Entwicklung eines Verfahrens zur quantitativen Bestimmung ... · Der Stamm der Cyanophyta enthält...
92
1 Entwicklung eines Verfahrens zur quantitativen Bestimmung von Algengruppen mit Hilfe computergestützter Auswertung spektralaufgelöster Fluoreszenzanregungsspektren Diplomarbeit der Mathematisch-Naturwissenschaftlichen Fakultät der Christian-Albrechts-Universität zu Kiel vorgelegt von Martin Beutler Kiel, Juni 1998
Transcript of Entwicklung eines Verfahrens zur quantitativen Bestimmung ... · Der Stamm der Cyanophyta enthält...

1
Entwicklung eines Verfahrens zur quantitativen Bestimmung vonAlgengruppen mit Hilfe computergestützter Auswertung
spektralaufgelöster Fluoreszenzanregungsspektren
Diplomarbeitder Mathematisch-Naturwissenschaftlichen Fakultät
der Christian-Albrechts-Universitätzu Kiel
vorgelegt vonMartin BeutlerKiel, Juni 1998

2
INHALTSVERZEICHNIS
KAPITEL...........................................................................................................................SEITE
1 VERZEICHNIS VERWENDETER ABKÜRZUNGEN UND BEGRIFFE............................4
2 EINLEITUNG..........................................................................................................................5
3 BIOPHYSIKALISCHE GRUNDLAGEN...............................................................................83.1 Algenstämme und -klassen..............................................................................................8
3.1.1 Cyanophyta, Cyanobakterien, Blaualgen...............................................................93.1.2 Glaucophyta.........................................................................................................103.1.3 Rhodophyta, Rotalgen .........................................................................................103.1.4 Heterokontophyta ................................................................................................103.1.5 Haptophyta...........................................................................................................123.1.6 Cryptophyta .........................................................................................................123.1.7 Dinophyta ............................................................................................................123.1.8 Chlorophyta .........................................................................................................13
3.2 Photosynthese................................................................................................................143.3 Die PS-II-Chlorophyll-a-Fluoreszenz ...........................................................................18
3.3.1 Das zeitliche Verhalten der PS-II-Chlorophyll-Fluoreszenz ...............................213.4 Photosysteme der einzelnen Algen................................................................................23
3.4.1 Innere Antennen des PS II ...................................................................................233.4.2 Äußere Antennen des PS II..................................................................................23
4 VORVERSUCHE..................................................................................................................314.1 Aufbau eines Fluorometers ...........................................................................................31
4.1.1 Das Anregungsmeßlicht.......................................................................................314.1.2 Die Photodetektoreinheit .....................................................................................324.1.3 Anordnung des Anregungsmeßlichtes und Photodetektoreinheit........................324.1.4 Ansteuerung des Meßlichtes und Signalverarbeitung .........................................344.1.5 Test der Linearität des Fluoreszenzsignals ..........................................................38
4.2 Aufnahme von Anregungsspektren verschiedener Algenkulturen................................404.2.1 Durchführung.......................................................................................................404.2.2 Ergebnisse............................................................................................................41
5 VERFAHREN ZUR ANPASSUNG MIT NORMKURVEN................................................465.1 Theorie des Fitverfahrens..............................................................................................465.2 Struktur des Rechenprogramms ....................................................................................505.3 Test des Normkurvenfitverfahrens mit simulierten Daten ............................................52

3
6 AUTOMATISIERUNG DES FLUOROMETERS................................................................556.1 Aufbau der Leuchtdiodenansteuerung...........................................................................556.2 Leuchtdioden-Takt ........................................................................................................56
7 TEST DES AUTOMATISIERTEN FLUOROMETERS ......................................................607.1 Verdünnungsreihen .......................................................................................................60
7.1.1 Künstliche Algenblüten .......................................................................................607.1.2 Verdünnung bei gleichbleibenden Verhältnissen der Algengruppen ..................62
7.2 Fluoreszenzverhalten gealterter Algenkulturen.............................................................667.2.1 Fluoreszenzverhalten der Grünalgen während der Alterung ...............................687.2.2 Fluoreszenzverhalten der Blaualgen während der Alterung................................727.2.3 Fluoreszenzverhalten der Kieselalgen während der Alterung .............................76
7.3 Messung an natürlichen Algenpopulationen der Schlei ................................................79
8 DISKUSSION UND AUSBLICK .........................................................................................83
9 ZUSAMMENFASSUNG ......................................................................................................87
10 LITERATURVERZEICHNIS .............................................................................................89

4
1 Verzeichnis verwendeter Abkürzungen und Begriffe
[A]:................................................................................................................ ElektronenakzeptorADP: ............................................................................................................AdenosindiphosphatATP:............................................................................................................ Adenosintriphosphat[Chl]:................................................................................................................ AntennenpigmentCyt b6/f-Komplex.:............................................................Cytochrom b6/Cytochrom f-KomplexD-Latch.: .....................................................................................................................Data-Latche-:.....................................................................................................................................Elektronf0:...................................................................................................................... Grundfluoreszenzfm: .................................................................................................................MaximalfluoreszenzGAL: ............................................................................................................Generic Array LogicH+: ...................................................................................................................................... ProtonHPLC: ......................................................................High Performance Liquid ChromatographykD, kF, kPC, kt, kT, kf, kd: ...................................................ReaktionsgeschwindigkeitskonstantenLED:..........................................................................................................................LeuchtdiodeLHCI: .............................................................................................. Light-Harvesting-Complex ILHCII: ............................................................................................Light-Harvesting-Complex IINADP: .................................................................... Nicotinamid-Adenin-Dinucleotid-PhosphatPQH2: ..............................................................................................................PlastohydrochinonPQ: ........................................................................................................................... PlastochinonPS I:........................................................................................................................ Photosystem IPS II: .....................................................................................................................Photosystem IIP 680 ................................................................................ Reaktionszentrum des Photosystem IIP 700 ..................................................................................Reaktionszentrum des Photosystem IQA, QB ..............................................................................Chinone des PS II (Primärakzeptoren)[R]: ....................................................................... photoaktives Pigment des ReaktionszentrumsWC-Nährmedium, WC-Medium: .......................................Modifiziertes-Woods-Hole-Medium

2 Einleitung
5
2 Einleitung
In den letzten Jahrzehnten entstand im Zusammenhang mit steigendem Kohlendioxidgehalt inder Erdatmosphäre ein stärkeres Interesse an der Primärproduktion von Biomasse durchPflanzen, die mit CO2-Aufnahme und Sauerstoffabgabe einhergeht und somit die Zusammen-setzung der Erdatmosphäre prägt. Eine der wichtigsten Gruppen der Primärproduzenten sinddie planktischen Algen in den Gewässern dieser Welt. Daher gewinnt die Detektion vonPhytoplankton-Konzentrationen zunehmend an Bedeutung. Als einfachstes und ältestes Ver-fahren ist das Auszählen von Phytoplanktern mit Hilfe eines Mikroskops zu nennen. GrößereProbenmengen werden durch Sedimentation in Zählkammern konzentriert und dann analy-siert. Dies Verfahren ermöglicht zwar eine sehr artenspezifische Analyse, ist aber verständ-licherweise sehr zeitaufwendig. Es erreicht aufgrund der doch recht kleinen Mengen ausge-zählter Zellen selten statistische Sicherheit, die sicherlich auch vom Fleiß des Mikroskopie-renden abhängt.Anfang der sechziger Jahre begann man die Konzentration des photosynthetischen PigmentsChlorophyll-a in den zu untersuchenden Wasserproben zu bestimmen. Hierbei fand man Pro-portionalitäten zwischen den Zellzahlen der Phytoplankter und der Chlorophyll-a-Konzentrationen in den Proben. Deshalb wurde im UNESCO-Monograph „Determination ofphotosynthetic pigments in seawater“ (1966) erstmalig ein standardisiertes Verfahren zurChlorophyll-a-Konzentrationsbestimmung entwickelt. Hierbei handelte es sich um ein soge-nanntes naßchemisches Verfahren, bei dem die Phytoplankter von dem umgebenden Wasserdurch Filtration getrennt werden. Anschließend wird das Filtrat in einem organischen Lö-sungsmittel aufgelöst und der Chlorophyllgehalt der Lösung dann durch Messung der opti-schen Absorption bei wenigen diskreten Wellenlängen bestimmt. Im Laufe der Jahre wurdenbis heute viele Abwandlungen dieser Methode entwickelt (Jeffrey und Humphrey 1975). Aus-serdem wurden fluorometrische in-vitro-Messungen eingeführt, die statt einer Absorptions-messung die Fluoreszenzemission der gelösten Probe ausnutzten (Yentsch und Menzel 1963;Holm-Hansen 1965). Der Nachteil all dieser naßchemischen Methoden liegt in dem großenZeitaufwand einer Bestimmung. Zudem muß während des Abfiltrierens und beim Lösungs-vorgang höchste Sorgfalt gewährleistet sein, da Chlorophyll hierbei leicht zerfällt. Weiterhinexistieren inzwischen derartig viele Abwandlungen dieser Methoden, die alle einen gewissenAnspruch auf Absolutheit erheben, so daß für diese Bestimmungsmöglichkeiten in nächsterZeit ein großer Diskussionsbedarf besteht.

2 Einleitung
6
Parallel dazu entwickelte sich aus chromatographischen Methoden die HPLC-Analyse (HighPerformance Liquid Chromatography) (Mantoura et al. 1997). Bei diesem Verfahren werdendie Pigmente aus den Phytoplanktern in Lösungsmittel gelöst und im Gegensatz zur normalenSäulenchromatographie unter hohem Druck durch die Säulen gepumpt. Je nach Retentions-zeiten lassen sich die Pigmente dann durch Messung der optischen Absorption oder Fluores-zenz-emission detektieren. Dieses ist bislang das präziseste Verfahren zur Chlorophyllbe-stimmung, da auch die Abbauprodukte des Chlorophylls korrekt bestimmt werden können.Die Aufbereitung der Proben erfordert jedoch viel Erfahrung, und die Analytik erfordert vielZeit.Eine weitere Methode der Chlorophyll-a-Konzentrationsbestimmung ist die in-situ-Messungder 685 nm Chlorophyll-a-Fluoreszenzemission des Photosystems II der Phytoplankter. Da-durch können auch Erkenntnisse über die photosynthetische Aktivität des Phytoplanktons ge-wonnen werden (Genty et al. 1989; Schreiber 1994; Moldaenke et al. 1995). Da man die ent-nommenen Proben nicht aufbereiten muß, steht hiermit eine praktische Methode für Feldver-suche und Meßstationen zur Verfügung, die es erlaubt, zu einer Chlorophyllgehaltsbestim-mung zu gelangen.In zunehmendem Maße gewinnen Phytoplankter an Bedeutung als Gewässergüte-Indikatoren,da sie durch ihr großes Oberflächen/Volumenverhältnis und durch ihre kurzen Regenerations-zeiten sehr stark und schnell auf die Inhaltsstoffe eines Gewässers reagieren. Dabei ist dieErforschung ungleichgewichtiger biologischer Systeme vonnöten, z.B. giftige Algenblüten ineinem künstlich eutrophierten Gewässer, obwohl hier auch natürliche toxische Algenblütenbekannt sind. Die Auswirkungen können jedoch in beiden Fällen verheerend sein. So kann derVerzehr von Austern und Muscheln aus einem Gebiet, das von einer sog. „roten Tide“, beste-hend aus Dinoflagellaten, betroffen war, für den Menschen toxisch sein, da die Muscheln dieGifte der von ihnen verdauten Dinoflagellaten anreichern. Im Sommer 1997 kam es in derOstsee vor den Küsten Kopenhagens und Fehmarns zu einer giftigen Cyanobakterien- (Blau-algen)-blüte der Gattungen Nodularia und Aphanizonemon, wobei erst seit kurzem bekanntist, daß die Cyanobakterien auch in den Meeren (und nicht nur in Süß- und Brackwasser) vonso großer Bedeutung sind. Durch ihre extrem geringe Größe sind die marinen Cyanobakterienbisher den Beobachtungen entgangen (Hoek 1993).Daher ist es von Interesse, eine Meßmethode zu entwickeln, die nicht nur Aufschluß über dieMenge des Phytoplanktons im Gewässer gibt, sondern auch die Zusammensetzung bestimmt.Das Auszählen unter dem Mikroskop wäre das Verfahren, welches am meisten Aufschlußüber die Zusammensetzung der Arten des Phytoplanktons liefert. Dieser Ansatz zur Bestim-mung erfordert jedoch eine beträchtliche Artenkenntnis.

2 Einleitung
7
Da die Klassifizierung von Phytoplanktern zu einem Großteil durch ihren Gehalt an photo-synthetischen Pigmenten, die zusätzlich zum Chlorophyll für die Lichtabsorption zuständigsind, geschieht, wird versucht, über HPLC-Analysen den Gehalt dieser photosynthetischenPigmente zu bestimmen und daraus Schlüsse über die Zusammensetzung des Phytoplanktonszu ziehen (Jeffrey 1997). Hierbei bereiten aber gelegentlich Cyanobakterien Schwierigkeiten,da diese als charakteristische Pigmente Phycobiliproteine enthalten, die bisher nicht bei einerHPLC-Analyse zu erfassen sind. Daher wird ihr Vorkommen über ein spezielles Carotinoid(Zeaxanthin) bestimmt, das jedoch nicht in allen Spezies vorkommt.Durch spezifische Pigmentierung der verschiedenen Algengruppen ergeben sich auch unter-schiedliche Fluoreszenzanregungsspektren für die Chlorophyll-a-Fluoreszenzemission. Diesführt bei der Messung der Chlorophyll-a-Fluoreszenz zum einen zu Fehlern in der Bestim-mung der Chlorophyll-a-Konzentration, eröffnet aber zum anderen Möglichkeiten zur Cha-rakterisierung der verschiedenen Algengruppen durch ihre Anregungsspektren. Versuche, An-regungsspektren des gesamten optischen Bereiches aufzunehmen, zeigten, daß dies möglichist (Yentsch und Yentsch 1979; Yentsch und Phinney 1985). Feldversuche machen es abererforderlich, daß möglichst häufig gemessen wird bzw. die Dauer einer Einzelmessung sehrkurz ist. Deshalb wird in dieser Arbeit zu überprüfen sein, inwieweit ein Anregungsspektrummit nur wenigen Wellenlängen ausreicht, eine Algengruppe zu bestimmen, und welcheSchwankungen dabei in den einzelnen Gruppen auftreten können. Weiterhin soll untersuchtwerden, ob sich daraus ein Verfahren zur Algenpopulationsbestimmung entwickeln läßt.

3 Biophysikalische Grundlagen
8
3 Biophysikalische Grundlagen
3.1 Algenstämme und -klassen
Als Grundlage für die Entwicklung eines Meßverfahrens, das Algen als Meßobjekte verwen-det, soll hier ein Umriß gegeben werden, was eigentlich Algen sind und wie sie eingeteiltwerden. Übersichten sind in der Literatur häufig zu finden (z.B. Hoek et al. 1995; Bold undWynne 1985). Grundsätzlich sind Algen Organismen ohne Wurzeln oder beblätterte Stengel,die zur oxygenen Photosynthese fähig sind. Ihr Lebensraum erstreckt sich auf Gewässer,feuchte Böden, aber sie sind auch in Wüsten oder auf Gletschern zu finden. In Gewässernkönnen benthische Formen, d.h. auf dem Grund oder im Sediment wachsende Algen, undplanktische, im Wasser frei schwebende Formen, vorkommen. Die Einteilung aller Algenerfolgt über Stämme (-phyta) und deren Untergruppen in Klassen (-phyceae). Die taxonomi-sche Einteilung der Algen erfolgt dabei nach Zusammensetzung der Photosynthesepigmente(d.h. Chlorophylle, Carotinoide und Phycobiliproteine), die auch die Farbgebung der Algenbestimmt. Außerdem spielen die chemische Zusammensetzung der Reservestoffe und der Bauder Zellwände eine wesentliche Rolle bei der Einteilung in Abteilungen und Klassen. EineÜbersicht über die verschiedenen Stämme mit ihren dazugehörigen Klassen ist in Tab. 3.1gegeben.

3 Biophysikalische Grundlagen
9
Stamm (Phyta) Klasse
Cyanophyta (Blaualgen) • Cyanophyceae
Glaucophyta • Glaucophyceae
Rhodophyta (Rotalgen) • Bangiophyceae• Florideophyceae
Heterokontophyta • Chrysophyceae (Goldalgen)• Xanthophyceae (Gelbgrüne Algen)• Bacillariophyceae (Kieselalgen)• Raphidophyceae• Phaeophyceae (Braunalgen)
Haptophyta • Haptophyceae
Cryptophyta • Cryptophyceae
Dinophyta • Dinophyceae
Chlorophyta (Grünalgen) • Prasinophyceae• Chlorophyceae• Charophyceae
Tab. 3.1: Übersicht der Algenstämme und deren Klassen (Hoek et al. 1995)
3.1.1 Cyanophyta, Cyanobakterien, Blaualgen
Der Stamm der Cyanophyta enthält nur die Klasse der Cyanophyceae. Die wichtigsten Photo-synthesepigmente sind Chlorophyll-a, „blaues“ Phycocyanin, Allophycocyanin und Phycoe-rythrin. Die Cyanophyta enthalten keine Chloroplasten und keinen eukaryotischen Zellkern(prokaryotische Zellen). Die akzessorischen Pigmente liegen in Reihen oder kugeligen Kör-pern vor, den Phycobilisomen (ähnliche Anordnung wie in den Chloroplasten der Rhodophytaund Glaucophyta).Die verschiedenen Arten sind im Salzwasser, Süßwasser und an Land zu finden. Am häufig-sten besiedeln sie stehendes oder leicht bewegtes, eutrophes Süßwasser.Es gibt einzellige, koloniebildende, fädige, aber keine begeißelten Blaualgen. Einzellige Blau-algen verfügen über eine Art „Schwimmbläschen“ (Gasvesikel). Bei der höheren Lichtinten-sität an der Wasseroberfläche kollabieren die Vesikel, und daraus folgt ein Absinken der Zel-len. Häufig bilden Cyanobakterien einen Film an der Oberfläche eines Gewässers. Einige Ar-ten z.B. Microcystis können giftige Substanzen (Microcystin) produzieren, so daß eine Algen-blüte toxische Auswirkung auf die Fauna (und die Trinkwasserqualität) haben kann. Die Zel-len vieler im Meer lebender Blaualgen mit nur einer Größe von 0,2 µm - 2 µm im Durchmes-

3 Biophysikalische Grundlagen
10
ser werden zum Picoplankton gezählt. Durch ihre geringe Größe wurde ihr Vorkommen imMeer lange Zeit übersehen.
3.1.2 Glaucophyta
Auch bei den Glaucophyta existiert nur eine Klasse, die der Glaucophyceae. Die Pigmentzu-sammensetzung ist denen der Blaualgen ganz ähnlich. Der Aufbau dieser eukaryotischen Zel-len wird durch ein evolutionäres „Verschlucken“ (Ingestion) einer Blaualge erklärt. Ihre Chlo-roplasten ähneln dem Aufbau einer Cyanobakterie. Die Zellen der Glaucophyceae sind begei-ßelt. Die Verbreitung dieses Stammes ist sehr gering, so daß er in dieser Arbeit nicht berück-sichtigt wird.
3.1.3 Rhodophyta, Rotalgen
Die Rotalgen unterteilen sich in zwei Klassen: die Bangiophyceae und Florideophyceae. IhrePigmentkomposition ist sehr ähnlich wie bei den Blaualgen. Dabei überwiegt jedoch das rotePhycoerythrin, und die Farbstoffe liegen in Chloroplasten vor. Es gibt 5000-5500 Arten; 150Arten davon kommen im Süßwasser und der Rest davon kommt im Salzwasser vor. Fast alleRhodophyta wachsen festgeheftet auf dem Grund an felsigen Meeresküsten. Die meistenRhodophyta sind Mehrzeller, und es gibt nur sehr wenige planktische Exemplare, die seltenervorkommen. Für einige Kulturkreise sind Rotalgen als Nahrungsmittel von Bedeutung. DieChloroplasten ähneln den Strukturen der Blaualgen und denen der Glaucophyta.
3.1.4 Heterokontophyta
Alle Klassen der Heterokontophyta besitzen als gemeinsames Merkmal den Gehalt an Chlo-rophyll-a, -c1, und -c2 und einem Xanthophyll Fucoxanthin (bei Chrysophyceae, Bacillario-phyceae, Phaeophyceae) oder Vaucheriaxanthin (bei Xanthophyceae). Dieses verleiht denHeterokontophyta die charakteristische bräunlich-gelbe Farbe. Die Zellen der begeißeltenVertreter sind heterokont, d.h. sie besitzen eine nach vorn gerichtete, lange Flimmergeißel undeine kurze nach hinten gerichtete Geißel ohne Flimmern.
3.1.4.1 Chrysophyceae, Goldalgen
Diese Fucoxanthin enthaltende Klasse der Heterokontophyta umfaßt ca. 1000 Arten, von de-nen die meisten im Süßwasser leben. Einige Arten bevölkern aber auch die Ozeane. Die Süß-wasserarten dominieren in oligotrophen Seen mit schwach saurem oder neutralem Wasser.Ein typischer Vertreter ist z.B. Ochromonas.

3 Biophysikalische Grundlagen
11
3.1.4.2 Xanthophyceae, Gelbgrüne Algen
Die Xanthophyceae erhalten ihre gelbgrüne Farbe durch das Chlorophyll-a. Zudem enthaltensie sehr geringe Mengen Chlorophyll-c1, -c2 und Vaucheriaxanthin. Fucoxanthin fehlt in die-ser Klasse.Sie sind hauptsächlich im Süßwasser verbreitet. Aber auch hier sind sie eher seltener anzutref-fen. Des weiteren existieren viele terrestrische Arten, die auf feuchter Erde leben. Außerdemgibt es einzellige und koloniebildende Arten.
3.1.4.3 Bacillariophyceae (Diatomeen, Kieselalgen)
Bei dieser weitverbreiteten Klasse findet man die Pigmente Chlorophyll-a, -c1,-c2, manchmal -c3 statt -c2. Ihre braune Farbe erhalten sie vornehmlich, wie bei den meisten Heterokonto-phyta, durch Fucoxanthin. Insgesamt leben ca. 100000 Arten im Süßwasser, im Meer und auffeuchten oder trockenen Böden, in gemäßigten bis kalten Gebieten der Weltmeere, besondersaber in nährstoffreichen Gewässern.Die Zellen sind mit einer Kieselsäureschale umgeben. Zum Aufbau der Kieselsäureschalebenötigen sie eine silikathaltige Umgebung. Einige Arten besitzen begeißelte Zellen, hierbeisind nur die männlichen Zellen begeißelt. Durch ihre große Anzahl und weite Verbreitunghaben die Kieselalgen große Bedeutung für die Primärproduktion. Die Klasse der Bacillario-phyceae besteht aus zwei Ordnungen (Pennales und Centrales), die sich hauptsächlich in derSymmetrie der Zellen unterscheiden.
3.1.4.4 Raphidophyceae
Diese Klasse ist hauptsächlich im marinen Bereich aufzufinden. Auch sie enthalten das farb-gebende Fucoxanthin. Fünf Gattungen kommen in saurem Süßwasser, wie kleinen Tümpeln,auf Sand oder im Moor, vor. Im Meer, z.B. in Meeresbuchten an der japanischen Küste, ver-ursachen Raphidophyceae toxische „rote Tiden“. Hieraus ergibt sich ein erheblicher Schadenfür dort ansässige Fischfarmen. Toxische „rote Tiden“ sind von vielen anderen Meeren be-kannt, werden aber meistens von Arten der Dinophyta erzeugt. (Okaichi et al. 1989)
3.1.4.5 Phaeophyceae, Braunalgen
Diese Klasse besteht aus 265 Gattungen mit 1500-2000 Arten und besiedelt ähnliche Lebens-räume wie die Rhodophyta, z.B. felsige Meeresküsten. Ihre braune Farbe entsteht durch dasPigment Fucoxanthin. Außerdem besitzen Phaeophyceae zahlreiche lichtbrechende Bläschen,die mit Tannin gefüllt sind. Das Tannin wird bei Oxidation dunkelbraun, so daß an Land ge-schwemmte, abgestorbene Algen eine schwarzbraune Farbe erhalten. Alle Arten der Klassesind Mehrzeller. Obwohl einige mikroskopische kleine, stark verzweigte Arten vorkommen,sind sie für das Phytoplankton eher unbedeutend.

3 Biophysikalische Grundlagen
12
3.1.5 Haptophyta
Dieser Algenstamm enthält nur eine Klasse, die Haptophyceae. Die Pigmentzusammenset-zung dieses Stammes ähnelt sehr der Pigmentzusammensetzung der Heterokontophyta, da sieChlorophyll-a, -c1 oder -c3 und -c2 sowie durch Fucoxanthin maskiert. Bei einigen Arten sindauch 19`-Hexanoxyloyfucoxanthin und 19`-Butanoyloxyfucoxanthin zu finden.Die Haptophyta spielen besonders in offenen Weltmeeren eine Rolle. Außerdem gibt es Al-genblüten in der südlichen Nordsee; 1988 wurde von einer riesigen Algenblüte von Chry-sochromulina polylepis an der Westküste Schwedens und der Südküste Norwegens berichtet.Diese Algenblüte war im höchsten Maße toxisch.
3.1.6 Cryptophyta
Der Stamm besteht aus einer Klasse, den Cryptophyceae, und umfaßt zwölf Gattungen mit100 Süßwasserarten und 100 marinen Arten. Die Süßwasserarten findet man manchmal inTümpeln, Pfützen, aber auch in großen, oligotrophen Seen. Die Salzwasserarten sind inBrackwasserpfützen und Gezeitentümpeln, und einige sind auch im offenen Meer zu finden.Die Art Mesodinium rubrum ensteht regelmäßig mit vom Grund aufsteigendem Wasser anden Küsten Perus und Baja California, da dort nährstoffreiches Tiefenwasser an die Oberflä-che gelangt. Dieses führt dann zur Algenblüte. Fast alle Cryptophyceae sind einzellige Fla-gellaten.Durch Ihre Pigmentzusammensetzung sind Cryptophyta eine Besonderheit. Sie besitzen Chlo-rophyll-a und -c2 und Phycocyanine und Phycoerythrine. Die Chloroplasten (1-2 pro Zelle)sind olivgrün, braun oder gelbbraun gefärbt. Die Farbe entsteht aufgrund der Maskierung vonChlorophyll-a mit (Cryptophyceen)-Phycocyanin, (Cryptophyceen)-Phycoerythrin. Das Phy-cocyanin und das Phycoerythrin liegt im Unterschied zu anderen Abteilungen im Innern derThyllakoide vor.
3.1.7 Dinophyta
Die Dinophyta besitzen als wichtigstes Pigment Chlorophyll-a, -c2 und das Xanthophyll Peri-dinin. 90% aller Dinophyta sind planktisch und marin. 50% aller marinen Dinophyta tragenkeine Chloroplasten und sind somit farblos. Dinophyta besitzen eine dicke Zellwand, die ausZellulose besteht. Bei einigen Arten ist ein vergleichbares Pigmentmuster wie bei den Hetero-kontophyta, und bei anderen Arten ist sogar eine Verwandtschaft zu den Ciliaten feststellbar.Diesen Vertretern fehlt das Peridinin.

3 Biophysikalische Grundlagen
13
3.1.8 Chlorophyta
Der Stamm der Chlorophyta umfaßt neben Algen auch Moose und Gefäßpflanzen. Chloro-phytina (Grünalgen) sind eine nur Algen umfassende Untergruppe der Chlorophyta. Die Chlo-rophytina bestehen aus drei Klassen: Den Chlorophyceae, Prasinophyceae und Charophyceae.Da sich diese nur durch wenige strukturelle Merkmale unterscheiden, werden im folgendendie Pigmentkombinationen und die Verbreitung des gesamten Stammes beschrieben. Grünal-gen sind am häufigsten im Süßwasser zu finden, trotzdem gibt es auch zahlreiche marine undim Brackwasser lebende Arten. Die Grünalgen bestehen aus 500 Gattungen und ca. 8000 Ar-ten. Es gibt einzellige oder koloniebildende, planktische Algen, aber auch viele mehrzellige,makroskopische Formen. Chlorophyceae ist die größte Klasse, früher wurden fast alle Chloro-phytina ihr zugerechnet.Die charakteristischen Pigmente der Grünalgen sind Chlorophyll-a und -b sowie das Caroti-noid Lutein, wodurch sie eine kräftig grüne Farbe erhalten. Die Grünalgen besitzen Chlo-roplasten, die eine sehr ähnliche Struktur besitzen wie die der höheren Pflanzen.

3 Biophysikalische Grundlagen
14
3.2 Photosynthese
Die Photosynthese ist ein komplexer endergonischer Prozeß, bei dem mit Hilfe absorbierterStrahlungsenergie organische Substanz in Form von Kohlenhydrat aus CO2 und Wasser gebil-det wird (Lawlor 1990). Dabei wird bei fast allen photosynthetisch aktiven Organismen (höhe-re Pflanzen, Algen und photosynthetisch aktive Bakterien) molekularer Sauerstoff (mit Aus-nahme einiger Schwefelbakterien) gebildet. Die Photosynthese ist also der für die Primärpro-duktion auf der Erde zuständige Prozeß, der die Strahlungsenergie der Sonne in eine auf derErde langlebigere Energieform, den chemischen Bindungen des gebildeten Kohlenhydrates,umsetzt.Der chemische Gesamtprozeß kann mit folgender Formel umschrieben werden:
6 CO2+ 12 H2O → C6 H12O6 + 6 O2 + 6 H2O
Dieser Gesamtprozeß besteht aus vielen Einzelschritten, die man in eine Licht- oder Hellreak-tion und Dunkelreaktionen (Calvin-Zyklus) zusammenfassen kann (Buschmann und Grum-bach 1985). Während der Hellreaktion werden aus ADP (Adenosindiphosphat) und NADP(Nicotinamid-Adenin-Dinucleotid-Phosphat) die energiereicheren Stoffe ATP (Adenosintri-phosphat) und NADP-H+H+ unter Absorption von Licht gebildet. Diese werden während derDunkelreaktion zur Bildung von Kohlenhydraten benötigt. In der Hellreaktion werden durchein im folgenden beschriebenen Mechanismus die Treibstoffe für den Calvin-Zyklus ATP undNADPH/H+ erzeugt.Sie werden in der Dunkelreaktion wie folgt verbraucht:
6CO2+18ATP+12NADPH/H+→C6H12O6+18ADP+18Pi+12NADP++6H2OPi: Phosphatgruppe
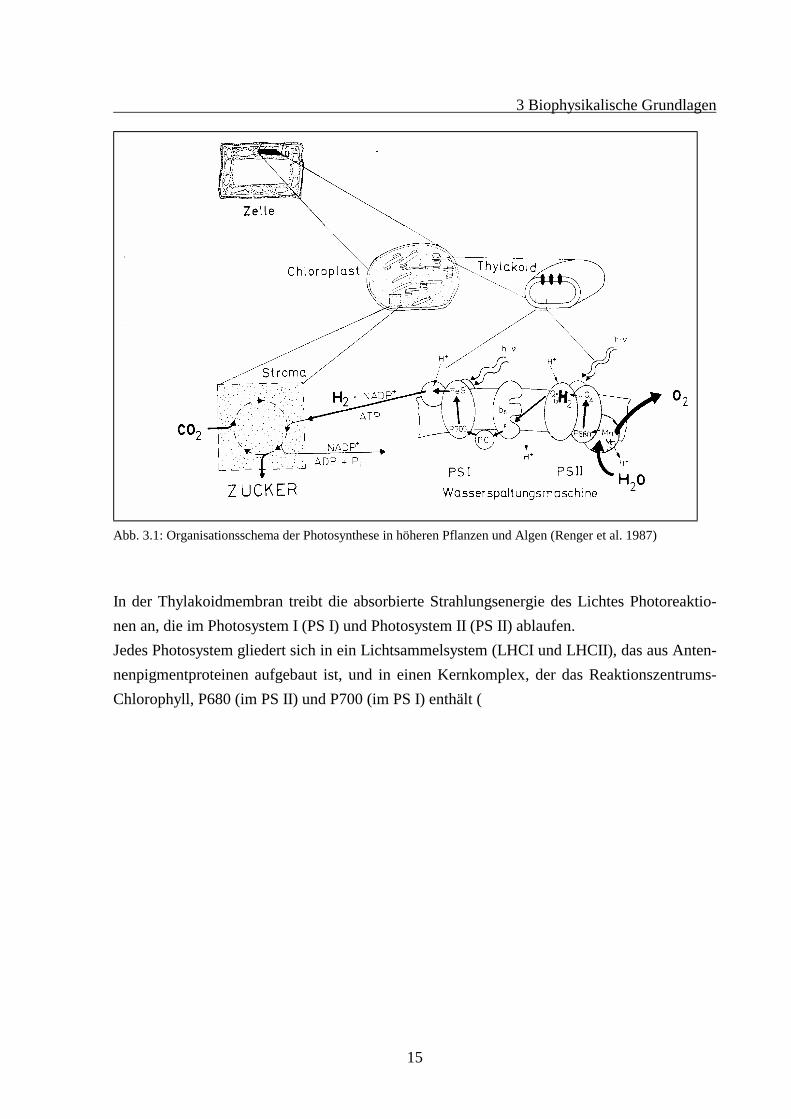
Das photochemische Geschehen vollzieht sich an abgeflachten Vesikeln, den Thylakoiden, dievon der Thylakoidmembran, einer charakteristischen Funktionsmembran umschlossen ist. Sietrennt das Lumen (innen, wäßrig) vom Stroma (außen, plasmatisch). Die Thylakoiden der hö-heren Pflanzen und Algen (mit Ausnahme der Cyanobakterien), befinden sich innerhalb derChloroplasten der einzelnen Zellen. Die Chloroplasten sind durch eine Doppelmembran vomübrigen Zellraum abgegrenzt und bilden einen eigenständigen Reaktionsraum (Abb. 3.1).
3 Biophysikalische Grundlagen
15
Abb. 3.1: Organisationsschema der Photosynthese in höheren Pflanzen und Algen (Renger et al. 1987)
In der Thylakoidmembran treibt die absorbierte Strahlungsenergie des Lichtes Photoreaktio-nen an, die im Photosystem I (PS I) und Photosystem II (PS II) ablaufen.Jedes Photosystem gliedert sich in ein Lichtsammelsystem (LHCI und LHCII), das aus Anten-nenpigmentproteinen aufgebaut ist, und in einen Kernkomplex, der das Reaktionszentrums-Chlorophyll, P680 (im PS II) und P700 (im PS I) enthält (

3 Biophysikalische Grundlagen
16
Abb. 3.2).
Abb. 3.2: Schematischer Aufbau der zwei Photosysteme (PS II und PS I)
Werden nun Lichtquanten von den Lichtsammelsystemen absorbiert, wird die aufgenommeneEnergie strahlungslos über die Kernkomplexe zum Reaktionszentrum weitergeleitet.Das angeregte Chlorophyll-a kann eine Ladungstrennung durchführen. Es besitzt nach einerEnergieaufnahme durch Lichteinstrahlung ein niedrigeres Redoxpotential und gibt an denPrimärelektronenakzeptor Phaeophytin ein Elektron ab. Die im Reaktionszentrum fehlendeLadung wird aus der Wasserspaltung an einem Mangankomplex geliefert. Jede Wasserspal-tung setzt vier Elektronen frei. Jeweils eins kann von einem P+680 aufgenommen werden. Die
685nmPSII-Fluoreszenz
Anregungs-Photon
720 nmPS I-Fluoreszenz
Elektronen-Transfer
Anregungs-Photon
ÄußereAntenne
ÄußereAntenne
Reaktions-Zentrum
Reaktions-Zentrum
EnergieTransfer
EnergieTransfer
Kern-Komplex
PS I
PS II
3 Biophysikalische Grundlagen
17
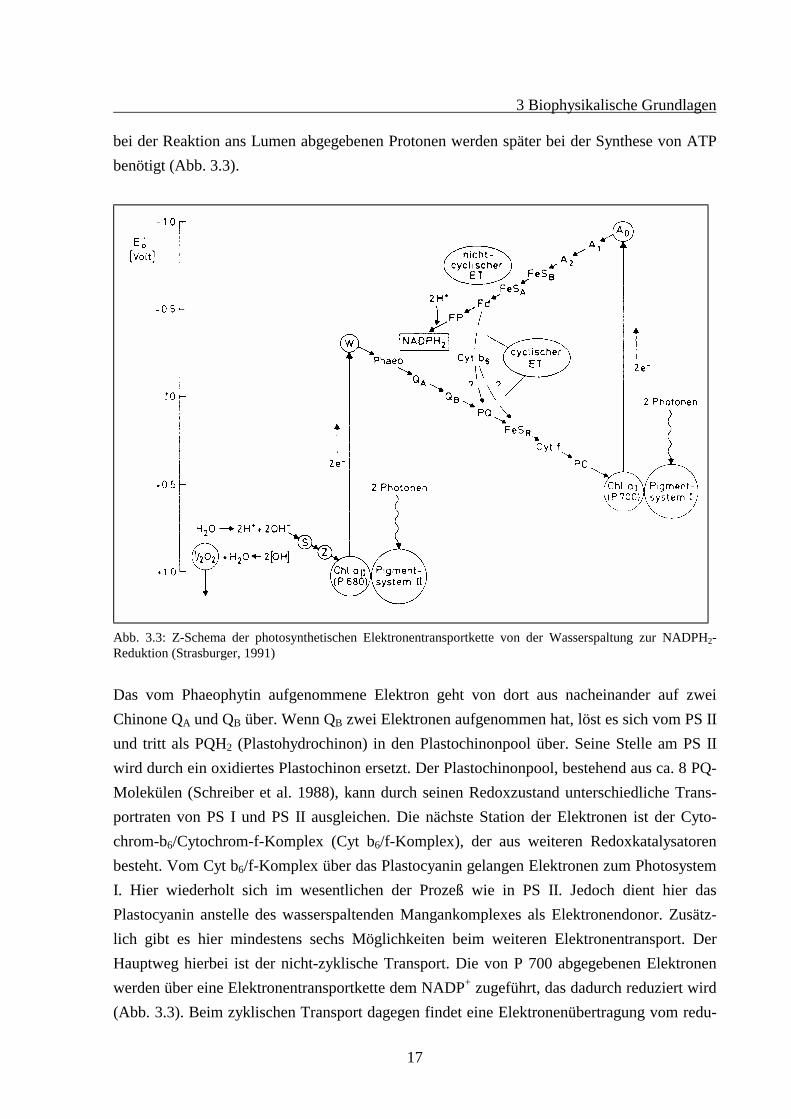
bei der Reaktion ans Lumen abgegebenen Protonen werden später bei der Synthese von ATPbenötigt (Abb. 3.3).
Abb. 3.3: Z-Schema der photosynthetischen Elektronentransportkette von der Wasserspaltung zur NADPH2-Reduktion (Strasburger, 1991)
Das vom Phaeophytin aufgenommene Elektron geht von dort aus nacheinander auf zweiChinone QA und QB über. Wenn QB zwei Elektronen aufgenommen hat, löst es sich vom PS IIund tritt als PQH2 (Plastohydrochinon) in den Plastochinonpool über. Seine Stelle am PS IIwird durch ein oxidiertes Plastochinon ersetzt. Der Plastochinonpool, bestehend aus ca. 8 PQ-Molekülen (Schreiber et al. 1988), kann durch seinen Redoxzustand unterschiedliche Trans-portraten von PS I und PS II ausgleichen. Die nächste Station der Elektronen ist der Cyto-chrom-b6/Cytochrom-f-Komplex (Cyt b6/f-Komplex), der aus weiteren Redoxkatalysatorenbesteht. Vom Cyt b6/f-Komplex über das Plastocyanin gelangen Elektronen zum PhotosystemI. Hier wiederholt sich im wesentlichen der Prozeß wie in PS II. Jedoch dient hier dasPlastocyanin anstelle des wasserspaltenden Mangankomplexes als Elektronendonor. Zusätz-lich gibt es hier mindestens sechs Möglichkeiten beim weiteren Elektronentransport. DerHauptweg hierbei ist der nicht-zyklische Transport. Die von P 700 abgegebenen Elektronenwerden über eine Elektronentransportkette dem NADP+ zugeführt, das dadurch reduziert wird(Abb. 3.3). Beim zyklischen Transport dagegen findet eine Elektronenübertragung vom redu-

3 Biophysikalische Grundlagen
18
zierten Ferredoxin über den Cytochrom-b6-Komplex und Plastochinon zurück auf P+700 statt(Abb. 3.3).Für die Synthese von ATP, das im Calvin-Zyklus neben NADPH/H+ zur Kohlenstoffassimi-lation benötigt wird, ist ein pH-Gradient über der Thylakoidmembran notwendig. Er treibt diesogenannte ATPase (Witt 1979). Der Gradient entsteht hauptsächlich durch den Elektronen-fluß durch die lineare Elektronentransportkette mit der Stöchiometrie 2H+/2e-. Je 2H+ pro 2e-
werden bei der Wasserspaltung ins Lumen abgegeben und beim NADPH/2H+ aus dem Stromaaufgenommen. Die anderen 2H+/2e- transportiert PQH2 vom Stroma ins Lumen. Von den We-gen zu den anderen Elektronenakzeptoren des PS I ist der zyklische Weg besonders effektiv.Durch das im zyklischen Weg energetisierte Protonenpumpen im Cytochrom-b6-Komplexwird der Aufbau des Protonengradienten im linearen Weg stark unterstützt.
3.3 Die PS-II-Chlorophyll-a-Fluoreszenz
Von dem Photosyntheseapparat wird nicht die gesamte von den Lichtsammelsystemen absor-bierte Strahlungsenergie in chemische Energie umgewandelt. Ein Teil der von den Antennen-pigmenten aufgenommenen Lichtenergie wird in Form von Wärme und Fluoreszenz durch dieChlorophyll-a-Moleküle der Antennen emittiert. Dieses wird als die „fundamentale Alternati-ve“ bezeichnet (Lavorel und Joliot 1972). An dieser Stelle werden nur die Energietransport-prozesse im Photosystem II behandelt, da die bei dem Photosystem I auftretende Fluoreszenznicht wie die PS-II-Fluoreszenz vom Zustand der Elektronentransportkette abhängt und somitzeitlich konstant ist (Baker und Webber 1987).Ein vom LHC II aufgenommenes Lichtquant bildet einen Anregungszustand. Dieser wirdhauptsächlich zum Reaktionszentrum weitergeleitet (Geschwindigkeitskonstante kT). Ein Teilwird aber bereits hier in Form von Wärme (kD) oder Fluoreszenzemission (kF) an die Umge-bung abgegeben (Bolhar-Nordenkampf und Öquist 1993). Das von dem Reaktionszentrumaufgenommene Exciton kann entweder in die Elektronentransportkette eingespeist (kPC), indie Antenne zurückwandern (kt), als Fluoreszenz (kf) oder als Wärme (kd) abgestrahlt werden.Die Energietransferprozesse mit ihren Geschwindigkeitskonstanten sind schematisch in Abb.3.4 abgebildet.

3 Biophysikalische Grundlagen
19
Äußere Antenne des PS IIim Grundzustand
[CHL]
Äußere Antenne des PS IIim angeregten Zustand
[CHL]*
Reaktions-Zentrumim Grundzustand
[R]
Reaktions-Zentrumim angeregten Zustand
[R]*
Elektronentransportkette
kdkPC[A]
kT
ktIa hνννν
Ia hνννν
kD
kF
kf
685nmPSII-Fluoreszenz
Abb. 3.4: Energietransportprozesse im Photosystem II (Butler 1978), darin bedeuten:Ia: Zahl der pro Zeiteinheit absorbierten Photonen, [Chl]: Antennenpigment, [R]: photoaktives Pigment des Re-aktionszentrums, [A]: Elektronenakzeptor für stabile Ladungsseparierung, k: Geschwindigkeitskonstanten fürFluoreszenz (kF), strahlungslosen Abbau (kd, kD ), Anregungsenergietransfer (kt, kT ) und photochemische Reak-tion (kPC)
Hieraus resultiert ein gekoppeltes Differentialgleichungssystem (siehe Gleichung [ 3.1] undGleichung [ 3.2]; Renger et al. 1987). Dabei wurden zur Vereinfachung der weiteren Be-trachtungen von R (da für jedes R weit mehr als 100 Antennenpigmentmoleküle existieren, istdie direkte Absorptionswahrscheinlichkeit von R, dem Reaktionszentrum, sehr gering) derAnregungstransfer zwischen verschiedenen Antennenkomplexen vernachlässigt.
[ 3.1]
[ 3.2]
mit: Ia: Zahl der pro Zeiteinheit absorbierten Photonen[Chl*]: Anzahl der Antennenpigmente im angeregten Zustand[R*]: Anzahl der angeregten photoaktiven Pigmente des Reaktionszen-
trumsA: auf eins normierte Anzahl der Elektronenakzeptoren für stabile La-
dungsseparierungk: Geschwindigkeitskonstanten für: Fluoreszenz: kF
strahlungslosen Abbau: kd, kDAnregungsenergietransfer: kt, kTphotochemische Reaktion kPC
d[Chl*]dt = Ia - (kT + kF + kD) [Chl*] + kt[R*]
d[R*]dt = kT [Chl*] - (kt + kf + kd + kPC [A]) [R*]

3 Biophysikalische Grundlagen
20
Das Differentialgleichungssystem ist für den stationären Fall leicht zu lösen. Für die Fluores-zenzquantenausbeute gilt allgemein:
[ 3.3]
Da der Betrag von [R*] vernachlässigbar klein ist, ergibt sich dann aus den Gleichungen[ 3.1], [ 3.2] und [ 3.3] für den stationären Fall:
[ 3.4]
Dabei bedeuten:
ΨΨΨΨF = kF
(kT+kF+kD)Fluoreszenzquantenausbeute der Antenne
ΨΨΨΨT = kT
(kT+kF+kD)Anregungsenergietransferwahrscheinlich-keit aus der Antenne in das Reaktions-zentrum
ϕϕϕϕt = kt
(kF+kt+kd+kPC[A])Anregungsenergietransferwahrscheinlich-keit aus dem Reaktionszentrum in die An-tenne
ΦF ist also unter anderem vom Funktionszustand der Elektronentransportkette abhängig. Beivollständiger Blockierung etwa durch ein sättigendes Hintergrundlicht wird die Elektronen-transportkette komplett ausgelastet, und damit wird [A] = 0 und die Fluoreszenzquantenaus-beute maximal. Bei ganz schwachen Anregungslichtintensitäten würde die absorbierte Energiemaximal durch die Elektronentransportkette abfließen können, und damit würde [A]=1 undΦF minimal werden.Für die Fluoreszenzintensität bei einer festen Anregungswellenlänge finden wir dann dieGleichung [ 3.5] (Schmidt 1993).
[ 3.5]
mit κλ: Absorptionskoeffizient der Antennec[Chl]: Anzahl der AntennenpigmentmoleküleIλ: eingestrahlte LichtintensitätΦF: Fluoreszenzquantenausbeute aus Gleichung [ 3.4]
ΦΦΦΦF = kFIa
([Chl*] + [R*])
ΦΦΦΦF = ΨΨΨΨF
(1-ϕϕϕϕtΨΨΨΨΤΤΤΤ))))
Fλλλλ= Il (1-e(κκκκλ λ λ λ c[Chl]))ΦΦΦΦF
3 Biophysikalische Grundlagen
21
Aus der Gleichung [ 3.5] folgt, daß das Emissionssignal Fλ proportional zur eingestrahltenQuantenzahl Iλ und der Fluoreszenzquantenausbeute ΦF ist, aber nicht zur Anzahl der Anten-nenpigmente. Aber aus einer Taylorentwicklung für Fλ, diese kann für kleine c[Chl] nach derersten Potenz abgebrochen werden ( e-y = 1-y ), ergibt sich Gleichung [ 3.6].
[ 3.6]
Würde die Fluoreszenzquantenausbeute konstant werden, so ergäbe sich die Fluoreszenzin-tensität lediglich als eine Funktion der Anzahl der Antennenpigmentmoleküle, des spektralenAbsorptionskoeffizienten und der Anregungslichtintensität.
3.3.1 Das zeitliche Verhalten der PS-II-Chlorophyll-Fluoreszenz
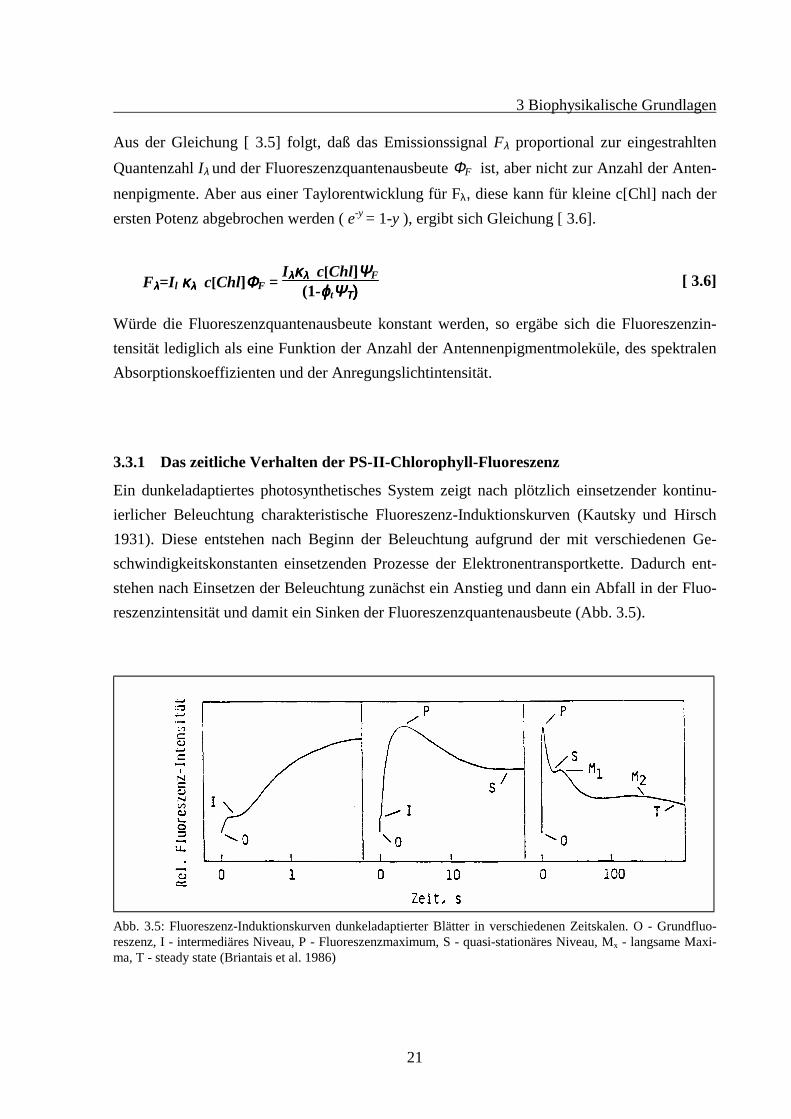
Ein dunkeladaptiertes photosynthetisches System zeigt nach plötzlich einsetzender kontinu-ierlicher Beleuchtung charakteristische Fluoreszenz-Induktionskurven (Kautsky und Hirsch1931). Diese entstehen nach Beginn der Beleuchtung aufgrund der mit verschiedenen Ge-schwindigkeitskonstanten einsetzenden Prozesse der Elektronentransportkette. Dadurch ent-stehen nach Einsetzen der Beleuchtung zunächst ein Anstieg und dann ein Abfall in der Fluo-reszenzintensität und damit ein Sinken der Fluoreszenzquantenausbeute (Abb. 3.5).
Abb. 3.5: Fluoreszenz-Induktionskurven dunkeladaptierter Blätter in verschiedenen Zeitskalen. O - Grundfluo-reszenz, I - intermediäres Niveau, P - Fluoreszenzmaximum, S - quasi-stationäres Niveau, Mx - langsame Maxi-ma, T - steady state (Briantais et al. 1986)
Fλλλλ=Il κκκκλλλλ c[Chl]ΦΦΦΦF = Iλλλλκκκκλλλλ c[Chl]ΨΨΨΨF
(1-ϕϕϕϕtΨΨΨΨΤΤΤΤ))))

3 Biophysikalische Grundlagen
22
Der zeitliche Verlauf der Kurve läßt sich in verschiedene Phasen einteilen. Im Dunkeln voreiner Beleuchtung ist die Fluoreszenzausbeute nicht gleich null. Mit einem schwachen Meß-licht würde eine Grundfluoreszenz f0 gemessen werden können. Nach dem Beginn der Be-leuchtung mit einem hellen Reaktionslicht steigt die Fluoreszenzausbeute über ein Zwischen-plateau I zum Maximum P. Hatte das Reaktionslicht sättigende Wirkung auf den Photosynthe-seapparat ist der Primärakzeptor QA vollständig reduziert. Durch Einsetzen des Calvin-Zyklusbeginnt eine Reoxidation von QA, was zu einem Sinken der Fluoreszenzausbeute führt. Etwaim Minutenbereich liegt die letzte Phase T, die sog. Steady-State-Phase. Ziel wird es später inden Versuchen sein, durch ausreichende Adaptation mit nichtsättigenden Lichtintensitäten diePhotosysteme (lebende Algen) in die Steady-State-Phase zu bringen, um für zeitlich konstanteFluoreszenzquantenausbeute zu sorgen.

3 Biophysikalische Grundlagen
23
3.4 Photosysteme der einzelnen Algen
Die Pigmentproteinkomplexe der Algen, die die Sonnenenergie aufnehmen und umwandeln,sind zusammen mit anderen Faktoren verantwortlich für die Anpassung der marinen und imSüßwasser lebenden Algen an ihre Lebensräume (Anderson und Barrett 1986). Bei aquati-schen Algen ist die Photosynthese stark beeinflußt durch die veränderliche Lichtqualität imWasser, die sich mit zunehmender Wassertiefe nicht nur quantitativ ändert. Zudem ändert sichdie Intensität des eingestrahlten Sonnenlichtes auch mit den verschiedenen Jahreszeiten. ImGegensatz zu Landpflanzen haben im Wasser lebende Algen während ihrer Evolution eineVielfalt von Photosystemen entwickelt, die sich in ihren optischen Absorptionseigenschaftenunterscheiden. Dadurch können sie an verschiedene Lichtqualitäten im Wasser unterschied-lichste räumliche und jahreszeitliche Nischen besetzen. Im weiteren Verlauf dieses Kapitelssoll versucht werden, eine Systematik zu erstellen, um Gemeinsamkeiten und Unterschiede inden Pigmentzusammenstellungen der Antennensysteme der einzelnen Algengruppen zu fin-den, da die Absorptionseigenschaften der Antennen ausschlaggebend für die Chlorophyll-a-Fluoreszenzanregungsspektren sind.
3.4.1 Innere Antennen des PS II
Die Pigmentproteinkomplexe der inneren Antennen des PS II haben sich bei den verschiede-nen Algengruppen während der Evolution kaum verändert, da diese Komplexe nur wenig zurAbsorption von Lichtenergie aus dem Sonnenlichtspektrum beitragen und eher für den Ener-gietransfer zum Reaktionszentrum zuständig sind. Bislang wurden hier in allen AlgengruppenChlorophyll-a/β-Carotin-Komplexe (gelegentlich α-Carotin) gefunden (Delepelaire und Chua1979).
3.4.2 Äußere Antennen des PS II
Für die vorliegende Arbeit spielen die äußeren Antennen der Algen eine wichtige Rolle, da dieunterschiedlichen Absorptionsspektren der äußeren Antennen verschiedener AlgengruppenUnterschiede in den Fluoreszenzanregungsspektren Gleichung [ 3.6] hervorrufen.Bei den für die Lichtabsorption zuständigen Pigmentproteinkomplexen der in dem Kapitel 3.1beschriebenen Algenstämme und -klassen lassen sich fünf verschiedene spektrale Gruppenbilden, die sich in ihren Absorptiosspektren unterscheiden. Zunächst sind hier die Xantho-phyll-Chlorophyll-a/b-Proteine der Grünalgen und grünalgenähnlichen Algen zu finden, diePhycobilisomen, reich an Phycocyanin, der Blaualgen und blaualgenähnlichen Algen, die

3 Biophysikalische Grundlagen
24
Xanthophyll-Chlorophyll-a/c-Proteine der bräunlichen Heterokontophyta und heterokonto-phytaähnlichen Algen, die Chlorophyll-a/c-Phycobiliprotein-Antennen der Cryptophyta, sowiedie Phycobilisomen, reich an Phycoerythrin, der Rotalgen. Da die Rotalgen hauptsächlich alsbenthische Algen vorkommen und im Phytoplankton nur selten zu finden sind, werden sie beiden weiteren Untersuchungen nicht berücksichtigt. Abb. 3.6 zeigt die Zuordnung der ver-schiedenen Algenstämme zu den spektralen Gruppen.
Chlorophyll-a/b-Xanthophyll
Chlorophyll-a/c-Phycobiliprotein
Phycobilisomen(Phycoerythrin)
Chlorophyll-a/c-Xanthophyll
Phycobilisomen(Phycocyanin)
Chlorophyta CryptophytaRhodophytaHeterokontophyta
HaptophytaDinophyta
CyanophytaGlaucophyta
GrünespektraleGruppe
GemischtespektraleGruppe
Rote spektraleGruppe
BraunespektraleGruppe
Blaue spektraleGruppe
Farbe derGruppe
ÄußereAntennen-Komplexe
Zur Gruppegehörende
Stämme
Abb. 3.6: Übersicht über die Einteilung der verschiedenen Algenstämme in spektrale Algengruppen. Oben:Spektrale Gruppe; Mitte: Dazugehörige äußere Antennen-Komplexe; Unten: Dazugehöriger Algenstamm
3.4.2.1 Spektrale Gruppe der Grünalgen
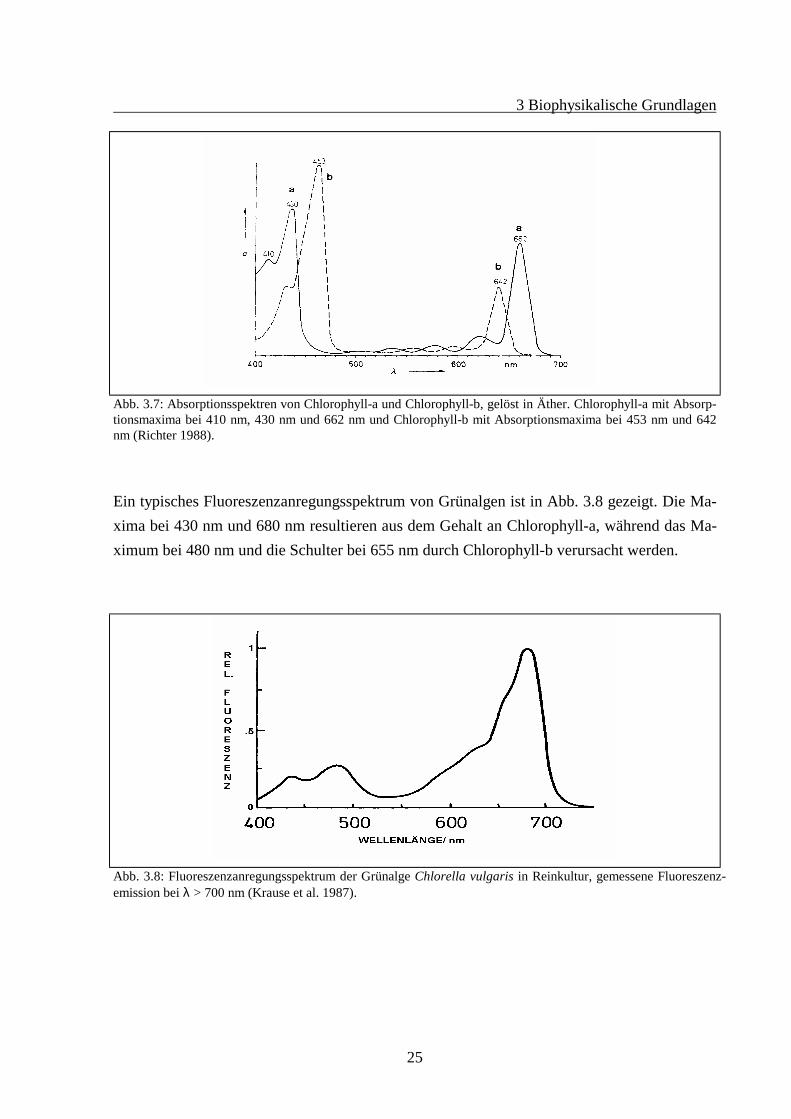
Zu dieser Gruppe gehören die Chlorophyta und Euglenophyta. Alle Angehörigen dieser Grup-pe besitzen Chlorophyll-a und Chlorophyll-b sowie ein Xanthophyll in den äußeren Antennendes PS II. Am häufigsten ist hier das Xanthophyll Lutein anzutreffen. In Abb. 3.7 sind dieAbsorptionsspektren von Chlorophyll-a und Chlorophyll-b, gelöst in Äther, aufgetragen. DiePhotosysteme dieser Gruppe sind eng verwandt mit denen der höheren Pflanzen.Bei der Lichtabsorption haben die Xanthophylle eine untergeordnete Rolle, so daß in einemFluoreszenzanregungsspektrum mit einer Emissionswellenlänge, die dem Chlorophyll-a imPS II entspricht, die Maxima bei den Absorptionsmaxima von Chlorophyll-a und Chlorophyll-b liegen (Diese sind in-vivo allerdings um ca. 15 nm in den roten Wellenlängenbereich ver-schoben).
3 Biophysikalische Grundlagen
25
Abb. 3.7: Absorptionsspektren von Chlorophyll-a und Chlorophyll-b, gelöst in Äther. Chlorophyll-a mit Absorp-tionsmaxima bei 410 nm, 430 nm und 662 nm und Chlorophyll-b mit Absorptionsmaxima bei 453 nm und 642nm (Richter 1988).
Ein typisches Fluoreszenzanregungsspektrum von Grünalgen ist in Abb. 3.8 gezeigt. Die Ma-xima bei 430 nm und 680 nm resultieren aus dem Gehalt an Chlorophyll-a, während das Ma-ximum bei 480 nm und die Schulter bei 655 nm durch Chlorophyll-b verursacht werden.
Abb. 3.8: Fluoreszenzanregungsspektrum der Grünalge Chlorella vulgaris in Reinkultur, gemessene Fluoreszenz-emission bei λ > 700 nm (Krause et al. 1987).

3 Biophysikalische Grundlagen
26
3.4.2.2 Spektrale Gruppe der Blaualgen
Cyanobakterien und Glaucophyta besitzen im Unterschied zu den meisten Algen stattXanthophyll/Chlorophyll-Proteinen in den äußeren Antennen der Photosysteme sog. Phycobi-lisomen (Ausnahme ist hier z.B. die Abteilung der Rhodophyceae, die einen sehr ähnlichenAufbau der Photosysteme besitzen wie die Cyanobakterien). Das PS II sitzt zusammen mitdem PS I in der Thylakoidmembran. Die Phycobilisomen befinden sich auf der Oberflächevon Stromathylakoiden. Granathylakoidstapel sind in diesen Organismen nicht zu finden(Gantt 1986). Die Phycobilisomen bestehen hauptsächlich aus den Phycobiliproteinen Phy-cocyanin, Phycoerythrin und Allophycocyanin. Abb. 3.9 zeigt den schematischen Aufbau derPhotosysteme von Blaualgen. Wahrscheinlich sind die Phycobilisomen über ein sog. Termi-nal-Pigment mit dem PS II gekoppelt. Bei den Blaualgen besitzen die PS I keine eigene äußereAntenne.
Abb. 3.9: Schematische Darstellung von Phycobilisomen typisch für viele hemidiskoidale Phycobilisomen, linksund rechts ein Querschnitt durch einen komplexeren halbkugelförmigen Phycobilisomen (Gantt 1986)
Auf der Thylakoidmembran sind nach außen hin Allophycocyanin, Phycocyanin und ganzaußen befindlich Phycoerythrin angeordnet. Der Energietransport erfolgt in umgekehrter Rei-henfolge von außen nach innen, zum Terminal Pigment (Lawlor 1990; Abb. 3.10). Cya-nobakterien besitzen im Vergleich zu Rhodophyceae ein höheres Phycocyanin-zu-Phycoerythrin-Verhältnis.

3 Biophysikalische Grundlagen
27
Abb. 3.10: Energietransport innerhalb eines typischen Phycobilisomen einer Cyanobakterie (Lawlor 1990)
Phycocyanin, Phycoerythrin und Allophycocyanin haben unterschiedliche Spektren in deroptischen Absorption. Die aus diesen drei Biliproteinen aufgebauten Phycobilisomen habeneine charakteristische Fluoreszenzemissionswellenlänge von 675 nm (in-vivo 660 nm). DieAbsorptions- und Fluoreszenzemissionsspektren isolierter Phycobilisomen sind in Abb. 3.11gezeigt.
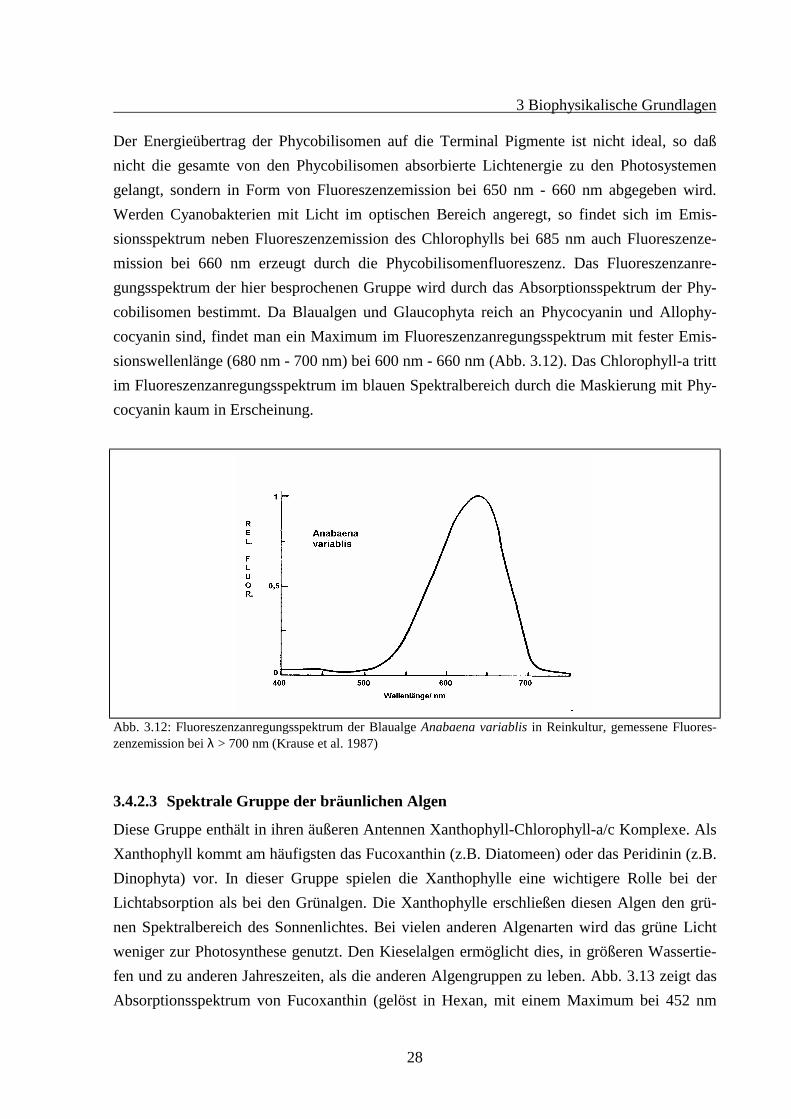
Abb. 3.11:Linke Seite: Die Absorptionsspektren von Phycobilisomen der Rotalge Porphyridium cruentum (( ) mitMaxima von Phycoerythrin bei ca. 545 - 565 nm, Phycocyanin bei 620 nm und Allophycocyanin bei 650 nm) undder Cyanobakterie Anacystis nidulans ((------) mit Phycocyanin und Allophycocyanin (Schulter bei 650 nm) alsdie dominierenden Pigmente).Rechte Seite: Die Fluoreszenzemission bei 675 nm für beide Arten. Von den Phycobilisomen absorbierte Energiewird in beiden Fällen nur über das Allophycocyanin abgegeben. (Gantt 1986)
3 Biophysikalische Grundlagen
28
Der Energieübertrag der Phycobilisomen auf die Terminal Pigmente ist nicht ideal, so daßnicht die gesamte von den Phycobilisomen absorbierte Lichtenergie zu den Photosystemengelangt, sondern in Form von Fluoreszenzemission bei 650 nm - 660 nm abgegeben wird.Werden Cyanobakterien mit Licht im optischen Bereich angeregt, so findet sich im Emis-sionsspektrum neben Fluoreszenzemission des Chlorophylls bei 685 nm auch Fluoreszenze-mission bei 660 nm erzeugt durch die Phycobilisomenfluoreszenz. Das Fluoreszenzanre-gungsspektrum der hier besprochenen Gruppe wird durch das Absorptionsspektrum der Phy-cobilisomen bestimmt. Da Blaualgen und Glaucophyta reich an Phycocyanin und Allophy-cocyanin sind, findet man ein Maximum im Fluoreszenzanregungsspektrum mit fester Emis-sionswellenlänge (680 nm - 700 nm) bei 600 nm - 660 nm (Abb. 3.12). Das Chlorophyll-a trittim Fluoreszenzanregungsspektrum im blauen Spektralbereich durch die Maskierung mit Phy-cocyanin kaum in Erscheinung.
Abb. 3.12: Fluoreszenzanregungsspektrum der Blaualge Anabaena variablis in Reinkultur, gemessene Fluores-zenzemission bei λ > 700 nm (Krause et al. 1987)
3.4.2.3 Spektrale Gruppe der bräunlichen Algen
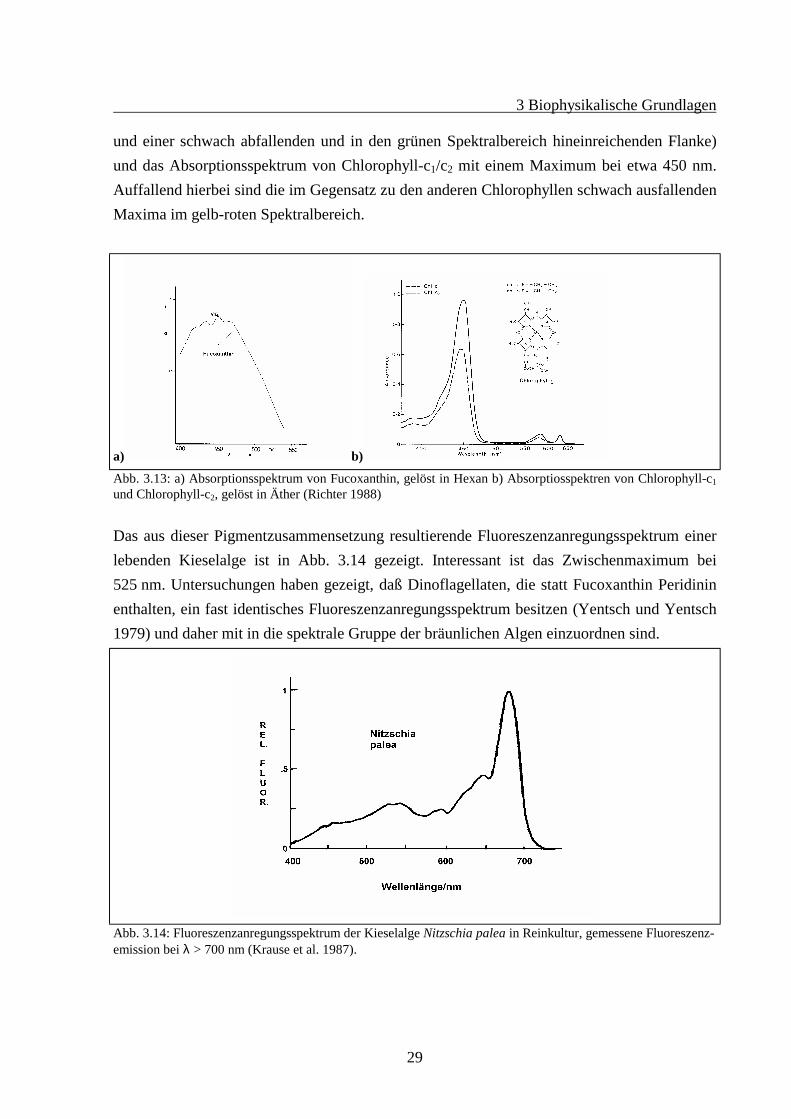
Diese Gruppe enthält in ihren äußeren Antennen Xanthophyll-Chlorophyll-a/c Komplexe. AlsXanthophyll kommt am häufigsten das Fucoxanthin (z.B. Diatomeen) oder das Peridinin (z.B.Dinophyta) vor. In dieser Gruppe spielen die Xanthophylle eine wichtigere Rolle bei derLichtabsorption als bei den Grünalgen. Die Xanthophylle erschließen diesen Algen den grü-nen Spektralbereich des Sonnenlichtes. Bei vielen anderen Algenarten wird das grüne Lichtweniger zur Photosynthese genutzt. Den Kieselalgen ermöglicht dies, in größeren Wassertie-fen und zu anderen Jahreszeiten, als die anderen Algengruppen zu leben. Abb. 3.13 zeigt dasAbsorptionsspektrum von Fucoxanthin (gelöst in Hexan, mit einem Maximum bei 452 nm
3 Biophysikalische Grundlagen
29
und einer schwach abfallenden und in den grünen Spektralbereich hineinreichenden Flanke)und das Absorptionsspektrum von Chlorophyll-c1/c2 mit einem Maximum bei etwa 450 nm.Auffallend hierbei sind die im Gegensatz zu den anderen Chlorophyllen schwach ausfallendenMaxima im gelb-roten Spektralbereich.
a) b)
Abb. 3.13: a) Absorptionsspektrum von Fucoxanthin, gelöst in Hexan b) Absorptiosspektren von Chlorophyll-c1und Chlorophyll-c2, gelöst in Äther (Richter 1988)
Das aus dieser Pigmentzusammensetzung resultierende Fluoreszenzanregungsspektrum einerlebenden Kieselalge ist in Abb. 3.14 gezeigt. Interessant ist das Zwischenmaximum bei525 nm. Untersuchungen haben gezeigt, daß Dinoflagellaten, die statt Fucoxanthin Peridininenthalten, ein fast identisches Fluoreszenzanregungsspektrum besitzen (Yentsch und Yentsch1979) und daher mit in die spektrale Gruppe der bräunlichen Algen einzuordnen sind.
Abb. 3.14: Fluoreszenzanregungsspektrum der Kieselalge Nitzschia palea in Reinkultur, gemessene Fluoreszenz-emission bei λ > 700 nm (Krause et al. 1987).

3 Biophysikalische Grundlagen
30
3.4.2.4 Spektrale Gruppe der Cryptophyta
Die äußeren Antennen der Cryptophyta bestehen aus Chlorophyll-a/c2 und einem zusätzlicheneinzelnen Phycobiliprotein; das kann Phycoerythrin oder aber Phycocyanin sein (Hill undRowan 1989). Durch das Vorkommen von jeweils nur einer Art eines Phycobiliproteins (Phy-cocyanin oder Phycoerythrin) ist diese Gruppe farblich recht gemischt. Dieses dürfte sich auchin stark unterschiedlichen Fluoreszenzanregungsspektren widerspiegeln. In dieser Arbeit wur-de sich jedoch auf die phycoerythrinhaltige Gattung Cryptomonas beschränkt, da diese inheimischem Süßwasser und Brackwasser häufiger vorkommt. Im marinen Bereich ist der ge-samte Stamm in unseren Breiten eher unbedeutend. Außerdem ist Cryptomonas ein schönesBeispiel für phycoerythrinhaltige Algen. In Abb. 3.15 ist ein Fluoreszenzanregungsspektrumeiner Cryptomonas S2 Kultur dargestellt. Deutlich ist ein Maximum bei 450 nm (Chlorophyll-c2) und ein Maximum bei 565 nm (Phycoerythrin) zu erkennen.
Abb. 3.15: Fluoreszenzanregungsspektrum von Cryptomonas S2 : ( ): Emissionswellenlänge λ=685 nm;(-----): Emissionswellenlänge λ=695 nm; (-⋅-⋅-⋅-): Emissionswellenlänge λ=715 nm (Schimek 1994)

4 Vorversuche
31
4 Vorversuche
4.1 Aufbau eines Fluorometers
Für Vorversuche sollte ein Fluorometer aufgebaut werden, das es ermöglichte, Anregungs-spektren lebender Zellen verschiedener Algengruppen zu messen. Basierend auf den mit die-sem Gerät gewonnenen Daten sollte die Möglichkeit der Unterscheidbarkeit dieser Gruppengeprüft werden. Außerdem sollte herausgefunden werden, ob und in welchem Maße Schwan-kungen in den Pigmentzusammensetzungen der Antennensysteme innerhalb einer spektralenGruppe auftreten können. Dabei wurde ein „Küvetten-Fluorometer“ der Firma „bbe Moldaen-ke“ für diese Zwecke erweitert.Um ein Fluorometer zu bauen, das Anregungsspektren von Algengruppen aufnimmt und Un-terschiede in der Zusammensetzung der Antennenpigmente detektiert, war es notwendig, dieFluoreszenzemission von Chlorophyll-a des PS II bei 680 nm - 700 nm für verschiedene An-regungswellenlängen zu bestimmen. Es konnte daher bei fester Emissionswellenlänge gemes-sen werden. Da es sich um eine Meßmethode handeln sollte, die Fluoreszenzanregungsspek-tren kontinuierlich und schnell bestimmt, war es zweckmäßig, nur ein diskretes Anregungs-spektrum mit nur wenigen Anregungswellenlängen zu verwenden, da dieses Konzept Meß-und Rechenzeit spart. Bei den Messungen sollten lebende Algenproben verwendet werden.Daher bot sich eine Messung in einer zweckmäßigerweise in unserem Meßbereich nicht fluo-reszierenden Quarzglasküvette an, so daß es möglich war, Proben direkt (ohne chemischeAufbereitung) zu untersuchen.
4.1.1 Das Anregungsmeßlicht
Da nur ein diskretes Anregungsspektrum benötigt wurde, konnten mehrere Lichtquellen mitfester Wellenlänge verwendet werden. Diese waren auf die Absorptionsspektren der Anten-nenpigmente der äußeren Antennen der verschiedenen Algengruppen abgestimmt. Um statio-näre Messungen zu ermöglichen, die nicht durch Erschöpfung oder Auffüllung metabolischerPools gestört werden, sollte Anregungslicht geringer Intensität benutzt werden.Mit Korrelationsmethoden sollte das Signal-Rausch-Verhältnis der Meßergebnisse verbessertwerden. Deshalb wurde das Anregungsmeßlicht moduliert. Die Auslösung von Induktionski-netiken während eines Meßlichtimpulses sollte verhindert werden, daher mußte die Anzeitund Auszeit des Meßlichtes gegenüber den Zeitkonstanten der relevanten Photosynthesereak-

4 Vorversuche
32
tionen klein sein (Dau 1994). Hieraus ergab sich eine Modulationsfrequenz des Meßlichtes,die größer sein mußte als 1kHz. Dafür eignen sich Leuchtdioden, die seit einigen Jahren mitverschiedenen Emissionswellenlängen, ausreichenden Lichtintensitäten und guter Modulier-barkeit erhältlich sind. Es wurden fünf Leuchtdioden passend zu den Absorptionsmaxima derPhotosynthesepigmente Chlorophyll-a, -b, -c, Fucoxanthin bzw. Peridinin sowie Phycobili-proteine mit den Wellenlängen 450 nm, 525 nm, 570 nm, 590 nm und 610 nm ausgewählt. Dadie 450-nm-Leuchtdiode laut Herstellerangabe ein sehr breites Emissionsspektrum besitzt,wurde diese sowohl mit jeweils 2 Farbfiltern (HT 121 und HT058 von LEE Filters) als auchohne Filter betrieben, so daß wir hier jeweils drei Wellenlängencharakteristika zur Verfügunghatten (bzw. insgesamt sieben verschiedene Wellenlängen). Zusätzlich schneidet jeweils eineeinen Tiefpaß bildende Filterkombination (bbe-FK630) die Spektren aller Leuchtdioden über630 nm ab. Aufgrund der gemessenen schwachen Lichtintensität der 570-nm-Leuchtdiode von1,3 µE/m-2s-1 sollten hiervon zwei Leuchtdioden parallel betrieben werden. Um am Meßpunktdie emittierende Fläche der Leuchtdioden abzubilden, wurde hinter den Filtern eine Sammel-linse mit einer Brennweite von f=40mm und einem Durchmesser von 25 mm installiert.
4.1.2 Die Photodetektoreinheit
An der Photodetektoreinheit des „Küvetten-Fluorometers“ wurde nichts verändert. Als Photo-detektor wurde für die Messungen ein Photomultiplier (Hamamatsu H-578) verwendet, dadieser sich gegenüber Photodioden durch ein größeres Signal-Rausch-Verhältnis, einer höhe-ren Empfindlichkeit und geringerem Dunkelrauschen auszeichnet. Um sicherzustellen, daßnur die Fluoreszenzemission von Chlorophyll-a in einem Wellenlängenintervall 690 nm - 730 nm detektiert wurde und kein Streulicht der Leuchtdioden einfiel, wurde eine Filterkom-bination (bbe-FK690) verwendet. Eine Sammellinse soll das emittierte Fluoreszenzlicht aufdas Fenster des Photomultiplier fokussieren.
4.1.3 Anordnung des Anregungsmeßlichtes und Photodetektoreinheit
Die Photodetektoreinheit und das Anregungsmeßlicht wurden so angeordnet, daß ihre opti-schen Achsen einen Winkel von 90° bildeten. Abb. 4.1 zeigt die Skizze der Anordnung. In derMitte befand sich bei der Messung die die Probe enthaltende Quarzglasküvette (Fassungsver-mögen 25ml, Maße: Länge×Breite×Höhe=25mm×25mm×70mm). In die Küvette konnte beieiner Messung ein magnetischer Rührer eingefügt werden. Dieser konnte vor einer Messungdurch die sich unter der Küvette befindlichen magnetischen Rührer-Platte betrieben werden.Dies verhinderte eine Sedimentation der Algen während der Messung und sorgte für einegleichmäßige Durchmischung der Probe.

4 Vorversuche
33
Aus Platzgründen wurden die Anregungsleuchtdioden des Meßlichtes links und rechts von derKüvette angeordnet, so daß sich auf der linken Seite der Küvette die drei 450-nm-Leuchtdioden (zwei davon mit Farbfiltern bestückt) und die 590-nm-Leuchtdiode befanden.Auf der rechten Seite der Küvette befanden sich die beiden 570-nm-, die 525-nm- und die610-nm-Leuchtdioden. Zwischen Leuchtdioden und Küvette wurden jeweils die optischenFilter und die Abbildungslinsen angeordnet. Diese bildeten das Licht der Leuchtdioden je-weils in der Mitte der Küvette ab.
Abb. 4.1: Skizze zur Anordnung des Anregungsmeßlichtes und der Photodetektoreinheit
Hinter der Küvette in einem rechten Winkel zu dem einfallenden Licht der Leuchtdioden be-fand sich der Photomultiplier mit den optischen Filtern und der Fokussierlinse ausgerüstet.Der gesamte Meßaufbau war in einer lichtdichten, schwarzen Meßkammer angeordnet, so daßAußenlicht keine störende Auswirkung auf den Meßvorgang hatte.
Magnetische-Rührer-Platte
900900
Quarzglas-Küvette
Photomultiplier
Filtersatz
Linse zur Fokussierungder Fluoreszenz auf den
Photomutiplier
4kHz modulierteAnregungs-leuchtdiode
4kHz modulierteAnregungsleucht-diode
Filtersatz mitAbbildungslinse
Filtersatz mitAbbildungslinse
Magnet.Rührer

4 Vorversuche
34
4.1.4 Ansteuerung des Meßlichtes und Signalverarbeitung
Eine Übersicht über die Ansteuerung der Leuchtdioden und die Verarbeitung der gemessenenSignale gibt Abb. 4.2. Die Leuchtdioden wurden mit einem 4-kHz-Rechteck-Signal mit einen1:1 Taktverhältnis angesteuert. Dieses Rechteck wurde von einem Mikrocontroller (Katce68332-System) erzeugt, das danach von einer Stromreglerstufe auf eine Spitzenstromstärkevon Iss=40 mA eingestellt wurde.
K üvette m itA lgen
suspens ion C hlorop hyll-aF luo reszenz-em ission
525nm 610nm450nm
445nm590nm
A btas t-S igna l
2*570n m455nm
M anuellerS cha lter
P hoto m ultip lierm it
V erstä rkerstu fe
kH z-S ignal
A nregun gs-Leu chtd io den
M ikro-C ontro ller
S trom -R egle r
A D -W an dler
Abb. 4.2: Übersicht über die Ansteuerungsvorgänge bei einem Meßablauf. Der Mikrocontroller gibt ein 4-kHz-Rechteck-Signal (1:1 Taktverhältnis) über einen Stromregler an die durch den manuellen Schalter ausgewählteLeuchtdiode aus. Das mit einer Frequenz von 4 kHz modulierte Anregungslicht der Leuchtdiode löst die jetztebenfalls 4-kHz-modulierte Chlorophyll-a-Fluoreszenz der Algensuspension aus. Das Fluoreszenzsignal wirdvom Photomultiplier detektiert und über eine Verstärkerstufe an einen AD-Wandler geleitet, der wiederum vomMikrocontroller mit einem 128-kHz-Abtastimpuls ausgelesen wird.
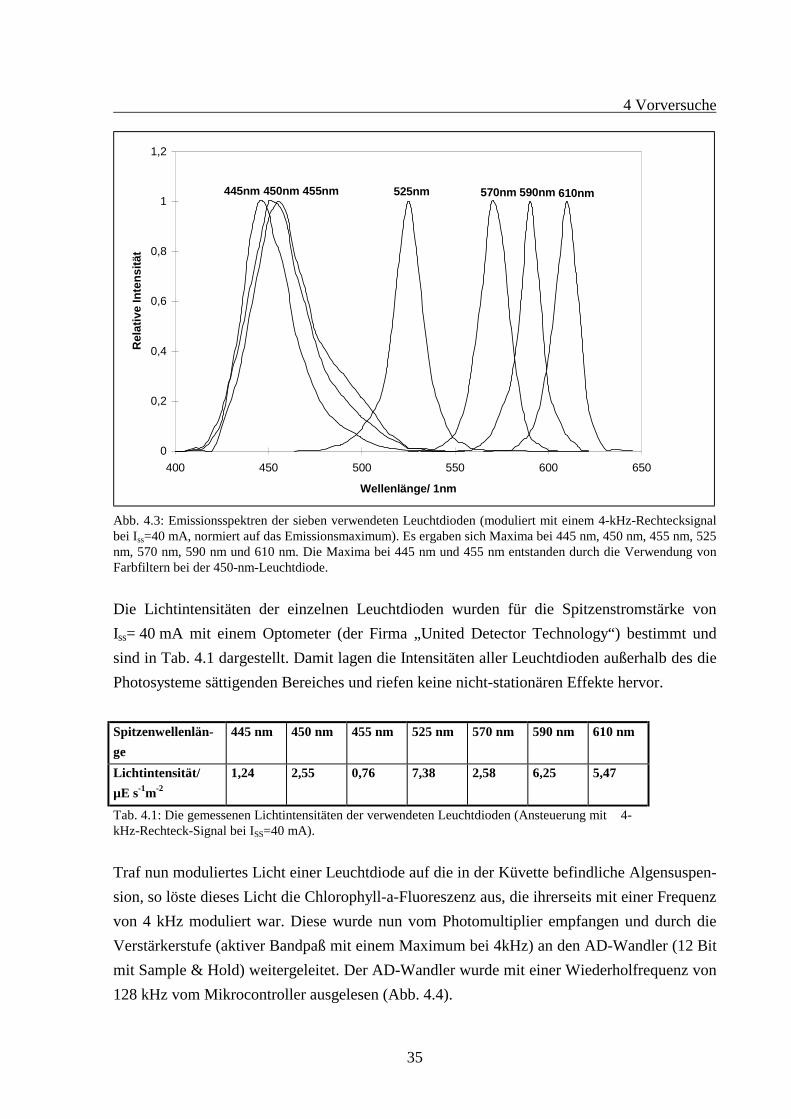
Die Auswahl der Leuchtdioden erfolgte in den Vorversuchen über einen Schalter. Hierausergab sich die Möglichkeit, jeweils eine Leuchtdiode einzeln betreiben zu können. Die Emis-sionsspektren der Leuchtdioden wurden mit einem Kontron-Spektralfluorometer gemessenund sind in Abb. 4.3 dargestellt. Nach diesen Messungen lagen also die sieben Anregungs-peakwellenlängen bei 445 nm, 450 nm, 455 nm, 525 nm, 570 nm, 590 nm und 610 nm.
4 Vorversuche
35
0
0,2
0,4
0,6
0,8
1
1,2
400 450 500 550 600 650
Wellenlänge/ 1nm
Rel
ativ
e In
tens
ität
445nm 450nm 455nm 525nm 570nm 590nm 610nm
Abb. 4.3: Emissionsspektren der sieben verwendeten Leuchtdioden (moduliert mit einem 4-kHz-Rechtecksignalbei Iss=40 mA, normiert auf das Emissionsmaximum). Es ergaben sich Maxima bei 445 nm, 450 nm, 455 nm, 525nm, 570 nm, 590 nm und 610 nm. Die Maxima bei 445 nm und 455 nm entstanden durch die Verwendung vonFarbfiltern bei der 450-nm-Leuchtdiode.
Die Lichtintensitäten der einzelnen Leuchtdioden wurden für die Spitzenstromstärke vonIss= 40 mA mit einem Optometer (der Firma „United Detector Technology“) bestimmt undsind in Tab. 4.1 dargestellt. Damit lagen die Intensitäten aller Leuchtdioden außerhalb des diePhotosysteme sättigenden Bereiches und riefen keine nicht-stationären Effekte hervor.
Spitzenwellenlän-ge
445 nm 450 nm 455 nm 525 nm 570 nm 590 nm 610 nm
Lichtintensität/µE s-1m-2
1,24 2,55 0,76 7,38 2,58 6,25 5,47
Tab. 4.1: Die gemessenen Lichtintensitäten der verwendeten Leuchtdioden (Ansteuerung mit 4-kHz-Rechteck-Signal bei ISS=40 mA).
Traf nun moduliertes Licht einer Leuchtdiode auf die in der Küvette befindliche Algensuspen-sion, so löste dieses Licht die Chlorophyll-a-Fluoreszenz aus, die ihrerseits mit einer Frequenzvon 4 kHz moduliert war. Diese wurde nun vom Photomultiplier empfangen und durch dieVerstärkerstufe (aktiver Bandpaß mit einem Maximum bei 4kHz) an den AD-Wandler (12 Bitmit Sample & Hold) weitergeleitet. Der AD-Wandler wurde mit einer Wiederholfrequenz von128 kHz vom Mikrocontroller ausgelesen (Abb. 4.4).

4 Vorversuche
36
Abb. 4.4: Taktdiagramm zur Leuchtdiodenansteuerung und Abtastung - oben: Das Ansteuerungssignal derLeuchtdiode - unten Der Abtastimpuls, während einer Periode des Leuchtdiodenansteuerungstaktes wird der AD-Wandler 32-mal ausgelesen.
So ergab sich pro Periode des Leuchtdiodenansteuerungssignals eine Folge von 32 Wertenund insgesamt über die Meßzeit eine Anzahl von l= 128 kHz*Meßzeit Meßwerten. Die Folgevon Meßwerten ytn (siehe Gleichung [ 4.1]) setzt sich aus dem Fluoreszenzsignal f(t) und dem
bei der Messung auftretendem Rauschen R(t) zusammen.
[ 4.1]
mit f(t): FluoreszenzsignalR(t): Meßrauschenδ(t-tn): Normierte Abtastfunktion = 1 für t = tn
= 0 sonstIn einer Korrelationsrechnung wurden die abgetasteten Signale dann verwendet, um die Inten-sität der Fluoreszenz zu bestimmen. Dabei wurden jeweils die Werte einer Anzeit des Leucht-diodensignals und die Werte einer Auszeit getrennt aufsummiert. Die beiden Summen wurdendann voneinander subtrahiert und als ein Wert abgespeichert. Da die Leuchtdiode und dieReglerstufe eine Einschwingzeit besaßen, wurden die ersten drei abgetasteten Werte einer An-bzw. Auszeit bei der Berechnung nicht berücksichtigt (Lincke 1994). Nach Beendigung derMeßzeit wurden alle pro Periode abgespeicherten Werte noch einmal aufsummiert und durch
4 k H z -L e u c h td io d en -A n steu er u n g
H i
L o
H i
L o
A b ta sts ig n a lfa= 1 2 8 k H z
n = 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 19 2 0 2 1 2 2 2 3 2 4 2 5 2 6 2 7 2 8 29 3 0 3 1 3 2
ytn = ∑n=1
l = 128 kHz*Meßzeit
{f(t)+R(t)}*δδδδ(t-tn)

4 Vorversuche
37
die Anzahl der Perioden des Leuchtdiodenansteuerungssignals l geteilt, so daß sich der Mit-telwert X ergab (Gleichung [ 4.2])
[ 4.2]
Durch Ausführen der Multiplikation mit der normierten Abtastfunktion δ(t-tn) ergibt sich:
X =
∑k=0
l=4000*Meßzeit
∑
n=(k*32)+2
(k*32)+14{f(tn)+R(tn)} -
∑
n=(k*32)+19
(k*32)+30{f(tn)+R(tn)}
l
Dieses führt nach Vertauschen der Summanden zu:
=
∑k=0
l=4000*Meßzeit
∑
n=(k*32)+2
(k*32)+14f(tn) -
∑
n=(k*32)+19
(k*32)+30f(tn)
l +
∑k=0
l=4000*Meßzeit
∑
n=(k*32)+2
(k*32)+14R(tn) -
∑
n=(k*32)+19
(k*32)+30R(tn)
l
→→→→ 0 für l→→→→∞∞∞∞
Die Anteile des Meßrauschens werden bei zunehmender Meßzeit verschwindend gering, da esangenommen werden kann, daß es sich bei R(tn) um eine Funktion handelt, die gleichmäßigpositive wie negative Anteile besitzt.Folglich ergibt sich mit X ein Wert, der proportional zur Differenz der Fluoreszenzintensitätenwährend einer An- und Auszeit der Leuchtdiode ist. Weil aber in der Auszeit keine Fluores-zenz auftritt, ergibt sich X als ein Maß für die Fluoreszenzintensität während einer Leucht-dioden-Anzeit, also gilt:
X∼∼∼∼ Fluoreszenzantwort (Anzeit)
Der Mikrocontroller gab die gemessenen X-Werte in Digits aus. Dabei entsprachen 100 Digitseiner Spannung von U=1V am Photomultiplier.
X =
∑k=0
l=4000*Meßzeit
∑
n=(k*32)+2
(k*32)+14{f(t)+R(t)}*δδδδ(t-tn) -
∑
n=(k*32)+19
(k*32)+30{f(t)+R(t)}*δδδδ(t-tn)
l

4 Vorversuche
38
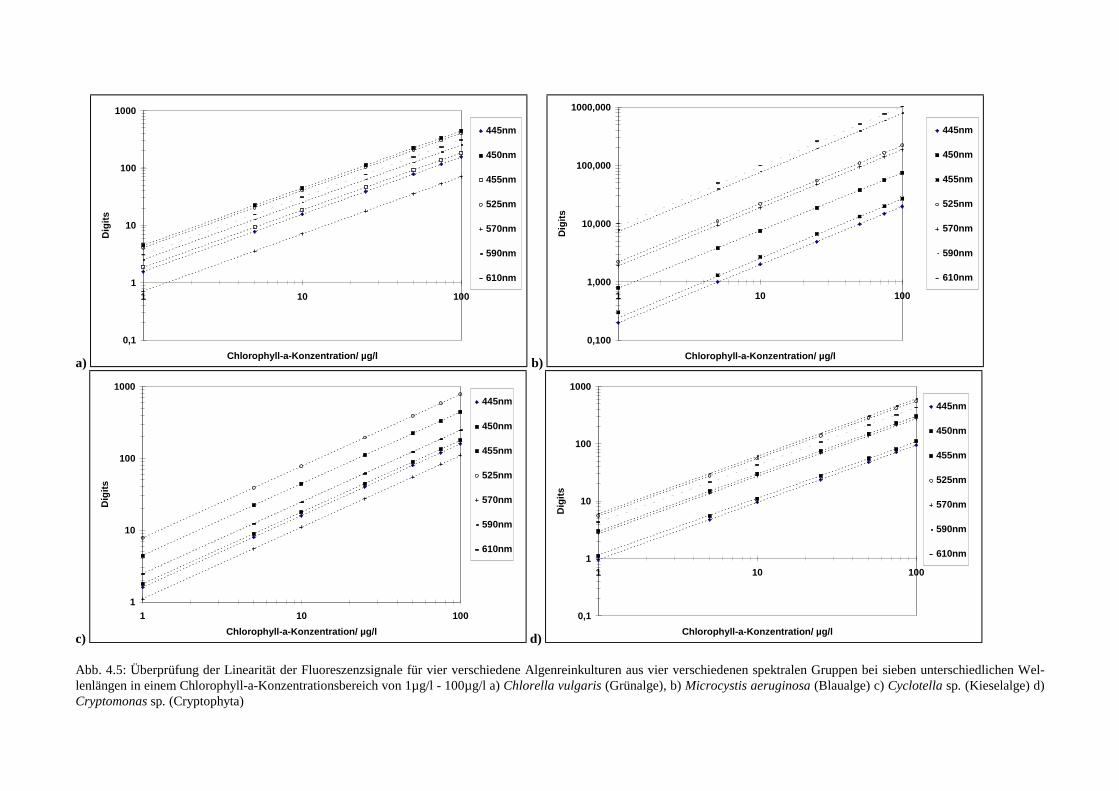
4.1.5 Test der Linearität des Fluoreszenzsignals
Für die weiteren Untersuchungen war es notwendig sicherzustellen, daß die verwendeten Pro-ben der Algenkulturen Konzentrationen besaßen, die zu einer Fluoreszenzintensität führten,welche im linearen Bereich der Fluoreszenzsignale lag, d.h. die Fluoreszenzintensität sollteproportional zur Chlorophyll-a-Konzentration sein. Zum Test der Linearität der Fluoreszenz-signale verschiedener Algenreinkulturen, erzeugt durch Anregung mit den sieben zur Verfü-gung stehenden Wellenlängen, wurden vier verschiedene Algenarten (jeweils ein Vertretereiner spektralen Gruppe) in verschiedenen Chlorophyll-a-Konzentrationen untersucht. Hierzuwurde jeweils der Chlorophyll-a-Gehalt einer hochkonzentrierten Stammkultur spektropho-tometrisch (Nusch 1980) bestimmt und diese auf Konzentrationen zwischen 1µg/l und100µg/l Chlorophyll-a verdünnt. Zur Anzucht und Verdünnung der Proben bei der Messungwurde WC-Nährmedium (Schlösser 1994) verwendet. Als Algenkulturen wurden benutzt:Chlorella vulgaris (Grünalge), Microcystis aeruginosa (Cyanobakterie), Cyclotella sp. (Kie-selalge) und Cryptomonas sp. (Cryptophyta).Vor jeder Messung wurde die Probe mit demMeßlicht 90s beleuchtet, um die Photosysteme in den Steady-State zu bringen. Dann wurdebei einer Meßzeit von 9s gemessen. Dieser Vorgang wurde jeweils dreimal wiederholt. Vorjeder Messung wurde die Probe 10s lang gerührt. Dieses verhinderte das Herabsinken der Al-genzellen. Die aus den drei Messungen gewonnenen Meßwerte (Fluoreszenzintensitäten)wurden für jede Leuchtdiode gemittelt und in Digits gegenüber dem spektrophotometrischbestimmten Chlorophyll-a-Gehalt in Abb. 4.5 aufgetragen. Daraus ist zu entnehmen, daß allespektralen Algengruppen mit Chlorophyll-a-Konzentrationen von 1-100µg/l im linearen Meß-bereich liegen. In den nun folgenden Experimenten sollte in diesen Konzentrationsbereichengearbeitet werden.
a)
0,1
1
10
100
1000
1 10 100
Chlorophyll-a-Konzentration/ µg/l
Dig
its
445nm
450nm
455nm
525nm
570nm
590nm
610nm
b)
0,100
1,000
10,000
100,000
1000,000
1 10 100
Chlorophyll-a-Konzentration/ µg/l
Dig
its
445nm
450nm
455nm
525nm
570nm
590nm
610nm
c)
1
10
100
1000
1 10 100Chlorophyll-a-Konzentration/ µg/l
Dig
its
445nm
450nm
455nm
525nm
570nm
590nm
610nm
d)
0,1
1
10
100
1000
1 10 100
Chlorophyll-a-Konzentration/ µg/l
Dig
its
445nm
450nm
455nm
525nm
570nm
590nm
610nm
Abb. 4.5: Überprüfung der Linearität der Fluoreszenzsignale für vier verschiedene Algenreinkulturen aus vier verschiedenen spektralen Gruppen bei sieben unterschiedlichen Wel-lenlängen in einem Chlorophyll-a-Konzentrationsbereich von 1µg/l - 100µg/l a) Chlorella vulgaris (Grünalge), b) Microcystis aeruginosa (Blaualge) c) Cyclotella sp. (Kieselalge) d)Cryptomonas sp. (Cryptophyta)

4 Vorversuche
40
4.2 Aufnahme von Anregungsspektren verschiedener Algenkulturen
Mit Hilfe des in Kapitel 4.1 beschriebenen Fluorometers sollten die Chlorophyll-a-Fluoreszenzanregungsspektren der vier spektralen Gruppen, Grünalgen, Blaualgen, bräunlichespektrale Gruppe und Cryptophyta, untersucht werden. Dabei sollte auch herausgefundenwerden, welchen Schwankungen diese Spektren unterliegen. Weiter sollte geklärt werden, obes möglich ist, aus der Form und der Größe des Fluoreszenzanregungsspektrums Rückschlüs-se auf die Art der Algen und die Chlorophyll-a-Konzentration einer Wasserprobe zu ziehen.Die wichtigste Bedingung dabei ist sicherlich, daß die Zusammensetzung der Antennenpig-mente innerhalb einer spektralen Gruppe ähnlich ist und nur wenig schwankt. Dabei gibt eseinige Faktoren, die die Zusammensetzung der Antennenpigmente beeinflussen könnten. Zu-nächst wurde der wichtigste Faktor betrachtet. Dieses sind die Schwankungen der Zusammen-setzung der Antennenpigmente bei verschiedenen Arten einer spektralen Gruppe.
4.2.1 Durchführung
Für die Messungen standen 34 Algentaxa, davon 17 Blaualgen, 9 Grünalgen, 3 Kieselalgen, 2Dinoflagellaten und 3 Cryptophyta zur Verfügung (die verwendeten Algenarten sind in An-hang A aufgeschlüsselt). Da bei den Blaualgen die größten Schwankungen in dem Aufbau derAntennenpigmente zu vermuten waren, ist eine möglichst große Anzahl von Blaualgenartenuntersucht worden. Von den in der Natur dem Vorkommen nach selteneren Cryptophyta stan-den nur drei Kulturen zur Verfügung. Durch die Untersuchung von Kieselalgen und Dinofla-gellaten sollte herausgefunden werden, inwieweit sich diese Stämme ähneln oder ob eine Un-terscheidung dieser Klassen durch die Fluoreszenzanregungsspektren möglich ist.Um bei dieser Messung möglichst nur Unterschiede in den Antennenpigmenten zu bestim-men, die algenartenspezifisch sind, wurden Anzuchtbedingungen und -dauer bei allen Kultu-ren identisch gehalten. Alle Kulturen besaßen ein Kulturalter von 14 Tagen. Sämtliche unter-suchte Kulturen wurden in WC-Nährmedium gezogen. Bei allen Messungen ist der Chloro-phyll-a-Gehalt der Probe durch eine HPLC-Analyse (Wiltshire und Schroeder 1994) ermitteltworden.

4 Vorversuche
41
Da einige Kulturen sehr hohe Chlorophyll-a-Konzentrationen besaßen, wurden sie auf denverifizierten linearen Meßbereich (1 µg/l - 100 µg/l Chlorophyll-a, Abb. 4.5) der Fluorozenz-signale mit WC-Nährmedium verdünnt. Vor der Messung wurden die Algen 90 Sekundenlang an das Meßlicht adaptiert, d.h. mit dem Licht der bei der Messung als Anregungslicht-quelle verwendeten Leuchtdiode vorbeleuchtet. Bei einer Meßdauer von 9 Sekunden wurdedanach die Fluoreszenzintensität bei dieser Wellenlänge bestimmt. Dieser Wert wurde jeweilsdreifach bestimmt und dann gemittelt.
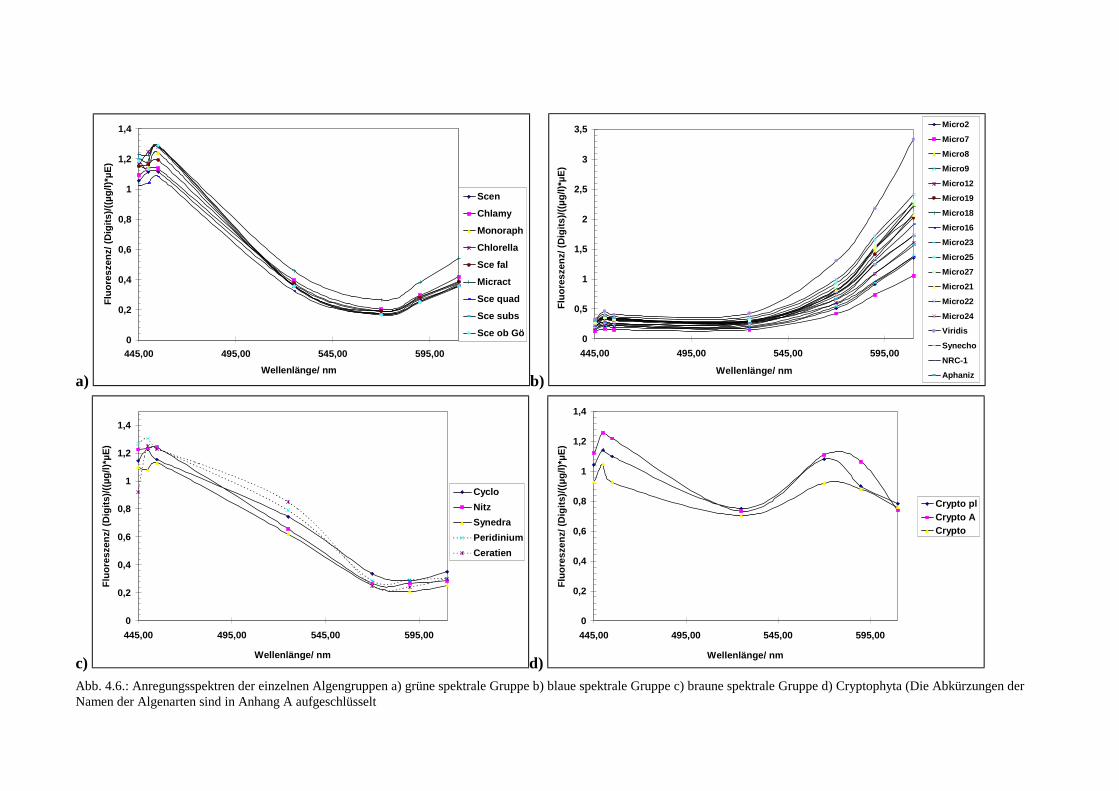
4.2.2 Ergebnisse
Zur Auswertung wurden die Fluoreszenzintensitäten (in Digits) bei jeder der sieben Wellen-längen auf den Chlorophyll-a-Gehalt der Probe und die Lichtintensität bei der jeweiligenWellenlänge normiert. Abb. 4.6 zeigt eine Übersicht der einzelnen Spektren der vier spektra-len Gruppen. Obwohl nur sieben diskrete Punkte des Fluoreszenzanregungsspektrums aufge-nommen wurden, sind diese Punkte im Diagramm durch Linien miteinander verbunden wor-den. Dieses soll weniger eine Aussage über das kontinuierliche Spektrum sein, sondern denVerlauf eines Spektrums einer einzelnen Algenart verdeutlichen.Bei den Grünalgen findet sich das Maximum bei 455 nm, verursacht durch die Chlorophyll-a/b-Antenne. In dieser Gruppe scheint es im Vergleich zu den anderen Gruppen die geringstenSchwankungen innerhalb einer Gruppe zu geben.Die Angehörigen der bräunlichen spektralen Gruppe, die Kieselalgen und Dinoflagellaten,haben ein sehr ähnliches Spektrum. Das Maximum des Spektrums liegt im Blauen bei Anre-gung mit 450 nm und ein hohes Signal liegt bei 525 nm, verursacht durch Fucoxanthin bzw.Peridinin. Bei den Dinoflagellaten scheint der Verlauf bei 525 nm etwas höher zu sein als beiden Kieselalgen. Eine Unterscheidung von Dinoflagellaten und Kieselalgen anhand der Spek-tren dürfte jedoch Probleme bereiten, da die Unterschiede deutlich zu gering sind und in derVariationsbandbreite der Algen, besonders wenn andere Algenarten hinzukämen, untergehenwürden. Die Unterschiede der Spektren dieser Gruppe werden hervorgerufen durch Fucoxan-thin und Peridinin und haben eine höhere Streuung der Meßwerte in dieser Gruppe bei Anre-gung mit 525 nm zur Folge.Die Cyanobakterien haben das Maximum im Fluoreszenzanregungsspektrum bei 610 nm; hierist jedoch auch die prozentuale Standardabweichung der Meßwerte am höchsten. Dieses Er-gebnis war zu erwarten, da bekannt ist, daß Phycobilisomen im Vergleich zu den Antennensy-stemen der anderen Algenarten recht variabel in ihrer Größe sein können (Gantt 1986). Dasausgeprägte Maximum bei 610 nm ist jedoch so charakteristisch, daß diese Gruppe von denanderen unterscheidbar sein wird.

4 Vorversuche
42
Bei den Cryptophyta befindet sich das Maximum des Spektrums im Blauen bei einer Wellen-länge von 450 nm, und bei 570 nm ist ein Nebenmaximum festzustellen. Dieses wird durchdas Phycoerythrin erzeugt. Hohe Schwankungen konnten nicht festgestellt werden, da auchnur drei Arten zur Verfügung standen.Abb. 4.7 zeigt eine Übersicht über die mittleren Spektren der untersuchten Algengruppen undderen aus den Unterschieden der einzelnen Arten resultierenden Standardabweichungen.Daraus läßt sich erkennen, daß die grüne und bräunliche Gruppe sich am stärksten bei 525 nmunterscheiden. Außerdem ist bei dem Vergleich der beiden Gruppen auffällig, daß die grüneGruppe bei Anregung mit einer Wellenlänge von 610 nm stärker fluoresziert. Dies liegt andem Gehalt von Chlorophyll-b der Grünalgen. Die Gruppe der Blaualgen setzt sich durch ih-ren Gehalt an Phycocyanin mit dem daraus resultierenden Maximum im Fluoreszenzanre-gungsspektrum bei 610 nm von den anderen Gruppen am deutlichsten ab. Weiterhin ist indieser Gruppe charakteristisch, daß es nur eine relativ schwache Fluoreszenzantwort bei An-regung im Blauen gibt.Die drei untersuchten Cryptophyta zeigen das Maximum bei 450 nm. Dies liegt an dem Chlo-rophyll-a/c-Komplex. Einzigartig ist jedoch das zweite Maximum bei 570 nm, mit dem sichdiese Gruppe von den anderen absetzt. Eine Aussage über die Schwankungen der Spektreninnerhalb dieser Gruppe war angesichts der wenigen untersuchten Kulturen nicht sinnvoll.Obwohl bei den Grünalgen das Maximum des Spektrums bei 455 nm (Chlorophyll-b) liegt, istder Gebrauch dieser Wellenlänge sowie der Wellenlänge 445 nm nicht zwingend notwendig,da die Algengruppen im 5-nm-Abstand der Fluoreszenzanregungsspektren keine großen Un-terschiede zeigen. Deshalb konnte bei den weiteren Untersuchungen auf diese Wellenlängenverzichtet werden.
a)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
445,00 495,00 545,00 595,00Wellenlänge/ nm
Fluo
resz
enz/
(Dig
its)/(
(µg/
l)*µE
)
Scen
Chlamy
Monoraph
Chlorella
Sce fal
Micract
Sce quad
Sce subs
Sce ob Gö
b)
0
0,5
1
1,5
2
2,5
3
3,5
445,00 495,00 545,00 595,00
Wellenlänge/ nm
Fluo
resz
enz/
(Dig
its)/(
(µg/
l)*µE
)
Micro2
Micro7Micro8Micro9
Micro12Micro19
Micro18Micro16Micro23
Micro25Micro27Micro21
Micro22
Micro24Viridis
Synecho
NRC-1Aphaniz
c)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
445,00 495,00 545,00 595,00
Wellenlänge/ nm
Fluo
resz
enz/
(Dig
its)/(
(µg/
l)*µE
)
CycloNitzSynedraPeridiniumCeratien
d)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
445,00 495,00 545,00 595,00
Wellenlänge/ nm
Fluo
resz
enz/
(Dig
its)/(
(µg/
l)*µE
)
Crypto plCrypto ACrypto
Abb. 4.6.: Anregungsspektren der einzelnen Algengruppen a) grüne spektrale Gruppe b) blaue spektrale Gruppe c) braune spektrale Gruppe d) Cryptophyta (Die Abkürzungen derNamen der Algenarten sind in Anhang A aufgeschlüsselt
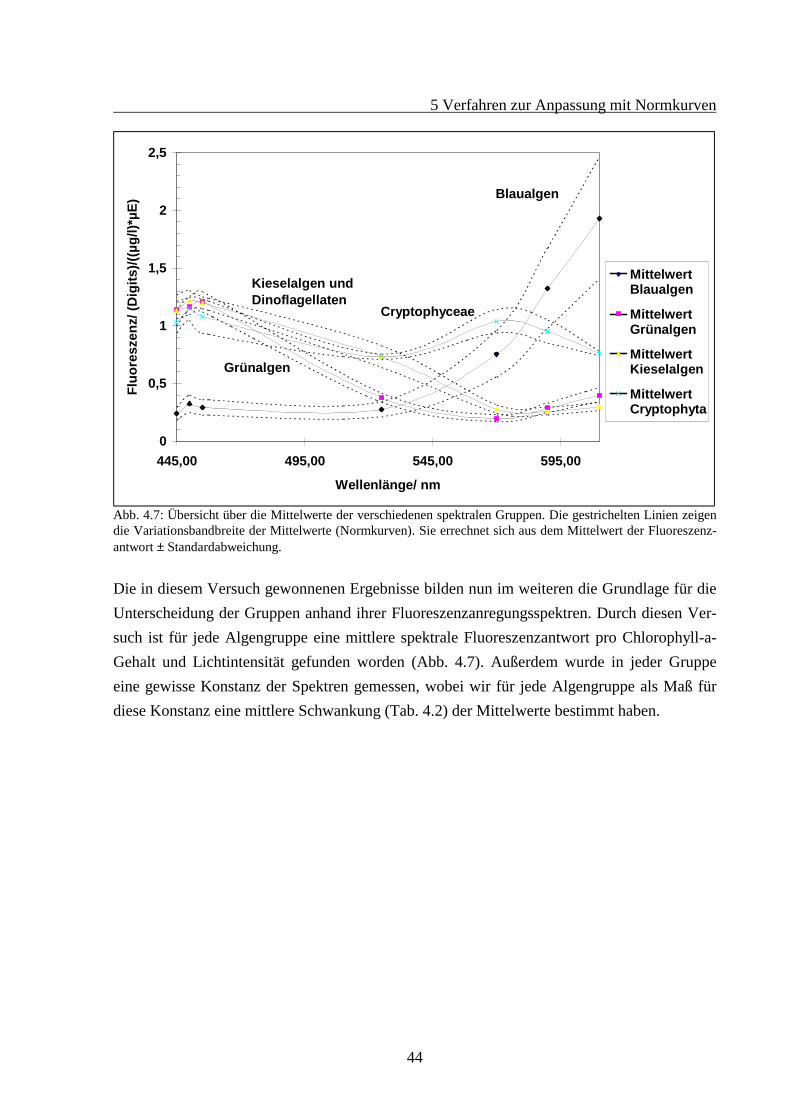
5 Verfahren zur Anpassung mit Normkurven
44
0
0,5
1
1,5
2
2,5
445,00 495,00 545,00 595,00
Wellenlänge/ nm
Fluo
resz
enz/
(Dig
its)/(
(µg/
l)*µE
)
MittelwertBlaualgen
MittelwertGrünalgen
MittelwertKieselalgen
MittelwertCryptophyta
Blaualgen
Cryptophyceae
Grünalgen
Kieselalgen und Dinoflagellaten
Abb. 4.7: Übersicht über die Mittelwerte der verschiedenen spektralen Gruppen. Die gestrichelten Linien zeigendie Variationsbandbreite der Mittelwerte (Normkurven). Sie errechnet sich aus dem Mittelwert der Fluoreszenz-antwort ± Standardabweichung.
Die in diesem Versuch gewonnenen Ergebnisse bilden nun im weiteren die Grundlage für dieUnterscheidung der Gruppen anhand ihrer Fluoreszenzanregungsspektren. Durch diesen Ver-such ist für jede Algengruppe eine mittlere spektrale Fluoreszenzantwort pro Chlorophyll-a-Gehalt und Lichtintensität gefunden worden (Abb. 4.7). Außerdem wurde in jeder Gruppeeine gewisse Konstanz der Spektren gemessen, wobei wir für jede Algengruppe als Maß fürdiese Konstanz eine mittlere Schwankung (Tab. 4.2) der Mittelwerte bestimmt haben.
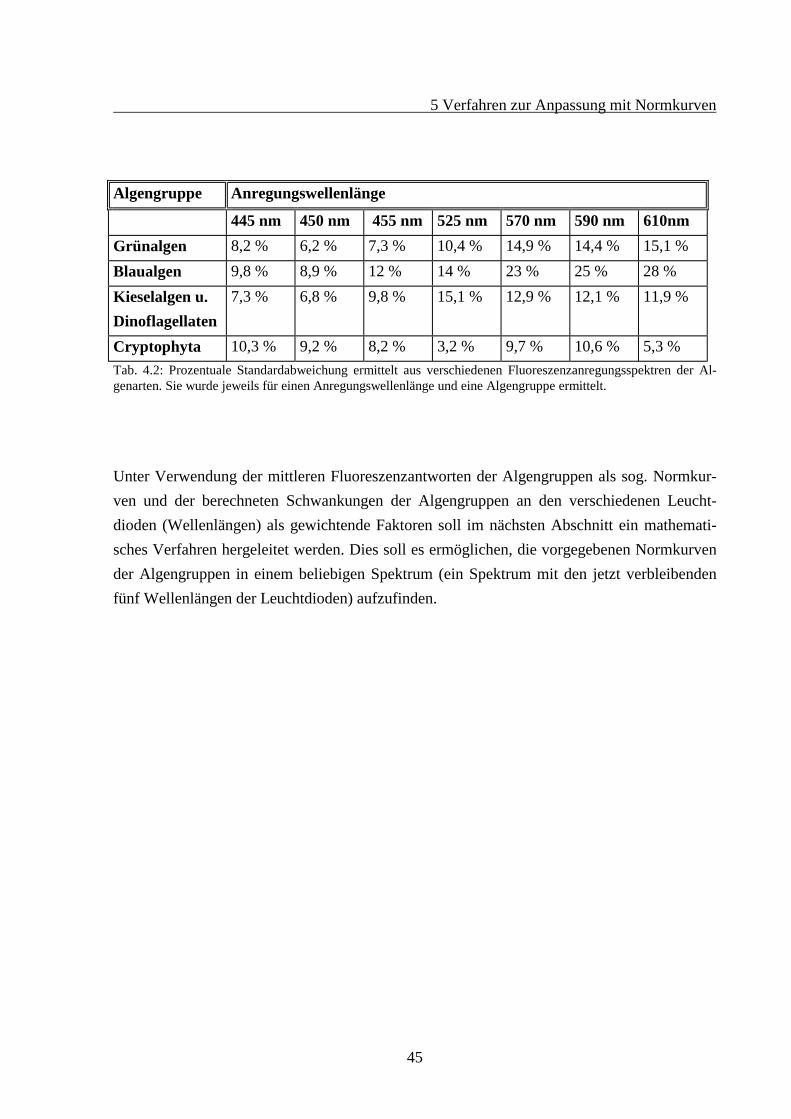
5 Verfahren zur Anpassung mit Normkurven
45
Algengruppe Anregungswellenlänge
445 nm 450 nm 455 nm 525 nm 570 nm 590 nm 610nmGrünalgen 8,2 % 6,2 % 7,3 % 10,4 % 14,9 % 14,4 % 15,1 %Blaualgen 9,8 % 8,9 % 12 % 14 % 23 % 25 % 28 %Kieselalgen u.Dinoflagellaten
7,3 % 6,8 % 9,8 % 15,1 % 12,9 % 12,1 % 11,9 %
Cryptophyta 10,3 % 9,2 % 8,2 % 3,2 % 9,7 % 10,6 % 5,3 %Tab. 4.2: Prozentuale Standardabweichung ermittelt aus verschiedenen Fluoreszenzanregungsspektren der Al-genarten. Sie wurde jeweils für einen Anregungswellenlänge und eine Algengruppe ermittelt.
Unter Verwendung der mittleren Fluoreszenzantworten der Algengruppen als sog. Normkur-ven und der berechneten Schwankungen der Algengruppen an den verschiedenen Leucht-dioden (Wellenlängen) als gewichtende Faktoren soll im nächsten Abschnitt ein mathemati-sches Verfahren hergeleitet werden. Dies soll es ermöglichen, die vorgegebenen Normkurvender Algengruppen in einem beliebigen Spektrum (ein Spektrum mit den jetzt verbleibendenfünf Wellenlängen der Leuchtdioden) aufzufinden.

5 Verfahren zur Anpassung mit Normkurven
46
5 Verfahren zur Anpassung mit Normkurven
In Kapitel 4.2.2 ergab sich für jede der vier untersuchten spektralen Gruppen ein mittleres, fürjede Gruppe charakteristisches, Fluoreszenzanregungsspektrum (Normkurve), bezogen aufeine Chlorophyll-a-Konzentration und die verwendete Lichtintensität. Unter Zuhilfenahme derNormkurven soll versucht werden, aus einem Fluoreszenzanregungsspektrum eines Algenge-misches unterschiedlicher spektraler Gruppen und unter Verwendung der Normkurven dieChlorophyll-a-Konzentration des Gemisches zu bestimmen. Außerdem soll jeder spektralenGruppe ein Chlorophyll-a-Gehalt zugeordnet werden.
5.1 Theorie des Fitverfahrens
Die erste Anwendung von Curve-fitting Verfahren im Gebiet der Photosynthese führtenKolbowski et al. (1990) durch, um aus komplexen Antworten die Anteile vonEinzelkomponenten herauszufinden. Dabei zerlegten sie die photoakustische Druckwelle nacheinem Lichtimpuls in die thermischen Verluste, O2-Entwicklung und CO2-Aufnahme.Kolbowski wandte damals die in Kiel entwickelten Programme (Willkomm 1987) zumNormkurvenfit an. Hierbei wird das gemessene Signal mit einer gewichteten Summe vonNormkurven genähert. Im Falle der photoakustischen Signale wurde das Verfahren in derDissertation von Tabrizi (1997), durch einen Streckfaktor für die Normkurve der CO2-Aufnahme ergänzt (Tabrizi et al. 1998), dargestellt.Das Verfahren des Normkurvenfits soll auf die Anregungsspektren der Algen übertragenwerden; dazu folgende Betrachtungen:Die Fluoreszenzintensität ist proportional zu der in den Pigmenten der Antennen absorbiertenund zum Chlorophyll-a geleiteten Lichtenergie. Die effektive Absorption einer Algengruppeist durch die in Abb. 4.7 gezeigten Normkurven beschrieben. Damit stellen sich diegemessenen Fluoreszenzintensitäten als gewichtete Summe dieser Normkurven dar.

5 Verfahren zur Anpassung mit Normkurven
47
[ 5.1]
mit: yi: die Gesamtfluoreszenzintensität des Chlorophyll-a’sder Algen aller vier spektraler Gruppen bei Anregungmit der Leuchtdiode ’i’
xki: die Fluoreszenzintensität der Normkurve der spek-tralen Algengruppe ‘k’ bei Anregung mit der Leucht-diode ’i’
ak : Chlorophyll-a-Konzentration der spektralen Algen-gruppe ‘k’
Da die Normkurven auf den Chlorophyll-a-Gehalt normiert sind, geben die Koeffizienten ak
auch den Chlorophyll-a-Gehalt pro Algengruppe an. Die Summe der ak ist derGesamtchlorophyllgehalt.Die Programme zum Normkurvenfit der Photoakustik-Kurven (Tabrizi 1997) konnten allerdingsnicht übernommen werden, weil die Streuung der Kurven in Abb. 4.7 als Gewichtsfaktoren be-rücksichtigt werden sollten.Die Berücksichtigung der Streuung führte zu einer Gewichtung des Fehlers mit σσσσi (Willkomm1987; Press 1989). Statt dessen kann σσσσi auch in den Nenner gezogen werden (Glei-chung [ 5.2]).
[ 5.2]
mit σi : die Meßungenauigkeit der Fluoreszenzintensität bei An-regung mit der Leuchtdiode ‘i’
Für den Fall, daß σσσσi konstant und bekannt ist, liegt das Minimum der χ2 -Funktion an derStelle, an der die partielle Ableitung gleich null wird. Dies führt zu Gleichung [ 5.3].
0 = ∂∂∂∂χχχχ2
∂∂∂∂an =
∂∂∂∂ ∂∂∂∂an
∑i=1
5
yi - ∑
k=1
4
ak xki
σσσσi
2
n = 1,..., 4
= 2∑i=1
5
xin ∑
k=1
4
xik ak - yi
σσσσi2
yi = ∑k=1
4ak xki i = 1,.., 5
χχχχ2 = ∑i=1
5
yi - ∑
k=1
4 ak xki
σσσσi
2

5 Verfahren zur Anpassung mit Normkurven
48
[ 5.3]
Die Definition des Lösungsvektors a lautet a =
a1
a2 a3 a4
[ 5.4]
Der Vektor b in Gleichung [ 5.5] gibt die gemessenen Fluoreszenzintensitäten bezüglich derLeuchtdiode ‘i’, dividiert durch die Meßungenauigkeit σσσσi an
[ 5.5]
Unter Bildung der Matrix der Normkurven der einzelnen spektralen Algengruppen, dividiertdurch die Meßungenauigkeit σσσσi bezüglich der Leuchtdiode ‘i’ (Gleichung [ 5.6]),
[ 5.6]
ergibt sich für die Koeffizienten cnk des Matrix Produkts ATA : cnk = ∑i=1
5
xin xik
σσσσi2 .
Außerdem gleicht ∑i=1
5
xin yi
σσσσi2 der n-ten Komponente des Produkts ATb .
= 2
∑k=1
4
ak ∑i=1
5
xin xik
σσσσi2 - 2
∑
i=1
5
xin yi
σσσσi2
b =
y1 σσσσ1
y2 σσσσ2
y3 σσσσ3
y4 σσσσ4
y5 σσσσ5
A =
x11
σσσσ1
x12
σσσσ1
x13
σσσσ1
x14
σσσσ1
x21
σσσσ1
x22
σσσσ2
x23
σσσσ2
x24
σσσσ2
x31
σσσσ3
x32
σσσσ3
x33
σσσσ3
x34
σσσσ3
x41
σσσσ4
x42
σσσσ4
x43
σσσσ4
x44
σσσσ4
x51
σσσσ5
x52
σσσσ5
x53
σσσσ5
x54
σσσσ5

5 Verfahren zur Anpassung mit Normkurven
49
Dies bedeutet, daß :
[ 5.7]
(bekannt als „Gaußsche Normalgleichung“ (Bronstein und Semendjajew 1991))Nach a (dem Vektor der Chlorophyll-a-Konzentrationen) aufgelöst, ergibt sich dann die Glei-chung [ 5.8].
[ 5.8]
Für die auftretenden Meßungenauigkeiten σσσσi, hatte sich in Kapitel 4.2.2 ergeben, daß jedeNormkurve einer Algengruppe eine für jede Anregungswellenlänge spezifische Schwankungaufweist. Damit erhalten wir für die Meßungenauigkeiten nicht nur einen mit der Anzahl derverwendeten Leuchtdioden dimensionierten Vektor, sondern eine Matrix mit den Dimensio-nen: Anzahl der Leuchtdioden × Anzahl der Algengruppen. Dieses würde uns bei der Auf-stellung der χ2-Funktion zu einem nicht-linearen Fall führen:
[ 5.9]
mit σσσσi 2 = ∑
k=1
4
( ) ak σσσσkj 2
In Gleichung [ 5.9] ist der Zusammenhang zwischen den Fitparametern ak und dem Fehlernicht-linear. Damit ist eine Lösung mit der Gaußschen Normalgleichung ausgeschlossen.
( )AT A a = AT b
a = ( )AT A -1 AT b
χχχχ2 = ∑i=1
5
yi - ∑
k=1
4 ak xki
σσσσi
2
= ∑
j=1
5
yj - ∑
k=1
4 ak xkj
∑k=1
4
( ) ak σσσσkj 2
2

5 Verfahren zur Anpassung mit Normkurven
50
Nicht-lineare Lösungsmethoden wie z.B. der Simplex-Algorithmus benötigen sehr viele Itera-tionsschritte, was für eine on-line Bestimmung zuviel Rechenzeit kostet.Da die Gewichtung mit Meßungenauigkeiten nur kleinere Korrekturen der Fitergebnisse be-wirkt, kann hier ein vereinfachtes Verfahren angewandt werden.Im ersten Schritt dieses iterativen Verfahrens wird ein sinnvoller Schätzwert für die Streuun-gen σσσσi gewählt. Dieser Vektor mit der Dimension der Anzahl der Meßpunkte (Leuchtdioden)geht von gleichen Algenkonzentrationen aus, so daß das σσσσi
2 die Summe der Meßungenauig-keiten aller Algengruppen bei jeder Leuchtdiode ist (Gleichung [ 5.10] ).
[ 5.10]
Damit sind die σσσσi bekannt, und die ak (d.h., die Konzentrationen der einzelnen Algengruppenkönnen durch Lösen der Gaußschen Normalgleichung (Gleichung [ 5.8]) bestimmt werden).Nun werden die Meßungenauigkeiten σσσσi neu berechnet. Diese ergeben sich jetzt aus der Mul-tiplikation der zuvor berechneten Konzentration einer Algengruppe ak mit der zur Algengrup-pe dazugehörigen Meßungenauigkeit. Diese werden dann für alle Algengruppen aufsummiert
und als neuer Wert für die Meßungenauigkeit eingesetzt (σσσσi 2 = ∑
k=1
4
( ) ak σσσσkj 2). Mit dieser jetzt
neu entstandenen Gewichtung werden die Konzentrationen der Algengruppen durch Anwen-dung der Gaußschen Normalgleichung (Gleichung [ 5.8]) im zweiten Iterationsschritt neu be-rechnet. Daraus ergeben sich neue ak’s. In Probeläufen zeigte sich, daß bei einer dritten Itera-tion die ak’s sich nur noch in der 5. Stelle hinter dem Komma ändern. Deshalb wurde nach der2. Iterationsstufe der Algorithmus gestoppt.Das zugehörige Computerprogramm wird im nächsten Abschnitt besprochen.
5.2 Struktur des Rechenprogramms
Zur Lösung des im vorigen Kapitel beschriebenen Algorithmus ist ein Pascal-Computerprogramm (Pascal-Quelltext siehe Anhang B) entworfen worden, das die GaußscheNormalgleichung aufstellt und löst, sowie die Iteration mit den Meßungenauigkeiten durch-führt. Hierbei wurden teilweise Programmroutinen aus der Literatur übernommen (Press1989).In einem ersten Schritt werden die zur Lösung von a = ( )AT A -1 AT b benötigten Matrizen
und Vektoren A, AT und b aufgestellt. Die dazu benötigten Meßungenauigkeiten σσσσi für jede
σσσσi2 = ∑
k=1
4 σσσσkj
2

5 Verfahren zur Anpassung mit Normkurven
51
Leuchtdiode ‘i’ berechnet sich wie in Gleichung [ 5.10] beschrieben. Die Matrixinversion vonAAT wird durch eine Gauss-Jordan-Elimination (Press 1989) gelöst. Durch diese Schritte er-hält man den Lösungsvektor a.Bei dieser Lösungsmethode können ak’s auftreten, die einen negativen Wert besitzen. Dieseswürde aber bedeuten, daß die betreffende Algengruppe eine negative Konzentration besäße,was natürlich nicht möglich ist. Anschaulich gesehen würde eine Normkurve, die nur negativin das aufgenommene diskrete Spektrum paßt, dem diskreten Spektrum am wenigsten ent-sprechen und daher nicht vorhanden sein. Deshalb wird bei Auftreten von negativen Elemen-ten in a das kleinste Element von a (falls mehrere Elemente kleiner als null sind) gleich nullgesetzt, und die Lösung von a = ( )AT A -1 AT b wird mit einem niederdimensionaleren Vektorfür a wiederholt. Dieses wiederholt sich so oft, bis a keine negativen Elemente mehr enthält.Mit Hilfe des Lösungsvektors a können jetzt die Meßungenauigkeiten bei den fünf Leucht-dioden berechnet werden. Dazu berechnen sich die σσσσi wie in Kapitel 5.1 beschrieben zu
σσσσi 2 = ∑
k=1
4
( ) ak σσσσkj 2 . Diese Werte werden jetzt wieder am Anfang des Programmes eingesetzt.
Erster Fit mit denMittelwerten der
Standardabweichungenals Gewichtsfaktoren
DiskretesAnregungspektrum der
Algen bei fünfWellenlängen
Lösungsvektor a
Neuer Fit, wenn einElement von akleiner als 0 ist(dieses Element
wird dann auf nullgesetzt)
Berechnung derneuen
Gewichtsfaktoren
Neuer Fit mit denneuen
Gewichtsfaktoren
Lösungsvektor a
Neuer Fit, wenn einElement von akleiner als 0 ist(dieses Element
wird dann auf nullgesetzt)
Abb. 5.1: Schema zum Rechenprogrammablauf: Von einem diskreten Anregungsspektrum ausgehend, wird zu-nächst ein Lösungsvektor a (dabei wird wie in (Gleichung [ 5.10]) beschrieben gewichtet) berechnet. DieserLösungsvektor a wird auf negative Elemente überprüft. Sofern ein negatives Element auftritt, wird dieses auf nullgesetzt, und Vektor a wird mit einer Dimension weniger neu gefittet. Danach werden die Gewichtsfaktoren durchden Lösungsvektor a neu bestimmt und der Fit wird erneut ausgeführt. Hieraus ergibt sich ein neuer Lösungs-vektor a.
Als Abschluß wird die Summe über alle Elemente von a gebildet. Dieses ergibt die Ge-samtchlorophyll-a-Konzentration. Die gesamte Programmstruktur ist noch einmal oben inAbb. 5.1 schematisch dargestellt.Mit diesem Rechenprogramm ist es nun möglich, Fluoreszenzanregungsspektren von Algen-gemischen bezüglich der Zusammensetzung aus verschiedenen Algengruppen und dem Chlo-rophyll-a-Gehalt zu interpretieren.

5 Verfahren zur Anpassung mit Normkurven
52
5.3 Test des Normkurvenfitverfahrens mit simulierten Daten
In einem „Trockendurchlauf“ sollten die in Abschnitt 4.2.2 gewonnenen Normkurven undSchwankungen der spektralen Algengruppen bei unterschiedlichen Wellenlängen benutztwerden, um aus theoretisch zusammengesetzten Anregungsspektren der Algen die gesuchtenParameter zu berechnen.Hierzu wurde zunächst die Normkurve einer Algengruppe benutzt und die mittleren Schwan-kungen dieser Normkurven addiert bzw. subtrahiert. So ergaben sich zwei neue Kurven, diejetzt mit Fehlern behaftet waren (im weiteren sind diese beiden Kurven mit Fehlerplus beiAddition und Fehlerminus bei Subtraktion der mittleren Schwankungen der Normkurven be-zeichnet. Diese Kurven sind als Ränder der Fehlerschläuche in Abb. 4.7 (Seite 44) schon ein-mal zu sehen gewesen). Fehlerplus und Fehlerminus stellen ein mögliches gemessenes Spek-trum dar.Das Einsetzen von Fehlerplus und -minus als Anregungsspektrum sollte die Fluoreszenzant-wort einer Algenreinkultur bei einer Konzentration von 1µg/l Chlorophyll-a simulieren, diemit natürlichen Meßfehlern (hier Abweichung der Spektren der Algen, die durch veränderli-che Antennenpigmentzusammensetzungen variieren) behaftet sind. Es sollte insbesondereuntersucht werden, ob die Näherung von Fehlerplus und Fehlerminus das Vorhandensein an-derer Algengruppen vortäuscht.Durch Einsetzen von Fehlerplus und -minus in den Rechenalgorithmus aus dem vorigen Ab-schnitt ergab sich eine Chlorophyll-a-Konzentration und eine Zuordnung zu den spektralenGruppen. Dabei sollte die Berechnung einmal mit Gewichtung der mittleren Schwankungendurchgeführt werden und einmal ohne, d.h. daß die Matrix der mittleren Schwankungen anallen Stellen auf eins gesetzt wurde.
Die Rechnungen wurden für Grünalgen, Blaualgen und die bräunliche Gruppe durchgeführt.Bei allen Bestimmungen der Zusammensetzung der spektralen Gruppen wurde die Gruppe zu100% richtig erkannt. Die Unempfindlichkeit gegen Schwankungen der charakteristischenSpektren ist sehr wichtig für die Zuverlässigkeit des Gerätes. Schwankungen gab es dagegenbei der Bestimmung der Chlorophyll-a-Konzentrationen, die jedoch bei Durchführung einer
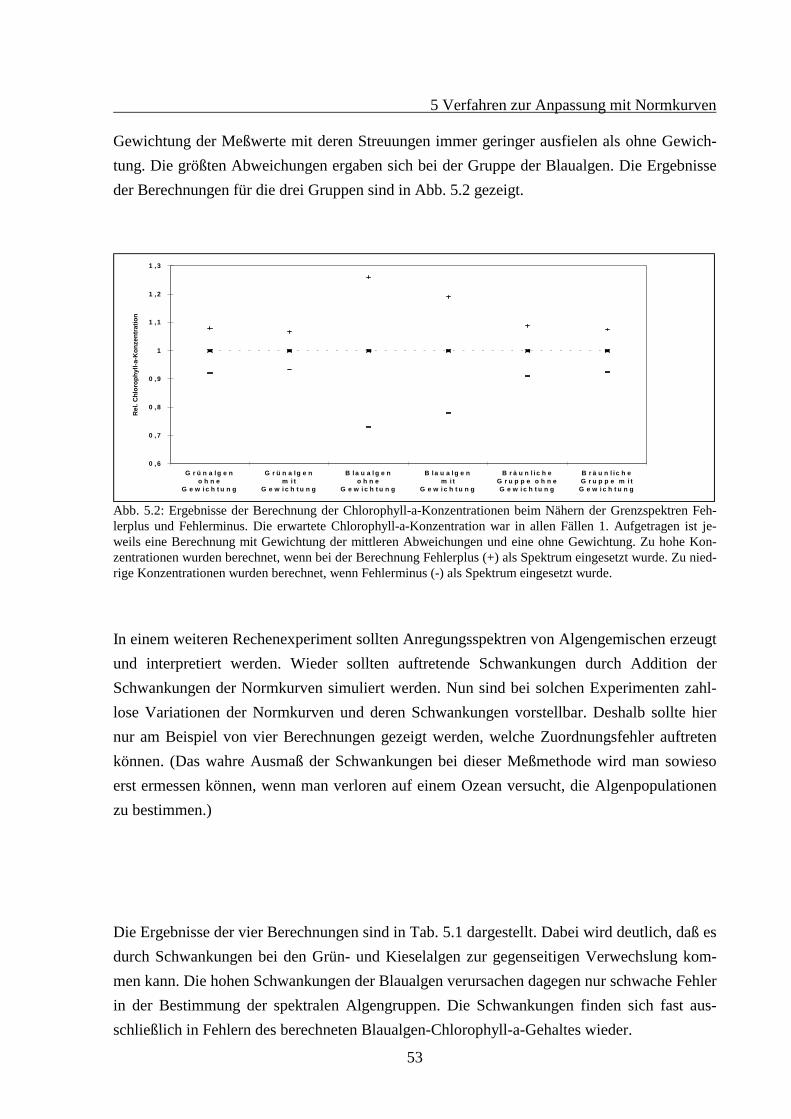
5 Verfahren zur Anpassung mit Normkurven
53
Gewichtung der Meßwerte mit deren Streuungen immer geringer ausfielen als ohne Gewich-tung. Die größten Abweichungen ergaben sich bei der Gruppe der Blaualgen. Die Ergebnisseder Berechnungen für die drei Gruppen sind in Abb. 5.2 gezeigt.
0 ,6
0 ,7
0 ,8
0 ,9
1
1 ,1
1 ,2
1 ,3
G r ü n a lg e no h n e
G e w i c h t u n g
G r ü n a lg e nm i t
G e w i c h t u n g
B la u a l g e no h n e
G e w i c h t u n g
B la u a l g e nm i t
G e w i c h t u n g
B r ä u n l ic h eG r u p p e o h n eG e w i c h t u n g
B r ä u n l ic h eG r u p p e m i t
G e w i c h t u n g
Rel
. Chl
orop
hyll-
a-K
onze
ntra
tion
Abb. 5.2: Ergebnisse der Berechnung der Chlorophyll-a-Konzentrationen beim Nähern der Grenzspektren Feh-lerplus und Fehlerminus. Die erwartete Chlorophyll-a-Konzentration war in allen Fällen 1. Aufgetragen ist je-weils eine Berechnung mit Gewichtung der mittleren Abweichungen und eine ohne Gewichtung. Zu hohe Kon-zentrationen wurden berechnet, wenn bei der Berechnung Fehlerplus (+) als Spektrum eingesetzt wurde. Zu nied-rige Konzentrationen wurden berechnet, wenn Fehlerminus (-) als Spektrum eingesetzt wurde.
In einem weiteren Rechenexperiment sollten Anregungsspektren von Algengemischen erzeugtund interpretiert werden. Wieder sollten auftretende Schwankungen durch Addition derSchwankungen der Normkurven simuliert werden. Nun sind bei solchen Experimenten zahl-lose Variationen der Normkurven und deren Schwankungen vorstellbar. Deshalb sollte hiernur am Beispiel von vier Berechnungen gezeigt werden, welche Zuordnungsfehler auftretenkönnen. (Das wahre Ausmaß der Schwankungen bei dieser Meßmethode wird man sowiesoerst ermessen können, wenn man verloren auf einem Ozean versucht, die Algenpopulationenzu bestimmen.)
Die Ergebnisse der vier Berechnungen sind in Tab. 5.1 dargestellt. Dabei wird deutlich, daß esdurch Schwankungen bei den Grün- und Kieselalgen zur gegenseitigen Verwechslung kom-men kann. Die hohen Schwankungen der Blaualgen verursachen dagegen nur schwache Fehlerin der Bestimmung der spektralen Algengruppen. Die Schwankungen finden sich fast aus-schließlich in Fehlern des berechneten Blaualgen-Chlorophyll-a-Gehaltes wieder.
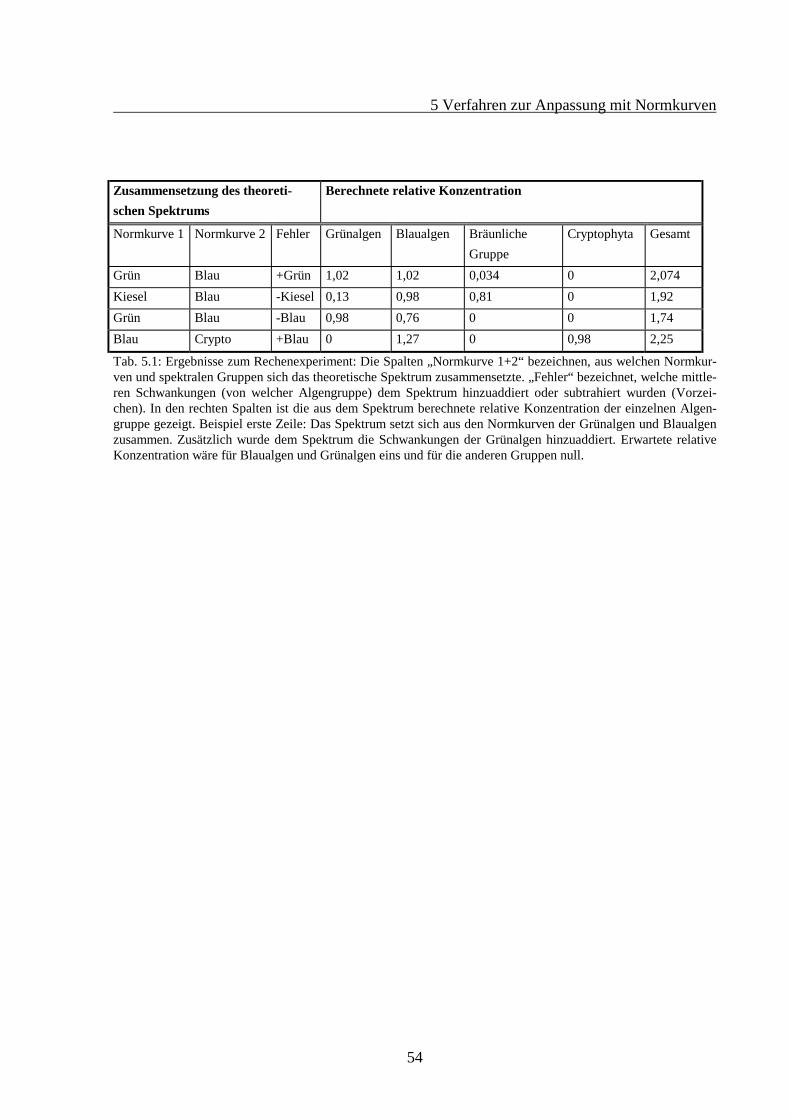
5 Verfahren zur Anpassung mit Normkurven
54
Zusammensetzung des theoreti-schen Spektrums
Berechnete relative Konzentration
Normkurve 1 Normkurve 2 Fehler Grünalgen Blaualgen BräunlicheGruppe
Cryptophyta Gesamt
Grün Blau +Grün 1,02 1,02 0,034 0 2,074Kiesel Blau -Kiesel 0,13 0,98 0,81 0 1,92Grün Blau -Blau 0,98 0,76 0 0 1,74Blau Crypto +Blau 0 1,27 0 0,98 2,25
Tab. 5.1: Ergebnisse zum Rechenexperiment: Die Spalten „Normkurve 1+2“ bezeichnen, aus welchen Normkur-ven und spektralen Gruppen sich das theoretische Spektrum zusammensetzte. „Fehler“ bezeichnet, welche mittle-ren Schwankungen (von welcher Algengruppe) dem Spektrum hinzuaddiert oder subtrahiert wurden (Vorzei-chen). In den rechten Spalten ist die aus dem Spektrum berechnete relative Konzentration der einzelnen Algen-gruppe gezeigt. Beispiel erste Zeile: Das Spektrum setzt sich aus den Normkurven der Grünalgen und Blaualgenzusammen. Zusätzlich wurde dem Spektrum die Schwankungen der Grünalgen hinzuaddiert. Erwartete relativeKonzentration wäre für Blaualgen und Grünalgen eins und für die anderen Gruppen null.

6 Automatisierung des Fluorometers
55
6 Automatisierung des Fluorometers
Zur automatischen Meßwertaufnahme ist in das in Kapitel 1 beschriebene Fluorometer eineAnsteuerungselektronik eingebaut worden, die es ermöglichte, alle verbleibenden fünfLeuchtdioden (mit den Wellenlängen 450 nm, 525 nm, 570 nm, 590 nm und 610 nm) alternie-rend zu betreiben und dabei die auftretende Fluoreszenzintensität dieser Leuchtdiode zuzu-ordnen. Im wesentlichen wurde hierzu der in Abb. 4.2 (Seite 34) gezeigte Schalter durch einein der Regelungseinheit integrierte Leuchtdiodenansteuerungsplatine ersetzt (Eine Photogra-phie des Fluorometers ist in Anhang E gezeigt).
6.1 Aufbau der Leuchtdiodenansteuerung
Der Aufbau zur Leuchtdiodenansteuerung ist schematisch in Abb. 6.1 dargestellt. Eine vomRechner ausgegebene 3-Bit-Adresse und 1 Datenbit zum Setzen und Löschen der Adressewird an einen GAL ausgegeben. Jeder der fünf Leuchtdioden ist eine Adresse zugeordnet. DerGAL ist als D-Latch programmiert (GAL-Programmierung siehe Anhang D), d.h. er setzt dieAdresse an seinen Ausgängen, bis sie vom Rechner wieder ausgeschaltet wird.
Mikro-controller
Multi-plexer
GalvanischeTrennung
Latch(GAL)
Adressefür LED's
Adressefür LED's
Adressefür LED's
Sollwert
Istwert
4kHz-Signal
610nm
450nm525nm
570nm590nm
Referenz-Leuchtdiode
Photodiode
Abb. 6.1: Schematischer Aufbau der Leuchtdiodenansteuerung: Die vom Rechner ausgegebene Adresse einerLeuchtdiode gelangt über ein D-Latch und eine galvanische Trennung zu einem Multiplexer, der die betreffendeLeuchtdiode durchschaltet. Diese wird durch einen Istwert/Sollwert-Vergleich auf die vorgegebene Stromstärkeeingestellt (Schaltplan dazu siehe Anhang C).

6 Automatisierung des Fluorometers
56
Durch hinter den GAL geschaltete Optokoppler wird eine galvanische Trennung der Signalevollzogen. Diese gelangen dann an einen Multiplexer. Der Multiplexer erhält die Adresse vomGAL und schaltet die betreffende Leuchtdiode durch, bis sie am Ausgang des GAL gelöschtwird. Die betreffende Leuchtdiode ist in Reihe mit einer Referenzleuchtdiode geschaltet. DieHelligkeit dieser Leuchtdiode wird überprüft und mit dem Sollwert verglichen und auf dieseneingestellt.
6.2 Leuchtdioden-Takt
Bei dem Betrieb der Leuchtdioden wurde jeweils eine Leuchtdiode einzeln angesteuert. DasTaktdiagramm hierzu ist in Abb. 6.2 aufgetragen. Die Leuchtdioden sind mit einem 4kHz-Signal synchronisiert worden. Bei Beginn einer Messung wurde zunächst die 450-nm-Leuchtdiode durchgeschaltet. Sie war genau während einer Anzeit des 4kHz-Rechteck-Signalseingeschaltet und wurde danach wieder ausgeschaltet. Bei der nächsten Anzeit des 4kHz-Signals wurde die 525-nm-Leuchtdiode eingeschaltet, die jetzt auch wieder nur eine Anzeitdes 4kHz-Signals betrieben wurde. So wurden in der Reihenfolge 450 nm, 525 nm, 570 nm,590 nm und 610 nm alle Leuchtdioden einmal durchgeschaltet. Dieser Vorgang wiederholtesich solange, bis die Meßzeit beendet war.Dabei wurden die Meßwerte der auftretenden Fluoreszenz jeweils für eine Leuchtdiode ge-speichert und dann der gleichen Korrelationsrechnung unterzogen, wie in Kapitel 4.1.4 be-schrieben. Nach der Messung ergab sich für die Fluoreszenzantworten bei den fünf verschie-denen Wellenlängen jeweils ein Wert pro Wellenlänge.

6 Automatisierung des Fluorometers
57
Hi4kHz-
Lo LeuchtdiodenansteuerungHi
450nmLo
Hi525nm
Lo
Hi570nm
Lo
Hi590nm
Lo
Hi610nm
Lo
Abb. 6.2: Taktdiagramm zur Ansteuerung der Leuchtdioden. Oben: Das vom Rechner ausgegebene 4kHz-Signal.Darunter: An- und Auszeiten der fünf Leuchtdioden
Um den Meßaufbau und die Ansteuerung der Leuchtdioden zu testen, wurden die Fluores-zenzantworten bei Anregung mit der oben beschriebenen alternierenden Beleuchtung durchdie fünf Leuchtdioden von vier Algenreinkulturen mit einer Art aus einer spektralen Gruppeuntersucht. Dazu wurde die Spannung, die durch die Fluoreszenz entstand, am Photomulti-plierausgang mit einem Oszilloskop gemessen. Die Chlorophyll-a-Konzentration lag in allenProben bei 20 µg/l Chlorophyll-a (spektrophotometrische Bestimmung, Nusch 1980). Abb.6.3 zeigt die Messungen der vier Algenkulturen. Deutlich sind für alle verschiedenen Artenverschiedene, reproduzierbare „Spektren“ zu erkennen.
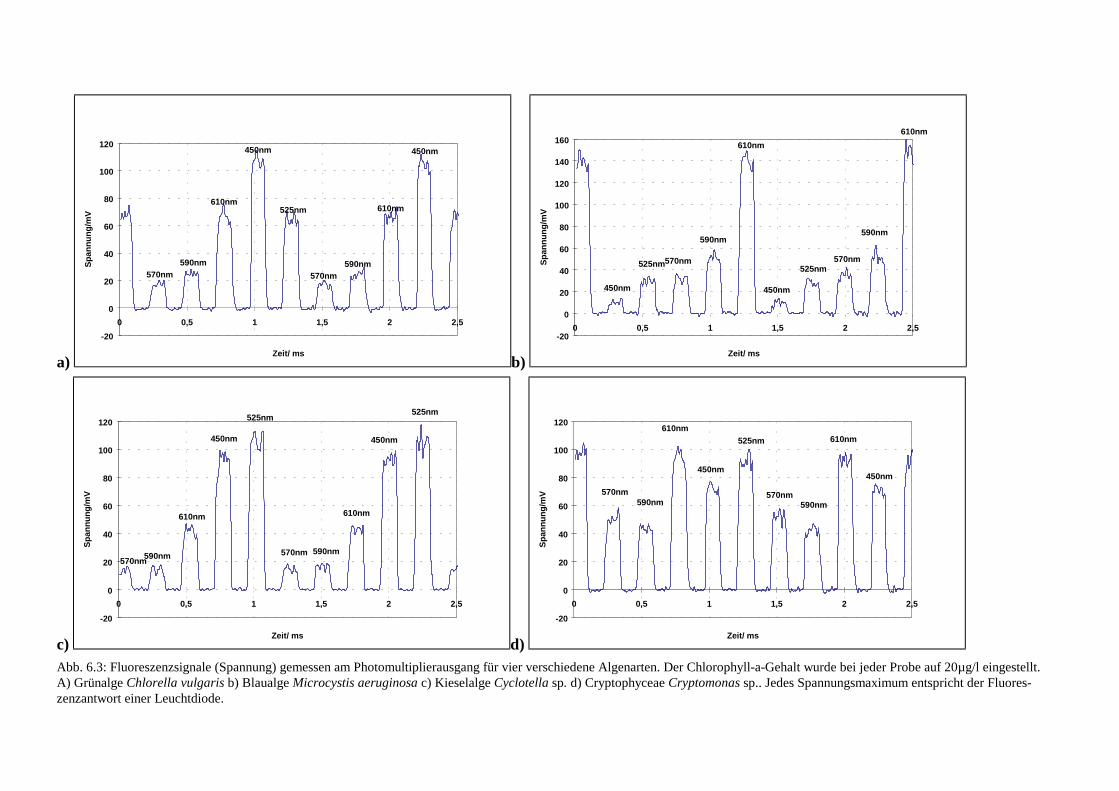
a)
-20
0
20
40
60
80
100
120
0 0,5 1 1,5 2 2,5
Zeit/ ms
Span
nung
/mV
450nm
525nm
570nm590nm
610nm
570nm590nm
610nm
450nm
b)
-20
0
20
40
60
80
100
120
140
160
0 0,5 1 1,5 2 2,5
Zeit/ ms
Span
nung
/mV
450nm
525nm570nm
590nm
610nm
570nm
590nm
610nm
450nm
525nm
c)
-20
0
20
40
60
80
100
120
0 0,5 1 1,5 2 2,5
Zeit/ ms
Span
nung
/mV
450nm
525nm
570nm590nm
610nm
570nm 590nm
610nm
450nm
525nm
d)
-20
0
20
40
60
80
100
120
0 0,5 1 1,5 2 2,5
Zeit/ ms
Span
nung
/mV
450nm
525nm
570nm590nm
610nm
570nm590nm
610nm
450nm
Abb. 6.3: Fluoreszenzsignale (Spannung) gemessen am Photomultiplierausgang für vier verschiedene Algenarten. Der Chlorophyll-a-Gehalt wurde bei jeder Probe auf 20µg/l eingestellt.A) Grünalge Chlorella vulgaris b) Blaualge Microcystis aeruginosa c) Kieselalge Cyclotella sp. d) Cryptophyceae Cryptomonas sp.. Jedes Spannungsmaximum entspricht der Fluores-zenzantwort einer Leuchtdiode.
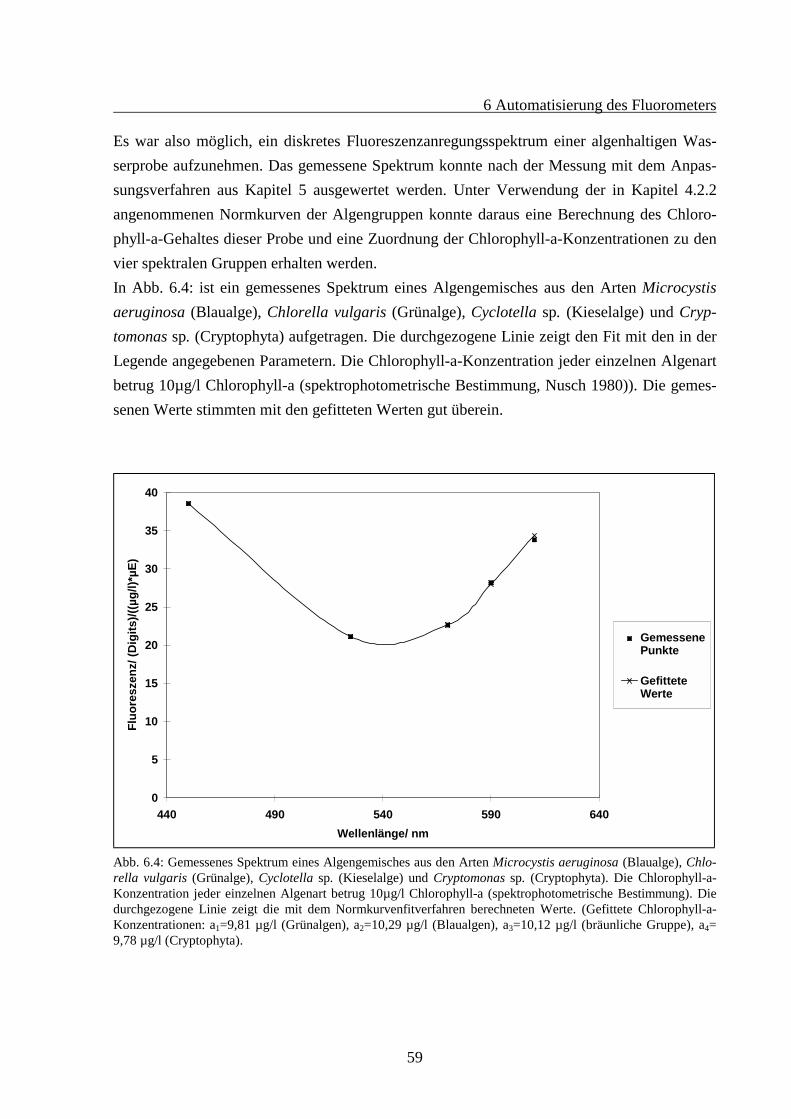
6 Automatisierung des Fluorometers
59
Es war also möglich, ein diskretes Fluoreszenzanregungsspektrum einer algenhaltigen Was-serprobe aufzunehmen. Das gemessene Spektrum konnte nach der Messung mit dem Anpas-sungsverfahren aus Kapitel 5 ausgewertet werden. Unter Verwendung der in Kapitel 4.2.2angenommenen Normkurven der Algengruppen konnte daraus eine Berechnung des Chloro-phyll-a-Gehaltes dieser Probe und eine Zuordnung der Chlorophyll-a-Konzentrationen zu denvier spektralen Gruppen erhalten werden.In Abb. 6.4: ist ein gemessenes Spektrum eines Algengemisches aus den Arten Microcystisaeruginosa (Blaualge), Chlorella vulgaris (Grünalge), Cyclotella sp. (Kieselalge) und Cryp-tomonas sp. (Cryptophyta) aufgetragen. Die durchgezogene Linie zeigt den Fit mit den in derLegende angegebenen Parametern. Die Chlorophyll-a-Konzentration jeder einzelnen Algenartbetrug 10µg/l Chlorophyll-a (spektrophotometrische Bestimmung, Nusch 1980)). Die gemes-senen Werte stimmten mit den gefitteten Werten gut überein.
0
5
10
15
20
25
30
35
40
440 490 540 590 640Wellenlänge/ nm
Fluo
resz
enz/
(Dig
its)/(
(µg/
l)*µE
)
GemessenePunkte
GefitteteWerte
Abb. 6.4: Gemessenes Spektrum eines Algengemisches aus den Arten Microcystis aeruginosa (Blaualge), Chlo-rella vulgaris (Grünalge), Cyclotella sp. (Kieselalge) und Cryptomonas sp. (Cryptophyta). Die Chlorophyll-a-Konzentration jeder einzelnen Algenart betrug 10µg/l Chlorophyll-a (spektrophotometrische Bestimmung). Diedurchgezogene Linie zeigt die mit dem Normkurvenfitverfahren berechneten Werte. (Gefittete Chlorophyll-a-Konzentrationen: a1=9,81 µg/l (Grünalgen), a2=10,29 µg/l (Blaualgen), a3=10,12 µg/l (bräunliche Gruppe), a4=9,78 µg/l (Cryptophyta).

7 Test des automatisierten Fluorometers
60
7 Test des automatisierten Fluorometers
7.1 Verdünnungsreihen
Durch die Verdünnungsreihen (hier ist die Messung der Anregungsspektren von Algengemi-schen unterschiedlicher Zusammensetzung und Konzentrationen gemeint) sollte herausgefun-den werden, ob es mit dem vorliegenden Meßaufbau praktisch möglich ist, den Chlorophyll-a-Gehalt einer Wasserprobe zu bestimmen und diesen den spektralen Algengruppen richtig zu-zuordnen. Dazu wurde das dominierende Auftreten einer Algenspezies (Algenblüte) simuliert.Außerdem sollten die linearen Meßbereiche der Methode untersucht werden, die nach untenhin, bei geringen Konzentrationen, durch die Auflösung des Fluorometers gegeben sind. Beihöheren Chlorophyll-a-Konzentrationen können Vorgänge wie Selbstabsorption von Fluores-zenzstrahlung durch die Algen die Messung und damit die Meßbereiche beeinflussen. In die-sen Meßbereichen gilt nicht mehr die angenommene Proportionalität der Fluoreszenzsignalezur Chlorophyll-a-Konzentration.
7.1.1 Künstliche Algenblüten
Im Labor sollten Kulturen untersucht werden, die in der Zelldichte einer Blaualgenblüte sowieeiner Grünalgenblüte entsprachen. Dazu wurde ein Gemisch aus drei Algenreinkulturen mitdefinierten Chlorophyll-a-Konzentrationen hergestellt. Dem Gemisch wurden dann stufenwei-se bekannte Mengen einer vierten Algenreinkultur hinzugefügt. Dies kam einem Anwachseneiner Algenart gleich, da die Konzentrationen der anderen Algenarten konstant gehalten wur-den. Dieses Vorgehen simulierte eine Algenblüte in natürlichen Gewässern.In einem ersten Versuch sollte eine künstliche Blaualgenblüte detektiert werden. Dazu wurdeein Gemisch aus Grünalgen (Chlorophyll-a-Konzentration: 5 µg/l), Kieselalgen (Chlorophyll-a-Konzentration: 7 µg/l) und Cryptophyceae (Chlorophyll-a-Konzentration: 2 µg/l; die Chlo-rophyllgehalte wurden durch spektrophotometrische Bestimmung (Nusch 1980) ermittelt)hergestellt. Von dem Gemisch wurde ein Fluoreszenzanregungsspektrum aufgenommen undunter Anwendung des Normkurvenfitverfahrens der Chlorophyll-a-Gehalt berechnet und dieZuordnung zu den Algengruppen getroffen. Danach wurden verschiedene Mengen einer Blau-algenkultur (Microcystis aeruginosa) dem Gemisch hinzugefügt, so daß die Wasserproben
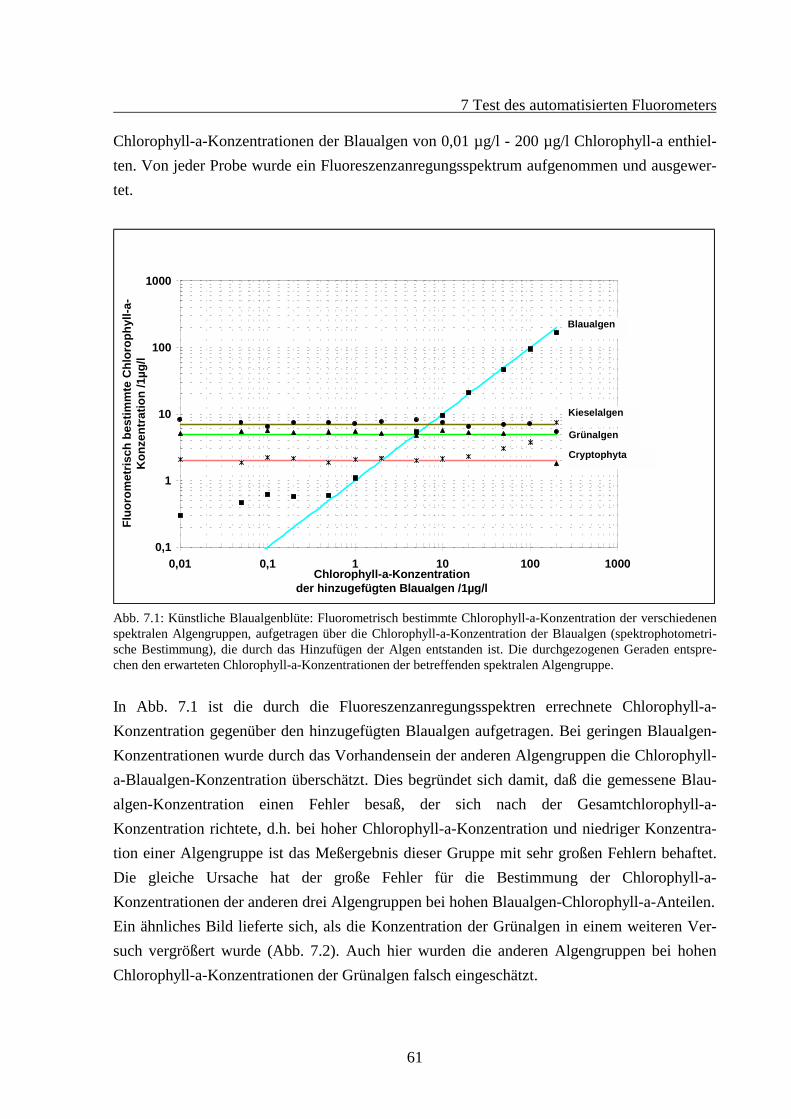
7 Test des automatisierten Fluorometers
61
Chlorophyll-a-Konzentrationen der Blaualgen von 0,01 µg/l - 200 µg/l Chlorophyll-a enthiel-ten. Von jeder Probe wurde ein Fluoreszenzanregungsspektrum aufgenommen und ausgewer-tet.
0,1
1
10
100
1000
0,01 0,1 1 10 100 1000Chlorophyll-a-Konzentration
der hinzugefügten Blaualgen /1µg/l
Fluo
rom
etris
ch b
estim
mte
Chl
orop
hyll-
a-K
onze
ntra
tion
/1µg
/l
Blaualgen
Kieselalgen
Grünalgen
Cryptophyta
Abb. 7.1: Künstliche Blaualgenblüte: Fluorometrisch bestimmte Chlorophyll-a-Konzentration der verschiedenenspektralen Algengruppen, aufgetragen über die Chlorophyll-a-Konzentration der Blaualgen (spektrophotometri-sche Bestimmung), die durch das Hinzufügen der Algen entstanden ist. Die durchgezogenen Geraden entspre-chen den erwarteten Chlorophyll-a-Konzentrationen der betreffenden spektralen Algengruppe.
In Abb. 7.1 ist die durch die Fluoreszenzanregungsspektren errechnete Chlorophyll-a-Konzentration gegenüber den hinzugefügten Blaualgen aufgetragen. Bei geringen Blaualgen-Konzentrationen wurde durch das Vorhandensein der anderen Algengruppen die Chlorophyll-a-Blaualgen-Konzentration überschätzt. Dies begründet sich damit, daß die gemessene Blau-algen-Konzentration einen Fehler besaß, der sich nach der Gesamtchlorophyll-a-Konzentration richtete, d.h. bei hoher Chlorophyll-a-Konzentration und niedriger Konzentra-tion einer Algengruppe ist das Meßergebnis dieser Gruppe mit sehr großen Fehlern behaftet.Die gleiche Ursache hat der große Fehler für die Bestimmung der Chlorophyll-a-Konzentrationen der anderen drei Algengruppen bei hohen Blaualgen-Chlorophyll-a-Anteilen.Ein ähnliches Bild lieferte sich, als die Konzentration der Grünalgen in einem weiteren Ver-such vergrößert wurde (Abb. 7.2). Auch hier wurden die anderen Algengruppen bei hohenChlorophyll-a-Konzentrationen der Grünalgen falsch eingeschätzt.
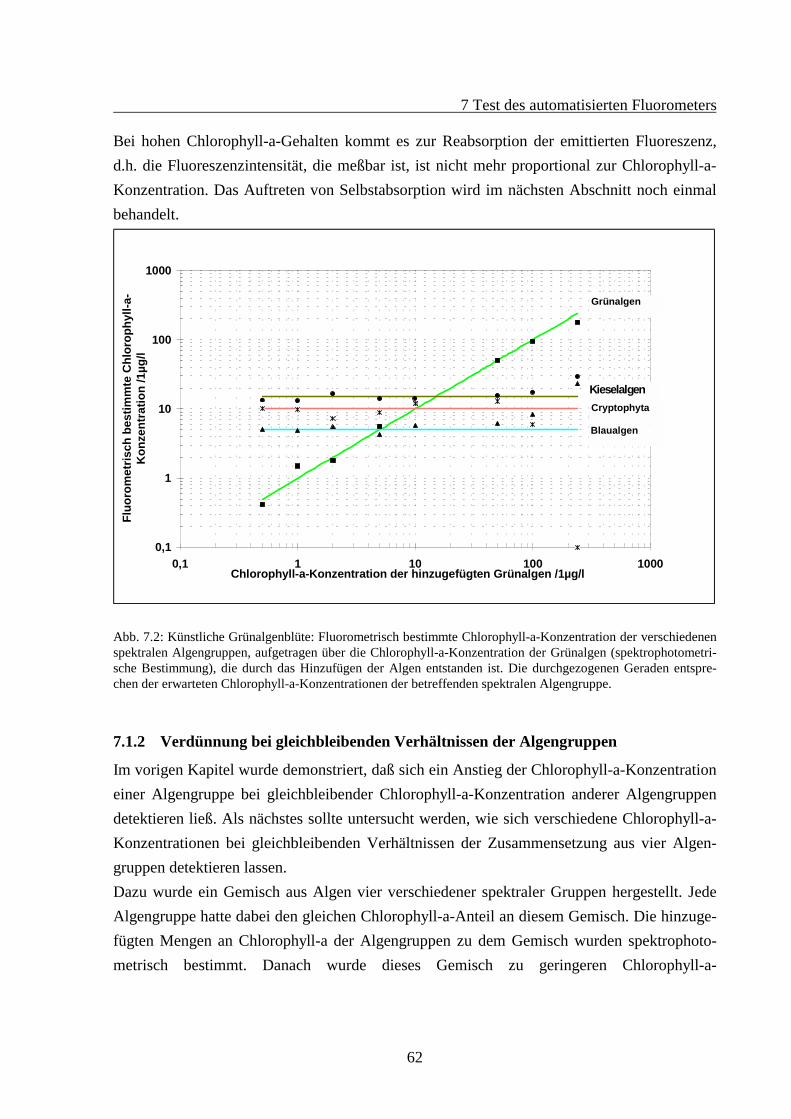
7 Test des automatisierten Fluorometers
62
Bei hohen Chlorophyll-a-Gehalten kommt es zur Reabsorption der emittierten Fluoreszenz,d.h. die Fluoreszenzintensität, die meßbar ist, ist nicht mehr proportional zur Chlorophyll-a-Konzentration. Das Auftreten von Selbstabsorption wird im nächsten Abschnitt noch einmalbehandelt.
0,1
1
10
100
1000
0,1 1 10 100 1000Chlorophyll-a-Konzentration der hinzugefügten Grünalgen /1µg/l
Fluo
rom
etris
ch b
estim
mte
Chl
orop
hyll-
a-K
onze
ntra
tion
/1µg
/l
Grünalgen
KieselalgenCryptophyta
Blaualgen
Abb. 7.2: Künstliche Grünalgenblüte: Fluorometrisch bestimmte Chlorophyll-a-Konzentration der verschiedenenspektralen Algengruppen, aufgetragen über die Chlorophyll-a-Konzentration der Grünalgen (spektrophotometri-sche Bestimmung), die durch das Hinzufügen der Algen entstanden ist. Die durchgezogenen Geraden entspre-chen der erwarteten Chlorophyll-a-Konzentrationen der betreffenden spektralen Algengruppe.
7.1.2 Verdünnung bei gleichbleibenden Verhältnissen der Algengruppen
Im vorigen Kapitel wurde demonstriert, daß sich ein Anstieg der Chlorophyll-a-Konzentrationeiner Algengruppe bei gleichbleibender Chlorophyll-a-Konzentration anderer Algengruppendetektieren ließ. Als nächstes sollte untersucht werden, wie sich verschiedene Chlorophyll-a-Konzentrationen bei gleichbleibenden Verhältnissen der Zusammensetzung aus vier Algen-gruppen detektieren lassen.Dazu wurde ein Gemisch aus Algen vier verschiedener spektraler Gruppen hergestellt. JedeAlgengruppe hatte dabei den gleichen Chlorophyll-a-Anteil an diesem Gemisch. Die hinzuge-fügten Mengen an Chlorophyll-a der Algengruppen zu dem Gemisch wurden spektrophoto-metrisch bestimmt. Danach wurde dieses Gemisch zu geringeren Chlorophyll-a-

7 Test des automatisierten Fluorometers
63
Konzentrationen verdünnt. Hierbei wurde von jeder Mischung zehnmal hintereinander eindiskretes Spektrum aufgenommen. Daraus wurden wieder der Chlorophyll-a-Gesamtgehaltund die Konzentration der einzelnen vier spektralen Algengruppen bestimmt. Durch die zehn-fache Bestimmung war es möglich, die Standardabweichung jeder Messung zu bestimmen.Die Ergebnisse der Bestimmung der Chlorophyll-a-Konzentration der jeweiligen Algengruppesind in Abb. 7.3 aufgetragen. Dabei ist zu erkennen, daß die Standardabweichung der Meßer-gebnisse bei allen Algengruppen prozentual gesehen bei niedrigen Chlorophyll-a-Konzentrationen anwuchs. Dieses war ein Fehler, der durch den Meßaufbau bedingt war. Erwurde z.B. durch das Dunkelrauschen des Photomultipliers bestimmt.Bei größeren Konzentrationen verringerte sich die Streuung der Meßwerte zunehmend. Beisehr hohen Konzentrationen wurde die von den Algen emittierte Fluoreszenzstrahlung merk-lich reabsorbiert und es kam zu Fehlbestimmungen der Algengruppen.Dabei wird die Fluoreszenz dann vornehmlich von den Photosystem-I-Antennen absorbiertwerden, da diese den langwelligeren Lichtbereich abdecken. Aufgrund der unterschiedlichenAbsorptionsspektren der verschiedenen Algengruppen findet die Reabsorption der Fluores-zenz nicht bei allen Gruppen gleichmäßig statt. Vermutlich verändern sich durch die Reab-sorption bei höheren Chlorophyll-a-Konzentrationen auch die Fluoreszenzanregungsspektren.Dieses wäre denkbar; denn durch die verstärkte Anregung der PS I-Systeme findet der sog.„Emerson-Effekt“ (Emerson 1958) statt. Hierbei wird unter Einstrahlung von Rotlicht und derdamit verstärkten Anregung der PS I-Systeme die Photosyntheserate pro absorbiertem Licht-quant gesteigert. Dadurch würde die gesteigerte Photosyntheserate die Fluoreszenzquanten-ausbeute senken. Dieser Effekt würde bei den verschiedenen Algengruppen ebenfalls unter-schiedlich starke Auswirkungen haben. Daher wäre es an dieser Stelle recht spekulativ, überdie genaue Auswirkung der Selbstabsorption und ihre damit verbundenen Effekte zu diskutie-ren.Aus den Messungen wird ersichtlich, daß bei hohen Konzentrationen in unserem Fall einerGesamtkonzentration von 400 µg/l Chlorophyll-a der Gehalt an Grünalgen und Cryptophy-ceae erheblich unterschätzt wurde.Bei Betrachtung des gemessenen Chlorophyll-a-Gesamtgehalts durch die Fluorezenzspektrenwährend der Verdünnungsreihe (Abb. 7.4) ergab sich ein ganz ähnliches Bild. Bei niedrigenKonzentrationen nahm die Standardabweichung der Meßwerte erheblich zu und bei hohenKonzentrationen (400 µg/l Chlorophyll-a) entstand eine Unterschätzung des tatsächlichenChlorophyll-a-Gesamtgehaltes (spektrophotometrische Bestimmung).
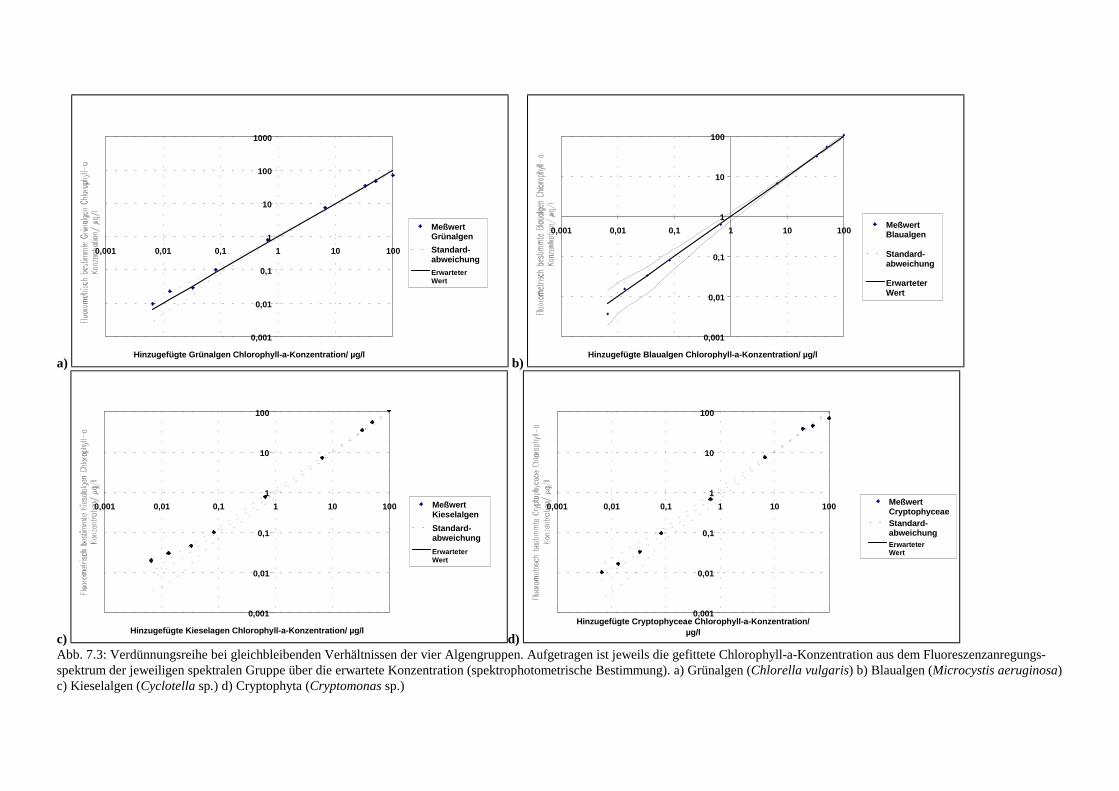
a)
0,001
0,01
0,1
1
10
100
1000
0,001 0,01 0,1 1 10 100
Hinzugefügte Grünalgen Chlorophyll-a-Konzentration/ µg/l
MeßwertGrünalgenStandard-abweichungErwarteter Wert
b)
0,001
0,01
0,1
1
10
100
0,001 0,01 0,1 1 10 100
Hinzugefügte Blaualgen Chlorophyll-a-Konzentration/ µg/l
MeßwertBlaualgen
Standard-abweichung
Erwarteter Wert
c)
0,001
0,01
0,1
1
10
100
0,001 0,01 0,1 1 10 100
Hinzugefügte Kieselagen Chlorophyll-a-Konzentration/ µg/l
MeßwertKieselalgenStandard-abweichungErwarteter Wert
d)
0,001
0,01
0,1
1
10
100
0,001 0,01 0,1 1 10 100
Hinzugefügte Cryptophyceae Chlorophyll-a-Konzentration/ µg/l
MeßwertCryptophyceaeStandard-abweichungErwarteter Wert
Abb. 7.3: Verdünnungsreihe bei gleichbleibenden Verhältnissen der vier Algengruppen. Aufgetragen ist jeweils die gefittete Chlorophyll-a-Konzentration aus dem Fluoreszenzanregungs-spektrum der jeweiligen spektralen Gruppe über die erwartete Konzentration (spektrophotometrische Bestimmung). a) Grünalgen (Chlorella vulgaris) b) Blaualgen (Microcystis aeruginosa)c) Kieselalgen (Cyclotella sp.) d) Cryptophyta (Cryptomonas sp.)

7 Test des automatisierten Fluorometers
65
0,001
0,01
0,1
1
10
100
1000
0,01 0,1 1 10 100 1000
Hinzugefügte Gesamtchlorophyll-a-Konzentration/ µg/l
Fluo
rom
etris
ch b
estim
mte
Ges
amt-C
hlor
ophy
ll-a-
Kon
zent
ratio
n/ µ
g/l
Gemessener Gesamtgehalt
Standard-abweichung
Erwarteter Gesamtgehalt
Abb. 7.4: Verdünnungsreihe bei gleichbleibenden Verhältnissen der vier Algengruppen. Aufgetragen ist jeweilsdie gefittete Gesamtchlorophyll-a-Konzentration aus dem Fluoreszenzanregungsspektrum über die erwarteteGesamtchlorophyll-a-Konzentration (spektrophotometrische Bestimmung).

7 Test des automatisierten Fluorometers
66
7.2 Fluoreszenzverhalten gealterter Algenkulturen
In den vorangegangenen Versuchen war untersucht worden, ob sich definierte Mengen vonAlgengruppen in einem Gemisch detektieren lassen. Dabei wurden Normkurven und derenSchwankungen dazu benutzt, die Chlorophyll-a-Konzentrationen der einzelnen Algengruppenaus dem diskreten Fluoreszenzanregungsspektrum zu bestimmen. Zur Ermittlung der Norm-kurven (Abschnitt 4.2) wurden jeweils verschiedene Vertreter aus den spektralen Gruppenuntersucht. Dabei wurden Algenkulturen verwendet, die unter identischen Anzuchtbedingun-gen kultiviert worden waren und eine identische Anzuchtdauer besaßen. Anzuchtbedingungenund Anzuchtdauer waren so gehalten, daß keine Streßbedingungen, also Mangel an Nährstof-fen oder zu geringe Lichtintensität, herrschten.In weiteren Versuchen sollte untersucht werden, welche Auswirkungen unterschiedliche An-zuchtdauer und damit einhergehende Anzuchtbedingungen auf die detektierten Fluoreszenzan-regungsspektren der Algen haben können.Als Musterbeispiel wurden drei Algenarten untersucht. Es handelte sich dabei um eineGrünalge (Scenedesmus acutus), eine Blaualge (Synechococcus sp.) und eine Kieselalge (Cy-clotella sp.).Von den drei Algenarten wurden jeweils drei 300 ml Kulturen (insgesamt neun Kulturen) an-gesetzt. Als Nährmedium wurde WC-Medium verwendet. Eine Leuchtstoffröhre (Lichtinten-sität 150 µE m-2s-1) sorgte für kontinuierliche Beleuchtung. Um Sedimentation am Gefäßbo-den zu verhindern und um für einen gleichmäßigen Kohlendioxid und Sauerstoffgehalt in derKultur zu sorgen, wurden die Kulturen permanent gerührt. Der Versuchszeitraum erstrecktesich über 36 Tage. An neun Tagen während dieser Zeit wurde jeder Kultur eine Probe von 10ml entnommen. 5ml davon wurden für eine HPLC-Analyse (Wiltshire und Schroeder 1994)verwendet, mit der der Chlorophyll-a-Gehalt der Probe und der Gehalt an Carotinen undXanthophyllen bestimmt wurde (Die Zusammensetzung der Phycobilisomen der Blaualgenkonnte mit einer HPLC-Analyse aufgrund der Wasserlöslichkeit und der besonderen Polaritätdieser Pigmente noch nicht untersucht werden bzw. war mir dieses unbekannt). Von der ande-ren Hälfte der entnommenen Probe wurde das diskrete Fluoreszenzanregungsspektrum aufge-nommen. Bei zu hohen Konzentrationen der Probe wurde diese mit WC-Nährmedium ent-sprechend verdünnt.
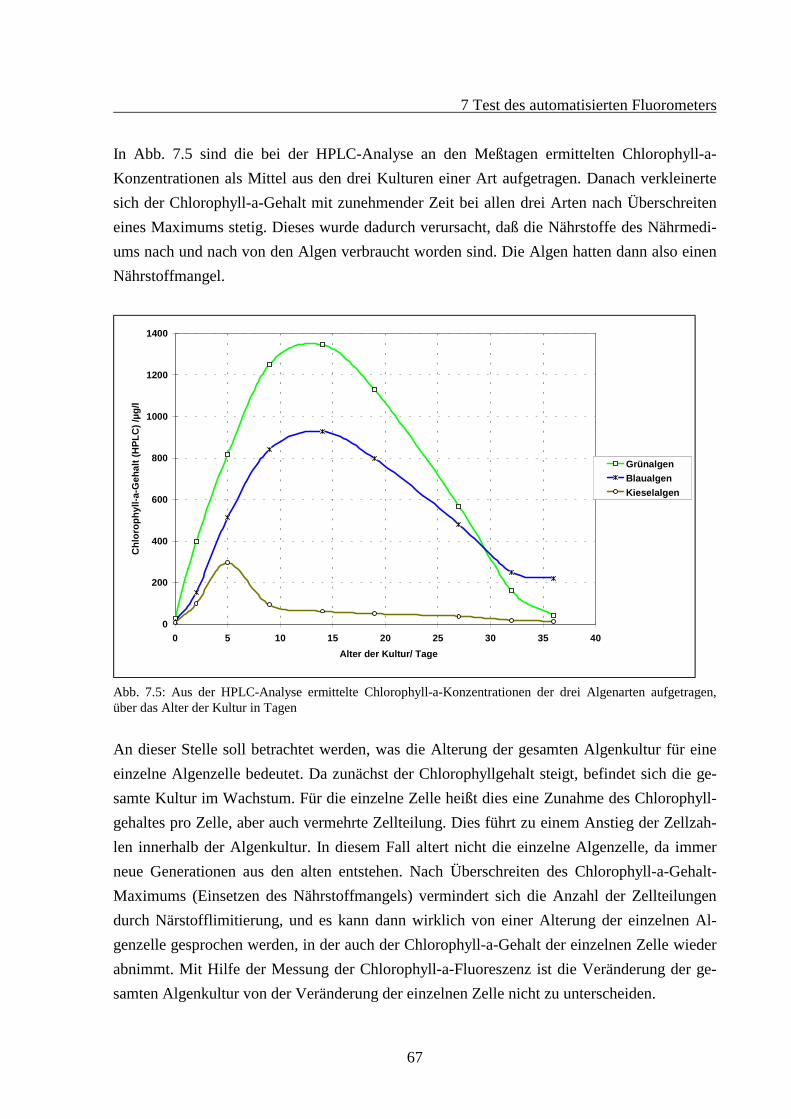
7 Test des automatisierten Fluorometers
67
In Abb. 7.5 sind die bei der HPLC-Analyse an den Meßtagen ermittelten Chlorophyll-a-Konzentrationen als Mittel aus den drei Kulturen einer Art aufgetragen. Danach verkleinertesich der Chlorophyll-a-Gehalt mit zunehmender Zeit bei allen drei Arten nach Überschreiteneines Maximums stetig. Dieses wurde dadurch verursacht, daß die Nährstoffe des Nährmedi-ums nach und nach von den Algen verbraucht worden sind. Die Algen hatten dann also einenNährstoffmangel.
0
200
400
600
800
1000
1200
1400
0 5 10 15 20 25 30 35 40Alter der Kultur/ Tage
Chl
orop
hyll-
a-G
ehal
t (H
PLC
) /µg
/l
GrünalgenBlaualgenKieselalgen
Abb. 7.5: Aus der HPLC-Analyse ermittelte Chlorophyll-a-Konzentrationen der drei Algenarten aufgetragen,über das Alter der Kultur in Tagen
An dieser Stelle soll betrachtet werden, was die Alterung der gesamten Algenkultur für eineeinzelne Algenzelle bedeutet. Da zunächst der Chlorophyllgehalt steigt, befindet sich die ge-samte Kultur im Wachstum. Für die einzelne Zelle heißt dies eine Zunahme des Chlorophyll-gehaltes pro Zelle, aber auch vermehrte Zellteilung. Dies führt zu einem Anstieg der Zellzah-len innerhalb der Algenkultur. In diesem Fall altert nicht die einzelne Algenzelle, da immerneue Generationen aus den alten entstehen. Nach Überschreiten des Chlorophyll-a-Gehalt-Maximums (Einsetzen des Nährstoffmangels) vermindert sich die Anzahl der Zellteilungendurch Närstofflimitierung, und es kann dann wirklich von einer Alterung der einzelnen Al-genzelle gesprochen werden, in der auch der Chlorophyll-a-Gehalt der einzelnen Zelle wiederabnimmt. Mit Hilfe der Messung der Chlorophyll-a-Fluoreszenz ist die Veränderung der ge-samten Algenkultur von der Veränderung der einzelnen Zelle nicht zu unterscheiden.

7 Test des automatisierten Fluorometers
68
7.2.1 Fluoreszenzverhalten der Grünalgen während der Alterung
Die reinen Grünalgenkulturen entwickelten die höchsten Chlorophyll-a-Konzentrationen derAlgenkulturen während der Meßzeit. Dabei war die Chlorophyll-a-Konzentration in den Pro-ben zwischen dem zehnten und fünfzehnten Tag am höchsten.Die Fluoreszenzspektren wurden wie in Abschnitt 7.1 mit dem Normkurvenfit analysiert. DieFits ergaben den Gesamtchlorophyll-a-Gehalt und die Aufspaltung in die vier Algengruppen.Da es sich um eine Grünalgenkultur handelte, sollte das Fitergebnis 100% Grünalgen und 0%für die anderen Algengruppen sein. Abweichungen sind ein Maß für die Treffsicherheit desVerfahrens. Der aus der Fluoreszenzmessung berechnete Chlorophyll-a-Gehalt und der Anteilder vier Algengruppen daran während der 36-Tage-Meßzeit ist in Abb. 7.6 gezeigt. Dabei istzu erkennen, daß den Grünalgen immer mehr als 92 Prozent des Gesamtchlorophyll-a-Gehaltes zugeordnet wird.
0
200
400
600
800
1000
1200
1400
0 2 5 9 14 19 27 32 36
Alter der Kultur/ Tage
Chl
orop
hyll-
a-K
onze
ntra
tion/
µg/
l
Berechneter Chlorophyll-a-Gehalt der Kieselalgen Berechneter Chlorophyll-a-Gehalt der Grünalgen
Abb. 7.6: Chlorophyll-a-Gehaltsbestimmung aus den Fluoreszenzanregungsspektren der Grünalgenkulturen überdie 36-Tage-Meßzeit, die verschiedene Färbung gibt den gefitteten Anteil der verschiedenen spektralen Gruppenan dem Gesamtgehalt an. (Hier Grünalgen und Kieselalgen)
Als nächstes sollte überprüft werden, ob die Berechnung des Gesamtchlorophyll-a-Gehaltesmit den Ergebnissen aus der HPLC-Analyse übereinstimmten.Die aus den aufgenommenen Fluoreszenzspektren errechneten Chlorophyll-a-Konzentrationensind in Abb. 7.7 aufgetragen.
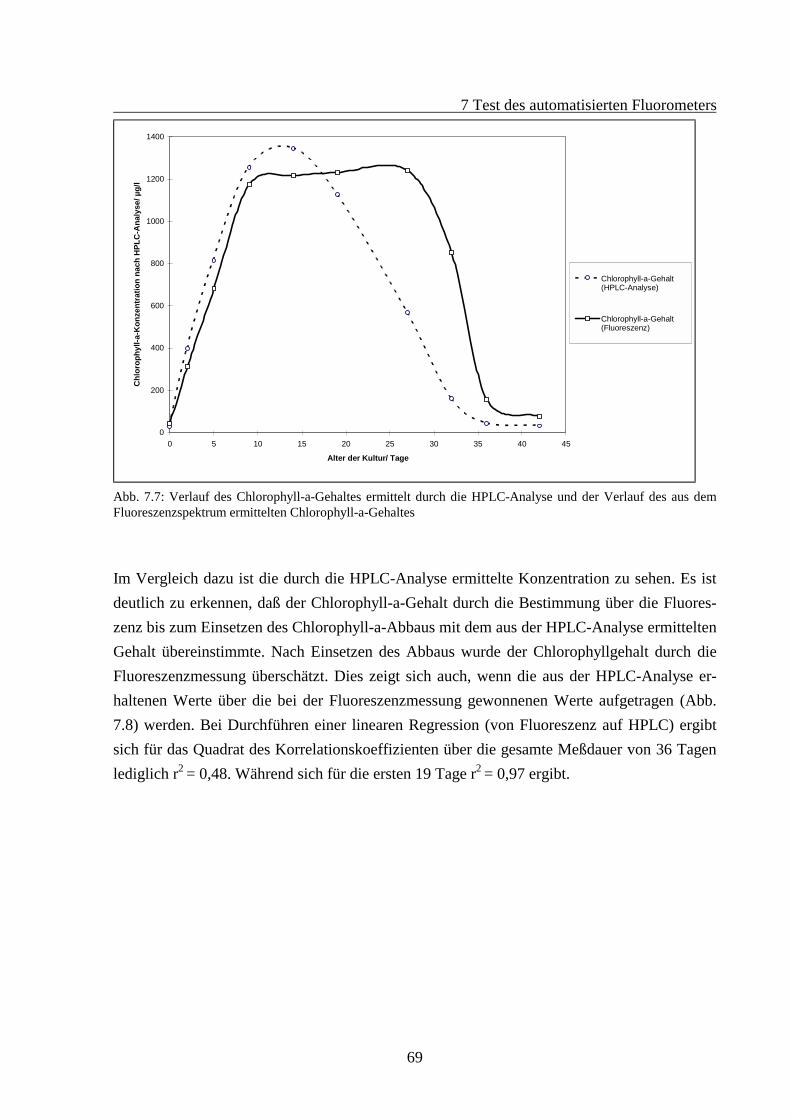
7 Test des automatisierten Fluorometers
69
0
200
400
600
800
1000
1200
1400
0 5 10 15 20 25 30 35 40 45
Alter der Kultur/ Tage
Chl
orop
hyll-
a-K
onze
ntra
tion
nach
HPL
C-A
naly
se/ µ
g/l
Chlorophyll-a-Gehalt (HPLC-Analyse)
Chlorophyll-a-Gehalt (Fluoreszenz)
Abb. 7.7: Verlauf des Chlorophyll-a-Gehaltes ermittelt durch die HPLC-Analyse und der Verlauf des aus demFluoreszenzspektrum ermittelten Chlorophyll-a-Gehaltes
Im Vergleich dazu ist die durch die HPLC-Analyse ermittelte Konzentration zu sehen. Es istdeutlich zu erkennen, daß der Chlorophyll-a-Gehalt durch die Bestimmung über die Fluores-zenz bis zum Einsetzen des Chlorophyll-a-Abbaus mit dem aus der HPLC-Analyse ermitteltenGehalt übereinstimmte. Nach Einsetzen des Abbaus wurde der Chlorophyllgehalt durch dieFluoreszenzmessung überschätzt. Dies zeigt sich auch, wenn die aus der HPLC-Analyse er-haltenen Werte über die bei der Fluoreszenzmessung gewonnenen Werte aufgetragen (Abb.7.8) werden. Bei Durchführen einer linearen Regression (von Fluoreszenz auf HPLC) ergibtsich für das Quadrat des Korrelationskoeffizienten über die gesamte Meßdauer von 36 Tagenlediglich r2 = 0,48. Während sich für die ersten 19 Tage r2 = 0,97 ergibt.

7 Test des automatisierten Fluorometers
70
R2 = 0,4849
R2 = 0,9674
0
200
400
600
800
1000
1200
1400
0 200 400 600 800 1000 1200 1400
Chlorophyll-a-Konzentration nach HPLC-Analyse/ µg/l
Chl
orop
hyll-
a-K
onze
ntra
tion
erre
chne
t aus
der
Chl
orop
hyll-
a Fl
uore
szen
z/
µg/l
Linerare Regression über die ersten 19 Tage
Linerare Regression über die gesamte Meßdauer
Abb. 7.8: Die fluorometrisch ermittelten Chlorophyll-a-Gehalte der alternden Grünalgenkultur aufgetragen überdie bei der HPLC-Analyse ermittelten Chlorophyll-a-Gehalte. Die gestrichelte Linie verdeutlicht den zeitlichenVerlauf.
Welche Erklärungsmöglichkeiten stehen hierfür zur Verfügung?Prinzipiell gäbe es für die Überschätzung der Chlorophyll-a-Konzentration während der Ab-bauphase drei Erklärungen.Als erstes ist bei einsetzendem Abbau von Chlorophyll-a sicherlich eine störende Wirkungvon Chlorophyll-a-Abbauprodukten, den Chlorophylliden und Phaeophorbiden, auf das Fluo-reszenzanregungsspektrum zu vermuten. Diese können ein ähnliches Fluoreszenzverhaltenwie Chlorophyll-a besitzen (Doerffer 1986). Die Abbauprodukte hätten aber bei der HPLC-Analyse detektiert werden können und wären bestimmbar gewesen. Diese Stoffe waren abernicht nachweisbar. Daher konnte dies kein Grund für die Meßergebnisse sein.Eine zweite Erklärung für das Zustandekommen der Meßergebnisse wäre eine erhebliche Ver-änderung der äußeren Antennen des Photosystem II. Zeitlich gesehen ist zum Ende der Meß-zeit hin eine Erhöhung des Chlorophyll-b-zu-Chlorophyll-a-Verhältnisses festzustellen. DerVerlauf des Chlorophyll-b-zu-Chlorophyll-a-Verhältnisses während der Alterung der Grünal-genkultur ist in Abb. 7.9 gezeigt. Deutlich ist an den letzten drei Meßtagen ein Anstieg desVerhältnisses zu beobachten.

7 Test des automatisierten Fluorometers
71
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 5 10 15 20 25 30 35 40Alter der Kultur/ Tage
Rel
ativ
es C
hlor
ophy
ll-b/
Chl
orop
hyll-
a-Ve
rhäl
tnis
Abb. 7.9: Verlauf des relativen Chlorophyll-b-zu-Chlorophyll-a-Verhältnisses (Bestimmung durch die HPLC-Analyse) während der Alterung der Grünalgenkultur
Trägt man nun die Fluoreszenzantwort einer bestimmten Leuchtdiode, geteilt durch den ausder HPLC-Analyse gewonnenen Chlorophyll-a-Gehalt über dieses Pigmentverhältnis auf, soerhält man einen linearen Zusammenhang (Abb. 7.10). Eine Veränderung der äußeren Anten-nen scheint also eine mögliche Erklärung für die Überschätzung des Chlorophyll-a-Gehaltesdurch die Fluoreszenzmessung an den letzten Meßtagen des Alterungsversuches.

7 Test des automatisierten Fluorometers
72
0
5
10
15
20
25
30
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Relatives Chlorophyll-b/Chlorophyll-a-Verhältnis
Fluo
resz
enzi
nten
sitä
t/1µg
/l C
hlor
ophy
ll-a
450nm
525nm
570nm
590nm
610nm
Abb. 7.10: Abhängigkeit der Fluoreszenzintensität vom Chlorophyll-b-zu-Chlorophyll-a-Verhältnis bei konstan-tem Chlorophyll-a-Gehalt. Hierbei ist die von einer LED angeregte Fluoreszenz auf den aus der HPLC-Analysebestimmten Chlorophyll-a-Gehalt normiert. Die durchgezogenen Geraden sind durch Regression für jeweils eineLeuchtdiode (siehe Kästchen) bestimmt worden.
Eine weitere Möglichkeit, die zu einer Vergrößerung der Fluoreszenz pro Chlorophyll-a-Gehalt führen könnte, ist die Veränderung der photosynthetischen Aktivität der Photosysteme.Bei einer absterbenden Algenkultur, die unter Nährstoffmangel leidet, ist es denkbar, daß diePhotosysteme nicht mehr voll funktionsfähig sind. Daher könnte die von den äußeren Anten-nen der Alge aufgenommene Lichtenergie nicht mehr in dem Maße von den Photosystemenumgewandelt werden, wie es bei einer gesunden Alge der Fall wäre. Daher würde sich dieFluoreszenzquantenausbeute erhöhen und die Fluoreszenzintensität pro Chlorophyll-a-Konzentration zunehmen. Dieses ist ein bekanntes Phänomen. Da die Fluoreszenzquanten-ausbeute durch eine Messung der maximalen Fluoreszenz (fm) bestimmbar ist (Genty et al.1989; Moldaenke et al. 1995), wäre die Erhöhung der Fluoreszenzquantenausbeute und damitverursachte Fehler bei der Chlorophyll-a-Gehaltsbestimmung korrigierbar. Dies ist für einNachfolgegerät geplant.
7.2.2 Fluoreszenzverhalten der Blaualgen während der Alterung
Der Verlauf der Chlorophyll-a-Konzentration in den alternden Blaualgenkulturen zeigte eineähnliche Form wie bei den Grünalgen, wenngleich ein niedrigeres Maximum vorhanden warund der Abfall nach Überschreiten des Chlorophyll-a-Gehalt-Maximums nicht ganz so steil

7 Test des automatisierten Fluorometers
73
verläuft. Bei einem Vergleich der durch die HPLC-Analyse ermittelten Chlorophyll-a-Konzentrationen mit den aus der Fluoreszenz berechneten Chlorophyll-a-Konzentrationen(Abb. 7.11), zeigt sich, daß der Chlorophyll-a-Gehalt zum Ende der Messung hin unterschätztwurde.
0
100
200
300
400
500
600
700
800
900
1000
0 5 10 15 20 25 30 35 40
Alter der Kultur/ Tage
Chlorophyll-a- Konzentration(HPLC)
Chlorophyll-a- Konzentration(Fluoreszenz)
Abb. 7.11: Verlauf der aus der Fluoreszenz berechneten Chlorophyll-a-Konzentrationen über die 36-Tage-Meßzeit, im Vergleich dazu die durch die HPLC-Analyse ermittelten Chlorophyll-a-Konzentrationen.
Die auftretenden Fehler bei der Chlorophyll-a-Gehaltsbestimmung durch die Fluoreszenz las-sen sich wie bei den Grünalgen durch drei Faktoren erklären, die bei einer Alterung auftretenkönnen:Das Auftreten von Chlorophyll-a-Abbauprodukten, die Veränderung der äußeren Antennenund eine Variabilität in der photosynthetischen Aktivität der Photosysteme. Doch wieder wa-ren Abbauprodukte des Chlorophylls nicht nachweisbar.Als nächster Punkt wurde wieder eine Veränderung in den äußeren Antennen der Blaualgen,den Phycobilisomen, in Erwägung gezogen. Obwohl die Phycobiliproteine der Phycobiliso-men in der HPLC-Analyse nicht nachweisbar sind, müßte sich eine Veränderung der äußerenAntennen, bzw. speziell des Phycocyanin-zu-Chlorophyll-a-Verhältnisses, in einer Verände-rung des Verhältnisses der Fluoreszenzantworten von 450 nm und 610 nm Anregungswellen-
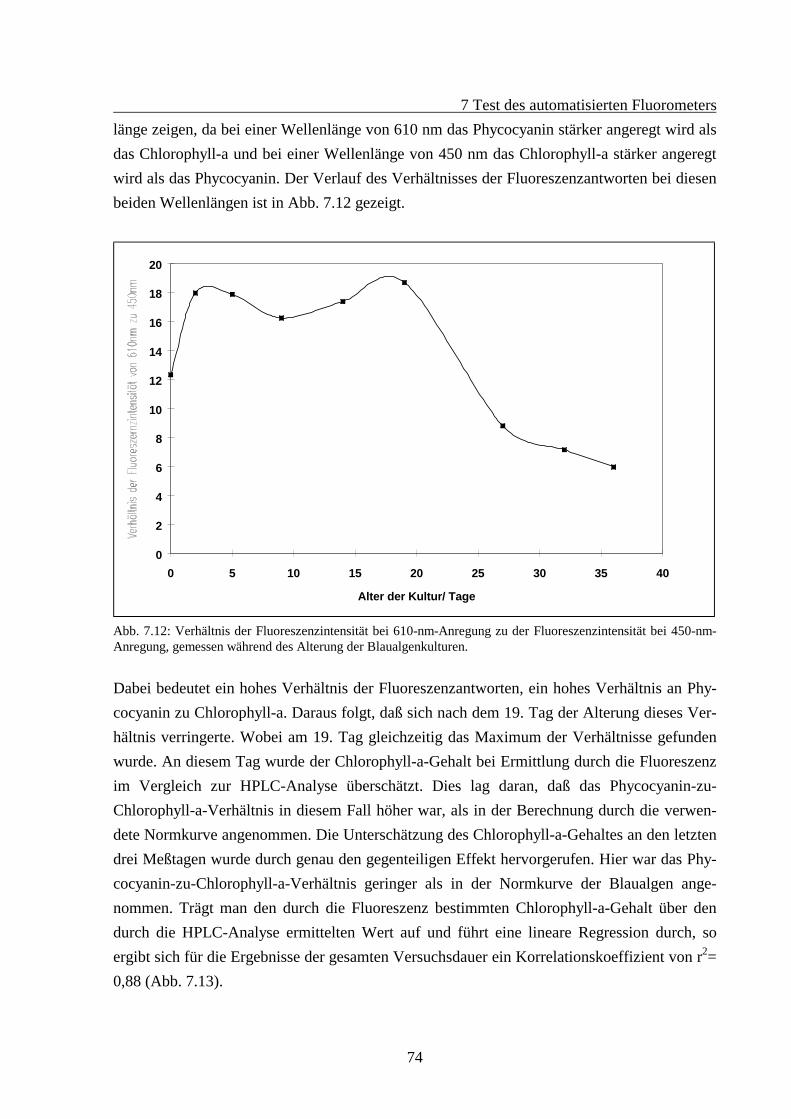
7 Test des automatisierten Fluorometers
74
länge zeigen, da bei einer Wellenlänge von 610 nm das Phycocyanin stärker angeregt wird alsdas Chlorophyll-a und bei einer Wellenlänge von 450 nm das Chlorophyll-a stärker angeregtwird als das Phycocyanin. Der Verlauf des Verhältnisses der Fluoreszenzantworten bei diesenbeiden Wellenlängen ist in Abb. 7.12 gezeigt.
0
2
4
6
8
10
12
14
16
18
20
0 5 10 15 20 25 30 35 40
Alter der Kultur/ Tage
Abb. 7.12: Verhältnis der Fluoreszenzintensität bei 610-nm-Anregung zu der Fluoreszenzintensität bei 450-nm-Anregung, gemessen während des Alterung der Blaualgenkulturen.
Dabei bedeutet ein hohes Verhältnis der Fluoreszenzantworten, ein hohes Verhältnis an Phy-cocyanin zu Chlorophyll-a. Daraus folgt, daß sich nach dem 19. Tag der Alterung dieses Ver-hältnis verringerte. Wobei am 19. Tag gleichzeitig das Maximum der Verhältnisse gefundenwurde. An diesem Tag wurde der Chlorophyll-a-Gehalt bei Ermittlung durch die Fluoreszenzim Vergleich zur HPLC-Analyse überschätzt. Dies lag daran, daß das Phycocyanin-zu-Chlorophyll-a-Verhältnis in diesem Fall höher war, als in der Berechnung durch die verwen-dete Normkurve angenommen. Die Unterschätzung des Chlorophyll-a-Gehaltes an den letztendrei Meßtagen wurde durch genau den gegenteiligen Effekt hervorgerufen. Hier war das Phy-cocyanin-zu-Chlorophyll-a-Verhältnis geringer als in der Normkurve der Blaualgen ange-nommen. Trägt man den durch die Fluoreszenz bestimmten Chlorophyll-a-Gehalt über dendurch die HPLC-Analyse ermittelten Wert auf und führt eine lineare Regression durch, soergibt sich für die Ergebnisse der gesamten Versuchsdauer ein Korrelationskoeffizient von r2=0,88 (Abb. 7.13).

7 Test des automatisierten Fluorometers
75
R2 = 0,8762
0
100
200
300
400
500
600
700
800
900
1000
0 100 200 300 400 500 600 700 800 900 1000Chlorophyll-a-Konzentration (HPLC)/ µg/l
Abb. 7.13: Blaualgenalterung; Chlorophyll-a-Gehalte, ermittelt aus den Fluoreszenzanregungsspektren an denneun Meßtagen, aufgetragen über die Chlorophyll-a-Gehalte aus der HPLC-Analyse.
Die Veränderungen in den Pigmenten der äußeren Antennen der reinen Blaualgenkultur wäh-rend des Alterungsversuches spiegelten sich auch in der Zuordnung zu den einzelnen spektra-len Gruppen aufgrund der Fluoreszenzanregungsspektren der Proben wider (Abb. 7.14). Da-nach wurden an den letzten drei Meßtagen die Chlorophyll-a-Konzentrationen auch denGrünalgen zugeordnet. Dabei war die höchste Fehleinschätzung ein 15-prozentiger Grünal-genanteil am letzten Meßtag.Da am 19. Tag das Phycocyanin-zu-Chlorophyll-a-Verhältnis am höchsten war, läßt sich dieÜberschätzung des Chlorophyll-a-Gehaltes an diesem Tag erklären. Bei der Berechnung derspektralen Algengruppen dieses Tages erfolgt eine Zuordnung des Chlorophyll-a-Gehaltes nurzu den Blaualgen.An Tagen, an denen das Chlorophyll-a-zu-Phycocyanin-Verhältnis niedriger war, als in derNormkurve der Blaualgen angenommen, folgte, daß bei der Zuordnung zu den AlgengruppenGrünalgen mitberechnet wurden. So wird klar, daß bei einem zu großen Phycocyanin-zu-Chlorophyll-a-Verhältnis die Blaualgen in einer Monokultur zwar korrekt bestimmt wurden,aber bei Zugabe von Grünalgen zu dieser Kultur wäre die Chlorophyll-a-Konzentration derGrünalgen unterschätzt worden.
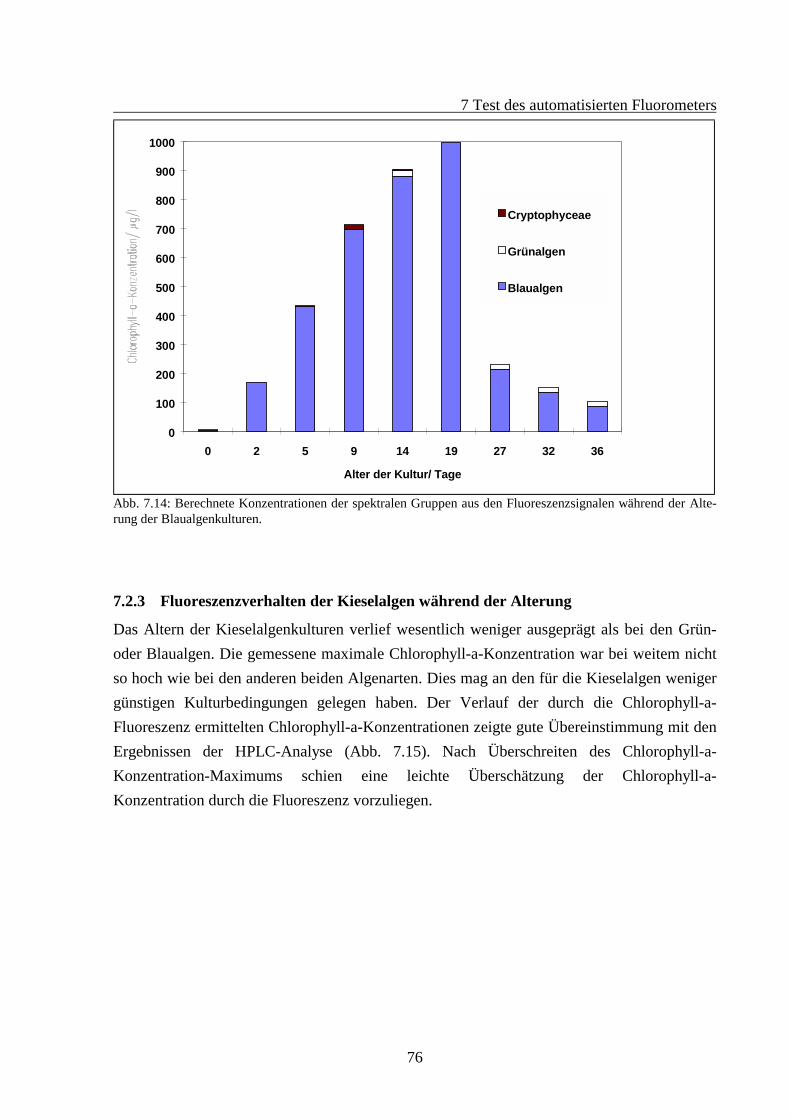
7 Test des automatisierten Fluorometers
76
0
100
200
300
400
500
600
700
800
900
1000
0 2 5 9 14 19 27 32 36
Alter der Kultur/ Tage
Cryptophyceae
Grünalgen
Blaualgen
Abb. 7.14: Berechnete Konzentrationen der spektralen Gruppen aus den Fluoreszenzsignalen während der Alte-rung der Blaualgenkulturen.
7.2.3 Fluoreszenzverhalten der Kieselalgen während der Alterung
Das Altern der Kieselalgenkulturen verlief wesentlich weniger ausgeprägt als bei den Grün-oder Blaualgen. Die gemessene maximale Chlorophyll-a-Konzentration war bei weitem nichtso hoch wie bei den anderen beiden Algenarten. Dies mag an den für die Kieselalgen wenigergünstigen Kulturbedingungen gelegen haben. Der Verlauf der durch die Chlorophyll-a-Fluoreszenz ermittelten Chlorophyll-a-Konzentrationen zeigte gute Übereinstimmung mit denErgebnissen der HPLC-Analyse (Abb. 7.15). Nach Überschreiten des Chlorophyll-a-Konzentration-Maximums schien eine leichte Überschätzung der Chlorophyll-a-Konzentration durch die Fluoreszenz vorzuliegen.
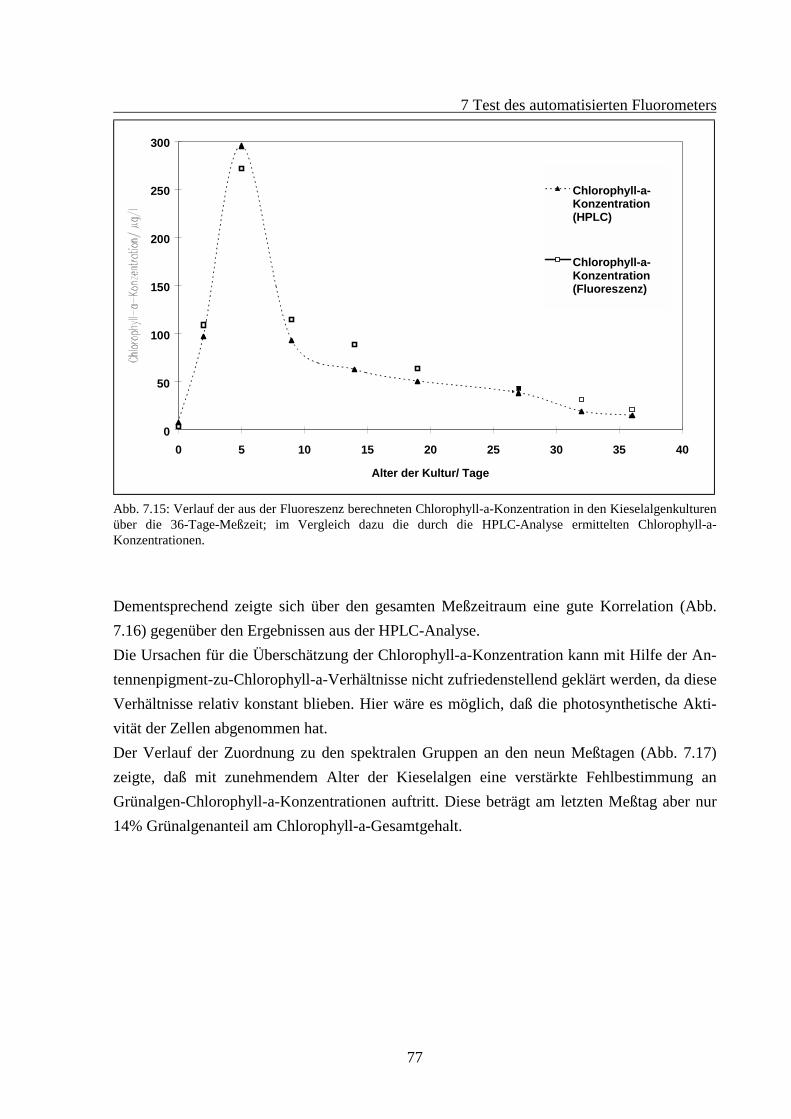
7 Test des automatisierten Fluorometers
77
0
50
100
150
200
250
300
0 5 10 15 20 25 30 35 40
Alter der Kultur/ Tage
Chlorophyll-a- Konzentration (HPLC)
Chlorophyll-a- Konzentration(Fluoreszenz)
Abb. 7.15: Verlauf der aus der Fluoreszenz berechneten Chlorophyll-a-Konzentration in den Kieselalgenkulturenüber die 36-Tage-Meßzeit; im Vergleich dazu die durch die HPLC-Analyse ermittelten Chlorophyll-a-Konzentrationen.
Dementsprechend zeigte sich über den gesamten Meßzeitraum eine gute Korrelation (Abb.7.16) gegenüber den Ergebnissen aus der HPLC-Analyse.Die Ursachen für die Überschätzung der Chlorophyll-a-Konzentration kann mit Hilfe der An-tennenpigment-zu-Chlorophyll-a-Verhältnisse nicht zufriedenstellend geklärt werden, da dieseVerhältnisse relativ konstant blieben. Hier wäre es möglich, daß die photosynthetische Akti-vität der Zellen abgenommen hat.Der Verlauf der Zuordnung zu den spektralen Gruppen an den neun Meßtagen (Abb. 7.17)zeigte, daß mit zunehmendem Alter der Kieselalgen eine verstärkte Fehlbestimmung anGrünalgen-Chlorophyll-a-Konzentrationen auftritt. Diese beträgt am letzten Meßtag aber nur14% Grünalgenanteil am Chlorophyll-a-Gesamtgehalt.
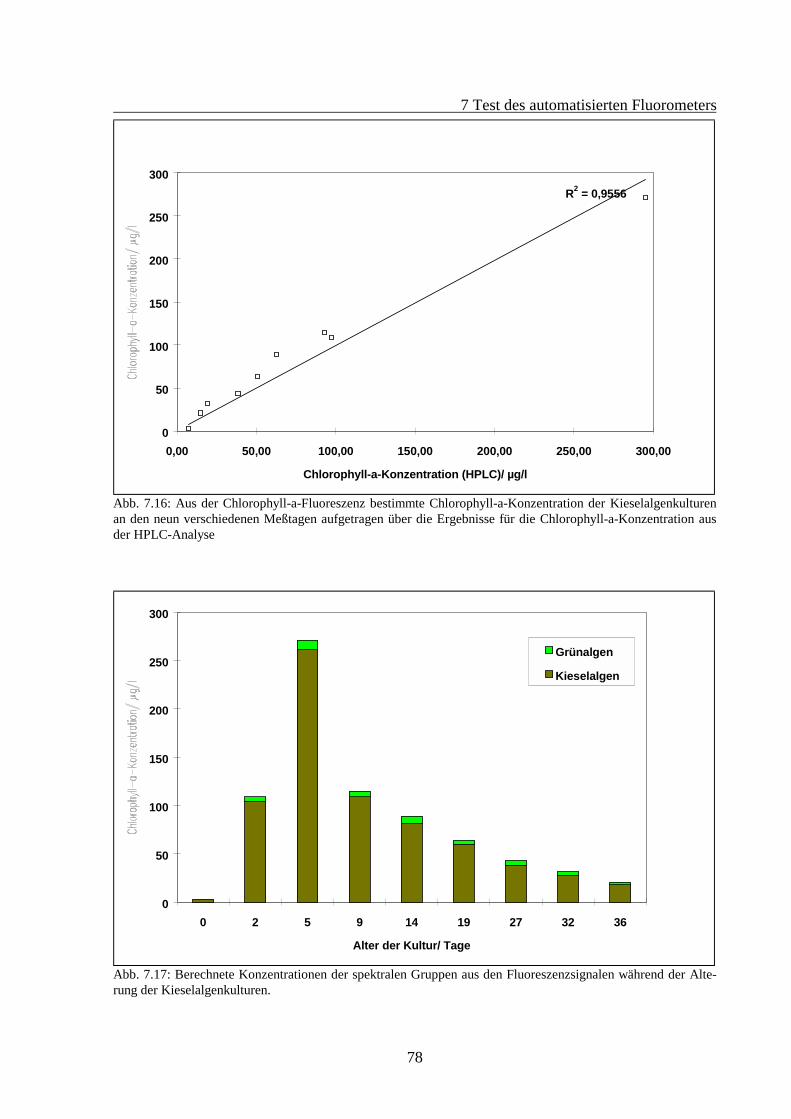
7 Test des automatisierten Fluorometers
78
R2 = 0,9556
0
50
100
150
200
250
300
0,00 50,00 100,00 150,00 200,00 250,00 300,00
Chlorophyll-a-Konzentration (HPLC)/ µg/l
Abb. 7.16: Aus der Chlorophyll-a-Fluoreszenz bestimmte Chlorophyll-a-Konzentration der Kieselalgenkulturenan den neun verschiedenen Meßtagen aufgetragen über die Ergebnisse für die Chlorophyll-a-Konzentration ausder HPLC-Analyse
0
50
100
150
200
250
300
0 2 5 9 14 19 27 32 36
Alter der Kultur/ Tage
Grünalgen
Kieselalgen
Abb. 7.17: Berechnete Konzentrationen der spektralen Gruppen aus den Fluoreszenzsignalen während der Alte-rung der Kieselalgenkulturen.

7 Test des automatisierten Fluorometers
79
7.3 Messung an natürlichen Algenpopulationen der Schlei
Zum Abschluß des experimentellen Teils dieser Arbeit wurden Fluoreszenzanregungsspektrenvon Proben aus natürlichen Gewässern aufgenommen, daraus mit dem Normkurvenfitverfah-ren der Chlorophyll-a-Gehalt berechnet und die Zuordnung zu den spektralen Algengruppengetroffen.Dazu wurden während einer Ausfahrt im März ‘98 mit dem Forschungsschiff „Littorina“ aufder Schlei fünf Proben von verschiedenen Orten (in der Sprache der Meereskundler „Statio-nen“; Schleimünde, Bienebek, Ulsnis, Große Breite und Fahrdorf, siehe Abb. 7.18) untersuchtund mit einer HPLC-Analyse verglichen.
Abb. 7.18: Lageorte der fünf „Stationen“ auf der Schlei
Dabei wurden neben dem Chlorophyll-a-Gehalt auch die Mengen der Carotine und Xantho-phylle durch die HPLC-Analyse (Meyerhöfer 1994) bestimmt, um einen Hinweis auf die Zu-sammensetzung der Algenpopulationen zu erhalten. Die Proben wurden bei Schleimünde auseiner Wassertiefe von vier Metern und in der Schlei aus einer Tiefe von einem Meter ent-nommen. Abb. 7.19 zeigt den Chlorophyll-a-Gehalt an den unterschiedlichen Meßstationen,ermittelt durch das Fluoreszenzanregungsspektrum und durch die HPLC-Analyse.
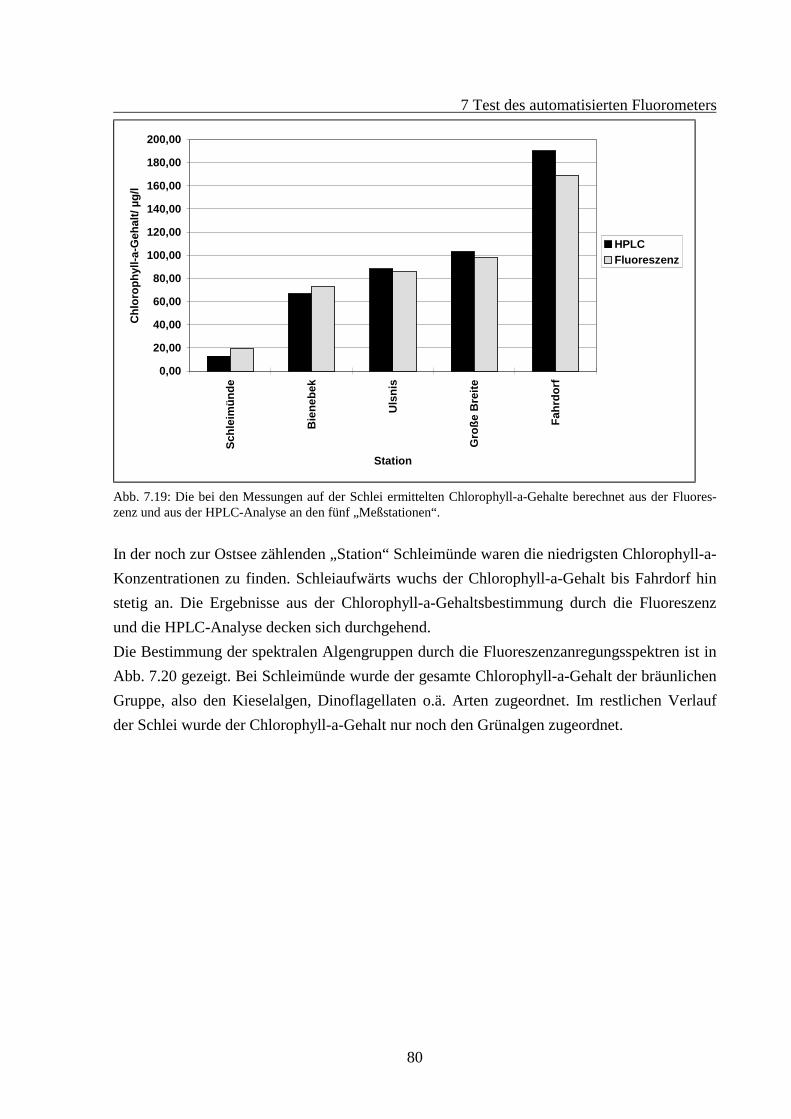
7 Test des automatisierten Fluorometers
80
0,00
20,00
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
200,00
Schl
eim
ünde
Bie
nebe
k
Uls
nis
Gro
ße B
reite
Fahr
dorf
Station
Chl
orop
hyll-
a-G
ehal
t/ µg
/l
HPLCFluoreszenz
Abb. 7.19: Die bei den Messungen auf der Schlei ermittelten Chlorophyll-a-Gehalte berechnet aus der Fluores-zenz und aus der HPLC-Analyse an den fünf „Meßstationen“.
In der noch zur Ostsee zählenden „Station“ Schleimünde waren die niedrigsten Chlorophyll-a-Konzentrationen zu finden. Schleiaufwärts wuchs der Chlorophyll-a-Gehalt bis Fahrdorf hinstetig an. Die Ergebnisse aus der Chlorophyll-a-Gehaltsbestimmung durch die Fluoreszenzund die HPLC-Analyse decken sich durchgehend.Die Bestimmung der spektralen Algengruppen durch die Fluoreszenzanregungsspektren ist inAbb. 7.20 gezeigt. Bei Schleimünde wurde der gesamte Chlorophyll-a-Gehalt der bräunlichenGruppe, also den Kieselalgen, Dinoflagellaten o.ä. Arten zugeordnet. Im restlichen Verlaufder Schlei wurde der Chlorophyll-a-Gehalt nur noch den Grünalgen zugeordnet.

7 Test des automatisierten Fluorometers
81
������������������������������0
20
40
60
80
100
120
140
160
180
Schl
eim
ünde
Bie
nebe
k
Uls
nis
Gro
ße B
reite
Fahr
dorf
Station
Chl
orop
hyll-
a-G
ehal
t
Grünalgen�������� Bräunliche
GruppeCryptophyceae
Abb. 7.20: Chlorophyll-a-Gehalte der vier spektralen Gruppen ermittelt aus den Fluoreszenzanregungsspektrender Proben von den fünf „Stationen“ in µg Chl./l
Die Auswertungsergebnisse der bei der HPLC-Analyse gemessenen Photosynthesepigmente(Abb. 7.21) zeigten Übereinstimmung zu den Ergebnissen aus der Fluoreszenz. Bei Schlei-münde war relativ zum Chlorophyll-a gesehen hauptsächlich Fucoxanthin, Chlorophyll-c1, -c2, und -c3 zu finden. Dieses sind die akzessorischen Pigmente der bräunlichen Gruppe. ImVerlauf der Schlei war ein hoher Gehalt an Chlorophyll-b und Lutein feststellbar, welcher einHinweis auf Grünalgen ist. Spezielle charakteristische Markercarotinoide der Blaualgen warennicht zu beobachten. Das gefundene Alloxanthin läßt auf einige Cryptophyceen schließen, derAnteil am Chlorophyll-a-Gesamtgehalt dürfte jedoch an allen Stationen unter 3% liegen.Damit ist nicht nur die richtige Bestimmung des Gesamtchlorophyllgehalts durch das Fluoro-meter belegt, sondern auch die Aufschlüsselung auf die Pigmentgruppen durch die HPLC-Messung bestätigt.
7 Test des automatisierten Fluorometers
82
����������������������
��������������
����������
��������������
��������������
���������������������������������������������
�����������������������������������
������������������������������
����������������������������������������������������������������
������������������������������������������
��������������������������������������������������������
������������������������������������������
��������������������������������������������������������
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Schleimünde Bienebek Ulsnis Große Breite Fahrdorf
Pigm
ent-z
u-C
hlor
ophy
ll-a-
Verh
ältn
is
Chlorophyll c3
Chlorophyll c1+2
Fucoxanthin�����
Alloxanthin�����
Lutein����������Chlorophyll-b
Abb. 7.21: In der HPLC-Analyse bestimmte Pigment-zu-Chlorophyll-a-Verhältnisse an den fünf Meßstationen.

8 Diskussion und Ausblick
83
8 Diskussion und Ausblick
In dieser Arbeit zeigte sich, daß die Verwendung von nur wenigen, auf die Absorptionsmaxi-ma der verschiedenen Antennenpigmente der Algengruppen abgestimmten Anregungswel-lenlängen, eine sehr schnelle Auswertung der aufgenommenen Anregungsspektren mit nurwenig Rechenleistung ermöglicht. Dadurch wurde die Berechnung des Chlorophyll-a-Gehaltes einer Wasserprobe und die Berechnung der Anteile von vier spektralen Algengrup-pen daran möglich.Durch die Berücksichtigung der natürlichen Schwankungsbreite der Anregungsspektren beider Ausführung des Rechenalgorithmus erfolgt eine genauere Einschätzung der gesuchtenParameter. Die üblichen Chlorophyll-a-Fluoreszenz-Meßverfahren, die mit monochromati-scher Anregung arbeiten, können eine Fehleinschätzung des zu bestimmenden Chlorophyll-a-Gehaltes einer Wasserprobe liefern. Dies wird durch die unterschiedlichen Anregungsspektrender spektralen Algengruppen verursacht. Durch die multispektrale Anregung werden Algen-gruppen richtig erkannt und die Fehleinschätzung damit beseitigt.Im wesentlichen hat das Verfahren sehr gute Ergebnisse (10% Fehlbestimmung) geliefert. Daseinzige nennenswerte Problem stellen die in Abb. 7.7 und Abb. 7.11 auftretenden Fehler inder Chlorophyll-a-Gehaltsbestimmung bei den gealterten Grün- und Blaualgenkulturen dar.Dies wurde hauptsächlich durch die Veränderung in der Zusammensetzung der Antennenpig-mente, aber vermutlich auch durch eine Verminderung der photosynthetischen Aktivität derabsterbenden Algenzellen verursacht. Dadurch war die Fluoreszenzintensität pro Chlorophyll-a-Gehalt eine andere als in den Normkurven angenommen, was dann letztendlich zu den Feh-lern führte. Da dies Effekte waren, die in Laborkulturen durch hohe Nährstoffkonzentrationenentstanden sind (Chlorophyllkonzentrationen bis zu 1500 µg/l), wird man diese Effekte ver-mutlich in der Natur nur seltener und in geringerem Ausmaße wahrnehmen können. Dennochsollte bei Messungen in stark eutrophierten Gewässern bei beginnendem Altern einer Algen-blüte die Möglichkeit der Messung eines zu hohen Chlorophyllgehaltes in Betracht gezogenwerden. Bei Einsetzen der Algenblüte sind jedoch noch keine Fehlbestimmungen zu erwarten.Weiterhin ist in nicht-eutrophierten Gewässern mit diesen Effekten nicht zu rechnen.Es besteht aber auch die Möglichkeit, bei einer Weiterentwicklung des Verfahrens die im Zu-sammenhang mit der Grünalgenalterung auftretende Veränderung des Chlorophyll-a-zu-Chlorophyll-b-Verhältnisses zu berücksichtigen. Würde dieses Verhältnis bekannt sein, sokönnte die korrekte Normkurve für eine Grünalge mit diesem speziellen Chlorophyll-a-zu-Chlorophyll-b-Verhältnis errechnet werden. Dies wäre durch die Ergänzung um eine weitere

8 Diskussion und Ausblick
84
für Chlorophyll-b spezifischere Anregungswellenlänge durchführbar, so daß man bereits ausdem Anregungsspektrum einer unbekannten Probe dieses Verhältnis ermitteln könnte. Gün-stig wäre hier eine Anregungswellenlänge von 470 nm - 490 nm. In diesem Bereich sind aberim Moment noch keine lichtstarken Leuchtdioden erhältlich. Eine weitere Möglichkeit ist eineAnregung bei 650 nm. Aufgrund der Breitbandigkeit von Leuchtdioden würde die notwendigestarke Filterung zur Verhinderung eines Einfalls in den Detektorfilter die Intensität zu starkschwächen.Bei der Alterung der Blaualgenkulturen haben sich Schwankungen im Phycocyanin-zu-Chlorophyll-a-Verhältnis ergeben. Das Phycocyanin ist in der äußeren Antenne der Blaualgen,den Phycobilisomen, eingebunden. Diese können Fluoreszenzlicht bei etwa 650 nm emittie-ren. In einem Emissionsspektrum von Blaualgen ist deutlich eine Schulter bei 650 nm zu se-hen (Abb. 8.1). Würde nun mit einem geeigneten Aufbau die Fluoreszenzemission bei 650 nmgemessen werden, etwa mit einem zweiten Detektor und entsprechenden Filtern, so könnteunter Zuhilfenahme des Anregungsspektrums der Phycocyaningehalt und der Gehalt der ande-ren Phycobiliproteine bestimmt werden, so daß auch hier für verschiedene Phycocyanin-zu-Chlorophyll-a-Verhältnisse unterschiedliche Normkurven errechnet werden könnten.
0
0,2
0,4
0,6
0,8
1
1,2
600 650 700 750 800Emissionswellenlänge/ nm
Rel
. Flu
ores
zenz
inte
nsitä
t
BlaualgenGrünalgen
Abb. 8.1: Fluoreszenzemissionsspektrum einer Blaualgenkultur (Microcystis aeruginosa) und im Vergleich dazudas Emissionsspektrum einer Grünalgenkultur (Chlorella vulgaris)Beide Messungen wurden bei einer Anregungswellenlänge von 500 nm aufgenommen. Bei beiden Algenartenliegt das Maximum bei einer Anregungswellenlänge von 685 nm, verursacht durch das Chlorophyll-a. Zusätzlichentsteht bei den Blaualgen eine Schulter bei 650 nm, erzeugt durch die Fluoreszenz der Phycobilisomen.
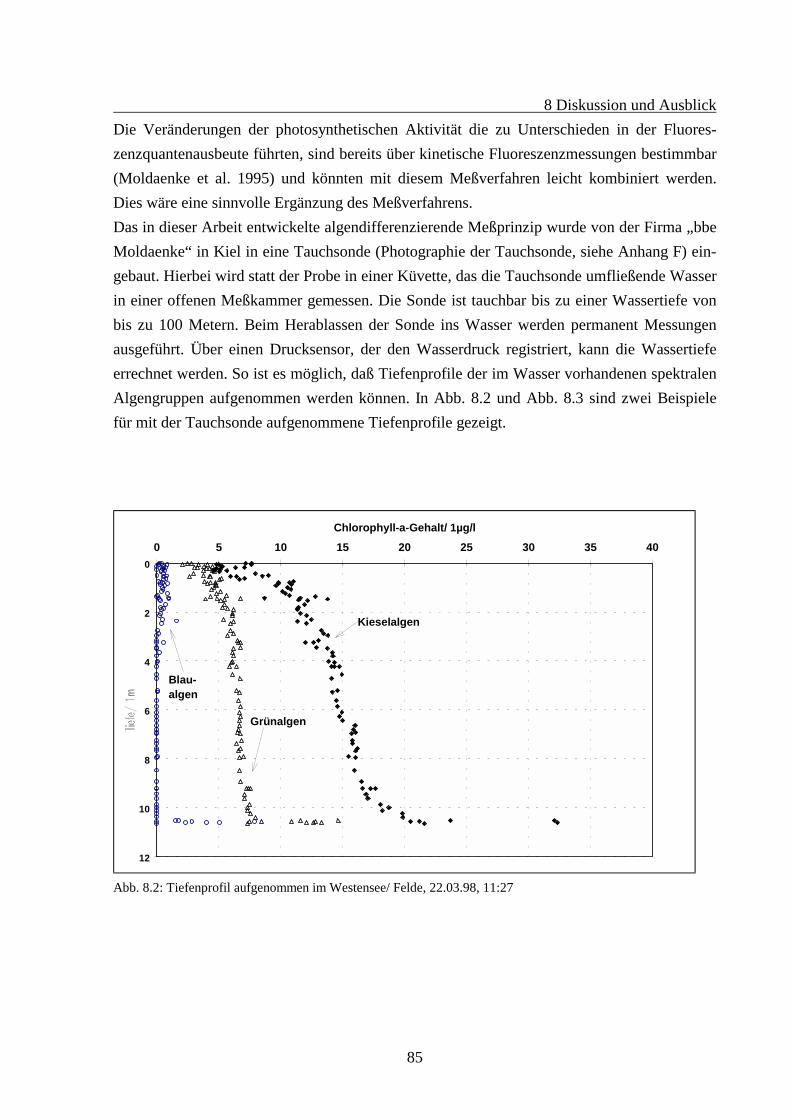
8 Diskussion und Ausblick
85
Die Veränderungen der photosynthetischen Aktivität die zu Unterschieden in der Fluores-zenzquantenausbeute führten, sind bereits über kinetische Fluoreszenzmessungen bestimmbar(Moldaenke et al. 1995) und könnten mit diesem Meßverfahren leicht kombiniert werden.Dies wäre eine sinnvolle Ergänzung des Meßverfahrens.Das in dieser Arbeit entwickelte algendifferenzierende Meßprinzip wurde von der Firma „bbeMoldaenke“ in Kiel in eine Tauchsonde (Photographie der Tauchsonde, siehe Anhang F) ein-gebaut. Hierbei wird statt der Probe in einer Küvette, das die Tauchsonde umfließende Wasserin einer offenen Meßkammer gemessen. Die Sonde ist tauchbar bis zu einer Wassertiefe vonbis zu 100 Metern. Beim Herablassen der Sonde ins Wasser werden permanent Messungenausgeführt. Über einen Drucksensor, der den Wasserdruck registriert, kann die Wassertiefeerrechnet werden. So ist es möglich, daß Tiefenprofile der im Wasser vorhandenen spektralenAlgengruppen aufgenommen werden können. In Abb. 8.2 und Abb. 8.3 sind zwei Beispielefür mit der Tauchsonde aufgenommene Tiefenprofile gezeigt.
0
2
4
6
8
10
12
0 5 10 15 20 25 30 35 40Chlorophyll-a-Gehalt/ 1µg/l
Kieselalgen
Grünalgen
Blau-algen
Abb. 8.2: Tiefenprofil aufgenommen im Westensee/ Felde, 22.03.98, 11:27
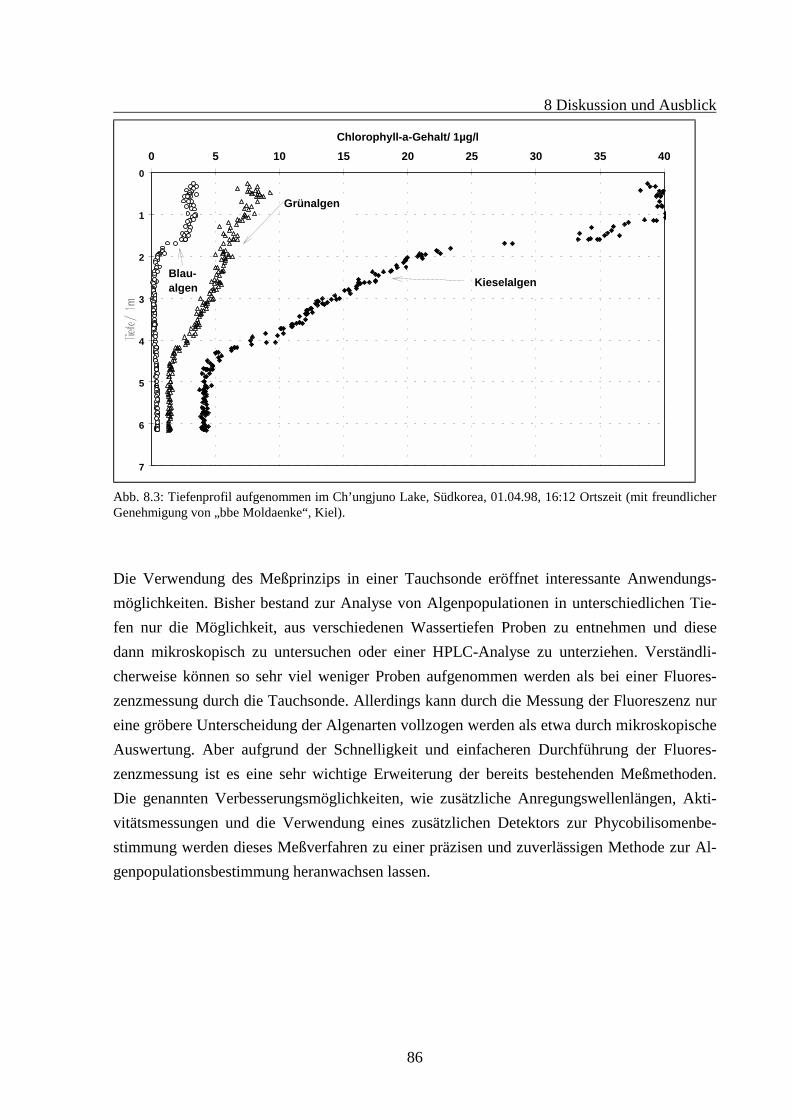
8 Diskussion und Ausblick
86
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30 35 40Chlorophyll-a-Gehalt/ 1µg/l
Kieselalgen
Grünalgen
Blau-algen
Abb. 8.3: Tiefenprofil aufgenommen im Ch’ungjuno Lake, Südkorea, 01.04.98, 16:12 Ortszeit (mit freundlicherGenehmigung von „bbe Moldaenke“, Kiel).
Die Verwendung des Meßprinzips in einer Tauchsonde eröffnet interessante Anwendungs-möglichkeiten. Bisher bestand zur Analyse von Algenpopulationen in unterschiedlichen Tie-fen nur die Möglichkeit, aus verschiedenen Wassertiefen Proben zu entnehmen und diesedann mikroskopisch zu untersuchen oder einer HPLC-Analyse zu unterziehen. Verständli-cherweise können so sehr viel weniger Proben aufgenommen werden als bei einer Fluores-zenzmessung durch die Tauchsonde. Allerdings kann durch die Messung der Fluoreszenz nureine gröbere Unterscheidung der Algenarten vollzogen werden als etwa durch mikroskopischeAuswertung. Aber aufgrund der Schnelligkeit und einfacheren Durchführung der Fluores-zenzmessung ist es eine sehr wichtige Erweiterung der bereits bestehenden Meßmethoden.Die genannten Verbesserungsmöglichkeiten, wie zusätzliche Anregungswellenlängen, Akti-vitätsmessungen und die Verwendung eines zusätzlichen Detektors zur Phycobilisomenbe-stimmung werden dieses Meßverfahren zu einer präzisen und zuverlässigen Methode zur Al-genpopulationsbestimmung heranwachsen lassen.

9 Zusammenfassung
87
9 Zusammenfassung
Ziel der Arbeit war die Konstruktion einer Apparatur, die in der Lage ist, in einer unbehan-delten Wasserprobe den Chlorophyllgehalt und die Zusammensetzung der spektralen Algen-gruppen zu bestimmen. Dazu wurde ein mit diskreten Anregungswellenlängen arbeitendesFluoreszenzspektrometer aufgebaut. Vorversuche zeigten, daß kein kontinuierliches Spektrumausgewertet werden muß, sondern daß die Messung an fünf diskreten Wellenlängen ausreicht.Fünf Leuchtdioden mit Peakwellenlängen von 450 nm, 525 nm, 570 nm, 590 nm und 610nmerzeugten das Licht. In diesen Vorversuchen wurden anhand reiner Kulturen für vier verschie-dene spektrale Algengruppen - Grünalgen, Blaualgen, bräunliche spektrale Gruppe und Cryp-tophyceae - typische „Spektren“ und ihre Streuung bestimmt. Die „Normspektren“ dientendazu, in einem gemessenen Anregungsspektrum die jeweiligen Anteile der vier Algengruppenin einem „Normkurvenfit“ zu bestimmen. Dazu wurde das Gesamtspektrum durch eine ge-wichtete Summe der „Normspektren“ genähert. Ein besonderes Problem war die Einführungvon Gewichtsfaktoren, die die Ungenauigkeit aufgrund der Streuung der „Normspektren“ be-rücksichtigen sollte. Hier war die Schwierigkeit, daß die Streuung der Gesamtkurve wiederumeine gewichtete Summe der Normkurvenstreuung ist. Die Gewichtsfaktoren sind dabei genaudie Koeffizienten, die im Fit bestimmt werden sollen. Damit tauchen die Fitparameter auch im
Nenner der χ2-Funktion auf, und das Problem wird nicht-linear.
Da ein nicht-lineares Fitprogramm, wie z. B der Simplex, zu viel Zeit für eine on-line Analysebenötigt hätte, wurde ein 2-Schritt-Verfahren eingeführt. In einem ersten Schritt wurde eineGleichverteilung aller Algengruppen angenommen, so daß die Streuung die Summe der Ein-zelstreuungen war. Hiermit wurden dann in einem linearen, gewichteten Gauss-Jordan-Fit dieKoeffizienten bestimmt. Diese Koeffizienten dienten dann dazu, die Streuung erneut zu be-rechnen und den Fit mit den korrigierten Streuungen durchzuführen. Es zeigte sich, daß beieinem folgenden Iterationsschritt die Anteile nur noch in der 5. Stelle hinter dem Komma ab-wichen. Deshalb wurde im endgültig implementierten Algorithmus nach dem 2. Schritt abge-brochen. Eine Ansteuerungselektronik ermöglichte die automatische Meßwertaufnahme.Der Test des Gerätes geschah mit Hilfe von Verdünnungsreihen bekannter Algengemischeund in natürlichen Gewässern, wobei die Ergebnisse durch HPLC-Analysen überprüft wurden.In simulierten Algenblüten und bei einer Verdünnungsreihe eines Algengemisches mit gleich-bleibenden Anteilen vier spektraler Algengruppen am Gesamtchlorophyll-a-Gehalt konnte

9 Zusammenfassung
88
nachgewiesen werden, daß es anhand der Fluoreszenzanregungsspektren möglich ist, denChlorophyll-a-Gehalt vier verschiedener spektraler Algengruppen zu berechnen.Durch Alterungsversuche dreier verschiedener Algenarten (Scenedesmus acutus, Synechococ-cus sp. und Cyclotella sp.), wurde gezeigt, daß trotz einhergehender Veränderung der Anten-nensysteme der Algen die richtige Zuordnung zu den spektralen Gruppen aufgrund der Fluo-reszenzanregungsspektren getroffen werden konnte.Die Ergebnisse der Bestimmung von natürlichen Algenpopulationen der Schlei mit der ent-wickelten Methode zeigten Übereinstimmung mit Resultaten aus einer parallel unternomme-nen HPLC-Pigmentanalyse.

10 Literaturverzeichnis
89
10 Literaturverzeichnis
Anderson, J.M., Barrett, J., 1986. Light-harvesting pigment-protein complexes of algae. In:Staehelin, L.A. und Arntzen, C.J. (eds.), Photosynthesis III. Encyclopedia of Plant Physi-ology. New Ser. V. 5-6, 19. Springer-Verlag, Berlin, pp 269 - 285
Baker, N., Webber, A., 1987. Interactions between photosystems. Advances in Botanical Re-search 13: 2 - 56
Bold, H. C., Wynne, J. W., 1985. Introduction to the algae. Prentice-Hall, Inc., New Jersey
Bolhar-Nordenkampf, H.R., Öquist, G., 1993. Chlorophyll fluorescence as a tool in photo-synthesis research. In: Hall, D.O. (ed.), Photosynthesis and Production in a changing En-vironment: Field and Laboratory Manual. Chapman & Hall, London, pp 193- 206
Briantais, J.M., Vernotte, C., Krause, G.H., Weis, E., 1986. Chlorophyll a fluorescence ofhigher plants: chloroplasts and leaves. In: Govindjee, J Amesz, D.C. Fork ( eds), LightEmission by Plants and Bacteria. Academic Press, Orlando, pp 539 - 586
Bronstein, I.N., Semendjajew, K.A., 1991. Taschenbuch der Mathematik, 25.Aufl, B.G.Teubner, Thun, Stuttgart, Leipzig
Buschmann, C., Grumbach, K., 1985. Physiologie der Photosynthese. Springer-Verlag, Berlin
Butler, W.L., 1978. Energy distribution in the photochemical apparatus of photosynthesis.Ann. Rev. Plant Physiol. 29: 345 - 378
Dau, H., 1994. Short-term adaption of plants to changing light intensities and its relation toPhotosystem II photochemistry and fluorescence emission. Journal of Photochem. andPhotobiol. B: Biology 26: 3 - 27
Delepelaire, P., Chua, N.H., 1979. Lithium dodecylsulfate/polyacrylamide gel electrophoresisof thylakoid membranes at 4 °C. Proc Natl Acad Sci USA 76: 111 - 115
Doerffer, R., 1986. The effect of short term illumination changes on the sun light stimulatedfluorescence of phytoplankton. In: The Use of Chlorophyll Fluorescence Measurementsfrom Space for Separating Constitutents of Sea Water. Volume I Summary Report, ESAContract No. RFQ3-5059/84/NL/MD
Emerson, R., 1958. The quantum yield of photosynthesis. Ann. Rev. Plant Physiol. 9: 1 - 24
Gantt E, 1986. Phycobilisomes. In: Staehelin L.A., Arntzen C.J. (eds), Photosynthesis III. En-cyclopedia of Plant Physiology. New Ser., V. 5-6, 19. Springer-Verlag, Berlin, pp 260 -268

10 Literaturverzeichnis
90
Genty, B., Briantais, J.-M., Baker, N., R., 1989. The relationship between the quantum yieldof photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochem.Biophys. Acta 990: 87 - 92
Hill, D.R.A., Rowan, K.S., 1989. The biliproteins of the cryptophyceae. Phycologia 28: 455 -463
Hoek, C.v.d., 1993. Algen. 3. Aufl., Thieme, Stuttgart
Hoek, C.v.d., Mann, D.G., Jahns, H.M., 1995. Algae: An introduction to phycology. Univer-sity Press, Cambridge
Holm-Hansen, O., 1965. Fluorometric determination of chlorophyll. J. Cons. Perm. Int. Ex-plor. Mer, 30: 3 - 15
Jeffrey, S.W., 1997. Application of pigment methods to oceanography. In: Phytoplanktonpigments in oceanography: Guidelines to modern methods. Monographs on Oceano-graphic Methodology. SCOR UNESCO, pp. 127 - 166
Jeffrey, S.W., Humphrey, G.F., 1975. New spectrometric equations for determining chloro-phylls a, b, c1 and c2 in algae, phytoplankton and higher plants. Biochem. Physiol. Planz.B.P.P., 167: 191 - 194
Kautsky, H., Hirsch, A., 1931. Neue Versuche zur Kohlensäureassimilation. Naturwiss. 19:964 -975
Kolbowski, J., Reising, H. Schreiber, U., 1990. Computer controlled pulse modulation systemfor analysis of photoacoustic signals in the time domain. Photosynth. Res. 25: 309 - 316
Krause, H., Dirnhofer, P., Gerhardt, V., 1987. Anwendung der verzögerten Fluoreszenz vonPhytoplankton in der Ökologie - The application of delayed fluorescence of phytoplanktonin ecology. Arch. Hydrobiol. Beih. Ergebn. Limnol. 29: 55 - 62
Lavorel, J., Joliot, P., 1972. A connected model of the photosynthetic unit. Biophys. J. 12: 815- 831
Lawlor, DW, 1990. Photosynthese: Stoffwechsel-Kontrolle-Physiologie. Thieme, Stuttgart
Lincke, M., 1994. Entwicklung und Erprobung eines signalprozessorgesteuerten digitalenKorrelators zur Unterdrückung des Phasenrauschens bei mechanischen Choppern.Diplomarbeit Univ. Kiel.
Mantoura, R.F.C., Barlow, R.G., Head, J. H., 1997. Simple isocratic methods for chlorophyllsand their degradation products. In: Phytoplankton pigments in oceanography: Guidelinesto modern methods. Monographs on Oceanographic Methodology. SCOR UNESCO, pp.383 - 428

10 Literaturverzeichnis
91
Meyerhöfer, M., 1994. Plankton-Pigmente und deren Abbauprodukte als Biomarker zur Be-schreibung und Abschätzung der Phytoplankton-Sukzession und Sedimentation im Nord-atlantik. Berichte aus dem IfM, Kiel, Nr. 251
Moldaenke, C., Vanselow, K.H., Hansen, U.-P., 1995. The 1-Hz fluorometer: A new approachto fast and sensitive long-term studies of active chlorophyll and environmental influences.Helgol. Meeresunters. 49: 785 - 796
Nusch, E.A., 1980. Comparison of different methods for chlorophyll and phaeopigment de-termination. Arch. Hydrobiol. Beih. Ergebn. Limnol.14: 14 - 36
Okaichi, T., Anderson, D.M., Nemoto, T. (eds.), 1989, Red Tides, Biology, EnvironmentalScience and Toxicology. Elsevier, Amsterdam
Press W.H., 1989. Numerical Recipes in Pascal. University Press, Cambridge
Renger, G., Dohnt, G., Kayed, A., Voss, M., Gräber, P., 1987. Die Chlorophyll-a-Fluoreszenzals Indikator des Funktionszustandes des Photosyntheseapparats in Pflanzen. TheoretischeGrundlagen und praktische Anwendungen [The fluorescence of chlorophyll a - an indica-tor of the functional state of the photosynthetic apparatus in plants.]. Arch. Hydrobiol.Beih. Ergebn. Limnol. 29: 1 - 24
Richter,G., 1988. Stoffwechselphysiology der Pflanzen. 5. Aufl., Thieme, Stuttgart,
Schimek, C., 1994. Der Photosyntheseapparat der Cryptophyceen. Diss. Univ. Marburg
Schlösser, U. G., 1994. SAG - Sammlung von Algenkulturen at the University of Göttingen -Catalogue of Strains. Bot. Acta 107: 113 - 186
Schmidt, W., 1993. Optische Spektroskopie: Eine Einführung für Naturwissenschaftler undTechniker. Verlag Chemie, Weinheim; New York
Schreiber, U., Klughammer, C., Neubauer C., 1988. Measuring P700 absorbance changesaround 830nm with a new type of pulse modulation system. Z. Naturforsch. 43c: 686 -698
Schreiber, U., 1994. New emitter-detector-cuvette assembly for measuring modulated chloro-phyll fluorescence of high diluted suspensions in conjunction with the standard PAMfluorometer. Z. Naturforsch. 49c: 646 - 656
Strasburger, E., 1991. Lehrbuch der Botanik, 33.Aufl. Gustav-Fischer-Verlag, Stuttgart
Tabrizi, H., 1997. Komponentenzerlegung photoakustischer Impulsantworten aus dem Photo-syntheseapparat sowie ihr Einsatz bei der Untersuchung des transthylakoiden Protonen-flusses und beim Nachweis der Mehlerreaktion. Diss. Univ. Kiel
Tabrizi, H., Schinner, K., Spors, J. and Hansen, U.P., 1998. Deconvolution of the three com-ponents of the photoacoustic signal by curve fitting and the relationship of CO2 uptake toproton fluxes. Photosynth. Res. (in press)

10 Literaturverzeichnis
92
UNESCO, 1966. Determination of photosynthetic pigments. In: Seawater.-Rep.SCOR/UNESCO Working Group 17. Monographs on Oceanographic Methology.UNESCO, Paris, pp. 1 - 69.
Willkomm, R., 1987. Rausch- und Impedanzmessungen zur Untersuchung von Kontrollme-chanismen des Membrantransportes von Characeen. Diss. Univ. Kiel
Wiltshire, K.H., Schroeder, F., 1994. Pigment patterns in suspended matter from the Elbe es-tuary, Northern Germany. Netherlands Journal of Aquatic Ecology 28 (3 - 4), 255 - 265
Witt, H.T., 1979. Energy conversion in the functional membrane photosynthesis. Analysis bylight pulse and electric pulse methods. Biochim. Biophys. Acta 104: 355 - 427
Yentsch, C.S., Menzel, D.W., 1963. A method for the determination of phytoplankton chloro-phyll by fluorescence. Deep Sea Res. 10: 1221 - 1231
Yentsch, C.S., Phinney, D.A., 1985. Spectral fluorescence: A taxonomic tool for studying thestructure of phytoplankton populations. J. Plankt. Res.7, 617 - 632
Yentsch, C.S., Yentsch, C.M., 1979. Fluorescence spectral signatures: The characterization ofphytoplankton populations by the use of excitation and emission spectra. J. Mar. Res. 37:471 - 483