
Die Wirkung von Nikotin auf trigeminal exprimierte Ionenkanäle · Der Nervus mandibularis enthält...
153
Die Wirkung von Nikotin auf trigeminal exprimierte Ionenkanäle Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Biologie und Biotechnologie an der Internationalen Graduiertenschule Biowissenschaften der Ruhr-Universität Bochum angefertigt im Lehrstuhl für Zellphysiologie Prof. Dr. Dr. Dr. Hanns Hatt vorgelegt von Benjamin Sven Philip Schreiner aus Essen Bochum Februar 2013
Transcript of Die Wirkung von Nikotin auf trigeminal exprimierte Ionenkanäle · Der Nervus mandibularis enthält...

Die Wirkung von Nikotin
auf trigeminal exprimierte Ionenkanäle
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie und Biotechnologie
an der Internationalen Graduiertenschule Biowissenschaften
der Ruhr-Universität Bochum
angefertigt im
Lehrstuhl für Zellphysiologie
Prof. Dr. Dr. Dr. Hanns Hatt
vorgelegt von
Benjamin Sven Philip Schreiner
aus
Essen
Bochum
Februar 2013

The Effect of Nicotine
on Trigeminal Expressed Ion Channels
Dissertation to obtain the degree
of Doctor Rerum Naturalium
at the Faculty of Biology and Biotechnology,
International Graduate School of Biosciences
Ruhr-University Bochum
prepared at the
Department of Cell Physiology,
Prof. Dr. Dr. Dr. Hanns Hatt
submitted by
Benjamin Sven Philip Schreiner
from
Essen
Bochum,
February 2013

Referent: Prof. Dr. Dr. Dr. Hanns Hatt
Korreferent: PD Dr. Matthias Schmidt

Erklärung
Hiermit erkläre ich, dass ich die Arbeit selbstständig verfasst und bei keiner anderen Fakultät
eingereicht und dass ich keine anderen als die angegebenen Hilfsmittel verwendet habe. Es
handelt sich bei der heute von mir eingereichten Dissertation um sechs in Wort und Bild völlig
übereinstimmende Exemplare.
Weiterhin erkläre ich, dass digitale Abbildungen nur die originalen Daten enthalten und in
keinem Fall inhaltsverändernde Bildbearbeitung vorgenommen wurde.
Bochum, den
- Unterschrift -

Inhaltsverzeichnis
Seite
1. Einleitung 1
1.1 Der Nervus trigeminus 1
1.1.1 Anatomie des Nervus trigeminus 2
1.1.2 Der trigeminale Sinn 3
1.2 Liganden-gesteuerte Ionenkanäle 5
1.2.1 Cys-loop-Kanäle 6
1.2.2 Ionotrope Glutamatrezeptoren 8
1.2.3 P2X-Rezeptoren 9
1.3 TRP-Kanäle 9
1.4 Nikotin 11
1.5 Transkriptomanalyse 13
1.6 Zielsetzung der Arbeit 17
2. Material und Methoden 19
2.1 Material 19
2.1.1 Medien 19
2.1.2 Kits 20
2.1.3 Enzyme 20
2.1.4 Oligonukleotide 20
2.1.5 DNA-Längenstandard 20
2.1.6 Plasmide 21
2.1.7 Pharmakologisch wirksame Substanzen 21
2.1.8 Sonstige Chemikalien 22
2.1.9 Verbrauchsmaterialien 22
2.1.10 Versuchstiere 23
2.1.11 Geräte 23
2.1.12 Software 24

Seite
2.2 Methoden 24
2.2.1 Entnahme von Oozyten aus Xenopus laevis 24
2.2.2 Aufbereitung der Oozyten und Injektion der RNA 25
2.2.3 Die Zwei-Elektroden-Voltage-Clamp Technik 26
2.2.4 Trigeminus-Primärzellkultur 28
2.2.5 Die Calcium-Imaging Methode 28
2.2.6 Isolation einzelner Neurone aus der trigeminalen Zellkultur 29
2.2.7 Molekularbiologische Methoden 30
2.2.7.1 Quantitative Polymerase Kettenreaktion (qPCR) 30
2.2.7.2 Agarosegelelektrophorese 32
2.2.7.3 Transformation 32
2.2.7.4 Anzucht von Bakterien 32
2.2.7.5 Präparative DNA-Aufreinigung 32
2.2.7.6 Ethanol/Natriumacetat-Fällung 32
2.2.7.7 Quantifizierung von DNA und RNA 33
2.2.7.8 In vitro-Transkription 33
2.2.7.9 Ammonium-Acetat-Fällung 33
2.2.7.10 Elution von DNA aus Agarosegelen 33
2.2.7.11 DNA-Sequenzierung 34
2.2.7.12 Restriktion von Plasmiden 34
2.2.7.13 RNA-Isolation 34
2.2.7.14 Transkriptom-Amplifikation und cDNA-Synthese bei Einsatz kleinster 35
Mengen RNA
2.2.7.15 Next Generation Sequencing-Verfahren 35
2.2.7.16 Transkriptomanalyse 35
3. Ergebnisse 40
3.1 Elektrophysiologische Untersuchung der Nikotin-Responsivität 40
rekombinanter Ionenkanäle
3.1.1 Antagonismus von Nikotin an humanen 5-HT3-Rezeptoren 40
3.1.2 Aktivierung humaner nikotinischer Azetylcholinrezeptoren 44
durch (+)-Nikotin

Seite
3.1.3 Untersuchung der Wirkung von Nikotin an humanen 47
α1 Glycinrezeptoren (GlyR α1)
3.1.4 Untersuchung der Wirkung von Nikotin an humanen GABAA-Rezeptoren 48
3.1.5 Untersuchung der Wirkung von Nikotin am humanen P2X-Rezeptor 50
3.1.6 Untersuchung der Wirkung von Nikotin an humanen Glutamatrezeptoren 51
3.1.7 Die Wirkung von (+)-Nikotin auf den TRPA1-Kanal 54
3.1.8 Untersuchung der Wirkung von Nikotin an den Vanilloid-Rezeptoren TRPV1-3 56
3.1.9 Untersuchung der Wirkung von Nikotin am humanen 58
Melastatinrezeptor TRPM8
3.1.10 Zusammenfassung der elektrophysiologischen Untersuchungen 60
3.2 Untersuchungen der Wirkung von Nikotin auf trigeminale Neurone der Maus 61
3.2.1 Stereoselektive Responsivität trigeminaler Ganglienzellen auf Nikotin 61
3.2.2 Suche nach weiteren Rezeptoren für Nikotin in trigeminalen Neuronen 62
3.2.2.1 Responsivität trigeminaler Neurone für (-)-Nikotin unter pharmakologischer 62
Inhibition von nAChR und TRPA1
3.2.2.2 Responsivität trigeminaler Neurone für (+)-Nikotin 65
3.2.3 Einfluss extrazellulären Calciums auf Nikotin-Responsivität trigeminaler 69
Neurone
3.2.4 Inhibitorischer Einfluss von Nikotin auf 5-HT3-vermittelte 70
Calcium-Signale trigeminaler Neurone
3.3 Transkriptomanalyse Nikotin-responsiver trigeminaler Neurone 72
3.3.1 Vergleich der Transkriptome neuronaler Gewebe und einzelner Nikotin- 78
responsiver Neurone
3.3.1.1 Expression von housekeeping-Genen 78
3.3.1.2 Expression der Cys-loop-Rezeptoren 80
3.3.1.3 Expression der TRP-Kanäle 83
3.3.1.4 Expression der KCNK-Kanäle 85
3.3.2 Differentielle Expressionen zwischen einzelnen Nikotin-responsiven Zellen 86
4. Diskussion 91
4.1 Charakterisierung der stereospezifischen Wirkung von Nikotin 91
auf nAChR und TRPA1
4.2 Die Wirkung von Nikotin auf 5-HT3-Rezeptoren 95
4.3 Nikotinblock an NMDA-Kanälen 97

Seite
4.4 Keine Wirkung von Nikotin auf weitere untersuchte Ionenkanäle 99
4.5 Chemosensorisch relevante Nikotinkonzentrationen 100
4.6 Calcium-Imaging der trigeminalen Zellkultur – neue Nikotinrezeptoren 101
4.7 Vergleichende Transkriptomanalyse 104
4.7.1 Die Qualität der Sequenzierung 106
4.7.2 Die Expression der bekannten Nikotinrezeptoren nAChR und TRPA1 107
4.7.3 Die Expression der weiteren Cys-loop-Rezeptoren 109
4.7.4 Die Expression weiterer chemosensorisch relevanter Rezeptoren 110
4.7.5 Validierung der Sequenzierergebnisse 111
4.7.6 Differentielle Expression der einzelnen Nikotin-responsiven Neurone 113
4.7.7 Die Bedeutung der chemosensorischen Wahrnehmung von Nikotin 115
5. Zusammenfassung 117
5. Summary 120
6. Anhang 122
6.1 Literaturverzeichnis 122
6.2 Abbildungsverzeichnis 138
6.3 Tabellenverzeichnis 140
6.4 Abkürzungsverzeichnis 141
6.5 Publikationsliste 143

1
Einleitung
1. Einleitung
1.1 Der Nervus trigeminus
Die Wahrnehmung chemischer Signale aus der Umwelt ist für das Überleben jedes Lebewesens
essentiell. Der chemische Sinn ist demnach auch einer der ursprünglichsten Sinne im Tierreich. Bei
unterschiedlichsten tierischen Organismen vom Einzeller bis zu hochentwickelten Vertebraten
haben sich Mechanismen entwickelt, um Moleküle aus der Umgebung als Reiz für zielgerichtetes
Verhalten zu nutzen.
Der Geschmackssinn vermittelt Sinneseindrücke, die Lebewesen in die Lage versetzen, qualitative
Eigenschaften von Nahrung zu bewerten, zu der sie direkten Kontakt haben, weshalb der
Geschmackssinn auch als Nahsinn bezeichnet wird. Nicht immer muss dafür bei Vertebraten das
ausschließliche Sinnesorgan die Zunge sein, jedoch werden alle Geschmacksorgane in direkter
Berührung mit der Nahrung benutzt. Es handelt sich bei den wahrgenommenen Reizen um nicht
flüchtige, aber wasserlösliche Substanzen. Es werden die fünf Geschmacksrichtungen „süß“,
„sauer“, „salzig“, „bitter“ und „umami“ unterschieden. Geschmackseindrücke enthalten nicht nur
Informationen über den Nährstoffgehalt der Nahrung, Bittereindrücke weisen in der Regel auf
Giftstoffe, wie beispielsweise Pflanzenalkaloide, in der Nahrung hin.
Der Geruchssinn ist ein Fernsinn. Über ihn können leicht flüchtige Substanzen wahrgenommen
werden, die beispielsweise das Vorhandensein einer Nahrungsquelle sowie die Anwesenheit von
Fressfeinden oder Artgenossen anzeigen. Makrosmaten wie die Ratte richten einen Großteil ihres
Verhaltens an olfaktorischen Signalen aus (Doty, 1986; Wallace et al., 2002). Auch beim Menschen
ist der Geruchssinn bedeutend, obwohl er für das eigene Überleben nicht mehr essenziell ist
(Hatt, 1996). Trotzdem kann der Mensch eine Vielzahl verschiedener Geruchsqualitäten
unterscheiden, jedoch besitzt der Mensch nur ~350 funktionelle olfaktorische Rezeptorgene,
wohingegen bei Nagern wie der Maus ~1000 Gene funktionell sind (Niimura and Nei, 2005; Zhang
and Firestein, 2002; Zhang et al., 2004; Zozulya et al., 2001). Die Rezeptoren sind auf den Zilien
der Rezeptorneurone exprimiert, die in den Schleim des olfaktorischen Epithels ragen. Jedes
olfaktorische Rezeptorneuron exprimiert dabei nur einen Rezeptortyp (Chess et al., 1994; Ishii et
al., 2001; Serizawa et al., 2004). Ein Duft kann mehrere Rezeptortypen aktivieren (Hatt, 2004;
Malnic et al., 1999). Die Vielfalt unterschiedlicher Geruchsqualitäten wird durch spezifische
Aktivierungsmuster verschiedener olfaktorischer Rezeptoren für jeden Geruch kodiert
(Mombaerts, 2006; Mori et al., 2006). Der Vermeidung schädlicher chemischer Umwelteinflüsse
verschiedener Art dient insbesondere der allgemeine chemische Sinn, den der Nervus trigeminus

2
Einleitung
vermittelt. Reize, die mit diesem System wahrgenommen werden, können eine schmerzhafte
Empfindung, wie „Brennen“ und „Stechen“ erzeugen, die Vermeidungsverhalten oder aversive
Reflexe, wie „Nießen“ oder kurzfristigen Atemstopp nach sich ziehen. Daneben vermittelt der
Trigeminus Empfindungen wie Adstringenz, und „Prickeln“. Außerdem werden
Temperatureindrücke über den Trigeminus vermittelt. Diese Sensibilität des Trigeminus für Hitze
und Kälte sowie für mechanische Reize, die auch schmerzhafte Empfindungen erzeugen kann,
dient ebenfalls zur Vermeidung schädlicher Umwelteinflüsse.
1.1.1 Anatomie des Nervus trigeminus
Aufgrund seiner Gliederung in drei Hauptäste erhielt der fünfte Hirnnerv der Vertebraten, die
Bezeichnung Nervus trigeminus (lateinisch für Drillingsnerv). Er wird unterteilt in die Nervi
ophtalmicus, maxillaris und mandibularis. Der ausschließlich sensible Nervus ophtalmicus, teilt
sich wiederum in weitere Nerven, welche im Bereich der Augen unter anderem die Orbita,
Hornhaut, Bindehaut, Tränendrüsen und obere Bereiche der Gesichtshaut sowie die Stirn
innervieren (Abb. 1.1). Im Bereich der Nasenhöhle innerviert der Nervus ophtalmicus zudem die
Schleimhaut von Nasenscheidewand und Siebbein sowie die Keilbeinhöhle.
Abbildung 1.1: Schema des Nervus trigeminus des Menschen. Sensible Fasern der Hauptäste Nervus
opthalmicus (1), Nervus maxillaris (2) und Nervus mandibularis (3) haben ihren Ursprung im Ganglion
gasseri (Semilunarganglion). Aus “Gray’s Anatomy: The Anatomical Basis of Medicine and Surgery” (Gray,
2004).

3
Einleitung
Der ebenfalls ausschließlich sensible Nervus maxillaris innerviert im Bereich der Augen einen Teil
der Orbita und der Augenlider. Zudem innerviert er die Nasenschleimhaut und im Mundraum das
Gaumenepithel, Zahnfleisch sowie die Zähne des Oberkiefers. Außerdem werden weite Bereiche
der Gesichtshaut vom Nervus maxillaris innerviert.
Der Nervus mandibularis enthält sowohl motorische Fasern, welche vor allem die Kaumuskulatur
innervieren, als auch sensible Fasern. Diese innervieren die vorderen zwei Drittel der Zunge, die
Wangenschleimhaut, die Zähne und Zahnfleisch des Unterkiefers sowie untere Bereiche der
Gesichtshaut. Die sensiblen Afferenzen des Nervus trigeminus stammen von Neuronen, die im
Ganglion gasseri (auch Semilunarganglion) residieren. Diese Neurone erster Ordnung projizieren
in die zentralen Trigeminuskerne des Hirnstammes (Trepel, 1999). Das Ganglion gasseri ist als Sitz
der Neurone der sensiblen Afferenz den Ganglien der Dorsalwurzeln seriell homolog (Lazarov,
2002).
1.1.2 Der trigeminale Sinn
Die pseudounipolaren Neurone des Ganglion gasseri des trigeminalen chemosensorischen
Systems erstrecken ihre Afferenzen als freie Nervenendigungen in das innervierte Zielgewebe, zu
denen auch die Schleimhäute im Mund-, Nasen- und Rachenraum gehören (Trepel, 1999).
Chemische Reize werden vor allem über schwach myelinisierte Aδ- und unmyelinisierte C-Fasern
mit geringer Geschwindigkeit in den Hirnstamm geleitet (Anton and Peppel, 1991; Hensel et al.,
1974; Heppelmann et al., 2001; Jessel and Basbaum, 2000; Sekizawa and Tsubone, 1994),
wohingegen die stark myelinisierten Aα-Fasern mechanische Stimuli mit höherer Geschwindigkeit
weiterleiten (Jessel and Basbaum, 2000). Aδ-Fasern sind an der Vermittlung von Signalen beteiligt,
die zu einer kurzen, stechenden Schmerzempfindung führen (Mackenzie et al., 1975; Torebjork
and Hallin, 1973). C-Fasern vermitteln Signale, die brennenden, lang anhaltenden Schmerz zur
Folge haben (Mackenzie et al., 1975; Torebjork and Hallin, 1970).
Der Trigeminus kann jedoch auch einen Anteil zur Geschmacks- und Geruchsempfindung
beitragen. Bekannt ist, dass praktisch alle Duftstoffe, die in niedriger Konzentration das
olfaktorische System aktivieren, in höherer Konzentration eine trigeminale Komponente
vermitteln und damit bisweilen begrenztes Riechvermögen im Falle einer Anosmie
aufrechterhalten können (Cain, 1976; Doty et al., 1978; Silver et al., 1985; Hummel & Livermore,
2002). Stoffe, die eine starke trigeminale Empfindung auslösen, haben oft eine hohe
Lipidlöslichkeit, die es erst möglich macht, freie Nervenendigungen zu erreichen (Cometto-Muniz
et al., 1998). Zwischen olfaktorischem und trigeminalen System ist eine Interaktion bekannt, die

4
Einleitung
durch trigeminale Kollateralen zu olfaktorischen Rezeptorzellen in der Schleimhaut aber auch in
den Bulbus olfactorius entsteht (Schaefer et al., 2002). Es konnte daher an olfaktorischen
Rezeptorzellen des Frosches eine Modulation der Aktivität durch elektrische Stimulation des
ophtalmischen Astes des Trigeminus beobachtet werden (Bouvet et al., 1987). Eine Interaktion
konnte jedoch auch auf einer zentralen Ebene beobachtet werden. In einem Kerngebiet des
Thalamus der Ratte, dem Nucleus mediodorsalis, konnten Inokuchi et al. (1993) Aktivität bei
olfaktorischer Stimulation feststellen, die sich durch simultane Stimulation durch trigeminale
Reize verringert. In psychophysischen Studien konnten Cain und Murphy (1980), sowie Kobal und
Hummel (1988) zeigen, dass trigeminale Reize die empfundene Intensität verschiedener
Geruchsstoffe verringern, wenn beide Reize simultan appliziert werden. Auch der trigeminal
vermittelte Schmerz wurde gleichzeitig als geringer empfunden als bei alleiniger Applikation des
trigeminalen Reizes ohne Duftstimulus. Allerdings konnten Jacquot et al. (2004), ebenfalls in
psychophysischen Studien, ein Herabsenken der Wahrnehmungsschwelle und somit eine
Sensibilisierung für die Duftstoffe Phenylethylalkohol und Butanol nach vorangegangener
Stimulation mit dem trigeminalen Reiz Allylisothiocyanat beobachten. An schlafenden
Versuchspersonen, die durch einen trigeminalen Stimulus CO2 geweckt wurden, konnte zudem
beobachtet werden, dass eine simultane olfaktorische Stimulation mit Schwefelwasserstoff das
CO2-induzierte Erwachen fördert (Stuck et al., 2011). Offenbar kann sowohl durch trigeminale
Stimuli die olfaktorische als auch durch olfaktorische die trigeminale Wahrnehmung positiv wie
negativ moduliert werden. Ein gewisser Anteil dieser Modulation scheint auf der Ebene direkter
Zellkontakte zwischen olfaktorischen Rezeptorneuronen und trigeminalen Nervenendigungen
stattzufinden.
Trigeminale Neurone exprimieren eine Vielzahl von Rezeptoren, an die Stoffe verschiedenster
Substanzklassen binden können. Es wurde für unterschiedliche Klassen ionotroper Rezeptoren die
Expression im Nervus trigeminus nachgewiesen. Aus der Familie der transient receptor potential
Kanäle werden beispielsweise TRPV1-4, TRPM8 (Caterina et al., 1997; McKemy et al., 2002; Silver
et al., 2006; Xu et al., 2002; Yamamoto et al., 2009) und TRPA1 (Story et al., 2003; Kobayashi et
al., 2005) exprimiert. Aus der Familie der Cys-loop-Kanäle ist die Expression nikotinischer
Acetylcholinrezeptoren (Alimohammadi and Silver, 2000; Flores et al., 1996; Keiger and Walker,
2000; Liu et al., 1998; Liu et al., 1996; Liu et al., 1993), 5-HT3- (Hu et al., 2004; Tecott et al., 1993)
und GABA-Rezeptoren (Durkin et al., 1999; Hayasaki et al., 2006; Kondo et al., 1994) bekannt. Es
kommen zudem Glutamat- (Chun et al., 2008; Gu and Huang, 1994; Sahara et al., 1997) und
Purinozeptoren (Kuroda et al., 2012; Xiang et al., 1998; Spehr et al., 2004) vor. Aus der Familie der
KCNK Kanäle wurde die Expression von KCNK2, KCNK10 und KCNK18 nachgewiesen

5
Einleitung
(Lafreniere et al., 2010; Yamamoto et al., 2009). Daneben werden auch G-Protein gekoppelte
Rezeptoren, beispielsweise metabotrope Serotoninrezeptoren (Bonaventure et al., 1998a;
Bonaventure et al., 1998b) oder Opioidrezeptoren (Zhu et al., 1998), exprimiert. Alle diese
Rezeptoren könnten potentiell die sensorische Wahrnehmung eines chemischen Reizes direkt
oder modulatorisch beeinflussen, jedoch ist bei Weitem noch nicht die gesamte trigeminale
exprimierte Rezeptorvielfalt erfasst.
1.2 Liganden-gesteuerte Ionenkanäle
Zur Familie der „klassischen“ Liganden-gesteuerten Ionenkanäle rechnet man die Cys-loop-
Rezeptoren, die ionotropen Glutamatrezeptoren und die ionotropen Purinozeptoren. In die
Gruppe der Cys-loop-Rezeptoren fallen neben den nikotinischen Acetylcholinrezeptoren auch die
5-HT3-, die GABA-, die Glycinrezeptoren und die zuletzt bekannt gewordenen ZAC-Kanäle (Zn2+-
activated ion channel). GABA- und Glycinrezeptoren sind Cl--Ionenkanäle, nACh- und 5-HT3-
Rezeptoren sowie ZAC-Kanäle hingegen unspezifische Kationenkanäle. Glutamatrezeptoren und
Purinozeptoren sind ebenfalls Kationenkanäle. Untereinheiten aus den drei erwähnten
Rezeptorfamilien unterscheiden sich strukturell.
Eine Untereinheit der Cys-loop-Rezeptoren besteht aus vier Transmembrandomänen (Abb. 1.2
Mitte), dem extrazellulären N-Terminus - auf dem auch die Ligandenbindestelle liegt - und dem
ebenfalls extrazellulären C-Terminus (Corringer et al., 2000). Die zweite Transmembrandomäne
(TM2) bildet die Kanalpore. Das namensgebende Merkmal, der Cys-loop, liegt am N-Terminus der
α-Untereinheiten und ist an der Bildung der Ligandenbindestelle beteiligt (Brejc et al., 2001).
Eine Untereinheit ionotroper Glutamatrezeptoren besteht aus drei Transmembrandomänen und
einer Membranschlaufe auf zytoplasmatischer Seite, die die Kanalpore bildet (Abb. 1.2 rechts).
Der N-Terminus liegt extra-, der C-Terminus intrazellulär (Bennett und Dingledine, 1995;
Hollmann et al., 1994; Wo und Oswald, 1994; 1995; Wood et al., 1995).
Eine Untereinheit der P2X-Rezeptoren besteht aus zwei Transmembrandomänen, die durch eine
extrazelluläre Schlaufe verbunden sind (Abb. 1.2 links). C- und N-Terminus befinden sich
dementsprechend intrazellulär. P2X-Rezeptoren sind leitfähig für monovalente Kationen und Ca2+-
Ionen, der Selektivitätsfilter wird durch beide Transmembrandomänen, die Kanalpore durch TM2
gebildet (Egan and Khakh, 2004; Migita et al., 2001; Samways and Egan, 2007; Samways et al.,
2008).

6
Einleitung
Abbildung. 1.2: Schema der Untereinheiten Liganden-gesteuerter Ionennkanäle. Verändert nach Khakh
(2001).
1.2.1 Cys-loop-Kanäle
Liganden-gesteuerte Ionenkanäle aus der Superfamilie der Cys-loop-Kanäle bilden homo- oder
heteropentamere funktionelle Ionenkanalproteine. Die in dieser Gruppe zusammengefassten
Rezeptorfamilien haben einen gemeinsamen entwicklungsgeschichtlichen Ursprung (Ortells und
Lunt, 1995).
Von der Subfamilie der nikotinischen Acetylcholinrezeptoren wurden bisher 17 verschiedene
Untereinheiten identifiziert. In fester Stöchiometrie bilden die Untereinheiten (α1)2, β1, δ und ε
die Rezeptorkanäle der neuromuskulären Synapse. Die Untereinheit γ, kommt nur in
Jugendstadien vor und wird später durch die stabilere ε-UE ersetzt. Alle weiteren Untereinheiten
treten vorwiegend in neuronalem Gewebe auf. Die Untereinheiten α7-9 können Homomere
bilden oder Heteromere untereinander sowie mit α10 bilden. Bei den Untereinheiten α2-6 und
β2-4 sind verschiedene Heteromultimere in unterschiedlicher Stöchiometrie möglich
(Albuquerque et al., 2009; Millar und Gotti, 2009;). Im trigeminalen Ganglion der Ratte ist eine
relative Expression der Untereinheiten in der Reihenfolge α7 ≈ α3 > α6 > α4 ≈ α5 > α9 ≥ α2, und
β2 ≈ β3 > β4 festgestellt worden (Liu et al., 1998).
Die α2-Untereinheit, die bei Nagern sehr niedrig exprimiert wird, ist bei Primaten weit höher
exprimiert. Die Expression dieser Untereinheit ist in Primaten der von α4 in Stärke und Expression
sehr ähnlich, wobei bei Nagern die verbreitete Untereinheitenkombination α4β2 ist, bei Primaten
bei ähnlichem Expressionsmuster daneben auch α2β4 vorkommt (Han et al., 2000). Zumindest für
die Untereinheitenkombinationen α4β2 und α3β4 konnte die Expression in freien trigeminalen
Nervenendigungen festgestellt werden (Flores et al., 1996). Boyd et al. (1991) beobachteten eine

7
Einleitung
nAChR-abhängige Modulation primärer Neurone sensorischer Ganglien durch cholinerge Fasern,
welche keine fokussierten synaptischen Strukturen bilden, sondern Acetylcholin diffus
ausschütten.
Für Serotoninrezeptoren des 5-HT3-Typs sind die Untereinheiten A-E bekannt. Für eine
funktionelle Expression müssen die Untereinheiten B-E mit 5-HT3A heteromultimerisieren (Davies
et al., 1999; Niesler et al., 2007). Die Aktivierung von 5-HT3-Rezeptoren wird im
Zentralnervensystem mit der Vermittlung des Brechreizes in Verbindung gebracht. In zwei der
dafür verantwortlichen Strukturen, der Area postrema und dem Nucleus tractus solitarii, über den
auch gustatorische Informationen umgeschaltet werden, ist eine deutliche Expression von 5-HT3-
Rezeptoren bekannt (Kilpatrick et al., 1989; Pratt and Bowery, 1989). So werden spezifische 5-HT3-
Antagonisten als Antiemetika genutzt (Hsu, 2010). Obwohl 5-HT3-Rezeptoren in vielen Strukturen
des Zentralnervensystems vorkommen, konnte für diese Rezeptoren auch eine Funktion auf
primären afferenten Neuronen bei der Schmerzempfindung festgestellt werden. Freie nozizeptive
Nervenendigungen peripherer sensorischer Ganglien können durch Serotonin und 5-HT3-
spezifische Agonisten schmerzhafte Eindrücke erzeugen (Richardson, 1990), die durch spezifische
5-HT3-Antagonisten gehemmt werden können (Richardson et al., 1985).
Im zentralen Nervensystem, insbesondere im Gehirn der Vertebraten, ist GABA der wichtigste
inhibitorische Neurotransmitter (Somogyi et al., 1998). Die Gruppe der ionotropen GABA-
Rezeptoren, die GABAA-Rezeptoren, trägt demnach zur Steuerung vielfältiger
neurophysiologischer Prozesse bei (Hevers und Luddens, 1998) und sind bedeutende
pharmakologische Ziele für Anästhesie des Zentralnervensystems (Reynolds et al., 2003; et al,
2010). Es sind 19 GABAA-Untereinheiten (α1-6, β1-3, γ1-3, δ, ε, π, θ sowie ρ1-3) bekannt (Simon et
al., 2004). Die bis vor einigen Jahren noch als GABAC-Rezeptoren bekannten Untereinheiten ρ1-3
wurden hier, wie von Barnard et al. (1998) vorgeschlagen, zur Gruppe der GABAA-Rezeptoren
gezählt. Im trigeminalen Ganglion der Ratte konnte die Expression von α1-6, β1-3, γ1-3, und delta
festgestellt werden. Hayasaki et al. (2006) und Kondo et al. (1994) spekulieren, dass GABA nicht
synaptisch fokussiert, sondern diffus im trigeminalen Ganglion ausgeschüttet wird, um auf eine
große Anzahl von Neuronen modulatorisch zu wirken. Durch die Expression von Glutamat-
Decarboxylase, einem bei der Produktion von GABA notwendigen Enzym, in trigeminalen
Neuronen schließen die Autoren darauf, dass der Transmitter von den Neuronen selbst
ausgeschüttet wird.
Glycinrezeptoren kommen im Zentralnervensystem der Vertebraten vor allem im Rückenmark,
der Medulla oblongata und der Retina vor (Gisselmann et al., 2002; Lynch, 2009) und dienen an
inhibitorischen Synapsen als Rezeptorkanäle mit Cl--Leitfähigkeit. Bei willkürlicher oder

8
Einleitung
reflexartiger Kontraktion von Skelettmuskeln sorgen sie beispielsweise für die Relaxation
antagonistischer Muskelgruppen (West, 1985). Es gibt die Untereinheiten α1-4 (Grenningloh et
al., 1987; Lynch, 2004) und eine β-Untereinheit (Grenningloh et al., 1990). Die α1-3-
Untereinheiten können funktionelle Homopentamere oder Heteropentamere mit der β-
Untereinheit bilden (Lynch, 2009). Die α4-Untereinheit ist beim Menschen nicht funktionell
(Simon et al., 2004). Heteromere vor allem aus α1β aber auch aus α3β sind die am häufigsten
exprimierten Glycinrezeptoren (Malosio et al., 1991). Es werden einerseits Stöchiometrien von
(α)2(β)3 (Grudzinska et al., 2005), aber auch von (α)3(β)2 (Becker et al., 1988; Burzomato et al.,
2003; Kuhse et al., 1993) vermutet. Im trigeminalen Ganglion konnte die Expression von
Glycinrezeptoren bisher nicht nachgewiesen werden, jedoch in den Nuclei der Hirnnerven und
somit auch in den zentralen Trigeminuskernen des Hirnstammes (Zarbin et al., 1981).
1.2.2 Ionotrope Glutamatrezeptoren
Die Familie der ionotropen Glutamatrezeptoren wird in die Subtypen AMPA- (α-amino-3-hydroxy-
5-methyl-4-isoxazolepropionic acid), Kainat- und die NMDA (N-Methyl-D-Aspartat)-Rezeptoren
unterteilt, die nach Substanzen benannt wurden, die neben Glutamat auch spezifisch aktivierend
auf einen dieser Ionenkanäle wirken. Es sind vier Untereinheiten von AMPA- (GluR1-4), fünf vom
Kainat- (GluR5-7, KA-1,2) und sechs vom NMDA-Typ (NR1, NR2A-D, NR3) bekannt (Dingledine et
al., 1999). Von AMPA-Rezeptoren existieren je Untereinheit zwei unterschiedliche splice-
Varianten „flip“ und „flop“, die sich um die Expression eines Exons unterscheiden. In adulten
Säugern dominiert die „flop“-Variante (Monyer et al., 1991; Sommer et al., 1990). Vier
Untereinheiten bilden ein funktionelles Ionenkanalprotein (Behe et al., 1995; Dingledine et al.,
1999). AMPA- und Kainatrezeptoren können miteinander heteromultimerisieren (Dingledine et
al., 1999), NMDA-Rezeptoren bestehen stets aus NR1 und weiteren NMDA-Untereinheiten (Behe
et al., 1995). Bisher wurden die AMPA-Rezeptoren GluR1, 2 und 3 (Chun et al., 2008), die
Kainatrezeptoren GluR5 und KA-2 und schwächer GluR6 (Sahara et al., 1997), sowie NMDA-
Rezeptoren (Gu und Huang, 1994) im Trigeminus festgestellt.
Seit langer Zeit ist die Bedeutung von Glutamatrezeptoren bei Lernvorgängen im Zusammenhang
mit synaptischer Plastizität insbesondere im Hippocampus bekannt. Die Leitfähigkeit von NMDA-
Rezeptoren für Ca2+-Ionen führen dabei zu einer intrazellulären Regulationskaskade, die eine
Potenzierung stark aktivierter Synapsen zur Folge hat. Ein spannungsabhängiger Block durch
Mg2+-Ionen sorgt jedoch dafür, dass eine Aktivierung von NMDA-Rezeptoren nur an aktiven,
bereits depolarisierten Synapsen stattfindet. Die Depolarisation ist meist auf die Koexpression von

9
Einleitung
AMPA-Kanälen zurückzuführen (Asztely und Gustafsson, 1996; Maren und Baudry, 1995; Nowak
et al., 1984).
1.2.3 P2X-Rezeptoren
Rezeptoren der P2X-Familie sind ionotrope Purinozeptoren, die durch den natürlichen Liganden
ATP aktiviert werden. Es sind sieben verschiedene Untereinheiten (P2X1-7) bekannt. Ein
funktionelles Kanalprotein besteht aus drei Untereinheiten, die sowohl homo- als auch heteromer
auftreten (Collo et al., 1996; Lambrecht, 2000; Nicke et al., 1998; Soto et al., 1997). Eine
bedeutende Funktion von P2X-Rezeptoren ist die Beteiligung an der Vermittlung von
Schmerzeindrücken bei Entzündungsprozessen (Barclay et al., 2002).
In primären sensorischen Nervenenden kommen alle Subtypen von P2X-Rezeptoren vor.
Insbesondere im Nervus trigeminus tritt die P2X2-Untereinheit sowohl homomer in nicht
nozizeptiven Neuronen (Spehr et al., 2004) als auch heteromer mit P2X3 auf nozizeptiven
Nervenendigungen auf (Chen et al., 1995). So erregt die Applikation von ATP auch Capsaicin-
sensitive Fasern auf der Zunge der Ratte (Rong et al., 2000). Zudem löst die Applikation von ATP
auf beschädigte Hautpartien eine schmerzhafte Empfindung aus. Nach mechanischen
Gewebeschäden kann ATP sowohl aus sympathischen Nervenendigungen als auch aus Zellen der
Haut freigesetzt werden (Burnstock, 2007). Da ohnehin ein Großteil trigeminaler Neurone ATP-
sensitiv ist (>80%), konnten trigeminale Ganglienzellen identifiziert werden, die sowohl auf
Capsaicin, ATP und Nikotin reagieren (Xu et al., 2010).
1.3 TRP-Kanäle
Die Bezeichnung der Familie der transient receptor potential Kanäle wurde durch die
Eigenschaften des zuerst entdeckten Kanals geprägt. Dieser vermittelte im Auge von Drosophila
melanogaster eine transiente Antwort auf einen Lichtstimulus (Minke et al., 1975; Montell et al.,
1985). TRP-Kanäle gibt es in vielen Organismen, von Hefen über Würmern, Taufliegen und auch in
Säugern wie Mäusen und Primaten inklusive dem Menschen (Venkatachalam und Montell, 2007).
Die Kanäle werden in sieben Subfamilien eingeteilt, von denen sechs auch bei Mäusen und
Menschen vorkommen. Beim Menschen werden 27 Kanäle unterschieden. Es gibt sieben
Mitglieder der „klassischen“ TRPC- (canonical) Familie, sechs in der TRPV- (vanilloid), acht in der
TRPM- (melastatin), TRPA1 (ankyrin-repeat), drei TRPML- (mucolipin) und drei in der TRPP-
(polycystic kidney disease-related protein) Familie (Clapham, 2003; Abb. 1.3).

10
Einleitung
Eine TRP-Kanaluntereinheit beinhaltet sechs Transmembrandomänen und eine Poren bildende
Schlaufe zwischen den Domänen 5 und 6. Beide Termini liegen auf der zytoplasmatischen Seite
(Ramsey et al., 2006). Vier Untereinheiten bilden ein funktionelles Kanalprotein (Moiseenkova-
Bell et al., 2008), welches als Homo- und Heteromer gebildet sein kann (Clapham, 2003). Die TRP-
Domäne, ein weiteres Merkmal von Kanälen der TRPC und M-Subfamilien, ist eine sehr
konservierte hydrophobe Region C-terminal der 6. Transmembrandomäne, die aus 23-25
Aminosäuren besteht (Montell et al., 2002). Obwohl die Funktion der TRP-Box weitgehend unklar
ist, haben Rohacs et al. (2005) herausgefunden, dass phosphatidylinositol 4,5-bisphosphate (PIP2)
an diese Region binden kann und so modulierend auf den Kanal wirken könnte. Die N-terminalen
Enden insbesondere von TRPA1, aber auch bei den Subfamilien TRPV und TRPC beinhalten
Ankyrinsequenzen, die als Proteininteraktionssequenz vor allem auch der Verankerung am
Zytoskelett dienen (Kohl et al., 2003). TRP-Kanäle sind kationenselektiv, einige wie TRPV1
insbesondere für Ca2+- und Mg2+-Ionen, V5 und V6 ausschließlich für Ca2+-Ionen (Caterina et al.,
1997; Montell, 2001; Venkatachalam und Montell, 2007).
Abbildung 1.3: TRP-Kanaluntereinheiten verschiedener Subfamilien. Ankyrinsequenzen (A) sind grün, coiled coil-Domäne (cc) ist rosa dargestellt. Verändert nach Montell (2005).
Thermo-TRP-Kanäle
Die TRP-Kanäle, die im Trigeminus insbesondere exprimiert sind, dienen unter anderem als Sensor
für Temperatur. Zu den Thermo-TRP-Kanälen gehören TRPV1-4, TRPM8 und TRPA1 (Patapoutian
et al., 2003). TRPV3 und V4 werden aktiviert durch Wärme im Temperaturbereich von 33-40°C
(V3; Peier et al., 2002b; Smith et al., 2002; Xu et al., 2002) und 25-34°C (V4; Guler et al., 2002;
Watanabe et al., 2002b). Durch Hitze ab einer Temperatur von 43°C werden TRPV1 (Caterina et
al., 1997) und ab 52°C TRPV2 (Caterina et al., 1999) aktiviert. Durch niedrige Temperaturen
unterhalb von 25°C wird TRPM8 (McKemy et al., 2002; Peier et al., 2002a) unterhalb von 17°C
wird TRPA1 (Karashima et al., 2009; Sawada et al., 2007; Story et al., 2003) aktiviert.

11
Einleitung
Die genannten Kanäle sind jedoch durch multiple Reizmodi aktivierbar. So ist beispielsweise
TRPV4 auch als Mechanosensor bekannt (Liedtke und Friedman, 2003; Liedtke et al., 2003; Vriens
et al., 2004). Eine gemeinsame Eigenschaft der Thermo-TRP-Kanäle ist jedoch, dass sie auch durch
chemische Stimuli aktivierbar sind (Venkatachalam und Montell, 2007). Oft sind es
Pflanzeninhaltsstoffe, die als Agonisten wirken. Capsaicin, die Substanz, die Schärfe von Chili-
Schoten auslöst, ist ein bekanntes Beispiel für einen TRPV1-Agonisten (Caterina et al., 1997).
TRPV2 wird durch Inhaltsstoffe der Hanfpflanze (Cannabis sativa), wie Cannabidiol und
Tetrahydrocannabinol, aktiviert (Qin et al., 2008). Für TRPV3 wurden die Terpenoide Thymol und
Carvacrol (Vogt-Eisele et al., 2007), insbesondere aus dem Echten Thymian (Thymus vulgaris) und
für TRPV4 Phorbolester, aus dem Milchsaft einiger Wolfsmilchgewächse (Watanabe et al., 2002a)
als Agonisten bekannt. Menthol aus verschiedenen Minzpflanzen aktiviert TRPM8 (Peier et al.,
2002a) und weniger stark auch TRPV3 (Macpherson et al., 2006). Allylisothiocyanat, die Schärfe
gebende Substanz aus dem Schwarzen Senf (Brassica nigra), aktiviert TRPA1 (Jordt et al., 2004).
Die Aktivierung der genannten Kanäle durch einen chemischen Reiz vermittelt ein Signal, das wie
ein thermischer Reiz in einem Temperatureindruck resultiert. Die Wahrnehmung von Hitze und
starker Kälte ist mit einem schmerzhaften Eindruck verbunden. Die Aktivierung der
entsprechenden Thermosensoren wie TRPV1 für Hitze und TRPA1 für Kälte mittels eines
chemischen Agonisten geht dementsprechend mit Empfindungen wie „brennend“ bei Aufnahme
von Chili und „stechend“ bei Aufnahme von Senf einher (Caterina et al., 1997; Jordt et al., 2004).
Es sind zahlreiche weitere Substanzen, sowohl Natur- als auch synthetische Stoffe bekannt, die
durch eine Wirkung auf Thermo-TRP-Kanäle eine trigeminale Empfindung hervorrufen (Viana,
2011).
1.4 Nikotin
Nikotin gehört zu den Suchtmitteln, deren Genuss eine sehr große weltweite Verbreitung
erfahren hat. Es gibt weltweit ein bis zwei Milliarden Tabak-Raucher. Durch den Genuss von
Nikotin in Form von Tabakkonsum sterben durch damit verbundene Krankheitsbilder etwa 5
Millionen Menschen pro Jahr (Hatsukami et al., 2008). Das Pflanzenalkaloid Nikotin ist ein
sekundärer Pflanzeninhaltsstoff insbesondere der Gattung der Tabakpflanzen (Nicotiana spec.)
und dient diesen als Fraßschutz gegen Herbivoren (Ames, 1983; Davis et al., 1991; Kuhn, 1964). Da
das Molekül ein stereogenes Zentrum besitzt, existiert Nikotin in zwei Stereoisomeren (Barlow
und Hamilton, 1965; Pictet und Rotschy, 1900; Abb. 1.4).

12
Einleitung
Abbildung 1.4: Strukturformel beider Nikotin-Enantiomere. Links: (-)-Nikotin; rechts: (+)-Nikotin. Das
markierte C-Atom (*) ist das stereogene Zentrum.
Das (-)-Enantiomer wird von Pflanzen bei Weitem häufiger synthetisiert als die (+)-Isoform
(Armstrong et al., 1998; Perfetti and Coleman, 1998; Perfetti et al., 1998). Der Anteil des (+)-
Enantiomers an der Nikotingesamtmenge liegt im Rohtabak nur bei etwa 1%, jedoch findet
während der Pyrolyse im Verbrennnungsprozess eine Razemisierung statt, bei der sich der Gehalt
von (+)-Nikotin auf 2-3% erhöht (Baker und Bishop, 2004; Clayton et al., 2010). Bei Tabakkonsum
wird Nikotin über die Schleimhaut von Nase, Mund und Lunge aufgenommen und gelangt in die
Blutbahn.
Es sind bisher verschiedene Rezeptorproteine für Nikotin bekannt. Nikotinische Acetylcholin-
rezeptoren und TRPA1 werden durch Nikotin aktiviert, TRPV1 positiv moduliert und 5-HT3- und
spannungsgesteuerte Natriumkanäle sowie IH-Kanäle geblockt.
Die Wirkung von Nikotin im Zentralnervensystem ist seit geraumer Zeit erforscht (Aceto und
Martin, 1982; Domino, 1998; Rand, 1989; Zevin et al., 1998). Die maßgeblichen Rezeptoren, die
diese Wirkung vermitteln, sind nikotinische Acetylcholinrezeptoren (Dani und Bertrand, 2007;
Laviolette und van der Kooy, 2004). Griguoli et al. (2010) fanden ferner an Zellen des
Hippocampus der Maus heraus, dass Nikotin außerdem hyperpolarisationsaktivierte
IH-Kanäle inhibiert.
Die Wirkung von Nikotin auf chemosensorische Systeme hingegen ist bisher noch nicht
erschöpfend untersucht worden, wobei sich hierbei Ergebnisse mehren, die neben der Beteiligung
von nAChR auf eine größere Vielfalt verschiedener Rezeptoren deuten. In Abhängigkeit von der
Konzentration kann Nikotin verschiedene chemosensorische Eindrücke vermitteln. Beide
Enantiomere können sowohl das olfaktorische als auch das trigeminale System aktivieren
(Hummel et al., 1992a; Hummel et al., 1992b; Thuerauf et al., 2006). In niedrigen Konzentrationen
ruft Nikotin einen Geruchseindruck, in höheren Konzentrationen zudem sowohl brennende als
auch stechende Eindrücke hervor (Thuerauf et al., 1999; Walker et al., 1996). Aus einer Arbeit von
Thuerauf et al. (2006) geht hervor, dass trigeminale Chemorezeption in der Nase mit dem nAChR
Antagonisten Mecamylamin inhibiert werden kann. Der von Nikotin hervorgerufene

13
Einleitung
Geruchseindruck wird dadurch jedoch nicht verhindert und demnach nicht mit Beteiligung von
nAChR vermittelt. Die Autoren vermuten deshalb, dass olfaktorische Rezeptoren für die
Wahrnehmung niedriger Konzentrationen von Nikotin verantwortlich sind. Zudem konnte in
psychophysischen Studien festgestellt werden, dass das trigeminale System stereoselektiv auf
Nikotin reagiert (Thuerauf et al., 1999; Thuerauf et al., 2000; Walker et al., 1996). Für das (-)-
Enantiomer wurde dabei eine niedrigere Reizschwelle und stärkere subjektive Empfindungsstärke
festgestellt als für das (+)-Enantiomer. An Elektroolfaktogrammen am olfaktorischen Epithel des
Krallenfrosches wurde hingegen keine Stereoselektivität festgestellt (Thuerauf et al., 1995). Es
konnte mit dem TRPA1-Kanal von Mensch und Maus bereits ein weiterer trigeminal exprimierter
Rezeptor für (-)-Nikotin identifiziert werden (Talavera et al., 2009). Neben diesem und den nAChR
als direkt aktivierten Kanalklassen wurden mittlerweile antagonistische oder modulatorische
Wirkungen von (-)-Nikotin an einigen weiteren Ionenkanälen bekannt. Liu et al. (2004) konnten
eine positive Modulation der Capsaicin-induzierten Antworten am TRPV1-Kanal der Ratte
feststellen. Außerdem beobachteten sie eine Inhibition menschlicher Spannungsgesteuerter
Natriumkanäle, die sie als Ursache für eine analgetische Wirkung von Nikotin ansahen. Diese
Wirkung wurde beispielsweise in Schmerzvermeidungstest an Ratten festgestellt (Carstens et al.,
2001). An homomeren 5-HT3-Rezeptoren der Maus konnten Gurley und Lanthorn (1998) ferner
einen kompetitiven Antagonismus für beide Nikotin-Enantiomere feststellen. In Radioliganden-
Bindungsstudien am Gehirn der Ratte konnte eine Bindung von (-)-Nikotin an native 5-HT3-
Rezeptoren nachgewiesen werden (Drisdel et al., 2008).
Olfaktorische Rezeptoren für Nikotin wurden bisher nicht identifiziert. Die bisher bekannten
Rezeptoren, die durch Nikotin aktiviert, inhibiert oder moduliert werden können, sind nicht
ausreichend, um die Wahrnehmung von Nikotin auf zellulärer Ebene vollständig erklären zu
können.
1.5 Transkriptomanalyse
Das Transkriptom ist die Gesamtheit exprimierter RNA einer betrachteten biologischen Struktur,
wie beispielsweise eines Organs, eines Gewebetyps oder einer einzelnen Zelle. Es beinhaltet eine
Vielzahl verschiedener RNA-Klassen, von denen die mRNA mit etwa 1% den Anteil darstellt, von
dem Information bei der Translation in Proteine umgesetzt wird (He und Hannon, 2004). Da jedes
Organ, Gewebe oder jede einzelne Zelle charakteristische Transkriptome hat, die Rückschlüsse auf
die jeweilige Proteinausstattung zulassen, wurden schon seit langer Zeit Anstrengungen
unternommen, Transkriptome möglichst komplett zu analysieren, um qualitative, wie quantitative

14
Einleitung
Aussagen über die Expression der Gene treffen zu können. Dabei wurden in den letzten Jahren
technische Fortschritte insbesondere bei der DNA-Sequenziertechnik erreicht. Zu einer
Transkriptomanalyse auf diesem Wege muss die RNA des Transkriptoms, zunächst in cDNA
umgeschrieben werden. Trotzdem wird diese Sequenzierung üblicherweise als „RNA-Seq“
bezeichnet (Wang et al., 2009). Mit Hilfe der in den letzten Jahren entstandenen Techniken kann
eine wesentlich größere Anzahl von DNA-Fragmenten zu geringerem Preis sequenziert werden,
als es mit der Technik nach Sanger möglich ist. Eines dieser Verfahren, die auch unter dem Begriff
next generation sequencing zusammengefasst werden, ist das Illumina-Solexa
Sequenzierverfahren, das seinen Vorteil in einer sehr hohen Sequenzierleistung hat (bis zu 600
Gigabasen). Damit kann eine sehr große Anzahl von über 300 Millionen vergleichsweise kurzen
Fragmenten (35-150 bp) sequenziert werden (Liu et al., 2012). Das Verfahren ist in dieser Arbeit
zur Transkriptomanalyse genutzt worden.
Zunächst besteht bei dieser Technik der erste wichtige Schritt in der Generierung der „cDNA-
Library“. Dazu wird zunächst – wie erwähnt – eine cDNA-Synthese durchgeführt und entstandene
cDNA dann in Fragmente mit einer Länge von 200-300 Basenpaaren zerlegt. Einzelsträngige
Endabschnitte werden im nächsten Schritt aufgefüllt und an die Enden anschließend eine
Adaptersequenz ligiert (Abb. 1.5 A-D). An die Oberfläche der Reaktionskammer (flow cell) sind
nun Ankerfragmente gebunden, die zu den Adaptern komplementäre Sequenzen haben, sodass
im zweiten Schritt die einzelsträngigen DNA-Fragmente bei der Beschickung der Kammer an den
Ankerfragmenten haften. Um jedes Fragment wird anschließend mit der PCR-basierten
sogenannten „Bridge-Amplifizierung“ ein Cluster identischer DNA-Moleküle auf der
Reaktionsfläche der Kammer erzeugt (Abb. 1.5 E-G). Im dritten Schritt folgt die eigentliche
Sequenzierung, wobei eine Polymerase an den anhaftenden Fragmenten einen komplementären
Strang fortschreibt. Dazu sind die vier Nukleotide mit verschiedenen Fluoreszenzmarkierungen
versehen und werden über die Kammer gespült. Das jeweils passende Nukleotid wird von der
Polymerase eingebaut. Durch einen Syntheseblock stoppt die Synthese nach dem Einbau jedes
Nukleotids und die Fluoreszenz kann gemessen werden. Nach dem Entfernen des Syntheseblocks
kann das nächste Nukleotid fortgeschrieben werden. Die Reihenfolge detektierter
Fluoreszenzfarben gibt die komplementäre Sequenz der Fragmente eines Clusters wieder (Abb.
1.5 H-K).
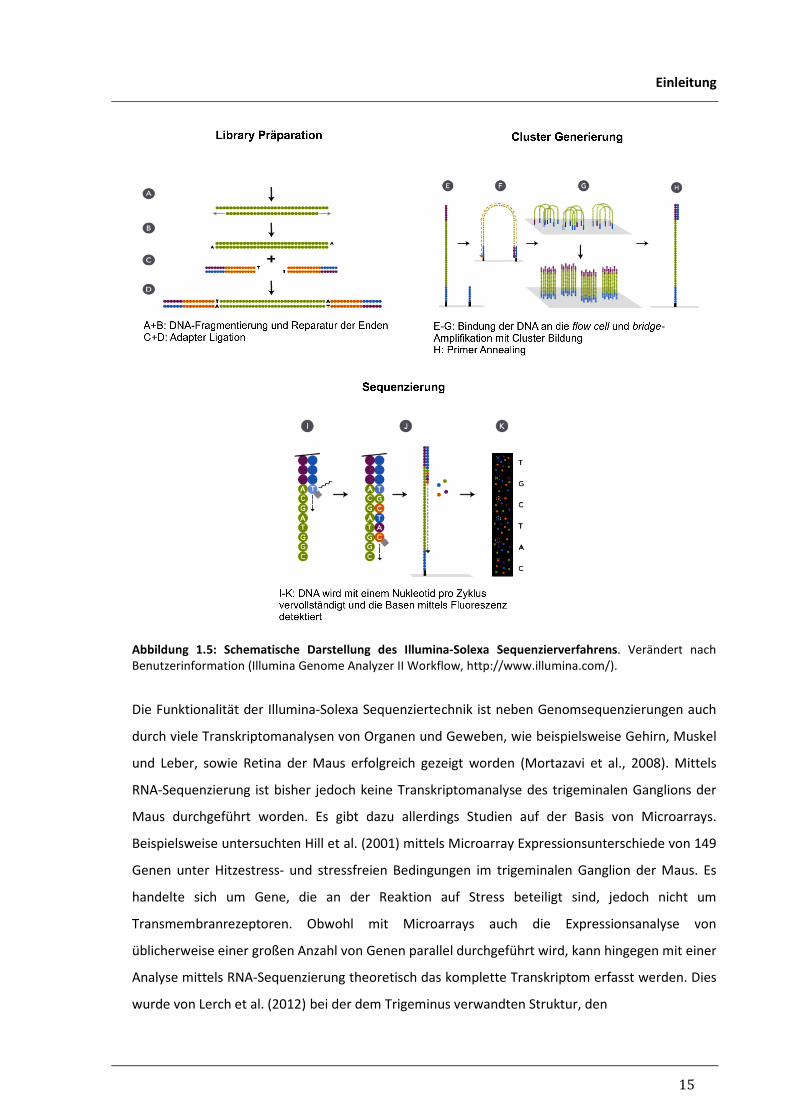
15
Einleitung
Abbildung 1.5: Schematische Darstellung des Illumina-Solexa Sequenzierverfahrens. Verändert nach
Benutzerinformation (Illumina Genome Analyzer II Workflow, http://www.illumina.com/).
Die Funktionalität der Illumina-Solexa Sequenziertechnik ist neben Genomsequenzierungen auch
durch viele Transkriptomanalysen von Organen und Geweben, wie beispielsweise Gehirn, Muskel
und Leber, sowie Retina der Maus erfolgreich gezeigt worden (Mortazavi et al., 2008). Mittels
RNA-Sequenzierung ist bisher jedoch keine Transkriptomanalyse des trigeminalen Ganglions der
Maus durchgeführt worden. Es gibt dazu allerdings Studien auf der Basis von Microarrays.
Beispielsweise untersuchten Hill et al. (2001) mittels Microarray Expressionsunterschiede von 149
Genen unter Hitzestress- und stressfreien Bedingungen im trigeminalen Ganglion der Maus. Es
handelte sich um Gene, die an der Reaktion auf Stress beteiligt sind, jedoch nicht um
Transmembranrezeptoren. Obwohl mit Microarrays auch die Expressionsanalyse von
üblicherweise einer großen Anzahl von Genen parallel durchgeführt wird, kann hingegen mit einer
Analyse mittels RNA-Sequenzierung theoretisch das komplette Transkriptom erfasst werden. Dies
wurde von Lerch et al. (2012) bei der dem Trigeminus verwandten Struktur, den

16
Einleitung
Dorsalwurzelganglien (DRG), durchgeführt. Die oben erwähnten Einzelnachweise für die
Expression von Liganden-gesteuerten Ionen- (1.1.2, 1.2.1-1.2.3) und TRP-Kanälen (1.1.2, 1.3) im
Trigeminus eignen sich zwar nicht für einen quantitativen Vergleich mit diesen Ergebnissen,
jedoch ist in den Dorsalwurzelganglien eine ähnlich große Vielfalt an Ionenkanälen exprimiert. Die
Vielfalt exprimierter nikotinischer Acetylcholinrezeptoren ist in den DRG nach Lerch et al. (2012)
zwar geringer als im Trigeminus, jedoch scheint die Reihenfolge der Expressionsstärke ähnlich zu
sein (α3 > α7 > α4; β2 > β4). Auch in den DRG ist 5-HT3A exprimiert, 5-HT3B jedoch gemäß diesen
Ergebnissen nicht. Es werden ebenfalls weniger verschiedene GABAA-Rezeptoruntereinheiten
exprimiert (α1 >> α2 > α3; β3 >> β1 ≈ β3; γ2 > γ1 ≈ δ). Zudem sind Glycinrezeptoren (β >> α2),
TRP-Kanäle (V1 > M8 > M2 ≈ M4 > A1 > M7 > M3) und KCNK-Kanäle (3 > 12 > 2 ≈ 13 > 1 > 10 > 9 ≈
4) exprimiert. Auch Purinozeptoren und ionotrope Glutamatrezeptoren werden exprimiert. Da
das trigeminale Ganglion und die Dorsalwurzelganglien seriell homologe Strukturen sind, die auch
für ihren jeweiligen Innervationsbereich eine vergleichbare Funktion haben, ist zumindest eine
ähnliche Rezeptorausstattung zu erwarten.
Obwohl RNA-Sequenzierungen zur Transkriptomanalyse von Organen oder Gewebeverbänden
mittlerweile verbreitet sind, stellt dies auf der Ebene einzelner Zellen immer noch eine technische
Herausforderung dar. Analysen des Transkriptoms einzelner Zellen können immer dann von
Bedeutung sein, wenn man innerhalb eines Gewebes verschiedene Zellpopulationen mit sehr
heterogener Proteinausstattung erwartet. Handelt es sich um eine seltene Zellpopulation, so kann
auf herkömmlichem Weg nicht genug RNA isoliert werden, um eine ausreichende Probe für eine
RNA-Sequenzierung zu gewinnen. Einzelne Säugerzellen enthalten durchschnittlich etwa 10 pg
RNA (~ 0,1 pg mRNA; Gonzalez-Roca et al., 2010; Sambrook und Russell, 2001), für eine RNA-
Sequenzierung sind jedoch Mengen im Bereich von mehreren hundert ng bis wenigen µg
notwendig. Um diesen Anforderungen zu entsprechen, wird zur Transkriptomanalyse einzelner
Zellen ohne eine verlustreiche RNA-Aufreinigung, oftmals die cDNA-Synthese direkt im Zelllysat
vollzogen und eine anschließende Amplifikation durchgeführt. Diese kann im Wesentlichen auf
zwei verschiedenen Techniken basieren. Zum einen kann eine lineare Amplifikation mittels in-
vitro Transkription, zum anderen eine PCR-basierte exponentielle Amplifikation durchgeführt
werden (Tang et al., 2011). Da letztere größere Mengen cDNA ergeben, wurde in dieser Arbeit mit
dem „TransPlex Complete Whole Transcriptome Amplification System“ genau ein solches
Verfahren gewählt. Mit dieser Technik wurde bereits gezeigt, dass bei einem Einsatz von 100 pg
teilweise degradierter Gesamt-RNA bei der TransPlex Amplifikation und einer anschließenden
Analyse mittels Microarray reproduzierbar vergleichbare Expressionsmuster festgestellt werden
können wie durch quantitative PCR und ohne Amplifikation (Tomlins et al., 2006).

17
Einleitung
Gonzalez-Roca et al. (2010) konnten dies ebenfalls mit Microarrays bei einem Einsatz von zehn
Zellen bei der Amplifikation grundsätzlich bestätigen, was zeigt, dass die TransPlex Amplifikation
prinzipiell auch bei geringen Mengen eingesetzter RNA ein Amplikon erzeugen kann, das relative
Expressionsverhältnisse wiederspiegelt. In ihrer Studie haben sie jedoch auch gezeigt, dass in
Einzelfällen Expressionsunterschiede, die mittels qPCR festgestellt wurden, nach der Amplifikation
nicht erkannt werden konnten. In einer weiteren Studie konnte jedoch gezeigt werden, dass die
Kombination von PCR-basierter Amplifikation und der Transkriptomanalyse mittels next
generation sequencing das Erstellen von Expressionsprofilen aber auch auf Einzelzellebene
ermöglicht. Embryonale Stammzellen der Maus konnten mit Hilfe der Illumina Sequenziertechnik
durch die Expression von RNA für bekannte Markerproteine verschiedenen Zellpopulationen
zugeordnet werden (Islam et al., 2011).
1.6 Zielsetzung der Arbeit
Das Ziel dieser Arbeit ist die Untersuchung der direkten oder modulatorischen Wirkung von
Nikotin auf Membranrezeptoren, die in primär afferenten chemosensorischen Neuronen des
Nervus trigeminus exprimiert werden. Die alte Erkenntnis, dass die chemosensorische
Wahrnehmung von Nikotin durch nikotinische Acetylcholinrezeptoren vermittelt wird, wurde vor
einiger Zeit ergänzt durch die Feststellung, dass der TRPA1-Kanal durch Nikotin aktiviert werden
und somit an der Vermittlung des sensorischen Eindrucks von Nikotin beteiligt sein kann. Der
Mensch ist in der Lage beim olfaktorischen wie auch trigeminal vermittelten sensorischen
Eindruck zwischen (+)- und (-)-Nikotin zu unterscheiden. Die unterschiedlichen Qualitäten dieses
Eindruckes führen zu der Vermutung, dass weitere, noch unbekannte Transmembranrezeptoren
an der chemosensorischen Wahrnehmung von Nikotin beteiligt sind. Solche neuen „Targets“ für
Nikotin sollen im Rahmen dieser Arbeit gefunden werden. Die trigeminalen afferenten Neurone
weisen eine Vielfalt von Rezeptoren auf. Es sind Liganden-gesteuerte Ionenkanäle, TRP-Kanäle
sowie G-Protein-gekoppelte Rezeptoren exprimiert. Unter diesen sind mit 5-HT3A und TRPV1
bereits „Targets“ bekannt, von denen eine Modulation durch (-)-Nikotin gezeigt wurde. Die
Entdeckung unbekannter modulatorischer Einflüsse von Nikotin auf Transmembranrezeptoren ist
weiteres Ziel dieser Arbeit. Eine systematische Studie, in der trigeminal exprimierte
Membranrezeptoren aus den Familien der klassischen Liganden-gesteuerten Ionenkanäle und der
TRP-Kanäle in ihrem Antwortverhalten auf Nikotin untersucht wurden, ist bisher nicht
durchgeführt worden. Um ein besseres Verständnis der physiologischen Basis der sensorischen
Wahrnehmung von Nikotin zu bekommen, soll die Wirkung des (+)- und (-)-Enantiomers auf

18
Einleitung
heterolog exprimierte humane Rezeptoren untersucht werden. Als Expressionssystem sollen
Oozyten von Xenopus laevis genutzt werden, um typische Vertreter humaner Cys-loop-
Rezeptoren, ionotroper Glutamatrezeptoren, ionotroper Purinozeptoren sowie TRP-Kanäle
verschiedener Unterfamilien zu exprimieren. Mittels der Voltage-Clamp Methode soll eine
mögliche pharmakologische Wirkung von (+)- und (-)-Nikotin auf diese Rezeptoren
elektrophysiologisch untersucht werden.
Um zunächst die Ergebnisse des ersten Projektteils zu bestätigen, soll in einem zweiten Ansatz die
Wirkung beider Nikotinenantiomere auf kultivierte trigeminale Neurone der Maus unter
Zuhilfenahme der Calcium-Imaging Technik analysiert werden. Im Weiteren sollen die bekannten
Nikotinrezeptoren nAChR und TRPA1 auf pharmakologischem Weg blockiert und persistierende
Antworten weiter untersucht werden.
Im dritten Teil dieser Arbeit sollen in einer Transkriptomanalyse die Expressionsprofile Nikotin-
responsiver Neurone miteinander verglichen werden, um Hinweise auf mögliche weitere
Rezeptoren für Nikotin zu finden. Es sollen dazu Neurone mit der erwähnten persistierenden
Nikotinantwort unter Blockerbedingungen für nAChR und TRPA1 im Calcium-Imaging identifiziert
und deren Transkriptom isoliert werden. Mittels Next-Generation Sequencing ist eine
Einzelzelltranskriptomanalyse schon mit verhältnismäßig wenigen Zellen möglich.
Membranrezeptoren, die insbesondere in Nikotin-responsiven Neuronen exprimiert sind, können
eine physiologische Bedeutung für die chemosensorische Wahrnehmung von Nikotin haben.

19
Material und Methoden
2. Material und Methoden
2.1 Material
2.1.1 Medien
Extrazellulär-Medium 140 mM NaCl, 5 mM KCl, 2 mM CaCl2,
1 mM MgCl2, 10 mM HEPES, pH 7.4
(NaOH/HCl), 300-310 mOsmol (Glucose)
Hochkalium-Lösung 100 mM NaCl, 45 mM KCl, 2 mM CaCl2,
1 mM MgCl2, 10 mM HEPES, pH 7.4
(NaOH/HCl), 300-310 mOsmol (Glucose)
Ca2+-freies Extrazellulär-Medium 140 mM NaCl, 5 mM KCl, 1 mM MgCl2,
5 mM EGTA, 10 mM HEPES, pH 7.4
(NaOH/HCl), 300-310 mOsmol (Glucose)
ND96-Lösung 96 mM NaCl, 2 mM KCl, 1,8 mM CaCl2,
1 mM MgCl2, 5 mM HEPES, 100U/ml
Penicillin, 100 U/ml Streptomycin,
pH 7,2 (NaOH/HCl)
Ca2+-freie Barth-Lösung 88 mM NaCl, 1 mM KCl, 2,4 mM NaHCO3,
7,5 mM Tris-HCl, 0,82 mM MgSO4, pH 7,4
Standard Barth-Lösung 88 mM NaCl, 1 mM KCl, 2,4 mM NaHCO3,
7,5 mM Tris-HCl, 0,82 mM MgSO4,
0,41 mM CaCl2, 0,33 mM Ca(NO3)2 pH 7,4
Standard Frosch-Ringer (NFR) 115 mM NaCl, 2,5 mM KCl, 1,8 mM CaCl2,
10 mM HEPES, pH 7,2
Mg2+ Frosch-Ringer 115 mM NaCl, 2,5 mM KCl, 1,8 mM MgCl2,
10 mM HEPES, pH 7,2
Medien der Zellkultur
Leibovitz Medium (L15) Invitrogen “Dulbeccos´s Modified Eagle Medium”
(DMEM/F-12)
Invitrogen
“Minimum Essential Medium” (MEM) Invitrogen
PBS+/+ Invitrogen
“Fetal Bovine Serum” (FBS) Invitrogen

20
Material und Methoden
2.1.2 Kits
cDNA-Synthese iScript™ cDNA Synthesis Kits, Bio-Rad
Laboratories
In vitro Transkription von cRNA AmpliCap-T7 Message Maker Kit, Epicentre
Präparative DNA-Aufreinigung Pure Yield Maxiprep System, Promega PCR PCR- SYBR-Green-Mix Peglab
PCR-Aufreinigung und Gel-Extraktion QIAquick PCR Purification Kit, Qiagen
Wizard® SV Gel and PCR Clean-Up System,
Promega
Transkriptom-Amplifikation Message Booster cDNA-synthesis from cell
lysates Kit, Epicentre
WT-Ovation One-Direct RNA-Amplification
System, NuGen
Transplex Complete WTA2 Kit, Sigma-Aldrich
RNA-Isolation RNeasy Micro-, Mini-Kit, Qiagen RNA-Aufreinigung RNA Clean & Concentrator, Zymo Research
2.1.3 Enzyme
Taq-DNA-Polymerase Invitrogen
Collagenase Typ 1 Sigma-Aldrich
Worthington Biochemical Corporation
DNase I Qiagen
Reverse Transcriptase SuperScript® III, Invitrogen
Restriktionsendonukleasen MBI Fermentas
New England Biolabs
2.1.4 Oligonukleotide
GAPDHmmfw TGTGTCCGTCGTGGATCTGA
GAPDHmmRV CCTGCTTCACCACCTTCTTGA RPL29mmfw TTGCCAAGAAGCACAACAAG
RPL29mmrv TGTCTTCACACTGGCAGGAG
BS_mmB3T-1555-fw GTCCACTTGGCTCTGTCCTC
BS_mmB3T-1646-rv CCCCCGAATATAAACACAACC
BS_mmCNPAse-2698-fw ACCCTGAGCTGGCAAGAGTA
BS_mmCNPAse-2802-rv CCAGATTCTCGGGTGACAAC
BS_mmACTB-fw AAGGTGACAGCATTGCTTCTG
BS_mmACTB-rv CCTGGGCCATTCAGAAATTA
2.1.5 DNA-Längenstandard
Für die Bestimmung der Länge von DNA-Fragmenten nach Agarosegelelektrophorese wurde der
Längenstandard „GeneRuler™ 1 kb DNA Ladder“ der Firma „Fermentas - Thermo Fisher Scientific“
verwendet.

21
Material und Methoden
2.1.6 Plasmide
Vektor Insert Restriktionsenzym
zur Linearisierung
pSGEM 5-HT3A SacI
GlyR α1 PacI
GABAA α1-2, β1-3, γ2, ρ1 PacI
GluR1(Q)flop, GluK2(Q), NR1-3a,
NR2B
NheI
TRPA1 PacI
TRPV1-3 PacI
TRPM8 PacI
pcDNA3 5-HT3B SmaI
P2X2 XbaI
pCRBluntIITopo nAChR α2 XbaI
nAChR β4 NotI
pOTB7 nAChR α7 XhoI
2.1.7 Pharmakologisch wirksame Substanzen
Substanz Hersteller
2-APB (2-Aminoethoxydiphenylborat) Sigma-Aldrich
Acetylcholin Sigma-Aldrich
AITC (Allylisothiocyanat) Fluka
ATP (Adenosintriphosphat) Roche
Borneol Sigma-Aldrich
Capsaicin Calbiochem
mCPBG (m-Chlorophenylbiguanidhydrochlorid)
Tocris
Edelfosin Tocris
GABA (γ-Aminobuttersäure) Sigma-Aldrich
Gentamycin Sigma-Aldrich
Glutamat Sigma-Aldrich
Glycin Roth
HC030031 Tocris
Hexamethoniumchlorid Sigma-Aldrich
Mecamylamin Sigma-Aldrich
Menthol Henkel (-)-Nikotin Sigma-Aldrich
(+)-Nikotin Dr. Berthold Renner,
Universität Erlangen
Rutheniumrot Sigma-Aldrich
Serotonin Sigma-Aldrich
U-73122 Sigma-Aldrich
Zinkchlorid Riedel-de Haёn

22
Material und Methoden
2.1.8 Sonstige Chemikalien
Substanz Hersteller
Ethyl-3-Aminobenzoat, Sigma-Aldrich
Poly-L-Lysin Sigma-Aldrich
Penicillin / Streptomycin, PAA Laboratories GmbH
Fura-2AM Invitrogen
Ethidiumbromid Serva/AppliChem
Agarose „ultra pure“ Gibco BRL Mineralöl 1:1 hoch-/niederviskos Sigma-Aldrich
SYBR Green Sigma-Aldrich
Puffer Rezeptur
TBE-Puffer 89 mM TRIS,2 mM EDTA-Na2-Salz, 89 mM
Borsäure
Laufpuffer 20% Ficoll (w/v), 100 mM EDTA,
0,025% Bromphenolblau (w/v),
0,025% Xylencyanol (w/v)
2.1.9 Verbrauchsmaterialien
Glaskapillaren, filamentiert GB 150 EFT-10 1.17 x 1.50 x 100 mm
with filament, Science Products GmbH
Glaskapillaren, ohne Filament Glass capillaries Nanoliter 2000; 3,5
inches, 1.14 ID, 0.5 mm OD, World
Precesion Instruments
Glasplättchen für Zellkultur Borosilikatdeckgläser 30 mm, Thermo
Scientific Homogenisator-Röhrchen Precellys-Keramik Kit, 1,4 mm PeqLab
Nahtmaterial MonoPlus®, resorbierbar, B. Braun
Aesculap
Pasteurpipetten Brand
PCR-Platte (96-wells) Thermo Scientific
PCR-Gefäße Low profil Thermo Strip, Thermo
Scientific
Reaktionsgefäße 15 / 50 ml, Sarstedt,
1,5 / 2 ml, Bio-Rad,
DNA LoBind® tube, Eppendorf Sterilfilter Schleicher & Schüll
Zellsieb 70 µm Cell Strainer, BD Falcon
Zellkulturplatten, 24-well plates Tissue culture testplates, gamma
sterilized, Orange Scientific
Zellkulturschälchen, 35 mm Sarstedt

23
Material und Methoden
2.1.10 Versuchstiere
Weibliche Frösche der Art Xenopus laevis wurden von der Firma „Xenopus Express France“
bezogen.
Zudem wurden frisch präparierte Xenopus laevis Oozyten von der Firma „EcoCyte Bioscience“
verwendet.
Alle verwendeten Mäuse entstammen der Zuchtlinie „Crl:CD1(ICR)“ und wurden von „Charles
River Laboratories“ bezogen.
2.1.11 Geräte
Analog-digitale Schnittstelle HEKA LIH 1600
Analysenwaage Sartorius
Applikationssystem 7-in-1 System aus lehrstuhleigener
Entwicklung
Automatische Pipette Eppendorf Research Pro
Brutschränke Heraeus Kelvitron K
Thermo Scientific Hera Cell 150
CCD-Kamera Zeiss Photometrics, Axiocam MRM Dyn Mag-Spin Invitrogen
Fluoreszenzlampe Lambda DG4 Sutter
Instrument Company
Geldokumentationsanlage Biozym Multiimage Light Cabinet
Gewebe-Homogenisator Bertin PreCellys 24
Kondensor Voltacraft
Magnetventile Lee Company 3-Way Ventil
Mikromanipulatoren Narishige Japan
Merzheuser
Monochromator T.I.L.L. Photonics Nanoinjektor WPI Nanoliter 2000 + Micro 4 Syringe
Pump Controler
Objektiv UplanApo (20x/0.75) Olympus
Photometer Peqlab ND-1000
Shutter Driver Uniblitz Vmm-D1
Stereomikroskop Zeiss Axiovert 200
Thermocycler Eppendorf realplex2 epgradient S
Tischheizblock Eppendorf Thermomixer comfort
Tischzentrifuge Eppendorf Mini-Spin Plus Vertikales Elektrodenziehgerät List Medical LM-2P-a
Voltage-Clamp Verstärker NPI Electronic Instruments Turbo Tec-03
Warmluftschüttler B. Braun Biotech international Centromat
Zentrifuge Sorvall RC 6+

24
Material und Methoden
2.1.12 Software
Elektrophysiologische Daten wurden mit dem Programm „Pulse“ der Firma „HEKA“ aufgezeichnet
und ausgewertet. Konzentrations-Wirkungs-Kurven wurden mit der 3-Parameter Hill Gleichung in
dem Programm „SigmaPlot“ (Systat Software Inc.) berechnet.
Die Aufzeichnung der Daten im Calcium-Imaging wurde mit dem Programm „SlideBook“ (3I
Imaging) geleistet. Zur Auswertung wurde die Software „IGOR Pro“ (WaveMetrics) herangezogen.
Analysen einzelner Nukleinsäuresequenzen wurden mit den Programmen „EditSeq, MapDraw und
SeqMan“ (DNA-Star) durchgeführt.
Für die Transkriptomanalyse wurden die Programme
- „TopHat” (Trapnell et al., 2009),
- „Cufflinks“ und „CuffDiff“ (Trapnell et al., 2010)
benutzt, die am “Institute of Genetic Medicine, Johns Hopkins University”, den “Departments of
Mathematics“ und “Molecular and Cell Biology, University of California” und dem “Department of
Stem Cell and Regenerative Biology, Harvard University” entwickelt worden sind.
Für statistische Auswertungen wurde „MS Excel“ (Microsoft), für graphische Darstellungen „Corel
Draw X4“ (Corel Corporation) benutzt.
2.2 Methoden
2.2.1 Entnahme von Oozyten aus Xenopus laevis
Zur Vorbereitung der Operation wurden die Frösche zunächst in einer 1%-igen 3-Ethyl-3-
Aminobenzoat-Lösung (in NFR, Sigma-Aldrich) narkotisiert. Nach völliger Erschlaffung der
Muskulatur wurde das Tier auf Eis gelagert, bedeckt und für 30 Minuten dort belassen, sodass der
Stoffwechsel stark verlangsamte. Die eigentliche Operation fand ebenfalls auf Eis statt. Es wurde
hierzu lediglich die Bauchseite des Frosches mit dem Operationsbereich vom Eis befreit. Haut und
Muskelschicht wurden ventral etwa 1,5 cm bis 3 cm lateral der Medianlinie und etwa 2 cm cranial
der Hautfalte, zwischen Abdomen und Oberschenkel über eine Länge von circa 0,5 cm bis 1 cm
eingeschnitten. Dadurch wurde der Bauchraum eröffnet und der Ovariallappen wurde
entnommen.

25
Material und Methoden
Dieser wurde in Ca2+-freie Barth-Lösung überführt und anschließend wurden die
Bauchmuskelschicht sowie die Haut separat vernäht um die Öffnung der Bauchhöhle zu
verschließen. Nach ca. zwei bis vier Stunden Quarantäne wurden die Tiere wieder in die
Tierhaltung verbracht.
2.2.2 Aufbereitung der Oozyten und Injektion der RNA
In den entnommenen Ovariallappen kommen neben den Oozyten weitere Zelltypen vor, die die
Oozyten umgeben und den Gewebsverband des Ovariums bilden. Es handelt sich dabei in erster
Linie um Bindegewebe, Blutgefäße sowie Follikelzellen, die jede einzelne Oozyte umhüllen.
Insbesondere die Follikelschicht beeinflusst elektrophysiologische Messungen wegen der
Expression endogener K+-Kanäle und isoliert zudem die Zellmembran der Oozyte von applizierten
pharmakologisch wirksamen Substanzen (Arellano et al., 1996; Miledi und Woodward, 1989a; b).
Um diese umgebenden Zellschichten zu entfernen, wurden die Ovariallappen für 1,5 bis 2
Stunden bei Raumtemperatur unter sanftem Schütteln mit 0,02% Collagenase I in Ca2+-freier
Barth-Lösung behandelt. Nach erfolgter Defollikulierung wurde die Collagenase schrittweise durch
Barth-Lösungen mit aufsteigenden Ca2+-Konzentrationen ausgewaschen. Dadurch wurden die
Oozyten schonend in Ca2+-haltige Standard Barth-Lösung überführt und im Brutschrank bei 17°C
gelagert.
Etwa nach 24 Stunden nach der enzymatischen Dissoziierung der Oozyten erfolgte die Injektion
von cRNA zur Expression der zu untersuchenden Transmembranproteine. Dazu wurden äußerlich
einwandfreie Oozyten der Stadien IV, V und VI (Dumont, 1972) ausgewählt und in eine
Kulturschale mit eingefügtem Plastik-Gitternetz (0,7 x 0,7 mm Maschengröße) überführt, welches
die Oozyten für die Injektion ausreichend fixierte. Die cRNA wurde mit einem Nanoliter-
Injektionsgerät (WPI Nanoliter 2000 + Micro 4 Syringe Pump Controler) Mikromanipulator-
gesteuert (Merzheuser) über eine Glaskapillare in die Oozyten injiziert. Die dafür notwendigen
Glaspipetten (ohne Filament) wurden mit einem Vertikalelektrodenziehgerät (List Medical LM-2P-
a) zu einer feinen Spitze ausgezogen und durch Abschneiden manuell geöffnet. Die Glaspipette
wurde mit Mineralöl (Sigma-Aldrich) befüllt und mit Hilfe des Injektors wurden 1 – 2 µl cRNA
aufgezogen. Pro Oozyte wurden etwa 10–30 ng cRNA injiziert. Die Oozyten wurden nach der
Injektion in 1 ml antibiotikahaltige ND96-Lösung (Penicillin/Streptomycin, je 100 U/ml) überführt
und bei 18°C in 24-well Zellkulturplatten (Tissue culture testplates, gamma sterilized, Orange
Scientific) gelagert. Die Expression spezifischer Transmembranproteine wurde nach etwa 2-5
Tagen mit dem Zwei-Elektroden-Voltage-Clamp Verfahren funktionell überprüft.

26
Material und Methoden
2.2.3 Die Zwei-Elektroden-Voltage-Clamp Technik
Mit Hilfe der Zwei-Elektroden-Voltage-Clamp Methode lassen sich in Oozyten exprimierte
Transmembranproteine elektrophysiologisch untersuchen. Gemäß dem Ohmschen Gesetz (U = R x
I) steigt an einer Zellmembran der über die Membran fließende Strom (I) bei konstanter Spannung
(U) mit sinkendem elektrischem Widerstand (R). Im Allgemeinen wird die elektrische Leitfähigkeit
an Zellmembranen im Bereich der gewählten Klemmspannungen von -40 - -80 mV maßgeblich
durch die Ionenleitfähigkeit von Transmembranproteinen vorgegeben. In Xenopus laevis Oozyten
sind dies nach der cRNA-Injektion vorwiegend heterolog exprimierte Ionenkanäle. Werden diese
Kanäle aktiviert, so verringert sich der Widerstand der Membran. Dadurch ändert sich der über
die Membran fließende Strom und führt zu einer Änderung der Membranspannung. Bei dem
Verfahren der Spannungsklemme wird über einen elektrischen Regelkreis das Potential an der
Membran auf einen konstanten Wert eingestellt und gehalten (-40 - -80 mV). Es werden dazu
zwei Elektroden, eine Strom- und eine Spannungselektrode in die Oozyte eingestochen. Die
Spannungselektrode dient zur Messung der an der Membran anliegenden Spannung, wobei eine
extrazelluläre Badelektrode als Referenz dient. Die Stromelektrode liefert genau die Menge
Strom, die zur Aufrechterhaltung des eingestellten Potentials an der Membran notwendig ist.
Dieser Haltestrom ist proportional zu der über die Membran fließenden Ladung, welche durch
Ionenkanalöffnung hervorgerufen wird. Dieser Regelkreis wird mit Hilfe eines Voltage-Clamp
Verstärkers (Turbo Tec-03, NPI Electronic Instruments) gesteuert und die Änderung des
Haltestroms als Maß für die Kanalöffnung wird über eine analog-digitale Schnittstelle (HEKA LIH
1600) mit dem Programm Pulse (HEKA) aufgezeichnet (Abb. 2.1).

27
Material und Methoden
Abbildung 2.1: Schematische Darstellung des Voltage-Clamp Messaufbaus. EI=intrazelluläres
Potential, ER=Referenzpotential, UM=Messspannung, UH=Haltespannung, IK=Kompensationsstrom,
EE=Erdpotential
Die Oozyten wurden während der Messung in eine spezielle Messkammer gelegt. Um ein
ungewolltes Bewegen der Oozyte während der Messung zu vermeiden, wurde diese in einer
Vertiefung fixiert. Zwischen den Applikationen pharmakologisch wirksamer Substanzen wurde die
Oozyte mit NFR oder Mg2+-Frosch Ringer gespült, um diese zu entfernen. Pro Applikation wurden
in der Regel 100 µl Testlösung mit einer automatischen Pipette (Eppendorf Research Pro) für eine
Dauer von 20 s appliziert und ein Stromverlauf von meist zwei Minuten aufgezeichnet. Die für die
Erstellung von Konzentrations-Wirkungsbeziehungen notwendigen Applikationen verschiedener
Substanzkonzentrationen wurden stets in aufsteigender Konzentrationsfolge vorgenommen. Für
die Aufzeichnung von Strom-Spannungsbeziehungen wurde mit dem Programm „Pulse“ eine
Spannungsrampe (-150 bis +50 mV) aufgezeichnet. Die Messung der Amplituden erfolgte mit
„Pulse“, die graphische Darstellung von Stromspuren mit „Corel Draw“ (X4, Corel Corporation).
Eine Prüfung auf statistische Signifikanz wurde mit dem Student’s t-Test (MS Excel) durchgeführt,
wobei p < 0,05 als Signifikanzlevel gewählt wurde. Konzentrations-Wirkungs-Kurven und
halbmaximale Agonisten-konzentrationen (EC50) wurden durch „SigmaPlot“ (Systat Software Inc.)
mit Hilfe der 3 Parameter Hill Gleichung berechnet.

28
Material und Methoden
2.2.4 Trigeminus-Primärzellkultur
Trigeminale Neurone wurden aus dem Ganglion gasseri zwei bis fünf Tage alter Mäuse (P2-P5)
präpariert. Dazu wurden die Mäuse dekapitiert, die Kopfhaut sagittal eingeschnitten, entfernt und
der kaum ossifizierte Schädel horizontal von occipital nach frontal eröffnet. Nach dem Entfernen
des Gehirns konnten beide trigeminalen Ganglien der Schädelbasis entnommen, in Phosphat
gepufferter Salzlösung (PBS+/+, Invitrogen) gewaschen und in eisgekühltes Leibovitz Medium (L15,
Invitrogen) überführt werden. Die gesammelten Ganglien wurden anschließend in 2 ml Minimum
Essential Medium (MEM, Invitrogen) und 0.025% Collagenase (T-1A, Sigma-Aldrich) für 45
Minuten in einen Brutschrank bei 37°C, 95% Luftfeuchtigkeit und 6% CO2 inkubiert, um den
Gewebsverband enzymatisch zu dissoziieren. Im Anschluss erfolgte eine mechanische Trituration
mit einer feuerpolierten Pasteurpipette (Brand). Die Zellsuspension wurde dann durch ein Zellsieb
mit 70 µM Maschenweite (BD Falcon) gegeben, um eventuelle nicht vollständig dissoziierte
Gewebeteile aus der Suspension zu entfernen. Anschließend wurde die Suspension für vier
Minuten bei 1000 rpm zentrifugiert, der Überstand verworfen und das erhaltene Zellpellet im
Kulturmedium (Dulbeccos´s Modified Eagle Medium (DMEM/F12, Invitrogen), 10% fötales
Kälberserum (FBS, Invitrogen), 1% Penicillin/Streptomycin (PAA Laboratories GmbH)
resuspendiert. Die Zellsuspension wurde auf Poly-L-Lysin (Sigma-Aldrich) beschichtete 30 mm
Deckgläschen in 35 mm Zellkulturschalen mit etwa 50 µl Suspension pro Deckgläschen
aufgetragen. Die Kulturschalen wurden dann zunächst für eine Stunde in den Brutschrank bei
37°C, 95% Luftfeuchtigkeit und 6% CO2 verbracht, um den Zellen zu ermöglichen, in der
Suspension abzusinken und der Beschichtung anzuhaften. Im Anschluss wurden die
Zellkulturschalen mit 2 ml Kulturmedium aufgefüllt und für mindestens vier Stunden, maximal vier
Tage bis zum Gebrauch im Brutschrank unter vorher genannten Bedingungen gelagert.
2.2.5 Die Calcium-Imaging Methode
Der Fluoreszenzfarbstoff „Fura-2/AM“ (Invitrogen) dient im Calcium-Imaging als Reporter-
Farbstoff für die intrazelluläre Calcium-Konzentration. Bei einer Bindung von Ca2+-Ionen an den
Farbstoff kommt es zu einer Konformationsänderung des Moleküls, die eine Verschiebung des
elektromagnetischen Absorptionsspektrums bewirkt. Das Ca2+-gesättigte Molekül hat ein
Absorptionsmaximum bei 340 nm Wellenlänge, freies „Fura-2/AM“ hingegen bei 380 nm. Bei
einer Messung im Calcium-Imaging wird ein mit „Fura-2/AM“ beladene Zelle mit Licht beider
Wellenlängen hintereinander angeregt. Die Mit Hilfe der Emissionswellenlänge von 510 nm
werden die Fluoreszenzintensitäten (f) gemessen und der Quotient (f340/f380), der ein Maß der

29
Material und Methoden
intrazellulären Calcium-Konzentration ist, aus diesen beiden gebildet. Zelluläre
Signaltransduktionen in deren Verlauf es zu Ca2+-Konzentrationsänderung kommt, können durch
die resultierende Fluoreszenzveränderung untersucht werden.
Zur Beladung wurden die Zellen der trigeminale Zellkultur mit 3 µM des Farbstoffes für 45-60
Minuten vor dem Experiment im Brutschrank bei 37°C inkubiert. Anschließend wurden die
Deckgläschen den Kulturschalen entnommen, in eine Messkammer aus rostfreiem Edelstahl
gelegt und diese mit Extrazellulär-Medium befüllt. Die Messungen wurden mit einem inversen
Stereomikroskop (Zeiss Axiovert 200) und einem 20-fach Objektiv (UplanApo 20x/0.75, Olympus)
durchgeführt. Die Zellen wurden bei einer Frequenz von 1 Hz für 100-200 ms bei 340 und 380 nm
Wellenlänge angeregt (Lambda DG4 Fluoreszenzlampe, Sutter Instrument Company, Uniblitz
Vmm-D1 Shutter Driver und Voltakraft Kondensor). Emittiertes Licht wurde bei 510 nm
Wellenlänge mit einer CCD-Kamera („charge-coupled device“, Zeiss Photometrics Axiocam MRM)
registriert und mit dem Programm „SlideBook“ (3I Imaging) aufgezeichnet. Die Auswertung
erfolgte mit „IGOR Pro“ (WaveMetrics), graphische Darstellung mit „Corel Draw X4“( Corel
Corporation).
Während des Experimentes wurde durch Flüssigkeitszufuhr und -absaugung ein permanenter
Austausch des Extrazellulär-Mediums in der Messkammer gewährleistet. Zudem wurde mittels
eines eigens entwickelten Applikationssystems, bei dem sieben separate Bevorratungsgefäße in
einen Applikationskopf münden, ein konstanter Flüssigkeitsstrom von Extrazellulär-Medium
angelegt, der bei gleicher Strömungsgeschwindigkeit übergangslos gegen die zu applizierende
Testlösung umgeschaltet werden konnte. Am Ende jeden Experiments wurde für 5 s Hochkalium-
Lösung appliziert, um gemessene Zellen als vitale Neurone zu verifizieren und somit von Gliazellen
unterscheiden zu können.
2.2.6 Isolation einzelner Neurone aus trigeminaler Zellkultur
Um die Rezeptortranskripte der zuvor im Calcium-Imaging charakterisierten trigeminalen Neurone
zu untersuchen, musste die identifizierten Zellen einzeln aus der Kultur herausgelöst werden. Für
diesen Zweck wurde ein Mikromanipulator gesteuertes (Merzheuser sowie Narishige Japan)
Nanoliter-Injektionsgerät (WPI Nanoliter 2000 + Micro 4 Syringe Pump Controler) benutzt,
welches für Aufnahme und Abgabe von Volumina im Nanoliterbereich geeignet ist. Um einzelne
Neurone aus dem Zellverband herauszulösen wurde das Gerät mit einer ölbefüllten Glaspipette
(ohne Filament, Glass capillaries Nanoliter 2000; 3,5 inches, 1.14 ID, 0.5 mm OD, World Precision
Instruments) bestückt, die zuvor mit einem Vertikalelektrodenziehgerät (List Medical LM-2P-a) zu

30
Material und Methoden
einer feinen Spitze ausgezogen wurde. Unter optischer Kontrolle mit Hilfe eines optischen
Eichgitters (100 µM Gitterweite, Neubauer-Zählkammer) wurde der Spitzendurchmesser auf 30-
60 µm eingestellt und anschließend feuerpoliert.
Vitale Neurone in der trigeminalen Zellkultur vernetzen sich in der Regel über zelluläre Ausläufer
mit den nächstgelegenen Zellen. Durch gezieltes Ansteuern und Aufsetzen der Pipettenspitze auf
das Zellkulturgläschen wurden zu isolierende Zellen zunächst vereinzelt, anschließend vom Glas
gelöst und mit 5 nl Extrazellulär-Lösung aufgesaugt. Dabei konnte eine Verunreinigung der Probe
durch benachbarte Zellen oder deren Fragmente durch dieses geringe Aufnahmevolumen
zuverlässig verhindert werden. Die aufgenommene Zelle wurde im Anschluss in ein
Reaktionsgefäß (0,5 ml, DNA LoBind® Tube, Eppendorf) mit RNA-stabilisierendem Lysis-Puffer zur
Transkriptom-Amplifikation überführt und bei -80°C bis zur Amplifikation gelagert.
2.2.7 Molekularbiologische Methoden
2.2.7.1 Quantitative Polymerase Kettenreaktion (qPCR)
Die PCR oder polymerase chain reaction ist eine Methode, mit der sich Teile einer DNA-Matrize –
auch Template genannt – vervielfältigen und damit nachweisen lassen können (Saiki et al., 1988).
Die Vervielfältigung wird dabei mit einer DNA-Polymerase erreicht, die entlang eines DNA-
Einzelstranges einen komplementären Strang aus freien Nukleotiden neu synthetisiert. Das Enzym
braucht dabei jedoch einen kurzen doppelsträngigen Abschnitt, um an die DNA-Matrize binden zu
können. Zu diesem Zweck werden künstlich erzeugte Oligonukleotide (engl. Primer) mit einer
Länge von 20-25 Basenpaaren verwendet, die jeweils an die 5‘-Enden beider komplementärer
Stränge des zu amplifizierenden Abschnittes spezifisch binden. Dieser Vorgang wird als Annealing
bezeichnet und kann sich bedingt durch die Oligonukleotidsequenz in seinem
Temperaturoptimum beträchtlich unterscheiden. Nach diesem Schritt folgt die Elongation, wobei
die Polymerase das Template in 3‘-Richtung fortschreibt. Dieser Schritt erfolgt am
Temperaturoptimum des Enzyms, was in der Regel 72°C beträgt. Im Anschluss folgt die
Denaturierung, wobei die Temperatur bis auf 94°C erhöht wird und der Doppelstrang wieder in
zwei Einzelstränge zerfällt. Da die auf diese Weise entstanden Stränge jeweils wieder als Matrize
dienen, kann der Vorgang in vielen Zyklen (Annealing, Elongation, Denaturierung) wiederholt
werden und so die Molekülanzahl des zu amplifizierenden Fragmentes exponentiell erhöht
werden.
Im Rahmen einer quantitativen PCR (realtime qPCR) wird diese exponentielle Erhöhung während
der Entstehung doppelsträngigen Amplifikats in der Elongation quantitativ erfasst. Dazu wird ein

31
Material und Methoden
Fluoreszenzfarbstoff (SYBR Green, Sigma-Aldrich) zum Reaktionsansatz gegeben, der in
doppelsträngige DNA interkaliert, was zu einem Anstieg an Fluoreszenz führt. Im Laufe der qPCR
gibt es einen Zeitpunkt ab dem die spezifische Fluoreszenz der Probe erstmalig über die Intensität
der Hintergrundfluoreszenz steigt und fortan in steilem exponentiellem Verlauf anwächst. Die
Anzahl der PCR-Zyklen, die in diesem Zeitpunkt bereits stattgefunden hat, wird bei einer qPCR als
CT-Wert (cycle treshold) bezeichnet und ist ein relatives Maß für die eingesetzte Anzahl von DNA-
Matrizen. Je mehr zu Anfang eingesetzt wurde, desto eher ist der cycle treshold erreicht; ein
niedriger CT-Wert bedeutet also eine hohe Anzahl eingesetzter Matrizen.
Jede Reaktion wurde aus demselben Ansatz in drei separaten Gefäßen (als Triplet) mit je 25 µl
Reaktionsansatz durchgeführt.
Dabei beinhaltete jeder Ansatz von 75 µl Gesamtvolumen:
37,5 μl SYBR Green Mix (Peqlab)
1,7 μl Template (20 ng)
1,7 μl forward-Primer (17 pmol)
1,7 μl reverse-Primer (17 pmol)
32,4 μl A.dest.
Das Standardprotokoll einer qPCR:
1. 95°C für 3 min, initiale Denaturierung
2. 95°C 45 s, Denaturierung im Zyklus
60-65°C 45 s, Annealing im Zyklus
72°C 1 Minute, Elongation im Zyklus
Es wurden 40 Zyklen je PCR durchgeführt.
3. 72°C 2 Minuten, terminale Elongation
4. 95°C 15 s, Temperaturrampe über 20 Minuten von 60°C bis 95°C, Schmelzkurve
5. 12°C dauerhaft, Stopp der PCR.
Die nach der terminalen Elongation durchgeführte Schmelzkurve diente einer Überprüfung der
Spezifität der durchgeführten Reaktion. Bei Amplifikaten verschiedener Sequenz setzt die
Denaturierung bei unterschiedlichen Temperaturen ein. Die Denaturierung setzt gebundenen
Fluoreszenzfarbstoff wieder frei, was bei einer spezifischen Temperatur zu punktuell starkem
quantifizierbarem Absinken der Fluoreszenz führt. Das benutzte PCR-Gerät (Thermocycler
Eppendorf realplex2) berechnet CT-Werte und spezifische Schmelztemperaturen automatisch.

32
Material und Methoden
2.2.7.2 Agarosegelelektrophorese
Die Auftrennung der DNA erfolgte unter Standardbedingungen in einem etwa 0,5-1,0%-igem
Agarosegel (Sambrook et al., 1989). Die fotografische Dokumentation erfolgte mit dem
„MultiImage Light Cabinet“ (Biozym).
2.2.7.3 Transformation
Um Fremd-DNA in Form eines Plasmids zu vervielfältigen, wurden Bakterien (E. coli XL1-Blue,
Stratagene) mit abgewandelter Rubidium-Chlorid-Methode nach Hanahan und Meselson (1983)
chemisch transformiert.
2.2.7.4 Anzucht von Bakterien
Transformierte Bakterien wurden in Abhängigkeit von der Plasmid-vermittelten
Antibiotikaresistenz auf Kanamycin- oder Ampicillin-haltigen (jeweils 100 mg/ml, PAA)
Agarplatten über Nacht bei 37°C im Brutschrank (Heraeus) angezogen (Sambrook et al., 1989).
Bakterien einer Einzelkolonie wurden in 200 ml LB-Medium ebenfalls mit entsprechendem
Antibiotikum über Nacht bei 37°C und 250 rpm im Warmluftschüttler (B. Braun) vermehrt.
2.2.7.5 Präparative DNA-Aufreinigung
Mit Hilfe des “Pure Yield Maxiprep Systems” (Promega) wurde die Plasmid-DNA aus den
Bakterienkulturen (siehe 2.2.7.4) nach dem Prinzip der alkalischen Lyse isoliert (Birnboim und
Doly, 1979).
2.2.7.6 Ethanol/Natriumacetat-Fällung
Die Aufreinigung und Ankonzentrierung der DNA wurde mittels Ethanol/Natriumacetat-Fällung
durchgeführt. Bei dieser Methode macht man sich zu Nutze, dass Nukleinsäuren in Salzlösungen
unter Zugabe von Ethanol oder Isopropanol auszufallen. Dafür wurden der Probe 0,1 Volumina
3 M Natriumacetat und 0,7 Volumina Isopropanol hinzugefügt, der Ansatz für 30 Minuten bei -
20°C inkubiert und im Anschluss 10 Minuten mit 13.000 rpm zentrifugiert (Eppendorf Mini-Spin
Plus). Der Überstand wurde abgenommen und verworfen. Dem Pellet wurden 400 µl Ethanol (70
%) zugegeben, und die Probe erneut für 20 Minuten mit 13.000 rpm zentrifugiert. Der Überstand
wurde abgenommen und verworfen, das Pellet für fünf Minuten bei Raumtemperatur und

33
Material und Methoden
offenem Laborgefäß belassen, um restliches Ethanol zu entfernen (Evaporation). Im Anschluss
wurde die DNA in RNase-freiem, sterilem H2O aufgenommen und bei -20°C gelagert.
2.2.7.7 Quantifizierung von DNA und RNA
Zur Konzentrationsbestimmung von Nukleinsäurelösungen wurde die Extinktion photometrisch
bei 260 nm Wellenlänge ermittelt. Die Messung erfolgte mit dem „Nano-Drop 2000“ Photometer
(Peqlab), das die optische Dichte kleiner Volumina im Bereich von 1-2 µl messen kann.
2.2.7.8 In vitro-Transkription
Zur Synthese von cRNA, die zur heterologen Expression von Proteinen in Xenopus Oozyten
benötigt wird, wurde isolierte linearisierte Plasmid-DNA als Matrize für eine RNA-abhängige
Polymerase genutzt. Dafür besitzen alle benutzten Plasmide die Promotorsequenz des T7-Phagen
und wurden mit der sehr effizienten T7-RNA-Polymerase transkribiert. Das linearisierte Plasmid
beinhaltet hinter der Promotorregion das Insert, welches die kodierende Sequenz für zu
exprimierende Ionenkanäle enthält. Zur in vitro-Transkription wurde das „AmpliCap-T7 Message
Maker Kit“ (Epicentre) nach Herstellerprotokoll benutzt. Im Anschluss wurde die synthetisierte
cRNA in einer Ammonium-Acetat-Fällung aufgereinigt.
2.2.7.9 Ammonium-Acetat-Fällung
Nukleinsäuren können aus einer wässrigen Lösung mit Ammonium-Acetat gefällt werden. Im
Gegensatz zur Natriumacetat Fällung (siehe 2.2.7.6) werden bei dieser Methode keine
kurzkettigen Oligonukleotide gefällt, so dass ein hoch reines Produkt entsteht. Die in der in vitro-
Transkription synthetisierte cRNA wurde nach diesem Prinzip aufgereinigt. Dabei wurden einer
Probe 0,5 Volumina NH4-Acetat (10 M) zugegeben, 15 Minuten auf Eis inkubiert und im Anschluss
für 15 Minuten bei 4°C mit 14.000 rpm zentrifugiert (Eppendorf Mini-Spin Plus). Der Überstand
wurde abgezogen und verworfen, danach mit Ethanol (70%) gewaschen und die RNA in RNase-
freiem, sterilem H2O aufgenommen und bei -80°C gelagert.
2.2.7.10 Elution von DNA aus Agarosegelen
Um DNA, die mittels Gelelektrophorese aufgetrennt wurde, aus dem Agarose-Gel zu eluieren,
wurde die entsprechende unter UV-Licht identifizierte Bande mit einem Skalpell aus dem Gel

34
Material und Methoden
ausgeschnitten und in Lysis-Puffer des „Wizard® SV Gel and PCR Clean-Up Systems“ (Promega)
eluiert.
2.2.7.11 DNA-Sequenzierung
Zur Sequenzierung von Plasmid-DNA und PCR-Produkten wurden durch den Service des
Lehrstuhls für Rezeptorbiochemie der Ruhr-Universität Bochum durchgeführt. Der Service nutzt
den Sequenzierautomaten „3130xl Genetic Analyzer“ (Applied Biosystems), der mit einem
Sequenzierverfahren nach Sanger (1975) analysiert.
2.2.7.12 Restriktion von Plasmiden
Mit Hilfe von Restriktionsendonukleasen können spezifische doppelsträngige DNA-
Sequenzabschnitte erkannt und durch Hydrolyse an den Phosphodiesterbindungen aufgetrennt
werden. Neben der Erkennungssequenz sind auch Reaktionsbedingungen (Temperatur und
Puffer) spezifisch für diese Endonukleasen. Die Reaktion wurde hier genutzt, um den
Expressionsvektor für die anschließende In vitro-Transkription zu linearisieren. Dazu wird ein
Enzym ausgewählt, dessen Erkennungssequenz am 3‘-Ende der zu synthetisierenden Sequenz liegt
(2.1.6). Restriktionsenzyme wurden von Fermentas und New England Biolabs bezogen und unter
den vom Hersteller angegebenen Optimalbedingungen benutzt. Dabei wurden 10 µg zu
schneidendes Plasmid, 2 µl Restriktionspuffer (10x), 1 µl Enzym (10 U/µl) mit sterilem H2O auf 20
µl Gesamtvolumen aufgefüllt und für 60 Minuten inkubiert. Es folgte eine Aufreinigung wie unter
2.2.7.10 beschrieben.
2.2.7.13 RNA-Isolation
Mit Hilfe des „RNeasy-Systems“ (Qiagen) kann Total-RNA aus Gewebeproben isoliert und über
eine Silica-Säule aufgereinigt werden. Dabei wurden sowohl das Micro- als auch das Mini-Format
des Kits, in Abhängigkeit von der eingesetzten Gesamtprobenmasse benutzt. Dabei wurde stets
der zusätzliche Schritt eines säulengebundenen DNase-I-Verdaus durchgeführt. Das Kit wurde
nach Herstellerprotokoll für Gewebeaufschluss benutzt. Isolierte RNA wurde nach Elution in
sterilem RNase-freiem H2O bei -80°C gelagert.

35
Material und Methoden
2.2.7.14 Transkriptom-Amplifikation und cDNA-Synthese bei Einsatz kleinster Mengen RNA
Nach der Isolation einzelner trigeminaler Neurone zum Zweck der Einzelzelltranskriptomanalyse
muss das gesamte Transkriptom – also die Gesamtheit aller in der Zelle vorliegenden Transkripte
– in gleichbleibendem Verhältnis amplifiziert werden, denn aus einzelnen Zellen kann nicht genug
RNA gewonnen werden, um die für das gewählte Sequenzierverfahren erforderliche
Mindestmenge von 1 µg einsetzen zu können. Diese Amplifikation wurde mit Hilfe des „TransPlex
Complete WTA2-Systems“ (Sigma-Aldrich) durchgeführt. Die Amplifikation erfolgt in zwei
Schritten: 1. eine reverse Transkription, welche eine cDNA-Bibliothek aus zufälligen,
sequenzüberlappenden Fragmenten einer Länge von 100-1000 Basenpaaren erzeugt und dabei
den Fragmenten ein universales Sequenzende angefügt und 2. einer PCR-basierten
repräsentativen Vervielfältigung, bei der das universelle Sequenzende als Primerbindestelle dient.
Das System wurde nach Herstellerprotokoll benutzt, wobei die Probe bei Bedarf nach der
reversen Transkription bis zur PCR-basierten Amplifikation bei -20°C gelagert wurde. Nach dem
gesamten Prozess wurde die erzeugte cDNA-Bibliothek aufgereinigt (2.2.7.10) und deren
Konzentration bestimmt (2.2.7.7).
2.2.7.15 Next Generation Sequencing-Verfahren
Zur Analyse amplifizierter Transkriptome einzelner isolierter Neurone mussten die wie unter
2.2.7.14 beschrieben gewonnenen cDNA-Proben zunächst sequenziert werden. Ein geeignetes
Verfahren muss dazu insbesondere eine hohe Anzahl paralleler Ansätze bieten können. Die
Transkriptomsequenzierungen wurden nach dem „Illumina Solexa“ Verfahren durchgeführt, was
durch den Service des „Cologne Center for Genomics“ geleistet wurde. Sequenzierergebnisse
wurden als Dateien des „fastq“-Formats übermittelt, wofür frei zugängliche Software zur
Datenanalyse von der homepage des „Center for Bioinformatics & Computational Biology“
(http://www.cbcb.umd.edu/) bezogen wurde.
2.2.7.16 Transkriptomanalyse
Eine Transkriptomanalyse nach einer Methode der Massensequenzierung beginnt zunächst mit
einem systematischen Vergleich der Sequenzierergebnisse mit einem Referenzgenom (Trapnell et
al., 2009). Zu diesem Zweck wurde das Referenzgenom „mm9“ (Mus musculus) verwendet. Um
die Sequenzierdaten an das Genom zu ‚alignen‘ wurde das Programm „TopHat“ benutzt, welches
auf den „Bowtie“-Algorithmus zurückgreift (Langmead et al., 2009). Dieser Algorithmus kann

36
Material und Methoden
neben den Übereinstimmungen mit dem verwendeten Referenzgenom auch Intron/Exon-
überspannende Transkripte zuordnen. Nach dieser ersten Zuordnung folgt die Analyse der
Expressionsstärke mit dem Programm „Cufflinks“ (Trapnell et al., 2010), wobei das
Referenztranskriptom „mm9refseq“ als Vergleich dient. Die relative Expressionsstärke eines
Transkriptes wird in „fragments per kilobase of exon per million mapped reads“ (FPKM)
angegeben. Dabei wird die Anzahl einem Gen zugeordneter reads wegen ihrer für gewöhnlich
unterschiedlichen Länge auf 1000 bp normiert und diese normierte Anzahl als Anteil an einer
Million reads ausgedrückt. Zum Vergleich der Expression können Sequenzierergebnisse anderer
Gewebe aus der NCBI SRA-Datenbank (www.ncbi.nlm.nih.gov/) herangezogen werden, um mit
dem Programm „CuffDiff“ gewebsspezifische Transkripte zu identifizieren. Außerdem ermöglicht
„CuffDiff“ die Untersuchung von Expressionsschwankungen in verschiedenen Sequenzierläufen
des gleichen Gewebes, was eine Analyse der physiologischen Expressionsvariabilität ermöglicht.
TopHat
Mit Hilfe des „Bowtie“-Algorithmusses vergleicht „TopHat“ kurze Nukleinsäure-Sequenzen, die im
„Next-Generation-Sequencing“ als sogenannte reads bezeichnet werden, mit dem
Referenzgenom (Mus musculus/mm9) und identifiziert so Exon-Exon-splice-junctions. Zunächst
findet das Programm potentielle Exone und legt eine splice-junction Datenbank an. Über einen
Abgleich mit den tatsächlichen reads werden die splice-junctions bestätigt. „TopHat“ unterteilt
reads in kleinere Sequenzfragmente, ordnet diese stückweise der Referenzsequenz zu und fügt
diese anschließend wieder als Exon zusammen (Trapnell et al., 2009). Das Softwarepacket zur
Transkriptomanalyse wurde in einer Linux-basierten Version benutzt. Eine Befehlszeile zur
Datenanalyse besteht beispielsweise aus folgenden Kommandokomponenten:
o Tophat –o mmTG1 –r 128 –solexa1.3-quals –F 0 -G /home/ubuntu/ngs/gtf/mm9refseq.gtf/home/ubuntu/ngs/ bowtie/idexes/mm9 mTG1_s_3_1.fq mTG1_s_3_2.fq

37
Material und Methoden
Option Funktion
TopHat Auswahl des Programms -o Speicherort der Analyseergebnisse
-r 128 Innere Distanz der Sequenzen auf dem
Genom bei paired end Sequenzierung
Solexa.1.3-quals Qualitätsbewertung der Analysen
-F 0 Isoform-Filter wurde deaktiviert
-G Referenzgenom
Anschließend folgt der Pfad der zu analysierenden Zieldateien. In diesem Beispiel wurde eine
paired end Sequenzierung durchgeführt.
SAMtools
Im Anschluss werden Ergebnis-Dateien der „TopHat“-Analyse mit dem Programm „SAMtools“
sortiert und indiziert. Mit dem Befehl „samtools sort accepted_hits.bam Sorted“ wird die
Sortierung, mit „samtools index sorted.bam“ die Indizierung gestartet.
Cufflinks
Das „Cufflinks“-Programm assembliert Transkripte und ermittelt deren Expressionsstärke in der
zugrunde liegenden Probe unter Berücksichtigung aller möglichen Isoformen. Das Programm
berechnet für jedes bekannte Transkript die FPKM. Sollte die Sequenzierung als paired-end
Experiment durchgeführt worden sein, existieren für jedes Transkript zwei reads, was „Cufflinks“
bei der Berechnung der zugeordneten reads berücksichtigt. Sollten einzelne reads mehreren
Positionen des Genoms zugeordnet werden, berechnet „Cufflinks“ die Expression an jedem
Genort anteilig (Mortazavi et al., 2008). Eine Befehlszeile zur Datenanalyse besteht beispielsweise
aus folgenden Kommandokomponenten:
o Cufflinks -v -u -o cloutput --min-frags-per-transfrag 1 -G /home/ubuntu/ngs/gtf/mm9refseq.gtf -b/home/ubuntu/ngs/bowtie/idexes/mm9.fa sorted.bam

38
Material und Methoden
Option Funktion
Cufflinks Auswahl des Programms -v Status updates und Diagnoseinformationen
-u Bewertet Zuordnung der reads die an
mehreren Stellen im Genom mappen
-o Speicherort der Analyseergebnisse
--min-frags-per-transfrag
Anzahl der reads wird auf 1 gesetzt, damit
das Programm pro read ein Transkript
zuordnet (niedrige Transkripte werden
detektiert)
-G Referenzgenom
-b Referenzsequenz zur Berechnung des Standardfehlers und Korrektur-Algorithmus
Anschließend folgt der Pfad der zu analysierenden Zieldateien.
CuffDiff
Mit dem „Cufflinks“-Unterprogramm „CuffDiff“ können Expressionslevel verschiedener
Sequenzierdurchläufe miteinander verglichen und auf Signifikanz untersucht werden. Dies macht
man es sich bei der vergleichenden Analyse verschiedener Gewebe oder zur Identifizierung
spezifischer Genregulationen zunutze. Eine Befehlszeile zur Datenanalyse besteht beispielsweise
aus folgenden Kommandokomponenten:
o Cuffdiff -o cuffdiff_Trig1_Trig2 -b /home/ubuntu/ngs/bowtie/indexes/mm9.fa -c 1 /home/ubuntu /ngs/gtf/mm9refseq.gtf/home/ubuntu/benjamin/DatenSeq/mmTG1/ accepted_hits.bam/home/benjamin/DatenSeq/mmTG1/mmTG2/ accepted_hits.bam
Option Funktion
CuffDiff Auswahl des Programms
-o Speicherort der Analyseergebnisse
-b Referenzsequenz zur Berechnung des
Standardfehlers und Korrektur-Algorithmus
-c Schwellendifferenzlevel der Expression ab dem das Programm Signifikanz prüft; in
Anzahl der alignments

39
Material und Methoden
Anschließend wird hier ebenfalls der Pfad der zu analysierenden Zieldateien angehängt.
Ergebnisse nach „Cufflinks“- oder „CuffDiff“-Analysen können im Weiteren in Excel überführt und
mit diesem Programm weiter verwaltet werden. Da die bisherigen Programme die Identität jedes
Gens mit der NM-Nummer darstellen, muss diesen im Anschluss eine gene-id zugeordnet werden,
die von der UCSC-Datenbank stammt (http://genome.ucsc.edu/). Die Differenz der Genexpression
wird als fold change (Änderungsfaktor) berechnet und logarithmisch ausgegeben. „CuffDiff“ führt
verschiedene statistische Analysen durch, um Signifikanz zu ermitteln
(http://cufflinks.cbcb.umd.edu/howitworks.html). Die Berechnung der Signifikanz wird nach
Benjamini Hochberg korrigiert (FDR > 0.05).

40
Ergebnisse
3. Ergebnisse
3.1 Elektrophysiologische Untersuchungen der Nikotin-Responsivität rekombinanter
Ionenkanäle
Im ersten Teil dieser Arbeit wurde der Einfluss von Nikotin auf humane Liganden-gesteuerte
Ionenkanäle und TRP-Kanäle untersucht. Es wurden dazu Subfamilien gewählt, deren Expression
und zumeist auch deren funktionelle Bedeutung im Trigeminus bekannt sind. Aus der Klasse der
Liganden-gesteuerten Ionenkanäle wurden die Subfamilien der 5-HT3-, nACh-, Glycin- und GABA-
Rezeptoren untersucht. Zudem wurden P2X-Rezeptoren und ionotrope Glutamatrezeptoren aus
den Subfamilien der AMPA-, Kainat- und NMDA-Rezeptoren untersucht. Die Bedeutung der
Thermo-TRP-Kanäle für die trigeminale Sensorik ist seit Längerem bekannt. Aus der Klasse der
TRP-Kanäle wurden daher TRPA1, TRPV1-3 und TRPM8 untersucht. Zu untersuchende
Ionenkanäle wurden heterolog in Xenopus laevis Oozyten exprimiert (2.2.2) und im Zwei-
Elektroden Voltage-Clamp Verfahren elektrophysiologisch charakterisiert (2.2.3). Zunächst wurde
dazu eine saturierende Konzentration des Agonisten des jeweiligen Ionenkanals appliziert, um
dessen Expression nachzuweisen. Im Anschluss wurde 1 mM (+)- oder (-)-Nikotin appliziert, um
einen direkten Agonismus von Nikotin auf die exprimierten Ionenkanäle festzustellen. Konnte
keine direkte Aktivierung festgestellt werden, wurde im Anschluss die konzentrationsabhängige
Wirkung des Agonisten sowie des Agonisten bei Koapplikation von (+)- oder (-)-Nikotin
untersucht. Dazu wurden je Zelle alternierend für beide Bedingungen Lösungen ansteigender
Agonistenkonzentrationen appliziert. Die Konzentration des Nikotins betrug dabei stets 1 mM.
Durch die Bildung des Quotienten aus absoluten Stromamplituden und dem maximal gemessenen
Strom an der Oozyte bei sättigender Agonistenkonzentration (I/I (max)), wurden die Antworten
normiert und konnten in Konzentrations-Wirkungs-Beziehungen aufgetragen werden. Im
Anschluss wurde die Agonistenkonzentration bei halbmaximaler Antwort (EC50) und der Hill-
Koeffizient (nH) ermittelt.
3.1.1 Antagonismus von Nikotin an humanen 5-HT3-Rezeptoren
Von den ionotropen Serotoninrezeptoren (5-HT3) wurde zunächst die humane alpha-Untereinheit
5-HT3A untersucht, da diese zur Bildung eines funktionellen homo- wie heteropentameren
Serotoninrezeptoren essentiell ist (Davies et al., 1999). In einer vorangegangenen Studie konnte
bereits eine antagonistische Wirkung von Nikotin auf den 5-HT3A-Rezeptor der Maus
nachgewiesen werden (Gurley und Lanthorn, 1998), jedoch wurde die Wirkung des

41
Ergebnisse
(+)-Nikotin-Enantiomers und somit eine mögliche Stereoselektivität bzw. mögliche Spezies-
Unterschiede nicht untersucht. Die alleinige Applikation der beiden Nikotin-Enantiomere auf
heterolog exprimierte 5-HT3A-Rezeptoren führte zu keiner nennenswerten Stromamplitude. Es
konnte an keiner Oozyte bei keinem der beiden Enantiomere ein größerer Strom als 20 nA
gemessen werden (Imax[(-/+)-Nikotin] < 0,5% Imax[Serotonin]; n = 8). Durch den natürlichen
Agonisten Serotonin vermittelte Ströme erreichten hingegen bei saturierenden Konzentrationen
Stromamplituden von über 5 µA. Um eine modulatorische Wirkung des Nikotins auf die
exprimierten Kanäle zu untersuchen, wurden Konzentrationen von 0,3 µM – 100 µM Serotonin
mit und ohne Nikotin beider Enantiomere appliziert und Konzentrations-Wirkungs-Kurven
aufgetragen. Es konnte eine starke Reduktion der Ströme bei Serotoninkonzentrationen bis 10 µM
durch (-)-Nikotin (1 mM) beobachtet werden. Der inhibitorische Effekt von (-)-Nikotin wurde bei
Serotoninkonzentrationen von 30 µM und 100 µM geringer. Der EC50-Wert von Serotonin betrug
dabei 2,22 ± 0,55 µM (nH = 1,97 ± 0,16), wohingegen der EC50-Wert bei Koapplikation von 1 mM
(-)-Nikotin bei 17,7 ± 2,05 µM (nH = 3,08 ± 0,34) lag. Es konnte somit eine signifikante
Rechtsverschiebung der Konzentrations-Wirkungs-Kurve beobachtet werden (p < 0,001; n = 8).
Dies ist vergleichbar mit dem von Gurley und Lanthorn (1998) beobachteten kompetitiven
Wirkmechanismus von Nikotin auf den 5-HT3A-Rezeptor der Maus, wenngleich am menschlichen
Rezeptor die Maximalwirkung bei Koapplikation von (-)-Nikotin etwas niedriger bleibt als bei der
Kontrolle (Abb. 3.1 A, B).
Zur Untersuchung der Wirksamkeit des zweiten Stereoisomers wurden die Versuche mit (+)-
Nikotin wiederholt. Da dies an anderen Oozyten durchgeführt wurde und so auch die
Kontrollversuche ohne Nikotin wiederholt werden mussten, wurde hier eine zweite
Konzentrations-Wirkungskurve aufgetragen und für Serotonin ohne Nikotin erneut ein EC50-Wert
ermittelt. Hier konnte lediglich eine schwache Inhibition des Rezeptors beobachtet werden. Es
wurde für Serotonin ein EC50-Wert von 2,57 ± 0,28 µM (nH = 2,2 ± 0,27) festgestellt. Der EC50-Wert
für Serotonin unter Koapplikation von (+)-Nikotin betrug 3,76 ± 0,50 µM (nH = 2,17 ± 0,4; p =
0,031; n = 13). Für (+)-Nikotin wurde ebenfalls ein, wenngleich schwacher, doch signifikanter
Effekt der Rechtsverschiebung der Konzentrations-Wirkung-Kurve beobachtet. Der untersuchte
humane 5-HT3A-Rezeptor wird somit von Nikotin stark stereospezifisch inhibiert (Abb. 3.1 C, D).
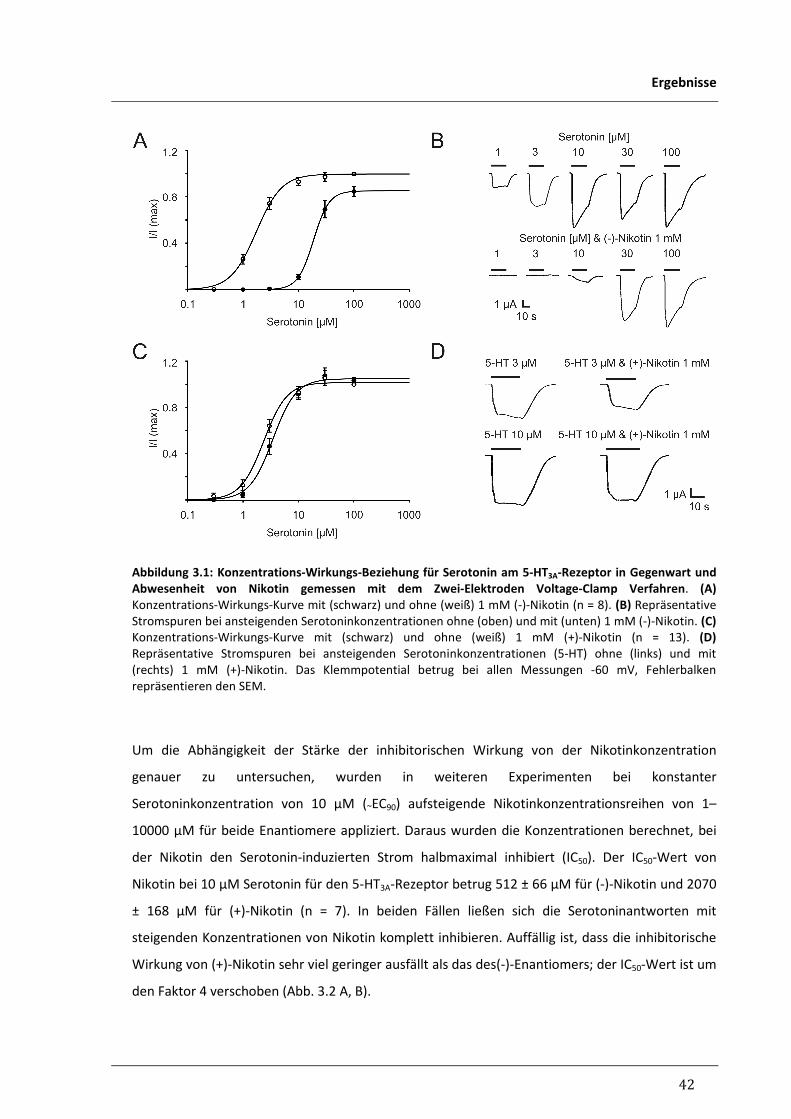
42
Ergebnisse
Abbildung 3.1: Konzentrations-Wirkungs-Beziehung für Serotonin am 5-HT3A-Rezeptor in Gegenwart und
Abwesenheit von Nikotin gemessen mit dem Zwei-Elektroden Voltage-Clamp Verfahren. (A)
Konzentrations-Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (-)-Nikotin (n = 8). (B) Repräsentative
Stromspuren bei ansteigenden Serotoninkonzentrationen ohne (oben) und mit (unten) 1 mM (-)-Nikotin. (C)
Konzentrations-Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (+)-Nikotin (n = 13). (D)
Repräsentative Stromspuren bei ansteigenden Serotoninkonzentrationen (5-HT) ohne (links) und mit
(rechts) 1 mM (+)-Nikotin. Das Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken
repräsentieren den SEM.
Um die Abhängigkeit der Stärke der inhibitorischen Wirkung von der Nikotinkonzentration
genauer zu untersuchen, wurden in weiteren Experimenten bei konstanter
Serotoninkonzentration von 10 µM (~EC90) aufsteigende Nikotinkonzentrationsreihen von 1–
10000 µM für beide Enantiomere appliziert. Daraus wurden die Konzentrationen berechnet, bei
der Nikotin den Serotonin-induzierten Strom halbmaximal inhibiert (IC50). Der IC50-Wert von
Nikotin bei 10 µM Serotonin für den 5-HT3A-Rezeptor betrug 512 ± 66 µM für (-)-Nikotin und 2070
± 168 µM für (+)-Nikotin (n = 7). In beiden Fällen ließen sich die Serotoninantworten mit
steigenden Konzentrationen von Nikotin komplett inhibieren. Auffällig ist, dass die inhibitorische
Wirkung von (+)-Nikotin sehr viel geringer ausfällt als das des(-)-Enantiomers; der IC50-Wert ist um
den Faktor 4 verschoben (Abb. 3.2 A, B).
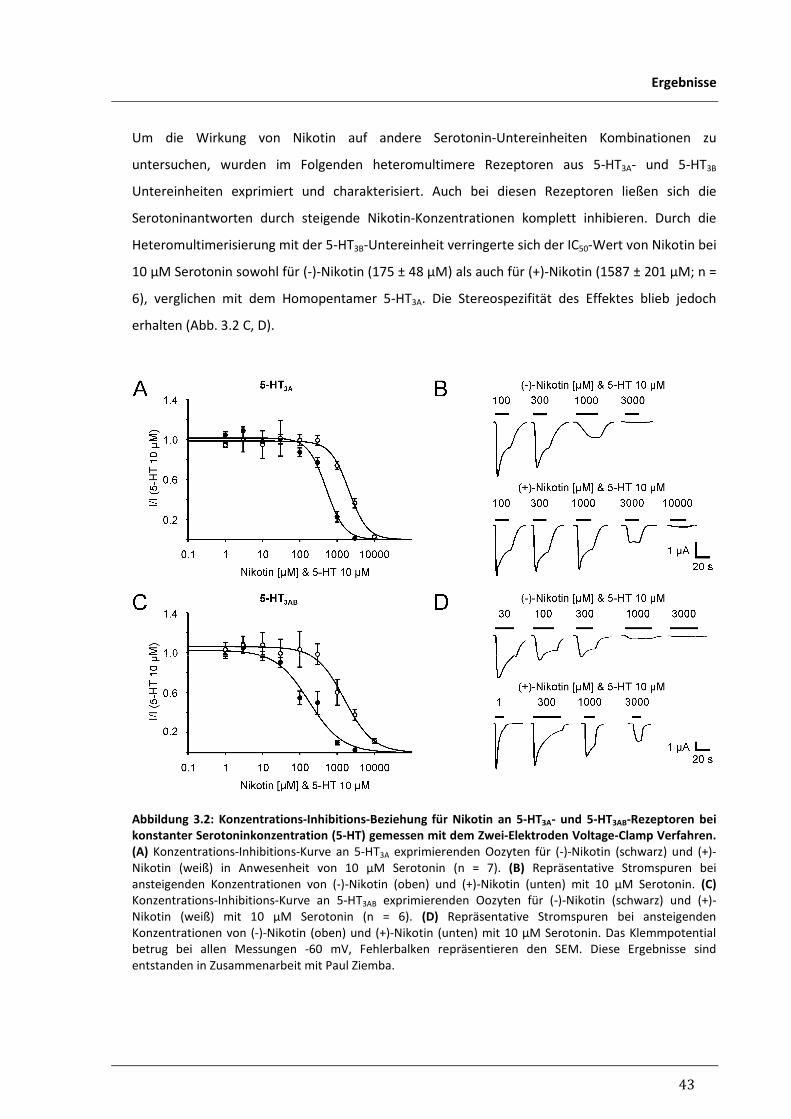
43
Ergebnisse
Um die Wirkung von Nikotin auf andere Serotonin-Untereinheiten Kombinationen zu
untersuchen, wurden im Folgenden heteromultimere Rezeptoren aus 5-HT3A- und 5-HT3B
Untereinheiten exprimiert und charakterisiert. Auch bei diesen Rezeptoren ließen sich die
Serotoninantworten durch steigende Nikotin-Konzentrationen komplett inhibieren. Durch die
Heteromultimerisierung mit der 5-HT3B-Untereinheit verringerte sich der IC50-Wert von Nikotin bei
10 µM Serotonin sowohl für (-)-Nikotin (175 ± 48 µM) als auch für (+)-Nikotin (1587 ± 201 µM; n =
6), verglichen mit dem Homopentamer 5-HT3A. Die Stereospezifität des Effektes blieb jedoch
erhalten (Abb. 3.2 C, D).
Abbildung 3.2: Konzentrations-Inhibitions-Beziehung für Nikotin an 5-HT3A- und 5-HT3AB-Rezeptoren bei
konstanter Serotoninkonzentration (5-HT) gemessen mit dem Zwei-Elektroden Voltage-Clamp Verfahren.
(A) Konzentrations-Inhibitions-Kurve an 5-HT3A exprimierenden Oozyten für (-)-Nikotin (schwarz) und (+)-
Nikotin (weiß) in Anwesenheit von 10 µM Serotonin (n = 7). (B) Repräsentative Stromspuren bei
ansteigenden Konzentrationen von (-)-Nikotin (oben) und (+)-Nikotin (unten) mit 10 µM Serotonin. (C)
Konzentrations-Inhibitions-Kurve an 5-HT3AB exprimierenden Oozyten für (-)-Nikotin (schwarz) und (+)-
Nikotin (weiß) mit 10 µM Serotonin (n = 6). (D) Repräsentative Stromspuren bei ansteigenden
Konzentrationen von (-)-Nikotin (oben) und (+)-Nikotin (unten) mit 10 µM Serotonin. Das Klemmpotential
betrug bei allen Messungen -60 mV, Fehlerbalken repräsentieren den SEM. Diese Ergebnisse sind
entstanden in Zusammenarbeit mit Paul Ziemba.

44
Ergebnisse
3.1.2 Aktivierung humaner nikotinischer Acetylcholinrezeptoren durch (+)-Nikotin
Eine Aktivierung nikotinischer Acetylcholinrezeptoren (nAChR) durch (+)-Nikotin wurde zunächst
an der nAChR Untereinheit α7 untersucht, da diese funktionale Homopentamere bilden und die
am häufigsten vorkommenden nAChR im Trigeminus sind (Drisdel and Green, 2000; Liu et al.,
1998). Die direkte Aktivierung des nAChR α7 durch Nikotin wurde in einer vorhergehenden Studie
bereits untersucht (Peng et al., 1994). Die Stereoselektivität nikotinischer AChR ist seit langem
bekannt und wurde mehrfach anhand von Radioliganden-Bindungsstudien nachgewiesen
(Ikushima et al., 1982; Sloan et al., 1985; Zhang and Nordberg, 1993). Mit einer
elektrophysiologischen Messmethode im rekombinanten System wurde dies bei nAChR des
Haushuhns untersucht, wobei erwartungsgemäß (+)-Nikotin als ein schwächerer Agonist
nachgewiesen werden konnte (Amar et al., 1993). Eine elektrophysiologische Untersuchung der
Stereoselektivität eines humanen nAChR ist jedoch nicht bekannt. Um die Stereoselektivität von
Nikotin auf den humanen nAChR α7 zu untersuchen, wurden aufsteigende Nikotin-
Konzentrationen von (+)-Nikotin appliziert. Aus den normierten, relativen Stromamplituden
wurde eine Konzentrations-Wirkungs-Beziehung mit einem EC50 von 519 ± 189 µM (nH = 2,53 ±
1,4; n = 7-9) erstellt. Bei dieser Charakterisierung der Direktwirkung von (+)-Nikotin auf α7 wurden
durchschnittliche Ströme von 46 ± 16 nA (100 µM) bis 196 ± 44 nA (3 mM) gemessen. Ein
Vergleich der normierten Stromamplituden (I/I(ACh)) aller drei untersuchten Agonisten bei einer
sättigenden Konzentration von 1 mM zeigt, dass der natürliche Agonist Acetylcholin die höchste
gemittelte Stromamplitude hervorruft. Die durch (-)-Nikotin hervorgerufene Amplitude ist mit
0,77 ± 0,1 etwas geringer, jedoch unterscheiden sich beide Amplituden nicht signifikant
voneinander (p = 0,69). Die von (+)-Nikotin hervorgerufene Stromantwort beträgt lediglich 0,12 ±
0,02 des durch ACh induzierten Stroms (p < 0,001). Nikotin besitzt als Agonist am nAChR α7 in
Form des (-)-Enantiomers eine etwa 6,4-fach höhere Maximalwirkung als (+)-Nikotin (Abb. 3.3). Im
Vergleich zu Acetylcholin ist Nikotin in beiden Enantiomeren am nAChR α7 ein partieller Agonist.
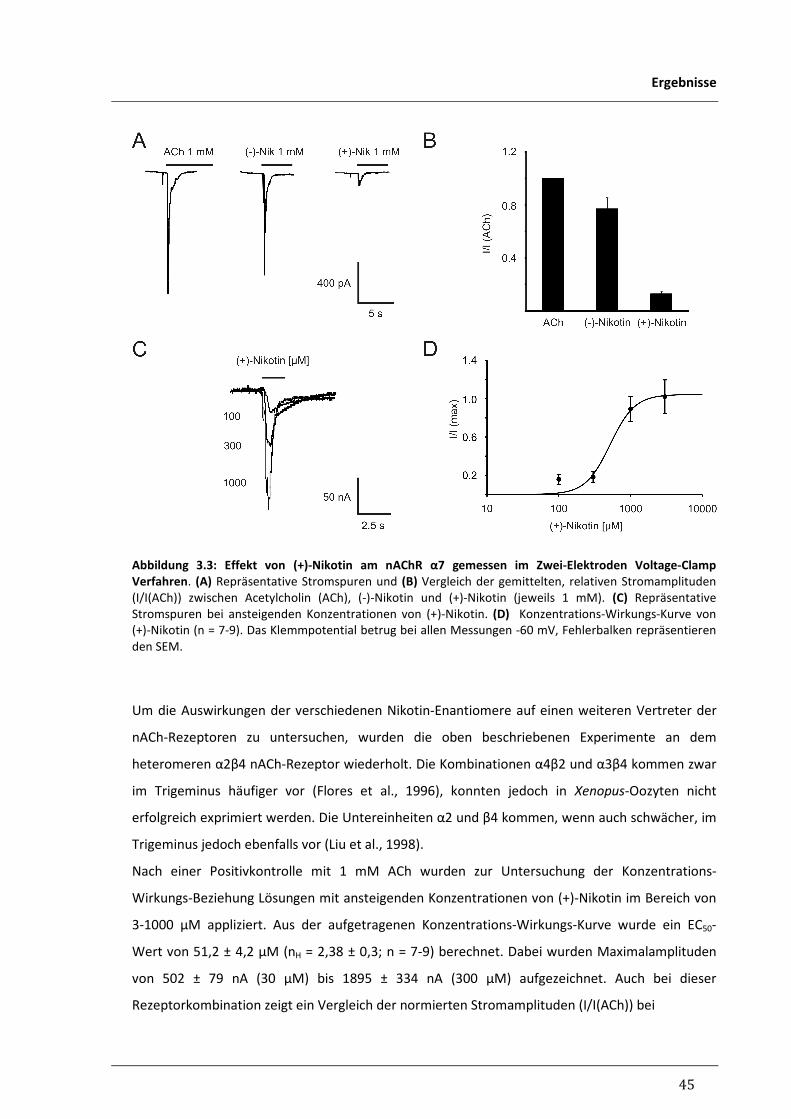
45
Ergebnisse
Abbildung 3.3: Effekt von (+)-Nikotin am nAChR α7 gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren. (A) Repräsentative Stromspuren und (B) Vergleich der gemittelten, relativen Stromamplituden
(I/I(ACh)) zwischen Acetylcholin (ACh), (-)-Nikotin und (+)-Nikotin (jeweils 1 mM). (C) Repräsentative
Stromspuren bei ansteigenden Konzentrationen von (+)-Nikotin. (D) Konzentrations-Wirkungs-Kurve von
(+)-Nikotin (n = 7-9). Das Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken repräsentieren
den SEM.
Um die Auswirkungen der verschiedenen Nikotin-Enantiomere auf einen weiteren Vertreter der
nACh-Rezeptoren zu untersuchen, wurden die oben beschriebenen Experimente an dem
heteromeren α2β4 nACh-Rezeptor wiederholt. Die Kombinationen α4β2 und α3β4 kommen zwar
im Trigeminus häufiger vor (Flores et al., 1996), konnten jedoch in Xenopus-Oozyten nicht
erfolgreich exprimiert werden. Die Untereinheiten α2 und β4 kommen, wenn auch schwächer, im
Trigeminus jedoch ebenfalls vor (Liu et al., 1998).
Nach einer Positivkontrolle mit 1 mM ACh wurden zur Untersuchung der Konzentrations-
Wirkungs-Beziehung Lösungen mit ansteigenden Konzentrationen von (+)-Nikotin im Bereich von
3-1000 µM appliziert. Aus der aufgetragenen Konzentrations-Wirkungs-Kurve wurde ein EC50-
Wert von 51,2 ± 4,2 µM (nH = 2,38 ± 0,3; n = 7-9) berechnet. Dabei wurden Maximalamplituden
von 502 ± 79 nA (30 µM) bis 1895 ± 334 nA (300 µM) aufgezeichnet. Auch bei dieser
Rezeptorkombination zeigt ein Vergleich der normierten Stromamplituden (I/I(ACh)) bei

46
Ergebnisse
sättigenden Konzentrationen (jeweils 1 mM), dass Acetylcholin die höchste gemittelte
Stromamplitude hervorruft, während (-)-Nikotin mit 0,88 ± 0,07 ein starker Agonist ist. Die
Maximalwirkung von (-)-Nikotin unterscheidet sich nicht signifikant von der Wirkung von
Acetylcholin (p = 0,18). Mit 0,13 ± 0,03 des Acetylcholin-induzierten Maximalstromes ist (+)-
Nikotin ein schwacher partieller Agonist (p < 0,001). Als Agonist am nAChR α2β4 hat das (-)-
Enantiomer somit etwa eine 6,7-fach höhere Maximalwirkung als (+)-Nikotin. Beim Betrachten
der Kinetik wird deutlich, dass diese Untereinheitenkombination in Gegenwart von (+)-Nikotin
verglichen mit dem vollen Agonisten Acetylcholin und (-)-Nikotin weniger stark desensitisiert.
(Abb. 3.4).
Abbildung 3.4: Effekt von (+)-Nikotin am nAChR α2β4 gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren. (A) Repräsentative Stromspuren und (B) Vergleich der gemittelten, relativen Stromamplituden
(I/I(ACh)) bei Acetylcholin (ACh), (-)-Nikotin und (+)-Nikotin (jeweils 1 mM). (C) Repräsentative Stromspuren
bei ansteigenden Konzentrationen von (+)-Nikotin. (D) Konzentrations-Wirkungs-Kurve von (+)-Nikotin (n =
7-9). Das Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken repräsentieren den SEM.
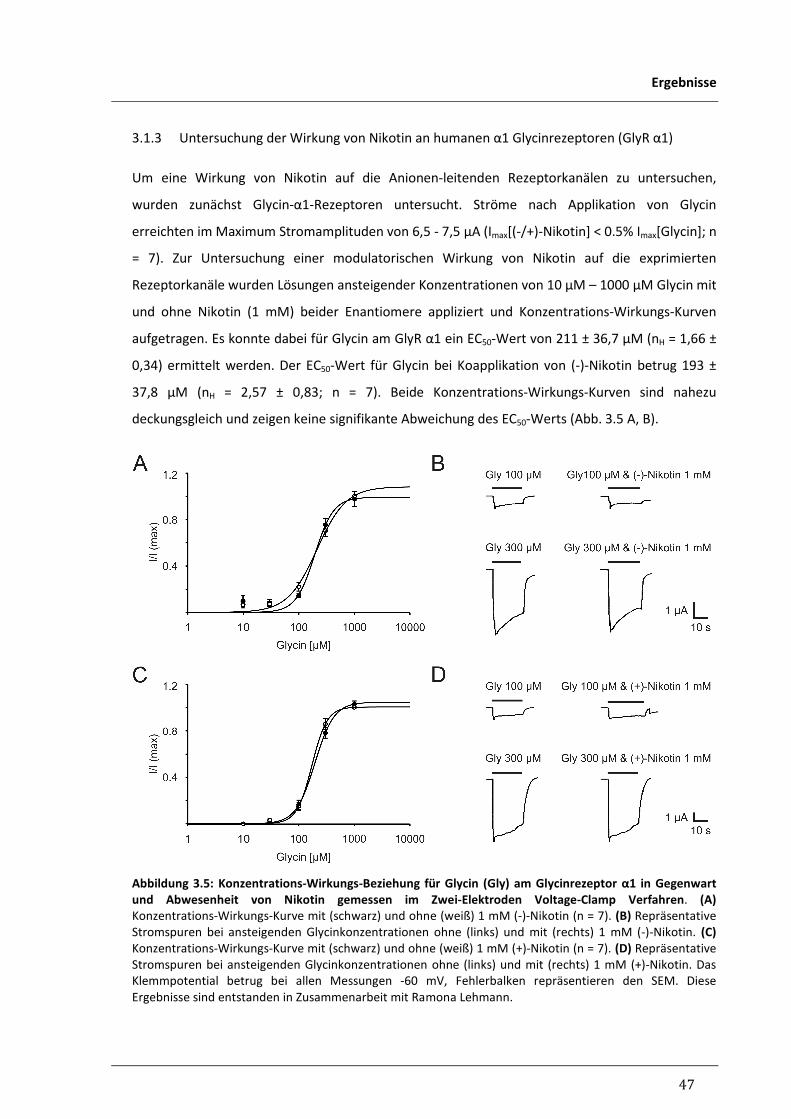
47
Ergebnisse
3.1.3 Untersuchung der Wirkung von Nikotin an humanen α1 Glycinrezeptoren (GlyR α1)
Um eine Wirkung von Nikotin auf die Anionen-leitenden Rezeptorkanälen zu untersuchen,
wurden zunächst Glycin-α1-Rezeptoren untersucht. Ströme nach Applikation von Glycin
erreichten im Maximum Stromamplituden von 6,5 - 7,5 µA (Imax[(-/+)-Nikotin] < 0.5% Imax[Glycin]; n
= 7). Zur Untersuchung einer modulatorischen Wirkung von Nikotin auf die exprimierten
Rezeptorkanäle wurden Lösungen ansteigender Konzentrationen von 10 µM – 1000 µM Glycin mit
und ohne Nikotin (1 mM) beider Enantiomere appliziert und Konzentrations-Wirkungs-Kurven
aufgetragen. Es konnte dabei für Glycin am GlyR α1 ein EC50-Wert von 211 ± 36,7 µM (nH = 1,66 ±
0,34) ermittelt werden. Der EC50-Wert für Glycin bei Koapplikation von (-)-Nikotin betrug 193 ±
37,8 µM (nH = 2,57 ± 0,83; n = 7). Beide Konzentrations-Wirkungs-Kurven sind nahezu
deckungsgleich und zeigen keine signifikante Abweichung des EC50-Werts (Abb. 3.5 A, B).
Abbildung 3.5: Konzentrations-Wirkungs-Beziehung für Glycin (Gly) am Glycinrezeptor α1 in Gegenwart
und Abwesenheit von Nikotin gemessen im Zwei-Elektroden Voltage-Clamp Verfahren. (A)
Konzentrations-Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (-)-Nikotin (n = 7). (B) Repräsentative
Stromspuren bei ansteigenden Glycinkonzentrationen ohne (links) und mit (rechts) 1 mM (-)-Nikotin. (C)
Konzentrations-Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (+)-Nikotin (n = 7). (D) Repräsentative
Stromspuren bei ansteigenden Glycinkonzentrationen ohne (links) und mit (rechts) 1 mM (+)-Nikotin. Das
Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken repräsentieren den SEM. Diese
Ergebnisse sind entstanden in Zusammenarbeit mit Ramona Lehmann.

48
Ergebnisse
Für das (+)-Enantiomer wurden vergleichbare Ergebnisse festgestellt. Da Messungen für (+)- und
(-)-Enantiomer an verschiedenen Oozyten durchgeführt wurden, musste für diese zweite
Messreihe aus den entsprechenden Kontrollen erneut ein EC50-Wert für Glycin separat ermittelt
werden. Der EC50-Wert für Glycin betrug hier 172 ± 17,8 µM (nH 3,17 ± 0,22), der für Glycin unter
Koapplikation von (+)-Nikotin betrug 195 ± 16,7 µM (nH = 2,62 ± 0,15; n = 7) Die EC50-Werte und
der Hill-Koeffizient
unterscheiden sich nicht signifikant voneinander (Abb. 3.5 C, D). Die oben aufgeführten
Ergebnisse zeigen deutlich, dass beide Nikotin-Enantiomere keine Wirkung auf humane GlyR α1
Rezeptoren haben.
3.1.4 Untersuchung der Wirkung von Nikotin an humanen GABAA-Rezeptoren
Als weiterer Vertreter der ligandengesteuerter Ionenkanäle wurden GABAA-Rezeptoren
untersucht. Dazu wurden fünf verschiedene Untereinheiten-Kombinationen untersucht, drei
Heteropentamere (α1β2γ2, α2β1γ2, α2β3γ2) und zwei Homopentamere (β3, ρ1). Von den
untersuchten Kombinationen gehören insbesondere α1β2γ2, α2β3γ2 zu den am weitesten
verbreiteten Kombinationen gehören (McKernan and Whiting, 1996; Nutt, 2006; Pirker et al.,
2000). Die einzelnen Untereinheiten mit Ausnahme von α2 und ρ1 gehören im Trigeminus zudem
zu den stark exprimierten GABA-Rezeptoren (Hayasaki et al., 2006).
Die Wirkung von Nikotin wurde mit dem (-)-Nikotin-Enantiomer charakterisiert. Um eine direkte
Wirkung auf GABAA-Rezeptoren zu untersuchen, wurde 1 mM (-)-Nikotin appliziert, worauf
Stromantworten im Bereich von 15 – 30 nA registriert werden konnten. Verglichen mit der
Applikation des Agonisten GABA, welcher in den verschiedenen Rezeptorkombinationen sich stark
unterscheidende Maximalstromamplituden hervorrief, waren die durch (-)-Nikotin induzierten
Amplituden vernachlässigbar gering (Imax[Nikotin] < 0.5% Imax[GABA]; n = 4-5). Im Anschluss
wurden durch ansteigende GABA-Konzentrationen (1 µM – 1000 µM) mit und ohne 1 mM
(-)-Nikotin Konzentrations-Wirkungs-Kurven erstellt.
Für GABA ergab diese Konzentrations-Wirkungs-Kurve im Falle des α1β2γ2 GABAA-Rezeptors ein
EC50-Wert von 52,3 ± 28,6 µM (nH = 1,62 ± 0,25), bei Koapplikation von (-)-Nikotin einen EC50-Wert
von 32,2 ± 6,81 µM (nH = 1,74 ± 0,15; n = 4). Die Koapplikation von Nikotin und GABA führte dabei
zu keiner signifikanten Veränderung der Konzentrations-Wirkungs-Beziehung (p = 0,45; Abb. 3.6
A).
Im Folgenden wurden α2β1γ2 GABAA-Rezeptoren wie oben beschrieben untersucht. Dabei
ergaben sich EC50-Wert für GABA von 53,3 ± 8,45 µM (nH = 1,43 ± 0,15) und GABA + (-)-Nikotin von

49
Ergebnisse
63,5 ± 7,14 µM (nH = 1,65 ± 0,13; n = 5), welche sich nicht signifikant voneinander unterscheiden
(p = 0,12; Abb. 3.6 B).
Des Weiteren wurde für den α2β3γ2 GABAA-Rezeptor ein EC50-Wert von 119 ± 1,74 µM (nH = 3,43
± 0,64) für den Agonisten GABA und 117 ± 2,87 µM (nH = 3,1 ± 0,33; n = 5) für die Koapplikation
von GABA und (-)-Nikotin ermittelt, welche sich ebenfalls nicht signifikant voneinander
unterscheiden (p = 0,78; Abb. 3.6 C).
Abbildung 3.6: Der Einfluss von (-)-Nikotin auf die Konzentrations-Wirkungs-Beziehung für GABA an
GABAA-Rezeptoren gemessen im Zwei-Elektroden Voltage-Clamp Verfahren. Konzentrations-Wirkungs-
Kurven am GABAA-Rezeptor (A) α1β2γ2 (n = 4), (B) α 2β1γ2 (n = 5), (C) α 2β3γ2 (n = 5), (D) β3 (n = 5) und (E)
ρ1 (n = 5) mit (schwarz) und ohne (weiß) 1 mM (-)-Nikotin, sowie exemplarische Membranstromspuren
ohne (links) und mit (rechts) 1 mM (-)-Nikotin. Das Klemmpotential betrug bei allen Messungen -60 mV,
Fehlerbalken repräsentieren den SEM.

50
Ergebnisse
Der GABAA-Rezeptor β3 kann homomer ohne eine α-Untereinheit funktionell exprimiert werden
(Saras et al., 2008). Für diesen homopentameren Rezeptor konnte ein GABA EC50-Wert von 29,9 ±
13,7 µM (nH = 2,35 ± 0,87) und 14,4 ± 2,55 µM (nH = 1,63 ± 0,4; n = 5) für GABA + (-)-Nikotin
festgestellt werden. Beide Werte unterscheiden sich ebenfalls nicht signifikant (p = 0,31; Abb. 3.6
D). Darüber hinaus wurde die GABAA-Untereinheit ρ1 untersucht, welche nativ sowohl
homopentamer als auch heteropentamer mit anderen ρ-Untereinheiten vorkommt (Cutting et al.,
1991). Der für GABA ermittelte EC50-Wert beträgt für diesen Rezeptor 0,88 ± 0,11 µM (nH = 1,32 ±
0,23), für GABA + (-)-Nikotin 1,51 ± 0,3 µM (nH = 0,93 ± 0,18; n = 5). Die Unterschiede in den
normierten Stromamplituden wie auch der Unterschied in den EC50-Werten sind nicht signifikant
(p = 0,21; Abb. 3.6 E). Es konnte für (-)-Nikotin an keinem der untersuchten GABAA-Rezeptoren
eine Wirkung nachgewiesen werden.
3.1.5 Untersuchung der Wirkung von Nikotin am humanen P2X-Rezeptor
Aus der Familie der Purinozeptoren wurde die Wirkung von Nikotin am P2X2-Rezeptor untersucht.
Die Applikation des Agonisten ATP führt zu Maximalstromamplituden von etwa 4,5-6,5 µA
(Imax[(-/+)-Nikotin] < 0.5% Imax[ATP]; n = 6). Zur Untersuchung einer modulatorischen Wirkung von
Nikotin auf P2X wurden Lösungen ansteigender Konzentrationen von ATP im Bereich von 0,1 µM
– 100 µM mit und ohne Nikotin (1 mM) beider Enantiomere appliziert. Es konnten aus den
normierten relativen Stromamplituden Konzentrations-Wirkungs-Kurven aufgetragen und EC50-
Werte berechnet werden. Für ATP wurde ein Wert von 13,7 ± 0,56 µM (nH = 2,37 + 0,19) und für
ATP mit (-)-Nikotin ein Wert von 14,2 ± 0,3 µM (nH = 2,03 + 0,07; n = 6) bestimmt. Beide
aufgetragenen Konzentrations-Wirkungs-Kurven verlaufen nahezu deckungsgleich. Die EC50-Werte
unterscheiden sich nicht signifikant voneinander (Abb. 3.7 A, B).
Für das (+)-Enantiomer wurden vergleichbare Experimente durchgeführt. Da Messungen für (+)-
und (-)-Enantiomer an verschiedenen Oozyten durchgeführt wurden, musste für diese zweite
Messreihe aus den entsprechenden Kontrollen erneut ein EC50-Wert für ATP separat ermittelt
werden. Es konnten ebenfalls Konzentrations-Wirkungs-Kurven aufgetragen und EC50-Werte
berechnet werden. Für ATP wurde hierbei ein Wert von 13,9 ± 0,2 µM (nH = 2,0 ± 0,1), für ATP und
(+)-Nikotin ein Wert von 9,64 ± 1,1 µM (nH = 2,51 + 0,87; n = 9) berechnet. Die Kurven verlaufen
nahezu deckungsgleich. Die EC50-Werte unterscheiden sich nicht signifikant (Abb. 3.7 C, D). Es
konnte für keines der beiden Nikotinenantiomere eine direkte oder modulatorische Wirkung am
humanen P2X-Rezeptor nachgewiesen werden.
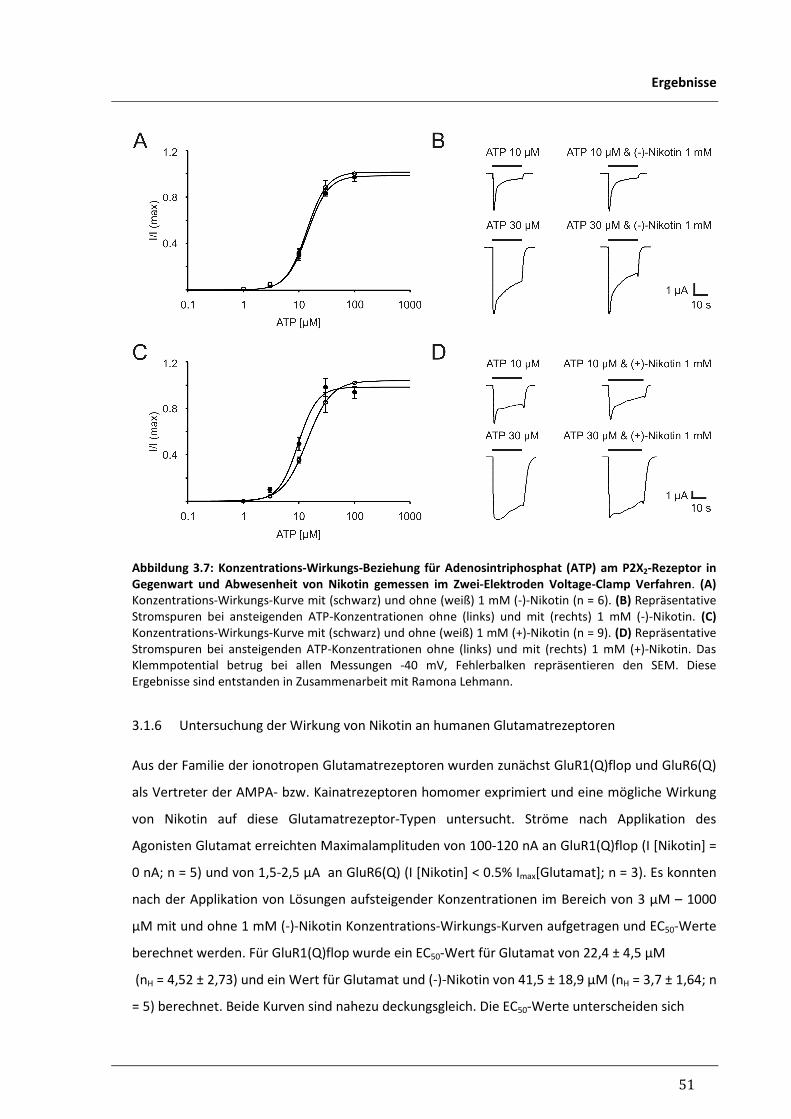
51
Ergebnisse
Abbildung 3.7: Konzentrations-Wirkungs-Beziehung für Adenosintriphosphat (ATP) am P2X2-Rezeptor in
Gegenwart und Abwesenheit von Nikotin gemessen im Zwei-Elektroden Voltage-Clamp Verfahren. (A)
Konzentrations-Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (-)-Nikotin (n = 6). (B) Repräsentative
Stromspuren bei ansteigenden ATP-Konzentrationen ohne (links) und mit (rechts) 1 mM (-)-Nikotin. (C)
Konzentrations-Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (+)-Nikotin (n = 9). (D) Repräsentative
Stromspuren bei ansteigenden ATP-Konzentrationen ohne (links) und mit (rechts) 1 mM (+)-Nikotin. Das
Klemmpotential betrug bei allen Messungen -40 mV, Fehlerbalken repräsentieren den SEM. Diese
Ergebnisse sind entstanden in Zusammenarbeit mit Ramona Lehmann.
3.1.6 Untersuchung der Wirkung von Nikotin an humanen Glutamatrezeptoren
Aus der Familie der ionotropen Glutamatrezeptoren wurden zunächst GluR1(Q)flop und GluR6(Q)
als Vertreter der AMPA- bzw. Kainatrezeptoren homomer exprimiert und eine mögliche Wirkung
von Nikotin auf diese Glutamatrezeptor-Typen untersucht. Ströme nach Applikation des
Agonisten Glutamat erreichten Maximalamplituden von 100-120 nA an GluR1(Q)flop (I [Nikotin] =
0 nA; n = 5) und von 1,5-2,5 µA an GluR6(Q) (I [Nikotin] < 0.5% Imax[Glutamat]; n = 3). Es konnten
nach der Applikation von Lösungen aufsteigender Konzentrationen im Bereich von 3 µM – 1000
µM mit und ohne 1 mM (-)-Nikotin Konzentrations-Wirkungs-Kurven aufgetragen und EC50-Werte
berechnet werden. Für GluR1(Q)flop wurde ein EC50-Wert für Glutamat von 22,4 ± 4,5 µM
(nH = 4,52 ± 2,73) und ein Wert für Glutamat und (-)-Nikotin von 41,5 ± 18,9 µM (nH = 3,7 ± 1,64; n
= 5) berechnet. Beide Kurven sind nahezu deckungsgleich. Die EC50-Werte unterscheiden sich
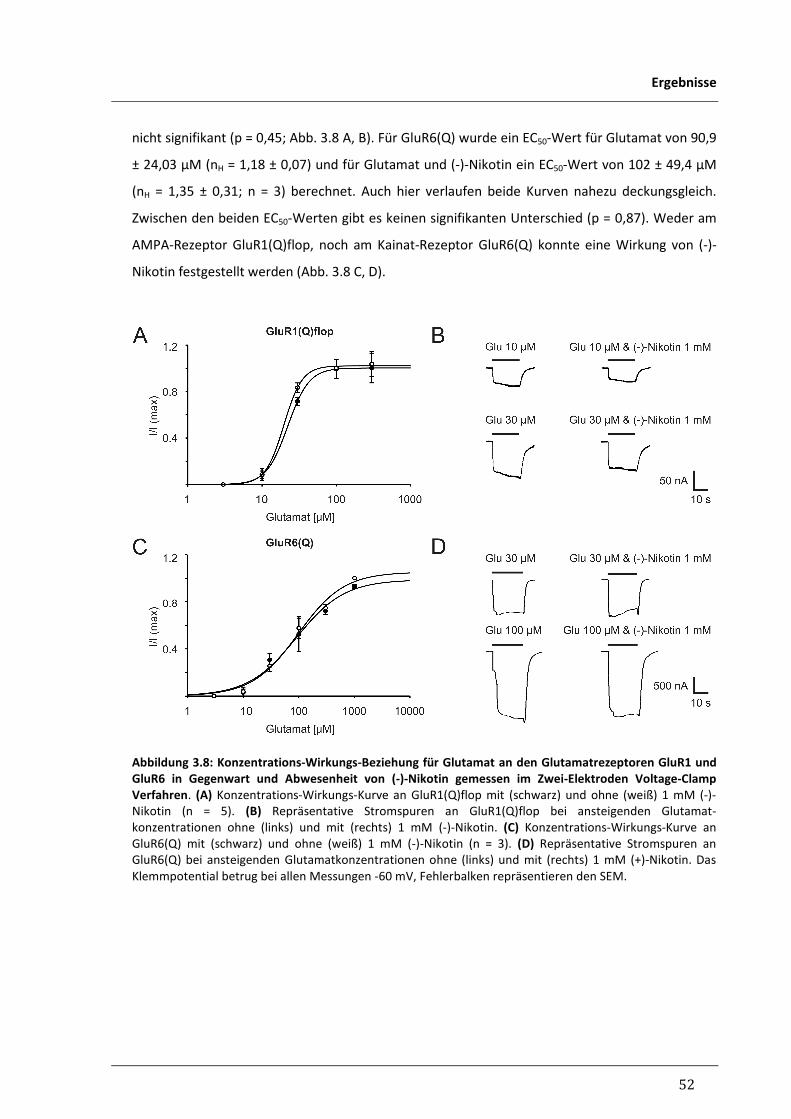
52
Ergebnisse
nicht signifikant (p = 0,45; Abb. 3.8 A, B). Für GluR6(Q) wurde ein EC50-Wert für Glutamat von 90,9
± 24,03 µM (nH = 1,18 ± 0,07) und für Glutamat und (-)-Nikotin ein EC50-Wert von 102 ± 49,4 µM
(nH = 1,35 ± 0,31; n = 3) berechnet. Auch hier verlaufen beide Kurven nahezu deckungsgleich.
Zwischen den beiden EC50-Werten gibt es keinen signifikanten Unterschied (p = 0,87). Weder am
AMPA-Rezeptor GluR1(Q)flop, noch am Kainat-Rezeptor GluR6(Q) konnte eine Wirkung von (-)-
Nikotin festgestellt werden (Abb. 3.8 C, D).
Abbildung 3.8: Konzentrations-Wirkungs-Beziehung für Glutamat an den Glutamatrezeptoren GluR1 und
GluR6 in Gegenwart und Abwesenheit von (-)-Nikotin gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren. (A) Konzentrations-Wirkungs-Kurve an GluR1(Q)flop mit (schwarz) und ohne (weiß) 1 mM (-)-
Nikotin (n = 5). (B) Repräsentative Stromspuren an GluR1(Q)flop bei ansteigenden Glutamat-
konzentrationen ohne (links) und mit (rechts) 1 mM (-)-Nikotin. (C) Konzentrations-Wirkungs-Kurve an
GluR6(Q) mit (schwarz) und ohne (weiß) 1 mM (-)-Nikotin (n = 3). (D) Repräsentative Stromspuren an
GluR6(Q) bei ansteigenden Glutamatkonzentrationen ohne (links) und mit (rechts) 1 mM (+)-Nikotin. Das
Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken repräsentieren den SEM.

53
Ergebnisse
Als weiterer Glutamatrezeptor wurde aus der Familie der NMDA-Rezeptoren ein Heteromer aus
NR1-3a und NR2B untersucht. Nach Applikation von Glutamat konnten Maximalströme von etwa
1,5-2 µA gemessen werden, nach Applikation von 1 mM (-)-Nikotin konnte hingegen an keiner
Oozyte wurde ein nennenswerter Strom (Imax[(-)-Nikotin] < 0.5% Imax[Glutamtat]; n = 5) gemessen
werden, weshalb eine direkte Aktivierung durch (-)-Nikotin ausgeschlossen werden konnte. Bei
einer Klemmspannung von -60 mV wurden nach Applikation ansteigender
Glutamatkonzentrationen (0,1 µM – 100 µM) mit und ohne 1 mM (-)-Nikotin Konzentrations-
Wirkungs-Kurven aufgetragen. Aus diesen konnte ein EC50-Wert von 3,88 ± 0,77 µM (nH = 1,92 ±
0,47) für Glutamat und 4,8 ± 3,74 µM (nH = 1,0 ± 0,12) für Glutamat und (-)-Nikotin berechnet
werden. Die gemittelte Stromamplitude ist nach jeder Applikation mit (-)-Nikotin geringer im
Vergleich zur Glutamat Kontrolle. Diese Unterschiede sind jedoch nicht signifikant und
beeinflussen den EC50-Wert nicht (p = 0,13; Abb. 3.9 A, B).
Für NMDA-Rezeptoren ist ein Block durch Mg2+-Ionen beschrieben, der von der
Membranspannung abhängig ist (Nowak et al., 1984). Dabei werden positiv geladene Mg2+-Ionen
von einer negativen Membranspannung (unter -40 bis -20 mV) angezogen und setzen sich in die
Kanalpore, was zu einer Blockierung des Ionenflusses und damit des NMDARs führt. Um diesen
möglichen Mg2+-Block auszuschließen, wurden die Messungen unter Mg2+-freien Bedingungen
durchgeführt. Interessanterweise konnte bei Applikation von 10 µM Glutamat und 100 µM – 10
mM (-)-Nikotin ein spannungsabhängiger Block beobachtet werden, wenn die Klemmspannung
von -40 mV auf -100 mV verringert wurde. Es zeigte sich, dass dieser Block stärker wurde je
negativer das gewählte Klemmpotential war und weist damit starke Ähnlichkeiten mit dem Mg2+-
Block auf (Abb. 3.9 C). Eine signifikante Verringerung des Stroms durch diesen Block tritt erst bei
Potentialen negativer als -80 mV bei Koapplikation von 10 mM (-)-Nikotin mit 10 µM Glutamat (p
= 0,016) auf. Durch eine Verringerung der Nikotin-Konzentration auf 1 mM (-)-Nikotin bei -80 mV
konnte keine signifikante Verringerung des Stroms mehr registriert werden; durch eine
Erniedrigung der Klemmspannung auf -100 mV zeigt sich jedoch ein signifikanter Block (p =
0,0039) bei gleichbleibender Nikotin Konzentration. Im Vergleich zum Mg2+-Block (1,8 mM), der
bei einem Klemmpotential von -60 mV den Glutamat-induzierten Strom nahezu vollständig
verhinderte, konnte der spannungsabhängige Nikotin-Block selbst bei hohen (-)-Nikotin-
Konzentrationen (10 mM) und einem Klemmpotential von -100 mV nur etwa 70% des Glutamat-
induzierten Stromes verhindern (Abb. 3.9 D).
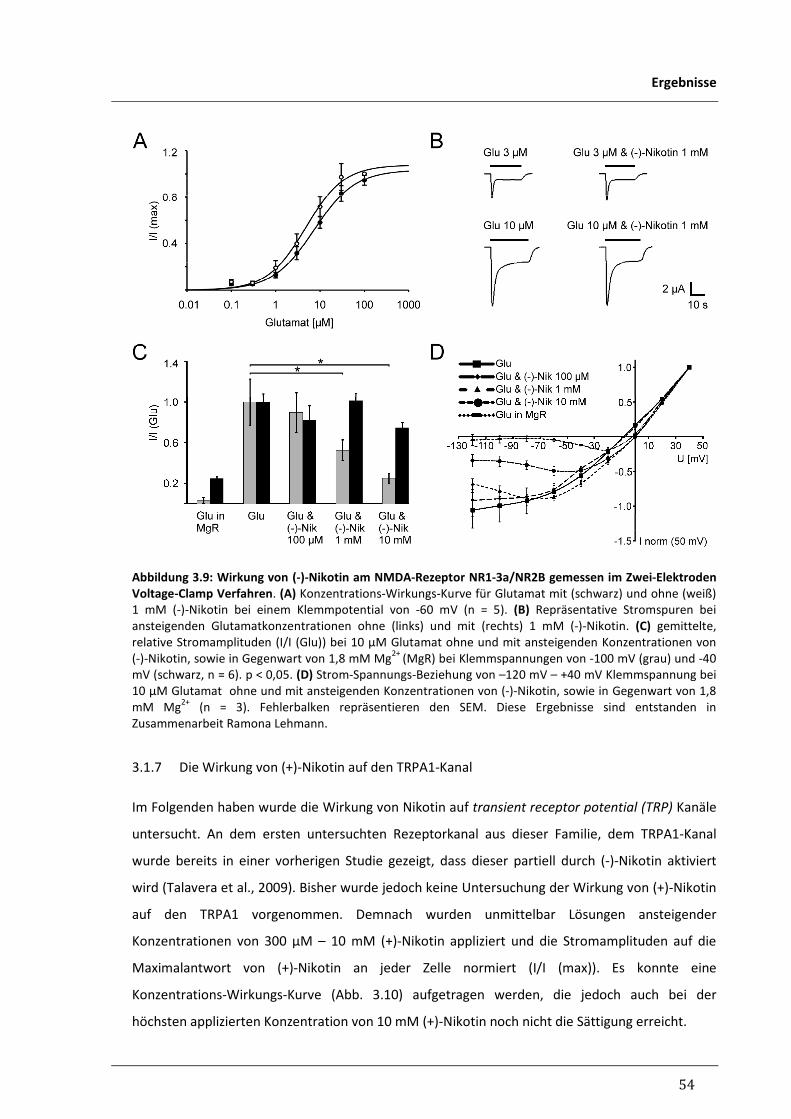
54
Ergebnisse
Abbildung 3.9: Wirkung von (-)-Nikotin am NMDA-Rezeptor NR1-3a/NR2B gemessen im Zwei-Elektroden
Voltage-Clamp Verfahren. (A) Konzentrations-Wirkungs-Kurve für Glutamat mit (schwarz) und ohne (weiß)
1 mM (-)-Nikotin bei einem Klemmpotential von -60 mV (n = 5). (B) Repräsentative Stromspuren bei
ansteigenden Glutamatkonzentrationen ohne (links) und mit (rechts) 1 mM (-)-Nikotin. (C) gemittelte,
relative Stromamplituden (I/I (Glu)) bei 10 µM Glutamat ohne und mit ansteigenden Konzentrationen von
(-)-Nikotin, sowie in Gegenwart von 1,8 mM Mg2+
(MgR) bei Klemmspannungen von -100 mV (grau) und -40
mV (schwarz, n = 6). p < 0,05. (D) Strom-Spannungs-Beziehung von –120 mV – +40 mV Klemmspannung bei
10 µM Glutamat ohne und mit ansteigenden Konzentrationen von (-)-Nikotin, sowie in Gegenwart von 1,8
mM Mg2+
(n = 3). Fehlerbalken repräsentieren den SEM. Diese Ergebnisse sind entstanden in
Zusammenarbeit Ramona Lehmann.
3.1.7 Die Wirkung von (+)-Nikotin auf den TRPA1-Kanal
Im Folgenden haben wurde die Wirkung von Nikotin auf transient receptor potential (TRP) Kanäle
untersucht. An dem ersten untersuchten Rezeptorkanal aus dieser Familie, dem TRPA1-Kanal
wurde bereits in einer vorherigen Studie gezeigt, dass dieser partiell durch (-)-Nikotin aktiviert
wird (Talavera et al., 2009). Bisher wurde jedoch keine Untersuchung der Wirkung von (+)-Nikotin
auf den TRPA1 vorgenommen. Demnach wurden unmittelbar Lösungen ansteigender
Konzentrationen von 300 µM – 10 mM (+)-Nikotin appliziert und die Stromamplituden auf die
Maximalantwort von (+)-Nikotin an jeder Zelle normiert (I/I (max)). Es konnte eine
Konzentrations-Wirkungs-Kurve (Abb. 3.10) aufgetragen werden, die jedoch auch bei der
höchsten applizierten Konzentration von 10 mM (+)-Nikotin noch nicht die Sättigung erreicht.

55
Ergebnisse
Bei der Applikation höherer Konzentrationen setzten trotz der Spannungsklemme des
Messsystems starke Schwankungen der Membranspannung der Oozyte ein, sodass Messungen
nur noch schwer möglich waren. Aus diesem Grund konnte kein EC50-Wert berechnet werden. Als
Kontrolle der Expression von TRPA1 wurde nach jeder Nikotin Konzentrations-Wirkungs-Kurve der
TRPA1-Agonist Allylisothiocyanat (AITC) in einer Konzentration von 100 µM appliziert.
Abbildung 3.10: Effekt von (+)-Nikotin am TRPA1 gemessen im Zwei-Elektroden Voltage-Clamp Verfahren.
(A) Repräsentative Stromspuren und (B) Vergleich der gemittelten, relativen Stromamplituden (I/I(AITC)) für
Allylisothiocyanat (AITC), (-)-Nikotin und (+)-Nikotin (jeweils 10 mM; n = 4-7). (C) Repräsentative
Stromspuren bei ansteigenden Konzentrationen von (+)-Nikotin. (D) Konzentrations-Wirkungs-Kurve für
(+)-Nikotin (n = 5-7). Das Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken repräsentieren
den SEM.
Zum Vergleich der relativen Effektivität beider Nikotin-Enantiomere wurden in weiteren
zusätzlichen Experimenten 10 mM (+)-Nikotin, 10 mM (-)-Nikotin und im Anschluss ein sättigender
Stimulus von 100 µM AITC (Bandell et al., 2004) appliziert (Abb. 3.10). Für (+)-Nikotin konnten
Maximalstromamplituden von 300-570 nA gemessen werden. Für (-)-Nikotin lagen diese bei etwa
bei 250-1800 nA, die Applikation von AITC hingegen führte zu Maximalströmen von 590-9300 nA
(n = 4-7). Ein Vergleich der normierten Stromamplituden (I/I(AITC)) aller drei applizierten

56
Ergebnisse
Agonisten zeigt, dass AITC der Agonist mit der bei Weitem höchsten Maximalwirkung ist. Die bei
(-)-Nikotin gemessene Amplitude beträgt etwa 16 ± 12%, die für (+)-Nikotin etwa 17 ± 9% des
AITC-induzierten Maximalstromes. Die Werte der beiden Nikotin-Enantiomere unterscheiden sich
nicht signifikant voneinander (p = 0,41). Am humanen TRPA1 wirkt demnach nicht nur das
(-)-Enantiomer, sondern auch (+)-Nikotin als partieller Agonist mit vergleichbar hoher
Maximalwirkung. Der TRPA1-Kanal weist den Ergebnissen zufolge in Bezug auf Nikotin keine
Stereoselektivität auf.
3.1.8 Untersuchung der Wirkung von Nikotin an den Vanilloid-Rezeptoren TRPV1-3
Als weitere Rezeptoren der Familie der TRP-Kanäle wurden TRPV1, TRPV2 und TRPV3 aus der
Unterfamilie der Vanilloidrezeptoren untersucht. Liu et al., (2004) zeigten, dass der TRPV1-Kanal
der Spezies Rattus norvegicus positiv durch (-)-Nikotin moduliert wird. Aufbauend auf diesen
Ergebnissen wurde die modulatorische und die direkte Wirkung von beiden Nikotin-Enantiomeren
auf den humanen TRPV1 untersucht. Bei der Applikation von 1 mM Nikotin beider Enantiomere
zeigte sich, dass Nikotin keine direkte Aktivierung des Kanals hervorruft, da keine Stromantwort
registriert werden konnte (Imax[(-/+)-Nikotin] = 0; n = 8-10). Als Positivkontrolle wurde der Agonist
Capsaicin (Caterina et al., 1997) verwendet, nach dessen Applikation Maximalstromamplituden
von 250-900 nA gemessen wurden. Um die modulatorische Wirkung auf TRPV1-Kanäle zu
untersuchen, wurden Lösungen ansteigender Konzentrationen von 10 nM – 10 µM Capsaicin mit
und ohne Nikotin (1 mM) beider Enantiomere appliziert. Die normierten, relativen
Stromamplituden (I/I(max)) wurden als Konzentrations-Wirkungs-Kurve aufgetragen. Die EC50-
Verschiebung für die Modulation der Capsaicin Aktivierung durch Nikotin konnte nicht berechnet
werden, da mit den verwendeten Konzentrationen keine Sättigung erreicht wurde. Die beiden
Nikotin-Enantiomere zeigten keine unterschiedliche Wirkung auf den TRPV1 Rezeptor, da sich der
Mittelwert der Stromamplituden bei keiner Capsaicinkonzentration signifikant voneinander
unterscheiden (Abb. 3.11 A, B). Es konnte am humanen TRPV1 bei keinem der beiden Nikotin-
Enantiomere eine direkte oder modulatorische Wirkung von Nikotin festgestellt werden.
Des Weiteren wurden die Vanilloidrezeptoren TRPV2 und TRPV3 in Hinblick auf ihre Modulation
durch Nikotin untersucht. Nach der Applikation von 1mM und Nikotin beider Enantiomere
konnten an diesen Kanälen keine Stromantwort festgestellt werden (Imax[(-/+)-Nikotin] = 0 nA; n =
5).
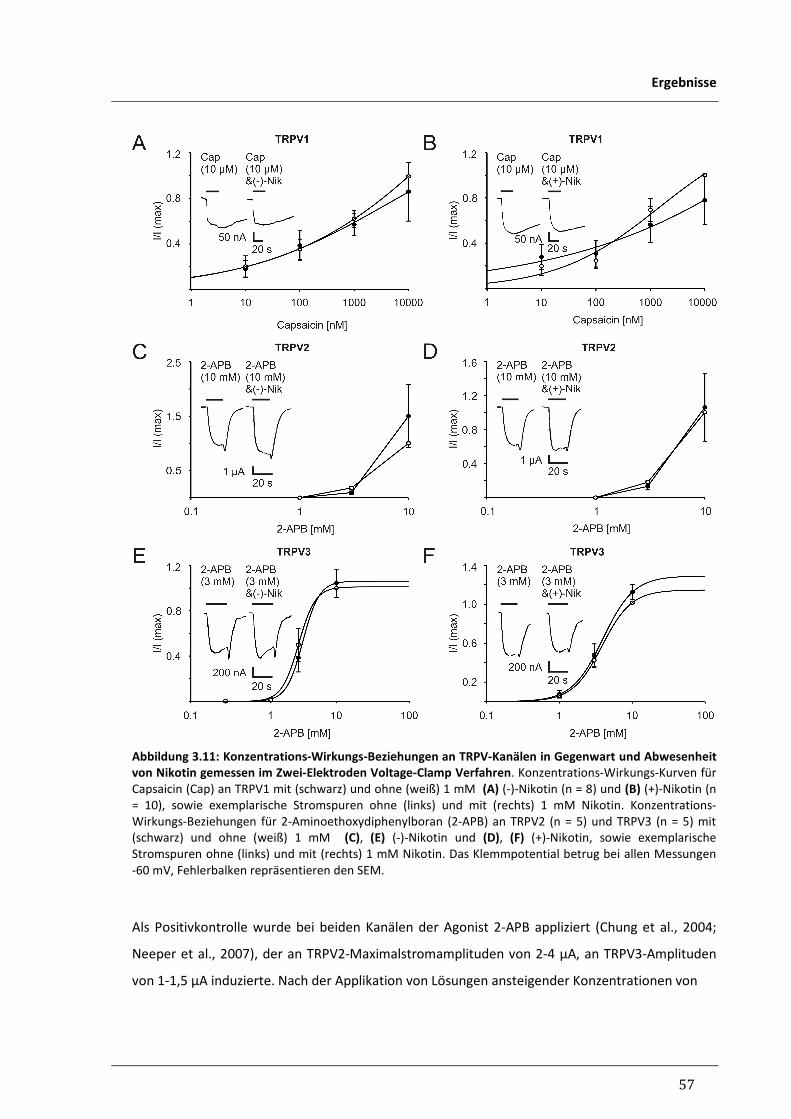
57
Ergebnisse
Abbildung 3.11: Konzentrations-Wirkungs-Beziehungen an TRPV-Kanälen in Gegenwart und Abwesenheit
von Nikotin gemessen im Zwei-Elektroden Voltage-Clamp Verfahren. Konzentrations-Wirkungs-Kurven für
Capsaicin (Cap) an TRPV1 mit (schwarz) und ohne (weiß) 1 mM (A) (-)-Nikotin (n = 8) und (B) (+)-Nikotin (n
= 10), sowie exemplarische Stromspuren ohne (links) und mit (rechts) 1 mM Nikotin. Konzentrations-
Wirkungs-Beziehungen für 2-Aminoethoxydiphenylboran (2-APB) an TRPV2 (n = 5) und TRPV3 (n = 5) mit
(schwarz) und ohne (weiß) 1 mM (C), (E) (-)-Nikotin und (D), (F) (+)-Nikotin, sowie exemplarische
Stromspuren ohne (links) und mit (rechts) 1 mM Nikotin. Das Klemmpotential betrug bei allen Messungen
-60 mV, Fehlerbalken repräsentieren den SEM.
Als Positivkontrolle wurde bei beiden Kanälen der Agonist 2-APB appliziert (Chung et al., 2004;
Neeper et al., 2007), der an TRPV2-Maximalstromamplituden von 2-4 µA, an TRPV3-Amplituden
von 1-1,5 µA induzierte. Nach der Applikation von Lösungen ansteigender Konzentrationen von

58
Ergebnisse
1 mM - 10 mM 2-APB mit und ohne Nikotin (1 mM) beider Enantiomere zeigte sich keine
signifikante Änderung der normierten Stromamplituden. (Abb. 3.11 C, D).
Für TRPV3 wurde ein EC50-Wert von 3026 + 324 µM (nH = 3,75 ± 0,29) für 2-APB und 3522 ± 319
µM (nH = 3,85 ± 0,01; n = 5) für 2-APB und (-)-Nikotin über die Auftragung der Messdaten in einer
Konzentrations-Wirkungs-Beziehung errechnet. Die oben beschrieben Experimente wurden für
das (+)-Enantiomer wiederholt. Da Messungen für (+)- und (-)-Enantiomer an verschiedenen
Oozyten durchgeführt wurden, musste für diese zweite Messreihe aus den entsprechenden
Kontrollen erneut ein EC50-Wert für 2-APB ohne Nikotin separat ermittelt werden. Dabei betrug
der EC50-Wert für 2-APB 4059 ± 530 µM (nH = 2,42 ± 0,27), der für 2-APB und (+)-Nikotin betrug
3367 ± 375 µM (nH = 2,34 ± 0,17; n = 5). Die berechneten EC50-Werte unterscheiden sich nicht
signifikant (p [-] = 0,13; p [+] = 0,35; Abb. 3.11 E, F). Aus diesen Ergebnissen geht hervor, dass
weder (-)- noch (+)-Nikotin eine direkte oder modulatorische Wirkung auf TRPV2 oder TRPV3
haben.
3.1.9 Untersuchung der Wirkung von Nikotin am humanen Melastatinrezeptor TRPM8
Als letzter Kanal aus der Familie der TRP-Kanäle wurde TRPM8 untersucht. Nach Applikation des
Agonisten Menthol (McKemy et al., 2002) wurden Maximalstromamplituden im Bereich von 3-6,5
µA registriert, nach der Applikation von 1 mM Nikotin beider Enantiomere wurde hingegen keine
Stromantwort festgestellt (Imax[(-/+)-Nikotin] < 0,5% Imax[Menthol]). Eine mögliche modulatorische
Wirkung von Nikotin wurde durch die Applikation von Lösungen ansteigender Konzentration von
10 µM – 3 mM Menthol mit und ohne Nikotin (1 mM) beider Enantiomere untersucht. Diese
Messungen ergaben für Menthol einen EC50-Wert von 511 ± 125 µM (nH = 0,89 ± 0,1), für Menthol
und (-)-Nikotin ein Wert von 415 ± 122 µM (nH = 0,93 ± 0,14) und für Menthol und (+)-Nikotin ein
Wert von 291 ± 23,9 µM (nH = 1,02 ± 0,06; n = 7). Die berechneten EC50-Werte unterscheiden sich
nicht signifikant (Abb. 3.12). Es konnte anhand der Ergebnisse weder eine direkte noch eine
modulatorische Wirkung von Nikotin auf den humanen Melastatinrezeptor TRPM8 festgestellt
werden.

59
Ergebnisse
Abbildung 3.12: Der Einfluss von Nikotin auf die Konzentrations-Wirkungs-Beziehung für Menthol (Men)
am TRPM8-Kanal gemessen im Zwei-Elektroden Voltage-Clamp Verfahren. (A) Konzentrations-Wirkungs-
Kurve mit (schwarz) und ohne (weiß) 1 mM (-)-Nikotin (n = 7). (B) Repräsentative Stromspuren bei
ansteigenden Mentholkonzentrationen ohne (links) und mit (rechts) 1 mM (-)-Nikotin. (C) Konzentrations-
Wirkungs-Kurve mit (schwarz) und ohne (weiß) 1 mM (+)-Nikotin (n = 7). (D) Repräsentative Stromspuren
bei ansteigenden Mentholkonzentrationen ohne (links) und mit (rechts) 1 mM (+)-Nikotin. Das
Klemmpotential betrug bei allen Messungen -60 mV, Fehlerbalken repräsentiert den SEM.
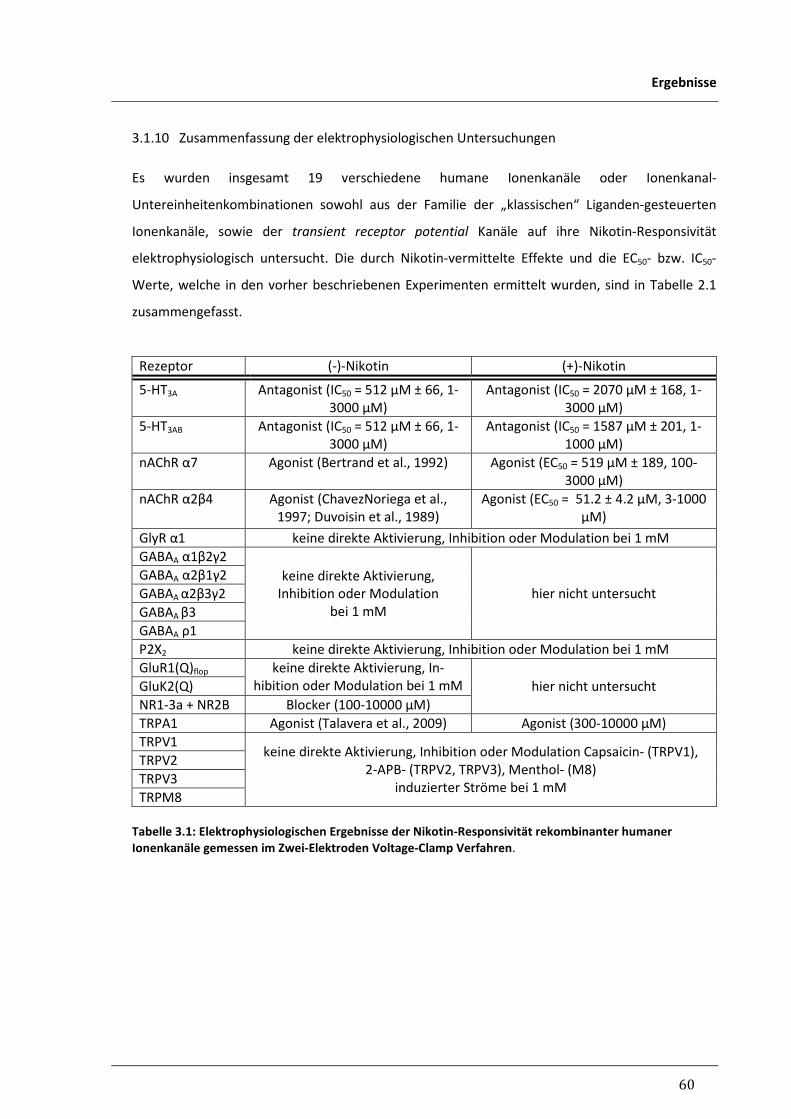
60
Ergebnisse
3.1.10 Zusammenfassung der elektrophysiologischen Untersuchungen
Es wurden insgesamt 19 verschiedene humane Ionenkanäle oder Ionenkanal-
Untereinheitenkombinationen sowohl aus der Familie der „klassischen“ Liganden-gesteuerten
Ionenkanäle, sowie der transient receptor potential Kanäle auf ihre Nikotin-Responsivität
elektrophysiologisch untersucht. Die durch Nikotin-vermittelte Effekte und die EC50- bzw. IC50-
Werte, welche in den vorher beschriebenen Experimenten ermittelt wurden, sind in Tabelle 2.1
zusammengefasst.
Rezeptor (-)-Nikotin (+)-Nikotin
5-HT3A Antagonist (IC50 = 512 µM ± 66, 1-
3000 µM)
Antagonist (IC50 = 2070 µM ± 168, 1-
3000 µM)
5-HT3AB Antagonist (IC50 = 512 µM ± 66, 1-
3000 µM)
Antagonist (IC50 = 1587 µM ± 201, 1-
1000 µM)
nAChR α7 Agonist (Bertrand et al., 1992) Agonist (EC50 = 519 µM ± 189, 100-
3000 µM)
nAChR α2β4 Agonist (ChavezNoriega et al.,
1997; Duvoisin et al., 1989)
Agonist (EC50 = 51.2 ± 4.2 µM, 3-1000
µM)
GlyR α1 keine direkte Aktivierung, Inhibition oder Modulation bei 1 mM
GABAA α1β2γ2
keine direkte Aktivierung, Inhibition oder Modulation
bei 1 mM
hier nicht untersucht
GABAA α2β1γ2
GABAA α2β3γ2
GABAA β3
GABAA ρ1
P2X2 keine direkte Aktivierung, Inhibition oder Modulation bei 1 mM
GluR1(Q)flop keine direkte Aktivierung, In-
hibition oder Modulation bei 1 mM hier nicht untersucht GluK2(Q)
NR1-3a + NR2B Blocker (100-10000 µM)
TRPA1 Agonist (Talavera et al., 2009) Agonist (300-10000 µM)
TRPV1 keine direkte Aktivierung, Inhibition oder Modulation Capsaicin- (TRPV1),
2-APB- (TRPV2, TRPV3), Menthol- (M8)
induzierter Ströme bei 1 mM
TRPV2
TRPV3
TRPM8
Tabelle 3.1: Elektrophysiologischen Ergebnisse der Nikotin-Responsivität rekombinanter humaner
Ionenkanäle gemessen im Zwei-Elektroden Voltage-Clamp Verfahren.

61
Ergebnisse
3.2 Untersuchungen der Wirkung von Nikotin auf trigeminale Neurone der Maus
Da freie Nervenendigungen trigeminaler Neurone auch Schleimhäute des Mund- und
Rachenraumes innervieren, sind die Neurone bei Rauchern regelmäßig einem nikotinhaltigem
Aerosol ausgesetzt. Zur Untersuchung des Einflusses von Nikotin auf trigeminale Neurone der
Maus wurden zunächst Trigeminus-Primärzellkulturen angelegt (2.2.4). Als
Untersuchungsmethode wurde die Calcium-Imaging Methode verwendet (2.2.5), da sie es erlaubt
die Wirkung pharmakologisch aktiver Substanzen auf eine Vielzahl von Zellen gleichzeitig zu
beobachten. Dabei diente die Auswahl applizierter spezifischer Agonisten und Antagonisten dem
Zweck, die Gesamtheit Nikotin-responsiver Neurone in der trigeminalen Zellkultur bestimmten
pharmakologischen Antwortprofilen zuzuordnen. Diese Antwortprofile ermöglichten Rückschlüsse
auf die funktionelle Expression bestimmter Membranrezeptoren. Als abschließende Kontrolle
wurde stets eine Hochkalium-Lösung appliziert, welche nur vitale Neurone aktiviert. Mit diesem
Test ist es möglich, funktionale Neurone von anderen Zellen in der Trigeminuskultur zu
unterscheiden.
3.2.1 Stereoselektive Responsivität trigeminaler Ganglienzellen auf Nikotin
Um zu untersuchen, ob trigeminale Neurone stereoselektiv auf Nikotin antworten, wurden im
ersten Ansatz 250 µM (-)-Nikotin und nach einer Auswaschphase (> 80 s) 250 µM (+)-Nikotin
appliziert. Um eine Beeinflussung von (-)-Nikotin auf die Antwort von (+)-Nikotin auszuschließen,
wurden die beiden Enantiomere in der Hälfte der durchgeführte Experimente vice versa appliziert.
Bei diesen Experimenten wurden 83 Neurone identifiziert, die auf einen K+-Stimulus mit einem
Calciumtransienten reagierten, wovon 66 keine Nikotin-Responsivität zeigten. Es reagierten 7
Neurone mit einem Calciumtransienten auf beide Nikotinenantiomere, 9 ausschließlich auf (+)-
Nikotin und lediglich 1 Neuron reagierte ausschließlich auf (-)-Nikotin (Abb. 3.13). Diese ersten
Ergebnisse zeigen, dass zum einen die Rezeptorausstattung von trigeminalen Neuronen
heterogen ist, da drei verschiedene Populationen anhand ihrer Nikotin-Responsivität identifiziert
wurden und zum anderen exprimiert demnach etwa die Hälfte der Nikotin-responsiven Neurone
einen Rezeptor, der selektiv auf (+)-Nikotin reagiert. Ein solcher Rezeptor ist bisher unbekannt.
Auch mit den elektrophysiologischen Untersuchungen, welche unter 3.1 beschrieben sind, wurde
kein Rezeptor identifiziert, der stereoselektiv auf (+)-Nikotin reagiert.

62
Ergebnisse
Abbildung 3.13: Calcium-Imaging Signale trigeminaler Neurone auf Stimuli von Nikotin beider
Enantiomere. (A) Exemplarische Messspuren eines (+)-Nikotin- (hellgrau), (-)-Nikotin- (schwarz) und (+/-)-
Nikotin- (dunkelgrau) responsiven Neurons. (B) Prozentualer Anteil der Populationen gemessener Neurone.
Diese Ergebnisse sind entstanden in Zusammenarbeit mit Ulrike Thiel.
3.2.2 Suche nach weiteren Rezeptoren für Nikotin in trigeminalen Neuronen
3.2.2.1 Responsivität trigeminaler Neurone für (-)-Nikotin unter pharmakologischer Inhibition von
nAChR und TRPA1
Für den Nachweis weiterer Membranrezeptoren welche auf Nikotin reagieren, mussten im
Calcium-Imaging der trigeminalen Zellkultur die bekannten Nikotin-induzierten Antworten von
nAChR und TRPA1-Kanälen auf pharmakologischem Wege ausgeschlossen werden. Dazu wurde
die Substanz Mecamylamin eingesetzt, die bei Konzentrationen höher als 1 mM sowohl
nikotinische Acetylcholinrezeptoren als auch TRPA1-Kanäle inhibiert (Talavera et al., 2009). In
weiteren Experimenten wurde Hexamethonium, ein Blocker neuronaler nAChRs (Bertrand et al.,
1992; Charnet et al., 1992; Liu et al., 1993) und HC030031, ein spezifischer TRPA1-Blocker
(McNamara et al., 2007), koappliziert, um Nikotin-induzierte Antworten dieser beiden Rezeptoren
zu inhibieren. Zuvor wurden diese Blocker einzeln eingesetzt. Dabei konnte durch 1 mM
Hexamethonium ein Großteil der durch 250 µM (-)-Nikotin-induzierte Antworten blockiert
werden. Antworten von 154 der 219 (70%) responsiven Neurone wurden inhibiert. Bei einer
Gesamtanzahl vitaler Neuronen von 727 beträgt der inhibierte Anteil etwa 21% (Abb. 3.14 A, B).
Der TRPA1 spezifische Blocker HC030031 (30 µM) konnte 46 von 94 (49%) der (-)-Nikotin-
induzierten Antworten inhibieren. Bei einer Gesamtanzahl gemessener vitaler Neurone von 435
beträgt der inhibierte Anteil hier etwa 11%. Dass es sich bei dieser Zellpopulation um TRPA1
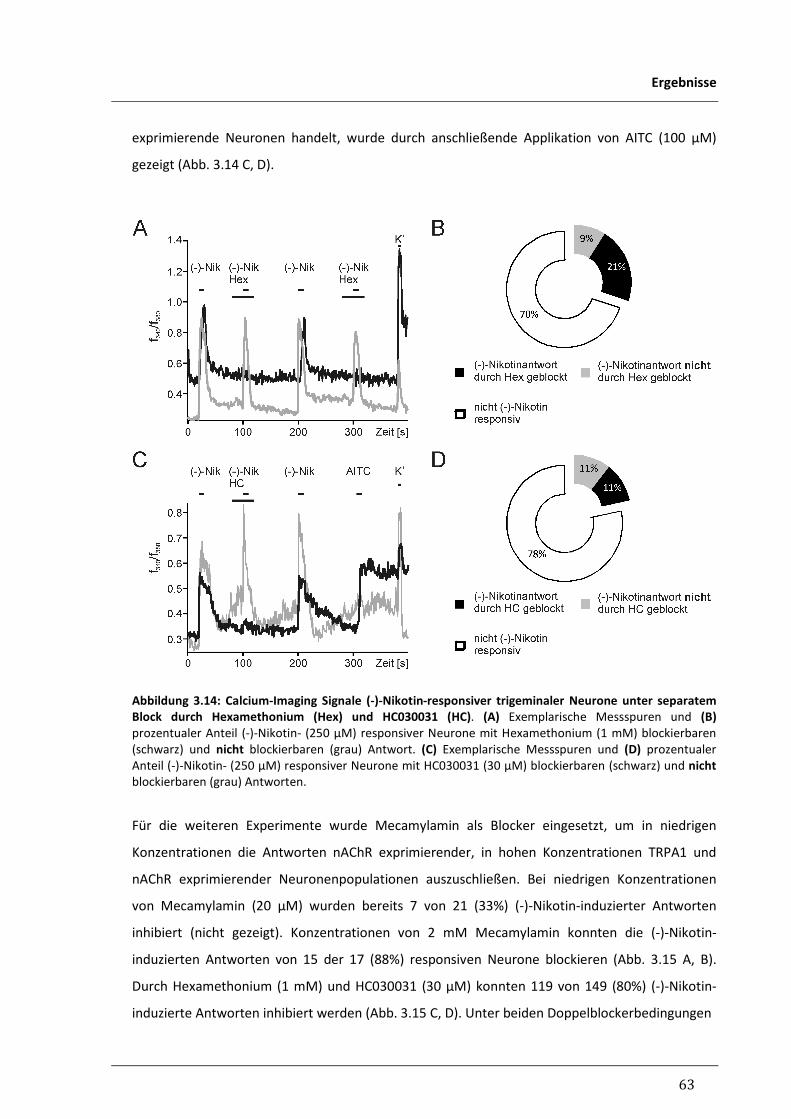
63
Ergebnisse
exprimierende Neuronen handelt, wurde durch anschließende Applikation von AITC (100 µM)
gezeigt (Abb. 3.14 C, D).
Abbildung 3.14: Calcium-Imaging Signale (-)-Nikotin-responsiver trigeminaler Neurone unter separatem
Block durch Hexamethonium (Hex) und HC030031 (HC). (A) Exemplarische Messspuren und (B)
prozentualer Anteil (-)-Nikotin- (250 µM) responsiver Neurone mit Hexamethonium (1 mM) blockierbaren
(schwarz) und nicht blockierbaren (grau) Antwort. (C) Exemplarische Messspuren und (D) prozentualer
Anteil (-)-Nikotin- (250 µM) responsiver Neurone mit HC030031 (30 µM) blockierbaren (schwarz) und nicht
blockierbaren (grau) Antworten.
Für die weiteren Experimente wurde Mecamylamin als Blocker eingesetzt, um in niedrigen
Konzentrationen die Antworten nAChR exprimierender, in hohen Konzentrationen TRPA1 und
nAChR exprimierender Neuronenpopulationen auszuschließen. Bei niedrigen Konzentrationen
von Mecamylamin (20 µM) wurden bereits 7 von 21 (33%) (-)-Nikotin-induzierter Antworten
inhibiert (nicht gezeigt). Konzentrationen von 2 mM Mecamylamin konnten die (-)-Nikotin-
induzierten Antworten von 15 der 17 (88%) responsiven Neurone blockieren (Abb. 3.15 A, B).
Durch Hexamethonium (1 mM) und HC030031 (30 µM) konnten 119 von 149 (80%) (-)-Nikotin-
induzierte Antworten inhibiert werden (Abb. 3.15 C, D). Unter beiden Doppelblockerbedingungen
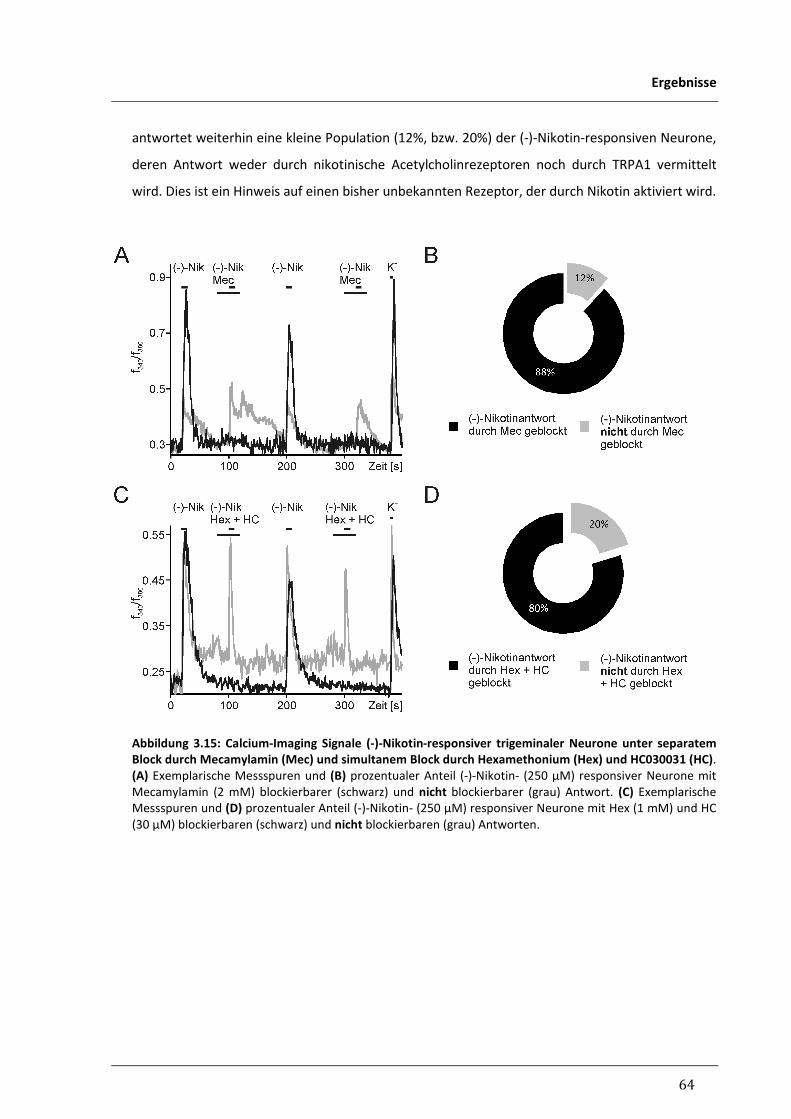
64
Ergebnisse
antwortet weiterhin eine kleine Population (12%, bzw. 20%) der (-)-Nikotin-responsiven Neurone,
deren Antwort weder durch nikotinische Acetylcholinrezeptoren noch durch TRPA1 vermittelt
wird. Dies ist ein Hinweis auf einen bisher unbekannten Rezeptor, der durch Nikotin aktiviert wird.
Abbildung 3.15: Calcium-Imaging Signale (-)-Nikotin-responsiver trigeminaler Neurone unter separatem
Block durch Mecamylamin (Mec) und simultanem Block durch Hexamethonium (Hex) und HC030031 (HC).
(A) Exemplarische Messspuren und (B) prozentualer Anteil (-)-Nikotin- (250 µM) responsiver Neurone mit
Mecamylamin (2 mM) blockierbarer (schwarz) und nicht blockierbarer (grau) Antwort. (C) Exemplarische
Messspuren und (D) prozentualer Anteil (-)-Nikotin- (250 µM) responsiver Neurone mit Hex (1 mM) und HC
(30 µM) blockierbaren (schwarz) und nicht blockierbaren (grau) Antworten.

65
Ergebnisse
3.2.2.2 Responsivität trigeminaler Neurone für (+)-Nikotin
Unter 3.2.1 wurde bereits gezeigt, dass Populationen trigeminaler Neurone stereoselektiv auf
Nikotin reagieren. Anschließend sollte die Beteiligung verschiedener Membranrezeptoren an
(+)-Nikotin-induzierten Antworten der Neurone untersucht werden. Um die beteiligten
Rezeptoren einzugrenzen, wurden für die folgenden Experimente Rutheniumrot als Blocker
eingesetzt, da diese Substanz verschiedene TRP-Kanäle (TRPA1, TRPV und verschiedene
Calciumkanäle) blockiert (Clapham et al., 2005). Es wurden zuerst (+)-Nikotin (250 µM) und
(-)-Nikotin (250 µM) appliziert. Anschließend folgte die Koapplikation von (+)-Nikotin mit
Rutheniumrot (10 µM). Zuletzt wurde der TRPA1-Agonist AITC (100 µM) appliziert. Von 292
untersuchten trigeminalen Neuronen waren 85 (29%) Nikotin-responsiv. Die Neurone konnten
aufgrund ihrer Antwortmuster in verschiedene Populationen eingeteilt werden: 1) 24 (8%) Zellen
wurden identifiziert, die lediglich auf (-)-Nikotin reagierten, 2) 20 (7%) der Zellen reagierten auf
alle drei applizierten Agonisten. Die (+)-Nikotinantworten dieser Zellen konnten nicht durch
Rutheniumrot geblockt werden. 3) 25 (9%) Zellen antworteten auf beide Nikotinenantiomere,
jedoch nicht auf AITC. Die (+)-Nikotinantworten dieser Zellen konnten ebenfalls nicht geblockt
werden. 4) 16 (5%) Zellen antworteten ausschließlich auf (+)-Nikotin, wobei die Antworten durch
Rutheniumrot geblockt werden konnten (Abb. 3.16). Die Antworten dieser Zellen wurden nicht
durch TRPA1 vermittelt, denn diese zeigten keine Sensitivität für AITC. Außerdem wirkt
Rutheniumrot nicht auf nAChR. So können nAChR an diesen geblockten Antworten nicht beteiligt
sein. Die gemessene Antwort wird demnach offenbar von einem weiteren unbekannten
(+)-Nikotinrezeptor vermittelt.

66
Ergebnisse
Abbildung 3.16: Calcium-Imaging Antwortprofile Nikotin-responsiver trigeminaler Neurone bei Block
durch Rutheniumrot (RR). (A) Prozentualer Anteil der Populationen gemessener Neurone. (B), (C)
Exemplarische Messspuren verschiedener Neurone. Legende unter (A) ist gleichermaßen für (B) und (C)
gültig. Es wurden 250 µM beider Nikotinenantiomere, 10 µM Rutheniumrot und 100 µM AITC appliziert.
In einem weiteren Experiment wurde mit Hilfe des spezifischen Antagonisten Gentamycin (Nagata
et al., 2005) die TRPA1 vermittelte (+)-Nikotinantwort geblockt. Dabei wurde zuerst (+)-Nikotin
(250 µM) appliziert, worauf die Koapplikation von (+)-Nikotin und Gentamycin (30 µM) folgte.
Anschließend wurden sukzessiv (+)-Nikotin und (-)-Nikotin (250 µM) appliziert. Von 165
untersuchten Neuronen wurden 44 (27%) als Nikotin-responsiv identifiziert. Anhand der
unterschiedlichen Antwortprofile wurden diese in unterschiedliche Populationen eingeteilt: 1) 20
(12%) ausschließlich (-)-Nikotin-responsive Neurone wurden charakterisiert. 2) Weitere 8 (5%)
Neurone waren responsiv für beide Enantiomere, wobei die (+)-Nikotinantwort durch Gentamycin
geblockt werden konnte. 3) Bei weiteren 6 (4%) Neuronen konnte die (+)-Nikotinantwort geblockt
werden, jedoch waren die Zellen nicht (-)-Nikotin-responsiv. Der Block der (+)-Nikotinantwort der
beiden Populationen 2) und 3) weist auf die Beteiligung des TRPA1-Kanals hin. Zudem wurden

67
Ergebnisse
zwei weitere Populationen festgestellt, deren Antwort auf (+)-Nikotin nicht geblockt werden
konnte: 4) Davon waren zum einen 5 Zellen (3%) auch (-)-Nikotin-responsiv, 5) zum anderen
weitere 5 Neurone (3%) ausschließlich (+)-Nikotin-responsiv (Abb. 3.17). Es ist möglich, dass die
Antworten Ersterer auf eine Aktivität nikotinischer Acetylcholinrezeptoren zurückgehen. Da bei
Letzteren jedoch Antworten auf (-)-Nikotin ausbleiben, scheint offenbar bei etwa 3% der Neurone
die Expression eines unbekannten Rezeptors für (+)-Nikotin vorzuliegen.
Abbildung 3.17: Calcium-Imaging Antwortprofile Nikotin-responsiver trigeminaler Neurone bei Block
durch Gentamycin (A) Prozentualer Anteil der Populationen gemessener Neurone. (B), (C) Exemplarische
Messspuren verschiedener Neurone. Legende unter (A) ist gleichermaßen für (B) und (C) gültig. Es wurden
250 µM Nikotin bei beiden Enantiomeren und 30 µM Gentamycin appliziert.
In einem weiteren Experiment sollten die Ergebnisse des vorangegangenen Experimentes mit
Borneol, einem weiteren Blocker für TRPA1 (lehrstuhleigene Ergebnisse, Korrespondenz mit Azhar
Sherkheli) verifiziert werden. Zusätzlich wurden die nikotinischen Acetylcholinrezeptoren mit
Hexamethonium blockiert (Abb. 3.18).

68
Ergebnisse
Abbildung 3.18: Calcium-Imaging Antwortprofile Nikotin-responsiver trigeminaler Neurone bei Block
durch Hexamethonium und Borneol. (A) Prozentualer Anteil der Populationen gemessener Neurone. (B),
(C) Exemplarische Messspuren verschiedener Neurone. Legende unter (A) ist gleichermaßen für (B) und (C)
gültig. Es wurden 250 µM Nikotin bei beiden Enantiomeren, 1 mM Hexamethonium und 1 mM Borneol
appliziert.
Zunächst wurden (+)-Nikotin (250 µM) und (-)-Nikotin (250 µM) appliziert. Anschließend folgte die
Koapplikation von (+)-Nikotin mit Hexamethonium (1 mM) und/oder Borneol (1 mM); darauf
folgte eine weitere Applikation von (+)-Nikotin. Von 392 untersuchten Neuronen wurden 135
(35%) als Nikotin-responsiv klassifiziert, die wieder anhand ihrer Antwortprofile in verschiedene
Klassen eingeordnet werden konnten: 1) 26 Neurone (7%) ausschließlich (-)-Nikotin-responsive
Neurone wurden identifiziert. 2) Weiterhin antworteten 14 (4%) Neurone auf beide Enantiomere,
wobei die (+)-Nikotinantwort durch das Antagonistengemisch geblockt wurde. 3) Bei einer
weiteren Population von 52 Neuronen (13%) konnte die (+)-Nikotinantwort geblockt werden,
jedoch waren die Zellen nicht (-)-Nikotin-responsiv. Hinzu kamen zwei Populationen, deren
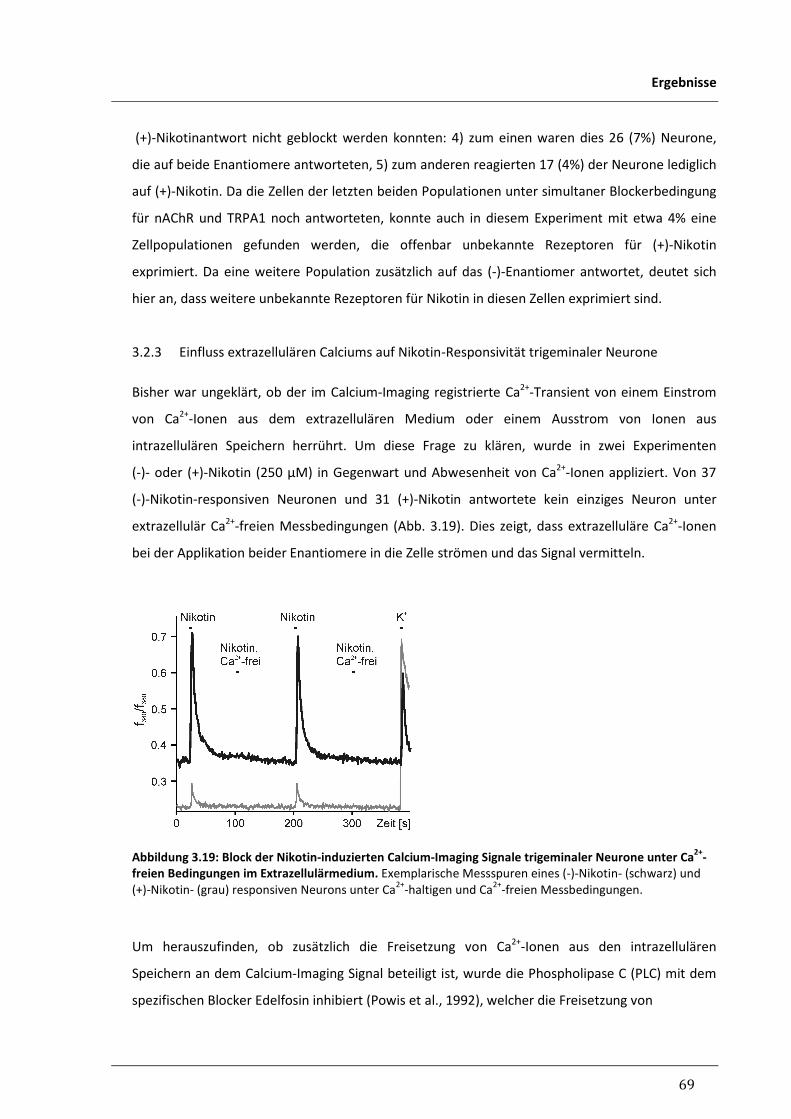
69
Ergebnisse
(+)-Nikotinantwort nicht geblockt werden konnten: 4) zum einen waren dies 26 (7%) Neurone,
die auf beide Enantiomere antworteten, 5) zum anderen reagierten 17 (4%) der Neurone lediglich
auf (+)-Nikotin. Da die Zellen der letzten beiden Populationen unter simultaner Blockerbedingung
für nAChR und TRPA1 noch antworteten, konnte auch in diesem Experiment mit etwa 4% eine
Zellpopulationen gefunden werden, die offenbar unbekannte Rezeptoren für (+)-Nikotin
exprimiert. Da eine weitere Population zusätzlich auf das (-)-Enantiomer antwortet, deutet sich
hier an, dass weitere unbekannte Rezeptoren für Nikotin in diesen Zellen exprimiert sind.
3.2.3 Einfluss extrazellulären Calciums auf Nikotin-Responsivität trigeminaler Neurone
Bisher war ungeklärt, ob der im Calcium-Imaging registrierte Ca2+-Transient von einem Einstrom
von Ca2+-Ionen aus dem extrazellulären Medium oder einem Ausstrom von Ionen aus
intrazellulären Speichern herrührt. Um diese Frage zu klären, wurde in zwei Experimenten
(-)- oder (+)-Nikotin (250 µM) in Gegenwart und Abwesenheit von Ca2+-Ionen appliziert. Von 37
(-)-Nikotin-responsiven Neuronen und 31 (+)-Nikotin antwortete kein einziges Neuron unter
extrazellulär Ca2+-freien Messbedingungen (Abb. 3.19). Dies zeigt, dass extrazelluläre Ca2+-Ionen
bei der Applikation beider Enantiomere in die Zelle strömen und das Signal vermitteln.
Abbildung 3.19: Block der Nikotin-induzierten Calcium-Imaging Signale trigeminaler Neurone unter Ca2+
-
freien Bedingungen im Extrazellulärmedium. Exemplarische Messspuren eines (-)-Nikotin- (schwarz) und
(+)-Nikotin- (grau) responsiven Neurons unter Ca2+
-haltigen und Ca2+
-freien Messbedingungen.
Um herauszufinden, ob zusätzlich die Freisetzung von Ca2+-Ionen aus den intrazellulären
Speichern an dem Calcium-Imaging Signal beteiligt ist, wurde die Phospholipase C (PLC) mit dem
spezifischen Blocker Edelfosin inhibiert (Powis et al., 1992), welcher die Freisetzung von

70
Ergebnisse
Ca2+-Ionen aus intrazellulären Speichern verhindert. Trigeminale Zellkulturen wurden mit
Edelfosin (10 µM) für 20 Minuten präinkubiert und unter Gegenwart von Edelfosin im Calcium-
Imaging auf Nikotin-Responsivität untersucht. Dabei wurden (-)-Nikotin (250 µM) und die Blocker
HC030031 (30 µM) und Hexamethonium (1 mM) koappliziert, um zusätzlich TRPA1 und nAChR
vermittelte Antworten zu inhibieren. Die Applikation wurde in einem Experiment zwei weitere
Male wiederholt. Von den 56 untersuchten Neuronen waren 7 (12%) unter den beschriebenen
Blockerbedingungen (-)-Nikotin-responsiv (Abb. 3.20). Der (-)-Nikotin-induzierte Calciumtransient,
der hier beobachteten Population, scheint demnach offenbar nicht über eine PLC-gekoppelte
Signaltransduktion vermittelt zu werden.
Abbildung 3.20: Einfluss des Phospholipase C-Blockers Edelfosin auf (-)-Nikotin-induzierte Calcium-
Imaging Signale trigeminaler Neurone (A) Repräsentative Messspur mit 10 µM Edelfosin inkubierter,
(-)-Nikotin- (250 µM) responsiver Neurone unter simultanem Block durch Hexamethonium (1 mM) und
HC030031 (30 µM). (B) Prozentualer Anteil der Populationen unter Edelfosineinfluss gemessener Neurone.
3.2.4 Inhibitorischer Einfluss von Nikotin auf 5-HT3 vermittelte Calcium-Signale trigeminaler
Neurone
Die Serotonin-vermittelte Stromantwort rekombinant exprimierter 5-HT3-Rezeptoren konnte im
Voltage-Clamp Verfahren durch Koapplikation von Nikotin inhibiert werden. Im Folgenden wurde
nun mittels Calcium-Imaging untersucht, ob dieser Antagonismus auch an trigeminalen Neuronen
nachgewiesen werden kann. Dazu wurde 10 µM Serotonin appliziert. Da sowohl ionotrope wie
metabotrope Serotoninrezeptoren durch den Liganden aktiviert werden, wurde anschließend
1-(m-Chlorophenyl)-biguanid (mCPBG, 100 µM; Kilpatrick et al., 1990), ein spezifischer Agonist
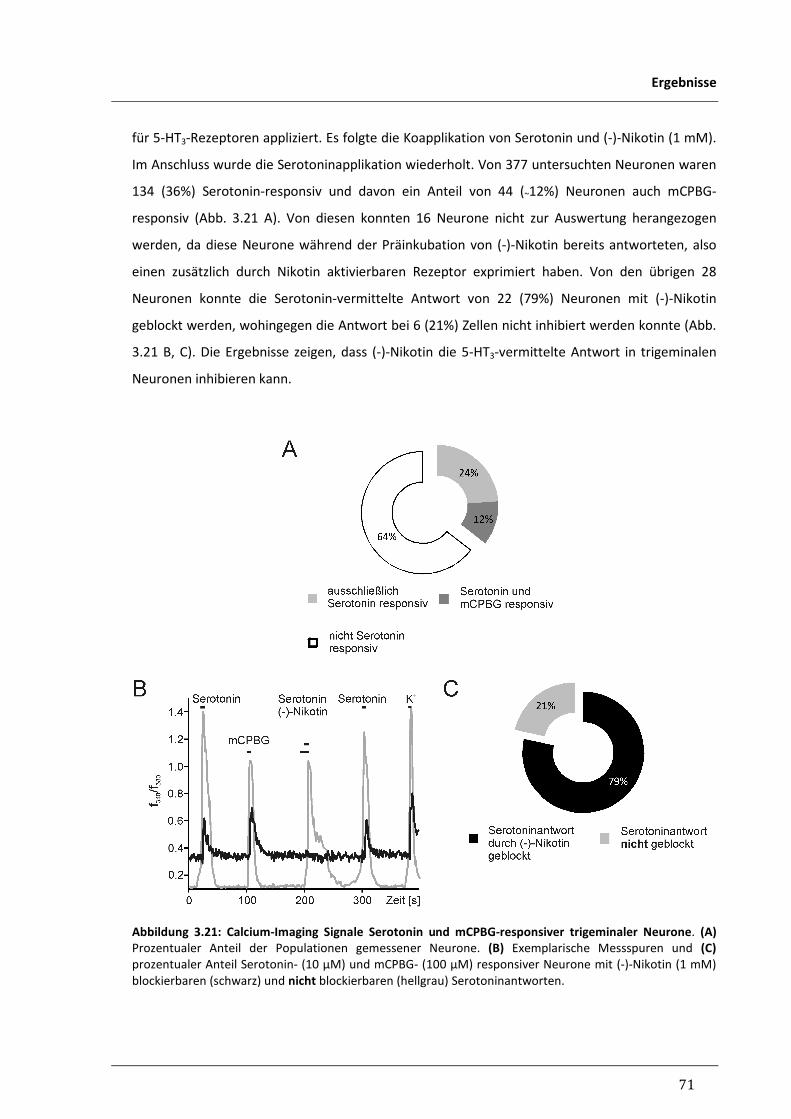
71
Ergebnisse
für 5-HT3-Rezeptoren appliziert. Es folgte die Koapplikation von Serotonin und (-)-Nikotin (1 mM).
Im Anschluss wurde die Serotoninapplikation wiederholt. Von 377 untersuchten Neuronen waren
134 (36%) Serotonin-responsiv und davon ein Anteil von 44 (~12%) Neuronen auch mCPBG-
responsiv (Abb. 3.21 A). Von diesen konnten 16 Neurone nicht zur Auswertung herangezogen
werden, da diese Neurone während der Präinkubation von (-)-Nikotin bereits antworteten, also
einen zusätzlich durch Nikotin aktivierbaren Rezeptor exprimiert haben. Von den übrigen 28
Neuronen konnte die Serotonin-vermittelte Antwort von 22 (79%) Neuronen mit (-)-Nikotin
geblockt werden, wohingegen die Antwort bei 6 (21%) Zellen nicht inhibiert werden konnte (Abb.
3.21 B, C). Die Ergebnisse zeigen, dass (-)-Nikotin die 5-HT3-vermittelte Antwort in trigeminalen
Neuronen inhibieren kann.
Abbildung 3.21: Calcium-Imaging Signale Serotonin und mCPBG-responsiver trigeminaler Neurone. (A)
Prozentualer Anteil der Populationen gemessener Neurone. (B) Exemplarische Messspuren und (C)
prozentualer Anteil Serotonin- (10 µM) und mCPBG- (100 µM) responsiver Neurone mit (-)-Nikotin (1 mM)
blockierbaren (schwarz) und nicht blockierbaren (hellgrau) Serotoninantworten.

72
Ergebnisse
3.3 Transkriptomanalyse Nikotin-responsiver trigeminaler Neurone
Im Rahmen der vergleichenden Transkriptomanalyse wurden Ergebnisse verschiedener
Massensequenzierungen ausgewertet. Dies waren zum einen Sequenzierungen von Gesamt-RNA
verschiedener Gewebe der Maus (Trigeminus, Dorsalwurzelganglien, Gehirn), wobei die
Ergebnisse für das Gehirn aus dem Datensatz einer bereits publizierten Studie stammen
(SRA001030; Mortazavi et al., 2008). Zum anderen waren dies Ergebnisse der Sequenzierungen
von Amplikons, die vom Transkriptom einzelner Zellen amplifiziert wurden, sowie einem
Amplikon, amplifiziert von der Gesamt-RNA des Trigeminus (RNA-Standard). Für die Gewinnung
der Proben wurde zum einen Gesamt-RNA aus ganzen trigeminalen Ganglien und
Dorsalwurzelganglien (weiterhin bezeichnet als Explantate) adulter Mäuse isoliert (2.2.7.13). Zum
anderen wurden die Amplikons der Transkriptome einzelner Neurone von trigeminalen
Primärkulturen amplifiziert, die sich vorher in Calcium-Imaging Experimenten als Nikotin-
responsiv erwiesen hatten. Dazu wurde ein Messprotokoll wie in Abbildung 3.15 durchgeführt, bei
der die Nikotinantwort der Neurone mit den spezifischen Blockern für nAChR (Hexamethonium,
Hex) und TRPA1 (HC030031, HC) in blockierbar und nicht blockierbar eingeordnet wird. Es wurden
sowohl blockierbare als auch nicht blockierbare Neurone aus der Zellkultur herausgelöst. Im
Anschluss wurden Transkriptome von je zwei Neuronen beider Populationen amplifiziert
(2.2.7.14), um für die nachfolgende Amplikonsequenzierung eine ausreichend große cDNA-Probe
zu erhalten. Dabei waren die (-)-Nikotinantworten der Zellen 5-1 und 6-1 nicht durch Hex und HC
blockierbar, jene von Zelle 2-1 und 2-2 waren blockierbar. Zellen 5-1 und 2-2, sowie Zellen 6-1 und
2-1 stammen aus derselben Charge kultivierter Zellen. Außerdem wurde mit dem Einsatz von
isolierter Gesamt-RNA (100 pg) aus trigeminalen Ganglien über die Amplifikationsreaktion ein
Amplikon (weiterhin als RNA-Standard bezeichnet) erzeugt, dass ebenfalls der
Massensequenzierung zugeführt wurde. Beim Vergleich dieser Probe mit denen der trigeminalen
Gesamt-RNA lässt sich beurteilen, ob der gesamte Prozess der Amplifikation die
Expressionsergebnisse beeinflusst.
Abbildung 3.22 zeigt ein Schema der Amplifikationsreaktion. Zunächst wurde an der RNA
einzelner Zellen beziehungsweise an trigeminaler Gesamt-RNA mit einem randomisierten
Primergemisch eine cDNA-Synthese vorgenommen, bei der Sequenzfragmente über die gesamte
Länge aller Transkripte entstehen können. Die erzeugte cDNA wird vom Hersteller des Systems als
„OmniPlex Library“ bezeichnet. Die Enden der darin enthaltenen Fragmente wurden bei der
cDNA-Synthese mit einer Sequenz versehen, die als Bindestelle für die Primer der anschließenden
PCR-basierten Amplifikation dient.

73
Ergebnisse
Abbildung 3.22: Schematische Darstellung der Transkriptomamplifikation. Mit dem Amplifikationssystem
„TransPlex Complete WTA2“ (Sigma-Aldrich) wurde in einer PCR-basierten Reaktion ein Amplikon des
gesamten Transkriptoms erzeugt. Verändert nach Benutzerinformation (WTA2 Procedure,
http://www.sigmaaldrich.com).
Die auf diese Weise erzeugten Amplikons des Transkriptoms der vier Neurone und der RNA-
Standard sowie die Proben aus Dorsalwurzel- und trigeminalen Ganglien wurden durch den
Service des „Cologne Center for Genomics“ im Next Generation Sequencing-Verfahren (2.2.7.15)
sequenziert. Die Daten der sieben Transkriptome sowie die Daten eines bereits publizierten
Datensatzes vom Gehirn der Maus wurden im Anschluss mit den - unter 2.2.7.16 genannten -
Algorithmen bioinformatisch prozessiert, wobei jedes einzelne sequenzierte Fragment (read)
einem Abschnitt des Referenzgenoms der Maus (mm9) zugeordnet wurde. Dieser Prozess ergab
ein Maß für relative Stärke der Expression eines Transkriptes innerhalb des untersuchten
Transkriptoms und wird als FPKM (fragments per kilobase of exon per million mapped reads)
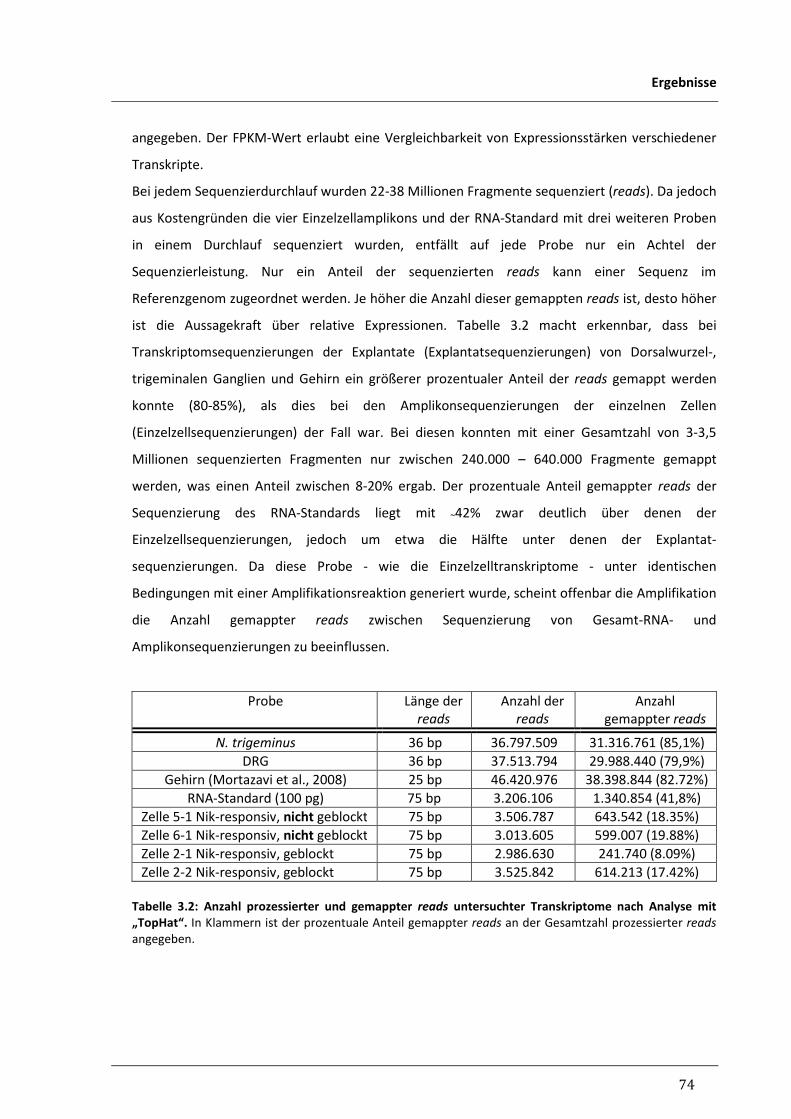
74
Ergebnisse
angegeben. Der FPKM-Wert erlaubt eine Vergleichbarkeit von Expressionsstärken verschiedener
Transkripte.
Bei jedem Sequenzierdurchlauf wurden 22-38 Millionen Fragmente sequenziert (reads). Da jedoch
aus Kostengründen die vier Einzelzellamplikons und der RNA-Standard mit drei weiteren Proben
in einem Durchlauf sequenziert wurden, entfällt auf jede Probe nur ein Achtel der
Sequenzierleistung. Nur ein Anteil der sequenzierten reads kann einer Sequenz im
Referenzgenom zugeordnet werden. Je höher die Anzahl dieser gemappten reads ist, desto höher
ist die Aussagekraft über relative Expressionen. Tabelle 3.2 macht erkennbar, dass bei
Transkriptomsequenzierungen der Explantate (Explantatsequenzierungen) von Dorsalwurzel-,
trigeminalen Ganglien und Gehirn ein größerer prozentualer Anteil der reads gemappt werden
konnte (80-85%), als dies bei den Amplikonsequenzierungen der einzelnen Zellen
(Einzelzellsequenzierungen) der Fall war. Bei diesen konnten mit einer Gesamtzahl von 3-3,5
Millionen sequenzierten Fragmenten nur zwischen 240.000 – 640.000 Fragmente gemappt
werden, was einen Anteil zwischen 8-20% ergab. Der prozentuale Anteil gemappter reads der
Sequenzierung des RNA-Standards liegt mit ~42% zwar deutlich über denen der
Einzelzellsequenzierungen, jedoch um etwa die Hälfte unter denen der Explantat-
sequenzierungen. Da diese Probe - wie die Einzelzelltranskriptome - unter identischen
Bedingungen mit einer Amplifikationsreaktion generiert wurde, scheint offenbar die Amplifikation
die Anzahl gemappter reads zwischen Sequenzierung von Gesamt-RNA- und
Amplikonsequenzierungen zu beeinflussen.
Probe Länge der
reads
Anzahl der
reads
Anzahl
gemappter reads
N. trigeminus 36 bp 36.797.509 31.316.761 (85,1%)
DRG 36 bp 37.513.794 29.988.440 (79,9%)
Gehirn (Mortazavi et al., 2008) 25 bp 46.420.976 38.398.844 (82.72%)
RNA-Standard (100 pg) 75 bp 3.206.106 1.340.854 (41,8%)
Zelle 5-1 Nik-responsiv, nicht geblockt 75 bp 3.506.787 643.542 (18.35%)
Zelle 6-1 Nik-responsiv, nicht geblockt 75 bp 3.013.605 599.007 (19.88%)
Zelle 2-1 Nik-responsiv, geblockt 75 bp 2.986.630 241.740 (8.09%)
Zelle 2-2 Nik-responsiv, geblockt 75 bp 3.525.842 614.213 (17.42%)
Tabelle 3.2: Anzahl prozessierter und gemappter reads untersuchter Transkriptome nach Analyse mit
„TopHat“. In Klammern ist der prozentuale Anteil gemappter reads an der Gesamtzahl prozessierter reads
angegeben.

75
Ergebnisse
Die Gesamtzahl exprimierter Gene beträgt bei den Explantaten etwa 14000-16000, beim RNA-
Standard wurden in etwa 11000, bei den einzelnen Zellen zwischen 2000-5000 exprimierte Gene
festgestellt. Die geringe Anzahl festgestellter Gene bedeutet nicht zwangsläufig eine tatsächlich
geringere Anzahl exprimierter Gene in den einzelnen Zellen. Sie könnte aber dadurch begründet
sein, dass die Wahrscheinlichkeit sehr gering ist, dass unter wesentlich weniger prozessierten
reads bei Amplikonsequenzierungen im Vergleich zu den Explantatsequenzierungen, Transkripte
sehr schwach exprimierter Gene überhaupt festgestellt werden. In einer ersten näheren
Betrachtung wurden für die untersuchten Transkriptome die exprimierten Gene in Stufen der
Expressionsstärke eingeteilt, die von niedrigen FPKM-Werten von 0,1 bis sehr hohen im Bereich
zwischen 3000-10000 reichen. Mit einer Auftragung der Genanzahl gegen die zugehörige
Expressionsstärke wurde diese Klassifizierung in Abbildung 3.23 dargestellt.
Abbildung 3.23: Einteilung der Transkriptome in Expressionsklassen. Jede FPKM-Klasse stellt ein Intervall
dar, dem die Anzahl von Genen zugeordnet wurde, die dieser Expressionsstärke entspricht.
Der Darstellung entnimmt man, dass die meisten Gene bei den niedrigen Expressionsstärken
gelistet werden, wobei insbesondere bei den Transkriptomen von Trigeminus, Gehirn und den

76
Ergebnisse
Einzelzelltranskriptomen zwischen 3-10 FPKM viele Gene exprimiert sind. Bei den Transkriptomen
der Dorsalwurzelganglien und trigeminalem RNA-Standard fallen die meisten Gene in die Klasse
von 0,1-1 FPKM. Es fällt auf, dass der Anteil der sehr niedrig exprimierten Gene von 0,1-1 FPKM
bei den Transkriptomen der einzelnen Neurone sehr gering ist, bei den anderen Transkriptomen
hat er hingegen einen großen Anteil. Dies kann daran liegen, dass bei den
Einzelzelltranskriptomen bedingt durch die niedrige Gesamtzahl gemappter reads, einzelne
gemappte reads für ein Transkript bereits in einem FPKM von 1-2 resultieren können.
Da FPKM-Werte relative Expressionsanteile von 1.000.000 gemappter reads darstellen, wirft die
wesentlich geringere Anzahl gemappter reads und exprimierter Gene der Einzelzelltranskriptome
die Frage auf, inwieweit die FPKM-Werte von Explantat- und Einzelzelltranskriptomen
vergleichbar sind. Hohe FPKM-Werte können bei einer geringen Gesamtzahl gemappter reads auf
wenige sequenzierte Fragmente eines Transkriptes zurückgehen. So repräsentiert ein einzelnes
gemapptes Fragment bei angenommener Exonlänge von 1000 Basenpaaren im Transkriptom der
untersuchten Explantate einen FPKM von etwa 0,03-0,66. Bei den Einzelzelltranskriptomen ergibt
sich hingegen ein FPKM von etwa 1,6-4,1. Gleiche FPKM-Werte bei Explantat- und
Einzelzelltranskriptomen gehen somit aus deutlich verschiedenen Anzahlen absoluter gemappter
reads hervor. In dieser Arbeit wurden zusätzlich zu der Berechnung der relativen FPKM-Werte die
alignments sequenzierter Fragmente an das Genom für die Einzelzelltranskriptome bei
ausgewählten Genen überprüft und die absolute Anzahl der tatsächlichen gemappten Fragmente
ermittelt. Tang et al. (2011) kommen in einer Übersichtsarbeit zu dem Schluss, dass heutige
technische Möglichkeiten der Amplikonsequenzierung von Transkriptomen einzelner Zellen
quantitative Aussagen über Expressionsmuster prinzipiell zulassen. Allerdings sind die
resultierenden FPKM-Werte bisher nur bedingt vergleichbar mit denen der Sequenzierungen von
Gesamt-RNA, denn bei den geringen Mengen RNA, die bei der vorangehenden
Amplifikationsreaktion eingesetzt werden, ist es sehr wahrscheinlich, dass die Expression sehr
schwach exprimierter Gene sich nicht verhältnisgenau im Amplikon wiederspiegelt, wenn nur sehr
wenige oder gar nur ein Transkript pro Zelle vorliegt. Andererseits können stark exprimierte Gene
im Amplikon überrepräsentiert sein. Die Generierung des Amplikons geht somit mit einer
gewissen Verzerrung des Transkriptoms einher, was einen direkten Vergleich von FPKM-Werten
erschwert.
In Abbildung 3.24 sind graphische Darstellungen zweier exemplarischer alignments gezeigt, die
mit dem Programm „Integrative Genomics Viewer“ (www.broadinstitute.org/igv/) aus den
Amplikonsequenzierungen der einzelnen Zellen und der Sequenzierung von trigeminalem Gewebe
erzeugt wurden.

77
Ergebnisse
Abbildung 3.24: Exemplarische alignments sequenzierter Transkripte aus Amplikonsequenzierungen
einzelner Zellen und der Trigeminus-Explantatsequenzierung an das Genom der Maus (mm9). Mit Gapdh
ist ein Beispiel für ein stark exprimiertes, mit Ldha ein Beispiel für ein schwächer exprimiertes
housekeeping-Gen dargestellt. Dick blaue Abschnitte markieren Exone des entsprechenden Gens im
Genom. Vereinzelt sind die alignments so tief gestapelt, dass nur ein Teil der Fragmente abgebildet werden
kann.
Im linken Teil ist mit Gapdh ein Beispiel für ein stark exprimiertes housekeeping-Gen gezeigt, für
das bei den Einzelzelltranskriptomen zwischen 88 und 321 Fragmente gemappt werden konnten,
die FPKM-Werte zwischen 118 und 1080 ergaben. Bei der Trigeminussequenzierung wurden
einige tausend reads gemappt, es ergab sich ein FPKM-Wert von 22,5. Dunkelgraue
Sequenzabschnitte stellen gemappte Fragmente dar, die einem Exon im Genom, blau dargestellt,
zugeordnet wurden. Bei stark exprimierten Genen konnten meist an allen Exonen Fragmente
gemappt werden. Auf der rechten Seite ist mit Ldha ein Beispiel für ein in den Einzelzelltranskrip-
tomen schwach exprimiertes housekeeping-Gen dargestellt. Für dieses Gen wurden bei den
einzelnen Zellen zwischen 1 und 37 Fragmente gemappt, die FKPM-Werte zwischen 1 und 38

78
Ergebnisse
ergaben. Es konnten hier nicht an allen Exonen Fragmente gemappt werden. Bei der Trigeminus-
Explantatsequenzierung wurden wieder einige tausend reads gemappt. Durch die ebenfalls sehr
hohe Gesamtanzahl gemappter reads bei dieser Sequenzierung ergab sich hier ein FPKM-Wert
von etwa 33, der in ähnlicher Größenordnung liegt, wie bei den Amplikonsequenzierungen. Dies
verdeutlicht die Abhängigkeit des FPKM als relatives Expressionsmaß von der Gesamtzahl der
gemappten reads.
3.3.1 Vergleich der Transkriptome neuronaler Gewebe und einzelner Nikotin-responsiver
Neurone
Um die große Menge an Expressionsdaten aus acht verschiedenen Transkriptomen betrachten zu
können, wurden FPKM-Werte in Expressionsklassen dargestellt, die bei aufsteigender
Expressionsstärke mit zunehmenden Farbintensitäten kodiert sind. Es wurden damit heatmaps
aufgetragen, die sowohl einen direkten Vergleich der Expressionen zwischen verschiedenen
Genen innerhalb eines Transkriptoms als auch zwischen verschiedenen Transkriptomen - bezogen
auf ein Gen - ermöglichen. Bei den Einzelzelltranskriptomen wurden zusätzlich die absoluten
Zahlen gemappter Transkripte angegeben.
Die Expression einer Vielzahl Liganden-gesteuerter Ionenkanäle, TRP-Kanäle und KCNK-Kanäle im
Trigeminus ist bekannt. Viele dieser Kanäle werden mit chemosensorischen Prozessen in
Verbindung gebracht. Daher wurde insbesondere die Expression der Gene dieser drei
Ionenkanalfamilien in Trigeminus, DRG und Gehirn im Vergleich zu den einzelnen Zellen in der
Darstellung als heatmaps betrachtet.
3.3.1.1 Expression von housekeeping-Genen
Vor der Beurteilung der Expression erwähnter Ionenkanäle wurde zunächst die Expression
sogenannter housekeeping-Gene betrachtet (Abb. 3.25). Diese Gene sind dafür bekannt, dass ihre
Expression über verschiedene Gewebetypen hinweg verhältnismäßig geringen Schwankungen
unterliegt, da ihre Genprodukte in der Regel allgemeine Funktionen für den Zellstoffwechsel
haben (Barber et al., 2005; Kouadjo et al., 2007; Zhou et al., 2010). Die Gene Actb, Gapdh und
Ppia sind bei den Einzelzelltranskriptomen besonders stark exprimiert. Dies trifft bei den Proben
der Explantate und dem RNA-Standard insbesondere auch auf Actb zu. Gapdh und Ppia sind auch
in DRG und Trigeminus, im Gehirn jedoch wesentlich schwächer exprimiert. Ldha und Ubc sind
allgemein schwächer, jedoch in den meisten Proben in ähnlicher Größenordnung exprimiert. Die
Gene Rpl5, Rpl19 und Rpl29 sind in den Einzelzellen stärker exprimiert.

79
Ergebnisse
Insbesondere Rpl19 und Rpl29 scheinen in den Explantaten kaum exprimiert zu sein. Im
Gegensatz dazu ist Hprt in den Explantaten hoch, jedoch in drei der vier Einzelzellen gar nicht und
in Zelle 2-2 nur schwach exprimiert.
Mit Tubb3 (Beta-3-Tubulin) ist ein Gen dargestellt, dass in Neuronen stark exprimiert wird, die
gerade Ausläufer bilden (Jiang et al., 1994a; b; Kost und Oblinger, 1993). Dieses Gen wird in allen
Explantaten exprimiert, jedoch konnte in den Einzelzellproben kein Fragment gefunden werden.
Cnp (CNPase) wird spezifisch in Oligodendrozyten und Schwann-Zellen exprimiert (Chandross et
al., 1999; Nishizawa et al., 1985; Radtke et al., 2011; Sprinkle, 1989; Trapp et al., 1988). Da erstere
im Gehirn und letztere in den DRG und dem Trigeminus vorkommen, wird das Gen in den
entsprechenden Proben exprimiert. In drei der vier Einzelzelltranskriptome ist Cnp
erwartungsgemäß nicht exprimiert. In Zelle 6-1 konnte lediglich 1 Fragment festgestellt werden.
Dies ist ein starker Hinweis, dass keine Kontamination der isolierten Proben durch RNA aus
Schwann-Zellen der Zellkultur auftrat. In der trigeminalen Zellkultur finden sich zumeist neben
intakten Zellen auch Zelltrümmer und vermutlich auch RNA zerstörter Schwann-Zellen, die jedoch
während der Messungen vom permanenten Flüssigkeitsstrom des Extrazellulärmediums stark
verdünnt, beziehungsweise fortgespült werden. Offenbar hat hier keine Kontamination der Probe
mit freier RNA untergegangener Zellen stattgefunden.
Zusammengefasst ist die Expression der housekeeping-Gene in Trigeminus und Einzelzell-
transkriptomen tendenziell ähnlich, jedoch wurden zwischen den einzelnen Neuronen größere
Schwankungen festgestellt als zwischen den verschiedenen Geweben. Ein nennenswerter
Expressionsunterschied zwischen blockierbaren und nicht blockierbaren Neuronen konnte nicht
festgestellt werden.

80
Ergebnisse
Abbildung 3.25: Heatmap der
Expressionsstärken untersuchter Trans-
kriptome für eine Auswahl von
housekeeping-Genen.
Die acht Expressionsklassen wurden durch
FPKM-Intervalle festgelegt. Bei den
Einzelzelltranskriptomen ist zusätzlich für
jedes Gen die absolute Anzahl gemappter
Transkripte angegeben. Zellen 5-1 und 6-1
sind Nikotin-responsiv, nicht Hex, HC
geblockt, Zellen 2-1 und 2-2 sind Nikotin-
responsiv, Hex, HC geblockt.
Gapdh: Glycerinaldehyd-3-Phosphat-
Dehydrogenase; Tubb3: Beta-3-Tubulin;
Ldha: Lactatdehydrogenase A; Actb: Beta-
Actin; Ubc: Ubiquitin C; Ppia: Peptidyl-
Prolyl-Isomerase; Tbp: TATA-Box
bindendes Protein; Hprt: Hypoxanthin-
Guanin-Phosphoribosyltransferase; Rpl:
Ribosomales Protein L; Cnp: Cyclic-
nucleotide-phosphodiesterase. Die
Transkriptomanalysen von DRG und
Trigeminus-Explantat erfolgten in
Zusammenarbeit mit Ramona Lehmann.
3.3.1.2 Expression der Cys-loop-Rezeptoren
In einer ersten Betrachtung der Expression der Subfamilien der Cys-loop Rezeptoren wurde
festgestellt, dass eine Vielzahl der 16 untersuchten Untereinheiten (α1-7, α9, 10, β1-4, γ, δ, ε) der
nikotinischen Acetylcholinrezeptoren in allen untersuchten Geweben exprimiert wird (Abb. 3.25
links). Im Gehirn sind α2-7 exprimiert, davon α4 am stärksten. Im Trigeminus und den DRG sind
α3, 4, 6 und 7 exprimiert, davon in den DRG α6 und 7 deutlich. Im Trigeminus ist die α7
Untereinheit am stärksten exprimiert, was sich auch im trigeminalen RNA-Standard
wiederspiegelt. Erstaunlicherweise wurden beim Trigeminus auch Fragmente der muskulären α1
Untereinheit festgestellt. In allen Proben der Explantate konnten β1-4 nachgewiesen werden,
wobei β2 am stärksten exprimiert wird. Auch hier wurden Fragmente der muskulären β1
Untereinheit festgestellt, jedoch konnte keine Expression der muskulären γ, δ, ε Untereinheiten
festgestellt werden.

81
Ergebnisse
Die Nikotin-responsiven Neurone unterschieden sich im Calcium-Imaging in der Blockierbarkeit
ihrer Antwort durch ein Blockergemisch mit dem nAChR-Blocker Hexamethonium. Die Antwort
der Zellen 2-1 und 2-2 ließ sich blocken, was auf die Expression von nAChR hinweist. Dies konnte
mit einer Expression von α4, β2 und β4 zumindest für die Zelle 2-1 festgestellt werden. Bei den
nicht blockierbaren Zellen 5-1 und 6-1 erwartet man hingegen keine Expression nikotinischer
Acetylcholinrezeptoren. In beiden Zellen wurde jedoch eine sehr schwache Expression von β4, in
Zelle 5-2 zusätzlich α4 - sehr schwach exprimiert - festgestellt. In jeder einzelnen Zelle wurde
außerdem die muskuläre Untereinheit α1, in Zelle 5-1 auch β1 nachgewiesen, wohingegen γ, δ
und ε nicht vorkamen.
Von 5-HT3-Rezeptoren ist die Expression der Untereinheiten A und B im Trigeminus bekannt. Die
Expression beider Untereinheiten wurde im Gehirn und in den DRG festgestellt, jedoch ist die
Expression im Trigeminus wesentlich höher (Abb. 3.26 links). In keiner der einzelnen Neurone
konnten 5-HT3-Rezeptoren nachgewiesen werden.
Für die meisten GABA-Rezeptoruntereinheiten (α1-6, β1-3, γ1-3 und δ) wurde deutliche
Expression im Gehirn festgestellt, ε, θ und ρ2 sind schwach exprimiert (Abb. 3.25 rechts). In den
DRG und dem Trigeminus ist die exprimierte Vielfalt geringer, es sind α1, 2, β3 und γ2 deutlich
exprimiert. Zusätzlich konnten im Trigeminus α3 und 5, sowie β1, β2 und y1 nachgewiesen
werden. Im trigeminalen RNA-Standard spiegelt sich die hohe Expression dieser Untereinheiten
wieder. Ebenfalls sind α2 und 5 β3 und γ2 stark exprimiert, denn insbesondere die Untereinheiten
α2, β3 und γ2 sind an Bildung vieler Untereinheitenkombinationen von GABA-Rezeptoren
beteiligt. So wird auch α2 in vier der Einzelzelltranskriptome, β2 in den Zellen 2-1 und 2-2, sowie
γ2 auch in Zelle 2-1 nachgewiesen. In dieser kommen auch zusätzlich α3 und ρ3 vor. Lediglich eine
der vier Einzelzelltranskriptome (Zelle 2-1) zeigt für GABA-Rezeptoren tendenziell ähnliche
Expressionsschwerpunkte, wie man sie im Trigeminus-Explantat feststellen kann.
Glycin ist als inhibitorischer Transmitter insbesondere im Rückenmark bekannt. Glycinrezeptoren
werden aber auch im Gehirn exprimiert (Abb. 3.26 rechts). Die Expressionsdaten bestätigen dies
hier für alle fünf Untereinheiten, wobei α1 und 2, sowie die β-Untereinheit besonders stark
exprimiert sind. In den DRG wurde neben einer sehr schwachen Expression von α4 keine weitere
α-Untereinheit festgestellt, β hingegen ist stark exprimiert. Für den Trigeminus konnte bisher die
Expression einer α-Untereinheit der Glycinrezeptoren nicht nachgewiesen werden.

82
Ergebnisse
Abbildung 3.26: Heatmap der Expressionsstärken untersuchter Transkriptome für die Cys-loop-Kanäle.
Die sechs Expressionsklassen wurden durch FPKM-Intervalle festgelegt. Bei den Einzelzelltranskriptomen ist
zusätzlich für jedes Gen die absolute Anzahl gemappter Transkripte angegeben. Zellen 5-1 und 6-1 sind
Nikotin-responsiv, nicht Hex, HC geblockt, Zellen 2-1 und 2-2 sind Nikotin-responsiv, Hex, HC geblockt. Die
Transkriptomanalysen von DRG und Trigeminus-Explantat erfolgten in Zusammenarbeit mit Ramona
Lehmann.

83
Ergebnisse
Die Expressionsergebnisse bestätigen diese Abwesenheit mit der Ausnahme einer sehr schwachen
Expression von α2. β hingegen wird sowohl in den DRG als auch im Trigeminus deutlich
exprimiert, was durch den trigeminalen RNA-Standard bestätigt wird. In den
Einzelzelltranskriptomen von Zelle 2-1 und 2-2 konnten β-Untereinheiten festgestellt werden,
jedoch keine Expression einer α-Untereinheit bestätigt werden. Über die untersuchten
Transkriptome hinweg wird ein deutlicher Überschuss der β-Untereinheit deutlich.
3.3.1.3 Expression der TRP-Kanäle
Bei den TRP-Kanälen ist insbesondere die Expression der Thermo-TRPs von Interesse. Für den
durch Nikotin aktivierbaren TRPA1-Kanal ist die Expression in chemosensorischen afferenten
Ganglien - wie den DRG und dem trigeminalen Ganglion - bekannt. Die Expressionsdaten dieser
beiden Transkriptome bestätigen dies (Abb. 3.27). In den DRG und im Trigeminus gehört TRPA1 zu
den deutlich exprimierten TRP-Kanälen. Für das Gehirn konnte keine Expression festgestellt
werden. Bei zwei der einzelnen Zellen, Zellen 2-1 und 2-2, konnten im vorangegangenen Calcium-
Imaging die Nikotinantworten durch ein Blockergemisch aus Hexamethonium und dem
spezifischen TRPA1-Blocker HC030031 geblockt werden. Dieser Block der Antwort wäre durch die
Expression des TRPA1-Kanals erklärbar. In keinem der vier Einzelzelltranskriptome konnte jedoch
ein TRPA1-Transkript festgestellt werden.
Bei den TRPM-Kanälen 1-8 fällt insbesondere die starke Expression des TRPM8 im Trigeminus ins
Auge. Obwohl TRPM2-4 und TRPM7 im Trigeminus exprimiert sind, scheint TRPM8 der bei
Weitem am stärksten exprimierte TRP-Kanal im Trigeminus zu sein. In den DRG wurden ebenfalls
M2-4 und M7 festgestellt, M8 wird auch in den DRG exprimiert, fehlt jedoch im Gehirn. In den
Einzelzelltranskriptomen konnte bei keiner Zelle ein TRPM8-Fragment gemappt werden,
allerdings wurde in Zelle 2-1 eine sehr starke Expression von TRPM2 und M3 festgestellt.
Von den Vanilloid-Rezeptoren TRPV1-6 sind insbesondere V1 und V2 in den DRG und dem
Trigeminus stark exprimiert, wohingegen V3-6 sehr schwach bis gar nicht exprimiert wurde. V2
und V4 werden im Gehirn deutlich exprimiert. Für V1 konnte jedoch kein Fragment gemappt
werden. In den Einzelzelltranskriptomen konnte keine Expression für Vanilloidrezeptoren
festgestellt werden.
Die TRPC-Kanäle 1-7 sind grundsätzlich alle im Gehirn exprimiert, wobei C1, C3 und C7 am
deutlichsten exprimiert sind. In den DRG und im Trigeminus sind C6 und ebenfalls C1 und C3
deutlich exprimiert, C2, C4, C5 und C7 sehr schwach bis gar nicht. Bei den
Einzelzelltranskriptomen konnte lediglich ein Transkript für TRPC1 in Zelle 2-1 gemappt werden,

84
Ergebnisse
ansonsten wurde kein TRPC exprimiert. Allgemein zeigt der trigeminale RNA-Standard eine
wesentlich geringere Expression von TRP-Kanälen.
Deutlich exprimiert waren jedoch TRPM3, M7, M8 und TRPV2 (Abb. 3.27).
Abbildung 3.27: Heatmap der
Expressionsstärken untersuchter
Transkriptome für die TRP-Kanäle. Die
sechs Expressionsklassen wurden durch
FPKM-Intervalle festgelegt. Bei den
Einzelzelltranskriptomen ist zusätzlich für
jedes Gen die absolute Anzahl gemappter
Transkripte angegeben. Zellen 5-1 und 6-1
sind Nikotin-responsiv, nicht Hex, HC
geblockt, Zellen 2-1 und 2-2 sind Nikotin-
responsiv, Hex, HC geblockt. Die
Transkriptomanalysen von DRG und
Trigeminus-Explantat erfolgten in
Zusammenarbeit mit Ramona Lehmann.
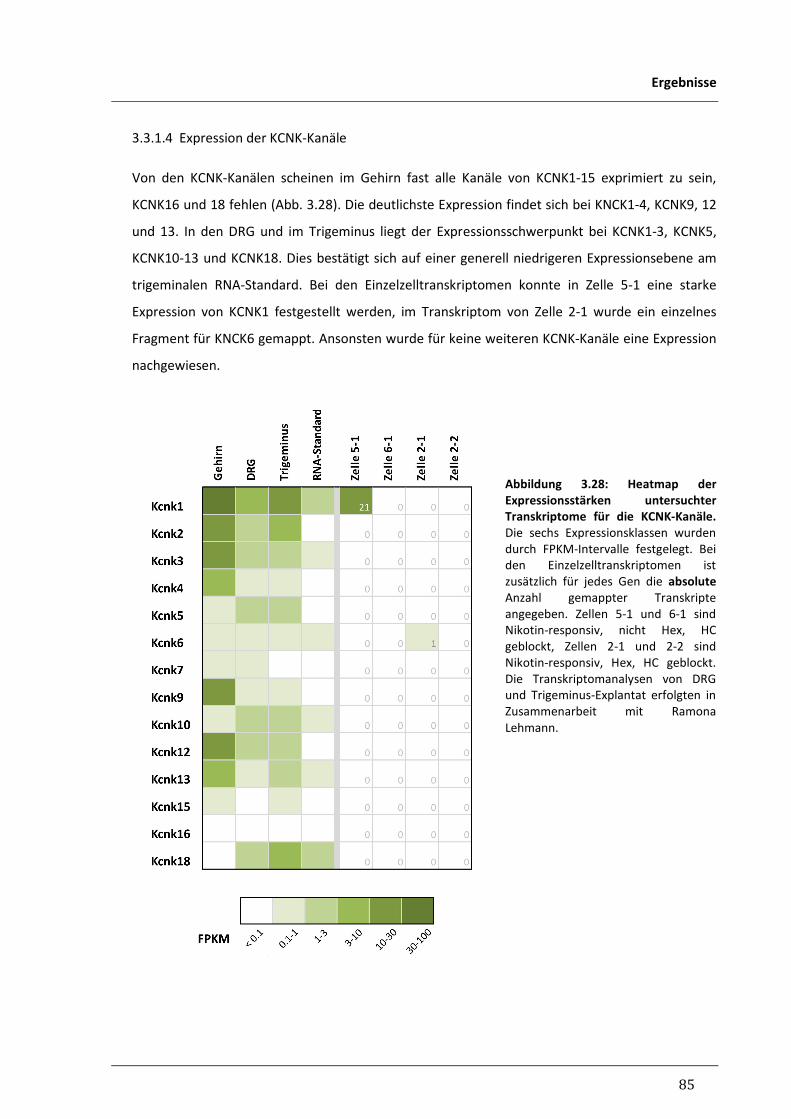
85
Ergebnisse
3.3.1.4 Expression der KCNK-Kanäle
Von den KCNK-Kanälen scheinen im Gehirn fast alle Kanäle von KCNK1-15 exprimiert zu sein,
KCNK16 und 18 fehlen (Abb. 3.28). Die deutlichste Expression findet sich bei KNCK1-4, KCNK9, 12
und 13. In den DRG und im Trigeminus liegt der Expressionsschwerpunkt bei KCNK1-3, KCNK5,
KCNK10-13 und KCNK18. Dies bestätigt sich auf einer generell niedrigeren Expressionsebene am
trigeminalen RNA-Standard. Bei den Einzelzelltranskriptomen konnte in Zelle 5-1 eine starke
Expression von KCNK1 festgestellt werden, im Transkriptom von Zelle 2-1 wurde ein einzelnes
Fragment für KNCK6 gemappt. Ansonsten wurde für keine weiteren KCNK-Kanäle eine Expression
nachgewiesen.
Abbildung 3.28: Heatmap der
Expressionsstärken untersuchter
Transkriptome für die KCNK-Kanäle.
Die sechs Expressionsklassen wurden
durch FPKM-Intervalle festgelegt. Bei
den Einzelzelltranskriptomen ist
zusätzlich für jedes Gen die absolute
Anzahl gemappter Transkripte
angegeben. Zellen 5-1 und 6-1 sind
Nikotin-responsiv, nicht Hex, HC
geblockt, Zellen 2-1 und 2-2 sind
Nikotin-responsiv, Hex, HC geblockt.
Die Transkriptomanalysen von DRG
und Trigeminus-Explantat erfolgten in
Zusammenarbeit mit Ramona
Lehmann.
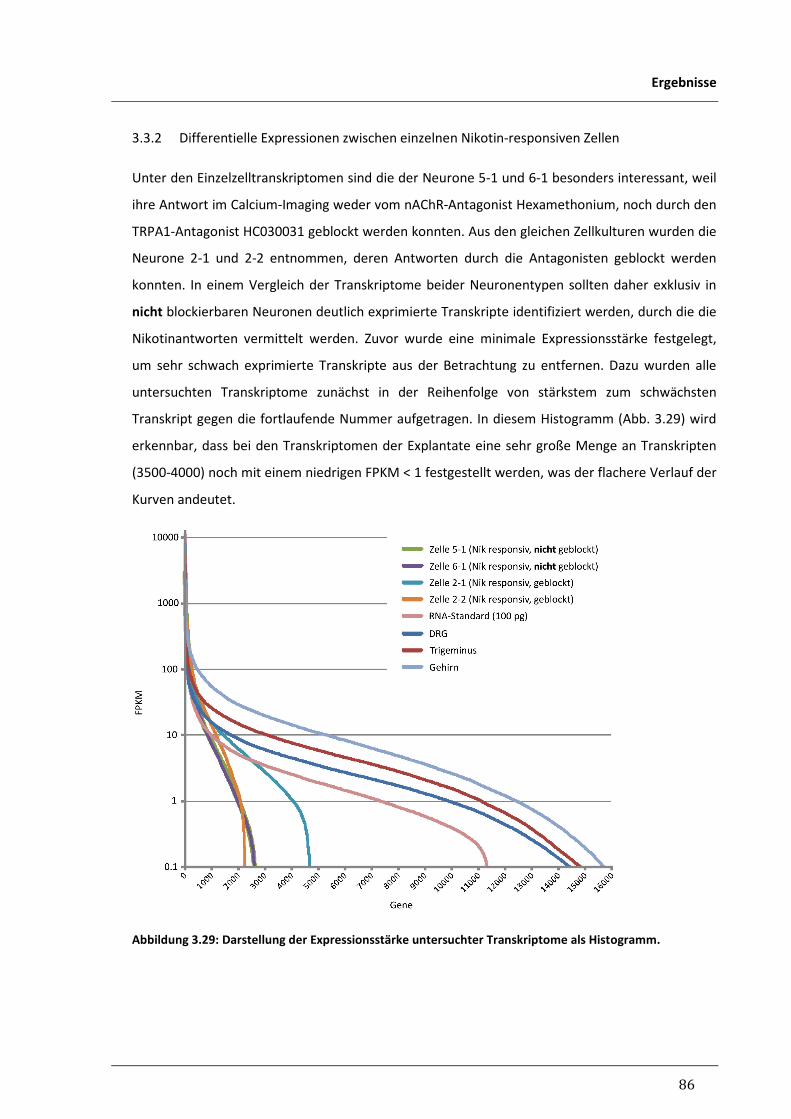
86
Ergebnisse
3.3.2 Differentielle Expressionen zwischen einzelnen Nikotin-responsiven Zellen
Unter den Einzelzelltranskriptomen sind die der Neurone 5-1 und 6-1 besonders interessant, weil
ihre Antwort im Calcium-Imaging weder vom nAChR-Antagonist Hexamethonium, noch durch den
TRPA1-Antagonist HC030031 geblockt werden konnten. Aus den gleichen Zellkulturen wurden die
Neurone 2-1 und 2-2 entnommen, deren Antworten durch die Antagonisten geblockt werden
konnten. In einem Vergleich der Transkriptome beider Neuronentypen sollten daher exklusiv in
nicht blockierbaren Neuronen deutlich exprimierte Transkripte identifiziert werden, durch die die
Nikotinantworten vermittelt werden. Zuvor wurde eine minimale Expressionsstärke festgelegt,
um sehr schwach exprimierte Transkripte aus der Betrachtung zu entfernen. Dazu wurden alle
untersuchten Transkriptome zunächst in der Reihenfolge von stärkstem zum schwächsten
Transkript gegen die fortlaufende Nummer aufgetragen. In diesem Histogramm (Abb. 3.29) wird
erkennbar, dass bei den Transkriptomen der Explantate eine sehr große Menge an Transkripten
(3500-4000) noch mit einem niedrigen FPKM < 1 festgestellt werden, was der flachere Verlauf der
Kurven andeutet.
Abbildung 3.29: Darstellung der Expressionsstärke untersuchter Transkriptome als Histogramm.

87
Ergebnisse
Auch beim trigeminalen RNA-Standard verläuft die Kurve ähnlich. Es werden insgesamt weniger
Transkripte festgestellt, jedoch trotzdem noch viele (~3500) mit einem FPKM < 1. Die Kurven der
Einzelzelltranskriptome enden abrupter. Unterhalb des FPKM-Wertes von 1 knickt der Verlauf bei
den Zellen 2-1 und 2-2 stark, bei Zellen 5-1 und 6-1 leicht ab, was zeigt, dass ein FPKM < 1 in den
Einzelzelltranskriptomen für wesentlich weniger (zwischen 200-800) Transkripte festgestellt wird.
Die minimale Expressionsstärke wurde daher bei einem FPKM von 1 festgelegt und somit die am
schwächsten exprimierten 200-800 Gene aus der Betrachtung ausgenommen.
Da die Zellen 5-1 und 2-2 sowie die Zellen 6-1 und 2-1 jeweils eine nicht geblockte und eine
geblockte Zelle aus demselben Versuchsdurchlauf sind, wurden sie miteinander verglichen. Mit
einem FPKM > 1 wurde bei einer Gesamtzahl von 3101 exprimierten Genen in Zelle 5-1 eine
Anzahl von 2425 exklusiv exprimierter Gene festgestellt. Bei Zelle 6-1 wurde mit 1895
exprimierten Genen mit einem FPKM > 1 eine Anzahl von 1244 exklusiv exprimierter Gene
festgestellt (Abb. 3.30 oben). Ein Großteil dieser Gene kodiert für Proteine des Zellstoffwechsels
und Strukturproteine. Gene, die für G-Protein-gekoppelte Rezeptoren und Ionenkanäle kodieren,
sind in Tabelle 3.3 in absteigender Expressionsstärke gelistet. Die Liste zeigt, dass in den
untersuchten Zellen eine große Vielfalt Transmembranproteine vorliegt, zu denen in beiden Zellen
auch stark exprimierte olfaktorische Rezeptoren (OR) gehören. Unter diesen Genen kann sich
auch ein weiterer unbekannter Rezeptor für Nikotin befinden, was jedoch schwer zu überprüfen
ist. Mit 75 gemappten reads ist in Zelle 5-1 Olfr293 das stärkste exklusiv exprimierte
Transmembranprotein. Tabelle 3.4 zeigt die Expression aller olfaktorischer Rezeptoren in allen
untersuchten einzelnen Zellen. Zum Vergleich sind für jeden exprimierten Rezeptor auch die
FPKM-Werte des RNA-Standards und des Trigeminus-Explantats angegeben. Es wurden einige
Rezeptoren als nicht exprimiert gewertet, deren Expression auf der Zuordnung sequenzierter
Fragmente an repetitive Sequenzabschnitte beruht. Da solche Sequenzabschnitte mehrfach im
Genom der Maus vorkommen, ist eine eindeutige Zuordnung des Fragments zu einem einzigen
Exon im Genom nicht möglich. Jede Zelle hat offenbar ein individuelles Expressionsprofil
unterschiedlicher olfaktorischer Rezeptoren. Dabei sind bei Zelle 2-1 mit 26 OR besonders viele
exprimiert, Zelle 5-1 exprimiert 6 OR, Zelle 6-1 und 2-2 nur einen OR. In Zelle 5-1 übertrifft
allerdings die Expression des Olfr293 die der anderen bei Weitem. Die Expression dieses
Rezeptors sowie der olfaktorischen Rezeptoren in Zellen 2-2 und 6-1 konnten entweder mit den
Ergebnissen für RNA-Standard oder für Trigeminus-Explantat zumindest auf niedriger
Expressionsebene bestätigt werden. Bei Zelle 2-1 ist kein Rezeptor besonders stark exprimiert
oder übertrifft die anderen stark in der Expression. Es wird auch lediglich die Expression zweier
Rezeptoren (Olfr1340, Olfr341) im Trigeminus-Explantat festgestellt.

88
Ergebnisse
Es wurde daneben eine gemeinsame Expression von 767 Genen zwischen den blockierbaren
Zellen 5-1 und 6-1 ermittelt (Abb. 3.30 unten). Diese beiden Zellen haben damit höhere
Übereinstimmungen im Transkriptom als zwischen 5-1 und 2-2 sowie 6-1 und 2-1 vorliegen. Es
wurde unter diesen gemeinsam exprimierten Genen ebenfalls nach G-Protein-gekoppelten
Rezeptoren und Ionenkanälen gesucht und mit Gpr137 lediglich ein Gen für einen G-Protein-
gekoppelten Rezeptor gefunden. Dieser wird jedoch in beiden Zellen von vielen anderen
Rezeptoren in der Expressionsstärke bei Weitem übertroffen.
Abbildung 3.30: Vergleich der Anzahl exprimierter Gene der Einzelzelltranskriptome. Es wurden
ausschließlich Gene gewertet, die mit einem FPKM > 1 exprimiert wurden. Schnittmengen zeigen die Anzahl
der Gene, die in beiden verglichenen Zellen exprimiert wurden. Zusätzlich ist die Gesamtzahl in einer Zelle
exprimierter Gene (Summe von Schnittmenge und jeweiliger Restmenge der exklusiv exprimierten Gene) zu
jeder Zelle angegeben.

89
Ergebnisse
gene_id Gen FPKM Zelle
5-1
Frag-
mente
gene_id Gen FPKM
Zelle 6-1
Frag-
mente
NM_001011752 Olfr293 114.8 75 NM_008310 Htr1f 25.17 30
NM_025791 Tmem223 84.03 40 NM_001005227 Olfr1200 24.66 13
NR_028418 Tmem179b 67.02 15 NM_008963 Ptgds 17.85 7
NM_147029 Olfr1120 53.89 28 NM_026582 Wls 14.75 24
NM_144534 Tmem38a 45.86 31 NM_001177362 Gpr137 7.46 6
NM_025460 Tmem126a 45.74 19 NM_013582 Lhcgr 5.85 9
NM_023671 Clns1a 32.07 28 NM_007937 Epha5 3.62 18
NM_133189 Cacng7 29.74 12 NM_001081405 Vmn2r28 3.61 6
NM_011695 Vdac2 27.33 27
NM_009601 Chrnb1 25.59 31
NM_008536 Tm4sf1 21.79 24
NM_146982 Olfr1257 21.61 11
NM_026211 Tmed9 17.13 15
NM_207567 Olfr1198 15.78 8
NM_019631 Tmem45a 14.25 17
NM_008430 Kcnk1 14.23 21
NM_010098 Opn3 14.08 14
NM_175502 Tmem74 13.47 12
NM_009630 Adora2a 10.37 14
NM_026145 Kctd10 9.84 19
NM_001011867 Olfr538 9.78 5
NM_001159392 Tnfaip1 8.93 22
NM_009144 Sfrp2 7.92 10
NM_001033271 Tmem55b 7.81 21
NR_033648 Vmn2r-ps129 7.78 11
NM_146498 Olfr490 7.69 4
NM_008169 Grin1 7.55 16
NM_178751 Orai2 6.63 17
NM_172671 Lgr4 5.85 17
NM_175519 Kctd8 5.29 10
NM_018733 Scn1a 5.16 37
NM_133683 Tmem19 5.07 10
NM_025781 Tmem170 4.91 6
NM_001008497 P2ry14 4.85 5
NM_176996 Smo 4.14 11
NM_032397 Kcnn1 4.07 11
NM_144830 Tmem106a 4.06 6
NM_001044741 Cacnb3 3.46 6
NM_144936 Tmem45b 3.11 4
NM_033444 Clic1 2.98 3
NM_010934 Npy1r 2.54 5
NM_029722 Gje1 2.42 4
NM_010348 Grik1 1.69 4
NM_153059 Tmem5 1.21 1
NM_009219 Sstr4 1.2 1
NM_001177362 Gpr137 1.16 1
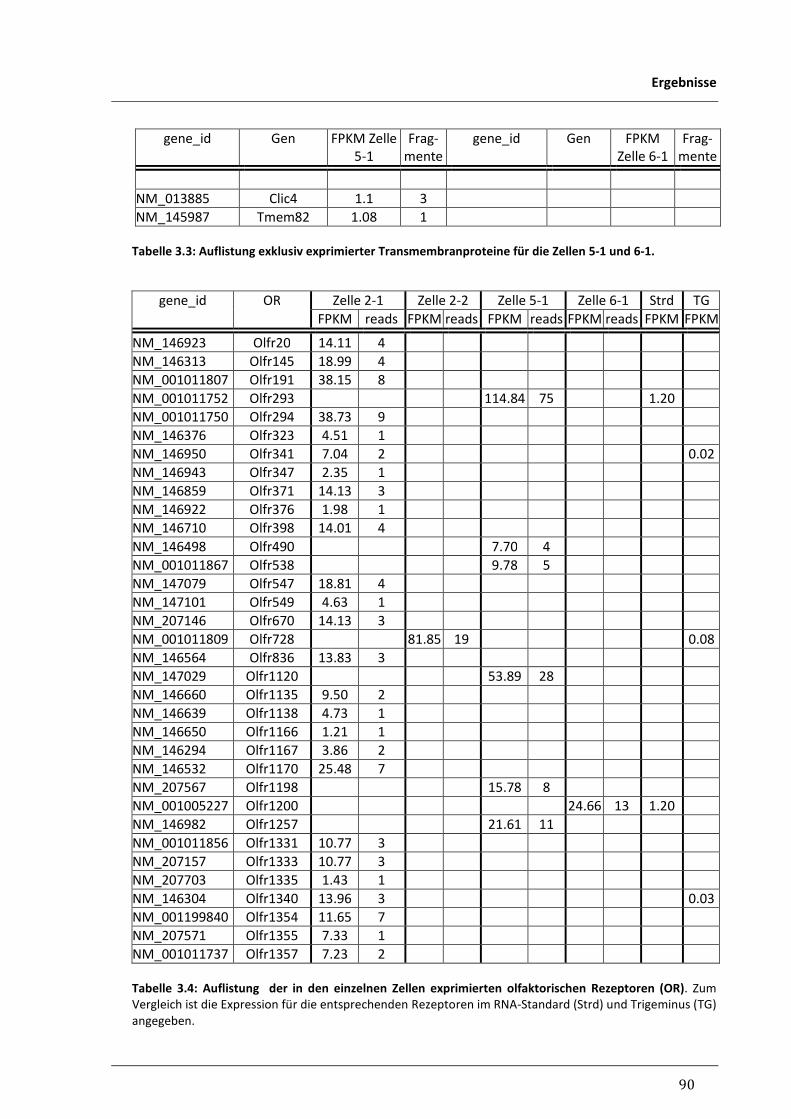
90
Ergebnisse
gene_id Gen FPKM Zelle
5-1
Frag-
mente
gene_id Gen FPKM
Zelle 6-1
Frag-
mente
NM_013885 Clic4 1.1 3
NM_145987 Tmem82 1.08 1
Tabelle 3.3: Auflistung exklusiv exprimierter Transmembranproteine für die Zellen 5-1 und 6-1.
gene_id OR Zelle 2-1 Zelle 2-2 Zelle 5-1 Zelle 6-1 Strd TG
FPKM reads FPKM reads FPKM reads FPKM reads FPKM FPKM
NM_146923 Olfr20 14.11 4
NM_146313 Olfr145 18.99 4
NM_001011807 Olfr191 38.15 8
NM_001011752 Olfr293 114.84 75 1.20
NM_001011750 Olfr294 38.73 9
NM_146376 Olfr323 4.51 1
NM_146950 Olfr341 7.04 2 0.02
NM_146943 Olfr347 2.35 1
NM_146859 Olfr371 14.13 3
NM_146922 Olfr376 1.98 1
NM_146710 Olfr398 14.01 4
NM_146498 Olfr490 7.70 4
NM_001011867 Olfr538 9.78 5
NM_147079 Olfr547 18.81 4
NM_147101 Olfr549 4.63 1
NM_207146 Olfr670 14.13 3
NM_001011809 Olfr728 81.85 19 0.08
NM_146564 Olfr836 13.83 3
NM_147029 Olfr1120 53.89 28
NM_146660 Olfr1135 9.50 2
NM_146639 Olfr1138 4.73 1
NM_146650 Olfr1166 1.21 1
NM_146294 Olfr1167 3.86 2
NM_146532 Olfr1170 25.48 7
NM_207567 Olfr1198 15.78 8
NM_001005227 Olfr1200 24.66 13 1.20
NM_146982 Olfr1257 21.61 11
NM_001011856 Olfr1331 10.77 3
NM_207157 Olfr1333 10.77 3
NM_207703 Olfr1335 1.43 1
NM_146304 Olfr1340 13.96 3 0.03
NM_001199840 Olfr1354 11.65 7
NM_207571 Olfr1355 7.33 1
NM_001011737 Olfr1357 7.23 2
Tabelle 3.4: Auflistung der in den einzelnen Zellen exprimierten olfaktorischen Rezeptoren (OR). Zum
Vergleich ist die Expression für die entsprechenden Rezeptoren im RNA-Standard (Strd) und Trigeminus (TG)
angegeben.

91
Diskussion
4. Diskussion
4.1 Charakterisierung der stereospezifischen Wirkung von Nikotin auf nAChR und TRPA1
Im Rahmen dieser Arbeit habe ich mit elektrophysiologischen Experimenten die Wirkung von
Nikotin in seinen beiden stereoisomeren Erscheinungsformen auf verschiedene ionotrope
Rezeptoren untersucht. Im Fokus dieser Untersuchungen standen insbesondere die Rezeptoren
der Klasse der Liganden-gesteuerten Ionenkanäle und der TRP-Kanäle, die im Trigeminus
exprimiert sind. Bisher sind zwei ionotrope Rezeptortypen bekannt, die durch Nikotin aktiviert
werden können. Zum einen sind dies nikotinische Acetylcholinrezeptoren, zum anderen der
TRPA1-Kanal. Es ist bekannt, dass diese einerseits als maßgebliche Rezeptoren bei der
chemosensorischen Wahrnehmung von Nikotin fungieren, aber andererseits auch rekombinant
exprimiert in elektrophysiologischen Experimenten durch Nikotin aktiviert werden können
(Talavera et al., 2009; Thuerauf et al., 1999; Walker et al., 1996).
In der Familie der nAChR gibt es – wie in der Einleitung erwähnt – zwei verschiedene
Rezeptortypen: homo- und heteromere Rezeptoren. Es wurden deshalb α7 als Beispiel eines
homomeren Rezeptors mit starker Expression im Trigeminus (Liu et al., 1998) und α2β4 als
Beispiel eines heteromeren Rezeptors untersucht (3.1.2). Für die Untersuchung heteromerer
nAChR wären α4β2 und α3β4 interessantere Kandidaten gewesen, da diese Kombinationen sehr
deutlich im Trigeminus nachgewiesen worden sind (Flores et al., 1996). Die Expression dieser
Rezeptoren in Xenopus-Oozyten schlug jedoch fehl.
Bei Radioliganden-Bindungsstudien wurde hinsichtlich der Aktivierung von nAChR eine gewisse
Stereoselektivität für das (-)-Enantiomer von Nikotin festgestellt (Khan et al., 1994; Sloan et al.,
1985). Allerdings wurde eine Aktivierung heterolog exprimierter menschlicher nAChR durch (+)-
Nikotin mit elektrophysiologischen Methoden noch nicht gezeigt. Im Beispiel des nAChR α7 des
Haushuhns wurde eine Aktivierung durch (+)-Nikotin jedoch von Amar et al. (1993)
elektrophysiologisch im Voltage-Clamp Verfahren nachgewiesen. Die Autoren stellten allerdings
mit EC50-Werten von 24 ± 0,7 µM für (-)-Nikotin und 45 ± 1 µM für (+)-Nikotin eine nur geringe
Stereoselektivität fest. Ich habe in dieser Arbeit die Stereoselektivität für (+/-)-Nikotin beim
menschlichen α7 nAChR untersucht und einen EC50 von 519 ± 189 µM ermittelt. Die von
ChavezNoriega et al. (1997) publizierten Werte für (-)-Nikotin (113 µM) und Acetylcholin (180 µM)
wurden ebenfalls mittels Voltage-Clamp an Xenopus-Oozyten ermittelt und sind deshalb mit
meinen Ergebnissen vergleichbar. Ich habe erwartungsgemäß festgestellt, dass der EC50 für
(+)-Nikotin höher ist. Aus beiden letztgenannten Studien wird außerdem klar, dass (-)-Nikotin für

92
Diskussion
α7 beider Spezies eine höhere pharmakologische Potenz besitzt als Acetylcholin. Amar et al.
(1993) stellten zudem auch bei (+)-Nikotin für α7 eine höhere Potenz als Acetylcholin fest, was
den Ergebnissen dieser Arbeit entsprechend nicht für den menschlichen nAChR α7 gilt. In meinen
Ergebnissen wird zusätzlich deutlich, dass die Maximalwirkung für (-)-Nikotin am menschlichen
nAChR α7 über 6-mal größer ist als die von (+)-Nikotin. Die Autoren der letztgenannten Studie
fanden diesbezüglich beim nAChR α7 des Haushuhns hingegen keinen nennenswerten
Unterschied. Die Stereoselektivität für (-)-Nikotin scheint bei den nAChR α7 von Huhn und
Mensch unterschiedlich stark ausgeprägt zu sein. Zwischen den Extrazellulärdomänen von
menschlichem α7 und jenem der Ratte besteht eine Gemeinsamkeit von 91-99% (ChavezNoriega
et al., 1997). Galzi et al. (1991) zeigten, dass in diesem Bereich schon der Austausch einer einzigen
Aminosäure zu massiven Veränderungen der Pharmakologie führt. Die festgestellte
Stereoselektivität am menschlichen nAChR α7 passt näherungsweise zu der aus Radioliganden-
Bindungsstudien am Gehirn der Ratte bekannten 10-fach höheren Bindungsaffinität für (-)-
Nikotin, obwohl dabei eine Zuordnung zu einem Rezeptortyp nicht möglich ist (Sloan et al., 1985).
Die für den nAChR α7 im Voltage-Clamp aufgezeichneten Stromantworten zeigten eine schnelle
Desensitisierung. Hierbei geht die Stromantwort noch während der Applikation des Agonisten
zurück, was die Messung der Maximalstromamplitude erschwert. Die von ChavezNoriega et al.
(1997) aufgezeichneten α7-vermittelten Ströme und die für den nAChR α7 des Haushuhns (Amar
et al., 1993) publizierten, zeigten ebenso eine starke Desensitisierung. Auch diese Konzentrations-
Wirkungsbeziehungen wurden mit ähnlich stark desensitisierenden Stromantworten erstellt,
wobei die Autoren darauf hinwiesen, dass Maximalamplituden möglicherweise zu klein gemessen
werden. Nach Wagner et al. (2004) ist mit einer Unterschätzung der Maximalamplitude
grundsätzlich im Zwei-Elektroden-Voltage-Clamp an Xenopus-Oozyten zu rechnen, da die
Konzentration des Agonisten während der Applikation nur langsam bis zur Zielkonzentration
ansteigt. Bei der benutzten manuellen Applikationsmethode wird ein Austausch der gesamten
Messkammerflüssigkeit in einem Zeitraum von maximal einer Sekunde erreicht. Innerhalb dieses
Zeitraumes erreicht auch die Agonistenkonzentration die maximale Konzentration. Während
dieser Zeit setzt aber bereits eine Desensitisierung ein, die dazu führen kann, dass die gemessene
Stromamplitude kleiner als die tatsächliche Maximalamplitude ist und dabei ermittelte
pharmakologische Kenngrößen wie den EC50 und den maximalen Effekt beeinflusst. Dieser
desensitisierende Effekt ist von der pharmakologischen Affinität des Liganden zum Rezeptor und
der applizierten Agonistenkonzentration abhängig. Bei Agonisten mit niedriger Affinität zum
Rezeptor fällt der Effekt nicht stark ins Gewicht. Mit steigender Konzentration steigt auch die
Desensitisierung und damit der Messfehler im Verlauf der Konzentrations-Wirkungskurve.

93
Diskussion
Diesen Effekt kann man nur mit Einzelkanalmessungen gänzlich ausschließen. Die für (+)-Nikotin
festgestellten Stromamplituden bei nAChR α7 sind jedoch wesentlich kleiner als die für (-)-Nikotin
und ACh festgestellten Amplituden, sodass ausgeschlossen werden kann, dass der Effekt auf
Desensitisierung zurückgeht. Die Differenz der Maximalstromantworten von ACh und (+)-Nikotin
im Gegensatz zu ACh und (-)-Nikotin wurde als signifikant festgestellt.
Bei nikotinischen Acetylcholinrezeptoren sind auch β-Untereinheiten an der Bildung der
Agonistenbindestelle beteiligt (Corringer et al., 2000) und beeinflussen pharmakologische
Eigenschaften, wie die Spezifität für cholinerge Agonisten (Luetje and Patrick, 1991). Die Autoren
letzterer Studie haben gezeigt, dass eine Spezifität für Acetylcholin und Nikotin von der
Untereinheitenkombination abhängt. Dabei war α2β4 eine der Kombinationen mit höherer
Spezifität für Nikotin. Als Beispiel eines heteromeren Rezeptors habe ich in dieser Arbeit die
Kombination α2β4 mit Blick auf eine Stereoselektivität für Nikotin untersucht. Ähnlich den
Ergebnissen bei α7 wurde hier festgestellt, dass die Maximalwirkung für (-)-Nikotin beim
menschlichen nAChR α2β4 über 6-mal größer ist als von (+)-Nikotin. Auch für nAChR α2β4 wurde
in dieser Arbeit keine Konzentrations-Wirkungs-Beziehung für (-)-Nikotin und Acetylcholin
ermittelt. Da die von ChavezNoriega et al. (1997) publizierten EC50-Werte für (-)-Nikotin (20,7 µM)
und Acetylcholin (82,5 µM) ebenfalls im Voltage-Clamp Verfahren an Xenopus-Oozyten gemessen
wurden, konnten sie mit dem in dieser Arbeit ermittelten Wert für
(+)-Nikotin (51 µM) verglichen werden. Es zeigt sich, dass α2β4 stereoselektiv für (-)-Nikotin ist
und in diesem Fall sogar (+)-Nikotin wie das (-)-Enantiomer eine höhere pharmakologische Potenz
hat als der natürliche Agonist Acetylcholin.
Die Reihenfolgen der pharmakologischen Potenz für beide untersuchten menschlichen
nikotinischen Acetylcholinrezeptoren lautet somit für die nAChR α7: (-)-Nik > ACh > (+)-Nik und für
die Untereinheitenkombination α2β4: (-)-Nik > (+)-Nik > ACh.
Vor einiger Zeit haben Talavera et al. (2009) (-)-Nikotin als einen Agonisten vom TRPA1-Kanal
identifiziert. Dies ist bis jetzt nach dem nAChR die zweite Entdeckung eines Ionenkanals, der durch
Nikotin aktiviert wird, jedoch untersuchten die Autoren nicht die Wirksamkeit des
(+)-Enantiomers und ließen somit die Frage offen, ob TRPA1 in Bezug auf Nikotin stereoselektiv
ist. Diese Frage habe ich in dieser Arbeit untersucht. In letztgenannter Studie wurden
rekombinant exprimierte menschliche TRPA1-Kanäle mit elektrophysiologischen Experimenten in
chinese hamster ovary-Zellen (CHO- Zellen) untersucht. Die festgestellte Maximalwirkung von (-)-
Nikotin entspricht dabei etwa einem Viertel der Maximalwirkung des Agonisten AITC. Um eine
relative Stromantwort in etwa dieser Höhe hervorzurufen, habe ich in dieser Arbeit am
menschlichen TRPA1-Kanal eine Konzentration von 10 mM beider Nikotinenantiomere appliziert.

94
Diskussion
Sowohl (-)- als auch (+)-Enantiomer sind partielle Agonisten, wobei sie bei der applizierten hohen
Konzentration die gleiche Wirkung haben, der Kanal also nicht stereoselektiv antwortet (3.1.7). Da
allerdings in der vorliegenden Arbeit keine Konzentrations-Wirkungs-Kurve bis zur Sättigung
vorgelegt werden konnte, ist ein Vergleich der pharmakologischen Potenz von (+)-Nikotin mit dem
von Talavera et al. (2009) ermittelten EC50-Wert (20-30 µM) für (-)-Nikotin nur bedingt möglich.
Allerdings zeigt schon die Tatsache, dass die Stromantworten bei einer Konzentration von 1000
µM (+)-Nikotin weniger als ein Drittel der Stromantworten bei 10 mM beträgt, dass der EC50-Wert
für (+)-Nikotin wesentlich größer als 1000 µM sein muss und sich somit stark von dem publizierten
unterscheidet. Der pharmakologische Unterschied beruht jedoch nicht nur in einer Verschiebung
der Konzentrations-Wirkungs-Kurve. Talavera et al. (2009) stellen für (-)-Nikotin bei
Konzentrationen größer als 1000 µM einen Block des Kanals fest, sodass bei 10 mM (-)-Nikotin
bereits keine Stromantwort mehr registriert werden kann. In dieser Arbeit konnte dieser Befund
nicht bestätigt werden. Im Gegenteil dazu werden bei dieser Konzentration für beide Enantiomere
die größten Stromantworten gemessen. Da es sich in beiden Fällen um den menschlichen TRPA1
handelt, kommen Spezies bezogene Unterschiede nicht in Frage, somit könnte die Begründung
des beobachteten pharmakologischen Unterschiedes vor allem in unterschiedlicher Ausstattung
der Expressionssysteme Xenopus-Oozyten und CHO-Zellen liegen. Beispielsweise wurde ein bis zu
100-fach höherer EC50 beim TRPM8 des Hundes in Xenopus-Oozyten im Vergleich zu HEK-Zellen
für den Agonisten Icillin festgestellt (Liu et al., 2006). Die Tatsache, dass in dieser Arbeit keine
Stereospezifität des TRPA1 für ein Nikotinenantiomer festgestellt werden konnte, ist eine wichtige
Feststellung für die Interpretation psychophysischer Studien (Thuerauf et al., 1999; Thuerauf et
al., 2000; Walker et al., 1996). Die Ergebnisse machen klar, dass TRPA1 einen Beitrag zur
trigeminalen Empfindung von Nikotin leistet, jedoch kann der Kanal an der Vermittlung
unterschiedlicher trigeminaler Empfindungsstärken der beiden Nikotinenantiomere nicht beteiligt
sein.
Mit den Ergebnissen dieser Arbeit habe ich gezeigt, dass (+)-Nikotin den menschlichen nAChR im
rekombinanten System aktiviert. Obschon dies im Vergleich zu (-)-Nikotin mit reduzierter
pharmakologischer Potenz und Maximalwirkung auftritt, könnte durch (+)-Nikotin eine
trigeminale nAChR vermittelte Empfindung hervorrufen werden, denn trotz der für (+)-Nikotin
niedrigeren Potenz sind nAChR immerhin 10-100-mal sensitiver für (+)-Nikotin als der menschliche
TRPA1-Kanal im gleichen Expressionssystem.

95
Diskussion
In der Studie von Talavera et al. (2009) wurde jedoch auch in vivo die Bedeutung von TRPA1 für
Irritationen durch Nikotin nachgewiesen. Die Autoren haben festgestellt, dass TRPA1-knockout-
Mäuse weniger Nikotin-induzierte Atemwegsreflexe zeigen als Wildtyptiere. Zudem haben die
Autoren in Calcium-Imaging Experimenten 23% Nikotin-responsiver trigeminaler Neurone
festgestellt, wobei 72% dieser Neurone ebenfalls auf den TRPA1-Agonisten AITC antworteten.
In dieser Arbeit habe ich ebenfalls im Calcium-Imaging Nikotinantworten trigeminaler Neurone
untersucht und konnte mit 20-35% Nikotin-responsiver Neurone eine ähnliche Größenordnung
feststellen. Es wurden hier etwa 49% Nikotin-induzierter Signale mit einem spezifischen TRPA1-
Blocker geblockt und somit durch diesen Kanal vermittelt (3.2.2.1). In weiteren Experimenten
wurden mit dem spezifischen nAChR-Blocker Hexamethonium 70% der Nikotin-induzierten
Signale geblockt und so gezeigt, dass nAChR einen großen Anteil der trigeminalen Nikotinantwort
vermitteln. Die Nikotinantworten wurden mit 250 µM bei einer niedrigeren Konzentration
beobachtet, als die Minimalkonzentration die bei rekombinanten TRPA1-Kanälen eine
Stromantwort erzeugte. Die Empfindlichkeit für Nikotin ist somit bei den TRPA1-exprimierenden
Neuronen wesentlich höher als im heterologen Expressionssystem.
Zusammenfassend kann man anhand der elektrophysiologischen Experimenten sagen, dass es bei
nAChR und TRPA1 die pharmakologische Voraussetzung für die trigeminale Wahrnehmung von
Nikotin beider Enantiomere gibt. An trigeminalen Neuronen konnte gezeigt werden, dass
Nikotinantworten über beide Rezeptoren vermittelt werden.
4.2 Die Wirkung von Nikotin auf 5-HT3-Rezeptoren
Neben der direkten Aktivierung habe ich die Modulation bzw. Inhibition ionotroper Rezeptoren
durch Nikotin untersucht und dabei zunächst rekombinant exprimierte 5-HT3-Rezeptoren
untersucht. 5-HT3-Rezeptoren treten als homomere 5-HT3A- und heteromere 5-HT3AB-Rezeptoren
auf, die sich sowohl pharmakologisch als auch in den Kanaleigenschaften unterscheiden. Aufgrund
des sehr geringen Leitwertes (< 1 pS) der Homomere vermuteten bereits Maricq et al. (1991) bei
der ersten Charakterisierung rekombinanter 5-HT3A-Rezeptoren die Expression weiterer
Untereinheiten, die in einem Heteromultimer mit 5-HT3A einen höheren Leitwert von
16 pS verursachen. Die Koexpression von homo-, wie heteromeren 5-HT3-Rezeptoren sind in
Serotonin-sensitiven Neuronen der Regelfall. Homo- und Heteromere haben unterschiedliche
pharmakologische Eigenschaften, da diese durch die A- und B-Untereinheit beeinflusst werden,
denn die Ligandenbindestelle wird an den Kontaktstellen einer A- mit einer weiteren A- oder einer
B-Untereinheit gebildet (Boyd et al., 2002). Für homomere 5-HT3A-Rezeptoren der Maus wurden

96
Diskussion
(+)- und (-)-Nikotin als kompetitive Antagonisten Serotonin-induzierter Stromantworten
festgestellt (Gurley und Lanthorn, 1998). Dabei wurde eine stereoselektiv stärkere
antagonistische Wirkung für das (-)-Enantiomer beobachtet. Die 5-HT3B-Untereinheit konnte in
letztgenannter Studie nicht untersucht werden, da sie erst ein Jahr später kloniert wurde (Davies
et al., 1999; Dubin et al., 1999). Eine Wirkung von Nikotin auf die heteromeren 5-HT3AB-
Rezeptoren wurde bisher nicht publiziert. In dieser Arbeit wurde deshalb die Wirkung von Nikotin
auf 5-HT3AB untersucht und gezeigt, dass in Xenopus-Oozyten exprimierte menschliche 5-HT3A- und
5-HT3AB-Rezeptoren sowohl durch (+)- als auch (-)-Nikotin geblockt werden (3.1.1). Von Gurley und
Lanthorn (1998) wurde am homomeren 5-HT3A der Maus ein kompetitiver Block mit niedrigeren
IC50-Werten ((-)-Nikotin: 32 µM; (+)-Nikotin: 107 µM) festgestellt. Allerdings wurden in diesem Fall
die Stromantworten auch durch niedrigere Serotoninkonzentrationen hervorgerufen (1 µM
anstatt 10 µM), was die höheren IC50-Werte in dieser Arbeit ((-)-Nikotin: 512 µM; (+)-Nikotin:
2070 µM) erklärt. Mit Radioliganden-Bindungsstudien zeigten Drisdel et al. (2008) die Bindung
von (-)-Nikotin auch an native 5-HT3-Rezeptoren aus dem Gehirn der Ratte. Es gibt viele
Substanzen, die Bindungsaffinitäten sowohl zu nAChR als auch zu 5-HT3-Rezeptoren besitzen,
jedoch nicht auf beide Rezeptorfamilien die gleiche Wirkung haben. Die Autoren letztgenannter
Studie zeigten beispielsweise in elektrophysiologischen Experimenten, dass das - wie Nikotin - als
cholinerger Agonist bekannte Epibatidin ebenfalls ein 5-HT3-Antagonist ist. Zudem ist das als
spezifischer 5-HT3-Antagonist eingesetzte Tropisetron ein starker partieller Agonist von nAChR α7
(Macor et al., 2001). Diese Tatsachen passen auch dazu, dass die zuerst für nAChR bekannt
gewordene Stereoselektivität für das (-)-Enantiomer von Nikotin auch bei 5-HT3-Rezeptoren
auftritt. In dieser Arbeit konnte diese Stereospezifität menschlicher 5-HT3-Rezeptoren sowohl für
Hetero- als auch Homomere gezeigt werden.
Die Bedeutung dieses Blocks für chemosensorische Prozesse an trigeminalen Neuronen wurde im
Rahmen dieser Arbeit mittels Calcium-Imaging untersucht, wobei ich zeigen konnte, dass 79%
Serotonin-induzierter Signale trigeminaler Neurone durch (-)-Nikotin inhibiert wurden (3.2.4).
Mittels der 5-HT3-spezifischen Agonisten mCPBG konnte bestätigt werden, dass es sich bei dieser
Population um 5-HT3-exprimierende Neurone handelt. Im Trigeminus werden neben den
ionotropen 5-HT3-Rezeptoren auch verschiedene metabotrope Serotoninrezeptoren exprimiert
(Amrutkar et al., 2012), was die Population der Zellen mit nicht inhibierten Serotoninsignalen
erklärt. Es wurde in diesen Experimenten zwar mit 1 mM Nikotin eine hohe Konzentration
appliziert, jedoch zeigt die Konzentrations-Inhibitionskurve insbesondere für 5-HT3AB bereits bei
100 µM (-)-Nikotin eine starke Inhibition des Serotonin-induzierten (10 µM) Stromes. Eine
Aktivierung von 5-HT3-Rezeptoren auf nozizeptiven

97
Diskussion
Fasern führt zu einem schmerzhaften Eindruck (Richardson, 1990). Nikotin kann somit einen
5-HT3-induzierten schmerzhaften Eindruck vermindern. 5-HT3-Rezeptoren können somit auch die
Wahrnehmung von Nikotin beeinflussen; die beschriebene Inhibition könnte aber auch im
Allgemeinen beim Vorliegen einer ausreichend hohen Nikotinkonzentration einen
modulatorischen Einfluss auf die Funktion des trigeminalen sensorischen Systems haben. Dies gilt
allerdings auch für das gustatorische System. Beim gustatorischen System wurde zum einen eine
Signalübertragung von gustatorischen Rezeptorzellen an die afferenten Nervenendigungen mit
ATP gezeigt (Finger et al., 2005), es konnte jedoch zum anderen auch die Ausschüttung von
Serotonin festgestellt werden (Huang et al., 2005). Kaya et al. (2004) konnten eine Expression von
5-HT3-Rezeptoren in gustatorischen Rezeptorzellen der Ratte zeigen. Durch den Block 5-HT3-
vermittelter Signale kann Nikotin einen gustatorischen Eindruck vermindern.
4.3 Nikotinblock an NMDA-Kanälen
Von der Familie der ionotropen Glutamatrezeptoren ist eine Beteiligung an chemosensorischen
Prozessen bekannt. Eine Funktion als olfaktorischer Rezeptor ist beispielsweise für die Taufliege
Drosophila melanogaster bekannt, bei der Glutamatrezeptoren in den Antennen exprimiert
werden (Abuin et al., 2011; Benton et al., 2009). Die bei Vertebraten vorkommenden
Glutamatrezeptoren werden in die drei Subfamilien der AMPA-, Kainat- und NMDA-Rezeptoren
unterteilt. Die Expression von NMDA- im olfaktorischen Epithel des Menschen (Borgmann-Winter
et al., 2009) und die Koexpression von AMPA- und NMDA-Rezeptoren im olfaktorischen System
der Maus sowohl in der Mucosa als auch im olfaktorischen Bulbus wurden bereits nachgewiesen
(Rash et al., 2005). Erst kürzlich wurde eine NMDA-Rezeptor-abhängige Potenzierung der
Duftstoff-induzierten Aktivität von Mitralzellen gezeigt, die durch NMDA-Rezeptor-Antagonisten
verhindert werden kann (Lethbridge et al., 2012). Es liegt die Vermutung nahe, dass durch die
Aktivierung ionotroper Glutamatrezeptoren somit im olfaktorischen System von Säugern eine
Modulation chemosensorischer Wahrnehmung stattfinden könnte. Da auch alle ionotropen
Glutamatrezeptoren in trigeminalen Neuronen exprimiert werden (Gu und Huang, 1994; Sahara
et al., 1997), könnte auch hier eine Modulation die trigeminale Empfindung beeinflussen. Aus
diesem Grund wurden in dieser Arbeit von jeder Subfamilie ionotroper Rezeptoren, die alle auch
im Trigeminus exprimiert werden, ein Rezeptor exemplarisch untersucht, um eine Wirkung von
Nikotin auf diese Rezeptoren zu identifizieren.

98
Diskussion
Es wurden in dieser Arbeit für die non-NMDA-Glutamat-Rezeptoren der AMPA- und
Kainatunterfamilie (GluR1(Q)flop, GluR6(Q)) Konzentrations-Wirkungskurven für den jeweiligen
Agonisten mit und ohne (-)-Nikotin einer Konzentration von 1 mM erstellt (3.1.6), wobei keine
Wirkung von (-)-Nikotin feststellt wurde.
Es konnte jedoch in dieser Arbeit das erste Mal ein Konzentrations-abhängiger Block rekombinant
exprimierter menschlicher NMDA-Rezeptoren durch (-)-Nikotin gezeigt werden. Die zunächst an
NMDA-Rezeptor-exprimierenden Xenopus-Oozyten durchgeführten Experimente zur
Konzentrations-Wirkungs-Beziehung ergaben bei der üblichen Klemmspannung von -60 mV
keinen signifikanten Effekt bei Koapplikation von (-)-Nikotin. Es wurden sodann Experimente zur
Strom-Spannungs-Beziehung durchgeführt. Der dabei entdeckte Block zeigt die gleiche
Spannungsabhängigkeit wie der von Nowak et al. (1984) für NMDA-Rezeptoren beschriebene
Mg2+-Block, bei dem das Mg2+-Ion im Inneren der Kanalpore den Durchfluss anderer Ionen
verhindert. Es handelt sich um einen spannungsabhängigen nicht-kompetitiven Block. Für NMDA-
Rezeptoren ist eine Vielzahl solcher Kanalblocker bekannt, die vor allem erforscht wurden, um
Substanzen wie beispielsweise Memantin zu finden, die mit hoher Spezifität aber niedriger
Affinität binden, um NMDA-Rezeptor vermittelte Excitotoxizität zu unterbinden, aber normale
physiologische Aktivität zu erlauben (Chen et al., 1992). Es konnte vor einiger Zeit gezeigt werden,
dass NMDA-vermittelte Excitotoxizität bei kultivierten cortikalen Neuronen der Ratte in der
Gegenwart von (-)-Nikotin verhindert wird (Akaike et al., 1994). Kürzlich wurde dies auch bei der
Gabe von Epibatidin, einem starken nAChR-Agonisten, auf retinales Gewebe festgestellt (Sieber,
2012). Die Autoren beider Studien vermuten eine Koexpression nikotinischer
Acetylcholinrezeptoren in den untersuchten Zellen, die der schädlichen Wirkung einer verstärkten
NMDA-Rezeptor-Aktivierung entgegenwirken. Allerdings könnten cholinerge Agonisten auch
direkt an der Inhibition von NMDA-Rezeptoren beteiligt sein. Es wurde eine große Anzahl von
Substanzen, wie beispielsweise Dextrorphan (Damaj et al., 2005; Wong et al., 1988) oder das oben
genannte Memantin gefunden, die sowohl als NMDA-Rezeptor-Blocker fungieren aber auch eine
Wirkung als Antagonist nikotinischer Acetylcholinrezeptoren haben. Dazu passt der in dieser
Arbeit dargestellte Block von (-)-Nikotin auf NMDA-Rezeptoren (3.1.6). Da dieser Effekt jedoch
erst bei Spannungen negativer als -80 mV und bei Konzentrationen von über 1 mM auftritt, hat er
vermutlich keine systemische Relevanz, kann höchstwahrscheinlich nicht bei NMDA-Rezeptor-
exprimierenden Mitralzellen im olfaktorischen Bulbus stattfinden, ist aber für experimentelle
Bedingungen, bei denen Zellen einer ausreichend hohen Konzentration von Nikotin ausgesetzt
werden, durchaus zu bedenken. Zudem könnte Nikotin in der Schleimhaut des
chemosensorischen Epithels durch die verschiedenen Wege der Selbstverabreichung höhere

99
Diskussion
Konzentrationen erreichen und so auch die trigeminale Chemosensorik über eine NMDA-
Rezeptor-vermittelte Modulation beeinflussen.
4.4 Keine Wirkung von Nikotin auf weitere untersuchte Ionenkanäle
In der umfangreichen systematischen Untersuchung zur Wirkung von Nikotin auf Liganden-
gesteuerter Ionenkanäle und TRP-Kanäle wurden im Rahmen dieser Arbeit viele weitere
ionotrope Rezeptoren untersucht, für die sich herausstellte, dass Nikotin auf diese keine Wirkung
hat. Dazu gehörten bei den klassischen Liganden-gesteuerten Ionenkanälen alle untersuchten
GABAA-Rezeptoren (α1β2γ2, α2β1γ2, α2β3γ2, β3, ρ1; 3.1.4), α1 Glycin-Rezeptoren (3.1.3) und die
Purinozeptoren P2X2 (3.1.5). Wie in der Einleitung erwähnt, ist die Expression von GABAA- und
P2X-Rezeptoren im Trigeminus gezeigt. Es konnte vor kurzen bei einem Großteil trigeminaler
Neurone GABA-induzierte Cl--Ausströme gezeigt werden (Schöbel et al., 2012).
Für P2X-Rezeptoren ist eine Beteiligung bei der Schmerzempfindung gezeigt, die durch ein
Vorkommen auf nozizeptiven Nervenendigungen begründet ist (Barclay et al., 2002; Chen et al.,
1995). Für bestimmte chemische Stimuli ist eine Modulation P2X-vermittelter Signale bei
trigeminalen Neuronen der Ratte festgestellt worden (Spehr et al., 2004). Von diesen
untersuchten Rezeptoren wurde bei den GABAA-Rezeptoren ausschließlich eine Wirkung des (-)-
Enantiomers untersucht. Aus der Klasse der TRP-Kanäle wurden zudem die Vanilloidrezeptoren
V1-3 (3.1.8) und der Melastatinrezeptor M8 (3.1.9) untersucht. Auch für diese Kanäle war keine
Wirkung von Nikotin festzustellen.
Es wurden bei diesen Experimenten Konzentrations-Wirkungskurven für den jeweiligen Agonisten
mit und ohne Nikotin einer Konzentration von 1 mM erstellt. Für 1 mM ist eine aktivierende
Wirkung bei nAChR und TRPA1 gezeigt worden, aber auch eine inhibierende Wirkung an 5-HT3-
und NMDA-Rezeptoren. Die Applikation auch einer 10-fach höheren Nikotinkonzentration wäre
problemlos möglich, bei noch höheren Konzentrationen setzen starke Potentialschwankungen der
Oozyten ein. Da jedoch eine Nikotinkonzentration von 10 mM auch am chemosensorischen
Epithel sehr unwahrscheinlich ist, wäre ein Effekt, der bei dieser Konzentration erst einsetzt,
physiologisch irrelevant.
Die Überschneidung des Ligandenspektrums, die für viele Rezeptoren unterschiedlichster
Rezeptorfamilien gezeigt wurde, ist neben der vielseitigen Expression der Rezeptorfamilien im
Trigeminus ein Grund für die umfassende Untersuchung in dieser Arbeit. Bereits an nAChR und 5-
HT3-Rezeptoren zeigt sich, dass innerhalb einer Rezeptorklasse (Cys-loop-Rezeptoren) eine
Substanz als Ligand für unterschiedliche Unterfamilien auftritt, allerdings auch mit gegensätzlicher

100
Diskussion
Wirkung. Dies tritt auch bei TRP-Kanälen auf. Beispielsweise ist Menthol als TRPM8-Agonist
bekannt, was jedoch auch für bestimmte Spezies in geringerem Maß auf TRPA1 zutrifft. TRPV1 ist
ein Rezeptor für Piperidin, was in schwächerem Maß auch auf TRPA1 zutrifft. TRPA1 ist hingegen
als Rezeptor für Allylisothiocyanat, Allicin und Dialllyldisulfid, schwefelhaltigen Inhaltsstoffen von
Knoblauch und Senf, bekannt. Jedoch aktivieren diese im Gegenzug in schwachem Maße auch
TRPV1 (Viana, 2011). Liu et al. (2004) zeigten für den TRPV1-Kanal der Ratte, eine positive
Modulation Capsaicin-induzierter Stromantworten durch 1 mM (-)-Nikotin. Diese Beobachtung
konnte ich jedoch in dieser Arbeit am menschlichen TRPV1 nicht bestätigen. Dies könnte zum
einen in der unterschiedlichen Ausstattung verschiedener Expressionssysteme beruhen, denn die
Autoren der genannten Studie nutzten CHO-Zellen (chinese hamster ovary), wohingegen die
Ergebnisse dieser Arbeit an Xenopus-Oozyten entstanden. Zum anderen könnten Spezies-
bezogene Unterschiede der Rezeptoren die Ursache sein. Es wurden beispielsweise für den
menschlichen TRPA1 und den TRPA1 der Maus unterschiedliche Aktivierungskinetiken durch
Menthol gezeigt (Karashima et al., 2007; Story et al., 2003). Ansonsten ist bisher für keinen der
anderen genannten Rezeptoren weder eine Bindung, noch eine pharmakologische Wirkung von
Nikotin bekannt.
4.5 Chemosensorisch relevante Nikotinkonzentrationen
Im Zusammenhang mit der Konzentration bestimmter Agonisten bei Experimenten im
heterologen Expressionssystem aber auch in Experimenten mit kultivierten Neuronen stellt sich
die Frage, ob die gewählten Konzentrationen der Applikationslösungen der physiologischen
Realität entsprechen. Generell überstiegen die Konzentrationen der hier applizierten Lösungen
die maximalen Blutplasmakonzentrationen von Nikotin bei Weitem. In dieser Arbeit wurde der
Einfluss von Nikotin bei allen Rezeptoren mit einer Konzentration von 1 mM getestet. Im
Blutplasma von Rauchern wurde bisher in Abhängigkeit vom Zigarettenkonsum eine
Konzentration im Bereich von 0,5–2 µM Nikotin gemessen (Pidoplichko et al., 1997; Zevin et al.,
1998). Die Nikotinkonzentration wurde allerdings auch bereits im Speichel von Rauchern
gemessen. Die Speichelkonzentration überstieg die des Plasmas in der Regel um etwa das 10-
fache auf bis zu 25 µM (Hukkanen et al., 2005; Teneggi et al., 2002). Für die Gewinnung der Probe
wurde dazu allerdings der Speichelfluss angeregt und die Konzentrationen des Flüssigkeitsfilms
der Schleimhaut dadurch verdünnt. Die tatsächliche Konzentration von Nikotin am
chemosensorischen Epithel könnte über die Bereiche der Schleimhaut im Mund- und Rachenraum
sowie in der Nasenhöhle stark variieren, da das lokale Volumen von Schleim oder Speichel, die als

101
Diskussion
Lösungsmittel fungieren, unterschiedlich sein kann. Konzentrationsmessungen über eine im
Mundraum gesammelte Speichelprobe können die verschiedenen Konzentrationsmaxima in der
Schleimhaut nicht repräsentieren. Ginzkey et al. (2012) betrachten deshalb eine Konzentration
von 1 mM Nikotin als sinnvoll für den Einsatz in physiologischen Experimenten, da millimolare
Konzentrationen von Nikotin in der Mucosa erreicht werden können. Die in dieser Arbeit
beschriebenen Nikotin-vermittelten Effekte bei 5-HT3-, TRPA1- und NMDA-Rezeptoren haben
vermutlich keine Relevanz auf der Ebene des Zentralnervensystems, jedoch könnten sie die
chemosensorische Wahrnehmung auf der Ebene des sensorischen Epithels beeinflussen.
4.6 Calcium-Imaging der trigeminalen Zellkultur – neue Nikotinrezeptoren
Neben den elektrophysiologischen Experimenten wurde in dieser Arbeit die Wirkung von Nikotin
auf kultivierte trigeminale Neurone charakterisiert. Auf diesem Wege konnten Hinweise auf
weitere Rezeptoren sowohl für (-)- als auch für (+)-Nikotin gewonnen werden. Ein Großteil
Nikotin-vermittelter Antworten geht auf nAChR und TRPA1-Kanäle zurück, wie bereits unter 4.1
dargestellt wurde. Mit den unter 3.2.2.1 zunächst einzeln eingesetzten Blockern für nikotinische
Acetylcholinrezeptoren (Hexamethonium, Hex) und TRPA1-Kanäle (HC030031, HC) konnte
erwartungsgemäß nur ein Teil der Nikotin-induzierten Signale geblockt werden. Es wurde
anschließend unter Einsatz beider Blocker, also simultanen Blockerbedingungen für nAChR und
TRPA1, ein noch größerer Anteil der Antworten geblockt als mit den einzeln eingesetzten
Blockern. Jedoch war stets eine Population von Zellen vorhanden, die nicht geblockt werden
konnte. In einem vergleichbaren Experiment, bei dem beide Kanalfamilien durch hohe
Konzentrationen von Mecamylamin geblockt wurden, konnte erneut der größte Anteil Nikotin-
vermittelter Signale geblockt werden und wiederum bleibt ein Anteil der Signale unter
Blockerbedingungen bestehen. Der Anteil der Nikotin-sensitiven Neurone, deren Signale auf einen
der bekannten Nikotinrezeptoren nAChR und TRPA1 zurückgehen, wurde demnach zwischen 80
und 88% ermittelt. Dies erklärt auf zellulärer Ebene, dass Mecamylamin die sensorische
Wahrnehmung von Nikotin teilweise inhibiert, wie Thuerauf et al. (2006) in psychophysischen
Tests mit Nikotin-haltigem Aerosol feststellte. Allerdings folgern die Autoren, dass die Wirkung
des Blockers ausschließlich auf der Inhibition nikotinischer Acetylcholinrezeptoren beruht. Die von
Thuerauf et al. (2006) in den psychophysischen Experimenten applizierte Konzentration von
Mecamylamin (> 2 mM) ist jedoch ausreichend, um gleichermaßen TRPA1 zu inhibieren.
Tatsächlich wurde sowohl durch Talavera et al. (2009) als auch in dieser Arbeit (3.2.2.1) gezeigt,
dass ein nicht unerheblicher Anteil Nikotin-sensitiver trigeminaler Neurone ebenfalls AITC-sensitiv

102
Diskussion
ist, beziehungsweise durch HC inhibiert werden kann. Ich habe in dieser Arbeit unter dem Einfluss
des nAChR- und in hoher Konzentration auch TRPA1-Blockers Mecamylamin sowie unter den
erwähnten simultanen Blockerbedingungen mit Hex und HC allerdings auch zwischen 12-20%
persistierende Nikotinsignale festgestellt, für die eine Signalvermittlung durch nAChR oder TRPA1
nicht in Frage kommt. Demnach wurde auf der Ebene sensorischer Neurone gezeigt, dass an der
trigeminalen Wahrnehmung von (-)-Nikotin ein weiterer, noch unbekannter Rezeptor beteiligt ist.
Wie einleitend erwähnt, reagiert das trigeminale System stereoselektiv auf Nikotin (Thuerauf et
al., 1999; Thuerauf et al., 2000; Walker et al., 1996). Von Thuerauf et al. (1999) wird dabei sowohl
für den brennenden als auch für den stechenden Anteil der Empfindung in psychophysischen
Versuchen eine niedrigere Schwelle und eine höhere Intensität für (-)-Nikotin als für (+)-Nikotin
beschrieben. Die allerdings unterschiedlich stark ausgeprägte Stereoselektivität zwischen beiden
Empfindungen erklären die Autoren durch die Ausstattung von C- und Aδ-Fasern mit
verschiedenen Untereinheitenkombinationen nikotinischer Acetylcholinrezeptoren. Talavera et al.
(2009) weisen nach, dass TRPA1, der die stechende Wirkung des Senfölinhaltsstoffes AITC
vermittelt durch Nikotin aktivierbar ist. Gemäß den elektrophysiologischen Ergebnissen in dieser
Arbeit ist es jedoch unwahrscheinlich, dass TRPA1 für eine stereospezifisch stechende
Empfindung verantwortlich ist. Thuerauf et al. (1999) vermuten, dass Subpopulationen
trigeminaler Neurone spezifisch auf ein Nikotinenantiomer antworten könnten. Das Vorkommen
solcher Nikotin-stereoselektiven trigeminalen Neurone konnte in dieser Arbeit mit einem
weiteren Experiment im Calcium-Imaging nachgewiesen werden (3.2.1), was die Vermutung von
Thuerauf et al. (1999) bestätigt. Ein großer Teil der Zellen reagiert auf beide Enantiomere. Es ist
wahrscheinlich, dass diese Zellen einen der bekannten Nikotinrezeptoren nAChR oder TRPA1
exprimieren. Die Fähigkeit beider Ionenkanalfamilien, durch beide Nikotinenantiomere aktiviert
zu werden, wurde bereits gezeigt. Trotz der größeren pharmakologischen Potenz von (-)-Nikotin
bei nAChR reicht die applizierte Konzentration von (+)-Nikotin (250 µM) aus – wie in dieser Arbeit
exemplarisch für homo- wie heteromere nAChR gezeigt wurde, um auch nAChR-vermittelte
(+)-Nikotinsignale hervorzurufen.
Ein weiterer großer Teil der untersuchten trigeminalen Neurone antwortet ausschließlich auf
(+)-Nikotin. Das Ausbleiben einer (-)-Nikotinantwort zeigt, dass der diese Antwort vermittelnde
Rezeptor weder ein TRPA1-Kanal noch ein nAChR sein kann. Neben der oben bereits gezeigten
Existenz eines unbekannten Typs von (-)-Nikotinrezeptoren scheint hier also ein weiterer
selektiver Rezeptortyp für (+)-Nikotin in trigeminalen Neuronen exprimiert zu sein. Ein solcher
Rezeptor ist bisher unbekannt.

103
Diskussion
In weiteren Experimenten im Calcium-Imaging wurde die Wirkung von (+)-Nikotin auf trigeminale
Neurone weiter untersucht, um den bisher nicht beschriebenen selektiven (+)-Nikotinrezeptor zu
identifizieren (3.2.2.2). Es wurde eine ausschließlich (+)-Nikotin-responsive Zellpopulation (5% der
untersuchten Neurone) identifiziert, deren (+)-Nikotinantwort bei Koapplikation von
Rutheniumrot, einem unspezifischen TRP-Kanalblocker, inhibiert werden konnte. Rutheniumrot
blockiert neben TRPA1 (Clapham et al., 2005) auch viele weitere Kanäle, wobei der Blocker nicht
auf nAChR wirkt (eigene Beobachtung, Daten nicht gezeigt). Die blockierbare Antwort kann also
einerseits nicht durch nAChR, andererseits aber auch nicht durch TRPA1 vermittelt worden sein,
denn die beschriebene Neuronenpopulation antwortet nicht auf AITC und nicht auf (-)-Nikotin.
Der Blocker muss in diesem Fall allerdings nicht direkt den gesuchten Rezeptor blockiert haben, es
könnte lediglich ein Ca2+-leitender Effektorkanal der pharmakologische Wirkort gewesen sein. In
einem weiteren Experiment wurde mit Gentamycin, einem spezifischen Blocker für TRPA1
(Nagata et al., 2005), eine kleine Neuronenpopulation (3%) nachgewiesen, deren Antwort nicht
blockiert werden konnte, die jedoch nur auf (+)- und nicht auf (-)-Nikotin antwortete. Eine
Signalvermittlung durch den TRPA1 wird hier ausgeschlossen, eine Beteiligung nikotinischer
Acetylcholinrezeptoren kann wegen der ausbleibenden (-)-Nikotinantwort ebenfalls
ausgeschlossen werden. Auch diese Ergebnisse deuten auf einen Rezeptor hin, der spezifisch (+)-
Nikotin-responsiv ist.
Diese Ergebnisse konnten mit einem weiteren Experiment bestätigt werden. Mit Borneol,
ebenfalls einem TRPA1-spezifischen Blocker (lehrstuhleigene Ergebnisse, Korrespondenz mit M.A.
Sherkheli) und Hexamethonium konnte unter simultanen Blockerbedingungen für nAChR und
TRPA1 eine ausschließlich (+)-Nikotin-responsive Population (4%) festgestellt werden. Bryant et al.
(2010) haben in olfaktorischen Rezeptorneuronen der Ratte ebenfalls Zellpopulationen
identifiziert, die selektiv auf (+)-Nikotin reagieren und vermuten, dass olfaktorische Rezeptoren an
dieser Antwort beteiligt sind. Die letztgenannten Autoren fanden, dass immerhin etwa 14%
Nikotin-responsiver olfaktorischer Rezeptorneurone ausschließlich auf (+)-Nikotin antworteten,
was den in dieser Arbeit ermittelten Anteil bei trigeminalen Neuronen nur geringfügig übertrifft.
Es konnte im Rahmen dieser Arbeit erstmals gezeigt werden, dass es im trigeminalen System
Neuronenpopulationen gibt, die einen unbekannten, ausschließlich für (+)-Nikotin-responsiven
Rezeptor funktionell exprimieren. Dies ist eine Erkenntnis, die zur Vervollständigung des
Verständnisses der sensorischen Wahrnehmung von Nikotin beiträgt.
Im Weiteren sollte in dieser Arbeit untersucht werden, ob die nachgewiesenen Rezeptoren für (-)-
und (+)-Nikotin zur Klasse der G-Protein-gekoppelten Rezeptoren gehören könnten. Verschiedene
G-Proteine können unterschiedliche second-messenger-Transduktionsprozesse einleiten.

104
Diskussion
Da es pharmakologische Substanzen gibt, die selektiv metabotrope Signaltransduktionskaskaden
inhibieren, kann mit weitergehenden Blockerexperimenten die Beteiligung bestimmter Kaskaden
ausgeschlossen werden. In dieser Arbeit wurde nun zunächst eine Beteiligung einer
Phospholipase C-gekoppelten Transduktion am Nikotin-induzierten Signal im Calcium-Imaging
untersucht. Das im Calcium-Imaging beobachtete Signal wird durch einen Anstieg der Ca2+-
Konzentration im Zellplasma verursacht, der entweder auf einem Einstrom von außen durch
Ionenkanäle der Zellmembran oder auf einer Ausschüttung aus intrazellulären Speichern durch
Ionenkanäle des endoplasmatischen Retikulums beruhen kann. In Bezug auf die Nikotinantworten
der trigeminalen Neurone würde der Einstrom von Ca2+-Ionen aus intrazellulären Speichern auf
eine Beteiligung einer Phospholipase C-gekoppelten Signaltransduktion deuten, wobei diese
durch Erzeugung eines second messengers IP3-gesteuerte Ionenkanäle aktiviert (Berridge, 1993).
In dieser Arbeit konnte gezeigt werden, dass alle im Calcium-Imaging messbaren
Nikotinantworten auf dem Einstrom extrazelluärer Ca2+-Ionen beruhen (3.2.3). Unter extrazellulär
Ca2+-freien Messbedingungen konnten weder (-)- noch (+)-Nikotin-induzierte Signale festgestellt
werden. Zudem konnten mit dem PLC-Blocker Edelfosin (Powis et al., 1992) und bei gleichzeitigen
Blockerbedingungen für nAChR und TRPA1 (Hex, HC) nicht alle (-)-Nikotinantworten geblockt
werden (3.2.3). Die hier persistierenden Nikotin-induzierten Signale sind offenbar nicht PLC-
gesteuert, wobei allerdings auch eine PLC-gesteuerte Signaltransduktion über den in der
Zellmembran verbleibenden second messenger Diacylglycerol (DAG) stattfinden kann. Dieser kann
beispielsweise TRPC2-Kanäle aktivieren (Zufall et al., 2005). Die Beteiligung weiterer ionotroper
oder metabotroper Prozesse bei der trigeminalen Nikotinwahrnehmung wie beispielsweise
zyklisch-Nukleotid-gesteuerte Signaltransduktionskaskaden konnten gegen Ende der Arbeit nicht
mehr untersucht werden, kommen jedoch weiterhin in Frage als Ursache der persistierenden
Nikotinantworten in trigeminalen Neuronen.
4.7 Vergleichende Transkriptomanalyse
Bisher wurde weder eine Transkriptomanalyse des Trigeminus noch der Dorsalwurzelganglien der
Maus mittels Next-generation-sequencing publiziert. Um die Expression verschiedener
Transmembranrezeptoren und Ionenkanäle in den genannten Geweben in umfangreichem
Maßstab zu untersuchen, wurde eine Analyse des Transkriptoms mittels NGS im Rahmen dieser
Arbeit durchgeführt. Es wurden zudem bereits publizierte Sequenzierergebnisse für das Gehirn in
die Analyse einbezogen. Die Ergebnisse dieser RNA-Sequenzierungen wurden mit einigen
Amplikon-Sequenzierungen einzelner Nikotin-responsiver Neurone verglichen.

105
Diskussion
Aufgrund von Experimenten im Calcium-Imaging wurden diese Zellen zwei Typen zugeordnet. Bei
zwei der Zellen wurde das Nikotinsignal durch Blocker der bekannten Nikotinrezeptoren nAChR
und TRPA1 inhibiert. Bei zwei weiteren Zellen konnten diese Blocker das Nikotinsignal nicht
verhindern. Bei den letzteren wurde die Expression eines weiteren Nikotinrezeptors
angenommen, für den eine Transkriptomanalyse weitere Hinweise liefern kann.
Expressionsnachweise werden mittlerweile nicht mehr nur rein qualitativ geführt. Neue
Methoden ermöglichen zuverlässige quantitative Aussagen zur Expressionsstärke und erfahren
zunehmende technische Optimierung. Dabei kann man zwischen Methoden zur
Expressionsanalyse weniger bekannter Gene und Massenansätzen unterscheiden. Bei ersteren ist
die quantitative realtime PCR (qPCR) die verbreitete Referenzmethode, die aber für die Analyse
ganzer Transkriptome ungeeignet ist. Zu den Methoden, mit denen die große Anzahl von
Transkripten, die in einem Transkriptom vorkommen kann, möglichst umfassend quantifiziert
werden kann, zählen neben DNA-Microarrays vor allem auch das Next-generation-sequencing-
Verfahren. Wang et al. (2009) stellen im einer Übersichtsarbeit die Vorteile von Next-generations-
sequencing gegenüber DNA-Microarrays heraus. Zum einen können mit RNA-Sequenzierungen
alle Transkripte innerhalb eines Transkriptoms erfasst werden, mit Hilfe von DNA-Microarrays
lediglich jene, für die Sonden auf dem Microarray aufgebracht sind. Zum anderen enthalten
Sequenzierergebnisse im Gegensatz zu DNA-Microarrays kein Hintergrundsignal, weshalb auch
niedrige Expressionsstärken festgestellt werden können. Ebenfalls können sehr hohe
Expressionsstärken festgestellt werden. Arrays zeigen Expressionsstärken mit einer geringeren
dynamischen Breite an als Sequenzierergebnisse; ab einer gewissen Expressionshöhe
unterscheiden sich Signalstärken verschiedener Expressionshöhen nur noch gering, was die
Genauigkeit von Arrays bei sehr hohen Expressionsstärken abnehmen lässt. Mit RNA-
Sequenzierungen können mehrere tausendfache Expressionsunterschiede festgestellt werden,
mit DNA-Arrays lediglich einige hundert. Diese Eigenschaften machen Next-generation-sequencing
zur bisher brauchbarsten Methode zur Transkriptomanalyse.
Die erwähnten Expressionsnachweise für TRP- und Liganden-gesteuerte Ionenkanäle im
trigeminalen System wurden bisher mit Methoden geführt, mit denen in der Regel ein qualitativer
Nachweis geleistet wird. So wurden beispielsweise die Expressionsverhältnisse der Transkripte
verschiedener nikotinischer Acetylcholinrezeptoren im Trigeminus von Liu et al. (1998) mittels RT-
PCR ermittelt, wobei die Bandenstärke nach der Gelelektrophorese als Hinweis der
Expressionsstärke genutzt wurde. Tecott et al. (1993) weisen mittels In-situ-Hybridisierung 5-HT3-
Rezeptor exprimierende trigeminale Neurone nach. Kobayashi et al. (2005) zeigen die Expression
von TRPA1, M8 und V1 im Trigeminus sowohl mittels RT-PCR als auch In-situ-Hybridisierung.

106
Diskussion
Aus diesen Ergebnissen lassen sich keine zuverlässigen relativen Expressionsstärken zwischen
verschiedenen Transkripten ableiten. Die bisherigen Expressionsnachweise erübrigen also
keinesfalls einen systematischen quantitativen Ansatz. Die einzelnen Next-generation-sequencing-
Verfahren sind zurzeit noch in einem starken Optimierungsprozess begriffen (Wang et al., 2009),
was unter anderem dazu geführt hat, dass die angebotenen Sequenzierlängen auch bei dem in
dieser Arbeit benutzten Illumina-Solexa Verfahren mittlerweile gestiegen sind. So sind die aus der
früheren Studie herangezogenen Ergebnisse einer RNA-Sequenzierung des Gehirns der Maus mit
Sequenzlängen von 25 bp entstanden (Mortazavi et al., 2008). Die im Rahmen dieser Arbeit
durchgeführten Sequenzierungen hatten Sequenzlängen von 35 und 75 bp (3.3). Um ein
sequenziertes Transkript an einen spezifischen Abschnitt des Genoms zu mappen, ist eine Länge
von 25 Basenpaaren generell ausreichend, allerdings kommen im gesamten Genom repetitive
Motive vor, die eine eindeutige Zuordnung eines sequenzierten Fragmentes erschweren. Mit
zunehmender Sequenzierlänge dürfte die Wahrscheinlichkeit einer eindeutigen Zuordnung
steigen. Es existieren weitere Next-generation-sequencing-Verfahren, die größere
Sequenzierlängen ergeben, jedoch wurde in dieser Arbeit eine Methode gewählt, die bei kürzerer
Sequenzierlänge eine wesentlich größere Anzahl von Sequenzierungen liefert, damit sich die
Berechnung der relativen Expressionsstärken auf eine große Datengesamtmenge stützt. Mit dem
in dieser Arbeit benutzten Illumina-Solexa Verfahren lassen sich mit Abstand die größten
Anzahlen sequenzierter Fragmente erzielen (Liu et al., 2012).
4.7.1 Die Qualität der Sequenzierung
Für den Trigeminus der Maus wurde in dieser Arbeit erstmals eine Transkriptomanalyse basierend
auf einer Sequenzierung nach dem Illumina-Solexa Verfahren durchgeführt. Ferner wurde auf
dem gleichen Weg auch das Transkriptom der Dorsalwurzelganglien der Maus untersucht. Der
Anteil gemappter reads an der gesamten Anzahl prozessierter reads dieser beiden untersuchten
Gewebe ist mit 80-85% in etwa so hoch, wie bei der von Mortazavi et al. (2008) durchgeführten
Sequenzierung. Es zeigt sich also, dass mit dem benutzten Verfahren reproduzierbar ein
vergleichbar großer Anteil von Sequenzierergebnissen verwerten lässt. Die restlichen 15-20%
prozessierter reads könnten unspezifisch während der cDNA-Synthese an Primerdimeren
entstanden sein. Diese Fragmente stören jedoch die Auswertung nicht, da sie wegen der nicht
möglichen spezifischen Zuordnung zum Genom der Maus keine falsch positiven Ergebnisse
erzeugen können. Allerdings wird ein gewisser Anteil der Sequenzierleistung durch diese
Fragmente beansprucht. Obwohl vermutlich in der Zukunft ein Ziel der technischen

107
Diskussion
Weiterentwicklung der Synthese der cDNA-Library die Verringerung unspezifischer cDNA-
Fragmente sein wird, ist die Einbuße der Sequenzierleistung durch nicht gemappte reads bei einer
Menge von etwa 30 Millionen gemappten reads bei den in dieser Arbeit sequenzierten
Transkriptomen von Trigeminus und DRG von geringer Bedeutung.
Bei den im Rahmen dieser Arbeit durchgeführten Amplikonsequenzierungen ist der Anteil
gemappter reads allerdings mit 8-42% deutlich geringer als bei den RNA-Sequenzierungen der
Explantate. Die für eine Sequenzierung des Transkriptoms einzelner Zellen notwendige
Amplifikation wurde in dieser Arbeit mit dem TransPlex Verfahren (3.3) durchgeführt. Wie in der
Einleitung dargestellt, kann mit der TransPlex Amplifikation prinzipiell auch bei geringen Mengen
eingesetzter RNA ein Amplikon erzeugt werden, dass relative Expressionsverhältnisse
wiederspiegelt. Neben den spezifisch vom Transkriptom amplifizierten Fragmenten scheint
allerdings bei dieser Reaktion auch ein großer Anteil unspezifischer Fragmente erzeugt zu werden,
die nicht an das Genom gemappt werden können. Es wird also bei der Amplikonsequenzierung
mit diesem System weit über die Hälfte der Sequenzierleistung eingebüßt, was zu einer starken
Verkleinerung der Datenmenge führt. Die Primersequenzen der PCR-basierten Amplifikation
dieses Verfahrens, die jedes Fragment des Amplikons flankieren, können für einen Teil der nicht
gemappten reads verantwortlich sein (Gonzalez-Roca et al., 2010). Auch die Methoden der
Amplikonerzeugung werden zurzeit stark weiterentwickelt. Die meisten optimierten Verfahren
starten die cDNA-Synthese am 3‘-Ende des Transkripts und erzeugen ein Amplikon, dass für eine
Analyse mit dafür optimierten DNA-Microarrays brauchbar ist (Caretti et al., 2008; Kurimoto et al.,
2007). Amplikonsequenzierungen nach derart 3‘-fokussierten Amplifikationsverfahren können
keine Sequenzierergebnisse über das gesamte Transkriptom hinweg erzeugen, was gegenüber
dem hier benutzten TransPlex Verfahren einen Nachteil darstellt. Die hierbei eingesetzten
randomisierten Primer ermöglichen einen cDNA-Synthesestart an jedem Abschnitt des
Transkriptoms.
4.7.2 Die Expression der bekannten Nikotinrezeptoren nAChR und TRPA1
In dieser Arbeit wurden die Transkriptome der erwähnten Gewebe Trigeminus und DRG mit den
amplifizierten Transkriptomen einzelner (-)-Nikotin-responsiver trigeminaler Neurone verglichen
(3.3). Bei zwei der untersuchten vier (-)-Nikotin-responsiven Zellen konnte die Antwort durch die
spezifischen nAChR- und TRPA1-Blocker Hex und HC (3.2.2.1) geblockt werden (Zellen 2.1 und
2.2). Bei zwei weiteren (Zellen 5-1 und 6-1) persistierte das (-)-Nikotinsignal auch bei
Koapplikation der Blocker, sodass angenommen werden konnte, dass die Antworten nicht von

108
Diskussion
diesen beiden Rezeptoren vermittelt wurden. Die erste Erwartung an das Transkriptom dieser
beiden Zellen ist somit das Fehlen von Transkripten dieser beiden Kanäle. Tatsächlich ist TRPA1 in
keiner der vier untersuchten Zellen exprimiert (3.3.1.3). Dies passt zu dem Anteil TRPA1
funktionell exprimierender trigeminaler Neurone von 7-11%, der in dieser Arbeit in Calcium-
Imaging Experimenten mit dem spezifischen TRPA1-Blocker HC und dem TRPA1-Agonisten AITC
ermittelt wurde.
Allerdings zeigen die Ergebnisse dieser Arbeit für die untersuchten Gewebe der Maus eine
durchaus deutliche Expression von TRPA1 in trigeminalen und Dorsalwurzelganglien und
bestätigen die Ergebnisse von Kobayashi et al. (2005), die ebenfalls keinen signifikanten
Unterschied der TRPA1-Expression zwischen Trigeminus und DRG bei der Ratte feststellten.
Für die Expression von nAChR wurden bei einer der beiden Zellen mit geblocktem (-)-Nikotinsignal
die Untereinheiten α4 und β2 festgestellt, die eine häufige Kombination in sensorischen Ganglien
miteinander bilden (Albuquerque et al., 2009). Das beobachtete Signal könnte demnach also
durch diesen Rezeptor vermittelt worden sein. Allerdings wurden in einer der beiden Zellen mit
nicht geblocktem (-)-Nikotinsignal ebenfalls neuronale nAChR, wenngleich auch sehr niedrig
exprimiert festgestellt. Hier ist der nAChR jedoch nicht für das Signal verantwortlich, denn dieses
würde durch Hexamethonium geblockt. Die Expression muskulärer nAChR α1 und β1 (3.3.1.2) ist
zwar erstaunlich, da sie nativ nur in Muskeln vorkommt, jedoch kann hier kein funktioneller
Rezeptor exprimiert worden sein, denn muskuläre nAChR bilden nativ nur in den beiden
Kombinationen α1β1γδ und α1β1δε funktionelle Rezeptoren (Millar und Gotti, 2009), die zum
einen hier nicht gebildet werden können und zum anderen wäre auch diese Antwort durch
Hexamethonium mit der applizierten Konzentration zu einem Großteil geblockt worden (Papke et
al., 2010).
Erstmals wurde im Rahmen dieser Arbeit die Reihenfolge der Expressionsstärke nikotinischer
Acetylcholinrezeptoren für ein Explantat des trigeminalen Ganglions der Maus mit Next–
generation-sequencing herausgefunden. Für die Reihenfolge der nAChR gilt: α7 > α6 > α3 > α4 >
α1 und β2 > β3 > β1 > β4, was in etwa den Ergebnissen von Liu et al. (1998) entspricht, die diese
Reihenfolge mittels PCR feststellten: α7 ≈ α3 > α6 > α4 ≈ α5 > α9 ≥ α2, und β2 ≈ β3 > β4. Die
geringfügigen Unterschiede zwischen den Ergebnissen könnten durch Differenzen zwischen Ratte
und Maus begründet, oder auch den unterschiedlichen technischen Verfahren geschuldet sein.
Das nAChR-Expressionsprofil entspricht zudem dem der Dorsalwurzelganglien, was die
Ähnlichkeiten von Trigeminus und DRG als sensorisch afferente Ganglien unterstreicht. Es ist
bisher weder klar, aus welchem Grund eine derart hohe Zahl verschiedener nAChR in den
untersuchten Ganglien exprimiert wird, noch ob sich diese Expression mit dem Alter des Tieres

109
Diskussion
stark verändert. Durch die Vielfalt an nAChR wird eine große Zahl verschiedener
Untereinheitenkombinationen verschiedener pharmakologischer Profile in trigeminalen Ganglien
möglich.
4.7.3 Die Expression der weiteren Cys-loop-Rezeptoren
Für die weiteren Cys-loop-Rezeptoren wurde im Trigeminus eine ähnlich große Vielfalt festgestellt
(3.3.1.2). Für 5-HT3-Rezeptoren konnte die von Hu et al. (2004) und Tecott et al. (1993)
festgestellte Expression im Trigeminus bestätigt werden. Auch hier ähnelt das trigeminale
Expressionsmuster dem der DRG. Die von Hayasaki et al. (2006) und Kondo et al. (1994)
berichtete Expression der GABAA-Rezeptoren α1-6, β1-3, γ1-3 im Trigeminus konnte bestätigt
werden, zusätzlich wurde mit diesen Ergebnissen Hinweise auf die Expression von δ und ρ2
gefunden. Die von Mortazavi et al. (2008) festgestellte Rezeptorvielfalt im Gehirn bestätigt die
von Nutt (2006) nachgewiesene Expression von α1-6, β1-3, γ1-3, δ, ε und θ und unterstreicht die
Funktion von GABA als wichtigsten inhibitorischen Neurotransmitter im Gehirn. Die im Gehirn
exprimierte Vielfalt der GABAA-Rezeptoren ist deutlich höher als die Vielfalt, die in den
sensorischen Ganglien exprimiert wird. Glycinrezeptoren werden, wie in der Einleitung
beschrieben, im Rückenmark zur Inhibition des motorischen Systems exprimiert. Im Gehirn sind
sie hingegen nicht die vorherrschenden Cl--Kanäle (Lynch, 2009; Somogyi et al., 1998). So sind vier
der sechs exprimierten GABAA-Rezeptor α-Untereinheiten im Gehirn deutlich stärker exprimiert
als die stärkste Glycinrezeptor α-Untereinheit α1. Für die Expression von Glycinrezeptoren im
Trigeminus ist β die am stärksten exprimierte Untereinheit. Dies ist insofern erstaunlich, als β
alleine keine funktionellen Rezeptoren bilden kann. Da die Expression von α2 nur einen Bruchteil
der Höhe von β erreicht, kann die Höhe der β-Expression auch nicht durch die Stöchiometrie von
(α)2(β)3 (Grudzinska et al., 2005) erklärt werden, weshalb der Grund der starken Expression der β-
Untereinheit hier unklar bleibt.
Die Expressionsprofile der Cys-loop-Rezeptoren der einzelnen Zellen spiegeln die Vielfalt der
Expression im Trigeminus nicht wieder. Für den Großteil der Cys-loop-Rezeptoren konnte keine
Expression festgestellt werden. Demnach kann man vermuten, dass die beobachtete
Rezeptorvielfalt im Trigeminus auf Zellpopulationen mit starker Expression eines individuellen
Teils der Rezeptorvielfalt beruht, die auf das Explantat bezogen eine durchschnittliche
Expressionsstärke zur Folge haben. Für GABAA-Rezeptoren widerspricht dem allerdings, dass im
Trigeminus der Ratte nahezu 100% der Neurone GABA-responsiv sind (Hayasaki et al., 2006;
Kondo et al., 1994).

110
Diskussion
Die als RNA-Standard bezeichnete Probe sollte als Amplikon, das vom trigeminalen Transkriptom
amplifiziert wurde, ein mit dem Trigeminus vergleichbares Expressionsprofil aufweisen. Dabei fällt
für die meisten Cys-loop-Rezeptoren auf, dass nur die im Trigeminus stark exprimierten Kanäle im
Amplikon festgestellt werden. Die Amplifizierung mit dem TransPlex Verfahren hat also offenbar
zur Folge, dass die schwach exprimierten Transkripte im Amplikon nicht repräsentiert werden,
was auch der Einschätzung von Gonzalez-Roca et al. (2010) entspricht.
4.7.4 Die Expression weiterer chemosensorisch relevanter Rezeptoren
Die Expression der weiteren TRP-Kanäle (TRPA1 wurde bereits erwähnt), TRPC, TRPM und TRPV,
konnte im Trigeminus bestätigt werden (3.3.1.3). Wie in der Einleitung dargestellt, sind die
Thermo-TRPs die Rezeptoren, die für die Temperaturwahrnehmung verantwortlich sind. Sie
vermitteln zudem die schmerzhaften Wirkungen von Irritantien. Die Expression von TRPV1-4 und
TRPM8 im Trigeminus ist nachgewiesen (Caterina et al., 1997; McKemy et al., 2002; Silver et al.,
2006; Xu et al., 2002; Yamamoto et al., 2009). Die Transkriptomanalyse zeigt Ähnlichkeiten des
Expressionsprofils von DRG und Trigeminus, die sich sehr deutlich nur durch die höhere
Expression von TRPM8 im Trigeminus unterscheiden. Dies entspricht den Ergebnissen von
Kobayashi et al. (2005), die in 35% trigeminaler Neurone und 22% DRG-Neuronen eine TRPM8-
Expression feststellten. Außerdem konnten letztgenannte Autoren keinen signifikanten
Unterschied der Expression von TRPA1 und TRPV1 zwischen Trigeminus und DRG feststellen. Dies
stimmt mit den Ergebnissen der Transkriptomanalyse überein. Von TRPC-Kanälen ist eine
vielseitige Expression über verschiedenste Gewebe hinweg bekannt. In einer Übersichtsarbeit
weist Montell (2005) auf eine starke Expression vor allem im Gehirn hin, wobei sie auch in nicht
neuronalem Gewebe auftreten. Zudem wurden alle TRPC-Kanäle in Dorsalwurzelganglien
festgestellt (Elg et al., 2007). Bei der hier durchgeführten Transkriptomanalyse wurden bis auf
TRPC7 alle Kanäle der Familie im Trigeminus und den DRG festgestellt. Bisher sind kaum
brauchbare spezifische Blocker für TRPC bekannt. Dies ist für eine Charakterisierung im nativen
System problematisch. Es sind vor allem intrazelluläre Modulationen bekannt. So ist für TRPC3, C6
und C7 eine Modulation durch Diacylglycerol bekannt, das durch die intrazelluläre Phospholipase
C gebildet wird (Hofmann et al., 1999; Okada et al., 1999). Es erscheint wahrscheinlich, dass diese
Kanäle bei chemosensorischen Prozessen des Trigeminus keine Funktion als ionotroper Rezeptor,
sondern als metabotroper Effektor ausüben.
Auch für KCNK-Kanäle 2, 4, 10, 18 ist eine Expression im Trigeminus bekannt (Lafreniere et al.,
2010; Yamamoto et al., 2009). Diese können als Rezeptoren für Reize von außen wirken und eine

111
Diskussion
irritierende Wirkung hervorrufen. Das durch den Inhaltsstoff Hydroxy-α-sanshool des
Szechuanpfeffers ausgelöste Prickeln wird durch KNCK-Kanäle vermittelt (Bautista et al., 2008;
Lennertz et al., 2010). Yamamoto et al. (2009) stellten eine Koexpression von KCNK-Kanälen und
vielen Thermo-TRP-Kanälen in kleineren trigeminalen Neuronen fest. Noel et al. (2009) konnten
nachweisen, dass durch KCNK-Kanäle die Schwellentemperatur, bei der ein thermischer Reiz ein
Vermeidungsverhalten auslöst, also eine Temperatur einen schmerzhaften Eindruck erzeugt,
durch KCNK-Kanäle bei Mäusen maßgeblich beeinflusst wird. Eine pharmakologisch wirksame
Substanz an KCNK-Kanälen kann also Schmerz- und Temperaturempfindung modulieren. Im
Rahmen einer weiteren Arbeit des Lehrstuhls wurde eine inhibierende Wirkung hoher
Konzentrationen von Nikotin auf die KCNK-Kanäle 2, 10 und 18 festgestellt (Daten hier nicht
gezeigt, persönliche Korrespondenz, L. Beltrán Márques ). Die Transkriptomanalyse bestätigt die
Expression von KCNK 1, 2, 10 und 18 im Trigeminus und den DRG und zeigt die Expression
weiterer zehn KCNK-Kanäle im Trigeminus (KCNK 4-6, 9, 12-14; 3.3.1.4).
4.7.5 Validierung der Sequenzierergebnisse
Im Rahmen der Transkriptomanalyse von Trigeminus und DRG in dieser Arbeit wurde auch die
Expression verschiedener housekeeping-Gene (Barber et al., 2005; Kouadjo et al., 2007; Zhou et
al., 2010) untersucht. Da dies Gene sind, die in der Regel keinen starken gewebespezifischen
Regulationen unterliegen, werden sie im Allgemeinen als Standard für Expressionsvergleiche
zwischen verschiedenen Strukturen genutzt. Zum einen zeigt beispielsweise die über die
verglichenen Gewebe hinweg starke Expression von Actb, Ubc und Ldha, mittelstarke Expression
von Tbp und schwache Expression ribosomaler Proteine Rpl5, 19 und 29, dass im Grunde
genommen das gewählte Sequenzierverfahren zur quantitativen Transkriptomanalyse geeignet ist
(3.3.1.1). In dieser Arbeit wurden housekeeping-Gene zum anderen als Positivkontrolle für eine
erfolgreiche Amplifikation genutzt. Um zu zeigen, dass mit einem per TransPlex Verfahren
erzeugten Amplikon ebenfalls eine zuverlässige quantitative Transkriptomanalyse möglich ist,
muss also für die housekeeping-Gene ein mit dem Trigeminus vergleichbares Expressionsprofil bei
den einzelnen Zellen festgestellt werden. Dies ist mit einigen Einschränkungen der Fall. Starke
Expression stimmt für Actb, deutliche bei Ldha, Ubc und Ppia im Wesentlichen überein. Mit Hprt
liegt allerdings ein Gen vor, dass im Trigeminus deutlich, in den einzelnen Zellen aber schwach
und teilweise gar nicht exprimiert wird, umgekehrt sind alle ribosomalen Proteine (RPL5, 19, 29)
im Trigeminus schwach, in den einzelnen Zellen jedoch stark exprimiert. Für den RNA-Standard
ergibt sich ein ähnliches Bild.

112
Diskussion
Dieses Amplikon wurde zum direkten Vergleich des trigeminalen Transkriptoms ohne und mit
Amplifikation generiert. Die Ähnlichkeit des Expressionsprofils stark exprimierter Gene und die
Abweichungen bei schwächer exprimierten Genen führt zu der Annahme, dass die von Gonzalez-
Roca et al. (2010) im Einzelfall festgestellten Unterschiede zwischen Expressionsergebnissen
mittels Sequenzierung nach TransPlex Amplifikation und der klassischen qPCR je stärker auftreten,
desto schwächer das Gen tatsächlich exprimiert ist.
Eine fehlende Expression von CNPase (bis auf einem Fragment im Amplikon von Zelle 6-1) in den
einzelnen Zellen wurde erwartet, da CNPase ein Marker für Schwann-Zellen ist und somit in
Amplikons trigeminaler Neurone nur als Kontamination auftreten kann. Im Transkriptom des
gesamten Ganglions kommt CNPase wegen der vorhandenen Schwann-Zellen vor (Chandross et
al., 1999; Nishizawa et al., 1985; Radtke et al., 2011; Sprinkle, 1989; Trapp et al., 1988). Beta-3-
Tubulin (Tubb3) ist für immunhistochemische Experimente ein allgemeiner Neuronenmarker.
Allerdings liegt die mRNA eher während der Wachstumsphasen von Zellausläufern in hohem
Maße vor (Jiang et al., 1994a; b; Kost und Oblinger, 1993). So könnte der Grund für die fehlende
Expression von Beta-3-Tubulin in den einzelnen Neuronen schlicht ein fehlendes Wachstum in der
Zellkultur sein.
Die bisher betrachteten Expressionsprofile der vier einzelnen Neurone ermöglichen einige
Folgerungen:
1. Die Expression der housekeeping-Gene zeigt, dass ein Transplex Amplikon eines Transkriptoms
mit einigen Einschränkungen für quantitative Expressionsanalyse geeignet ist. Einschränkungen
der Genauigkeit der Methode sind im Bereich der sehr niedrigen Expression zu erwarten.
2. Die fehlende Expression des Schwann-Zellmarker CNPase in den Transkriptomen der einzelnen
Neurone weist darauf hin, dass das Verfahren der Isolation einzelner Neurone aus der Zellkultur
kontaminationsfrei technisch durchführbar ist und die unbeabsichtigte Aufnahme von
Zellfragmenten oder freier mRNA vermeidbar ist.
Nur eines der beiden Neurone mit blockierbarem (-)-Nikotinsignal exprimiert einige der in
peripheren Ganglienzellen weit verbreiteten nAChR.
3. Es ist also offenbar mit Einschränkungen möglich, mit der TransPlex Amplifikation und
anschließender Amplikonsequenzierung, den für eine physiologische Eigenschaft
verantwortlichen Rezeptor zu identifizieren.
Im Gegensatz zu den housekeeping-Genen ist der Großteil der untersuchten Ionenkanäle nicht in
den einzelnen Neuronen exprimiert.

113
Diskussion
4. Die Vielfalt an ionotropen Rezeptoren im trigeminalen Transkriptom entsteht vermutlich nicht
durch weit verbreitete Expression, sondern durch verschiedene Zellpopulationen mit spezifischem
Expressionsprofil.
Allerdings zeigten Gonzalez-Roca et al. (2010) mittels qPCR, dass eine differentielle
Transkriptomanalyse zweier Amplikons des gewählten Systems bei einer derart geringen
eingesetzten RNA-Menge von etwa 10 pg (Sambrook und Russell, 2001), wie sie in einzelnen
Zellen enthalten ist, einen gewissen Anteil falsch negativer Ergebnisse ergibt. Die natürliche
Detektionsgrenze der Methode liegt bei einem RNA-Molekül. Allerdings ist sowohl die
Wahrscheinlichkeit gering, dass die Expression eines Genes mit einem einzelnen Transkript bei der
TransPlex Amplifikation repräsentiert, als auch bei der Sequenzierung festgestellt wird.
Vermutlich enthält demnach die Menge der gleichsam nicht exprimiert festgestellten Ionenkanäle
in den einzelnen Neuronen viele Gene, deren spezifische Expression aus technischen Gründen
nicht festgestellt werden konnten. Im Gegenzug ist es sehr wahrscheinlich, dass festgestellte
Expression auch tatsächlich auf einem ursprünglich vorliegenden Transkript beruht.
4.7.6 Differentielle Expression der einzelnen Nikotin-responsiven Neurone
Im Rahmen dieser Arbeit wurden die Transkriptome von vier (-)-Nikotin-responsiven trigeminalen
Neuronen untersucht. Bei zwei der Neuronen konnte das Nikotinsignal durch ein Gemisch aus
nAChR- und TRPA1-Blockern unterbunden werden. Da das Signal der anderen beiden Neurone in
der Gegenwart des Blockergemisches persistierte, ist zu vermuten, dass ein zusätzlicher, bisher
unbekannter Nikotinrezeptor für die Vermittlung des Signals verantwortlich ist. Aus diesem Grund
wurden die Transkriptome der Neurone mit nicht geblocktem Nikotinsignal mit denen der
Neurone mit geblocktem Signal aus jeweils demselben Versuchsdurchlauf verglichen, um
exprimierte Gene zu identifizieren, die exklusiv in den nicht geblockten Neuronen vorkommen
(3.3.2). Die nicht geblockten Zellen exprimierten 1244 (Zellen 6-1 vs 2-1) und 2425 (Zellen 5-1 vs
2-2) Gene exklusiv. Unter diesen befanden sich neben Strukturgenen auch 48 (Zelle 5-1)
beziehungsweise 8 (Zelle 6-1) Ionenkanäle und G-Protein-gekoppelte Rezeptoren. Der Vergleich
dieser beiden „Exklusivlisten“ zeigte, dass mit dem Gpr137 einer dieser Rezeptoren in beiden
nicht geblockten Zellen exprimiert wird. Ginge man davon aus, dass man das Transkript eines
Rezeptors, der im Calcium-Imaging ein deutliches Signal vermittelt, in der Transkriptomanalyse
ebenfalls zuverlässig nachweisen kann, so wäre mit dem Gpr137 ein sinnvoller Kandidat für
weitergehende Experimente zur pharmakologischen Charakterisierung identifiziert. Gpr137
codiert für den G-Protein-gekoppelten Rezeptor Latrophilin 1 (Vakonakis et al., 2008), der im

114
Diskussion
gesamten Nervensystem exprimiert wird (Magdaleno et al., 2006) und auch im olfaktorischen
Epithel vorkommt (persönliche Korrespondenz, N. Kanageswaran). Über die Funktion dieses
Rezeptors ist bisher nichts bekannt. Man weiß, dass das Spinnengift α-Latrotoxin an ihn bindet
(Grishin, 1998). Obwohl es denkbar wäre, dass dieser Rezeptor das (-)-Nikotinsignal der
trigeminalen Neurone vermittelt hat, ist die gemeinsame Expression des Rezeptors in den beiden
nicht geblockten Neuronen nur ein schwaches Indiz. Die bereits erwähnten Einschränkungen der
Einzelzelltranskriptomanalyse, die nicht auf das Sequenzierverfahren, sondern vielmehr auf der
Generierung des Amplikons beruhen, können Ursache dafür sein, dass exprimierte Rezeptoren in
Einzelfällen nicht festgestellt werden. Dies könnte auch bei den beiden geblockten Neuronen der
Fall sein. Aufgrund der Blockierbarkeit der Nikotinantwort durch die nAChR- und TRPA1-
spezifischen Antagonisten erwartet man, dass die Antwort durch einen dieser Kanäle vermittelt
wurde. Eine der Zellen zeigt eine Expression, der in sensorischen Ganglien verbreiteten nAChR
α4β2 (2.1), bei der anderen Zelle (2.2) ist kein neuronaler nAChR exprimiert. Zudem konnte auch
keine Expression von TRPA1 festgestellt werden. Da es aber bisher erst wenige Erkenntnisse
darüber gibt, wie hoch der Anteil der mittels Transkriptomanalyse eines TransPlex Amplikons
fälschlicherweise nicht festgestellten Gene bei minimalem RNA-Einsatz in der Amplifikation ist,
kann nicht ausgeschlossen werden, dass in den vier untersuchten Neuronen mehr
Transmembranproteine exprimiert werden, als festgestellt wurden. Zwischen den beiden
untersuchten nicht geblockten Zellen könnten mehr Transmembranproteine gemeinsam
exprimiert werden.
Es wurde zudem eine deutliche Expression olfaktorischer Rezeptoren in den untersuchten
Neuronen festgestellt. Mittlerweile wurden olfaktorische Rezeptoren außerhalb des
olfaktorischen Epithels schon in verschiedenen Geweben wie beispielsweise Muskel (Griffin et al.,
2009) und Hirngewebe der Maus (Otaki et al., 2004), sowie Leber (Yuan et al., 2001), Hoden
(Spehr et al., 2003) aber auch der Zunge von Mensch (Durzynski et al., 2005; Gaudin et al., 2001)
und Maus (Gaudin et al., 2006) nachgewiesen. Spehr et al. (2004) zeigten, dass im Trigeminus
keine olfaktorischen Rezeptoren exprimiert werden. Bei der im Rahmen dieser Arbeit
durchgeführten Transkriptomanalyse wurde die Expression einiger olfaktorischer Rezeptoren
festgestellt. Jedoch lag die Expressionsstärke weit unterhalb von 1 FPKM. Dies traf zudem auch
auf gustatorische Rezeptoren zu. Die in den nicht geblockten Neuronen teils deutlich exprimierten
olfaktorischen Rezeptoren wie Olfr293 und Olfr1200, für die bisher kein Ligand bekannt ist,
könnten ebenfalls in Betracht gezogen werden, für die Vermittlung des Nikotinsignals
verantwortlich zu sein. Ihre Expression wurde allerdings im Explantat des Trigeminus nicht
gefunden, was darauf hinweist, dass es sich bei den Zellen, die diese Rezeptoren exprimieren, um

115
Diskussion
eine sehr kleine Neuronenpopulation handelt. Der Anteil der Transkripte für diese Rezeptoren am
Transkriptom des gesamten Trigeminus ist offenbar zu gering, um beim Next-generation-
sequencing ins Gewicht zu fallen. Die Ergebnisse unterstützen die Vermutung der Existenz Nikotin-
sensitiver olfaktorischer Rezeptoren von Thuerauf et al. (2006) und kann die von Bryant et al.
(2010) festgestellte in Gegenwart von nAChR-Blockern Nikotin-responsive Population
olfaktorischer Rezeptorneurone erklären.
4.7.7 Die Bedeutung der chemosensorischen Wahrnehmung von Nikotin
Die chemosensorische Wahrnehmung von Nikotin ist maßgeblich an der suchtstillenden Wirkung
beteiligt, die Zigarettenrauch auf Raucher ausübt (Rose et al., 1985). Eine zusätzliche sensorische
Stimulation über den Mund- und Rachenraum verstärkt so auch die Effektivität von
Nikotinersatzprodukten wie Nikotinpflastern, verringert also die Rückfallwahrscheinlichkeit
(Westman et al., 1995). Nikotin ist ein wesentlicher Bestandteil des Aromas von Zigarettenrauch
(Pritchard et al., 1996), dementsprechend wirkt der erste sensorische Eindruck nikotinfreier
Tabakprodukte weniger suchtstillend als bei nikotinhaltigen (Naqvi und Bechara, 2005). Allerdings
stellten Rose et al. (1993) fest, dass Raucher unter ad libitum Versuchsbedingungen den Konsum
eines verdünnten Zigarettenaerosols so erhöhen, dass der gesteigerte Konsum die Verdünnung
wieder aufwiegt. Dies passierte bei einem Aerosol mit starkem sensorischem Eindruck aber mit
sehr niedrigem Nikotingehalt nicht. Der Konsum des Aerosols wurde also durch den sensorischen
Eindruck bestimmt, nicht durch die Nikotindosis. Sehr deutlich wird die Bedeutung des
chemosensorischen Eindrucks für das Rauchverhalten an Zusatzstoffen in Tabakwaren. Als ein
verbreiteter Aromastoff wird Zigaretten Menthol zugegeben. Neben der Aktivierung von TRPM8
hat Menthol eine antagonistische Wirkung beim TRPA1-Kanal (Macpherson et al., 2006). Uhl et al.
(2011) stellten fest, dass ein menschlicher TRPA1-Haplotyp mit einem missense-Polymorphismus
insbesondere bei starken Rauchern mit einer häufigeren Wahl von Mentholzigaretten einhergeht.
Der durch den Rauch hervorgerufene sensorische Gesamteindruck beeinflusst hier durch den
Nikotinrezeptor TRPA1 also maßgeblich die Wahl des Tabakprodukts. Allerdings wird der Kanal
durch weitere Inhaltsstoffe des Tabakrauchs aktiviert, wie beispielsweise Formaldehyd
(McNamara et al., 2007). Es wurde außerdem festgestellt, dass Raucher von Mentholzigaretten
Anzeichen einer tendenziell stärkeren Nikotinabhängigkeit zeigen, was dadurch passieren kann,
dass die Hemmung des TRPA1 die durch Irritantien hervorgerufene Verringerung der Inhalation
verhindert (Fagan et al., 2010). Menthol im Aerosol führt also zu einer tieferen Inhalation und zu
größerer Nikotinaufnahme. Die gezielte Zugabe von Zusätzen in

116
Diskussion
Tabakwaren kann also – vor allem durch ein Zusammenspiel chemosensorischer Vorgänge – eine
Entwöhnung vom Tabakkonsum erschweren. Es ist zu vermuten, dass weitere Zusammenhänge –
wie sie für die Wirkung von Nikotin und Menthol im Zigarettenrauch gefunden wurden – das
Verhalten von Tabakkonsumenten beeinflussen. Nikotin kann zu einem konditionierten Reiz
werden, der neben der pharmakologischen Wirkung im Zentralnervensystem auch als
chemosensorischer Stimulus deutlich das Suchtverhalten beeinflusst (Bryant et al., 2010). Bisher
sind nAChR und TRPA1 als die einzigen Rezeptoren bekannt, die direkt durch Nikotin aktiviert
werden. Es konnte in dieser Arbeit gezeigt werden, dass diese beiden Rezeptoren nicht
ausreichen, um die chemosensorische Wirkung von Nikotin zu erklären. Es geht außerdem aus
dieser Arbeit hervor, dass es sowohl für das (+)- als auch das (-)-Enantiomer je einen weiteren
stereoselektiven Rezeptor im Trigeminus geben muss. Die weitere Aufklärung der
chemosensorischen Signalwege zur Wahrnehmung von Nikotin ist von Bedeutung, um das
Verständnis der Suchtwirkung von Nikotin zu vervollständigen.

117
Zusammenfassung
5. Zusammenfassung
Nikotin ist ein weit verbreitetes Suchtmittel, dessen Konsum mit schweren gesundheitlichen
Folgen verbunden sein kann. Der von Nikotin hervorgerufene chemosensorische Eindruck trägt
einen Anteil zur Wirkung der Droge bei. Nikotin existiert in zwei Stereoisomeren, das (+)- und das
(-)-Nikotin, von denen (-)-Nikotin das wesentlich häufiger in der Natur vorkommende Enantiomer
ist. Die Wirkung der Droge wird im Zentralnervensystem durch nikotinische
Acetylcholinrezeptoren vermittelt. Nikotin ruft einen chemosensorischen Eindruck hervor, der
unter anderem einen olfaktorischen Eindruck und trigeminale Eindrücke wie Brennen und
Stechen einschließt. Bei der chemosensorischen Wirkung von Nikotin tritt eine gewisse
Stereoselektivität auf. Probanden können die Enantiomere olfaktorisch unterscheiden, die
trigeminalen Komponenten haben für beide Enantiomere unterschiedliche Schwellen. Für den
chemosensorischen Eindruck von Nikotin ist neben der Beteiligung nikotinischer
Acetylcholinrezeptoren auch eine Beteiligung des transient receptor potential A1-Kanals bekannt,
der auch den stechenden trigeminalen Eindruck von Senf vermittelt. Psychophysische
Experimente lassen vermuten, dass der chemosensorische Eindruck von Nikotin nicht vollständig
durch nAChR und TRPA1 erklärt werden kann.
In dieser Arbeit wurde die Wirkung von Nikotin auf im Trigeminus exprimierte ionotrope
Rezeptoren untersucht. Dazu wurde das Zwei-Elektroden-Voltage-Clamp-Verfahren zur
elektrophysiologischen Charakterisierung der Wirkung von (+)- und (-)-Nikotin auf heterolog
exprimierte Rezeptoren genutzt. Für heterolog exprimierte menschliche nAChR sowie den TRPA1-
Kanal wurde in dieser Arbeit erstmals eine elektrophysiologische Untersuchung der
Stereoselektivität der Rezeptoren durchgeführt. Für je ein Beispiel eines homo- (α7) und
heteromultimeren (α2β4) menschlichen nAChR konnte die Stereoselektivität für das
(-)-Enantiomer auf diesem Wege gezeigt werden. Beim TRPA1-Kanal wurde erstmals (+)-Nikotin
als Agonist festgestellt und zudem im Vergleich mit (-)-Nikotin keine Stereospezifität beobachtet.
Für die mit den nAChR verwandten humanen 5-HT3-Rezeptoren wurde festgestellt, dass Nikotin
als Antagonist wirkt, wobei für (-)-Nikotin eine stärkere Wirkung beobachtet wurde. Diese Effekte
wurden sowohl am homomeren menschlichen 5-HT3A- als auch am heteromeren 5-HT3AB-
Rezeptor gezeigt. Bei einer Untersuchung ionotroper Glutamatrezeptoren wurde ein (-)-Nikotin-
hervorgerufener (10 mM), spannungsabhängiger Block von NMDA-Rezeptoren (NR1-3a/NR2B)
festgestellt, der in seiner Erscheinung dem bekannten Mg2+-Block ähnelt, jedoch nicht die Stärke
erreicht. Für GABAA-, Glycin-, AMPA- (GluR1(Q)flop), Kainat- (GluR6(Q)), P2X2-Rezeptoren sowie
TRPV1-3- und TRPM8-Kanäle wurde keine Wirkung von Nikotin beobachtet.

118
Zusammenfassung
Die positive Modulation von (-)-Nikotin bei TRPV1-Rezeptoren der Ratte konnte am menschlichen
TRPV1 nicht bestätigt werden.
Ferner wurde im Rahmen der Arbeit die Wirkung von Nikotin auf trigeminale Neurone der Maus
im Calcium-Imaging untersucht und in Experimenten unter simultanen Blockerbedingungen für
die bekannten Nikotinrezeptoren nAChR und TRPA1 festgestellt, dass es eine kleine
Neuronenpopulation gibt, deren (-)-Nikotinantwort sich durch die spezifischen Blocker für diese
Kanäle nicht blockieren lässt. Damit konnte ich zeigen, dass es in trigeminalen Neuronen einen
weiteren Rezeptor für (-)-Nikotin geben muss, der wahrscheinlich an der Vermittlung des
chemosensorischen Eindrucks von Nikotin beteiligt ist. Erstmals wurde zudem eine (+)-Nikotin-
spezifische Population trigeminaler Neurone entdeckt, für die ein Rezeptor bisher nicht bekannt
ist. In weiteren Experimenten konnte unter Blockerbedingungen für nAChR und TRPA1-Kanälen
gezeigt werden, dass die Signale der (+)-Nikotin-spezifischen Neuronenpopulation nicht durch die
bekannten Nikotinrezeptoren vermittelt werden. Es wurde in dieser Arbeit also nachgewiesen,
dass es im Trigeminus einen spezifischen (+)-Nikotinrezeptor gibt. Da die nicht nAChR- oder
TRPA1-vermittelten Nikotinantworten sowohl für (+)- als auch (-)-Nikotin möglicherweise durch G-
Protein-gekoppelte Rezeptoren vermittelt werden können, wurde in dieser Arbeit die Beteiligung
metabotroper Signalwege untersucht. Es konnte gezeigt werden, dass weder (+)- noch
(-)-Nikotinsignale unter Ca2+-freien Messbedingungen auftreten, die Nikotinsignale sind also auf
Ca2+-Ionen von außen angewiesen. Es konnte außerdem erstmals gezeigt werden, dass der für
5-HT3-Rezeptoren bekannte Block durch Nikotin auch an nativen Rezeptoren in trigeminalen
Neuronen auftritt. Es wurde also mit 5-HT3-Rezeptoren ein drittes wichtiges Target für Nikotin im
Trigeminus nachgewiesen, das durch einen Nikotin-vermittelten Block trigeminales
Schmerzempfinden beeinflusst.
Mit einer systematischen Transkriptomanalyse von Trigeminus und Dorsalwurzelganglien mittels
Next-generation-sequencing wurden die bisher zumeist qualitativen Expressionsnachweise für die
chemosensorisch relevanten Cys-loop-, TRP- und KCNK-Kanäle im Trigeminus in einem
quantitativen Verfahren bestätigt und mit DRG verglichen. Mit Hilfe einer Amplikonsequenzierung
wurde das Transkriptom einiger einzelner im Calcium-Imaging unter den erwähnten simultanen
Blockerbedingungen identifizierten und aus der Zellkultur isolierten Neurone untersucht, um
Kandidaten eines unbekannten Rezeptors für (-)-Nikotin zu identifizieren. Dabei wurde eine
deutliche Expression olfaktorischer Rezeptoren in einzelnen Neuronen festgestellt. In einem der
zwei bei simultanem Block Nikotin-responsiven Neurone wurde der Olfr293, in dem anderen
Olfr1200 exprimiert. Aus der Menge bekannter Transmembranrezeptoren exprimierten diese
beiden Zellen nur den Latrophilinrezeptor Gpr137 gemeinsam.

119
Zusammenfassung
Die mit der Einzelzelltranskriptomanalyse gewonnen Expressionsergebnisse bieten weitere
Ansätze, unter den vielen im Trigeminus exprimierten Transmembranrezeptoren die gesuchten
unbekannten Rezeptoren für Nikotin zu finden.

120
Summary
5. Summary
The consumption of the widespread addictive drug nicotine can cause severe diseases. The
nicotine-induced sensory effect is known to contribute to the addictive effect. Nicotine exists in
two stereoisomers, (+)- and (-)-nicotine, of which the (-)-enantiomer is much more abundant in
nature. The drug’s central nervous effect is mediated by nicotinic acetylcholine receptors. Both,
the olfactory system as well as trigeminal sensations like “burning” and “stinging” contribute to
the chemosensory impression induced by nicotine. The chemosensory impressions are
stereoselective. Subjects can discriminate the two isomers by olfactory sensation; the trigeminal
effects appear at different detection thresholds for each enantiomer. Besides nicotinic
acetylcholine receptors, the transient receptor potential channel A1, that mediates the “stinging”
mustard oil impression, is known to contribute to the nicotine impression. Psychophysical studies
indicate that nAChR and TRPA1 cannot fully explain the nicotine-induced chemosensory
impression.
In this study, we investigated the effect of nicotine on trigeminal expressed ionotropic receptors.
We used the electrophysiological two-electrode-voltage-clamp technique to characterize the
effect of (+)- and (-)-nicotine on heterologously expressed receptors. For homo- (α7) and
heteromeric (α2β4) human nAChRs the stereoselectivity for (-)-nicotine could be shown. For the
first time, (+)-nicotine could be shown to be a TRPA1-agonist. Additionally, no stereospecifity was
observed. Further, nicotine was identified as an antagonist for human 5-HT3-receptors with a
stereospecifity for the (-)-enantiomer. These effects were observed for homomeric 5-HT3A- as well
as heteromeric 5-HT3AB-receptors. Experiments analyzing ionotropic glutamate receptors revealed
a voltage-dependent block of NMDA-receptors (NR1-3a/NR2B) induced by (-)-nicotine (10 mM),
which looks similar to the known voltage-dependent Mg2+-block, but remains incomplete. No
effect was observed at GABAA-, Glycine-, AMPA- (GluR1(Q)flop), Kainat- (GluR6(Q)), P2X2-
receptors as well as TRPV1-3- and TRPM8-channels. In our experiments with human TRPV1,
(-)-nicotine could not evoke the positive modulation, that is known for TRPV1 from rat.
Further, in this study the effect of nicotine on murine trigeminal neurons was investigated in
calcium-imaging experiments. Under nAChR and TRPA1 blocking conditions a certain population
of neurons was found, showing a persisting (-)-nicotine response. With these results I could prove
the existence of an unknown (-)-nicotine receptor, expressed in trigeminal neurons, that may
contribute to the chemosensory impression of nicotine. Further, a population of neurons was
identified, responding specifically to (+)-nicotine. A specific receptor for this enantiomer is not
known to date. In further experiments, again under nAChR and TRPA1 blocking conditions, we

121
Summary
could prove signals of specifically (+)-nicotine-responding neurons not to be mediated by both
known receptor families. Therefore, in this study the existence of a (+)-nicotine-specific receptor
expressed in trigeminal neurons was shown. As the unknown (+)- and (-)-nicotine-evoked signals
may be mediated by G-protein-coupled receptors, the involvement of metabotropic signal
pathways was investigated. Neither (+)- nor (-)-nicotine-induced signals were found under
Ca2+-free conditions, indicating that the observed nicotine-induced signals depend on extracellular
Ca2+-ions. Additionally, the blocking effect of nicotine on 5-HT3-receptors, for the first time, was
observed on native receptors in trigeminal neurons. 5-HT3-receptors were identified as the third
important target of nicotine in the trigeminal system with the capacity to influence nociception.
In a systematical approach, in this study, we analyzed the transcriptome of trigeminal and dorsal
root ganglia via next-generation-sequencing methods. We could quantify the trigeminal
expression of chemosensory relevant Cys-loop-, TRP- and KCNK-channels. In previous studies
these channels were detected by qualitative methods. Further, we tried to identify unknown
receptors of (+)- and (-)-nicotine by analyzing the amplicons of single cell transcriptomes, that
showed nicotine-signals independently from nAChRs and TRPA1. Transcriptome analysis of these
cells showed a clear expression of olfactory receptors in single neurons responsive under blocking
conditions. One neuron expressed Olfr293, the other Olfr1200. From the mass of known
transmembrane receptors, both cells shared just the expression of one single receptor, the
Latrophilin receptor Gpr137. Results gained from single-cell transcriptome analysis provide
further candidates of trigeminal expressed receptors that could be the unknown nicotine
receptors.

122
Anhang
6. Anhang
6.1 Literaturverzeichnis
Abuin, L., Bargeton, B., Ulbrich, M.H., Isacoff, E.Y., Kellenberger, S., Benton, R., 2011. Functional
Architecture of Olfactory Ionotropic Glutamate Receptors. Neuron 69, 44-60.
Aceto, M.D., Martin, B.R., 1982. Central Actions of Nicotine. Med Res Rev 2, 43-62.
Akaike, A., Tamura, Y., Yokota, T., Shimohama, S., Kimura, J., 1994. Nicotine-Induced Protection of
Cultured Cortical-Neurons against N-Methyl-D-Aspartate Receptor-Mediated Glutamate
Cytotoxicity. Brain Research 644, 181-187. Albuquerque, E.X., Pereira, E.F.R., Alkondon, M., Rogers, S.W., 2009. Mammalian Nicotinic
Acetylcholine Receptors: From Structure to Function. Physiol Rev 89, 73-120.
Alimohammadi, H., Silver, W.L., 2000. Evidence for nicotinic acetylcholine receptors on nasal
trigeminal nerve endings of the rat. Chem Senses 25, 61-66.
Amar, M., Thomas, P., Johnson, C., Lunt, G.G., Wonnacott, S., 1993. Agonist pharmacology of the
neuronal alpha 7 nicotinic receptor expressed in Xenopus oocytes. FEBS Lett 327, 284-
288.
Ames, B.N., 1983. Dietary Carcinogens and Anticarcinogens - Oxygen Radicals and Degenerative
Diseases. Science 221, 1256-1264.
Amrutkar, D.V., Ploug, K.B., Hay-Schmidt, A., Porreca, F., Olesen, J., Jansen-Olesen, I., 2012. mRNA expression of 5-hydroxytryptamine 1B, 1D, and 1F receptors and their role in controlling
the release of calcitonin gene-related peptide in the rat trigeminovascular system. Pain
153, 830-838.
Anton, F., Peppel, P., 1991. Central Projections of Trigeminal Primary Afferents Innervating the
Nasal-Mucosa - a Horseradish-Peroxidase Study in the Rat. Neuroscience 41, 617-628.
Arellano, R.O., Woodward, R.M., Miledi, R., 1996. Ion channels and membrane receptors in
follicle-enclosed Xenopus oocytes. Ion Channels 4, 203-259.
Armstrong, D.W., Wang, X.D., Ercal, N., 1998. Enantiomeric composition of nicotine in smokeless
tobacco, medicinal products, and commercial reagents. Chirality 10, 587-591.
Asztely, F., Gustafsson, B., 1996. Ionotropic glutamate receptors - Their possible role in the expression of hippocampal synaptic plasticity. Mol Neurobiol 12, 1-11.
Baker, R.R., Bishop, L.J., 2004. The pyrolysis of tobacco ingredients. J Anal Appl Pyrol 71, 223-311.
Bandell, M., Story, G.M., Hwang, S.W., Viswanath, V., Eid, S.R., Petrus, M.J., Earley, T.J.,
Patapoutian, A., 2004. Noxious cold ion channel TRPA1 is activated by pungent
compounds and bradykinin. Neuron 41, 849-857.
Barber, R.D., Harmer, D.W., Coleman, R.A., Clark, B.J., 2005. GAPDH as a housekeeping gene:
analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol Genomics
21, 389-395.
Barclay, J., Patel, S., Dorn, G., Wotherspoon, G., Moffatt, S., Eunson, L., Abdel'al, S., Natt, F., Hall,
J., Winter, J., Bevan, S., Wishart, W., Fox, A., Ganju, P., 2002. Functional Downregulation of P2X(3) receptor subunit in rat sensory neurons reveals a significant role in chronic
neuropathic and inflammatory pain. Journal of Neuroscience 22, 8139-8147.
Barlow, R.B., Hamilton, J.T., 1965. Stereospecificity of Nicotine. Brit J Pharm Chemoth 25, 206-
212.
Barnard, E.A., Skolnick, P., Olsen, R.W., Mohler, H., Sieghart, W., Biggio, G., Braestrup, C., Bateson,
A.N., Langer, S.Z., 1998. International Union of Pharmacology. XV. Subtypes of gamma-
aminobutyric acid(A) receptors: Classification on the basis of subunit structure and
receptor function. Pharmacological Reviews 50, 291-313.

123
Anhang
Bautista, D.M., Sigal, Y.M., Milstein, A.D., Garrison, J.L., Zorn, J.A., Tsuruda, P.R., Nicoll, R.A., Julius,
D., 2008. Pungent agents from Szechuan peppers excite sensory neurons by inhibiting two-pore potassium channels. Nat Neurosci 11, 772-779.
Becker, C.M., Hoch, W., Betz, H., 1988. Glycine Receptor Heterogeneity in Rat Spinal-Cord during
Postnatal-Development. Embo Journal 7, 3717-3726.
Behe, P., Stern, P., Wyllie, D.J.A., Nassar, M., Schoepfer, R., Colquhoun, D., 1995. Determination of
Nmda Nr1 Subunit Copy Number in Recombinant Nmda Receptors. P Roy Soc B-Biol Sci
262, 205-213.
Bennett, J.A., Dingledine, R., 1995. Topology profile for a glutamate receptor: three
transmembrane domains and a channel-lining reentrant membrane loop. Neuron 14,
373-384.
Benton, R., Vannice, K.S., Gomez-Diaz, C., Vosshall, L.B., 2009. Variant Ionotropic Glutamate Receptors as Chemosensory Receptors in Drosophila. Cell 136, 149-162.
Berridge, M.J., 1993. Inositol Trisphosphate and Calcium Signaling. Nature 361, 315-325.
Bertrand, D., Bertrand, S., Ballivet, M., 1992. Pharmacological Properties of the Homomeric Alpha-
7 Receptor. Neuroscience Letters 146, 87-90.
Birnboim, H.C., Doly, J., 1979. A rapid alkaline extraction procedure for screening recombinant
plasmid DNA. Nucleic Acids Res 7, 1513-1523.
Bonaventure, P., Voorn, P., Luyten, W.H.M.L., Jurzak, M., Schotte, A., Leysen, J.E., 1998a. Detailed
mapping of serotonin 5-HT1B and 5-HT1D receptor messenger RNA and ligand binding
sites in guinea-pig brain and trigeminal ganglion: Clues for function. Neuroscience 82,
469-484. Bonaventure, P., Voorn, P., Luyten, W.H.M.L., Leysen, J.E., 1998b. 5HT(1B) and 5HT(1D) receptor
mRNA differential co-localization with peptide mRNA in the guinea pig trigeminal
ganglion. Neuroreport 9, 641-645.
Borgmann-Winter, K.E., Rawson, N.E., Wang, H.Y., Wang, H., Macdonald, M.L., Ozdener, M.H.,
Yee, K.K., Gomez, G., Xu, J., Bryant, B., Adamek, G., Mirza, N., Pribitkin, E., Hahn, C.G.,
2009. Human Olfactory Epithelial Cells Generated in Vitro Express Diverse Neuronal
Characteristics. Neuroscience 158, 642-653.
Bouvet, J.F., Delaleu, J.C., Holley, A., 1987. Olfactory receptor cell function is affected by
trigeminal nerve activity. Neurosci Lett 77, 181-186.
Boyd, G.W., Low, P., Dunlop, J.I., Robertson, L.A., Vardy, A., Lambert, J.J., Peters, J.A., Connolly, C.N., 2002. Assembly and cell surface expression of homomeric and heteromeric 5-HT3
receptors: The role of oligomerization and chaperone proteins. Molecular and Cellular
Neuroscience 21, 38-50.
Boyd, R.T., Jacob, M.H., McEachern, A.E., Caron, S., Berg, D.K., 1991. Nicotinic acetylcholine
receptor mRNA in dorsal root ganglion neurons. J Neurobiol 22, 1-14.
Brejc, K., van Dijk, W.J., Klaassen, R.V., Schuurmans, M., van der Oost, J., Smit, A.B., Sixma, T.K.,
2001. Crystal structure of an ACh-binding protein reveals the ligand-binding domain of
nicotinic receptors. Nature 411, 269-276.
Bryant, B., Xu, J., Audige, V., Lischka, F.W., Rawson, N.E., 2010. Cellular Basis for the Olfactory
Response to Nicotine. Acs Chem Neurosci 1, 246-256. Burnstock, G., 2007. Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev
87, 659-797.
Burzomato, V., Groot-Kormelink, P.J., Sivilotti, L.G., Beato, M., 2003. Stoichiometry of
recombinant heteromeric glycine receptors revealed by a pore-lining region point
mutation. Receptor Channel 9, 353-361.

124
Anhang
Cain, W.S., 1976. Olfaction and the common chemical sense: some psychophysical contrasts. Sens
Processes 1, 57-67. Cain, W.S., Murphy, C.L., 1980. Interaction between Chemo-Receptive Modalities of Odor and
Irritation. Nature 284, 255-257.
Cano, J., Lobera, B., Rodriguezechandia, E.L., 1982. Influence of Innervation on the Levels of
Noradrenaline and Serotonin in the Circumvallate Papilla of the Rat. Journal of
Neurobiology 13, 1-7.
Caretti, E., Devarajan, K., Coudry, R., Ross, E., Clapper, M.L., Cooper, H.S., Bellacosa, A., 2008.
Comparison of RNA amplification methods and chip platforms for microarray analysis of
samples processed by laser capture microdissection. J Cell Biochem 103, 556-563.
Carstens, E., Anderson, K.A., Simons, C.T., Carstens, M.I., Jinks, S.L., 2001. Analgesia induced by
chronic nicotine infusion in rats - Differences by gender and pain test. Psychopharmacology 157, 40-45.
Caterina, M.J., Rosen, T.A., Tominaga, M., Brake, A.J., Julius, D., 1999. A capsaicin-receptor
homologue with a high threshold for noxious heat. Nature 398, 436-441.
Caterina, M.J., Schumacher, M.A., Tominaga, M., Rosen, T.A., Levine, J.D., Julius, D., 1997. The
capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389, 816-
824.
Chandross, K.J., Cohen, R.I., Paras, P., Gravel, M., Braun, P.E., Hudson, L.D., 1999. Identification
and characterization of early glial progenitors using a transgenic selection strategy.
Journal of Neuroscience 19, 759-774.
Charnet, P., Labarca, C., Cohen, B.N., Davidson, N., Lester, H.A., Pilar, G., 1992. Pharmacological and Kinetic-Properties of Alpha-4-Beta-2 Neuronal Nicotinic Acetylcholine-Receptors
Expressed in Xenopus Oocytes. J Physiol-London 450, 375-394.
ChavezNoriega, L.E., Crona, J.H., Washburn, M.S., Urrutia, A., Elliott, K.J., Johnson, E.C., 1997.
Pharmacological characterization of recombinant human neuronal nicotinic
acetylcholine receptors h alpha 2 beta 2, h alpha 2 beta 4, h alpha 3 beta 2, h alpha 3
beta 4, h alpha 4 beta 2, h alpha 4 beta 4 and h alpha 7 expressed in Xenopus oocytes.
Journal of Pharmacology and Experimental Therapeutics 280, 346-356.
Chen, C.C., Akopian, A.N., Sivilotti, L., Colquhoun, D., Burnstock, G., Wood, J.N., 1995. A P2x
Purinoceptor Expressed by a Subset of Sensory Neurons. Nature 377, 428-431.
Chen, H.S.V., Pellegrini, J.W., Aggarwal, S.K., Lei, S.Z., Warach, S., Jensen, F.E., Lipton, S.A., 1992. Open-Channel Block of N-Methyl-D-Aspartate (Nmda) Responses by Memantine -
Therapeutic Advantage against Nmda Receptor-Mediated Neurotoxicity. Journal of
Neuroscience 12, 4427-4436.
Chess, A., Simon, I., Cedar, H., Axel, R., 1994. Allelic Inactivation Regulates Olfactory Receptor
Gene-Expression. Cell 78, 823-834.
Chun, Y.H., Frank, D., Lee, J.S., Zhang, Y.P., Auh, Q.S., Ro, J.Y., 2008. Peripheral AMPA receptors
contribute to muscle nociception and c-fos activation. Neurosci Res 62, 97-104.
Chung, M.K., Lee, H., Mizuno, A., Suzuki, M., Caterina, M.J., 2004. 2-aminoethoxydiphenyl borate
activates and sensitizes the heat-gated ion channel TRPV3. Journal of Neuroscience 24,
5177-5182. Clapham, D.E., 2003. TRP channels as cellular sensors. Nature 426, 517-524.
Clapham, D.E., Julius, D., Montell, C., Schultz, G., 2005. International Union of Pharmacology. XLIX.
Nomenclature and structure-function relationships of transient receptor potential
channels. Pharmacol Rev 57, 427-450.
Clayton, P., Lu, A., Bishop, L., 2010. The pyrolysis of (-)-(S)-nicotine: racemization and
decomposition. Chirality 22, 442-446.

125
Anhang
Collo, G., North, R.A., Kawashima, E., MerloPich, E., Neidhart, S., Surprenant, A., Buell, G., 1996.
Cloning of P2X(5) and P2X(6) receptors and the distribution and properties of an extended family of ATP-gated ion channels. Journal of Neuroscience 16, 2495-2507.
Cometto-Muniz, J.E., Cain, W.S., Abraham, M.H., 1998. Nasal pungency and odor of homologous
aldehydes and carboxylic acids. Experimental Brain Research 118, 180-188.
Corringer, P.J., Le Novere, N., Changeux, J.P., 2000. Nicotinic receptors at the amino acid level.
Annu Rev Pharmacol 40, 431-458.
Cutting, G.R., Lu, L., Ohara, B.F., Kasch, L.M., Montroserafizadeh, C., Donovan, D.M., Shimada, S.,
Antonarakis, S.E., Guggino, W.B., Uhl, G.R., Kazazian, H.H., 1991. Cloning of the Gamma-
Aminobutyric-Acid (Gaba) Rho-1 Cdna - a Gaba Receptor Subunit Highly Expressed in the
Retina. P Natl Acad Sci USA 88, 2673-2677.
Damaj, M.I., Flood, P., Ho, K.K., May, E.L., Martin, B.R., 2005. Effect of dextrometorphan and dextrorphan on nicotine and neuronal nicotinic receptors: In vitro and in vivo selectivity.
Journal of Pharmacology and Experimental Therapeutics 312, 780-785.
Dani, J.A., Bertrand, D., 2007. Nicotinic acetylcholine receptors and nicotinic cholinergic
mechanisms of the central nervous system. Annu Rev Pharmacol Toxicol 47, 699-729.
Davies, P.A., Pistis, M., Hanna, M.C., Peters, J.A., Lambert, J.J., Hales, T.G., Kirkness, E.F., 1999. The
5-HT3B subunit is a major determinant of serotonin-receptor function. Nature 397, 359-
363.
Davis, R.A., Stiles, M.F., Debethizy, J.D., Reynolds, J.H., 1991. Dietary Nicotine - a Source of Urinary
Cotinine. Food Chem Toxicol 29, 821-827.
Dingledine, R., Borges, K., Bowie, D., Traynelis, S.F., 1999. The glutamate receptor ion channels. Pharmacological Reviews 51, 7-61.
Domino, E.F., 1998. Tobacco smoking and nicotine neuropsychopharmacology: Some future
research directions. Neuropsychopharmacol 18, 456-468.
Doty, R.L., 1986. Odor-guided behavior in mammals. Experientia 42, 257-271.
Doty, R.L., Brugger, W.E., Jurs, P.C., Orndorff, M.A., Snyder, P.J., Lowry, L.D., 1978. Intranasal
Trigeminal Stimulation from Odorous Volatiles - Psychometric Responses from Anosmic
and Normal Humans. Physiology & Behavior 20, 175-185.
Drisdel, R.C., Green, W.N., 2000. Neuronal alpha-bungarotoxin receptors are alpha 7 subunit
homomers. Journal of Neuroscience 20, 133-139.
Drisdel, R.C., Sharp, D., Henderson, T., Hales, T.G., Green, W.N., 2008. High affinity binding of epibatidine to serotonin type 3 receptors. Journal of Biological Chemistry 283, 9659-
9665.
Dubin, A.E., Huvar, R., D'Andrea, M.R., Pyati, J., Zhu, J.Y., Joy, K.C., Wilson, S.J., Galindo, J.E., Glass,
C.A., Luo, L., Jackson, M.R., Lovenberg, T.W., Erlander, M.G., 1999. The pharmacological
and functional characteristics of the serotonin 5-HT3A receptor are specifically modified
by a 5-MT3B receptor subunit. Journal of Biological Chemistry 274, 30799-30810.
Dumont, J.N., 1972. Oogenesis in Xenopus-Laevis (Daudin) .1. Stages of Oocyte Development in
Laboratory Maintained Animals. J Morphol 136, 153-179.
Durkin, M.M., Gunwaldsen, C.A., Borowsky, B., Jones, K.A., Branchek, T.A., 1999. An in situ
hybridization study of the distribution of the GABA(B2) protein mRNA in the rat CNS. Mol Brain Res 71, 185-200.
Durzynski, L., Gaudin, J.C., Myga, M., Szydlowski, J., Gozdzicka-Jozefiak, A., Haertle, T., 2005.
Olfactory-like receptor cDNAs are present in human lingual cDNA libraries. Biochem
Bioph Res Co 333, 264-272.
Duvoisin, R.M., Deneris, E.S., Patrick, J., Heinemann, S., 1989. The Functional Diversity of the
Neuronal Nicotinic Acetylcholine-Receptors Is Increased by a Novel Subunit - Beta-4.
Neuron 3, 487-496.

126
Anhang
Egan, T.M., Khakh, B.S., 2004. Contribution of calcium ions to P2X channel responses. J Neurosci
24, 3413-3420. Elg, S., Marmigere, F., Mattsson, J.P., Ernfors, P., 2007. Cellular subtype distribution and
developmental regulation a TRPC channel members in the mouse dorsal root ganglion.
Journal of Comparative Neurology 503, 35-46.
Fagan, P., Moolchan, E.T., Hart, A., Rose, A., Lawrence, D., Shavers, V.L., Gibson, J.T., 2010.
Nicotine dependence and quitting behaviors among menthol and non-menthol smokers
with similar consumptive patterns. Addiction 105, 55-74.
Finger, T.E., Danilova, V., Barrows, J., Bartel, D.L., Vigers, A.J., Stone, L., Hellekant, G., Kinnamon,
S.C., 2005. ATP signaling is crucial for communication from taste buds to gustatory
nerves. Science 310, 1495-1499.
Flores, C.M., DeCamp, R.M., Kilo, S., Rogers, S.W., Hargreaves, K.M., 1996. Neuronal nicotinic receptor expression in sensory neurons of the rat trigeminal ganglion: Demonstration of
alpha 3 beta 4, a novel subtype in the mammalian nervous system. Journal of
Neuroscience 16, 7892-7901.
Galzi, J.L., Bertrand, D., Devillersthiery, A., Revah, F., Bertrand, S., Changeux, J.P., 1991.
Functional-Significance of Aromatic-Amino-Acids from 3 Peptide Loops of the Alpha-7
Neuronal Nicotinic Receptor-Site Investigated by Site-Directed Mutagenesis. Febs
Letters 294, 198-202.
Gaudin, J.C., Breuils, L., Haertle, T., 2001. New GPCRs from a human lingual cDNA library. Chem
Senses 26, 1157-1166.
Gaudin, J.C., Breuils, L., Haertle, T., 2006. Mouse orthologs of human olfactory-like receptors expressed in the tongue. Gene 381, 42-48.
Ginzkey, C., Stueber, T., Friehs, G., Koehler, C., Hackenberg, S., Richter, E., Hagen, R., Kleinsasser,
N.H., 2012. Analysis of nicotine-induced DNA damage in cells of the human respiratory
tract. Toxicology Letters 208, 23-29.
Gisselmann, G., Galler, A., Friedrich, F., Hatt, H., Bormann, J., 2002. Cloning and functional
characterization of two glycine receptor alpha-subunits from the perch retina. Eur J
Neurosci 16, 69-80.
Gonzalez-Roca, E., Garcia-Albeniz, X., Rodriguez-Mulero, S., Gomis, R.R., Kornacker, K., Auer, H.,
2010. Accurate Expression Profiling of Very Small Cell Populations. Plos One 5, e14418.
Gray, H., 2004. Gray's Anatomy: The Anatomical Basis of Medicine and Surgery. 39. Edition: Churchill-Livingstone.
Grenningloh, G., Rienitz, A., Schmitt, B., Methfessel, C., Zensen, M., Beyreuther, K., Gundelfinger,
E.D., Betz, H., 1987. The Strychnine-Binding Subunit of the Glycine Receptor Shows
Homology with Nicotinic Acetylcholine-Receptors. Nature 328, 215-220.
Grenningloh, G., Schmieden, V., Schofield, P.R., Seeburg, P.H., Siddique, T., Mohandas, T.K.,
Becker, C.M., Betz, H., 1990. Alpha-Subunit Variants of the Human Glycine Receptor -
Primary Structures, Functional Expression and Chromosomal Localization of the
Corresponding Genes. Embo Journal 9, 771-776.
Griffin, C.A., Kafadar, K.A., Pavlath, G.K., 2009. MOR23 Promotes Muscle Regeneration and
Regulates Cell Adhesion and Migration. Dev Cell 17, 649-661. Griguoli, M., Maul, A., Nguyen, C., Giorgetti, A., Carloni, P., Cherubini, E., 2010. Nicotine Blocks the
Hyperpolarization-Activated Current I-h and Severely Impairs the Oscillatory Behavior of
Oriens-Lacunosum Moleculare Interneurons. Journal of Neuroscience 30, 10773-10783.
Grishin, E.V., 1998. Black widow spider toxins: The present and the future. Toxicon 36, 1693-1701.
Grudzinska, J., Schemm, R., Haeger, S., Nicke, A., Schmalzing, G., Betz, H., Laube, B., 2005. The
beta subunit determines the ligand binding properties of synaptic glycine receptors.
Neuron 45, 727-739.

127
Anhang
Gu, Y.P., Huang, L.Y.M., 1994. Modulation of Glycine Affinity for Nmda Receptors by Extracellular
Ca2+ in Trigeminal Neurons. Journal of Neuroscience 14, 4561-4570. Guler, A.D., Lee, H.S., Iida, T., Shimizu, I., Tominaga, M., Caterina, M., 2002. Heat-evoked
activation of the ion channel, TRPV4. Journal of Neuroscience 22, 6408-6414.
Gurley, D.A., Lanthorn, T.H., 1998. Nicotinic agonists competitively antagonize serotonin at mouse
5-HT3 receptors expressed in Xenopus oocytes. Neuroscience Letters 247, 107-110.
Han, Z.Y., Le Novere, N., Zoli, M., Hill, J.A., Champtiaux, N., Changeux, J.P., 2000. Localization of
nAChR subunit mRNAs in the brain of Macaca mulatta. Eur J Neurosci 12, 3664-3674.
Hanahan, D., Meselson, M., 1983. Plasmid Screening at High Colony Density. Method Enzymol
100, 333-342.
Hatsukami, D.K., Stead, L.F., Gupta, P.C., 2008. Tobacco addiction. Lancet 371, 2027-2038.
Hatt, H., 1996. Molecular mechanisms of olfactory processing in the mammalian olfactory epithelium. ORL J Otorhinolaryngol Relat Spec 58, 183-194.
Hatt, H., 2004. Molecular and cellular basis of human olfaction. Chem Biodivers 1, 1857-1869.
Hayasaki, H., Sohma, Y., Kanbara, K., Maemura, K., Kubota, T., Watanabe, M., 2006. A local
GABAergic system within rat trigeminal ganglion cells. Eur J Neurosci 23, 745-757.
He, L., Hannon, G.J., 2004. MicroRNAs: Small RNAs with a big role in gene regulation (vol 5, pg 522
2004). Nat Rev Genet 5, 522-531.
Hensel, H., Andres, K.H., During, M.V., 1974. Structure and Function of Cold Receptors. Pflug Arch
Eur J Phy 352, 1-10.
Heppelmann, B., Gallar, J., Trost, B., Schmidt, R.F., Belmonte, C., 2001. Three-dimensional
reconstruction of scleral cold thermoreceptors of the cat eye. Journal of Comparative Neurology 441, 148-154.
Hevers, W., Luddens, H., 1998. The diversity of GABA(A) receptors - Pharmacological and
electrophysiological properties of GABA(A) channel subtypes. Mol Neurobiol 18, 35-86.
Hill, J.M., Lukiw, W.J., Gebhardt, B.M., Higaki, S., Loutsch, J.M., Myles, M.E., Thompson, H.W.,
Kwon, B.S., Bazan, N.G., Kaufman, H.E., 2001. Gene expression analyzed by microarrays
in HSV-1 latent mouse trigeminal ganglion following heat stress. Virus Genes 23, 273-
280.
Hofmann, T., Obukhov, A.G., Schaefer, M., Harteneck, C., Gudermann, T., Schultz, G., 1999. Direct
activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature 397, 259-263.
Hollmann, M., Maron, C., Heinemann, S., 1994. N-glycosylation site tagging suggests a three transmembrane domain topology for the glutamate receptor GluR1. Neuron 13, 1331-
1343.
Hsu, E.S., 2010. A Review of Granisetron, 5-Hydroxytryptamine(3) Receptor Antagonists, and
Other Antiemetics. Am J Ther 17, 476-486.
Hu, W.P., Guan, B.C., Ru, L.Q., Chen, J.G., Li, Z.W., 2004. Potentiation of 5-HT3 receptor function
by the activation of coexistent 5-HT2 receptors in trigeminal ganglion neurons of rats.
Neuropharmacology 47, 833-840.
Huang, Y.J., Maruyama, Y., Lu, K.S., Pereira, E., Plonsky, I., Baur, J.E., Wu, D.Q., Roper, S.D., 2005.
Mouse taste buds use serotonin as a neurotransmitter. Journal of Neuroscience 25, 843-
847. Hukkanen, J., Jacob, P., 3rd, Benowitz, N.L., 2005. Metabolism and disposition kinetics of nicotine.
Pharmacol Rev 57, 79-115.
Hummel, T., Hummel, C., Pauli, E., Kobal, G., 1992a. Olfactory Discrimination of Nicotine-
Enantiomers by Smokers and Nonsmokers. Chem Senses 17, 13-21.
Hummel, T., Livermore, A., Hummel, C., Kobal, G., 1992b. Chemosensory Event-Related Potentials
in Man - Relation to Olfactory and Painful Sensations Elicited by Nicotine. Electroen Clin
Neuro 84, 192-195.

128
Anhang
Hummel, T., Livermore, A., 2002. Intranasal chemosensory function of the trigeminal nerve and
aspects of its relation to olfaction. Int Arch Occ Env Hea 75, 305-313. Ikushima, S., Muramatsu, I., Sakakibara, Y., Yokotani, K., Fujiwara, M., 1982. The effects of d-
nicotine and l-isomer on nicotinic receptors. J Pharmacol Exp Ther 222, 463-470.
Inokuchi, A., Kimmelman, C.P., Snow, J.B., 1993. Convergence of Olfactory and Nasotrigeminal
Inputs and Possible Trigeminal Contributions to Olfactory Responses in the Rat
Thalamus. Eur Arch Oto-Rhino-L 249, 473-477.
Ishii, T., Serizawa, S., Kohda, A., Nakatani, H., Shiroishi, T., Okumura, K., Iwakura, Y., Nagawa, F.,
Tsuboi, A., Sakano, H., 2001. Monoallelic expression of the odorant receptor gene and
axonal projection of olfactory sensory neurones (vol 6, pg 71, 2001). Genes Cells 6, 573-
573.
Islam, S., Kjallquist, U., Moliner, A., Zajac, P., Fan, J.B., Lonnerberg, P., Linnarsson, S., 2011. Characterization of the single-cell transcriptional landscape by highly multiplex RNA-seq.
Genome Res 21, 1160-1167.
Jacquot, L., Monnin, J., Brand, G., 2004. Influence of nasal trigeminal stimuli on olfactory
sensitivity. Cr Biol 327, 305-311.
Jessel, T., Basbaum, A., 2000. The Bodily Senses. In: Principles of Neural Science. (Kandel E,
Schwartz J, Jessel T, eds) New York: McGraw-Hill., pp 430-450.
Jiang, Y.Q., Pickett, J., Oblinger, M.M., 1994a. Comparison of Changes in Beta-Tubulin and Nf
Gene-Expression in Rat Drg Neurons under Regeneration-Permissive and Regeneration-
Prohibitive Conditions. Brain Research 637, 233-241.
Jiang, Y.Q., Pickett, J., Oblinger, M.M., 1994b. Long-Term Effects of Axotomy on Beta-Tubulin and Nf Gene-Expression in Rat Drg Neurons. J Neural Transp Plas 5, 103-114.
Jordt, S.E., Bautista, D.M., Chuang, H.H., McKemy, D.D., Zygmunt, P.M., Hogestatt, E.D., Meng,
I.D., Julius, D., 2004. Mustard oils and cannabinoids excite sensory nerve fibres through
the TRP channel ANKTM1. Nature 427, 260-265.
Karashima, Y., Damann, N., Prenen, J., Talavera, K., Segal, A., Voets, T., Nilius, B., 2007. Bimodal
action of menthol on the transient receptor potential channel TRPA1. J Neurosci 27,
9874-9884.
Karashima, Y., Talavera, K., Everaerts, W., Janssens, A., Kwan, K.Y., Vennekens, R., Nilius, B., Voets,
T., 2009. TRPA1 acts as a cold sensor in vitro and in vivo. P Natl Acad Sci USA 106, 1273-
1278. Kaya, N., Shen, T., Lu, S.G., Zhao, F.L., Herness, S., 2004. A paracrine signaling role for serotonin in
rat taste buds: expression and localization of serotonin receptor subtypes. Am J Physiol
Regul Integr Comp Physiol 286, R649-658.
Keiger, C.J.H., Walker, J.C., 2000. Individual variation in the expression profiles of nicotinic
receptors in the olfactory bulb and trigeminal ganglion and identification of alpha 2,
alpha 6, alpha 9, and beta 3 transcripts. Biochem Pharmacol 59, 233-240.
Khakh, B.S., 2001. Molecular physiology of P2X receptors and ATP signalling at synapses. Nat Rev
Neurosci 2, 165-174.
Khan, I.M., Yaksh, T.L., Taylor, P., 1994. Ligand Specificity of Nicotinic Acetylcholine-Receptors in
Rat Spinal-Cord - Studies with Nicotine and Cytisine. Journal of Pharmacology and Experimental Therapeutics 270, 159-166.
Kilpatrick, G.J., Butler, A., Burridge, J., Oxford, A.W., 1990. 1-(M-Chlorophenyl)-Biguanide, a
Potent High-Affinity 5-Ht3 Receptor Agonist. Eur J Pharmacol 182, 193-197.
Kilpatrick, G.J., Jones, B.J., Tyers, M.B., 1989. Binding of the 5-Ht3 Ligand, [H-3] Gr65630, to Rat
Area Postrema, Vagus Nerve and the Brains of Several Species. Eur J Pharmacol 159,
157-164.

129
Anhang
Kobal, G., Hummel, C., 1988. Cerebral Chemosensory Evoked-Potentials Elicited by Chemical-
Stimulation of the Human Olfactory and Respiratory Nasal-Mucosa. Electroen Clin Neuro 71, 241-250.
Kobayashi, K., Fukuoka, T., Obata, K., Yamanaka, H., Dai, Y., Tokunaga, A., Noguchi, K., 2005.
Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons
with A delta/C-fibers and colocalization with trk receptors. Journal of Comparative
Neurology 493, 596-606.
Kohl, A., Binz, H.K., Forrer, P., Stumpp, M.T., Pluckthun, A., Grutter, M.G., 2003. Designed to be
stable: Crystal structure of a consensus ankyrin repeat protein. P Natl Acad Sci USA 100,
1700-1705.
Kondo, E., Kiyama, H., Araki, T., Shida, T., Ueda, Y., Tohyama, M., 1994. Coexpression of Gaba(a)
Receptor Gamma-1 and Gamma-2 Subunits in the Rat Trigeminal Ganglion. Mol Brain Res 21, 363-367.
Kost, S.A., Oblinger, M.M., 1993. Immature Corticospinal Neurons Respond to Axotomy with
Changes in Tubulin Gene-Expression. Brain Res Bull 30, 469-475.
Kouadjo, K.E., Nishida, Y., Cadrin-Girard, J.F., Yoshioka, M., St-Amand, J., 2007. Housekeeping and
tissue-specific genes in mouse tissues. BMC Genomics 8, 127.
Kuhn, H., 1964. Tobacco alkaloids and their pyrolysis products in smoke. In Tobacco Alkaloids and
Related Compounds; van Euler, U. S., Ed. Proceedings of the Fourth Wenner-Gren
International Symposium; Macmillan: New York, 1964, 37-51.
Kuhse, J., Laube, B., Magalei, D., Betz, H., 1993. Assembly of the Inhibitory Glycine Receptor -
Identification of Amino-Acid-Sequence Motifs Governing Subunit Stoichiometry. Neuron 11, 1049-1056.
Kurimoto, K., Yabuta, Y., Ohinata, Y., Saitou, M., 2007. Global single-cell cDNA amplification to
provide a template for representative high-density oligonucleotide microarray analysis.
Nat Protoc 2, 739-752.
Kuroda, H., Shibukawa, Y., Soya, M., Masamura, A., Kasahara, M., Tazaki, M., Ichinohe, T., 2012.
Expression of P2X(1) and P2X(4) receptors in rat trigeminal ganglion neurons.
Neuroreport 23, 752-756.
Lafreniere, R.G., Cader, M.Z., Poulin, J.F., Andres-Enguix, I., Simoneau, M., Gupta, N., Boisvert, K.,
Lafreniere, F., McLaughlan, S., Dube, M.P., Marcinkiewicz, M.M., Ramagopalan, S.,
Ansorge, O., Brais, B., Sequeiros, J., Pereira-Monteiro, J.M., Griffiths, L.R., Tucker, S.J., Ebers, G., Rouleau, G.A., 2010. A dominant-negative mutation in the TRESK potassium
channel is linked to familial migraine with aura. Nat Med 16, 1157-U1501.
Lambrecht, G., 2000. Agonists and antagonists acting at P2X receptors: selectivity profiles and
functional implications. N-S Arch Pharmacol 362, 340-350.
Langmead, B., Trapnell, C., Pop, M., Salzberg, S.L., 2009. Ultrafast and memory-efficient alignment
of short DNA sequences to the human genome. Genome Biol 10, R25.
Laviolette, S.R., van der Kooy, D., 2004. The neurobiology of nicotine addiction: Bridging the gap
from molecules to behaviour. Nat Rev Neurosci 5, 55-65.
Lazarov, N.E., 2002. Comparative analysis of the chemical neuroanatomy of the mammalian
trigeminal ganglion and mesencephalic trigeminal nucleus. Prog Neurobiol 66, 19-59. Lennertz, R.C., Tsunozaki, M., Bautista, D.M., Stucky, C.L., 2010. Physiological Basis of Tingling
Paresthesia Evoked by Hydroxy-alpha-Sanshool. Journal of Neuroscience 30, 4353-4361.
Lerch, J.K., Kuo, F., Motti, D., Morris, R., Bixby, J.L., Lemmon, V.P., 2012. Isoform Diversity and
Regulation in Peripheral and Central Neurons Revealed through RNA-Seq. Plos One 7,
e30417.
Lethbridge, R., Hou, Q.L., Harley, C.W., Yuan, Q., 2012. Olfactory Bulb Glomerular NMDA
Receptors Mediate Olfactory Nerve Potentiation and Odor Preference Learning in the
Neonate Rat. Plos One 7.

130
Anhang
Liedtke, W., Friedman, J.M., 2003. Abnormal osmotic regulation in trpv4(-/-) mice. P Natl Acad Sci
USA 100, 13698-13703. Liedtke, W., Tobin, D.M., Bargmann, C.I., Friedman, J.M., 2003. Mammalian TRPV4 (VR-OAC)
directs behavioral responses to osmotic and mechanical stimuli in Caenorhabditis
elegans. P Natl Acad Sci USA 100, 14531-14536.
Liu, L., Chang, C.Q., Jiao, Y.Q., Simon, S.A., 1998. Neuronal nicotinic acetylcholine receptors in rat
trigeminal ganglia. Brain Research 809, 238-245.
Liu, L., Keiger, C.J.H., Simon, S.A., 1996. Nicotinic acetylcholine and capsaicin receptors in rat
trigeminal ganglia. Chem Senses 21, 635.
Liu, L., Li, Y.H., Li, S.L., Hu, N., He, Y.M., Pong, R., Lin, D.N., Lu, L.H., Law, M., 2012. Comparison of
Next-Generation Sequencing Systems. J Biomed Biotechnol 2012, 251364.
Liu, L., Pugh, W., Ma, H., Simon, S.A., 1993. Identification of Acetylcholine-Receptors in Adult-Rat Trigeminal Ganglion Neurons. Brain Research 617, 37-42.
Liu, L., Zhu, W., Zhang, Z.S., Yang, T., Grant, A., Oxford, G., Simon, S.A., 2004. Nicotine inhibits
voltage-dependent sodium channels and sensitizes vanilloid receptors. J Neurophysiol
91, 1482-1491.
Liu, Y., Lubin, M.L., Reitz, T.L., Wang, Y., Colburn, R.W., Flores, C.M., Qin, N., 2006. Molecular
identification and functional characterization of a temperature-sensitive transient
receptor potential channel (TRPM8) from canine. Eur J Pharmacol 530, 23-32.
Luetje, C.W., Patrick, J., 1991. Both Alpha- and Beta-Subunits Contribute to the Agonist Sensitivity
of Neuronal Nicotinic Acetylcholine-Receptors. Journal of Neuroscience 11, 837-845.
Lynch, J.W., 2004. Molecular structure and function of the glycine receptor chloride channel. Physiol Rev 84, 1051-1095.
Lynch, J.W., 2009. Native glycine receptor subtypes and their physiological roles.
Neuropharmacology 56, 303-309.
Mackenzie, R.A., Burke, D., Skuse, N.F., Lethlean, A.K., 1975. Fiber Function and Perception during
Cutaneous Nerve Block. J Neurol Neurosur Ps 38, 865-873.
Macor, J.E., Gurley, D., Lanthorn, T., Loch, J., Mack, R.A., Mullen, G., Tran, O., Wright, N., Gordon,
J.C., 2001. The 5-HT3 antagonist tropisetron (ICS 205-930) is a potent and selective alpha
7 nicotinic receptor partial agonist. Bioorganic & Medicinal Chemistry Letters 11, 319-
321.
Macpherson, L.J., Hwang, S.W., Miyamoto, T., Dubin, A.E., Patapoutian, A., Story, G.M., 2006. More than cool: Promiscuous relationships of menthol and other sensory compounds.
Molecular and Cellular Neuroscience 32, 335-343.
Magdaleno, S., Jensen, P., Brumwell, C.L., Seal, A., Lehman, K., Asbury, A., Cheung, T., Cornelius,
T., Batten, D.M., Eden, C., Norland, S.M., Rice, D.S., Dosooye, N., Shakya, S., Mehta, P.,
Curran, T., 2006. BGEM: An in situ hybridization database of gene expression in the
embryonic and adult mouse nervous system. Plos Biol 4, 497-500.
Malnic, B., Hirono, J., Sato, T., Buck, L.B., 1999. Combinatorial receptor codes for odors. Cell 96,
713-723.
Malosio, M.L., Marquezepouey, B., Kuhse, J., Betz, H., 1991. Widespread Expression of Glycine
Receptor Subunit Messenger-Rnas in the Adult and Developing Rat-Brain. Embo Journal 10, 2401-2409.
Maren, S., Baudry, M., 1995. Properties and mechanisms of long-term synaptic plasticity in the
mammalian brain: relationships to learning and memory. Neurobiol Learn Mem 63, 1-
18.
Maricq, A.V., Peterson, A.S., Brake, A.J., Myers, R.M., Julius, D., 1991. Primary Structure and
Functional Expression of the 5ht3 Receptor, a Serotonin-Gated Ion Channel. Science 254,
432-437.

131
Anhang
McKemy, D.D., Neuhausser, W.M., Julius, D., 2002. Identification of a cold receptor reveals a
general role for TRP channels in thermosensation. Nature 416, 52-58. McKernan, R.M., Whiting, P.J., 1996. Which GABA(A)-receptor subtypes really occur in the brain?
Trends Neurosci 19, 139-143.
McNamara, C.R., Mandel-Brehm, J., Bautista, D.M., Siemens, J., Deranian, K.L., Zhao, M., Hayward,
N.J., Chong, J.A., Julius, D., Moran, M.M., Fanger, C.M., 2007. TRPA1 mediates formalin-
induced pain. P Natl Acad Sci USA 104, 13525-13530.
Migita, K., Haines, W.R., Voigt, M.M., Egan, T.M., 2001. Polar residues of the second
transmembrane domain influence cation permeability of the ATP-gated P2X(2) receptor.
J Biol Chem 276, 30934-30941.
Miledi, R., Woodward, R.M., 1989a. Effects of Defolliculation on Membrane Current Responses of
Xenopus Oocytes. J Physiol-London 416, 601-621. Miledi, R., Woodward, R.M., 1989b. Membrane Currents Elicited by Prostaglandins, Atrial
Natriuretic Factor and Oxytocin in Follicle-Enclosed Xenopus Oocytes. J Physiol-London
416, 623-643.
Millar, N.S., Gotti, C., 2009. Diversity of vertebrate nicotinic acetylcholine receptors.
Neuropharmacology 56, 237-246.
Minke, B., Wu, C.F., Pak, W.L., 1975. Induction of Photoreceptor Voltage Noise in Dark in
Drosophila Mutant. Nature 258, 84-87.
Moiseenkova-Bell, V.Y., Stanciu, L.A., Serysheva, I.I., Tobe, B.J., Wensel, T.G., 2008. Structure of
TRPV1 channel revealed by electron cryomicroscopy. P Natl Acad Sci USA 105, 7451-
7455. Mombaerts, P., 2006. Axonal wiring in the mouse olfactory system. Annu Rev Cell Dev Bi 22, 713-
737.
Montell, C., 2001. An end in sight to a long TRP. Neuron 30, 3-5.
Montell, C., 2005. The TRP superfamily of cation channels. Sci STKE 2005, re3.
Montell, C., Birnbaumer, L., Flockerzi, V., Bindels, R.J., Brudorf, E.A., Caterina, M.J., Clapham, D.E.,
Harteneck, C., Heller, S., Julius, D., Kojima, I., Mori, Y., Penner, R., Prawitt, D.,
Scharenberg, A.M., Schultz, G., Shimizu, N., Zhu, M.X., 2002. A unified nomenclature for
the superfamily of TRP cation channels. Molecular Cell 9, 229-231.
Montell, C., Jones, K., Hafen, E., Rubin, G., 1985. Rescue of the Drosophila Phototransduction
Mutation Trp by Germline Transformation. Science 230, 1040-1043. Monyer, H., Seeburg, P.H., Wisden, W., 1991. Glutamate-Operated Channels - Developmentally
Early and Mature Forms Arise by Alternative Splicing. Neuron 6, 799-810.
Mori, K., Takahashi, Y.K., Igarashi, K.M., Yamaguchi, M., 2006. Maps of odorant molecular features
in the mammalian olfactory bulb. Physiol Rev 86, 409-433.
Mortazavi, A., Williams, B.A., Mccue, K., Schaeffer, L., Wold, B., 2008. Mapping and quantifying
mammalian transcriptomes by RNA-Seq. Nat Methods 5, 621-628.
Nagata, K., Duggan, A., Kumar, G., Garcia-Anoveros, J., 2005. Nociceptor and hair cell transducer
properties of TRPA1, a channel for pain and hearing. Journal of Neuroscience 25, 4052-
4061.
Naqvi, N.H., Bechara, A., 2005. The airway sensory impact of nicotine contributes to the conditioned reinforcing effects of individual puffs from cigarettes. Pharmacol Biochem
Be 81, 821-829.
Neeper, M.P., Liu, Y., Hutchinson, T.L., Wang, Y., Flores, C.M., Qin, N., 2007. Activation properties
of heterologously expressed mammalian TRPV2 - Evidence for species dependence.
Journal of Biological Chemistry 282, 15894-15902.
Nicke, A., Baumert, H.G., Rettinger, J., Eichele, A., Lambrecht, G., Mutschler, E., Schmalzing, G.,
1998. P2X1 and P2X3 receptors form stable trimers: a novel structural motif of ligand-
gated ion channels. EMBO J 17, 3016-3028.

132
Anhang
Niesler, B., Walstab, J., Combrink, S., Moller, D., Kapeller, J., Rietdorf, J., Bonisch, H., Gothert, M.,
Rappold, G., Bruss, M., 2007. Characterization of the novel human serotonin receptor subunits 5-HT3C, 5-HT3D, and 5-HT3E. Mol Pharmacol 72, 8-17.
Niimura, Y., Nei, M., 2005. Comparative evolutionary analysis of olfactory receptor gene clusters
between humans and mice. Gene 346, 13-21.
Nishizawa, Y., Kurihara, T., Masuda, T., Takahashi, Y., 1985. Immunohistochemical Localization of
2',3'-Cyclic Nucleotide 3'-Phosphodiesterase in Adult Bovine Cerebrum and Cerebellum.
Neurochem Res 10, 1107-1118.
Noel, J., Zimmermann, K., Busserolles, J., Deval, E., Alloui, A., Diochot, S., Guy, N., Borsotto, M.,
Reeh, P., Eschalier, A., Lazdunski, M., 2009. The mechano-activated K+ channels TRAAK
and TREK-1 control both warm and cold perception. Embo Journal 28, 1308-1318.
Nowak, L., Bregestovski, P., Ascher, P., Herbet, A., Prochiantz, A., 1984a. Magnesium gates glutamate-activated channels in mouse central neurones. Nature 307, 462-465.
Nutt, D., 2006. GABAA receptors: subtypes, regional distribution, and function. J Clin Sleep Med 2,
7-11.
Okada, T., Inoue, R., Yamazaki, K., Maeda, A., Kurosaki, T., Yamakuni, T., Tanaka, I., Shimizu, S.,
Ikenaka, K., Imoto, K., Mori, Y., 1999. Molecular and functional characterization of a
novel mouse transient receptor potential protein homologue TRP7 - Ca2+-permeable
cation channel that is constitutively activated and enhanced by stimulation of G protein-
coupled receptor. Journal of Biological Chemistry 274, 27359-27370.
Ortells, M.O., Lunt, G.G., 1995. Evolutionary History of the Ligand-Gated Ion-Channel Superfamily
of Receptors. Trends Neurosci 18, 121-127. Otaki, J.M., Yamamoto, H., Firestein, S., 2004. Odorant receptor expression in the mouse cerebral
cortex. Journal of Neurobiology 58, 315-327.
Papke, R.L., Wecker, L., Stitzel, J.A., 2010. Activation and Inhibition of Mouse Muscle and
Neuronal Nicotinic Acetylcholine Receptors Expressed in Xenopus Oocytes. Journal of
Pharmacology and Experimental Therapeutics 333, 501-518.
Patapoutian, A., Peier, A.M., Story, G.M., Viswanath, V., 2003. Thermotrp channels and beyond:
Mechanisms of temperature sensation. Nat Rev Neurosci 4, 529-539.
Peier, A.M., Moqrich, A., Hergarden, A.C., Reeve, A.J., Andersson, D.A., Story, G.M., Earley, T.J.,
Dragoni, I., McIntyre, P., Bevan, S., Patapoutian, A., 2002a. A TRP channel that senses
cold stimuli and menthol. Cell 108, 705-715. Peier, A.M., Reeve, A.J., Andersson, D.A., Moqrich, A., Earley, T.J., Hergarden, A.C., Story, G.M.,
Colley, S., Hogenesch, J.B., McIntyre, P., Bevan, S., Patapoutian, A., 2002b. A heat-
sensitive TRP channel expressed in keratinocytes. Science 296, 2046-2049.
Peng, X., Katz, M., Gerzanich, V., Anand, R., Lindstrom, J., 1994. Human Alpha-7-Acetylcholine
Receptor - Cloning of the Alpha-7-Subunit from the Sh-Sy5y Cell-Line and Determination
of Pharmacological Properties of Native Receptors and Functional Alpha-7-Homomers
Expressed in Xenopus-Oocytes. Mol Pharmacol 45, 546-554.
Perfetti, T.A., Coleman, W.M., 1998. Chiral-gas chromatography-selected ion monitoring-mass
selective detection analysis of tobacco materials and tobacco smoke. Beitr Tabakforsch
Int 18, 15-33. Perfetti, T.A., Coleman, W.M., Smith, W.S., 1998. Determination of mainstream and sidestream
cigarette smoke components for cigarettes of different tobacco types and a set of
reference cigarettes. Beitr Tabakforsch Int 18, 95-113.
Pictet, A., Rotschy, A., 1900. Ueber inactives Nicotin. Ber. Dtsch. chem. Ges. 33, 2353-2355.
Pidoplichko, V.I., DeBiasi, M., Williams, J.T., Dani, J.A., 1997. Nicotine activates and desensitizes
midbrain dopamine neurons. Nature 390, 401-404.

133
Anhang
Pirker, S., Schwarzer, C., Wieselthaler, A., Sieghart, W., Sperk, G., 2000. GABA(A) receptors:
Immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience 101, 815-850.
Powis, G., Seewald, M.J., Gratas, C., Melder, D., Riebow, J., Modest, E.J., 1992. Selective-Inhibition
of Phosphatidylinositol Phospholipase-C by Cytotoxic Ether Lipid Analogs. Cancer Res 52,
2835-2840.
Pratt, G.D., Bowery, N.G., 1989. The 5-Ht3 Receptor Ligand, [H-3] Brl 43694, Binds to Presynaptic
Sites in the Nucleus Tractus Solitarius of the Rat. Neuropharmacology 28, 1367-1376.
Pritchard, W.S., Robinson, J.H., Guy, T.D., Davis, R.A., Stiles, M.F., 1996. Assessing the sensory role
of nicotine in cigarette smoking. Psychopharmacology 127, 55-62.
Qin, N., Neeper, M.P., Liu, Y., Hutchinson, T.L., Lubin, M.L., Flores, C.M., 2008. TRPV2 is activated
by cannabidiol and mediates CGRP release in cultured rat dorsal root ganglion neurons. Journal of Neuroscience 28, 6231-6238.
Radtke, C., Sasaki, M., Lankford, K.L., Gallo, V., Kocsis, J.D., 2011. CNPase expression in olfactory
ensheathing cells. J Biomed Biotechnol 2011, 608496.
Ramsey, I.S., Delling, M., Clapham, D.E., 2006. An introduction to TRP channels. Annu Rev Physiol
68, 619-647.
Rand, M.J., 1989. Neuropharmacological Effects of Nicotine in Relation to Cholinergic
Mechanisms. Nicotinic Receptors in the Cns 79, 3-11.
Rash, J.E., Davidson, K.G.V., Kamasawa, N., Yasumura, T., Kamasawa, M., Zhang, C.B., Michaels, R.,
Restrepo, D., Ottersen, O.P., Olson, C.O., Nagy, J.I., 2005. Ultrastructural localization of
connexins (Cx36, Cx43, Cx45), glutamate receptors and aquaporin-4 in rodent olfactory mucosa, olfactory nerve and olfactory bulb. J Neurocytol 34, 307-341.
Reynolds, D.S., Rosahl, T.W., Cirone, J., O'Meara, G.F., Haythornthwaite, A., Newman, R.J., Myers,
J., Sur, C., Howell, O., Rutter, A.R., Atack, J., Macaulay, A.J., Hadingham, K.L., Hutson,
P.H., Belelli, D., Lambert, J.J., Dawson, G.R., McKernan, R., Whiting, P.J., Wafford, K.A.,
2003. Sedation and anesthesia mediated by distinct GABA(A) receptor isoforms. Journal
of Neuroscience 23, 8608-8617.
Richardson, B.P., 1990. Serotonin and Nociception. Ann Ny Acad Sci 600, 511-520.
Richardson, B.P., Engel, G., Donatsch, P., Stadler, P.A., 1985. Identification of Serotonin M-
Receptor Subtypes and Their Specific Blockade by a New Class of Drugs. Nature 316,
126-131. Rohacs, T., Lopes, C.M.B., Michailidis, I., Logothetis, D.E., 2005. PI(4,5)P-2 regulates the activation
and desensitization of TRPM8 channels through the TRP domain. Nat Neurosci 8, 626-
634.
Rong, W.F., Burnstock, G., Spyer, K.M., 2000. P2X purinoceptor-mediated excitation of trigeminal
lingual nerve terminals in an in vitro intra-arterially perfused rat tongue preparation. J
Physiol-London 524, 891-902.
Rose, J.E., Behm, F.M., Levin, E.D., 1993. Role of Nicotine Dose and Sensory Cues in the Regulation
of Smoke Intake. Pharmacol Biochem Be 44, 891-900.
Rose, J.E., Tashkin, D.P., Ertle, A., Zinser, M.C., Lafer, R., 1985. Sensory Blockade of Smoking
Satisfaction. Pharmacol Biochem Be 23, 289-293. Sahara, Y., Noro, N., Lida, Y., Soma, K., Nakamura, Y., 1997. Glutamate receptor subunits GluR5
and KA-2 are coexpressed in rat trigeminal ganglion neurons. Journal of Neuroscience
17, 6611-6620.
Saiki, R.K., Gelfand, D.H., Stoffel, S., Scharf, S.J., Higuchi, R., Horn, G.T., Mullis, K.B., Erlich, H.A.,
1988. Primer-Directed Enzymatic Amplification of DNA with a Thermostable DNA-
Polymerase. Science 239, 487-491.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning – A Laboratory Manual. New
York: Cold Spring Harbor Laboratory Press.

134
Anhang
Sambrook, J., Russell, D.W., 2001. Molecular Cloning - A Laboratory Manual. New York: Cold
Spring Harbor Laboratory Press. Samways, D.S., Egan, T.M., 2007. Acidic amino acids impart enhanced Ca2+ permeability and flux
in two members of the ATP-gated P2X receptor family. J Gen Physiol 129, 245-256.
Samways, D.S., Migita, K., Li, Z., Egan, T.M., 2008. On the role of the first transmembrane domain
in cation permeability and flux of the ATP-gated P2X2 receptor. J Biol Chem 283, 5110-
5117.
Sanger, F., 1975. The Croonian Lecture, 1975. Nucleotide sequences in DNA. Proc R Soc Lond B
Biol Sci 191, 317-333.
Saras, A., Gisselmann, G., Vogt-Eisele, A.K., Erlkamp, K.S., Kletke, O., Pusch, H., Hatt, H., 2008.
Histamine action on vertebrate GABA(A) receptors - Direct channel gating and
potentiation of GABA responses. Journal of Biological Chemistry 283, 10470-10475. Sawada, Y., Hosokawa, H., Horia, A., Matsumura, K., Kobayashi, S., 2007. Cold sensitivity of
recombinant TRPA1 channels. Brain Research 1160, 39-46.
Schaefer, M.L., Bottger, B., Silver, W.L., Finger, T.E., 2002. Trigeminal collaterals in the nasal
epithelium and olfactory bulb: A potential route for direct modulation of olfactory
information by trigeminal stimuli. Journal of Comparative Neurology 444,221-226.
Schöbel, N., Radtke, D., Lubbert, M., Gisselmann, G., Lehmann, R., Cichy, A., Schreiner, B.S.P.,
Altmuller, J., Spector, A.C., Spehr, J., Hatt, H., Wetzel, C.H., 2012. Trigeminal Ganglion
Neurons of Mice Show Intracellular Chloride Accumulation and Chloride-Dependent
Amplification of Capsaicin-Induced Responses. Plos One 7, e48005.
Sekizawa, S., Tsubone, H., 1994. Nasal Receptors Responding to Noxious Chemical Irritants. Resp Physiol 96, 37-48.
Sergeeva, O.A., Kletke, O., Kragler, A., Poppek, A., Fleischer, W., Schubring, S.R., Gorg, B., Haas,
H.L., Zhu, X.R., Lubbert, H., Gisselmann, G., Hatt, H., 2010. Fragrant Dioxane Derivatives
Identify beta 1-Subunit-containing GABA(A) Receptors. Journal of Biological Chemistry
285, 23985-23993.
Serizawa, S.U., Miyamichi, K., Sakano, H., 2004. One neuron-one receptor rule in the mouse
olfactory system. Trends Genet 20, 648-653.
Sieber, M., 2012. Neuroprotective Properties of Nicotine. Curr Med Chem 19, 292-297.
Silver, W.L., Clapp, T.R., Stone, L.M., Kinnamon, S.C., 2006. TrpV1 receptors and nasal trigeminal
chemesthesis. Chem Senses 31, A59-A59. Silver, W.L., Mason, J.R., Marshall, D.A., Maruniak, J.A., 1985. Rat Trigeminal, Olfactory and Taste
Responses after Capsaicin Desensitization. Brain Research 333, 45-54.
Simon, J., Wakimoto, H., Fujita, N., Lalande, M., Barnard, E.A., 2004. Analysis of the set of GABA(A)
receptor genes in the human genome. Journal of Biological Chemistry 279, 41422-
41435.
Sloan, J.W., Martin, W.R., Hernandez, J., Hook, R., 1985. Binding characteristics of (-)- and (+)-
nicotine to the rat brain P2 fraction. Pharmacol Biochem Behav 23, 987-993.
Smith, G.D., Gunthorpe, J., Kelsell, R.E., Hayes, P.D., Reilly, P., Facer, P., Wright, J.E., Jerman, J.C.,
Walhin, J.P., Ooi, L., Egerton, J., Charles, K.J., Smart, D., Randall, A.D., Anand, P., Davis,
J.B., 2002. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 418, 186-190.
Sommer, B., Keinanen, K., Verdoorn, T.A., Wisden, W., Burnashev, N., Herb, A., Kohler, M., Takagi,
T., Sakmann, B., Seeburg, P.H., 1990. Flip and Flop - a Cell-Specific Functional Switch in
Glutamate-Operated Channels of the Cns. Science 249, 1580-1585.
Somogyi, P., Tamas, G., Lujan, R., Buhl, E.H., 1998. Salient features of synaptic organisation in the
cerebral cortex. Brain Res Rev 26, 113-135.
Soto, F., Garcia-Guzman, M., Stuhmer, W., 1997. Cloned ligand-gated channels activated by
extracellular ATP (P2X receptors). J Membr Biol 160, 91-100.

135
Anhang
Spehr, J., Spehr, M., Hatt, H., Wetzel, C.H., 2004. Subunit-specific P2X-receptor expression defines
chemosensory properties of trigeminal neurons. Eur J Neurosci 19, 2497-2510. Spehr, M., Gisselmann, G., Poplawski, A., Riffell, J.A., Wetzel, C.H., Zimmer, R.K., Hatt, H., 2003.
Identification of a testicular odorant receptor mediating human sperm chemotaxis.
Science 299, 2054-2058.
Sprinkle, T.J., 1989. 2',3'-cyclic nucleotide 3'-phosphodiesterase, an oligodendrocyte-Schwann cell
and myelin-associated enzyme of the nervous system. Crit Rev Neurobiol 4, 235-301.
Story, G.M., Peier, A.M., Reeve, A.J., Eid, S.R., Mosbacher, J., Hricik, T.R., Earley, T.J., Hergarden,
A.C., Andersson, D.A., Hwang, S.W., McIntyre, P., Jegla, T., Bevan, S., Patapoutian, A.,
2003. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold
temperatures. Cell 112, 819-829.
Stuck, B.A., Baja, J., Lenz, F., Herr, R.M., Heiser, C., 2011. Co-Stimulation with an Olfactory Stimulus Increases Arousal Responses to Trigeminal Stimulation. Neuroscience 176, 442-
446.
Talavera, K., Gees, M., Karashima, Y., Meseguer, V.M., Vanoirbeek, J.A.J., Damann, N., Everaerts,
W., Benoit, M., Janssens, A., Vennekens, R., Viana, F., Nemery, B., Nilius, B., Voets, T.,
2009. Nicotine activates the chemosensory cation channel TRPA1. Nat Neurosci 12,
1293-U1214.
Tang, F.C., Lao, K.Q., Surani, M.A., 2011. Development and applications of single-cell
transcriptome analysis. Nat Methods 8, S6-S11.
Tecott, L.H., Maricq, A.V., Julius, D., 1993. Nervous-System Distribution of the Serotonin 5-Ht3
Receptor Messenger-Rna. P Natl Acad Sci USA 90, 1430-1434. Teneggi, V., Squassante, L., Iavarone, L., Milleri, S., Bye, A., Gomeni, R., 2002. Correlation and
predictive performances of saliva and plasma nicotine concentration on tobacco
withdrawal-induced craving. Brit J Clin Pharmaco 54, 407-414.
Thuerauf, N., Kaegler, M., Dietz, R., Barocka, A., Kobal, G., 1999. Dose-dependent stereoselective
activation of the trigeminal sensory system by nicotine in man. Psychopharmacology
142, 236-243.
Thuerauf, N., Kaegler, M., Renner, B., Barocka, A., Kobal, G., 2000. Specific sensory detection,
discrimination, and hedonic estimation of nicotine enantiomers in smokers and
nonsmokers: Are there limitations in replacing the sensory components of nicotine? J
Clin Psychopharm 20, 472-478. Thuerauf, N., Markovic, K., Braun, G., Bleich, S., Reulbach, U., Kornhuber, J., Lunkenheimer, J.,
2006. The influence of mecamylamine on trigeminal and olfactory chemoreception of
nicotine. Neuropsychopharmacol 31, 450-461.
Thuerauf, N., Renner, B., Kobal, G., 1995. Responses Recorded from the Frog Olfactory Epithelium
after Stimulation with R(+)- and S(-)- Nicotine. Chem Senses 20, 337-344.
Tomlins, S.A., Mehra, R., Rhodes, D.R., Shah, R.B., Rubin, M.A., Bruening, E., Makarov, V.,
Chinnaiyan, A.M., 2006. Whole transcriptome amplification for gene expression profiling
and development of molecular archives. Neoplasia 8, 153-162.
Torebjork, H.E., Hallin, R.G., 1970. C-fibre units recorded from human sensory nerve fascicles in
situ. A preliminary report. Acta Soc Med Ups 75, 81-84. Torebjork, H.E., Hallin, R.G., 1973. Perceptual Changes Accompanying Controlled Preferential
Blocking of a and C Fiber Responses in Intact Human Skin Nerves. Experimental Brain
Research 16, 321-332.
Trapnell, C., Pachter, L., Salzberg, S.L., 2009. TopHat: discovering splice junctions with RNA-Seq.
Bioinformatics 25, 1105-1111.

136
Anhang
Trapnell, C., Williams, B.A., Pertea, G., Mortazavi, A., Kwan, G., van Baren, M.J., Salzberg, S.L.,
Wold, B.J., Pachter, L., 2010. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat
Biotechnol 28, 511-515.
Trapp, B.D., Bernier, L., Andrews, S.B., Colman, D.R., 1988. Cellular and subcellular distribution of
2',3'-cyclic nucleotide 3'-phosphodiesterase and its mRNA in the rat central nervous
system. J Neurochem 51, 859-868.
Trepel, M., 1999. Peripheres Nervensystem. In: Neuroanatomie - Struktur und Funktion. (Trepel
M, ed): Urban & Fischer.
Uhl, G.R., Walther, D., Behm, F.M., Rose, J.E., 2011. Menthol Preference Among Smokers:
Association With TRPA1 Variants. Nicotine Tob Res 13, 1311-1315.
Vakonakis, I., Langenhan, T., Promel, S., Russ, A., Cambell, I.D., 2008. Solution structure and sugar-binding mechanism of mouse latrophilin-1 RBL: a 7TM receptor-attached lectin-like
domain. Structure 16, 944-953.
Venkatachalam, K., Montell, C., 2007. TRP channels. Annu Rev Biochem 76, 387-417.
Viana, F., 2011. Chemosensory Properties of the Trigeminal System. Acs Chem Neurosci 2, 38-50.
Vogt-Eisele, A.K., Keber, K., Sherkheli, M.A., Vielhaber, G., Panten, J., Gisselmann, G., Hatt, H.,
2007. Monoterpenoid agonists of TRPV3. Brit J Pharmacol 151, 530-540.
Vriens, J., Watanabe, H., Janssens, A., Droogmans, G., Voets, T., Nilius, B., 2004. Cell swelling,
heat, and chemical agonists use distinct pathways for the activation of the cation
channel TRPV4. P Natl Acad Sci USA 101, 396-401.
Wagner, D.A., Czajkowski, C., Jones, M.V., 2004. An arginine involved in GABA binding and unbinding but not gating of the GABA(A) receptor. Journal of Neuroscience 24, 2733-
2741.
Walker, J.C., KendalReed, M., Keiger, C.J., Bencherif, M., Silver, W.L., 1996. Olfactory and
trigeminal responses to nicotine. Drug Develop Res 38, 160-168.
Wallace, D.G., Gorny, B., Whishaw, I.Q., 2002. Rats can track odors, other rats, and themselves:
implications for the study of spatial behavior. Behav Brain Res 131, 185-192.
Wang, Z., Gerstein, M., Snyder, M., 2009. RNA-Seq: a revolutionary tool for transcriptomics. Nat
Rev Genet 10, 57-63.
Watanabe, H., Davis, J.B., Smart, D., Jerman, J.C., Smith, G.D., Hayes, P., Vriens, J., Cairns, W.,
Wissenbach, U., Prenen, J., Flockerzi, V., Droogmans, G., Benham, C.D., Nilius, B., 2002a. Activation of TRPV4 channels (hVRL-2/mTRP12) by phorbol derivatives. Journal of
Biological Chemistry 277, 13569-13577.
Watanabe, H., Vriens, J., Suh, S.H., Benham, C.D., Droogmans, G., Nilius, B., 2002b. Heat-evoked
activation of TRPV4 channels in a HEK293 cell expression system and in native mouse
aorta endothelial cells. Journal of Biological Chemistry 277, 47044-47051.
West, J.B., 1985. Best and Taylor’s Physiological Basis of Medical Practice. 11th edition Williams
and Wilkins, Baltimore.
Westman, E.C., Behm, F.M., Rose, J.E., 1995. Airway Sensory Replacement Combined with
Nicotine Replacement for Smoking Cessation - a Randomized, Placebo-Controlled Trial
Using a Citric-Acid Inhaler. Chest 107, 1358-1364. Wo, Z.G., Oswald, R.E., 1994. Transmembrane topology of two kainate receptor subunits revealed
by N-glycosylation. Proc Natl Acad Sci U S A 91, 7154-7158.
Wo, Z.G., Oswald, R.E., 1995. A topological analysis of goldfish kainate receptors predicts three
transmembrane segments. J Biol Chem 270, 2000-2009.
Wong, B.Y., Coulter, D.A., Choi, D.W., Prince, D.A., 1988. Dextrorphan and Dextromethorphan,
Common Antitussives, Are Antiepileptic and Antagonize N-Methyl-D-Aspartate in Brain-
Slices. Neuroscience Letters 85, 261-266.

137
Anhang
Wood, M.W., VanDongen, H.M., VanDongen, A.M., 1995. Structural conservation of ion
conduction pathways in K channels and glutamate receptors. Proc Natl Acad Sci U S A 92, 4882-4886.
Xiang, Z.H., Bo, X.N., Burnstock, G., 1998. Localization of ATP-gated P2X receptor
immunoreactivity in rat sensory and sympathetic ganglia. Neuroscience Letters 256,
105-108.
Xu, H.X., Ramsey, I.S., Kotecha, S.A., Moran, M.M., Chong, J.H.A., Lawson, D., Ge, P., Lilly, J., Silos-
Santiago, I., Xie, Y., DiStefano, P.S., Curtis, R., Clapham, D.E., 2002. TRPV3 is a calcium-
permeable temperature-sensitive cation channel. Nature 418, 181-186.
Xu, S.H., Ono, K., Inenaga, K., 2010. Electrophysiological and Chemical Properties in Subclassified
Acutely Dissociated Cells of Rat Trigeminal Ganglion by Current Signatures. J
Neurophysiol 104, 3451-3461. Yamamoto, Y., Hatakeyama, T., Taniguchi, K., 2009. Immunohistochemical colocalization of TREK-
1, TREK-2 and TRAAK with TRP channels in the trigeminal ganglion cells. Neuroscience
Letters 454, 129-133.
Yuan, T.T.T., Toy, P., McClary, J.A., Lin, R.J., Miyamoto, N.G., Kretschmer, P.J., 2001. Cloning and
genetic characterization of an evolutionarily conserved human olfactory receptor that is
differentially expressed across species. Gene 278, 41-51.
Zarbin, M.A., Wamsley, J.K., Kuhar, M.J., 1981. Glycine Receptor - Light Microscopic
Autoradiographic Localization with [Strychnine-H-3. Journal of Neuroscience 1, 532-547.
Zevin, S., Gourlay, S.G., Benowitz, N.L., 1998. Clinical pharmacology of nicotine. Clinics in
Dermatology 16, 557-564. Zhang, X., Nordberg, A., 1993. The competition of (-)-[3H]nicotine binding by the enantiomers of
nicotine, nornicotine and anatoxin-a in membranes and solubilized preparations of
different brain regions of rat. Naunyn Schmiedebergs Arch Pharmacol 348, 28-34.
Zhang, X.M., Firestein, S., 2002. The olfactory receptor gene superfamily of the mouse. Nat
Neurosci 5, 124-133.
Zhang, X.M., Rogers, M., Tian, H.K., Zhang, X.H., Zou, D.J., Jian, L., Ma, M.H., Shepherd, G.M.,
Firestein, S.J., 2004. High-throughput microarray detection of olfactory receptor gene
expression in the mouse. P Natl Acad Sci USA 101, 14168-14173.
Zhou, L.H., Lim, Q.E., Wan, G.Q., Too, H.P., 2010. Normalization with genes encoding ribosomal
proteins but not GAPDH provides an accurate quantification of gene expressions in neuronal differentiation of PC12 cells. BMC Genomics 11, 75.
Zhu, Y.X., Hsu, M.S., Pintar, J.E., 1998. Developmental expression of the mu, kappa, and delta
opioid receptor mRNAs in mouse. Journal of Neuroscience 18, 2538-2549.
Zozulya, S., Echeverri, F., Nguyen, T., 2001. The human olfactory receptor repertoire. Genome Biol
2, research0018.1-research0018.12.
Zufall, F., Ukhanov, K., Lucas, P., Leinders-Zufall, T., 2005. Neurobiology of TRPC2: from gene to
behavior. Pflug Arch Eur J Phy 451, 61-71.

138
Anhang
6.2 Abbildungsverzeichnis
Abbildung 1.1 Schema des Nervus trigeminus des Menschen (Gray, 2004).
Abbildung 1.2 Schema der Untereinheiten Liganden-gesteuerter Ionennkanäle
(Khakh, 2001).
Abbildung 1.3 TRP-Kanaluntereinheiten verschiedener Subfamilien (Montell, 2005).
Abbildung 1.4 Strukturformel beider Nikotin-Enantiomere.
Abbildung 1.5 Schematische Darstellung des Illumina-Solexa Sequenzierverfahrens
(Illumina Genome Analyzer II Workflow, http://www.illumina.com/).
Abbildung 2.1 Schematische Darstellung des Voltage-Clamp Messaufbaus.
Abbildung 3.1 Konzentrations-Wirkungs-Beziehung für Serotonin am 5-HT3A-Rezeptor in Gegenwart und Abwesenheit von Nikotin gemessen mit dem Zwei-Elektroden
Voltage-Clamp Verfahren.
Abbildung 3.2 Konzentrations-Inhibitions-Beziehung für Nikotin an 5-HT3A- und 5-HT3AB-
Rezeptoren bei konstanter Serotoninkonzentration (5-HT) gemessen mit dem
Zwei-Elektroden Voltage-Clamp Verfahren.
Ergebnisse sind entstanden in Zusammenarbeit mit Paul Ziemba.
Abbildung 3.3 Effekt von (+)-Nikotin am nAChR α7 gemessen im Zwei-Elektroden Voltage-
Clamp Verfahren.
Abbildung 3.4 Effekt von (+)-Nikotin am nAChR α2β4 gemessen im Zwei-Elektroden Voltage-
Clamp Verfahren. Abbildung 3.5 Konzentrations-Wirkungs-Beziehung für Glycin (Gly) am Glycinrezeptor α1 in
Gegenwart und Abwesenheit von Nikotin gemessen im Zwei-Elektroden
Voltage-Clamp Verfahren.
Ergebnisse sind entstanden in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.6 Der Einfluss von (-)-Nikotin auf die Konzentrations-Wirkungs-Beziehung für
GABA an GABAA-Rezeptoren gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren.
Abbildung 3.7 Konzentrations-Wirkungs-Beziehung für Adenosintriphosphat (ATP) am P2X2-
Rezeptor in Gegenwart und Abwesenheit von Nikotin gemessen im Zwei-
Elektroden Voltage-Clamp Verfahren. Ergebnisse sind entstanden in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.8 Konzentrations-Wirkungs-Beziehung für Glutamat an den Glutamatrezeptoren
GluR1 und GluR6 in Gegenwart und Abwesenheit von (-)-Nikotin gemessen im
Zwei-Elektroden Voltage-Clamp Verfahren.
Abbildung 3.9 Wirkung von (-)-Nikotin am NMDA-Rezeptor NR1-3a/NR2B gemessen im Zwei-
Elektroden Voltage-Clamp Verfahren.
Ergebnisse sind entstanden in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.10 Effekt von (+)-Nikotin am TRPA1 gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren.
Abbildung 3.11 Konzentrations-Wirkungs-Beziehungen an TRPV-Kanälen in Gegenwart und Abwesenheit von Nikotin gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren.
Abbildung 3.12 Der Einfluss von Nikotin auf die Konzentrations-Wirkungs-Beziehung für
Menthol (Men) am TRPM8-Kanal gemessen im Zwei-Elektroden Voltage-Clamp
Verfahren.
Abbildung 3.13 Calcium-Imaging Signale trigeminaler Neurone auf separate Stimuli von Nikotin
beider Enantiomere.
Ergebnisse sind entstanden in Zusammenarbeit mit Ulrike Thiel.

139
Anhang
Abbildung 3.14 Calcium-Imaging Signale (-)-Nikotin-responsiver trigeminaler Neurone unter
separatem Block durch Hexamethonium (Hex) und HC030031 (HC). Abbildung 3.15 Calcium-Imaging Signale (-)-Nikotin-responsiver trigeminaler Neurone unter
separatem Block durch Mecamylamin (Mec) und simultanem Block durch
Hexamethonium (Hex) und HC030031 (HC).
Abbildung 3.16 Calcium-Imaging Antwortprofile Nikotin-responsiver trigeminaler Neurone bei
Block durch Rutheniumrot (RR).
Abbildung 3.17 Calcium-Imaging Antwortprofile Nikotin-responsiver trigeminaler Neurone bei
Block durch Gentamycin.
Abbildung 3.18 Calcium-Imaging Antwortprofile Nikotin-responsiver trigeminaler Neurone bei
Block durch Hexamethonium und Borneol.
Abbildung 3.19 Block der Nikotin-induzierten Calcium-Imaging Signale trigeminaler Neurone unter Ca2+-freien Bedingungen im Extrazellulärmedium.
Abbildung 3.20 Einfluss des Phospholipase C-Blockers Edelfosin auf (-)-Nikotin-induzierte
Calcium-Imaging Signale trigeminaler Neurone.
Abbildung 3.21 Calcium-Imaging Signale Serotonin und mCPBG-responsiver trigeminaler
Neurone
Abbildung 3.22 Schematische Darstellung der Transkriptomamplifikation.
(Verändert nach Benutzerinformation WTA2 Procedure,
http://www.sigmaaldrich.com).
Abbildung 3.23 Einteilung der Transkriptome in Expressionsklassen.
Abbildung 3.24 Exemplarische alignments sequenzierter Transkripte aus Amplikon-
sequenzierungen einzelner Zellen und der Trigeminus-
Explantatsequenzierung an das Genom der Maus (mm9).
Abbildung 3.25 Heatmap der Expressionsstärken untersuchter Transkriptome für eine
Auswahl von housekeeping-Genen. Die Transkriptomanalysen von DRG und
Trigeminus-Explantat erfolgten in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.26 Heatmap der Expressionsstärken untersuchter Transkriptome für die Cys-loop- Kanäle. Die Transkriptomanalysen von DRG und Trigeminus-Explantat erfolgten
in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.27 Heatmap der Expressionsstärken untersuchter Transkriptome für die TRP-
Kanäle. Die Transkriptomanalysen von DRG und Trigeminus-Explantat erfolgten
in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.28 Heatmap der Expressionsstärken untersuchter Transkriptome für die KCNK-
Kanäle. Die Transkriptomanalysen von DRG und Trigeminus-Explantat erfolgten
in Zusammenarbeit mit Ramona Lehmann.
Abbildung 3.29 Darstellung der Expressionsstärke untersuchter Transkriptome als Histogramm. Abbildung 3.30 Vergleich der Anzahl exprimierter Gene der Einzelzelltranskriptome.

140
Anhang
6.3 Tabellenverzeichnis
Tabelle 3.1 Elektrophysiologischen Ergebnisse der Nikotin-Responsivität rekombinanter
humaner Ionenkanäle gemessen im Zwei-Elektroden Voltage-Clamp Verfahren.
Tabelle 3.2 Anzahl prozessierter und gemappter reads untersuchter Transkriptome nach
Analyse mit „TopHat“.
Tabelle 3.3 Auflistung exklusiv exprimierter Transmembranproteine für die Zellen 5-1 und
6-1.
Tabelle 3.4 Auflistung der in den einzelnen Zellen exprimierten olfaktorischen Rezeptoren
(OR).

141
Anhang
6.4 Abkürzungsverzeichnis
2-APB 2-Aminoethoxydiphenylborat
5-HT3A 5-Hydroxy-Tryptamin-Rezeptor Typ 3A
ACh Acetylcholin
AITC Allylisothiocyanat
AMPA α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
ATP Adenosintriphosphat
bp Basenpaare
CCD-Kamera charge-coupled device-Kamera
cDNA complementary desoxyribonucleic acid
CHO-Zellen Chinese hamster ovary Zellen
CNPase cyclic nucleotide phosphodiesterase
cRNA complementary ribonucleic acid
CT cycle treshold
DAG Diacylglycerol
DMEM/F-12 Dulbeccos´s Modified Eagle Medium
DRG dorsal root ganglia
EC50 half maximal effective concentration
EGTA ethylene glycol tetraacetic acid
FBS fetal bovine serum
FPKM fragments per kilobase of exon per million mapped reads
GABA γ-Aminobuttersäure
GABAAR γ-Aminobuttersäure-Rezeptor Typ A
GluR Glutamatrezeptor
Gly Glycin
HC HC030031
HEK Zellen Human embryonal kidney 293 Zellen
HEPES 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
Hex Hexamethonium
Hz Hertz I Strom
ICH-Kanäle hyperpolarization-activated, cyclic nucleotide-gated cation channel
KCNK Kanal Tandemporen-Kaliumkanal Typ K
L15 Leibovitz Medium 15
mCPBG m-Chlorophenylbiguanidhydrochlorid
Mec Mecamylamin
MEM minimum essential medium
mRNA messenger ribonucleic acid
nAChR nikotinische Acetylcholinrezeptoren
NFR normal frog ringer
Nik Nikotin
NMDA N-Methyl-D-Aspartat
NR N-Methyl-D-Aspartat-Rezeptor
OR olfaktorische Rezeptoren
P2X-Rezeptor Purinozeptor Typ 2X
PBS+/+ Phosphat gepufferte Salzlösung
PCR polymerase chain reaction
PIP2 Phosphatidylinositol 4,5-Bisphosphat

142
Anhang
PLC Phospholipase C
qPCR quantitative polymerase chain reaction rpm rounds per minute
RR Rutheniumrot
RT-PCR Reverse transcription polymerase chain reaction
Strd RNA-Standard
TBE-Puffer Tris/Borate/EDTA-Puffer
TG Trigeminus
TM2 Transmembrandomäne 2
TRPA1 transient receptor potential ankyrin-repeat 1
TRPC transient receptor potential canonical
TRPM transient receptor potential melastatin TRPML transient receptor potential mucolipin
TRPP transient receptor potential polycystic kidney disease-related protein
TRPV transient receptor potential vanilloid
ZAC Zn2+
-activated ion channel
UE Untereinheiten

143
Anhang
6.5 Publikationsliste
Veröffentlicht
Schöbel, N., Radtke, D., Lubbert, M., Gisselmann, G., Lehmann, R., Cichy, A., Schreiner, B.S.P.,
Altmüller, J., Spector, A.C., Spehr, J., Hatt, H., Wetzel, C.H., 2012. Trigeminal Ganglion Neurons of Mice Show Intracellular Chloride Accumulation and Chloride-Dependent Amplification of
Capsaicin-Induced Responses. Plos One 7, e48005.
Eingereicht
Schreiner, B.S.P., Lehmann, R., Thiel , U., Ziemba, P., Beltrán, L.R., Sherkheli, M.A., Jeanbourquin,
P., Hugi, A., Werner, M., Gisselmann, G., Hatt, H.
Direct action and modulating effect of (+)- and (–)-nicotine on ion channels expressed in
trigeminal sensory neurons.
In Vorbereitung
Kanageswaran, N., Demond, M., Schreiner, B.S.P., Baumgart, S., Nagel, M., Altmueller, J., Becker,
C., Dörner, J.F., Benecke, H., Wetzel, C.H., Hatt, H., Gisselmann, G.
Deep sequencing of the murine olfactory epithelium transcriptome.
Manteniotis, S., Lehmann, R., Flegel, C., Vogel, F., Hofreuter, A., Schreiner, B.S.P., Schöbel, N.,
Altmüller, J., Hatt, H., Gisselmann, G.
Comprehensive RNA-Seq expression analysis of sensory neurons with focus on trigeminal ganglia.

Lebenslauf
Zur Person
Name Benjamin Schreiner
Geboren am 12.02.1980
Geburtsort Essen
Schulbildung
1986-1990 Katholische Grundschule Hattingen-Niederwenigern
1990-1999 Gymnasium Waldstraße in Hattingen
Akademische Ausbildung
1999 – 2008 Studium der Biologie an der Ruhr-Universität Bochum
2007- 2008 Diplomarbeit am Lehrstuhl für Zellphysiologie der Ruhr-
Universität Bochum mit dem Titel „Die Funktion zyklisch-
Nukleotid gesteuerter Ionenkanäle bei der Chemorezeption von
Drosophila melanogaster“, betreut durch Prof. Dr. Dr. Dr. Hanns
Hatt

Danksagung
An dieser Stelle möchte ich mich bei Herrn Professor Dr. Dr. Dr. Hanns Hatt für die Überlassung
des interessanten Themas, die Bereitstellung des Arbeitsplatzes und die Betreuung bedanken.
Für die freundliche Übernahme des Korreferats danke ich Herrn PD Dr. Matthias Schmidt.
Mein besonderer Dank geht an Herrn Dr. Markus Werner und Herrn Dr. Günter Gisselmann,
die in vielen konstruktiven Gesprächen zum Gelingen dieser Arbeit beigetragen haben und in
deren wissenschaftlichem Erfahrungsschatz so mancher wertvolle Rat zu unerwarteten
Problemstellungen zu finden ist.
Für technische Unterstützung möchte ich Herrn Harry Bartel, Herrn Thomas Lichtleitner, Frau
Ute Müller und Frau Jasmin Gerkrath danken. Für die Unterstützung in Dingen der Bürokratie
danke ich Frau Ulrike Thomes.
Ein großes Dankeschön an alle Mitarbeiter des Lehrstuhls für Zellphysiologie für die grandiose
Atmosphäre. Besonders möchte ich mich bei aktuellen und ehemaligen Insassen vom „Raum
der Götter“ Sabrina Baumgart, Olaf Kletke, Lian Gelis, Stefan Kurtenbach, Sebastian Rasche und
Ivonne Wallrabenstein bedanken. Neben der Wissenschaft ging es mit Euch auch mal lustig zu.
Ihr habt meine Marotten ertragen. Es war eine gute Zeit. Ein weiteres herzliches Dankeschön
auch an die lieb gewonnenen Ehemaligen des Lehrstuhls Ainhara Aguado, Debbie Radtke,
Nicole Schöbel und Ramona Lehmann. Ohne Euch ist blöd.
Ganz besonders möchte ich meinen Eltern für die Unterstützung in jeder Hinsicht danken.
Mein ganzes Leben lang habt ihr mich noch nie im Regen stehen lassen.
Ein ganz großes Dankeschön an meine Familie, an Gott und an meine Freunde. Ihr habt mir viel
Verständnis entgegengebracht, selbst wenn ich mal irgendwie komisch geworden bin. Und
danke Oma für immer lecker Essen.


















