
Biogenese und Virusassembly des filamentösen Coliphagen M13 · Biogenese und Virusassembly des...
220
Biogenese und Virusassembly des filamentösen Coliphagen M13 Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften (Dr. rer. nat.) Fakultät Naturwissenschaften Universität Hohenheim Institut für Mikrobiologie vorgelegt von Martin Ploß aus Bietigheim-Bissingen 2012
Transcript of Biogenese und Virusassembly des filamentösen Coliphagen M13 · Biogenese und Virusassembly des...

Biogenese und Virusassembly des filamentösen
Coliphagen M13
Dissertation zur Erlangung des Doktorgrades
der Naturwissenschaften (Dr. rer. nat.)
Fakultät Naturwissenschaften
Universität Hohenheim
Institut für Mikrobiologie
vorgelegt von
Martin Ploß
aus Bietigheim-Bissingen
2012

Dekan: Prof. Dr. Heinz Breer
1. berichtende Person: Prof. Dr. Andreas Kuhn
2. berichtende Person: Prof. Dr. Artur Pfitzner
Eingereicht am: 08.05.2012
Mündliche Prüfung am: 17.10.2012
Die vorliegende Arbeit wurde am 18.07.2012 von der Fakultät Naturwissenschaften
der Universität Hohenheim als „Dissertation zur Erlangung des Doktorgrades der
Naturwissenschaften“ angenommen.

Gewidmet
meiner geliebten Frau Stefanie
und
meinem geliebten Sohn Florian


Inhaltsverzeichnis
2
Inhaltsverzeichnis
1 Einleitung 8
1.1 Der Bakteriophage M13 8
1.1.1 Klassifizierung des Bakteriophagen M13 8
1.1.2 Morphologie und Genom der Inoviridae 8
1.1.3 Gene, Genprodukte und ihre Funktionen 10
1.1.4 Struktur des Bakteriophagen M13 13
1.1.5 Der Zyklus des Bakteriophagen M13 14
1.2 Der gp1/11-Assembly-Komplex 21
1.3 Das Hüllprotein gp9 24
1.4 „Phage-Display“ 25
1.5 Proteintransport-Systeme 28
1.6 Aufgabenstellung der vorliegenden Arbeit 35
2 Material und Methoden 37
2.1 Material 37
2.1.1 Bakterien- und Bakteriophagenstämme 37
2.1.2 Plasmide 39
2.1.3 DNS-Standard für Agarose-Gelelektrophoresen 41
2.1.4 Enzyme 41
2.1.5 Antikörper 42
2.1.6 Proteinstandard 43
2.1.7 Aminosäuren, Zucker und Vitamine 44
2.2 Chemikalien und Verbrauchsmaterialien 44
2.3 Oligonukleotide 48
2.3.1 Sequenzierprimer 48
2.3.2 Oligonukleotide für ortsgerichtete Mutagenesen 48
2.3.3 Oligonukleotide für die Amplifikation von Genen 49
2.3.4 Oligonukleotide für die Insertion eines antigenen Epitops 49
2.4 Geräte 50
2.5 Verwendete Kits 52
2.6 Kulturmedien 52
2.6.1 Festmedien 52
2.6.2 Flüssigmedien 53
2.7 Puffer und Lösungen 54

Inhaltsverzeichnis
3
2.7.1 Agarosegelelektrophorese 54
2.7.2 Coomassiefärbung 55
2.7.3 DNS-Puffer 55
2.7.4 DNS-Sequenziergel 55
2.7.5 Herstellung von Phagenstammlösungen 56
2.7.6 Induktion von Expressionssystemen 56
2.7.7 Kompetente E. coli-Zellen 56
2.7.8 Nanogoldmarkierung von M13-Phagen 57
2.7.9 Negativkontrastierung von biologischen Proben für die Transmissions-
elektronenmikroskopie (TEM) 57
2.7.10 Proteinstandard für die präparative Gelfiltration 57
2.7.11 Radioaktive Markierung in vivo mit anschließender Immunpräzipitation
und Protease-Mapping 58
2.7.12 Reinigung der gp1/11-Assembly-Komplexe mittels präparativer Gel-
filtration 59
2.7.13 Reinigung der gp1/11-Assembly-Komplexe mittels Metallaffinitätschro-
matographie (IMAC) 59
2.7.14 SDS-Polyacrylamid-Gelelektrophorese (PAGE) 61
2.7.14.1 SDS-PAGE nach Laemmli (1970) 61
2.7.14.2 SDS-Tricin-Gelelektrophorese nach Schägger & Jagow (1987) 63
2.7.15 Stammlösungen 64
2.7.16 Dot-Blot / Western-Blot 65
2.8 Methoden 65
2.8.1 Herstellung von E. coli-Vorkulturen 65
2.8.2 Herstellung transformationskompetenter E. coli-Zellen 65
2.8.3 Transformation von E. coli 66
2.8.4 Transfektion von E. coli 66
2.8.5 Isolierung von Plasmid-DNS aus E. coli 67
2.8.6 Isolierung von M13 RF-Plasmid-DNS aus E. coli 67
2.8.7 Agarose-Gelelektrophorese 68
2.8.8 Restriktionsverdau von Plasmid-DNS und DNS-Fragmenten 68
2.8.9 Ligationsansätze 69
2.8.10 Hybridisierung von Oligonukleotiden zur Ligation 69
2.8.11 Ethanol-Fällung von Plasmid-DNS 70

Inhaltsverzeichnis
4
2.8.12 DNS-Sequenzierung 70
2.8.13 PCR-Reaktionen 71
2.8.14 Ortsgerichtete Mutagenese (QuikChange® Mutagenese) 72
2.8.15 Komplementationstest 73
2.8.16 Konstruktion spezifischer M13-Phagenstämme 75
2.8.17 Plasmidkonstrukte 76
2.8.18 „One-step growth“ Experiment 78
2.8.19 M13 Adsorptionsassay 79
2.8.20 Messung des Extrusionsverlaufs von M13-Phagen auf der Zellober-
fläche von E. coli 80
2.8.21 Herstellung von M13-Phagenstammlösungen 81
2.8.22 Phagentiterbestimmung 81
2.8.23 TCA-Fällung von Proteinen 82
2.8.24 Bestimmung der Bakterienzellzahl 83
2.8.25 Präparation transfizierter E. coli DH5α-Zellen für AFM und
TEM-Messungen 83
2.8.26 Präparation und Adsorption von E. coli-Zellen für AFM-Messungen 83
2.8.27 Präparation und Adsorption von E. coli-Zellen für TEM-Messungen 84
2.8.28 „Atomic Force Microscopy“ (AFM) 84
2.8.29 Transmissionselektronenmikroskopie (TEM) 85
2.8.30 Immunogold-Markierung von M13-Phagen 85
2.8.31 Immunogold-Markierung infizierter/uninfizierter E. coli DH5α-Zellen 86
2.8.32 Expression des gp1/11-Assembly-Komplexes 86
2.8.33 Proteinreinigung mittels Metallaffinitätschromatographie (IMAC) 87
2.8.34 Präparative Gelfiltration 88
2.8.35 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE) 90
2.8.36 Coomassiefärbung von SDS-Proteingelen 91
2.8.37 Western-Blot (semi-dry) und Membranbehandlung 91
2.8.38 Dot-Blot Analyse und Membranbehandlung 92
2.8.39 Pulse-labelling-Experimente, Immunpräzipitation und Protease-Mapping 93
2.8.40 In vivo Versuch zur YidC-abhängigen Membraninsertion von gp9T7 95
2.8.41 Quantifizierung radioaktiv markierter Proteine („Phosphorimaging“) 95
2.8.42 Präparation FPLC gereinigter gp1/11-Assembly-Komplexe für TEM-Auf-
nahmen 96

Inhaltsverzeichnis
5
3 Ergebnisse 97
3.1 Quantifizierung der M13-Phagenextrusion 98
3.1.1 Quantifizierung der wildtypischen M13-Phagenextrusion 98
3.1.2 Quantifizierung der M13mp19am8-Phagenextrusion während der
Komplementation durch das plasmidkodierte Haupthüllprotein gp8 101
3.1.3 M13-Adsorption an F-Pili in Abhängigkeit von der M.O.I. 104
3.2 Visualisierung M13-infizierter E. coli-Zellen 106
3.2.1 Immunogold-Markierung von M13-Phagen 107
3.2.2 TEM-Aufnahmen M13-sekretierenderE. coli DH5α-Zellen 108
3.2.3 AFM-Aufnahmen M13-sekretierender E. coli DH5α-Zellen 110
3.2.4 Veränderung der bakteriellen Zelloberfläche nach M13-Infektion 111
3.2.4.1 Extrusionsverlauf der M13-Phagen auf der Zelloberfläche von E. coli 111
3.3 Funktionelle Präsentation und Detektion von antigenen Epitopen am
N-Terminus des „Minor Coat“ Proteins gp9 116
3.3.1 Komplementationstest der gp9 Konstrukte 119
3.3.2 Expressionsnachweis der gp9 Mono-Tag Konstrukte in E. coli K38 121
3.3.3 Expressionsnachweis der gp9 Di-Tag Konstrukte in E. coli K38 122
3.3.4 Membraneinbau der Proteine gp9T7SD und gp9T7repSD in E. coli K38 124
3.3.5 Nachweis der exprimierten gp9 Konstrukte im M13-Phagen 126
3.3.5.1 Nachweis der exprimierten gp9-Konstrukte im M13-Phagen mit Hilfe
von Immun-Dot-Blots 126
3.3.5.2 Nachweis der exprimierten gp9 Doppel-Tag-Konstrukte im M13-Phagen
durch Markierung mit Protein-A konjugiertem Gold 129
3.3.6 Translokation des Konstrukts gp9T7SD in Abhängigkeit von der E. coli
Membraninsertase YidC 131
3.4 Massenbestimmung des gp1/11-Assembly-Komplexes und TEM-Auf-
nahmen der gereinigten Komplexe 135
3.4.1 Überexpression und Identifikation des M13 gp1/11-Assembly-Komplexes
in E. coli C41 135
3.4.2 Reinigung überexprimierter M13 gp1/11-Assembly-Komplexe aus E. coli
C41 mittels Metallaffinitätschromatographie (IMAC) 137
3.4.3 Massenbestimmung des gereinigten M13 gp1/11-Assembly-Komplexes
aus E. coli C41 durch präparative Gelfiltration 139

Inhaltsverzeichnis
6
3.4.4 Massenbestimmung des gereinigten M13 gp1/1-Assembly-Komplexes
aus infizierten E. coli K37 durch präparative Gelfiltration 142
3.4.5 Identifikation von Proteinbanden im IMAC-Eluat durch MALDI-TOF/MS 145
3.4.6 TEM-Aufnahmen FPLC gereinigter gp1/11-Assembly-Komplexe 147
4 Diskussion 149
4.1 Phagenextrusion aus infizierten E. coli-Zellen 149
4.2 M13-Sekretion aus infizierten E. coli-Zellen 155
4.2.1 TEM und AFM-Studien an infizierten E. coli DH5α-Zellen 155
4.2.2 Extrusionsverlauf der M13-Phagen auf der Zelloberfläche von E. coli 158
4.3 Funktionelle Präsentation antigener Epitope am N-Terminus des
„Minor Coat“ Proteins gp9 161
4.3.1 Die Translokation des Konstrukts gp9T7SD erfolgt durch die Membran-
insertase YidC 168
4.4 Massenbestimmung des gp1/11-Assembly-Komplexes 169
4.4.1 Identifikation der Proteine im IMAC-Eluat 173
4.4.2 Massenbestimmung des gereinigten gp1/11-Assembly-Komplexes aus
nicht infizierten E. coli-Zellen durch präparative Gelfiltration 175
4.4.3 Massenbestimmung des gereinigten gp1/11-Assembly-Komplexes aus
infizierten E. coli-Zellen durch präparative Gelfiltration 177
4.4.4 TEM-Aufnahmen FPLC gereinigter gp1/11-Assembly-Komplexe 179
5 Zusammenfassung 183
6 Literaturverzeichnis 187
7 Anhang 205
Abkürzungsverzeichnis 205
MALDI-TOF/MS Analysedaten 210
Spektrum 37 kDa gp1-Proteinbande 210
Spektrum 31 kDa gp1-Proteinbande 211
Spektrum 20 kDa gp1-Proteinbande 212
Sequenzen 213
gp1 DNA-Sequenz 213
gp1 AS-Sequenz 213
gp11 DNA-Sequenz 214
gp11 AS-Sequenz 214

Inhaltsverzeichnis
7
Publikationen 215
Danksagung 216

Einleitung
8
1 Einleitung
1.1 Der Bakteriophage M13
1.1.1 Klassifizierung des Bakteriophagen M13
Der Bakteriophage M13 wurde erstmals 1963 von Peter Hans Hofschneider be-
schrieben (Hofschneider, 1963). Wegen der einzelsträngigen DNA wird M13 taxo-
nomisch der Gruppe II, den Einzelstrang-DNA-Phagen (ssDNA-Phagen; ss, „single
strand“), zugeordnet. Charakteristisch für diese Gruppe ist die genomische Repli-
kation über ein doppelsträngiges DNA-Intermediat, die sogenannte replikative Form
(RF). Generell können Phagen morphologisch in sphärische und filamentöse Phagen
unterteilt werden, die sich sowohl in Form als auch im Grad ihrer verschachtelten
Gene voneinander unterscheiden (Klaus et al., 1992).
Innerhalb der einzelsträngigen DNA-Phagen wird der filamentöse Phage M13, wie
auch die Phagen fd und f1, zur Familie der Inoviridae, der sogenannten Ff-Gruppe
(„F“ für F-Pili, „f“ für filamentös) zugeordnet.
1.1.2 Morphologie und Genom der Inoviridae
Die Phagen dieser Familie zeichnen sich durch eine einzelsträngig zirkuläre DNA
aus, welche ausschließlich durch die helikale Anordnung von Hüllproteinen vor
äußeren Einflüssen geschützt wird. Hier ergibt sich aus der Kombination von
Anordnung der Hüllproteine und DNA das filamentöse Erscheinungsbild dieser
Phagen, deren Charakteristika flexible Virionen mit einem Durchmesser von ~ 7 nm
und einer Länge von ~ 900 nm sind (Abb. 1-1).
Das M13-Genom besteht aus ~ 6400 Nukleotiden und kodiert 11 verschiedene
Genprodukte (gp), die sowohl in der Kopienzahl als auch im Molekulargewicht stark
variieren. Das Genom enthält neben einer nicht-kodierenden intergenischen Region
(IR, „intergenic region“), welche den Replikationsursprung (ORI, „origin of
replication“) für die (+) und (– )-Strang-Synthese beinhaltet, eine morphogenetisch
cis-aktive Haarnadelschlaufe („hairpin packaging signal“), welche für die Assem-
blierung des Phagen essenziell ist und der DNA innerhalb der Proteinhülle eine
Orientierung verleiht (Dotto & Zinder, 1983, Russel & Model, 1989). Innerhalb der
Gene 1 und 2 befindet sich jeweils eine translationale Initiierungsstelle, durch welche
ein weiteres „in-frame“ Phagenprotein synthetisiert wird. In beiden Fällen sind die
„in-frame“ Produkte mit dem C-terminalen Drittel des jeweils vollständigen Gen-

Einleitung
9
produkts identisch. Von den 11 phagenkodierten Proteinen bilden fünf (gp3, gp6,
gp8, gp7, gp9) die Phagenhülle um das zentral positionierte DNA-Molekül, drei (gp2,
gp5, gp10) werden für die Synthese der ssDNA und weitere drei (gp1, gp11, gp4) für
die Assemblierung des Phagen benötigt (Tabelle 1-1). Die Synthese aller Phagen-
proteine erfolgt simultan, jedoch findet hier auf der Ebene der Promotoren sowie
-abhängiger Terminationssignale eine Regulierung der jeweiligen Transkriptions-
und Translationsraten statt, um ein stöchiometrisches Gleichgewicht zwischen den
einzelnen Proteinen zu gewährleisten.
Gen Genprodukt Funktion Lokalisierung Menge Größe(AS) MG(kDa)
2 (10) gp2
Repli-
kation
Nickase(4)
Zytoplasma 1000(1)
410 46,1
gp10 Zytoplasma 500(1)
111 12,7
5 gp5 ssDNAbp Zytoplasma 100000(1)
87 9,7
3 gp3
Hüllprotein
Virion Basis 5(2)
406 42,5
6 gp6 Virion Basis 5(2)
112 12,3
7 gp7 Virion Spitze 5(2)
33 3,6
8 gp8 Filament 2700(2)
50 5,2
9 gp9 Virion Spitze 5(2)
32 3,63
1 (11) gp1
Assem-
blierung
Assembly-
Komplex iM
(3)
348 38
gp11 108 12,4
4 gp4 Pore äM 405 43,5
Abb. 1-1: AFM (Atomic Force Microscope) Auf-nahme von M13wt-Phagen. Die Länge des Mess-balkens beträgt 500 nm.
Tabelle 1-1: Übersicht über die Gene und Genprodukte F-spezifischer filamentöser Phagen sowie deren Eigenschaften. Die Gene wurden hinsichtlich der Funktion ihrer Produkte geordnet. AS = Aminosäuren, ssDNAbp = Einzelstrang-DNA bindendes Protein, iM = innere Membran, äM = äußere Membran,
(1)Anzahl pro Zelle,
(2)Anzahl pro Phage,
(3)Anzahl unbekannt,
(4)Nickase: Endonuklease mit
Topoisomeraseaktivität (Daten: Rasched & Oberer, 1986, Marvin, 1998 und Russel, 2004)

Einleitung
10
1.1.3 Gene, Genprodukte und ihre Funktionen
Gene, durch welche die Replikation des Phagen gesteuert wird
Gen 2 kodiert sowohl gp2, eine spezifische Nickase (Endonuklease mit
Topoisomeraseaktivität), welche im „rolling circle“ Replikationsmechanismus
für die Bildung eines Einzelstrangbruchs in der replikativen Form der viralen
DNA verantwortlich ist, als auch gp10, welches im Leseraster von Gen 2
liegt, wodurch 111 Aminosäuren des gp10 mit dem des C-terminalen Endes
von gp2 identisch sind (Yen & Webster, 1981, Fulford & Model 1984). In
Abwesenheit von gp10 ist die DNA-Synthese stark beeinträchtigt, wodurch
diesem „in-frame“ Produkt eine essenzielle Rolle bei der DNA-Synthese des
Phagen zufällt (Fulford & Model 1984).
Gen 5 kodiert das DNA-Bindeprotein gp5, welches in β-Konformation als
Dimer an die einzelsträngige Phagen-DNA bindet. In vitro kristallisierte
Komplexe zeigen, dass pro Helix-Windung 6 dieser Dimere (McPherson et
al., 1980) gebunden werden. Durch die Assoziation von gp5 mit der einzel-
strängigen, replizierten Phagen-DNA, welche durch elektrostatische und
Abb. 1-2: Schematische Darstellung der Phagengene und deren Genprodukte. Gen 2 kodiert
zwei Genprodukte, gp2 und gp10. Gp2 besitzt eine Endonuklease- sowie Topoisomeraseaktivität und bindet an die intergenische Region der RF-DNA. Hier erzeugt es einen Strangbruch im (+)-Strang, wodurch die Replikation der Phagen-DNA durch die wirtseigenen Proteine initiiert wird. Gp10 bewirkt im späteren Verlauf der Infektion eine Akkumulation der ssDNA. Gen 5 kodiert das ssDNA-Binde-protein, welches zusammen mit der replizierten ssDNA ein flexibles Filament ausbildet. Die Gene 7 und 9 kodieren kleine „Minor Coat“ Proteine, die an dem Ende des Phagen lokalisiert sind, welches zu Beginn der Assemblierung die Wirtszelle als Erstes verlässt. Gen 8 kodiert das Haupthüllprotein (MCP, „Major Coat“ Protein), das mit ~ 2800 Kopien am Phagen präsent ist. Die Gene 3 und 6 kodieren weitere „Minor Coat“ Proteine, die an der Basis des Phagen lokalisiert sind, durch welche die Assemblierung terminiert und zu Beginn der Infektion die Adsorption des Phagen an den F-Pilus initiiert wird. Gen 1 kodiert gp1 und gp11, die für die Assemblierung des Phagen essenziell sind und in der inneren Membran (iM) einen Assembly-Komplex ausbilden. Gen 4 kodiert gp4, welches in der äußeren Membran (äM) eine homomultimere Pore ausbildet, über die der Phage das Wirtsbakterium verlässt. Die IR-Region („intergenic region“) ist ein nicht-kodierender Bereich, der den Replikations-ursprung (ORI, „origin of replication“) für die (+) und (-)-Strang-Synthese sowie eine morphogenetisch cis-aktive Haarnadelschlaufe („hairpin packaging signal“) beinhaltet.

Einleitung
11
hydrophobe Wechselwirkungen zustande kommt, wird die Synthese des
komplementären (-)-Stranges unterbunden und ein DNA-gp5-Komplex
gebildet, der einer linksdrehenden Helix entspricht (Stassen et al., 1995,
Konings et al., 1995, Guan et al., 1995, Olah et al., 1995). Zusätzlich kommt
dem gp5 eine regulatorische Funktion während des Replikationsprozesses
zu. Hohe Konzentrationen hemmen die Konvertierung des (+)-Stranges in
die RF-Form, wodurch der Übergang von der Replikations- zur Assembly-
phase neuer Phagen-Nachkommen eingeleitet wird. Zudem reguliert gp5
durch die Expressionskontrolle von gp2 und gp10 die Replikationsphase
(Salstrom & Pratt, 1971, Model et al., 1982).
Gene für die Synthese der Hüllproteine
Die Gene 7 und 9 kodieren die „Minor Coat“ Proteine gp7 bzw. gp9, die mit
jeweils ~ 5 Kopien eine Kappenstruktur an dem Phagenende ausbilden,
welches zu Beginn der Assemblierung des Phagen zuerst das Wirts-
bakterium über den Sekretionskanal verlässt. Beide Hüllproteine sind für die
Initiation der Assemblierung essenziell, da sie mit dem „hairpin packaging
signal“ der DNA interagieren (Russel & Model, 1989, Endemann & Model,
1995). Gp7 und gp9 gehören mit 33 bzw. 32 Aminosäuren zu den kleinsten
von Ribosomen translatierten Proteinen und werden ohne N-terminale
Signalsequenz synthetisiert.
Gen 8 kodiert das Haupthüllprotein (MCP, „Major Coat“ Protein) gp8 des
Phagen. Mit ~ 2800 Kopien bildet es eine schuppenförmig helikale Umman-
telung über die gesamte Phagenlänge. Gp8 interagiert mit seiner apolaren
Domäne über hydrophobe Wechselwirkungen mit anderen gp8 sowie „Minor
Coat“ Proteinen, wodurch die Phagenhülle stabilisiert wird (Rasched &
Oberer, 1986, Marvin, 1989). Die Synthese des Haupthüllproteins erfolgt als
73 Aminosäuren langes Präprotein („procoat“) mit einer N-terminal abspalt-
baren, 23 Aminosäuren langen, Leadersequenz. Diese wird nach der Trans-
lokation des Proteins durch die Translokase YidC in der inneren Membran
von Escherichia coli durch die Leaderpeptidase abgespalten und somit zum
maturen MCP umgewandelt (Kuhn et al., 1986, Kuhn & Troschel, 1992,
Samuelson et al., 2001).

Einleitung
12
Gen 3 und 6 kodieren die „Minor Coat“ Proteine gp3 bzw. gp6, die mit
jeweils ~ 5 Kopien eine Kappenstruktur an der Phagenbasis ausbilden,
welche die Assemblierung des Phagen terminiert und anschließend zu
Beginn der Infektion die Anheftung des Phagen an den bakteriellen F-Pilus
einleitet. Im Gegensatz zu gp6 wird gp3 mit einer 18 Aminosäuren langen
Signalsequenz synthetisiert, durch welche die Translokation über das
SecYEG-Translokasesystem erfolgt (Rapoza & Webster, 1993, Thie et al.,
2008). In Abwesenheit von gp3 erfolgt eine proteolytische Prozessierung des
gp6, was ein Indiz dafür ist, dass gp3 und gp6 vor ihrer Verwendung am
Phagenpartikel in der inneren Membran assemblieren. Dies wird dadurch
unterstützt, dass beide Proteine nur als Komplex isoliert werden können
(Gailus & Rasched, 1994, Rakonjak et al., 1999).
Für die Assemblierung und die Sekretion neuer Phagenpartikel essenzielle
Gene
Gen 1 kodiert sowohl das integrale Membranprotein gp1 als auch das
integrale Membranprotein gp11, welches im Leseraster von Gen 1 liegt,
wodurch 108 Aminosäuren des gp11 mit dem C-terminalen Ende von gp1
identisch sind. Gp1 wird ohne Signalsequenz synthetisiert, jedoch weist die
Insertion des Proteins eine Abhängigkeit vom Sec-Translokasesystem auf
(Rapoza & Webster, 1993). Gp1 interagiert mit dem Thioredoxin des Wirts,
welches in E. coli von dem Gen trxA kodiert wird (Russel & Model, 1983, Lim
et al., 1985). Zudem besitzt gp1 ein Nukleotid-Bindemotiv in der zytoplas-
matischen Domäne, was nahelegt, dass die Assemblierung des Phagen
unter ATP-Umsetzung stattfindet (Russel, 1991, 1993). Gp1 und gp11 sind
essenziell für die Assemblierung neuer Phagen-Nachkommen (Silver &
Wickner, 1980, Rapoza & Webster, 1995, Marvin 1998), jedoch führt eine
Überproduktion von gp1 zu einem raschen Tod der Wirtszelle (Horabin &
Webster, 1986). Gp1 und gp11 bilden in der inneren Membran der Wirtszelle
einen Assembly-Komplex aus, welcher mit der aus gp4-Monomeren
bestehenden Pore in der äußeren Membran zu einem 2-Komponenten-
Sekretionskomplex interagiert (Rapoza & Webster, 1995, Haigh & Webster,
1999).

Einleitung
13
Gen 4 kodiert das integrale Membranprotein gp4. Dies wird mit einer 21
Aminosäuren langen Signalsequenz synthetisiert und durch das SecYEG-
Translokasesystem in das Periplasma transloziert, wo es sich anschließend
in die äußere Membran des Wirts integriert und eine Sekretionspore aus
~ 12 - 14 Homomultimeren bildet (Brissette & Russel, 1990, Russel &
Kazmierczak, 1993, Rapoza & Webster, 1993, Kazmierczak et al., 1994).
Gp4, gp1 und gp11 interagieren zu einem Sekretionskomplex, der für das
Ausschleusen neugebildeter Phagen essenziell ist (Rapoza & Webster,
1995, Haigh & Webster, 1999).
1.1.4 Struktur des Bakteriophagen M13
Die Hülle des Phagen besteht anteilsmäßig im Wesentlichen aus dem Haupthüll-
protein gp8, welches mit ~ 2800 Kopien eine schuppenförmig helikale Ummantelung
über die gesamte Phagenlänge ausbildet (Abb. 1-3). Charakteristisch für gp8 ist eine
apolare Domäne, welche aus 19 Aminosäuren besteht und über deren hydrophoben
Wechselwirkungen mit anderen gp8-Molekülen sowie den intrinsischen Interaktionen
mit apolaren Domänen der „Minor Coat“ Proteine die Phagenhülle stabilisiert wird
(Marvin, 1989). Neben gp8 sind noch vier weitere, sogenannte „Minor Coat“ Proteine,
die mit jeweils ~ 5 Kopien pro Partikel vorhanden sind, an der Zusammensetzung
des Phagen beteiligt (Glucksman et al. 1992, Stopar & Spruijt 2003). Hier bilden die
„Minor Coat“ Proteine gp7 und gp9 eine Kappenstruktur an der Phagenspitze, welche
zu Beginn der Phagenassemblierung zuerst das Wirtsbakterium über den Sekretions-
kanal (ein gp4-Oligomer) verlässt. Gp6 und gp3 hingegen bilden abschließend eine
Kappenstruktur am entgegengesetzten Ende des Phagen, wodurch die Phagen-
assemblierung beendet wird und der Phage eine Orientierung erhält (Simons et al.,
1980, 1981, Grant et al., 1981, Endemann & Model, 1995). Die Adsorption des
Phagen an den F-Pilus der E. coli-Zelle erfolgt durch das nach außen hin frei
zugängliche „Minor Coat“ Protein gp3, wodurch die Infektion eingeleitet wird. Die
Unterteilung des gp3 erfolgt in 3 Segmente (N1, N2 und C), die jeweils über 2 flexible
Glycin-reiche Linker miteinander verbunden sind und denen während des Infektions-
vorgangs spezifische Funktionen zugeordnet werden können (Abb. 1-4). Durch das
Segment N2 des gp3 wird die Adsorption des Phagen an den F-Pilus initiiert,
während N1 im Verlauf der Infektion an den Korezeptor TolA in der Wirtsmembran

Einleitung
14
bindet. Über das Segment C ist das gp3 in die Proteinhülle des Phagen eingebettet
(Stengele et al., 1990, Click & Webster, 1997, Rakonjac et al., 1999).
1.1.5 Der Zyklus des Bakteriophagen M13
Alle, bis zum derzeitigen Stand, charakterisierten Phagen der Ff-Gruppe verwenden
lange, dünne Strukturen auf der Oberfläche des Wirtsbakteriums, sogenannte „Pili“,
als Rezeptoren für die Initiation der Wirtsinfektion. Von den filamentösen Phagen
M13, fd und f1 wird der F-Pilus (F+, F´: „fertility“), von Ike dagegen die N- und P-Pili
als spezifische Rezeptoren präferiert (Webster, 1996). Charakteristisch für alle Infek-
tionsverläufe aus dieser Gruppe ist, dass die Wirtsorganismen im Verlauf der
Phagenvermehrung nicht durch Lyse geschädigt werden. Vielmehr sind die infizierten
Wirte weiterhin befähigt zu wachsen und sich zu teilen, obwohl ein signifikanter Effekt
bei der Zunahme der Generationszeit zu beobachten ist (Hoffmann-Berling & Mazé,
1964, Ploss & Kuhn, 2010). Durch die veränderte Generationszeit sind auch
Abb. 1-3: Schematisierte Darstellung des M13-Phagen. Die Hülle des Phagen wird hauptsächlich durch das Hüllprotein gp8 gebildet, das die DNA über die gesamte Phagenlänge schuppenförmig umgibt. Gp3 und gp6 sind an dem Ende des Phagen lokalisiert, welches zu Beginn des Infektionsprozesses an den F-Pilus der Wirtszelle adsorbiert; gp7 und gp9 befinden sich am opposi-tionären Ende (Ref. Ploss & Kuhn, 2010)
Abb. 1-4: Schematisierte Darstellung der Segmentstruktur des „Minor Coat“ Proteins gp3. Das gp3 wird in 3 Segmente unterteilt, N1, N2 und C. Die einzelnen Segmente sind über die flexiblen, Glycin-reichen Linker L1 und L2 verbunden. Durch das Segment N2 wird die Adsorption des Phagen an den F-Pilus initiiert, während N1 im Verlauf der Infektion an den Korezeptor TolA in der Wirts-membran bindet. Über das Segment C ist das gp3 in die Proteinhülle des Phagen eingebettet. AS = Aminosäure (Daten: Rakonjak et al., 1999, verändert nach Russel, 2004).

Einleitung
15
filamentöse Phagen befähigt auf Festmedien „Plaques“ auszubilden, ohne lytisch zu
sein. Hier handelt es sich um lokale Bereiche, welche sich durch das verlangsamte
Bakterienwachstum und der hieraus resultierenden geringeren Zelldichte von dem
restlichen Bakterienrasen hervorheben.
Die infizierten Zellen sekretieren bereits 10 Minuten nach erfolgreicher Infektion neue
Phagen-Nachkommen in das umgebende Medium. Die Sekretion kommt erst dann
zum Erliegen, wenn die Wirtszelle in die stationäre Phase übergeht (Klaus et al.,
1992). Allgemein kann der Zyklus des Bakteriophagen M13 in die drei Phasen
Infektion, Reproduktion und Assembly gegliedert werden. Während der Infektions-
phase erfolgt der Kontakt zwischen Phage und Wirt sowie die Einschleusung der
genetischen Information des Phagen in die Wirtszelle.
Abb. 1-5: Modell des M13-Zyklus. (I.) Die Infektion des Wirtsbakteriums wird durch die Adsorption von M13 an die Spitze des F-Pilus eingeleitet. Durch den Abbau der Pilus-Segmente an der Pilus-Basis wird der gebundene Phage in Kontakt mit der Zelloberfläche gebracht. Über die Interaktion der Phagenhüllproteine gp3 mit dem zellulären TolQRA gelangen die Hüllproteine in die innere Membran und die Phagen-DNA in das Zytoplasma. (II.) Im Verlauf der Replikation wird aus dem einzel-strängigen (+)-Strang des Phagen durch wirtseigene Enzyme eine replikative Form (RF-Form) herge-stellt, die einerseits durch phagenkodierte Proteine (gp2, gp10) vervielfältigt wird und andererseits zur Bildung neuer (+)-Einzelstränge dient. (III.) Während der Proteinsynthese werden, ausgehend von 9 Genen, 11 verschiedene Genprodukte (gp) synthetisiert, wobei hier die RF-Form als Matrize dient. Gp2, gp10 und gp5 verbleiben als lösliche Proteine im Zytoplasma. Bei den 8 verbleibenden Produkten handelt es sich um integrale Membranproteine, von denen nur gp4 in die äußere Membran inseriert, während die restlichen in die innere Membran eingebaut werden. (IV.) Zusammen mit gp5-Dimeren bildet die einzelsträngige DNA ((+)-Strang) einen Komplex. (V.) Während des Assembly werden die synthetisierten Strukturuntereinheiten mit der replizierten Phagen-DNA an der inneren Membran der Wirtszelle zu neuen Phagen-Nachkommen zusammengebaut und verlassen diese, ohne sie zu lysieren (nach Webster, 1996, abgeändert).

Einleitung
16
Die Reproduktionsphase ist geprägt durch die Vermehrung der Phagen-DNA sowie
durch die Synthese der phagenkodierten Proteine. In der Assemblyphase erfolgt
schließlich der Zusammenbau der synthetisierten Strukturuntereinheiten mit der
replizierten Phagen-DNA zu neuen Phagen-Nachkommen sowie deren Aus-
schleusung aus der Wirtszelle (Abb. 1-5).
1. Infektionsphase
Der Bakteriophage M13 infiziert Gram-negative Escherichia coli-Zellen, die als
Rezeptoren F-Pili ausbilden. Die Infektion erfolgt hier in einem zweistufigen Prozess
(Model & Russel, 1988). Während der ersten Stufe adsorbiert der Phage mit Hilfe
des N2-Segments von gp3 (Abb. 1-4) an der Spitze des F-Pilus. Durch eine Depoly-
merisierung der Pilinuntereinheiten an der Pilus-Basis in der inneren Membran
verkürzt sich der Pilus, wodurch der gebundene Phage in Kontakt mit der Zellober-
fläche gebracht wird (Frost, 1993). Daraufhin bindet das N-terminale Segment N1
des gp3 an die Untereinheit TolAIII seines Korezeptors TolA. Die Proteine TolQ, TolR
und TolA sind in der inneren Membran von E. coli lokalisiert und für die Translokation
von Gruppe A Colicinen verantwortlich (Webster, 1991). TolQ ist hier durch drei, TolR
und TolA jeweils durch eine Transmembrandomäne in der inneren Membran
verankert (Kampfenkel & Braun, 1993, Vianney et al., 1994). Über die intermole-
kularen Wechselwirkungen der drei Proteine entsteht ein Komplex, durch den die
innere Membran mit der äußeren in Kontakt steht, wodurch unter anderem die
Stabilität der äußeren Membran bedingt wird. TolA besteht aus 3 Domänen: über die
N-terminale Domäne AI ist das Protein in der inneren Membran verankert und
interagiert hierdurch mit den Proteinen TolQ und TolR. Die Domäne AII weist eine
α-helikale Struktur auf, welche das Periplasma durchspannt und die C-terminale
Domäne AIII so positioniert, dass diese mit Komponenten der äußeren Membran
interagieren kann (Abb. 1-6). AIII dient sowohl den Colicinen als auch dem Segment
N1 des gp3 als Rezeptor (Webster, 1991, Raggett et al., 1998, Karlsson et al., 2003).
In der zweiten Stufe erfolgt nach einer Konformationsänderung der α-helikalen
Struktur des TolAII-Proteins die Integration der Phagenhüllproteine in die innere
Membran, wodurch die Translokation der Phagen-DNA in die Wirtszelle zustande
kommt (Abb. 1-6) (Marvin, 1998, Karlsson et al., 2003).

Einleitung
17
2. Reproduktionsphase
Direkt nach der Translokation der einzelsträngigen Phagen-DNA (das (+)-Strang
Genom hat die Polarität einer mRNA) in das Zytoplasma bindet die wirtseigene RNA-
Polymerase am origin in der nicht-kodierenden intergenischen Region (IR) und
synthetisiert einen kurzen Primer, über dessen 3´-Ende durch die wirtseigene DNA-
Polymerase III, sowie Topoisomerase, der Einzelstrang in eine doppelsträngige Form
konvertiert wird, die sogenannte „replikative Form“ (RF) (Horiuchi, 1997). Die RF-
Form dient sowohl bei der Proteinbiosynthese der viralen Proteine als auch bei der
Synthese weiterer (+)-Stränge als Matrize. Die Replikation der RF-Form erfolgt durch
den Mechanismus des „rolling circle“. Dabei spielt die, durch das M13-Phagengenom
kodierte, Endonuklease (gp2) eine wichtige Rolle, da sie durch das Erzeugen eines
Einzelstrangbruches im (+)-Strang der RF-DNA den Replikationsprozess einleitet
und durch die Ligation der offenen Enden des (+)-Stranges nach Vollendung der
Abb. 1-6: Modell des M13-Infektionsvorgangs an einer E. coli-Zelle. (1) Die Infektion des Wirts-bakteriums wird durch die Adsorption des N2-Segments des gp3-Hüllproteins an die Spitze des F-Pilus eingeleitet. (2) Durch den Abbau der Pilinuntereinheiten an der Pilus-Basis wird der gebundene Phage in Kontakt mit der Zelloberfläche und dem Korezeptor TolA gebracht. (3) Über die Interaktion des N1-Segments mit der TolAIII-Domäne und dessen Konformationsänderung werden die Hüllpro-teine schließlich in die innere Membran integriert und die Phagen-DNA in das Zytoplasma transloziert (nach Karlsson et al., 2003)

Einleitung
18
Replikation diese rezirkularisiert. Der DNA-Replikationsprozess erfolgt parallel zur
viralen Proteinbiosynthese. Zu Beginn der frühen Replikationsphase ist die Menge
des löslichen Einzelstrang-Bindeproteins gp5 sehr gering, was dazu führt, dass die
während der Replikation neu synthetisierten (+)-Stränge sofort in die RF-Form
konvertiert werden. Während der frühen Phase steigen jedoch die Mengen an DNA
und Phagenproteinen exponentiell an. Dabei werden die Proteine gp3, gp6, gp8,
gp7, gp9, welche die Phagenhülle bilden, sowie gp1 und gp11, die für die Assem-
blierung neuer Phagenpartikel benötigt werden, in die innere Membran des Wirts
transloziert. Der Bestimmungsort für das phagenkodierte gp4 ist die äußere
Membran, in der ~ 12 - 14 gp4-Monomere eine Sekretionspore ausbilden, durch die
neue Phagen in die äußere Umgebung sekretiert werden (Russel et al., 1997).
Aufgrund erhöhter gp5-Konzentrationen kommt es in der späteren Reproduktions-
phase zu Interaktionen von gp5-Dimeren mit den synthetisierten (+)-Strängen,
wodurch die Konvertierung in die RF-Form unterdrückt wird und vermehrt ssDNA-
gp5-Komplexe gebildet werden, welche das Substrat der Assemblyphase darstellen
(Webster & Cashman, 1973). Zu diesem Zeitpunkt erfolgt gleichzeitig eine Expres-
sionskontrolle von gp2 und gp10 durch das virale gp5, indem es deren Translation
reprimiert (Yen & Webster, 1982).
3. Assemblyphase
Der Assembly-Vorgang filamentöser Phagen ist ein sekretorischer Prozess, welcher
an speziellen, membranassoziierten Bereichen stattfindet, wo innere und äußere
Membran in engem Kontakt miteinander stehen (Lopez & Webster, 1985). Diese
Assembly-Stellen werden, unabhängig von den anderen Phagenproteinen, durch die
Interaktion der periplasmatischen Domänen der drei morphogenen Proteine gp1,
gp11 und gp4 gebildet. Der Vorgang der Phagenassemblierung wird in 5 Abschnitte
eingeteilt: Preinitiation, Initiation, Elongation, Pretermination und Termination (Russel
et al., 2004).
Preinitiation: Stellt mit der Ausbildung von Assembly-Stellen, die sich aus den
morphogenen Proteinen gp1, gp11 und gp4 zusammensetzen und mit Hilfe des
Elektronenmikroskops auf den Wirtsbakterien nachgewiesen werden können, den
ersten Abschnitt in der Phagenmorphogenese dar (Lopez & Webster, 1985, Feng et
al., 1999).

Einleitung
19
Initiation: Wenn sowohl die Assembly-Stelle mit den gebundenen membran-
assoziierten „Minor Coat“ Proteinen gp7 und gp9 als auch der ssDNA-gp5-Komplex
in der Zelle vorliegen, wird durch die Bindung des Nukleoproteinkomplexes an den
Preinitiationskomplex die Assemblierung des Phagen initiiert. Hier erfolgt eine Inter-
aktion zwischen der zytoplasmatischen Domäne von gp1 mit dem „packaging signal“
des ssDNA-gp5-Komplexes, welches aus diesem herausragt (Russel & Model,
1989). Durch diesen Prozess wird bereits die Spitze des Phagen gebildet. Obwohl
die Initiation keinerlei Redoxpartner bedarf, ist das zytoplasmatische Thioredoxin
Bestandteil des Initiationskomplexes und vermutlich an der Prozessivität der Elonga-
tionsreaktionen beteiligt.
Elongation: Nachdem durch die Initiation der Assemblierung die Phagenspitze
gebildet wurde, erfolgt nun der sukzessive Austausch der gp5-Dimere am ssDNA-
gp5-Komplex gegen das Haupthüllprotein gp8. Dieser Austausch wird durch die
C-terminale Domäne des gp1 katalysiert und erfolgt vermutlich unter ATP-Hydrolyse
(Russel, 1991). Hierdurch wird die ssDNA mit einer Phagenhülle aus ~ 2800 gp8-
Molekülen umgeben, wobei die umhüllte DNA während des Elongationsprozesses
über die homomultimere gp4-Sekretionspore in die äußere Umgebung transloziert
wird (Abb. 1-7). Während der Elongation ändern die gp8-Moleküle aufgrund positiv
geladener Aminosäuren an ihrer C-terminalen Domäne im gp1/11-Assembly-
Komplex ihre Position dahingehend, dass eine Interaktion mit dem negativ gela-
denen Phosphatrückgrat der ssDNA erfolgen kann (Rapoza & Webster, 1995).
Dieser Vorgang ist persistent bis das Ende der viralen DNA erreicht wird.
Pretermination und Termination: Die Pretermination der Assemblierung wird durch
den Einbau des „Minor Coat“ Proteinkomplexes gp3/gp6 an das Ende der viralen
Einzelstrang-DNA eingeleitet. Für die Termination der Assemblierung und die Ent-
lassung des fertigen Phagen in das umgebende Medium werden 93 Aminosäuren
des C-terminalen Segments von gp3 benötigt, wobei erst ab einer Länge von 132
Aminosäuren der C-terminalen Domäne stabile Phagenpartikel entstehen (Rakonjak
et al., 1999).

Einleitung
20
Abb. 1-7: Schematisierte Darstellung der M13-Phagenassemblierung während der Elongation. Die Assemblierung des Phagen erfolgt an einer speziellen Assembly-Stelle in der inneren Membran der Wirtszelle, welche durch die Interaktionen der periplasmatischen Domänen des gp1/11-Assembly-Komplexes und der homomultimeren gp4-Pore zustande kommt. Während der Elongation werden vermutlich unter ATP-Hydrolyse die an die ssDNA gebundenen gp5-Dimere abgelöst und durch das Haupthüllprotein gp8, das innerhalb der inneren Membran mit weiteren gp8-Molekülen interagiert, ersetzt. Aufgrund positiv (+) geladener Aminosäuren an der C-terminalen Domäne der gp8-Moleküle werden vermutlich ihre Positionen im gp1/11-Assembly-Komplex dahingehend verändert, dass eine Interaktion mit dem negativ geladenen Phosphatrückgrat der ssDNA erfolgen kann. Die umhüllte DNA wird während des Elongationsprozesses über die homomultimere gp4-Sekretionspore in die äußere Umgebung sekretiert (angepasst nach Webster, 1996).

Einleitung
21
1.2 Der gp1/11-Assembly-Komplex
Die Phagenassemblierung stellt einen sekretorischen Prozess an der inneren E. coli-
Membran dar. Dabei entsteht der Phage direkt durch die Umhüllung der einzel-
strängigen Phagen-DNA mit spezifischen Phagenhüllproteinen, welche nach ihrer
Synthese in die innere Membran transloziert und über den, für die Assemblierung
essenziellen, gp1/11-Assembly-Komplex herangeführt werden (Silver & Wickner,
1980, Rapoza & Webster, 1995, Marvin 1998).
Die Synthese von gp1 und gp11 erfolgt über eine polycistronische mRNA ohne
N-terminale Signalsequenz, jedoch scheint die Translokation des 348 Aminosäuren
langen gp1 vom SecYEG-System des Wirts abhängig zu sein (LaFarina & Model,
1983, Horabin & Webster, 1986, Rapoza & Webster, 1993). In infizierten Zellen sind
gp1 und gp11 mit einer Ratio von 1:1 bis 1:2 präsent. Die Proteine werden jeweils
über eine 20 Aminosäuren lange hydrophobe Domäne in der inneren Membran
verankert und besitzen eine 75 Aminosäuren lange C-terminale Domäne, welche im
Periplasma lokalisiert ist. Die 13 Aminosäuren lange N-terminale Domäne des gp11
bzw. die 253 Aminosäuren lange N-terminale Domäne des gp1 sind zytoplasmatisch
lokalisiert, wobei die N-terminale Domäne von gp1 sowohl im Bereich der Amino-
säure 142 mit dem Genprodukt des bakteriellen trxA interagiert (Russel & Model,
1983) als auch im Bereich der Aminosäuren 8-15 das Walker Nukleotid-Bindemotiv
GXXGXGKT aufweist (Abb. 1-8) (Walker et al., 1982). Die Rolle dieses Motivs im
Assembly-Komplex ist nicht ganz geklärt, jedoch handelt es sich hier um ein beson-
deres Merkmal, welches in vielen pro- und eukaryotischen Transportsystemen vor-
handen ist und nahelegt, dass die Translokation des Phagen während der Assem-
blierung vermutlich unter ATP-Hydrolyse stattfindet (Russel, 1991, 1993) und von der
protonenmotorischen Kraft („proton motiv force“) abhängig ist (Feng et al., 1997).
Über positiv geladene Aminosäuren der N-terminalen Domänen von gp1 und gp11
erfolgt die Interaktion zwischen dem Assembly-Komplex und der einzelsträngigen
Phagen-DNA (Abb. 1-7). Während der Elongation bedingen diese Ladungen wahr-
scheinlich eine Positionsänderung der gp8-Moleküle, sodass diese an das negativ
geladene Phosphatrückgrat der ssDNA binden können (Rapoza & Webster, 1995).
Jedoch sind Einzelheiten über die weiteren Vorgänge am Assembly-Komplex weit-
gehend unbekannt.

Einleitung
22
Die Aufgabe des gp11 im Assembly-Komplex ist immer noch ungeklärt, doch zeigt
sich bei Anwesenheit dieses Proteins eine Verringerung der proteolytischen
Prozessierung des gp1, was ein Indiz dafür ist, dass gp11 sowohl für die Stabilität
des Komplexes als auch für den Schutz des gp1 vor periplasmatischen Proteasen
verantwortlich ist (Feng et al., 1999).
Der 2-Komponenten-Sekretionskomplex
Gp1 und gp11 bilden vermutlich nach Spekulation von Russel (Russel et al., 2004) in
der inneren Membran des Wirts einen multimeren Komplex aus jeweils 5 - 6 der
einzelnen Moleküle, welcher ebenfalls eine Pore darstellt, da in Abwesenheit aller
weiteren Phagenproteine eine Überproduktion an gp1 und gp11 einen Zusammen-
bruch des Membranpotentials verursacht (Horabin & Webster, 1988). Die periplas-
matisch lokalisierte C-terminale Domäne der gp1-Moleküle des Assembly-Komplexes
interagieren ab den Aminosäuren an Position 275 - 320 mit dem periplasmatischen
N-terminalen Bereich der homomultimeren gp4-Pore in der äußeren Membran zu
einem Sekretionskomplex (Abb. 1-9) (Rapoza & Webster, 1995, Feng et al., 1999,
Haigh & Webster, 1999). Dieser Komplex wird unabhängig von anderen Phagen-
proteinen ausgebildet. Die homomultimere gp4-Sekretionspore setzt sich dabei aus
Abb. 1-8: Schematisierter Aufbau der gp1- und gp11-Moleküle. Die Einteilung der gp1- und gp11-Moleküle erfolgt in drei Bereiche: (1) Die zytoplasmatisch lokalisierte N-terminale Domäne mit einem positiv geladenen amphiphilen Bereich, der mit der Phagen-DNA und den Hüllproteinen interagiert. (2) Eine hydrophobe Domäne, über die gp1 und gp11 in der inneren Membran des Wirts verankert sind und (3) die periplasmatisch lokalisierte C-terminale Domäne (nach Daten von Guy-Caffey et al., 1991 und Webster, 1996)
gp11 gp1
AS 8-15: ATP-Bindemotiv
AS 142: Interaktion mit Thioredoxin
(1) N-terminale Domäne, Zytoplasma:
gp1 AS 1-253
gp11 AS 241-253
(2) hydrophobe Domäne, innere Membran:
AS 254-273
(3) C-terminale Domäne, Periplasma:
AS 274-348

Einleitung
23
~ 12 - 14 gp4-Monomeren zusammen, welche eine fassähnliche Struktur mit einem
Durchmesser von 13,5 nm und einer Länge von 24 nm ausbildet. Der Innen-
durchmesser der Pore variiert zwischen 6 nm – 8,8 nm und ist an der Basis der
N-terminalen Domäne geschlossen, was nahelegt, dass die gp4-Sekretionspore
während der Phagenassemblierung einer Konformationsänderung unterworfen ist
(Linderoth et al., 1997, Opalka et al., 2003).
Die C-terminale Domäne des gp4 hat mit einer Anzahl von 212 Aminosäuren eine
77 %ige Sequenzübereinstimmung mit dem homologen Protein PulD aus Klebsiella,
welches zu der Kategorie der Sekretine gezählt wird (Pugsley, 1993). Bakterielle
Sekretine sind zusammen mit weiteren Proteinen in der äußeren Membran lokalisiert,
wo diese im Fall der Typ II-Sekretion die Ausschleusung hydrolytischer sowie
toxischer Proteine in das Periplasma vermitteln, die anschließend über die äußere
Membran in das umgebende Milieu transloziert werden. Im Fall der Typ III-Sekretion
vermitteln Sekretine den Transport von Kinasen und Phosphatasen über zwei
bakterielle und eine eukaryotische Membran in tierische oder pflanzliche Zellen
(Galan & Collmer, 1999). Abgesehen von den Sekretinen besitzen sowohl die Typ II-
als auch die Typ III-Sekretionssysteme, wie auch der Sekretionskomplex von M13,
innere Membran
Periplasma
Murein
äußere Membran
C-Terminus
gp1/11-Assembly-Komplex
gp4-Multimer
Abb. 1-9: Schematisierter gp1/11-gp4-Sekretionskomplex. Die C-terminale Domäne des gp1/11-Assembly-Komplexes interagiert mit der N-terminalen Domäne der 13,5 nm großen gp4-Pore zu einem Sekretionskomplex, wodurch das gp1 vor einer proteolytischen Prozessierung geschützt wird. Während der Assemblierung neuer Phagenpartikel durch den Assembly-Komplex werden die naszier-enden Phagen durch die gp4-Pore in der äußeren Membran in das umgebende Milieu abgegeben (nach Webster, 1996).
13,5 nm
N
C
N
gp11 gp1 gp4
C
C
N
Zytoplasma

Einleitung
24
eine ATP-bindende Komponente. Verglichen mit den genannten Sekretionstypen
stellt der Sekretionskomplex von M13 einen eigenen, speziell an die Phagenassem-
blierung angepassten, Sekretionstyp dar.
1.3 Das Hüllprotein gp9
Gp9 wird, wie die Proteine gp7, gp3 und gp6, zu den „Minor Coat“ Proteinen gezählt,
da es mit nur ~ 5 Kopien im Phagenpartikel präsent ist (Makowski, 1992). Es bildet
zusammen mit gp7 eine Kappenstruktur an der Phagenspitze, welche zu Beginn der
Phagenassemblierung zuerst das Wirtsbakterium über den Sekretionskanal verlässt.
In Deletionsversuchen am „hairpin packaging signal“ konnte gezeigt werden, dass
Mutationen in gp7 und gp9 in der Lage sind, die Veränderungen im „hairpin
packaging signal“ zu kompensieren. Diese Ergebnisse zeigen, dass beide „Minor
Coat“ Proteine essenziell für den Initiationsvorgang der Assemblierung sind (Russel
& Model, 1989, Endemann & Model, 1995). Gp9 ist mit 32 Aminosäuren das kleinste
translatierte Protein von insgesamt 5 Hüllproteinen (Russel et al., 2004) und wird, wie
gp7 und gp6, ohne N-terminale Signalsequenz synthetisiert (Peters et al., 1994).
Basierend auf der Erkenntnis, dass gp9 nach der Insertion in die innere Membran
weiterhin eine N-terminale Formylgruppe aufweist, wurde vermutet, dass die Trans-
lokation entweder sofort nach oder während der Translation erfolgt (Simons et al.,
1980).
Wie die Proteinanalyse in Abbildung 1-10 zeigt, weist gp9 eine dem Haupthüllprotein
gp8 ähnliche Struktur auf. Wie gp8 besitzt es eine hydrophobe Domäne mit einem
anschließenden hydrophilen C-terminalen Bereich mit positiv geladenen Seiten-
gruppen (Marvin, 1998). Die N-terminale Domäne dagegen, die bei gp8 in der
inneren Membran lokalisiert ist, fehlt.
Abb. 1-10: Gegenüberstellung der Aminosäuresequenzen von gp9 und gp8, sowie deren Positionierung in der inneren Membran von E. coli. Das „Minor Coat“ Protein gp9 weist eine mit dem gp8 vergleichbare Struktur auf. Gp9 besitzt wie gp8 eine hydrophobe Domäne (schwarz hinter-legt), die von einem positiv geladenen hydrophilen C-terminalen Bereich gefolgt wird. Die Ladungen der Aminosäurereste sind durch (+) und (-) gekennzeichnet (nach Marvin, 1998).

Einleitung
25
Die Orientierung des gp9 nach Translokation in die innere Membran erfolgt dabei so,
dass der N-Terminus dem Periplasma zugewendet und der C-Terminus im Zyto-
plasma ist (Simons et al., 1981, Ploss & Kuhn, 2011). Durch FTIR und CD-Unter-
suchungen konnte für das gp9 eine einfache α-helikale Struktur in künstlich herge-
stellten Vesikeln aus DOPG (1,2 Dioleyl-sn-glycerol-3-phosphoglycerol) nachge-
wiesen werden (Makowski, 1992, Houbiers et al., 1999).
1.4 „Phage-Display“
Das „Phage-Display“ wurde 1985 von Smith (Smith, 1985) eingeführt und stellt eine
biotechnologische Methode dar, bei der auf genetischer Ebene kodierende Se-
quenzen von Peptiden oder Proteinteilen wie z. B. Antikörperfragmenten bzw. kom-
pletten Proteinen, nach einer Insertion in ein Phagenhüllgen funktionell auf der Ober-
fläche von Bakteriophagen präsentiert werden. Hierdurch entspricht der Genotyp
dem vorliegenden Phänotyp. Mit Hilfe des „Phage-Display“ können unter anderem
Protein-Protein Interaktionen untersucht werden. Aufgrund der geringen Genom-
größe des wildtypischen Phagen M13 sowie dessen komplett analysierter Genom-
sequenz besteht die Möglichkeit, gezielt heterologe Hüllproteine auf der Phagen-
oberfläche zu erzeugen. Jedoch kommt es hier zu einer Limitierung, da die Insertion
sowohl von für E. coli toxischen als auch von zu langen und hydrophoben Proteinen
den Phagenzyklus beeinträchtigen können. Das „Minor Coat“ Protein gp3 weist eine
höhere Toleranz gegenüber N-terminalen Insertionen als das Haupthüllprotein gp8
auf. Während gp3 Insertionen von zusätzlichen 6 - 38 Aminosäuren in die N-termi-
nale Domäne sowohl an Position 198 als auch 249 toleriert (Scott & Smith, 1990, Kay
et al., 1993, Russel, 2004), ist die Insertion in die N-terminale Domäne von gp8 auf
8–10 Aminosäuren limitiert (Kishchenko et al., 1994, Iannolo et al., 1995). Jedoch ist
auch im Fall des gp3 die Insertion limitiert, da hier nur Peptide fusioniert werden
können, welche die Infektivität des Phagen nicht negativ beeinflussen. Weiterhin
können einzelne Aminosäuren und zusätzliche positive Ladungen der inserierten
Peptide das „Phage-Display“ dahingehend beeinträchtigen, dass hierdurch sowohl
die Translokation des Sec-abhängigen gp3, als auch die Assemblierung der Phagen-
partikel durch veränderte gp8-Moleküle gestört wird (Yamane & Mizushima, 1988,
Andersson & Heijne, 1991, Schuenemann et al., 1999, Rodi et al., 2002). Außer an
gp3 und gp8 besteht des Weiteren die Möglichkeit, ein „Phage-Display“ an den
„Minor Coat“ Proteinen gp7 und gp9 durchzuführen (Gao et al., 1999, Sweeney et al.,

Einleitung
26
2006, Løset et al., 2011). In gp9 konnten erfolgreich bis zu 34 zusätzliche Amino-
säuren in die N-terminale Domäne inseriert werden, ohne den Infektionszyklus des
Phagen zu beeinträchtigen (Ploss & Kuhn, 2011). Wie in Abbildung 1-11 dargestellt,
besteht hierdurch die Möglichkeit zur Insertion und Präsentation von Peptiden an drei
unterschiedlichen Positionen des Phagen, wodurch mono-, bi- und trifunktionale
Phagen generiert werden können (Huang et al., 2005, Guo et al., 2010).
Monovalentes-/Polyvalentes „Phage-Display“
Für die funktionelle Präsentation von Fremdpeptiden auf der Phagenoberfläche, bei
der der Phänotyp dem Genotyp entspricht, stehen zwei Methoden zur Verfügung
(Lowman, 1997). Bei einer direkten Verwendung des Phagenvektors werden die
Nukleotidsequenzen des zu präsentierenden Peptids „in-frame“ in das Gen des
entprechenden Hüllproteins inseriert. In einer anschließenden Infektion werden hier-
durch, während der Assemblierung des Phagen in E. coli, ausschließlich Phagen-
partikel erzeugt, welche nur die entsprechenden heterologen Hüllproteine präsen-
tieren, wodurch ein polyvalentes „Phage-Display“ generiert wird (Parmley & Smith,
Abb. 1-11: Schematische Darstellung der an M13 möglichen „Phage-Display“-Positionen. Durch die Insertion unterschiedlicher Peptidsequenzen in die Gene der für M13 kodierenden Hüll-proteine, besteht die Möglichkeit zur Generierung mono-, bi- und trifunktionaler Phagen.

Einleitung
27
1988). Eine alternative Methode führt über ein Phagemid-System. Hier handelt es
sich um ein Hybridvektor-System, welches sich aus einem M13-Helferphagen und
einem zusätzlichen kleinen Plasmid, dem sogenannten Phagemid, zusammensetzt
(Russel et al., 1986). Der dabei verwendete Helferphage ist jedoch dahingehend
modifiziert, dass durch Mutationen im „ori“ der intergenischen Region des Phagen die
„Verpackungseffizienz“ des Phagengenoms in neue Phagenpartikel stark beein-
trächtigt ist. Im Gegenzug besitzt das Phagemid einen wildtypischen „Ff-ori“, über
den das Defizit in der „Verpackungseffizienz“ des Phagen ausgeglichen werden
kann. Durch eine Infektion von phagemidtransformierten Zellen kommt es während
der Replikationsphase zu einer homologen Rekombination zwischen Phagemid und
Helferphagengenom. Parallel zu den wildtypischen Hüllproteinen erfolgt während der
Proteinbiosynthese auch die Herstellung des phagemidkodierten, heterologen Hüll-
proteins, welches ebenfalls für die Assemblierung neuer Phagen verwendet werden
kann. Durch die Rekombination entstehen während der Replikationsphase sowohl
wildtypische als auch rekombinante Genome mit der doppelten Genanzahl eines
Hüllproteins und wildtypischem „ori“. Da letztere eine höhere „Verpackungseffizienz“
der einzelsträngigen DNA in neue Phagenpartikel aufweisen, erfolgt während der
Assemblierung hauptsächlich die Generierung von Hybridphagen. Hierdurch entsteht
ein monovalentes „Phage-Display“, bei dem auf den Phagen sowohl wildtypische als
auch die heterologe Variante des entsprechenden Hüllproteins präsentiert wird
(Russel et al., 2004). Ein großer Vorteil dieser Methode besteht darin, dass mit Hilfe
eines kleinen Vektors mit geringem Aufwand die Präsentation heterologer Hüll-
proteine auf der Phagenoberfläche möglich ist, ohne die komplexe Struktur des
Phagengenoms beeinflussen zu müssen. Ein Nachteil dieser Methode ist, dass nur
bis zu 10 % der neugebildeten Phagen mindestens ein heterologes Hüllprotein auf-
weisen (Russel et al., 2004). Über eine dritte Methode, die Komplementation, besteht
ebenfalls die Möglichkeit zur funktionellen, polyvalenten Präsentation heterologer
Fusionsproteine. Hier entspricht jedoch der Phänotyp nicht dem Genotyp, da die
Expression des genomisch kodierten Hüllproteins zuvor über eine Punktmutation
unterbunden und anschließend durch eine induzierbare Expression, von einem
sogenannten Helferplasmid aus, für die Assemblierung des Phagen bereitgestellt
wird (Ploss & Kuhn, 2011). In Abbildung 1-12 sind die gängigen Methoden für poly-
und monovalentes „Phage-Display“ am Bakteriophagen M13 in einer Übersicht dar-
gestellt.

Einleitung
28
1.5 Proteintransport-Systeme
In allen Zellsystemen erfolgt sowohl die Synthese der organismenspezifischen als
auch, im Hinblick einer Infektion von E. coli mit Bakteriophagen, die Synthese viraler
Proteine im Zytoplasma. Im Anschluss müssen diese, um ihrer Bestimmung nach-
kommen zu können, zu individuellen Kompartimenten der Zellen transportiert
werden. Infolge dessen kann der Transport eukaryotischer Proteine zu verschie-
denen Zellorganellen erfolgen wie z. B. dem Zellkern, dem endoplasmatischen Reti-
kulum, den Peroxisomen, den Mitochondrien oder den Chloroplasten. Prokaryo-
tische, wie auch virale Proteine hingegen können aus dem Zytoplasma in die innere
Membran, ins Periplasma oder in die äußere Membran transportiert werden
(Brissette & Russel, 1990, Rapoza & Webster, 1995, Wang & Dalbey, 2011). Um
diese Translokationen zu ermöglichen, stehen in Mitochondrien, Chloroplasten und
Bakterien unterschiedliche Protein Import- und Export-Systeme zur Verfügung (Abb.
1-13).
Abb. 1-12: Allgemeines Schema von poly- und monovalenten „Phage-Display“ an M13. Hier sind jeweils am Beispiel des „Minor Coat“ Proteins gp3 die Unterschiede zwischen den einzelnen „Phage-Display“-Systemen verdeutlicht. Die Insertion der Peptidsequenzen erfolgt für das „Display“ hinter die N-terminale Signalsequenz von gp3. Im Helferphagen-System ist sowohl im Phagemid als auch in der ssDNA des Helferphagen ein „Ff-ori“ vorhanden, der jedoch im Fall des Helferphagen durch Mutationen in der Replikations- und Verpackungseffizienz der ssDNA gemindert ist. Das Komplementationssystem unterscheidet sich von den anderen zwei Systemen dahingehend, dass hier der Phänotyp nicht identisch mit dem Genotyp ist (nach Russel et al., 2004, verändert).

Einleitung
29
Abb. 1-13: Darstellung der identifi-zierten Protein Import- und Export-Systeme eu- und prokaryotischer Zellen am Beispiel des Mitochondriums, des Chloroplasten und des Bakteriums. Mitochondrium: TOM = translocase of the outer membrane, TIM = translocase of the inner membrane, SAM/TOB = sorting and assembly machinery / translocase of the outer mitochondrial membrane, MIA = mitochondrial intermembrane space import and assembly, Oxa1 = oxidase assembly. Chloroplast: TOC = translocase of the outer envelope membrane of chloroplast, TIC = translocase of the inner envelope membrane of chloroplast, TAT = twin arginine translocation, Sec = secretion, Alb3 = albino. Bakterium: BAM = β-barrel assembly machinery, Sec = secretion, Tat = twin arginine translocation (nach Wang & Dalbey, 2011)

Einleitung
30
Eukaryotische Proteintransport-Systeme
In eukaryotischen Zellen werden Proteine, deren Bestimmungsort das Mitochondrium
ist, als signalsequenztragendes Präprotein im Zytoplasma synthetisiert und an-
schließend über den TOM-Komplex („translocase of the outer membrane“) unter
ATP-Verbrauch und mit Hilfe des Membranpotentials durch die äußere Membran in
den Intermembranraum (IMS, „inter membrane space“) transloziert. Von hier aus
kann der Transport der Proteine in drei weitere Organellkompartimente erfolgen, die
mitochondriale Matrix, die äußere sowie die innere Mitochondrienmembran. Aus-
gehend vom Intermembranraum werden Proteine mit β-Faltblatt-Strukturmotiven,
deren endgültiger Bestimmungsort die äußere Mitochondrienmembran ist, über den
SAM/TOB-Komplex („sorting and assembly machinery“ / „translocase of the outer
mitochondrial membrane“) transloziert (Kozjak et al., 2003, Paschen et al., 2003).
Proteine mit konservierten Cysteinresten hingegen werden über den, in der inneren
Mitochondrienmembran lokalisierten, Mia40/Erv-Komplex („mitochondrial intermem-
brane space import and assembly“ / „essential for respiration and vegetative
growth“), unter Ausbildung essenzieller Disulfidbrücken gefaltet, wodurch das Ener-
gieniveau der zu translozierenden Proteine herabgesetzt und ihr Verbleib im Inter-
membranraum erzielt wird („folding trap“-Mechanismus, Lutz et al., 2003, Herrmann
& Köhl, 2007). Die Translokation von Proteinen, deren endgültige Lokalisation die
innere Mitochondrienmembran ist, kann auf drei Wege geschehen. Ein Weg der
Translokation erfolgt über den TIM22-Komplex („translocase of the inner
membrane“). Die hier zu translozierenden Proteine werden mit einer Signalsequenz
synthetisiert, welche nach der Translokation in den IMS mit den Chaperonen dieses
Komplexes interagieren, um anschließend über den TIM22-Komplex in die innere
Membran transloziert zu werden. Membranproteine mit einer α-helikalen Trans-
membrandomäne hingegen werden bevorzugt über den TIM23-Komplex in die innere
Membran transloziert (Glick et al.,1992). Alternativ kann die Translokation auch über
Oxa1 („oxidase assembly“), dessen Analogon im bakteriellen YidC zu finden ist,
katalysiert werden. Über diesen Weg werden die Transmembranproteine zunächst
über den TIM23-Komplex aus dem IMS in die mitochondriale Matrix importiert, um
daraufhin über Oxa1 in die innere Membran transloziert zu werden. Durch Oxa1
werden weiterhin die in der mitochondrialen Matrix translatierten Membranproteine in
die innere Mitochondrienmembran inseriert (Bonnefoy et al., 2009).

Einleitung
31
Chloroplastenspezifische Proteine werden im Zytoplasma der pflanzlichen Zelle
ebenfalls als Präproteine mit spezifischen Signalsequenzen synthetisiert. Im Unter-
schied zu den mitochondrialen Proteinen ist hier der Transport zu sechs Organell-
kompartimenten möglich. Hierzu zählen die äußere, die innere und die Thylakoid-
membran sowie der Intermembranraum, das Stroma und das Thylakoidlumen. Die
Translokation der über die äußere Membran zu importierenden Proteine wird durch
den TOC-Komplex („translocase of the outer envelope membrane of chloroplast“),
die der über die innere Membran zu importierenden Proteine durch den TIC-Komplex
(„translocase of the inner envelope membrane of chloroplast“) vermittelt. Stroma-
assoziierte Peptide verbleiben nach Abspaltung der Signalsequenz durch Stroma-
spezifische Peptidasen in diesem Kompartiment, wohingegen Thylakoidlumen-
spezifische Proteine entweder über den Sec- („secretion“) oder Tat- („twin arginine
translocation“) abhängigen Transport in das Lumen exportiert werden (Cline & Theg,
2007). Diese Proteine werden mit einer zweiteiligen N-terminalen Signalsequenz
synthetisiert, welche sich aus einem „Stroma-targeting“ und einem Thylakoidlumen-
Signal zusammensetzt, die Übereinstimmungen mit bakteriellen Signalpeptiden
zeigen. Das Tat-Signalpeptid unterscheidet sich vom Sec-Signalpeptid insofern, dass
das Tat-Signalpeptid ein sogenanntes RR-Motiv (Arginin-Arginin) vor der hydro-
phoben Transmembrandomäne aufweist (Berks, 1996). Über das Sec-Translokase-
system können ungefaltete Proteine entweder in die Thylakoidmembran transloziert
oder in das Thylakoidlumen exportiert werden, wohingegen über das Tat-Translo-
kasesystem der Export gefalteter Proteine in das Lumen erfolgt (Mould et al.,1997).
Chloroplastenkodierte Proteine wie z. B. die Chlorophyll a- und b-Bindeproteine
besitzen ein stark konserviertes, aus 18 Aminosäuren bestehendes Motiv (L18-Motiv,
VDPLYPGGSFDPLGLADD), wodurch diese mittels des chloroplasteneigenen
cpSRP-Systems („chloroplast signal recognition particle“) cotranslational über cpFtsY
(„chloroplast filamentous temperature-sensitive Y“) an die Thylakoidmembran geleitet
und nach GTP-Hydrolyse über das Alb3-System inseriert werden (Tu et al., 1999,
2000, Henry et al., 2007).

Einleitung
32
Prokaryotische Proteintransport-Systeme
Ähnlich den Sec- und Tat-Translokasesystemen der Chloroplasten, sind die
bakteriellen Analoga gleichfalls für die Proteintranslokation über die innere Bakterien-
membran in das Periplasma verantwortlich. Wie bei den Mitochondrien- und Chloro-
plastenproteinen werden sowohl bakterielle als auch virale M13-Proteine wie gp3,
gp8 und gp4, welche zu anderen Zellkompartimenten delegiert werden müssen,
ebenfalls als signalsequenztragende Präproteine synthetisiert. Für die Sekretion von
ungefalteten Proteinen in das Periplasma ist die SecYEG-Translokase das bevor-
zugte System, während nur eine sehr geringe Anzahl an gefalteten und mit Metall-
Kofaktoren assoziierten Proteinen über das Tat-Translokasesystem in das Peri-
plasma von E. coli exportiert wird. Nach der Translokation werden Proteine mit
β-Faltblatt-Strukturmotiven, deren endgültiger Bestimmungsort die äußere Membran
ist, durch den BAM-Komplex („β-barrel assembly machinery“), der zu dem
SAM/TOB-Komplex der Mitochondrien analog ist, transloziert (Knowles et al., 2009).
Der Export bakterieller Lipoproteine in die äußere Membran hingegen wird über das
bakterienspezifische Lol-System, in dem die Proteinkomponenten LolABCDE invol-
viert sind, vermittelt (Tokuda & Matsuyama, 2004). Die Insertion membranasso-
ziierter Proteine kann in E. coli entweder Sec-abhängig kotranslational über den
SRP-Sec-Weg („signal recognition protein“) oder Sec-unabhängig über den YidC-
Weg erfolgen (Ulbrandt et al., 1997).
Translokationssysteme in Escherichia coli
Die Delegation der bakteriellen Proteine und die Bestimmung des in Folge dafür
benötigten Transportwegs erfolgt während der Proteinsynthese am Ribosom. Einer
der wichtigsten Translokationswege wird von den Proteinen SecA und SecB ver-
mittelt und als Sec-abhängige Translokation bzw. Sekretion bezeichnet (Abb. 1-14
b)). Während der Proteinsynthese erfolgt die Bindung eines ribosomenassoziierten
„Trigger Factor“ (TF) an eine N-terminale Sec-Signalsequenz der naszierenden
Peptidkette. Nach Vollendung der Synthese wird das ungefaltete Präprotein post-
translational von SecB gebunden und über elektrostatische Wechselwirkungen mit
dem membranassoziierten SecA an die innere Membran geführt (Wickner & Leonard,
1996). Durch die Interaktion der Präprotein-Signalsequenz mit SecA wird das SecB-
assoziierte Präprotein auf SecY übertragen und anschließend unter ATP-Hydrolyse
und mit Hilfe der protonenmotorischen Kraft (PMF, „proton motive force“) durch die

Einleitung
33
SecY-Pore in das Periplasma sekretiert. Während dieses Sekretionsvorgangs wird
die N-terminale Signalsequenz durch die membrangebundene Leaderpeptidase
prozessiert (Watanabe et al., 1988). Über diesen Weg werden Außenmembran-
proteine wie OmpA („outer membrane protein“), die alkalische Phosphatase PhoA,
das periplasmatische Maltose Bindeprotein MBP und das M13-kodierte gp4, welches
in der äußeren Membran eine homomultimere Pore bildet, sekretiert (Lecker et al.,
1990, Bassford, 1990, Brissette & Russel, 1990).
Eine weitere Sec-abhängige Translokation stellt der SRP-Sec-Weg dar, über den
Membranproteine, deren endgültiger Bestimmungsort die innere Membran ist, trans-
loziert werden (Abb.1-14 A)). Weist die während der Proteinsynthese aus dem Ribo-
som heraustretende Peptidkette eine Signalsequenz mit hydrophobem (von Heijne,
1985) und helikalem Charakter (Bruch et al., 1989) auf, so bindet das ribosomen-
assoziierte SRP den Ribonukleoproteinkomplex. Durch die Bindung des SRPs ent-
steht ein RNC/SRP-Komplex („ribosome nascent chain“), der anschließend mit dem
membranassoziierten SRP-Rezeptor FtsY („filamentous temperature-sensitive Y“) an
der inneren Membran interagiert. Unter GTP-Hydrolyse dissoziiert der FtsY/SRP-
Komplex, wodurch die Peptidkette an die Translokase SecYEG übertragen wird.
Anschließend erfolgt die Membraninsertion des Proteins unter Mitwirkung von YidC,
das für die richtige Faltung des Proteins verantwortlich ist. Als Sec-abhängige Subs-
trate wurden die ATP-Synthase Untereinheit a (FOa) (Yi et al., 2003, 2004), die
Zytochrom-bo3-Oxidase-Untereinheit CyoA (Celebi et al., 2006, Bloois et al., 2006,
Plessis et al., 2006,), die NADH Dehydrogenase I Untereinheit (Price & Driessen,
2010), als auch das „Minor Coat“ Protein gp3 des Bakteriophagen M13 charak-
terisiert (Rapoza & Webster, 1993, Thie et al., 2008).
Die Insertion integraler Membranproteine kann jedoch auch Sec-unabhängig über
den YidC-Weg erfolgen (Abb. 1-14 C)). Dieses System wird aber nur von wenigen
Proteinen wie z. B. dem Hüllprotein gp8 des Phagen M13, dem Hüllprotein des
Phagen Pf3, der c-Untereinheit der F1FO-ATP-Synthase und dem mechanosensitiven
Kanalprotein MscL genutzt (Samuelson et al., 2000, Chen et al., 2002, Van der Laan
et al., 2004, Facey et al., 2007,). Während SRP am kotranslationalen Targeting von
MscL und der c-Untereinheit der F1FO-ATP-Synthase beteiligt ist, erfolgt der Trans-
port des Hüllproteins von Pf3 als auch der des Präproteins gp8 des Phagen M13
unter Ausschluss weiterer Proteine (van Bloois et al., 2004, Facey et al., 2007). Nach
der Bindung des Substrates an die Insertase YidC und der darauf folgenden Faltung
Einleitung
34
des zu inserierenden Proteins wird dieses in die innere Membran abgegeben und,
wie im Fall des gp8-Präproteins, anschließend die N-terminale Leader-Signalse-
quenz durch die Leaderpeptidase abgespalten (Chang et al., 1978).
SecYEG
YidC
SRP
FtsY
YidC
GTP
Lep
3.
Transfer an YidC
1. SRP-vermitteltes Targeting an
FtsY
Ribosom
mRNA
2. Translokation durch SecY
4. Membraninsertion
ATP +
PMF
Lep
Ribosom
mRNA
SecA SecA
SecYEG
SecB
SecB
TF
naszierendes Polypeptid 1.
Translation
2. SecB-vermitteltes post-translationales
Targeting
3. Sekretion durch
SecY
4. Abspaltung des
Signalpeptids
Ribosom
mRNA
Sec-Signalpeptid TF
1.
Translation
naszierendes Polypeptid
YidC
Lep
FtsY
SRP
2a. post-
translationales Targeting
3. Translokation durch YidC + PMF
4. Membraninsertion
5. Abspaltung des Signalpeptids
Tat
C Tat
B
Tat A
Tat
C Tat A Oligo-mere
Tat
B
Lep
Tat
C Tat
B
Tat A Oligo-mere
Insertion Sekretion Insertion durch YidC
Sec-abhängiger Proteintransport Sec-unabhängiger
Proteintransport
naszierendes Polypeptid
Ribosom
mRNA
Tat-Signalpeptid TF
1. Translation
KF
KF 2. Nach Faltung und Targeting des Proteins
durch Kofaktoren (KF)
3. Bindung des Signalpeptids
an TatC
4. Oligomerisation der TatA-Untereinheiten
PMF
6. Abspaltung des
Signalpeptids
5. Translokation durch TatA + PMF
Insertion/Sekretion durch Tat
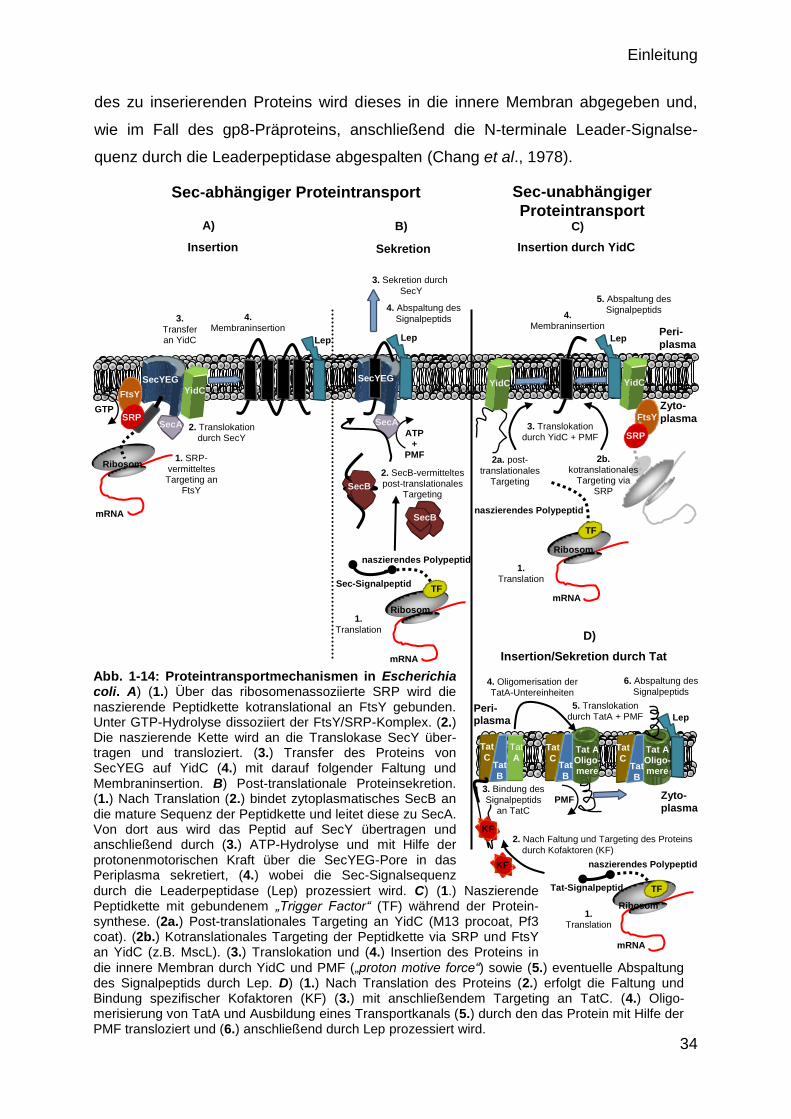
Abb. 1-14: Proteintransportmechanismen in Escherichia coli. A) (1.) Über das ribosomenassoziierte SRP wird die naszierende Peptidkette kotranslational an FtsY gebunden. Unter GTP-Hydrolyse dissoziiert der FtsY/SRP-Komplex. (2.) Die naszierende Kette wird an die Translokase SecY über-tragen und transloziert. (3.) Transfer des Proteins von SecYEG auf YidC (4.) mit darauf folgender Faltung und Membraninsertion. B) Post-translationale Proteinsekretion. (1.) Nach Translation (2.) bindet zytoplasmatisches SecB an die mature Sequenz der Peptidkette und leitet diese zu SecA. Von dort aus wird das Peptid auf SecY übertragen und anschließend durch (3.) ATP-Hydrolyse und mit Hilfe der protonenmotorischen Kraft über die SecYEG-Pore in das Periplasma sekretiert, (4.) wobei die Sec-Signalsequenz
durch die Leaderpeptidase (Lep) prozessiert wird. C) (1.) Naszierende Peptidkette mit gebundenem „Trigger Factor“ (TF) während der Protein-synthese. (2a.) Post-translationales Targeting an YidC (M13 procoat, Pf3 coat). (2b.) Kotranslationales Targeting der Peptidkette via SRP und FtsY an YidC (z.B. MscL). (3.) Translokation und (4.) Insertion des Proteins in
A) B) C)
D)
Peri-
plasma
Zyto-
plasma
Zyto-
plasma
Peri-plasma
die innere Membran durch YidC und PMF („proton motive force“) sowie (5.) eventuelle Abspaltung des Signalpeptids durch Lep. D) (1.) Nach Translation des Proteins (2.) erfolgt die Faltung und Bindung spezifischer Kofaktoren (KF) (3.) mit anschließendem Targeting an TatC. (4.) Oligo-merisierung von TatA und Ausbildung eines Transportkanals (5.) durch den das Protein mit Hilfe der PMF transloziert und (6.) anschließend durch Lep prozessiert wird.
2b. kotranslationales
Targeting via SRP

Einleitung
35
Ein weiterer, Sec-unabhängiger, Proteintransportweg stellt das, zu den Chloroplasten
homologe, Tat-System dar (Robinson et al, 2001) (Abb. 1-14 D)). Über dieses
System werden bereits gefaltete Präproteine, welche ein N-terminales Doppel-
Arginin Motiv aufweisen, entweder in die innere Membran inseriert oder ins Peri-
plasma transloziert (Berks et al., 2000). Das Tat-System besteht aus einem TatABC-
Komplex, in dem wahrscheinlich multimere TatA-Proteine einen Transportkanal in
der Zytoplasmamembran ausbilden (Line & Mori, 2001). Durch die Bindung des
Substrates an die Untereinheit TatC des Rezeptorelements TatBC, wird die Poly-
merisation der TatA-Untereinheiten zu einem Transportkanal initiiert, woraufhin das
zu transportierende Protein auf TatA übertragen und mit Hilfe der protonen-
motorischen Kraft (PMF, „proton motive force“) entweder in die Membran inseriert
oder in das Periplasma transloziert wird (Yahr & Wickner, 2001, Alami et al.,2003,
Cline & Theg, 2007, Tarry et al., 2009). Nach dem Proteintransport erfolgt abschlie-
ßend die Prozessierung der Peptide, in welcher das N-terminale Tat-Signalpeptid
durch die, in der Zytoplasmamembran lokalisierte, Leaderpeptidase entfernt wird. Zu
den Tat-Substraten zählen vornehmlich Proteine mit Redoxeigenschaften wie z. B.
die Trimethylamin-N-Oxid-Reduktase (TorA) (Sargent et al., 1998), die -Untereinheit
der Formiatdehydrogenase N (FdnG) (Wexler et al., 1998) und die kleine Unter-
einheit Hya der Hydrogenase 1 (Mori & Cline, 1998). Weitere Tat-Substrate sind
unter anderem die -Lactamase (Nivière at al., 1992), die P2-Domäne der Leader
Peptidase (Cristobal et al., 1999) und die Nitratreduktase (Weiner et al., 1998).
1.6 Aufgabenstellung der vorliegenden Arbeit
In dieser Arbeit soll durch die Untersuchung vier essenzieller Bereiche des M13-
Phagenzyklus, welche die Abschnitte Infektion, Proteininsertion, Assembly sowie die
Phagensekretion betreffen, neue Erkenntnisse über die Biogenese und die Assem-
blierung des filamentösen Coliphagen M13 erhalten werden. Mit Hilfe einer Quantifi-
zierung der M13-Phagenextrusion sollen anhand detaillierter „one-step growth“
Experimente neue Einblicke sowohl in die wildtypisch verlaufende Infektion als auch
in dem Verlauf einer induzierten Sekretion erhalten werden. Über die Quantifi-
zierungen können anschließend Rückschlüsse auf die Dynamik der Phagen-Wirts-
bindung, die Anzahl der sekretierten Phagen-Nachkommenschaften pro Zelle und
den Infektionsverlauf in infizierten Zellen gezogen werden. Diese Erkenntnisse sollen
im Verlauf dieser Arbeit dazu verwendet werden, die Sekretion neu gebildeter

Einleitung
36
Phagenpartikel auf der Bakterienoberfläche, sowohl mit elektronenmikroskopischen
(TEM, „transmission electron microscope“) als auch mit rasterkraftmikroskopischen
(RKM = AFM, „Atomic Force Microscope“) Methoden, näher untersuchen zu können.
Auch sind noch nicht für alle Phagenhüllproteine, die während des Assemblierungs-
vorgangs für die Sekretion neuer Phagen in der inneren Wirtsmembran zur Ver-
fügung stehen, deren wirtsspezifisch verwendeten Proteintransportsysteme identi-
fiziert. Mit Hilfe der funktionellen Präsentation von Fremdpeptiden am „Minor Coat“
Protein gp9 soll durch die Detektion antigener Epitope Aufklärung über dessen
Insertion in die innere Wirtsmembran erhalten werden.
Über den phagenkodierten gp1/11-Assembly-Komplex konnten hinsichtlich seiner
Bedeutung für den M13-Phagenzyklus sowie dessen Interaktion mit phagen-
kodierten als auch wirtsspezifischen Proteinen in der Vergangenheit fundamentale
Erkenntnisse erhalten werden. Trotz der eingehenden Untersuchungen um die
Bedeutung des Komplexes für die Assemblierung, ist es noch nicht gelungen,
dessen Größe und Molekulargewicht zu erörtern bzw. diesen zu visualisieren. Eine
Überexpression der Phagenproteine gp1/11 in E. coli-Zellen sollte es ermöglichen,
die Masse des Komplexes zu ermitteln und erste strukturelle Charakterisierungen
des gereinigten gp1/11-Assembly-Komplexes durchzuführen.

Material und Methoden
37
2 Material und Methoden
2.1 Material
2.1.1 Bakterien- und Bakteriophagenstämme
Bakterienstämme
Escherichia coli C41 (DE3) (Miroux & Walker, 1996)
F– ompT gal dcm lon hsdSB(rB- mB
-) λ(DE3 [lacI lacUV5-T7 gene 1 ind1 sam7 nin5])
Dieser Stamm stellt eine Spontanmutation des E. coli-Stammes BL21(DE3) dar. Er
wird zur Überexpression rekombinanter Proteine in Expressionsvektoren mit T7
Promotoren verwendet (Sambrook et al., 1989).
Escherichia coli DH5α (Hanahan, 1985)
F- gyrA96 (Nalr) recA1 relA1 endA1 thi-1 hsdR17 (rk-, mk
+) glnV44 deoR ∆(lacZYA-
argF) U169 [Φ80d ∆(lacZ) M15]
Dieser Bakterienstamm bildet weder F-Pili noch Fimbrien aus und eignet sich
aufgrund dieser Eigenschaften für die Messungen der M13-Phagensekretion mit dem
AFM und TEM. Zur Infektion muss der Bakterienstamm mit dem RF-Plasmid des
M13-Phagen transfiziert werden.
Escherichia coli JS7131 (Samuelson et al., 2000)
MC1060 ΔyidC attB::R6Kori ParaBADyidC+ (Specr)
Bei diesem Bakterienstamm handelt es sich um einen YidC-Depletionsstamm. Die
genomische YidC-Expression wird durch einen araBAD Promotor kontrolliert wo-
durch die Expression in diesen Zellen durch Arabinose (Expression) bzw. Glukose
(keine Expression) reguliert werden kann.
Escherichia coli K37 (Hines & Ray, 1980)
HfrC supD32 relA1 pit-10 spoT1 tonA22 ompF627 phoA4 T2R λ-
Dieser Bakterienstamm ist ein Wirt des Bakteriophagen M13. Als Suppressor-Stamm
kann dieser sowohl für M13wt als auch für Phagen mit Ambermutation zur Eta-
blierung von Phagenstammlösungen verwendet werden. Er kann ebenfalls zum Aus-
plattieren bei Spot- und Plaque-Tests eingesetzt werden.
Escherichia coli K38 (Garen et al., 1965, Lyons & Zinder, 1972)
HfrC T2R relA1 pit-10 spoT1 tonA22 ompF627 phoA4 λ-

Material und Methoden
38
Dieser Bakterienstamm ist ein Wirt des Bakteriophagen M13wt. Als Nicht-Sup-
pressor-Stamm führen Ambermutation in Genen zur Termination der Translation. Er
wird sowohl zur Identifikation von M13-Ambermutanten als auch in Komplemen-
tationstests, in denen die Translation genomischer Gene des M13-Phagen zuvor
durch Insertion von Ambermutantionen unterbunden und nur anhand hilfsplasmid-
kodierter Gene kompensiert werden kann, verwendet.
Escherichia coli MC4100 (Casadaban, 1976)
F- [araD139]B/r Δ(argF-lac)169* lambda- e14- flhD5301 Δ(fruK-yeiR)725 (fruA25)‡
relA1 rpsL150(strR) rbsR22 Δ(fimB-fimE)632(::IS1) deoC1
Aufgrund des fehlenden F-Plasmids ist dieser Stamm kein natürlicher Wirt des
Bakteriophagen M13. Der Stamm wurde für die Quantifizierung der unspezifischen
Adsorption von M13-Phagen an Bakterien verwendet.
Escherichia coli XL1-blue (Bullock et al., 1987)
supE44 hsdR17 recA1 endA1 gyrA46 thi, relA1 lac- F´[proAB+ laclq lacZ∆M15
Tn10(tetr)]
Dieser Bakterienstamm wird allgemein wegen seines Rekombinationsdefizits zur
Transformation und Isolierung von Vektoren verwendet. Durch den ΔlacZ-Genotypen
ist eine Blau-Weiß-Selektion mit X-Gal (Sambrook et al., 1989) möglich. Der Bakte-
rienstamm wird wegen der F-Pili wie E. coli K37 als Suppressor-Stamm zur Eta-
blierung von Phagenstammlösungen und zum Ausplattieren bei Spot- und Plaque-
Tests eingesetzt.
Bakteriophagenstämme:
M13mp19
Entspricht dem M13wt, in dessen Genom zusätzlich ein lacZ-Gen inseriert wurde,
welches für das N-terminale α-Fragment der β-Galactosidase kodiert und zwischen
den Kodons 6-7 eine „Multiple Cloning Site“ aufweist. Dadurch ist eine Blau-Weiß-
Selektion möglich.
M13amp
Ursprünglicher Phagenstamm ist M13mp19. In dessen „Multiple Cloning Site“ wurde
das β-Lactamase-Gen aus pGZamp inseriert, wodurch nach IPTG-Induktion die
infizierten E. coli-Zellen eine Resistenz gegen das Antibiotikum Ampicillin erhalten.

Material und Methoden
39
M13am9-M7
Ursprünglicher Phagenstamm ist M13mp19. In das Gen 9 wurde an der Position +7
durch eine ortsgerichtete Mutagenese eine Ambermutation inseriert.
M13mp19-am8-M2
Ursprünglicher Phagenstamm ist M13mp19. Zusätzlich wurde in das Gen 8 eine
Ambermutation an der Position +2 inseriert.
2.1.2 Plasmide
pETg1/11
Der Ursprungsvektor ist das Plasmid pET-16b (Novagen), welcher eine durch den
T7-Promotor kontrollierte „Multiple Cloning Site“ besitzt. Der „Multiple Cloning Site“ ist
eine kodierende Sequenz für einen 10x Histidin-Tag vorangestellt, so dass durch die
Klonierung von Genen in diesen Bereich nach Expression Fusionsproteine mit
N-terminalen 10x Histidin-Tag entstehen. Zur Reinigung und Charakterisierung des
gp1/11-Assembly-Komplexes wurde das Gen 1 mittels PCR-Methode amplifiziert und
anschließend zwischen die Schnittstellen XhoI und Bpu1102I inseriert.
pGEMT-easy (Promega)
Das offene Plasmid mit T-Überhängen an den 3´-Enden der Plasmid-DNS diente zur
Insertion des mittels PCR-Amplifikation erhaltenen Fragments der Gene 7 und 9.
pGEMg7/9
Vektor mit T7-Promotor. Die Gene 7 und 9 des Bakteriophagen M13 wurden über
A-Überhänge in die „Multiple Cloning Site“ ligiert.
pGZamp
Der Ursprungsvektor ist das Plasmid pGZ119EH (Lessl et al., 1992), in welches das
β-Lactamase-Gen aus pUC18 in die SmaI-Schnittstelle inseriert wurde.
pMSg1/11
Der Ursprungsvektor ist das Plasmid pMS119EH (Strack et al., 1992), welches eine
durch den Tac-Promotor kontrollierte „Multiple Cloning Site“ besitzt. Zur Expression
des gp1/11-Assembly-Komplexes mit 10x N-terminalem Histidin-Tag in E. coli-
Stämmen, welche keine T7-Polymerase exprimieren, wurde das Gen aus pETg1/11
zwischen die Schnittstellen XbaI und HindIII inseriert.

Material und Methoden
40
pGZg804
Der Ursprungsvektor ist das Plasmid pGZ119HE (Lessl et al.,1992), in welches das
M13-Gen 8 aus pJF804 (Fürste et al., 1986) zwischen die Schnittstellen SalI und
EcoRI inseriert wurde.
pMSg7/9
Der Ursprungsvektor ist das Plasmid pMS119EH (Strack et al., 1992), welches eine
durch den Tac-Promotor kontrollierte „Multiple Cloning Site“ besitzt. Die M13-Gene 7
und 9 wurden zur genetischen Manipulation sowie für Expressions- und Komplemen-
tationsstudien aus dem Vektor pGEMg7/9 zwischen die Schnittstellen PaeI und SacI
inseriert.
pMSg9MunI
Entspricht pMSg7/9, jedoch wurde in Gen 9 eine MunI-Schnittstelle an Position +3
inseriert.
pMSg9T7
Entspricht pMSg7/9, jedoch wurde in Gen 9 an Position +3 die kodierende Sequenz
des T7-Tags inseriert.
pMSg9T7rep
Entspricht pMSg9T7, jedoch wurde in Gen 9 an Position +22 die kodierende Start-
sequenz +1 und +2 von gp9 an das Ende des kodierenden T7-Tags eingefügt, um
die wildtypische Aminosäuresequenz wiederherzustellen.
pMSg9HA
Entspricht pMSg7/9, jedoch wurde in Gen 9 an Position +3 die kodierende Sequenz
des HA-Tags inseriert.
pMSg9HArep
Entspricht pMSg9HA, jedoch wurde in Gen 9 an Position +20 die kodierende Start-
sequenz +1 und +2 von gp9 an das Ende des kodierenden HA-Tags eingefügt, um
die wildtypische Aminosäuresequenz wiederherzustellen.
pMSg9T7SD
Entspricht pMSg9T7, jedoch mit der zusätzlichen, 8 bp vor dem Startcodon von Gen
9 eingefügten, E. coli Shine-Dalgarno-Sequenz AGGAGG.

Material und Methoden
41
pMSg9DT7SD
Entspricht pMSg9T7SD, jedoch mit einem zusätzlichen T7-Tag an Position +20.
pMSg9T7repSD
Entspricht pMSg9T7rep, jedoch mit der zusätzlichen, 8 bp vor dem Startcodon von
Gen 9 eingefügten, E. coli Shine-Dalgarno-Sequenz AGGAGG.
pMSg9DT7repSD
Entspricht pMSg9T7repSD, jedoch mit einem zusätzlichen T7-Tag an Position +20.
pMSg9HASD
Entspricht pMSg9HA, jedoch mit der zusätzlichen, 8 bp vor dem Startcodon von Gen
9 eingefügten, E. coli Shine-Dalgarno-Sequenz AGGAGG.
pMSg9DHASD
Entspricht pMSg9HASD, jedoch mit einem zusätzlichen HA-Tag an Position +18.
pMSg9HArepSD
Entspricht pMSg9HArep, jedoch mit der zusätzlichen, 8 bp vor dem Startcodon von
Gen 9 eingefügten, E. coli Shine-Dalgarno-Sequenz AGGAGG.
pMSg9DHArepSD
Entspricht pMSg9HArepSD, jedoch mit einem zusätzlichen HA-Tag an Position +18.
pUC18 (Fermentas)
Aus diesem Plasmid entstammt das β-Lactamase-Gen, welches im Vektor pGZamp
unter der Kontrolle des induzierbaren Tac-Promotors steht.
2.1.3 DNS-Standard für Agarose-Gelelektrophoresen
DNA Ladder, 1 kb (N3232S), 100 bp (N3231S ) von New England Biolabs GmbH,
Frankfurt, D
2.1.4 Enzyme
DNase II, Sigma-Aldrich Chemie GmbH, Steinheim, D
Lysozym, Serva, Hamburg, D
Proteinase K, Sigma-Aldrich Chemie GmbH, Steinheim, D

Material und Methoden
42
Restriktionsendonukleasen
Alle in dieser Arbeit eingesetzten Restriktionsendonukleasen wurden von der Firma
Fermentas Life Sciences GmbH (St. Leon-Rot, D) bezogen.
DNS-Ligase
Die in dieser Arbeit für Ligationsreaktionen verwendete T4 DNA-Ligase wurde von
Fermentas Life Sciences GmbH (St. Leon-Rot, D) bezogen.
DNS modifizierende Enzyme
Die folgenden Enzyme der Firma Fermentas Life Sciences GmbH (St. Leon-Rot, D)
wurden zur Modifikation von Plasmiden und DNS-Fragmenten verwendet:
Klenow-Fragment (DNA-Polymerase I)
T4 DNA-Polymerase
Shrimp Alkaline Phosphatase
T4 Polynukleotidkinase
DNS-Polymerasen
Für QuikChange® Mutagenesen und PCR-Amplifikationen verwendete Enzyme:
PfuUltraII™ Fusion HS, Stratagene, La Jolla, USA
GoTaq® Flexi DNA-Polymerase, Promega Corp., Madison,
WI, USA
2.1.5 Antikörper
Primäre Antikörper:
Anti-M13 Coat (AII), Western/Dot-Blot 1:10000 in 1x TBS, Immunpräzipitation
8 µl Serum, Kaninchenserum, Institutsbestand
Anti-M13 pI/XI (C-terminaler Antikörper), Western-Blot 1:10000 in 1x TBS, Kanin-
chenserum. Der polyklonale Antikörper wurde mit Hilfe des synthetischen Peptids
CVSIKKGNSNEIVK hergestellt, GenScript corp., Scotch Plains, NJ, USA
Anti-T7-Tag, Western-Blot 1:5000 in 1x TBS, Immunpräzipitation 3 µl Serum,
monoklonaler Antikörper, Novagen, USA
Anti-HA-Tag, Western-Blot 1:1000 in 1x TBS, Immunpräzipitation 5 µl Serum,
monoklonaler Antikörper, Roche Pharma AG, Grenzach-Wyhlen, D
Anti-YidC, Western-Blot 1:10000 in 1x TBS, C-terminaler Antikörper, Kanin-
chenserum, Institutsbestand

Material und Methoden
43
Anti-GroEL, Western-Blot 1:10000 in 1x TBS, Immunpräzipitation 1 µl Serum,
Kaninchenserum, Sigma-Aldrich Chemie GmbH, Steinheim, D
Anti-OmpA, Western-Blot 1:10000 in 1x TBS, Immunpräzipitation 1 µl Serum,
Kaninchenserum, Institutsbestand
Sekundäre Antikörper:
Serum gegen primäre Antikörper aus Kaninchen, monoklonaler anti-Rabbit IgG
aus Ziege mit konjugierter Peroxidase, Sigma-Aldrich GmbH, Steinheim, D
Serum gegen primäre Antikörper aus Mäusen, monoklonaler anti-mouse IgG aus
Schaf mit konjugierter Meerrettichperoxidase, Amersham, GE Healthcare,
Buckinghamshire, UK
Detektion und Visualisierung gebundener Erstantikörper an M13-Phagen mit Hilfe
von Protein-A konjugiertem Gold mit 20 nm Durchmesser, P6855, Sigma-Aldrich
Chemie GmbH, Steinheim, D
Protein-A Zysorbin™ Immunoadsorbent für die Isolierung von Proteinen mit
primär gebundenen Antikörpern während der Immunpräzipitation, ZYTOMED
GmbH, Berlin, D
2.1.6 Proteinstandard
PageRuler™ Prestained Protein Ladder PR0671, Fermentas Life Sciences GmbH,
St. Leon-Rot, D
PageRuler™ Prestained
170 kDa
130 kDa
100 kDa
70 kDa
55 kDa
40 kDa
35 kDa
25 kDa
15 kDa
10 kDa
Tabelle 2-1: Größenverteilung der Proteine im Proteinstandardgemisch nach gelelektrophoretischer
Auftrennung.

Material und Methoden
44
2.1.7 Aminosäuren, Zucker und Vitamine
Aminosäuren:
[35S]-L-Methionin, MP Biomedicals, Irvine, USA
Alle anderen in dieser Arbeit verwendeten Aminosäuren stammen von BIOMOL
Feinchemikalien GmbH, Hamburg, D.
Zucker:
Arabinose, USB, Cleveland, USA
Glukose, USB, Cleveland, USA
Vitamine:
Vitamin B1, Merck, Darmstadt, D
2.2 Chemikalien und Verbrauchsmaterialien
A
Aceton Carl Roth KG, Karlsruhe, D
Acrylamid Carl Roth KG, Karlsruhe, D
AFM Messspitzen NPS-20 Brucker AXS, Palaiseau, F
Agar GERBU Biotechnik GmbH, Gaiberg, D
Agarose Ultrapure USB Corporation, Cleveland, USA
Alamethicin (aus Trichoderma viride) Sigma-Aldrich Chemie GmbH, Steinheim, D
Ampicillin USB Corporation, Cleveland, USA
Ammoniumchlorid, NH4Cl Merck, Darmstadt, D
Ammoniumperoxodisulfat (APS) Merck, Darmstadt, D
B
Bisacrylamid Sigma-Aldrich Chemie GmbH, Steinheim, D
Borsäure Merck, Darmstadt, D
Bromphenolblau Bio Rad Laboratories, CA, USA
C
Carbonylcyanid-m-Chlorophenyl- Merck, Darmstadt, D
hydrazon (CCCP)
Caesiumchlorid, CsCl USB, Cleveland, OH, USA
Calciumchlorid, CaCl2 Dihydrat Sigma-Aldrich Chemie GmbH, Steinheim, D
Chloramphenicol Sigma-Aldrich Chemie GmbH, Steinheim, D

Material und Methoden
45
Coomassie Brilliant Blue R250 Serva Electrophoresis GmbH, Heidelberg, D
D
2’-Desoxy-Nukleosid-5’-Triphosphat USB, Cleveland, OH, USA
(dNTP)
Di-2’-Desoxy-Nukleosid-5’-Triphos- USB, Cleveland, OH, USA
phat (ddNTP)
Dimethylsulfoxid (DMSO) Merck, Darmstadt, D
Dinatriumhydrogenphosphat, J. T. Baker, Deventer, Nl
Na2HPO4
1,4-Dithiotreitol, DTT GERBU Biotechnik GmbH, Gaiberg, D
Dodecylphosphocholin Anatrace Inc., Maumee, OH, USA
(Fos-cholin12 )
E
ECL Immobilon Western Detection Millipore GmbH, Schwalbach/Taunus, D
Reagens
Eisessig Merck, Darmstadt, D
Eisen(III)-citrat tribasisch Hydrat Merck, Darmstadt, D
Entwickler Eukobrom TETENAL AG & Co. KG,
Norderstedt, D
Ethanol Merck, Darmstadt, D
Ethidiumbromid Merck, Darmstadt, D
Essigsäure (100 %) Merck, Darmstadt, D
Ethylendiamintetraessigsäure USB Corporation, Cleveland, OH, USA
(EDTA)
F
Film (Valmex Blue sensitive X-Ray Valmex photographische Produkte,
VA711B) Augsburg, D
Filterpapier (1 mm, 3 mm) Whatman GB 004 Gel Blotting paper GmbH,
Schleicher & Schuell, Dassel D
Fixierer Superfix Plus, Eukobrom TETENAL AG &
Co. KG, Norderstedt, D
G
Gelfiltrationssäule (XK 16/40) Amersham, GE Healthcare,
Buckinghamshire, UK

Material und Methoden
46
Glaswaren Schott AG, Jena, D
Glycerin Fluka Chemie GmbH, Buchs, CH
Glycin USB, United States Biochemical Corporation,
Cleveland, OH, USA
H
Harnstoff, Urea Sigma-Aldrich Chemie GmbH, Steinheim, D
HClkonz Merck, Darmstadt, D
Hefeextrakt Oxoid LTD., Basingstone, Hampshire, UK
I
Imidazol Sigma-Aldrich Chemie GmbH, Steinheim, D
Isopropanol Fluka Chemie GmbH, Buchs, CH
Isopropyl-β-D-galactopyranosid Gerbu Biotechnik GmbH, Gaiberg, D
(IPTG)
K
Kaliumchlorid, KCl Sigma-Aldrich Chemie GmbH, Steinheim, D
Kaliumdihydrogenphosphat, KH2PO4 Fluka Chemie GmbH, Buchs, CH
Kaliumhydroxid, KOH Merck, Darmstadt, D
Kupfernetzchen (Athene 200) Plano, Wetzlar, D
L
Long RangerTM Gel Lösung FMC BioProducts, Rockland, ME, USA
M
Magermilchpulver Heirler Cenovis GmbH, Radolfzell, D
Magnesiumchlorid, MgCl2 6 H2O Merck, Darmstadt, D
Magnesiumsulfat, MgSO4 Merck, Darmstadt, D
Mangan(II)-chlorid, MnCl2 4 H2O Sigma-Aldrich Chemie GmbH, Steinheim, D
Membran für Western-Blot Hybond Nitrocellulose Membrane, Amersham
ECL Bioscience, Buckinghamshire, UK
Minisäule BD PlastipakTM, 5 ml-Spritze, Becton
Dickinson, Heidelberg, D
Monomerlösung Carl Roth GmbH & Co., Karlsruhe, D
(30% Acrylamid, 0,8% Bisacrylamid)
Muscovite Mica V-1 Quality Electron Microscopy Sciences, München, D

Material und Methoden
47
N
Natriumchlorid, NaCl USB Corporation, Cleveland, OH, USA
Natriumdodecylsulfat (SDS) Serva Electrophoresis GmbH, Heidelberg, D
Natriumhydroxid, NaOH Merck, Darmstadt, D
Ni-NTA-Agarose, Ni-NTA-Superflow, Qiagen, Hilden, D
Bindungskapazität 50 mg/ml
Nitrocellulose Transfere Membrane Whatman GmbH, Dassel, D
PROTRAN®
Nukleotidmix, 4dNTP Fermentas GmbH, St. Leon-Rot, D
P
Phenylmethansulfonsäurefluorid Sigma-Aldrich Chemie GmbH,
(PMSF) Steinheim, D
Polyethylenglycol, PEG 6000 Applichem GmbH, Darmstadt, D
Ponceau S Sigma-Aldrich Chemie GmbH, Steinheim, D
Primer Biomers, Ulm, D
R
Rubidiumchlorid, RbCl Sigma-Aldrich Chemie GmbH, Steinheim, D
Reaktionsgefäße (1,5 ml, 2 ml) Sarstedt, Nümbrecht, D
S
Schwefelsäure, H2SO4 Merck, Darmstadt, D
Sephadex G10 Sigma-Aldrich Chemie GmbH, Steinheim, D
Superdex 200TM Amicon, Schorndorf, D
Spritzen BD PlastipakTM, 50 ml, Becton Dickinson,
Heidelberg, D
Spritzenfilter, ø 25 mm, 0,22 µm Fisherbrand, Syringe Filter, Fisher Scientific
GmbH, Schwerte, D
T
Tetramethylethylendiamin (TEMED) Sigma-Aldrich Chemie GmbH, Steinheim, D
Thermo SequenaseTM Cycle USB, Cleveland, OH, USA
Sequencing Kit
Trichloressigsäure, TCA Merck, Darmstadt, D
Tricin Sigma-Aldrich Chemie GmbH, Steinheim, D
Tris (hydroxymethyl)-aminomethan USB, Cleveland, OH, USA
TritonX-100 Sigma-Aldrich Chemie GmbH, Steinheim, D

Material und Methoden
48
Trypton BitTekTM Oxoid LTD., Basingstone, Hampshire, UK
2.3 Oligonukleotide
Der Bezug sowohl von fluoreszenzmarkierten Sequenzierprimern als auch von
Primern für die Durchführung ortsgerichteter Mutagenesen in Gen 8 und 9 des
Phagen M13 erfolgte über die Firma Eurofins MWG Operon (Ebersberg, D). Primer
für die Amplifizierung und Durchführung ortsgerichteter Mutagenesen in Gen 1 des
Phagen M13 erfolgte über die Firma Biomers.net GmbH (Ulm, D).
2.3.1 Sequenzierprimer Annealing Temperatur
Procoat Start: 5´-ATTACGTATTTTACCCG-3´ 47°C
Procoat Back: 5´-GGAGCCTTTATTTGTATCGGT-3´ 59°C
Tac Forward: 5´-GACAATTAATCATCGGCTCG 56°C
Tac Back: 5´-GGCTGAAAATCTTCTCTCATCCG-3´ 59°C
T7 Forward: 5´-TAATACGACTCACTATAGGG-3´ 55°C
T7 Back: 5´-CGCTGAGATAGGTGCCTCAC-3´ 53°C
Universal reverse: 5´-TTGTGAGCGGATAACAATTTC-3´ 52°C
2.3.2 Oligonukleotide für ortsgerichtete Mutagenesen
Insertion einer Ambermutation in Gen 8 an Position +3:
(A): 5´-CCTCATGAAATAGTACTTAGTCCTCAAAGC-3´ Tm 68°C
(B): 5´-GCTTTGAGGACTAAGTACTATTTCATGAGG-3´
Insertion einer Ambermutation in Gen 9 an Position +7:
(A): 5´-GTTTTAGTGTATTAGTTCGCCTCTTTCG-3´ Tm 66°C
(B): 5´-CGAAAGAGGCGAACTAATACACTAAAAC-3´
Insertion einer MunI-Schnittstelle in Gen 9 an Position +2:
(A): 5´-GGGTCAAAGATGAGTCAATTGGTTTTAGTGTATTC-´3 Tm 70°C
(B): 5´-GAATACACTAAAACCAATTGACTCATCTTTGACCC-´3
Insertion der Shine-Dalgarno-Sequenz AGGAGG vor das Startcodon
von Gen 9 an Position -8:
(A): 5´-GCTTGGTATAATCGCAGGAGGTCAAAGATGAGTC-3´ Tm 74°C
(B): 5´-GACTCATCTTTGACCTCCTGCGATTATACCAAGC-3´

Material und Methoden
49
Reparatur der Startsequenzen in Gen 9 durch Insertion des Bereichs +1 und +2 von
gp9 an Position +20 (gp9HArep) bzw +22 (gp9T7rep):
gp9HA
(A): 5´-GATTATGCACAATTGATGAGTGTTTTAGTGTATTCTTTTG-3´ Tm 70°C
(B): 5´-CAAAAGAATACACTAAAACACTCATCAATTGTGCATAATC-3´
gp9T7
(A): 5´-GGTTCAGCGCAATTGATGAGTGTTTTAGTGTATTCTTTTG-3´ Tm 74°C
(B): 5´-CAAAAGAATACACTAAAACACTCATCAATTGCGCTGAACC-3´
2.3.3 Oligonukleotide für die Amplifikation von Genen
Gen 7/9:
(A): 5´-TGTACACCGTTCATCTGTCC-3´ Tm 60°C
(B): 5´-TCATGAGGAAGTTTCCATTAAACG-3´ Tm 59°C
Gen 1:
(A): 5´-CCGCCTCGAGATGGCTGTTTATTTTGTAACTGGC-3´ Tm 68°C
(B): 5´-CCGCTCAGCTTAATTACATTTAACAATTTCATTTGAATTACC-3´ Tm 66°C
2.3.4 Oligonukleotide für die Insertion eines antigenen Epitops
T7-Tag:
(A): 5´-AATTAATGGCGTCCATGACCGGTGGTCAACAGATGGGTTCAGGTTCA
GCGC-3´ Tm 85°C
(B): 5´-AATTGCGCTGAACCTGAACCCATCTGTTGACCACCGGTCATGGACGC
CATT-3´ Tm 85°C
HA-Tag:
(A): 5´-AATTAGCGTCAGGTTCATATCCGTATGACGTTCCGGATTATGCAC-3´
Tm 79°C
(B): 5´-AATTGTGCATAATCCGGAACGTCATACGGATATGAACCTGACGCT-3´
Tm 79°C

Material und Methoden
50
2.4 Geräte
AFM („Atomic Force Microscope“)
Nanoscope IIIa, Digital Instruments, USA
Agarose-Gelelektrophoresekammer
Easy Electrophoresis Systems, Model #B1A, Owl Scientific Inc., USA
Äkta
Basic Chromatographie System pH/C-900, UV-900, P-900, Amersham Biosciences,
Freiburg, D
Autoklaven
H+P Varioklav® 75S, H+P Labortechnik GmbH, Oberschleißheim, D
H+P Varioklav® 135S, H+P Labortechnik GmbH, Oberschleißheim, D
Bildaufnahmesysteme (Aufnahme von Gelen und LB-Medienplatten)
Digitales Bildaufnahmesystem, Darkroom Evo III mit Software Diana/95 Camera
Control v. 1.6, Raytest Isotopenmessgeräte, Straubengardt, D
Bio-IMAGING Analyser BAS-1500 und Geldokumentationssystem mit Software
AIDA, Raytest Isotopenmessgeräte, Straubengardt, D
Scanner UMAX PowerLook III mit Software Adobe Photoshop 5.0, Amersham,
GE Healthcare, Buckinghamshire, UK
Blotkammer (Western-Blot)
semi-dry Blotkammer, Owl Separation Systems, Schleicher & Schuell, Dassel, D
Brutschränke
Heraeus B6030, Heraeus Sepatech GmbH, Osterode, D
Heraeus T6200, Heraeus Sepatech GmbH, Osterode, D
DNS-Sequenzierer
DNA-Sequencer LI-COR® Modell 4200, Software e-Seq v3.0, LI-COR® Inc., Lincoln,
USA
Drehrad
Gerlinde Kisker, Produkte für die Biotechnologie, Mühlhausen, D
Eismaschine
Scotsman® AF20, Scotsman Ice Systems, Vernon Hills, Illinois, USA
Geltrockner
BioRad Gel Dryer Model 583, Bio Rad Laboratories, Hercules, CA, USA

Material und Methoden
51
Heizblock
Unitek HB-130, Miyachi Unitek, Monrovia, CA, USA
Magnetrührer
Magnetrührer MR 2000, Heidolph Elektro GmbH & Co. KG, Kelheim, D
Mikroskop
Standard 18, Carl Zeiss, Oberkochen, D
Netzgeräte
Power Pac 300, BioRad Laboratories, Inc., Hercules, CA, USA
Power Pac 3000, BioRad Laboratories, Inc., Hercules, CA, USA
pH-Messgerät
Metrohm 620 pH-Meter, Metrohm, Herisau, CH
Photometer
Cell Density Meter CO 8000, WPA Biowave, Cambridge, UK
Lambda 15 UV/VIS Spectrophotometer, Perkin-Elmer Inc., Waltham, MA, USA
SDS-PAGE
Hoefer Electrophoresis Unit SE600 Series, Amersham Biosciences, Freiburg, D
Sterilisation
gebrauchsfertige Filtereinheit, Filtration Life Science Diagnostic Components,
Schleicher & Schuell, Dassel, D
Sterilbank
BDK Luft- und Reinraumtechnik GmbH, Sonnenbühl-Genkingen, D
Thermoblock/Mixer
Thermoblock, Unitek HB-130, Biozym, Hermeln, D
Thermomixer 5436, Eppendorf AG, Hamburg, D
Thermocycler
T3 Thermocycler und TGradient, Biometra GmbH, Goettingen, D
Tischschüttler
Vortex genie 2, Scientific Industries, bender & Hobein AG, Zürich, CH
Transmissionselektronenmikroskop (TEM)
LEO 912AB mit Omega Filter für EFTEM, 80kV, Punkt zu Punkt Auflösung 0,37 nm,
Carl Zeiss NTS GmbH, Oberkochen, D

Material und Methoden
52
Vakuumkonzentration
SpeedVac Concentrator, Savant Instruments Inc. Model RH40-11 mit Refrigerated
Condensation Trap (Kühlfalle) und High Vacuum Pump, Bachofer Laboratoriums-
geräte, Reutlingen, D
Waagen
Sartorius 1264MP, Sartorius Werke GmbH, Göttingen, D
Sartorius Competence CP 224 S, Sartorius Werke GmbH, Göttingen, D
Wasseraufbereitungsanlage (zur Erzeugung von H2Obidest)
Milli-Q® Water Purification System, Q-Garde® 2, Millipore GmbH, Eschborn, D
Wasserbad
Wasserbadschüttler HT, Infors AG, Bottmingen, CH
Zellaufschlusssystem
French Pressure Cell Press, American Instrument Company, MA, USA
Zentrifugen
Tischzentrifuge Z 200 M/H, HERMLE Labortechnik GmbH, Wehingen, D
Tischkühlzentrifuge Z 233 MK, HERMLE Labortechnik GmbH, Wehingen, D
Kühlzentrifuge Model J2-21, Zentrifuge mit den Rotoren JA14, JA20, Beckman
Instruments GmbH, USA
Kühlzentrifuge Model Avanti J25, Zentrifuge mit den Rotoren JLA25.50,
JLA10.500, JLA16.250, Beckman Coulter, Inc. Fullerton, CA, USA
Ultrazentrifuge Optima LE-80K, Zentrifuge mit den Rotoren 50Ti, 60Ti, SW28,
Beckman Instruments GmbH, Fullerton, CA, USA
2.5 Verwendete Kits
ZippyTM Plasmid Miniprep Kit, Zymo Research, Irvine, CA, USA
ZymocleanTM, Gel DNS Recovery Kit, Zymo Research, Irvine, CA, USA
GFXTM PCR DNS and Gel Band purification Kit, Amersham Inc., Cleveland, OH,
USA
ECL PlusWestern Blotting Reagent Pack, Amersham Inc., Cleveland, OH, USA
2.6 Kulturmedien
2.6.1 Festmedien
LB-Agarplatten:
LB-Medium, 1,5 % (w/v) Agar, pH 7,5 (NaOH), autoklaviert

Material und Methoden
53
LB-Agarplatten mit Ampicillin:
LB-Medium, 1,5 % (w/v) Agar, pH 7,5 (NaOH), autoklaviert, Ampicillin (100 µg/ml)
sterilfiltriert
LB-Topagar:
LB-Medium, 0,7 % (w/v) Agar, pH 7,5 (NaOH), autoklaviert
2.6.2 Flüssigmedien
M9 Minimalmedium (10 x):
300 mM Na2HPO4 x 2 H2O, 220 mM KH2PO4, 180 mM NH4Cl, 80 mM NaCl
„Pulse“-Medium:
10 % (v/v) M9 Minimalmedium (10 x), 0,1 % (v/v) 1 M MgSO4, 0,1 % (v/v) 1 M CaCl2,
0,5 % (v/v) Vitamin B1-Lösung (1 mg/ml), 0,5 % (v/v) Eisencitrat (0,1 %), 0,5 % (v/v)
Glukoselösung (40 %), Aminosäuremix ohne L-Methionin 10 % (v/v).
Für Pulsversuche mit araBAD Promotor wird zur Induktion plasmidkodierter Proteine
die Glukoselösung durch eine Arabinoselösung ersetzt.
Aminosäuremix ohne L-Methionin:
Für die Herstellung werden alle Aminosäuren aus Tabelle 2-2 getrennt eingewogen
und, mit Ausnahme der Asparaginsäure und Tyrosin, in ddH2O gelöst.
Tabelle 2-2: Molarität der einzelnen Aminosäurelösungen für den Aminosäuremix ohne
L-Metionin. *Lösen in 1,5 ml 0,5 N HCl, **Lösen in 10 ml 0,1 N NaOH
Aminosäure Molarität [mM] Alanin 100 Arginin 50
Asparagin 100 Asparaginsäure 100*
Cystein 100 Glutamin 100
Glutaminsäure 50 Glycin 100 Histidin 100
Isoleucin 100 Leucin 100 Lysin 100
Phenylalanin 100 Prolin 100 Serin 100
Threonin 100 Tryptophan 50
Tyrosin 5**
Valin 100

Material und Methoden
54
Hiervon werden 2 ml Tyrosinlösung, 200 µl Argininlösung, 200 µl Glutaminsäure-
lösung, 200 µl Tryptophanlösung und jeweils 100 µl der verbleibenden Aminosäure-
lösungen mit 6 ml ddH2O gemischt und mit 10 µl HClkonz neutralisiert.
LB-Medium:
1 % (w/v) Trypton, 0,5 % (w/v) NaCl, 0,5 % (w/v) Hefeextrakt, pH 7,5 (NaOH),
autoklaviert
LB-Medium mit Ampicillin:
1 % (w/v) Trypton, 0,5 % (w/v) NaCl, 0,5 % (w/v) Hefeextrakt, pH 7,5 (NaOH),
autoklaviert, Ampicillin (100 µg/ml) sterilfiltriert
2.7 Puffer und Lösungen
2.7.1 Agarosegelelektrophorese
10x TBE-Puffer (pH 8,3):
Substanz Menge
Tris 2,2 M
Borsäure 1,8 M
EDTA pH 8,0 50 mM
4x DNS-Probenpuffer:
Substanz Menge
Glycerol 50 % (w/v)
Bromphenolblau 0,4 % (w/v)
EDTA 1 mM
10x Ethidiumbromidlösung:
Substanz Menge
Ethidiumbromid 10 mg/ml

Material und Methoden
55
2.7.2 Coomassiefärbung
Färbelösung:
Substanz Menge Coomassie Brilliant blue
R250 0,1 % (w/v)
Ethanol 40 % (v/v)
Essigsäure 100 % 10 % (v/v)
Entfärbelösung:
Substanz Menge
Essigsäure 100 % 10 % (v/v)
2.7.3 DNS-Puffer
10x TE-Puffer (pH 8 mit HCl):
Substanz Menge
Tris 0,1 M
EDTA 10 mM
2.7.4 DNS-Sequenziergel
Stammlösungen:
10x TBE (pH 8,3)
Substanz Menge
Tris 2,2 M
Borsäure 1,8 M
EDTA 50 mM
sterilfiltrieren
Sequenziergel
Substanz Menge
Harnstoff 42 % (w/v)
Acrylamidlösung LongRanger
TM
12 % (v/v)
10 x TBE 12 % (v/v)
10 % (w/v) APS 0,005 % (v/v)
TEMED 0,0005 % (v/v)

Material und Methoden
56
2.7.5 Herstellung von Phagenstammlösungen
Präzipitationspuffer:
Substanz Menge
PEG 6000 20 % (w/v)
NaCl 2,5 M
sterilfiltrieren
2.7.6 Induktion von Expressionssystemen
1 M IPTG:
Substanz Menge
IPTG 23,8 % (w/v)
sterilfiltrieren
Arabinose:
Substanz Menge
Arabinose 25 % (w/v)
sterilfiltrieren
2.7.7 Kompetente E. coli-Zellen
TFB1-Puffer (pH 5,8 mit Essigsäure):
Substanz Menge
RbCl 100 mM
MnCl2 50 mM
Kaliumacetat 30 mM
CaCl2 10 mM
Glycerin 15 % (w/v)
sterilfiltrieren
TFB2-Puffer (pH 7,0 mit KOH):
Substanz Menge
MOPS 10 mM
RbCl 10 mM
CaCl2 75 mM
Glycerin 15 % (w/v)
sterilfiltrieren

Material und Methoden
57
2.7.8 Nanogoldmarkierung von M13-Phagen
10x TBS-Puffer (pH 7,4):
Substanz Menge
Tris 200 mM
NaCl 1,5 M
KCl 27 mM
autoklavieren
Block-Lösung:
Substanz Menge
10x TBS 10 % (v/v)
BSA 0,1 % (w/v)
Staph-A Nanogold-Lösung:
Substanz Menge
10x TBS 10 % (v/v)
BSA 0,1 % (w/v)
Staph-A Nanogoldlösung 0,05 % (v/v)
2.7.9 Negativkontrastierung von biologischen Proben für die Transmissions-
elektronenmikroskopie (TEM)
Substanz Menge
Phosphorwolframsäure 5 % (w/v)
Ethanol 50 % (w/v)
Der pH-Wert der Lösung wird mit Natronlauge auf pH 7 eingestellt.
2.7.10 Proteinstandard für die präparative Gelfiltration
Substanz Menge
Apoferritin 25 mg/ml
β-Amylase 5 mg/ml
BSA 5 mg/ml
Carboanhydrase 3 mg/ml
Tris/HCl pH 7,5 20 mM
NaCl 50 mM
Fos-cholin 12 0,8 % (v/v)

Material und Methoden
58
2.7.11 Radioaktive Markierung in vivo mit anschließender Immunpräzipitation
und Protease-Mapping
Stammlösungen:
Substanz Menge
EDTA pH 8 100 mM
IPTG 1 M
[35
S]-L-Methionin 10 µCi/µl
Proteinase K 20 mg/ml
Protein-A
Lysozym 0,5 mg/ml
Fixierlösung für radioaktive SDS-Gele:
Substanz Menge
Ethanol 40 % (v/v)
Essigsäure 100 % 10 % (v/v)
Probenaufbereitung:
Substanz Menge
Aceton
TCA 100 % (w/v)
Pufferlösungen:
TEN-Puffer
Substanz Menge
Tris/HCl pH 8 10 mM
NaCl 150 mM
EDTA 1 mM
TENTX-Puffer
Substanz Menge
Tris/HCl pH 8 10 mM
NaCl 150 mM
EDTA 1 mM
Triton X-100 2 % (v/v)

Material und Methoden
59
Spheroplastenpuffer
Substanz Menge
Saccharose 40 % (w/v)
Tris/HCl pH 8 33 mM
2.7.12 Reinigung der gp1/11-Assembly-Komplexe mittels präparativer Gelfil-
tration
Puffer für Säule XK 16/40
Substanz Menge
Tris/HCl pH 7,5 20 mM
NaCl 50 mM
Fos-cholin 12 0,8 % (v/v)
2.7.13 Reinigung der gp1/11-Assembly-Komplexe mittels Metallaffinitätschro-
matographie (IMAC)
Aufbewahrungspuffer für E. coli-Zellen:
Substanz Menge
Tris/HCl pH 7,5 20 mM
Saccharose 25 % (w/v)
Zellen können über mehrere Monate bei -80 °C gelagert werden.
Stammlösungen für den Zellaufschluß im „French-Press“ Verfahren:
Substanz Menge
DNase II 25 mg/ml
PMSF in DMSO 100 mM
MgCl2 1 M
CaCl2 20 mg/ml
Stammlösungen für die Metallaffinitätschromatographie (IMAC):
Substanz Menge
Imidazol/HCl pH 7 1 M
NaCl 5 M
Tris/HCl pH 7,5 1 M
Fos-cholin 12 10 %

Material und Methoden
60
Puffer für die Metallaffinitätschromatographie (IMAC):
Equilibrierungspuffer für Ni-NTA-Säulenmatrix
Substanz Menge
Imidazol 10 mM
Tris/HCl pH 7,5 20 mM
NaCl 300 mM
Solubilisierungspuffer
Substanz Menge
Imidazol 10 mM
Tris/HCl pH 7,5 20 mM
NaCl 300 mM
Fos-cholin 12 2 % (v/v)
Säulenwaschpuffer 1
Substanz Menge
Imidazol 15 mM
Tris/HCl pH 7,5 20 mM
NaCl 300 mM
Fos-cholin 12 0,8 % (v/v)
Säulenwaschpuffer 2
Substanz Menge
Imidazol 40 mM
Tris/HCl pH 7,5 20 mM
NaCl 300 mM
Fos-cholin 12 0,8 % (v/v)
Elutionspuffer
Substanz Menge
Imidazol 300 mM
Tris/HCl pH 7,5 20 mM
NaCl 300 mM
Fos-cholin 12 0,8 % (v/v)

Material und Methoden
61
2.7.14 SDS-Polyacrylamid-Gelelektrophorese (PAGE)
Probenaufbereitung/Stammlösungen:
Substanz Menge
Aceton
APS 10 % (w/v)
SDS 25 % (w/v)
TEMED
TCA 100 % (w/v)
SDS-Probenpuffer:
Lösung 1
Substanz Menge
Tris/HCl pH 7,0 200 mM
EDTA/HCl pH 7,0 20 mM
Lösung 2
Substanz Menge
Tris/HCl pH 7,0 83,3 mM
Glycerin 29,2 % (v/v)
SDS 8,8 % (w/v)
Bromphenolblau 0,14 % (w/v)
1 M DDT
Die Lösungen werden im folgenden Verhältnis vor der Verwendung miteinander
gemischt: Lösung 1 : Lösung 2 : 1 M DTT
5 : 4 : 1 ( v/v)
2.7.14.1 SDS-PAGE nach Laemmli (1970)
10x SDS-Laufpuffer:
Substanz Menge
Tris 3 % (w/v)
Glycin 14,4 % (w/v)
SDS 1 % (w/v)

Material und Methoden
62
SDS-Gele ohne Harnstoff
Monomerlösung:
Substanz Menge
Acrylamid 30 % (w/v)
Bisacrylamid 0,8 % (w/v)
Zusammensetzung von Sammel- und Trenngel (18 x 14 cm):
Sammelgel (3 %)
Substanz Menge
1 M Tris/HCl pH 6,8 1,75 ml
Monomerlösung 1,33 ml
ddH2O 7,3 ml
25 % (w/v) SDS 40 µl
10 % (w/v) APS 50 µl
TEMED 5 µl
Trenngel (12 %)
Substanz Menge
2 M Tris/HCl pH 8,8 2,81 ml
Monomerlösung 6,0 ml
ddH2O 6,0 ml
25 % (w/v) SDS 60 µl
10 % (w/v) APS 75 µl
TEMED 5 µl
SDS-Gele mit Harnstoff
Lösungen:
Substanz Menge
Acrylamid 45 % (w/v)
Bisacrylamid 2 % (w/v)
Zusammensetzung von Sammel- und Trenngel (18 x 14 cm):
Sammelgel (3 %)
Substanz Menge
Harnstoff 3,6 g
1 M Tris/HCl pH 6,8 0,6 ml
45 % (w/v) Acrylamid 1,1 ml

Material und Methoden
63
2 % (w/v) Bisacrylamid 0,65 ml
ddH2O 4,85 ml
25 % (w/v) SDS 40 µl
10 % (w/v) APS 100 µl
TEMED 8 µl
Trenngel (22 %)
Substanz Menge
Harnstoff 7,2 g
3,3 M Tris/HCl pH 8,7 2,5 ml
45 % (w/v) Acrylamid 9,8 ml
2 % (w/v) Bisacrylamid 0,8 ml
25 % (w/v) SDS 100 µl
10 % (w/v) APS 70 µl
TEMED 10 µl
2.7.14.2 SDS-Tricin-Gelelektrophorese nach Schägger & Jagow (1987)
10x Kathodenpuffer (- Pol) (pH 8,25 mit HCl):
Substanz Menge
Tris 1 M
Tricin 1 M
SDS 1 % (w/v)
10x Anodenpuffer (+ Pol) (pH 8,9 mit HCl):
Substanz Menge
Tris 2 M
Monomerlösung:
Substanz Menge
Acrylamid 48 % (w/v)
Bisacrylamid 1,5 % (w/v)
Gel-Pufferlösung:
Substanz Menge
Tris/HCl pH 8,9 3 M
SDS 0,3 % (w/v)

Material und Methoden
64
Zusammensetzung von Sammel-, Spacer- und Trenngel (18 x 14 cm):
Sammelgel (4 %)
Substanz Menge
Monomerlösung 480 µl
Gel-Pufferlösung 1,49 ml
ddH2O 4,03 ml
10 % (w/v) APS 30 µl
TEMED 3 µl
„Spacer“-Gel (10 %)
Substanz Menge
Monomerlösung 0,61 ml
Gel-Pufferlösung 1 ml
ddH2O 1,39 ml
10 % (w/v) APS 10 µl
TEMED 2 µl
Trenngel (16,5 %)
Substanz Menge
Monomerlösung 2,66 ml
Gel-Pufferlösung 2,66 ml
Glycerol 1,06 g
ddH2O 1,62 ml
10 % (w/v) APS 30 µl
TEMED 3 µl
2.7.15 Stammlösungen
Substanz Menge
Ampicillin 20 mg/ml
Chloramphenicol 25 mg/ml
Glukose 40 % (w/v)
NaCl 5 M

Material und Methoden
65
2.7.16 Dot-Blot / Western-Blot
10x TBS-Puffer (pH 7,4):
Substanz Menge
Tris 200 mM
NaCl 1,5 M
KCl 27 mM
10x Transferpuffer:
Substanz Menge
Tris 500 mM
Glycin 3 M
Ponceau S-Lösung:
Substanz Menge
Ponceau S-Farbstoff 0,2 % (w/v)
Essigsäure 5 % (v/v)
Block-Lösung:
Substanz Menge
10x TBS 10 % (v/v)
Milchpulver 5 % (v/v)
„Stripping“-Puffer (pH 2,3) zur Entfernung gebundener Antikörper:
Substanz Menge
Glycin 200 mM
NaCl 500 mM
2.8 Methoden
2.8.1 Herstellung von E. coli-Vorkulturen
2 ml LB-Medium wurde mit einer E. coli-Einzelkolonie beimpft und über Nacht bei
37°C geschüttelt. Bei Transformanten erfolgte der Zusatz des entsprechenden Anti-
biotikums zum Medium.
2.8.2 Herstellung transformationskompetenter E. coli-Zellen
Zum Beimpfen einer 50 ml LB-Kultur im Verhältnis 1:100 wurde eine frische Über-
nachtvorkultur des gewünschten E. coli-Stamms verwendet. Die Inkubation der

Material und Methoden
66
Zellkultur erfolgte anschließend unter Schütteln bei 37°C bis zu einer optischen
Dichte (OD600) von 0,5. Daraufhin wurde die Kultur auf Eis gestellt, in ein steriles
Zentrifugengefäß überführt und zentrifugiert (5000 g, 10 min, 4°C). Der Überstand
wurde verworfen, das Zellpellet in 30 ml TFB1-Puffer (4°C) aufgenommen und
60 Minuten auf Eis gestellt. Es erfolgte eine erneute Zentrifugation, nach welcher der
Überstand abermals verworfen und das Pellet nun in 2 ml TFB2-Puffer aufge-
nommen wurde. Die Aliquotierung der Zellen erfolgte zu 50 µl in auf Eis vorgekühlte
1,5 ml Plastikreaktionsgefäße, welche direkt in flüssigem Stickstoff eingefroren und
bei -80°C gelagert wurden.
2.8.3 Transformation von E. coli
50 µl transformationskompetenter Bakterienzellen wurden auf Eis aufgetaut und
anschließend mit 1-2 µl Plasmid-DNS versetzt. Hierauf folgte eine 30-minütige
Inkubation der Zellen auf Eis mit einem anschließenden Hitzeschock (42°C, 90 s)
und Reinkubation der Zellen auf Eis (2 min). Die transformierten Zellen wurden
anschließend auf LB-Agarplatten mit entsprechendem Antibiotikum ausgebracht und
über Nacht bei 37°C inkubiert.
2.8.4 Transfektion von E. coli
50 µl transformationskompetenter Bakterienzellen wurden auf Eis aufgetaut und
anschließend mit 1-2 µl RF-Plasmid-DNS versetzt. Hierauf folgte eine 30-minütige
Inkubation der Zellen auf Eis mit einem anschließenden Hitzeschock (42°C, 90 s)
und Reinkubation der Zellen auf Eis (2 min). Im Fall des M13-Phagenstamms mit
β-Lactamase-Gen wurden die transfizierten Zellen auf LB-Agarplatten mit Ampicillin
(100 µg/ml) und IPTG (1 mM) ausgebracht und bei 37°C über Nacht inkubiert. Für
Phagenstämme ohne β-Lactamase-Gen wurden die transfizierten Zellen nach dem
Hitzeschock max. 30 Minuten bei 37°C inkubiert und mit diesen dann eine
Verdünnungsreihe bis 1:100000 angefertigt. Von den Verdünnungsstufen wurden je
100 µl in, mit Plattierbakterien versetzten, Top-Agar (350 µl E. coli F+, 46°C) auf LB-
Agarplatten ausgebracht und nach Verfestigung des Top-Agars über Nacht bei 37°C
inkubiert.

Material und Methoden
67
2.8.5 Isolierung von Plasmid-DNS aus E. coli
Minipräparation
a) Anzucht einer E. coli-Flüssigkultur
Für die Minipräparation wurden die Plasmide von Interesse zuvor in E. coli-Zellen
transformiert (Kap. 2.8.3). Mit einer Einzelkolonie erfolgte dann die Beimpfung von
2 ml LB-Medium, zu dem entsprechend benötigtes Antibiotikum zugesetzt wurde. Die
Flüssigkultur wurde anschließend über Nacht bei 37°C inkubiert.
b) Isolierung von Plasmid-DNS
Die Plasmidisolierung wurde nach dem Protokoll des ZippyTM Plasmid Miniprep Kits
von Zymo Research durchgeführt.
2.8.6 Isolierung von M13 RF-Plasmid-DNS aus E. coli
a1) Minipräparation nach Infektion
Für die Minipräparation wurde eine, bei 37°C gewachsene, E. coli K37-Kultur (2 ml)
bei einer optischen Dichte (OD600) von 0,2 mit dem M13-Phagenstamm von Interesse
mit einer M.O.I. („Multiplicity of infection“) von 10 infiziert. Anschließend wurde die
Kultur bei 37°C über Nacht inkubiert.
a2) Minipräparation nach Transfektion
Für die Minipräparation wurde das RF-Plasmid des Phagenstamms von Interesse in
kompetente E. coli-Zellen transformiert (Kap. 2.8.4) und diese anschließend in Top-
Agar auf LB-Kulturplatten ausplattiert. Von den gebildeten Plaques auf dem Bakte-
rienrasen wurde eine Einzelplaque mit Hilfe einer Pasteurpipette isoliert und in 500 µl
sterilem ddH2O aufgenommen und anschließend für mindestens 30 Minuten bei RT
inkubiert. 250 µl dieser Phagenlösung wurde dann für die Infektion einer bei 37°C
gewachsene E. coli K37-Kultur (2 ml, siehe a1)) verwendet.
Alternativ kann die isolierte Einzelplaque direkt in 2 ml LB-Medium aufgenommen
und anschließend über Nacht bei 37°C inkubiert werden.
b) Isolierung von M13 RF-Plasmid-DNS
Die Plasmidisolierung wurde nach dem Protokoll des ZippyTM Plasmid Miniprep Kits
von Zymo Research durchgeführt.

Material und Methoden
68
2.8.7 Agarose-Gelelektrophorese
Zu 40 ml 1x TBE-Puffer wurden, entsprechend der zu trennenden DNS-Fragmente
(Tabelle 2-3), die Mengen (w/v) Agarose zugegeben und aufgekocht. Nach dem
Abkühlen auf circa 50°C wurden 20 µg Ethidiumbromid zu der flüssigen Agarose-
schmelze zugesetzt und anschließend in die Agarose-Gelelektrophorese-Vorrichtung
„Easy Electrophoresis Systems“ gegossen. Als Laufpuffer diente 1x TBE. Zu 10 µl
DNS-Probe wurden 5 µl 4x DNS-Probenpuffer zugegeben und diese wurden dann
bei einer konstanten Spannung von 90 V aufgetrennt. Die Abschätzung der DNS-
Fragmentgrößen erfolgte anhand aufgetragener DNS-Standards (1 kb DNA Ladder,
100 bp DNA Ladder, New England BioLabs Inc.)
Fragmentlänge Agarosekonzentration
(w/v)
1 bis 30 kb 0,5 %
0,8 bis 12 kb 0,7 %
0,5 bis 7 kb 1,0 %
0,4 bis 6 kb 1,2 %
0,2 bis 3 kb 1,5 %
0,1 bis 2 kb 2,0 %
2.8.8 Restriktionsverdau von Plasmid-DNS und DNS-Fragmenten
Zur Herstellung der in dieser Arbeit vorgestellten Plasmidkonstrukte wurden folgende
Verdaustrategien angewendet:
a) Einfachverdau (10 µl Gesamtvolumen)
1 µl 10x Enzymreaktionspuffer
1 µl Restriktionsenzym
350 – 400 ng Plasmid-DNS
mit x µl ddH2O steril auf 10 µl ergänzen
Der Ansatz wurde abhängig vom verwendeten Enzym 2 h bei 37°C oder 30°C
inkubiert. Zur Entfernung des Restriktionsenzyms wurde die verdaute DNS über das
ZymocleanTM, Gel DNA Recovery Kit von Zymo Research gereinigt.
Tabelle 2-3: Agarosekonzentrationen und die jeweiligen Fragmentlängen-Trennbereiche

Material und Methoden
69
b) Doppelverdau in geeignetem Enzymreaktionspuffer (15 µl Gesamtvolumen)
1,5 µl 10x Enzymreaktionspuffer
1 µl Restriktionsenzym A
1 µl Restriktionsenzym B
350 – 400 ng Plasmid-DNS
mit x µl ddH2O steril auf 15 µl ergänzen
Der Ansatz wurde abhängig von den verwendeten Enzymen 2 h bei 37°C oder 30°C
inkubiert. Zur Entfernung der Restriktionsenzyme und zur Isolierung des DNS-
Fragments von Interesse wurde die verdaute DNS über eine Agarosegelelektro-
phorese aufgetrennt, das DNS-Fragment isoliert und mit Hilfe des ZymocleanTM Gel
DNA Recovery Kit von Zymo Research nach Vorschrift gereinigt.
C) Doppelverdau ohne geeigneten Enzymreaktionspuffer
Bei Doppelverdauen mit Restriktionsenzymen, welche keinen geeigneten gemein-
samen Reaktionspuffer besaßen, wurden nacheinander Einfachverdaue wie unter a)
beschrieben durchgeführt.
2.8.9 Ligationsansätze
Ligationen wurden mit T4 DNA-Ligase in einem Endvolumen von 10 µl entsprechend
der Konzentrationen und Bedingungen laut Herstellerangaben (MBI-Fermentas)
durchgeführt.
2.8.10 Hybridisierung von Oligonukleotiden zur Ligation
Phosphorylierung der Oligonukleotide
Für je ein Oligonukleotid wurde ein Ansatz hergestellt:
1 µg Oligonukleotid
3 µl 5x T4 Polynukleotidkinasepuffer
1 µl 10 mM ATP
2 µl T4 Polynukleotidkinase
mit x µl ddH2O steril auf 15 µl ergänzen
Die Ansätze wurden für 1 h bei 37°C und anschießend für 20 Minuten bei 65°C
inkubiert.

Material und Methoden
70
Hybridisierung der Oligonukleotide
Zur Generierung von hybridisierten Oligonukleotiden wurden jeweils die 15 µl
Ansätze der phosphorylierten Einzeloligonukleotide vereinigt.
15 µl Oligonukleotid A
15 µl Oligonukleotid B
4 µl 10x Annealing Puffer (200 mM Tris/HCl (pH 7,4), 20 mM MgCl2,
500 mM NaCl)
6 µl ddH2O
Der Ansatz wurde für 10 Minuten auf 95°C erhitzt, langsam auf 50°C abgekühlt,
0,1 µg – 0,5 µg MunI linearisierter pMSg9MunI Vektor hinzugefügt und anschließend
weiter langsam auf Raumtemperatur abgekühlt. Der Ansatz wurde daraufhin mit
Ethanol gefällt (Kap. 2.8.11), eine Ligation (Kap. 2.8.9) durchgeführt und in E. coli-
Zellen transformiert (Kap. 2.8.3).
2.8.11 Ethanol-Fällung von Plasmid-DNS
Zur Fällung wurde 1/10 des Ausgangsvolumens an Natriumacetat (3 M, pH 5,5)
sowie das 2,5-fache Volumen an Ethanol (100 %) zur DNS-Probe zugegeben und
gemixt. Der Ansatz wurde dann 1 h bei -80°C inkubiert und anschließend 20 Minuten
bei 4°C mit 16 000 g zentrifugiert. Der Überstand wurde vorsichtig abgenommen und
verworfen. Als Waschschritt erfolgte die Zugabe von 1 ml eiskaltem, 80 %igem
Ethanol zu der pelletierten DNS. Die Probe wurde dann 5 Minuten bei 4°C mit 16 000
g zentrifugiert, der Überstand abgenommen und verworfen. Zur Entfernung des Rest-
ethanols aus der DNS wurde die Probe im Speed Vac Concentrator getrocknet und
anschließend in 10 µl 1x TE-Puffer aufgenommen.
2.8.12 DNS-Sequenzierung
Die in dieser Arbeit durchgeführten Sequenzierungen wurden nach der Methode von
Sanger (Sanger et al., 1977) unter Verwendung des Thermo SequenaseTM Cycle
Sequencing Kits (USB, Cleveland, OH, USA) und fluoreszenzmarkierter Primer
(Kapitel 2.3) nach folgendem Programm im Thermocycler durchgeführt:

Material und Methoden
71
Anzahl der Zyklen Temperatur Zeit
Vorheizen 1 95°C 2 min
Denaturierung
30
95°C 30 s
Annealing Primer TM – 10°C 30 s
Elongation 72°C 15 s / 1 kb
Final Elongation 1 72°C 3 min
Ende 4°C ∞
Sequenziergel
Der Zusammenbau der Gelapparatur erfolgte nach Vorschrift des Herstellers. Für ein
Sequenziergel wurden die Substanzen aus Kapitel 2.7.4 in 10 ml ddH2O gelöst, an-
schließend zu 50 ml mit ddH2O ergänzt und entgast. Aus dieser 50 ml Lösung
wurden sodann 3 ml entnommen, mit 4 µl TEMED und 32 µl 10 %iger APS-Lösung
versetzt und zum Abdichten der Unterseite der Gelapparatur verwendet. Nach
dessen Polymerisation wurde nach Zugabe von 25 µl TEMED und 250 µl 10 %iger
APS-Lösung zu der Restmenge mit diesem das Sequenziergel gegossen und ein
Vorkamm an der Oberseite der Gelapparatur eingesteckt. Nach vollständiger Poly-
merisation des Gelmaterials (1 h) wurde der Vorkamm gegen einen Haifischzahn-
kamm ausgetauscht und die Gelapparatur in das Sequenziergerät installiert.
2.8.13 PCR-Reaktionen
Die Amplifikation von Genfragmenten durch PCR („polymerase chain reaktion“) zur
anschließenden Klonierung in Plasmidvektoren erfolgte mit Hilfe der GoTaq® Flexi
DNA-Polymerase von Promega und speziellen Oligonukleotiden (Kap. 2.3) nach
folgendem Protokoll:
a) PCR-Mix (50 µl)
10 µl 5x Green GoTaq Flexi-Buffer
1 µl dNTP-Mix (10 mM each)
100 – 150 ng Templat DNS
4 µl MgCl2 25 mM
3 µl Primer A (10 pM/µl)
3 µl Primer B (10 pM/µl)
x µl dd H2O steril auf 49,5 µl ergänzen
0,5 µl GoTaq® Flexi DNA-Polymerase

Material und Methoden
72
b) PCR-Reaktionsprotokoll:
Anzahl der Zyklen Temperatur Zeit
Initiale Denaturierung 1 95°C 2 min
Denaturierung
30
95°C 30 s
Annealing Primer TM – 10°C 30 s
Elongation 72°C 1 min / 1 kb
Final Elongation 1 72°C 5 min
Ende 4°C ∞
Nach der PCR wurde der Mix über eine Agarose-Gelelektrophorese aufgetrennt und
das Amplifikat mit Hilfe des ZymocleanTM Gel DNA Recovery Kit von Zymo Research
gereinigt.
2.8.14 Ortsgerichtete Mutagenese (QuikChange® Mutagenese)
Die ortsspezifische Manipulation von Gensequenzen zur Herstellung der gp9 Kon-
strukte in Kapitel 2.8.17 sowie der M13-Phagen mit Ambermutationen in den Genen
8 und 9 in Kapitel 2.8.16 erfolgte mit Hilfe spezifischer Oligonukleotide (Kap. 2.3) und
der Polymerase PfuUltraII™ Fusion HS von Stratagene nach folgendem Protokoll:
a) QuikChange®-Ansatz (50 µl)
5 µl 10x PfuUltraII™ Fusion HS Reaction-Buffer
1 µl dNTP-Mix (10 mM each)
30 ng Templat DNS
2 µl Primer A (100 ng/µl)
2 µl Primer B (100 ng/µl)
x µl dd H2O steril auf 49 µl ergänzen
1 µl PfuUltraII™ Fusion HS DNA-Polymerase
b) PCR-Reaktionsprotokoll:
Anzahl der Zyklen Temperatur Zeit
Initiale Denaturierung 1 95°C 2 min
Denaturierung 19
95°C 20 s
Annealing Primer TM – 10°C 20 s

Material und Methoden
73
Elongation 72°C 15 s / 1 kb
Final Elongation 1 72°C 3 min
Ende 4°C ∞
Nach Ablauf der Reaktion wurden 2 µl des Restriktionsenzyms DpnI (5 u/µl) zu dem
Reaktionsansatz hinzugegeben und für 2 h bei 37°C inkubiert. Danach wurde die
Plasmid-DNS durch eine Ethanol-Fällung (Kap. 2.8.11) gereinigt und in E. coli XL1-
blue transformiert (Kap. 2.8.3).
2.8.15 Komplementationstest
Mit Hilfe des Komplementationstest wurde ein induziertes „one-step growth“
Experiment zur Quantifizierung der M13-Phagenextrusion durchgeführt und die
Funktionalität der hergestellten gp9-Konstrukte überprüft.
Das Prinzip des Komplementationstest besteht in einer Überprüfung der Funk-
tionalität eines auf genetischer Ebene modifizierten Genes und die hieraus resul-
tierende Beeinflussung des Phagenzyklus. Hierzu werden einmal Bakterienstämme
benötigt, welche als Nicht-Suppressoren bezeichnet werden und nicht in der Lage
sind eine spezielle „nonsense“ Mutation, die Ambermutation, während der Trans-
lation selbst wiederherzustellen. Als weiteres wird ein M13-Phagenstamm benötigt,
welcher in dem Gen des Genprodukts, welches untersucht werden soll, eine Amber-
mutation besitzt und daher nicht in der Lage ist, dieses Genprodukt zu bilden. Durch
diese Konstellation kommt es bei einer Infektion von E. coli Nicht-Suppressor-
stämmen wie z.B. E. coli K38 zu keiner Vermehrung des Phagen. Besitzt jedoch die
Zelle durch eine vorausgegangene Transformation ein Plasmid, welches für das
mutationsfreie Genprodukt kodiert, kann dieses exprimiert, für die Phagenassem-
blierung verwendet und somit die Funktionalität des plasmidkodierten Genprodukts
untersucht und hierdurch ebenfalls der Assembly-Vorgang kontrolliert werden.
a) Induziertes „one-step growth“ Experiment
Phagenstamm M13mp19am8-M2: Damit die Phagensekretion aus E. coli induzierbar
und somit kontrollierbar war, wurde die genetische Erbsubstanz des wildtypischen
M13mp19-Phagen durch eine Ambermutation im Gen 8 verändert, wodurch kein
funktionsfähiges Haupthüllprotein gebildet wird.

Material und Methoden
74
Nicht-Suppressorstamm E. coli K38: In diesen Bakterienstamm wurden zur Induzier-
barkeit der Assemblierung von M13mp19am8-M2 das Plasmid pGZg8wt (Kap. 2.1.2)
transformiert (Kap. 2.8.3) und eine Vorkultur (Kap. 2.8.1) hergestellt.
Die Durchführung entspricht der des Kapitels 2.8.18.
b) Komplementationstest der gp9-Konstrukte
Phagenstamm M13am9-M7: Um die Funktionalität der plasmidkodierten Konstrukte
untersuchen zu können, wurden die Komplementationsexperimente mit diesem
Phagenstamm durchgeführt, dessen genetische Erbsubstanz durch Insertion einer
Ambermutation im Gen 9 verändert wurde und der somit kein funktionsfähiges gp9
mehr bildet.
Suppressorstamm E. coli K37: In diesem Bakterienstamm werden die Genprodukte
von Genen mit Ambermutation in nahezu wildtypischer Expressionsrate hergestellt.
Dieser Bakterienstamm stellte somit den Referenzwert für die wildtypische Ver-
mehrung des Phagenstamms M13am9-M7 dar.
Nicht-Suppressorstamm E. coli K38: In diesen Bakterienstamm wurden zur Funktio-
nalitätsüberprüfung die plasmidkodierten gp9-pMS-Konstrukte aus Kapitel 2.1.2
transformiert (Kap. 2.8.3). Weiterhin diente der nicht transformierte Stamm als
Kontrolle, um die Anzahl der M13-Revertanten bestimmen zu können.
Für Komplementationsexperimente wurde von der M13am9-M7 Phagenstamm-
lösung (Phagentiter 1 x 109 pfu/ml) eine Verdünnungsreihe so hergestellt, dass in
den einzelnen Verdünnungsstufen Phagentiter von 106 pfu/ml bis 101 pfu/ml
vorlagen. Damit die Komplementationsexperimente als Spot-Test (Kap. 2.8.22 a))
durchführbar waren wurden von allen transformierten pMSgp9-Konstrukten in E. coli
K38 je 2 ml LBamp Vorkulturen (Kap. 2.8.1) hergestellt und diese anschließend bei
37°C bis zu einer Zelldichte von 4 x 108 Bakterien/ml kultiviert. Wie in Kapitel 2.8.22
a) beschrieben, wurden dann für jedes Konstrukt Spot-Tests durchgeführt, bei denen
jeweils abwechselnd die einzelnen plasmidtragenden E. coli K38-Kulturen als
Plattierbakterien verwendet wurden. Zur Kontrolle wurde eine E. coli K37-Kultur ohne
Plasmid und eine E. coli K38-Kultur ohne Plasmid zum Ausplattieren verwendet. Zu
allen Komplementationsexperimenten wurde IPTG (1 mM Endkonzentration) zur
Induktion der plasmidkodierten gp9-Konstrukte zum Top-Agar hinzugegeben. Nach
Verfestigung des Top-Agars wurden jeweils 10 µl der zuvor hergestellten Verdün-

Material und Methoden
75
nungsstufen von M13am9-M7, 106 pfu/ml bis 101 pfu/ml auf die Oberseite der
einzelnen LB-Platten aufgebracht und bei RT getrocknet. Die LB-Platten wurden
dann über Nacht bei 37°C inkubiert und daraufhin die Plaque-Bildung am Folgetag
analysiert.
2.8.16 Konstruktion spezifischer M13-Phagenstämme
M13amp
Dieser Phagenstamm wurde zur Selektion infizierter Bakterienzellen generiert.
Hierfür wurde das β-Lactamase-Gen durch Restriktionsverdau (Kap. 2.8.8) mit dem
Enzym PagI aus dem Plasmid pUC18 isoliert und das ~1 kb große Fragment nach
enzymatischer Modifikation seiner 5´- und 3´-Überhänge mit Hilfe des Klenow-
Fragments (nach Anleitung MBI-Fermentas) in die SmaI-Position des Vektors
pGZ119EH ligiert, wodurch der Vektor pGZamp hervorging. Anschließend erfolgte
die Subklonierung des β-Lactamase-Gens aus dem Vektor pGZamp über die Restrik-
tionsschnittstellen EcoRI und PstI (Kap. 2.8.8) in die „Multiple Cloning Site“ des
Phagenstamms M13mp19. Die generierten Konstrukte wurden durch DNA-Sequen-
zierung mit Hilfe des „Universal reverse“-Primers (Kap. 2.3.1) überprüft.
M13am9-M7
Dieser Phagenstamm wurde zur Überprüfung der Funktionsfähigkeit der in dieser
Arbeit hergestellten gp9-Konstrukte generiert. Über eine ortsgerichtete Mutagenese
(Kap. 2.8.14) wurde mit Hilfe eines modifizierten Primerpaares (Kap. 2.3.2) in Gen 9
des Phagenstamms M13mp19 eine Ambermutation (TAG) an Aminosäureposition +7
inseriert und mit Hilfe des „Procoat back“-Primers (Kap. 2.3.1) durch DNA-Sequen-
zierung überprüft.
M13mp19-am8-M2
Dieser Phagenstamm wurde zur Realisierung eines induzierbaren Phagensekretions-
systems generiert. Über eine ortsgerichtete Mutagenese (Kap. 2.8.14) wurde mit
Hilfe eines modifizierten Primerpaares (Kap. 2.3.2) in Gen 8 des Phagenstamms
M13mp19 eine Ambermutation (TAG) an Aminosäureposition +2 inseriert und mit
Hilfe des „Procoat Start“-Primers (Kap. 2.3.1) durch DNA-Sequenzierung überprüft.

Material und Methoden
76
2.8.17 Plasmidkonstrukte
Gen 9-Konstrukte
pGEMg7/9
Die Gene 7 und 9 wurden durch PCR unter Verwendung des M13mp19 RF-Plasmids
als „Template“ mit Hilfe des Primerpaares aus Kapitel 2.3.3 amplifiziert und über
A-Überhänge in den Vektor pGEMT-easy (Promega) ligiert (Kap. 2.8.9) und mit Hilfe
des „T7 Forward“-Primers (Kap. 2.3.1) durch DNA-Sequenzierung überprüft.
pMSg7/9
Sowohl zur weiteren genetischen Modifikation als auch für Expressions- und Komple-
mentationsstudien wurden die Gene 7 und 9 des M13-Phagen aus dem Vektor
pGEMg7/9 über die Restriktionsschnittstellen PaeI und SacI (Kap. 2.8.8) in den
Vektor pMS119HE ligiert (Kap. 2.8.9). Das generierte gp9-Konstrukt wurde mit Hilfe
des „Tac Forward“-Primers (Kap. 2.3.1) durch DNA-Sequenzierung überprüft.
pMSg9MunI
Über eine ortsgerichtete Mutagenese (Kap. 2.8.14) erfolgte mit Hilfe eines modi-
fizierten Primerpaares (Kap. 2.3.2) die Insertion einer MunI-Schnittstelle (CAATTG)
an die Aminosäureposition +2 des Gens 9 im Vektor pMSg7/9. Das generierte gp9-
Konstrukt wurde mit Hilfe des „Tac Forward“-Primers (Kap. 2.3.1) durch DNA-
Sequenzierung überprüft.
pMSg9T7 und pMSg9HA
Die Insertion der antigenen T7- und HA-Epitope in den N-terminalen Bereich des
Gens 9 erfolgte durch die Ligation (Kap. 2.8.9) der zuvor hybridisierten Oligo-
nukleotide (Kap. 2.8.10) aus Kapitel 2.3.4 in die MunI-Schnittstelle des Vektors
pMSg9MunI. Die generierte gp9-Konstrukte wurde mit Hilfe des „Tac Forward“-
Primers (Kap. 2.3.1) durch DNA-Sequenzierung überprüft.
pMSg9T7rep und pMSg9HArep
Zur Wiederherstellung der wildtypischen Aminosäuresequenz nach den Sequenzen
der zuvor eingefügten antigenen Epitope wurde in Gen 9 des pMSg9T7- bzw.
pMSg9HA-Vektors an Position +20 (HA) bzw. +22 (T7) die Startsequenz ATGAGT
des wildtypischen gp9 über eine ortsgerichtete Mutagenese (Kap. 2.8.14) mit Hilfe
modifizierter Primerpaare (Kap. 2.3.2) eingefügt. Die generierten gp9-Konstrukte

Material und Methoden
77
wurden mit Hilfe des „Tac Forward“-Primers (Kap. 2.3.1) durch DNA-Sequenzierung
überprüft.
pMSg9T7SD, pMSg9HASD, pMSg9T7repSD und pMSg9HArepSD
Damit bei der Expression der gp9-pMS-Konstrukte in den E. coli-Zellen höhere
Proteinmengen erhalten werden konnten, wurden in die Konstrukte über eine orts-
gerichtete Mutagenese (Kap. 2.8.14) mit Hilfe modifizierter Primerpaare (Kap. 2.3.2)
8 bp vor dem Startcodon von Gen 9 die Shine-Dalgarno-Sequenz AGGAGG von
E. coli inseriert. Die generierten gp9-Konstrukte wurden anschließend mit Hilfe des
„Tac Forward“-Primers (Kap. 2.3.1) durch DNA-Sequenzierung überprüft.
pMSg9DT7SD, pMSg9DHASD, pMSg9DT7repSD und pMSg9DHArepSD
Durch eine weitere Insertion der zuvor hybridisierten Oligonukleotide (Kap. 2.8.10)
aus Kapitel 2.3.4 in die MunI-Schnittstelle (Kap. 2.8.8) der Vektoren pMSg9T7SD,
pMSg9HASD, pMSg9T7repSD und pMSg9HArepSD konnten durch die anschlie-
ßende Ligation (Kap. 2.8.9) jeweils ein weiteres antigenes T7- bzw. HA-Epitop in den
N-terminalen Bereich des Gens 9 eingefügt werden. Die generierten gp9-Konstrukte
wurden mit Hilfe des „Tac Forward“-Primers (Kap. 2.3.1) durch DNA-Sequenzierung
überprüft.
Gen 1-Konstrukte
pETg1/11
Zur Reinigung und Charakterisierung des gp1/11-Assembly-Komplexes wurde das
Gen 1 durch PCR unter Verwendung des M13mp19 RF-Plasmids als Templat mit
Hilfe des Primerpaares aus Kapitel 2.3.3 amplifiziert, anschließend in die Schnitt-
stellen XhoI und Bpu1102I des Vektors pET-16b (Novagen) inseriert (Kap. 2.8.8) und
das Konstrukt mit Hilfe des „T7 Forward“-Primers (Kap. 2.3.1) durch DNA-Sequen-
zierung überprüft.
pMSg1/11
Zur Expression des gp1/11-Assembly-Komplexes mit 10x N-terminalem Histidin-Tag
in E. coli-Stämmen, welche keine T7-Polymerase exprimieren, wurde das Gen 1 aus
pETg1/11 über die Restriktionsschnittstellen XbaI und HindIII (Kap. 2.8.8) in den
Vektor pMS119EH subkloniert. Hierbei wurde darauf geachtet, dass die Ribosomen-
bindestelle aus pET-16b Bestandteil des zu klonierenden Fragments war.

Material und Methoden
78
2.8.18 „One-step growth“ Experiment
Die Quantifizierung der Phagensekretion aus E. coli stellt eine zeitliche Veränderung
der Phagenmenge im umgebenden Medium nach erfolgter Infektion einer Bakterien-
kultur dar. Da die Phagenproteinbiosynthese unmittelbar von der Stoffwechsel-
leistung des Wirtes abhängig ist, kann diese unter anderem durch die Variation der
Kulturmedientemperatur gesteuert werden.
Phagenstamm M13mp19: Zur Quantifizierung der wildtypischen Phagensekretion
aus E. coli K38 wurde dieser, dem M13 Wildtyp entsprechende, Phagenstamm
verwendet.
Bakterienstamm E. coli K38: In diesem Stamm kann sich M13mp19 wildtypisch
vermehren.
Zur Durchführung des „one-step growth“ Experiments wurde ein steriler Erlenmeyer-
kolben mit 50 ml LB-Flüssigmedium 1/100 mit einer E. coli K38-Vorkultur (Kap 2.8.1)
bzw. im Fall des induzierbaren Experiments (Kap. 2.8.15 a)) mit einer mit pGZg8wt
transformierten E. coli K38 Vorkultur beimpft. Die Zellen wurden bei 37°C bis zu einer
Zelldichte von circa 2 x 108 Bakterien/ml kultiviert und anschließend im Eiswasserbad
abgekühlt. Daraufhin folgte eine 5-minütige Infektion der Zellkultur mit M13mp19
(M.O.I 100) bzw. mit M13mp19am8 (M.O.I. 10) im Eiswasserbad. Zur Abtrennung
des phagenhaltigen Kulturmediums wurde die Zellkultur zentrifugiert (7500 g, 4°C,
5 min), der Überstand verworfen und die Zellen in 5 ml frischem LB-Flüssigmedium
(4°C) aufgenommen und in einen neuen, mit 50 ml LB-Flüssigmedium vorge-
wärmten, Erlenmeyerkolben (37°C) überführt. Anschließend erfolgte eine erneute
Inkubation der Zellen bei 37°C. Dies stellt den Zeitpunkt t = 0 min dar. Im Fall des
induzierten „one-step growth“ Experiments erfolgte an dieser Stelle die Zugabe von
IPTG (1 mM Endkonzentration). Zu definierten Zeiten (Kap. 3.1.1 bzw. 3.1.2) wurden
Proben zu 500 µl aus der infizierten Kultur entnommen und in ein steriles 1,5 ml
Reaktionsgefäß überführt und zentrifugiert (16 000 g, 4°C, 2 min). Der Überstand
wurde in ein neues steriles 1,5 ml Reaktionsgefäß überführt und auf Eis gelagert. Zur
Bestimmung der Veränderung des Phagentiters im Kulturmedium wurden anschlie-
ßend von den Überständen Verdünnungsreihen bis zu einer Verdünnungsstufe von
1/108 angefertigt und mit diesen ein Spot-Test (Kap. 2.8.22 a)) und daraufhin, zur

Material und Methoden
79
eindeutigen Ermittlung der Phagentiter, ein Plaque-Test (Kap. 2.8.22 b)) durchge-
führt. Die Tests wurden 3-mal wiederholt.
Im Fall des induzierten „one-step growth“ Experiments aus Kapitel 2.8.15 a) wurde
als Kontrolle die Veränderung des Phagentiters in einer zweiten infizierten, jedoch
nicht induzierten, Zellkultur ermittelt.
Die Veränderung des Bakterientiters während des wildtypisch verlaufenden „one-step
growth“ Experiments wurde durch die Entnahme von 100 µl Proben zu definierten
Zeiten (Kap. 3.1.1) mittels Lichtmikroskopie bestimmt (Kap. 2.8.23).
2.8.19 M13 Adsorptionsassay
Phagenstamm M13mp19: Zur Quantifizierung der wildtypischen Phagenadsorption
an E. coli K38 wurde dieser, dem M13 Wildtyp entsprechende, Phagenstamm ver-
wendet.
Bakterienstamm E. coli K38: Dieser Stamm besitzt für den erfolgreichen Infektions-
vorgang von M13 F-Pili.
Bakterienstamm E. coli MC4100: Dieser Stamm besitzt keine F-Pili für die Adsorption
von M13-Phagen auf der Bakterienoberfläche und stellt somit die Kontrolle dar.
Zur Durchführung des Adsorptionsassays wurde ein steriler Erlenmeyerkolben mit
50 ml LB-Flüssigmedium 1/100 mit einer E. coli K38-Vorkultur (Kap 2.8.1) bzw. ein
zweiter mit einer E. coli MC4100-Vorkultur beimpft. Die Zellen wurden bei 37°C bis
zu einer Zelldichte von circa 2 x 108 Bakterien/ml kultiviert und anschließend im Eis-
wasserbad abgekühlt. Daraufhin folgte eine 5-minütige Infektion der Zellkulturen mit
M13mp19 auf Eis. Damit die Anzahl der gebundenen Phagen anhand der Abnahme
des eingesetzten Phagentiters bestimmt werden konnte, wurde einmal eine Probe zu
500 µl kurz nach der Zugabe der entsprechenden Phagenmenge und ein weiteres
Mal nach Beendigung der 5-minütigen Infektion entnommen und in ein steriles 1,5 ml
Reaktionsgefäß überführt und zentrifugiert (16 000 g, 4°C, 2 min). Der Überstand
wurde in ein neues steriles 1,5 ml Reaktionsgefäß überführt und auf Eis gelagert. Zur
Bestimmung der Veränderung des Phagentiters im Kulturmedium wurden anschlie-
ßend von den Überständen Verdünnungsreihen bis zu einer Verdünnungsstufe von
1/109 angefertigt und mit diesen ein Spot-Test (Kap. 2.8.22 a)) und daraufhin, zur

Material und Methoden
80
eindeutigen Ermittlung der Phagentiter, ein Plaque-Test (Kap. 2.8.22 b)) durchge-
führt. Sowohl der Spot- als auch der Plaque-Test wurden 3-mal wiederholt.
Im Fall der Kontrolle E. coli K38 wurde dieser Adsorptionsassay mit den M.O.I.-
Werten 5, 10, 20, 52 und 90 durchgeführt.
Im Fall der Negativkontrolle E. coli MC4100 wurde für die Ermittlung der unspezi-
fischen Adsorption von M13-Phagen an Glasgeräte und Bakterien dieser Adsorp-
tionsassay mit den M.O.I.-Werten 10, 50 und 100 durchgeführt.
2.8.20 Messung des Extrusionsverlaufs von M13-Phagen auf der Zellober-
fläche von E. coli
Phagenstamm M13mp19: Zur Quantifizierung der wildtypischen Phagensekretion
aus E. coli XL1-blue wurde dieser, dem M13 Wildtyp entsprechende, Phagenstamm
verwendet.
Bakterienstamm E. coli XL1-blue: In diesem Stamm kann sich M13mp19 wildtypisch
vermehren.
Um den zeitlichen Verlauf des Sekretionsvorgangs auf der Zelloberfläche mit Hilfe
des AFM („Atomic Force Microscope“) messen zu können, wurde ein steriler
Erlenmeyerkolben mit 50 ml LB-Flüssigmedium 1/100 mit einer E. coli XL1-blue-
Vorkultur (Kap 2.8.1) beimpft. Die Zellen wurden bei 37°C bis zu einer Zelldichte von
circa 2 x 108 Bakterien/ml kultiviert und anschließend im Eiswasserbad abgekühlt.
Daraufhin folgte die Infektion (M.O.I. 20) der Zellkultur für 5 Minuten mit M13mp19.
Anschließend wurde die Batch-Kultur sofort wieder bei 37°C geschüttelt. Für die
Messungen der einzelnen Sekretionsstadien am AFM wurden aus der Batch-Kultur
Probenmengen von 500 µl entnommen und in sterile 1,5 ml Reaktionsgefäße über-
führt, die sowohl Alamethicin (5 µM Endkonzentration) als auch CCCP (50 µM End-
konzentration) enthielten und auf Eis gelagert. Die Probenentnahmen erfolgten kurz
vor Infektion sowie direkt nach erneutem Schütteln bei 37°C zu den Zeitpunkten
t = 0, 2, 4, 5, 6, 7, 8, 9, 10, 12, 14 und 16 Minuten. Die Proben wurden daraufhin, wie
in Kapitel 2.8.26 beschrieben, für die Messungen am AFM vorbereitet und gemessen
(Kap. 2.8.28). Die Messungen wurden 3-mal wiederholt.

Material und Methoden
81
2.8.21 Herstellung von M13-Phagenstammlösungen
a) Phagenvermehrung
Für die Phagenvermehrung wurden 100 ml LB-Medium mit einer frischen Vorkultur
von E. coli K37 im Verhältnis 1/100 beimpft. Die Kultur wurde anschließend bis zu
einer optischen Dichte (OD600) von 0,2 bei 37°C geschüttelt. Daraufhin erfolgte die
Infektion der Bakterienkultur für 5 Minuten mit einer M.O.I. von 5 bei Raumtemperatur
ohne Schütteln. Anschließend wurde die Kultur über Nacht bei 37°C geschüttelt.
b) Präzipitation der Phagen
Die infizierte Übernachtkultur wurde in ein steriles Zentrifugengefäß überführt und
zentrifugiert (8000 g, 15 min). Anschließend erfolgte die vorsichtige Überführung des
phagenhaltigen Überstands in ein neues steriles Gefäß. Zum Überstand wurde 1/6
des Ausgangsvolumens an Präzipitationspuffer (Kap. 2.7.5) hinzugegeben und für
4 h bei 6°C gemischt. Anschließend wurde die Suspension erneut bei 8000 g für
15 Minuten zentrifugiert. Das pelletierte Phagenpräzipitat wurde vorsichtig vom Über-
stand abgetrennt und das trübe Phagenpellet in 1 ml 1x TE-Puffer aufgenommen.
Zur Entfernung von kopräzipitierten Bakterienzellen wurde das eluierte Phagenpellet
in ein 2 ml Reaktionsgefäß überführt und für 2 Minuten bei 16 000 g zentrifugiert. Der
Überstand wurde dann vorsichtig in ein neues steriles 1,5 ml Reaktionsgefäß über-
führt und der Phagentiter bestimmt.
2.8.22 Phagentiterbestimmung
a) Spot-Test
Dieses Verfahren stellt eine schnelle Methode zur Ermittlung der Phagenkon-
zentration in einer Phagenlösung dar. Es handelt sich hierbei jedoch nicht um eine
genaue Methode, sie dient vielmehr zur Ermittlung einer sinnvollen Verdünnungs-
stufe aus einer Phagenlösung anhand derer dann eine genaue Phagentiterbe-
stimmung mit Hilfe des Plaque-Test Verfahrens durchgeführt werden kann. Als Basis
für den Test wird der zellfreie Überstand einer E. coli-Flüssigkultur verwendet. Hierfür
wird eine Probe von 500 µl aus der Kultur entnommen und zentrifugiert (16 000 g,
3 min). Der Überstand wird in ein neues steriles 1,5 ml Reaktionsgefäß überführt und
hieraus eine Verdünnungsreihe bis 1/1010 hergestellt.
Für den Spot-Test wurden 3 ml geschmolzener Top-Agar in ein steriles Reagenzglas
gegeben und dieses in einem Thermoblock auf 47°C temperiert. Anschließend
wurden zum geschmolzenen Top-Agar 350 µl Plattierbakterien (E. coli, F+, z.B. K37

Material und Methoden
82
oder XL1-blue) zugegeben und gemischt. Die Suspension wurde auf eine LB-Agar-
platte ausgebracht, woraufhin sie sich nach 5 Minuten verfestigte. Auf den Top-Agar
wurden dann jeweils 10 µl der einzelnen Phagenverdünnungen pipettiert. Man lässt
die Tropfen in den Agar einwirken bis keine Flüssigkeit auf der Oberfläche mehr zu
sehen ist. Danach wurde die LB-Agarplatte bei 37°C über Nacht im Brutschrank
inkubiert. Am nächsten Tag erfolgte die Auswertung der gebildeten Plaques.
b) Plaque-Test
Durch diese Methode erfolgt eine genaue Bestimmung der Phagenpartikelanzahl pro
Volumeneinheit in einer Flüssigkeit.
Für den Plaque-Test wurden 3 ml geschmolzener Top-Agar in ein steriles Reagenz-
glas gegeben und dieses in einem Thermoblock auf 47 °C temperiert. Anschließend
wurden zum geschmolzenen Top-Agar 350 µl der Plattierbakterien (E. coli, F+, z.B.
K37 oder XL1-blue) und 100 µl einer angemessenen Phagenverdünnung aus a)
zugegeben und gemischt. Die Suspension wurde dann auf eine LB-Agarplatte
ausgebracht, woraufhin sie sich nach 5 Minuten verfestigte. Danach wurde die LB-
Agarplatte bei 37°C über Nacht im Brutschrank inkubiert. Am nächsten Tag erfolgten
die Auswertung der gebildeten Plaques und die Bestimmung des Phagentiters in der
vorliegenden Phagenstammlösung.
Der Phagentiter in „plaque forming units“ (pfu) pro Milliliter lässt sich wie folgt
errechnen:
2.8.23 TCA-Fällung von Proteinen
Zur Konzentrierung und Aufbereitung von Proteinen für SDS-Polyacrylamid-
Gelelektrophoresen sowie für SDS-Tricin-Gelelektrophoresen wurden diese zuvor mit
Hilfe von Trichloressigsäure (TCA) gefällt. Hierfür wurden die Proben mit TCA
versetzt (12 % Endkonzentration), kurz gemischt, 60 Minuten auf Eis inkubiert und
anschließend zentrifugiert (10 min, 4°C und 16 000 g). Der Überstand wurde vor-
sichtig abgenommen und das Pellet zweimal mit 1 ml eiskaltem Aceton gewaschen
und zentrifugiert (10 min, 4°C und 16 000 g). Abschließend wurde das Pellet bei
50°C getrocknet, in Probenpuffer aufgenommen und kurz vor der Verwendung für die
SDS-Gelelektrophorese 5 Minuten bei 95°C mit Hilfe eines Thermoblocks gekocht.

Material und Methoden
83
2.8.24 Bestimmung der Bakterienzellzahl
Die Bestimmung der Bakterienzellzahl von E. coli-Flüssigkulturen erfolgte mit Hilfe
der Thoma-Zählkammer am Lichtmikroskop. Hierfür wurde aus der Kultur steril eine
Probe von 200 µl entnommen und diese nach Bedarf verdünnt bis bei einer 400-
fachen Vergrößerung am Mikroskop die Bakterienzellen mit der Thoma-Zählkammer
auszählbar waren. Zur Bestimmung der Bakterienzellzahl wurden die Werte aus vier
ausgezählten Kleinquadraten gemittelt und wie folgt die Zellzahl pro Milliliter
errechnet:
2.8.25 Präparation transfizierter E. coli DH5α-Zellen für AFM und TEM-
Messungen
Kompetente E. coli DH5α-Zellen (Kapitel 2.8.2) wurden, wie in Kapitel 2.8.4
beschrieben, mit dem RF-Plasmid des Phagenstamms M13amp (Kapitel 2.1.1) trans-
fiziert und auf eine LB-Agarplatte mit Ampicillin (Kap. 2.6.1) und IPTG (1 mM End-
konzentration) ausplattiert. Die LB-Agarplatte wurde über Nacht bei 37°C inkubiert
und am Folgetag mit einer Einzelkolonie eine Vorkultur hergestellt (Kap. 2.8.1). Vor
der Messung der transfizierten E. coli-Zellkultur wurde der Überstand mit Hilfe des
Spot-Tests (Kap. 2.8.22 a)) auf die Präsenz von M13-Phagen hin untersucht und
dann wie in den Kapiteln 2.8.26 und 2.8.27 für die Untersuchungen aufbereitet.
Als Kontrollen wurden nicht infizierte E. coli DH5α-Zellen verwendet.
2.8.26 Präparation und Adsorption von E. coli-Zellen für AFM-Messungen
Für die Messungen von Bakterien am AFM wurde als Ausgangsmaterial eine
Bakterienflüssigkultur in LB-Medium verwendet (Kapitel 2.8.1). Die Zellen wurden bei
Abb. 2-1: Schematischer Aufbau des Zählfeldes bei einer Thoma-Zählkammer. Kleinquadrate sind in grau hervorgehoben.

Material und Methoden
84
37°C bis zu einer Zelldichte von 2 x 108 Bakterien/ml kultiviert. Aus dieser Kultur
erfolgte dann die Entnahme von 500 µl, die sodann in ein steriles 1,5 ml Reaktions-
gefäß überführt und vorsichtig zentrifugiert (7500 g, 3 min) wurden. Der Überstand
wurde verworfen. Daraufhin folgten drei Waschschritte in denen das Zellpellet
vorsichtig in sterilem ddH2O aufgenommen und abermals zentrifugiert wurde (7500 g,
3 min). Abschließend wurden die Zellen in 500 µl sterilem ddH2O aufgenommen.
Von den 500 µl der Zellsuspension wurden anschließend 10 µl auf die neu abge-
zogene Glimmeroberfläche (Muscovite Mica, V-1 Quality), welche zuvor mit 2-Kom-
ponentenklebstoff auf die Oberfläche eines AFM-Probenträgers befestigt wurde,
aufgebracht und adsorbiert (4°C, 15 min). Danach wurde die Restflüssigkeit mit einer
Pipette abgenommen, die AFM-Probenoberfläche 2-mal mit 15 µl ddH2O gewaschen
und 5 Minuten bei Raumtemperatur getrocknet.
2.8.27 Präparation und Adsorption von E. coli-Zellen für TEM-Messungen
Die Präparation der Proben erfolgte wie unter Kapitel 2.8.26 beschrieben.
Von der gereinigten Zellsuspension wurden 10 µl auf ein zuvor plasmagereinigtes
(Benada & Pokorný, 1990) Kupfernetzchen (Athene 200) aufgebracht und für
5 Minuten bei Raumtemperatur adsorbiert. Die Flüssigkeit wurde anschließend mit
Hilfe eines Filterpapiers von der Oberfläche abgesaugt. Die Oberfläche wurde dann
durch 2-maliges eintauchen des Netzchens in 2 Tropfen ddH2O á 50 µl gewaschen,
die Restflüssigkeit mit Hilfe eines Filterpapiers von der Oberfläche des Netzchens
entfernt und für 5 Minuten bei Raumtemperatur getrocknet. Zur Negativkontras-
tierung der Proben wurden dann 5 µl einer 5 %igen Phosphorwolframsäure (pH 7)
auf die Netzchenoberfläche aufgebracht, nach 30 s mit Hilfe eines Filterpapiers von
der Oberfläche entfernt und für 10 Minuten bei Raumtemperatur getrocknet.
2.8.28 „Atomic Force Microscopy“ (AFM)
Alle Messungen wurden mit dem kommerziell erhältlichen Nanoscope IIIa von Digital
Instruments aufgenommen. Das Gerät war mit einem J-Scanner mit 150 µm Scan-
Weite ausgestattet. Die Messung der leicht feuchten biologischen Proben erfolgte im
Kontakt-Modus mit Hilfe einer goldbedampften Siliziumnitritmesssonde („Cantilever“),
die eine Federkonstante von k = 0,06 N/m (NP-S) besaß. Die Geschwindigkeit, mit
welcher die Probenoberfläche gerastert wurde, betrug 3,52 Hz und der Rasterwinkel

Material und Methoden
85
90° zur Achse der Messsonde. Die Auflagekraft der Messsonde während der
Probenmessungen betrug weniger als 1 nN. Alle AFM-Messdaten, welche als soge-
nannte „Deflection“-Bilder in dieser Arbeit gezeigt sind, wurden mit dem frei erhält-
lichen Programm WSxM v.4.0 Develop 12.4 der Firma Nanotec Electronica ver-
arbeitet.
2.8.29 Transmissionselektronenmikroskopie (TEM)
Alle Messungen der hier gezeigten TEM-Aufnahmen erfolgten mit einem LEO 912AB
der Firma Zeiss GmbH mit Omega Filter für EFTEM und einer Punkt zu Punkt
Auflösung von 0,37 nm. Die Messungen wurden mit einer Beschleunigungs-
spannung von 80 kV betrieben; als Kathode wurde eine Lanthanhexaborid (LaB6) –
Kristallelektrode mit pyramidenförmiger Spitze als Elektronenquelle verwendet. Die
Bildaufnahme erfolgte durch eine 2k x 2k CCD-Kamera der Firma Olympus und die
Bildverarbeitung durch das Programm iTEM der Firma Olympus Soft Imaging
Solutions.
2.8.30 Immunogold-Markierung von M13-Phagen
Für die Markierung wurde die M13-Phagenstammlösung (Kap. 2.8.21) zu einer
Endkonzentration von 5 x 109 pfu/ml entsprechend mit Wasser verdünnt. Aus dieser
Verdünnung wurden 10 µl auf einem zuvor plasmagereinigten (Benada & Pokorný,
1990) Kupfernetzchen (Athene 200) aufgebracht und für 1 Minute bei Raum-
temperatur adsorbiert. Die Flüssigkeit wurde anschließend mit Hilfe eines Filter-
papiers von der Oberfläche abgesaugt. Die Oberfläche wurde dann durch 2-maliges
Eintauchen des Netzchens in 2 Tropfen ddH2O á 50 µl gewaschen, die Rest-
flüssigkeit mit Hilfe eines Filterpapiers von der Oberfläche des Netzchens entfernt
und für 5 Minuten bei Raumtemperatur getrocknet. Zur Vermeidung unspezifischer
Bindungen des Erstantikörpers an das Trägermaterial des Kupfernetzchens sowie
den Phagen wurde die Probe mit 50 µl 1x TBS/0,1 % BSA (Kap. 2.7.8) für 10
Minuten inkubiert. Der überschüssige Puffer wurde anschließend mit Hilfe eines
Filterpapiers von der Oberfläche entfernt, direkt durch 20 µl Erstantikörperlösung
(1:5000 in 1x TBS) ersetzt und für 30 Minuten bei Raumtemperatur inkubiert.
Anschließend erfolgte die Entfernung der Antikörperlösung wieder mit Filterpapier
und die Probe wurde mit 50 µl 1x TBS für 3 Minuten inkubiert. Nach Entfernen des
Puffers mit Filterpapier wurden 20 µl einer 1:20 Verdünnung in 1x TBS des Protein-A
konjugierten Goldes (Protein-A – 20 nm kolloidales Gold, Sigma Aldrich) auf die

Material und Methoden
86
Netzchenoberfläche aufgebracht und für weitere 30 Minuten inkubiert. Abschließend
wurde die Probe 1-mal mit 50 µl 1x TBS/0,1% BSA für 3 Minuten und 2-mal mit 50 µl
ddH2O für 2 s gewaschen. Überschüssiges Wasser wurde mit Hilfe von Filterpapier
entfernt und das Netzchen bei Raumtemperatur getrocknet.
Für die Messungen am TEM wurde die Probe, wie in Kapitel 2.8.27 beschrieben,
negativ kontrastiert.
Als Kontrolle wurde eine Probe nicht mit Primärantikörpern vorbehandelt.
2.8.31 Immunogold-Markierung infizierter/uninfizierter E. coli DH5α-Zellen
Für die Markierung wurden transfizierte E. coli DH5α-Zellen wie in Kapitel 2.8.25
hergestellt und wie in Kapitel 2.8.25 präpariert. Im Unterschied dazu wurden die
transfizierten Zellen nach dem letzten Waschschritt in 500 µl 1x TBS/0,1 % BSA
(Kap. 2.7.8) aufgenommen und 10 Minuten bei Raumtemperatur inkubiert.
Anschließend wurden die Zellen zentrifugiert (7500 g, 3 min) und in 50 µl Primär-
antikörperlösung (1:5000 in 1x TBS) aufgenommen und 30 Minuten auf einem
Drehrad bei Raumtemperatur inkubiert. Die Zellen wurden daraufhin 2-mal mit 500 µl
1x TBS gewaschen und zentrifugiert (7500 g, 3 min). Nach dem letzten Waschschritt
erfolgte dann die Aufnahme der Zellen in 50 µl Protein-A konjugierter Goldlösung,
welche zuvor 1:20 in 1x TBS verdünnt wurde. Die Probe wurde für weitere 30
Minuten bei Raumtemperatur auf einem Drehrad inkubiert. Abschließend wurden die
Zellen 2-mal mit 500 µl ddH2O gewaschen und zentrifugiert (7500 g, 3 min) und dann
in 100 µl ddH2O aufgenommen.
Für die Messungen am TEM wurde die Probe, wie in Kapitel 2.8.27 beschrieben,
präpariert.
Als Kontrolle wurde eine Probe nicht infizierter E. coli DH5α dieser Prozedur unter-
zogen.
2.8.32 Expression des gp1/11-Assembly-Komplexes
Expression in E. coli C41
Für die Expression und Reinigung der gp1/11-Assembly-Komplexe wurde das
Plasmid pETg1/11 (Kap. 2.8.17), wie in Kapitel 2.8.3 beschrieben, in E. coli C41
transformiert und eine Vorkultur hergestellt (Kap. 2.8.1.). Die Vorkultur wurde
anschließend dazu verwendet, um Hauptkulturen (2 ml, 2 L, 4 L) im Verhältnis 1:100
beimpfen zu können. Die Hauptkulturen wurden bei 37°C bis zu einer optischen
Dichte (OD600) von 0,5 kultiviert. Die Expression des plasmidkodierten gp1/11 wurde

Material und Methoden
87
dann durch die Zugabe von IPTG (1 mM Endkonzentration) induziert. Nach
2,5 h wurden die Großkulturen zentrifugiert (7500 g, 4°C, 10 min) und in 2 ml pro
Gramm Zellpellet Tris/Sucrose Puffer (2.7.13) aufgenommen. Die 2 ml Kleinkulturen
wurden 3 Minuten bei 16 000 g zentrifugiert und TCA gefällt (Kap. 2.8.23).
Expression in E. coli K37
Für die Expression und Reinigung der gp1/11-Assembly-Komplexe wurde das
Plasmid pMSg1/11 (Kap. 2.8.17), wie in Kapitel 2.8.3 beschrieben, in E. coli K37
transformiert und eine Vorkultur hergestellt (Kap. 2.8.1). Die Vorkultur wurde
anschließend dazu verwendet, um die Hauptkultur (4 L) im Verhältnis 1:100 beimpfen
zu können. Die Hauptkultur wurde bei 37°C bis zu einer optischen Dichte (OD600) von
0,5 kultiviert. Die Expression des plasmidkodierten gp1/11 wurde dann durch die
Zugabe von IPTG (1 mM Endkonzentration) induziert und die Zellkultur mit einer
M.O.I. von 5 mit M13mp19-Phagen infiziert. Nach 2,5 h wurden die Großkulturen
zentrifugiert (7500 g, 4°C, 10 min) und in 2 ml pro Gramm Zellpellet Tris/Sucrose
Puffer (Kap. 2.7.13) aufgenommen.
2.8.33 Proteinreinigung mittels Metallaffinitätschromatographie (IMAC)
Die Reinigung des gp1/11-Assembly-Komplexes mittels IMAC erfolgte aus nicht
infizierten E. coli C41 und infizierten E. coli K37 (Kap. 2.8.32). Um die in der inneren
Membran von E. coli lokalisierten gp1/11-Assembly-Komplexe reinigen zu können,
wurden die in Tris/Sucrose Puffer (Kap. 2.7.13) aufgenommen Zellen mit PMSF
(2,5 mM Endkonzentration), MgCl2 (15 mM Endkonzentration) und DNAse II (8 µg
Endkonzentration) versetzt und im „French Press“-Verfahren (Milner et al., 1950) bei
8000 Psi aufgeschlossen. Die Suspension wurde bei 4000 g für 10 Minuten bei 4°C
zentrifugiert und das Pellet verworfen. Zur Pelletierung der inneren Zellmembranen
im resultierenden Überstand erfolgte eine Zentrifugation bei 177 614 g für 60 Minuten
bei 4°C, der Überstand wurde anschließend verworfen, das Pellet in 20 ml eiskaltem
Solubilisierungspuffer (Kap. 2.7.13) aufgenommen und mittels Dounce Homogeni-
sators homogenisiert. Die Solubilisierung der Membranen erfolgte für 2 h unter
Rühren bei 4°C. Zur Abtrennung nicht solubilisierbarer Bestandteile wurde die
Suspension nochmals bei 177 614 g für 60 Minuten bei 4°C zentrifugiert und der
Überstand in ein steriles 50 ml Reaktionsgefäß überführt. Zu dieser Lösung wurde
1 ml Ni-NTA-Agarose zugegeben, welche zuvor 3-mal mit 15 ml ddH2O gewaschen
und anschließend 15 Minuten mit 10 ml Äquilibrierungspuffer (Kap 2.7.13) inkubiert

Material und Methoden
88
wurde. Nach Zugabe der Ni-NTA-Agarose zur solubilisierten Fraktion folgte eine
Inkubationsphase für 2 h unter Rotation bei 4°C. Die darauf folgenden Prozeduren
wurden bei 4°C durchgeführt. Nach der Inkubationsphase wurde die Suspension in
eine Minisäule (BD PlastipakTM, 5 ml, Becton Dickinson) überführt und die Ni-NTA-
Agarose im „Gravity flow“-Verfahren einmal mit 10 ml Säulenwaschpuffer 1 (Kap.
2.7.13) und ein weiteres Mal mit 10 ml Säulenwaschpuffer 2 (Kap. 2.7.13) gereinigt.
Abschließend wurde die Ni-NTA-Agarose zur Entfernung der gebundenen Proteine
mit 1,5 ml Elutionspuffer (Kap. 2.7.13) für 1 h bei 4°C inkubiert und dann die
Elutionsfraktion bei 1500 g (2 min) in ein steriles 15 ml Reaktionsgefäß überführt. Zur
Aufbewahrung der eluierten Proteine wurden diese in flüssigem Stickstoff tiefgefroren
und bei -80°C gelagert.
2.8.34 Präparative Gelfiltration
Kalibrierung des Trennsystems
Durch die präparative Gelfiltration werden Proteine aufgrund ihrer hydrodynamischen
Eigenschaften über eine Säulenmatrix getrennt. Für die Auftrennung des Protein-
gemisches in der Elutionsfraktion aus der Metallaffinitätschromatographie in Kapitel
2.8.33 wurde als Säulenmatrix SuperdexTM 200 mit einem Trennbereich von 10 – 600
kDa in einer XK 16/40 Säule (Amersham, GE Healthcare) gewählt. Um das Trenn-
verhalten der Säule zu bestimmen und den daraus erhaltenen Fraktionsvolumina
entsprechende Molekulargewichte zuordnen zu können, wurde eine Kalibrierung der
Säule mit Hilfe von 2 ml des Proteinstandards aus Kapitel 2.7.10 durchgeführt,
welcher Proteine mit bekannten molekularen Massen enthielt. Die Kalibrierung
erfolgte mit Hilfe des Äkta Basic Chromatographie Systems (Amersham Biosciences)
mit einer konstanten Flussrate von 0,5 ml/min. Die eluierten Proteine wurden in
Fraktionen zu je 2 ml gesammelt. Die molekularen Massen der eluierten Standard-
proteine wurden logarithmisch gegen das Elutionsvolumen (Ve) aufgetragen (Tabelle
2-4) und hieraus eine Standardkurve für die XK 16/40-Säule mit SuperdexTM 200
Säulenmatrix ermittelt (Abb. 2-2). Durch den Vergleich dieser Werte mit dem
Elutionsverhalten der Proteine aus der Elutionsfraktion der Metallaffinitätschromato-
graphie (Kap. 2.8.33) wurden deren Massen ermittelt.

Material und Methoden
89
Tabelle 2-4: Proteine des Standardgemisches mit den zugehörigen Massen in kDa und
Elutionsvolumina (Ve) zur Kalibrierung des Trennverhaltens der XK 16/40 Säule mit SuperdexTM
200 Säulenmatrix.
Protein Ve Masse
Apoferritin 20,4 ml 440 kDa
beta-Amylase 24,2 ml 200 kDa
BSA Dimer 27,7 ml 132 kDa
BSA Monomer 31,5 ml 66,5 kDa
Carboanhydrase 38,9 ml 30 kDa
Abb. 2-2: Ermittelte Standardkurve der durch Gelfiltration getrennten Standardproteine. Die
molekularen Massen der eluierten Standardproteine wurden logarithmisch gegen das Elutions-
volumen (Ve) aufgetragen.
Gelfiltration der Elutionsfraktionen aus der Metallaffinitätschromatographie
Die Auftrennung des Elutionsgemisches aus Kapitel 2.8.33 erfolgte durch das Äkta
Basic Chromatographie System (Amersham Biosciences) mit einer XK 16/40-Säule
und SuperdexTM 200 als Säulenmatrix. Für die Prozedur wurde der Puffer aus Kapitel
2.7.12 verwendet. Die Gelfiltration wurde mit einer konstanten Durchflussrate von
0,5 ml/min durchgeführt und die eluierten Fraktionen zu je 2 ml gesammelt. Zur
Identifikation der Proteine in den jeweiligen Elutionsfraktionen wurden aus diesen
150 µl Proben entnommen und mit TCA gefällt (Kap. 2.8.23). Anschließend erfolgte
die Auftrennung der Proben über ein 12 %iges SDS-Gel (Kap. 2.8.35). Das Protein-
bandenmuster wurde, wie in Kapitel 2.8.36 beschrieben, mit Coomassie sichtbar
20 40 60 80 10010
100
1000
BSA Monomer 66,5 kDa
BSA Dimer 132 kDa
Carboanhydrase 30 kDa
Apoferritin 440 kDa
beta-Amylase 200 kDa
Masse [
kD
a]
Ve [ml]

Material und Methoden
90
gemacht. Die Zuordnung der in den Elutionsfraktionen vorherrschenden Massen
erfolgte anhand der für die Säule XK 16/40 ermittelten Standardkurve und der
Elutionsvolumina (Ve) der einzelnen Fraktionen.
2.8.35 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE)
Zur Auftrennung des kleinen Hüllproteins gp9 des Bakteriophagen M13 wurden
sowohl SDS-Polyacrylamid-Gele mit Harnstoff (22 %) nach Laemmli (1970) als auch
SDS-Tricin-Gele (16,5 %) nach Schägger und Jagow (1987) verwendet. Die Auf-
trennung der gp1/11-Assembly-Komplexe erfolgte durch 12 % SDS- Polyacrylamid-
Gele ohne Harnstoffanteile.
Probenvorbereitung
Nach TCA-Fällung der Proben (Kap. 2.8.23) wurden diese in SDS-Probenpuffer
(Kap. 2.7.14) aufgenommen, 5 Minuten bei 95°C inkubiert und anschließend in die
vorgesehenen Probentaschen der SDS-Gele pipettiert.
SDS Polyacrylamid-Gelelektrophorese ohne Harnstoff (12 %)
Für das Trenngel wurden die Substanzen aus Kapitel 2.7.14.1 gelöst, entgast und
mit SDS, APS und TEMED versetzt, zwischen zwei Gelplatten gegossen und mit
Isopropanol überschichtet. Nach der Polymerisation des Trenngels wurde das
Isopropanol mit dH2O entfernt, die Lösung für das Sammelgel aus Kapitel 2.7.14.1
gelöst, auf das Trenngel gegossen und der Kamm für die Probentaschen
eingesteckt. Nach 15-minütiger Polymerisation wurde der Kamm entfernt, die
Taschen mit dH2O gespült und das Gel in die dafür vorgesehene Apparatur installiert
(Laufzeit bei 25 mA ~ 3-4 h).
SDS Polyacrylamid-Gelelektrophorese mit Harnstoff (22 %)
Für das Trenngel wurden die Substanzen aus Kapitel 2.7.14.1 gelöst, entgast und
mit SDS, APS und TEMED versetzt, zwischen zwei Gelplatten gegossen und mit
Isopropanol überschichtet. Nach der Polymerisation des Trenngels wurde das
Isopropanol mit dH2O entfernt, die Lösung für das Sammelgel aus Kapitel 2.7.14.1
gelöst, auf das Trenngel gegossen und der Kamm für die Probentaschen
eingesteckt. Nach 15-minütiger Polymerisation wurde der Kamm entfernt, die
Taschen mit dH2O gespült und das Gel in die dafür vorgesehene Apparatur installiert
(Laufzeit bei 25 mA ~ 5,5-6,5 h).

Material und Methoden
91
SDS Tricin-Gelelektrophorese (16,5 %)
Für das Trenngel wurden die Substanzen aus Kapitel 2.7.14.2 gelöst, mit APS und
TEMED versetzt, zwischen zwei Gelplatten gegossen und mit Isopropanol über-
schichtet. Nach der Polymerisation des Trenngels wurde das Isopropanol mit dH2O
entfernt, die Lösung für das Spacer-Gel aus Kapitel 2.7.14.2 gelöst, auf das Trenngel
gegossen und mit Isopropanol überschichtet. Nach der Polymerisation des Spacer-
Gels wurde das Isopropanol mit dH2O entfernt, die Lösung für das Sammelgel aus
Kapitel 2.7.14.2 gelöst, auf das Spacer-Gel gegossen und der Kamm für die Proben-
taschen eingesteckt. Nach der Polymerisation wurde der Kamm entfernt, die
Taschen mit dH2O gespült und das Gel in die dafür vorgesehene Apparatur installiert
(Laufzeit bei 25 mA ~ 6,5-7,5 h).
2.8.36 Coomassiefärbung von SDS-Proteingelen
Die Visualisierung der durch SDS-PAGE aufgetrennten Proteinbandenmuster
erfolgte mit Hilfe der Färbe- und Entfärbelösung aus Kapitel 2.7.2. Nach Beendigung
der SDS-PAGE wurde das Gel aus der Apparatur entfernt, in eine Schale mit Färbe-
lösung gelegt und 2 x 3 Minuten in der Mikrowelle bei 600 Watt gefärbt. Zur
Entfärbung wurde das Gel 3 x 5 Minuten in der Mikrowelle bei 600 W in Entfärbe-
lösung erhitzt.
2.8.37 Western-Blot (semi-dry) und Membranbehandlung
Durch SDS-PAGE aufgetrennte Proteine können mit dem Western-Blot Verfahren
durch elektrischen Strom auf eine proteinbindende Membran übertragen und
anschließend mit geeigneten Antikörpern nachgewiesen werden. Für die Durch-
führung eines semi-dry Western-Blots wurden auf die Kathodenseite zuerst zwei mit
1x Transferpuffer getränkte Filterpapiere gelegt. Auf die Filter wurden blasenfrei das
SDS-Gel, die Nitrocellulose Transfermembran und noch 2 mit Transferpuffer
getränkte Filterpapiere gelegt. Auf diese Schichtung wurde der Deckel mit der Anode
aufgesetzt, verschraubt und die Elektroden angeschlossen. Die Stromdichte beim
semi-dry Western-Blot betrug 1,0-1,5 mA pro cm2 Gelfläche. Bei einem Minigel
entspricht das einer Stromstärke von 50 mA, bei einem Maxigel (10 cm x 18 cm)
einer Stromstärke von 250 mA. Der Blot war nach 75 Minuten beendet.

Material und Methoden
92
Immundetektion der Proteine und ECL
Proteine, welche durch das Western-Blot Verfahren auf eine proteinbindende
Membran übertragen wurden, können durch spezifische Antikörper detektiert und
anschließend durch ECL (Enhanced Chemoluminescence) nachgewiesen werden.
Nach dem Transfer wurde die Nitrocellulose Membran für 2 Minuten in eine 0,2 %ige
Ponceau-S-Lösung gegeben, um die Proteine zu fixieren. Die Membran wurde unter
fließendem Wasser entfärbt und über Nacht in 1x TBS/5 % Magermilchpulver bei 4°C
geschüttelt. Am Folgetag wurde die Membran 3-mal 5 Minuten mit 1x TBS ge-
waschen. Danach erfolgte die Zugabe des 1. Antikörpers (Kap. 2.1.5) auf die
Membran und die Inkubation (2 h) bei 4°C unter Schütteln. Die Membran wurde dann
2-mal 5 Minuten mit 1x TBS gewaschen und anschließend mit dem 2. Antikörper
(Kap. 2.1.5) inkubiert (90 min) und 3-mal 5 Minuten mit 1x TBS gewaschen.
Das Proteinbandenmuster wurde dann mit dem ECL Immobilon Western Detection
Reagents (Millipore) nach Anleitung auf einen Röntgenfilm übertragen.
2.8.38 Dot-Blot Analyse und Membranbehandlung
Die Detektion der plasmidkodierten gp9-Konstrukte am Phagen erfolgte mit Hilfe von
Dot-Blot Analysen. Hierfür wurden zuvor die plasmidkodierten gp9-Konstrukte in
E. coli K38-Zellen transformiert (Kap. 2.8.3). Von den Konstrukten wurde je eine
Einzelkolonie für die Herstellung von Vorkulturen verwendet (Kap. 2.8.1). Die
Vorkulturen dienten anschließend zur Beimpfung von 50 ml LB-Batchkulturen
(1:100). Nach Beimpfung der Batch-Kulturen erfolgte die Zugabe von IPTG (1 mM
Endkonzentration) und die Inkubation für 15 Minuten bei 37°C unter Schütteln.
Hierauf wurden die Batch-Kulturen jeweils mit dem Phagenstamm M13am9-M7 bei
Raumtemperatur für 5 Minuten ohne Schütteln infiziert (M.O.I. 5). Anschließend
erfolgte die Kultivierung der infizierten Kulturen über Nacht bei 37°C unter Schütteln.
Am Folgetag wurden aus den 50 ml Batch-Kulturen, wie in Kapitel 2.8.21 b)
beschrieben, Phagenstammlösungen hergestellt und anschließend deren Phagen-
titer bestimmt (Kap. 2.8.22).
Als Kontrollen bei den Dot-Blot Analysen dienten die induzierten, jedoch nicht infi-
zierten, Übernacht-Batch-Kultur Varianten der hier transformierten Zellen. Diese
Kulturen wurden ebenfalls, wie in Kapitel 2.8.21 b) und 2.8.22 beschrieben, präzi-
pitiert und anschließend deren Phagentiter bestimmt.

Material und Methoden
93
Von den hergestellten Phagenstammlösungen wurden Verdünnungsreihen so ange-
fertigt, dass in diesen nahezu gleiche Mengen an Phagen pro 400 µl vorlagen
(106 pfu bis 108 pfu). Die Kontrollen wurden entsprechend den Verdünnungsstufen
der Phagenstammlösungen verdünnt (10-1 bis 10-3).
In die Dot-Blot Apparatur (MiniFold®) wurden zwei Filterpapiere (1 mm, Whatmann)
und auf diese eine Nitrocellulose Transfermembran (PROTRAN®), die zuvor
zusammen 5 Minuten in 1x Transferpuffer (Kap. 2.7.16) inkubiert wurden, aufgelegt
und eingespannt. Die Apparatur wurde mit einer Vakuumpumpe verbunden und unter
Vakuum gesetzt, woraufhin aus den zuvor hergestellten Verdünnungsstufen jeweils
400 µl der Phagenlösungen mit Hilfe der Dot-Blot Apparatur auf die Nitrocellulose
Transfermembran aufgebracht wurden.
Immundetektion der gp9-Konstrukte und ECL
Nach dem Transfer der Phagen wurde die Membran für 2 Minuten in eine 0,2 %ige
Ponceau-S-Lösung gegeben, um die Phagen zu fixieren. Die Membran wurde unter
fließendem Wasser entfärbt und über Nacht in 1x TBS/5 % Magermilchpulver bei
4°C geschüttelt. Am Folgetag wurde die Membran 3-mal 5 Minuten mit 1x TBS
gewaschen. Danach erfolgte die Zugabe des 1. Antikörpers (Kap. 2.1.5) auf die
Membran und die Inkubation (2 h) bei 4°C unter Schütteln. Die Membran wurde dann
2-mal 5 Minuten mit 1x TBS gewaschen und anschließend mit dem 2. Antikörper
(Kap. 2.1.5) inkubiert (90 min) und 3-mal 5 Minuten mit 1x TBS gewaschen.
Die Signale wurden dann mit dem ECL Immobilon Western Detection Reagents
(Millipore) nach Anleitung auf einen Röntgenfilm übertragen.
2.8.39 Pulse-labelling-Experimente, Immunpräzipitation und Protease-Mapping
„Pulse-labelling“-Experimente
Für „Pulse-labelling“-Experimente wurden die entsprechenden Plasmide mit den zu
untersuchenden gp9-Konstrukten in E. coli K38 transformiert (Kap. 2.8.3) und
daraufhin Vorkulturen hergestellt (Kap. 2.8.1). Mit den Vorkulturen wurden jeweils
2 ml „Pulse“-Medium (Kap. 2.6.2) 1:10 beimpft und bei 37°C geschüttelt. Ab einer
Zellzahl von 2 x 108 Zellen/ml wurden die Zellen zentrifugiert (16 000 g, 2 min), in
500 µl neuem „Pulse“-Medium aufgenommen und mit IPTG (1 mM Endkonzentration)
induziert. Nach 10 Minuten wurden die gebildeten Proteine durch Zugabe von 20 µCi
[35S]-Methionin radioaktiv markiert und nach weiteren 10 Minuten die Proben mit TCA
präzipitiert, anschließend mit Aceton gereinigt und abschließend getrocknet (Kap.

Material und Methoden
94
2.8.23). Die resultierenden Präzipitate wurden in 50 µl Tris/SDS-Puffer (10 mM
Tris/HCl pH 8.0, 2 % SDS) aufgenommen, 5 Minuten bei 95°C im Thermoblock
erhitzt und immunpräzipitiert.
Immunpräzipitation
Für eine Immunpräzipitation wurden zu den Proben 1 ml TENTX–Puffer (Kap. 2.7.11)
und 15 µl Protein-A Substrat (ZysorbinTM) zugegeben, für 30 Minuten bei 4°C auf
einem Drehrad inkubiert und anschließend bei 16 000 g für 15 Minuten zentrifugiert.
Der Überstand wurde in ein neues Reaktionsgefäß überführt, mit 1-8 µl Primär-
antikörper versetzt (Kap. 2.1.5) und über Nacht bei 4°C auf dem Drehrad inkubiert.
Dann erfolgte die Zugabe von 20 µl der Protein-A Suspension zu den Proben sowie
eine weitere Inkubation für eine Stunde auf dem Drehrad mit anschließender
Zentrifugation (14 000 g, 4°C, 30 sec) der Proben. Das Pellet wurde zweimal mit
1 ml TENTX und einmal mit 1 ml TEN-Puffer (Kap. 2.7.11) gewaschen, in 20 µl SDS-
Probenpuffer (Kap. 2.7.14) aufgenommen, 5 Minuten bei 95°C inkubiert und
gelelektrophoretisch (22 % SDS-Gel mit Harnstoff Kap. 2.7.14.1; 16,5 % SDS-Tricin
Gel Kap. 2.7.14.2) analysiert. Nach der gelelektrophoretischen Auftrennung (Kap.
2.8.35) wurden die Proteine durch eine 10-minütige Inkubation des Gels in
Fixierlösung (Kap. 2.7.11) fixiert und anschließend mit Hilfe einer Geltrocknungs-
apparatur (80°C, 4 h) auf 1 mm Filterpapier (Whatman®) aufgebracht.
Protease-Mapping
Für das Protease-Mapping der zu analysierenden gp9-Konstrukte wurden die Zellen
wie im vorherigen Abschnitt kultiviert und die nach Induktion gebildeten Proteine für
10 Minuten mit [35S]-Methionin radioaktiv markiert. Nach dem 10-minütigen „Pulse-
labelling“ wurden die Proben zentrifugiert (12 000 g, 4°C, 3 min), der radioaktive
Überstand verworfen, die Zellen in 500 µl eiskaltem Spheroplastenpuffer (Kap.
2.7.11) resuspendiert, 5 µl EDTA (0,1 M Stammlösung), 5 µl Lysozym (0,5 mg/ml
Stammlösung) zugegeben und für 15 Minuten auf Eis inkubiert. Die Zellsuspensionen
wurden anschließend in je 3 Ansätze (1x 100 µl, 2 x 200 µl) aliquotiert. Ein 200 µl
Aliquot wurde unmittelbar mit TCA präzipitiert (Kap. 2.8.23), zu dem zweiten 200 µl
Aliquot wurden 7,5 µl Proteinase K (20 mg/ml Stammlösung), zu dem dritten Aliquot
mit 100 µl sowohl 7,5 µl Proteinase K als auch 30 µl Triton X-100 (10 % Stamm-
lösung) zugegeben und für 1 h auf Eis inkubiert. Nach der Inkubation wurde der
Proteaseverdau durch die Präzipitation der Proben mit TCA (Kap. 2.8.23) gestoppt.

Material und Methoden
95
Die Proben wurden anschließend, wie im obigen Abschnitt beschrieben, immun-
präzipitiert unter der Verwendung von anti-OmpA, anti-GroEL, anti-HA, anti-T7 und
anti-M13 Coat Seren.
2.8.40 In vivo Versuch zur YidC-abhängigen Membraninsertion von gp9T7
Zur Untersuchung der YidC-abhängigen Membraninsertion des gp9T7-Konstruktes
wurde das Plasmid pMSgp9T7SD (Kap. 2.1.2) in E. coli JS7131 (Kap. 2.1.1)
transformiert (Kap. 2.8.3) und eine Vorkultur hergestellt (Kap. 2.8.1). Die Vorkultur
diente zur Beimpfung (1:100) von zwei 2 ml LB-Medien mit Arabinose (0,2 %
Endkonzentration). Die Kulturen wurden bei 37°C bis zu einer Zellzahl von 1x 108
Zellen/ml geschüttelt, dann dreimal mit 1x M9-Minimalmedium gewaschen und
zentrifugiert (12 000 g, 3 min). Daraufhin wurden die Zellen einmal in 2 ml „Pulse“-
Medium (Kap. 2.6.2) mit Arabinose (0,2 % Endkonzentration) sowie ein weiteres Mal
in 2 ml „Pulse“-Medium mit Glukose (0,2 % Endkonzentration) aufgenommen und für
weitere 2 h bei 37°C geschüttelt.
Zum Nachweis der YidC Depletion wurden sowohl aus der Arabinose- als auch aus
der Glukose-Kultur Proben von 100 µl entnommen, TCA präzipitiert (Kap. 2.8.23) und
über eine 12 %ige SDS-PAGE (Kap. 2.8.35) mit anschließendem Western-Blot (Kap.
2.8.37) unter Verwendung eines anti-YidC Serums (Kap. 2.1.5) analysiert.
Zur Analyse der YidC-abhängigen Membraninsertion von gp9T7 wurden die Kulturen,
wie in Kapitel 2.8.39 für das „Pulse-labelling“ beschrieben, induziert, radioaktiv
behandelt und anschließend ein Protease-Mapping durchgeführt. Daraufhin folgte die
Immunpräzipitation der Proben unter der Verwendung der anti-OmpA, anti-GroEL
und anti-T7 Seren. Die Proben wurden anschließend gelelektrophoretisch über ein
16,5 % SDS-Tricin-Gel (Kap. 2.7.14.2) analysiert. Nach der gelelektrophoretischen
Auftrennung (Kap. 2.8.35) wurden die Proteine durch eine 10-minütige Inkubation
des Gels in Fixierlösung (Kap. 2.7.11) fixiert und anschließend mit Hilfe einer
Geltrocknungsapparatur (80°C, 4 h) auf 1 mm Filterpapier (Whatman®) aufgebracht.
2.8.41 Quantifizierung radioaktiv markierter Proteine („Phosphorimaging“)
Die Quantifizierung des Bandenmusters der radioaktiv markierten und über SDS-
PAGE aufgetrennten Proteine als Autoradiogram wurde nach Trocknung der Gele mit
der Software AIDA, Raytest, Fuji-Film (Straubenhardt, D) durchgeführt. Die Auto-
radiographie erfolgte für mindestens 5 Tage.

Material und Methoden
96
2.8.42 Präparation FPLC gereinigter gp1/11-Assembly-Komplexe für TEM-Auf-
nahmen
Aus der Elutionsfraktion, über FPLC gereinigter Assembly-Komplexe, wurden 10 µl
entnommen, auf ein Kupfernetzchen (Athene 200) aufgebracht und für 2 Minuten bei
Raumtemperatur adsorbiert. Überschüssige Flüssigkeit wurde mit Hilfe eines Filter-
papiers von der Oberfläche entfernt und danach das Kupfernetzchen 5 Minuten bei
Raumtemperatur getrocknet. Zur Negativkontrastierung der Proben wurden dann 5 µl
einer 5 %igen Phosphorwolfransäure (pH 7) auf die Netzchenoberfläche aufgebracht,
diese nach 30 Sekunden mit Hilfe eines Filterpapiers von der Oberfläche entfernt und
das Kupfernetzchen für 10 Minuten bei Raumtemperatur getrocknet.

Ergebnisse
97
3 Ergebnisse
Der filamentöse Coliphage M13 ist einer der bestuntersuchten Objekte in der Mikro- und
Molekularbiologie. Jedoch sind trotz aller bisher durchgeführten Untersuchungen noch
einige grundlegende Aspekte der Biogenese des Phagen ungeklärt. Um die Biogenese
und die Phagenassemblierung des filamentösen Coliphagen M13 besser verstehen zu
können, wurden folgende Versuche durchgeführt:
Abschnitt 1:
Quantifizierung der M13-Phagenextrusion aus infizierten Escherichia coli-Zellen: Die
Quantifizierung erfolgte anhand eines detaillierten „one-step growth“ Experiments einer
wildtypisch verlaufenden Infektion (Kap. 3.1.1). Neben der wildtypischen Infektion wurde
ein induzierbares Phagenextrusionssystem für eine weitere Quantifizierung verwendet
(Kap. 3.1.2), bei dem das Haupthüllprotein gp8 durch ein induzierbares Plasmid in trans
bereitgestellt wird (Kuhn und Wickner, 1985). Außerdem wurden Adsorptions-
experimente von M13mp19-Phagen an F-Pili von E. coli K38-Zellen durchgeführt, um
die Bindungseffizienz des Phagen bei verschieden hohen Werten der „Multiplicity of
infection“ (M.O.I.) zu erörtern (Kap. 3.1.3).
Abschnitt 2:
Visualisierung M13-infizierter Escherichia coli-Zellen: Die Visualisierung erfolgte mit
Hilfe des Transmissionselektronenmikroskops (TEM) und des „Atomic Force Micros-
cope“ (AFM) (Kap. 3.2). Hierzu wurden die aus Versuchsabschnitt 1 erhaltenen Infor-
mationen herangezogen, um den Extrusionsverlauf von M13-Phagen direkt auf der
Zelloberfläche von E. coli, in Abhängigkeit von der Zeit nach erfolgter Infektion, mit dem
AFM messen zu können (Kap. 3.2.4).
Abschnitt 3:
Funktionelle Präsentation und Detektion von Fremdpeptiden am „Minor Coat“ Protein
gp9: Ein „Phage-Display“ am N-Terminus des „Minor Coat“ Proteins gp9 wurde dazu
verwendet, um eine Abhängigkeitsuntersuchung der gp9-Translokation von der E. coli
Membraninsertase YidC (Kap. 3.3.1 - 3.3.6) durchführen zu können.
Abschnitt 4:
Überexpression der Phagenproteine gp1/11 in Escherichia coli-Zellen: Die Proteine gp1
und gp11 bilden zusammen durch intermolekulare Wechselwirkungen in der Zyto-

Ergebnisse
98
plasmamembran von E. coli einen Komplex, über den die M13-Phagen in die äußere
Umgebung sekretiert werden. Von gereinigten gp1/11-Assembly-Komplexen wurde die
Masse ermittelt und TEM-Aufnahmen angefertigt (Kap. 3.4).
Abschnitt 1:
3.1 Quantifizierung der M13-Phagenextrusion
Damit neue Daten über den M13-Infektionsverlauf erhalten werden konnten, wurde in
diesem Abschnitt die Infektion des Wirtsbakteriums Escherichia coli K38 mit M13-
Phagen anhand von Adsorptionsexperimenten untersucht und anschließend die
Sekretion neuer M13-Phagen-Nachkommen mit Hilfe von „one-step growth“ Experi-
menten analysiert.
3.1.1 Quantifizierung der wildtypischen M13-Phagenextrusion
Für die Quantifizierung der M13-Phagenextrusion wurde ein „one-step growth“
Experiment mit M13-infizierten E. coli-Zellen durchgeführt (Kap. 2.8.18). Für diesen
Versuch kam der Bakterienstamm E. coli K38 zur Verwendung. In diesem können sich
M13 Wildtyp-Phagen normal vermehren, während er für Phagen mit Ambermutation
einen Suppressor-Stamm darstellt. Hierdurch sollte die Quantifizierung der induzierten
Sekretion anhand von M13mp19am8 in Kapitel 3.1.2 mit der wildtypischen vergleichbar
sein. Um die Phagenextrusion quantifizieren zu können, wurde eine Batch-Kultur
exponentiell wachsender E. coli K38-Zellen bei 37°C bis zu einer Zelldichte von
2 x 108 Bakterien/ml geschüttelt, anschließend in einem Eiswasserbad abgekühlt und
daraufhin mit M13mp19-Phagen mit einer M.O.I. von 100 infiziert. Die Adsorption der
Phagen an die F-Pili der E. coli-Zellen erfolgte für 5 Minuten im Eiswasserbad, wodurch
die Aufnahme der Phagen-DNA in das Wirtsbakterium durch Herabsetzung des
bakteriellen Stoffwechsels verhindert wurde. Ungebundene Phagen konnten durch
behutsames Zentrifugieren bei 7500 g, um das Abscheren der Pili zu verhindern, von
den infizierten Bakterien abgetrennt werden. Der phagenhaltige Überstand wurde
verworfen und das Zellpellet in neuem vorgewärmten LB-Flüssigmedium aufge-
nommen. Anschließend erfolgte eine erneute Inkubation der Zellen bei 37°C, um die
Aufnahme der Phagen-DNA in die E. coli-Zellen zu ermöglichen. Dies stellt den Zeit-
punkt t = 0 min in Abbildung 3-1 dar. Zu definierten Zeitpunkten wurden Proben
entnommen, zentrifugiert und mit Hilfe des phagenhaltigen Kulturüberstands die Verän-
derung des Phagentiters durch „Plaque-Assay“ (Kap. 2.8.22 b)) und die Veränderung
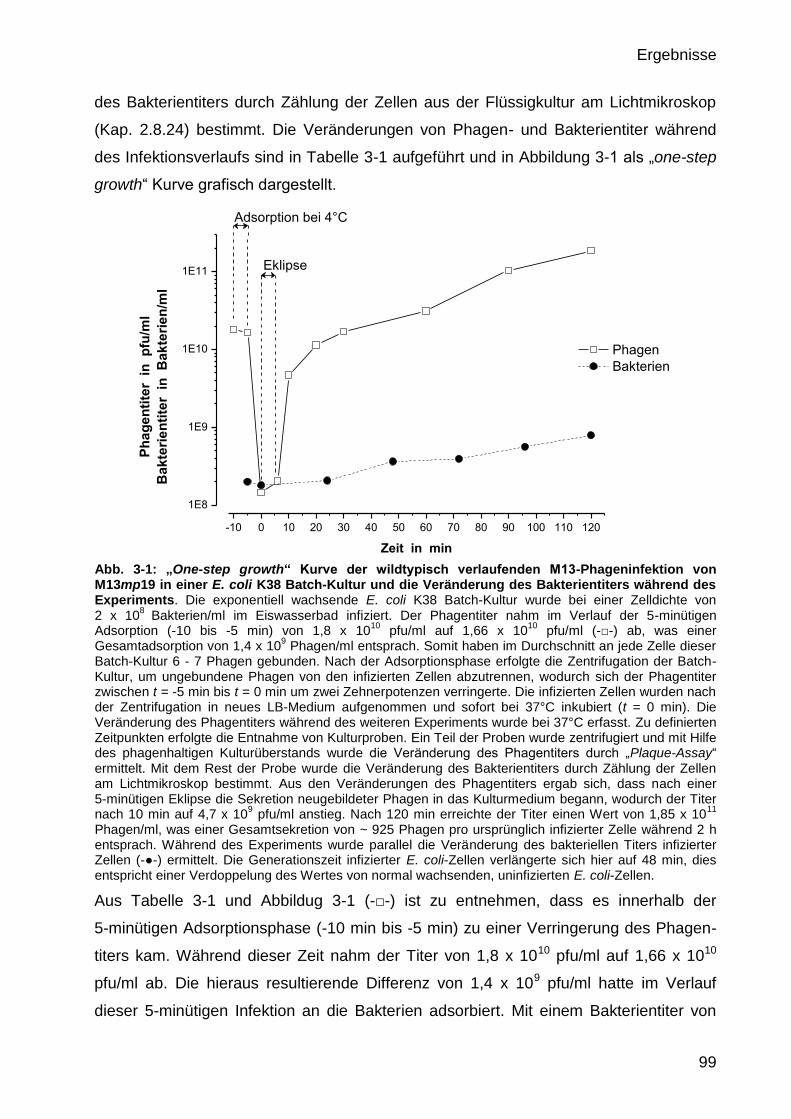
Ergebnisse
99
des Bakterientiters durch Zählung der Zellen aus der Flüssigkultur am Lichtmikroskop
(Kap. 2.8.24) bestimmt. Die Veränderungen von Phagen- und Bakterientiter während
des Infektionsverlaufs sind in Tabelle 3-1 aufgeführt und in Abbildung 3-1 als „one-step
growth“ Kurve grafisch dargestellt.
Aus Tabelle 3-1 und Abbildug 3-1 (-□-) ist zu entnehmen, dass es innerhalb der
5-minütigen Adsorptionsphase (-10 min bis -5 min) zu einer Verringerung des Phagen-
titers kam. Während dieser Zeit nahm der Titer von 1,8 x 1010 pfu/ml auf 1,66 x 1010
pfu/ml ab. Die hieraus resultierende Differenz von 1,4 x 109 pfu/ml hatte im Verlauf
dieser 5-minütigen Infektion an die Bakterien adsorbiert. Mit einem Bakterientiter von
-10 0 10 20 30 40 50 60 70 80 90 100 110 120
1E8
1E9
1E10
1E11
Adsorption bei 4°C
Eklipse
Phagen
Bakterien
Ph
ag
en
tite
r in
p
fu/m
l
Ba
kte
rie
nti
ter
in
B
ak
teri
en
/ml
Zeit in min
Abb. 3-1: „One-step growth“ Kurve der wildtypisch verlaufenden M13-Phageninfektion von M13mp19 in einer E. coli K38 Batch-Kultur und die Veränderung des Bakterientiters während des Experiments. Die exponentiell wachsende E. coli K38 Batch-Kultur wurde bei einer Zelldichte von 2 x 10
8 Bakterien/ml im Eiswasserbad infiziert. Der Phagentiter nahm im Verlauf der 5-minütigen
Adsorption (-10 bis -5 min) von 1,8 x 1010
pfu/ml auf 1,66 x 1010
pfu/ml (-□-) ab, was einer Gesamtadsorption von 1,4 x 10
9 Phagen/ml entsprach. Somit haben im Durchschnitt an jede Zelle dieser
Batch-Kultur 6 - 7 Phagen gebunden. Nach der Adsorptionsphase erfolgte die Zentrifugation der Batch-Kultur, um ungebundene Phagen von den infizierten Zellen abzutrennen, wodurch sich der Phagentiter zwischen t = -5 min bis t = 0 min um zwei Zehnerpotenzen verringerte. Die infizierten Zellen wurden nach der Zentrifugation in neues LB-Medium aufgenommen und sofort bei 37°C inkubiert (t = 0 min). Die Veränderung des Phagentiters während des weiteren Experiments wurde bei 37°C erfasst. Zu definierten Zeitpunkten erfolgte die Entnahme von Kulturproben. Ein Teil der Proben wurde zentrifugiert und mit Hilfe des phagenhaltigen Kulturüberstands wurde die Veränderung des Phagentiters durch „Plaque-Assay“ ermittelt. Mit dem Rest der Probe wurde die Veränderung des Bakterientiters durch Zählung der Zellen am Lichtmikroskop bestimmt. Aus den Veränderungen des Phagentiters ergab sich, dass nach einer 5-minütigen Eklipse die Sekretion neugebildeter Phagen in das Kulturmedium begann, wodurch der Titer nach 10 min auf 4,7 x 10
9 pfu/ml anstieg. Nach 120 min erreichte der Titer einen Wert von 1,85 x 10
11
Phagen/ml, was einer Gesamtsekretion von ~ 925 Phagen pro ursprünglich infizierter Zelle während 2 h entsprach. Während des Experiments wurde parallel die Veränderung des bakteriellen Titers infizierter Zellen (-●-) ermittelt. Die Generationszeit infizierter E. coli-Zellen verlängerte sich hier auf 48 min, dies entspricht einer Verdoppelung des Wertes von normal wachsenden, uninfizierten E. coli-Zellen.

Ergebnisse
100
2 x 108 Zellen/ml ergibt sich eine durchschnittliche Bindung von 7 Phagen pro
Bakterienzelle. Nach der Adsorptionsphase folgte die Zentrifugation der infizierten
Batch-Kultur, um ungebundene Phagen von den infizierten Zellen abzutrennen.
Dadurch nahm der Phagentiter zwischen t = -5 min und 0 min um zwei Zehnerpotenzen
ab. Nach der Aufnahme der infizierten Zellen in neues LB-Medium und der daraufhin
folgenden Inkubation bei 37°C (Abb.3-1, t = 0) folgte eine Eklipse, welche 6 Minuten
anhielt.
Während dieser Zeit wurden keine Phagen-Nachkommen sekretiert. Die Sekretion der
ersten Phagen-Nachkommen in das LB-Medium der Batch-Kultur begann 6 Minuten
nach Inkubation der Zellen bei 37°C. Dies wurde durch den Anstieg des Phagentiters
zwischen der 6. und 10. Minute in Abbildung 3-1 (-□-) dokumentiert. In dieser kurzen
Zeit erhöhte sich der Phagentiter von 2 x 108 pfu/ml auf 4,7 x 109 pfu/ml. Dies stellt eine
Vermehrung der Ausgangsmenge um das circa 30-fache dar, während der Bakterien-
titer in dieser Zeit nahezu konstant blieb. Daraus ergab sich, dass in dieser Zeitspanne
von 4 Minuten jede infizierte Zelle im Mittel 6 neue Phagen pro Minute in das Medium
sekretiert hatte. Erst nach 20 Minuten verlangsamte sich der Anstieg des Phagentiters
auf durchschnittlich 4 Phagen/min für jede infizierte Zelle. Nach insgesamt 120 Minuten
erreichte der Phagentiter, nach Abzug des Phagenhintergrunds zu t = 0 min, einen Wert
von 1,85 x 1011 pfu/ml. Hieraus ergab sich innerhalb dieser 120 Minuten eine durch-
schnittliche Gesamtsekretion von ~ 925 Phagen pro ursprünglich infizierter Zelle. Mit
einem Bakterientiter von 7,9 x 108 Bakterien/ml, welcher im Verlauf dieses zweistün-
digen Experiments erreicht wurde, erhielt man eine gemittelte Sekretion von 300 neuen
Phagen pro Bakterienzelle. Der ursprüngliche Phagentiter zu t = 0 min konnte während
dieser 120 Minuten um das 1200-fache erhöht werden. Das Experiment zeigte auch,
dass sich die Generationszeit für die infizierten E. coli-Zellen veränderte. Analysiert man
Zeit in min
Phagentiter in pfu/ml
Zeit in min
Bakterientiter in Bakterien/ml
-10 1,80 x 1010
-5 2,0 x 108
-5 1,66 x 1010
0 1,8 x 108
0 1,44 x 108 24 2,07 x 10
8
6 2,04 x 108 48 3,64 x 10
8
10 4,70 x 109 72 3,93 x 10
8
20 1,14 x 1010
96 5,61 x 108
30 1,70 x 1010
120 7,92 x 108
60 3,10 x 1010
90 1,04 x 1011
120 1,85 x 1011
Tabelle 3-1: Veränderung des Phagen- und Bakterientiters während des wildtypischen „one-step growth“ Experiments bei einer M.O.I von 100. Die Bestimmung des Phagentiters [pfu/ml] zu den Entnahmezeiten erfolgte durch „Plaque-Assay“; die Anzahl der infizierten Bakterien wurde durch Auszählung am Lichtmikroskop bestimmt. Zur Bestimmung des Bakterientiters wurde nach dem Zeitpunkt t = 0 min ein Probenintervall von 24 Minuten gewählt, welches der Generationszeit von E. coli entspricht.

Ergebnisse
101
den Verlauf des Bakterientiters in Tabelle 3-1, so ergab sich für infizierte E. coli-Zellen
eine Generationszeit von ~ 48 Minuten. Dies stellte eine Verdoppelung der Gene-
rationszeit gegenüber uninfizierten Zellen dar, da diese während der exponentiellen
Wachstumsphase üblicherweise nur 24 Minuten für eine Zellteilung benötigen.
3.1.2 Quantifizierung der M13mp19am8-Phagenextrusion während der Komple-
mentation durch das plasmidkodierte Haupthüllprotein gp8
Bei diesem „one-step growth“ Experiment war die Phagensekretion aus E. coli durch die
Regulation der gp8-Expression induzierbar. Es sollte der Verlauf der Phagensekretion
mit dem der wildtypischen Sekretion verglichen werden. Zunächst wurde ein induzier-
bares Expressionssystem konstruiert (Kap. 2.8.15 a)). Das Gen 8, welches für das
Haupthüllprotein kodiert, wurde in das Plasmid pGZ119HE kloniert. In diesem Plasmid
stand die gp8-Expression unter der Kontrolle des induzierbaren tac Promotors. Das
resultierende Plasmid pGZg8wt wurde in E. coli K38 transformiert. Für die Infektion der
transformierten Zellen kam der Phagenstamm M13mp19am8 zur Verwendung, in dem
die Aminosäure Lysin an Position 3 des Gens 8 in ein Amber-Stoppcodon verändert
war. Für die Quantifizierung wurden zwei Batch-Kulturen exponentiell wachsender
E. coli K38-Zellen mit transformiertem pGZg8wt verwendet, welche zuvor bei 37°C bis
zu einem Titer von 2 x 108 Bakterien/ml kultiviert, dann in einem Eiswasserbad abge-
kühlt und anschließend mit M13mp19am8-Phagen infiziert wurden (Kap 2.8.18).
Während einer 5-minütigen Adsorptionsphase erfolgte die Bindung der Phagen an die
F-Pili der E. coli-Zellen. Durch behutsame Zentrifugation bei 7500 g konnten anschlie-
ßend ungebundene Phagen von den infizierten Bakterien getrennt werden. Die phagen-
haltigen Überstände wurden verworfen und die Zellpellets in neuen vorgewärmten LB-
Flüssigmedien aufgenommen. Hierauf folgte eine erneute Inkubation der Zellsuspen-
sionen bei 37°C, um die Aufnahme der Phagen-DNA in die E. coli-Zellen zu ermög-
lichen. Sodann erfolgte die Induktion einer der Batch-Kulturen durch Zugabe von IPTG
(1mM Endkonzentration) und damit die Bereitstellung des Haupthüllproteins gp8 in
trans für die Phagenassemblierung. Dies stellt den Zeitpunkt t = 0 min in Abbildung 3-2
dar. Die zweite Batch-Kultur blieb als Referenz uninduziert. In Intervallen wurden
Proben zu definierten Zeitpunkten aus beiden Batch-Kulturen entnommen, zentrifugiert
und mit Hilfe der phagenhaltigen Kulturüberstände durch „Plaque-Assay“ (Kap. 2.8.22
b)) der Verlauf der Phagentiter verfolgt. Die Veränderungen der Phagentiter sind in
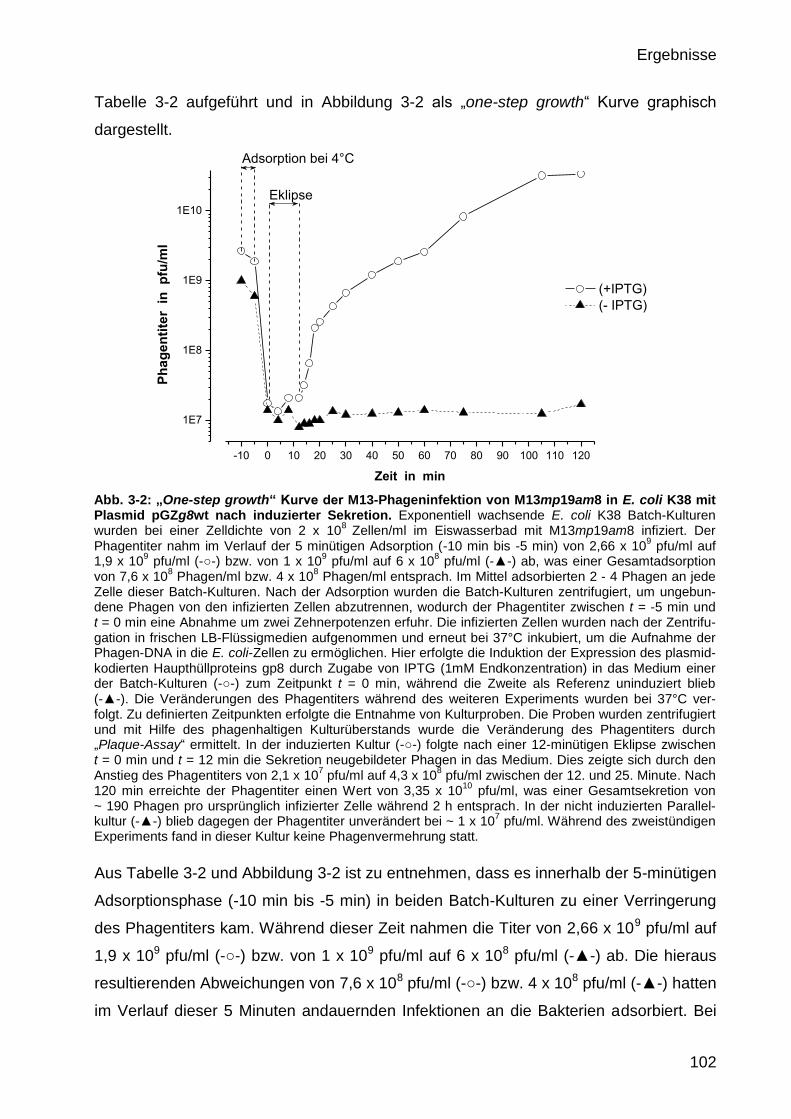
Ergebnisse
102
Tabelle 3-2 aufgeführt und in Abbildung 3-2 als „one-step growth“ Kurve graphisch
dargestellt.
Aus Tabelle 3-2 und Abbildung 3-2 ist zu entnehmen, dass es innerhalb der 5-minütigen
Adsorptionsphase (-10 min bis -5 min) in beiden Batch-Kulturen zu einer Verringerung
des Phagentiters kam. Während dieser Zeit nahmen die Titer von 2,66 x 109 pfu/ml auf
1,9 x 109 pfu/ml (-○-) bzw. von 1 x 109 pfu/ml auf 6 x 108 pfu/ml (-▲-) ab. Die hieraus
resultierenden Abweichungen von 7,6 x 108 pfu/ml (-○-) bzw. 4 x 108 pfu/ml (-▲-) hatten
im Verlauf dieser 5 Minuten andauernden Infektionen an die Bakterien adsorbiert. Bei
-10 0 10 20 30 40 50 60 70 80 90 100 110 120
1E7
1E8
1E9
1E10
Eklipse
Ph
ag
en
tite
r in
p
fu/m
l
Zeit in min
(+IPTG)
(- IPTG)
Adsorption bei 4°C
Abb. 3-2: „One-step growth“ Kurve der M13-Phageninfektion von M13mp19am8 in E. coli K38 mit Plasmid pGZg8wt nach induzierter Sekretion. Exponentiell wachsende E. coli K38 Batch-Kulturen wurden bei einer Zelldichte von 2 x 10
8 Zellen/ml im Eiswasserbad mit M13mp19am8 infiziert. Der
Phagentiter nahm im Verlauf der 5 minütigen Adsorption (-10 min bis -5 min) von 2,66 x 109 pfu/ml auf
1,9 x 109 pfu/ml (-○-) bzw. von 1 x 10
9 pfu/ml auf 6 x 10
8 pfu/ml (-▲-) ab, was einer Gesamtadsorption
von 7,6 x 108 Phagen/ml bzw. 4 x 10
8 Phagen/ml entsprach. Im Mittel adsorbierten 2 - 4 Phagen an jede
Zelle dieser Batch-Kulturen. Nach der Adsorption wurden die Batch-Kulturen zentrifugiert, um ungebun-dene Phagen von den infizierten Zellen abzutrennen, wodurch der Phagentiter zwischen t = -5 min und t = 0 min eine Abnahme um zwei Zehnerpotenzen erfuhr. Die infizierten Zellen wurden nach der Zentrifu-gation in frischen LB-Flüssigmedien aufgenommen und erneut bei 37°C inkubiert, um die Aufnahme der Phagen-DNA in die E. coli-Zellen zu ermöglichen. Hier erfolgte die Induktion der Expression des plasmid-kodierten Haupthüllproteins gp8 durch Zugabe von IPTG (1mM Endkonzentration) in das Medium einer der Batch-Kulturen (-○-) zum Zeitpunkt t = 0 min, während die Zweite als Referenz uninduziert blieb (-▲-). Die Veränderungen des Phagentiters während des weiteren Experiments wurden bei 37°C ver-folgt. Zu definierten Zeitpunkten erfolgte die Entnahme von Kulturproben. Die Proben wurden zentrifugiert und mit Hilfe des phagenhaltigen Kulturüberstands wurde die Veränderung des Phagentiters durch „Plaque-Assay“ ermittelt. In der induzierten Kultur (-○-) folgte nach einer 12-minütigen Eklipse zwischen t = 0 min und t = 12 min die Sekretion neugebildeter Phagen in das Medium. Dies zeigte sich durch den Anstieg des Phagentiters von 2,1 x 10
7 pfu/ml auf 4,3 x 10
8 pfu/ml zwischen der 12. und 25. Minute. Nach
120 min erreichte der Phagentiter einen Wert von 3,35 x 1010
pfu/ml, was einer Gesamtsekretion von ~ 190 Phagen pro ursprünglich infizierter Zelle während 2 h entsprach. In der nicht induzierten Parallel-kultur (-▲-) blieb dagegen der Phagentiter unverändert bei ~ 1 x 10
7 pfu/ml. Während des zweistündigen
Experiments fand in dieser Kultur keine Phagenvermehrung statt.

Ergebnisse
103
einem Bakterientiter von 2 x 108 Zellen/ml ergab sich hier eine durchschnittliche
Bindung von 2 bis 4 Phagen pro Bakterienzelle.
Nach der Adsorptionsphase folgte die Zentrifugation der infizierten Batch-Kulturen, um
ungebundene Phagen von den infizierten Zellen abzutrennen. Dadurch nahmen die
Phagentiter zwischen t = -5 min und t = 0 min um zwei Zehnerpotenzen ab. Nach Auf-
nahme der infizierten Zellen in frisches LB-Medium und unter Schütteln bei 37°C (Abb.
3-2, t = 0 min) folgte in der induzierten Kultur (-○-) eine Eklipse, welche circa 12 Minuten
anhielt. Während dieser Zeit wurden keine Phagen-Nachkommen sekretiert. Die
Sekretion der ersten Phagen-Nachkommen in das LB-Medium der induzierten Batch-
Kultur erfolgte 12 Minuten nach Inkubation der Zellen. Hierdurch stieg der Phagentiter
bis zur 18. Minute von 2,10 x 107 pfu/ml auf 2,10 x 108 pfu/ml an. In dieser kurzen
Zeitspanne verzehnfachte sich der Phagentiter gegenüber der Ausgangsmenge. Nach
35 Minuten erreichte der Titer einen Wert von 6,7 x 108 pfu/ml, was einer Sekretion von
6,5 x 108 neuer Phagen pro ml entsprach. Somit wurden von jeder infizierten Bakterien-
zelle ~ 3 Phagen sekretiert. Nach 120 Minuten erreichte der Phagentiter einen Endwert
von 3,35 x 1010 pfu/ml, was einer Gesamtsekretion von ~ 190 Phagen pro ursprünglich
infizierter Zelle innerhalb von 2 h entsprach. Während dieser zweistündigen Infektion
Zeit in min
-IPTG Phagentiter in
pfu/ml
+ IPTG Phagentiter in
pfu/ml
-10 1,00 x 109 2,66 x 109 -5 6,00 x 108 1,90 x 109 0 1,40 x 107 1,75 x 107 4 1,00 x 107 1,35 x 107 8 1,40 x 107 2,10 x 107 12 8,00 x 106 2,10 x 107 14 9,00 x 106 3,20 x 107 16 9,00 x 106 6,60 x 107 18 1,00 x 107 2,10 x 108 20 1,00 x 107 2,55 x 108 25 1,35 x107 4,30 x 108 30 1,20 x 107 6,70 x 108 40 1,25 x 107 1,20 x 109 50 1,30 x 107 1,90 x 109 60 1,40 x 107 2,60 x 109 75 1,30 x 107 8,30 x 109
105 1,25 x 107 3,20 x 1010 120 1,70 x 107 3,35 x 1010
Tabelle 3-2: Veränderung des Phagentiters mit und ohne Induktion des plasmidkodierten Haupthüllproteins gp8 in infizierten E. coli K38 Batch-Kulturen. Die Bestimmung der Phagentiter zu den Entnahmezeiten erfolgte durch „Plaque-Assay“. Zum Zeitpunkt t = 0 min wurde die Expression des Haupthüllproteins gp8 durch Zugabe von IPTG zu einer der Batch-Kulturen induziert, während die Zweite als Referenz uninduziert blieb.

Ergebnisse
104
konnte bei der induzierten Kultur die Ausgangsmenge der Phagen zu t = 0 min um das
1900-fache erhöht werden. Der Verlauf des Phagentiters in der nicht induzierten Kultur
(-▲-) hingegen blieb während des gesamten Experiments unverändert bei ~ 1 x 107
pfu/ml. Dies zeigte, dass keine neugebildeten Phagen in das Medium sekretiert wurden.
3.1.3 M13-Adsorption an F-Pili in Abhängigkeit von der M.O.I.
In diesem Adsorptionsexperiment wurde die maximal mögliche Anzahl von Phagen
ermittelt, die während einer 5-minütigen Adsorptionsphase pro E. coli-Zelle an F-Pili
gebunden werden können.
Für dieses Adsorptionsexperiment wurden Batch-Kulturen exponentiell wachsender
E. coli K38-Zellen bei einer Zelldichte von 2 x 108 Zellen/ml mit M13mp19-Phagen infi-
ziert (Kap. 2.8.19). Hier wurden Adsorptionsverläufe für die M.O.I.-Werte von 5, 10, 20,
52 und 90 erstellt. Die Adsorption der Phagen an die F-Pili der E. coli-Zellen erfolgte für
5 Minuten im Eiswasserbad. Die Analyse der Anzahl an gebundenen Phagen pro Zelle
erfolgte anhand der Abnahmen der eingesetzten Phagentiter während der Adsorptions-
phase. Hierfür wurde einmal der Phagentiter direkt zum Zeitpunkt der Phagenzugabe
zur Batch-Kultur (t = 0 min) und nochmals nach einer 5-minütigen Adsorptionsphase
(t = 5 min) mit Hilfe des „Plaque-Assay“ (Kap. 2.8.22 b)) bestimmt. Aus den sich hieraus
ergebenden Abweichungen in den Phagentitern wurden anschließend die Phagenad-
sorptionen pro Minute für die 5 Minuten andauernden Adsorptionsphasen ermittelt.
Parallel dazu wurde der Wert für die unspezifische Bindung der M13mp19-Phagen an
Glasgeräte und unpilierte Bakterien als Kontrolle bestimmt (Kap. 2.8.19). Hier wurden
Batch-Kulturen exponentiell wachsender E. coli MC4100 verwendet, welche keine F-Pili
besitzen. Unter gleichen Bedingungen wie bei E. coli K38 erfolgten Adsorptionsverläufe
für die M.O.I.-Werte 10, 50 und 100. Alle durchgeführten Experimente wurden 2-mal
wiederholt. Dabei konnte eine ~ 8 %ige Standardabweichung zwischen den Mess-
werten der Einzelmessungen ermittelt werden, welche in den Darstellungen der Adsorp-
tionskurven in Abbildung 3-3 in Form von Fehlerbalken berücksichtigt wurde. Die
ermittelten Werte für die unspezifischen Bindungen sind in Tabelle 3-4, die an E. coli
K38 in Tabelle 3-3 wiedergegeben. Die gemittelten Werte wurden zusätzlich graphisch
als Adsorptionskurven in Abbildung 3-3 dargestellt.

Ergebnisse
105
Tabelle 3-3: Abnahme des Phagentiters in E. coli K38 Batch-Kulturen während der Phagen-Adsorptionsphase: Die Abnahme des Phagentiters während der 5-minütigen Adsorptionsphase wurde für verschiedene M.O.I.-Werte durchgeführt. Hierfür erfolgte die Ermittlung der Phagentiter direkt zum Zeitpunkt der Phagenzugabe zur Batch-Kultur und nochmals nach 5-minütiger Adsorptionsphase mit Hilfe des „Plaque-Assay“. Ab M.O.I. 20 kam es bei einem Wert von ~ -2,8 x 10
8 pfu/min zu einer
Sättigung von gebundenen Phagen an E. coli K38.
Tabelle 3-4: Abnahme des Phagentiters in E. coli MC4100 Batch-Kulturen während der Phagen-Adsorptionsphase: Die Abnahme des Phagentiters während der 5-minütigen Adsorptionsphase wurde für verschiedene M.O.I.-Werte durchgeführt. Hier erfolgte die Bestimmung der Phagentiter direkt zum Zeitpunkt der Phagenzugabe zur Batch-Kultur und nochmals nach 5-minütiger Adsorptionsphase mit Hilfe des „Plaque-Assay“. Aus den Versuchen ergab sich ein Wert von -1 x 10
8 pfu/min für die unspezifische
Bindung während der 5-minütigen Adsorptionsphase in Eiswasserbad, unabhängig von der eingesetzten M.O.I.
M.O.I. Abnahme PhagentiterΔt=5min
pfu/min
5 - 8.00 x 107
10 - 1.52 x 108
20 - 2.70 x 108
52 - 2.74 x 108
90 - 2.80 x 108
M.O.I. Abnahme PhagentiterΔt=5min
[pfu/min]
10 - 1 x 108
50 - 1 x 108
100 - 1 x 108
0 10 20 30 40 50 60 70 80 90 100 110
0,0
-5,0x107
-1,0x108
-1,5x108
-2,0x108
-2,5x108
-3,0x108
-3,5x108
spezifische Bindung
unspezifische Bindung
Ab
na
hm
e P
ha
ge
nti
ter
in
p
fu/m
in
M.O.I.
Abb. 3-3: Adsorption der M13mp19-Phagen in E. coli Batch-Kulturen in Abhängigkeit der ver-wendeten M.O.I. Batch-Kulturen exponentiell wachsender E. coli K38 (-■-) bzw. E. coli MC4100 (-○-) Zellen wurden bei einer Zelldichte von 2 x 10
8 Zellen/ml mit unterschiedlichen M.O.I.-Werten im
Eiswasserbad infiziert. Die Abnahmen der Phagentiter während der 5-minütigen Adsorptionsphasen wurden durch Probenentnahme zur Infektionszeit t = 0 min und nach t = 5 min durch „Plaque-Assay“ ermittelt. Aus den sich hieraus ergebenden Abweichungen in den Phagentitern wurden anschließend die Phagenadsorptionen pro Minute für die 5-minütige Adsorptionsphase ermittelt. Für die Adsorptionsex-

Ergebnisse
106
Die Abnahmen der Phagentiter in den einzelnen Adsorptionsexperimenten sind
graphisch in Abbildung 3–3 gegen die eingesetzte M.O.I. aufgetragen. Die ermittelten
Adsorptionswerte in den E. coli K38 Batch-Kulturen (-■-) wurden hier mit dem
ermittelten Wert für die unspezifische Adsorption der Phagen (-○-) bereinigt. Die
Adsorptionskurve zeigt eine ansteigende Bindung von Phagen an die Zellen während
der 5-minütigen Adsorptionsphase mit Zunahme der eingesetzten M.O.I. im Bereich
zwischen M.O.I. 5 – 20. Ab M.O.I. 20 erreichte die Phagenbindung eine gemittelte
Sättigung von ~ -3 x 108 Phagen/Minuten, darüber hinaus konnten durch weitere
Erhöhungen der M.O.I. keine weiteren Phagen während der 5-minütigen Adsorptions-
phase an die F-Pili der Zellen gebunden werden. Bezogen auf die Anzahl der maximal
infizierbaren E. coli-Zellen in den Batch-Kulturen während dieser 5 Minuten, ergab sich
somit ein maximales Verhältnis von 1:7, d.h. nach dieser 5-minütigen Adsorptionsphase
konnten durchschnittlich ~ 7 Phagen pro E. coli-Zelle gebunden werden.
Die Adsorptionsexperimente zur Ermittlung der unspezifischen Bindung an E. coli
MC4100 (Abb. 3-3, -○-) sowie an verwendete Glasgeräte (Erlenmeyerkolben, Glas-
pipetten) ergaben einen konstanten Wert von durchschnittlich -1 x 108 pfu/min nach der
5-minütigen Adsorptionsphase, welcher unabhängig von der verwendeten M.O.I. war.
Abschnitt 2:
3.2 Visualisierung M13-infizierter E. coli-Zellen
Zur Visualisierung neu gebildeter Phagenpartikel auf der Bakterienoberfläche wurden in
diesem Abschnitt M13-infizierte Escherichia coli DH5α-Zellen mit Hilfe des Transmissi-
onselektronenmikroskops (TEM) und des Rasterkraftmikroskops (RKM = AFM, „Atomic
Force Microscope“) untersucht.
perimente mit E. coli K38 wurden M.O.I.-Werte von 5, 10, 20, 52 und 90 verwendet und die Anzahl adsorbierter Phagen/min dargestellt (-■-). Alle durchgeführten Experimente wurden 2-mal wiederholt. Die ermittelte Standardabweichung von ~ 8 % zwischen den Messwerten der Einzelmessungen ist jeweils in den Werten der Adsorptionskurven als Fehlerbalken dargestellt. Die maximale Abnahme lag hier bei ~ -2,8 x 10
8 pfu/min. Dabei trat eine Adsorptionssättigung ab einer M.O.I. von 20 auf. Dies
entsprach einer Adsorption von ~ 7 Phagen pro Zelle nach 5 Minuten. Für die unspezifische Bindung von M13-Phagen an Glasgeräte und Bakterien wurden Adsorptionsexperimente mit E. coli MC4100 durchgeführt, welche keine F-Pili besitzen. Hier wurden M.O.I.-Werte von 10, 50 und 100 eingesetzt. Die Anzahl der adsorbierten Phagen pro Minute an die Zellen und Glasgeräte in den 5-minütigen Adsorptionsphasen wurden in Abhängigkeit von der M.O.I. dargestellt (-○-). Der konstante Wert von -1 x 10
8 pfu/min zeigt, dass die unspezifische Bindung nicht von der eingesetzten M.O.I. abhängig ist.
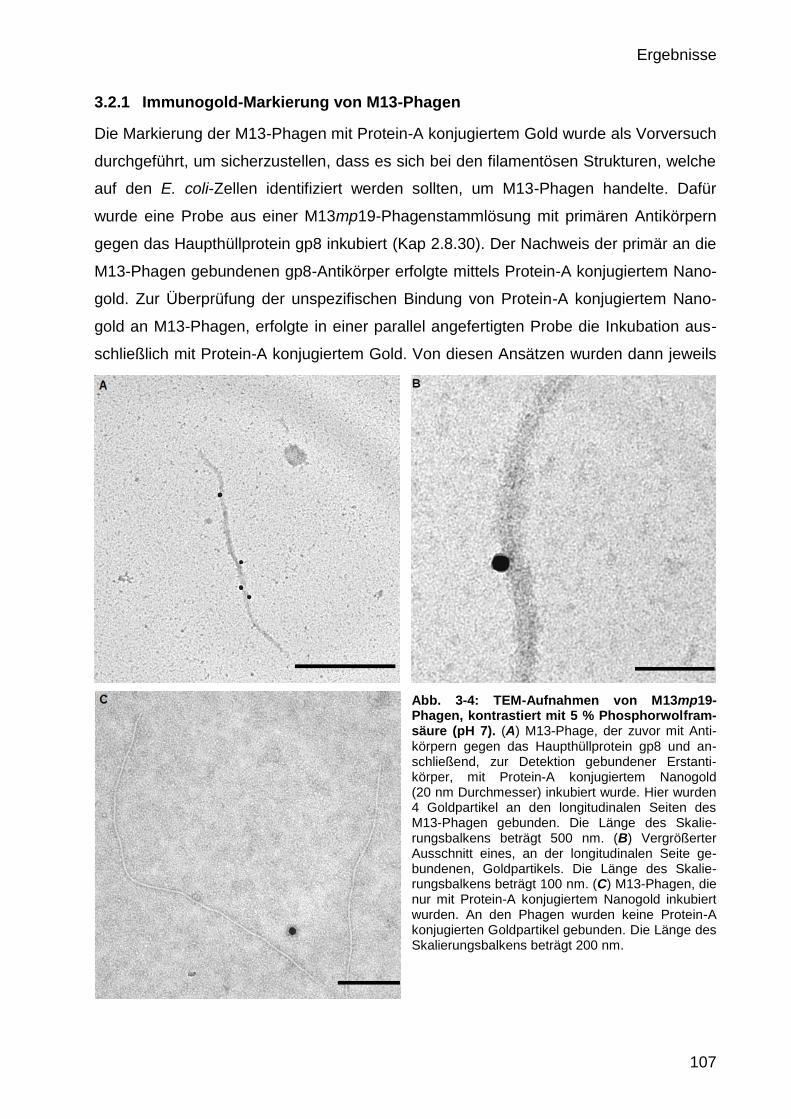
Ergebnisse
107
3.2.1 Immunogold-Markierung von M13-Phagen
Die Markierung der M13-Phagen mit Protein-A konjugiertem Gold wurde als Vorversuch
durchgeführt, um sicherzustellen, dass es sich bei den filamentösen Strukturen, welche
auf den E. coli-Zellen identifiziert werden sollten, um M13-Phagen handelte. Dafür
wurde eine Probe aus einer M13mp19-Phagenstammlösung mit primären Antikörpern
gegen das Haupthüllprotein gp8 inkubiert (Kap 2.8.30). Der Nachweis der primär an die
M13-Phagen gebundenen gp8-Antikörper erfolgte mittels Protein-A konjugiertem Nano-
gold. Zur Überprüfung der unspezifischen Bindung von Protein-A konjugiertem Nano-
gold an M13-Phagen, erfolgte in einer parallel angefertigten Probe die Inkubation aus-
schließlich mit Protein-A konjugiertem Gold. Von diesen Ansätzen wurden dann jeweils
Abb. 3-4: TEM-Aufnahmen von M13mp19- Phagen, kontrastiert mit 5 % Phosphorwolfram-säure (pH 7). (A) M13-Phage, der zuvor mit Anti-körpern gegen das Haupthüllprotein gp8 und an-schließend, zur Detektion gebundener Erstanti-körper, mit Protein-A konjugiertem Nanogold (20 nm Durchmesser) inkubiert wurde. Hier wurden 4 Goldpartikel an den longitudinalen Seiten des M13-Phagen gebunden. Die Länge des Skalie-rungsbalkens beträgt 500 nm. (B) Vergrößerter Ausschnitt eines, an der longitudinalen Seite ge-bundenen, Goldpartikels. Die Länge des Skalie-rungsbalkens beträgt 100 nm. (C) M13-Phagen, die nur mit Protein-A konjugiertem Nanogold inkubiert wurden. An den Phagen wurden keine Protein-A konjugierten Goldpartikel gebunden. Die Länge des Skalierungsbalkens beträgt 200 nm.

Ergebnisse
108
10 µl auf Trägernetzchen aus Kupfer adsorbiert und eine Negativkontrastierung mit
Phosphorwolframsäure (5 %, pH 7) durchgeführt (Kap. 2.8.27). Die kontrastierten
Proben wurden anschließend am Transmissionselektronenmikroskop (TEM) untersucht
(Kap. 2.8.29). Die Ergebnisse der mit Immunogold markierten Phagen sind in Abbildung
3-4 (A – C) zu sehen. Die Abbildung 3-4 (A) zeigt einen M13-Phagen, der zuvor mit
gp8-Antikörpern inkubiert wurde, bevor die Detektion mit Protein-A konjugiertem Nano-
gold erfolgte. Hier ist ein M13-Phage mit einer Länge von ~ 1 µm zu sehen, an dem vier
Goldpartikel gebunden wurden. In Abbildung 3-4 (B) ist ein vergrößerter Ausschnitt
eines, an der longitudinalen Seite des M13-Phagen, gebundenen Goldpartikels mit
20 nm Durchmesser zu sehen. Die Abbildung 3-4 (C) zeigt eine TEM-Aufnahme der
Probe, die nicht mit dem primären Antikörper gegen gp8 sondern nur mit Protein-A kon-
jugiertem Nanogold inkubiert wurde. An diesen M13-Phagen wurden keine Goldpartikel
gebunden.
3.2.2 TEM-Aufnahmen M13-sekretierenderE. coli DH5α-Zellen
Für TEM-Aufnahmen M13-sekretierender E. coli-Zellen wurden unpilierte E. coli DH5α-
Zellen verwendet, da nur so sichergestellt werden konnte, dass es sich bei filamen-
tösen Strukturen auf den Zelloberflächen um M13-Phagen handelte. Dieser Stamm ver-
fügt über keine F-Pili (F-), sodass die Infektion nur über eine Transfektion mit dem RF-
Plasmid von M13 erfolgen konnte (Kap. 2.8.25). Zur Überprüfung der Transfektion von
E. coli DH5α wurde der Phagenstamm M13amp konstruiert (Kap. 2.8.16), der das
β-Lactamase Gen unter der Kontrolle des lac-Promotors in der „Multiple Cloning Site“
von M13mp19 trug. Mit Hilfe des Phagenstamms M13amp konnten, nach Induktion mit
IPTG, transfizierte Zellen auf ampicillinhaltigen LB-Medien selektiert werden. Nach
Transfektion wurden mehrere gewachsene Einzelkolonien von Selektivplatten in
ampicillin- und IPTG-haltiges LB-Flüssigmedium überführt und über Nacht bei 37°C
unter Schütteln kultiviert. Die anschließenden Bestimmungen der Phagentiter in den
Überständen der einzelnen Kulturen erfolgte mit Hilfe des „Plaque-Assay“ (Kap. 2.8.22
b)). Für Aufnahmen von phagensekretierenden Zellen wurde eine infizierte E. coli
DH5α-Flüssigkultur verwendet, in deren Überstand eindeutig M13-Phagen nachge-
wiesen werden konnten. Damit die filamentösen Oberflächenstrukturen auf den Zellen
eindeutig als M13-Phagen zu identifizieren waren, erfolgte bei diesen Zellen die gleiche
Markierungsprozedur wie bei den Immunogold markierten M13-Phagen in Kap. 3.2.1.
Zur Überprüfung der unspezifischen Bindungen von gp8-Antikörpern und Protein-A

Ergebnisse
109
konjugiertem Nanogold an nicht transfizierte E. coli DH5α-Zellen, wurde parallel eine
nicht infizierte Kultur der gleichen Prozedur unterzogen; diese diente als Kontrolle. Von
den Ansätzen wurden, wie in Kapitel 2.8.27 beschrieben, jeweils 10 µl auf Kupfer-
netzchen adsorbiert und daraufhin mit Phosphorwolframsäure (5 %, pH 7) negativ
kontrastiert. Anschließend folgte die Untersuchung der Proben am TEM (Kap. 2.8.29).
Die Aufnahmen der Immunogold markierten E. coli DH5α-Zellen sind in Abbildung 3-5
(A – C) zu sehen.
Die transfizierten E. coli DH5α (F-)-Zellen in Abbildung 3-5 (A) und (B) zeigten
filamentöse Strukturen an deren Polenden, die an den longitudinalen Seiten Protein-A
konjugierte Nanogoldpartikel von 20 nm Durchmesser gebunden hatten. Die filamen-
tösen Strukturen hatten Längen von ~ 1 µm. Die nicht transfizierte Kontrolle in
Abb. 3-5: TEM-Aufnahmen von gebundenem Protein-A konjugiertem Gold an extrudierenden M13-Phagen. M13amp transfizierte E. coli DH5α- Zellen (F
-) wurden zuerst mit Antikörpern gegen das
Haupthüllprotein gp8 und anschließend mit Protein-A konjugiertem Nanogold behandelt. Negativkon-trastierung der Proben erfolgte mit 5 % Phosphor-wolframsäure (pH 7). (A), (B) Bakterienoberflächen transfizierter Zellen mit filamentösen Strukturen an den polaren Enden, die Längen von ~ 1 µm be-saßen. An den longitudinalen Seiten der Filamente waren Protein-A konjugierte Nanogoldpartikel mit 20 nm Durchmesser gebunden. (C) Nicht transfi-zierte Zelle, die mit gp8-Antikörpern und Protein-A konjugiertem Nanogold inkubiert wurde. Die Zelle zeigte weder filamentöse Strukturen noch gebun-dene Goldpartikel auf der Zelloberfläche. Die Län-gen der Skalierungsbalken betragen 500 nm.

Ergebnisse
110
Abbildung 3-5 (C) hingegen, welche ebenfalls mit gp8-Antikörpern und Protein-A
konjugiertem Nanogold inkubiert wurde, zeigte weder filamentöse Strukturen mit asso-
ziiertem Gold noch gebundene Goldpartikel auf der Bakterienoberfläche.
3.2.3 AFM-Aufnahmen M13-sekretierender E. coli DH5α-Zellen
Das „Atomic Force Microscope“ (AFM) stellt ein geeignetes Instrument dar, um bio-
logische Proben zu messen. Mit Hilfe des AFM ist es möglich, Oberflächenstrukturen
biologischer Proben detailgetreu messen und darstellen zu können. Über ein piezo-
elektrisches Verfahren wird eine Messsonde („Cantilever“), welche eine sehr feine
Spitze („Tip“) besitzt, über eine Probenoberfläche gerastert. Während des Rastervor-
gangs werden die Wechselwirkungskräfte („van der Waals Kräfte“) zwischen der Probe
und der Messsonde zur topographischen Darstellung der Probenoberfläche ausgenutzt.
Für die Messungen M13-sekretierender Zellen am AFM kam die transfizierte E. coli
DH5α-Flüssigkultur aus Kapitel 3.2.2 zur Verwendung. Die infizierte Kultur wurde, wie in
Kapitel 2.8.25 beschrieben, bei 37°C geschüttelt und bis zu einer Zelldichte von
2 x 108 Bakterien/ml inkubiert. Anschließend erfolgte die Aufbereitung der Zellen für die
Messung am AFM (Kap. 2.8.26). Eine 10 µl Probe der infizierten Kultur wurde auf eine
Glimmeroberfläche gegeben, damit die Bakterien auf der Oberfläche des Probenträgers
adsorbieren konnten. Parallel wurde eine Probe nicht infizierter E. coli DH5α-Zellen, die
als Kontrolle diente, auf einem AFM-Probenträger präpariert. Anschließend folgten die
Messungen der hergestellten Proben mit dem AFM im Kontakt-Modus (Kap. 2.8.28).
Die Auflagekraft der Messsonde auf der Probenoberfläche war < 1 nN. Die Ergebnisse
der Messungen sind in Abbildung 3-6 (A – C) als AFM „deflection“-Aufnahmen darge-
stellt. Dabei handelt es sich nicht um topographische Bilder, sondern um Bilder, die im
sogenannten „deflection“-Modus aufgenommen wurden und eine Ableitung der topo-
grafischen Bildinformation darstellen. Dadurch können kleinere Strukturen, wie Phagen,
dargestellt und von größeren Strukturen bei großen Höhendifferenzen besser unter-
schieden werden.
Die Abbildung 3-6 zeigt E. coli DH5α-Zellen mit Strukturierungen auf den Zellober-
flächen. Bei diesen Zellstrukturen handelt es sich um Eintrocknungsartefakte, welche
durch die Aufbereitung der Bakterienzellen für die AFM-Messungen entstehen. Die
Messungen der Zelloberflächen transfizierter Zellen in Abbildung 3-6 (A) und (B) zeigten
filamentöse Strukturen, welche seitlich an den Zellen über die gesamte Oberfläche

Ergebnisse
111
verteilt waren. Diese Strukturen haben Längen zwischen 1,3 – 1,4 µm. Die Messung
der nicht transfizierten Kontrolle (C) hingegen zeigte keine filamentösen Strukturen.
3.2.4 Veränderung der bakteriellen Zelloberfläche nach M13-Infektion
Die aus den Kapiteln 3.1.1 und 3.1.3 in Versuchsabschnitt 1 erhaltenen Ergebnisse
wurden hier verwendet, um in diesem 2. Abschnitt den Extrusionsverlauf von M13-
Phagen direkt auf der Zelloberfläche von E. coli-Zellen in Abhängigkeit von der Zeit,
nach erfolgter Infektion, mit dem „Atomic Force Microscope“ (AFM) messen zu können.
3.2.4.1 Extrusionsverlauf der M13-Phagen auf der Zelloberfläche von E. coli
Aus den vorangegangenen Versuchen in Abschnitt 1 ging hervor, dass die Zeitspanne
zwischen Infektion und Sekretion neuer M13-Phagen aus den Wirtszellen sehr kurz
Abb. 3-6: AFM-Messungen von E. coli DH5α-Zellen. (A), (B) E. coli DH5α (F
-) Zellen wurden mit
dem RF-Plasmid des Phagenstamms M13amp transfiziert. Die Zellen zeigten filamentöse Struk-turen mit Längen von 1,3 – 1,4 µm, welche sich über die gesamte Zelloberfläche verteilten. Die Länge des Skalierungsbalkens in (A) beträgt 1 µm, in (B) 2 µm. (C) Nicht transfizierte Zellen zeigen keine filamentösen Strukturen auf der Zellober-fläche. Die Länge des Skalierungsbalkens beträgt 2 µm.

Ergebnisse
112
war. Damit eine zeitliche Abfolge der einzelnen Sekretions- bzw. Extrusionsstadien
nach Infektion mit dem AFM gemessen werden konnte, kamen hier das Antibiotikum
Alamethicin (aus Trichoderma viride), welches durch Oligomerisierung von 4 – 6 Mole-
külen einen Ionenkanal in Zellmembranen ausbildet (Jones et al., 1980, Fox &
Richards, 1982), und das Ionophor Carbonylcyanid-m-Chlorophenylhydrazon (CCCP)
zur Anwendung. Bei diesen Substanzen handelt es sich um Ionophore, die in der
Membran von E. coli oligomerisieren und die protonenmotorische Kraft, über welche die
Zelle Energie für ihre Synthesevorgänge gewinnt, rasch zusammenbrechen lassen
(Cavari et al., 1966, Gášková et al., 1999). Hierdurch war die Möglichkeit gegeben, die
Zellen in dem Stadium zu konservieren, in welchem sie sich zum Zeitpunkt der Proben-
entnahme befanden. Dadurch erhielt man eine Momentaufnahme der zellulär intakten
E. coli-Oberflächen.
Für die AFM-Messungen des Extrusionsverlaufs der M13-Phagen auf der Zellober-
fläche von E. coli wurde eine Batch-Kultur exponentiell wachsender E. coli XL1-blue-
Zellen, wie in Kapitel 2.8.20 beschrieben, bei einer Zelldichte von 2 x 108 Zellen/ml mit
M13mp19-Phagen (M.O.I. 20) infiziert. Die Adsorption der Phagen an die F-Pili der
E. coli-Zellen erfolgte für 5 Minuten im Eiswasserbad. Anschließend wurde die Batch-
Kultur sofort wieder bei 37°C geschüttelt, um die Aufnahme der Phagen-DNA in die
Zellen zu ermöglichen. Für die Messungen der einzelnen Stadien am AFM wurden aus
der Batch-Kultur Proben entnommen und diesen Alamethicin (5 µM Endkonzentration)
sowie CCCP (50 µM Endkonzentration) zugesetzt. Probenentnahmen erfolgten kurz vor
Infektion sowie direkt nach erneutem Schütteln bei 37°C zu den Zeitpunkten t = 0, 2, 4,
5, 6, 7, 8, 9, 10, 12, 14 und 16 Minuten. Diese Intervallabstände waren das Ergebnis,
welches sich aus dem wildtypisch verlaufenden „one-step growth“ Experiment aus
Kapitel 3.1 ergab. Die einzelnen Proben wurden daraufhin, wie in Kapitel 2.8.26
beschrieben, für die Messungen am AFM vorbereitet. Auf die Glimmeroberfläche des
AFM-Probenträgers wurden 10 µl der aufbereiteten Proben gegeben, damit die
Bakterien an die Oberfläche des Probenträgers adsorbieren konnten. Anschließend
erfolgten die Messungen der Probenoberflächen mit dem AFM im Kontakt-Modus mit
der Trockenzelle (Kap. 2.8.28). Die Auflagekraft der Messsonde auf der Probenober-
fläche war < 1 nN. Die Messergebnisse der Zelloberflächen von infizierten E. coli XL1-
blue-Zellen sind in Abbildung 3-7, Abbildung 3-8 und Abbildung 3-9 als AFM
„deflection“-Aufnahmen dargestellt. Sie dokumentieren die zeitliche Abfolge des Extru-
sionsverlaufs der M13-Phagen auf der Zelloberfläche von E. coli XL1-blue.
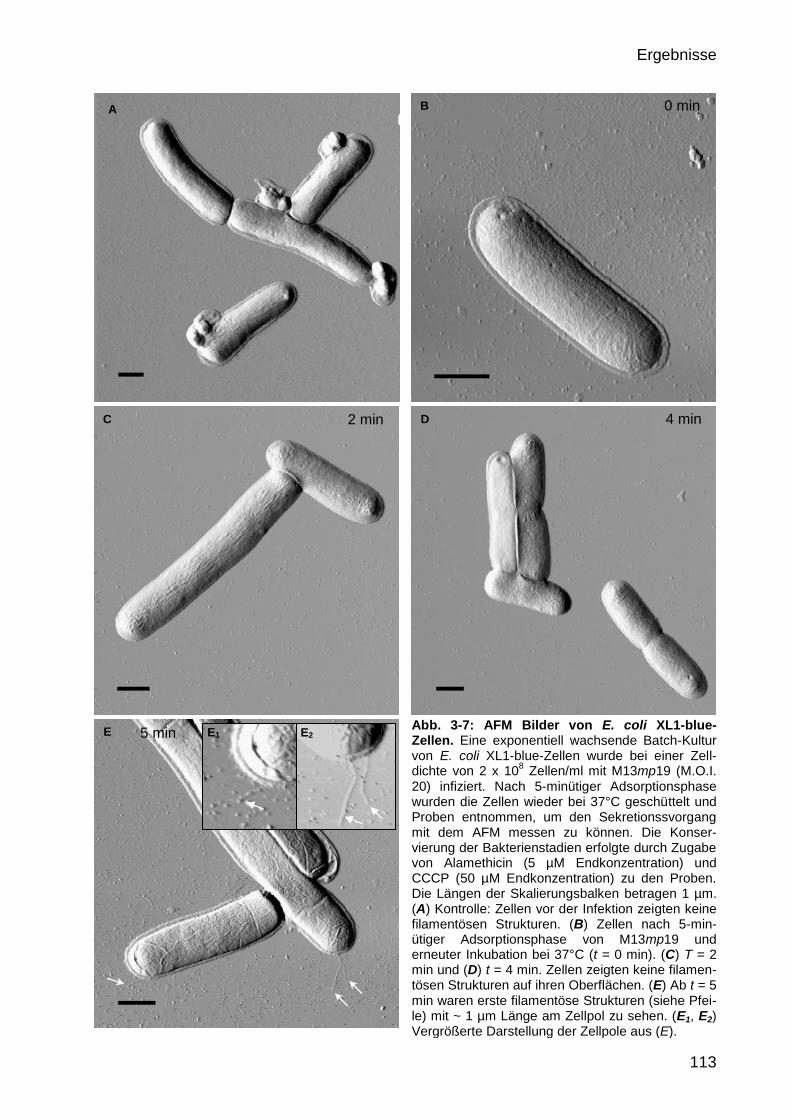
Ergebnisse
113
E
B
C D
Abb. 3-7: AFM Bilder von E. coli XL1-blue-Zellen. Eine exponentiell wachsende Batch-Kultur von E. coli XL1-blue-Zellen wurde bei einer Zell-dichte von 2 x 10
8 Zellen/ml mit M13mp19 (M.O.I.
20) infiziert. Nach 5-minütiger Adsorptionsphase wurden die Zellen wieder bei 37°C geschüttelt und Proben entnommen, um den Sekretionssvorgang mit dem AFM messen zu können. Die Konser-vierung der Bakterienstadien erfolgte durch Zugabe von Alamethicin (5 µM Endkonzentration) und CCCP (50 µM Endkonzentration) zu den Proben. Die Längen der Skalierungsbalken betragen 1 µm. (A) Kontrolle: Zellen vor der Infektion zeigten keine filamentösen Strukturen. (B) Zellen nach 5-min-ütiger Adsorptionsphase von M13mp19 und erneuter Inkubation bei 37°C (t = 0 min). (C) T = 2 min und (D) t = 4 min. Zellen zeigten keine filamen-tösen Strukturen auf ihren Oberflächen. (E) Ab t = 5 min waren erste filamentöse Strukturen (siehe Pfei-le) mit ~ 1 µm Länge am Zellpol zu sehen. (E1, E2) Vergrößerte Darstellung der Zellpole aus (E).
A
E1 E2
2 min 4 min
5 min
0 min

Ergebnisse
114
A
G
H I
J
F
Abb. 3-8: AFM Bilder M13-sekretierender E. coli XL1-blue-Zellen. Eine exponentiell wachsende Batch-Kultur von E. coli XL1-blue-Zellen wurde bei einer Zelldichte von 2 x 10
8 Zellen/ml mit M13mp19
(M.O.I. 20) infiziert. Nach 5-minütiger Adsorptions-phase wurden die Zellen wieder bei 37°C ge-schüttelt und Proben entnommen, um den Sekre-tionsvorgang mit dem AFM messen zu können. Die Konservierung der Bakterienstadien erfolgte durch Zugabe von Alamethicin (5 µM Endkonzentration) und CCCP (50 µM Endkonzentration) zu den Proben. Die Längen der Skalierungsbalken betra-gen 1 µm. (F) T = 6 min; vermehrt zeigen sich fila-mentöse Phagenstrukturen von ~ 1 µm Länge am Zellpol. (G) T = 7 min, filamentöse Strukturen auch am Zellpolrand. (H) T = 8 und (I) t = 9 min. Auf-treten filamentöser Strukturen in geringer Anzahl an den longitudinalen Seiten der Zellen. (J) T = 10 min, filamentöse Strukturen in noch zählbarer Anzahl auf der gesamten Zelloberfläche.
8 min
6 min 7 min
9 min
10 min

Ergebnisse
115
Der Nachweis, dass E. coli XL1-blue-Zellen außer F-Pili keinerlei filamentöse Strukturen
auf ihren Zelloberflächen besaßen, erfolgte durch die AFM-Messung der Probe, welche
kurz vor der Infektion der Batch-Kultur entnommen wurde. Die Abbildung 3-7 (A) zeigt
die Messung der Kontrolle. Die Zellen besaßen keine filamentösen Strukturen auf ihren
Zelloberflächen. Die Messungen der Proben t = 0 min bis t = 4 min in Abbildung 3-7 (B)
– (D) ließen weiterhin keine filamentösen Strukturen auf den Bakterienoberflächen
erkennen. Erst die Messung der Probe t = 5 min in Abbildung 3-7 (E) zeigte erstmalig
filamentöse Strukturen mit Längen von ~ 1 µm an den Zellpolen der E. coli-Zellen
(weiße Pfeile). Nach einer weiteren Minute (Abb. 3-7 (F); t = 6 min) war die Anzahl der
Filamente mit ~ 1 µm Länge am Zellpol angestiegen. Ab t = 7 min (Abb. 3-8 (G)) traten
filamentöse Strukturen an beiden Zellpolen auf. Zwischen den Zeitpunkten t = 8 min bis
L
M
K
Abb. 3-9: AFM Bilder M13-sekretierender E. coli XL1-blue-Zellen. Eine exponentiell wachsende Batch-Kultur von E. coli XL1-blueZellen wurde bei einer Zelldichte von 2 x 10
8 Zellen/ml mit M13mp19
(M.O.I. 20) infiziert. Nach 5-minütiger Adsorptions-phase wurden die Zellen wieder bei 37°C ge-schüttelt und Proben entnommen, um den Sekre-tionsvorgang mit dem AFM messen zu können. Die Konservierung der Bakterienstadien erfolgte durch Zugabe von Alamethicin (5 µM Endkonzentration) und CCCP (50 µM Endkonzentration) zu den Proben. Die Länge der Skalierungsbalken beträgt 1 µm. (K) T = 12 min; Anzahl filamentöser Struk-turen über die gesamte Zelloberfläche nimmt zu. (L) T = 14 min und (M) t = 16 min; die Anzahl der Filamente auf den Zelloberflächen ist nicht mehr zählbar.
12 min 14 min
16 min

Ergebnisse
116
t = 10 min (Abb. 3-8 (H), (I) und (J)) traten die Strukturen mit zunehmender Anzahl auch
an den longitudinalen Seiten der Bakterienoberfläche auf. Die Bilder in Abbildung 3-9
(K) – (M) zeigen die Messungen der Zeitpunkte t = 12 min bis t = 16 min. Die Anzahl der
Filamente nahm in dieser Zeitspanne auf den gesamten Zelloberflächen weiterhin zu.
Ab der 14. Minute (Abb. 3-9 (L)) kam es zu einer Sättigung der bakteriellen Oberflächen
mit filamentösen Strukturen. Die Messung der Bakterienoberflächen für t = 16 min in
Abbildung 3-9 (M) wies keine Veränderungen der Oberflächenbelegung mit Filamenten
zur 14. Minute auf; die Mengen waren nicht mehr zählbar.
Abschnitt 3:
In diesem Abschnitt konnte das wirtsspezifische Translokationssystem identifiziert wer-
den, welches das „Minor Coat“ Protein gp9 in die Zytoplasmamembran von E. coli inse-
riert und somit für die Assemblierung neuer Phagen-Nachkommen zur Verfügung steht.
3.3 Funktionelle Präsentation und Detektion von antigenen Epitopen am
N-Terminus des „Minor Coat“ Proteins gp9
In diesem Versuchsabschnitt konnte unter anderem erörtert werden, ob antigene Epi-
tope am N-terminalen Ende des „Minor Coat“ Proteins gp9 funktionell präsentiert
werden können, ohne die wildtypische Sekretion der M13-Phagen zu beeinträchtigen. In
einem weiteren Schritt wurde dann die Abhängigkeit der Translokation des Hüllproteins
gp9 in die Zytoplasmamembran von E. coli durch die Membraninsertase YidC erörtert.
Das „Minor Coat“ Protein gp9 ist ein sehr kleines Protein, das am Phagen mit nur ~ 5
Kopien präsent ist und sich aus 32 Aminosäuren zusammensetzt. Im direkten Vergleich
mit dem Präprotein des Haupthüllproteins gp8 weist es keine abspaltbare Signal-
sequenz („Leadersequenz“) auf. Der sehr kurze N-terminale Bereich des gp9 befindet
sich in der Zytoplasmamembran und ist periplasmatisch orientiert. Um eine ausreichend
lange und abspaltbare Peptidsequenz im N-terminalen Bereich generieren zu können,
wurde ein „Phage-Display“ am „Minor Coat“ Protein gp9 entwickelt. Funktionell präsen-
tierte antigene Epitope können mit Hilfe spezifischer primärer Antikörper detektiert und
daraufhin über eine sekundäre Reaktion, vermittelt durch Protein-A aus Staphylococcus
aureus, über eine Immunpräzipitation isoliert oder mit Protein-A konjugiertem Nanogold
markiert werden. Gleichzeitig wird durch die Insertion antigener Epitope der N-terminale
Bereich des gp9 verlängert, wodurch eine Zugänglichkeit dieser inserierten Epitope für
Peptidasen, wie z. B. Proteinase K, möglich sein sollte.
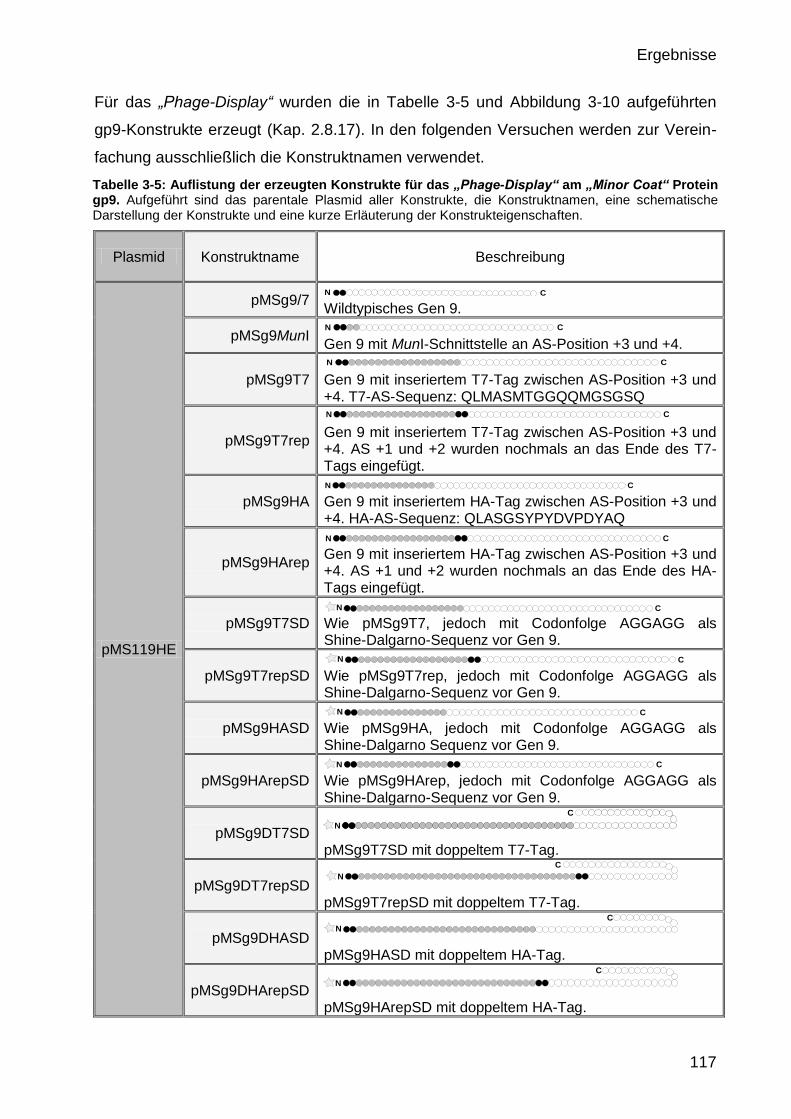
Ergebnisse
117
N
C
Für das „Phage-Display“ wurden die in Tabelle 3-5 und Abbildung 3-10 aufgeführten
gp9-Konstrukte erzeugt (Kap. 2.8.17). In den folgenden Versuchen werden zur Verein-
fachung ausschließlich die Konstruktnamen verwendet.
Plasmid Konstruktname Beschreibung
pMS119HE
pMSg9/7 Wildtypisches Gen 9.
pMSg9MunI Gen 9 mit MunI-Schnittstelle an AS-Position +3 und +4.
pMSg9T7 Gen 9 mit inseriertem T7-Tag zwischen AS-Position +3 und +4. T7-AS-Sequenz: QLMASMTGGQQMGSGSQ
pMSg9T7rep
Gen 9 mit inseriertem T7-Tag zwischen AS-Position +3 und +4. AS +1 und +2 wurden nochmals an das Ende des T7-Tags eingefügt.
pMSg9HA Gen 9 mit inseriertem HA-Tag zwischen AS-Position +3 und +4. HA-AS-Sequenz: QLASGSYPYDVPDYAQ
pMSg9HArep
Gen 9 mit inseriertem HA-Tag zwischen AS-Position +3 und +4. AS +1 und +2 wurden nochmals an das Ende des HA-Tags eingefügt.
pMSg9T7SD Wie pMSg9T7, jedoch mit Codonfolge AGGAGG als Shine-Dalgarno-Sequenz vor Gen 9.
pMSg9T7repSD Wie pMSg9T7rep, jedoch mit Codonfolge AGGAGG als Shine-Dalgarno-Sequenz vor Gen 9.
pMSg9HASD Wie pMSg9HA, jedoch mit Codonfolge AGGAGG als Shine-Dalgarno Sequenz vor Gen 9.
pMSg9HArepSD Wie pMSg9HArep, jedoch mit Codonfolge AGGAGG als Shine-Dalgarno-Sequenz vor Gen 9.
pMSg9DT7SD pMSg9T7SD mit doppeltem T7-Tag.
pMSg9DT7repSD pMSg9T7repSD mit doppeltem T7-Tag.
pMSg9DHASD pMSg9HASD mit doppeltem HA-Tag.
pMSg9DHArepSD pMSg9HArepSD mit doppeltem HA-Tag.
C
N
N
C
N
C
N
C
C
N
N
C
N
C
C
N
N
C
N
C
N
C
C
N
N
C
Tabelle 3-5: Auflistung der erzeugten Konstrukte für das „Phage-Display“ am „Minor Coat“ Protein gp9. Aufgeführt sind das parentale Plasmid aller Konstrukte, die Konstruktnamen, eine schematische Darstellung der Konstrukte und eine kurze Erläuterung der Konstrukteigenschaften.

Ergebnisse
118
Abb. 3-10: Schematische Darstellung der erzeugten Konstrukte für das „Phage-Display“ am „Minor Coat“ Protein gp9. Ausgehend von der Wildtypsequenz gp9wt wurden die in den Darstellungen der einzelnen Konstrukte aufgezeigten Veränderungen in gp9 eingefügt. In die Wildtypsequenz wurde eine MunI-Schnittstelle inseriert (gp9MunI), welche dazu diente, kurze, für einen T7- oder HA-Tag kodie-rende Sequenzen, einzubringen. Hierdurch entstanden die Konstrukte gp9T7, gp9T7rep, gp9HA und gp9HArep. Anschließend wurde in die MunI-Schnittstelle dieser Konstrukte jeweils eine weitere kodier-ende Sequenz eines T7- bzw. HA-Tags eingebracht. Hierdurch entstanden die Konstrukte gp9DT7, gp9DT7rep, gp9DHA und gp9DHArep. In den rep-Konstrukten wurde nach den eingefügten Tags die gp9-Wildtypsequenz (schwarz hinterlegt) durch Insertion der 1. und 2. AS wiederhergestellt (schwarz unterstrichen). Die Proteine sind im 1-Buchstabencode dargestellt.
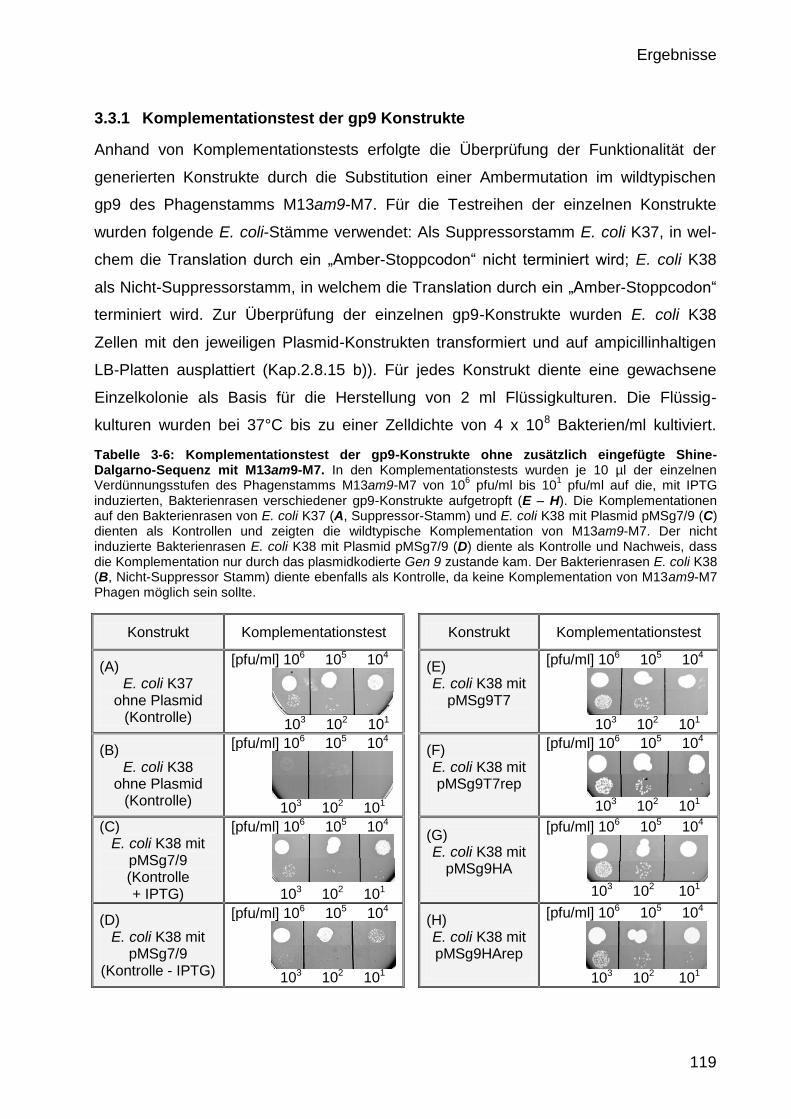
Ergebnisse
119
Tabelle 3-6: Komplementationstest der gp9-Konstrukte ohne zusätzlich eingefügte Shine-Dalgarno-Sequenz mit M13am9-M7. In den Komplementationstests wurden je 10 µl der einzelnen Verdünnungsstufen des Phagenstamms M13am9-M7 von 10
6 pfu/ml bis 10
1 pfu/ml auf die, mit IPTG
induzierten, Bakterienrasen verschiedener gp9-Konstrukte aufgetropft (E – H). Die Komplementationen auf den Bakterienrasen von E. coli K37 (A, Suppressor-Stamm) und E. coli K38 mit Plasmid pMSg7/9 (C) dienten als Kontrollen und zeigten die wildtypische Komplementation von M13am9-M7. Der nicht induzierte Bakterienrasen E. coli K38 mit Plasmid pMSg7/9 (D) diente als Kontrolle und Nachweis, dass die Komplementation nur durch das plasmidkodierte Gen 9 zustande kam. Der Bakterienrasen E. coli K38 (B, Nicht-Suppressor Stamm) diente ebenfalls als Kontrolle, da keine Komplementation von M13am9-M7 Phagen möglich sein sollte.
3.3.1 Komplementationstest der gp9 Konstrukte
Anhand von Komplementationstests erfolgte die Überprüfung der Funktionalität der
generierten Konstrukte durch die Substitution einer Ambermutation im wildtypischen
gp9 des Phagenstamms M13am9-M7. Für die Testreihen der einzelnen Konstrukte
wurden folgende E. coli-Stämme verwendet: Als Suppressorstamm E. coli K37, in wel-
chem die Translation durch ein „Amber-Stoppcodon“ nicht terminiert wird; E. coli K38
als Nicht-Suppressorstamm, in welchem die Translation durch ein „Amber-Stoppcodon“
terminiert wird. Zur Überprüfung der einzelnen gp9-Konstrukte wurden E. coli K38
Zellen mit den jeweiligen Plasmid-Konstrukten transformiert und auf ampicillinhaltigen
LB-Platten ausplattiert (Kap.2.8.15 b)). Für jedes Konstrukt diente eine gewachsene
Einzelkolonie als Basis für die Herstellung von 2 ml Flüssigkulturen. Die Flüssig-
kulturen wurden bei 37°C bis zu einer Zelldichte von 4 x 108 Bakterien/ml kultiviert.
Konstrukt Komplementationstest Konstrukt Komplementationstest
(A) E. coli K37
ohne Plasmid (Kontrolle)
[pfu/ml] 106 105 104
103 102 101
(E) E. coli K38 mit
pMSg9T7
[pfu/ml] 106 105 104
103 102 101
(B) E. coli K38
ohne Plasmid (Kontrolle)
[pfu/ml] 106 105 104
103 102 101
(F) E. coli K38 mit pMSg9T7rep
[pfu/ml] 106 105 104
103 102 101
(C) E. coli K38 mit
pMSg7/9 (Kontrolle + IPTG)
[pfu/ml] 106 105 104 103 102 101
(G) E. coli K38 mit
pMSg9HA
[pfu/ml] 106 105 104
103 102 101
(D) E. coli K38 mit
pMSg7/9 (Kontrolle - IPTG)
[pfu/ml] 106 105 104
103 102 101
(H) E. coli K38 mit pMSg9HArep
[pfu/ml] 106 105 104
103 102 101

Ergebnisse
120
Tabelle 3-7: Komplementationstest der gp9-Konstrukte mit zusätzlich eingefügter Shine-Dalgarno-Sequenz und M13am9-M7. In den Komplementationstests wurden je 10 µl der einzelnen Verdünnungs-stufen des Phagenstamms M13am9-M7 von 10
6 pfu/ml bis 10
1 pfu/ml auf die, mit IPTG induzierten, Bak-
terienrasen verschiedener gp9-Konstrukte (I – P) aufgetropft. Bis auf E. coli mit pMSg9HASD (K) und E. coli mit pMSg9HArepSD (L) konnten alle Konstrukte das, durch Ambermutation ausgeschaltete, gp9 des Phagenstamms M13am9-M7 bis zu einer Verdünnungsstufe von 10
1 pfu/ml komplementieren.
Die gewachsenen Kulturen wurden dann zur Generierung von Bakterienrasen für Spot-
Tests verwendet, auf denen Verdünnungsreihen des Phagenstamms M13am9-M7 auf-
getropft wurden (Kap.2.8.22 a)). Die Induktion der gp9-Konstruktexpression in den ver-
wendeten Bakterienrasen erfolgte durch Zugabe von IPTG. Nach Inkubation bei 37°C
über Nacht, folgte die Auswertung der Plaquebildung auf den einzelnen Bakterien-
rasen.
Die Tabelle 3-6 und Tabelle 3-7 zeigen die Ergebnisse, welche in den durchgeführten
Komplementationstests auf den einzelnen Bakterienrasen der jeweiligen Konstrukte
erhalten wurden. Bis auf pMSg9HASD (K), pMSg9HArepSD (L), pMSg9DHASD (O) und
pMSg9DHArepSD (P) (Tabelle 3-7) konnten alle Konstrukte das, durch Ambermutation
ausgeschaltete, gp9 des Phagenstamms M13am9-M7 bis zu einer Verdünnungsstufe
von 101 pfu/ml komplementieren. Dies entsprach der wildtypischen Komplementation
des Amber 9-Phagen auf dem Bakterienrasen des Suppressorstamms E. coli K37 (A) in
Tabelle 3-6. Auf den Bakterienrasen der Konstrukte pMSg9HASD (K), pMSg9HArepSD
(L), pMSg9DHASD (O) und pMSg9DHArepSD (P) gab es nur Komplementationen bis
zu einer Verdünnung von 102 pfu/ml. Verglichen mit dem Komplementationstest auf
dem Bakterienrasen des Nicht-Suppressorstamms E. coli K38 (B), der hier als Kontrolle
Konstrukt Komplementationstest Konstrukt Komplementationstest
(I) E. coli K38 mit
pMSg9T7 SD
[pfu/ml] 106 105 104
103 102 101
(M) E. coli K38 mit
pMSg9DT7 SD
[pfu/ml] 106 105 104
103 102 101
(J) E. coli K38 mit pMSg9T7rep
SD
[pfu/ml] 106 105 104
103 102 101
(N) E. coli K38 mit pMSg9DT7rep
SD
[pfu/ml] 106 105 104
103 102 101
(K) E. coli K38 mit
pMSg9HA SD
[pfu/ml] 106 105 104
103 102 101
(O) E. coli K38 mit pMSg9DHA
SD
[pfu/ml] 106 105 104
103 102 101
(L) E. coli K38 mit pMSg9HArep
SD
[pfu/ml] 106 105 104
103 102 101
(P) E. coli K38 mit pMSg9DHArep
SD
[pfu/ml] 106 105 104
103 102 101

Ergebnisse
121
diente, haben alle hergestellten Konstrukte die Befähigung das fehlende gp9 des
M13am9-M7-Phagen funktionell zu ersetzen. Dass die Komplementationen auf das
plasmidlokalisierte Gen 9 zurückzuführen waren, zeigte der Vergleich der induzierten
(C) und nicht induzierten (D) Plasmid-Kontrolle pMSg7/9 in Tabelle 3-6. Bei der nicht
induzierten Kontrolle (D) kam es bis zur Verdünnungsstufe von 103 pfu/ml zu Plaque-
bildungen, welche durch Revertanten verursacht wurden. Bei der induzierten Kontrolle
(C) hingegen erfolgte die Komplementation bis zur Verdünnungsstufe von 101 pfu/ml.
3.3.2 Expressionsnachweis der gp9 Mono-Tag Konstrukte in E. coli K38
Anhand der durchgeführten Komplementationstests in Kapitel 3.3.1 konnte gezeigt
werden, dass die Konstrukte mit N-terminalem Mono-Tag imstande waren, das fehlende
gp9 in M13am9-M7 funktionell zu substituieren. Mit dem Expressionsnachweis sollte
gezeigt werden, dass eine Translation der gp9-Konstrukte mit Mono-Tag in den Zellen
erfolgt. Bei gp9 handelt es sich, trotz der fusionierten antigenen Epitope, um ein sehr
kleines Protein mit geringer Expressionsrate. Deshalb wurden die Konstruktexpres-
sionen in vivo radioaktiv mit Hilfe von „Pulse“-Experimenten (Kap. 2.8.39) nachge-
wiesen.
Hierfür erfolgte die Kultivierung von E. coli K38-Flüssigkulturen der Komplemen-
tationstests in Tabelle 3-7 in M9-Minimalmedium bis zur frühen logarithmischen Wachs-
tumsphase bei 37°C. Die Expression der gp9-Konstrukte wurde durch Zugabe von
IPTG (1mM Endkonzentration) zum Kulturmedium induziert. Nach 10 Minuten erfolgte
die Markierung der in der Synthese befindlichen Proteine für weitere 10 Minuten durch
Zusatz von 20 µCi [35S]-Methionin („Pulse“). Die „Pulse“-Reaktionen wurden anschlie-
ßend durch Präzipitation der Proben mit TCA gestoppt. Nach TCA-Fällung der radio-
aktiv markierten Proben erfolgten, unter Verwendung von anti-T7- bzw. anti-HA-Serum,
Immunpräzipitationen (Kap. 2.8.39) zum Nachweis exprimierter gp9T7- und gp9HA-
Konstrukte aus den Zelllysaten. Die immunpräzipitierten Proteine wurden mittels SDS-
PAGE (mit Harnstoff) (Kap. 2.8.35) aufgetrennt und die radioaktiv markierten Proteine
anschließend durch Phosphorimaging (Kap. 2.8.41) als Autoradiogramme detektiert. Als
Kontrolle für die T7- als auch für die HA-Konstruktexpression wurde eine mit dem
Ausgangsvektor pMS119EH transformierte Kultur jeweils mit den entsprechend
spezifischen Antiseren nach Induktion immunpräzipitiert und das Bandenmuster auf das
Vorkommen der zusätzlichen Proteinbande der Konstrukte verglichen.

Ergebnisse
122
Die Autoradiogramme in Abbildung 3-11 zeigen die einzelnen „Pulse“-Analysen der
in vivo Konstruktexpressionen. Für die Konstrukte gp9T7SD (A), gp9T7repSD (B),
gp9HASD und gp9HArepSD (C) wurden die Expressionen der Proteine ( ) in E. coli
K38 nachgewiesen. Aus dem Vergleich des Konstrukts gp9T7repSD in (B) mit der
Markerbande (M) ( ), welche das Haupthüllprotein gp8 mit ~ 5,2 kDa darstellt, kann
gefolgert werden, dass die gp9-Konstrukte ein Molekulargewicht von ~ 5,2 kDa –
5,5 kDa besitzen müssen, da diese sich untereinander nur in der Summe von 2 – 4
Aminosäuren unterscheiden. Aus den radioaktiv markierten Kulturen konnten bei der
Immunpräzipitation, mit Hilfe der spezifischen Antiseren gegen das T7- und HA-Epitop,
gp9 Mono-Tag-Konstrukte nachgewiesen werden, wohingegen aus den Kontrollkulturen
(K) mit pMS119EH keine vergleichbaren Proteine präzipitiert wurden.
3.3.3 Expressionsnachweis der gp9 Di-Tag Konstrukte in E. coli K38
Anhand der durchgeführten Komplementationstests in Kapitel 3.3.1 konnte gezeigt
werden, dass die Konstrukte mit N-terminalem Doppel-Tag imstande waren, das ausge-
schaltete gp9 in M13am9-M7 funktionell zu substituieren. Mit dem Expressionsnachweis
konnte gezeigt werden, dass eine Translation der gp9-Konstrukte mit Doppel-Tag in
den Zellen erfolgte. Der Nachweis der Konstruktexpressionen in vivo erfolgte, wie in
Abb. 3-11: „Pulse“-Analysen der gp9 Mono-Tag-Konstruktexpressionen in E. coli K38. Für den Expressionsnachweis der gp9 Mono-Tag-Konstrukte wurden die transformierten E. coli K38-Kulturen der Komplementationstests in M9-Minimalmedium bei 37°C bis zu einem Bakterientiter von 2 x 10
8 Zellen/ml
kultiviert. Die Expressionen der plasmidkodierten Mono-Tag-Konstrukte erfolgten durch Zugabe von IPTG (1 mM Endkonzentration) zu den Kulturmedien. Nach 10-minütiger Induktion wurden die Kulturen mit 20 µCi [
35S]-Methionin für 10 Minuten radioaktiv markiert („Pulse“) und anschließend mit TCA (12 % End-
konzentration) gefällt. Nach den Immunpräzipitationen folgte die Auftrennung der radioaktiv markierten Proben durch eine 22 %ige SDS-Urea-PAGE mit anschließendem Phosphorimaging. Als Kontrolle (K) wurde eine mit dem Ausgangsvektor pMS119EH transformierte Kultur mit anti-T7 bzw. anti-HA Serum immunpräzipitiert. Die Markerbande (M) ( ) in (B) stellt das Haupthüllprotein gp8 mit einem Molekular-gewicht von ~ 5,2 kDa dar. In den Proben gp9T7 (A), gp9T7rep (B), gp9HA und gp9HArep (C) konnten mit Hilfe der Immunpräzipitationen die einzelnen gp9 Mono-Tag-Konstrukte ( ) aus den Zelllysaten nachgewiesen werden. Zur Identifikation der gp9-Konstrukte wurden die Proben mit den Kontrollen (K) verglichen. Aus dem Vergleich mit der Markerbande (M) konnte gefolgert werden, dass die gp9 Mono-Tag-Konstrukte ein Molekulargewicht von ~ 5,2 kDa – 5,5 kDa besitzen.
M
(A) (B) (C)
~ 5,2
kDa
gp9T7 repSD
K gp9HA SD
K gp9HA repSD
gp9T7SD
K
Antikörper: anti-T7: + + - + + - - -
Antikörper: anti-HA: - - - - - + + +
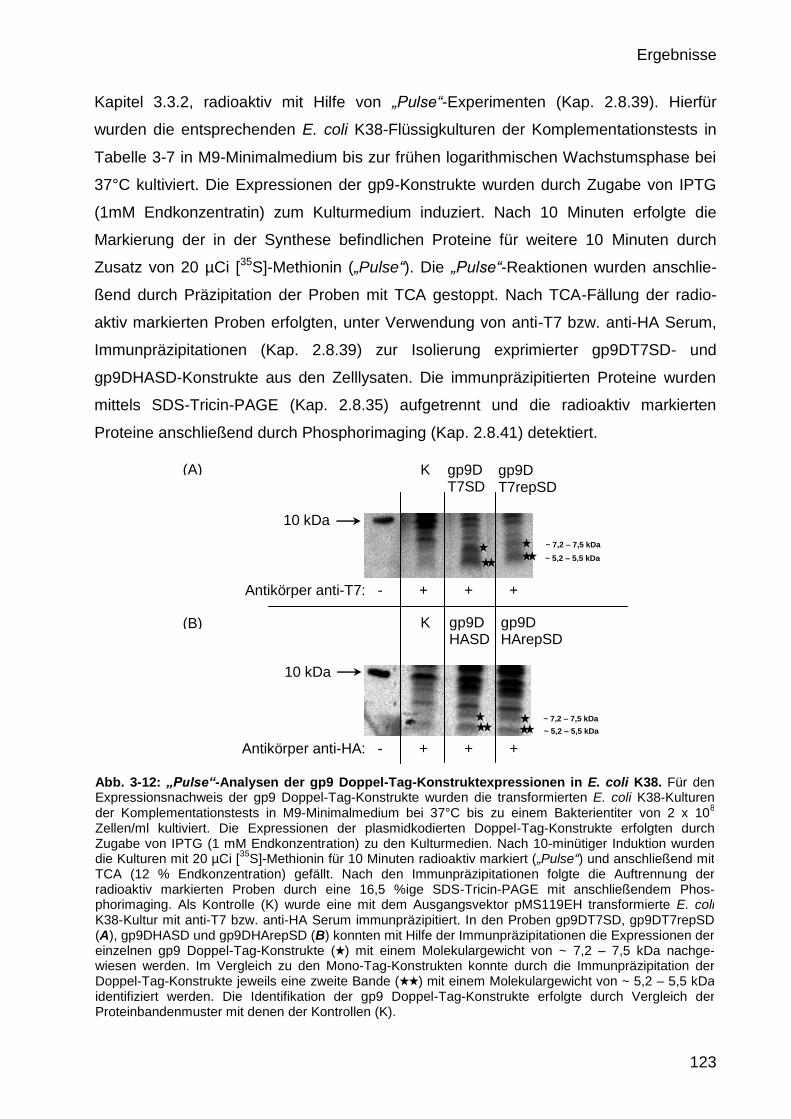
Ergebnisse
123
Kapitel 3.3.2, radioaktiv mit Hilfe von „Pulse“-Experimenten (Kap. 2.8.39). Hierfür
wurden die entsprechenden E. coli K38-Flüssigkulturen der Komplementationstests in
Tabelle 3-7 in M9-Minimalmedium bis zur frühen logarithmischen Wachstumsphase bei
37°C kultiviert. Die Expressionen der gp9-Konstrukte wurden durch Zugabe von IPTG
(1mM Endkonzentratin) zum Kulturmedium induziert. Nach 10 Minuten erfolgte die
Markierung der in der Synthese befindlichen Proteine für weitere 10 Minuten durch
Zusatz von 20 µCi [35S]-Methionin („Pulse“). Die „Pulse“-Reaktionen wurden anschlie-
ßend durch Präzipitation der Proben mit TCA gestoppt. Nach TCA-Fällung der radio-
aktiv markierten Proben erfolgten, unter Verwendung von anti-T7 bzw. anti-HA Serum,
Immunpräzipitationen (Kap. 2.8.39) zur Isolierung exprimierter gp9DT7SD- und
gp9DHASD-Konstrukte aus den Zelllysaten. Die immunpräzipitierten Proteine wurden
mittels SDS-Tricin-PAGE (Kap. 2.8.35) aufgetrennt und die radioaktiv markierten
Proteine anschließend durch Phosphorimaging (Kap. 2.8.41) detektiert.
Abb. 3-12: „Pulse“-Analysen der gp9 Doppel-Tag-Konstruktexpressionen in E. coli K38. Für den Expressionsnachweis der gp9 Doppel-Tag-Konstrukte wurden die transformierten E. coli K38-Kulturen der Komplementationstests in M9-Minimalmedium bei 37°C bis zu einem Bakterientiter von 2 x 10
8
Zellen/ml kultiviert. Die Expressionen der plasmidkodierten Doppel-Tag-Konstrukte erfolgten durch Zugabe von IPTG (1 mM Endkonzentration) zu den Kulturmedien. Nach 10-minütiger Induktion wurden die Kulturen mit 20 µCi [
35S]-Methionin für 10 Minuten radioaktiv markiert („Pulse“) und anschließend mit
TCA (12 % Endkonzentration) gefällt. Nach den Immunpräzipitationen folgte die Auftrennung der radioaktiv markierten Proben durch eine 16,5 %ige SDS-Tricin-PAGE mit anschließendem Phos-phorimaging. Als Kontrolle (K) wurde eine mit dem Ausgangsvektor pMS119EH transformierte E. coli K38-Kultur mit anti-T7 bzw. anti-HA Serum immunpräzipitiert. In den Proben gp9DT7SD, gp9DT7repSD (A), gp9DHASD und gp9DHArepSD (B) konnten mit Hilfe der Immunpräzipitationen die Expressionen der einzelnen gp9 Doppel-Tag-Konstrukte ( ) mit einem Molekulargewicht von ~ 7,2 – 7,5 kDa nachge-wiesen werden. Im Vergleich zu den Mono-Tag-Konstrukten konnte durch die Immunpräzipitation der Doppel-Tag-Konstrukte jeweils eine zweite Bande ( ) mit einem Molekulargewicht von ~ 5,2 – 5,5 kDa identifiziert werden. Die Identifikation der gp9 Doppel-Tag-Konstrukte erfolgte durch Vergleich der Proteinbandenmuster mit denen der Kontrollen (K).
(A)
(B)
Antikörper anti-T7: - + + +
Antikörper anti-HA: - + + +
K gp9D T7SD
gp9D T7repSD
10 kDa
10 kDa
gp9D HASD
K gp9D HArepSD
~ 5,2 – 5,5 kDa
~ 5,2 – 5,5 kDa
~ 7,2 – 7,5 kDa
~ 7,2 – 7,5 kDa

Ergebnisse
124
Als Kontrolle für die DT7SD- als auch für die DHASD-Konstruktexpression wurde eine
mit dem Ausgangsvektor pMS119EH transformierte E. coli K38-Kultur jeweils mit den
entsprechend spezifischen Antiseren nach Induktion immunpräzipitiert und mit den
Proteinbandenmustern der gp9-Konstrukte verglichen.
Die Autoradiogramme in Abbildung 3-12 zeigen die einzelnen „Pulse“-Analysen der in
vivo Konstruktexpressionen. Aus den radioaktiv markierten Kulturen der Konstrukte
gp9DT7SD, gp9DT7repSD (A), gp9DHASD und gp9DHArepSD (B) konnten, während
der Immunpräzipitation mit dem jeweiligen Antiserum gegen das T7- oder HA-Epitop,
Doppel-Tag tragende gp9 Proteine ( ) mit Molekulargewichten zwischen ~ 7,2 – 7,5
kDa in E. coli K38 nachgewiesen werden. In den Kontrollkulturen (K) wurden keine
vergleichbaren Produkte nachgewiesen. Im Vergleich zu den Mono-Tag-Konstrukten in
Abbildung 3-11 konnte durch die Immunpräzipitationen der Doppel-Tag-Konstrukte
jeweils ein weiteres Produkt als zweite Bande ( ) nachgewiesen werden, welches mit
einem Molekulargewicht von ~ 5,2 – 5,5 kDa mit dem der gp9 Mono-Tag-Konstrukte
vergleichbar war.
3.3.4 Membraneinbau der Proteine gp9T7SD und gp9T7repSD in E. coli K38
In den Kapiteln 3.3.2 und 3.3.3 konnten die Expressionen der Mono- und Doppel-Tag-
Konstrukte in vivo nachgewiesen werden. Nachfolgend sollte der Membraneinbau der
Proteine gp9T7SD und gp9T7repSD in die Zytoplasmamembran von E. coli K38
überprüft werden. Die Translokation des T7-Epitops in das Periplasma von E. coli K38
wurde in vivo radioaktiv mit Hilfe von „Pulse“-Experimenten (Kap. 2.8.40) mit anschlie-
ßendem Protease-Mapping überprüft. Der N-Terminus des wildtypischen gp9 ist peri-
plasmatisch orientiert, so dass die fusionierten T7-Epitope für eine proteolytische
Prozessierung durch Proteinase K zugänglich sein sollten. Bei Abspaltungen der anti-
genen Epitope sollten sich die Konstrukte während der Immunpräzipitation nicht mehr
aus den präzipitierten Zelllysaten isolieren lassen.
Für die Translokationsnachweise wurden die mit pMSg9T7SD und pMSg9T7repSD
transformierten E. coli K38-Flüssigkulturen (Tabelle 3-7) bis zur frühen logarithmischen
Wachstumsphase in M9-Minimalmedium bei 37°C kultiviert. Die Expressionen der gp9-
Konstrukte wurden durch Zugabe von IPTG (1mM Endkonzentration) zum Kultur-
medium induziert. Nach 10 Minuten erfolgte die Markierung der in der Synthese befind-
lichen Proteine für weitere 10 Minuten durch Zusatz von 20 µCi [35S]-Methionin
(„Pulse“). Die Proben wurden dann, wie in Kapitel 2.8.39 beschrieben, dem Protease-

Ergebnisse
125
Mapping unterzogen, in welchem die radioaktiv markierten Proteine in den Sphero-
plasten gp9-exprimierender Zellen mit Proteinase K einer Proteolyse unterzogen
wurden. Nach Proteinase K Verdau erfolgte die Präzipitation der Proben mit TCA und
anschließend die Immunpräzipitation (Kap. 2.8.39) unter Verwendung von anti-T7-
Serum, um die g9T7-Proteine in den Zelllysaten nachzuweisen. Die immunpräzipitierten
Proteine wurden mittels SDS-Tricin-PAGE (Kap. 2.8.35) aufgetrennt und die radioaktiv
markierten Proteine anschließend durch Phosphorimaging (Kap. 2.8.41) detektiert. Als
Kontrolle (K) wurden Zellen, die nur den Ausgangsvektor pMS119EH enthielten,
ebenfalls mit anti-T7 Serum immunpräzipitiert und dann das Proteinbandenmuster
verglichen. Das äußere Membranprotein OmpA sowie das zytosolische Chaperon
GroEL der E. coli-Zellen dienten bei dem Protease-Mapping als interne Kontrollen, um
Abb. 3-13: Protease-Mapping der Konstrukte gp9T7SD und gp9T7repSD in E. coli K38. Für den Translokationsnachweis der gp9T7SD- und gp9T7repSD-Konstrukte wurden die transformierten E. coli K38 Kulturen in M9-Minimalmedium bei 37°C bis zu einem Bakterientiter von 2 x 10
8 Zellen/ml kultiviert.
Die Expressionen der plasmidkodierten Konstrukte erfolgten durch Zugabe von IPTG (1 mM Endkonzen-tration) zu den Kulturmedien. Nach 10 min Induktion wurden die Kulturen mit 20 µCi [
35S]-Methionin für
10 min radioaktiv markiert („Pulse“) und anschließend einem Protease-Mapping unterzogen. Nach Proteinase K Verdau und Immunpräzipitation folgte die Auftrennung der radioaktiv markierten Proben über eine 16,5 %ige SDS-Tricin-PAGE mit anschließendem Phosphorimaging. Als Kontrolle (K) wurde eine mit dem Ausgangsvektor pMS119EH transformierte E. coli K38 Kultur mit anti-T7 Serum immun-präzipitiert. In den nicht mit Proteinase K behandelten Proben (Proteinase K -) von gp9T7SD in (A) und gp9T7repSD in (B) konnten mit Hilfe der Immunpräzipitation exprimierte gp9T7-Proteine ( ) aus den Zell-lysaten isoliert werden. Dass es sich jeweils um die gp9-Konstrukte handelte, ergab sich aus dem Ver-gleich mit den Kontrollen (K), welche auf Höhe der gp9-Bande keine Proteinbanden zeigten. Die mit Pro-teinase K behandelten Proben (Proteinase K +) wiesen ebenfalls auf dieser Höhe keine Proteinbanden auf, was auf eine proteolytische Prozessierung der T7-Epitope hinwies. Dass die Integrität der Sphero-plasten gegeben und die Proteinase funktionsfähig war, ergab sich aus der, trotz Proteinase K Verdau, unprozessierten Kontrolle des zytosolischen GroEL und der abgebauten Kontrolle OmpA der äußeren Membran. (C) Schematische Darstellung der T7-Tag-Prozessierung durch Proteinase K während des Protease-Mappings.
K
(C)
(A)
(B)
OmpA GroEL gp9T7 SD
K
Proteinase K: - + - - + - +
Proteinase K: - + - + - +
gp9T7repSD
OmpA GroEL

Ergebnisse
126
die Integrität der hergestellten Spheroplasten bestätigen zu können. Aus den präzipi-
tierten Proben wurden 10 % der Proben jeweils mit anti-OmpA und anti-GroEL Serum
immunpräzipitiert. In Spheroplasten kann das zytosolische GroEL nicht durch Prote-
inase K proteolytisch abgebaut werden, während OmpA in der äußeren Membran durch
die Protease vollkommen abgebaut wird.
Die Autoradiogramme in Abbildung 3-13 zeigen die aufgetrennten, immunpräzipitierten
Proben der radioaktiven Protease-Mappings. Aus den Proben der Konstrukte gp9T7SD
in (A) wie auch gp9T7repSD in (B), die nicht mit Proteinase K (Proteinase K -)
behandelt wurden, konnten gp9 Proteine mit T7-Epitopen ( ) nachgewiesen werden.
Aus den Proben in (A) und (B), welche mit Proteinase K (Proteinase K +) behandelt
wurden, war keine entsprechende Proteinbande detektierbar. Dass es sich jeweils um
die exprimierten gp9T7-Konstrukte handelte, ergab sich aus dem Vergleich mit den anti-
T7 behandelten Kontrollen (K) in Abbildung 3-13 (A, B), welche ebenfalls keine Banden
auf dieser Höhe zeigten. Somit konnte gezeigt werden, dass die gp9T7-Konstrukte in
die Zytoplasmamembran von E. coli K38 transloziert wurden. Dass die Unversehrtheit
der Spheroplasten gegeben sowie die Protease aktiv war, ergab sich aus der, trotz Pro-
teinase K Verdau, ungeschnittenen zytosolischen Kontrolle GroEL und dem Verdau der
Kontrolle OmpA, welche in der äußeren Membran von E. coli lokalisiert ist.
3.3.5 Nachweis der exprimierten gp9 Konstrukte im M13-Phagen
Nachdem in Komplementations- (Kap. 3.3.1), Expressions- (Kap. 3.3.2; 3.3.3) und
Translokationsexperimenten (Kap. 3.3.4) gezeigt werden konnte, dass die gp9-Kon-
strukte in der E. coli-Zelle synthetisiert und für die Assemblierung neuer M13-Phagen-
Nachkommen in der Zytoplasmamembran vorhanden waren, sollte der Einbau der
modifizierten gp9 Proteine über die Detektion der antigenen Epitope am Phagen nach-
gewiesen werden. Der Nachweis erfolgte zuerst durch Immun-Dot-Blot von, auf Nitro-
zellulosemembran, immobilisierten und komplementierten M13am9-M7-Phagen mittels
spezifischer Seren gegen die T7- und HA-Epitope. In einem ergänzenden Experiment
sollten die antigenen Epitope an den M13am9-M7-Phagen mit Hilfe von Protein-A
konjugiertem Gold markiert und anschließend am TEM visualisiert werden.
3.3.5.1 Nachweis der exprimierten gp9-Konstrukte im M13-Phagen mit Hilfe von
Immun-Dot-Blots
Der Nachweis, dass die konstruierten gp9 Proteine in die Hülle der M13-Phagen
eingebaut wurden, sollte durch Detektion der antigenen Epitope in der N-terminalen

Ergebnisse
127
Domäne der gp9-Proteine erfolgen. Um dies zu ermöglichen wurden zuerst, wie in
Kapitel 2.8.38 beschrieben, Komplementationen von M13am9-M7-Phagen durchge-
führt. Die Batch-Kulturen der transformierten E. coli K38-Zellen (Tabelle 3-7) wurden
nach IPTG-Induktion (1mM Endkonzentration) mit M13am9-M7-Phagen infiziert (M.O.I.
5) und über Nacht bei 37°C kultiviert. Die Abtrennung der komplementierten M13-
Phagen von den Bakterien erfolgte durch die Zentrifugation der Batch-Kulturen. Von
den phagenhaltigen Überständen wurden dann, wie in Kapitel 2.8.38 beschrieben,
Verdünnungsreihen so hergestellt, dass in 400 µl Entnahmevolumen eine Anzahl von
108 pfu bis 106 pfu vorhanden waren. Damit ausgeschlossen werden konnte, dass die
exprimierten Proteine in den E. coli-Zellen keine Signalverfälschung während der
Detektion der Epitope an den Phagen verursachten, wurden von den Überständen der
proteinexprimierenden, nicht infizierten Übernachtkulturen, den Phagenstammlösungen
entsprechend, Verdünnungsstufen (10-1 – 10-3) hergestellt. Anschließend wurden je
400 µl der verdünnten Lösungen mit Hilfe einer Dot-Blot Apparatur auf Nitrozellulose-
membranen adsorbiert. Die Membranen wurden anschließend, wie in Kapitel 2.8.38
erläutert, zuerst mit spezifischen primären und dann mit Meerrettichperoxidase gekop-
pelten Sekundärantikörpern zur Detektion der T7- und HA-Epitope inkubiert. Der
Nachweis der Epitope in den Phagen erfolgte anschließend mittels Chemolumineszenz.
In Abbildung 3-14 sind die Ergebnisse der durchgeführten Immun-Dot-Blots zu sehen.
Um ausschließen zu können, dass es sich bei den Detektionen der antigenen Epitope in
den komplementierten M13am9-M7-Phagen um Signale bzw. Signalverfälschungen von
unspezifisch gebundenen Primärantikörpern handelte, erfolgte zuerst die Inkubation des
Phagenstamms M13am9-M7 mit anti-T7 und anti-HA Serum. Die Immun-Dot-Blots der
adsorbierten Verdünnungsreihen von 108 pfu/400 µl bis 106 pfu/400 µl in (A), welche mit
anti-T7 und anti-HA Serum behandelt wurden, zeigten, dass die Inkubation mit diesen
Seren keine unspezifischen Bindungen bewirkte. Dass in den verwendeten Verdünnun-
gen Phagen vorhanden waren, zeigte der Nachweis des Haupthüllproteins gp8. Mit
Hilfe des anti-gp8 Serums konnten hier bis zu einer Verdünnung von 106 pfu/400 µl
M13am9-M7-Phagen detektiert werden.
Dass in den Verdünnungsreihen der darauf folgenden spezifischen Epitopdetektionen
der Phagenkonstrukte Phagen vorhanden waren, zeigte der Immun-Dot-Blot in
Abbildung 3-14 (B1). Hier erfolgte der Nachweis der Phagen ebenfalls durch Detektion
des Haupthüllproteins gp8 in den jeweils verwendeten Verdünnungen mit Hilfe von anti-
gp8 Serum.
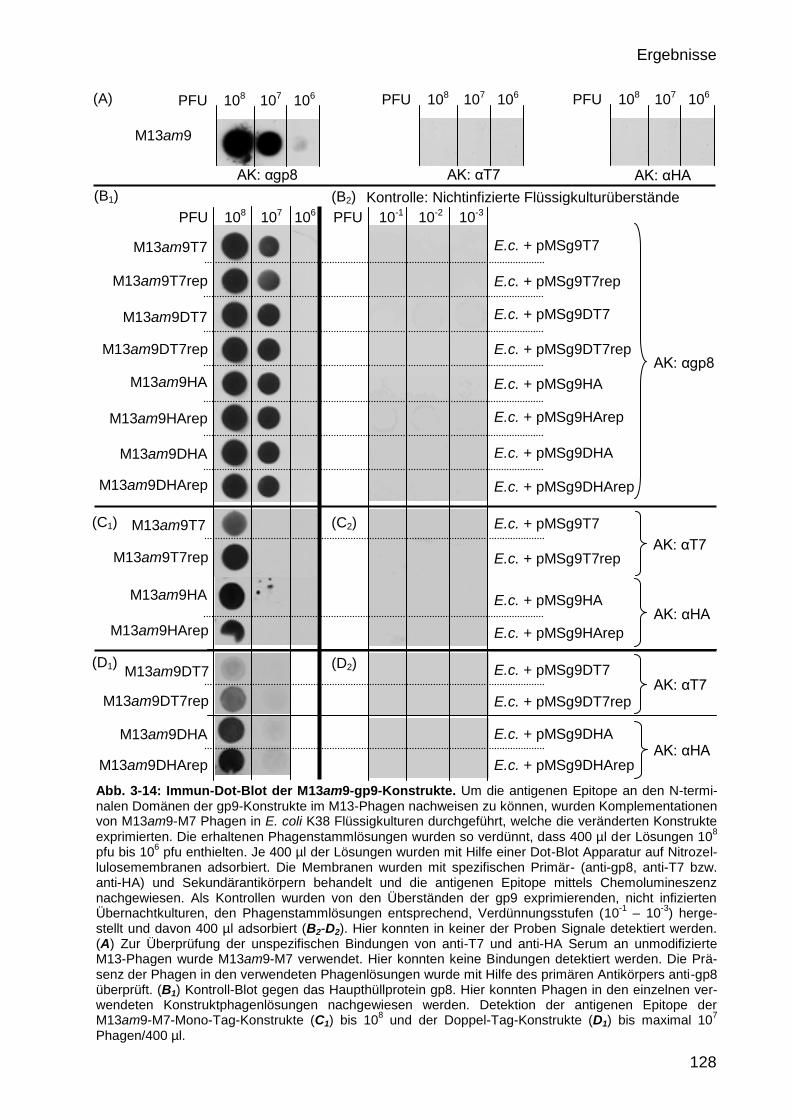
Ergebnisse
128
Abb. 3-14: Immun-Dot-Blot der M13am9-gp9-Konstrukte. Um die antigenen Epitope an den N-termi-nalen Domänen der gp9-Konstrukte im M13-Phagen nachweisen zu können, wurden Komplementationen von M13am9-M7 Phagen in E. coli K38 Flüssigkulturen durchgeführt, welche die veränderten Konstrukte exprimierten. Die erhaltenen Phagenstammlösungen wurden so verdünnt, dass 400 µl der Lösungen 10
8
pfu bis 106 pfu enthielten. Je 400 µl der Lösungen wurden mit Hilfe einer Dot-Blot Apparatur auf Nitrozel-
lulosemembranen adsorbiert. Die Membranen wurden mit spezifischen Primär- (anti-gp8, anti-T7 bzw. anti-HA) und Sekundärantikörpern behandelt und die antigenen Epitope mittels Chemolumineszenz nachgewiesen. Als Kontrollen wurden von den Überständen der gp9 exprimierenden, nicht infizierten Übernachtkulturen, den Phagenstammlösungen entsprechend, Verdünnungsstufen (10
-1 – 10
-3) herge-
stellt und davon 400 µl adsorbiert (B2-D2). Hier konnten in keiner der Proben Signale detektiert werden. (A) Zur Überprüfung der unspezifischen Bindungen von anti-T7 und anti-HA Serum an unmodifizierte M13-Phagen wurde M13am9-M7 verwendet. Hier konnten keine Bindungen detektiert werden. Die Prä-senz der Phagen in den verwendeten Phagenlösungen wurde mit Hilfe des primären Antikörpers anti-gp8 überprüft. (B1) Kontroll-Blot gegen das Haupthüllprotein gp8. Hier konnten Phagen in den einzelnen ver-wendeten Konstruktphagenlösungen nachgewiesen werden. Detektion der antigenen Epitope der M13am9-M7-Mono-Tag-Konstrukte (C1) bis 10
8 und der Doppel-Tag-Konstrukte (D1) bis maximal 10
7
Phagen/400 µl.
M13am9
M13am9T7
M13am9T7rep
M13am9DT7
M13am9DT7rep
M13am9HA
M13am9HArep
M13am9DHA
M13am9DHArep
AK: αgp8
AK: αT7
AK: αHA
M13am9T7
M13am9T7rep
M13am9HA
M13am9HArep
(A)
(B1)
(C1)
(D1)
PFU 10-1 10-2 10-3
Kontrolle: Nichtinfizierte Flüssigkulturüberstände
E.c. + pMSg9T7
E.c. + pMSg9T7rep
E.c. + pMSg9DT7
E.c. + pMSg9DT7rep
E.c. + pMSg9HA
E.c. + pMSg9HArep
E.c. + pMSg9DHArep
E.c. + pMSg9DHA
E.c. + pMSg9T7
E.c. + pMSg9T7rep
E.c. + pMSg9HA
E.c. + pMSg9HArep
E.c. + pMSg9DT7
E.c. + pMSg9DT7rep
E.c. + pMSg9DHA
E.c. + pMSg9DHArep
(B2)
(C2)
(D2)
M13am9DT7rep
M13am9DT7
M13am9DHA
M13am9DHArep
AK: αT7
AK: αHA
PFU 108 107 106
AK: αgp8 AK: αT7
PFU 108 107 106
AK: αHA
PFU 108 107 106
PFU 108 107 106

Ergebnisse
129
In den verwendeten Phagenverdünnungen konnten bis zu einer Verdünnung von
106 pfu/400 µl Phagen nachgewiesen werden. Die adsorbierten, den Verdünnungen der
Phagenstammlösungen entsprechenden, Verdünnungsreihen der Überstände induzier-
ter, jedoch nicht infizierter Übernachtkulturen in Abbildung 3-14 (B2), zeigten keine
Signale. Hierdurch konnte ausgeschlossen werden, dass die verwendeten Kulturen mit
wildtypischen M13-Phagen infiziert waren. Bei den adsorbierten M13am9-M7-Phagen
mit Mono-Tag-Konstrukten (C1) konnten T7- und HA-Epitope maximal nur bis zu einer
Anzahl von 108 pfu/400 µl nachgewiesen werden. Im Vergleich hierzu ließen sich, bis
auf das Konstrukt M13am9DT7, in den M13am9-M7-Phagen mit Doppel-Tag-
Konstrukten die Epitope noch schwach bis zu einer Anzahl von 107 pfu/400 µl detek-
tieren. Dass die in Abbildung 3-14 (C1) und (D1) erhaltenen Signale nicht durch die
Präparationsmethodik der jeweiligen gp9-exprimierenden E. coli-Zellen verursacht
wurden, zeigten die adsorbierten Überstände der verwendeten Flüssigkulturen in
Abbildung 3-14 (C2) und (D2), welche keine Signale mit den jeweils verwendeten
primären Antikörpern aufwiesen.
3.3.5.2 Nachweis der exprimierten gp9 Doppel-Tag-Konstrukte im M13-Phagen
durch Markierung mit Protein-A konjugiertem Gold
In Kapitel 3.3.5.1 konnte gezeigt werden, dass alle, über Komplementation herge-
stellten, M13am9T7- und HA-Konstrukte detektierbare Epitope im Phagen besaßen. Mit
Hilfe von Protein-A konjugiertem Gold sollten hier, wie in Kapapitel 3.2.1 für das Haupt-
hüllprotein gp8, die Doppel-T7- und Doppel-HA-Epitope durch Markierung der M13-
Phagen detektiert und am Transmissionselektronenmikroskop (TEM) visualisiert werden
(Kap. 2.8.29). Dafür wurden 100 µl aus den Phagenstammlösungen aus Kapitel 3.3.5.1,
wie in Kapitel 2.8.30 beschrieben, mit 3 µl primären Antikörpern, entweder mit anti-T7
oder anti-HA, für 1 Stunde inkubiert. Der Nachweis der an die M13-Phagen gebun-
denen Primärantikörper erfolgte mittels 1/20 verdünntem Protein-A konjugiertem Nano-
gold (Kap. 2.8.30). Zur Überprüfung der unspezifischen Bindung von Protein-A konju-
giertem Nanogold an M13am9-M7-Phagen erfolgte in einer parallel angefertigten Probe
die Inkubation ausschließlich mit Protein-A konjugiertem Nanogold. Von diesen
Ansätzen wurden dann jeweils 10 µl auf Trägernetzchen aus Kupfer adsorbiert und eine
Negativkontrastierung mit Phosphorwolframsäure (5 %, pH 7) durchgeführt (Kap.
2.8.27). Die kontrastierten Proben wurden anschließend am TEM untersucht. Die
Ergebnisse der Immunogold-Markierungen sind in Abbildung 3-15 zu sehen.
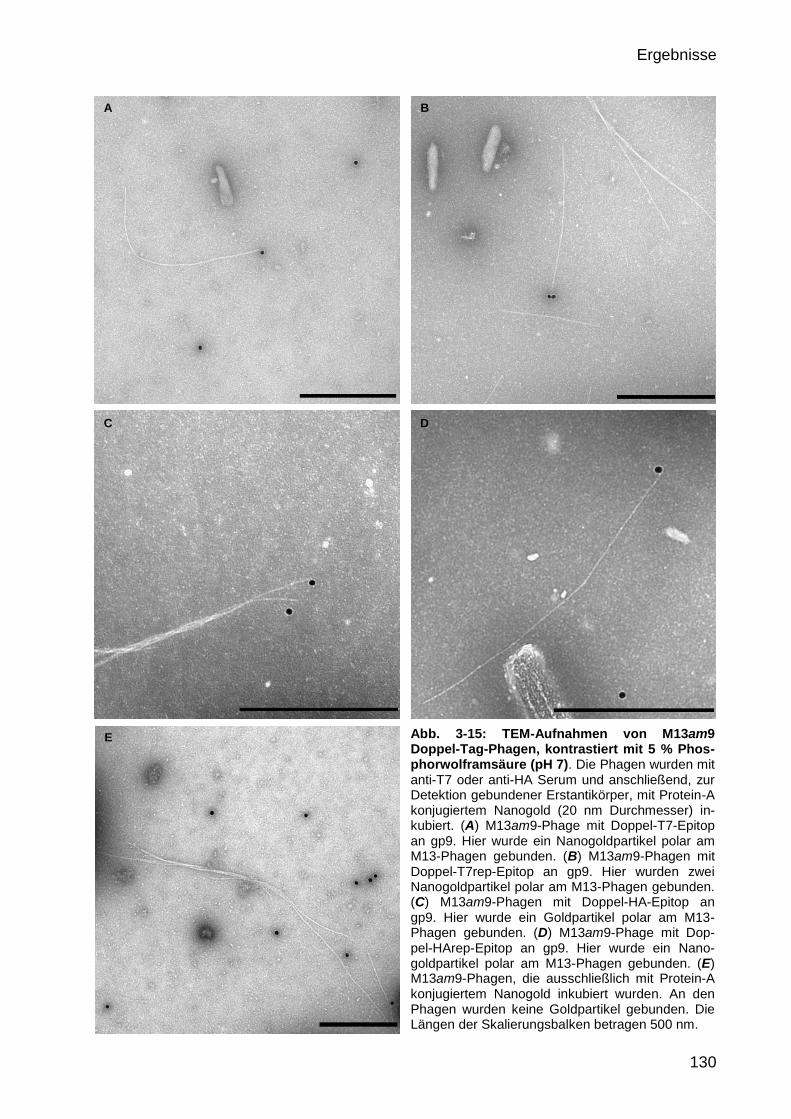
Ergebnisse
130
Abb. 3-15: TEM-Aufnahmen von M13am9 Doppel-Tag-Phagen, kontrastiert mit 5 % Phos-phorwolframsäure (pH 7). Die Phagen wurden mit anti-T7 oder anti-HA Serum und anschließend, zur Detektion gebundener Erstantikörper, mit Protein-A konjugiertem Nanogold (20 nm Durchmesser) in-kubiert. (A) M13am9-Phage mit Doppel-T7-Epitop an gp9. Hier wurde ein Nanogoldpartikel polar am M13-Phagen gebunden. (B) M13am9-Phagen mit Doppel-T7rep-Epitop an gp9. Hier wurden zwei Nanogoldpartikel polar am M13-Phagen gebunden. (C) M13am9-Phagen mit Doppel-HA-Epitop an gp9. Hier wurde ein Goldpartikel polar am M13-Phagen gebunden. (D) M13am9-Phage mit Dop-pel-HArep-Epitop an gp9. Hier wurde ein Nano-goldpartikel polar am M13-Phagen gebunden. (E) M13am9-Phagen, die ausschließlich mit Protein-A konjugiertem Nanogold inkubiert wurden. An den Phagen wurden keine Goldpartikel gebunden. Die Längen der Skalierungsbalken betragen 500 nm.
A B
C D
E

Ergebnisse
131
Die Abbildung 3-15 (A – D) zeigt TEM-Aufnahmen von M13am9-M7-Phagen mit
Doppel-Tag-gp9-Konstrukten. Die Längen der dargestellten Skalierungsbalken in den
hier gezeigten Aufnahmen betragen 500 nm. Phagen mit T7-Tags wurden zuvor mit
anti-T7 und Phagen mit HA-Tags mit anti-HA Serum inkubiert, bevor die Inkubation mit
Protein-A konjugiertem Nanogold erfolgte. In Abbildung 3-15 (A) ist ein ~ 1 µm langer
M13am9-M7-Phage mit Doppel-T7-Epitop zu sehen, an dem polar ein Nanogoldpartikel
mit 20 nm Durchmesser gebunden wurde. In Abbildung 3-15 (B) sind M13am9-M7-
Phagen mit Doppel-T7rep-Epitop zu sehen, hier wurden an einem M13am9-M7-Phagen
zwei Nanogoldpartikel polar gebunden. Abbildung 3-15 (C) zeigt M13am9-M7-Phagen
mit Doppel-HA-Epitop. An einem Phagen wurde polar ein Nanogoldpartikel gebunden.
In Abbildung 3-15 (D) ist ein ~ 1 µm langer M13am9-M7-Phage mit Doppel-HArep-
Epitop zu sehen, an dem ebenfalls polar ein Nanogoldpartikel gebunden werden
konnte. Die TEM-Aufnahme der Probe in Abbildung 3-15 (E) zeigt M13am9-M7-
Phagen, die nicht mit primären Antikörpern sondern nur mit Protein-A konjugiertem
Nanogold inkubiert wurden. An diesen M13-Phagen wurden keine Nanogoldpartikel
unspezifisch gebunden.
3.3.6 Translokation des Konstrukts gp9T7SD in Abhängigkeit von der E. coli
Membraninsertase YidC
Das YidC-Protein von E. coli ist eine, in der Zytoplasmamembran lokalisierte, Membran-
insertase, welche die Translokation essenzieller E. coli eigener Membranproteine
katalysiert. YidC kann eigenständig oder zusammen mit dem SecYEG Translokations-
kanal der Sec-Translokase Proteine in die Zytoplasmamembran inserieren. Für das
M13-Haupthüllprotein gp8 konnte gezeigt werden, dass dessen Insertion in die Zyto-
plasmamembran nicht durch den bakteriellen Sec-Translokaseapparat katalysiert wird
(Kuhn, 1988), sondern vom Membranpotential und YidC abhängig ist. Das „Minor Coat“
Protein gp9 ist mit einer Länge von 32 Aminosäuren das kleinste Protein unter den
M13-Hüllproteinen. Im Vergleich zum Haupthüllprotein gp8 besitzt es keine N-terminal
abspaltbare Signalsequenz („Leadersequenz“).
Hier sollte, mit Hilfe des durch Proteinase K abspaltbaren antigenen T7-Epitops, die
Abhängigkeit der gp9 Membraninsertion von der Membraninsertase YidC in vivo radio-
aktiv mit Hilfe von „Pulse“-Experimenten (Kap. 2.8.40) und anschließendem Protease-
Mapping überprüft werden. Wie in Kapitel 3.3.4 gezeigt wurde, lassen sich die inse-
rierten Epitope in den gp9-Konstrukten durch Proteinase K proteolytisch abspalten.

Ergebnisse
132
Durch die Verwendung des Bakterienstamms E. coli JS7131, in dem die genomische
YidC-Expression unter Kontrolle des araBAD Promotors steht, kann die YidC-
Expression in diesen Zellen durch Arabinose (Expression) bzw. Glukose (keine
Expression) reguliert werden. Falls die gp9T7-Insertion eine von YidC abhängige
Translokation aufweist, sollte während des Protease-Mappings, bei Abwesenheit von
YidC, keine Prozessierung des N-terminalen antigenen Epitops erfolgen und dadurch
das Protein sich anhand einer Immunpräzipitation aus den Zelllysaten isolieren und
nachweisen lassen. Für den YidC abhängigen Translokationsnachweis wurden E. coli
JS7131-Zellen mit dem Konstrukt pMSg9T7SD transformiert und eine Einzelkolonie in
M9-Minimalmedium bis zur frühen logarithmischen Wachstumsphase bei 37°C kultiviert.
Die gewachsene E. coli-Kultur wurde dann zur Herstellung zweier Flüssigkulturen
verwendet. Zum Medium der ersten Kultur wurde Arabinose (Ara, 0,2 % Endkon-
zentration) zugesetzt, um YidC wildtypisch in den Zellen zu exprimieren.
Der zweiten Kultur wurde Glukose (Glc, 0,2 % Endkozentration) zum Medium zuge-
setzt, um die YidC-Expression für 120 Minuten zu depletieren. Nach Erreichen des
Bakterientiters von 2 x 108 Bakterien/ml erfolgte, wie in Kapitel 2.8.40 beschrieben, eine
Probenentnahme aus den Kulturen für den Nachweis der YidC-Expression. Die Proben
wurden dann durch TCA-Zugabe präzipitiert. Anschließend erfolgte die Auftrennung der
Proben über eine SDS-PAGE (Kap. 2.8.35). Die YidC-Expression in den Kulturen wurde
mit Hilfe eines Western-Blots (Kap. 2.8.37) nachgewiesen (Abb. 3-16). Als Kontrolle (K)
diente gereinigtes YidC, das den gefällten Proben gegenüber gestellt wurde.
Für den Translokationsnachweis wurden in den restlichen Proben die Expression der
gp9-Konstrukte durch Zugabe von IPTG (1mM Endkonzentration) zum Kulturmedium
induziert. Nach 10 Minuten erfolgte die Markierung der in der Synthese befindlichen
Proteine für weitere 10 Minuten durch Zugabe von 20 µCi [35S]-Methionin („Pulse“).
E. coli JS7131 K
Ara Glc
YidC-10xHis
YidC
Abb. 3-16: Western-Blot Analyse der YidC-Expressionen in E. coli JS7131. Die TCA-präzipitierten Proben wurden durch SDS-PAGE (12 %) aufgetrennt und exprimiertes YidC durch Western-Blot und anti-YidC Serum nachgewiesen. Gereinigtes YidC mit 10x His-Tag diente als Kontrolle (K). In der in 0,2 % Arabinose (Ara) kultivierten Kultur wurde YidC exprimiert; in der in 0,2 % Glukose (Glc) kultivierten Kultur konnte keine YidC-Expression nachgewiesen werden.

Ergebnisse
133
Die Proben wurden dann, wie in Kapitel 2.8.39 beschrieben, einem Protease-Mapping
unterzogen, in welchem die radioaktiv markierten Proteine in den Spheroplasten der
gp9-exprimierenden Zellen, durch Zugabe von Proteinase K, einer proteolytischen
Prozessierung ausgesetzt wurden. Nach der Proteolyse erfolgte die Präzipitation der
Proben mit TCA und darauf folgender Immunpräzipitation (Kap. 2.8.39) unter Verwen-
dung von anti-T7 Serum zur Isolierung der T7-Konstrukte aus den Zelllysaten. Die
immunpräzipitierten Proteine wurden mittels SDS-Tricin-PAGE (Kap. 2.8.35) aufge-
trennt und die radioaktiv markierten Proteine anschließend durch Phosphorimaging
(Kap. 2.8.41) detektiert (Abb. 3-17). Als Kontrolle (K) wurden mit pMS119EH trans-
formierte E. coli JS7131-Zellen ebenfalls radioaktiv markiert und anschließend mit anti-
T7 Serum immunpräzipitiert. Das äußere Membranprotein OmpA sowie das zyto-
solische Chaperon GroEL der E. coli-Zelle dienten bei diesem Translokationsversuch
als interne Kontrolle, um die Funktionalität der hergestellten Spheroplasten und die
Aktivität der Protease überprüfen zu können.
Glc
Ara
Proteinase K: - + - - + - +
Proteinase K: - + - + - +
gp9T7SD K OmpA GroEL
Abb. 3-17: Protease-Mapping des Konstrukts gp9T7SD in E. coli JS7131. Für den Translokations-nachweis des gp9T7SD Konstrukts wurden transformierte E. coli JS7131-Kulturen in M9-Minimalmedium mit 0,2 % Arabinose (Ara) bzw. mit 0,2 % Glukose (Glc; 120 min Depletion) bei 37°C bis zu einem Bakte-rientiter von 2 x 10
8 Zellen/ml kultiviert. Die Expression des plasmidkodierten Konstrukts erfolgte durch
Zugabe von IPTG (1 mM Endkonzentration). Nach 10-minütiger Induktion wurden die Kulturen mit 20 µCi [35
S]-Methionin für 10 Minuten radioaktiv markiert („Pulse“) und anschließend einem Protease-Mapping unterzogen. Nach Proteinase K Verdau und Immunpräzipitation mit anti-T7 Serum folgte die Auftrennung der radioaktiv markierten Proben über eine SDS-Tricin-PAGE (16,5 %) mit anschließendem Phosphor-imaging. Als Kontrolle (K) wurde eine mit pMS119EH transformierte E. coli JS7131-Kultur mit anti-T7 Serum immunpräzipitiert. In den in Glukose kultivierten Zellen (A) konnte gp9T7SD ( ) sowohl in der nicht mit Proteinase K behandelten als auch in der mit Proteinase K behandelten Probe nachgewiesen werden. Im direkten Vergleich konnte in den in Arabinose kultivierten Zellen (B) nur in der nicht mit Proteinase K behandelten Probe exprimiertes gp9T7SD ( ) nachgewiesen werden. Dies zeigte, dass unter wildtypischen Bedingungen (YidC-Expression) das gp9T7SD in die Zytoplasmamembran trans-loziert und hierfür YidC benötigt wird. Dass es sich jeweils um die gp9-Konstrukte handelte, ergab sich aus der Kontrolle (K), welche auf Höhe des gp9 keine Proteinbanden zeigte. Dass die Integrität der Spheroplasten gegeben war ergab sich aus der, trotz Proteinase K Verdau, unprozessierten Kontrolle GroEL. Die Funktionalität der Protease zeigte der Abbau der OmpA-Kontrolle.
(A)
(B)

Ergebnisse
134
Aus den präzipitierten Proben wurden 10 % des Probenvolumens entnommen und mit
anti-OmpA und anti-GroEL Serum koimmunpräzipitiert, um die Integrität der Sphero-
plasten überprüfen zu können. Bei intakten Spheroplasten kann das zytosolische GroEL
nicht durch Proteinase K proteolytisch prozessiert werden, während OmpA in der
äußeren Membran vollständig abgebaut wird.
Die Western-Blot Analyse der YidC-Expressionen in E. coli JS7131 (Abb. 3-16) zeigte,
dass die in 0,2 % Arabinose (Ara) kultivierten Zellen YidC exprimierten (siehe Pfeil),
während die Expression von YidC in den in 0,2 % Glukose (Glc) gewachsenen Zellen
depletiert war und kein YidC detektiert werden konnte. Die Autoradiogramme in
Abbildung 3-17 zeigen die aufgetrennten, immunpräzipitierten Proben der radioaktiven
Protease-Mappings. Aus den in Glukose kultivierten Zellen (Abb. 3-17 (A)) konnte das
Konstrukt gp9T7SD ( ) sowohl in der mit Proteinase K behandelten, als auch in der
unbehandelten Probe mit Hilfe von anti-T7 Serum während der Immunpräzipitation
nachgewiesen werden. Das T7-Epitop wurde in beiden Proben nicht proteolytisch
prozessiert. Im Vergleich hierzu konnte in den in Arabinose kultivierten Zellen (Abb.
3-17 (B)) das gp9T7SD-Konstrukt nur in der nicht mit Proteinase K behandelten Probe
mit anti-T7 Serum nachgewiesen werden ( ). Die Prozessierung des T7-Epitops in
der mit Proteinase K behandelten Probe zeigte, dass gp9T7SD in Anwesenheit von
YidC in die Zytoplasmamembran von E. coli JS7131 eingebaut wurde. Dass es sich
jeweils um die exprimierten gp9T7SD-Proteine handelte, ergab sich aus dem Vergleich
mit der anti-T7 behandelten Kontrolle (K) (Abb. 3-17 (A)), welche ebenfalls keine
Banden auf dieser Höhe zeigte. Dass die Unversehrtheit der Spheroplasten gegeben
sowie die Protease aktiv war, ergab sich aus der, trotz Proteinase K Verdau, unge-
schnittenen zytosolischen Kontrolle GroEL und dem Abbau der OmpA-Kontrolle, welche
in der äußeren Membran von E. coli lokalisiert ist. Das Protease-Mapping in Abbildung
3-17 (B) zeigte zudem wiederholt in Übereinstimmung mit Kapitel 3.3.4, dass auch hier
das gp9 mit inseriertem T7-Epitop in die Zytoplasmamembran von E. coli eingebaut
wurde.
Abschnitt 4:
Das Gen 1 des Phagen M13 kodiert sowohl das integrale Membranprotein gp1 als auch
das integrale Membranprotein gp11. Nach Spekulationen von Russel et al. (2004)
bilden diese Proteine vermutlich in der inneren Wirtsmembran einen multimeren Kom-
plex aus jeweils 5 – 6 der einzelnen Moleküle. Dieser Komplex ist für die Assemblierung

Ergebnisse
135
neuer Phagen-Nachkommen essenziell (Silver & Wickner, 1980, Rapoza & Webster,
1995, Marvin 1998). Da weder das Gewicht noch die Größe dieses Assembly-
Komplexes bekannt sind, sollte in diesem Abschnitt zur Gewinnung neuer Daten, mit
Hilfe einer Überexpression der Phagenproteine gp1/11 in E. coli-Zellen, die Masse des
Komplexes durch eine präparative Gelfiltration bestimmt und die gereinigten Komplexe
durch TEM-Aufnahmen visualisiert und analysiert werden.
3.4 Massenbestimmung des gp1/11-Assembly-Komplexes und TEM-Auf-
nahmen der gereinigten Komplexe
Bei den Proteinen gp1 und gp11 handelt es sich um integrale Membranproteine, die in
der Zytoplasmamembran von E. coli den M13 Assembly-Komplex bilden. Dieser
Komplex bildet zusammen mit dem gp4-Porenkomplex in der äußeren Membran von
E. coli einen gp1/11-gp4-Sekretionskomplex (Rapoza & Webster, 1995, Feng et al.,
1999, Haigh & Webster, 1999), durch welchen die M13-Phagen in die äußere Umge-
bung sekretiert werden. Für die Bestimmung der Masse des gp1/11-Assembly-Kom-
plexes sollten diese Proteine in E. coli C41-Zellen überexprimiert und anschließend aus
den Zelllysaten gereinigt werden. Um ausreichende Mengen an gp1/11-Komplexen zu
erhalten, wurde der Bakterienstamm E. coli C41 in Kombination mit dem Vektor pET-
16b gewählt. Mit diesem Expressionssystem können durch Überexpression größere
Proteinmengen synthetisiert und anschließend gereinigt werden. Vor der „Multiple
Cloning Site“ (MCS) enthält das Plasmid pET-16b einen T7 RNA-Polymerase Promotor
und eine ϕ10 Shine-Dalgarno-Sequenz, so dass in speziellen E. coli-Stämmen wie C41,
welche die T7 RNA-Polymerase exprimieren, gezielt klonierte Gene mit hoher Effizienz
translatiert und so Proteine auf diesem Weg überexprimiert werden können. Zusätzlich
enthält die MCS eine 10x His-Tag Sequenz, welche nach Klonierung N-terminale His-
Tag Fusionsprodukte liefert. Diese Fusionsprodukte können dann anhand immobili-
sierter Metallaffinitätschromatographie (IMAC) aus den Zelllysaten isoliert werden.
3.4.1 Überexpression und Identifikation des M13 gp1/11-Assembly-Komplexes in
E. coli C41
Zur Konstruktion des gp1/11-Expressionssystems wurde das für gp1/11 kodierende
Gen 1 durch PCR aus dem Phagengenom amplifiziert und anschließend in die MCS
von pET-16b kloniert. Aus dieser Klonierung ging das Konstrukt pETgp1/11 hervor
(Kap. 2.8.17). Daraufhin erfolgte die Überprüfung der gp1/11-Expression in den E. coli-

Ergebnisse
136
Zellen sowie die Ermittlung der nach der Induktion optimalen Kultivierungsdauer,
während der die höchste Menge an Proteinen erhalten wurde (Kap. 2.8.32). Hierfür
erfolgte die Transformation des Vektors pETgp1/11 in E. coli C41. Von den auf ampicil-
linhaltigen LB-Agarplatten gewachsenen Transformanten wurde eine Einzelkolonie zur
Beimpfung von 2 ml Flüssigkulturen verwendet. Die Kultivierung der Flüssigkulturen
erfolgte bei 37°C unter Schütteln bis zu einem Bakterientiter von 2 x 108 Bakterien/ml,
woraufhin die Expression der Proteine durch Zugabe von IPTG (1mM Endkonzen-
tration) zu den Flüssigkulturen induziert wurde. Nach weiterer Kultivierung bei 37°C
erfolgten die Präzipitationen der Kulturen durch Zugabe von TCA nach 2 h, 4 h und 5 h.
Anschließend wurden die Proben durch SDS-PAGE (Kap. 2.8.35) aufgetrennt und die
exprimierten gp1- und gp11-Proteine mit Hilfe eines Western-Blots (Kap. 2.8.37), unter
Verwendung von anti-gp1/11 Serum gegen die C-terminale Domäne der Proteine,
identifiziert. Als Kontrollen wurden Kulturen von nicht transformierten E. coli C41-Zellen,
sowie mit pET-16b und pETgp1/11 (nicht induziert) transformierten Zellen ebenfalls mit
TCA präzipitiert.
Mit Hilfe der Western-Blot Analyse in Abbildung 3-18 konnte, unter Verwendung des
anti-gp1/11 Serums, die Expression von gp1 ( ) und gp11 ( ) in E. coli C41 nachge-
wiesen werden. In E. coli C41 (K1), E. coli C41 mit pET-16b Vektor (K2) und E. coli C41
mit pETgp1/11 Vektor, jedoch nicht induziert (K3), wurden keine Expressionen von gp1
und gp11 nachgewiesen. Der Vergleich der nach 2 h, 4 h und 5 h präzipitierten Proben
zeigte, dass die Probe mit 2-stündiger Kultivierung nach erfolgter Induktion bei 37°C die
höchsten Mengen an gp1 und gp11 aufwies. Nach 4 h bzw. 5 h Expression nahmen die
Proteinmengen deutlich ab.
K1 K2 K3 2 h 4 h 5 h Abb. 3-18: Western-Blot Nachweis von exprimiertem gp1 und gp11 in E. coli C41: Mit pETgp1/11 transformierte E. coli C41-Zellen wurden bei 37°C bis zu einem Bakterientiter von 2 x 10
8 Zellen/ml kultiviert. Die Expression von gp1 und gp11
erfolgte durch Zugabe von IPTG (1 mM Endkonzentration) zu den 2 ml Flüssigkulturen. Nach 2 h, 4 h und 5 h Expression wurden die Kulturen durch TCA präzipitiert und anschließend über eine 12 %ige SDS-PAGE aufgetrennt. Der Nachweis von exprimiertem gp1 und gp11 erfolgte mit Hilfe eines Western-Blots unter Ver-wendung von anti-gp1/11 Serum gegen die C-terminale Domäne der Proteine. Als Kontrollen wurden Kulturen von nicht trans-formierten E. coli C41-Zellen (K1), sowie mit pET-16b (K2) und pETgp1/11 (K3; nicht induziert) transformierten Zellen aufge-tragen. In den Proben mit 2 h, 4 h und 5 h Expression konnte sowohl gp1 ( ) als auch gp11 ( ) nachgewiesen werden. Im Vergleich dazu erfolgte in den Kontrollen K1 – K3 keine gp1 und gp11-Expression.
130 100 70 55 40 35
25
15
10

Ergebnisse
137
3.4.2 Reinigung überexprimierter M13 gp1/11-Assembly-Komplexe aus E. coli
C41 mittels Metallaffinitätschromatographie (IMAC)
In Kapitel 3.4.1 wurde gezeigt, dass in E. coli C41-Zellen, welche zuvor mit dem Kon-
strukt pETg1/11 transformiert wurden, die Expression von gp1 und gp11 durch IPTG
induzierbar war und nach 2-stündiger Kultivierung die höchsten Proteinmengen erhalten
wurden. Nach erfolgreicher Expression folgte nun die Reinigung der gp1/11-Assembly-
Komplexe aus E. coli C41. Durch die Klonierung des Gens 1 in die „Multiple cloning
site“ (MCS) von pET-16b entstehen bei der Expression von gp1-Fusionsproteine mit
N-terminalem 10x His-Tag. Gp11 ist das Produkt einer internen Initiierung innerhalb des
Gens 1 und bildet dadurch keine Fusionsproteine mit N-terminalem His-Tag. Während
der Reinigung der exprimierten Komplexe aus der Zytoplasmamembran, mittels immo-
bilisierter Metallaffinitätschromatographie (IMAC), können die Komplexe anhand des
N-terminalen His-Tags der gp1-Fusionsproteine isoliert werden.
Für die Überexpression (Kap. 2.8.32) und anschließende Reinigung der Komplexe
(Kap. 2.8.33) wurde das Konstrukt pETg1/11 in E. coli C41 transformiert. Die Reinigung
erfolgte zuerst aus einer 2 L und anschließend nochmals aus einer 4 L LB-Batch-Kultur,
um die Menge an exprimierten gp1/11-Assembly-Komplexen zu erhöhen. Bei allen
durchgeführten Expressionen wurden die Zellen bei 37°C bis zu einer Zelldichte von
2 x 108 Bakterien/ml kultiviert. Die Expression des plasmidkodierten gp1 und gp11
erfolgte durch Zugabe von IPTG (1mM Endkonzentration) zum Kulturmedium. Die
Zellen wurden nach Induktion für weitere 2 h bei 37°C kultiviert und nach anschlie-
ßender Zentrifugation in Tris-Sucrose Lösung (25% Sucrose in 20 mM Tris pH 7,5)
aufgenommen.
Um die in der Zytoplasmamembran lokalisierten gp1/11-Assembly-Komplexe reinigen
zu können, wurden die Zellen im „French Press“-Verfahren aufgeschlossen und die
Zellmembranen unter Verwendung von 3,5 % des zwitterionischen Detergenzes Fos-
cholin 12 (n-Dodecylphosphocholin) solubilisiert. Die Reinigung des gp1/11-Assembly-
Komplexes erfolgte anschließend über den N-terminal fusionierten His-Tag der gp1-
Fusionsproteine mit Hilfe einer Ni-NTA-Säule. Die solubilisierte Proteinfraktion wurde
nach Zugabe von Ni-NTA-Agarose für 2 h unter Rotation bei 7°C inkubiert, damit die
Interaktion zwischen den His-Tags und der Ni-NTA-Matrix erfolgen konnte. Nach Inku-
bation wurde die Ni-NTA-Agarose als Säulenmaterial in eine Spritzenhülse gegeben
und eine Ni-NTA-Säule hergestellt. Im „Gravity flow“-Verfahren wurde anschließend das
Säulenmaterial mit Hilfe ansteigender Imidazolkonzentrationen in den Waschpuffern

Ergebnisse
138
von unspezifisch gebundenen Proteinen gereinigt. Danach folgte die Elution gebun-
dener Proteine von der Ni-NTA-Matrix mit einer Imidazolkonzentration von 300 mM. Im
Verlauf der Reinigungen wurden Proben (à 50 µl) entnommen, um den Verbleib der
gp1- und gp11-Proteine während der einzelnen Reinigungsphasen überprüfen zu
können. Die Proben wurden durch Zugabe von TCA präzipitiert und anschließend durch
SDS-PAGE aufgetrennt (Kap. 2.8.35). Die Identifikation der gp1- und gp11-Proteine
erfolgte über eine Western-Blot Analyse (Kap. 2.8.37) unter Verwendung von anti-
gp1/11 Serum. Um den einzelnen Proteinbanden ein Molekulargewicht zuordnen zu
können, wurde ein Proteinstandard aufgetragen.
Das Coomassie gefärbte SDS-Gel in Abbildung 3-19 (A) zeigte, dass in der Elutions-
fraktion (E) während der Metallaffinitätschromatographie drei Proteine aus der solubili-
sierten Proteinfraktion gereinigt werden konnten. Mit Hilfe des Proteinstandards (M)
konnten für die eluierten Proteine näherungsweise die Molekulargewichte ermittelt
werden. Das Molekulargewicht der oberen Bande ( ) betrug ~ 39 kDa, das der mittleren
( ) ~ 37 kDa und das der unteren ( ) ~ 31 kDa. Auffällig war, dass die Intensität
der ~ 39 kDa Bande ( ) mit der ~ 31 kDa Bande ( ) vergleichbar war.
Abb. 3-19: SDS-PAGE und Western-Blot Analyse der gp1/11-Assembly-Komplex Reinigung aus E. coli C41 (2 L Flüssigkultur). Gp1/11-exprimierende E. coli C41-Zellen wurden mittels „French Press“-Verfahren aufgeschlossen und die Membranen unter Verwendung von 3,5 % des zwitterionischen Detergenzes Fos-cholin 12 (n-Dodecyl-phosphocholin) solubilisiert. Die Reinigung des gp1/11-Assembly-Komplexes erfolgte anschließend über den N-terminalen His-Tag der gp1-Fusionsproteine mit Hilfe einer Ni-NTA-Säule. Im Verlauf der Reinigung wurden 50 µl Proben entnommen, um den Verbleib der gp1- und gp11-Proteine während der einzelnen Reinigungsschritte überprüfen zu können. Die Proben wurden dann nach TCA-Fällung über eine 12 %ige SDS-PAGE aufgetrennt. M = Proteinstandard (Marker), SP = solubilisiertes Pellet, ÜUZ = Überstand Ultrazentrifugation, DS = Durchlauf Ni-NTA-Säule, W1 und W2 = Waschschritt, E = Elution der gebundenen Proteine. (A) Coomassie gefärbtes, 12 %iges SDS-Gel der aufgetrennten Proben. In der Elution waren Banden dreier Proteine zu sehen. Eine Bande bei ~ 39 kDa ( ), eine zweite bei ~ 37 kDa ( ) und eine dritte bei ~ 31 kDa ( ). (B) Western-Blot der Proben aus den Reinigungsschritten. Die Identifikation von exprimierten gp1 und gp11 erfolgte mit anti-gp1/11 Serum. In den Proben des solubilisierten Pellets, des Überstandes nach Ultrazentrifugation und auch im Durchlauf der Säule konnte gp1 ( ) bei ~ 39 kDa und gp11 ( ) bei ~ 12 kDa identifiziert werden. In der Elution wurde ausschließlich gp1 nachgewiesen.
130 100 70
55 40
35
25
15
10
kDa SP ÜUZ Ds W1 W2 E
130 100 70
55 40
35
25
15
10
kDa M SP Üuz Ds W1 W2 E
(A) (B)

Ergebnisse
139
In den Western-Blot Analysen (Abb. 3-18 und Abb. 3-19 (B)) konnte die ~ 39 kDa
Bande ( ) als gp1 identifiziert werden. In den Proben des solubilisierten Pellets (SP,
Abb. 3-19 (B)), des Überstandes nach Ultrazentrifugation (ÜUZ) und auch im Durchlauf
der Säule (DS) konnte bei ~ 12 kDa ( ) gp11 identifiziert werden. In der Elution
hingegen konnte ausschließlich nur gp1 nachgewiesen werden. Das starke gp1-Signal
in der Probe (ÜUZ) des Immun-Blots zeigte außerdem, dass die Membranen mit Fos-
cholin 12 ausreichend solubilisierbar waren. Das gp1-Signal im Durchlauf der Ni-NTA-
Säule (DS) hingegen wies darauf hin, dass ein Teil des gp1 und gp11 während der
2-stündigen Inkubation nicht an die Ni-NTA-Matrix gebunden werden konnte.
Das Coomassie gefärbte Gel in Abbildung 3-20 zeigt die aufgetrennte Probe der
Elutionsfraktion (E) sowie die Waschschritte (W1) und (W2), welche aus der Metall-
affinitätschromatographie von 4 L gp1/11-exprimierenden E. coli C41-Zellen erhalten
wurden. Im Vergleich zur Elutionsfraktion (E) in Abbildung 3-19 (A) konnte in dieser
Elutionsfraktion mit Hilfe der Coomassiefärbung eine vierte Bande bei ~ 20 kDa ( )
und eine fünfte bei ~ 12 kDa ( ) identifiziert werden. Die 12 kDa Bande ent-
sprach, dem Molekulargewicht nach, der des gp11 ( ) im Western-Blot in Abbildung
3-19 (B). Das Bandenmuster des Waschschrittes (W2) hingegen zeigte, dass ab einer
Imidazolkonzentration von 75 mM geringe Mengen gp1 und gp11 von der Säulenmatrix
eluiert wurden, da diese Proteinbanden ebenfalls in der Elutionsfraktion vorhanden
waren. Diese waren jedoch nicht im Waschschritt (W1) zu sehen, welcher mit einer
Imidazolkonzentration von 40 mM durchgeführt wurde.
3.4.3 Massenbestimmung des gereinigten M13 gp1/11-Assembly-Komplexes aus
E. coli C41 durch präparative Gelfiltration
Mit Hilfe präparativer Gelfiltration können Proteine aufgrund ihrer hydrodynamischen
Eigenschaften über eine Säulenmatrix getrennt werden. Je kleiner das hydrody-
Abb. 3-20: SDS-PAGE der gp1/11-Komplexreinigung aus E. coli C41 (4 L Flüssigkultur). Der Reinigungsablauf entsprach dem der 2 L Flüssigkultur. Eine Probe aus der Elutionsfraktion wurde nach TCA-Fällung über eine 12 %ige SDS-PAGE aufgetrennt und anschließend mit Coomassie gefärbt. In der Elution konnten mit Hilfe der Comassiefärbung fünf Proteinbanden sichtbar gemacht werden: Eine Bande bei ~ 39 kDa ( ), eine zweite bei ~ 37 kDa ( ), eine dritte bei ~ 31 kDa ( ) eine vierte bei ~ 20 kDa ( ) und eine fünfte bei ~ 12 kDa ( ). Das Banden-muster des Waschschrittes W2 zeigte, dass ab einer Konzentration von 75 mM Imidazol im Waschpuffer geringe Mengen der Proteine, welche in der Elutionsfraktion vorhanden waren, eluiert wurden. Diese waren jedoch nicht im Waschschritt W1 zu sehen.
kDa E W1 W2
130 100 70
55 40 35
25
15
10

Ergebnisse
140
namische Volumen eines Proteins ist, desto langsamer passiert es die Matrix, große
Proteine bzw. Proteinkomplexe hingegen passieren aufgrund des größeren hydrody-
namischen Volumens die Matrix schneller. Für eine Bestimmung der Masse, des in
E. coli C41 überexprimierten gp1/11-Assembly-Komplexes, sollte das aus der IMAC-
Reinigung erhaltene Proteingemisch der Elutionsfraktion durch FPLC („fast protein
liquid chromatography“) aufgetrennt werden (Kap. 2.8.34). Damit eine möglichst genaue
Auftrennung des Proteingemisches erfolgen konnte, wurde für die FPLC als Säulen-
matrix SuperdexTM 200 verwendet, da diese einen Trennbereich von 10 – 600 kDa
besitzt. Um in den gesammelten Fraktionen der FPLC die entsprechenden Molekular-
gewichte zuordnen zu können, wurde, wie in Kapitel 2.8.34 beschrieben, zuvor eine
Kalibrierung der Säule durchgeführt. Hierfür erfolgte die Auftrennung eines Gemisches
aus Proteinen (Apoferritin, β-Amylase, BSA, Carboanhydrase) mit bekannten Mole-
kulargewichten zwischen 30 kDa – 440 kDa über die SuperdexTM 200 Matrix. Aus den
Verhältnissen von Elutionsvolumina zu eluierten Molekulargewichten wurde eine Eich-
gerade erstellt. Mit Hilfe dieser Eichgeraden sollten anschließend die Bestimmungen
der Massen in den Elutionsfraktionen der FPLC erfolgen.
Für die Bestimmung der Masse des gp1/11-Assembly-Komplexes wurde das 2 ml
Elutionskonzentrat aus der IMAC in Abbildung 3-20 über die zuvor geeichte XK 16/40
Säule mit SuperdexTM 200 Matrix aufgetrennt. Um die Massen eindeutig zuordnen zu
können, erfolgte, wie bei der Eichung, die Auftrennung des Proteingemisches in deter-
genzhaltigem Laufpuffer (20 mM Tris/HCl pH 7, 50 mM NaCl, 0,8 % Fos-cholin 12) bei
einer Flussrate von 0,35 ml/min. Während der FPLC wurden Elutionsfraktionen von 2 ml
Probenvolumen gesammelt. Die Detektion der eluierten Proteine während der FPLC
erfolgte bei 280 nm. Um die Proteine in den gesammelten Fraktionen identifizieren zu
können, wurden aus diesen jeweils 100 µl entnommen, mit TCA präzipitiert und
anschließend über eine 12 %ige SDS-PAGE aufgetrennt (Kap. 2.8.35) und Coomassie
gefärbt (Kap. 2.8.36). Aufgrund der Säulenkalibrierung und der daraus resultierenden
Eichgeraden (Kap. 2.8.34) konnten den über SDS-PAGE aufgetrennten Einzelfrak-
tionen entsprechende Massen zugeordnet werden.
Das Chromatogramm in Abbildung 3-21 (A) zeigt den Proteindetektionsverlauf sowie
die Anzahl der gesammelten 2 ml Fraktionen während der Gelfiltration. Nach 20 ml
Elutionsvolumen wurde, durch den Anstieg der Absorption auf 10 mAU, die Elution der
ersten Proteine nachgewiesen. Anhand der Kalibrierung der Säule konnten den hier
eluierten Proteinen Molekulargewichte zwischen ~ 350 kDa – 290 kDa zugeordnet
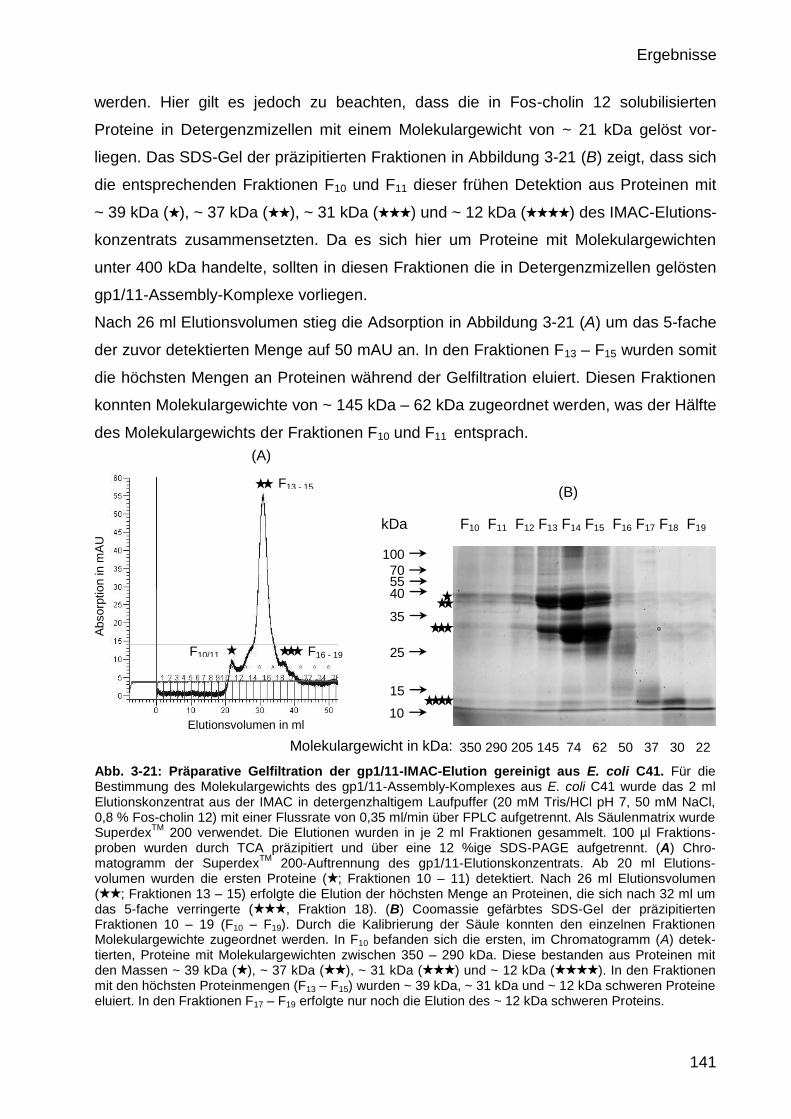
Ergebnisse
141
werden. Hier gilt es jedoch zu beachten, dass die in Fos-cholin 12 solubilisierten
Proteine in Detergenzmizellen mit einem Molekulargewicht von ~ 21 kDa gelöst vor-
liegen. Das SDS-Gel der präzipitierten Fraktionen in Abbildung 3-21 (B) zeigt, dass sich
die entsprechenden Fraktionen F10 und F11 dieser frühen Detektion aus Proteinen mit
~ 39 kDa ( ), ~ 37 kDa ( ), ~ 31 kDa ( ) und ~ 12 kDa ( ) des IMAC-Elutions-
konzentrats zusammensetzten. Da es sich hier um Proteine mit Molekulargewichten
unter 400 kDa handelte, sollten in diesen Fraktionen die in Detergenzmizellen gelösten
gp1/11-Assembly-Komplexe vorliegen.
Nach 26 ml Elutionsvolumen stieg die Adsorption in Abbildung 3-21 (A) um das 5-fache
der zuvor detektierten Menge auf 50 mAU an. In den Fraktionen F13 – F15 wurden somit
die höchsten Mengen an Proteinen während der Gelfiltration eluiert. Diesen Fraktionen
konnten Molekulargewichte von ~ 145 kDa – 62 kDa zugeordnet werden, was der Hälfte
des Molekulargewichts der Fraktionen F10 und F11 entsprach.
100
70 55 40
35
25
15
10
kDa F10 F11 F12 F13 F14 F15 F16 F17 F18 F19
350 290 205 145 74 62 50 37 30 22 Molekulargewicht in kDa:
(B)
Abb. 3-21: Präparative Gelfiltration der gp1/11-IMAC-Elution gereinigt aus E. coli C41. Für die Bestimmung des Molekulargewichts des gp1/11-Assembly-Komplexes aus E. coli C41 wurde das 2 ml Elutionskonzentrat aus der IMAC in detergenzhaltigem Laufpuffer (20 mM Tris/HCl pH 7, 50 mM NaCl, 0,8 % Fos-cholin 12) mit einer Flussrate von 0,35 ml/min über FPLC aufgetrennt. Als Säulenmatrix wurde Superdex
TM 200 verwendet. Die Elutionen wurden in je 2 ml Fraktionen gesammelt. 100 µl Fraktions-
proben wurden durch TCA präzipitiert und über eine 12 %ige SDS-PAGE aufgetrennt. (A) Chro-matogramm der Superdex
TM 200-Auftrennung des gp1/11-Elutionskonzentrats. Ab 20 ml Elutions-
volumen wurden die ersten Proteine ( ; Fraktionen 10 – 11) detektiert. Nach 26 ml Elutionsvolumen ( ; Fraktionen 13 – 15) erfolgte die Elution der höchsten Menge an Proteinen, die sich nach 32 ml um das 5-fache verringerte ( , Fraktion 18). (B) Coomassie gefärbtes SDS-Gel der präzipitierten Fraktionen 10 – 19 (F10 – F19). Durch die Kalibrierung der Säule konnten den einzelnen Fraktionen Molekulargewichte zugeordnet werden. In F10 befanden sich die ersten, im Chromatogramm (A) detek-tierten, Proteine mit Molekulargewichten zwischen 350 – 290 kDa. Diese bestanden aus Proteinen mit den Massen ~ 39 kDa ( ), ~ 37 kDa ( ), ~ 31 kDa ( ) und ~ 12 kDa ( ). In den Fraktionen mit den höchsten Proteinmengen (F13 – F15) wurden ~ 39 kDa, ~ 31 kDa und ~ 12 kDa schweren Proteine eluiert. In den Fraktionen F17 – F19 erfolgte nur noch die Elution des ~ 12 kDa schweren Proteins.
Elutionsvolumen in ml
Ab
so
rptio
n in
mA
U
(A)
F10/11
F13 - 15
F16 - 19

Ergebnisse
142
Auf dem SDS-Gel in Abbildung 3-21 (B) zeigte sich, dass in diesen Fraktionen Proteine
mit den Massen von ~ 39 kDa ( ), ~ 37 kDa ( ), ~ 31 kDa ( ) und ~ 12 kDa
( ) der IMAC-Elution vorkamen.
In Abbildung 3-21 (A) erfolgte zwischen den Elutionsvolumina 36 ml – 38 ml ( ) die
Elution der niedermolekularen Proteine. Den Fraktionen F16 – F19 wurden Massen
zwischen ~ 50 kDa und 22 kDa zugeordnet. Das SDS-Gel in Abbildung 3-21 (B) zeigte,
dass in diesen Fraktionen hauptsächlich die Elution des ~ 12 kDa schweren Proteins
( ) erfolgte.
3.4.4 Massenbestimmung des gereinigten M13 gp1/11-Assembly-Komplexes aus
infizierten E. coli K37 durch präparative Gelfiltration
Das Chromatogramm der FPLC-Auftrennung des gp1/11-Elutionkonzentrats aus Kapitel
3.4.3 zeigte, dass die gp1/11-Assembly-Komplexe nach 20 ml Säulenvolumen, das
einem Molekulargewicht von ~ 350 kDa – 290 kDa entspricht, eluiert wurden. Jedoch
war die Proteinmenge an eluierten Komplexen zu gering. Um eine mögliche Beteiligung
weiterer Phagenproteine identifizieren zu können, wurde eine weitere Reinigung des
gp1/11-Assembly-Komplexes aus infizierten E. coli K37-Zellen durchgeführt. Dieser
Bakterienstamm besitzt F-Pili und ist ein natürlicher Wirt des M13-Phagen. Hier sollten
die gp1/11-exprimierenden Zellen zum Zeitpunkt der induzierten Proteinexpression mit
M13mp19-Phagen zusätzlich infiziert werden. Durch die Infektion sollte eine Degra-
dierung des Komplexes in der Zytoplasmamembran aufgrund seiner aktiven Verwen-
dung während der Phagensekretion verringert werden. Durch die zusätzliche Infektion
der Zellen entstehen erwartungsgemäß heterologe Komplexe, die sich wohl aus wild-
typischen und His-Tag fusionierten gp1 zusammensetzen. Über den His-Tag sollten
sich daher auch diese Komplexe mit Hilfe der IMAC reinigen und anschließend über
FPLC auftrennen lassen. Damit eine Reinigung mit anschließender Massenbestimmung
erfolgen konnte, wurde das gp1/11-Fragment, wie in Kapitel 2.8.17 beschrieben, mit
N-terminalem His-Tag aus dem Vektor pETg1/11 in den Vektor pMS119EH kloniert, aus
dem das Konstrukt pMSg1/11 hervorging. Hier stand die Expression der gp1- und gp11-
Proteine unter der Kontrolle des induzierbaren tac Promotors. Diese Umklonierung
wurde durchgeführt, da der Bakterienstamm E. coli K37 keine T7 RNA-Polymerase
exprimiert, die für die Proteinexpression in pET-klonierter Gene benötigt wird.
Für die Reinigung des Komplexes wurden E. coli K37-Zellen mit dem Vektor pMSg1/11
transformiert. Von auf ampicillinhaltigen LB-Agarplatten gewachsenen Transformanten
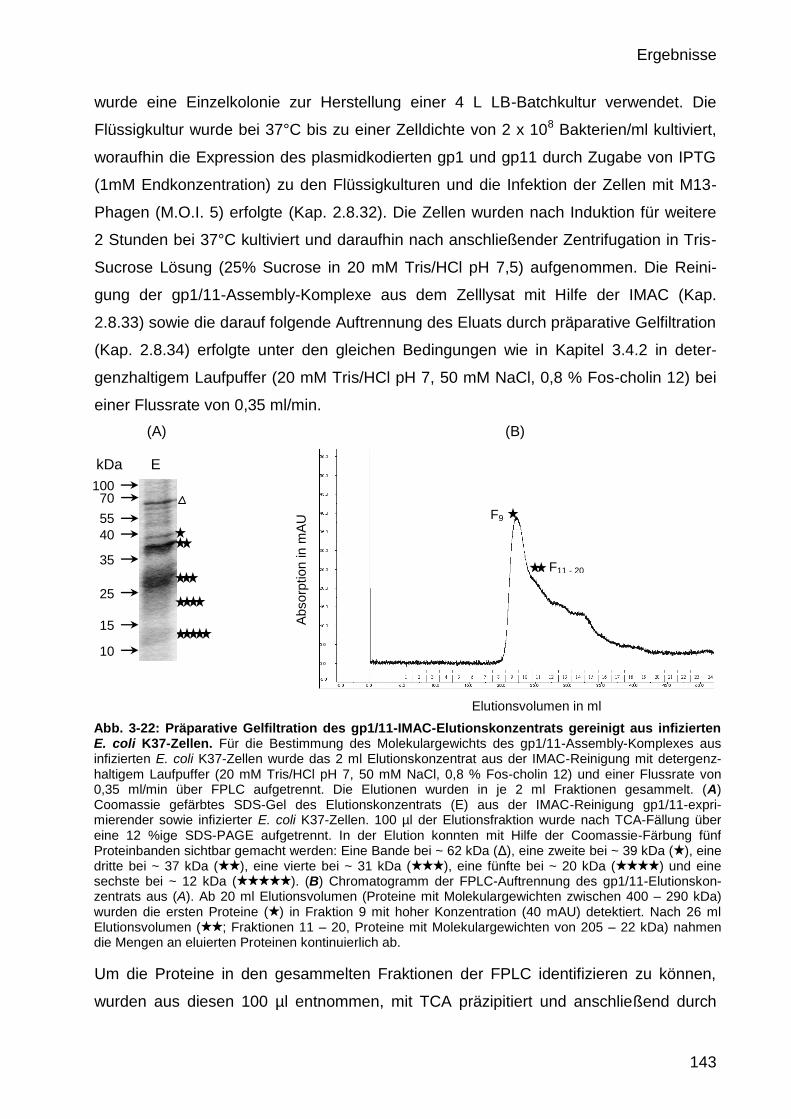
Ergebnisse
143
wurde eine Einzelkolonie zur Herstellung einer 4 L LB-Batchkultur verwendet. Die
Flüssigkultur wurde bei 37°C bis zu einer Zelldichte von 2 x 108 Bakterien/ml kultiviert,
woraufhin die Expression des plasmidkodierten gp1 und gp11 durch Zugabe von IPTG
(1mM Endkonzentration) zu den Flüssigkulturen und die Infektion der Zellen mit M13-
Phagen (M.O.I. 5) erfolgte (Kap. 2.8.32). Die Zellen wurden nach Induktion für weitere
2 Stunden bei 37°C kultiviert und daraufhin nach anschließender Zentrifugation in Tris-
Sucrose Lösung (25% Sucrose in 20 mM Tris/HCl pH 7,5) aufgenommen. Die Reini-
gung der gp1/11-Assembly-Komplexe aus dem Zelllysat mit Hilfe der IMAC (Kap.
2.8.33) sowie die darauf folgende Auftrennung des Eluats durch präparative Gelfiltration
(Kap. 2.8.34) erfolgte unter den gleichen Bedingungen wie in Kapitel 3.4.2 in deter-
genzhaltigem Laufpuffer (20 mM Tris/HCl pH 7, 50 mM NaCl, 0,8 % Fos-cholin 12) bei
einer Flussrate von 0,35 ml/min.
Um die Proteine in den gesammelten Fraktionen der FPLC identifizieren zu können,
wurden aus diesen 100 µl entnommen, mit TCA präzipitiert und anschließend durch
Abb. 3-22: Präparative Gelfiltration des gp1/11-IMAC-Elutionskonzentrats gereinigt aus infizierten E. coli K37-Zellen. Für die Bestimmung des Molekulargewichts des gp1/11-Assembly-Komplexes aus infizierten E. coli K37-Zellen wurde das 2 ml Elutionskonzentrat aus der IMAC-Reinigung mit detergenz-haltigem Laufpuffer (20 mM Tris/HCl pH 7, 50 mM NaCl, 0,8 % Fos-cholin 12) und einer Flussrate von 0,35 ml/min über FPLC aufgetrennt. Die Elutionen wurden in je 2 ml Fraktionen gesammelt. (A) Coomassie gefärbtes SDS-Gel des Elutionskonzentrats (E) aus der IMAC-Reinigung gp1/11-expri-mierender sowie infizierter E. coli K37-Zellen. 100 µl der Elutionsfraktion wurde nach TCA-Fällung über eine 12 %ige SDS-PAGE aufgetrennt. In der Elution konnten mit Hilfe der Coomassie-Färbung fünf Proteinbanden sichtbar gemacht werden: Eine Bande bei ~ 62 kDa (Δ), eine zweite bei ~ 39 kDa ( ), eine dritte bei ~ 37 kDa ( ), eine vierte bei ~ 31 kDa ( ), eine fünfte bei ~ 20 kDa ( ) und eine sechste bei ~ 12 kDa ( ). (B) Chromatogramm der FPLC-Auftrennung des gp1/11-Elutionskon-zentrats aus (A). Ab 20 ml Elutionsvolumen (Proteine mit Molekulargewichten zwischen 400 – 290 kDa) wurden die ersten Proteine ( ) in Fraktion 9 mit hoher Konzentration (40 mAU) detektiert. Nach 26 ml Elutionsvolumen ( ; Fraktionen 11 – 20, Proteine mit Molekulargewichten von 205 – 22 kDa) nahmen die Mengen an eluierten Proteinen kontinuierlich ab.
(A)
100 70
55
40
35
25
15
10
kDa E
(B)
F9
F11 - 20
Absorp
tion
in
mA
U
Elutionsvolumen in ml

Ergebnisse
144
eine 12 %ige SDS-PAGE aufgetrennt (Kap. 2.8.35) und Coomassie gefärbt (Kap.
2.8.36). Aufgrund der Säulenkalibrierung und der daraus resultierenden Eichgeraden
(Kap. 2.8.34) konnten den aufgetragenen Fraktionen entsprechende Massen zuge-
ordnet werden.
Das Chromatogramm in Abbildung 3-22 (B) zeigt sowohl den Proteindetektionsverlauf
als auch die Anzahl der gesammelten 2 ml Fraktionen während der präparativen Gel-
filtration. Nach 20 ml Elutionsvolumen wurde, durch den Anstieg der Absorption auf
40 mAU, die Elution der ersten Proteine in Fraktion 9 angezeigt. Anhand der Kali-
brierung der Säule konnten den hier eluierten Proteinen Molekulargewichte zwischen
~ 350 – 290 kDa zugeordnet werden. Hier gilt es jedoch zu beachten, dass die in Fos-
cholin 12 solubilisierten Proteine in Detergenzmizellen mit einem Molekulargewicht von
~ 21 kDa gelöst vorliegen. Das SDS-Gel der präzipitierten Fraktionen in Abbildung 3-23
zeigte, dass sich die Fraktion F9 hauptsächlich aus Proteinen mit der Masse von ~ 37
kDa ( ; gp1 wt) sowie aus Proteinen mit den Massen von ~ 62 kDa (Δ; unbek.), ~ 39
kDa ( ; gp1-His), ~ 31 kDa ( ; gp1p) und ~ 20 kDa ( ; gp1 Dimer?) des IMAC-
Elutionskonzentrats zusammensetzten.
Abb. 3-23: SDS-PAGE der aufgetrennten Fraktionen aus der präparativen Gelfiltration des gp1/11-IMAC-Elutionskonzentrats gereinigt aus infizierten E. coli K37-Zellen. Die Gelfiltration der gp1/11-Assembly-Komplexe erfolgte in detergenzhaltigem Laufpuffer (20 mM Tris/HCl pH 7, 50 mM NaCl, 0,8 % Fos-cholin 12) bei einer Flussrate von 0,35 ml/min. Während der FPLC wurden die Elutionen in je 2 ml Fraktionen gesammelt. Aus den Fraktionen wurden 100 µl mit TCA präzipitiert und über eine 12 %ige SDS-PAGE aufgetrennt. F9 – F18 = Elutionsfraktionen. Ab 20 ml Elutionsvolumen wurden die ersten Proteine in Fraktion 9 mit hoher Konzentration (40 mAU) detektiert. Die Auftrennung der Probe auf dem SDS-Gel zeigte, dass sich diese Fraktion mit Massen zwischen ~ 350 kDa bis 290 kDa hauptsächlich aus dem ~ 37 kDa ( ; gp1 wt) schweren Protein des Elutionskonzentrats der IMAC sowie einem ~ 62 kDa (Δ; unbekannt), dem ~ 39 kDa ( ; gp1-His), dem ~ 31 kDa ( ; gp1 p) und dem ~ 20 kDa ( ; gp11 Dimer?) schweren Protein zusammensetzte. Erst nach 31 ml Elutionsvolumen (Fraktionen 14 – 18) erfolgte die Elution des 12 kDa ( ; gp11 wt) schweren Proteins. Durch Kalibrierung der Säule konnten den einzelnen Fraktionen die entsprechenden Molekulargewichte zugeordnet werden.
350 290 205 145 74 62 50 37 30 22 Molekulargewicht in kDa:
F9 F10 F11 F12 F13 F14 F15 F16 F17 F18 kDa
Unbek.
gp1-His
gp1p
gp11 (Dimer?)
gp11 wt
35
70
55
40
15
10
100
25
gp1 wt

Ergebnisse
145
Da es sich hier um Proteine mit Molekulargewichten unter 400 kDa handelte, sollten,
wie in Abbildung 3-21 (B), in dieser Fraktion in Detergenzmizellen gelöste gp1/11-
Assembly-Komplexe vorhanden sein. Im Verlauf der aufgetragenen Elutionen nahm
zwischen F10 und F12 die Menge eines eluierten Proteins mit ~ 62 kDa (Δ) auffällig zu,
welches ebenfalls in der Elutionsfraktion in Abbildung 3-22 (A) und sehr schwach in den
Western-Blot Analysen in Abbildung 3-18 und Abbildung 3-19 (B) zu sehen war. Im
Vergleich zur Fraktion F9 in Abbildung 3-21 (A) hatte die Fraktion F9 in Abbildung 3-23
mit einer Absorption von 40 mAU eine 40-mal höhere Proteinkonzentration. Somit
konnte durch die Infektion der E. coli K37 Batch-Kultur die Menge an gp1/11-Assembly-
Komplexen zugunsten der Komplexe hin verschoben werden. Jedoch war hier im
Vergleich zu Abbildung 3-21 (B), in der Fraktion F9 kein Protein mit der Masse von ~ 12
kDa zu erkennen. In den Fraktionen F10 – F14 (Massen mit 290 kDa - 62 kDa) nehmen
die Mengen der ~ 39 kDa ( ), ~ 37 kDa ( ), ~ 31 kDa ( ) und ~ 20 kDa ( )
schweren Proteine kontinuierlich ab. Nach 31 ml Elutionsvolumen (F14 – F18; ~ 62 kDa -
22 kDa) erfolgte die Elution eines ~ 12 kDa schweren Proteins.
3.4.5 Identifikation von Proteinbanden im IMAC-Eluat durch MALDI-TOF/MS
In Kapitel 3.4.1und 3.4.2 konnte mit Hilfe des anti-gp1/11 Serums die Proteinbande mit
einem Molekulargewicht von ~ 39 kDa als gp1 und die Proteinbande mit ~ 12 kDa als
gp11 identifiziert werden. Sowohl in den IMAC-Eluaten in Abbildung 3-20 bzw.
Abbildung 3-22, als auch in den FPLC-Elutionsfraktionen in Abbildung 3-23 konnten
weitere Proteine koeluiert werden. Hierbei galt das Interesse vor allem der ~ 37 kDa,
der ~ 31 kDa und der ~ 20 kDa schweren Proteinbanden, da diese kontinuierlich in den
IMAC-Elutionen präsent waren. Zur Identifizierung wurden diese Banden aus den
Coomassie gefärbten SDS-Gelen isoliert und mittels MALDI-TOF/MS („Matrix Assisted
Laser Desorption/Ionization – Time Of Flight / Mass-Spectrometry“) analysiert. Bei
dieser Methode werden zuerst Proteine mittels Trypsinverdau in Peptidfragmente abge-
baut, deren Einzelmassen für jedes Protein einzigartig sind und durch MALDI-TOF/MS-
Analyse bestimmt werden können. Mit Hilfe eines anschließenden Vergleichs des
hieraus entstandenen Peptid-Massen-Fingerprints (PMF), mit den in Datenbanken
hinterlegten PMFs (Mascot, http://www.matrixscience.com), können dann die jeweiligen
Proteine identifiziert werden.
Die MALDI-TOF/MS-Analysen in Abbildung 3-24 (B-D) identifizierten die aus dem SDS-
Gel isolierten Banden als gp1-Proteinfragmente des filamentösen Phagen M13 (s. An-
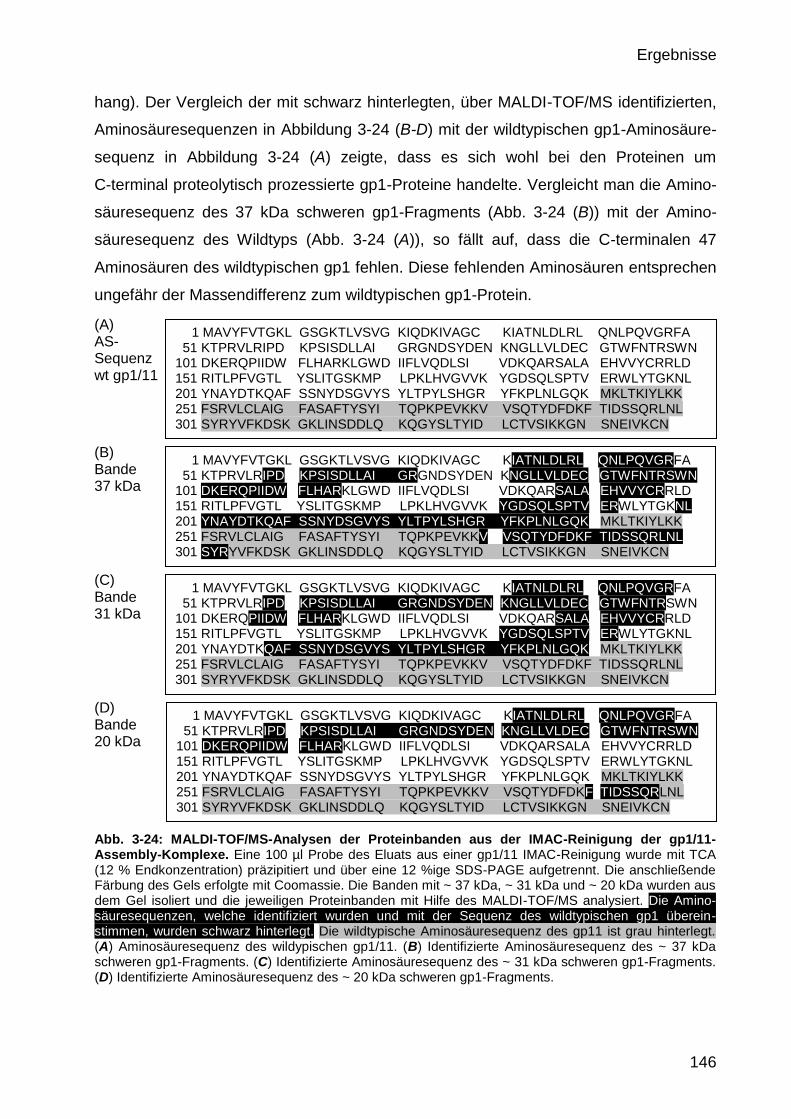
Ergebnisse
146
hang). Der Vergleich der mit schwarz hinterlegten, über MALDI-TOF/MS identifizierten,
Aminosäuresequenzen in Abbildung 3-24 (B-D) mit der wildtypischen gp1-Aminosäure-
sequenz in Abbildung 3-24 (A) zeigte, dass es sich wohl bei den Proteinen um
C-terminal proteolytisch prozessierte gp1-Proteine handelte. Vergleicht man die Amino-
säuresequenz des 37 kDa schweren gp1-Fragments (Abb. 3-24 (B)) mit der Amino-
säuresequenz des Wildtyps (Abb. 3-24 (A)), so fällt auf, dass die C-terminalen 47
Aminosäuren des wildtypischen gp1 fehlen. Diese fehlenden Aminosäuren entsprechen
ungefähr der Massendifferenz zum wildtypischen gp1-Protein.
1 MAVYFVTGKL GSGKTLVSVG KIQDKIVAGC KIATNLDLRL QNLPQVGRFA 51 KTPRVLRIPD KPSISDLLAI GRGNDSYDEN KNGLLVLDEC GTWFNTRSWN 101 DKERQPIIDW FLHARKLGWD IIFLVQDLSI VDKQARSALA EHVVYCRRLD 151 RITLPFVGTL YSLITGSKMP LPKLHVGVVK YGDSQLSPTV ERWLYTGKNL 201 YNAYDTKQAF SSNYDSGVYS YLTPYLSHGR YFKPLNLGQK MKLTKIYLKK 251 FSRVLCLAIG FASAFTYSYI TQPKPEVKKV VSQTYDFDKF TIDSSQRLNL 301 SYRYVFKDSK GKLINSDDLQ KQGYSLTYID LCTVSIKKGN SNEIVKCN
1 MAVYFVTGKL GSGKTLVSVG KIQDKIVAGC KIATNLDLRL QNLPQVGRFA 51 KTPRVLRIPD KPSISDLLAI GRGNDSYDEN KNGLLVLDEC GTWFNTRSWN 101 DKERQPIIDW FLHARKLGWD IIFLVQDLSI VDKQARSALA EHVVYCRRLD 151 RITLPFVGTL YSLITGSKMP LPKLHVGVVK YGDSQLSPTV ERWLYTGKNL 201 YNAYDTKQAF SSNYDSGVYS YLTPYLSHGR YFKPLNLGQK MKLTKIYLKK 251 FSRVLCLAIG FASAFTYSYI TQPKPEVKKV VSQTYDFDKF TIDSSQRLNL 301 SYRYVFKDSK GKLINSDDLQ KQGYSLTYID LCTVSIKKGN SNEIVKCN
1 MAVYFVTGKL GSGKTLVSVG KIQDKIVAGC KIATNLDLRL QNLPQVGRFA 51 KTPRVLRIPD KPSISDLLAI GRGNDSYDEN KNGLLVLDEC GTWFNTRSWN 101 DKERQPIIDW FLHARKLGWD IIFLVQDLSI VDKQARSALA EHVVYCRRLD 151 RITLPFVGTL YSLITGSKMP LPKLHVGVVK YGDSQLSPTV ERWLYTGKNL 201 YNAYDTKQAF SSNYDSGVYS YLTPYLSHGR YFKPLNLGQK MKLTKIYLKK 251 FSRVLCLAIG FASAFTYSYI TQPKPEVKKV VSQTYDFDKF TIDSSQRLNL 301 SYRYVFKDSK GKLINSDDLQ KQGYSLTYID LCTVSIKKGN SNEIVKCN
1 MAVYFVTGKL GSGKTLVSVG KIQDKIVAGC KIATNLDLRL QNLPQVGRFA 51 KTPRVLRIPD KPSISDLLAI GRGNDSYDEN KNGLLVLDEC GTWFNTRSWN 101 DKERQPIIDW FLHARKLGWD IIFLVQDLSI VDKQARSALA EHVVYCRRLD 151 RITLPFVGTL YSLITGSKMP LPKLHVGVVK YGDSQLSPTV ERWLYTGKNL 201 YNAYDTKQAF SSNYDSGVYS YLTPYLSHGR YFKPLNLGQK MKLTKIYLKK 251 FSRVLCLAIG FASAFTYSYI TQPKPEVKKV VSQTYDFDKF TIDSSQRLNL 301 SYRYVFKDSK GKLINSDDLQ KQGYSLTYID LCTVSIKKGN SNEIVKCN
(A) AS-Sequenz wt gp1/11
(B) Bande 37 kDa
(C) Bande 31 kDa
(D) Bande 20 kDa
Abb. 3-24: MALDI-TOF/MS-Analysen der Proteinbanden aus der IMAC-Reinigung der gp1/11- Assembly-Komplexe. Eine 100 µl Probe des Eluats aus einer gp1/11 IMAC-Reinigung wurde mit TCA (12 % Endkonzentration) präzipitiert und über eine 12 %ige SDS-PAGE aufgetrennt. Die anschließende Färbung des Gels erfolgte mit Coomassie. Die Banden mit ~ 37 kDa, ~ 31 kDa und ~ 20 kDa wurden aus dem Gel isoliert und die jeweiligen Proteinbanden mit Hilfe des MALDI-TOF/MS analysiert. Die Amino-säuresequenzen, welche identifiziert wurden und mit der Sequenz des wildtypischen gp1 überein-stimmen, wurden schwarz hinterlegt. Die wildtypische Aminosäuresequenz des gp11 ist grau hinterlegt. (A) Aminosäuresequenz des wildypischen gp1/11. (B) Identifizierte Aminosäuresequenz des ~ 37 kDa schweren gp1-Fragments. (C) Identifizierte Aminosäuresequenz des ~ 31 kDa schweren gp1-Fragments. (D) Identifizierte Aminosäuresequenz des ~ 20 kDa schweren gp1-Fragments.

Ergebnisse
147
Dies gilt ebenfalls für das isolierte ~ 31 kDa schwere gp1-Fragment (Abb. 3-24 (C)). Die
Differenz der fehlenden C-terminalen Aminosäuren zum wildtypischen gp1 entspricht
auch hier ungefähr der Massendifferenz. Eine Ausnahme stellt jedoch das isolierte
~ 20 kDa schwere gp1-Fragment dar (Abb. 3-24 (D)), da hier die nachgewiesenen
Peptidfragmente nur einem ~ 10 kDa schweren entsprechen. Zusätzlich fällt auf, dass
hier ein 8 Aminosäuren langes Fragment nachgewiesen wurde, welches dem Bereich
der Aminosäuren 290 - 297 des wildtypischen gp1 (Abb. 3-24 (A)) entspricht und
ansonsten nur noch in der analysierten 37 kDa schweren Bande, nicht aber in der
analysierten 31 kDa schweren Bande, vorkommt. Hier könnte es sich wahrscheinlich
um 2 Fragmente des gp1 handeln, welche einmal die N- und einmal die C-terminal
prozessierten Form darstellen.
3.4.6 TEM-Aufnahmen FPLC gereinigter gp1/11-Assembly-Komplexe
Für die Visualisierung der gp1/11-Assembly-Komplexe wurden 10 µl Proben aus der
Elutionsfraktion F9 in Abbildung 3-23 auf Trägernetzchen aus Kupfer adsorbiert und
eine Negativkontrastierung mit Phosphorwolframsäure (5 %, pH 7) durchgeführt (Kap.
2.8.42). Die kontrastierten Proben wurden anschließend am Transmissionselektronen-
mikroskop (TEM) untersucht (Kap. 2.8.29).
Die TEM-Aufnahmen in Abbildung 3-25 (A) und (B) zeigen, dass in den adsorbierten
Proben aus Fraktion F9, in die während der FPLC-Auftrennung Komplexe mit Moleku-
largewichten zwischen ~ 400 kDa und 290 kDa eluiert wurden, eine große Anzahl
gleichförmiger und gleichgroßer Ringstrukturen vorhanden waren. In der Abbildung 3-25
(C1) und (D1) sind einzelne Komplexstrukturen zu erkennen, die einen Innendurch-
messer von ~ 7 – 8 nm und einen Außendurchmesser von ~ 11 – 12 nm aufweisen. Mit
Hilfe mathematischer Bildprozessierung konnten in den Abbildungen 3-25 (C2) und (D2)
die ringförmigen Strukturen aus (C1) und (D1) verbessert dargestellt werden.

Ergebnisse
148
Abb. 3-25: TEM-Aufnahmen FPLC gereinigter gp1/11-Assembly-Komplexe. Der gp1/11-Assembly-Komplex wurde in E. coli K37-Zellen mit Hilfe des Vektors pMSg1/11 überexprimiert. Durch eine parallel verlaufende Infektion der Zellen mit M13mp19-Phagen wurde die Degradierung des Assembly-Komplexes durch zelluläre Proteasen verringert. Aus den Zelllysaten wurden anschließend über eine IMAC-Reinigung die Komplexe durch einen N-terminal fusionierten His-Tag isoliert und über FPLC gereinigt. Die TEM- Aufnahmen zeigen Proben aus Fraktion F9, welche zuvor mit 5 % Phosphorwolframsäure (pH 7) negativ kontrastiert wurden und Komplexe mit Molekulargewichten von ~ 400 kDa bis 290 kDa besaßen. (A, B) Aufnahmen, welche ringartige Strukturen zeigen, die einen Innendurchmesser von ~ 7 – 8 nm und einen Außendurchmesser von ~ 11 – 12 nm aufweisen. (C1, D1) Einzeldarstellungen der Ringstrukturen aus (A). (C2, D2) Durch Software verbesserte Darstellungen der Einzelpartikel aus C1 und D1. Die Einzelpartikel zeigen deutlich ringförmige Strukturen. Die Längen der Skalierungsbalken betragen 10 nm.
A B C1
D1
C2
D2

Diskussion
149
4 Diskussion
4.1 Phagenextrusion aus infizierten E. coli-Zellen
Charakteristisch für die filamentösen Phagen M13, fd und f1 sind ihre nicht-lytischen
Infektionsverläufe in den Wirtsorganismen. Jedoch ist der zeitliche Ablauf der Wirts-
infektion und die Synthese neuer Phagen-Nachkommen zwischen den Stämmen
filamentöser Phagen variabel und abhängig von den verwendeten Wirtsorganismen
und den äußeren Bedingungen (Panter & Symons, 1966, Minashima et al., 1968).
Vereinfacht kann ein Infektionsverlauf in einer E. coli-Wirtszelle mit den oben
genannten filamentösen Phagen durch die Phasen Adsorption, Infektion, Eklipse und
die Freisetzung neuer Phagen-Nachkommen durch die Wirtszelle beschrieben
werden. Dabei stellt die Eklipse das Stadium zwischen der Aufnahme der Phagen-
DNA in die Wirtszelle und der Freisetzung neuer Phagen-Nachkommen dar
(Delbrück, 1940).
Damit der zeitliche Verlauf einer M13-Phageninfektion speziell für das Wirts-
bakterium E. coli K38 ermittelt werden konnte, wurde eine Quantifizierung der M13-
Phagenextrusion aus infizierten E. coli K38-Kulturen anhand zweier „one-step
growth“ Modellversuche durchgeführt. Diese Modelle sollten anschließend als Basis
dienen, um den Extrusionsverlauf von M13-Phagen auf der Zelloberfläche, in Abhän-
gigkeit von der Zeit nach Infektion, mit dem AFM messen zu können. Im ersten
Modellversuch erfolgte die Quantifizierung anhand der wildtypisch verlaufenden M13-
Infektion in E. coli K38 durch eine Infektion mit M13mp19. Für den zweiten Modell-
versuch wurde ein induzierbares Phagenextrusionssystem hergestellt, in welchem
die Sekretion neugebildeter M13am8 Phagen über das, in trans bereitgestellte,
Haupthüllprotein gp8 kontrolliert werden kann (Kuhn & Wickner, 1985). Mit beiden
Systemen wurden „one-step growth“ Experimente mit infizierten E. coli K38 Batch-
Kulturen durchgeführt.
Bei einer M.O.I. von 20 können während einer 5-minütigen Adsorptionsphase
~ 7 Phagen pro Bakterienzelle binden
Um vergleichbare Infektionsverläufe aus den Experimenten zu erhalten, erfolgten die
Adsorptionen der Phagen an die E. coli-Zellen bei 4°C. Hierdurch wurde die Auf-
nahme der Phagen DNA in die Zellen unterdrückt (Tzagoloff & Pratt, 1964) und
sichergestellt, dass die Aufnahme der Phagen-DNA in beiden Experimenten unter

Diskussion
150
gleichen Bedingungen erfolgen konnte. Aus den erhaltenen „one-step growth“
Kurven beider Experimente ging hervor, dass sich während der 5-minütigen
Adsorption der Phagen an E. coli K38 die Phagentiter verringerten. Dies konnte
ebenfalls von Tzagoloff und Pratt für die Adsorption von M13-Phagen an E. coli 112-
12 sowie von O’ Callaghan (O´ Callaghan et al., 1973) an E. coli K-12 gezeigt
werden. Die Phagen, die sich aus den jeweiligen Differenzen der Phagentiter
ergaben, hatten während dieser 5 Minuten an die F-Pili der Zellen gebunden. Aus
den M13-Adsorptionsversuchen in Kapitel 3.1.3, in welchem die Bindung der Phagen
an die F-Pili in Abhängigkeit der eingesetzten M.O.I. untersucht wurde, ergab sich
eine Adsorptionskonstante von 3 x 108 pfu/min. In diesem ermittelten Wert ist die
Phagenmenge von 1 x 108 pfu/min berücksichtigt, welche unspezifisch an Bakterien
und Glaswaren bindet (Kap. 3.1.3). Tzagoloff und Pratt berechneten für die
Adsorption von M13-Phagen an E. coli 112-12 eine Adsorptionskonstante von
3 x 10-11 pfu/min. Hier muss jedoch beachtet werden, dass die von Tzagoloff und
Pratt angewendeten Methoden, die zu diesem Adsorptionswert führten, weder eine
unspezifische Adsorption der M13-Phagen noch das Abscheren der F-Pili berück-
sichtigten, das während der Zentrifugation und der anschließenden Wiederaufnahme
der Bakterienkultur in frisches Kulturmedium vor der unmittelbaren Adsorptions-
messung der M13-Phagen auftreten kann. Für die wildtypische „one-step growth“
Kurve ergab sich in dem hier durchgeführten Versuch eine durchschnittliche
Adsorption von 6 – 7 Phagen pro Zelle im Verlauf der 5-minütigen Adsorptionsphase
bei einer M.O.I. von 100. Das induzierte „one-step growth“ Experiment ergab eine
durchschnittliche Adsorption von 2 – 4 Phagen pro Zelle bei einer M.O.I. von 10.
Dass diese in den „one-step growth“ Kurven erhaltenen Adsorptionswerte realisierbar
sind, wurde von Ann Jacobson (Jacobson, 1972) bei ihren Adsorptionsmessungen
des Bakteriophagen f1 an E. coli K-12 gezeigt. Sie konnte in ihren Untersuchungen
feststellen, dass die Anzahl der F-Pili pro E. coli-Zelle limitiert ist und diese bis zu 12
F-Pili ausbilden. Die Adsorptionsexperimente in Kap. 3.1.3 zeigten außerdem, dass
ab einer M.O.I. von 20 ein Adsorptionsmaximum von ~ 7 Phagen pro Bakterienzelle
erreicht wurde. Trotz Erhöhung der M.O.I. konnte während dieser 5-minütigen
Adsorptionsphasen keine Steigerung an gebundenen Phagen erreicht werden.
Hierdurch lassen sich auch die unterschiedlichen Adsorptionswerte in den „one-step
growth“ Experimenten dieser Arbeit erklären. Während bei einer M.O.I. von 10
durchschnittlich nur 50 % der F-Pili mit Phagen besetzt waren, wurde bei einer M.O.I.

Diskussion
151
von 100 nahezu eine Bindungssättigung an den F-Pili erreicht. Zudem zeigt die
Kurve des Adsorptionsexperiments in Kap. 3.1.3, dass erst ab einer M.O.I. größer als
5 aussagekräftige Werte für die Adsorption erhalten wurden. Bei M.O.I.-Werten
kleiner als 5 kommt es, aufgrund der Höhe der unspezifischen Bindung, zu unge-
nauen Werten, die keine Berechnung der Adsorption mehr zulässt.
Die Eklipse der wildtypischen Infektion beträgt 5 Minuten, die der induzierten
Sekretion 12 Minuten
Für die Evaluierung des zeitlichen Verlaufs einer wildtypischen Infektion sowie die
einer induzierten Sekretion der Phagen aus E. coli K38, wurden die Kulturen zuvor
mit M13-Phagen infiziert. Im Fall des wildtypischen Verlaufs erfolgte die Infektion der
Bakterienkultur mit einer M.O.I. von 100, im Fall der induzierten Sekretion wurden die
Zellen nur mit einer M.O.I. von 10 für 5 Minuten infiziert, wobei aber trotzdem davon
ausgegangen werden konnte, dass mindestens ein Phage pro Bakterienzelle
gebunden und diese somit auch infiziert wird. Nach der Infektion wurden die
ungebundenen Phagen durch Zentrifugation der Zellkulturen entfernt und die
infizierten E. coli K38-Zellen in neues LB-Medium aufgenommen und bei 37°C
rekultiviert, wodurch sich die 100-fache Abnahme des Phagentiters der „one-step
growth“ Kurve der wildtypisch verlaufenden Infektion und der induzierten Sekretion in
Abbildung 4-1 zwischen t = -5 und t = 0 erklärt. Im Vergleich zur wildtypischen „one-
step growth“ Kurve sind die Phagentiter der induzierten „one-step growth“ Kurve
10-mal geringer, was wohl in diesem Fall auf den 10-mal geringeren M.O.I-Wert der
Infektion dieser Kultur zurückzuführen ist. Trotz des 10-fachen Unterschiedes der
Phagentiter in den erstellten „one-step growth“ Kurven, zeigen diese innerhalb des
120 Minuten andauernden Infektions-Assays einen dennoch vergleichbaren Verlauf.
Diese „one-step growth“ Kurven weisen zudem Übereinstimmungen mit der von
Brown und Dowell (Brown & Dowell, 1968) für die synchronisierte M13-Infektion einer
E. coli S-26 Batch-Kultur erstellte „one-step growth“ Kurve auf. Sowohl in der wild-
typischen, der induzierten als auch in der von Brown und Dowell gezeigten „one-step
growth“ Kurve steigt der Phagentiter nach der M13-Infektion der Bakterienkultur
innerhalb von 20 Minuten um das ~ 100-fache an, woraus gefolgert werden kann,
dass die Sekretion neuer Phagen-Nachkommen durch die infizierten Zellen sehr
schnell erfolgt. Vergleicht man die wildtypische und die induzierte Sekretion (Abb.
4-1), so stellt sich heraus, dass der wildtypische Infektionsverlauf eine Eklipse von

Diskussion
152
-10 0 10 20 30 40 50 60 70 80 90 100 110 120
1E7
1E8
1E9
1E10
1E11
Zugabe von IPTG
Eklipsendauer bei Induktion
Adsorption bei 4°C
Eklipsendauer bei wildtypischer Infektion
Phagensekretion wt nach
Infektion mit M.O.I. 100
Induzierte Phagensekretion
nach Infektion mit M.O.I. 10
Ph
ag
en
tite
r in
p
fu/m
l
Zeit in min
Abb. 4-1: Grafische Gegenüberstellung der wildtypischen und induzierten Phagensekretion aus infizierten E. coli-Zellen. Im Fall der wildtypischen Sekretion wurde die E. coli Flüssigkultur mit einer M.O.I. von 100 mit M13-Phagen infiziert, im Fall der induzierten Sekretion erfolgte die Infektion mit einer M.O.I von 10. Der hellgrau schraffierte Bereich markiert die 5-minütige Eklipse des wild-typischen Infektionsverlauf, der dunkelgrau schraffierte Bereich die 12-minütige Eklipse der indu-zierten Phagensekretion nach Zugabe von IPTG zu t = 0
5 Minuten nach Aufnahme in frisches Kulturmedium (t = 0) und die induzierte
Sekretion eine Eklipse von 12 Minuten nach Zugabe von IPTG zum Kulturmedium
(t = 0) aufweist.
Im Vergleich zu der von Brown und Dowell publizierten „one-step growth“ Kurve,
welche eine 10-minütige Eklipse während des M13-Infektionsverlaufs in E. coli S26
aufwies, zeigt die wildtypische „one-step growth“ Kurve der M13-Phagen in einer
E. coli K38-Kultur eine um 5 Minuten verkürzte Eklipse. Ursache für die unter-
schiedlichen Eklipsenlängen könnte die Abweichung vom verwendeten Bakterien-
stamm sein. Eine schlüssigere Erklärung für die verkürzte Eklipse der wildtypischen
M13-Phagensekretion aus E. coli K38 könnte jedoch die Vortemperierung des Kultur-
mediums sein, wodurch der bakterielle Stoffwechsel und damit die Proteinsynthese
schneller zur ursprünglichen Ausgangsleistung gefunden hat. Möglicherweise ist dies
sogar ein Resultat aus der Kombination von vortemperiertem Kulturmedium und der
Abweichung vom E. coli-Stamm. Da die von Brown und Dowell verwendete E. coli
S26-Kultur nach Zugabe der M13-Phagen bei 0°C erst auf 37°C erwärmt wurde, kam
es wohl hierdurch zu einer Verzögerung der Phagenprotein-Biosynthese, aus der

Diskussion
153
sich eine Verlängerung der Eklipse ergab. Im Vergleich dazu wurde die E. coli K38-
Kultur direkt nach der Adsorptionsphase und Zentrifugation in frisches, bei 37°C vor-
gewärmtes, Kulturmedium überführt, wodurch sich eine Eklipsenlänge von nur
5 Minuten ergab. Während dieser Eklipse erfolgte die Aufnahme der Phagen-DNA
sowie die Synthese der phagenkodierten Proteine durch die Wirtszelle (Jacobson,
1972).
Extrusion neugebildeter Phagen erfolgt 5 Minuten nach Infektion
Während im Verlauf der wildtypischen Infektion nach 5 Minuten neue Phagen-
Nachkommen in das Kulturmedium abgegeben wurden, erfolgte bei der induzierten
Phagenextrusion erst 12 Minuten nach Zugabe von IPTG zum Kulturmedium zu t = 0
die Sekretion neuer Phagen (Abb. 4-1). Ursache für die Differenz von 7 Minuten zum
Wildtyp könnte die Menge des in trans bereitgestellten Haupthüllproteins gp8 sein.
Für die Sekretion werden pro Phage ~ 2800 Moleküle des Haupthüllproteins gp8
benötigt (Stopar et al., 2003). Da es sich bei dem Vektor pGZ119 (Kuhn, 1986, Lessl,
1992) jedoch um ein „medium-copy“ Plasmid handelt, entstand vermutlich durch die
Bereitstellung des plasmidkodierten Haupthüllproteins in trans nach Induktion zu t = 0
über das Konstrukt pGZg8wt eine anfängliche Unterversorgung. Hieraus kann
gefolgert werden, dass erst ~ 7 Minuten nach Induktion ausreichend gp8 in den
Zellen synthetisiert wurde, damit neue Phagen sekretiert werden konnten.
Eine infizierte E. coli-Zelle kann in 115 Minuten bis zu 925 neue Phagen
sekretieren
Nach den Eklipsen folgte, wie auch in dem „one-step growth“ Experiment von Brown
und Dowell für eine M13-infizierte E. coli S26-Kultur (Brown & Dowell, 1968), die
exponentielle Vermehrung der Phagen. In der wildtypisch verlaufenden Infektion
erhöhte sich während dieser Zeitspanne der Phagentiter um das 30-fache aufgrund
einer Sekretion von durchschnittlich 6 neuen Phagen/min pro infizierter Zelle. Bei der
induzierten Sekretion stieg der Phagentiter nur um das 10-fache an, so dass sich
hierfür eine Sekretion von 3 neuen Phagen/min pro infizierter Zelle ergab. Die
Differenz in den Phagensekretionsraten wird wahrscheinlich auch hier durch eine zu
schwache Expression des plasmidkodierten Haupthüllproteins gp8 bei der indu-
zierten Sekretion verursacht. Nach 120 Minuten konnte der Phagentiter in der
Bakterienkultur der wildtypisch verlaufenden Phageninfektion im Vergleich zum
Zeitpunkt t = 0 um das 1200-fache an Phagen erhöht werden. Das bedeutet, dass im

Diskussion
154
Durchschnitt während der 115-minütigen Sekretionssphase jede ursprünglich
infizierte Zelle 925 neue Phagen gebildet hatte. Bezieht man die Sekretionsrate auf
die durchschnittliche Gesamtmenge an Bakterienzellen in dieser Bakterienkultur
während dieses Versuchszeitraums, so ergibt sich eine gemittelte Sekretion von 440
Phagen pro Zelle, was nahezu der Menge entspricht, die Hoffmann-Berling und Mazé
(Hoffmann-Berling & Mazé, 1964) für fd-Phagen-infizierte E. coli K-12-Zellen nachge-
wiesen haben. Ein Grund, weshalb hier keine absoluten Sekretionsraten pro Zelle
über den gesamten Zeitraum angegeben werden können, liegt daran, dass in jeder
E. coli-Kultur die Zellteilungen sowie die Zellstoffwechsel der Bakterien asynchron
verlaufen und dies somit nur eine gemittelte Berechnung der sekretierten Phagen
erlaubt.
Die Menge neugebildeter Phagen wird bei der induzierten Sekretion durch die
in trans bereitgestellten Proteine limitiert
In der Bakterienkultur, in welcher die Sekretion neuer M13-Phagen induziert wurde,
konnte der ursprüngliche Ausgangsphagentiter zu t = 0 nach 120 Minuten um das
1900-fache angereichert werden. Das bedeutet, dass im Durchschnitt durch jede
ursprünglich infizierte sowie induzierte Zelle, während der 115-minütigen Sekretions-
phase, 170 neue Phagen gebildet wurden. Bezieht man die Sekretionsrate auf die
durchschnittliche Gesamtmenge an Bakterienzellen in dieser Bakterienkultur wäh-
rend dieses Versuchszeitraums, so ergibt sich eine gemittelte Sekretion von 80
Phagen pro Zelle. Somit entspricht die Expressionsrate des plasmidkodierten gp8
ungefähr einem Fünftel der Menge, die während der wildtypischen Infektion gebildet
wird. Hier erweist sich ebenfalls das Plasmid pGZg8wt als limitierender Faktor der
Sekretionsrate. Mit Hilfe eines „high-copy“ Plasmids könnte sich eventuell, durch die
Erhöhung der Syntheserate von gp8, diese Limitierung aufheben und eine wild-
typische Sekretion erreichen lassen. Vergleicht man die Steigerungen der Phagen-
titer zwischen der wildtypischen und der induzierten Phagensekretion, so fällt auf,
dass die Steigerungsrate des Ausgangsphagentiters in der induzierten Sekretion um
das 1,5 fache höher ist als in der wildtypischen Sekretion. Da sich aber die Werte
hier auf das Verhältnis von Ausgangsphagentiter zu Endphagentiter beziehen und
die Ausgangsphagentiter davon abhängig sind, wie vollständig die phagenhaltigen
Überstände abgenommen wurden bevor die Aufnahme der infizierten Zellen in neues

Diskussion
155
LB-Medium zu t = 0 erfolgte, sind diese Werte, verglichen mit denen aus der expo-
nentiellen Phagenvermehrungsphase, nur beschränkt aussagekräftig.
M13-Infektionen bewirken eine Veränderung der Generationszeit des Wirtes
Aufgrund der Bestimmungen der Bakterienzellzahlen während des wildtypischen
Infektionsverlaufs in der E. coli K38 Batch-Kultur konnte, wie schon von Hoffmann-
Berling und Mazé (Hoffmann-Berling & Mazé, 1964) für fd-Phagen-infizierte E. coli
K-12 sowie von Brown und Dowell (Brown & Dowell, 1968) für M13-infizierte E. coli
S26 beschrieben, gezeigt werden, dass die Infektion eine deutliche Auswirkung auf
die Generationszeit der E. coli-Zellen hatte. Hier ergab sich für M13-infizierte E. coli
K38-Zellen eine Verdoppelung der Generationszeit von 24 auf 48 Minuten. Da sich
jedoch Phageninfektionen unterschiedlich auf die Generationszeiten verschiedener
Bakterienstämme auswirken können (Panter & Symons, 1966; Minashima et al.,
1968), kann die hier ermittelte Generationszeit von 48 Minuten nur auf M13-infizierte
E. coli K38-Kulturen bezogen werden.
4.2 M13-Sekretion aus infizierten E. coli-Zellen
4.2.1 TEM und AFM-Studien an infizierten E. coli DH5α-Zellen
Infizierte E. coli-Zellen sekretieren M13 über die gesamte Zelloberfläche
Die visuelle Darstellung erfolgte anhand zweier unterschiedlicher Mikroskopie-
verfahren, um jeweils die Vorteile der angewandten Methoden nutzen zu können.
Hierdurch sollten detaillierte und aussagekräftige Ergebnisse von M13-infizierten
E. coli-Zellen erhalten werden. Mit Hilfe des Transmissionselektronenmikroskops
(TEM) und des „Atomic Force Microscope“ (AFM) wurden M13-infizierte E. coli-Zellen
untersucht. Zur eindeutigen Bestimmbarkeit von sekretierten M13-Phagen auf den
Zelloberflächen wurde hier der Bakterienstamm DH5α verwendet. Dieser Bakterien-
stamm besitzt, wie in einer elektronenmikroskopischen Aufnahme einer nicht
infizierten Zelle gezeigt wurde, weder F-Pili noch Fimbrien auf seiner Zelloberfläche.
Dies war ein essenzielles Kriterium, da eine Unterscheidung zwischen diesen
zellulären Oberflächenstrukturen und M13-Phagen aufgrund ihrer ähnlichen Gestalt
und Größe nur schwer durchführbar ist. Zwar bilden Zellen nach einer Infektion durch
M13 keine F-Pili mehr aus (Jacobson, 1972), jedoch gilt dies nicht für Fimbrien,
welche zur Adsorption der E. coli-Zellen an Oberflächen dienen. Des Weiteren
wurden die Zellen mit dem speziell für die Visualisierung infizierter Zellen herge-

Diskussion
156
stellten Phagenstamm M13amp transfiziert. Mit Hilfe dieses Phagenstamms waren
infizierte Zellen resistent gegenüber dem Antibiotikum Ampicillin, wodurch eine
Selektion auf und in selektiven Nährmedien möglich war. Da E. coli DH5α keine F-Pili
besitzt, können diese Zellen ausschließlich über eine Transfektion infiziert werden.
Zur eindeutigen Bestimmbarkeit infizierter Zellen wurden diese auf ampicillinhaltigen
Kulturmedien gehalten. Da mit Hilfe des TEM eine bessere Darstellung unter-
schiedlich dichter Materialien wie z.B. elektronendichtem Nanogoldpartikel und mit
Schwermetallen negativ kontrastierten schwach elektronenstreuenden biologischen
Proben möglich ist, wurde aus einer, in ampicillinhaltigem Medium gewachsenen,
E. coli-Kultur eine Probe infizierter Zellen entnommen und diese für die Unter-
suchung am TEM präpariert. Der eindeutige Nachweis der M13-Phagen an den
E. coli-Zellen sollte hier mit Hilfe eines primären Antikörpers gegen das Haupt-
hüllprotein gp8 mit anschließender Bindung eines Sekundärantikörpers erfolgen,
welcher mit Protein-A konjugiertem Nanogold assoziiert war. Die präparierten Zellen
wurden sodann am TEM untersucht. Durch dieses Verfahren konnten während der
TEM-Untersuchungen an E. coli DH5α-Zellen filamentöse Strukturen mit gebun-
denen Nanogoldpartikeln mit Längen zwischen 1,3 – 1,4 µm dargestellt werden (Abb.
3-5, (A), (B), S. 108). Die visualisierten Strukturen waren zum Teil länger als
wildtypische M13-Phagen, was darauf zurückzuführen ist, dass die hier für die
Infektion verwendeten M13amp-Phagen, aufgrund des im Phagengenom inserierten
β-Lactamase-Gens, ein größeres Genom besitzen als die wildtypischen M13-
Phagen. Da sich die Genomgröße direkt auf die Länge des M13-Phagen auswirkt,
sind M13amp-Phagen um ~ 400 – 500 nm länger als wildtypische M13-Phagen. Die
Spezifität der Detektion des Haupthüllproteins gp8 durch das anti-gp8 Serum sowie
der Sekundärdetektion des primär an die M13-Phagen gebundenen Antikörpers mit
Hilfe von Protein-A konjugiertem Nanogold wurde zuvor experimentell in Kap. 3.2.1
anhand von Phagenstammlösungen überprüft. Hier konnten die longitudinalen Seiten
der M13-Phagen, welche sich aus gp8 Proteinen zusammensetzen, mit Nanogold
markiert werden (Abb. 3-4 (A), (B), S.106). Jedoch waren nicht alle Positionen
entlang der longitudinalen Seiten der Phagen mit den 20 nm großen Nanogold-
partikeln besetzt. Im Vergleich hierzu erreichte Guo et al. (2010) durch die Fusion
eines Goldbindepeptids an die N-terminale Domäne des Haupthüllproteins gp8 eine
nahezu völlige Bedeckung der Phagenoberfläche mit 10 nm großen Nanogold-
partikel. Ursachen für diesen hohen Adsorptionsgrad könnten geringere sterische

Diskussion
157
Beeinträchtigungen durch die 10 nm großen Goldpartikel sowie eine erhöhte Affinität
der Goldbindepeptide für Goldpartikel sein. Weiterhin zeigten die in Kapitel 3.2.1 mit
Immunogold markierten und mit Phosphorwolframsäure negativ kontrastierten
Phagen eine Abweichung des Phagendurchmessers von 7 nm auf 20 nm. Während
der Immunogoldmarkierung der Phagen wurden sie zur Verringerung unspezifischer
Bindungen mit bovinem Serumalbumin (BSA) präinkubiert. Aus der Arbeit von
Rozamond et al. (2006) ist bekannt, dass bei BSA-behandelten und mit Phosphor-
wolframsäure kontrastierten M13-Phagen Abweichungen in deren Durchmessern
beobachtet werden. Sie führten die Ursache dieser Abweichung auf eine, durch die
Verwendung von BSA zur Verringerung unspezifischer Antikörperbindungen,
gebildete Schicht auf der Phagenoberfläche zurück, durch die sich eine Verdickung
der filamentösen Strukturen ergibt.
Das AFM stellt eine weitere Mikroskopietechnik dar, um biologische Proben zu
messen. Mit Hilfe des AFM können Oberflächenstrukturen biologischer Proben ohne
aufwändige Vorbehandlungen mit nahezu atomarer Auflösung detailgetreu ge-
messen und dargestellt werden. Für die AFM-Studien an M13-sekretierenden Zellen
diente eine transfizierte E. coli DH5α-Flüssigkultur, wie sie schon bei den Unter-
suchungen mit dem Transmissionselektronenmikroskop verwendet wurde. Die AFM-
Messungen in Kapitel 3.2.3 zeigten Zelloberflächen mit filamentösen Strukturen
deren Längen 1,3 – 1,4 µm betrugen und somit vergleichbar mit denen aus der TEM-
Studie waren. Im Vergleich zu den TEM Untersuchungen waren hier die sekretierten
M13-Phagen nicht nur auf die Zellpole beschränkt. In der AFM-Studie konnten
infizierte Zellen gemessen werden, welche M13-Phagen über die gesamte Zellober-
fläche sekretierten. Dieser Unterschied zu der TEM-Studie war wahrscheinlich ein
Effekt, der auf die probenschonendere Präparationsmethodik für AFM-Messungen
zurück zu führen ist. Während die Proben für die TEM-Untersuchungen, aufgrund der
Antikörper- und Protein-A Nanogold-Markierungen, mehrmaligen Waschschritten
unterzogen wurden, in denen vermutlich sekretierte Phagen auf den Zelloberflächen
abgeschert wurden, konnten diese während der Probenaufbereitung für die AFM-
Messungen durch die weniger aufwändige Präparationsmethode auf den Zellober-
flächen bewahrt werden. Hier wurden die infizierten Zellen maximal 3-mal mittels
Zentrifugation bei geringen Zentrifugalkräften gewaschen, um Salze und Verunreini-

Diskussion
158
gungen aus den Kulturmedien zu entfernen, die sich sonst störend auf die AFM-
Messungen ausgewirkt hätten.
Durch die AFM-Messung uninfizierter E. coli DH5α-Zellen konnte sichergestellt
werden, dass dieser Stamm keine filamentösen Strukturen auf der Zelloberfläche
besaß.
4.2.2 Extrusionsverlauf der M13-Phagen auf der Zelloberfläche von E. coli
Alamethicin und das Ionophor Carbonylcyanid m-Chlorphenylhydrazon (CCCP)
eignen sich zur Konservierung der Stadien infizierter E. coli-Zellen
Mit Hilfe der aus Kap. 3.1 erhaltenen Informationen über den Infektions- bzw. Sekre-
tionsverlauf sollte der Extrusionsverlauf der M13-Phagen direkt auf der Zellober-
fläche von E. coli XL1-blue, in Abhängigkeit von der Zeit nach erfolgter Infektion, mit
dem AFM gemessen und visualisiert werden. Aus den Ergebnissen, welche für den
wildtypischen Sekretionsverlauf in Kapitel 3.1.1 erhalten wurden, ging hervor, dass
die Sekretion erster Phagen-Nachkommen zwischen der 5. und 6. Minute nach
Rekultivierung der infizierten Zellen bei 37°C erfolgte. Zwischen der 8. und 10.
Minute fand die exponentielle Vermehrung der Phagen statt. Dies zeigte, dass die
Sekretion neuer Phagen aus den E. coli-Zellen ein sehr schnell ablaufender Prozess
war. Damit jedoch eine zeitliche Abfolge der einzelnen Extrusionsstadien nach
Infektion mit dem AFM gemessen werden konnte, wurde zu den, nach Infektion und
Rekultivierung, aus der E. coli XL1 Kultur entnommenen Proben Alamethicin und
Carbonylcyanid m-Chlorophenylhydrazon (CCCP) zugegeben. Bei diesen Sub-
stanzen handelt es sich um Ionophore, die in der Zytoplasmamembran von E. coli
Poren bilden (Jones et al., 1980, Fox & Richards, 1982), welche die protonen-
motorische Kraft rasch zusammenbrechen lassen (Cavari et al., 1966, Gášková et
al., 1999). Hierdurch war die Möglichkeit gegeben, eine Konservierung der zellulären
Oberflächen von E. coli-Zellen zum Probenentnahmezeitpunkt zu erhalten, da der
Sekretionsprozess von M13 ATP-abhängig ist (Feng et al., 1997). Für diesen
Versuch wurde der E. coli-Stamm XL1-blue verwendet, da dieser F-Pili besitzt, über
welche die Zellen mit M13 infiziert werden können. Hierdurch konnte der wildtypische
Infektionsverlauf aus Kap. 3.1.1 auf diesen Bakterienstamm übertragen werden. Die
F-Pili der Zellen wirkten sich hier nicht störend auf die AFM-Messungen aus, da
diese nach M13-Infektion nicht mehr ausgebildet werden (O´Callaghan et al., 1973).
Im Vergleich zu E. coli K38 aus Kap. 3.1.1, besitzt E. coli XL1-blue keine, durch ihr

Diskussion
159
Aussehen und Längen störenden, Fimbrien. Wie aus der AFM-Messung uninfizierter
E. coli XL1-blue Zellen in Kapitel 3.2.4.1 hervorging, waren auf den Oberflächen der
Bakterien keinerlei filamentöse Strukturen zu erkennen. Die Abwesenheit der F-Pili
könnte hier eine Folge der Probenpräparation sein. Während der Präparation
könnten durch die einzelnen Waschschritte Abscherungen der labilen F-Pili verur-
sacht worden sein.
Phagensekretion beginnt nach 5 Minuten an den Zellpolen von E. coli
Aus den AFM-Messungen der Proben in Kapitel 3.2.4.1 ging hervor, dass nahezu in
Übereinstimmung mit der „one-step growth“ Kurve aus Kapitel 3.1.1 nach 5 Minuten
erste filamentöse Strukturen an den Zellpolen zu sehen waren. Da diese Strukturen
in Aussehen und Länge (1 – 1,4 µm) den Phagenstrukturen der elektronenmikros-
kopischen Aufnahmen in Kapitel 3.2.2 und der AFM-Messungen M13-infizierter
E. coli DH5α-Zellen in Kapitel 3.2.3 entsprechen, kann davon ausgegangen werden,
dass es sich auch bei den hier am Zellpol visualisierten filamentösen Strukturen um
M13-Phagen handelt. Eine zusätzliche Identifikation der Strukturen über eine
Immunogoldmarkierung, wie es für die TEM-Studien in Kapitel 3.2 angewendet
wurde, wäre hier aufgrund des AFM-Funktionsprinzips sowie dem zu geringen
Größenunterschied zwischen den Nanogoldpartikeln und dem Hintergrund des AFM-
Probenträgers, welcher durch auskristallisierende Salze verursacht wird, nicht
möglich gewesen.
Phagensekretion breitet sich innerhalb von 9 Minuten über die gesamte
Zelloberfläche aus
In der Zeit zwischen der 6. und 7. Minute nahm die Anzahl der Phagen zu den
longitudinalen Seiten der Zelle hin zu. Ab der 8. Minute zeigte die „one-step growth“
Kurve in Kapitel 3.1.1 den Beginn der exponentiellen Phagenvermehrung. Die AFM-
Messungen der entsprechenden Proben ab der 8. Minute zeigten, dass die Sekretion
der Phagen-Nachkommen nun nicht mehr nur auf die Zellpole beschränkt war,
sondern sich jetzt über die gesamte Zelloberfläche erstreckte. Innerhalb der 10. bis
14. Minute steigerte sich exponentiell die Anzahl der sekretierten Phagen, so dass
diese über die gesamten Bakterienoberflächen sekretiert wurden. Nach der 14.
Minute waren die Zelloberflächen soweit mit Phagen belegt, dass sich zu den
Messungen der Bakterien zum Zeitpunkt der 16. Minute keine Veränderungen mehr
ergaben. Wie schon aus der wildtypischen „one-step growth“ Kurve hervorging,

Diskussion
160
erfolgt die Sekretion neuer Phagen durch den infizierten Wirt innerhalb eines kurzen
Zeitraums. Die AFM-Messungen zeigten, dass die Sekretion zunächst an den
Zellpolen beginnt und sich daraufhin sehr schnell und mit zunehmender Anzahl über
die gesamte Zelloberfläche hin ausbreitet. Der Anlass für den Beginn der
Phagensekretion an den Zellpolen ist noch unklar. In der Arbeit von Edgar et al.
(Edgar et al., 2008) konnte mittels Quantum-Dot markierter Phagen gezeigt werden,
dass sowohl nach der Bindung von T4 an die äußeren Membranproteine OmpF und
OmpC (Hashemolhosseini et al., 1994) als auch nach der Bindung des Phagen λ an
den Maltoporinrezeptor LamB (Ryter et al., 1975) die adsorbierten Phagen zu den
Zellpolen transportiert werden und dort die Aufnahme der Phagen-DNA in die Zellen
erfolgt. Außerdem konnten sie durch Fluoreszenzmarkierung des mannosespezi-
fischen Enzyms IIC ManY, welches bei der Aufnahme der λ-Phagen-DNA essenziell
ist (Scandella & Arber, 1974), und der Protease FtsH, welche über den lysogenen
oder lytischen Weg des Phagen λ bestimmt (Herman et al., 1997), zeigen, dass diese
Proteine an den Zellpolen lokalisiert sind. M13 hingegen nutzt für die Infektion von
E. coli-Zellen den F-Pilus (Marvin & Hohn, 1969). Nach Adsorption an den F-Pilus
verkürzt sich dieser (Jacobson, 1972), wodurch der Phage mit dem bakteriellen
TolQRA in der Zytoplasmamembran in Kontakt kommt, welches für die Aufnahme der
Phagen-DNA in die Zelle verantwortlich ist (Click & Webster, 1997, 1998). TolQRA ist
für die Aufnahme des bakteriellen Colicin A in die Zytoplasmamembran verant-
wortlich (Davies & Reeves, 1975a, 1975b), in der es spannungsabhängige Kanäle
bildet (Bourdineaud et al., 1990). Die Aufnahme des Colicin A erfolgt über die
Bindung an BtuB und OmpF (Cavard & Lazdunski, 1981) in der äußeren Membran
von E. coli. Aus den Ergebnissen von Edgar, Cavard und Lazdunski könnte
geschlossen werden, dass das bakterielle TolQRA, welches das OmpF gebundene
Colicin A in die Membran transloziert, sich ebenfalls am Zellpol befindet und daraus
resultierend die Aufnahme der M13-DNA auch dort erfolgt. Von Trenkner et al. und
Smilowitz (Trenkner et al., 1967, Smilowitz, 1974) konnte gezeigt werden, dass bei
einer M13-Infektion die in die Zytoplasmamembran inserierten Hüllproteine wieder in
neugebildete Phagen eingebaut werden. Dies bedeutet, dass durch die in der
Zytoplasmamembran verbleibenden Hüllproteine aus der Infektionsphase eine
anfänglich präferierte Sekretionsstelle an den Zellpolen vorhanden wäre und die
Sekretion hierdurch an den Zellpolen beginnen könnte.

Diskussion
161
4.3 Funktionelle Präsentation antigener Epitope am N-Terminus des „Minor
Coat“ Proteins gp9
Insertion antigener Epitope am N-Terminus von gp9
Das „Minor Coat“ Protein gp9 ist mit 32 Aminosäuren eines der kleinsten von
Ribosomen translatierten Proteine (Russel et al., 2004). Verglichen mit dem „Minor
Coat“ Protein gp3 und dem Haupthüllprotein gp8 wird gp9 ohne N-terminale
Signalsequenz (Kuhn et al., 1986, Peters et al., 1994) synthetisiert. Von Endemann
und Model (Endemann & Model, 1995) konnte gezeigt werden, dass eine an die
N-terminale Domäne inserierte Glutathion-S-Transferase (GST) als antigenes Epitop
verwendet werden kann. Jedoch war das über 35 kDa schwere gp9-GST-Fusions-
protein aufgrund seiner Größe nicht geeignet, einen M13-Phagen mit Ambermutation
in Gen 9 in trans zu komplementieren. Während die Translokation des „Minor Coat“
Proteins gp3 über die Translokase SecYEG (Thie et al., 2008) und die des gp8 über
YidC (Samuelson et al. 2000) erfolgt, ist der Translokationsweg des gp9 in die
Zytoplasmamembran noch nicht eindeutig dargelegt worden. In der Arbeit von
Houbiers (Houbiers et al. 2001), in welcher die Translokation der gp9 Proteine in vitro
in künstlich hergestellte Liposomen aus DOPC (1,2-Dioleoyl-sn-glycero-3-phospho-
cholin) und DOPG (1,2-Dioleoyl-sn-glycero-3-phosphoglycerol) untersucht wurde,
wurde die These aufgestellt, dass dieses Protein aufgrund seiner geringen Größe
spontan in die Membran inseriert. Aus der Arbeit von Endemann und Model
hingegen ging hervor, dass die Fusion eines großen Proteins an die N-terminale
Domäne des gp9 nicht funktional ist. Deshalb sollte ähnlich der Arbeit von Facey und
Kuhn (Facey & Kuhn, 2003), in welcher über N-terminale T7- und HA-Epitope gezeigt
werden konnte, dass das Sensor-Kinaseprotein KdpD unabhängig von den Trans-
lokasen SecYEG und YidC in die Cytoplasmamembran transloziert wird, hier eben-
falls mit Hilfe funktionell präsentierter N-terminaler Epitope am „Minor Coat“ Protein
gp9 die Assemblierung am M13-Phagen verfolgt und die Translokation untersucht
werden. Hierfür wurde das Gen 9 in den Expressionsvektor pMS119EH kloniert und
über die QuikChange® Methode eine MunI-Schnittstelle in die N-terminale Domäne
eingefügt. Über diese Schnittstelle wurden Oligonukleotide inseriert, welche 17, 19,
32, und 36 Aminosäuren lange Peptide kodierten (Abb. 4-2). Auf diese Weise
konnten sowohl T7- als auch HA-Epitope mit unterschiedlichen Längen an die
N-terminale Domäne von gp9 fusioniert werden. Hierdurch sollte der insgesamt
hydrophobe Charakter des N-terminalen Bereichs erhalten bleiben und durch eine

Diskussion
162
Verlängerung der N-terminalen Domäne eine verbesserte Detektion der antigenen
Epitope mit Hilfe spezifischer Antikörper erreicht werden. Des Weiteren erfolgte in
den Konstrukten gp9T7rep, gp9DT7rep, gp9HArep und gp9DHArep die Wiederher-
stellung der wildtypischen Aminosäurenfolge des gp9 (Abb. 4-2) nach den kodier-
enden Sequenzen der inserierten Epitope. Durch die wildtypische Proteinstruktur an
den Enden der Epitope sollte eine eventuelle Interaktion zwischen gp9 und gp7
wiederhergestellt werden. Außerdem wurde vor das Gen 9 der jeweiligen Konstrukte
eine verbesserte „Shine-Dalgarno“-Sequenz (AGGAGG; Shine & Dalgarno 1974)
eingefügt, durch die eine gesteigerte Expression der Fusionsproteine erreicht werden
sollte.
Abb. 4-2: Schematische Darstellung der erzeugten gp9-Konstrukte mit Angabe der zusätzlich inserierten Aminosäuren. Die Anzahl der zusätzlich an die N-terminale Domäne von gp9 inserierten Aminosäuren ist grau hinterlegt. Die dem Wildtyp entsprechende Originalsequenz ist in den rep-Konstrukten schwarz hinterlegt (Ref.: Ploss & Kuhn, 2011)

Diskussion
163
Gp9-Konstrukte mit T7- und HA-Epitopen sind befähigt einen M13-Phagen mit
Ambermutation in Gen 9 in trans zu komplementieren
Die Komplementation stellt eine Methode dar, um die Funktionalität der genetisch
veränderten Proteine während der Phagenreproduktion in vivo zu überprüfen.
Deshalb wurden die hergestellten gp9-Konstrukte in Tabelle 3-5 in Komplemen-
tationsassays dahingehend untersucht, ob die in trans exprimierten Proteine befähigt
waren, an dieser Stelle eine Ambermutation in Gen 9 eines M13-Phagen funktionell
ersetzen zu können.
Im Gegensatz zum Komplementationstest von Endemann und Model (Endemann &
Model, 1995), welcher mit einem gp9-GST-Fusionsprotein durchgeführt wurde, zeigte
sich durch Plaquebildungen in den Komplementationstests in Kapitel 3.3.1, dass
sowohl alle gp9 Mono-Tag- als auch alle gp9 Doppel-Tag-Konstrukte befähigt waren
die Ambermutation des M13am9-Phagen in trans zu ersetzen. Im direkten Vergleich
mit der wildtypischen M13am9-Vermehrung auf dem Suppressorstamm E. coli K37
zeigten die Komplementationen aller untersuchten gp9-Konstrukte eine nahezu
wildtypische Vermehrung. Dass die Komplementationen durch die gp9-Konstrukte
zustande kamen, konnte anhand des Vergleichs zwischen uninduziertem nicht-
Suppressorstamm E. coli K38 mit transformierten pMSg7/9 und dem Komplemen-
tationstest auf E. coli K38 mit induzierten gp9T7- und HA-Konstrukten gezeigt
werden. Die auf dem uninduzierten Bakterienrasen von E. coli K38 + pMSg7/9
gebildeten Plaques, welche bis zu einer Verdünnungsstufe von 103 pfu/ml zu sehen
sind, wurden durch Revertanten und eine geringe Transkriptionsrate des gp9
verursacht und stellen somit den unspezifischen Hintergrund der Komplemen-
tationstests dar. Dass in der Phagenstammlösung von M13am9 Revertanten vor-
handen waren, ging auch aus dem Komplementationstest der M13am9-Phagen auf
dem Bakterienrasen des nicht-Suppressorstamms E. coli K38 hervor, da hier in der
Verdünnungsstufe von 106 pfu/ml eine geringe Plaquebildung zustande kam. Im
Vergleich zu den HA-Tag-Konstrukten konnten nur die T7-Tag-Konstrukte eine
wildtypische Vermehrung bis zur Verdünnungsstufe von 101 pfu/ml erreichen. Die
HA-Tag-Konstrukte zeigten hier eine Vermehrung bis zur Verdünnungsstufe von
102 pfu/ml, was eine Potenz niedriger als bei den T7-Tags ist. Der Grund hierfür ist
unklar, jedoch könnten die negativen Ladungen in den antigenen HA-Epitopen dieser
Konstrukte, welche hier durch vier Aspartate verursacht werden, dafür verantwortlich
sein. Aufgrund dieser Ladungen ist der Translokationsvorgang über die Zytoplasma-

Diskussion
164
membran energieaufwendiger (Facey & Kuhn, 2004). Hierdurch könnte es dazu
kommen, dass mengenmäßig mehr gp9T7-Konstrukte innerhalb einer kurzen Zeit-
spanne in die Membran transloziert werden als gp9HA-Konstrukte, wodurch eine
Limitierung während der Phagensekretion zustande käme und mehr Phagen mit T7-
Konstrukten gebildet werden könnten. Im Gegensatz zu dem von Endemann und
Model (Endemann & Model, 1995) verwendeten antigenen GST-Epitop (Glutathion-
S-Transferase) an der N-terminalen Domäne von gp9 konnten hier, durch die Inser-
tion kürzerer N-terminaler Peptide mit Längen von 17-36 Aminosäuren, nahezu wild-
typische Titer des M13am9-Phagen erhalten werden.
N-terminale T7- und HA-Epitope haben keine negativen Effekte auf die gp9-
Expression
Endemann und Model (Endemann & Model 1995) war es möglich mit Hilfe eines
30 kDa schweren GST-Proteins, welches N-terminal an gp9 fusioniert wurde, mittels
eines spezifischen anti-GST Serums die Expression des plasmidkodierten Fusions-
proteins in E. coli-Zellen durch Western-Blot Analyse nachzuweisen. Vergleichsweise
konnte hier in den Kapiteln 3.3.2 und 3.3.3 ebenfalls die Expression des plasmid-
kodierten gp9 mit Hilfe von N-terminal fusionierten antigenen T7- und HA-Epitopen in
E. coli K38 nachgewiesen werden. Die Autoradiogramme zeigten, dass sich sowohl
die gp9 Mono-Tag-Konstrukte mit zusätzlichen 17 (HA-Tag) bzw. 19 (T7-Tag) Amino-
säuren als auch die gp9 Doppel-Tag-Konstrukte mit zusätzlichen 32 (HA-Tag) bzw.
36 (T7-Tag) Aminosäuren in den E. coli-Zellen in trans exprimieren ließen. Durch den
Vergleich des exprimierten gp9T7repSD-Konstrukts mit dem Haupthüllprotein gp8,
welches ein Molekulargewicht von 5,2 kDa besitzt, konnte dem T7-Fusionsprotein ein
Molekulargewicht von ~ 5,2 kDa bis 5,5 kDa zugeordnet werden. Aufgrund der
Insertion des antigenen, 19 Aminosäuren langen, T7-Epitops war dieses Konstrukt
~ 1,9 kDa schwerer als das wildtypische gp9, welches laut Literatur nur ~ 3,6 kDa
schwer ist (Rasched & Oberer, 1986). Da die weiteren, über Immunpräzipitation
nachgewiesenen, gp9T7-, HA- und HArep-Mono-Tag-Konstrukte in Kapitel 3.3.2 nur
2 – 4 Aminosäuren weniger besitzen als gp9T7repSD, kann davon ausgegangen
werden, dass auch diese Konstrukte ~ 5,2 kDa bis 5,5 kDa schwer sind.
Im Gegensatz zu den gp9 Mono-Tag-Konstrukten konnte bei den Expressionsstudien
der gp9 Doppel-Tag-Konstrukte mit Hilfe der Immunpräzipitationen in den Autoradio-
grammen jeweils ein weiteres, ~ 2 kDa leichteres, Protein in den Proben der

Diskussion
165
Konstrukte nachgewiesen werden. Eine eindeutige Aussage, um welches Protein es
sich jeweils handelt, kann hier nicht gemacht werden. Eine mögliche Erklärung
könnte jedoch sein, dass durch periplasmatische Proteasen an den verlängerten
N-terminalen Domänen der Konstrukte eine proteolytische Abspaltung eines
antigenen Epitops stattgefunden haben könnte und die Doppel-Tag-Konstrukte so zu
Mono-Tag ähnlichen Formen prozessiert wurden (Abb. 4-3). Anhand der verblie-
benen antigenen Epitope könnten durchaus mit Hilfe der Immunpräzipitation Mono-
Tag-Formen nachgewiesen werden. Dies würde bedeuten, dass in den 20 Minuten,
in denen die Expression der Proteine induziert und diese radioaktiv markiert wurden,
ein Abbau der N-terminalen Domäne im Periplasma erfolgte.
N-terminale T7- und HA-Epitope zeigen keine negativen Effekte auf die
Membraninsertion
Die Membraninsertion der gp9-Konstrukte mit N-terminalem T7- bzw. HA-Tag wurde
durch radioaktive Markierungen der in E. coli exprimierten gp9-Konstrukte mit an-
schließenden Protease-Mappings untersucht. Hier konnten in Kapitel 3.3.2 sowohl für
das gp9T7-Mono-Tag- als auch für das gp9HA-Mono-Tag-Konstrukt durch die pro-
teolytische Abspaltung der antigenen Epitope der Einbau der gp9-Konstrukte in die
Zytoplasmamembran nachgewiesen werden. Während aus den Proteinase K behan-
delten Proben keine gp9-Konstrukte durch Immunpräzipitation nachgewiesen werden
konnten, war es möglich gp9-Konstrukte aus den Proteinase K unbehandelten
Abb. 4-3: Schematische Darstellung der proteo-lytischen Prozessierung von gp9 Doppel-Tag- zu gp9 Mono-Tag-Konstrukten durch periplasmatische
Proteasen.

Diskussion
166
Proben mit Hilfe von T7- und HA-Antiseren durch Immunpräzipitation nachzuweisen.
Die Abspaltungen der Epitope zeigten, dass der N-Terminus der gp9-Konstrukte peri-
plasmatisch lokalisiert und für Proteasen zugänglich war. Zudem belegte dies, dass
die Insertion eines antigenen Epitops mit zusätzlichen negativen Ladungen in die
N-terminale Domäne von gp9, wie im Fall des HA-Tags, keine unmittelbaren Auswir-
kungen auf die Membraninsertion hatte und somit für die Assemblierung neuer M13-
Phagen zur Verfügung stehen sollte. Die Unversehrtheit der hergestellten Sphero-
plasten in den durchgeführten Protease-Versuchen ging aus dem proteolytisch
unprozessierten GroEL und die proteolytische Aktivität der Proteinase K aus dem
Abbau des OmpA hervor.
N-terminal inserierte Epitope können mit Hilfe spezifischer Antikörper am M13-
Phagen detektiert werden
In Komplementationstests (Kap. 3.3.1), Expressionsnachweisen (Kap. 3.3.2, 3.3.3)
und Membraninsertionsnachweisen (Kap 3.3.4) konnte gezeigt werden, dass die
hergestellten gp9-Konstrukte funktionsfähig waren und theoretisch für die Phagen-
assemblierung in der Zytoplasmamembran zur Verfügung standen. Durch Infektion
der konstruktexprimierenden E. coli K38-Zellen mit M13am9 wurden, in Folge der
Substitution des wildtypischen gp9 in trans durch T7-, HA-Mono- bzw. T7-, HA-
Doppel-Tag-gp9, Phagen hergestellt, welche theoretisch die gp9-Konstrukte in der
Phagenhülle aufweisen sollten. Nach Titration der Phagenstammlösungen konnte mit
Hilfe von Dot-Blot Analysen, T7 und HA spezifischer Antikörper, der Nachweis der
antigenen Epitope am gp9 in den einzelnen Tag präsentierenden Phagen mittels
Chemolumineszenz nachgewiesen werden. Die ECL-Signale auf den Röntgenfilmen
bestätigten, dass die hier in gp9 inserierten Peptide durch Antikörper detektierbar
waren und die gp9-Konstrukte während der Assemblierung neuer M13-Phagen an
Stelle des wildtypischen gp9 eingebaut wurden. Der Vergleich der Dot-Blot Analysen
von Mono- und Doppel-Tag präsentierenden Phagen in Kapitel 3.3.5.1 zeigte, dass
die Detektion der antigenen Epitope der gp9 Doppel-Tag-Phagen besser war, da
noch bei der geringen Anzahl von 107 Phagen schwache Chemolumineszenz-
Signale nachgewiesen werden konnten. Bei den Mono-Tag-Konstrukten konnten
Chemolumineszenz-Signale nur bis 108 Phagen detektiert werden. Wegen der
Präsenz von zwei antigenen Epitopen besteht wahrscheinlich bei den Doppel-Tag-
Konstrukten die Möglichkeit, eine verbesserte Detektion durch die Bindung der

Diskussion
167
doppelten Anzahl von Antikörpern zu erreichen. Jedoch stellt die Dot-Blot Analyse
nur eine quantitative Methode dar, wodurch es schwierig ist, qualitative Aussagen zu
treffen. Aufgrund statistischer Pipettierfehler in der Größenordnung von ~ 10 % kann
davon ausgegangen werden, dass nicht alle hergestellten Verdünnungsstufen die
gleichen Phagenmengen besaßen. Während bei den T7-Konstrukten mit der
Zunahme der Epitopanzahl die Intensität der Detektionen an den Virionen abnahm,
konnte diese im Fall der HA-Konstrukte gesteigert werden. Da sich in den Expres-
sionsnachweisen der gp9 Doppel-Tag-Konstrukte in Kapitel 3.3.3 durch die Immun-
präzipitationen der Konstrukte ein weiteres, ~ 2 kDa leichteres, Protein identifizieren
ließ, könnte davon ausgegangen werden, dass diese Konstrukte nicht sehr stabil
sind und während der Assemblierung neuer Phagen die gp9 Doppel-Tag-Konstrukte
durch periplasmatische Proteasen zu Mono-Tag ähnlichen Formen prozessiert und in
die Phagenhülle eingebaut werden. Hierdurch würde sich die Anzahl der antigenen
Epitope und in Folge das ECL-Signal bei einer Dot-Blot Analyse verringern.
Zusätzlich müssen hier auch die unterschiedlichen Nachweisqualitäten der hier
verwendeten Erstantikörper berücksichtigt werden, da beide vermutlich nicht
vergleichbar gut an ihre antigenen Epitope binden.
Der ECL-Signalunterschied zwischen gp8 und den gp9-Konstrukten liegt, außer an
den Erstantikörper-Nachweisqualitäten, vermutlich an der Ratio von 135 gp8 zu
1 gp9 im Phagen (Endemann & Model, 1995). Dadurch werden mehr primäre sowie
Peroxidase gekoppelte sekundäre Antikörper an gp8 gebunden und folglich während
der ECL-Reaktion stärkere Signale erhalten.
Antigene Epitope der gp9-Konstrukte sind am Phagen für Antikörper
zugänglich und ermöglichen die Bindung von Goldnanopartikeln
Die Visualisierung phagenpräsentierter gp9-Tag-Konstrukte erfolgte mit Hilfe von
Protein-A konjugiertem Nanogold und anschließender elektronenmikroskopischer
Untersuchungen. Hier wurden die gp9-Konstrukt präsentierenden Phagen zuerst mit
primären Antikörpern gegen das jeweils vorhandene Epitop und anschließend mit
Protein-A konjugiertem Nanogold inkubiert. Diese Methode des Nachweises
inserierter antigener Epitope wurde ebenfalls von Gao et al. (Gao et al., 1999), nach
Insertion der variablen Region der schweren und leichten Kette eines Kokain-
Antikörpers in die M13-Phagenhüllproteine gp7 und gp9, und von Løset et al. (Løset
et al., 2011), nach Insertion eines Fragments des T-Zell Rezeptors (scTCR) in die

Diskussion
168
M13-Phagenhüllproteine gp7 und gp9, angewendet. Für das Display der antigenen
Epitope verwendeten beide Arbeitsgruppen ein Helferphagensystem, durch welches
ein monovalentes „Phage-Display“ zustande kommt, bei dem weniger als 10 % der
veränderten gp9-Konstrukte in die Phagenhülle inseriert werden (Russel et al., 2004).
Trotz der geringen Insertionsrate konnten beide Arbeitsgruppen durch die Bindung
von Protein-A konjugiertem Nanogold an die Phagenspitze die inserierten Epitope in
den Hüllproteinen gp7 und gp9 nachweisen. Im Vergleich hierzu wurden die hier
konstruierten gp9T7- und HA-Konstrukte, nach Infektion der konstruktexprimierenden
E. coli K38-Zellen mit M13am9, in trans für die Assemblierung bereitgestellt und in
Folge der Substitution des wildtypischen gp9 durch die Konstrukte Phagen herge-
stellt, welche theoretisch ausschließlich gp9-Konstrukte in der Phagenhülle auf-
weisen sollten. Durch die elektronenmikroskopischen Untersuchungen und den
Nachweis polar gebundener Nanogoldpartikel an den jeweiligen gp9T7- und HA-
Konstrukt präsentierenden Phagen konnte gezeigt werden, dass die Präsentation
antigener Epitope am „Minor Coat“ Protein gp9 auch unabhängig von einer zusätz-
lichen N-terminalen Leadersequenz und eines Helferphagensystems erfolgen kann.
4.3.1 Die Translokation des Konstrukts gp9T7SD erfolgt durch die Membran-
insertase YidC
Für das Sensor-Kinaseprotein KdpD konnte von Facey und Kuhn (Facey & Kuhn,
2003) mit Hilfe eines N-terminal lokalisierten HA-Epitops nachgewiesen werden, dass
KdpD unabhängig von den Translokasen Sec und YidC in die Zytoplasmamembran
von E. coli transloziert wird. Vergleichbar wie für KdpD, sollte auch hier für das „Minor
Coat“ Protein gp9 die Translokation des Proteins in Abhängigkeit von YidC anhand
eines T7-Epitops in der N-terminalen Domäne untersucht werden. Hierfür erfolgte die
Expression des Konstrukts gp9T7SD in E. coli JS7131 in Abwesenheit von YidC, was
durch eine Western-Blot Analyse bestätigt wurde. Anhand eines „Pulse“-Experiments
und Protease-Mappings konnte gezeigt werden, dass in Abwesenheit von YidC keine
Abspaltung des N-terminalen T7-Epitops durch Proteinase K erfolgte, wodurch sich
gp9T7 durch Immunpräzipitation nachweisen ließ. Im Vergleich hierzu erfolgte in
Anwesenheit von YidC der Abbau des N-terminalen T7-Epitops durch Proteinase K,
wodurch gp9T7 durch Immunpräzipitation nicht mehr nachweisbar war. Dies zeigte,
dass die Translokation von gp9T7SD, ebenso wie die des M13-Haupthüllproteins
gp8 (Samuelson et al., 2001, Stiegler et al., 2011), von YidC abhängig ist. Die

Diskussion
169
Unversehrtheit der im Protease-Mapping hergestellten Spheroplasten ging aus dem
unverdauten GroEL und die proteolytische Aktivität der Proteinase K aus dem
abgebauten OmpA hervor. Von Houbiers et al. (Houbiers et al., 2001) wurden mit
Hilfe überexprimiertem und anschließend gereinigtem gp9 in vitro Insertionsstudien
durchgeführt. Nach einer Reinigung des Proteins aus E. coli erfolgte die Inkubation
von gp9-Molekülen mit künstlich hergestellten Lipid-Monolayern aus DOPC und
DOPG. Die Insertion von gp9 in die Lipidschichten wurde über die Veränderung des
Oberflächendrucks dieser Monolayer verfolgt. Aufgrund der Beobachtung einer
Druckzunahme auf den Oberflächen der Monolayer nach Zugabe von gereinigtem
gp9, postulierte diese Arbeitsgruppe, dass gp9 wegen seiner geringen Größe
spontan in Lipidmembranen inseriert und dies wohl auch in vivo der Fall sein könnte.
4.4 Massenbestimmung des gp1/11-Assembly-Komplexes
Die Phagenassemblierung stellt einen sekretorischen Prozess an der inneren E. coli-
Membran dar. Dabei entsteht der Phagenpartikel direkt durch die Umhüllung der
einzelsträngigen Phagen-DNA mit spezifischen Phagenhüllproteinen, welche nach
ihrer Synthese in die innere Membran transloziert und über den, für die Assem-
blierung essenziellen, gp1/11-Assembly-Komplex herangeführt werden (Silver &
Wickner, 1980, Rapoza & Webster, 1995, Marvin 1998). Das Leseraster des Gens 1
kodiert für zwei Genprodukte, gp1 und gp11 (Russel & Model 1989, Guy-Caffey et
al., 1992, Russel 1993). Die Synthese von gp1 und gp11 erfolgt über eine polycis-
tronische mRNA ohne N-terminale Signalsequenz. Die in der inneren Membran
lokalisierten Genprodukte sind essenziell für die Assemblierung des Phagen. Von
Russel (Russel, 1991) konnte gezeigt werden, dass der zytoplasmatische Bereich
des gp1 ein Nukleotid-Bindemotiv besitzt und von Feng (Feng et al., 1997), dass die
Phagenassemblierung ATP benötigt. Weiter konnte Kazmierczak (Kazmierczak et al.,
1994) nachweisen, dass die periplasmatisch lokalisierten Termini von gp1 und gp4
miteinander interagieren. Über diesen 2-Komponenten-Komplex werden die neu
gebildeten M13-Phagen aus den E. coli-Zellen in die äußere Umgebung sekretiert
(Rapoza & Webster, 1995, Haigh & Webster 1999). Eine weitere und essenzielle
Komponente des gp1/11-Assembly-Komplexes ist das E. coli-Protein Thioredoxin
(Russel & Model, 1985, 1986), dessen Rolle im Assembly-Komplex noch ungeklärt
ist.

Diskussion
170
Gp1 und gp11 liegen im Verhältnis von ~ 1:1,4 in exprimierenden E. coli C41-
Zellen vor
Damit ausreichende Mengen an gp1/11-Assembly-Komplexen für eine Quantifi-
zierung ihrer Masse und ihrer anschließenden Visualisierung mittels TEM erhalten
werden konnten, wurde das Gen I aus dem Genom des M13-Phagen mittels PCR
amplifiziert und in den Vektor pET-16b kloniert. Die Expression erfolgte anschließend
in E. coli C41-Zellen. Mit Hilfe eines, gegen die C-terminale Domäne gerichteten,
Antikörpers ließ sich die Expression von gp1 und gp11 in den Zellen durch eine
Western-Blot Analyse nachweisen. Hier konnte ein Protein mit einer Masse von
~ 39 kDa als gp1 und ein weiteres mit einer Masse von ~ 12 kDa als gp11 identifiziert
werden, die mit den Massen übereinstimmen, die Guy-Caffey für die Genprodukte 1
und 11 des filamentösen Phagen f1 aus infizierten E. coli-Zellen beschrieben hatte
(Guy-Caffey et al., 1992). Ein anschließender Versuch einer Quantifizierung eines
Western-Blots zur Ermittlung des Expressionsverhältnisses von plasmidexprimiertem
gp1 und gp11 in E. coli C41 durch den Vergleich von drei gp1-Banden mit der von
drei gp11-Banden ergab, dass über das pET-Vektor-System ein gemitteltes
Verhältnis von einem gp1 zu ~ 1,5 gp11 (Abb. 4-4) in den Zellen vorlag. Jedoch ist
bei solch einer Quantifizierung zu beachten, dass allgemein die erzeugten Signale in
antikörpervermittelten Proteindetektionen von den Qualitäten und Spezifitäten der
verwendeten Erst- und Zweitantikörper abhängig sind und hierdurch nur beschränkt
aussagekräftige Ergebnisse erhalten werden. Dass es sich in der Western-Blot
Analyse um gp1 und gp11 handelte, ging aus der antikörperspezifischen Detektion
und aus dem direkten Vergleich mit den Negativkontrollen hervor. Guy-Caffey et al.
(1992) konnten für f1-infizierte E. coli-Zellen anhand einer Western-Blot Analyse
zeigen, dass hier die Proteine ein Verhältnis von einem gp1 zu 1 - 2 gp11 in den
Zytoplasmamembranen aufwiesen. Dies zeigt, dass auch in einem E. coli-
Expressionssystem, in welchen gp1/11 über ein pET-Vektor-System unabhängig von
anderen M13-Proteinen exprimiert wird, die Proteine in nahezu wildtypischem
Verhältnis synthetisiert werden können. Bei den durchgeführten gp1/11-Expressions-
experimenten in Kap. 3.5.1 war außerdem aus den Western-Blot Analysen
ersichtlich, dass sich die Mengen von gp1 und gp11 nach zweistündiger Expression
verringerten. Diese Mengenminderung ist wahrscheinlich ein Effekt, der auf eine
Hemmung des Bakterienwachstums durch die verstärkte Expression des gp1/11-
Assembly-Komplexes zurückzuführen ist, wie dies auch schon von Horabin &
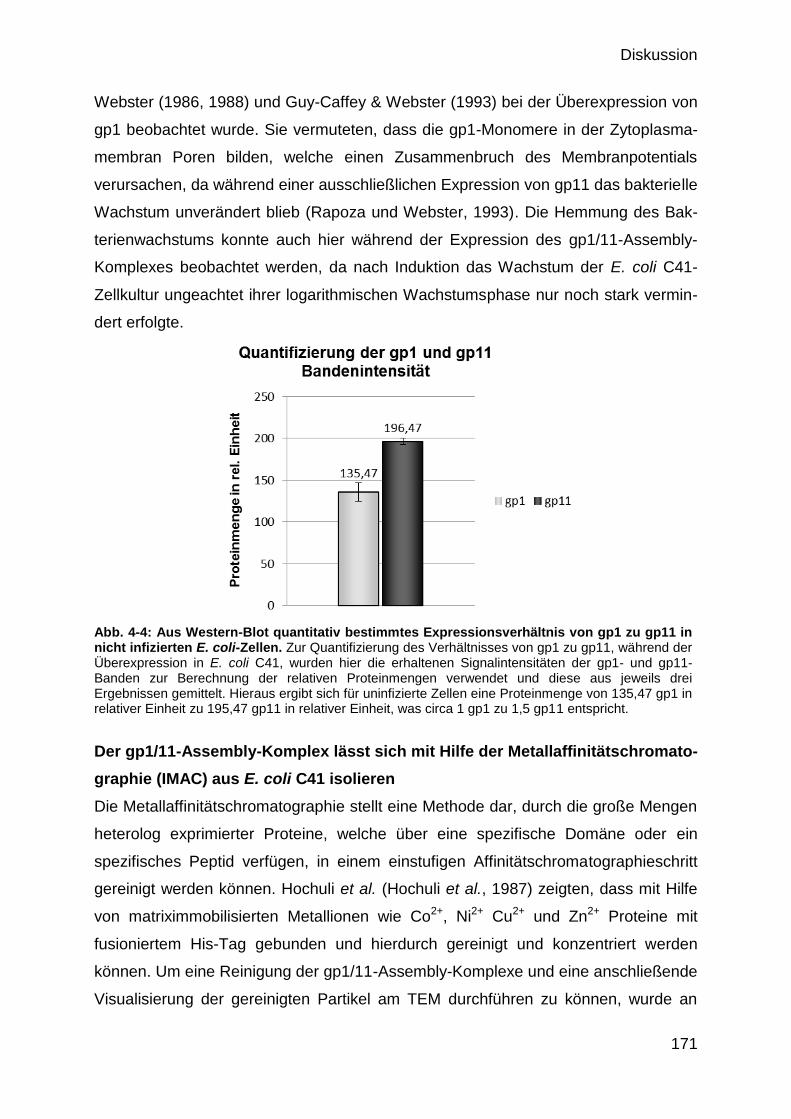
Diskussion
171
Webster (1986, 1988) und Guy-Caffey & Webster (1993) bei der Überexpression von
gp1 beobachtet wurde. Sie vermuteten, dass die gp1-Monomere in der Zytoplasma-
membran Poren bilden, welche einen Zusammenbruch des Membranpotentials
verursachen, da während einer ausschließlichen Expression von gp11 das bakterielle
Wachstum unverändert blieb (Rapoza und Webster, 1993). Die Hemmung des Bak-
terienwachstums konnte auch hier während der Expression des gp1/11-Assembly-
Komplexes beobachtet werden, da nach Induktion das Wachstum der E. coli C41-
Zellkultur ungeachtet ihrer logarithmischen Wachstumsphase nur noch stark vermin-
dert erfolgte.
Der gp1/11-Assembly-Komplex lässt sich mit Hilfe der Metallaffinitätschromato-
graphie (IMAC) aus E. coli C41 isolieren
Die Metallaffinitätschromatographie stellt eine Methode dar, durch die große Mengen
heterolog exprimierter Proteine, welche über eine spezifische Domäne oder ein
spezifisches Peptid verfügen, in einem einstufigen Affinitätschromatographieschritt
gereinigt werden können. Hochuli et al. (Hochuli et al., 1987) zeigten, dass mit Hilfe
von matriximmobilisierten Metallionen wie Co2+, Ni2+ Cu2+ und Zn2+ Proteine mit
fusioniertem His-Tag gebunden und hierdurch gereinigt und konzentriert werden
können. Um eine Reinigung der gp1/11-Assembly-Komplexe und eine anschließende
Visualisierung der gereinigten Partikel am TEM durchführen zu können, wurde an
Abb. 4-4: Aus Western-Blot quantitativ bestimmtes Expressionsverhältnis von gp1 zu gp11 in nicht infizierten E. coli-Zellen. Zur Quantifizierung des Verhältnisses von gp1 zu gp11, während der Überexpression in E. coli C41, wurden hier die erhaltenen Signalintensitäten der gp1- und gp11-Banden zur Berechnung der relativen Proteinmengen verwendet und diese aus jeweils drei Ergebnissen gemittelt. Hieraus ergibt sich für uninfizierte Zellen eine Proteinmenge von 135,47 gp1 in relativer Einheit zu 195,47 gp11 in relativer Einheit, was circa 1 gp1 zu 1,5 gp11 entspricht.

Diskussion
172
den N-Terminus von gp1 ein His-Tag aus zehn Histidinen fusioniert. Die Reinigung
der gp1/11-Assembly-Komplexe erfolgte zunächst aus einer 2 L und anschließend
aus einer 4 L E. coli C41 Batch-Kultur. In den durchgeführten Reinigungen wurden
die gp1/11-Assembly-Komplexe mit Hilfe des zwitterionischen Detergenzes Fos-
cholin-12 (n-Dodecylphosphocholin) aus den Membranen solubilisiert.
Im Verlauf der Reinigungen wurden Proben entnommen und anschließend über
SDS-PAGE aufgetrennt, um den Verbleib der gp1/11-Assembly-Komplexe über-
prüfen zu können. Die mit Coomassie gefärbten SDS-Gele in Kapitel 3.4.2 zeigten
durch die Präsenz mehrerer Proteine in den Elutionsfraktionen, dass sowohl aus der
2 L als auch aus der 4 L E. coli C41-Batch-Kultur, nach zweistündiger gp1/11-
Expression, mit Hilfe der Metallaffinitätschromatographie Assembly-Komplexe gerei-
nigt werden konnten. Mit Hilfe eines spezifischen Antikörpers gegen die C-terminale
Domäne von gp1 konnte durch Western-Blot Analyse sowohl in den solubilisierten,
den ultrazentrifugierten als auch in den Elutionsproben gp1 nachgewiesen werden.
Mit 39 kDa Molekulargewicht war das gp1 um ~ 1 kDa schwerer als das wildtypische
gp1. Diese Abweichung kann vermutlich auf den fusionierten 10 x His-Tag zurück-
geführt werden. Von Feng et al. wurde nach der Fusion eines His-Tags an die
C-terminale Domäne von gp1 eine ähnliche Veränderung des apparenten Molekular-
gewichtes nach erfolgter SDS-PAGE beobachtet (Feng et al., 1999).
In der aus 2 L Bakterienkultur durchgeführten IMAC-Reinigung konnte mit Hilfe einer
Western-Blot Analyse, unter Verwendung des gp1 Antikörpers gegen die C-terminale
Domäne, außerdem in der solubilisierten Probe und der Überstandsprobe nach
Zentrifugation das ~ 12 kDa schwere gp11 nachgewiesen werden. In der Elutions-
fraktion war jedoch kein gp11 detektierbar. Dieser Verlust des gp11 könnte durch
eine Störung der intramolekularen Wechselwirkung zwischen gp1 und gp11, verur-
sacht durch das Detergenz Fos-Cholin-12, oder durch eine mangelhafte Detektion
des gp11 durch den verwendeten Erstantikörper erklärt werden. Zudem zeigte die
Probe des Säulendurchlaufs, dass ein großer Anteil der gp1/11-Assembly-Komplexe
nicht gebunden werden konnte und hierdurch eine ~ 50 %ige Verringerung der
Gesamtproteinmenge zustande kam.
Mit der anschließenden Erhöhung der Bakterienkultur auf 4 L konnte in der darauf-
folgenden IMAC-Reinigung die Menge an gebundenen und eluierten gp1/11-
Assembly-Komplexen gesteigert werden. Während in der Elutionsfraktion der 2 L
Reinigung kein gp11 detektierbar war, konnte in der IMAC-Elutionsfraktion der 4 L

Diskussion
173
Reinigung deutlich ein Protein mit einem Molekulargewicht von ~ 12 kDa identifiziert
werden. Weiterhin konnten in dieser Elutionsfraktion Proteine mit Molekulargewichten
von ~ 39 kDa, ~ 37 kDa, ~ 31 kDa und ~ 20 kDa nachgewiesen werden. Feng et al.
konnten nach IMAC-Reinigungen von plasmidexprimierten gp1 mit C-terminal fusio-
nierten His-Tags gp1-Abbaufragmente mit ~ 30 kDa und ~ 31 kDa (Feng et al.,
1999), unter Verwendung eines spezifischen Antikörpers gegen die N-terminale
Domäne von gp1, durch Western-Blot Analyse nachweisen und mit Hilfe eines
spezifischen Antikörpers gegen die C-terminale Domäne nach weiterer Western-Blot
Analyse diese als proteolytische Abbauprodukte der C-terminalen Domäne identi-
fizieren. Im Weiteren war es ihnen möglich, durch den Vergleich einer wildtypischen
mit einer f1-Phageninfektion, bei der kein funktionsfähiges gp4 über das Phagen-
genom gebildet werden konnte, zu zeigen, dass die gp1-Prozessierung durch gp4
verhindert werden konnte. Außerdem konnten sie durch die Expression von plasmid-
kodierten gp1 in Bakterienstämmen, in denen gezielt Proteasegene deletiert wurden,
ausschließen, dass für die Prozessierungen das periplasmatisch lokalisierte OmpT
und DegP verantwortlich war.
4.4.1 Identifikation der Proteine im IMAC-Eluat
Zur Identifikation der koeluierten Proteinbanden im IMAC-Eluat aus Kapitel 3.4.2
wurden diese mit Hilfe von MALDI-TOF/MS (Matrix Assisted Laser Desorption /
Ionization–Time Of Flight / Mass-Spectrometry) analysiert. Durch diese Methode
können unbekannte Proteine nach Trypsinverdau, aufgrund der resultierenden
Peptidfragmentgrößen, identifiziert werden. Die entstehenden Fragmente sind für
jedes Protein spezifisch und ermöglichen durch den hieraus erzeugten Peptid-
Massen-Fingerprint (PMF) einen Vergleich mit den in Datenbanken hinterlegten
PMFs bekannter Proteine, wodurch diese identifiziert und zugeordnet werden
können.
Anhand der durchgeführten MALDI-TOF/MS Analysen der aus dem SDS-Gel
isolierten ~ 37 kDa, ~ 31 kDa und ~ 20 kDa schweren Proteine wurden diese, wie
auch im Anhang zu entnehmen ist, durch Vergleich mit in der Proteindatenbank
„Mascot“ (http://www.matrixscience.com) hinterlegten PMFs, als gp1-Proteine des
filamentösen Phagen M13 identifiziert. Aus den identifizierten Peptidfragmenten ging
hervor, dass es sich um gp1-Abbauprodukte handelte, welche sich nur in der Länge
ihrer C-terminalen Domänen von dem wildtypischen gp1 mit N-terminal fusioniertem

Diskussion
174
His-Tag unterschieden (Abb. 4-5) während die N-terminalen Domänen bei allen drei
zu identifizierenden Proteinen unverändert blieben. Wegen des His-Tags besteht
ebenfalls die Möglichkeit, in dem hier verwendeten Expressionssystem, proteolytisch
verkürzte C-terminale gp1-Fragmente durch eine Metallaffinitätschromatographie zu
reinigen. Aufgrund der verkürzten C-terminalen Domäne konnten wahrscheinlich
diese gp1-Abbauprodukte unter Verwendung des C-terminalen Antikörpers nicht in
den Western-Blot Analysen der IMAC-Reinigung in Kapitel 3.4.2 und der Expres-
sionsanalyse in Kapitel 3.4.1 nachgewiesen werden. Durch einen sukzessiven
proteolytischen Abbau der C-terminalen Domäne des gp1 wäre das antigene Epitop,
gegen das der C-terminale Antikörper gerichtet ist, in kürzeren gp1-Fragmenten nicht
mehr vorhanden und so auch nicht mehr detektierbar.
Die MALDI-TOF/MS Analysen zeigten weiterhin, dass im Vergleich zum wildty-
pischen gp1 mit N-terminalem His-Tag, die C-terminale Domäne des ~ 37 kDa
schweren gp1-Fragments um ~ 15 % und die des ~ 31 kDa schweren gp1-Fragments
um ~ 33 % verkürzt war. Da in der MALDI-TOF/MS Analyse des ~ 20 kDa schweren
gp1-Fragments ein 8 Aminosäuren langer Abschnitt (AS 290 – 297) der C-terminalen
Domäne identifiziert wurde, der nur im Fall des um ~ 15 % verkürzten ~ 37 kDa
schweren gp1-Fragments nachweisbar war, kann hier keine endgültige Aussage
Abb. 4-5: Schematische Darstellung der isolierten und durch MALDI-TOF/MS identifizierten gp1-Fragmente im Vergleich zum His-Tag fusionierten gp1 Wildtyp-Protein. Die Verkürzungen der einzelnen C-terminalen Domänen sind prozentual im Vergleich zum His-Tag fusionierten gp1 Wildtyp-Protein (~ 39 kDa, His-Tag grau hinterlegt) dargestellt. Die Bildung der Fragmente ist wahrscheinlich auf eine proteolytische Prozessierung der C-terminalen Domäne durch periplas-matische Proteasen zurückzuführen. Im Fall des ~ 20 kDa schweren Fragments könnte es sich um ein 70 % verkürztes gp1 handeln, denkbar wäre auch eine dimere Form aus prozessiertem gp1 mit ~10 kDa und gp11 mit ~10 kDa.

Diskussion
175
getroffen werden, ob es sich hier wirklich um ein ~ 70 % verkürztes gp1-Fragment
handelt. Eine denkbare Möglichkeit wäre, dass hier eventuell ein Dimer aus
prozessiertem gp1 mit ~ 10 kDa und einem gp 11 mit ebenfalls ~ 10 kDa vorliegt, da
der identifizierte Abschnitt von Aminosäure 290 – 297 sich im Bereich von gp11
befindet (Abb. 4-5).
4.4.2 Massenbestimmung des gereinigten gp1/11-Assembly-Komplexes aus
nicht infizierten E. coli-Zellen durch präparative Gelfiltration
Durch die Größenausschlußchromatographie erfolgt die Auftrennung eines Substrat-
gemisches über eine stationäre Phase aus porösen Polymeren, wie z.B. Sepha-
dexTM, aufgrund des hydrodynamischen Volumens der Moleküle. Hierbei ist die
Effizienz, mit der die einzelnen Moleküle getrennt werden, von den Oberflächen der
eingesetzten porösen Matrizes abhängig. Mixed-Bed-Säulen, in denen Polymere mit
verschiedenen Porengrößen als stationäre Phase eingesetzt werden, trennen über
einen großen Massenbereich linear mit dem hydrodynamischen Volumen. Die hier
zur Massenbestimmung verwendete SuperdexTM 200 Matrix ermöglicht die Auftren-
nung von Stoffgemischen mit Massen von 10 kDa bis 600 kDa. Durch die Verwen-
dung von Proteinstandards mit geringen Molmassenverteilungen kann eine Kali-
brierung solcher Mixed-Bed-Säulen erfolgen. Aus der erhaltenen logarithmischen
Kalibrierungskurve lassen sich die Molmassen berechnen, da das Produkt von
Molmasse und intrinsischer Viskosität proportional zum hydrodynamischen Radius
der Moleküle ist.
Der gp1/11-Assembly-Komplex hat ein gemitteltes Molekulargewicht von circa
300 kDa
Mit Hilfe der Größenausschlußchromatographie konnte in Kapitel 3.4.3 die gp1/11-
Elutionsfraktion aus der Metallaffinitätschromatographie in Fraktionen mit unter-
schiedlichen Massen und Konzentrationen aufgetrennt werden. Durch die vorherige
Kalibrierung der Säule konnten den eluierten, detergenzassoziierten gp1/11-
Assembly-Komplexen entsprechend quantitativ Massen zugewiesen werden. Für das
verwendete Detergenz Fos-cholin 12 wurde für die gebildeten Detergenzmizellen,
unter Einberechnung der substratspezifischen Aggregationszahl, ein Molekular-
gewicht von ~ 21 kDa errechnet. Unter Berücksichtigung der Detergenzmizelle war
es möglich, dem eluierten Molekülkomplex ein gemitteltes reines Gewicht von
~ 300 kDa zuzuordnen.

Diskussion
176
Die Identifikation der Proteinkomponenten in Kapitel 3.4.3 mittels SDS-PAGE ergab,
dass sich die Elutionsfraktion der Größenausschlußchromatographie, welche die
Molekülkomplexe mit Massen von 330 – 270 kDa beinhaltete, überwiegend aus
Proteinen mit den Massen von ~ 39 kDa und ~ 37 kDa und geringfügig aus Proteinen
mit ~ 31 kDa und ~ 12 kDa zusammensetzte. In MALDI-TOF/MS Analysen wurden
die ~ 37 kDa und ~ 31 kDa schweren Proteine als gp1-Fragmente identifiziert,
welche sich vermutlich durch proteolytischen Abbau in der Länge ihrer C-terminalen
Domänen unterscheiden. Aufgrund der geringen Anteile dieser prozessierten gp1-
Fragmente im eluierten Molekülkomplex kann hier wahrscheinlich davon ausge-
gangen werden, dass in dieser durchgeführten Größenausschlußchromatographie,
unter Berücksichtigung der Detergenzmizelle, gp1/11-Assembly-Komplexe mit
Massen zwischen ~ 330 kDa - 270 kDa eluiert wurden. Hieraus lässt sich ein
gemitteltes reines Gewicht von ~ 300 kDa für den gp1/11-Assembly-Komplex
bestimmen.
Feng et al. (1999) konnten mit Hilfe eines Vergleichs von gp1-Proteinen aus einer mit
f1-Phagen-infizierten E. coli-Kultur, bei der kein funktionsfähiges gp4 über das
Phagengenom gebildet werden konnte, mit einer wildtypisch verlaufenden Infektion
in E. coli zeigen, dass eine Prozessierung des gp1 durch gp4 unterbunden werden
konnte. In der hier nach IMAC-Reinigung der überexprimierten gp1/11-Assembly-
Komplexe durchgeführten Größenausschlusschromatographie in Kapitel 3.4.3
konnten in den eluierten Komplexen durch SDS-PAGE gp1-Moleküle mit unter-
schiedlichen Massen identifiziert werden. Hier könnte es sich vermutlich um einen
gp1/11-Assembly-Komplex handeln, in dem, aufgrund des fehlenden gp4 in der
äußeren Membran von E. coli, die gp1-Moleküle nach Translokation in die Zyto-
plasmamembran durch periplasmatische Proteasen C-terminal prozessiert wurden.
Weiterhin war zu beobachten, dass in der Fraktion mit Proteinkomplexen von
145 kDa circa die 4-fache und in der mit 74 kDa circa die 10-fache Menge der zuvor
identifizierten Proteinkomponenten eluiert wurden. Dies könnte ein Hinweis sein,
dass der gp1/11-Assembly-Komplex selbst bei Funktionslosigkeit oder eventuell nach
einem kurzen Intervall einem proteolytischen Abbau unterliegt.

Diskussion
177
4.4.3 Massenbestimmung des gereinigten gp1/11-Assembly-Komplexes aus
infizierten E. coli-Zellen durch präparative Gelfiltration
Für die Assemblierung und die Sekretion neuer Phagen-Nachkommenschaften
werden drei, durch den Phagen kodierte, Proteine, gp4 (Brissette und Russel, 1990),
gp1 und gp11 (Guy-Caffey et al., 1992, Rapoza & Webster, 1995) sowie das von
E. coli synthetisierte Thioredoxin benötigt (Russel & Model, 1985; 1986). Zur Identi-
fikation neuer Proteinkomponenten am Assembly-Komplex und zur Verringerung des
proteolytischen gp1-Abbaus durch periplasmatische Proteasen (Feng et al., 1999)
wurde eine Reinigung des Assembly-Komplexes aus einer mit M13mp19-infizierten
E. coli K37 Batch-Kultur durchgeführt.
Durch Infektion gp1/11-exprimierender Zellen entstehen gp1/11-Hybrid-
komplexe
Der Vergleich der über SDS-PAGE aufgetrennten Elutionsproben aus nicht infizierten
E. coli-Zellen und infizierten E. coli-Zellen zeigt, dass trotz Infektion weiterhin die
gleichen Abbauprodukte in der Elution der Metallaffinitätschromatographie vorhan-
den waren. Auffällig ist jedoch, dass das IMAC-Elutionskonzentrat aus der gp1/11-
Reinigung der infizierten Kultur nur geringe Mengen an gp1 mit ~ 39 kDa aufweist,
aber dafür große Mengen an gp1 mit ~ 37 kDa. Durch eine zusätzliche Infektion der
gp1-His-Tag exprimierenden E. coli K37-Zellen mit M13-Phagen kommt es zur
Bildung von gp1/11-Hybrid-Assembly-Komplexen, welche sich aus wildtypischen und
His-Tag präsentierenden gp1-Proteinen zusammensetzen. Diese lassen sich an-
schließend ebenso über die Metallaffinitätschromatographie isolieren und reinigen.
Durch die Infektion und die hieraus entstehende additive Expression des gp1
verringert sich jedoch die Anzahl der His-Tag präsentierenden gp1-Moleküle mit
einem Molekulargewicht von ~ 39 kDa. Gleichzeitig kommt es in den Komplexen zu
einer Anreicherung des wildtypischen gp1, welches ein Molekulargewicht von
~ 37 kDa besitzt. Trotz der Infektion werden weiterhin die ~ 31 kDa schweren gp1-
Abbauprodukte gebildet. Eine Ursache hierfür könnte ein stöchiometrisches Un-
gleichgewicht zwischen dem additiv exprimierten gp1 und dem durch Infektion gebil-
deten gp4 sein. Vermutlich ist die bei einer Infektion gebildete Menge an gp4 nicht
ausreichend, damit alle synthetisierten gp1-Moleküle vor einem proteolytischen
Abbau im Periplasma geschützt sind. Dass ein proteolytischer Abbau eines plasmid-
exprimierten gp1 im Periplasma erfolgt, konnte von Feng et al. durch die Identi-

Diskussion
178
fikation eines ~ 31 kDa schweren gp1-Abbauprodukts (Feng et al., 1999) anhand
einer Western-Blot Analyse unter Verwendung spezifischer Antikörper gegen die
N-terminale Domäne von gp1 nachgewiesen werden. Mit Hilfe eines inserierten His-
Tags und einer Ambermutation in der C-terminalen Domäne von gp1 konnten Feng
et al. die proteolytische Spaltung auf den Bereich der Aminosäuren 275 bis 298
einschränken. Da dieser Bereich des gp1 vermutlich durch intermolekulare Wechsel-
wirkungen mit gp4 den gp1/11-gp4-Sekretionskomplex bildet, spekulierten sie, dass
gp1 möglicherweise hierdurch vor dem proteolytischen Abbau im Periplasma ge-
schützt wird.
Eine M13-Infektion gp1/11-exprimierender Zellen erhöht den Schutz des
Assembly-Komplexes vor proteolytischer Prozessierung
Das Chromatogramm der Größenausschlusschromatographie in Kapitel 3.4.4 zeigte,
dass die Reinigung des Elutionskonzentrats aus der IMAC-Reinigung infizierter
Zellen sich wie das Elutionskonzentrat aus der IMAC-Reinigung uninfizierter Zellen
auftrennen ließ. Auch hier ließen sich die gp1/11-Elutionsfraktionen aus der Metall-
affinitätschromatographie in Fraktionen mit unterschiedlichen Massen und Konzen-
trationen auftrennen. Im Vergleich zur Reinigung der gp1/11-Assembly-Komplexe
aus uninfizierten Zellen erfolgte hier die Elution hoher Molekulargewichte von 350 -
290 kDa in einer früheren Fraktion. Im Unterschied zu den uninfizierten Zellen war es
hier durch die Infektion wahrscheinlich möglich, eine Protektion des gp1/11-
Assembly-Komplexes zu bewirken, da die Menge an unprozessierten gp1/11-
Assembly-Komplexen um das circa 40-fache erhöht werden konnte. Dies deutet
darauf hin, dass mit Hilfe einer M13-Infektion von gp1/11-exprimierenden Zellen die
Reinigung zugunsten intakter Assembly-Komplexe verschoben werden konnte. Die
Identifikation der Proteinkomponenten in dieser frühen Elutionsfraktion mittels SDS-
PAGE ergab, dass sich die eluierten Assembly-Komplexe hauptsächlich aus gp1-
Molekülen mit einer Masse von ~ 37 kDa zusammensetzten, was ungefähr der
Masse des wildtypischen gp1 entspricht. Die geringe Präsenz des ~ 39 kDa
schweren gp1 zeigte, dass die hier eluierten Assembly-Komplexe auch gp1-Moleküle
mit His-Tag enthielten. Über diese konnten wahrscheinlich während der Metallaffini-
tätschromatographie die Bindungen der Komplexe an die Ni-NTA-Matrix erfolgen.
Zudem ist dies ein Hinweis, dass bei der hier angewendeten Reinigungsmethode nur
ein geringer Anteil von His-Tag fusionierten gp1 benötigt wird, um größere Mengen

Diskussion
179
an gp1/11-Assembly-Komplexen isolieren zu können. Die außerdem in dieser
Elutionsfraktion identifizierten ~ 31 kDa und ~ 20 kDa schweren gp1-Abbauprodukte
sind vermutlich eine Folge des stöchiometrischen Ungleichgewichts zwischen gp4,
welches durch die M13-Infektion gebildet wurde, und gp1. Durch dieses Ungleich-
gewicht könnte in den gp1/11-exprimierenden Zellen ein Gemisch aus intakten und
prozessierten Assembly-Komplexen entstehen, welche während der Metallaffinitäts-
chromatographie gereinigt und in der Größenausschlusschromatographie wegen der
zu geringen Massenunterschiede in die gleiche Fraktion eluiert wurden. Da jedoch,
wie aus der SDS-PAGE der Fraktion hervorgeht, die Mengen der gp1-Abbauprodukte
im Vergleich zu den wildtypischen gp1-Proteinen sehr gering waren, kann davon aus-
gegangen werden, dass in dieser frühen Elutionsfraktion 90 % der gereinigten
gp1/11-Assembly-Komplexe in ihrer nativen Form vorlagen. Für eine höhere Aus-
beute an unprozessierten Assembly-Komplexen könnte es essenziell sein, das
stöchiometrische Gleichgewicht zwischen gp1 und gp4 herzustellen. Dies würde
unter anderem dadurch erreicht werden, indem man das gp1/11-Expressionssystem
dahingehend verändert, dass die Expressionsrate von gp1/11 verringert oder die
gp4-Expressionsrate erhöht wird. Im Unterschied zur Größenausschlusschromato-
graphie der IMAC-Reinigung des Elutionskonzentrats aus infizierten Zellen, konnte in
der frühen Fraktion durch SDS-PAGE kein gp11 als Bestandteil des Assembly-
Komplexes identifiziert werden. Eine mögliche Ursache hierfür könnte eine zu
geringe Menge des Proteins sein. Da jedoch in den späteren Fraktionen das ~ 12
kDa schwere gp11 nachgewiesen wurde, kann davon ausgegangen werden, dass
dieses auch im Assembly-Komplex präsent war, jedoch in einer zu geringen
Konzentration.
4.4.4 TEM-Aufnahmen FPLC gereinigter gp1/11-Assembly-Komplexe
Der gp1/11-Assembly-Komplex bildet eine ringförmige Struktur mit einem
Durchmesser von ~ 11 – 12 nm
Die TEM-Aufnahmen der Proben aus der Elutionsfraktion mit Assembly-Komplexen
mit Molekulargewichten von 330 kDa - 270 kDa aus Kapitel 3.4.4, welche über
Größenausschlusschromatographie gereinigte gp1/11-Assembly-Komplexe aus infi-
zierten E. coli K37-Zellen enthielt, zeigten nach Negativkontrastierung mit 5 %iger
Phosphorwolframsäure deutlich ringförmige Strukturen. Mit Hilfe einer software-
gestützten Bildprozessierung konnte die Erkennbarkeit des ringförmigen Charakters

Diskussion
180
dieser Strukturen verbessert werden. Aus diesen Bildern lässt sich für den Außen-
durchmesser des Assembly-Komplexes eine Länge von ~ 11 – 12 nm und für den
Innendurchmesser eine Länge von ~ 7 – 8 nm ermitteln. Vergleicht man die durch
Opalka et al. ermittelten Größen für die homomultimere gp4-Pore in der äußeren
Membran von E. coli, welche mit dem gp1/11-Assembly-Komplex einen Sekretions-
komplex bildet, mit den hier ermittelten Größen des Assembly-Komplexes, so könnte
dieser in das Innere der 13,5 nm großen gp4-Pore (Opalka et al., 2003) hineinragen
(Abb. 4-6).
Durch diese intermolekulare Interaktion könnte die C-terminale Domäne des gp1 im
Bereich der Aminosäuren 275 – 320 (Feng et al., 1999) durch die N-terminale
Domäne der gp4-Pore vor einer proteolytischen Prozessierung geschützt werden.
Dabei interagieren nur die C-terminalen Domänen der gp1-Monomere mit den
N-terminalen Domänen der gp4-Monomere und bilden nach einer Hypothese von
Feng et al. einen gp1/11-gp4-Sekretionskomplex. Der Innendurchmesser des
gp1/11-Assembly-Komplexes wäre hier mit einem Innendurchmesser von 7 – 8 nm
ausreichend dimensioniert für die Sekretion neugebildeter M13-Phagen. Die hier
innere Membran
Periplasma
Murein
äußere Membran
13,5 nm
11 - 12 nm
gp1/11-Assembly-Komplex
gp4-Multimer
Abb. 4-6: Schematische Darstellung des gp1/11-gp4-Sekretionskomplexes. Aufgrund des gp1/11-Assembly-Komplex-Außendurchmessers von 11-12 nm besteht die Möglichkeit, dass der C-terminale Bereich des Assembly-Komplexes in den N-terminalen Bereich der 13,5 nm großen gp4-Pore hineinragt und hierdurch das gp1 vor einer proteolytischen Prozessierung geschützt ist (nach Webster, 1996, verändert).
gp11 gp1 gp4
N
N N
C C
C
Zytoplasma

Diskussion
181
verwendeten Proben enthielten, aufgrund der Analyse der Größenausschlusschro-
matographie, ausschließlich Moleküle mit Massen von ~ 330 – 270 kDa. Hier kann
somit davon ausgegangen werden, dass die aufgenommenen ringförmigen Struk-
turen Assembly-Komplexe darstellen, welche sich, wie aus der SDS-PAGE in Kapitel
3.4.4 hervorging, hauptsächlich aus gp1-Molekülen mit ~ 37 kDa und einem sehr
geringen Anteil aus gp11 mit ~ 12 kDa zusammensetzen.
Der gp1/11-Assembly-Komplex könnte ein Decamer oder Dodecamer sein
Auf der Basis des über Größenausschlußchromatographie ermittelten Molekular-
gewichts von gemittelten ~ 300 kDa für den Assembly-Komplex, den anschließend
durch SDS-PAGE identifizierten gp1 Fragmenten und deren ermittelten Einzel-
massen, könnte vermutet werden, dass es sich bei den hier gereinigten Assembly-
Komplexen um Multimere handelt. Aus den identifizierten Einzelmassen der gp1-
Fragmente in diesen Komplexen ergibt sich für diese ein gemitteltes Molekular-
gewicht von ~ 37 kDa. Aus einer angenommenen Ratio von einem gp1 mit ~ 37 kDa
zu einem gp11 mit ~ 12 kDa würde sich für ein theoretisches gp1/11-Dimer eine
Gesamtmasse von ~ 49 kDa errechnen lassen. Da dem hier gereinigten Assembly-
Komplex eine Masse von ~ 300 kDa zugeordnet werden konnte, könnte sich dieser
somit aus 5 - 6 gp1/11-Dimeren zusammensetzen und hieraus ein decamerer bzw.
ein dodecamerer Komplex resultieren. Dies ist jedoch rein hypothetisch, da in den
gereinigten Komplexen kein gp11 nachweisbar war. Dass sich der Assembly-
Komplex eventuell aus jeweils 5 - 6 gp1 Monomeren und 5 - 6 gp11 Monomeren
zusammensetzen könnte, wurde auch schon von Russel et al. spekuliert (Russel et
al., 2004). In Anbetracht der vorgegebenen Symmetrie durch die gp4-Pore, welche
aus ~ 12 – 14 gp4-Monomeren gebildet wird (Opalka et al., 2003), ergäbe sich im
Fall eines dodecameren gp1/11-Assembly-Komplexes bei einer intermolekularen
Interaktion mit der gp4-Sekretionspore eine plausible stöchiometrische Überein-
stimmung.
Wie aus der SDS-PAGE Analyse der Elution größenausschlußchromatographie-
gereinigter Assembly-Komplexe aus infizierten E. coli K37-Zellen in Kapitel 3.4.4
hervorging, entsprechen aufgrund des zu großen Anteils an prozessierten gp1-
Molekülen und der zu geringen Menge von gp11-Molekülen die gereinigten Kom-
plexe nicht zu 100 % der nativen Form.

Diskussion
182
Hypothetisch könnte, mit Hilfe der aus den TEM-Untersuchungen erhaltenen
Informationen, unter Berücksichtigung einer ringförmigen Symmetrie des Assembly-
Komplexes und des Durchmessers des deutlichsten Signals eines Monomers im
Komplex der vergrößerten TEM-Aufnahme in Abbildung 4-7 (A, siehe Pfeil), nach
einer theoretischen Positionierung von gedachten gp1- (rot) und gp11-Monomeren
(gelb) in (B) im Verhältnis 1:1, ein dodecamerer Assembly-Komplex modelliert
werden, dessen innerer Durchmesser ~ 7 – 8 nm und äußerer Durchmesser ~ 11 –
12 nm (C) beträgt.
Abb. 4-7: Schematische Darstellung des gp1/11-Assembly-Komplexes. (A) Vergrößerte Darstellung einer elektronenmikroskopischen Aufnahme eines ringförmigen gp1/11-Assembly-Komplexes. Das deutlichste Signal eines Monomers wird durch einen weißen Pfeil markiert. (B) Nach der theoretischen Positionierung virtueller gp1- (rot) und gp11-Monomere (gelb) im Verhältnis 1:1 ergibt sich ein Dodecamer mit einem Durchmesser von 11 – 12 nm. (C) Schematisches Modell des sich aus (B) ergebenden gp1/11-Assembly-Komplexes. Die Längen der Skalierungsbalken betragen 10 nm.
N
Ø 11 - 12 nm
C
gp1/11-Assembly-Komplex
gp11 gp1
(C)
N
C

Zusammenfassung/Summary
183
5 Zusammenfassung
Der Phage M13 ist taxonomisch den Einzelstrang-DNA-Phagen zugeordnet und
gehört zur Familie der Inoviridae. Für die Vermehrung verwendet M13 Gram-nega-
tive Escherichia coli-Zellen, die F-Pili ausbilden, als Wirt. Durch die Vermehrung wird
die Wirtszelle nicht geschädigt. In dieser Arbeit konnten neue Erkenntnisse über die
M13-Biogenese erhalten werden, welche den Bereich Infektionsprozess, die Assem-
blierung und die Phagensekretion betreffen.
Durch Adsorptionsexperimente, in denen das Wirtsbakterium E. coli K38 mit M13-
Phagen infiziert wurde, konnte gezeigt werden, dass innerhalb der ersten 5 Minuten
die Bindung der Phagen an die Zellen stattfindet und hier aufgrund einer Limitierung
der F-Pili pro Zelle maximal 7 Phagen pro E. coli-Zelle binden konnten.
Die Insertion des Phagenhüllproteins gp9 in die Zytoplasmamembran der Wirtszelle
wurde mit Hilfe antigener Epitope in der N-terminalen Domäne nachgewiesen und es
konnte gezeigt werden, dass das YidC-Protein von E. coli dafür benötigt wird.
Zudem wurde gezeigt, dass plasmidkodiertes gp9 mit zusätzlich eingebrachten
antigenen Epitopen in der N-terminalen Domäne keinen negativen Effekt auf die
Assemblierung neuer Phagen-Nachkommen hatte. Daher kann das gp9 durch die
Insertion kurzer Peptidsequenzen (17 – 36 Aminosäuren) für ein „Phage-Display“
verwendet werden.
Hinsichtlich der Phagenassemblierung war es möglich, den membranständigen
gp1/11-Assembly-Komplex, nach Überexpression in E. coli und Größenausschluß-
chromatographie, zu charakterisieren und ihm ein Molekulargewicht von ~ 300 kDa
zuzuordnen. Die transmissionselektronenmikroskopische (TEM) Untersuchung des
gereinigten gp1/11-Assembly-Komplexes zeigte ringförmige Strukturen mit einem
Innendurchmesser von ~ 7 – 8 nm und einem Außendurchmesser von ~ 11 – 12 nm.
Die Untersuchung der wildtypischen Phagensekretion zeigte, dass nach einer
5-minütigen Latenzphase (Eklipse) die Sekretion neuer Phagen-Nachkommen
erfolgte und eine infizierte E. coli-Zelle in einem Zeitraum von 115 Minuten bis zu 925
neue Phagen sekretieren kann, was einer durchschnittlichen Sekretion von 7 Phagen
pro Minute entspricht. Durch die M13-Infektion verlängerte sich die Generationszeit
der E. coli K38-Zellen von 24 auf 48 Minuten.

Zusammenfassung/Summary
184
Die Untersuchung der Phagensekretion und der Phagenreplikation genetisch
veränderter Phagen, deren Synthese des Haupthüllproteins gp8 durch das Wirts-
bakterium mit Hilfe einer Punktmutation im Phagengenom unterbunden wurde,
erfolgte in Wirtszellen mit zusätzlich plasmidkodiertem Gen 8 des Phagen. Es zeigte
sich, dass das plasmidkodierte gp8 limitiert war, was eine Verringerung der Phagen-
sekretion auf ~ 2 Phagen pro Minute verursachte und die Latenzzeit (Eklipse) auf
12 Minuten verzögerte.
Die Sekretion von M13-Phagen aus infizierten E. coli-Zellen konnte mit dem „Atomic
Force Microscope“ (AFM) dargestellt und die Phagen am TEM, nach Markierung mit
Protein-A konjugiertem Gold, nachgewiesen werden. Die Sekretion von M13-
Phagen-Nachkommen begann an den Zellpolen von E. coli und breitete sich im
Verlauf von 4 Minuten von diesen zu den Seiten der Zellen hin aus und erstreckte
sich nach 16 Minuten über die gesamte Zelloberfläche.

Zusammenfassung/Summary
185
5 Summary
Taxonomically, the bacteriophage M13 is assigned to the single-stranded DNA
phage and belongs to the family of Inoviridae. For propagation the Gram-negative
bacteria Escherichia coli with F-pili is required. The host cell is not lysed by the
phage. New findings about the M13 phage biogenesis are presented here within four
essential areas of the M13 phage cycle concerning the sections infection, assembly,
and phage secretion.
Phage adsorption experiments in which the host bacterium E. coli K38 was infected
by M13 phage showed that the phage adsorption to the cells takes place within the
first 5 minutes and because of a limitation of F-pili per cell a maximum of 7 phages
per cell were found to be adsorbed.
The insertion of the phage coat protein gp9 into the cytoplasmic membrane of the
host cell was verified by the periplasmic location of antigenic epitopes introduced into
the N-terminal domain of gp9. The membrane insertion of gp9 was found to depend
on the host protein YidC. Plasmid-encoded gp9 exhibiting antigenic epitopes at the
N-terminal domain did not interfere with the assembly of new progeny phage.
Therefore, the development of a phage display system with gp9 by introducing short
peptide sequences (17 – 36 amino acids) is feasible.
After overexpression of gp1/11 assembly complexes in E. coli and size exclusion
chromatography, respectively, the complex was characterized and a molecular
weight of ~ 300 kDa was assigned. Examinations of the purified gp1/11 assembly
complexes by transmission electron microscopy (TEM) revealed ring-like structures
with ~ 7 – 8 nm in inner diameter and ~ 11 – 12 nm in outer diameter.
The investigation of M13 wild-type infection showed that the secretion of new
progeny phage starts after a short lag period (eclipse). An infected E. coli cell
secreted upto 925 progeny in a time period of 115 minutes which corresponds to an
average of 7 secreted phages per minute. The generation time of the infected E. coli
K38 cells rose from 24 minutes to 48 minutes.
Experiments were carried out with genetically manipulated phages which were
hindered to synthesize the major coat protein gp8 in the host cell by a nonsense
mutation in the phage genome. Therefore, phage replication was only observed in

Zusammenfassung/Summary
186
host cells bearing plasmid encoding gene 8. Since the quantity of the protein was
limited the lag period (eclipse) was extended to 12 minutes and the efficiency of
phage secretion was decreased to about 2 phages per minute.
The M13 phage secretion from infected E. coli cells was visualized by atomic force
microscopy (AFM). The identity of the phage was verified by labeling with protein-A
conjugated gold and transmission electron microscopy (TEM). The secretion of M13
progeny was first observed at the cell poles of E. coli and then spreaded within
4 minutes along the cell surface. After 16 minutes the secretion was observed over
the entire cell surface.

Literaturverzeichnis
187
6 Literaturverzeichnis
Alami, M., Luke, I., Deitermann, S., Eisner, G., Koch, H. G., Brunner, J. & Müller, M.
(2003). Differential interactions between a twin-arginine signal peptide and its
translocase in Escherichia coli. Mol. Cell. 12(4), 937-46
Andersson, H. & von Heijne, G. (1991). A 30-residue-long ‘‘export initiation domain’’
adjacent to the signal sequence is critical for protein translocation across the inner
membrane of Escherichia coli. Proc. Natl. Acad. Sci. USA 88, 9751–4
Bassford, P. J. Jr. (1990). Export of the periplasmic maltose-binding protein of
Escherichia coli. J. Bioenerg. Biomembr. 22(3), 401-39
Benada, O. & Pokorný, V. (1990). Modification of the Polaron sputter-coater unit for
glow-discharge activation of carbon support films. J. Electron. Microsc. Tech. 16,
235-239
Berks, B. C. (1996). A common export pathway for proteins binding complex redox
cofactors? Mol. Microbiol. 22, 393–404
Berks, B. C. (1998). Targeting signals for a bacterial Sec-independent export system
direct plant thylakoid import by the delta pH pathway. FEBS Lett. 431, 339-42
Berks, B. C., Sargent, F. & Palmer, T. (2000). The Tat protein export pathway. Mol.
Microbiol. 35(2), 260-74
Bonnefoy, N., Fiumera, H. L., Dujardin, G. & Fox, T.D. (2009). Roles of Oxa1-related
inner-membrane translocases in assembly of respiratory chain complexes. Biochim.
Biophys. Acta. 1793, 60–70
Bourdineaud, J. P., Boulanger, P., Lazdunski, C. & Letellier, L. (1990). In vivo
properties of colicin A: channel activity is voltage dependent but translocation may be
voltage independent. Proc. Natl. Acad. Sci. USA 87, 1037-41
Brissette, J. L. & Russel, M. (1990). Secretion and membrane itegration of a
filamentous phage-encoded morphogenetic protein. J. Mol. Biol. 211, 565-80

Literaturverzeichnis
188
Brown, L. R. & Dowell, C. E. (1968). Replication of coliphage M13: effects on host
cells after synchronized infection. J. Virol. 2(11), 1290-5
Bruch, M. D., McKnight, C. J. & Gierasch, L. M. (1989). Helix formation and stability
in a signal sequence. Biochem. 28(21), 8554-61
Bullock, W. O., Fernandez, J. M. & Short, J. M. (1987). XL1-blue: a high efficiency
plasmid transforming recA Escherichia coli strain with beta-galactosidase section.
Biotechniques 5, 376-78
Casadaban, M. J. (1976). Transposition and fusion of the lac genes to selected
promoters in Escherichia coli using bacteriophage lambda and Mu. J. Mol. Biol. 104,
541-55
Cavard, D. & Lazdunski, C. (1981). Involvement of BtuB and OmpF proteins in
binding and uptake of colicin A. FEMS Microbiol. Lett. 12, 311-6
Cavari, B. Z., Avi-Dor, Y. & Grossowicz, N. (1966). Effect of Carbonyl Cyanide m-
Chlorophenylhydrazone on Respiration and Respiration-Dependent Phosphorylation
in Escherichia coli. Biochem. J. 103, 601-8
Celebi, N., Yi, L., Facey, S. J., Kuhn, A. & Dalbey, R. E. (2006). Membrane
biogenesis of subunit II of cytochrome bo oxidase: contrasting requirements for
insertion of N-terminal and C-terminal domains. J. Mol. Biol. 357, 1428-36
Chang, N. C., Blobel, G. & Model, P. (1978). Detection of prokaryotic signal
peptidase in an Escherichia coli membrane fraction: endoproteolytic cleavage of
nascent f1 pre-coat protein. Proc. Natl. Acad. Sci. USA 75(1), 361-5
Chen, M., Samuelson, J. C., Jiang, F., Muller, M., Kuhn, A. & Dalbey, R. E. (2002).
Direct interaction of YidC with the Sec-independent Pf3 coat protein during its
membrane protein insertion. J. Biol. Chem. 277, 7670–5
Click, E. M. & Webster, R. E. (1997). Filamentous phage infection: required
interactions with the TolA protein. J. Bacteriol. 179, 6464-71

Literaturverzeichnis
189
Click, E. M. & Webster, R. E. (1998). The TolQRA proteins are required for
membrane insertion of the major capsid protein of the filamentous phage f1 during
infection. J. Bacteriol. 180, 1723-8
Cline, K. & Theg, S. M. (2007). The Sec and protein translocation pathways in
chloroplast, In: Ross E. Dalbey, Carla M. Koehler, Fuyuhiko Tamanoi (Eds.),
Molecular Machines Involved in Protein Transport across Cellular Membranes, 3rd
Edition, The Enzymes, Vol 25, Academic Press, San Diego, 463–92
Cristobal, S., de Gier, J.-W., Nielsen, H., & von Heijne, G. (1999). Competition
between Sec- and TAT-dependent protein translocation in Escherichia coli. EMBO J.
18, 2982-90
Davies, J. K. & Reeves, P. (1975a). Colicin tolerance and map location of
conjugation-deficient mutants. J. Bacteriol. 123, 372-3
Davies, J. K. & Reeves, P. (1975b). Genetics of resistance to colicins in Escherichia
coli K-12: cross-resistance among colicins of group A. J. Bacteriol. 123, 102-17
Delbrück, M. (1942). Bacterial viruses (bacteriophages). Adv. Enzymol. 2, 1–32
Dotto, G. P. & Zinder, N. D. (1983). The morphogenic signal of bacteriophage f1.
Virol. 130(1), 252-6
du Plessis, D. J., Nouwen, N. & Driessen, A. J. (2006). Subunit a of cytochrome o
oxidase requires both YidC and SecYEG for membrane insertion. J. Biol. Chem. 281,
12248–52
Edgar, R., Rokney, A., Feeney, M., Semsey, S., Kessel, M., Goldberg, M. B., Adhya,
S. & Oppenheim, A. B. (2008). Bacteriophage infection is targeted to cellular poles.
Mol. Microbiol. 68(5), 1107–16
Endemann, H. & Model, P. (1995). Location of filamentous phage minor coat proteins
in phage and in infected cells. J. Mol. Biol. 250, 496–506
Facey, S. J. & Kuhn, A. (2003). The sensor protein KdpD inserts into the Escherichia
coli membrane independent of the Sec translocase and YidC. Eur. J. Biochem. 270,
1724–34

Literaturverzeichnis
190
Facey, S. J. & Kuhn, A. (2004). Membrane integration of E. coli model membrane
proteins. Biochim. et Biophys. Acta 1694, 55-66
Facey, S. J., Neugebauer, S. A., Krauss, S. & Kuhn, A. (2007). The
mechanosensitive channel protein MscL is targeted by the SRP to the novel YidC
membrane insertion pathway of Escherichia coli. J. Mol. Biol. 365, 995–1004
Feng, J. N., Russel, M. & Model, P. (1997). A permeabilized cell system that
assembles filamentous bacteriophage. Proc. Natl. Acad. Sci. USA 94(8), 4068-73
Feng, J. N., Model, P. & Russel, M. (1999). A trans-envelope protein complex
needed for filamentous phage assembly and export. Mol. Microbiol. 34, 745-755
Fox Jr., R. O. & Richards, F. M. (1982). A voltage-gated ion channel model inferred
from the crystal structure of alamethicin at 1.5-Å resolution. Nature 300, 325-30
Frost, L. S. (1993). Conjugative pili and pilus-specific phages. In: "Bacterial
Conjugation", D. B. Clewell, (ed.), 189-221. Plenum, New York.
Fröderberg, L., Houben, E. N., Baars, L., Luirink, J. & de Gier, J. W. (2004).
Targeting and translocation of two lipoproteins in Escherichia coli via the
SRP/Sec/YidC pathway. J. Biol. Chem. 279(30), 31026-32
Fulford, W. & Moder, P. (1984). Gene X of bacteriophage f1 is required for phage
DNA synthesis. J. Mol. Biol. 178, 137-53
Fürste, J. P., Pansegrau, W., Frank, R., Blocker, H., Scholz, P., Bagdasarian, M. &
Lanka, E. (1986). Molecular cloning of the plasmid RP4 primase region in a multi-
host–range tacP expression vector. Gene 48(1), 119-31
Galan, J. E. & Collmer, A. (1999). Type III secretion machines: bacterial devices for
protein delivery into host cells. Science 284(1), 322-28
Gaskova, D., Brodska, B., Holoubek, A. & Sigler, K. (1999). Factors and processes
involved in membrane potential build-up in yeast: diS-C3(3) assay. Int. J. Biochem. &
Cell. Biol. 31(5), 575-84

Literaturverzeichnis
191
Gao, C., Mao, S., Lo, C. H., Wirsching, P., Lerner, R. A. & Janda, K. D. (1999).
Making artificial antibodies: a format for phage display of combinatorial heterodimeric
arrays. Proc. Natl. Acad. Sci. USA 96(11), 6025–30
Garen, A., Garen, S. & Wilhelm, R. C. (1965). Suppressor genes for nonsense
mutations. I. The Su-1, Su-2, and Su-3 genes of Escherichia coli. J. Mol. Biol. 14(1),
167-78
Glick, B. S., Beasley, E. M. & Schatz, G. (1992). Protein sorting in mitochondria,
Trends. Biochem. Sci. 17, 453–59
Glucksman, M. J. & Bhattacharjee, S. (1992). Three-dimensional structure of a
cloning vector. J. Mol. Biol. 226, 455-70
Grant, R. A., Lin, T. C., Webster, R. E. & Konigsberg, W. (1981). Structure of the
filamentous bacteriophage: isolation, charcterization, and localization of the minor
coat proteins and orientation of the DNA, 413-428. In: M. DuBow (ed.),
Bacteriophage assembly. Progress in clinical and biological research, vol. 64, Alan R.
Liss, Inc., New York
Gray, C. W., Brown, R. S. & Marvin, D. A. (1981). Adsorption complex of filamentous
fd virus. J. Mol. Biol. 146(4), 621-7
Guan, Y., Zhang, H. & Wang, A. H. J. (1995). Electrostatic potential distribution of the
gene V protein from Ff phage facilitates cooperative DNA binding: a model of the
GVP-ssDNA complex. Protein Sci. 4(2), 187-97
Guo, Y., Liang, X., Zhou, Y., Zhang, Z., Wie, H., Men, D., Luo, M. & Zhang, X. E.
(2010). Construction of bifunctional phage display for biological analysis and
immunoassay. Anal. Biochem. 396, 155-7
Guy-Caffey, J. K., Rapoza, M. P., Jolley, K. A. & Webster, R. E. (1992). Membrane
localization and topology of a viral assembly protein. J. Bacteriol. 174, 2460-5
Guy-Caffey, J. K. & Webster, R. E. (1993). The membrane domain of a
bacteriophage assembly protein: membrane insertion and growth inhibition. J. Biol.
Chem. 268, 5496-503

Literaturverzeichnis
192
Haigh, N. G. & Webster, R. E. (1999). The pI and pVI assembly proteins serve
seperate and essential roles in filamentous phage assembly. J. Mol. Biol. 293, 1017-
27.
Hanahan, D. (1985). DNA Cloning: A Practical Approach. Glover, D. M. (ed.), Vol. 1,
p. 109, IRL Press, McLean, Virginia.
Hashemolhosseini, S., Holmes, Z., Mutschler, B. & Henning, U. (1994). Alterations of
receptor specificities of coliphages of the T2 family. J. Mol. Biol. 240, 105-10
Henry, R., Goforth, R. L. & Schunemann, D. (2007). Chloroplast SRP/FtsY and Alb3
in protein integration into the thylakoid membrane, in: Ross E. Dalbey, Carla M.
Koehler, Fuyuhiko Tamanoi (Eds.), Molecular Machines Involved in Protein Transport
across Cellular Membranes, 3rd Edition, The Enzymes, Vol. 25, Academic Press,
San Diego, CA, pp. 493–521
Herman, C., Thevenet, D., D’Ari, R., & Bouloc, P. (1997). The HflB protease of
Escherichia coli degrades its inhibitor l cIII. J. Bacteriol. 179, 358-63
Herrmann. J. M. & Köhl, R. (2007). Catch me if you can! Oxidative protein trapping in
the intermembrane space of mitochondria. J. Cell. Biol. 176(5), 559-63
Hines, J. C. & Ray, D. S. (1980). Construction and characterization of new coliphage
M13 cloning vectors. Gene 11(3-4), 207-18
Hochuli, E., Dobeli. H. & Schachner, A. (1987). New chelate adsorbant selective for
proteins and peptides containing neighbouring histidine residues. J. Chromatogr.
417, 177-184
Hoffmann-Berling, H. & Mazé, R. (1964). Release of male-mpecific bacteriophages
from surviving host bacteria. Virol. 22, 305-13
Hofschneider, P. H. (1963). Untersuchungen über "kleine" Bakteriophagen. Die
Isolierung und Eigenschaften der "kleinen" Bakteriophagen M12, M13, und M20. Z.
Naturforschg. 18b, 203-5

Literaturverzeichnis
193
Horabin, J. I. & Webster, R. E. (1986). Morphogenesis of f1 filamentous
bacteriophage. Increased expression of gene I inhibits bacterial growth. J. Mol. Biol.
188, 403-13.
Horiuchi, K. (1997). Initiation mechanism in replication of filamentous phage DNA.
Genes Cells 2(7), 425-32
Houbiers, M. C., Spruijt, R. B., Wolfs, C. J. A. M. & Hemminga, M. A. (1999).
Conformational and aggregational properties of the gene 9 minor coat protein of
bacteriophage M13 in membrane-mimicking systems. Biochem. 38, 1128-35
Houbiers, M. C., Spruijt, R. B., Demel, R. A., Hemminga, M. A. & Wolfs, C. J. A. M.
(2001). Spontaneous insertion of gene 9 minor coat protein of bacteriophage M13 in
model membranes. Biochem. Biophys. Acta 1511, 309-16
Huang, Y., Chiang, C. Y., Lee, S. K., Gao, Y., Hu, E. L., Yoreo, J. D. & Belcher, A. M.
(2005). Programmable assembly of nanoarchitectures using genetically engineered
viruses. Nano Letters 5(7), 1429-34
Iannolo, G., Minenkova, O., Petruzzelli, R. & Cesareni, G. (1995). Modifying
filamentous phage capsid: limits in the size of the major capsid protein. J. Mol. Biol.
248, 835-44
Jones, L. R., Maddock, S. W. & Besch Jr., H. R. (1980). Unmasking effect of
alamethicin on the (Na+,K+)-ATPase, beta-adrenergic receptor-coupled adenylate
cyclase, and cAMP-dependent protein kinase activities of cardiac sarcolemmal
vesicles. J. Biol. Chem. 255(20), 9971-80
Kampfenkel, K. & Braun, V. (1993). Membrane topologies of the TolQ and TolR
proteins of Escherichia coli: inactivation of TolQ by a missense mutation in the
proposed first transmembrane segment. J. Bacteriol. 175, 4485-91
Karlsson, F., Borrebaeck, C. A. K., Nilsson, N. & Malmborg-Hagert, A. C. (2003). The
mechanism of bacterial infections between TolA and phage protein 3 domains.
Microbiol. Rev. 185, 2628-34

Literaturverzeichnis
194
Kay, B. K., Adey, N. B., He, Y. S., Manfredi, J. P., Mataragnon, A. H. & Fowlkes, D.
M. (1993). An M13 phage library displaying random 38-amino-acid peptides as a
source of novel sequences with affinity to selected targets. Gene 128 (1), 59-65
Kazmierczak, B., Mielke, D. L., Russel, M. & Model, P. (1994). Filamentous phage
pIV forms a multimer that mediates phage export across the bacterial cell envelope.
J. Mol. Biol. 238, 187-98
Kishchenko, G., Batliwala, H. & Makowski, L. (1994). Structure of a foreign peptide
displayed on the surface of bacteriophage M13. J. Mol. Biol. 241, 208-13
Knowles, T. J., Scott-Tucker, A., Overduin, M. & Henderson, I. R. (2009). Membrane
protein architects: the role of the BAM complex in outer membrane protein assembly.
Nat. Rev. Microbiol. 7, 206–14
Konings, R. N. H., Folmer, R. H. A., Folkers, P. J. M., Nilges, M. & Hilbers, C. W.
(1995). Three-dimensional structure of the single-stranded DNA-binding protein
encoded by gene V of the filamentous bacteriophage M13 and a model of its
complex with single-stranded DNA. FEMS Microbiol. Rev. 17(1-2), 57-72
Kozjak, V., Wiedemann, N., Milenkovic, D., Lohaus, C., Meyer, H. E., Guiard, B.,
Meisinger, C. & Pfanner, N. (2003). An essential role of Sam50 in the protein sorting
and assembly machinery of the mitochondrial outer membrane, J. Biol. Chem. 278
48520-3
Kuhn, A., Kreil, G. & Wickner, W. (1986). Both hydrophobic domains of M13 procoat
are required to initiate membrane insertion. Embo J. 5(13), 3681-5
Kuhn, A. & Troschel, D. (1992). Distinct steps in the insertion pathway of
bacteriophage coat proteins. In: “Membrane Biogenesis and Protein Targeting” (W.
Newport, and R. Lill, eds.), pp. 33-47. Elsevier New York
LaFarina, M. & Model, P. (1983). Transcription in bacteriophage f1-infected
Escherichia coli: messenger populations in the infected cell. J. Mol. Biol. 164, 377-93
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 227(259), 680-5

Literaturverzeichnis
195
Lecker, S. H., Driessen, A. J. & Wickner, W. (1990). ProOmpA contains secondary
and tertiary structure prior to translocation and is shielded from aggregation by
association with SecB protein. EMBO J. 9(7), 2309-14
Lessl, M., Balzer, D., Lurz, R., Waters, V. L., Guiney, D. G. & Lanka, E. (1992).
Dissection of IncP conjugative plasmid transfer: definition of the transfer region Tra2
by mobilization of theTra1 region in trans. J. Bacteriol. 174, 2493-500
Lim, C. J., Haller, B. & Fuchs, J. A. (1985). Thioredoxin is the bacterial protein
encoded by fip that is required for filamentous phage f1 assembly. J. Bacteriol.
161(2), 799-802
Lopez, J. & Webster, R. E. (1985). Assembly site of bacteriophage f1 corresponds to
adhesion zones between the inner and outer membranes of the host cell. J.
Bacteriol. 163, 1270-4
Løset, G. A., Roos, N., Bogen, B. & Sandlie, I. (2011). Expanding the versatility of
phage display II: improved affinity selection of folded domains on protein VII and IX of
the filamentous phage. PLoS ONE 6(2), 1-10
Lowman, H. B. (1997). Bacteriophage display and discovery of peptide leads for drug
development. Annu. Rev. Biophys. Biomol. Struct. 26, 401-24
Lutz, T., Neupert, W. & Herrmann, J. M. (2003). Import of small Tim proteins into the
mitochondrial intermembrane space. EMBO J. 22, 4400-8
Lyons, L. B. & Zinder, N. D. (1972). The genetic map of the filamentous
bacteriophage f1. Virol. 49(1), 45-60
Makowski, L. (1992). Terminating a macromolecular helix. Structural model for the
minor proteins of bacteriophage M 13. J. Mol. Biol. 228, 885-92
Marvin, D. A & Hohn, B. (1969). Filamentous bacterial viruses. Bacteriol. Rev. 33,
172-209
Marvin, D. A. (1998). Filamentous phage strukture, infection and assembly. Curr.
Opin. Struct. Biol. 8, 150-8

Literaturverzeichnis
196
McPherson, A., Wang, A. H., Jurnak, F. A., Molineux, I., Kolpak, F. & Rich, A. (1980).
X-Ray diffraction studies on crystalline complexes of gene 5 DNA-unwinding protein
with deoxyoligonucleotides. J. Biol. Chem. 255(7), 3174-7
Milner, H.W., König, M.L. & Lawrence, M.S. (1950). Reactivation of dispersed
chloroplast material by reaggregation. Arch. Biochem. 28(2), 185-19
Minamishima, Y., Takeya, K. Ohnishi, Y. & Amako, K. (1968). Physicochemical and
biological properties of fibrous Pseudomonas bacteriophages. J. Virol. 2, 208-13
Miroux, B. & Walker, J. E. (1996). Over-production of proteins in Escherichia coli:
mutant hosts that allow synthesis of some membrane proteins and globular proteins
at high levels. J. Mol. Biol. 60, 289-98
Model, P., McGill, C., Mazur, B. & Fulford, W. D. (1982). The replication of
bacteriophage f1: gene 5 protein regulates the synthesis of gene 2 protein. Cell
29(2), 329-35
Model, P., & Russel, M. (1988). Filamentous bacteriophage, 375-456 In: "The
Bacteriophages" R. Calendar, (ed.), (2), Plenum, New York.
Mori, H. & Cline, K. (1998). A signal peptide that directs non-Sec transport in bacteria
also directs efficient and exclusive transport on the thylakoid ∆pH pathway. J. Biol.
Chem. 273, 11405-8
Mould, R. M., Knight, J. S., Bogsch, E. & Gray, J. C. (1997). Azide-sensitive thylakoid
membrane insertion of chimeric cytochrome f polypeptides imported by isolated pea
chloroplasts. Plant J. 11, 1051–58
Nivière, V., Wong, S.L., & Voordouw, G. (1992). Site-directed mutagenesis of the
hydrogenase signal peptide consensus box prevents export of a β-lactamase fusion
protein. J. Gen. Microbiol. 138, 2173-83
O’Callaghan, R., Bradley, R. & Paranchych, W. (1973). The effect of M13 phage
infection upon the F pili of E. coli. Virol. 64, 220-9
Olah, G. A., Gray, D. M., Gray, C. M., Kergil, D. L., Sosnick, T. R., Mark, B. L.,
Vaughan, M. R. & Trewhella, J. (1995). Structures of fd gene 5 protein-nucleic acid

Literaturverzeichnis
197
complexes: a combined solution scattering and electron microscopy study. J. Mol.
Biol. 249(3), 576-94
Opalka, N., Beckmann, R., Boisset, N., Simon, M. N., Russel, M. & Darst, S. A.
(2003). Structure of the filamentous phage pIV multimer by cryo-electron mivroscopy.
J. Mol. Biol. 325, 461-470
Panter, R. A. & Symons, R. H. (1966). Isolation and properties of a DNA-containing
rod-shaped bacteriophage. Australian J. Biol. Sci. 19, 565-73
Parmley, S. F. & Smith, G. P. (1988). Antibody-selectable filamentous fd phage
vectors: affinity purification of target genes. Gene 73, 305-318
Paschen, S. A., Waizenegger, T., Stan, T., Preuss, M., Cyrklaff, M., Hell, K.,
Rapaport, D. & Neupert, W. (2003). Evolutionary conservation of biogenesis of beta-
barrel membrane proteins. Nature 426, 862-6
Peters, E. A., Schatz, P. J., Johnson, S. S & Dower, W. J. (1994). Membrane
insertion defects caused by positive charges in the early mature region of protein plll
of filamentous phage fd can be corrected by prlA suppressors. J. Bact. 176(14),
4296-4305
Ploss, M. & Kuhn, A. (2010). Kinetics of filamentous phage assembly. Phys. Biol.
7(4), 045002
Ploss, M. & Kuhn, A. (2011). Membrane insertion and assembly of epitope-tagged
gp9 at the tip of the M13 phage. BMC Microbiol. 11(1), 211-9
Price, C. E. & Driessen, A. J. (2010). Conserved negative charges in the
transmembrane segments of subunit K of the NADH:ubiquinone oxidoreductase
determine its dependence on YidC for membrane insertion. J. Biol. Chem. 285, 3575-
81
Pugsley, A. P. (1993). The complete general secretary pathway in Gram-negative
bacteria. Microbiol. Rev. 57, 50-108

Literaturverzeichnis
198
Raggett, E. M., Bainbridge, L. J., Evans, J., Cooper, A. & Lakey, J. H. (1998).
Discovery of critical TolA-binding residues in the bactericidal toxin colicin N: a
biophysical approach. Mol. Microbiol. 28(6), 1335-43
Rakonjac, J., Feng, J. & Model, P. (1999). Filamentous phage are released from the
bacterial membrane by a two-step mechanism involving a short C-terminal fragment
of pIII. J. Mol. Biol. 289, 1253-65
Rapoza, M. P. & Webster, R. E. (1993). The filamentous bacteriophage assembly
proteins require the bacterial SecA protein for correct localization to the membrane.
J. Bacteriol. 175, 1856-9
Rapoza, M. P. & Webster, R. E. (1995). The products of gen I and the overlapping in-
frame gene XI are required for filamentous phage assembly. J. Mol. Biol. 248, 627-38
Rasched, I. & Oberer, E. (1986). Ff coliphages: structural and functional
relationships. Microbiol. Rev. 50, 401-27
Ray, D. S., Bscheider, H. P. & Hofschneider, P. H. (1966). Replication of the single-
stranded DNA of the male-specific bacteriophage M13. Isolation of intracellular forms
of phage-specific DNA. J. Mol. Biol. 21, 473-83
Robinson, C. & Bolhuis, A. (2001). Protein targeting by the twin-arginine translocation
pathway. Nat. Rev. Mol. Cell. Biol. 2(5), 350-356
Rodi, D. J., Soares, A. & Makowski, L. (2002). Quantitative assessment of peptide
sequence diversity in M13 combinatorial peptide phage display libraries. J. Mol. Biol.
322(5), 1039–52
Russel, M. & Model, P. (1983). A bacterial gene, fip, required for filamentous phage
f1 assembly. J. Bacteriol. 154(3), 1064-76
Russel, M. & Model, P. (1985). Thioredoxin is required for filamentous phage
assembly. Proc. Natl. Acad. Sci. USA 82, 29-33
Russel, M. & Model, P. (1986). The role of thioredoxin in filamentous phage
assembly. Construction, isolation, and characterization of mutant thioredoxins. J.
Biol. Chem. 261, 14997-15005

Literaturverzeichnis
199
Russel, M., Kidd, S. & Kelley, M. R. (1986). An improved filamentous helper phage
for generating singe-stranded plasmid DNA. Gene 45, 333-38
Russel, M. & Model, P. (1989). Genetic analysis of the filamentous bacteriophage
packaging signal and of the proteins that interact with it. J. Virol. 63, 3284-3295
Russel, M. (1991). Filamentous phage assembly. Mol. Microbiol. 5, 1607-1613
Russel, M. (1993). Protein-protein interactions during filamentous phage assembly. J.
Mol. Biol. 231, 687-689
Russel, M. & Kazmierczak, B. (1993). Analysis of the structure and subcellular
location of filamentous phage plV. J. Bacteriol. 175, 3998-4007
Russel, M., Linderoth, N. A. & Sali, A. (1997). Filamentous phage assembly: variation
on a protein export theme. Gene 192(1), 23-32
Russel, M., Lowman, H., B. & Clackson, T. (2004). Introduction to phage biology and
phage display, 1–26. In: Phage Display: A practical approach, Practical approach
series 266, Edited by Tim Clackson and Henry B. Lowman, Oxford University Press
Great Clarendon Street, Oxford OX2 6DP
Ryter, A., Shuman, H. & Schwartz, M. (1975). Intergration of the receptor for
bacteriophage lambda in the outer membrane of Escherichia coli: coupling with cell
division. J. Bacteriol. 122, 295-301
Salstrom, J. S. & Pratt, D. (1971). Role of coliphage M13 gene 5 in single-stranded
DNA production. J. Mol. Biol. 61(3), 489-501
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecular cloning, a laboratory
manual. 2nd edit, 3, Cold Spring Habor Laboratory Press, New York
Samuelson, J. C., Chen, M., Jiang, F., Moller, I., Wiedmann, M., Kuhn, A., Phillips, G.
J. & Dalbey, R. E. (2000). YidC mediates membrane protein insertion on bacteria.
Nature 406(6796), 637-41
Samuelson, J. C., Jiang, F., Yi, L., Chen, M., de Gier, J. W., Kuhn, A. & Dalbey, R. E.
(2001). Function of YidC for the insertion of M13 procoat protein in Escherichia coli.
J. Biol. Chem. 276, 34847-52

Literaturverzeichnis
200
Sanger, F., Nicklen, S. & Coulson, A. R. (1977). DNA sequencing with chain-
terminating inhibitors. Proc. Natl. Acad. Sci. USA 74(12), 5463-67
Sargent, F., Bogsch, E. G., Stanley, N. R., Wexler, M., Robinson, C., Berks, B. C., &
Palmer, T. (1998). Overlapping functions of components of a bacterial Sec-
independent protein export pathway. EMBO J. 17, 3640-50
Scandella, D., & Arber, W. (1974). An Escherichia coli mutant which inhibits the
injection of phage lambda DNA. Virol. 58, 504-13
Scott, J. K. & Smith, G. P. (1990). Searching for peptide ligands with an epitope
library. Science 249, 386-90
Schägger, H. & Jagow, G. (1987). Tricine-sodium dodecyl sulfate-polyacrylamide gel
electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal.
Biochem. 166(2), 368-79
Schuenemann, T. A., Delgado-Nixon, V. M. & Dalbey, R. E. (1999). Direct evidence
that the proton motive force inhibits membrane translocation of positively charged
residues within membrane proteins. J. Biol. Chem. 274, 6855–64
Shine, J., & Dalgarno, L. (1974). The 3′-terminal sequence of Escherichia coli 16s
ribosomal RNA: complementarity to nonsense triplets and ribosome binding sites.
Proc. Natl. Acad. Sci. USA 71, 1342-46
Silver, P. & Wickner, W. T. (1980). Role of M13 gene 1 in filamentous virus
assembly. J. Virol. 35(1), 256-8
Simons, G. R., Beintema, J., Duisterwinkel, F., Konings, R. & Schoenmakers, J.
(1980). Genes 6, 7 and 9 of bacteriophage M13: identification of their products as
minor capsid proteins, 413-428. In: M. DuBow (ed.), Bacteriophage assembly.
Progress in clinical and biological research, vol 64, Alan R. Liss, Inc., New York
Simons, G. R., Konings, R. & Schoenmakers, J. (1981). Genes VI, VII, and IX of
phage M13 code for minor capsid proteinsof the virion. Proc. Natl. Acad. Sci. USA
78(7), 4194-98

Literaturverzeichnis
201
Smilowitz, H. (1974). Bacteriophage f1 infection: fate of the parental major coat
protein. J. Virol. 13, 94-9
Smith, G. P. (1985). Filamentous fusion phage: novel expression vectors that display
cloned antigens on the virion surface. Science 228, 1315-17
Stassen, A. P. M., Folmer, R. H. A., Hilbers, C. W. & Konings, R. N. H. (1995).
Single-stranded DNA binding protein encoded by the filamentous bacteriophage
M13: structural and functional characteristics. Mol. Biol. Reports 20(3), 109-27
Stopar, D., Spruijt, R. B., Wolfs, C. J. A. M. & Hemminga, M. A. (2003) Protein-lipid
interactions of bacteriophage M13 major coat protein. Biochim. Biophys. Acta
1611(1-2), 5-15
Strack, B., Lessl, M., Calendar, M. & Lanka, E. (1992). Common sequence motif,
EGYATA, identified within the primase domains of plasmid-encoded I- and P-type
DNA primases and the alpha protein of the Escherichia coli satellite phage P4. J.
Biol. Chem. 267, 13062-72
Sweeney, R. Y., Park, E. Y., Iverson, B. L. & Georgiou G. (2006). Assembly of
multimeric phage nanostructures through leucine zipper interactions. Biotechnol.
Bioeng. 95(3), 539-45
Tarry, M. J., Schäfer, E., Chen, S., Buchanan, G., Greene, N. P., Lea, S. M., Palmer,
T., Saibil, H. R. & Berks, B. C. (2009). Structural analysis of substrate binding by the
TatBC component of the twin-arginine protein transport system. Proc. Natl. Acad. Sci.
USA 106(32), 13284-9
Thie, H., Schirrmann, T., Paschke, M., Dübel, S. & Hust, M. (2008). SRP and Sec
pathway leader peptides for antibody phage display and antibody fragment
production in E. coli. New Biotech. 25(1), 49-54
Tokuda, H. & Matsuyama, S. (2004). Sorting of lipoproteins to the outer membrane in
E.coli. Biochim. Biophys. Acta 1694(1-3), IN1-9
Trenkner, E., Bonhoeffer, F. & Gierer, A. (1967). The fate of the protein component of
bacteriophage fd during infection. Biochem. Biophys. Res. Comm. 28, 932-9

Literaturverzeichnis
202
Tu, C. J., Schuenemann, D. & Hoffman, N. E. (1999). Chloroplast FtsY, chloroplast
signal recognition particle, and GTP are required to reconstitute the soluble phase of
light-harvesting chlorophyll protein transport into thylakoid membranes. J. Biol.
Chem. 274, 27219–24
Tu, C. J., Peterson, E. C., Henry, R. & Hoffman, N. E. (2000). The L18 domain of
light-harvesting chlorophyll proteins binds to cpSRP43. J. Biol. Chem. 275, 13187–90
Tzagoloff, H. & Pratt, D. (1964). The initial steps in infection with coliphage M13.
Virol. 24, 373-80
Ulbrandt, N. D., Newitt, J. A. & Bernstein H. D. (1997). The E. coli signal recognition
particle is required for the insertion of a subset of innermembrane proteins. Cell 88,
187-96
van Bloois, E., Haan, G. J., de Gier, J. W., Oudega, B. & Luirink, J. (2004). F1FO
synthase subunit c is targeted by the SRP to YidC in the E. coli inner membrane.
FEBS Letters 576(1-2), 97-100
van Bloois, E., Haan, G. J., de Gier, J. W., Oudega, B. & Luirink, J. (2006). Distinct
requirements for translocation of the N-tail and C-tail of the Escherichia coli inner
membrane protein CyoA. J. Biol. Chem. 281, 10002-9
Van Der Laan, M., Bechtluft, P., Kol, S., Nouwen N. & Driessen, A. J. (2004). F1FO
ATP synthase subunit c is a substrate of the novel YidC pathway for membrane
protein biogenesis. J. Cell. Biol. 165, 213-22
Vianney, A., Lewin, T. M., Beyer, W. F., Jr., Lazzaroni, J. C., Portalier, R. & Webster,
R. E. (1994). Membrane topology and mutational analysis of the TolQ protein of
Escherichia coli required for the uptake of macromolecules and cell envelope
integrity. J. Bacteriol. 176, 822-9
von Heijne, G. (1985). Signal sequences. The limits of variation. J. Mol. Biol. 184(1),
99-105
Wang, P. & Dalbey, R. E. (2011). Inserting membrane proteins: the YidC/Oxa1/Alb3
machinery in bacteria, mitochondria, and chloroplasts. Biochim. Biophys. Acta 1808,
866-75

Literaturverzeichnis
203
Watanabe, T., Hayashi, S. & Wu, H. C. (1988). Synthesis and Export of the outer
membrane lipoprotein in Escherichia coli mutants defective in generalized protein
export. J. Bacteriol. 170(9), 4001-7
Webster, R. E. & Cashman, J. S. (1973). Abortive infection of E. coli with the
bacteriophage f1: cytoplasmic membrane proteins and the f1 DNA-gene 5 complex.
Virol. 55(1), 20-38
Webster, R. E. (1991). The tol gene products and the import of macromolecules into
Escherichia coli. Mol. Microbiol. 5, 1005-11
Webster, R. E. (1996). Biology of the filamentous bacteriophage, 1-16. In: “Phage
Display of Peptides and Proteins: A Laboratory Manual”, B. K. Kay, J. Winter, J.
McCafferty (ed.). Academic Press Inc., San Diego, California.
Weiner, J. H., Bilous, P.T., Shaw, G. M., Lubitz, S. P., Frost, L., Thomas, G. H., Cole,
J. & Turner, R. J. (1998). A novel and ubiquitous system for membrane targeting and
secretion of cofactor-containing proteins. Cell 93, 93-101
Wexler, M., Bogsch, E. G., Klösgen, R. B., Palmer, T., Robinson, C. & Berks, B. C.
(1998). Targeting signals for a bacterial Sec-independent export system direct plant
thylakoid import by the delta pH pathway. FEBS Lett. 431(3), 339-42
Wu, L. F., Ize, B., Chanal, A., Quentin, Y. & Fichant, G. (2000). Bacterial twin-
arginine signal peptide-dependent protein translocation pathway: evolution and
mechanism. J. Mol. Microbiol. Biotechnol. 2, 179-89
Yahr, T. L. & Wickner, W. T. (2001). Functional reconstitution of bacterial Tat
translocation in vitro. EMBO J. 20(10), 2472-9
Yamane, K. & Mizushima, S. (1988). Introduction of basic amino acid residues after
the signal peptide inhibits protein translocation across the cytoplasmic membrane of
Escherichia coli. J. Bio.l Chem. 263, 19690–96
Yen, T. S. B. & Webster, R. E. (1981). Bacteriophage f1 gene II and X proteins. J.
Biol. Chem. 256(21), 11259-65

Literaturverzeichnis
204
Yen, T. S. B. & Webster, R. E. (1982). Translational control of bacteriophage f1 gene
II and gene X proteins by gene V protein. Cell 29(2), 337-45
Yi, L., Jiang F., Chen, M., Cain, B., Bolhuis, A. & Dalbey, R. E. (2003). YidC is strictly
required for membrane insertion of subunits a and c of the F(1)F(0)ATP synthase and
SecE of the SecYEG translocase. Biochem. 42, 10537-44
Yi, L., Celebi, N., Chen, M. & Dalbey, R. E. (2004). Sec/SRP requirements and
energetics of membrane insertion of subunits a, b, and c of the Escherichia coli F1F0
ATP synthase. J. Biol. Chem. 279, 39260-7

Anhang
205
7 Anhang
Abkürzungsverzeichnis
A
A Adenin
Abb. Abbildung
AFM Atomic Force Microscope
Amp Ampicillin
APS Ammoniumpersulfat
Ara Arabinose
AS Aminosäure(n)
ATP Adenosintriphosphat
B
bp Basenpaare
bzw. Beziehungsweise
C
C Cystein
ca. circa
CaCl2 Calciumchlorid
cm Zentimeter
C-Terminus Carboxyl-Terminus
D
D Asparaginsäure
dATP Desoxyadenosintriphosphat
dCTP Desoxycytosintriphosphat
ddNTP Didesoxyribonukleotidtriphosphat, z. B. ddATP
ddH2O zweifach destilliertes Wasser
dGTP Desoxyguanidintriphosphat
dNTP Desoxyribonukleotidphosphat, z. B. dATP
dTTP Desoxythymidinphospha
d.h. das heißt
DNA Desoxyribonucleinsäure
DNase Desoxyribonuclease
ds doppelsträngig

Anhang
206
DMSO Dimethylsulfoxid
DTT Dithiothreitol
E. coli Escherichia coli
E
E Glutaminsäure
E Extinktion
ECL Enhanced chemoluminescence, verstärkte
Chemolumineszenz
EDTA Ethylendiamintetraacetat
et al et aliter; und andere
F
F Phenylalanin
F´ Fertility (Fruchtbarkeit)
G
G Glycin
g Vielfaches der Erdbeschleunigung
g Gramm
Glc Glukose
gp Genprodukt
H
H Histidin
h Stunde(n)
HCl Chlorwasserstoffsäure
His Histidin
Hz Hertz
I
I Isoleucin
IPTG Isopropyl-β-D-thiogalactopyranosid
K
K Lysin
Kap. Kapitel
kb Kilobasen
kDa Kilodalton
KCl Kaliumchlorid

Anhang
207
L
L Leucin
L Liter
LB Luria-Bertani
M
M Methionin
M Mol
mA Milliampére
MCS Multiple Clonig Site, multiple Klonierungsstelle
mg Milligramm
MgCl2 Magnesiumchlorid
MgSO4 Magnesiumsulfat
min Minute
ml Milliliter
MnCl2 Manganchlorid
M.O.I. Multiplicity of infection
MOPS 3-(N-Morpholino)-Propansulfonsäure
N
N Asparagin
NaCl Natriumchlorid
Na2CO3 Natriumcarbonat
µCi Mikro-Curie
µg Mikrogramm
µl Mikroliter
µM Mikromol
µm Mikrometer
NaOH Natriumhydroxid
nN Nanonewton
nm Nanometer
ng Nanogramm
O
OD Optische Dichte
P
P Prolin

Anhang
208
PAGE Polyacrylamidgelelektrophorese
PCR polymerase chain reaction,
Polymerasekettenreaktion
PEG Polyethylenglycol
pfu plaque forming units
pN Pikonewton
Q
Q Glutamin
R
R Arginin
Ref. Referenz
RF Replikative Form
RbCl Rubidiumchlorid
RNA Ribonucleinsäure
RT Raumtemperatur
RKM Rasterkraftmikroskop
RNase Restriktionsendonuclease
rpm rotations per minute
S
S Serin
S Seite
s Sekunde
s. siehe
SDS Sodiumdodecylsulfate
SRP Signal-Erkennungs-Partikel
ss single stranded, Einzelstrang
T
T Threonin
Tab. Tabelle
TBE Tris-Borsäure-EDTA
TBS Tris-buffered saline
TCA Trichloressigsäure
TE Tris-EDTA
TEMED N,N,N´,N´-Tetramethylendiamin

Anhang
209
Tris Tris(hydroxymethyl)aminomethan
U
u. a. unter anderem
UV Ultraviolett
V
V Valin
V Volt
v/v Volumen/Volumen
W
W Tryptophan
wt Wildtyp
w/v Gewicht/Voluen
X
X-Gal 5-Bromo 4-Chlor-indoxyl-β-D-galactosid
Y
Y Tyrosin
Z
z. B. zum Beispiel
Diverse Zeichen
Ø Durchmesser
λ Wellenlänge Lambda (nm)
~ circa

Anhang
210
MALDI-TOF/MS Analysedaten
Spektrum 37 kDa gp1-Proteinbande
Mit der gp1 AS-Sequenz übereinstimmende AS sind rot dargestellt:

Anhang
211
Spektrum 31 kDa gp1-Proteinbande
Mit der gp1 AS-Sequenz übereinstimmende AS sind rot dargestellt:
m/z 804.2520 1.0 805.0855 1.0 805.2674 1.0 805.7854 1.0 806.2630 1.0 821.2963 1.0 826.2472 1.0 827.3998 1.0 829.4222 1.0 842.2302 1.0 858.1888 1.0 887.6396 1.0 902.3538 1.0 915.5013 1.0 917.2537 1.0 1024.5669 1.0 1118.5679 1.0 1207.6451 1.0 1247.5983 1.0 1262.6629 1.0 1304.6133 1.0 1318.6193 1.0 1351.7017 1.0 1378.6950 1.0 1380.6159 1.0 1381.5103 1.0
m/z 1395.7302 1.0 1409.7034 1.0 1411.7314 1.0 1427.7209 1.0 1594.8936 1.0 1780.8429 1.0 1894.8778 1.0 1908.8965 1.0 2116.0044 1.0 2327.1174 1.0 2568.1682 1.0 2575.2975 1.0 2595.1471 1.0 2597.8727 1.0 2599.1066 1.0 2612.2274 1.0 2628.1731 1.0 2645.2213 1.0 2669.2267 1.0 2901.2248 1.0 2917.2928 1.0 2920.3233 1.0 2931.3118 1.0 2933.3349 1.0

Anhang
212
Spektrum 20 kDa gp1-Proteinbande
Mit der gp1 AS-Sequenz übereinstimmende AS sind rot dargestellt:

Anhang
213
Sequenzen
gp1 DNA-Sequenz
1 ATGGCTGTTTATTTTGTAACTGGCAAATTAGGCTCTGGAAAGACGCTCGTTA
53 GCGTTGGTAAGATTCAGGATAAAATTGTAGCTGGGTGCAAAATAGCAACTAA
105 TCTTGATTTAAGGCTTCAAAACCTCCCGCAAGTCGGGAGGTTCGCTAAAAC
156 GCCTCGCGTTCTTAGAATACCGGATAAGCCTTCTATATCTGATTTGCTTGCT
208 ATTGGGCGCGGTAATGATTCCTACGATGAAAATAAAAACGGCTTGCTTGTTC
260 TCGATGAGTGCGGTACTTGGTTTAATACCCGTTCTTGGAATGATAAGGAAAG
312 ACAGCCGATTATTGATTGGTTTCTACATGCTCGTAAATTAGGATGGGATATT
364 ATTTTTCTTGTTCAGGACTTATCTATTGTTGATAAACAGGCGCGTTCTGCATT
417 AGCTGAACATGTTGTTTATTGTCGTCGTCTGGACAGAATTACTTTACCTTTTG
470 TCGGTACTTTATATTCTCTTATTACTGGCTCGAAAATGCCTCTGCCTAAATTA
522 CATGTTGGCGTTGTTAAATATGGCGATTCTCAATTAAGCCCTACTGTTGAGC
575 GTTGGCTTTATACTGGTAAGAATTTGTATAACGCATATGATACTAAACAGGC
627 TTTTTCTAGTAATTATGATTCCGGTGTTTATTCTTATTTAACGCCTTATTTATC
681 ACACGGTCGGTATTTCAAACCATTAAATTTAGGTCAGAAGATGAAATTAACT
733 AAAATATATTTGAAAAAGTTTTCTCGCGTTCTTTGTCTTGCGATTGGATTTGC
786 ATCAGCATTTACATATAGTTATATAACCCAACCTAAGCCGGAGGTTAAAAAG
839 GTAGTCTCTCAGACCTATGATTTTGATAAATTCACTATTGACTCTTCTCAGCG
891 TCTTAATCTAAGCTATCGCTATGTTTTCAAGGATTCTAAGGGAAAATTAATTA
944 ATAGCGACGATTTACAGAAGCAAGGTTATTCACTCACATATATTGATTTATGT
997 ACTGTTTCCATTAAAAAAGGTAATTCAAATGAAATTGTTAAATGTAATTAA
gp1 AS-Sequenz
1 M A V Y F V T G K L G S G K T L V S V G K I Q D K I V A G C K I A T N L D L R
40 L Q N L P Q V G R F A K T P R V L R I P D K P S I S D L L A I G R G N D S Y D
80 E N K N G L L V L D E C G T W F N T R S W N D K E R Q P I I D W F L H A R K
119 L G W D I I F L V Q D L S I V D K Q A R S A L A E H V V Y C R R L D R I T L P
158 F V G T L Y S L I T G S K M P L P K L H V G V V K Y G D S Q L S P T V E R W
196 L Y T G K N L Y N A Y D T K Q A F S S N Y D S G V Y S Y L T P Y L S H G R Y
234 F K P L N L G Q K M K L T K I Y L K K F S R V L C L A I G F A S A F T Y S Y I
273 T Q P K P E V K K V V S Q T Y D F D K F T I D S S Q R L N L S Y R Y V F K D
309 S K G K L I N S D D L Q K Q G Y S L T Y I D L C T V S I K K G N S N E I V K C
348 N

Anhang
214
gp11 DNA-Sequenz
1 ATGAAATTAACTAAAATATATTTGAAAAAGTTTTCTCGCGTTCTTTGTCTTGC
54 GATTGGATTTGCATCAGCATTTACATATAGTTATATAACCCAACCTAAGCCGG
108 AGGTTAAAAAGGTAGTCTCTCAGACCTATGATTTTGATAAATTCACTATTGAC
161 TCTTCTCAGCGTCTTAATCTAAGCTATCGCTATGTTTTCAAGGATTCTAAGG
213 GAAAATTAATTAATAGCGACGATTTACAGAAGCAAGGTTATTCACTCACATAT
266 ATTGATTTATGTACTGTTTCCATTAAAAAAGGTAATTCAAATGAAATTGTTAA
318 ATGTAATTAA
gp11 AS-Sequenz
1 M K L T K I Y L K K F S R V L C L A I G F A S A F T Y S Y I T Q P K P E V K K V
41 V S Q T Y D F D K F T I D S S Q R L N L S Y R Y V F K D S K G K L I N S D D L
80 Q K Q G Y S L T Y I D L C T V S I K K G N S N E I V K C N

Anhang
215
Publikationen
Ploss, M. & Kuhn, A. (2010). Kinetics of filamentous phage assembly. Phys. Biol.
7(4), 045002
Ploss, M. & Kuhn, A. (2011). Membrane insertion and assembly of epitope-tagged
gp9 at the tip of the M13 phage. BMC Microbiol. 11(1), 211-9

Anhang
216
Danksagung
Bedanken möchte ich mich bei Herrn Prof. Dr. Andreas Kuhn für die freundliche
Überlassung des interessanten Themas, die Bereitstellung des Arbeitsplatzes und
die Ermöglichung dieser Arbeit. Vielen Dank für das Korrekturlesen des Manuskripts
sowie der hilfreichen Ratschläge.
Herrn Prof. Dr. Artur Pfitzner vom Institut für Genetik, Fg. Allgemeine Virologie,
danke ich für die freundliche Bereitschaft zur Übernahme des Koreferats.
Bedanken möchte ich mich auch bei Frau Dr. Sandra Facey für Ihre Hilfsbereitschaft,
den hilfreichen Diskussionen, konstruktiven Anregungen und der Unterstützung in
allen Lebenslagen sowie dem Lesen des Manuskripts; ich danke Dir!
Danken möchte ich auch allen Mitarbeitern des Instituts für Mikrobiologie für die
Zusammenarbeit und Hilfsbereitschaft.
Des Weiteren möchte ich mich herzlich bei meiner Frau Stefanie für die hilfreichen
und konstruktiven Diskussionen, das Manuskript betreffend, bedanken. Ich danke Dir
für das Korrekturlesen dieser Arbeit, den allerbesten Rückhalt, Deine Geduld und für
Deine Aufmunterungen. Ich bin froh, dass es Dich in meinem Leben gibt!!
Ein weiteres Dankeschön gilt meinem Sohn Florian, der mich während des
Schreibens des Manuskripts stets mit selbst gebrühten Kaffee versorgt hat, für seine
abwechslungsreichen Ablenkungen und die Ausflüge in den Garten, zum Sand-
kasten und auf den Spielplatz, wodurch ich immer wieder neue Kräfte sammeln
konnte. Flo ich bin dankbar, dass Du Teil meines Lebens bist!!
Meiner Schwiegermutter Ingrid möchte ich herzlich für das gute Essen, das
Korrekturlesen des Manuskripts und ihre stets herzliche Hilfe und Unterstützung
danken. Du bist die weltbeste Schwiegermutter die man sich als Schwiegersohn nur
wünschen kann.
Meiner Schwägerin Monika und meinem Schwager Gregor möchte ich für die Hilfe
bei der Klärung grammatikalischer Fragen bedanken. Gregor ich möchte mich bei Dir
recht herzlich für die Zeit, die Du Dir, trotz Deines engen Zeitplans, genommen hast
um meine „Summary“ zu lesen, bedanken.

















![ATHLETIC-TEAM WITTLICH Bestenliste.pdf · 12,09 Koller, Lars 04 Athletic-Team Wittlich 06.05.2017 Daun 3 x 800 m [M13 - Disziplinübersicht ] [ Klassenübersicht ] 8:46,05 Athletic-Team](https://static.fdokument.com/doc/165x107/5d5172cb88c993ad428b9e99/athletic-team-bestenlistepdf-1209-koller-lars-04-athletic-team-wittlich-06052017.jpg)

