
Grundlagen Mechanosensitivität -...
59
Biophysik der Zelle Grundlagen der zellulären Mechanosensitivität Carsten Grashoff MPI für Biochemie, Molekulare Mechanotransduktion [email protected]
Transcript of Grundlagen Mechanosensitivität -...

Biophysik der Zelle
Grundlagen der zellulären
Mechanosensitivität
Carsten Grashoff
MPI für Biochemie, Molekulare
Mechanotransduktion
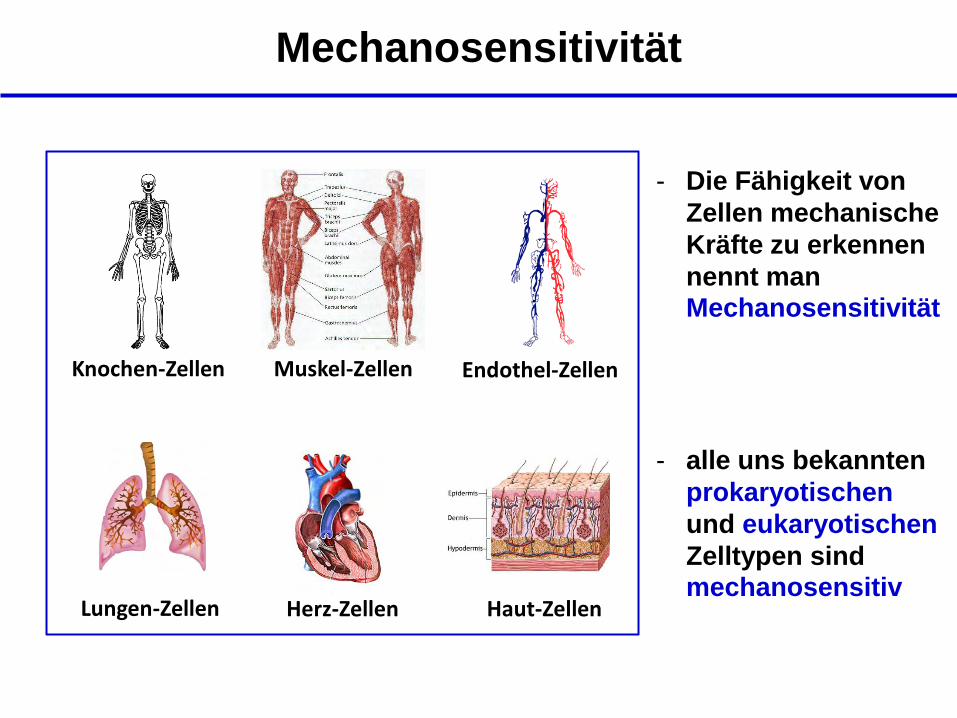
Mechanosensitivität
Knochen-Zellen Muskel-Zellen Endothel-Zellen
Lungen-Zellen Haut-Zellen Herz-Zellen
- Die Fähigkeit von
Zellen mechanische
Kräfte zu erkennen
nennt man Mechanosensitivität
- alle uns bekannten
prokaryotischen
und eukaryotischen
Zelltypen sind mechanosensitiv

1. Wie erkennt eine Zelle die
mechanische Umgebung ?
Die biophysikalische Fragestellung
2. Wie verwandelt eine Zelle mechanische
Information in biologische Prozesse ?
3. Wie kann ich diese biophysikalischen
Prozesse messen ?

1. Wie erkennt eine Zelle die
mechanische Umgebung ?
Die biophysikalische Fragestellung
Wodurch ist die Umgebung der Zelle
charakterisiert ?

Wodurch ist die Umgebung der Zelle charakterisiert?
- die mechanische
Umgebung der Zelle
ist durch die
extrazelluläre Matrix (EZM) charakterisiert
- die EZM diktiert die
biophysikalischen
und biochemischen
Eigenschaften des Gewebes
Die Extrazelluläre Matrix (EZM)
Die EZM ist aus vielen
Makromolekülen aufgebaut, die
von Zellen sezerniert werden
Man unterscheidet:
- Faserbildende Proteine (z.B.
Kollagene, Elastine)
- Glykosaminoglykane (GAGs)
- Proteoglykane
- Adhäsionsproteine (z.B.
Fibronektin, Laminin)
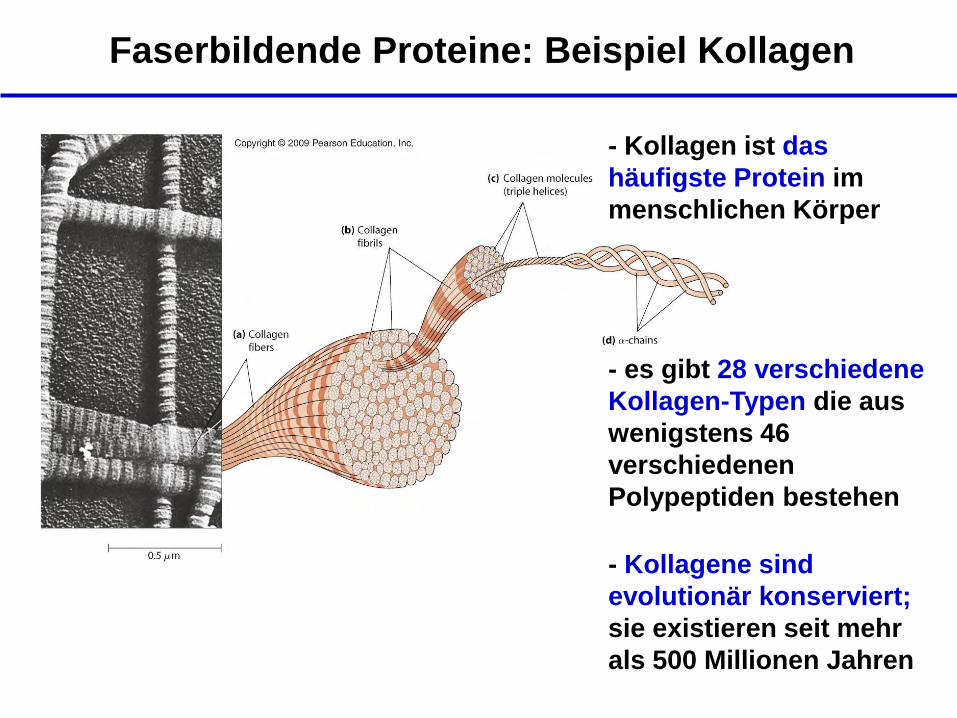
Faserbildende Proteine: Beispiel Kollagen
- Kollagen ist das
häufigste Protein im
menschlichen Körper
- Kollagene sind
evolutionär konserviert;
sie existieren seit mehr
als 500 Millionen Jahren
- es gibt 28 verschiedene
Kollagen-Typen die aus
wenigstens 46
verschiedenen
Polypeptiden bestehen
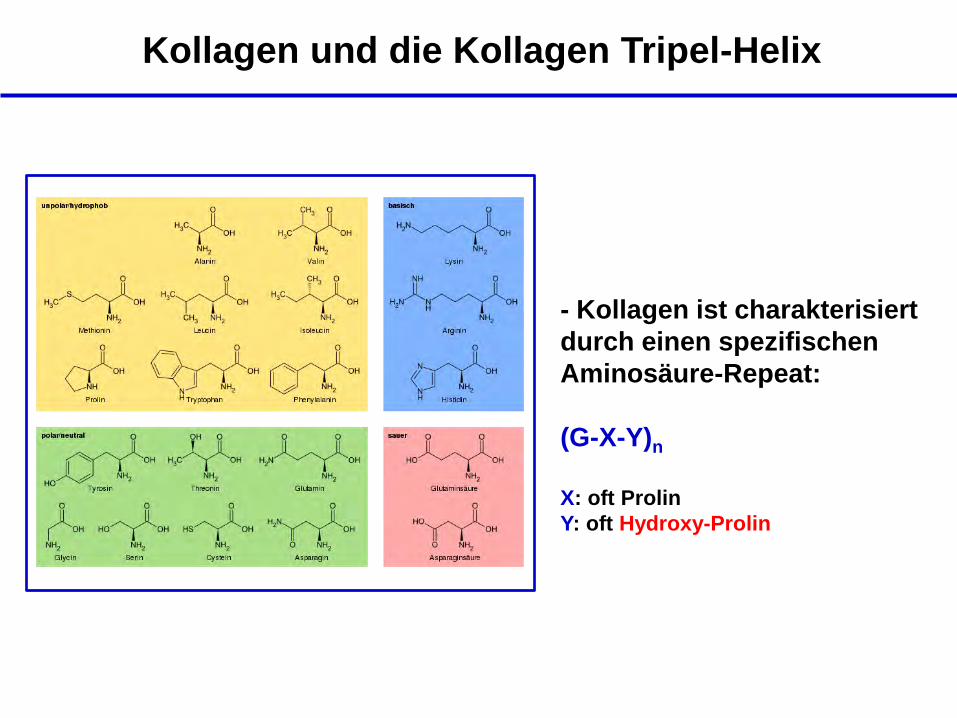
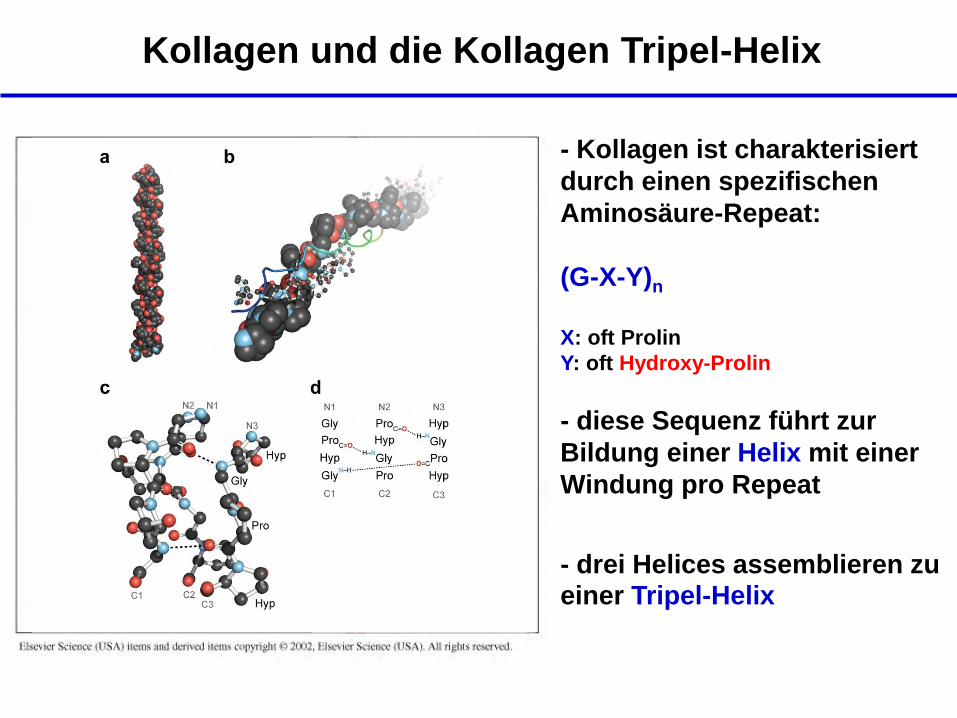
Kollagen und die Kollagen Tripel-Helix
- Kollagen ist charakterisiert
durch einen spezifischen
Aminosäure-Repeat:
(G-X-Y)n
X: oft Prolin
Y: oft Hydroxy-Prolin
Kollagen und die Kollagen Tripel-Helix
- Kollagen ist charakterisiert
durch einen spezifischen
Aminosäure-Repeat:
(G-X-Y)n
X: oft Prolin
Y: oft Hydroxy-Prolin
- diese Sequenz führt zur
Bildung einer Helix mit einer
Windung pro Repeat
- drei Helices assemblieren zu einer Tripel-Helix
Die Rolle des Hydroxyprolin im Kollagen
(G-X-Y)n
X: oft Prolin
Y: oft Hydroxy-Prolin
- Hydroxyprolin stabilisiert
die Tripel-Helix durch
stereo-elektronische
Effekte
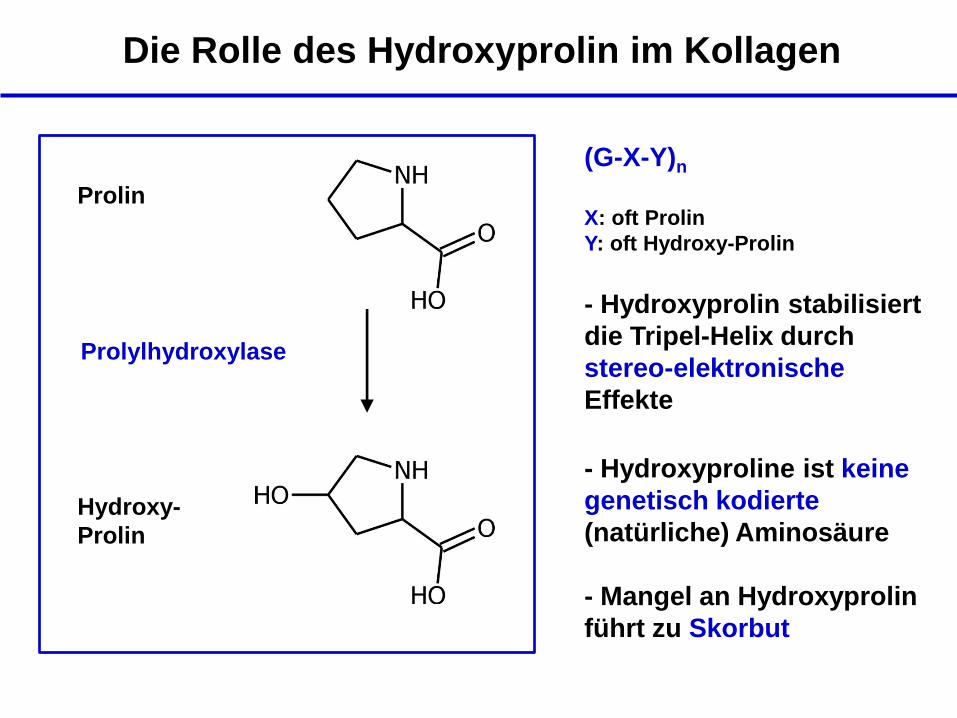
Prolin
Hydroxy-
Prolin
Prolylhydroxylase
+ Vitamin C
- Mangel an Hydroxyprolin
führt zu Skorbut
- Hydroxyproline ist keine
genetisch kodierte
(natürliche) Aminosäure
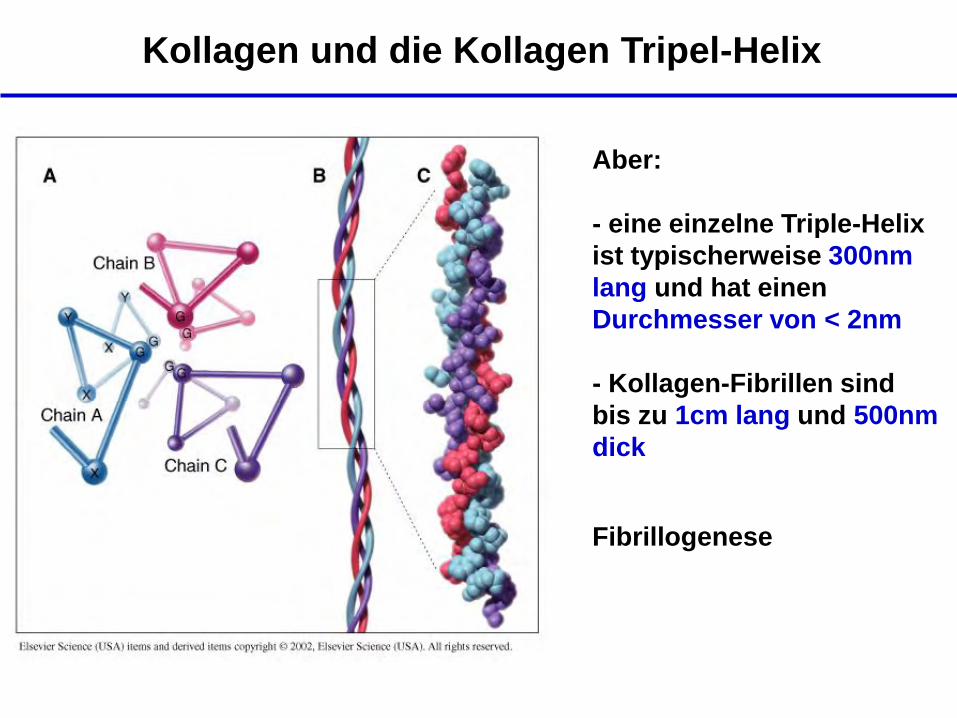
Kollagen und die Kollagen Tripel-Helix
Aber:
- eine einzelne Triple-Helix
ist typischerweise 300nm
lang und hat einen
Durchmesser von < 2nm
- Kollagen-Fibrillen sind
bis zu 1cm lang und 500nm
dick
Fibrillogenese
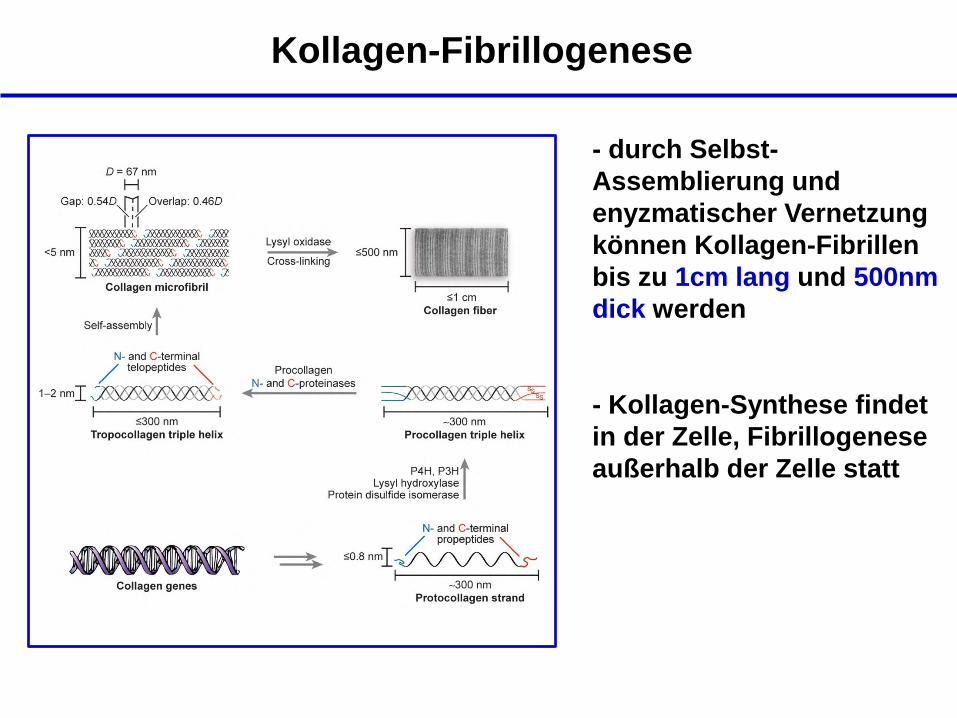
Kollagen-Fibrillogenese
- durch Selbst-
Assemblierung und
enyzmatischer Vernetzung
können Kollagen-Fibrillen
bis zu 1cm lang und 500nm
dick werden
- Kollagen-Synthese findet
in der Zelle, Fibrillogenese
außerhalb der Zelle statt
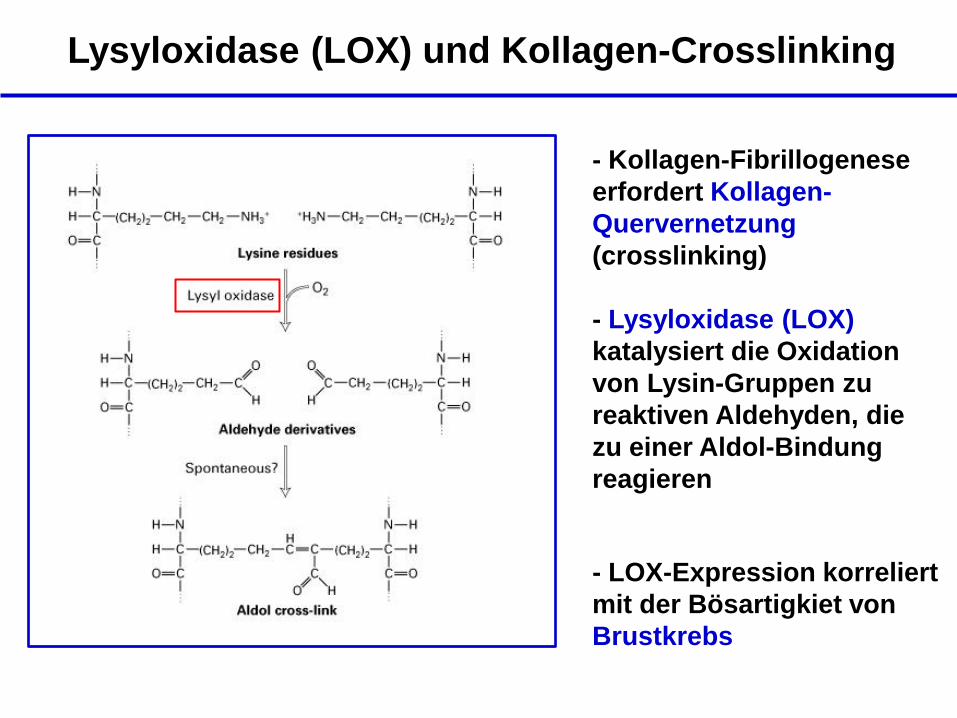
Lysyloxidase (LOX) und Kollagen-Crosslinking
- Kollagen-Fibrillogenese
erfordert Kollagen-
Quervernetzung
(crosslinking)
- LOX-Expression korreliert
mit der Bösartigkiet von
Brustkrebs
- Lysyloxidase (LOX)
katalysiert die Oxidation
von Lysin-Gruppen zu
reaktiven Aldehyden, die
zu einer Aldol-Bindung
reagieren

Es gibt eine Vielzahl von Kollagen-Netzwerken
- es gibt mindestens 28
verschiedene Kollagene,
die unterschiedlich stark
vernetzt werden können
- daher werden eine
Vielzahl von Strukturen
gebildet wie z.B. Fibrillen,
Netzwerke, Membranen,
etc.
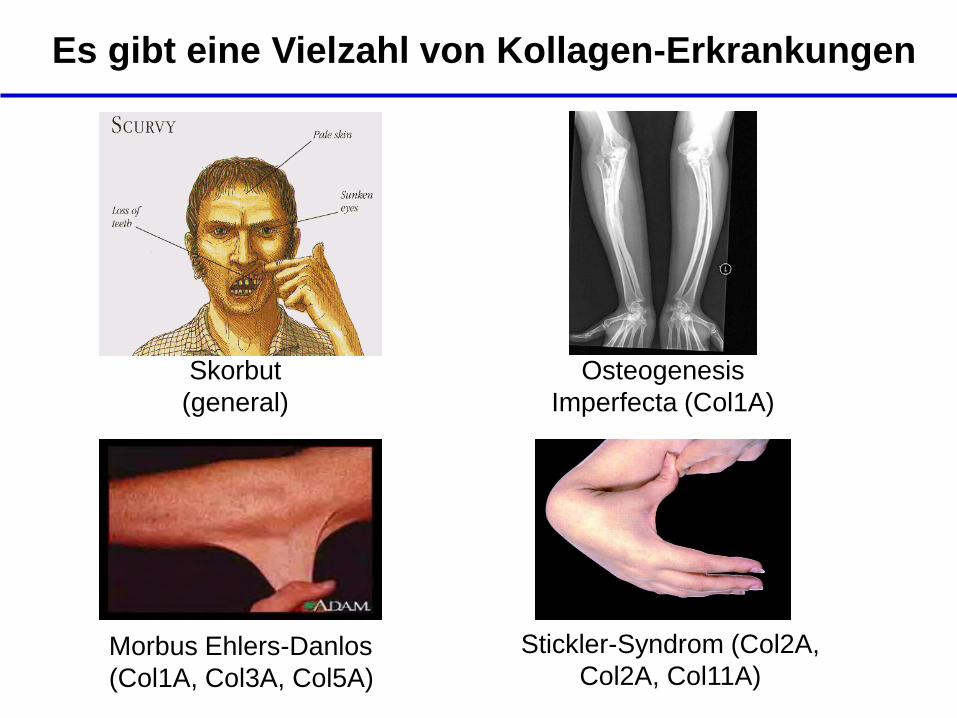
Es gibt eine Vielzahl von Kollagen-Erkrankungen
Osteogenesis
Imperfecta (Col1A)
Morbus Ehlers-Danlos
(Col1A, Col3A, Col5A)
Stickler-Syndrom (Col2A,
Col2A, Col11A)
Skorbut
(general)
Aber die EZM muss auch elastisch sein !
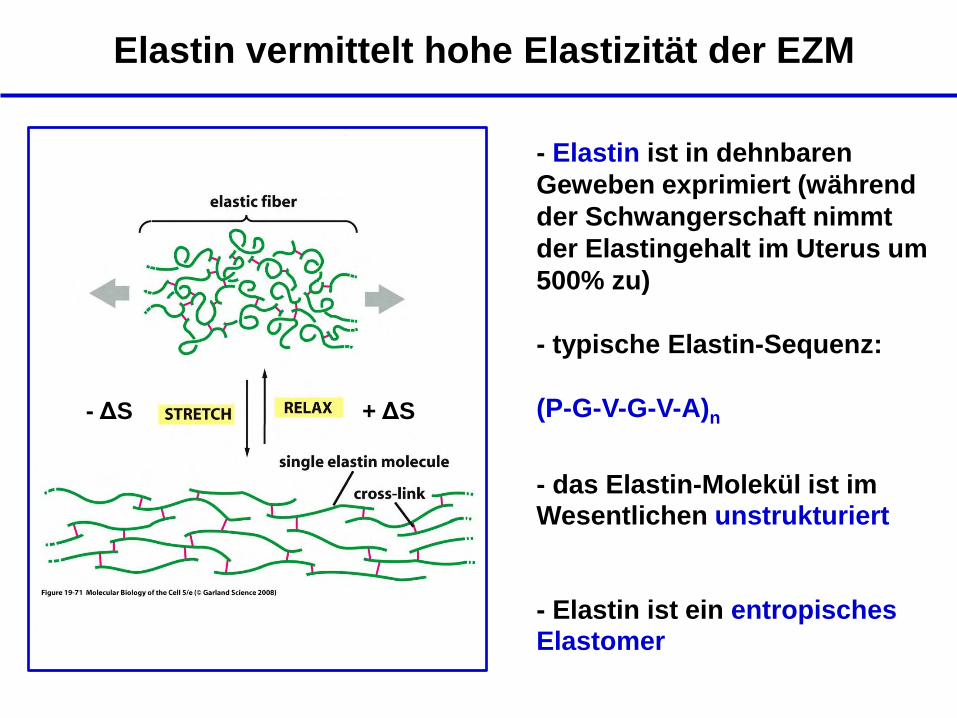
Elastin vermittelt hohe Elastizität der EZM
- Elastin ist in dehnbaren
Geweben exprimiert (während
der Schwangerschaft nimmt
der Elastingehalt im Uterus um
500% zu)
- typische Elastin-Sequenz:
(P-G-V-G-V-A)n
- das Elastin-Molekül ist im Wesentlichen unstrukturiert
+ ΔS - ΔS
- Elastin ist ein entropisches Elastomer
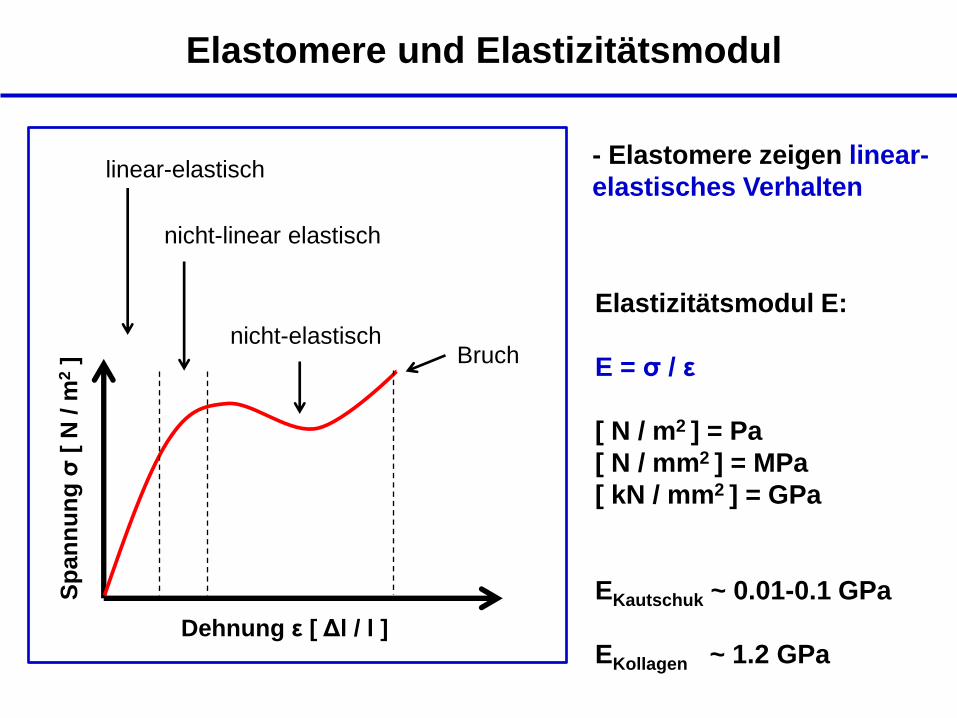
Elastomere und Elastizitätsmodul
- Elastomere zeigen linear-
elastisches Verhalten
nicht-elastisch
Dehnung ε [ Δl / l ]
Sp
an
nu
ng
σ [
N / m
2 ]
Bruch
linear-elastisch
nicht-linear elastisch
Elastizitätsmodul E:
E = σ / ε
[ N / m2 ] = Pa
[ N / mm2 ] = MPa
[ kN / mm2 ] = GPa
EKautschuk ~ 0.01-0.1 GPa
EKollagen ~ 1.2 GPa

Beispiele von natürlichen entropischen Elastomeren
Elastin
(P-G-V-G-V-A)n
Flagelliform
(G-P-G-G-A)n
Resilin
(G-G-R-P)n
E ~ 0.0011 GPa E ~ 0.003 GPa E ~ 0.002 GPa
EKautschuk ~ 0.01-0.1 GPa EKollagen ~ 1.2 GPa
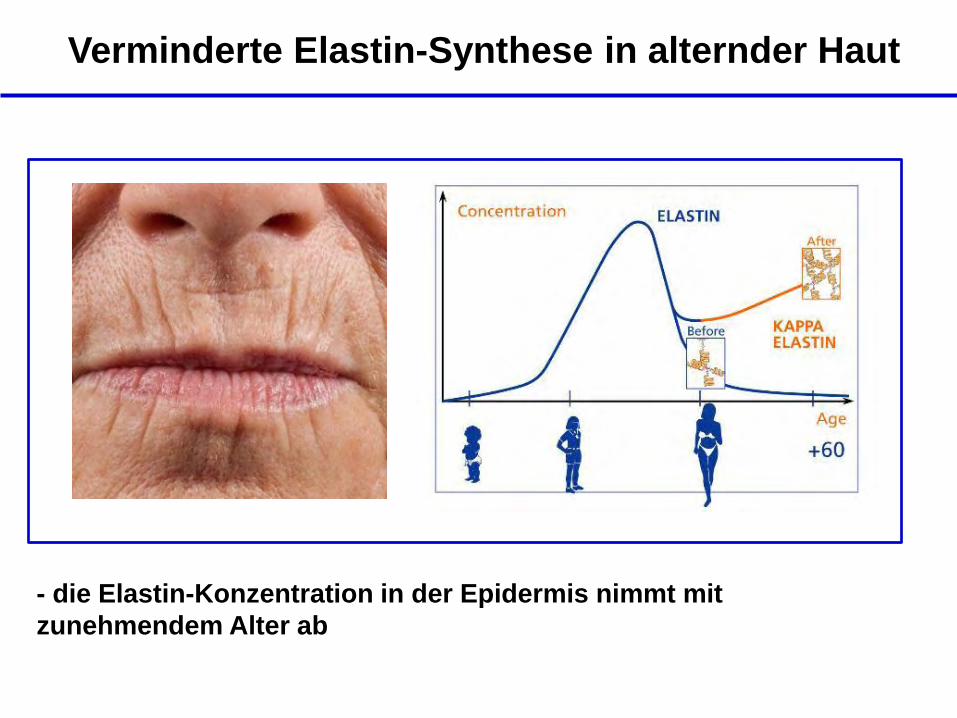
Verminderte Elastin-Synthese in alternder Haut
- die Elastin-Konzentration in der Epidermis nimmt mit
zunehmendem Alter ab
Was vermittelt Kompressionswiderstand ?
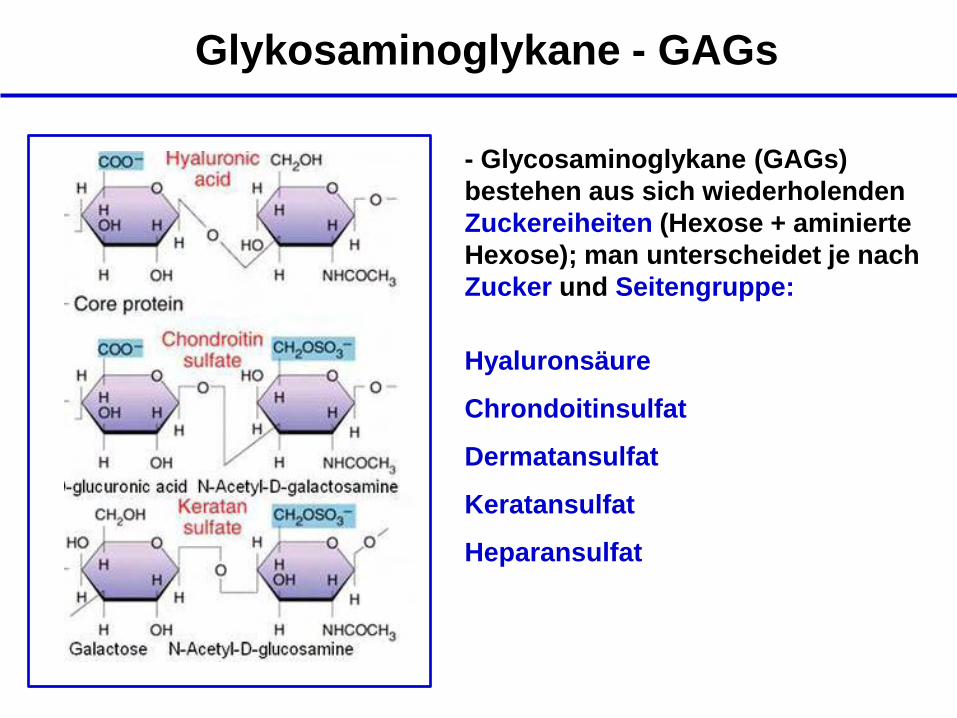
Glykosaminoglykane - GAGs
- Glycosaminoglykane (GAGs)
bestehen aus sich wiederholenden
Zuckereiheiten (Hexose + aminierte
Hexose); man unterscheidet je nach
Zucker und Seitengruppe:
Hyaluronsäure
Chrondoitinsulfat
Dermatansulfat
Keratansulfat
Heparansulfat
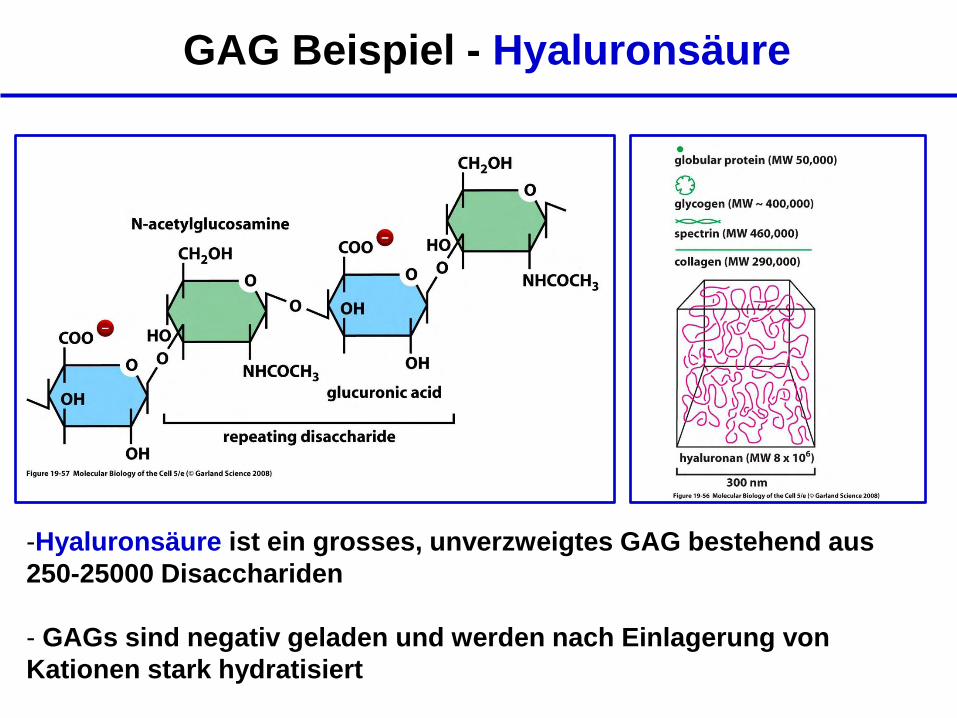
GAG Beispiel - Hyaluronsäure
-Hyaluronsäure ist ein grosses, unverzweigtes GAG bestehend aus
250-25000 Disacchariden
- GAGs sind negativ geladen und werden nach Einlagerung von
Kationen stark hydratisiert
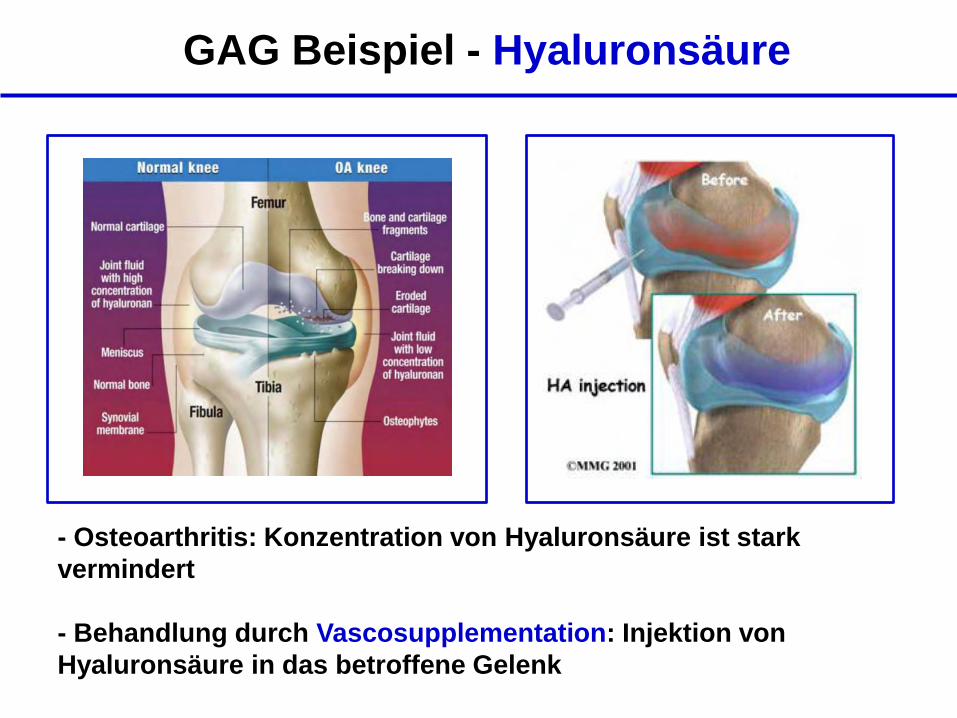
GAG Beispiel - Hyaluronsäure
- Osteoarthritis: Konzentration von Hyaluronsäure ist stark
vermindert
- Behandlung durch Vascosupplementation: Injektion von
Hyaluronsäure in das betroffene Gelenk
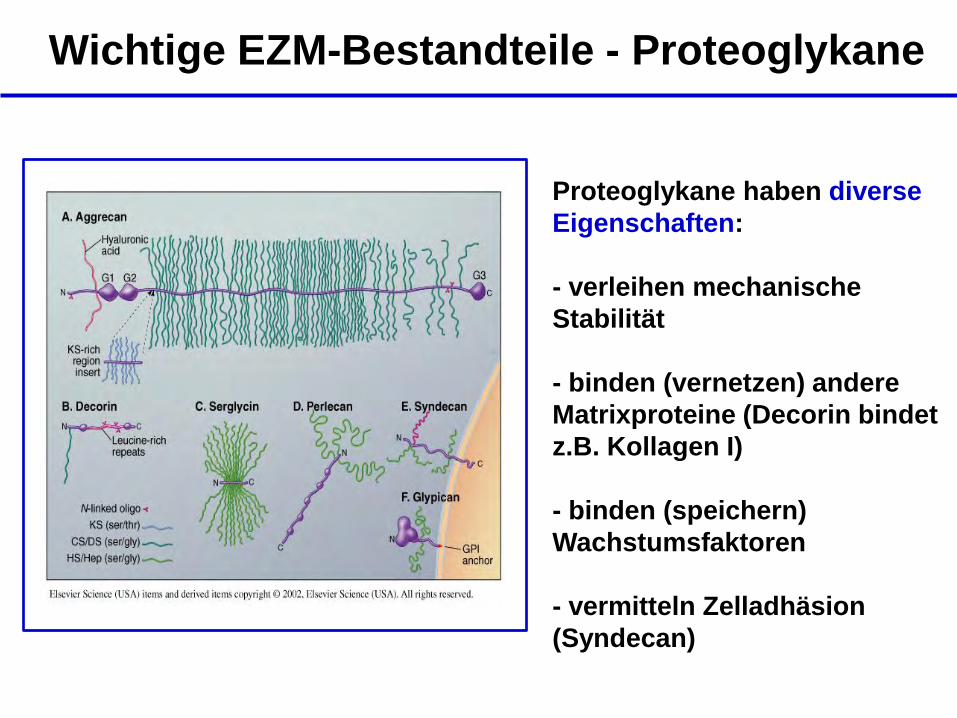
Wichtige EZM-Bestandteile - Proteoglykane
Proteoglykane haben diverse
Eigenschaften:
- verleihen mechanische
Stabilität
- binden (vernetzen) andere
Matrixproteine (Decorin bindet
z.B. Kollagen I)
- binden (speichern)
Wachstumsfaktoren
- vermitteln Zelladhäsion
(Syndecan)
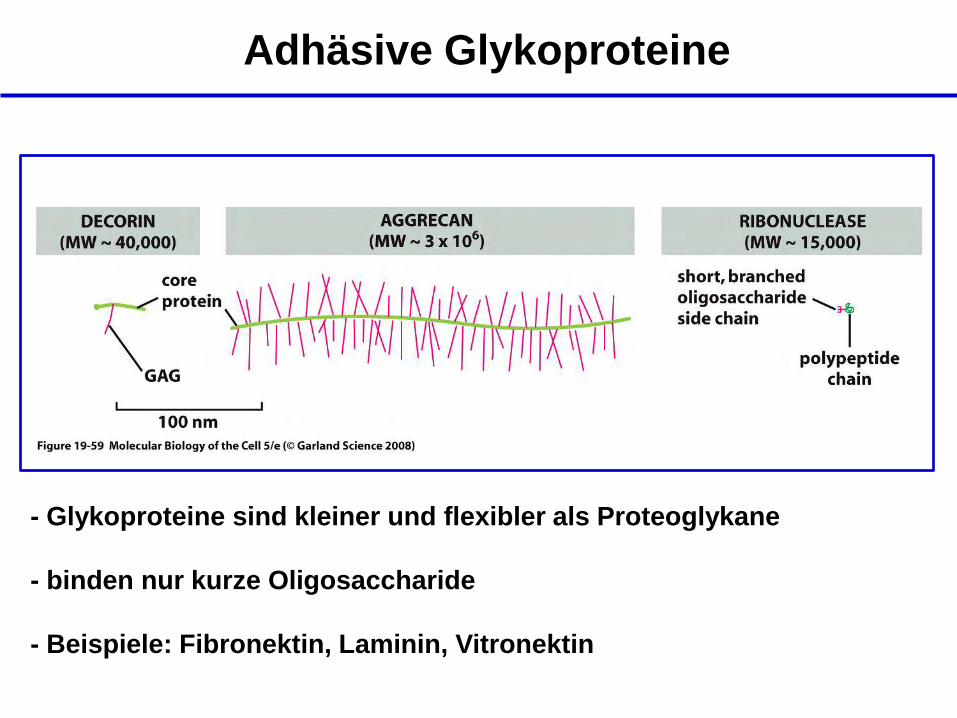
Adhäsive Glykoproteine
- Glykoproteine sind kleiner und flexibler als Proteoglykane
- binden nur kurze Oligosaccharide
- Beispiele: Fibronektin, Laminin, Vitronektin
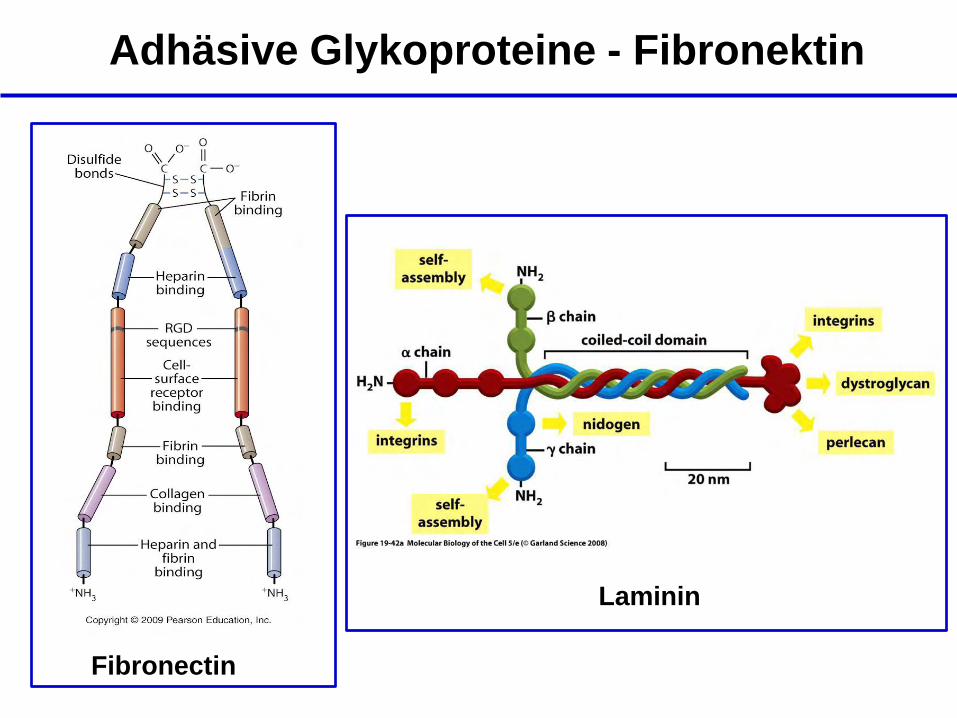
Adhäsive Glykoproteine - Fibronektin
Fibronectin
Laminin

Zusammenfassung - EZM
- die Zusammensetzung der EZM ist komplex; aus ihr
ergeben sich die mechanischen Eigenschaften des
Gewebes
- die EZM besteht aus Proteinen, Glykosaminoglykanen,
Proteoglykanen und Glykoproteinen
- Kollagene vermitteln hohe Zugfestigkeit
- Elastin vermittelt elastische Eigenschaften
- GAGs vermitteln hohen Kompressionswiderstand
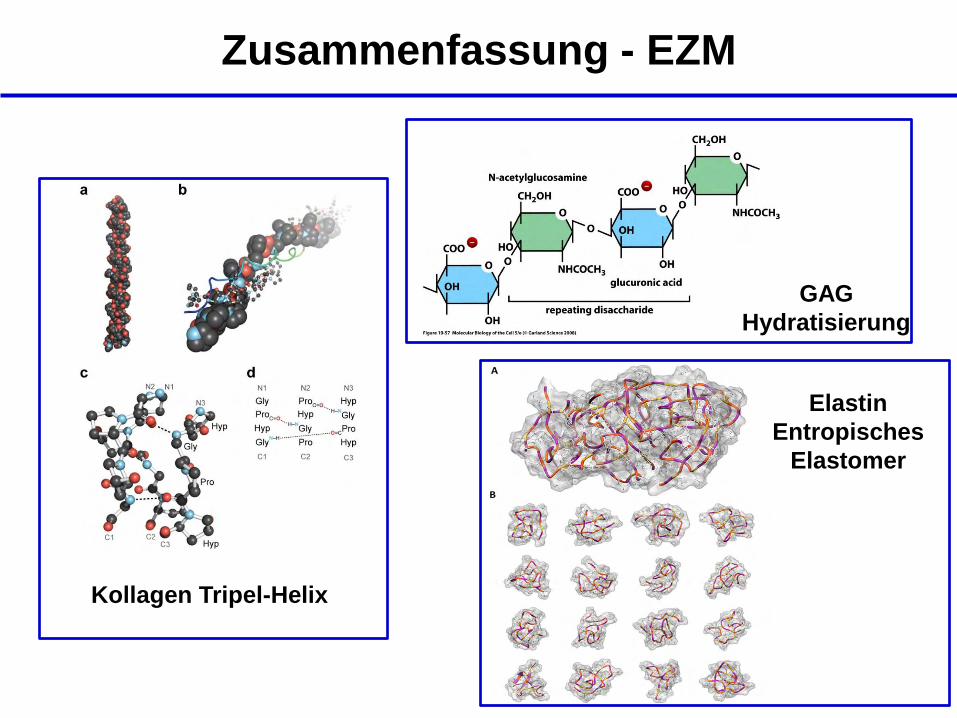
Zusammenfassung - EZM
Kollagen Tripel-Helix
Elastin
Entropisches
Elastomer
GAG
Hydratisierung

Zusammenfassung - EZM

1. Wie erkennt eine Zelle seine
mechanische Umgebung?
Die biophysikalische Fragestellung
Welche Moleküle erkennen die EZM ?
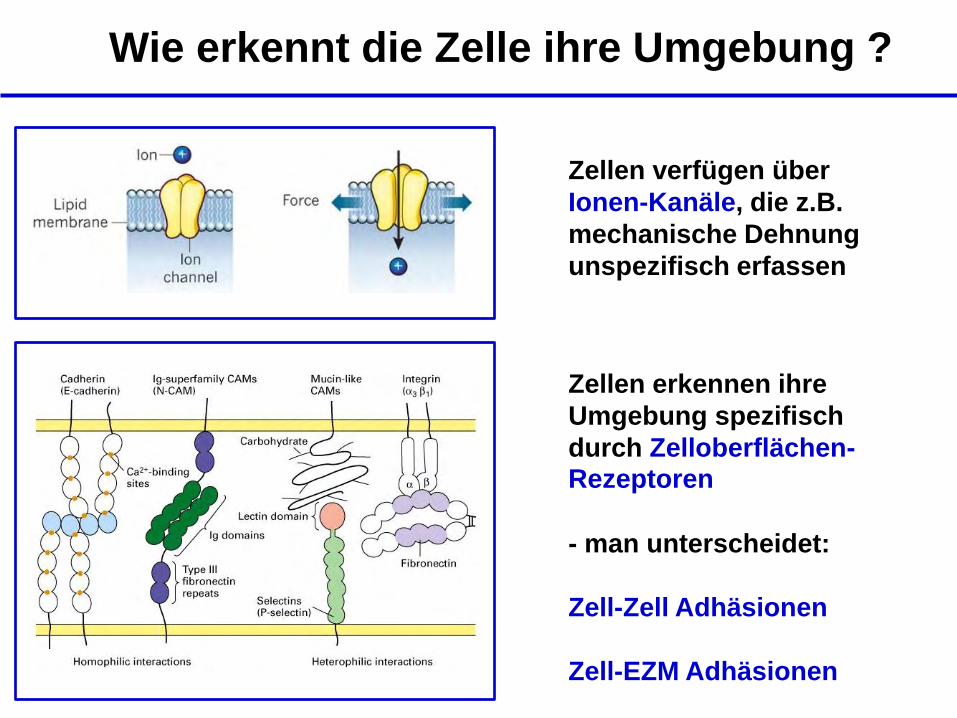
Wie erkennt die Zelle ihre Umgebung ?
Zellen verfügen über
Ionen-Kanäle, die z.B.
mechanische Dehnung
unspezifisch erfassen
Zellen erkennen ihre
Umgebung spezifisch
durch Zelloberflächen-Rezeptoren
- man unterscheidet:
Zell-Zell Adhäsionen
Zell-EZM Adhäsionen
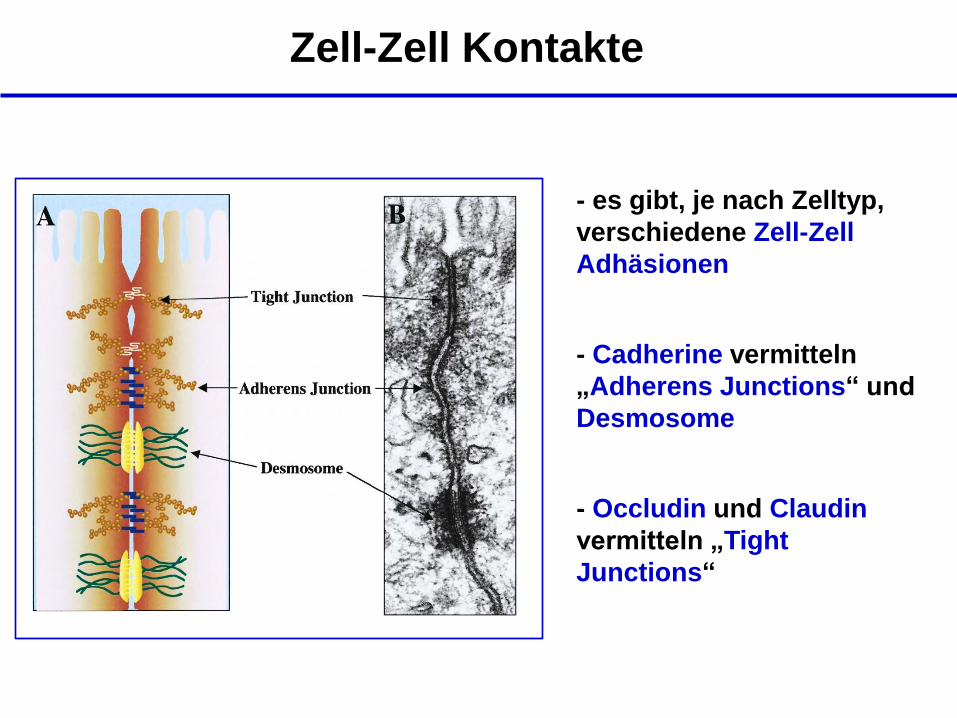
Zell-Zell Kontakte
- es gibt, je nach Zelltyp,
verschiedene Zell-Zell
Adhäsionen
- Cadherine vermitteln
„Adherens Junctions“ und
Desmosome
- Occludin und Claudin
vermitteln „Tight
Junctions“
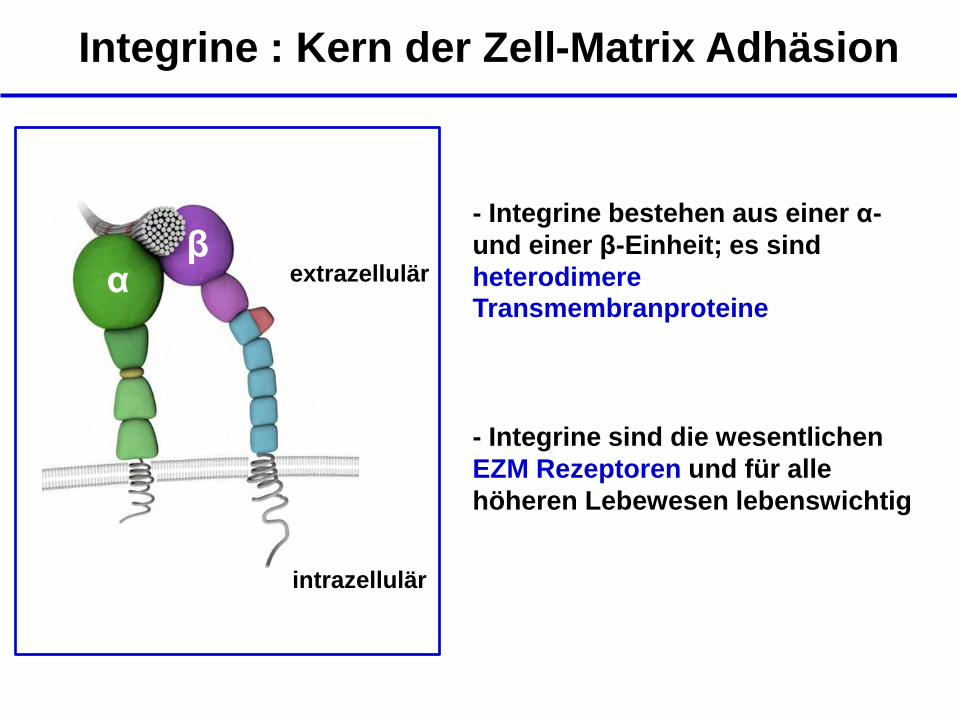
Integrine : Kern der Zell-Matrix Adhäsion
β α
intrazellulär
extrazellulär
- Integrine bestehen aus einer α-
und einer β-Einheit; es sind
heterodimere Transmembranproteine
- Integrine sind die wesentlichen
EZM Rezeptoren und für alle
höheren Lebewesen lebenswichtig
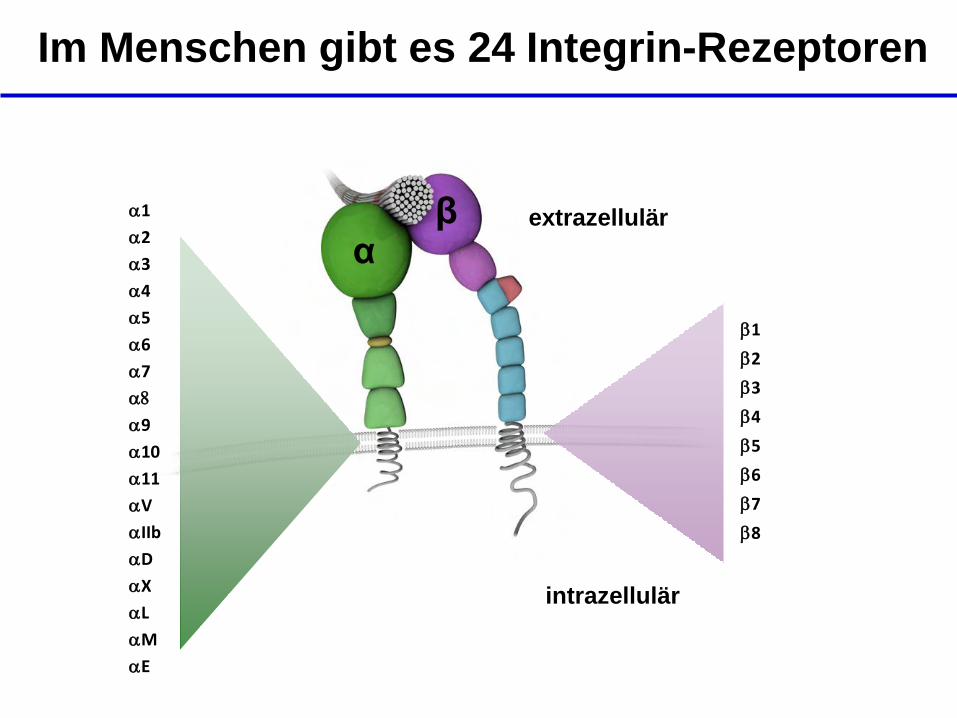
Im Menschen gibt es 24 Integrin-Rezeptoren
β α
b1
b2
b3
b4
b5
b6
b7
b8
a1
a2
a3
a4
a5
a6
a7
a8
a9
a10
a11
aV
aIIb
aD
aX
aL
aM
aE
intrazellulär
extrazellulär
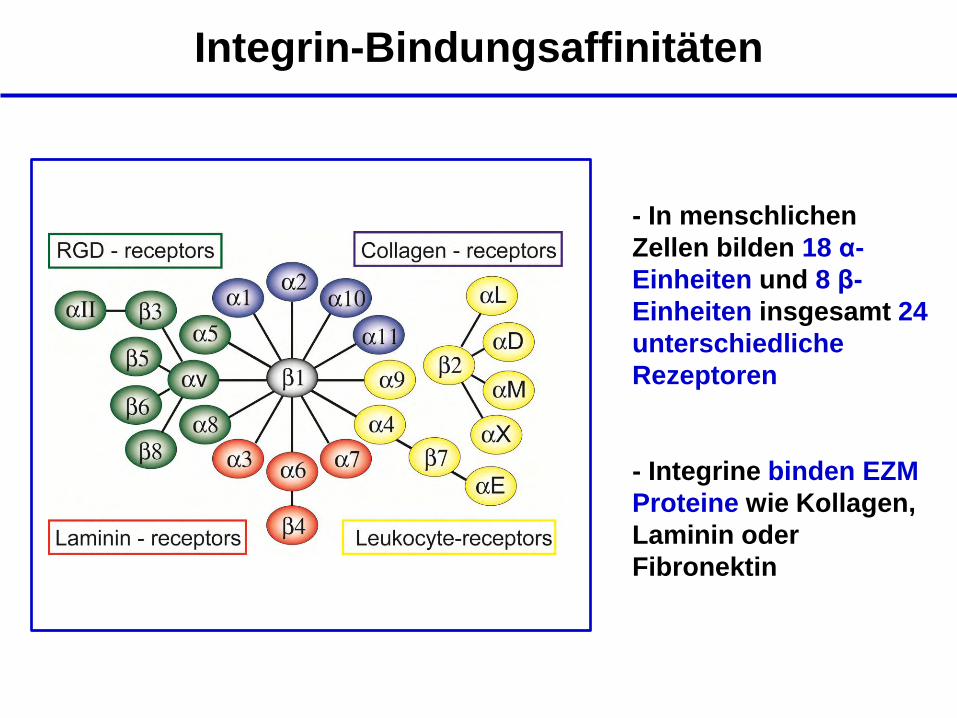
Integrin-Bindungsaffinitäten
- In menschlichen
Zellen bilden 18 α-
Einheiten und 8 β-
Einheiten insgesamt 24
unterschiedliche
Rezeptoren
- Integrine binden EZM
Proteine wie Kollagen,
Laminin oder
Fibronektin
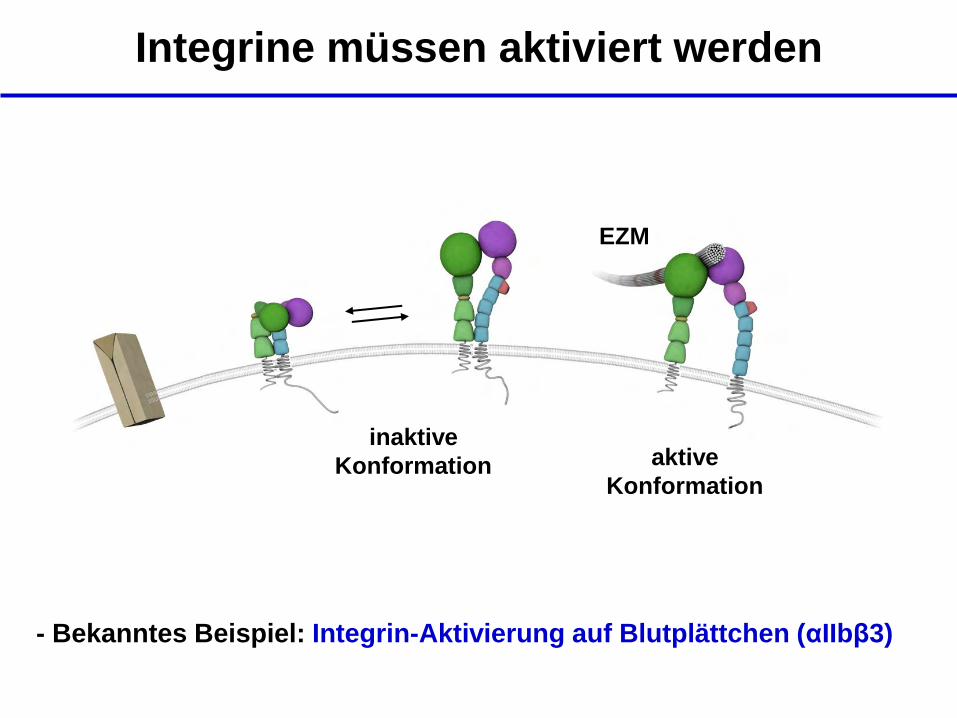
Integrine müssen aktiviert werden
EZM
inaktive
Konformation aktive
Konformation
- Bekanntes Beispiel: Integrin-Aktivierung auf Blutplättchen (αIIbβ3)
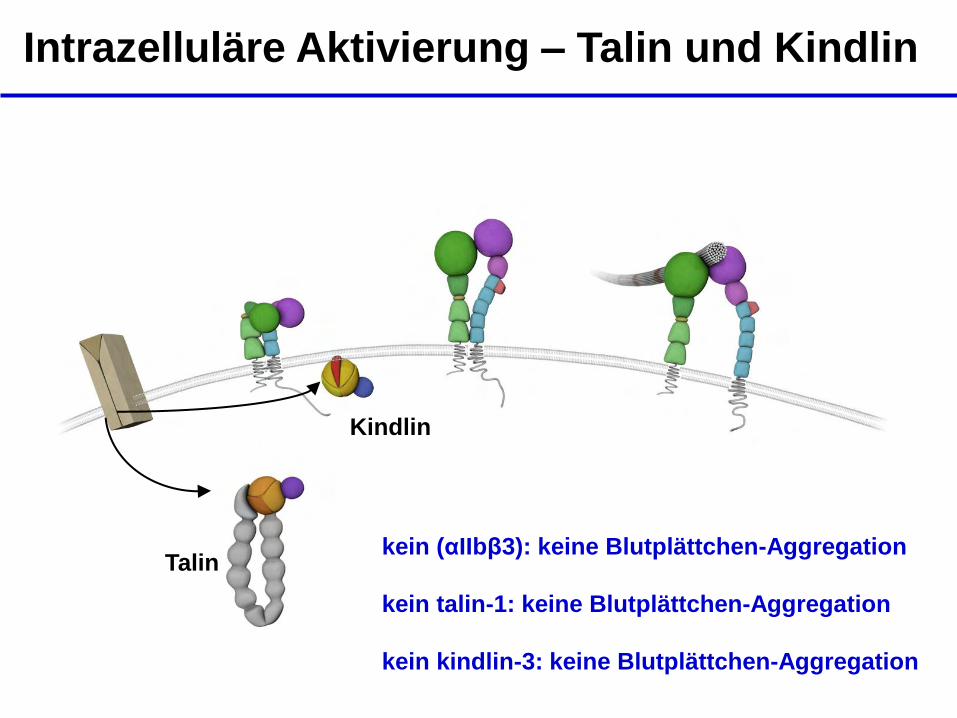
Intrazelluläre Aktivierung – Talin und Kindlin
Talin
Kindlin
kein (αIIbβ3): keine Blutplättchen-Aggregation
kein talin-1: keine Blutplättchen-Aggregation kein kindlin-3: keine Blutplättchen-Aggregation
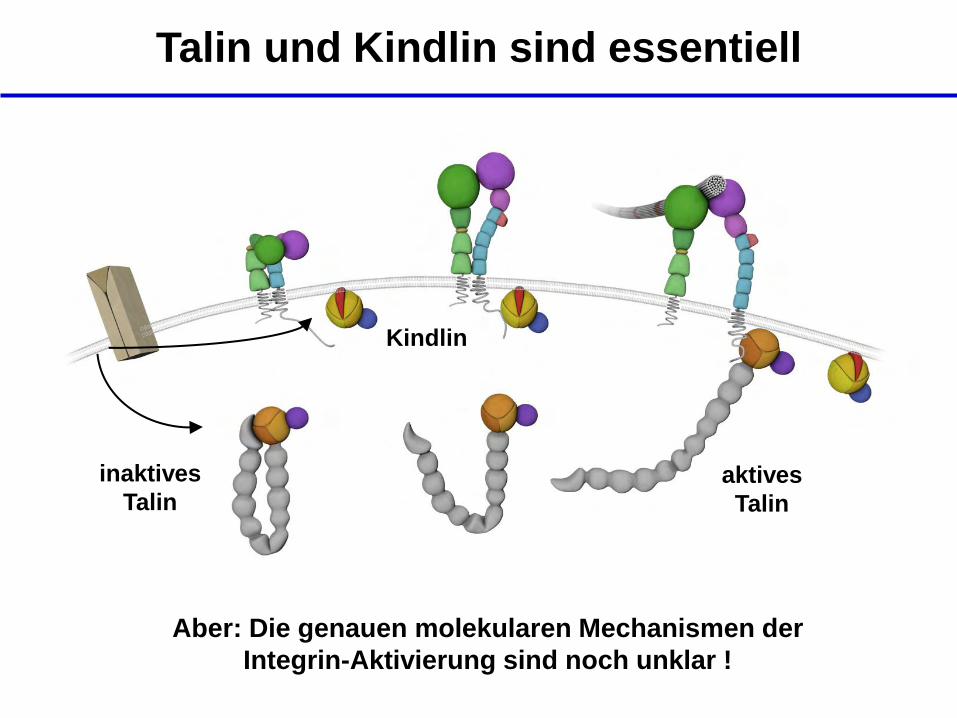
Talin und Kindlin sind essentiell
inaktives
Talin aktives
Talin
Kindlin
Aber: Die genauen molekularen Mechanismen der
Integrin-Aktivierung sind noch unklar !
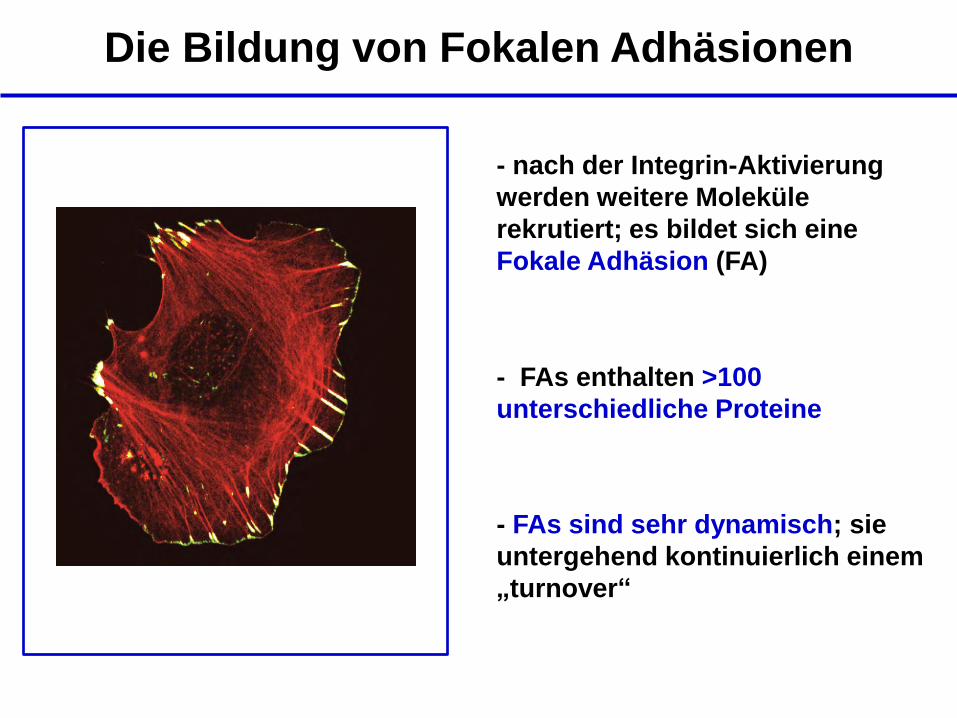
Die Bildung von Fokalen Adhäsionen
- nach der Integrin-Aktivierung
werden weitere Moleküle
rekrutiert; es bildet sich eine
Fokale Adhäsion (FA)
>100 Proteine
- FAs enthalten >100
unterschiedliche Proteine
- FAs sind sehr dynamisch; sie
untergehend kontinuierlich einem
„turnover“
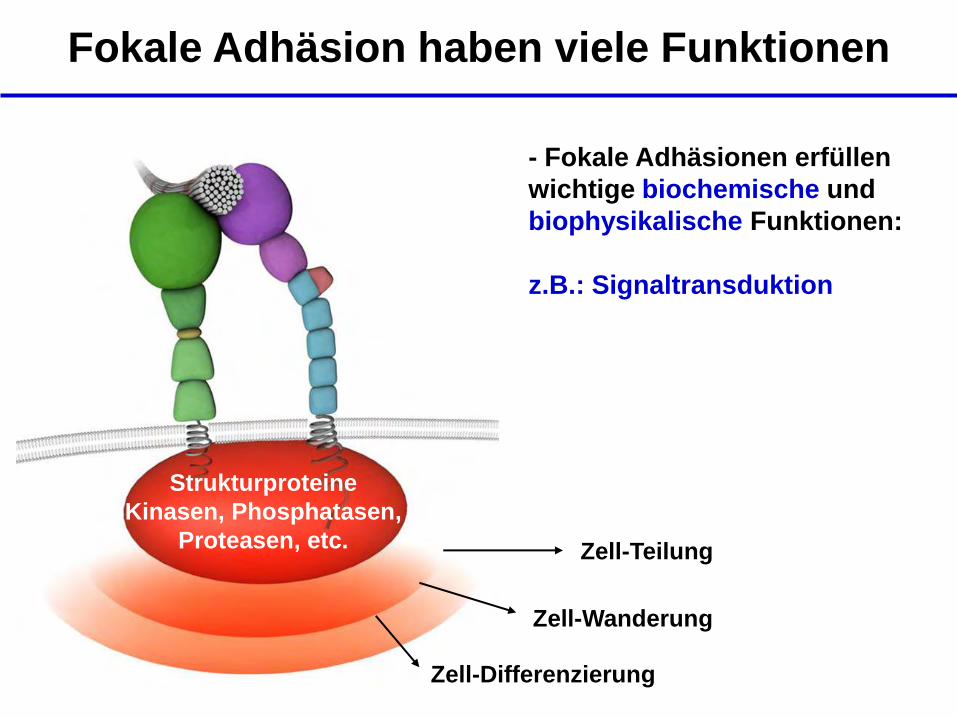
Fokale Adhäsion haben viele Funktionen
- Fokale Adhäsionen erfüllen
wichtige biochemische und
biophysikalische Funktionen:
z.B.: Signaltransduktion
Zell-Teilung
Zell-Wanderung
Zell-Differenzierung
Strukturproteine
Kinasen, Phosphatasen,
Proteasen, etc.

Zusammenfassung – Zell-Matrix Adhäsion
Zellen erfassen ihre Umgebung auf vielfache Weise:
- Dehnungsabhängige Ionen-Kanäle registrieren Spannung an der Plasma-Membran
- Cadherine vermitteln Zell-Zell Kontakte
- Integrine stellen spezifische Interaktionen zu EZM Proteinen her

Zusammenfassung – Integrine
- Integrine sind heterodimere Transmembranproteine
- zu der zytoplasmischen Integrin-Domäne werden
eine Vielzahl von Proteinen rekrutiert; diese bilden
eine fokale Adhäsion
- Integrin-Aktivierung ist Talin und Kindlin-abhängig

2. Wie verwandelt eine Zelle
physikalische Information in
biologische Prozesse ?
Die biophysikalische Fragestellung
Wie testen Integrine / fokale
Adhäsionen die mechanischen
Eigenschaften EZM ?
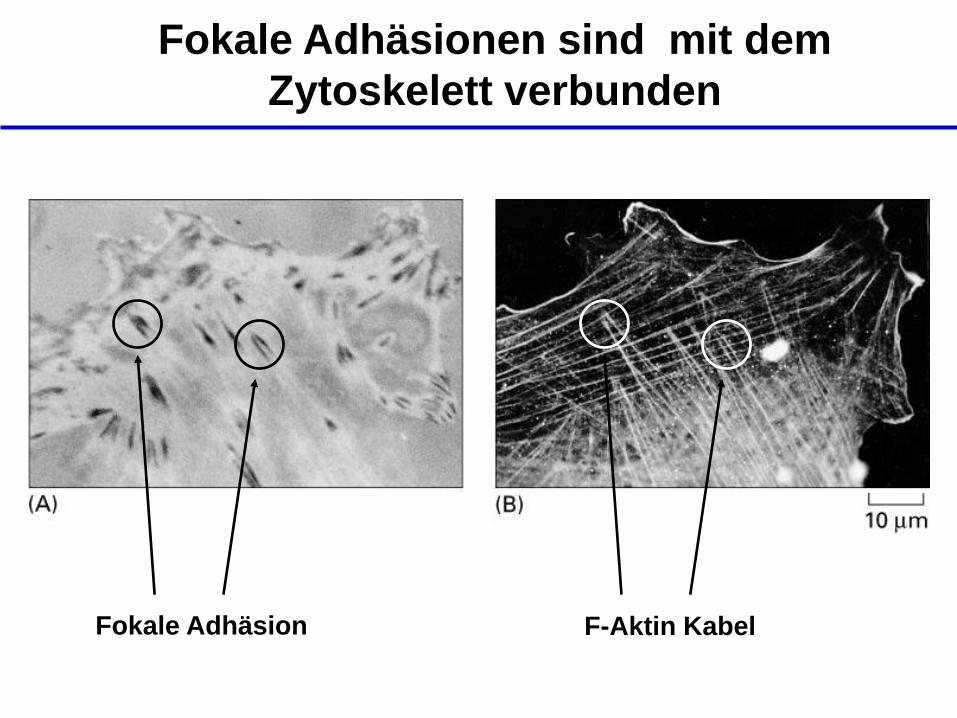
Fokale Adhäsionen sind mit dem
Zytoskelett verbunden
Fokale Adhäsion F-Aktin Kabel
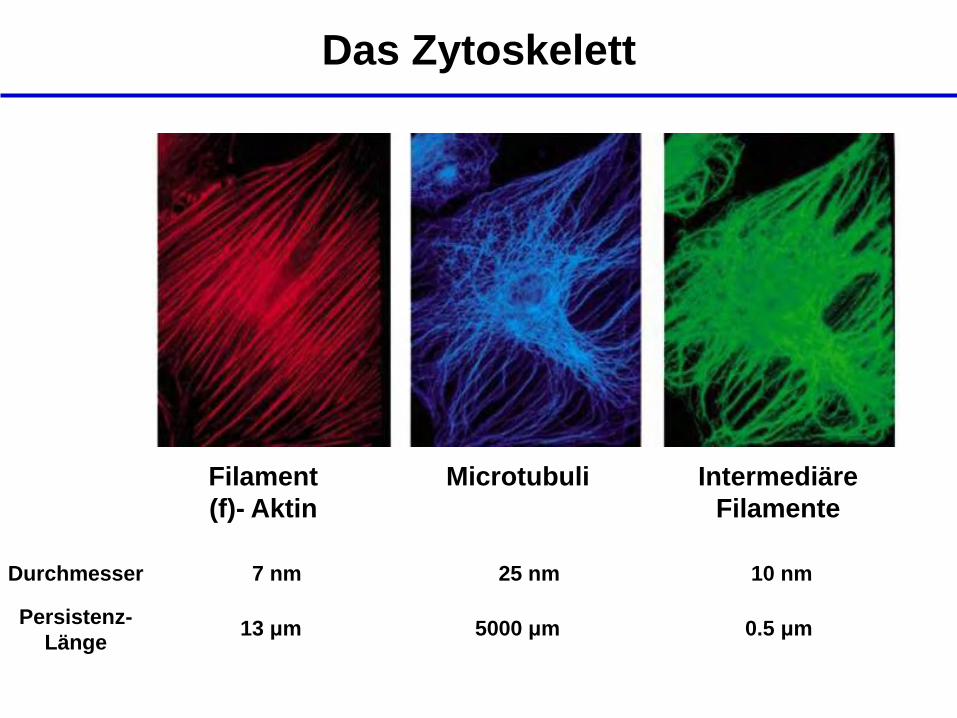
Das Zytoskelett
Filament
(f)- Aktin
Microtubuli Intermediäre
Filamente
Durchmesser 7 nm 25 nm 10 nm
Persistenz-
Länge 13 μm 5000 μm 0.5 μm
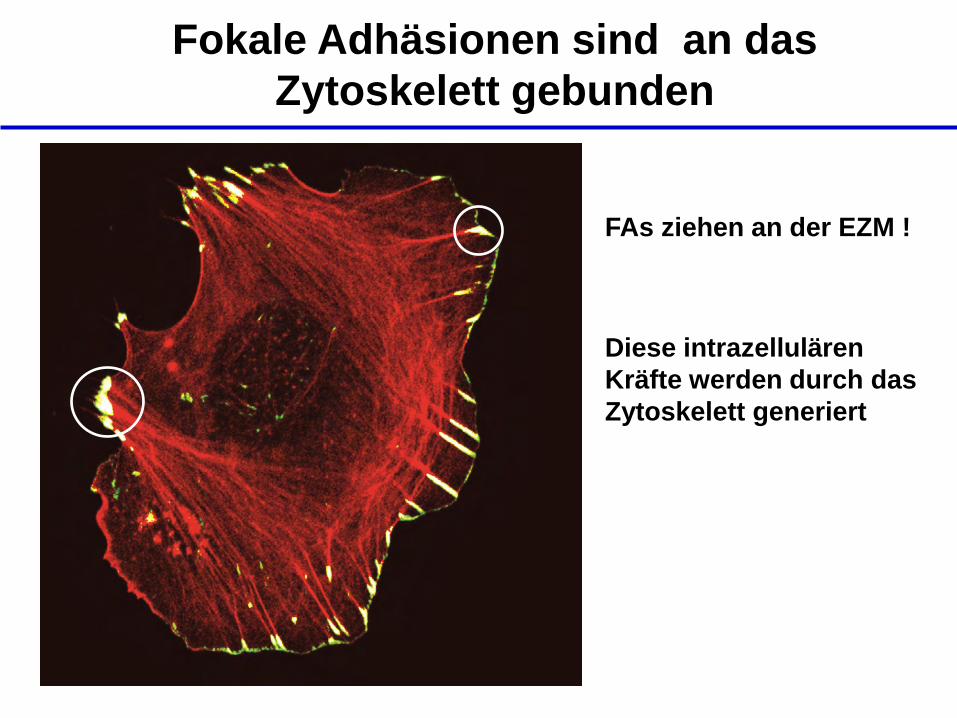
Fokale Adhäsionen sind an das
Zytoskelett gebunden
FAs ziehen an der EZM !
Diese intrazellulären
Kräfte werden durch das
Zytoskelett generiert
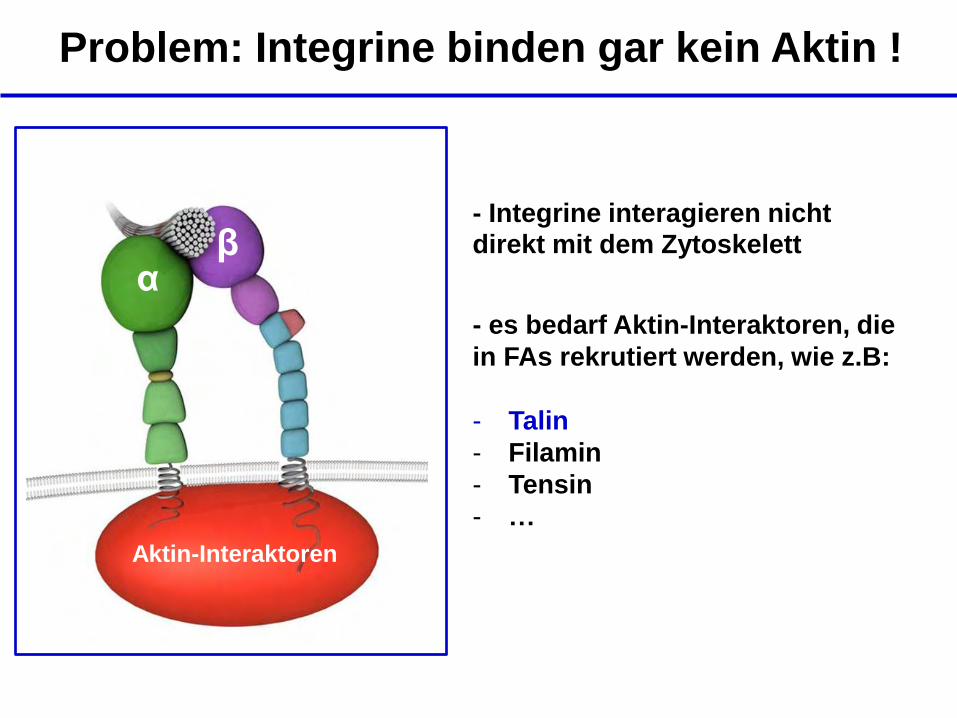
Problem: Integrine binden gar kein Aktin !
β α
- Integrine interagieren nicht direkt mit dem Zytoskelett
- es bedarf Aktin-Interaktoren, die
in FAs rekrutiert werden, wie z.B:
- Talin
- Filamin
- Tensin
- …
Aktin-Interaktoren
β α
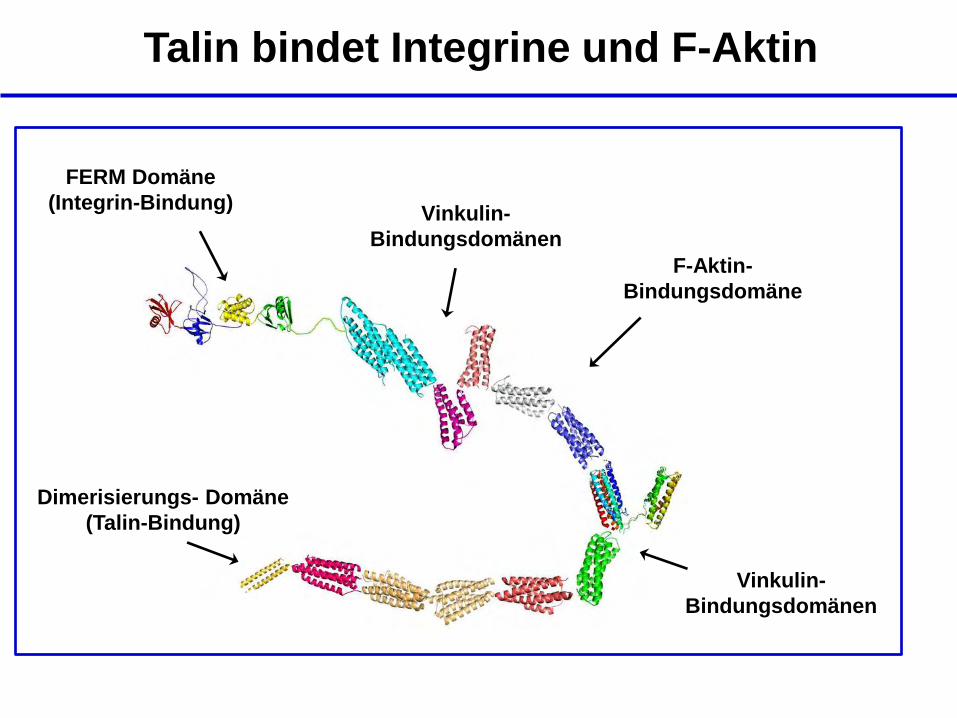
Talin bindet Integrine und F-Aktin
Dimerisierungs- Domäne
(Talin-Bindung)
FERM Domäne
(Integrin-Bindung)
F-Aktin-
Bindungsdomäne
Vinkulin-
Bindungsdomänen
Vinkulin-
Bindungsdomänen
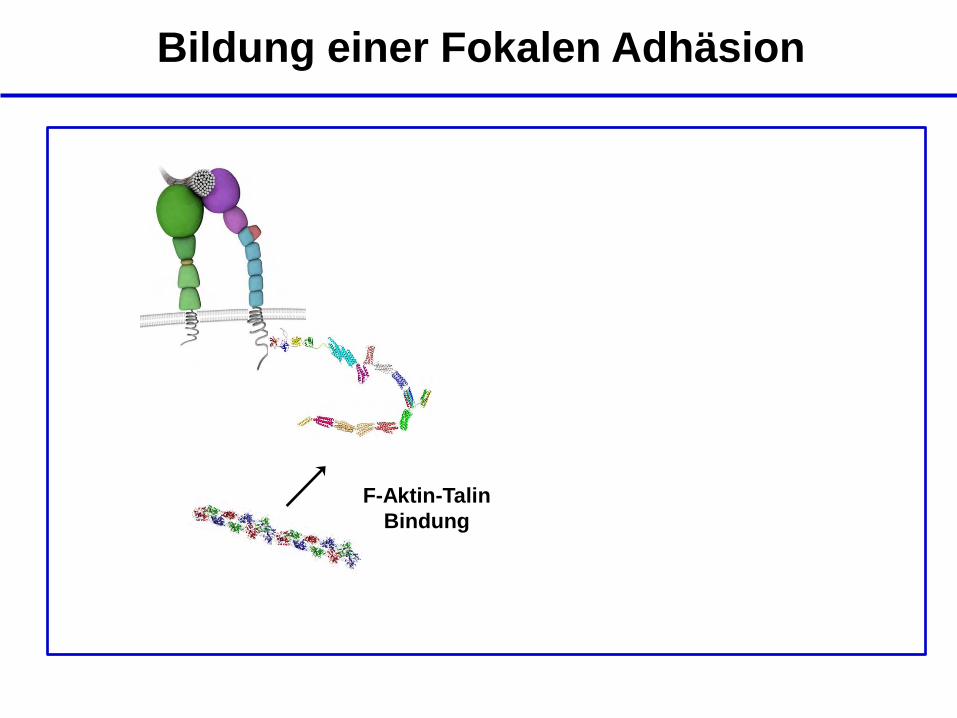
Bildung einer Fokalen Adhäsion
F-Aktin-Talin
Bindung
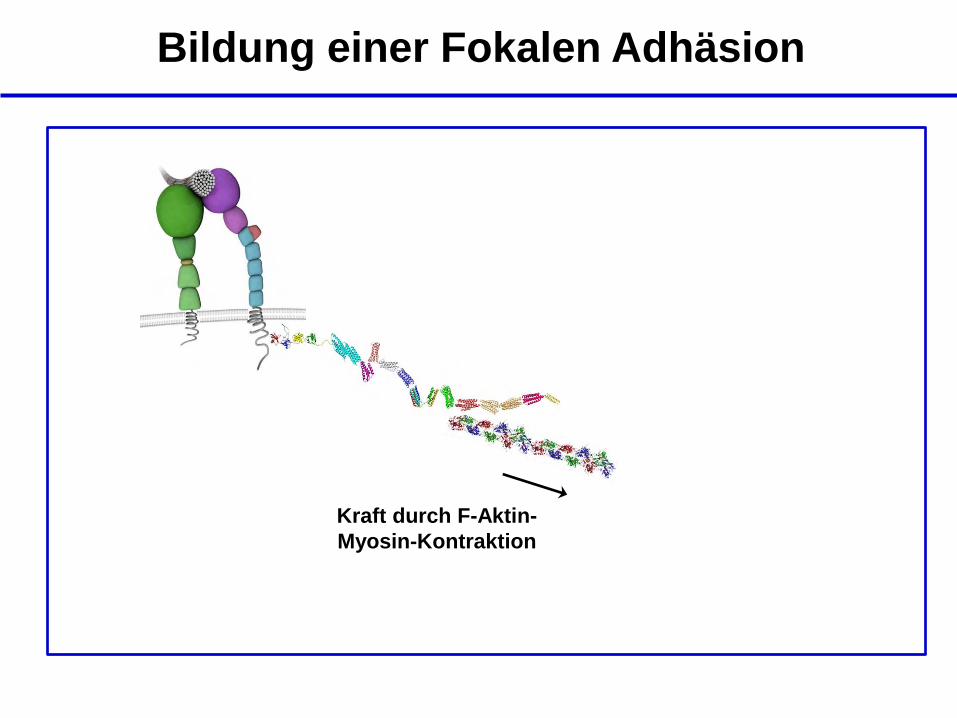
Kraft durch F-Aktin-
Myosin-Kontraktion
Bildung einer Fokalen Adhäsion
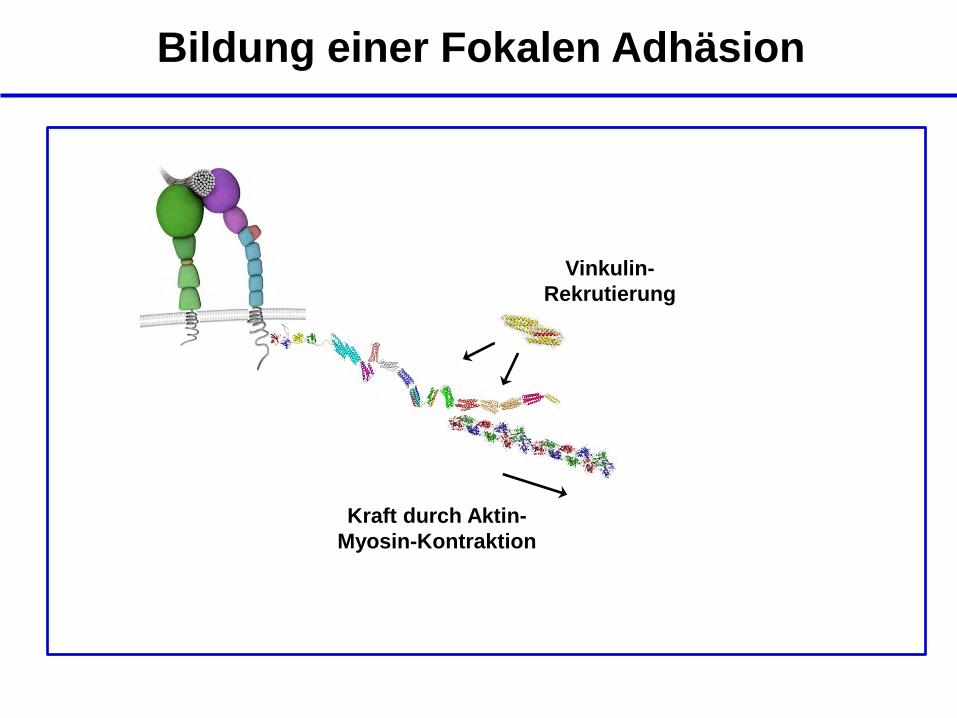
Kraft durch Aktin-
Myosin-Kontraktion
Vinkulin-
Rekrutierung
Bildung einer Fokalen Adhäsion
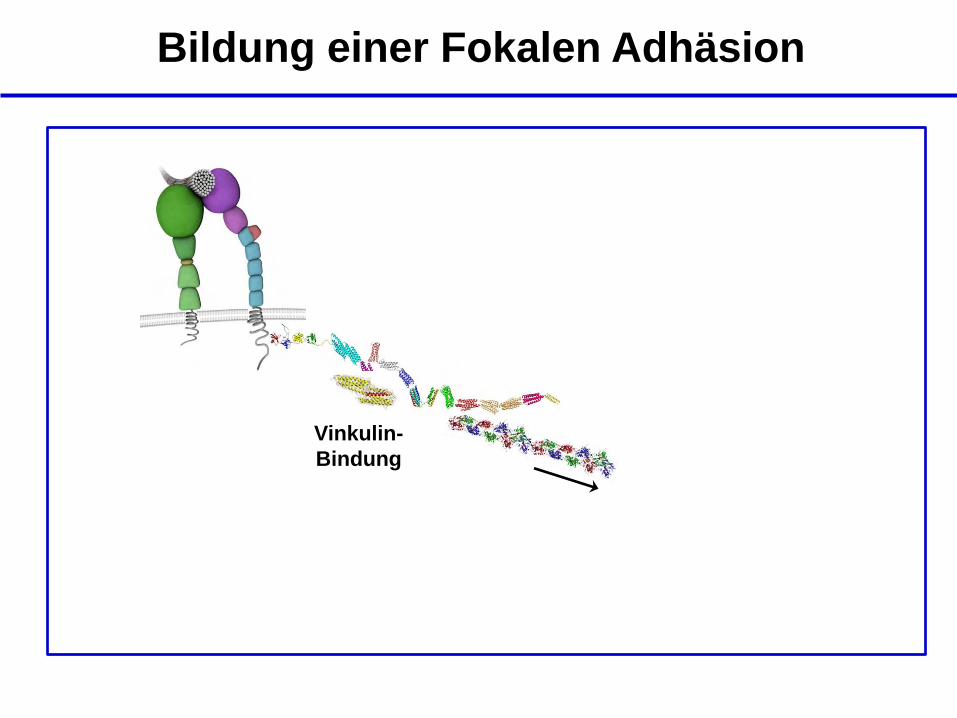
Vinkulin-
Bindung
Bildung einer Fokalen Adhäsion
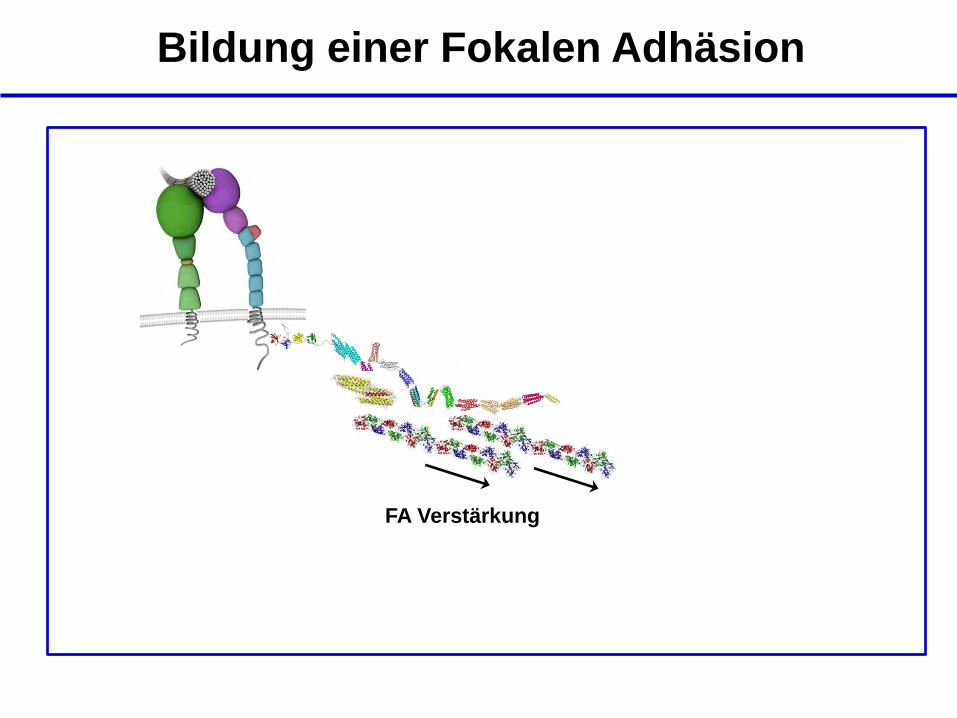
FA Verstärkung
Bildung einer Fokalen Adhäsion
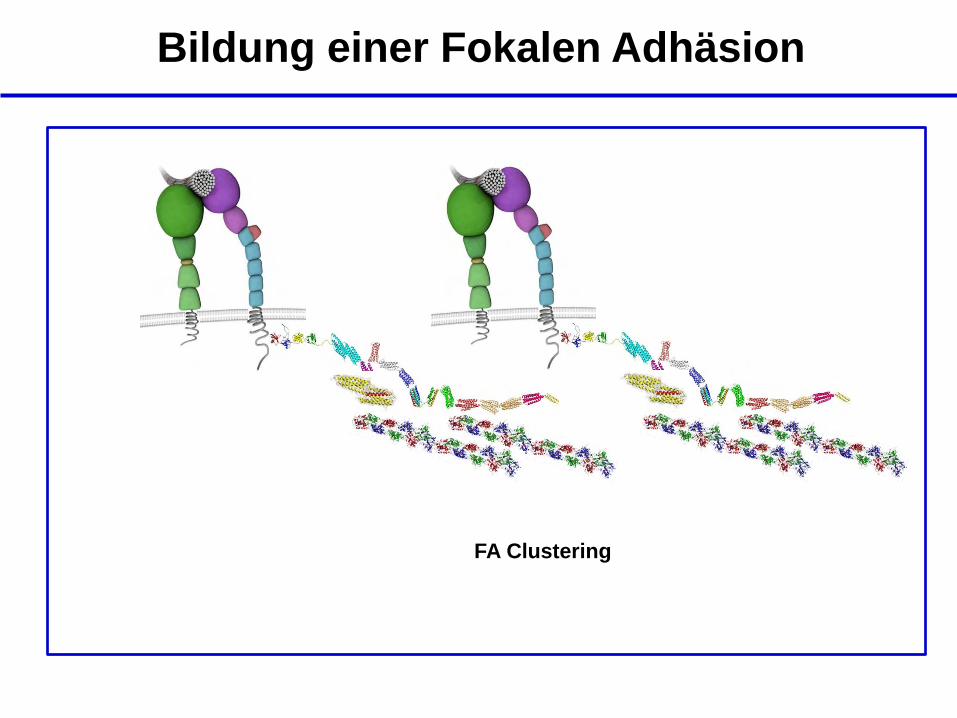
Bildung einer Fokalen Adhäsion
FA Clustering
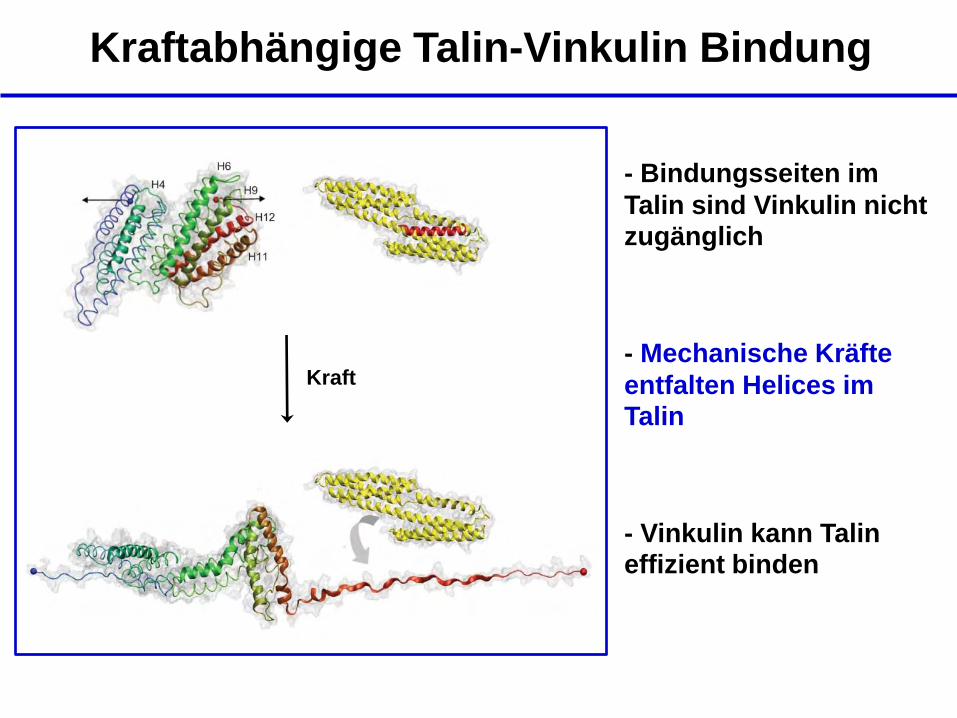
Kraftabhängige Talin-Vinkulin Bindung
- Bindungsseiten im
Talin sind Vinkulin nicht zugänglich
Kraft - Mechanische Kräfte
entfalten Helices im Talin
- Vinkulin kann Talin effizient binden
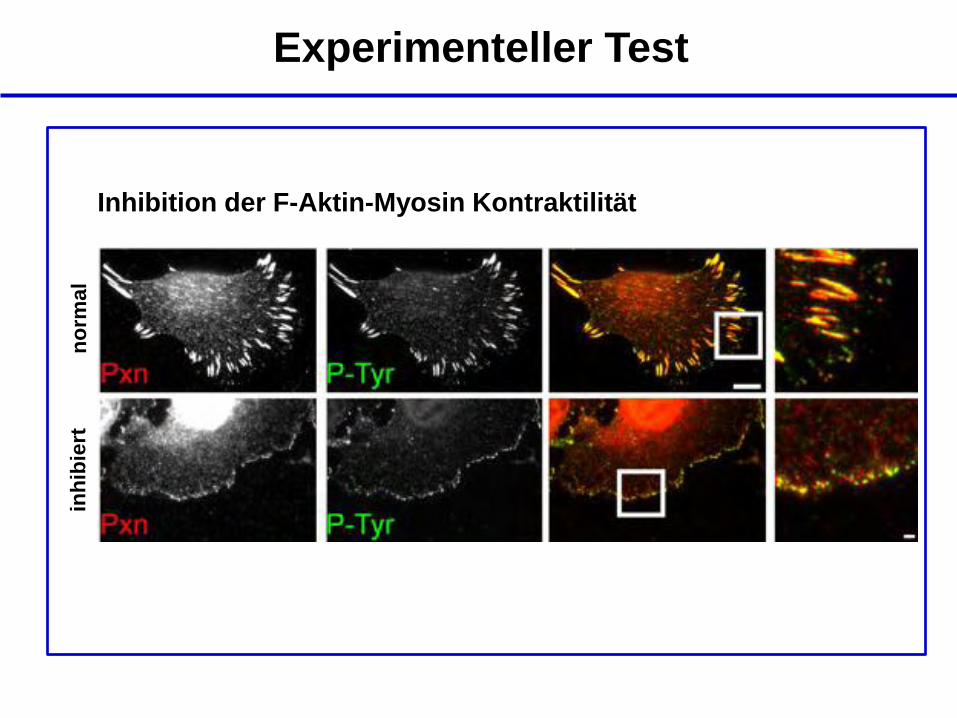
Experimenteller Test
Inhibition der F-Aktin-Myosin Kontraktilität
no
rmal
inh
ibie
rt
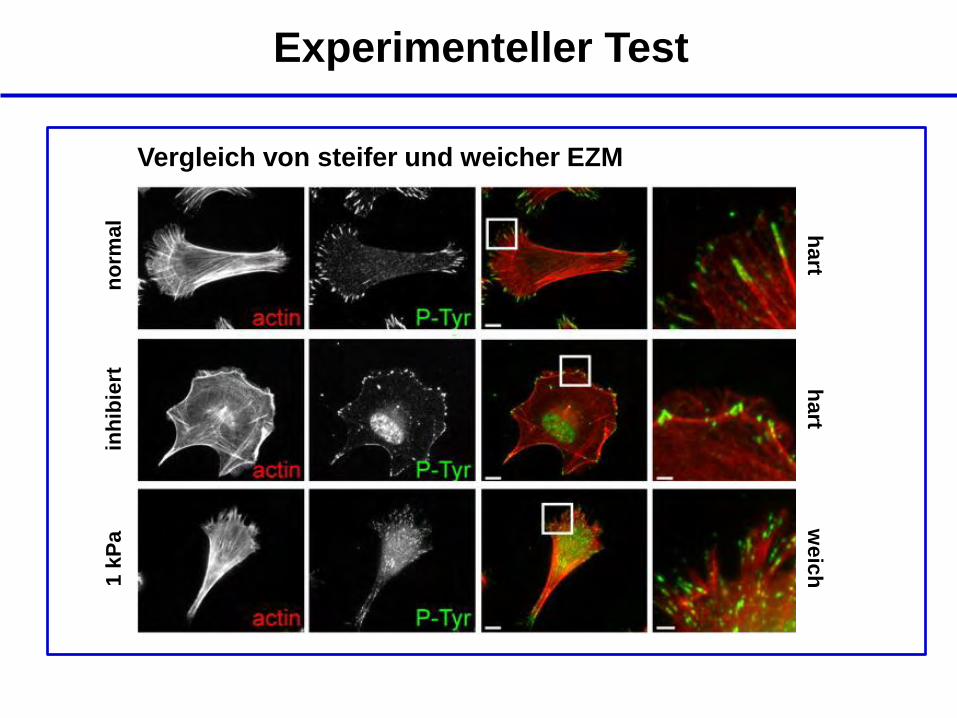
Experimenteller Test
Vergleich von steifer und weicher EZM
no
rmal
inh
ibie
rt
1 k
Pa
hart
hart
weic
h

Zusammenfassung –Integrin-Aktin Bindung
- Die Reifung Fokaler Adhäsionen sind Talin-abhängig
- Fokale Adhäsionen sind mechanosensitiv
- Talin vermittelt die Bindung von Integrinen an das F-Aktin Zytoskelett
- Fokale Adhäsionen erkennen intrazelluläre
Kontraktion sowie extrazelluläre Matrix-Rigidität


















