1 Allgemeine Physiologie und Zellphysiologie · 2013. 11. 7. · Im strengen Sinne semipermeabel...
14
1 Allgemeine Physiologie und Zellphysiologie C. Hick, J. Hartmann 1.1 Physiologische Maßeinheiten ......... 1 1.2 Osmose .......................... 2 1.3 Stofftransport ..................... 3 1.3.1 Stofftransport in Gasen und Flu ¨ ssigkeiten .... 3 1.3.2 Stofftransport durch Membranen .......... 3 Passiver Transport .................... 3 Aktiver Transport ..................... 5 1.3.3 Stofftransport in Zellen ................. 7 Mitochondriale ATP-Synthetase ........... 7 Intrazellula ¨rer Transport in Vesikeln ........ 7 Zytoskelett ......................... 8 Axonaler Transport ................... 8 1.3.4 Informationsu ¨ bermittlung .............. 9 1.4 Zellorganisation................... 9 1.4.1 Funktionelle Kompartimentierung ......... 9 1.4.2 Histokompatibilita ¨ ts-Antigene ........... 9 1.4.3 Intrazellula ¨ re Botenstoffe .............. 10 Die cAMP-Kaskade .................. 10 Die IP 3 -Kaskade .................... 10 NO als Signalstoff ................... 12 1.5 Elektrische Pha ¨ nomene an der Zelle ... 12 1.6 Energetik ........................ 12 1.7 Regelung und Steuerung ............ 12 Die physiologischen Funktionen des Organismus werden in den Kapiteln 2 bis 20 im Zusammenhang der verschiedenen Organsysteme besprochen. Wich- tige Grundpha ¨ nomene wie Osmose (+ Kap. 1.2), ak- tiver und passiver Stofftransport an Membranen (+ Kap. 1.3) oder die Grundlagen von Zellorganisa- tion und Zellidentita ¨ t (Histokompatibilita ¨t, + Kap. 1.4) sind jedoch fu ¨ r alle Organsysteme identisch. Das gilt auch fu ¨ r die Mechanismen der Informations- u ¨ bermittlung (+ Kap. 1.3.4 und 1.4.3) und allgemeine Regel- und Steuerungsprinzipien (+ Kap. 1.7). Diese elementaren physiologischen Mechanismen und Funktionen sollen deshalb im folgenden Kapitel nach einem kurzen U ¨ berblick zu physiologisch wich- tigen Maßeinheiten (+ Kap. 1.1) zusammenfassend besprochen werden. Erregungsvorga ¨ nge an Zellen (Ruhemembranpotenti- al und Aktionspotential) werden im Kapitel 12 darge- stellt. 1.1 Physiologische Maßeinheiten Die Physiologie beschreibt Austauschvorga ¨ nge zwi- schen Zellen und Zellverba ¨ nden in einer quantitati- ven Sprache. Praktisch wichtig sind die folgenden Maßeinheiten: Druck = Kraft pro Fla ¨che (Pa = N/m 2 ) Einheit des Drucks ist das Pascal (Pa), die Kraft wird in Newton (N), die Fla ¨ che in Quadratmetern (m 2 ) an- gegeben. Als a ¨ ltere Druckeinheiten werden in der Physiologie noch mmHg (Quecksilber) und cmH 2 O verwendet. Dabei gilt: 1 cmH 2 O = 0,098 kPa 1 mmHg = 0,133 kPa Arbeit (= Energie = Wa ¨ rmemenge) = Kraft mal Weg (J = N m) Die Einheit von Energie, Arbeit oder Wa ¨ rmemenge – diese drei Ausdru ¨ cke sind im physikalischen Sinne a ¨ quivalent – ist das Joule (J). Fu ¨ r die Umrechnung aus der a ¨ lteren Energieeinheit Kalorie (cal) gilt: 1 cal = 4,185 J Lernziel! physikalische Grundlagen physiologischer Prozesse Stofftransport im Ko ¨ rper Zellorganisation, -kommunikation und Signalkaskaden. 1
Transcript of 1 Allgemeine Physiologie und Zellphysiologie · 2013. 11. 7. · Im strengen Sinne semipermeabel...

1 Allgemeine Physiologie undZellphysiologie
C. Hick, J. Hartmann
1.1 Physiologische Maßeinheiten . . . . . . . . . 1
1.2 Osmose . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1.3 Stofftransport . . . . . . . . . . . . . . . . . . . . . 3
1.3.1 Stofftransport in Gasen und Flussigkeiten . . . . 3
1.3.2 Stofftransport durch Membranen . . . . . . . . . . 3Passiver Transport . . . . . . . . . . . . . . . . . . . . 3Aktiver Transport. . . . . . . . . . . . . . . . . . . . . 5
1.3.3 Stofftransport in Zellen. . . . . . . . . . . . . . . . . 7Mitochondriale ATP-Synthetase . . . . . . . . . . . 7Intrazellularer Transport in Vesikeln . . . . . . . . 7Zytoskelett . . . . . . . . . . . . . . . . . . . . . . . . . 8Axonaler Transport . . . . . . . . . . . . . . . . . . . 8
1.3.4 Informationsubermittlung . . . . . . . . . . . . . . 9
1.4 Zellorganisation. . . . . . . . . . . . . . . . . . . 9
1.4.1 Funktionelle Kompartimentierung . . . . . . . . . 9
1.4.2 Histokompatibilitats-Antigene . . . . . . . . . . . 9
1.4.3 Intrazellulare Botenstoffe . . . . . . . . . . . . . . 10Die cAMP-Kaskade . . . . . . . . . . . . . . . . . . 10Die IP3-Kaskade . . . . . . . . . . . . . . . . . . . . 10NO als Signalstoff . . . . . . . . . . . . . . . . . . . 12
1.5 Elektrische Phanomene an der Zelle . . . 12
1.6 Energetik. . . . . . . . . . . . . . . . . . . . . . . . 12
1.7 Regelung und Steuerung . . . . . . . . . . . . 12
Die physiologischen Funktionen des Organismuswerden in den Kapiteln 2 bis 20 im Zusammenhangder verschiedenen Organsysteme besprochen. Wich-tige Grundphanomene wie Osmose (+ Kap. 1.2), ak-tiver und passiver Stofftransport an Membranen(+ Kap. 1.3) oder die Grundlagen von Zellorganisa-tion und Zellidentitat (Histokompatibilitat, + Kap.1.4) sind jedoch fur alle Organsysteme identisch.Das gilt auch fur die Mechanismen der Informations-ubermittlung (+Kap. 1.3.4 und 1.4.3) und allgemeineRegel- und Steuerungsprinzipien (+ Kap. 1.7). Dieseelementaren physiologischen Mechanismen undFunktionen sollen deshalb im folgenden Kapitelnach einem kurzen Uberblick zu physiologisch wich-tigen Maßeinheiten (+ Kap. 1.1) zusammenfassendbesprochen werden.Erregungsvorgange an Zellen (Ruhemembranpotenti-al und Aktionspotential) werden im Kapitel 12 darge-stellt.
1.1 Physiologische Maßeinheiten
Die Physiologie beschreibt Austauschvorgange zwi-schen Zellen und Zellverbanden in einer quantitati-ven Sprache. Praktisch wichtig sind die folgendenMaßeinheiten:
Druck = Kraft pro Flache (Pa = N/m2)
Einheit des Drucks ist das Pascal (Pa), die Kraft wirdinNewton (N), die Flache in Quadratmetern (m2) an-gegeben. Als altere Druckeinheiten werden in derPhysiologie noch mmHg (Quecksilber) und cmH2Overwendet. Dabei gilt:
1 cmH2O = 0,098 kPa1 mmHg = 0,133 kPaArbeit (= Energie = Warmemenge) = Kraft mal Weg(J = N � m)
Die Einheit von Energie, Arbeit oder Warmemenge –diese drei Ausdrucke sind im physikalischen Sinneaquivalent – ist das Joule (J). Fur die Umrechnungaus der alteren Energieeinheit Kalorie (cal) gilt:
1 cal = 4,185 J
Lernziel!� physikalische Grundlagen physiologischer Prozesse� Stofftransport im Korper� Zellorganisation, -kommunikation und Signalkaskaden.
1

Merke!Von praktischer Bedeutung ist in der Physiologie,dass das Produkt aus Druck (N/m2) und Volumen(m3) ebenfalls Arbeit (N � m) ergibt (Druck-Volu-men-Arbeit). Dies ermoglicht z. B. die Berechnungder Herzarbeit (+ Kap. 3.3.4).
Klinik!Ein plotzlicher Blutdruckanstieg fuhrt zu einem ra-schen Anstieg der Druck-Volumen-Arbeit des Her-zens. Um diese Mehrarbeit leisten zu konnen, istdas Herz auf eine Steigerung der Energieversorgungdurch Sauerstoff angewiesen. Diese gesteigerteSauerstoffzufuhr wird uber eine Erhohung der Koro-nardurchblutung sichergestellt. Ist dies z. B. bei arte-riosklerotisch verengten Koronararterien nicht mog-lich, entsteht ein Sauerstoffmangel im Myokardge-webe, der beim Patienten einen Angina-pectoris-Schmerz in der Brust auslosen kann.
Leistung = Arbeit pro Zeit (W = J/s)
Einheit der Leistung ist dasWatt (W), das einem JouleArbeit (J) pro Sekunde (s) entspricht (+ Kap. 8, Ener-gie- und Warmehaushalt).
StoffmengeDie Stoffmenge einer Substanz wird in Mol (Symbol:mol) angegeben. Dabei gilt:
1 mol = 6,022 � 1023 Teilchen
Will man bei den Angaben der Stoffmenge dieWertig-keit der Substanz berucksichtigen, empfiehlt sich dieAngabe in Val:
1 val ¼ 1 mol �Wertigkeit
Beispiel: 1 val zweiwertiger Ca2+-Ionen entspricht 0,5mol Ca2+.
KonzentrationDie Konzentration einer Substanz kann auf drei ver-schiedene Weisen ausgedruckt werden:� Die Massenkonzentration gibt die Masse eines
Stoffes pro Volumeneinheit an. So betragt z. B.die Massenkonzentration von Hamoglobin imBlut beim Mann 15–16 g/100 ml.
� Die Stoffmengenkonzentration, auch als molareKonzentration bezeichnet, gibt die Stoffmengepro Volumen einer Losung an. Die Stoffmengen-konzentration von K+-Ionen im Blutplasma z.B.liegt bei 5 mmol/l.
� Diemolale Konzentration gibt die Stoffmenge proMasseneinheit eines Losungsmittels an. MolaleKonzentrationsangaben beziehen sich also nichtauf die gesamte Losung, sondern auf die Massedes Losungsmittels. Außerdem sind sie – im Ge-
gensatz zu volumenbezogenen molaren Konzen-trationsangaben – von Temperaturschwankungenund den hieraus resultierenden Volumenschwan-kungen unabhangig. Deshalb werden vor allemdie Konzentrationen osmotisch wirksamer Sub-stanzen (+ Kap. 1.2) besser in molalen und nichtin molaren Einheiten angegeben. Blutserum z.B.hat eine Osmolalitat von ca. 280–295 mosm/kgH2O (+ Kap. 2.3.1).
In physiologischen Flussigkeiten wie dem Blutplasmamachen die gelosten Bestandteile (vor allem Salzeund Eiweiße) bis zu etwa 7% des Gesamtvolumensder Losung aus. Molare Konzentrationsangaben,die sich auf dieses Volumen derGesamtlosung bezie-hen, konnen daher die Anzahl der tatsachlich in einerdefinierten Stoffmenge gelosten und damit fur einechemische Reaktion zur Verfugung stehenden Teil-chen einer Substanz nicht exakt angeben. Dahersind molale Konzentrationsangaben, die sich aufdie Masse des Losungsmittels beziehen, fur physiolo-gische Flussigkeiten im Allgemeinen praziser.
Merke!� Osmolaritat: osmotisch wirksame Stoffmenge proLiter Losung (osm/l)
� Osmolalitat: osmotisch wirksame Stoffmenge proKilogramm Losungsmittel (osm/kg).
1.2 Osmose
Definition der Osmose
Die Diffusion von Losungsmittel durch eine semiper-meable Membranwird als Osmose bezeichnet. Semi-permeable, d. h. halb-durchlassige Membranen, sindMembranen, die nur fur das Losungsmittel, nicht aberfur die in ihm gelosten Substanzen durchlassig sind.Wird beispielsweise eine Zuckerlosung von einerWasserlosung durch eine semipermeable Membrangetrennt, stromen die Wassermolekule entlang demKonzentrationsgefalle in die Zuckerlosung ein. Diegroßeren Zuckermolekule werden dagegen an der se-mipermeablen Membran zuruckgehalten. Durch die-sen Wasserzustrom steigt das Volumen der Zuckerlo-sung an (+ Abb. 1.1).
Definition des osmotischen Drucks
Auf der anderen Seite uben die nicht-diffusiblen Glu-cosemolekule einen Druck auf die semipermeableMembran aus, der als osmotischer Druck bezeichnetwird. Dieser osmotische Druck hangt nur von derAn-zahl der gelosten Teilchen ab, nicht von ihrer chemi-schen Beschaffenheit. Ist die Anzahl (n) der Teilchenbekannt, kann der an einer semipermeablen Mem-bran entstehende osmotische Druck (Posm) nach van’tHoff analog zur allgemeinen Gasgleichung (+ Kap.5.2) berechnet werden:
1 Allgemeine Physiologie und Zellphysiologie
2

1
Posm ¼ n
V� R � T
Der osmotische Druck (Posm) steigt also1. proportional zur Anzahl n der in einemVolumen V
gelosten Teilchen und2. proportional zur Temperatur T.R ist die allgemeine Gaskonstante.
Osmolaritat und Osmolalitat
Wird die osmotisch wirksame Stoffmengenkonzen-tration der gelosten Teilchen als Molaritat, also inmol/l Losung ausgedruckt, erhalt man die Osmolari-tat als Maß der osmotischen Aktivitat. Wird die Teil-chenzahl alsMolalitat ausgedruckt, also auf dieMassedes Losungsmittels bezogen, erhalt man die Osmola-litat der Losung. Osmolaritat undOsmolalitat sind di-rekt proportional zu dem durch die Losung an semi-permeablen Membranen entstehenden osmotischenDruck. Losungen, die den gleichen osmotischenDruck wie das Plasma aufweisen, werden als isotoneLosungen bezeichnet.Der osmotische Druck von Plasma oder physiologi-scher Kochsalzlosung betragt 745 kPa (+Kap. 2.3.1).Im strengen Sinne semipermeabel ist eine Membrannur, wenn sie fur bestimmte geloste Teile vollig un-durchlassig ist. Trifft dies nicht ganz zu, muss dievan’t-Hoff-Gleichung durch einen Reflexionskoeffi-zienten r erganzt werden.
Posm ¼ n
V� R � T � r
Der Reflexionskoeffizient liegt zwischen 0 (Membranvollig durchlassig, auch fur die gelosten Teile) und 1(Membran vollig undurchlassig fur die gelosten Teile).In manchen Fallen reißt der osmotische Losungsmit-telstrom auch kleinere geloste Teile (z. B. Elektrolyte)mit sich und transportiert sie durch die semipermea-ble Membran. Durch diesen Solvent-drag-Effektwerden beispielsweise im proximalen NierentubulusNa+-Ionen aus dem Primarharn ruckresorbiert(+ Kap.9.2.4).
Klinik!Losungen, die in den Blutkreislauf infundiert wer-den, mussen zum Plasma isoton sein, damit keineFlussigkeitsverschiebungen zwischen dem Intrava-salraum und dem umgebenden Gewebe auftreten.In bestimmten Fallen kann es jedoch erwunschtsein, Flussigkeit aus dem Gewebe zu mobilisierenund in den Blutkreislauf zu uberfuhren (z. B. beistarken Blutverlusten). Hierzu verwendet man Plas-maexpander, die einen hoheren kolloidosmotischenDruck haben als das Plasma und so Flussigkeit in dieGefaße „saugen“.
1.3 Stofftransport
1.3.1 Stofftransport in Gasen und Flussigkeiten
In Gasen und Flussigkeiten konnen sich die Teilchenfrei bewegen. Der Stoffaustausch folgt hierbei zweiKraften:� Konzentrationsunterschiede sind die treibende
Kraft beim Stofftransport durch Diffusion.� Temperatur- oder Druckdifferenzen sind die trei-
bende Kraft der Konvektion, d. h. des Stoffaus-tauschs durch Stromung eines Gases oder einerFlussigkeit.
1.3.2 Stofftransport durch Membranen
Membranen stellen fur den freien Stofftransport einHindernis dar. Sie bestehen aus einer 4–5 nm dickenLipiddoppelschicht. Die hydrophoben Fettsaurere-ste bilden im Inneren der Membran eine lipophile„Olphase“. Die hydrophilenKopfgruppen der Lipidesind auf der Innenseite der Membran dem Zellinne-ren und auf der Außenseite der Membran der Umge-bung der Zelle zugewandt. In die Lipiddoppelschichteingelassen sind Membranproteine, die als Ionenka-nale dienen konnen (+ Abb. 1.2).
Merke!Je großer der Gehalt an ungesattigten Fettsauren, de-sto flexibler ist die Membran.
Passiver Transport
Einfache Diffusion
Frei durch die Plasmamembran diffundieren konnengeloste Gase und kleine lipophile Substanzen (z. B.Harnstoff). Die Diffusion folgt den Gesetzmaßigkei-ten des Fick-Diffusionsgesetzes. Danach ist die proZeiteinheit durch Diffusion transportierte StoffmengeJ [mol/s] direkt proportional zur Diffusionsflache A[m2] und zur Konzentrationsdifferenz uber der Mem-
Abb. 1.1 Osmose. Die Wassermolekule wandern ihremKonzentrationsgefalle folgend auch gegen das Gravita-tionsgefalle uber die semipermeable Membran in dieGlucoselosung ein. Die großeren Glucosemolekule ( )konnen die Membran nicht passieren.
1.3 Stofftransport
3

bran Dc [mol/m3]. Zur Dicke der Membran d [m] istder Diffusionsfluss umgekehrt proportional:
J = � D � Ad� c
Dabei ist D der Diffusionskoeffizient, der von der dif-fundierenden Substanz, dem Losungsmittel und derTemperatur abhangig ist.
Klinik!Bei bestimmten Lungenerkrankungen (z. B. Lun-genfibrosen) kommt es zu einer Verdickung der Al-veolarmembran. Dadurch wird nach der obigenGleichung die Sauerstoffdiffusion uber diese Mem-bran behindert, da die Dicke der Diffusionsmem-bran zugenommen hat. Laborchemisch ist die Sauer-stoffsattigung des arteriellen Blutes vermindert, kli-nisch leidet der Patient unter Atemnot (+ Kap.5.9.3).
Der Diffusionskoeffizient (D) und die Dicke derMembran (d) werden oft zur Permeabilitat (P) fureine Substanz an einer bestimmten Membran zusam-mengefasst. Die Permeabilitat P [m/s] gibt an, wierasch eine bestimmte Substanz eine Membran passie-ren kann (+ Abb. 1.3).Vor allem geladene Substanzen konnen wegen ihrerelektrischen Ladung auch bei geringer Große eine Li-piddoppelmembran kaum passieren (z. B. Na+- oderK+-Ionen). Sie sind fur die Diffusion auf Ionenkanaleangewiesen, die von Membranproteinen gebildetwerden (+ Abb. 1.5a). Die treibende Kraft fur den Io-nentransport sind elektrochemische Potentialdiffe-renzen oder Konzentrationsgradienten uber der
Membran. Die Ionenkanale haben einen Durchmes-ser von weniger als 1 nm und sind durch die in ihrerWand enthaltenen Molekulstrukturen relativ spezi-fisch fur bestimmte Ionen. So lassen sich Kalium-, Na-trium- und Calciumkanale unterscheiden. Zur phy-siologischen Bedeutung der Ionenkanale + Kapitel12.1.1.Einfache Diffusionsvorgange, die keine Transport-proteine benotigen und direkt durch die Membranoder durch Ionenkanale ablaufen, folgen einer linea-ren Transportcharakteristik:Mit zunehmender Kon-zentration des zu transportierenden Molekuls steigtauch die Transportrate linear an (+ Abb. 1.4a).
Glykolipide
Phospholipid-kopfgruppe
Fettsäurereste
integralesMembranprotein
Intrazellulärraum
Ionenkanal
hydrophileSchichten
hydrophobeSchicht
ca. 5 nm
Extrazellulärraum
Abb. 1.2 Aufbau einer Plasmamembran.
Na+
K+
Cl–
Glucose
Tryptophan
Glycerin
Harnstoff
H2O
Permeabilität [m/s]10–14 10–10 10–8 10–6 10–4 10–210–12
Abb. 1.3 Permeabilitat einer typischen Lipiddoppelmem-bran. Die Membrandurchlassigkeit ist fur H2O am großtenund fur Na+ und K+ am niedrigsten.
1 Allgemeine Physiologie und Zellphysiologie
4

1
Erleichterte Diffusion
Von erleichterter Diffusion spricht man, wenn dieMembran fur den diffundierenden Stoff spezielleCar-rier-Proteine enthalt. Im Gegensatz zu Ionenkana-len, die jeweils eine bestimmte Ionengruppe passierenlassen, transportieren Carrier-Proteine kleine Mole-kule wie Glucose. Die treibende Kraft des Stofftrans-portes ist hierbei wie bei der einfachen Diffusion einKonzentrationsgradient. Der Organismus muss alsokeine Transportenergie aufwenden. Durch die erfor-derlichen Carrier-Proteine ist die erleichterte Diffusi-on jedoch substanzspezifisch – wie ein aktiver Trans-port (+ unten).Erfolgt die erleichterte Diffusion nur in einer Rich-tung, spricht man von einem Uniport. Typisches Bei-spiel fur einen solchen Uniport sind die Glucose-Transportproteine (GLUTs, + Kap. 10.6.1), durchdie Glucose in die Zellen gelangt.Da die erleichterte Diffusion auf nur in begrenzterZahl zur Verfugung stehende Transportproteine an-gewiesen ist, weist sie eine Sattigungscharakteristiknach der Michaelis-Menten-Kinetik auf: Mit zuneh-mender extrazellularer Konzentration [c] des zutransportierenden Stoffes nahert sich die Transportra-te JA einem nicht uberschreitbaren Maximalwert Jmax
(+ Abb. 1.4, Kurve b):
JA ¼ ðJmax � ½c�ÞðKm þ ½c�Þ
Die Michaelis-Konstante Km gibt die Affinitat des zutransportierenden Stoffes zu seinem Carrier wieder:Sie bezeichnet die extrazellulare Konzentration desStoffes, bei der die Halfte der Maximalgeschwindig-keit Jmax des Transportvorgangs erreicht ist.
Merke!Ein hoher Km-Wert bedeutet eine geringe Affinitat!
Aktiver Transport
Stoffe, fur die kein elektrisches oder chemisches Kon-zentrationsgefalle uber dieMembran besteht, oder dieentgegen einem bestehenden Konzentrationsgefalletransportiert werden mussen, sind auf aktive, Energieverbrauchende Transportvorgange angewiesen. Hier-bei unterscheidet man einen primar aktiven Trans-port von einem sekundar und einem tertiar aktivenTransport. Diese aktiven Transportformen unter-scheiden sich von einfachen passiven Diffusionspro-zessen durch drei Charakteristika.� Strukturspezifitat: Jedes Transportsystem ist auf
bestimmte Substanzen spezialisiert und kann nurdiese transportieren.
� Hemmbarkeit: Stoffe mit ahnlicher Struktur wiedie zu transportierenden Substanzen konnen dieTransportproteine besetzen und so den Stofftrans-port blockieren.
� Sattigung: Wegen der begrenzten Zahl der Trans-portproteine gibt es fur die Transportrate einenMaximalwert.
Primar aktiver Transport
In den Zellen liegt die Konzentration von K+-Ionenmit 155 mmol/l deutlich uber der extrazellularen Ka-liumkonzentration von 5 mmol/l. Umgekehrt ist ex-trazellular die Konzentration von Na+-Ionen mit145 mmol/l großer als ihre intrazellulare Konzentra-tion von 12 mmol/l (+ Tab. 1.1). Die Konzentrations-unterschiede wurden sich ohne aktive Gegenmaß-nahmen durch Diffusion und Osmose in kurzerZeit ausgleichen. Die Aufrechterhaltung der Konzen-trationsdifferenzen ist jedoch fur die Funktion derZellen unverzichtbar. Deshalb ist ein aktiver undEnergie verbrauchender Ionentransport uber die Zell-membran erforderlich, der diese Konzentrationsun-terschiede stabilisiert.Der wichtigste aktive Transportprozess ist die Na+-K+-Pumpe. Dieses spezialisierte Membranproteinist in allen Plasmamembranen zu finden und reicht
Transportrate
a)
b)
JA
Konzentration destransportierten Moleküls [c]
Jmax
passiver Transport(Diffusion)
aktiver Transport(Pumpe)
50% Jmax
Km
Abb. 1.4 Abhangigkeit der Transportrate von der Kon-zentration des zu transportierenden Molekuls. a: LineareTransportcharakteristik bei freier Diffusion durch dieMembran oder durch Membrankanale. b: Sattigungs-charakteristik bei erleichterter Diffusion uber Carrier oderbei aktiven Transportvorgangen durch Pumpen. Gestri-chelt ist die Ableitung der Michaelis-Menten-KonstanteKm gezeigt.
Tab. 1.1 Intra- und extrazellulare Ionenkonzentrationen
intrazellulareKonzentration[mmol/l]
extrazellulareKonzentration[mmol/l]
Na+ 12 145
K+ 155 5
Ca2+ 10–5–10–4 2,5
Cl– 4 120
HCO3 8 27
große Anionen 155 5
1.3 Stofftransport
5

durch die Lipiddoppelschicht hindurch. Biochemischist die Na+-K+-Pumpe eine ATPase, d. h. ein Enzym,das unter Energieverbrauch ATP in ADP und Phos-phat spaltet. Diese ATPase ist an der Innenseite derZellmembran lokalisiert. Durch die ATP-Spaltungwerden auf der Innenseite der Na+-K+-Pumpe Bin-dungsstellen fur drei Na+-Ionen aktiviert. Diese dreiNa+-Ionen werden entgegen dem Na+-Konzentrati-onsgradienten aus der Zelle heraus transportiert. ImGegenzug gelangen zwei K+-Ionen, ebenfalls gegenihren Konzentrationsgradienten, ins Innere der Zelle.Pro gespaltenem ATP-Molekul werden also drei posi-tive Ladungen aus der Zelle entfernt, wahrend nurzwei positive Ladungen in die Zelle gelangen(+ Abb. 1.5b).Die Na+-K+-Pumpe ist daher elektrogen, d. h. sie bautdurch ihre Pumparbeit nicht nur einen Konzentrati-onsgradienten, sondern auch einen elektrischen Gra-dienten uber der Zellmembran auf. Der Energieauf-wand fur die Na+-K+-Pumpe ist betrachtlich. Er be-tragt ein Drittel des gesamten Energieumsatzes derZelle. Neben der Na+-K+-Pumpe sind noch zwei wei-tere aktive Pumpmechanismen physiologisch von Be-deutung:� Protonen-Pumpen (H+-K+-ATPasen) transportie-
ren unter ATP-Verbrauch H+-Ionen uber eineMembran. Sie sind z. B. die Protonenlieferantenfur die Bildung der Salzsaure des Magens(+ Kap. 7.3.3).
� Calcium-Pumpen (Ca2+-ATPasen) konnen hoheCalcium-Konzentrationsdifferenzen aufbauen.Sie finden sich z. B. im sarkoplasmatischen Retiku-lum (dem Calciumspeicher der Zelle) oder in derZellmembran von Herzmuskelzellen (+ Kap.3.1.3).
Klinik!Diese primar aktiven Transporter sind Ansatzpunktefur verschiedene Medikamente:� DurchHerzglykoside (Digitalis) wird die Na+-K+-
ATPase blockiert. Uber Zwischenschritte (+Kap.3.1.3) kommt es zu einem intrazellularen Calci-um-Anstieg und einer kraftigeren Herzaktion.
� Protonenpumpeninhibitoren (PPI) blockierendie H+-K+-ATPase, was zu einer vermindertenMagensaureproduktion fuhrt und die AbheilungvonMagengeschwuren begunstigt (+ Kap. 7.3.3).
Sekundar aktiver Transport
Der durch dieNa+-K+-Pumpe unter Energieverbrauchaktiv aufgebaute elektrochemische Natrium-Konzen-trationsgradient dient als Motor fur weitere Trans-portmechanismen. Diese Transportmechanismen,die keine eigene Energiezufuhr benotigen und alsoohne ATP-Spaltung auskommen, werden als sekun-dar aktive Transportmechanismen bezeichnet. Manunterscheidet:� Antiport-Mechanismen (= Countertransporte):
Das wichtigste Antiport-System ist der Ca2+-Na+-
Antiport der Zellmembranen. Dabei liefern dreieinstromende Na+-Ionen die Energie fur den Aus-wartstransport eines Ca2+-Ions (+ Abb. 1.5c). AufdieseWeise wird die hoheKonzentrationsdifferenzfur Ca2+-Ionen uber der Zellmembran (innen10–5 mmol/l, außen 2,5 mmol/l) aufrechterhalten.
� Symport-Mechanismen (= Cotransporte): EinNa+-Ion stromt entlang seinem elektrochemischenKonzentrationsgradienten in die Zelle ein und lie-fert hierdurch die Energie fur den Transport einesweiteren Molekuls, das dann auch gegen seineneigenen Konzentrationsgradienten in die Zelletransportiert werden kann. Der Na+-Glucose-Symport der Darmschleimhaut ist der bekanntestedieser sekundar aktiven Symport-Transportmecha-nismen (+ Abb. 1.5d und Kap. 7.5.2). Auch dieAminosauren werden uber solche Symport-Me-chanismen im Verbund mit Na+-Ionen in dieDarmschleimhaut aufgenommen (+ Kap. 7.5.3).
Tertiar aktiver Transport
Ein tertiar aktiver Transport ist ein Transportvorgang,der seine Energie von einem sekundar aktiven Trans-portprozess erhalt. So wird ein H+-Dipeptid-Cotrans-porter im Dunndarm, der H+-Ionen und Di- oder Tri-peptide aus dem Darmraum aufnimmt, von einem se-kundar aktiven Na+-H+-Antiport angetrieben, der H+-Ionen ins Darmlumen sezerniert. Der hierdurch ent-stehende, ins Zellinnere gerichtete H+-Gradient istdie treibende Kraft dieses tertiar aktiven H+-Dipep-tid-Transportsystems. Der sekundar aktive Na+-H+-Antiport wiederum wird von einer an der Blutseiteder Darmzelle gelegenen primar aktiven Na+-K+-ATPase angetrieben.
Endozytose und Exozytose
Stoffe, die nicht durch die Plasmamembran diffundie-ren konnen und fur die keine Transportproteine exi-stieren, konnen durch Endozytose in die Zelle aufge-nommen werden. Hierbei handelt es sich z. B. umCholesterin oder um bestimmte Proteine. Bei der En-dozytose stulpt sich zuerst die Plasmamembran ein.Diese Einstulpung vertieft sich und es entsteht einVe-sikel, das sich von der Plasmamembran abschnurtund die endozytotisch aufgenommene Substanz miteiner aus der Plasmamembran gebildeten Lipiddop-pelschicht umhullt. Oft sind bestimmte Gebiete derZellmembran durch Rezeptorproteine fur eine indie Zelle aufzunehmende Substanz (z. B. fur Insulin,+ Kap. 10.6.1) oder fur Antigene besonders sensibel.Die Bindung der Substanz an das Rezeptorproteinlost dann den Vorgang der Endozytose aus.Uber den umgekehrten Vorgang der Exozytose wer-den Stoffe aus der Zelle entfernt. Hierbei kann essich z. B. um Hormone oder um Enzyme handeln.Die Vesikelmembran verschmilzt mit der Zellmem-bran und entlasst so die imVesikel gespeicherten Sub-stanzen in den Extrazellularraum.
1 Allgemeine Physiologie und Zellphysiologie
6

1.3.3 Stofftransport in Zellen
Auch innerhalb der Zellen finden aktive Transport-vorgange statt. Diese werden analog zu den Trans-portvorgangen an der außeren Zellmembran auchan den inneren Membranen des Zytosols beobachtet.Außerdem werden in der Zelle Syntheseprodukte inForm von Vesikeln transportiert. Fur diesen Trans-port sind kontraktile Vorgange imZytoskelett von Be-deutung.
Mitochondriale ATP-Synthetase
Ein besonders interessanter Transportmechanismusauf Membranebene findet sich in den Mitochondri-en.Die an der innerenMitochondrienmembran loka-lisierten Enzyme der Atmungskette transportierenH+-Ionen vom mitochondrialen Matrixraum in denSpalt zwischen innerer und außerer Mitochondrien-membran und bauen so einen Protonengradientenuber der inneren Mitochondrienmembran auf. DieserProtonengradient treibt eine in der inneren Mito-chondrienmembran lokalisierte ATP-Synthetase an:Der Energie freisetzende Ruckstrom der Ionen in
die mitochondriale Matrix ermoglicht so die Synthesevon ATP aus ADP. Diese protonengetriebene ATP-Synthetase der Mitochondrien kann als ruckwartslaufende Protonenpumpe aufgefasst werden(+ Abb. 1.6).
Klinik!Bei einer Blausaurevergiftung (z. B. durch Kalium-cyanid) werden Enzyme der Atmungskette inakti-viert, was zu einer Blockade des Sauerstoff- undElektronentransfers fuhrt: Es kommt zur inneren Er-stickung. Da das Gewebe den Sauerstoff nicht mehraufnehmen kann, weist das venose Blut einen demarteriellen Blut vergleichbar hohen O2-Gehalt auf.Dies erklart die typische Rotfarbung der Haut.
Intrazellularer Transport in Vesikeln
Intrazellulare Transportvorgange konnen auch uberVesikel erfolgen. Die Wande dieser Vesikel bestehenaus Lipiddoppelschichten, die einen ahnlichen Auf-bau wie die Zellmembran haben. Beispielsweise er-
außen
Membran
innen
b
c d
a
–
––
–
K+
K+-Kanal
K+ K+ATP ADP
Ca2+
Na+GlcNa+Na+
Na+
Na+Na+
Na+
Abb. 1.5 Transportmechanismen an Membranen. a: Passiver Transport von K+ durch Diffusion uber Ionenkanale. b:Primar aktiver Transport uber eine Na+-K+-Pumpe. c: Sekundar aktiver Ca2+-Na+-Antiport. d: Sekundar aktiver Na+-Glucose-Symport.
1.3 Stofftransport
7
1

halten die an den Ribosomen des rauen endoplasma-tischen Retikulums synthetisierten Proteine eine sol-che Lipidhulle vom endoplasmatischen Retikulum.Die entstehenden Vesikel schnuren sich vom endo-plasmatischen Retikulum ab, gelangen zum Golgi-Komplex und verschmelzen mit den Zisternen desGolgi-Apparates. Hier konnen noch Modifikationen,z. B. Glykosylierungen, am neu synthetisierten Prote-in vorgenommen werden. Wird die Zelle zur Protein-sekretion stimuliert, schnuren sich vom Golgi-Appa-rat Sekretvesikel ab, die zur Plamamembran wandernund die Proteine uber eine Exozytose in den Extrazel-lularraum abgeben.
Zytoskelett
Die Transportvesikel bewegen sich durch die Zelle,indem sie an den Mikrotubuli des Zytoskeletts ent-langgleiten. Außer den Mikrotubuli enthalt das Zyto-skelett auch Mikrofilamente, die uberwiegend ausdem Protein F-Aktin bestehen, das sich auch inden Aktinfilamenten vonMuskelzellen findet. Mikro-filamente und Mikrotubuli sind im Zytosol von Dy-nein- und Myosin-Molekulen umgeben. Myosinkommt – in Faserform – auch in Muskelzellen vor(+ Kap. 13.1.1). Unter ATP-Einsatz sind Energie ver-brauchende Verschiebungen im Zytoskelett moglich,die fur Transportvorgange, aber auch fur Formande-rungen der Zellen oder fur Zilienschlage verantwort-lich sind. Der grundlegendeMechanismus dieser Ver-schiebungen beruht hierbei analog zu den Vorgangenin der Muskelzelle auf einer Wechselwirkung vonAktin und Myosin (+ Kap. 13.1 und 13.2) bzw.beim Zilienschlag auf Dynein-Dynein-Wechselwir-kungen. Da Aktin und Myosin in der Muskelzellein geordneter Faserform und nicht als lockere, zytoso-lische Proteinaggregate vorkommen, ist die Kraftent-
wicklung der Muskelfaser effektiver und gerichteterals die Aktin-Myosin-Interaktionen im Zytoskelettder normalen Zellen es sind.
Klinik!Die zur Therapie der Gicht eingesetzte SubstanzColchicin hemmt die Bewegung der Mikrotubuli.Ihre klinische Wirkung beim Gichtanfall erklartsich dadurch, dass durch diese Hemmung der Mi-krotubuli die Phagozytosefahigkeit der Makropha-gen (+ Kap. 2.5.1) im entzundeten Gelenk gehemmtwird. Die Phagozytose der Harnsaurekristalle in derGelenkflussigkeit ist Ausloser der schmerzhaftenEntzundungsreaktionen im Gelenk.Auch viele in der Krebstherapie eingesetzte Che-motherapeutika hemmen die Bewegungen des Zy-toskeletts. Krebszellen teilen sich schneller als nor-male Zellen und sind daher auf rasche Umbauvor-gange im Zytoskelett angewiesen. Wenn diese Um-bauvorgange gehemmt werden, sind die sich schnellteilenden Krebszellen besonders stark betroffen. Al-lerdings werden auch korpereigene Zellen, die einehohe Teilungsrate haben, mitgeschadigt, vor allemHaarzellen, Zellen der Darm- und Mundschleim-haut, Knochenmarkzellen (Immunsuppression)und Keimzellen.
Axonaler Transport
Am deutlichsten sind intrazellulare Transportvorgan-ge in den bis uber 1 m langen Zellfortsatzen von Ner-venzellen, den Axonen, zu beobachten. Der schnelleaxonale Transport schreitet mit einer Geschwindig-keit von etwa 12 mm/Stunde vom Zellkorper in Rich-tung Peripherie voran. Der schnelle retrograde axo-
Zytoplasma
äußereMitochondrienmembran
hohe H+-KonzentrationH+H+H+
H+
2e–
H+ H+
2e– ADP + Pi
H+2H+ 1/2O2
H2O
energiereicheElektronen
(Citratzyklus)
ATP-Synthe-
tase
innereMitochondrien-
membran
H+
H+-Poolim Matrixraum
ATP
NADH
Abb. 1.6 ATP-Gewinnung in den Mitochondrien: Die energiereichen Elektronen aus dem Citratzyklus bauen einen H+-Gradienten uber der inneren Mitochondrienmembran auf. Hierdurch wird die ATP-Synthetase angetrieben. [1]
1 Allgemeine Physiologie und Zellphysiologie
8

nale Transport bewegt sich mit etwa der Halfte dieserGeschwindigkeit von der Peripherie zum Zellkorperhin. Die schnellen axonalen Transporte bedienen sichals Transportmedium der Vesikel und Organellen derZelle (z. B. der Mitochondrien) und verbrauchenEnergie (ATP).
Klinik!Durch die retrograden axonalen Transportmecha-nismen gelangen z. B.Herpes- oder Poliomyelitisvi-ren von ihrer peripheren Infektionsstelle in die Zell-korper der Nervenzellen. Auch das Tetanustoxinwird von den peripheren Axonen im Wundgebietaufgenommen und retrograd in die Nervenzellkor-per transportiert. Das Tollwut-Virus gelangt uber re-trograden Transport in das ZNS und breitet sich an-schließend durch anterograden Transport in die Or-gane (Speicheldruse, Niere etc.) aus.
Neben diesen beiden schnellen axonalen Transport-mechanismen gibt es auch noch langsame axonaleTransporte von nur 1–5 mm pro Tag. Tubulin undAktin bewegen sich mit dieser Geschwindigkeit; siekonnen verschiedene Enzyme und Proteine mitneh-men. Die Geschwindigkeit des langsamen axonalenTransportes entspricht der Regenerationsgeschwin-digkeit eines geschadigten Nervs.
1.3.4 Informationsubermittlung
Information kann zwischen Zellen auf verschiedenenWegen ubertragen werden:� durch direkte Kommunikation uber Gap juncti-
ons: Durch diese Poren in den Zellmembranensind die Einzelzellen zu einem funktionellen Syn-zytium verbunden. Bei pH-Abfall oder Ca++-An-stieg (z. B. bei Zellschadigung oder Sauerstoffman-gel) sinkt die Offen-Wahrscheinlichkeit der Gapjunctions. Die geschadigte Zelle wird so von ihrerUmgebung isoliert und ein Ausbreiten der Schadi-gung verhindert.
� durchKommunikation uberNervenfasern (+Kap.12, 14, 15, 16 und 20).
� durch die Bildung von Botenstoffen, den Hormo-nen. Die physiologischen Funktionen der einzel-nen Hormone werden im Detail in Kapitel 10 be-sprochen.
1.4 Zellorganisation
1.4.1 Funktionelle Kompartimentierung
Der Innenraum der Zelle enthalt verschiedene Orga-nellen, deren Arbeitsgebiete durch Membranstruktu-ren getrennt sind; man spricht von funktionellerKompartimentierung. Physiologisch wichtig sind:
� endoplasmatisches Retikulum (in der Muskelzel-le: sarkoplasmatisches Retikulum): Ort der Cal-ciumspeicherung (+ Kap. 3.1.3 und 13.1.1)
� Mitochondrien: aerobe Energiegewinnung durchdie Enzyme der Atmungskette („Kraftwerke derZelle“)
� Ribosomen: Proteinsynthese nach den Anweisun-gen der im Zellkern produzierten mRNA (messen-ger RNA)
� Golgi-Apparat: Glykosylierung der Proteine undBereitstellung von Lipidmembranen fur die Sekret-vesikeln
� Lysosomen: bauen uber Protonenpumpen ein sau-res Milieu auf, das den sauren Hydrolasen ein op-timales Aktivitatsniveau zum Abbau der phagozy-tierten Proteine garantiert
� Peroxisomen: oxidieren phagozytierte Substanzenmithilfe von Peroxiden (aus molekularem Sauer-stoff gewonnen) und machen sie so unschadlich.Zur Entgiftung dieser fur die korpereigenen Zell-strukturen toxischen Peroxide dient das EnzymKatalase, das Peroxide zu Wasser umwandelnkann.
1.4.2 Histokompatibilitats-Antigene
Die Zelloberflache enthalt die sog. Histokompatibi-litats-Antigene (MHC-Antigene). Dies sind auf deraußeren Oberflache der Zellmembranen angesiedelteMolekule, die durch einen speziellen Genkomplexkodiert werden (Major histocompatibility complex,MHC). Dieser Genkomplex zeichnet sich durcheine große Vielfalt (Polymorphismus) der beteiligtenGene aus. Durch diese Vielfalt haben die MHC-Anti-gene, die in drei Klassen eingeteilt werden (MHC-I,-II und -III), bei jedem Individuum eine unterschied-liche Struktur. Genetisch verwandte Individuen stim-men in mehr MHC-Eigenschaften (MHC-Loci) uber-ein als genetisch nicht verwandte. MHC-Antigenekonnen daher zur Uberprufung vonVerwandtschafts-beziehungen eingesetzt werden.
Klinik!Eine entscheidende Voraussetzung fur den Erfolgvon Transplantationen ist eine moglichst großeUbereinstimmung der Histokompatibilitats-Antige-ne der Klasse I (Transplantationsantigene), dieauf allen kernhaltigen Zellen des Organismus undauf Thrombozyten vorkommen.
Histokompatibilitats-Antigene der Klasse II findensich als dimere, integrale Membranproteine vorwie-gend auf den Membranoberflachen von Phagozytenund B-Lymphozyten. Sie steuern u.a. die Interaktionder Antigen prasentierenden Makrophagen mit denT-Helfer-Lymphozyten (+ Kap. 2.5).
1.4 Zellorganisation
9
1

1.4.3 Intrazellulare Botenstoffe
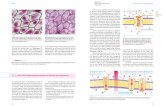
Innerhalb der Zelle werden Informationen uber Bo-tenstoffe ubertragen: Secondmessenger.Zwei grund-legende Mechanismen der Informationsubertragungdurch Peptidhormone sind� die cAMP-Kaskade (cAMP = zyklisches Adeno-
sinmonophosphat) und� die IP3-Kaskade (IP3 = Inositoltriphosphat).
Die cAMP-Kaskade
Peptidhormone (z. B. Adrenalin) sind hydrophil undkonnen die Plasmamembran nicht direkt durchdrin-gen. Sie wirken deshalb auf membranstandige Hor-monrezeptoren, die spezifisch fur die einzelnen Hor-mone sind. Durch die Bindung des Hormons an sei-nen Rezeptor wird eine Kette von biochemischen Pro-zessen ausgelost. Im Einzelnen unterscheidet man inder cAMP-Kaskade die folgenden funf Schritte(+ Abb. 1.7):1. Durch die Bindung des Hormons an seinen Rezep-
tor andert dieser seine Konformation.2. Hierdurchwird ein an der Innenseite der Zellmem-
bran lokalisiertes G-Protein aktiviert. Diese Akti-vierung besteht darin, dass ein an das G-Protein ge-bundenes Molekul GDP (Guanosindiphosphat)durch seine energiereichere TriphosphatformGTP ersetzt wird.
3. Das durch GTP aktivierte G-Protein reagiert miteiner ebenfalls an der Innenseite der Membran lo-kalisierten Adenylatcyclase (AC). Adenylatcycla-sen bilden cAMP aus ATP.
4. Dieses cAMP dient als zweiter Botenstoff (Secondmessenger) und bindet sich an die im Zytosol ge-
legene Proteinkinase A (P-A) und aktiviert sie da-durch. cAMP vermittelt also im Zellinneren dieWirkung eines Hormons, das selbst nicht die Zell-membran durchdringen kann.
5. Der aktive Komplex aus cAMP und Proteinki-nase A ist das eigentliche ausfuhrende Enzymder cAMP-Kaskade: Er phosphoryliert Proteine,welche dann die jeweils spezifischen Hormonwir-kungen in der Zelle vermitteln (im Falle des Adre-nalins z. B. die Freisetzung vonGlucose durchGly-kogenolyse).
Der Second messenger cAMP wird durch die Spal-tung der Phosphodiesterbindung deaktiviert, wobeieinfaches 5’-AMP entsteht. Das hierfur verantwortli-che Protein, die Phosphodiesterase, wird durchTheophyllin und Koffein gehemmt. Die „Zellaktivie-rung“ durch Theophyllin oder Koffein lasst sich alsoauf einen durch diese Substanzen bewirkten Anstiegdes intrazellularen cAMP-Spiegels zuruckfuhren.Die cAMP-Kaskade lauft bei den unterschiedlichstenhydrophilen Peptidhormonen (+ Kap. 10.1.1) in glei-cher Weise ab. Die G-Proteine konnen dabei in zweiVarianten vorkommen:� Stimulierende G-Proteine (GS-Proteine) aktivie-
ren die Adenylatcyclase und steigern damit dieHormonwirkung.
� Hemmende G-Proteine (Gi-Proteine) bremsendie Adenylatcyclase und die Hormonwirkung(+ Tab. 1.2).
Klinik!Das Choleratoxin verhindert die hydrolytische In-aktivierung des GTP-aktivierten stimulierenden G-Proteins, indem es eine Untereinheit dieses G-Pro-teins ribosyliert. Hieraus resultiert eine Dauerakti-vierung der Adenylatcyclase, was zu einer Offnungvon Chloridkanalen in den luminalen Membranendes Ileums fuhrt. Der pathologische Chloridaus-strom zieht Wasser mit sich und bewirkt so denenormen Wasserverlust von mehreren Litern proTag (sekretorische Diarrho).Das Pertussistoxin blockiert in ahnlicher Weise einhemmendes G-Protein. Auch hierdurch wird zuvielcAMP gebildet: Eine pathologisch vermehrte Sekre-tion von NaCl und H2O im Trachealepithel ist dieFolge.
Die IP3-Kaskade
Ein zweiter intrazellularer Botenstoff, der von vielenHormonen (z. B. Adrenalin am a1-Rezeptor, Dop-amin am D2-Rezeptor, Acetylcholin an muskariner-gen M-Rezeptoren) zur Signalubermittlung benutztwird, ist das Inositoltriphosphat (IP3). Auch hierlauft die Ubermittlung der Hormonwirkung in Stufenab (+ Abb. 1.8):1. Das Hormon bindet sich an den Hormonrezeptor,
der daraufhin seine Konformation andert.
–+ –
–
–
+
(2)(3)
(1)erregendes
Hormonsignal
(1)hemmendes
Hormonsignal
GiGS
ACGTP
ATP
GDP + P
Pertussis-toxin
GTP
GDP + PProtein + P
(inaktiv)(5)cAMP + P-A
(4)
Protein-P(aktiv)
Hormon-wirkung
AMPPhosphodiesterase
Theophyllin,Koffein
GTP
Cholera-toxin
RiRS
Abb. 1.7 Die cAMP-Kaskade. RS = stimulierender Rezeptor(z. B. b1-Rezeptoren), Ri = inhibierende Rezeptoren (z. B.a2-Rezeptoren), GS = stimulierendes G-Protein, AC =Adenylatcyclase, P-A = Proteinkinase A, P = Phosphat. [2]
1 Allgemeine Physiologie und Zellphysiologie
10

2. Hierdurch wird ein G-Protein an der Innenseiteder Plasmamembran durch Bindung von GTP ak-tiviert.
3. Dieses aktivierte G-Protein aktiviert seinerseits dasebenfalls an der Innenseite der Plasmamembran lo-kalisierte Enzym Phospholipase C.
4. Die Phospholipase C spaltet das in den Plasma-membranen enthaltene Phosphatidylinosindi-phosphat in IP3 und Diacylglycerin (DG).
5. IP3 setzt Ca2+ aus dem endoplasmatischen Retiku-lum frei und aktiviert hierdurch eine Proteinki-nase.
6. DG aktiviert eine in der Plasmamembran liegendeC-Kinase.
7. Die aktivierte Proteinkinase (durch IP3) und dieaktivierte C-Kinase (durch DG) bilden die letzteStufe der Kaskade und phosphorylieren Funkti-onsproteine, welche die spezifische Hormonant-wort auslosen.
Im Gegensatz zur cAMP-Kaskade gibt es in der IP3-Kaskade keine hemmenden G-Proteine (+ Tab.1.3).
Tab. 1.2 cAMP-vermittelte Wirkungen (Auswahl)
Botenstoff Rezeptor
uber stimulierendeGs-Proteine
Adenosin A2A, A2b
Adiuretin (= Vasopressin) V2
Adrenalin/Noradrenalin b1, b2
Dopamin D1, D5
Histamin H2
Serotonin 5-HT4, 5-HT7
Calcitonin, FSH, Glucagon, Sekretin, VIP, TRH und TSH vers.
uber inhibitorischeGi-Proteine
Acetylcholin M2, M4
Adenosin A1, A3
Adrenalin/Noradrenalin a2
Dopamin D2, D3, D4
GABA GABAB
Glutamat mGLU2–4 und mGLU6–8
Serotonin 5-HT1
Angiotensin II, Melatonin, Neuropeptid c, Opioide,Somatostatin u.a.
vers.
(2) (3)
+
(1)Hormonsignal
R
PIP2 (6)G
PLCGTP
GDP
Inositol-triphosphatase
IP2IP3
(4)
(7)Protein(inaktiv)
Hormon-wirkung
(7)
Protein(inaktiv)
PKCa2+
(5)
endoplasmatisches Retikulum
CK
DG
Protein-P(aktiv)
Abb. 1.8 Die IP3-Kaskade. R = Rezeptor, G = G-Protein, PLC= Phospholipase C, IP3 = Inositoltriphosphat, CK = C-Ki-nase, DG = Diacylglycerin, PIP2 = Phosphatidylinosindi-phosphat, PK = Proteinkinase, PLC = Phospholipase C. [2]
Tab. 1.3 IP3-vermittelte Wirkungen (Auswahl)
Botenstoff Rezeptor
uber sti-mulieren-de Gq-Pro-teine
Acetylcholin M1, M3
Adiuretin (= Vasopressin) V1
Adrenalin / Noradrenalin a1
Glutamat mGLU1,mGLU5
Histamin H1
Serotonin 5-HT2
Bradykinin, CCK, Endothelin,Gastrin, Oxytocin, TRH undTSH
vers.
1.4 Zellorganisation
11
1

NO als Signalstoff
Auch das kurzlebige Radikal Stickstoffmonoxid (NO)mit einer Halbwertszeit von wenigen Sekunden dientder Signalubertragung zwischen Zellen. In Gefaßendo-thelzellen und in einigen Neuronen konnen bestimmteReize (Ca2+-Anstieg, Schubspannung im Endotheldurch vorbeifließendes Blut) eine Anregung der NO-Synthase auslosen (vermittelt uber den Ca2+-Calmodu-lin-Komplex). Dadurch wird Arginin zu Citrullin umge-wandelt und es entsteht einNO-Molekul. DasNOkannaus der Endothelzelle herausdiffundieren. Im Gefaßlu-men bewirkt es eine Hemmung der Thrombozytenag-gregation. Im Zytosol von benachbarten glatten Gefaß-muskelzellen aktiviert es eine losliche Guanylatcyclase,die GTP zu cGMP umbaut. Hierdurch wird eine Pro-teinkinase G aktiviert, wodurch die intrazellulareCa2+-Konzentration abfallt. Es resultiert eine Vasodila-tation (+ Abb. 1.9), die besonders fur die metabolischeKontrolle der Durchblutung wichtig ist (+ Kap. 4.4.2).
Klinik!Die bei Angina pectoris oder Herzinfarkt zur Sen-kung der arteriellen Pumplast des Herzens verwen-deten Nitrate (z. B. Nitroglycerin) bewirken uber dieFreisetzung von NO eine Vasodilatation. Durch diegeringere Pumplast reduziert sich der Sauerstoffver-brauch des Herzens (+ Kap. 3.4.2).
Merke!Nicht uber cAMP- oder IP3-Kaskade wirkt das Hor-mon Insulin. Die Bindung an den Insulinrezeptorlost unmittelbar – ohne zwischengeschalteten Se-cond messenger – die Aktivierung einer Proteinki-nase aus. Lipophile Hormone (Steroidhormoneund Thyroxin) passieren die Zellmembranen undbinden sich an spezifische intrazellulare Hormonre-zeptoren, sind also ebenfalls nicht auf die Second-messenger-Systeme angewiesen (+ Kap. 10.1.2).
1.5 Elektrische Phanomene an der Zelle
Die Phanomene des Membranruhepotentials unddes Erregungsablaufs an Zellen werden zusammen-hangend inKapitel 12.1 und 12.2 besprochen, die spe-ziellen Veranderungen des Ruhe- und Aktionspoten-tials am Herzmuskel in Kapitel 3.1.1.
1.6 Energetik
Die Prinzipien aerober und anaerober Energiegewin-nung, von Energiespeicherung und Energieverbrauchwerden in Kapitel 6 „Arbeits- und Leistungsphysiolo-gie“ und in Kapitel 8 „Energie- und Warmehaushalt“im Zusammenhang dargestellt.
1.7 Regelung und Steuerung
Auf physiologischer Ebene ist der Organismus in allenBereichen bemuht, einen Gleichgewichtszustand alsBasis fur ungestorte, „normale“ Organfunktionen zugewahrleisten. Die Physiologie versucht solche dyna-mischen Strukturen, die ein ebenso dynamischesGleichgewicht anstreben, als Regelkreise zu be-schreiben. Der Begriff Regelkreis erweitert den star-ren Reflexbegriff der klassischen Physiologie. Regel-kreise unterscheiden sich von linearen Reflexen inzwei Punkten:� Das Ergebnis der Reizung wirkt auf den Reiz zu-
ruck.� Der Informationsfluss vom Reizergebnis auf den
Reiz ist kontinuierlich und kein einmaliges Ereig-nis.
Blutfluss
NOArginin
CitrullinEndothelzelle glatte Gefäßmuskelzelle
ZellmembranZellmembran
NO-Synthase
Ca2+
Schub-spannung
Guanylat-cyclase
GTP cGMP
Proteinkinase G
[Ca2+]iVasodilatation
Ca2+-Calmodulin-Komplex
Abb. 1.9 NO als Signalstoff.
1 Allgemeine Physiologie und Zellphysiologie
12

In Regelkreisen kann das Reizergebnis auf zweierleiWeise auf den Reiz zuruckwirken:� Das Ergebnis der Reizung hemmt das Auftreten
weiterer Reize: negative Ruckkopplung. Beispiel:Blutdruckanstieg fuhrt uber die Aktivierung desPressosensorenreflexes reflektorisch zur Gefaßer-weiterung und damit zur Reduktion des erhohtenBlutdrucks.
� Das Ergebnis der Reizung fordert das Auftretenweiterer Reize: positive Ruckkopplung. Beispiel:Steigerung der LH-Freisetzung durch Ostradiolkurz vor der Ovulation (LH steigert physiologi-scherweise die Ostradiol-Freisetzung; + Kap.11.2).
Durch diese eingebauten Ruckkopplungsschleifenunterscheiden sich Regelkreise von einfachen Steue-rungssystemen, bei denen der Zielwert fest vorgege-ben wird (Beispiel: Frontalvorlesung mit Abschluss-klausur). Regelkreise konnen daher erheblich flexi-bler auf Außeneinflusse reagieren als starre Steue-rungssysteme (Beispiel: Seminar mit „Feedback“ derTeilnehmer).Ein besonders gut erforschter Regelkreis ist der Pres-sosensorenreflex (+ Kap. 4.2.3). An ihm lassen sichdie einzelnen Komponenten eines Regelkreises ver-anschaulichen:� Die Regelgroße ist der mittlere arterielle Blut-
druck.� Das Regelziel, die Fuhrungsgroße oder der Soll-
wert der Regelgroße ist die Hohe des optimalen,situationsgerechten Blutdruckwertes.
� Die Regelstrecke ist das Hochdrucksystem desBlutkreislaufs.
� Der Regler liegt in den medullaren Kreislaufarea-len des ZNS, in denen alle Informationen zur Blut-drucksteuerung zusammenlaufen. Dies sind Infor-mationen aus
� den Fuhlern, d. h. den Pressorezeptoren in der A.carotis und der Aorta. Diese Rezeptoren lieferndem medullaren Kreislaufregler den
� Istwert, d. h. die aktuelle Entladungsfrequenz derPressosensoren.
� Storgroßen des Blutdruckwertes, die durch denRegelkreis ausgeglichenwerdenmussen, entstehenz. B. durch Anderungen der Korperlage: Blut-druckabfall beim Aufstehen durch „Versacken“des Blutes in den Gefaßen der unteren Korper-halfte.
� Stellglieder des Pressosensorenreflexes sind dieje-nigen Organe, die in der Lage sind, die RegelgroßeBlutdruck zu modifizieren, d. h. das Herz und dieWiderstandsgefaße des großen Kreislaufs.
� Stellgroßen sind diejenigen physiologischen Para-meter, welche die Aktivitat der Stellglieder unmit-telbar beeinflussen, also in diesem Beispiel die Ent-ladungsfrequenzen der sympathischen und para-sympathischen Innervation des Herzens und derWiderstandsgefaße.
Der Regelkreis des Pressosensorenreflexes halt denBlutdruck auf einem konstanten Niveau. Man sprichtvon einem Halteregler. Kann der Sollwert beliebigverstellt werden und folgt der Regelkreis diesen Soll-wertverstellungen und stabilisiert sie, arbeitet der Re-gelkreis als Folgeregler (Servoregler).Das Beispiel der Blutdruckregulation zeigt aber auch,dass selbst dieses relativ komplexe Regelkreismodellzur Blutdruckregulation noch nicht alle Einflussgro-ßen adaquat berucksichtigt. So spielen neben denuber den Pressorezeptorenreflex vermittelten kurzfri-stigen Regulationsmechanismen auch weitere mittel-und langfristige Einflussgroßen, zentrale Kontrollme-chanismen und außere Umgebungsfaktoren einewichtige Rolle (+ Kap. 4.2.3).
1.7 Regelung und Steuerung
13
1