

CELL-SPECIFIC REGULATION OF PTX3 BY GLUCOCORTICOID ... · CELL-SPECIFIC REGULATION OF PTX3 BY...
26
CELL-SPECIFIC REGULATION OF PTX3 BY GLUCOCORTICOID HORMONES IN HEMATOPOIETIC AND NON-HEMATOPOIETIC CELLS* Andrea Doni ¶ , Giovanna Mantovani ƒ , Chiara Porta ¶ , Jan Tuckermann φ , Holger M. Reichardt ϕ , Anna Kleiman φ , Marina Sironi ¶ , Luca Rubino ¶ , Fabio Pasqualini ¶ , Manuela Nebuloni ξ , Stefano Signorini ß , Giuseppe Peri ¶ , Antonio Sica ¶ , Paolo Beck-Peccoz ƒ , Barbara Bottazzi ¶ , and Alberto Mantovani ¶,† from ¶ Istituto Clinico Humanitas (ICH), IRCCS, Department of Immunology and Inflammation, Rozzano, Milan, Italy ƒ Department of Medical Sciences, Endocrine Unit, University of Milan, Fondazione Policlinico, IRCCS, Milan, Italy φ Molecular Biology of Tissue specific Hormone Action, Leibniz Institute for Age Research Fritz-Lipmann, Jena, Germany ϕ Department of Cellular and Molecular Immunology, University of Goettingen, Humboldtallee, Goettingen, Germany ß Department of Laboratory Medicine, Hospital of Desio, Desio, Milan, Italy ξ Pathology Unit, Luigi Sacco Department of Clinical Sciences, University of Milan, Milan, Italy † Institute of General Pathology, University of Milan, Milan, Italy Runnung Title: PTX3 regulation by glucocorticoids Address correspondence to: Alberto Mantovani, Istituto Clinico Humanitas (ICH), via Manzoni, 56 20089 Rozzano, Milan, Italy. Tel. 0039-0282242445; Fax. 0039-0282245101; E-mail address: [email protected] The prototypic long pentraxin 3 (PTX3) is a fluid phase pattern recognition receptor which plays non-redundant roles in the resistance against diverse pathogens, in the assembly of a hyaluronic acid-rich extracellular matrix and in female fertility. Inflammatory signals induce production of PTX3 in diverse cell types including myeloid dendritic cells (DC), fibroblasts and endothelial cells (EC). The present study was designed to explore the effect of glucocorticoid hormones (GC) on PTX3 production in different cellular contexts. In myeloid DC GC inhibited the PTX3 production. In contrast, in fibroblasts and EC GC alone induced and, under inflammatory conditions, enhanced and extended PTX3 production. In vivo administration of GC augmented the blood levels of PTX3 in mice and humans. Moreover, patients with Cushing’s syndrome had increased levels of circulating PTX3 whereas PTX3 levels were decreased in subjects affected by iatrogenic hypocortisolism. In non-hematopoietic cells, GC receptor (GR) functioned as a ligand- dependent transcription factor (dimerization- dependent) to induce PTX3 gene expression. In contrast, in hematopoietic cells, GR repressed PTX3 gene transcription by interfering (dimerization-independent) with the action of other signalling pathways, likely NFκB and AP-1. Thus, divergent effects of GC were found to be due to different GR mechanisms. The results presented here indicate that GC have divergent effects on PTX3 production in hematopoietic (DC and macrophages) and non-hematopoietic (fibroblasts and EC) cells. The divergent effects of GC on PTX3 production is likely to reflect the different functions of this multifunctional molecule in innate immunity and in the construction of the extracellular matrix. Pentraxins are a superfamily of conserved proteins usually characterized by a cyclic multimeric structure (1-4). The classic short pentraxins C-reactive protein (CRP) and serum amyloid P component (SAP) are acute phase proteins in man and mouse, respectively. They 1 http://www.jbc.org/cgi/doi/10.1074/jbc.M805631200 The latest version is at JBC Papers in Press. Published on August 14, 2008 as Manuscript M805631200 Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on April 30, 2020 http://www.jbc.org/ Downloaded from
Transcript of CELL-SPECIFIC REGULATION OF PTX3 BY GLUCOCORTICOID ... · CELL-SPECIFIC REGULATION OF PTX3 BY...

CELL-SPECIFIC REGULATION OF PTX3 BY GLUCOCORTICOID HORMONES IN HEMATOPOIETIC AND NON-HEMATOPOIETIC
CELLS* Andrea Doni ¶, Giovanna Mantovani ƒ, Chiara Porta ¶, Jan Tuckermann φ, Holger M. Reichardt ϕ, Anna Kleiman φ, Marina Sironi ¶, Luca Rubino ¶,
Fabio Pasqualini ¶, Manuela Nebuloni ξ, Stefano Signorini ß, Giuseppe Peri ¶, Antonio Sica ¶, Paolo Beck-Peccoz ƒ, Barbara Bottazzi ¶, and Alberto
Mantovani ¶,†
from ¶ Istituto Clinico Humanitas (ICH), IRCCS, Department of Immunology and Inflammation, Rozzano, Milan, Italy
ƒ Department of Medical Sciences, Endocrine Unit, University of Milan, Fondazione Policlinico, IRCCS, Milan, Italy
φ Molecular Biology of Tissue specific Hormone Action, Leibniz Institute for Age Research Fritz-Lipmann, Jena, Germany
ϕ Department of Cellular and Molecular Immunology, University of Goettingen, Humboldtallee, Goettingen, Germany
ß Department of Laboratory Medicine, Hospital of Desio, Desio, Milan, Italy ξ Pathology Unit, Luigi Sacco Department of Clinical Sciences, University of Milan, Milan, Italy
† Institute of General Pathology, University of Milan, Milan, Italy Runnung Title: PTX3 regulation by glucocorticoids
Address correspondence to: Alberto Mantovani, Istituto Clinico Humanitas (ICH), via Manzoni, 56 20089 Rozzano, Milan, Italy. Tel. 0039-0282242445; Fax. 0039-0282245101; E-mail address: [email protected] The prototypic long pentraxin 3 (PTX3) is a fluid phase pattern recognition receptor which plays non-redundant roles in the resistance against diverse pathogens, in the assembly of a hyaluronic acid-rich extracellular matrix and in female fertility. Inflammatory signals induce production of PTX3 in diverse cell types including myeloid dendritic cells (DC), fibroblasts and endothelial cells (EC). The present study was designed to explore the effect of glucocorticoid hormones (GC) on PTX3 production in different cellular contexts. In myeloid DC GC inhibited the PTX3 production. In contrast, in fibroblasts and EC GC alone induced and, under inflammatory conditions, enhanced and extended PTX3 production. In vivo administration of GC augmented the blood levels of PTX3 in mice and humans. Moreover, patients with Cushing’s syndrome had increased levels of circulating PTX3 whereas PTX3 levels were decreased in subjects affected by iatrogenic hypocortisolism. In non-hematopoietic cells, GC receptor (GR) functioned as a ligand-
dependent transcription factor (dimerization-dependent) to induce PTX3 gene expression. In contrast, in hematopoietic cells, GR repressed PTX3 gene transcription by interfering (dimerization-independent) with the action of other signalling pathways, likely NFκB and AP-1. Thus, divergent effects of GC were found to be due to different GR mechanisms. The results presented here indicate that GC have divergent effects on PTX3 production in hematopoietic (DC and macrophages) and non-hematopoietic (fibroblasts and EC) cells. The divergent effects of GC on PTX3 production is likely to reflect the different functions of this multifunctional molecule in innate immunity and in the construction of the extracellular matrix.
Pentraxins are a superfamily of conserved proteins usually characterized by a cyclic multimeric structure (1-4). The classic short pentraxins C-reactive protein (CRP) and serum amyloid P component (SAP) are acute phase proteins in man and mouse, respectively. They
1
http://www.jbc.org/cgi/doi/10.1074/jbc.M805631200The latest version is at JBC Papers in Press. Published on August 14, 2008 as Manuscript M805631200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

are produced in the liver in response to inflammatory signals, most prominently IL-6. CRP and SAP bind different ligands, in a calcium dependent manner, and they are involved in the innate resistance to microbes and the scavenging of cellular debris and extracellular matrix components (5).
The prototypic long pentraxin PTX3 shares similarities with the classical short pentraxins. However it has an unrelated long N-terminal domain coupled to the C-terminal pentraxin domain, and differs in gene organization, cellular source, inducing stimuli and ligands recognized (3,4,6). PTX3 is rapidly produced and released by several cell types, in particular by mononuclear phagocytes, myeloid DC, fibroblasts, epithelial and EC in response to primary inflammatory signals (e.g. TLR engagment, TNFα, IL-1β) (3,7-11) and its production is amplified by IL-10 (12). PTX3 binds the complement component C1q, apoptotic cells, the extracellular matrix component TSG6 and selected microorganisms, including Aspergillus fumigatus and Pseudomonas aeruginosa (13-19).
Recent studies in gene-modified mice have shown that PTX3 plays complex non-redundant functions in vivo, ranging from innate immunity against diverse microorganisms to the assembly of a hyaluronic acid-rich extracellular matrix and female fertility (3,6,14,16,17,20-22). PTX3 acts as a fluid phase pattern recognition receptor which plays a non-redundant role in resistance against selected pathogens and has properties similar to antibodies. Its production is induced by pathogen recognition, it recognizes microbial moieties, activates the classical pathway of complement and facilitates recognition by macrophages and DC. Thus, PTX3 behaves as a bona fide ante-antibody (3,6,14,17,18).
PTX3 is also a constituent of the extracellular matrix which plays a non-redundant role in the assembly of the hyaluronan-rich viscoelastic matrix of the cumulus oophorus and hence in female fertility (3,6,14,16,21,23). PTX3 binds TSG6, a hyaluronic acid-binding protein and may act as a focal point of the assembly of hyaluronic acid-rich matrices (3,6,16).
In addition, PTX3 has a tuning function under inflammatory conditions at least in part limiting tissue damage by regulating apoptotic cell clearance and acting as a third-party agent
between microbial stimuli and dying cells (3,19,22,24-27).
GC have a complex regulatory activity on components of innate and adaptive immunity (28-30). As PTX3 is a multifunctional fluid phase pattern recognition receptor which links innate immunity, inflammation and matrix deposition, and shows a unique pattern of regulation (3,12), we explored the GC effect on the PTX3 production. The present study reports that exogenous, as well as endogenous GC, regulate PTX3 production in a cell-type specific manner and through different actions of GR. The divergent effect of GC on PTX3 production in hematopoietic (DC and macrophages) and non-hematopoietic (fibroblasts and EC) cells mirrors the diverse functions of this molecule in innate immunity and in the assembly of the extracellular matrix.
EXPERIMENTAL PROCEDURES
Media and reagents The following reagents were used for tissue culture: pyrogen-free saline (S. A. L., Bergamo, Italy), RPMI 1640 medium (Biochrom, Berlin, Germany), M199 medium (Biochrom, Berlin, Germany), 200mM L-glutamine (Biochrom, Berlin, Germany), aseptically collected fetal calf serum (FCS) (Hyclone, Laboratories, Logan, UT) and PBS (Cambrex Bio Science, Belgium). Heparin and collagenase were from SIGMA-Aldrich. Human recombinant granulocyte macrophage colony-stimulation factor (GM-CSF) was a gift from Novartis (Milan, Italy) and human IL-13 was a gift from A. Minty (Sanofi Elf Bio Recherches, Labège, France). Recombinant TNFα and IL-1β were purchased by Peprotech (London, GB) and LPS from E. Coli serotype R515 (RE) (>99.9% of purity) was obtained from Alexis. Diverse Dex (Dexamethasone-21-phosphate disodium salt) preparations from MP Biomedicals (Germany) and Sigma-Aldrich were used. RU486 (Mifepristone) and PGE2 were obtained from SIGMA-Aldrich. Vitamin D3 (VitD3) was a gift from Luciano Adorini (Bioxell, Milan, Italy). Reagents were controlled to exclude endotoxin contamintion by LAL test (BioWhittaker Inc.-Cambrex, MD). DC and Macrophages Monocyte-derived DC were generated as previously described (8). Briefly, blood monocytes were obtained from fresh buffy coats of healthy donors (courtesy of the Centro
2
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Trasfusionale, Ospedale di Desio, Milan, Italy) by Ficoll (SIGMA-Aldrich) and Percoll (Amersham, Uppsala, Sweden) and after discard of non-adherent cells. Purified monocytes were cultured for 6 days at 1 x 106 cells/mL in 6-well tissue culture plates (BD Biosciences) in RPMI 1640 medium supplemented with 2mM/L L-glutamine and 10% FCS, 50 ng/mL GM-CSF and 20 ng/mL IL-13. RAW 264.7 murine macrophage cell line was obtained from ATCC. Bone marrow-derived macrophages (BMM) were obtained from WT and GR dimerization knock-out (GRdim/dim) mice as previously described (31). Cells were then cultured for 24 h in RPMI 1640 medium supplemented with 2mM/L L-glutamine and 2% FCS at 1 x 106 cells/mL in 24-well tissue culture plates (BD Biosciences) in the presence of LPS, 1 and 100ng/ml, Dex, ranging from 10-5 to 10-7M and RU486 10-5M. RU486 and Dex were added 1h prior to the challenge with LPS. Fibroblasts and EC Human normal lung fibroblasts, WI38, and the human fibrosarcoma cell line, 8387, were obtained from ATCC. WI38 and 8387 cell lines were cultured in RPMI supplemented with 10% FCS and 2mM/L L-glutamine until stimulation. Mouse embryonic fibroblasts (MEF) from WT and GRdim/dim mice were obtained as previously described (32) and prepared according to standard techniques. Cells were plated at a density of 1 x 105 cells and expanded in 6-well plates in RPMI 1640 medium supplemented with 2mM/L L-glutamine, 10% FCS and β-mercaptoethanol. The number of MEF at confluence was 5 x 105 cells per well. Cells were stimulated at passage 7-9. Human umbilical vein endothelial cells (HUVEC) were isolated from umbilical cord vein by digestion with 0.5% collagenase as previously described (33). HUVEC were then plated at a density of 1 x 105 cells per well in gelatin-coated 24-well plates (BD Biosciences) and grown to confluence in medium 199/DMEM (1:1) supplemented with 20% FCS, 2mM/L L-glutamine, 100µg/mL streptomycin, 100U/mL penicillin, 5µg/mL endothelial cell growth factor and 10µg/ml heparin. Cultured medium was refreshed every other day. The number of HUVEC at confluence was 5 x 105 cells per well. Confluent cells were then cultured for the reported periods in 24-well tissue culture plates (BD Biosciences) in medium supplemented with
2mM/L L-glutamine, 2% FCS in the presence of 20ng/mL TNFα or murine TNFα, 20ng/mL IL-1β, Dex ranging from 10-5 to 10-8M and RU486 10-5M. RU486 and Dex were added 1h before to challenge with proinflammatory stimuli. Plasmid Construction, Transfection and Luciferase assay A 1.37-kilobase EcoRI-PvuII genomic fragment from lP2 phage (10) which spans nucleotides 1 to 1374 of human PTX3 promoter (accession X97748) was used as DNA template for polymerase chain reaction (PCR) with the following oligonucleotides: Forward primer (5’-AAAAAGGTACCCCGGATCTCCCTTCTAAC-3’) was designed in antisense orientation on PTX3 promoter from nucleotide 7 to nucleotide 25 with an additional 11 nucleotides at the 5’ end; 6 of them (underlined) give rise to a KpnI restriction site; Revers primer (5’-TTTCCACAGCTGGCGGGAGGAGACTCTCAA -3’) was designed in sense orientation on the PTX3 promoter from nucleotides 1351 to 1380, it includes (underlined) a PvuII site. The PCR reaction products were subcloned in pGEMT easy vector, next human PTX3 promoter fragment was excised by KpnI-PvuII digestion and subcloned in SmaI-KpnI digested pGV-B2 vector. The luciferase reporter clones containing the human PTX3 promoter were sequenced (Primm) and subsequently used in functional assays. Cells transfections were performed using lipofectamine 2000TM according to the manufacturer’s instructions (Invitrogen, Life Technologies). Luciferase assay was performed using the Luciferase Assay Kit by Promega (Cat. # E1500). Briefly, cells were stimulated at various times as described, washed in PBS and resuspended in Cell Lysis Buffer (Promega Kit). Cell Lysate proteins were quantified using Comassie method (Biorad). In order to perform luciferase assay, 50 μL of luciferase substrate, reconstituted following the manufacturer’s instructions, were added to 15µg of total proteins. Signals was read on Luminometer LB 9507 (Berthold Inc.) RNA For Northern Blot analysis total RNA was extracted by TRIzol method in according to manufacturer’s instructions (Invitrogen, Life Technologies), blotted and hybridized as described (8). PTX3 mRNA densitometric analysis was performed by quantification of relative optical density (ROD) x area using an
3
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Image Analysis Software (AIS) (Image Research, Inc., Ontario). For gene expression analysis by quantitative PCR, according to the manufacturer’s instructions, total RNA was reverse transcribed using cDNA Archive kit (Applied Biosystem, NJ, USA), then real time PCR was performed with Power Syber Green PCR Master Mix (Applied Biosystem) and detected by 7900HT Fast Realtime System (Applied Biosystem). Data was processed using the SDS2.2.2 software (Applied Biosystem). All results were normalized to the expression of the housekeeping gene, β-actin, in the PCR reactions and expressed as folds change in mRNA expression with respect to the untreated cells. Data represented are from 3 independent experiments done in triplicate. Chromatin Immunoprecipitation Assay (ChIP) ChIP assays were performed in HUVEC, WI38 and DC as previously described (34). Chromatin was sheared by sonication to give fragments 500-1000bp length. Chromatin immunoprecipitation was performed overnight at 4°C with 4µg of a rabbit polyclonal anti-RNA Pol II antibody (N-20, #B0408), a rabbit polyclonal anti-GR (H-300, #B0604) and a control antisera (Santa Cruz Biotechnologies, CA). Immunoprecipitated DNA was amplified by Real-time PCR with PTX3 promoter-specific primers spanning PTX3 promoter from nucleotide 1231 to nucleotide 1380. The ChIP results were normalized to input and expressed as fold induction with respect to the control cells. Data are 3 independent experiments representative done in triplicate. Proteins Human PTX3 (PTX3) was measured using an ELISA previously described (35). IL-6 was measured by ELISA in according to manufacturer’s instructions (R&D Systems, Minneapolis, MN). Murine PTX3 (mPTX3) was measured by ELISA based on mAbs obtained by immunization of PTX3-/- mice (14). In brief, 96-well ELISA plates (Nunc, Roskilde, Denmark) were coated with 100ng of anti-mPTX3 monoclonal antibody (mAb) MnmE1. Non-specific binding sites were saturated by 300µl of 5% dry milk, prior to the addition of purified recombinant mPTX3 standards (from 0.1 to 10ng/mL) and unknown samples, diluted in PBS (Cambrex Bio Science, Belgium) plus 2% of bovine serum albumin BSA (SIGMA-Aldrich). After incubation, 25ng of biotin-conjugated anti-
mPTX3 mAb MNmE2 were added. Streptavidin-peroxidase was then added followed by the addition of TMB liquid Substrate System (SIGMA-Aldrich). After each step, plates were extensively washed with PBS (Cambrex Bio Science, Belgium) containing 0.05% Tween 20 (Merk, Germany), also used for reagent diluent. Immunohistochemistry Skin samples (1 normal and 1 with Cushing syndrome) were collected and fixed in 4% formalin. Paraffin embedded tissue were cut at 4 μm, and incubated with 2% H2O2 in methanol (to eliminate endogenous peroxidase activity). Sections were pre-incubated with PBS + 1% BSA + 1% normal human serum + 0.02% NP40 (SIGMA-Aldrich) to block non specific sites for 10 minutes at room temperature and than incubated at room temperature for 2 hours with an anti-PTX3 affinity-purified rabbit polyclonal (0.25µg/mL). A purified rabbit serum was also used as negative control. After extensive wash sections were incubated with secondary antibodies, Super Sensitive IHC Detection Systems (BioGenex, San Ramon, USA) was used according to the manufacturer’s instructions. Staining reactions were performed with 3,3-diaminobenzidine (DAB, BioGenex) as substrate. Sections were counterstained with hematoxylin and mounted with Eukitt. Mice PTX3 levels were measured in male, adult (8 weeks), C57/BL6 mice (Charles River Lab., Calco, Italy). Dex was prepared as a suspension in PBS (Cambrex Bio Science, Belgium) at 5mg/mL and was administrated, at 30mg/Kg, via i. p. injection prior to blood collection at 24h. Control mice received i. p. injections of vehicle. Six animals were examined for each experimental group. Patients The study included 16 patients with hypercortisolism and 8 patients with hypocortisolism admitted to the Endocrine Unit, Fondazione Policlinico, IRCCS. All patients with hypercortisolism (5 males and 11 females, aged 34 to 72) were affected by ACTH-dependent Cushing syndrome due to either a pituitary adenoma or to ectopic ACTH syndrome. All patients had elevated 24h urinary cortisol excretion and lack of suppression of cortisol levels after low dose Dex test. Patients with hypocortisolism (4 males and 5 females, aged 40 to 67) had a history of Cushing disease due to a pituitary adenoma surgically removed
4
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

by the transsphenoidal route. Three months after surgery, all patients had reduced 24h urinary cortisol excretion and lack of induction of cortisol levels after a low dose (1μg) ACTH stimulation test. The patients were evaluated after adequate cortisone acetate treatment withdrawal. Serum PTX3 and cortisol levels were measured in 7 normal subjects, both under basal conditions and after an overnight treatment with 1 mg Dex. PTX3 was measured by ELISA as described. Serum cortisol levels were measured by EIA method (Nichols Institute, San Juan Capistrano, USA). The control cohort was from the same unit. Informed consent was obtained in all cases, after project approval by the local Ethic Committee.
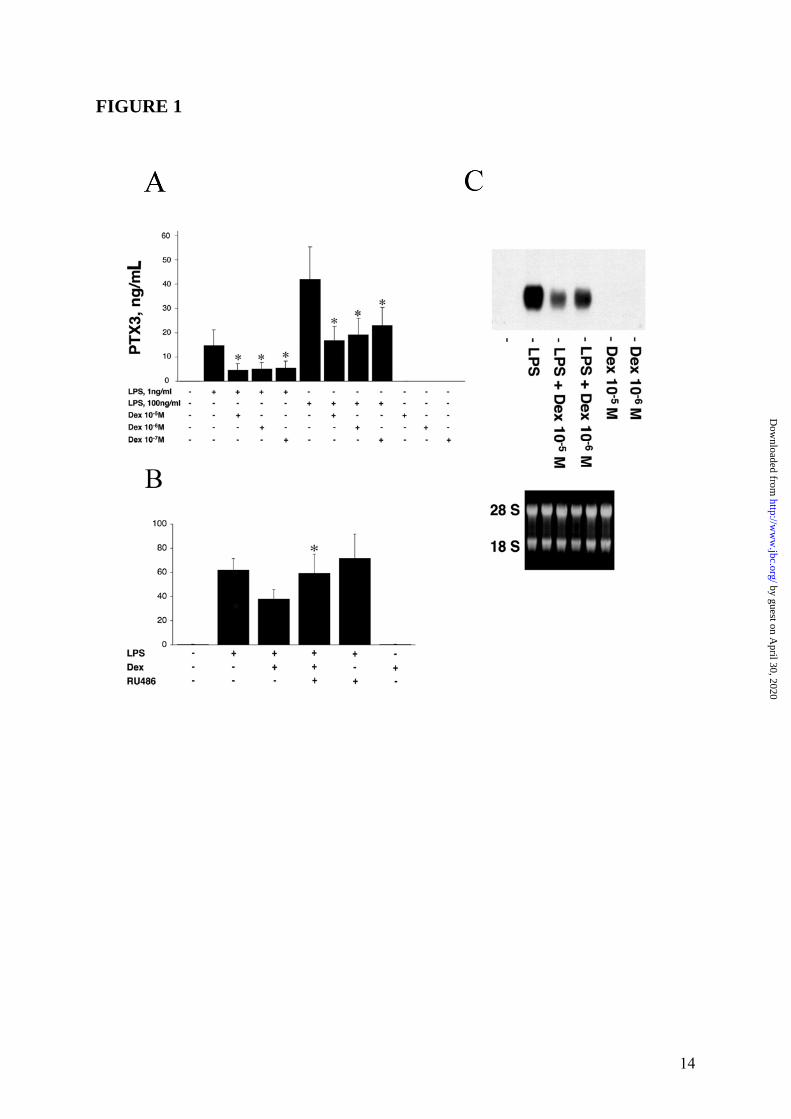
RESULTS Regulation of PTX3 production by GC in monocyte-derived DC In a first series of experiments, we evaluated the effect of GC on monocyte-derived DC and their capability to regulate the PTX3 production. Figure 1 Panel A summarizes results of a series of 12 experiments where monocyte-derived DC were treated with LPS (1 and 100ng/mL), Dex (10-5-10-7M) and combinations thereof. As expected, LPS induced PTX3 production in DC (8) and resulted in release of 14.8 ± 6.7ng/mL and 42.7 ± 12.0 ng/mL (mean ± SD) when DC were challenged for 24h with either 1 or 100ng/mL of LPS respectively. GC alone did not induce PTX3 in DC. In all experiments performed, Dex caused a strong inhibition of the LPS-induced PTX3 production. As shown in Figure 1 Panel A, when Dex (10-5M) was combined to either 1 or 100ng/mL LPS, the inhibition of PTX3 production was 63.0 ± 14.5% or 59.5 ± 12.0%, respectively. The inhibitory effect of GC on PTX3 production by monocyte-derived DC was dose-related (range of 10-5-10-
7M) (Figure 1 Panel A). When RU486 (mifepristone), a GR antagonist, was combined to LPS and Dex, the regulatory effect of GC on PTX3 production by myeloid DC was blocked. As shown in Figure 1 Panel B, in a series of 4 experiments, PTX3 production induced by LPS (61.1 ± 4.2ng/mL) was inhibited by the addition of Dex (37.0 ± 3.4ng/mL), but not when RU486 was added to the culture (57.1 ± 12.1ng/mL). RU486 did not induce PTX3 and did not alter the LPS-induced PTX3 production by DC (65.1 ± 13.2ng/mL) (Figure 1 Panel B).
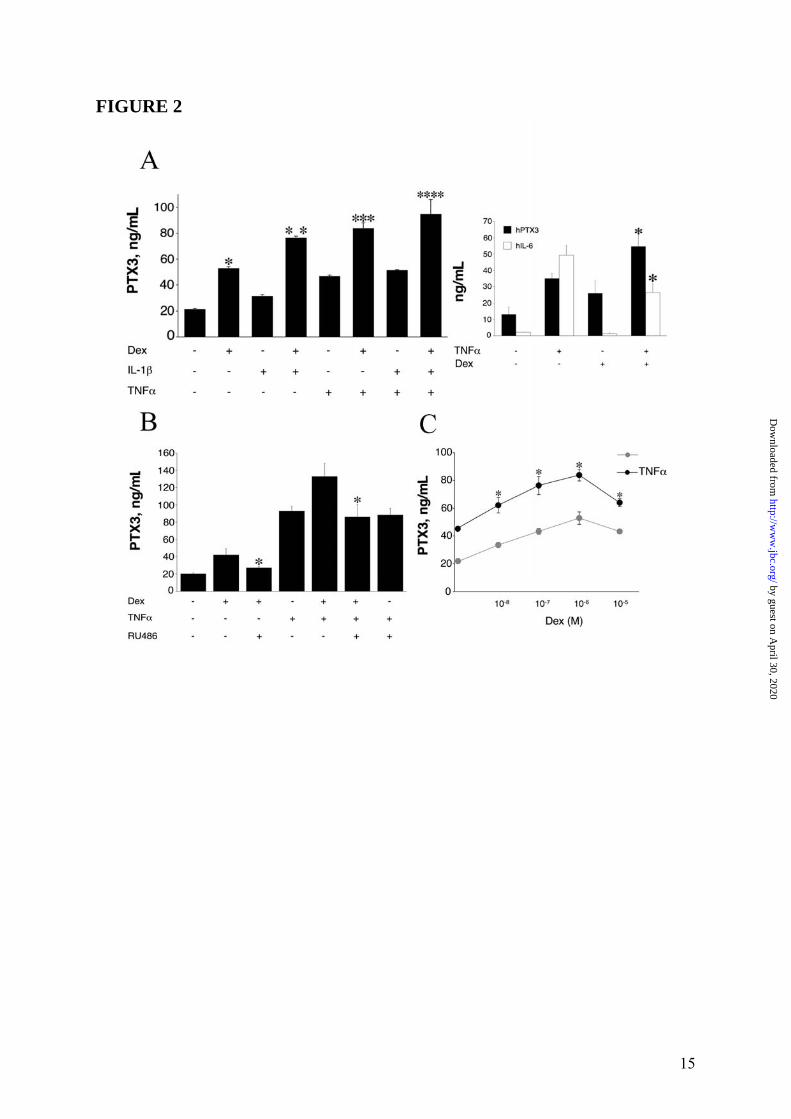
Thus, the inhibitory effect of Dex on LPS-induced PTX3 production by myeloid DC is GR-dependent. PTX3 transcript levels were assessed by Northern Blot analysis and, as shown in Figure 1 Panel C, Dex was found to cause a marked inhibition of PTX3 mRNA expression induced by LPS in DC after 8h of culture. Densitometric analysis revealed that a combined treatment with LPS and Dex used at 10-5M or 10-6M, caused an inhibition of transcript levels corresponding to 64% or 59% of the response to LPS, respectively. GC also inhibited the PTX3 production by LPS-treated monocytes and macrophages (data not shown). Regulation of PTX3 production by GC in non-hematopoietic cells As PTX3 is produced by several cell types, including fibroblasts and EC (3), we evaluated also the ability of GC to regulate the PTX3 production in a human fibrosarcoma (8387), normal human fibroblasts (WI38) and HUVEC. In all experiments performed (n = 4), unexpectedly Dex alone induced production of PTX3 in 8387 cells (52.9 ± 1.1ng/mL) after 24h of culture (Figure 2 Panel A, left part). PTX3 induction was observed following challenge with IL-1β (31.5 ± 1.0ng/mL) and TNFα (46.9 ± 0.8ng/mL). When Dex was combined with IL-1β or TNFα, an additive or synergistic enhancement of PTX3 production was observed in 8387 (142.7 ± 3.9%, 78.9 ± 9.1%) (Figure 2 Panel A). As expected, the TNFα-induced IL-6 was significantly down-regulated by GC both in myeloid DC (data not shown) and in 8387 (series of 3 experiments, Figure 2 Panel A, right part). It was important to ascertain whether the production of PTX3 by GC in cells of fibroblastic origin was receptor-dependent. In a series of 3 experiments (Figure 2 Panel B), PTX3 production by GC-stimulated 8387 (42.0 ± 7.1ng/mL) was significantly inhibited when cells were cultured in the presence of RU486 (26.1 ± 1.6ng/mL, p < 0.05). As expected, TNFα-induced PTX3 production (93.1 ± 5.4ng/mL) by 8387 was markedly augmented in combination with Dex (132.8 ± 15.0ng/mL). The augmentation by Dex was inhibited by the addition of RU486 (86.0 ± 14.6ng/mL). RU486 did not inhibit the TNFα-induced PTX3 production (88.4 ± 7.6ng/mL) (Figure 2 Panel
5
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

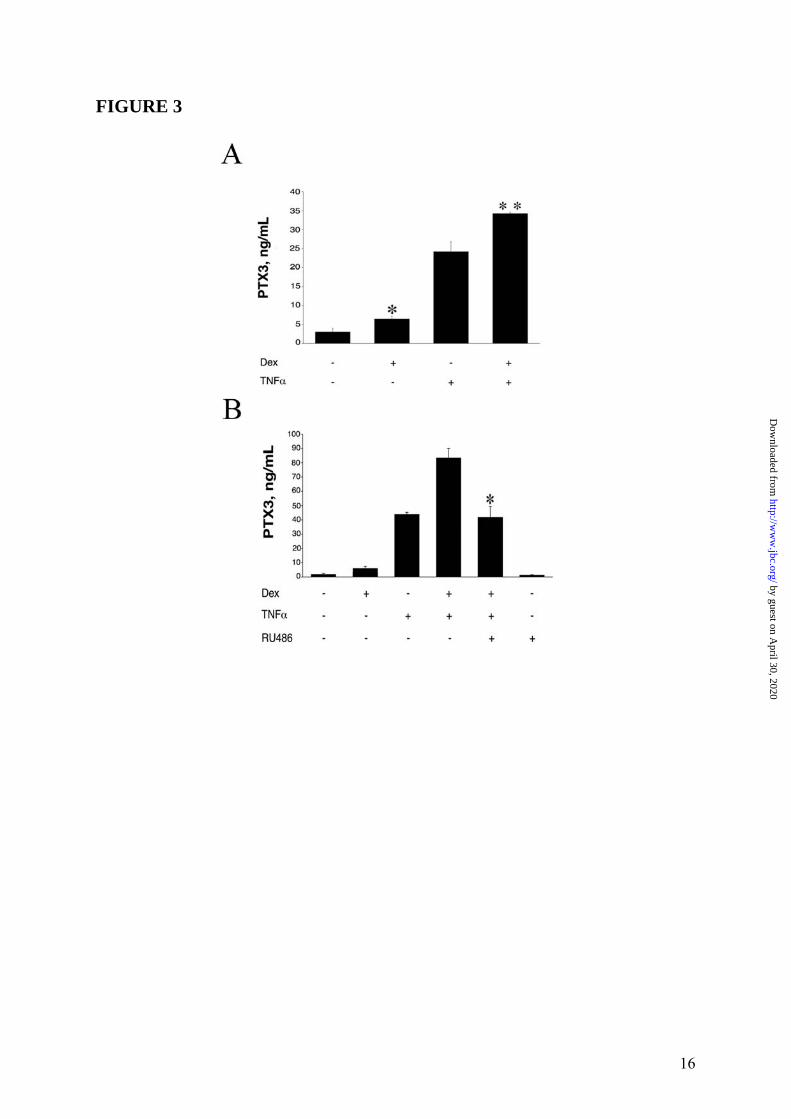
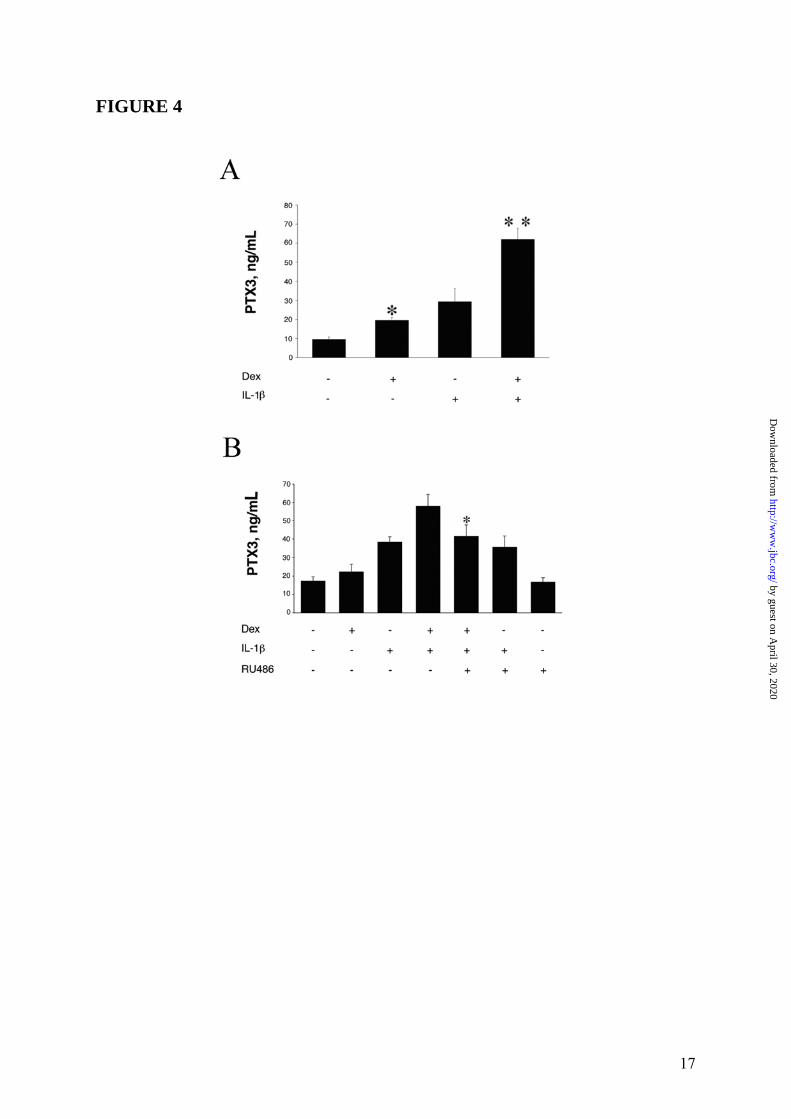
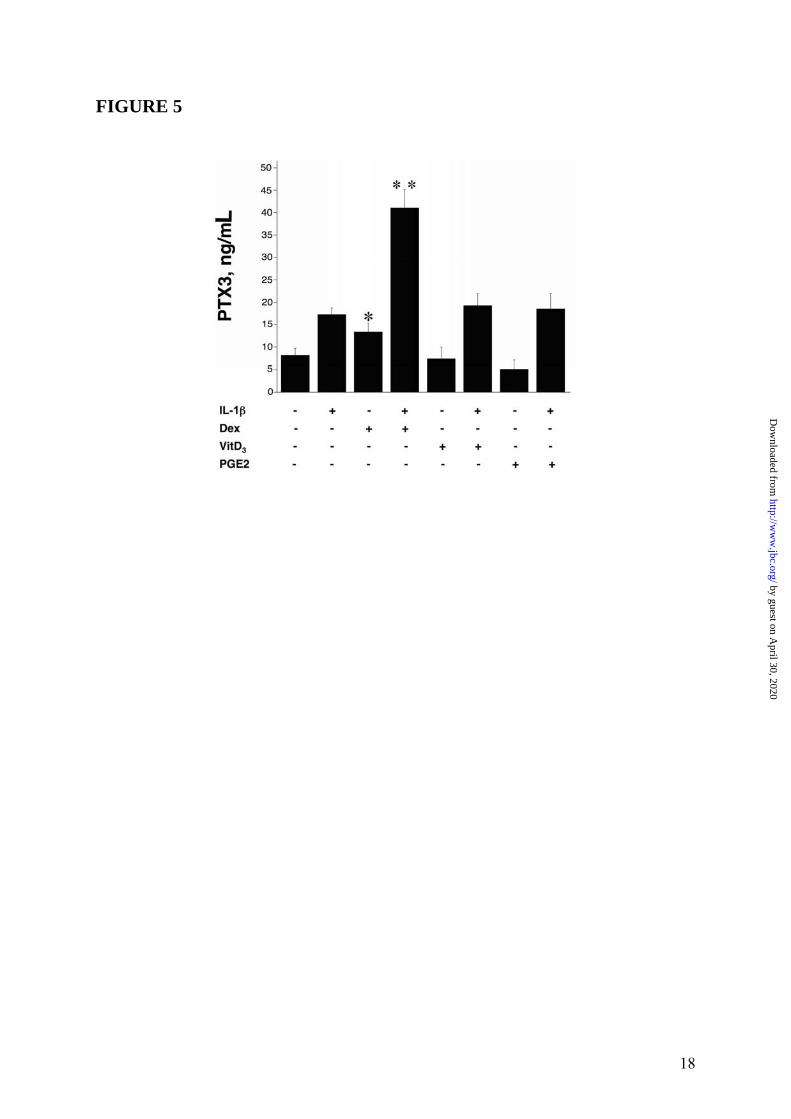
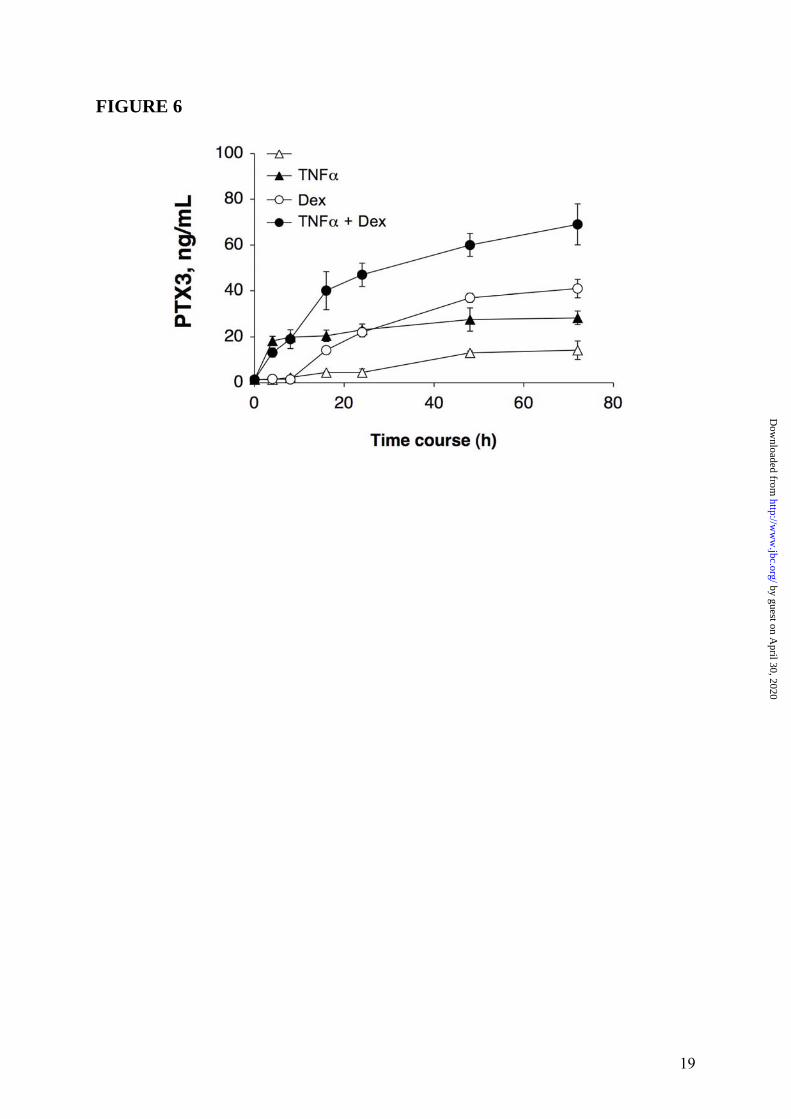
B). Thus, RU486 blocked the effects of GC on PTX3 production both in fibroblasts and in DC. Similar results were obtained when the WI38 normal fibroblast cell line was used (4 experiments performed, a representative shown in Figure 3 Panel A and Panel B). EC are a major source of PTX3 (3). Dex induced PTX3 production in HUVEC (19.7 ± 1.7ng/mL, in the experiment shown in Figure 4 Panel A, representative of 4 performed), although less efficiently than IL-1β (28.7 ± 6.2ng/mL). Dex superinduced PTX3 in combination with IL-1β, corresponding to 164.7 ± 11.2% of the IL-1β response (63.3 ± 5.2ng/mL) (Figure 4 Panel A). RU486 completely blocked the production of PTX3 by Dex but did not alter the IL-1β-induced PTX3 production (Figure 4 Panel B). The effect of GC on PTX3 production by 8387 fibroblasts was dose-related (10-5M-10-8M) (Figures 2 Panel C). VitD3 and PGE2, which inhibit the LPS-induced PTX3 production in myeloid DC (12), did not induce and did not alter PTX3 production in HUVEC (Figure 5) and in fibroblasts (data not shown). Stimulation with different hormones (e. g. T3 and T4), which, as GC, engage receptors belonging to superfamily of nuclear receptors, did not affect PTX3 production by DC and fibroblasts (data not shown). The time course of PTX3 production in non-hematopoietic cells was different in response to TNFα and GC (Figure 6, 3 experiments performed). PTX3 induction in response to TNFα was already evident after 8h whereas induction by GC required 16h. GC-induced PTX3 production was sustained up to 48h. Sustained production over time was also observed when TNFα and GC were combined (Figure 6). GC were found to induce PTX3 mRNA and to enhance the induction by TNFα and IL-1β in fibroblasts and EC (range of 5h-10h). Addition of RU486 inhibited the GR-mediated augmentation of PTX3 expression (Figure 7). Mechanism of PTX3 regulation by GC It was important to explore the molecular mechanism of the divergent PTX3 regulation by GC. PTX3 promoter was found to contain binding sites for the transcription factors NFκB and AP-1, which confer responsiveness to IL-1β, LPS and TNFα (36,37). We investigated whether GC directly modulate the PTX3 gene expression. The analysis of human PTX3 gene
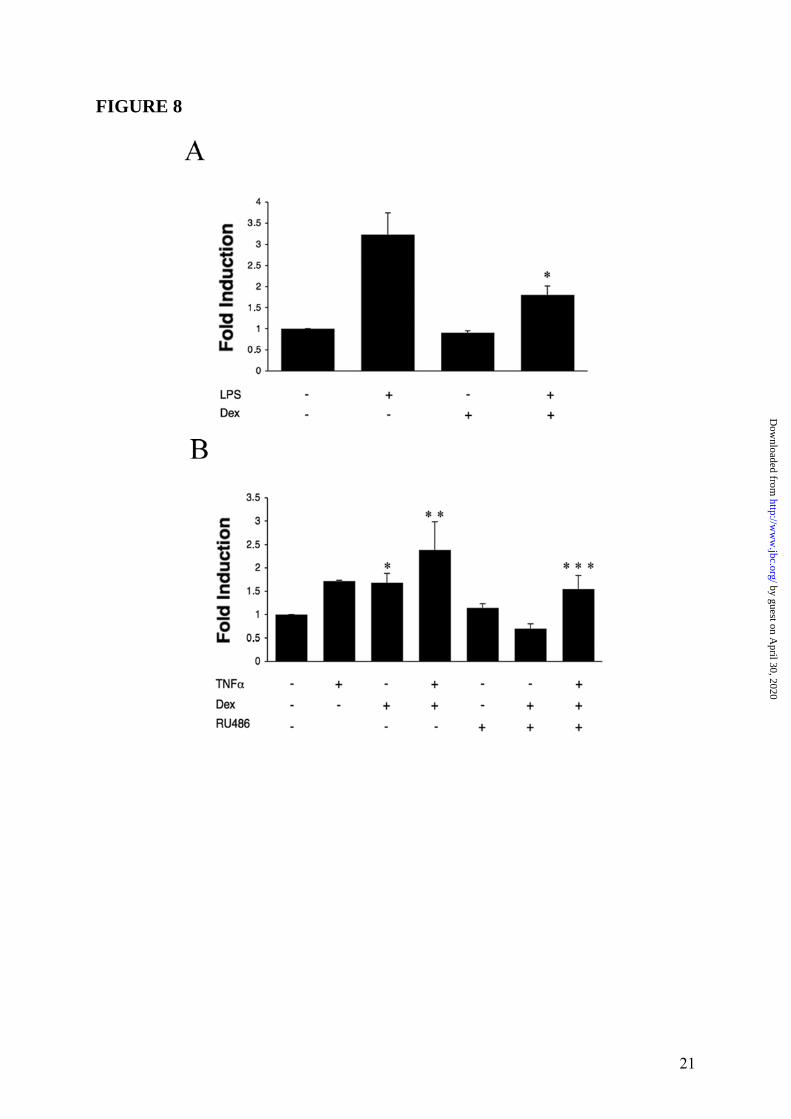
promoter (GenBank accession no. X97748) by Signal Scan software (38) allowed us to identify putative GR responsive regions. Thus, to evaluate GC regulation of PTX3 gene expression in both fibroblasts and macrophages, a reporter plasmid (pGV-B2) carrying human PTX3 promoter linked to luciferase gene was generated and used to transiently transfect RAW 264.7 and 8387 cell lines. Therefore we evaluated the transcriptional regulation of the PTX3 promoter by GC in fibroblasts and macrophages. In RAW 264.7 cell line PTX3 promoter activity significantly increased (3.2-fold, p = 0.009) following a 16h challenge with LPS (10ng/mL) compared to untreated cells. Treatment with Dex (10-6M) counteracted (-1.79-fold, p = 0.039) the transcriptional activation of PTX3 promoter induced by LPS (a representative experiment of 3 performed is shown in Figure 8 Panel A). In RAW 264.7 cells Dex did not induce PTX3 promoter activity. The GC-mediated PTX3 gene repression was also observed when cells were also treated for 4 and 8h (not shown). Figure 8 Panel A shows one out of three experiments performed, with the other two shown in the Supplemental Data (Figure S1). In this cellular context, Dex indeed inhibited NFκB and AP-1 activation, as shown by Western Blot and EMSA analysis (Figure S2, Supplemental Data). The 8387 cell line was treated with TNFα (20ng/mL), Dex (10-6M) and RU486 (10-5M) for different times after 16h of transient transfection with PTX3 promoter-pGV-B2 reporter plasmid. Figure 8 Panel B shows one representative experiment at 16h out of three performed (see Supplemental Data Figure S3, for the two experiments not shown). The PTX3 promoter responded to both TNFα (1.73-fold, p = 0.003) and Dex (1.72-fold, p = 0.013) used alone, and its activity was further enhanced following a combined treatment (2.34-fold, p = 0.017). The luciferase activity induced by GC was repressed when cells were incubated in the presence of the antagonist RU486 (Figure 8 Panel B). Similar results were obtained at 4h and 8h (data not shown).
Following binding to the GR, GC exert diverse regulatory activities through complex mechanisms. The GR either directly engages GC response elements (GRE) in the regulatory regions of genes, or indirectly, via interactions with other transcription factors (30,39). GR dimerization deficient (GR dim/dim) mice provide a tool to differ the mechanism of action of GC,
6
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

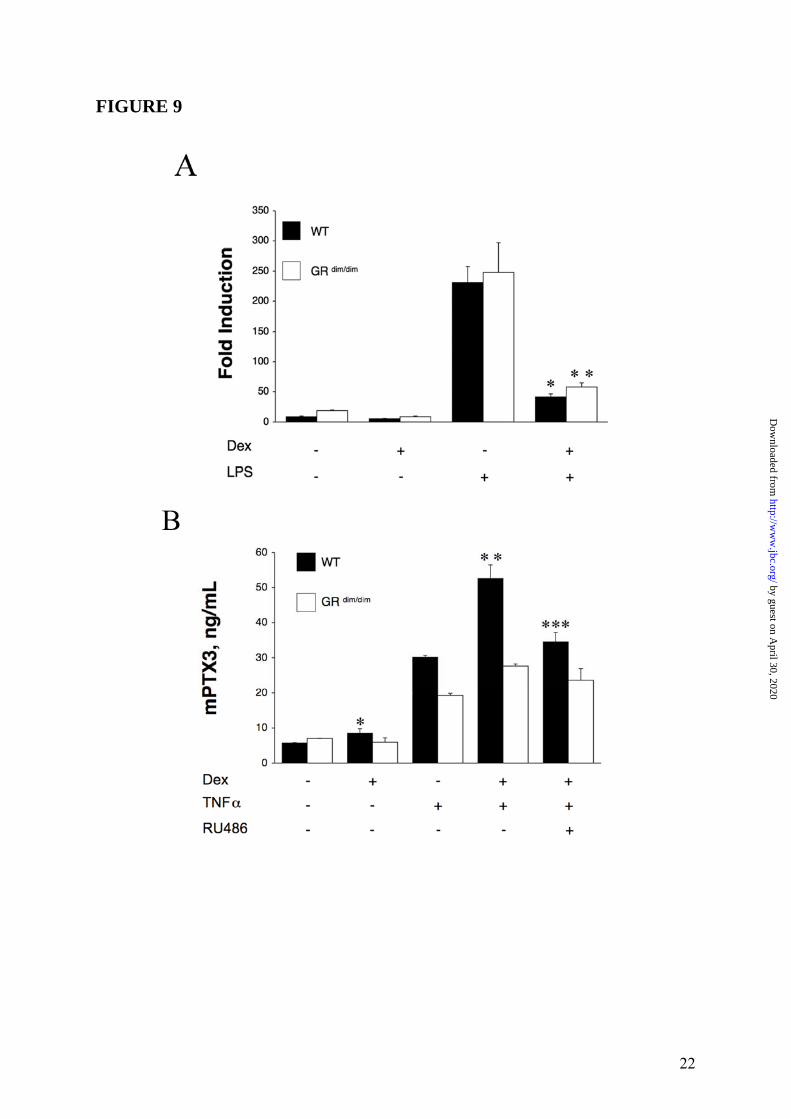
in which GRE-mediated gene activation, which is entirely dependent on GR dimerization, is removed but GR interactions with NFκB and AP-1, which are independent of dimerization, are still possible (32). Therefore, we ascertained the effect of GC on PTX3 expression in bone marrow-derived macrophages (BMM) and MEF from WT or GR dim/dim mice. First, we employed a quantitative PCR to explore the changes in regulation of LPS-induced expression of PTX3 mRNA in BMM from WT and GR dim/dim mice. RNA from macrophages was collected following treatment with LPS (10ng/mL), Dex (10-6M) or LPS plus Dex for different times. One representative experiment at 8h out of three performed is shown in Figure 9 Panel A and the other two are presented in Supplemental Data Figure S4. Dex alone was able to appreciably modify the steady state levels of PTX3 mRNA in macrophages from WT and GR dim/dim mice, whereas LPS caused a 20-fold increase in transcript in treated macrophages derived from WT or GR dim/dim mice. PTX3 expression was strikingly down-regulated by Dex both in WT (p = 0.0098) and GR dim/dim (p = 0.0040) macrophages (Figure 9 Panel A). Similar results were obtained at 2, 4 and 24h (data not shown). Then, MEF from GR dim/dim mice were assessed for their ability to produce PTX3 in response to GC. A series of 3 experiments were performed (Figure 9 Panel B). As expected, MEF from WT mice produced 8.4 ± 1.34ng/mL (p = 0.003 vs untreated), 30.1 ± 0.6ng/mL and 52.6 ± 3.8ng/mL (p = 0.002 vs TNFα) of PTX3, in response to Dex, TNFα and to the combined treatment respectively. PTX3 induction and superinduction by Dex corresponded to 28.0% and 174.6% of the response to TNFα (Figure 9 Panel B). In contrast, MEF from GR dim/dim mice did to not produce PTX3 in response to Dex (5.9 ± 1.3ng/mL) compared to 7.0 ± 0.1ng/mL for untreated cells. In GR dim/dim mice no significant superinduction of the response to TNFα by Dex was observed (Figure 9 Panel B). Overall these results indicate that in hematopoietic cells, GC-GR complex binding to NFκB and AP-1 transcription factors leads to inhibition of PTX3 gene expression, while in non-hematopoietic cells GC-GR complexes directly activate PTX3 transcription by GR binding to the PTX3 promoter. To definitely assess the importance of GC-GR complexes in
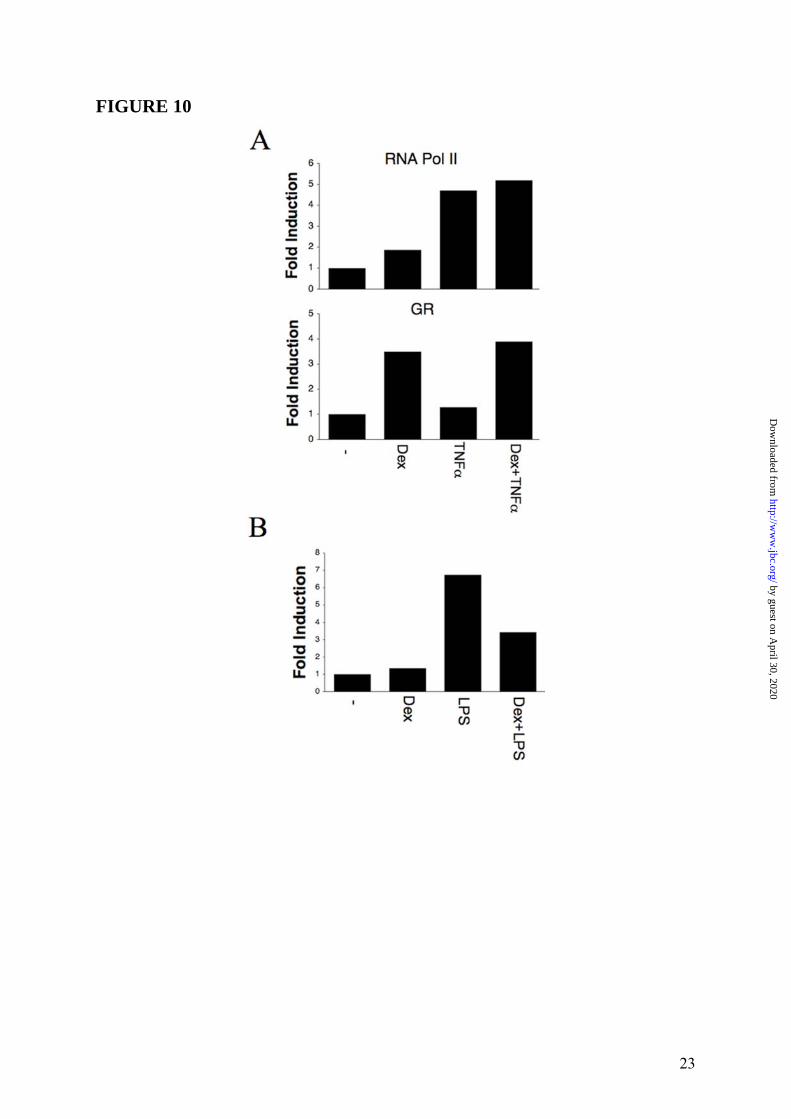
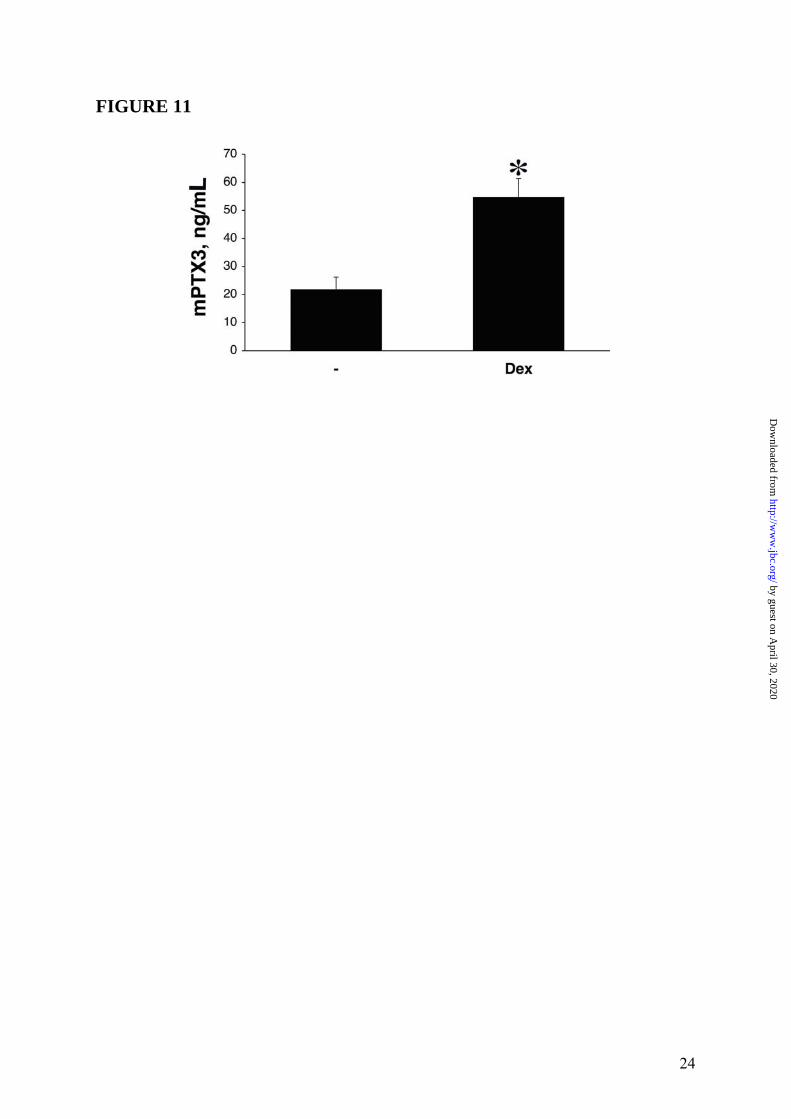
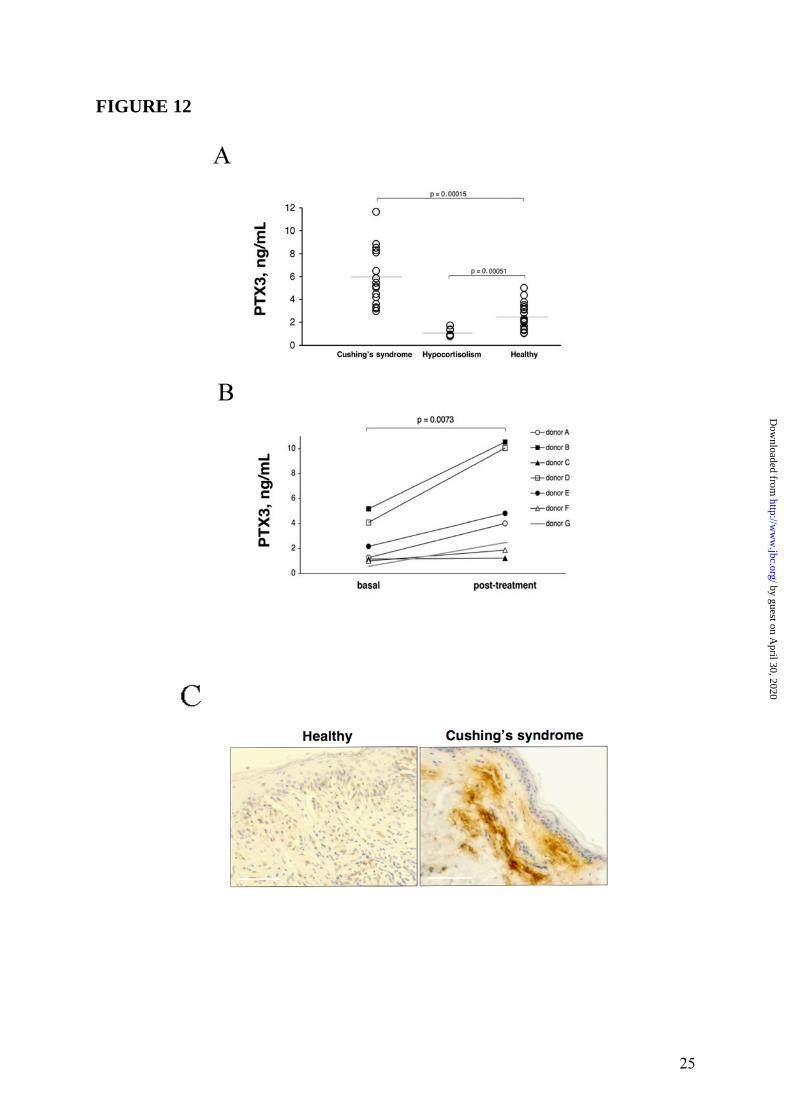
the transcriptional activation of PTX3 gene, ChIP analysis was conducted in WI38 (n = 3) and HUVEC (n = 2) with RNA pol II and GR antibodies. In agreement with gene expression analysis, results obtained in WI38 (one representative experiment is shown in Figure 10 Panel A, upper part) and HUVEC (Supplemental Data Figure S5) showed that binding of RNA pol II to PTX3 promoter was induced upon stimulation (2h) with Dex (10-6M), TNFα (20ng/mL) and the combination of two stimuli. Furthermore, Dex, but not TNFα, stimulation was able to induce GR recruitment to the PTX3 promoter (Figure 10 Panel A, lower part), thus demonstrating that GC-GR complexes play a role in the transcriptional activation of PTX3 gene in WI38 and HUVEC. In contrast, ChIP analysis performed in hematopoietic cells (DC) (n = 2, one experiment shown) showed that the LPS-induced binding of RNA pol II to the PTX3 promoter was inhibited by Dex (Figure 10 Panel B). Effect of GC on PTX3 levels in mice and humans It was of interest to ascertain the in vivo effect of exogenous GC on PTX3 levels in mice. Figure 11 shows a typical experiment, of 3 performed, in which C57/BL6 mice following a 24h treatment with Dex (30mg/Kg) markedly showed increased circulating levels of PTX3 (53.7 ± 6.8ng/mL, n = 6, p < 0.05) compared to control animals (22.0 ± 4.4ng/mL, n = 6). Circulating levels of PTX3 were then measured in patients affected by Cushing’s syndrome (hypercortisolism) and hypocortisolism in order to evaluate a regulatory effect of endogenous GC hormones on PTX3 in humans. As shown in Figure 12 Panel A, the PTX3 serum concentration in 18 normal donors was 2.6 ± 0.3ng/mL, similar to that observed in other studies (33,38). On the other hand, PTX3 levels were augmented in patients affected by Cushing’s syndrome (6.0 ± 0.6ng/mL, n = 16, p = 0.00015 vs. control group) and significantly, decreased in patients affected by hypocortisolism (1.1 ± 0.1ng/mL, n = 9, p = 0.00051 vs. control group). In 7 normal subjects circulating levels of PTX3 were measured 24h after treatment with a low dose of Dex (1mg). PTX3 was found to be augmented after Dex administration from 2.2 ± 0.7ng/mL to 5.0 ± 1.6ng/mL, p = 0.0073 (Figure 12 Panel B). When hystological analysis were conducted in skin of a patient affected by severe Cushing’s
7
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

syndrome, strong interstitial staining for PTX3 was observed in the derma (Figure 12 Panel C).
DISCUSSION
Glucocorticoid hormones (GC) have
profound effects on innate and adaptive immune responses and are a major tool in the armamentarium of anti inflammatory/immunosuppressive agents (28-30). In view also of the use of PTX3 as a novel marker of inflammation (3,40-42) it was important to assess the effect of GC on this key component of humoral innate immunity. Unexpectedly, we found that GC have divergent effects on PTX3 production in hematopoietic (macrophages and DC) and non-hematopoietic (fibroblasts and EC) cells, with inhibition in the former and induction in the latter.
Cell context-dependent regulation was also observed when the PTX3 promoter was tested in a reporter gene assay with divergent regulation in macrophages (inhibition) and fibroblastic cells (induction). Inhibition and induction are both GR-mediated, as revealed by a specific antagonist (RU486), but ordered by different mechanisms of action.
GC receptor (GR) exerts regulatory activity through complex mechanisms: it functions as ligand-dependent transcription factor through direct (dimerization-dependent) DNA binding or it represses gene transcription through a protein-protein (dimerization-independent) interference with the action of other signalling pathways (30,39,43,44). Here we report that GC-mediated down-regulation of PTX3 in hematopoietic cells is independent of GR dimerization since LPS-induced PTX3 expression was efficiently down-regulated in macrophages from GR dim/dim mice. On the other hand, PTX3 induction was observed in fibroblasts from WT mice but not GR dim/dim mice. GC exert many of anti-inflammatory actions by interfering with other signalling pathways and GR interactions with NFκB and AP-1 occur in GR dim/dim mice, in which GR-mediated gene activation is impaired (32,45). The human and murine PTX3 promoters have been both characterized and NFκB plays a pivotal role in PTX3 gene regulation (36,37). More recently, PTX3 has been shown to be regulated by the FUS/CHOP oncogene (46,47). Here we report that the human PTX3 promoter contains putative GR responsive elements, that in non-
hematopoietic cells the PTX3 promoter is activated by GC and that the stimulation of PTX3 gene expression and production is dimerization-dependent. In contrast, GC-mediated suppression of PTX3 expression in cells of hemopoietic origin is dimerization-independent and likely mediated by interference with the NFκB pathway. Chromatin immunoprecipitation experiments confirmed the transcriptional activation of the PTX3 gene by Dex in non-hematopoietic (fibroblasts and endothelial cells), but not in myeloid cells. The results offer new unexpected vistas on cell context-dependent PTX3 gene regulation.
The finding that the effect of GC on PTX3 production is cell context dependent is hardly surprising. Data presented here establish that PTX3 production is subjected to regulatory constraints that differ according to cell type. The phenomenon of GC-mediated opposite cell-specific effects on the apoptotic pathway has been demonstrated by different authors. For instance, while GC mediate pro-apoptotic events in the majority of hematological cells, GC-induced anti-apoptotic signaling has been identified in cells of epithelial origin, and the underlying mechanisms have not been fully understood yet (48). Moreover, cell type-specific opposite regulation of the same gene has been recently described for thyroid receptor (TR), a nuclear receptor highly homologous to GR, that has been shown to positively regulate type 1 iodothyronine deiodinase gene in COS-7 and HEK-293 cells, and suppress the same gene expression in the JEG-3 choriocarcinoma cell line. This negative regulation was demonstrated to require both TR-DNA binding and the presence of a JEG-3 specific transcription factor, not present in COS-7 or HEK-293 cells (49).
GC are more than simple inhibitors of inflammation and immunity. Indeed GC have been shown to enhance the expression of scavenger receptors (e. g. MARCO, CD163) as well as complement components (e.g. C1q, C3, C5), TLR family members, SAA family proteins, Fibrinogen, the collectin MBL and surfactant proteins SP-A and SP-D (50-56). Interestingly, PTX3 binds C1q and activates the classic pathway of complement activation and both are augmented by GC. SAA, MBL, Fibrinogen, SP-A and SP-D play key roles in the extracellular matrix, are components of the humoral arm of innate immunity produced by non-hematopoietic cells and share with PTX3 the property to be augmented by GC.
8
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

PTX3 is a prototypic long pentraxin structurally related to the short pentraxins CRP and SAP (3,4,6). It was previously shown that induction of the acute phase proteins (e.g. SAA) is impaired in the liver of GR dim/dim mice (57). GC selectively induce regulate acute-phase and extracellular matrix proteins such as SAA family proteins, Fibrinogen, surfactant proteins SP-A and SP-D, but not CRP and SAP. In addition, GC oppositely regulate IL-6 (inhibition) and IL-10 (induction), key inducers of CRP/SAP and PTX3 (1,3).
PTX3 binds TSG-6 and acts as a focal point in the assembly of hyaluronic acid rich extracellular matrices. PTX3 levels are augmented in humans after GC administration and are elevated in patients affected by Cushing’s syndrome. Interestingly, PTX3 is found to localize in the derma of these patients. Patients with Cushing’s syndrome have alterations in the extracellular matrix with loss
of connective tissue, leading to fragility and atrophy of the skin, thinning of the stratum corneum and loss of subcutaneous fat. All this explains skin easy bruising after minimal trauma, the frequent presence of extensive ecchymoses and the formation of typical striae rubrae on the stretched, fragile skin. It remains to be ascertained whether unbalanced production of PTX3 by fibroblasts and endothelial cells induced by GC plays a role in these manifestations of Cushing’s syndrome.
The results presented here indicate the regulation of PTX3 by GC is cell context-dependent, with inhibition in hematopoietic cells (macrophages and DC) and induction in non-hematopoietic cells (fibroblasts and endothelial cells). The divergent effects of GC on PTX3 production in diverse cellular contexts is likely to reflect the diverse roles of this multifunctional protein in innate immunity, inflammation, and in the assembly of the extracellular matrix.
REFERENCES
1. Pepys, M. B., and Baltz, M. L. (1983) Adv Immunol 34, 141-212 2. Baumann, H., and Gauldie, J. (1994) Immunol Today 15(2), 74-80 3. Garlanda, C., Bottazzi, B., Bastone, A., and Mantovani, A. (2005) Annu Rev Immunol 23,
337-366 4. Mantovani, A., Garlanda, C., Doni, A., and Bottazzi, B. (2008) J Clin Immunol 28(1), 1-13 5. Du Clos, T. W., and Mold, C. (2004) Immunol Res 30(3), 261-277 6. Bottazzi, B., Garlanda, C., Salvatori, G., Jeannin, P., Manfredi, A., and Mantovani, A. (2006)
Curr Opin Immunol 18(1), 10-15 7. Alles, V. V., Bottazzi, B., Peri, G., Golay, J., Introna, M., and Mantovani, A. (1994) Blood
84(10), 3483-3493 8. Doni, A., Peri, G., Chieppa, M., Allavena, P., Pasqualini, F., Vago, L., Romani, L., Garlanda,
C., and Mantovani, A. (2003) Eur J Immunol 33(10), 2886-2893 9. Lee, G. W., Lee, T. H., and Vilcek, J. (1993) J Immunol 150(5), 1804-1812 10. Breviario, F., d'Aniello, E. M., Golay, J., Peri, G., Bottazzi, B., Bairoch, A., Saccone, S.,
Marzella, R., Predazzi, V., Rocchi, M., and et al. (1992) J Biol Chem 267(31), 22190-22197 11. Han, B., Mura, M., Andrade, C. F., Okutani, D., Lodyga, M., dos Santos, C. C., Keshavjee,
S., Matthay, M., and Liu, M. (2005) J Immunol 175(12), 8303-8311 12. Doni, A., Michela, M., Bottazzi, B., Peri, G., Valentino, S., Polentarutti, N., Garlanda, C., and
Mantovani, A. (2006) J Leukoc Biol 79(4), 797-802 13. Bottazzi, B., Vouret-Craviari, V., Bastone, A., De Gioia, L., Matteucci, C., Peri, G.,
Spreafico, F., Pausa, M., D'Ettorre, C., Gianazza, E., Tagliabue, A., Salmona, M., Tedesco, F., Introna, M., and Mantovani, A. (1997) J Biol Chem 272(52), 32817-32823
14. Garlanda, C., Hirsch, E., Bozza, S., Salustri, A., De Acetis, M., Nota, R., Maccagno, A., Riva, F., Bottazzi, B., Peri, G., Doni, A., Vago, L., Botto, M., De Santis, R., Carminati, P., Siracusa, G., Altruda, F., Vecchi, A., Romani, L., and Mantovani, A. (2002) Nature 420(6912), 182-186.
15. Nauta, A. J., Bottazzi, B., Mantovani, A., Salvatori, G., Kishore, U., Schwaeble, W. J., Gingras, A. R., Tzima, S., Vivanco, F., Egido, J., Tijsma, O., Hack, E. C., Daha, M. R., and Roos, A. (2003) Eur J Immunol 33(2), 465-473
9
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

16. Salustri, A., Garlanda, C., Hirsch, E., De Acetis, M., Maccagno, A., Bottazzi, B., Doni, A., Bastone, A., Mantovani, G., Beck Peccoz, P., Salvatori, G., Mahoney, D. J., Day, A. J., Siracusa, G., Romani, L., and Mantovani, A. (2004) Development 131(7), 1577-1586
17. Diniz, S. N., Nomizo, R., Cisalpino, P. S., Teixeira, M. M., Brown, G. D., Mantovani, A., Gordon, S., Reis, L. F., and Dias, A. A. (2004) J Leukoc Biol 75(4), 649-656
18. Jeannin, P., Bottazzi, B., Sironi, M., Doni, A., Rusnati, M., Presta, M., Maina, V., Magistrelli, G., Haeuw, J. F., Hoeffel, G., Thieblemont, N., Corvaia, N., Garlanda, C., Delneste, Y., and Mantovani, A. (2005) Immunity 22(5), 551-560
19. Rovere, P., Peri, G., Fazzini, F., Bottazzi, B., Doni, A., Bondanza, A., Zimmermann, V. S., Garlanda, C., Fascio, U., Sabbadini, M. G., Rugarli, C., Mantovani, A., and Manfredi, A. A. (2000) Blood 96(13), 4300-4306
20. Dias, A. A., Goodman, A. R., Dos Santos, J. L., Gomes, R. N., Altmeyer, A., Bozza, P. T., Horta, M. F., Vilcek, J., and Reis, L. F. (2001) J Leukoc Biol 69(6), 928-936
21. Varani, S., Elvin, J. A., Yan, C., DeMayo, J., DeMayo, F. J., Horton, H. F., Byrne, M. C., and Matzuk, M. M. (2002) Mol Endocrinol 16(6), 1154-1167
22. Souza, D. G., Soares, A. C., Pinho, V., Torloni, H., Reis, L. F., Teixeira, M. M., and Dias, A. A. (2002) Am J Pathol 160(5), 1755-1765
23. Bottazzi, B., Bastone, A., Doni, A., Garlanda, C., Valentino, S., Deban, L., Maina, V., Cotena, A., Moalli, F., Vago, L., Salustri, A., Romani, L., and Mantovani, A. (2006) J Leukoc Biol 79(5), 909-912
24. Ravizza, T., Moneta, D., Bottazzi, B., Peri, G., Garlanda, C., Hirsch, E., Richards, G. J., Mantovani, A., and Vezzani, A. (2001) Neuroscience 105(1), 43-53
25. Baruah, P., Dumitriu, I. E., Peri, G., Russo, V., Mantovani, A., Manfredi, A. A., and Rovere-Querini, P. (2006) J Leukoc Biol 80(1), 87-95
26. Baruah, P., Propato, A., Dumitriu, I. E., Rovere-Querini, P., Russo, V., Fontana, R., Accapezzato, D., Peri, G., Mantovani, A., Barnaba, V., and Manfredi, A. A. (2006) Blood 107(1), 151-158
27. Salio, M., Chimenti, S., De Angelis, N., Molla, F., Maina, V., Nebuloni, M., Pasqualini, F., Latini, R., Garlanda, C., and Mantovani, A. (2008) Circulation 117(8), 1055-1064
28. Chrousos, G. P. (1995) N Engl J Med 332(20), 1351-1362 29. Franchimont, D. (2004) Ann N Y Acad Sci 1024, 124-137 30. Sternberg, E. M. (2006) Nat Rev Immunol 6(4), 318-328 31. Abraham, S. M., Lawrence, T., Kleiman, A., Warden, P., Medghalchi, M., Tuckermann, J.,
Saklatvala, J., and Clark, A. R. (2006) J Exp Med 203(8), 1883-1889 32. Reichardt, H. M., Kaestner, K. H., Tuckermann, J., Kretz, O., Wessely, O., Bock, R., Gass,
P., Schmid, W., Herrlich, P., Angel, P., and Schutz, G. (1998) Cell 93(4), 531-541 33. Napoleone, E., Di Santo, A., and Lorenzet, R. (1997) Blood 89(2), 541-549 34. Schioppa, T., Uranchimeg, B., Saccani, A., Biswas, S. K., Doni, A., Rapisarda, A.,
Bernasconi, S., Saccani, S., Nebuloni, M., Vago, L., Mantovani, A., Melillo, G., and Sica, A. (2003) J Exp Med 198(9), 1391-1402
35. Muller, B., Peri, G., Doni, A., Torri, V., Landmann, R., Bottazzi, B., and Mantovani, A. (2001) Crit Care Med 29(7), 1404-1407
36. Basile, A., Sica, A., d'Aniello, E., Breviario, F., Garrido, G., Castellano, M., Mantovani, A., and Introna, M. (1997) J Biol Chem 272(13), 8172-8178
37. Altmeyer, A., Klampfer, L., Goodman, A. R., and Vilcek, J. (1995) J Biol Chem 270(43), 25584-25590
38. Prestridge, D. S. (1991) Comput Appl Biosci 7(2), 203-206 39. Webster, J. I., and Sternberg, E. M. (2004) J Endocrinol 181(2), 207-221 40. Peri, G., Introna, M., Corradi, D., Iacuitti, G., Signorini, S., Avanzini, F., Pizzetti, F.,
Maggioni, A. P., Moccetti, T., Metra, M., Cas, L. D., Ghezzi, P., Sipe, J. D., Re, G., Olivetti, G., Mantovani, A., and Latini, R. (2000) Circulation 102(6), 636-641
41. Latini, R., Maggioni, A. P., Peri, G., Gonzini, L., Lucci, D., Mocarelli, P., Vago, L., Pasqualini, F., Signorini, S., Soldateschi, D., Tarli, L., Schweiger, C., Fresco, C., Cecere, R., Tognoni, G., and Mantovani, A. (2004) Circulation 110(16), 2349-2354
10
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

42. Inoue, K., Sugiyama, A., Reid, P. C., Ito, Y., Miyauchi, K., Mukai, S., Sagara, M., Miyamoto, K., Satoh, H., Kohno, I., Kurata, T., Ota, H., Mantovani, A., Hamakubo, T., Daida, H., and Kodama, T. (2007) Arterioscler Thromb Vasc Biol 27(1), 161-167
43. Aranda, A., and Pascual, A. (2001) Physiol Rev 81(3), 1269-1304 44. Liberman, A. C., Refojo, D., Druker, J., Toscano, M., Rein, T., Holsboer, F., and Arzt, E.
(2007) Faseb J 21(4), 1177-1188 45. De Bosscher, K., Vanden Berghe, W., and Haegeman, G. (2003) Endocr Rev 24(4), 488-522 46. Willeke, F., Assad, A., Findeisen, P., Schromm, E., Grobholz, R., von Gerstenbergk, B.,
Mantovani, A., Peri, S., Friess, H. H., Post, S., von Knebel Doeberitz, M., and Schwarzbach, M. H. (2006) Eur J Cancer 42(15), 2639-2646
47. Kuroda, M., Ishida, T., Horiuchi, H., Kida, N., Uozaki, H., Takeuchi, H., Tsuji, K., Imamura, T., Mori, S., Machinami, R., and et al. (1995) Am J Pathol 147(5), 1221-1227
48. Herr, I., Gassler, N., Friess, H., and Buchler, M. W. (2007) Apoptosis 12(2), 271-291 49. Kim, S. W., Hong, S. J., Kim, K. M., Ho, S. C., So, E. C., Harney, J. W., and Larsen, P. R.
(2004) Mol Endocrinol 18(12), 2924-2936 50. Thorn, C. F., Lu, Z. Y., and Whitehead, A. S. (2003) Eur J Immunol 33(9), 2630-2639 51. Silverstein, R., and Johnson, D. C. (2003) J Leukoc Biol 73(4), 417-427 52. Imasato, A., Desbois-Mouthon, C., Han, J., Kai, H., Cato, A. C., Akira, S., and Li, J. D.
(2002) J Biol Chem 277(49), 47444-47450 53. Galon, J., Franchimont, D., Hiroi, N., Frey, G., Boettner, A., Ehrhart-Bornstein, M., O'Shea,
J. J., Chrousos, G. P., and Bornstein, S. R. (2002) Faseb J 16(1), 61-71 54. Vizzardelli, C., Pavelka, N., Luchini, A., Zanoni, I., Bendickson, L., Pelizzola, M., Beretta,
O., Foti, M., Granucci, F., Nilsen-Hamilton, M., and Ricciardi-Castagnoli, P. (2006) Eur J Immunol 36(6), 1504-1515
55. Ashwell, J. D., Lu, F. W., and Vacchio, M. S. (2000) Annu Rev Immunol 18, 309-345 56. Duan, H. O., and Simpson-Haidaris, P. J. (2006) J Biol Chem 281(18), 12451-12457 57. Reichardt, H. M. (2004) Curr Pharm Des 10(23), 2797-2805
FOOTNOTES * This work was supported by Associazione Italiana per la Ricerca sul Cancro (AIRC), Ministero Istruzione, Università e Ricerca (MIUR), and European Commission, Mugen (005203), Muvapred (503240), DC-Thera (512074). We are grateful to Pietro Ghezzi (Mario Negri Institute of Milan, Milan, Italy) for helpful discussion. Andrea Doni and Giovanna Mantovani have equally contributed to the work. The abbreviations used are: Acute-phase protein / Pentraxins / PTX3 / Glucocorticoid hormones / Innate immunity / Inflammation / Pattern recognition receptor / Extracellular matrix / Tissue remodelling
FIGURE LEGENDS FIGURE 1: Inhibition by GC of PTX3 production in myeloid DC. Panel A summarizes results of a series of 12 experiments. Results refer to mean ± SD. ∗ = p < 0.01 vs. LPS. Panel B shows a series of 4 experiments performed in which the GC antagonist RU486 is used. LPS (100ng/mL), Dex (10-6M) and RU486 (10-5M) are used. Results refer to mean ± SD. ∗ = p < 0.05 vs. LPS plus Dex. Panel C, Northern blot analysis; the lower part is the ethidium bromide-stained membrane. FIGURE 2: Induction of PTX3 production by GC in 8387 fibrosarcoma cells. Panel A, left part, shows a series of 4 experiments with Dex (10-6M), IL-1β (20ng/mL) and TNFα (20ng/mL), and combinations thereof. Results refer to mean ± SD of a triplicate. ∗ = p < 0.01 vs. unstimulated; ∗∗ = p < 0.01 vs. IL-1β; ∗∗∗ = p < 0.05 vs. TNFα; ∗∗∗∗ = p < 0.01 vs. IL-1β + TNFα. Right part, TNFα
11
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

(20ng/mL) and Dex (10-6M) are used. ∗ = p < 0.01 vs. TNFα. Panel B, inhibition by RU486. A series of 3 experiments is shown. TNFα (20ng/mL), Dex (10-6M) and RU486 (10-5M) are used. Results refer to mean ± SD. ∗ = p < 0.05 vs. Dex or Dex plus TNFα. Panel C, different concentrations of Dex; TNFα (20ng/mL) is used. Results are mean ± SD of a triplicate. ∗ = p < 0.01 vs. TNFα. FIGURE 3: Regulation of PTX3 production by GC in WI38 fibroblast. Panel A summarizes a series of 4 experiments performed with Dex (10-6M) and TNFα (20ng/mL). Results refer to mean ± SD. ∗ = p < 0.05 vs. unstimulated. ∗∗ = p < 0.01 vs. TNFα. Panel B refers to a series of 3 experiments. TNFα (20ng/mL), Dex (10-6M) and RU486 (10-5M) are used. Results refer to mean ± SD. ∗ = p < 0.01 vs. Dex plus TNFα. FIGURE 4: Regulation of PTX3 production by GC in HUVEC. Panel A refers to a representative experiment out of 3 with Dex (10-6M) and IL-1β (20ng/mL). Results refer to mean ± SD. ∗ = p < 0.01 vs. unstimulated. ∗∗ = p < 0.01 vs. IL-1β. Panel B. Effect of RU486 (3 experiments). IL-1β (20ng/mL), Dex 10-6M and RU486 10-5M are used. Results refer to mean ± SD. ∗ = p < 0.05 vs. Dex plus IL-1β. FIGURE 5: Lack of effect of VitD3 and PGE2 on PTX3 regulation in HUVEC. Summary of 3 experiments performed with IL-1β (20ng/mL), Dex (10-6M), VitD3 (10-6M) and PGE2 (10-5M) and combinations thereof. Results represent mean ± SD. ∗ = p < 0.05 vs. unstimulated. ∗∗ = p < 0.01 vs. IL-1β. FIGURE 6: Kinetics of PTX3 induction by GC. Results are for a series of 3 different experiments performed. 8387 fibrosarcoma cells are stimulated with TNFα (20ng/mL), Dex (10-6M) and combinations thereof. Results are mean ± SD. FIGURE 7: Induction of PTX3 mRNA by GC in fibroblasts and endothelial cells. WI38 fibroblasts (left part) and HUVEC (right part) were used. WI38 and HUVEC mRNA were obtained after 5h of stimulation with TNFα (20ng/mL), IL-1β (20ng/mL), Dex (10-6M), RU486 (10-5M) and combinations thereof. The lower parts are the ethidium bromide-stained membrane. FIGURE 8: Transcriptional regulation of the PTX3 promoter by GC. Reporter gene assays were conducted as described in Experimental Procedures. PTX3 promoter activity is measured after 16h of stimulation with LPS (10ng/mL), TNFα (20ng/mL), Dex (10-6M), RU486 (10-5M), or the combined treatments. Similar results have been obtained at 4 and 8h. Results are relative to basal level and refer to mean ± SD of a triplicate (1 experiment of 3 performed). Panel A. Raw 264.7 macrophage cell line; ∗ = p < 0.05 vs. LPS. Panel B. 8387 fibrosarcoma cell line; ∗ = p < 0.02 vs. unstimulated, ∗∗ = p < 0.02 vs. TNFα, ∗∗∗ = p < 0.05 vs. TNFα + Dex. FIGURE 9: Regulation of PTX3 in GR dim/dim mice-derived cells. Panel A. Representative Quantitative RT-PCR are shown. RNA from macrophages was extracted and collected 8h after stimulation. LPS (10ng/mL) and Dex (10-6M) are used as stimuli. Results represent mean ± SD. ∗ = p < 0.01 vs. LPS. ∗∗ = p < 0.005 vs. LPS. Panel B, MEF from WT and GR dim/dim mice (representative of 4 experiments performed). TNFα (20ng/mL), Dex (10-6M) and RU486 (10-5M) were used as stimuli. Results represent mean ± SD. ∗ = p < 0.005 vs. unstimulated, ∗∗ = p < 0.005 vs. TNFα, ∗∗∗ = p < 0.01 vs. TNFα + Dex. FIGURE 10: GC-GR complexes directly activate PTX3 transcription by GR binding to PTX3 promoter. Dex (10-6M), TNFα (20ng/mL) and LPS (100ng/mL) are used. Panel A, ChIP analysis in WI38. Polyclonal RNA pol II (upper part) and GR (lower part) antibodies are used. Panel B, ChIP analysis in DC. A polyclonal RNA pol II antibody is used. Chromatin immunoprecipitation was conducted as described in Experimental Procedures. RT-PCR results are normalized to input and
12
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

expressed as fold induction with respect to the control cells. Data represents 3 independent experiments of several performed (WI38 n = 3, HUVEC n = 2, DC n = 2) and mean of a triplicate. FIGURE 11: Effect of GC on PTX3 blood levels in mice. The figure presents a typical in vivo experiment of 3 performed. C57/BL6 mice (n = 6 per group) were given Dex (30mg/Kg) i. p. or vehicle 24h before sacrifice. Results are mean ± SD. ∗ = p < 0.05 vs. control group. FIGURE 12: Regulation of peripheral blood PTX3 levels by GC in humans. Panel A, PTX3 levels in Cushing’s syndrome patients (n = 16), patients affected by hypocortisolism (n = 9) and healthy donors (n = 18). In Panel B PTX3 levels in 7 normal subjects treated with Dex (1mg) before obtaining peripheral blood (overnight treatment). Panel C, hystological analysis of PTX3 localization in a skin of a patient affected by Cushing’s syndrome and healthy donor are shown. PTX3 staining is performed as described in Experimental Procedures. PTX3 = brown; Bar = 50µm.
13
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Bottazzi and Alberto MantovaniNebuloni, Stefano Signorini, Giuseppe Peri, Antonio Sica, Paolo Beck-Peccoz, Barbara
Reichardt, Anna Kleiman, Marina Sironi, Luca Rubino, Fabio Pasqualini, Manuela Andrea Doni, Giovanna Mantovani, Chiara Porta, Jan Tuckermann, Holger M.
non-hematopoietic cellsCell-specific regulation of PTX3 by glucocorticoid hormones in hematopoietic and
published online August 14, 2008J. Biol. Chem.
10.1074/jbc.M805631200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/08/19/M805631200.DC1
by guest on April 30, 2020
http://ww
w.jbc.org/
Dow
nloaded from








![Analysis of the erythropoietin of a Tibetan Plateau ...Erythropoietin (EPO) is a glycoprotein hormone that ... hematopoietic progenitor cells [21]. EPO:EPOR engage-ment leads to the](https://static.fdokument.com/doc/165x107/60fa5d2bf20891506e1ddeac/analysis-of-the-erythropoietin-of-a-tibetan-plateau-erythropoietin-epo-is.jpg)









