
Charakterisierung von FLT3 assoziierten ... · Charakterisierung von FLT3 assoziierten...
93
Aus der Medizinischen Klinik und Poliklinik III - Großhadern der Ludwig-Maximilians-Universität München Direktor: Prof. Dr. med. Wolfgang Hiddemann Charakterisierung von FLT3 assoziierten Signaltransduktionsproteinen und TET2 als epigenetischer Regulator in der akuten myeloischen Leukämie Dissertation zum Erwerb des Doktorgrades der Medizin an der Medizinischen Fakultät der Ludwig-Maximilians-Universität München vorgelegt von Judith Hecker aus Karlsruhe 2017
Transcript of Charakterisierung von FLT3 assoziierten ... · Charakterisierung von FLT3 assoziierten...

Aus der
Medizinischen Klinik und Poliklinik III - Großhadern
der Ludwig-Maximilians-Universität München
Direktor: Prof. Dr. med. Wolfgang Hiddemann
Charakterisierung von FLT3 assoziierten Signaltransduktionsproteinen und TET2 als epigenetischer
Regulator in der akuten myeloischen Leukämie
Dissertation zum Erwerb des Doktorgrades der Medizin
an der Medizinischen Fakultät der
Ludwig-Maximilians-Universität München
vorgelegt von
Judith Hecker
aus Karlsruhe
2017

II
Mit Genehmigung der Medizinischen Fakultät
der Universität München
Berichterstatter: Prof. Dr. med. Karsten Spiekermann
Mitberichterstatter: Prof. Dr. Irmela Jeremias, Prof Dr. Heinrich Leonhardt
Mitbetreuung durch den promovierten Mitarbeiter: Dr. rer. nat. Harald Polzer
Dekan: Prof. Dr. med. dent. Reinhard Hickel
Tag der mündlichen Prüfung: 29.06.2017

III
Eidesstattliche Versicherung Hecker, Judith
Ich erkläre hiermit an Eides statt,
dass ich die vorliegende Dissertation mit dem Thema
Charakterisierung von FLT3 assoziierten Signaltransduktionsproteinen und TET2 als epigenetischer Regulator in der akuten myeloischen Leukämie
selbständig verfasst, mich außer der angegebenen keiner weiteren Hilfsmittel bedient und
alle Erkenntnisse, die aus dem Schrifttum ganz oder annähernd übernommen sind, als
solche kenntlich gemacht und nach ihrer Herkunft unter Bezeichnung der Fundstelle
einzeln nachgewiesen habe.
Ich erkläre des Weiteren, dass die hier vorgelegte Dissertation nicht in gleicher oder in
ähnlicher Form bei einer anderen Stelle zur Erlangung eines akademischen Grades
eingereicht wurde.
München, August 2016
Judith Hecker

IV
Abkürzungsverzeichnis
α Anti, Alpha
Abb. Abbildung
ad auffüllen auf
AKT Protein-Kinase-B
ALL akute lymphatische Leukämie
AML akute myeloische Leukämie
CBL engl. casitas B-lineage lymphoma protein
CC coiled-coil
CIN85 engl. CBL-interacting protein of 85kDa
CML chronische myeloische Leukämie
CMML chronische myelomonozytäre Leukämie
DNA engl. desoxyribonucleic acid
DMEM Dulbecco´s Modified Eagle Medium
EDTA Ethylendiamintetraessigsäure
EGF engl. epidermal growth factor
EGFR engl. epidermal growth factor receptor
FAB French-American-British
FACS engl. fluorescence activated cell sorter
FKS fötales Kälberserum
FL FLT3-Ligand
FLT3 engl. FMS-like tyrosin kinase 3
GRB2 engl. growth factor receptor-bound protein 2
h Stunde
HBS engl. hepes-buffered saline solution
IL-3 Interleukin-3
ITD engl. internal tandem duplication
JM juxtamembranös
kb Kilobasenpaar
kDa Kilodalton
l Liter
MAPK engl. mitogen-activated protein kinase
MDS Myelodysplastisches Syndrom
mg Milligramm
min Minute

V
MIG pMSCV-IRES-eGFP
MIY pMSCV-IRES-eYFP
MPN myeoloproliferative Neoplasien
MLL engl. mixed-lineage leukemia
NPM1 Nucleophosmin Member 1
OS engl. overall survival
PBS engl. phosphat buffer saline
PCR engl. polymerase chain reaction
PI3K Phosphatidylinositol-3 Kinase
PIP2 Phosphatidylinositol-4,5-Bisphosphat
PIP3 Phosphatidylinositol-3,4,5-Trisphosphat
PTK Proteintyrosinkinase
RING engl. really interesting new gene
rpm engl. rounds per minute
RT Raumtemperatur
RTK Rezeptor-Tyrosinkinase
s Sekunde
SB1 engl. SETA binding protein 1
SCF engl. stem cell factor
SDS Sodiumdodecylsulfat
SH2 engl. SRC-homology 2
SH3 engl. SRC-homology 3
SNP engl. Single Nucleotide Polymorphism
STAT5 engl. signal transducer and activator of transcription 5
TET Tet-Methylcytosin-Dioxygenase
TKB Tyrosinkinasebindung
TKD Tyrosinkinasedomäne
TKI Tyrosinkinaseinhibitor
U engl. Unit
VCM engl. virus containing medium
VEGF engl. vascular endothelial growth factor
VEGFR engl. vascular endothelial growth factor receptor
WHO engl. World Health Organization
WT Wildtyp
μg Mikrogramm
μl Mikroliter
°C Grad Celsius

VI
5-hmC 5-Hydroxy-Methylcytosin
5-mC 5-Methylcytosin

VII
Inhaltsverzeichnis
1 Einleitung ................................................................................................................ 1 1.1 Die akute myeloische Leukämie ...................................................................................... 1 1.2 Die Rezeptor-Tyrosinkinase FLT3 ................................................................................... 3
1.2.1 Struktureller Aufbau und Expression der Rezeptor-Tyrosinkinase FLT3 ................... 4 1.2.2 Signalkaskaden und Mutationen des FLT3-Rezeptors .............................................. 5 1.2.3 Funktion von FLT3 in der malignen Hämatopoese .................................................... 6
1.3 CBL, CIN85 und SB1 in der Regulation von Rezeptor-Tyrosinkinasen ............................ 8 1.4 TET2 als epigenetischer Regulator ................................................................................ 13 1.5 Zielsetzung der vorliegenden Arbeit ............................................................................... 15 2 Material und Methoden ........................................................................................ 17 2.1 Material.......................................................................................................................... 17
2.1.1 Inhibitoren, Enzyme, Zytokine und Chemikalien ...................................................... 17 2.1.2 Software ................................................................................................................. 19 2.1.3 Kits ......................................................................................................................... 20 2.1.4 Laborausstattung .................................................................................................... 20 2.1.5 Zelllinien ................................................................................................................. 21 2.1.6 Antikörper ............................................................................................................... 22 2.1.7 Plasmide ................................................................................................................. 23 2.1.8 Oligonukleotide ....................................................................................................... 24 2.1.9 Puffer, Medien, Lösungen ....................................................................................... 25
2.2 Methoden ...................................................................................................................... 26 2.2.1 Zellkultur ................................................................................................................. 26 2.2.1.1 Zellkultur von adhärenten Suspensionszellen ...................................................... 26 2.2.1.2 Einfrieren und Auftauen von Zellen ...................................................................... 27 2.2.1.3 Transiente Transfektion ....................................................................................... 27 2.2.1.4 Stabile Ba/F3-Zell-Transduktion ........................................................................... 27 2.2.1.5 Lebendzellzahl-Bestimmung ................................................................................ 28 2.2.1.6 Apoptosemessung mit Annexin-V-/7-AAD-Färbung ............................................. 28 2.2.1.7 Expressionsanalyse von Oberflächenantigenen mittels Durchflusszytometrie ...... 29 2.2.1.8 Hungern der Zellen und Stimulation vor der Zelllyse ............................................ 29 2.2.1.9 Immunfluoreszenz von adhärenten Zellen (Immunostaining) ............................... 29 2.2.2 Proteinbiochemische Methoden .............................................................................. 30 2.2.2.1 Proteinlysate ........................................................................................................ 30 2.2.2.2 Proteinkonzentrationsbestimmung nach Bradford ................................................ 30 2.2.2.3 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE) .......................................... 31 2.2.2.4 Western-Blot ........................................................................................................ 31 2.2.2.5 Strippen der Nitrozellulosemembran .................................................................... 31 2.2.3 Bakterienkultur ........................................................................................................ 32 2.2.3.1 Transformation kompetenter E.coli Bakterien ....................................................... 32 2.2.3.2 Plasmid-DNA-Präparation .................................................................................... 32 2.2.3.2.1 Mini-Präparation................................................................................................ 32 2.2.3.2.2 Maxi-Präparation ............................................................................................... 32 2.2.4 Molekularbiologische Methoden .............................................................................. 33 2.2.4.1 DNA-Verdau durch Restriktionsenzyme ............................................................... 33 2.2.4.2 Ligation von DNA-Fragmenten ............................................................................. 33 2.2.4.3 Isolierung von mRNA ........................................................................................... 34 2.2.4.4 Die Synthese von cDNA durch reverse Transkription ........................................... 34 2.2.4.5 Extraktion genomischer DNA ............................................................................... 34 2.2.4.6 Polymerasekettenreaktion (PCR) ......................................................................... 34 2.2.4.7 Ortsgerichtete Mutagenese .................................................................................. 35 2.2.4.8 Sequenzierung ..................................................................................................... 35 2.2.5 Präparation der Patientenproben ............................................................................ 35

VIII
2.2.5.1 Ficoll .................................................................................................................... 35 2.2.5.2 MACS Zellseparation ........................................................................................... 36
3 Ergebnisse ............................................................................................................ 37 3.1 Funktionelle Charakterisierung von CIN85 und SB1 in rezeptor-exprimierenden Zellen 37
3.1.1 Endogene Expression von CIN85 in AML-Zelllinien ................................................ 38 3.1.2 Expressionsnachweis von FLT3-WT, CIN85-WT und SB1-WT und Mutanten in einzeln bzw. doppelt-transduzierten Ba/F3-Zelllinien .......................................................... 39 3.1.3 Untersuchung einer möglichen Induktion von IL-3-unabhängigem Ba/F3-Zellwachstum durch SB1-Mutanten .................................................................................... 41 3.1.4 Untersuchung einer möglichen Apoptoseresistenz nach Zytokinentzug durch SB1-Mutanten ............................................................................................................................ 44 3.1.5 Die Internalisierung des FLT3-Rezeptors wird durch die Koexpression der CIN85 bzw. SB1-Konstrukte nicht beeinflusst ................................................................................ 45 3.1.6 Detektion der Lokalisation von SB1-WT und SB1-Mutanten mittels Immunostaining.......... ........................................................................................................ 47 3.1.7 Zusammenfassung der Ergebnisse der funktionellen Charakterisierung von CIN85 und SB1 in rezeptor-exprimierenden Zellen ........................................................................ 49
3.2 Vergleich des transformierenden Potentials der AKT1-E17K-Mutante in FLT3-exprimierenden Zellen mit der FLT3-ITD-Mutante W51 .......................................................... 49
3.2.1 Stabile Expression der FLT3- und AKT1–Konstrukte in den Ba/F3-Zelllinien .......... 50 3.2.2 Vergleich des pro-proliferativen Phänotyps der AKT1-E17K-Mutante und FLT3-ITD-Mutante W51 in den Ba/F3-Zelllinien und Inhibition des AKT1-E17K-induzierten proliferativen Phänotyps mittels AKT- und FLT3-Inhibitoren ............................................... 51 3.2.3 Einfluss der AKT1-E17K-Mutante auf die Apoptose nach IL-3-Entzug .................... 54 3.2.4 Untersuchung der Signalwege STAT5, AKT und MAPK in den Ba/F3-FLT3/AKT-Zelllinien ............................................................................................................................. 56 3.2.5 Zusammenfassung der Ergebnisse des biologischen Phänotyps von AKT1-E17K und FLT3-ITD im Vergleich ................................................................................................. 58
3.3 Der epigenetische Regulator TET2 ................................................................................ 59 3.3.1 Funktionelle Charakterisierung von hTET2-WT in Ba/F3-Zellen .............................. 59 3.3.2 Screening von CMML-Patienten auf TET2-Mutationen ........................................... 60 3.3.3 Zusammenfassung der Ergebnisse des epigenetischen Regulators TET2 .............. 62
4 Diskussion ............................................................................................................ 63 4.1 Charakterisierung der neu detektierten Punktmutationen in SB1 ................................... 63 4.2 Der biologische Phänotyp von AKT1-E17K und FLT3-ITD im Vergleich ........................ 66 4.3 Mutationen des epigenetischen Regulators TET2 in CMML-Patienten .......................... 69 5 Zusammenfassung............................................................................................... 72
6 Literaturverzeichnis ............................................................................................. 74
7 Anhang .................................................................................................................. 82 7.1 Tabellen......................................................................................................................... 82 7.2 Abbildungen .................................................................................................................. 83 8 Danksagung ......................................................................................................... 85

1
1 Einleitung
1.1 Die akute myeloische Leukämie
Bei akuten Leukämien handelt es sich um hochmaligne Stammzellerkrankungen mit klonaler
Expansion hämatopoetischer Vorläuferzellen, die unbehandelt innerhalb weniger Wochen zum
Tod führen [1]. Durch maligne Transformation von Stammzellen kommt es zur Vermehrung
eines leukämischen Zellklons, auch Blast genannt, welcher neben einem unkontrollierten
Wachstum auch einen apoptotischen und Differenzierungsblock aufweist, der auf
verschiedenen Ebenen der Zellreifung entstehen kann. Die autonome Proliferation einer
malignen Zellpopulation mit Akkumulation unreifer und funktionsloser Blasten führt zur
Verdrängung der normalen Hämatopoese mit konsekutiver Anämie, Thrombozytopenie und
Neutropenie als Folge der Knochenmarkinsuffizienz. In Abhängigkeit der eingeschlagenen
Differenzierungsrichtung unterscheidet man die akute lymphatische Leukämie, die durch
unkontrollierte Proliferation von Vorläuferzellen der Lymphopoese gekennzeichnet ist, von der
akuten myeloischen Leukämie, bei der es sich um eine Neoplasie der Myelopoese mit
variabler Beteiligung myeloischer Zellen, insbesondere der Granulozyten und Monozyten,
handelt. Selten gibt es biphänotypische Formen [2].
Die akute myeloische Leukämie ist mit 80 % die häufigste Unterform der akuten Leukämien im
Erwachsenenalter. Die Inzidenz der AML liegt bei 3-4/100.000, im Alter kontinuierlich
zunehmend mit einem mittleren Erkrankungsalter von ca. 65 Jahren [1, 3, 4]. Ab einem
Infiltrationsgrad des Knochenmarks mit mehr als 20 % unreifer Vorläuferzellen ist laut WHO
(„World-Health-Organization“)-Klassifikation eine akute Leukämie gegeben [5].
Neben der Einteilung der AML nach zytomorphologischen Gesichtspunkten wie in der FAB
(„French-American-British“)-Klassifikation aus dem Jahr 1976, die auf einer Analyse von
Knochenmark- oder peripheren Blutausstrichen durch zytomorphologische und zytochemische
Methoden basiert, erlaubt die neue WHO-Klassifikation von 2008 (siehe Tab. 1.1) durch die
Kombination von verschiedenen additiven Techniken wie Immunphänotypisierung, Zytogenetik
sowie Molekulargenetik eine genaue Definition von Subtypen in der AML und damit eine
präzise Diagnosestellung [2, 5].

2
Tabelle 1.1: Einteilung der akuten myeloischen Leukämie. Einteilung nach der WHO 2008-
Klassifizierung [2, 5]. Tabelle aus [2].
Akute myeloische Leukämie AML mit wiederkehrenden zytogenetischen Abnormalitäten
- mit t(8;21)(q22;q22), (AML1/ETO) - mit inv(16)(p13;q22) oder t(16;16)(p13;q22), (CBFß/MYH11) - akute Promyelozytenleukämie: mit t(15;17)(q22;q12), (PML/RARα) und
Varianten - mit 11q23 (MLL) Abnormalitäten
AML mit Dysplasie mehrerer Zellreihen (multilineär) - mit MDS-Vorphase - ohne MDS-Vorphase
AML und MDS, therapiebedingt - nach alkylierenden Substanzen - nach Topoisomerase-Inhibitoren
AML ohne andere Einordnungsmöglichkeit - mit minimaler Ausreifung - ohne Ausreifung - akute myelomonozytäre Leukämie - akute monoblastische und monozytäre Leukämie - akute Erythroleukämie - akute Megakaryoblastenleukämie - akute Basophilenleukämie - akute Panmyelosis mit Myelofibrose - myeloisches Sarkom - akute biphänotypische Leukämie
Vor allem Zyto- und Molekulargenetik sind für die Wahl der Therapie, aber auch für die
Prognoseeinschätzung unerlässlich, gezielte Therapieansätze rücken immer mehr in den
Fokus moderner Behandlungskonzepte. Rund 60 % der AML-Patienten weisen in der nicht
vorbehandelten zytogenetischen Analyse einen abnormalen Karyotyp auf, der je nach
zytogenetischer Entität, wie in Abbildung 1.1 zu sehen, unterschiedlichen Einfluss auf das
Gesamtüberleben der AML-Patienten hat [6, 7].
Auch in der Gruppe der AML-Patienten mit normalem Karyotyp (rund 40 %) wurde durch
molekulare Analysen eine prognostische Signifikanz und Relevanz der sehr heterogenen
genetischen Alterationen entdeckt. Zu diesen gehören beispielsweise Mutationen des Gens
für die Rezeptor-Tyrosinkinase „fibroblast-macrophage stimulating factor receptor (FMS)-
related tyrosine kinase 3“ (FLT3), Mutationen das Nucleophosmin Member 1 (NPM1)-Gens,
aber auch partielle Tandem-Duplikationen des myeloid/lymphoid oder mixed-lineage leukemia
(MLL)- sowie Überexpression des BAALC-Gens [8].
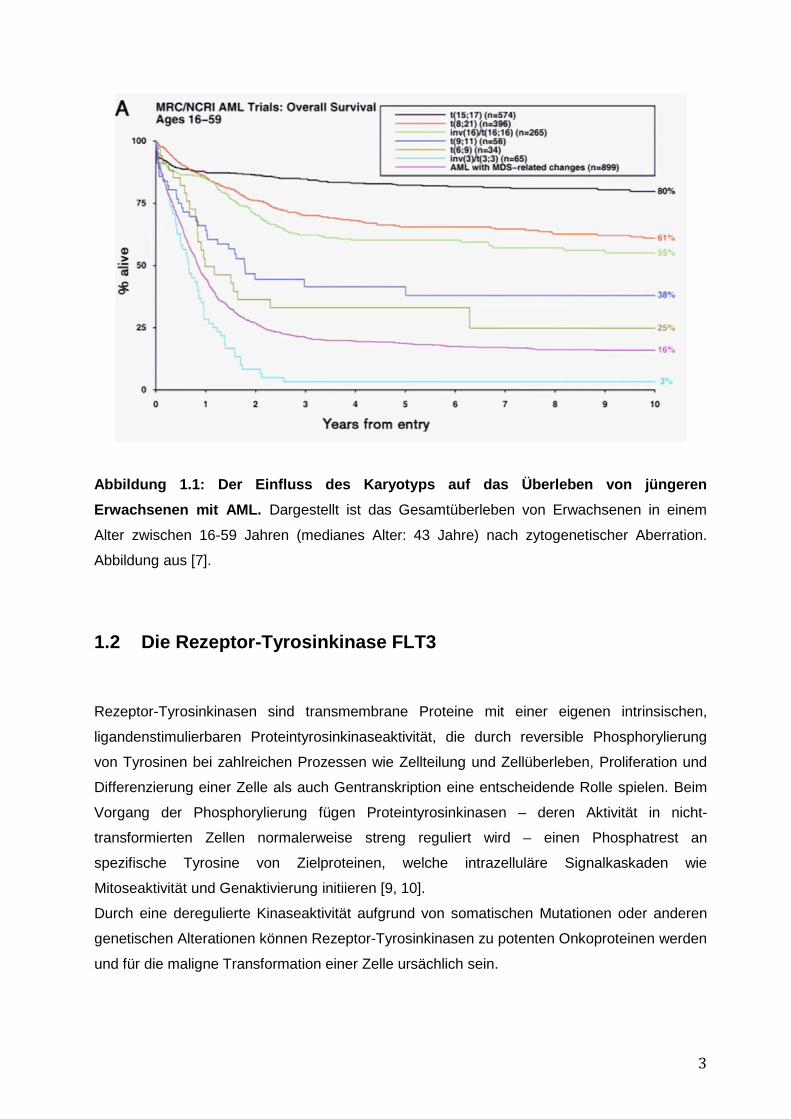
3
Abbildung 1.1: Der Einfluss des Karyotyps auf das Überleben von jüngeren Erwachsenen mit AML. Dargestellt ist das Gesamtüberleben von Erwachsenen in einem
Alter zwischen 16-59 Jahren (medianes Alter: 43 Jahre) nach zytogenetischer Aberration.
Abbildung aus [7].
1.2 Die Rezeptor-Tyrosinkinase FLT3 Rezeptor-Tyrosinkinasen sind transmembrane Proteine mit einer eigenen intrinsischen,
ligandenstimulierbaren Proteintyrosinkinaseaktivität, die durch reversible Phosphorylierung
von Tyrosinen bei zahlreichen Prozessen wie Zellteilung und Zellüberleben, Proliferation und
Differenzierung einer Zelle als auch Gentranskription eine entscheidende Rolle spielen. Beim
Vorgang der Phosphorylierung fügen Proteintyrosinkinasen – deren Aktivität in nicht-
transformierten Zellen normalerweise streng reguliert wird – einen Phosphatrest an
spezifische Tyrosine von Zielproteinen, welche intrazelluläre Signalkaskaden wie
Mitoseaktivität und Genaktivierung initiieren [9, 10].
Durch eine deregulierte Kinaseaktivität aufgrund von somatischen Mutationen oder anderen
genetischen Alterationen können Rezeptor-Tyrosinkinasen zu potenten Onkoproteinen werden
und für die maligne Transformation einer Zelle ursächlich sein.

4
Das humane Genom umfasst 58 Rezeptor-Tyrosinkinasen (RTK), die in 20 Familien unterteilt
sind. Mehr als die Hälfte dieser sind mit humanen malignen Erkrankungen assoziiert [11]. Alle
Rezeptor-Tyrosinkinasen verbindet ihre ähnliche molekulare Architektur, wobei vor allem die
extrazelluläre liganden-bindende Domäne die größte Varianz aufweist. Neben der Ligand-
bindenden extrazellulären Domäne bestehen RTKs aus der darauffolgenden
transmembranösen Domäne mit einer singulären transmembranösen Helix und zudem der
zytoplasmatischen Region, die die juxtamembranöse Domäne und die Kinasedomäne
beinhaltet [12].
1.2.1 Struktureller Aufbau und Expression der Rezeptor-Tyrosinkinase FLT3
FLT3 (Fms-like tyrosine kinase 3), auch FLK-2 (fetal liver kinase-2) oder STK-1 (human stem
cell kinase-1) genannt, wurde erstmals 1991 von zwei unabhängigen Gruppen isoliert und das
humane FLT3-Gen auf Chromosom 13q12 lokalisiert [13-15].
FLT3 kommt eine wichtige Rolle in der Entwicklung pluripotenter Stammzellen und B-Zellen zu
und wird größtenteils in hämatopoetischen Stammzellen, frühen, unreifen Vorläuferzellen und
dendritischen Zellen exprimiert.
Das humane FLT3-Protein mit 993 Aminosäuren kommt in der Zelle als ein posttranslational
N-linked glykosyliertes Protein mit rund 160 kDa, welches in der Plasmamembran lokalisiert
ist, sowie als schwächer glykosyliertes Protein mit etwa 140 kDa vor [16, 17].
Mit Progression der Zelldifferenzierung in der Hämatopoese nimmt die Expression des
humanen FLT3 (hFLT3) ab. Die Expression des hFLT3 ist auf Stammzellen und
Vorläuferzellen wie beispielsweise frühe lymphoide Progenitorzellen, myeloische und
granulozytär / makrophagische Progenitorzellen beschränkt [13, 18].
Der FLT3-Rezeptor zählt zu den Klasse-III-Rezeptor-Tyrosinkinasen. Sie sind durch große
Homologie in ihrer Sequenz, aber auch in ihrer Struktur gekennzeichnet. Sie zeigen eine
extrazelluläre Domäne bestehend aus fünf immunglobulin-ähnlichen Domänen, eine
transmembranöse Domäne und eine zytoplasmatische Domäne, welche aus
juxtamembranöser Domäne und zwei Proteintyrosinkinasedomänen, die durch eine
Interkinasedomäne voneinander getrennt sind, besteht [16, 19]. Neben FLT3 besteht die
Familie der Klasse-III-Rezeptor-Tyrosinkinasen (siehe auch Abbildung 1.2.1) aus den
Rezeptoren „stem cell factor receptor“ (KIT), platelet-derived growth factor receptor (PDGFR)
A und B, „colony-stimulating-factor 1“-Rezeptor (CSF1R) sowie „macrophage colony-
stimulating factor receptor“ (FMS) [12, 20].

5
Abb. 1.2.1: Einige in maligne hämatologische Erkrankungen involvierte Rezeptor-Tyrosinkinase-Familien und deren Aufbau. Unterschiedliche Rezeptorfamilien mit ihren
extrazellulären, transmembranösen und intrazellulären Domänen; in Gelb sind die
intrazellulären Kinasedomänen dargestellt. Abbildung nach [21].
1.2.2 Signalkaskaden und Mutationen des FLT3-Rezeptors Der inaktive FLT3-Rezeptor liegt als monomere, unphosphorylierte Form mit einer inaktiven
Kinasedomäne in der Membran vor. Die Bindung seines spezifischen Liganden FLT3-Ligand
(FL) führt zur Aktivierung und Konformationsänderung des Rezeptors, was durch Freilegung
der Dimerisierungsdomäne die Dimerisierung von Rezeptormonomeren induziert. Nach
Aktivierung der zytoplasmatischen Kinasedomänen kommt es zur Phosphorylierung von
Tyrosinresten intrazellulärer Rezeptoranteile. Die durch die Tyrosinphosphorylierung
geschaffenen Erkennungsmotive für Src-Homologie-2 (SH2)-Domänen anderer
zytoplasmatischer Proteine führen dazu, dass diese mit der Kinasedomäne interagieren und
ebenfalls an Tyrosinresten phosphoryliert werden [22, 23].

6
Zu den Adapterproteinen des Komplexes der Protein-Protein-Interaktionen, die an der
intrazellulären Domäne mit dem aktivierten FLT3-Rezeptor stattfinden, gehören beispielsweise
Casitas B-lineage lymphoma (CBL), CBLB (CBLB-related protein), SHIP, GRB2 und GRB2-
associated binder 2 (GAB2) [22, 24-27].
Die Kaskade an Phosphorylierungen resultiert in der Aktivierung von Signalkaskaden wie dem
„mitogen-activated protein“ (MAP)-Kinase-, Protein-Kinase-B (AKT) / „Phosphatidylinositol-3“
(PI3) - Kinase-Signalweg und dem „signal transducer and activator of transcription 5“ (STAT)-
Signalweg, die im Zellkern die transkriptionelle Regulierung von Genen beeinflussen, die
Zellüberleben, Proliferation, Apoptose und Differenzierung spezifischer Vorläuferpopulationen
steuern [28].
FLT3 kann als Protoonkogen Zellen transformieren und das Überleben und die Proliferation
leukämischer Blasten fördern [28]. Das Wildtyp (WT)-FLT3-Protein wird in vielen
hämatologischen Tumorerkrankungen exprimiert. Vor allem in der AML mit 70 bis zu 100 %
der Patienten und in der ALL finden sich hohe Expressionslevel von WT-FLT3, aber auch in
der fortgeschrittenen Krankheitsphase der CML, in der Blastenkrise, wurde die Expression
nachgewiesen [16, 29].
1.2.3 Funktion von FLT3 in der malignen Hämatopoese
Zudem zeigten Analysen von leukämischen Blasten in AML-Patienten somatische Mutationen
im FLT3-Gen. Die 1996 von Nakao et al. zuerst beschriebenen „internal tandem duplication“
(ITD)-Mutationen des FLT3-Gens stellen mit beispielsweise 20 - 30 % in der AML die
häufigsten FLT3-Mutationen dar [16, 30]. Durch in-frame Duplikationen eines Fragments der
juxtamembranösen Domäne, die in ihrer Länge von 2 bis 68 Aminosäuren variieren, resultiert
eine konstitutive Autophosphorylierung und damit Aktivierung des FLT3-Rezeptors aufgrund
von gestörter Autoinhibition [31]. FLT3-ITD-Mutationen korrelieren mit schlechter Prognose,
erhöhter Rezidivwahrscheinlichkeit, erniedrigter Remissionsdauer und erniedrigtem
Gesamtüberleben [32-34].
Mit bis zu 7 % in der AML ist die zweite Tyrosinkinasedomäne (TKD) des FLT3-Rezeptors
betroffen und stellt eine weitere häufige FLT3-Mutation dar. Hierbei kommt es durch
Punktmutationen zum Austausch des Aspartats 835 meist gegen ein Tyrosin (D835Y),
seltener gegen ein Histidin, Valin oder Glutamat oder aber durch kleine Deletionen und
Insertionen zu konstitutiver ligandenunabhängiger Rezeptoraktivierung. Eine prognostische
Auswirkung der TKD-Mutationen konnte bisher nicht nachgewiesen werden und bleibt
ungewiss [35-37].
Des Weiteren wurden Punktmutationen wie beispielsweise in der juxtamembranösen Domäne

7
(FLT3-JM-PMs) und Längenmutationen wie FLT3-ITD-W51, FLT3-ITD-NPOS und FLT3-ITD-
W78 beschrieben [38-42].
Auch durch Mutationen verursachte konstitutive Aktivierung nachgeschalteter Signalwege von
RTKs wie zum Beispiel im PI3K/AKT-Signalweg stellt ein bekanntes Charakteristikum der AML
und anderer humaner Tumorerkrankungen dar [43, 44]. Zu Beginn des PI3K/AKT-Signalwegs
steht die Aktivierung der Phosphoinositid-Kinase (PI3K). Sie kann durch mehrere
Mechanismen stattfinden: beispielsweise durch direkte Bindung an phosphorylierte RTKs oder
assoziierte Tyrosinkinasen oder über die membrangebundene GTPase Ras. Aktivierte PI3K
bindet an der Membraninnenseite Phosphatidylinositol-4,5-Bisphosphat (PIP2) und
phosphoryliert dieses zu Phosphatidylinositol-3,4,5-Trisphosphat (PIP3) [45]. Als sekundärer
Botenstoff lokalisiert PIP3 die Proteinkinase B (AKT), welche zu den Serin-Threonin-Kinasen
gehört, an die Plasmamembran. AKT wird dort mittels Phosphorylierung am Threonin 308
durch die 3-phosphoinositid-abhängige Kinase-1 (PDK1) sowie mittels Phosphorylierung am
Serin 473 durch eine weitere Kinase aktiviert [46]. Die Downstream-Wirkungen von AKT - wie
Blockade der Apoptose, Aktivierung der Translation oder Verhinderung von
zellteilungshemmender Wirkung - sind vielfältig. Über die Aktivierung multipler Effektoren und
Substrate generiert AKT-Signale, die in die Regulation der Apoptose und Proliferation
eingreifen [23, 47, 48].
Auch die deregulierte „signal transducer and activator of transcription 5“ (STAT5)-
Signalübertragung und konstitutive Aktivierung von STAT5, was zu erhöhter Transkription von
antiapoptotischen und pro-proliferativen Genen führt, wurde in leukämischen Blasten von
AML-Patienten nachgewiesen [49, 50]. Der STAT5-Signalweg wird entweder direkt von RTKs,
über JAK-Kinasen oder über SRC family-Kinasen (SFK) aktiviert [51]. Phosphorylierte STATs
dimerisieren und gelangen als STAT-Dimere in den Zellkern, in dem sie durch Bindung an
DNA die Expression von Zielgenen verändern. Die aberrante Aktivierung des STAT-
Signalwegs bei FLT3-ITDs induziert zu einem entscheidenden Teil die maligne Transformation
[50].
Die aberrant aktivierte FLT3-Kinase ist ein attraktiver Angriffspunkt zielgerichteter Therapien.
Mehrere FLT3-Tyrosinkinase-Inhibitoren (TKIs) befinden sich in präklinischen und klinischen
Studien, wobei sich in den meisten AML-Patienten nicht anhaltende oder eingeschränkte
Effektivität der Inhibitoren zeigt. Ein weiteres Problem stellt Resistenzbildung dar, sodass eine
Kombination von TKI mit beispielsweise konventioneller Chemotherapie als neue
Therapieoption evaluiert wird [52-54].

8
1.3 CBL, CIN85 und SB1 in der Regulation von Rezeptor-Tyrosinkinasen Die Identifikation des Protoonkogens CBL erfolgte erstmals 1989 in mit dem murinen
Leukämie-Virus Cas-NS1 infizierten Mäusen, welcher zur Induktion von pro-B-Lymphomen
und myeloischen Leukämien führt [55].
CBL, auch Casitas B-lineage lymphoma genannt, ist eine E3-Ubiquitin-Ligase, 120 kDa groß
und hoch konserviert. Als zur CBL-Familie der Säugetiere zugehörig konnten bisher die drei
Proteine CBL (c-CBL), CBL-b und CBL-3 charakterisiert werden. CBL gilt unter anderem als
Negativregulator, der das Proliferationssignal von Rezeptor-Tyrosinkinasen wie FLT3 und KIT
abschwächt, indem er zum Beispiel deren lysosomalen Abbau und Ubiquitinierung initiiert [56-
60].
Das Protein CBL besteht aus einer N-terminalen Tyrosinkinase-Bindungsdomäne (TKB) zur
Bindung phosphorylierter Tyrosine von aktivierten RTKs und PTKs [61]. Diese ist gefolgt von
der LINKER- und der RING-Finger-Domäne. Die RING-Finger-Domäne ist für die Rekrutierung
von E2-Enzymen und die Vermittlung des Ubiquitintransfers auf Zielproteine verantwortlich
[62]. Die LINKER-Domäne spielt eine entscheidende Rolle in der Positionierung der TKB-
Domäne zur RING-Finger-Domäne und ist entscheidend an der Effektivität des
Ubiquitintransfers beteiligt [63]. C-terminal des RING-Fingers folgt eine Prolin-reiche Region
(PXXP), C-terminale Phosphorylierungsstellen, die „ubiquitin-assoziierte“ (UBA)-Domäne und
eine Leuzin-Zipper (LZ)-Domäne, wodurch CBL als Multi-Adapter-Protein fungiert als auch die
Dimerisierung von CBL ermöglicht wird [64-66].
CBL ist als Multi-Adapter-Protein und Ubiquitin-Ligase in eine Vielfalt von zellulären Prozessen
involviert. Abbildung 1.3.1 zeigt einen Ausschnitt der Komplexität, mit wie vielen bisher
bekannten Proteinen CBL interagiert bzw. wie viele Proteine durch CBL reguliert werden. Eine
wichtige Rolle in der Rezeptor-Endozytose spielen beispielsweise die Adaptermoleküle CIN85
und CD2AP [64].

9
Abb. 1.3.1: Das CBL-Interaktom. Die Darstellung des Osprey-Diagramms zeigt die
momentan bekannten, mit CBL interagierenden Proteine in einer Übersicht. Die Gruppierung
erfolgte entsprechend der CBL-Bindungsstelle mittels farbiger Punkte. Abbildung aus [64].
Bereits Im Jahr 2002 wurde für CBL-b und 2003 für c-CBL eine atypische Poly-Prolin-Arginin-
Region (PXXXPR), die spezifisch von den SH3-Domänen von CIN85 erkannt wird, identifiziert
[67-69].
CIN85 (CBL-interacting protein of 85kDa), auch als RUK (regulator of ubiquitous kinase),
SH3KBP1 (SH3 (Src homology 3)-domain kinase binding protein 1) oder SETA (SH3 domain-
containing gene expressed in tumorigenic astrocytes) bekannt, agiert als zytoplasmatisches
Multi-Adapter-Protein. Über die C-terminale Region PXXXPR von c-CBL (auch bei CBL-b und
einigen anderen Proteinen) erfolgt die Bindung von CIN85 an c-CBL [59, 67-70]. Es entsteht
ein CBL-CIN85-Endophilin-Komplex, der mit Induktion einer negativen Membrankurvatur
entscheidend für die Invagination der Plasmamembran und somit entscheidend für die frühe
Phase der Endozytose und für die Rezeptor-Internalisierung ist [64, 71, 72]. Auch eine
Bindung von CIN85 und dessen Homolog CD2AP (CD2-associated protein) an adaptor-

10
protein-2 (AP-2) als Alternativ-Signalweg zur clathrin-vermittelten Endozytose ist möglich [64].
Die Rolle von CBL und CIN85 am Beispiel der EGFR-Endozytose zeigt Abbildung 1.3.2.
Abb. 1.3.2: Die Rolle von CBL und CIN85 in der EGFR-Endozytose. Darstellung der
Induktion einer negativen Membrankurvatur und Invagination der Plasmamembran durch die
Komplexbildung von EGFR, CBL, CIN85 und die Rekrutierung von Endophilinen. Abbildung
nach [73, 74].
CIN85 beeinflusst des Weiteren die Apoptose und ist zudem durch verschiedene
Mechanismen in die Negativregulation von CBL-Funktionen involviert. Beispielsweise induziert
EGF-Stimulation die Synthese von Sprouty2, welches an der Plasmamembran phosphoryliert
wird und die CBL-Bindungsstelle am EGFR imitiert. Die vermehrte Bindung von CBL an
Sprouty2 verringert in der Folge die Bindung von CBL am aktivierten EGFR, sodass die
Downregulation von EGFR inhibiert wird. Dies ist nur Sprouty-Isoformen möglich, die eine
CIN85-Bindedomäne (PXXXPR) besitzen [64, 75].
Im Jahr 2010 wurden von J Adélaíde et al. 63 MDS-Patienten und 53 CMML-Patienten
analysiert. Sowohl bei einem der MDS- (1,6 %) als auch bei einem der CMML-Patienten (1,9
%) wurden mit dem Zugewinn von jeweils genau einer zusätzlichen Kopie des CIN85-Gens
Alterationen des Gens, das für ein mit CBL interagierendes Protein kodiert, identifiziert.
Aufgrund der Tatsache, dass weder bei Mamma- noch bei Kolonkarzinomen eine Amplifikation
dieses Gens detektiert werden konnte, spricht diese Alteration für eine Assoziation mit den

11
Erkrankungen MDS und CMML. Eine ähnliche Rolle wie die von CBL-Mutationen in
myeloischen Erkrankungen scheint denkbar [60].
Es sind verschiedene CIN85-Isoformen bekannt; die längste besteht aus drei N-terminalen
SH3-Domänen, die die CBL-Bindung vermitteln, gefolgt von einer zentralen Prolin-reichen
Domäne und C-terminal einer coiled-coil (CC)-Domäne (siehe Abb. 1.3.3) [73, 76].
Aufgrund der neu gefundenen dritten SH3-Domäne im Jahr 2000 gelang mittels Yeast-two-
hybrid-Klonierung die Isolation eines neuen, mit CIN85 assoziierten Gens: SB1, auch Seta
binding protein 1 oder SHKBP1 genannt. Wie in Abbildung 1.3.4 dargestellt ist es SB1 in vitro
möglich, an alle 3 SH3-Domänen von CIN85 zu binden. Außerdem konnte eine starke
Interaktion von SB1 mit GRB2 als auch eine schwache Interaktion von CIN85 mit GRB2
nachgewiesen werden, was auf einen möglichen intrazellulären Komplex hinweisen könnte
[76]. Eine Überexpression von SHKBP1 zeigt anti-apoptotische Effekte in Zelllinien, was auf
eine Rolle als mutmaßliches Protoonkogen schließen lassen könnte [77].
Über Transkriptom-Sequenzierung einer 69jährigen de-novo-AML-Patientin, initial bei
Erstdiagnose des Knochenmarks und der Remissionskontrolle im peripheren Blut nach
Leukozytenregeneration, konnte durch unsere Arbeitsgruppe erstmals eine somatische SB1-
Mutation beschrieben werden. Die Patientin hatte mit 46, XX[20] einen normalen Karyotyp;
eine FLT3-ITD, NPM1-Mutation als auch partielle Tandem-Duplikation des MLL-Gens wurden
ausgeschlossen. Die gefundene tumorspezifische Einzelnukleotid-Variante resultierte in der
Missense-Mutation in SB1 (V89I). Aufgrund dieser Identifikation wurde über
Kapillarsequenzierung die genomische DNA weiterer 95 zytogenetisch normaler AML-
Patienten analysiert. Hierbei konnten zwei weitere Missense-Mutationen von SB1 (R454Q und
R672Q) identifiziert werden, die in Kombination mit NPM1- und FLT3-Mutationen vorlagen
[78].
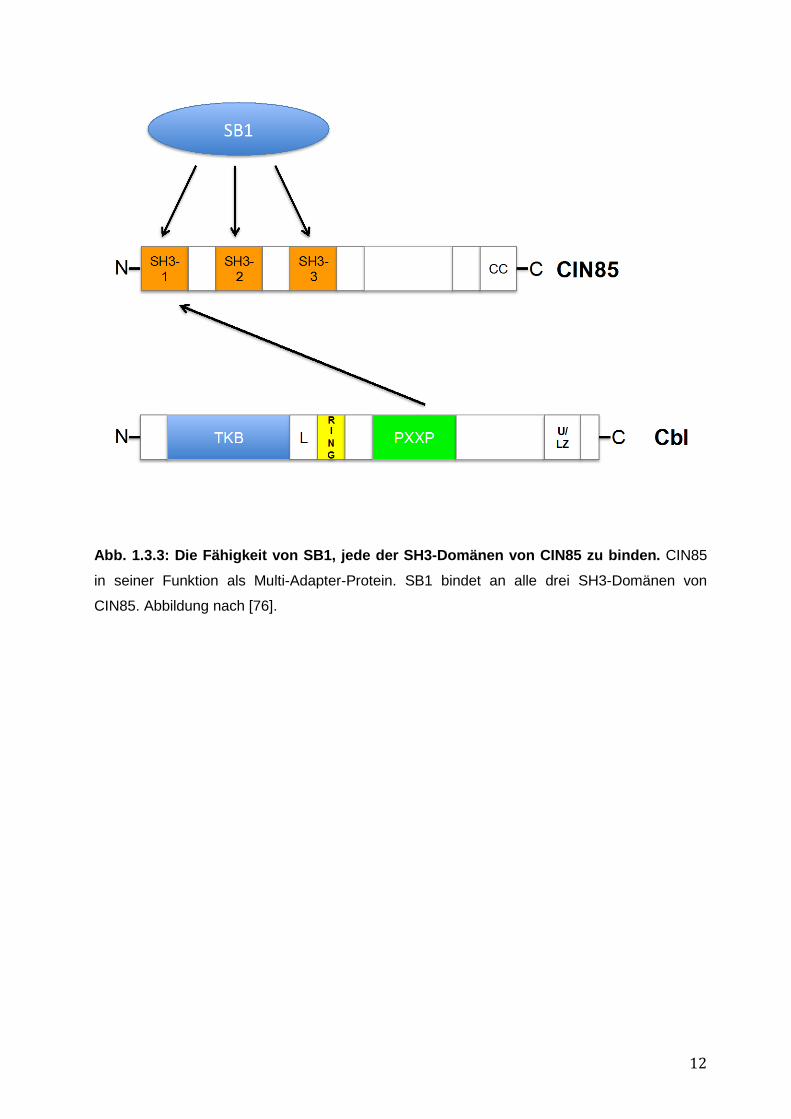
12
Abb. 1.3.3: Die Fähigkeit von SB1, jede der SH3-Domänen von CIN85 zu binden. CIN85
in seiner Funktion als Multi-Adapter-Protein. SB1 bindet an alle drei SH3-Domänen von
CIN85. Abbildung nach [76].

13
1.4 TET2 als epigenetischer Regulator
Die Identifikation von Mutationen in Genen involviert in epigenetische Regulation, wie
beispielsweise die des Ten-Eleven Translocation oncogene family member 2 (TET2), hat zur
deutlichen Erweiterung des Verständnisses der Leukämieentstehung beigetragen.
Das Gen TET2 - auf Chromosom 4q24 lokalisiert - wurde über SNP-arrays initial eine
mögliche Rolle als Tumor-Suppressor-Gen in myeloischen Neoplasien zugewiesen [79, 80].
Mutiertes oder deletiertes TET2 wurde anschließend in verschiedenen myeloischen
Erkrankungen wie in myelodysplastischem Syndrom (MDS), in myeloproliferativen Neoplasien
(MPN), in der AML, sekundärer AML und in chronisch myelomonozytärer Leukämie (CMML)
beschrieben [79, 81]. Die Gesamtfrequenz der Mutationen beträgt in der AML um die 10-20 %,
eine besonders hohe Mutationsfrequenz mit bis zu 50 % wurde in der CMML beschrieben [81].
Aufgrund des Nachweises der Konversion von 5-Methylcytosin (5-mC) zu 5-Hydroxy-
Methylcytosin (5-hmC) in DNA durch humanes TET2 bestätigte sich die Rolle von TET2 im
Rahmen der epigenetischen Expressionskontrolle (siehe Abb. 1.4) [82]. TET2 katalysiert die
Oxidation von 5-mC, was zu einer Demethylierung der DNA führt. Cytosin-Methylierung
ermöglicht in Eukaryonten eine epigenetische Regulation, die die Expression des jeweiligen
Gens supprimiert (= “Silencing“) [83]. Eine eingeschränkte katalytische Aktivität bei TET2-
Mutationen konnte Ko et al. 2010 in Knochenmarksproben von TET2-mutierten Patienten
nachweisen, die durchweg erniedrigte 5-hmC-Level in genomischer DNA im Vergleich zu den
gesunden Kontrollproben aufwiesen [84].
Kosmider et al. beschrieb eine Assoziation von TET2-Mutationen mit einem kürzen
Gesamtüberleben („overall survival“, OS) bei CMML-Patienten. Andere klinische Studien
zeigten entweder keine signifikante Korrelation einer TET2-Mutation mit dem Überleben bei
AML-Patienten oder längeres OS bei AML-Patienten, sodass die prognostische Relevanz
letztlich umstritten bleibt [81, 85-88].
Unumstritten ist die außerordentlich wichtige Rolle von TET2 in der normalen als auch
malignen Hämatopoese sowie seine Rolle in der Entwicklung hämatologischer Neoplasien, die
zukünftig mit Sicherheit Fokus weiterer Charakterisierungen sein wird.

14
Abb. 1.4: Mechanismen der TET2-induzierten Genexpression. A: TET2 fungiert als Katalysator von 5-mC zu 5-hmC in DNA mit α–KG als essentiellen
Kofaktor. 5-hmC ist ein Zwischenprodukt des Demethylierungs-Signalwegs. B: Durch TET2-
Mutationen wird 5-mC in geringerem Maße hydroxyliert, wodurch Gene der Negativregulation
der Zellselbsterneuerung und Gene der Förderung der Zelldifferenzierung nicht aktiviert
werden. Abbildung aus ([89], Referenz verändert).

15
1.5 Zielsetzung der vorliegenden Arbeit
Aufgrund von neuen Sequenzierungsmethoden wie beispielsweise der Transkriptom-
Sequenzierung wurden innerhalb der letzten Jahre viele neue Mutationen in Genen
identifiziert.
Ein Ziel dieser Dissertation besteht darin, das mutmaßliche onkogene Potential der in AML-
Patienten neu gefundenen SB1-Mutationen SB1-V89I, SB1-R454Q und SB1-R672Q
funktionell zu charakterisieren und in einem ersten Schritt Effekte auf Proliferation und
Apoptose zu untersuchen. Durch die nachgewiesene Bindung von SB1 an die SH3-Domänen
von CIN85, welches wiederum mit CBL interagiert, lässt sich auf eine potentielle Rolle von
SB1 in der Regulation von Rezeptor-Tyrosinkinasen und deren Endozytose, Degradierung und
Ubiquitinierung schließen. Des Weiteren zeigte eine Überexpression von SB1 in Zelllinien anti-
apoptotische Effekte.
Die zentrale Bedeutung der aktivierenden Mutationen von Rezeptor-Tyrosinkinasen wie FLT3
in der Leukämieentstehung ist seit Langem bekannt. FLT3 liegt bei rund 30 % der AML-
Patienten mutiert vor, speziell FLT3-ITD-Mutationen sind in der AML mit schlechter Prognose
und erhöhter Rezidivwahrscheinlichkeit assoziiert. Des Weiteren sollte in der vorliegenden
Dissertation das transformierende Potential der aktivierenden FLT3-ITD-Mutation W51 mit
dem der AKT1-E17K-Mutante in Form von Proliferations-, Apoptose- als auch Signaling-
Untersuchungen verglichen werden.
Auch die bedeutende Rolle von TET2 als epigenetischer Regulator in der normalen und
malignen Hämatopoese wurde innerhalb der letzten Jahre zunehmend besser verstanden. In
dieser Dissertation erfolgte das Screening einer CMML-Patientengruppe auf TET2-Mutationen
als auch die Isolation von RNA, cDNA und Proteinlysaten aus einer fast vollständig reinen
Monozytenpopulation der CMML-Patienten sowie die funktionelle Charakterisierung von
hTET2-WT in Ba/F3-Zellen.
Abbildung 1.5 zeigt die in der vorliegenden Doktorarbeit untersuchten Aspekte vereinfacht
dargestellt im Überblick.

16
Abb. 1.5: Überblick der untersuchten Aspekte der vorliegenden Doktorarbeit. Die
Aktivierung des FLT3-Rezeptors resultiert in einer Vielzahl von Signalkaskaden, welche in die
Regulation von Apoptose und Proliferation einer Zelle eingreifen. TET2 ist in die epigenetische
Regulation involviert. Abbildung nach [22, 78, 90].

17
2 Material und Methoden
2.1 Material
2.1.1 Inhibitoren, Enzyme, Zytokine und Chemikalien
Tabelle 2.1.1.1: Inhibitoren Inhibitor Bemerkung Bezugsquelle AC220 FLT3-Inhibitor SYNthesis med chem
(Cambridge, UK)
MK2206 AKT-Inhibitor Selleck Chemicals
(Houston, TX, USA)
Tabelle 2.1.1.2: Enzyme Enzym Bezugsquelle DyNAzyme-II-DNA-Polymerase Finnzymes, Espoo, Finnland Herculase-II-Fusion-DNA-Polymerase Agilent Technologies, Santa Clara, CA Phusion High-Fidelity-DNA-Polymerase Finnzymes, Espoo, Finnland Restriktionsenzyme New England Biolabs, Frankfurt Taq-DNA-Polymerase Roche, Mannheim T4-DNA-Ligase New England Biolabs, Frankfurt;
Fermentas, St. Leon-Rot
Tabelle 2.1.1.3: Zytokine Zytokin Bemerkung Bezugsquelle FLT3-Ligand (FL) rekombinant human Promocell, Heidelberg;
Immunotools, Friesoythe Interleukin-3 (IL-3) rekombinant murin Biosource, Camarillo, CA,
USA;
Immunotools, Friesoythe
18
Tabelle 2.1.1.4: Chemikalien Chemikalie Bezugsquelle Acrylamid/Bisacrylamid 30 % Roth, Karlsruhe Agarose Sigma, Taufkirchen Ammoniumperoxidisulfat (APS) BioRad, München Aprotinin Sigma, Taufkirchen Bio-Rad Protein Assay Dye Reagenz BioRad, München Bovines Serumalbumin (BSA) Fluka, Buchs, Schweiz BM Cyclin Roche, Mannheim Coulter Clenz Cleaning Agent Beckman Coulter, Krefeld Coulter Isoton II Diluent Beckman Coulter, Krefeld Dimethylsulfoxid (DMSO) Sigma, Taufkirchen DNA-Leiter Promega, Madison, WI, USA DNAse I Invitrogen, Karlsruhe dNTPs Invitrogen, Karlsruhe ECL Detektionsreagenz Amersham Pharmacia, Freiburg Ethanol Merck, Darmstadt Ethidiumbromid Fluka, Buchs, Schweiz Fötales Kälberserum (FKS) Sigma, Taufkirchen Glycerin Sigma, Taufkirchen HEPES 2-(4-(2-Hydroxyethyl)-1-
piperazinyl)-ethansulfonsäure
Sigma, Taufkirchen
High Performance Chemiluminescence Film Amershan Pharmacia, Freiburg Leupeptin A Sigma, Taufkirchen 2-Mercaptoethanol Sigma, Taufkirchen Methanol Merck, Darmstadt Milchpulver Merck, Darmstadt Natriumacetat Sigma, Taufkirchen Natriumazid Roth, Karlsruhe Natriumcitrat Sigma, Taufkirchen Natriumfluorid Sigma, Taufkirchen tetra-Natriumdiphosphatdekahydrat Sigma, Taufkirchen Natriumvanadat Sigma, Taufkirchen N-Ethylmaleimid Sigma, Taufkirchen Nitrocellulose-Membran Schleicher & Schüll, Dassel Orthovanadat Sigma, Taufkirchen

19
Paraformaldehyd 4 % (PFA) Carl Roth, Karlsruhe Penicillin-Streptomycin Invitrogen, Darmstadt peqGold Protein Marker VI Peqlab, Erlangen Phenylmethylsulfonylfluorid (PMSF) Sigma, Taufkirchen Ponceau Sigma, Taufkirchen Polyfect Transfektionsreagenz Qiagen, Hilden Polybren Sigma, Taufkirchen 2-Propanol Roth, Karlsruhe Propidiumiodid Sigma, Taufkirchen Rainbow Marker Amersham Pharmacia, Freiburg Sodiumdodecylsulfat (SDS) BioRad, München Tetramethylethylendiamin (TEMED) Serva, Heidelberg Tris(hydroxymethyl)aminomethan (Tris) Merck, Darmstadt Triton-X-100 Sigma, Taufkirchen; AppliChem, Darmstadt Trypan Blau Invitrogen, Darmstadt; Sigma, Taufkirchen
Trypsin-EDTA Invitrogen, Darmstadt Tween 20 Merck, Darmstadt
2.1.2 Software
Tabelle 2.1.2: Software Software Hersteller Adobe Illustrator Adobe Systems, Unterschleißheim BioEdit 7.0 ISIS Pharmaceutica, Carlsbad, USA Cellquest 3.3 Beckton Dickinson, Heidelberg E-capt 15.06 Vilber Lourmat EndNote 6.0.2 Thompson ISI, Carlsbad, CA, USA Microsoft Office Redmond, WA, USA Sigma Plot 11.0 SPSS Incorporated, Chicago, USA Vector NTI Advance 10 Invitrogen, Darmstadt WinMDI 2.9 Joseph Trotter

20
2.1.3 Kits
Tabelle 2.1.3: Kits Bezeichnung Bezugsquelle Annexin V-PE Apoptosis Detection Kit I Beckton Dickinson, Heidelberg BigDye Terminator Cycle Sequencing Kit Applied Biosystems, Darmstadt Endofree Plasmid Maxi Kit Qiagen, Hilden InFusion Advantage PCR Cloning Kit Clontech, CA MycoAlert Mycoplasma Detection Kit Lonza, Rockland, ME, USA Qiagen Endofree Plasmid Maxi Kit Qiagen, Hilden QIAquick Gel Extraction Kit Qiagen, Hilden QIAspin Mini Kit Qiagen, Hilden QuikChange II XL Site-Directed Mutagenesis
Kit Stratagene, Waldbronn
RevertAid Premium First Strand cDNA Fermentas, St. Leon-Rot Rneasy MiniKit Synthesis Kit Qiagen, Hilden
2.1.4 Laborausstattung
Tabelle 2.1.4: Laborausstattung Gerät Hersteller Bakterien-Brutschrank Heraeus, Osterode Blotting Kammer BioRad, München CO
2-Inkubator Binder, Tuttlingen
DNA-ThermoCycler Peqlab, Erlangen Eismaschine ZBE 70-35 Ziegra, Isernhagen Entwicklermaschine Kodak, Stuttgart Expositionskassetten Amersham, Pharmacia, Freiburg FACS Calibur Beckton Dickinson, Heidelberg FACS Sorter Beckton Dickinson, Heidelberg Fluoreszenzmikroskop Carl Zeiss, Jena Gelelektrophoresekammer BioRad, München GS Gene Linker UV Chamber Bio Rad, Kalifornien, USA Heizblock BT 130-2 Heap Labor Consult, Bovenden Heiz Drehrad Innova, Göllheim

21
Konfokales Mikroskop TCS-SP2, Leica, Heidelberg Kühlzentrifuge Eppendorf, Hamburg Mikroskope Carl Zeiss, Jena Pipettierer Accu Jet Brand, Wertheim Proteingelelektrophoresekammer Invitrogen, Darmstadt QuadroMACS Separator Miltenyi Biotec, Bergisch Gladbach Sequenzierer, ABI Prism 310
Genetic Analyzer Applied Biosystems, Darmstadt
Spektophotometer, Nanodrop 1000 Peqlab, Erlangen Sterilbank BDK, Luft und Reinraumtechnik Bio Flow Technik, Meckenheim Sysmex = Leukozytenzähler F820 Sysmex Deutschland Tischzentrifuge Rotanta Hettich, Tuttlingen Ultrazentrifuge LTF, Labortechnik Vi-Cell XR, Vi-Cell AS Beckman Coulter, Krefeld Vortexer Cenco, Breda, NL Wasserbad Haske, Karlsruhe
Zentrifuge Eppendorf, Heraeus
Zytozentrifuge MA 1677 E 9810 Shandon, Hettich, Tuttlingen
2.1.5 Zelllinien
Tabelle 2.1.5: Zelllinien Zelllinie Bemerkung Ba/F3 murine pro-B-Zelllinie, IL-3-abhängig Eol-1 humane AML-Zelllinie, aus dem peripheren Blut eines 33-jährigen Mannes
mit AML FAB M4eo [91] HEK 293 humane, embryonale Nierenepithel-Zelllinie [92]
HL60 humane AML-Zelllinie, aus peripherem Blut einer 35-jährigen Frau mit AML
FAB M2 [93] Kasumi-1 humane AML-Zelllinie, aus dem peripheren Blut eines 7-jährigen
japanischen Jungen mit AML FAB M2 [94] KG-1a humane AML-Zelllinie, aus dem Knochenmark eines 59-jährigen Mannes,
Variante der KG-1-AML-Zelllinie [95, 96] MM6 humane MonoMac6-Zelllinie, aus dem peripheren Blut eines 64-jährigen
Mannes mit AML FAB M5 [97]
22
MOLM-13 humane AML-Zelllinie, aus peripherem Blut eines 20-jährigen Mannes mit
AML FAB M5a [98] MV4-11 humane akute monozytäre Leukämie-Zelllinie, aus peripherem Blut eines
10- jährigen Jungen mit AML FAB M5 [99] NOMO1 humane AML-Zelllinie, aus derm Knochenmark einer 31-jährigen Frau mit
AML FAB M5a [100] OCI-AML3 humane AML-Zelllinie, aus dem peripheren Blut eines 57-jährigen Mannes
mit AML FAB M4 [101] Phoenix Eco humane, embryonale Nierenepithel-Zelllinie THP-1 humane AML-Zelllinie, aus dem peripheren Blut eines 1-jährigen Jungen
[102]
2.1.6 Antikörper
Tabelle 2.1.6: Antikörper Antikörper Bemerkung Herkunft α-β-Actin (AC-15) (A5441) Maus, monoklonal Sigma (Taufkirchen) α-AKT (#9272) Kaninchen, polyklonal Cell Signaling
(NEB, Frankfurt) CD14-FITC (IgG2a) (A07765) Maus Beckman Coulter (Krefeld) CD14 MicroBeads human Miltenyi Biotec
(Bergisch Gladbach) CD135-PE (IgG1) (IM2234U) Maus Immunotech (Marseille, FRA) α-CIN85 clone 179.1.E1
(#05-731) Maus, monoklonal Merck Millipore (Darmstadt)
α-FLT3 (S-18) (sc-480) Kaninchen, polyklonal Santa Cruz (CA, USA) α-HA tag (ab9110) Kaninchen, polyklonal Abcam (Cambridge,
Vereinigtes Königreich)
IgG1-PE Isotyp Kontrolle
(A07796) Maus Immunotech (Marseille, FRA)
IgG1-PE Isotyp Kontrolle
(#555749) Maus BD Pharmingen
(San Diego, USA) IgG2a-PE Isotyp Kontrolle
(#559319)
Maus BD Pharmingen
(San Diego, USA) IgG2b-PE Isotyp Kontrolle Maus BD Pharmingen
23
(#555743) (San Diego, USA) α-Kaninchen IgG-HRP (GAR) Ziege Sigma (Taufkirchen) α-44/42 MAP-Kinase (#9102) Kaninchen, polyklonal Santa Cruz (CA, USA) α-Maus IgG-HRP (GAM) Ziege Santa Cruz (CA, USA) α-pAKT (Ser473) (D9E)
(#4060S) Kaninchen, monoklonal Cell Signaling
(NEB, Frankfurt) α-Phospho-p44/42-MAPK
(Thr202/Tyr204) (#4370)
Kaninchen, monoklonal Cell Signaling
(NEB, Frankfurt) α-pSTAT5 (Tyr694) (#9351) Kaninchen, polyklonal Cell Signaling
(NEB, Frankfurt) α–Rabbit IgG-Cy3 (GAR) Ziege, polyklonal Dianova (Hamburg) α-STAT5 (C-17) (sc-835) Kaninchen, polyklonal Santa Cruz (CA, USA)
2.1.7 Plasmide Tabelle 2.1.7. Plasmide Bezeichnung Eigenschaften Herkunft
pMSCV-IRES-EGFP retroviraler Expressionsvektor R.K. Humphries, (Vancouver,
CAN) pMSCV-IRES-EYFP retroviraler Expressionsvektor R.K. Humphries, (Vancouver,
CAN) MIG AKT1 WT retroviraler Expressionsvektor AG Spierkermann,
KKG Leukämie MIG AKT1 E17K retroviraler Expressionsvektor AG Spierkermann,
KKG Leukämie pcDNA6/V5-His CIN85
XNC(123)cc
Expressionsvektor L. Gibson (Houston, USA)
MIG CIN85 WT retroviraler Expressionsvektor diese Arbeit MIY CIN85 WT retroviraler Expressionsvektor diese Arbeit MIY FLT3 WT retroviraler Expressionsvektor AG Spierkermann,
KKG Leukämie MIY FLT3 W51 retroviraler Expressionsvektor AG Spierkermann,
KKG Leukämie pCMV-HA SB1 WT Expressionsvektor YU-Yang Li
(Shanghai, China)

24
MIG SB1 WT retroviraler Expressionsvektor diese Arbeit MIG SB1 V89I retroviraler Expressionsvektor diese Arbeit MIG SB1 R454Q retroviraler Expressionsvektor diese Arbeit MIG SB1 R672Q retroviraler Expressionsvektor diese Arbeit pCMV6-Entry TET2 WT Expressionsvektor Jon Licht (Chicago, USA) MIY TET2 WT retroviraler Expressionsvektor diese Arbeit
2.1.8 Oligonukleotide
Tabelle 2.1.8.1:Oligonukleotide zur Sequenzierung von hTET2 Bezeichnung Sequenz Primer Sequenzieren TET2 1F 5´-TGGAGACACCAAGTGGCACTCTTT-3´
Primer Sequenzieren TET2 2F 5´-CACAAAGCTAGCGTCTGGTGAAGA-3´ Primer Sequenzieren TET2 3F 5´-AATCAAGGGCAGTCCCAAGGTACA-3´ Primer Sequenzieren TET2 4F 5´-ACAGGAGCAGAAGTCACAACAAGC-3´ Primer Sequenzieren TET2 5F 5´-AGCAGGTCCTAATGTGGCAGCTAT-3´ Primer Sequenzieren TET2 6F 5´-TGAGTTTGGGAGTGTGGAAGCTCA-3´ Primer Sequenzieren TET2 7F 5´-ATGCAGGGAGATGGTTTCAGCAGT-3´ Primer Sequenzieren TET2 8F mit Vektor 5´-AAGCCCGTGAGAAAGAGGAAGAGT-3´ Primer Sequenzieren TET2 Reverse 5´-ACACACACATGGTGAACTCCTGGA-3´ Tabelle 2.1.8.2: Oligonukleotide hTET2 WT PCR Bezeichnung Sequenz Primer Sequenzieren TET2 1F 5´-TGGAGACACCAAGTGGCACTCTTT-3´ Primer Sequenzieren TET2 Reverse 5´-ACACACACATGGTGAACTCCTGGA-3´
Tabelle 2.1.8.3: Oligonukleotide zur Sequenzierung von CIN85 (der Ratte) Bezeichnung Sequenz Forward Primer 1 (=Seq_rat SH3KBP1 1F) 5´-ACAGATCAACGGCAGGAGAGGTTT-3´
Forward Primer 2 (Seq_rat SH3KBP1 2F) 5´-AGTTACCTCCAGCTACATCAACCC-3´
Forward Primer 3 (Seq_rat SH3KBP1 3F) 5´-CAAAGATTGACTTGGCAGGCAGCA-3´
Reverse Primer (=Seq_rat SH3KBP1 R) 5´-AAAGCTGCTCATCCTGGGAAATGC-3´ Tabelle 2.1.8.4: Oligonukleotide zur Sequenzierung von hSB1 Bezeichnung Sequenz Forward Primer 1 (=Seq_human SHKBP1 1F) 5´-ACGCATCTCGACGCTGAAAGATGA-3´

25
Forward Primer 2 (=Seq_human SHKBP1 2F) 5´-AACATGACAAGATGGTGGCAGCAG-3´
Forward Primer 3 (=Seq_human SHKBP1 3F) 5´-AAGATCATGCTGTCGGAGAAGCAC-3´
Forward Primer 4 (=Seq_human SHKBP1 4F) 5´-AGCTGATGGAACAGCTGGAACACT-3´
Reverse Primer (=Seq_human SHKBP1 R) 5´-TGAGCCCATAGAACTGGGCTTCAT-3´
Tabelle 2.1.8.5: Oligonukleotide zur Mutagenese von hSB1 Bezeichnung Sequenz SB1 V89I Mutagenese-Primer
Sense 5'-GTTGGATCCCAGGGGTATCCACGGTTCCAG-3' Antisense 5'-CTGGAACCGTGGATACCCCTGGGATCCAAC-3'
SB1 R454Q Mutagenese-Primer
Sense 5'-GACAACAACCACGTGCAGACATGGTCTGTGACT-3' Antisense 5'-AGTCACAGACCATGTCTGCACGTGGTTGTTGTC-3'
SB1 R672Q Mutagenese-Primer
Sense 5'-CAGGAACTGGTGCAGAGTGGGCCAGAC-3' Antisense 5'-GTCTGGCCCACTCTGCACCAGTTCCTG-3'
2.1.9 Puffer, Medien, Lösungen
Tabelle 2.1.9: Puffer, Medien, Lösungen Material Zusammensetzung Agarosegel 1 % 1g Agarose in 100 ml TAE-Puffer
Calciumchlorid 2 M CaCl2 DAPI 1 µg DAPI in 1 ml PBS
G-NET NET mit 2,5g Gelatine GTE-Puffer 25 mM Tris/HCl pH8, 50 mM Glucose, 10 mM EDTA pH 8
HBS-Puffer 0,133 g Na2HPO3x2 H2O, 5,96 g HEPES, 8,16 g NaCl,
ad H2O dest. (50ml)
5 x KCM 5 ml 3 M KCl, 4,5 ml 1 M CaCl2, 7,5 ml 1 M MgCl2, ad H2O dest. (13ml) Lämmli-Puffer 187.5 mM Tris, 30 % Glyzerin, 6 % SDS, Spatelspitze Bromphenolblau LB-Agar 32 g LB Agar, ad 1000 ml H2O dest.
LB-Amp-Platten LB-Agar mit 100 μg/ml Ampicillin Ligase-Puffer 10fach NEB
Lysispuffer 15 ml 5 M NaCl, 2,5 ml 200 mM EGTA, 25 ml 1 M Hepes pH 7,5, 100 ml
Glycerol 50%, 5 ml Triton X-100, 2,1g NaF,
2,23 g Na4P2O7x10 H2O (500ml) MACS-Puffer PBS, BSA, EDTA, 0.09 % azide, pH 7,2
Natriumazidpuffer PBS, 0,1 % Natriumazid

26
10 x NET 438.3 g NaCl, 93.6 g EDTA, 802.9 g Tris, 25 g Triton-X 100, ad 5000 ml,
pH 7.7 Ponceau-Lösung 0.5 g Ponceau-S, 1 ml Eisessig, ad 100 ml H2O dest. Stripping-Puffer 0.1 M 2-Mercaptoethanol, 62.5 mM Tris pH 6.8,
2 % SDS (Natriumdodecylsulfat)
TAE-Puffer 57,1 ml Essigsäure 100 %, 242 g Tris, 18,61 g EDTA ad H2O dest. (5l)
10 x TBS 12,11 g Tris, 87,66 g NaCl, ad H2O dest., pH 8,0 (5l)
TBST 0,1 % Tween20 mit 1xTBS
Tris-HCl pH 6,8 908,55 g Tris, ad H2O dest., pH 6,8 (5l) Tris-HCl pH 8,8 908,55 g Tris, ad H2O dest., pH 8,8 (5l)
2.2 Methoden
2.2.1 Zellkultur
2.2.1.1 Zellkultur von adhärenten Suspensionszellen
Das Kultivieren der unter 2.1.5 angegebenen Zelllinien erfolgte in einem Inkubator bei 37 °C,
95 % relativer Luftfeuchtigkeit sowie in Gegenwart von 5 % CO2.
Die murinen Ba/F3-Zellen wurden in RPMI-1640-Medium unter Zugabe von 10 % fötalem
Kälberserum (FKS), 50 μg/ml Streptomycin, 50 U/ml Penicillin als auch 10 % WEHI-3B-
konditioniertem Medium als Produzent des murinen IL-3 gehalten. Das Splitten der Zellen
erfolgte alle 72 Stunden im Verhältnis 1:20.
Die AML-Zelllinien wurden in RPMI-Medium unter Zugabe von 20 % FKS, 50 μg/ml
Streptomycin und 50 U/ml Penicillin kultiviert und alle 3 Tage im Verhältnis 1:3 gesplittet.
Die adhärenten Zelllinien HEK-293 und Phoenix-Eco wurden in DMEM-Medium mit 10 % FKS
und 50 μg/ml Streptomycin als auch 50 U/ml Penicillin kultiviert. Alle 72 Stunden erfolgte die
Verdünnung im Verhältnis 1:5. Lösen der Zellen von ihrer Kulturflasche erfolgte durch
einmalige Waschung mit DPBS, anschließend fünfminütiger Inkubation mit Trypsin-EDTA bei
37 °C und Verdünnung mit DMEM-Vollmedium. Nach einmaliger Zentrifugation konnte das
Zellpellet erneut in frischem DMEM-Vollmedium aufgenommen und replatiert werden.

27
2.2.1.2 Einfrieren und Auftauen von Zellen
Es wurden 6 x 106 Suspensionszellen bzw. 3 x 106 adhärente Zellen in 1 ml Einfriermedium,
welches aus 90 % FKS und 10 % DMSO besteht, aufgenommen, in 1,5 ml Einfriergefäße
(Nunc-Cryotube) transferiert und in einem speziellen Freezing-Container (Typ Cryo 1 °C,
Nalgene, Deutschland) bei -80 °C eingefroren.
Durch Erwärmung der Zellsuspension in den Einfriergefäßen im 37 °C-Wasserbad,
Resuspension in Medium, Abzentrifugation und anschließend erneute Resuspension in
frischem Medium und Kultivierung konnten die Zellen aufgetaut und nach drei Tagen für
Experimente verwendet werden.
2.2.1.3 Transiente Transfektion
Die transiente Transfektion dient der Produktion von Virus enthaltendem Medium (VCM
englisch für virus containing medium), welches anschließend für die Transduktion von Ba/F3-
Zellen verwendet werden kann. Die adhärenten Phoenix-Eco- oder HEK293-Zelllinien
produzierten über Ca3(PO4)2-DNA-Transfektion den gewünschten retroviralen Überstand.
Hierzu wurde die cDNA in die multiple cloning site des pMSCV-IRESEGFP/EYFP-Vektors
kloniert. An Tag 1 wurden 6 x 106 Zellen in einer 10 cm Kulturschale für adhärente Zellen
ausgesät. An Tag 2 bei um die 70-80 %iger Zellkonfluenz erfolgte vier Stunden vor der
Transfektion erneut vorsichtiger Mediumwechsel und das Zugeben von 13 μg Plasmid-DNA zu
sterilem H2O dest. zu einem Gesamtvolumen von 450 μl. Zu den 450 μl wurden 50 μl 2 M
CaCl2-Lösung zugegeben, gemischt und einem HBS-Puffer tröpfchenweise beigegeben.
Inkubation bei RT für 3-4 Minuten und anschließend vorsichtiges Tropfen möglichst ohne
Irritation auf die zu transfizierenden Zellen. Einige Stunden danach erfolgte erneut vorsichtiger
Mediumwechsel. 2-3 Tage nach Transfektion wurde der VCM abgenommen, die
Transfektionseffizienz via Fluoreszenzmikroskop überprüft und die Zellen lysiert.
2.2.1.4 Stabile Ba/F3-Zell-Transduktion
Für die stabile Transduktion der Ba/F3-Zelllinien mit dem gewünschten retroviralen Überstand
wurden 1,5 x 106 Ba/F3-Zellen in 3 ml RPMI-Vollmedium aufgenommen, die Zellsuspension
mit einer Polybren-Lösung (Konzentration 8 μg/ml) versetzt und gemischt und mit derselben
Menge an VCM versehen. Anschließend wurde die Gesamtsuspension gleichmäßig auf zwei
Kammern einer 6-well-Schale verteilt und für 90 Minuten bei 2500 rpm und 32 °C in einer

28
Tischzentrifuge von Hettich zentrifugiert. Daraufhin erfolgte eine 2,5-stündige Inkubation der
Zellen bei 37 °C im Brutschrank und Mediumaustausch. Die Zellen wurden expandiert und 4-5
Tage nach Transduktion via YFP- oder GFP-Fluoreszenz im FACS („Fluorescence
Activated Cell Sorter“) gesortet. Anschließend erfolgte erneute Expansion der YFP/GFP-
positiven Zellen sowie eine zweite Sortierung anhand der Fluoreszenz nach etwa einer
Woche, wodurch eine zu über 95 % reine Zellpopulation erhalten wurde.
2.2.1.5 Lebendzellzahl-Bestimmung
Der Trypan-Blau-Exklusions-Test dient der Lebendzellzahl-Bestimmung. Aufgrund von
poröser Zellmembran kann bei toten Zellen der Farbstoff Trypan-Blau in die Zellen eindringen,
bei lebenden Zellen ist ein Passieren der Zellmembran nicht möglich. Dadurch erscheinen tote
Zellen im Lichtmikroskop blau, lebende Zellen sind farblos. Durch den Vi-Cell-Zellzähler von
Beckman Coulter konnte so in einem definierten Verhältnis von Zellsuspension und
Trypanblau die Lebendzellzahl bestimmt werden.
Für Ba/F3-Proliferationsexperimente wurden 4 x 104 Zellen pro ml Medium ausgesät und eine
festgelegte Menge an Inhibitoren und Zytokinen zugegeben. Nach 72h erfolgte die
Lebendzellzahlbestimmung.
2.2.1.6 Apoptosemessung mit Annexin-V-/7-AAD-Färbung
Zur Bestimmung früh- und spät-apoptotischer Zellen diente das Annexin V-PE Apoptosis
Detection Kit I von Beckton Dickinson. Die Zellen werden mit Annexin V-PE (Annexin V
konjugiert mit Phycoerythrin) und 7-AAD (7-Amino-actinomycin D) entsprechend dem
Herstellerprotokoll doppelgefärbt und via Durchflusszytometrie analysiert. Annexin V detektiert
Zellen in frühapoptotischem Stadium, 7-AAD Zellen in spätapoptotischem.
Für Apoptoseexperimente erfolgte das Kultivieren von 2 x 105 Ba/F3-Zellen pro ml ohne IL-3
und von 4 x 104 Ba/F3-Zellen pro ml mit IL-3 für 72 Stunden, nachdem die Zellen zweimalig
gewaschen und in RPMI mit 10 % FKS, 50 U/ml Penicillin und 50 μg/ml Streptomycin
aufgenommen wurden.

29
2.2.1.7 Expressionsanalyse von Oberflächenantigenen mittels Durchflusszytometrie
Zur Analyse der Expression von Oberflächenantigenen diente ebenfalls die
Durchflusszytometrie. Es wurden 1 x 106 Zellen mit PBS gewaschen, für 15 min bei RT mit
spezifischem Antikörper als auch dessen Isotyp-Kontrolle (z.B. CD135-PE und murines IgG1-
PE für FLT3) inkubiert, anschließend erneut zweimal gewaschen und in 250 μl FACS-Puffer
überführt. Die jeweilige Fluoreszenz der Zellen wurde via FACS-Calibur-Durchflusszytometer
detektiert. Daraufhin erfolgte die Auswertung der Expressionsstärke via WinMDI 2.9.
Zur Analyse der Internalisierung des FLT3-Rezeptors wurden pro Ansatz 3 x 105 Zellen in 100
μl PBS mit 5 % FKS aufgenommen, mit 100 ng FL für die verschiedenen Zeitintervalle von 5,
10 bis zu 30 Minuten inkubiert. Als 0 Minuten-Wert galt ein Ansatz, welcher gar nicht stimuliert
wurde. Nach dem jeweiligen Zeitintervall wurde die Internalisierung mittels Zugabe von 1 ml
PBS mit 0,1 % Natriumazid gestoppt. Nach dreimaligem Waschen mit Natriumazidpuffer
erfolgte die Aufnahme in 100 μl PBS und die Färbung mittels Oberflächenantikörper CD135-
PE als auch Analyse via FACS-Calibur und WinMDI 2.9 (siehe oben).
2.2.1.8 Hungern der Zellen und Stimulation vor der Zelllyse
Um spezifische phosphorylierte Signalproteine in Signalkaskaden der transduzierten Ba/F3-
Zelllinien analysieren zu können, wurden die Zellen vor der Herstellung von Zelllysaten
gehungert. Dafür wurden die Zellen zweimal mit PBS gewaschen und anschließend in
Hungermedium, d.h. RPMI mit 0,3 % FKS, 50 U/ml Penicillin und 50 μg/ml Streptomycin, für
24 h im Inkubator kultiviert.
Zudem erfolgte die Stimulation der Zellen vor der Zelllyse mit dem jeweiligen Liganden oder
Zytokin, beispielsweise um die Induktion der Signalkaskaden durch Zytokine zu untersuchen.
2.2.1.9 Immunfluoreszenz von adhärenten Zellen (Immunostaining)
Um die intrazelluläre Lokalisation eines Proteins genauer zu untersuchen, wurden HEK 293-
Zellen auf Deckgläschen kultiviert und mit SB1-WT oder Mutanten transfiziert. Bei rund 80
%iger Konfluenz der Zellen nach in etwa 24-48 h wurden diese mit PBS gewaschen und mit 2
% - 4 % Paraformaldehyd (PFA) in PBS für 10 min fixiert. Anschließend wurden die Zellen
erneut mit PBS gewaschen und mittels 0,1 % Triton X in PBS (3 x 5 min) permeabilisiert und
daraufhin 3 x 5 min mittels 10 % FKS in PBS blockiert. Es erfolgte die Inkubation im Dunkeln

30
mit dem ersten Antikörper α-HA (Abcam) bei 4°C über Nacht. Nach erneutem Waschen mit
PBS wurde der Sekundär-Antikörper α-Cy3 für eine Stunde im Dunkeln bei RT hinzugefügt.
Nach erneuten Waschschritten wurde mit 1 μg DAPI in 1 ml PBS für 2 min bei RT zur DNA-
Kernfärbung inkubiert. Daraufhin wurden die Deckgläschen auf Objektträgern fixiert und
mittels Konfokalem Mikroskop (TCS-SP2, Leica, Heidelberg) analysiert. Die Bilderverarbeitung
erfolgte mit Hilfe von Adobe Illustrator.
2.2.2 Proteinbiochemische Methoden
2.2.2.1 Proteinlysate
Um die Expression von zellulären Proteinen zu analysieren, erfolgte die Herstellung von
Proteinlysaten. Für die Proteinextraktion wurden 3 x 107 Ba/F3-Zellen in kaltem PBS
gewaschen und nach Zentrifugation in 500 μl Lysispuffer lysiert. Nach Resuspension wurden
die Lysate bei 4°C 30 min auf einem Drehrad (9 rpm) inkubiert, die Zelldebris anschließend
über Zentrifugation mit 13000 rpm und bei 4°C pelletiert. Der Proteinüberstand wurde bei
-20°C gelagert.
2.2.2.2 Proteinkonzentrationsbestimmung nach Bradford
Die genaue Bestimmung der Proteinkonzentration in den Proteinlysaten wurde mittels
Methode nach Bradford photometrisch ermittelt [103]. Hierzu wurden 980 μl von 1:5
verdünnter Farbstofflösung (Bradford-Reagenz, BioRad, München) mit 20 μl von 1:10
verdünntem Proteinlysat gemischt und 10 Minuten bei RT inkubiert. Im Spektralphotometer
erfolgte die Messung der Absorption bei 595 nm gegen die blank-Kontrolle (Bradford-Reagenz
mit Zusatz von Wasser). Mittels Verdünnungsreihe mit unterschiedlichen Konzentrationen an
BSA und deren Messung wurde eine Eichkurve erstellt, um die absolute Konzentration
bestimmen zu können.

31
2.2.2.3 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE)
Die Auftrennung der Proteine entsprechend ihres Molekulargewichts erfolgte mittels SDS-
Polyacrylamid-Gelelektrophorese (SDS-PAGE). Hierzu wurden Trenngele zwischen 7,5 % und
18 % verwendet, je nach Molekulargewicht der Proteine. Die Trenngele wurden gegossen und
mit Wasser überschichtet. Nach dem Auspolymerisieren wurde das Trenngel mit 4 %igem
Sammelgel überschichtet, welches durch Einbringen eines Kamms zum Beladen der Taschen
mit den Lysaten befähigt. Die Proteinlysate mit gleicher Menge an Protein wurden 1:1 mit 2x
Lämmli-Puffer versetzt und zur Denaturierung 5 Minuten bei 95°C erhitzt. Anschließend
wurden die präparierten Lysate als auch ein Protein-Größenstandard (Rainbow-Marker) in die
Geltaschen pipettiert und bei 80mA in einer mit Elektrophoresepuffer befüllten Gelkammer
aufgetrennt.
2.2.2.4 Western-Blot
Der Transfer der Proteine auf eine Nitrocellulose-Membran erfolgte mittels „Semidry-Blot-
Verfahren“ [104]. Dies geschah in Transblot-Puffer über 90 Minuten bei 250 mA. Anschließend
wurde die Membran mit Ponceau reversibel angefärbt, um den erfolgreichen Transfer und den
äquivalenten Transfer der Proteine grob zu überprüfen. Zur Entfärbung und Blockade
unspezifischer Proteinbindestellen wurde die Membran dreimal 20 Minuten mit G-NET
blockiert. Über Nacht wurde die Nitrozellulose-Membran mit entsprechend den
Herstellerangaben verdünnter Primärantikörper-Lösung bei 4°C inkubiert, am nächsten Tag
dreimal für 10 Minuten mit TBST gewaschen und daraufhin für 60 Minuten mit dem
entsprechenden Sekundärantikörper bei RT inkubiert. Als letzte Schritte wurde erneut dreimal
für 10 Minuten mit TBST und kurz mit PBS ausgewaschen und die Proteine mittels
Chemolumineszenz-Reagenz (ECL Detektionsreagenz) auf einem Hyperfilm (Amersham,
Freiburg) mit unterschiedlicher Expositionszeit detektiert.
2.2.2.5 Strippen der Nitrozellulosemembran
Um die Nitrocellulose-Membran von zuvor verwendeten Antikörpern zu reinigen und zur
Detektion eines anderen Proteins verwenden zu können, wird diese durch Strippen der
Membran entfernt. Dazu erfolgte die Inkubation der Membran in Stripping-Puffer für 40 min bei
56°C auf einem Drehrad. Anschließend wurde die Membran dreimal 20 min mit TBST
ausgewaschen als auch erneut dreimal 20 min mit G-NET blockiert.

32
2.2.3 Bakterienkultur
2.2.3.1 Transformation kompetenter E.coli Bakterien
Für die Bakterien-Transformation wurde ein Aliquot von kompetenten Zellen auf Eis aufgetaut.
Ein 50 μl-Ansatz aus 5 μl 5xKCM (500 mM KCl, 150 mM CaCl2, 250 mM MgCl2), 1 μg Plasmid-
DNA oder 10 μl Ligationsansatz wurde mit Aqua dest. auf 25 μl aufgefüllt und 25 μl
kompetente Bakterien zugegeben. Das 5xKCM sorgt für kurze Durchlässigkeit der Bakterien-
Zellmembran. Anschließend erfolgte die Inkubation für 20 min auf Eis sowie 10 min bei RT,
daraufhin die Zugabe von 250 μl LB- oder S.O.C.S.-Medium und das Schütteln über eine
Stunde bei 37°C. Nach Ausplattieren von 100 μl dieses Ansatzes auf einer meist Ampicillin-
versetzten LB-Agar-Platte erfolgte erneut die Inkubation über Nacht bei 37°C. Am nächsten
Tag konnte durch Isolation der gewachsenen Einzelkolonien und Präparation von Plasmid-
DNA eine weitere Expansion stattfinden.
2.2.3.2 Plasmid-DNA-Präparation
Für die Isolation der Plasmid-DNA aus Bakterien wurde das Prinzip der alkalischen Lyse
angewendet [105].
2.2.3.2.1 Mini-Präparation
Für die Isolation von Plasmid-DNA in kleinerer Menge (Mini-Präparation) wurden gewachsene
Einzelkolonien der transformierten Bakterien mit einer sterilen Pipettenspitze gepickt, über
Nacht bei 37°C mit 2 ml LB-Medium versetzt und unter Zugabe des jeweiligen Antibiotikums
bei 225 rpm im Bakterienschüttler inkubiert. Die DNA-Präparation erfolgte mittels QIAprep-
Spin-Miniprep-Kit (Qiagen, Hilden). Dies erfolgte meist um nachzuweisen, dass mittels
Ligation und Transformation eine Integration der Plasmid-DNA stattgefunden hat.
2.2.3.2.2 Maxi-Präparation
Zur Gewinnung größerer Plasmid-DNA-Mengen wurde eine Einzelkolonie gepickt, in 2 ml
Vorkultur für acht Stunden 37°C im Bakterienschüttler inkubiert und anschließend in 100ml LB-

33
Medium mit jeweiligem Antibiotikum überführt und erneut über Nacht inkubiert. Am nächsten
Tag konnte mittels Endofree-Plasmid-Maxi-Kit oder QIAgen-Plasmid-Maxi-Kit (Qiagen, Hilgen)
die Extraktion der Plasmid-DNA nach Herstellerangaben durchgeführt werden. Die Messung
der DNA-Konzentration erfolgte mittels Spektralphotometer Nanodrop 1000 (Peqlab,
Erlangen).
2.2.4 Molekularbiologische Methoden
2.2.4.1 DNA-Verdau durch Restriktionsenzyme
Durch Restriktionsenzyme der Firma New England Biolabs (Frankfurt) konnte die Plasmid-
DNA, welche durch Mini- oder Maxipräparation erhalten wurde, auf ihre Identität überprüft
werden. Hierzu wurden die Bedingungen entsprechend den Angaben des Herstellers gewählt.
Ein analytischer Verdau erfolgte über zwei Stunden in einem Reaktionsvolumen von 20 μl. Ein
präparativer Verdau erfolgte über drei Stunden in einem Reaktionsvolumen von 50 μl. Bei der
blunt-end Klonierung erfolgte zudem nach Inaktivierung der Enzyme bei 65°C für 20 min die
Zugabe von alkalischer Phosphatase (CIAP), CIAP-Puffer und EDTA und Inkubation für 30
min bei 37, wodurch eine Religation des Vektors durch Dephosphorylierung der Phosphatreste
am 5´-Ende des Vektors verhindert wurde. Der jeweilige Restriktionsansatz konnte
anschließend auf ein 1 %iges Agarosegel aufgetragen und über Ethidiumbromid oder Syber-
Safe detektiert und die DNA ggf. mit Hilfe des „Gel Extraction Kits“ von Qiagen
protokollgerecht aufgereinigt und isoliert werden.
2.2.4.2 Ligation von DNA-Fragmenten
Um Insert und Vektor zu ligieren, wurden meist pro Versuch drei Ansätze gewählt: ein Vektor-
zu Insert-DNA-Verhältnis von 3:1, 5:1 und 10:1 als auch eine Reaktion ohne Insert als
Religationskontrolle. Beide DNA-Fragmente wurden mit Ligasepuffer, T4-Ligase (NEB,
Frankfurt am Main) und H2O in einem Reaktionsvolumen von 20 μl über Nacht bei einer
Tempertatur von 16°C ligiert. Vor Transformation in Bakterien erfolgte die Inaktivierung der
Enzyme für 20 min bei 65°C.

34
2.2.4.3 Isolierung von mRNA
Die Isolierung von mRNA wurde mit Hilfe des RNeasy MiniKits (Qiagen, Hilden) nach
Herstellerangaben durchgeführt. Das Produkt wurde in 20-50 μl RNase-freiem Wasser
aufgenommen.
2.2.4.4 Die Synthese von cDNA durch reverse Transkription
Mittels RevertAid-Premium-First-Strand-cDNA-Synthesis-Kit (Fermentas, St. Leon-Rot)
erfolgte nach Herstellerangaben durch reverse Transkription der isolierten RNA die Synthese
der komplementären DNA (cDNA). Die entstandene cDNA wurde bei -80°C gelagert.
2.2.4.5 Extraktion genomischer DNA
Um genomische DNA aus Zellen zu extrahieren, wurde mittels DNAzol (Invitrogen, Darmstadt)
und nach Herstellerangaben aus rund 2 x 107 Zellen pro Ansatz die DNA gewonnen.
2.2.4.6 Polymerasekettenreaktion (PCR)
In der Polymerasekettenreaktion (PCR) können über Denaturierung, Annealing und Elongation
mittels Polymerase und zweier komplementärer Primer DNA-Menge hochspezifisch
amplifiziert werden. Dies eignet sich für eine Vielzahl an Untersuchungen, beispielsweise
Expressionsnachweise oder Amplifikation von Mutantenfragmenten für die anschließende
Klonierung. Ein Ansatz aus 10 μM 5´Primer, 10 μM 3´Primer, Taq-Polymerase, 25 mM MgCl2,
10 mM sNTPs, 10x Low Salt Buffer und Matrize wurde auf 25 μl H2O aufgefüllt und nach 5-
minütiger Denaturierung bei 95°C wie folgt in einem T3 Thermocycler (Biometra) durch 30
Zyklen des folgenden Programms amplifiziert:
30 Sekunden bei 95°C, 30 Sekunden bei der Primer-spezifischen Annealing-Temperatur, 30
Sekunden bis sechs Minuten (30 Sekunden pro 500 bp eines Fragments) bei 72°C. Nach
erneuten 5-10 Minuten bei 72°C erfolgte das Abkühlen des Ansatzes als auch Auftragen auf
ein Agarosegel.

35
2.2.4.7 Ortsgerichtete Mutagenese
Das Einfügen der einzelnen Mutanten erfolgte in einem PCR-basiertem Verfahren nach
Herstellerangaben mittels QuikChange-II-XL-Site-Directed-Mutagenesis-Kit von Stratagene
(Waldbronn). Anschließend wurden 20 μl des PCR-Produkts in Bakterien transformiert und
mittels Mini-Präparation isoliert. Die Plasmid-DNA wurde final vollständig durchsequenziert,
um die gewünschte Mutation zu detektieren als auch andere ungewollte Mutationen
auszuschließen.
2.2.4.8 Sequenzierung
Alle in dieser Arbeit durchgeführten Sequenzierungen wurden durch die Firma Sequiserve
(Vaterstetten) durchgeführt.
2.2.5 Präparation der Patientenproben
2.2.5.1 Ficoll
Die CMML-Patientenproben (erhalten aus der Probenbank des Leukämie-Diagnostiklabors
des Universitäts-Klinikums München als auch von dem niedergelassenen Hämato-Onkologen
Dr. Siegfried Völkl in München und Dr. Ole Maywald, Onkologische Praxis Ingolstadt) wurden
in 10-20 ml peripherem EDTA-Blut abgenommen und sofort weiterverarbeitet. Die Trennung
und Isolierung humaner mononukleärer Zellen aus dem peripheren Blut erfolgte mit Hilfe des
Ficoll-Gradienten. Hierfür wurden in ein Falcon 15 ml Biocoll Separating Solution gegeben.
Das erhaltene Blutprobenröhrchen wurde mehrfach gedreht und anschließend in ein zweites
Falcon gegeben und dieses nun auf 30 ml bei bis zu 15 ml Material, auf 50 ml bei über 15 ml
Material mit PBS Dulbecco aufgefüllt und erneut mehrmals vorsichtig gedreht. Anschließend
wurde der gesamte Inhalt des zweiten Falcons auf die 15 ml Biocoll Separating Solution des
ersten Falcons vorsichtig und langsam mittels Accu-Jet aufgeschichtet und bei 2270 rpm für
30 min bei 20°C ohne Bremse zentrifugiert. Nach der Zentrifugation wurde der mononukleäre
Zellring abgenommen, in ein neues Falcon gegeben und auf 50 ml mit PBS Dulbecco
aufgefüllt. Danach für 10 min bei 1440 rpm und 4°C zentrifugiert, der Überstand verworfen und
erneut in 1 ml PBS Dulbecco resuspendiert. Die Zellzahl der Probe wurde mittels Sysmex
bestimmt und unmittelbar weiterverarbeitet.

36
2.2.5.2 MACS Zellseparation
Um nach der Ficoll-Methode aus den mononukleären Zellen eine möglichst reine
Monozytenpopulation zu erhalten, erfolgte anschließend die Magnetic Cell Separation, auch
Magnetic-Activated Cell Sorting oder MACS genannt, über bestimmte Oberflächenstrukturen
der Zellen. Hierfür wurde pro Ansatz 2,5 x 107 Zellen in 450 μl MACS-Puffer aufgenommen
und mit 50 μl CD14 MicroBeads für 15 min auf Eis inkubiert. Nach Inkubation erfolgte die
Zugabe von 7,5 ml MACS-Puffer und eine fünfminütige Zentrifugation bei RT und 1200 rpm.
Der Überstand wurde abgenommen und verworfen, das Zellpellet in 1 ml MACS-Puffer neu
gelöst. Die MACS Separation Columns wurden in der magnetischen Halterung des
QuadroMACS Separators befestigt und jede Säule einmal mit 3 ml MACS-Puffer gewaschen.
Anschließend wurde die Probe nach der Antikörperinkubation auf die Säule pipettiert,
nachdem diese durchgelaufen war erfolgte das dreimalige Waschen der Säule mit je 3 ml
MACS-Puffer. Dann wurde die Säule aus der magnetischen Halterung genommen, 5 ml
MACS-Puffer auf die Säule pipettiert und mittels mitgeliefertem Stempel in ein Falcon
gedrückt. Die Suspension wurde bei 1200 U/min zentrifugiert, 2,5 ml des Überstands
verworfen und mit den restlichen 2,5 ml resuspendiert. Mittels Sysmex erfolgte dann die
Bestimmung der Zellzahl der über das Zellseparation isolierten Monozyten-Population. Über
Durchflusszytometrie wurde der Anteil CD14-positiver Zellen mittels des Antikörpers CD14-
FITC vor dem Ficoll, nach dem Ficoll und nach erfolgter Zellseparation mittels MACS
bestimmt. Aus der fast vollständig reinen CD14-positiven Zellpopulation wurden RNA, cDNA
und Proteinlysate hergestellt.

37
3 Ergebnisse
3.1 Funktionelle Charakterisierung von CIN85 und SB1 in rezeptor-exprimierenden Zellen
CIN85 ist ein Multi-Adapter-Protein und in die Internalisierung und in andere regulatorische
Prozesse von Rezeptor-Tyrosinkinasen involviert. Auch SB1 ist eines von vielen mit CIN85-
interagierenden Proteinen, dessen Überexpression in Zelllinien bereits anti-apoptotische
Effekte zeigte, sodass eine Rolle als mögliches Protoonkogen denkbar ist.
Zur Untersuchung des biologischen Phänotyps von SB1-WT und den SB1-Mutanten SB1-
V89I, SB1-R454Q sowie SB1-R672Q wurden Ba/F3-Zelllinien hergestellt, die FLT3 als
Rezeptor sowie die verschiedenen SB1-Konstrukte exprimieren. Eine schematische
Darstellung des SB1- und CBL-Proteins mit Mutationen zeigt Abbildung 3.1. Zusätzlich wurde
die endogene Expression von CIN85 untersucht. Bei der Ba/F3-Zelllinie handelt es sich um
murine pro-B-Zellen, die aufgrund ihrer Interleukin-3 (IL-3)-Abhängigkeit geeignet ist, um
Onkogene im zellulären Kontext zu charakterisieren. Nur die Transduktion transformierender
Gene ermöglicht ihnen IL-3-faktorunabhängiges Zellwachstum [106, 107].
Abb. 3.1: Schematische Darstellung des SB1- und CBL-Proteins mit Mutationen. N-
terminal die BR-C, ttk und bab-Domäne (BTB) des SB1-Proteins, gefolgt von drei WD40-

38
Domänen. Zudem Darstellung der im Folgenden untersuchten SB1-Mutationen SB1-V89I,
SB1-R454Q und SB1-R672Q. Das CBL-Protein besteht N-terminal aus der
Tyrosinkinasebindungs-Domäne (TKB), gefolgt von der Linker-Region (L), RING-Finger-
Region sowie einer Prolin-reichen Region (PXXP). C-terminal die Ubiquitin-assoziierte (U) und
Leuzin-zipper Domäne (LZ). Zudem Darstellung der häufigen humanen CBL-Mutationen CBL-
L380P, CBL-Y371H, CBL-C384R, CBL-C404Y, CBL-R420Q sowie der Deletionen CBL-Δ366-
408.
3.1.1 Endogene Expression von CIN85 in AML-Zelllinien
Zunächst wurden Ganzzell-Lysate von den verschiedenen AML-Zelllinien MM6, MOLM-13,
OCI-AML3, KG1alpha, Kasumi-1, Eol-1, MV4-11, HL60, THP-1 und NOMO1 auf ihre
endogene Expression von CIN85 untersucht. Dies geschah über Immunoblotting mittels
CIN85-spezifischem Antikörper (siehe Abbildung 3.1.1). Die endogene Expression von CIN85
zeigte sich in jeder der aufgeführten Zelllinien, die geringste Expression mit einer nur sehr
schwachen Bande wies die Zelllinie THP-1 auf. Zudem zeigte sich eine endogene Expression
auch in nativen Ba/F3-Zellen, wohingegen der Rezeptor FLT3 nicht in nativen Ba/F3-Zellen
exprimiert wird (s. Abb. 3.1.2.1 und 3.1.2.2). Der Nachweis über die endogene Expression von
SB1 gelang mangels fehlender Funktionalität der kommerziellen SB1-Antikörper nicht und
konnte daher nicht dargestellt werden.
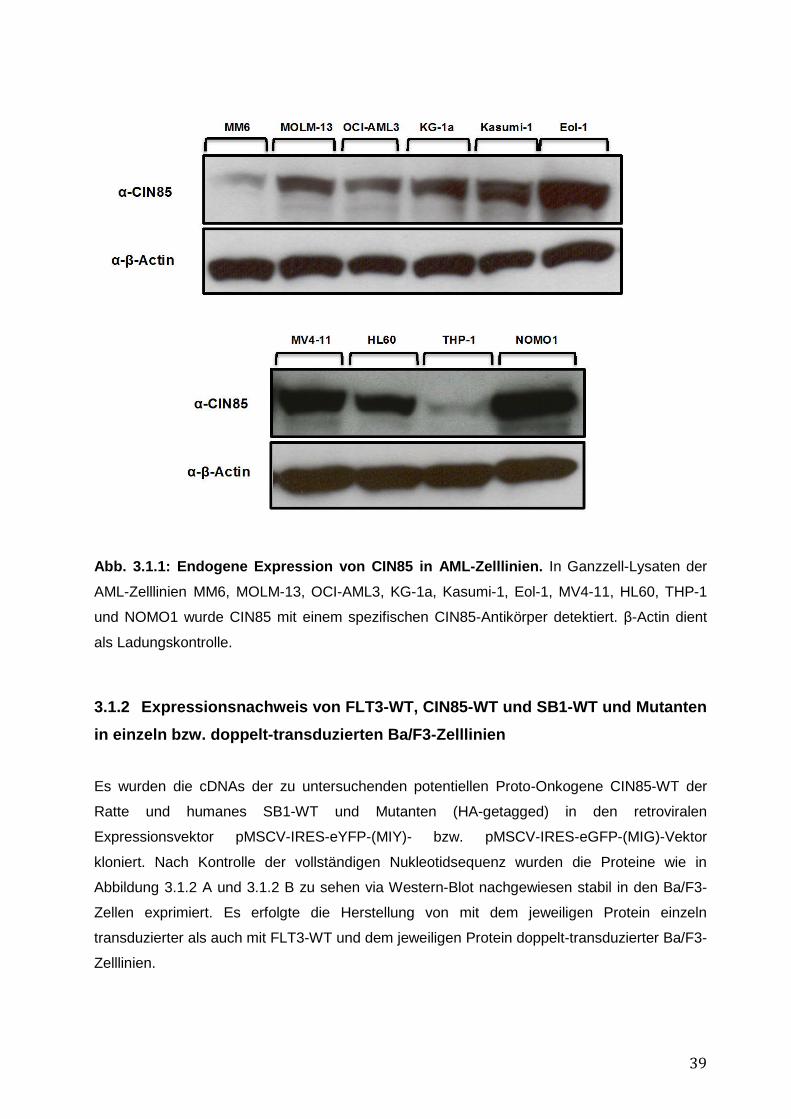
39
Abb. 3.1.1: Endogene Expression von CIN85 in AML-Zelllinien. In Ganzzell-Lysaten der
AML-Zelllinien MM6, MOLM-13, OCI-AML3, KG-1a, Kasumi-1, Eol-1, MV4-11, HL60, THP-1
und NOMO1 wurde CIN85 mit einem spezifischen CIN85-Antikörper detektiert. β-Actin dient
als Ladungskontrolle.
3.1.2 Expressionsnachweis von FLT3-WT, CIN85-WT und SB1-WT und Mutanten in einzeln bzw. doppelt-transduzierten Ba/F3-Zelllinien
Es wurden die cDNAs der zu untersuchenden potentiellen Proto-Onkogene CIN85-WT der
Ratte und humanes SB1-WT und Mutanten (HA-getagged) in den retroviralen
Expressionsvektor pMSCV-IRES-eYFP-(MIY)- bzw. pMSCV-IRES-eGFP-(MIG)-Vektor
kloniert. Nach Kontrolle der vollständigen Nukleotidsequenz wurden die Proteine wie in
Abbildung 3.1.2 A und 3.1.2 B zu sehen via Western-Blot nachgewiesen stabil in den Ba/F3-
Zellen exprimiert. Es erfolgte die Herstellung von mit dem jeweiligen Protein einzeln
transduzierter als auch mit FLT3-WT und dem jeweiligen Protein doppelt-transduzierter Ba/F3-
Zelllinien.
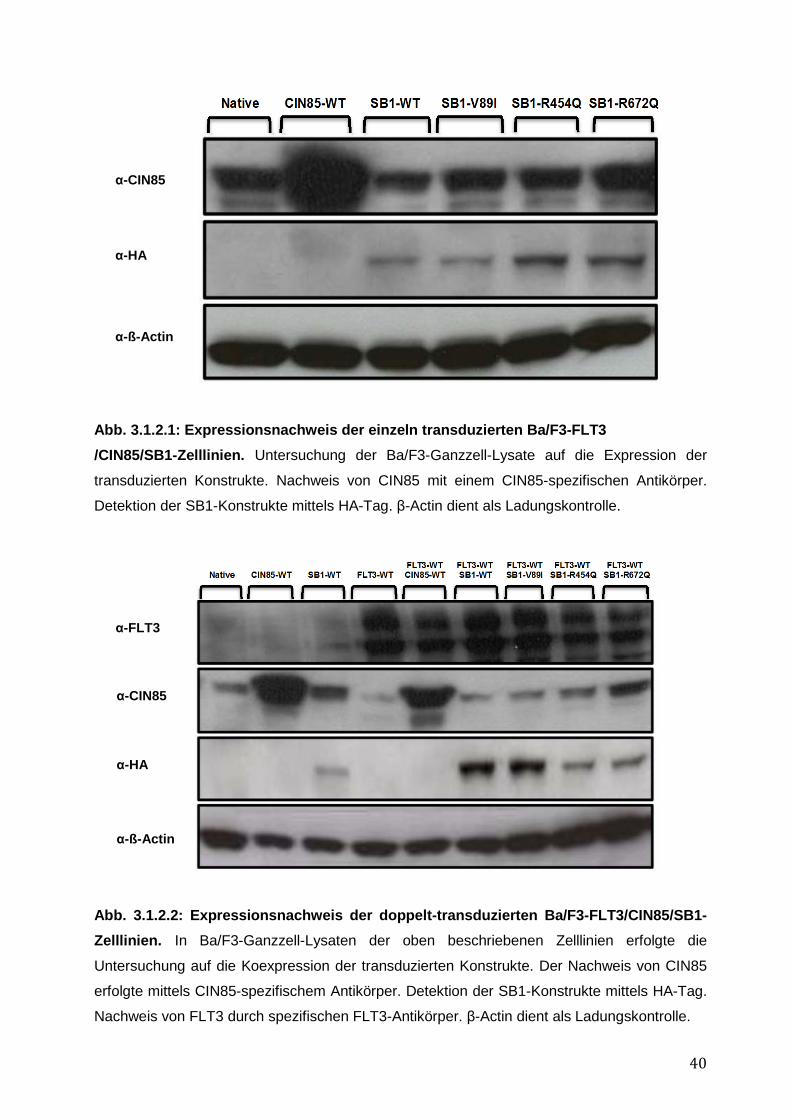
40
Abb. 3.1.2.1: Expressionsnachweis der einzeln transduzierten Ba/F3-FLT3 /CIN85/SB1-Zelllinien. Untersuchung der Ba/F3-Ganzzell-Lysate auf die Expression der
transduzierten Konstrukte. Nachweis von CIN85 mit einem CIN85-spezifischen Antikörper.
Detektion der SB1-Konstrukte mittels HA-Tag. β-Actin dient als Ladungskontrolle.
Abb. 3.1.2.2: Expressionsnachweis der doppelt-transduzierten Ba/F3-FLT3/CIN85/SB1-Zelllinien. In Ba/F3-Ganzzell-Lysaten der oben beschriebenen Zelllinien erfolgte die
Untersuchung auf die Koexpression der transduzierten Konstrukte. Der Nachweis von CIN85
erfolgte mittels CIN85-spezifischem Antikörper. Detektion der SB1-Konstrukte mittels HA-Tag.
Nachweis von FLT3 durch spezifischen FLT3-Antikörper. β-Actin dient als Ladungskontrolle.
α-CIN85
α-HA
α-ß-Actin
α-CIN85
α-HA
α-ß-Actin
α-FLT3
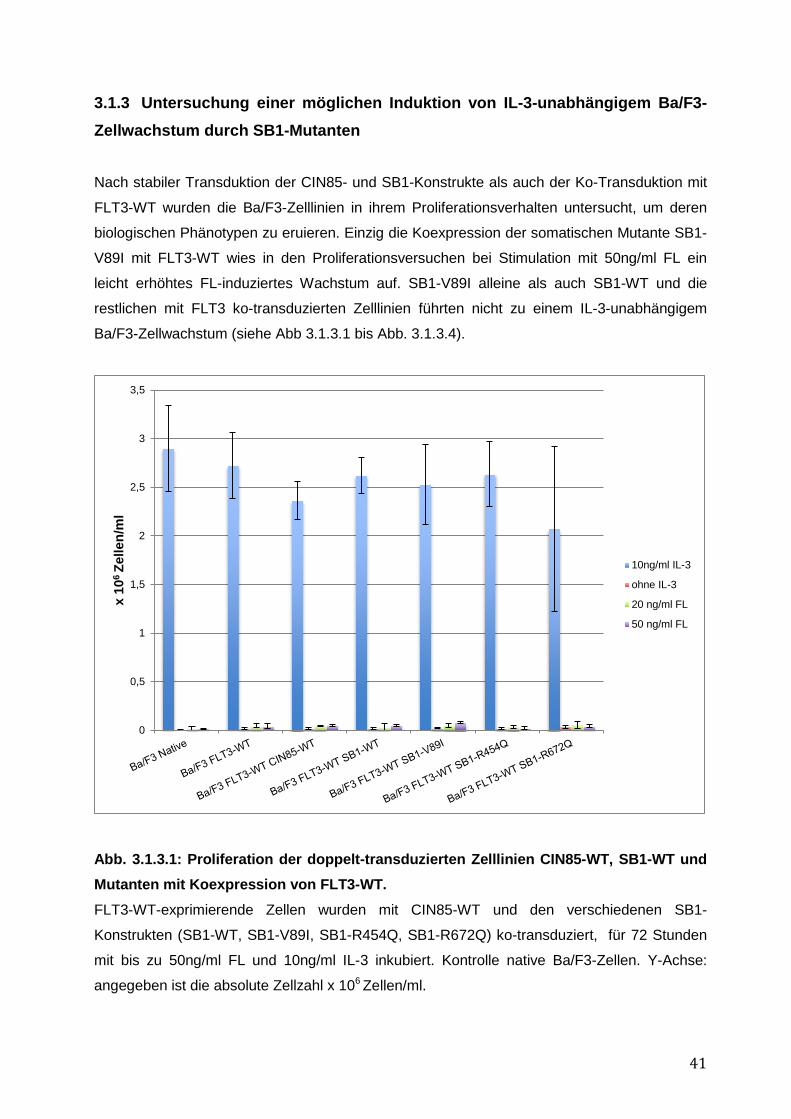
41
3.1.3 Untersuchung einer möglichen Induktion von IL-3-unabhängigem Ba/F3-Zellwachstum durch SB1-Mutanten
Nach stabiler Transduktion der CIN85- und SB1-Konstrukte als auch der Ko-Transduktion mit
FLT3-WT wurden die Ba/F3-Zelllinien in ihrem Proliferationsverhalten untersucht, um deren
biologischen Phänotypen zu eruieren. Einzig die Koexpression der somatischen Mutante SB1-
V89I mit FLT3-WT wies in den Proliferationsversuchen bei Stimulation mit 50ng/ml FL ein
leicht erhöhtes FL-induziertes Wachstum auf. SB1-V89I alleine als auch SB1-WT und die
restlichen mit FLT3 ko-transduzierten Zelllinien führten nicht zu einem IL-3-unabhängigem
Ba/F3-Zellwachstum (siehe Abb 3.1.3.1 bis Abb. 3.1.3.4).
Abb. 3.1.3.1: Proliferation der doppelt-transduzierten Zelllinien CIN85-WT, SB1-WT und Mutanten mit Koexpression von FLT3-WT. FLT3-WT-exprimierende Zellen wurden mit CIN85-WT und den verschiedenen SB1-
Konstrukten (SB1-WT, SB1-V89I, SB1-R454Q, SB1-R672Q) ko-transduziert, für 72 Stunden
mit bis zu 50ng/ml FL und 10ng/ml IL-3 inkubiert. Kontrolle native Ba/F3-Zellen. Y-Achse:
angegeben ist die absolute Zellzahl x 106 Zellen/ml.
0
0,5
1
1,5
2
2,5
3
3,5
x 10
6 Ze
llen/
ml
10ng/ml IL-3
ohne IL-3
20 ng/ml FL
50 ng/ml FL
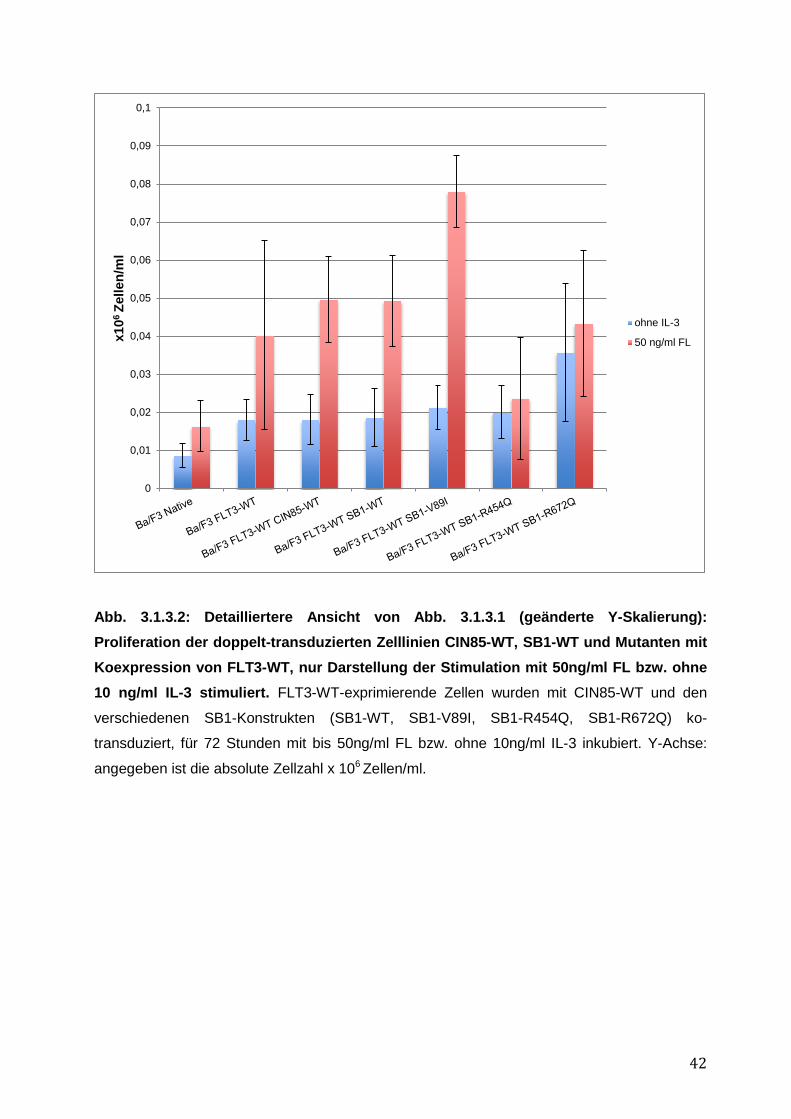
42
Abb. 3.1.3.2: Detailliertere Ansicht von Abb. 3.1.3.1 (geänderte Y-Skalierung): Proliferation der doppelt-transduzierten Zelllinien CIN85-WT, SB1-WT und Mutanten mit Koexpression von FLT3-WT, nur Darstellung der Stimulation mit 50ng/ml FL bzw. ohne 10 ng/ml IL-3 stimuliert. FLT3-WT-exprimierende Zellen wurden mit CIN85-WT und den
verschiedenen SB1-Konstrukten (SB1-WT, SB1-V89I, SB1-R454Q, SB1-R672Q) ko-
transduziert, für 72 Stunden mit bis 50ng/ml FL bzw. ohne 10ng/ml IL-3 inkubiert. Y-Achse:
angegeben ist die absolute Zellzahl x 106 Zellen/ml.
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,1x1
06 Z
elle
n/m
l
ohne IL-3
50 ng/ml FL
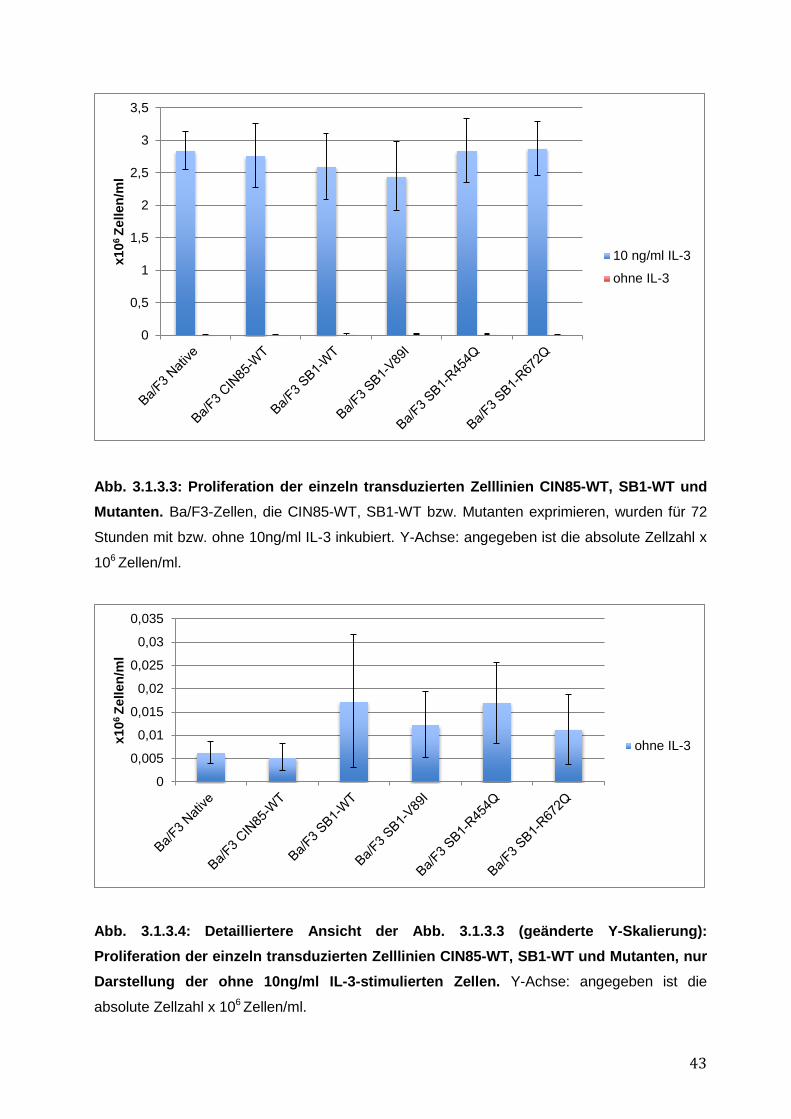
43
Abb. 3.1.3.3: Proliferation der einzeln transduzierten Zelllinien CIN85-WT, SB1-WT und Mutanten. Ba/F3-Zellen, die CIN85-WT, SB1-WT bzw. Mutanten exprimieren, wurden für 72
Stunden mit bzw. ohne 10ng/ml IL-3 inkubiert. Y-Achse: angegeben ist die absolute Zellzahl x
106 Zellen/ml.
Abb. 3.1.3.4: Detailliertere Ansicht der Abb. 3.1.3.3 (geänderte Y-Skalierung): Proliferation der einzeln transduzierten Zelllinien CIN85-WT, SB1-WT und Mutanten, nur Darstellung der ohne 10ng/ml IL-3-stimulierten Zellen. Y-Achse: angegeben ist die
absolute Zellzahl x 106 Zellen/ml.
0
0,5
1
1,5
2
2,5
3
3,5x1
06 Z
elle
n/m
l
10 ng/ml IL-3
ohne IL-3
0
0,005
0,01
0,015
0,02
0,025
0,03
0,035
x106
Zel
len/
ml
ohne IL-3
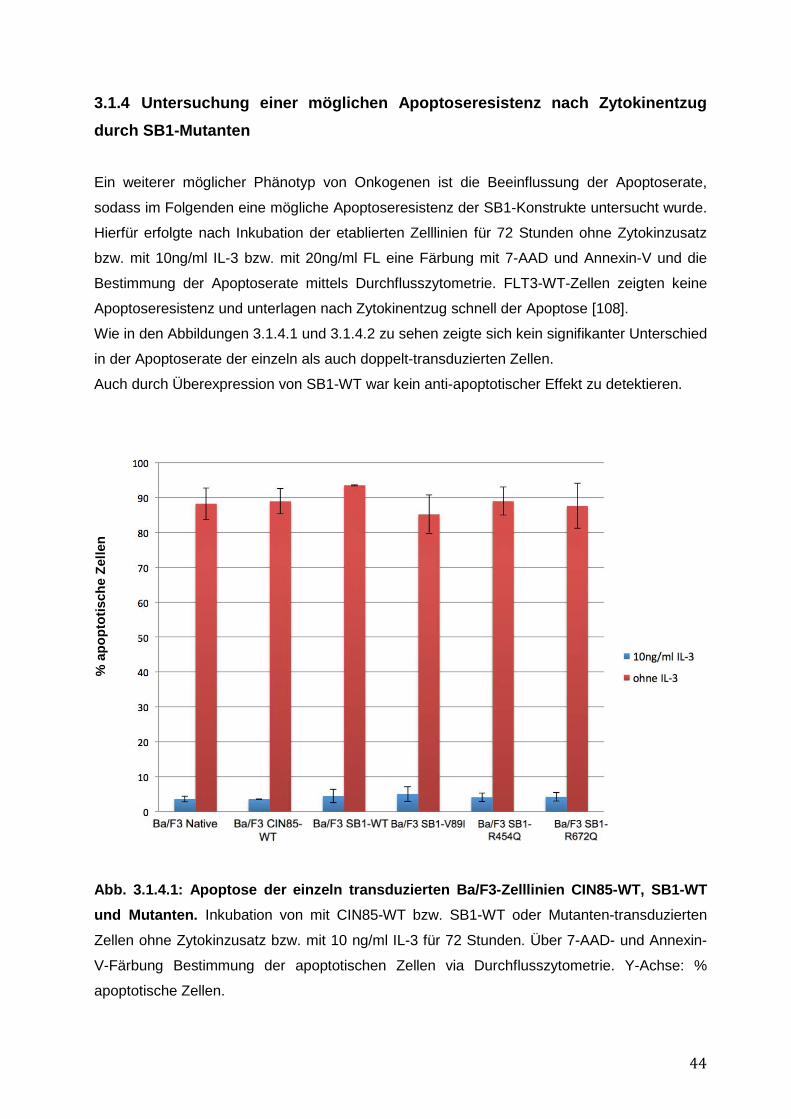
44
3.1.4 Untersuchung einer möglichen Apoptoseresistenz nach Zytokinentzug durch SB1-Mutanten
Ein weiterer möglicher Phänotyp von Onkogenen ist die Beeinflussung der Apoptoserate,
sodass im Folgenden eine mögliche Apoptoseresistenz der SB1-Konstrukte untersucht wurde.
Hierfür erfolgte nach Inkubation der etablierten Zelllinien für 72 Stunden ohne Zytokinzusatz
bzw. mit 10ng/ml IL-3 bzw. mit 20ng/ml FL eine Färbung mit 7-AAD und Annexin-V und die
Bestimmung der Apoptoserate mittels Durchflusszytometrie. FLT3-WT-Zellen zeigten keine
Apoptoseresistenz und unterlagen nach Zytokinentzug schnell der Apoptose [108].
Wie in den Abbildungen 3.1.4.1 und 3.1.4.2 zu sehen zeigte sich kein signifikanter Unterschied
in der Apoptoserate der einzeln als auch doppelt-transduzierten Zellen.
Auch durch Überexpression von SB1-WT war kein anti-apoptotischer Effekt zu detektieren.
Abb. 3.1.4.1: Apoptose der einzeln transduzierten Ba/F3-Zelllinien CIN85-WT, SB1-WT und Mutanten. Inkubation von mit CIN85-WT bzw. SB1-WT oder Mutanten-transduzierten
Zellen ohne Zytokinzusatz bzw. mit 10 ng/ml IL-3 für 72 Stunden. Über 7-AAD- und Annexin-
V-Färbung Bestimmung der apoptotischen Zellen via Durchflusszytometrie. Y-Achse: %
apoptotische Zellen.
% a
popt
otis
che
Zelle
n
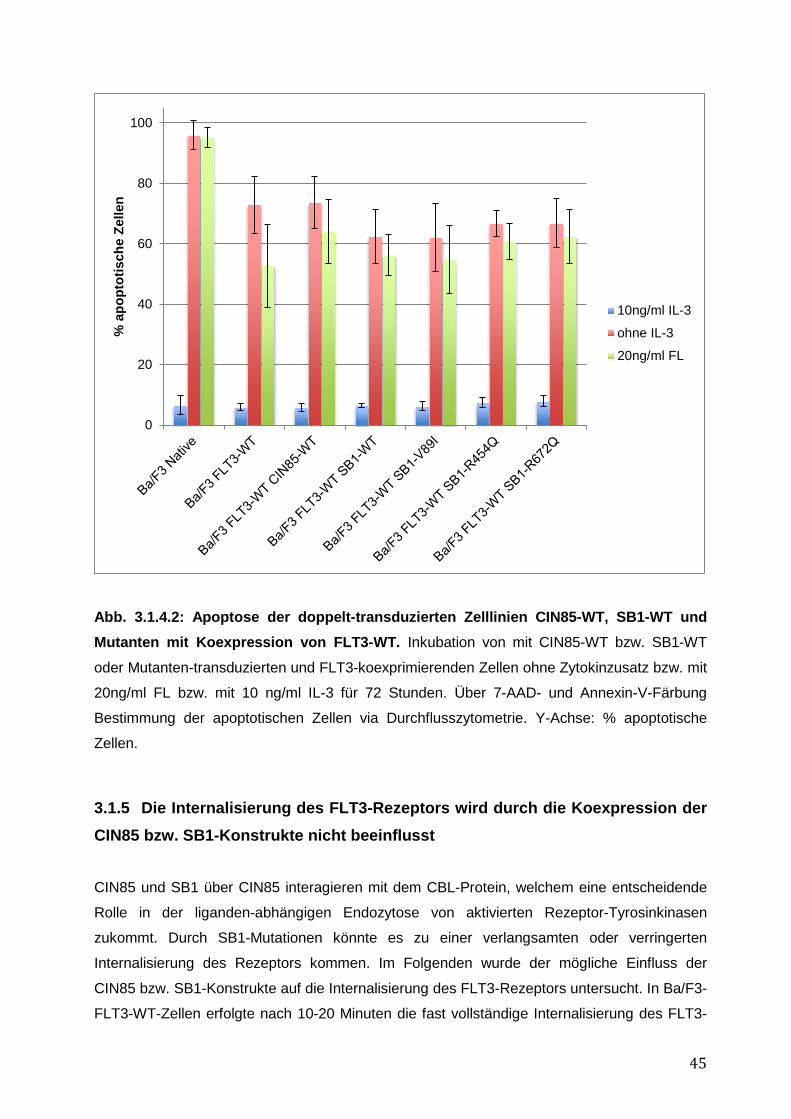
45
Abb. 3.1.4.2: Apoptose der doppelt-transduzierten Zelllinien CIN85-WT, SB1-WT und Mutanten mit Koexpression von FLT3-WT. Inkubation von mit CIN85-WT bzw. SB1-WT
oder Mutanten-transduzierten und FLT3-koexprimierenden Zellen ohne Zytokinzusatz bzw. mit
20ng/ml FL bzw. mit 10 ng/ml IL-3 für 72 Stunden. Über 7-AAD- und Annexin-V-Färbung
Bestimmung der apoptotischen Zellen via Durchflusszytometrie. Y-Achse: % apoptotische
Zellen.
3.1.5 Die Internalisierung des FLT3-Rezeptors wird durch die Koexpression der CIN85 bzw. SB1-Konstrukte nicht beeinflusst
CIN85 und SB1 über CIN85 interagieren mit dem CBL-Protein, welchem eine entscheidende
Rolle in der liganden-abhängigen Endozytose von aktivierten Rezeptor-Tyrosinkinasen
zukommt. Durch SB1-Mutationen könnte es zu einer verlangsamten oder verringerten
Internalisierung des Rezeptors kommen. Im Folgenden wurde der mögliche Einfluss der
CIN85 bzw. SB1-Konstrukte auf die Internalisierung des FLT3-Rezeptors untersucht. In Ba/F3-
FLT3-WT-Zellen erfolgte nach 10-20 Minuten die fast vollständige Internalisierung des FLT3-
0
20
40
60
80
100%
apo
ptot
isch
e Ze
llen
10ng/ml IL-3
ohne IL-3
20ng/ml FL
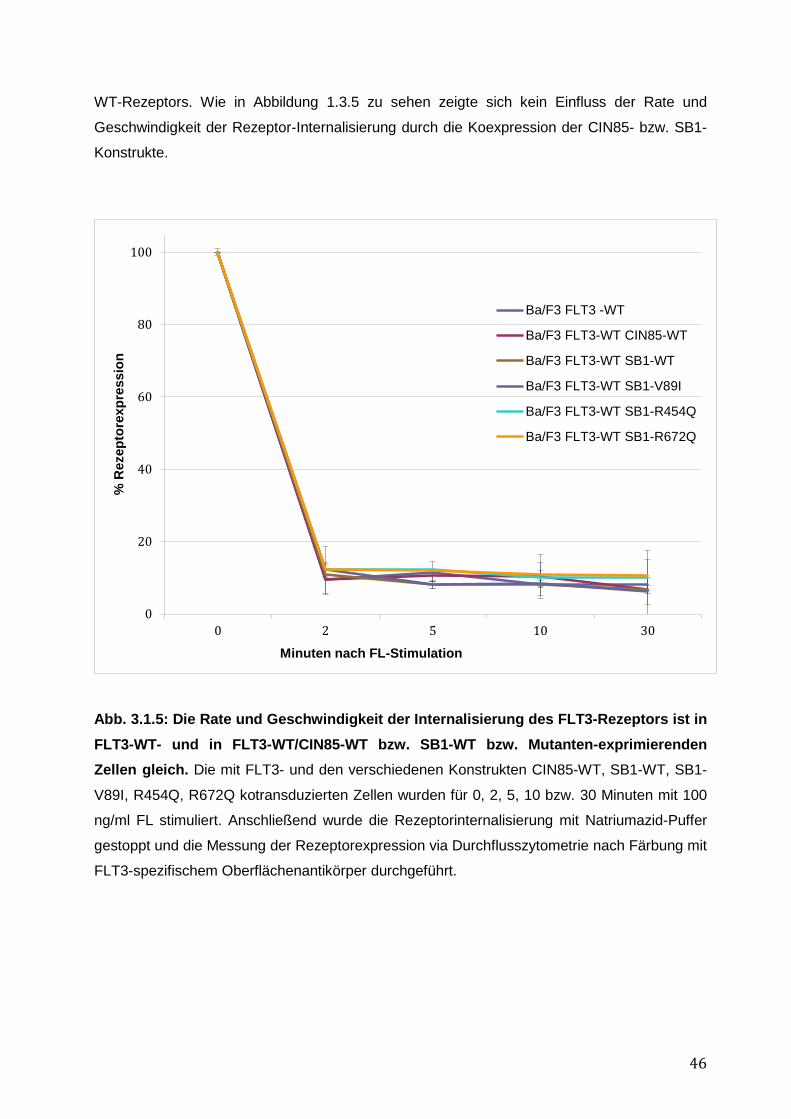
46
WT-Rezeptors. Wie in Abbildung 1.3.5 zu sehen zeigte sich kein Einfluss der Rate und
Geschwindigkeit der Rezeptor-Internalisierung durch die Koexpression der CIN85- bzw. SB1-
Konstrukte.
Abb. 3.1.5: Die Rate und Geschwindigkeit der Internalisierung des FLT3-Rezeptors ist in FLT3-WT- und in FLT3-WT/CIN85-WT bzw. SB1-WT bzw. Mutanten-exprimierenden Zellen gleich. Die mit FLT3- und den verschiedenen Konstrukten CIN85-WT, SB1-WT, SB1-
V89I, R454Q, R672Q kotransduzierten Zellen wurden für 0, 2, 5, 10 bzw. 30 Minuten mit 100
ng/ml FL stimuliert. Anschließend wurde die Rezeptorinternalisierung mit Natriumazid-Puffer
gestoppt und die Messung der Rezeptorexpression via Durchflusszytometrie nach Färbung mit
FLT3-spezifischem Oberflächenantikörper durchgeführt.
0
20
40
60
80
100
0 2 5 10 30
% R
ezep
tore
xpre
ssio
n
Minuten nach FL-Stimulation
Ba/F3 FLT3 -WT
Ba/F3 FLT3-WT CIN85-WT
Ba/F3 FLT3-WT SB1-WT
Ba/F3 FLT3-WT SB1-V89I
Ba/F3 FLT3-WT SB1-R454Q
Ba/F3 FLT3-WT SB1-R672Q
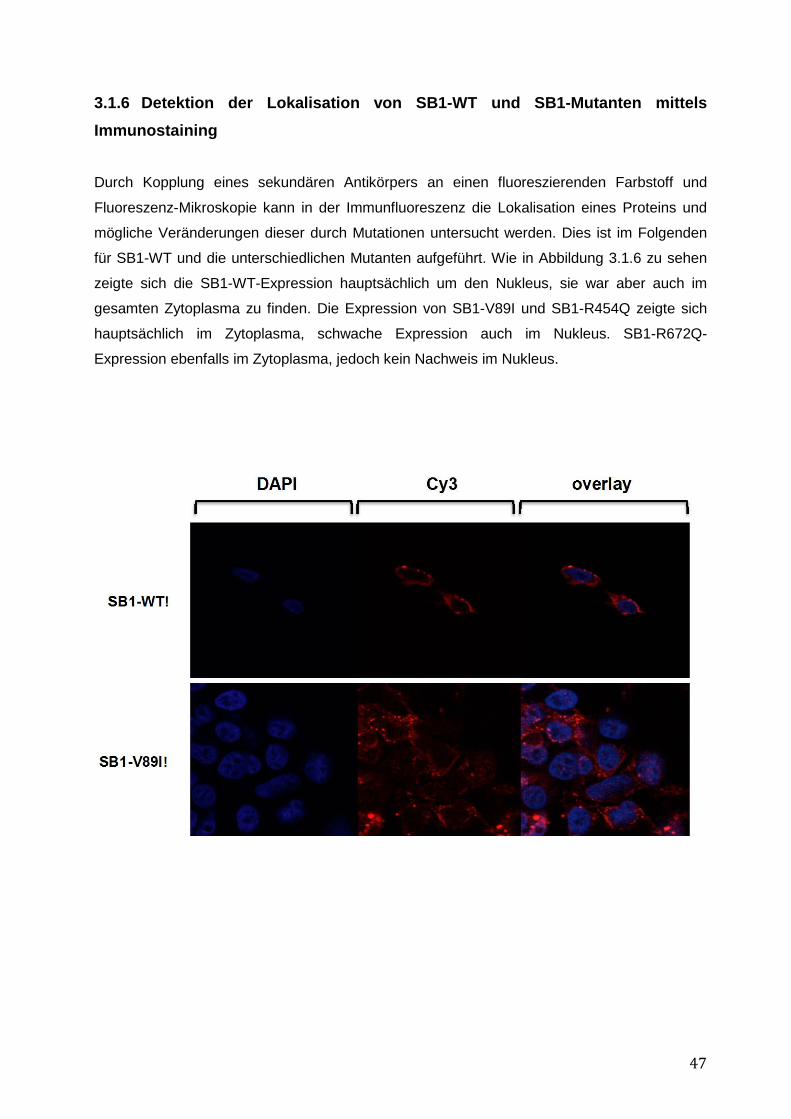
47
3.1.6 Detektion der Lokalisation von SB1-WT und SB1-Mutanten mittels Immunostaining
Durch Kopplung eines sekundären Antikörpers an einen fluoreszierenden Farbstoff und
Fluoreszenz-Mikroskopie kann in der Immunfluoreszenz die Lokalisation eines Proteins und
mögliche Veränderungen dieser durch Mutationen untersucht werden. Dies ist im Folgenden
für SB1-WT und die unterschiedlichen Mutanten aufgeführt. Wie in Abbildung 3.1.6 zu sehen
zeigte sich die SB1-WT-Expression hauptsächlich um den Nukleus, sie war aber auch im
gesamten Zytoplasma zu finden. Die Expression von SB1-V89I und SB1-R454Q zeigte sich
hauptsächlich im Zytoplasma, schwache Expression auch im Nukleus. SB1-R672Q-
Expression ebenfalls im Zytoplasma, jedoch kein Nachweis im Nukleus.
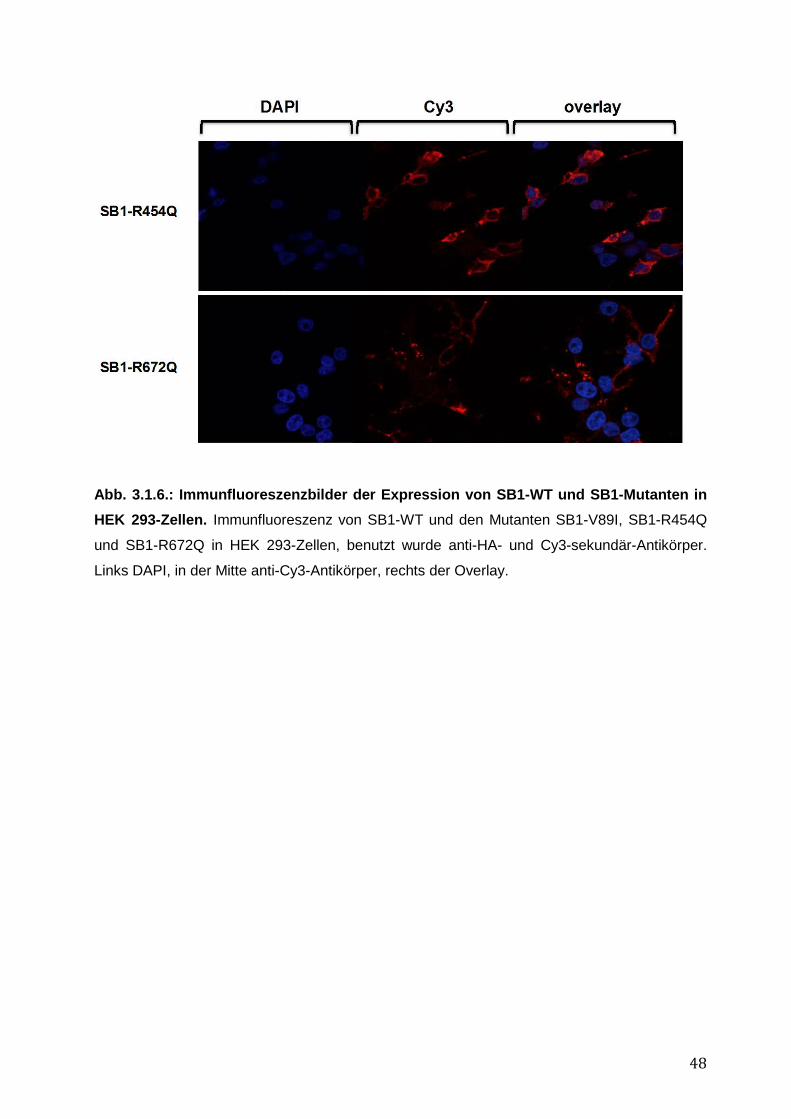
48
Abb. 3.1.6.: Immunfluoreszenzbilder der Expression von SB1-WT und SB1-Mutanten in HEK 293-Zellen. Immunfluoreszenz von SB1-WT und den Mutanten SB1-V89I, SB1-R454Q
und SB1-R672Q in HEK 293-Zellen, benutzt wurde anti-HA- und Cy3-sekundär-Antikörper.
Links DAPI, in der Mitte anti-Cy3-Antikörper, rechts der Overlay.

49
3.1.7 Zusammenfassung der Ergebnisse der funktionellen Charakterisierung von CIN85 und SB1 in rezeptor-exprimierenden Zellen
Nach Darstellung der endogenen Expression von CIN85 in AML-Zelllinien sowie Nachweis
einer stabilen Expression der transduzierten Konstrukte der einzeln bzw. doppelt-
transduzierten Ba/F3-FLT3/CIN85/SB1-Zelllinien konnte durch die Koexpression der
somatischen Mutante SB1-V89I mit FLT3-WT in den Proliferationsversuchen bei Stimulation
mit 50ng/ml FL ein leicht erhöhtes FL-induziertes Wachstum detektiert werden. SB1-V89I
alleine als auch SB1-WT und die restlichen mit FLT3 ko-transduzierten Zelllinien führten nicht
zu einem IL-3-unabhängigem Ba/F3-Zellwachstum. In der Untersuchung einer möglichen
Apoptoseresistenz nach Zytokinentzug durch SB1-Mutanten wiesen weder die einzeln
transduzierten Zelllinien CIN85-WT, SB1-WT und Mutanten noch die doppelt-transduzierten
Zelllinien CIN85-WT, SB1-WT und Mutanten mit Koexpression von FLT3-WT einen anti-
apoptotischen Effekt auf. Ebenso zeigte sich kein Einfluss auf die Rate und Geschwindigkeit
der FLT3-Rezeptor-Internalisierung durch die Koexpression der CIN85 bzw. SB1-Konstrukte.
In der Fluoreszenz-Mikroskopie konnte die SB1-WT-Expression hauptsächlich um den
Nukleus, ebenso schwach im Zytoplasma detektiert werden. Dieses Verhältnis verschob sich
bei den SB1-Mutanten SB1-V89I und SB1-R454Q zugunsten des Zytoplasmas, außerdem
zeigte sich eine schwache Expression auch im Nukleus. Für die SB1-Mutane R672Q konnte
keine Expression im Nukleus detektiert werden.
3.2 Vergleich des transformierenden Potentials der AKT1-E17K-Mutante in FLT3-exprimierenden Zellen mit der FLT3-ITD-Mutante W51
FLT3-ITD-Mutationen sind in der AML mit Leukozytose sowie schlechter Prognose als auch
erhöhter Rezidivrate assoziiert und bewirken eine konstitutive Autophosphorylierung und damit
Aktivierung des Rezeptors [31, 33, 109, 110].
Als wichtige Komponenten der Signaltransduktion sind die dem FLT3-Rezeptor
nachgeordneten MAPK-, AKT- und STAT5-Signalwege bekannt. Vor allem das Signalprotein
STAT5, aber auch die Aktivierung des AKT- und MAPK-Signalwegs spielt eine entscheidende
Rolle in der Transformation durch FLT3-ITD-Mutationen und damit in der AML-Entstehung
[111, 112].
Es wurden im Folgenden die Aktivierung dieser drei Signalproteine durch Phosphorylierung

50
als auch Proliferation, Inhibition und Apoptose für FLT3-exprimierende Zellen mit der AKT1-
E17K-Mutante im Vergleich zu der FLT3-ITD-Mutante W51 untersucht.
3.2.1 Stabile Expression der FLT3- und AKT1–Konstrukte in den Ba/F3-Zelllinien
Um das transformierende Potential des AKT-Signalwegs zu zeigen und dieses mit dem der
FLT3-ITD-Mutante zu vergleichen, erfolgte die Kotransduktion von AKT1-WT und der
konstitutiv aktivierten AKT1-E17K-Mutante mit jeweils FLT3-WT bzw. der FLT3-ITD-Mutante
W51. Die Mutation AKT1-E17K konnte in der T-ALL sowie in soliden Tumoren beispielsweise
des Harntrakts, der Brust oder der Haut detektiert werden, Mausmodelle wiesen einen
leukämischen Phänotyp auf [113, 114].
In der Abbildung 3.2.1 sind die Expressionsnachweise der transduzierten Zelllinien mit den
jeweiligen FLT3- und AKT1-Konstrukten, die eine deutliche Überexpression aufweisen, zu
sehen.
Abbildung 3.2.1: Expressionsnachweis der Ba/F3-FLT3/AKT1-Zelllinien. Untersuchung
der Ba/F3-Ganzzell-Lysate auf die Expression der transduzierten Konstrukte. Nachweis des
FLT3-Rezeptors mit einem FLT3-spezifischen Antikörper. Detektion der AKT1-Konstrukte
mittels spezifischem AKT-Antikörper. β-Actin dient als Ladungskontrolle.

51
3.2.2 Vergleich des pro-proliferativen Phänotyps der AKT1-E17K-Mutante und FLT3-ITD-Mutante W51 in den Ba/F3-Zelllinien und Inhibition des AKT1-E17K-induzierten proliferativen Phänotyps mittels AKT- und FLT3-Inhibitoren
Um den biologischen Phänotyp der AKT1-E17K-Mutante zu analysieren und mit dem der
FLT3-ITD-Mutante W51 vergleichen zu können, wurde die Proliferation mit und ohne
Inhibitoren als auch die Apopotoserate der verschiedenen Zellreihen untersucht.
In der Abbildung 3.2.2.1 ist zu sehen, dass sowohl die alleinige Expression von FLT3-W51 als
auch die alleinige Expression von AKT1-E17K in den Ba/F3-Zellen im Vergleich zu den FLT3-
WT- als auch AKT1-WT-Zellen zu bereits deutlich erhöhtem Zellwachstum führte. Eine
Annäherung des pro-proliferativen Phänotyps von AKT1-E17K zu den FLT3-W51-Zellen zeigte
sich durch dessen Koexpression mit dem FLT3-Wildtyp-Rezeptor (siehe Abbildung 3.2.2.2).
Eine leicht erhöhte Proliferation zeigte sich ebenso in der doppelt-transduzierten Zelllinie
FLT3-WT/AKT1-WT im Vergleich zu den nur FLT3-WT-exprimierenden Ba/F3-Zellen.
Wie gleichzeitig in den Abbildungen 3.2.2.1, 3.2.2.2 und 3.2.2.3 zu sehen erfolgte die
Hemmung der Zelllinien mittels spezifischer Inhibitoren gegen PI3K/AKT (MK2206) und FLT3
(AC220) in verschiedenen Konzentrationen und in Gegenwart von 20 ng/ml FL. Hier zeigte
sich die vollständige Hemmung der Ba/F3-FLT3-W51- und Ba/F3-FLT3-W51/AKT1-WT-
Zelllinien durch den FLT3-Tyrosinkinase-Inhibitor (FLT3-TKI) AC220. Auch die FLT3-
W51/AKT1-E17K-Zellen ließen sich durch AC220 auf das Proliferationsniveau der alleinigen
Expression der AKT1-E17K-Mutante hemmen. Der PI3K/AKT-Inhibitor MK2206 hingegen
hemmte das Wachstum aller drei FLT3-W51-exprimierenden Zelllinien nicht, was auf die
Unabhängigkeit des pro-proliferativen Phänotyps der FLT3-ITD-Mutation W51 von dem AKT-
Signalweg und auf die größere Bedeutung des STAT5-Signalwegs hindeutet.
Die Zelllinie FLT3-WT mit Koexpression der AKT1-E17K-Mutante als auch die Zelllinie mit der
alleinigen Expression von AKT1-E17K ließ sich durch MK2206 vollständig inhibieren, was die
Abhängigkeit des proliferativen Phänotyps von dem AKT-Signalweg bestätigt. Mittels AC220
war keine Inhibition des Ba/F3-AKT1-E17K-Zellwachstums zu verzeichnen.
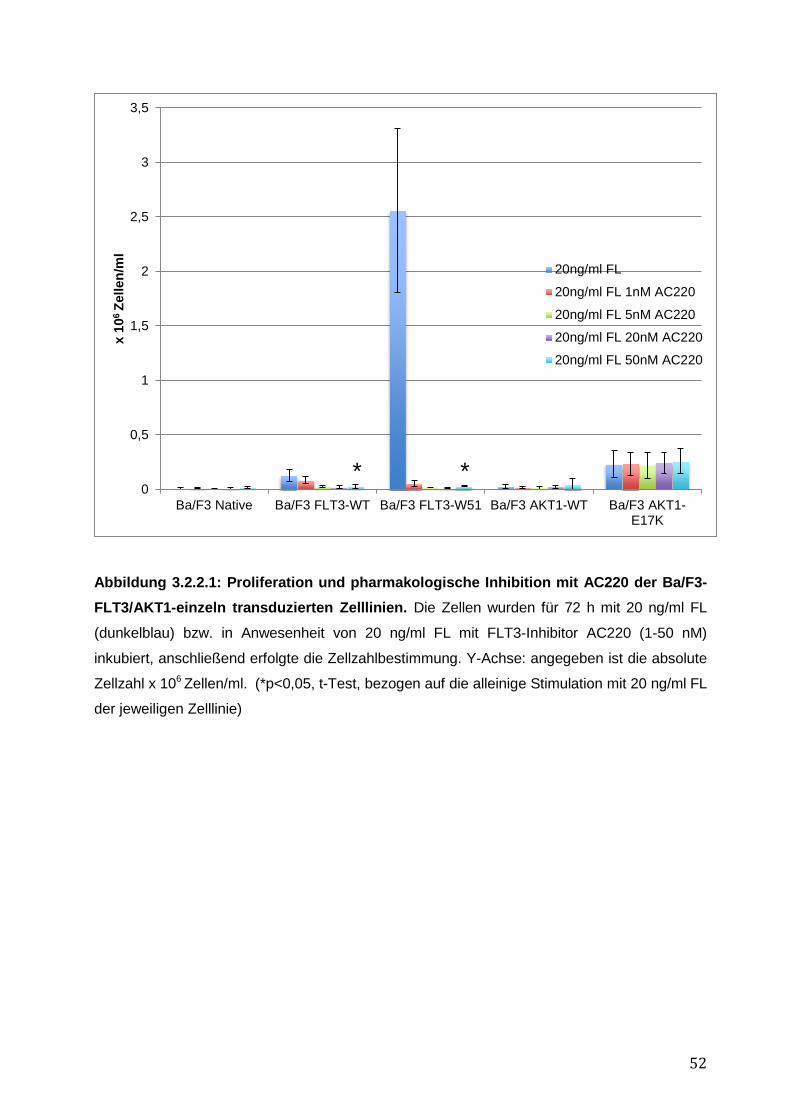
52
Abbildung 3.2.2.1: Proliferation und pharmakologische Inhibition mit AC220 der Ba/F3-FLT3/AKT1-einzeln transduzierten Zelllinien. Die Zellen wurden für 72 h mit 20 ng/ml FL
(dunkelblau) bzw. in Anwesenheit von 20 ng/ml FL mit FLT3-Inhibitor AC220 (1-50 nM)
inkubiert, anschließend erfolgte die Zellzahlbestimmung. Y-Achse: angegeben ist die absolute
Zellzahl x 106 Zellen/ml. (*p<0,05, t-Test, bezogen auf die alleinige Stimulation mit 20 ng/ml FL
der jeweiligen Zelllinie)
0
0,5
1
1,5
2
2,5
3
3,5
Ba/F3 Native Ba/F3 FLT3-WT Ba/F3 FLT3-W51 Ba/F3 AKT1-WT Ba/F3 AKT1-E17K
x 10
6 Ze
llen/
ml
20ng/ml FL
20ng/ml FL 1nM AC220
20ng/ml FL 5nM AC220
20ng/ml FL 20nM AC220
20ng/ml FL 50nM AC220
* *
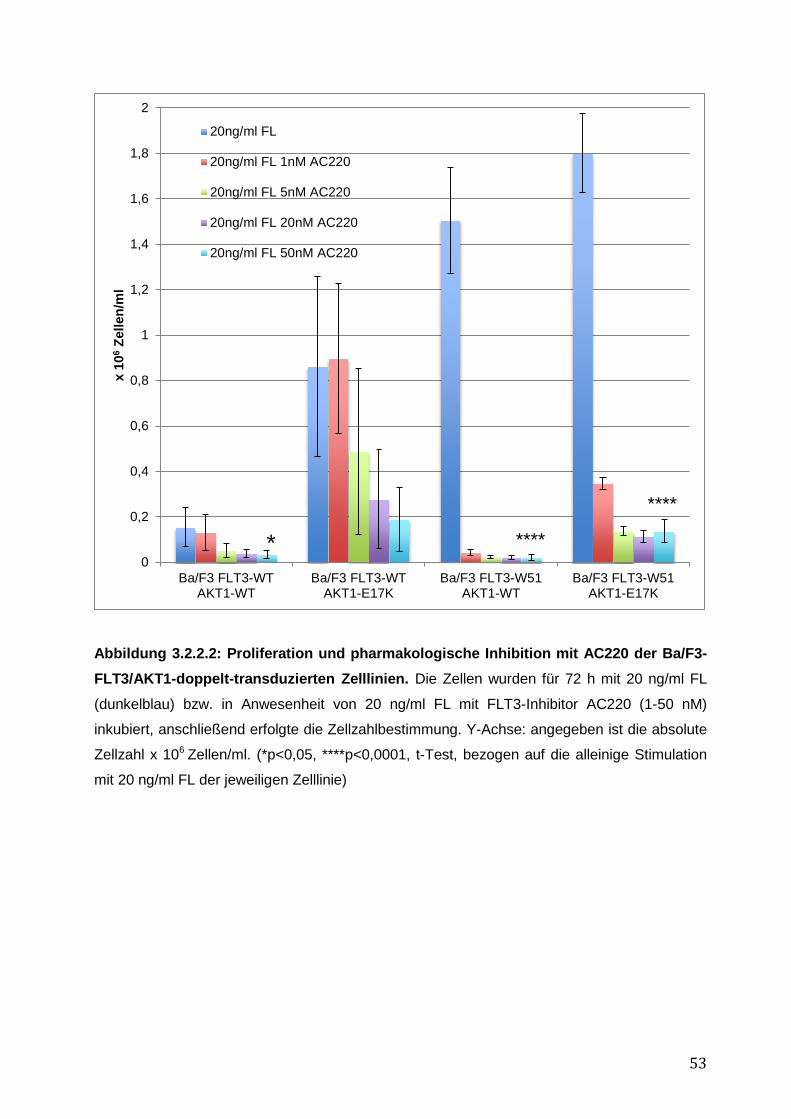
53
Abbildung 3.2.2.2: Proliferation und pharmakologische Inhibition mit AC220 der Ba/F3-FLT3/AKT1-doppelt-transduzierten Zelllinien. Die Zellen wurden für 72 h mit 20 ng/ml FL
(dunkelblau) bzw. in Anwesenheit von 20 ng/ml FL mit FLT3-Inhibitor AC220 (1-50 nM)
inkubiert, anschließend erfolgte die Zellzahlbestimmung. Y-Achse: angegeben ist die absolute
Zellzahl x 106 Zellen/ml. (*p<0,05, ****p<0,0001, t-Test, bezogen auf die alleinige Stimulation
mit 20 ng/ml FL der jeweiligen Zelllinie)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
Ba/F3 FLT3-WTAKT1-WT
Ba/F3 FLT3-WTAKT1-E17K
Ba/F3 FLT3-W51AKT1-WT
Ba/F3 FLT3-W51AKT1-E17K
x 10
6 Zel
len/
ml
20ng/ml FL
20ng/ml FL 1nM AC220
20ng/ml FL 5nM AC220
20ng/ml FL 20nM AC220
20ng/ml FL 50nM AC220
* ****
****
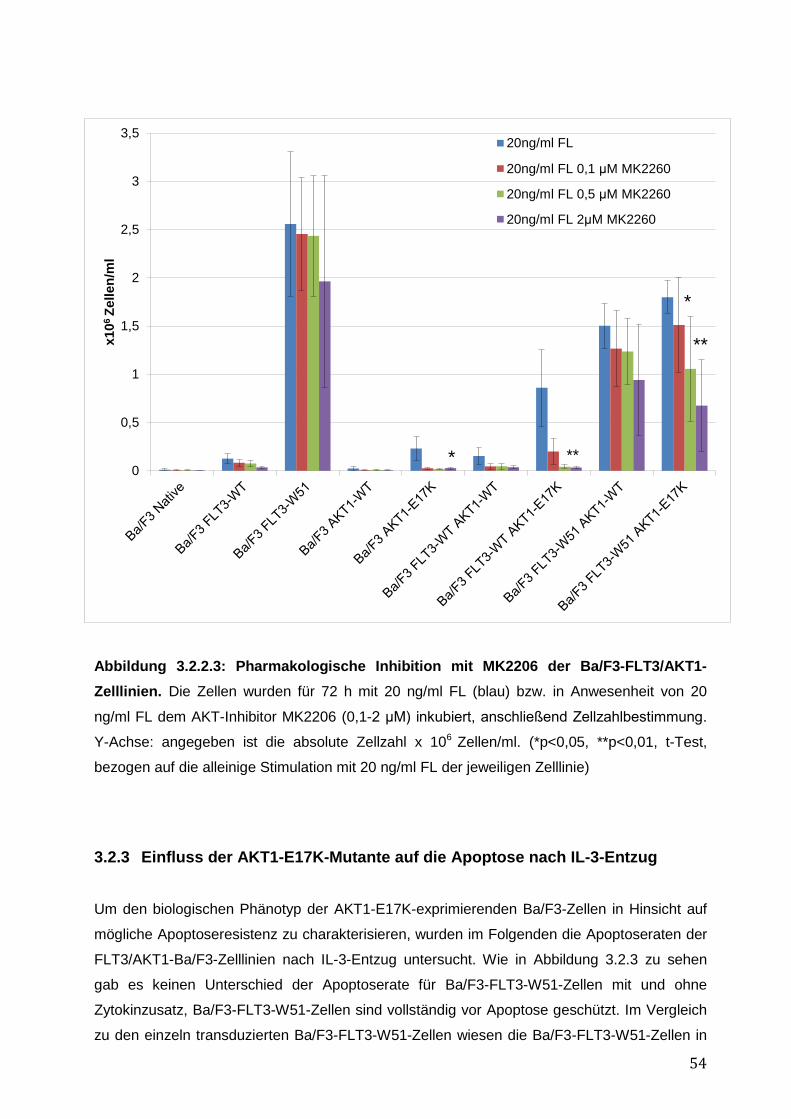
54
Abbildung 3.2.2.3: Pharmakologische Inhibition mit MK2206 der Ba/F3-FLT3/AKT1-Zelllinien. Die Zellen wurden für 72 h mit 20 ng/ml FL (blau) bzw. in Anwesenheit von 20
ng/ml FL dem AKT-Inhibitor MK2206 (0,1-2 μM) inkubiert, anschließend Zellzahlbestimmung.
Y-Achse: angegeben ist die absolute Zellzahl x 106 Zellen/ml. (*p<0,05, **p<0,01, t-Test,
bezogen auf die alleinige Stimulation mit 20 ng/ml FL der jeweiligen Zelllinie)
3.2.3 Einfluss der AKT1-E17K-Mutante auf die Apoptose nach IL-3-Entzug
Um den biologischen Phänotyp der AKT1-E17K-exprimierenden Ba/F3-Zellen in Hinsicht auf
mögliche Apoptoseresistenz zu charakterisieren, wurden im Folgenden die Apoptoseraten der
FLT3/AKT1-Ba/F3-Zelllinien nach IL-3-Entzug untersucht. Wie in Abbildung 3.2.3 zu sehen
gab es keinen Unterschied der Apoptoserate für Ba/F3-FLT3-W51-Zellen mit und ohne
Zytokinzusatz, Ba/F3-FLT3-W51-Zellen sind vollständig vor Apoptose geschützt. Im Vergleich
zu den einzeln transduzierten Ba/F3-FLT3-W51-Zellen wiesen die Ba/F3-FLT3-W51-Zellen in
0
0,5
1
1,5
2
2,5
3
3,5
x106
Zel
len/
ml
20ng/ml FL
20ng/ml FL 0,1 μM MK2260
20ng/ml FL 0,5 μM MK2260
20ng/ml FL 2μM MK2260
* **
*
**
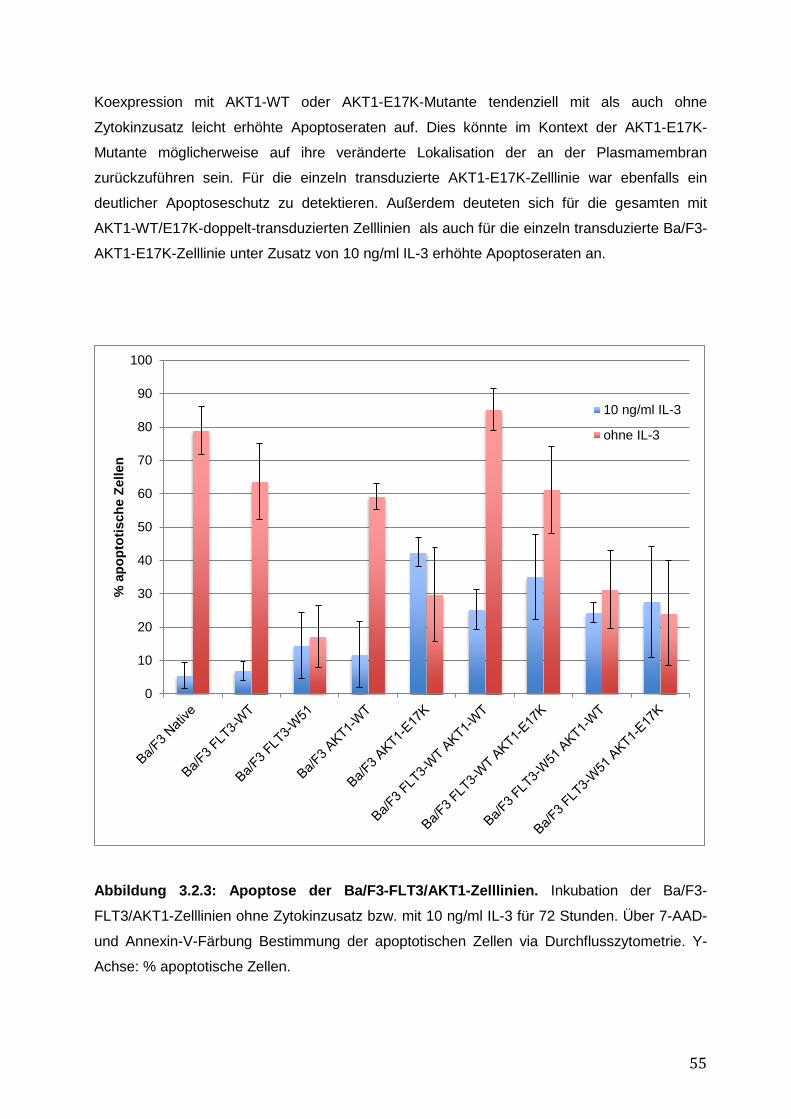
55
Koexpression mit AKT1-WT oder AKT1-E17K-Mutante tendenziell mit als auch ohne
Zytokinzusatz leicht erhöhte Apoptoseraten auf. Dies könnte im Kontext der AKT1-E17K-
Mutante möglicherweise auf ihre veränderte Lokalisation der an der Plasmamembran
zurückzuführen sein. Für die einzeln transduzierte AKT1-E17K-Zelllinie war ebenfalls ein
deutlicher Apoptoseschutz zu detektieren. Außerdem deuteten sich für die gesamten mit
AKT1-WT/E17K-doppelt-transduzierten Zelllinien als auch für die einzeln transduzierte Ba/F3-
AKT1-E17K-Zelllinie unter Zusatz von 10 ng/ml IL-3 erhöhte Apoptoseraten an.
Abbildung 3.2.3: Apoptose der Ba/F3-FLT3/AKT1-Zelllinien. Inkubation der Ba/F3-
FLT3/AKT1-Zelllinien ohne Zytokinzusatz bzw. mit 10 ng/ml IL-3 für 72 Stunden. Über 7-AAD-
und Annexin-V-Färbung Bestimmung der apoptotischen Zellen via Durchflusszytometrie. Y-
Achse: % apoptotische Zellen.
0
10
20
30
40
50
60
70
80
90
100
% a
popt
otis
che
Zelle
n
10 ng/ml IL-3
ohne IL-3

56
3.2.4 Untersuchung der Signalwege STAT5, AKT und MAPK in den Ba/F3-FLT3/AKT-Zelllinien
Die drei wichtigsten von FLT3 aktivierten Signalkaskaden sind der STAT5-, AKT- und MAPK-
Signalweg, welche in FLT3-ITDs konstitutiv aktiviert sind. Im Folgenden wurde die Aktivierung
durch Phosphorylierung dieser Signalproteine mittels phosphospezifischer Antikörper in FLT3-
exprimierenden Zellen mit der AKT1-E17K-Mutante im Vergleich zu der FLT3-ITD-Mutante
W51 untersucht. Wie in der Abbildung 3.2.4 zu sehen zeigte pSTAT5 in allen drei FLT3-W51-
Zelllinien einen deutlich erhöhten Grad der Aktivierung sowohl in stimulierten als auch
unstimulierten Zellen. Es zeigte sich ein gleichbleibend erhöhter Aktivierungsgrad für die
FLT3-W51-Zellen unabhängig von der Koexpression mit AKT1-WT oder AKT1-E17K.
Für AKT zeigte sich eine konstitutive Aktivierung in AKT1-WT- und AKT1-E17K-
exprimierenden Zellen mit teilweise leichter Verstärkung durch FL-Stimulation in FLT3-WT-
koexprimierenden Zellen. Wie in der Abbildung 3.2.4 zu sehen fand eine starke AKT-
Phosphorylierung nur in Anwesenheit der FLT3-ITD-Mutante statt. Diese erhöhte AKT-
Aktivierung war in den BA/F3-FLT3-W51-doppelt-transduzierten Zelllinien sowohl mit als auch
ohne Stimulation deutlich verstärkt. Ein Unterschied der Aktivierung war eher zugunsten der
Ba/F3-FLT3-W51-AKT1-WT-Zelllinie festzustellen.
Die MAPK war in allen drei AKT1-WT-exprimierenden Zelllinien stärker aktiviert als in der
AKT1-E17K-Mutante. In den FLT3-WT/W51-Zelllinien zeigte sich jedoch kein Unterschied in
der Aktivierung der MAPK zwischen den jeweils nur rezeptor-exprimierenden oder AKT1-WT-
koexprimierenden Zellen.
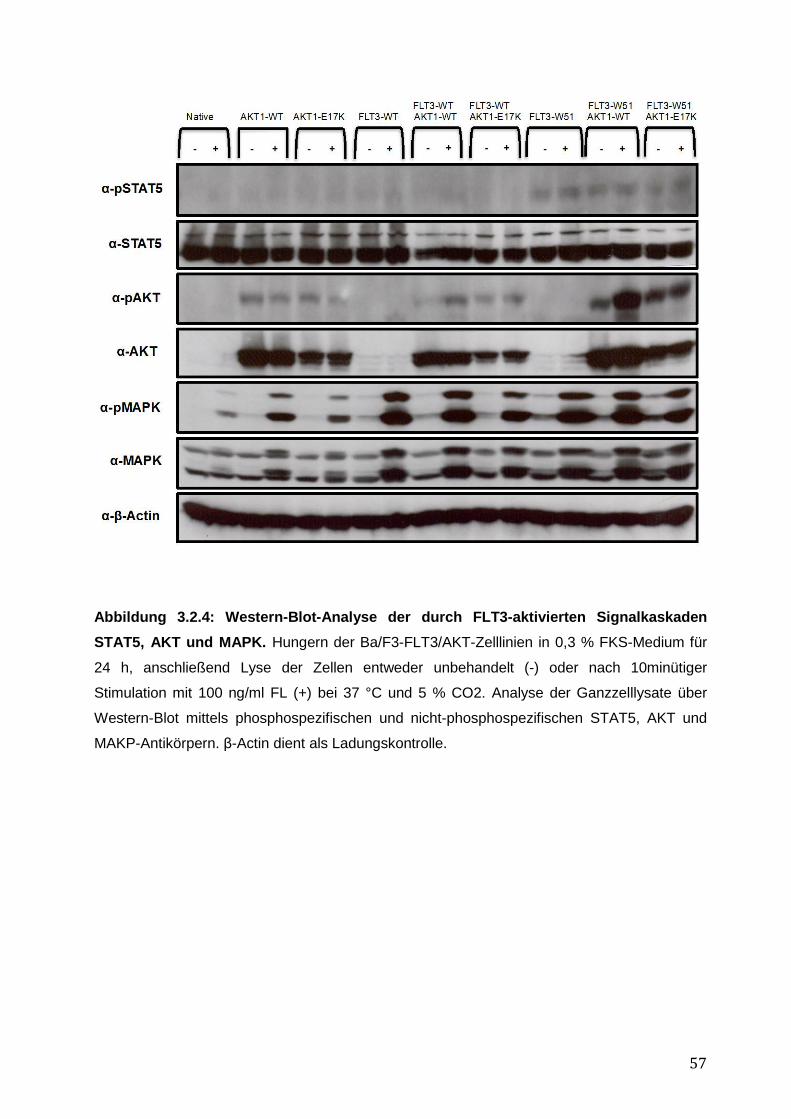
57
Abbildung 3.2.4: Western-Blot-Analyse der durch FLT3-aktivierten Signalkaskaden STAT5, AKT und MAPK. Hungern der Ba/F3-FLT3/AKT-Zelllinien in 0,3 % FKS-Medium für
24 h, anschließend Lyse der Zellen entweder unbehandelt (-) oder nach 10minütiger
Stimulation mit 100 ng/ml FL (+) bei 37 °C und 5 % CO2. Analyse der Ganzzelllysate über
Western-Blot mittels phosphospezifischen und nicht-phosphospezifischen STAT5, AKT und
MAKP-Antikörpern. β-Actin dient als Ladungskontrolle.

58
3.2.5 Zusammenfassung der Ergebnisse des biologischen Phänotyps von AKT1-E17K und FLT3-ITD im Vergleich
Nach Sicherstellung der stabilen Expression der transduzierten Konstrukte der einzeln bzw.
doppelt-transduzierten Ba/F3-FLT3/AKT1-Zelllinien erfolgte die Analyse des Phänotyps der
AKT1-E17K-Mutante im Vergleich zu FLT3-ITD in Proliferations- und Apoptose-
Untersuchungen. Die alleinige Expression von FLT3-W51 als auch die alleinige Expression
von AKT1-E17K im Vergleich zu den FLT3-WT- als auch AKT1-WT-Zellen führte bereits zu
deutlich erhöhtem, IL-3-unabhängigem Zellwachstum. Eine Annäherung des pro-proliferativen
Phänotyps von AKT1-E17K zu den FLT3-W51-Zellen zeigte sich durch dessen Koexpression
mit dem FLT3-Wildtyp-Rezeptor. Durch den FLT3-Tyrosinkinase-Inhibitor AC220 wurde eine
vollständige Hemmung der ligand-induzierten Proliferation der Ba/F3-FLT3-W51- und Ba/F3-
FLT3-W51/AKT1-WT-Zelllinien gezeigt. Auch die FLT3-W51/AKT1-E17K-Zellen ließen sich
durch AC220 hemmen, allerdings nur auf das Proliferationsniveau der alleinigen Expression
der AKT1-E17K-Mutante. Mittels AC220 war keine Inhibition des Ba/F3-AKT1-E17K-
Zellwachstums zu verzeichnen.
Durch Inhibition mit dem PI3K/AKT-Inhibitor MK2206 hingegen konnte das Wachstum der
Zelllinie mit der alleinigen Expression von AKT1-E17K als auch der Zelllinie FLT3-WT mit
Koexpression der AKT1-E17K-Mutante vollständig gehemmt werden.
In der Untersuchung einer möglichen Apoptoseresistenz nach 72 h Zytokin-Entzug wies die
AKT1-E17K-Zelllinie als auch die Ba/F3-FLT3-W51-Zelllinie einen hohen Apoptoseschutz auf.
Hinsichtlich der Signalübertragung wurde für pSTAT5 in allen drei FLT3-W51-Zelllinien ein
deutlich erhöhter Grad der Aktivierung sowohl in stimulierten als auch unstimulierten Zellen
deutlich. Es zeigte sich ein gleichbleibend erhöhter Aktivierungsgrad für die FLT3-W51-Zellen
unabhängig von der Koexpression mit AKT1-WT oder AKT1-E17K. AKT zeigte in AKT1-WT-
und AKT1-E17K-exprimierenden Zellen mit teilweise leichter Verstärkung durch FL-Stimulation
in FLT3-WT-koexprimierenden Zellen eine konstitutive Aktivierung. Eine starke AKT-
Phosphorylierung fand nur in Anwesenheit der FLT3-ITD-Mutante statt.
MAPK wurde nach Stimulation in allen drei AKT1-WT-exprimierenden Zelllinien stärker
aktiviert als in der AKT1-E17K-Mutante.

59
3.3 Der epigenetische Regulator TET2
TET2 spielt als epigenetischer Regulator eine wichtige Rolle in der Hämatopoese und
Entstehung von hämatologischen Neoplasien. Neben Mutationen in SB1 wurden innerhalb der
letzten Jahre viele TET2-Mutationen in Leukämien identifiziert. In der Literatur ist eine
besonders hohe TET2-Mutationsrate von ca. 50% in der CMML beschrieben, aber auch in der
AML wurde mutiertes oder deletiertes TET2 mit einer Gesamtfrequenz von um die 10-20%
detektiert [79, 81]. Ein Ziel dieses Teilprojekts der vorliegenden Doktorarbeit ist es, weitere
CMML-Patienten auf Rate und Art von TET2-Mutationen zu untersuchen. Des Weiteren sollte
hTET2 in der murinen pro-B-Zelllinie Ba/F3 funktionell charakterisiert werden.
3.3.1 Funktionelle Charakterisierung von hTET2-WT in Ba/F3-Zellen Es erfolgte im Folgenden die Klonierung der cDNA von humanem TET2-WT in den
retroviralen Expressionsvektor pMSCV-IRES-eYFP (MIY). Durch vollständige Sequenzierung
wurde die hTET2-WT-Nukleotidsequenz verifiziert. Das Konstrukt wurde anschließend stabil in
Ba/F3-Zellen exprimiert (siehe Abbildung 3.3.1.1).
Außerdem wurden Proliferations- als auch Apoptose-Assays von Ba/F3-hTET2-WT
durchgeführt, in denen kein pro-proliferativer oder anti-apoptotischer Phänotyp von hTET2-WT
zu sehen war (siehe Abbildung 3.3.1.2).
Abbildung 3.3.1.1: Expressionskontrolle von hTET2-WT in Ba/F3-Zellen. PCR mit hTET2-
spezifischen Primern aus der cDNA der Zelllinien Ba/F3-pMIY (als Negativkontrolle), Ba/F3
MIY-hTET2-WT, außerdem pMIY-hTET2-WT (als Positivkontrolle) als auch d2H2O zur
Kontrolle der Expression von hTET2-WT. Zu erwartende Größe der Bande: 534 bp. Mangels
fehlender Funktionalität der kommerziellen TET2-Antikörper erfolgte die TET2-
Expressionskontrolle mittels Detektion von RNA-Expression.
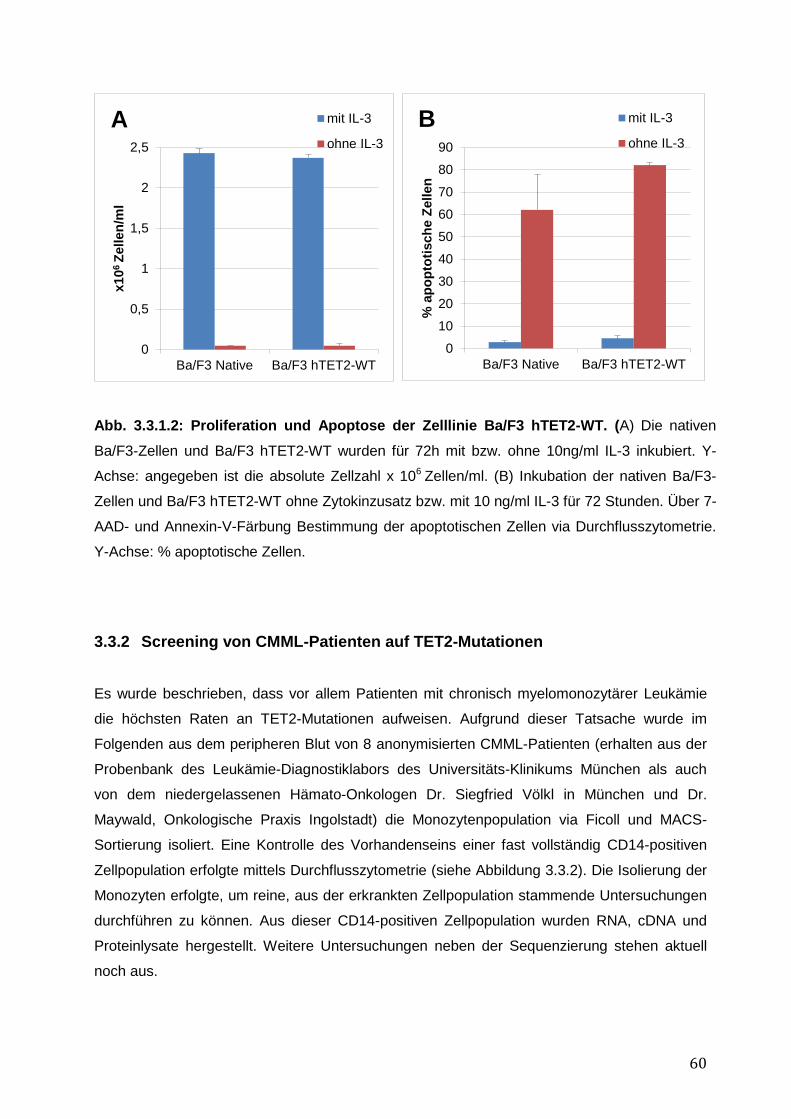
60
Abb. 3.3.1.2: Proliferation und Apoptose der Zelllinie Ba/F3 hTET2-WT. (A) Die nativen
Ba/F3-Zellen und Ba/F3 hTET2-WT wurden für 72h mit bzw. ohne 10ng/ml IL-3 inkubiert. Y-
Achse: angegeben ist die absolute Zellzahl x 106 Zellen/ml. (B) Inkubation der nativen Ba/F3-
Zellen und Ba/F3 hTET2-WT ohne Zytokinzusatz bzw. mit 10 ng/ml IL-3 für 72 Stunden. Über 7-
AAD- und Annexin-V-Färbung Bestimmung der apoptotischen Zellen via Durchflusszytometrie.
Y-Achse: % apoptotische Zellen.
3.3.2 Screening von CMML-Patienten auf TET2-Mutationen
Es wurde beschrieben, dass vor allem Patienten mit chronisch myelomonozytärer Leukämie
die höchsten Raten an TET2-Mutationen aufweisen. Aufgrund dieser Tatsache wurde im
Folgenden aus dem peripheren Blut von 8 anonymisierten CMML-Patienten (erhalten aus der
Probenbank des Leukämie-Diagnostiklabors des Universitäts-Klinikums München als auch
von dem niedergelassenen Hämato-Onkologen Dr. Siegfried Völkl in München und Dr.
Maywald, Onkologische Praxis Ingolstadt) die Monozytenpopulation via Ficoll und MACS-
Sortierung isoliert. Eine Kontrolle des Vorhandenseins einer fast vollständig CD14-positiven
Zellpopulation erfolgte mittels Durchflusszytometrie (siehe Abbildung 3.3.2). Die Isolierung der
Monozyten erfolgte, um reine, aus der erkrankten Zellpopulation stammende Untersuchungen
durchführen zu können. Aus dieser CD14-positiven Zellpopulation wurden RNA, cDNA und
Proteinlysate hergestellt. Weitere Untersuchungen neben der Sequenzierung stehen aktuell
noch aus.
0
0,5
1
1,5
2
2,5
Ba/F3 Native Ba/F3 hTET2-WT
x106
Zel
len/
ml
A mit IL-3
ohne IL-3
0102030405060708090
Ba/F3 Native Ba/F3 hTET2-WT
% a
popt
otis
che
Zelle
n
B mit IL-3
ohne IL-3
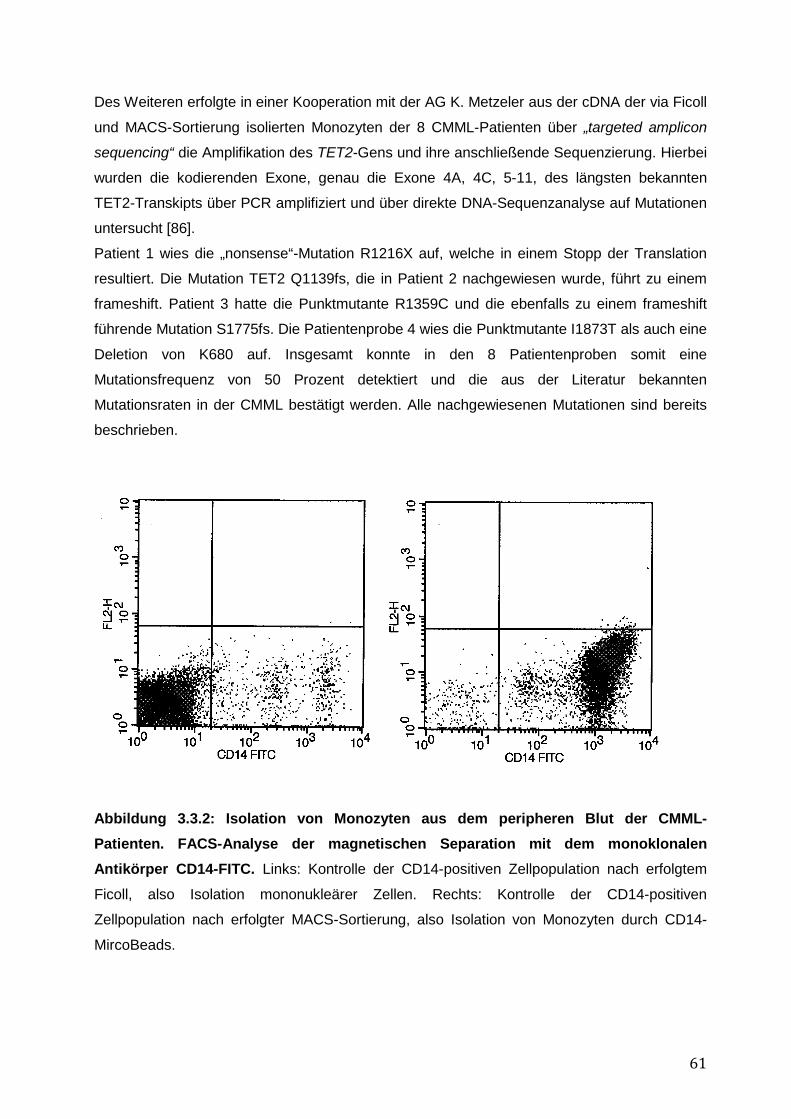
61
Des Weiteren erfolgte in einer Kooperation mit der AG K. Metzeler aus der cDNA der via Ficoll
und MACS-Sortierung isolierten Monozyten der 8 CMML-Patienten über „targeted amplicon
sequencing“ die Amplifikation des TET2-Gens und ihre anschließende Sequenzierung. Hierbei
wurden die kodierenden Exone, genau die Exone 4A, 4C, 5-11, des längsten bekannten
TET2-Transkipts über PCR amplifiziert und über direkte DNA-Sequenzanalyse auf Mutationen
untersucht [86].
Patient 1 wies die „nonsense“-Mutation R1216X auf, welche in einem Stopp der Translation
resultiert. Die Mutation TET2 Q1139fs, die in Patient 2 nachgewiesen wurde, führt zu einem
frameshift. Patient 3 hatte die Punktmutante R1359C und die ebenfalls zu einem frameshift
führende Mutation S1775fs. Die Patientenprobe 4 wies die Punktmutante I1873T als auch eine
Deletion von K680 auf. Insgesamt konnte in den 8 Patientenproben somit eine
Mutationsfrequenz von 50 Prozent detektiert und die aus der Literatur bekannten
Mutationsraten in der CMML bestätigt werden. Alle nachgewiesenen Mutationen sind bereits
beschrieben.
Abbildung 3.3.2: Isolation von Monozyten aus dem peripheren Blut der CMML-Patienten. FACS-Analyse der magnetischen Separation mit dem monoklonalen Antikörper CD14-FITC. Links: Kontrolle der CD14-positiven Zellpopulation nach erfolgtem
Ficoll, also Isolation mononukleärer Zellen. Rechts: Kontrolle der CD14-positiven
Zellpopulation nach erfolgter MACS-Sortierung, also Isolation von Monozyten durch CD14-
MircoBeads.

62
3.3.3 Zusammenfassung der Ergebnisse des epigenetischen Regulators TET2
Nach erfolgreichem Nachweis einer stabilen Expression von hTET2-WT in Ba/F3-Zellen
erfolgten sowohl Proliferations- als auch Apoptose-Assays von Ba/F3-hTET2-WT, in denen
kein pro-proliferativer oder anti-apoptotischer Phänotyp von hTET2-WT zu sehen war. Zudem
wurde von 8 anonymisierten CMML-Patienten die Monozytenpopulation isoliert, aus dieser
CD14-positiven Zellpopulation wurden RNA, cDNA und Proteinlysate hergestellt. Des
Weiteren fand über „targeted amplicon sequencing“ die Amplifikation der Exone 4A, 4C, 5-11
des TET2-Gens aus der cDNA der 8 CMML-Patienten und ihre anschließende Sequenzierung
statt. Hierbei konnten die bereits beschriebenen Mutationen TET2 R1216X, TET2 Q1139fs,
TET2 R1359C, TET2 S1775fs, TET2 I1873T und TET2 K680del nachgewiesen und eine
Mutationsfrequenz von 50% detektiert werden.

63
4 Diskussion
4.1 Charakterisierung der neu detektierten Punktmutationen in SB1
Durch neue Sequenzierungsmethoden wie beispielsweise der Transkriptom-Sequenzierung
konnten innerhalb der letzten Jahre eine Vielzahl von neuen Genmutationen in Tumoren
identifiziert werden. Diese neu entdeckten Mutationen untermauern die Komplexität und
Vielfalt der zellulären Prozesse, die eine Rolle in der Initiation und Propagation der Leukämie
spielen können. Um den Einfluss der neu identifizierten Mutationen auf die maligne
Transformation oder Tumorentstehung zu verstehen, müssen diese funktionell untersucht
werden. Ein Ziel dieser Arbeit ist es, die ebenfalls über Transkriptom-Sequenzierung neu
detektierten Punktmutationen in SB1, nämlich SB1-V89I, SB1-R454Q und SB1-R672Q,
funktionell zu charakterisieren.
Nach der Identifikation des Gens SB1, auch Seta binding protein 1 oder SHKBP1 genannt, im
Jahr 2000 von Borinstein et al. konnte im Jahr 2011 durch unsere Arbeitsgruppe erstmals eine
somatische SB1-Mutation SB1-V89I in der AML beschrieben werden [76, 78]. Zudem gelang
über Kapillarsequenzierung genomischer DNA weiterer AML-Patienten die Identifikation zwei
weiterer Missense-Mutationen von SB1 (R454Q und R672Q). Wie der Name bereits impliziert
bindet SB1 an Seta (=CIN85), welches als Multi-Adapter-Protein fungiert und unter anderem
über CBL in die Internalisierung und Regulation von Rezeptor-Tyrosinkinasen und in die
Apoptose involviert ist [64, 75]. Zudem wurden anti-apoptotische Effekte in Zelllinien mit einer
Überexpression von SB1 gezeigt, was darauf hindeutet, dass es sich bei SB1 um ein
Protoonkogen handelt [77]. Außerdem wird SHKBP1 aufgrund seiner Zugehörigkeit zu einem
Netzwerk von Inflammations-Proteinen eine potentielle prognostische Rolle nach einem
Herzinfarkt zugesprochen [115].
Um den biologischen Phänotyp von SB1 genauer charakterisieren zu können, erfolgten
Proliferations-, Apoptose-, Internalisierungs- als auch Immunfärbungs-Untersuchungen von
SB1-WT und SB1-Mutanten. Hierfür wurde das Ba/F3-Zellmodell, eine murine, IL-3-abhängige
pro-B-Zelllinie, verwendet. In unterschiedlichen AML-Zelllinien als auch den Ba/F3-Zellen
konnte eine endogene Expression von CIN85 unterschiedlicher Stärke nachgewiesen werden,
was ein ubiquitäres Vorhandensein von CIN85 vermuten lässt. Mangels fehlender
Funktionalität der kommerziellen SB1-Antikörper konnte die endogene Expression für SB1
nicht dargestellt werden. Aufgrund der IRES-Sequenz des retroviralen Vektors pMSCV-IRES-

64
EGFP - in welchen die Klonierung der SB1-Konstrukte erfolgte - wird gewährleistet, dass sich
das EGFP und das jeweilige Insert auf derselben mRNA befinden, was somit gleichzeitig eine
hohe Wahrscheinlichkeit auf die stabile Expression der SB1-Inserts sicherstellt. Zudem wurde
für weitere Expressionsuntersuchungen mittels HA-Tag der SB1-Konstrukte der
Expressionsnachweis gewährleistet. Aufgrund der endogenen Expression von CBL als auch
CIN85 in unserem Zellmodell konnte nach Doppelt-Transduktion der Ba/F3-Zellen mit dem
FLT3-Rezeptor als auch mit den unterschiedlichen SB1-Konstrukten (SB1-WT und SB1-
Mutanten) von der Vollständigkeit der Signalkaskade FLT3-CBL-CIN85-SB1 ausgegangen
werden.
Die Bindung von CIN85 an c-CBL spielt in der Endozytose von Rezeptor-Tyrosinkinasen
sowie deren Degradation eine entscheidende Rolle. Die Interaktion findet zwischen dem
Prolin-Arginin-Motiv (PXXXPR) von CBL und den SH3-Domänen von CIN85 statt. Die Isoform
von CIN85 mit drei SH3-Domänen häuft über die verschiedenen SH3-Domänen mehrere CBL-
Moleküle an, was einen kritischen Schritt für eine effiziente Degradation von RTKs darstellt.
CIN85 interagiert als Multi-Adapter-Protein neben CBL mit einer Vielzahl von weiteren
Proteinen wie beispielweise GRB2 oder dem in der vorliegenden Doktorarbeit näher
untersuchten SB1. Die Formation eines solchen Protein-Netzwerks ist in die Kontrolle von
Rezeptor-Tyrosinkinasen entscheidend miteingebunden [69]. Wie in dem Paper von Borinstein
et al. gezeigt wurde, ist SB1 in vitro in der Lage jede der drei SH3-Domänen von CIN85 zu
binden, wobei die stärkste Bindung in der SH3-1-Domäne detektiert wurde (siehe Abbildung
1.3.3). Das Paper von Feng et al. zeigt, dass CBL und SB1 um die Bindung an CIN85
konkurrieren und dass SB1 die Formierung des CBL-CIN85-Komplexes beeinflusst. Des
Weiteren wurde gezeigt, dass SB1 die Degradation des EGF-Rezeptors (engl. für Epidermal
Growth Factor Receptor) verzögert und dessen Signalwege verstärken kann [116].
Aufgrund dieser Daten sollte in der vorliegenden Doktorarbeit der Einfluss der in der AML neu
detektierten SB1-Mutanten untersucht sowie der Effekt der alleinigen Überexpression von
SB1-WT in unserem Zellmodell überprüft werden.
In unseren durchgeführten Untersuchungen zeigte die Koexpression von FLT3-WT mit CIN85-
bzw. den unterschiedlichen SB1-Konstrukten keinen Einfluss auf die FLT3-Internalisierung.
Der FLT3-Rezeptor wurde durch die Koexpression von CIN85-WT oder SB1-WT bzw. SB1-
Mutanten weder in der Geschwindigkeit noch bzgl. der Rate signifikant abweichend
internalisiert. Es wurde eine ubiquitäre endogene Expression der in der Signalkaskade
involvierten Proteine CBL und CIN85 in unserer Arbeitsgruppe sichergestellt. Durch Ko-
Transduktion des nicht endogen exprimierten Konstrukts FLT3-WT sowie der zu
untersuchenden SB1-WT- und SB1-Mutanten-Konstrukte wurde die Vollständigkeit des FLT3-
CBL-CIN85-SB1-Signalwegs sichergestellt. Dennoch könnten andere, in unserem Zellmodell
möglicherweise fehlende Proteine für den Signalweg eine wichtige Rolle spielen. Zudem

65
scheint auch die Verwendung von Konstrukten unterschiedlicher Spezies als möglicher
Störfaktor der Interaktion der verschiedenen Proteine denkbar.
In Hinblick auf einen möglichen Einfluss der Überexpression von SB1-WT oder Mutanten auf
die Zellproliferation oder –apoptose durch Beeinflussung der FLT3-CBL-CIN85-SB1-
Signalkaskade folgten weiterführende in-vitro-Ansätze. Nach stabiler Expression der
unterschiedlichen Konstrukte in den Ba/F3-Zelllinien zeigte sich bezüglich eines möglichen IL-
3-unabhänigigen Zellwachstums der SB1-Mutanten kein pro-proliferativer Phänotyp. Einzig die
Koexpression der somatischen Mutante SB1-V89I mit FLT3-WT wies bei Stimulation mit
50ng/ml FL ein minimal erhöhtes FL-induziertes Wachstum auf.
Auch in den Apoptoseexperimenten konnte keine Apoptoseresistenz der Ba/F3-Zelllinien
durch SB1-Mutanten, ebenso wenig durch die alleinige Überexpression von hSB1-WT wie
durch das Paper Liu et al. beschrieben gezeigt werden. In der Studie von Liu et al. zeigte sich
ein vermindertes TNF-induziertes Apoptoselevel durch Überexpression von humanem SB1 in
der ovariellen Zelllinie OV-90 [77].
Bei der Immunfärbung zeigte sich die SB1-WT-Expression hauptsächlich um den Nukleolus
und deutlich schwächer im Zytoplasma, was sich bei SB1-V89I und SB1-R454Q leicht und bei
SB1-R672Q gänzlich zugunsten des Zytoplasmas verschob.
In den Untersuchungen unserer Arbeitsgruppe ließ sich kein transformierender Phänotyp der
neu identifizierten SB1-Mutanten detektieren. Auch die alleinige Überexpression von SB1-WT
zeigte in unserem verwendeten Zellmodell keinen Phänotyp. Es bleibt offen, ob das
Verwenden eines anderen Zellmodells wie beispielsweise lentivirale Transduktion der
verschiedenen SB1-Konstrukte in AML-Zelllinien möglicherweise eine Relevanz für die
Detektion eines Phänotyps haben könnte. Zudem wurde in dem murinen Ba/F3-Zellmodell mit
dem CIN85-Konstrukt der Ratte und humanen SB1-Konstrukten gearbeitet. CIN85 der Ratte
zeichnet sich zwar durch eine hohe Homologie der Sequenzen und vor allem einer hohen
Homologie innerhalb der SH3-Domänen zu humanem CIN85 aus, trotzdem könnte dies ein
möglicher Störfaktor für die Interaktionskette FLT3-CBL-CIN85-SB1 darstellen. Des Weiteren
könnten die Ergebnisse dieses Teilprojekts dafür sprechen, dass es sich bei den über
Transkriptom-Sequenzierung neu detektierten SB1-Mutationen SB1-V89I, SB1-R454Q und
SB1-R672Q um Passenger- und nicht um Driver-Mutationen handelt, welche keine
funktionelle Konsequenz haben, oft während der Zellteilung auftreten und somit weder einen
Wachstumsvorteil aufweisen noch zur Krebsentstehung beitragen [117]. SB1-Mutationen
wurden inzwischen in einer Vielzahl von Krebserkrankungen verschiedenster Gewebe wie
beispielsweise der Speiseröhre, der Schilddrüse, der Haut, der Niere, der Leber und der
Lunge - um nur einige zu nennen – nachgewiesen. Ihre Rolle in der Krebsentstehung bleibt
aktuell weiterhin unklar.

66
4.2 Der biologische Phänotyp von AKT1-E17K und FLT3-ITD im Vergleich
Die Rezeptor-Tyrosinkinase FLT3 hat eine entscheidende Rolle in der normalen als auch
malignen Hämatopoese und ist als eines der am häufigsten mutierten Gene in der AML
bekannt [16]. Vor allem die aktivierenden FLT3-ITD-Mutationen, welche zu 20 - 30 % in der
AML detektiert werden, sind mit einer schlechteren Prognose, mit erniedrigtem
Gesamtüberleben und erhöhter Rezidivwahrscheinlichkeit assoziiert [16, 32-34]. Die aberrant
aktivierte FLT3-Kinase stellt einen attraktiven Angriffspunkt von zielgerichteten Therapien dar,
allerdings zeigt sich bei mehreren FLT3-Tyrosinkinase-Inhibitoren in präklinischen und
klinischen Studien in AML-Patienten entweder eingeschränkte oder nicht anhaltende
Effektivität sowie Resistenzentwicklung gegenüber den TKIs [52-54]. Auch die Induktion von
maligner Transformation durch konstitutive Aktivierung nachgeschalteter Signalwege von
RTKs wie beispielsweise die der STAT5- oder PI3K/AKT-Signalübertragung spielt sowohl in
der AML als auch in anderen humanen Tumorerkrankungen eine entscheidende Rolle [43,
44].
Der PI3K/AKT-Signalweg ist in viele biologische Prozesse wie beispielsweise der Regulation
der Apoptose, des Zellzyklus, der Proliferation und der Differenzierung involviert. Eine
Deregulation und aktivierende Mutationen in diesem Signalweg sind häufig in humanen
Tumorerkrankungen vorhanden, womit dem AKT-Signalweg eine kritische Rolle in der
Entstehung von malignen Grunderkrankungen zugesprochen wird [118-120]. Eine dieser
aktivierenden Mutationen ist die der AKT1-Kinase AKT1-E17K [121]. Im Gegensatz zu der
häufigen FLT3-ITD-Mutation wird diese in hämatologischen Erkrankungen nur selten detektiert
[120, 122], der AKT-Signalweg allerdings liegt in mehr als der Hälfte der Blasten von AML-
Patienten als konstitutiv aktiviert, d.h. phosphoryliert, vor [43, 123, 124]. Die Mutation AKT1-
E17K weist in in-vivo-Mausmodellen einen leukämischen Phänotyp auf und konnte in der T-
ALL sowie hauptsächlich in soliden Tumoren, beispielsweise des Harntrakts oder des
Endometriums, detektiert werden [113, 114]. In hämatologischen Neoplasien wie zum Beispiel
der Myelofibrose oder dem Multiplen Myelom wurden weitere AKT1-Mutationen wie AKT1-
M306T oder AKT1-R376C nachgewiesen [125, 126]. In Zusammenschau dieser Daten
könnten AKT-Mutationen oder die konstitutive Aktivierung des AKT-Signalwegs eine
entscheidende Rolle nicht nur in der Induktion von maligner Transformation, sondern auch in
der Pharmakoresistenz gegenüber zielgerichteter Therapie auch in der akuten myeloischen
Leukämie spielen. Um dieses transformierende Potential nachzuweisen, wurde die AKT1-
E17K-Mutante verwendet, welche konstitutiv aktiv ist.

67
AKT1-E17K ist eine Punktmutation in der Pleckstrin-Homologie-Domäne, die zu einer
verstärkten Affinität zu den Membranlipiden PIP2 und PIP3 führt. Daraus resultiert sowohl die
konstitutive Aktivierung der mutierten Kinase als auch eine veränderte, nämlich stärkere
Lokalisation der AKT1-Mutante an der Plasmamembran [113, 121].
Eine Fragestellung dieses Teilprojekts der vorliegenden Doktorarbeit war, ob ein aktivierter
AKT-Signalweg die Pharmakoresistenz gegenüber FLT3-Tyrosinkinase-Inhibitoren in
Patienten erklären kann. Hierzu wurde der biologische Phänotyp der häufig in der AML
detektierten FLT3-ITD-Mutante mit dem der konstitutiv aktiven AKT1-E17K-Mutante im FLT3-
WT-Hintergrund dargestellt, verglichen und mit spezifischen Inhibition wie dem FLT3-Inhibitor
AC220 sowie einem PI3K/AKT-Inhibitor MK2206 behandelt.
Die in dieser Arbeit durchgeführte Western-Blot-Analyse der Signalweg-Aktivierung bestätigte
die deutliche Phosphorylierung und damit Aktivierung von STAT5 aller mit FLT3-ITD (W51)-
transduzierten Zelllinien sowohl im FL-stimulierten als auch im unstimulierten Zustand. In den
anderen, nicht FLT3-ITD transduzierten Zelllinien konnte kein verstärktes pSTAT5-Signal
festgestellt werden. Dies untermauert die große Bedeutung und Abhängigkeit des
transformierenden Potentials der FLT3-ITD von dem STAT5-Signalweg. Die Aktivierung des
STAT5-Signalwegs durch FLT3-ITD-Mutationen wurde bereits in vielen Studien beschrieben
[107, 108, 112, 127, 128]. Zudem zeigte sich ebenfalls einzig in den mit FLT3-W51- und den
unterschiedlichen AKT1-Konstrukten doppelt-transduzierten Zelllinien eine starke AKT-
Aktivierung – auch in unstimuliertem Zustand. In AKT1-WT- und AKT1-E17K-einzeln-
transduzierten als auch mit FLT3-WT-doppelt-transduzierten Zellen stellte sich nur ein
schwaches pAKT-Signal dar. Die Anwesenheit von FLT3-ITD im Kontext einer Überexpression
von AKT1 oder der aktivierenden Mutante AKT1-E17K scheint deren Phosphorylierung und
damit Aktivierung sowohl mit als auch ohne FL-Stimulation deutlich zu verstärken. Die MAPK-
Aktivierung war in der AKT1-E17K-Mutanten-Zelllinie weniger stark als in den AKT1-WT-
exprimierenden Zelllinien, was möglicherweise auf einem sogenannten „Crosstalk“, also einer
Interaktion, zwischen dem Raf-AKT-Signalweg beruhen könnte [129].
In Proliferationsversuchen wuchsen Ba/F3-Zellen, die mit der AKT1-E17K-Mutante
transduziert waren, IL-3-unabhängig. Dies zeigt die deutliche Potenz von AKT in der Induktion
einer malignen Transformation einer Zelle. Auch die Überexpression von AKT1-WT in FLT3-
WT-koexprimierenden Zellen führte zu leicht erhöhten Proliferationsraten. Durch
Kotransduktion der AKT1-E17K-Mutante mit dem FLT3-WT-Rezeptor zeigten die Ba/F3-Zellen
eine weitere Annäherung des pro-proliferativen Phänotyps an den der FLT3-W51-Zellen,
welche das deutlichste IL-3-unabhängige Wachstum aufwiesen.
Der FLT3-Inhibitor AC220 erzielte die vollständige Hemmung der Ba/F3-FLT3-W51- und
Ba/F3-FLT3-W51/AKT1-WT-Zelllinien. Die Ba/F3-Zellen, kotransduziert mit FLT3-W51 und der
AKT-Mutante, konnten durch die Zugabe des FLT3-Inhibitors nur auf das Proliferationsniveau

68
der mit AKT1-E17K-transduzierten Zellen gehemmt werden und wuchsen weiterhin IL-3-
unabhängig. Dies könnte eine mögliche Erklärung für Resistenzen gegenüber FLT3-
Tyrosinkinase-Inhibitoren in AML-Patienten darstellen.
Der PI3K/AKT-Inhibitor MK2206 hingegen war in der Lage, den pro-proliferativen Phänotyp
der Zelllinie FLT3-WT mit Koexpression der AKT1-E17K-Mutante als auch die Zelllinie mit der
alleinigen Expression von AKT1-E17K vollständig zu inhibieren, was die alleinige Abhängigkeit
dieses Phänotyps vom AKT-Signalweg demonstriert. Die FLT3-W51-exprimierenden Zellen
dagegen wurden durch MK2206 nicht inhibiert, was die Unabhängigkeit dieses pro-
proliferativen Phänotyps vom AKT-Signalweg erneut untermauert.
Auch in der Untersuchung der Apoptoseraten nach IL-3-Entzug zeigte die einzeln-
transduzierte Zelllinie AKT1-E17K einen deutlichen Apoptoseschutz, wodurch das onkogene
Potential erneut bekräftigt wurde. Unter IL-3-Zusatz deuteten sich für alle mit AKT1-WT bzw.
AKT1-E17K-doppelt-transduzierten Zelllinien erhöhte Apoptoseraten an. Die angedeutete,
allerdings nicht immer signifikant erniedrigte Apoptoserate von mit AKT1-WT-einzeln- oder
doppelt-transduzierten Zelllinien unter IL-3-Zugabe im Vergleich zu den mit AKT1-E17K-
einzeln- oder doppelt-transduzierten Zelllinien könnte auf die veränderte, nämlich stärkere
Lokalisation der AKT1-Mutante an der Plasmamembran zurückzuführen sein [113, 121].
Die verschiedenen Untersuchungen zu Proliferation, Apoptose und Signalwegen verdeutlichen
ein starkes transformierendes Potential des AKT-Signalwegs hier dargestellt anhand der
AKT1-E17K-Mutante im FLT3-WT-Hintergrund. Das transformierende Potential scheint
annähernd so potent wie das einer FLT3-ITD-Mutation. Der aktivierte AKT-Signalweg
kooperiert mit FLT3-WT in der AML. Ba/F3-Zellen, welche mit der AKT1-E17K-Mutante und
FLT3-WT kotransduziert wurden, wuchsen IL-3-unabhängig. Einzeln transduziert wiesen die
AKT1-E17K-Zellen einen deutlichen Apoptoseschutz und ebenfalls IL-3-unabhängiges
Wachstum auf, was die Rolle von AKT als Schlüsselprotein erneut verdeutlicht.
Unsere Ergebnisse zeigen, dass eine Deregulation des AKT-Signalwegs durch eine
konstitutive Aktivierung von AKT in FLT3-WT exprimierenden Zellen ein starkes
transformierendes Potential vermittelt, allerdings schwächer als in FLT3-ITD exprimierenden
Zellen vermittelt durch STAT5. Es wurde des Weiteren nachgewiesen, dass die Mutation
AKT1-E17K zu einer partiellen Resistenz gegenüber FLT3-Tyrosinkinase-Inhibitoren führt.
Eine Kombination aus einem FLT3-Tyrosinkinase-Inhibitor mit einem PI3K/AKT-Inhibitor
könnte eine solche Resistenzbildung überkommen. Gezielte Inhibition von Signalproteinen des
PI3K/AKT-Signalwegs könnte vielversprechend für therapeutische Ansätze sein. AKT bleibt
aufgrund seiner vielfältigen Funktionen und aufgrund der Tatsache, dass der PI3K/AKT-
Signalweg einer der am häufigsten aberrant regulierten Signalwege in humanen Tumoren ist,
ein sehr interessantes Forschungsthema und möglicher therapeutischer Angriffspunkt [130].

69
4.3 Mutationen des epigenetischen Regulators TET2 in CMML-Patienten
2009 wurden erstmals TET2-Mutationen oder -Deletionen in hämatologischen Erkrankungen
wie dem myelodysplastischen Syndrom (MDS), den myeloproliferativen Neoplasien (MPN),
der akuten myeloischen Leukämie (AML), der chronischen myelomonozytären Leukämie
(CMML) und der sekundären AML detektiert [79]. Es wurde gezeigt, dass TET2-Mutationen
bereits in den primitiven CD34+CD38--Zellen nachweisbar sind [79]. Die DNA-Methylierung hat
entscheidende Konsequenzen in der Regulierung von Genexpression, die Methylierung von
Cytosin in der Promotorregion eines Gens resultiert in seiner Inaktivierung ( = “Silencing“). Der
Nachweis der Konversion von 5-Methylcytosin zu 5-Hydroxy-Methylcytosin in der DNA über
Oxidation durch TET2, was zu einer Demethylierung der DNA und somit zu einer Aktivierung
führt, machte dessen entscheidende Rolle in der epigenetischen Regulation deutlich [82].
2011 wurde des Weiteren demonstriert, dass die TET-Proteinfamilie, d.h. TET1, TET2 und
TET3, auch in der Lage ist, die weitere Oxidierung von 5-hmC zu 5-Formylcytosin (5-fC) und
5-Carboxylcytosin (5-caC) zu katalysieren [131, 132]. Inzwischen wurde TET2 in einer Reihe
von hämatologischen Erkrankungen beschrieben und erlaubt, wie beispielsweise in der AML
in einigen der durchgeführten klinischen Studien, eine prognostische Einordnung. TET2 hat
damit eine wichtige Funktion in der normalen und malignen Hämatopoese [133].
Bereits 2010 konnte Ko et al. eine eingeschränkte katalytische Aktivität in
Knochenmarksproben von TET2-mutierten Patienten nachweisen, welche im Vergleich zu
gesunden Kontrollproben analog erniedrigte 5-hmC-Level in genomischer DNA aufwiesen
[84]. Diese erniedrigten 5-hmC-Level in TET2-mutierten Patienten mit hämatologischen
Erkrankungen wurden unmittelbar nach Entdeckung der enzymatischen Funktion von TET2
bzw. der TET-Familie durch mehrere Arbeitsgruppen bestätigt. Außerdem konnte in mutierten
AML-Patienten eine erhöhte Promotor-Methylierung nachgewiesen werden [134].
Die genaue Rolle von TET2 in der Pathogenese von hämatologischen Erkrankungen ist
allerdings noch nicht verstanden und die Literatur hierzu teilweise widersprüchlich. Generell
scheint ein Funktionsverlust von TET2, beispielsweise durch eine TET2-Mutation, zu
erniedrigten 5-hmC-Leveln und demzufolge zu Akkumulation von 5-mC zu führen, welches mit
Genregression assoziiert ist [135].
In vivo-Studien weisen TET2 als einen kritischen Regulator sowohl der Differenzierung als
auch der Selbsterneuerung der hämatopoetischen Stammzellen (HSCs) aus. Bei TET2-/TET2-
-Mäusen konnte nach ca. 10 Wochen eine deutlich erhöhte Zellzahl im Knochenmark
nachgewiesen werden. Zudem wurde eine erhöhte Anzahl von lineage-negativen (lin-), c-
Kit+Sca-1+ (LSK) - Zellen als auch Lin-c-Kit+Sca-1- (LK) - Zellen detektiert, was eine regressive

70
Funktion von TET2 in der Entwicklung der hämatopoetischen Stamm- und Vorläuferzellen
impliziert [82]. Durch Figueroa et al. wurde zudem gezeigt, dass eine Reduktion der TET2-
Expression zu einer Inhibition der normalen myeloischen Differenzierung führt [134]. Mehrere
Mausmodelle zeigen, dass genetische Deletion von TET2 in zunehmender HSC-
Selbsterneuerung, deutlicher Ausweiterung des hämatopoetischen Stamm- und Progenitor-
Kompartments sowie Entwicklung einer proliferativen hämatologischen Erkrankung ähnlich der
CMML resultiert [136-138]. 2015 zeigte Rasmussen et al. in einem TET2-abhängigen
leukämischen Mausmodell einen Hypermethylierungs-Phänotyp von Enhancern, welcher in
AML-Patienten mit TET2-Mutation bestätigt wurde und für eine Induktion der Leukämogenese
durch Verlust der TET2-Funktion spricht [139]. Das Mausmodell von Shih et al. zeigt, dass
Mutationen in TET2 und FLT3 gemeinschaftlich die Propagation zur Induktion einer AML
durch DNA-Hypermethylierung sowie Genexpressionsveränderungen bewirken [140].
Aufgrund der hohen TET2-Mutationsfrequenz von bis zu 50 % in Patienten mit chronisch
myelomonozytärer Leukämie erfolgte in der vorliegenden Doktorarbeit aus dem peripheren
Blut von 8 anonymisierten CMML-Patienten die Isolation der Monozytenpopulation. Mittels
„targeted amplicon sequencing“ wurde anschließend die Amplifikation und Sequenzierung der
Exone 4A, 4C, 5-11 des TET2-Gens aus der cDNA der 8 CMML-Patienten durchgeführt.
Hierbei konnte in unserem CMML-Patientenkollektiv ebenfalls eine Mutationsfrequenz von 50
% detektiert werden. Diese Daten bestätigen somit die in der Literatur bisher bereits
beschriebene Mutationsrate von TET2 in der CMML [81]. Die von uns in den Patienten
detektierten Mutationen sind TET2 R1216X, TET2 Q1139fs, TET2 R1359C, TET2 S1775fs,
TET2 I1873T und TET2 K680del. Die Hälfte der vier TET2-mutierten Patienten wiesen zwei
TET2-Mutationen auf. Die gefundenen Mutationen sind alle bereits beschrieben und führen
beispielsweise zu Deletion, frameshift oder Stopp der Translation. Weitere Investigationen aus
dieser CD14-positiven Zellpopulation, aus der zudem RNA, cDNA und Proteinlysate
hergestellt wurden, stehen aktuell noch aus.
Durch unsere Sequenzierung konnten keine neuen Mutationen detektiert werden, das
Ergebnis untermauert jedoch die hohe Mutationsfrequenz und den Stellenwert von TET2 in
myeloischen Erkrankungen. Durch Erhöhung der Patientenzahl wie beispielsweise durch
Erweiterung des Patientenkollektivs hätten möglicherweise neue, noch nicht beschriebene
Mutationen nachgewiesen werden können. Jedoch ist für keine andere Erkrankung neben der
CMML in der Literatur eine so hohe Mutationsfrequenz von TET2 beschrieben. Die CMML ist
mit 1-3 Neuerkrankungen pro Jahr pro 100 000 Einwohner eine sehr seltene Erkrankung, was
die Rekrutierung von Patientenproben erschwert und limitiert hat.
In diesem Teilprojekt der vorliegenden Doktorarbeit wurden außerdem Apoptose- und
Proliferationsversuche der Ba/F3-hTET2-WT-Zellen durchgeführt. In dem Ba/F3-Zellmodell

71
war kein pro-proliferativer oder anti-apoptotischer Phänotyp festzustellen. Möglicherweise
wäre ein Phänotyp nicht durch alleinige Überexpression von TET2-WT, sondern durch
Klonierung und Transduktion von TET2-Mutanten zu detektieren gewesen. Auch die
Verwendung eines anderen Zellmodells wie beispielsweise AML-Zelllinien und lentivirales
Einbringen unterschiedlicher TET2-Konstrukte könnte von Nöten sein, um Effekte nachweisen
zu können.
Umfassende Projekte beschäftigten sich seit der Erstbeschreibung von TET2 mit dessen
Funktion, prognostischer Relevanz und klinischer Korrelation [81-89]. Trotz deutlichen
Erkenntniszugewinns bleiben viele Fragen offen, die Wichtigkeit weiterführender
Investigationen in TET2 wird durch unsere Ergebnisse untermauert.

72
5 Zusammenfassung
Die AML ist eine heterogene Erkrankung, bei der durch die Kombination aus
Immunphänotypisierung, Zytogenetik sowie Molekulargenetik eine immer präzisere
Diagnosestellung als auch Prognoseeinschätzung gelingt. Zytogenetische als auch
molekulargenetische Aberrationen führen zu Fehlregulationen der Zelle, was in konstitutiver
Aktivierung von onkogenen Signalwegen resultieren kann. Neue Sequenzierungsmethoden
identifizierten innerhalb der letzten Jahre eine Vielzahl von neuen Mutationen in Genen, deren
Rolle in der Initiation und Propagation der AML es einzuordnen gilt.
In dieser Doktorarbeit erfolgte die funktionelle Charakterisierung der im Jahr 2011 in der AML
durch Transkriptom-Sequenzierung detektierten Punktmutationen in SB1. SB1 (=Seta binding
protein1) bindet an Seta (=CIN85), das als Multi-Adapter-Protein u.a. über CBL in die
Internalisierung von RTKs sowie deren Degradation als auch in die Apoptose involviert ist. Um
zu klären, ob die identifizierte SB1-Mutation einen Phänotyp induziert, haben wir diese
funktionell charakterisiert. Nach Nachweis einer ubiquitären CIN85-Expression konnte in den
mit den unterschiedlichen SB1-Konstrukten transduzierten Ba/F3-Zelllinien eine stabile
Expression der Konstrukte nachgewiesen werden, sodass von der Vollständigkeit der
Signalkaskade FLT3-CBL-CIN85-SB1 ausgegangen werden konnte. Der FLT3-Rezeptor
wurde durch die Koexpression von SB1-WT bzw. SB1-Mutanten weder in der Geschwindigkeit
noch Rate signifikant abweichend internalisiert. Die Überexpression von SB1-WT oder
Mutanten zeigte weder einen pro-proliferativer Phänotyp noch eine Apoptoseresistenz. In der
Immunfärbung wurde SB1-WT vorwiegend um den Nukleolus exprimiert, was sich bei SB1-
V89I bzw. -R454Q minimal und bei SB1-R672Q gänzlich zugunsten des Zytoplasmas
verschob. Es konnte kein transformierender Phänotyp der neu identifizierten SB1-Mutanten
detektiert werden, wofür das Verwenden von Konstrukten unterschiedlicher Spezies trotz
hoher Sequenzhomologie einen möglichen Störfaktor der Interaktionskette FLT3-CBL-CIN85-
SB1 darstellen oder aber das Zellmodell an sich eine Rolle spielen könnte. Die neu
detektierten SB1-Mutationen könnten außerdem lediglich Passenger-Mutationen darstellen.
Des Weiteren wurde der transformierende Phänotyp der prognostisch entscheidenden FLT3-
ITD-Mutation mit der konstitutiv aktivierenden Mutation der AKT-Kinase AKT1-E17K
verglichen. Der FLT3-Rezeptor ist eines der am häufigsten mutierten Gene der AML, die ITD-
Mutationen stellen einen attraktiven, allerdings aktuell nur eingeschränkt wirksamen
Angriffspunkt zielgerichteter Therapien dar. Auch die konstitutive Aktivierung nachgeschalteter
Signalwege wie der PI3K/AKT-Signalweg spielt in der AML eine entscheidende Rolle. In
dieser Arbeit konnte gezeigt werden, dass sowohl AKT1-E17K als auch FLT3-ITD-
exprimierende Ba/F3-Zellen ein deutlich erhöhtes IL-3-unabhängiges Wachstum aufwiesen

73
sowie nach IL-3-Entzug vor Apoptose geschützt waren. Eine deutlich stärkere AKT-
Phosphorylierung war einzig in allen mit FLT3-ITD- und den unterschiedlichen AKT1-
Konstrukten doppelt-transduzierten Zelllinien feststellbar. Die pharmakologische Inhibition
verdeutlichte AKT als kritischen Mediator maligner Transformation. Durch AC220 ließen sich
die Ba/F3-Zellen, kotransduziert mit FLT3-W51 und der AKT-Mutante, nur auf das
Proliferationsniveau der mit AKT1-E17K-transduzierten Zellen hemmen. Der PI3K/AKT-
Inhibitor MK2206 hingegen war in der Lage, die Zelllinie FLT3-WT mit Koexpression der
AKT1-E17K-Mutante als auch mit der alleinigen Expression von AKT1-E17K vollständig zu
inhibieren. Diese Ergebnisse deuten darauf hin, dass AKT-Aktivierung zu partieller
Apoptoseresistenz gegenüber Tyrosinkinase-Inhibitoren in AML-Patienten führen könnte
sowie der PI3K/AKT-Signalweg als einer der am häufigsten aberrant regulierten Signalwege in
humanen Tumoren einen interessanten therapeutischen Angriffspunkt darstellt.
Im dritten Teil der Doktorarbeit wurden CMML-Patientenproben nach Isolation der CD14-
positiven Zellpopulation auf TET2-Mutationen untersucht. Der Nachweis der Konversion von
5-Methylcytosin zu 5-Hydroxy-Methylcytosin in der DNA über Oxidation durch TET2
verdeutlichte dessen entscheidende Rolle als epigenetischer Expressionsregulator. Die in der
Literatur beschriebene Mutationsfrequenz von bis zu 50% mit Nachweis der bekannten TET2-
Mutationen TET2 R1216X, TET2 Q1139fs, TET2 R1359C, TET2 S1775fs, TET2 I1873T und
TET2 K680del konnte bestätigt werden. Die Ba/F3-hTET2-WT-Zellen zeigten keinen pro-
proliferativen oder anti-apoptotischen Phänotyp. Weitere Untersuchungen mit der
hergestellten RNA, cDNA und Proteinlysaten stehen aktuell noch aus.
Individuelle und somit zielgerichtete Therapieansätze rücken zunehmend in den Fokus
moderner Medizin. Die Identifikation und genaue Charakterisierung von möglichen
Zielmolekülen innerhalb einer Erkrankung bleibt dabei unerlässlich und weiterhin eine große
Herausforderung.

74
6 Literaturverzeichnis
1. Berger DP, D.J., Engelhardt M, Engelhardt R, Henß H, Mertelsmann R,
Hämatologische Neoplasien in Das Rote Buch, Hämatologie und internistische Onkologie. Ecomed Medizin, 5. Überarbeitete Auflage, 2014: p. 636-645.
2. Siegenthaler W, B.H., Akute Leukämien in Klinische Pathophysiologie. New York, Thieme Verlag, 9. Auflage, 2006: p. 463-65.
3. Löwenberg B, D.J., Burnett A., Acute myeloid leukemia. N Engl J Med, 1999. 341(14): p. 1051–62.
4. Zenhäusern R, Z.C., Solenthaler M, Fey MF, Tobler A, Akute Leukämie beim Erwachsenen. Curriculum, Inselspital Bern, Schweiz Med Forum, 2003. Nr. 29/30.
5. Vardiman, J.W., et al., The 2008 revision of the World Health Organization (WHO) classification of myeloid neoplasms and acute leukemia: rationale and important changes. Blood, 2009. 114(5): p. 937-51.
6. D., G., Impact of cytogenetics on clinical outcome in AML. Acute Myelogenous Leukemia, Contemporary Hematology, 2007: p. 177-192.
7. Grimwade D, H.R., Independent prognostic factors for AML outcome. Hematology, 2009: p. 385-95.
8. Mrózek K, M.G., Paschka P, Whitman SP, Bloomfield CD., Clinical relevance of mutations and gene-expression changes in adult acute myeloid leukemia with normal cytogenetics: are we ready for a prognostically prioritized molecular classification? Blood, 2007. 109(2): p. 431-448.
9. Plattner H, H.J., Signale von außen an den Zellkern in Zellbiologie. Thieme, 4. Auflage, 2010: p. 427-29.
10. T, H., The phosphorylation of proteins on tyrosine: its role in cell growth and disease. The Croonian Lecture, 1997: p. 583-585.
11. Blume-Jensen, P.a.T.H., Oncogenic kinase signalling. Nature, 2001. 411(6835): p. 355-65.
12. Lemmon MA, S.J., Cell signaling by receptor-tyrosine kinases. Cell, 2010. 141(7): p. 1117–1134.
13. Mathews W, J.C., Wiegand GW, Pardoll D, Lemischka IR, A receptor tyrosine kinase specific to hematopoietic stem and progenitor cellenriched populations. Cell, 1991. 65: p. 1143-1152.
14. Rosnet O, M.S., deLapeyriere O, Birnbaum D, Murine Flt3, a gene encoding a novel tyrosine kinase receptor of the PDGFR/CSF1R family. Oncogene, 1991. 6: p. 1641-1650.
15. Rosnet O, M.M., Marchetto S, Birnbaum D, Isolation and chromosomal localization of a novel FMS-like tyrosine kinase gene. Genomics, 1991. 9: p. 380-385.
16. Gilliland DG, G.J., The roles of FLT3 in hematopoiesis and leukemia. Blood, 2002. 100: p. 1532-42.
17. Lyman, S., et al., Characterization of the protein encoded by the flt3 (flk2) receptorlike tyrosine kinase gene. Oncogene, 1993. 8(4): p. 815-22.
18. Kikushige, Y., et al., Human Flt3 is expressed at the hematopoietic stem cell and the granulocyte/macrophage progenitor stages to maintain cell survival. J Immunol, 2008. 180(11): p. 7358-67.
19. Rosnet, O., et al., Human FLT3/FLK2 receptor tyrosine kinase is expressed at the surface of normal and malignant hematopoietic cells. Leukemia, 1996. 10(2): p. 238-48.

75
20. Hannum, C., et al., Ligand for FLT3/FLK2 receptor tyrosine kinase regulates growth of haematopoietic stem cells and is encoded by variant RNAs. Nature, 1994. 368(6472): p. 643-8.
21. Toffalini, F., et al., New insights into the mechanisms of hematopoietic cell transformation by activated receptor tyrosine kinases. Blood, 2010. 116(14): p. 2429-2437.
22. Meshinchi S, A.F., Structural and functional alterations of FLT3 in acute myeloid leukemia. Clin Cancer Res, 2009. 15(13): p. 4263-69.
23. Wagener C, M.O., Signalwege der Tumorentstehung in Molekulare Onkologie, Entstehung, Progression, klinische Aspekte. Thieme, 3.Auflage: p. 207-18.
24. Dosil M, W.S., Lemischka IR, Mitogenic signalling and substrate specificity of the Flk2/Flt3 receptor tyrosine kinase in fibroblasts and interleukin 3-dependent hematopoietic cells. Mol Cell Biol, 1993. 13(10): p. 6572-85.
25. Zhang S, B.H., Flt3 ligand induces tyrosine phosphorylation of gab1 and gab2 and their association with shp-2, grb2, and PI3 kinase. Biochem Biophys Res Commun, 2000. 277(1): p. 195-9.
26. Marchetto S, F.E., Beslu N, et al., SHC and SHIP phosphorylation and interaction in response to activation of the FLT3 receptor. Leukemia, 1999. 13(9): p. 1374-82.
27. Lavagna-Sévenier C, M.S., Birnbaum D,Rosnet O, FLT3 signaling in hematopoietic cells involves CBL, SHC and an unknown P115 as prominent tyrosine-phosphorylated substrates. Leukemia, 1998. 12(3): p. 301-10.
28. Parcells BW, I.A., Simms-Waldrip T, Moore TB, Sakamoto KM, FMS-like tyrosine kinase 3 in normal hematopoiesis and acute myeloid leukemia. Stem Cells, 2006. 24(5): p. 1174-84.
29. Birg F, C.M., Rosnet O, Bardin F, Pebusque MJ, Marchetto S, Tabilio A, Mannoni P, Birnbaum D, Blood Expression of the FMS/KIT-like gene FLT3 in human acute leukemias of the myeloid and lymphoid lineages, 1992. 80(10): p. 2584-93.
30. Nakao M, Y.S., Iwai T, et al., Internal tandem duplication of the flt3 gene found in acute myeloid leukemia. Leukemia, 1996. 10(12): p. 1911-1918.
31. Vempati, S., et al., Arginine 595 is duplicated in patients with acute leukemias carrying internal tandem duplications of FLT3 and modulates its transforming potential. Blood, 2007. 110(2): p. 686-94.
32. Thiede C, S.C., Mohr B, Schaich M, Schakel U, Platzbecker U, Wermke M, Bornhauser M, Ritter M, Neubauer A, Ehninger G, Illmer T, Analysis of FLT3-activating mutations in 979 patients with acute myelogenous leukemia: association with FAB subtypes and identification of subgroups with poor prognosis. Blood 2002. 99(12): p. 4326-35.
33. Kiyoi H, T.M., Yokota S, Hamaguchi M, Ohno R, Saito H, Naoe T, Internal tandem duplication of the FLT3 gene is a novel modality of elongation mutation which causes constitutive activation of the product. Leukemia, 1998. 12(9): p. 1333-37.
34. Kottaridis PD, G.R., Frew ME, Harrison G, Langabeer SE, Belton AA, Walker H, Wheatley K, Bowen DT, Burnett AK, Goldstone AH, Linch DC, The presence of a FLT3 internal tandem duplication in patients with acute myeloid leukemia (AML) adds important prognostic information to cytogenetic risk group and response to the first cycle of chemotherapy: analysis of 854 patients from the United Kingdom Medical Research Council AML 10 and 12 trials. Blood, 2001. 98(6): p. 1752-59.
35. Abu-Duhier, F., et al., Identification of novel FLT-3 Asp835 mutations in adult

76
acute myeloid leukaemia. Br J Haematol, 2001. 113(4): p. 983-8. 36. Yamamoto, Y., et al., Activating mutation of D835 within the activation loop of
FLT3 in human hematologic malignancies. Blood, 2001. 97(8): p. 2434-9. 37. Mead AJ, L.D., Hills RK, Wheatley K, Burnett AK and Gale RE, FLT3 tyrosine
kinase domain mutations are biologically distinct from and have a significantly more favorable prognosis than FLT3 internal tandem duplications in patients with acute myeloid leukemia. Blood, 2007. 110(4): p. 1262-70.
38. Reindl C, B.K., Vempati S, Schnittger S, Ellwart J, et al., Point mutations in the juxtamembrane domain of FLT3 define a new class of activating mutations in AML. Blood, 2006. 107: p. 3700-07.
39. Kindler T, B.F., Kasper S, Estey E, Giles F, et al., Identification of a novel activating mutation (Y842C) within the activation loop of FLT3 in patients with acute myeloid leukemia (AML). Blood, 2005. 105(1): p. 335-340.
40. Schittenhelm MM, Y.K., Tyner JW, McGreevey L, Haley AD, et al., FLT3 K663Q is a novel AMLassociated oncogenic kinase: Determination of biochemical properties and sensitivity to Sunitinib (SU11248). Leukemia, 2006. 20: p. 2008-14.
41. Jiang J, P.J., Lee JC, Bo R, Stone RM, et al., Identifying and characterizing a novel activating mutation of the FLT3 tyrosine kinase in AML. Blood, 2004. 104(6): p. 1855-8.
42. Spiekermann K, B.K., Schoch C, Haferlach T, Hiddemann W, et al., A new and recurrent activating length mutation in exon 20 of the FLT3 gene in acute myeloid leukemia. Blood, 2002. 100(9): p. 3423-5.
43. Martelli AM, T.P., Evangelisti C, Chiarini F, Blalock WL, Billi AM, Manzoli L, McCubrey JA, Cocco L, Targeting the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin module for acute myelogenous leukemia therapy: from bench to bedside. Curr Med Chem, 2007. 14(19): p. 2009-23.
44. Martelli AM, E.C., Chiarini F, McCubrey JA, The phosphatidylinositol 3-kinase/Akt/mTOR signaling network as a therapeutic target in acute myelogenous leukemia patients. Oncotarget, 2010. 1(2): p. 89-103.
45. Martelli AM, N.M., Tabellini G, Bortul R, Tazzari PL, Evangelisti C and Cocco L, Phosphoinositide 3-kinase/Akt signaling pathway and its therapeutical implications for human acute myeloid leukemia. Leukemia, 2006. 20(6): p. 911-28.
46. Ni Y, S.-S.J., Young SH, Rozengurt E, PKD1 Mediates Negative Feedback of PI3K/Akt Activation in Response to G Protein-Coupled Receptors. PLoS One, 2013. 8(9): p. e73149.
47. Engelman, J., Luo J, Cantley LC, The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat Rev Genet, 2006. 7(8): p. 606-19.
48. Altman JA, S.A., Platanias LC, Targeting mTOR for the treatment of AML. New agents and new directions. Oncotarget, 2011. 2(6): p. 510-7.
49. Coffer PJ, K.L., de Groot RP, The role of STATs in myeloid differentiation and leukemia. Oncogene, 2000. 19(21): p. 2511-22.
50. Bar-Natan M, N.E., Xiang M, Frank DA, STAT signaling in the pathogenesis and treatment of myeloid malignancies. JAKSTAT, 2012. 1(2): p. 55-64.
51. Choudhary C, B.C., Schwable J, Tickenbrock L, Sargin B, Ueker A, Böhmer FD, Berdel WE, Müller-Tidow C, Serve H., Activation mechanisms of STAT5 by oncogenic Flt3-ITD. Blood, 2007. 110(1): p. 370-4.
52. Pratz KW, L.M., Bench to bedside targeting of FLT3 in acute leukemia. Curr

77
Drug Targets, 2010. 11(7): p. 781-9. 53. Kayser S, L.M., FLT3 tyrosine kinase inhibitors in acute myeloid leukemia:
clinical implications and limitations. Leuk Lymphoma, 2014. 55(2): p. 243-55. 54. Fischer, T., et al., Phase IIB trial of oral Midostaurin (PKC412), the FMS-like
tyrosine kinase 3 receptor (FLT3) and multi-targeted kinase inhibitor, in patients with acute myeloid leukemia and high-risk myelodysplastic syndrome with either wild-type or mutated FLT3. J Clin Oncol, 2010. 28(28): p. 4339-45.
55. Langdon, W.Y., et al., v-cbl, an oncogene from a dual-recombinant murine retrovirus that induces early B-lineage lymphomas. Proc Natl Acad Sci U S A, 1989. 86(4): p. 1168-72.
56. Keane, M., et al., Cloning and characterization of cbl-b: a SH3 binding protein with homology to the c-cbl proto-oncogene. Oncogene, 1995. 10(12): p. 2367-77.
57. Keane, M., et al., cbl-3: a new mammalian cbl family protein. Oncogene, 1999. 18(22): p. 3365-75.
58. Rao, N., et al., Negative regulation of Lck by Cbl ubiquitin ligase. Proc Natl Acad Sci U S A, 2002. 99(6): p. 3794-9.
59. Swaminathan G, T.A., The Cbl family proteins: ring leaders in regulation of cell signaling. J Cell Physiol, 2006. 209(1): p. 21-43.
60. Adélaíde, J., et al., Gain of CBL-interacting protein, a possible alternative to CBL mutations in myeloid malignancies. Leukemia, 2010. 24: p. 1539–1541.
61. Bonita, D., et al., Phosphotyrosine binding domain-dependent upregulation of the platelet-derived growth factor receptor alpha signaling cascade by transforming mutants of Cbl: implications for Cbl's function and oncogenicity. Mol Cell Biol, 1997. 17(8): p. 4597-610.
62. Joazeiro, C.A., et al., The tyrosine kinase negative regulator c-Cbl as a RING-type, E2-dependent ubiquitin-protein ligase. Science, 1999. 286(5438): p. 309-12.
63. Zheng, N., et al., Structure of a c-Cbl-UbcH7 complex: RING domain function in ubiquitin-protein ligases. Cell, 2000: p. 533-9.
64. Schmidt, M., Dikic, I, The Cbl interactome and its functions. Nat Rev Mol Cell Biol, 2005. 6(12): p. 907-18.
65. Blake, T., et al., The sequences of the human and mouse c-cbl proto-oncogenes show v-cbl was generated by a large truncation encompassing a proline-rich domain and a leucine zipper-like motif. Oncogene, 1991: p. 653-7.
66. Bartkiewicz M, H.A., Baron R, Leucine zipper-mediated homodimerization of the adaptor protein c-Cbl. A role in c-Cbl's tyrosine phosphorylation and its association with epidermal growth factor receptor. J Biol Chem, 1999. 274(43): p. 30887-95.
67. Szymkiewicz I, K.K., Soubeyran P, Dinarina A, Lipkowitz S, Dikic I., CIN85 participates in Cbl-b-mediated down-regulation of receptor tyrosine kinases. J Biol Chem, 2000. 277(42): p. 39666–39672.
68. Kurakin AV, W.S., Bredesen DE, Atypical recognition consensus of CIN85/SETA/Ruk SH3 domains revealed by target-assisted iterative screening. J Biol Chem, 2003. 278(36): p. 34102–34109.
69. Kowanetz K, S.I., Haglund K, Kowanetz M, Husnjak K, Taylor JD, Soubeyran P, Engstrom U, Ladbury JE, Dikic I, Identification of a novel proline-arginine motif involved in CIN85-dependent clustering of Cbl and down-regulation of epidermal growth factor receptors. J Biol Chem, 2003. 278(41): p. 39735–39746.
70. Take H, W.S., Takeda K, Yu ZX, Iwata N, Kajigaya S, Cloning and

78
characterization of a novel adaptor protein, CIN85, that interacts with c-Cbl. Biochem Biophys Res Commun, 2000. 268(2): p. 321–328.
71. Petrelli, A., et al., The endophilin–CIN85–Cbl complex mediates ligand-dependent downregulation of c-Met. Nature, 2002. 416: p. 187–190.
72. Soubeyran P, K., K, Szymkiewicz, I, Langdon, WY, Dikic, I, Cbl-CIN85-endophilin complex mediates ligand-induced downregulation of EGF receptors. Nature, 2002. 416: p. 183–187.
73. Dikic, I., Mechanisms controlling EGF receptor endocytosis and degradation. Biochem Soc Trans, 2003. 31: p. 1178-81.
74. Oved S, Y.Y., Signal transduction: molecular ticket to enter cells. Nature, 2002. 416(6877): p. 133-6.
75. Haglund K, S.M., Wong ES, Guy GR, Dikic I, Sprouty2 acts at the Cbl/CIN85 interface to inhibit epidermal growth factor receptor downregulation. EMBO Rep, 2005. 6(7): p. 635–641.
76. Borinstein, S., et al., SETA is a multifunctional adapter protein with three SH3 domains that binds Grb2, Cbl, and the novel SB1 proteins. Cellular Signalling, 2000. 12(11-12): p. 769-79.
77. Liu JP, L.N., Yuan HY, Guo Q, Lu H, Li YY, Human homologue of SETA binding protein 1 interacts with cathepsin B and participates in TNF-Induced apoptosis in ovarian cancer cells. Mol Cell Biochem, 2006. 292(1-2): p. 189-195.
78. Greif, P., et al., Identification of recurring tumor-specific somatic mutations in acute myeloid leukemia by transcriptome sequencing. Leukemia, 2011. 25(5): p. 821-7.
79. Delhommeau F, D.S., Della Valle V, et al, Mutation in TET2 in myeloid cancers. N Engl J Med, 2009. 360: p. 2289-2301.
80. Langemeijer SM, K.R., Berends M, et al, Acquired mutations in TET2 are common in myelodysplastic syndromes. Nat Genet, 2009. 41: p. 838- 842.
81. Kosmider O, G.-B.V., Ciudad M, Racoeur C, Jooste V, Vey N, et al., TET2 gene mutation is a frequent and adverse event in chronic myelomonocytic leukemia. Haematologica, 2009. 94: p. 1676–81.
82. Ko M, B.H., An J, et al., Ten‐Eleven‐Translocation 2 (TET2) negatively regulates homeostasis and differentiation of hematopoietic stem cells in mice. Proc Natl Acad Sci U S A, 2011. 108(35): p. 14566–14571.
83. Law, J., Jacobsen, SE, Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet, 2010. 11(3): p. 204–220.
84. Ko M, H.Y., Jankowska AM, et al., Impaired hydroxylation of 5-methylcytosine in myeloid cancers with mutant TET2. Nature, 2010. 468: p. 839-843.
85. Chou WC, C.S., Liu CY, et al., TET2 mutation is an unfavorable prognostic factor in acute myeloid leukemia patients with intermediate‐risk cytogenetics. Blood, 2011. 118(14): p. 3803–3810.
86. Metzeler KH, M.K., Radmacher MD, et al., TET2 mutations improve the new European LeukemiaNet risk classification of acute myeloid leukemia: a Cancer and Leukemia Group B study. J Clin Oncol, 2011. 29(10): p. 1373–1381.
87. Nibourel O, K.O., Cheok M, et al., Incidence and prognostic value of TET2 alterations in de novo acute myeloid leukemia achieving complete remission. Blood, 2010. 116: p. 1132-1135.
88. Kosmider O, G.-B.V., Cheok M, et al., TET2 mutation is an independent favorable prognostic factor in myelodysplastic syndromes (MDSs). Blood, 2009. 114: p. 3285-3291.
89. Chan, S., et al., Role of DNMT3A, TET2, and IDH1/2 mutations in pre-leukemic

79
stem cells in acute myeloid leukemia. Int J Hematol, 2013. 98: p. 648–657. 90. Vainchenker, W., et al., New mutations and pathogenesis of myeloproliferative
neoplasms. Blood, 2011. 118(7): p. 1723-35. 91. Saito, H., et al., Establishment and characterization of a new human
eosinophilic leukemia cell line. Blood, 1985. 66(6): p. 1233-40. 92. Graham, F., et al., Characteristics of a human cell line transformed by DNA
from human adenovirus type 5. J Gen Virol, 1977. 36(1): p. 59-74. 93. Collins SJ, G.R., Gallagher, RE, Continuous growth and differentiation of
human myeloid leukaemic cells in suspension culture. Nature, 1977. 270(5635): p. 347-9.
94. Asou, H., et al., Estabilshment of a human acute myeloid leukemia cell line (Kasumi-1) with 8;21 chromosome translocation. Blood, 1991. 77(9): p. 2031-36.
95. Koeffler HP, G.D., Acute myelogenous leukemia: a human cell line responsive to colony-stimulating activity. Science, 1978. 200: p. 1153-1154.
96. Koeffler, H., et al., An undifferentiated variant derived from the human acute myelogenous leukemia cell line (KG-1). Blood, 1980. 56: p. 265-273.
97. Ziegler-Heitbrock, H., et al., Establishment of an human cell line (Mono Mac 6) with characteristics of mature monocytes. International journal of cancer, 1988. 41(3): p. 456-61.
98. Matsuo, Y., et al., Two acute monocytic leukemia (AML-M5a) cell lines (MOLM-13 and MOLM-14) with interclonal phenotypic heterogeneity showing MLL-AF9 fusion resulting from an occult chromosome insertion, ins(11;9)(q23;p22p23). Leukemia, 1997. 11(9): p. 1469-77.
99. Lange, B., et al., Growth factor requirements of childhood acute leukemia: establishment of GM-CSF-dependent cell lines. Blood, 1987. 70(1): p. 192-9.
100. Kato, Y., et al., Establishment of peroxidase positive, human monocytic leukemia cell line (NOMO-1) and ist characteristics. Acta Haematol, 1986. 49: p. 277.
101. Wang, C., et al., Expression of a retinoic acid receptor gene in myeloid leukemia cells. Leukemia, 1989. 3(4): p. 264-69.
102. Tsuchiya, S., et al., Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). International journal of cancer, 1980. 26(2): p. 171-76.
103. Bradford, M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 1976. 72: p. 248-54.
104. Towbin, H., et al., Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A, 1979. 76(9): p. 4350-4.
105. Birnboim, H., A rapid alkaline extraction method for the isolation of plasmid DNA. Methods Enzymol, 1983. 100: p. 243-55.
106. Palacios, R., et al., Interleukin-3 supports growth of mouse pre-B-cell clones in vitro. Nature, 1984. 309(5964): p. 126-31.
107. Hayakawa, F., et al., Tandem-duplicated Flt3 constitutively activates STAT5 and MAP kinase and introduces autonomous cell growth in IL-3-dependent cell lines. Oncogene, 2000. 19(5): p. 624-31. .
108. Mizuki, M., et al., Flt3 mutations from patients with acute myeloid leukemia induce transformation of 32D cells mediated by the Ras and STAT5 pathways. Blood, 2000. 96: p. 3907-14.
109. Gale, R., et al., Relationship between FLT3 mutation status, biologic

80
characteristics, and response to targeted therapy in acute promyelocytic leukemia. Blood, 2005. 106(12): p. 3768-76. .
110. Gale, R., et al., No evidence that FLT3 status should be considered as an indicator for transplantation in acute myeloid leukemia (AML): an analysis of 1135 patients, excluding acute promyelocytic leukemia, from the UK MRC AML10 and 12 trials. Blood, 2005. 106(10): p. 3658-65. .
111. Gouilleux-Gruart, V., et al., STAT-related transcription factors are constitutively activated in peripheral blood cells from acute leukemia patients. Blood, 1996. 87(5): p. 1692-7.
112. Spiekermann K, B.K., Schwab R, Schmieja K,Hiddemann W, Overexpression and constitutive activation of FLT3 induces STAT5 activation in primary acute myeloid leukemia blast cells. Clin Cancer Res, 2003. 9: p. 2140-50.
113. Landgraf KE, P.C., Falke JJ, Molecular mechanism of an oncogenic mutation that alters membrane targeting: Glu17Lys modifies the PIP lipid specificity of the AKT1 PH domain. Biochemistry, 2008. 47(47): p. 12260-9.
114. Gutierrez, A., et al., High frequency of PTEN, PI3K, and AKT abnormalities in T-cell acute lymphoblastic leukemia. Blood, 2009. 114(3): p. 647-50.
115. Azuaje, F.J., et al., Information encoded in a network of inflammation proteins predicts clinical outcome after myocardial infarction. BMC Med Genomics, 2011. 14;4: p. 59.
116. Feng, L., et al., SH3KBP1-binding protein 1 prevents epidermal growth factor receptor degradation by the interruption of c-Cbl-CIN85 complex. Cell Biochem Funct., 2011. 29(7): p. 589-96.
117. Stratton, M.R., et al., The cancer genome. Nature, 2009. 458: p. 719-724. 118. Ligresti, G., et al., PIK3CA mutations in human solid tumors: role in sensitivity to
various therapeutic approaches. Cell Cycle, 2009. 8(9): p. 1352-8. 119. Samuels, Y., et al., High frequency of mutations of the PIK3CA gene in human
cancers. Science, 2004. 304(5670): p. 554. 120. Kim, M., et al., Mutational analysis of oncogenic AKT E17K mutation in common
solid cancers and acute leukaemias. Br J Cancer, 2008. 98(9): p. 1533-5. 121. Carpten, J., et al., A transforming mutation in the pleckstrin homology domain of
AKT1 in cancer. Nature, 2007. 448(7152): p. 439-44. 122. Luo, J., et al., Possible dominant-negative mutation of the SHIP gene in acute
myeloid leukemia. Leukemia, 2003. 17(1): p. 1-8. 123. Grandage, V., et al., PI3-kinase/Akt is constitutively active in primary acute
myeloid leukaemia cells and regulates survival and chemoresistance via NFkappaB, Mapkinase and p53 pathways. Leukemia, 2005. 19(4): p. 586-94.
124. Min, Y., et al., Constitutive phosphorylation of Akt/PKB protein in acute myeloid leukemia: its significance as a prognostic variable. Leukemia, 2003. 17(5): p. 995-7.
125. Lundberg, P., et al., Clonal evolution and clinical correlates of somatic mutations in myeloproliferative neoplasms. Blood, 2014. 123(14): p. 2220-8.
126. Leich, E., et al., Multiple myeloma is affected by multiple and heterogeneous somatic mutations in adhesion- and receptor tyrosine kinase signaling molecules. Blood Cancer, 2013. 3: p. e102.
127. Choudhary C, S.J., Brandts C, Tickenbrock L, Sargin B, et al, AML-associated Flt3 kinase domain mutations show signal transduction differences compared with Flt3 ITD mutations. Blood, 2005. 106: p. 265–273.
128. Grundler R, M.C., Thiede C, Peschel C, Duyster J, FLT3-ITD and tyrosine kinase domain mutants induce 2 distinct phenotypes in a murine bone marrow transplantation model. Blood, 2005. 105: p. 4792–4799.

81
129. Moelling, K., et al., Regulation of Raf-Akt Cross-talk. J Biol Chem, 2002. 277(34): p. 31099-106.
130. Yuan TL, C.L., PI3K pathway alterations in cancer: variations on a theme. Oncogene, 2008. 27(41): p. 5497-510.
131. He, Y.F., et al., Tet‐mediated formation of 5‐carboxylcytosine and its excision by TDG in mammalian DNA. Science, 2001. 333(6047): p. 1303-7.
132. Ito, S., et al., Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science, 2011. 333(6047): p. 1300-3.
133. Tian, X., et al., TET2 gene mutation is unfavorable prognostic factor in cytogenetically normal acute myeloid leukemia patients with NPM1+ and FLT3-ITD-mutations. Int J Hematol, 2014. 100(1): p. 96-104.
134. Figueroa, M.E., et al., Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell, 2010. 18(6): p. 553-567.
135. Tahiliani, M., et al., Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Sciene, 2009. 324(5929): p. 930-5.
136. Quivoron, C., et al., TET2 inactivation results in pleiotropic hematopoietic abnormalities in mouse and is a recurrent event during human lymphomagenesis. Cancer Cell, 2011. 20(1): p. 25-38.
137. Moran-Crusio, K., et al., Tet2 loss leads to increased hematopoietic stem cell self-renewal and myeloid transformation. Cancer Cell, 2011. 20(1): p. 11-24.
138. Abdel-Wahab, O., et al., Mutations in epigenetic modifiers in the pathogenesis and therapy of acute myeloid leukemia. Blood, 2013. 121(18): p. 3563-72.
139. Rasmussen, K.D., et al., Loss of TET2 in hematopoietic cells leads to DNA hypermethylation of active enhancers and induction of leukemogenesis. Genes Dev, 2015. 29(9): p. 910-22.
140. Shih, A.H., et al., Mutational Cooperativity Linked to Combinatorial Epigenetic Gain of Function in Acute Myeloid Leukemia. Cancer Cell, 2015. 27(4): p. 502-15.

82
7 Anhang
7.1 Tabellen
Tabelle 1.1: Einteilung der akuten myeloischen Leukämie. ...................................................... 2
Tabelle 2.1.1.1: Inhibitoren........................................................................................................17
Tabelle 2.1.1.2: Enzyme ......................................................................................................... 17
Tabelle 2.1.1.3: Zytokine ........................................................................................................ 17
Tabelle 2.1.1.4: Chemikalien .................................................................................................. 18
Tabelle 2.1.2: Software ........................................................................................................... 19
Tabelle 2.1.3: Kits ................................................................................................................... 20
Tabelle 2.1.4: Laborausstattung ............................................................................................. 20
Tabelle 2.1.5: Zelllinien ........................................................................................................... 21
Tabelle 2.1.6: Antikörper ........................................................................................................ 22
Tabelle 2.1.7: Plasmide .......................................................................................................... 23
Tabelle 2.1.8.1: Oligonukleotide zur Sequenzierung von hTET2 ............................................ 24
Tabelle 2.1.8.2: Oligonukleotide hTET2 WT PCR ................................................................... 24
Tabelle 2.1.8.3: Oligonukleotide zur Sequenzierung von CIN85 (der Ratte) ........................... 24
Tabelle 2.1.8.4: Oligonukleotide zur Sequenzierung von hSB1 .............................................. 24
Tabelle 2.1.8.5: Oligonukleotide zur Mutagenese von hSB1 ................................................... 25
Tabelle 2.1.9: Puffer, Medien, Lösungen ................................................................................ 25

83
7.2 Abbildungen
Abbildung 1.1: Der Einfluss des Karyotyps auf das Überleben von jüngeren Erwachsenen mit
AML .................................................................................................................................. 3
Abbildung 1.2.1: Einige in maligne hämatologische Erkrankungen involvierte Rezeptor-
Tyrosinkinase-Familien und deren Aufbau ........................................................................ 5
Abbildung 1.3.1: Das CBL-Interaktom ...................................................................................... 9
Abbildung 1.3.2: Die Rolle von CBL und CIN85 in der EGFR-Endozytose .............................. 10
Abbildung 1.3.3: Die Fähigkeit von SB1, jede der SH3-Domänen von CIN85 zu binden ......... 12
Abbildung 1.4: Mechanismen der TET2-induzierten Genexpression. ...................................... 14
Abbildung 1.5: Überblick der untersuchten Aspekte der vorliegenden Doktorarbeit................. 16
Abbildung 3.1: Schematische Darstellung des SB1- und CBL-Proteins mit Mutationen .......... 37
Abbildung 3.1.1: Endogene Expression von CIN85 in AML-Zelllinien ..................................... 39
Abbildung 3.1.2.1: Expressionsnachweis der einzeln transduzierten Ba/F3-FLT3/CIN85/SB1-
Zelllinien ......................................................................................................................... 40
Abbildung 3.1.2.2: Expressionsnachweis der doppelt-transduzierten Ba/F3-FLT3/CIN85/SB1-
Zelllinien ......................................................................................................................... 40
Abbildung 3.1.3.1: Proliferation der doppelt-transduzierten Zelllinien CIN85-WT, SB1-WT und
Mutanten mit Koexpression von FLT3-WT ...................................................................... 41
Abbildung 3.1.3.2: Detailliertere Ansicht der Ab. 3.1.3.1 ......................................................... 42
Abbildung 3.1.3.3: Proliferation der einzeln transduzierten Zelllinien CIN85-WT, SB1-WT und
Mutanten ........................................................................................................................ 43
Abbildung 3.1.3.4: Detailliertere Ansicht der Ab. 3.1.3.3 ......................................................... 43
Abbildung 3.1.4.1: Apoptose der einzeln transduzierten Ba/F3-Zelllinien CIN85-WT, SB1-WT
und Mutanten .................................................................................................................. 44
Abbildung 3.1.4.2: Apoptose der doppelt-transduzierten Zelllinien CIN85-WT, SB1-WT und
Mutanten mit Koexpression von FLT3-WT ...................................................................... 45
Abbildung 3.1.5: Die Rate und Geschwindigkeit der Internalisierung des FLT3-Rezeptors ist in
FLT3-WT- und in FLT3-WT/CIN85-WT bzw. SB1-WT bzw. Mutanten-exprimierenden
Zellen gleich ................................................................................................................... 46
Abbildung 3.1.6: Immunfluoreszenzbilder der Expression von SB1-WT und SB1-Mutanten in
HEK 293-Zellen .............................................................................................................. 47
Abbildung 3.2.1: Expressionsnachweis der Ba/F3-FLT3/AKT-Zelllinien .................................. 50
Abbildung 3.2.2.1: Proliferation und pharmakologische Inhibition mit AC220 der Ba/F3-
FLT3/AKT-einzeln transduzierten Zelllinien..................................................................... 52
Abbildung 3.2.2.2: Proliferation und pharmakologische Inhibition mit AC220 der Ba/F3-
FLT3/AKT-doppelt-transduzierten Zelllinien .................................................................... 53

84
Abbildung 3.2.2.3: Pharmakologische Inhibition mit MK2206 der Ba/F3-FLT3/AKT-Zelllinien
..................................................................................................................................... ..54
Abbildung 3.2.3: Apoptose der Ba/F3-FLT3/AKT-Zelllinien ..................................................... 55
Abbildung 3.2.4: Western-Blot-Analyse der durch FLT3-aktivierten Signalkaskaden STAT5,
AKT und MAPK .............................................................................................................. 57
Abbildung 3.3.1.1: Expressionskontrolle von hTET2-WT in Ba/F3-Zellen ............................... 59
Abbildung 3.3.1.2: Proliferation und Apoptose der Zelllinie Ba/F3 hTET2-WT ........................ 60
Abbildung 3.3.2: Isolation von Monozyten aus dem peripheren Blut der CMML-Patienten.
FACS-Analyse der magnetischen Separation mit dem monoklonalen Antikörper CD14-
FITC ............................................................................................................................... 61

85
8 Danksagung Mein herzlichster Dank geht an ..... Prof. Dr. med. W. Hiddemann, der es mir ermöglichte, die Promotionsarbeit an der
Medizinischen Klinik III des Klinikums Großhadern durchzuführen.
Prof. Dr. med. Karsten Spiekermann, der mir die Promotionsarbeit in seiner Arbeitsgruppe bei
einem tollen Klima, großartiger Betreuung als auch stetiger Unterstützung ermöglichte.
Dr. rer. nat. Harald Polzer, den ich als einen der zuverlässigsten Menschen kennenlernen
durfte und der stets auf alle Fragen eine Antwort hatte.
Dr. med. Siegfried Völkl und Dr. med. Ole Maywald als auch dem Team des Leukämie-
Diagnostik-Labors, die mich beim Rekrutieren der Patientenproben unterstützt haben.
Dr. med. Philipp Greif, Dr. med. Klaus Metzeler und Dr. med. Friederike Schneider für die
Unterstützung bzgl. SB1 bzw. TET2.
Alle Laborkollegen und Mitarbeiter der KKG Leukämie Diana Schmid, Jutta Sturm, Anna
Vetter, Daniel Reichart, Stephanie Fischbach-Scholz, Nadine Sandhöfer, Verena Weth,
Sayantanee Dutta, Hanna Janke, Belay Tizazu und Bianka Ksienzyk für die tolle Zeit, eure
Unterstützung und Hilfe.
Meine Eltern und meine Oma, denen ich Alles zu verdanken habe.


















