3 Blut, Herz und Kreislauf - qnap.e3.physik.tu-dortmund.de · 3 Blut, Herz und Kreislauf 5 l Blut...
26
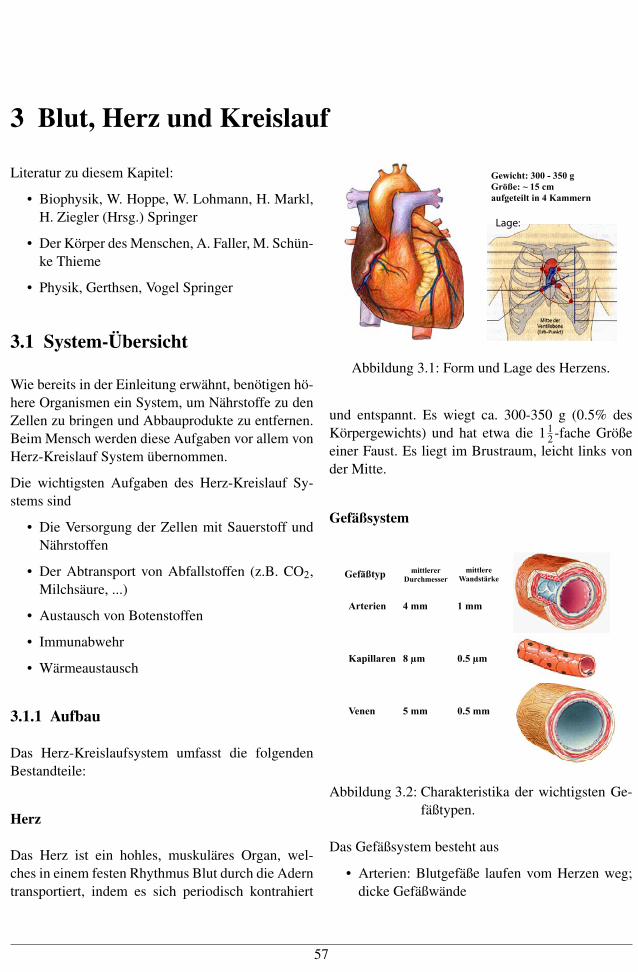
3 Blut, Herz und Kreislauf Literatur zu diesem Kapitel: • Biophysik, W. Hoppe, W. Lohmann, H. Markl, H. Ziegler (Hrsg.) Springer • Der Körper des Menschen, A. Faller, M. Schün- ke Thieme • Physik, Gerthsen, Vogel Springer 3.1 System-Übersicht Wie bereits in der Einleitung erwähnt, benötigen hö- here Organismen ein System, um Nährstoffe zu den Zellen zu bringen und Abbauprodukte zu entfernen. Beim Mensch werden diese Aufgaben vor allem von Herz-Kreislauf System übernommen. Die wichtigsten Aufgaben des Herz-Kreislauf Sy- stems sind • Die Versorgung der Zellen mit Sauerstoff und Nährstoffen • Der Abtransport von Abfallstoffen (z.B. CO 2 , Milchsäure, ...) • Austausch von Botenstoffen • Immunabwehr • Wärmeaustausch 3.1.1 Aufbau Das Herz-Kreislaufsystem umfasst die folgenden Bestandteile: Herz Das Herz ist ein hohles, muskuläres Organ, wel- ches in einem festen Rhythmus Blut durch die Adern transportiert, indem es sich periodisch kontrahiert Gewicht: 300 - 350 g Größe: ~ 15 cm aufgeteilt in 4 Kammern Lage: Abbildung 3.1: Form und Lage des Herzens. und entspannt. Es wiegt ca. 300-350 g (0.5% des Körpergewichts) und hat etwa die 1 1 2 -fache Größe einer Faust. Es liegt im Brustraum, leicht links von der Mitte. Gefäßsystem !"#$%&'( )*&&+",", -.,/0)"11", )*&&+"," 23451&$,6" 7,&",*"4 8 )) 9 )) :3(*++3,"4 ; ) <=> ) ?"4"4 > )) <=> )) Abbildung 3.2: Charakteristika der wichtigsten Ge- fäßtypen. Das Gefäßsystem besteht aus • Arterien: Blutgefäße laufen vom Herzen weg; dicke Gefäßwände 57
Transcript of 3 Blut, Herz und Kreislauf - qnap.e3.physik.tu-dortmund.de · 3 Blut, Herz und Kreislauf 5 l Blut...
3 Blut, Herz und Kreislauf
Literatur zu diesem Kapitel:
• Biophysik, W. Hoppe, W. Lohmann, H. Markl,H. Ziegler (Hrsg.) Springer
• Der Körper des Menschen, A. Faller, M. Schün-ke Thieme
• Physik, Gerthsen, Vogel Springer
3.1 System-Übersicht
Wie bereits in der Einleitung erwähnt, benötigen hö-here Organismen ein System, um Nährstoffe zu denZellen zu bringen und Abbauprodukte zu entfernen.Beim Mensch werden diese Aufgaben vor allem vonHerz-Kreislauf System übernommen.
Die wichtigsten Aufgaben des Herz-Kreislauf Sy-stems sind
• Die Versorgung der Zellen mit Sauerstoff undNährstoffen
• Der Abtransport von Abfallstoffen (z.B. CO2,Milchsäure, ...)
• Austausch von Botenstoffen
• Immunabwehr
• Wärmeaustausch
3.1.1 Aufbau
Das Herz-Kreislaufsystem umfasst die folgendenBestandteile:
Herz
Das Herz ist ein hohles, muskuläres Organ, wel-ches in einem festen Rhythmus Blut durch die Aderntransportiert, indem es sich periodisch kontrahiert
Gewicht: 300 - 350 gGröße: ~ 15 cmaufgeteilt in 4 Kammern
Lage:
Abbildung 3.1: Form und Lage des Herzens.
und entspannt. Es wiegt ca. 300-350 g (0.5% desKörpergewichts) und hat etwa die 1 1
2 -fache Größeeiner Faust. Es liegt im Brustraum, leicht links vonder Mitte.
Gefäßsystem
!"#$%&'( )*&&+",",-.,/0)"11",
)*&&+","23451&$,6"
7,&",*"4 8 )) 9 ))
:3(*++3,"4 ; ) <=> )
?"4"4 > )) <=> ))
Abbildung 3.2: Charakteristika der wichtigsten Ge-fäßtypen.
Das Gefäßsystem besteht aus
• Arterien: Blutgefäße laufen vom Herzen weg;dicke Gefäßwände
57

3 Blut, Herz und Kreislauf
• Kapillaren: Haargefäße in denen der Stoffaus-tausch stattfindet
• Venen: Blutgefäße führen zum Herz; dünnwan-dig
• Lymphgefäße: transportieren Flüssigkeit undAbwehrstoffe
Blut
Blut ist eine komplexe Flüssigkeit, bestehend ausdem Blutplasma und lebenden Zellen. Es trägt et-wa 8% zum gesamten Körpergewicht bei, d.h. einMensch mit 80 kg Körpergewicht besitzt ca. 6.4 kgBlut. Die Produktion der Blutkörperchen erfolgt imKnochenmark.
Regelung
Das gesamte Herz-Kreislauf System wird durch ei-ne Reihe von Regelmechanismen gesteuert, welchez.B. Herzfrequenz und Gefäßwiderstand dem aktu-ellen Bedarf anpassen. Die Regelungsmechanismenverwenden sowohl nervliche wie auch chemische In-formationsübertragung.
3.1.2 Kreislauf: Übersicht
obere Körperhälfte
untere Körperhälfte
Lymphgefäßsystem
Lunge
Herz
Darm
Leber
Niederdrucksystem
(Kapazitätsystem)
arterielles System
(Widerstandssystem)
Lunge
Gehirn
Herz
Leber Darm
Nieren
Haut, Muskel, Skelett
Abbildung 3.3: Überblick über arteriellen und venö-sen Kreislauf.
Abbildung 3.3 zeigt einen Überblick über das Herz-und Gefäßsystem. Sauerstoffgesättigtes, “arteriel-les” Blut ist rot gezeichnet, die Teilkreisläufe mitteilweise entsättigtem, “venösen” Blut sind blau ge-zeichnet, und das Lymphsystem, ein zusätzlichesTransportsystem des Körpers, ist grün eingefärbt.Auf Grund der starken Verzweigung der Blutgefäßeergibt sich eine Gesamtlänge von mehreren 10000km.
Auf der rechten Seite der Abbildung ist das arteri-elle System mit relativ starken und relativ starrenWänden dicker eingezeichnet. Das venöse Systembesitzt relativ schwache und stark dehnbare Wän-de und wird als Niederdrucksystem zusammenge-fasst. Der Lungenkreislauf, auch kleiner Kreislaufbenannt, liegt mit dem rechten Herzventrikel alsPumpe in Serie mit dem großen Körper- oder Organ-kreislauf. Die prozentuale Verteilung des Blutvolu-mens auf die einzelnen Organe und die Druckver-hältnisse im Herz-Kreislaufsystem wird später ge-nauer diskutiert. Das arterielle Blut ist zu rund 97%mit Sauerstoff gesättigt, das venöse zu rund 73%.
Bei Jugendlichen pumpt das Herz pro Minute etwa5 l Blut, bei 70-jährigen sinkt diese Menge auf etwa2.5 l.
3.1.3 Zusammensetzung des Blutes
Blut ist eine komplexe Flüssigkeit, welche eine Viel-zahl von gelösten Stoffen enthält (Salze, Protei-ne, Kohlenwasserstoffe etc.), wie auch suspendierteStoffe (v. A. rote und weisse Blutkörperchen). Wiein Abbildung 3.4 gezeigt, macht das Blutplasma gutdie Hälfte des Volumens aus, der Rest besteht zumgrößten Teil aus roten, zum kleineren Teil aus wei-ßen Blutkörperchen (Zellen). Das Blutplasma ent-hält neben Wasser vor allem Proteine (ca. 7%) undrund 1% Elektrolyte und kleine organische Molekü-le.
Den prozentualen Anteil der roten Blutkörperchen(Erythrozyten) am Gesamtvolumen bezeichnet manals Hämatokrit. Er liegt bei Männern bei etwa 47%,bei Frauen bei 43%.
Die roten Blutkörperchen sind für den Transport vonSauerstoff verantwortlich, die weißen für die Im-
58

3 Blut, Herz und Kreislauf
5 l Blut 2.8 l Plasma 190 g Protein
Abbildung 3.4: Zusammensetzung des menschli-chen Bluts.
munabwehr. Das Blutplasma transportiert Nährstof-fe und Stoffwechselendprodukte von den Zellen zuden Ausscheidungsorganen (z.B. Nieren), Kohlendi-oxid, Botenstoffe (Hormone) zwischen unterschied-lichen Organen, und Wärme für den Temperaturaus-gleich zwischen unterschiedlich aktiven Organen.
3.1.4 Erythrozyten
Die Erythrozyten oder roten Blutkörperchen sind imWesentlichen für den Sauerstofftransport im Blutverantwortlich.
Abbildung 3.5: Rote Blutkörperchen
Die roten Blutkörperchen sind ca. 7.5 µm im Durch-messer und etwa 2µm dick.
Damit haben sie eine große spezifische Oberflächedurch die sie effizient Sauerstoff mit ihrer Umge-bung austauschen können. Sie sind außerdem gutverformbar, was ihnen erlaubt, auch in sehr kleinenKapillaren noch durchzukommen.
Insgesamt besitzt ein erwachsener Mensch rund 3 ·1013 Erythrozyten. Ihre Lebensdauer beträgt ca. 120
Erythrozytendurchmesser / µm
Häufigkeit/%
Abbildung 3.6: Der mittlere Erythrozytendurchmes-ser beträgt ca. 7,5µm.
Tage, danach werden sie in Leber und Milz abge-baut. Damit die Zahl konstant bleibt müssen dement-sprechend
N =3 ·1013
120 ·24 ·3600s≈ 3 ·106 s−1
Erythrozyten neu gebildet werden. Dies geschieht imroten Knochenmark.
Rote Blutkörperchen haben interessante mechani-sche Eigenschaften, welche in erster Linie aufden Aufbau der Zellmembran zurückgeführt wer-den können. Diese besteht aus einer Doppel-schicht von Phospholipid-Molekülen. Diese Lipid-Doppelschicht wird auf der Innenseite der Zelledurch ein Protein-Netzwerk verstärkt. Das Netzwerkbesteht aus Spectrin und Actin und ist mit der Mem-bran über Ankerpunkte verknüpft. Dadurch könnenrote Blutkörperchen leicht geschert werden, aber dieOberfläche kann nur schwer lokal vergrößert oderverkleinert werden.
3.2 Eigenschaften von Flüssigkeiten
3.2.1 Viskosität und innere Reibung
Bei einer Flüssigkeit lassen sich die Moleküle ge-genseitig verschieben. Die Gestalt einer Flüssigkeitist daher beliebig, das Volumen ist aber bestimmt.Im Gegensatz zu Gasen wirken noch erheblicheKräfte zwischen den Molekülen (Kohäsionskräfte).In weiten Grenzen gilt Volumenelastizität, nach derEntlastung einer Kompression stellt sich wieder dasAnfangsvolumen ein.
59
3 Blut, Herz und Kreislauf
Eine reine Formänderung bedarf nur dann einer
Kraft, wenn sie schnell ausgeführt werden soll. In
diesem Fall führt die innere Reibung zu einem Wi-
derstand gegen die Scherung. Auf molekularer Ebe-
ne können diese Reibungskräfte auf van der Waals
Bindungen zwischen den Molekülen zurückgeführt
werden, welche eine gewisse (schwache) Kohäsion
zwischen benachbarten Flüssigkeitsvolumina erzeu-
gen.
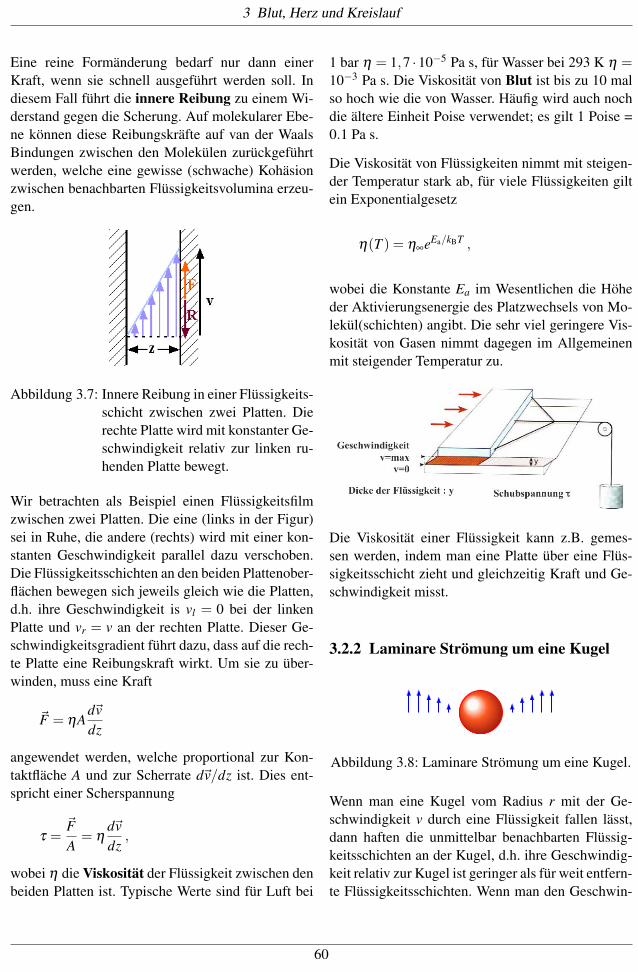
Abbildung 3.7: Innere Reibung in einer Flüssigkeits-
schicht zwischen zwei Platten. Die
rechte Platte wird mit konstanter Ge-
schwindigkeit relativ zur linken ru-
henden Platte bewegt.
Wir betrachten als Beispiel einen Flüssigkeitsfilm
zwischen zwei Platten. Die eine (links in der Figur)
sei in Ruhe, die andere (rechts) wird mit einer kon-
stanten Geschwindigkeit parallel dazu verschoben.
Die Flüssigkeitsschichten an den beiden Plattenober-
flächen bewegen sich jeweils gleich wie die Platten,
d.h. ihre Geschwindigkeit is vl = 0 bei der linken
Platte und vr = v an der rechten Platte. Dieser Ge-
schwindigkeitsgradient führt dazu, dass auf die rech-
te Platte eine Reibungskraft wirkt. Um sie zu über-
winden, muss eine Kraft
�F = ηAd�vdz
angewendet werden, welche proportional zur Kon-
taktfläche A und zur Scherrate d�v/dz ist. Dies ent-
spricht einer Scherspannung
τ =�FA= η d�v
dz,
wobei η die Viskosität der Flüssigkeit zwischen den
beiden Platten ist. Typische Werte sind für Luft bei
1 bar η = 1,7 ·10−5
Pa s, für Wasser bei 293 K η =10
−3Pa s. Die Viskosität von Blut ist bis zu 10 mal
so hoch wie die von Wasser. Häufig wird auch noch
die ältere Einheit Poise verwendet; es gilt 1 Poise =
0.1 Pa s.
Die Viskosität von Flüssigkeiten nimmt mit steigen-
der Temperatur stark ab, für viele Flüssigkeiten gilt
ein Exponentialgesetz
η(T ) = η∞eEa/kBT ,
wobei die Konstante Ea im Wesentlichen die Höhe
der Aktivierungsenergie des Platzwechsels von Mo-
lekül(schichten) angibt. Die sehr viel geringere Vis-
kosität von Gasen nimmt dagegen im Allgemeinen
mit steigender Temperatur zu.
Die Viskosität einer Flüssigkeit kann z.B. gemes-
sen werden, indem man eine Platte über eine Flüs-
sigkeitsschicht zieht und gleichzeitig Kraft und Ge-
schwindigkeit misst.
3.2.2 Laminare Strömung um eine Kugel
Abbildung 3.8: Laminare Strömung um eine Kugel.
Wenn man eine Kugel vom Radius r mit der Ge-
schwindigkeit v durch eine Flüssigkeit fallen lässt,
dann haften die unmittelbar benachbarten Flüssig-
keitsschichten an der Kugel, d.h. ihre Geschwindig-
keit relativ zur Kugel ist geringer als für weit entfern-
te Flüssigkeitsschichten. Wenn man den Geschwin-
60

3 Blut, Herz und Kreislauf
digkeitsgradienten durch dvdz ≈
vr abschätzt, dann er-
hält man für die Reibungskraft
FR =−η dvdz
·4πr2 =−4πηvr .
Eine genaue, sehr aufwändige Rechnung ergibt dasStokes-Gesetz
FR =−6πηvr .
Dieses Gesetz wird bei dem Kugelfallviskosimeterdazu benutzt, die Viskosität einer Flüssigkeit zu be-stimmen.
v
Zu messende
Flüssigkeit
Abbildung 3.9: Kugelfallviskosimeter.
Bestimmt man die Sinkgeschwindigkeit v einer Ku-gel mit einem Radius r in einer Flüssigkeit der Dich-te ρFl , dann kann man daraus die unbekannte Vis-kosität η dieser Flüssigkeit mit Hilfe des Stokes-Gesetzes berechnen. Die Sinkgeschwindigkeit istgegeben aus dem Gleichgewicht zwischen der Rei-bungskraft FR und der um die Auftriebskraft vermin-derten Gewichtskraft der Kugel:
FR = FG −FA =43
πr3g(ρK −ρFl) .
Die dynamische Viskosität ist also
η =2gr2(ρK −ρFl)
9v.
Ein anderes Gerät zur Viskositätsbestimmung istdas Kapillarviskosimeter, das auf dem Hagen-Poiseuilleschen Gesetz beruht (siehe 3.3.4).
!"#$%%&'$ ()*(+
!"#$%,-&../.0
1$2'3.4,"#$ 567,,809$8'
(86&'&.'$,56/8(
:,$/(3-6&,'8,"#$, 56/8(
;8.0#&<=-6&,'8,"#$, 56/8(
Abbildung 3.10: Viskosität von nicht-NewtonschenFluiden
3.2.3 Nicht-Newtonsche Flüssigkeiten
Die Proportionalität zwischen Geschwindigkeit undReibungskraft gilt jedoch nicht bei allen Fluiden. Indiesen Fällen gilt häufig eine verallgemeinerte Be-ziehung zwischen der Scherrate und der Scherspan-nung. Man kann hier jeweils eine differenzielle Vis-kosität definieren, als die Ableitung dτ
d�v/dz der Scher-spannung nach der Scherrate.
Spezielle Fälle sind
• dilatante Fluide: In Suspensionen (z.B. Quarz-kugeln) wird durch zunehmende Reibung mitwachsender Scherrate mehr Entropie erzeugt.
• Bingham-plastisches Fluid: In Suspensionenaus nicht-sphärischen Teilchen können sich dieTeilchen mit zunehmender Scherung ausrich-ten.
• Pseudoplastische Flüssigkeiten: BiologischeFlüssigkeiten wie z.B. Blut; die Scherung kannzu Aggregation und Orientierungsphänomenenführen, sowie zu Deformation der Komponen-ten, in Abhängigkeit von der Scherrate.
In einigen Fällen existiert auch für verschwindendeScherrate eine Scherspannung. Hier handelt es sichalso in einem weiteren Sinn um Festkörper.
3.2.4 Suspensionen
Suspensionen führen nicht nur zu einer Änderungdes Strömungswiderstandes; umgekehrt beeinflusst
61
3 Blut, Herz und Kreislauf
auch die Strömung die Eigenschaften der Suspensi-on. So können sich suspendierte Teilchen unter demEinfluss einer Scherung ausrichten oder aggregieren.Dabei sind Orientierung, Aggregation und Deforma-tion abhängig von der Scherrate dv/dz.
In einfachen Fällen kann man die Änderung der Vis-kosität durch die suspendierten Teilchen additiv be-schreiben:
η = ηL(1+ηspec).
Hier stellt ηL die Viskosität des reinen Lösungsmit-tels dar. Die relative Änderung ist gegeben durch dieEinstein-Relation
ηspec = 2.5Vrel
mit
Vrel =VolumenderTeilchen
GesamtvolumenderSuspension.
Diese Näherung gilt nur für geringe Konzentratio-nen, Vrel � 1.
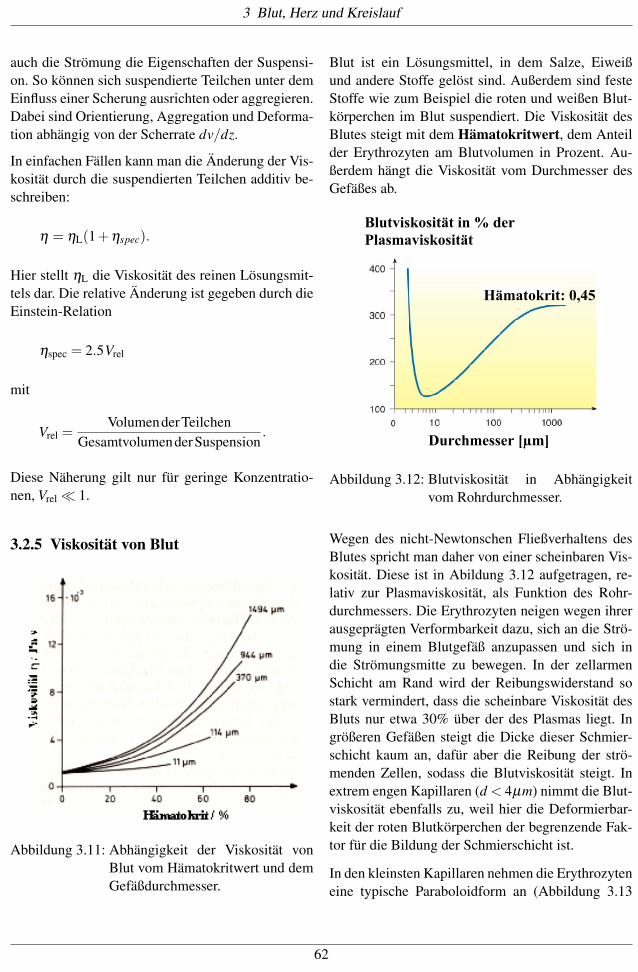
3.2.5 Viskosität von Blut
Abbildung 3.11: Abhängigkeit der Viskosität vonBlut vom Hämatokritwert und demGefäßdurchmesser.
Blut ist ein Lösungsmittel, in dem Salze, Eiweißund andere Stoffe gelöst sind. Außerdem sind festeStoffe wie zum Beispiel die roten und weißen Blut-körperchen im Blut suspendiert. Die Viskosität desBlutes steigt mit dem Hämatokritwert, dem Anteilder Erythrozyten am Blutvolumen in Prozent. Au-ßerdem hängt die Viskosität vom Durchmesser desGefäßes ab.
Blutviskosität in % der
Plasmaviskosität
Hämatokrit: 0,45
Durchmesser [µm]
Abbildung 3.12: Blutviskosität in Abhängigkeitvom Rohrdurchmesser.
Wegen des nicht-Newtonschen Fließverhaltens desBlutes spricht man daher von einer scheinbaren Vis-kosität. Diese ist in Abildung 3.12 aufgetragen, re-lativ zur Plasmaviskosität, als Funktion des Rohr-durchmessers. Die Erythrozyten neigen wegen ihrerausgeprägten Verformbarkeit dazu, sich an die Strö-mung in einem Blutgefäß anzupassen und sich indie Strömungsmitte zu bewegen. In der zellarmenSchicht am Rand wird der Reibungswiderstand sostark vermindert, dass die scheinbare Viskosität desBluts nur etwa 30% über der des Plasmas liegt. Ingrößeren Gefäßen steigt die Dicke dieser Schmier-schicht kaum an, dafür aber die Reibung der strö-menden Zellen, sodass die Blutviskosität steigt. Inextrem engen Kapillaren (d < 4µm) nimmt die Blut-viskosität ebenfalls zu, weil hier die Deformierbar-keit der roten Blutkörperchen der begrenzende Fak-tor für die Bildung der Schmierschicht ist.
In den kleinsten Kapillaren nehmen die Erythrozyteneine typische Paraboloidform an (Abbildung 3.13
62

3 Blut, Herz und Kreislauf
Abbildung 3.13: Änderung der Erythrozytenformin Gefäßen mit unterschiedlichemDurchmesser.
oben), als Anpassung an die lokalen Strömungsver-hältnisse. In geringfügig größeren Gefäßen (Abbil-dung 3.13 unten) unterliegen sie ständigen Formän-derungen.
Auch wenn der Durchmesser des Blutgefäßes dieGröße der Erythrozyten überschreitet, wirken wegendes inhomogenen Geschwindigkeitsfeldes Scher-kräfte auf die Zellmembran. Je nach Viskosität desBlutes und der Zellflüssigkeit, sowie der Scherra-te erzeugen diese Kräfte eine Rotation der Zel-le oder eine Rotation der Zellmembran um dasZellinnere.[16]
3.2.6 Reibung in laminaren Strömungen
Bei der laminaren Strömung ist das Verhalten derFlüssigkeit durch die innere Reibung bestimmt. Da-bei gleiten selbst sehr dünne Flüssigkeitsschichtenparallel zueinander, ohne sich zu mischen.
Wir betrachten in einer laminaren Strömung dieKräfte, die auf ein Volumenelement der Flüssigkeitwirken. An jedem (gedachten) Würfel dxdydz wir-ken Spannungen auf die Seiten des Würfels. Dieskönnen Scherspannungen oder Normalspannungensein.
Wir betrachten zunächst den Einfluss einer Scher-spannung, d.h. einer Reibungskraft. Diese kommtdadurch zustande, dass die Flüssigkeit sich mit va-riabler Geschwindigkeit v in y-Richtung bewegt. Auf
dy
dx
dF1
dF2
Abbildung 3.14: Vorbeigleiten vonFlüssigkeitsschichten.
der linken Seite schreiben wir für die Beziehungzwischen der Kraft (= Scherspannung mal Flächedydz) und der Scherrate
dF1 =−η�
∂v∂x
�
ldydz .
Der entsprechende Ausdruck für die Kraft auf derrechten Seite des Flüssigkeitselementes lautet
dF2 = η�
∂v∂x
�
rdydz = η(
�∂v∂x
�
l+
∂ 2v∂x2 dx)dydz .
Die gesamte Reibungskraft, die auf das Flüssigkeits-element wirkt, beträgt somit
dFr = dF1 +dF2 = η ∂ 2v∂x2 dV
(Der Index r steht hier für Reibung). Somit ist dieKraft nur dann von Null verschieden, wenn das Ge-schwindigkeitsprofil gekrümmt ist.
Berücksichtigt man die Beiträge aus allen Raumdi-mensionen, so erhält man
dFr = η�
∂ 2v∂x2 +
∂ 2v∂y2 +
∂ 2v∂ z2
�dV = η ∆vdV .
63

3 Blut, Herz und Kreislauf
Da die Kraft proportional zur Größe des Volumen-
elementes ist, können wir sie auch als Kraftdichte
schreiben:
�fr =d�Fr
dV= η∆�v .
Hier wurde auch berücksichtigt, dass die Bewegung
nicht unbedingt parallel zur y-Achse erfolgt.
3.2.7 Druckkraft
Nachdem wir bisher nur die Scherspannung berück-
sichtig haben, berechnen wir jetzt den Einfluss der
Normalspannung, welche auf zwei gegenüberliegen-
de Flächen wirkt.
Abbildung 3.15: Druckkraft auf ein Volumenele-
ment.
In einer Flüssigkeit ist die Normalspannung gleich
dem Druck. Somit wirkt auf das Flächenelement
dydz bei x die Kraft
dF1 = p(x)dydz
in x-Richtung. Auf das gleich große Flächenelement
bei x+dx wirkt entsprechend
dF2 = −p(x+dx)dydz
= −(p(x)+∂ p∂x
dx)dydz .
In beiden Fällen haben wir nur den Außendruck be-
rücksichtigt. Die gesamte Druckkraft in x-Richtung
auf das Volumenelement wird damit
dFp = dF1 +dF2 =−(∂ p∂x
)dV .
Die gleiche Überlegung gilt für die y- und z-
Richtung. Somit erhalten wir in 3 Dimensionen
d�Fp =−dV �∇p .
Als Kraftdichte kann dies wiederum als
d�fp =d�Fp
dV=−�∇p
geschrieben werden.
3.2.8 Die Grundgleichungen der
Hydrodynamik
Kann man äußere Felder (z.B. die Schwerkraft) ver-
nachlässigen, so sind die Druck- und Reibungskraft
die einzigen Kräfte, die auf das Flüssigkeitselement
wirken. Aus der Newtonschen Gleichung folgt für
die Beschleunigung dieses Masseelementes
DDt
(�vρ dV ) = −dV �∇p + η ∆�vdV .
Für inkompressible Flüssigkeiten ist die Dichte kon-
stant und kann vor die Ableitung gesetzt werden:
ρ DDt
�v =−�∇p + η ∆�v .
Die “substantielle Ableitung”DDt eines Vektorfeldes
�u in einem bewegten Medium (wie einer Flüssig-
keit) entspricht im Wesentlichen der vollständigen
Ableitung. Sie muss zum einen die zeitliche Ände-
rung der Geschwindigkeit berücksichtigen, zum an-
dern die Änderung, welche durch den Transport an
eine andere Stelle des Vektorfeldes erfolgt. Die ge-
samte Ableitung ist deshalb
DDt
(�u) =∂�u∂ t
+ (�v ·�∇)�u .
Der zweite Term wird als Konvektionsterm bezeich-
net. Er entfällt wenn das Medium sich nicht bewegt.
Setzen wir dies in die obige Gleichung für inkom-
pressible Flüssigkeiten ein, erhalten wir
ρ�
∂�v∂ t
+(�v ·�∇) ·�v�
= −�∇p + η ∆�v . (3.1)
64

3 Blut, Herz und Kreislauf
Dies ist die Navier-Stokes’sche Gleichung der Hy-
drodynamik für inkompressible Flüssigkeiten. Esist eine nichtlineare Differenzialgleichung, dahergibt es die Möglichkeit von Turbulenz. Wichtig inder Medizin ist dies z. B. bei einem Herzinfarkt, aus-gelöst durch eine Stenose.
Eine andere wichtige Grundgleichung der Hydrody-namik ist die Kontinuitätsgleichung
∂ρ(�r)∂ t
+ �∇ · (ρ(�r)�v(�r)) = 0 .
Sie stellt die Massenerhaltung dar: die Änderung derDichte an einem Ort ist gegeben durch die Divergenzdes Flusses an diesem Ort, d.h. durch die Differenzder ein- und ausfließenden Massenströme. Der Be-weis folgt der Kontinuitätsgleichung der Elektrosta-tik.
Aus der Kontinuitätsgleichung folgt unmittelbar,dass in inkompressiblen Flüssigkeiten (ρ = const.)das Geschwindigkeitsfeld quellenfrei ist, �∇ ·�v = 0.
3.3 Laminare und turbulente
Strömungen
3.3.1 Laminar vs. turbulent
In der Strömungsdynamik unterscheidet man zwi-schen zwei grundsätzlich unterschiedlichen Artenvon Strömung: laminarer und turbulenter Strömung.Bei einer laminaren Strömung fließt die Flüssigkeitin Schichten, die sich nicht miteinander vermischen.Bei turbulente Strömung treten Wirbel auf.
Ob eine Strömung laminar oder turbulent wird hängtvor allem von den drei Faktoren
• Viskosität
• Geschwindigkeit
• geometrische Abmessungen
ab. Im Falle von laminaren Strömungen kann mandas Strömungsfeld analytisch exakt beschreiben. ImFall von turbulenten Strömungen kann man nur das
Laminar
Turbulent
Innere Reibung
dominiert
Typisch für Blutzirkulation
Abbildung 3.16: Beispiel für das gemeinsame Auf-treten von laminarer und turbulen-ter Strömung.
globale Verhalten voraussagen, nicht aber die De-tails. Abb. 3.16 zeigt, dass beide Arten von Strö-mung gleichzeitig auftreten können und dass ein Sy-stem von einem zum anderen Typ wechseln kann.
Abbildung 3.17: Laminarer v. turbulenter Blutfluss.
Im Blutkreislauf findet man vor allem laminare Strö-mung. Dies hat einen entscheidenden Vorteil: dieTransportkapazität eines Blutgefäßes ist bei lamina-rer Strömung wesentlich größer als bei turbulenterStrömung. Turbulenter Blutfluss tritt vor allem aufbei krankhaften Veränderungen, wie z.B. Vereng-ungen und Verletzungen, und kann dann zu schwe-ren Schäden führen. Bei laminarem Blutfluss ist das(Minuten-)Blutvolumen proportional zur Druckdif-ferenz, Vl ∝ ∆p, bei turbulenter Strömung (nur) zurWurzel daraus, Vt ∝
√∆p. Dies hängt damit zusam-
men, dass bei turbulenten Strömungen ein größererTeil der Druck-Energie nicht für den Transport, son-dern für die Erzeugung von Wirbeln verwendet wird.
65

3 Blut, Herz und Kreislauf
Wir beginnen die Diskussion mit laminaren Strö-
mungen. Ausgangspunkt für die Beschreibung ist die
Navier-Stokes Gleichung. In einer laminaren Strö-
mung fällt der Konvektionsterm weg, (�v ·�∇) ·�v = 0.
Ist die Strömung auch stationär, so reduziert sich die
Navier-Stokes-Gleichung auf
0 = �∇p − η ∆�v . (3.2)
3.3.2 Laminare Strömung zwischen zweifesten Platten
Die Lösung dieser Gleichung hängt entscheidend
von den Randbedingungen ab. Diese definieren ins-
besondere den Widerstand, welcher auf die Flüssig-
keit wirkt.
v
z
x
d
0
x
0
V
Abbildung 3.18: Laminare Strömung zwischen zwei
Platten (links) und zugehöriges Ge-
schwindigkeitsprofil (rechts).
Wir betrachten als einfaches Beispiel einen linea-
ren Fluss in z-Richtung zwischen zwei Platten, die
senkrecht auf dei x-Achse stehen. Wir suchen statio-
näre Lösungen �v(�r) = vz(x) der hydrodynamischen
Gleichungen, die unabhängig von y und z sind. Ei-
ne Druckvariation in x-Richtung würde durch einen
entsprechenden Fluss ausgeglichen und verschwin-
det deshalb im Gleichgewicht. Somit bleibt für die
Druckabhängigkeit �∇p = ∂ p∂ z .
Damit erhalten wir für die Bewegungsgleichung
(3.2)
η ∂ 2vz
∂x2=
∂ p∂ z
. (3.3)
Hier stellt die linke Seite die Reibung dar, die rech-
te Seite die Druckkraft, welche damit im Gleich-
gewicht ist. Damit wir einen Fluss in positiver z-
Richtung erhalten, muss der Druckgradient nega-
tiv sein, ∂ p/∂ z < 0. Die Reibung entsteht durch
die Krümmung des Geschwindigkeitprofils und wird
minimal, wenn diese konstant ist. Dadurch wird das
Geschwindigkeitsprofil quadratisch. Die Krümmung
ist gegeben durch den Quotienten aus dem Druck-
gradienten und der Viskosität.
Integration der Bewegungsgleichung (3.3) ergibt
vz(x) =x2
2η∂ p∂ z
+ c1x + c2 .
Aus der Symmetrie des Problems folgt, dass c1 = 0.
Damit wird die Fließgeschwindigkeit in der Mit-
te maximal, v(0) = vmax = c2. Außerdem muss die
Fließgeschwindigkeit auf den Platten verschwinden,
vz(±d2) = 0.
Damit muss das Profil die Form
v(x) = vmax(1−4x2
d2)
haben. Einsetzen in die Bewegungsgleichung (3.3)
ergibt
∂ 2vz
∂x2= −8
vmax
d2=
1
η∂ p∂ z
.
Wir lösen auf nach
vmax =− d2
8η∂ p∂ z
.
3.3.3 Laminare Rohrströmung
Wir betrachten jetzt eine laminare Strömung in ei-
nem Rohr, wobei wir die Gravitationskraft vernach-
lässigen. Aus Symmetriegründen erwarten wir, dass
die Geschwindigkeit nur vom Radius (nicht vom Po-
larwinkel) abhängt. Aufgrund der Massenerhaltung
verschwindet die Abhängigkeit von z (für konstanten
66

3 Blut, Herz und Kreislauf
Abbildung 3.19: Laminare Rohrströmung. [9]
Rohrdurchmesser). Die Druckkraft auf einen Flüs-sigkeitszylinder mit Radius r ist
Fp = πr2(p1 − p2) .
p2 stellt hier den Druck am unteren Ende des Zylin-ders dar, p1 den am oberen Ende.
Die Reibungskraft, welche auf den gleichen Zylinderwirkt, ist
Fr = 2πrlη ∂v∂ r
.
Hier stellt l die Länge des Flüssigkeitszylinders dar.
Für eine stationäre Strömung müssen die beidenKräfte sich gegenseitig aufheben, Fp +Fr = 0. So-mit
πr2(p1 − p2) =−2πrlη ∂v∂ r
oder
∂v∂ r
= − 12η
(p1 − p2)
lr < 0.
Man erhält somit wiederum ein parabolisches Ge-schwindigkeitsprofil. Wir schreiben die Druckdiffe-renz in differenzieller Form, p1−p2
l = p(z)−p(z+l)l →
− ∂ p∂ z und erhalten
∂v∂ r
=+1
2η∂ p∂ z
r.
Mit der Randbedingung, dass die Geschwindigkeitam Rand des Rohres verschwindet, v(R) = 0, undv(0) = vmax, erhalten wir das Geschwindigkeitsprofil
v(r) = vmax
�1− r2
R2
�.
mit
vmax =1
4η(p1 − p2)
lR2 = −∂ p
∂ z1
4ηR2.
Auch in einem Rohr haftet die Flüssigkeit an derWand und strömt in der Mitte am schnellsten, undes ergibt sich wieder ein parabolisches Profil.
3.3.4 Durchfluss und Strömungswiderstand
Der Volumenstrom Q hat die Dimension Volu-men/Zeit oder Fläche·Geschwindigkeit. Den Volu-menstrom durch ein Flächenelement dA ist dQ =dA · v. Für einen Ring mit innerem Radius r und äu-ßerem Radius r + dr ist die Fäche 2πr dr und derVolumenstrom
dQ(r) = 2πr dr · vmax(1− r2/R2) .
Durch Integration über den Radius r erhält man, dassder Volumenstrom in diesem Fall proportional zurvierten Potenz des Zylinderradius ist:
Q = vmax2π� R
0r dr
�1− r2
R2
�
= vmax2π�
R2
2− R4
4
�=
π(p1 − p2)
8η lR4 .
Dies wird als Hagen-Poiseuillesches Gesetz be-zeichnet.
Der Volumenstrom ist somit auch proportional zurDruckdifferenz. In Analogie zum Ohmschen Gesetzder Elektrizitätslehre gilt
Stromungswiderstand =Druckdifferenz
Stromstarke.
67

3 Blut, Herz und Kreislauf
Für ein zylindrisches Rohr beträgt der Strömungswi-derstand somit
W =∆pQ
=8η lπR4 .
Eine Änderung der Durchflussmenge kann somitentweder über die Druckdifferenz oder über denStrömungswiderstand erreicht werden. Im Falle desmenschlichen Gefäßsystems ist die Druckdifferenzrelativ konstant: in den Arterien herrscht ein Druckvon 80-120 mm Hg = 10-16 kPa, während der Druckim Venensystem etwa 3 mm Hg = 0.4 kPa beträgt. Dadie Durchflussmenge je nach Bedarf um mehr als ei-ne Größenordnung variieren kann, wird diese überden Strömungswiderstand angepasst.
Diese Änderung erfolgt vor allem in den Kapillarge-fäßen. Dort ist auf Grund der R4-Abhängigkeit derStrömungswiderstand am größten, so dass sich Än-derungen am stärksten auswirken. Eine Änderungdes Gefäßdurchmessers um 19% führt bereits zu ei-ner Verdoppelung des Durchflusses (1.194 = 2). Au-ßerdem ist dadurch eine bedarfsgerechte und unab-hängige Steuerung für einzelne Organe möglich.
3.3.5 Größenordnungen und Längenskalen
Ausgehend von den Navier-Stokes-Gleichungenkann man verschiedene Strömungstypen unterschei-den. Betrachtet werden soll eine stationäre Strö-mung, für die ∂�v
∂ t = 0 gilt. Die Navier-Stokes-Gleichung lautet dann
ρ(�v ·�∇) ·�v =−�∇p+η�∇2�v.
Schätzt man die Größenordnungen der einzelnenTerme ab, so bekommt man
ρv2
l1����Beschleunigung
≈ pl2����
Druck
+ηvl23����
Reibung
,
wobei l1,2,3 die charakteristischen Längen für dielongitudinale Geschwindigkeits- und Druckände-rung sowie die transversale Geschwindigkeitsände-rung (bei der Reibungskraft) sind. Interessant ist
hier, dass der Reibungsterm mit dem Quadrat derLängenskala abnimmt, während der Druckterm mit1/l skaliert. Bei kleinen Längenskalen wird deshalbdie Viskosität wichtiger gegenüber dem Druck.
Ist die Reibung klein, ηvl23� p
l2, dann ist p ≈ 1
2 ρv2,wenn die beiden charakteristischen Längen für dieBeschleunigung und den Druck etwa gleich großsind. Der so erhaltene Druck ist der Staudruck undist für die Medizinphysik von untergeordneter Be-deutung.
Im Fall der laminaren Strömung ist die longitudinaleGeschwindigkeitsänderung klein, ρv2
l1� p
l2, und es
gilt �∇p = η�∇2�v .
Eine wichtige Größe ist das Verhältnis aus der Träg-heitskraft ρv2
l1und der Reibungskraft ηv
l23
:
Re :=ρvlη
.
Hier wurden die beiden Längenskalen gleich gesetzt,l1 ≈ l3 = l und stellen die charakteristische Längens-kala für die entsprechende Geometrie dar. Re wirdals Reynoldszahl bezeichnet. Wichtig ist u.a., dassdie Reynoldszahl proportional ist zur Längenskalaund indirekt proportional zur Viskosität. In Blutge-fäßen mit kleinem Querschnitt (Kapillaren) oder fürkleine Lebewesen (Bakterien, Amöben) wird des-halb die Reynoldszahl sehr klein. Die gleichen Ver-hältnisse für uns würden bedeuten, dass das Mediumdie Viskosität von Honig hätte.
Analog kann man das Verhältnis der Trägheits- undder Druckkraft bilden und erhält die Eulerzahl Eu :=
pρv2 . Beide Größen spielen unter anderem eine Rollebei Ähnlichkeitsbetrachtungen von Strömungen.
3.3.6 Turbulente Strömungen
Am wichtigsten für die Medizin(physik) ist die la-minare Strömung und die turbulente Strömung. Ei-ne laminare Strömung kann durch eine Störung oderzu große Geschwindigkeiten in eine turbulente Strö-mung umschlagen.
Ob eine Strömung turbulent wird, hängt wesentlichvom Verhältnis aus Trägheitskraft zu Reibungskraft
68

3 Blut, Herz und Kreislauf
Abbildung 3.20: Umschlagen von laminarer Strö-mung in turbulente Strömung an ei-ner Verengung.
ab, welches durch die Reynoldszahl Re = ρvl
η quan-tifiziert wird. Bei Re > 400 treten an Arterienveren-gungen oder Verzweigungen lokale Wirbel in denRandschichten der Strömung auf, bei Re > 2000kann die Strömung vollkommen turbulent werden.Bei turbulenten Strömungen steigt der Widerstandsprunghaft an.
In Blutgefäßen findet man sehr unterschiedlicheReynoldszahlen, wobei die wichtigste Variable dieLängenskala ist: in Kapillaren sind die Dimensio-nen im Bereich von wenigen µm und der Fluss istvollständig laminar. In der Aorta und der Vena ca-va sind die Dimensionen im Bereich von cm unddie Reynoldszahl kann in den Bereich von turbu-lenter Strömung gelangen. Allerdings sind die Strö-mungen im Körper nicht stationär, sondern gepulst,und die Gefäße sind stark verzweigt. Außerdem exi-stieren einige Mechanismen, welche Turbulenz un-terdrücken.
Abbildung 3.21: Simulation einer turbulenten Blut-strömung. [2]
Figur 3.21 zeigt das Auftreten einer Turbulenz an derVerzweigung einer Arterie, wenn diese an einer Stel-le verengt ist. Das Fließprofil wurde durch numeri-
sche Simulation mit der Technik finiter Elemente be-rechnet.
Im Allgemeinen werden Reynoldszahlen von >2000nur in den großen Arterien bei maximaler Geschwin-digkeit kurzzeitig überschritten. In den Blutgefäßenwird die laminare Strömung stabilisiert, indem zumBeispiel ein Teil der kinetischen Energie des systo-lisch ausgeworfenen Blutes kurzzeitig in potentiel-le Energie (Dehnung der Aorta) umgewandelt wird.Während der Diastole wird die potentielle Energiewieder in kinetische umgewandelt. Diese Möglich-keit der Dehnbarkeit des arteriellen Systems wirdals Windkesselfunktion bezeichnet. Mit diesem Ener-giespeicher können Turbulenzen unterdrückt wer-den. Es kann zum Herzflimmern kommen, wenn diesnicht mehr funktioniert (zum Beispiel im Alter).
Turbulenz führt zu wesentlich reduziertem Blut-durchfluss und wird deshalb im Körper so weit wiemöglich vermieden. Sie tritt deshalb praktisch nur inpathologischen Situationen auf, wie z.B. bei fehler-haften Aortaklappen.
3.3.7 Ähnlichkeit von Strömungen
Die Reynoldszahl ist nicht nur ein dimensionsloserParameter einer Strömung, die als Kriterium für denÜbergang zwischen laminarer und turbulenter Strö-mung nützlich ist. Sie gibt außerdem die Ähnlichkeitvon Strömungen an, denn Strömungen verhalten sichähnlich, wenn ihre Reynoldszahlen Re =
ρvl
η über-einstimmen.
Dieses Ähnlichkeitsgesetz ist die Grundlage für dieMessung von Modellen in Windkanälen. Ein verklei-nertes (oder auch vergrößertes) Modell M stimmtströmungsmäßig mit dem Original O überein, wennes ähnliche Geometrie wie das Original besitzt, dieStrömung die gleiche Reynoldszahl hat,
ReM ≈ ReO,
und auch die gleiche Euler-Zahl hat:
pM
ρMv2M
≈ pO
ρOv2O
.
69
3 Blut, Herz und Kreislauf
(pM: Druck des Fluids im Modellversuch; genauer:Differenz Druck minus Dampfdruck, ρM: Dichte desFluids im Modellversuch, vM: Geschwindigkeit desFluids im Modellversuch).
Abbildung 3.22: NASA-Windkanal mit Flugzeug-modell. [3]
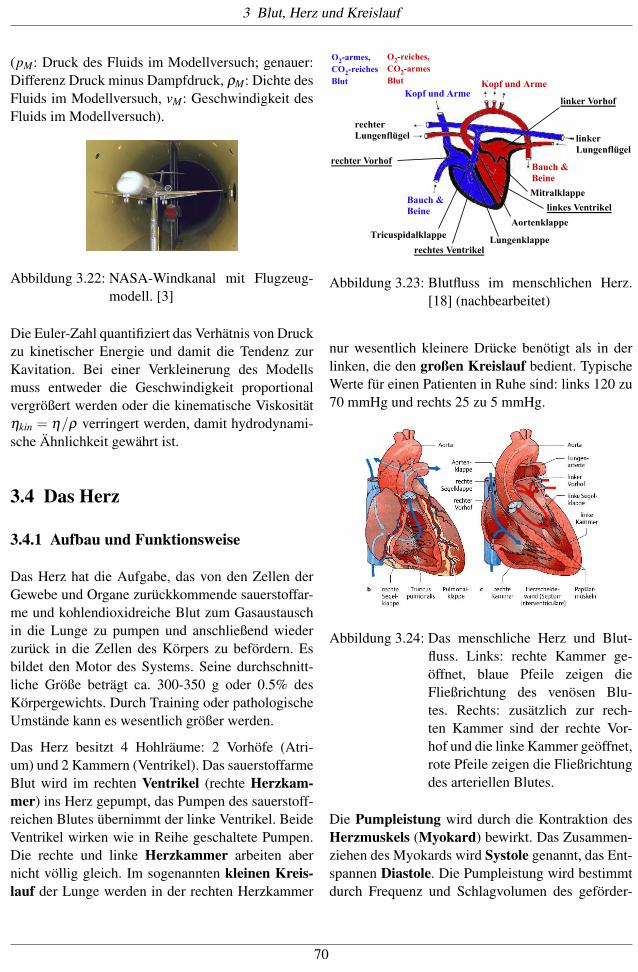
Die Euler-Zahl quantifiziert das Verhätnis von Druckzu kinetischer Energie und damit die Tendenz zurKavitation. Bei einer Verkleinerung des Modellsmuss entweder die Geschwindigkeit proportionalvergrößert werden oder die kinematische Viskositätηkin = η/ρ verringert werden, damit hydrodynami-sche Ähnlichkeit gewährt ist.
3.4 Das Herz
3.4.1 Aufbau und Funktionsweise
Das Herz hat die Aufgabe, das von den Zellen derGewebe und Organe zurückkommende sauerstoffar-me und kohlendioxidreiche Blut zum Gasaustauschin die Lunge zu pumpen und anschließend wiederzurück in die Zellen des Körpers zu befördern. Esbildet den Motor des Systems. Seine durchschnitt-liche Größe beträgt ca. 300-350 g oder 0.5% desKörpergewichts. Durch Training oder pathologischeUmstände kann es wesentlich größer werden.
Das Herz besitzt 4 Hohlräume: 2 Vorhöfe (Atri-um) und 2 Kammern (Ventrikel). Das sauerstoffarmeBlut wird im rechten Ventrikel (rechte Herzkam-
mer) ins Herz gepumpt, das Pumpen des sauerstoff-reichen Blutes übernimmt der linke Ventrikel. BeideVentrikel wirken wie in Reihe geschaltete Pumpen.Die rechte und linke Herzkammer arbeiten abernicht völlig gleich. Im sogenannten kleinen Kreis-
lauf der Lunge werden in der rechten Herzkammer
!"#$ %&' ()*+
)+,-.+)/%&0+&$120+1 13&4+)
/%&0+&$120+1
56%,- 75+3&+
!"#$ %&' ()*+
56%,- 75+3&+
8)3,%9#3'61416##+
:3.)61416##+
/%&0+&416##+
(").+&416##+13&4+9 ;+&.)34+1
)+,-.+9 ;+&.)34+1
)+,-.+) ;")-"$
13&4+) ;")-"$
!"#$%&'()*!"#%'+,-'(./01
!"#%'+,-'()*!"#$%&'(./01
Abbildung 3.23: Blutfluss im menschlichen Herz.[18] (nachbearbeitet)
nur wesentlich kleinere Drücke benötigt als in derlinken, die den großen Kreislauf bedient. TypischeWerte für einen Patienten in Ruhe sind: links 120 zu70 mmHg und rechts 25 zu 5 mmHg.
Abbildung 3.24: Das menschliche Herz und Blut-fluss. Links: rechte Kammer ge-öffnet, blaue Pfeile zeigen dieFließrichtung des venösen Blu-tes. Rechts: zusätzlich zur rech-ten Kammer sind der rechte Vor-hof und die linke Kammer geöffnet,rote Pfeile zeigen die Fließrichtungdes arteriellen Blutes.
Die Pumpleistung wird durch die Kontraktion desHerzmuskels (Myokard) bewirkt. Das Zusammen-ziehen des Myokards wird Systole genannt, das Ent-spannen Diastole. Die Pumpleistung wird bestimmtdurch Frequenz und Schlagvolumen des geförder-
70

3 Blut, Herz und Kreislauf
ten Blutes. Die Frequenz in Ruhe liegt bei etwa 70
Schlägen pro Minute und kann bei Belastung stark
gesteigert werden. Das Schlagvolumen kann durch
die Verkleinerung des systolischen Volumens der
linken Herzkammer durch Steigerung der Kontrak-
tion des Herzmuskels beeinflusst werden. Normaler-
weise werden nur ungefähr 60ml von 130ml Blut
in der Herzkammer in die Aorta entleert. Dies ent-
spricht pro Minute ca. 5 l. Bei entsprechender kör-
perlicher Leistung kann das Herz-Minutenvolumen
bis auf ca. 20-30 l gesteigert werden.
3.4.2 Der Herzzyklus
p[kPa]
i[l/s]
Anfang Aorta
Linker Ventrikel
Anfang Aorta
A- und P-Klappe
M- und T-Klappe
AnspannungSystole
Austreibung
p[kPa]
i[l/s]
EntspannungDiastole
p[kPa]
i[l/s]
Füllungszeit
p[kPa]
i[l/s]
Füllungszeit
p[kPa]
i[l/s]
p[kPa]
Rechter Ventrikel
Pulmonararterie
p[kPa]
i[l/s]
Abbildung 3.25: Herzzyklus: Druck im linken Ven-
trikel und am Anfang der Aorta
(oben), Blutstrom i am Anfang der
Aorta (Mitte) und Druck im rech-
ten Ventrikel bzw. dem Anfang der
Pulmonararterie (unten). Ganz un-
ten ist angegeben, welche Herz-
klappen während der einzelnen
Herzphasen geschlossen (schwarz)
oder geöffnet sind (weiß).
In der Abbildung 3.25 sind die Druckverhältnisse im
linken Ventrikel und am Anfang der Aorta zu se-
hen, der Blutstrom in die Aorta sowie die Druck-
verhältnisse im linken Ventrikel und am Anfang der
Lungenarterie. In der ersten Phase des Herzzyklus,
der Anspannungsphase, sind alle Herzklappen ge-
schlossen. Der Druck steigt in dem linken Ventrikel
stark an, bis sich auf Grund des höheren Drucks im
Vergleich zu dem in der Aorta die Aortenklappe öff-
net.
Nun beginnt die Austreibungsphase, in welcher
der Druck im linken Ventrikel langsamer ansteigt
und ein Maximum durchläuft. Wegen der geöffneten
Aortenklappe folgt der Druck am Anfang der Aorta
dem Druck des linken Ventrikels. Der Blutstrom zu
Beginn der Aorta steigt steil an und fällt dann wieder
ab. Am Ende der Austreibungsphase sinkt der Ven-
trikeldruck schnell ab, aber in der elastisch dehnba-
ren Arterie ist er noch höher, daher strömt etwas Blut
zurück ins Herz und die Aortenklappe schließt sich
wieder. Diese beiden beschriebenen Phasen gehören
zur Systole, in der die Ventrikel ihre Hauptarbeit lei-
sten.
Die sich anschließende Diastole ist im Wesentlichen
passiv und beginnt mit einer Entspannung der Ven-
trikelwand. Dadurch sinkt der Druck im linken Ven-
trikel rasch ab. Wenn er unter denjenigen im lin-
ken Vorhof gesunken ist, dann öffnet sich die Mi-
tralklappe, und es strömt Blut vom Vorhof in die
Kammer (Füllungszeit). Dies wird durch die ein-
setzende Vorhofkontraktion aktiv unterstützt. Wegen
der Füllung der erschlafften Kammer erhöht sich der
Druck dort bis zum sogenannten enddiastolischenFüllungsdruck. Danach beginnt ein neuer Herzzy-
klus.
3.4.3 Leistung des Herzens
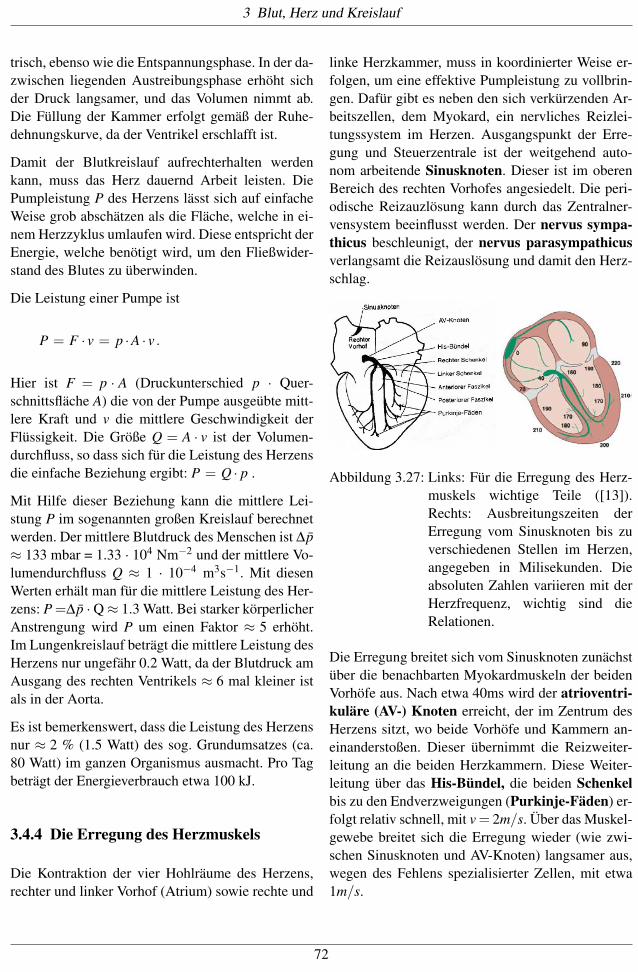
Abbildung 3.26: Pumpzyklus des Herzens. [18]
Der Pumpzyklus kann auch als pV-Diagramm dar-
gestellt werden. In der Abbildung 3.26 ist das pV-
Diagramm des linken Ventrikels idealisiert darge-
stellt. Zu Beginn der Systole (A) herrscht im Ven-
trikel der enddiastolische Füllungsdruck. Die Druck-
erhöhung der Anspannungsphase erfolgt isovolume-
71
3 Blut, Herz und Kreislauf
trisch, ebenso wie die Entspannungsphase. In der da-zwischen liegenden Austreibungsphase erhöht sichder Druck langsamer, und das Volumen nimmt ab.Die Füllung der Kammer erfolgt gemäß der Ruhe-dehnungskurve, da der Ventrikel erschlafft ist.
Damit der Blutkreislauf aufrechterhalten werdenkann, muss das Herz dauernd Arbeit leisten. DiePumpleistung P des Herzens lässt sich auf einfacheWeise grob abschätzen als die Fläche, welche in ei-nem Herzzyklus umlaufen wird. Diese entspricht derEnergie, welche benötigt wird, um den Fließwider-stand des Blutes zu überwinden.
Die Leistung einer Pumpe ist
P = F · v = p ·A · v .
Hier ist F = p · A (Druckunterschied p · Quer-schnittsfläche A) die von der Pumpe ausgeübte mitt-lere Kraft und v die mittlere Geschwindigkeit derFlüssigkeit. Die Größe Q = A · v ist der Volumen-durchfluss, so dass sich für die Leistung des Herzensdie einfache Beziehung ergibt: P = Q · p .
Mit Hilfe dieser Beziehung kann die mittlere Lei-stung P im sogenannten großen Kreislauf berechnetwerden. Der mittlere Blutdruck des Menschen ist ∆p≈ 133 mbar = 1.33 · 104 Nm−2 und der mittlere Vo-lumendurchfluss Q ≈ 1 · 10−4 m3s−1. Mit diesenWerten erhält man für die mittlere Leistung des Her-zens: P =∆p · Q ≈ 1.3 Watt. Bei starker körperlicherAnstrengung wird P um einen Faktor ≈ 5 erhöht.Im Lungenkreislauf beträgt die mittlere Leistung desHerzens nur ungefähr 0.2 Watt, da der Blutdruck amAusgang des rechten Ventrikels ≈ 6 mal kleiner istals in der Aorta.
Es ist bemerkenswert, dass die Leistung des Herzensnur ≈ 2 % (1.5 Watt) des sog. Grundumsatzes (ca.80 Watt) im ganzen Organismus ausmacht. Pro Tagbeträgt der Energieverbrauch etwa 100 kJ.
3.4.4 Die Erregung des Herzmuskels
Die Kontraktion der vier Hohlräume des Herzens,rechter und linker Vorhof (Atrium) sowie rechte und
linke Herzkammer, muss in koordinierter Weise er-folgen, um eine effektive Pumpleistung zu vollbrin-gen. Dafür gibt es neben den sich verkürzenden Ar-beitszellen, dem Myokard, ein nervliches Reizlei-tungssystem im Herzen. Ausgangspunkt der Erre-gung und Steuerzentrale ist der weitgehend auto-nom arbeitende Sinusknoten. Dieser ist im oberenBereich des rechten Vorhofes angesiedelt. Die peri-odische Reizauzlösung kann durch das Zentralner-vensystem beeinflusst werden. Der nervus sympa-
thicus beschleunigt, der nervus parasympathicus
verlangsamt die Reizauslösung und damit den Herz-schlag.
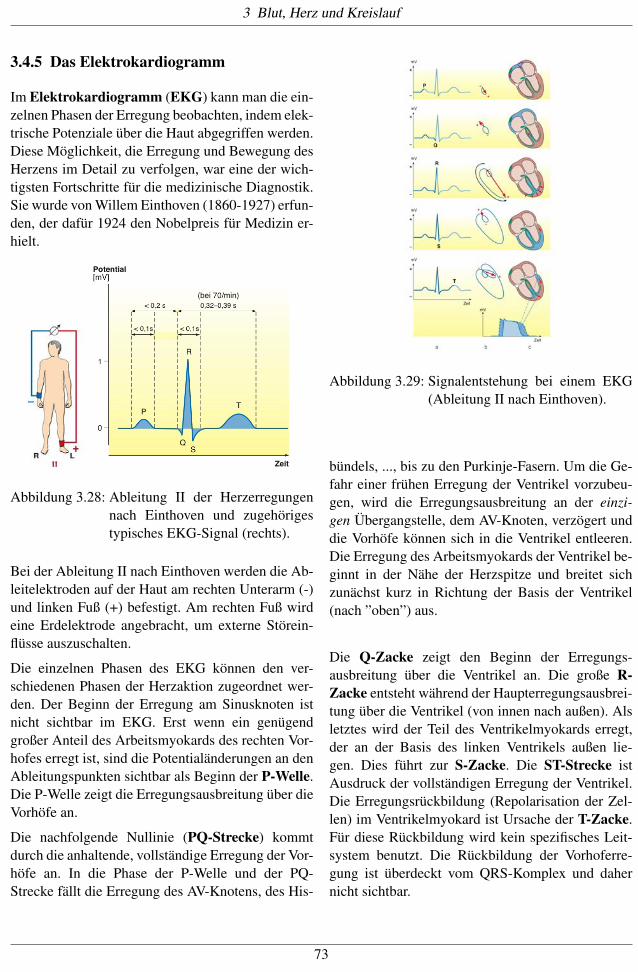
Abbildung 3.27: Links: Für die Erregung des Herz-muskels wichtige Teile ([13]).Rechts: Ausbreitungszeiten derErregung vom Sinusknoten bis zuverschiedenen Stellen im Herzen,angegeben in Milisekunden. Dieabsoluten Zahlen variieren mit derHerzfrequenz, wichtig sind dieRelationen.
Die Erregung breitet sich vom Sinusknoten zunächstüber die benachbarten Myokardmuskeln der beidenVorhöfe aus. Nach etwa 40ms wird der atrioventri-
kuläre (AV-) Knoten erreicht, der im Zentrum desHerzens sitzt, wo beide Vorhöfe und Kammern an-einanderstoßen. Dieser übernimmt die Reizweiter-leitung an die beiden Herzkammern. Diese Weiter-leitung über das His-Bündel, die beiden Schenkel
bis zu den Endverzweigungen (Purkinje-Fäden) er-folgt relativ schnell, mit v= 2m/s. Über das Muskel-gewebe breitet sich die Erregung wieder (wie zwi-schen Sinusknoten und AV-Knoten) langsamer aus,wegen des Fehlens spezialisierter Zellen, mit etwa1m/s.
72
3 Blut, Herz und Kreislauf
3.4.5 Das Elektrokardiogramm
Im Elektrokardiogramm (EKG) kann man die ein-zelnen Phasen der Erregung beobachten, indem elek-trische Potenziale über die Haut abgegriffen werden.Diese Möglichkeit, die Erregung und Bewegung desHerzens im Detail zu verfolgen, war eine der wich-tigsten Fortschritte für die medizinische Diagnostik.Sie wurde von Willem Einthoven (1860-1927) erfun-den, der dafür 1924 den Nobelpreis für Medizin er-hielt.
Abbildung 3.28: Ableitung II der Herzerregungennach Einthoven und zugehörigestypisches EKG-Signal (rechts).
Bei der Ableitung II nach Einthoven werden die Ab-leitelektroden auf der Haut am rechten Unterarm (-)und linken Fuß (+) befestigt. Am rechten Fuß wirdeine Erdelektrode angebracht, um externe Störein-flüsse auszuschalten.
Die einzelnen Phasen des EKG können den ver-schiedenen Phasen der Herzaktion zugeordnet wer-den. Der Beginn der Erregung am Sinusknoten istnicht sichtbar im EKG. Erst wenn ein genügendgroßer Anteil des Arbeitsmyokards des rechten Vor-hofes erregt ist, sind die Potentialänderungen an denAbleitungspunkten sichtbar als Beginn der P-Welle.Die P-Welle zeigt die Erregungsausbreitung über dieVorhöfe an.
Die nachfolgende Nullinie (PQ-Strecke) kommtdurch die anhaltende, vollständige Erregung der Vor-höfe an. In die Phase der P-Welle und der PQ-Strecke fällt die Erregung des AV-Knotens, des His-
Abbildung 3.29: Signalentstehung bei einem EKG(Ableitung II nach Einthoven).
bündels, ..., bis zu den Purkinje-Fasern. Um die Ge-fahr einer frühen Erregung der Ventrikel vorzubeu-gen, wird die Erregungsausbreitung an der einzi-gen Übergangstelle, dem AV-Knoten, verzögert unddie Vorhöfe können sich in die Ventrikel entleeren.Die Erregung des Arbeitsmyokards der Ventrikel be-ginnt in der Nähe der Herzspitze und breitet sichzunächst kurz in Richtung der Basis der Ventrikel(nach ”oben”) aus.
Die Q-Zacke zeigt den Beginn der Erregungs-ausbreitung über die Ventrikel an. Die große R-Zacke entsteht während der Haupterregungsausbrei-tung über die Ventrikel (von innen nach außen). Alsletztes wird der Teil des Ventrikelmyokards erregt,der an der Basis des linken Ventrikels außen lie-gen. Dies führt zur S-Zacke. Die ST-Strecke istAusdruck der vollständigen Erregung der Ventrikel.Die Erregungsrückbildung (Repolarisation der Zel-len) im Ventrikelmyokard ist Ursache der T-Zacke.Für diese Rückbildung wird kein spezifisches Leit-system benutzt. Die Rückbildung der Vorhoferre-gung ist überdeckt vom QRS-Komplex und dahernicht sichtbar.
73

3 Blut, Herz und Kreislauf
3.4.6 Regulierung und Störungen
Es gibt mehrere autoregulierende Mechanismen, dieeine autonome Teilregulation übernehmen können,so dass ein gestörter Erregungsablauf vom orga-nischen Herzschrittmacher, dem Sinusknoten, nichtkomplettes Herzversagen bedeutet. Die Herzleistungist in diesen Fällen allerdings im Allgemeinen starkeingeschränkt. Bei Ausfall des Sinusknotens über-nimmt zum Beispiel der AV-Knoten die Schritt-macherfunktion, wenn auch mit einer wesentlich ge-ringeren Frequenz von 30 bis 40 Schlägen pro Mi-nute. Wenn die Überleitung der Erregung von denVorhöfen auf den Ventrikel vollkommen blockiertist, dann spricht man von einem totalen Herzblock.Als Ersatzschrittmacher fungiert ein tertiäres Zen-trum im Reizleitungssystem der Herzkammern mitetwa 30 Erregungen pro Minute. Vorhöfe und Herz-kammern schlagen dann unkoordiniert, die Herzlei-stung ist stark herabgesetzt.
Die Abnahme der Herzfrequenz wird ganz allgemeinals Bradykardie, eine Zunahme als Tachykardie
bezeichnet. Es ändert sich nicht nur die Frequenz,sondern auch die Form der Erregung. Besonders ge-fährlich sind Tachykardien, die zu einer ungeregel-ten hochfrequenten Dauererregung des Herzmuskelsführen, dem sogenannten Herzflimmern.
Abbildung 3.30: Oben: Vorhofflimmern mit abso-luter Arrhythmie (nicht unmittel-bar lebensbedrohlich); unten: Kam-merflimmern (führt unbehandeltzum Tode).
Dieses führt zu einem Stillstand des Kreislaufs undunbehandelt zum Tode. Abhilfe bringt (neben Me-dikamenten in einigen besonderen Fällen) dann nurdie Defibrillation.
Abbildung 3.31: Oben: Normaler Herzrhythmus;unten: Totaler Herzblock.
Herzschrittmacher werden eingesetzt um pathologi-sche Störungen im Ablauf der Herzreizleitung zu be-handeln. Ein Beispiel ist der totale Herzblock, beidem es zu keiner Überleitung vom Vorhof zum Ven-trikel kommt.
3.5 Hydrodynamik der
Blutkreislaufs
3.5.1 Grundlagen
Abbildung 3.32: Übersicht über den menschlichenBlutkreislauf. [7]
Die Blutströmung durch die Blutgefäße wird durchDruckdifferenzen zwischen den einzelnen Gefäßab-schnitten bewirkt. Diese müssen eingestellt und imGleichgewicht gehalten werden. Dafür steuert dieNatur
74
3 Blut, Herz und Kreislauf
• Strömung, Druck und Widerstand im Gefäßssy-stem
• Herzzeitvolumen
• Organdurchblutung
• Blutzirkulation in den Kapillaren
• venöser Rückstrom zum Herzen
Steigt z.B. der Bedarf an Blut in einem Muskel,so wird einerseits die Leistung des Herzens erhöht;andererseits wird der Gefäßdurchmesser in diesemMuskel erhöht und dadurch der Strömungswider-stand reduziert. Eine Erweiterung des Gefäßdurch-messers wird z.B. durch Sauerstoffmangel induziert.
Abbildung 3.33: Prinzip des Blutkreislaufs. [13]
Bei einer laminaren Strömung hängt der Strömungs-widerstand nur von den Gefäßabmessungen und denEigenschaften des Blutes ab, aber nicht von des-sen Stromfluss. Im Kreislaufsystem können funk-tionell zwei Teilkreisläufe unterschieden werden:ein großer Körperkreislauf und ein kleiner Lungen-kreislauf. Die Organe sind parallel “geschaltet”, so-dass eine gleichmäßige Versorgung der Körperorga-ne und Kapillaren mit hoch oxygeniertem Blut si-chergestellt ist.
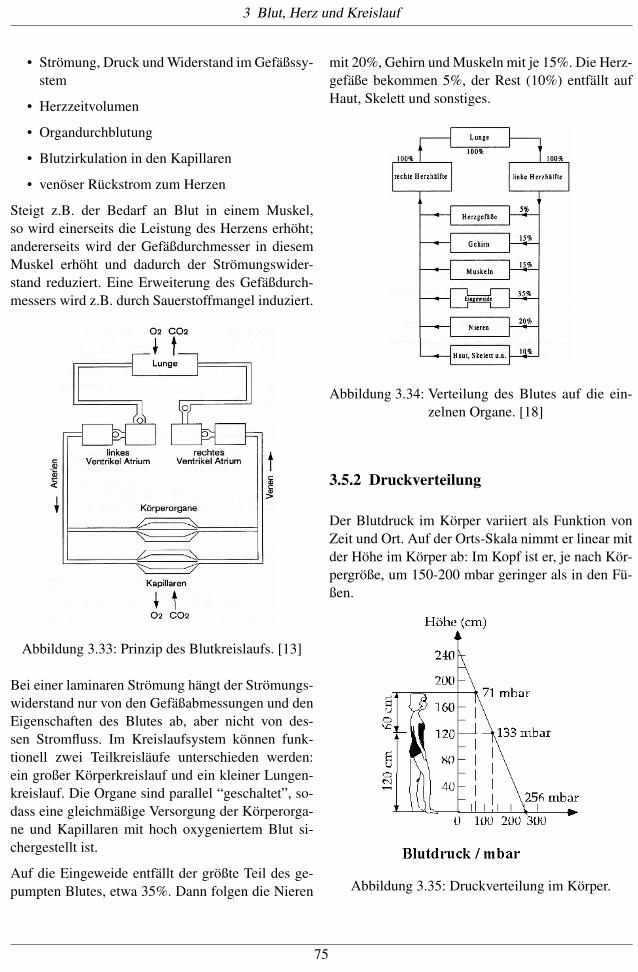
Auf die Eingeweide entfällt der größte Teil des ge-pumpten Blutes, etwa 35%. Dann folgen die Nieren
mit 20%, Gehirn und Muskeln mit je 15%. Die Herz-gefäße bekommen 5%, der Rest (10%) entfällt aufHaut, Skelett und sonstiges.
Abbildung 3.34: Verteilung des Blutes auf die ein-zelnen Organe. [18]
3.5.2 Druckverteilung
Der Blutdruck im Körper variiert als Funktion vonZeit und Ort. Auf der Orts-Skala nimmt er linear mitder Höhe im Körper ab: Im Kopf ist er, je nach Kör-pergröße, um 150-200 mbar geringer als in den Fü-ßen.
Abbildung 3.35: Druckverteilung im Körper.
75

3 Blut, Herz und Kreislauf
Außerdem hängt der Druck vom Organ ab, vomBlutkreislauf, der Sauerstoffversorgung, und derZeit. Im kleinen Kreislauf herrscht ein geringe-rer Strömungswiderstand, daher sind die systoli-schen/diastolischen Drücke gering (25/5 mm Hg),während sie im großen Kreislauf in Ruhe 120/70 mmHg betragen (siehe Abbildung 3.33). Im Kreislaufsy-stem wird das Blut vom linken Ventrikel über Arte-rien, die sich in Arteriolen verzweigen, in die Kapil-laren gepumpt. Die vom Herzen kommende Druck-welle wird in den Arteriolen sehr stark gedämpft,weil dort der Strömungswiderstand hoch ist.
Blutdruck
[mmHg]
Gefäßquerschnitte
Verteilung des
Blutvolumens
Geschw. [cm/s] Geschw.
Querschnitt
[cm2]Querschnitt
Niederdruck-
system
arterielles
System
Abbildung 3.36: Druckverteilung, Blutvolumenver-teilung und Geschwindigkeit in denverschiedenen Gefäßen.
Das führt zu einem kontinuierlichen Blutstrom inden Kapillaren, wo der Gasaustausch mit dem Ge-webe stattfindet. Da sich mehr als 70% des Blutvolu-mens im venösen System befinden, wird dieses auchals Kapazitätssystem bezeichnet.
3.5.3 Widerstand und Fließgeschwindigkeit
Wir diskutieren den Widerstand unter der Annah-me einer laminaren Strömung. Die treibende Kraftist die Druckdifferenz, die Reibungskraft wird durchdie Viskosität, die Fläche und den Geschwindig-keitsgradienten bestimmt (siehe Kapitel 3.2). In derRealität sind Abweichungen von diesem einfachenModell zu erwarten, da die Viskosität des Blutes
nicht konstant ist, und das Blut aus Plasma mit sus-pensierten Zellen (u.a. weiße und rote Blutkörper-chen) besteht. In der Aorta und der Arteria femo-ralis gilt, dass die Ausbreitungsgeschwindigkeit desPulses gleich der Strömungsgeschwindigkeit ist. DerEinfluss der Elastizität der Blutgefäße kann durchdas Hookesche Gesetz für die Spannung (Dehnung)der Blutgefäße berücksichtigt werden. Da Blutge-fäße jedoch auch aktive Komponenten in Form vonMuskeln enthalten, ist die Realität schwierig durcheinfache Modelle zu beschreiben. Im nächsten Ka-pitel werden die Ergebnisse eines einfachen Modellsfür die pulsatorischen Vorgänge im Arteriensystemvorgestellt.
Der mittlere Blutdruck in der Aorta beträgt 13kPa.Der wesentliche Strömungswiderstand kommt vonden Arteriolen und Kapillaren, er wird auch als peri-pherer Widerstand bezeichnet. Für den Menschenerhält man mit dem zeitlichen Mittelwert der Strom-stärke des Blutes (Herzminutenvolumen = Schlagvo-lumen · Herzfrequenz),
Q ≈ 70ml ·70/min ≈ 5l/min ≈ 83 ·10−3 l/s
und der Differenz der mittleren Drucke in Aorta undrechtem Vorhof (< prechterVorhof >≈ 1kPa):
Rper =�pAorta�−�prechterVorhof�
Q
≈ 1,5 ·108 Pas
m3 .
Die Strömungsgeschwindigkeit in der herznahenAorta beträgt etwa vm ≈ 0.25m/s. Da die Quer-schnittsfläche A der Kapillaren insgesamt etwa 500mal größer ist als die der Aorta, und Q = vmA ist,beträgt die Geschwindigkeit in den Kapillaren nurvk ≈ 0,25m/s
500 = 0.5mm/s. Diese Verlangsamung derStrömungsgeschwindigkeit, sowie die große Ober-fläche ermöglichen einen effektiven Austausch vonSauerstoff mit dem Gewebe.
Dieselben Beziehungen gelten auch für das Lungen-gefäßsystem, allerdings beträgt der periphere Wider-stand des Lungengefäßsystems nur etwa 10% desobigen Wertes, da der mittlere Druck in den Lun-genvenen bei etwa 0.7 − 1kPa liegt, und der mitt-lere Druck der Pulmonararterie etwa 2kPa beträgt.
76

3 Blut, Herz und Kreislauf
Der zeitliche Mittelwert der Stromstärke ist kon-stant. Der Gesamtquerschnitt der Lungenkapillarenist 100 bis 150 mal so hoch wie der Querschnitt derPulmonararterie, daher ergibt sich eine mittlere Strö-mungsgeschwindigkeit von 2mm/s in den Lungen-kapillaren.
3.5.4 Druckwelle
Die Beschreibung der pulsatorischen Vorgänge istwesentlich komplizierter als die der zeitlichen Mit-telwerte. Zur vollständigen Beschreibung werdenKontinuitätsgleichung und die Navier-Stokes Glei-chung
ρ�
∂�v∂ t
+(�v�∇)�v�=−�∇p+η�∇2�v
benötigt. Hier stellt die linke Seite die lokale Im-pulsänderung dar (∼ dρv/dt), die rechte Seite dieKräfte aufgrund des Druckgradienten und der Rei-bung. Zusätzlich werden die Randbedingungen (Ge-fäßwände) benötigt. Im Gegensatz zu unseren ver-einfachenden Annahmen sind die Gefäße in Wirk-lichkeit nicht starr, sondern elastisch. Dieses voll-ständige Gleichungssystem ist nicht analytisch zubehandeln. Lösungen existieren nur unter vereinfa-chenden Annahmen und für Spezialfälle.
Es wird im Folgenden angenommen, dass die pulsie-rende Strömung in einem starren, zylindrischen Rohr(Innenradius Ri) erfolgt, und dass nur Geschwin-digkeiten in longitudinaler (axialer) Richtung auf-treten, �v = v�ez. Dann hat man es mit einer lamina-ren Strömung in einem starren Rohr zu tun, äuße-re Kräfte wie die Schwerkraft werden nicht berück-sichtigt. Der Konvektionsterm in der Navier-StokesGleichung verschwindet und es bleibt
ρ ∂v∂ t
=−∂ p∂ z
+η∆v .
Der Laplace-Operator lautet in Zylinderkoordinaten
∆ =1r
∂∂ r
�r
∂∂ r
�+
1r2
∂ 2
∂ϕ2 +∂ 2
∂ z2 .
Aus Symmetriegründen gilt v = v(r), so dass
ρ ∂v∂ t
=−∂ p∂ z
+η�
1r
∂∂ r
�r
∂v∂ r
��.
Für nichtstationäre, pulsierende Strömungen im ein-geschwungenen Zustand wird ein sinusförmigerDruckgradient in axialer Richtung angenommen,∂ p∂ z = p�zeiωt , der zu einem sinusförmigen Ablauf derGeschwindigkeiten führt: v = vzeiωt . Damit wird
iωρvz =−p�z +η�
1r
∂∂ r
�r
∂vz
∂ r
��.
Mit den dimensionslosen Größen y := r/Ri und α :=Ri�
ωρ/η erhält man daraus eine Besselsche Diffe-renzialgleichung
d2vz
dy2 +1y
dvz
dy− iα2vz =−p�z
R2i
η.
Unter Berücksichtigung der Randbedingung, dassdie Flüssigkeit an der Rohrwand haftet (vz = 0 beiy = 1) bekommt man als Lösung
v = vzeiωt =p�zR2
iiηα2
�1− J0(αy
√−i)
J0(α√−i)
�eiωt .
J0 ist die Besselfunktion erster Art und nullter Ord-nung.
Abbildung 3.37 zeigt drei Beispiele für Geschwin-digkeitsprofile oszillatorischer Strömungen. Die Ge-schwindigkeitsumkehr beginnt stets in den wandna-hen Zonen, und im mittleren Rohrteil eilt die Pha-se der Geschwindigkeit derjenigen in den Randzo-nen nach. Mit steigendem Parameter α =Ri
�ωρ/η
wird das Profil flacher, und die zentrale Flüssigkeits-säule schwingt fast wie ein kompakter Körper.
3.5.5 Reales Verhalten
Es existieren verschiedene Möglichkeiten für dieMessung des Blutflusses. Eine relativ einfache ist diegepulste Ultraschall-Doppler-Strömungsmessung.Sie erlaubt es, die Geschwindigkeitsprofile inArterien in vivo zu messen.
77

3 Blut, Herz und Kreislauf
Abbildung 3.37: Geschwindigkeitsprofile für α =3,6,9. Zu sehen ist jeweils die Mo-mentangeschwindigkeit als Funkti-on des Ortes für eine vollständigeSchwingung (0◦ −360◦). [10]
Abbildung 3.38: Gemessenes Geschwindigkeitspro-fil und zugehörige Pulswelle (un-ten). [10]
Die Ergebnisse zeigen Gemeinsamkeiten mit denoben diskutierten analytischen Lösungen, aber auchUnterschiede. Diese hängen u.a. damit zusammen,dass es sich in der Realität nicht um einen sinusför-migen Verlauf des Druckgradienten und starre Ge-fäßwände handelt: In einem starren Blutgefäß wirdbei Druckanstieg die gesamte Blutmenge beschleu-nigt. Durch die Dehnbarkeit der Wände der Blutge-fäße wird jedoch ein Teil der Druckschwankungenaufgefangen.
Die Ausdehnung der Blutgefäße folgt in erster Nähe-rung dem Hookeschen Gesetz. Bei genauerer Mes-sung findet man jedoch Abweichungen. Die Blut-gefäße sind aus elastischem und kollagenem Bin-degewebe sowie glatter Muskulatur aufgebaut. Dar-aus resultiert ein relativ komplexes Verhalten und es
Abbildung 3.39: Elastizität der Arterien: Druck unfDurchmesser als Funktion der Zeit.[10]
wird eine Hysterese beobachtet, D = D(p). Dies ent-spricht qualitativ einem gedämpften System.
3.5.6 Ausbreitungsgeschwindigkeit inelastischen Blutgefäßen
In starren Röhren kann sich eine inkompressibleFlüssigkeit nur als ganzes bewegen; die Ausbrei-tungsgeschwindigkeit eines Druckpulses ist damitunendlich. Wir erhalten eine realistischere Beschrei-bung, wenn wir die Elastizität mit berücksichtigen.
Wir betrachten dafür die Ausbreitung der Druckpul-se in einem elastischen Blutgefäß mit Hilfe eineseindimensionalen Modells, bei dem wir nur die Län-genkoordinate (z) berücksichtigen. Im Gegensatz zuAbschnitt 3.5.4 vernachlässigen wir somit die radia-le Abhängigkeit der Geschwindigkeit. Ebenso ver-nachlässigen wir Reibungskräfte und Nichtlinearitä-ten. Aus den Navier-Stokes-Gleichungen (3.1) wer-den so die Euler’sch(en) Gleichung(en):
ρ ∂v∂ t
=−∂ p∂ z
.
Für die folgende Diskussion bringen wir diese Glei-chung in eine andere Form: Wir definieren eine ef-fektive Masse M := ρ
A . Damit erhalten wir
M∂Q∂ t
=−∂ p∂ z
, (3.4)
78

3 Blut, Herz und Kreislauf
wobei Q = Av die Stromstärke darstellt. Die zuge-
hörige Kontinuitätsgleichung lautet
− ∂Q∂ z
=∂A∂ t
=C∂ p∂ t
, (3.5)
wobei C := dAd p die Kapazität oder Nachgiebigkeit
(Compliance) pro Länge der Röhre ist. Sie kann z.B.
aus Fig. 3.39 abgelesen werden. Wir leiten Glei-
chung (3.4) nach z ab und Gl. (3.5) nach der Zeit:
∂ 2Q∂ t ∂ z
=− 1
M∂ 2 p∂ z2
=−C∂ 2 p∂ t2
.
Man erhält somit eine Wellengleichung, welche die
Ausbreitung einer Druckwelle beschreibt. Analog
kann man auch eine Wellengleichung für die Strom-
stärke Q verwendet werden:
∂ 2 p∂ t2
= c2∂ 2 p∂ z2
,∂ 2Q∂ t2
= c2∂ 2Q∂ z2
.
Harmonische Lösungen sind z.B.
p(z, t) = p0ei(wt±kz).
Die Ausbreitungsgeschwindigkeit ist dabei
c2 =ω2
k2=
1
CM.
Die Wellengleichung wird gelöst durch vorwärts-
laufende, antegrade Wellen p∼ = p0 · f (t − z/c)und durch rückwärtslaufende, retrograde Wellenmit p∼ = p0 · f (t + z/c). In den Arterien werden die
antegraden Wellen durch das Herz erzeugt, während
retrograde Wellen durch Reflexionen entstehen.
3.5.7 Wellengeschwindigkeit undWellenimpedanz
Wir berechnen die Wellengeschwindigkeit aus
c2 =1
CM=
d pdA
Aρ
(3.6)
=d p
2πRidRi
πR2
iρ
=d pdRi
Ri
2ρ.
Die Änderung dRi des Blutgefässradius wird be-
stimmt durch die Druckänderung d p und die Elasti-
zität des Blutgefässes. Wir nehmen an, dass die Län-
ge der Blutgefässe konstant sei, dass ihr Radius je-
doch durch den Druck elastisch geändert wird. Dies
entspricht einer relativen Längenänderung in tangen-
tialer Richtung um
dε =dRi
Ri→ dRi = Ri dε. (3.7)
Die Änderung dε der Dehnung können wir über
den differenziellen Elastizitätsmodul E auf die Span-
nungsänderung dσ zurückführen:
dσ = E dε.
Die Zugspannung in der Gefäßwand ist, bei einer
Wanddicke d,
σ = pRi
d→ dσ =
Ri
dd p.
Wir können also die Druckänderung d p schreiben
als
d p = dσ dRi
= E dε dRi. (3.8)
Mit (3.7) und (3.8) erhalten wir für die Ableitung
d pdRi
=E dε d
Ri
1
Ri dε= E
dR2
i.
Setzt man dies in Gleichung (3.6) für die Wellenge-
schwindigkeit ein, so erhält man
c =
�Ed
2Riρ. (3.9)
Wie in elektrischen Leitungen treten auch bei Flüs-
sigkeiten Reflexionen auf. Das Auftreten von Re-
flexionen kann man relativ leicht über den Wellen-
widerstand berechnen. Dieser ist gegeben durch die
Impedanz
Z =p∼Q∼
=p∼
v∼A.
79

3 Blut, Herz und Kreislauf
Hier drückt der Index ∼ aus, dass jweils nur die Ab-weichung vom Gleichgewichtswert zu berücksichti-gen ist. Setzt man
Q = Q0 f (t ± z/c) , p∼ = p0 f (t ± z/c)
in die Euler-Gleichung (3.4) ein, dann bekommt manfür antegrade Wellen (t − z/c):
MQ0 f =p0
cf → Q0 =
p0
M c
und damit
Z =p0
Q0= Mc =
ρcA. (3.10)
3.5.8 Reflexionen
An den Stellen, an denen sich Z ändert, gibt es Re-flexionen. Der Reflexionsfaktor r ist das Verhältnisder Druckamplitude der reflektierten zu derjenigender einlaufenden Welle. Er kann berechnet werdenzu
r =Z2 −Z1
Z2 +Z1,
wenn Z2 der Wellenwiderstand hinter der Reflexi-onsstelle ist und Z1 derjenige davor. Der Druck amReflexionsort ist eine Überlagerung aus ankommen-der (e) und reflektierter (r) Welle:
pd = pe + pr = pe(1+ r).
Für die reflektierte Welle ist p∼ > 0 und Q∼ < 0.
Im Arteriensystem überlagern sich hin- und zurück-laufende Wellen in weiten Bereichen, daher stimmtder Verlauf des (resultierenden) Drucks nicht mitdem Verlauf der (resultierenden) Stromstärke über-ein. Bereits zu Beginn der Aorta unterscheiden sichdie beiden Formen erheblich (siehe Abbildung 3.25).Der Druckpuls beginnt zeitgleich mit dem Strom-puls, aber die beiden Maxima stimmen nicht überein.Der Unterschied ist besonders groß in der Diasto-le, die hier in der arteriellen Dynamik nicht nur dieEntspannungs- und Füllungszeit, sondern auch nochdie Anspannungszeit des nächsten Zyklus umfasst.
Die Puls-Wellengeschwindigkeit nimmt mit demAbstand zum Herzen zu, da d/Ri anwächst, und, laufGleichung (3.9) damit auch die Wellengeschwindig-keit. Die Aortengeschwindigkeit beträgt ≈ 4 m/s, inden Extremitäten ist c bis zu 8 m/s, zum Teil (amEnde der Beinarterien) bis über 10 m/s.
Die Wellenimpedanz Z = ρcA (3.10) wächst mit
dem abnehmenden Querschnitt A eines Gefäßes (mitzunehmender Zahl von Verzweigungen) in dista-ler Richtung. Der Blutkreislauf kann daher nichtals eine homogene Leitung aufgefasst werden. DieZunahme von Z passiert kontinuierlich, aber auchsprunghaft, und es kommt zu vielfachen Reflexio-nen innerhalb des Systems. Die zurücklaufendenund an der Aortenklappe reflektierten Wellen sindzum Beispiel der Grund dafür, dass während derAustreibungsphase des Herzens das Druckmaximumspäter auftritt als das Strömungsmaximum (sieheAbb.3.25).
Die zahlreichen Reflexionen im Verlauf und am En-de des Arteriensystems bewirken, dass die Druck-amplitude mit dem Abstand vom Herzen zunimmt(siehe Abb. 3.36). Wegen einer (hier nicht genauerdiskutieren) ω-abhängigen Dämpfung runden sichdie Pulse mit wachsendem Abstand vom Herzenab. Zusätzliche, hier nicht behandelte, Komplikatio-nen sind die zum Teil eingeschränkten Anwendungs-möglichkeiten der Näherungen der Kontinuumsme-chanik, da der Durchmesser der Arteriolen mit demDurchmesser der roten Blutkörperchen vergleichbarist.
3.5.9 Kreislaufkontrolle
Reguliert werden folgende Größen des Blutkreis-laufs:
• Herzzeitvolumen
• Durchblutung der einzelnen Organe
• arterieller Druck
Das Herzzeitvolumen kann durch Erhöhung derHerzfrequenz und das Schlagvolumen beeinflusstwerden, wobei das Schlagvolumen einen geringenEinfluss hat. Die Durchblutung wird durch Ge-fäßerweiterung (Vasodilatation) erhöht, zum Bei-
80
3 Blut, Herz und Kreislauf
spiel wenn der Partialdruck von CO2 steigt oder dervon Sauerstoff fällt, ect.
Die Steuerung des arteriellen Blutdrucks kann mandurch einen Regelkreis beschreiben.
Regler
Kreislaufzentrum
Pressorezeptoren Stellglieder:
Herzzeitvolumen
Periph. Widerst.
Gefäß-
system
Steuersignale
Störgrößen
Sollwert
IstwertGeregeltes System
Abbildung 3.40: Regelkreis des Blutdrucks.
Das Kreislaufzentrum im Hirnstamm erhält Infor-mationen über den aktuellen Blutdruck von Pres-sorezeptoren im Aortenbogen sowie an der Auftei-lungsstelle der Halsschlagadern (Kopfschlagadern).Außerdem werden Informationen über den Füllstanddes Gefäßsystems von speziellen Dehnungsrezepto-ren im Bereich der oberen und unteren Hohlvene so-wie den beiden Vorhöfen des Herzens und der lin-ken Herzkammer geliefert. Die Steuerung des Herz-zeitvolumens und des peripheren Widerstandes wirdvom Kreislaufzentrum geregelt. Zusätzliche Stör-größen wie plötzlicher Blutverlust fließen ebenfallsdiesem Regelkreis zu.
3.5.10 Bluttransport in Venen
Innerhalb des venösen Systems unterscheidet manein oberflächliches, zwischen Muskelfaszie undHaut eingelagertes Venennetz und ein System tiefergelegener Venen. Das gesamte Venensystem ist nacheinem grundsätzlich anderen Prinzip aufgebaut alsdie Arterien. Der Druck, den das Herz aufgebaut hat,ist praktisch vollständig in den Kapillaren abgefallenund trägt deshalb kaum mehr zum Transport in denVenen bei.
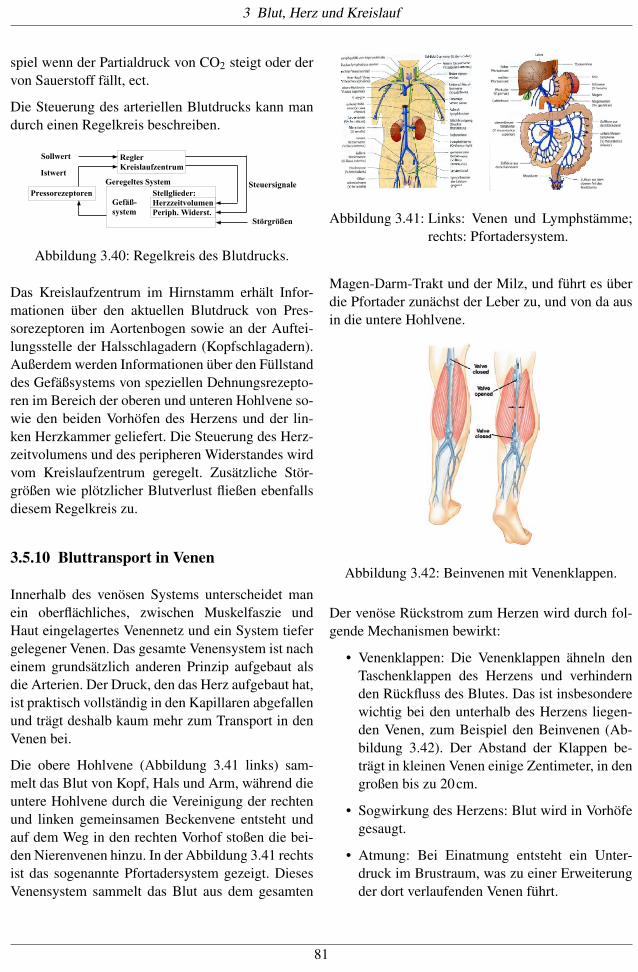
Die obere Hohlvene (Abbildung 3.41 links) sam-melt das Blut von Kopf, Hals und Arm, während dieuntere Hohlvene durch die Vereinigung der rechtenund linken gemeinsamen Beckenvene entsteht undauf dem Weg in den rechten Vorhof stoßen die bei-den Nierenvenen hinzu. In der Abbildung 3.41 rechtsist das sogenannte Pfortadersystem gezeigt. DiesesVenensystem sammelt das Blut aus dem gesamten
Abbildung 3.41: Links: Venen und Lymphstämme;rechts: Pfortadersystem.
Magen-Darm-Trakt und der Milz, und führt es überdie Pfortader zunächst der Leber zu, und von da ausin die untere Hohlvene.
Abbildung 3.42: Beinvenen mit Venenklappen.
Der venöse Rückstrom zum Herzen wird durch fol-gende Mechanismen bewirkt:
• Venenklappen: Die Venenklappen ähneln denTaschenklappen des Herzens und verhindernden Rückfluss des Blutes. Das ist insbesonderewichtig bei den unterhalb des Herzens liegen-den Venen, zum Beispiel den Beinvenen (Ab-bildung 3.42). Der Abstand der Klappen be-trägt in kleinen Venen einige Zentimeter, in dengroßen bis zu 20cm.
• Sogwirkung des Herzens: Blut wird in Vorhöfegesaugt.
• Atmung: Bei Einatmung entsteht ein Unter-druck im Brustraum, was zu einer Erweiterungder dort verlaufenden Venen führt.
81

3 Blut, Herz und Kreislauf
• Arteriovenöse Kopplung: Neben den Arteri-en liegen in der Peripherie meist zwei Venen.Durch Bindegewebe sind Arterien und Venenzu einem Gefäßbündel zusammengefasst. DiePulswelle in den Arterien presst die eng anlie-genden Venen zusammen, und wegen der Ve-nenklappen kann das venöse Blut nur in Rich-tung des Herzens fließen.
• Muskelpumpe: Der Druck der Skelettmuskula-tur bewirkt ebenfalls in Kombination mit Ve-nenklappen einen Rücktransport des venösenBlutes.
Beim stehenden Menschen in Ruhe reicht der nied-rige Blutdruck fast nicht aus, um das venöse Blutvon den Füßen zurück ins Herz zu leiten. Als Fol-ge davon sammelt sich Blut in den Venen der Beinean und erhöht den Druck in den Kapillaren, was einAnschwellen der Beine bewirken kann. Beim Gehenhingegen wird durch die Kontraktion der Muskeln inden Waden der Rückfluss des Blutes begünstigt.
3.5.11 Lymphsystem
Die Lymphstämme sind zusammen mit den wichtig-sten zentralen Venen in Abbildung 3.41 gezeigt.
In den Kapillaren erfolgt der Stoffaustausch mitdem umliegenden zwischenzelligen Raum, dem In-terstitium. Pro Tag werden rund 20 l Flüssigkeit indas Interstitium abfiltriert. Etwa 90% davon wer-den wieder resorbiert, die restlichen 10% werdenüber das Lymphgefäßsystem als Lymphflüssigkeitabtransportiert. Die Proteine, die ins Bindegewebegelangt sind, können ausschließlich von den Lymph-kapillaren in das Blut zurückgeführt werden. Drai-nage ist also eine wichtige Aufgabe des Lymphge-fäßsystems.
Das Lymphgefäßsystems enthält auch Lymphkno-ten, welche die Funktion von biologischen Filternwahrnehmen. In ihnen findet zum Beispiel die Aus-einandersetzung von Antigenen und Abwehrzellenstatt. Nach ihrer Vermehrung verlassen die Lym-phozyten (Teil der Leukozyten, der “weißen Blut-körperchen”) die Lymphknoten, und gelangen überdas Lymphgefäßsystem zurück ins Blut und andereTeile des Körpers. Die Immunzellen werden in den
Abbildung 3.43: Stofftransport und Lymphsystem.
primären lymphatischen Organen, im Thymus undKnochenmark, gebildet und wandern in die sekun-dären lymphatischen Organe: die Milz, die Lymph-knoten und das lymphatische System der Schleim-häute (z.B. Mandeln).
82