
Feeding ecology of the adult Himalayan Salamander Tylototriton
11
HERPETOZOA 9 (1/2): 19 - 29 Wien, 30. Juni 1996 Feeding ecology of the adult Himalayan Salamander Tylototriton verrucosus ANDERSON, 1871 (Caudata: Salamandridae) Nahrungsökologie des adulteri Krokodilmolches Tylototriton verrucosus ANDERSON, 1871 (Caudata: Salamandridae) Rrrwnc DASGUPTA KURZFASSUNG In der Regenzeit (Juni bis September), in der Tylototriton verrucosus ANDERSON, 1871 überwiegend aquatisch lebt und zur Fortpflanzung schreitet, bestand die Nahrung adulter Salamander in Darjeeling (Indien) in beiden Geschlechtern sowohl aus aquatischen als auch terrestrischen Nahrungselementen. Da die Männchen bereits im August und September vermehrt wieder das Land aufsuchen, wiesen sie im Vergleich zu den Weib- chen, die länger im Wasser bleiben, eine mehr aus terrestrischen Objekten zusammengesetzte Nahrung auf. Die Nahrung adulter Salamander bestand überwiegend aus unspezialisierten Elementen (Lumbriciden, Dipte- ren-Larven, Coleopteren-Larven, Lepidopteren-Larven, Odonaten-Larven, adulte Dytisciden, Rhacophoriden- Larven, konspezifische Gelege). Daneben fanden sich spezialisierte Nahrungsobjekte (Rhacophoriden-Schaum- nester, Dipteren-Puppen und Lepidopteren-Puppen) sowie sich rasch bewegende terrestrische Invertebraten. Magenfüllungs-Indices, Nahrungsnischenbreite und -Überlappung zwischen den Geschlechtern waren am höchsten während der stärksten Regenperioden, der Zeit größter Primärproduktion und größten Beutedruckes. ABSTRACT In Darjeeling (India), Tylototriton verrucosus ANDERSON, 1871 lives predominantly aquatic in the mon- soon months June through September. During this time of reproduction, the diet of adult male and female salamanders was found to comprise both aquatic and terrestrial elements. Males started to become terrestrial already in August/September and therefore had a more terrestrial diet than females which stayed longer in the water. Adult diet comprised mainly Lumbricidae, larval Diptera, larval Coleoptera, larval Lepidoptera, larval Odonata, adult dytiscid beetles, larval Rhacophoridae and cannibalized eggs while foam nests of Rhacopho- ridae, Diptera pupae, Lepidoptera pupae and fast moving terrestrial invertebrates were rare. Digestive tract fill-indices, food niche breadth and niche overlap between sexes, were highest during periods of high rainfall and high primary productivity in the ponds. KEY WORDS Tylototriton verrucosus; feeding ecology, behaviour; India, Himalayas INTRODUCTION Knowledge on feeding ecology of the sent in the diet of larvae of stage IV and Himalayan Salamander, Tylototriton verru- V, although they engorged themselves with cosus ANDERSON, 1871 is fragmentary. aquatic insects and attacked syntopic crabs DASGUPTA (1983) kept adult Salamanders (JPotamon potamiscus sikkimense). Adult alive in aquaria by feeding them with diet was found to be highly variable depen- earthworms and noted (1988) that larval ding on the study site. During reproductive stages of this salamander fed upon micro- peaks, adults preyed on a mixed diet of crustaceans, chironomids, mosquito larvae, terrestrial and aquatic components, and various benthic prey while post- The results of further work are pre- metamorphic juveniles preyed on small sented here in view of the paucity of de- bivalves (Sphaerium indicum). According tailed knowledge on food and feeding hab- to KUZMIN & al. (1994), transition from its of T. verrucosus, and lack of informa- feeding on endogenous yolk to exogenous tion on food niche breadth, food electivity diet occurred within stage II (sensu DAS- and digestive tract fill index in relation to GUPTA 1983). Microcrustaceans were ab- abiotic and biotic environmental factors. ©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
Transcript of Feeding ecology of the adult Himalayan Salamander Tylototriton

HERPETOZOA 9 (1/2): 19 - 29Wien, 30. Juni 1996
Feeding ecology of the adult Himalayan SalamanderTylototriton verrucosus ANDERSON, 1871
(Caudata: Salamandridae)
Nahrungsökologie des adulteri KrokodilmolchesTylototriton verrucosus ANDERSON, 1871
(Caudata: Salamandridae)
Rrrwnc DASGUPTA
KURZFASSUNG
In der Regenzeit (Juni bis September), in der Tylototriton verrucosus ANDERSON, 1871 überwiegendaquatisch lebt und zur Fortpflanzung schreitet, bestand die Nahrung adulter Salamander in Darjeeling (Indien)in beiden Geschlechtern sowohl aus aquatischen als auch terrestrischen Nahrungselementen. Da die Männchenbereits im August und September vermehrt wieder das Land aufsuchen, wiesen sie im Vergleich zu den Weib-chen, die länger im Wasser bleiben, eine mehr aus terrestrischen Objekten zusammengesetzte Nahrung auf.Die Nahrung adulter Salamander bestand überwiegend aus unspezialisierten Elementen (Lumbriciden, Dipte-ren-Larven, Coleopteren-Larven, Lepidopteren-Larven, Odonaten-Larven, adulte Dytisciden, Rhacophoriden-Larven, konspezifische Gelege). Daneben fanden sich spezialisierte Nahrungsobjekte (Rhacophoriden-Schaum-nester, Dipteren-Puppen und Lepidopteren-Puppen) sowie sich rasch bewegende terrestrische Invertebraten.
Magenfüllungs-Indices, Nahrungsnischenbreite und -Überlappung zwischen den Geschlechtern waren amhöchsten während der stärksten Regenperioden, der Zeit größter Primärproduktion und größten Beutedruckes.
ABSTRACT
In Darjeeling (India), Tylototriton verrucosus ANDERSON, 1871 lives predominantly aquatic in the mon-soon months June through September. During this time of reproduction, the diet of adult male and femalesalamanders was found to comprise both aquatic and terrestrial elements. Males started to become terrestrialalready in August/September and therefore had a more terrestrial diet than females which stayed longer in thewater. Adult diet comprised mainly Lumbricidae, larval Diptera, larval Coleoptera, larval Lepidoptera, larvalOdonata, adult dytiscid beetles, larval Rhacophoridae and cannibalized eggs while foam nests of Rhacopho-ridae, Diptera pupae, Lepidoptera pupae and fast moving terrestrial invertebrates were rare.
Digestive tract fill-indices, food niche breadth and niche overlap between sexes, were highest duringperiods of high rainfall and high primary productivity in the ponds.
KEY WORDS
Tylototriton verrucosus; feeding ecology, behaviour; India, Himalayas
INTRODUCTION
Knowledge on feeding ecology of the sent in the diet of larvae of stage IV andHimalayan Salamander, Tylototriton verru- V, although they engorged themselves withcosus ANDERSON, 1871 is fragmentary. aquatic insects and attacked syntopic crabsDASGUPTA (1983) kept adult Salamanders (JPotamon potamiscus sikkimense). Adultalive in aquaria by feeding them with diet was found to be highly variable depen-earthworms and noted (1988) that larval ding on the study site. During reproductivestages of this salamander fed upon micro- peaks, adults preyed on a mixed diet ofcrustaceans, chironomids, mosquito larvae, terrestrial and aquatic components,and various benthic prey while post- The results of further work are pre-metamorphic juveniles preyed on small sented here in view of the paucity of de-bivalves (Sphaerium indicum). According tailed knowledge on food and feeding hab-to KUZMIN & al. (1994), transition from its of T. verrucosus, and lack of informa-feeding on endogenous yolk to exogenous tion on food niche breadth, food electivitydiet occurred within stage II (sensu DAS- and digestive tract fill index in relation toGUPTA 1983). Microcrustaceans were ab- abiotic and biotic environmental factors.
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

20 R. DASGUPTA
Monsoon rain is the most importantabiotic environmental factor in the Indiansubcontinent. Amphibians and other am-phibious organisms start to deposit eggswith the coming of monsoon rains (MAL-
LICK & al. 1980). The level of primaryproductivity increases with rainfall and thelevel of prédation pressure on T. verruco-sus increases with increasing primary pro-ductivity (DASGUPTA 1995).
MATERIAL AND METHODS
Material
The present investigations were donefrom June to September 1994, at two studysites (26°27'10u N/ 87°59'30" E; 27° 13'04" N/ 88°53'OO" E), at altitudes of 1350m and 1200 m a.s.l., respectively, in Dar-jeeling Himalayas, India.
93 salamander specimens were sam-pled with the help of a throwing net duringthe entire study period (June: 42, July: 20;August: 18; September: 13).
Study site
At the head waters of Beltar (a trib-utary of the Balasone river) an elongatednatural depression within the stream chan-nel constitutes a small fluvio-lacustrinebasin (5000 m2) at an altitude of 1350 m.The actual water surface is confined bysteep inward facing terraces. The super-ficial water supply is controlled by firstand second order streams and fluctuatesthroughout the year. Two spring-initiatedchannels are mainly responsible for per-ennial water conditions during the drypost-monsoon periods. The second studysite (Namthing lake, 1200 m a.s.l., 7700m 2 ) is a typical structure of elongatedvalleys which are the results of faults; thelake is confined by a north and a southfacing slope. Rain contributes to the waterregime of this natural depression althoughground water seepage is the main source ofwater; there is no natural outlet. With theend of monsoon rains, the water leveldrops, mainly due to decreasing groundwater input. The landform characterized bymulti-tiered terraces resulting from tec-tono-fluvial agencies. Submerged vegeta-tion at both study sites comprise Poa sp.,and Hydrocotyle asiatica while semi-sub-merged vegetation includes Polygonumglabrum, P. aula, Cyperus cephalotes, C.rotundus, Scirpus articulât us, Acorus cala-mus, Cautleya hirta.
T. verrucosus lives predominantlyterrestrial in its non-reproductive period,and aquatic in its reproductive period.During June-July (courtship and egg-lay-ing period) both sexes migrated into thewater. In the post-breeding period (Au-gust-September) males moved back to landwhile females stayed in water. As a con-sequence, males were encountered moreoften on land, than females in August-Sep-tember. Females were larger in length thanmales by 10-20%.
For the present paper, it turned out tobe practical to subdivide the four monsoonmonths into a period of heavy rainfall(June and July, total precipitation in bothmonths was 865 mm) and in a period withlight rainfall (August and September, totalprecipitation in both months was 430 mm).
Methods
S t o m a c h f l u s h i n g . In thefield, the broad end of a water filled pi-pette was inserted into the mouth of T. ver-rucosus. Water was pumped into the stom-ach by squeezing the rubber bulb gentlythree times, till the food items were dis-placed along the sides of the pipette andcame to rest in the mouth. There was nooverflow of water from the pipette outsidethe oral cavity. Items so stuck in the mouthand esophagus were removed gently with apair of blunt forceps. This process was re-peated three times for every specimen ex-amined. The efficiency of this method wasnot checked by comparing its results withthat of stomach contents obtained by dis-section, because killing of this animal isprohibited in India. Similar methods ofamphibian stomach flushing were usedsuccessfully in the former Soviet Unionfollowing recommendations given by Bu-LAKOV (1976) and PlSARENKO & VORONIN(1976). Caudata do not suffer any ill-ef-fects after such stomach flushing (GRIF-FITH 1986; JOLY 1987).
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Feeding ecology of Tylototnton verrucosus ANDERSON, 1871 21
The stomach contents were placed ona waxed graph paper for measuring up tothe nearest mm, prior to drying, sievingthrough a dry cheese cloth and weighing ina petri dish on a single pan balance in thefield. The sieved sample was then fixedand preserved in 5% neutral formalin asrecommended by JOLY (1987).
It was not possible to identify each ofthese stomach-flushed items up to genericlevel and so only the family names arementioned. This is so, as the details of thefauna of the region still remain inade-quately explored. E. g., gammarids whichare usually common in lentie waters arestill unknown from most of these habitats.
V o l u m e of f o o d i t e m swas calculated using the formulae des-cribed by GRIFFITH (1986). The formulawas 2 TI a b2 for cylindrical objects (Lum-bricidae, Megascolecidae, adult Hymen-optera, larvae of the orders Diptera, Cole-optera, Trichoptera, Odonata and Ephe-meroptera, pupae of Diptera and Lepid-optera, teleost hatchling); 7i a b c for ellip-tical-cylindrical forms (Orthoptera nymphs,crab legs, Arachnida legs, adult Gastropo-da without shells, Isopoda); (47t/3)a2b foroblate spheroid forms (Bivalvia, adult Ga-stropoda with shells, aquatic Coleoptera,Hemiptera nymphs); and (47i/3)ab2 forprolate spheroid forms (eggs of Caudata,larval Anura), where 2a = length, 2b =width, c = depth of the prey item.
S t o m a c h t r a c t - f i l l i n d e x(J) in adults, is given as per mille of thestomach contents' wet weight (m) in thebody weight of the salamander (M) with-out food (KUZMIN 1985, 1986).
J = 1000 m / (M-m)Total body wet weight of salaman-
ders was taken prior to stomach flushing.F o o d n i c h e b r e a d t h (A)
was calculated according to the methods ofLEVINS (1968) by the formula
A = l / £ ( P x i ) 2
where pxi=percentage of the i-th compo-nent in the diet of predator species x (T.verrucosus). [1 ^ A <, +°°]
F o o d n i c h e o v e r l a p : Themethod of SCHOENER (1970) was used tocalculate food niche overlap between sexesby the formula
Co=l - [0 .5£ (px i -p y i ) ]
where pix, piy=percentage of the i-th com-ponent in the diet of predator species x(male) and y (female), respectively. [0 <,C o ^ l ]
S a m p l i n g o f a v a i l a b l ef o o d . The above methods for calculationof food niche breadth, food niche overlapand food electivity were selected, as it wasnot practical to estimate the abundance ofterrestrial prey items.
For determination of food electivity,food available in ponds was sampled witha playwood box with open ends, lm2 insize. The box was placed gently at twelvestations (six along the water edge, six inthe middle of the lake) with minimum dis-turbance of the water. All larger animalsencountered were scooped out with a teastrainer (72 meshes /cm2) and preserved in5% neutral formalin.
F o o d e l e c t i v i t y . Themethod of IVLEV (1961) was used to assessfood electivity (E) for aquatic prey itemsby the formula
E=(r, - pi) / (n + pi)where n=percentage of the j-th prey com-ponent in the diet and pi=percentage of thei-th prey component in the environment(water body). [-1 <> E <, +1]
This method indicates differences bet-ween relative frequency of prey categoriesin a predator's diet and that in the environ-ment by giving clearer upper and lowerlimits as compared to the electivity indexof STRAUSS (1979).
P r i m a r y p r o d u c t i v i t y .The method followed for the estimation ofprimary productivity (P) was the light anddark bottle method recommended by SAN-TANAM & al. (1989). Primary productivitywas estimated by measuring the amounts ofdissolved oxygen (DO) in two 250 mlreagent bottles. One of these bottles wasdark and the other was transparent. Thedifference in DO between the transparentand dark bottle was used to estimateprimary productivity (P) by converting theobserved difference (A) to the equivalentof inorganic carbon that had been incor-porated as organic carbon by multiplyingwith 0.375. This carbon equivalent wasdivided by the product of the photosyn-thetic quotient (PQ= 1.2) and the period inhours (t) for which the bottle was kept
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Table 1 : Percentages of taxonomic categories according to number (n) and volume (v) found in stom-achs of adult Tylototriton verrucosus during the monsoon months June through September. T - terrestrial taxa,A - aquatic taxa, (1) - legs only, (2) - hatchling, (3) -slough, (4) - eggs.
Tab. 1: Prozentueller Anteil verschiedener Taxa hinsichtlich Anzahl (n) und Volumen (v) am Magen-inhalt adulter Tylototriton verrucosus während der Monsunmonate Juni bis September. T - terrestrischeTaxa,A - aquatische Taxa, (1) -nur Extremitäten, (2) -Schlüpfling, (3) -Häutung, (4) -Laich.
TTTTTTTTTTTTTTTTTTTTTTTT
AAAAAAAAAAAAAAAAAAAAAAAAA
Gastropoda
Clitellata
CrustaceaArachnida (1)
Insecta
BivalviaCrustacea (1)Insecta
OsteichthyesAmphibia
Taxon
StylommatophoraStylommatophora indet.Oligochaeta
IsopodaAraneae
Orthoptera
Diptera (larvae)
Diptera (pupae) indet.Hymenoptera (imagines)
Coleoptera (imagines)Lépidoptère (larvae)
Lépidoptère (pupae)
EulamellibranchiataDecapodaDiptera (imagines)
Trichoptera (imagines)Odonata (larvae)
Coleoptera (larvae)Coleoptera (imagines)
TeleosteiAnura (larvae)Urodela
Arionidae
LumbricidaeMegascolecidaeOniscidaeTetragnathidaeCyclosidaeTheridiidaeGryllidaeTettigoniidaeDrosophilidaeSciaridaeTephritidaeLonchopteridaePhoridaeAsilidae
BraconidaeFormicidaeLagriidaeBombycidaeNoctuidaeSaturniidaeSatyridae
Euglesiidae, PisidiidaePotamonidaeSyrphidaeTabanidaeTipulidaeChironomidaeCulicidaeChaoboridaeDolichopodidaeEphydridaeCeratopogonidaePhryganeidaeAeshnidaeGomphidaePseudagrionidaeCrocothemidaeDytiscidaeGyrinidaeLaccophilidaeHydaticidaeDytiscidaeSiluridae (2)RhacophoridaeT. verrucosus (3)T. verrucosus (4)
%(n)
1.120.563.351.120.561.120.560.561.670.564.463.353.913.911.681.121.120.560.560.562.232.242.231.12
0.565.591.671.120.569.507.820.564.472.232.794.471.682.231.121.101.120.561.121.791.000.564.470.561.12
%(v)
0.4210.834.531.010.490.130.030.030.570.060.030.020.030.030.010.010.010.060.010.082.366.687.0011.50
1.396.291.170.380.031.380.440.010.250.280.040.500.490.150.790.790.190.330.450.690.190.0336.780.000.04
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
Feeding ecology of Tylototriton verrucosus ANDERSON, 1871 23
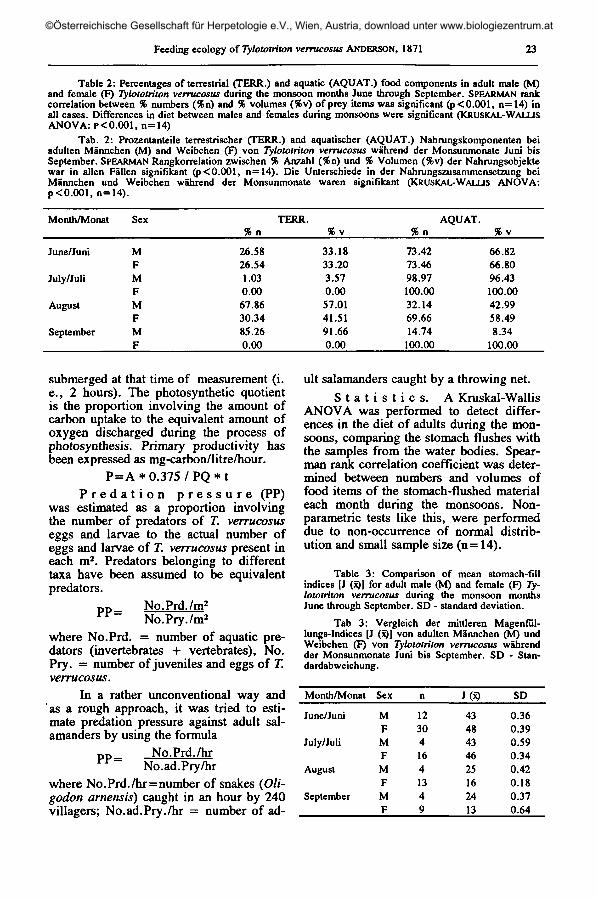
Table 2: Percentages of terrestrial (TERR.) and aquatic (AQUAT.) food components in adult male (M)and female (F) Tylototriton verrucosus during the monsoon months June through September. SPEARMAN rankcorrelation between % numbers (%n) and % volumes (%v) of prey items was significant (p<0.001, n=14) inall cases. Differences in diet between males and females during monsoons were significant (KRUSKAL-WALLISANOVA: P< 0.001, n=14)
Tab. 2: Prozentanteile terrestrischer (TERR.) and aquatischer (AQUAT.) Nahrungskomponenten beiadulten Männchen (M) and Weibchen (F) von Tylototriton verrucosus während der Monsunmonate Juni bisSeptember. SPEARMAN Rangkorrelation zwischen % Anzahl (%n) und % Volumen (%v) der Nahrungsobjektewar in allen Fällen signifikant (p<0.001, n=14). Die Unterschiede in der Nahrungszusammensetzung beiMännchen und Weibchen während der Monsunmonate waren signifikant (KRUSKAL-WALUS ANOVA:p <0.001, n= 14).
Month/Monat Sex TERR.% n %v % n
AQUAT.%v
June/Juni
July/Juli
August
September
MFMFMFMF
26.5826.541.030.0067.8630.3485.260.00
33.1833.203.570.0057.0141.5191.660.00
73.4273.4698.97100.0032.1469.6614.74
100.00
66.8266.8096.43100.0042.9958.498.34
100.00
submerged at that time of measurement (i.e., 2 hours). The photosynthetic quotientis the proportion involving the amount ofcarbon uptake to the equivalent amount ofoxygen discharged during the process ofphotosynthesis. Primary productivity hasbeen expressed as mg-carbon/litre/hour.
P = A * 0.375 / P Q * tP r é d a t i o n p r e s s u r e (PP)
was estimated as a proportion involvingthe number of predators of T. verrucosuseggs and larvae to the actual number ofeggs and larvae of T. verrucosus present ineach m2. Predators belonging to differenttaxa have been assumed to be equivalentpredators.
No.Prd./m2
No.Pry./m2
where No.Prd. = number of aquatic pre-dators (invertebrates + vertebrates), No.Pry. = number of juveniles and eggs of T.verrucosus.
In a rather unconventional way andas a rough approach, it was tried to esti-mate prédation pressure against adult sal-amanders by using the formula
p p _ No.Prd./hrrv~ No.ad.Pry/hr
where No.Prd./hr=number of snakes (Oli-godon arnensis) caught in an hour by 240villagers; No.ad.Pry./hr = number of ad-
PP=
ult salamanders caught by a throwing net.
S t a t i s t i c s . A Kruskal-WallisANOVA was performed to detect differ-ences in the diet of adults during the mon-soons, comparing the stomach flushes withthe samples from the water bodies. Spear-man rank correlation coefficient was deter-mined between numbers and volumes offood items of the stomach-flushed materialeach month during the monsoons. Non-parametric tests like this, were performeddue to non-occurrence of normal distrib-ution and small sample size (n= 14).
Table 3: Comparison of mean stomach-fillindices [J (x)] for adult male (M) and female (F) Ty-lototriton verrucosus during the monsoon monthsJune through September. SD - standard deviation.
Tab 3: Vergleich der mittleren Magenföl-lungs-Indices [J (x)] von adulten Männchen (M) undWeibchen (F) von Tylototriton verrucosus währendder Monsunmonate Juni bis September. SD - Stan-dardabweichung .
Month/Monat
June/Juni
July/Juli
August
September
Sex
MFMFMFMF
n
123041641349
J(x)
4348434625162413
SD
0.360.390.590.340.420.180.370.64
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

24 R. DASGUPTA
Table 4: Food niche breadth (A) in adult male(M) and female (F) Tylototriton verrucosus calculatedaccording to LEVINS (1964), based on number (n)and volume (v) of prey items.
Tab 4: Nahrungsnischenbreite (A) bei adultenMännchen (M) und Weibchen (F) von Tylototritonverrucosus auf Grundlage der Anzahl (n) und desVolumens (v) der Nahrungsobjekte; berechnet nachder Methode von LEVINS (1964).
Table S: Food niche overlap between sexes(Co) of adult Tylototriton verrucosus, according tonumber [Co (n)] and volume [Co (v)] of food items,during the monsoon months June through September.Indices calculated according to SCHOENER (1970).
Tab 5: Überlappung der Nahrungsnischen derGeschlechter (Co) bei adulten Tylototriton verrucosusauf Grundlage der Anzahl [Co (n)] und des Volu-mens [Co (v)] der Nahrungsobjekte, während der
Month/Monat
June/Juni
July/Juli
August
September
Sex
MFMFMFMF
A(n)
0.0520.0550.0520.0550.0300.0310.0150.016
A(v)
0.0530.0560.0530.0560.0330.0340.0160.018
Monsunmonaie jum DIS aepiemDer; oder Methode von SCHOENER (1970).
Month/Monat
June/JuniJuly/JuliAugustSeptember
RESULTS
Co(n)
0.69620.34810.02010.0201
lerecnnet nacn
Co(v)
0.63190.12650.01960.0201
Primary productivity
Primary productivity was high (max-imum 26.23 mg/l/hr) during June and July(rainfall 582 mm and 283 mm, respect-ively) at 1350 m. With progress of mon-soons, a decline in primary productivity(maximum 24.32 mg/l/hr) in August andSeptember (rainfall 222 mm and 208 mm,respectively) was recorded at 1350 m. At1200 m, the highest levels of primaryproductivity recorded were 26.23 mg/l/hrfor June-July and 26.00/1/hr for August-September.
Food during different monthsof the monsoon period
(tables 1 & 2)
In the monsoon period, food itemsobtained by stomach flushing comprisedboth aquatic and terrestrial elements; plane-tonic crustaceans and other microarthro-pods were absent in the stomach-flushedmaterial. Aquatic prey taxa predominatedin the stomachs during times of heavyrainfall (June-July), and concomitant highlevels of primary productivity.
During periods of low rainfall (endof monsoon rains, August-September), ter-restrial prey taxa predominated in thestomach-flushed samples obtained frommales and aquatic prey taxa in that obtainedfrom females. Compared to the females'diet, the diet of males was more terrestrialat any time and included Lepidoptera (lar-
vae and pupae of Noctuidae, Satumidae,Bombycidae) and larval Diptera. Food offemales included rhacophorid foam nests.
In males, diet was mainly aquatic inJune, almost exclusively aquatic in July,mainly terrestrial in August and predomi-nantly terrestrial in September. In females,diet was mainly aquatic in June, exclu-sively aquatic in July, predominantly aqua-tic in August and exclusively aquatic inSeptember.
Differences in the bulk, number andcomposition of the diet during differentmonths of the rainy season was significant(P< 0.001; Kruskal-Wallis ANOVA).
Stomach-fill index(table 3)
Stomach-fill indices were highestduring periods of heavy rainfall (June: 43-48 %o; July: 43-46%o) and subsequent highlevels of primary productivity. Stomach-
Table 6: Prédation pressure (PP) on adults,eggs and larvae of Tylototriton verrucosus in themonsoon months June through September.
Tab 6: Beutedruck (PP) auf adulte, Eier undLarven von Tylototriton verrucosus in den Monsun-monaten Juni bis September.
S tage/Stadium
Adults/ AdulteOva + Larvae
June /Juni
0.050.17
July/Juli
0.030.05
PP
August
0.000.03
Sep-tember
0.000.03
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Feeding ecology of Tylototriton verrucosus ANDERSON, 1871 25
fill indices were lower when rainfall waslow (August: 16-25 %o; September: 13-24%o). Females had higher stomach-fill in-dices than males during the egg-layingperiod (June-July), males had higher stom-ach-fill indices than females during thepost-reproductive period (August-Septem-ber.
Food niche breadth(table 4)
Food niche breadth was largely thesame in males and females. Indices weretwice as high in August and Septemberthan in June and July. During the post-re-productive period (August-September) fe-males were mostly sampled from water andmales from land.
Food niche overlapbetween sexes
(table 5)
Food niche overlap between sexes(Co) was clearly expressed during repro-ductive peaks when both sexes were in thewater (June-July, beginning of strong mon-soon rains, high primary productivity).
Overlap in diet was almost absentduring August-September (periods of lowrainfall) in the post-reproductive periodwhen males were leaving or had alreadyleft the water and primary productivity waslow.
Prédation pressure(table 6)
Prédation pressure on adults, eggsand larvae of T. verrucosus varied sea-sonally. Prédation pressure decreased fromJune through September in response togradual lowering of primary productivityand rainfall with progress of monsoons.
Food niche overlap and breadth,stomach-fill index, and their relation to
rainfall and primary productivity(table 7)
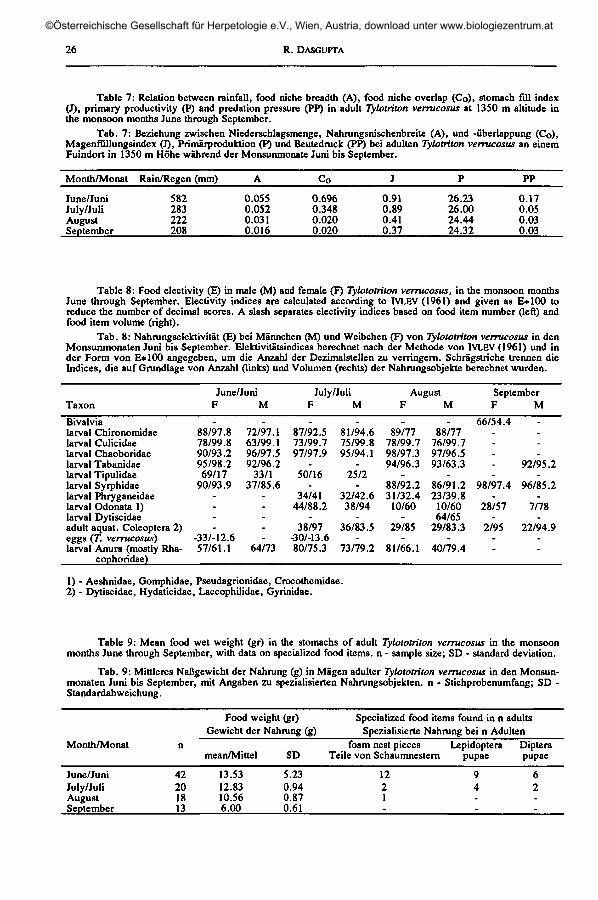
Stomach-fill indices, food niche over-lap and breadth between sexes were highestduring periods of heavy rainfall, coincidingwith the time of oviposition, high level ofprimary productivity and high prédationpressure.
Food electivity(table 8)
Despite the great abundance of T.verrucosus eggs in the ponds during Juneand July, conspecific eggs contributed onlyvery little (1.12 %, negative electivity in-dices) to the female stomach contents andwere never found in males. A teleosthatchling (Neomacheilus sp.) was detectedin the stomach of a female in June (periodof maximum rainfall) (table 1).
In June, aquatic dipterous larvae (Chi-ronomidae, Culicidae, Chaoboridae, Taba-nidae, Tipulidae, Syrphidae) and larvalAnura were present in the stomach con-tents, while aquatic Coleoptera, Bivalvia,Trichoptera and Odonata nymphs were ab-sent. Cannibalized eggs were obtained onlyfrom female stomachs.
In July, Odonata nymphs (Aeshnidae,Gomphidae, Pseudagrionidae, Crocothe-midae), Chironomidae, Tipulidae, Culici-dae, Chaoboridae, adult aquatic Coleoptera(Dytiscidae, Hydaticidae, Laccophilidae,Gyrinidae), and larval Anura were thedominating prey taxa in the stomach con-tents, while larval Tabanidae and Syrphi-dae were absent.
In August, Phryganeidae (Trichop-tera), Tabanidae, Syrphidae, and other lar-val Diptera, Odonata nymphs, larval An-ura, adult and larval Dytiscidae were pres-ent in the stomach flushes while Tipulidaewere not detected.
In September, Tabanidae, Syrphidae,Bivalvia, adult aquatic Coleoptera, andOdonata nymphs dominated in stomach-flushed samples, whereas Chironomidae,Culicidae, Chaoboridae, Tipulidae, and lar-val Anura were absent.
Differences in food electivity valuesby numbers and volumes are due to varia-tion in the relative size and proportion ofindividual food items at different altitudesduring different monsoon periods. This isreflective of differences in trophic struc-ture and spatial distribution of the prey atdifferent altitudes during different mon-soon months.
Food quantity (table 9)
There were clear differences betweenmonths concerning the mean quantity offood found in the stomachs of adult T. ver-
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
26 R. DASGUPTA
Table 7: Relation between rainfall, food niche breadth (A), food niche overlap (Co), stomach fill index(J), primary productivity (P) and prédation pressure (PP) in adult Tylotriton verrucosus at 1350 m altitude inthe monsoon months June through September.
Tab. 7: Beziehung zwischen Niederschlagsmenge, Nahrungsnischenbreite (A), und -Überlappung (Co),Magenfullungsindex (J), Primärproduktion (P) und Beutedruck (PP) bei adulten Tylotriton verrucosus an einemFuindort in 1350 m Höhe während der Monsunmonate Juni bis September.
Month/Monat
June/JuniJuly/JuliAugustSeptember
Rain/Regen (mm)
582283222208
A
0.0550.0520.0310.016
Co
0.6960.3480.0200.020
J
0.910.890.410.37
P
26.2326.0024.4424.32
PP
0.170.050.030.03
Table 8: Food electivity (E) in maie (M) and female (F) Tylototriton verrucosus, in the monsoon monthsJune through September. Electivity indices are calculated according to IVLEV (1961) and given as E+100 toreduce the number of decimal scores. A slash separates electivity indices based on food item number (left) andfood item volume (right).
Tab. 8: Nahrungselektivität (E) bei Männchen (M) und Weibchen (F) von Tylototriton verrucosus in denMonsunmonaten Juni bis September. Elektivitätsindices berechnet nach der Methode von IVLEV (1961) und inder Form von E*100 angegeben, um die Anzahl der Dezimalstellen zu verringern. Schrägstriche trennen dieIndices, die auf Grundlage von Anzahl (links) und Volumen (rechts) der Nahrungsobjekte berechnet wurden.
Taxon
Bivalvialarval Chironomidaelarval Culicidaelarval Chaoboridaelarval Tabanidaelarval Tipulidaelarval Syrphidaelarval Phryganeidaelarval Odonata 1)larval Dytiscidaeadult aquat. Coleoptera 2)eggs (T. verrucosus)larval Anura (mostly Rha-
cophoridae)
June/JuniF
_88/97.878/99.890/93.295/98.269/17
90/93.9----
-33/-12.657/61.1
M
_72/97.163/99.196/97.592/96.2
33/137/85.6
-----
64/73
July/JuliF
87/92.573/99.797/97.9
50/16-
34/4144/88.2
-38/97
-30/-13.680/75.3
M
.81/94.675/99.895/94.1
25/2-
32/42.638/94
-36/83.5
-73/79.2
AugustF
89/7778/99.798/97.394/96.3
-88/92.231/32.4
10/60-
29/85-
81/66.1
M
.88/77
76/99.797/96.593/63.3
-86/91.223/39.8
10/6064/65
29/83.3-
40/79.4
SeptemberF
66/54.4
-98/97.4
-28/57
-2/95
--
M
_
92/95.2-
96/85.2-
7/78-
22/94.9--
1) - Aeshnidae, Gomphidae, Pseudagrionidae, Crocothemidae.2) - Dytiscidae, Hydatictdae, Laccophilidae, Gyrinidae.
Table 9: Mean food wet weight (gr) in the stomachs of adult Tylototriton verrucosus in the monsoonmonths June through September, with data on specialized food items, n - sample size; SD - standard deviation.
Tab. 9: Mittleres Naßgewicht der Nahrung (g) in Mägen adulter Tylototriton verrucosus in den Monsun-monaten Juni bis September, mit Angaben zu spezialisierten Nahrungsobjekten, n - Stichprobenumfang; SD -Standardabweichung.
Month/Monat
Food weight (gr)Gewicht der Nahrung (g)
Specialized food items found in n adultsSpezialisierte Nahrung bei n Adulten
mean/Mittel SDfoam nest pieces
Teile von SchaumnesternLepidoptera
pupaeDipterapupae
June/JuniJuly/JuliAugustSeptember
42201813
13.5312.8310.566.00
5.230.940.870.61
1221-
94--
62--
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Feeding ecology of Tylototriton verrucosus ANDERSON, 1871 27
rucosus (P< 0.0001; Kruskal-Wallis AN-OVA) indicating increased food ingestion
by both sexes during the reproductiveperiod in June and July.
DISCUSSION
Diversity of food items
SATTMANN (1989) found that the dietof Triturus alpestris comprised aquatic andterrestrial prey. The diet was predominant-ly aquatic during the aquatic breedingphase of this newt. Towards the end of itsreproductive period, food became increas-ingly composed of terrestrial prey, espe-cially in males. He further noted negativefood electivity indices for cannibalizedeggs.
As was found earlier as well as in thepresent study, the diet of adult T. verruco-sus comprised both aquatic and terrestrialelements. In stomach contents at differenthabitats in Darjeeling, KUZMIN & al.(1994) found Lumbricidae (16.7% -85.7%), Chironomidae (Cryptochironomusand Tanytarsus, Yl.1% - 95.2%), aquaticDiptera other than Chironomidae (7.1% -66.7%), imagines of Hydrophilidae (19% -25%), terrestrial gastropods (16.7%), andeggs of Amphibia (25% - 66.7%). Terres-trial prey items included larval Diptera.Furthermore, these authors mentioned thepresence of low percentages of puparia ofaquatic Diptera (4.8%), larval T. verrucosus of 13 mm SVL (4.8%), shed skin of T.verrucosus (4.8%) and anuran larvae(11.8%). They further reported on larvalT. verrucosus attacking syntopic crabs (Po-tamon potamiscus sikkimense).
Arthropod remains obtained fromstomach flushed material during the pres-ent study additionally comprised Onisci-dae, Drosophilidae, Sciaridae; Tephritidae,Lonchopteridae, Asilidae, Phoridae, Lagri-idae, Bombycidae, Noctuidae and Saturni-idae. Crabs were not likely to be swal-lowed entirely, since only crab legs (Pota-mon potamiscus sikkimense) were flushedfrom the stomachs of T. verrucosus. Mol-lusk remains included Arionidae and otherStylommatophora.
A hatchling of Neomacheilus sp. ob-tained from stomach flushed material dur-ing the present study represents an unusualinstance of ingestion of a lotie organism bya lentie caudate; possibly after the formerhad either accidentally strayed or drifted
(monsoon rain) from an adjoining streaminto the lentie habitat occupied by the adultT. verrucosus during its breeding period.There appears to be no earlier record of T.verrucosus feeding on teleost hatchlings.
Different sensory systems in larvaeand adults used for prey detection resultedin selection for larger objects in the latter(KUZMIN 1986). Small prey items (micro-arthropods) were absent from the diet ofadult T. verrucosus (KUZMIN & al. 1994;this paper) but occurred in the diet of lar-vae and metamorphs (KUZMIN & al.1994). Adults may ingest such items acci-denally.
According to KUZMIN (1991), stom-ach contents of caudate amphibians did notonly allow for indication of the local in-vertebrate fauna but also served as anestimator of its spatial distribution. Theresults of stomach flushes obtained fromthe present study have added Tephritidae(Diptera) to the faunal list of the studysite. The absence of gammarids in stomachcontents in the previous (KUZMIN & al.1994) and in the present study, suggeststhat Gammarids might not occur in thehabitats under study.
Sex-specific oophagy
There is a number of observationspublished concerning oophagy in salaman-ders: Intrauterine cannibalism of the oo-phagous type viz., epitheliophagy andadelphophagy of Salamandra s. terrestrisembryos was thought to be responsible fordeposition of larger but fewer young byfemales of this species at higher altitudesas compared to females from the foothills(JOLY 1961). WOOD & GOODWIN (1954)reported on Notophthalmus viridescens fee-ding on conspecific eggs; larval and adultN. viridescens and larval Ambystoma opa-cum fed on eggs of several anuran andcaudate species (WALTERS 1975); KAPLAN& SHERMAN (1980) recorded Tarichapreying on conspecific eggs, and KUZMIN& al. (1994) reported on egg cannibalismin T. verrucosus. According to DUELLMAN& TRUEB (1986), newt eggs are hidden
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

28 R. DASGUPTA
singly or in clumps amidst aquatic vege-tation because adults are known to con-sume eggs with moving embryos. Sala-manders which guard their clutches do notforage actively during this time, neverthe-less, eggs could serve as an important en-ergy source for these adults (HARRIS &GILL 1980; POLIS & MYERS 1985; SOLA-NO 1987). Attending female Desmognathusochrophaeus fed on dead or infected eggsfrom their clutches (FORESTER 1979,1986). Oophagy in newts was consideredindiscriminate cannibalism (DUELLMAN &TRUEB 1986) because individuals may notrecognize their own eggs. The sameauthority considered preying on con-specific eggs to represent just exploitationof an available food resource. AmongstDendrobatidae, feeding on eggs laid byother females was interpreted as an ex-ample of competition (SUMMERS 1989).MARSHALL & al. (1990) reported that onlymales cannibalized eggs in Bombino orien-talis and only females in case of Tarichatorosa. KUZMIN (1989, 1991a) listed 59amphibian species that were oophagousand noted (1991a) that cannibalism was asimple manifestation of polyphagy.
In the present study, only female T.verrucosus cannibalized eggs. Detection ofcannibalized eggs in females at times ofhigh levels of food availibility (June, July)suggests that oophagy in this species wasnot a consequence of depletion of foodresources. The high abundance of eggsappeared to have induced oophagy. Thus,oophagy in case of this species is in ac-cordance with the optimal foraging theory:the electivity of polyphages increases instep with improvement in feeding con-ditions.
Food niche breadthand overlap
There appears to be little divergencein the diet between sexes which is not un-common and is known in case of somespecies of Triturus, where male-female dietoverlap is sometimes high. According to
GRIFFITHS (1987) a high degree of foodniche overlap is known to occur e.g. bet-ween Triturus vulgaris and Triturus hel-veticus and such co-existence betweenspecies with nearly identical food niches isexplained by unlimited food resources orextremely high rates of prédation.
Female T. verrucosus were larger inlength than males by 10-20%. So it is notsurprising that different sexes actually con-sumed prey of different mean size andquantity.
Stomach-fill index
In both sexes, stomach fill indiceswere higher in June-July (period of heavi-est rainfall and highest levels of primaryproductivity) than in the post-breeding pe-riod (August-September).
Males had lower stomach fill indicesthroughout the breeding period (June-July) and higher stomach fill indices dur-ing the post-breeding period (August-Sep-tember), as compared to the females. Thismay be explained by the females' higherreproductive expenses in June-July (Kuz-MIN 1989a) and by the males' more terres-trial life in August-September.
During the post-breeding period tillhatching of the young, the females ap-parently did not forage actively.
Prey composition in relation toprecipitation
Seasonal differences in diet reflectprey availibility and selectivity (TOFT1985).
In adult T. verrucosus, stomach-fillindices, food niche breadth and overlapbetween sexes were highest, largest andwidest respectively, during the peaks ofrainfall, primary productivity and pre-dation-pressure, thus, coinciding with thebreeding period in June-July when bothsexes migrated into the water.
In the post-breeding period (August-September), males moved away from thewater. This was reflected in their having amore terrestrial diet than females.
ACKNOWLEDGMENTS
Thanks are due to S. L. KUZVON for supply ofliterature and criticism of the present manuscript, andto M. DUTTA, T. MITRA and various other ento-mologists and malacologists from the Z. S. I., India,
for helping in identification of invertebrate remains.The British High Commission, New Delhi is great-fully acknowledged for financing the present studyby way of a "Radhakrishnan award".
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Feeding ecology of Tylototriton verrucosus ANDERSON, 1871 29
REFERENCES
BULAKOV, V. L. (1976): Methods of food in-vestigation in live amphibians.- Voropsy stepnogoLesovedeniya i Okhrany Prirody; Dnepropetrovsk;(6): 146-156.
DASGUPTA, R. (1983): Observations on theHimalayan newt.- J. Bengal. Nat. Hist. Soc.; (N. S.)2(2): 59-60.
DASGUPTA, R. (1988): Young of the Himala-yan newt.- J. Bengal. Nat. Hist. Soc.; (N. S.) 7 ( 1 ):3-18.
DASGUPTA, R. (1995): Trophic structure andphysical characteristics of some lentic-habitats inDarjeeling.- J. Bengal. Nat. Hist. Soc.; (N. S.) 14(1): 11-40.
DUELLMAN, W. E. & TRUEB, L. (1986): Bio-logy of amphibians. New York, St. Louis, San Fran-cisco (McGraw Hill).
FORESTER, D. C. (1979): The adaptiveness ofparental care in Desmognathus ochrophaeus COPE(Urodela: Plethodontidae).- Copeia; 1979: 332.
FORESTER, D. C. (1986): The recognition anduse of chemical signals by a nesting salamander. In:DUVALL, D. & MULLER-SCHWARSE, D. & SlL-VERSTEIN, R. M. (eds.): Chemical signals of verte-brates.
GRIFFITHS, R. A. (1986): Feeding niche over-lap and food selection in smooth and palmate newts,Triturus vulgaris and T. helveticus at a pond in Mid-Wale s.-J. Animal Ecology; 55: 201-214.
GRIFFITHS, R. A. (1987): Micohabitat andseasonal niche dynamics of smooth and palmatenewts, Triturus vulgaris and T. cristatus, at a pond inMid-Wales.- J. Animal Ecology; 56: 441-451.
HARRIS, R. N. & GILL, D. E. (1980): Com-munal nesting, brooding behaviour, and embryonicsurvival of the four toed salamander, Hemidactyliumscutatum.- Herpetologica; 36 (2): 141-144.
IVLEV, V. S (1961): Experimental ecology ofthe feeding of fishes. New Haven (Yale UniversityPress).
JOLY, P. (1961): Le cycle sexuel biennal chezla femelle de Salamandra s. quadrivirgata dans lesHautes-Pyrenees.- Compt. Rend. Avad. Sci., Paris;252 (20): 3145-3147.
JOLY, P. (1987): Le regime alimentaire desamphibiens: méthodes d'étude.- Alytes; 6 (1-2): 11-17.
KAPLAN, R. H. & SHERMAN, P. w . (1980):Intraspecific oophagy in California newts.- J.Herpetol.; 14: 183-185.
KUZMIN, S. L. (1985): Rate of food con-sumption and prey size in the Siberian newt.-Ecologiya; 15 (5): 265-271. [in Russian]
KUZMIN, S. L. (1986): Elective feeding habitsand feeding behaviour of Siberian Angle Tooth lar-vae.- Ecologiya; 16 (5): 282-286. [in Russian]
KUZMIN, S. L. (1989): Populational aspects ofamphibian trophology. In: Problemy populatsinnoiekologii zemnovodnykh i presmykayuszczikhsyua.Moscow, pp. 52-100. [in Russian]
KUZMIN, S. L. (1989a): On the sex differ-ences in amphibian feeding. In: Voprosy herpe-
tologii. Kiev (Naukova Dumka). [in Russian]KUZMIN, S. L. (1991): A review of studies on
amphibian and reptilian feeding ecology in theUSSR.- Herpetozoa, Wien; 4 (3/4): 99-115.
KUZMIN, S. L. (1991a): The ecology andevolution of amphibian cannibalism.- J- Bengal Nat.Hist. Soc.; (N. S.) 10 (2): U-27.
KUZMIN, S. L. & DASGUPTA, R. & SMIRNA,E. (1994): Ecology of the Himalayan newt (Tylo-totriton verrucosus) in Darjeeling Himalayas, India.-Russian J. Herpetology; 1 ( 1 ) : 69-76.
LEVINS, R. (1968): Evolution in changingenvironments.- Princeton, U.S.A (Princeton Uni-versity press).
MALUCK, P. K. & MALUCK, S. C. & DAS,D. K. (1980) Spawning behaviour of four salientiansof West Bengal.- Indian Biol. J.; 2 (1): 25-28.
MARSHALL, C. J. & DOVALE, L. S. & KAP-LAN, R. H. (1990): Intraspecific and sex specificoophagy in a salamander and frog; Reproductiveconvergence of Taricha torosa and Bombino orienta-lis.- Herpetologica; 64 (4): 395-399.
PlSARENKO; S. S. & VORONIN, A. A. (1976):Method for feeding study of live amphibians.- Eko-logiya, Sverdlovsk; (2): 106. On Russian).
POLIS, G. A. & MYERS, C. A. (1985): A sur-vey of intraspecific prédation among reptiles and am-phibians.-J. Herpetol.; 19 (1): 99-107.
SANTANAM, R. & VELAYUTHAM, P. &JAGATHEESAN, S. (1989): A manual of fresh-waterecology: an aspect of fishery environment. New Del-hi (Daya publishing House).
SATTMANN, H. (1989): Über die Nahrung desBergmolches Triturus alpestris (LAURENTI, 1768), inder aquatischen Phase (Caudata: Salamandridae).-Herpetozoa, Wien; 2. (1/2): 31-94.
SCHOENER, T. W. (1910): The evolution ofbill size differences amongst sympatric congenericspecies of birds.-Evolution; 19: 189-213.
SOLANO, H. (1987): Algunos aspectos de labiologia reproductiva del sapito silbador Leptodac-tylus fiiscus SCHNEIDER (Amphibia: Leptodactyli-dae).- Amphibia-Reptilia; ( (2): 111-128.
STRAUSS, R. E. (1979): Reliability estimatesfor Ivlev's index, the forage ratio, and a proposedindex of linear food-selection.- Trans. Amer. Fisher-ies Soc.; 108: 344-352.
SUMMERS, K. (1989): Sexual selection and intra-female competition in the green poison-dart frogDendrobates auratus.- Animal Behaviour; 37 (5):797-805.
TOFT.C. A. (1985): Resource partitioningamongst amphibians and reptiles.- Copeia; 1985 (1):1-21.
WALTERS, B. L. (1975): Studies on interspe-cific prédation within an amphibian community.- J.Herpetol.; 9: 267-280.
WOOD, J. T. & GOODWIN, O. K. (1954):Observations on the abundance, food, and feedingbehaviour of the newt, Notophthalmus v. viridescens(RAFINESQUE) in Virginia.- J. Elisha Mitchell Sci.Soc.; 70: 27-30.
DATE OF SUBMISSION: November 1st, 1995 Corresponding editor: Heinz Grillitsch
AUTHOR: Dr. RJTWlK DASGUPTA, Postgraduate Department of Zoology, Darjeeling GovernmentCollege, Darjeeling 734 101, West Bengal, India.
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at


















