
Histaminintoleranz: Ursache, Diagnostik und Therapie · PDF fileHISTAMINE$ Neuro transmier$$...
25
Histaminintoleranz: Ursache, Diagnostik und Therapie Prof. Dr. med. Yurdagül Zopf Medizinische Klinik 1, Universitätsklinik Erlangen
Transcript of Histaminintoleranz: Ursache, Diagnostik und Therapie · PDF fileHISTAMINE$ Neuro transmier$$...

Histaminintoleranz:
Ursache, Diagnostik und
Therapie
Prof. Dr. med. Yurdagül Zopf
Medizinische Klinik 1, Universitätsklinik Erlangen
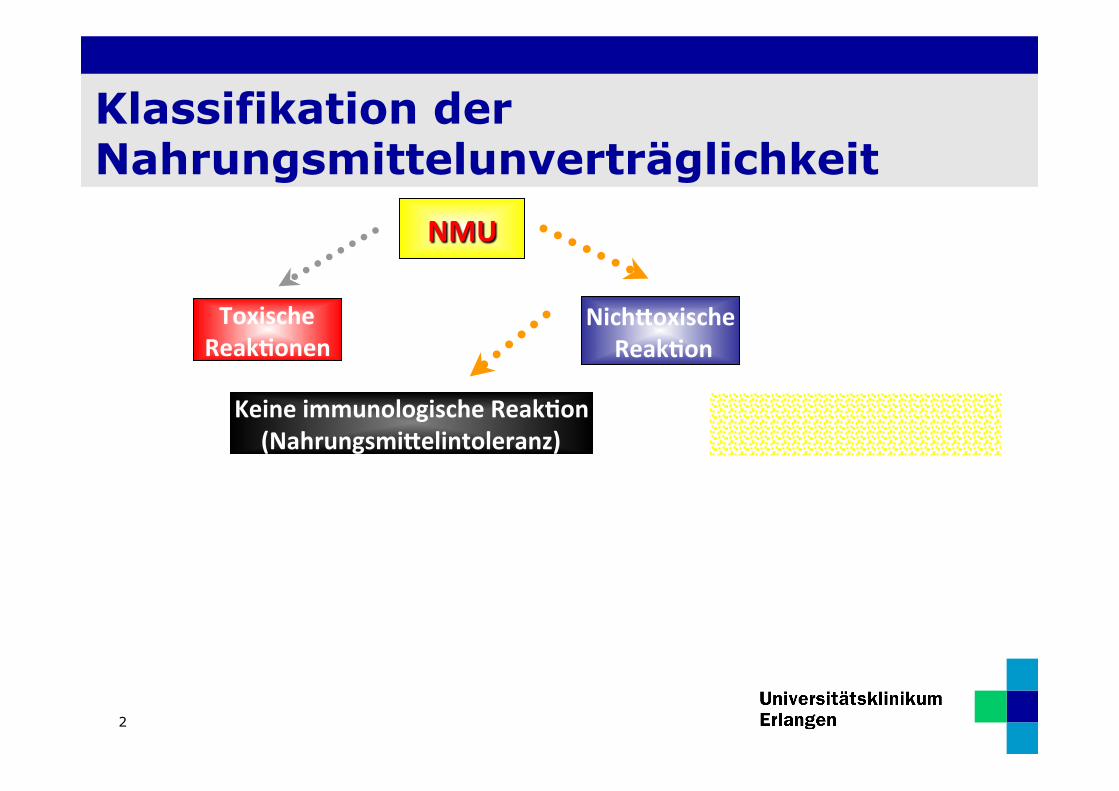
Klassifikation der Nahrungsmittelunverträglichkeit
2
NMU
Toxische Reak0onen
Keine immunologische Reak0on (Nahrungsmi9elintoleranz)
Nich9oxische Reak0on
Immunologische Reak0on Nahrungsmi9elallergie
sons0ge enzyma0sch
Histamin Intoleranz Laktose Intoleranz
Nicht IgE vermi9elt IgE-‐vermi9elt Aufnahmestörung
Fruktose Malabsorp0on
EAACI Position Paper. Allergy 1995; 50: 623-635)
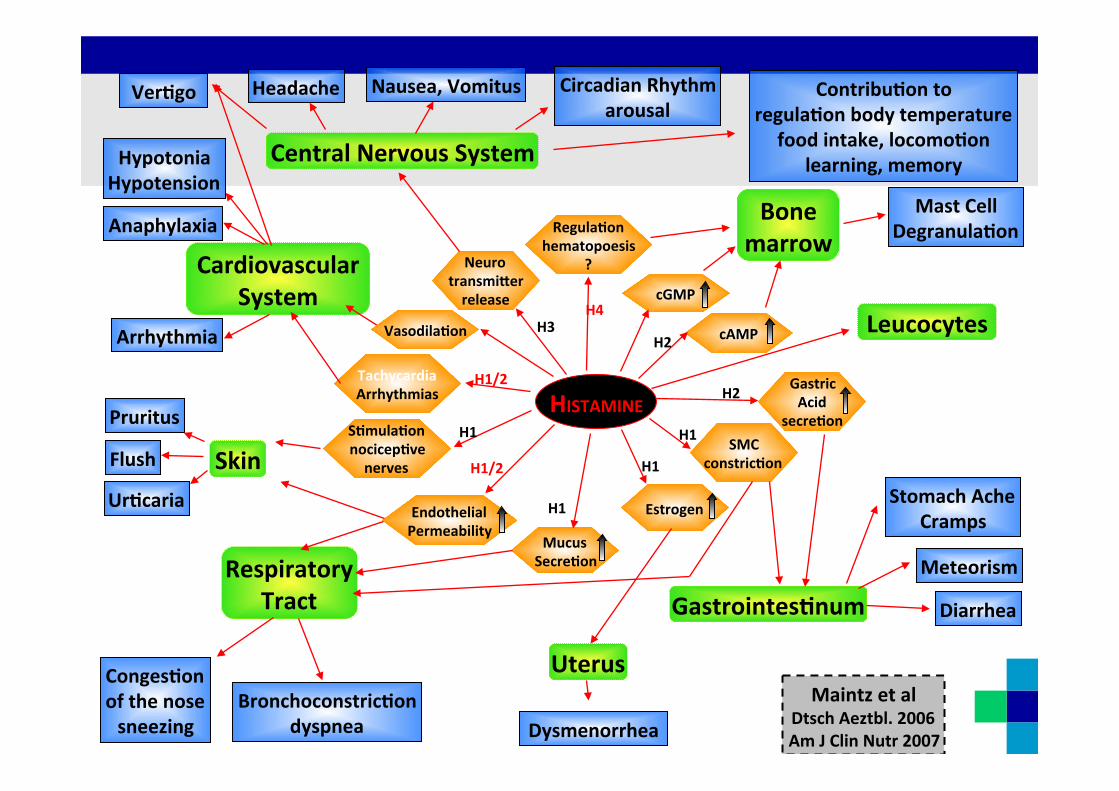
HISTAMINE
Neuro transmi9er release
H3
Regula0on hematopoesis
?
H4
Tachycardia Arrhythmias
Endothelial Permeability
H1/2
H1/2
S0mula0on nocicep0ve nerves
H1
Mucus Secre0on
Estrogen
SMC constric0on
H1
H1
H1
cAMP
Gastric Acid
secre0on
H2
H2
Central Nervous System
Ver0go Contribu0on to regula0on body temperature
food intake, locomo0on learning, memory
Circadian Rhythm arousal
Nausea, Vomitus Headache
Cardiovascular System
Anaphylaxia
Hypotonia Hypotension
Arrhythmia
Skin Flush
Pruritus
Ur0caria
Uterus
Dysmenorrhea
Respiratory Tract
Conges0on of the nose sneezing
Bronchoconstric0on dyspnea
Gastrointes0num Meteorism
Stomach Ache Cramps
Diarrhea
cGMP
Vasodila0on Leucocytes
Bone marrow
Mast Cell Degranula0on
Maintz et al Dtsch Aeztbl. 2006 Am J Clin Nutr 2007

Histaminintoleranz
4
n Gemeinsamer Nenner für Beschwerden nach
Histamin-‐reicher Nahrung
• Ungleichgewicht von Diaminoxidase und
Histamin in der Nahrung
• Diagnose über klinische Parameter schwierig
• Valide laborchemische Tests fehlen
Food-‐induced histaminosis as an epidemiological problem: plasma histamine eleva0on and haemodynamic altera0ons aaer oral histamine administra0on and blockade of diamine oxidase (DAO). Sa9ler J et. al, Agents Ac0ons. 1988 Apr;23(3-‐4):361-‐5.
HN
NH2
Histamin
HNNH
2HistaminHypotonie
Rhinits Anaphylaxie
Ur0karia Asthma
Diarrhoe

Histamin: ein biogener Amin
5
Hauptanteil der biogenen Amine stellen neben Histamin die biogenen Amine Cadaverin, Putrescin und Tyramin.
Freundlicherweise von Herrn Missbichler zur Verfügung gestellt

6
PROTEIN
Glutamin
Lysin
Tyrosin
Histidin Tryptophan
Cadaverin Histamin Tryptamin
Ornithin
Putrescin
Tyramin
Spermidin
Dopamin
Biogene Amine
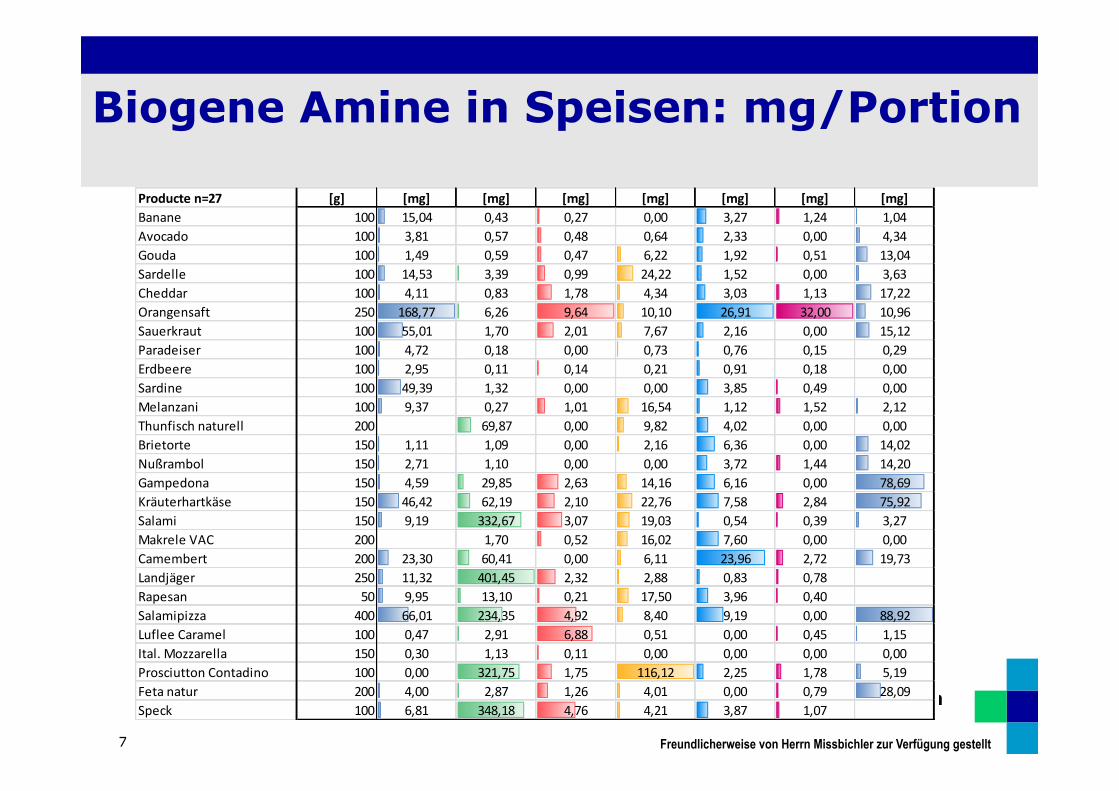
Biogene Amine in Speisen: mg/Portion
7
Producte n=27 [g] [mg] [mg] [mg] [mg] [mg] [mg] [mg]Banane 100 15,04 0,43 0,27 0,00 3,27 1,24 1,04Avocado 100 3,81 0,57 0,48 0,64 2,33 0,00 4,34Gouda 100 1,49 0,59 0,47 6,22 1,92 0,51 13,04Sardelle 100 14,53 3,39 0,99 24,22 1,52 0,00 3,63Cheddar 100 4,11 0,83 1,78 4,34 3,03 1,13 17,22Orangensaft 250 168,77 6,26 9,64 10,10 26,91 32,00 10,96Sauerkraut 100 55,01 1,70 2,01 7,67 2,16 0,00 15,12Paradeiser 100 4,72 0,18 0,00 0,73 0,76 0,15 0,29Erdbeere 100 2,95 0,11 0,14 0,21 0,91 0,18 0,00Sardine 100 49,39 1,32 0,00 0,00 3,85 0,49 0,00Melanzani 100 9,37 0,27 1,01 16,54 1,12 1,52 2,12Thunfisch naturell 200 69,87 0,00 9,82 4,02 0,00 0,00Brietorte 150 1,11 1,09 0,00 2,16 6,36 0,00 14,02Nußrambol 150 2,71 1,10 0,00 0,00 3,72 1,44 14,20Gampedona 150 4,59 29,85 2,63 14,16 6,16 0,00 78,69Kräuterhartkäse 150 46,42 62,19 2,10 22,76 7,58 2,84 75,92Salami 150 9,19 332,67 3,07 19,03 0,54 0,39 3,27Makrele VAC 200 1,70 0,52 16,02 7,60 0,00 0,00Camembert 200 23,30 60,41 0,00 6,11 23,96 2,72 19,73Landjäger 250 11,32 401,45 2,32 2,88 0,83 0,78Rapesan 50 9,95 13,10 0,21 17,50 3,96 0,40Salamipizza 400 66,01 234,35 4,92 8,40 9,19 0,00 88,92Luflee Caramel 100 0,47 2,91 6,88 0,51 0,00 0,45 1,15Ital. Mozzarella 150 0,30 1,13 0,11 0,00 0,00 0,00 0,00Prosciutton Contadino 100 0,00 321,75 1,75 116,12 2,25 1,78 5,19Feta natur 200 4,00 2,87 1,26 4,01 0,00 0,79 28,09Speck 100 6,81 348,18 4,76 4,21 3,87 1,07
Freundlicherweise von Herrn Missbichler zur Verfügung gestellt
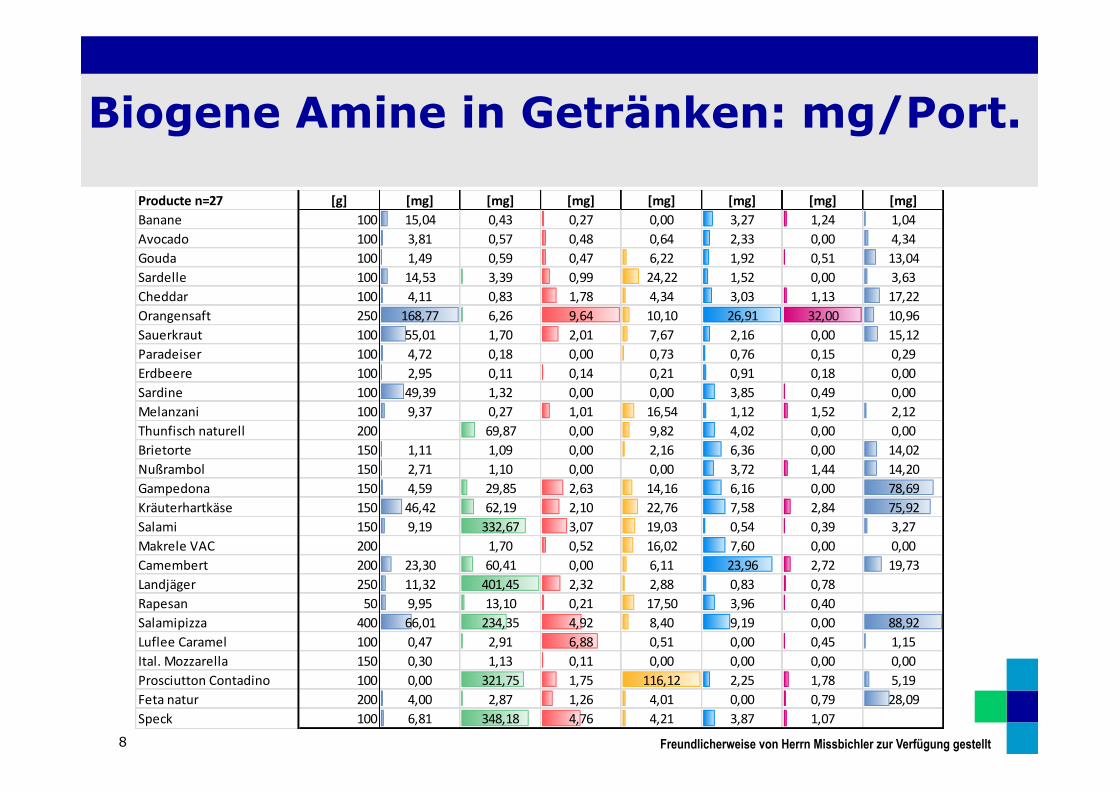
Biogene Amine in Getränken: mg/Port.
8
Producte n=27 [g] [mg] [mg] [mg] [mg] [mg] [mg] [mg]Banane 100 15,04 0,43 0,27 0,00 3,27 1,24 1,04Avocado 100 3,81 0,57 0,48 0,64 2,33 0,00 4,34Gouda 100 1,49 0,59 0,47 6,22 1,92 0,51 13,04Sardelle 100 14,53 3,39 0,99 24,22 1,52 0,00 3,63Cheddar 100 4,11 0,83 1,78 4,34 3,03 1,13 17,22Orangensaft 250 168,77 6,26 9,64 10,10 26,91 32,00 10,96Sauerkraut 100 55,01 1,70 2,01 7,67 2,16 0,00 15,12Paradeiser 100 4,72 0,18 0,00 0,73 0,76 0,15 0,29Erdbeere 100 2,95 0,11 0,14 0,21 0,91 0,18 0,00Sardine 100 49,39 1,32 0,00 0,00 3,85 0,49 0,00Melanzani 100 9,37 0,27 1,01 16,54 1,12 1,52 2,12Thunfisch naturell 200 69,87 0,00 9,82 4,02 0,00 0,00Brietorte 150 1,11 1,09 0,00 2,16 6,36 0,00 14,02Nußrambol 150 2,71 1,10 0,00 0,00 3,72 1,44 14,20Gampedona 150 4,59 29,85 2,63 14,16 6,16 0,00 78,69Kräuterhartkäse 150 46,42 62,19 2,10 22,76 7,58 2,84 75,92Salami 150 9,19 332,67 3,07 19,03 0,54 0,39 3,27Makrele VAC 200 1,70 0,52 16,02 7,60 0,00 0,00Camembert 200 23,30 60,41 0,00 6,11 23,96 2,72 19,73Landjäger 250 11,32 401,45 2,32 2,88 0,83 0,78Rapesan 50 9,95 13,10 0,21 17,50 3,96 0,40Salamipizza 400 66,01 234,35 4,92 8,40 9,19 0,00 88,92Luflee Caramel 100 0,47 2,91 6,88 0,51 0,00 0,45 1,15Ital. Mozzarella 150 0,30 1,13 0,11 0,00 0,00 0,00 0,00Prosciutton Contadino 100 0,00 321,75 1,75 116,12 2,25 1,78 5,19Feta natur 200 4,00 2,87 1,26 4,01 0,00 0,79 28,09Speck 100 6,81 348,18 4,76 4,21 3,87 1,07
Freundlicherweise von Herrn Missbichler zur Verfügung gestellt

Biogen-Amin-Abbau
9
D ü n n d a r m
Rezeptoren
DAO
Histamin Putrescin
Cadaverin
Tyramin
Serotonin
Biogen
e Am
ine
Abbaukapazität abhängig von der oralen Aufnahme
SYMPTOME

Laborchemische Messungen
10
D ü n n d a r m
Rezeptoren
DAO
Histamin Putrescin
Cadaverin
Tyramin
Serotonin
Biogen
e Am
ine
Abbaukapazität abhängig von der oralen Aufnahme
SYMPTOME
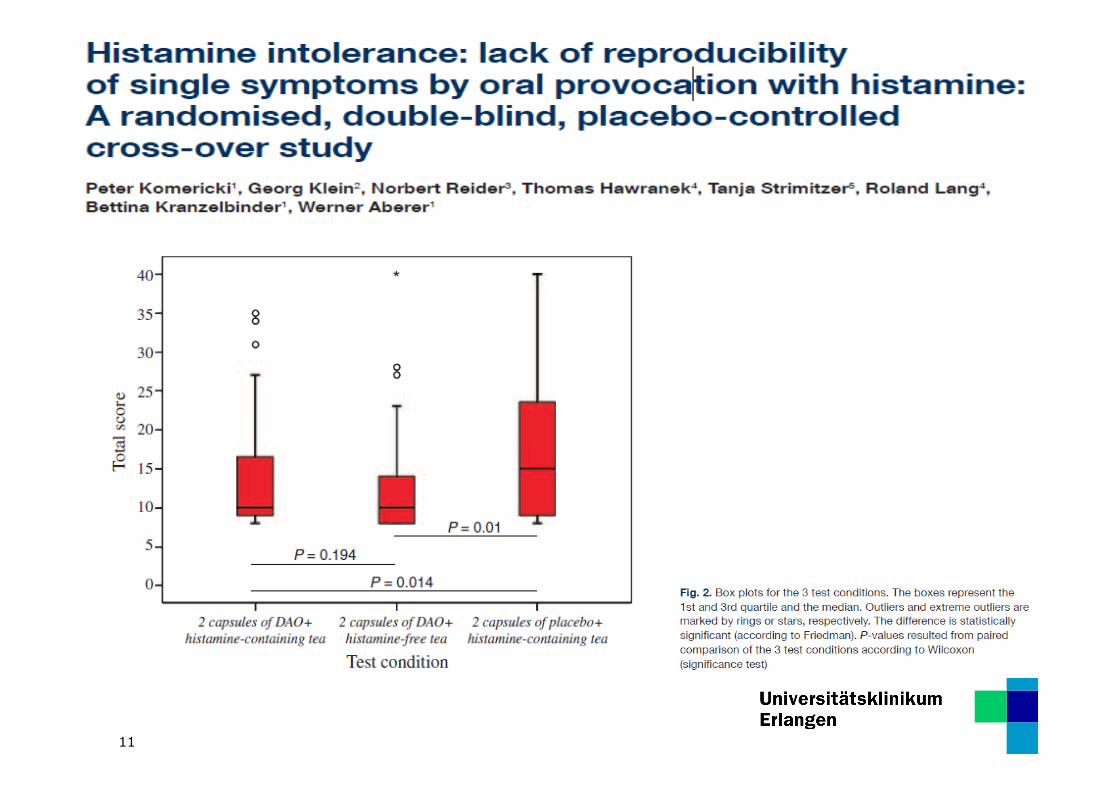
11

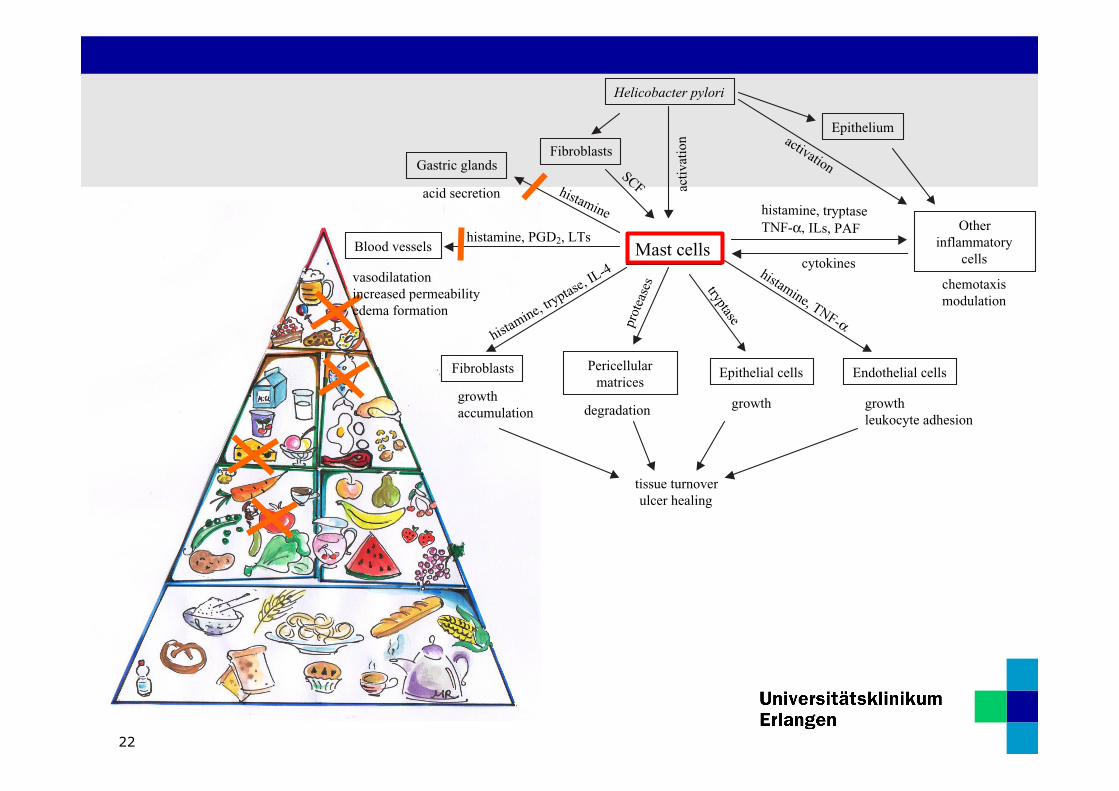
Therapie: Histaminarme Kost
12
• Rotwein, Sekt
• Thunfisch
• Hart-‐ und Räucherkäse
• Salami und andere Hartwürste
• Tomaten, Ketchup
• Schokolade
• Sauerkraut
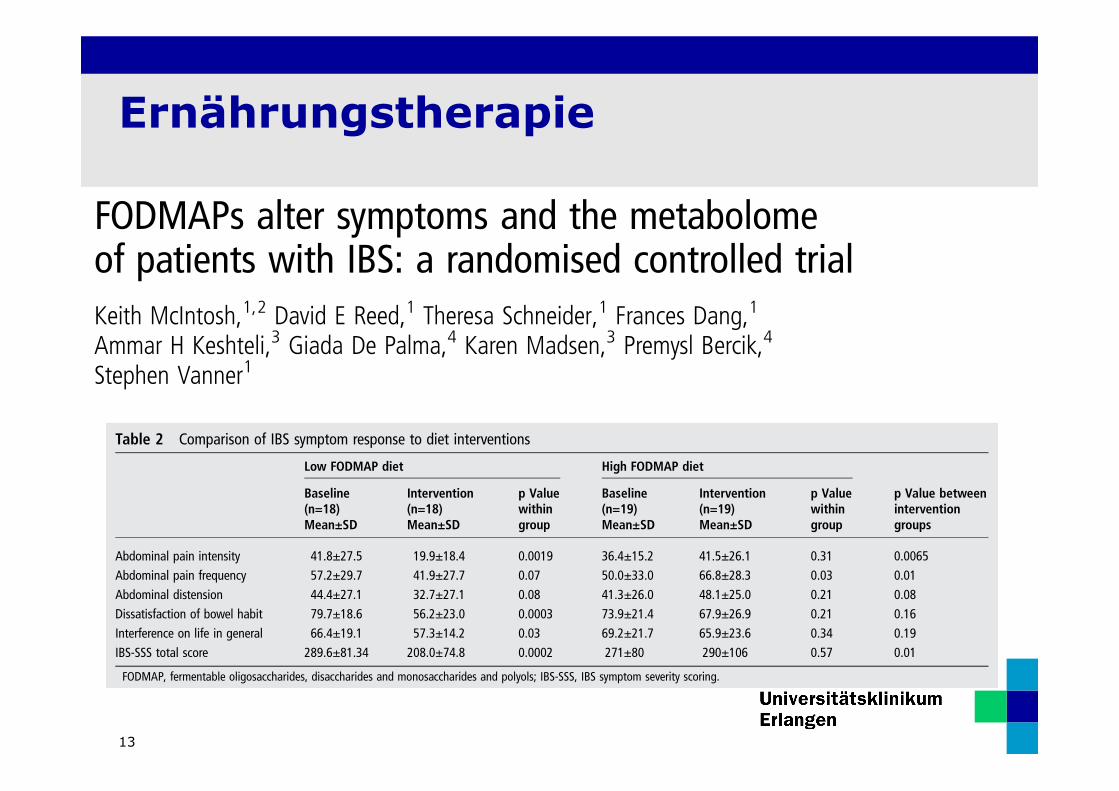
Ernährungstherapie
13
ORIGINAL ARTICLE
FODMAPs alter symptoms and the metabolomeof patients with IBS: a randomised controlled trialKeith McIntosh,1,2 David E Reed,1 Theresa Schneider,1 Frances Dang,1
Ammar H Keshteli,3 Giada De Palma,4 Karen Madsen,3 Premysl Bercik,4
Stephen Vanner1
▸ Additional material ispublished online only. To viewplease visit the journal online(http://dx.doi.org/10.1136/gutjnl-2015-311339).1GI Diseases Research Unit,Queen’s University,Kingston General Hospital,Kingston, Ontario, Canada2St. Joseph’s Hospital,Western University, London,Ontario, Canada3University of Alberta,Edmonton, Alberta, Canada4Farmcombe Institute,McMaster University, Hamilton,Ontario, Canada
Correspondence toDr Stephen Vanner, GIDiseases Research Unit,Kingston General Hospital,76 Stuart St., Kingston,Ontario, Canada K7L 2V7;[email protected]
Received 22 December 2015Revised 4 February 2016Accepted 17 February 2016
To cite: McIntosh K,Reed DE, Schneider T, et al.Gut Published Online First:[please include Day MonthYear] doi:10.1136/gutjnl-2015-311339
ABSTRACTObjective To gain mechanistic insights, we comparedeffects of low fermentable oligosaccharides,disaccharides and monosaccharides and polyols(FODMAP) and high FODMAP diets on symptoms, themetabolome and the microbiome of patients with IBS.Design We performed a controlled, single blind studyof patients with IBS (Rome III criteria) randomised to alow (n=20) or high (n=20) FODMAP diet for 3 weeks.Symptoms were assessed using the IBS symptom severityscoring (IBS-SSS). The metabolome was evaluated usingthe lactulose breath test (LBT) and metabolic profiling inurine using mass spectrometry. Stool microbiotacomposition was analysed by 16S rRNA gene profiling.Results Thirty-seven patients (19 low FODMAP; 18high FODMAP) completed the 3-week diet. The IBS-SSSwas reduced in the low FODMAP diet group (p<0.001)but not the high FODMAP group. LBTs showed a minordecrease in H2 production in the low FODMAP comparedwith the high FODMAP group. Metabolic profiling ofurine showed groups of patients with IBS differedsignificantly after the diet (p<0.01), with threemetabolites (histamine, p-hydroxybenzoic acid, azelaicacid) being primarily responsible for discriminationbetween the two groups. Histamine, a measure ofimmune activation, was reduced eightfold in the lowFODMAP group (p<0.05). Low FODMAP diet increasedActinobacteria richness and diversity, and high FODMAPdiet decreased the relative abundance of bacteriainvolved in gas consumption.Conclusions IBS symptoms are linked to FODMAPcontent and associated with alterations in themetabolome. In subsets of patients, FODMAPs modulatehistamine levels and the microbiota, both of which couldalter symptoms.Trial registration number NCT01829932.
INTRODUCTIONThere is increasing evidence that interactionsbetween food components and the microbiotacould provide mechanistic insights into the patho-physiology of IBS.1–3 In particular, a number ofstudies suggest that fermentable oligosaccharides,disaccharides and monosaccharides and polyols(FODMAPs) can induce IBS symptoms.3–8 Dietaryinterventions using low FODMAP diets showedmarked improvement in symptoms, benefiting upto 70% of patients in some studies.3 Furthermore,a recent study suggested that the reduction of
FODMAPs in a gluten-free diet could explain, atleast in part, the benefit observed with glutenavoidance in patients with IBS who do not havecoeliac disease.4 Despite this growing enthusiasmfor the low FODMAP diet, mechanisms underlyingthe diet are unclear. Furthermore, there are
Significance of this study
What is already known on this subject?▸ Meals trigger symptoms in many patients with
IBS and specific diets can improve symptoms.▸ Low fermentable oligosaccharides,
disaccharides and monosaccharides and polyols(FODMAP) diets appear to improve symptomsbut the specific role of FODMAPs is unclear.
▸ Metabolomics can identify metabolites of thehost-microbial interactions and providemechanistic insights.
What are the new findings?▸ Metabolic profiling of urine showed diets
caused significant separation of metabolome.▸ There was an eightfold reduction in urinary
histamine in the low FODMAP group.▸ Low FODMAP diet increased Actinobacteria
richness and diversity and high FODMAP dietdecreased the relative abundance of bacteriainvolved in gas consumption, likely contributingto symptoms.
▸ The lactulose breath test did not predictresponders of a low FODMAP diet.
How might this impact on clinical practice inthe foreseeable future?▸ Low FODMAP diets have a specific action that
is beneficial in patients with IBS but furtherstudies are needed to identify the specificFODMAPs that produce this benefit.
▸ There is evidence that FODMAPs modulateimmune activation in subsets of patients andhistamine may be an importantpathophysiological mediator.
▸ Changes in gut microbiota may play a role insymptom generation in a subset of patientswith IBS; however, a low FODMAP diet mightinduce potential ‘unhealthy’ changes at themicrobial level but requires long-term studies.
McIntosh K, et al. Gut 2016;0:1–11. doi:10.1136/gutjnl-2015-311339 1
Neurogastroenterology Gut Online First, published on March 14, 2016 as 10.1136/gutjnl-2015-311339
Copyright Article author (or their employer) 2016. Produced by BMJ Publishing Group Ltd (& BSG) under licence.
group.bmj.com on March 18, 2016 - Published by http://gut.bmj.com/Downloaded from
a more uniform group, all with some diarrhoea) we observedhigher bacterial richness (corrected p=0.047) in the lowFODMAP group compared with the high FODMAP group(figure 6A). Specifically, Firmicutes, Clostridiales andActinobacteria richness was higher (all corrected p=0.029,0.023 and 0.029, respectively), and Actinobacteria bacterialdiversity was increased (corrected p=0.013). Analysis of thetaxonomic composition at the highest assigned taxonomic level(genus level) revealed that bacteria from the genus Adlercreutzia(Coriobacteriaceae), Dorea (Lachnospiraceae) and the family ofActinomycetaceae were lower after a high FODMAP diet(uncorrected p=0.02, 0.05 and 0.044, respectively) (figure 6B).
When analysing the paired data, we observed thatPropionibacteriaceae (Actinomycetales) decreased (uncorrected
p=0.043) while unclassified genera within the Clostridialesfamily XIII Incertae sedis (uncorrected p=0.008) andPorphyromonas (uncorrected p=0.01) increased after the lowFODMAP diet. Interestingly, the high FODMAP diet inducedmore changes as several bacterial groups decreased after thehigh FODMAP diet, but the family Bifidobacteriaceae (uncor-rected p=0.041) and unclassified genera within the familyLachnospiraceae increased (uncorrected p=0.036).
Correlating microbial changes with diet, symptoms andmetabolomeSeveral abundant genera were negatively correlated with increas-ing FODMAP dietary content, including two genera belongingto the Lachnospiraceae family (Clostridium and Roseburia)
Table 2 Comparison of IBS symptom response to diet interventions
Low FODMAP diet High FODMAP diet
Baseline(n=18)Mean±SD
Intervention(n=18)Mean±SD
p Valuewithingroup
Baseline(n=19)Mean±SD
Intervention(n=19)Mean±SD
p Valuewithingroup
p Value betweeninterventiongroups
Abdominal pain intensity 41.8±27.5 19.9±18.4 0.0019 36.4±15.2 41.5±26.1 0.31 0.0065Abdominal pain frequency 57.2±29.7 41.9±27.7 0.07 50.0±33.0 66.8±28.3 0.03 0.01Abdominal distension 44.4±27.1 32.7±27.1 0.08 41.3±26.0 48.1±25.0 0.21 0.08Dissatisfaction of bowel habit 79.7±18.6 56.2±23.0 0.0003 73.9±21.4 67.9±26.9 0.21 0.16Interference on life in general 66.4±19.1 57.3±14.2 0.03 69.2±21.7 65.9±23.6 0.34 0.19IBS-SSS total score 289.6±81.34 208.0±74.8 0.0002 271±80 290±106 0.57 0.01
FODMAP, fermentable oligosaccharides, disaccharides and monosaccharides and polyols; IBS-SSS, IBS symptom severity scoring.
Figure 2 Global IBS symptoms improved on a low fermentable oligosaccharides, disaccharides and monosaccharides and polyols (FODMAP) dietbut not a high FODMAP diet. (A) Individual scores showing almost all patients improved in the low FODMAP group whereas most patients in thehigh FODMAP group were unchanged or worse. (B) Increases in dietary FODMAP content based on a FODMAP scoring system (see text andsupplementary table S1), independent of diet assignment, positively correlated with increasing symptoms in patients with IBS. IBS-SSS, IBS symptomseverity scoring.
McIntosh K, et al. Gut 2016;0:1–11. doi:10.1136/gutjnl-2015-311339 5
Neurogastroenterology
group.bmj.com on March 18, 2016 - Published by http://gut.bmj.com/Downloaded from

14
symptoms, regardless of diet assignment. This also showed sig-nificant correlation between symptoms and FODMAP content,further attesting to the conclusion that FODMAPs are linked toIBS symptoms.
There is rapidly growing interest in the study of food metabo-lomics to understand the complex interaction between diet andpatients.15 This metabolome has two different fractions,endogenous metabolome and food metabolome, and arederived from the digestion of food, their absorption and bio-transformation by the human donor and the microbiota. Ourstudy identified major changes in specific metabolites and asso-ciations between these metabolites and the microbiota inresponse to the dietary interventions, as shown in table 4. Thesereflect changes in both the endogenous and food metabolomeand while the full significance of these changes has yet to be elu-cidated, the implications of our metabolomic data are that mul-tiple pathways could lead to symptoms in IBS. For example, this
could include signalling by active and inert (eg, gases) metabo-lites due to fermentation and degradation by specific bacteria.Furthermore, dietary modulation of the microbiota could eitherincrease the abundance of specific bacteria or their metabolites,thereby amplifying abnormal signalling by these pathways.
Our analysis of the urinary metabolome identified three spe-cific metabolites that were altered by the diet, histamine, pHBAand azelaic acid. The reduction in histamine in the lowFODMAP group and increase in the high FODMAP group insubsets of patients is of immediate interest given the strong evi-dence that histamine is an important signalling molecule under-lying IBS symptoms. It is widely recognised that histamine isincreased in the mucosa of many patients with IBS,25–27 is asso-ciated with increased mast cell activation, and sensitises nocicep-tive and enteric neurons.28–30 The mast cell stabiliser ketotifenincreased threshold for patients with discomfort IBS with vis-ceral hypersensitivity as well as improved IBS symptoms.31
Table 3 Concentrations (mmol/mmol creatine) of the three major urinary metabolites that discriminate between the two diets
Low FODMAP diet High FODMAP dietp Valuebetweeninterventiongroups *
Baseline(n=16)Mean±SD
Intervention(n=16)Mean±SD
p Valuewithingroup1
Baseline(n=18)Mean±SD
Intervention(n=18)Mean±SD
p Valuewithingroup†
VIPscore
p-Hydroxybenzoic acid 0.8106±1.1212 1.4754±1.8456 0.43 0.7817±1.4526 0.2480±0.454 0.10 0.02 2.14Histamine 0.0061±0.0117 0.0008±0.0011 0.05 0.0094±0.1860 0.0085±0.0158 0.61 0.04 1.86Azelaic acid 3.7947±3.1003 5.6972±2.9191 0.14 3.7086±3.2912 3.5870±2.8614 0.98 0.04 1.52
*Obtained from analysis of covariance and adjusted for the baseline metabolite level, age, gender and IBS subtype.†Obtained from Wilcoxon signed-rank test.FODMAP, fermentable oligosaccharides, disaccharides and mono-saccharides and polyols; VIP, variable importance in projection.
Figure 5 Low fermentable oligosaccharides, disaccharides and monosaccharides and polyols (FODMAP) diet significantly decreases urine histaminelevels. (A) Urine histamine levels remained unchanged after 21 days of a high FODMAP diet (normalised to creatine) whereas there was a significantreduction in urine histamine in the low FODMAP group. *p<0.05. B. Individual patients that had elevated urine histamine (normalised to creatine)on day 0, day 21 or both in the high FODMAP group (left panel) and the low FODMAP group (right panel).
McIntosh K, et al. Gut 2016;0:1–11. doi:10.1136/gutjnl-2015-311339 7
Neurogastroenterology
group.bmj.com on March 18, 2016 - Published by http://gut.bmj.com/Downloaded from
symptoms, regardless of diet assignment. This also showed sig-nificant correlation between symptoms and FODMAP content,further attesting to the conclusion that FODMAPs are linked toIBS symptoms.
There is rapidly growing interest in the study of food metabo-lomics to understand the complex interaction between diet andpatients.15 This metabolome has two different fractions,endogenous metabolome and food metabolome, and arederived from the digestion of food, their absorption and bio-transformation by the human donor and the microbiota. Ourstudy identified major changes in specific metabolites and asso-ciations between these metabolites and the microbiota inresponse to the dietary interventions, as shown in table 4. Thesereflect changes in both the endogenous and food metabolomeand while the full significance of these changes has yet to be elu-cidated, the implications of our metabolomic data are that mul-tiple pathways could lead to symptoms in IBS. For example, this
could include signalling by active and inert (eg, gases) metabo-lites due to fermentation and degradation by specific bacteria.Furthermore, dietary modulation of the microbiota could eitherincrease the abundance of specific bacteria or their metabolites,thereby amplifying abnormal signalling by these pathways.
Our analysis of the urinary metabolome identified three spe-cific metabolites that were altered by the diet, histamine, pHBAand azelaic acid. The reduction in histamine in the lowFODMAP group and increase in the high FODMAP group insubsets of patients is of immediate interest given the strong evi-dence that histamine is an important signalling molecule under-lying IBS symptoms. It is widely recognised that histamine isincreased in the mucosa of many patients with IBS,25–27 is asso-ciated with increased mast cell activation, and sensitises nocicep-tive and enteric neurons.28–30 The mast cell stabiliser ketotifenincreased threshold for patients with discomfort IBS with vis-ceral hypersensitivity as well as improved IBS symptoms.31
Table 3 Concentrations (mmol/mmol creatine) of the three major urinary metabolites that discriminate between the two diets
Low FODMAP diet High FODMAP dietp Valuebetweeninterventiongroups *
Baseline(n=16)Mean±SD
Intervention(n=16)Mean±SD
p Valuewithingroup1
Baseline(n=18)Mean±SD
Intervention(n=18)Mean±SD
p Valuewithingroup†
VIPscore
p-Hydroxybenzoic acid 0.8106±1.1212 1.4754±1.8456 0.43 0.7817±1.4526 0.2480±0.454 0.10 0.02 2.14Histamine 0.0061±0.0117 0.0008±0.0011 0.05 0.0094±0.1860 0.0085±0.0158 0.61 0.04 1.86Azelaic acid 3.7947±3.1003 5.6972±2.9191 0.14 3.7086±3.2912 3.5870±2.8614 0.98 0.04 1.52
*Obtained from analysis of covariance and adjusted for the baseline metabolite level, age, gender and IBS subtype.†Obtained from Wilcoxon signed-rank test.FODMAP, fermentable oligosaccharides, disaccharides and mono-saccharides and polyols; VIP, variable importance in projection.
Figure 5 Low fermentable oligosaccharides, disaccharides and monosaccharides and polyols (FODMAP) diet significantly decreases urine histaminelevels. (A) Urine histamine levels remained unchanged after 21 days of a high FODMAP diet (normalised to creatine) whereas there was a significantreduction in urine histamine in the low FODMAP group. *p<0.05. B. Individual patients that had elevated urine histamine (normalised to creatine)on day 0, day 21 or both in the high FODMAP group (left panel) and the low FODMAP group (right panel).
McIntosh K, et al. Gut 2016;0:1–11. doi:10.1136/gutjnl-2015-311339 7
Neurogastroenterology
group.bmj.com on March 18, 2016 - Published by http://gut.bmj.com/Downloaded from
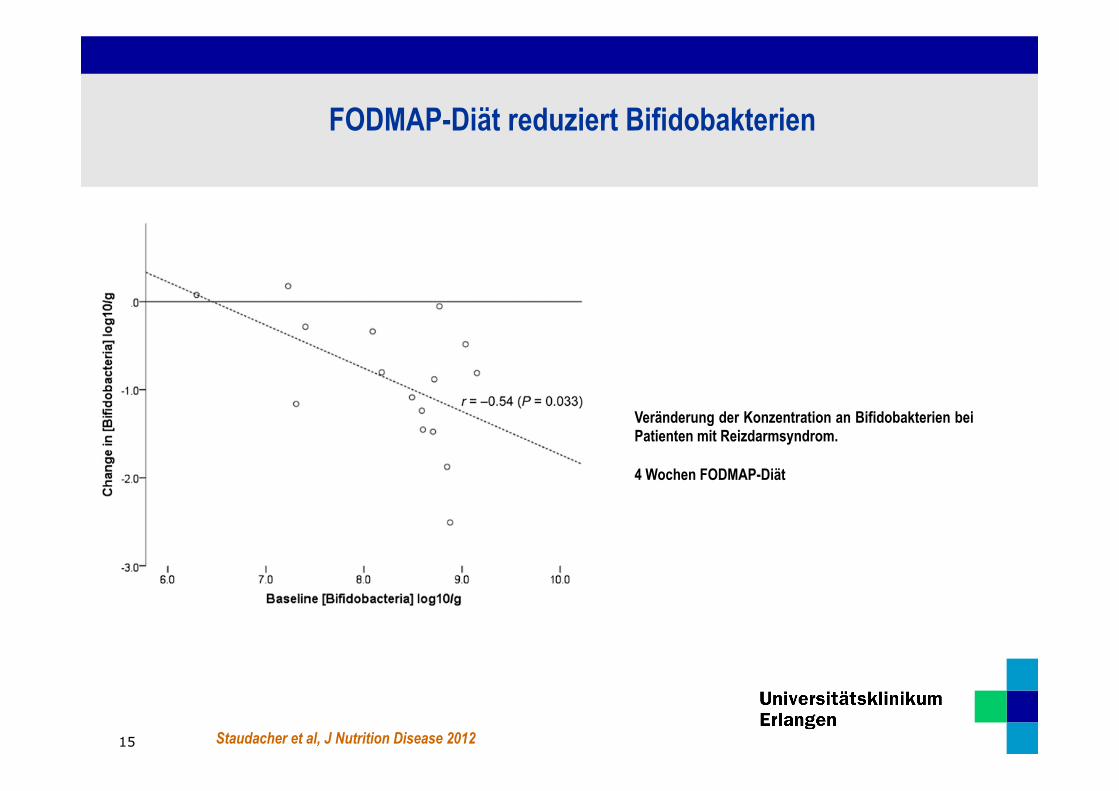
Staudacher et al, J Nutrition Disease 2012
Veränderung der Konzentration an Bifidobakterien bei Patienten mit Reizdarmsyndrom. 4 Wochen FODMAP-Diät
FODMAP-Diät reduziert Bifidobakterien
15

Faktoren, die das Mikrobiom beeinflussen
Meyer et al, European J Clin Nutrition 2009
Stress
Alter
Genetik
Medikamente
Infektionen
Diät
16
17
H.PYLORI INFECTION AND DEGRANULATION OF
MAST CELLS
As indicated in our previous study,3 mast cells undergodegranulation in H. pylori-infected gastric mucosa,meaning that they actively release chemical mediatorssuch as histamine and tryptase in such tissue.3 Themechanisms that trigger degranulation in mast cells arenot precisely understood. Kurose et al. demonstratedmast cell degranulation in the vicinity of rat mesentericvenules treated with H. pylori water extract under real-time microscopic observation,12 while Yamamoto et al.showed that H. pylori water extract directly induceddegranulation in rat peritoneal mast cells.13 In contrast,Bechi et al., Masini et al. and Lutton et al. found thatH. pylori components did not induce mast cell degranu-lation directly, but rather affected degranulation.14–16
Bechi et al. and Masini et al. reported that a cellcomponent ofH. pylori enhancedmast cell degranulationstimulated by compound 48/80, calcium ionophoreA23187 or bile acid,14, 15 while Lutton et al. reportedthat a cell component of H. pylori reduced mast celldegranulation stimulated by calcium ionophoreA23187, compound 48/80, concanavalin A or anti-IgE.16 These differences may partly depend on the
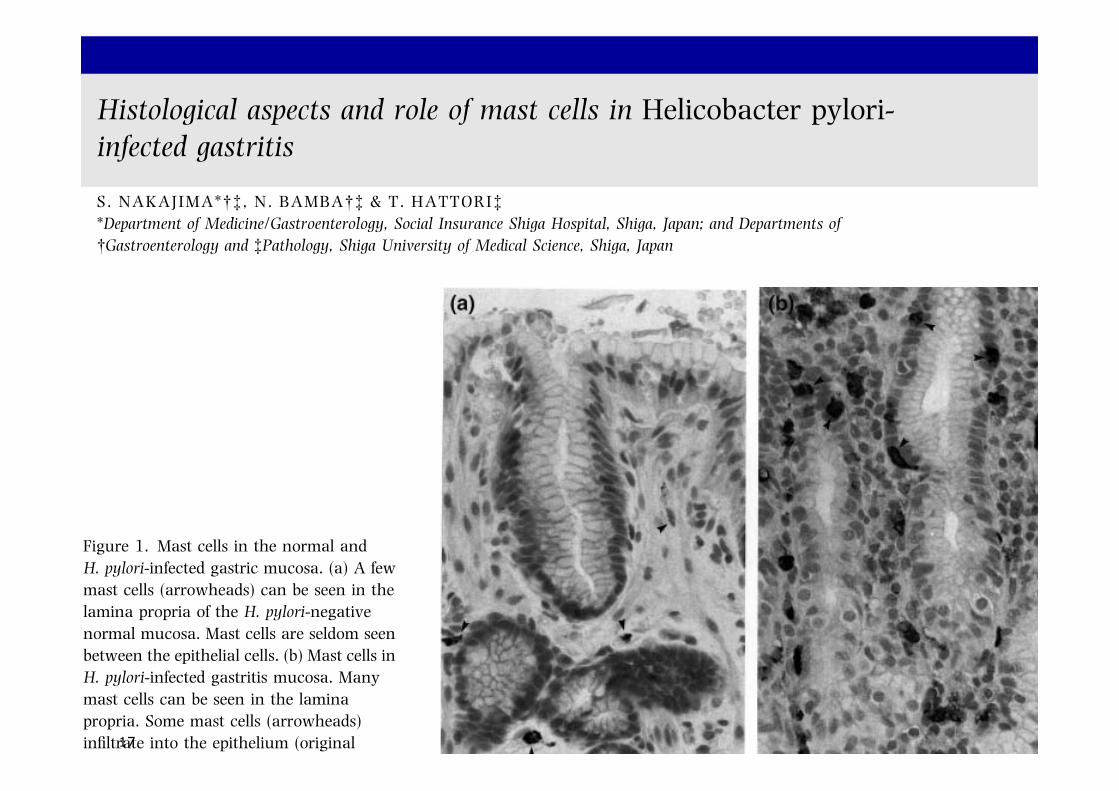
Figure 1. Mast cells in the normal andH. pylori-infected gastric mucosa. (a) A fewmast cells (arrowheads) can be seen in thelamina propria of the H. pylori-negativenormal mucosa. Mast cells are seldom seenbetween the epithelial cells. (b) Mast cells inH. pylori-infected gastritis mucosa. Manymast cells can be seen in the laminapropria. Some mast cells (arrowheads)infiltrate into the epithelium (originalmagnification ·400). Reprinted fromNakajima S et al., Gastroenterology 1997;113: 746–54,3 with kind permission fromThe American Gastroenterological Associ-ation.
Figure 2. Electron micrograph of a mast cell adjacent to plasmacell in the lamina propria of H. pylori-infected gastric mucosa.The cytoplasm of the mast cell show degranulation withmultiple ghost vacuoles (arrows) and targetoid-type granules(open arrows). Original magnification ·8800. Reprinted fromNakajima S et al., Gastroenterology 1997; 113: 746–54,3 withkind permission from The American GastroenterologicalAssociation.
MAST CELLS IN H. PYLORI GASTRITIS 167
! 2004 Blackwell Publishing Ltd, Aliment Pharmacol Ther 20 (Suppl. 1), 165–170
Histological aspects and role of mast cells in Helicobacter pylori-infected gastritis
S. NAKAJIMA*!" , N. BAMBA!" & T. HATTORI"*Department of Medicine/Gastroenterology, Social Insurance Shiga Hospital, Shiga, Japan; and Departments of!Gastroenterology and "Pathology, Shiga University of Medical Science, Shiga, Japan
SUMMARY
Background: Mast cells are one of the main pro-inflammatory cells, while their knowledge in Helicobac-ter pylori infection has not been summarized.Methods: We reviewed studies on mast cells in H. pyloriinfection, and summarized the histological aspects androles.Results: The density of mast cells is greater in H. pylori-infected than in non-infected subjects. Increased mastcell density in infected gastritis significantly decreasesafter eradication. On electron microscopy, mast cells ininfected gastric mucosa show degranulation. Someexperimental studies demonstrate that mast cells aredegranulated with H. pylori-derived products.
Conclusions: Mast cells are actively involved in thepathogenesis of H. pylori-infected gastritis. The possibleroles are to initiate and promote the formation ofoedema through degranulated and secreted mediators,and to release multiple chemotactic factors, whichinduce inflammatory cells to infiltrate to the site ofoedema, showing acute inflammatory changes. Mastcells also stimulate the degradation of pericellularmatrices and the growth of cells in their vicinity, andthereby promote tissue turnover. In chronic H. pyloriinfection, these reactions continue until the bacteria areeradicated. Mast cells may act both to maintain gastritisand to repair tissue damage in H. pylori-infected chronicgastritis.
INTRODUCTION
Mast cells are one of a number of inflammatory cellsderived from bone marrow. They are present not only ininflamed tissue, but also in normal tissue in preparationfor forthcoming tissue damage or inflammation.Because mast cells release various cytokines or chemicalmediators at the initiation of inflammation, they areconsidered to be a type of proinflammatory cell. Until1966, it was believed that mast cells were not present inthe mucosa of the gastrointestinal tract because theyare not stained by ordinary histological staining withhaematoxylin and eosin in tissue fixed with formalde-
hyde. In 1966, Enerback discovered that mast cells werepresent in the mucosa of the gastrointestinal tract usingtissue fixed with Carnoy’s fixative and stained withtoluidine blue.1 Since then, mast cells have beenclassified histochemically into two subpopulations:connective tissue-type and mucosal-type mast cells.The former is the classic type of mast cell, which stainswith toluidine blue or alcian blue in tissue fixed withformaldehyde, while the latter is the type found byEnerback. In 1990, Walls et al. demonstrated thatmucosal-type mast cells were immunohistochemicallystained with anti-tryptase antibody even in tissue fixedwith formaldehyde.2 In 1997, we demonstratedmucosal mast cells in normal and Helicobacter pylori-infected gastric mucosa using an immunohistochemicaltechnique and showed the relationship of mast cellswith H. pylori gastritis.3 The present review describes
Correspondence to: Dr S. Nakajima, Department of Medicine/Gastroenter-ology, Social Insurance Shiga Hospital, 16-1 Fujimidai, Otsu, Shiga 520-0846, Japan.E-mail: [email protected]
Aliment Pharmacol Ther 2004; 20 (Suppl. 1): 165–170.
! 2004 Blackwell Publishing Ltd 165
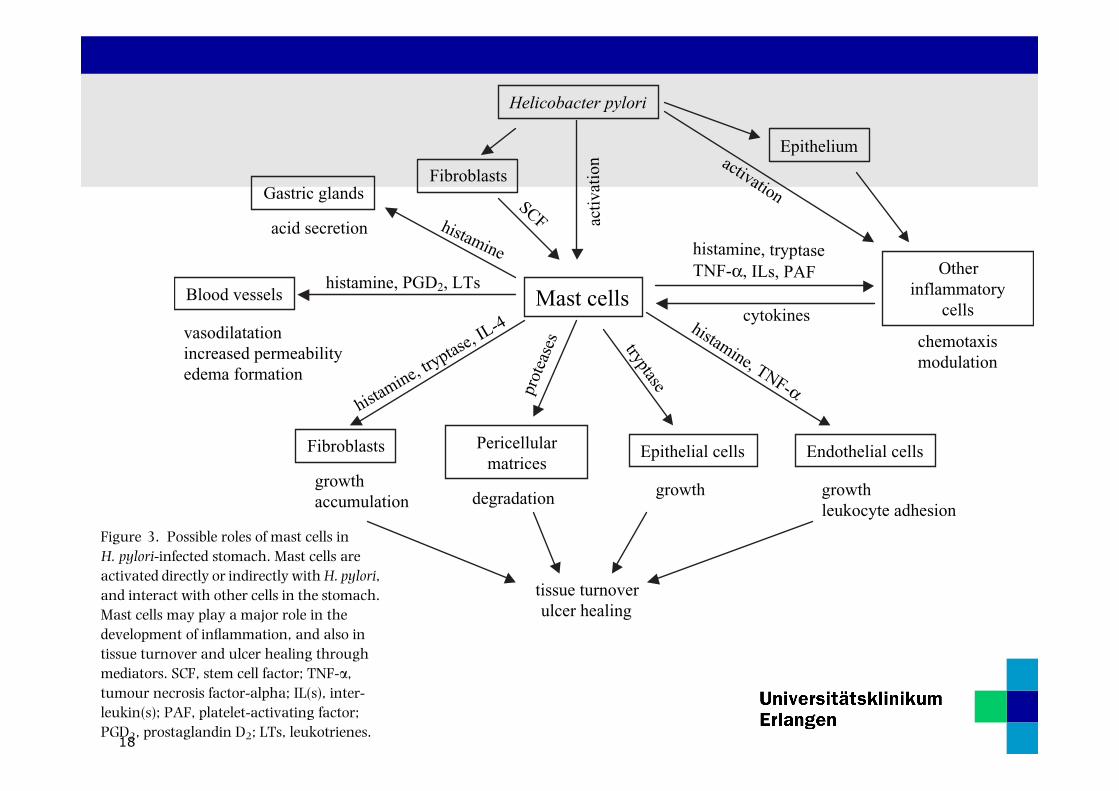
18
through H1 receptors.31 Interleukin-4, which is secreted
from activated mast cells32 stimulates fibroblasts toaccumulate at the site of inflammation.33 Tryptase,which is released with degranulation of mast cells,activates matrix metalloproteinases, especially matrixmetalloproteinase-3.34 Tryptase also stimulates thegrowth of epithelial cells and fibroblasts.35, 36 Thesemediators may act together to stimulate tissue and cellturnover in the vicinity of mast cells after the onset ofinflammation (Figure 3).It is uncertain whether histamine derived from mast
cells is involved in gastric acid secretion through H2
receptors; however, mast cell-derived histamine may beinvolved in a gastrin-independent mechanism of acidsecretion.37
POSSIBLE ROLE OF MAST CELLS
IN H. PYLORI-INFECTED CHRONIC GASTRITIS
A summary of the possible roles of mast cells in H.pylori-infected gastric mucosa is shown in Figure 3.Mast cells initiate and promote the formation of oedemathrough degranulated and secreted chemical mediators.They also release multiple chemotactic factors, whichinduce inflammatory cells to infiltrate to the site ofoedema. Mast cells and other inflammatory cells engagein complex and cooperative interplay with variouschemical mediators. They also stimulate the degrada-tion of pericellular matrices in H. pylori-infected gastricmucosa and the growth of cells in their vicinity, andpromote tissue turnover.
The phenomena described above are reactions in acuteand the subsequent phases of gastritis. Because H. pyloriinfection usually induces chronic inflammation throughchronic infection, these reactions continue in chronicH. pylori-infected gastritis until the bacteria are eradi-cated. Therefore mast cells in H. pylori-infected chronicgastritis may act both to maintain gastritis and to repairtissue damage.
REFERENCES
1 Enerback L. Mast cells in rat gastrointestinal mucosa. 2. Dye-binding and metachromatic properties. Acta Pathol MicrobiolScand 1966; 66: 303–12.
2 Walls AF, Jones DB, Williams JH, Church MK, Holgate ST.Immunohistochemical identification of mast cells in formalde-hyde-fixed tissue using monoclonal antibodies specific fortryptase. J Pathol 1990; 162: 119–26.
3 Nakajima S, Krishnan B, Ota H, et al. Mast cell involvement ingastritis with or without Helicobacter pylori infection. Gastro-enterology 1997; 113: 746–54.
4 Enerback L. Mast cell heterogeneity: The evolution of theconcept of a specific mucosal mast cell. In: Befus, AD, Bie-nenstock, J, Denburg, JA, eds. Mast Cell Differentiation andHeterogeneity. New York: Raven Press, 1986: 1–26.
5 Enerback L, Miller HRP, Mayrhofer G. Methods for the iden-tification and characterization of mast cells by light micro-scopy. In: Befus, AD, Bienenstock, J, Denburg, JA, eds. MastCell Differentiation and Heterogeneity. New York: RavenPress, 1986: 405–17.
6 Nakajima S, Arizono N, Hattori T, Bamba T. Increase inmucosal and connective tissue-type mast cells in thestomach with acetic acid-induced ulcer in rat. APMIS 1996;104: 19–29.
Mast cells
Helicobacter pylori
Epithelial cells
Other inflammatory
cells
Gastric glands
Blood vessels
Fibroblasts Pericellular matrices
Endothelial cells
activ
atio
n
histamineacid secretion
vasodilatationincreased permeabilityedema formation
histamine, try
ptase, IL-4
growthaccumulation
prot
ease
s tryptase
degradation growth
histamine, TNF-!
growthleukocyte adhesion
histamine, tryptaseTNF-!, ILs, PAF
cytokines
chemotaxismodulation
histamine, PGD2, LTs
tissue turnoverulcer healing
Epitheliumactivation
Fibroblasts
SCF
Figure 3. Possible roles of mast cells inH. pylori-infected stomach. Mast cells areactivated directly or indirectly withH. pylori,and interact with other cells in the stomach.Mast cells may play a major role in thedevelopment of inflammation, and also intissue turnover and ulcer healing throughmediators. SCF, stem cell factor; TNF-a,tumour necrosis factor-alpha; IL(s), inter-leukin(s); PAF, platelet-activating factor;PGD2, prostaglandin D2; LTs, leukotrienes.
MAST CELLS IN H. PYLORI GASTRITIS 169
! 2004 Blackwell Publishing Ltd, Aliment Pharmacol Ther 20 (Suppl. 1), 165–170
through H1 receptors.31 Interleukin-4, which is secreted
from activated mast cells32 stimulates fibroblasts toaccumulate at the site of inflammation.33 Tryptase,which is released with degranulation of mast cells,activates matrix metalloproteinases, especially matrixmetalloproteinase-3.34 Tryptase also stimulates thegrowth of epithelial cells and fibroblasts.35, 36 Thesemediators may act together to stimulate tissue and cellturnover in the vicinity of mast cells after the onset ofinflammation (Figure 3).It is uncertain whether histamine derived from mast
cells is involved in gastric acid secretion through H2
receptors; however, mast cell-derived histamine may beinvolved in a gastrin-independent mechanism of acidsecretion.37
POSSIBLE ROLE OF MAST CELLS
IN H. PYLORI-INFECTED CHRONIC GASTRITIS
A summary of the possible roles of mast cells in H.pylori-infected gastric mucosa is shown in Figure 3.Mast cells initiate and promote the formation of oedemathrough degranulated and secreted chemical mediators.They also release multiple chemotactic factors, whichinduce inflammatory cells to infiltrate to the site ofoedema. Mast cells and other inflammatory cells engagein complex and cooperative interplay with variouschemical mediators. They also stimulate the degrada-tion of pericellular matrices in H. pylori-infected gastricmucosa and the growth of cells in their vicinity, andpromote tissue turnover.
The phenomena described above are reactions in acuteand the subsequent phases of gastritis. Because H. pyloriinfection usually induces chronic inflammation throughchronic infection, these reactions continue in chronicH. pylori-infected gastritis until the bacteria are eradi-cated. Therefore mast cells in H. pylori-infected chronicgastritis may act both to maintain gastritis and to repairtissue damage.
REFERENCES
1 Enerback L. Mast cells in rat gastrointestinal mucosa. 2. Dye-binding and metachromatic properties. Acta Pathol MicrobiolScand 1966; 66: 303–12.
2 Walls AF, Jones DB, Williams JH, Church MK, Holgate ST.Immunohistochemical identification of mast cells in formalde-hyde-fixed tissue using monoclonal antibodies specific fortryptase. J Pathol 1990; 162: 119–26.
3 Nakajima S, Krishnan B, Ota H, et al. Mast cell involvement ingastritis with or without Helicobacter pylori infection. Gastro-enterology 1997; 113: 746–54.
4 Enerback L. Mast cell heterogeneity: The evolution of theconcept of a specific mucosal mast cell. In: Befus, AD, Bie-nenstock, J, Denburg, JA, eds. Mast Cell Differentiation andHeterogeneity. New York: Raven Press, 1986: 1–26.
5 Enerback L, Miller HRP, Mayrhofer G. Methods for the iden-tification and characterization of mast cells by light micro-scopy. In: Befus, AD, Bienenstock, J, Denburg, JA, eds. MastCell Differentiation and Heterogeneity. New York: RavenPress, 1986: 405–17.
6 Nakajima S, Arizono N, Hattori T, Bamba T. Increase inmucosal and connective tissue-type mast cells in thestomach with acetic acid-induced ulcer in rat. APMIS 1996;104: 19–29.
Mast cells
Helicobacter pylori
Epithelial cells
Other inflammatory
cells
Gastric glands
Blood vessels
Fibroblasts Pericellular matrices
Endothelial cells
activ
atio
n
histamineacid secretion
vasodilatationincreased permeabilityedema formation
histamine, try
ptase, IL-4
growthaccumulation
prot
ease
s tryptase
degradation growth
histamine, TNF-!
growthleukocyte adhesion
histamine, tryptaseTNF-!, ILs, PAF
cytokines
chemotaxismodulation
histamine, PGD2, LTs
tissue turnoverulcer healing
Epitheliumactivation
Fibroblasts
SCF
Figure 3. Possible roles of mast cells inH. pylori-infected stomach. Mast cells areactivated directly or indirectly withH. pylori,and interact with other cells in the stomach.Mast cells may play a major role in thedevelopment of inflammation, and also intissue turnover and ulcer healing throughmediators. SCF, stem cell factor; TNF-a,tumour necrosis factor-alpha; IL(s), inter-leukin(s); PAF, platelet-activating factor;PGD2, prostaglandin D2; LTs, leukotrienes.
MAST CELLS IN H. PYLORI GASTRITIS 169
! 2004 Blackwell Publishing Ltd, Aliment Pharmacol Ther 20 (Suppl. 1), 165–170
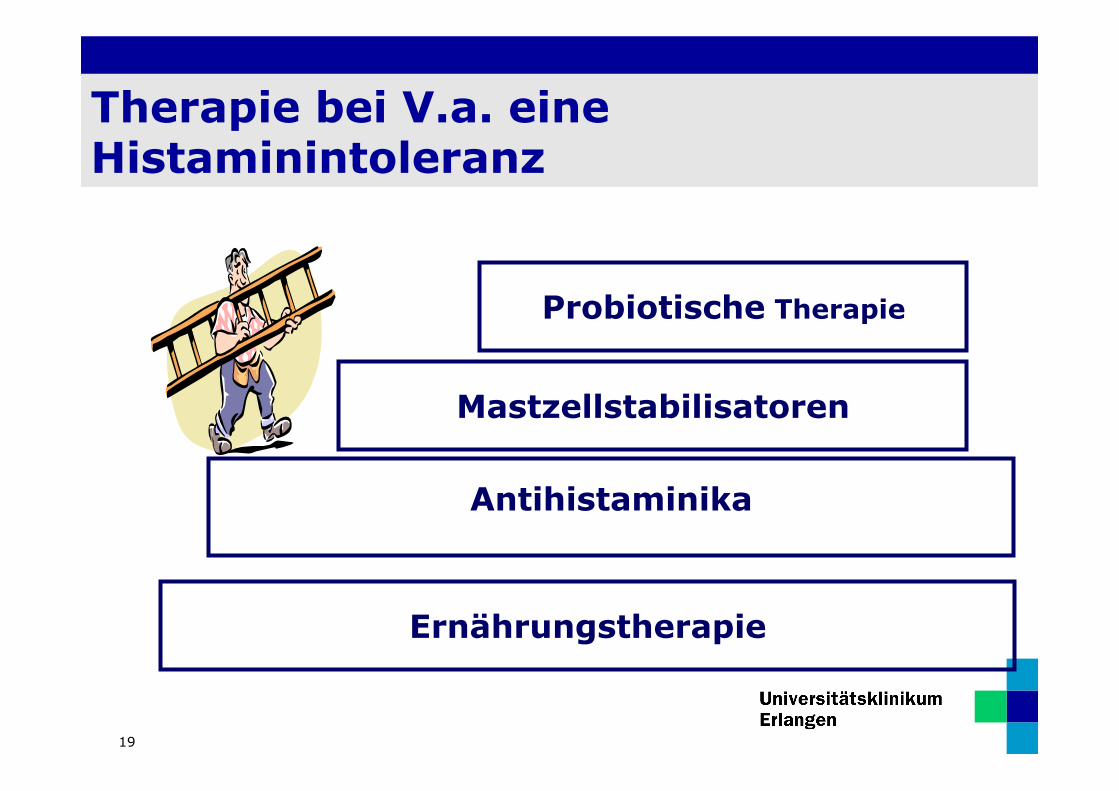
Therapie bei V.a. eine Histaminintoleranz
19
Antihistaminika
Mastzellstabilisatoren
Probiotische Therapie
Ernährungstherapie

20

Therapie
21
Antihistaminika
Mastzellstabilisatoren
Probiotische Therapie
Ernährungstherapie
22
through H1 receptors.31 Interleukin-4, which is secreted
from activated mast cells32 stimulates fibroblasts toaccumulate at the site of inflammation.33 Tryptase,which is released with degranulation of mast cells,activates matrix metalloproteinases, especially matrixmetalloproteinase-3.34 Tryptase also stimulates thegrowth of epithelial cells and fibroblasts.35, 36 Thesemediators may act together to stimulate tissue and cellturnover in the vicinity of mast cells after the onset ofinflammation (Figure 3).It is uncertain whether histamine derived from mast
cells is involved in gastric acid secretion through H2
receptors; however, mast cell-derived histamine may beinvolved in a gastrin-independent mechanism of acidsecretion.37
POSSIBLE ROLE OF MAST CELLS
IN H. PYLORI-INFECTED CHRONIC GASTRITIS
A summary of the possible roles of mast cells in H.pylori-infected gastric mucosa is shown in Figure 3.Mast cells initiate and promote the formation of oedemathrough degranulated and secreted chemical mediators.They also release multiple chemotactic factors, whichinduce inflammatory cells to infiltrate to the site ofoedema. Mast cells and other inflammatory cells engagein complex and cooperative interplay with variouschemical mediators. They also stimulate the degrada-tion of pericellular matrices in H. pylori-infected gastricmucosa and the growth of cells in their vicinity, andpromote tissue turnover.
The phenomena described above are reactions in acuteand the subsequent phases of gastritis. Because H. pyloriinfection usually induces chronic inflammation throughchronic infection, these reactions continue in chronicH. pylori-infected gastritis until the bacteria are eradi-cated. Therefore mast cells in H. pylori-infected chronicgastritis may act both to maintain gastritis and to repairtissue damage.
REFERENCES
1 Enerback L. Mast cells in rat gastrointestinal mucosa. 2. Dye-binding and metachromatic properties. Acta Pathol MicrobiolScand 1966; 66: 303–12.
2 Walls AF, Jones DB, Williams JH, Church MK, Holgate ST.Immunohistochemical identification of mast cells in formalde-hyde-fixed tissue using monoclonal antibodies specific fortryptase. J Pathol 1990; 162: 119–26.
3 Nakajima S, Krishnan B, Ota H, et al. Mast cell involvement ingastritis with or without Helicobacter pylori infection. Gastro-enterology 1997; 113: 746–54.
4 Enerback L. Mast cell heterogeneity: The evolution of theconcept of a specific mucosal mast cell. In: Befus, AD, Bie-nenstock, J, Denburg, JA, eds. Mast Cell Differentiation andHeterogeneity. New York: Raven Press, 1986: 1–26.
5 Enerback L, Miller HRP, Mayrhofer G. Methods for the iden-tification and characterization of mast cells by light micro-scopy. In: Befus, AD, Bienenstock, J, Denburg, JA, eds. MastCell Differentiation and Heterogeneity. New York: RavenPress, 1986: 405–17.
6 Nakajima S, Arizono N, Hattori T, Bamba T. Increase inmucosal and connective tissue-type mast cells in thestomach with acetic acid-induced ulcer in rat. APMIS 1996;104: 19–29.
Mast cells
Helicobacter pylori
Epithelial cells
Other inflammatory
cells
Gastric glands
Blood vessels
Fibroblasts Pericellular matrices
Endothelial cells
activ
atio
n
histamineacid secretion
vasodilatationincreased permeabilityedema formation
histamine, try
ptase, IL-4
growthaccumulation
prot
ease
s tryptase
degradation growth
histamine, TNF-!
growthleukocyte adhesion
histamine, tryptaseTNF-!, ILs, PAF
cytokines
chemotaxismodulation
histamine, PGD2, LTs
tissue turnoverulcer healing
Epitheliumactivation
Fibroblasts
SCF
Figure 3. Possible roles of mast cells inH. pylori-infected stomach. Mast cells areactivated directly or indirectly withH. pylori,and interact with other cells in the stomach.Mast cells may play a major role in thedevelopment of inflammation, and also intissue turnover and ulcer healing throughmediators. SCF, stem cell factor; TNF-a,tumour necrosis factor-alpha; IL(s), inter-leukin(s); PAF, platelet-activating factor;PGD2, prostaglandin D2; LTs, leukotrienes.
MAST CELLS IN H. PYLORI GASTRITIS 169
! 2004 Blackwell Publishing Ltd, Aliment Pharmacol Ther 20 (Suppl. 1), 165–170

Therapie
23
Antihistaminika
Mastzellstabilisatoren
Probiotische Therapie
Ernährungstherapie

24
through H1 receptors.31 Interleukin-4, which is secreted
from activated mast cells32 stimulates fibroblasts toaccumulate at the site of inflammation.33 Tryptase,which is released with degranulation of mast cells,activates matrix metalloproteinases, especially matrixmetalloproteinase-3.34 Tryptase also stimulates thegrowth of epithelial cells and fibroblasts.35, 36 Thesemediators may act together to stimulate tissue and cellturnover in the vicinity of mast cells after the onset ofinflammation (Figure 3).It is uncertain whether histamine derived from mast
cells is involved in gastric acid secretion through H2
receptors; however, mast cell-derived histamine may beinvolved in a gastrin-independent mechanism of acidsecretion.37
POSSIBLE ROLE OF MAST CELLS
IN H. PYLORI-INFECTED CHRONIC GASTRITIS
A summary of the possible roles of mast cells in H.pylori-infected gastric mucosa is shown in Figure 3.Mast cells initiate and promote the formation of oedemathrough degranulated and secreted chemical mediators.They also release multiple chemotactic factors, whichinduce inflammatory cells to infiltrate to the site ofoedema. Mast cells and other inflammatory cells engagein complex and cooperative interplay with variouschemical mediators. They also stimulate the degrada-tion of pericellular matrices in H. pylori-infected gastricmucosa and the growth of cells in their vicinity, andpromote tissue turnover.
The phenomena described above are reactions in acuteand the subsequent phases of gastritis. Because H. pyloriinfection usually induces chronic inflammation throughchronic infection, these reactions continue in chronicH. pylori-infected gastritis until the bacteria are eradi-cated. Therefore mast cells in H. pylori-infected chronicgastritis may act both to maintain gastritis and to repairtissue damage.
REFERENCES
1 Enerback L. Mast cells in rat gastrointestinal mucosa. 2. Dye-binding and metachromatic properties. Acta Pathol MicrobiolScand 1966; 66: 303–12.
2 Walls AF, Jones DB, Williams JH, Church MK, Holgate ST.Immunohistochemical identification of mast cells in formalde-hyde-fixed tissue using monoclonal antibodies specific fortryptase. J Pathol 1990; 162: 119–26.
3 Nakajima S, Krishnan B, Ota H, et al. Mast cell involvement ingastritis with or without Helicobacter pylori infection. Gastro-enterology 1997; 113: 746–54.
4 Enerback L. Mast cell heterogeneity: The evolution of theconcept of a specific mucosal mast cell. In: Befus, AD, Bie-nenstock, J, Denburg, JA, eds. Mast Cell Differentiation andHeterogeneity. New York: Raven Press, 1986: 1–26.
5 Enerback L, Miller HRP, Mayrhofer G. Methods for the iden-tification and characterization of mast cells by light micro-scopy. In: Befus, AD, Bienenstock, J, Denburg, JA, eds. MastCell Differentiation and Heterogeneity. New York: RavenPress, 1986: 405–17.
6 Nakajima S, Arizono N, Hattori T, Bamba T. Increase inmucosal and connective tissue-type mast cells in thestomach with acetic acid-induced ulcer in rat. APMIS 1996;104: 19–29.
Mast cells
Helicobacter pylori
Epithelial cells
Other inflammatory
cells
Gastric glands
Blood vessels
Fibroblasts Pericellular matrices
Endothelial cells
activ
atio
n
histamineacid secretion
vasodilatationincreased permeabilityedema formation
histamine, try
ptase, IL-4
growthaccumulation
prot
ease
s tryptase
degradation growth
histamine, TNF-!
growthleukocyte adhesion
histamine, tryptaseTNF-!, ILs, PAF
cytokines
chemotaxismodulation
histamine, PGD2, LTs
tissue turnoverulcer healing
Epitheliumactivation
Fibroblasts
SCF
Figure 3. Possible roles of mast cells inH. pylori-infected stomach. Mast cells areactivated directly or indirectly withH. pylori,and interact with other cells in the stomach.Mast cells may play a major role in thedevelopment of inflammation, and also intissue turnover and ulcer healing throughmediators. SCF, stem cell factor; TNF-a,tumour necrosis factor-alpha; IL(s), inter-leukin(s); PAF, platelet-activating factor;PGD2, prostaglandin D2; LTs, leukotrienes.
MAST CELLS IN H. PYLORI GASTRITIS 169
! 2004 Blackwell Publishing Ltd, Aliment Pharmacol Ther 20 (Suppl. 1), 165–170

25
Zusammenfassung
1. Diagnose der Histaminintoleranz – im Zusammenschau der Klinik
2. Eliminationsdiät
3. Re-Exposition bei unklarer Klinik
4. Kombinierte medikamentöse Therapie
5. Ggf. zusätzliche probiotische Therapie


















![Computation of the permeability of porous materials from their microstructure by … · 2013. 1. 21. · images of many porous materials [24, 8]. There is great interest in performing](https://static.fdokument.com/doc/165x107/60aef016c07a5904946638d7/computation-of-the-permeability-of-porous-materials-from-their-microstructure-by.jpg)