
· 2008. 5. 27. · Methodenentwicklung zur gaschromatographisch-massenspektrometrischen Analyse...
126
Methodenentwicklung zur gaschromatographisch- massenspektrometrischen Analyse freier faecaler Sterole und Gallensäuren im Single Ion Monitoring; Einfluss von Nahrungssupplementen auf die Exkretion Dissertation zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) vorgelegt dem Rat der Biologisch-Pharmazeutischen Fakultt der Friedrich-Schiller-Universitt Jena von Diplomchemikerin Sylvia Keller geboren am 10. November 1974 in Mühlhausen
Transcript of · 2008. 5. 27. · Methodenentwicklung zur gaschromatographisch-massenspektrometrischen Analyse...

Methodenentwicklung zur gaschromatographisch-
massenspektrometrischen Analyse freier faecaler Sterole und
Gallensäuren im Single Ion Monitoring;
Einfluss von Nahrungssupplementen auf die Exkretion
Dissertation
zur Erlangung des akademischen Grades
doctor rerum naturalium (Dr. rer. nat.)
vorgelegt dem Rat der Biologisch-Pharmazeutischen Fakultät
der Friedrich-Schiller-Universität Jena
von Diplomchemikerin Sylvia Keller
geboren am 10. November 1974 in Mühlhausen

Inhaltsverzeichnis
Inhaltsverzeichnis 1 EINLEITUNG UND ZIELSTELLUNG 1
2 ALLGEMEINE BETRACHTUNGEN 3
2.1 Chemie, Physiologie und Analytik faecaler Steroide 3
2.1.1 Faecale Sterole 3
2.1.1.1 Struktur und Physiologie 3
2.1.1.2 Darmflora und bakterielle Metabolisierung von Cholesterol 4
2.1.1.3 Analytische Voraussetzungen zur Sterolbestimmung 7
2.1.2 Gallensäuren 9
2.1.2.1 Struktur und Physiologie 9
2.1.2.2 Modifikation der Gallensäuren durch die Intestinalflora 10
2.1.2.3 Analytische Voraussetzungen zur Gallensäurenbestimmung 13
2.2 Wechselwirkungen der faecalen Steroide - Einfluss der
Ernährung 19
2.2.1 Pathogene Wirkung und Cholesterolmetabolismus 19
2.2.1.1 Toxizität 19
2.2.1.2 Cholesterol- und Gallensäurenmetabolismus 21
2.2.2 Einfluss von Nahrungsfaktoren auf die Steroidexkretion 23
2.2.2.1 Calcium 23
2.2.2.2 Probiotika 24
3 ENTWICKLUNG DER ANALYSENMETHODEN 27
3.1 Faecale Sterole 27
3.1.1 Probenvorbereitung 27
3.1.2 Analyse mit GC/MS QP 5000 27
3.1.2.1 Probleme der gaschromatographischen Analyse 27
3.1.2.2 Probleme der massenspektrometrischen Detektion 28
3.1.2.3 Messparameter 31
3.1.3 Präzision und Richtigkeit 33

Inhaltsverzeichnis
3.1.4 Vergleich der Ergebnisse zwischen derivatisierten und
freien Sterolen 34
3.1.5 Kalibration faecaler Sterole 35
3.1.6 Interner Standard: 5α-Cholestan 35
3.2 Faecale Gallensäuren 37
3.2.1 Probenvorbereitung 37
3.2.2 Analyse mit GC/MS QP 5000 37
3.2.2.1 Probleme der gaschromatographischen Analyse 37
3.2.2.2 Probleme der massenspektrometrischen Detektion 38
3.2.2.3 Messparameter 41
3.2.3 Präzision und Richtigkeit 43
3.2.4 Kalibration der Gallensäuren 44
3.2.5 Interner Standard: Hyodeoxycholsäure 45
4 HUMANSTUDIEN 46
4.1 Supplementation von Calciumphosphaten 46
4.2 Supplementation von probiotischer Rohwurst 48
4.3 Supplementation von probiotischem Joghurt 48
4.4 Stuhlproben und in der Auswertung berücksichtigte
Parameter 49
4.5 Statistik 50
5 ERGEBNISSE 51
5.1 Studie 1 - Supplementation von Calciumphosphaten 51
5.2 Studie 2 - Supplementation von probiotischer Rohwurst 54
5.3 Studie 3 - Supplementation von probiotischem Joghurt 57
5.4 Studie 4 - Supplementation von probiotischem Joghurt 63
5.5 Studienübergreifende Ergebnisse 69

Inhaltsverzeichnis
6 DISKUSSION 70
6.1 Analysenmethoden 70
6.2 Steroidgehalte in der Trockensubstanz und die tägliche
Exkretion 77
6.3 Der Einfluss von Calcium und Probiotika auf die Steroid-
ausscheidung 83
6.4 Serumcholesterol und faecale Steroide 90
7 SCHLUSSFOLGERUNGEN 93
8 ZUSAMMENFASSUNG 95
Quellenverzeichnis I - XVI
Selbständigkeitserklärung
Lebenslauf
Danksagung

Abbildungsverzeichnis
Abbildungsverzeichnis Abbildung 1 Grundgerüst und Struktur bedeutender faecaler Sterole 3
Abbildung 2 Metabolisierung von Cholesterol durch die Intestinalflora 6
Abbildung 3 Grundgerüst und Struktur bedeutender Gallensäuren 9
Abbildung 4 Metabolisierungsreaktionen an Gallensäuren durch die
Darmflora 12
Abbildung 5 Alkylesterbildung an der Carboxylgruppe freier Gallensäuren 14
Abbildung 6 Silylierungsreaktion an Hydroxygruppen des Steroidgerüstes 15
Abbildung 7 Cholesterol- und Gallensäurenmetabolismus 22
Abbildung 8 Fällung von Gallensäuren und Cholesterol durch Calcium-
phosphat-Komplexe 23
Abbildung 9 Probiotikawirkung im Colon 25
Abbildung 10 Chromatogramme eines Sterolstandardgemisches 28
Abbildung 11 Massenspektren faecaler Sterole 29
Abbildung 12 Separation von Cholesterol und Coprostanon im SIM- und
TIC-Modus 30
Abbildung 13 Temperaturverlauf zur Gaschromatographie faecaler Sterole 31
Abbildung 14 Sterolchromatogramm einer Faecesprobe 32
Abbildung 15 Vergleich der Ergebnisse von derivatisiertem und freiem
Cholesterol in einer internationalen Laborvergleichsunter-
suchung mit 20 Teilnehmern nach DGF [2001] 34
Abbildung 16 Beziehung zwischen dem Analysenergebnis und dem
Analysenwert von Cholesterol 36
Abbildung 17 Übereinanderliegende Darstellung der Chromatogramme
von 3-Oxo-LCA und 12-Oxo-DCA 38
Abbildung 18 Chromatogramm eines Gallensäurenstandardgemisches 38
Abbildung 19 Massenspektren faecaler Gallensäuren 39
Abbildung 20 Chromatogramm (MIC) von Cholsäurestandard und Chol-
säure in den Faeces 40

Abbildungsverzeichnis
Abbildung 21 Vermutliche Isomere der DCA 41
Abbildung 22 Gallensäurenchromatogramm einer Faecesprobe 42
Abbildung 23 Beziehung zwischen dem Analysenergebnis und dem
Analysenwert von DCA bzw. internem Standard 45
Abbildung 24 Struktur einer Studienperiode 49
Abbildung 25 Steroidexkretion der Probanden 24 und 25 in Studie 1 52
Abbildung 26 Sterolkonzentrationen in der Gefriertrockensubstanz 52
Abbildung 27 Tägliche Steroidexkretion in Abhängigkeit von der
Verzehrsreihenfolge der Joghurts 59
Abbildung 28 Zusammenhänge zwischen den Konzentrationen an Serum-
cholesterol und primärer Gallensäuren im Stuhl 61
Abbildung 29 Vergleich der täglichen Gallensäurenexkretion und Serum-
cholesterolkonzentrationen unter Berücksichtigung der
Verzehrsreihenfolge der Joghurts und in Abhängigkeit vom
Fettstoffwechsel 62
Abbildung 30 Tägliche faecale Steroidexkretion in Abhängigkeit von der
Verzehrsreihenfolge der Joghurts 66
Abbildung 31 Zusammenhänge zwischen den Konzentrationen an Serum-
cholesterol und primärer bzw. sekundärer Gallensäuren
im Stuhl 67
Abbildung 32 Beziehung zwischen Gehalten an Cholesterol und Copros-
tanol in der Stuhltrockensubstanz 69
Abbildung 33 TIC und MIC eines Sterolstandardgemisches 70
Abbildung 34 Chromatogramm faecaler Steroide nach BATTA et al. [1999] 73
Abbildung 35 Chromatogramm faecaler Gallensäuren nach SETCHELL et al.
[1983] 74
Abbildung 36 Fragmentierung der Sterole und Gallensäuren 75
Abbildung 37 Änderungen im Cholesterol- und Gallensäurenmetabolismus 91

Tabellenverzeichnis
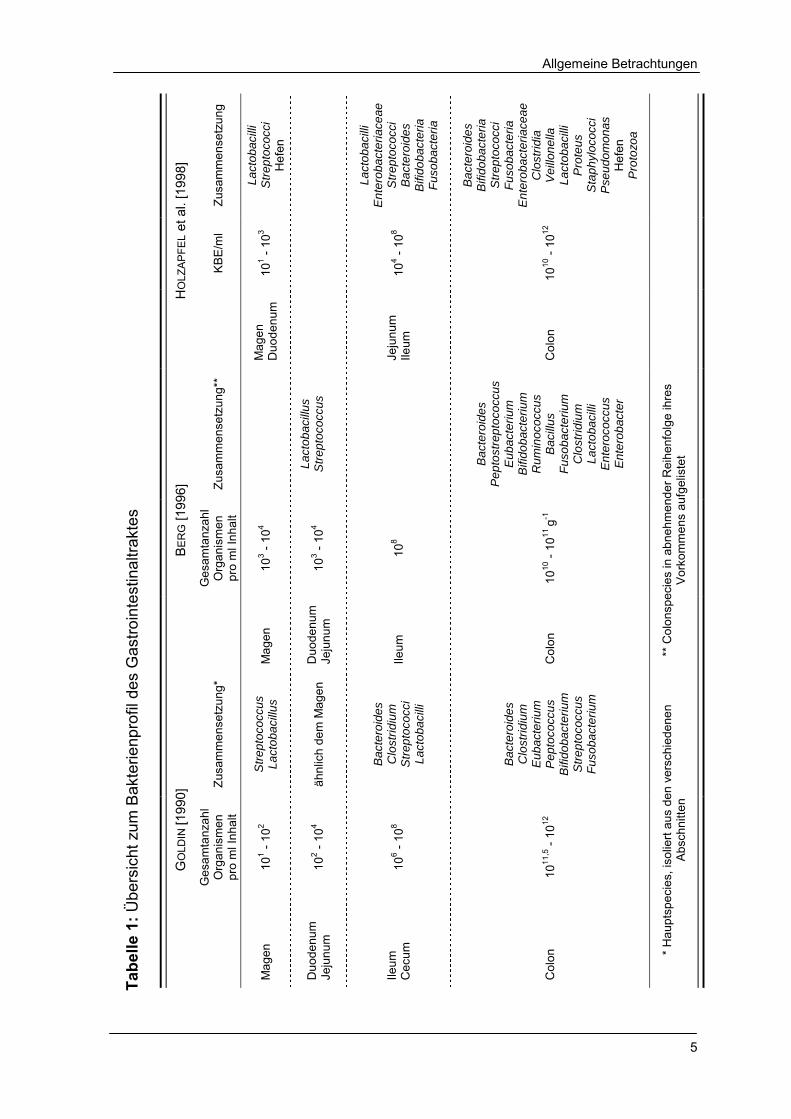
Tabellenverzeichnis Tabelle 1 Übersicht zum Bakterienprofil des Gastrointestinaltraktes 5
Tabelle 2 Parameter der Gallensäurenanalyse - Literaturübersicht 15
Tabelle 3 Elutionsreihenfolgen von Gallensäuren verschiedenen
Ursprungs 16
Tabelle 4 Molare Massen freier und derivatisierter Gallensäuren 17
Tabelle 5 Interne Standards zur Gallensäurenquantifizierung 18
Tabelle 6 Auswahl spezifischer Fragmente zur SIM-Detektion der
Sterole 30
Tabelle 7 Retentionszeiten der faecalen Sterole 32
Tabelle 8 Wiederholungsmessungen zur Präzision der Sterolanalyse 33
Tabelle 9 Parameter der Kalibration faecaler Sterole 35
Tabelle 10 Auswahl spezifischer Fragmente zur SIM-Detektion der
Gallensäuren 40
Tabelle 11 Retentionszeiten der Gallensäuren 42
Tabelle 12 Wiederholungsmessungen zur Präzision der Gallensäuren-
analyse 43
Tabelle 13 Parameter der Kalibration faecaler Gallensäuren 44
Tabelle 14 Berücksichtigte Humanstudien des Lehrstuhls
Ernährungsphysiologie im Überblick 47
Tabelle 15 Mittlere Stuhlausscheidung in Studie 1 51
Tabelle 16 Faecale Exkretion an Sterolen und Gallensäuren in Studie 1 53
Tabelle 17 Mittlere Stuhlausscheidung in Studie 2 54
Tabelle 18 Faecale Exkretion an Sterolen und Gallensäuren in Studie 2 55
Tabelle 19 Tägliche Gallensäurenexkretion unter Berücksichtigung der
Fettaufnahme und Trockensubstanzexkretion 56
Tabelle 20 Mittlere Stuhlausscheidung in Studie 3 57
Tabelle 21 Faecale Exkretion an Sterolen und Gallensäuren in Studie 3 58

Tabellenverzeichnis
Tabelle 22 Gallensäurenausscheidung bei normierter Fettaufnahme
der verschieden cholesterolämischen Probanden 60
Tabelle 23 Mittlere Stuhlausscheidung in Studie 4 63
Tabelle 24 Faecale Exkretion an Sterolen und Gallensäuren in Studie 4 65
Tabelle 25 Steroidexkretion der Frauen und Männer 68
Tabelle 26 Qualitative Bestimmung faecaler Sterole nach verschie-
denen Analysenmethoden 72
Tabelle 27 Übersicht zum Sterolgehalt in der Stuhltrockensubstanz 78
Tabelle 28 Übersicht zur täglichen Sterolexkretion 78
Tabelle 29 Übersicht zum Gallensäurengehalt in der Stuhltrocken-
substanz 80
Tabelle 30 Übersicht zur täglichen Gallensäurenexkretion 81
Tabelle 31 Minimal- und Maximalgehalte von Steroiden im Stuhl;
Studien 1 - 4 82
Tabelle 32 Einfluss von Calcium auf den Steroidgehalt in der Stuhl-
trockensubstanz 84
Tabelle 33 Mögliche Ursachen einer Änderung des faecalen Sterol-
profiles 86

Abkürzungsverzeichnis
Abkürzungsverzeichnis Ac Acetat
But Butylester
ca. circa
CA Cholsäure
CDCA Chenodeoxycholsäure
Chol Cholesterol
DC Dünnschichtchromatographie
DCA Deoxycholsäure
df Filmdicke
DGE Deutsche Gesellschaft für Ernährung
DGF Deutsche Gesellschaft für Fettforschung
DNS Desoxyribonucleinsäure
eV Elektronenvolt
Fa. Firma
GC Gaschromatographie
GCA Glycocholsäure
griech. griechisch
GS Gallensäure
GTS Gefriertrockensubstanz
HCA Hyocholsäure
HDCA Hyodeoxycholsäure
HDL High Density Lipoprotein
KBE Koloniebildende Einheiten
L. Lactobacillus
LCA Lithocholsäure
lat. lateinisch
LDL Low Density Lipoprotein
Max Maximum
MCA Muricholsäure
Metab Cholesterolmetabolite
Meth Methylester

Abkürzungsverzeichnis
MIC Multi Ion Chromatogram
Min Minimum
MS Massenspektroskopie
Mw Mittelwert
n. d. nicht detektiert
NWG Nachweisgrenze
o. A. ohne Angabe
prim. primär
QP Quadrupol
Rel. Int. Relative Intensität
Rel. Std.abw. Relative Standardabweichung
sek. sekundär
SIM Single Ion Monitoring
Std.abw. Standardabweichung
subsp. subspecies
TIC Total Ion Chromatogram, Total Ion Current
TMS Trimethylsilylether
TS Trockensubstanz
u. a. und andere; unter anderem
UDCA Ursodeoxycholsäure
vgl. vergleiche
vs. versus

Einleitung und Zielstellung
1
1 EINLEITUNG UND ZIELSTELLUNG
„0,02 g Cholsäure werden in 0,5 g Alkohol gelöst und mit 1 ccm n/10 Jodlösung
versetzt. Beim vorsichtigen Verdünnen mit Wasser erstarrt die Flüssigkeit zu
einem Kristallbrei von Nadeln, die im durchfallenden Licht blaue Färbung
zeigen.“
Die Anfänge der Analytik von neutralen und sauren Steroiden beruhen auf nass-
chemischen Methoden wie der hier dargestellte Nachweis von Cholsäure aus dem
Jahr 1887 von MYLIUS. Viele saure Steroide wurden erstmals aus
Gallenflüssigkeiten bzw. Gallensteinen verschiedener Tiere, wie z. B. von Pavian,
Kabeljau, Warzenschwein oder Eisbär, isoliert. Noch heute gebräuchliche
Trivialnamen wie Ursodeoxycholsäure (ursus [lat.]: Bär) oder Lithocholsäure (litho
[griech.]: Stein) machen diesen Ursprung deutlich.
Durch die Beobachtung des Reaktionsverhaltens konnten chemische Strukturen
erkannt [WIELAND 1928] und die Rolle der Gallensäuren als Emulgatoren bei der
Fettverdauung des Menschen geklärt werden [LETTRÉ 1954]. In der Mitte des
20. Jahrhunderts wurden vor allem Titrations- oder Spektroskopieverfahren zur
Bestimmung genutzt [GORDON et al. 1957, MOSBACH et al. 1954], wobei Letztere
im nahen Infrarot-Bereich noch heute angewendet werden [NAKAMURA et al. 1998].
Mit der Entwicklung der Gaschromatographie im Jahre 1944 durch Erika CREMER
[1976] konnten in den folgenden Jahrzehnten Steroide aus komplexen Matrizes
wie humanen Faeces chromatographisch getrennt und quantitativ bestimmt
werden [GRUNDY et al. 1965]. Durch die Kopplung an Massenspektrometer sind
außerdem Aussagen über Strukturelemente der Verbindungen ermöglicht worden
[ENEROTH et al. 1966]. Trotz moderner Technologie ist zur Bestimmung neutraler
und saurer Steroide in den Faeces auch heute eine aufwändige
Probenvorbereitung (Festphasenextraktion, Hydrolyse, Dünnschichtchromato-
graphie, flüssig/flüssig-Extraktion, Derivatisierung) zur gaschromatographischen
Analyse notwendig [PANDA und BROITMAN 2001, SETCHELL 1982].
Mit den erweiterten Möglichkeiten der Analytik hat sich ein breites Spektrum an
Untersuchungen zum Einfluss faecaler Steroide bei der Entstehung und
Vorbeugung von Erkrankungen des Gastrointestinaltraktes, der Leber oder bei

Einleitung und Zielstellung
2
Atherosklerose entfaltet. So werden Untersuchungen u. a. hinsichtlich der Gallen-
steinformation sowie zu den Effekten der Gallensäuren bei Familiärer Polyposis,
Ulcerativer Colitis, Durchfallerkrankungen, Coloncarcinogenese oder Hyper-
lipidämie durchgeführt [SETCHELL et al. 1988, WELLS et al. 2000]. Da die Ernährung
des Menschen hierbei ursächlich und vorbeugend wirken kann, erfolgte die
Durchführung einer Vielzahl von Studien zur Beeinflussung der Exkretion faecaler
Steroide durch Ballaststoffe [MIETTINEN 1987], Proteine [DUANE 1999], Fette
[ELLEGARD et al. 2000], Vitamine [DUANE und HUTTON 1983], Mineralien [VAN DER
MEER et al. 1991], Milchprodukte [HOLT 1999], Präbiotika und Probiotika [MARTEAU
2001].
Im Allgemeinen ist eine Veränderung des faecalen Steroidprofils entweder durch
Modifizierung der intestinalen Mikroflora, z. B. aufgrund verstärkter Fermentation
bei erhöhter Ballaststoffzufuhr, oder durch Änderung physiko-chemischer
Parameter im Darmlumen (pH-Wert, Löslichkeit, Adsorption) möglich.
Ziel der Arbeit ist es, effiziente Methoden zur gaschromatographisch-
massenspektrometrischen Analyse von faecalen Sterolen und Gallensäuren zu
erarbeiten, wobei die Vorteile der massenspektrometrischen Detektion hinsichtlich
Spezifität und Sensitivität in der Realisierung der Aufgabenstellung genutzt
werden. Weiterhin sollen die Methoden zur Analyse realer Proben aus
Humanstudien verschiedener Fragestellungen Anwendung finden und mögliche
Einflüsse von supplementiertem Calciumphosphat, probiotischer Rohwurst und
probiotischem Joghurt auf die Exkretion der Steroide ermittelt werden.

Allgemeine Betrachtungen
3
2 ALLGEMEINE BETRACHTUNGEN 2.1 Chemie, Physiologie und Analytik faecaler Steroide 2.1.1 Faecale Sterole
2.1.1.1 Struktur und Physiologie
Die zu den neutralen Steroiden gehörenden Sterole werden häufig mit der
Bezeichnung �Sterine� dargestellt. Man unterscheidet nach ihrem Vorkommen in
Zoosterole (Tiere), Phytosterole (Pflanzen) und Mykosterole (Pilze). Sie leiten sich
vom cyclischen, hydrierten Cyclopentanophenanthren ab, welches das typische
Grundgerüst der Steroide bildet. Zwischen den Kohlenstoffatomen 5/6 oder 7/8
und in der Seitenkette enthalten sie eine oder mehrere Doppelbindungen. Die
gesättigten Abkömmlinge enden in der trivialen Nomenklatur auf �-stanol� (Abb. 1).
Funktionelle Strukturelemente wie Hydroxy- oder Oxogruppen sind an der
Position 3 des Kohlenstoffgerüstes lokalisiert und die Seitenkette kann an C24 mit
Methyl- oder Ethylgruppen substituiert sein. Daher wird die Variabilität der Sterole
durch die Anzahl und Position der Doppelbindungen und die Struktur der
Seitenkette geprägt, hingegen ist die Vielfalt der Gallensäuren vor allem auf die
funktionellen Gruppen zurückzuführen (vgl. 2.1.2.1). Unter dem Begriff �faecale
Sterole� werden häufig die ungesättigten und gesättigten Vertreter tierischer und
pflanzlicher Herkunft zusammengefasst.
Abbildung 1: Grundgerüst und Struktur bedeutender faecaler Sterole
R1 Konfiguration A+B/
Position des H-Atoms an C5
∆5 Substituent an C24 Bezeichnung
H trans/5α - - 5α-Cholestan
β-OH - + - Cholesterol
β-OH - + -C2H5 Sitosterol
β-OH - + -CH3 Campesterol
β-OH cis/5β - - Coprostanol
β-OH trans/5α - - Cholestanol
=O cis/5β - - Coprostanon
=O trans/5α - - Cholestanon
A B
C D
R1
1
2
3
45
6
7
8
9
10
11
12
13
1415
16
1718
19
2021 22
23
24
25
26
27

Allgemeine Betrachtungen
4
Beim Menschen wird zwischen endogenem und exogenem Cholesterol
unterschieden, das entweder in der Leber des Menschen aus Isoprenbausteinen
über Squalen synthetisiert oder aus der Nahrung im Jejunum absorbiert werden
kann. Cholesterol ist ein bedeutender Bestandteil in den Membranen des
Menschen und ist der Ausgangsstoff für die Bildung von Vitamin D,
Sexualhormonen und Gallensäuren. Freies Cholesterol wird über die Galle in den
Darm abgegeben, rückresorbiert oder durch Darmbakterien des Colon
metabolisiert und ausgeschieden [BIESALSKI 1995].
Bei den aus Faecesproben isolierten Sterolen handelt es sich vorwiegend um
Cholesterol und dessen bakterielle Transformationsprodukte Coprostanol und
Coprostanon. Außerdem werden weitere Sterolabbauprodukte und mit der
Nahrung aufgenommene Phytosterole wie Sitosterol oder Campesterol
nachgewiesen.
2.1.1.2 Darmflora und bakterielle Metabolisierung von Cholesterol
Der Intestinaltrakt weist eine sehr differenzierte Besiedlung mit Mikroorganismen
auf. So befinden sich im Magen und oberen Abschnitt des Darmes vorwiegend
säuretolerante, mikroaerophile Lactobacillen und fakultative Streptococcen mit
einem Gesamtgehalt von 101 - 103 Organismen pro ml Inhalt. Der ileale bis cecale
Bereich bildet den Übergang zum Colon und ist sowohl durch Bakterien des
gastrisch, duodenalen Abschnittes als auch des Dickdarmes besiedelt. Der
Mikroorganismengehalt in diesem Bereich verdeutlicht ebenfalls das Übergangs-
milieu. Im anaeroben Colon ist die Artenvielfalt mit 400 - 500 Species und die
Anzahl an Organismen mit über 1010 KBE/ml Inhalt am höchsten (Tab. 1) [KELLER
und JAHREIS 2001].
Cholesterol kann durch anaerobe, gram-positive Bakterien [MIDTVEDT et al. 1990]
zu 80 - 100 % [PONZ DE LEON et al. 1987] in Coprostanol transformiert werden,
wobei vor allem Eubacterium-Stämme (z. B. Eubacterium coprostanoligenes
ATCC 51222) reduktiv wirksam sind [MACDONALD et al. 1983a].
Allgemeine Betrachtungen
5
Zusa
mm
ense
tzun
g
Lact
obac
illi
Stre
ptoc
occi
H
efen
Lact
obac
illi
Ente
roba
cter
iace
ae
Stre
ptoc
occi
Ba
cter
oide
s Bi
fidob
acte
ria
Fuso
bact
eria
Bact
eroi
des
Bifid
obac
teria
St
rept
ococ
ci
Fuso
bact
eria
En
tero
bact
eria
ceae
C
lost
ridia
Ve
illon
ella
La
ctob
acill
i Pr
oteu
s St
aphy
loco
cci
Pseu
dom
onas
H
efen
Pr
otoz
oa
KBE/
ml
101 -
103
104 -
108
1010
- 10
12
HO
LZAP
FEL
et a
l. [1
998]
Mag
en
Duo
denu
m
Jeju
num
Ile
um
Col
on
Zusa
mm
ense
tzun
g**
Lact
obac
illus
St
rept
ococ
cus
Bact
eroi
des
Pept
ostre
ptoc
occu
s Eu
bact
eriu
m
Bifid
obac
teriu
m
Rum
inoc
occu
s Ba
cillu
s Fu
soba
cter
ium
C
lost
ridiu
m
Lact
obac
illi
Ente
roco
ccus
En
tero
bact
er
Ges
amta
nzah
l O
rgan
ism
en
pro
ml I
nhal
t
103 -
104
103 -
104
108
1010
- 10
11 g-1
B ER
G [1
996]
Mag
en
Duo
denu
m
Jeju
num
Ileum
Col
on
** C
olon
spec
ies
in a
bneh
men
der R
eihe
nfol
ge ih
res
Vork
omm
ens
aufg
elis
tet
Zusa
mm
ense
tzun
g*
Stre
ptoc
occu
s La
ctob
acill
us
ähnl
ich
dem
Mag
en
Bact
eroi
des
Clo
strid
ium
St
rept
ococ
ci
Lact
obac
illi
Bact
eroi
des
Clo
strid
ium
Eu
bact
eriu
m
Pept
ococ
cus
Bifid
obac
teriu
m
Stre
ptoc
occu
s Fu
soba
cter
ium
Ges
amta
nzah
l O
rgan
ism
en
pro
ml I
nhal
t
101 -
102
102 -
104
106 -
108
1011
,5 -
1012
GO
LDIN
[199
0]
Mag
en
Duo
denu
m
Jeju
num
Ileum
C
ecum
Col
on * H
aupt
spec
ies,
isol
iert
aus
den
vers
chie
dene
n Ab
schn
itten
Tabe
lle 1
: Übe
rsic
ht z
um B
akte
rienp
rofil
des
Gas
troin
test
inal
trakt
es

Allgemeine Betrachtungen
6
Mit Hilfe von radiomarkierten Wasserstoff- und Kohlenstoffatomen im
Cholesterolmolekül werden zwei verschiedene Wege zur Umwandlung in
Coprostanol nachgewiesen (Abb. 2). Der erste Mechanismus ist die direkte
Reduktion der ∆5-Doppelbindung [ROSENFELD et al. 1954]. Der zweite Weg, mit
anfänglicher Oxidation der Hydroxygruppe, Isomerisierung der Doppelbindung und
anschließender Reduktion zu Coprostanon bzw. Coprostanol, wird als vorwiegend
ablaufender Mechanismus angesehen [MACDONALD et al. 1983a, REN et al. 1996].
Abbildung 2: Metabolisierung von Cholesterol durch die Intestinalflora
1: direkte Umwandlung; 2a - c: indirekte Umwandlung
Cholesterol
Coprostanol Coprostanon
Cholest-4-en-3-on
1
2a
2b
2c
A B
1
2
3
45
6
7
8
9
10
19
HO
A B
1
2
3
45
6
7
8
9
10
19
HOA B
1
2
3
45
6
7
8
9
10
19
O
A B
1
2
3
45
6
7
8
9
10
19
O
R1
R2
R2 R2
R2
R1R1
R1

Allgemeine Betrachtungen
7
2.1.1.3 Analytische Voraussetzungen zur Sterolbestimmung
Probenvorbereitung
Sterole liegen durch ihre geringe Polarität im Lipidextrakt des Stuhles vor
(vgl. 2.1.2.3). Die Trennung von anderen Steroiden, die Separation in freie und
veresterte Sterole und die Verteilung in einzelne Sterolklassen (Oxo-Sterole,
5β-Sterole, 5α-Sterole) kann mit chromatographischen Methoden an Gelen
[BARKER et al. 1993, KORPELA 1982] oder auf Dünnschichtplatten [ARCA et al.
1983] erfolgen.
Zur Abspaltung der Fettsäuren aus Sterolestern wird in nahezu allen Methoden
eine alkalische Hydrolyse in alkoholischen Medien durchgeführt [HOVING 1995].
Dabei werden methanolische bzw. ethanolische Natron- oder Kalilaugen ver-
wendet [BÖRJESSON et al. 1998, KAIBARA et al. 1983, LINDENTHAL et al. 2001,
VERBEEK et al. 1995]. Mit unpolaren Lösungsmitteln wie Hexan oder Cyclohexan
ist eine Extraktion der Sterole aus der polaren Reaktionslösung möglich [AUSMAN
et al. 1993, CLIFFORD et al. 1986].
Gaschromatographische Analyse
In den meisten Anwendungen wird die Hydroxygruppe an Position 3 des
Kohlenstoffgerüstes in Alkylsilylether, Acetate oder Haloacetate überführt
[GÖRÖG 1983]. Für Phytosterole sind auch Methoden ohne Derivatisierung be-
schrieben [SUPELCO 2002]. Für freie, faecale Sterole werden Koelutionen von
Cholestanol/Cholesterol [HOVING 1995] und Coprostanon/Cholesterol [OWEN et al.
1987] bei Verwendung unpolarer gepackter Säulen beobachtet. Die Trennung von
Cholestanol und Cholesterol wird durch eine optimierte Probenvorbereitung
(Dünnschichtchromatographie) [TURJMAN und NAIR 1984] und/oder durch die
Modifizierung des gaschromatographischen Verfahrens erreicht. Dies kann
entweder durch Trimethylsilylierung der Hydroxygruppe mit Applikation auf
unpolare Kapillarsäulen [MIETTINEN 1982], durch eine Erhöhung der Polarität der
stationären Phase der gepackten Säulen [ARCA et al. 1983] oder durch duales
Analysieren mit zwei gepackten Säulen [GERHARDT et al. 1977] geschehen.

Allgemeine Betrachtungen
8
Eine Separation von underivatisiertem Coprostanon und Cholesterol ist bis heute
nur durch die Probenaufarbeitung möglich bzw. es wird eine Überquantifizierung
des Cholesterol in den Faeces hingenommen [OWEN et al. 1987].
Die beschriebenen Alternativen führen zu einem höheren Zeitaufwand in der
Probenvorbereitung [SHIMADA et al. 2001], zu möglichen Substanzverlusten
besonders an 3-Oxo-Sterolen (vgl. 2.1.2.3) oder zum Einsatz gepackter Säulen,
die heute in der Regel durch Kapillarsäulen ersetzt werden.
Massenspektrometrische Detektion und interner Standard
Obwohl 5α-Cholestan keine funktionelle Gruppe am Steroidgerüst trägt, wird es in
den meisten Anwendungen als interner Standard eingesetzt [ARCA et al. 1983,
BÖRJESSON et al. 1998, KORPELA 1982, LINDENTHAL et al. 2001, PONZ DE LEON et al.
1987, VERBEEK et al. 1995]. Als Alternativen werden 3α-Hydroxy-5β-Cholestan,
Stigmasterol oder radiomarkiertes Cholesterol verwendet [HOVING 1995].
Massenspektrometrische Daten zur quantitativen Bestimmung im Modus des
Single Ion Monitoring (SIM) sind fast ausschließlich für die Serumcholesterol-
analyse dokumentiert [GAMBERT et al. 1979, WOLTHERS et al. 1980]. Zur Analyse
von freiem Cholesterol wird das Fragment m/z = 386 erwähnt, welches dem
Molekülion der Verbindung entspricht [BJÖRKHEM et al. 1974]. Außerdem werden
Trimethylsilylether oxidierter Pflanzensterole im Serum oder Epididymissterole der
Ratte mittels SIM quantifiziert [LINDENTHAL et al. 2001, PLAT et al. 2001]. In der
Analytik weiterer Sterole wird die Fragmentierung zur Identifikation der Analyten
angewendet, jedoch nicht zur Quantifizierung [MATUSIK et al. 1988, SONG et al.
2000].

Allgemeine Betrachtungen
9
2.1.2 Gallensäuren
2.1.2.1 Struktur und Physiologie
Die Struktur der Gallensäuren basiert auf dem Steroidgerüst, das am Ende der
Seitenkette eine Carboxylgruppe trägt. Deshalb werden sie den sauren Steroiden
zugeordnet. Neben den C24-Hauptvertretern sind auch natürliche Abkömmlinge mit
einer Anzahl von 21 Kohlenstoffatomen dokumentiert [LUND et al. 1991,
MURI-BOBERG et al. 1990]. Im Unterschied zu den faecalen Cholesterol-
metaboliten sind die A- und B-Ringe hauptsächlich in cis-Konfiguration verknüpft,
was eine gewinkelte räumliche Struktur der Moleküle zur Folge hat. Die Vielzahl
der verschiedenen Gallensäuren ist durch Variationen in Anzahl, Position und
Lage von Hydroxy- und Oxogruppen (α: Hydroxygruppe unterhalb der
Molekülebene, β: Hydroxygruppe oberhalb der Molekülebene) am Steroidgerüst
bedingt (Abb. 3).
R1 R2 R3 R4 Trivialname (Abkürzung)
H H H H 5ß-Cholansäure
α-OH H H H Lithocholsäure (LCA)
β-OH H H H iso-Lithocholsäure (iso-LCA)
α-OH H H α-OH Deoxycholsäure (DCA)
β-OH H H α-OH iso-Deoxycholsäure (iso-DCA)
α-OH α-OH H H Hyodeoxycholsäure (HDCA)
α-OH H β-OH H Ursodeoxycholsäure (UDCA)
α-OH H α-OH H Chenodeoxycholsäure (CDCA)
α-OH H α-OH α-OH Cholsäure (CA)
α-OH H H =O 12-Oxo-Deoxycholsäure (12-Oxo-DCA)
Abbildung 3: Grundgerüst und Struktur bedeutender Gallensäuren
Beim Säuger werden primäre und sekundäre Gallensäuren unterschieden. Die
primären Gallensäuren Cholsäure (CA) und Chenodeoxycholsäure (CDCA)
werden in den Hepatozyten des Menschen aus Cholesterol mittels
7α-Hydroxylasen gebildet, mit Taurin oder Glycin an der Säuregruppe verestert
A B
C D
O
OH
R1
R2
R3
1
2
3
45
6
7
8
9
10
11
12
13
1415
16
1718
19
2021 22
23
24R4

Allgemeine Betrachtungen
10
und über die Gallenblase oder direkt in das Duodenum sezerniert. Die
Mizellenbildung ist die Voraussetzung zur Absorption hydrophober Moleküle (z. B.
Fette) im wässrigen Medium des Darmes. Sie bilden ein Mehrkomponen-
tensystem, bestehend aus Gallensäuren (40 � 100 %), Lecithin (0 - 60 %) und
Cholesterol (0 - 15 %). Neben den Nahrungsbestandteilen werden über 90 % der
Gallensäuren im Ileum reabsorbiert [LÖFFLER und PETRIDES 1998]. Der Durchtritt
kann sowohl passiv als auch aktiv erfolgen. Trotzdem erreichen konjugierte
Gallensäuren das Colon und werden durch die anaerobe Mikroflora metabolisiert
und anschließend als sekundäre Gallensäuren Lithocholsäure (LCA) und
Deoxycholsäure (DCA) erneut absorbiert oder mit den Faeces ausgeschieden.
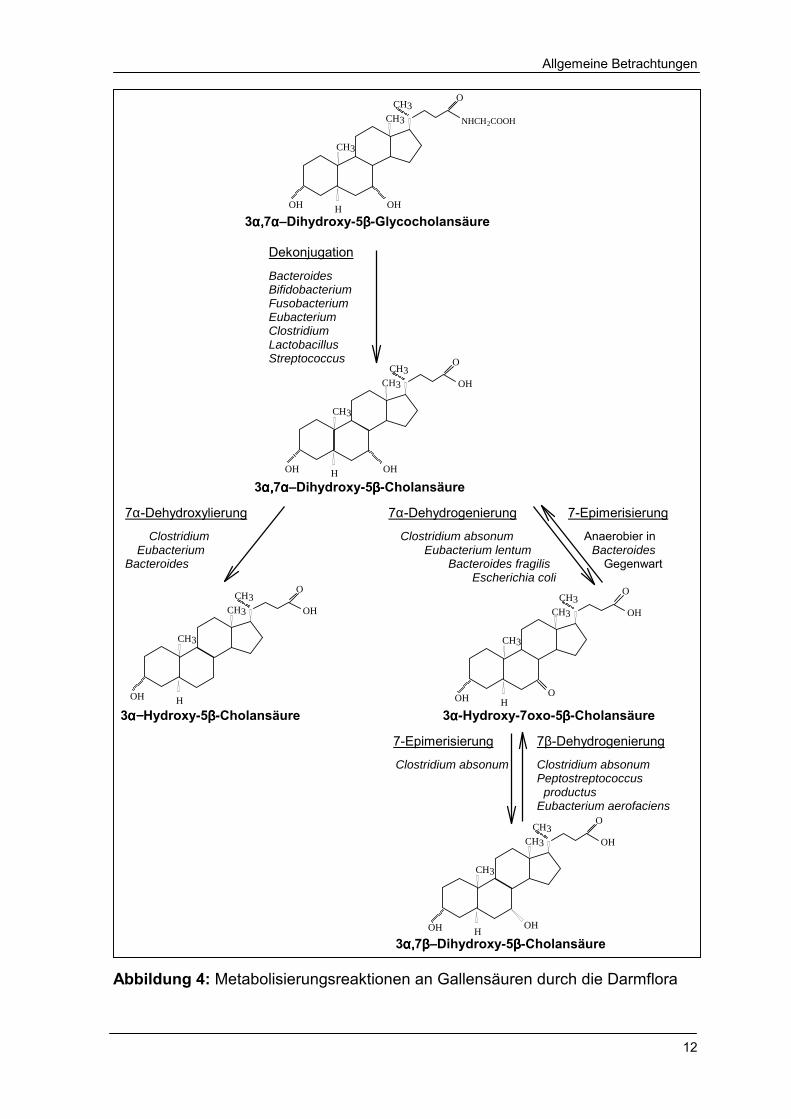
2.1.2.2 Modifikation der Gallensäuren durch die Intestinalflora
Die Bakterien des Colon sind in der Lage, 15 bis 20 Metabolite aus Gallensäuren
zu formen [VAN ELDERE 1999]. Dabei sind die Hydrolyse in freie Gallensäuren, die
Dehydroxylierung der 7α-Hydroxygruppe und die Epimerisierung der Hydroxy-
gruppen am Steroidgerüst die vorwiegenden Reaktionen (Abb. 4).
Die Hydrolyse der Peptidbindung zwischen den Gallensäuren und Taurin bzw.
Glycin ist die Initialtransformation durch die intestinale Mikroflora. Sie ist nahezu
vollständig, so dass in den Faeces hauptsächlich freie Gallensäuren vorliegen
[MACDONALD et al. 1983a]. Hydrolaseaktivität entfalten viele Bakterien, so konnte
bei Bacteroides-, Bifidobacterium-, Fusobacterium-, Eubacterium-, Clostridium-,
Lactobacillus- und Streptococcus-Species die Fähigkeit zur Dekonjugation gezeigt
werden [HYLEMON und GLASS 1983, SHIMADA et al. 1969, VAN ELDERE 1999].
Die Dehydroxylierung an der C7-Position der Gallensäuren bewirkt die
Umwandlung der primären Gallensäuren in die sekundären Gallensäuren. Zu den
7α-dehydroxylierenden Bakterien zählen Clostridium [WELLS und HYLEMON 2000],
Eubacterium und Bacteroides [EDENHARDER 1984]. Bei dieser Metabolisierung
werden die Gallensäuren zuerst mittels Adenosindiphosphat aktiviert [COLEMAN et
al. 1987] und anschließend zu 3-Oxo-4-Cholensäure oxidiert [BJÖRKHEM et al.
1989]. Weiterhin erfolgt eine trans-Eliminierung von Wasser, so dass ein ∆6 unge-
sättigtes Intermediat entsteht [MACDONALD et al. 1983a].

Allgemeine Betrachtungen
11
In Kombination dieser Thesen müsste ein ∆4, ∆6 konjugiertes System entstehen,
das zur 7-Dehydroxy-Gallensäure gesättigt wird. Die Reaktion ist irreversibel, das
heißt, die Umkehrreaktion von z. B. DCA zu CA ist nicht möglich [VAN ELDERE
1999].
Ein weiteres bakterielles Enzym zur Transformation der Gallensäuren ist die
α-Dehydrogenase. Sie bewirkt die Epimerisierung von α-ständigen Hydroxy-
gruppen in β-Epimere über Oxo-Intermediate. Dabei können Hydroxygruppen an
den Positionen 3, 6, 7 und 12 des Steroidgerüstes umgewandelt werden. Die
Epimerisierung an der Position 7 tritt in verstärkter Konkurrenz zu der irreversiblen
Dehydroxylierung auf. 7α-Dehydrogenaseaktivität wurde in Bacteroides [SHERROD
und HYLEMON 1977], Eubacterium lentum [HIRANO und MASUDA 1981], Escherichia
coli [MACDONALD et al. 1973] und Clostridium absonum [MACDONALD et al. 1983b,
MACDONALD und ROACH 1981] gezeigt. 7β-Dehydrogenaseaktivität wurde ebenfalls
in Clostridium absonum [MACDONALD et al. 1983b, MACDONALD und ROACH 1981]
aber auch in Eubacterium aerofaeciens und Peptostreptococcus productus
gefunden [HIRANO und MASUDA 1982]. EDENHARDER und KNAFLIC [1981]
beschreiben die Epimerisierung von CDCA in UDCA über das 7-Oxo-Intermediat
durch aus den Faeces isolierten Lecithinase-Lipase-negativen Clostridien,
verweisen aber darauf, dass eine Rückreaktion nicht erfolgt.
Allgemeine Betrachtungen
12
3α,α,α,α,7αααα�Dihydroxy-5ββββ-Glycocholansäure
Dekonjugation
Bacteroides Bifidobacterium Fusobacterium Eubacterium Clostridium Lactobacillus Streptococcus
3α,α,α,α,7αααα�Dihydroxy-5ββββ-Cholansäure
7α-Dehydroxylierung 7α-Dehydrogenierung 7-Epimerisierung
Clostridium Clostridium absonum Anaerobier in Eubacterium Eubacterium lentum Bacteroides
Bacteroides Bacteroides fragilis Gegenwart Escherichia coli
3α−α−α−α−Hydroxy-5ββββ-Cholansäure 3αααα-Hydroxy-7oxo-5ββββ-Cholansäure
7-Epimerisierung 7β-Dehydrogenierung
Clostridium absonum Clostridium absonum Peptostreptococcus productus
Eubacterium aerofaciens
3α,α,α,α,7ββββ�Dihydroxy-5ββββ-Cholansäure Abbildung 4: Metabolisierungsreaktionen an Gallensäuren durch die Darmflora
CH3
CH3
H
CH3
OH
O
OH
NHCH2COOH
CH3
CH3
H
CH3
OH
O
OH
OH
CH3
CH3
H
CH3
OH
O
OH
CH3
CH3
H
CH3
OH
O
OH
O
CH3
CH3
H
CH3
OH
O
OH
OH

Allgemeine Betrachtungen
13
2.1.2.3 Analytische Voraussetzungen zur Gallensäurenbestimmung
Probenvorbereitung
Den meisten Aufarbeitungsprozeduren aus homogenisierten Faeces geht eine
Extraktion der Lipide voraus. Dabei werden Soxhlet-Extraktionen mit Petrolether
[OWEN et al. 1987], Methanol/Dichlorethan [ALI et al. 1966], Methanol/Chloroform
[Eneroth et al. 1968] oder Ethanol [PANDA und BROITMAN 2001, HORI et al. 1998]
angewendet. Weitere Möglichkeiten wie sequenzielles Extrahieren am Rückfluss
mit 90 % Ethanol, 80 % Ethanol und Chloroform/Wasser [SETCHELL et al. 1983]
oder flüssig/flüssig-Extraktion mit Toluol nach Erhitzen mit Eisessig [EVRARD und
JANSSEN 1968] werden ebenfalls beschrieben. Allerdings ist eine
Probenaufarbeitung auch direkt aus der Probe möglich [CHILD et al. 1987, GRUNDY
et al. 1965].
Die folgenden Schritte der Probenaufarbeitung dienen der Reinigung und
Separation der interessierenden Steroide und variieren je nach analytischer
Fragestellung. So können durch Ionen-Austausch-Chromatographie mit Amberlyst
A-26 [OKISHIO und NAIR 1967] neutrale und saure Steroide oder mit Lipidex-DEAP
freie, glycin- und taurinkonjugierte Gallensäuren separiert werden [SETCHELL et al.
1983]. Eine Vielzahl chromatographischer Verfahren mit Gelen (Sephadex LH-20,
Ammoniumform), Reversed-Phase C18, Aluminiumoxid, Papier, Kieselgel auf
Platten u. a. kommen zum Einsatz [ENEROTH und SJÖVALL 1971].
In der Reihenfolge der Anwendungen ist die alkalische oder enzymatische
Hydrolyse der konjugierten Gallensäuren der entscheidende Schritt. Da in
Faecesproben vorwiegend freie Gallensäuren vorliegen und bei großem
Probenumfang eine mehrtägige Aufarbeitung einer einzelnen Probe ineffektiv ist,
kann eine Separation der neutralen Steroide mittels flüssig/flüssig-Extraktion zur
Analyse freier Gallensäuren zufriedenstellende Ergebnisse liefern. So werden von
CZUBAYKO et al. [1991] nach Entfernen der neutralen Komponenten eine alkalische
Hydrolyse durchgeführt, die Gallensalze durch Ansäuern mit Salzsäure in freie
Säuren überführt, extrahiert und derivatisiert.
Bei Hydrolyse in Gegenwart von Natronlauge ist allerdings ein Verlust an Oxo-
Gallensäuren möglich. Die Ursache hierfür ist vermutlich eine basekatalysierte

Allgemeine Betrachtungen
14
Keto-Enol-Tautomerisierung, die eine Artefaktbildung zur Folge hat. Dies gilt
besonders für Oxogruppen am Kohlenstoffatom 3 des Steroidgerüstes [SJÖVALL et
al. 1971]. Da jedoch in den Faeces die 12-Oxo-Verbindungen die quantitativ
Bedeutendsten sind [LAWSON und SETCHELL 1988], dient die Aufarbeitung nach
CZUBAYKO et al. [1991] als Basis in der vorliegenden Arbeit.
Gaschromatographische Analyse
Aufgrund der hohen Siedepunkte der Gallensäuren ist eine Derivatisierung der
Carboxyl-, Hydroxy- und eventuell der Oxogruppen erforderlich, die in aufeinander
folgenden Schritten realisiert wird. Zur Veresterung der Carboxylgruppe können
Butanol [BATTA et al. 1999], Methanol/Dimethoxypropan [BRYDON et al. 1979,
HAYASHI et al. 1986] oder Diazomethan [KELSEY et al. 1980, TANIDA et al. 1981]
eingesetzt werden (Abb. 5).
Abbildung 5: Alkylesterbildung an der Carboxylgruppe freier Gallensäuren
Da Oxogruppen in Acetylierungsreaktionen Enolester und in Silylierungs-
reaktionen Enolether bilden können [SJÖVALL et al. 1971] wird in einigen Fällen
eine Umwandlung der Oxogruppen in Methyloxime vor die Derivatisierung der
Hydroxygruppen gesetzt [TANDON 1984]. Diese werden abschließend in Acetate
[NAKAMURA et al. 1992, SZCZEPANIK et al. 1976], Trifluoracetate [HYLLA et al. 1998,
REDDY et al. 1988] oder Trimethylsilylether [CRONHOLM et al. 1972, OWEN et al.
1984, SUMMERFIELD et al. 1976] überführt. Als Silylierungsmittel hat sich ein
Gemisch aus Hexamethyldisilazan, Trimethylchlorsilan und Pyridin bewährt [IIDA et
al. 2001] (Abb. 6).
O
OH17
2021 22
23
24
R
O
17
2021 22
23
24
ROC4H9 +
O
17
2021 22
23
24
ROCH3
a: H2SO4, C4H9OHb: HCl, CH3OH, (CH3)2C(OCH3)2c: CH2N2
a b, c

Allgemeine Betrachtungen
15
Trotz der Vielfalt der Derivatisierungsmöglichkeiten ist die Bildung von
Trimethylsilylether-Methylester am weitesten verbreitet.
Abbildung 6: Silylierungsreaktion an Hydroxygruppen des Steroidgerüstes
Zur Trennung der Gallensäuren werden vorwiegend unpolare stationäre Phasen in
gepackten Säulen oder Kapillarsäulen verschiedener Längen verwendet [SCALIA
1995]. Es wird ein programmierter Temperaturverlauf bevorzugt, der sich über
einen weiten Bereich erstrecken kann [ALME et al. 1977, BARKER et al. 1994,
BATTA et al. 1999, SETCHELL et al. 1983, SRIKUMAR et al. 1998] (Tab. 2).
Tabelle 2: Parameter der Gallensäurenanalyse - Literaturübersicht
Referenz stationäre Phase Säulenlänge [m]
Temperatur [°C] (Start - Ende) Derivate
ALME et al. [1977] 1,5 % SE 30 2,5 210 - 240 Meth/TMS
BARKER et al. [1994] DB-1 25 o. A. Meth/TMS
BATTA et al. [1999] CP-Sil-5CB 25 100 - 278 But/TMS
SETCHELL et al. [1983] OV-1 25 220 - 285 Meth/TMS
SRIKUMAR et al. [1998] DB-5 30 50 - 310 But/Ac
Meth: Methylester; But: Butylester; TMS: Trimethylsilylether; Ac: Acetat; o. A.: ohne Angabe
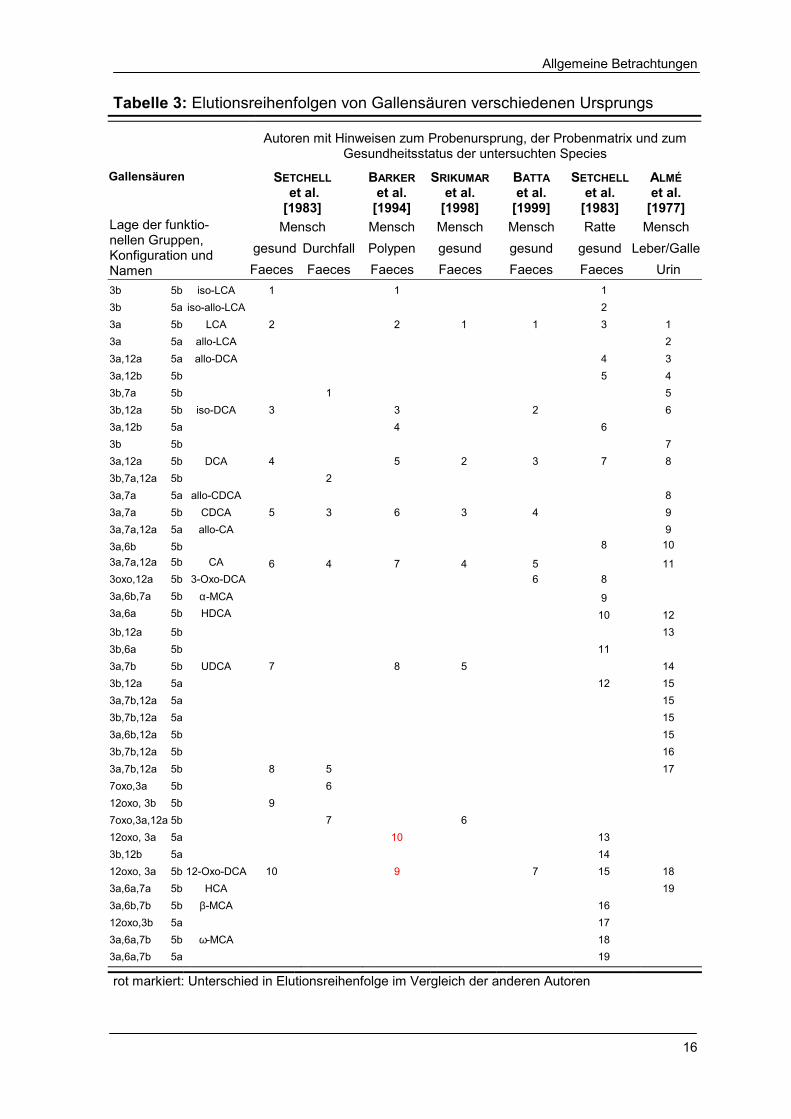
Trotz unterschiedlicher Messparameter zeigen die Chromatogramme der
verschiedenen Autoren die gleichen Elutionsreihenfolgen der jeweils analysierten
Gallensäuren. Allerdings erscheinen bei BARKER et al. [1994] die cis/trans-Isomere
der 12-Oxo-DCA in umgekehrter Reihenfolge (Tab. 3).
+5
Pyridin
HO
A
1
2
3
4
+ HX5
Hexamethyldisilazan (HMDS):Trimethylchlorsilan (TMCS):
X = NHSi(CH3)3X = Cl
CH3
Si
CH3
H3C XA
1
2
3
4
CH3
Si
CH3
H3C O
Allgemeine Betrachtungen
16
Tabelle 3: Elutionsreihenfolgen von Gallensäuren verschiedenen Ursprungs
Autoren mit Hinweisen zum Probenursprung, der Probenmatrix und zum
Gesundheitsstatus der untersuchten Species Gallensäuren SETCHELL
et al. [1983]
BARKER et al.
[1994]
SRIKUMAR et al.
[1998]
BATTA et al.
[1999]
SETCHELL et al.
[1983]
ALMÉ et al.
[1977] Mensch Mensch Mensch Mensch Ratte Mensch
gesund Durchfall Polypen gesund gesund gesund Leber/Galle
Lage der funktio- nellen Gruppen, Konfiguration und Namen Faeces Faeces Faeces Faeces Faeces Faeces Urin 3b 5b iso-LCA 1 1 1 3b 5a iso-allo-LCA 2 3a 5b LCA 2 2 1 1 3 1 3a 5a allo-LCA 2 3a,12a 5a allo-DCA 4 3 3a,12b 5b 5 4 3b,7a 5b 1 5 3b,12a 5b iso-DCA 3 3 2 6 3a,12b 5a 4 6 3b 5b 7 3a,12a 5b DCA 4 5 2 3 7 8 3b,7a,12a 5b 2 3a,7a 5a allo-CDCA 8 3a,7a 5b CDCA 5 3 6 3 4 9 3a,7a,12a 5a allo-CA 9 3a,6b 5b 8 10 3a,7a,12a 5b CA 6 4 7 4 5 11 3oxo,12a 5b 3-Oxo-DCA 6 8 3a,6b,7a 5b α-MCA 9 3a,6a 5b HDCA 10 12 3b,12a 5b 13 3b,6a 5b 11 3a,7b 5b UDCA 7 8 5 14 3b,12a 5a 12 15 3a,7b,12a 5a 15 3b,7b,12a 5a 15 3a,6b,12a 5b 15 3b,7b,12a 5b 16 3a,7b,12a 5b 8 5 17 7oxo,3a 5b 6 12oxo, 3b 5b 9 7oxo,3a,12a 5b 7 6 12oxo, 3a 5a 10 13 3b,12b 5a 14 12oxo, 3a 5b 12-Oxo-DCA 10 9 7 15 18 3a,6a,7a 5b HCA 19 3a,6b,7b 5b β-MCA 16 12oxo,3b 5a 17 3a,6a,7b 5b ω-MCA 18 3a,6a,7b 5a 19
rot markiert: Unterschied in Elutionsreihenfolge im Vergleich der anderen Autoren

Allgemeine Betrachtungen
17
Die Variabilität der renalen konjugierten Gallensäuren scheint im Vergleich zur
faecalen Ausscheidung beim Menschen größer zu sein [BATTA und SALEN 1999].
Das Gallensäurenprofil der Ratte unterscheidet sich vom Menschen durch
Vertreter der Muricholsäure (MCA), die Hydroxygruppen an Position 6 des
Steroidgerüstes tragen.
Massenspektrometrische Detektion und interner Standard
Generelle Fragmentierungen der Trimethylsilylether-Methylester der Gallensäuren
sind die Abspaltung der Seitenkette (M - 115), der Trimethylsilanole (M - 90) oder
angularer Methylgruppen (M - 15) (Tab. 4). Weiterhin treten Ringbrüche in A, B, C
und D auf [LAWSON und SETCHELL 1988]. Je höher die Energie des
Elektronenstoßes ist, um so stärker ist das Ausmaß der Fragmentierung. Bei
niedrigen Energien (20 eV) werden Fragmente mit einem höheren
Informationsgehalt zur Struktur der Gallensäuren erhalten [REIMENDAL und
SJÖVALL 1973]. Der Elektronenfluss in Massenspektrometern mit
Elektronenstoßionisation ist in vielen Geräten auf 70 eV standardisiert
[ANDEREGG 1990].
Tabelle 4: Molare Massen freier und derivatisierter Gallensäuren
Anzahl und Art der funktionellen Gruppen
Molare Masse (frei) [g/mol]
Molare Masse (Meth und TMS) [g/mol]
1 �OH 376 462
2 �OH 392 550
3 �OH 408 638 1 �OH + 1 =O 390 476
Meth: Methylester; TMS: Trimethylsilylether
Quantitative Analysen können sowohl mit dem Totalionenstrom (TIC, SCAN) der
gebildeten Fragmente als auch im SIM realisiert werden. Das Arbeiten mit dem
gesamten Spektrum bietet eine größere Anzahl von Fragmenten zur Analyse,
weshalb DE WEERDT et al. [1980] dieses Verfahren der Analyse im SIM vorziehen.
Da sich bei der Bestimmung aller gebildeten Fragmente die Empfindlichkeit der
Messung verringert und bei großem Probenumfang die erfasste Datenmenge

Allgemeine Betrachtungen
18
bedeutend vergrößert ist, zeigt die Quantifizierung im SIM einige Vorteile. So
werden bei SCALIA et al. [2000] UDCA-Gehalte im Plasma mit einer
Nachweisgrenze von 0,12 µmol/l (0,05 ppm) detektiert.
Zur Minimierung von Fehlern innerhalb der Probenvorbereitung oder der
Messmethode sind verschiedene interne Standards zur Bestimmung faecaler
Gallensäuren in der Literatur beschrieben (Tab. 5).
Tabelle 5: Interne Standards zur Gallensäurenquantifizierung
Referenz Interner Standard
IMRAY et al. [1992] nor-DCA; 3H-CA
JENKINS et al. [1999] 5β-Cholansäure ROY et al. [1999] HDCA
SETCHELL et al. [1983] Coprostanol, 14C-CA, 14C-CDCA
SUBBIAH [1973] HCA WESTSTRATE et al. [1999] 7α,12α-Dihydroxy-5β-Cholansäure

Allgemeine Betrachtungen
19
2.2 Wechselwirkungen der faecalen Steroide - Einfluss der Ernährung 2.2.1 Pathogene Wirkung und Cholesterolmetabolismus
2.2.1.1 Toxizität
Da der Verzehr von viel Fett und wenig Ballaststoffen mit einem erhöhten Risiko
der Coloncarcinogenese assoziiert wird [SLATTERY et al. 2000], sind Unter-
suchungen zu toxischen Wirkungen der Gallensäuren durchgeführt worden. So
wirken sie indirekt, indem sie die Fecapentaen-Biosynthese beeinflussen, die als
Mutagene in den Faeces bekannt sind [VAN TASSELL et al. 1982]. Außerdem wird
bei fettreicher Ernährung die Abgabe des Gallensaftes stimuliert, wodurch
anaerobe Bacteroides-Species im Darm gefördert werden, die Fecapentaene
bilden können [MOORE und MOORE 1995].
In genotoxischen Untersuchungen verändert LCA in vitro embryonale Hamster-
zellen und verursacht zum einen DNS-Strangbrüche in L1210-Zellen und zum
anderen steigert es die Reparaturmechanismen nach DNS-Strangbrüchen im
Rattengewebe [NAIR 1988]. In einer Studie von VENTURI et al. [1997] ist
Faeceswasser von 35 gesunden Probanden auf Genotoxizität untersucht worden.
Dabei konnte bei elf Personen ein stark erhöhter Wert ermittelt werden. Weitere
Untersuchungen haben ergeben, dass die Schädigungen der DNS vermutlich
durch oxidativen Einfluss und nicht durch Deoxycholat oder Lithocholat verursacht
worden ist. CRAVEN et al. [1987] postulieren, dass Gallensäuren einen Anstieg
reaktiver Sauerstoffspecies im Colonepithel induzieren. Generell haben
konjugierte Gallensalze ein geringeres genotoxisches Potenzial als freie Mono-
und Dihydroxygallensäuren und darüber hinaus liegt freie LCA durch die geringe
Löslichkeit im Faeceswasser nur in geringen Konzentrationen vor [NAGENGAST et
al. 1995].
Die Zytotoxizität der Gallensäuren ist stark von ihrer chemischen Struktur
abhängig. Allgemein gilt: je hydrophober eine Gallensäure ist, umso zytotoxischer
wirkt sie. Als Monohydroxygallensäure entwickelt LCA in vitro die höchste
Toxizität. Die Dihydroxygallensäuren DCA und CDCA sind stark hydrophob und
wirken verstärkt zytotoxisch. Aufgrund der räumlichen Lage der Alkoholgruppen ist

Allgemeine Betrachtungen
20
UDCA mit ebenfalls zwei Hydroxygruppen weniger hydrophob. CA ist ein
Intermediat und wirkt nur in hohen Konzentrationen als Zellgift [HOFMANN 1999,
LATTA et al. 1993].
Konjugierte Gallensäuren haben keine toxische Wirkung auf Zellen ohne
Gallensäuretransporter. Steigt die extrazelluläre Konzentration stark an, können
Membranschäden durch unterschiedliche Mechanismen (Prostaglandin E2, Proteinkinasen) auftreten [DE RUPERTIS et al. 1984, NISHIZUKA 1984]. Die
schädigende Konzentration ist der kritischen Mizellenkonzentration ähnlich. Da
CDCA und DCA im Vergleich zu CA geringe kritische Mizellenkonzentrationen
aufgrund ihrer hydrophoben Natur aufweisen, wirken sie verstärkt toxisch
[NAGENGAST et al. 1995].
Eine cholesterolreiche Diät erhöht den CDCA-Pool im Körper und steigende
Gehalte an CDCA und LCA im Colon können folgen [CHAPLIN 1998]. Außerdem
scheint der pH-Wert im Faeceswasser Einfluss auf die extrazelluläre Zytotoxizität
der Gallensäuren zu nehmen [DE KOK et al. 1999]. Bei Störungen der Integrität der
Zellmembranen stimuliert der Zellverlust die Erneuerung durch die mukosale
Proliferation. Dieses scheint der Schlüsselschritt hinsichtlich einer Tumorbildung
im Colon zu sein, da eine hyperproliferierende Mukosa sensibler auf Carcinogene
reagiert [NAGENGAST et al. 1995].
Im Gallentrakt und im Dünndarm bei Gesunden ist die Zytotoxizität vernach-
lässigbar, da bei der Verdauung von Lipiden mit konjugierten Gallensäuren
gemischte Mizellen gebildet werden, deren Konzentrationen unterhalb der
schädigenden Wirkung liegen [HOFMANN 1999]. Neben der extrazellulären
Zytotoxizität erfolgen weiterhin Untersuchungen zu intrazellulären Veränderungen
durch Gallensäuren [LATTA et al. 1993].
Die Rolle der neutralen Sterole in der Coloncarcinogenese ist nicht geklärt. So
deklarieren WEISBURGER et al. [1983] neutrale Sterole nicht als Tumorpromotoren,
obwohl gezeigt wird, dass Vegetarier als Gruppe mit einem niedrigen Risiko zur
Darmkrebsinzidenz einen signifikant verminderten Gehalt an bakteriellen
Cholesterolabbauprodukten und geringere oder unveränderte Konzentrationen an
Cholesterol in den Faeces im Vergleich zu Omnivoren oder Ovo-Lacto-Vegetariern
aufweisen [NAIR et al. 1984]. Im Gegensatz dazu zeigen NAIR und TURJMAN [1983],
dass das Darmkrebsrisiko mit niedrigen Konversionsraten des Cholesterols

Allgemeine Betrachtungen
21
korreliert, d. h. hohe Konzentrationen an Cholesterol und geringe Gehalte an
Coprostanol im Stuhl.
Es kann geschlussfolgert werden, dass eine fettreiche Ernährung die anaerobe
Darmflora beeinflussen kann [MOORE und MOORE 1995], was u. a. eine Erhöhung
von Enzymaktivitäten (Nitroreduktase, Azoreduktase, Glucuronidase, De-
hydroxylase) zur Folge hat [GOLDIN und GORBACH 1976]. Außerdem kommt es zu
einer vermehrten Gallensekretion. Dadurch sind die Konzentrationen an
Gallensäuren im Stuhl erhöht [REDDY et al. 1983] und infolge der Enzymaktivität
findet eine verstärkte Metabolisierung zu zytotoxischen, sekundären Gallensäuren
statt [STADLER et al. 1988]. Dies kann zu einer erhöhten Darmkrebsinzidenz
führen.
2.2.1.2 Cholesterol- und Gallensäurenmetabolismus
Gallensäuren wirken als Regulatoren für den Cholesterolmetabolismus in
Leberzellen. Sie sind Abbauprodukte des Cholesterols und hemmen die
Biosynthese (Abb. 7). Die Fütterung von CA und CDCA an Ratten und Hamster
unterdrückt die hepatische Cholesterolsynthese. Die Gehalte an Cholesterolestern
und LDL-Cholesterolkonzentrationen steigen an [CHIANG et al. 1998, SPADY et al.
1986].
Zur Gallensäurensynthese sind zwei Wege bekannt [VLAHCEVIC et al. 1992]. In
dem vorwiegend stattfindenden Prozess gilt die 7α-Hydroxylase als
Schlüsselenzym. Es wird hauptsächlich CA und weiterhin CDCA gebildet.
Alternativ kann eine Seitenkettenoxidation am Cholesterol als Initialreaktion
auftreten. Es wird 27-Hydroxycholesterol gebildet, welches die Cholesterol-
synthese nach unten reguliert. In diesem Fall ist CDCA das Hauptprodukt [CHAPLIN
1998, DOWLING und MURPHY 1990].
Durch Resorption im Darm gelangen primäre und sekundäre Gallensäuren erneut
in die Leber. Die Konzentration und Zusammensetzung des Gallensäurenpools
beeinflussen den Metabolismus des hepatischen Cholesterols. Ein hoher Gallen-
säurenpool induziert die Exkretion von Cholesterol in die Galle und vermindert die
CA-Synthese.

Allgemeine Betrachtungen
22
GS-Pool
Durch hydrophobe Gallensäuren wie CA, CDCA und DCA wird die
12α-Hydroxylaseaktivität in Ratten vermindert. Hydrophile Gallensäuren wie
UDCA und HCA erhöhen die Spezifität. Die Fütterung von Cholesterol unterdrückt
die 12α-Hydroxylaseaktivität und fördert die 7α-Hydroxylaseaktivität [VLAHCEVIC et
al. 2000].
Die Tarahumara-Indianer in Mexiko weisen lebenslang sehr geringe Cholesterol-
konzentrationen im Plasma auf, und sie haben eine höhere Exkretion faecaler
Gallensäuren. Die intestinale Absorption von Cholesterol ist im Vergleich zu
Amerikanern und Australiern gering, die endogene Biosynthese ist im Gegensatz
dazu erhöht [MCMURRY et al. 1985].
Chol Chol
CA, CDCA
CA,CDCA DCA, iso-~ LCA, iso-~ UDCA, Chol
Metab
Chol, Metab CA, CDCA
sek. GS
Abbildung 7: Cholesterol- und Gallensäurenmetabolismus
Chol: Cholesterol; GS: Gallensäuren; Metab: Cholesterolmetabolite
Chol.ester Acetyl-Co A HMG-Co A-R • -
Chol
CA
CDCA Leber
LDL: Chol, 27OH-Chol HDL: Chol
Serum
GalleChol CA
CDCA sek. GS
27-H
ydro
xyla
se
7-H
ydro
xyla
se
Ileum
Jej
unum
D
uode
num
F
aece
s
Col
on
Förderung Hemmung Gallensäuren � � Cholesterol + -
���
� +

Allgemeine Betrachtungen
23
2.2.2 Einfluss von Nahrungsfaktoren auf die Steroidexkretion
2.2.2.1 Calcium
Untersuchungen zur Verminderung der zytotoxischen Wirkung der Gallensäuren
im Colon beziehen sich auf den Einfluss von Calcium. Oral aufgenommenes
Calcium soll die oberflächenaktiven Gallensäuren inaktivieren. Konjugierte
Gallensäuren weisen eine höhere Löslichkeit bei niedrigem pH-Wert und eine
größere Stabilität gegen Fällungsmittel wie Calcium auf. Daher sind keine
Änderungen im Ileum zu beobachten [GOVERS und VAN DER MEER 1993]. Durch
den Angriff von Hydrolasen der Colonbakterien werden die Gallensäuren
dekonjugiert. Calcium kann entweder als freies Ion mit der ionisierten Gallensäure
ein Salz bilden, oder es verbindet sich mit ebenfalls aus der Nahrung
aufgenommenen anorganischen Phosphaten im Lumen zu einem Komplex, der
eine Fällung der Gallensäuren und eine Mitfällung von Cholesterol bewirkt, wobei
der letztere Mechanismus wirksam sein soll [VAN DER MEER et al. 1991, VAN DER
MEER et al. 1997] (Abb. 8).
Faeceswasser CaPi + freie Gallensäuren + Cholesterol CaPi ··· freie Gallensäuren ↓ + Cholesterol ↓
Abbildung 8: Fällung von Gallensäuren und Cholesterol durch Calciumphosphat-
Komplexe
So zeigt BOVEE-OUDENHOVEN et al. [1999] an Ratten eine Verminderung der
Gallensäurenkonzentration mit einhergehender Verminderung der Zytotoxizität
des Faeceswassers. LUPTON et al. [1996] stellen in einer Studie mit 22 Probanden,
deren Nahrung mit Calcium supplementiert wurde, keine signifikante
Verminderung des Gesamtgallensäurengehaltes, aber eine Verminderung an
primären Gallensäuren im Faeceswasser fest. Eine weitere Humanstudie mit
kontrolliertem Milchverzehr ergibt eine erhöhte faecale Exkretion an Gallensäuren,
der Gehalt im Faeceswasser ist jedoch vermindert [GOVERS et al. 1996].

Allgemeine Betrachtungen
24
Im Gegensatz dazu hat die Supplementation von Calciumcarbonat keinen Einfluss
auf die vermehrte Tumorbildung bei Nitrosomethylharnstoff-exponierten Ratten.
Sie führt zu einer erhöhten Cholesterol- und unveränderten Coprostanolexkretion
[QUILLIOT et al. 1999]. In einer prospektiven Kohortenstudie mit ca. 10.000
Probanden und einer Langzeitstudie über drei Jahre mit 116 Probanden zeigen
sich keine Korrelationen zwischen der Calciumaufnahme und dem Risiko an
Darmkrebs zu erkranken [HOFSTAD et al. 1998, JÄRVINEN et al. 2001].
In Studien zu Veränderungen der Darmflora durch den Einfluss von Calcium
werden in Ratten die Salmonelleninfektionen vermindert und intestinale
Lactobacillen stimuliert [BOVEE-OUDENHOVEN et al. 1999, BOVEE-OUDENHOVEN und
VAN DER MEER 1997]. Milchsäure wirkt auf gram-negative Bakterien in vitro
antimikrobiell und erhöht durch Entfernen von Lipopolysacchariden die
Permeabilität der Bakterienmembranen [ALAKOMI et al. 2000].
2.2.2.2 Probiotika
Den Probiotika werden eine Vielzahl von gesundheitsfördernden Wirkungen
zugeschrieben. Neben Untersuchungen zur Lactoseintoleranz, zur Stimulation des
Immunsystems und zur Senkung der Cholesterolkonzentrationen im Serum sind
Studien zur Prävention von Darmkrebs durchgeführt worden (Abb. 9). Dabei wird
u. a. festgestellt, dass die enzymatische Aktivität von Enzymen des Dickdarmes,
die zur Bildung von Carcinogenen führen, vermindert wird. Dies konnte für
β-Glucuronidase, Nitroreduktase, Glycocholsäurehydrolase und Azoreduktase
gezeigt werden [BEZKOROVAINY 2001]. Neben den erwähnten Enzymen sind die
7α-Dehydroxylase und die 7α-Dehydrogenase von Interesse, da LCA und DCA
als mögliche Promotoren einer Tumorbildung gelten (vgl. 2.2.1.1). So werden die
Dekonjugation und Dehydroxylierung von Gallensalzen als unerwünschte
physiologische Leistungen von Milchsäurebakterien dargestellt [HAMMES 1998].
Der gezielte Einsatz diätetischer Erzeugnisse (Ballaststoffe, fermentierte Milch-
produkte, Milchsäurebakterien) sollte eine Absenkung des pH-Wertes aufgrund
kurzkettiger, organischer Säuren hervorrufen [GARROTE et al. 2000], so dass
dieser nicht mehr im Aktivitätsoptimum krebsrelevanter Enzyme, z. B. der
7α-Steroid-Dehydrogenase liegt [MACDONALD et al. 1978].

Allgemeine Betrachtungen
25
Eine Verminderung der Aktivität müsste einen Abfall des Gehaltes an sekundären
Gallensäuren in den Faeces zur Folge haben. In einer Humanstudie von VAN
MUNSTER et al. [1994] wird 14 gesunden Probanden eine mit resistenter Stärke
angereicherte, standardisierte Diät verabreicht. Die Exkretion an DCA vermindert
sich um 50 % und die Ausscheidung an kurzkettigen Fettsäuren steigt um 35 %
an. In vitro wird eine verminderte Umwandlung von CA und CDCA in ihre
sekundären Verbindungen durch Darmbakterien bei pH = 6 im Vergleich zu einem
neutralen Wert (pH = 7) beschrieben [CHRISTL et al. 1996]. Weiterhin wird eine
verminderte bakterielle Umwandlung von CDCA bei Zusatz von Lactobacillus
acidophilus zum faecalen Medium beobachtet [FERNANDES und SHAHANI 1990].
Daraus schlussfolgernd soll eine Erniedrigung des pH-Wertes im Colon eine
Verminderung an sekundären Gallensäuren zur Folge haben.
Im Kontrast dazu führt in einer randomisierten Doppelblindstudie von MARTEAU et
al. [1995] die Supplementation eines mit Bifidobacterium und Lactobacillus
acidophilus fermentierten Milchproduktes bei acht Ileostomiepatienten zu einer
signifikanten Erhöhung freier und sekundärer Gallensäuren, aufgrund einer hohen
Metabolisierungsaktivität der verabreichten Kulturen. In Untersuchungen zu
beiden Stämmen in vitro wird eine schnelle und umfassende Dekonjugation von
Gallensalzen beobachtet, jedoch keine Dehydroxylierung.
Senkung der Serumcholesterolkonzentration Verminderung des Darmkrebsrisikos Cholesterolassimilation Dekonjugation von GS Bindung von GS β-Glucuronidase ↓
an Zellwand Nitroreduktase ↓ Azoreduktase ↓ (GCA-Hydrolase)
Löslichkeit von GS ↓ (7-Dehydroxylase) (7-Dehydrogenase)
Fällung von Cholesterol + GS
Neusynthese an GS
Verminderung an Cholesterol Verminderung an Gallensäuren Verminderte Enzymaktivitäten
Abbildung 9: Probiotikawirkung im Colon (GCA: Glycocholsäure)
sek. GS

Allgemeine Betrachtungen
26
Lactobacillen können auf verschiedenen Wegen den Gehalt an Taurocholsäure
vermindern. Dabei erfolgt beim Abbau durch L. casei subsp. casei TMC 0409
keine Substanzumwandlung, so dass vermutlich die konjugierte Gallensäure an
die Zellwand gebunden wird und ein physikalischer Prozess die Ursache für eine
Verminderung ist. Beim Abbau durch L. reuteri TMC 4405 hingegen werden
chromatographisch neben der Taurocholsäure auch andere Substanzen detektiert.
Nach massenspektrometrischer Analyse der unbekannten Stoffe wird davon
ausgegangen, dass eine Oxidation und Dekonjugation zur Verminderung der
Taurocholsäure führt [HASHIMOTO et al. 2000].
Keine signifikanten Änderungen in der faecalen Gallensäuren- und Sterolexkretion
werden in einer Humanstudie von BARTRAM et al. [1994] bei Gabe eines mit
Bifidobacterium longum supplementierten Joghurts beobachtet.
Weiterhin ist die Gallensalz-Hydrolase zur Dekonjugation der Gallensalze in
Hinblick auf die cholesterolsenkende Wirkung der Probiotika von besonderer
Bedeutung. Die Gallensalz-Hydrolase ist ein wesentlicher Faktor zur Vermin-
derung von Cholesterol in vitro durch Lactobacillus reuteri [TARANTO et al. 1997].
Durch die Bildung freier Gallensäuren mit geringer Löslichkeit bei einem niedrigen
pH-Wert soll ein Ausfällen der Gallensäuren unter Kopräzipitation des
Cholesterols folgen. Andere Studien beschreiben die Assimilation und eine daraus
folgende Verminderung von Cholesterol durch Lactobacillus acidophilus und
Bifidobacterium longum [GILLILAND et al. 1985, DAMBEKODI und GILLILAND 1998]
oder eine ansteigende Exkretion dekonjugierter Gallensäuren infolge erhöhter
Hydrolaseaktivität im Colon durch Lactobacillus plantarum [DE SMET et al. 1994,
DE SMET et al. 1998] und damit eine verstärkte hepatische Neusynthese von
Gallensäuren aus Cholesterol. Dieser Weg wurde in den letzten Jahren verstärkt
kritisiert, da eine daraus resultierende Erhöhung sekundärer Gallensäuren im
Colon aufgrund der engen Beziehung zur Coloncarcinogenese unerwünscht ist
[SANDERS 2000].

Entwicklung der Analysenmethoden
27
3 ENTWICKLUNG DER ANALYSENMETHODEN 3.1 Faecale Sterole 3.1.1 Probenvorbereitung
Die aus den jeweiligen Perioden der Humanstudien gesammelten individuellen
Stuhlproben werden gepoolt und Aliquote lyophilisiert. Zur Doppelbestimmung
werden zwei Proben der gepulverten Faeces (je 50 mg) in Gefäße mit 250 µg
internem Standard eingewogen. Aufgrund des stabilen chemischen Verhaltens,
der einfachen Handhabung und der in der Literatur beschriebenen, positiven
Erfahrung wird 5α-Cholestan eingesetzt.
50 µl destilliertes Wasser werden zum Quellen der Probe zugegeben. Nach einer
milden alkalischen Hydrolyse mit 1 M ethanolischer Natronlauge (90 %) bei 70 °C
für 1 h und Zugabe von 0,5 ml destilliertem Wasser erfolgt viermal die Extraktion
der freien Sterole mit je 1 ml Cyclohexan. Die vereinigten Extrakte werden unter
einem Stickstoffstrom eingeengt, in 500 µl Decan aufgenommen und ohne weitere
Derivatisierung in den Gaschromatographen GC/MS QP 5000 (Fa. SHIMADZU)
injiziert.
3.1.2 Analyse mit GC/MS QP 5000
3.1.2.1 Probleme der gaschromatographischen Analyse
Die gaschromatographische Analyse eines Standardgemisches mit einer
unpolaren Kapillarsäule (100 %-Dimethylpolysiloxan, Optima 1 ms) und Detektion
des Totalionenstromes zeigt, dass eine Separation von freiem Cholesterol und
Cholestanol möglich ist. Nach Zugabe von Coprostanon erscheint es zusammen
mit Cholesterol unter einem Peak (vgl. 2.1.1.3). Bei der Optimierung des
Temperaturprogramms durch einen verminderten Temperaturanstieg erfolgt keine
Änderung.
Die geringe Erhöhung der Säulenpolarität durch einen Anteil von 5 %
Phenylsubstituenten (95 %-Dimethylpolysiloxan, DB 5) im Trägermaterial bewirkt

Entwicklung der Analysenmethoden
28
zwar eine erhöhte Auflösung der beiden Substanzen, jedoch ist diese für eine
Quantifizierung nicht ausreichend (Abb. 10). Eine weitere Steigerung der Polarität
der stationären Phase durch einen 20 bzw. 50 %igen Gehalt an Phenylgruppen
zeigt keine zufriedenstellende Separation. Kapillarsäulen höherer Polarität mit
Cyanopropylsubstituenten oder auf Polyethylenglycolbasis können aufgrund der
geringeren thermischen Stabilität nicht berücksichtigt werden. Eine Separation der
zu untersuchenden Komponenten ist allein durch die gaschromatographische
Trennung nicht zu realisieren.
Abbildung 10: Chromatogramme eines Sterolstandardgemisches
1: Coprostanol; 2: Cholesterol; 3: Coprostanon; 4: Cholestanol I + II: Optima 1 ms; 50 m; 0,2 mm; 0,2 µm df
III: DB 5; 50 m; 0,2 mm; 0,2 µm df
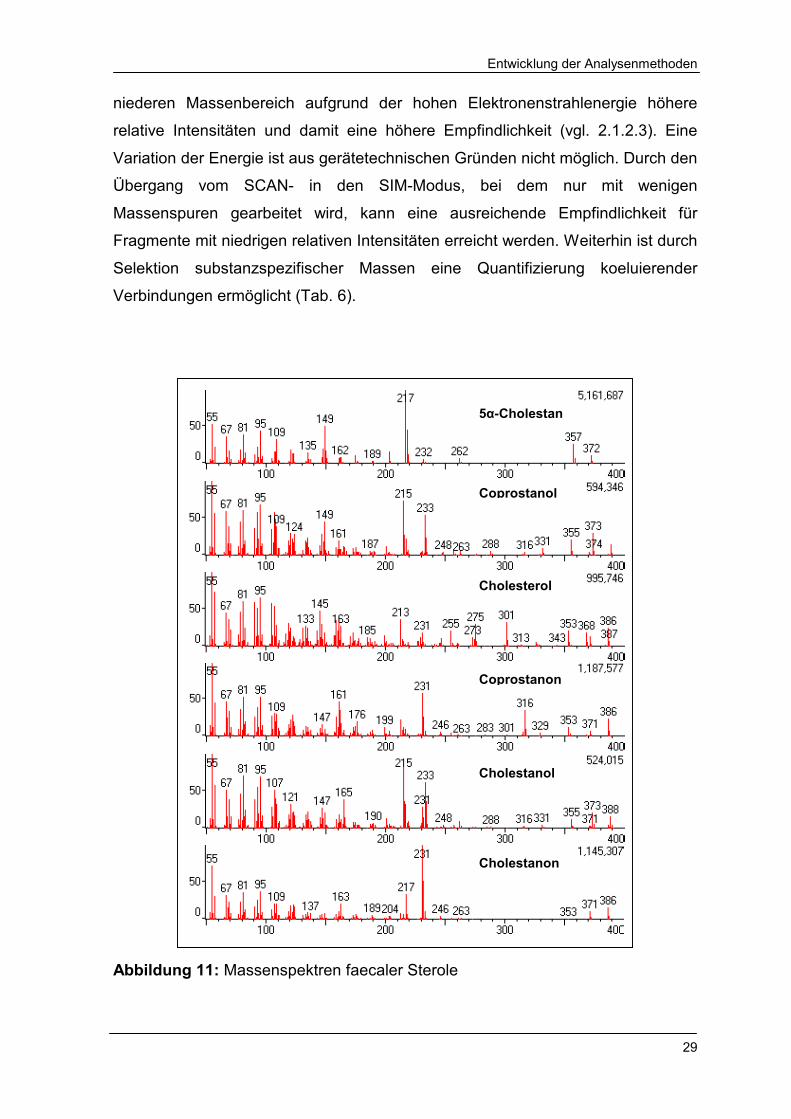
3.1.2.2 Probleme der massenspektrometrischen Detektion
Mit Hilfe von Einzelstandardlösungen sind die Massenspektren von 5α-Cholestan
(M = 372 g/mol), Coprostanol (M = 388 g/mol), Cholesterol (M = 386 g/mol),
Coprostanon (M = 386 g/mol), Cholestanol (M = 388 g/mol) und Cholestanon
(M = 386 g/mol) bei einer Energie des Elektronenstromes von 70 eV
aufgenommen worden (Abb. 11). In den Spektren aller Verbindungen werden die
Molekülionen nachgewiesen, allerdings haben die Fragmente im mittleren bis
1
2
4 2
2 4
4
1
1 3
3
III
II
I
Entwicklung der Analysenmethoden
29
niederen Massenbereich aufgrund der hohen Elektronenstrahlenergie höhere
relative Intensitäten und damit eine höhere Empfindlichkeit (vgl. 2.1.2.3). Eine
Variation der Energie ist aus gerätetechnischen Gründen nicht möglich. Durch den
Übergang vom SCAN- in den SIM-Modus, bei dem nur mit wenigen
Massenspuren gearbeitet wird, kann eine ausreichende Empfindlichkeit für
Fragmente mit niedrigen relativen Intensitäten erreicht werden. Weiterhin ist durch
Selektion substanzspezifischer Massen eine Quantifizierung koeluierender
Verbindungen ermöglicht (Tab. 6).
Abbildung 11: Massenspektren faecaler Sterole
5α-Cholestan
Coprostanol
Cholesterol
Coprostanon
Cholestanol
Cholestanon
Entwicklung der Analysenmethoden
30
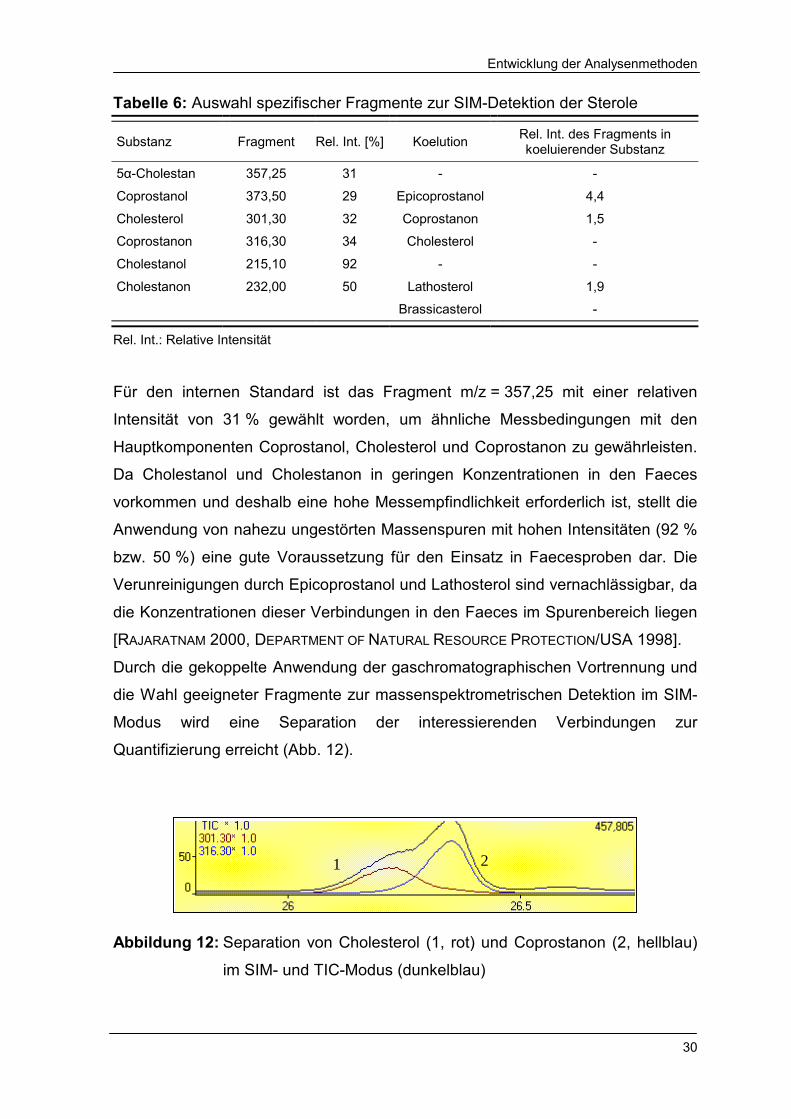
Tabelle 6: Auswahl spezifischer Fragmente zur SIM-Detektion der Sterole
Substanz Fragment Rel. Int. [%] Koelution Rel. Int. des Fragments in koeluierender Substanz
5α-Cholestan 357,25 31 - -
Coprostanol 373,50 29 Epicoprostanol 4,4
Cholesterol 301,30 32 Coprostanon 1,5 Coprostanon 316,30 34 Cholesterol -
Cholestanol 215,10 92 - -
Cholestanon 232,00 50 Lathosterol 1,9 Brassicasterol -
Rel. Int.: Relative Intensität
Für den internen Standard ist das Fragment m/z = 357,25 mit einer relativen
Intensität von 31 % gewählt worden, um ähnliche Messbedingungen mit den
Hauptkomponenten Coprostanol, Cholesterol und Coprostanon zu gewährleisten.
Da Cholestanol und Cholestanon in geringen Konzentrationen in den Faeces
vorkommen und deshalb eine hohe Messempfindlichkeit erforderlich ist, stellt die
Anwendung von nahezu ungestörten Massenspuren mit hohen Intensitäten (92 %
bzw. 50 %) eine gute Voraussetzung für den Einsatz in Faecesproben dar. Die
Verunreinigungen durch Epicoprostanol und Lathosterol sind vernachlässigbar, da
die Konzentrationen dieser Verbindungen in den Faeces im Spurenbereich liegen
[RAJARATNAM 2000, DEPARTMENT OF NATURAL RESOURCE PROTECTION/USA 1998].
Durch die gekoppelte Anwendung der gaschromatographischen Vortrennung und
die Wahl geeigneter Fragmente zur massenspektrometrischen Detektion im SIM-
Modus wird eine Separation der interessierenden Verbindungen zur
Quantifizierung erreicht (Abb. 12).
Abbildung 12: Separation von Cholesterol (1, rot) und Coprostanon (2, hellblau)
im SIM- und TIC-Modus (dunkelblau)
1 2
Entwicklung der Analysenmethoden
31
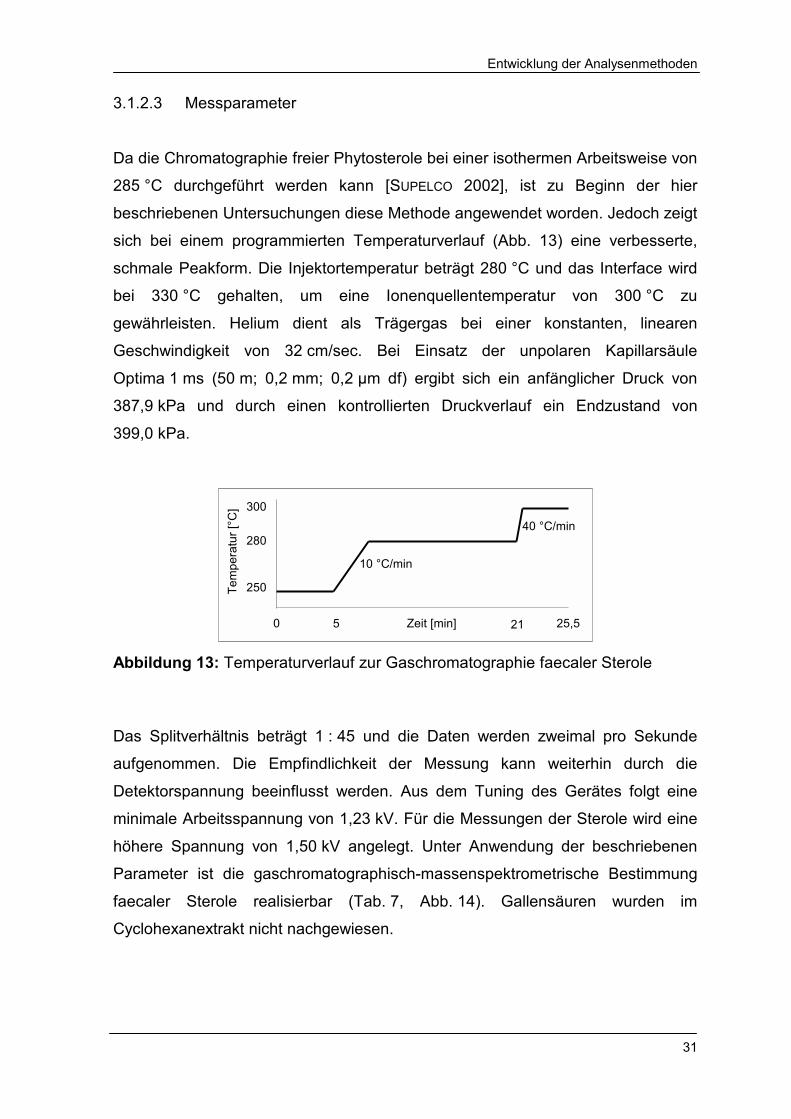
3.1.2.3 Messparameter
Da die Chromatographie freier Phytosterole bei einer isothermen Arbeitsweise von
285 °C durchgeführt werden kann [SUPELCO 2002], ist zu Beginn der hier
beschriebenen Untersuchungen diese Methode angewendet worden. Jedoch zeigt
sich bei einem programmierten Temperaturverlauf (Abb. 13) eine verbesserte,
schmale Peakform. Die Injektortemperatur beträgt 280 °C und das Interface wird
bei 330 °C gehalten, um eine Ionenquellentemperatur von 300 °C zu
gewährleisten. Helium dient als Trägergas bei einer konstanten, linearen
Geschwindigkeit von 32 cm/sec. Bei Einsatz der unpolaren Kapillarsäule
Optima 1 ms (50 m; 0,2 mm; 0,2 µm df) ergibt sich ein anfänglicher Druck von
387,9 kPa und durch einen kontrollierten Druckverlauf ein Endzustand von
399,0 kPa.
Abbildung 13: Temperaturverlauf zur Gaschromatographie faecaler Sterole
Das Splitverhältnis beträgt 1 : 45 und die Daten werden zweimal pro Sekunde
aufgenommen. Die Empfindlichkeit der Messung kann weiterhin durch die
Detektorspannung beeinflusst werden. Aus dem Tuning des Gerätes folgt eine
minimale Arbeitsspannung von 1,23 kV. Für die Messungen der Sterole wird eine
höhere Spannung von 1,50 kV angelegt. Unter Anwendung der beschriebenen
Parameter ist die gaschromatographisch-massenspektrometrische Bestimmung
faecaler Sterole realisierbar (Tab. 7, Abb. 14). Gallensäuren wurden im
Cyclohexanextrakt nicht nachgewiesen.
300
280
250
0 25,5 Zeit [min]
10 °C/min
40 °C/min
Tem
pera
tur [
°C]
5 21

Entwicklung der Analysenmethoden
32
Tabelle 7: Retentionszeiten der faecalen Sterole
Substanz Absolute Retentionszeit [min] Relative Retentionszeit [min]
5α-Cholestan 12,8 1,00
Coprostanol 16,7 1,30
Cholesterol 17,6 1,38
Coprostanon 17,6 1,38 Cholestanol 17,8 1,39
Cholestanon 18,7 1,46
Abbildung 14: Sterolchromatogramm einer Faecesprobe
1: 5α-Cholestan; 2: Coprostanol; 3: Cholesterol (rot, oben);
4: Coprostanon (blau); 5: Cholestanol; 6: Cholestanon
1
6
4 3
2
5
Entwicklung der Analysenmethoden
33
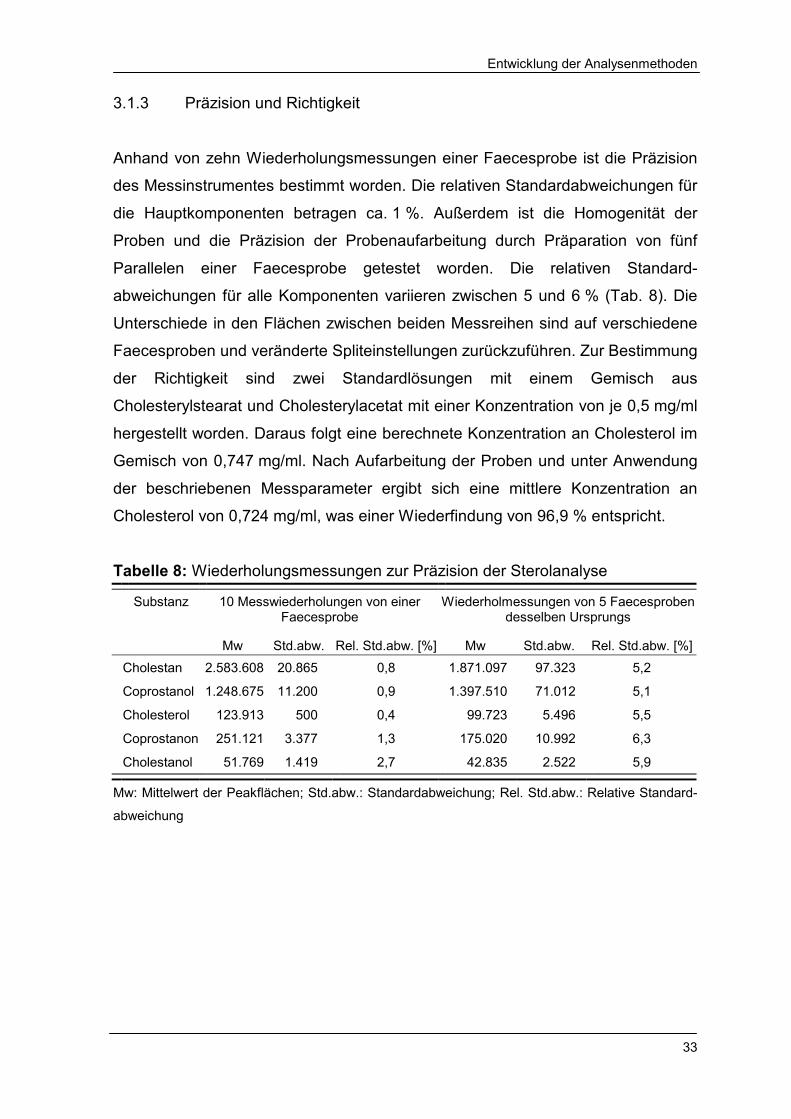
3.1.3 Präzision und Richtigkeit
Anhand von zehn Wiederholungsmessungen einer Faecesprobe ist die Präzision
des Messinstrumentes bestimmt worden. Die relativen Standardabweichungen für
die Hauptkomponenten betragen ca. 1 %. Außerdem ist die Homogenität der
Proben und die Präzision der Probenaufarbeitung durch Präparation von fünf
Parallelen einer Faecesprobe getestet worden. Die relativen Standard-
abweichungen für alle Komponenten variieren zwischen 5 und 6 % (Tab. 8). Die
Unterschiede in den Flächen zwischen beiden Messreihen sind auf verschiedene
Faecesproben und veränderte Spliteinstellungen zurückzuführen. Zur Bestimmung
der Richtigkeit sind zwei Standardlösungen mit einem Gemisch aus
Cholesterylstearat und Cholesterylacetat mit einer Konzentration von je 0,5 mg/ml
hergestellt worden. Daraus folgt eine berechnete Konzentration an Cholesterol im
Gemisch von 0,747 mg/ml. Nach Aufarbeitung der Proben und unter Anwendung
der beschriebenen Messparameter ergibt sich eine mittlere Konzentration an
Cholesterol von 0,724 mg/ml, was einer Wiederfindung von 96,9 % entspricht.
Tabelle 8: Wiederholungsmessungen zur Präzision der Sterolanalyse
Substanz
10 Messwiederholungen von einer Faecesprobe
Wiederholmessungen von 5 Faecesproben desselben Ursprungs
Mw Std.abw. Rel. Std.abw. [%] Mw Std.abw. Rel. Std.abw. [%]
Cholestan 2.583.608 20.865 0,8 1.871.097 97.323 5,2
Coprostanol 1.248.675 11.200 0,9 1.397.510 71.012 5,1
Cholesterol 123.913 500 0,4 99.723 5.496 5,5
Coprostanon 251.121 3.377 1,3 175.020 10.992 6,3
Cholestanol 51.769 1.419 2,7 42.835 2.522 5,9
Mw: Mittelwert der Peakflächen; Std.abw.: Standardabweichung; Rel. Std.abw.: Relative Standard-
abweichung
Entwicklung der Analysenmethoden
34
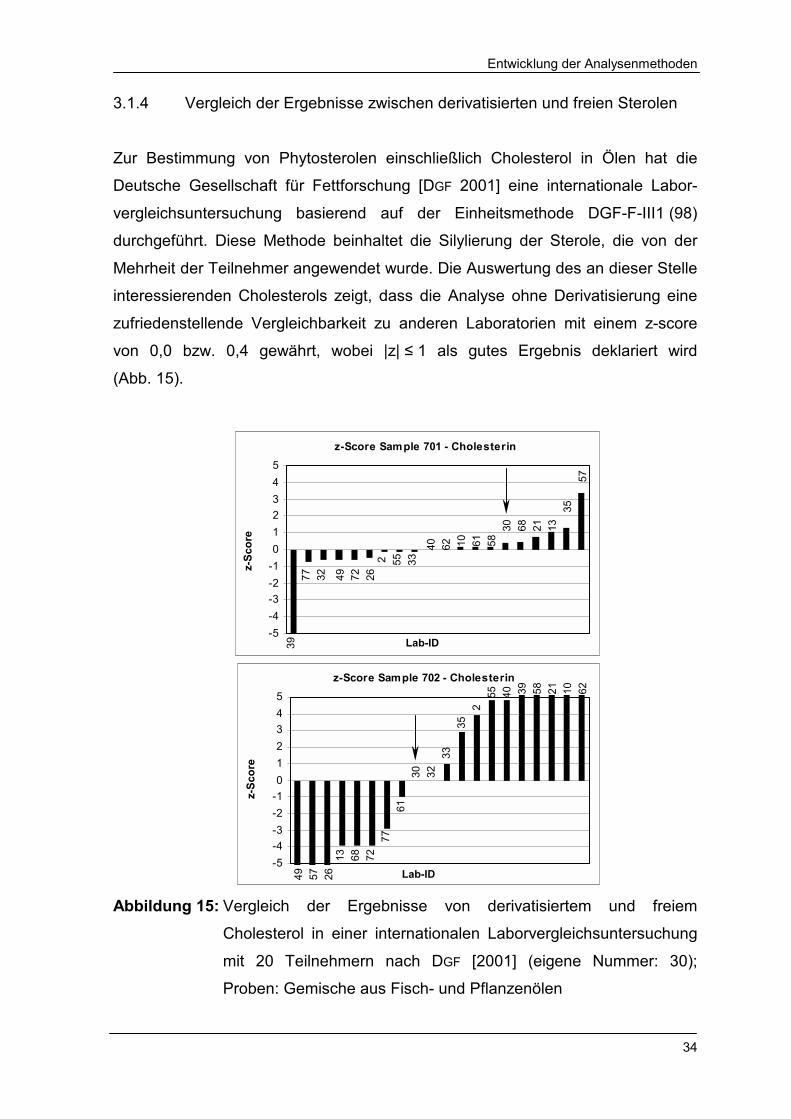
3.1.4 Vergleich der Ergebnisse zwischen derivatisierten und freien Sterolen
Zur Bestimmung von Phytosterolen einschließlich Cholesterol in Ölen hat die
Deutsche Gesellschaft für Fettforschung [DGF 2001] eine internationale Labor-
vergleichsuntersuchung basierend auf der Einheitsmethode DGF-F-III1 (98)
durchgeführt. Diese Methode beinhaltet die Silylierung der Sterole, die von der
Mehrheit der Teilnehmer angewendet wurde. Die Auswertung des an dieser Stelle
interessierenden Cholesterols zeigt, dass die Analyse ohne Derivatisierung eine
zufriedenstellende Vergleichbarkeit zu anderen Laboratorien mit einem z-score
von 0,0 bzw. 0,4 gewährt, wobei |z| ≤ 1 als gutes Ergebnis deklariert wird
(Abb. 15).
Abbildung 15: Vergleich der Ergebnisse von derivatisiertem und freiem
Cholesterol in einer internationalen Laborvergleichsuntersuchung
mit 20 Teilnehmern nach DGF [2001] (eigene Nummer: 30);
Proben: Gemische aus Fisch- und Pflanzenölen
z-Score Sample 701 - Cholesterin
-5-4-3-2-1012345
z-Sc
ore
57
26
68
72
77
61
32 33
62
35
55
39
58 21
10
49
13
30
2
40
z-Score Sample 702 - Cholesterin
-5-4-3-2-1012345
z-Sc
ore
77
49
57
26 13
68
72
33
61
30
32
35 2
40
39
21
10
58
62
55
Lab-ID
Lab-ID
Entwicklung der Analysenmethoden
35
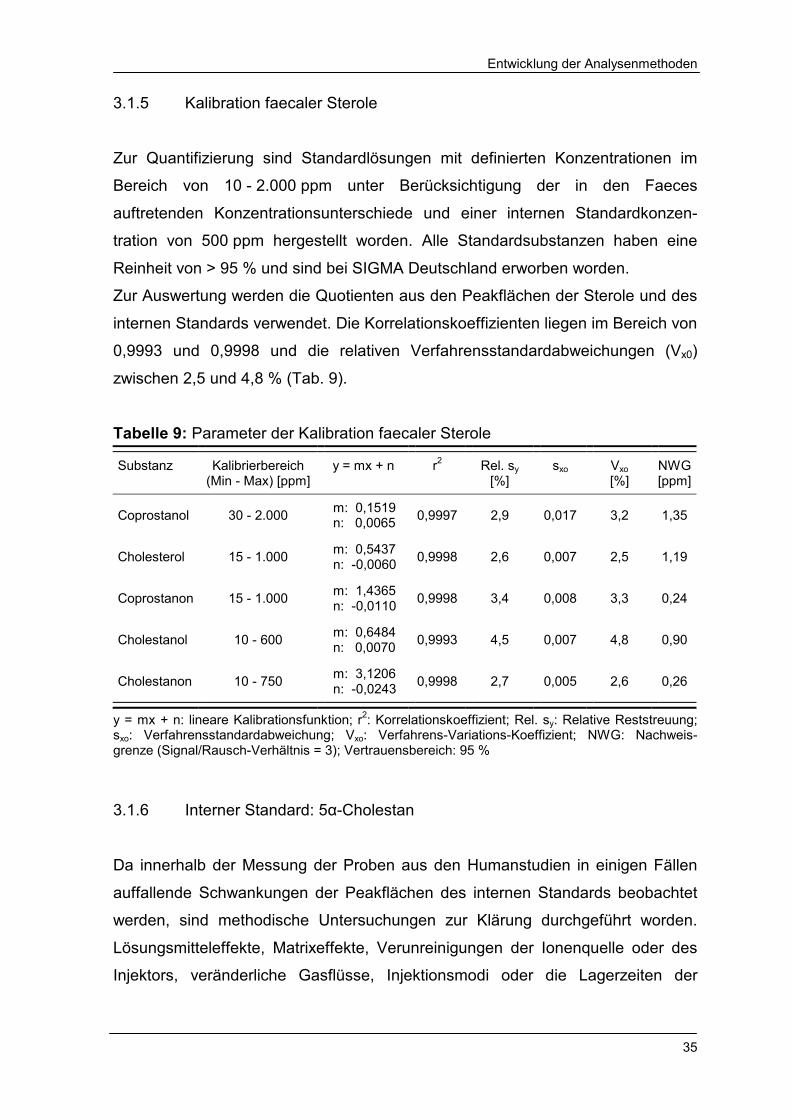
3.1.5 Kalibration faecaler Sterole
Zur Quantifizierung sind Standardlösungen mit definierten Konzentrationen im
Bereich von 10 - 2.000 ppm unter Berücksichtigung der in den Faeces
auftretenden Konzentrationsunterschiede und einer internen Standardkonzen-
tration von 500 ppm hergestellt worden. Alle Standardsubstanzen haben eine
Reinheit von > 95 % und sind bei SIGMA Deutschland erworben worden.
Zur Auswertung werden die Quotienten aus den Peakflächen der Sterole und des
internen Standards verwendet. Die Korrelationskoeffizienten liegen im Bereich von
0,9993 und 0,9998 und die relativen Verfahrensstandardabweichungen (Vx0)
zwischen 2,5 und 4,8 % (Tab. 9).
Tabelle 9: Parameter der Kalibration faecaler Sterole
Substanz
Kalibrierbereich (Min - Max) [ppm]
y = mx + n
r2
Rel. sy
[%] sxo
Vxo [%]
NWG [ppm]
Coprostanol 30 - 2.000 m: 0,1519 n: 0,0065 0,9997 2,9 0,017 3,2 1,35
Cholesterol 15 - 1.000 m: 0,5437 n: -0,0060 0,9998 2,6 0,007 2,5 1,19
Coprostanon 15 - 1.000 m: 1,4365 n: -0,0110 0,9998 3,4 0,008 3,3 0,24
Cholestanol 10 - 600 m: 0,6484 n: 0,0070 0,9993 4,5 0,007 4,8 0,90
Cholestanon 10 - 750 m: 3,1206 n: -0,0243 0,9998 2,7 0,005 2,6 0,26
y = mx + n: lineare Kalibrationsfunktion; r2: Korrelationskoeffizient; Rel. sy: Relative Reststreuung; sxo: Verfahrensstandardabweichung; Vxo: Verfahrens-Variations-Koeffizient; NWG: Nachweis-grenze (Signal/Rausch-Verhältnis = 3); Vertrauensbereich: 95 % 3.1.6 Interner Standard: 5α-Cholestan
Da innerhalb der Messung der Proben aus den Humanstudien in einigen Fällen
auffallende Schwankungen der Peakflächen des internen Standards beobachtet
werden, sind methodische Untersuchungen zur Klärung durchgeführt worden.
Lösungsmitteleffekte, Matrixeffekte, Verunreinigungen der Ionenquelle oder des
Injektors, veränderliche Gasflüsse, Injektionsmodi oder die Lagerzeiten der
Entwicklung der Analysenmethoden
36
Proben können als Ursachen ausgeschlossen werden. Eine weitere mögliche
Ursache kann ein unterschiedlicher Fragmentierungsgrad aufgrund Verschleiß-
erscheinungen (Oberflächendeformierung) der Filamente des Massen-
spektrometers sein.
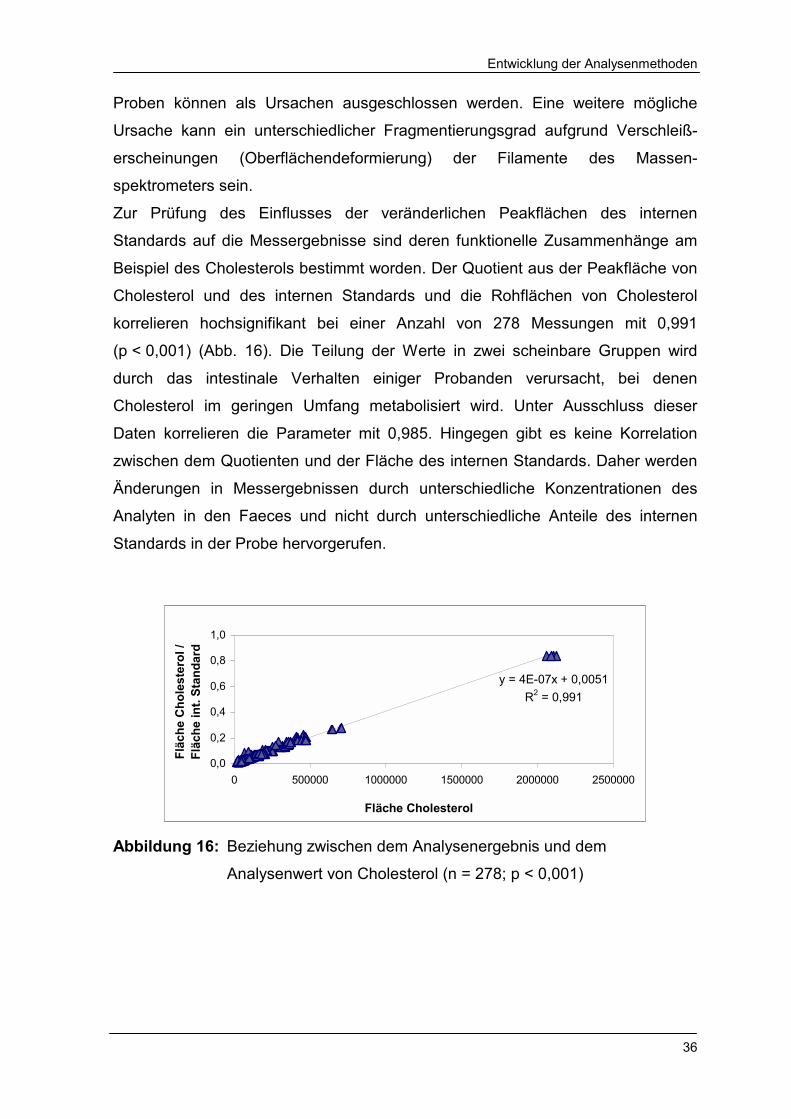
Zur Prüfung des Einflusses der veränderlichen Peakflächen des internen
Standards auf die Messergebnisse sind deren funktionelle Zusammenhänge am
Beispiel des Cholesterols bestimmt worden. Der Quotient aus der Peakfläche von
Cholesterol und des internen Standards und die Rohflächen von Cholesterol
korrelieren hochsignifikant bei einer Anzahl von 278 Messungen mit 0,991
(p < 0,001) (Abb. 16). Die Teilung der Werte in zwei scheinbare Gruppen wird
durch das intestinale Verhalten einiger Probanden verursacht, bei denen
Cholesterol im geringen Umfang metabolisiert wird. Unter Ausschluss dieser
Daten korrelieren die Parameter mit 0,985. Hingegen gibt es keine Korrelation
zwischen dem Quotienten und der Fläche des internen Standards. Daher werden
Änderungen in Messergebnissen durch unterschiedliche Konzentrationen des
Analyten in den Faeces und nicht durch unterschiedliche Anteile des internen
Standards in der Probe hervorgerufen.
Abbildung 16: Beziehung zwischen dem Analysenergebnis und dem
Analysenwert von Cholesterol (n = 278; p < 0,001)
y = 4E-07x + 0,0051R2 = 0,991
0,0
0,2
0,4
0,6
0,8
1,0
0 500000 1000000 1500000 2000000 2500000
Fläche Cholesterol
Fläc
he C
hole
ster
ol /
Fläc
he in
t. St
anda
rd

Entwicklung der Analysenmethoden
37
3.2 Faecale Gallensäuren 3.2.1 Probenvorbereitung
Aufgrund der Ähnlichkeit zu den Analyten, einfacher Zugänglichkeit und der
zentralen Lage im Chromatogramm wird als interner Standard zur Gallensäuren-
bestimmung Hyodeoxycholsäure (HDCA) gewählt.
Die wässrige Phase der Sterolextraktion wird zur Isolierung der Gallensäuren mit
200 µl 10 M Natronlauge versetzt und 2 h bei 120 °C gehalten. Die entstandenen
Natriumsalze werden durch Zugabe von Salzsäure (25 %) bis pH = 1 in die freien
Säuren überführt. Nach vierfacher Extraktion mit je 1 ml Diethylether, Vereinigung
der Etherextrakte und der Zugabe von 125 µg HDCA wird das Lösungsmittel unter
einem Stickstoffstrom bis zur Trockne eingedampft. Die Methylierung erfolgt durch
Zugabe von 650 µl Dimethoxypropan, 950 µl Methanol und 50 µl methanolischer
Salzsäure (3 N) bei einer Temperatur von 50 °C für 45 min. Die Reaktionslösung
wird bis zur Trockne eingedampft, in 150 µl Sylon (Hexamethyldi-
silazan : Trimethylchlorsilan : Pyridin = 3 : 1 : 9, Fa. SUPELCO, Deutschland)
aufgenommen und eine Stunde bei 90 °C silyliert. Nach Abdampfen unter einem
Stickstoffstrom wird der Rückstand in 250 µl Decan gelöst, 10 min geschüttelt und
20 min bei 3.000 Umdrehungen/min zentrifugiert. Der klare Überstand wird
abgenommen und 1 µl wird in den Gaschromatographen injiziert. 3.2.2 Analyse mit GC/MS QP 5000
3.2.2.1 Probleme der gaschromatographischen Analyse
Zur gaschromatographischen Analyse werden von iso-LCA, LCA, iso-DCA, DCA,
CDCA, CA, HDCA, HCA, UDCA, 12-Oxo-DCA, 3-Oxo-LCA, 3,7-Dioxo-CDCA und
3,7,12-Trioxo-CA einzelne Chromatogramme im SCAN-Modus aufgenommen. Im
Gegensatz zu 12-Oxo-DCA bilden Gallensäuren mit einer 3-Oxogruppe infolge der
Probenvorbereitung Artefakte, die im Chromatogramm sichtbar werden (vgl.
2.1.2.3, Abb. 17).

Entwicklung der Analysenmethoden
38
Abbildung 17: Übereinanderliegende Darstellung der Chromatogramme von
3-Oxo-LCA (blau) und 12-Oxo-DCA (grün)
Unter Ausschluss der instabilen Verbindungen kann eine zufriedenstellende
Separation der Gallensäuren in einem Standardgemisch mit einer unpolaren
Kapillarsäule (ZB 5; 30 m; 0,25 mm; 0,25 µm df) erreicht werden (Abb. 18).
Abbildung 18: Chromatogramm (SCAN) eines Gallensäurenstandardgemisches 3.2.2.2 Probleme der massenspektrometrischen Detektion
Da die humane Stuhlprobe eine komplexe Matrix mit vielen möglichen
Verunreinigungen darstellt, wird trotz zufriedenstellender Separation der
interessierenden Gallensäuren eine Quantifizierung im SIM-Modus bevorzugt, um
Fehler durch Peakunreinheiten weitestgehend auszuschließen. Daher sind die
Massenspektren bei 70 eV von iso-LCA, LCA, iso-DCA, DCA, CDCA, CD, HDCA,
UDCA und 12-Oxo-DCA zur Auswahl geeigneter Fragmente registriert worden
(Tab. 10). Molekülionen können nur für 12-Oxo-DCA nachgewiesen werden, alle
anderen Gallensäuren fragmentieren in kleinere Bruchstücke (Abb. 19).
UD
CA
HC
A
12-O
xo-
DC
A HD
CA
CD
CA
CA
DC
A is
o-D
CA
LCA
iso-
LCA
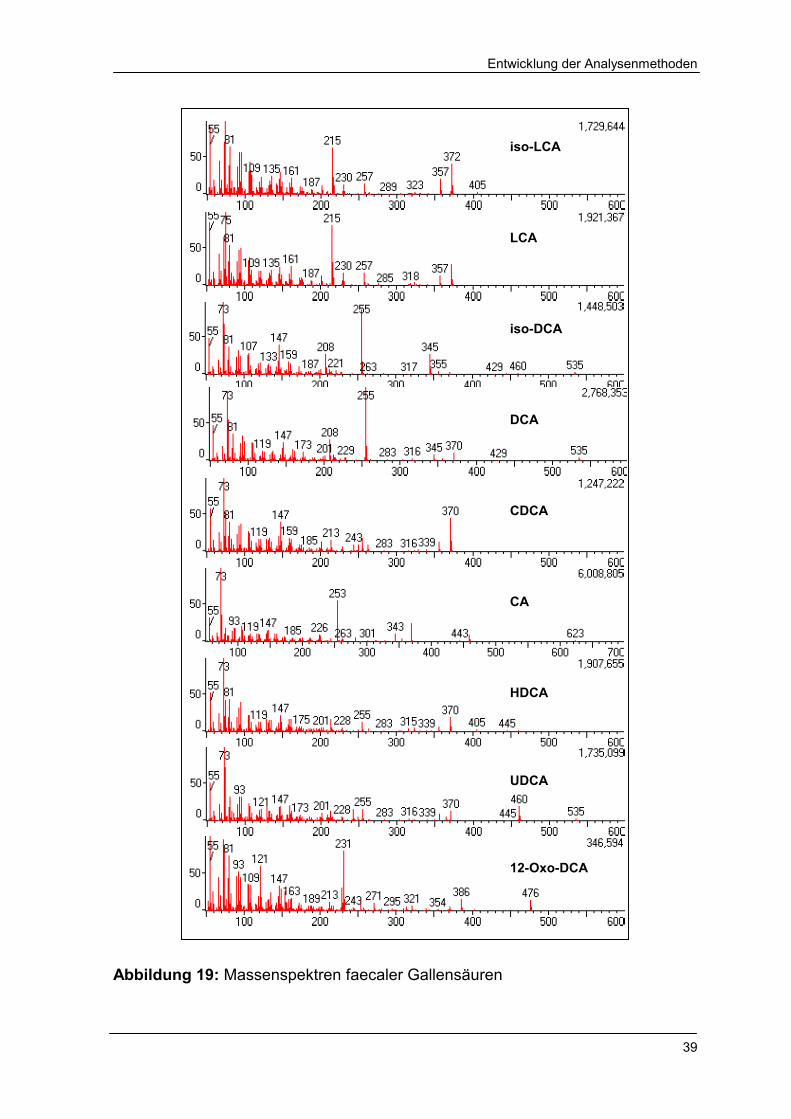
Entwicklung der Analysenmethoden
39
Abbildung 19: Massenspektren faecaler Gallensäuren
12-Oxo-DCA
iso-LCA
LCA
iso-DCA
DCA
CDCA
CA
HDCA
UDCA

Entwicklung der Analysenmethoden
40
Tabelle 10: Auswahl spezifischer Fragmente zur SIM-Detektion der Gallensäuren
Substanz Fragment Relative Intensität [%]
iso-LCA 215,25 31
LCA 215,25 58
iso-DCA 75,10 91 DCA 255,30 93
CDCA 73,10 100
CA 253,20 44 HDCA 81,15 45
UDCA 460,00 14
12-Oxo-DCA 231,25 52
Um eine größtmögliche Peakreinheit in Faecesproben zu sichern, sind weiterhin
Multi Ion Chromatogramme (MIC) von Standard- und Faecesproben erstellt
worden. Dabei hat sich eine Koelution einer weiteren Steroidverbindung mit
Cholsäure in Faecesproben gezeigt, die im SCAN-Modus nicht zu erkennen ist.
Während im Standard die relativen Intensitäten der Fragmente m/z = 253,20 und
m/z = 255,30 ein Verhältnis von 25 : 1 aufweisen, zeigt sich in der Faecesprobe
ein Verhältnis von ca. 1 : 1 und eine Verschiebung der Peakmaxima (Abb. 20).
Abbildung 20: Chromatogramm (MIC) von Cholsäurestandard (I) und Cholsäure
in den Faeces (II)
I
II
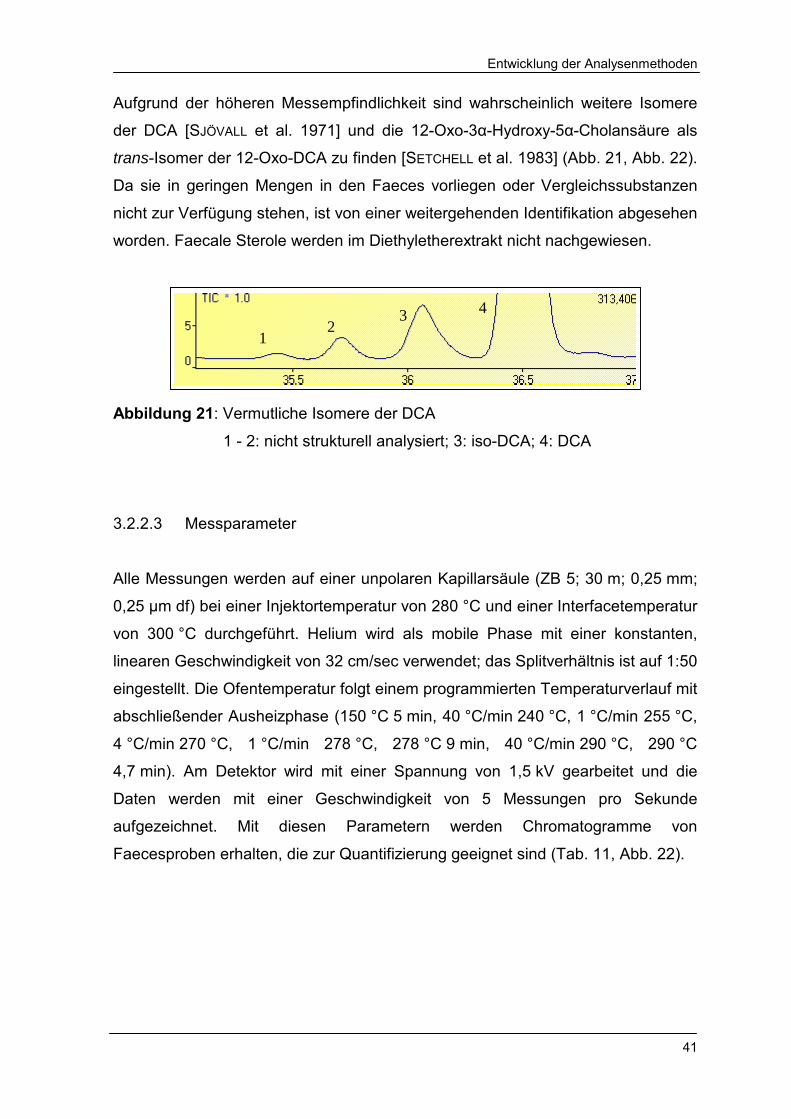
Entwicklung der Analysenmethoden
41
Aufgrund der höheren Messempfindlichkeit sind wahrscheinlich weitere Isomere
der DCA [SJÖVALL et al. 1971] und die 12-Oxo-3α-Hydroxy-5α-Cholansäure als
trans-Isomer der 12-Oxo-DCA zu finden [SETCHELL et al. 1983] (Abb. 21, Abb. 22).
Da sie in geringen Mengen in den Faeces vorliegen oder Vergleichssubstanzen
nicht zur Verfügung stehen, ist von einer weitergehenden Identifikation abgesehen
worden. Faecale Sterole werden im Diethyletherextrakt nicht nachgewiesen.
Abbildung 21: Vermutliche Isomere der DCA
1 - 2: nicht strukturell analysiert; 3: iso-DCA; 4: DCA
3.2.2.3 Messparameter
Alle Messungen werden auf einer unpolaren Kapillarsäule (ZB 5; 30 m; 0,25 mm;
0,25 µm df) bei einer Injektortemperatur von 280 °C und einer Interfacetemperatur
von 300 °C durchgeführt. Helium wird als mobile Phase mit einer konstanten,
linearen Geschwindigkeit von 32 cm/sec verwendet; das Splitverhältnis ist auf 1:50
eingestellt. Die Ofentemperatur folgt einem programmierten Temperaturverlauf mit
abschließender Ausheizphase (150 °C 5 min, 40 °C/min 240 °C, 1 °C/min 255 °C,
4 °C/min 270 °C, 1 °C/min 278 °C, 278 °C 9 min, 40 °C/min 290 °C, 290 °C
4,7 min). Am Detektor wird mit einer Spannung von 1,5 kV gearbeitet und die
Daten werden mit einer Geschwindigkeit von 5 Messungen pro Sekunde
aufgezeichnet. Mit diesen Parametern werden Chromatogramme von
Faecesproben erhalten, die zur Quantifizierung geeignet sind (Tab. 11, Abb. 22).
1 2
3 4
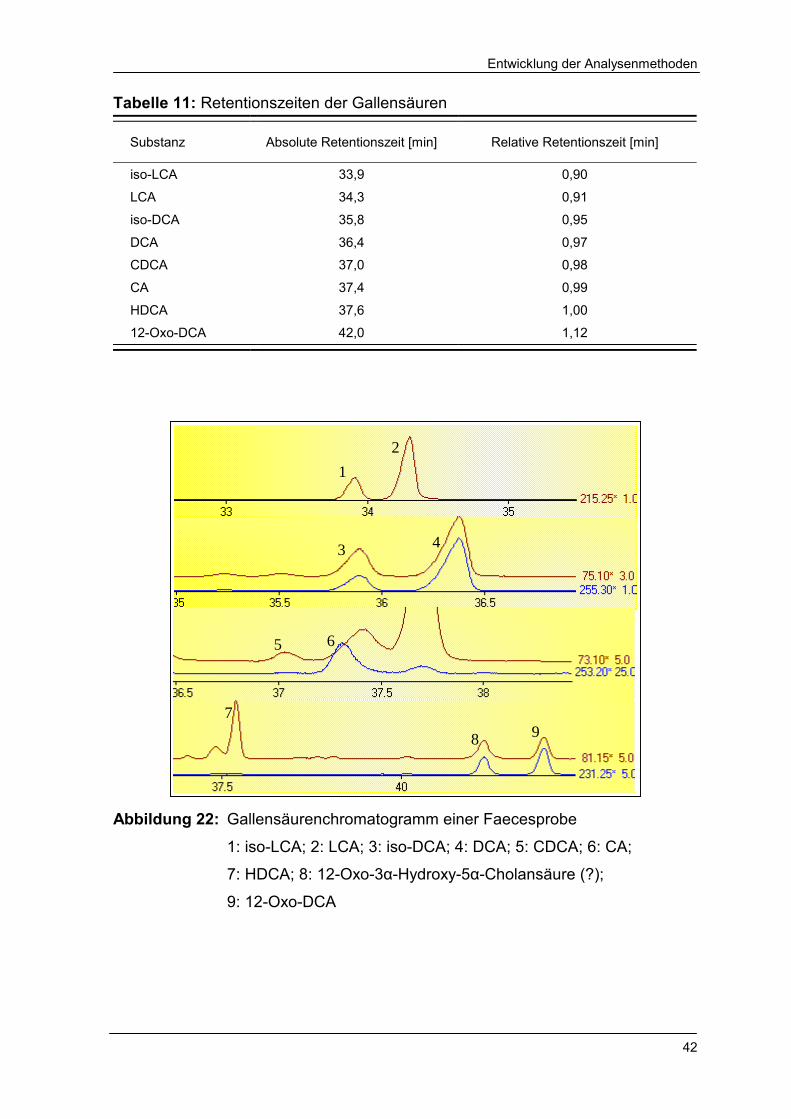
Entwicklung der Analysenmethoden
42
Tabelle 11: Retentionszeiten der Gallensäuren
Substanz Absolute Retentionszeit [min] Relative Retentionszeit [min]
iso-LCA 33,9 0,90
LCA 34,3 0,91
iso-DCA 35,8 0,95
DCA 36,4 0,97 CDCA 37,0 0,98
CA 37,4 0,99
HDCA 37,6 1,00 12-Oxo-DCA 42,0 1,12
Abbildung 22: Gallensäurenchromatogramm einer Faecesprobe 1: iso-LCA; 2: LCA; 3: iso-DCA; 4: DCA; 5: CDCA; 6: CA;
7: HDCA; 8: 12-Oxo-3α-Hydroxy-5α-Cholansäure (?);
9: 12-Oxo-DCA
1 2
3 4
5 6
7
8 9
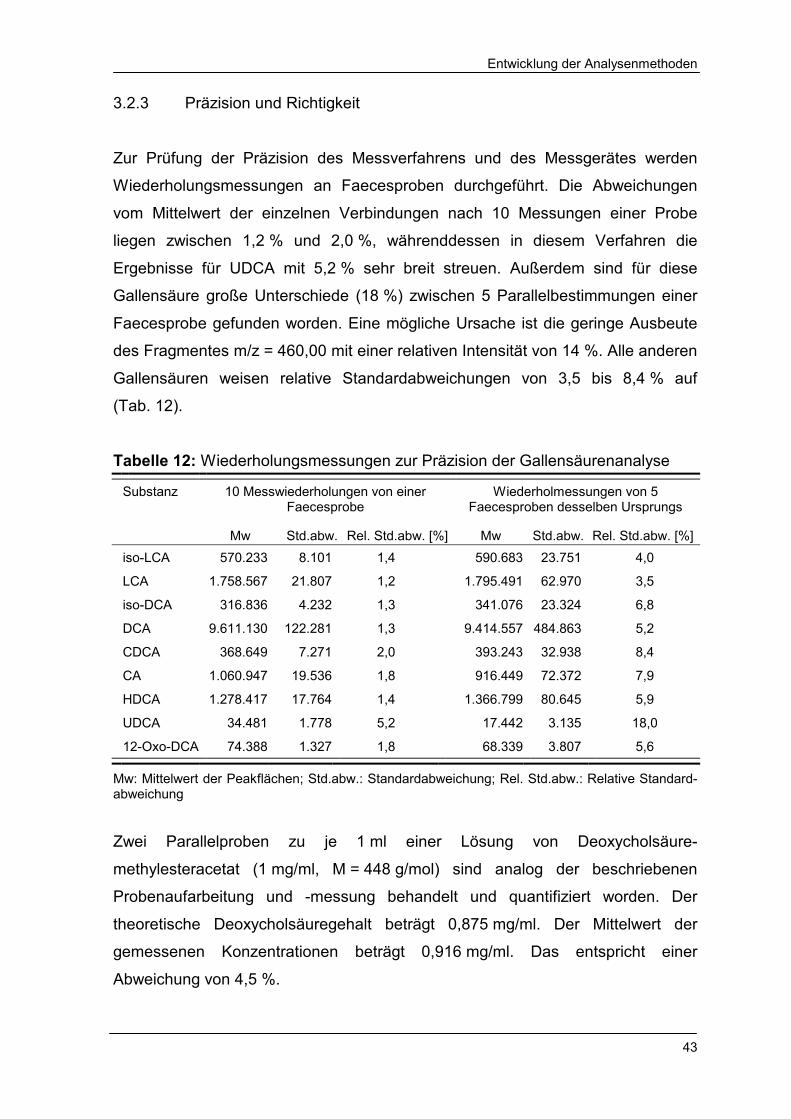
Entwicklung der Analysenmethoden
43
3.2.3 Präzision und Richtigkeit
Zur Prüfung der Präzision des Messverfahrens und des Messgerätes werden
Wiederholungsmessungen an Faecesproben durchgeführt. Die Abweichungen
vom Mittelwert der einzelnen Verbindungen nach 10 Messungen einer Probe
liegen zwischen 1,2 % und 2,0 %, währenddessen in diesem Verfahren die
Ergebnisse für UDCA mit 5,2 % sehr breit streuen. Außerdem sind für diese
Gallensäure große Unterschiede (18 %) zwischen 5 Parallelbestimmungen einer
Faecesprobe gefunden worden. Eine mögliche Ursache ist die geringe Ausbeute
des Fragmentes m/z = 460,00 mit einer relativen Intensität von 14 %. Alle anderen
Gallensäuren weisen relative Standardabweichungen von 3,5 bis 8,4 % auf
(Tab. 12).
Tabelle 12: Wiederholungsmessungen zur Präzision der Gallensäurenanalyse
Substanz
10 Messwiederholungen von einer Faecesprobe
Wiederholmessungen von 5 Faecesproben desselben Ursprungs
Mw Std.abw. Rel. Std.abw. [%] Mw Std.abw. Rel. Std.abw. [%]
iso-LCA 570.233 8.101 1,4 590.683 23.751 4,0
LCA 1.758.567 21.807 1,2 1.795.491 62.970 3,5
iso-DCA 316.836 4.232 1,3 341.076 23.324 6,8
DCA 9.611.130 122.281 1,3 9.414.557 484.863 5,2
CDCA 368.649 7.271 2,0 393.243 32.938 8,4
CA 1.060.947 19.536 1,8 916.449 72.372 7,9
HDCA 1.278.417 17.764 1,4 1.366.799 80.645 5,9
UDCA 34.481 1.778 5,2 17.442 3.135 18,0
12-Oxo-DCA 74.388 1.327 1,8 68.339 3.807 5,6
Mw: Mittelwert der Peakflächen; Std.abw.: Standardabweichung; Rel. Std.abw.: Relative Standard-abweichung Zwei Parallelproben zu je 1 ml einer Lösung von Deoxycholsäure-
methylesteracetat (1 mg/ml, M = 448 g/mol) sind analog der beschriebenen
Probenaufarbeitung und -messung behandelt und quantifiziert worden. Der
theoretische Deoxycholsäuregehalt beträgt 0,875 mg/ml. Der Mittelwert der
gemessenen Konzentrationen beträgt 0,916 mg/ml. Das entspricht einer
Abweichung von 4,5 %.
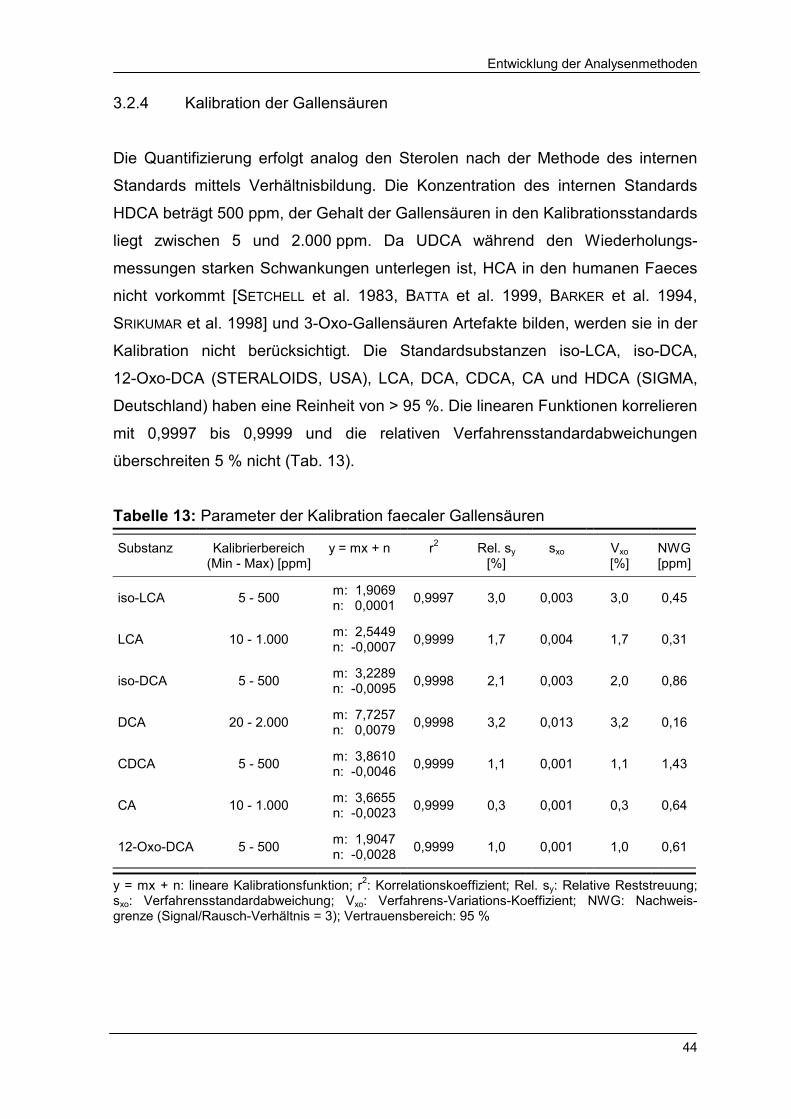
Entwicklung der Analysenmethoden
44
3.2.4 Kalibration der Gallensäuren
Die Quantifizierung erfolgt analog den Sterolen nach der Methode des internen
Standards mittels Verhältnisbildung. Die Konzentration des internen Standards
HDCA beträgt 500 ppm, der Gehalt der Gallensäuren in den Kalibrationsstandards
liegt zwischen 5 und 2.000 ppm. Da UDCA während den Wiederholungs-
messungen starken Schwankungen unterlegen ist, HCA in den humanen Faeces
nicht vorkommt [SETCHELL et al. 1983, BATTA et al. 1999, BARKER et al. 1994,
SRIKUMAR et al. 1998] und 3-Oxo-Gallensäuren Artefakte bilden, werden sie in der
Kalibration nicht berücksichtigt. Die Standardsubstanzen iso-LCA, iso-DCA,
12-Oxo-DCA (STERALOIDS, USA), LCA, DCA, CDCA, CA und HDCA (SIGMA,
Deutschland) haben eine Reinheit von > 95 %. Die linearen Funktionen korrelieren
mit 0,9997 bis 0,9999 und die relativen Verfahrensstandardabweichungen
überschreiten 5 % nicht (Tab. 13).
Tabelle 13: Parameter der Kalibration faecaler Gallensäuren
Substanz
Kalibrierbereich (Min - Max) [ppm]
y = mx + n
r2
Rel. sy
[%] sxo
Vxo [%]
NWG [ppm]
iso-LCA 5 - 500 m: 1,9069 n: 0,0001 0,9997 3,0 0,003 3,0 0,45
LCA 10 - 1.000 m: 2,5449 n: -0,0007 0,9999 1,7 0,004 1,7 0,31
iso-DCA 5 - 500 m: 3,2289 n: -0,0095 0,9998 2,1 0,003 2,0 0,86
DCA 20 - 2.000 m: 7,7257 n: 0,0079 0,9998 3,2 0,013 3,2 0,16
CDCA 5 - 500 m: 3,8610 n: -0,0046 0,9999 1,1 0,001 1,1 1,43
CA 10 - 1.000 m: 3,6655 n: -0,0023 0,9999 0,3 0,001 0,3 0,64
12-Oxo-DCA 5 - 500 m: 1,9047 n: -0,0028 0,9999 1,0 0,001 1,0 0,61
y = mx + n: lineare Kalibrationsfunktion; r2: Korrelationskoeffizient; Rel. sy: Relative Reststreuung; sxo: Verfahrensstandardabweichung; Vxo: Verfahrens-Variations-Koeffizient; NWG: Nachweis-grenze (Signal/Rausch-Verhältnis = 3); Vertrauensbereich: 95 %
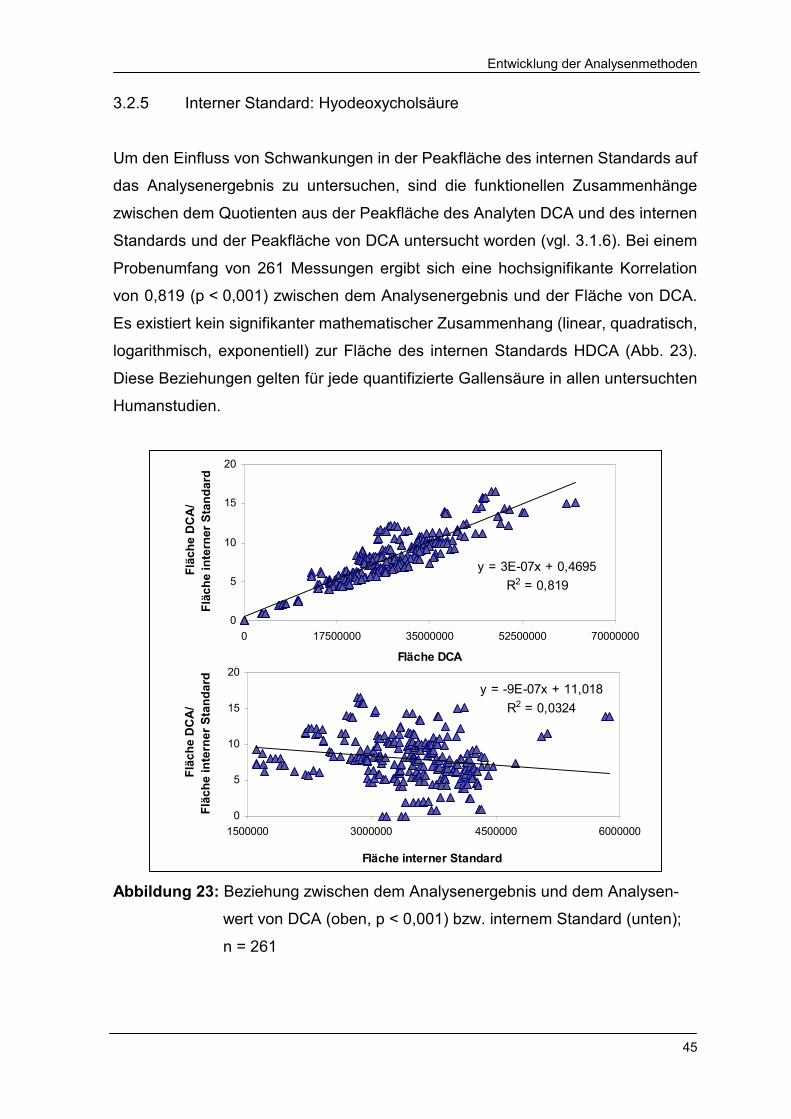
Entwicklung der Analysenmethoden
45
3.2.5 Interner Standard: Hyodeoxycholsäure
Um den Einfluss von Schwankungen in der Peakfläche des internen Standards auf
das Analysenergebnis zu untersuchen, sind die funktionellen Zusammenhänge
zwischen dem Quotienten aus der Peakfläche des Analyten DCA und des internen
Standards und der Peakfläche von DCA untersucht worden (vgl. 3.1.6). Bei einem
Probenumfang von 261 Messungen ergibt sich eine hochsignifikante Korrelation
von 0,819 (p < 0,001) zwischen dem Analysenergebnis und der Fläche von DCA.
Es existiert kein signifikanter mathematischer Zusammenhang (linear, quadratisch,
logarithmisch, exponentiell) zur Fläche des internen Standards HDCA (Abb. 23).
Diese Beziehungen gelten für jede quantifizierte Gallensäure in allen untersuchten
Humanstudien.
Abbildung 23: Beziehung zwischen dem Analysenergebnis und dem Analysen-
wert von DCA (oben, p < 0,001) bzw. internem Standard (unten);
n = 261
y = 3E-07x + 0,4695R2 = 0,819
0
5
10
15
20
0 17500000 35000000 52500000 70000000
Fläche DCA
Fläc
he D
CA
/Fl
äche
inte
rner
Sta
ndar
d
y = -9E-07x + 11,018R2 = 0,0324
0
5
10
15
20
1500000 3000000 4500000 6000000
Fläche interner Standard
Fläc
he D
CA
/Fl
äche
inte
rner
Sta
ndar
d

Humanstudien
46
4 HUMANSTUDIEN
Alle Studien sind vor ihrem Beginn von der Ethikkommission der Medizinischen
Fakultät der Friedrich-Schiller-Universität Jena auf ihre Durchführbarkeit geprüft
und genehmigt worden. Die Probanden erklärten ihre freiwillige Teilnahme.
4.1 Supplementation von Calciumphosphaten
In einer Humanstudie (Studie 1) mit 11 männlichen Probanden wird eine Diät mit
einer gesteigerten Zufuhr von Calcium und Phosphor im Verhältnis Ca : P = 1 : 1,3
verzehrt [GRIMM 2000]. Nach einer dreiwöchigen Basisperiode, in der die
Probanden ein 7-Tage-Ernährungsprotokoll erstellen, folgen zwei Zulageperioden
mit einer Dauer von je zwei Wochen. Auf der Basis der Ergebnisse aus den
Verzehrsdaten wird eine Standarddiät mit einem Calciumgehalt von 1.231 mg/d
und einem Phosphorgehalt von 1.600 mg/d erstellt. In der ersten Zulageperiode
werden täglich 609 mg Calcium und 793 mg Phosphor in Form von Tabletten
supplementiert, was einer gesteigerten Zufuhr von 50 % entspricht. Die Tabletten
setzen sich aus Tricalciumphosphat (41,6 %), Mononatriumphosphat (29,7 %),
Dinatriumphosphat (25,2 %) und Tablettierzusatz (3,5 %) zusammen. In der
zweiten Zulageperiode wird die Dosis verdoppelt, so dass die tägliche
Gesamtaufnahme an Calcium 2.462 mg und an Phosphor 3.200 mg beträgt
(Tab. 14).

Humanstudien
47
Stud
ie 4
Lact
obac
illus
acid
ophi
lus
145
Bifid
obac
teriu
m s
peci
es 4
20
Jogh
urt
13 M
änne
r; 13
Fra
uen
-
25 ±
3
13
3
3; 5
; 5
cros
s-ov
er
Stan
dard
jogh
urt
Stud
ie 3
Lact
obac
illus
acid
ophi
lus
145
Bifid
obac
teriu
m lo
ngum
913
Jogh
urt
29 F
raue
n
14 F
raue
n m
it m
oder
ater
H
yper
chol
este
rolä
mie
34 ±
9
21
3
7; 7
; 7
cros
s-ov
er
Stan
dard
jogh
urt
Stud
ie 2
Lact
obac
illus
para
case
i LTH
257
9
Roh
wur
st
10 M
änne
r; 10
Fra
uen
-
33 ±
8
10
2 5; 5
para
llel
Roh
wur
st
Stud
ie 1
Ca
: P =
1 :
1,3
Tabl
ette
11 M
änne
r
-
25 ±
5
7 3
3; 2
; 2
para
llel
ohne
Zul
age
Supp
lem
ent
Med
ium
Prob
ande
n
Beso
nder
heite
n
Alte
r [a]
Stud
iend
auer
[w]
Perio
dena
nzah
l
Perio
dend
auer
[w]
Stud
iend
esig
n
Ver
lauf
Kon
trolle
Tabe
lle 1
4: B
erüc
ksic
htig
te H
uman
stud
ien
des
Lehr
stuh
les
Ernä
hrun
gsph
ysio
logi
e im
Übe
rblic
k

Humanstudien
48
4.2 Supplementation von probiotischer Rohwurst
In einer Interventionsstudie (Studie 2) wird Lactobacillus paracasei LTH 2579
(108 KBE/g) an 10 männliche und 10 weibliche gesunde Probanden verabreicht
[JAHREIS et al. 2002]. Die Probanden durchlaufen zwei Versuchsperioden, in
denen täglich 50 g Rohwurst verzehrt wird. In der ersten Periode erhalten alle
Teilnehmer eine konventionelle und in der zweiten eine probiotische Wurst. Beide
Versuchsphasen dauern je 5 Wochen und innerhalb der letzten 9 Tage wird
jeweils eine vollständig vorgegebene Diät bereitgestellt, die aus den Ergebnissen
von zuvor erfassten Ernährungsprotokollen der Probanden zusammengestellt ist
(Tab. 14).
4.3 Supplementation von probiotischem Joghurt In randomisierten, placebo-kontrollierten Doppelblindstudien (Studien 3 und 4)
werden die Effekte verschiedener, mit probiotischen Bakterien supplementierter
Joghurts untersucht. Der Testjoghurt der dritten Studie enthält Lactobacillus
acidophilus 145 (106-108 KBE/g) und Bifidobacterium longum 913 (105 KBE/g)
[KIEßLING et al. 2002]. Die Studie ist in drei 7-wöchige Perioden strukturiert, wobei
die erste eine Adaptationsphase mit konventionellem Joghurt darstellt. Die
Starterkulturen zur Fermentation sind Streptococcus thermophilus und
Lactobacillus lactis.
Die 29 normocholesterolämischen oder moderat hypercholesterolämischen
Frauen sind auf zwei Gruppen verteilt. Sie verzehren anschließend über
7 Wochen entweder 300 g konventionellen Joghurt oder Testjoghurt und das
Supplement wechselt in den folgenden 7 Wochen.
In der vierten Studie erhalten je 13 männliche und weibliche Probanden täglich
300 g mit Lactobacillus acidophilus 742 (107 KBE/ml) und Bifidobacterium species
420 (104 KBE/ml) versetzten Joghurt [KLEIN et al. 2002]. Im Gegensatz zu Studie 3
wird während der ersten Phase kein Kontrolljoghurt gegeben, als Starterkulturen
werden Streptococcus thermophilus und Lactobacillus bulgaricus eingesetzt und
die beiden Studienphasen dauern je 5 Wochen (Tab. 14).

Humanstudien
49
4.4 Stuhlproben und in der Auswertung berücksichtigte Parameter Die Stuhlproben werden in jeder Studie auf die gleiche Weise erfasst. Während
den letzten 7 - 9 Tagen der Basis-, Kontroll-, Adaptations- oder Zulageperioden
wird jedem Proband die gesamte Diät auf der Basis von zuvor ausgewerteten
Ernährungsprotokollen vorgegeben. Nach einer zweitägigen Ausschleusphase
wird der Stuhl über 5 bzw. 7 Tage quantitativ gesammelt, tiefgefroren, am Ende
der Studie homogenisiert und gepoolt (Abb. 24). Aus den Proben werden Aliquote
lyophilisiert und ihr Sterol- und Gallensäurengehalt analysiert (vgl. 3).
Abbildung 24: Struktur einer Studienperiode
Neben der Bestimmung der faecalen Steroide werden weitere Erhebungen aus
den Studien berücksichtigt und statistisch ausgewertet.
• Studie 1: Frisch- und Gefriertrockensubstanzexkretion
pH-Wert im Stuhl
• Studie 2: Frisch- und Trockensubstanzexkretion
pH-Wert im Stuhl
Keimzahlbestimmung der Lactobacillen im Stuhl
Fettaufnahme
• Studie 3: Frisch- und Gefriertrockensubstanzexkretion
pH-Wert im Stuhl
Fettaufnahme
1 - 6 Wochen 2 Tage 5 - 7 Tage Basis-, Kontroll-, Ausschleusphase Sammelphase Adaptations- oder Zulageperiode
Standardisierte Diät

Humanstudien
50
Gesamtcholesterolkonzentration im Serum
LDL-Cholesterolkonzentration im Serum
HDL-Cholesterolkonzentration im Serum
Triglyceridkonzentration im Serum
• Studie 4: Frisch- und Gefriertrockensubstanzexkretion
pH-Wert im Stuhl
Fettaufnahme
LDL-Cholesterolkonzentration im Serum
HDL-Cholesterolkonzentration im Serum
4.5 Statistik Zur Auswertung aller erhobenen Parameter wird die Computer-Software SPSS
10.0 unter Anwendung des allgemeinen linearen Modells für Messwiederholungen
eingesetzt. Diese Statistik berücksichtigt die Änderungen der abhängigen
Variablen bei wiederholter Erhebung von Daten zur Untersuchung des
interessierenden Einflusses (Calcium, Probiotika, Joghurt) im Einzelfall. Es
beschreibt die Unterschiede der mittleren Differenz geschätzter Randmittel im
untersuchten Einfluss. Dadurch werden inter-individuelle Unterschiede weniger
gewichtet als bei dem Vergleich von Mittelwerten mittels t-Test und F-Test.
Eventuelle Einflüsse der faecalen pH-Werte (Studie 1), der Keimzahlen an
Lactobacillen im Stuhl (Studie 2), der Serumkonzentrationen an Cholesterol
(Studie 3), der Reihenfolge der Supplementation (Studie 3, 4) und geschlechts-
spezifische Unterschiede (Studie 4) haben in diesem mathematischen Modell als
Zwischensubjektfaktoren Anwendung gefunden. Bei einem Auftreten von
signifikanten Unterschieden in diesen Faktoren werden die Mittelwerte zwischen
den jeweiligen Gruppen (normocholesterolämische vs. hypercholesterolämische
Probanden, Frauen vs. Männer) mit Hilfe des Modells der einfaktoriellen ANOVA
statistisch geprüft. In allen Studien werden sowohl die jeweiligen Gehalte in der
Stuhltrockensubstanz als auch die tägliche Exkretion der Sterole bzw.
Gallensäuren ausgewertet.

Ergebnisse
51
5 ERGEBNISSE 5.1 Studie 1 - Supplementation von Calciumphosphaten In allen drei Perioden sind weder in der Exkretion an Frischsubstanz noch an
Gefriertrockensubstanz Änderungen zu verzeichnen (Tab. 15). In der ersten
Zulageperiode ist eine signifikante Verminderung des pH-Wertes in den Faeces
festzustellen, obwohl keine Änderungen in den Konzentrationen an kurzkettigen,
flüchtigen Fettsäuren im Stuhl aufgetreten sind [GRIMM 2000].
In der Auswertung der faecalen Exkretion wird ein Proband (Nr. 24)
ausgeschlossen, da dieser sowohl in der Ausscheidung der Sterole als auch der
Gallensäuren ein inverses Steroidprofil aufweist (Abb. 25). Das bedeutet, dass die
Gehalte der Ausgangsverbindungen (Cholesterol, primäre Gallensäuren) größer
oder gleich den Gehalten der bakteriellen Metabolisierungsprodukte (Coprostanol,
sekundäre Gallensäuren) sind (vgl. 6.2).
Tabelle 15: Mittlere Stuhlausscheidung in Studie 1
Periode I Periode II Periode III
Gefriertrockensubstanz [g/d] 39,2 ± 6,6 40,0 ± 13,4 45,3 ±14,0
Frischsubstanz [g/d] 164 ± 42 164 ± 63 180 ± 80
pH 6,9 ± 0,2 6,6 ± 0,2a 6,8 ± 0,2
a: Signifikanz zu Periode I (p < 0,05)
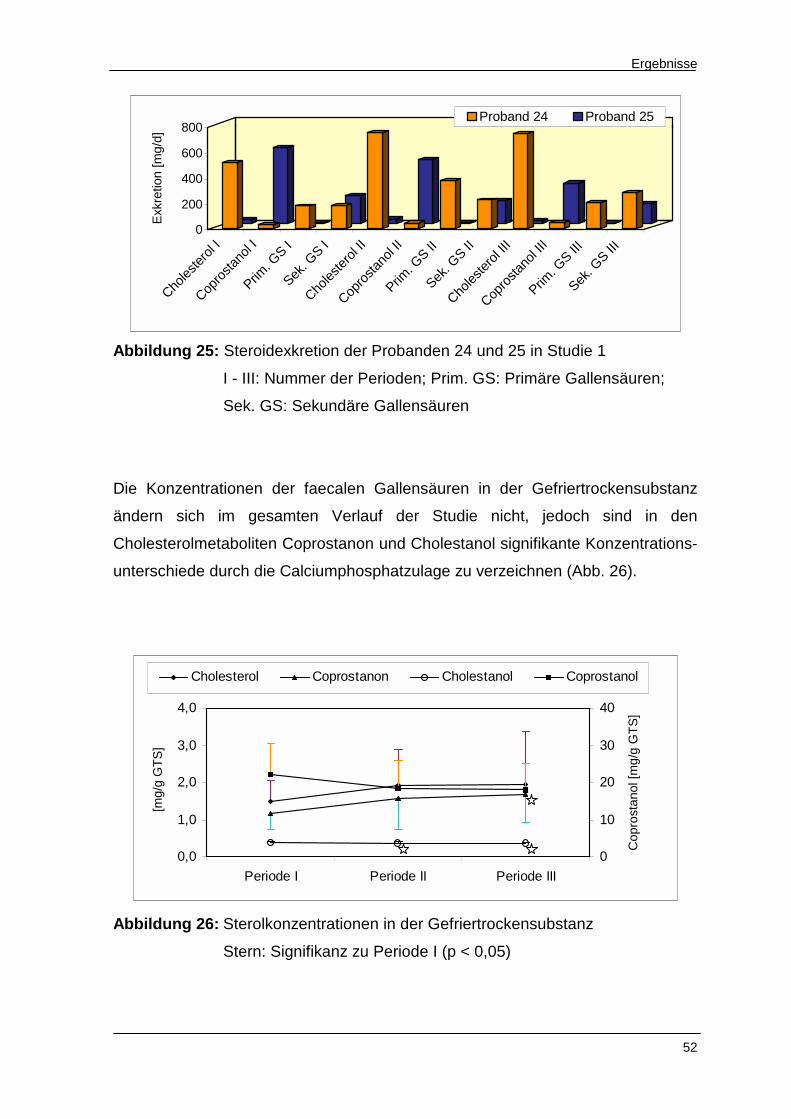
Ergebnisse
52
Abbildung 25: Steroidexkretion der Probanden 24 und 25 in Studie 1
I - III: Nummer der Perioden; Prim. GS: Primäre Gallensäuren;
Sek. GS: Sekundäre Gallensäuren
Die Konzentrationen der faecalen Gallensäuren in der Gefriertrockensubstanz
ändern sich im gesamten Verlauf der Studie nicht, jedoch sind in den
Cholesterolmetaboliten Coprostanon und Cholestanol signifikante Konzentrations-
unterschiede durch die Calciumphosphatzulage zu verzeichnen (Abb. 26).
Abbildung 26: Sterolkonzentrationen in der Gefriertrockensubstanz
Stern: Signifikanz zu Periode I (p < 0,05)
0
200
400
600
800
Exkr
etio
n [m
g/d]
Choles
terol
I
Copros
tanol
I
Prim. G
S I
Sek. G
S I
Choles
terol
II
Copros
tanol
II
Prim. G
S II
Sek. G
S II
Choles
terol
III
Copros
tanol
III
Prim. G
S III
Sek. G
S III
Proband 24 Proband 25
0,0
1,0
2,0
3,0
4,0
Periode I Periode II Periode III
[mg/
g G
TS]
0
10
20
30
40
Cop
rost
anol
[mg/
g G
TS]
Cholesterol Coprostanon Cholestanol Coprostanol
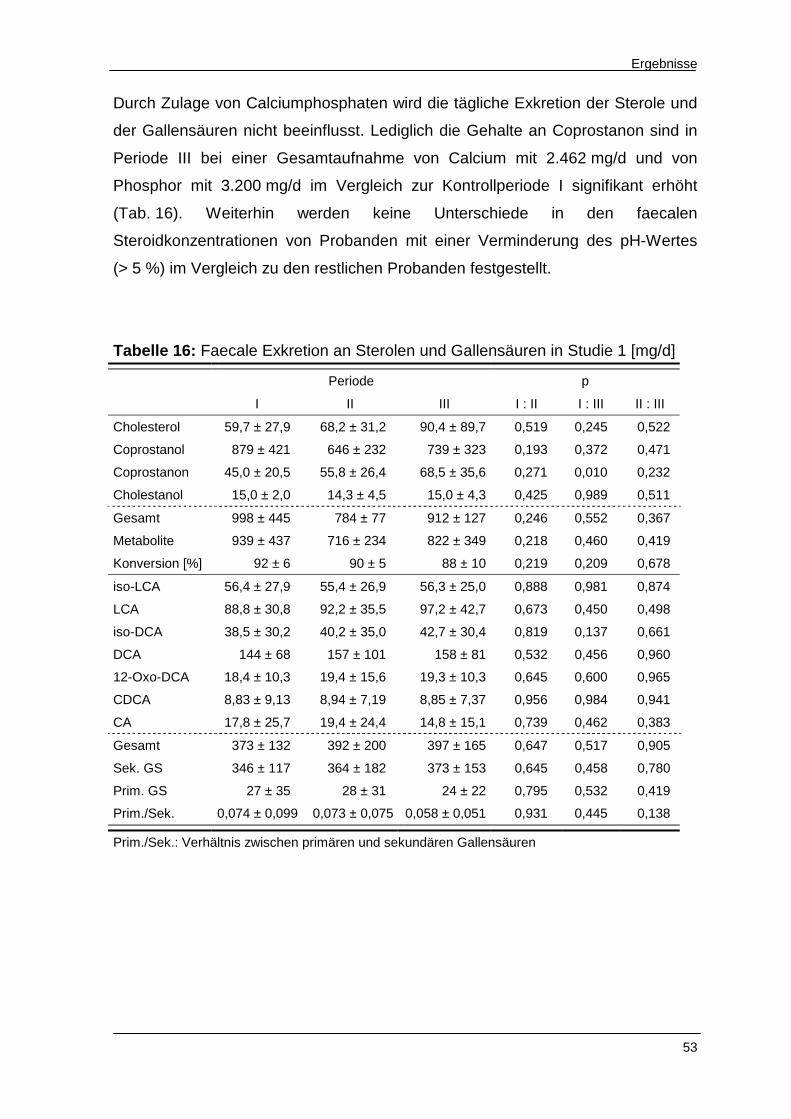
Ergebnisse
53
Durch Zulage von Calciumphosphaten wird die tägliche Exkretion der Sterole und
der Gallensäuren nicht beeinflusst. Lediglich die Gehalte an Coprostanon sind in
Periode III bei einer Gesamtaufnahme von Calcium mit 2.462 mg/d und von
Phosphor mit 3.200 mg/d im Vergleich zur Kontrollperiode I signifikant erhöht
(Tab. 16). Weiterhin werden keine Unterschiede in den faecalen
Steroidkonzentrationen von Probanden mit einer Verminderung des pH-Wertes
(> 5 %) im Vergleich zu den restlichen Probanden festgestellt.
Tabelle 16: Faecale Exkretion an Sterolen und Gallensäuren in Studie 1 [mg/d]
Periode p
I II III I : II I : III II : III
Cholesterol 59,7 ± 27,9 68,2 ± 31,2 90,4 ± 89,7 0,519 0,245 0,522
Coprostanol 879 ± 421 646 ± 232 739 ± 323 0,193 0,372 0,471
Coprostanon 45,0 ± 20,5 55,8 ± 26,4 68,5 ± 35,6 0,271 0,010 0,232
Cholestanol 15,0 ± 2,0 14,3 ± 4,5 15,0 ± 4,3 0,425 0,989 0,511
Gesamt 998 ± 445 784 ± 77 912 ± 127 0,246 0,552 0,367
Metabolite 939 ± 437 716 ± 234 822 ± 349 0,218 0,460 0,419
Konversion [%] 92 ± 6 90 ± 5 88 ± 10 0,219 0,209 0,678
iso-LCA 56,4 ± 27,9 55,4 ± 26,9 56,3 ± 25,0 0,888 0,981 0,874
LCA 88,8 ± 30,8 92,2 ± 35,5 97,2 ± 42,7 0,673 0,450 0,498
iso-DCA 38,5 ± 30,2 40,2 ± 35,0 42,7 ± 30,4 0,819 0,137 0,661
DCA 144 ± 68 157 ± 101 158 ± 81 0,532 0,456 0,960 12-Oxo-DCA 18,4 ± 10,3 19,4 ± 15,6 19,3 ± 10,3 0,645 0,600 0,965
CDCA 8,83 ± 9,13 8,94 ± 7,19 8,85 ± 7,37 0,956 0,984 0,941
CA 17,8 ± 25,7 19,4 ± 24,4 14,8 ± 15,1 0,739 0,462 0,383
Gesamt 373 ± 132 392 ± 200 397 ± 165 0,647 0,517 0,905
Sek. GS 346 ± 117 364 ± 182 373 ± 153 0,645 0,458 0,780
Prim. GS 27 ± 35 28 ± 31 24 ± 22 0,795 0,532 0,419 Prim./Sek. 0,074 ± 0,099 0,073 ± 0,075 0,058 ± 0,051 0,931 0,445 0,138
Prim./Sek.: Verhältnis zwischen primären und sekundären Gallensäuren
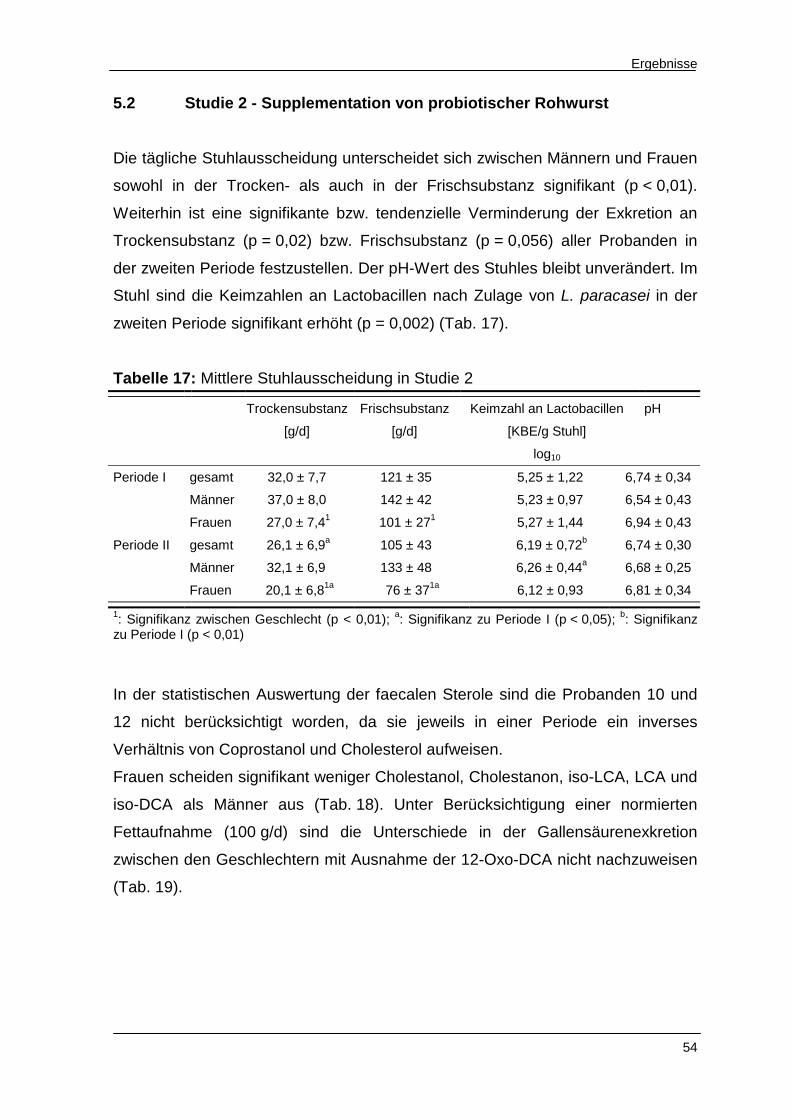
Ergebnisse
54
5.2 Studie 2 - Supplementation von probiotischer Rohwurst
Die tägliche Stuhlausscheidung unterscheidet sich zwischen Männern und Frauen
sowohl in der Trocken- als auch in der Frischsubstanz signifikant (p < 0,01).
Weiterhin ist eine signifikante bzw. tendenzielle Verminderung der Exkretion an
Trockensubstanz (p = 0,02) bzw. Frischsubstanz (p = 0,056) aller Probanden in
der zweiten Periode festzustellen. Der pH-Wert des Stuhles bleibt unverändert. Im
Stuhl sind die Keimzahlen an Lactobacillen nach Zulage von L. paracasei in der
zweiten Periode signifikant erhöht (p = 0,002) (Tab. 17).
Tabelle 17: Mittlere Stuhlausscheidung in Studie 2
Trockensubstanz Frischsubstanz Keimzahl an Lactobacillen pH
[g/d] [g/d] [KBE/g Stuhl]
log10
Periode I gesamt 32,0 ± 7,7 121 ± 35 5,25 ± 1,22 6,74 ± 0,34 Männer 37,0 ± 8,0 142 ± 42 5,23 ± 0,97 6,54 ± 0,43
Frauen 27,0 ± 7,41 101 ± 271 5,27 ± 1,44 6,94 ± 0,43
Periode II gesamt 26,1 ± 6,9a 105 ± 43 6,19 ± 0,72b 6,74 ± 0,30 Männer 32,1 ± 6,9 133 ± 48 6,26 ± 0,44a 6,68 ± 0,25
Frauen 20,1 ± 6,81a 76 ± 371a 6,12 ± 0,93 6,81 ± 0,34
1: Signifikanz zwischen Geschlecht (p < 0,01); a: Signifikanz zu Periode I (p < 0,05); b: Signifikanz zu Periode I (p < 0,01)
In der statistischen Auswertung der faecalen Sterole sind die Probanden 10 und
12 nicht berücksichtigt worden, da sie jeweils in einer Periode ein inverses
Verhältnis von Coprostanol und Cholesterol aufweisen.
Frauen scheiden signifikant weniger Cholestanol, Cholestanon, iso-LCA, LCA und
iso-DCA als Männer aus (Tab. 18). Unter Berücksichtigung einer normierten
Fettaufnahme (100 g/d) sind die Unterschiede in der Gallensäurenexkretion
zwischen den Geschlechtern mit Ausnahme der 12-Oxo-DCA nicht nachzuweisen
(Tab. 19).

Ergebnisse
55
< 10
6 [KBE
/g]
27,3
± 2
7,6
573
± 26
6
63,4
± 4
5,8
11,1
± 1
,5
3,07
± 0
,56
706
± 26
9
651
± 27
4
94 ±
5
42,8
± 1
4,0
53,7
± 2
0,4c
24,4
± 1
5,8
145
± 69
16,8
± 7
,8
3,99
± 5
,99
4,83
± 8
,64
291
± 13
3
282
± 10
5
9 ±
18
0,03
1 ±
0,04
8
Keim
zahl
an
Lact
obac
illen
> 10
6 [KBE
/g]
55,5
± 2
7,8
619
± 26
7
50,0
± 4
6,2
11,9
± 1
,7
3,32
± 0
,58
772
± 27
0
684
± 27
6
91 ±
5
56,7
± 1
3,9
85,2
± 2
0,5
40,5
± 1
5,9
166
± 71
12,7
± 7
,4
7,74
± 6
,04
10,1
± 8
,7
379
± 11
3
361
± 10
7
18 ±
14
0,05
5 ±
0,04
6
Frau
en
29,0
± 3
1,7
504
± 30
8
32,5
± 5
2,9
10,2
± 1
,9b
2,64
± 0
,72b
603
± 28
8
549
± 31
7
93 ±
6
36,1
± 1
6,8b
51,8
± 2
4,6b
20,2
± 1
8,9b
135
± 85
12,1
± 9
,3
5,04
± 7
,30
7,57
± 1
0,41
273
± 13
61
260
± 12
92
13 ±
17
0,05
2 ±
0,05
7
G
esch
lech
t
Män
ner
53,8
± 3
3,0
688
± 32
0
80,9
± 5
5,2
12,8
± 2
,0
3,75
± 0
,70
874
± 32
3
786
± 33
0
93 ±
6
63,3
± 1
7,4
87,2
± 2
5,3
44,8
± 1
9,6
177
± 88
17,3
± 9
,8
6,70
± 7
,61
7,32
± 1
0,79
398
± 14
1
384
± 13
4
14 ±
18
0,03
4 ±
0,06
0
I
I
39,7
± 5
0,3
543
± 36
0
50,5
± 4
5,4
9,7
± 2,
8a
2,71
± 0
,83a
671
± 36
8
606
± 37
0
93 ±
9
44,7
± 2
3,5
70,0
± 3
6,6
27,3
± 1
7,9a
122
± 86
a
10,4
± 1
0,5a
5,34
± 1
1,10
5,97
± 1
5,31
276
± 15
8a
264
± 14
7a
11 ±
26
0,04
0 ±
0,06
1
Perio
de
I
43,1
± 3
4,6
649
± 36
4
62,9
± 7
1,3
13,3
± 3
,3
13,3
± 3
,3
806
± 39
5
729
± 39
1
93 ±
7
54,7
± 2
0,5
83,9
± 3
0,1
37,7
± 2
4,4
190
± 10
3
19,1
± 1
4,8
6,39
± 8
,40
8,91
± 1
1,79
395
± 15
8
380
± 15
2
15 ±
20
0,04
7 ±
0,07
4
Cho
lest
erol
Cop
rost
anol
Cop
rost
anon
Cho
lest
anol
Cho
lest
anon
Ges
amt
Met
abol
ite
Konv
ersi
on [%
]
iso-
LCA
LCA
iso-
DC
A
DC
A
12-O
xo-D
CA
CD
CA
CA
Ges
amt
Sek.
GS
Prim
. GS
Prim
./Sek
.
a : Sig
nifik
anz
zwis
chen
Per
iode
n (p
< 0
,05)
; b : Sig
nifik
anz
zwis
chen
Ges
chle
cht;
c : Sig
nifik
anz
zwis
chen
Pro
band
en m
it Ke
imza
hlen
grö
ßer b
zw. k
lein
er 1
06 KBE
/g
Stuh
l; 1 : p
= 0
,068
; 2 : p =
0,0
58; P
rim./S
ek.:
Verh
ältn
is z
wis
chen
prim
ären
und
sek
undä
ren
Gal
lens
äure
n
Tabe
lle 1
8: F
aeca
le E
xkre
tion
an S
tero
len
und
Gal
lens
äure
n in
Stu
die
2 [m
g/d]

Ergebnisse
56
Die Zulage von L. paracasei bewirkt keine Änderung der Konversionsrate von
Cholesterol in die bakteriellen Metabolite und beeinflusst nicht das Verhältnis von
primären zu sekundären Gallensäuren. Allerdings zeigt sich eine geringere
Konzentration der Gesamtgallensäuren in den Faeces, die durch eine verminderte
Exkretion sekundärer Gallensäuren verursacht wird. Weiterhin sind die Probanden
mit einer Keimzahl an Lactobacillen in der zweiten Periode über und unter
106 KBE/g Stuhl differenziert betrachtet worden. Eine signifikant erhöhte tägliche
Ausscheidung an LCA ist in Probanden mit hohen Keimzahlen zu beobachten,
außerdem haben sie tendenziell erhöhte Konzentrationen an Cholesterol, iso-LCA
und iso-DCA im Stuhl (Tab. 18).
Wird die signifikant verminderte Stuhlausscheidung in der zweiten Periode
berücksichtigt, und die Steroidexkretion auf eine Trockensubstanzexkretion von
30 g/d bezogen, treten keine Unterschiede nach Zulage von L. paracasei in den
faecalen Gallensäurengehalten auf (Tab. 19). Dieses Ergebnis wird durch
unveränderte Konzentrationen an Gallensäuren in der faecalen Trockensubstanz
bestätigt. Eine tendenzielle Verminderung der DCA im Stuhl ist weiter vorhanden
(p = 0,085).
Tabelle 19: Tägliche Gallensäurenexkretion [mg/d] unter Berücksichtigung der
Fettaufnahme und Trockensubstanzexkretion
Normierte Fettaufnahme (100 g/d) Normierte TS-Exkretion (30 g/d) Männer Frauen p Periode I Periode II p
Cholesterol 69,4 ± 64,4 41,4 ± 18,4 0,104 47,7 ± 26,3 53,6 ± 40,8 0,539
Coprostanol 671 ± 268 679 ± 411 0,943 640 ± 329 688 ± 412 0,429
iso-LCA 57,7 ± 21,0 53,7 ± 20,2 0,677 52,5 ± 24,4 57,1 ± 25,7 0,193
LCA 80,9 ± 34,1 72,7 ± 32,9 0,602 73,7 ± 23,5 72,0 ± 28,3 0,785
iso-DCA 42,1 ± 24,8 27,4 ± 23,9 0,205 34,7 ± 24,4 32,4 ± 19,6 0,533
DCA 167 ± 101 186 ± 97 0,684 174 ± 72 148 ± 62 0,085
12-Oxo-DCA 11,1 ± 12,3 23,6 ± 11,8 0,093 16,6 ± 10,3 13,4 ± 16,5 0,479
CDCA 6,74 ± 9,52 6,61 ± 9,17 0,975 6,04 ± 8,00 6,06 ± 10,51 0,996
CA 7,38 ± 13,32 9,88 ± 12,82 0,682 8,73 ± 12,41 6,85 ± 14,72 0,629
Gesamt 373 ± 180 379 ± 173 0,935 366 ± 120 336 ± 115 0,209
Sek. GS 359 ± 166 363 ± 159 0,954 352 ± 116 323 ± 109 0,179
Prim. GS 14 ± 22 16 ± 22 0,817 14 ± 20 13 ± 25 0,781
TS: Trockensubstanz
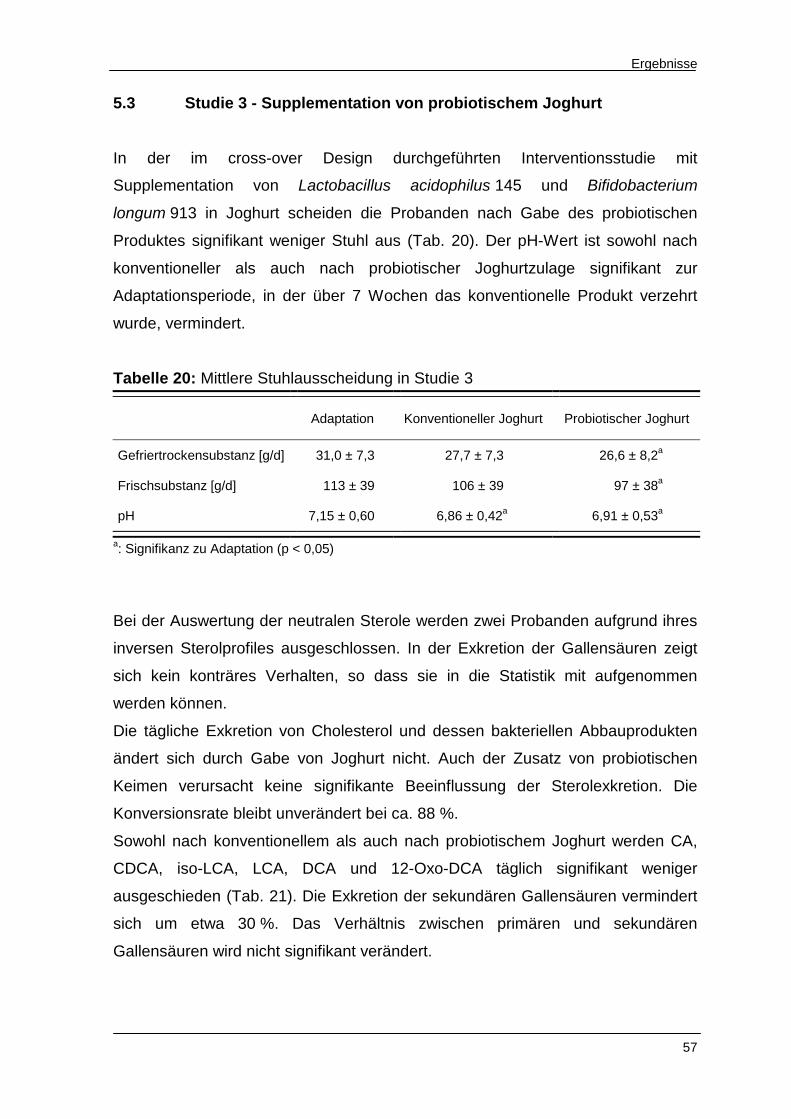
Ergebnisse
57
5.3 Studie 3 - Supplementation von probiotischem Joghurt
In der im cross-over Design durchgeführten Interventionsstudie mit
Supplementation von Lactobacillus acidophilus 145 und Bifidobacterium
longum 913 in Joghurt scheiden die Probanden nach Gabe des probiotischen
Produktes signifikant weniger Stuhl aus (Tab. 20). Der pH-Wert ist sowohl nach
konventioneller als auch nach probiotischer Joghurtzulage signifikant zur
Adaptationsperiode, in der über 7 Wochen das konventionelle Produkt verzehrt
wurde, vermindert.
Tabelle 20: Mittlere Stuhlausscheidung in Studie 3
Adaptation Konventioneller Joghurt Probiotischer Joghurt
Gefriertrockensubstanz [g/d] 31,0 ± 7,3 27,7 ± 7,3 26,6 ± 8,2a
Frischsubstanz [g/d] 113 ± 39 106 ± 39 97 ± 38a
pH 7,15 ± 0,60 6,86 ± 0,42a 6,91 ± 0,53a
a: Signifikanz zu Adaptation (p < 0,05)
Bei der Auswertung der neutralen Sterole werden zwei Probanden aufgrund ihres
inversen Sterolprofiles ausgeschlossen. In der Exkretion der Gallensäuren zeigt
sich kein konträres Verhalten, so dass sie in die Statistik mit aufgenommen
werden können.
Die tägliche Exkretion von Cholesterol und dessen bakteriellen Abbauprodukten
ändert sich durch Gabe von Joghurt nicht. Auch der Zusatz von probiotischen
Keimen verursacht keine signifikante Beeinflussung der Sterolexkretion. Die
Konversionsrate bleibt unverändert bei ca. 88 %.
Sowohl nach konventionellem als auch nach probiotischem Joghurt werden CA,
CDCA, iso-LCA, LCA, DCA und 12-Oxo-DCA täglich signifikant weniger
ausgeschieden (Tab. 21). Die Exkretion der sekundären Gallensäuren vermindert
sich um etwa 30 %. Das Verhältnis zwischen primären und sekundären
Gallensäuren wird nicht signifikant verändert.

Ergebnisse
58
Prob
iotis
cher
Jo
ghur
t
2,1
3 ±
0,95
18,4
± 5
,9
1,46
± 0
,68
0,52
± 0
,05
22,5
± 6
,2
20,4
± 6
,1
88,4
± 7
,2
1,44
± 0
,66b
2,06
± 0
,71b
1,55
± 0
,85
4,46
± 1
,68c
0,28
± 0
,21a
0,21
± 0
,08
0,12
± 0
,07a
10,1
± 3
,0c
9,8
± 3,
0c
0,3
± 0,
1a
0,03
5 ±
0,01
6
Konv
entio
nelle
r Jo
ghur
t
2,05
± 1
,14
18,9
± 7
,1
1,43
± 0
,68
0,51
± 0
,05
22,9
± 7
,4
20,9
± 7
,4
88,9
± 7
,8
1,39
± 0
,56c
2,03
± 0
,79c
1,61
± 0
,89
4,09
± 1
,34c
0,32
± 0
,27a
0,20
± 0
,08
0,09
± 0
,05b
9,7
± 2,
6c
9,4
± 2,
6c
0,3
± 0,
1a
0,03
3 ±
0,01
6
mg/
g G
efrie
rtroc
kens
ubst
anz
Adap
tatio
n
2,16
± 1
,21
17,8
± 5
,3
1,50
± 1
,18
0,51
± 0
,04
22,0
± 6
,4
19,8
± 6
,0
88,4
± 6
,7
1,79
± 0
,78
2,63
± 1
,23
1,57
± 1
,06
5,95
± 2
,26
0,43
± 0
,42
0,25
± 0
,16
0,23
± 0
,23
12,9
± 4
,4
12,4
± 4
,4
0,5
± 0,
4
0,04
4 ±
0,04
2
Prob
iotis
cher
Jo
ghur
t
57,2
± 2
8,8
491
± 20
6
38,3
± 1
9,5
13,6
± 3
,2
600
± 22
8
543
± 21
7
88,4
± 7
,2
39,4
± 2
0,7b
55,3
± 2
4,6c
43,9
± 2
9,6
122
± 60
c
8,0
± 7,
3b
5,46
± 2
,18a
3,10
± 1
,80b
277
± 12
2c
268
± 12
1c
9 ±
4b
0,03
5 ±
0,01
6
Konv
entio
nelle
r Jo
ghur
t
55,6
± 2
6,9
520
± 19
9
38,9
± 1
8,8
14,0
± 2
,5
629
± 21
0
573
± 20
8
88,9
± 7
,8
39,2
± 1
8,3b
56,2
± 2
6,9c
45,0
± 2
7,1
114
± 48
c
8,6
± 7
,9a
5,26
± 1
,71a
2,58
± 1
,28b
271
± 10
2c
263
± 10
1c
8 ±
3b
0,03
3 ±
0,01
6
mg/
d
Adap
tatio
n
62,7
± 3
8,6
519
± 18
9
42,7
± 2
9,2
14,6
± 2
,8
639
± 21
7
577
± 20
4
88,4
± 6
,7
53,2
± 2
8,3
78,2
± 3
8,5
46,3
± 3
2,1
176
± 73
12,8
± 1
2,1
7,62
± 5
,25
7,04
± 6
,92
382
± 14
7
367
± 14
4
15 ±
12
0,04
4 ±
0,04
2
Cho
lest
erol
Cop
rost
anol
Cop
rost
anon
Cho
lest
anol
Ges
amt
Met
abol
ite
Konv
ersi
on [%
]
iso-
LCA
LCA
iso-
DC
A
DC
A
12-O
xo-D
CA
CD
CA
CA
Ges
amt
Sek.
GS
Prim
. GS
Prim
./Sek
.
Tabe
lle 2
1: F
aeca
le E
xkre
tion
an S
tero
len
und
Gal
lens
äure
n in
Stu
die
3
a : Sig
nifik
anz
zu A
dapt
atio
n (p
< 0
,05)
; b : Sig
nifik
anz
zu A
dapt
atio
n (p
< 0
,01)
; c : Sig
nifik
anz
zu A
dapt
atio
n (p
< 0
,001
); P
rim./S
ek.:
Verh
ältn
is z
wis
chen
prim
ären
und
sek
undä
ren
Gal
lens
äure
n
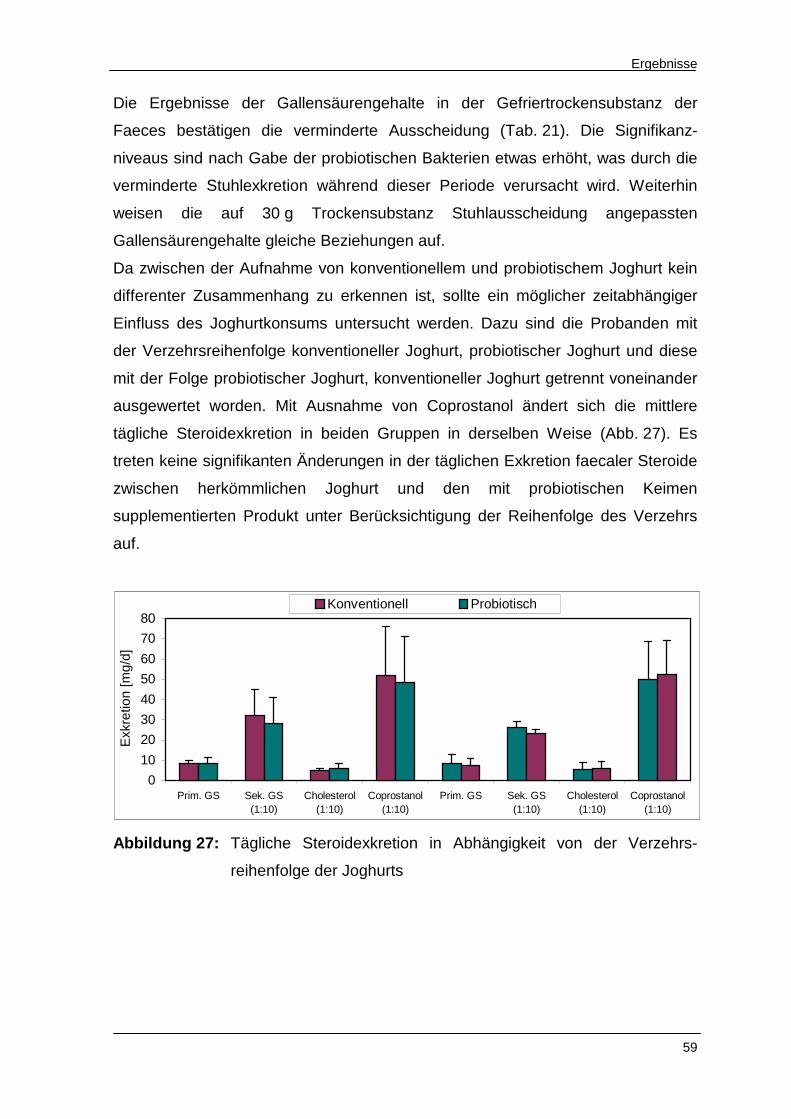
Ergebnisse
59
Die Ergebnisse der Gallensäurengehalte in der Gefriertrockensubstanz der
Faeces bestätigen die verminderte Ausscheidung (Tab. 21). Die Signifikanz-
niveaus sind nach Gabe der probiotischen Bakterien etwas erhöht, was durch die
verminderte Stuhlexkretion während dieser Periode verursacht wird. Weiterhin
weisen die auf 30 g Trockensubstanz Stuhlausscheidung angepassten
Gallensäurengehalte gleiche Beziehungen auf.
Da zwischen der Aufnahme von konventionellem und probiotischem Joghurt kein
differenter Zusammenhang zu erkennen ist, sollte ein möglicher zeitabhängiger
Einfluss des Joghurtkonsums untersucht werden. Dazu sind die Probanden mit
der Verzehrsreihenfolge konventioneller Joghurt, probiotischer Joghurt und diese
mit der Folge probiotischer Joghurt, konventioneller Joghurt getrennt voneinander
ausgewertet worden. Mit Ausnahme von Coprostanol ändert sich die mittlere
tägliche Steroidexkretion in beiden Gruppen in derselben Weise (Abb. 27). Es
treten keine signifikanten Änderungen in der täglichen Exkretion faecaler Steroide
zwischen herkömmlichen Joghurt und den mit probiotischen Keimen
supplementierten Produkt unter Berücksichtigung der Reihenfolge des Verzehrs
auf.
Abbildung 27: Tägliche Steroidexkretion in Abhängigkeit von der Verzehrs-
reihenfolge der Joghurts
01020304050607080
Prim. GS Sek. GS(1:10)
Cholesterol(1:10)
Coprostanol(1:10)
Prim. GS Sek. GS(1:10)
Cholesterol(1:10)
Coprostanol(1:10)
Exkr
etio
n [m
g/d]
Konventionell Probiotisch
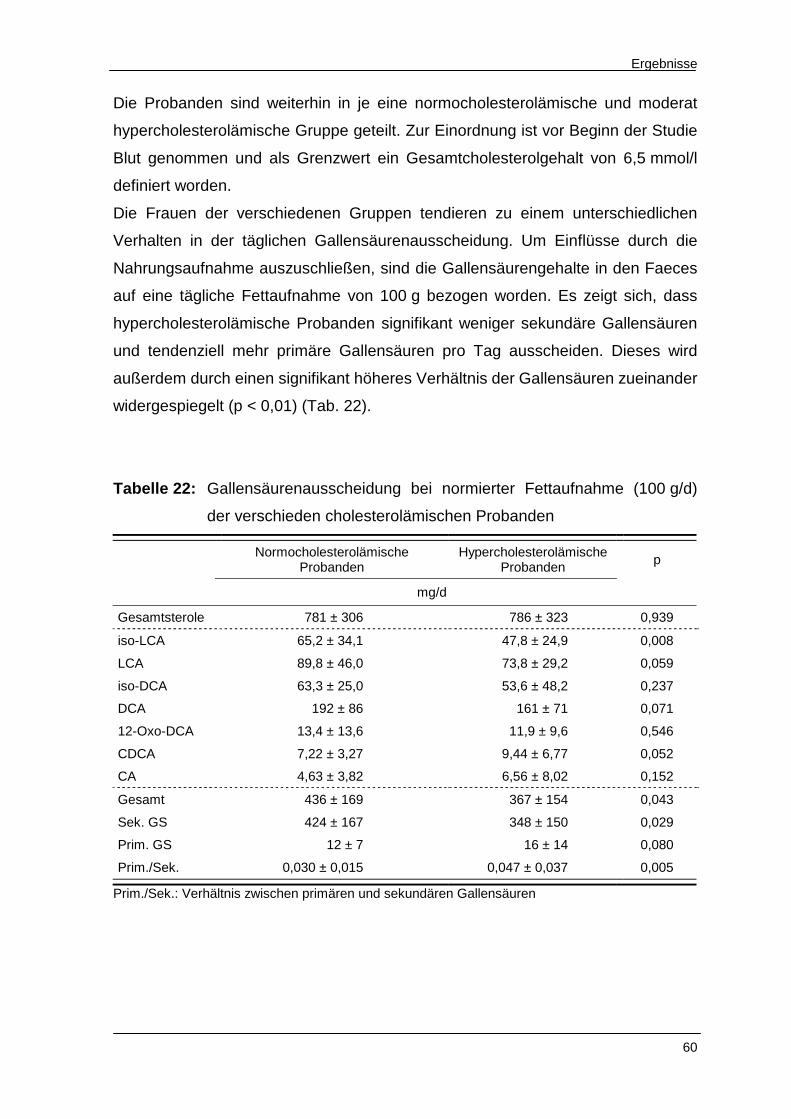
Ergebnisse
60
Die Probanden sind weiterhin in je eine normocholesterolämische und moderat
hypercholesterolämische Gruppe geteilt. Zur Einordnung ist vor Beginn der Studie
Blut genommen und als Grenzwert ein Gesamtcholesterolgehalt von 6,5 mmol/l
definiert worden.
Die Frauen der verschiedenen Gruppen tendieren zu einem unterschiedlichen
Verhalten in der täglichen Gallensäurenausscheidung. Um Einflüsse durch die
Nahrungsaufnahme auszuschließen, sind die Gallensäurengehalte in den Faeces
auf eine tägliche Fettaufnahme von 100 g bezogen worden. Es zeigt sich, dass
hypercholesterolämische Probanden signifikant weniger sekundäre Gallensäuren
und tendenziell mehr primäre Gallensäuren pro Tag ausscheiden. Dieses wird
außerdem durch einen signifikant höheres Verhältnis der Gallensäuren zueinander
widergespiegelt (p < 0,01) (Tab. 22).
Tabelle 22: Gallensäurenausscheidung bei normierter Fettaufnahme (100 g/d)
der verschieden cholesterolämischen Probanden
Normocholesterolämische Probanden
Hypercholesterolämische Probanden p
mg/d
Gesamtsterole 781 ± 306 786 ± 323 0,939
iso-LCA 65,2 ± 34,1 47,8 ± 24,9 0,008
LCA 89,8 ± 46,0 73,8 ± 29,2 0,059
iso-DCA 63,3 ± 25,0 53,6 ± 48,2 0,237 DCA 192 ± 86 161 ± 71 0,071
12-Oxo-DCA 13,4 ± 13,6 11,9 ± 9,6 0,546
CDCA 7,22 ± 3,27 9,44 ± 6,77 0,052 CA 4,63 ± 3,82 6,56 ± 8,02 0,152
Gesamt 436 ± 169 367 ± 154 0,043
Sek. GS 424 ± 167 348 ± 150 0,029 Prim. GS 12 ± 7 16 ± 14 0,080
Prim./Sek. 0,030 ± 0,015 0,047 ± 0,037 0,005
Prim./Sek.: Verhältnis zwischen primären und sekundären Gallensäuren
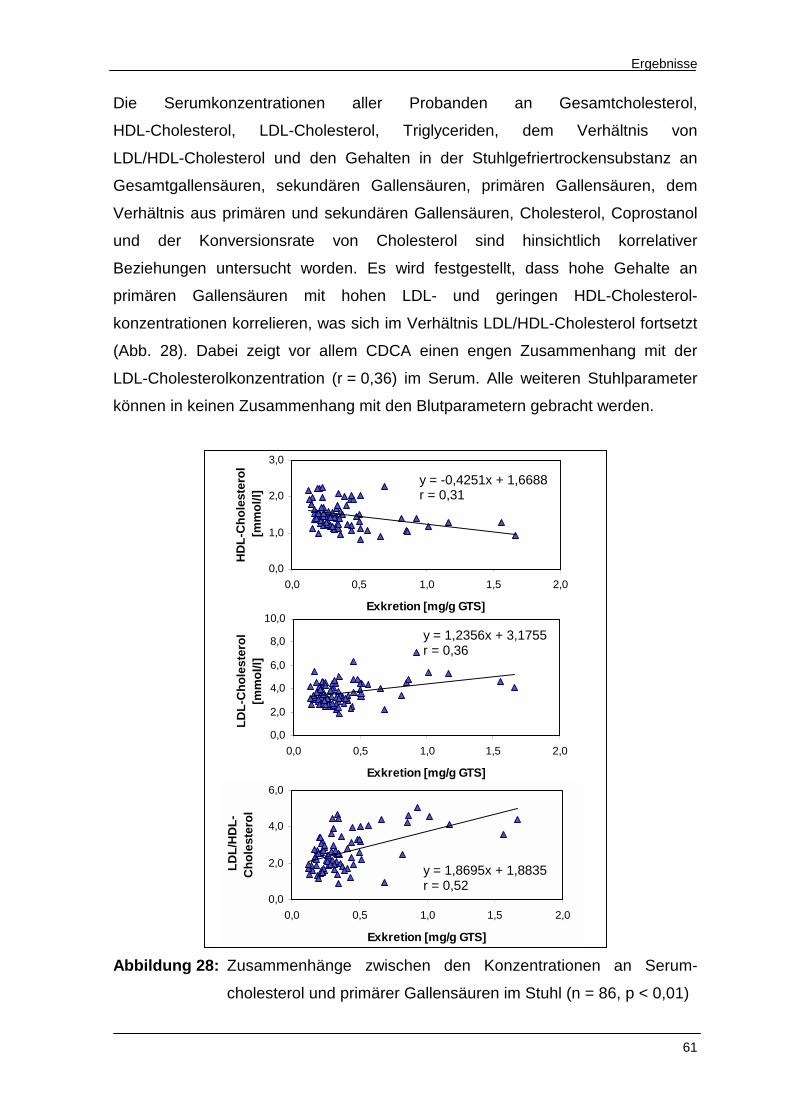
Ergebnisse
61
Die Serumkonzentrationen aller Probanden an Gesamtcholesterol,
HDL-Cholesterol, LDL-Cholesterol, Triglyceriden, dem Verhältnis von
LDL/HDL-Cholesterol und den Gehalten in der Stuhlgefriertrockensubstanz an
Gesamtgallensäuren, sekundären Gallensäuren, primären Gallensäuren, dem
Verhältnis aus primären und sekundären Gallensäuren, Cholesterol, Coprostanol
und der Konversionsrate von Cholesterol sind hinsichtlich korrelativer
Beziehungen untersucht worden. Es wird festgestellt, dass hohe Gehalte an
primären Gallensäuren mit hohen LDL- und geringen HDL-Cholesterol-
konzentrationen korrelieren, was sich im Verhältnis LDL/HDL-Cholesterol fortsetzt
(Abb. 28). Dabei zeigt vor allem CDCA einen engen Zusammenhang mit der
LDL-Cholesterolkonzentration (r = 0,36) im Serum. Alle weiteren Stuhlparameter
können in keinen Zusammenhang mit den Blutparametern gebracht werden.
Abbildung 28: Zusammenhänge zwischen den Konzentrationen an Serum-
cholesterol und primärer Gallensäuren im Stuhl (n = 86, p < 0,01)
0,0
2,0
4,0
6,0
0,0 0,5 1,0 1,5 2,0
Exkretion [mg/g GTS]
LDL/
HD
L-C
hole
ster
ol
0,0
1,0
2,0
3,0
0,0 0,5 1,0 1,5 2,0
Exkretion [mg/g GTS]
HD
L-C
hole
ster
ol [m
mol
/l]
0,0
2,0
4,0
6,0
8,0
10,0
0,0 0,5 1,0 1,5 2,0
Exkretion [mg/g GTS]
LDL-
Cho
lest
erol
[mm
ol/l]
y = 1,8695x + 1,8835 r = 0,52
y = -0,4251x + 1,6688 r = 0,31
y = 1,2356x + 3,1755 r = 0,36
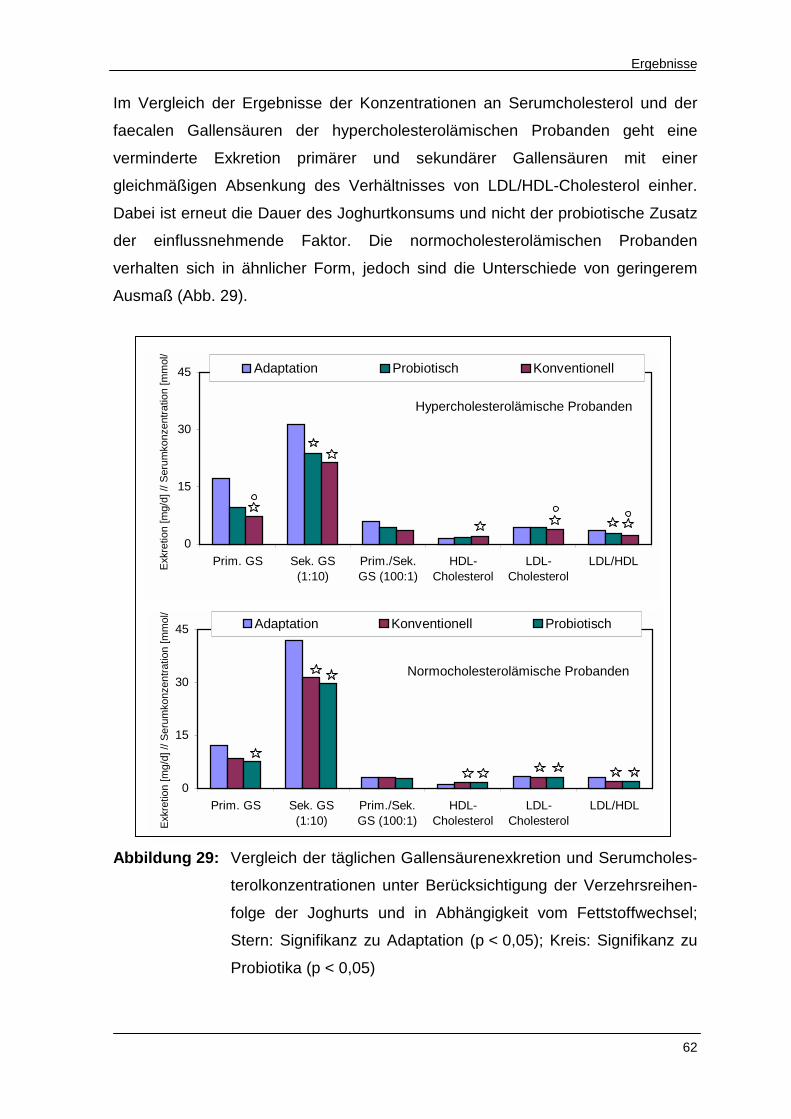
Ergebnisse
62
0
15
30
45
Prim. GS Sek. GS(1:10)
Prim./Sek.GS (100:1)
HDL-Cholesterol
LDL-Cholesterol
LDL/HDLExkr
etio
n [m
g/d]
// S
erum
konz
entra
tion
[mm
ol/
Adaptation Probiotisch Konventionell
0
15
30
45
Prim. GS Sek. GS(1:10)
Prim./Sek.GS (100:1)
HDL-Cholesterol
LDL-Cholesterol
LDL/HDL
Exkr
etio
n [m
g/d]
// S
erum
konz
entra
tion
[mm
ol/
Adaptation Konventionell Probiotisch
Im Vergleich der Ergebnisse der Konzentrationen an Serumcholesterol und der
faecalen Gallensäuren der hypercholesterolämischen Probanden geht eine
verminderte Exkretion primärer und sekundärer Gallensäuren mit einer
gleichmäßigen Absenkung des Verhältnisses von LDL/HDL-Cholesterol einher.
Dabei ist erneut die Dauer des Joghurtkonsums und nicht der probiotische Zusatz
der einflussnehmende Faktor. Die normocholesterolämischen Probanden
verhalten sich in ähnlicher Form, jedoch sind die Unterschiede von geringerem
Ausmaß (Abb. 29).
Abbildung 29: Vergleich der täglichen Gallensäurenexkretion und Serumcholes-
terolkonzentrationen unter Berücksichtigung der Verzehrsreihen-
folge der Joghurts und in Abhängigkeit vom Fettstoffwechsel;
Stern: Signifikanz zu Adaptation (p < 0,05); Kreis: Signifikanz zu
Probiotika (p < 0,05)
Hypercholesterolämische Probanden
Normocholesterolämische Probanden
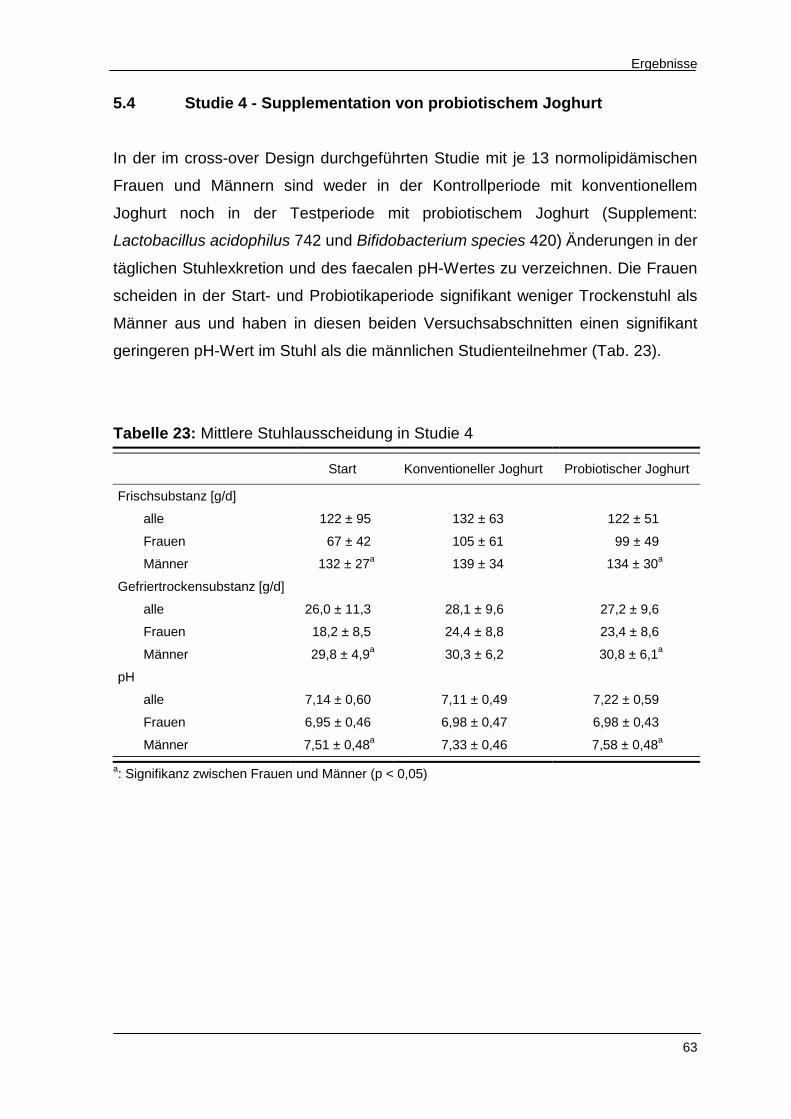
Ergebnisse
63
5.4 Studie 4 - Supplementation von probiotischem Joghurt
In der im cross-over Design durchgeführten Studie mit je 13 normolipidämischen
Frauen und Männern sind weder in der Kontrollperiode mit konventionellem
Joghurt noch in der Testperiode mit probiotischem Joghurt (Supplement:
Lactobacillus acidophilus 742 und Bifidobacterium species 420) Änderungen in der
täglichen Stuhlexkretion und des faecalen pH-Wertes zu verzeichnen. Die Frauen
scheiden in der Start- und Probiotikaperiode signifikant weniger Trockenstuhl als
Männer aus und haben in diesen beiden Versuchsabschnitten einen signifikant
geringeren pH-Wert im Stuhl als die männlichen Studienteilnehmer (Tab. 23).
Tabelle 23: Mittlere Stuhlausscheidung in Studie 4
Start Konventioneller Joghurt Probiotischer Joghurt
Frischsubstanz [g/d]
alle 122 ± 95 132 ± 63 122 ± 51
Frauen 67 ± 42 105 ± 61 99 ± 49 Männer 132 ± 27a 139 ± 34 134 ± 30a
Gefriertrockensubstanz [g/d]
alle 26,0 ± 11,3 28,1 ± 9,6 27,2 ± 9,6 Frauen 18,2 ± 8,5 24,4 ± 8,8 23,4 ± 8,6
Männer 29,8 ± 4,9a 30,3 ± 6,2 30,8 ± 6,1a
pH alle 7,14 ± 0,60 7,11 ± 0,49 7,22 ± 0,59
Frauen 6,95 ± 0,46 6,98 ± 0,47 6,98 ± 0,43
Männer 7,51 ± 0,48a 7,33 ± 0,46 7,58 ± 0,48a
a: Signifikanz zwischen Frauen und Männer (p < 0,05)

Ergebnisse
64
Drei Probanden (Nummer: 12, 22, 26) werden aufgrund ihres inversen Sterol- und
Gallensäurenprofils in der statistischen Auswertung nicht berücksichtigt.
Der Konsum von konventionellem Joghurt führt zu signifikant erhöhten Gehalten
an Cholesterol und Cholestanol in der Gefriertrockensubstanz im Vergleich zu den
Ausgangswerten. Daraus folgt eine verminderte Konversionsrate des
Cholesterols. Nach Verzehr des mit probiotischen Keimen supplementierten
Produktes sind die Konzentrationen von Cholesterol und den Metaboliten in der
Gefriertrockensubstanz signifikant zu den Kontrollgehalten vermindert. Die
Konzentrationen an Gallensäuren in der Gefriertrockensubstanz sind im gesamten
Verlauf der Studie unverändert (Tab. 24).
Die tägliche Exkretion der Sterole ist nach konventioneller Joghurtgabe hinsichtlich
der Startbedingungen erhöht. Mit Ausnahme des Coprostanols tendieren die
Sterolexkretionen pro Tag nach der probiotischen Periode ebenfalls zu erhöhten
Werten. Die Ausscheidung der sekundären Gallensäure DCA ist nach dem
Verzehr von Joghurt im Vergleich zu den Ausgangswerten und unabhängig von
der Joghurtart angestiegen. Weiterhin ist eine signifikant erhöhte tägliche
Exkretion an der primären Gallensäure CDCA nach Gabe von probiotischem
Joghurt im Vergleich zu den Basisdaten zu beobachten. Da die Probanden (vor
allem die Frauen) in der Startperiode geringe Mengen an Stuhl ausscheiden, ist
ein Einfluss durch ansteigende Stuhlmassen nach den Joghurtperioden auf die
höhere tägliche Steroidexkretion nicht auszuschließen.
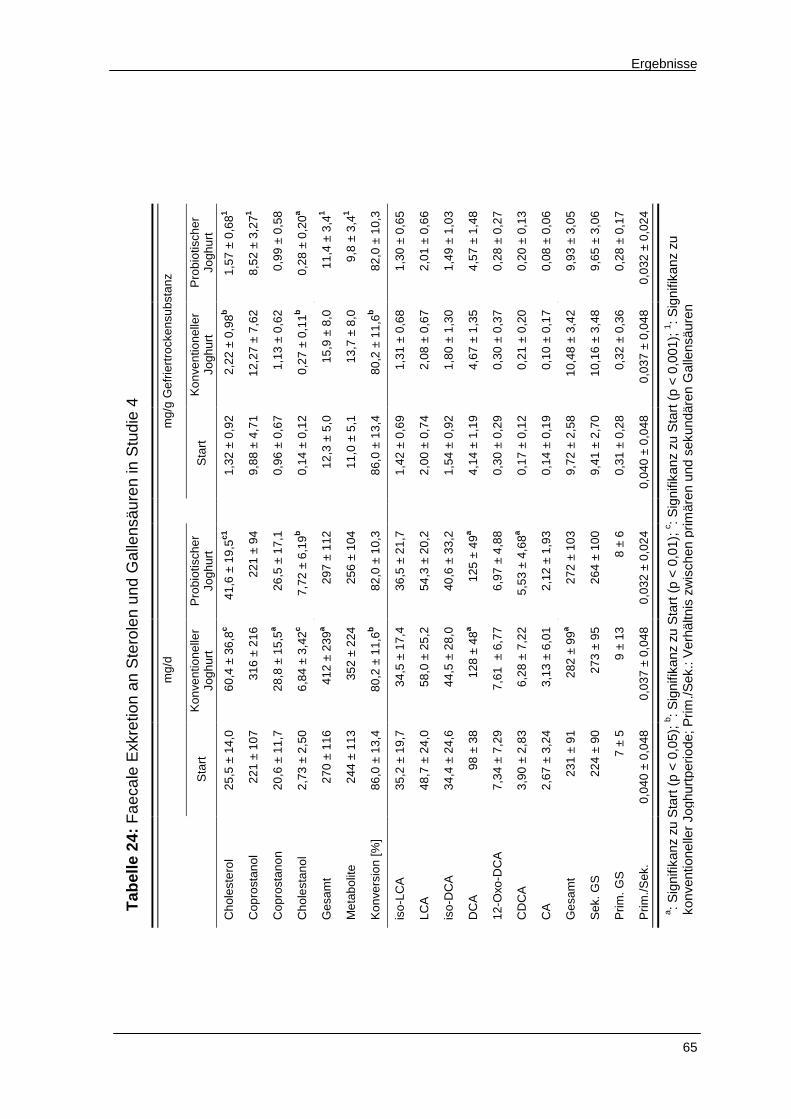
Ergebnisse
65
Prob
iotis
cher
Jo
ghur
t
1,57
± 0
,681
8,52
± 3
,271
0,99
± 0
,58
0,28
± 0
,20a
11,4
± 3
,41
9,8
± 3,
41
82,0
± 1
0,3
1,30
± 0
,65
2,01
± 0
,66
1,49
± 1
,03
4,57
± 1
,48
0,28
± 0
,27
0,20
± 0
,13
0,08
± 0
,06
9,93
± 3
,05
9,65
± 3
,06
0,28
± 0
,17
0,03
2 ±
0,02
4
Konv
entio
nelle
r Jo
ghur
t
2,22
± 0
,98b
12,2
7 ±
7,62
1,13
± 0
,62
0,27
± 0
,11b
15,9
± 8
,0
13,7
± 8
,0
80,2
± 1
1,6b
1,31
± 0
,68
2,08
± 0
,67
1,80
± 1
,30
4,67
± 1
,35
0,30
± 0
,37
0,21
± 0
,20
0,10
± 0
,17
10,4
8 ±
3,42
10,1
6 ±
3,48
0,32
± 0
,36
0,03
7 ±
0,04
8
mg/
g G
efrie
rtroc
kens
ubst
anz
Star
t
1,32
± 0
,92
9,88
± 4
,71
0,96
± 0
,67
0,14
± 0
,12
12,3
± 5
,0
11,0
± 5
,1
86,0
± 1
3,4
1,42
± 0
,69
2,00
± 0
,74
1,54
± 0
,92
4,14
± 1
,19
0,30
± 0
,29
0,17
± 0
,12
0,14
± 0
,19
9,72
± 2
,58
9,41
± 2
,70
0,31
± 0
,28
0,04
0 ±
0,04
8
Prob
iotis
cher
Jo
ghur
t
41,6
± 1
9,5c1
221
± 94
26,5
± 1
7,1
7,72
± 6
,19b
297
± 11
2
256
± 10
4
82,0
± 1
0,3
36,5
± 2
1,7
54,3
± 2
0,2
40,6
± 3
3,2
125
± 49
a
6,97
± 4
,88
5,53
± 4
,68a
2,12
± 1
,93
272
± 10
3
264
± 10
0
8 ±
6
0,03
2 ±
0,02
4
Konv
entio
nelle
r Jo
ghur
t
60,4
± 3
6,8c
316
± 21
6
28,8
± 1
5,5a
6,84
± 3
,42c
412
± 23
9a
352
± 22
4
80,2
± 1
1,6b
34,5
± 1
7,4
58,0
± 2
5,2
44,5
± 2
8,0
128
± 48
a
7,61
± 6
,77
6,28
± 7
,22
3,13
± 6
,01
282
± 99
a
273
± 95
9 ±
13
0,03
7 ±
0,04
8
mg/
d
Star
t
25,5
± 1
4,0
221
± 10
7
20,6
± 1
1,7
2,73
± 2
,50
270
± 11
6
244
± 11
3
86,0
± 1
3,4
35,2
± 1
9,7
48,7
± 2
4,0
34,4
± 2
4,6
98 ±
38
7,34
± 7
,29
3,90
± 2
,83
2,67
± 3
,24
231
± 91
224
± 90
7 ±
5
0,04
0 ±
0,04
8
Cho
lest
erol
Cop
rost
anol
Cop
rost
anon
Cho
lest
anol
Ges
amt
Met
abol
ite
Konv
ersi
on [%
]
iso-
LCA
LCA
iso-
DC
A
DC
A
12-O
xo-D
CA
CD
CA
CA
Ges
amt
Sek.
GS
Prim
. GS
Prim
./Sek
.
Tabe
lle 2
4: F
aeca
le E
xkre
tion
an S
tero
len
und
Gal
lens
äure
n in
Stu
die
4
a : Sig
nifik
anz
zu S
tart
(p <
0,0
5); b : S
igni
fikan
z zu
Sta
rt (p
< 0
,01)
; c : Sig
nifik
anz
zu S
tart
(p <
0,0
01);
1 : Sig
nifik
anz
zu
konv
entio
nelle
r Jo g
hurtp
erio
de; P
rim./S
ek.:
Verh
ältn
is z
wis
chen
prim
ären
und
sek
undä
ren
Gal
lens
äure
n

Ergebnisse
66
Die Auswertung der täglichen Steroidexkretion nach Separation in Gruppen mit
unterschiedlicher Reihenfolge des Joghurtverzehrs ergibt eine gleichförmige
Veränderung des Steroidprofiles mit Ausnahme von Coprostanol (Abb. 30). Die
mittlere Coprostanolausscheidung ist nach Konsum von herkömmlichen Joghurt
erhöht, vor allem bei den Probanden, denen in der ersten Studienperiode
probiotischer Joghurt verabreicht wurde.
In der Probandengruppe, die zuerst konventionellen Joghurt verzehrt hat, sind die
Änderungen in der Gallensäureexkretion im zeitlichen Verlauf verstärkt
ausgeprägt. Hingegen ist mit beginnender probiotischer Joghurtgabe die tägliche
Exkretion faecaler Sterole im Studienverlauf signifikant beeinflusst.
Abbildung 30: Tägliche faecale Steroidexkretion in Abhängigkeit von der
Verzehrsreihenfolge der Joghurts
Stern: Signifikanz zwischen Perioden (p < 0,05)
01020304050607080
Prim. GS Sek. GS(1:10)
Cholesterol(1:10)
Coprostanol(1:10)
Prim. GS Sek. GS(1:10)
Cholesterol(1:10)
Coprostanol(1:10)
Exkr
etio
n [m
g/d]
Konventionell Probiotisch

Ergebnisse
67
Zwischen dem LDL/HDL-Cholesterol-Verhältnis im Serum und der Konzentration
der faecalen Steroide in der Gefriertrockensubstanz bestehen lineare
Beziehungen, wobei der Gehalt primärer Gallensäuren im Stuhl positiv (p < 0,01)
und die Konzentrationen sekundärer Gallensäuren negativ (p < 0,01) mit dem
Quotienten korrelieren (Abb. 31). Keine mathematischen Zusammenhänge
können zu den Gehalten in der Stuhlgefriertrockensubstanz an Cholesterol,
Coprostanol und der Konversionsrate von Cholesterol nachgewiesen werden.
Abbildung 31: Zusammenhänge zwischen den Konzentrationen an Serum-
cholesterol und primärer bzw. sekundärer Gallensäuren im Stuhl
(n = 69, p < 0,01)
Sekundäre Gallensäuren
0,0
1,0
2,0
3,0
4,0
0,0 5,0 10,0 15,0 20,0 25,0
Exkretion [mg/g GTS]
LDL/
HD
L-C
hole
ster
ol
Primäre Gallensäuren
0,0
1,0
2,0
3,0
4,0
0,0 0,5 1,0 1,5
Exkretion [mg/g GTS]
LDL/
HD
L-C
hole
ster
ol
y = 0,8641x + 1,3638 r = 0,33
y = -0,0599x + 2,1751 r = 0,35
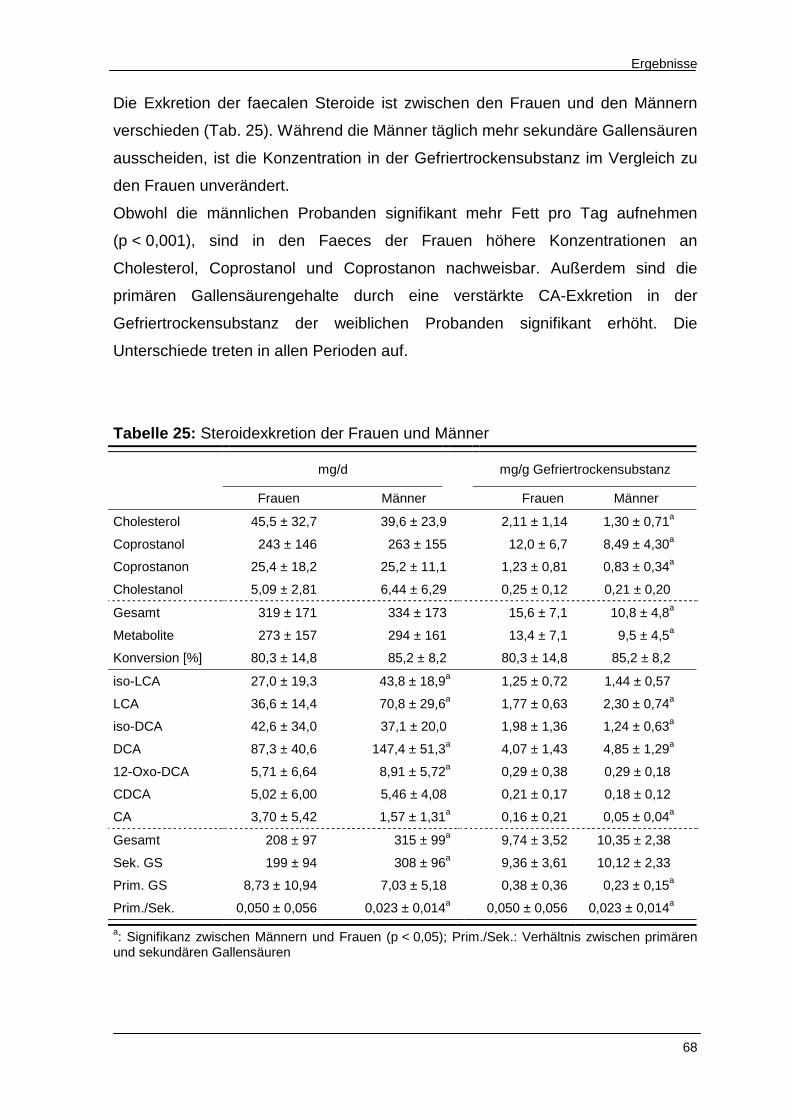
Ergebnisse
68
Die Exkretion der faecalen Steroide ist zwischen den Frauen und den Männern
verschieden (Tab. 25). Während die Männer täglich mehr sekundäre Gallensäuren
ausscheiden, ist die Konzentration in der Gefriertrockensubstanz im Vergleich zu
den Frauen unverändert.
Obwohl die männlichen Probanden signifikant mehr Fett pro Tag aufnehmen
(p < 0,001), sind in den Faeces der Frauen höhere Konzentrationen an
Cholesterol, Coprostanol und Coprostanon nachweisbar. Außerdem sind die
primären Gallensäurengehalte durch eine verstärkte CA-Exkretion in der
Gefriertrockensubstanz der weiblichen Probanden signifikant erhöht. Die
Unterschiede treten in allen Perioden auf.
Tabelle 25: Steroidexkretion der Frauen und Männer
mg/d
mg/g Gefriertrockensubstanz
Frauen Männer Frauen Männer
Cholesterol 45,5 ± 32,7 39,6 ± 23,9 2,11 ± 1,14 1,30 ± 0,71a
Coprostanol 243 ± 146 263 ± 155 12,0 ± 6,7 8,49 ± 4,30a Coprostanon 25,4 ± 18,2 25,2 ± 11,1 1,23 ± 0,81 0,83 ± 0,34a
Cholestanol 5,09 ± 2,81 6,44 ± 6,29 0,25 ± 0,12 0,21 ± 0,20
Gesamt 319 ± 171 334 ± 173 15,6 ± 7,1 10,8 ± 4,8a
Metabolite 273 ± 157 294 ± 161 13,4 ± 7,1 9,5 ± 4,5a Konversion [%] 80,3 ± 14,8 85,2 ± 8,2 80,3 ± 14,8 85,2 ± 8,2
iso-LCA 27,0 ± 19,3 43,8 ± 18,9a 1,25 ± 0,72 1,44 ± 0,57
LCA 36,6 ± 14,4 70,8 ± 29,6a 1,77 ± 0,63 2,30 ± 0,74a iso-DCA 42,6 ± 34,0 37,1 ± 20,0 1,98 ± 1,36 1,24 ± 0,63a
DCA 87,3 ± 40,6 147,4 ± 51,3a 4,07 ± 1,43 4,85 ± 1,29a
12-Oxo-DCA 5,71 ± 6,64 8,91 ± 5,72a 0,29 ± 0,38 0,29 ± 0,18 CDCA 5,02 ± 6,00 5,46 ± 4,08 0,21 ± 0,17 0,18 ± 0,12
CA 3,70 ± 5,42 1,57 ± 1,31a 0,16 ± 0,21 0,05 ± 0,04a
Gesamt 208 ± 97 315 ± 99a 9,74 ± 3,52 10,35 ± 2,38
Sek. GS 199 ± 94 308 ± 96a 9,36 ± 3,61 10,12 ± 2,33 Prim. GS 8,73 ± 10,94 7,03 ± 5,18 0,38 ± 0,36 0,23 ± 0,15a
Prim./Sek. 0,050 ± 0,056 0,023 ± 0,014a 0,050 ± 0,056 0,023 ± 0,014a
a: Signifikanz zwischen Männern und Frauen (p < 0,05); Prim./Sek.: Verhältnis zwischen primären und sekundären Gallensäuren
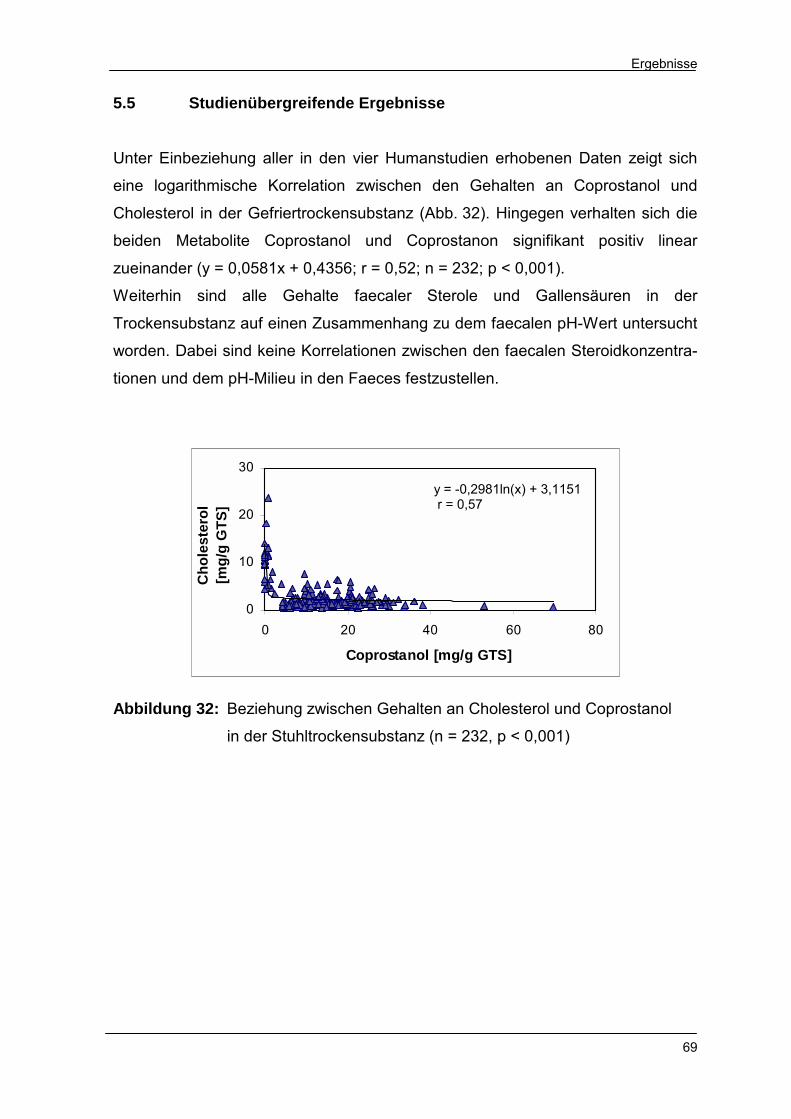
Ergebnisse
69
5.5 Studienübergreifende Ergebnisse Unter Einbeziehung aller in den vier Humanstudien erhobenen Daten zeigt sich
eine logarithmische Korrelation zwischen den Gehalten an Coprostanol und
Cholesterol in der Gefriertrockensubstanz (Abb. 32). Hingegen verhalten sich die
beiden Metabolite Coprostanol und Coprostanon signifikant positiv linear
zueinander (y = 0,0581x + 0,4356; r = 0,52; n = 232; p < 0,001).
Weiterhin sind alle Gehalte faecaler Sterole und Gallensäuren in der
Trockensubstanz auf einen Zusammenhang zu dem faecalen pH-Wert untersucht
worden. Dabei sind keine Korrelationen zwischen den faecalen Steroidkonzentra-
tionen und dem pH-Milieu in den Faeces festzustellen.
Abbildung 32: Beziehung zwischen Gehalten an Cholesterol und Coprostanol
in der Stuhltrockensubstanz (n = 232, p < 0,001)
0
10
20
30
0 20 40 60 80
Coprostanol [mg/g GTS]
Cho
lest
erol
[mg/
g G
TS]
y = -0,2981ln(x) + 3,1151 r = 0,57
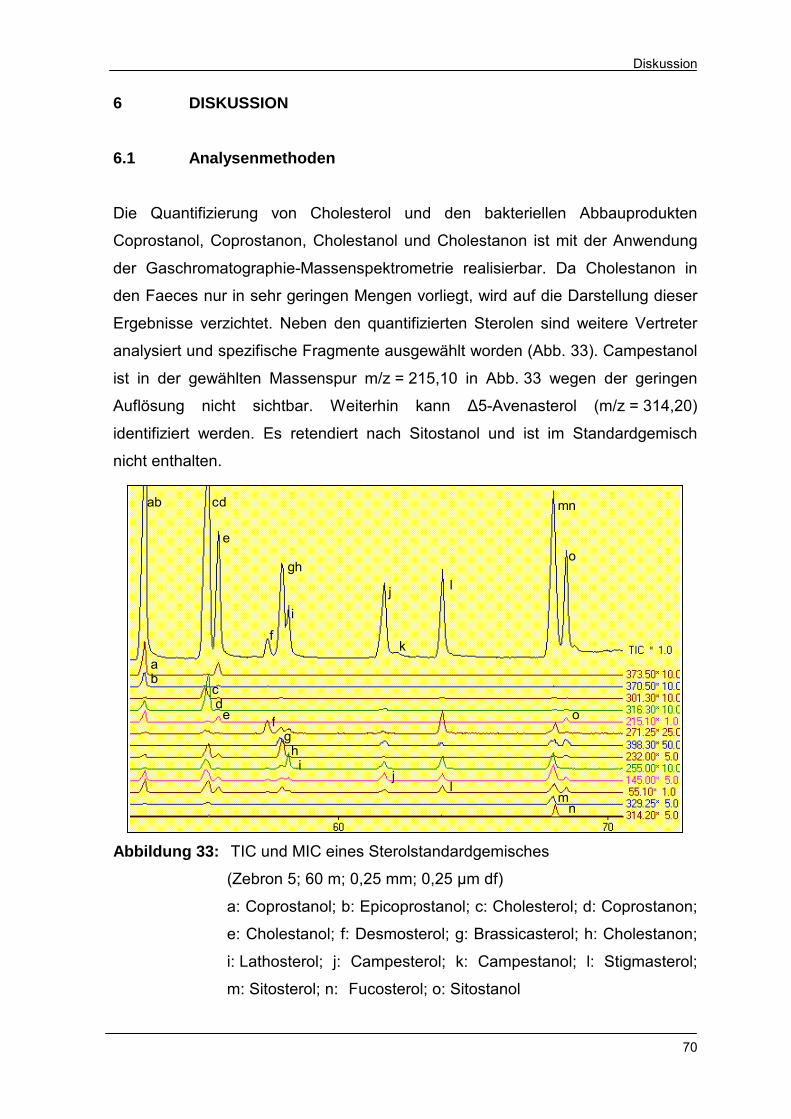
Diskussion
70
6 DISKUSSION
6.1 Analysenmethoden Die Quantifizierung von Cholesterol und den bakteriellen Abbauprodukten
Coprostanol, Coprostanon, Cholestanol und Cholestanon ist mit der Anwendung
der Gaschromatographie-Massenspektrometrie realisierbar. Da Cholestanon in
den Faeces nur in sehr geringen Mengen vorliegt, wird auf die Darstellung dieser
Ergebnisse verzichtet. Neben den quantifizierten Sterolen sind weitere Vertreter
analysiert und spezifische Fragmente ausgewählt worden (Abb. 33). Campestanol
ist in der gewählten Massenspur m/z = 215,10 in Abb. 33 wegen der geringen
Auflösung nicht sichtbar. Weiterhin kann ∆5-Avenasterol (m/z = 314,20)
identifiziert werden. Es retendiert nach Sitostanol und ist im Standardgemisch
nicht enthalten.
Abbildung 33: TIC und MIC eines Sterolstandardgemisches
(Zebron 5; 60 m; 0,25 mm; 0,25 µm df)
a: Coprostanol; b: Epicoprostanol; c: Cholesterol; d: Coprostanon;
e: Cholestanol; f: Desmosterol; g: Brassicasterol; h: Cholestanon;
i: Lathosterol; j: Campesterol; k: Campestanol; l: Stigmasterol;
m: Sitosterol; n: Fucosterol; o: Sitostanol
a
b
b
c
c d
e
e
f i
g
g
h
i j
j
k
l
l m
m n
n
o
o
a
fd
h

Diskussion
71
Im Literaturvergleich zeigt sich, dass die beschriebene Analysenmethode zur
Bestimmung von Sterolen tierischen Ursprungs und deren intestinalen
Abbauprodukten sowie von Phytosterolen mit etablierten Methoden vergleichbar
ist (Tab. 26). Die Analyse von Abbauprodukten der Pflanzensterole mit dieser
Methode ist aufgrund nicht zur Verfügung stehender Referenzsubstanzen nicht
möglich gewesen. Für zukünftige Untersuchungen bei denen der Gehalt
pflanzlicher Sterole in den Faeces von Interesse sein kann, wird die Erweiterung
des Sterolprofiles in der Analyse mit den Abbauprodukten der Phytosterole
angestrebt und weiterhin ist die Anwendung eines radiomarkierten Sterols mit
einer Hydroxygruppe an Position 3 des Steroidgerüstes als interner Standard zu
prüfen.
Die Methoden von BARKER et al. [1993], CZUBAYKO et al. [1991] und MIETTINEN
[1982] beinhalten die Analyse von Coprostanon � eines der Hauptabbauprodukte
des Cholesterols � ohne auf die Artefaktbildung von 3-Oxo-Steroiden nach einer
Trimethylsilylierung einzugehen. In eigenen Voruntersuchungen zeigt sich, dass in
reinen Standardsubstanzen die Trimethylsilylierung des Coprostanon frei von
Nebenprodukten möglich ist. Hingegen werden durch mögliche Einflüsse der
komplexen Faecesmatrix in realen Proben Artefakte des Coprostanons gefunden.
Lediglich CHILD et al. [1987] verweisen auf das instabile Verhalten dieser
Steroidgruppe und führen Coprostanon in der Sterolbestimmung nicht auf.
Die Hydrolyse mit ethanolischer Natronlauge ist mit der beschriebenen Methode
weiterhin notwendig, um die GC-Kapillarsäule vor hochsiedenden Verun-
reinigungen zu schützen. Aus diesem Grund ist nur die Analyse unveresterter
Sterole möglich. BATTA et al. [1999] beschreiben eine simultane Sterol- und
Gallensäurenanalysenmethode, bei der die Methylierung und Silylierung direkt in
der gepulverten Trockenfaecesprobe durchgeführt und die Steroide anschließend
extrahiert werden. Das Chromatogramm zeigt viele unidentifizierte Peaks und
Coprostanon sowie Cholestanol werden nicht qualitativ angegeben. Im Bereich
der Gallensäuren wird keine Basislinientrennung von iso-DCA und DCA erreicht,
iso-LCA scheint nicht enthalten zu sein, dafür zeigt der LCA-Peak im unteren
Bereich eine leichte Verbreiterung, die auf eine Überlagerung deuten kann
(Abb. 34).
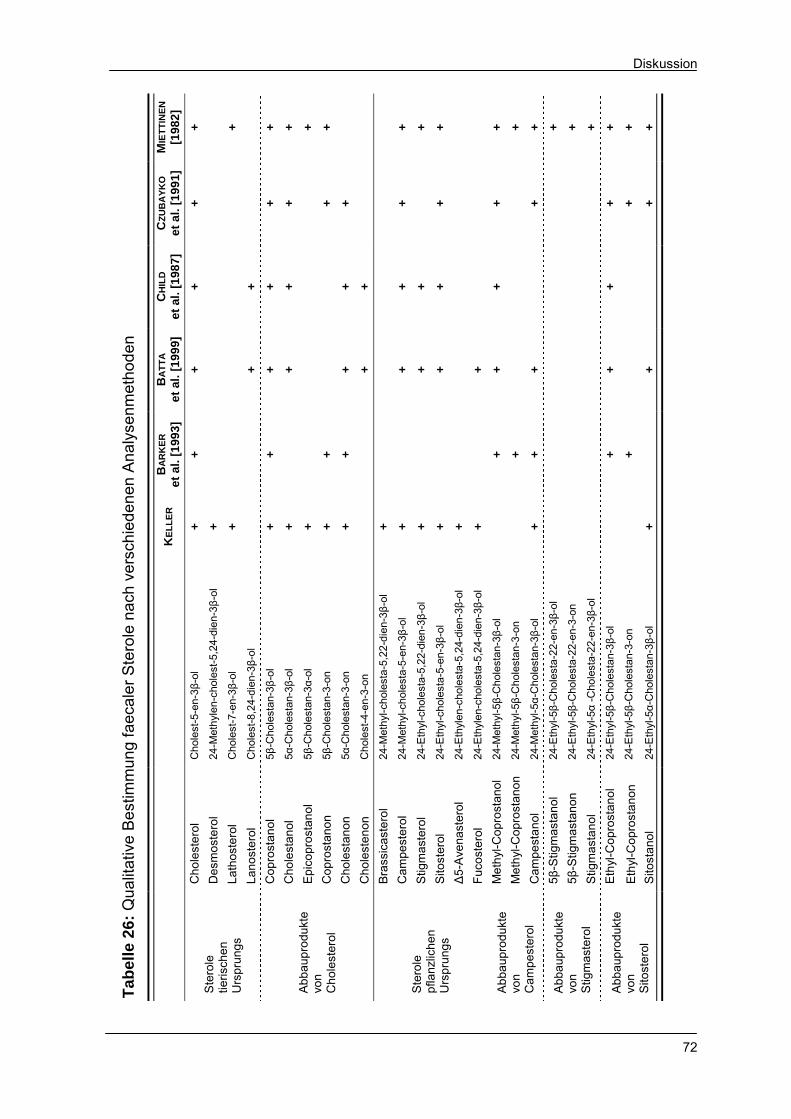
Diskussion
72
MIE
TTIN
EN
[198
2]
+ + + + + + + + + + + + + + + + + +
CZU
BAY
KO
et
al.
[199
1]
+ + + + + + + + + + + +
CH
ILD
et
al.
[198
7]
+ + + + + + + + + + +
BAT
TA
et a
l. [1
999]
+ + + + + + + + + + + + + +
BAR
KER
et
al.
[199
3]
+ + + + + + + + +
KEL
LER
+ + + + + + + + + + + + + + + +
Cho
lest
-5-e
n-3β
-ol
24-M
ethy
len-
chol
est-5
,24-
dien
-3β-
ol
Cho
lest
-7-e
n-3β
-ol
Cho
lest
-8,2
4-di
en-3β-
ol
5β-C
hole
stan
-3β-
ol
5α-C
hole
stan
-3β-
ol
5β-C
hole
stan
-3α-
ol
5β-C
hole
stan
-3-o
n
5α-C
hole
stan
-3-o
n
Cho
lest
-4-e
n-3-
on
24-M
ethy
l-cho
lest
a-5,
22-d
ien-
3β-o
l
24-M
ethy
l-cho
lest
a-5-
en-3β-
ol
24-E
thyl
-cho
lest
a-5,
22-d
ien-
3β-o
l
24-E
thyl
-cho
lest
a-5-
en-3β-
ol
24-E
thyl
en-c
hole
sta-
5,24
-die
n-3β
-ol
24-E
thyl
en-c
hole
sta-
5,24
-die
n-3β
-ol
24-M
ethy
l-5β-
Cho
lest
an-3β-
ol
24-M
ethy
l-5β-
Cho
lest
an-3
-on
24-M
ethy
l-5α-
Cho
lest
an-3β-
ol
24-E
thyl
-5β-
Cho
lest
a-22
-en-
3β-o
l
24-E
thyl
-5β-
Cho
lest
a-22
-en-
3-on
24-E
thyl
-5α
-Cho
lest
a-22
-en-
3β-o
l
24-E
thyl
-5β-
Cho
lest
an-3β-
ol
24-E
thyl
-5β-
Cho
lest
an-3
-on
24-E
thyl
-5α-
Cho
lest
an-3β-
ol
Cho
lest
erol
D
esm
oste
rol
Lath
oste
rol
Lano
ster
ol
Cop
rost
anol
C
hole
stan
ol
Epic
opro
stan
ol
Cop
rost
anon
C
hole
stan
on
Cho
lest
enon
Br
assi
cast
erol
C
ampe
ster
ol
Stig
mas
tero
l Si
tost
erol
∆5
-Ave
nast
erol
Fu
cost
erol
M
ethy
l-Cop
rost
anol
M
ethy
l-Cop
rost
anon
C
ampe
stan
ol
5β-S
tigm
asta
nol
5β-S
tigm
asta
non
Stig
mas
tano
l Et
hyl-C
opro
stan
ol
Ethy
l-Cop
rost
anon
Si
tost
anol
Ster
ole
tieris
chen
U
rspr
ungs
Abba
upro
dukt
e vo
n C
hole
ster
ol
Ster
ole
pfla
nzlic
hen
Urs
prun
gs
Abba
upro
dukt
e vo
n C
ampe
ster
ol
Abba
upro
dukt
e vo
n St
igm
aste
rol
Abba
upro
dukt
e vo
n Si
tost
erol
Tabe
lle 2
6: Q
ualit
ativ
e Be
stim
mun
g fa
ecal
er S
tero
le n
ach
vers
chie
dene
n An
alys
enm
etho
den
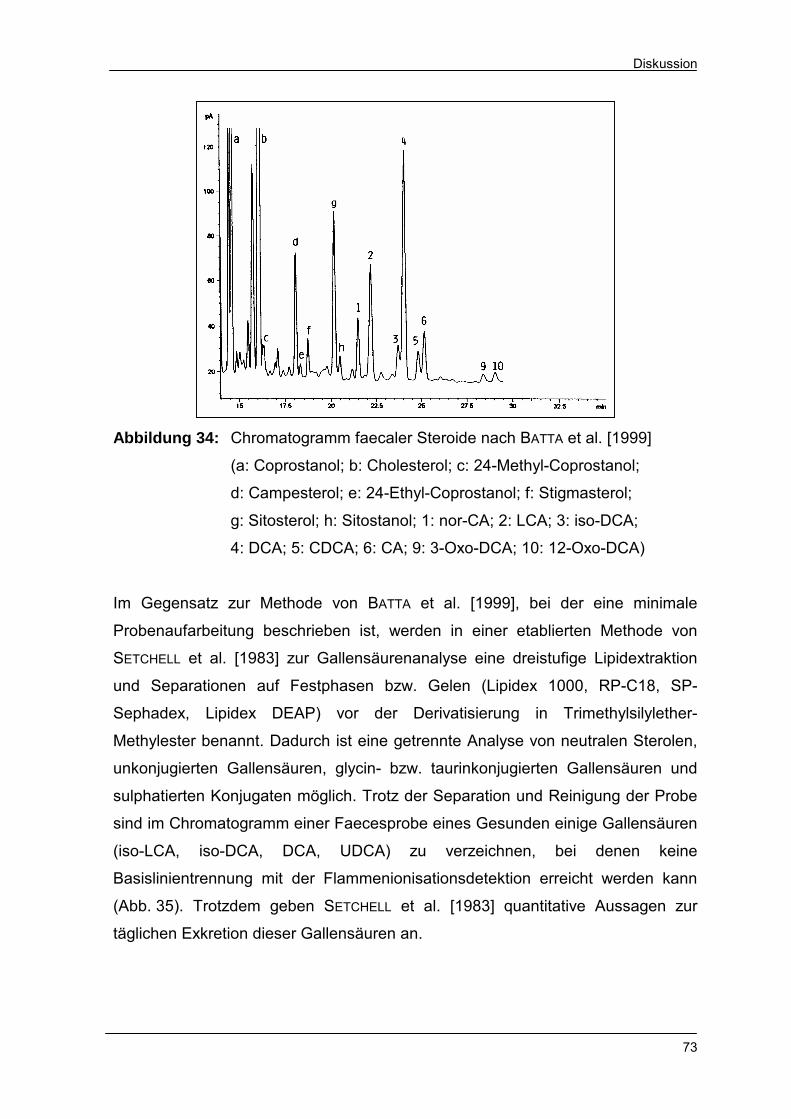
Diskussion
73
Abbildung 34: Chromatogramm faecaler Steroide nach BATTA et al. [1999]
(a: Coprostanol; b: Cholesterol; c: 24-Methyl-Coprostanol;
d: Campesterol; e: 24-Ethyl-Coprostanol; f: Stigmasterol;
g: Sitosterol; h: Sitostanol; 1: nor-CA; 2: LCA; 3: iso-DCA;
4: DCA; 5: CDCA; 6: CA; 9: 3-Oxo-DCA; 10: 12-Oxo-DCA)
Im Gegensatz zur Methode von BATTA et al. [1999], bei der eine minimale
Probenaufarbeitung beschrieben ist, werden in einer etablierten Methode von
SETCHELL et al. [1983] zur Gallensäurenanalyse eine dreistufige Lipidextraktion
und Separationen auf Festphasen bzw. Gelen (Lipidex 1000, RP-C18, SP-
Sephadex, Lipidex DEAP) vor der Derivatisierung in Trimethylsilylether-
Methylester benannt. Dadurch ist eine getrennte Analyse von neutralen Sterolen,
unkonjugierten Gallensäuren, glycin- bzw. taurinkonjugierten Gallensäuren und
sulphatierten Konjugaten möglich. Trotz der Separation und Reinigung der Probe
sind im Chromatogramm einer Faecesprobe eines Gesunden einige Gallensäuren
(iso-LCA, iso-DCA, DCA, UDCA) zu verzeichnen, bei denen keine
Basislinientrennung mit der Flammenionisationsdetektion erreicht werden kann
(Abb. 35). Trotzdem geben SETCHELL et al. [1983] quantitative Aussagen zur
täglichen Exkretion dieser Gallensäuren an.

Diskussion
74
Abbildung 35: Chromatogramm faecaler Gallensäuren nach SETCHELL et al.
[1983]
(1: iso-LCA; 2: LCA; 3: iso-DCA; 4: DCA; 5: CDCA; 6: CA;
7: UDCA; 8: 7β-CA; 9: 12-Oxo-iso-DCA; 10: 12-Oxo-DCA)
Beim Übergang zur massenspektrometrischen Detektion ist die vollständige
Separation mit Ausnahme von UDCA realisierbar. Wahrscheinlich ist die relative
Intensität des gewählten Fragments m/z = 460,00 mit 14 % zu gering um
reproduzierbare Ergebnisse zu erhalten (vgl. 3.2.3). Außerdem kann diese
Massenspur eventuell durch andere Steroide gestört sein, da bei SETCHELL et al.
[1983] im Zeitbereich der UDCA des Chromatogramms eine Peakhäufung zu
erkennen ist. Die Wahl eines anderen Fragmentes bzw. die Anwendung eines
Targetfragmentes zur Peakreinheitskontrolle (Prüfung eines konstanten
Verhältnisses zweier Fragmente der UDCA) in der Routineanalyse sollten
zukünftig geprüft werden.

Diskussion
75
Mit Ausnahme der iso-DCA (m/z = 75,10), CDCA (m/z = 73,10) und HDCA
(m/z = 81,15) sind Strukturangaben zu den analysierten Fragmenten aller anderen
Sterole und Gallensäuren möglich (Abb. 36).
M [g/mol] Fragment m/z M
[g/mol] Fragment m/z
Cholestan 372 M � (15) 357 iso-LCA 462 M � (90 + 115 + 42) 215
Coprostanol 388 M � (15) 373 LCA 462 M � (90 + 115 + 42) 215
Cholesterol 386 M � (15 + 70) 301 iso-DCA 550 - 75
Coprostanon 386 M � (70) 316 DCA 550 M � (2 � 90 + 115) 255
Cholestanol 388 M � (113 + 42 + 18) 215 12-Oxo-DCA 476 M � (90 + 115 + 40) 231
Cholestanon 386 M � (113 + 41) 232 CDCA 550 - 73
CA 638 M � (3 � 90 + 115) 253
HDCA 550 - 81
Abbildung 36: Fragmentierung der Sterole und Gallensäuren
Durch den Verlust einer Methylgruppe werden bei Cholestan und Coprostanol
Fragmente gebildet, deren Masse um m/z = 15,00 geringer ist als die
Molekülmasse. Hierbei werden wahrscheinlich angulare Methylgruppen des
Steroidgerüstes abgespalten, wie LAWSON und SETCHELL [1988] es für die
strukturähnlichen Gallensäuren beschreiben.
OM - (70)
M - (113)
M - (113 + 41)M - (113 + 42)
M - (70)
M - (90)
M - (115)
M - (115 + 40)M - (115 + 42)
O
O
OTMS
Meth
Sterole Gallensäuren

Diskussion
76
Sowohl bei den Sterolen als auch bei den Gallensäuren ist die Abspaltung der
Seitenketten und das Aufbrechen des Cyclopentanringes aufgetreten. Dabei
können die Protonen entweder am Steroidgerüst verbleiben oder mit der
abgespaltenen Gruppe aus dem Molekül austreten. Dadurch sind Fragmente mit
M - (113 + 42), M - (113 + 41) bzw. M - (113 + 40) möglich. Die verschiedenen
Fragmentierungsprodukte werden übereinstimmend bei SJÖVALL et al. [1971]
beschrieben.
Die Trimethylsilylgruppen werden mit einem Proton des Steroidgerüstes als
Alkohol abgespalten, so dass Fragmente mit M - (n � 90) gebildet werden
(n = Anzahl der Trimethylsilylgruppen im Molekül). Die Abgangsgruppen werden
von IIDA et al. [2001] auch bei tertiär hydroxylierten Steroiden detektiert. SJÖVALL
et al. [1971] verweisen weiterhin auf den Austritt von Trimethylsiloxygruppen bei
dem Fragmente mit M - (n � 89) gebildet werden. Diese Fragmente sind in den in
dieser Arbeit erhaltenen Massenspektren � jedoch von geringer Intensität �
nachweisbar.
Für das bei Coprostanon gebildete Fragment M - (70) sind zwei Fragmentierungen
möglich. Zum einen kann ein Bruch in der Seitenkette erfolgen [SJÖVALL et al.
1971], zum anderen ist die Spaltung des Cyclohexanringes A möglich [LAWSON
und SETCHELL 1988, MATUSIK et al. 1988].

Diskussion
77
6.2 Steroidgehalte in der Trockensubstanz und die tägliche Exkretion
Unter Berücksichtigung aller Probanden aus allen Perioden der vier Humanstudien
(n = 232, 86 Probanden) ergibt sich ein mittlerer Gesamtsterolgehalt von
19,4 ± 9,1 mg/g Trockensubstanz, der im Bereich der von PEROGAMBROS et al.
[1982] und REDDY et al. [1989] beschriebenen Konzentrationen von
21,8 ± 3,7 mg/g bzw. 23,1 ± 10,9 mg/g Trockensubstanz liegt (Tab. 27). Die
Konversionsrate des Cholesterols in die Metabolite ist mit 87 ± 10 % höher als bei
BARKER et al. [1993] und KORPELA und ADLERCREUTZ [1985]. Sowohl die
Omnivoren als auch die Vegetarier in der Studie von KORPELA und ADLERCREUTZ
[1985] weisen höhere Gehalte an Cholesterol in den Faeces auf. Die von BARKER
et al. [1993] beschriebenen Ergebnisse der einzelnen Sterole sind im Vergleich zu
den hier dargestellten Werten und den Literaturangaben [KORPELA und
ADLERCREUTZ 1985, PEROGAMBROS et al. 1982, REDDY et al. 1989, WESTSTRATE et
al. 1999] deutlich niedriger, was auf die Angabe des Median und nicht des
arithmetischen Mittels zurückzuführen sein kann.
Die Angaben für die tägliche Exkretion faecaler Sterole sind in der Literatur
differenziert. Diese Unterschiede können durch ungleich abgesetzte Stuhlmengen
der Probanden hervorgerufen werden (Tab. 28). Lediglich BARTRAM et al. [1991]
berücksichtigen die tägliche Exkretion an Trockenstuhl. Sie ermitteln eine höhere
Stuhlmasse und damit eine gesteigerte tägliche Gesamtsterolexkretion im
Vergleich zu den Probanden der vorliegenden Arbeit.
Die in der Literatur beschriebene Cholesterolexkretion variiert zwischen
43 ± 10 mg/d und 287 ± 248 mg/d [NAIR et al. 1984, NAKAMURA et al. 1992]. Die
ermittelte tägliche Ausscheidung an Cholesterol der Probanden ohne inversem
Sterolprofil aus den untersuchten Humanstudien beträgt 54 ± 32 mg. Unter
Berücksichtigung aller Probanden werden im Mittel 86 ± 111 mg Cholesterol pro
Tag mit den Faeces abgegeben. Die tägliche Coprostanolausscheidung ist im
Vergleich mit den in der Literatur aufgeführten Studien erhöht. Dies führt zu einer
hohen Konversionsrate von 87 ± 10 %, die den Ergebnissen von PONZ DE LEON et
al. [1987] mit einem Cholesterolumsatz von 89 ± 10 % entspricht.
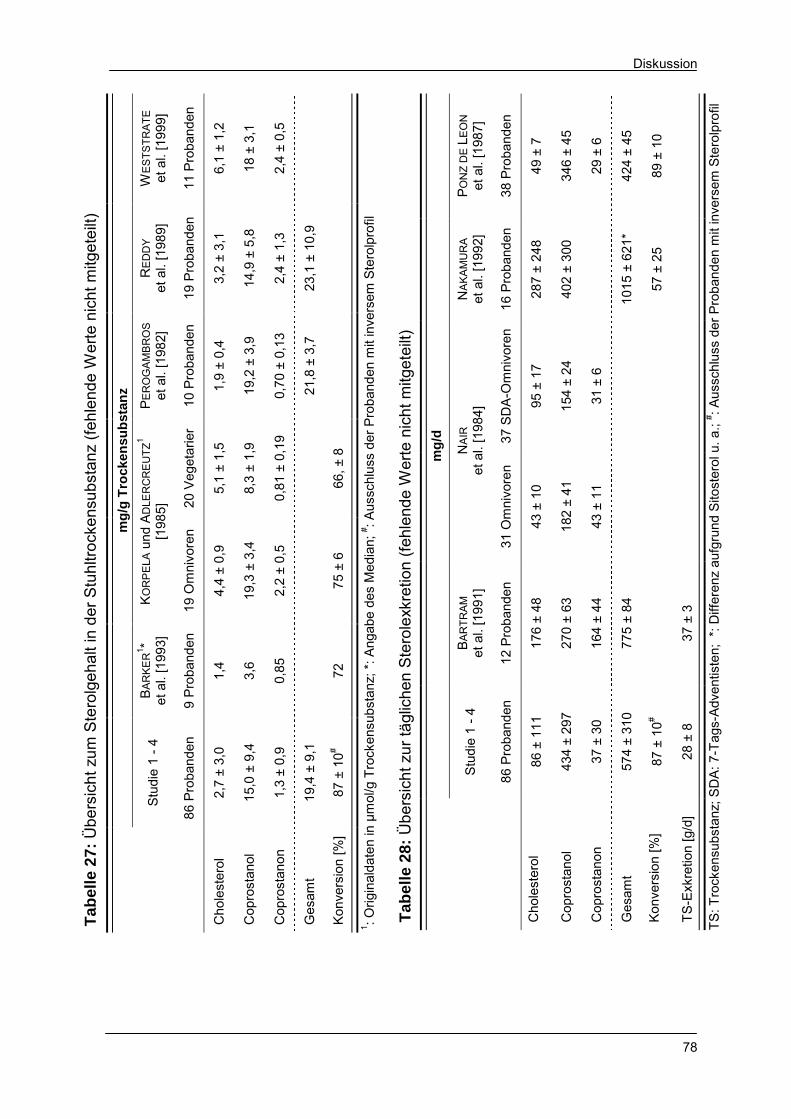
Diskussion
78
WES
TSTR
ATE
et a
l. [1
999]
11 P
roba
nden
6,1
± 1,
2
18 ±
3,1
2,4
± 0,
5
RED
DY
et a
l. [1
989]
19 P
roba
nden
3,2
± 3,
1
14,9
± 5
,8
2,4
± 1,
3
23,1
± 1
0,9
P ER
OG
AMBR
OS
et a
l. [1
982]
10 P
roba
nden
1,9
± 0,
4
19,2
± 3
,9
0,70
± 0
,13
21,8
± 3
,7
20 V
eget
arie
r
5,1
± 1,
5
8,3
± 1,
9
0,81
± 0
,19
66, ±
8
K OR
PELA
und
AD
LER
CR
EUTZ
1 [1
985]
19 O
mni
vore
n
4,4
± 0,
9
19,3
± 3
,4
2,2
± 0,
5
75 ±
6
B AR
KER
1 * et
al.
[199
3]
9 Pr
oban
den
1,4
3,6
0,85
72
mg/
g Tr
ocke
nsub
stan
z
Stud
ie 1
- 4
86 P
roba
nden
2,7
± 3,
0
15,0
± 9
,4
1,3
± 0,
9
19,4
± 9
,1
87 ±
10#
Cho
lest
erol
Cop
rost
anol
Cop
rost
anon
Ges
amt
Konv
ersi
on [%
]
P ON
Z D
E L E
ON
et
al.
[198
7]
38 P
roba
nden
49 ±
7
346
± 45
29 ±
6
424
± 45
89 ±
10
NAK
AMU
RA
et a
l. [1
992]
16 P
roba
nden
287
± 24
8
402
± 30
0
1015
± 6
21*
57 ±
25
37 S
DA-
Om
nivo
ren
95 ±
17
154
± 24
31 ±
6
NAI
R
et a
l. [1
984]
31 O
mni
vore
n
43 ±
10
182
± 41
43 ±
11
B AR
TRAM
et
al.
[199
1]
12 P
roba
nden
176
± 48
270
± 63
164
± 44
775
± 84
37 ±
3
mg/
d
Stud
ie 1
- 4
86 P
roba
nden
86 ±
111
434
± 29
7
37 ±
30
574
± 31
0
87 ±
10#
28 ±
8
Cho
lest
erol
Cop
rost
anol
Cop
rost
anon
Ges
amt
Konv
ersi
on [%
]
TS-E
xkre
tion
[g/d
]
Tabe
lle 2
7: Ü
bers
icht
zum
Ste
rolg
ehal
t in
der S
tuhl
trock
ensu
bsta
nz (f
ehle
nde
Wer
te n
icht
mitg
etei
lt)
Tabe
lle 2
8: Ü
bers
icht
zur
tägl
iche
n St
erol
exkr
etio
n (fe
hlen
de W
erte
nic
ht m
itget
eilt)
TS: T
rock
ensu
bsta
nz; S
DA:
7-T
ags-
Adve
ntis
ten;
*: D
iffer
enz
aufg
rund
Sito
ster
ol u
. a.;
# : Aus
schl
uss
der P
roba
nden
mit
inve
rsem
Ste
rolp
rofil
1 : Orig
inal
date
n in
µm
ol/g
Tro
cken
subs
tanz
; *: A
ngab
e de
s M
edia
n; # : A
ussc
hlus
s de
r Pro
band
en m
it in
vers
em S
tero
lpro
fil

Diskussion
79
Der mittlere Gesamtgallensäurengehalt in der Trockensubstanz aller erhobenen
Daten aus den Humanstudien entspricht mit 10,6 ± 3,5 mg/g den in der Literatur
aufgeführten Konzentrationen, die zwischen 4,0 ± 0,5 bis 9,5 ± 6,4 mg/g
Trockensubstanz variieren [REDDY et al. 1989, HOFSTAD et al. 1998] (Tab. 29). Die
etwas erhöhte Tendenz innerhalb der Studien 1 - 4 im Vergleich zur Literatur ist
u. a. darauf zurückzuführen, dass die Autoren weniger Gallensäuren analysiert
haben.
Bei den Teilnehmern der im Rahmen dieser Arbeit beschriebenen Studien liegen
ähnliche Konzentrationen an LCA und DCA in der Faecestrockensubstanz vor, wie
bei Probanden der Studien von REDDY et al. [1989], SPANHAAK et al. [1998],
WESTSTRATE et al. [1999] und den Patienten der Studie von HOFSTAD et al. [1998].
Zwischen den Studien unterscheiden sich die Gehalte der primären Gallensäuren
im Bereich von 0,00 bis 0,61 ± 1,51 mg/g Trockensubstanz. Auch innerhalb der
Studien von KAIBARA et al. [1983], HOFSTAD et al. [1998], SPANHAAK et al. [1998]
und den hier dargestellten Werten streuen die Gehalte an CDCA und CA in der
Trockensubstanz teilweise über 100 %.
Die Gehalte der 12-Oxo-DCA in der Trockensubstanz sind mit 0,37 ± 0,35 mg den
bei REDDY et al. [1989] und KAIBARA et al. [1983] dokumentierten Gehalten von
0,34 ± 0,02 mg bzw. 0,47 ± 0,29 mg vergleichbar. Es ist weiter festzustellen, dass
in früheren Arbeiten durchschnittlich geringere Konzentrationen an sekundären
und größere Mengen an primären Gallensäuren in den Faeces nachgewiesen
werden [KAIBARA et al. 1983, MOWER et al. 1979]. So sind die von KAIBARA et al.
[1983] ermittelten Gehalte an faecaler iso-LCA und iso-DCA niedriger als die
entsprechenden Ergebnisse der Probanden dieser Arbeit. Auch die von
WESTSTRATE et al. [1999] dargestellten Werte an iso-DCA im Stuhl liegen unter
dem mittleren Gehalt der 86 Teilnehmer der angeführten Humanstudien; iso-LCA
ist nicht quantifiziert worden. Unter Berücksichtigung der inter-individuellen
Streuung sind die Ergebnisse vergleichbar.
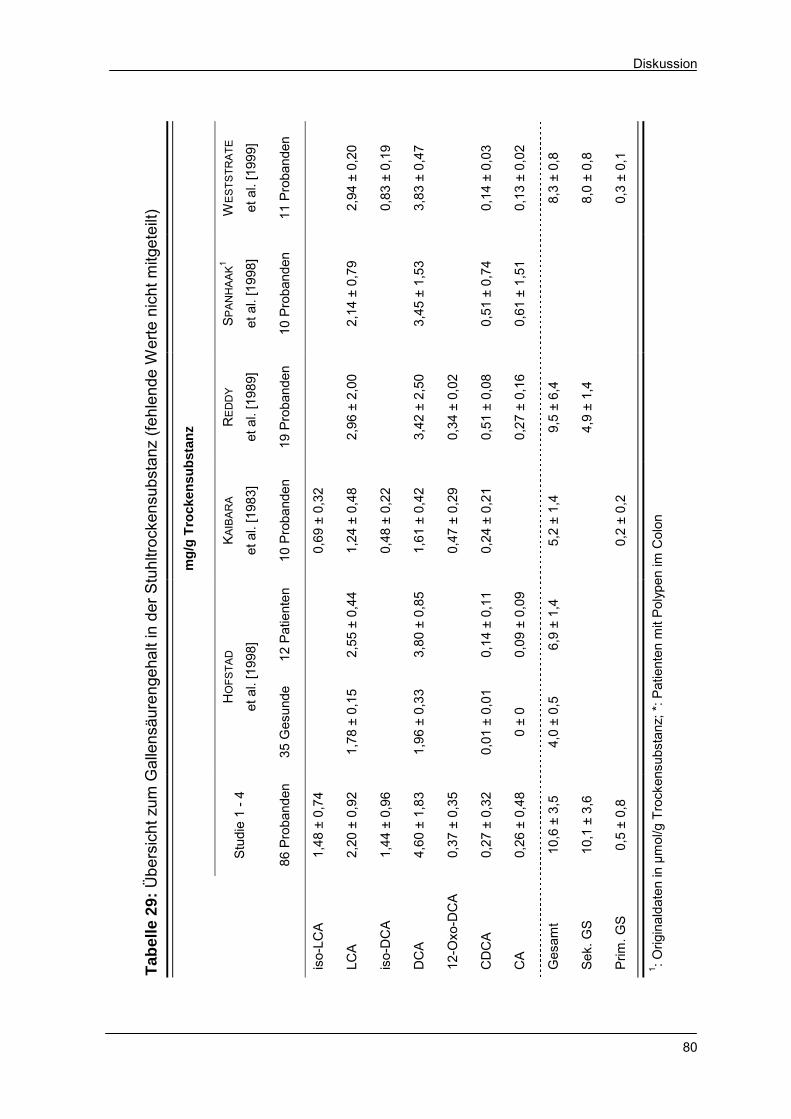
Diskussion
80
WES
TSTR
ATE
et a
l. [1
999]
11 P
roba
nden
2,94
± 0
,20
0,83
± 0
,19
3,83
± 0
,47
0,14
± 0
,03
0,13
± 0
,02
8,3
± 0,
8
8,0
± 0,
8
0,3
± 0,
1
S PAN
HAA
K1
et a
l. [1
998]
10 P
roba
nden
2,14
± 0
,79
3,45
± 1
,53
0,51
± 0
,74
0,61
± 1
,51
RED
DY
et a
l. [1
989]
19 P
roba
nden
2,96
± 2
,00
3,42
± 2
,50
0,34
± 0
,02
0,51
± 0
,08
0,27
± 0
,16
9,5
± 6,
4
4,9
± 1,
4
K AIB
ARA
et a
l. [1
983]
10 P
roba
nden
0,69
± 0
,32
1,24
± 0
,48
0,48
± 0
,22
1,61
± 0
,42
0,47
± 0
,29
0,24
± 0
,21
5,2
± 1,
4
0,2
± 0,
2
12 P
atie
nten
2,55
± 0
,44
3,80
± 0
,85
0,14
± 0
,11
0,09
± 0
,09
6,9
± 1,
4
HO
FSTA
D
et a
l. [1
998]
35 G
esun
de
1,78
± 0
,15
1,96
± 0
,33
0,01
± 0
,01
0 ±
0
4,0
± 0,
5
mg/
g Tr
ocke
nsub
stan
z
Stud
ie 1
- 4
86 P
roba
nden
1,48
± 0
,74
2,20
± 0
,92
1,44
± 0
,96
4,60
± 1
,83
0,37
± 0
,35
0,27
± 0
,32
0,26
± 0
,48
10,6
± 3
,5
10,1
± 3
,6
0,5
± 0,
8
iso-
LCA
LCA
iso-
DC
A
DC
A
12-O
xo-D
CA
CD
CA
CA
Ges
amt
Sek.
GS
Prim
. GS
Tabe
lle 2
9: Ü
bers
icht
zum
Gal
lens
äure
ngeh
alt i
n de
r Stu
hltro
cken
subs
tanz
(feh
lend
e W
erte
nic
ht m
itget
eilt)
1 : Orig
inal
date
n in
µm
ol/g
Tro
cken
subs
tanz
; *: P
atie
nten
mit
Poly
pen
im C
olon
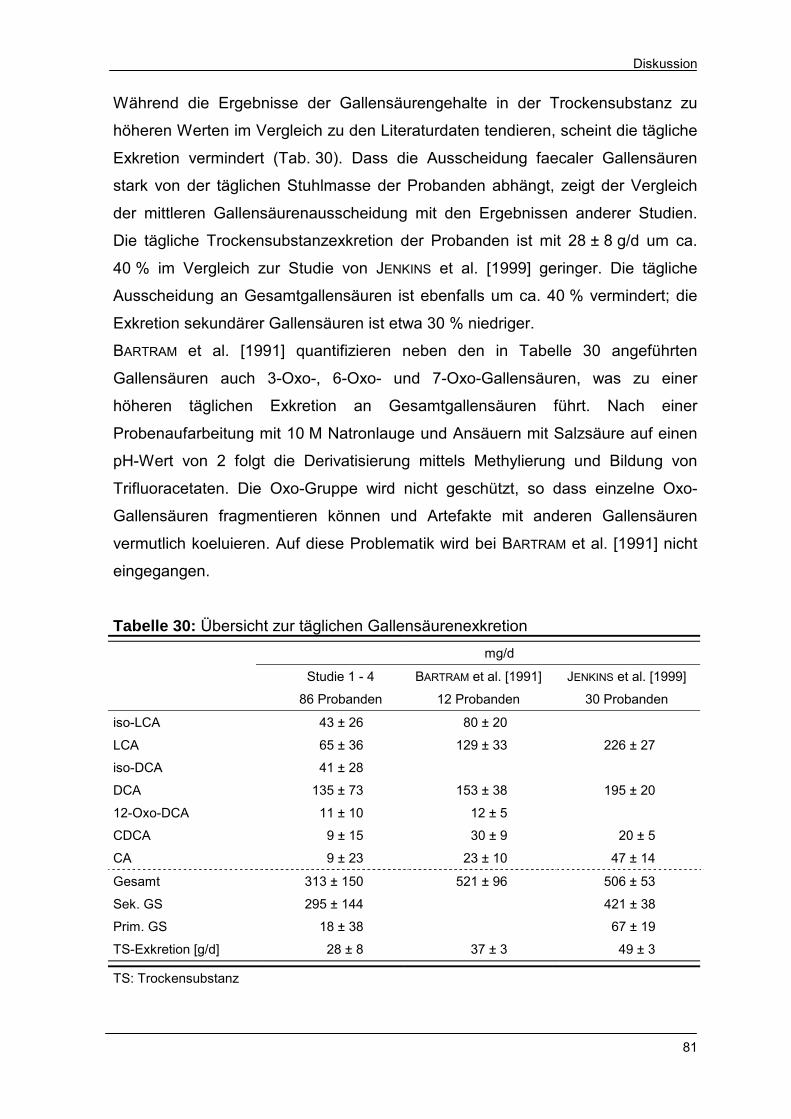
Diskussion
81
Während die Ergebnisse der Gallensäurengehalte in der Trockensubstanz zu
höheren Werten im Vergleich zu den Literaturdaten tendieren, scheint die tägliche
Exkretion vermindert (Tab. 30). Dass die Ausscheidung faecaler Gallensäuren
stark von der täglichen Stuhlmasse der Probanden abhängt, zeigt der Vergleich
der mittleren Gallensäurenausscheidung mit den Ergebnissen anderer Studien.
Die tägliche Trockensubstanzexkretion der Probanden ist mit 28 ± 8 g/d um ca.
40 % im Vergleich zur Studie von JENKINS et al. [1999] geringer. Die tägliche
Ausscheidung an Gesamtgallensäuren ist ebenfalls um ca. 40 % vermindert; die
Exkretion sekundärer Gallensäuren ist etwa 30 % niedriger.
BARTRAM et al. [1991] quantifizieren neben den in Tabelle 30 angeführten
Gallensäuren auch 3-Oxo-, 6-Oxo- und 7-Oxo-Gallensäuren, was zu einer
höheren täglichen Exkretion an Gesamtgallensäuren führt. Nach einer
Probenaufarbeitung mit 10 M Natronlauge und Ansäuern mit Salzsäure auf einen
pH-Wert von 2 folgt die Derivatisierung mittels Methylierung und Bildung von
Trifluoracetaten. Die Oxo-Gruppe wird nicht geschützt, so dass einzelne Oxo-
Gallensäuren fragmentieren können und Artefakte mit anderen Gallensäuren
vermutlich koeluieren. Auf diese Problematik wird bei BARTRAM et al. [1991] nicht
eingegangen.
Tabelle 30: Übersicht zur täglichen Gallensäurenexkretion
mg/d
Studie 1 - 4 BARTRAM et al. [1991] JENKINS et al. [1999]
86 Probanden 12 Probanden 30 Probanden
iso-LCA 43 ± 26 80 ± 20
LCA 65 ± 36 129 ± 33 226 ± 27 iso-DCA 41 ± 28
DCA 135 ± 73 153 ± 38 195 ± 20
12-Oxo-DCA 11 ± 10 12 ± 5 CDCA 9 ± 15 30 ± 9 20 ± 5
CA 9 ± 23 23 ± 10 47 ± 14
Gesamt 313 ± 150 521 ± 96 506 ± 53
Sek. GS 295 ± 144 421 ± 38 Prim. GS 18 ± 38 67 ± 19
TS-Exkretion [g/d] 28 ± 8 37 ± 3 49 ± 3
TS: Trockensubstanz
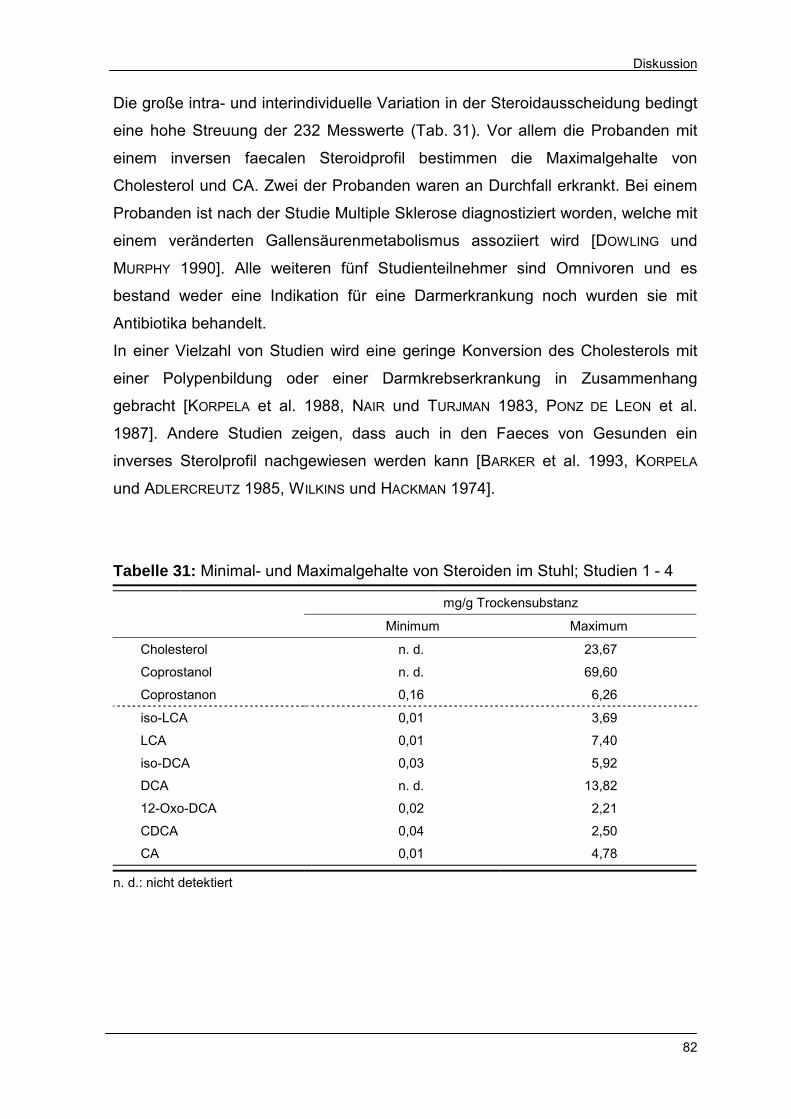
Diskussion
82
Die große intra- und interindividuelle Variation in der Steroidausscheidung bedingt
eine hohe Streuung der 232 Messwerte (Tab. 31). Vor allem die Probanden mit
einem inversen faecalen Steroidprofil bestimmen die Maximalgehalte von
Cholesterol und CA. Zwei der Probanden waren an Durchfall erkrankt. Bei einem
Probanden ist nach der Studie Multiple Sklerose diagnostiziert worden, welche mit
einem veränderten Gallensäurenmetabolismus assoziiert wird [DOWLING und
MURPHY 1990]. Alle weiteren fünf Studienteilnehmer sind Omnivoren und es
bestand weder eine Indikation für eine Darmerkrankung noch wurden sie mit
Antibiotika behandelt.
In einer Vielzahl von Studien wird eine geringe Konversion des Cholesterols mit
einer Polypenbildung oder einer Darmkrebserkrankung in Zusammenhang
gebracht [KORPELA et al. 1988, NAIR und TURJMAN 1983, PONZ DE LEON et al.
1987]. Andere Studien zeigen, dass auch in den Faeces von Gesunden ein
inverses Sterolprofil nachgewiesen werden kann [BARKER et al. 1993, KORPELA
und ADLERCREUTZ 1985, WILKINS und HACKMAN 1974].
Tabelle 31: Minimal- und Maximalgehalte von Steroiden im Stuhl; Studien 1 - 4
mg/g Trockensubstanz
Minimum Maximum
Cholesterol n. d. 23,67
Coprostanol n. d. 69,60
Coprostanon 0,16 6,26
iso-LCA 0,01 3,69
LCA 0,01 7,40
iso-DCA 0,03 5,92 DCA n. d. 13,82
12-Oxo-DCA 0,02 2,21
CDCA 0,04 2,50 CA 0,01 4,78
n. d.: nicht detektiert

Diskussion
83
6.3 Der Einfluss von Calcium und Probiotika auf die Steroid-ausscheidung
Da von den faecalen Steroiden vor allem die Gallensäuren aufgrund ihres
toxischen Verhaltens im Faeceswasser und nicht in der Stuhltrockensubstanz
hinsichtlich einer Calciumsupplementation untersucht werden, sind wenige Daten
in der Literatur zum Sterolprofil dokumentiert. In Studie 1 sind durch die Zulage
von anorganischem Calciumphosphat keine signifikanten Änderungen im
Gallensäurengehalt der Trockensubstanz und in der täglichen Exkretion zu
verzeichnen. Der Gehalt an Coprostanon in der Trockensubstanz steigt nach einer
täglichen Zufuhr von 2.462 mg Calcium und 3.200 mg Phosphor signifikant an. Es
ergibt sich ein paralleler Verlauf der Cholesterol- und Coprostanongehalte. Die
Cholesterolexkretion ist aufgrund der starken Streuung nicht signifikant verändert.
Die an der Position 3 hydrogenierten Abbauprodukte Coprostanol und Cholestanol
tendieren zu geringeren Gehalten. Es ist an dieser Stelle zu erwähnen, dass keine
Aussage zum Gallensäurenprofil im Faeceswasser möglich ist. Differenzen durch
ein Calcium-bedingtes Fällen der sauren Steroide werden in der Analyse der
Trockensubstanz nicht berücksichtigt. Die Untersuchung der wässrigen Phase ist
für spätere Studien vorbehalten.
Hinsichtlich der Sterolausscheidung in den Faeces verweisen QUILLIOT et al.
[1999] nach einer 8-monatigen Studie an Ratten auf eine signifikant erhöhte
wöchentliche Cholesterol- und unveränderte Coprostanolexkretion (Tab. 32). Die
Basisdiät enthält 24 % Sonnenblumenöl. In 100 g von diesem Pflanzenöl sind
etwa 200 - 450 mg Phytosterole enthalten [VLAHAKIS und HAZEBROEK 2000].
Phytosterole führen zu einer verminderten Cholesterolabsorption und einer
verstärkten Cholesterolexkretion [NTANIOS et al. 2001]. Die Exkretion an Sitosterol
und Cholesterol ist sowohl in der Kontrollgruppe als auch in der Testgruppe der
Ratten angestiegen. Bei Zulage von Calciumcarbonat sind die wöchentlichen
Ausscheidungen beider Sterole im Vergleich zur Kontrollgruppe signifikant höher.
In einer Studie an Ratten [APPLETON et al. 1992] in der über 2 Wochen
Calciumlactat verabreicht worden ist, werden keine signifikanten Unterschiede in
der Konzentration an Cholesterol und dem Gesamtsterolgehalt in der
Trockensubstanz festgestellt.
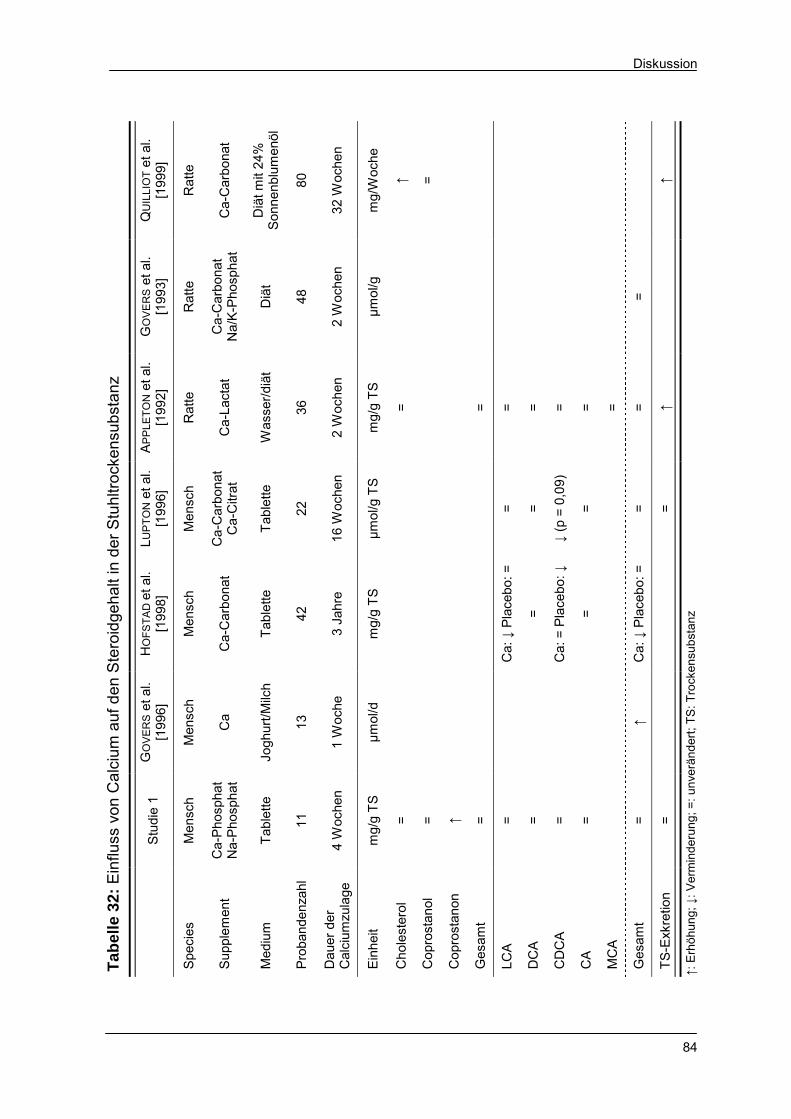
Diskussion
84
QU
ILLI
OT
et a
l. [1
999]
Rat
te
Ca-
Car
bona
t
Diä
t mit
24%
So
nnen
blum
enöl
80
32 W
oche
n
mg/
Woc
he
↑ = ↑
GO
VER
S et
al.
[199
3]
Rat
te
Ca-
Car
bona
t N
a/K-
Phos
phat
Diä
t
48
2 W
oche
n
µmol
/g
=
A PPL
ETO
N e
t al.
[199
2]
Rat
te
Ca-
Lact
at
Was
ser/d
iät
36
2 W
oche
n
mg/
g TS
= = = = = = = = ↑
L UPT
ON
et a
l. [1
996]
Men
sch
Ca-
Car
bona
t C
a-C
itrat
Tabl
ette
22
16 W
oche
n
µmol
/g T
S
= =
↓ (p
= 0
,09)
= = =
HO
FSTA
D e
t al.
[199
8]
Men
sch
Ca-
Car
bona
t
Tabl
ette
42
3 Ja
hre
mg/
g TS
Ca:
↓ P
lace
bo: =
=
Ca:
= P
lace
bo: ↓
=
Ca:
↓ P
lace
bo: =
GO
VER
S et
al.
[199
6]
Men
sch
Ca
Jogh
urt/M
ilch
13
1 W
oche
µmol
/d
↑
Stud
ie 1
Men
sch
Ca-
Phos
phat
N
a-Ph
osph
at
Tabl
ette
11
4 W
oche
n
mg/
g TS
= = ↑ = = = = = = =
Spec
ies
Supp
lem
ent
Med
ium
Prob
ande
nzah
l
Dau
er d
er
Cal
cium
zula
ge
Einh
eit
Cho
lest
erol
Cop
rost
anol
Cop
rost
anon
Ges
amt
LCA
DC
A
CD
CA
CA
MC
A
Ges
amt
TS-E
xkre
tion
Tabe
lle 3
2: E
influ
ss v
on C
alci
um a
uf d
en S
tero
idge
halt
in d
er S
tuhl
trock
ensu
bsta
nz
↑: E
rhöh
ung;
↓: V
erm
inde
rung
; =: u
nver
ände
rt; T
S: T
rock
ensu
bsta
nz

Diskussion
85
Durch Aufnahme eines mit Bifidobacterium und Lactobacillus supplementierten
Joghurts ist in Studie 4 ein kontinuierlicher Anstieg der Cholesterolexkretion zu
verzeichnen. Nach einem 10-wöchigen Verzehr von Joghurt hat sich die tägliche
Cholesterolexkretion unabhängig von der Probiotikazulage zum Ausgangswert ca.
verdreifacht. Der Vergleich zu den Werten ohne Joghurtkonsum ist in Studie 3
nicht möglich, da der erste Messwert die Cholesterolexkretion nach 7 Wochen
Adaptation an konventionellen Joghurt repräsentiert. In dieser Studie ist sowohl
nach 14-wöchigem Joghurtkonsum als auch nach 21 Wochen ein der Adaptation
vergleichbarer Cholesterolgehalt nachweisbar.
Da die Effekte auf den faecalen Cholesterolgehalt unabhängig von der Joghurtart
auftreten und in Studie 2 nach Gabe von probiotischen Keimen ohne
Calciumsupplementation keine Änderungen im Sterolgehalt zu verzeichnen sind,
scheint die Calciumzufuhr einen erhöhten Cholesterolgehalt in den Faeces zu
bewirken.
In Studie 4 wird nach probiotischer Joghurtgabe im Vergleich zur konventionellen
Studienphase ein signifikant verminderter faecaler Coprostanolgehalt in der
Trockensubstanz gefunden. Dies führt zu einer herabgesetzten Gesamtsterol-
konzentration und durch den erhöhten Cholesterolgehalt zu einer verminderten
Konversionsrate. Die Coprostanolexkretion der Probanden in den Studien 2 und 3
ist in allen Perioden in der Stuhltrockensubstanz unverändert, tendiert aber in der
Studie 3 nach Probiotikasupplementation zu einer verminderten täglichen
Exkretion.
Es ist zu berücksichtigen, dass der Joghurtkonsum zu einer erhöhten
Cholesterolaufnahme führt und damit eine erhöhte Cholesterolexkretion zur Folge
haben kann. In einer Humanstudie von BARTRAM et al. [1994] werden nach
3-wöchigem Joghurtkonsum (500 ml/d) keine Änderungen in der Cholesterol- und
Coprostanolexkretion unabhängig von einer Probiotikasupplementation ermittelt.
Weiterhin beschreiben SPANHAAK et al. [1998] keine Änderungen der faecalen
Cholesterol- und Coprostanolgehalte nach 4-wöchiger Aufnahme von 100 ml mit
L. casei Shirota fermentierter Milch. Hingegen sind in einer Studie an Hamstern,
denen Sojamilch bzw. mit Bifidobacterium breve fermentierte Sojamilch auf der
Basis einer cholesterolfreien bzw. -angereicherten Diät verabreicht worden ist, die
Cholesterolgehalte in den Faeces nach Cholesterolzulage signifikant erhöht.

Diskussion
86
Weder die Sojamilch noch das fermentierte Produkt beeinflussen die faecalen
Cholesterolgehalte [KIKUCHI-HAYAKAWA et al. 1998].
Da sowohl in der Studie an Ratten von QUILLIOT et al. [1999] als auch in Studie 1
die Cholesterolaufnahmen zwischen den Perioden nicht signifikant verschieden
sind, können die höheren Gehalte an Cholesterol in der Trockensubstanz nicht
durch eine verstärkte Zufuhr exogenen Cholesterols verursacht werden. Daher
lassen sich mögliche Thesen ableiten (Tab. 33):
1. der Cholesterolfluss in das Colon ist erhöht
2. der Cholesterolgehalt in den Faeces ist durch veränderte Konversion durch
Darmbakterien erhöht
Tabelle 33: Mögliche Ursachen einer Änderung des faecalen Sterolprofiles
Cholesterolfluss in das Colon erhöht Cholesterolfluss in das Colon
unverändert
Gehalte in den Faeces Gehalte in den Faeces
Konversionsrate Cholesterol Metabolite Gesamt Cholesterol Metabolite Gesamt
gleich ↑ ↑ ↑ = = = erhöht ↓ / = / ↑ ↑ ↑ ↓ ↑ =
vermindert ↑ ↓/ = / ↑ ↑ ↑ ↓ =
↑: Erhöhung; ↓: Verminderung; =: unverändert
Für beide Thesen sind Hinweise zu finden. VAN DER MEER et al. [1991, 1997]
postulieren, dass Calcium im Colon die Fällung der Gallensäuren und eine
Mitfällung von Cholesterol bewirkt, was zu einer erhöhten Cholesterol-
ausscheidung führen kann (vgl. 2.2.2.1). Dies geschieht nach GOVERS und VAN
DER MEER [1993] nicht im jejunal-ilealen Bereich, da konjugierte Gallensäuren eine
höhere Löslichkeit bei niedrigem pH-Wert und eine größere Stabilität gegen
Fällungsmittel aufweisen. Weder das Lösungsverhalten von freiem noch von
verestertem Cholesterol in diesem Darmabschnitt unter Calciumeinfluss wird
diskutiert.

Diskussion
87
Unter Phytosterolsupplementation wird die Cholesterolabsorption im Dünndarm
vermindert und die Cholesterolexkretion steigt [NTANIOS et al. 2001]. Da der
Cholesterolmetabolismus eng mit dem der Gallensäuren verbunden ist, kann die
Exkretion an Gallensäuren beeinflusst werden [CHAPLIN 1998] (Abb. 37). UCHIDA et
al. [1984] beschreiben endogenes Cholesterol als Präkursor für faecale CA-
Gehalte und exogenes Cholesterol als Präkursor für CDCA-Gehalte.
Probanden einer Humanstudie von WESTSTRATE et al. [1999] weisen nach
Phytosterolsupplementation höhere Gehalte an Cholesterol und geringere Gehalte
an primären Gallensäuren in den Faeces im Vergleich zu einer Placebogruppe
auf. Die Gehalte sekundärer Gallensäuren bleiben unverändert. HOFSTAD et al.
[1998] und LUPTON et al. [1996] beschreiben in den Langzeitstudien mit
Calciumsupplementation ebenfalls verminderte Gehalte primärer Gallensäuren,
vor allem der CDCA. Da die sekundäre Gallensäure LCA durch Dehydroxylierung
der primären Gallensäure CDCA gebildet wird, ist eine Verminderung des
Gehaltes, wie HOFSTAD et al. [1998] nachweisen, möglich. DE RODAS et al. [1996]
stellen nach Calciumsupplementation eine schnellere Verminderung der Gehalte
an Gesamtcholesterol und Gallensäuren im Serum von Schweinen fest, die
wahrscheinlich aus einer Beeinflussung des enterohepatischen Kreislaufes
resultieren.
Es ist weiter anzumerken, dass die Tiere in der Studie von QUILLIOT et al. [1999]
aus der Gruppe mit supplementiertem Calcium und erhöhter Cholesterolexkretion
langsamer wuchsen. Bei BEHLING et al. [1990] wird eine verminderte
Wachstumsrate bei Ratten, deren Futter mit Calciumcabonat supplementiert
worden ist, nachgewiesen. Die Calciumzulage führt aufgrund einer geringen
intestinalen Absorption zu einer verstärkten Lipidexkretion.
Neben den erhöhten Cholesterolgehalten im Dickdarm ist durch Veränderungen
der intestinalen Mikroflora eine verminderte Konversionsrate des Cholesterols
möglich, die zu einem erhöhten Cholesterolgehalt in den Faeces führen können.
BOVEE-OUDENHOVEN und VAN DER MEER [1997] sowie BOVEE-OUDENHOVEN et al.
[1999] zeigen, dass durch den Einfluss von Calcium die intestinalen Lactobacillen
in Ratten stimuliert werden.

Diskussion
88
Diese Bakterien haben nicht die Fähigkeit Cholesterol in Coprostanol zu
transformieren [NORIN et al. 1991]. Die gebildete Milchsäure wirkt weiterhin auf
gram-negative Bakterien in vitro antimikrobiell [ALAKOMI et al. 2000].
Die Umwandlung von Cholesterol in die faecalen Metabolite wird vor allem durch
gram-positive Stämme wie Eubacterium, Clostridium und Bifidobacterium
hervorgerufen. Aber auch gram-negative Species wie Escherichia coli oder
Bacteroides � ein Hauptvertreter der Colonflora � haben die Fähigkeit zur
Metabolisierung [Eyssen et al. 1973, Mott et al. 1980, Owen et al. 1978, Tenneson
et al. 1977]. Eine mögliche Änderung des Bakterienprofiles durch Calciumzulage
und zusätzlicher Gabe von Lactobacillen kann zu einer Wandlung des
Sterolprofiles in den Faeces führen, wobei der Sterolgesamtgehalt sich nicht
verändert. Die Ergebnisse der Studie 4 stützen diese These. In der probiotischen
Periode dieser Studie ist die tägliche Cholesterolexkretion im Vergleich zu den
Ausgangswerten signifikant erhöht, die Coprostanolausscheidung ist unverändert.
Bei Gabe von konventionellem Joghurt zeigt der Coprostanolgehalt eine steigende
Tendenz.
Weiterhin scheint die Expositionsdauer beeinflussend zu wirken, da keine
Änderungen im Steroidprofil nach einer Calciumsupplementation von
1 - 2 Wochen auftreten [APPLETON et al. 1992, GOVERS et al. 1993]. Ab einer
Calciumzulage von 4 Wochen werden Änderungen in der Steroidexkretion
verzeichnet [Studie 1, Studie 3, Studie 4, HOFSTADT et al. 1998, LUPTON et al.
1996, QUILLIOT et al. 1999].
Eine verminderte CDCA-Exkretion unter erhöhter Calciumzufuhr scheint im
Widerspruch zur erhöhten täglichen CDCA-Ausscheidung nach Joghurtkonsum
der Studie 4 zu stehen. Da bei hoher Zufuhr Nahrungscholesterol in der Leber und
Geweben zu 27-Hydroxycholesterol umgewandelt und dieses zu CDCA
transformiert wird [LEE et al. 1996], kann der kontinuierliche Joghurtkonsum zu
erhöhten faecalen CDCA-Gehalten führen. Die tägliche Exkretion an sekundären
Gallensäuren ist nach Joghurtgabe tendenziell bzw. signifikant erhöht. Da dies
sowohl in der konventionellen als auch in der probiotischen Phase zu beobachten
ist, scheint kein Einfluss aus dem Dekonjugationsvermögen der probiotischen
Keime zu bestehen, vielmehr kann eine höhere Stuhlausscheidung bzw. eine
gesteigerte Exkretion primärer Gallensäuren ursächlich sein.

Diskussion
89
In Studie 3 sind nach einem Joghurtkonsum von 14 Wochen nahezu alle
Gallensäuren in den Faeces signifikant vermindert und bleiben nach 21 Wochen
auf einem verminderten Niveau. Die faecalen pH-Werte in diesen Perioden sind im
Vergleich zur Adaptation signifikant vermindert. Da eine Absenkung des faecalen
pH-Wertes mit einer sinkenden Enzymaktivität der Intestinalflora in
Zusammenhang gebracht wird, kann die Dehydroxylierung der primären
Gallensäuren beeinflusst werden [DGE 2000; MACDONALD et al. 1978]. Weiterhin
ist eine verminderte Hydrolyse der Konjugate möglich, was zu einer verminderten
Exkretion freier Gallensäuren führen kann. Da die Probenaufarbeitung vorwiegend
diese berücksichtigt, können die aus konjugierten Verbindungen stammenden
Gallensäuren nicht in die Analysenwerte eingegangen sein.
Aus den Ergebnissen der Studien 3 und 4 lässt sich schließen, dass sich durch die
Aufnahme von Joghurt anfangs das faecale Steroidprofil ändert. Mit zunehmender
Konsumdauer stabilisiert sich das Sterolprofil und die Gehalte unkonjugierter
Gallensäuren in den Faeces werden vermindert. Die Supplementation von
probiotischen Keimen wirkt nicht gegensätzlich (Studie 2).

Diskussion
90
6.4 Serumcholesterol und faecale Steroide Aus Studie 4 ist ersichtlich, dass die Probanden mit einer höheren faecalen
Exkretion sekundärer Gallensäuren ein geringeres LDL/HDL-Cholesterol-
verhältnis im Serum aufweisen. Weiterhin ist der Lipoproteinquotient bei
Probanden mit einer geringen Ausscheidung primärer Gallensäuren vermindert.
Diese Korrelation ist ebenfalls bei den Probanden der Studie 3 zu beobachten. Die
hypercholesterolämischen Probanden scheiden weniger sekundäre bzw. mehr
primäre Gallensäuren aus als die normocholesterolämischen Probanden. Die
Frauen der Studie 4 haben im Vergleich zu den männlichen Probanden höhere
Gehalte an Cholesterol im Blut [KLEIN 2002] und verminderte Gehalte sekundärer
bzw. erhöhte Gehalte primärer Gallensäuren im Stuhl.
In einer Studie mit 30 Patienten, die an Atherosklerose erkrankt sind, ist die
Exkretion sekundärer Gallensäuren hochsignifikant im Vergleich zu einer
gesunden Kontrollgruppe vermindert. Die HDL-Cholesterolgehalte sind in der
Patientengruppe niedriger [CHARACH et al. 1998].
Eine verringerte Gallensäurenausscheidung kann aus einem geringen
Gallensäurenpool im Körper (These 1, Abb. 37) oder aus einer erhöhten
Resorption sekundärer Gallensäuren im Colon folgen. Dadurch vergrößert sich der
Gallensäurenpool im Körper (These 2, Abb. 37), der durch Beeinflussung des
hepatischen Cholesterols einen erhöhten Cholesterolfluss in die Galle bzw. in das
Blut zur Folge hat [NAITO et al. 1996]. In der Leber, an Blutgefässwänden und
durch Makrophagen wird Cholesterol zu 27-Hydroxycholesterol oxidiert, was als
Substrat für den mitochondrialen, alternativen Weg zur Gallensäurensynthese gilt,
wobei vorwiegend CDCA gebildet wird [VLAHCEVIC et al. 2000, REISS et al. 1997,
JAVITT et al. 1994.]
In einer Studie von PULLINGER et al. [2002] wird eine verminderte Gesamtgallen-
säurenexkretion und ein erhöhter faecaler CDCA-Gehalt an einem Probanden mit
hypercholesterolämischen LDL-Cholesterolgehalten und verminderter
7α-Hydroxylaseaktivität in der Leber nachgewiesen. Die Autoren sehen als
Ursache eine Hochregulierung der alternativen Gallensäurensynthese und einen
verminderten Gallensäurenpool.
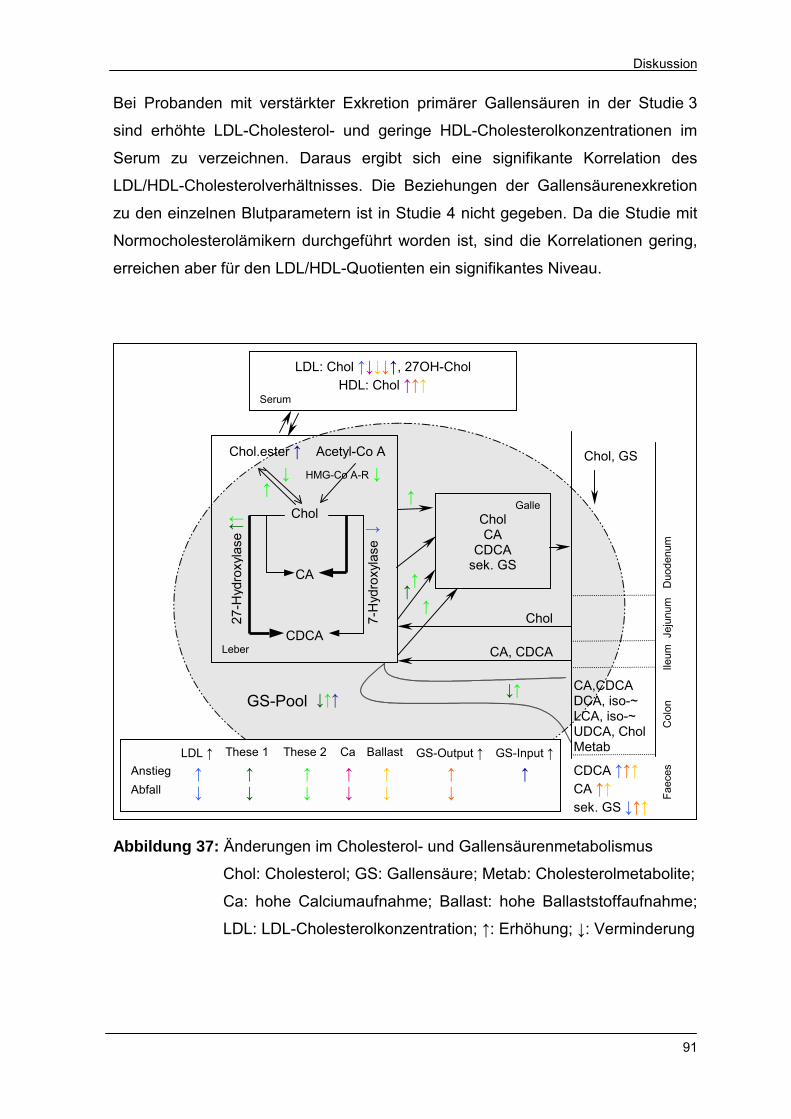
Diskussion
91
GS-Pool
Bei Probanden mit verstärkter Exkretion primärer Gallensäuren in der Studie 3
sind erhöhte LDL-Cholesterol- und geringe HDL-Cholesterolkonzentrationen im
Serum zu verzeichnen. Daraus ergibt sich eine signifikante Korrelation des
LDL/HDL-Cholesterolverhältnisses. Die Beziehungen der Gallensäurenexkretion
zu den einzelnen Blutparametern ist in Studie 4 nicht gegeben. Da die Studie mit
Normocholesterolämikern durchgeführt worden ist, sind die Korrelationen gering,
erreichen aber für den LDL/HDL-Quotienten ein signifikantes Niveau.
Chol, GS Chol
CA, CDCA
CA,CDCA DCA, iso-~ LCA, iso-~ UDCA, Chol
Metab
CDCA ↑↑↑ CA ↑↑
sek. GS ↓↑↑
Abbildung 37: Änderungen im Cholesterol- und Gallensäurenmetabolismus
Chol: Cholesterol; GS: Gallensäure; Metab: Cholesterolmetabolite;
Ca: hohe Calciumaufnahme; Ballast: hohe Ballaststoffaufnahme;
LDL: LDL-Cholesterolkonzentration; ↑: Erhöhung; ↓: Verminderung
Chol.ester ↑ Acetyl-Co A
HMG-Co A-R
Chol
CA
CDCA Leber
LDL: Chol ↑↓↓↓↑, 27OH-Chol HDL: Chol ↑↑↑
Serum
GalleChol CA
CDCA sek. GS
27-H
ydro
xyla
se
7-H
ydro
xyla
se
Ileum
Jej
unum
D
uode
num
F
aece
s
Col
on
LDL ↑ These 1 These 2 Ca Ballast GS-Output ↑ GS-Input ↑ Anstieg ↑ ↑ ↑ ↑ ↑ ↑ ↑ Abfall ↓ ↓ ↓ ↓ ↓ ↓
↑↓↓↑
↑↑
↑↑↓ ↓
↓ ↑ ↑
↑
↑

Diskussion
92
Die These der erhöhten Resorption von Gallensäuren wird durch die Arbeiten von
GYLLING et al. [2002] unterstützt. Dabei wird in Hypercholesterolämikern eine
Mangelabsorption an Gallensäuren induziert, die eine Verminderung des
LDL-Cholesterols und eine Erhöhung des HDL-Cholesterols zur Folge hat. Die
gleichen Änderungen der Blutparameter werden nach Gabe von Koriandersamen
von CHITHRA und LEELAMMA [1997] bei Anstieg der faecalen Gallensäurenexkretion
beobachtet. In einer Tierstudie mit Ratten von USMAN und HOSONO [2000] wird ein
hypocholesterolämischer Effekt von L. gasseri SBT0274 durch eine verminderte
Reabsorption von Gallensäuren im enterohepatischen Kreislauf und eine erhöhte
faecale Exkretion beschrieben. Die Fütterung von primären Gallensäuren führt
hingegen zu erhöhten LDL-Cholesterolkonzentrationen in Ratten und Hamstern
bei einem zunehmenden Gallensäurenpool [CHIANG et al. 1998, SPADY et al. 1986]
(Abb. 37).
In Studie 3 sind durch 6-monatigen Joghurtkonsum sowohl bei den normo- als
auch bei den hypercholesterolämischen Probanden die HDL-Cholesterol-
konzentrationen angestiegen, die LDL-Cholesterolkonzentrationen gesunken und
das Verhältnis von LDL/HDL-Cholesterol vermindert sich. Die Ausscheidung an
unkonjugierten primären und sekundären Gallensäuren ist in beiden
Probandengruppen vermindert. Während das Verhältnis der primären und
sekundären Gallensäuren in der normocholesterolämischen Probandengruppe
unverändert bleibt, ist es in der hypercholesterolämischen Gruppe mit
zunehmenden Joghurtkonsum tendenziell vermindert. In einer Studie an Ratten
wurde durch Calciumzulage die LDL-Cholesterolkonzentration vermindert und die
HDL-Cholesterolkonzentration erhöht, dies führt zu einem Anstieg im
HDL/LDL-Cholesterolquotienten und demzufolge in eine Verminderung des
LDL/HDL-Cholesterolverhältnisses [VASKONEN et al. 2002] (Abb. 37).

Schlussfolgerungen
93
7 SCHLUSSFOLGERUNGEN » Die gaschromatographisch-massenspektrometrische Analyse von faecalem
Cholesterol und dessen bakteriellen Metaboliten Coprostanol, Coprostanon,
Cholestanol und Cholestanon zeigt, dass sie für quantitative Aussagen
geeignet ist.
» Die Separation von Cholesterol und Cholestanol wird durch die Anwendung
der Gaschromatographie mit üblicher Detektion erreicht. Hingegen ist zur
Separation von Cholesterol und Coprostanon die spezifische Detektion im
Single Ion Monitoring erforderlich.
» Mit der Analyse underivatisierter Sterole können sowohl die Artefaktbildung
des Hauptabbauproduktes Coprostanon durch Trimethylsilylierung als auch
Substanzverluste durch eine unvollständige Reaktion verhindert werden.
» Die neue Methode erlaubt hinsichtlich des Zeit- und Chemikalienaufwandes
eine vereinfachte Probenaufarbeitung.
» Die quantitative gaschromatographische Analyse von faecaler iso-Deoxy-
cholsäure, Deoxycholsäure, iso-Lithocholsäure, Lithocholsäure, Cheno-
deoxycholsäure, Cholsäure und 12-Oxo-Deoxycholsäure ist realisierbar.
» Mittels massenspektrometrischer Detektion im Modus des Single Ion
Monitorings sind Peaküberlagerungen durch koeluierende Substanzen
vermindert, wie es am Beispiel der Cholsäure gezeigt werden kann.
» Die Analyse von 3-Oxo-Gallensäuren ist aufgrund ihres instabilen
chemischen Verhaltens innerhalb der Probenvorbereitung nicht möglich.
» Die entwickelten Methoden zur Sterol- und Gallensäurenanalyse sind für
einen hohen Probendurchsatz geeignet.

Schlussfolgerungen
94
» Die tägliche Aufnahme von bis zu 2.460 mg anorganischem Calcium und
3.200 mg Phosphor ist ohne Einfluss auf die faecale Exkretion von
Gesamtgallensäuren. Durch die Calciumphosphatzulage ist der Gehalt an
Coprostanon in der Trockensubstanz signifikant erhöht; die Cholesterol-
konzentrationen sind aufgrund starker Streuung unverändert (Studie 1).
» Der Verzehr von Joghurt über 6 Monate vermindert die faecale Exkretion
unkonjugierter primärer und sekundärer Gallensäuren, wobei die
Sterolausscheidung unverändert bleibt (Studie 3).
» Die probiotischen Stämme Lactobacillus paracasei LTH 2579, Lactobacillus
acidophilus 145, Bifidobacterium longum 913, Lactobacillus acidophilus 742
und Bifidobacterium species 420 bewirken keine Änderung des faecalen
Gallensäurenprofiles. Bei Supplementation von L. paracasei ist eine
Tendenz zu verminderten Deoxycholsäuregehalten erkennbar. Die in
Joghurt verabreichten Stämme führen zu verminderten faecalen
Coprostanolkonzentrationen (Studien 2 - 4).
» In allen vier Studien konnten Probanden mit einem inversen Sterolprofil
nachgewiesen werden, wobei in fünf von acht Fällen kein Hinweis auf eine
Darmerkrankung zu erkennen war.
» Hypercholesterolämische Probanden scheiden weniger sekundäre Gallen-
säuren aus als normocholesterolämische Probanden und tendieren zu
erhöhten Gehalten primärer Gallensäuren in den Faeces (Studie 3).
» Die tägliche Steroidexkretion von 86 erfassten Probanden und deren
Gehalte an Sterolen und Gallensäuren in der Stuhltrockensubstanz sind mit
den in der Literatur zitierten Werten vergleichbar.
» Die Gehalte an Cholesterol und Coprostanol in der Trockensubstanz stehen
in einem korrelativen Zusammenhang.

Zusammenfassung
95
8 ZUSAMMENFASSUNG
Ziel der Arbeit war es, analytische Methoden zur Bestimmung faecaler Sterole und
Gallensäuren zu erstellen. Weiterhin sollten unter Anwendung der Methoden
Einflüsse einer Supplementation der Nahrung mit Calciumphosphaten, Probiotika
und Joghurt auf die Exkretion dieser Steroide eruiert werden.
Etablierte gaschromatographische Methoden zur Sterolanalytik mit Flammen-
ionisationsdetektion erfordern eine Derivatisierung zur Trennung der Analyten. Mit
Einsatz der massenspektrometrischen Detektion ist es möglich, spezifische
Fragmente zu bestimmen und dadurch eine Separation koeluierender Substanzen
zu gewährleisten. Auf der Basis dieser analytischen Komponenten ist die
quantitative Analyse underivatisierter Sterole (Cholesterol, Coprostanol,
Coprostanon, Cholestanol, Cholestanon) realisierbar. Dazu werden die Sterole
hydrolysiert und anschließend mit Cyclohexan extrahiert. Nach Vereinigung der
Extrakte und Einengen bis zur Trockne werden die Sterole in Decan gelöst und in
den Gaschromatographen injiziert. Als interner Standard wird 5α-Cholestan
eingesetzt. Unter Auswertung der Massenspektren sind folgende Fragmente zur
Quantifizierung ausgewählt worden: 5α-Cholestan (m/z = 357,25), Coprostanol
(m/z = 373,50), Cholesterol (m/z = 301,30), Coprostanon (m/z = 316,30),
Cholestanol (m/z = 215,10) und Cholestanon (m/z = 232,00).
Durch den Einsatz des Single Ion Monitoring in der faecalen Gallensäurenanalytik
ist es möglich, Peaküberlagerungen durch die komplexe und individuelle
Faecesmatrix vor allem bei hohen Probendurchsätzen im Vergleich zur Detektion
mittels Flammenionisationsdetektion zu verringern. Zur Probenvorbereitung wird
die wässrige Phase der Sterolextraktion mit Natronlauge hydrolysiert,
anschließend mit Salzsäure auf einen pH-Wert von 1 eingestellt und die freien
Gallensäuren werden mit Diethylether separiert. Nach Methylierung und
Silylierung wird 1 µl in den Gaschromatographen injiziert. Zur quantitativen
Analyse sind die Fragmente für iso-LCA (m/z = 215,25), LCA (m/z = 215,25), iso-
DCA (m/z = 75,10), DCA (m/z = 255,30), CDCA (m/z = 73,10), CA (m/z = 253,20),
HDCA (interner Standard, m/z = 81,15), 12-Oxo-DCA (m/z = 231,25) aus den
Massenspektren ausgewählt worden.

Zusammenfassung
96
Um einen möglichen Einfluss von Calciumphosphat, Probiotika und Joghurt auf
die Steroidexkretion zu bestimmen, werden gefriergetrocknete Faecesproben aus
vier Humanstudien analysiert. Diese sind am Institut für Ernährungswissenschaft,
Lehrstuhl Ernährungsphysiologie, der Friedrich-Schiller-Universität Jena zwischen
1997 und 2001 durchgeführt worden.
In Studie 1 verzehren 11 männlichen Probanden eine Diät mit einer gesteigerten
Zufuhr von Calciumphosphat im Verhältnis Ca : P = 1 : 1,3. Die Studie ist in drei
Perioden gegliedert. In der Kontrollperiode nehmen die Probanden auf der Basis
von erhobenen Ernährungsprotokollen täglich 1.231 mg Calcium und 1.600 mg
Phosphor auf. In der ersten Zulageperiode werden täglich 609 mg Calcium und
793 mg Phosphor in Form von Tabletten supplementiert. Die Dosis wird in der
zweiten Zulageperiode verdoppelt.
Die Probanden setzen in allen Perioden vergleichbare Stuhlmassen ab. Eine
gesteigerte Zufuhr von anorganischem Calcium und Phosphor beeinflusst die
tägliche faecale Gallensäurenexkretion und die Konzentrationen in der
Trockensubstanz nicht. Durch Calciumphosphat (Ca: 1.218 mg/d bzw.
P: 1.586 mg/d) sind die täglichen Coprostanonausscheidungen und die Gehalte im
Stuhl signifikant erhöht. Die Änderungen der mittleren Konzentrationen an
Coprostanon und Cholesterol in der Trockensubstanz verlaufen gleichförmig. Die
Konzentrationsunterschiede des Cholesterols sind aufgrund einer starken
Streuung statistisch nicht gesichert.
Der Einfluss von Lactobacillus paracasei LTH 2579 (108 KBE/g) auf die faecale
Steroidexkretion wird an je 10 männlichen und weiblichen Probanden in Studie 2
untersucht. Dabei werden über zwei Versuchsperioden täglich 50 g Rohwurst mit
bzw. ohne Probiotikum verzehrt.
Die Frauen der Studie 2 setzen signifikant weniger Stuhl ab als die Männer. Bei
allen Probanden ist eine verringerte Stuhlexkretion in der zweiten Periode
festzustellen. Die Keimzahlen an Lactobacillen in den Faeces sind nach Zulage in
der zweiten Periode erhöht. Weibliche Probanden scheiden weniger Sterole und
Gallensäuren aus als die männlichen. Unter Berücksichtigung der Fettaufnahme
sind die geschlechtsspezifischen Unterschiede in der Gallensäurenexkretion nicht

Zusammenfassung
97
mehr nachzuweisen. Die gesamte tägliche Gallensäurenausscheidung ist in der
zweiten Periode signifikant vermindert. Allerdings bleiben unabhängig vom
Probiotikum die Gehalte an sauren Steroiden in der Trockensubstanz und nach
Standardisierung auf 30 g Trockensubstanzexkretion mit Ausnahme tendenziell
verminderter Deoxycholsäurewerte unbeeinflusst. Die Diskrepanz wird durch
verschiedene Stuhlmassen zwischen den Versuchsperioden hervorgerufen.
In Studie 3 wird der Einfluss eines mit Lactobacillus acidophilus 145
(106 - 108 KBE/g) und Bifidobacterium longum 913 (105 KBE/g) supplementierten
Joghurts auf Änderungen der faecalen Steroidexkretion von 29 normo-
cholesterolämischen bzw. moderat hypercholesterolämischen Frauen untersucht.
Die Studie ist im cross-over Design in drei 7-wöchige Perioden strukturiert, wobei
die erste eine Adaptationsphase mit konventionellem Joghurt darstellt.
In Analogie zu Studie 2 scheiden die Probanden nach Gabe des probiotischen
Produktes weniger Stuhl aus. Der faecale pH-Wert ist sowohl nach
konventionellem als auch nach probiotischem Joghurtverzehr signifikant zur
Adaptationsperiode vermindert. Die Konversionsrate des Cholesterols bleibt in
allen Perioden unverändert bei ca. 88 %. Die tägliche Exkretion der sekundären
Gallensäuren vermindert sich um etwa 30 % unabhängig vom Probiotika-
supplement im Vergleich zu den Adaptationswerten. Weiterhin ist die
Ausscheidung an primären Gallensäuren ohne Einfluss der zugesetzten Bakterien
verringert. Die Ergebnisse der Gallensäurengehalte in der Gefriertrockensubstanz
der Faeces bestätigen die verminderte Exkretion. Hypercholesterolämische
Probanden scheiden signifikant weniger sekundäre Gallensäuren und tendenziell
mehr primäre Gallensäuren pro Tag aus.
In der vierten Studie wird ein mit Lactobacillus acidophilus 742 (107 KBE/ml) und
Bifidobacterium species 420 (104 KBE/ml) versetzter Joghurt an je 13 männliche
und weibliche Probanden verabreicht. In den im cross-over Design durchgeführten
Untersuchungen dauern die beiden Studienphasen jeweils 5 Wochen und in der
Startperiode wird kein Joghurt verzehrt.
Im Verlauf der Studie sind keine signifikanten Änderungen in der täglichen
Stuhlexkretion und des faecalen pH-Wertes aufgetreten. Die Frauen scheiden in

Zusammenfassung
98
der Basisperiode mit 18 g/d Trockenstuhl bzw. 67 g/d Frischsubstanz geringe
Mengen aus. Im Studienverlauf ändern sich die Gallensäurenkonzentrationen in
der Gefriertrockensubstanz nicht. Nach konventioneller Joghurtgabe sind im
Vergleich zu den Basisdaten die Gehalte an Cholesterol erhöht und die
Konversionsrate vermindert. Die Gesamtsterol-, Coprostanol- und
Cholesterolkonzentrationen sind nach Konsum von probiotischem Joghurt im
Vergleich zu dem konventionellen Produkt vermindert. Die tägliche Ausscheidung
an Cholesterol, Cholestanol, Coprostanon und Deoxycholsäure ist nach dem
Verzehr von Joghurt im Vergleich zu den Ausgangswerten und unabhängig von
der Joghurtart angestiegen. Ein steigender Effekt durch erhöhte Stuhlmassen
nach den Joghurtperioden ist nicht auszuschließen. Im Stuhl der weiblichen
Probanden liegen höhere Konzentrationen an Cholesterol, Coprostanol,
Coprostanon und primären Gallensäuren vor als in dem der männlichen
Probanden. Diese scheiden hingegen mehr Litho- und Deoxycholsäure aus.
An allen vier Studien nahmen Probanden mit einem inversem Sterolprofil teil,
wobei in einigen Fällen kein Hinweis auf eine Darmerkrankung zu erkennen war.
Der Verzehr von Joghurt über 6 Monate verringert die faecale Ausscheidung freier
primärer und sekundärer Gallensäuren, was durch eine Verminderung des
faecalen pH-Wertes und einer folgenden Modifikation von Enzymaktivitäten im
Colon verursacht sein kann. Die supplementierten Lactobacillen und
Bifidobakterien bewirken keine Änderung des faecalen Gallensäurenprofiles,
führen aber in Kombination mit Joghurt zu verminderten Coprostanol-
konzentrationen. Bei Supplementation von L. paracasei ist eine Tendenz zu
verminderten Deoxycholsäuregehalten in den Faeces erkennbar.
Dass ein enger Zusammenhang zwischen dem Cholesterol- und dem
Gallensäurenkreislauf im Menschen besteht, wird durch ein differentes faecales
Gallensäurenprofil moderat hypercholesterolämischer Probanden im Vergleich zu
Gesunden bestätigt. Ein dauerhafter Joghurtkonsum kann die Gehalte freier
Gallensäuren in den Faeces vermindern und die Cholesterolgehalte im Blut positiv
beeinflussen. Geringe Gallensäurenkonzentrationen im Colon sind weiterhin unter
dem Aspekt der Carcinogenese von Vorteil, da freie Gallensäuren aufgrund ihres
toxischen Verhaltens als Promotoren einer Tumorgenese gelten.

Quellenverzeichnis
I
Quellenverzeichnis ALAKOMI HL, SKYTTÄ E, SAARELA M, MATTILA-SANDHOLM T, LATVA-KALA K, HELANDER IM: Lactic acid permeabilizes gram-negative bacteria by disruption the outer membrane. Appl Environ Microbiol 66 (2000) 2001-2005 ALI SS, KUKSIS A, BEVERIDGE JMR: Excretion of bile acids by three men on a fat-free diet. Can J Biochem 44 (1966) 957-969 ALMÉ B, BREMMELGAARD A, SJÖVALL J, THOMASSEN P: Analysis of metabolic profiles of bile acids in urine using a lipophilic anion exchanger and computerized gas-liquid chromatography-mass spectrometry. J Lipid Res 18 (1977) 339-362 ANDEREGG RJ: Mass spectrometry: an introduction. In: Suelter CH, Watson JT (Hrsg.) Methods of Biochemical Analysis. Volume 34: Biomedical Applications of Mass Spectrometry. New York/Chichester, John Wiley and Sons, 1990, 1-73 APPLETON GVN, OWEN RW, WILLIAMSON RCN: The effect of dietary calcium supplementation on intestinal lipid metabolism. J Steroid Biochem Molec Biol 42 (1992) 383-387 ARCA M, MONTALI A, CIOCCA S, ANGELICO F, CANTAFORA A: An improved gas-liquid chromatographic method for the determination of fecal neutral sterols. J Lipid Res 24 (1983) 332-335 AUSMAN LM, JOHNSON JA, GUIDRY C, NAIR PP: Fecal bile acids and neutral sterols in the cotton-top tamarin (Saguinus Oedipus). Comp Biochem Physiol 105B (1993) 655-663 BARKER GM, RADLEY S, DAVIS A, SETCHELL KDR, O'CONNELL N, DONOVAN IA, KEIGHLEY MRB, NEOPTOLEMOS JP: Analysis of faecal neutral sterols in patients with familial adenomatous polyposis by gas chromatography-mass spectrometry. Int J Colorect Dis 8 (1993) 188-192 BARKER GM, RADLEY S, DAVIS A, IMRAY CHE, SETCHELL KDR, O�CONNELL N, DONOVAN IA, KEIGHLEY MRB, NEOPTOLEMOS JP: Unconjugated faecal bile acids in familial adenomatous polyposis analysed by gas-liquid chromatography and mass spectrometry. Br J Surg 81 (1994) 739-742 BARTRAM H-P, SCHEPPACH W, HEID C, FABIAN C, KASPER H: Effect of starch malabsorption on fecal bile acids and neutral sterols in humans: Possible implications for colonic carcinogenesis. Cancer Res 51 (1991) 4238-4242 BARTRAM H-P, SCHEPPACH W, GERLACH S, RUCKDESCHEL G, KELBER E, KASPER H: Does yogurt enriched with Bifidobacterium longum affect colonic microbiology and fecal metabolites in healthy subjects? Am J Clin Nutr 59 (1994) 428-432 BATTA AK, SALEN G: Gas chromatography of bile acids. J Chromatogr B 723 (1999) 1-16

Quellenverzeichnis
II
BATTA AK, SALEN G, RAPOLE KR, BATTA M, BATTA P, ALBERTS D, EARNEST D: Highly simplified method for gas-liquid chromatographic quantitation of bile acids and sterols in human stool. J Lipid Res 40 (1999) 1148-1154 BEHLING AR, KAUP SM, CHOQUETTE LL, GREGER JL: Lipid absorption and intestinal tumour incidence in rats fed on varying levels of calcium and butterfat. Br J Nutr 64 (1990) 505-513 BERG RD: The indigenous gastrointestinal microflora. Trends Microbiol 4 (1996) 430-435 BEZKOROVAINY A: Probiotics: Determinants of survival and growth in the gut. Am J Clin Nutr 73 (2001) 399-405 BIESALSKI HK: Fette. In: Biesalski HK, Fürst P, Kasper H, Kluthe R, Pölert W, Puchstein C, Stähelein HB (Hrsg.) Ernährungsmedizin. Georg Thieme, Stuttgart/New York, 1995, 58-77 BJÖRKHEM I, BLOMSTRAND R, SVENSSON L: Serum cholesterol determination by mass fragmentographic. Clin Chim Acta 54 (1974) 185-193 BJÖRKHEM I, EINARSSON K, MELONE P, HYLEMON P: Mechanism of intestinal formation of deoxycholic acid from cholic acid in humans: Evidence for a 3-oxo-∆4-steroid intermediate. J Lipid Res 30 (1989) 1033-1039 BOVEE-OUDENHOVEN IM, VAN DER MEER R: Protective effects of dietary lactulose and calcium phosphate against Salmonella infection. Scand J Gastroenterol 32 (1997) 112-114 BOVEE-OUDENHOVEN IM, WISSINK ML, WOUTERS JT, VAN DER MEER R: Dietary calcium phosphate stimulates intestinal Lactobacilli and decreases the severity of a Salmonella infection in rats. J Nutr 129 (1999) 607-612 BÖRJESSON E, SUNDIN A, LEEMING R, TORSTENSSON L: New method for determination of fecal sterols in urine using non-chlorinated solvents. J Chromatogr B 713 (1998) 438-442 BRYDON WG, TADESSE K, SMITH DM, EASTWOOD MA: Gas chromatographic procedure for the measurement of bile acids in rat intestine. J Chromatogr 172 (1979) 450-452 CHAPLIN MF: Bile acids, fibre and colon cancer: The story unfolds. J Roy Soc Health 118 (1998) 53-61 CHARACH G, RABINOVICH PD, KONIKOFF FM, GROSSKOPF I, WEINTRAUB MS: Decreased fecal bile acid output in patients with coronary atherosclerosis. J Med 29 (1998) 125-136

Quellenverzeichnis
III
CHIANG MT, CHEN YC, HUANG AL : Plasma lipoprotein cholesterol levels in rats fed a diet enriched in cholesterol and cholic acid. Int J Vitam Nutr Res 68 (1998) 328-334 CHILD P, ALOE M, MEE D: Separation and quantitation of fatty acids, sterols, and bile acids in feces by gas chromatography as their butyl ester-acetate derivatives. J Chromatogr 415 (1987) 13-26 CHITHRA V, LEELAMMA S: Hypolipidemic effect of coriander seeds (Coriandrum sativum): Mechanism of action. Plant Foods Hum Nutr 51 (1997) 167-172 CHRISTL SU, BARTRAM HP, PAUL A, KLEBER E, SCHEPPACH W, KASPER H: Bile acid metabolism by colonic bacteria in continuous culture: Effects of starch and pH. Ann Nutr Metab 41 (1997) 45-51 CLIFFORD AJ, SMITH LM, CREVELING RK, HAMBLIN CL, CLIFFORD CK: Effects of dietary triglycerides on serum and liver lipids and sterol excretion of rats. J Nutr 116 (1986) 944-956 COLEMAN JP, WHITE WB, EGESTAD B, SJÖVALL J, HYLEMON PB: Biosynthesis of a novel bile acid nucleotide and mechanism of 7α-dehydroxylation by an intestinal Eubacterium species. J Biol Chem 262 (1987) 4701-4707 CRAVEN PA, PFANSTIEL J, DERUPERTIS FR: Role of activation of protein kinase C in the stimulation of colonic epithelial proliferation and reactive oxygen formation by bile acids. J Clin Invest 79 (1987) 532-541 CREMER E: Über die Wanderungsgeschwindigkeit der Zonen bei der chromatographischen Analyse. Nachdruck: Chromatographia 9 (1976) 363-366 CRONHOLM T, MAKINO I, SJÖVALL J: Steroid metabolism in rats given [1-2H2] ethanol. Biosynthesis of bile acids and reduction of 3-keto-5b-cholanoic acid. Eur J Biochem 24 (1972) 507-519 CZUBAYKO F, BEUMERS B, LAMMSFUSS S, LÜTJOHANN D, VON BERGMANN K: A simplified micro-method for quantification of fecal excretion of neutral and acidic sterols for outpatient studies in humans. J Lipid Res 32 (1991) 1861-1867 DAMBEKODI PC, GILLILAND SE: Incorporation of cholesterol into the cellular membrane of Bifidobacterium longum. J Dairy Sci 81 (1998) 1818-1824 DE KOK TMCM, VAN FAASSEN A, GLINGHAMMER B, PACHEN DMFA, ENG M, RAFTER JJ, BAETEN CGMI, ENGELS LGJB, KLEINJANS JCS: Bile acid concentrations, cytotoxicity, and pH of fecal water from patients with colorectal adenomas. Dig Dis Sci 44 (1999) 2218-2225

Quellenverzeichnis
IV
DEPARTMENT OF NATURAL RESOURCE PROTECTION (1998) Implementation of a chemical method for differentiating human and animal fecal impacts in surface waters and sediments. http://www.co.broward.fl.us/moi00708.pdf (Stand 24.06.2002) Adobe Acrobat Dokument, 34 Seiten DE RODAS B Z, GILLILAND SE, MAXWELL CV: Hypocholesterolemic action of Lactobacillus acidophilus ATCC 43121 and calcium in swine with hypercholesterolemia induced by diet. J Dairy Sci 79 (1996) 2121-2128 DE RUPERTIS FR, CRAVEN PA, SAITO R: Bile salt stimulation of colonic epithelial proliferation. Evidence for involvement of lipoxygenase products. J Clin Invest 74 (1984) 1614-1624 DE SMET I, VAN HOORDE L, DE SAEYER N, VANDE WOESTYNE M, VERSTRAETE M: In vitro study of bile salt hydrolase (BSH) activity of BSH isogenic Lactobacillus plantarum 80 strains and estimation of cholesterol lowering through enhanced BSH activity. Microb Ecol Health Dis 7 (1994) 315-329 DE SMET I, DE BOEVER P, VERSTRAETE W: Cholesterol-lowering in pigs through enhanced bile salt hydrolase activity. Br J Nutr 79 (1998) 185-194 DE WEERDT GA, BEKE R, VERDIEVEL H, BARBIER F: Quantitative determination of fecal bile acids as their methyl ether methyl ester by the repetitive scan technique. Biomed Mass Spectrom 7 (1980) 515-520 DGE, Deutsche Gesellschaft für Ernährung: Ernährungsbericht 2000. Frankfurt/Main, Deutsche Gesellschaft für Ernährung e. V., 2000, 263-274 DGF, Deutsche Gesellschaft für Fettwissenschaft: 7th Proficiency test, Final report, 2001 DOWLING RH, MURPHY GM: Bile acids and acquired disease: Old hypothesis, new concepts. In: Cohen RD, Lewis B, Alberti KGMM, Denman AM (Hrsg.) The Metabolic and Molecular Basis of Acquired Disease. Baillière Tindall, London/Philadelphia, 1990, 1231-1245 DUANE WC, HUTTON SW: Lack of effect of experimental ascorbic acid deficiency on bile acid metabolism, sterol balance, and biliary lipid composition in man. J Lipid Res 24 (1983) 1186-1195 DUANE WC: Effects of soybean protein and very low dietary cholesterol on serum lipids, biliary lipids, and fecal sterols in humans. Metabolism 48 (1999) 489-494 EDENHARDER R, KNAFLIC T: Epimerization of chenodeoxycholic acid to ursodeoxycholic acid by human intestinal lecithinase-lipase-negative Clostridia. J Lipid Res 22 (1981) 652-658 EDENHARDER R: Dehydroxylation of cholic acid at C12 and epimerization at C5 and C7 by Bacteroides species. J Steroid Biochem 21 (1984) 413-420

Quellenverzeichnis
V
ELLEGARD L, BOSAEUS I, ANDERSSON H: Will recommended changes in fat and fibre intake affect cholesterol absorption and sterol excretion? An ileostomy study. Eur J Clin Nutr 54 (2000) 306-313 ENEROTH P, GORDON B, SJÖVALL J: Characterization of trisubstituted cholanoic acids in human feces. J Lipid Res 7 (1966) 524-530 ENEROTH P, HELLSTRÖM K, SJÖVALL J: A method for quantitative determination of bile acids in human feces. Acta Chem Scand 22 (1968) 1729-1744 ENEROTH P, SJÖVALL J: Extraction, purification, and chromatographic analysis of bile acids in biological materials. In: Nair PP, Kritchevsky D (Hrsg.) The Bile Acids. Chemistry, Physiology, and Metabolism. Volume 1: Chemistry. New York/London, Plenum Press, 1971, 121-172 EVRARD E, JANSSEN G: Gas-liquid chromatographic determination of human fecal bile acids. J Lipid Res 9 (1968) 226-236 EYSSEN HJ, PARMENTIER GG, COMPERNOLLE FC, DE PAUW G, PIESSENS DENEF M: Biohydrogenation of sterols by Eubacterium ATCC 21, 408 - nova species. Eur J Biochem 36 (1973) 411-421 FERNANDES CF, SHAHANI KM: Anticarcinogenic and immunological properties of dietary lactobacilli. J Food Prot 53 (1990) 704-710 GAMBERT P, LALLEMANT C, ARCHAMBAULT A, MAUME BF, PADIEU P: Assessment of serum cholesterol by two methods: gas-liquid chromatography on a capillary column and chemical ionization-mass fragmentography with isotopic dilution of [3,4-13C] cholesterol as internal standard. J Chromatogr 162 (1979) 1-6 GARROTE GL, ABRAHAM AG, DE ANTONI GL: Inhibitory power of kefir: The role of organic acids. J Food Prot 63 (2000) 364-369 GERHARDT KO, GEHRKE CW, ROGERS IT, FLYNN MA, HENTGES DJ: Gas-liquid chromatography of fecal neutral steroids. J Chromatogr 135 (1977) 341-349 GILLILAND SE, NELSON CR, MAXWELL C: Assimilation of cholesterol by Lactobacillus acidophilus. Appl Environ Microbiol 49 (1985) 377-381 GOLDIN BR, GORBACH SL: The relationship between diet and rat fecal bacterial enzymes implicated in colon cancer. J Natl Cancer Inst 57 (1976) 371-375 GOLDIN BR: Intestinal microflora: Metabolism of drugs and carcinogens. Ann Med 22 (1990) 43-48 GORDON H, LEWIS B, EALES L, BROCK JF: Dietary fat and cholesterol metabolism. Faecal elimination of bile acids and other lipids. Lancet 2 (1957) 1299-1306

Quellenverzeichnis
VI
GOVERS MJAP, VAN DER MEER R: Effects of dietary calcium and phosphate on the intestinal interactions between calcium, phosphate, fatty acids, and bile acids. Gut 34 (1993) 365-370 GOVERS MJAP, TERMONT DSML, LAPRÉ JA, KLEIBEUKER JH, VONK RJ, VAN DER MEER R: Calcium in milk products precipitates intestinal fatty acids and secondary bile acids and thus inhibits colonic cytotoxicity in humans. Cancer Res 56 (1996) 3270-3275 GÖRÖG S: Cholesterol and related compounds. In: Pungor E, Simon W, Inczédy J (Hrsg.) Studies in Analytical Chemistry. 5. Quantitative Analysis of Steroids. Budapest, Akadémiai Kiadó, 1983, 247-289 GRIMM M: Einfluss einer exzessiven Phosphatzufuhr beim Menschen auf Knochenmarker und Endokrinum sowie anti-/genotoxische Effekte. Dissertation, Friedrich Schiller Universität, Jena, 2000 GRUNDY SM, AHRENS JR. EH, MIETTINEN TA: Quantitative isolation and gas-liquid chromatographic analysis of total fecal bile acids. J Lipid Res 6 (1965) 397-410 GYLLING H, MIETTINEN TA: LDL cholesterol lowering by bile acid malabsorption during inhibited synthesis and absorption of cholesterol in hypercholesterolemic coronary subjects. Nutr Metab Cardivasc 12 (2002) 19-23 HAMMES WP: Bewertung der gesundheitlichen Unbedenklichkeit von Milchsäurebakterien und Probiotika. Monatsschr Kinderheilkd 146 (1998) 31-38 HASHIMOTO H, KAWASE M, HOSODA M, HE F, MORITA H, HOSONO A: Binding, deconjugation and oxidation of taurocholic acid with lactobacilli cells. Milchwiss 55 (2000) 316-319 HAYASHI E, AMURO Y, ENDO T, YAMAMOTO H, MIYAMOTO M, KISHIMOTO S: Fecal bile acids and neutral sterols in rats with spontaneous colon cancer. Int J Cancer 37 (1986) 629-632 HIRANO S, MASUDA N: Transformation of bile acids by Eubacterium lentum. Appl Environ Microbiol 42 (1981) 912-915 HIRANO S, MASUDA N: Characterization of NADP-dependent 7β-hydroxysteroid dehydrogenases from Peptostreptococcus productus and Eubacterium aerofaciens. Appl Environ Microbiol 43 (1982) 1057-1063 HOFMANN AF: The continuing importance of bile acids in liver and intestinal disease. Arch Intern Med 159 (1999) 2647-2657 HOFSTAD B, VATN MH, ANDERSEN SN, OWEN RW, LARSEN S, OSNES M: The relationship between faecal bile acid profile with or without supplementation with calcium and antioxidants on recurrence and growth of colorectal polyps. Eur J Cancer Prev 7 (1998) 287-294

Quellenverzeichnis
VII
HOLT RH: Dairy foods and prevention of colon cancer: Human studies. J Am Coll Nutr 18 (1999) 379-391 HOLZAPFEL WH, HABERER P, SNEL J, SCHILLINGER U, HUIS IN�T VELD JHJ : Overview of gut flora and probiotics. Int J Food Microbiol 41 (1998) 85-101 HORI T, MATSUMOTO K, SAKAITANI Y, SATO M, MOROTOMI M: Effect of dietary deoxycholic acid and cholesterol on fecal steroid concentration and its impact on the colonic crypt cell proliferation in azoxymethane-treated rats. Cancer Lett124 (1998) 79-84 HOVING EB: Chromatographic methods in the analysis of cholesterol and related lipids. J Chromatogr B 671 (1995) 341-362 HYLEMON PB, GLASS TL: Biotransformation of bile acids and cholesterol by the intestinal microflora. In: Hentges DJ (Hrsg.) Human Intestinal Microflora in Health and Disease. New York/London, Academic Press, 1983, 189-213 HYLLA S, GOSTNER A, DUSEL G, ANGER H, BARTRAM HP, CHRISTEL SU, KASPER H, SCHEPPACH W: Effects of resistant starch on the colon in healthy volunteers: Possible implications for cancer prevention. Am J Clin Nutr 67 (1998) 136-142 IIDA T, HIKOSAKA M, GOTO J, NAMBARA T: Capillary gas chromatographic behaviour of tert.-hydroxylated steroids by trialkylsilylation. J Chromatogr A 937 (2001) 97-105 IMRAY CHE, MINOURA T, DAVIS A, RADLEY S, NEWBOLD KM, LAVELLE-JONES M, LAWSON AM, BAKER PR, NEOPTOLEMOS JP: Comparability of hamster with human faecal unconjugated bile acids in a model of colorectal cancer. Anticancer Res 12 (1992) 553-558 JAHREIS G, VOGELSANG H, KIESSLING G, SCHUBERT R, BUNTE C, HAMMES WP: Influence of probiotic sausage (Lactobacillus paracasei) on blood lipids and immunological parameters of healthy volunteers. Food Res Int 35 (2002) 133-138 JAVITT NB: Bile acid synthesis from cholesterol � regulatory and auxiliary pathways. FASEB J 8 (1994) 1308-1311 JÄRVINEN R, KNEKT P, HAKULINEN T, AROMAA A: Prospective study on milk products, calcium and cancers of the colon and rectum. Eur J Clin Nutr 55 (2001) 1000-1007 JENKINS DJA, KENDALL CWC, MEHLING CC, PARKER T, RAO AV, AGARWAL S, NOVOKMET R, JONES PJH, RAEINI M, STORY IA, FURUMOTO E, VIDGEN E, GRIFFIN LC, CUNNANE SC, RYAN MA, CONNELLY PW: Combined effect of vegetable protein (soy) and soluble fiber added to a standard cholesterol-lowering diet. Metabolism 48 (1999) 809-816 KAIBARA N, SASAKI T, IKEGUCHI M, KOGA S, IKAWA S: Faecal bile acids and neutral sterols in Japanese with large bowel carcinoma. Oncology 40 (1983) 255-258

Quellenverzeichnis
VIII
KELLER S, JAHREIS G: Gallensäuren und Einflussfaktoren auf ihre Modifikation und Exkretion. Ernährungs-Umschau 48 (2001) 142-147 KELSEY MI, MOLINA JE, HUANG SKS, HWANG KK: The identification of microbial metabolites of sulfolithocholic acid. J Lipid Res 21 (1980) 751-759 KIEßLING G, SCHNEIDER J, JAHREIS G: Long-term consumption of fermented dairy products over 6 months increases HDL cholesterol. Eur J Clin Nutr (2002), in press KIKUCHI-HAYAKAWA H, ONODERA N, MATSUBARA S, YASUDA E, SHIMAKAWA Y, ISHIKAWA F: Effects of soya milk and Bifidobacterium-fermented soya milk on plasma and liver lipids, and faecal steroids in hamsters fed on a cholesterol-free or cholesterol-enriched diet. Br J Nutr 79 (1998) 97-105 KLEIN A, KIEßLING G, JAHREIS G: Einfluss von probiotischen Keimen auf den Lipidstoffwechsel. 11. Atherosklerose Symposium Erfurt (2002) KLEIN A: unveröffentlichte Ergebnisse (2002) KORPELA JT: Capillary gas-liquid chromatography of faecal free and esterified neutral sterols. Scand J Clin Invest 42 (1982) 529-534 KORPELA JT, ADLERCREUTZ H: Fecal neutral sterols in omnivorous and vegetarian women. Scand J Gastroenterol 20 (1985) 1180-1184 KORPELA JT, ADLERCREUTZ H, TURUNEN MJ: Fecal free and conjugated bile acids and neutral sterols in vegetarians, omnivores, and patients with colorectal cancer. Scand J Gastroenterol 23 (1988) 277-283 LATTA RK, FIANDER H, ROSS NW, SIMPSON C, SCHNEIDER H: Toxicity of bile acids to colon cancer cell lines. Cancer Lett 70 (1993) 167-173 LAWSON AM, SETCHELL KDR: Mass spectrometry of bile acids. In: Setchell KDR, Kritchevsky D, Nair PP (Hrsg.) The Bile Acids. Chemistry, Physiology, and Metabolism. Volume 4: Methods and Applications. New York/London, Plenum Press, 1988, 167-268 LEE C, MARTIN KO, JAVITT NB: Bile acid synthesis: 7α-Hydroxylation of intermediates in the sterol 27-hydroxylase metabolic pathway. J Lipid Res 37 (1996) 1356-1362 LETTRÉ H: Über Sterine und Gallensäuren. In: Lettré H, Inhoffen HH, Tschesche R (Hrsg.) Über Sterine, Gallensäuren und verwandte Naturstoffe. Erster Band. Stuttgart, Ferdinand Enke, 1954, 1-164 LINDENTHAL B, ALDAGHLAS TA, KELLEHER JK, HENKEL SM, TOLBA R, HAIDL G, VON BERGMANN K: Neutral sterols of rat epididymis: High concentrations of dehydrocholesterol in rat caput epididymidis. J Lipid Res 42 (2001) 1089-1095

Quellenverzeichnis
IX
LÖFFLER G, PETRIDES PE (Hrsg) Biochemie und Pathobiochemie. Berlin/Heidelberg, Springer, 1998, 1001-1003 LUND E, MURI-BOBERG K, BYSTROM S, CARLSTROM K, OLUND J, BJÖRKHEM I: Formation of novel C21 bile acids from cholesterol in the rat. J Biol Chem 266 (1991) 4929-4937 LUPTON JR, STEINBACH G, CHANG WC, O�BRIEN BC, WIESE S, STOLTZFUS CL, GLOBER GA, WARGOVICH MJ, MCPHERSON RS, WINN RJ: Calcium supplementation modifies the relative amounts of bile acids in bile and affects key aspects of human colon physiology. J Nutr 126 (1996) 1421-1428 MACDONALD IA, WILLIAMS CN, MAHONY DE: 7α-Hydroxysteroid dehydrogenase from Escherichia coli B: Preliminary studies. Biochim Biophys Acta 309 (1973) 243-253 MACDONALD IA, SINGH G, MAHONY DE, MEIER CE: Effect of pH on bile salt degradation by mixed fecal cultures. Steroids 32 (1978) 245-256 MACDONALD IA, ROACH PD: Bile salt induction of 7α- and 7β-hydroxysteroid dehydrogenases in Clostridium absonum. Biochim Biophys Acta 665 (1981) 262-269 MACDONALD IA, BOKKENHEUSER VD, WINTER J, MCLERNON AM, MOSBACH EH: Degradation of steroids in the human gut. J Lipid Res 24 (1983a) 675-700 MACDONALD IA, WHITE BA, HYLEMON PB: Separation of 7α- and 7β-hydroxysteroid dehydrogenase activities from Clostridium absonum ATCC 27555 and cellular response of this organism to bile acid inducers. J Lipid Res 24 (1983b) 1119-1136 MARTEAU P, GERHARDT MF, MYARA A, BOUVIER E, TRIVIN F, RAMBAUD JC: Metabolism of bile salts by alimentary bacteria during transit in the human small intestine. Microb Ecol Health Dis 8 (1995) 151-157 MARTEAU P: Prebiotics and probiotics for gastrointestinal health. Clin Nutr 20 (2001) 41-45 MATUSIK JE, HOSKIN GP, SPHON JA: Gas chromatographic/mass spectrometric confirmation of identity of coprostanol in Mercenaria mercenaria (Bivalvia) taken from sewage-polluted water. J Assoc Off Anal Chem 71 (1988) 994-999 MCMURRY MP, CONNOR WE, LIN DS, CERQUEIRA MT, CONNOR SL: The absorption of cholesterol and the sterol balance in the Tarahumara Indians of Mexico fed cholesterol-free and high cholesterol diets. Am J Clin Nutr 41 (1985) 1289-1298 MIDTVEDT T, LINGAAS E, CARLSTEDT-DUKE B, HÖVERSTAD T, MIDTVEDT AC, SAXERHOLT H, STEINBAKK M, NORIN KE: Intestinal microbial conversion of cholesterol in man. Influence of antibiotics. APMIS 98 (1990) 839-844

Quellenverzeichnis
X
MIETTINEN TA: Gas-liquid chromatographic determination of fecal neutral sterols using a capillary column. Clin Chim Acta 124 (1982) 245-248 MIETTINEN TA: Dietary fiber and lipids. Am J Clin Nutr 45 (1987) 1237-1242 MOORE WEC, MOORE LH: Intestinal floras of populations that have a high risk of colon cancer. Appl Environ Microbiol 61 (1995) 3202-3207 MOSBACH EH, KALINSKY HJ, HALPERN E, KENDALL FE: Determination of deoxycholic and cholic acids in bile. Arch Biochem Biophys 51 (1954) 402-410 MOTT GE, BRINKLEY AW, MERSINGER CL: Biochemical characterization of cholesterol-reducing Eubacterium. Appl Environ Microbiol 40 (1980) 1017-1022 MOWER HF, RAY RM, SHOFF R, STEMMERMANN GN, NOMURA A, GLOBER GA, KAMIYAMA S, SHIMADA A, YAMAKAWA H: Fecal bile acids in two Japanese populations with different colon cancer risks. Cancer Res 39 (1979) 328-331 MURI-BOBERG K, LUND E, OLUND J, BJÖRKHEM I: Formation of C21 bile acids from plant sterols in the rat. J Biol Chem 265 (1990) 7967-7975 MYLIUS F: Ueber die blaue Jodstärke und die blaue Jodcholsäure. Hoppe Seylers Physiol Chem 11 (1887) 306-347 NAGENGAST FM, GRUBBEN MJA, VAN MUNSTER IP: Role of bile acids in colorectal carcinogenesis. Eur J Cancer 31A (1995) 1067-1070 NAIR P, TURJMAN N: Role of bile acids and neutral sterols in familial cancer syndromes of the colon. Dis Colon Rectum 00 (1983) 629-632 NAIR P, TURJMAN N, GOODMAN GT, GUIDRY C, CALKINS BM: Diet, nutrition intake, and metabolism in populations at high and low risk for colon cancer. Am J Clin Nutr 40 (1984) 931-936 NAIR PP: Role of bile acids and neutral sterols in carcinogenesis. Am J Clin Nutr 48 (1988) 768-774 NAITO T, KUROKI S, CHIJIIWA K, TANAKA M: Bile acid synthesis and biliary hydrophobicity during obstructive jaundice in rats. J Surg Res 65 (1996) 70-76 NAKAMURA T, KIKUCHI H, TAKEBE K, KUDOH K, TERADA A, TAN-DOH Y, YAMADA N: Faecal lipid extraction levels in normal Japanese females on an unrestricted diet and a fat-restricted diet measured by simultaneous analysis of faecal lipids. J Int Med Res 20 (1992) 461-466 NAKAMURA T, TAKEUCHI T, TERADA A, TANDO Y, SUDA T: Near-infrared spectrometry analysis of fat, neutral sterols, bile acids, and short-chain fatty acids in the feces of patients with pancreatic maldigestion and malabsorption. Int J Pancreatol 23 (1998) 137-143

Quellenverzeichnis
XI
NISHIZUKA Y: The role of protein kinase C in cell surface signal transduction and tumor promotion. Nature 308 (1984) 693-698 NORIN E, PERSSON A-K, SAXERHOLT H, MIDTVEDT T: Establishment of Lactobacillus and Bifidobacterium species in germfree mice and their influence on some microflora-associated characteristics. Appl Environ Microbiol 57 (1991) 1850-1852 NTANIOS F: Plant sterol-ester-enriched spreads as an example of a new functional food. Eur J Lipid Sci Technol 103 (2001) 102-106 OKISHIO T, NAIR PP: An improved column for gas-liquid chromatography of substituted cholanic acids. Anal Biochem 15 (1966) 360-363 OWEN RW, TENNESON ME, BILTON RF, MASON AN: The degradation of cholesterol by Escherichia coli isolated from human feces. Biochem Soc Trans 6 (1978) 377-379 OWEN RW, THOMPSON MH, HILL MJ: Analysis of metabolic profiles of steroids in faeces of healthy subjects undergoing chenodeoxycholic acid treatment by liquid-gel chromatography and gas-liquid chromatography-mass spectrometry. J Steroid Biochem 21 (1984) 593-600 OWEN RW, DODO M, THOMPSON MH, HILL MJ: Fecal steroids and colorectal cancer. Nutr Cancer 9 (1987) 73-80 PANDA SK, BROITMAN SA : An analytical method for the measurement of fecal neutral and acid sterols from rats with induced large bowel tumorigenesis. Anal Chim Acta 432 (2001) 317-324 PEROGAMBROS A, PAPAVASSILIOU J , LEGAKIS NJ: Faecal neutral sterols in patients with colon cancer. Oncology 39 (1982) 274-278 PLAT J, BRZEZINKA H, LÜTJOHANN D, MENSINK RP, VON BERGMANN K: Oxidized plant sterols in human serum and lipid infusions as measured by combined gas-liquid chromatography-mass spectrometry. J Lipid Res 42 (2001) 2030-2038 PONZ DE LEON M, RONCUCCI L, DI DONATO P, SACCHETTI C, PEZCOLLER C, ANNONI C, BERTANI C, REBECCHI P, BALLI F, GALLI D, CARULLI N: Fecal neutral steroids in normal conditions and in patients with polyps or cancer of the large bowel. Cancer Res 47 (1987) 305-310 PULLINGER CR, ENG C, SALEN G, SHEFER S, BATTA AK, ERICKSON SK, VERHAGEN A, RIVERA CR, MULVIHILL SJ, MALLOY MJ, KANE JP: Human cholesterol 7α-hydroxylase (CYP7A1) deficiency has a hypercholesterolemic phenotype. J Clin Invest 110 (2002) 109-117 QUILLIOT D, BELBRAOUET S, PELLETIER X, GUEANT JL, FLOQUET J, DEBRY G: Influence of a high-calcium carbonate diet on the incidence of experimental colon cancer in rats. Nutr Cancer 34 (1999) 213-219

Quellenverzeichnis
XII
RAJARATNAM R (2000) Impact of serum squalene, postprandial lipoproteins, and cholesterol metabolism on coronary artery disease in postmenopausal women. http://ethesis.helsinki.fi/julkaisut/laa/kliin/vk/rajaratnam/impactof.pdf (Stand: 24.06. 2002) Adobe Acrobat Dokument, 94 Seiten REDDY BS, EKELUND G, BOHE M, ENGLE A, DOMELLOF L: Metabolic epidemiology of colon cancer: Dietary pattern and fecal sterol concentrations of three populations. Nutr Cancer 5 (1983) 34-40 REDDY BS, ENGLE A, SIMI B, O'BRIEN LT, BARNARD RJ, PRITIKIN N, WYNDER EL: Effect of low-fat, high-carbohydrate, high-fiber diet on fecal bile acid and neutral sterols. Prev Med 17 (1988) 432-439 REDDY B, ENGLE A, KATSIFIS S, SIMI B, BARTRAM H-P, PERRINO P, MAHAN C: Biochemical epidemiology of colon cancer: Effect of types of dietary fiber on fecal mutagens, acid, and neutral sterols in healthy subjects. Cancer Res 49 (1989) 4629-4635 REDDY S, SANDERS TAB, OWEN RW, THOMPSON MH: Faecal pH, bile acid and sterol concentrations in premenopausal Indian and white vegetarians compared with white omnivores. Br J Nutr 79 (1998) 495-500 REIMENDAL R, SJÖVALL JB: Computer evaluation of gas chromatographic-mass spectrometric analyses of steroids from biological materials. Anal Chem 45 (1973) 1083-1089 REISS AB, MARTIN KO, ROJER DE, IYER S, GROSSI EA, GALLOWAY AC, JAVITT NB: Sterol 27-hydroxylase: Expression in human arterial endothelium. J Lipid Res 38 (1997) 1254-1260 REN D, LI L, SCHWABACHER AW, YOUNG JW, BEITZ DC: Mechanism of cholesterol reduction to coprostanol by Eubacterium coprostanoligenes ATCC 51222. Steroids 61 (1996) 33-40 ROSENFELD RS, FUKUSHIMA DK, HELLMANN L, GALLAGHER TF: The transformation of cholesterol to coprostanol. J Biol Chem 211 (1954) 301-311 ROY P, OWEN RW, FAIVRE J, SCHEPPACH W, SALDANHA MH, BECKLY DE, BOUTRON MC: Faecal neutral sterols and bile acids in patients with adenomas and large bowel cancer: An ECP case-control study. Eur J Cancer Prev 8 (1999) 409-415 SANDERS ME: Considerations for use of probiotic bacteria to modulate human health. J Nutr 130 (2000) 384-390 SCALIA S: Bile acid separation. J Chromatogr B 671 (1995) 299-317

Quellenverzeichnis
XIII
SCALIA S, SCAGLIARINI R, PAZZI P: Evaluation of ursodeoxycholic acid bioavailability from immediate- and sustained release preparations using gas chromatography-mass spectrometry and high-performance liquid chromatography. Arzneim Forsch/Drug Res 50 (2000) 129-134 SETCHELL KDR: Liquid-solid extraction, lipophilic gel chromatography and capillary column gas chromatography in the analysis of bile acids from biological samples. In: Barbara L, Dowling RH, Hofman AF, Roda E (Hrsg.) Bile Acids in Gastroenterology. Lancaster/Boston, MTP Press Limited Falcon House, 1982, 1-17 SETCHELL KDR, LAWSON AM, TANIDA N, SJÖVALL J: General methods for the analysis of metabolic profiles of bile acids and related compounds in feces. J Lipid Res 24 (1983) 1085-1100 SETCHELL KDR, STREET JM, SJÖVALL J: Fecal bile acids. In: Setchell KDR, Kritchevsky D, Nair PP (Hrsg.) The Bile Acids. Chemistry, Physiology, and Metabolism. Volume 4: Methods and Applications. New York/London, Plenum Press, 1988, 441-570 SHERROD JA, HYLEMON PB: Partial purification and characterization of NAD-dependent 7α-hydroxysteroid dehydrogenase from Bacteroides thetaiotaomicron. Biochim Biophys Acta 486 (1977) 351-358 SHIMADA K, BRICKNELL KS, FINEGOLD SM: Deconjugation of bile acids by intestinal bacteria: Review of literature and additional studies. J Infect Dis 119 (1969) 273-281 SHIMADA K, MITAMURA K, HIGASHI T: Gas chromatography and high-performance liquid chromatography of natural steroids. J Chromatogr A 935 (2001) 141-172 SJÖVALL J, ENEROTH P, RYHAGE R: Mass spectra of bile acids. In: Nair PP, Kritchevsky D (Hrsg.) The Bile Acids. Chemistry, Physiology, and Metabolism. Volume 1: Chemistry. New York/London, Plenum Press, 1971, 209-248 SLATTERY ML, POTTER JD, MA KN, CAAN BJ, LEPPERT M, SAMOWITZ W: Western diet, family history of colorectal cancer, NAT2, GSTM-1 and risk of colon cancer. Cancer Cause Control 11 (2000) 1-8 SONG YS, JIN C, PARK EH: Identification of metabolites of phytosterols in rat feces using GC/MS. Arch Pharm Res 23 (2000) 599-604 SPADY DK, STANGE EF, BILHARTZ LE, DIETSCHY JM: Bile acids regulate hepatic low density lipoprotein receptor activity in the hamster by altering cholesterol flux across the liver. Proc Natl Acad Sci USA 83 (1986) 1916-1920 SPANHAAK S, HAVENAAR R, SCHAAFSMA G: The effect of consumption of milk fermented by Lactobacillus casei strain Shirota on the intestinal microflora and immune parameters in humans. Eur J Clin Nutr 52 (1998) 899-907

Quellenverzeichnis
XIV
SRIKUMAR TS, WEZENDONK B, VAN DOKKUM W: Analysis of fecal bile acids and neutral steroids using gas-liquid chromatography. Ann Nutr Metab 42 (1998) 231-236 STADLER J, STERN HS, YEUNG KS, MCGUIRE V, FURRER R, MARCON N, BRUCE WR: Effect of high fat consumption on cell proliferation activity of colorectal mucosa and on soluble faecal bile acids. Gut 29 (1988) 1326-1331 SUBBIAH MTR: Methods of measuring bile acids in bile and feces. Ann Clin Lab Sci 3 (1973) 362-368 SUMMERFIELD JA, BILLING BH, SHACKLETON CHL: Identification of bile acids in the serum and urine in cholestasis. Biochem J 154 (1976) 507-516 SUPELCO (2002) Application. Note 37. http://infonew.sigma-aldrich.com/Graphics/Supelco/objects/4600/4584.pdf (Stand 23.06.2002), Adobe Acrobat Dokument, 2 Seiten SZCZEPANIK PA, HACHEY DL, KLEIN PD: Characterization of bile acid methyl ester acetate derivatives using gas-liquid chromatography, electron impact, and chemical ionization mass spectrometry. J Lipid Res 17 (1976) 314-334 TANDON R: Selective liquid chromatographic isolation and gas chromatographic-mass spectrometric analysis of ketonic bile acids in faeces. J Chromatogr 302 (1984) 1-14 TANIDA N, HIKASA Y, HOSOMI M, SATOMI M, OOHAMA I, SHIMOYAMA T: Fecal bile acid analysis in healthy Japanese subjects using lipophilic anion exchanger, capillary column gas chromatography and mass spectrometry. Gastroenterol Jpn 16 (1981) 363-371 TARANTO MP, SESMA F, PESCE DE RUIZ HOLGADO A, DE VALDEZ GF: Bile salts hydrolase plays a key role on cholesterol removal by Lactobacillus reuteri. Biotech Lett 19 (1997) 845-847 TENNESON ME, OWEN RW, MASON AN: The anaerobic side-chain cleavage of bile acids by Escherichia coli isolated from human feces. Biochem Soc Trans 5 (1977) 1758-1760 TURJMAN N, NAIR PP : Diet, nutrition intake, and metabolism in populations at high and low risk for colon cancer. In situ bromination for the separation of the cholesterol-cholestanol pair in human fecal extracts. Am J Clin Nutr 40 (1984) 952-956 UCHIDA K, TAKASE H, NOMURA Y, TAKEDA K, TAKEUCHI N: Changes in biliary and fecal bile acids in mice after treatments with diosgenin and ß-Sitosterol. J Lipid Res 25 (1984) 236-245

Quellenverzeichnis
XV
USMAN, HOSONO A: Effect of administration of Lactobacillus gasseri on serum lipids and fecal steroids in hypercholesterolemic rats. J Dairy Sci 83 (2000) 1705-1711 VAN DER MEER R, TERMONT DSML, DE VRIES HT: Differential effects of calcium ions and calcium phosphate on cytotoxicity of bile acids. Am J Physiol 260 (1991) G142-G147 VAN DER MEER R, LAPRÉ JA, GOVERS MJAP, KLEIBEUKER JH: Mechanisms of the intestinal effects of dietary fats and milk products on colon carcinogenesis. Cancer Lett 114 (1997) 75-83 VAN ELDERE J: Chemical transformations of bile salts by the intestinal microflora. In: Tannock GW (Hrsg.) Medical Importance of the Normal Microflora. Dordrecht/Boston, Kluwer Academic Publishers, 1999, 312-337 VAN MUNSTER IP, TANGERMAN A, NAGENGAST FM: Effect of resistant starch on colonic fermentation, bile acid metabolism, and mucosal proliferation. Dig Dis Sci 39 (1994) 834-842 VAN TASSELL RL, MACDONALD DK, WILKINS TD: Stimulation of mutagen production in human feces by bile and bile acids. Mutat Res 103 (1982) 233-239 VASKONEN T, MERVAALA E, SUMUVUORI V, SEPPANEN-LASKO T, KARPPANEN H: Effects of calcium and plant sterols on serum lipids in obese Zucker rats on a low-fat diet. Br J Nutr 87 (2002) 239-245 VENTURI M, HAMBLY RJ, GLINGHAMMAR B, RAFTER JJ, ROWLAND IR: Genotoxic activity in human faecal water and the role of bile acids: A study using the alkaline comet assay. Carcinogenesis 18 (1997) 2353-2359 VERBEEK MJF, DE DECKERE EAM, TIJBURG LBM, VAN AMELSVOORT JMM, BEYNEN AC: Influence of dietary retrograded starch on the metabolism of neutral steroids and bile acids in rats. Br J Nutr 74 (1995) 807-820 VLAHAKIS C, HAZEBROEK J: Phytosterol accumulation in canola, sunflower, and soybean oils: Effects of genetics, planting location, and temperature. JAOCS 77 (2000) 49-53 VLAHCEVIC ZR, PANDAK WM, HEUMAN DM, HYLEMON PB: Function and regulation of hydroxylases involved in the bile acid biosynthesis pathway. Semin Liver Dis 12 (1992) 403-419 VLAHCEVIC ZR, EGGERTSEN G, BJÖRKHEM I, HYLEMON PB, REDFORD K, PANDAK WM: Regulation of sterol 12a-hydroxylase and cholic acid biosynthesis in the rat. Gastroenterology 118 (2000) 599-607 WEISBURGER JH, REDDY BS, BARNES WS WYNDER EL: Bile acids, but not neutral sterols, are tumor promoters in the colon in man and in rodents. Environ Health Perspect 50 (1983) 101-107

Quellenverzeichnis
XVI
WELLS JE, BERR F, THOMAS LA, DOWLING RH, HYLEMON PB: Isolation and characterization of cholic acid 7a-dehydroxylating fecal bacteria from cholesterol gallstone patients. J Hepatol 32 (2000) 4-10 WELLS JE, HYLEMON PB: Identification and characterization of a bile acid 7a-dehydroxylating operon in Clostridium sp. strain TO-931, a highly active 7a-dehydroxylating strain isolated from human feces. Appl Environ Microbiol 66 (2000) 1107-1113 WESTSTRATE JA, AYESH R, BAUER-PLANK C, DREWITT PN: Safety evaluation of phytosterol esters. Part 4. Faecal concentrations of bile acids and neutral sterols in healthy normolipidaemic volunteers consuming a controlled diet either with or without a phytosterol ester-enriched margarine. Food Chem Toxicol 37 (1999) 1063-1071 WIELAND HO (1928) The chemistry of the bile acids. Nobel Lecture. http://www.nobel.se/chemistry/laureates/1927/wieland-lecture.html (Stand 23.06. 2002) Adobe Acrobat Dokument, 9 Seiten WILKINS TD, HACKMAN AS: Two patterns of neutral steroid conversion in the feces of normal North Americans. Cancer Res 34 (1974) 2250-2254 WOLTHERS BG, HINDRIKS FR, MUSKIET FA, GROEN A: Mass fragmentographic analysis of total cholesterol in serum using a heptadeuterated internal standard. Clin Chim Acta 103 (1980) 305-315

Selbständigkeitserklärung
Selbständigkeitserklärung Hiermit erkläre ich, dass ich die Arbeit selbständig verfasst und keine anderen als
die angegebenen Quellen und Hilfsmittel benutzt habe.
Die Arbeit wurde bisher keiner anderen Prüfungsbehörde vorgelegt und auch nicht
veröffentlicht.
Jena, 28. Oktober 2002
Sylvia Keller

Danksagung
Danksagung
Mein Dank gilt Herrn Prof. G. Jahreis, der es mir ermöglichte, die Freude an der
praktischen Arbeit der Gaschromatographie, die Neugier an den physiologischen
Prozessen im Menschen und die Vorliebe für Mathematik in der Aufgabenstellung
zur Steroidanalyse und deren Anwendung zu verbinden. Er verfolgte meine teils
auch unkonventionellen Ideen stets mit Aufmerksamkeit und unterstützte mich bei
der Umsetzung in eine praktizierbare Anwendung.
Weiterhin möchte ich den derzeitigen und ehemaligen Mitarbeitern des
Lehrbereichs Ernährungsphysiologie danken. Sie wurden nie müde meine häufigen
und wohl manchmal drängenden Fragen zur Physiologie und Ernährung des
Menschen zu beantworten. Innerhalb von vier Jahren meiner Arbeit gab es auch
Momente, in denen sich das Gesetz von Murphy bewahrheiten wollte. Jedoch der
humorvolle Geist meiner Kollegen erzeugte immer wieder die innere
Herausforderung sich das scheinbare Eigenleben der Gerätetechnik zu Nutze zu
machen.
Der Arbeit von Frau Andrea Klein, Frau Dr. Manuela Grimm und Frau Dr. Grit
Kießling bei der Durchführung der Humanstudien ist es zu verdanken, dass es mir
nie an Probenmaterial mangelte und mir alle notwendigen Informationen zur
Verfügung standen.
Die Gesprächsbereitschaft der Mitarbeiter der Firma SHIMADZU über Themen des
Handbuchs „GC-MS QP 5000“ hinaus, verschaffte mir theoretische und praktische
Hilfe.
Abschließend danke ich meiner Familie für die vielen Stunden des „Fezes“.


















