
Entwicklung einer Methode zur Identifizierung von ... · zur Identifizierung von Mikroorganismen...
100
Entwicklung einer Methode zur Identifizierung von Mikroorganismen über Raman-Spektroskopie Masterarbeit zur Erlangung des Master of Science an der Fakultät Life Sciences der Hochschule Rhein-Waal, Kleve Studiengang: Lebensmittelwissenschaften vorgelegt von Thomas Johann Tewes Kleve, Januar, 2019
Transcript of Entwicklung einer Methode zur Identifizierung von ... · zur Identifizierung von Mikroorganismen...

Entwicklung einer Methode
zur Identifizierung von Mikroorganismen
über Raman-Spektroskopie
Masterarbeit
zur
Erlangung des Master of Science
an der
Fakultät Life Sciences
der
Hochschule Rhein-Waal, Kleve
Studiengang: Lebensmittelwissenschaften
vorgelegt von
Thomas Johann Tewes
Kleve, Januar, 2019

1. Prüfer: Herr Prof. Dr.-Ing Frank Platte
2. Prüfer: Herr Prof. Dr. Dirk Bockmühl

I
Danksagung
Mein besonderer Dank gilt meinen beiden Betreuern Prof. Dr. Frank Platte, der sich sehr
viel Zeit für meine Einarbeitung in das Raman-System und die Software MATLAB nahm,
sowie Herrn Prof. Dr. Dirk Bockmühl, der es mir überhaupt erst ermöglichte an diesem
hochinteressanten Thema zu arbeiten.
Ebenfalls sehr dankbar bin ich:
Frau Britta Brands und Herrn Ralf Lucassen für die Bereitstellung der Mikroorganismen
und der Materialien, sowie für Ihre beispiellose Hilfsbereitschaft und dem Interesse an
meiner Arbeit.
Sowie meinen Eltern und meiner Familie, die mich auf verschiedenste Art und Weise un-
terstützten.

II
Inhaltsverzeichnis
Kurzzusammenfassung ................................................................................................ IV
Abstract ........................................................................................................................... V
Abbildungsverzeichnis ................................................................................................. VI
Tabellenverzeichnis .................................................................................................... VIII
Abkürzungsverzeichnis ................................................................................................ IX
1 Einleitung ...........................................................................................................1
1.1 Theoretischer Hintergrund ................................................................................3
1.1.1 Physiologie der Mikroorganismen ....................................................................3
1.1.2 Methoden zur Identifizierung von Mikroorganismen ........................................6
1.1.3 Raman-Spektroskopie ......................................................................................8
1.1.3.1 Theorie des Raman-Effekts .....................................................................8
1.1.3.2 Aufbau des Raman-Messsystems ........................................................10
1.1.3.3 Informationsreiche spektrale Regionen von Mikroorganismen ............10
1.1.3.4 Einflüsse auf die Qualität eines Raman-Spektrums .............................13
1.1.4 Datenvorbehandlung und Vorhersagemodelle ...............................................18
1.1.5 Einblick in Ergebnisse bisheriger Studien ......................................................19
1.2 Zielsetzung ......................................................................................................21
2 Material und Methoden ...................................................................................23
2.1 Material ............................................................................................................23
2.1.1 Geräte .............................................................................................................23
2.1.2 Software ..........................................................................................................24
2.1.3 Mikroorganismen.............................................................................................24
2.1.4 Verbrauchsmaterialien ....................................................................................24
2.2 Methoden ........................................................................................................25
2.2.1 Anzucht der Mikroorganismen ........................................................................25
2.2.2 Probenpräparation ..........................................................................................25
2.2.3 Vorbereitung des Raman-Messsystems ........................................................26
2.2.4 Sammeln der Raman-Spektren ......................................................................26
2.2.5 Datenvorbehandlung .......................................................................................27
2.2.5.1 Interpolierung .........................................................................................27
2.2.5.2 Basislinienkorrektur und Glättung .........................................................28
2.2.5.3 Normalisierung.......................................................................................29
2.2.6 Hauptkomponentenanalyse ............................................................................31
2.2.7 Modellentwicklung zur Identifizierung von Mikroorganismen durch verschiedene Klassifikationsverfahren ...........................................................34
2.2.7.1 Quadratische Diskriminanzanalyse .......................................................34
2.2.7.2 Nächster-Nachbar-Algorithmus .............................................................36
2.2.7.3 Support Vector Maschine ......................................................................37

III
2.2.8 Validierung der Vorhersagemodelle ...............................................................38
2.2.8.1 Kreuzvalidierung ....................................................................................38
2.2.8.2 Praktische Validierung ...........................................................................39
3 Ergebnisse .......................................................................................................40
3.1 Spektrenaufzeichnung ....................................................................................40
3.1.1 Störung durch Umgebungslicht ......................................................................40
3.1.2 Störung durch Fluoreszenz .............................................................................41
3.1.3 Einfluss der Probenpräparation auf ein Spektrum .........................................41
3.1.4 Einfluss der Kultivierungsart auf ein Spektrum ..............................................43
3.1.5 Einfluss der Vorbestrahlung auf ein Spektrum ...............................................45
3.2 Modellentwicklung ...........................................................................................47
3.2.1 Datenvorbehandlung .......................................................................................47
3.2.2 Hauptkomponentenanalyse ............................................................................50
3.2.3 Klassifikationsverfahren und Kreuzvalidierung ..............................................52
3.2.3.1 Quadratische Diskriminanzanalyse .......................................................52
3.2.3.2 Nächster-Nachbar-Algorithmus .............................................................53
3.2.3.3 Support Vector Maschine ......................................................................54
3.3 Praktische Validierung ....................................................................................55
4 Diskussion .......................................................................................................61
4.1 Spektrenaufzeichnung ....................................................................................61
4.2 Datenvorbehandlung .......................................................................................62
4.3 Beurteilung der Leistung und mögliche Limitationen der entwickelten Methode ..........................................................................................................64
4.4 Möglichkeiten zur Vermeidung falsch positiver Vorhersagen ........................65
4.5 Ausblick ...........................................................................................................66
Anhang ...........................................................................................................................67
5 Literaturverzeichnis ........................................................................................82
Eidesstattliche Versicherung .......................................................................................88

IV
Kurzzusammenfassung
Zielstellung: Das Ziel dieser Arbeit war es, eine Methode zur Identifizierung von Mikroor-
ganismen über Mikro-Raman-Spektroskopie zu entwickeln. Dafür musste zunächst eine
Methode gefunden werden, um Spektren verschiedener Mikroorganismen mit ausreichen-
der Qualität zu erhalten. Im Anschluss sollte die Datensammlung erfolgen, bei der möglichst
viele repräsentative Spektren gesammelt werden. Mit den Daten mussten danach Modelle
zur Vorhersage unbekannter Raman-Spektren entwickelt werden, was eine entsprechende
Datenvorbehandlung voraussetzte. Um die Praxistauglichkeit der Modelle zu prüfen, sollten
diese auf Spektren von Proben angewandt werden, die nicht in den Kalibrationssets ent-
halten waren. Neben der Entwicklung der Methode und der Vorhersagemodelle, sollten
auch verschiedene Einflüsse der Analysebedingungen berücksichtigt werden.
Methoden: Neun verschiedene Mikroorganismen, die aus kryokonservierten Reinkulturen
stammten, wurden bei 30 °C für 24 Stunden auf Trypton Soja Agar (TSA) bebrütet. Die
Spektrenaufzeichnung erfolgte bei einer Anregung von 633 nm über ein konfokales Raman-
Mikroskop der Firma Renishaw. Sowohl die Datenvorbehandlung (Basislinienkorrektur,
Glättung, Normalisierung, Hauptkomponentenanalyse (PCA)), als auch die Modellentwick-
lung, erfolgte über die Software MATLAB. Es wurden verschiedene Klassifikationsverfahren
angewandt, um ein Modell mit möglichst guter Leistung zu finden.
Ergebnisse: Für die Entwicklung der Kalibrationsmodelle wurden insgesamt 2953 Raman-
Spektren von neun verschiedenen Mikroorganismen gesammelt. Störungen durch Fluores-
zenz wurden durch Vorbestrahlung von bis zu 15 Minuten ausgebrannt. Die Modelle basie-
rend auf der Quadratischen Diskriminanzanalyse (QDA), dem k-Nächster-Nachbar-Algo-
rithmus (kNN) und der Support Vector Maschine (SVM), wurden mit verschiedener Anzahl
an Hauptkomponenten und einer fünffachen Kreuzvalidierung erstellt. Die theoretischen
Vorhersagegüten belaufen sich bei den genauesten Modellen auf 99,0 % (QDA), 99,1 %
(kNN) und 99,7 % (SVM). Für die praktische Validierung wurden für jeden Mikroorganismus
100 weitere „unbekannte“ Spektren gesammelt. Die Vorhersagegenauigkeiten in der Praxis
belaufen sich auf 98,1 % (QDA), 97,1 % (kNN) und 97,0 % (SVM).
Diskussion: Eine zuverlässige Identifizierung der in dieser Arbeit untersuchten Mikroorga-
nismen konnte über alle verwendeten Klassifikationsverfahren erreicht werden. Auf Stamm-
ebene erzielt die SVM in der Praxis die genauesten Ergebnisse, jedoch nicht bei den rest-
lichen Mikroorganismen. Die QDA erzielt in der Praxis etwas genauere Vorhersagen als
das kNN-Modell, jedoch basiert das kNN-Modell mit der besten Leistung auf den wenigsten
Hauptkomponenten. Dies kann als Vorteil angesehen werden, da weniger Dimensionen zur
erfolgreichen Klassifizierung in der Regel robustere Modelle in der Praxis bedeuten.

V
Abstract
Objective: The objective of this work was to develop a method for the identification of
microorganisms via micro-Raman spectroscopy. First, a method was established to obtain
spectra of sufficient quality for various microorganisms. Subsequently, the data collection
was performed, in which as many representative spectra as possible were collected. After-
wards, models for the prediction of unknown Raman spectra were developed with the data,
which required a corresponding data pre-treatment. To test the practicability of the models,
they were applied to spectra of samples that were not included in the calibration sets. In
addition to the development of the method and the predictive models, different influences
of the analysis conditions should also be considered.
Methods: Nine different microorganisms from cryopreserved pure cultures were incubated
at 30 °C for 24 hours on tryptone soy agar (TSA). The spectra were obtained at 633 nm
excitation via a confocal Raman microscope from the company Renishaw. Both the data
pre-treatment (baseline correction, smoothing, normalization, principal component analysis
(PCA)) and the model development were carried out using the MATLAB software. Various
classification methods were used to determine the model with the best possible perfor-
mance.
Results: For the development of the calibration models 2953 Raman spectra of nine diffe-
rent microorganisms were collected. Disturbances by fluorescence were bleached out by
pre-irradiation for up to 15 minutes. The models based on quadratic discriminant analysis
(QDA), the k-nearest neighbor algorithm (kNN), and support vector machine (SVM) were
created with different numbers of principal components and using a five-fold cross-valida-
tion. The theoretical prediction accuracies for the most accurate models are 99.0 % (QDA),
99.1 % (kNN) and 99.7 % (SVM). For practical validation, 100 more "unknown" spectra
were collected for each microorganism. The accuracies of the prediction in practice are
98.1 % (QDA), 97.1 % (kNN) and 97.0 % (SVM).
Discussion: All classification methods used in this thesis identified reliably the investigated
microorganisms. At strain-level, SVM achieves the most accurate results in practice, but not
with the remaining microorganisms. In practice, QDA achieves slightly more accurate pre-
dictions than the kNN-model. However, the kNN-model with the best performance is based
on the lowest number of principal components. This can be considered as an advantage,
because using less dimensions for successful classification usually means more robust mo-
dels in practice.

VI
Abbildungsverzeichnis
Abbildung 1: Schema einer prokaryotischen Zelle (A) und einer eukaryotischen Zelle (B) (Fritsche, 2016) ................................................................................................4
Abbildung 2: Schematischer Aufbau einer grampositiven Zellwand (A) und einer gramnegativen Zellwand (B) (Fuchs et al., 2007) .................................................6
Abbildung 3: Schematische Darstellung der Energieniveaus bei der Rayleigh-, Stokes- und Anti-Stokes-Streuung links und fiktives Raman-Spektrum rechts (modifiziert nach (Kim et al., 2017)) ......................................................................9
Abbildung 4: Schematischer Aufbau des konfokalen Raman Mikroskops inVia (Renishaw) (modifiziert nach (Nataf, 2016) und (Vargis et al., 2012)) ...............10
Abbildung 5: Beispiel eines mit erweitertem Wellenzahlbereich erfassten Raman-Spektrums von Saccharomyces cerevisiae (A) und hervorgehobene informationsreiche Region (B) .............................................................................11
Abbildung 6: Normalschwingungen eines dreiatomigen Moleküls (z.B. Wasser) (modifiziert nach (Hasegawa, 2017)) ..................................................................12
Abbildung 7: Raman-Spektrum von E. coli K12 mit Zuordnung des biochemischen Ursprungs der Banden nach (De Gelder, 2008; Maquelin et al., 2002) .............12
Abbildung 8: Raman-Spektren zweier Bakterien mit starken Fluoreszenzstörungen ....14
Abbildung 9: Auftreten von „Cosmic spikes“ in Raman-Spektren markiert durch Pfeile 16
Abbildung 10: Raman-Spektrum eines Glasobjektträgers (A) und Raman-Spektrum von getrocknetem Nährboden (TSA) (B) .............................................................18
Abbildung 11: Konfokales Raman-Mikroskop inVia Renishaw.......................................23
Abbildung 12: Prinzip eines Tiefpassfilters zur Glättung eines Stufenspektrums ..........29
Abbildung 13: Schrittweises Schema der Datenvorbehandlung anhand dreier Spektren verschiedener Mikroorganismen; (A) Interpolierte Spektren, (B) Basislinienkorrektur und Glättung, (C) Normalisierung .......................................31
Abbildung 14: Vereinfachtes Schema einer Dimensionsreduktion durch PCA (modifiziert nach (Powell & Lehe, 2014)) ............................................................33
Abbildung 15: Beispiel zweier Datenklassen mit markiertem Klassenschwerpunkt und Projektion der Daten um die größte Varianz zu erzielen (A) und Projektion der Daten in Diskriminanzrichtung (B) (Hastie et al., 2009) ......................................35
Abbildung 16: Daten aus drei verschiedenen Klassen mit linearen Entscheidungsgrenzen durch LDA (A), quadratische Entscheidungsgrenzen, welche durch Auffinden linearer Grenzen in einem höherdimensionalem Raum erhalten wurden, zurückgeführt auf den ursprünglichen Raum (B), mittels QDA gefundene quadratische Entscheidungsgrenzen (C) (modifiziert nach (Hastie et al., 2009)) .........................................................................................................36
Abbildung 17: Grafische Erklärung von kNN anhand normalisierter Intensitäten zweier ausgewählter Wellenzahlen aus Raman-Spektren von drei verschiedenen Mikroorganismen .................................................................................................37
Abbildung 18: Schematische Darstellung des Funktionsprinzips der SVM mit den Ausgangsdaten (A), dem höherdimensionalem Raum (B) und resultierenden Entscheidungsgrenzen bei zurücktransformieren der Daten (C); Schwarze Linien sind trennende Hyperebenen, gestrichelte Linien „Support Vectors“ und Entscheidungsgrenze im höherdimensionalem Raum als graues Dreieck hervorgehoben (modifiziert nach (Van den Burg & Groenen, 2016)) .................38
Abbildung 19: Vergleich von Raman-Spektren bei verschiedenen Umgebungslichtbedingungen ohne fokussierte Probe bei einem Prozent Laserleistung und 40 Sekunden Belichtungsdauer ............................................40

VII
Abbildung 20: Einfluss der Vorbestrahlungsdauer auf eine Probe E. coli K12 (Counts normalisiert) .........................................................................................................41
Abbildung 21: Beispiel zweier Koloniematerialprobenschichten auf Glasobjektträgern zur Einschätzung der Schichtdicken (a= dünn; b= mittel; c= dick; d= ungeeigneter, noch flüssiger Bereich) .................................................................42
Abbildung 22: Einfluss der Probenschichtdicke auf das Raman-Spektrum einer Sty TA98-Probe bei zwei Minuten Vorbestrahlung; (A) unbehandelte Daten, (B) normalisierte Spektren (Eine Einschätzung der Begriffe mittlere, dünne und dicke Schicht kann anhand Abbildung 21 erfolgen).....................................42
Abbildung 23: Vergleich der Raman-Spektren aller Mikroorganismen von Kultur 1 (blau) und Kultur 2 (rot) mit fett hervorgehobenen Mittelwertspektren .45
Abbildung 24: (A) Darstellung mehrerer hintereinander aufgezeichneter Spektren von ein und demselben Bereich einer dünnen Probenschicht Micrococcus luteus (DSM-Nr. 20030) mit t als Anregungsdauer in Minuten; (B) Gleiche Spektren nach Basislinienkorrektur, Glättung und Normalisierung ....................................46
Abbildung 25: Auswirkung längerer Bestrahlung auf Peaks im Wellenzahlbereich von 600 bis 900 cm-1 bei drei verschiedenen gramnegativen Bakterien; Blau hervorgehoben ein Spektrum mit kurzer Bestrahlungsdauer und Rot ein Spektrum mit langer Bestrahlungsdauer (Die Spektren wurden basislinienkorrigiert, geglättet und normalisiert) ..................................................47
Abbildung 26: Spektren vor (A) und nach Basislinienkorrektur, Glättung und Normalisierung (B) (Zur besseren Darstellung wurden in dieser Abbildung auch die unvorbehandelten Spektren normalisiert) ............................................48
Abbildung 27: Alle zur Kalibrationsentwicklung verwendete Spektren der 9 untersuchten Mikroorganismen nach Basislinienkorrektur, Glättung und Normalisierung in Grau und farblich hervorgehobene Mittelwertspektren (n= Anzahl der Spektren) ...........................................................................................49
Abbildung 28: Abnehmende erklärende Varianz der PCs nach PCA mit vorbehandelten Daten .........................................................................................50
Abbildung 29: Darstellung der ersten drei PCs der Hauptkomponentenanalyse von allen Kalibrations-Spektren mit und ohne Vorbehandlung aus jeweils zwei verschiedenen Betrachtungswinkeln ...................................................................51
Abbildung 30: Vorhersagematrices bei Verwendung der quadratischen Diskriminanzanalyse von Modellen mit 8 und 15 PCs mit der durchschnittlichen Vorhersagegüte in Prozent ...................................................52
Abbildung 31: Vorhersagematrices bei Verwendung des Nächste-Nachbar-Algorithmus von Modellen mit 8 und 15 PCs mit der durchschnittlichen Vorhersagegüte in Prozent ..................................................................................53
Abbildung 32: Vorhersagematrices bei Verwendung der Support Vector Maschine von Modellen mit 8 und 15 PCs mit der durchschnittlichen Vorhersagegüte in Prozent .................................................................................................................54
Abbildung 33: Darstellung der ersten drei Hauptkomponenten einer PCA der Kalibrierungsspektren (markiert als Punkte n= 2953) und Validierungsspektren (markiert als Quadrate n= 900) aus zwei Ansichten (A und B) ..........................55
Abbildung 34: Durchschnittliche Vorhersagegüte bei Anwendung aller entwickelten Modelle auf die Validierungsspektren aller 9 Mikroorganismen mit Standardabweichung ...........................................................................................56
Abbildung 35: Absolute Anzahl der richtig vorhergesagten Spektren bei der Validierung dreier Klassifizierungsverfahren bei variierender Anzahl PCs ........57

VIII
Tabellenverzeichnis
Tabelle 1: Relative Häufigkeit von Makromolekülen in einer Bakterienzelle (Madigan et al., 2015) ............................................................................................................5
Tabelle 2: Banden im Raman-Spektrum und Zuordnung des biochemischen Ursprungs nach (De Gelder et al., 2007; S. Kumar et al., 2016; Munchberg et al., 2014; Sil et al., 2017; Teng et al., 2016) .......................................................13
Tabelle 3: Verwendete Mikroorganismen, Kürzel und DSM-Nr......................................24
Tabelle 4: Anzahl der zur Kalibrationsentwicklung erfassten Raman-Spektren pro Kulturtyp (Summe= 2953)....................................................................................44
Tabelle 5: Validierungsvorhersagen des QDA-Modells mit 20 PCs (Vorhersagegüte 98,1 %) ....................................................................................58
Tabelle 6:Validierungsvorhersagen des kNN-Modells mit 15 PCs (Vorhersagegüte 97,1 %) ....................................................................................59
Tabelle 7: Validierungsvorhersagen des SVM-Modells mit 15 PCs (Vorhersagegüte 96,9 %) und 20 PCs (Vorhersagegüte 97,0 %) ......................60

IX
Abkürzungsverzeichnis
Einheit Bezeichnung
a.u. Unskalierte Einheit
Engl. arbitrary unit
cm Zentimeter
g Gramm
Hz Hertz
l/mm Linienzahl pro Millimeter
min Minuten
mW Milliwatt
nm Nanometer
rpm Umdrehungen pro Minute
Engl. rounds per minute
s Sekunden
Abkürzung Bezeichnung
A Adenin
C Cytosin
CCD Engl. „Charge-coupled Device“
Def. Deformationsschwingung(en)
DNA Desoxyribonukleinsäure
DUVRRS Tief-UV-Resonanz-Raman-Spektroskopie
Engl. deep UV resonance Raman spectroscopy
EMD Empirische Bandzerlegung
Engl. empirical mode decomposition
engl. Englisch
EP Europäisches Arzneibuch
EPS Extrazelluläre polymere Substanzen
FDA-BAM Engl. Food and Drug Administration-Bacteriological
Analytical Manual
FT Fouriertransformation
G Guanin
glyk. Verb. Glykosidische Verbindung
HCA Hierarchische Clusteranalyse
HeNe Helium-Neon
IR Infrarot
ISO Internationale Organisation für Normung
JP Japanische Pharmakopöe
kNN k-Nächster-Nachbar
LDA Lineare Diskriminanzanalyse
LPS Lipopolysaccharid(e)
n Anzahl Elemente/ Beobachtungen
NaCl Natriumchlorid
Nd:YAG Neodym-dotierter Yttrium-Aluminium-Granat-Laser
NGS Engl. Next Generation Sequencing
NIR Nahinfrarot
PC(s) Hauptkomponenten
Engl. principal component(s)

X
PCA Hauptkomponentenanalyse
Engl. pricipal component analysis
PCR Polymerase-Kettenreaktion
Engl. polymerase chain reaction
PHB Polyhydroxybutyrat
Phe Phenylalanin
QDA Quadratische Diskriminanzanalyse
RNA Ribonukleinsäure
SD Standardabweichung
Engl. Standard Derivation
SERDS Engl. Shifted-excitation Raman difference
spectroscopy
SERS Oberflächenverstärkte Raman-Spektroskopie
Engl. Surface-enhanced Raman Spectroscopy
Si Silizium
spp. species pluralis
Str. Streckschwingung(en)
SVD Singulärwertzerlegung
Singular Value Decomposition
SVM Engl. Support Vector Machine
t Dauer/ Zeit
TSA Trypton Soja Agar
TXT Textdatei (Dateiformat)
Tyr Tyrosin
U Uracil
USP United States Pharmakopöe
UV Ultraviolett
WDF WiRE Data File (Dateiformat)

Einleitung
1
1 Einleitung
Mikroorganismen sind mikroskopisch kleine einzellige Organismen, die uns praktisch über-
all umgeben. In der Natur stößt man unweigerlich auf Mikroorganismen; egal ob im Wasser,
im Boden, in der Luft, auf und im Menschen aber auch in extremen Umgebungen wie in
heißen Quellen oder Gletschereis. (Madigan et al., 2015)
Mikroorganismen haben die Entwicklung aller anderen Organismen ermöglicht, sind hoch-
gradig an ihre Umwelt angepasst und weisen die größte Vielfalt aller Lebewesen auf. Der
Mensch kann ohne Mikroorganismen nicht überleben, umgekehrt sind Mikroorganismen
jedoch ohne höhere Organismen überlebensfähig. (Cypionka, 2010)
Neben den vielen nützlichen Funktionen, die Mikroorganismen in unserem Körper haben,
wie beispielsweise die Produktion von Vitaminen, und der Aufschluss von Nahrungsbe-
standteilen durch Bakterien in der Darmflora (Fritsche, 2016) werden sie auch vielseitig in
der Industrie eingesetzt. Mikroorganismen dienen beispielsweise zur Herstellung und Ver-
edelung diverser Lebensmittel und Nahrungsmittelbestandteilen und zur Herstellung von
pharmakologisch wirksamen Substanzen wie Antibiotika oder Insulin. (Fuchs et al., 2007)
Es gibt allerdings auch Mikroorganismen, die für die Vergiftung oder den Verderb von Le-
bensmitteln und das Auslösen von Krankheiten verantwortlich sind. Es gibt sowohl Krank-
heitserreger, bei denen alle Arten einer Gattung pathogen sind, wie Shigella und Yersinia,
als auch Gattungen, bei denen nur einige Arten pathogen sind, wie beispielsweise bei der
Gattung Streptococcus die Art pneumoniae oder bei der Gattung Bacillus die Art anthracis,
deren Namen hier bereits auf die auslösenden Krankheiten schließen lassen. Besonders
bedeutsam sind Bakterien, die in einer einzigen Art harmlose und pathogene Stämme auf-
weisen, wie z.B. der ungefährliche Escherichia coli K12 und der pathogene E. coli O157:H7.
(Fuchs et al., 2007)
Nach dem Ausbruch einer Krankheit ist es in der Medizin unerlässlich, den Erreger richtig
und schnell zu ermitteln um die beste Therapie zu wählen (Fuchs et al., 2007). Prüfvor-
schriften für pharmazeutische Produkte machen den Nachweis bestimmter Keime ebenfalls
erforderlich. Für die Beurteilung des Hygienezustands bestimmter Gegenstände oder Ober-
flächen sowie für die Beurteilung der Prozesshygiene in der Industrie oder im Handwerk,
ist der alleinige Nachweis von Mikroorganismen kein Anzeichen für einen Mangel; es müs-
sen spezifische Hygieneindikatoren herangezogen werden. (Krämer, 2007)
Für diese und viele weitere Fälle sind Methoden zur Differenzierung von Mikroorganismen
erforderlich.

Einleitung
2
Um Mikroorganismen zu differenzieren bzw. zu identifizieren gibt es einfache taxonomische
Verfahren, mit denen die Identität eines Mikroorganismus immer weiter eingekreist werden
kann. (Steinbüchel et al., 2012)
Molekularbiologische Methoden wie die Polymerase-Ketten-Reaktion (PCR) (Cypionka,
2010) oder Methoden wie die Fluoreszenzmikroskopie (Ivnitski et al., 1999) und Massen-
spektroskopie (Sogawa et al., 2012) bilden leistungsfähige Alternativen zu klassischen Ver-
fahren. Als geeignet haben sich auch spektroskopische Verfahren, wie die Fourier-Trans-
formations-Infrarot- (FT-IR) Spektroskopie (Ngo-Thi et al., 2003) und die Raman-Spektro-
skopie erwiesen (Harz et al., 2005; Maquelin et al., 2002; Rösch et al., 2005; Strola et al.,
2014).
Die Identifizierung eines Mikroorganismus anhand eines elektromagnetischen Spektrums,
beruhend auf der chemischen Zusammensetzung dieses Organismus, erfolgt vereinfacht
durch den Abgleich des unbekannten Spektrums mit einer Datenbank bekannter Spektren.
Spektroskopische Methoden in Verbindung mit chemometrischen Verfahren zur Entwick-
lung leistungsfähiger Vorhersagemodelle, ermöglichen eine zerstörungsfreie, schnelle und
einfache Identifizierung von Mikroorganismen.

Einleitung
3
1.1 Theoretischer Hintergrund
Da die zu entwickelnde Methode auf der Wechselwirkung von Licht und der chemischen
Zusammensetzung von Mikroorganismen basiert, soll zunächst ein Überblick der relevan-
ten theoretischen Hintergründe zu Mikroorganismen und der Raman-Spektroskopie erfol-
gen. Um einen Vergleich der Raman-Spektroskopie mit anderen Verfahren zur Identifizie-
rung von Mikroorganismen zu ermöglichen, sollen einige gängige Verfahren ebenfalls kurz
beschrieben werden. Wichtig ist neben der Theorie der Raman-Spektroskopie die Theorie
hinter multivariaten Vorhersagemodellen, welche im Methoden-Teil der Arbeit näher be-
schrieben werden. Ein Einblick in Ergebnisse bisheriger Studien zur gleichen Thematik er-
möglicht die Einordnung der in dieser Arbeit erreichten Resultate und eine Einschätzung
des momentanen Forschungsstands.
1.1.1 Physiologie der Mikroorganismen
Mikroorganismus ist ein Überbegriff für Einzeller, die man zunächst grob in Prokaryoten
und Eukaryoten einteilen kann. Prokaryoten sind einfach gebaute einzellige Organismen
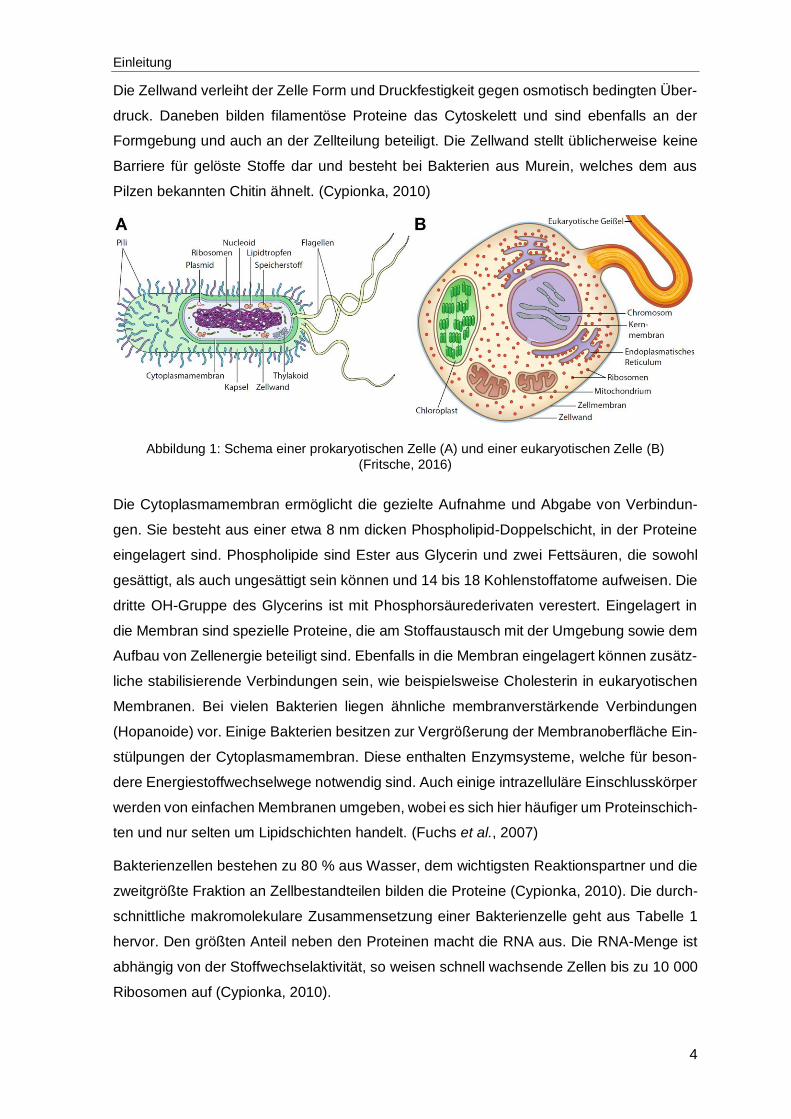
ohne abgegrenzten Zellkern (siehe Abbildung 1 A). Dazu zählen die Bakterien und Archaea.
Eukaryoten sind Organismen mit echtem Zellkern, zu denen auch Pflanzen und Tiere zäh-
len; in der Mikrobiologie ist jedoch von Pilzen, Algen und Protozoen die Rede. Eukaryoten
enthalten Organellen wie Mitochondrien, endoplasmatische Retikuli und Chloroplasten
(siehe Abbildung 1 B), welche bei Prokaryoten nur in wenigen Ausnahmefällen zu finden
sind. (Cypionka, 2010)
Die für diese Arbeit relevanten Mikroorganismen stellen die Bakterien und Hefen dar (ein-
zellige Pilze). Mittels dünnen filamentösen Zellanhängen, den sogenannten Flagellen oder
Geißeln, können sich einige Bakterien frei bewegen. Andere Zellanhängsel wie Fimbrien
(Typ-I-Pili) sind dünner und kürzer als Flagellen, dienen in der Regel nicht zur Fortbewe-
gung und sind in größerer Anzahl als Flagellen vorhanden. Sie bestehen aus Protein und
ermöglichen die Anheftung der Bakterien an Oberflächen. Spezielle Pili, wie der F-Pili die-
nen zur Herstellung von Zell-Zell-Kontakten, die den Austausch genetischer Information
ermöglichen. (Fuchs et al., 2007)
Die Zellhülle umschließt das Cytoplasma. In der Regel ist die Zellhülle aus einer äußeren
Wand und einer Membran zusammengesetzt. Die Membran ist an Transportprozessen und
am Energiestoffwechsel beteiligt. Das Cytoplasma enthält die genetische Information auf
einem Chromosom bzw. in einem Nucleoid (Kernäquivalent) und bei Bakterien häufig
extrachromosomale ringförmige DNA (Plasmide), sowie Ribosomen, Proteine, Coenzyme
und alle weiteren Bestandteile aufgenommener Stoffe. (Cypionka, 2010)
Einleitung
4
Die Zellwand verleiht der Zelle Form und Druckfestigkeit gegen osmotisch bedingten Über-
druck. Daneben bilden filamentöse Proteine das Cytoskelett und sind ebenfalls an der
Formgebung und auch an der Zellteilung beteiligt. Die Zellwand stellt üblicherweise keine
Barriere für gelöste Stoffe dar und besteht bei Bakterien aus Murein, welches dem aus
Pilzen bekannten Chitin ähnelt. (Cypionka, 2010)
Abbildung 1: Schema einer prokaryotischen Zelle (A) und einer eukaryotischen Zelle (B)
(Fritsche, 2016)
Die Cytoplasmamembran ermöglicht die gezielte Aufnahme und Abgabe von Verbindun-
gen. Sie besteht aus einer etwa 8 nm dicken Phospholipid-Doppelschicht, in der Proteine
eingelagert sind. Phospholipide sind Ester aus Glycerin und zwei Fettsäuren, die sowohl
gesättigt, als auch ungesättigt sein können und 14 bis 18 Kohlenstoffatome aufweisen. Die
dritte OH-Gruppe des Glycerins ist mit Phosphorsäurederivaten verestert. Eingelagert in
die Membran sind spezielle Proteine, die am Stoffaustausch mit der Umgebung sowie dem
Aufbau von Zellenergie beteiligt sind. Ebenfalls in die Membran eingelagert können zusätz-
liche stabilisierende Verbindungen sein, wie beispielsweise Cholesterin in eukaryotischen
Membranen. Bei vielen Bakterien liegen ähnliche membranverstärkende Verbindungen
(Hopanoide) vor. Einige Bakterien besitzen zur Vergrößerung der Membranoberfläche Ein-
stülpungen der Cytoplasmamembran. Diese enthalten Enzymsysteme, welche für beson-
dere Energiestoffwechselwege notwendig sind. Auch einige intrazelluläre Einschlusskörper
werden von einfachen Membranen umgeben, wobei es sich hier häufiger um Proteinschich-
ten und nur selten um Lipidschichten handelt. (Fuchs et al., 2007)
Bakterienzellen bestehen zu 80 % aus Wasser, dem wichtigsten Reaktionspartner und die
zweitgrößte Fraktion an Zellbestandteilen bilden die Proteine (Cypionka, 2010). Die durch-
schnittliche makromolekulare Zusammensetzung einer Bakterienzelle geht aus Tabelle 1
hervor. Den größten Anteil neben den Proteinen macht die RNA aus. Die RNA-Menge ist
abhängig von der Stoffwechselaktivität, so weisen schnell wachsende Zellen bis zu 10 000
Ribosomen auf (Cypionka, 2010).

Einleitung
5
Die hauptsächlich in den Membranen vorkommenden Lipide machen etwa 9,1 % des Tro-
ckengewichts aus. Polysaccharide und Lipopolysaccharide (LPS) gemeinsam 8,4 % und
die DNA, in Form eines einzelnen Moleküls oder als Plasmide 3,1 %.
Tabelle 1: Relative Häufigkeit von Makromolekülen in einer Bakterienzelle (Madigan et al., 2015)
Makromolekül Prozent des Trockengewichts Beispiele für Zellbestandteile
Protein 55,0 % Membranproteine, Flagellen,
Ribosome, Enzyme
Lipid 9,1 % Membrane, Vesikel
Polysaccharid 5,0 % Zellwand, Energiespeicher
Lipopolysaccharid 3,4 % Membrane
DNA 3,1 % Kernäquivalent, Plasmid
RNA 20,5 % Ribosome
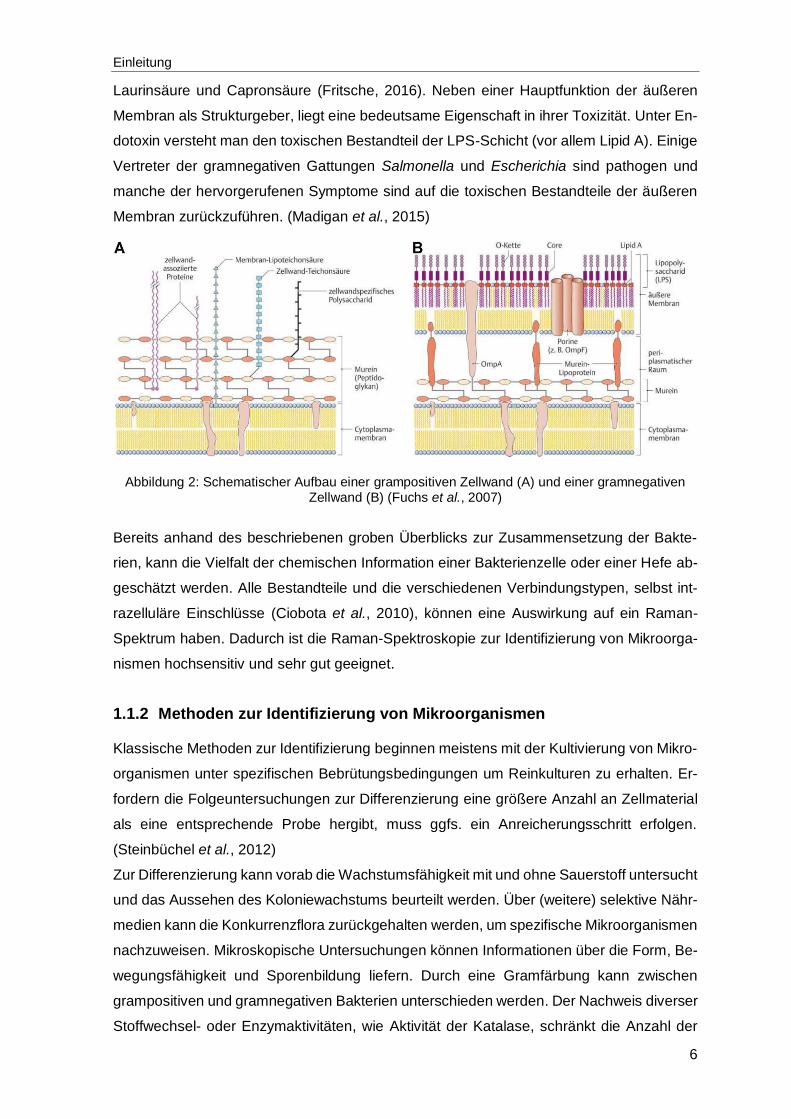
Ein sehr wichtiges Unterscheidungsmerkmal bei Bakterien beruht auf der Gramfärbung.
Abhängig von der Zellwandstruktur können über eine Färbungsreaktion Bakterien in gram-
positive und gramnegative eingeteilt werden (siehe Abbildung 2). Die gramnegative Zell-
wand besteht aus mindestens zwei Schichten und die grampositive Zellwand ist dicker und
besteht hauptsächlich aus einem einzigen Molekültyp. (Madigan et al., 2015)
Die Zellwände der Bakterien verfügen über eine starre Schicht aus Peptidoglykan, einem
Polysaccharid, das aus N-Acetylglucosamin, N-Acetylmuraminsäure und einigen wenigen
Aminosäuren besteht. Diese Bestandteile verbinden sich zu einer wiederholenden Struktur
(Glykantetrapeptid). Lange nebeneinanderliegende Peptidoglykanstränge umgeben die
Zelle als Schicht. Aminosäuren quervernetzen die Stränge. Glykosidische Bindungen, die
Zucker in den Glykansträngen verbinden, verleihen der Struktur nur in einer Richtung Fes-
tigkeit. Die Festigkeit in die andere Richtung bzw. die Quervernetzung entsteht bei den
gramnegativen durch Bildung einer Peptidbindung und bei grampositiven entsteht die Quer-
vernetzung durch kurze Interpeptidbrücken, mit je nach Spezies variierender Art und Anzahl
an Aminosäuren. (Madigan et al., 2015)
Die Zellwand grampositiver Bakterien (Abbildung 2 A) besteht zu 90 % aus Peptidoglykan.
In die Zellwand verankert sind Teichonsäuren. An Membranlipide gebundene Teichonsäu-
ren werden Lipoteichonsäuren genannt. Beide ragen nach außen hervor. Bei gramnegati-
ven Bakterien ist der Peptidoglykan-Anteil sehr viel geringer und eine zweite äußere Memb-
ran macht den größten Teil der Zellwand aus. Diese Membran besteht nicht nur aus einer
Lipiddoppelschicht und Proteinen, sondern beinhaltet auch Polysaccharide, die mit den Li-
piden zu einem Komplex verknüpft sind, weshalb die äußere Membran auch LPS-Schicht
genannt wird (siehe Abbildung 2 B). (Madigan et al., 2015)
Beim Lipid A sind die Fettsäuren im Gegensatz zu den Phospholipiden nicht an Glycin ge-
bunden, sondern an ein Disaccharid aus Glucosaminphosphat. Zu den Fettsäuren zählen
hier Palmitinsäure, Stearinsäure und auch kurzkettige Fettsäuren wie Myristinsäure,
Einleitung
6
Laurinsäure und Capronsäure (Fritsche, 2016). Neben einer Hauptfunktion der äußeren
Membran als Strukturgeber, liegt eine bedeutsame Eigenschaft in ihrer Toxizität. Unter En-
dotoxin versteht man den toxischen Bestandteil der LPS-Schicht (vor allem Lipid A). Einige
Vertreter der gramnegativen Gattungen Salmonella und Escherichia sind pathogen und
manche der hervorgerufenen Symptome sind auf die toxischen Bestandteile der äußeren
Membran zurückzuführen. (Madigan et al., 2015)
Abbildung 2: Schematischer Aufbau einer grampositiven Zellwand (A) und einer gramnegativen Zellwand (B) (Fuchs et al., 2007)
Bereits anhand des beschriebenen groben Überblicks zur Zusammensetzung der Bakte-
rien, kann die Vielfalt der chemischen Information einer Bakterienzelle oder einer Hefe ab-
geschätzt werden. Alle Bestandteile und die verschiedenen Verbindungstypen, selbst int-
razelluläre Einschlüsse (Ciobota et al., 2010), können eine Auswirkung auf ein Raman-
Spektrum haben. Dadurch ist die Raman-Spektroskopie zur Identifizierung von Mikroorga-
nismen hochsensitiv und sehr gut geeignet.
1.1.2 Methoden zur Identifizierung von Mikroorganismen
Klassische Methoden zur Identifizierung beginnen meistens mit der Kultivierung von Mikro-
organismen unter spezifischen Bebrütungsbedingungen um Reinkulturen zu erhalten. Er-
fordern die Folgeuntersuchungen zur Differenzierung eine größere Anzahl an Zellmaterial
als eine entsprechende Probe hergibt, muss ggfs. ein Anreicherungsschritt erfolgen.
(Steinbüchel et al., 2012)
Zur Differenzierung kann vorab die Wachstumsfähigkeit mit und ohne Sauerstoff untersucht
und das Aussehen des Koloniewachstums beurteilt werden. Über (weitere) selektive Nähr-
medien kann die Konkurrenzflora zurückgehalten werden, um spezifische Mikroorganismen
nachzuweisen. Mikroskopische Untersuchungen können Informationen über die Form, Be-
wegungsfähigkeit und Sporenbildung liefern. Durch eine Gramfärbung kann zwischen
grampositiven und gramnegativen Bakterien unterschieden werden. Der Nachweis diverser
Stoffwechsel- oder Enzymaktivitäten, wie Aktivität der Katalase, schränkt die Anzahl der

Einleitung
7
infrage kommenden Mikroorganismen, genauso wie Tests zum Nachweis spezifischer
Oberflächenantigene weiter ein. All diese Untersuchungen können insgesamt sehr zeitin-
tensiv sein und erfordern sowohl einen großen Platz- als auch Materialbedarf. Einige Sys-
teme wie „VITEK 2“ ermöglichen durch eine Vielzahl biochemischer Reaktionen die Identi-
fizierung mehrerer Proben, je nach Spezies innerhalb von 3 bis 17 Stunden (Ling et al.,
2003; Spanu et al., 2003).
Molekularbiologische Methoden zur Identifizierung von Mikroorganismen sind beispiels-
weise die PCR, mit der spezifische DNA-Abschnitte aus zuvor isolierter DNA vervielfältigt
und dann mittels Gelelektrophorese nachgewiesen werden können. Einige Nachteile hier-
bei liegen darin, dass gezielt mit entsprechenden Primern nach einer spezifischen Sequenz
gesucht werden muss und relativ viele Labor-Schritte notwendig sind. (Cypionka, 2010)
Beachtet werden sollte, dass es modernere PCR-Varianten wie die Echtzeit-PCR und die
Multiplex-PCR gibt. Bei der Echtzeit-PCR wird der Abschluss jedes DNA-Vermehrungszyk-
lus durch einen Fluoreszenzfarbstoff angezeigt, was Rückschlüsse über den Faktor der
DNA-Vervielfältigung zulässt. Bei der Multiplex-PCR werden mehrere Primerpaare einge-
setzt. (Fritsche, 2016)
Große Fortschritte wurden auch bei der DNA-Sequenzierung gemacht. Seit einigen Jahren
können ganze Genome oder spezifische DNA-Abschnitte vergleichsweise schnell und ein-
fach sequenziert werden, wobei die Kosten mit jedem Jahr um den Faktor 2 bis 3 sinken
(Pettersson et al., 2009). Mit „Next Generation Sequencing“ (NGS) sind Erregernachweise
in Proben möglich, bestimmte Einschränkungen der PCR können überwunden werden und
bisher nicht identifizierte noch unbekannte Organismen, können näher erforscht werden.
(Andrusch et al., 2018)

Einleitung
8
1.1.3 Raman-Spektroskopie
Die meisten spektroskopischen Verfahren beruhen auf den Wechselwirkungen zwischen
elektromagnetischer Strahlung mit chemischen Verbindungen. Der Physiker C. V. Raman
entdeckte 1928, dass sich die Wellenlänge eines geringen Teils, der von bestimmten Mo-
lekülen ausgehenden Streustrahlung von der des einfallenden Strahls unterscheidet und,
dass die Verschiebungen der Wellenlänge von der Struktur der streuenden Moleküle ab-
hängig sind (Skoog et al., 2013).
Wird die Lösung einer Substanz mit monochromatischem Licht bestrahlt, so tritt ein Teil der
Strahlung ungehindert hindurch, ein geringer Teil des Lichts wird in alle Raumrichtungen
gestreut und ein noch geringerer Teil tritt ebenfalls als Streustrahlung aus, besitzt jedoch
eine andere Frequenz als das eingestrahlte Licht. Diese Frequenzverschiebung entsteht
durch Absorption und Reemission in Verbindung mit Schwingungsanregung oder Schwin-
gungslöschung. Diese Streustrahlung kann spektral zerlegt und mittels eines photoelektri-
schen Detektors registriert werden. Die Differenz zwischen der Frequenz der eingestrahlten
Linie und einer Raman-Linie ist die Frequenz der dazugehörigen Schwingung. (Hesse et
al., 2008)
Das Raman-Spektrum ist ein Emissions-Spektrum. Die Frequenzen der Raman-Linien oder
-Banden können größer oder kleiner sein als die Anregungsfrequenz (Rayleigh-Linie). Cha-
rakteristisch für ein Molekül sind die Differenzen der Raman-Frequenzen von der Anre-
gungsfrequenz. Sie sind unabhängig von der Anregungsfrequenz und können auch im IR-
Spektrum wiedergefunden werden. (Hesse et al., 2008)
Die Differenz der Wellenlängen der einfallenden und gestreuten Strahlung (Raman-Ver-
schiebung; engl. Raman shift) entspricht den Wellenlängen im mittleren Infrarotbereich. Das
Raman-Streuspektrum und das IR-Absorptionsspektrum einer Verbindung ähneln sich häu-
fig sehr. Dennoch gibt es ausreichend Unterschiede zwischen speziell IR- und Raman-ak-
tiven Gruppen, sodass sich beide Verfahren ergänzen. Vorteile der Raman-Spektroskopie
gegenüber der IR-Spektroskopie sind, dass Wasser keinen starken störenden Einfluss auf
die Messung hat und die Probenvorbereitung gewöhnlich sehr viel einfacher ist.
(Skoog et al., 2013)
1.1.3.1 Theorie des Raman-Effekts
Der Raman-Effekt unterscheidet sich von der Absorption und Emission elektromagneti-
scher Strahlung von Materie. Vereinfacht kann der Raman-Effekt als Stoßprozess zwischen
Molekülen der Materie und den Photonen der elektromagnetischen Strahlung erklärt wer-
den. Bei der Rayleigh-Streuung haben das eingestrahlte Licht und das gestreute Licht die
gleiche Frequenz (elastischer Stoß). Die Energiezustände von Molekül und Photon haben
sich durch den Stoß nicht geändert; die Energie des Streulichts entspricht der Energie des

Einleitung
9
eingestrahlten Lichts (siehe Abbildung 3). Die Intensität der Rayleigh-Streuung beträgt ca.
ein Prozent der Ausgangsintensität der elektromagnetischen Welle. (Lechner, 2016)
Bei Stokes’schen Linien hat die gestreute elektromagnetische Welle eine kleinere Frequenz
als die eingestrahlte Welle (Rotverschiebung). Beim Stoß überträgt das Photon Energie auf
das Molekül und wird selbst um diesen Energiebetrag ärmer (inelastischer Stoß). Das
Streulicht ist energieärmer als das eingestrahlte Licht (siehe Abbildung 3). Ebenfalls dem
inelastischen Stoß entsprechend hat die gestreute elektromagnetische Welle bei den Anti-
stokes’schen Linien eine größere Frequenz als die eingestrahlte elektromagnetische Welle
(Blauverschiebung). Das sich im angeregten Zustand befindende Molekül überträgt beim
Stoß Energie auf das Photon, was bedeutet, dass das Streulicht energiereicher wird (siehe
Abbildung 3). (Lechner, 2016)
Abbildung 3: Schematische Darstellung der Energieniveaus bei der Rayleigh-, Stokes- und Anti-Stokes-Streuung links und fiktives Raman-Spektrum rechts (modifiziert nach (Kim et al., 2017))
Die Wellenzahl der Rayleigh-Linie aus Abbildung 3 rechts entspricht der Wellenzahl der
Anregungsstrahlung. Somit beträgt die Wellenzahl der Rayleigh-Linie bei einer Anregungs-
wellenlänge von 633 nm 15798 cm-1. Für eine einfachere Vergleichbarkeit von Raman-
Spektren mit IR-Spektren wird der Wert der Rayleigh-Linie auf null gesetzt, wobei der Be-
trag der Raman-Verschiebung für alle Anregungswellenlängen gleich ist (Salzer et al.,
2017). Die Verteilung der Stokes- und Anti-Stokes-Linien ist beidseitig der Rayleigh-Linie
symmetrisch, jedoch sind die Stokes-Linien im Vergleich zu den Anti-Stokes-Linien intensi-
ver, weshalb in der Raman-Spektroskopie häufig nur die Stokes-Seite eines Spektrums ge-
messen und ohne negatives Vorzeichen dargestellt wird. (Teixeira-Dias, 2017)
Einleitung
10
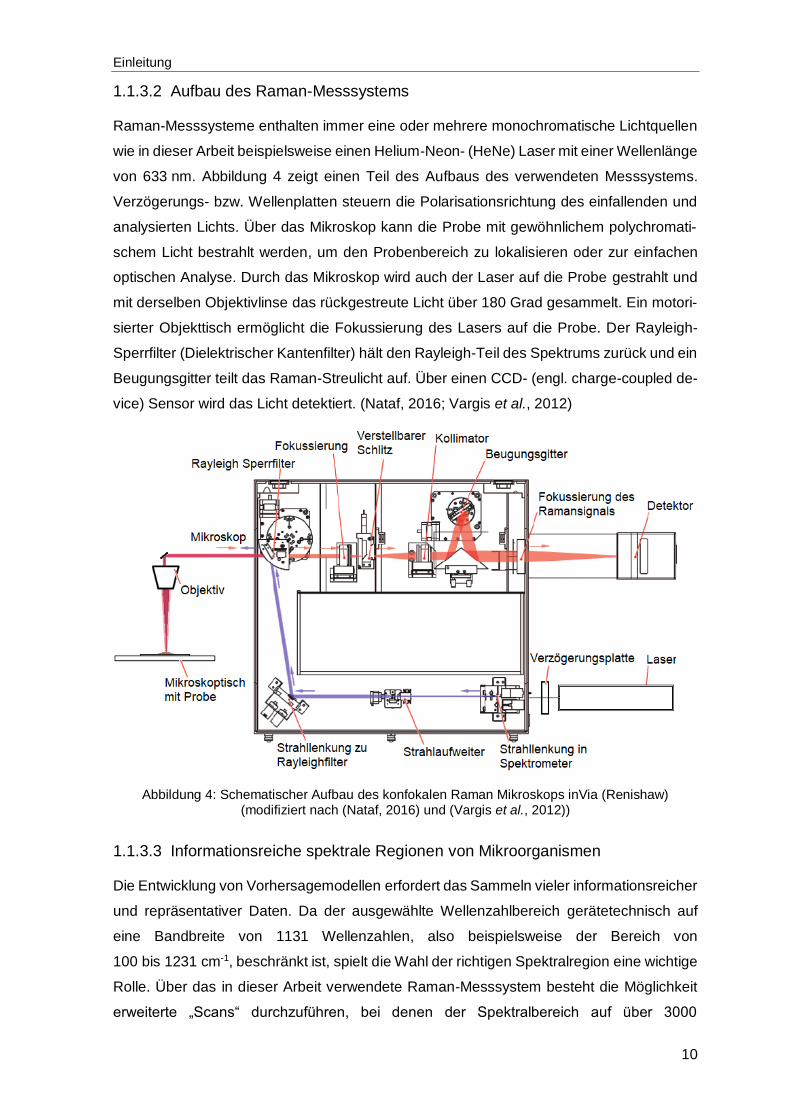
1.1.3.2 Aufbau des Raman-Messsystems
Raman-Messsysteme enthalten immer eine oder mehrere monochromatische Lichtquellen
wie in dieser Arbeit beispielsweise einen Helium-Neon- (HeNe) Laser mit einer Wellenlänge
von 633 nm. Abbildung 4 zeigt einen Teil des Aufbaus des verwendeten Messsystems.
Verzögerungs- bzw. Wellenplatten steuern die Polarisationsrichtung des einfallenden und
analysierten Lichts. Über das Mikroskop kann die Probe mit gewöhnlichem polychromati-
schem Licht bestrahlt werden, um den Probenbereich zu lokalisieren oder zur einfachen
optischen Analyse. Durch das Mikroskop wird auch der Laser auf die Probe gestrahlt und
mit derselben Objektivlinse das rückgestreute Licht über 180 Grad gesammelt. Ein motori-
sierter Objekttisch ermöglicht die Fokussierung des Lasers auf die Probe. Der Rayleigh-
Sperrfilter (Dielektrischer Kantenfilter) hält den Rayleigh-Teil des Spektrums zurück und ein
Beugungsgitter teilt das Raman-Streulicht auf. Über einen CCD- (engl. charge-coupled de-
vice) Sensor wird das Licht detektiert. (Nataf, 2016; Vargis et al., 2012)
Abbildung 4: Schematischer Aufbau des konfokalen Raman Mikroskops inVia (Renishaw) (modifiziert nach (Nataf, 2016) und (Vargis et al., 2012))
1.1.3.3 Informationsreiche spektrale Regionen von Mikroorganismen
Die Entwicklung von Vorhersagemodellen erfordert das Sammeln vieler informationsreicher
und repräsentativer Daten. Da der ausgewählte Wellenzahlbereich gerätetechnisch auf
eine Bandbreite von 1131 Wellenzahlen, also beispielsweise der Bereich von
100 bis 1231 cm-1, beschränkt ist, spielt die Wahl der richtigen Spektralregion eine wichtige
Rolle. Über das in dieser Arbeit verwendete Raman-Messsystem besteht die Möglichkeit
erweiterte „Scans“ durchzuführen, bei denen der Spektralbereich auf über 3000

Einleitung
11
Wellenzahlen vergrößert werden kann, allerdings auf Kosten der spektralen Auflösung und
der Messdauer. Der erweiterte Scan ist jedoch eine gute Möglichkeit Proben vorab grob zu
untersuchen, um informationsreiche Regionen ausfindig zu machen. Das Spektrum eines
Vorversuchs aus Abbildung 5 zeigt ein erweitertes Raman-Spektrum von Saccharomyces
cerevisiae. Die Banden im Bereich von 600 bis 1800 cm-1 (hervorgehoben in Abbildung 5
B) sind im Verhältnis zur Bande zwischen 2840 bis 3000 cm-1 schwächer ausgeprägt aber
liegen in der sogenannten „fingerprint“-Region, welche zur Identifizierung von Mikroorga-
nismen besonders gut geeignet ist (Sato & Martinho, 2018; Teng et al., 2016). Die Region
zwischen 1750 cm-1 und 2800 cm-1 besitzt keinerlei sich vom Grundrauschen abhebenden
Signaturen.
Abbildung 5: Beispiel eines mit erweitertem Wellenzahlbereich erfassten Raman-Spektrums von
Saccharomyces cerevisiae (A) und hervorgehobene informationsreiche Region (B)
Wie in Kapitel 1.1.1 beschrieben, bilden Lipide die äußere Membran der Mikroorganismen
und intrazelluläre Begrenzungen. Lipide verursachen mehrere Banden im Bereich zwischen
2800 und 3000 cm-1. Zurückzuführen sind diese auf asymmetrische Streckschwingungen
(siehe Abbildung 6 A) von CH3- und CH2-Gruppen der Fettsäuren in den Phospholipiden
(J. Kneipp, 2001).
Die Energie der asymmetrischen Schwingungen der Moleküle besitzen eine höhere Ener-
gie als die zugehörigen symmetrischen Schwingungen (siehe Abbildung 6 B). Deformati-
onsschwingungen (siehe Abbildung 6 C) von CH3 und CH2 befinden sich im Bereich von
1450 cm-1 und 1470 cm-1. Große Moleküle wie Proteine besitzen ein komplexes Schwin-
gungsspektrum und weisen daher eine Vielzahl von Banden auf. Beispielsweise die haupt-
sächlich durch C=O-Streckschwingungen der sekundären Amidstrukturen im Peptidrück-
grat hervorgerufene Amid I-Bande. Nukleinsäuren in Form von DNA und RNA weisen auf-
grund ihres Zucker-Phosphat-Rückgrats P=O-Schwingungen auf, welche in Banden im

Einleitung
12
Bereich zwischen 900 cm-1 und 1250 cm-1 resultieren. C=O-, C=N-, oder C=C-Schwingun-
gen der einzelnen Basen der DNA und RNA, liegen im Bereich von 1600 cm-1 bis
1700 cm-1. Diese Banden werden jedoch häufig von den ebenfalls in dieser Region liegen-
den Amid I-Banden der anteilmäßig stärker vorhandenen Proteine (siehe Tabelle 1) über-
lagert. Die weniger Energie besitzenden Ringschwingungen der Nukleinsäuren befinden
sich zwischen 1300 cm-1 und 1500 cm-1. Der Spektralbereich zwischen 600 und 900 cm-1
enthält äußerst spezifische spektrale Informationen, welche sich besonders aus den inten-
sitätsschwachen Ringschwingungen verschiedener Aromaten zusammensetzt (Tyrosin
(Tyr), Tryptophan (Trp), Nukleotide). (J. Kneipp, 2001)
Abbildung 6: Normalschwingungen eines dreiatomigen Moleküls (z.B. Wasser)
(modifiziert nach (Hasegawa, 2017))
Abbildung 7 zeigt ein geglättetes, ansonsten unverändertes Spektrum von E. coli K12 und
die bereits beschriebene Zuordnung einiger Banden zu bestimmten Biomolekülen und typi-
schen chemischen Verbindungen. Besonders auffällig ist der scharfe, intensive Peak bei
1003 cm-1, der durch Deformationsschwingungen von Kohlenstoffring-Verbindungen des
Phenylalanins (Phe) entsteht.
A= Adenin; C= Cytosin; G= Guanin; U= Uracil;
Def.= Deformationsschwingungen; Str.= Streckschwingungen; glyk.= glykosidische (Verbindung)
Abbildung 7: Raman-Spektrum von E. coli K12 mit Zuordnung des biochemischen Ursprungs der
Banden nach (De Gelder, 2008; Maquelin et al., 2002)

Einleitung
13
Aus Tabelle 2 gehen detaillierte Informationen zu den Banden und des biochemischen Ur-
sprungs innerhalb eines typischen Raman-Spektrums von Mikroorganismen hervor.
Tabelle 2: Banden im Raman-Spektrum und Zuordnung des biochemischen Ursprungs
nach (De Gelder et al., 2007; S. Kumar et al., 2016; Munchberg et al., 2014; Sil et al., 2017; Teng et al., 2016)
Bande [cm-1] Biochemischer Ursprung, Verbindungen, Schwingungstypen
622 Phenylalanin
637 Tyrosin
724 Adenin
783 Thymin, Cytosin, Uracil, DNA (PO2 bei DNA; Str.)
829 Protein (Fermi-Dublett)
853
895 CN, CON; Str./ CCH; Def.
960 C=C; Def.
1003 Phenylalanin (CC Ring; Def.)
1034 Phenylalanin
1087 Kohlenhydrate (C-C, C-O, -C-OH)
1128 Proteine (C-N, C-C; Str.)
1157 Amid III
1173 Phospholipide
1208 Phenylalanin, Tyrosin
1236 Amid III
1336 Tryptophan
1451 Lipide, Aminosäureseitenketten (CH2; Def.)
1557 Tryptophan (Indolring)
1582 Nukleinsäuren
1605 Tyrosin, Phenylalanin, Tryptophan
1667 Amid I (C=O; Str.)
2820-3050 Diverse Kohlenwasserstoffe (CH2, CH3, -C-H, =C-H; Str.)
(Str.= Streckschwingung, Def.= Deformationsschwingung)
1.1.3.4 Einflüsse auf die Qualität eines Raman-Spektrums
1.1.3.4.1 Störung durch Fluoreszenz
Zur Fluoreszenz kommt es, wenn ein angeregtes Molekül wieder in den Grundzustand zu-
rückfällt und dabei Strahlung bei einer Frequenz aussendet, die dem Übergang zwischen
dem angeregten und dem Grundzustand entspricht (Banwell & MacCash, 1999). Eigenflu-
oreszenz, verursacht von der biologischen Probe selbst oder von Verunreinigungen, kann
deutlich stärker als das eigentliche Raman-Signal sein, weshalb Fluoreszenz als „Offset“
auftritt, welcher das eigentliche Raman-Signal überdecken kann (Leon-Bejarano et al.,
2016).
Verschiedene Mikroorganismen verursachen unterschiedlich starke Fluoreszenzstörungen.
Pseudomonaden produzieren beispielsweise Siderophore wie Pyoverdine. Diese fluores-
zierenden Verbindungen, die vom Bakterium ausgeschieden werden, um das gering lösli-
che Eisen zu binden und aufzunehmen, sorgen beispielsweise bei P. fluorescens für starke

Einleitung
14
Fluoreszenzstörungen im Raman-Spektrum. Auch andere Bakterien produzieren Stoffe,
welche bei der „konventionellen“ Raman-Spektroskopie zu Problemen mit Fluoreszenz füh-
ren wie beispielsweise das Enterobactin von E. coli oder Salmonella Typhimurium.
(Kahlon, 2016)
Abbildung 8 zeigt exemplarisch stark durch Fluoreszenz beeinträchtigte Raman-Spektren
von P. fluorescens und B. subtilis. Die Art der Störung ist nicht identisch, sondern wirkt sich
bei B. subtilis vor allem als gleichmäßig abfallender Offset aus und bei P. fluorescens als
einen starken „Fluoreszenzhügel“ zwischen 600 und 900 cm-1 und einem Anstieg ab
1200 cm-1.
Abbildung 8: Raman-Spektren zweier Bakterien mit starken Fluoreszenzstörungen
Für die Fluoreszenzbeseitigung gibt es sowohl instrumentelle als auch mathematische An-
sätze (Leon-Bejarano et al., 2016). Wirkt sich die Fluoreszenz hauptsächlich auf die Basis-
linie eines Spektrums aus und hat nur einen geringen Einfluss auf die Banden selbst, so
können die Auswirkungen störender Fluoreszenz durch einfache mathematische Verfahren
beseitigt werden (siehe Methodik in Kapitel 2.2.5.2).
Die instrumentellen Ansätze umfassen beispielweise die Anpassung der Anregungswellen-
länge in den Nahinfraroten (NIR) Bereich, wie durch die Verwendung eines Neodym-dotier-
ten Yttrium-Aluminium-Granat- (Nd:YAG) Lasers mit einer üblichen Wellenlänge von
1064 nm. Aufgrund der geringen Quantenausbeute von Detektoren für den NIR-Spektral-
bereich und ihres hohen thermischen Rauschens, können jedoch keine kurzen Messzeiten
realisiert werden, weshalb eher im sichtbaren oder im ultravioletten (UV) Bereich angeregt
wird. Da UV-Strahlung für die meisten biologischen Materialien schädlich ist, hat sich die
Anregung im roten bzw. der Übergang in den NIR-Spektralbereich (785 nm) bewährt. Bei
785 nm Anregungswellenlänge ist der Fluoreszenzhintergrund niedrig und die

Einleitung
15
Quantenausbeute von CCD-Detektoren auf Siliziumbasis akzeptabel, weshalb sie sich als
geeignet für die Analyse biologischer Proben erwiesen hat. (Gebrekidan et al., 2016)
Bereits in zahlreichen wissenschaftlichen Veröffentlichungen wurde die oberflächenver-
stärkte Raman-Spektroskopie (SERS) (engl. Surface-enhanced Raman Spectroscopy) als
Methode zur Differenzierung von Bakterien angewandt. Bei der SERS wird die Raman-
Streuung von Molekülen in unmittelbarer Nähe einer nanostrukturierten metallischen Ober-
fläche enorm verstärkt. Die meisten SERS-Studien mit Bakterien wurden entweder mit Sil-
berkolloiden oder mit nanostrukturierten Silberoberflächen durchgeführt. (Sivanesan et al.,
2014)
Das verstärkte Signal wird durch die Kombination von elektromagnetischer und chemischer
Verstärkung erklärt. Die elektromagnetische Verstärkung ergibt sich aus der Resonanz des
angelegten Feldes mit Oberflächenplasmonen der metallischen Nanostrukturen. Die che-
mische Verstärkung bezieht sich auf den Ladungstransfer zwischen einem Substrat und
einem adsorbierten Molekül. Die elektromagnetische Verstärkung kann theoretisch Fakto-
ren von 103 bis 1011 erreichen, während für den chemischen Verstärkungsfaktor bis zu 103
berechnet wurde. Die enorme Steigerung der Raman-Streuung kann die Beobachtung ein-
zelner Moleküle ermöglichen, hat den Effekt den Fluoreszenzhintergrund zu beseitigen und
das Signal-Rausch-Verhältnis zu verbessern. (Witkowska et al., 2017)
Weitere instrumentelle Möglichkeiten sind beispielsweise die „Shifted-excitation Raman Dif-
ference Spectroscopy“ SERDS bei der mit zwei Lasern leicht unterschiedlicher Wellenlänge
zwei Spektren entstehen, von denen ein Differenzspektrum ohne Fluoreszenzstörungen
berechnet werden kann (Gebrekidan et al., 2016) und „Time-Gated Raman Spectroscopy“
bei der zeitgesteuerte Vorrichtungen kurze, intensive Laserimpulse abgeben und die Pro-
benantwort gleichzeitig mit den Impulsen aufgezeichnet wird. Die Ankunftszeit und die
Energie jedes Photons werden detektiert und durch Unterschiede in der Fluoreszenzle-
bensdauer und der „Lebensdauer“ des Raman-Signals, kann der Fluoreszenzhintergrund
eliminiert werden. (Lipiainen et al., 2018)
Da in der Praxis häufig nicht ohne Weiteres mit der Anregungswellenlänge variiert oder das
Verfahren grundlegend verändert werden kann, können auch einfache Waschschritte mit
sterilem deionisiertem Wasser (Kusic et al., 2015; Sengupta et al., 2006) oder Kochsalzlö-
sung zu einer Verringerung der Fluoreszenz führen, insbesondere wenn die Fluoreszenz
durch Pyoverdine ausgelöst wird. (Willemse-Erix et al., 2010)
Ein weiterer simpler Ansatz ist das Ausbrennen von Fluoreszenz durch Laserlicht, im eng-
lischen auch unter „Photobleaching“ bekannt. Beim „Photobleaching“ wird die Probe mit
dem Laser für einen bestimmten Zeitraum intensiv vorbestrahlt. Dies führt zu einem aus-
bleichen von Fluorophoren und somit zur Reduzierung des Fluoreszenzhintergrunds.
(Golcuk et al., 2006)

Einleitung
16
Das Ausbrennen der Fluoreszenz, hat allerdings den Nachteil, dass Proben verändert bzw.
beschädigt werden können. Pigmente, Farbstoffe und andere photochemisch empfindli-
chen Verbindungen können beispielsweise durch Anregung im sichtbaren Bereich zerstört
werden. (Schrader et al., 2000)
Veränderungen von Bakterienzellen durch elektromagnetische Strahlung haben Auswir-
kungen auf das Raman-Spektrum, welche eine Differenzierung verschiedener Mikroorga-
nismen und eine Standardisierung der Methode erschweren könnten.
1.1.3.4.2 Störung durch kosmische Strahlung und Umgebungslicht
Ein bedeutsamer Störfaktor in der Raman-Spektroskopie, sind gelegentlich auftretende un-
spezifische Peaks hoher Intensität. Diese sehr schmalen Peaks („Cosmic spikes“) entste-
hen durch kosmische Strahlung und werden von den empfindlichen CCD-Detektoren regis-
triert. (X. Zhang et al., 2017)
Abbildung 9 zeigt das mehrfache Auftreten solcher Störungen bei den Spektren zweier Mik-
roorganismen. Aufgrund ihres möglichen Einflusses auf multivariate Vorhersagemodelle,
dürfen solche Spektren nicht zur Modellentwicklung verwendet werden oder die Störungen
müssen zuvor entfernt werden. Besonders problematisch sind „Cosmic spikes“, wenn diese
sehr hohe Intensitäten aufweisen, wie beispielsweise der Peak bei ca. 960 cm-1 von B. sub-
tilis aus Abbildung 9. „Spikes“ die eine größere Bandbreite haben, als Banden mit spezifi-
scher chemometrischer Bedeutung oder „Spikes“, die direkt auf informationsreichen Regi-
onen auftreten, sind ebenfalls sehr problematisch. (L. Zhang & Henson, 2007)
Abbildung 9: Auftreten von „Cosmic spikes“ in Raman-Spektren markiert durch Pfeile

Einleitung
17
Zwar gibt es verschiedene Verfahren, um diese Störungen zu eliminieren, was vor allem in
der In-Prozess-Analytik eine wichtige Rolle spielt, jedoch werden bei Auftreten von „Cosmic
spikes“ bei den in dieser Arbeit gesammelten Spektren, keine Verfahren zur Eliminierung
angewandt, sondern betroffene Spektren verworfen.
Umgebungslicht hat insbesondere bei Verwendung von sehr schwachen Laserleistungen
einen Einfluss auf das aufgenommene Raman-Spektrum. Bestimmte Glühbirnen oder
Leuchtstoffröhren können sogar Peaks im Raman-Spektrum erzeugen, die leicht fehlinter-
pretiert werden könnten. Aus diesem Grund sollten Raman-Spektren immer in einer abge-
dunkelten Umgebung aufgezeichnet werden.
1.1.3.4.3 Einfluss der Probenpräparation
Das in dieser Arbeit vorliegende Mikro-Raman-Messsystem liefert eine Technik zur Analyse
von Oberflächen. Dennoch haben die Schichtdicken der Proben eine Auswirkung auf das
Messergebnis. Der Einfluss der Dicke des Probenfilms legt nahe, dass eine ideale Proben-
präparation gefunden werden muss, um möglichst deutliche Signale bei allen Proben zu
erhalten. Das Problem einer solchen standardisierten Probenschicht liegt darin, dass eine
Methode entwickelt werden soll, die für eine Vielzahl von Mikroorganismen geeignet ist.
Jedoch unterscheiden sich einige der hier analysierten Mikroorganismen so stark vonei-
nander, dass sich eine exakt definierte Probenpräparation, die für alle Probentypen geeig-
net ist, als schwierig erweisen könnte. Deshalb sollen vielmehr durch eine robuste Kalibrie-
rung mögliche Einflüsse durch kleinere Unterschiede in der Probenpräparation abgedeckt
werden. Im Ergebnisteil dieser Arbeit sollen auch die Auswirkungen der Probenpräparation
untersucht werden.
Neben der Schichtdicke spielt auch der Trocknungsgrad einer Probe eine Rolle. Allgemein
gilt in der Schwingungsspektroskopie, dass bei flüssigen Proben breitere, weniger scharfe
Peaks auftreten. Gase hingegen zeigen scharfe Banden, da dort die Moleküle mit einem
charakteristischem Drehimpuls frei rotieren können. Bei festen Proben treten im Vergleich
zu flüssigen Proben zusätzliche Banden aufgrund von Torsionsschwingungen auf. (Larkin,
2011)
Durch eine standardisierte Trocknungsdauer kann der Einfluss von Restfeuchte auf ein
Spektrum jedoch nahezu ausgeschlossen werden.
Kein oder nur ein sehr geringer Einfluss, könnte durch den verwendeten Glasobjektträger
entstehen (siehe Abbildung 10 A), wohingegen Rückstände des Nährmediums aufgrund
der darin enthaltenen Zucker, Proteine, Peptide und Aminosäuren durchaus einen Einfluss
haben. So zeigen sich bei einem Raman-Spektrum von getrocknetem Nährboden diverse
Peaks in informationsreichen Spektralgegenden (siehe Abbildung 10 B).

Einleitung
18
Abbildung 10: Raman-Spektrum eines Glasobjektträgers (A) und Raman-Spektrum von getrockne-tem Nährboden (TSA) (B)
1.1.4 Datenvorbehandlung und Vorhersagemodelle
Jede Datenvorbehandlung bedeutet zunächst einen Informationsverlust. Jedoch gibt es oft
Information in einem Spektrum, die für eine Klassifikation unwichtig oder sogar störend sein
können. So kann die Intensität eines Spektrums beispielsweise viel über die Leistung des
Lasers und die Anregbarkeit des Probenmaterials aussagen. Rauschen kann Rückschlüsse
auf die Sensitivität des Detektors erlauben und selbst Basislinienverschiebungen können
charakteristisch für bestimmte Umgebungsparameter sein. Doch die Art dieser Informatio-
nen sind für die Vorhersage eines unbekannten Mikroorganismus störend. Für die Entwick-
lung von Vorhersagemodellen gibt es deshalb diverse mathematische Verfahren, um die
für eine bestimmte Aufgabenstellung maximale Information aus einem Spektrum zu gewin-
nen. Viel wichtiger ist allerdings vorab die richtige Spektren-Aufnahme zu gewährleisten.
Dazu gehört beispielsweise die Wahl des für die Aufgabe geeigneten Wellenzahlbereichs,
die Standardisierung des Versuchsaufbaus oder des Prozesses und die optimalen Geräte-
Messeinstellungen. Unter Umständen kann mit einem optimierten Prozess die Datenvorbe-
handlung auf ein Minimum reduziert werden, dennoch ist sie speziell bei der Problemstel-
lung dieser Arbeit von großer Wichtigkeit. Die zuvor beschrieben Einflüsse auf ein Raman-
Spektrum, speziell Störungen durch Fluoreszenz aber auch mögliche Intensitätsunter-
schiede in Spektren von Mikroorganismen gleicher Art, Rauschen und Multikollinearität von
spektralen Daten, erfordern eine Datenvorbehandlung. Möglichkeiten zur Datenvorbehand-
lung und die Art der nötigen Verfahren werden in Kapitel 2.2.5 beschrieben.
Raman-Spektren bestehen je nach Messsystem aus vielen einzelnen Messpunkten (Vari-
ablen). Das erste Ziel der multivariaten Datenanalyse ist die Datenreduktion. Variablen mit
gleicher Information werden in sogenannte Hauptkomponenten zusammengefasst. Jedes
Objekt kann dann anstatt durch viele einzelne Variablen mit wenigen Hauptkomponenten
beschrieben werden. Ein weiterer Effekt der multivariaten Analyse ist, dass beim Berechnen
der Hauptkomponenten Variablen mit Information von Variablen ohne Information getrennt

Einleitung
19
werden. Dadurch wird auch gewährleistet, dass Rauschen nicht als Signal fehlinterpretiert
wird. Wurde die relevante Information aus einer großen Datenmenge herausgefunden,
kann daraus ein Modell erstellt werden, wie beispielsweise ein Klassifizierungsmodell zur
Vorhersage eines bestimmten Mikroorganismus aus neuen unbekannten Daten.
(Kessler, 2007)
Es gibt diverse Methoden zur Klassifizierung wie die lineare Diskriminanzanalyse (LDA)
oder die k-Nächste-Nachbarn-Klassifikation (kNN) bis hin zu komplexeren Methoden wie
„Support Vector Maschinen“ (SVM) oder künstliche neuronale Netze. Bei der Entwicklung
eines Modells sollte immer berücksichtigt werden, dass nicht nur die Vorhersageleistung
hoch ist, sondern dass die Modelle auch robust gegenüber unbekannten Daten und nicht
zu komplex sind, sodass sie für den Anwender oder zumindest den Entwickler interpretier-
bar bleiben. Um ein bestmögliches Modell zu finden, werden in dieser Arbeit drei verschie-
dene Klassifizierungsverfahren gegenübergestellt und deren Leistung verglichen.
1.1.5 Einblick in Ergebnisse bisheriger Studien
Bakterien konnten neben der Raman-Spektroskopie bereits erfolgreich über FT-IR-Spekt-
roskopie identifiziert werden (Helm et al., 1991; Ngo-Thi et al., 2003). Die Probenvorberei-
tung ist allerdings aufwendiger, als bei der Raman-Spektroskopie und durch konfokale
Mikro-Raman-Spektroskopie können bei der Analyse von Mikroorganismen anstelle meh-
rerer hundert Zellen, wie sie für die IR-Spektroskopie nötig sind, sogar einzelne Bakterien-
zellen innerhalb kurzer Zeit analysiert werden (Rösch et al., 2005).
2004 haben Lopez-Diez et al. über Tief-UV-Resonanz-Raman-Spektroskopie (engl. deep
UV resonance Raman spectroscopy, DUVRRS) verschiedene endosporenbildende Bakte-
rien der Gattungen Bacillus und Brevibacillus analysiert. Bei der Anregung im tiefen UV-
Bereich bei 244 nm findet ein Resonanzeffekt statt, welcher die Raman-Banden verstärkt.
Bei Anregung unter 260 nm Wellenlänge, werden die Messungen auch nicht von Fluores-
zenz gestört. Über Hauptkomponentenanalyse (Principal Component Analysis, PCA), Dis-
kriminanzanalyse und Hierarchische Clusteranalyse (HCA) wurde bestätigt, dass die
DUVRRS zur Differenzierung eng verwandter Endosporenbildner verwendet werden kann.
(Lopez-Diez & Goodacre, 2004)
Von wissenschaftlichen Untersuchungen zur Identifizierung von Mikroorganismen bei einer
Anregungswellenlänge von 532 nm wurde ebenfalls mehrfach berichtet, jedoch handelt es
sich dort meist um Analysen einzelner Bakterienzellen (Harz et al., 2005; Strola et al., 2014).
Raman-Spektren von einzelnen Bakterienzellen zeigen bei Anregung mit 532 nm im Ver-
gleich zu Spektren von Bakterienfilmen weniger Fluoreszenzstörung und zusätzliche Sig-
nale (Harz et al., 2005).

Einleitung
20
Möglichkeiten das Problem auftretender Fluoreszenz zu beseitigen und das schwache Ra-
man-Signal zu verstärken, wurden bereits in Kapitel 1.1.3.4.1 beschrieben. So wurde bei-
spielsweise SERS auch erfolgreich mit Goldpartikeln eingesetzt, um einzelne Bakterienzel-
len bei Anregung im NIR-Bereich zu analysieren. Das dadurch verstärkte Raman-Signal
ermöglicht im Vergleich zur herkömmlichen Raman-Messung in sehr kurzer Zeit umfang-
reiche Informationen über die Zelle und kleine chemische Veränderungen wie z.B. Ände-
rungen in den DNA-Banden zu registrieren. (K. Kneipp et al., 2002)
Eine deutliche Differenzierung von Bakterienarten und -stämmen mittels Raman-Spektro-
skopie, wurde bei Verwendung von nicht überwachten Klassifikationsverfahren wie der
Clusteranalyse nur erzielt, wenn die Kultivierungsbedingungen der Bakterien standardisiert
wurden, da verschiedene Wachstumsbedingungen einen starken Einfluss auf das Raman-
Spektrum haben. (Harz et al., 2005; Hutsebaut et al., 2004)
Baritaux et al. berichten über Raman-spektroskopische-Methoden zur Identifizierung von
Bakterien in Umweltproben. Es wurde eine Datenbank mit Raman-Spektren von Bakterien
eingerichtet, die verschiedenen Umgebungsbedingungen ausgesetzt waren. Dieser Daten-
satz wurde dazu verwendet die Möglichkeit einer Klassifizierung über eine SVM bei Mes-
sungen unter nicht idealen Bedingungen zu prüfen. Für Umweltproben sind Modelle mit
breiter Abdeckung wünschenswert, da dort die genauen Bedingungen der Bakterien nicht
kontrolliert werden können. (Baritaux et al., 2016)
Forschung mit direktem Bezug auf die Lebensmittelindustrie wurde unter anderem für die
Detektion der Bakteriengattung Brucella in Milch betrieben. Die Ergebnisse weisen darauf
hin, dass Mikro-Raman-Spektroskopie in Verbindung mit SVM eine vielversprechende Al-
ternative zur Identifizierung von Brucella spp. darstellen könnte, sowohl auf Agar-Platten,
als auch direkt in der Milch. Es kann eine Identifizierung auf Einzelzellebene innerhalb von
zwei Stunden ohne die Notwendigkeit einer Vorkultivierung erreicht werden.
(Meisel et al., 2012)
Mittels SERS und PCA konnte eine leistungsfähige Methode zum Nachweis und zur Iden-
tifizierung pathogener Bakterien in Lebensmittelproben entwickelt werden, welche als alter-
native Methode in die ISO-Normen aufgenommen werden könnte. Die Analysezeit könnte
von sechs Tagen, wie sie bei vielen gängigen Methoden nötig ist, auf zwei Tage reduziert
werden. Bei der Methode wurde die ansonsten zeitaufwendige Inkubation verkürzt und di-
rekte SERS-Analysen von auf selektiven Nährböden gewachsenen Bakterienkolonien
durchgeführt. Mit der Methode konnten in Lachs, Eiern, Säuglingsnahrung (Pulver), Milch
und Kräutermischungen gewachsene Bakterien (Salmonella enterica, Listeria monocytoge-
nes, Cronobacter sakazakii) mit 98-prozentiger Genauigkeit innerhalb von 48 Stunden iden-
tifiziert werden. (Witkowska et al., 2017)

Einleitung
21
1.2 Zielsetzung
Es wurde bereits in vielen wissenschaftlichen Veröffentlichungen nachgewiesen, dass eine
Identifizierung von Mikroorganismen über Raman-Spektroskopie sehr gute Resultate erzie-
len kann (siehe Kapitel 1.1.5). Zwar gibt es Spektren-Datenbanken, um Spektren einfacher
zu interpretieren und bestimmte Biomoleküle innerhalb eines Raman-Spektrums wiederzu-
finden, jedoch gibt es noch keine offiziellen und frei verfügbare Datenbanken, welche Ra-
man-Spektren vieler verschiedener Mikroorgansimen enthalten. Ein schneller oder automa-
tisierter Abgleich eines aufgenommenen Raman-Spektrums von einem Mikroorganismus
mit einer freien Datenbank ist bislang nicht möglich.
Neben der Auswahl des entsprechenden Wellenzahlbereichs liegt ein weiteres Problem bei
der Übertragung von bereits entwickelten Vorhersagemodellen von einem System auf ein
anderes darin, dass in der Forschung unterschiedlichste Geräte zur Erfassung der Daten
verwendet werden und jedes optische Element eines Messsystems einen Einfluss auf ein
Spektrum hat. Mit variierender Anregungswellenlänge verändern sich zwar nicht die Positi-
onen der Peaks, jedoch die Intensitäten entsprechender Banden. Auch die Umgebungsbe-
dingungen wie Licht und Temperatur unter welchen die Referenzspektren gesammelt wer-
den oder Unterschiede in der Probenpräparation, haben einen Einfluss auf ein Raman-
Spektrum. Diese Effekte können, zusammen mit diversen Variationen in den Kulturbedin-
gungen, dazu führen, dass bereits entwickelte Modelle bei einem anderen Versuchsaufbau
ungeeignet sind. Dennoch könnte eine universelle Datenbank bei entsprechender Robust-
heit und Vereinheitlichung, insbesondere bei der Datenvorbehandlung, möglich sein.
Die Hochschule Rhein-Waal in Kleve besitzt ein Mikro-Raman-Spektroskopie-System, wel-
ches eine Differenzierung von Mikroorganismen möglich machen könnte. In den meisten
Veröffentlichungen zur gleichen Thematik liegt die Anregungswellenlänge im NIR-Bereich
bei 785 nm und 1064 nm. Auch der UV-Bereich oder der sichtbare Bereich bei 532 nm
kommt häufiger zum Einsatz. Die in dieser Arbeit verwendete Anregungswellenlänge von
633 nm ist für organische Proben daher eher untypisch.
Das Ziel ist es, mit dem vorliegenden Messsystem eine zur Identifizierung einer Vielzahl
von Mikroorganismen geeigneten Methode zu entwickeln. Pathogene oder gentechnisch
veränderte Mikroorganismen werden in dieser Arbeit nicht analysiert. Es soll geprüft wer-
den, ob und welche Vorhersagemodelle die genauesten Ergebnisse erzielen und ob die
Methode in der Lage ist, Mikroorganismen auch auf Stammebene zu differenzieren. Um
diese Ziele zu erreichen muss zunächst eine Methode gefunden werden, um verwertbare
Spektren verschiedener Mikroorganismen zu erhalten. Anschließend erfolgt der Schritt der
Daten-Sammlung, bei welchem möglichst viele repräsentative Spektren verschiedener Mik-
roorganismen gesammelt werden. Mit den gesammelten Daten werden Modelle zur

Einleitung
22
Vorhersage entwickelt, was eine geeignete Datenvorbehandlung voraussetzt. Um zu über-
prüfen, wie die Modelle in der Praxis abschneiden, müssen diese auf Spektren von Proben
angewandt werden, die nicht im Kalibrationsset enthalten sind. Aus diesem Grund werden
zusätzliche Spektren für eine praktische Validierung der Modelle gesammelt. Neben dem
Schwerpunkt, nämlich der Entwicklung der Methode und der Vorhersagemodelle, sollen
auch verschiedene Einflüsse der Analysebedingungen berücksichtigt werden. Dazu gehö-
ren Einflüsse auf Raman-Spektren bei leichten Unterschieden in der Kultivierung, Neben-
effekte nach längerer Vorbestrahlung zur Fluoreszenzreduzierung und Auswirkungen der
Probenpräparation auf ein Spektrum.
Ein weiterer Fokus soll auf der späteren Anwenderfreundlichkeit der Methode liegen, was
eine einfache Durchführungsweise und robuste Vorhersagemodelle voraussetzt. Deshalb
soll ein kompakter Leitfaden zur Vorgehensweise bei der Identifizierung unbekannter Mik-
roorganismen entworfen werden. Die Ergebnisse dieser Arbeit können als Basis neuer
hochschulinternen Forschungsziele für den Einsatz der Raman-Spektroskopie in der Mik-
robiologie dienen.

Material und Methoden
23
2 Material und Methoden
2.1 Material
2.1.1 Geräte
• Brutschrank, HettCube 200 R (Hettich AG)
• Vortexschüttler, Fisherbrand ZX3 (Thermo Scientific)
• Zentrifuge, Heraeus Fresco 17 (Thermo Scientific)
• Raman Spektroskopie System, Konfokales Raman-Mikroskop inVia (Renishaw)
o Laser: HeNe-Laser, 17 mW, luftgekühlt
Plasmafilter für 633 nm, Gitter mit 1800 l/mm
o Anregungswellenlänge: 633 nm
o Spektrale Auflösung: 0,82 cm-1
o Mikroskop: adaptiertes Leica DM2700 (100X-Objektiv)
o Detektor: CCD-Chip
Abbildung 11 zeigt den Hauptteil des Raman-Messsystems. Er besteht aus dem Mikroskop,
dem Laser und dem Gehäuse unter welchem sich, wie aus Abbildung 4 zu entnehmen,
verschiedene optische Elemente sowie der Detektor befinden. Die Hardware ist mit einem
Computer verbunden und ein Trackball dient neben der mechanischen Möglichkeit zur Fo-
kussierung und Bewegung des Objekttisches, als zusätzliches Eingabegerät.
• Videokamera
• Wahlschalter
(Wechsel zwischen Lichtquelle
des Mikroskops und Laser)
• HeNe-Laser
• Objektiv(e)
• Motorisierter Mikroskoptisch
• CCD-Detektor
Abbildung 11: Konfokales Raman-Mikroskop inVia Renishaw

Material und Methoden
24
2.1.2 Software
• Bedienersoftware des Raman-Messsystems, WiRE Version 4.3 (Renishaw)
• Konvertierungssoftware, WiRE Batch Coverter (Renishaw)
• Software zur Datenverarbeitung, Datenvorbehandlung und zur Erstellung der Vor-
hersagemodelle, MATLAB R2017b Version 9.3.0.713579 (The MathWorks, Inc.)
o Spectr-O-Matic Toolbox for MATLAB Version 2.0 (Petar H. Lambrev)
o MATLAB Classification Learner, Statistics and Machine Learning Toolbox
Version 11.2 (The MathWorks, Inc.)
2.1.3 Mikroorganismen
Tabelle 3 enthält die in dieser Arbeit verwendeten Mikroorganismen. Die aufgelisteten Kür-
zel aus Tabelle 3 werden in nachfolgenden Tabellen, Abbildungen und Texten als Bezeich-
nung der Mikroorganismen verwendet.
Tabelle 3: Verwendete Mikroorganismen, Kürzel und DSM-Nr.
Kürzel Mikroorganismus DSM-Nr.
Bdi 7234 Brevundimonas diminuta 7234
Bsu 10 Bacillus subtilis 10
K12 423 Escherichia coli K12 423
MLB 28269 Micrococcus luteus 28269
Mlu 1790 Micrococcus luteus 1790
Mlu 20030 Micrococcus luteus 20030
Pfl 50090 Pseudomonas fluorescens 50090
Sce 1334 Saccharomyces cerevisiae 1334
Sty TA981 Salmonella Thyphimurium TA98 -
2.1.4 Verbrauchsmaterialien
• Trypton Soja Agar nach EP, USP, JP, ISO und FDA-BAM GranuCult (Merck KGaA)
• Impfschlingen, steril aus Polystyrol 10 µl (Sarstedt)
• Reaktionsgefäße mit Deckel, 1,5 ml
• Objektträger mit geschnittenen Kanten und Mattrand (VWR International)
1 TA98 Salmonella Thyphimurium-Stamm mit natürlicher Mutation, welche zur Apathogenität führt.

Material und Methoden
25
2.2 Methoden
2.2.1 Anzucht der Mikroorganismen
Für die Herstellung der Nährböden werden 40 g der TSA-Fertigmischung mit 1000 ml deio-
nisiertem Wasser bei 121 °C autoklaviert. Im Anschluss werden die Platten mit dem noch
flüssigen Agar gegossen und bei Raumtemperatur gelagert.
Zur Anzucht werden Mikroorganismen aus stickstoffgekühlten Reinkulturen (Suspensionen
gelagert bei -80 °C mit Glycerin als Frostschutzmittel) verwendet. Die aufgetauten Keim-
suspensionen werden mittels sterilen Einweg-Impfösen auf die TSA-Petrischalen ausge-
strichen. Die Bebrütung erfolgt bei 30 °C für 24 Stunden. Nach der Bebrütung wird ein Teil
des gewachsenen Koloniematerials erneut auf einer weiteren TSA-Platte ausgestrichen.
Zur Analyse werden sowohl die erste, als auch die zweite Kultur herangezogen. Nachfol-
gend werden in dieser Arbeit die Begriffe Kultur 1 und Kultur 2 dafür verwendet.
2.2.2 Probenpräparation
In 1,5 ml Reaktionsgefäße werden mit einer Kolbenhubpipette 1000 µl 0,9% NaCl-Lösung
vorgelegt. Mit einer Impfschlinge wird Koloniematerial von der Agarplatte in das Reaktions-
gefäß gegeben. Es wird so viel Material verwendet, wie mit drei vollgestrichenen Impfösen
aufgenommen werden kann. Beim Überführen des Koloniematerials wird beachtet, den
Nährboden nicht zu beschädigen und keine Bestandteile davon mitaufzunehmen.
Die Mischung aus Mikroorganismus und NaCl-Lösung wird solange auf einem Vortexer ge-
mischt, bis eine gleichmäßige Suspension entsteht. Mit einer Zentrifuge werden die Sus-
pensionen für drei Minuten bei 5000 Umdrehungen pro Minute (rpm) zentrifugiert. Der Über-
stand wird abpipettiert und verworfen. Das Material kann im Anschluss mittels Pipette oder
einem sterilen Kunststoffstab aus dem Reaktionsgefäß auf den Objektträger überführt wer-
den, wo die Masse dann für ein bis zwei Stunden bei Raumtemperatur getrocknet wird.
Je nach Bakterium bzw. Hefe ist die mit der Impföse aufgenommene Probenmenge variabel
und auch der Rest an NaCl-Waschlösung kann variieren, woraus kleinere Unterschiede in
der Probendarbietung auf dem Objektträger folgen können. Auch die Beschaffenheit des
Mikroorganismus selbst hat einen Einfluss auf die Trocknungsdauer und die Probendarbie-
tung. Klebriges Koloniematerial zerfließt weniger stark, wohingegen gut suspendierbare
Proben einfacher einen dünnen Film auf dem Objektträger bilden. Unabhängig von der Art
der Probe gilt, dass das Material gewöhnlich von außen nach innen durchtrocknet. Da für
die Kalibration sehr viele Spektren pro Probe gesammelt werden und es mehrere Stunden
dauert, kann dies ausgenutzt werden, indem zuerst Spektren der trockenen Randbereiche
gesammelt werden und während der Durchführung dieser Messungen die Probe im

Material und Methoden
26
Zentrum weiter trocknet. Es muss allerdings berücksichtigt werden, dass das Probenmate-
rial an keiner Stelle zu feucht ist, da die Gefahr besteht, dass Probenmaterial durch Kapil-
larkräfte auf das Objektiv gelangen könnte.
2.2.3 Vorbereitung des Raman-Messsystems
Das Raman-Messsystem wird mit folgenden Schritten in Betriebsbereitschaft gesetzt:
• Einschalten des Lasers
• Einschalten des Spektrometers
• Einschalten der Lichtquelle des Mikroskops
• Einschalten des Computers
• Starten der Software WiRE 4.3
Nach diesen ersten Schritten wird vor jedem Versuchstag über die Software WiRE 4.3 die
„Quick-Calibration“ initiiert. Diese beinhaltet die Messung eines Silizium- (Si) Standards.
Der Si-Standard wird auf dem Objekttisch fixiert und mittels Grob- und Feintrieb des Mikro-
skops wird die Oberfläche scharf gestellt. Mit dem Trackball kann die korrekte Fokussierung
überprüft werden, indem damit der Objekttisch bzw. die Probe bewegt wird und die Bewe-
gung am Bildschirm mitverfolgt wird. Über das Trackball-Eingabegerät kann die Fokussie-
rung auch nachjustiert werden.
2.2.4 Sammeln der Raman-Spektren
Pro Versuchstag werden von maximal drei verschiedenen Mikroorganismen Spektren ge-
sammelt. Der Objektträger mit der getrockneten Probe wird auf dem Objekttisch fixiert. Mit
dem 100X-Objektiv wird eine Stelle fokussiert. Danach erfolgt die Messung mit folgenden
Einstellungen:
• Wellenzahlzentrum: 1200 cm-1 (Wellenzahlbereich: ca. 605 cm-1 bis 1737 cm-1)
• Belichtungsdauer: 3 s
• Laserleistung: 100 %
• Anzahl der Einzelspektren2 für ein Spektrum: 15
Die Spektrenaufzeichnung erfolgt bei inaktiver Raumbeleuchtung und geschlossenen Ja-
lousien. Von der fokussierten Stelle werden 5 bis 10 Spektren hintereinander aufgezeich-
net. Zwischen den Messungen wird der Fokus überprüft und unter Umständen nachjustiert.
Nach dem Aufzeichnen dieser Spektren von ein und derselben Stelle, wird eine andere
Stelle auf dem Objektträger fokussiert, welche sich möglichst weit entfernt von der letzten
Stelle befindet. Von der neuen Stelle werden nun wieder 5 bis 10 Spektren aufgezeichnet
2 Durch Akkumulation mehrerer Einzelspektren wird das Rauschen reduziert

Material und Methoden
27
und danach wird die Stelle nach demselben Prinzip gewechselt. Dies wird so oft wiederholt,
bis an einem Versuchstag pro Mikroorganismus etwa 100 bis 200 Spektren entstehen. Bei
der Wahl der Stellen ist zu beachten, dass möglichst viele Variationen, die in der Praxis
vorkommen können, abgedeckt werden. Ist das Probenmaterial beim Trocknungsvorgang
beispielsweise so zerflossen, dass bestimmte Bereiche eine etwas geringere Schichtdicke
aufweisen als andere, so werden von allen Bereichen Spektren gesammelt. Die Anzahl der
Aufzeichnungen von Spektren bei variierenden Schichthöhen sollten in etwa gleichmäßig
gewichtet sein.
Die Messdauer eines Raman-Spektrums beträgt bei 3 Sekunden Belichtungszeit und
15 Einzelspektren entsprechend 45 Sekunden. Proben die starke Störung durch Fluores-
zenz aufweisen werden allerdings solange vorbestrahlt, bis für Mikroorganismen typische
Signale sichtbar werden. Sollten nach etwa zehn Minuten Vorbestrahlung keine typischen
Peaks zu sehen sein, wird die zu analysierende Stelle auf dem Objektträger gewechselt.
Es werden Spektren von Kultur 1, die direkt aus der Tiefkühlung einen Tag lang auf TSA
bebrütet werden, erfasst und Spektren von Kultur 2, bei der Koloniematerial von der ersten
Kultur überführt und ebenfalls einen Tag lang auf TSA bebrütet wird.
Um die gesammelten Daten anschließend einfach und effizient verarbeiten zu können, wer-
den die Dateinamen der Spektren nach der folgenden Logik gewählt:
• Jahr, Monat, Tag, fortlaufende Nummerierung innerhalb des Versuchstages begin-
nend mit 001, Kürzel des Mikroorganismus (z.B. 20180831 216 Sty TA98)
Mit dem WiRE Batch Converter werden die Spektren in das TXT-Format konvertiert.
Dadurch wird jedes Spektrum als eine Datei aus zwei Spalten mit der Wellenzahl und der
entsprechenden Intensität bzw. den „Counts“ abgespeichert. Jedes Spektrum hat 1015 Da-
tenpunkte bzw. 1015 Wellenzahlen (Raman shifts) mit den dazugehörigen Intensitäten
(Counts). Die originalen Dateien im sogenannten WDF-Format können mit der WiRE-Soft-
ware geöffnet werden und enthalten zusätzlich alle Informationen zu den Mess- und Gerä-
teeinstellungen.
2.2.5 Datenvorbehandlung
2.2.5.1 Interpolierung
Zunächst müssen die Spektren jedes Mikroorganismus in MATLAB eingelesen werden.
Trotz Kalibrierung des Raman-Messsystems mit dem Si-Standard, gibt es bei verschiede-
nen Versuchseinheiten kleinere Abweichungen im Wellenzahlbereich der Spektren. Diese
Unterschiede würden eine weitere Datenvorbehandlung nicht möglich machen oder könn-
ten Fehler bei der späteren Modellentwicklung hervorrufen. Aus diesem Grund werden die
Daten interpoliert. Dies hat zur Folge, dass alle Spektren auf den exakt gleichen

Material und Methoden
28
Wellenzahlbereich vereinheitlicht werden und den Vorteil, dass die Abstände zwischen den
1015 Datenpunkten äquidistant werden.
Für das weitere prozessieren der Spektren müssen die Daten als Matrix konvertiert werden.
Das Einlesen, das Interpolieren und das Transformieren als Datenmatrix, erfolgt über die
MATLAB-Befehle aus der Spectr-O-Matic Toolbox von Petar H. Lambrev. Die Vorgehens-
weise in MATLAB ist im Leitfaden zur Identifizierung von Mikroorganismen über Raman-
Spektroskopie (Anhang 7) beschrieben.
2.2.5.2 Basislinienkorrektur und Glättung
Wie in Kapitel 1.1.3.4 beschrieben, haben verschiedene Einflüsse einen Effekt auf die Qua-
lität eines Raman-Spektrums. Der bedeutendste Einfluss bei biologischen Proben stellt der
Fluoreszenzhintergrund dar.
Die vollständige Entfernung von Fluoreszenzstörung in Raman-Spektren ist aufgrund der
komplexen Merkmale biologischer Raman-Spektren eine Herausforderung.
Bei rechnerischen Verfahren zur Entfernung des Fluoreszenzhintergrunds bzw. zur Basis-
linienkorrektur, geht es gewöhnlich darum eine Basislinie zu schätzen und diese vom ge-
messenen Raman-Spektrum abzuziehen. Somit spielt die Art des Verfahrens zur Schät-
zung bzw. zur Angleichung einer Basislinie an das Spektrum eine entscheidende Rolle für
die Leistung einer Basislinienkorrektur. (Guo et al., 2016)
Rechnerische Verfahren zur Basislinienkorrektur sind neben der Bildung der ersten und
zweiten Ableitung eines Spektrums beispielsweise eine Polynomanpassung (Lieber &
Mahadevan-Jansen, 2003), Fourier Transformation (Schulze et al., 2005), Wavelet Trans-
formation und Empirische Bandzerlegung (EMD) (Leon-Bejarano et al., 2016).
Die Glättung eines Spektrums dient der Rauschverminderung. Bei Rauschen handelt es
sich um ein Störsignal, welches vom Spektrometer selbst verursacht wird. Je schwächer
das spektroskopische Signal ist, desto stärker wird das Rauschen. Zur Beseitigung oder
Eindämmung des Rauschens ist ein sehr einfacher Ansatz die Polynomglättung, bei der
nach Wahl einer Intervallgröße, ähnlich wie bei der Basislinienkorrektur, ein Polynom gefit-
tet wird. (Kessler, 2007)
Die in dieser Arbeit eingesetzten Methoden zur Basislinienkorrektur und Glättung basieren
auf einem Tiefpassfilter. Ein Tiefpass hat generell die Funktion tiefe Frequenzen eines
Spektrums passieren zu lassen bzw. höhere Frequenzen abzuschneiden (Runkler, 2016).
Ein Tiefpassfilter kann problemlos auf diverse Arten von Spektren oder Signale angewandt
werden.

Material und Methoden
29
Am ehesten kann die Funktion der Glättung durch den Tiefpassfilter anhand eines Stufen-
spektrums verdeutlicht werden. Die Kanten entsprechen den hohen Frequenzkomponen-
ten, die entfernt werden, während tiefe Frequenzen passieren können (siehe Abbildung 12).
Abbildung 12: Prinzip eines Tiefpassfilters zur Glättung eines Stufenspektrums
Bei elektromagnetischen Wellen ergibt sich die Frequenz aus der Wellenzahl multipliziert
mit der Lichtgeschwindigkeit (im Vakuum). Der Zusammenhang der Frequenz mit der Wel-
lenzahl, der Wellenlänge und der Lichtgeschwindigkeit bei elektromagnetischen Wellen
kann wie folgt beschrieben werden:
𝜈 =1
𝜆=
𝑓
𝑐
(1)
�̃� = Wellenzahl
𝜆 = Wellenlänge
𝑓 = Frequenz [1
𝑠] = [𝐻𝑧]
𝑐 = Lichtgeschwindigkeit im Vakuum ≈ 2,998 ∙ 1010 𝑐𝑚
𝑠
2.2.5.3 Normalisierung
Die Spektren gleicher Mikroorganismen unterscheiden sich nur geringfügig in ihren eigent-
lichen Signaturen, was die späteren Vorhersagen anhand unbekannter Spektren überhaupt
erst möglich macht, jedoch gibt es sehr starke Unterschiede in der Intensität der Spektren.
Insbesondre mit zunehmender Bestrahlungsdauer sinkt die Intensität eines Spektrums.
Diese Unterschiede verhindern die Entwicklung von Kalibrations-Modellen mit hoher Vor-
hersagegüte.
Eine Normalisierung gleicht systematische Veränderungen in der Intensität aus. Spektren,
bei denen die Signale das gleiche Verhältnis zueinander haben, aber unterschiedliche ma-
ximale Intensitäten, werden durch die Normalisierung identisch. Prinzipiell verändert sich
das Aussehen der Spektren durch die Normalisierung nicht. (Kessler, 2007)

Material und Methoden
30
Die Spektren werden in dieser Arbeit normalisiert, indem das Minimum eines Spektrums
als null und das Maximum als eins definiert wird. Aus Formel (2) geht die rechnerische
Vorgehensweise hervor, um die Intensität bei einer Wellenzahl eines Spektrums zu norma-
lisieren. Das entsprechende MATLAB-Skript um die Intensitäten jeder Wellenzahl aller
Spektren automatisiert zu normalisieren, befindet sich in Anhang 7.
𝐼𝑛𝑜𝑟𝑚 = 𝜇 ∙ (𝐼 − 𝐼𝑚𝑖𝑛) + 𝑖𝑚𝑖𝑛 mit 𝜇 =𝑖𝑚𝑎𝑥 − 𝑖𝑚𝑖𝑛
𝐼𝑚𝑎𝑥 − 𝐼𝑚𝑖𝑛
(2)
𝐼𝑛𝑜𝑟𝑚 = Normalisierte Intensität bei gewählter Wellenzahl des Ausgangsspektrums
𝜇 = Mittlere normalisierte Intensität des Ausgangsspektrums
𝐼 = Intensität bei gewählter Wellenzahl des Ausgangsspektrums
𝐼𝑚𝑖𝑛 = Minimalintensität des Ausgangsspektrums
𝐼𝑚𝑎𝑥 = Maximalintensität des Ausgangsspektrums
𝑖𝑚𝑖𝑛 = Definiertes Minimum = 0
𝑖𝑚𝑎𝑥 = Definiertes Maximum = 1
Abbildung 13 zeigt anhand dreier exemplarischer Spektren verschiedener Mikroorganis-
men den schrittweisen Effekt der einzelnen Vorbehandlungsarten. Die interpolierten aber
ansonsten noch unbehandelten Spektren aus Abbildung 13 A, weisen große Unterschiede
in der Intensität auf und K12 423 (blau) einen Anstieg im Bereich von 600 bis 950 cm-1.
Nach Basislinienkorrektur und Glättung (Abbildung 13 B) liegen die Spektren nahezu paral-
lel zur Wellenzahlachse, weisen geringeres Rauschen auf und die spezifischen Signaturen
werden deutlicher sichtbar. Nach Normalisierung (Abbildung 13 C) bewegen sich die Inten-
sitäten der Spektren zwischen null und eins und weisen keine negativen Intensitäten mehr
auf.

Material und Methoden
31
Abbildung 13: Schrittweises Schema der Datenvorbehandlung anhand dreier Spektren verschiede-
ner Mikroorganismen; (A) Interpolierte Spektren, (B) Basislinienkorrektur und Glättung, (C) Normalisierung
2.2.6 Hauptkomponentenanalyse
Eines der wichtigsten Ziele der Hauptkomponentenanalyse, im englischen Principal Com-
ponent Analysis (PCA) genannt, ist die Daten- bzw. Dimensionsreduktion. Viele einzelne
Variablen, wie in dieser Arbeit 1015 verschiedene Intensitäten für jedes Ramanspektrum,
werden zu wenigen Hauptkomponenten (PCs) zusammengefasst, welche für die Beschrei-
bung der Mikroorganismen ausreichen sollen. (Kessler, 2007)
Die PCA ist besonders aufgrund der hohen Multikollinearität bei spektroskopischen Daten
wichtig. Multikollinearität bedeutet, dass einzelne Intensitäten innerhalb eines Spektrums
stark miteinander korrelieren. Aufgrund der relativ hochauflösenden Abtastung benachbar-
ter Wellenzahlen sind dort die Intensitäten häufig nahezu identisch und korrelieren daher
sehr stark.
Dies bedeutet zum einen Redundanz in den Daten und somit weniger Information und zum
anderen lassen sich die vorhandenen Informationen nicht mehr eindeutig den Variablen
zuordnen (Backhaus et al., 2015). Zudem kann bei Heranziehen vieler oder aller Variablen
das Rauschen fehlerhafterweise als Signal interpretiert werden. Multikollinearität führt bei

Material und Methoden
32
vielen Klassifizierungsverfahren zu numerischen Problemen, denen eine vorangehende
PCA entgegenwirken kann (Jolliffe, 2002).
Mithilfe der PCA, werden aus den Ausgangsvariablen, also den Intensitäten der jeweiligen
Wellenzahlen, neue, latente Variablen berechnet, die sogenannten Faktoren bzw. Haupt-
komponenten. Die PCs sind mathematisch eine Linearkombination der Ausgangsvariablen.
Das heißt sie setzten sich aus einer linearen Summe der unterschiedlich gewichteten Ur-
sprungsvariablen zusammen. Zur Berechnung der PCs gibt es mehrere mathematische
Vorgehensweisen. Bei einer quadratischen Ausgangsmatrix X können die PCs als Eigen-
vektoren und zugehörige Eigenwerte der Datenmatrix X betrachtet und über einen Algorith-
mus zur Eigenwertberechnung bestimmt werden. Hierfür wird die Datenmatrix X zuerst in
die Korrelationsmatrix bzw. in die Kovarianzmatrix übertragen, damit sie quadratisch wird.
Mit der quadratischen Matrix wird die Eigenwertberechnung durchgeführt, die im englischen
Singular Value Decomposition genannt wird (SVD). (Kessler, 2007)
Jeder Eigenwert mit dem zugehörigem Eigenvektor bildet einen Faktor bzw. eine PC. Diese
PCs bilden die Faktormatrix P. Ein Faktor bildet je eine Spalte der Matrix P. Die Zeilenzahl
von P entspricht der Anzahl der Elemente pro Faktor und wird durch die Anzahl der Spalten
(Variable) in der Ausgangsmatrix bestimmt. Bei der Berechnung der Eigenwerte findet noch
keine Dimensionsreduktion statt. Die Elemente der Spalten in P werden Faktorladungen
(engl. Loadings) genannt. Der Eigenwert bestimmt hierbei den Anteil des Faktors an der
Gesamtvarianz der Ursprungsdaten. Je höher der Eigenwert ist, umso größer ist die erklärte
Gesamtvarianz und umso wichtiger ist der Faktor zur Beschreibung der Ursprungsdaten.
(Kessler, 2007)
Die Faktoren können auch bestimmt werden, indem die Richtung der maximalen Varianz
mit einem geeigneten Algorithmus in den Ausgangsdaten gesucht wird. Die hiermit gefun-
denen Faktoren stellen ein neues Koordinatensystem dar, welches die Daten besser be-
schreibt. Dann kann durch den Verzicht auf höhere Koordinatenachsen eine Datenreduk-
tion erfolgen. Es wird auf Koordinatenachsen verzichtet, die nur einen geringen Teil zur
Gesamtvarianz der Daten beitragen; somit wird nur ein Unterraum der Ausgangsdaten be-
trachtet. (Kessler, 2007)
Da die Daten durch das neue Faktorenkoordinatensystem beschrieben werden sollen, müs-
sen nach der Berechnung der PCs die Ursprungsdaten in den neuen Faktorenraum trans-
formiert werden. Um für jede Beobachtung die Koordinaten im neuen Faktorenraum zu be-
rechnen, muss jedes Objekt auf jeden der Faktoren abgebildet werden. Die dadurch erhal-
tenen Koordinaten werden Faktorenwerte bzw. mit dem englischen Begriff Scores genannt.
Für alle Objekte und jeden Faktor wird ein Scorewert berechnet, die dann die Matrix T bil-
den. Die Anzahl der Zeilen von T entsprechen der Anzahl der Objekte der Originalmatrix X

Material und Methoden
33
und die Anzahl der Spalten entsprechen der Zahl der verwendeten PCs bzw. der Dimension
des neuen Faktorenkoordinatensystems. (Kessler, 2007)
Abbildung 14 verdeutlicht das Prinzip der PCA grafisch anhand zweier Variablen (X und Y)
und fünf Beobachtungen. Das Koordinatensystem wird gedreht, damit die Beobachtungen
innerhalb der ersten Hauptachse die größte Varianz aufweisen. Das Beispiel aus Abbildung
14 verdeutlicht, dass bei Weglassen der zweiten Hauptkomponente (PC2) nur wenig Infor-
mation verloren geht. PC1 reicht also aus um die zweidimensionalen Daten mit nur einer
Dimension zu beschreiben. Genau nach diesem Prinzip können Raman-Spektren, mit je
nach Messsystem über 1000 Dimensionen, durch wenige PCs beschrieben werden.
Abbildung 14: Vereinfachtes Schema einer Dimensionsreduktion durch PCA
(modifiziert nach (Powell & Lehe, 2014))
Betrachtet man nur einen Unterraum der Originaldaten, also eine geringere Anzahl an PCs
als die Anzahl der Variablen zulassen würde, dann gibt es eine Matrix E, welche die Dimen-
sion der Originalmatrix X hat und den Anteil der Originaldaten, der nicht durch die PCs
erklärt wurde, die sogenannten Residuen enthält. Je mehr Faktoren berechnet werden,
desto kleiner werden die Werte in Matrix E. Bei Verwendung aller PCs für den Faktoren-
raum werden die Elemente der Matrix E null, womit die Berechnung der PCA beendet ist.
(Kessler, 2007)
Es sollte berücksichtigt werden, dass die ersten PCs, bzw. die Hauptkomponenten mit den
größeren Eigenwerten, nicht notwendigerweise die relevanteste Information für die Klassi-
fizierung enthalten (Chang, 1983). In dieser Arbeit soll vor jedem Klassifizierungsverfahren
eine PCA durchgeführt werden und zur Modellentwicklung eine unterschiedliche Anzahl an
PCs herangezogen werden. Jedes Klassifizierungsmodell wird jeweils unter Verwendung
von 8, 12, 15 und 20 PCs generiert.

Material und Methoden
34
2.2.7 Modellentwicklung zur Identifizierung von Mikroorganismen durch
verschiedene Klassifikationsverfahren
Ein Modell ist im Allgemeinen eine mathematische Funktion, mit welcher Vorhersagen über
unbekannte Daten gemacht werden können. Ein einfaches Beispiel eines Modells ist eine
Kalibrierungsgrade, wie sie in der quantitativen Chemie häufig verwendet wird. Zu Stan-
dards mit bekannter Konzentration werden Detektorwerte (z.B. Peakflächen bei Hochleis-
tungsflüssigkeitschromatographie) erzeugt. Werden die Konzentrationen in einem Koordi-
natensystem auf die entsprechenden Detektorwerte aufgetragen, kann eine Regressions-
gerade berechnet werden. Durch Umstellen der Geradengleichung können aus Detektor-
signalen unbekannter Proben, die unter denselben Bedingungen erfasst wurden, Konzent-
rationen berechnet werden. Einfache Vorhersagemodelle für qualitative Analysen funktio-
nieren nach dem gleichen Prinzip. Anhand einer detektierbaren Variable können Vorhersa-
gen über eine andere Eigenschaft oder Klasse gemacht werden. Die Zuordnung eines Mik-
roorganismus egal ob in Gattung, Art oder Stamm, anhand nur eines einzigen Merkmals,
wie das Gramverhalten, ist nicht möglich. Für solche Vorhersagen sind multivariate Modelle
notwendig, die mehrere beschreibende Variablen berücksichtigen. In dieser Arbeit sollen
drei gängige Klassifizierungsverfahren angewandt und Resultate verglichen werden. Hier-
bei handelt es sich um die quadratische Diskriminanzanalyse (QDA), kNN und SVM, welche
nachfolgend skizziert werden.
2.2.7.1 Quadratische Diskriminanzanalyse
Die Diskriminanzanalyse ermöglicht die Untersuchung der Unterschiedlichkeit mehrerer
Gruppen hinsichtlich einer bestimmten Anzahl von Variablen. (Backhaus et al., 2015)
Bei der LDA wird die Vorhersage unbekannter Daten erreicht, indem die Varianz innerhalb
einer Klasse (Mikroorganismus) minimiert und die Varianz zwischen verschiedenen Klas-
sen maximiert wird. (Hair et al., 2010)
Dadurch entsteht ähnlich wie bei der PCA ein neues Koordinatensystem bzw. eine Linear-
kombination. Sowohl mit der PCA, als auch über die LDA wird eine Dimensionsreduktion
erzielt. Die PCA zielt jedoch darauf ab die größte Varianz der Daten zu finden (siehe Abbil-
dung 15 A) und die LDA darauf die Trennung der bekannten Klassen zu maximieren (Ab-
bildung 15 B). Eine neue Beobachtung wird der Klasse mit dem nächstgelegenen Daten-
zentrum (Klassenschwerpunkt) zugeordnet. (Hastie et al., 2009)

Material und Methoden
35
Abbildung 15: Beispiel zweier Datenklassen mit markiertem Klassenschwerpunkt und Projektion
der Daten um die größte Varianz zu erzielen (A) und Projektion der Daten in Diskriminanzrichtung (B) (Hastie et al., 2009)
Die Diskriminanzfunktion wird auch kanonische Diskriminanzfunktion genannt und die Dis-
kriminanzvariable kanonische Variable. Der Begriff „kanonisch“ beschreibt, dass eine Line-
arkombination von Variablen vorgenommen wird. (Backhaus et al., 2015)
Die Formel der kanonischen Diskriminanzfunktion setzt sich wie folgt zusammen (Hair et
al., 2010):
𝑍𝑗𝑘 = 𝑎 + 𝑊1𝑋1𝑘 + 𝑊2𝑋2𝑘 + ⋯ + 𝑊𝑛𝑋𝑛𝑘 (3)
𝑍𝑗𝑘 = Diskriminanzvariable 𝑗 für Objekt 𝑘
𝑎 = Konstantes Glied (Achsenabschnitt)
𝑊𝑖 = Diskriminanzkoeffizient für Merkmalsvariable 𝑖
𝑋𝑖𝑘 = Merkmalsvariable 𝑖 für Objekt 𝑘
Häufig trennen lineare Entscheidungsgrenzen die Klassen nicht ausreichend. Vor allem
wenn viele Beobachtungen vorliegen, können komplexere Entscheidungsgrenzen zu ge-
naueren Vorhersagen führen. Die QDA ermöglicht im Gegensatz zur LDA quadratische
Entscheidungsgrenzen. LDA verwendet einen einzelnen Klassenschwerpunkt sowie eine
gemeinsame Kovarianzmatrix, um die Verteilung der Daten in jeder Klasse zu beschreiben.
In vielen Situationen sind mehrere Schwerpunkte geeigneter. Falls ungleiche Kovarianz-
Matrizen vorliegen, kann dies durch Anwendung einer QDA berücksichtigt werden
(Backhaus et al., 2015). Bei zu vielen korrelierten Prädiktoren, beispielsweise bei entspre-
chenden Wellenzahlen (Raman shifts), könnte die LDA zu viele Parameter verwenden, die
mit hoher Varianz geschätzt werden, worunter die Leistung des Modells leidet. (Hastie et
al., 2009)
Abbildung 16 A zeigt einige fiktive Daten aus drei verschiedenen Klassen, wobei lineare
Entscheidungsgrenzen durch LDA ermittelt wurden. Quadratische Entscheidungsgrenzen,
welche durch Auffinden linearer Grenzen in einem höherdimensionalen Raum erhalten wur-
den, gehen aus Abbildung 16 B hervor. Lineare Ungleichungen in einem entsprechend hö-
herdimensionalen Raum sind quadratische Ungleichungen im ursprünglichen Raum.

Material und Methoden
36
Abbildung 16 C zeigt die mittels QDA gefundenen quadratischen Entscheidungsgrenzen,
deren Unterschiede zum entsprechenden Fall aus Abbildung 16 B gewöhnlich gering sind.
(Hastie et al., 2009)
Die in dieser Arbeit angewandte Form der Diskriminanzanalyse ist die QDA.
Abbildung 16: Daten aus drei verschiedenen Klassen mit linearen Entscheidungsgrenzen durch LDA (A), quadratische Entscheidungsgrenzen, welche durch Auffinden linearer Grenzen in einem höherdimensionalem Raum erhalten wurden, zurückgeführt auf den ursprünglichen Raum (B), mit-
tels QDA gefundene quadratische Entscheidungsgrenzen (C) (modifiziert nach (Hastie et al., 2009))
2.2.7.2 Nächster-Nachbar-Algorithmus
Der k-Nächster-Nachbar-Algorithmus (kNN) beruht auf dem Abstand einer Beobachtung zu
seinen „k-nächsten“ Nachbarn. Es gibt hierfür verschiedene mathematische Abstands-
maße, wobei das am häufigsten verwendete der euklidische Abstand ist. Im Wesentlichen
ist dieser ein Maß für die Länge einer zwischen zwei Objekten gezogenen geraden Linie.
(Hair et al., 2010)
Vereinfacht wird kNN in Abbildung 17 anhand eines Beispiels erklärt. Zwei unbekannte Da-
tenpunkte werden mit k= 5 und k= 1 als jeweils einer der drei im Trainings-Datenset enthal-
tenen Mikroorganismen klassifiziert. Abbildung 17 macht besonders bei Datenpunkt „Unbe-
kannt 2“ deutlich, dass in Grenzregionen zweier Cluster eines Datenraums bei Wahl eines
kleinen k-Wertes eine Anfälligkeit gegenüber Ausreißern bestehen kann. Zu große k-Werte
führen allerdings zu Ungenauigkeit, insbesondere wenn die Zahl der Beobachtungen pro
Mikroorganismus stark variieren sollte.

Material und Methoden
37
Abbildung 17: Grafische Erklärung von kNN anhand normalisierter Intensitäten zweier ausgewähl-
ter Wellenzahlen aus Raman-Spektren von drei verschiedenen Mikroorganismen
Mit steigendem k-Wert existieren häufig mehr Nachbarn mit großem als mit kleinem Ab-
stand, weshalb die Vorhersage unbekannter Daten durch weit entfernt liegende Nachbarn
dominiert werden kann. Um dort entgegen zu wirken, können weiter entfernte Nachbarn so
gewichtet werden, dass sie geringeren Einfluss auf das Ergebnis haben. (Ertel, 2016)
Die Wahl der idealen Anzahl an nächsten Nachbarn könnte mittels Kreuzvalidierung be-
stimmt werden, indem Modelle mit verschiedenen k-Werten trainiert werden und die Ergeb-
nisse der Kreuzvalidierung verglichen werden. In dieser Arbeit wird die Anzahl der Nach-
barn k= 1 gewählt, womit auch keine Gewichtung erfolgt.
2.2.7.3 Support Vector Maschine
Das erste Ziel einer SVM zur Klassifizierung von Daten ist es eine Hyperebene zu finden,
die für alle enthaltenen Klassen einen möglichst großen datenfreien Bereich hat. Die der
Hyperebene am nächsten liegenden Datenpunkte werden „Support Vectors“ (Stützvekto-
ren) genannt und haben alle den gleichen Abstand zur Trenngeraden. (Ertel, 2016)
Objekte im Datenraum können bei der Problemstellung dieser Arbeit nicht linear voneinan-
der getrennt werden. Um eine nichtlineare Klassentrennung zu erzielen, können über den
sogenannten Kernel-Trick die Daten in einen höherdimensionalen Raum überführt werden,
sodass die Struktur der Daten für eine lineare Trennung geeignet wird. Werden die Daten
zurücktransformiert, wird aus der linearen eine nichtlineare Hyperebene. In vielen Fällen
wird die lineare Entscheidungsgrenze nur in einem sehr hochdimensionalen Merkmalsraum
oder sogar in einem unendlich dimensionalen Merkmalsraum erhalten, was ohne Kernel-
Trick sehr viel rechenintensiver oder unmöglich sein kann (Iba, 2018). (Runkler, 2016)
Abbildung 18 zeigt das allgemeine Prinzip der SVM anhand eines zweidimensionalen Da-
tensatzes mit drei verschiedenen Klassen, die durch variierende Symbole und Farben ge-
kennzeichnet sind. Die Originaldaten sind in Abbildung 18 A dargestellt und Abbildung 18 B

Material und Methoden
38
zeigt die Zuordnung der Daten im höherdimensionalen Raum, nachdem eine Kernel-Funk-
tion angewendet und die optimale Trennung ermittelt wurde. Die Entscheidungsgrenzen in
diesem Raum werden als graues Dreieck dargestellt. Abbildung 18 C zeigt die resultieren-
den Grenzen, die auf den ursprünglichen Datenraum zurückgebildet wurden. Der Abstand
von der Hyperebene zu den „Support Vectors“ (gestrichelte Linien) wird im englischen als
„Margin“ bezeichnet und bildet die maximale Trennspanne zwischen zwei Klassen. (Van
den Burg & Groenen, 2016)
Abbildung 18: Schematische Darstellung des Funktionsprinzips der SVM mit den Ausgangsdaten (A), dem höherdimensionalem Raum (B) und resultierenden Entscheidungsgrenzen bei zurück-transformieren der Daten (C); Schwarze Linien sind trennende Hyperebenen, gestrichelte Linien „Support Vectors“ und Entscheidungsgrenze im höherdimensionalem Raum als graues Dreieck
hervorgehoben (modifiziert nach (Van den Burg & Groenen, 2016))
2.2.8 Validierung der Vorhersagemodelle
2.2.8.1 Kreuzvalidierung
Ein Problem bei der Modellentwicklung, basierend auf multivariaten Daten ist das „Overfit-
ting“. „Overfitting“ bedeutet, dass eine Überanpassung des Modells innerhalb der soge-
nannten Trainingsdaten stattfindet (Runkler, 2016). Die theoretische Vorhersagegüte wird
mit zunehmender Variablenzahl innerhalb des Modells besser, jedoch nicht die praktische
Vorhersagegüte bei Anwenden des Modells auf unbekannte Daten. Werden für ein Modell
zu viele Hauptkomponenten bzw. zu viele Variablen herangezogen, steigt die Wahrschein-
lichkeit des Übertrainierens, da auch zufälliges Rauschen modelliert wird. (Kessler, 2007)
Ein solches „Overfitting“ kann durch eine Kreuzvalidierung vermieden werden. Bei der
Kreuzvalidierung wird jede Beobachtung, die zur Modellentwicklung verwendet wird, zur
Kalibrierung und zur Validierung eingesetzt. Dazu werden die Daten nach zufälliger Mi-
schung in Testdaten und Trainingsdaten aufgeteilt. Mit den Trainingsdaten wird das Modell
errechnet, mit welchem für den Testsatz Vorhersagen getroffen werden, um die Leistung
des Modells zu bewerten. Dieser Vorgang wird so oft wiederholt, bis jede Beobachtung
einmal weggelassen und mit dem Modell der anderen Daten vorhergesagt wurde. Aus dem
Vergleich der vorhergesagten Klasse (Mikroorganismus) mit dem tatsächlichen Wert, kann

Material und Methoden
39
der Validierfehler berechnet werden. (Runkler, 2016)
Um die auszulassenden Proben zu bestimmen, gibt es verschiedene Möglichkeiten. Bei
einer vollständigen Kreuzvalidierung wird jede Beobachtung einmal weggelassen. Bei
2000 Spektren würden demnach 2000 Kalibriermodelle erstellt und jedes Mal würde nur
eine Probe weggelassen werden. Bei großen Datensets werden gewöhnlich mehrere Be-
obachtungen auf einmal weggelassen. Die k-fache-Kreuzvalidierung (engl. k-fold-cross va-
lidation) teilt die Daten in „k“ Teile auf. Bei 2000 Beobachtungen und k= 5 werden die Daten
in Sets mit jeweils 400 Beobachtungen eingeteilt. Es werden 5 separate Trainings durch-
geführt, indem jeweils ein Set ausgelassen und als Testset verwendet wird. Die Vorhersa-
gegüte der jeweiligen Modelle (in Prozent) werden gemittelt, um die endgültige Vorhersa-
gegüte zu bestimmen. (Ertel, 2016; Perner & Petrou, 1999)
In dieser Arbeit wird für jedes Modell eine 5-fache Kreuzvalidierung durchgeführt.
2.2.8.2 Praktische Validierung
Eine Kreuzvalidierung ist nur der erste Schritt, um die Leistung eines Vorhersagemodells
zu überprüfen und „Overfitting“ entgegenzuwirken. Zwar steigt die Wahrscheinlichkeit einer
richtigen Identifizierung einer unbekannten Probe mit zunehmender Probenzahl innerhalb
des Kalibrationssets, jedoch kann die Repräsentativität gegenüber völlig unbekannten Da-
ten nicht gewährleistet werden. Für die externen Validierungen werden deshalb von neuen
Proben jedes der 9 Mikroorganismen 100 weitere Spektren gesammelt, was etwa einem
Drittel der Datenmenge, die für das Kalibrationsset verwendet werden, entspricht. Die Vali-
dierungsspektren bestehen zur Hälfte aus Spektren von Proben einer ersten Kultur und zur
anderen Hälfte aus Spektren einer zweiten Kultur (siehe Kapitel 2.2.1). Es werden für die
praktische Validierung keine Daten herangezogen, die in den entwickelten Modellen ent-
halten sind.
Die Probenpräparation, das Vorbereiten des Messsystems und das Sammeln der Validie-
rungsspektren, werden nach den exakt gleichen Methoden wie in Kapitel 2.2.2 bis 2.2.4
beschrieben, durchgeführt. Da in dieser Arbeit drei verschiedene Klassifizierungsverfahren
unter Verwendung von 8, 10, 12, 15 und 20 PCs gegenübergestellt werden, sind es insge-
samt 15 Modelle, die erstellt werden. Jedes Kalibrationsmodell wird für einen Vergleich der
Modelle untereinander auf alle 900 Validierungsspektren angewandt.

Ergebnisse
40
3 Ergebnisse
3.1 Spektrenaufzeichnung
3.1.1 Störung durch Umgebungslicht
Ein Effekt von Umgebungslicht bei Verwendung sehr geringer Laserleistungen geht aus
Abbildung 19 hervor. Dort wurden Spektren mit 1 % Laserleistung und einem 100X-Objektiv
erfasst, ohne eine Probe zu fokussieren. Abbildung 19 zeigt, dass es zu einer leichten Ab-
sorption bei ca. 1240 cm-1 kommt und bei aktiven Leuchtstoffröhren im Raum zu deutlichen
Peaks bei 710 cm-1, 1660 cm-1 und 1700 cm-1. Wird um den Laserstrahl komplett verdun-
kelt, so zeigen sich weder Absorption noch Peaks. Da diese Effekte bei höheren Laserleis-
tungen und insbesondere bei Vorhandensein einer fokussierten Probe nicht mehr auftreten,
reicht es aus, die Spektren bei dieser Arbeit wie in Kapitel 2.2.4 beschrieben bei geschlos-
senen Jalousien und inaktiver Raumbeleuchtung aufzuzeichnen.
Abbildung 19: Vergleich von Raman-Spektren bei verschiedenen Umgebungslichtbedingungen ohne fokussierte Probe bei einem Prozent Laserleistung und 40 Sekunden Belichtungsdauer
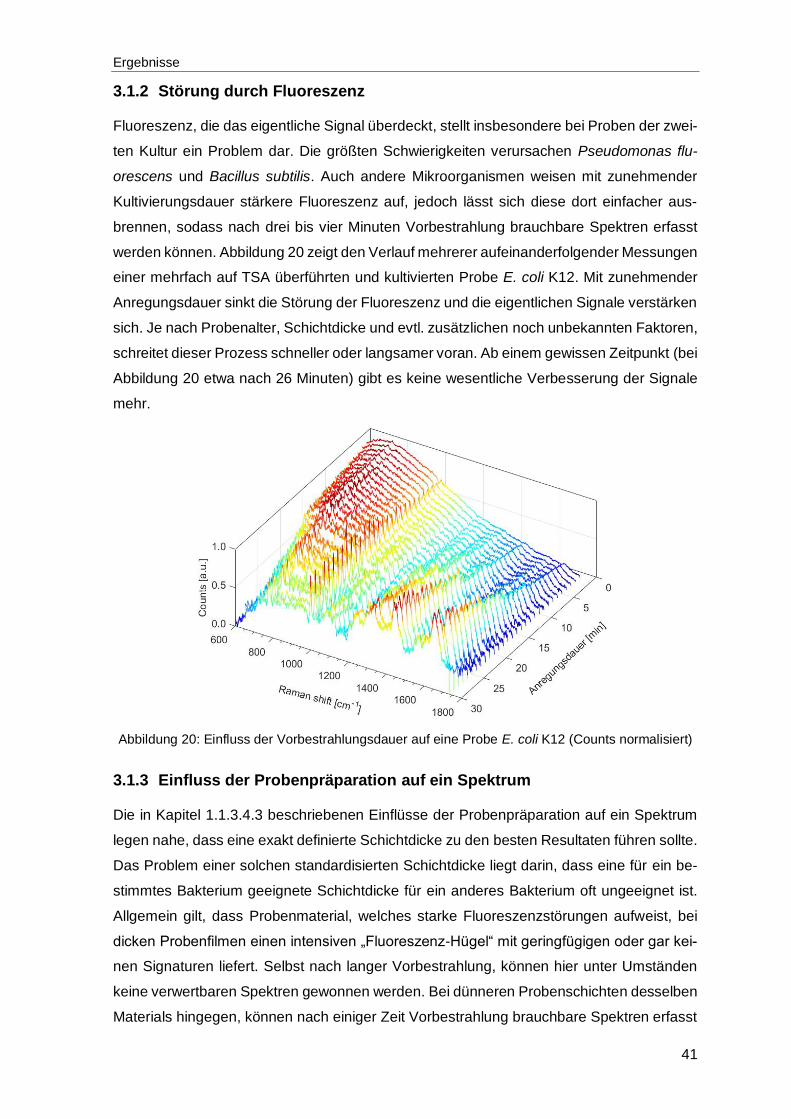
Ergebnisse
41
3.1.2 Störung durch Fluoreszenz
Fluoreszenz, die das eigentliche Signal überdeckt, stellt insbesondere bei Proben der zwei-
ten Kultur ein Problem dar. Die größten Schwierigkeiten verursachen Pseudomonas flu-
orescens und Bacillus subtilis. Auch andere Mikroorganismen weisen mit zunehmender
Kultivierungsdauer stärkere Fluoreszenz auf, jedoch lässt sich diese dort einfacher aus-
brennen, sodass nach drei bis vier Minuten Vorbestrahlung brauchbare Spektren erfasst
werden können. Abbildung 20 zeigt den Verlauf mehrerer aufeinanderfolgender Messungen
einer mehrfach auf TSA überführten und kultivierten Probe E. coli K12. Mit zunehmender
Anregungsdauer sinkt die Störung der Fluoreszenz und die eigentlichen Signale verstärken
sich. Je nach Probenalter, Schichtdicke und evtl. zusätzlichen noch unbekannten Faktoren,
schreitet dieser Prozess schneller oder langsamer voran. Ab einem gewissen Zeitpunkt (bei
Abbildung 20 etwa nach 26 Minuten) gibt es keine wesentliche Verbesserung der Signale
mehr.
Abbildung 20: Einfluss der Vorbestrahlungsdauer auf eine Probe E. coli K12 (Counts normalisiert)
3.1.3 Einfluss der Probenpräparation auf ein Spektrum
Die in Kapitel 1.1.3.4.3 beschriebenen Einflüsse der Probenpräparation auf ein Spektrum
legen nahe, dass eine exakt definierte Schichtdicke zu den besten Resultaten führen sollte.
Das Problem einer solchen standardisierten Schichtdicke liegt darin, dass eine für ein be-
stimmtes Bakterium geeignete Schichtdicke für ein anderes Bakterium oft ungeeignet ist.
Allgemein gilt, dass Probenmaterial, welches starke Fluoreszenzstörungen aufweist, bei
dicken Probenfilmen einen intensiven „Fluoreszenz-Hügel“ mit geringfügigen oder gar kei-
nen Signaturen liefert. Selbst nach langer Vorbestrahlung, können hier unter Umständen
keine verwertbaren Spektren gewonnen werden. Bei dünneren Probenschichten desselben
Materials hingegen, können nach einiger Zeit Vorbestrahlung brauchbare Spektren erfasst

Ergebnisse
42
werden. Mit abnehmender Schichtdicke werden entsprechende charakteristischen Peaks
eines Mikroorganismus schwächer und ab einer bestimmten Höhe sind sie mit der gewähl-
ten Methode nicht mehr sichtbar. Da es sich bei der nachfolgenden Einteilung der Proben-
schichten in „dick“, „dünn“ und „mittel“ um keine exakten Größen handelt, kann Abbildung
21 eine etwaige Einschätzung dieser Begrifflichkeiten ermöglichen.
Abbildung 21: Beispiel zweier Koloniematerialprobenschichten auf Glasobjektträgern zur Einschät-
zung der Schichtdicken (a= dünn; b= mittel; c= dick; d= ungeeigneter, noch flüssiger Bereich)
Aus Abbildung 22 sind die Spektren einer Sty TA98-Probe (2. Kultur) bei variierender
Schichtdicke zu entnehmen. Die Spektren wurden nach zwei Minuten Vorbestrahlung er-
zeugt. Abbildung 22 A zeigt im direkten Vergleich der Spektren die steigende Intensität mit
zunehmender Schichtdicke. Die normalisierten Spektren aus Abbildung 22 B verdeutlichen
die starken Fluoreszenzstörung bei dicker Probenschicht und die geringer ausgeprägten
Peaks bei einem dünneren Probenbereich. Die mittlere Probenschicht liefert die deutlichs-
ten Peaks.
Abbildung 22: Einfluss der Probenschichtdicke auf das Raman-Spektrum einer Sty TA98-Probe bei
zwei Minuten Vorbestrahlung; (A) unbehandelte Daten, (B) normalisierte Spektren (Eine Einschät-zung der Begriffe mittlere, dünne und dicke Schicht kann anhand Abbildung 21 erfolgen)

Ergebnisse
43
3.1.4 Einfluss der Kultivierungsart auf ein Spektrum
Kommen Mikroorganismen aus einer gefrorenen Reinkultur und werden einen Tag lang auf
TSA bebrütet (Kultur 1), so zeigen sie bei der Analyse nur geringe Störung durch Fluores-
zenz. Sind die Bakterien hingegen bereits auf dem Nährmedium gewachsen und werden
erneut auf TSA überführt und einen weiteren Tag bebrütet (Kultur 2), werden die Störungen
durch Fluoreszenz deutlich stärker. Bei der Analyse von Proben der zweiten Kultur muss
somit durchschnittlich länger vorbestrahlt werden, um Spektren mit deutlichen Signaturen
zu erhalten. Da die Mikroorganismen in der Praxis nicht aus einer tiefgekühlten Reinkultur
kommen werden und je nach durchführender Person kleinere Unterschiede in der Proben-
präparation entstehen, kann es sein, dass der Anwender Probleme hat, überhaupt ein ge-
eignetes Spektrum zu erhalten. Oft sind die „Counts“ bei stark fluoreszierenden Proben
auch so hoch, dass das Raman-Spektrum mit den ausgewählten Messeinstellungen aus
Kapitel 2.2.4 nicht einmal angezeigt werden kann. Hier empfiehlt sich in der Software
Wire 4.3 ein zweites Messfenster zu öffnen, bei welchem die Belichtungsdauer von 3 auf
0,25 Sekunden verringert wird und eine hohe Anzahl an Einzelspektren eingestellt wird
(z.B. 500). Die Akkumulationen sollen in diesem Fall nicht den eigentlich gewünschten Ef-
fekt der Rauschverminderung haben, zumal ab einer bestimmten Anzahl an Einzelspektren
keine sichtliche Besserung mehr erfolgt (Bei der vorliegenden Methode etwa nach 30 Ein-
zelspektren), sondern dazu dienen, die Messung aufrecht zu erhalten und die Entwicklung
des Raman-Spektrums zu beobachten. Die 500 Akkumulationen müssen somit nicht kom-
plett abgeschlossen werden, sondern können beispielsweise nach 60 Sekunden gestoppt
und direkt danach neu initiiert werden. Dieser Vorgang kann mehrfach wiederholt werden,
um mögliche Verbesserungen zu registrieren. Durch diese Vorgehensweise können sehr
viel schneller geeignete Probenbereiche lokalisiert werden und die Fluoreszenz effektiver
ausgebrannt werden. Sobald typische Signaturen und eine Fluoreszenzverminderung er-
sichtlich sind, kann auf die eigentliche Methode zurückgewechselt werden. Die genaue Vor-
gehensweise, um bei problematischen Proben geeignete Spektren zu erhalten, werden im
Leitfaden zur Identifizierung von Mikroorganismen über Raman-Spektroskopie (Anhang 7)
beschrieben.

Ergebnisse
44
Da für Kultur 2 durchschnittlich länger vorbestrahlt wurde, konnten dort pro Versuchstag
etwas weniger Spektren aufgezeichnet werden als von Kultur 1. Insgesamt wurden für die
Kalibrationsentwicklung 2953 Raman-Spektren erfasst, von denen 1602 aus Kultur 1 und
1351 aus Kultur 2 stammen (siehe Tabelle 4).
Tabelle 4: Anzahl der zur Kalibrationsentwicklung erfassten Raman-Spektren pro Kulturtyp
(Summe= 2953)
Mikroorganismus Kultur 1 Kultur 2
Bdi 7234 206 161
Bsu 10 163 126
K12 423 170 140
MLB 28269 191 178
Mlu 1790 208 198
Mlu 20030 151 178
Pfl 50090 130 100
Sce 1334 169 125
Sty TA98 214 145
Abbildung 23 zeigt ausgebleicht im Hintergrund alle aufgenommenen Spektren von Kultur 1
in Blau und Kultur 2 in Rot. Die Spektren sind normalisiert, ansonsten unbehandelt und die
Mittelwertspektren sind fett hervorgehoben. Mit Ausnahme von MLB 28269 sind überall die
bei Kultur 2 stärker ausgeprägten Fluoreszenzhintergründe zwischen ca. 600 bis 900 cm-1
zu erkennen.
Die Auswirkungen der beiden Kultivierungsarten sind auf Sty TA98 eher gering; dort sind
die Mittelwertspektren nahezu identisch und Fluoreszenz der Kultur 2 konnte schneller aus-
gebrannt werden als bei den anderen Proben. Effekte auf die Basislinie vieler Spektren sind
nicht nur auf die Eigenfluoreszenz der Mikroorganismen, sondern auf kleinere Variationen
in der Probenschichthöhe zurückzuführen. Die größte Streuung liegt bei Bsu 10 vor. Insbe-
sondere bei Kultur 2 sind bei Bsu 10 nur durch längeres Vorbestrahlen aussagekräftige
Spektren zu erzeugen. Die Spektren von Pfl 50090 leiden sowohl bei Kultur 2, als auch bei
Kultur 1 am stärksten unter der Fluoreszenz. Die Spektren zeigen dort nur schwach ausge-
prägte Signaturen, die sich teilweise nur gering vom Grundrauschen abheben.

Ergebnisse
45
Abbildung 23: Vergleich der Raman-Spektren aller Mikroorganismen von Kultur 1 (blau) und Kul-tur 2 (rot) mit fett hervorgehobenen Mittelwertspektren
3.1.5 Einfluss der Vorbestrahlung auf ein Spektrum
Zwar führt eine längere Bestrahlung im Normalfall zu einer Intensivierung verschiedener
Peaks (siehe Abbildung 20), allerdings kann es insbesondere bei sehr dünnen Proben-
schichten zu einem Rückgang bestimmter Peaks kommen. Abbildung 24 A zeigt mehrere
Spektren, die von ein und derselben Stelle einer dünnen M. luteus-Probenschicht gesam-
melt wurden. Es ist ersichtlich, dass die charakteristischen Peaks bei 1156 cm-1 und
1528 cm-1 mit zunehmender Bestrahlung stark an Intensität verlieren. Dies hat auch zur
Folge, dass bei Vorbehandlung der Daten bestimmte Bereiche stark gestaucht werden, was
zu Fehlinterpretationen führen könnte (siehe Abbildung 24 B).
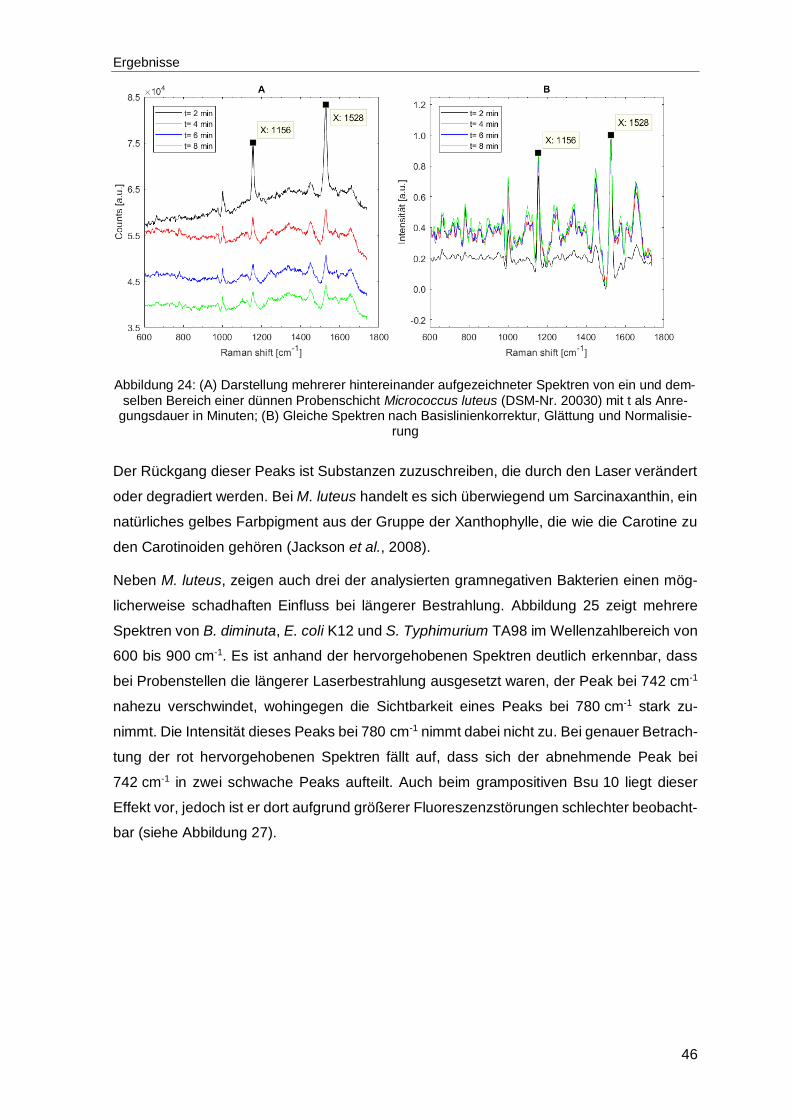
Ergebnisse
46
Abbildung 24: (A) Darstellung mehrerer hintereinander aufgezeichneter Spektren von ein und dem-
selben Bereich einer dünnen Probenschicht Micrococcus luteus (DSM-Nr. 20030) mit t als Anre-gungsdauer in Minuten; (B) Gleiche Spektren nach Basislinienkorrektur, Glättung und Normalisie-
rung
Der Rückgang dieser Peaks ist Substanzen zuzuschreiben, die durch den Laser verändert
oder degradiert werden. Bei M. luteus handelt es sich überwiegend um Sarcinaxanthin, ein
natürliches gelbes Farbpigment aus der Gruppe der Xanthophylle, die wie die Carotine zu
den Carotinoiden gehören (Jackson et al., 2008).
Neben M. luteus, zeigen auch drei der analysierten gramnegativen Bakterien einen mög-
licherweise schadhaften Einfluss bei längerer Bestrahlung. Abbildung 25 zeigt mehrere
Spektren von B. diminuta, E. coli K12 und S. Typhimurium TA98 im Wellenzahlbereich von
600 bis 900 cm-1. Es ist anhand der hervorgehobenen Spektren deutlich erkennbar, dass
bei Probenstellen die längerer Laserbestrahlung ausgesetzt waren, der Peak bei 742 cm-1
nahezu verschwindet, wohingegen die Sichtbarkeit eines Peaks bei 780 cm-1 stark zu-
nimmt. Die Intensität dieses Peaks bei 780 cm-1 nimmt dabei nicht zu. Bei genauer Betrach-
tung der rot hervorgehobenen Spektren fällt auf, dass sich der abnehmende Peak bei
742 cm-1 in zwei schwache Peaks aufteilt. Auch beim grampositiven Bsu 10 liegt dieser
Effekt vor, jedoch ist er dort aufgrund größerer Fluoreszenzstörungen schlechter beobacht-
bar (siehe Abbildung 27).

Ergebnisse
47
Abbildung 25: Auswirkung längerer Bestrahlung auf Peaks im Wellenzahlbereich von 600 bis 900 cm-1 bei drei verschiedenen gramnegativen Bakterien; Blau hervorgehoben ein Spektrum mit kurzer Bestrahlungsdauer und Rot ein Spektrum mit langer Bestrahlungsdauer (Die Spektren wur-
den basislinienkorrigiert, geglättet und normalisiert)
3.2 Modellentwicklung
3.2.1 Datenvorbehandlung
Abbildung 26 A enthält alle zur Modellentwicklung gesammelten unbehandelten Raman-
Spektren. Zur besseren Darstellung wurden die Spektren normalisiert. Die hauptsächlich
der Fluoreszenz zuzuschreibenden „Offsets“ variieren, wobei die Auswirkungen häufiger
auf dem niedrigeren Wellenzahlbereich zu beobachten sind. Nach Datenvorbehandlung
durch Basislinienkorrektur, Glättung und Normalisierung (Abbildung 26 B), werden die spe-
zifischen Banden deutlich sichtbar. Auch die beiden Banden, die bei den unbehandelten
Spektren kaum ersichtlich sind (bei etwa 740 cm-1 und 780 cm-1), sind nach Datenvorbe-
handlung bei allen gramnegativen Bakterien klar zu erkennen. Mit Ausnahme der drei
Micrococcus luteus-Stämme haben die Banden bei 1003 cm-1 (charakteristisch für Phe-
nylalanin) und 1450 cm-1 (charakteristisch für Lipide) am häufigsten die Maximalintensität.
Die für Carotinoide typischen Banden bei 1156 cm-1 und 1528 cm-1 sind das auffälligste
Merkmal aller Micrococcus luteus-Stämme.
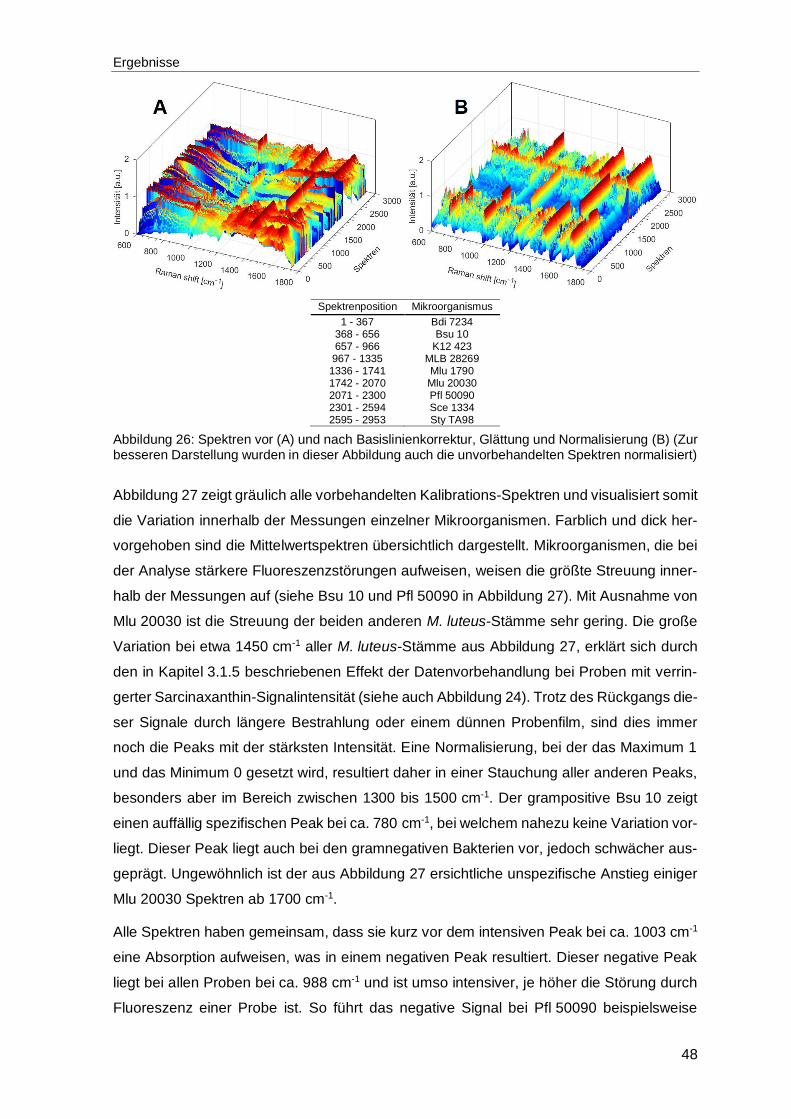
Ergebnisse
48
Spektrenposition Mikroorganismus
1 - 367 Bdi 7234 368 - 656 Bsu 10 657 - 966 K12 423 967 - 1335 MLB 28269 1336 - 1741 Mlu 1790 1742 - 2070 Mlu 20030 2071 - 2300 Pfl 50090 2301 - 2594 Sce 1334 2595 - 2953 Sty TA98
Abbildung 26: Spektren vor (A) und nach Basislinienkorrektur, Glättung und Normalisierung (B) (Zur besseren Darstellung wurden in dieser Abbildung auch die unvorbehandelten Spektren normalisiert)
Abbildung 27 zeigt gräulich alle vorbehandelten Kalibrations-Spektren und visualisiert somit
die Variation innerhalb der Messungen einzelner Mikroorganismen. Farblich und dick her-
vorgehoben sind die Mittelwertspektren übersichtlich dargestellt. Mikroorganismen, die bei
der Analyse stärkere Fluoreszenzstörungen aufweisen, weisen die größte Streuung inner-
halb der Messungen auf (siehe Bsu 10 und Pfl 50090 in Abbildung 27). Mit Ausnahme von
Mlu 20030 ist die Streuung der beiden anderen M. luteus-Stämme sehr gering. Die große
Variation bei etwa 1450 cm-1 aller M. luteus-Stämme aus Abbildung 27, erklärt sich durch
den in Kapitel 3.1.5 beschriebenen Effekt der Datenvorbehandlung bei Proben mit verrin-
gerter Sarcinaxanthin-Signalintensität (siehe auch Abbildung 24). Trotz des Rückgangs die-
ser Signale durch längere Bestrahlung oder einem dünnen Probenfilm, sind dies immer
noch die Peaks mit der stärksten Intensität. Eine Normalisierung, bei der das Maximum 1
und das Minimum 0 gesetzt wird, resultiert daher in einer Stauchung aller anderen Peaks,
besonders aber im Bereich zwischen 1300 bis 1500 cm-1. Der grampositive Bsu 10 zeigt
einen auffällig spezifischen Peak bei ca. 780 cm-1, bei welchem nahezu keine Variation vor-
liegt. Dieser Peak liegt auch bei den gramnegativen Bakterien vor, jedoch schwächer aus-
geprägt. Ungewöhnlich ist der aus Abbildung 27 ersichtliche unspezifische Anstieg einiger
Mlu 20030 Spektren ab 1700 cm-1.
Alle Spektren haben gemeinsam, dass sie kurz vor dem intensiven Peak bei ca. 1003 cm-1
eine Absorption aufweisen, was in einem negativen Peak resultiert. Dieser negative Peak
liegt bei allen Proben bei ca. 988 cm-1 und ist umso intensiver, je höher die Störung durch
Fluoreszenz einer Probe ist. So führt das negative Signal bei Pfl 50090 beispielsweise
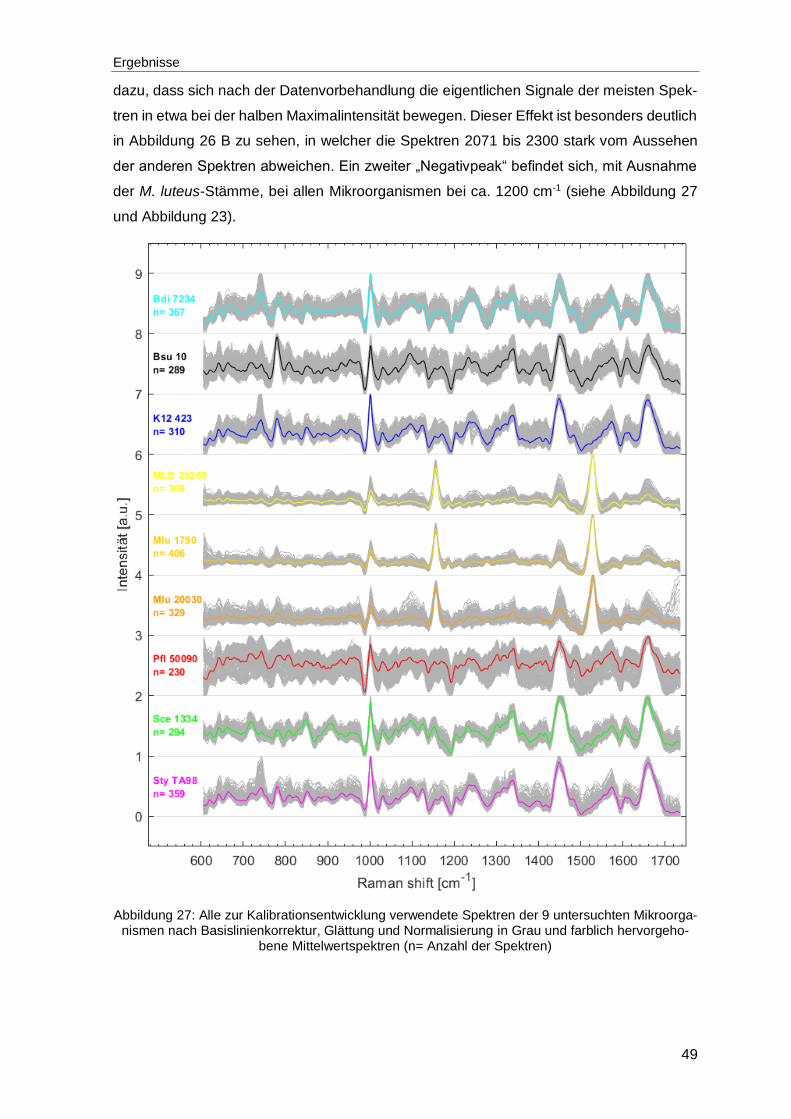
Ergebnisse
49
dazu, dass sich nach der Datenvorbehandlung die eigentlichen Signale der meisten Spek-
tren in etwa bei der halben Maximalintensität bewegen. Dieser Effekt ist besonders deutlich
in Abbildung 26 B zu sehen, in welcher die Spektren 2071 bis 2300 stark vom Aussehen
der anderen Spektren abweichen. Ein zweiter „Negativpeak“ befindet sich, mit Ausnahme
der M. luteus-Stämme, bei allen Mikroorganismen bei ca. 1200 cm-1 (siehe Abbildung 27
und Abbildung 23).
Abbildung 27: Alle zur Kalibrationsentwicklung verwendete Spektren der 9 untersuchten Mikroorga-nismen nach Basislinienkorrektur, Glättung und Normalisierung in Grau und farblich hervorgeho-
bene Mittelwertspektren (n= Anzahl der Spektren)

Ergebnisse
50
3.2.2 Hauptkomponentenanalyse
Eine PCA wurde sowohl mit den unbehandelten, als auch mit den behandelten Daten
durchgeführt. Die PCA nach Vorbehandlung der Daten kann mit den ersten drei Hauptkom-
ponenten 93,1 % der Varianz erklären. Die erklärte Varianz der ersten 10 PCs geht aus
Abbildung 28 hervor. Um 95 % der Varianz zu erklären, müssen die ersten fünf PCs heran-
gezogen werden.
Abbildung 28: Abnehmende erklärende Varianz der PCs nach PCA mit vorbehandelten Daten
Abbildung 29 stellt aus zwei Betrachtungswinkeln die ersten drei PCs nach der Hauptkom-
ponentenanalyse dar. Die unvorbehandelten Daten zeigen bei Darstellung der ersten drei
PCs keine deutlichen Muster. Datenpunkte der nicht behandelten Spektren, die nahezu auf
einer exakten Geraden hintereinander aufgereiht sind, erklären sich durch die bis zu zehn-
fache Mehrfachmessung von ein und derselben Stelle einer Probe was häufig zu Spektren
führt, die sich in ihren Signaturen kaum unterscheiden, jedoch stark in der Intensität. Be-
sonders deutlich sind solche Muster bei den unbehandelten Pfl 50090 Daten zu erkennen.
Aus der Darstellung der PCA mit den vorbehandelten Daten aus Abbildung 29, geht die
deutliche Aufteilung in drei Hauptcluster hervor. Die sich grundlegend in ihrem Aufbau von
Bakterien unterscheidenden Hefen (Sce 1334), weisen die größte Distanz zum Zentrum der
Daten auf. Die drei M. luteus-Stämme, welche sich durch die Besonderheit des enthaltenen
Carotinoids auszeichnen, liegen dicht zusammen und sind ebenfalls weit vom Zentrum ent-
fernt. Die Daten von Bdi 7234, Bsu 10, K12 423, Pfl 50090 und Sty TA98 überschneiden
sich zwar alle, sind jedoch größtenteils separat angehäuft und ohne größere Ausreißer.
Auffällig ist die Aufteilung der Pfl 50090-Daten in zwei Untergruppen, von denen sich eine
nahezu im Zentrum der Daten befindet (Kultur 2) und die andere weiter außerhalb (Kul-
tur 1). Eine Aufteilung der Pfl 50090-Spektren in zwei Bereiche, ist auch bei den unbehan-
delten Daten in Abbildung 29 zu erkennen.
71,32
19,32
2,45 1,55 0,74 0,55 0,51 0,45 0,25 0,210
20
40
60
80
1 2 3 4 5 6 7 8 9 10
Erk
lärt
e V
ari
an
z [%
]
PC

Ergebnisse
51
Abbildung 29: Darstellung der ersten drei PCs der Hauptkomponentenanalyse von allen Kalibrati-ons-Spektren mit und ohne Vorbehandlung aus jeweils zwei verschiedenen Betrachtungswinkeln

Ergebnisse
52
3.2.3 Klassifikationsverfahren und Kreuzvalidierung
Jedes Modell aller Klassifizierungsverfahren wurde mit dimensionsreduzierten Daten er-
stellt. Es wurden Modelle nach 8, 10, 12, 15 und 20 PCs generiert. Nachfolgend werden die
Wahrheits- bzw. Vorhersagematrices der Modelle nach 5-facher Kreuzvalidierung bei Ver-
wendung von 8 und 15 PCs dargestellt. Die restlichen Vorhersagematrices befinden sich in
Anhang 1 (QDA) Anhang 2 (kNN) und Anhang 3 (SVM). Eine Wahrheitsmatrix zeigt die
wahren Werte, also die zu klassifizierenden Mikroorganismen und die kreuzvalidierten Vor-
hersagewerte des Modells. Daraus kann eine erste Einschätzung der Vorhersagegüte der
Modelle erfolgen und es kann nach Mustern bei falsch positiven Vorhersagen gesucht wer-
den.
3.2.3.1 Quadratische Diskriminanzanalyse
Abbildung 30 zeigt die Vorhersagematrices zweier QDA-Modelle unter Verwendung von 8
und 15 PCs. Die Vorhersagegüte bewegt sich zwischen 95 und 98 %. Bei beiden Modellen
liegen die meisten falschen Vorhersagen bei den drei M. luteus-Stämmen vor, wobei
MLB 28269 am schlechtesten und Mlu 20030 am genauesten vorhergesagt wird. Beim Mo-
dell mit 15 PCs liegen kaum noch fehlerhafte Vorhersagen außerhalb der Stammebene vor,
wohingegen beim Modell mit 8 PCs falsche Vorhersagen stärker verteilt sind. Pfl 50090
wird dort 210 Mal richtig vorhergesagt und 20 Mal als Sty TA98, was einer Genauigkeit von
91,3 % entspricht. Bei Sty TA98 gibt es bei 8 PCs falsche Vorhersagen für Pfl 50090,
K12 423 und Bdi 7234. Bei all diesen falschen Vorhersagen handelt es sich, wie auch bei
Sty TA98 selbst, um gramnegative Bakterien.
8 PCs (94,9 %) 15 PCs (98,3 %)
Wa
hr
Vorhersagen Vorhersagen
Abbildung 30: Vorhersagematrices bei Verwendung der quadratischen Diskriminanzanalyse von
Modellen mit 8 und 15 PCs mit der durchschnittlichen Vorhersagegüte in Prozent

Ergebnisse
53
3.2.3.2 Nächster-Nachbar-Algorithmus
Sowohl die richtigen als auch fehlerhaften Vorhersagen aus den kNN-Modellen von Abbil-
dung 31 zeigen ähnliche Muster wie QDA-Modelle aus Abbildung 30. Wobei die Vorhersa-
gen außerhalb der Stammebene bei dem kNN-Modell mit 15 PCs etwas genauer sind als
die Vorhersagen des QDA-Modells. Nur eine einzige Vorhersage liegt bei kNN mit 15 PCs
außerhalb der Stammebene, womit dieses Modell die genauesten Vorhersagen auf Art-
Ebene erzielt. Auffallend ist, dass Mlu 20030 besonders bei nur 8 PCs deutlich ungenauer
vorhergesagt wird (mehr als 10 % falsche Vorhersagen) als bei den QDA-Modellen.
8 PCs (94,4 %) 15 PCs (98,4 %)
Wa
hr
Vorhersagen Vorhersagen
Abbildung 31: Vorhersagematrices bei Verwendung des Nächste-Nachbar-Algorithmus von Model-
len mit 8 und 15 PCs mit der durchschnittlichen Vorhersagegüte in Prozent

Ergebnisse
54
3.2.3.3 Support Vector Maschine
Prozentual gesehen liefert die SVM bei Verwendung von 8 PCs etwa 2 % genauere Vor-
hersagen als entsprechende kNN- und QDA-Modelle. Auch das SVM Modell mit 15 PCs
zeigt eine um 1 % genauere Vorhersage als die Modelle mit 15 PCs aus Abbildung 30 und
Abbildung 31. Besonders auf Stammebene scheint nach Abbildung 32 die SVM deutlich
besser abzuschneiden als die vergleichbaren Modelle der beiden anderen Klassifizierungs-
verfahren. Bei der SVM (15 PCs) wird jedoch zum ersten Mal eines der 329 Spektren eines
M. luteus-Stamm fälschlicherweise als Bsu 10 vorhergesagt. Zwar ist dies prozentual ge-
sehen immer noch sehr viel genauer als die anderen Modelle, jedoch handelt es sich bei
Bsu 10 um ein Bakterium, welches sich stark von Mlu 20030 unterscheidet, was die falsche
Vorhersage schwerer nachvollziehbar macht, als falsche Vorhersagen innerhalb der
Stammebene von M. luteus.
8 PCs (96,5 %) 15 PCs (99,4 %)
Wa
hr
Vorhersagen Vorhersagen
Abbildung 32: Vorhersagematrices bei Verwendung der Support Vector Maschine von Modellen mit 8 und 15 PCs mit der durchschnittlichen Vorhersagegüte in Prozent

Ergebnisse
55
3.3 Praktische Validierung
Die aus Kapitel 3.2.3 hervorgehenden Wahrheitsmatrices liefern eine erste Einschätzung
über die Genauigkeit der Vorhersagemodelle. Allerdings kann es trotz Kreuzvalidierung zu
„Overfitting“ kommen. Aus diesem Grund ist es unerlässlich die Modelle auf völlig unbe-
kannte Daten anzuwenden, also auf Spektren, welche nicht im Kalibrationsset enthalten
sind und aus anderen Proben der gleichen Mikroorganismen stammen.
Für die Validierung der entwickelten Kalibrationsmodelle wurden insgesamt 900 weitere
Spektren gesammelt. Die Validierungssets aller Mikroorganismen bestehen jeweils aus 100
Spektren, von denen 50 aus einer ersten Kultur und 50 aus einer zweiten Kultur stammen.
Aufgrund schlechten Wachstums der Pfl 50090-Kultur 1, wurden dort keine Spektren auf-
genommen, dafür 100 Validierungsspektren von einer Kultur 2.
Eine PCA, berechnet aus den Kalibrations- und Validierungsspektren, dargestellt in Abbil-
dung 33, zeigt sowohl die Kalibrierungsspektren (Punkte) als auch die Validierungsspektren
(Quadrate). Insgesamt decken die Kalibrationsspektren den größten Teil der Validierungs-
spektren ab. Eine Zuordnung der Validierungsspektren von M. luteus (auf Art-Ebene) und
S. cerevisiae, könnte bei Darstellung der ersten drei PCs mit dem bloßen Auge erfolgen.
Dennoch sind Unterschiede erkennbar. Einige Validierungsspektren von Sce 1334 sind
stärker in die PC3-Dimension gestreut und werden nicht vom Kalibrationsset abgedeckt.
Auch bei M. luteus bilden einige Validierungsspektren die äußersten Grenzen des entspre-
chenden Clusters. Jedoch beschrieb dort derselbe Stamm (Mlu 20030) zuvor bereits die
äußerste Grenze des Clusters in die gleiche Richtung, womit die Daten noch am ehesten
diesem Stamm zuzuordnen sind.
Abbildung 33: Darstellung der ersten drei Hauptkomponenten einer PCA der Kalibrierungsspektren
(markiert als Punkte n= 2953) und Validierungsspektren (markiert als Quadrate n= 900) aus zwei Ansichten (A und B)
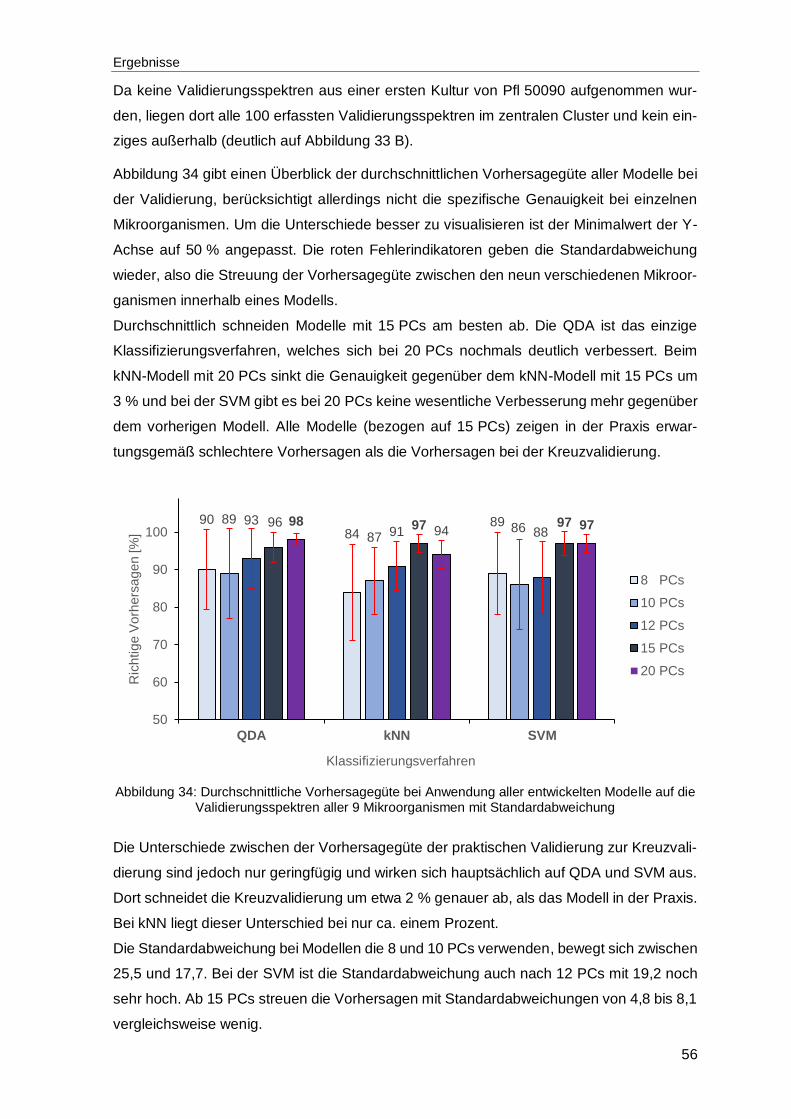
Ergebnisse
56
Da keine Validierungsspektren aus einer ersten Kultur von Pfl 50090 aufgenommen wur-
den, liegen dort alle 100 erfassten Validierungsspektren im zentralen Cluster und kein ein-
ziges außerhalb (deutlich auf Abbildung 33 B).
Abbildung 34 gibt einen Überblick der durchschnittlichen Vorhersagegüte aller Modelle bei
der Validierung, berücksichtigt allerdings nicht die spezifische Genauigkeit bei einzelnen
Mikroorganismen. Um die Unterschiede besser zu visualisieren ist der Minimalwert der Y-
Achse auf 50 % angepasst. Die roten Fehlerindikatoren geben die Standardabweichung
wieder, also die Streuung der Vorhersagegüte zwischen den neun verschiedenen Mikroor-
ganismen innerhalb eines Modells.
Durchschnittlich schneiden Modelle mit 15 PCs am besten ab. Die QDA ist das einzige
Klassifizierungsverfahren, welches sich bei 20 PCs nochmals deutlich verbessert. Beim
kNN-Modell mit 20 PCs sinkt die Genauigkeit gegenüber dem kNN-Modell mit 15 PCs um
3 % und bei der SVM gibt es bei 20 PCs keine wesentliche Verbesserung mehr gegenüber
dem vorherigen Modell. Alle Modelle (bezogen auf 15 PCs) zeigen in der Praxis erwar-
tungsgemäß schlechtere Vorhersagen als die Vorhersagen bei der Kreuzvalidierung.
Abbildung 34: Durchschnittliche Vorhersagegüte bei Anwendung aller entwickelten Modelle auf die Validierungsspektren aller 9 Mikroorganismen mit Standardabweichung
Die Unterschiede zwischen der Vorhersagegüte der praktischen Validierung zur Kreuzvali-
dierung sind jedoch nur geringfügig und wirken sich hauptsächlich auf QDA und SVM aus.
Dort schneidet die Kreuzvalidierung um etwa 2 % genauer ab, als das Modell in der Praxis.
Bei kNN liegt dieser Unterschied bei nur ca. einem Prozent.
Die Standardabweichung bei Modellen die 8 und 10 PCs verwenden, bewegt sich zwischen
25,5 und 17,7. Bei der SVM ist die Standardabweichung auch nach 12 PCs mit 19,2 noch
sehr hoch. Ab 15 PCs streuen die Vorhersagen mit Standardabweichungen von 4,8 bis 8,1
vergleichsweise wenig.
9084
8989
8786
9391 88
96 97 979894 97
50
60
70
80
90
100
QDA kNN SVM
Ric
htig
e V
orh
ers
ag
en
[%
]
Klassifizierungsverfahren
8 PCs
10 PCs
12 PCs
15 PCs
20 PCs

Ergebnisse
57
Um nachzuvollziehen, welche falschen Vorhersagen hauptsächlich für größere Stan-
dardabweichungen verantwortlich sind und um Modelle nach der Vorhersagegüte spezifi-
scher Mikroorganismen einschätzen zu können, ist es wichtig, die Anzahl korrekt hervorge-
sagter Spektren jedes Modells einzeln zu betrachten.
Abbildung 35 zeigt für alle drei angewandten Klassifizierungsverfahren „Heatmaps“, welche
die Anzahl der richtig vorhergesagten Spektren bei entsprechender Anzahl PCs darstellen.
Ungenaue Vorhersagen mit weniger als 50 % Trefferquote, werden als rötlich dargestellt.
Vorhersagen mit über 90 % Richtigkeit werden als grünlich dargestellt. Werte dazwischen
bewegen sich im orangefarbenen bis gelben Bereich.
QDA kNN SVM
PCs PCs PCs
Abbildung 35: Absolute Anzahl der richtig vorhergesagten Spektren bei der Validierung dreier Klas-sifizierungsverfahren bei variierender Anzahl PCs
Besonders deutlich gehen aus Abbildung 35 die ungenauen Vorhersagen für K12 423 her-
vor. Die 100 Spektren dieses Bakteriums werden bei allen drei Klassifizierungsverfahren
bei Verwendung von bis zu 12 PCs mit etwa weniger als der Hälfte richtig vorhergesagt. Mit
steigender Anzahl an PCs werden die Vorhersagen jedoch genauer, sodass mit dem kNN-
Modell mit 15 PCs alle 100 Spektren von K12 423 richtig vorhergesagt werden. Aus den
Wahrheitsmatrices der Kreuzvalidierung in Kapitel 3.2.3 und den aus den Anhängen 1 bis
3 geht hervor, dass die Genauigkeit der Modelle mit zunehmender Anzahl an Dimensionen
steigt. Die Validierung zeigt allerdings, dass sich die Modelle von kNN und SVM ab 20 PCs
wieder verschlechtern. Auf die ideale Anzahl an PCs, die für ein Modell herangezogen wer-
den sollten, kann daraus jedoch noch nicht klar geschlossen werden, da es vom Mikroor-
ganismus abhängig ist. So liefert beispielsweise die SVM bei 20 PCs die genauesten Vor-
hersagen für die drei M. luteus-Stämme, jedoch werden die Vorhersagen für Pfl 50090 um
13 % ungenauer als beim SVM-Modell mit nur 10 PCs. Auch bei den kNN-Modellen werden
23 % der Pfl 50090-Spektren erst ab 20 PCs falsch vorhergesagt. Bei den Modellen mit
weniger als 20 PCs liegen dort die Fehlerquoten zwischen null und einem Prozent.

Ergebnisse
58
Für eine solide Einschätzung über die Leistung der Modelle ist es nicht nur erforderlich zu
wissen, welche Mikroorganismen wie häufig richtig bzw. falsch vorhergesagt werden, son-
dern auch die falsch positiven Vorhersagen zu kennen. So zeigt sich beispielsweise, dass
nahezu alle falsch vorhergesagten K12 423 Spektren als Sty TA98 vorhergesagt wurden;
einem Bakterium, das mit K12 423 eng verwandt ist.
Die nachfolgenden Tabellen 5 bis 7 zeigen die Validierungsvorhersagen des jeweils genau-
esten Modells jedes Klassifizierungsverfahrens im Detail. Bei der SVM ist das Modell mit
20 PCs um 0,11 % genauer als das Modell mit 15 PCs, dennoch könnte das 20 PC-Modell
aufgrund des starken Verlusts der Genauigkeit bei Pfl 50090, als das schlechtere Modell
eingestuft werden. Aus diesem Grund werden die Vorhersagen beider SVM-Modelle dar-
gestellt. Die spezifischen Vorhersagen der restlichen Modelle befinden sich in den Anhän-
gen 4 bis 6.
Die aus Tabelle 5 hervorgehenden Vorhersagen zeigen, dass die fünf falschen Identifizie-
rungen für K12 423 Treffer für Sty TA98 sind. Die falschen Vorhersagen bei M. luteus liegen
innerhalb der Stammebene, wobei alle Mlu 20030 Spektren richtig vorhergesagt wurden.
Tabelle 5: Validierungsvorhersagen des QDA-Modells mit 20 PCs (Vorhersagegüte 98,1 %)
QDA 20 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wahr
Bdi 7234 100 Bsu 10 100
K12 423 95 5 MLB 28269 97 3 Mlu 1790 7 91 Mlu 20030 100 Pfl 50090 100 Sce 1334 100 Sty TA98 100
Das genauste kNN-Modell liefert mit 95 bis 100 % sehr gute Vorhersagen bei allen Mikro-
organismen mit Ausnahme der M. luteus-Stämme MLB 28269 und Mlu 1790 (siehe Tabelle
6). Dort liegt die Vorhersagegüte bei 92 bzw. 87 %. Beim kNN-Modell werden im Gegensatz
zum QDA-Modell alle K12 423 richtig vorhergesagt, jedoch werden genau komplementär
dazu fünf Sty TA98 Spektren als K12 423 interpretiert. Mit einer durchschnittlichen Vorher-
sagegüte von 97,1 % ist das kNN-Modell, mit geringem Abstand das in der Praxis leistungs-
fähigste Modell, aller auf 15 PCs basierenden Modelle.

Ergebnisse
59
Tabelle 6:Validierungsvorhersagen des kNN-Modells mit 15 PCs (Vorhersagegüte 97,1 %)
kNN 15 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 100 Bsu 10 100
K12 423 100 MLB 28269 92 7 1 Mlu 1790 13 87 Mlu 20030 100 Pfl 50090 100 Sce 1334 100 Sty TA98 5 95
Aus Tabelle 7 geht hervor, dass die SVM bei Verwendung von 15 PCs die genausten Vor-
hersagen auf Stammebene der M. luteus Proben liefert. Jedoch ist das genannte SVM-
Modell schlecht geeignet, um Vorhersagen für K12 423 zu treffen, da dort 20 % der Spek-
tren fälschlicherweise als Sty TA98 identifiziert wurden.
Das SVM-Modell basierend auf 20 PCs schneidet bei der Validierung geringfügig besser
ab als das 15 PC-Modell. Die Verbesserung auf Stammebene ist vernachlässigbar klein,
jedoch sinkt die Fehlerrate bei K12 423 von 20 % auf 9 %. Bei den falschen Vorhersagen
handelt es sich hierbei wieder um den eng verwandten Sty TA98. Auffällig ist, dass bei der
SVM mit 20 PCs Pfl 50090 acht Mal als StyTA98 und fünf Mal als Sce 1334 vorhergesagt
wird. Diese falschen Vorhersagen sind aufgrund der größeren spektralen Unterschiede die-
ser Mikroorganismen schwer nachvollziehbar. Aufgrund der Komplexität einer trennenden
Hyperebene der SVM in einem 20-dimensionalen Raum, sind fehlerhafte Vorhersagen oh-
nehin schwerer zu interpretieren als beispielsweise bei kNN.
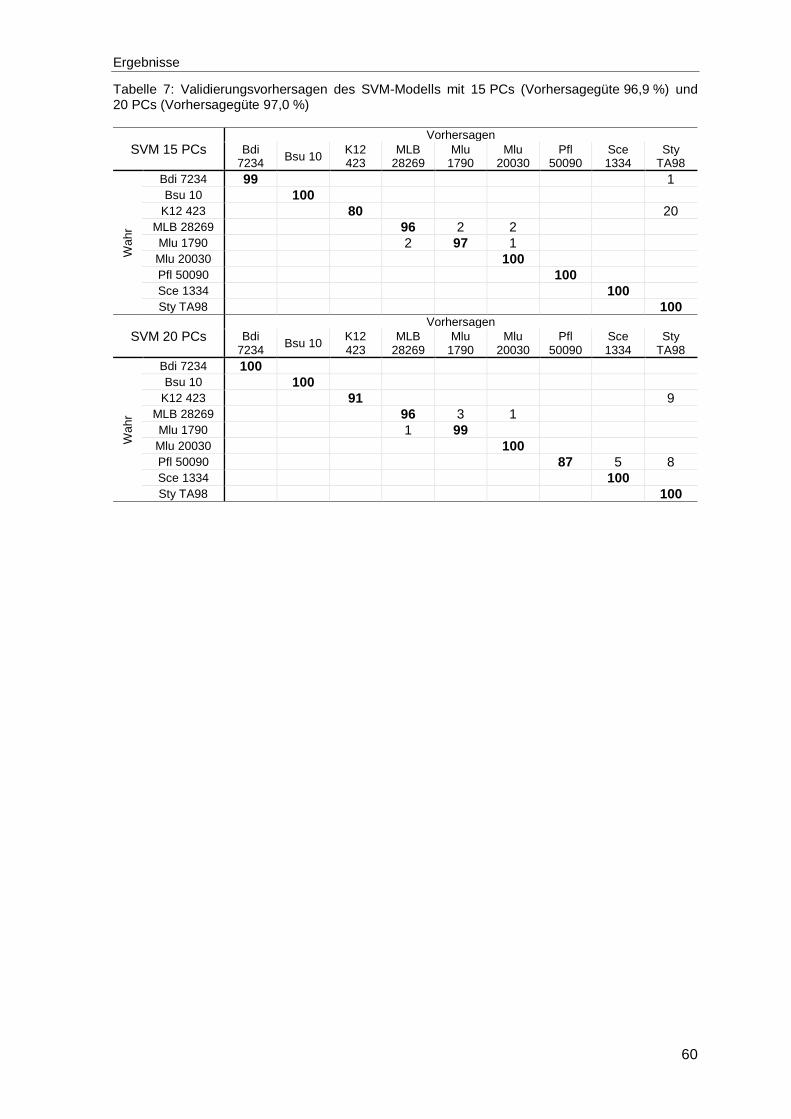
Ergebnisse
60
Tabelle 7: Validierungsvorhersagen des SVM-Modells mit 15 PCs (Vorhersagegüte 96,9 %) und 20 PCs (Vorhersagegüte 97,0 %)
SVM 15 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 99 1 Bsu 10 100
K12 423 80 20 MLB 28269 96 2 2 Mlu 1790 2 97 1 Mlu 20030 100 Pfl 50090 100 Sce 1334 100 Sty TA98 100
SVM 20 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 100 Bsu 10 100
K12 423 91 9 MLB 28269 96 3 1 Mlu 1790 1 99 Mlu 20030 100 Pfl 50090 87 5 8 Sce 1334 100 Sty TA98 100

Diskussion
61
4 Diskussion
4.1 Spektrenaufzeichnung
Zwar können bei gleichbleibender Laserleistung (100 %) und längeren Belichtungszeiten
bei einigen Mikroorganismen Spektren besserer Qualität gewonnen werden (z.B. bei
Bdi 7234 und Sty TA98), jedoch weisen Spektren bei Belichtungszeiten über 3 Sekunden
bei vielen Mikroorganismen zu hohe „Counts“ auf und können nicht mehr detektiert werden
(z.B. Bsu 10, Pfl 50090, K12 423). Die in der Methodik aufgelisteten Messeinstellungen
(Kapitel 2.2.4), insbesondere die Belichtungszeit von 3 Sekunden und die Akkumulation
von 15 Einzelspektren, erweisen sich als guter Kompromiss zwischen Messdauer, Qualität
des Spektrums und Wirksamkeit auf die hier verwendeten Mikroorganismen.
Störende Fluoreszenz konnte bei allen Proben, mit Ausnahme von Pfl 50090, effektiv aus-
gebrannt werden. Bei problematischen Proben wie Pfl 50090 können zusätzliche Wasch-
schritte des Probenmaterials mit NaCl-Lösung zu besseren Resultaten führen (Willemse-
Erix et al., 2010). Allerdings müsste dies im Kalibrationsset mit entsprechenden Referenz-
spektren berücksichtigt werden. Eine zuverlässige Differenzierung verschiedener Pseu-
domonaden-Arten ist mit der bislang entwickelten Methode ohne weitere Maßnahmen zur
Fluoreszenzverringerung eher unwahrscheinlich, da auch andere Pseudomonaden wie
P. aeruginosa störende Pyoverdine produzieren (Kahlon, 2016).
Eine Fluoreszenzverminderung durch Chemikalien wie Wasserstoffperoxid (H2O2), kann
bei einigen biologischen Proben wie Knochengewebe erfolgreich erreicht werden, wobei
die Zeitspanne zwischen Fluoreszenzreduzierung und der Beschädigung der Probe sehr
gering ist (Golcuk et al., 2006; Penel et al., 1998). Aufgrund der Toxizität von H2O2 gegen-
über vielen Mikroorganismen, schon bei geringer Konzentration, scheidet dieses als Mittel
zur Fluoreszenzverringerung sicherlich aus.
Eine noch längere Vorbestrahlung der Proben (z.B. mehr als 30 Minuten) kann zu aussa-
gekräftigeren Spektren führen. Hierbei besteht aber zum einen das Risiko das Material noch
schwerer zu beschädigen und zum anderen wäre der große Vorteil der schnellen Analyse
ab einem gewissen Punkt nicht mehr gegeben. Auch die Modellentwicklung würde sehr viel
länger dauern, wenn die Aufnahmezeit der Referenzspektren verlängert wird.
Die Auswirkungen längerer Vorbestrahlung auf das Spektrum von M. luteus legen nahe,
dass es zu einer Schädigung des enthaltenen Sarcinaxanthins kommt. Carotinoide dienen
dem Bakterium unter anderem zur Stärkung der Membranintegrität und als Schutz vor ioni-
sierender Strahlung. Nicht alle Carotinoide sind stark photoinstabil und die Diversität der

Diskussion
62
Carotinoide machen sie bei einigen Bakterien selbst auf Stammebene zu einem idealen
Unterscheidungsmerkmal. (B. N. V. Kumar et al., 2015)
Aus Abbildung 25 geht der Effekt des Rückgangs der Peaks bei ca. 742 cm-1 bei längerer
Laserbestrahlung dreier gramnegativer Bakterien hervor. Die Bande bei dieser Wellenzahl
könnte von Peptidoglycan stammen (Wang et al., 2017), jedoch sollte sie dann bei gram-
positiven deutlicher erkennbar sein, als bei gramnegativen und wäre weder photoinstabil
noch besonders empfindlich gegenüber einer möglichen Erhitzung durch den Laser. In der
Literatur, die für Tabelle 2 und Abbildung 7 herangezogen wurden, sind für Mikroorganis-
men keine spezifischen Banden im Bereich von 742 cm-1 aufgeführt. Die Bande könnte in
dieser Arbeit durch Rückstände des Nährmediums verursacht worden sein. Abbildung 10
zeigt bei einem Raman-Spektrum von getrocknetem TSA ebenfalls einen Peak bei ca.
742 cm-1, was diesen Verdacht erhärtet. Es könnte sich allerdings auch um extrazelluläre
polymere Substanzen (EPS) handeln, die durch die Energie des Laserbeschusses nach
und nach verdrängt werden. Kanematsu et al. berichten von Raman-Peaks bei 743 cm-1,
welche den Polysacchariden in EPS des Biofilms von E. coli zuzuschreiben sind
(Kanematsu et al., 2016).
Bei dem besonders stark ausgeprägten Peak von Bacillus subtilis bei 780 cm-1, handelt es
sich um ein für DNA/RNA typisches Signal (Ringschwingungen von Cytosin und Uracil). Da
das Genom von B. subtilis nicht größer als beispielsweise das Genom von E. coli ist, könn-
ten diese Signale aufgrund der unterschiedlich beschaffenen Zellwand bei grampositiven
leichter zu gewinnen sein als bei gramnegativen. Es könnte allerdings auch mit der Fähig-
keit der Sporenbildung von B. subtilis zusammenhängen; so enthalten Sporen drei bis vier
Mal so viel RNA als DNA (P. Zhang et al., 2011) und somit auch mehr Uracil, welches bei
etwa 780 cm-1 das charakteristische Signal zeigt.
4.2 Datenvorbehandlung
Trotz stark variierender Auswirkungen verschiedener Messbedingungen auf die Basislinie
(siehe Abbildung 23), kann mit der verwendeten MATLAB-Routine eine gleichmäßige
Transformierung der Spektren erzielt werden (siehe Abbildung 27). Anhand der PCA nach
Datenvorbehandlung wird eine klare Auftrennung der Daten in spezifische Cluster sichtbar.
Der ungewöhnliche Anstieg einiger Spektren von Mlu 20030 nach Datenvorbehandlung
(Abbildung 27) liegt an etwa 10 Spektren, welche von einer sehr dünnen Bakterienschicht
aufgenommen wurden. Hier verlaufen die Spektren, ähnlich wie das Spektrum des Glasob-
jektträgers (Abbildung 10) als ansteigende Gerade mit gering ausgeprägten Signaturen.
Teilweise erkennbar sind diese Spektren in den ausgebleichten Hintergrundspektren aus
Abbildung 23 (ab 1600 cm-1). Bei solchen Fällen, in denen die letzten Datenpunkte die

Diskussion
63
höchsten Intensitäten aufweisen, führt das angewandte Verfahren zur Basislinienkorrektur
zu einem entsprechenden Anstieg an dieser Stelle. Die betroffenen Spektren aus dem Ka-
librationsset zu entfernen, könnte zu einer leichten Verbesserung der Modelle führen.
Als besonders bedeutsam kann die auftretende Absorption nahezu aller Spektren bei etwa
988 cm-1 und 1200 cm-1 angesehen werden (siehe Abbildung 23). Mit zunehmender Fluo-
reszenz steigt besonders bei 988 cm-1 die Absorption. Dieser negative Peak verleiht dem
Spektrum nach Vorbehandlung einen gleichmäßigen „Offset“ aller Banden (siehe Abbildung
27). Möglicherweise beruhen die sehr genauen Vorhersagen zu einem großen Teil auf ge-
nau diesem Umstand. Mlu 20030 ist der Stamm, welcher durchweg am genauesten vorher-
gesagt wird, selbst bei Verwendung von nur 8 PCs wird er mit allen drei Klassifizierungs-
verfahren zu durchschnittlich 97,3 % richtig identifiziert (siehe Abbildung 35). Die beiden
anderen M. luteus-Stämme, die einen geringeren „Offset“ aufweisen, werden bei 8 PCs mit
allen Klassifizierungsverfahren durchschnittlich nur zu 79 % (MLB 28269) bzw. 90 %
(Mlu 1790) richtig identifiziert. Um zu untersuchen ob diese „Negativpeaks“ für genauere
Vorhersagen sorgen, müssten neue Modelle erstellt werden, bei denen der ausgewählte
Wellenzahlbereich vor der Basislinienkorrektur entsprechend angepasst wird. Sollte der
durch die Absorption verliehene „Offset“ für die gute Differenzierbarkeit verantwortlich sein,
wäre dies nicht zwingend nachteilig, da diese Auswirkungen auf die Spektren charakteris-
tisch und reproduzierbar sind. Allerdings wären die Modelle dann ggfs. nur beschränkt auf
externe Daten anwendbar, die ein solches Verhalten nicht aufweisen. Der „Negativpeak“
bei 988 cm-1 wird in diesem Ausmaß in der Literatur nicht vorgefunden, jedoch ist in vielen
Publikationen bei Raman-Spektren von Bakterien ein Minimum oder zumindest eine deutli-
che „Einkerbung“ innerhalb der Spektren bei 1200 cm-1 zu beobachten (De Gelder, 2008;
Van de Vossenberg et al., 2013).
Neben der Basislinienkorrektur, Glättung und der Normalisierung, stellt auch die PCA eine
Datenvorbehandlungsmethode dar. Es ist, wie in Kapitel 2.2.6 beschrieben, möglich, dass
die Wahl bzw. die Reihenfolge der PCs nicht die relevantesten Eigenschaften zur Vorher-
sage wiedergeben (Chang, 1983). Mögliche Verbesserungen der Modellleistungen, basie-
rend auf der Datenvorbehandlung, könnten unter Umständen durch eine stärkere Gewich-
tung ausgewählter spektraler Bereiche erzielt werden.
Ein sehr einfacher Ansatz der ggfs. zu weiteren Verbesserungen der Modelle führen könnte,
ist das Bilden von Ableitungen der Spektren zur Basislinienkorrektur. Vorversuche in die-
sem Bereich erzielten zwar schlechtere Resultate, jedoch könnte dies hauptsächlich mit der
Reihenfolge der einzelnen Vorbehandlungsschritte zusammenhängen. Ableitungsverfah-
ren neigen in Umgebungen mit hohem Rauschen dazu, schlechtere Ergebnisse zu erzielen
(Prakash & Wei, 2011). Es sollte also vor der Bildung der Ableitung die Glättung der

Diskussion
64
Spektren durchgeführt werden. Ein Nachteil dieser Vorbehandlungsmethode besteht darin,
dass die Spektren aufgrund der Veränderungen der Peaks schwerer zu interpretieren sind.
4.3 Beurteilung der Leistung und mögliche Limitationen der
entwickelten Methode
Die Ergebnisse dieser Arbeit zeigen, dass mit dem verwendeten Raman-Messsystem eine
zuverlässige Differenzierung der 9 untersuchten Mikroorganismen bei einer Anregungswel-
lenlänge von 633 nm möglich ist. Die Vorhersagegüten der leistungsfähigsten Modelle bei
praktischer Validierung liegen bei 98,1 % (QDA 20 PCs), 97,1 % (kNN 15 PCs) und 97,0 %
(SVM 20 PCs). Somit zeigt keines der Klassifizierungsverfahren einen deutlichen Vorteil
gegenüber den anderen, vielmehr Ergänzen sich die Verfahren. Das ausgewählte QDA-
Modell liefert beispielsweise auf Stammebene genauere Vorhersagen als das kNN-Modell,
jedoch basiert das kNN-Modell nur auf 15 PCs, was als Vorteil angesehen werden kann.
Weniger Dimensionen zur erfolgreichen Klassifizierung, bedeuten in der Regel robustere
Modelle in der Praxis. Die SVM-Modelle mit 15 und 20 PCs sind auf Stammebene deutlich
genauer als die QDA- und kNN-Modelle, schneiden aber bei ansonsten gut zu identifizie-
renden Bakterien (K12 423 und Pfl 50090) schlecht ab. Dies könnte zumindest bei
Pfl 50090 damit zusammenhängen, dass das Kalibrationsset aufgrund der zwei verschie-
denen Kultivierungsbedingungen zwei Cluster enthält, die leicht widersprüchlich sind. Eine
Vergrößerung des Kalibrationssets, oder eine Anpassung der Methode auf die Beschrän-
kung nur einer einzigen Kultivierungsart, könnte zu besseren Resultaten führen. Jedoch
sind diese Maßnahmen aufgrund der großen Störungen durch Fluoreszenz bei Pfl 50090
nicht ratsam. Die guten Vorhersagen für Pfl 50090 beruhen hauptsächlich auf dem großen
Unterschied zu den anderen Raman-Spektren, welche allerdings nicht direkt der chemi-
schen Zusammensetzung der Zelle zuzuschreiben sind, sondern der daraus resultierenden
Signalstörung. Da diese starke Störung bei nur einem einzigen Mikroorganismus innerhalb
des Kalibrationssets vorkommt, ermöglicht das schlechte Signal die gute Vorhersage.
Modelle für die weniger als 15 PCs herangezogen wurden, zeigen starke Probleme bei der
Unterscheidung von K12 423 von StyTA98 (siehe Abbildung 35). Die Ursache für die fal-
sche Identifizierung von K12 423 als Sty TA98 und nicht umgekehrt, könnte mit dem leicht
größeren Datenumfang der Sty TA98-Spektren zusammenhängen. Dies weist darauf hin,
dass sich eine gleichmäßige Datengewichtung eng verwandter Mikroorganismen innerhalb
eines Kalibrationsmodells positiv auf die Vorhersageleistung auswirken könnte.
Die sehr genauen Vorhersagen auf Stammebene von M. luteus müssen vorsichtiger be-
trachtet werden als die Vorhersagen bei den anderen Mikroorganismen. Erste Modelle, die
für M. luteus jeweils noch 100 Referenzspektren weniger enthielten als die finalen Modelle,

Diskussion
65
zeigten angewandt auf 200 völlig unbekannte Spektren Vorhersagegüten von 60 %
(MLB 28269), 70 % (Mlu 1790) und 99 % (Mlu 20030). Aufgrund der geringen Vorhersage-
güte bei den ersten beiden Stämmen, wurde das Kalibrationsset um jeweils 100 zufällig
gewählte Referenzspektren aus diesen 200 Spektren aufgestockt. Die restlichen 100 Vali-
dierungsspektren für M. luteus sind deshalb zwar nicht im Kalibrationsset enthalten, jedoch
wurden sie am gleichen Tag von demselben Probenmaterial erfasst. Da eine Mehrfachauf-
nahme von 5 bis 10 Spektren pro Stelle erfolgte, können im Kalibrationsset ähnliche Spek-
tren vorliegen wie bei der Validierung. Dies kann möglicherweise zu leicht überbestimmten
Resultaten geführt haben. Dennoch zeigt dies, dass mit robusten Referenzdatenbanken,
anhand vieler Proben aus mehreren Versuchstagen, immer genauere Vorhersagen für un-
bekannte Daten gemacht werden können. Die PCA aus Abbildung 33, welche sowohl die
Kalibrations-, als auch die Validierungsspektren enthält, verdeutlicht dies. Werden bei-
spielsweise nachträglich die Validierungsspektren in das Kalibrationsset mitaufgenommen,
würden insbesondere bei Sce 1334 und Mlu 20030 eine breitere Abdeckung erfolgen.
Mögliche Limitationen bei der Anwendung der Methode sind bei Mikroorganismen denkbar,
die besonders lichtempfindlich sind oder dunkles Koloniematerial aufweisen. Dunkles oder
farbiges Koloniematerial absorbiert einen größeren Anteil des Laserlichts, was zu einem
Verbrennen der Probe führen könnte (Banwell & MacCash, 1999).
4.4 Möglichkeiten zur Vermeidung falsch positiver Vorhersagen
Die entwickelten Kalibrationsmodelle basieren auf teilweise sehr komplexen Entschei-
dungsgrenzen in einem definierten Datenraum. Diese Grenzen führen dazu, dass ein
Spektrum eines unbekannten Mikroorganismus entweder der einen oder der anderen
Klasse zugeordnet wird. Unbekannte Spektren werden mit den entwickelten Modellen also
dem spektral ähnlichsten Mikroorganismus „zugeordnet“, selbst wenn die Unterschiede
sehr groß sein sollten. Bei einer Identifizierung pathogener Stämme müssten Kalibrations-
modelle auch das Ergebnis „unbekannt“ ausgeben können, oder einen Wert zur Beurteilung
der Wahrscheinlichkeit, dass ein Messergebnis korrekt ist. In der Medizin könnte eine sol-
che Vorhersage für das gesundheitliche Wohl eines Menschen entscheidend sein.
Unter anderem spricht man in der medizinischen Diagnostik von Sensitivität und Spezifität
eines Tests. Die Sensitivität ist ein Maß für das Erkennen eines oder mehrerer bestimmter
Merkmale. Führt das Vorhandensein dieser Merkmale immer zu einem positiven Test, so
beträgt die Sensitivität 100 %. Übertragen bedeutet dies, dass bei entsprechenden Intensi-
täten innerhalb eines definierten Wellenzahlbereichs ein Mikroorganismus vorhergesagt
wird. Die Spezifität ist hingegen ein Maß für das Erkennen, dass bestimmte Merkmale nicht
vorliegen. Werden alle negativen Proben, also Mikroorganismen, die nicht im Kalibratiosset
enthalten sind stets als „negativ“ vorhergesagt, so beträgt die Spezifität 100 %. Erst durch

Diskussion
66
Kombination von hoher Sensitivität und Spezifität werden Testergebnisse verlässlich.
(Suerbaum et al., 2016)
Für die hier verwendeten Klassifikationsverfahren gibt es verschiedene Ansätze, mit denen
die Zuverlässigkeit einer Vorhersage näher beschrieben werden könnte. Für den kNN-Al-
gorithmus wurden Methoden zur Berechnung der Wahrscheinlichkeit einer korrekten Klas-
sifizierung entwickelt, die über das sogenannte „Bootstrapping“-Verfahren erreicht werden
(Holmes & Adams, 2002; Villa et al., 2008). Bei der QDA können vergleichsweise einfach
über das Bayes-Theorem A-posteriori-Wahrscheinlichkeiten bestimmt werden, über welche
die Vertrauenswürdigkeit von Vorhersagen abgeschätzt werden können (Backhaus et al.,
2015). SVMs bieten solche Klassifizierungswahrscheinlichkeiten nicht direkt, jedoch gibt es
auch dort Ansätze, um A-posteriori-Wahrscheinlichkeiten zu erhalten. Es kann eine Schätz-
funktion bestimmt werden, die als trennende Hyperebene zwischen den Kalibrationsdaten
und abweichenden unbekannten Daten fungiert. (Platt, 1999; Schölkopf et al., 2001)
4.5 Ausblick
Aufgrund der nachgewiesenen Eignung des Messsystems für die Identifizierung der neun
verwendeten Mikroorganismen, ist eine Vielzahl weiterer Experimente denkbar. Die we-
sentlichsten neuen Projekte könnten unter anderem die Untersuchung weiterer Mikroorga-
nismen sein. Besonders Versuche mit Mikroorganismen, die photosensitive Pigmente oder
dunkles Koloniematerial aufweisen, könnten weitere Aufschlüsse über den Eignungsbe-
reich der Methode geben. Eine Untersuchung zur Auswirkung einer variierenden Anzahl an
Waschschritten auf die Fluoreszenzstörung bestimmter Mikroorganismen wäre ebenfalls
ein interessanter Schritt für die Weiterentwicklung der Methode.
Die Einschätzung der Zuverlässigkeit vorhergesagter Mikroorganismen mit den jetzigen
Daten könnte am einfachsten bei einem QDA-Modell umgesetzt werden. Dort bietet die
Software MATLAB die Möglichkeit neben der Vorhersage der Klasse auch A-posteriori-
Wahrscheinlichkeiten auszugeben. Allerdings können die Modelle hierfür nicht mit dem
Classification-Learner-Tool erstellt werden, sondern müssen manuell über die Kommando-
zeile programmiert werden.
Besonders interessant ist das Anpassen der entwickelten Methode, um Untersuchungen
zum Nachweis der Produktion bestimmter Substanzen durch Mikroorganismen zu ermögli-
chen. Über die Mikro-Raman-Spektroskopie können solche Analysen stark vereinfacht wer-
den. Beispielsweise ist das von einigen Bakterien produzierbare Polyhydroxybutyrat (PHB)
für die industrielle Produktion von biologisch abbaubaren Kunststoffen von großer Bedeu-
tung. Die Raman-Spektroskopie kann als schnelles und nichtinvasives Verfahren zum
Nachweis und zur Bestimmung des Gehalts von PHB in Bakterien dienen. (Gelder et al.,
2008; Samek et al., 2016)

Anhang
67
Anhang
Anhangsverzeichnis
Anhang 1: Vorhersagematrices QDA ..............................................................................68
Anhang 2: Vorhersagematrices kNN...............................................................................69
Anhang 3: Vorhersagematrices SVM ..............................................................................70
Anhang 4: Validierungsvorhersagen der restlichen QDA-Modelle .................................71
Anhang 5: Validierungsvorhersagen der restlichen kNN-Modelle ..................................72
Anhang 6: Validierungsvorhersagen der restlichen SVM-Modelle .................................73
Anhang 7: Leitfaden zur Identifizierung von Mikroorganismen über Raman-Spektroskopie ......................................................................................................74
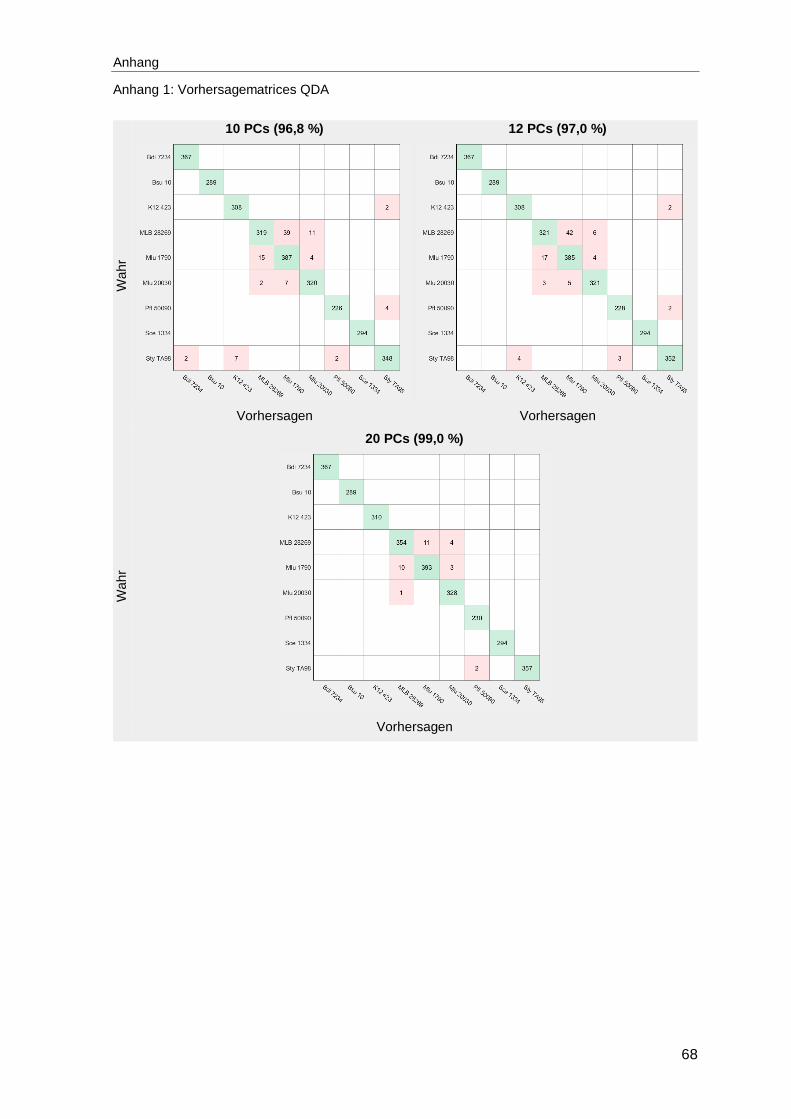
Anhang
68
Anhang 1: Vorhersagematrices QDA
10 PCs (96,8 %) 12 PCs (97,0 %) W
ah
r
Vorhersagen Vorhersagen
20 PCs (99,0 %)
Wa
hr
Vorhersagen

Anhang
69
Anhang 2: Vorhersagematrices kNN
10 PCs (96,9 %) 12 PCs (97,5 %) W
ah
r
Vorhersagen Vorhersagen
20 PCs (99,1 %)
Wa
hr
Vorhersagen

Anhang
70
Anhang 3: Vorhersagematrices SVM
10 PCs (98,3 %) 12 PCs (98,5 %) W
ah
r
Vorhersagen Vorhersagen
20 PCs (99,7 %)
Wa
hr
Vorhersagen

Anhang
71
Anhang 4: Validierungsvorhersagen der restlichen QDA-Modelle
QDA 8 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 100 Bsu 10 100
K12 423 1 34 3 62 MLB 28269 86 11 3 Mlu 1790 8 90 2 Mlu 20030 1 1 98 Pfl 50090 100 Sce 1334 100 Sty TA98 98
QDA 10 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 98 2 Bsu 10 100
K12 423 26 3 71 MLB 28269 94 3 3 Mlu 1790 6 91 3 Mlu 20030 3 97 Pfl 50090 100 Sce 1334 100 Sty TA98 1 99
QDA 12 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wahr
Bdi 7234 99 1 Bsu 10 100
K12 423 51 3 46 MLB 28269 96 3 1 Mlu 1790 4 93 3 Mlu 20030 1 99 Pfl 50090 100 Sce 1334 100 Sty TA98 2 98
QDA 15 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wahr
Bdi 7234 100 Bsu 10 100
K12 423 76 24 MLB 28269 97 3 Mlu 1790 6 91 3 Mlu 20030 100 Pfl 50090 100 Sce 1334 100 Sty TA98 100

Anhang
72
Anhang 5: Validierungsvorhersagen der restlichen kNN-Modelle
kNN 8 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 100 Bsu 10 99 1
K12 423 1 22 77 MLB 28269 67 23 10 Mlu 1790 12 86 2 Mlu 20030 4 1 95 Pfl 50090 99 1 Sce 1334 1 99 Sty TA98 1 7 4 88
kNN 10 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 100 Bsu 10 98 2
K12 423 46 54 MLB 28269 78 11 11 Mlu 1790 16 79 5 Mlu 20030 4 96 Pfl 50090 1 99 Sce 1334 1 99 Sty TA98 3 7 90
kNN 12 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wahr
Bdi 7234 100 Bsu 10 100
K12 423 59 41 MLB 28269 93 4 3 Mlu 1790 9 84 7 Mlu 20030 3 3 94 Pfl 50090 100 Sce 1334 100 Sty TA98 9 91
kNN 20 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wahr
Bdi 7234 100 Bsu 10 100
K12 423 95 5 MLB 28269 88 10 2 Mlu 1790 6 94 Mlu 20030 1 99 Pfl 50090 1 77 16 6 Sce 1334 1 1 98 Sty TA98 2 98

Anhang
73
Anhang 6: Validierungsvorhersagen der restlichen SVM-Modelle
SVM 8 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 99 1 Bsu 10 100
K12 423 32 68 MLB 28269 84 15 1 Mlu 1790 5 94 1 Mlu 20030 1 99 Pfl 50090 3 95 2 Sce 1334 4 96 Sty TA98 2 98
SVM 10 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wa
hr
Bdi 7234 83 17 Bsu 10 100
K12 423 26 74 MLB 28269 77 17 6 Mlu 1790 3 96 1 Mlu 20030 100 Pfl 50090 100 Sce 1334 6 94 Sty TA98 1 99
SVM 12 PCs Vorhersagen
Bdi 7234
Bsu 10 K12 423
MLB 28269
Mlu 1790
Mlu 20030
Pfl 50090
Sce 1334
Sty TA98
Wahr
Bdi 7234 83 17 Bsu 10 99 1
K12 423 39 61 MLB 28269 85 9 6 Mlu 1790 6 93 1 Mlu 20030 1 1 98 Pfl 50090 100 Sce 1334 5 95 Sty TA98 3 97

Anhang
74
Anhang 7: Leitfaden zur Identifizierung von Mikroorganismen über Raman-Spektroskopie
1. Allgemeines
• Befolgen Sie die allgemeinen und raumspezifischen Sicherheitsvorschriften.
• Beachten Sie bei der Verwendung des Raman-Mikroskopie-Systems inVia von Renishaw
das im Labor ausliegende Dokument „Vor jeder Messung“.
• Bisher (Stand Januar 2019) können folgende Mikroorganismen identifiziert werden:
Kürzel Mikroorganismus DSM-Nr. Geschätzte Vorhersagegüte [%]*
QDA kNN SVM
Bdi 7234 Brevundimonas diminuta 7234 100 100 100
Bsu 10 Bacillus subtilis 10 100 100 100
K12 423 Escherichia coli K12 423 95 100 91
MLB 28269 Micrococcus luteus 28269 97 92 96
Mlu 1790 Micrococcus luteus 1790 91 87 99
Mlu 20030 Micrococcus luteus 20030 100 100 100
Pfl 50090 Pseudomonas fluorescens 50090 100 100 87
Sce 1334 Saccharomyces cerevisiae 1334 100 100 100
Sty TA98 Salmonella Thyphimurium TA98 - 100 95 100
* basierend auf dem Ergebnis einer Validierung mit jeweils 100 unbekannten Spektren und Modellen mit 20 Hauptkomponenten (QDA und SVM) und 15 Hauptkomponenten (kNN)
2. Anzucht der Mikroorganismen und Probenvorbereitung
• Machen Sie von dem zu identifizierenden Koloniematerial einen Ausstrich auf TSA-Nähr-
boden und bebrüten Sie diesen bei 30 °C für 24 h.
• Legen Sie 1000 µl sterile NaCl-Lösung (0,9 %) in ein 1,5 ml-Reaktionsgefäß vor.
• Überführen Sie so viel Koloniematerial in das Reaktionsgefäß, wie Sie etwa mit drei Abstri-
chen einer 10 µl Impfschlinge aufnehmen können.
• Mischen Sie den Inhalt durch einen Vortex-Mi-
scher, bis eine gleichmäßige Suspension entsteht.
• Zentrifugieren Sie das Material für 3 Minuten bei
5000 rpm und verwerfen Sie anschließend den
Überstand.
• Überführen Sie das Zellmaterial auf einen Glasob-
jektträger (mittels Pipette oder Kunststoffstab).

Anhang
75
• Lassen Sie das Koloniematerial trocknen
(Bei Raumtemperatur ca. 1 bis 2 Stunden).
• Bereiten Sie das Raman-Messsystem nach den
Vorgaben des Dokuments „Vor jeder Messung“ vor
(Kalibrierung mit Si-Standard).
• Platzieren Sie die getrocknete Probe auf dem Ob-
jekttisch und fokussieren Sie diese mit dem 100X-
Objektiv.
3. Spektrenaufzeichnung
• Gehen Sie sicher, dass die Jalousien des Labors geschlossen sind und die Raumbeleuch-
tung inaktiv ist.
• Verwenden Sie folgende Messeinstellungen:
Wellenzahlzentrum: 1200 cm-1
Belichtungsdauer: 3 Sekunden
Laserleistung: 100 %
Akkumulationen: 15
• Starten Sie eine Messung und ordnen Sie das Resultat in eine der nachfolgenden
Kategorien A bis D ein.
A1
A2
B
C1
C2
D
• Zur Einschätzung der Begriffe „dünn“, „dick“, „mittel“ bei Probenschichten kann nachfol-
gende Abbildung eine Einschätzung ermöglichen:
Beispiel zweier Kolonie-
materialprobenschich-
ten auf Glasobjektträ-
gern zur Einschätzung
der Schichtdicken
(a= dünn; b= mittel;
c= dick; d= ungeeignet,
noch flüssig

Anhang
76
Weiteres Vorgehen bei Kategorie A (A1 und A2)
• Diese Spektren sind bereits verwertbar (Zeichnen Sie am besten mehrere Spektren an die-
ser Stelle auf, wechseln Sie anschließend die fokussierte Position und zeichnen Sie ggfs.
weitere Spektren auf).
• Speichern Sie Spektren unter einem Dateinamen ab, der einer bestimmten Logik folgt: z.B.
Jahr, Monat, Tag, fortlaufende Nummerierung, Kürzel des Mikroorganismus oder der
Probe (z.B. 20180831 001 K12 423).
Weiteres Vorgehen bei Kategorie B
• Ihre Probenschicht ist vermutlich zu dünn oder das Koloniematerial so blass, dass ein gro-
ßer Teil des Laserlichts hindurchtritt. Wechseln Sie die fokussierte Stelle oder sorgen Sie
für einen dickeren Probenfilm und versuchen Sie es erneut.
Weiteres Vorgehen bei Kategorie C (C1 und C2)
• Ihre Probenschicht ist vermutlich zu dick; Bestrahlen Sie die Probe länger vor, indem Sie
mit denselben Messeinstellungen nacheinander Messungen starten.
• Sie können die Belichtungszeit an dieser Stelle auch verringern und die Akkumulationen
erhöhen. Dadurch müssen Sie nicht so häufig eine neue Messung initiieren und die Aus-
wirkung der Vorbestrahlung ist schneller ersichtlich.
• Bestrahlen Sie insgesamt etwa 5 bis 15 Minuten vor und beobachten Sie ob Signaturen
erkennbar werden. Werden in diesem Zeitraum Peaks erkennbar, wechseln Sie ggfs. zu-
rück zu den 15 Akkumulationen und den 3 Sekunden Belichtungszeit und zeichnen Sie ein
Spektrum auf.
• Raman-Spektren die von „mittleren“ oder „dicken“ Probenschichten nach Vorbestrahlung
erhalten werden, entwickeln sich häufig wie das Beispiel A2. Diese Spektren sind verwert-
bar. Verfahren Sie dann nach Kategorie A weiter.
Sieht das Spektrum nach etwa 15 Minuten Vorbestrahlung noch aus wie bei C, sollten Sie
eine dünnere Probenschicht fokussieren.
Weiteres Vorgehen bei D:
• Die Counts sind zu hoch. Es wird kein Spektrum angezeigt. Dies kann bei Proben passie-
ren, die starke Eigenfluoreszenz aufweisen, insbesondere bei dicken Probenschichten.
• Untersuchen Sie die Probe mit kürzerer Belichtungszeit z.B. 0,25 Sekunden und stellen
Sie eine hohe Akkumulation ein z.B. 500. Bestrahlen Sie die Probe 2 Minuten vor und initi-
ieren Sie die Messung mit verkürzter Belichtungszeit erneut.
• Wiederholen Sie dies einige Male, bis die Fluoreszenz soweit ausgebrannt wurde, dass
auch bei 3 Sekunden Belichtungszeit ein Spektrum angezeigt wird.
• Prüfen Sie nach spätestens 15 Minuten Vorbestrahlung, ob das Spektrum in Kategorie A,
B, oder C fällt und verfahren Sie entsprechend weiter.

Anhang
77
4. Datenverarbeitung
• Konvertieren Sie die WDF-Dateien mit dem WiRE Batch Converter in das TXT-Format.
• Erstellen Sie auf dem Computer, auf dem die Software MATLAB (R2017b) installiert ist,
einen neuen Ordner.
• Kopieren Sie folgende Dateien in diesen Ordner:
o f = Wellenzahlachse
o lpf = Tiefpassfilter
o specdata = Tool-Sammlung
o Alle Modelle = Sammlung der entwickelten Vorhersagemodelle
o Ihre konvertierten TXT-Files der Spektren
• Starten Sie MATLAB und wählen Sie den entsprechenden Ordner aus.
• Nachfolgende Befehle müssen in das Kommandofenster eingegeben werden.
Was in der Kommandozeile nach einem %-Zeichen geschrieben wird, hat keinen Einfluss,
sondern dient als Kommentar. Die Schriftart Courier weist darauf hin, dass der Text di-
rekt in MATLAB kopiert und ausgeführt werden könnte.

Anhang
78
% Erstellen Sie ein Datenpaket, das Ihre Spektren enthält. Sie können das
Paket benennen wie Sie wollen (es empfiehlt sich eine kurze Bezeichnung).
In diesem Beispiel wird das Paket als „Bdi“ bezeichnet. Der Befehl
„specdata“ erlaubt eine Einschränkung der Suchkriterien durch „*“
Bdi=specdata.load('2018*Bdi*.txt')
%Standardisieren Sie die Wellenzahlachse (Interpolieren)
Bdi_x=Bdi.setx(606:1.114:1736)
%Konvertieren Sie die Spektren als Datenmatrix
Bdi_x_mat=Bdi_x.xymat
• Nun müssen Sie die erste Spalte ihrer Datenmatrix löschen (Wellenzahlen) und die Daten
anschließend transponieren, sodass die 1015 Wellenzahlen die Spalten ergeben und die
(in diesem Beispiel 50) Spektren die Zeilen.
• Kopieren Sie in Ihren „Workspace“ die Wellenzahlachse „f“
%Führen Sie die Basislinienkorrektur und Glättung aus.
dim = 50 %Anzahl Ihrer Spektren
for i = 1:1*dim
Bdi_x_mat_treat(i,:) = Bdi_x_mat(i,:) - lpf(f, Bdi_x_mat(i,:), 20) ;
Bdi_x_mat_treat(i,:) = lpf(f, Bdi_x_mat_treat(i,:),3) ;
end

Anhang
79
% Die Spektren werden nun normalisiert, also das Minimum 0 und das Maximum
als 1 definiert
imax = 1;
imin = 0;
% Zeilenweise Bestimmung
Bdi_x_mat_treat_min = min(Bdi_x_mat_treat,[],2);
Bdi_x_mat_treat_max = max(Bdi_x_mat_treat,[],2);
y = zeros(50,1015); %(50,1015)Entspricht dem Format der Datenmatrix
for i=1:length(Bdi_x_mat_treat_max)
Mittel = (imax - imin) / (Bdi_x_mat_treat_max(i) - Bdi_x_mat_treat_min(i));
% jede Zeile einzeln normieren
Bdi_x_mat_treat_norm (i,:) = Mittel*( Bdi_x_mat_treat (i,:) -
Bdi_x_mat_treat_min(i)) + imin;
end
• Die blau hinterlegten Dateien werden nicht weiter benötigt. Alle anderen sollten Sie abspei-
chern. Prüfen Sie Ihre Daten an dieser Stelle auf Plausibilität, indem Sie die Spektren in
MATLAB „plotten“ (plot(f, Bdi_x_mat_treat_norm))
• Kopieren Sie die vorbehandelte Datenmatrix und fügen Sie diese in Microsoft Excel ein,
wobei sie in Excel die erste Spalte frei lassen. Speichern Sie die Excel-Datei in Ihren er-
stellten Ordner.

Anhang
80
• Importieren Sie in MATLAB eine Datentabelle basierend auf Ihrer Excel-Datei und benen-
nen Sie diese entsprechend. Sorgen Sie dafür, dass auch die leere Spalte 1 importiert wird
(VarName1). Die 1015 Wellenzahlen werden automatisch als VarName2 bis Var-
Name1016 bezeichnet, was für die Vorhersage mit den entwickelten Modellen erforderlich
ist.
• Wählen Sie eines der Modelle aus dem Paket aus und fügen Sie es in Ihren „Workspace“
ein. Hier wird das kNN-Modell, basierend auf 15 Hauptkomponenten gewählt
(model5_KNN_15PC)

Anhang
81
• Nun erfolgt die Vorhersage der vorbehandelten unbekannten Spektren über das ausge-
wählte Modell.
% Geben Sie eine Bezeichnung für die Vorhersage ein (Vorhersage). Wählen
Sie den Namen des entsprechenden Modells (model5_KNN_15PC). Den Befehl zur
Vorhersage (.predictFcn) und den Namen der Datenmatrix für die Sie eine
Vorhersage machen wollen (Bdi_vali).
Vorhersage = model5_KNN_15PC.predictFcn(Bdi_vali)
• Sie erhalten nun eine Liste, in der mit dem ausgewählten Modell für jedes Raman-Spekt-
rum eine Vorhersage gemacht wurde.

Anhang
82
5 Literaturverzeichnis
Andrusch, A., Dabrowski, P. W., Klenner, J., Tausch, S. H., Kohl, C., Osman, A. A., . . . Nitsche, A. (2018). PAIPline: pathogen identification in metagenomic and clinical next generation sequencing samples. Bioinformatics, 34(17), S. i715-i721. doi: 10.1093/bioinformatics/bty595
Backhaus, K., Erichson, B., Plinke, W., & Weiber, R. (2015). Multivariate Analysemethoden: Eine anwendungsorientierte Einführung: Springer Berlin Heidelberg. S. 107, 216, 221, 250, 263, 268
Banwell, C. N., & MacCash, E. M. (1999). Molekülspektroskopie: Ein Grundkurs: Oldenbourg. S. 164, 165
Baritaux, J. C., Simon, A. C., Schultz, E., Emain, C., Laurent, P., & Dinten, J. M. (2016). A study on identification of bacteria in environmental samples using single-cell Raman spectroscopy: feasibility and reference libraries. Environ Sci Pollut Res Int, 23(9), S. 8184-8191. doi: 10.1007/s11356-015-5953-x
Chang, W.-C. (1983). On using principal components before separating a mixture of two multivariate normal distributions. Applied Statistics, 32, S. 267-275.
Ciobota, V., Burkhardt, E. M., Schumacher, W., Rösch, P., Kusel, K., & Popp, J. (2010). The influence of intracellular storage material on bacterial identification by means of Raman spectroscopy. Anal Bioanal Chem, 397(7), S. 2929-2937. doi: 10.1007/s00216-010-3895-1
Cypionka, H. (2010). Grundlagen der Mikrobiologie: Springer Berlin Heidelberg. S. 1, 2, 4, 15, 16, 19, 27, 30, 94, 95
De Gelder, J. (2008). Raman spectroscopy as a tool for studying bacterial cell compounds. Ghent University.
De Gelder, J., De Gussem, K., Vandenabeele, P., & Moens, L. (2007). Reference database of Raman spectra of biological molecules. Journal of Raman Spectroscopy, 38(9), S. 1133-1147. doi: 10.1002/jrs.1734
Ertel, W. (2016). Grundkurs Künstliche Intelligenz: Eine praxisorientierte Einführung: Springer Fachmedien Wiesbaden. S. 212, 232, 233, 298
Fritsche, O. (2016). Mikrobiologie: Springer Berlin Heidelberg. S. 6, 10, 11, 21, 326
Fuchs, G., Eitinger, T., & Schlegel, H. G. (2007). Allgemeine Mikrobiologie: Thieme. S. 26, 86, 130-133, 135-136, 138-142, 581, 612, 623
Gebrekidan, M. T., Knipfer, C., Stelzle, F., Popp, J., Will, S., & Braeuer, A. (2016). A shifted-excitation Raman difference spectroscopy (SERDS) evaluation strategy for the efficient isolation of Raman spectra from extreme fluorescence interference. Journal of Raman Spectroscopy, 47(2), S. 198-209. doi: 10.1002/jrs.4775
Gelder, J. D., Willemse-Erix, D., Scholtes, M. J., Sanchez, J. I., Maquelin, K., Vandenabeele, P., . . . Vos, P. D. (2008). Monitoring poly(3-hydroxybutyrate) production in cupriavidus necator DSM 428 (H16) with raman spectroscopy. Anal Chem, 80(6), S. 2155-2160. doi: 10.1021/ac702185d
Golcuk, K., Mandair, G. S., Callender, A. F., Sahar, N., Kohn, D. H., & Morris, M. D. (2006). Is photobleaching necessary for Raman imaging of bone tissue using a green laser? Biochim Biophys Acta, 1758(7), S. 868-873. doi: 10.1016/j.bbamem.2006.02.022

Anhang
83
Guo, S., Bocklitz, T., & Popp, J. (2016). Optimization of Raman-spectrum baseline correction in biological application. Analyst, 141(8), S. 2396-2404. doi: 10.1039/c6an00041j
Hair, J. F., Black, W. C., & Babin, B. J. (2010). Multivariate Data Analysis: A Global Perspective: Pearson Education. S. 340, 506, 507, 530
Harz, M., Rösch, P., Peschke, K. D., Ronneberger, O., Burkhardt, H., & Popp, J. (2005). Micro-Raman spectroscopic identification of bacterial cells of the genus Staphylococcus and dependence on their cultivation conditions. Analyst, 130(11), S. 1543-1550. doi: 10.1039/b507715j
Hasegawa, T. (2017). Quantitative Infrared Spectroscopy for Understanding of a Condensed Matter: Springer Japan. S. 5
Hastie, T., Tibshirani, R., & Friedman, J. (2009). The Elements of Statistical Learning: Data Mining, Inference, and Prediction, Second Edition: Springer New York. S. 103, 111, 116, 438, 439
Helm, D., Labischinski, H., Schallehn, G., & Naumann, D. (1991). Classification and identification of bacteria by Fourier-transform infrared spectroscopy. J Gen Microbiol, 137(1), S. 69-79. doi: 10.1099/00221287-137-1-69
Hesse, M., Meier, H., & Zeeh, B. (2008). Spectroscopic methods in organic chemistry: Thieme. S. 69, 70, 71
Holmes, C. C., & Adams, N. M. (2002). A probabilistic nearest neighbour method for statistical pattern recognition. Journal of the Royal Statistical Society: Series B (Statistical Methodology), 64(2), S. 295-306. doi: 10.1111/1467-9868.00338
Hutsebaut, D., Maquelin, K., De Vos, P., Vandenabeele, P., Moens, L., & Puppels, G. J. (2004). Effect of culture conditions on the achievable taxonomic resolution of Raman spectroscopy disclosed by three Bacillus species. Anal Chem, 76(21), S. 6274-6281. doi: 10.1021/ac049228l
Iba, H. (2018). Evolutionary Approach to Machine Learning and Deep Neural Networks: Neuro-Evolution and Gene Regulatory Networks: Springer Singapore. S. 37
Ivnitski, D., Abdel-Hamid, I., Atanasov, P., & Wilkins, E. (1999). Biosensors for detection of pathogenic bacteria. Biosensors and Bioelectronics, 14(7), S. 599-624. doi: 10.1016/S0956-5663(99)00039-1
Jackson, H., Braun, C. L., & Ernst, H. (2008). The chemistry of novel xanthophyll carotenoids. Am J Cardiol, 101(10A), S. 50D-57D. doi: 10.1016/j.amjcard.2008.02.008
Jolliffe, I. T. (2002). Principal Component Analysis: Springer. S. 167
Kahlon, R. S. (2016). Pseudomonas: Molecular and Applied Biology: Springer International Publishing. S. 48, 49, 146
Kanematsu, H., Kudara, H., Kanesaki, S., Kogo, T., Ikegai, H., Ogawa, A., & Hirai, N. (2016). Application of a Loop-Type Laboratory Biofilm Reactor to the Evaluation of Biofilm for Some Metallic Materials and Polymers such as Urinary Stents and Catheters. Materials (Basel), 9(10). doi: 10.3390/ma9100824
Kessler, W. (2007). Multivariate Datenanalyse: für die Pharma, Bio- und Prozessanalytik: Wiley. S. 21-36, 53, 156, 187
Kim, Y.-i., Jeong, S., Jun, B.-H., Lee, Y.-S., Lee, Y.-S., Jeong Dae, H., & Lee Dong, S. (2017). Endoscopic imaging using surface-enhanced Raman scattering. In European Journal of Nanomedicine (Vol. 9, pp. 91).

Anhang
84
Kneipp, J. (2001). Fourier-Transform-Infrarotmikrospektroskopische Charakterisierung transmissibler spongiformer Enzephalopathien. Freie Universität Berlin, https://docplayer.org/8549739-Fourier-transform-infrarotmikrospektroskopische-charakterisierung-transmissibler-spongiformer-enzephalopathien.html Abgerufen am 2.11.2018.
Kneipp, K., Haka, A. S., Kneipp, H., Badizadegan, K., Yoshizawa, N., Boone, C., . . . Feld, M. S. (2002). Surface-Enhanced Raman Spectroscopy in Single Living Cells Using Gold Nanoparticles. Applied Spectroscopy, 56(2), S. 150-154. doi: 10.1366/0003702021954557
Krämer, J. (2007). Lebensmittel-Mikrobiologie: 49 Tabellen: Ulmer. S. 364, 365
Kumar, B. N. V., Kampe, B., Rösch, P., & Popp, J. (2015). Characterization of carotenoids in soil bacteria and investigation of their photodegradation by UVA radiation via resonance Raman spectroscopy. Analyst, 140(13), S. 4584-4593. doi: 10.1039/c5an00438a
Kumar, S., Verma, T., Mukherjee, R., Ariese, F., Somasundaram, K., & Umapathy, S. (2016). Raman and infra-red microspectroscopy: towards quantitative evaluation for clinical research by ratiometric analysis. Chem Soc Rev, 45(7), S. 1879-1900. doi: 10.1039/c5cs00540j
Kusic, D., Kampe, B., Ramoji, A., Neugebauer, U., Rösch, P., & Popp, J. (2015). Raman spectroscopic differentiation of planktonic bacteria and biofilms. Anal Bioanal Chem, 407(22), S. 6803-6813. doi: 10.1007/s00216-015-8851-7
Larkin, P. (2011). Infrared and Raman Spectroscopy: Principles and Spectral Interpretation: Elsevier Science. S. 55-57
Lechner, M. D. (2016). Einführung in die Quantenchemie: Aufbau der Atome und Moleküle, Spektroskopie: Springer Berlin Heidelberg. S. 93
Leon-Bejarano, M., Dorantes-Mendez, G., Ramirez-Elias, M., Mendez, M. O., Alba, A., Rodriguez-Leyva, I., & Jimenez, M. (2016). Fluorescence background removal method for biological Raman spectroscopy based on empirical mode decomposition. Conf Proc IEEE Eng Med Biol Soc, 2016, S. 3610-3613. doi: 10.1109/EMBC.2016.7591509
Lieber, C. A., & Mahadevan-Jansen, A. (2003). Automated method for subtraction of fluorescence from biological Raman spectra. Appl Spectrosc, 57(11), S. 1363-1367. doi: 10.1366/000370203322554518
Ling, T. K., Liu, Z. K., & Cheng, A. F. (2003). Evaluation of the VITEK 2 system for rapid direct identification and susceptibility testing of gram-negative bacilli from positive blood cultures. J Clin Microbiol, 41(10), S. 4705-4707.
Lipiainen, T., Pessi, J., Movahedi, P., Koivistoinen, J., Kurki, L., Tenhunen, M., . . . Strachan, C. J. (2018). Time-Gated Raman Spectroscopy for Quantitative Determination of Solid-State Forms of Fluorescent Pharmaceuticals. Anal Chem, 90(7), S. 4832-4839. doi: 10.1021/acs.analchem.8b00298
Lopez-Diez, E. C., & Goodacre, R. (2004). Characterization of microorganisms using UV resonance Raman spectroscopy and chemometrics. Anal Chem, 76(3), S. 585-591. doi: 10.1021/ac035110d
Madigan, M. T., Martinko, J. M., Stahl, D. A., & Clark, D. P. (2015). Brock Mikrobiologie kompakt: Pearson Studium. S. 2, 8, 44-47, 81
Maquelin, K., Kirschner, C., Choo-Smith, L. P., van den Braak, N., Endtz, H. P., Naumann, D., & Puppels, G. J. (2002). Identification of medically relevant microorganisms by vibrational spectroscopy. J Microbiol Methods, 51(3), S. 255-271.

Anhang
85
Meisel, S., Stockel, S., Elschner, M., Melzer, F., Rösch, P., & Popp, J. (2012). Raman spectroscopy as a potential tool for detection of Brucella spp. in milk. Appl Environ Microbiol, 78(16), S. 5575-5583. doi: 10.1128/AEM.00637-12
Munchberg, U., Rösch, P., Bauer, M., & Popp, J. (2014). Raman spectroscopic identification of single bacterial cells under antibiotic influence. Anal Bioanal Chem, 406(13), S. 3041-3050. doi: 10.1007/s00216-014-7747-2
Nataf, G. F. (2016). New approaches to understand conductive and polar domain walls by Raman spectroscopy and low energy electron microscopy. (Doctoral dissertation), Université Paris-Saclay, https://tel.archives-ouvertes.fr/tel-01412787/document Abgerufen am 11.10.2018.
Ngo-Thi, N. A., Kirschner, C., & Naumann, D. (2003). Characterization and identification of microorganisms by FT-IR microspectrometry. Journal of Molecular Structure, 661-662, S. 371-380. doi: 10.1016/j.molstruc.2003.08.012
Penel, G., Leroy, G., & Bres, E. (1998). New Preparation Method of Bone Samples for Raman Microspectrometry (Vol. 52).
Perner, P., & Petrou, M. (1999). Machine Learning and Data Mining in Pattern Recognition: First International Workshop: Springer Berlin Heidelberg. S. 169, 170
Pettersson, E., Lundeberg, J., & Ahmadian, A. (2009). Generations of sequencing technologies. Genomics, 93(2), S. 105-111. doi: 10.1016/j.ygeno.2008.10.003
Platt, J. (1999). Probabilistic Outputs for Support Vector Machines and Comparisons to Regularized Likelihood Methods. 10.
Powell, V., & Lehe, L. (2014). Principal Component Analysis Explained Visually. http://setosa.io/ev/principal-component-analysis/ Abgerufen am 19.11.2018.
Prakash, B. D., & Wei, Y. C. (2011). A fully automated iterative moving averaging (AIMA) technique for baseline correction. Analyst, 136(15), S. 3130-3135. doi: 10.1039/c0an00778a
Rösch, P., Harz, M., Schmitt, M., Peschke, K. D., Ronneberger, O., Burkhardt, H., . . . Popp, J. (2005). Chemotaxonomic identification of single bacteria by micro-Raman spectroscopy: application to clean-room-relevant biological contaminations. Appl Environ Microbiol, 71(3), S. 1626-1637. doi: 10.1128/AEM.71.3.1626-1637.2005
Runkler, T. A. (2016). Data Analytics: Models and Algorithms for Intelligent Data Analysis: Springer Fachmedien Wiesbaden. S. 31, 79-81, 99-102
Salzer, R., Thiele, S., & Suemmchen, L. (2017). Schwingungsspektroskopie - kompakt. http://www.chemgapedia.de/vsengine/vlu/vsc/de/ch/3/anc/ir_spek/spek_kompakt.vlu/Page/vsc/de/ch/3/anc/ir_spek/schwspek/methoden/raman_spek/ramanspektrum_m21ht0502.vscml/Supplement/2.html Abgerufen am 2.12.2018.
Samek, O., Obruca, S., Siler, M., Sedlacek, P., Benesova, P., Kucera, D., . . . Zemanek, P. (2016). Quantitative Raman Spectroscopy Analysis of Polyhydroxyalkanoates Produced by Cupriavidus necator H16. Sensors (Basel), 16(11). doi: 10.3390/s16111808
Sato, E. T., & Martinho, H. (2018). First-principles calculations of Raman vibrational modes in the fingerprint region for connective tissue. Biomed Opt Express, 9(4), S. 1728-1734. doi: 10.1364/BOE.9.001728
Schölkopf, B., Platt, J. C., Shawe-Taylor, J., Smola, A. J., & Williamson, R. C. (2001). Estimating the support of a high-dimensional distribution. Neural Comput, 13(7), S. 1443-1471. doi: 10.1162/089976601750264965
Schrader, B., Schulz, H., Andreev, G. N., Klump, H. H., & Sawatzki, J. (2000). Non-destructive NIR-FT-Raman spectroscopy of plant and animal tissues, of food and works of art. Talanta, 53(1), S. 35-45.

Anhang
86
Schulze, G., Jirasek, A., Yu, M. M., Lim, A., Turner, R. F., & Blades, M. W. (2005). Investigation of selected baseline removal techniques as candidates for automated implementation. Appl Spectrosc, 59(5), S. 545-574. doi: 10.1366/0003702053945985
Sengupta, A., Mujacic, M., & Davis, E. J. (2006). Detection of bacteria by surface-enhanced Raman spectroscopy. Anal Bioanal Chem, 386(5), S. 1379-1386. doi: 10.1007/s00216-006-0711-z
Sil, S., Mukherjee, R., Kumar, N. S., S, A., Kingston, J., & Singh, U. K. (2017). Detection and classification of Bacteria using Raman Spectroscopy Combined with Multivariate Analysis. Defence Life Science Journal, 2(4). doi: 10.14429/dlsj.2.12275
Sivanesan, A., Witkowska, E., Adamkiewicz, W., Dziewit, L., Kaminska, A., & Waluk, J. (2014). Nanostructured silver-gold bimetallic SERS substrates for selective identification of bacteria in human blood. Analyst, 139(5), S. 1037-1043. doi: 10.1039/c3an01924a
Skoog, D. A., Holler, F. J., Crouch, S. R., Föllner, B., Möhring, H. J., & Niessner, R. (2013). Instrumentelle Analytik: Grundlagen - Geräte - Anwendungen: Springer Berlin Heidelberg. S. 321-333
Sogawa, K., Watanabe, M., Sato, K., Segawa, S., Miyabe, A., Murata, S., . . . Nomura, F. (2012). Rapid identification of microorganisms by mass spectrometry: improved performance by incorporation of in-house spectral data into a commercial database. Anal Bioanal Chem, 403(7), S. 1811-1822. doi: 10.1007/s00216-011-5656-1
Spanu, T., Sanguinetti, M., Ciccaglione, D., D'Inzeo, T., Romano, L., Leone, F., & Fadda, G. (2003). Use of the VITEK 2 system for rapid identification of clinical isolates of Staphylococci from bloodstream infections. J Clin Microbiol, 41(9), S. 4259-4263.
Steinbüchel, A., Oppermann-Sanio, F. B., Ewering, C., & Pötter, M. (2012). Mikrobiologisches Praktikum: Versuche und Theorie: Springer Berlin Heidelberg. S. 39, 40, 333
Strola, S. A., Baritaux, J. C., Schultz, E., Simon, A. C., Allier, C., Espagnon, I., . . . Dinten, J. M. (2014). Single bacteria identification by Raman spectroscopy. J Biomed Opt, 19(11), S. 111610. doi: 10.1117/1.JBO.19.11.111610
Suerbaum, S., Burchard, G. D., Kaufmann, S. H. E., & Schulz, T. F. (2016). Medizinische Mikrobiologie und Infektiologie: Springer Berlin Heidelberg. S. 147
Teixeira-Dias, J. J. C. (2017). Molecular Physical Chemistry: A Computer-based Approach using Mathematica® and Gaussian: Springer International Publishing. S. 176
Teng, L., Wang, X., Wang, X., Gou, H., Ren, L., Wang, T., . . . Xu, J. (2016). Label-free, rapid and quantitative phenotyping of stress response in E. coli via ramanome. Sci Rep, 6, S. 34359. doi: 10.1038/srep34359
Van de Vossenberg, J., Tervahauta, H., Maquelin, K., Blokker-Koopmans, C. H. W., Uytewaal-Aarts, M., van der Kooij, D., . . . van der Gaag, B. (2013). Identification of bacteria in drinking water with Raman spectroscopy. Analytical Methods, 5(11), S. 2679-2687. doi: 10.1039/C3AY40289D
Van den Burg, G., & Groenen, P. (2016). A Generalized Multiclass Support Vector Machine. Journal of Machine Learning Research, 17.
Vargis, E., Tang, Y. W., Khabele, D., & Mahadevan-Jansen, A. (2012). Near-infrared Raman Microspectroscopy Detects High-risk Human Papillomaviruses. Transl Oncol, 5(3), S. 172-179.

Anhang
87
Villa, J. L., Boqué, R., & Ferré, J. (2008). Calculation of the probability of correct classification in probabilistic bagged k-Nearest Neighbours. Chemometrics and Intelligent Laboratory Systems, 94(1), S. 51-59. doi: 10.1016/j.chemolab.2008.06.007
Wang, P., Pang, S., Pearson, B., Chujo, Y., McLandsborough, L., Fan, M., & He, L. (2017). Rapid concentration detection and differentiation of bacteria in skimmed milk using surface enhanced Raman scattering mapping on 4-mercaptophenylboronic acid functionalized silver dendrites. Anal Bioanal Chem, 409(8), S. 2229-2238. doi: 10.1007/s00216-016-0167-8
Willemse-Erix, D. F., Jachtenberg, J. W., Schut, T. B., van Leeuwen, W., van Belkum, A., Puppels, G., & Maquelin, K. (2010). Towards Raman-based epidemiological typing of Pseudomonas aeruginosa. J Biophotonics, 3(8-9), S. 506-511. doi: 10.1002/jbio.201000026
Witkowska, E., Korsak, D., Kowalska, A., Ksiezopolska-Gocalska, M., Niedziolka-Jonsson, J., Rozniecka, E., . . . Kaminska, A. (2017). Surface-enhanced Raman spectroscopy introduced into the International Standard Organization (ISO) regulations as an alternative method for detection and identification of pathogens in the food industry. Anal Bioanal Chem, 409(6), S. 1555-1567. doi: 10.1007/s00216-016-0090-z
Zhang, L., & Henson, M. J. (2007). A practical algorithm to remove cosmic spikes in Raman imaging data for pharmaceutical applications. Appl Spectrosc, 61(9), S. 1015-1020. doi: 10.1366/000370207781745847
Zhang, P., Kong, L., Wang, G., Setlow, P., & Li, Y. Q. (2011). Monitoring the wet-heat inactivation dynamics of single spores of Bacillus species by using Raman tweezers, differential interference contrast microscopy, and nucleic acid dye fluorescence microscopy. Appl Environ Microbiol, 77(14), S. 4754-4769. doi: 10.1128/AEM.00194-11
Zhang, X., Chen, S., Ling, Z., Zhou, X., Ding, D. Y., Kim, Y. S., & Xu, F. (2017). Method for Removing Spectral Contaminants to Improve Analysis of Raman Imaging Data. Sci Rep, 7, S. 39891. doi: 10.1038/srep39891

Eidesstattliche Versicherung
88
Eidesstattliche Versicherung
Ich versichere an Eides statt, dass ich die vorliegende Arbeit selbständig angefertigt und
mich fremder Hilfe nicht bedient habe.
Alle Stellen, die wörtlich oder sinngemäß veröffentlichtem oder nicht veröffentlichtem
Schrifttum entnommen sind, habe ich als solche kenntlich gemacht.
_____________________ ____________________________________
Ort, Datum Unterschrift
![Vorexperimente zur koh˜arent ˜uberh ˜ohten Raman-Streuung ... · B Spektroskopie des Zustands E2§ ... Stimulated Raman Scattering, SRS, [2{4,7,9{11]). Dabei wird in Molek˜ulen](https://static.fdokument.com/doc/165x107/5e2007194fe028629f270f1e/vorexperimente-zur-kohoearent-oeuberh-oeohten-raman-streuung-b-spektroskopie.jpg)

















