Identifizierung und Quantifizierung der Transforming ... · Abstract Regula Jens Ulrich...
158
Der Doktorvater ist Mitglied der Fakultät: Aus der Medizinischen Klinik und Poliklinik der Berufsgenossenschaftlichen Kliniken Bergmannsheil - Universitätsklinik - der Ruhr-Universität Bochum Direktor: Prof. Dr. med. H. Schatz Identifizierung und Quantifizierung der Transforming Growth Factor beta Rezeptor-Isotypen 1 und 2 sowie von Betaglykan in humanen Knochenbi- opsien und primären, humanen Knochenzellkulturen Inaugural-Dissertation zur Erlangung des Doktorgrades der Medizin einer Hohen Medizinischen Fakultät der Ruhr-Universität Bochum Vorgelegt von: Jens Ulrich Regula aus Heidelberg 2003
Transcript of Identifizierung und Quantifizierung der Transforming ... · Abstract Regula Jens Ulrich...

Der Doktorvater ist Mitglied der Fakultät:
Aus der Medizinischen Klinik und Poliklinik
der Berufsgenossenschaftlichen Kliniken Bergmannsheil - Universitätsklinik -
der Ruhr-Universität Bochum
Direktor: Prof. Dr. med. H. Schatz
Identifizierung und Quantifizierung der Transforming Growth Factor beta
Rezeptor-Isotypen 1 und 2 sowie von Betaglykan in humanen Knochenbi-
opsien und primären, humanen Knochenzellkulturen
Inaugural-Dissertation
zur
Erlangung des Doktorgrades der Medizin
einer
Hohen Medizinischen Fakultät
der Ruhr-Universität Bochum
Vorgelegt von:
Jens Ulrich Regula
aus Heidelberg
2003

AbstractRegula
Jens Ulrich
Identifizierung und Quantifizierung der Transforming Growth Factor ß Rezeptor-Isotypen
1 und 2 sowie von Betaglykan in humanen Knochenbiopsien und primären, humanen
Knochenzellkulturen
Problem: Transforming Growth Factor beta (TGF-ß) ist ein multifunktionaler, für den
Knochenstoffwechsel wichtiger Wuchs- und Wachstumsfaktor. TGF-ß fördert Proliferati-
on und Differenzierung der Zellen im Knochen. Die durch TGF-ß generierten Signale
werden über die sogenannten TGF-ß Typ I- und II- Rezeptoren (TßR), die Serin-/
Threonin-Kinasen sind, weitergeleitet. Der TGF-ß Typ III-Rezeptor (im Knochen Betagly-
kan) nimmt nicht direkt an Signalübermittlung teil. Dieser besitzt eine große extrazelluläre
Region, mit der die TGF-ß-Isoformen gebunden werden, und nur eine kurze intrazellulä-
re, nicht an der Signalübermittlung beteiligte Region.
Methode: Es werden humane Knochenproben und humane, primäre Knochenzell-
kulturen auf ihre TßR-I- bis -III-mRNA-Expression hin untersucht. Wegen der geringen
Mengen des Knochenmaterials wird ein Verfahren mit RT und anschließender quantita-
tiver „real-time“-PCR entwickelt, welches in der Lage ist, hochsensitiv und zuverlässig die
TßR zu detektieren. Anhand eines Probandenkollektives werden diverse Einflußgrößen
(Alter, Geschlecht, Mobilisation, Steroidbehandlung, BMI, Entnahmeort und präoperative
Diagnose) auf Unterschiede in der TßR-Expression untersucht. Zusätzlich wird ein Ver-
gleich mit der TGF-ß-mRNA-Expression und dem TGF-ß-Gehalt im Knochen und im Se-
rum durchgeführt. In Knochenzellkulturen wird die differenzierungsabhängige TßR-Ex-
pression und der Einfluß von Dexamethason (abhängig von Dosierung und Inkubations-
zeit) auf verschieden differenzierte Knochenzellen untersucht.
Ergebnisse und Diskussion: Signifikante Expressionsunterschiede ergeben sich für
TßR-II bei Osteoarthrose und zwischen Tibia und Beckenkamm, für den TßR-I zwischen
Patienten mit BMI größer oder kleiner als 25 kg/m^2. Hochsignifikante Korrelationen aller
TßR untereinander werden nachgewiesen. Expressionsunterschiede einzelner TßR tre-
ten im weiteren abhängig vom Geschlecht, vom Hormonstatus, vom Entnahmeort, von
der Mobilisation und von einer Steroidbehandlung auf. Steroide scheinen ihre Wirkung
auf die TßR in Abhängigkeit von der Inkubationszeit, Dosishöhe und Differenzierungs-
grad der Knochenzellen zu entfalten. Modulationen der TßR-Expression durch diverse
Faktoren (Steroide, Geschlecht, Lokalisation, Diagnose, BMI) werden nachgewiesen.

Dekan: Prof. Dr. med. G. Muhr
Referent: PD Dr. med. S. Hering
Koreferent: Prof. Dr. med. J. Krämer
Tag der mündlichen Prüfung: 27.01.2004

Meinen Eltern gewidmet

Inhaltsverzeichnis
Inhaltsverzeichnis1. Einleitung.................................................................................................................14
1.1 Der Knochen und seine Funktionen im Körper: Ein Überblick..........................14
1.2 Zelluläre und molekulare Aspekte des Knochenstoffwechsels.........................15
1.2.1 Biochemische Bestandteile des Knochens...............................................15
1.2.2 Der Knochenumbau..................................................................................15
1.2.3 Physiologische Einflüsse auf den Knochenstoffwechsel...........................16
1.2.3.1 Hormonelle Steuerung des Knochenstoffwechsels...........................16
1.2.3.2 Differenzierung der Knochenzellen....................................................17
1.2.3.3 Einfluß der Geschlechtshormone auf den Knochenstoffwechsel......18
1.2.3.4 Auswirkung von Bewegung auf den Knochenstoffwechsel...............18
1.2.3.5 Wirkung von Leptin im Knochenstoffwechsel....................................19
1.2.3.6 Weitere Faktoren mit Einfluß auf den Knochen.................................19
1.2.4 Veränderungen und Störungen des Knochenstoffwechsels.....................20
1.2.4.1 Altersbedingte Knochenveränderungen............................................20
1.2.4.2 Steroidinduzierte Knochenveränderungen........................................20
1.2.4.3 Weitere Einflußfaktoren und Erkrankungen.......................................21
1.3 Transforming Growth Factor beta (TGF-ß).......................................................23
1.3.1 Die TGF-ß-Superfamilie.............................................................................23
1.3.2 Aufbau, Struktur und Regulation von TGF-ß.............................................23
1.3.3 Physiologische Wirkung von TGF-ß..........................................................24
1.3.4 Wirkung von TGF-ß am Knochen..............................................................25
1.3.5 Funktion der „Bone Morphogenic Proteins“...............................................26
1.3.6 Krankheiten, die mit TGF-ß assoziiert sind................................................26
Seite 5

Inhaltsverzeichnis
1.4 Die Transforming Growth Factor-ß-Rezeptoren................................................28
1.4.1 Die TGF-ß-Rezeptoren beim Menschen....................................................28
1.4.2 Transforming Growth Factor ß Receptor Typ I..........................................28
1.4.3 Transforming Growth Factor ß Receptor Typ II.........................................29
1.4.4 Transforming Growth Factor ß Receptor Typ III........................................30
1.4.5 Bindungsverhalten der TßR.......................................................................31
1.4.6 Einfluß der TßR auf die Differenzierung....................................................32
1.4.7 Krankheiten, die mit Veränderungen der TßR assoziiert sind...................34
1.4.7.1 Osteoarthrose....................................................................................34
1.4.7.2 Weitere Erkrankungen.......................................................................34
1.5 Signalübertragung durch die TGF-ß-Rezeptoren..............................................35
1.6 Die „Smads“......................................................................................................38
1.6.1 Einteilung, Aufbau und Funktion...............................................................38
1.6.2 Signalübertragung durch die „Smads“.......................................................39
2. Fragestellung...........................................................................................................41
3. Methoden und Material............................................................................................43
3.1 Herstellung der Proben.....................................................................................43
3.1.1 Probengewinnung.....................................................................................43
3.1.2 Probenaufbereitung...................................................................................43
3.2 Zellkultur............................................................................................................44
3.3 RNA-Extraktion.................................................................................................45
3.3.1 RNA-Extraktion mit „TRIzol“.......................................................................45
3.3.2 RNA-Extraktion nach Chomczynski und Sacchi........................................46
3.3.3 RNA-Messung im Photometer...................................................................47
3.4 Reverse Transkription.......................................................................................47
Seite 6

Inhaltsverzeichnis
3.5 RNA-Gelelektrophorese....................................................................................48
3.6 Die Polymerase-Kettenreaktion........................................................................49
3.6.1 Prinzip der Polymerase-Kettenreaktion.....................................................49
3.6.2 Primerdesign.............................................................................................51
3.7 Quantitative PCR..............................................................................................52
3.7.1 Grundlagen................................................................................................52
3.7.2 Quantifizierungsstrategien.........................................................................52
3.7.3 Fluoreszenzfarbstoff SYBR Green I..........................................................55
3.7.4 Schmelzkurvenanalyse..............................................................................55
3.8 Grundlagen und Anwendung des „Light-Cycler“...............................................57
3.9 Grundlagen und Anwendung des „TaqMan“.....................................................60
3.10 Verwendung der Uracil-DNA-Glycosylase......................................................62
3.11 Agarose-Gelelektrophorese............................................................................63
3.12 Gel-Elution......................................................................................................63
3.13 Standardreihenherstellung..............................................................................64
3.14 Statistik...........................................................................................................64
3.15 Primer und Produkte.......................................................................................66
3.15.1 PDH.........................................................................................................66
3.15.2 TßR-I.......................................................................................................66
3.15.3 TßR-II.......................................................................................................66
3.15.4 Betaglykan...............................................................................................66
3.15.5 Weitere Primer........................................................................................66
3.16 Geräte.............................................................................................................67
3.16.1 PCR-Geräte.............................................................................................67
3.16.2 Gelelektrophorese...................................................................................67
Seite 7

Inhaltsverzeichnis
3.16.3 Zentrifugen..............................................................................................67
3.16.4 Computer und Software..........................................................................67
3.16.5 Sonstige Geräte......................................................................................68
3.17 Verwendete Chemikalien und Artikel..............................................................68
3.17.1 Einzelchemikalien....................................................................................68
3.17.2 Verwendete Reagenziensätze................................................................69
3.17.3 Enzyme....................................................................................................69
3.17.4 DNA-Größenstandards für Agarose-Gelelektrophorese..........................69
3.18 Puffer und Lösungen......................................................................................70
4. Ergebnisse...............................................................................................................73
4.1 Charakterisierung des Knochenkollektivs.........................................................73
4.2 PCR-Etablierung...............................................................................................77
4.2.1 Statistischer Vergleich der „real-time“-PCR-Geräte...................................77
4.2.1.1 Interassayvarianz...............................................................................77
4.2.1.2 Intraassayvarianz...............................................................................78
4.2.1.3 Sensitivität und Spezifität...................................................................78
4.2.1.4 Amplifikationseffektivität....................................................................81
4.2.1.5 Zuverlässigkeit der Standardreihen...................................................82
4.3 Datenanalyse der Knochenstudie.....................................................................84
4.3.1 Allgemeines Verteilungsmuster.................................................................84
4.3.2 Geschlechtsspezifische Verteilungsmuster...............................................85
4.3.3 Verteilungsmuster bezüglich Hormonstatus..............................................85
4.3.4 Altersabhängigkeit der TßR-Expression....................................................86
4.3.5 Diagnosespezifische Unterschiede der TßR-Expression..........................87
4.3.6 Entnahmeortspezifische Expression der TßR-Expression........................88
Seite 8

Inhaltsverzeichnis
4.3.7 Mobilisationsspezifische Expression der TßR...........................................89
4.3.8 Einfluß der Steroidmedikation auf die TßR-Expression.............................90
4.3.9 Expression der TßR in Abhängigkeit vom „Body Mass Index“...................91
4.3.10 Korrelationen zwischen TGF-ß-Isoformen und TGF-ß-Rezeptoren.........92
4.3.11 Korrelation zwischen den einzelnen TßR-Isoformen...............................93
4.3.12 Weitere Ergebnisse der Knochenstudie..................................................96
4.4 Analyse der TßR in humanen, primären Knochenzellkulturen..........................98
4.4.1 Einfluß der TßR auf die Differenzierung....................................................98
4.4.2 Einfluß der Steroide auf primäre Knochenzellkulturen..............................99
5. Diskussion.............................................................................................................102
5.1 Wertigkeit der angewandten Methoden..........................................................102
5.2 Einflüsse auf die PDH-Expression bei relativer Quantifizierung.....................106
5.3 Diskussion der Ergebnisse der Knochenstudie...............................................107
5.3.1 Generelle Aspekte...................................................................................107
5.3.2 Einfluß des Geschlechts auf die TßR-Expression...................................108
5.3.3 Einfluß des Alters auf die TßR-Expression..............................................110
5.3.4 TßR-Expression bei pathologischen Veränderungen des Knochens......112
5.3.5 Einfluß des Entnahmeortes auf die TßR-Expression..............................114
5.3.6 Einfluß der Mobilisation auf die TßR-Expression.....................................116
5.3.7 TßR-Expression unter längerfristiger Glukokortikoidbehandlung............118
5.3.8 Einfluß des BMI auf die TßR-Expression.................................................119
5.3.9 Korrelation zwischen den TßR-Isoformen...............................................121
5.3.10 Weitere Ergebnisse der Knochenstudie................................................122
5.4 Diskussion der Ergebnisse der humanen, primären Knochenzellkulturen......123
5.4.1 Einfluß der TßR auf die Differenzierung..................................................123
Seite 9

Inhaltsverzeichnis
5.4.2 Einfluß von Glukokortikoiden auf primäre Knochenzellkulturen..............124
6. Zusammenfassung................................................................................................128
7. Literatur.................................................................................................................130
8. Danksagung..........................................................................................................157
9. Lebenslauf.............................................................................................................158
Seite 10

Abkürzungen
Abkürzungen
A Adenosin
Ac Acetat
AP Alkalische Phosphatase
ATCC „American Tissue Cell Culture and Tissue Collection“
ATP Adenosintriphosphat
BAP „Bone Alkaline Phosphatase“ (engl.: knochenspezifische AP)
BMI „Body Mass Index“ (engl., BMI=kg Körpergewicht / (Größe in m)^2)
BMP „Bone Morphogenic Protein“ (engl.)
bp Basenpaare
c centi =10^-2
C Cytosin
ca. cirka
Cp „Crossing point“ (engl.)
CR „Collagenase Released“
Ct „threshold cycle“ (engl.)
CTP Cytosintriphosphat
Da Dalton
dATP Desoxyadenosintriphosphat
dCTP Desoxycytosintriphosphat
DEPC Di-Ethyl-Pyro-Carbonat
dGTP Desoxyguanosintriphosphat
dN6 Desoxynukleotide, aus 6 Basen bestehend
DNA Desoxyribonukleinsäure (engl.: desoxyribonukleic acid)
dNTP Desoxynukleotidtriphosphat
dTTP Desoxythymidintriphosphat
ECM Extrazelluläre Matrix
EDTA Ethylendiamintetraessigsäure
EGF „Epidermal Growth Factor“
engl. englisch
et al. und andere
EtBr Ethidiumbromid
f femto=10^-15
Seite 11

Abkürzungen
FGF „Fibroblast Growth Factor“
x g „gravity“ (engl.), Vielfaches der Erdanziehungskraft
g Gramm
G Guanosin
GAPDH Glycerinaldehyd-3-Phosphat-Dehydrogenase
GE Gelelektrophorese
GC Glukokortikoide
GRE „Glucocorticoid-response-element“
GTP Guanosintriphosphat
h Stunde(n)
HHT Hereditäre Hämorrhagische Teleangiektasie
HPLC Hochdruck-Flüssigkeits-Chromatographie
IGF „Insulin-like growth factor“
IGFBP „Insulin-like growth factor binding protein“
IL Interleukin
i.S. im Serum
J. Jahre
k kilo=1000
kb Kilo-Basen
l Liter
LTBP „Latent Transforming Growth Factor beta Binding Protein“
m milli =10^-3
m Meter
m männlich
µ mikro =10^-6
M Molar
M Mega =1Million
mA milli-Ampere
MADH „Mother Against Decapentaplegic Homolog“
min Minute(n)
M-Mulv „Moloney-Murine Leukemia Virus“
mRNA Boten-Ribonukleinsäure (engl.: „messenger ribonucleic acid“)
MW Mittelwert
n nano =10^-9
n Probenanzahl
Seite 12
Abkürzungen
nt Nukleotide
OC Osteocalcin
p pico =10^-12
p___ Protein mit dem Gewicht ___ kDa
PBS „Phosphate Buffered Saline“
PCR Polymerase-Kettenreaktion (engl.: „polymerase chain
reaction“)
PDGF „Platelet-Derived Growth Factor“
PDH Pyruvatdehydrogenase
pH pondus Hydrogenii
PTH Parathormon
PTHrP „PTH related protein“
RNA Ribonukleinsäure (engl.: „ribonucleic acid“)
RNAse Ribonuklease
RNAsin RNAse Inhibitor
RT Reverse Transkriptase
s Sekunde(n)
SD Standardabweichung (engl.: „standard deviation“)
Smad „Similar MAD“ („Mother Against Decapentaplegic“)
SZ Stromale Zellen
T Thymidin
TßR „Transforming Growth Factor beta Receptor“
TE „Treated Explants“
TGF-ß „Transforming Growth Factor beta“
TGIF „TGF-Interacting Factor“
Tm Schmelzpunkt
TNF Tumor Nekrose Faktor
TRAP „TGF-ß Receptor associated protein“
TRIP „TGF-ß Receptor interacting protein“
TTP Thymidintriphosphat
U Unit (Enzym-Aktivitäts-Einheit)
UV Ultraviolett
w weiblich
°C Grad Celsius
Seite 13

Kapitel 1. Einleitung
1. Einleitung
1.1 Der Knochen und seine Funktionen im Körper: Ein Überblick
Das Stützgerüst des menschlichen Körpers wird überwiegend von Knochen gebildet und
ermöglicht Sitzen und körperliche Bewegung. Auf molekularer Ebene besteht der
Knochen hauptsächlich aus Kalzium, Phosphat, verschiedenen Kollagenen und Matrix-
proteinen. Er besitzt einen hohen Anteil am Körpergewicht der Menschen und wird meist
als starres System betrachtet. Tatsächlich gehört er zu den mit am stärksten im Umbau
befindlichen und am besten regulierten Organen des menschlichen Körpers. Viele Hor-
mone wie Parathormon, Vitamin D, Calcitonin und andere sind am Erhalt, Auf- und Ab-
bau des Knochens beteiligt. Diese Hormone üben ihre Hauptwirkung auf die Osteo-
blasten und Osteoklasten im Knochen über eine Steuerung des Kalksalzgehaltes aus.
Weitere im Knochen enthaltenen Zellen sind die Osteozyten, von Knochenmatrix einge-
mauerte Osteoblasten, und die den Knochen von den anderen Geweben abgrenzenden
„lining cells“ (Junqueira et al., 1995). Des weiteren haben viele Wachstums- und Wuchs-
faktoren einen Einfluß auf den Knochenstoffwechsel und regeln die Speicherung von
Kalzium und Phosphat. Viele Umweltgifte werden in ihm abgelagert und verzögert in den
Kreislauf abgegeben. Der Knochen enthält neben am Knochenstoffwechsel beteiligten
Zellen das Knochenmark, das für die Hämatopoese und Synthese diverser Wachstums-
faktoren und Zytokine verantwortlich ist (Schölmerich et al., 1996).
Seite 14

Kapitel 1. Einleitung
1.2 Zelluläre und molekulare Aspekte des Knochenstoffwechsels
1.2.1 Biochemische Bestandteile des Knochens
Mineralbestandteile - vorwiegend Kalziumapatit (Kalziumphosphatsalze) - und das
Osteoid - die organische Knochenmatrix, zu 90 % aus Kollagen Typ I bestehend – bilden
die Hauptbestandteile des Knochens. Die Osteoblasten synthetisieren den Großteil der
Kollagene, das Osteocalcin und Osteonektin. Zu einem kleinen Anteil kommen in der
Knochenmatrix Proteine (z. B. Albumin, α-2-HS-Glykoprotein, ß-2-Mikroglobulin und
Transferrin), Enzyme (z.B. Kollagenase), Wuchs- und Wachstumsfaktoren wie „Fibro-
blast Growth Factor“ (FGF), „Platelet Derived Growth Factor“ (PDGF) Transforming
Growth Factor beta“ (TGF-ß) und Interleukin (IL) -1 und -6 vor (Blümcke et al., 1995).
1.2.2 Der Knochenumbau
Das Konzept des „bone remodeling“ spielt eine wichtige Rolle beim heutigen Verständnis
des Knochenstoffwechsels und seiner pathologischen Veränderungen. Das „remodeling”
vollzieht sich herdförmig in Kompakta und Spongiosa. Die konzertierte Aktion von Osteo-
blasten und Osteoklasten, durch ein genaues Zusammenspiel von parakrinen, auto-
krinen und endokrinen Hormonen gesteuert, bewirkt eine ständige Knochenresorption
und -neubildung. Dieses Ineinandergreifen von Knochenumbauvorgängen, das soge-
nannte „coupling“, wird durch einige Wuchsfaktoren geregelt, TGF-ß wird als einer der
maßgeblichsten angesehen. Ein „uncoupling”, eine Dissoziation der osteoblastären und
osteoklastären Aktivität, scheint mitverantwortlich für den bei Krankheiten auftretenden
Knochenverlust zu sein (Erlebacher et al., 1998; Schölmerich et al., 1996).
Primär ruhen lokalisationsabhänig etwa 80 % bis 90 % der Knochenzellen. Spontan
Seite 15

Kapitel 1. Einleitung
sammeln sich Prä-Osteoklasten, fusionieren und beginnen, Knochen zu resorbieren. La-
tentes TGF-ß wird aus der Knochenmatrix durch die Osteoklasten freigesetzt und ak-
tiviert. TGF-ß hemmt die osteoklastäre Aktivität stark, fördert die Apoptose der
Osteoklasten und bewirkt an den Osteoklasten einen Stopp der Knochenresorption. Die
Osteoblasten wandern durch chemotaktische Vorgänge, vermittelt durch Knochenmatrix-
bestandteile, Kollagenfragmente, Osteocalcin oder Wuchsfaktoren wie TGF-ß oder
PDGF, in die Lakunen. Knochenzellen fangen an, Mitogene wie „Insulin-like Growth Fac-
tor“ (IGF) oder TGF-ß, die die Proliferation von Osteoblastenvorläuferzellen fördern, zu
produzieren. Nach der Proliferation und der Synthese der extrazellulären Matrix (ECM)
folgt die Differenzierung der Osteoblasten, die wahrscheinlich durch „Bone Morphogenic
Proteins“ (BMP), 1,25-(OH)2 Vitamin D3, Fluoride und Vitamin A gefördert wird. Aktive
Vitamin D-Metabolite, die eine intramembranäre Ossifikation verursachen, oder BMPs
könnten die Mineralisierung der Matrix bewirken (Mundy et al., 1995; Guise et al., 1998).
1.2.3 Physiologische Einflüsse auf den Knochenstoffwechsel
1.2.3.1 Hormonelle Steuerung des Knochenstoffwechsels
Die hormonelle Steuerung im Knochen beruht auf zwei, eng miteinander verknüpften
Systemen: die Regelung des Kalksalzgehaltes und der Matrixsynthese. Der Kalksalzge-
halt der Knochenmatrix ist durch ein kontrolliertes Zusammenspiel der drei Hormone –
Calcitonin, Parathormon und Vitamin D - in vier Organen – Knochen, Nebenschilddrüse,
Niere und Verdauungstrakt – sowie der Knochensalze Kalzium und Phosphat reguliert.
Dies sorgt für einen sehr konstanten Kalziumspiegel im Blut, dessen Bilanz gleich null
sein soll. In die Knochenmatrix sind viele Wachstums- und Wuchsfaktoren eingebettet,
vor allem TGF-ß und IGF-2. Diese Faktoren werden durch die dauernd tätigen
Osteoklasten freigesetzt, die ihrerseits den Knochenaufbau fördern (Guise et al., 1998).
Folgende Übersicht zeigt die hormonellen Einflußfaktoren auf Osteoblasten und -klasten
bezüglich der Wirkung am Knochen (Mundy et al., 1996):
Seite 16

Kapitel 1. Einleitung
➔ Resorption inhibierende und/oder Anbau aktivierende Regulatoren:
Calcitonin, Östrogen, Progesteron, Androgene, „Transforming Growth Faktor-ß“ (TGF-ß),
„Insulin like Growth Faktor“-1 und -2 (IGF), „Bone Morphogenetic Protein“ (BMP), „Fibro-
blast Growth Faktor“ (FGF), Wachstumshormon (GH, engl.: „Growth Hormone“)
➔ Resorption aktivierende und/oder Anbau inhibierende Regulatoren:
Glukokortikoide, „Colony Stimulating Factors“ (CSF), Interleukine (vor allem IL-1, -6, -11),
„Tumor Necrosis Factor-α“ (TNF-α), Prostaglandin E2 (PG-E2)
➔ Resorption und Anbau aktivierende Regulatoren:
Parathormon (PTH), Vitamin D, „Platelet-Derived Growth Factor“ (PDGF)
1.2.3.2 Differenzierung der Knochenzellen
Die Produktion von Kollagen Typ I findet in einer frühen Phase der osteoblastären Diffe-
renzierung statt. Danach folgt die sequentielle Expression von alkalischer Phosphatase
und Osteocalcin. Am Übergang von Proliferation zur Differenzierung werden die verant-
wortlichen Gene der Proliferation und der Kollagensynthese herunterreguliert, die der
Differenzierung und Matrixreifung in ihrer Expression verstärkt und dies erlaubt die Mine-
ralisation. Tetranektin ist ein in der Mineralisationsphase des Knochens vermehrt ge-
bildetes Protein. Osteoblasten zeigen ein streng geregeltes, sequentielles Expressions-
muster mit einer reziproken, funktional gekoppelten Beziehung zwischen Differenzierung
und Proliferation (Kassem et al., 2000; Takeuchi et al., 1996). Folgende Marker sind
während Entwicklung und Reifung erhöht: 1.) c-fos während der Proliferationsphase, 2.)
Alkalische Phosphatase (AP) während der frühen Osteoblastendifferenzierung und 3.)
Osteocalcin (OC) in der späten Differenzierungsphase (Kostenuik et al., 1997).
Seite 17

Kapitel 1. Einleitung
1.2.3.3 Einfluß der Geschlechtshormone auf den Knochenstoffwechsel
Die Entdeckung der Östrogenrezeptoren auf Osteoblasten zeigt, daß die Osteoblasten
direkte Angriffspunkte der Östrogene sind. Studien zeigen wegen der Uneinheitlichkeit
der untersuchten Zellpopulationen sehr heterogene Ergebnisse. Östrogene stimulieren
die AP-Aktivität, unterdrücken die Osteocalcin-Expression und reduzieren die Zellprolife-
ration dosisabhängig. Die Kollagen-Expression wird nur gering durch Östrogene be-
einflußt (Robinson et al., 1997). Der androgenbedingte Knochenverlust über Androgen-
rezeptoren ist, genauso wie der östrogenbedingte über Östrogenrezeptoren, letztlich IL-6
vermittelt. IL-6 bewirkt eine Steigerung der Osteoklastogenese (Bellido et al., 1995).
Androgene übertragen ihre Effekte auf den Knochen über mindestens zwei Systeme,
eines wurde als das TGF-ß-System identifiziert (Kasperk et al., 1990). Adrenale und go-
nadale Androgene stimulieren die Knochenbildung und sind wichtig für die Erhaltung der
Knochenmasse. Androgene stimulieren die AP-Expression direkt und die Mitose über
Androgenrezeptoren (Kasperk et al., 1997).
1.2.3.4 Auswirkung von Bewegung auf den Knochenstoffwechsel
Kostenuik und Mitarbeiter zeigten die Belastungsabhängigkeit der Osteoblastendiffe-
renzierung in einem Rattenmodell mit Hinterbeinentlastung. Die Proliferation und weitere
Differenzierung der Osteoblasten wurde unterdrückt und ging mit reduzierter AP-Aktivität
und geringerer Matrixmineralisation einher (Kostenuik et al., 1997). Harris et al. be-
schrieben leichte, nicht-signifikante Unterschiede in der Expression von IL-1, IL-6 und
TGF-ß in fetalen, humanen Osteoblastenkulturen, die entweder in Schwerelosigkeit oder
auf der Erde kultiviert wurden. In vivo scheint es mehr auf die dynamischen, auf den
Knochen wirkenden Kräfte, als auf die statischen, in Schwerelosigkeit verminderten Kräf-
te anzukommen (Harris et al., 2000). Mechanische Belastung begünstigt die Prolifera-
tionsphase der Osteoblasten über eine verlängerte Wachstumsphase oder durch eine
Hemmung der Zelldifferenzierung. Die Zahl der Osteoblasten nimmt unter Belastung zu.
Es kommt zu verstärkter Kollagenbildung, zur Abnahme der AP-Aktivität und des Osteo-
Seite 18

Kapitel 1. Einleitung
calcingehaltes. Eine intensivierte Proliferationsphase führt zu einer besser an me-
chanische Reize angepaßten, stabileren Knochenmatrix (Kasper et al., 2000). Zhang et
al. führten Versuche mit hinterteilentlasteten Ratten durch und zeigten einen deutlichen
Rückgang der TGF-ß2-Expression in Osteozyten und im Periost bei Skelettentlastung
mittels in-situ-Hybridisierung. Da TGF-ß2 ortsspezifisch im Periost großteils von Prä- und
Osteoblasten exprimiert wird und da die TGF-ß2-Expression in den Knochen-
marksstromazellen bei immobiliserten Ratten reduziert ist, scheint es, daß TGF-ß2 im
Prozeß des „bone remodeling” an der Transduktion mechanischer Signalen beteiligt ist
(Zhang et al., 1999).
1.2.3.5 Wirkung von Leptin im Knochenstoffwechsel
Leptin ist ein 16 kDa großes, von weißem Fettgewebe synthetisiertes Peptidhormon. Es
ist das Produkt des obese-Gens (ob). Leptin kommt eine wesentliche Rolle bei der Rege-
lung der Essensaufnahme und des Energieverbrauches zu. Des weiteren wirkt Leptin
auf Angiogenese, Knochenbildung und Immunsystem (Huang et al., 2000). Seine
Wirkung besteht in der Förderung der Differenzierung von Knochenmarksstammzellen in
Richtung Osteoblasten und nicht in Richtung Adipozyt. Es hemmt die Knochenneubil-
dung über eine hypothalamische Achse (Kume et al., 2002), und wirkt mit den BMPs
fördernd auf die Differenzierung der Osteoblasten (Gori et al., 1999).
1.2.3.6 Weitere Faktoren mit Einfluß auf den Knochen
Die FGFs sind starke, knochenbildende Faktoren und fördern die Proliferation und Sti-
mulation der Osteoblastenvorläuferzellen (Dunstan et al., 1993). Knochenzellen
exprimieren PDGF und besitzen Rezeptoren für PDGF, über die eine starke
Knochenneubildung und eine erhöhte osteoklastäre Resorption initiiert wird (Graves et
al., 1989). IGF-1 und -2 haben eine positive Wirkung auf den Knochen in vivo und
fördern die Osteoklastentätigkeit (Mundy et al., 1995). Russell-Aulet et al. zeigten für GH
einen über die IGFs vermittelten, positiven Einfluß auf Knochendichte und -stoffwechsel
(Russell-Aulet et al., 1998).
Seite 19

Kapitel 1. Einleitung
1.2.4 Veränderungen und Störungen des Knochenstoffwechsels
1.2.4.1 Altersbedingte Knochenveränderungen
Mit zunehmendem Alter kommt es aufgrund verschiedener Faktoren zu einer
Verminderung der Knochenmasse. Darüber hinaus auftretende Verringerungen der
Knochendichte führen zu einem erhöhten Risiko des Auftretens von Frakturen. Die
pathologisch verminderte Knochenmasse ist als Osteopenie und das Unterschreiten fest-
gelegter Grenzwerte der Knochendichte oder das Auftreten von Frakturen bei noch de-
finitionsgemäß bestehender Osteopenie als Osteoporose definiert. Diese signalisieren
eine gestörte Mikroarchitektonik des Knochens und somit eine signifikante Verringerung
der ossären Stabilität. Ursachen können Alter, ungenügende körperliche Aktivität und
hormonelle Störungen sein (Allolio et al., 1996). Der Gipfel der Knochendichte, die „peak
bone mass“, ist am Ende des 3. Lebensjahrzehntes erreicht und läßt sich durch Bewe-
gung, Ernährung und Nikotinkarenz beeinflussen. Je höher die „peak bone mass“ in
jungen Jahren ist, desto geringer ist das Osteoporoserisiko im Alter (Eisman, 1999).
1.2.4.2 Steroidinduzierte Knochenveränderungen
Eine langdauernde und höher dosierte Steroidtherapie ist ein häufiger Grund für eine
Osteopenie oder Osteoporose. Steroide haben vielfältige Wirkungen auf den Knochen.
Sie erhöhen die PTH-induzierte Knochenresorption durch Osteoklasten sowie die renale
Kalziumausscheidung und reduzieren die intestinale Kalziumaufnahme. Die Sekretion
des Wachstumshormons, des Kalzitonins und gonadaler Steroide ist vermindert (Allolio
et al., 1996). Steroide verändern die Transkription verschiedentlich, sie beeinflussen An-
zahl und Funktion der Osteoblasten. Die Antwort der Osteoblasten auf Steroide hängt
stark von ihrer Differenzierung ab. Die Reifung von Osteoblasten wird gefördert, die Dif-
ferenzierung und Reifung von Vorläuferzellen zu Präosteoblasten gehemmt. Präosteo-
blasten sezernieren vermindert AP und Osteocalcin. Die reifen Osteoblasten synthe-
Seite 20

Kapitel 1. Einleitung
tisieren weniger Osteocalcin und Kollagen I α1. Kollagenase 3, die Kollagen Typ I ab-
baut, wird vermehrt sezerniert. Dem Knochen fehlen junge, proliferierende Osteoblasten
und die vorhandenen Osteoblasten funktionieren nur mäßig (Hulley et al., 1998). An
Knochenzellkulturen zeigten Hulley et al. eine Hemmung der Knochenreifung durch
Dexamethason im „Mitogen Activated Protein Kinase“-Signalweg (MAPK). Die Wachs-
tumshemmung durch Steroide zeigte eine Latenzzeit von 6 h. Daher vermuteten sie die
Regulation über eine veränderte Gentranskription (Hulley et al., 1998).
Dexamethason verzögert die Apoptose bei Osteoblasten durch eine Verhinderung der
Fas-Expression in den peripheren mononukleären Blutzellen. Die Gelenkzerstörung bei
Arthrose scheint über Fas-Liganden vermittelt zu werden, die eine gelenknahe Osteopo-
rose und eine vermehrte Apoptose bei Osteoblasten bewirken (Nakashima et al., 1998).
Pearce et al. behandelten Patienten ein halbes Jahr mit Steroiden. Bei den steroidthera-
pierten Patienten nahm in diesem Zeitraum die Knochendichte um 2 bis 4,5 % ab, dem
8- bis 16-fachen im Vergleich zu einem gesunden Kollektiv (0,25 % bis 0,5 % pro Jahr)
entsprechend. Osteocalcin und die knochenspezifische AP im Serum waren als Hinweis
auf eine Störung der Knochenneubildung erniedrigt (Pearce et al., 1998).
Iba et al. zeigten eine Reduktion der Dexamethason-induzierten Differenzierung und Te-
tranektin-Expression in osteoblastären Zellen durch TGF-ß1. Tetranektin wird durch
Dexamethason in einer durch das Simian Virus 40 (SV-40) immortalisierten Zellinie aus
fetalen, humanen Kalvariaknochenzellen (SV-HFO) hochreguliert. Dieser Anstieg wird
durch TGF-ß1 dosisabhängig gehemmt. Parallel zu der Tetranektin-Expression steigt die
AP. TGF-ß1 scheint die Proliferation, Differenzierung und ECM-Synthese zu initiieren
und weniger die späteren Phasen der Osteogenese zu beeinflussen (Iba et al., 1995).
1.2.4.3 Weitere Einflußfaktoren und Erkrankungen
Ein Polymorphismus des Vitamin D-Rezeptor-Gens scheint für Abweichungen von der
normalen Knochendichte verantwortlich zu sein (Audi et al., 1999). Ein Polymorphismus
im Intron 1 des Kollagen I α1-Gens geht mit Unterschieden in Knochendichte und Kör-
pergröße einher (Mann et al., 2001). Gewisse HLA-Phänotypen (Tsuji et al., 1998) und
chromosomale Regionen scheinen für eine erhöhte oder reduzierte Knochendichte zu
Seite 21

Kapitel 1. Einleitung
kodieren (Devoto et al., 1998). Eine multiple Allelie mit Einfluß auf die Knochendichte
existiert für die TGF-ß-Rezeptor-Gene und für TGF-ß3 (Han et al., 2001 und Gortz et al.,
2001).
Weitere Erkrankungen des Knochens und seines Stoffwechsels sind die Osteomalazie,
Rachitis, Osteopetrose, Ostitis deformans (M. Paget) und alle Formen des Hypo- und
Hyperparathyreodismus. Sie beruhen auf einer Störung im Makro- oder Mikrostoffwech-
sel des Knochens und bewirken eine Veränderung der Knochenstruktur (Schölmerich et
al., 1996).
Seite 22

Kapitel 1. Einleitung
1.3 Transforming Growth Factor beta (TGF-ß)
1.3.1 Die TGF-ß-Superfamilie
Der Wachstumsfaktor TGF-ß gehört zu einer Superfamilie von Polypeptiden, deren
Hauptaufgabe die Steuerung der Differenzierung und Entwicklung vieler Stamm- und
Vorläuferzellen ist. Zu dieser Familie gehören die TGF-ß-Isoformen, die BMPs, Aktivin
und Inhibin. Sie besitzen eine ausgeprägte phylogenetisch bedingte Sequenzhomologie
(Roberts et al., 1998).
1.3.2 Aufbau, Struktur und Regulation von TGF-ß
Bisher sind 23 TGF-ß-Isoformen beschrieben, bei den Säugetieren 5 und bei den Men-
schen 3, TGF-ß1, -2 und -3. Aus den transkribierten Vorläufer-Proteinen - zwischen 390
und 412 Aminosäuren lang – werden durch Endopeptidasen die bioaktiven oder latenten
TGF-ß-Isoformen gebildet (Linkhart et al., 1996). Die einzelnen TGF-ß-Isoformen sind
auf verschiedenen Chromosomen kodiert: TGF-ß1 auf 19q13.2, TGF-ß2 auf 1q41, TGF-
ß3 auf 14q24 (Quelle: http://www.ncbi.nlm.nih.gov/LocusLink).
Das bioaktive Molekül ist ein Homodimer mit 25 kDa und setzt sich aus zwei jeweils 12,5
kDa schweren, über Disulfidbindungen verbundenen Monomeren zusammen. Es können
auch Heterodimere gebildet werden. TGF-ß liegt normalerweise als latentes TGF-ß ge-
bunden an das 190 kDa schwere „Latent Transforming Growth Factor Binding Protein“
(LTBP) vor, das von Knochenzellen entweder in Granula gespeichert oder in die extra-
zelluläre Matrix sezerniert wird (Mundy et al., 1995). LTBP scheint die TGF-ß-Bioverfüg-
barkeit und darüber die TGF-ß-Wirkung zu beeinflussen (Dabovic et al., 2002). Latentes
TGF-ß wird durch diverse Enzyme aus diesem Komplex herausgelöst und bindet als bio-
Seite 23

Kapitel 1. Einleitung
aktives TGF-ß an die Matrix, α-Makroglobulin oder Decorin, durch die die Wirkung aufge-
hoben wird. Eine Bindung an Albumin oder IgG hebt die Wirkung nicht auf (Clark et al.,
1998; Dallas et al., 1994).
Die TGF-ß-Isoformen haben unterschiedliche Wirkungsstärken an gleichen Organen.
TGF-ß1 stimuliert das Osteoblastenwachstum und die Kollagensynthese 3- bis 5-fach
stärker als andere. TGF-ß2 inhibiert stärker als andere das Wachstum der Knochen-
markszellen. TGF-ß3 ist weniger fibrinogenbildend als TGF-ß1 und -2 und antagonisiert
leicht deren Wirkung (Clark et al., 1998).
Im TGF-ß1-Promotor findet sich ein „Glucocorticoid Response Element“ (GRE), welches
in Osteoblasten nicht aktiv zu sein scheint (Parrelli et al., 1998).
1.3.3 Physiologische Wirkung von TGF-ß
TGF-ß1 gilt als potenter Inhibitor der Proliferation von lymphoiden Zellen und Aktivator
der Zellproliferation bei der Gewebereparatur und -differenzierung (Sporn et al., 1999).
TGF-ß spielt eine wesentliche Rolle in Entwicklung und Wachstum von embryonalem
und adultem Gewebe, Entzündung, Wundheilung und Angiogenese. Seine Effekte
werden auf parakrine und autokrine Art vermittelt. Wachstum epithelialer und
endothelialer Zellen wird durch TGF-ß in vitro gehemmt, während mesenchymale Zellen
in ihrem Wachstum gefördert werden (Clark et al., 1998). TGF-ß3 steigert die Kollagen-
synthese. Im Zusammenspiel mit TGF-ß1 scheint TGF-ß3 eine inhibierende Wirkung auf
die Kollagensynthese und -ablagerung zu haben (Murata et al., 1997; Seitzer et al.,
1995; Centrella et al., 1992). Die Wirkung von TGF-ß im Gewebe ist von seiner Kon-
zentration abhängig. In niedrigen Konzentrationen bewirkt es eine Stimulierung, in hohen
Konzentrationen eine Hemmung der Zellproliferation. Verschiedene Mitglieder der TGF-
ß-Superfamilie rufen unterschiedliche Antworten im Körper hervor. TGF-ß beeinflußt die
„Cyclin Dependent Kinases“ (CDK) und die „Cyclin Dependent Kinase Inhibitors“ (CKI)
(Hill, 1996).
Steroide haben eine konträre Wirkung zu TGF-ß, erstere schwächen die Wundheilung,
während letzteres die Wundheilung fördert. Dexamethason blockiert die fibroseindu-
Seite 24

Kapitel 1. Einleitung
zierende Wirkung von TGF-ß durch die Hemmung der Kollagensynthese in Fibroblasten,
in denen sowohl die TGF-ß1-mRNA vermindert ist als auch weniger Proteine sezerniert
werden (Parrelli et al., 1998).
1.3.4 Wirkung von TGF-ß am Knochen
Die Knochenmatrix enthält die höchsten Konzentrationen von TGF-ß1 im Körper. Dort
stimuliert es die Kollagensynthese und beeinflußt die mRNA-Expression von pro-α1(I)-
Kollagen, Osteonektin, Osteopontin, Fibronektin und der AP in Osteoblasten und Fibro-
blasten. Es fördert die Matrixproteinsynthese und Knochenneubildung. Gehemmt wird
der Matrixabbau durch Verringerung matrixabbauender Enzyme und Vermehrung von
Proteaseinhibitoren (Guise et al., 1998). Direkte TGF-ß-Applikation auf die Schädelka-
lotte von Mäusen zeigt eine lokal vermehrte Knochenreifung und -bildung (Marcelli et al.,
1990).
Das Gleichgewicht zwischen Knochenbildung und Knochenresorption wird von Faktoren
wie TGF-ß, den IGFs und IL-6 gesteuert. TGF-ß1 wird vornehmlich von Osteoblasten ge-
bildet und in die Knochenmatrix inkorporiert. Es wird durch Plasmin und einen hohen
oder niedrigen pH-Wert aktiviert. Es wird während der Knochenresorption aus der
Knochenmatrix freigesetzt und durch den niedrigen pH in der Umgebung der
Osteoklasten aktiviert. TGF-ß1 scheint der wichtigste Mediator der Knochenumbaupro-
zesse zu sein, denn 1.) fördert es in den reifen Osteoklasten Apoptoseprozesse und
hemmt sie in ihrer knochenresorbierenden Funktion, 2.) hemmt es die Proliferation von
unreifen, mononukleären Vorläuferzellen der Osteoklasten in vitro, 3.) hemmt es die Fu-
sion von mononukleären Vorläuferzellen zu reifen Osteoklasten, 4.) fördert TGF-ß1 die
Proliferation und Differenzierung von Prä- und Osteoblasten und 5.) steigert es die Che-
motaxis der Osteoblasten. Es ist ein wesentlicher Vermittler des „coupling” (Parfitt et al.,
1979; Mundy et al., 1995). Langdahl et al. konnten zwei Sequenzvarianten im TGF-ß1-
Gen, die 783-8delC- und die C788T-Variante, identifizieren, die bei Osteoporose-Pati-
enten überrepräsentiert waren und mit erhöhten Knochenumbauparametern einher-
gingen. Beiden Varianten könnte ein erhöhter Knochenumbau durch Wegfallen der TGF-
ß1-vermittelten Osteoklastenhemmung zugrunde liegen (Langdahl et al., 1997).
Seite 25

Kapitel 1. Einleitung
Zugabe von TGF-ß1 zu einer Maus-Knochenmarkszellkultur reduziert die Bildung von
Osteoklasten-ähnlichen Zellen durch vermehrte Apoptose. Dies scheint teilweise über
die vermehrte Expression von Osteoprotegerin („osteoclastogenesis inhibitory factor“,
OCIF) vermittelt zu sein (Murakami et al., 1998).
Zhou et al. zeigten in einem Hasenmodell mit kontinuierlicher, subperiostaler TGF-ß-Infu-
sion nicht nur eine Wirkung an den Osteoblasten, sondern auch an den Osteoklasten.
Die Wirkung auf das „remodeling” des Knochens nach Defektsetzung durch TGF-ß setzt
in der Frühphase ein, indem primär die Knochenneubildung und später konsekutiv die
Knochenresorption gefördert wird (Zhou et al., 1995).
1.3.5 Funktion der „Bone Morphogenic Proteins“
„Bone Morphogenic Proteins“ (BMPs) gehören zu der TGF-ß-Superfamilie. Die BMPs
sind in der Knochenmatrix vorhanden und haben die einzigartige Fähigkeit, die ektope
Knochenbildung zu initiieren und zu stimulieren. Mit fortschreitender Osteoblastendiffe-
renzierung werden BMP-2, -3 und -4 zunehmend exprimiert (Harris et al., 1994). Exo-
genes BMP-2 fördert die Knochenmineralisation in vitro. Die Tatsache, daß bei Fehlen
von BMP-2 die Osteoblasten nicht differenzieren können, legt nahe, daß die BMPs auto-
krine Faktoren der Knochenzelldifferenzierung sind. Im BMP-2-Promotor findet sich ein
„BMP-2-response element“ und Elemente, die auf TGF-ß, p53 und Rb ansprechen
(Ghosh-Choudhury et al., 2000; Helvering et al., 2000).
1.3.6 Krankheiten, die mit TGF-ß assoziiert sind
In der Krebsentstehung wirkt TGF-ß im frühen Stadium antiproliferativ, später jedoch un-
terstützend bei Neoangiogenese, Zellausbreitung, Immunsuppression, Tumorzellinvasion
und Metastasierung (Guise et al., 1998).
Seite 26

Kapitel 1. Einleitung
Die Fibrosierung als Folge einer exzessiven Produktion und Ablagerung von extrazellulä-
ren Matrixproteinen wird bei Nierenfibrose oder diabetischer Retinopathie durch übermä-
ßige Stimulierung der TGF-ß-Aktivität oder eine aktivierende Mutation in diesem Signal-
weg hervorgerufen (Hill, 1999; Clark et al., 1995).
Eine inaktivierende Mutation in CDMP-1 der TGF-ß-Superfamilie führt zu akro-
mesomelischer Chondrodysplasie Typ Hunter-Thompson (Storm et al., 1994).
Seite 27

Kapitel 1. Einleitung
1.4 Die Transforming Growth Factor-ß-Rezeptoren
1.4.1 Die TGF-ß-Rezeptoren beim Menschen
Die Wirkung von TGF-ß wird über spezifische Rezeptoren vermittelt, deren Struktur und
Funktion erst in den letzten Jahren zunehmend entschlüsselt und charakterisiert worden
sind. Die TGF-ß Typ I und II Rezeptoren sind transmembranäre Serin-Threonin-Kinase-
Rezeptoren (Miyazono et al., 1994) und bilden einen heterooligomeren Signalkomplex.
Rezeptoren wie Betaglykan oder andere TGF-ß Typ III Rezeptoren binden TGF-ß, ohne
daß sie an der Signalübermittelung beteiligt sind (Massagué et al., 1992 und 1998). Die
Funktion wird in der Präsentation der TGF-ß-Isoformen für die TßR-I und -II gesehen. Für
TGF-ß sind bei dem Menschen drei Rezeptorgruppen beschrieben, die auf eigenen
Genen kodiert sind und durch verschiedene Mechanismen reguliert werden (Hill 1996).
1.4.2 Transforming Growth Factor ß Receptor Typ I
Der Transforming Growth Factor ß Receptor Type I (TßR-I) ist auf Chromosom 9q22 ko-
diert (Johnson et al., 1995). Seine Länge liegt zwischen 503 und 532 Aminosäuren. Das
Gewicht wird mit 53 kDa angegeben. Sieben verschiedene Typen konnten bisher identifi-
ziert werden und die Liganden sind großteils bekannt. Die Sequenzhomologie unterein-
ander ist hoch. Die extrazelluläre Domäne ist mit zehn konservierten Cystein-Resten
kürzer als die des TßR-II. Ein Cluster von 3 Cysteinen kommt analog im TßR-II vor. Der
Transmembrandomäne folgt die Juxtamembrandomäne. Die konservierte Signaturse-
quenz SGSGSGLP (GS-Region) des TßR-I liegt direkt vor der Serin/Threonin-Kinase-
Region, gefolgt vom wenige Aminosäuren langen C-terminale Ende. Der TßR-I ist durch
cytoplasmatische Kinasen konstitutiv phosphoryliert und an der Zelloberfläche als Homo-
dimer anzutreffen (Derynck et al., 1997). Die Promotorregion ist sehr GC-reich. In der
Seite 28

Kapitel 1. Einleitung
upstream-Region des TßR-I-Methionin-
Start-Codons liegen mindestens sieben
Sp1-Bindungsstellen, einige Sp3-
Elemente, AP2-Bindungssequenzen und
zwei CCAAT-Boxen. Ihr fehlt eine TATA-
oder CAAT-Box. Eine GC-reiche Region
bei -314 bis -229 bp ist für die transkrip-
tionelle Aktivität wichtig und enthält die
hypothetische Startsequenz. Die Sp1-
und Sp3-Bindungssequenzen scheinen
für die basale TßR-I-Expression verant-
wortlich zu sein. Für PEBP2/CBFα-Famili-
enmitglieder wurde eine Bindungsstelle
identifiziert, die für die Expression diffe-
renzierter Gene von großer Bedeutung zu
sein scheint (Bloom et al., 1996; Ji et al.,
1997).
1.4.3 Transforming Growth Factor ß Receptor Typ II
Der Genort des Transforming Growth Factor ß Receptor Type II (TßR-II) liegt auf Chro-
mosom 3p22 (Mathew et al., 1994). Sein Gewicht wird auf 73 kDa geschätzt. Die TßR-II
mRNA wird als einzelne Isoform von ca. 5,2 kb exprimiert. Die aus 7 Exons abgeleitete
567 Aminosäuren lange Polypeptidsequenz enthält eine 136 Aminosäuren lange N-gly-
kolisierte, cystein-reiche extrazelluläre Region, eine transmembranäre und eine cytoplas-
matische Domäne. Die cytoplasmatische Region besteht hauptsächlich aus einer Kina-
se-Domäne und liegt zwischen der juxtamembranären Region und dem C-terminalen
Ende. Das Signalpeptid ist in Exon 1, die cysteinreiche Extrazellulärregion in den Exons
Seite 29
Abbildung 1: Aufbau der TßR-I und -II (aus
Derynck et al., 1997)

Kapitel 1. Einleitung
2 und 3, die Transmembrandomäne in Exon 4, die Kinasedomäne in den Exons 4 bis 7
und das Stopsignal in Exon 7 codiert (Takenoshita et al., 1996).
Das C-terminale Ende ist serin- und threoninreich. Viele Cystein-Reste in der extrazellu-
lären Domäne lassen auf eine extensive Disulfid-Brückenbildung und -faltung für die kor-
rekte Ligandenbindung schließen. Die Kinase-Regionen der TßR-II-Isoformen sowie
einige Cysteinreste in Membrannähe sind zwischen den Spezies hochgradig konserviert.
Die extrazelluläre Region ist weniger gut konserviert und ermöglicht ein gewisses Maß
an Signalspezifität. Der Typ II-Rezeptor formt Homodimere, die sich aus zwei unter-
schiedlichen Typ II-Rezeptoren zusammensetzen können und eine Anlagerung von hete-
rodimeren Liganden ermöglichen. Die ligandenunabhängige Dimerisierung läßt eine kon-
stitutive Autophosphorylierung vermuten. Die Autophosphorylierung des TßR-II kann
durch andere Kinasen nur in geringerem Ausmaß in der cytoplasmatischen Region bei
kinase-deaktivierten Punktmutanten vorgenommen werden (Derynck et al., 1997).
In der Promotorstruktur des TßR-II finden sich keine TATA- oder CAAT-Box-Motive, aber
zwei Sp1- und zwei AP1-Motive. AP2-Elemente sind nicht enthalten. Die Promotorregion
ist nicht so GC-reich (ca. 70%) wie die des TßR-I (ca. 85%). Die proximale AP1-Sequenz
steigert die transkriptionelle Aktivität in Lungenfibroblasten. Da die Promotorregionen der
beiden TßR unterschiedlich sind, dürften auch unterschiedliche Transkriptionsfaktoren
für deren Regulation verantwortlich sein (Yu et al., 1996; Bloom et al., 1996). Sp1-
Elemente beeinflussen die Expression einiger Gene, die die Zelldifferenzierung und -pro-
liferation induzieren (Humphries et al., 1994).
1.4.4 Transforming Growth Factor ß Receptor Typ III
Abhängig von der Gewebeart werden unterschiedliche Transforming Growth Factor ß
Receptor Type III (TßR-III) exprimiert: Betaglykan in epithelialem, mesenchymalen und
neuralem Gewebe und Endoglin in hämatopoetischem Gewebe. Das genomische Äqui-
valent von Betaglykan liegt auf Chromosom 1p33-p32 und das von Endoglin auf Chro-
mosom 9q33-q34.1 (Johnson et al., 1995, Fernandez-Ruiz et al., 1993, Wang et al.,
1991). Von allen TßR-III-Isotypen wird Betaglykan im Knochen am meisten exprimiert
Seite 30

Kapitel 1. Einleitung
(Horner et al., 1998). Aufgrund der extensiven, posttranslationalen Modifikation der TßR-
III mit Kohlenhydraten, Glykosaminoglykanen und Heparan- und Chondroitinsulfaten
wandert Betaglykan in der Elektrophorese sehr heterogen zwischen 280 und 330 kDa,
obwohl das Kernprotein nur 100 bis 110 kDa wiegt. Das Protein ist 853 Aminosäuren
lang und besteht aus einem Signalpeptid, einer großen extrazellulären Domäne, einer
transmembranären Region und einer 41 Aminosäuren langen, cytoplasmatischen Do-
mäne. Die kurze intrazelluläre Region erlaubt es, einen löslichen, funktionsfähigen TßR-
III nachzuweisen, der mit den Signalrezeptoren TßR-I und -II in Konkurrenz tritt (Derynck
et al., 1997). Endoglin wird in vaskulären Endothelzellen und in einigen hämatopoe-
tischen Zellen gefunden. Die Länge des Polypeptides beträgt 561 Aminosäuren. Es wird
L- und S- Endoglin wegen der Struktur der C-terminalen Sequenz unterschieden. Endo-
glin kann mit dem TßR-I-TßR-II-Komplex assoziieren, und Betaglykan nur mit TßR-II.
Beide Moleküle haben eine Sequenzhomologie von 71 % und eine Identität von 63 % in
der extrazellulären Region und liegen als Homo-Oligomere, meist als Homodimere vor.
Die Dimerisierung ist unabhängig von der Ligandenbindung. Nicht-kovalente Kräfte
sorgen bei Betaglykan und Disulfid-Brücken bei Endoglin für den Zusammenhalt. Pro
Monomer oder Dimer wird ein TGF-ß-Molekül gebunden. Während lösliches Betaglykan
als kompetitiver Inhibitor der TGF-ß-Bindung an die TßR-I und -II dienen kann, ver-
vielfacht die membrangebundene Form die TGF-ß-Bindung an die TßR. Biologisch
wichtig scheint eine Reservoir-Funktion der TßR-III für verfügbares TGF-ß zu sein, wel-
ches hochaffin den Rezeptoren präsentiert wird (Derynck et al., 1997).
1.4.5 Bindungsverhalten der TßR
TßR-I und TßR-II besitzen eine größere Affinität zu TGF-ß1 und TGF-ß3 als zu TGF-ß2.
Alle drei Formen aktivieren den gleichen Signalweg. Zellen ohne exprimierte TßR-III
zeigen auf TGF-ß2 kein spezifisches „Antwortmuster“. Dagegen exprimieren Osteo-
blasten Betaglykan und können auf Stimulation mit TGF-ß2 typisch reagieren (Pfeilschif-
ter et al., 1990). Wenn Betaglykan mit TßR-II koexprimiert wird, verstärkt es die
Ligandenbindung an den TßR-II, besonders die des nur schwach an den TßR-II
bindenden TGF-ß2. TßR-II alleine bindet TGF-ß1 und -3 gut. TGF-ß2 wird nur in
Seite 31

Kapitel 1. Einleitung
Anwesenheit des TßR-I gebunden, da durch Zusammenlagerung eine neue Bindungs-
stelle für TGF-ß2 geschaffen wird. Betaglykan hat eine höhere Affinität zu TGF-ß2 und
kann dieses dem TßR-II gut präsentieren, was durch Koimmunopräzipitation von TßR-II
und -III in Anwesenheit von TGF-ß2 gezeigt wurde (Rodriguez et al., 1990).
Betaglykan bindet wegen des Proteingerüstes und unabhängig von den Modifikationen
alle TGF-ß-Isoformen mit einer ähnlichen Affinität. Endoglin bindet TGF-ß1 und -3 mit
deutlich höherer Affinität wie TGF-ß2. TßR-I bindet nur zusammen mit TßR-II Liganden.
Die Signalspezifität wird durch den TßR-II bestimmt. Differentielle Interaktionen zwischen
den verschiedenen Typ I- und Typ II-Rezeptoren tragen zur Signalspezifität bei und
erlauben eine Modulation der Antwortfähigkeit auf Liganden. In dem heteromeren Kom-
plex aus TßR-II und TßR-I wird der TßR-I durch TßR-II nach Ligandenbindung phospho-
ryliert und diese Phosphorylierung aktiviert die Kinasetätigkeit des TßR-I (Derynck et al.,
1997).
1.4.6 Einfluß der TßR auf die Differenzierung
Die Expression der TßR wird durch stimulierende und hemmende Cytokine während der
osteoblastären Differenzierung spezifisch reguliert. Die relative Menge des TßR-I in Form
von mRNA und Protein sowie die Promotoraktivität steigen im Verlauf der osteoblastären
Differenzierung parallel an, vermutlich über eine veränderte Sp1-Aktivität oder andere
Promotor geregelt (Ji et al., 1997). In undifferenziertem Mesenchym wird der TßR-II stark
exprimiert und im Laufe des Entwicklungsprozesses geht die Expression deutlich zurück
(Humphries et al., 1994). Die Differenzierung und Expression der TßR an der Zelloberflä-
che wird durch Interaktion mit Matrixkollagenen in osteoblastären Zellen reguliert. Da
TGF-ß die Differenzierung und Entwicklung von differenzierungsassoziierten Phä-
notypen wie die Expression der AP und Osteocalcin in osteoblastären Zellen hemmt,
müssen die TGF-ß-Effekte während der Differenzierung unterdrückt werden, damit die
Zellen von der Phase der Matrixsynthese in die der Mineralisation übergehen können.
Bei Fortbestehen der TGF-ß-Wirkung auf die Osteoblasten kommt es zu weiterer Matrix-
proteinakkumulation und die Matrix kann nicht mineralisiert werden. Die Erhöhung der
AP-Aktivität geht mit einer Reduzierung der TGF-ß-Aktivität einher, die auf einer
Seite 32

Kapitel 1. Einleitung
Verminderung der Bindung an die TßR zu beruhen scheint. Wird die Kollagensynthese
blockiert, bleibt die TßR-Expression auf hohem Niveau. Anhäufung von Matrixkollagen
verstärkt die Differenzierung der osteoblastären Zellen und unterdrückt die TGF-ß-Aktivi-
tät durch eine Verminderung der Rezeptorzahl. Diese Reduktion scheint nach heutigen
Vorstellungen einer der Mechanismen zu sein, mit dem die Zellen der TGF-ß-Kontrolle
„entkommen“ und in die Mineralisationsphase übergehen können. Die Verminderung der
TßR an der Zelloberfläche ist nicht durch eine Reduktion der Transkription bedingt, da
die mRNA sich nur wenig ändert. Die Ansprechbarkeit scheint durch eine veränderte in-
trazelluläre Kommunikation moduliert zu werden (Takeuchi et al., 1996).
Für die spezifischen Wirkungen der TGF-ß-Rezeptoren scheinen die relativen und nicht
die absoluten Verhältnisse der Rezeptoren zueinander wichtig zu sein. Bei überwiegend
osteoblasten-ähnlicher Aktivität tritt der TßR-I in den Vordergrund. Eine Reduktion der
Betaglykan-Expression scheint das TGF-ß-bedingte Wachstum besser zu regeln. Ein
Überfluß an Betaglykan kann TGF-ß-Moleküle abfangen und konservieren. Wird das
Verhältnis von TßR-I und TßR-II im Vergleich zu Betaglykan größer, so vermittelt TGF-ß
vermehrt mitogene Effekte. Wird das Verhältnis TßR-I zu TßR-II größer, kommt es zu
vermehrter Kollagensynthese und AP-Aktivität, einem der frühen Osteoblastendiffe-
renzierung entsprechendem Muster. Kortison verändert das Bindungsverhalten und
TGF-ß bindet vermehrt an Betaglykan und weniger an die TßR-I und -II. Dies zeigt einer-
seits die Wichtigkeit der Rezeptorstöchiometrie auf Knochenzellen, gibt andererseits je-
doch keine Erklärung für den TGF-ß-Funktionsverlust (Centrella et al., 1995). Behand-
lung von Knochenzellkulturen mit dem differenzierend wirkenden BMP-2 fördert die
Bindung von TGF-ß an TßR-I, während die Bindung an andere Rezeptoren in Abhängig-
keit von der Differenzierung mehr oder weniger stark abnimmt (Kessler et al., 2000).
Seite 33

Kapitel 1. Einleitung
1.4.7 Krankheiten, die mit Veränderungen der TßR assoziiert sind
1.4.7.1 Osteoarthrose
Die Expression eines verkürzten, Kinase-defizienten TßR-II in einem Mausmodell bewirkt
einen Differenzierungsschub der Chondrozyten, und führt zu einem Osteoarthrose-ähnli-
chen Phänotyp mit Gelenkknorpelaufsplitterung, Anhäufung und Proliferation der Chon-
drozyten, Osteophytenbildung und knorpeliger Metaplasie der Synovia. Der Verlust der
TGF-ß-Antwortfähigkeit bewirkt eine Verminderung der Proteoglykan-Synthese. Die Syn-
these von Kollagen Typ X - ein Marker der hypertrophen Chondrozytendifferenzierung,
der durch TGF-ß gehemmt wird - ist atypischerweise vermehrt, und könnte auf verzö-
gertem Einsetzen der TGF-ß-induzierten Apoptose beruhen. Endogenes TGF-ß scheint
für die Knorpelhomöostase und Chondrozytenreifung wichtig zu sein (Serra et al., 1997).
1.4.7.2 Weitere Erkrankungen
Eine inaktivierende Mutation mit fehlerhafter Funktion im Endoglin-Gen ist die Grundlage
für die autosomal-dominant vererbte hereditäre hämorrhagische Teleangiektasie Typ I
(HHT) oder Morbus Osler-Rendu (McAllister et al., 1994). Eine Mutation im humanen
„Activin Like Kinase“-1-Gen (ALK), einem Mitglied der TGF-ß-Superfamilie, führt zur
hereditären hämorrhagischen Teleangiektasie Typ II (Johnson et al., 1996).
Mutationen des Typ-II-Rezeptors für „Müllerian Inhibiting Substance“ (MIS), einem Mit-
glied der TGF-ß-Superfamilie, kann die unvollständige Regression des Müller-Ganges
und das Syndrom des persistierenden Müller-Ganges auslösen (Hata et al., 1998).
Sowohl die Gene als auch die Promotoren der TßR-I und -II können durch Rearrange-
ments, homozygote oder größere Deletionen, fehlende Transkripte, homozygote Genver-
luste, Transversions- und „frame-shift“- Mutationen für die Krebsentstehung verantwort-
lich sein (Vostriukhina et al., 1998; Reiss et al., 1999; Hu et al., 1998). Retinoblastom-
zellen sprechen nicht bei fehlender TßR-II-Expression auf TGF-ß an (Horie et al., 1998).
Seite 34

Kapitel 1. Einleitung
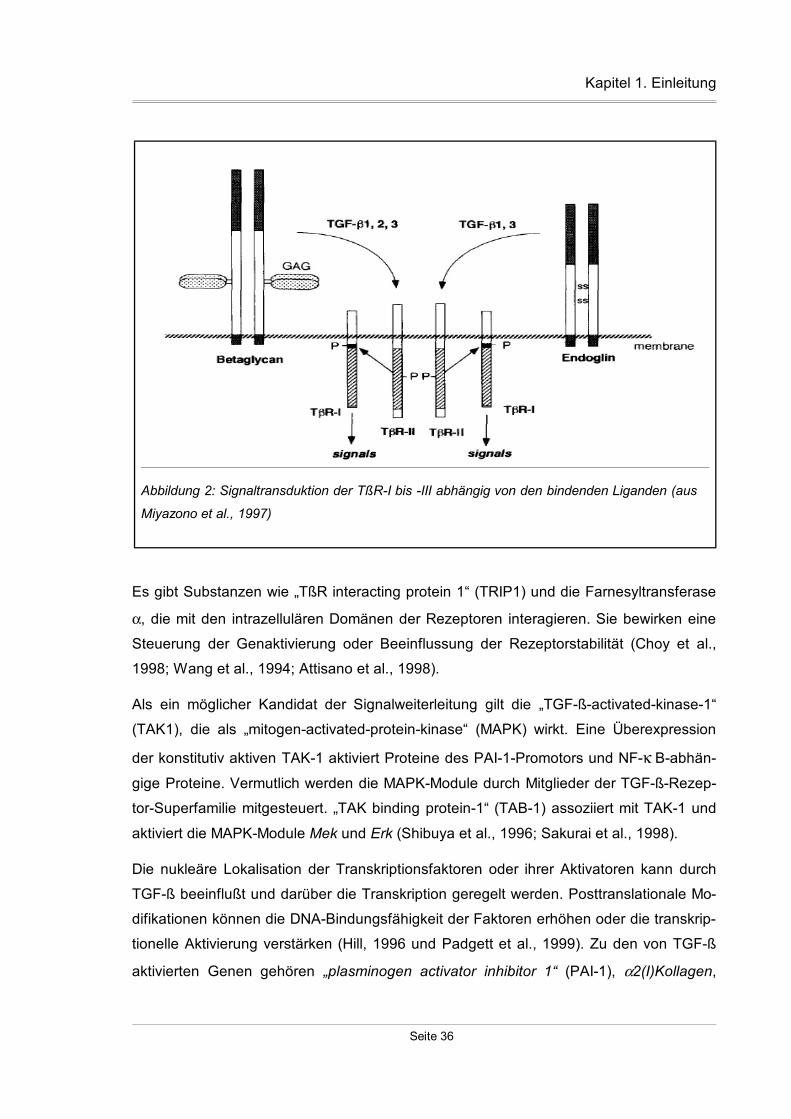
1.5 Signalübertragung durch die TGF-ß-Rezeptoren
Liganden binden an den TßR-II und bilden den Ligand-Rezeptor-Komplex. TßR-I bindet
normalerweise nur in Anwesenheit des TßR-II Liganden. Zur Signalübermittelung ist ein
Komplex aus mindestens zwei TßR-I und zwei TßR-II nötig. TßR-I und TßR-II können in
Abwesenheit von Liganden Komplexe bilden. Ligandenbindung an zuvor assoziierte he-
teromere Rezeptorkomplexe stabilisiert diese Komplexe. Der TßR-II ist konstitutiv aktiv.
Er autophosphoryliert sich unabhängig von der Ligandenbindung an Ser/Thr-Resten, die
das Ziel anderer Kinasen sein können. TßR-I erkennt den Ligand-Rezeptor-Komplex.
Phosphorylierungen durch den TßR-II an Thr- und Ser-Resten in der GS-Region des
TßR-I aktivieren die Kinaseaktivität, die ihrerseits weitere intrazelluläre Substrate phos-
phoryliert. Die Kinaseaktivität des TßR-I und die GS-Region sind von großer Bedeutung
für die intrazelluläre Signalweiterleitung. Es gibt Mutationen an den Ser/Thr-Resten, die
zu einem konstitutiv aktiven (Charng et al., 1996) oder zu einem kinase-defizienten,
funktionslosen Rezeptor (Brand et al., 1995) führen. Im Vergleich zu der aktiven Mutation
führt eine physiologische Ligandenaktivierung zu einer stärkeren Wirkung.
Der TßR-II ist für die Signalerkennung und Aktivierung des TßR-I zuständig und be-
stimmt, welche Liganden gebunden werden. Der TßR-I leitet das Signal weiter und ent-
scheidet, welche Proteine phosphoryliert werden. Das Phosphorylieren einer bestimmten
Aminosäure in der Kinase-Region des TßR-I durch den TßR-II führt immer zu der glei-
chen Zellantwort. Das Zellsignal ändert sich in Abhängigkeit von der phosphorylierten
Kinase-Region des TßR-I. Ein TßR-II kann mit verschiedenen TßR-I-Kinasedomänen in-
teragieren und eine Varianz der Zellantwort herbeiführen. Aufgrund der unterschiedli-
chen Rezeptoraffinitäten und Konzentrationen der Liganden werden verschiedene Gene
als Antwort aktiviert. Niedrige Ligandenkonzentrationen bewirken eine Aktivierung der
hochaffinen Promotoren. Andererseits führen hohe Konzentrationen der Liganden zu
starken Signalen, da Promotoren mit niedrigen Affinitäten aktiviert werden (Derynck et
al., 1997). Der TßR-III wirkt nicht an der Signalübermittelung mit, da er nur über eine
kurze intrazelluläre Region ohne Kinasefunktion verfügt. Eine Deletion der cytoplasma-
tischen Domäne bewirkt eine veränderte Phosphorylierung des TßR-II und veränderte
Präsentationsfähigkeit der TGF-ß-Isoformen (Blobe et al., 2001).
Seite 35
Kapitel 1. Einleitung
Es gibt Substanzen wie „TßR interacting protein 1“ (TRIP1) und die Farnesyltransferase
α, die mit den intrazellulären Domänen der Rezeptoren interagieren. Sie bewirken eine
Steuerung der Genaktivierung oder Beeinflussung der Rezeptorstabilität (Choy et al.,
1998; Wang et al., 1994; Attisano et al., 1998).
Als ein möglicher Kandidat der Signalweiterleitung gilt die „TGF-ß-activated-kinase-1“
(TAK1), die als „mitogen-activated-protein-kinase“ (MAPK) wirkt. Eine Überexpression
der konstitutiv aktiven TAK-1 aktiviert Proteine des PAI-1-Promotors und NF-κ B-abhän-
gige Proteine. Vermutlich werden die MAPK-Module durch Mitglieder der TGF-ß-Rezep-
tor-Superfamilie mitgesteuert. „TAK binding protein-1“ (TAB-1) assoziiert mit TAK-1 und
aktiviert die MAPK-Module Mek und Erk (Shibuya et al., 1996; Sakurai et al., 1998).
Die nukleäre Lokalisation der Transkriptionsfaktoren oder ihrer Aktivatoren kann durch
TGF-ß beeinflußt und darüber die Transkription geregelt werden. Posttranslationale Mo-
difikationen können die DNA-Bindungsfähigkeit der Faktoren erhöhen oder die transkrip-
tionelle Aktivierung verstärken (Hill, 1996 und Padgett et al., 1999). Zu den von TGF-ß
aktivierten Genen gehören „plasminogen activator inhibitor 1“ (PAI-1), α2(I)Kollagen,
Seite 36
Abbildung 2: Signaltransduktion der TßR-I bis -III abhängig von den bindenden Liganden (aus
Miyazono et al., 1997)

Kapitel 1. Einleitung
WAF1/Cip1/p21, der Transkriptionsfaktor NF κ B und TGF-ß selber. In den Promotoren
dieser Gene finden sich TGF-ß responsible Elemente. Transin enthält einen negativ
TGF-ß responsiblen Promotor. Direkte Ziele von TGF-ß sind die Transkriptionsfaktoren
CTF/NF1, AP1 und ATF. Bei TGF-ß-Stimulation werden c-fos, c-jun und junB aktiviert.
PAI-1 gilt als direktes, nukleäres Ziel von TGF-ß. Die Gene p16, p15 und CDK25a wirken
auf CyclinD, die CDK4 und 6, p21 und p27 auf CyclinE und CDK2 und bewirken eine
Steuerung der Zelle am Übergang der G1-Phase zur S-Phase des Zellzyklus (Hill, 1996).
Seite 37

Kapitel 1. Einleitung
1.6 Die „Smads“
1.6.1 Einteilung, Aufbau und Funktion
Die Smads sind Transkriptionsfaktoren, die die von den TßRs generierten Signale intra-
zellulär weiterverarbeiten. Smad steht für „Similar Mothers Against Decapentaplegic“
(MAD). MADs sind entwicklungsgeschichtlich konservierte, TßR-abhängige Proteine, die
zuerst in C. elegans und Drosophila melanogaster nachgewiesen wurden. Die Smads
werden durch die TßR-I-Kinase phosphoryliert und sind selber Kinasen, die andere
Proteine phosphorylieren. Eine Rezeptoraktivierung führt zu einer Anhäufung von spezi-
fischen Smads im Kern (Kawabata et al., 1999).
Smads bestehen aus verschiedenen Regionen, der MH1-, der Linker- und der MH2-Re-
gion. Die MH1-Domäne bindet unter anderem an den Vit-D-Rezeptor, während die MH2-
Domäne der meisten Smads mit einigen Transkriptionsfaktoren wie CBP, p300, TGIF,
„Forkhead Activin Signal Transducer“ (FAST) und CBFa1-3 assoziieren kann. FAST wird
durch Mitglieder der TGF-ß-Superfamilie aktiviert und für den DNA-Bindungskomplex von
Smad-2 und -4 benötigt (Verschueren et al., 1998).
Die „receptor associated“ Smads (R-Smads), das sind Smad-1, -2, -3, -5 und -8, bilden
nach Ligandenaktivierung einen Komplex mit TßR-I, durch dessen Kinasen die R-Smads
phosphoryliert werden. Diese bilden mit den „common mediator“ Smads (Co-Smads),
dessen einziger Vertreter Smad-4 ist, einen heterooligomeren Komplex und translozieren
in den Zellkern. Die anti-Smads (Smad-6 und -7) antagonisieren die Aktivität des R-
Smad–Co-Smad-Komplexes durch Verhinderung der Translokation des Komplexes in
den Kern. Smad-6 inhibiert den BMP-Signalweg, während Smad-7 den TGF-ß- und den
BMP-Signalweg inhibiert. Die anti-Smads lagern sich stabil am TßR-I an und verhindern
die Smad-2-Aktivierung. Sie konkurrieren mit Smad-4 kompetitiv um Smad-2. Freies
Smad-7 liegt im Zellkern vor, das die responsiven Gene vor Expression schützt. Die
Gene werden durch Rezeptoraktivierung angeschaltet. Als Anker für die Smads in der
Membran wirkt „smad anchor for receptor activation” (SARA) (Hill, 1999; Zhang et al.,
1999).
Seite 38

Kapitel 1. Einleitung
1.6.2 Signalübertragung durch die „Smads“
Nach Rezeptoraktivierung durch Ligandenbindung wird Smad-2 phosphoryliert, von
SARA gelöst und bildet mit Smad-4 einen Komplex, der in den Zellkern wandert. Ein
TßR-II kann mit mehreren TßR-I interagieren und unterschiedliche Wirkungen wegen der
Bindung verschiedener Smads entfalten. Der TßR-I scheint für die Signalspezifität
verantwortlich zu sein. Smads alleine besitzen nur eine geringe DNA-Bindungsaffinität
und benötigen weitere Transkriptionsfaktoren. Dies könnte die Zelltypspezifität durch ver-
schiedene TGF-ß-Familienmitglieder erklären: so leiten zum Beispiel Smad-2 und -3 die
Signale von TGF-ß und Smad-1 und -5 die Signale der BMPs weiter.
Seite 39
Abbildung 3: Überblick über den TGF-ß-Signalweg vom Rezeptor bis zum Zellkern (mit freundli-
cher Genehmigung der BioCarta Europe GmbH, Hamburg, nach: http://www.biocarta.com)

Kapitel 1. Einleitung
Unterschiede in den Affinitäten der einzelnen Faktoren gekoppelt mit der An- und
Abwesenheit von DNA-Bindungsstellen für Smads könnten die Aktivierung spezifischer
Gene durch unterschiedliche Konzentrationen der aktiven Smad-Komplexe erlauben.
Phosphorylierungen an unterschiedlichen Aminosäuren könnten ein anderes Signal
generieren. Ein anderer Ansatz erklärt die Wirkungsstärke der Signale im TGF-ß-Signal-
weg durch den relativen Mangel oder Überschuß von Ko-Aktivatoren über die Ko-Sup-
pressoren (Padgett, 1999). Aktivierung einzelner Smads durch den TßR-I kann nicht die
osteoblastäre Differenzierung bewirken, sondern dies können nur konstante Kombina-
tionen mehrerer Smads wie Smad-1 und Smad-5 im Zusammenspiel mit den BMPs
fördern. Smad-3 und -4 können entweder direkt an die DNA-Bindungsstelle mit der Folge
GTCT oder komplementär an das AGAC-Motiv der DNA binden, oder sie benötigen
DNA-Bindungsproteine wie c-jun und c-fos. PEBP2αA/CBFA1 scheint die Knochenbil-
dung durch direkten „Angriff“ an der DNA zu fördern, während PEBP2αB/CBFA2/AML1
die Hämatopoese nur durch Mithilfe von DNA-Bindungsproteinen zu stimulieren scheint.
Als transkriptioneller Ko-Aktivator dient p300 und bildet mit den R-Smads einen Komplex.
Die Transkription der Smads und anderer Faktoren wird durch p300 positiv beeinflußt.
p300 und „cAMP Response Elements Bindung Protein“ (CBP) sind funktionell und struk-
turell verwandt. Smads interagieren mit transkriptionellen Ko-Repressoren wie c-Ski oder
TGIF, die mit Smad-2 und -3 in Wechselwirkung treten. p300 und c-Ski verbindet ein di-
rekter Antagonismus auf die TGF-ß-Signalwirkung (Miyazono et al., 2000). „SKI related
novel gene“ (SnoN), ein c-Ski-ähnliches Protein, unterdrückt die TGF-ß-Zielgene und
TGF-ß-Promotoraktivität, indem es in Abwesenheit von TGF-ß nukleäres Smad-4 bindet.
Smad-3 bewirkt eine Degradation von SnoN. TGF-ß erhöht die SnoN-Expression und
sorgt für einen „negative-feedback“-Mechanismus (Stroschein et al., 1999). Andere Re-
zeptorsysteme beeinflussen die Smads. Der MAPK-Signalweg inhibiert die Smad-Si-
gnalübertragung über Smad-1. Tyrosin-Kinase-Rezeptoren reduzieren die Smad-Signal-
übertragung durch vermehrte Expression der inhibierend wirkenden Smad-6 und Smad-7
(Wrana et al., 2000). Interferon γ und EGF besitzen über Smad-6 und Smad-7 ant-
agonistische Wirkung auf die TGF-ß-Signalübertragung. Überexpression von Smad-3 ak-
tiviert sehr stark AP1-haltige Promotor (Attisano et al., 2000).
Seite 40

Kapitel 2. Fragestellung
2. Fragestellung
TGF-ß und seine Rezeptoren besitzen elementare Bedeutung für Aufbau, Erhalt, Funkti-
on und Differenzierung des Knochens. Bisherige Daten zeigen, daß eine Deregulation
oder Fehlsteuerung in diesem System deutliche Wirkungen und Veränderungen am
Knochen zur Folge hat. Verschiedene Einflüsse auf die TßR-Anzahl oder Aktivität durch
Stimuli wie Steroide, Kollagen-Interaktionen, BMP-2 oder Alter des Knochens sind be-
schrieben. Es gibt wenig Untersuchungen in humanem Knochen bezüglich der Expressi-
on von TGF-ß und seiner Rezeptoren, die über physiologische Veränderungen im
Rahmen des Alterungsprozesses, pathologische Veränderungen durch Immobilisation
oder längerfristige Steroidtherapie bei chronischer Lungenerkrankung Auskunft geben.
Aufgrunddessen wird in der durchgeführten Arbeit die TßR-Expression primär in hu-
manen Knochenproben analysiert. Über physiologische und pathologische Expressions-
profile der TßR an humanen Knochenzellen ist nur wenig bekannt. Bisher wurde nur die
Expression der TßR-I und -II untersucht, die von Betaglykan nicht.
Die Analyse der gereinigten, humanen Knochenproben soll Summationseffekte auf die
TßR-Expression darstellen und die Verhältnisse beleuchten, wie sie im Knochen ansatz-
weise zu finden sind. Nachteilig an den Knochenproben ist, daß eine Uneinheitlichkeit
der knöchernen Zellpopulation besteht. In ihr sind Zellen der Osteoblastogenese wie
Prä-Osteoblasten, Osteoblasten, Osteozyten, Zellen der Osteoklastogenese und weitere
enthalten. In dieser Arbeit werden hauptsächlich Zellen der Osteoblastogenese unter-
sucht, da Osteoblasten zu über 80 % die Knochenzellpopulation bilden. Die anderen, im
Knochen enthaltenen Zellen, sind von der Anzahl her in der Minderheit. Des weiteren
bilden diese Zellen im Vergleich zu osteoblastären Zellen deutlich weniger TGF-ß und
exprimieren deutlich weniger TßR. Daher wird angenommen, daß die TßR-Expression
fast ausschließlich auf osteoblastäre Zellen zurückgeht. Eine gründliche Vorreinigung
der Proben entfernt Knochenmarkszellen, die ebenfalls eine hohe TßR-Expression auf-
weisen.
Um tiefere Einblicke in die Regulation des Knochenstoffwechsels zu bekommen, wird zu-
sätzlich in primären, humanen Knochenzellkulturen, die direkt aus menschlichen
Knochenproben stammen, die TßR-Expression untersucht. Sie lassen sich nach ihrem
Seite 41

Kapitel 2. Fragestellung
Differenzierungsgrad einteilen. Um die Einflüsse auf die Zellpopulation und die unter-
schiedliche Differenzierung der Osteoblastenpopulation genauer zu erkennen, werden
primäre, humane Osteoblastenzellkulturen unterschiedlicher Differenzierung eingesetzt
und untersucht. Die Analyse solch genau definierter Zellpopulationen ermöglicht die dif-
ferenzierte Betrachtung der Expression der TßR in den einzelnen Subpopulationen.
Der Schwerpunkt dieser Arbeit liegt in der Entwicklung und Etablierung des mRNA-Ex-
pressionsnachweises der TGF-ß-Rezeptoren an humanen Knochenproben und in
primären Knochenzellkulturen mittels quantitativer „real-time“-PCR. Mit Hilfe herkömmli-
cher Knochenzellkulturen wird die PCR etabliert und optimiert. Diese Ergebnisse werden
auf den LightCycler7 und TaqMan7 für die quantitative PCR übertragen. Der anspruchs-
volle Nachweis mittels quantitativer PCR soll eine zuverlässige Detektion bei kleinen
Mengen an Ausgangsmaterial, sei es in Form der Knochenproben oder der primären hu-
manen Osteoblastenkulturen, ermöglichen. Des weiteren interessieren die Varianzen,
Nachweisgrenzen und Reproduzierbarkeit der „real-time“ PCR.
Zusammenfassend stellen sich folgende Fragen:
- Wird die TßR-Expression in humanen Knochenproben durch Glukokortikoide, Immobili-
sation, Alterung oder durch andere Parameter beeinflußt?
- Unterscheidet sich die TßR-Expression in unterschiedlich differenzierten Populationen
von primären, humanen Knochenzellkulturen? Wird die TßR-Expression durch Variation
der Steroidkonzentration und -einwirkungsdauer verändert?
- Wie genau arbeiten die „real-time“-PCR-Geräte und welche Grenzen besitzen sie?
Seite 42

Kapitel 3. Methoden und Material
3. Methoden und Material
3.1 Herstellung der Proben
3.1.1 Probengewinnung
Spongiöse Knochenproben wurden Patienten entnommen, die sich in den Berufsge-
nossenschaftlichen Kliniken Bergmannsheil der Ruhr-Universität Bochum einem
knochenchirurgischen Eingriff unterziehen mußten. Bei diesen Routineeingriffen
bestanden Operationsindikationen im Rahmen der Behandlung von arthrotischen Ge-
lenkveränderungen, einer direkten Traumaversorgung oder Knochenentnahmen am Be-
ckenkamm als Zusatzeingriff zur Spongiosagewinnung für die Frakturstabilisierung. Der
Entnahme der maximal 0,5 cm im Durchmesser großen Spongiosaanteile folgte eine so-
fortige Lagerung in flüssigem Stickstoff. Präoperativ wurde den Patienten zusätzlich Se-
rum entnommen und für spätere Analysen bei -80 °C eingefroren.
Die Studie wurde vor Beginn der zuständigen Ethikkommission vorgelegt und genehmigt.
3.1.2 Probenaufbereitung
Vor der weiteren Verarbeitung erfolgt eine chirurgische Säuberung der Spongiosa mit
Entfernung kompakter Knochenanteile, von Knorpelresten oder anhängendem Weichteil-
gewebe. Durch zweimaliges Waschen mit eisgekühlter physiologischer Kochsalzlösung
wird das trabekuläre Knochenmaterial von Knochenmarksbestandteilen getrennt und in
Spezialbehältern einer Schwingmühle (Fa. Retsch, Haan) mit sterilem Spülmedium ge-
säubert. Histologische Kontrollen während der Etablierung zeigten eine weitestgehende
Reinigung des Knochenmaterials von adhärenten Zellen.
Unter Stickstoffkühlung der Proben und Mahlkammern erfolgt die Knochenzerkleinerung
Seite 43

Kapitel 3. Methoden und Material
in einer Schwingmühle bei maximaler Leistung für 1 min. Das gewonnene Material wird
in mit „Solution D“ (siehe Puffer und Lösungen) gefüllte Röhrchen überführt und das Ge-
wicht über die Differenz vor und nach Knochenmehlzugabe ermittelt. Die RNA-Extraktion
erfolgt nach der Methode von Chomczynski und Sacchi (Chomczynski et al., 1987).
3.2 Zellkultur
Knochenzellkulturen der Linien HOS (ATCC: CRL-1543), HOS 58 (Siggelkow et al.,
1998) und MG 63 (Billiau et al., 1977) werden geführt und zur Etablierung der RNA-
Extraktion und PCR verwendet. Den HOS-Kulturen liegen humane Osteosarkom-Zellen
eines 13-jährigen Mädchens der kaukasischen Rasse zugrunde, die aus fibroblasten-
und epithel-ähnlichen Zellen bestehen. Die MG 63-Zellen sind von Billiau et al. erstmals
beschriebene Interferon-produzierende Osteosarkom-Zellen.
Primäre Knochenzellkulturen stellte freundlicherweise Dr. Achim Battmann aus Giessen
zur Verfügung und führte sie nach der Methode von Robey et al. (Robey et al., 1985).
Mit dieser Methode lassen sich aus einem Knochenfragment Zellen unterschiedlichen
Reife- und Differenzierungsgrades, die SZ-Zellen (Stroma-Zellen), die pluripotenten
Charakter haben, die CR-Zellen („collagenase released“), die (Prä-) Osteoblasten ent-
sprechen und die TE-Zellen („treated explants“), die osteozytär differenziert sind, extra-
hieren. Die Zellkulturen mit den Differenzierungsstufen SZ, CR und TE stammen jeweils
von einem Spender (? bedeutet Alter und Geschlecht unbekannt):
Tabelle 1: Aufstellung über die verwendeten primären Knochenzellkulturen, Alter und
Geschlecht
Zellkultur Alter (in Jahren) Geschlecht (M=Männ-
lich, W=Weiblich)738 60 M741 68 W767 73 W769 50 W776 62 W
Seite 44
Kapitel 3. Methoden und Material
Zellkultur Alter (in Jahren) Geschlecht (M=Männ-
lich, W=Weiblich)779 65 W780 76 M781 69 W782 87 W783 26 W797 ? ?
Vor der RNA-Extraktion der Zellkulturen mit TRIzol7 (Gibco Technologies, Gaithersburg,
USA) werden die Zellkulturen gründlich mit PBS (siehe Puffer und Lösungen) gespült.
3.3 RNA-Extraktion
3.3.1 RNA-Extraktion mit „TRIzol“
Für die RNA-Extraktion der Zellkulturplatten kommt TRIzol7 zum Einsatz, das eine modi-
fizierte und vereinfachte Variante der Extraktion nach Chomczynski und Sacchi ist. Nach
zweimaligem Waschen mit 1-fach PBS wird TRIzol7 abhängig von der Fläche und der
Zellzahl auf die Platten aufgetragen, bei den 6-“well“-Platten pro „well“ 1 ml TRIzol7.
Nach guter Spülung des Bodens mit TRIzol7 und Durchmischung der Lösung wird sie in
Eppendorfgefäße überführt, 5 min bei Raumtemperatur stehen gelassen, pro eingesetz-
tem ml TRIzol7 0,2 ml Chloroform hinzugefügt und kräftig geschüttelt. Vor weiterem
Zentrifugieren für 15 min bei 12000 x g und 4 °C inkubieren die Proben für 3 min bei
Raumtemperatur. Es folgt ein Abpipettieren der RNA-enthaltenden, obersten der drei
Schichten und Überführen in ein neues Gefäß. Nach Zugabe von 0,5 ml Isopropanol pro
ml TRIzol7 und erneuter Inkubation für 10 min bei Raumtemperatur werden die Proben
für 10 min bei 12000 x g und 4 °C zentrifugiert. Bei starker Verfärbung oder hohem Fett-
gehalt der Proben erfolgt eine Wiederholung dieses Fällungsschrittes. Ein RNA-Präzipi-
tat wird meist sichtbar. Nach einem ein- bis zweimal durchzuführenden Ethanolwasch-
schritt mit Zugabe von mindestens 1 ml 70 % Ethanol pro ml TRIzol7 und kräftigem
Seite 45

Kapitel 3. Methoden und Material
Schütteln wird bei 7500 x g und 4 °C für 5 min zentrifugiert. Der vorsichtigen Entfernung
der oberen Ethanolschicht folgt ein Zentrifugieren bei 7500 x g und 4 °C. Es wird gerade
so viel abpipettiert, daß die erhaltene RNA noch flüssigkeitsbedeckt ist. Die Proben
stehen bis zum fast vollständigen Verdunsten des Alkohols. In 50 bis 100 µl DMPC-
Wasser (siehe Puffer und Lösungen) löst sich die erhaltene, fast komplett getrocknete
RNA unter Anwärmung auf 60 °C für 10 min auf und lagert bei -80 °C bis zur weiteren
Verarbeitung.
3.3.2 RNA-Extraktion nach Chomczynski und Sacchi
Für die Extraktion der Gesamt-RNA aus dem Knochenmehl findet die Guanidinium-Thio-
cyanat-Methode nach Chomczynski et al. Anwendung (Chomczynski et al., 1987). Die
verwendeten Volumina sind für etwa 100 mg Gewebe berechnet. Während der ge-
samten RNA-Extraktion sollen die Proben durchgehend gekühlt werden. Nach Homo-
genisierung der Proben in einer Schwingmühle und Zugabe von 1 ml "Solution D" wird
die Suspension in ein 10 ml-Gefäß überführt, 100 µl 2 M Natriumacetat pH 4, 300 µl
Phenol und 200 µl Chloroform-Isoamylalkohol-Gemisch (49:1) hinzugegeben und nach
jeder Substanzzugabe gemischt. Der Lagerung auf Eis für 10 min folgt eine Zentrifugati-
on für 15 min bei 10000 x g und 4 °C. Nach Überführung der wäßrigen Phase in ein neu-
es Gefäß und Zugabe von 1 ml Isopropanol erfolgt eine Präzipitation bei -20 °C für 1 h.
Vorsichtigem Abgießen des Überstandes folgt eine Zugabe von 0,3 ml "Solution D".
Nach Auflösen des Pellets und nach erneuter Überführung in ein frisches Eppendorfge-
fäß werden 0,3 ml Isopropanol hinzu pipettiert. Es schließt sich eine Präzipitation für 1 h
bei -20 °C mit nachfolgender Zentrifugation für 15 min bei 10000 x g und 4 °C an. Nach
vorsichtigem Dekantieren des Überstandes wird das Pellet zweimal mit 0,6 ml 70% Etha-
nol gewaschen, anschließend bei 1000 x g und bei 4 °C für 5 min zentrifugiert, für etwa 4
min im Lyophilisator nicht ganz vollständig getrocknet und in 50 µl DMPC-Wasser
resuspendiert.
Seite 46

Kapitel 3. Methoden und Material
3.3.3 RNA-Messung im Photometer
Um näherungsweise die RNA-Konzentration nach der Extraktion zu erhalten und um
etwa gleiche absolute Mengen in die Reverse Transkription einzusetzen, wird 1 µl der
erhaltenen RNA-Lösung in ein neues Gefäß überführt und dieses mit DMPC-Wasser ad
100 µl aufgefüllt. Nach guter Durchmischung der Proben und gründlichster Reinigung
der Küvette werden 70 µl in die Meßküvette des Photometers gefüllt. Da Photometer-
messungen einer gewissen Schwankungsbreite unterworfen sind, erfolgt die Mittelwert-
berechnung nach dreimaliger Probenmessung gegen eine reine DMPC-Wasser-Probe.
Der Verdünnungsschritt erfordert eine Berechnung der ursprünglichen Konzentration.
3.4 Reverse Transkription
Die Reverse Transkription (RT) ist ein Verfahren zur Übersetzung von RNA in cDNA
(„copy“ oder „complementary“ DNA) mit Hilfe geeigneter Enzyme wie der M-Mulv-RT
(Moloney-Murine Leukemia Virus Reverse Transkriptase). Zur Denaturierung wird die
extrahierte RNA im Wasserbad für 5 min auf 80 °C erwärmt. Die eingesetzte RNA-
Menge beträgt standardmäßig 2 µg und wird mit DMPC-Wasser auf 25 µl verdünnt, um
gleiche Reaktionsbedingungen und -konzentrationen zu schaffen. Der Reaktionsansatz
wird als „Mastermix“ auf Eis hergestellt, gemischt, kurz abzentrifugiert und in die vorbe-
reiteten Gefäße hinzugegeben. Das Endvolumen beträgt 40 µl. Die RT wird für 1 h bei
42 °C durchgeführt und bei 80 °C für 10 min durch Hitzeinaktivierung beendet.
Zu dem Reaktionsansatz gehören:
Tabelle 2: Reaktionsansatz der Reversen Transkription
Reagenz Menge EndkonzentrationdN6-Primer (random primer) 6 µl 1,4 mM
dNTP-Mix (10mM jeweils/100µl) 1,5 µl 0,5 mM
M-Mulv-RT-Reaktionspuffer * 6 µl
Seite 47
Kapitel 3. Methoden und Material
Reagenz Menge EndkonzentrationRNAse-Inhibitor (26 U/µl) 0,5 µl 0,325 U/µl
M-Mulv-RT (20 U/µl) 2 µl 1U/µl
Summe: 15 µl
*: Verwendung findet der vom Hersteller mitgelieferte Puffer (Primer und M-Mulv-RT: Roche Bio-
chemica, Mannheim).
3.5 RNA-Gelelektrophorese
Die RNA-Gelelektrophorese verwendet einen „Formaldehyd gel running buffer“ (FGRB)
und einen „Formaldehyd gel loading buffer“ (siehe Puffer und Lösungen). Für ein 1,4 %-
iges Agarosegel mit 50 ml Volumen kommt folgender Ansatz zur Anwendung: Nach
Zugabe von 700 mg Agarose zu 31,22 ml DMPC-Wasser, vorsichtigem Erhitzen bis zum
Kochen und Abkühlung auf etwa 50 °C werden 8,92 ml Formaldehyd und 10 ml 5x
„FGRB“ dazugegeben. Einer kurzen Durchmischung der Lösung und dem Ausgießen in
die vorbereitete Gelkammer folgt das Aushärten des Gels bei Raumtemperatur. Der Gel-
träger und die Gelkammer werden vor dem Einsatz gründlichst mit Wasserstoffperoxid
und DMPC-Wasser gespült und die Kammer mit 1x „FGRB“ gefüllt.
Für die Probenvorbereitung werden 4,5 µl Probe mit 2 µl „FGRB“, 3,5 µl Formaldehyd,
10,0 µl Formamid und 1 µl Ethidiumbromid (1 mg/ml) zusammen gemischt, 15 min in-
kubiert und kurz anzentrifugiert. Die Zugabe von 2 µl „Formaldehyd gel loading buffer“
folgt. Nach dem Probenauftrag liegt an der Gelkammer eine Spannung von 80 V für 2 h
an.
Seite 48
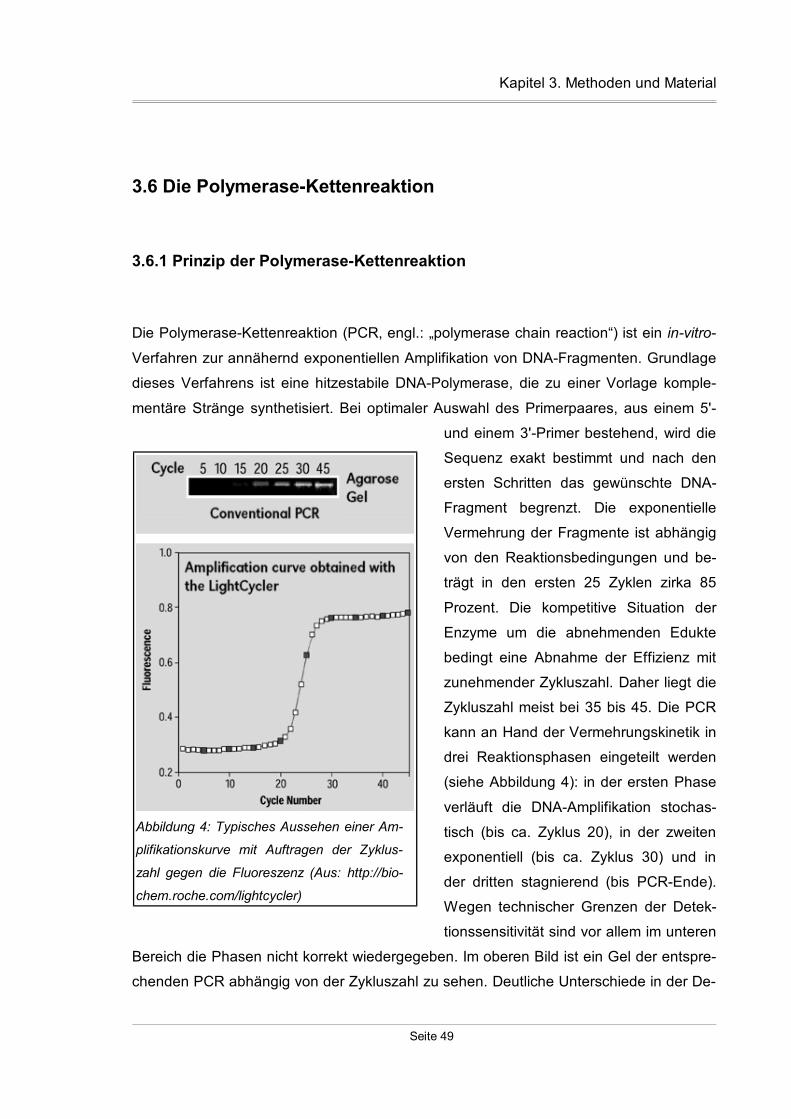
Kapitel 3. Methoden und Material
3.6 Die Polymerase-Kettenreaktion
3.6.1 Prinzip der Polymerase-Kettenreaktion
Die Polymerase-Kettenreaktion (PCR, engl.: „polymerase chain reaction“) ist ein in-vitro-
Verfahren zur annähernd exponentiellen Amplifikation von DNA-Fragmenten. Grundlage
dieses Verfahrens ist eine hitzestabile DNA-Polymerase, die zu einer Vorlage komple-
mentäre Stränge synthetisiert. Bei optimaler Auswahl des Primerpaares, aus einem 5'-
und einem 3'-Primer bestehend, wird die
Sequenz exakt bestimmt und nach den
ersten Schritten das gewünschte DNA-
Fragment begrenzt. Die exponentielle
Vermehrung der Fragmente ist abhängig
von den Reaktionsbedingungen und be-
trägt in den ersten 25 Zyklen zirka 85
Prozent. Die kompetitive Situation der
Enzyme um die abnehmenden Edukte
bedingt eine Abnahme der Effizienz mit
zunehmender Zykluszahl. Daher liegt die
Zykluszahl meist bei 35 bis 45. Die PCR
kann an Hand der Vermehrungskinetik in
drei Reaktionsphasen eingeteilt werden
(siehe Abbildung 4): in der ersten Phase
verläuft die DNA-Amplifikation stochas-
tisch (bis ca. Zyklus 20), in der zweiten
exponentiell (bis ca. Zyklus 30) und in
der dritten stagnierend (bis PCR-Ende).
Wegen technischer Grenzen der Detek-
tionssensitivität sind vor allem im unteren
Bereich die Phasen nicht korrekt wiedergegeben. Im oberen Bild ist ein Gel der entspre-
chenden PCR abhängig von der Zykluszahl zu sehen. Deutliche Unterschiede in der De-
Seite 49
Abbildung 4: Typisches Aussehen einer Am-
plifikationskurve mit Auftragen der Zyklus-
zahl gegen die Fluoreszenz (Aus: http://bio-
chem.roche.com/lightcycler)
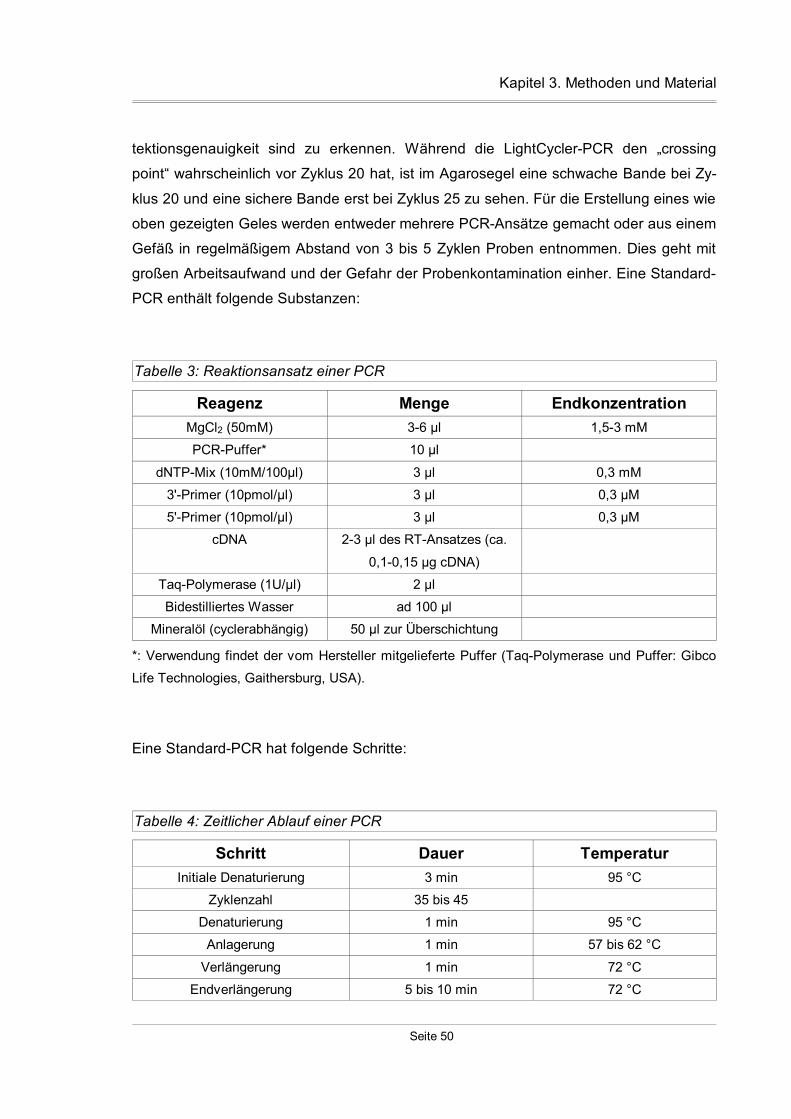
Kapitel 3. Methoden und Material
tektionsgenauigkeit sind zu erkennen. Während die LightCycler-PCR den „crossing
point“ wahrscheinlich vor Zyklus 20 hat, ist im Agarosegel eine schwache Bande bei Zy-
klus 20 und eine sichere Bande erst bei Zyklus 25 zu sehen. Für die Erstellung eines wie
oben gezeigten Geles werden entweder mehrere PCR-Ansätze gemacht oder aus einem
Gefäß in regelmäßigem Abstand von 3 bis 5 Zyklen Proben entnommen. Dies geht mit
großen Arbeitsaufwand und der Gefahr der Probenkontamination einher. Eine Standard-
PCR enthält folgende Substanzen:
Tabelle 3: Reaktionsansatz einer PCR
Reagenz Menge EndkonzentrationMgCl2 (50mM) 3-6 µl 1,5-3 mM
PCR-Puffer* 10 µl
dNTP-Mix (10mM/100µl) 3 µl 0,3 mM
3'-Primer (10pmol/µl) 3 µl 0,3 µM
5'-Primer (10pmol/µl) 3 µl 0,3 µM
cDNA 2-3 µl des RT-Ansatzes (ca.
0,1-0,15 µg cDNA)
Taq-Polymerase (1U/µl) 2 µl
Bidestilliertes Wasser ad 100 µl
Mineralöl (cyclerabhängig) 50 µl zur Überschichtung
*: Verwendung findet der vom Hersteller mitgelieferte Puffer (Taq-Polymerase und Puffer: Gibco
Life Technologies, Gaithersburg, USA).
Eine Standard-PCR hat folgende Schritte:
Tabelle 4: Zeitlicher Ablauf einer PCR
Schritt Dauer TemperaturInitiale Denaturierung 3 min 95 °C
Zyklenzahl 35 bis 45
Denaturierung 1 min 95 °C
Anlagerung 1 min 57 bis 62 °C
Verlängerung 1 min 72 °C
Endverlängerung 5 bis 10 min 72 °C
Seite 50

Kapitel 3. Methoden und Material
An die PCR können weitere Analyseschritte angeschlossen werden. Regelmäßig wurde
in dieser Arbeit die Agarosegelelektrophorese (siehe unten) im Anschluß durchgeführt.
Bei der quantitativen PCR kommt die Schmelzkurvenanalyse zum Einsatz (siehe unten).
Die PCR-Produkte werden im Anschluß mittels Sequenzierung auf ihre Spezifität hin un-
tersucht und mit den Ergebnissen der Gelelektrophorese verglichen.
3.6.2 Primerdesign
Die Primer werden anhand der von „GenBank“ – einer Datenbank zur Suche nach be-
kannten und öffentlich zugänglichen DNA-Sequenzen, betreut durch das „National In-
stitutes of Health“ (NIH), USA, - bezogenen Sequenzen unter Berücksichtigung von
Primerlänge, Zielsequenz, GC-Gehalt, Sekundär- und Tertiärstruktur mit Hilfe des
Primerdesignprogrammes Oligo 5.0 (National Biosciences, Plymouth, USA) ausgewählt.
Exonspezifische Primer werden gesucht, d.h. sie liegen auf unterschiedlichen Exons. Bei
Kontamination des PCR-Ansatzes mit genomischer DNA entstehen zusätzliche Banden
in der Gelelektrophorese durch längere PCR-Produkte, bedingt durch in genomischer
DNA enthaltene Introns. Dadurch ist eine Qualitätskontrolle der RNA-Extraktion möglich.
Die gewählten Primer erfüllen folgende Voraussetzungen: der GC-Gehalt der Primer liegt
bei etwa 50 bis 60 %, mehr als drei Nukleotide eines Typs in Folge werden vemieden
(Poly(G)-Bereiche) und es besteht keine Komplementarität zwischen den Primern.
Mehrere Formeln zur Berechnung des Schmelzpunktes Tm existieren (Von Ahsen et al.,
2001). In dieser Arbeit wird die „Nearest Neighbour“ Methode (SantaLucia, 1998) ange-
wendet. Der so bestimmte Tm des Primerpaares soll nicht um mehr als 5 oC voneinander
abweichen und in der Nähe von 60 °C liegen. Die Primer besitzen zwischen 18 und 24
Nukleotiden eine optimale Länge bei guter Spezifität. Längere Primer verursachen Pro-
bleme bei der Hybridisierung aufgrund von Sekundärstrukturausbildungen. Der Schmelz-
punkt Tm ist definiert als der Punkt, an dem die Hälfte der DNA-Stränge in Doppelhelix
sowie in Einzelstrang-Form vorliegt. Der Schmelzpunkt ist abhängig von der Salzkon-
zentration, der Konzentration von DMSO, SYBR Green I7 , dem GC-Gehalt, der Frag-
mentlänge, der Komplementarität sowie der Sekundär- und Tertiärstruktur (SantaLucia,
1998).
Seite 51

Kapitel 3. Methoden und Material
3.7 Quantitative PCR
3.7.1 Grundlagen
Das Prinzip der quantitativen PCR beruht auf dem Vergleich entweder der Amplifikation
nach einer bestimmten Reaktionszeit oder von Amplifikationskurven von bekannten und
unbekannten Proben. Die PCR ist eine Exponentialfunktion mit der Gleichung:
Y = X * (1+E)^n oder log y = log x + n * log (1+E),
wobei für Y = Menge an hergestelltem Amplifikat, X = Startkopienzahl und E = Effizienz
der PCR gilt.
Da die Primerauswahl und die Amplifikatlängen die PCR-Effizienz beeinflussen, werden
nur Amplifikate mit Längen zwischen 170 bp und 500 bp verwendet, da in diesem Be-
reich die PCR-Effizienz sich nicht wesentlich unterscheidet und bei 60 bis 80 % liegt.
3.7.2 Quantifizierungsstrategien
Es existiert die sogenannte „absolute“ und „relative Quantifizierung“. Ziel der „absoluten
Quantifizierung“ ist es, dem gemessenen Signal eine bestimmte Startkopienzahl oder
Konzentration zuzuordnen. Bei der absoluten Quantifizierung sollen Proben und Stan-
dard gleiche Sequenz und Amplifikatlängen haben, um Unterschiede der PCR-Effizienz
und mögliche Fehlerquellen auszuschließen. Die Quantifizierung kann endpunktge-
steuert durch Koamplifikation interner Standards mit den zu analysierenden Proben,
oder mit Echtzeitdatenaufnahme wie bei der „real-time“-PCR ablaufen. Letztere
ermöglicht eine genaue und reproduzierbare Quantifizierung durch Erfassung der PCR-
Seite 52
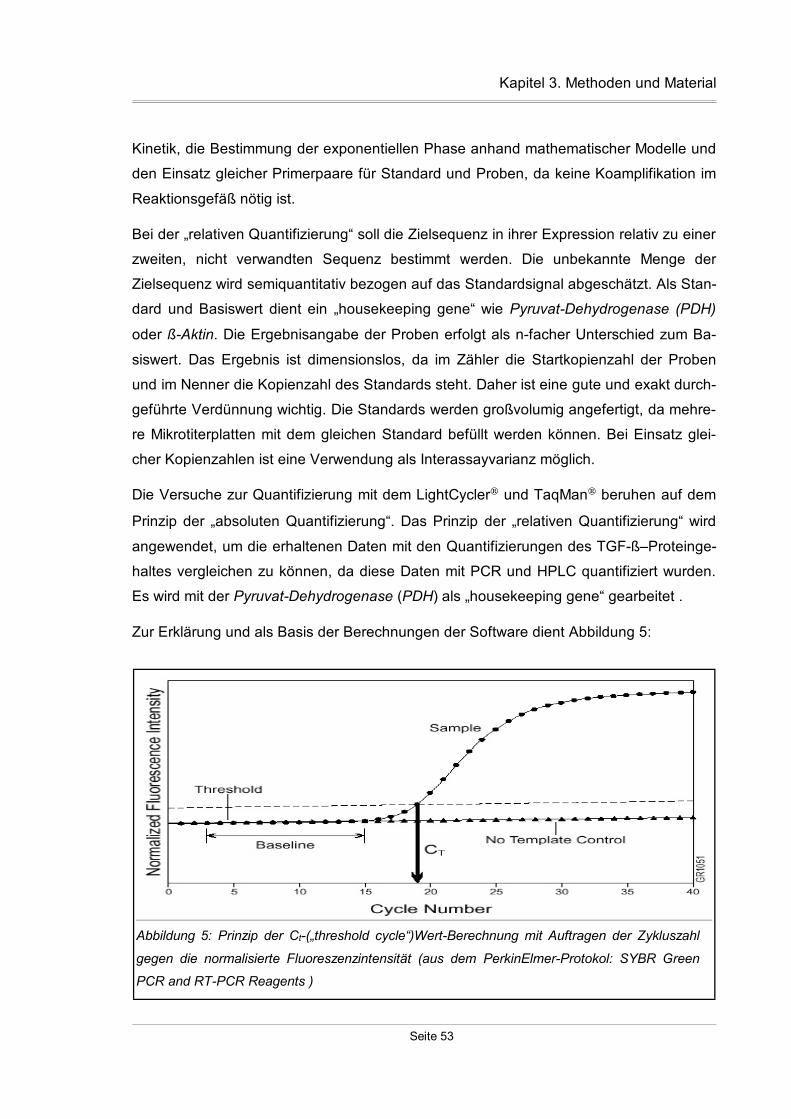
Kapitel 3. Methoden und Material
Kinetik, die Bestimmung der exponentiellen Phase anhand mathematischer Modelle und
den Einsatz gleicher Primerpaare für Standard und Proben, da keine Koamplifikation im
Reaktionsgefäß nötig ist.
Bei der „relativen Quantifizierung“ soll die Zielsequenz in ihrer Expression relativ zu einer
zweiten, nicht verwandten Sequenz bestimmt werden. Die unbekannte Menge der
Zielsequenz wird semiquantitativ bezogen auf das Standardsignal abgeschätzt. Als Stan-
dard und Basiswert dient ein „housekeeping gene“ wie Pyruvat-Dehydrogenase (PDH)
oder ß-Aktin. Die Ergebnisangabe der Proben erfolgt als n-facher Unterschied zum Ba-
siswert. Das Ergebnis ist dimensionslos, da im Zähler die Startkopienzahl der Proben
und im Nenner die Kopienzahl des Standards steht. Daher ist eine gute und exakt durch-
geführte Verdünnung wichtig. Die Standards werden großvolumig angefertigt, da mehre-
re Mikrotiterplatten mit dem gleichen Standard befüllt werden können. Bei Einsatz glei-
cher Kopienzahlen ist eine Verwendung als Interassayvarianz möglich.
Die Versuche zur Quantifizierung mit dem LightCycler7 und TaqMan7 beruhen auf dem
Prinzip der „absoluten Quantifizierung“. Das Prinzip der „relativen Quantifizierung“ wird
angewendet, um die erhaltenen Daten mit den Quantifizierungen des TGF-ß–Proteinge-
haltes vergleichen zu können, da diese Daten mit PCR und HPLC quantifiziert wurden.
Es wird mit der Pyruvat-Dehydrogenase (PDH) als „housekeeping gene“ gearbeitet .
Zur Erklärung und als Basis der Berechnungen der Software dient Abbildung 5:
Seite 53
Abbildung 5: Prinzip der Ct-(„threshold cycle“)Wert-Berechnung mit Auftragen der Zykluszahl
gegen die normalisierte Fluoreszenzintensität (aus dem PerkinElmer-Protokol: SYBR Green
PCR and RT-PCR Reagents )
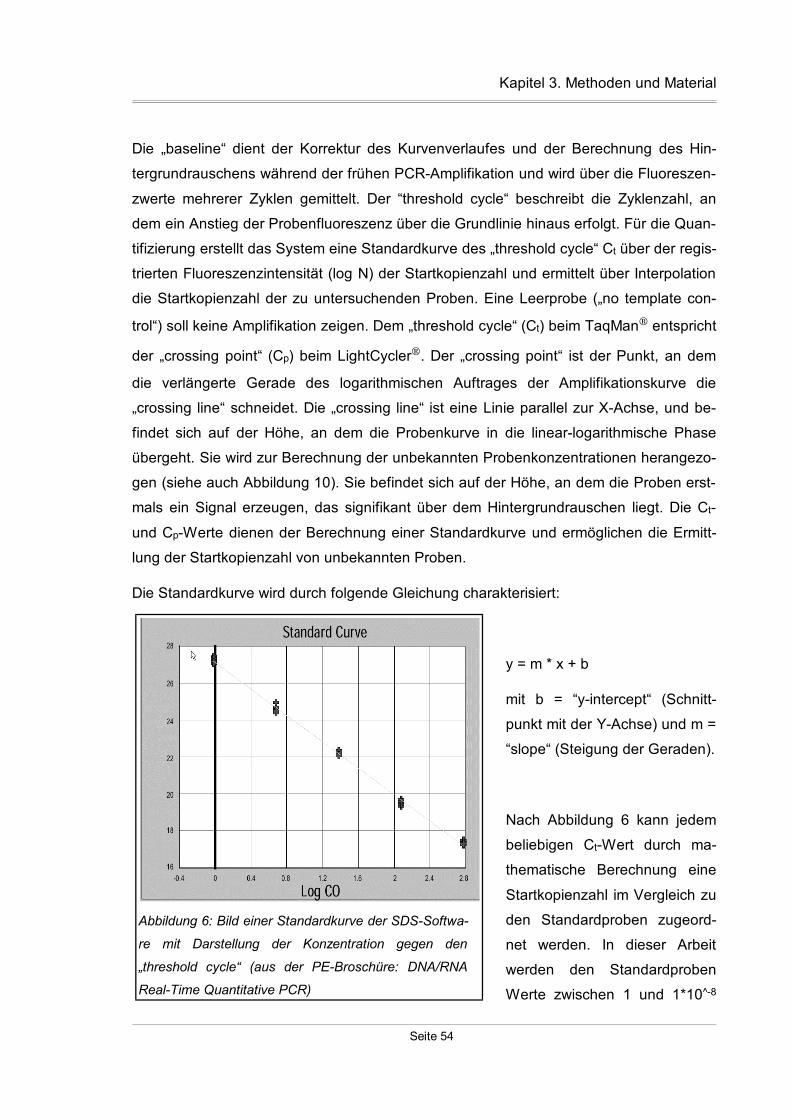
Kapitel 3. Methoden und Material
Die „baseline“ dient der Korrektur des Kurvenverlaufes und der Berechnung des Hin-
tergrundrauschens während der frühen PCR-Amplifikation und wird über die Fluoreszen-
zwerte mehrerer Zyklen gemittelt. Der “threshold cycle“ beschreibt die Zyklenzahl, an
dem ein Anstieg der Probenfluoreszenz über die Grundlinie hinaus erfolgt. Für die Quan-
tifizierung erstellt das System eine Standardkurve des „threshold cycle“ Ct über der regis-
trierten Fluoreszenzintensität (log N) der Startkopienzahl und ermittelt über Interpolation
die Startkopienzahl der zu untersuchenden Proben. Eine Leerprobe („no template con-
trol“) soll keine Amplifikation zeigen. Dem „threshold cycle“ (Ct) beim TaqMan7 entspricht
der „crossing point“ (Cp) beim LightCycler7. Der „crossing point“ ist der Punkt, an dem
die verlängerte Gerade des logarithmischen Auftrages der Amplifikationskurve die
„crossing line“ schneidet. Die „crossing line“ ist eine Linie parallel zur X-Achse, und be-
findet sich auf der Höhe, an dem die Probenkurve in die linear-logarithmische Phase
übergeht. Sie wird zur Berechnung der unbekannten Probenkonzentrationen herangezo-
gen (siehe auch Abbildung 10). Sie befindet sich auf der Höhe, an dem die Proben erst-
mals ein Signal erzeugen, das signifikant über dem Hintergrundrauschen liegt. Die Ct-
und Cp-Werte dienen der Berechnung einer Standardkurve und ermöglichen die Ermitt-
lung der Startkopienzahl von unbekannten Proben.
Die Standardkurve wird durch folgende Gleichung charakterisiert:
y = m * x + b
mit b = “y-intercept“ (Schnitt-
punkt mit der Y-Achse) und m =
“slope“ (Steigung der Geraden).
Nach Abbildung 6 kann jedem
beliebigen Ct-Wert durch ma-
thematische Berechnung eine
Startkopienzahl im Vergleich zu
den Standardproben zugeord-
net werden. In dieser Arbeit
werden den Standardproben
Werte zwischen 1 und 1*10^-8
Seite 54
Abbildung 6: Bild einer Standardkurve der SDS-Softwa-
re mit Darstellung der Konzentration gegen den
„threshold cycle“ (aus der PE-Broschüre: DNA/RNA
Real-Time Quantitative PCR)
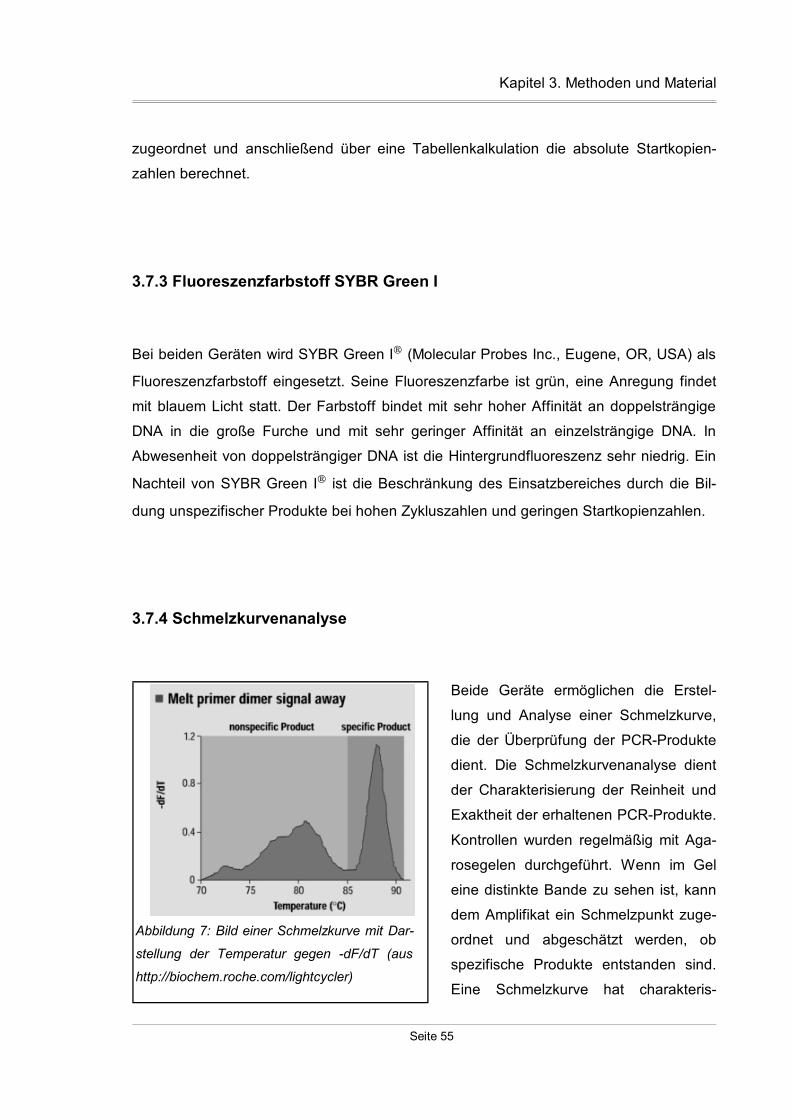
Kapitel 3. Methoden und Material
zugeordnet und anschließend über eine Tabellenkalkulation die absolute Startkopien-
zahlen berechnet.
3.7.3 Fluoreszenzfarbstoff SYBR Green I
Bei beiden Geräten wird SYBR Green I7 (Molecular Probes Inc., Eugene, OR, USA) als
Fluoreszenzfarbstoff eingesetzt. Seine Fluoreszenzfarbe ist grün, eine Anregung findet
mit blauem Licht statt. Der Farbstoff bindet mit sehr hoher Affinität an doppelsträngige
DNA in die große Furche und mit sehr geringer Affinität an einzelsträngige DNA. In
Abwesenheit von doppelsträngiger DNA ist die Hintergrundfluoreszenz sehr niedrig. Ein
Nachteil von SYBR Green I7 ist die Beschränkung des Einsatzbereiches durch die Bil-
dung unspezifischer Produkte bei hohen Zykluszahlen und geringen Startkopienzahlen.
3.7.4 Schmelzkurvenanalyse
Beide Geräte ermöglichen die Erstel-
lung und Analyse einer Schmelzkurve,
die der Überprüfung der PCR-Produkte
dient. Die Schmelzkurvenanalyse dient
der Charakterisierung der Reinheit und
Exaktheit der erhaltenen PCR-Produkte.
Kontrollen wurden regelmäßig mit Aga-
rosegelen durchgeführt. Wenn im Gel
eine distinkte Bande zu sehen ist, kann
dem Amplifikat ein Schmelzpunkt zuge-
ordnet und abgeschätzt werden, ob
spezifische Produkte entstanden sind.
Eine Schmelzkurve hat charakteris-
Seite 55
Abbildung 7: Bild einer Schmelzkurve mit Dar-
stellung der Temperatur gegen -dF/dT (aus
http://biochem.roche.com/lightcycler)
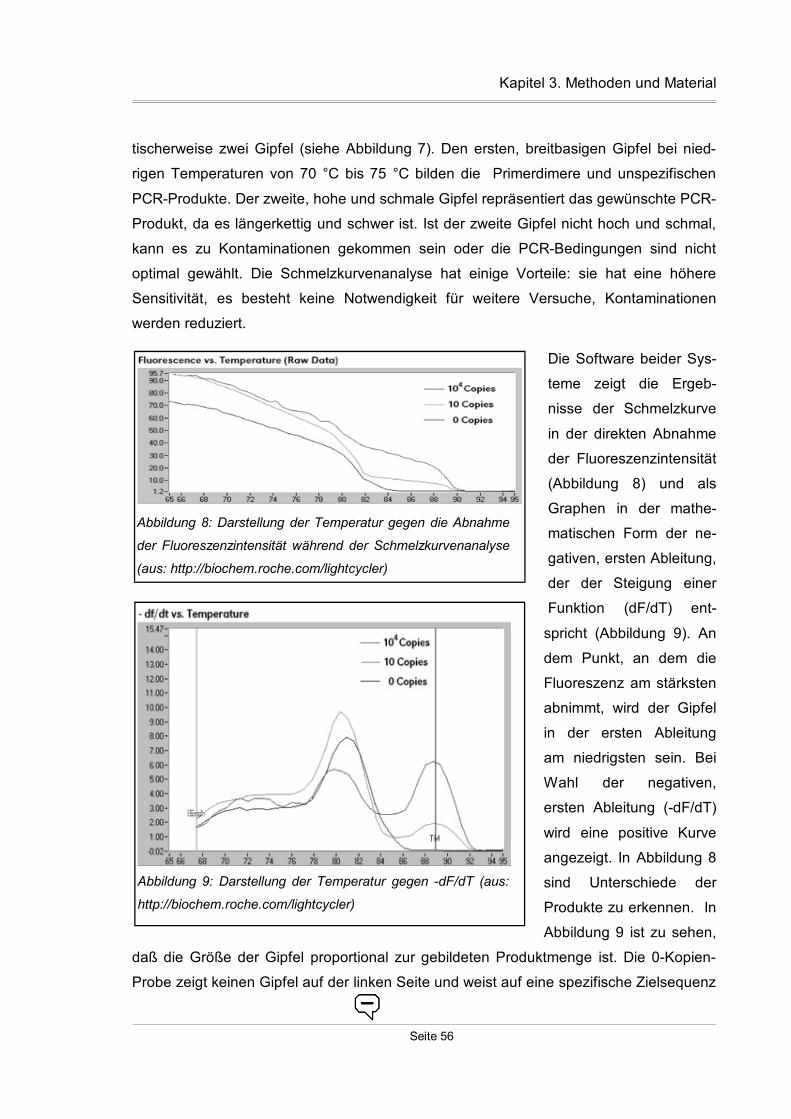
Kapitel 3. Methoden und Material
tischerweise zwei Gipfel (siehe Abbildung 7). Den ersten, breitbasigen Gipfel bei nied-
rigen Temperaturen von 70 °C bis 75 °C bilden die Primerdimere und unspezifischen
PCR-Produkte. Der zweite, hohe und schmale Gipfel repräsentiert das gewünschte PCR-
Produkt, da es längerkettig und schwer ist. Ist der zweite Gipfel nicht hoch und schmal,
kann es zu Kontaminationen gekommen sein oder die PCR-Bedingungen sind nicht
optimal gewählt. Die Schmelzkurvenanalyse hat einige Vorteile: sie hat eine höhere
Sensitivität, es besteht keine Notwendigkeit für weitere Versuche, Kontaminationen
werden reduziert.
Die Software beider Sys-
teme zeigt die Ergeb-
nisse der Schmelzkurve
in der direkten Abnahme
der Fluoreszenzintensität
(Abbildung 8) und als
Graphen in der mathe-
matischen Form der ne-
gativen, ersten Ableitung,
der der Steigung einer
Funktion (dF/dT) ent-
spricht (Abbildung 9). An
dem Punkt, an dem die
Fluoreszenz am stärksten
abnimmt, wird der Gipfel
in der ersten Ableitung
am niedrigsten sein. Bei
Wahl der negativen,
ersten Ableitung (-dF/dT)
wird eine positive Kurve
angezeigt. In Abbildung 8
sind Unterschiede der
Produkte zu erkennen. In
Abbildung 9 ist zu sehen,
daß die Größe der Gipfel proportional zur gebildeten Produktmenge ist. Die 0-Kopien-
Probe zeigt keinen Gipfel auf der linken Seite und weist auf eine spezifische Zielsequenz
Seite 56
Abbildung 8: Darstellung der Temperatur gegen die Abnahme
der Fluoreszenzintensität während der Schmelzkurvenanalyse
(aus: http://biochem.roche.com/lightcycler)
Abbildung 9: Darstellung der Temperatur gegen -dF/dT (aus:
http://biochem.roche.com/lightcycler)
Jens Regula
erratum: auf der rechten Seite

Kapitel 3. Methoden und Material
mit Tm=89 °C hin. Während der Schmelzkurvenerstellung erfolgt die Datenaufnahme der
optischen Einheit kontinuierlich. Vor der Durchführung der Schmelzkurvenaufnahme
werden die Proben zur Denaturierung auf 95 °C erhitzt. Für eine genaue Probenauf-
trennung nach dem Schmelzpunkt wird auf 60 °C abgekühlt und langsam auf 95 °C er-
hitzt. Nach der Schmelzkurvenaufnahme mit der Probenabkühlung ist die PCR beendet.
3.8 Grundlagen und Anwendung des „Light-Cycler“
Der LightCycler7 von Roche7 - bestehend aus Computer, Drucker und Cyclereinheit -
basiert auf der „rapid-cycle“-Technik, eine von Wittwer et al. entwickelte Technik der
DNA-Amplifikation, bei der in Glaskapillaren kleine Volumina (10-20 µl) mit Hilfe von
Heißluft und eines Ventilators rasch erhitzt und wieder abgekühlt werden (Wittwer et al.,
1989 und 1997). Der sichtbare Teil des Gerätes besteht aus einer Kammer mit
Probenkarussell, das 32 Glaskapillaren faßt, einem Ventilator, einem Temperaturfühler
und einer Linse, die das Licht auf die Proben leitet. In den Deckel sind Schlitze für die
Luftzufuhr und eine Heizspule eingebaut. In dem unteren Teil befinden sich die optische
Einheit, Temperatursensoren und die Optiksteuereinheit. Ein Ventilator sorgt in kürzester
Zeit für eine Temperaturangleichung in der Heizkammer. Die Angleichzeiten sind wegen
der Glaskapillaren kurz. Eine PCR mit Schmelzkurvenanalyse ist in 40 min durchführbar.
Die optische Einheit besteht aus einer Leuchtdiode, die blaues Licht mit der Wellenlänge
λ = 470 nm mit 1 mW Leistung emittiert, und einem Drei-Kanal-Fluorimeter, das die Fluo-
reszenz bei 530 nm, 640 nm und 710 nm mit einer Meßzeit von 20 ms für die einzelnen
Proben erfaßt. Die Gehäusekonstruktion läßt kein Störlicht von außen zu. Der Strah-
lengang geht von der blauen Diode über Spiegel- und Linsenkombinationen zur Spitze
der Glaskapillaren und regt das SYBR Green I7 zur Fluoreszenz an. Der „Rückweg“ des
Lichtes geht über Spiegel-, Filter- und Linsenkombinationen zu den detektierenden Pho-
tohybriden. Der LightCycler7 erhält während des einzelnen PCR-Zyklus eine spezifische
Amplifikationskurve „online“ als Aufzeichnung der Vermehrung des PCR-Produkts in
Anwesenheit des Fluorophoren, die erst produktspezifisch durch die anschließend durch-
geführte Schmelzkurvenanalyse wird. Die DNA-Amplifikation wird am Ende jeder
Verlängerungsphase gemessen.
Seite 57
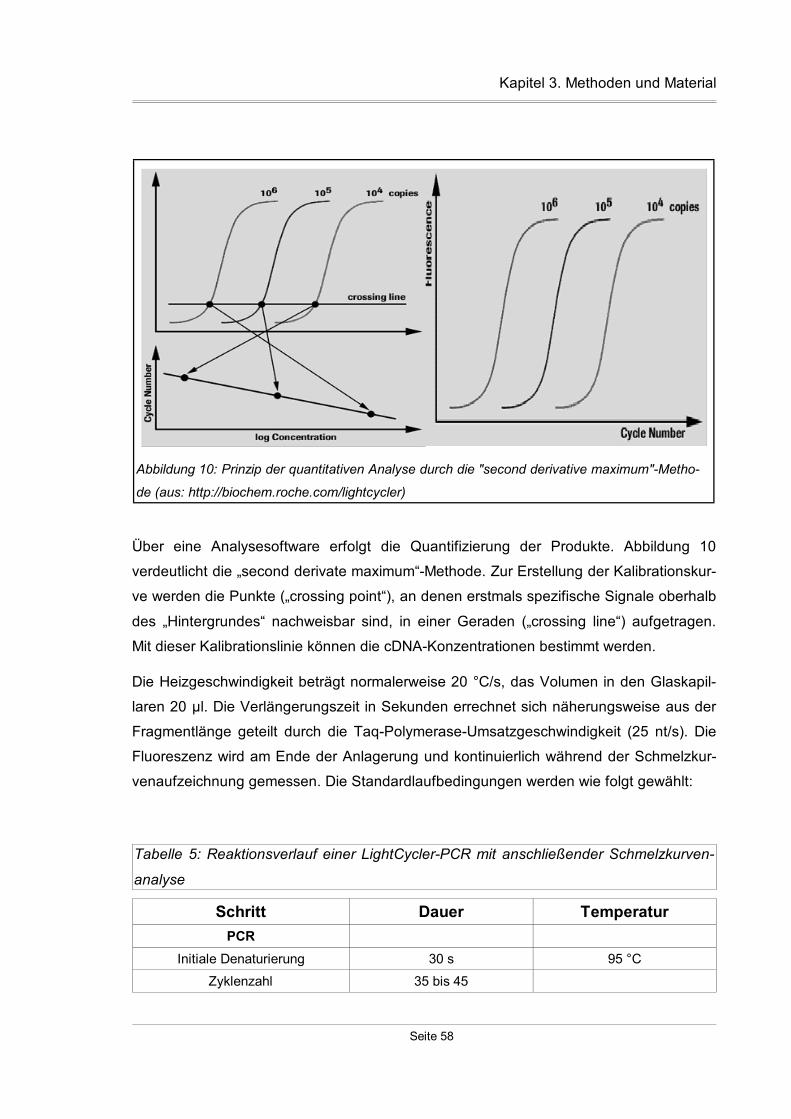
Kapitel 3. Methoden und Material
Über eine Analysesoftware erfolgt die Quantifizierung der Produkte. Abbildung 10
verdeutlicht die „second derivate maximum“-Methode. Zur Erstellung der Kalibrationskur-
ve werden die Punkte („crossing point“), an denen erstmals spezifische Signale oberhalb
des „Hintergrundes“ nachweisbar sind, in einer Geraden („crossing line“) aufgetragen.
Mit dieser Kalibrationslinie können die cDNA-Konzentrationen bestimmt werden.
Die Heizgeschwindigkeit beträgt normalerweise 20 °C/s, das Volumen in den Glaskapil-
laren 20 µl. Die Verlängerungszeit in Sekunden errechnet sich näherungsweise aus der
Fragmentlänge geteilt durch die Taq-Polymerase-Umsatzgeschwindigkeit (25 nt/s). Die
Fluoreszenz wird am Ende der Anlagerung und kontinuierlich während der Schmelzkur-
venaufzeichnung gemessen. Die Standardlaufbedingungen werden wie folgt gewählt:
Tabelle 5: Reaktionsverlauf einer LightCycler-PCR mit anschließender Schmelzkurven-
analyse
Schritt Dauer TemperaturPCR
Initiale Denaturierung 30 s 95 °C
Zyklenzahl 35 bis 45
Seite 58
Abbildung 10: Prinzip der quantitativen Analyse durch die "second derivative maximum"-Metho-
de (aus: http://biochem.roche.com/lightcycler)
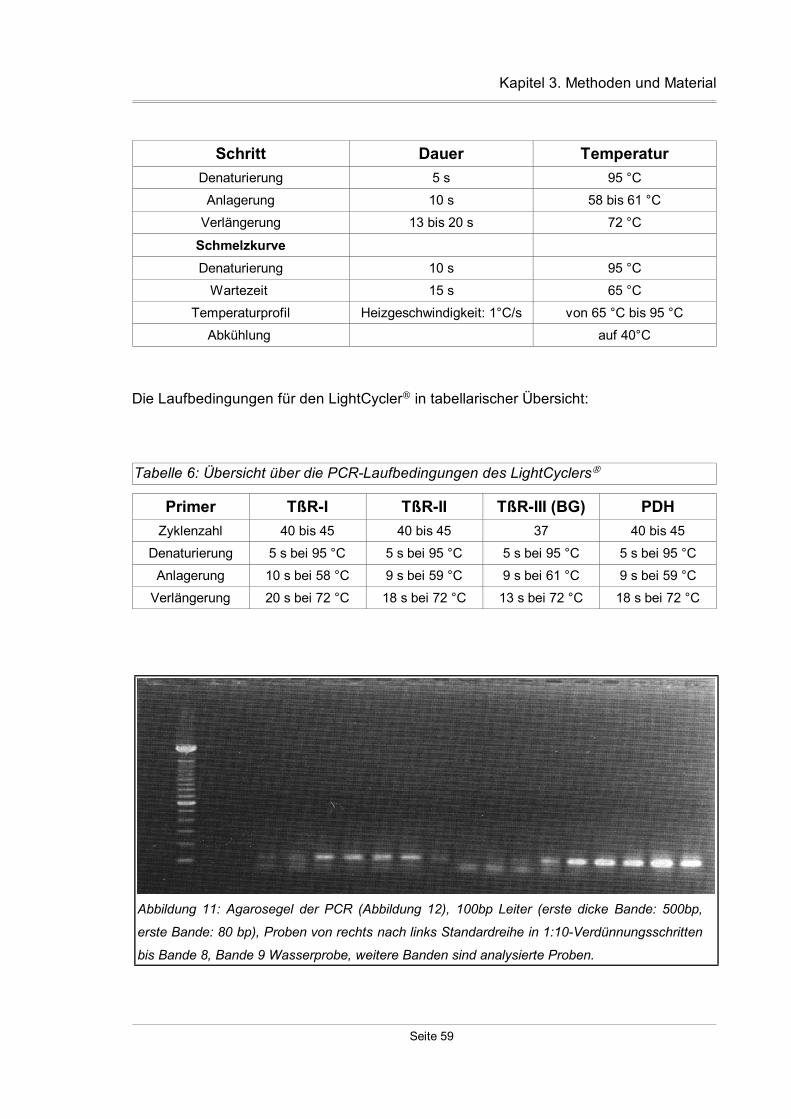
Kapitel 3. Methoden und Material
Schritt Dauer TemperaturDenaturierung 5 s 95 °C
Anlagerung 10 s 58 bis 61 °C
Verlängerung 13 bis 20 s 72 °C
SchmelzkurveDenaturierung 10 s 95 °C
Wartezeit 15 s 65 °C
Temperaturprofil Heizgeschwindigkeit: 1°C/s von 65 °C bis 95 °C
Abkühlung auf 40°C
Die Laufbedingungen für den LightCycler7 in tabellarischer Übersicht:
Tabelle 6: Übersicht über die PCR-Laufbedingungen des LightCyclers7
Primer TßR-I TßR-II TßR-III (BG) PDHZyklenzahl 40 bis 45 40 bis 45 37 40 bis 45
Denaturierung 5 s bei 95 °C 5 s bei 95 °C 5 s bei 95 °C 5 s bei 95 °C
Anlagerung 10 s bei 58 °C 9 s bei 59 °C 9 s bei 61 °C 9 s bei 59 °C
Verlängerung 20 s bei 72 °C 18 s bei 72 °C 13 s bei 72 °C 18 s bei 72 °C
Seite 59
Abbildung 11: Agarosegel der PCR (Abbildung 12), 100bp Leiter (erste dicke Bande: 500bp,
erste Bande: 80 bp), Proben von rechts nach links Standardreihe in 1:10-Verdünnungsschritten
bis Bande 8, Bande 9 Wasserprobe, weitere Banden sind analysierte Proben.
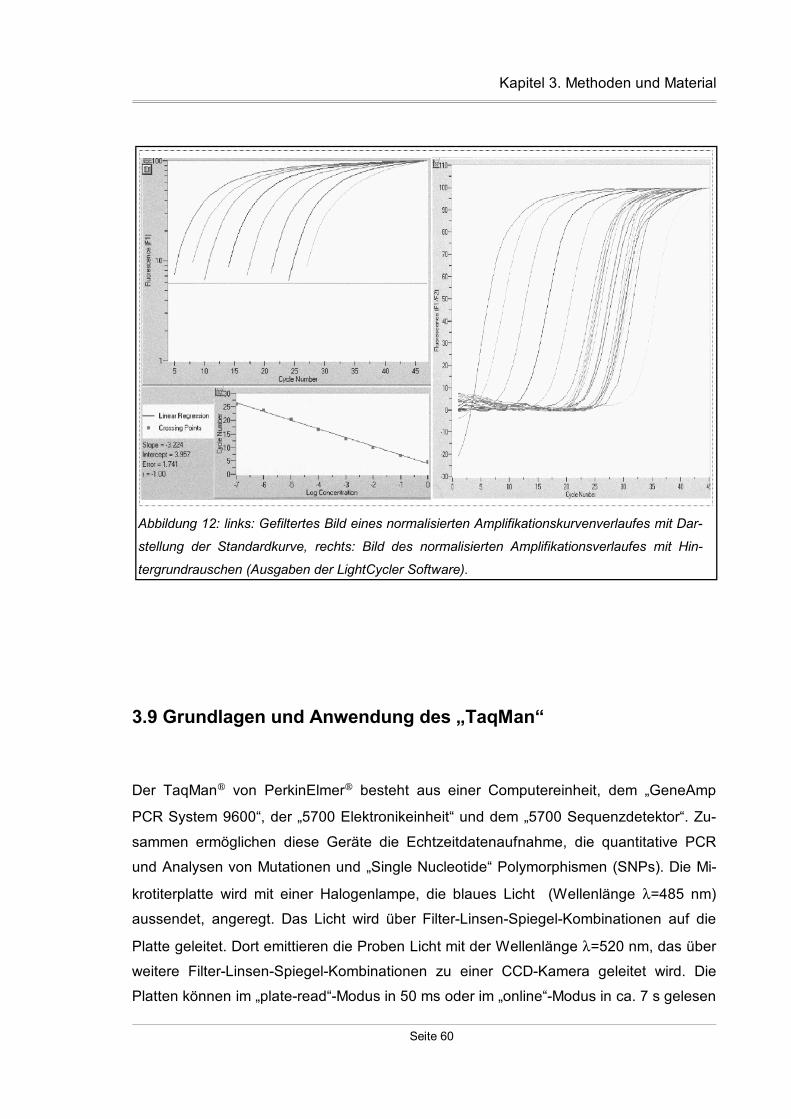
Kapitel 3. Methoden und Material
3.9 Grundlagen und Anwendung des „TaqMan“
Der TaqMan7 von PerkinElmer7 besteht aus einer Computereinheit, dem „GeneAmp
PCR System 9600“, der „5700 Elektronikeinheit“ und dem „5700 Sequenzdetektor“. Zu-
sammen ermöglichen diese Geräte die Echtzeitdatenaufnahme, die quantitative PCR
und Analysen von Mutationen und „Single Nucleotide“ Polymorphismen (SNPs). Die Mi-
krotiterplatte wird mit einer Halogenlampe, die blaues Licht (Wellenlänge λ=485 nm)
aussendet, angeregt. Das Licht wird über Filter-Linsen-Spiegel-Kombinationen auf die
Platte geleitet. Dort emittieren die Proben Licht mit der Wellenlänge λ=520 nm, das über
weitere Filter-Linsen-Spiegel-Kombinationen zu einer CCD-Kamera geleitet wird. Die
Platten können im „plate-read“-Modus in 50 ms oder im „online“-Modus in ca. 7 s gelesen
Seite 60
Abbildung 12: links: Gefiltertes Bild eines normalisierten Amplifikationskurvenverlaufes mit Dar-
stellung der Standardkurve, rechts: Bild des normalisierten Amplifikationsverlaufes mit Hin-
tergrundrauschen (Ausgaben der LightCycler Software).
Kapitel 3. Methoden und Material
werden. Das für die Quantifizierung relevante Bild wird am Ende der Anlagerungsphase
der PCR aufgezeichnet. Die Fluoreszenz jeder Vertiefung wird durch die SDS Software7
(Perkin Elmer7) getrennt verarbeitet und angezeigt. Eine „online“-Betrachtung der PCR
ist nicht möglich.
In die SDS-(„Sequence Detection System“) Software7 können die relevanten Daten für
die Mikrotiterplatte wie die verwendeten Primer, bei der Quantifizierung die Standardrei-
henkonzentration, Name und Art der Proben eingegeben werden. Für jede PCR kann ein
eigenes Temperaturprofil erstellt werden. Die Erstellung einer Schmelzkurve muß geson-
dert aktiviert werden. Alle wesentlichen Ergebnisse der PCR wie die Standardkurve mit
„slope“, „intercept“ und Korrelation, die Schmelzkurve, die Plattenbelegung, der Amplifi-
kationskurvenverlauf und ein Quantifizierungsbericht können angezeigt werden. Es gibt
eine Export-Funktion im Excel7-Format zur manuellen Nachbearbeitung.
Eine Bestückung der Cyclereinheit mit Mikrotiterplatten oder mit 0,2 ml Reaktionsge-
fäßen ist möglich. Wegen der optischen Einheit müssen lichtdurchlässige Deckel
verwendet werden. Der beheizte Deckel verhindert ein Verdampfen und Niederschlagen
am Probendeckel. Die Heizeinheit arbeitet nach dem Flüssigkeitsdurchstromprinzip, bei
dem eine wärmeleitende Flüssigkeit durch mehrere Röhren im Block die Heiz- und Kühl-
arbeit leistet.
Die PCR-Programmierung erfolgt nach einem 3-Schritt-PCR-Protokoll:
Tabelle 7: Reaktionsverlauf einer TaqMan-PCR
Reagenz Dauer TemperaturUracil-DNA-Glycosylase-Verdauung 2 min 50 °C
Initiale Denaturierung 10 min 95 °C
Zyklenzahl 35 bis 40
Denaturierung 50 s 95 °C
Anlagerung 50 s 60 bis 61 °C
Verlängerung 1 min 72 °C
Seite 61

Kapitel 3. Methoden und Material
Die Laufbedingungen für den TaqMan7 in tabellarischer Übersicht:
Tabelle 8: Übersicht über die Laufbedingungen des TaqMan7
Primer TßR-I TßR-II TßR-III (BG) PDHZyklenzahl 40 40 40 40
Denaturierung 50 s bei 95 °C 50 s bei 95 °c 50 s bei 95 °C 50 s bei 95 °C
Anlagerung 50 s bei 60 °C 50 s bei 60 °C 50 s bei 61 °C 50 s bei 61 °C
Verlängerung 1 min bei 72 °C 1 min bei 72 °C 1 min bei 72 °C 1 min bei 72 °C
Es folgt die einprogrammierte Schmelzkurvenaufnahme mit Beginn bei 60°C.
Das Probenvolumen beträgt 25 µl. In dem PCR Core Reagents Kit von PE Applied Bio-
systems ist dTTP gegen dUTP ausgetauscht, um mit der darin enthaltenen Uracil-DNA-
Glycosylase eine Kontamination zu verhindern. DMSO im Reagenziensatz vermindert
den „primer-mismatch“, die Fehlanlagerung von Primern an nicht-komplementäre Basen.
3.10 Verwendung der Uracil-DNA-Glycosylase
Der Einsatz der Uracil-DNA-Glycosylase (UNG) erfolgt mit dUTP, um in der PCR Konta-
minationen durch frühere Amplifikationen zu verringern. (Longo et al., 1990). Zu diesem
Zweck wird dTTP durch dUTP im Reaktionsansatz ersetzt und die entstehenden Produk-
te werden gegenüber der UNG empfindlich. Vor jeder PCR erfolgt eine Behandlung mit
UNG, um Uracil-enthaltende DNA abzubauen. In der DNA oder RNA bildet UTP wie TTP
Wasserstoffbrückenbindungen mit ATP aus. Uracil kommt natürlicherweise in der RNA
vor. Natürliche DNA oder cDNA enthält kein Uracil, so daß die Probe den UNG-Verdau
unbeschadet übersteht. Für den UNG-Verdau wird 1 U DNA-Glycosylase dem Re-
aktionsansatz hinzugefügt. Nach 10 min Inkubation bei 50 °C folgt die Inaktivierung der
UNG bei 95 °C für 2 bis 5 min. Gleichzeitig ist dies der erste PCR-Denaturierungsschritt.
Seite 62

Kapitel 3. Methoden und Material
3.11 Agarose-Gelelektrophorese
Die Agarose-Gelelektrophorese dient der Identifizierung der PCR-Produkte anhand ihrer
Größe und beinhaltet die Probenvorbereitung, die Agarosegelherstellung und den
Probenauftrag (für die Herstellung der Lösungen: siehe Puffer und Lösungen).
Für die Probenvorbereitung werden 20 µl des erhaltenen PCR-Ansatzes mit 5 µl Ficoll-
Farbmarker versetzt, gemischt und kurz abzentrifugiert.
Verwendung finden 1 %- bis 2 %-ige Agarosegele. Für den mittleren Gelträger werden 1-
2 g Agarose mit 100 ml 1-fach-TAE-Lösung versetzt und vorsichtig aufgekocht. Mit dem
auf 70 °C abgekühlten Agarose-TAE-Gemisch wird vom Rand her der zuvor vorbereitete
Gelträger aufgefüllt, so daß die eingelegten Kämme gut in die Gelschicht eintauchen.
Nach 5- bis 10-minütiger Abkühlung und Aushärtung werden die Kämme entfernt und
der Gelträger in die mit der 1-fach-TAE-Lösung gefüllte Gelkammer gestellt. Der vor-
sichtige Probenauftrag in die durch die Kämme entstandenen Löcher schließt sich an.
Neben den Proben wird in jeder Reihe ein Loch mit einem Größenmarker, der leiterartig
in gewissen Abständen von 100 bp bis 200 bp Vergleiche mit den PCR-Produkten
ermöglicht, befüllt. Eine Spannung zwischen 100 V und 125 V wird für 1,5 bis 2 h ange-
legt.
3.12 Gel-Elution
Die Gel-Elution erfolgt mit Hilfe des HighPure PCR-Product Purification Kit7 (Roche Bio-
chemica, Mannheim). Die zu eluierende Bande wird auf dem UV-Tisch ausgeschnitten,
in ein zuvor gewogenes steriles Eppendorfgefäß gelegt und anschließend erneut gewo-
gen. Pro 100 mg Agarosegel werden 300 ml Bindungspuffer (siehe Puffer und Lö-
sungen) hinzugegeben, kräftig geschüttelt und auf dem Thermomixer bei 60°C bis zum
vollständigen Auflösen des Gelstückes geschüttelt. Nach dem Auflösen folgt die Zugabe
von 150 ml Isopropanol pro 100 mg Agarosegel und ein gründliches Schütteln. Dem
Überführen in das HighPure-Isolationsgefäß folgt ein Zentrifugieren für 1 min bei voller
Seite 63

Kapitel 3. Methoden und Material
Leistung (12000 x g) zentrifugiert und ein Verwerfen des Durchlaufes. Der Zugabe von
500 µl Waschpuffer (siehe Puffer und Lösungen) in das obere Reservoir schließt sich ein
Zentrifugieren für 1 min bei 12000 x g und ein Verwerfen des Durchlaufs an. Nach einem
zweiten Waschschritt mit 200 µl Waschpuffer, Zentrifugieren und Abgießen des Durch-
laufs wird das Vlies-Gefäß in ein sauberes Eppendorfgefäß gesetzt. Die Elution der Nu-
kleinsäuren erfolgt mit 50 bis 100 µl bidestilliertem Wasser und erneutem Zentrifugieren
für 1 min bei voller Leistung. Die Konzentrationsbestimmung der im Eluat enthaltenen
DNA erfolgt mehrfach photometrisch gegen eine Leerprobe aus bidestilliertem Wasser.
3.13 Standardreihenherstellung
Die quantitative PCR erfordert die Herstellung von Standardreihen. Eine PCR wird mit
den für die Standardreihe benötigten Primern durchgeführt und das Ergebnis mittels
Agarosegel begutachtet. Bei Erhalt einer distinkten Bande folgt die Extraktion und pho-
tometrische Konzentrationsbestimmung nach oben beschriebenen Protokollen. Das Ori-
ginaleluat wird auf 1000 ml verdünnt und davon werden Verdünnungsreihen in 1:10
Schritten durch Zugabe von 100 µl der höheren Verdünnungsstufe zu 900 µl sterilem, bi-
destilliertem Wasser bis zu einer Verdünnung von 10^-7 hergestellt. Kurzes Mischen auf
dem Vortex und ein kurzes Anzentrifugieren schließt sich an. Mit beiden Verdünnungs-
reihen wird eine PCR durchgeführt und das Ergebnis mittels Agarosegel kontrolliert.
3.14 Statistik
Die Auswertung des Datenmaterials erfolgt mittels SPSS 10.0 sowie Lotus 1-2-3 für
Windows. Wenn möglich, wird mit dem chi-Quadrat-Test die Datenbasis auf signifikante
Unterschiede zwischen den Kollektiven untersucht. Ansonsten werden Korrelationen zwi-
schen den Basiswerten gesucht. Der Kolmogorov-Smirnov-Anpassungstest, bzw. der
Shapiro-Wilk-Test testet die Daten auf Normalverteilung. Da die Mehrheit der gebildeten
Seite 64

Kapitel 3. Methoden und Material
Gruppen keine Normalverteilung zeigt, werden Unterschiede in den einzelnen Gruppen
des Kollektivs mit Hilfe des nicht-parametrischen Mann-Whitney-U-Test auf Signifikanz
untersucht. Bei normalverteilten Werten kommt der t-Test zur Anwendung. Die Korrelati-
on nach Pearson vergleicht zwei metrische Größen miteinander, ansonsten finden Korre-
lationsuntersuchungen nach Spearman-Rho statt. Eine Wahrscheinlichkeit von p<0,05
wird primär als Signifikanzgrenze festgesetzt. Alle Daten sind als Mittelwerte +/- Stan-
dardabweichung angegeben und gegebenenfalls noch weitere statistische Größen
erwähnt.
Seite 65

Kapitel 3. Methoden und Material
3.15 Primer und Produkte
3.15.1 PDH
GenBank Zugangsnummer: D90086 (Koike et al., 1990)
upstream exon 2 5'-GGT-ATG-GAT-GAG-GAG-CTG-G-3' ( 4293-4312)
downstream exon 3 5'-CTT CCA CAG CCC TCG ACT A-3' (4627-4645)
DNA-Produktlänge: 185 bp
3.15.2 TßR-I
GenBank Zugangsnummer: NM_004612 (Franzen et al., 1993)
upstream 5'-ACG GCG TTA CAG TGT TTC TG-3' (170-190)
downstream 5'-GGC-GAT-CTA-ATG-AAG-GGT-C-3' (563-581)
Produktlänge: 411 bp
3.15.3 TßR-II
GenBank Zugangsnummer: M85079 (Lin et al., 1992)
upstream 5'-TCA-CCT-CCA-CAG-TGA-TCA-CA-3' (1412-1431)
downstream 5'-TAC-AGC-GAG-ATG-TCA-TTT-CC-3' (1700-1719)
Produktlänge: 307bp
3.15.4 Betaglykan
GenBank Zugangsnummer: NM_003243 (Wang et al., 1991; Lopez-Casillas et al., 1991)
upstream 5'-TTG-CTA-CTC-GGG-GAT-GGA-3' (1761-1778)
downstream 5'-GGT-CGG-GTG-AAC-AGG-GAA-G-3' (2018-2037)
Produktlänge: 276bp
3.15.5 Weitere Primer
Random-Primer: p(dN6), Roche Diagnostics GmbH, Mannheim
Seite 66

Kapitel 3. Methoden und Material
3.16 Geräte
3.16.1 PCR-Geräte
- Eppendorf Mastercycler gradient, Eppendorf GmbH, Hamburg
- bio med Thermocycler 60, bio med GmbH, Theres
- LightCycler, Roche, Mannheim
- TaqMan SDS 5700, Perkin Elmer Biosystems, Foster City, California, USA
3.16.2 Gelelektrophorese
- Consort E 425 Microcomputer electrophoresis power supply, Milford Haven, UK
- Consort E 452 Microcomputer electrophoresis power supply, Milford Haven, UK
- Serva Gelkammern, Heidelberg
3.16.3 Zentrifugen
- Eppendorf Centrifuge 5415 D und 5402, Hamburg
- Minifuge 2 und Megafuge 1.0 R, Heraeus Sepatech, Osterode
3.16.4 Computer und Software
- HP Vectra III mit LightCycler Software 3.39, Roche, Mannheim
- Dell P200MMX mit SDS-Software, Perkin Elmer, Foster City, Kalifornien, USA
- The Imager Software Version 2.04, Appligene Inc., San Diego, Kalifornien, USA
- Oligo 5.0, National Biosciences, Plymouth, USA
- StarOffice 5.2 für Windows, Sun Microsystems Inc., Palo Alto, USA
- OpenOffice 1.01. für Windows Sun Microsystems Inc., Palo Alto, USA
- SPSS 10.0 für Windows, SPSS Inc., Chicago, USA
- Align Plus for Windows 3.0 1997, Scientific&Educational Software, Durham, NC, USA
- Chromas 1.6, Technelysium Ltd., Helensvale, Australien
- Lotus 1-2-3 für Windows, Corel Corp., Cambridge, USA
- Endnote 3.0, Niles Software, Berkeley, Kalifornien, USA
- Siemens P200 MMX, München
Seite 67

Kapitel 3. Methoden und Material
- Windows 98 und Windows XP, Microsoft Corporation, Redmond, Washington, USA
3.16.5 Sonstige Geräte
- Schwingmühle MM 2000, Retsch, Haan
- Homogenisator UltraTurrax T25, Janke & Kunkel Labortechnik GmbH, Staufen i. Br.
- Vortex VF2, Janke & Kunkel Labortechnik GmbH, Staufen i. Br.
- Thermomixer compact, Eppendorf, Hamburg
- Waage MC1 Sartorius, Göttingen
- Reinstwasseranlage Milli-Q Plus, Millipore, Bedford, MA, USA
- Photometer GeneQuant DNA/RNA Calculator, Pharmacia Biotech, Freiburg
- pH-Meter CG 840, Schott-Geräte Hofheim a. Ts.
3.17 Verwendete Chemikalien und Artikel
3.17.1 Einzelchemikalien
- Ethanol absolut p.A:, Merck, Darmstadt
- Chloroform p.A., Merck, Darmstadt
- Phenol, Sigma-Aldrich, München
- Isoamylalkohol (Isomerengemisch), Merck, Darmstadt
- 2-Propanol, reinst, Merck, Darmstadt
- DMPC, Sigma-Aldrich, München
- Natriumacetat, wasserfrei p. A., Merck
- Morpholinopropansulfonsäure (MOPS), Serva Feinbiochemica GmbH, Heidelberg
- EDTA, Sigma-Aldrich, München
- Glycerol 87%, Merck, Darmstadt
- SeaKem LE agarose, for gel electrophoresis, FMC BioProducts, Rockland, Maine, USA
- Mineral Oil, for molecular biology, Sigma
- Formaldehyd, Sigma-Aldrich, München
- Formamid, Sigma-Aldrich, München
Seite 68

Kapitel 3. Methoden und Material
- Ethidiumbromid, Sigma-Aldrich, München
- Guanidiniumthiocyanat, for molecular biology, Fluka BioChemika, Buchs, Schweiz
- Tris(hydroxymethyl)-aminomethan p.A. Puffersubstanz, Merck, Darmstadt
- Eisessigsäure, Sigma-Aldrich, München
- PBS (50-fach)-Phosphatpuffer-Konzentrat pH 7,4, BAG, Lich
- Ficoll 400, Pharmacia Biotech, Uppsala, Sweden
- dATP 100mM, Promega, Madison, Wisconsin, USA
- dCTP 100mM, Promega, Madison, Wisconsin, USA
- dGTP 100mM, Promega, Madison, Wisconsin, USA
- dTTP 100mM, Promega, Madison, Wisconsin, USA
3.17.2 Verwendete Reagenziensätze
- High Pure PCR Product Purification Kit, Roche Biochemica, Mannheim
- TRIzol reg. (Total RNA Isolation) Reagent, Life Technologies, Gaithersburg, USA
- LightCycler - DNA Master SYBR Green 1, Roche Diagnostics GmbH, Mannheim
- SYBR Green PCR Core Reagents, PE Biosystems, Foster City, Kalifornien, USA
- DNA Sequencing Kit - Big Dye Terminator cycle sequencing ready reaction kit, PE Bio-
systems, Foster City, Kalifornien, USA
3.17.3 Enzyme
- Taq DNA Polymerase, recombinant, Gibco Life Technologies, Gaithersburg, USA
- M-MuLV Reverse Transkriptase, Roche Diagnostics GmbH, Mannheim
- porcine RNAguard RNase Inhibitor, Amersham Pharmacia Biotech, Uppsala, Schweden
- Uracil-DNA Glycosylase, heat-labile, Roche Diagnostics GmbH, Mannheim
3.17.4 DNA-Größenstandards für Agarose-Gelelektrophorese
- 100 bp DNA Ladder, Gibco Life Technologies, Gaithersburg, USA
- 1kB DNA Plus Ladder, Gibco Life Technologies, Gaithersburg, USA
Seite 69

Kapitel 3. Methoden und Material
3.18 Puffer und Lösungen
Für alle Enzymreaktionen werden, wenn nicht anders vermerkt, die vom Hersteller
gelieferten Puffer nach dessen Angaben verwendet.
Farbmarker für DNA-Gele:
0,1% Ponceau S
0,1% Orange G
0,1% Bromphenolblau
0,1% XyleneCyanol
1x Lade-Puffer für GE
Lade-Puffer für GE:
10 mM EDTA
25% Ficoll
Aqua dest. ad 100 ml
5x Formaldehyd gel running buffer (FGRB):
0,1M MOPS
40mM Natriumazetat
5 mM EDTA pH8
Formaldeyd gel loading buffer:
50% Glycerol
1mM EDTA
0,25% Bromphenolblau
0,25% Xylene Cyanol FF
TAE-Puffer (50x-Stock-Lösung):
242 g Tris
57,1 ml Eisessigsäure
100 ml 0,5 ml EDTA pH 8,0
ad 1000 ml Aqua dest.
Seite 70

Kapitel 3. Methoden und Material
1-fach TAE-Lösung (Agarosegelelektrophoresegebrauchslösung):
20 ml 50x TAE-Lösung
50 µl Ethidiumbromid (10mg/ml)
ad 1000 ml Aqua dest.
PBS-Puffer-Stock-Lösung:
8 g Natriumchlorid (NaCl)
0,2 g Kaliumchlorid (KCl)
1,44 g Natriumhydrogenphosphat ( Na2 HPO4)
0,24 g Kaliumdihydrogenphosphat ( K H2PO4)
auf pH 7,4 mit Salzsäure (HCl) einstellen und ad 1000ml Aqua dest.
3M Natriumazetat pH 5,2:
408,1 g NaAcetat x 3 H2O
auf pH 5,2 mit Eisessigsäureeinstellen und ad 1000ml Aqua dest.
dNTP-Mix (100µM jeweils):
10 µl 100mM dATP
10 µ 100mM dCTP
10 µl 100mM dGTP
10 µl 100mM dTTP
ad 100 µl mit DMPC-Wasser
Solution D (Denaturierungslösung):
4 M Guanidiniumthiocyanat
25 mM Natriumcitrat
0,5% Sarkosyl
0,1 mM 2-Mercaptoethanol
ad 337 ml Wasser
Bindungspuffer (HighPure PCR Purification Kit, Roche, Mannheim):
3M Guanidinium-Thiocyanat
10 mM Tris-HCl
5 % Ethanol (v/v)
pH 6,6 (25°C)
Seite 71

Kapitel 3. Methoden und Material
Waschpuffer (HighPure PCR Purification Kit, Roche, Mannheim):
20 mM NaCl
2 mM Tris-HCl
80 Vol-% Ethanol abs. p.A.
pH 7,5 (25°C)
DMPC-Wasser:
0,1 % Dimethylpyrocarbonat werden in Aqua bidest. gelöst, über Nacht geschüttelt und
danach zur Spaltung von DMPC in CO2 und Ethanol autoklaviert, im Gegensatz zur
Autoklavierung werden RNAsen durch das methylierende DMPC vollständig inaktiviert.
Seite 72
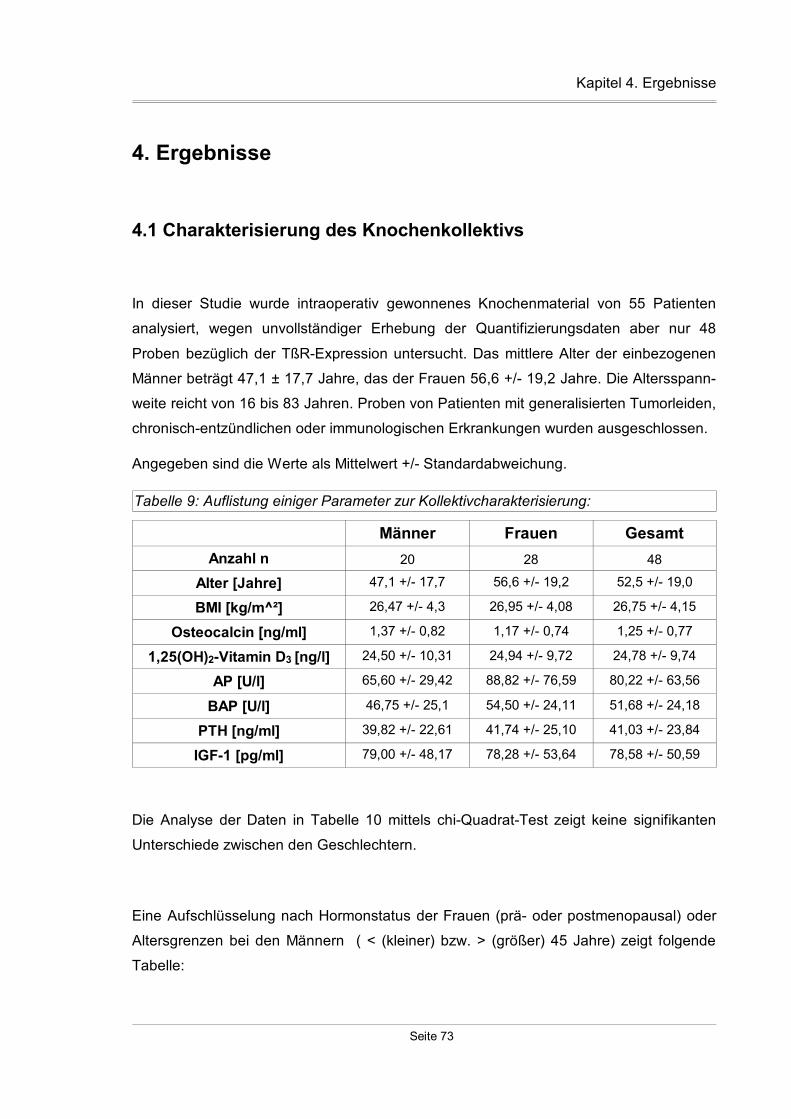
Kapitel 4. Ergebnisse
4. Ergebnisse
4.1 Charakterisierung des Knochenkollektivs
In dieser Studie wurde intraoperativ gewonnenes Knochenmaterial von 55 Patienten
analysiert, wegen unvollständiger Erhebung der Quantifizierungsdaten aber nur 48
Proben bezüglich der TßR-Expression untersucht. Das mittlere Alter der einbezogenen
Männer beträgt 47,1 ± 17,7 Jahre, das der Frauen 56,6 +/- 19,2 Jahre. Die Altersspann-
weite reicht von 16 bis 83 Jahren. Proben von Patienten mit generalisierten Tumorleiden,
chronisch-entzündlichen oder immunologischen Erkrankungen wurden ausgeschlossen.
Angegeben sind die Werte als Mittelwert +/- Standardabweichung.
Tabelle 9: Auflistung einiger Parameter zur Kollektivcharakterisierung:
Männer Frauen GesamtAnzahl n 20 28 48
Alter [Jahre] 47,1 +/- 17,7 56,6 +/- 19,2 52,5 +/- 19,0
BMI [kg/m^²] 26,47 +/- 4,3 26,95 +/- 4,08 26,75 +/- 4,15
Osteocalcin [ng/ml] 1,37 +/- 0,82 1,17 +/- 0,74 1,25 +/- 0,77
1,25(OH)2-Vitamin D3 [ng/l] 24,50 +/- 10,31 24,94 +/- 9,72 24,78 +/- 9,74
AP [U/l] 65,60 +/- 29,42 88,82 +/- 76,59 80,22 +/- 63,56
BAP [U/l] 46,75 +/- 25,1 54,50 +/- 24,11 51,68 +/- 24,18
PTH [ng/ml] 39,82 +/- 22,61 41,74 +/- 25,10 41,03 +/- 23,84
IGF-1 [pg/ml] 79,00 +/- 48,17 78,28 +/- 53,64 78,58 +/- 50,59
Die Analyse der Daten in Tabelle 10 mittels chi-Quadrat-Test zeigt keine signifikanten
Unterschiede zwischen den Geschlechtern.
Eine Aufschlüsselung nach Hormonstatus der Frauen (prä- oder postmenopausal) oder
Altersgrenzen bei den Männern ( < (kleiner) bzw. > (größer) 45 Jahre) zeigt folgende
Tabelle:
Seite 73
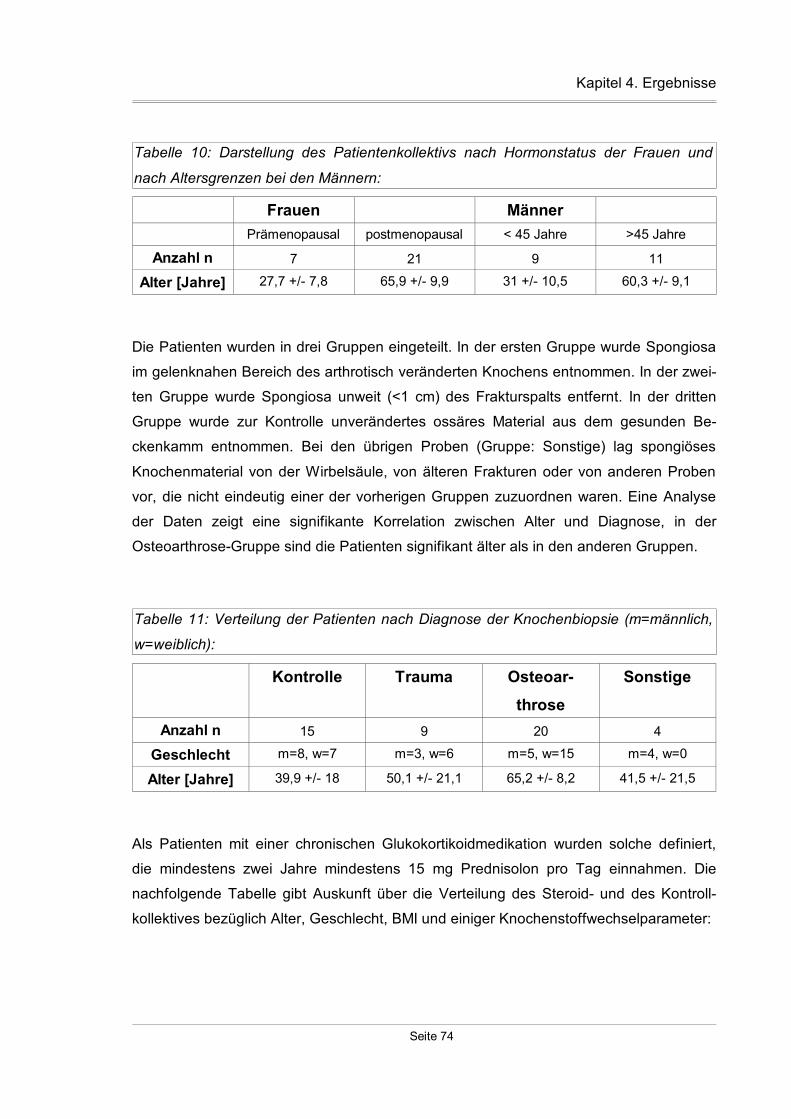
Kapitel 4. Ergebnisse
Tabelle 10: Darstellung des Patientenkollektivs nach Hormonstatus der Frauen und
nach Altersgrenzen bei den Männern:
Frauen MännerPrämenopausal postmenopausal < 45 Jahre >45 Jahre
Anzahl n 7 21 9 11
Alter [Jahre] 27,7 +/- 7,8 65,9 +/- 9,9 31 +/- 10,5 60,3 +/- 9,1
Die Patienten wurden in drei Gruppen eingeteilt. In der ersten Gruppe wurde Spongiosa
im gelenknahen Bereich des arthrotisch veränderten Knochens entnommen. In der zwei-
ten Gruppe wurde Spongiosa unweit (<1 cm) des Frakturspalts entfernt. In der dritten
Gruppe wurde zur Kontrolle unverändertes ossäres Material aus dem gesunden Be-
ckenkamm entnommen. Bei den übrigen Proben (Gruppe: Sonstige) lag spongiöses
Knochenmaterial von der Wirbelsäule, von älteren Frakturen oder von anderen Proben
vor, die nicht eindeutig einer der vorherigen Gruppen zuzuordnen waren. Eine Analyse
der Daten zeigt eine signifikante Korrelation zwischen Alter und Diagnose, in der
Osteoarthrose-Gruppe sind die Patienten signifikant älter als in den anderen Gruppen.
Tabelle 11: Verteilung der Patienten nach Diagnose der Knochenbiopsie (m=männlich,
w=weiblich):
Kontrolle Trauma Osteoar-
throse
Sonstige
Anzahl n 15 9 20 4
Geschlecht m=8, w=7 m=3, w=6 m=5, w=15 m=4, w=0
Alter [Jahre] 39,9 +/- 18 50,1 +/- 21,1 65,2 +/- 8,2 41,5 +/- 21,5
Als Patienten mit einer chronischen Glukokortikoidmedikation wurden solche definiert,
die mindestens zwei Jahre mindestens 15 mg Prednisolon pro Tag einnahmen. Die
nachfolgende Tabelle gibt Auskunft über die Verteilung des Steroid- und des Kontroll-
kollektives bezüglich Alter, Geschlecht, BMI und einiger Knochenstoffwechselparameter:
Seite 74
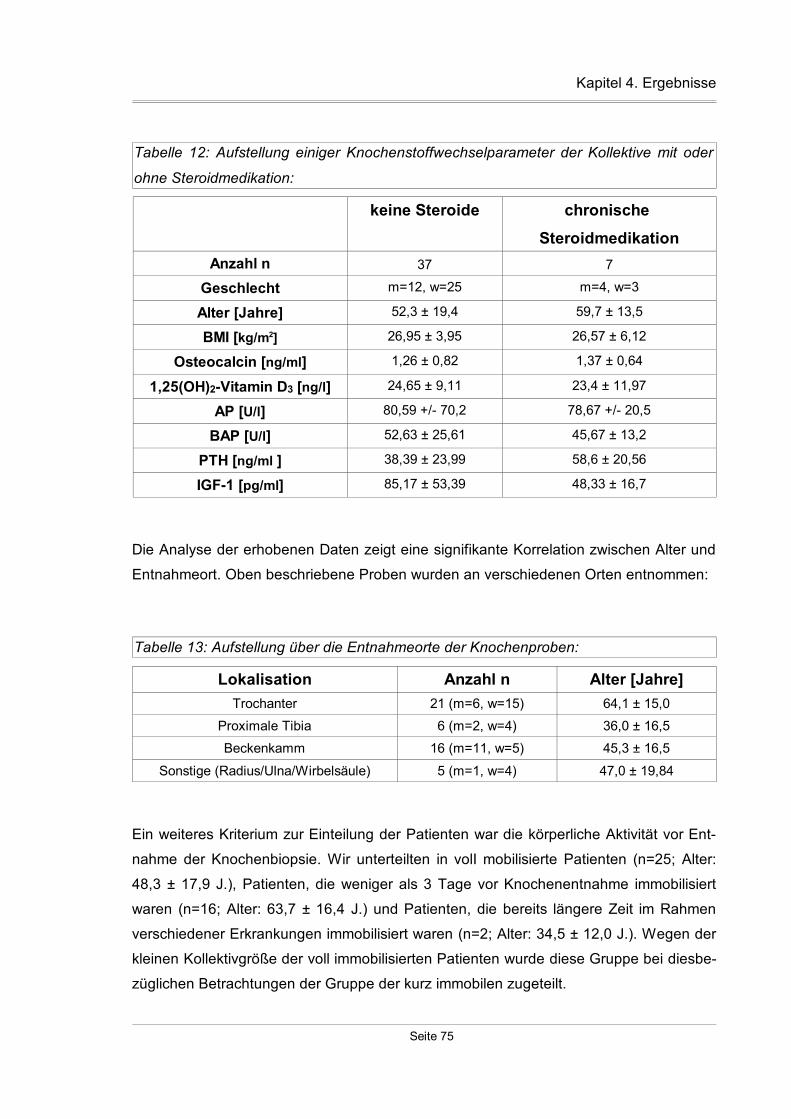
Kapitel 4. Ergebnisse
Tabelle 12: Aufstellung einiger Knochenstoffwechselparameter der Kollektive mit oder
ohne Steroidmedikation:
keine Steroide chronische
SteroidmedikationAnzahl n 37 7
Geschlecht m=12, w=25 m=4, w=3
Alter [Jahre] 52,3 ± 19,4 59,7 ± 13,5
BMI [kg/m2] 26,95 ± 3,95 26,57 ± 6,12
Osteocalcin [ng/ml] 1,26 ± 0,82 1,37 ± 0,64
1,25(OH)2-Vitamin D3 [ng/l] 24,65 ± 9,11 23,4 ± 11,97
AP [U/l] 80,59 +/- 70,2 78,67 +/- 20,5
BAP [U/l] 52,63 ± 25,61 45,67 ± 13,2
PTH [ng/ml ] 38,39 ± 23,99 58,6 ± 20,56
IGF-1 [pg/ml] 85,17 ± 53,39 48,33 ± 16,7
Die Analyse der erhobenen Daten zeigt eine signifikante Korrelation zwischen Alter und
Entnahmeort. Oben beschriebene Proben wurden an verschiedenen Orten entnommen:
Tabelle 13: Aufstellung über die Entnahmeorte der Knochenproben:
Lokalisation Anzahl n Alter [Jahre]Trochanter 21 (m=6, w=15) 64,1 ± 15,0
Proximale Tibia 6 (m=2, w=4) 36,0 ± 16,5
Beckenkamm 16 (m=11, w=5) 45,3 ± 16,5
Sonstige (Radius/Ulna/Wirbelsäule) 5 (m=1, w=4) 47,0 ± 19,84
Ein weiteres Kriterium zur Einteilung der Patienten war die körperliche Aktivität vor Ent-
nahme der Knochenbiopsie. Wir unterteilten in volI mobilisierte Patienten (n=25; Alter:
48,3 ± 17,9 J.), Patienten, die weniger als 3 Tage vor Knochenentnahme immobilisiert
waren (n=16; Alter: 63,7 ± 16,4 J.) und Patienten, die bereits längere Zeit im Rahmen
verschiedener Erkrankungen immobilisiert waren (n=2; Alter: 34,5 ± 12,0 J.). Wegen der
kleinen Kollektivgröße der voll immobilisierten Patienten wurde diese Gruppe bei diesbe-
züglichen Betrachtungen der Gruppe der kurz immobilen zugeteilt.
Seite 75
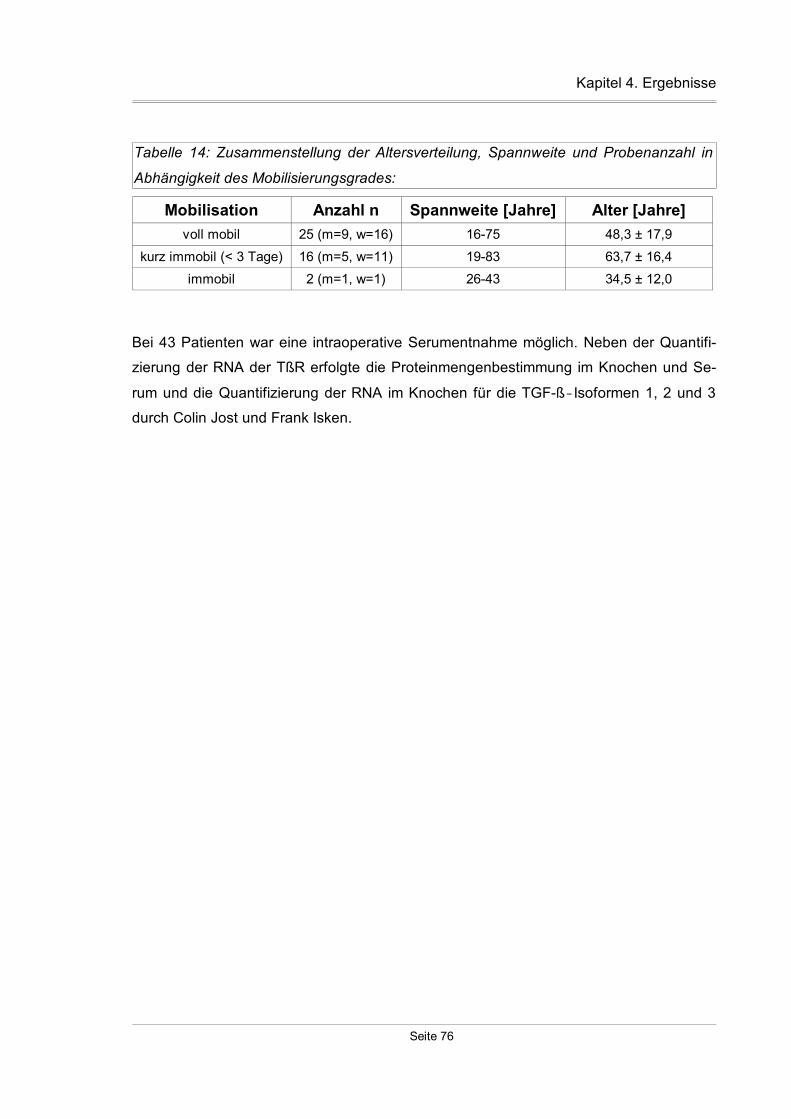
Kapitel 4. Ergebnisse
Tabelle 14: Zusammenstellung der Altersverteilung, Spannweite und Probenanzahl in
Abhängigkeit des Mobilisierungsgrades:
Mobilisation Anzahl n Spannweite [Jahre] Alter [Jahre]voll mobil 25 (m=9, w=16) 16-75 48,3 ± 17,9
kurz immobil (< 3 Tage) 16 (m=5, w=11) 19-83 63,7 ± 16,4
immobil 2 (m=1, w=1) 26-43 34,5 ± 12,0
Bei 43 Patienten war eine intraoperative Serumentnahme möglich. Neben der Quantifi-
zierung der RNA der TßR erfolgte die Proteinmengenbestimmung im Knochen und Se-
rum und die Quantifizierung der RNA im Knochen für die TGF-ß-Isoformen 1, 2 und 3
durch Colin Jost und Frank Isken.
Seite 76
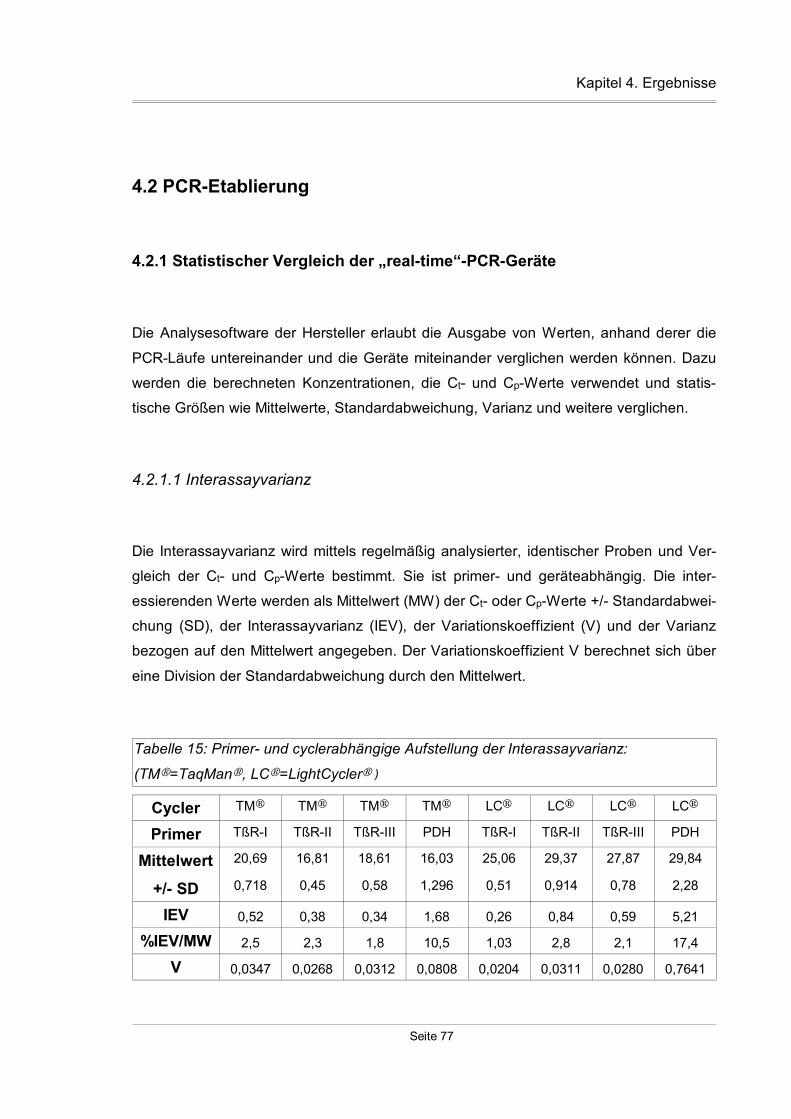
Kapitel 4. Ergebnisse
4.2 PCR-Etablierung
4.2.1 Statistischer Vergleich der „real-time“-PCR-Geräte
Die Analysesoftware der Hersteller erlaubt die Ausgabe von Werten, anhand derer die
PCR-Läufe untereinander und die Geräte miteinander verglichen werden können. Dazu
werden die berechneten Konzentrationen, die Ct- und Cp-Werte verwendet und statis-
tische Größen wie Mittelwerte, Standardabweichung, Varianz und weitere verglichen.
4.2.1.1 Interassayvarianz
Die Interassayvarianz wird mittels regelmäßig analysierter, identischer Proben und Ver-
gleich der Ct- und Cp-Werte bestimmt. Sie ist primer- und geräteabhängig. Die inter-
essierenden Werte werden als Mittelwert (MW) der Ct- oder Cp-Werte +/- Standardabwei-
chung (SD), der Interassayvarianz (IEV), der Variationskoeffizient (V) und der Varianz
bezogen auf den Mittelwert angegeben. Der Variationskoeffizient V berechnet sich über
eine Division der Standardabweichung durch den Mittelwert.
Tabelle 15: Primer- und cyclerabhängige Aufstellung der Interassayvarianz:
(TM7=TaqMan7, LC7=LightCycler7 )
Cycler TM7 TM7 TM7 TM7 LC7 LC7 LC7 LC7
Primer TßR-I TßR-II TßR-III PDH TßR-I TßR-II TßR-III PDH
Mittelwert
+/- SD
20,69
0,718
16,81
0,45
18,61
0,58
16,03
1,296
25,06
0,51
29,37
0,914
27,87
0,78
29,84
2,28
IEV 0,52 0,38 0,34 1,68 0,26 0,84 0,59 5,21
%IEV/MW 2,5 2,3 1,8 10,5 1,03 2,8 2,1 17,4
V 0,0347 0,0268 0,0312 0,0808 0,0204 0,0311 0,0280 0,7641
Seite 77
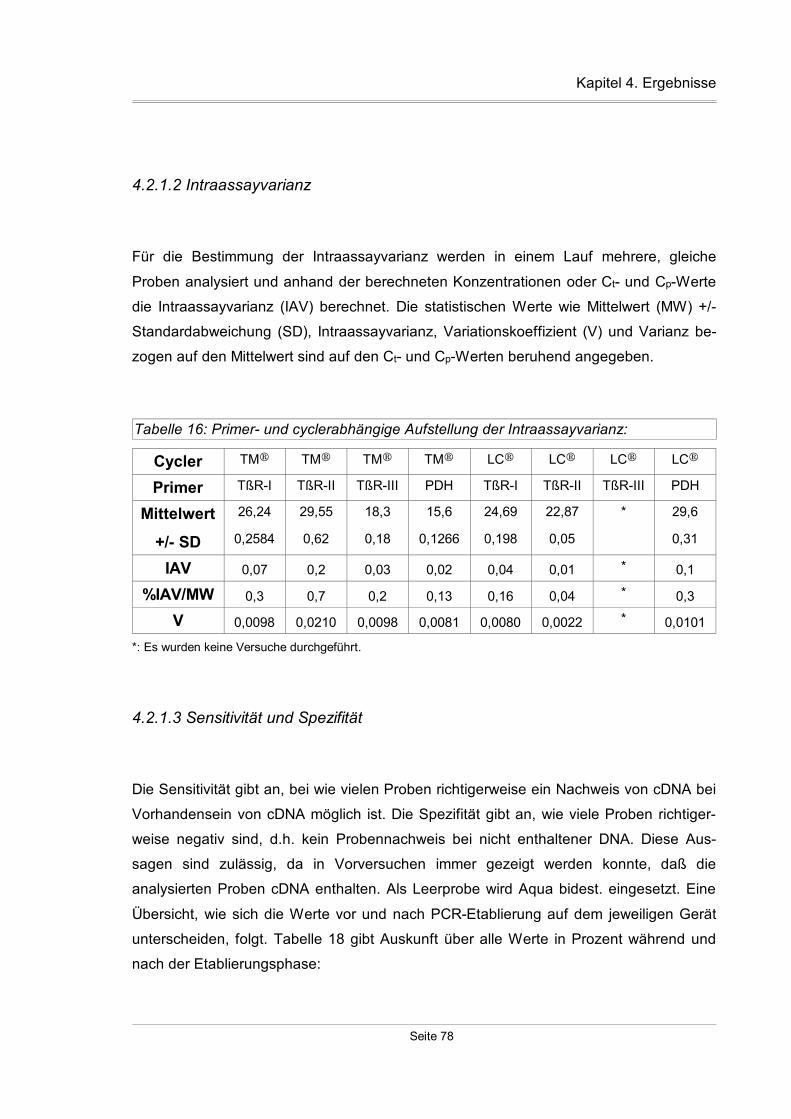
Kapitel 4. Ergebnisse
4.2.1.2 Intraassayvarianz
Für die Bestimmung der Intraassayvarianz werden in einem Lauf mehrere, gleiche
Proben analysiert und anhand der berechneten Konzentrationen oder Ct- und Cp-Werte
die Intraassayvarianz (IAV) berechnet. Die statistischen Werte wie Mittelwert (MW) +/-
Standardabweichung (SD), Intraassayvarianz, Variationskoeffizient (V) und Varianz be-
zogen auf den Mittelwert sind auf den Ct- und Cp-Werten beruhend angegeben.
Tabelle 16: Primer- und cyclerabhängige Aufstellung der Intraassayvarianz:
Cycler TM7 TM7 TM7 TM7 LC7 LC7 LC7 LC7
Primer TßR-I TßR-II TßR-III PDH TßR-I TßR-II TßR-III PDH
Mittelwert
+/- SD
26,24
0,2584
29,55
0,62
18,3
0,18
15,6
0,1266
24,69
0,198
22,87
0,05
* 29,6
0,31
IAV 0,07 0,2 0,03 0,02 0,04 0,01 * 0,1
%IAV/MW 0,3 0,7 0,2 0,13 0,16 0,04 * 0,3
V 0,0098 0,0210 0,0098 0,0081 0,0080 0,0022 * 0,0101
*: Es wurden keine Versuche durchgeführt.
4.2.1.3 Sensitivität und Spezifität
Die Sensitivität gibt an, bei wie vielen Proben richtigerweise ein Nachweis von cDNA bei
Vorhandensein von cDNA möglich ist. Die Spezifität gibt an, wie viele Proben richtiger-
weise negativ sind, d.h. kein Probennachweis bei nicht enthaltener DNA. Diese Aus-
sagen sind zulässig, da in Vorversuchen immer gezeigt werden konnte, daß die
analysierten Proben cDNA enthalten. Als Leerprobe wird Aqua bidest. eingesetzt. Eine
Übersicht, wie sich die Werte vor und nach PCR-Etablierung auf dem jeweiligen Gerät
unterscheiden, folgt. Tabelle 18 gibt Auskunft über alle Werte in Prozent während und
nach der Etablierungsphase:
Seite 78
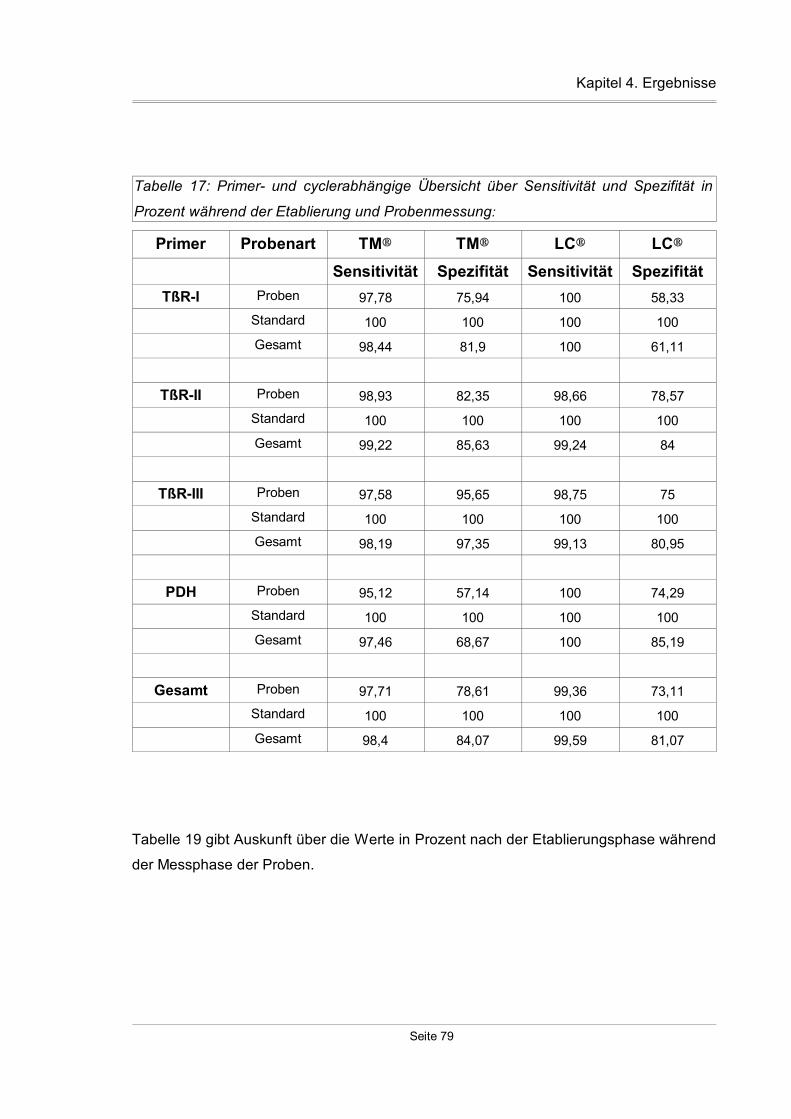
Kapitel 4. Ergebnisse
Tabelle 17: Primer- und cyclerabhängige Übersicht über Sensitivität und Spezifität in
Prozent während der Etablierung und Probenmessung:
Primer Probenart TM7 TM7 LC7 LC7
Sensitivität Spezifität Sensitivität SpezifitätTßR-I Proben 97,78 75,94 100 58,33
Standard 100 100 100 100
Gesamt 98,44 81,9 100 61,11
TßR-II Proben 98,93 82,35 98,66 78,57
Standard 100 100 100 100
Gesamt 99,22 85,63 99,24 84
TßR-III Proben 97,58 95,65 98,75 75
Standard 100 100 100 100
Gesamt 98,19 97,35 99,13 80,95
PDH Proben 95,12 57,14 100 74,29
Standard 100 100 100 100
Gesamt 97,46 68,67 100 85,19
Gesamt Proben 97,71 78,61 99,36 73,11
Standard 100 100 100 100
Gesamt 98,4 84,07 99,59 81,07
Tabelle 19 gibt Auskunft über die Werte in Prozent nach der Etablierungsphase während
der Messphase der Proben.
Seite 79
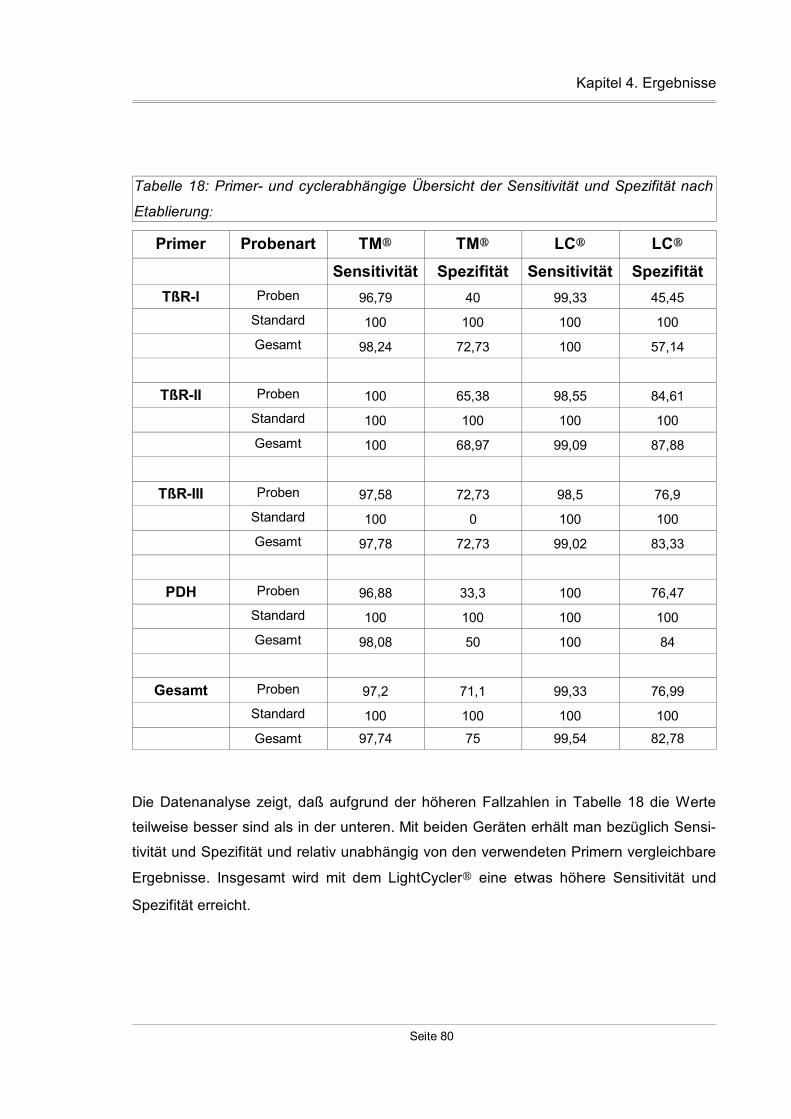
Kapitel 4. Ergebnisse
Tabelle 18: Primer- und cyclerabhängige Übersicht der Sensitivität und Spezifität nach
Etablierung:
Primer Probenart TM7 TM7 LC7 LC7
Sensitivität Spezifität Sensitivität SpezifitätTßR-I Proben 96,79 40 99,33 45,45
Standard 100 100 100 100
Gesamt 98,24 72,73 100 57,14
TßR-II Proben 100 65,38 98,55 84,61
Standard 100 100 100 100
Gesamt 100 68,97 99,09 87,88
TßR-III Proben 97,58 72,73 98,5 76,9
Standard 100 0 100 100
Gesamt 97,78 72,73 99,02 83,33
PDH Proben 96,88 33,3 100 76,47
Standard 100 100 100 100
Gesamt 98,08 50 100 84
Gesamt Proben 97,2 71,1 99,33 76,99
Standard 100 100 100 100
Gesamt 97,74 75 99,54 82,78
Die Datenanalyse zeigt, daß aufgrund der höheren Fallzahlen in Tabelle 18 die Werte
teilweise besser sind als in der unteren. Mit beiden Geräten erhält man bezüglich Sensi-
tivität und Spezifität und relativ unabhängig von den verwendeten Primern vergleichbare
Ergebnisse. Insgesamt wird mit dem LightCycler7 eine etwas höhere Sensitivität und
Spezifität erreicht.
Seite 80

Kapitel 4. Ergebnisse
4.2.1.4 Amplifikationseffektivität
Eine PCR läßt sich durch die Gleichung:
(1): N = N0 * 2^n
beschreiben, wobei N die Menge des hergestellten Amplifikates nach n Zyklen und N0
die Startkopienzahl ist. Der Term 2^n ist durch (1+E)^n mit E gleich der Amplifikations-
effektivität ersetzbar, da bei einer optimalen PCR E=1 ist.
(2): N = N0 * (1+E)^n
Die „real-time“-PCR-Geräte geben bei der Analyse eines PCR-Laufes die Steigung
„slope“ (s) und den y-Achsenschnittpunkt „intercept“ wieder. Zur Erstellung einer Stan-
dardkurve wird (2) logarithmiert und in die Form gebracht:
(3): log N = log N0 + n * log (1+E)
Die graphische Darstellung erfolgt folgendermaßen:
Aus Abbildung 13 geht hervor:
(4): s = -1/(log 1+E)
Mathematische Umformung er-
gibt Gleichung (5) zur Berech-
nung der Effektivität bei gege-
bener Steigung der Standard-
kurve:
(5): E = 10^(1/s) – 1
Mittels Formel (5) wird auf rech-
nerische Weise die Amplifika-
tionseffektivität bestimmt. Sie
liegt primer- und geräteabhän-
gig zwischen 56 % und 92 % (siehe Tabelle 20).
Seite 81
Abbildung 13: Graphische Darstellung der Amplifika-
tionseffektivität in halblogarithmischer Form.
Hohe Effizienz
Log c
Ct
Niedrige Effizienz
s=-1/log(1+E)
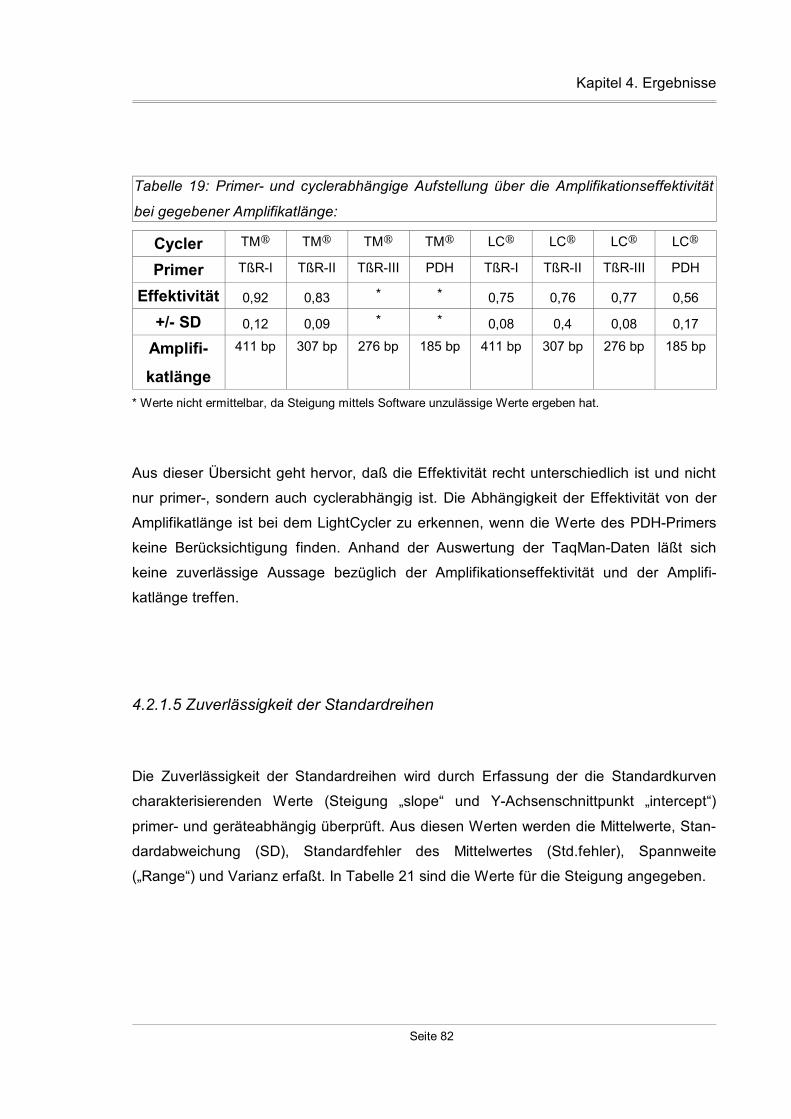
Kapitel 4. Ergebnisse
Tabelle 19: Primer- und cyclerabhängige Aufstellung über die Amplifikationseffektivität
bei gegebener Amplifikatlänge:
Cycler TM7 TM7 TM7 TM7 LC7 LC7 LC7 LC7
Primer TßR-I TßR-II TßR-III PDH TßR-I TßR-II TßR-III PDH
Effektivität 0,92 0,83 * * 0,75 0,76 0,77 0,56
+/- SD 0,12 0,09 * * 0,08 0,4 0,08 0,17
Amplifi-
katlänge
411 bp 307 bp 276 bp 185 bp 411 bp 307 bp 276 bp 185 bp
* Werte nicht ermittelbar, da Steigung mittels Software unzulässige Werte ergeben hat.
Aus dieser Übersicht geht hervor, daß die Effektivität recht unterschiedlich ist und nicht
nur primer-, sondern auch cyclerabhängig ist. Die Abhängigkeit der Effektivität von der
Amplifikatlänge ist bei dem LightCycler zu erkennen, wenn die Werte des PDH-Primers
keine Berücksichtigung finden. Anhand der Auswertung der TaqMan-Daten läßt sich
keine zuverlässige Aussage bezüglich der Amplifikationseffektivität und der Amplifi-
katlänge treffen.
4.2.1.5 Zuverlässigkeit der Standardreihen
Die Zuverlässigkeit der Standardreihen wird durch Erfassung der die Standardkurven
charakterisierenden Werte (Steigung „slope“ und Y-Achsenschnittpunkt „intercept“)
primer- und geräteabhängig überprüft. Aus diesen Werten werden die Mittelwerte, Stan-
dardabweichung (SD), Standardfehler des Mittelwertes (Std.fehler), Spannweite
(„Range“) und Varianz erfaßt. In Tabelle 21 sind die Werte für die Steigung angegeben.
Seite 82
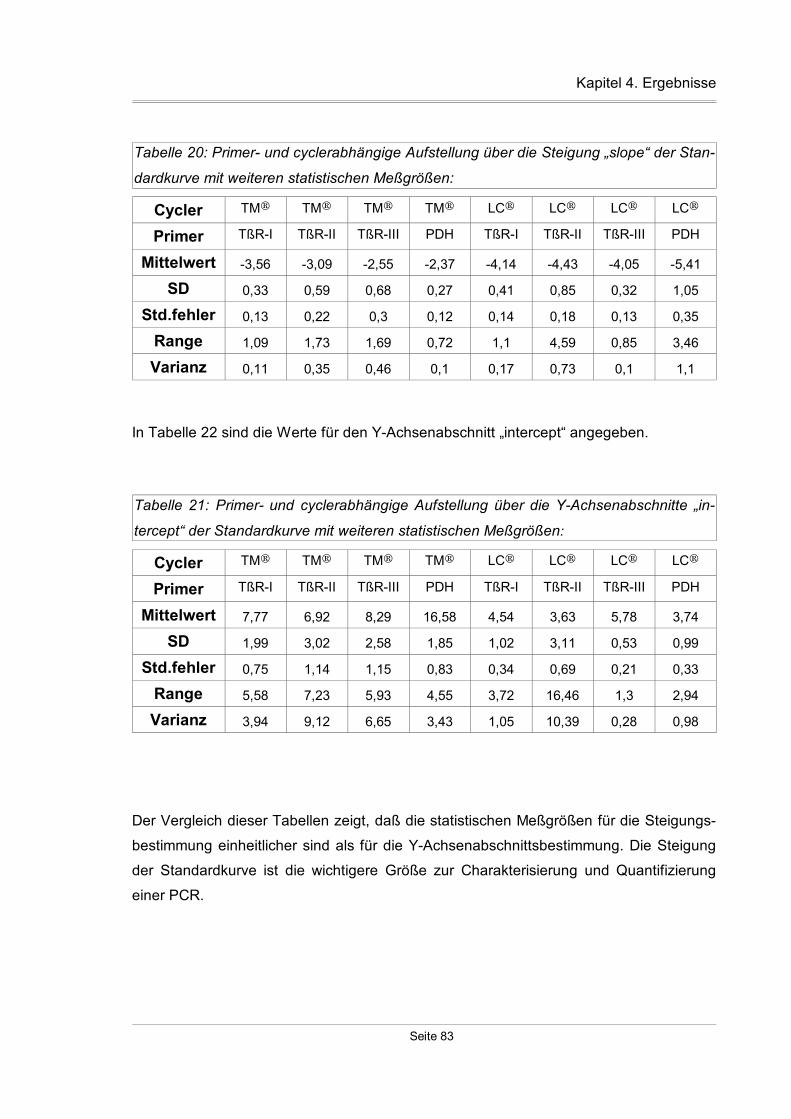
Kapitel 4. Ergebnisse
Tabelle 20: Primer- und cyclerabhängige Aufstellung über die Steigung „slope“ der Stan-
dardkurve mit weiteren statistischen Meßgrößen:
Cycler TM7 TM7 TM7 TM7 LC7 LC7 LC7 LC7
Primer TßR-I TßR-II TßR-III PDH TßR-I TßR-II TßR-III PDH
Mittelwert -3,56 -3,09 -2,55 -2,37 -4,14 -4,43 -4,05 -5,41
SD 0,33 0,59 0,68 0,27 0,41 0,85 0,32 1,05
Std.fehler 0,13 0,22 0,3 0,12 0,14 0,18 0,13 0,35
Range 1,09 1,73 1,69 0,72 1,1 4,59 0,85 3,46
Varianz 0,11 0,35 0,46 0,1 0,17 0,73 0,1 1,1
In Tabelle 22 sind die Werte für den Y-Achsenabschnitt „intercept“ angegeben.
Tabelle 21: Primer- und cyclerabhängige Aufstellung über die Y-Achsenabschnitte „in-
tercept“ der Standardkurve mit weiteren statistischen Meßgrößen:
Cycler TM7 TM7 TM7 TM7 LC7 LC7 LC7 LC7
Primer TßR-I TßR-II TßR-III PDH TßR-I TßR-II TßR-III PDH
Mittelwert 7,77 6,92 8,29 16,58 4,54 3,63 5,78 3,74
SD 1,99 3,02 2,58 1,85 1,02 3,11 0,53 0,99
Std.fehler 0,75 1,14 1,15 0,83 0,34 0,69 0,21 0,33
Range 5,58 7,23 5,93 4,55 3,72 16,46 1,3 2,94
Varianz 3,94 9,12 6,65 3,43 1,05 10,39 0,28 0,98
Der Vergleich dieser Tabellen zeigt, daß die statistischen Meßgrößen für die Steigungs-
bestimmung einheitlicher sind als für die Y-Achsenabschnittsbestimmung. Die Steigung
der Standardkurve ist die wichtigere Größe zur Charakterisierung und Quantifizierung
einer PCR.
Seite 83

Kapitel 4. Ergebnisse
4.3 Datenanalyse der Knochenstudie
4.3.1 Allgemeines Verteilungsmuster
Die Detektion der TGF-ß-Rezeptor mRNA Expression war in 48/55, bzw. 37/55 Knochen-
proben, die nach initialer grober Reinigung und vor der RNA-Extraktion ein Gewicht über
50 mg besaßen, möglich. Alle Knochenbiopsieproben wurden auf dem LightCycler7
analysiert. Die gemessene TßR-I mRNA Expression ist deutlich höher als die TßR-II und
-III mRNA Expression. Das Verhältnis TßR-I/TßR-II/TßR-III mRNA Expression beträgt je
nach Patient und Entnahmeort 200:1:2, hinweisend auf die differentielle Expression der
einzelnen TßR-Isoformen.
Allgemein sind die Angaben der absoluten TßR-Expression in Kopien pro ng. Die
Angaben der relativen Expression drücken das Verhältnis der TßR zu PDH als ein Vielfa-
ches der PDH-Expression aus und sind dimensionslos.
Tabelle 22: TGF-ß Typ I bis III Rezeptoren, Spannweite, Mittelwerte und Median:
Rezeptor Anzahl n Spannweite Mittelwert +/- SD MedianTßR-I [Kopien/ng] 48 0,216 - 4339,87 884,56 +/- 931,63 572,81
TßR-II [Kopien/ng] 37 0,005 - 21,3 4,22 +/- 4,38 2,39
TßR-III [Kopien/ng] 48 0,017 - 42,84 8,31 +/- 9,31 5,49
TßR-I/PDH 47 0,01 - 1445,16 221,18 +/- 286,12 134,26
TßR-II/PDH 36 0,002 – 4,286 0,817 +/- 1,00 0,33
TßR-III/PDH 47 0,002 – 28,906 2,74 +/- 4,77 1,06
Seite 84
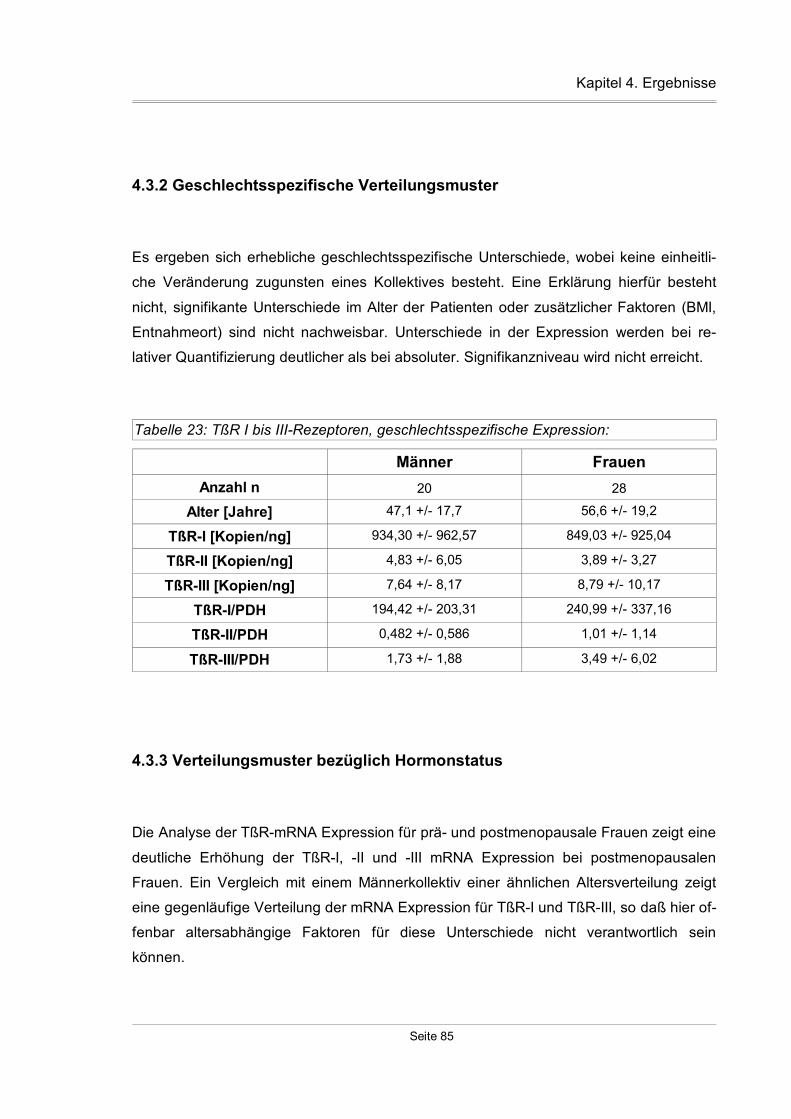
Kapitel 4. Ergebnisse
4.3.2 Geschlechtsspezifische Verteilungsmuster
Es ergeben sich erhebliche geschlechtsspezifische Unterschiede, wobei keine einheitli-
che Veränderung zugunsten eines Kollektives besteht. Eine Erklärung hierfür besteht
nicht, signifikante Unterschiede im Alter der Patienten oder zusätzlicher Faktoren (BMI,
Entnahmeort) sind nicht nachweisbar. Unterschiede in der Expression werden bei re-
lativer Quantifizierung deutlicher als bei absoluter. Signifikanzniveau wird nicht erreicht.
Tabelle 23: TßR I bis III-Rezeptoren, geschlechtsspezifische Expression:
Männer FrauenAnzahl n 20 28
Alter [Jahre] 47,1 +/- 17,7 56,6 +/- 19,2
TßR-I [Kopien/ng] 934,30 +/- 962,57 849,03 +/- 925,04
TßR-II [Kopien/ng] 4,83 +/- 6,05 3,89 +/- 3,27
TßR-III [Kopien/ng] 7,64 +/- 8,17 8,79 +/- 10,17
TßR-I/PDH 194,42 +/- 203,31 240,99 +/- 337,16
TßR-II/PDH 0,482 +/- 0,586 1,01 +/- 1,14
TßR-III/PDH 1,73 +/- 1,88 3,49 +/- 6,02
4.3.3 Verteilungsmuster bezüglich Hormonstatus
Die Analyse der TßR-mRNA Expression für prä- und postmenopausale Frauen zeigt eine
deutliche Erhöhung der TßR-I, -II und -III mRNA Expression bei postmenopausalen
Frauen. Ein Vergleich mit einem Männerkollektiv einer ähnlichen Altersverteilung zeigt
eine gegenläufige Verteilung der mRNA Expression für TßR-I und TßR-III, so daß hier of-
fenbar altersabhängige Faktoren für diese Unterschiede nicht verantwortlich sein
können.
Seite 85
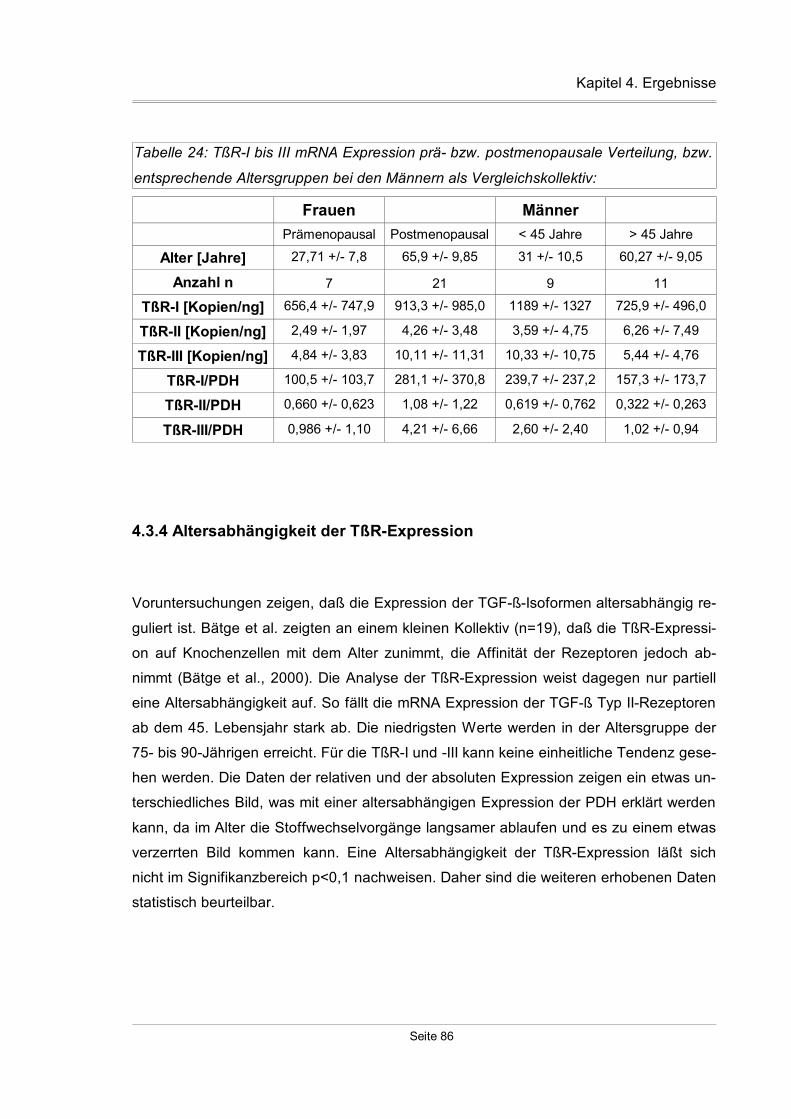
Kapitel 4. Ergebnisse
Tabelle 24: TßR-I bis III mRNA Expression prä- bzw. postmenopausale Verteilung, bzw.
entsprechende Altersgruppen bei den Männern als Vergleichskollektiv:
Frauen MännerPrämenopausal Postmenopausal < 45 Jahre > 45 Jahre
Alter [Jahre] 27,71 +/- 7,8 65,9 +/- 9,85 31 +/- 10,5 60,27 +/- 9,05
Anzahl n 7 21 9 11
TßR-I [Kopien/ng] 656,4 +/- 747,9 913,3 +/- 985,0 1189 +/- 1327 725,9 +/- 496,0
TßR-II [Kopien/ng] 2,49 +/- 1,97 4,26 +/- 3,48 3,59 +/- 4,75 6,26 +/- 7,49
TßR-III [Kopien/ng] 4,84 +/- 3,83 10,11 +/- 11,31 10,33 +/- 10,75 5,44 +/- 4,76
TßR-I/PDH 100,5 +/- 103,7 281,1 +/- 370,8 239,7 +/- 237,2 157,3 +/- 173,7
TßR-II/PDH 0,660 +/- 0,623 1,08 +/- 1,22 0,619 +/- 0,762 0,322 +/- 0,263
TßR-III/PDH 0,986 +/- 1,10 4,21 +/- 6,66 2,60 +/- 2,40 1,02 +/- 0,94
4.3.4 Altersabhängigkeit der TßR-Expression
Voruntersuchungen zeigen, daß die Expression der TGF-ß-Isoformen altersabhängig re-
guliert ist. Bätge et al. zeigten an einem kleinen Kollektiv (n=19), daß die TßR-Expressi-
on auf Knochenzellen mit dem Alter zunimmt, die Affinität der Rezeptoren jedoch ab-
nimmt (Bätge et al., 2000). Die Analyse der TßR-Expression weist dagegen nur partiell
eine Altersabhängigkeit auf. So fällt die mRNA Expression der TGF-ß Typ II-Rezeptoren
ab dem 45. Lebensjahr stark ab. Die niedrigsten Werte werden in der Altersgruppe der
75- bis 90-Jährigen erreicht. Für die TßR-I und -III kann keine einheitliche Tendenz gese-
hen werden. Die Daten der relativen und der absoluten Expression zeigen ein etwas un-
terschiedliches Bild, was mit einer altersabhängigen Expression der PDH erklärt werden
kann, da im Alter die Stoffwechselvorgänge langsamer ablaufen und es zu einem etwas
verzerrten Bild kommen kann. Eine Altersabhängigkeit der TßR-Expression läßt sich
nicht im Signifikanzbereich p<0,1 nachweisen. Daher sind die weiteren erhobenen Daten
statistisch beurteilbar.
Seite 86
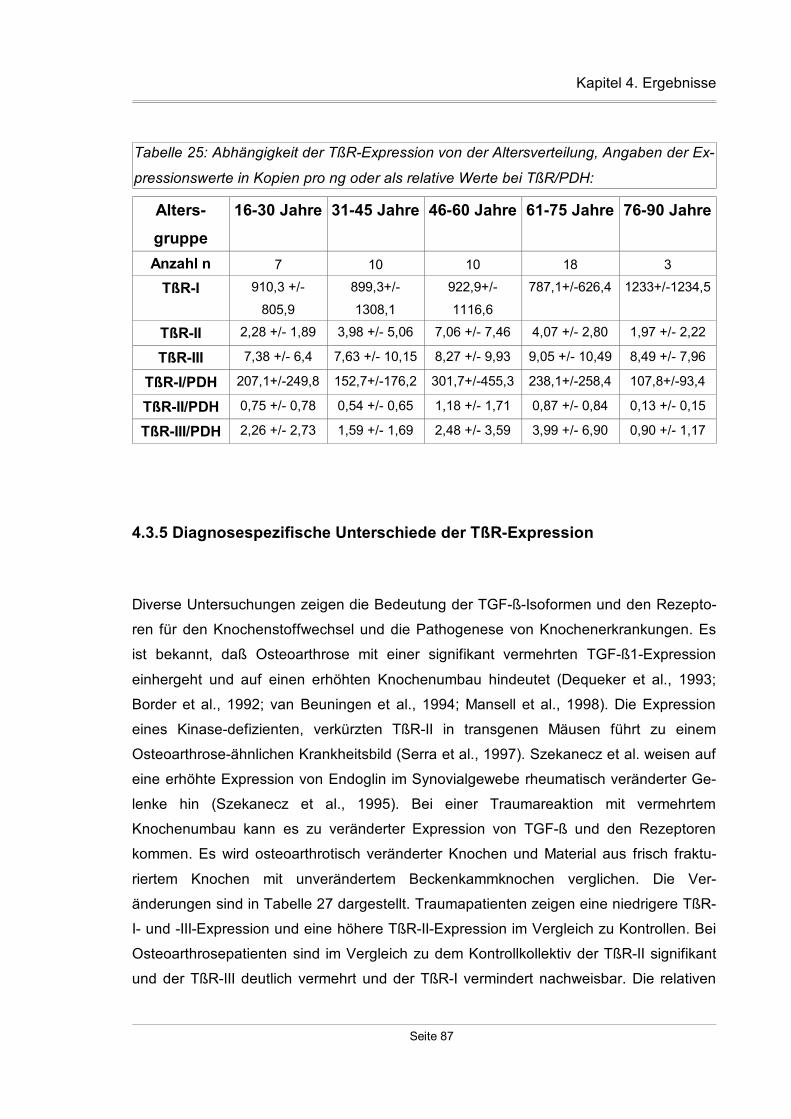
Kapitel 4. Ergebnisse
Tabelle 25: Abhängigkeit der TßR-Expression von der Altersverteilung, Angaben der Ex-
pressionswerte in Kopien pro ng oder als relative Werte bei TßR/PDH:
Alters-
gruppe
16-30 Jahre 31-45 Jahre 46-60 Jahre 61-75 Jahre 76-90 Jahre
Anzahl n 7 10 10 18 3
TßR-I 910,3 +/-
805,9
899,3+/-
1308,1
922,9+/-
1116,6
787,1+/-626,4 1233+/-1234,5
TßR-II 2,28 +/- 1,89 3,98 +/- 5,06 7,06 +/- 7,46 4,07 +/- 2,80 1,97 +/- 2,22
TßR-III 7,38 +/- 6,4 7,63 +/- 10,15 8,27 +/- 9,93 9,05 +/- 10,49 8,49 +/- 7,96
TßR-I/PDH 207,1+/-249,8 152,7+/-176,2 301,7+/-455,3 238,1+/-258,4 107,8+/-93,4
TßR-II/PDH 0,75 +/- 0,78 0,54 +/- 0,65 1,18 +/- 1,71 0,87 +/- 0,84 0,13 +/- 0,15
TßR-III/PDH 2,26 +/- 2,73 1,59 +/- 1,69 2,48 +/- 3,59 3,99 +/- 6,90 0,90 +/- 1,17
4.3.5 Diagnosespezifische Unterschiede der TßR-Expression
Diverse Untersuchungen zeigen die Bedeutung der TGF-ß-Isoformen und den Rezepto-
ren für den Knochenstoffwechsel und die Pathogenese von Knochenerkrankungen. Es
ist bekannt, daß Osteoarthrose mit einer signifikant vermehrten TGF-ß1-Expression
einhergeht und auf einen erhöhten Knochenumbau hindeutet (Dequeker et al., 1993;
Border et al., 1992; van Beuningen et al., 1994; Mansell et al., 1998). Die Expression
eines Kinase-defizienten, verkürzten TßR-II in transgenen Mäusen führt zu einem
Osteoarthrose-ähnlichen Krankheitsbild (Serra et al., 1997). Szekanecz et al. weisen auf
eine erhöhte Expression von Endoglin im Synovialgewebe rheumatisch veränderter Ge-
lenke hin (Szekanecz et al., 1995). Bei einer Traumareaktion mit vermehrtem
Knochenumbau kann es zu veränderter Expression von TGF-ß und den Rezeptoren
kommen. Es wird osteoarthrotisch veränderter Knochen und Material aus frisch fraktu-
riertem Knochen mit unverändertem Beckenkammknochen verglichen. Die Ver-
änderungen sind in Tabelle 27 dargestellt. Traumapatienten zeigen eine niedrigere TßR-
I- und -III-Expression und eine höhere TßR-II-Expression im Vergleich zu Kontrollen. Bei
Osteoarthrosepatienten sind im Vergleich zu dem Kontrollkollektiv der TßR-II signifikant
und der TßR-III deutlich vermehrt und der TßR-I vermindert nachweisbar. Die relativen
Seite 87
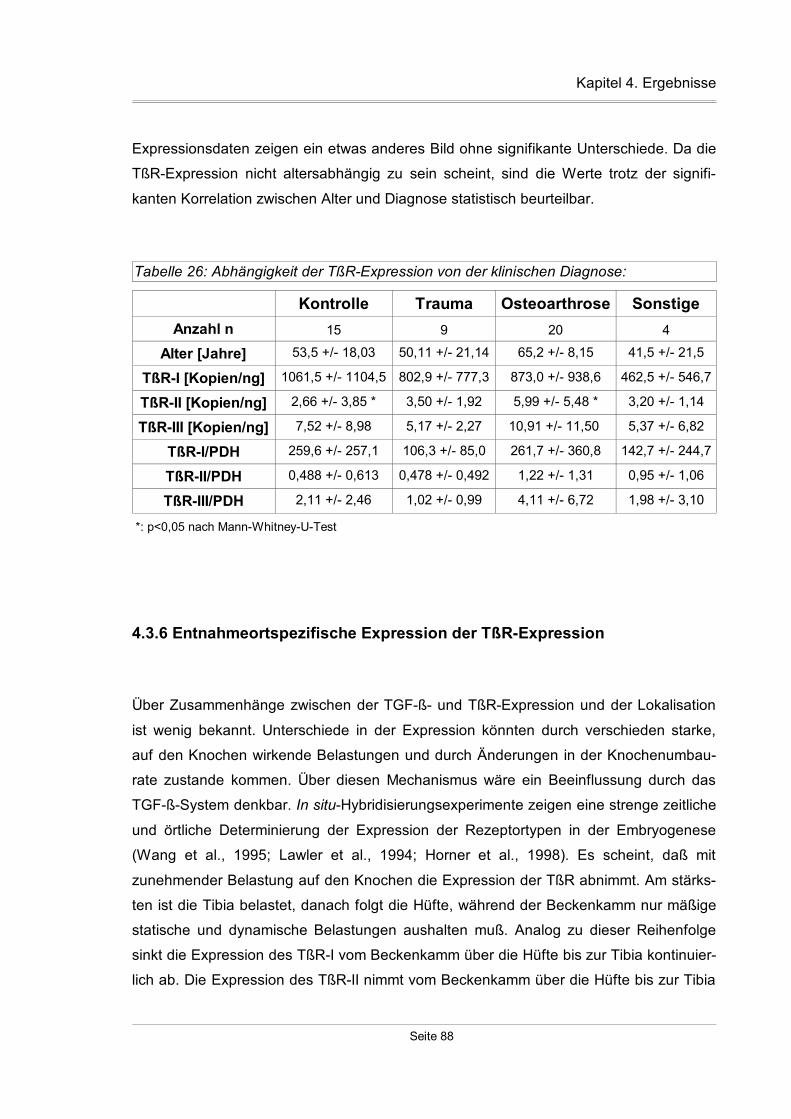
Kapitel 4. Ergebnisse
Expressionsdaten zeigen ein etwas anderes Bild ohne signifikante Unterschiede. Da die
TßR-Expression nicht altersabhängig zu sein scheint, sind die Werte trotz der signifi-
kanten Korrelation zwischen Alter und Diagnose statistisch beurteilbar.
Tabelle 26: Abhängigkeit der TßR-Expression von der klinischen Diagnose:
Kontrolle Trauma Osteoarthrose SonstigeAnzahl n 15 9 20 4
Alter [Jahre] 53,5 +/- 18,03 50,11 +/- 21,14 65,2 +/- 8,15 41,5 +/- 21,5
TßR-I [Kopien/ng] 1061,5 +/- 1104,5 802,9 +/- 777,3 873,0 +/- 938,6 462,5 +/- 546,7
TßR-II [Kopien/ng] 2,66 +/- 3,85 * 3,50 +/- 1,92 5,99 +/- 5,48 * 3,20 +/- 1,14
TßR-III [Kopien/ng] 7,52 +/- 8,98 5,17 +/- 2,27 10,91 +/- 11,50 5,37 +/- 6,82
TßR-I/PDH 259,6 +/- 257,1 106,3 +/- 85,0 261,7 +/- 360,8 142,7 +/- 244,7
TßR-II/PDH 0,488 +/- 0,613 0,478 +/- 0,492 1,22 +/- 1,31 0,95 +/- 1,06
TßR-III/PDH 2,11 +/- 2,46 1,02 +/- 0,99 4,11 +/- 6,72 1,98 +/- 3,10
*: p<0,05 nach Mann-Whitney-U-Test
4.3.6 Entnahmeortspezifische Expression der TßR-Expression
Über Zusammenhänge zwischen der TGF-ß- und TßR-Expression und der Lokalisation
ist wenig bekannt. Unterschiede in der Expression könnten durch verschieden starke,
auf den Knochen wirkende Belastungen und durch Änderungen in der Knochenumbau-
rate zustande kommen. Über diesen Mechanismus wäre ein Beeinflussung durch das
TGF-ß-System denkbar. In situ-Hybridisierungsexperimente zeigen eine strenge zeitliche
und örtliche Determinierung der Expression der Rezeptortypen in der Embryogenese
(Wang et al., 1995; Lawler et al., 1994; Horner et al., 1998). Es scheint, daß mit
zunehmender Belastung auf den Knochen die Expression der TßR abnimmt. Am stärks-
ten ist die Tibia belastet, danach folgt die Hüfte, während der Beckenkamm nur mäßige
statische und dynamische Belastungen aushalten muß. Analog zu dieser Reihenfolge
sinkt die Expression des TßR-I vom Beckenkamm über die Hüfte bis zur Tibia kontinuier-
lich ab. Die Expression des TßR-II nimmt vom Beckenkamm über die Hüfte bis zur Tibia
Seite 88
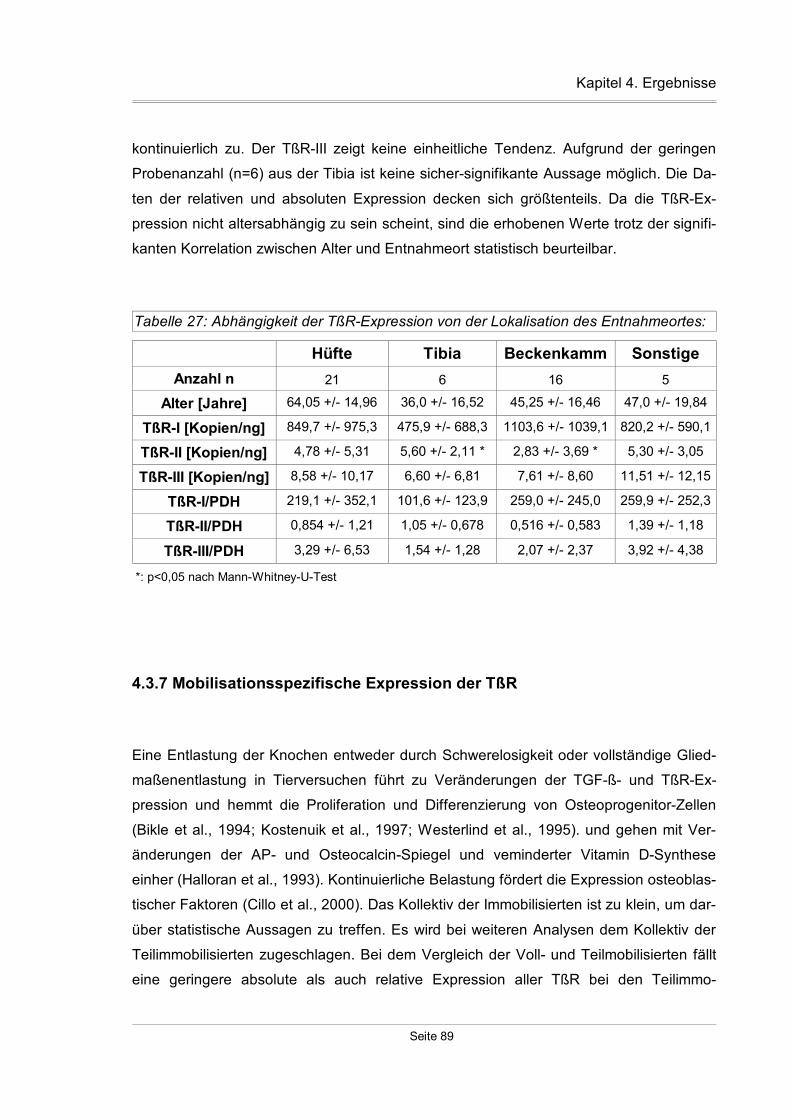
Kapitel 4. Ergebnisse
kontinuierlich zu. Der TßR-III zeigt keine einheitliche Tendenz. Aufgrund der geringen
Probenanzahl (n=6) aus der Tibia ist keine sicher-signifikante Aussage möglich. Die Da-
ten der relativen und absoluten Expression decken sich größtenteils. Da die TßR-Ex-
pression nicht altersabhängig zu sein scheint, sind die erhobenen Werte trotz der signifi-
kanten Korrelation zwischen Alter und Entnahmeort statistisch beurteilbar.
Tabelle 27: Abhängigkeit der TßR-Expression von der Lokalisation des Entnahmeortes:
Hüfte Tibia Beckenkamm SonstigeAnzahl n 21 6 16 5
Alter [Jahre] 64,05 +/- 14,96 36,0 +/- 16,52 45,25 +/- 16,46 47,0 +/- 19,84
TßR-I [Kopien/ng] 849,7 +/- 975,3 475,9 +/- 688,3 1103,6 +/- 1039,1 820,2 +/- 590,1
TßR-II [Kopien/ng] 4,78 +/- 5,31 5,60 +/- 2,11 * 2,83 +/- 3,69 * 5,30 +/- 3,05
TßR-III [Kopien/ng] 8,58 +/- 10,17 6,60 +/- 6,81 7,61 +/- 8,60 11,51 +/- 12,15
TßR-I/PDH 219,1 +/- 352,1 101,6 +/- 123,9 259,0 +/- 245,0 259,9 +/- 252,3
TßR-II/PDH 0,854 +/- 1,21 1,05 +/- 0,678 0,516 +/- 0,583 1,39 +/- 1,18
TßR-III/PDH 3,29 +/- 6,53 1,54 +/- 1,28 2,07 +/- 2,37 3,92 +/- 4,38
*: p<0,05 nach Mann-Whitney-U-Test
4.3.7 Mobilisationsspezifische Expression der TßR
Eine Entlastung der Knochen entweder durch Schwerelosigkeit oder vollständige Glied-
maßenentlastung in Tierversuchen führt zu Veränderungen der TGF-ß- und TßR-Ex-
pression und hemmt die Proliferation und Differenzierung von Osteoprogenitor-Zellen
(Bikle et al., 1994; Kostenuik et al., 1997; Westerlind et al., 1995). und gehen mit Ver-
änderungen der AP- und Osteocalcin-Spiegel und veminderter Vitamin D-Synthese
einher (Halloran et al., 1993). Kontinuierliche Belastung fördert die Expression osteoblas-
tischer Faktoren (Cillo et al., 2000). Das Kollektiv der Immobilisierten ist zu klein, um dar-
über statistische Aussagen zu treffen. Es wird bei weiteren Analysen dem Kollektiv der
Teilimmobilisierten zugeschlagen. Bei dem Vergleich der Voll- und Teilmobilisierten fällt
eine geringere absolute als auch relative Expression aller TßR bei den Teilimmo-
Seite 89
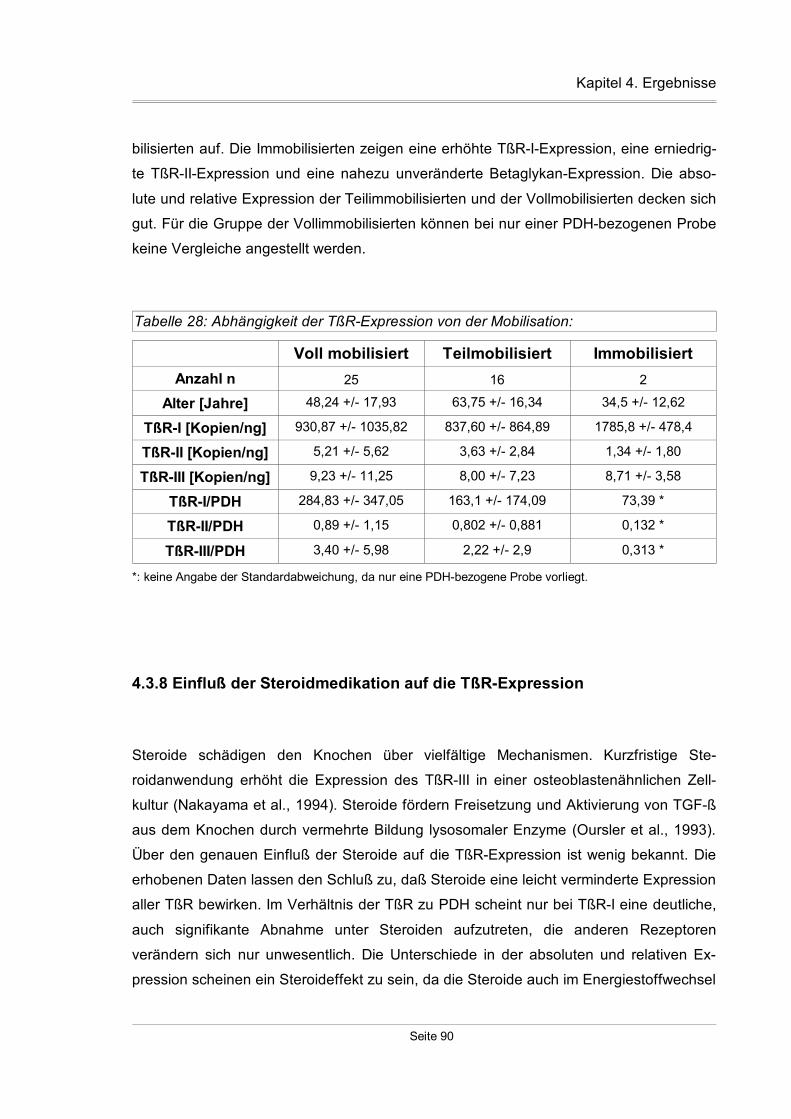
Kapitel 4. Ergebnisse
bilisierten auf. Die Immobilisierten zeigen eine erhöhte TßR-I-Expression, eine erniedrig-
te TßR-II-Expression und eine nahezu unveränderte Betaglykan-Expression. Die abso-
lute und relative Expression der Teilimmobilisierten und der Vollmobilisierten decken sich
gut. Für die Gruppe der Vollimmobilisierten können bei nur einer PDH-bezogenen Probe
keine Vergleiche angestellt werden.
Tabelle 28: Abhängigkeit der TßR-Expression von der Mobilisation:
Voll mobilisiert Teilmobilisiert ImmobilisiertAnzahl n 25 16 2
Alter [Jahre] 48,24 +/- 17,93 63,75 +/- 16,34 34,5 +/- 12,62
TßR-I [Kopien/ng] 930,87 +/- 1035,82 837,60 +/- 864,89 1785,8 +/- 478,4
TßR-II [Kopien/ng] 5,21 +/- 5,62 3,63 +/- 2,84 1,34 +/- 1,80
TßR-III [Kopien/ng] 9,23 +/- 11,25 8,00 +/- 7,23 8,71 +/- 3,58
TßR-I/PDH 284,83 +/- 347,05 163,1 +/- 174,09 73,39 *
TßR-II/PDH 0,89 +/- 1,15 0,802 +/- 0,881 0,132 *
TßR-III/PDH 3,40 +/- 5,98 2,22 +/- 2,9 0,313 *
*: keine Angabe der Standardabweichung, da nur eine PDH-bezogene Probe vorliegt.
4.3.8 Einfluß der Steroidmedikation auf die TßR-Expression
Steroide schädigen den Knochen über vielfältige Mechanismen. Kurzfristige Ste-
roidanwendung erhöht die Expression des TßR-III in einer osteoblastenähnlichen Zell-
kultur (Nakayama et al., 1994). Steroide fördern Freisetzung und Aktivierung von TGF-ß
aus dem Knochen durch vermehrte Bildung lysosomaler Enzyme (Oursler et al., 1993).
Über den genauen Einfluß der Steroide auf die TßR-Expression ist wenig bekannt. Die
erhobenen Daten lassen den Schluß zu, daß Steroide eine leicht verminderte Expression
aller TßR bewirken. Im Verhältnis der TßR zu PDH scheint nur bei TßR-I eine deutliche,
auch signifikante Abnahme unter Steroiden aufzutreten, die anderen Rezeptoren
verändern sich nur unwesentlich. Die Unterschiede in der absoluten und relativen Ex-
pression scheinen ein Steroideffekt zu sein, da die Steroide auch im Energiestoffwechsel
Seite 90

Kapitel 4. Ergebnisse
eine Rolle spielen. Steroide bewirken eine vermehrte Expression der PDH (Lehninger et
al., 2001). Somit stellt PDH in diesem Fall kein ideales „housekeeping gene“ dar, da es
selber Einflüssen unterworfen ist. Daher sind die relativen Expressionsdaten vorsichtig
zu interpretieren.
Tabelle 29: Abhängigkeit der TßR-Expression von der Steroidmedikation:
Steroidmedikation Keine SteroidmedikationAnzahl n 7 37
Alter [Jahre] 59,71 +/- 13,5 52,32 +/- 19,42
TßR-I [Kopien/ng] 871,32 +/- 913,22 932,69 +/- 973,16
TßR-II [Kopien/ng] 3,48 +/- 2,53 4,48 +/- 4,89
TßR-III [Kopien/ng] 7,65 +/- 10,93 8,76 +/- 9,40
TßR-I/PDH 169,48 +/- 186,09 239,95 +/- 308,14
TßR-II/PDH 0,89 +/- 1,05 0,788 +/- 1,02
TßR-III/PDH 2,99 +/- 4,53 2,78 +/- 5,04
4.3.9 Expression der TßR in Abhängigkeit vom „Body Mass Index“
Klinisch ist bekannt, daß Übergewicht die Entstehung einer Osteoarthrose fördert. Über
eine TGF-ß-Beteiligung in diesem Prozeß ist nichts Genaues bekannt. In der Literatur
finden sich keine Hinweise, welchen Einfluß das Gewicht und der BMI („Body Mass In-
dex“) auf TGF-ß, die Rezeptoren und die strukturelle Stabilität des Knochens hat. Ein hö-
herer BMI geht mit einem vermehrten Kalksalzgehalt des Knochens einher (Pettersson et
al., 2000). Bei adipösen Patienten mit einem BMI > 25 kg/m2 fällt eine deutliche Redukti-
on der TßR I- und III-Expression im Vergleich zu Normalgewichtigen auf. Der TßR-II ist
dagegen nur unwesentlich niedriger exprimiert. Die Daten der relativen Expression zu
PDH zeigen bei allen Rezeptoren eine Abnahme. Obwohl zwischen dem Alter der Pati-
enten und der Höhe des BMI eine signifikante Korrelation besteht, sind die Daten statis-
tisch beurteilbar, da die TßR-Expression nicht altersabhängig zu sein scheint.
Seite 91
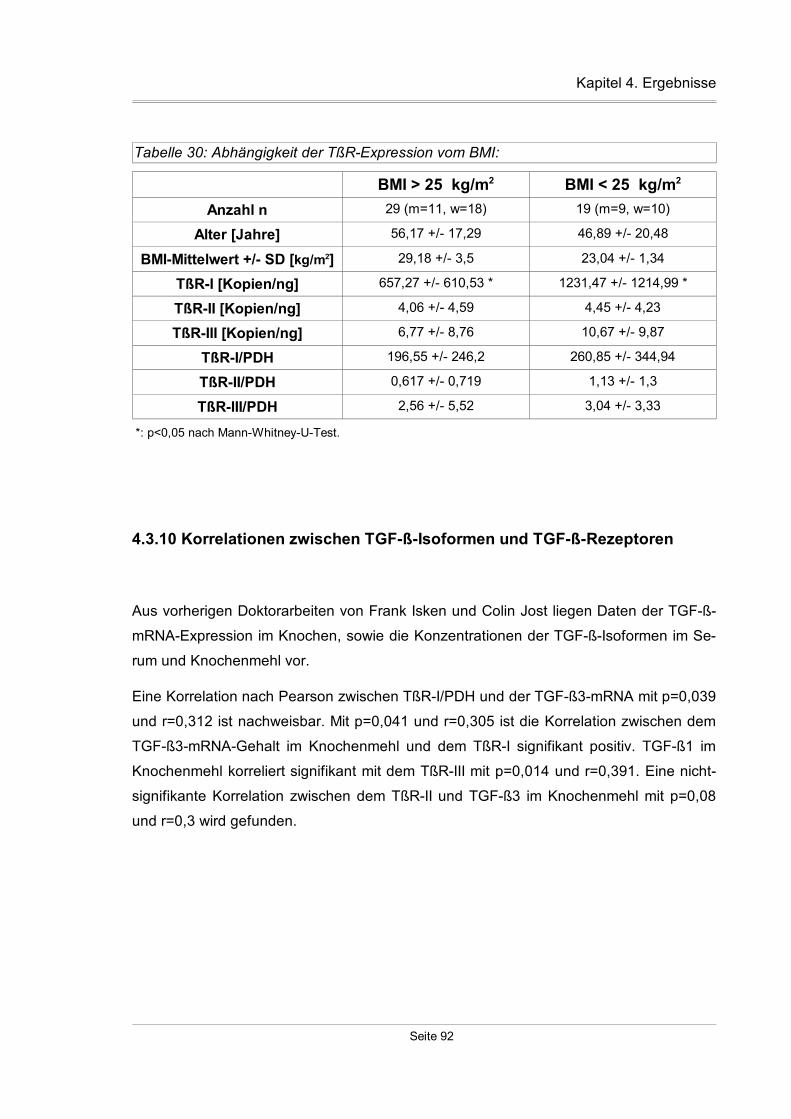
Kapitel 4. Ergebnisse
Tabelle 30: Abhängigkeit der TßR-Expression vom BMI:
BMI > 25 kg/m2 BMI < 25 kg/m2
Anzahl n 29 (m=11, w=18) 19 (m=9, w=10)
Alter [Jahre] 56,17 +/- 17,29 46,89 +/- 20,48
BMI-Mittelwert +/- SD [kg/m2] 29,18 +/- 3,5 23,04 +/- 1,34
TßR-I [Kopien/ng] 657,27 +/- 610,53 * 1231,47 +/- 1214,99 *
TßR-II [Kopien/ng] 4,06 +/- 4,59 4,45 +/- 4,23
TßR-III [Kopien/ng] 6,77 +/- 8,76 10,67 +/- 9,87
TßR-I/PDH 196,55 +/- 246,2 260,85 +/- 344,94
TßR-II/PDH 0,617 +/- 0,719 1,13 +/- 1,3
TßR-III/PDH 2,56 +/- 5,52 3,04 +/- 3,33
*: p<0,05 nach Mann-Whitney-U-Test.
4.3.10 Korrelationen zwischen TGF-ß-Isoformen und TGF-ß-Rezeptoren
Aus vorherigen Doktorarbeiten von Frank Isken und Colin Jost liegen Daten der TGF-ß-
mRNA-Expression im Knochen, sowie die Konzentrationen der TGF-ß-Isoformen im Se-
rum und Knochenmehl vor.
Eine Korrelation nach Pearson zwischen TßR-I/PDH und der TGF-ß3-mRNA mit p=0,039
und r=0,312 ist nachweisbar. Mit p=0,041 und r=0,305 ist die Korrelation zwischen dem
TGF-ß3-mRNA-Gehalt im Knochenmehl und dem TßR-I signifikant positiv. TGF-ß1 im
Knochenmehl korreliert signifikant mit dem TßR-III mit p=0,014 und r=0,391. Eine nicht-
signifikante Korrelation zwischen dem TßR-II und TGF-ß3 im Knochenmehl mit p=0,08
und r=0,3 wird gefunden.
Seite 92
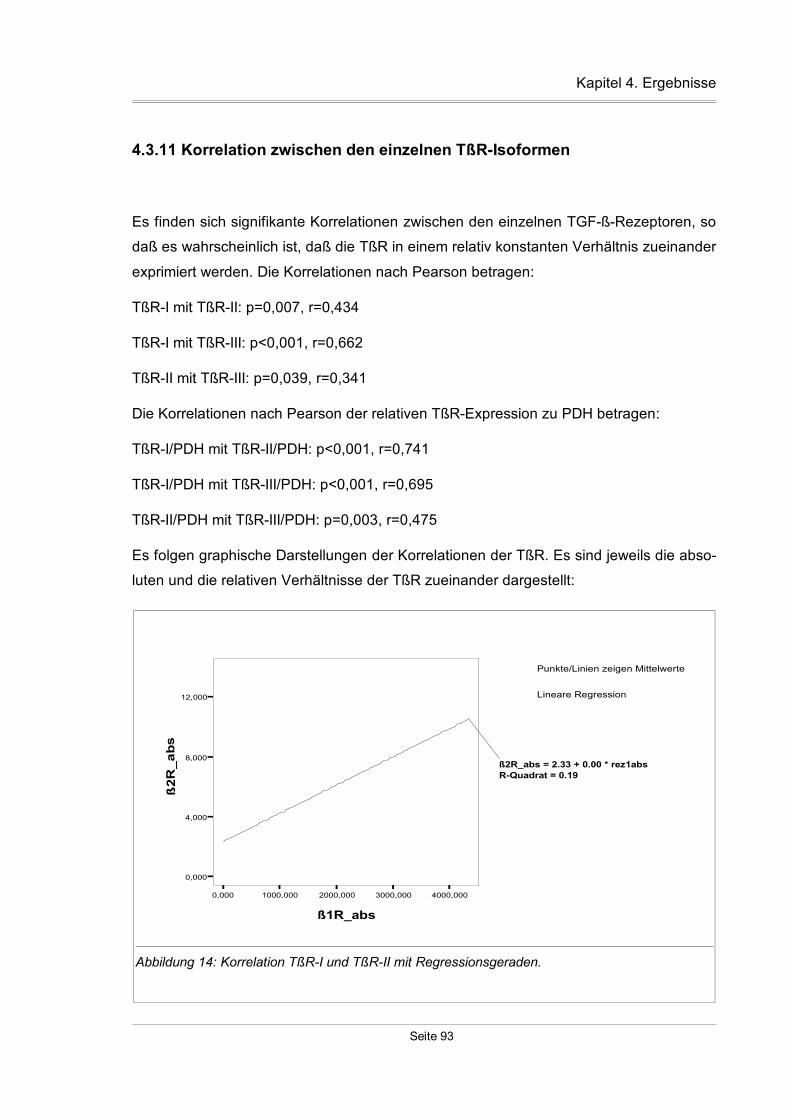
Kapitel 4. Ergebnisse
4.3.11 Korrelation zwischen den einzelnen TßR-Isoformen
Es finden sich signifikante Korrelationen zwischen den einzelnen TGF-ß-Rezeptoren, so
daß es wahrscheinlich ist, daß die TßR in einem relativ konstanten Verhältnis zueinander
exprimiert werden. Die Korrelationen nach Pearson betragen:
TßR-I mit TßR-II: p=0,007, r=0,434
TßR-I mit TßR-III: p<0,001, r=0,662
TßR-II mit TßR-III: p=0,039, r=0,341
Die Korrelationen nach Pearson der relativen TßR-Expression zu PDH betragen:
TßR-I/PDH mit TßR-II/PDH: p<0,001, r=0,741
TßR-I/PDH mit TßR-III/PDH: p<0,001, r=0,695
TßR-II/PDH mit TßR-III/PDH: p=0,003, r=0,475
Es folgen graphische Darstellungen der Korrelationen der TßR. Es sind jeweils die abso-
luten und die relativen Verhältnisse der TßR zueinander dargestellt:
Seite 93
Abbildung 14: Korrelation TßR-I und TßR-II mit Regressionsgeraden.
Punkte/Linien zeigen Mittelwerte
0,000 1000,000 2000,000 3000,000 4000,000
ß1R_abs
0,000
4,000
8,000
12,000
ß2R
_ab
s
�
��
�
���
�
��
�
�
�
�
�
�
�
�
�
�
�
��
��
�
�
�
�
�
�
�
�
�
�
�
ß2R_abs = 2.33 + 0.00 * rez1absR-Quadrat = 0.19
Lineare Regression
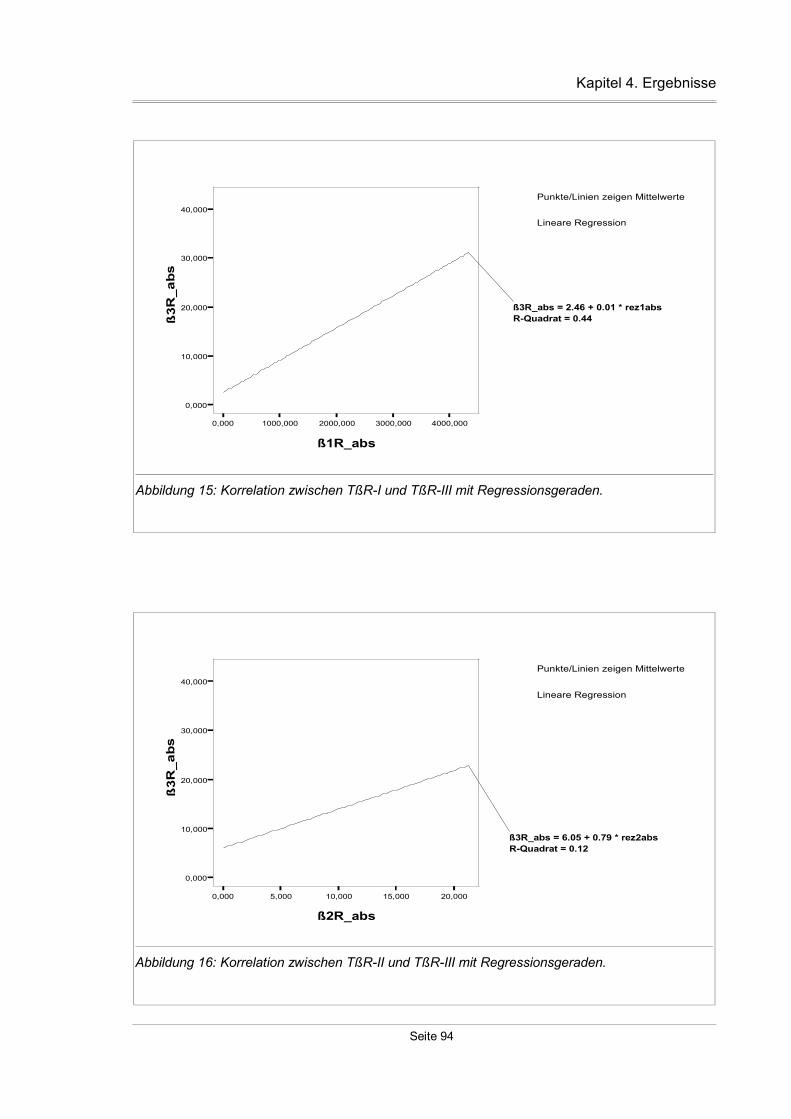
Kapitel 4. Ergebnisse
Seite 94
Abbildung 15: Korrelation zwischen TßR-I und TßR-III mit Regressionsgeraden.
Punkte/Linien zeigen Mittelwerte
0,000 1000,000 2000,000 3000,000 4000,000
ß1R_abs
0,000
10,000
20,000
30,000
40,000
ß3R
_ab
s
�
�
�
��������
��
�
��
���
�
�
���
�
�
�� �
�
�
�
�
�
��
�
��
�
�
�
�
�
�
�
�
ß3R_abs = 2.46 + 0.01 * rez1absR-Quadrat = 0.44
Lineare Regression
Abbildung 16: Korrelation zwischen TßR-II und TßR-III mit Regressionsgeraden.
Punkte/Linien zeigen Mittelwerte
0,000 5,000 10,000 15,000 20,000
ß2R_abs
0,000
10,000
20,000
30,000
40,000
ß3R
_ab
s
��
�
�
�
�
�
�
�
��
�
��
�
�
��
��
�
�
�
� � �
��
�
�
�
�
�
�
�
� ß3R_abs = 6.05 + 0.79 * rez2absR-Quadrat = 0.12
Lineare Regression
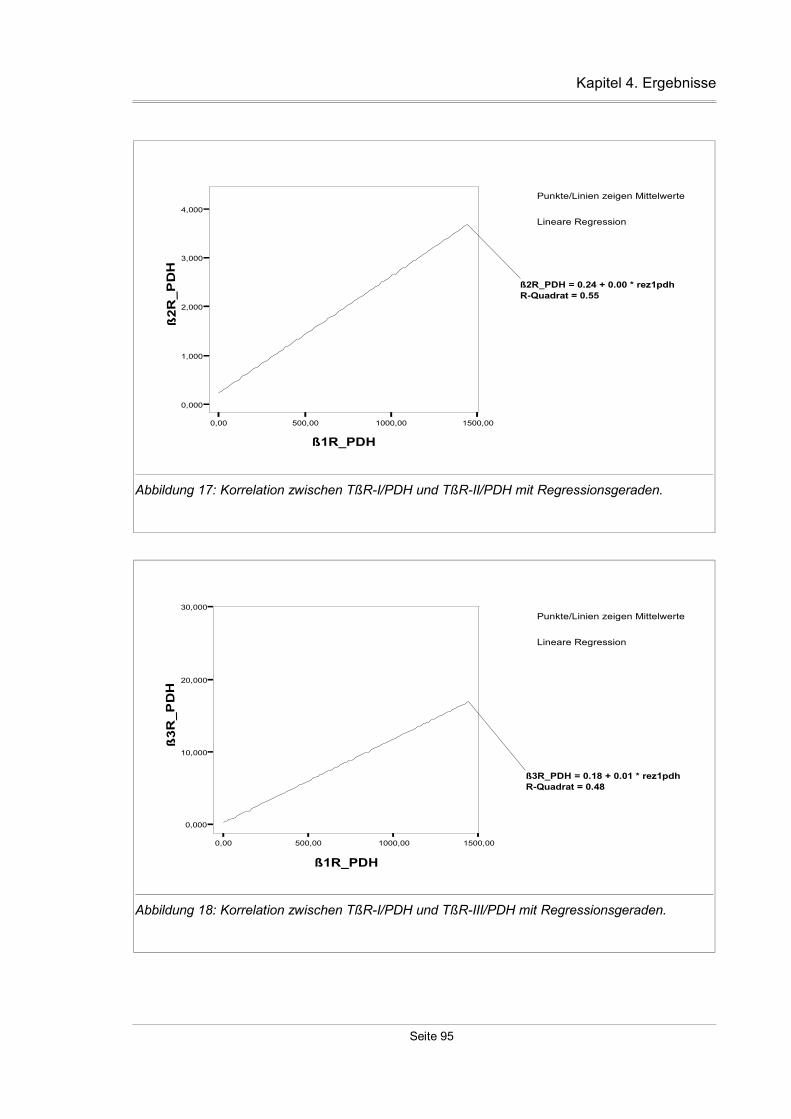
Kapitel 4. Ergebnisse
Seite 95
Abbildung 17: Korrelation zwischen TßR-I/PDH und TßR-II/PDH mit Regressionsgeraden.
Punkte/Linien zeigen Mittelwerte
0,00 500,00 1000,00 1500,00
ß1R_PDH
0,000
1,000
2,000
3,000
4,000
ß2R
_PD
H
�����
��������
�
�� �
�
�
�
���
�
�
�
�
�
�
�
�
�
�
�
�
�
ß2R_PDH = 0.24 + 0.00 * rez1pdhR-Quadrat = 0.55
Lineare Regression
Abbildung 18: Korrelation zwischen TßR-I/PDH und TßR-III/PDH mit Regressionsgeraden.
Punkte/Linien zeigen Mittelwerte
0,00 500,00 1000,00 1500,00
ß1R_PDH
0,000
10,000
20,000
30,000
ß3R
_PD
H
�������������������� �
��
���
�
�
����
�
��
�
� �
�
�
�
��
�
�
�
�
ß3R_PDH = 0.18 + 0.01 * rez1pdhR-Quadrat = 0.48
Lineare Regression
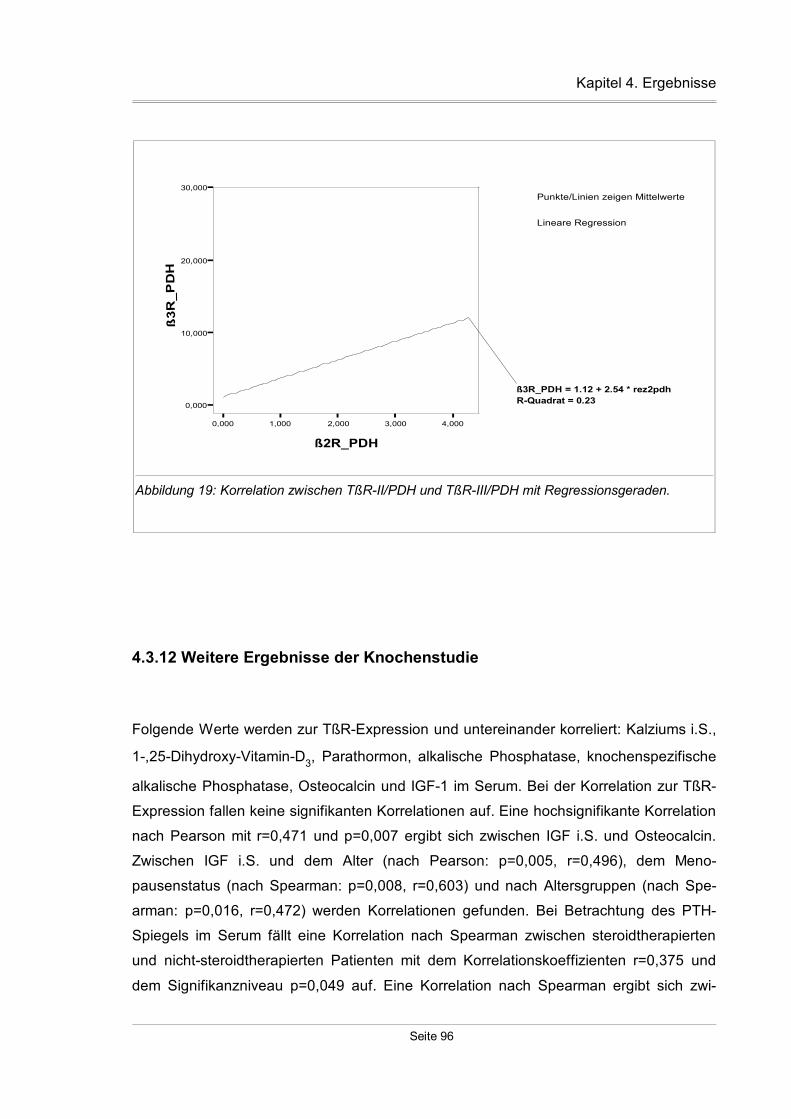
Kapitel 4. Ergebnisse
4.3.12 Weitere Ergebnisse der Knochenstudie
Folgende Werte werden zur TßR-Expression und untereinander korreliert: Kalziums i.S.,
1-,25-Dihydroxy-Vitamin-D3, Parathormon, alkalische Phosphatase, knochenspezifische
alkalische Phosphatase, Osteocalcin und IGF-1 im Serum. Bei der Korrelation zur TßR-
Expression fallen keine signifikanten Korrelationen auf. Eine hochsignifikante Korrelation
nach Pearson mit r=0,471 und p=0,007 ergibt sich zwischen IGF i.S. und Osteocalcin.
Zwischen IGF i.S. und dem Alter (nach Pearson: p=0,005, r=0,496), dem Meno-
pausenstatus (nach Spearman: p=0,008, r=0,603) und nach Altersgruppen (nach Spe-
arman: p=0,016, r=0,472) werden Korrelationen gefunden. Bei Betrachtung des PTH-
Spiegels im Serum fällt eine Korrelation nach Spearman zwischen steroidtherapierten
und nicht-steroidtherapierten Patienten mit dem Korrelationskoeffizienten r=0,375 und
dem Signifikanzniveau p=0,049 auf. Eine Korrelation nach Spearman ergibt sich zwi-
Seite 96
Abbildung 19: Korrelation zwischen TßR-II/PDH und TßR-III/PDH mit Regressionsgeraden.
Punkte/Linien zeigen Mittelwerte
0,000 1,000 2,000 3,000 4,000
ß2R_PDH
0,000
10,000
20,000
30,000
ß3R
_PD
H
�
�����������������
�
�
�
� ��
�
�
��
�
�
�
�
�
�
� �
ß3R_PDH = 1.12 + 2.54 * rez2pdhR-Quadrat = 0.23
Lineare Regression

Kapitel 4. Ergebnisse
schen Kalzium i. S. und der präoperativen Mobilisation mit r=0,311 und p=0,042. Zwi-
schen dem Hb-Gehalt, eingeteilt in Normo- und Hypohämoglobinämie und Ca i.S. wird
eine Korrelation nach Spearman mit den Werten r=0,349 und p=0,02 nachgewiesen.
Zwischen der Mobilität und der AP i.S. wird eine fast signifikante Korrelation nach Spe-
arman mit r=0,394 und p=0,052 gefunden. Eine Korrelation nach Pearson zeigt sich für
die AP- und BAP-Spiegel im Serum mit den Werten r=0,8 und p<0,001. Zwischen dem
Menopausenstatus und Osteocalcin i.S. findet sich eine Korrelation nach Spearman mit
p=0,034 und r=0,382.
Seite 97
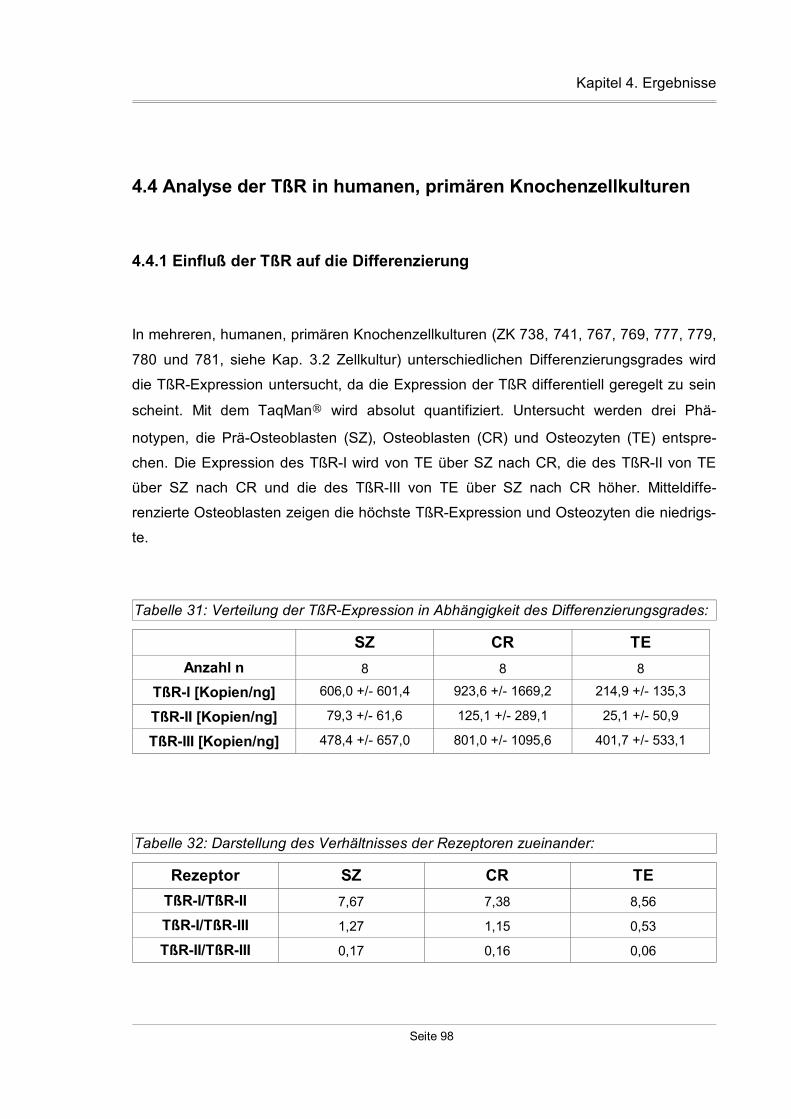
Kapitel 4. Ergebnisse
4.4 Analyse der TßR in humanen, primären Knochenzellkulturen
4.4.1 Einfluß der TßR auf die Differenzierung
In mehreren, humanen, primären Knochenzellkulturen (ZK 738, 741, 767, 769, 777, 779,
780 und 781, siehe Kap. 3.2 Zellkultur) unterschiedlichen Differenzierungsgrades wird
die TßR-Expression untersucht, da die Expression der TßR differentiell geregelt zu sein
scheint. Mit dem TaqMan7 wird absolut quantifiziert. Untersucht werden drei Phä-
notypen, die Prä-Osteoblasten (SZ), Osteoblasten (CR) und Osteozyten (TE) entspre-
chen. Die Expression des TßR-I wird von TE über SZ nach CR, die des TßR-II von TE
über SZ nach CR und die des TßR-III von TE über SZ nach CR höher. Mitteldiffe-
renzierte Osteoblasten zeigen die höchste TßR-Expression und Osteozyten die niedrigs-
te.
Tabelle 31: Verteilung der TßR-Expression in Abhängigkeit des Differenzierungsgrades:
SZ CR TEAnzahl n 8 8 8
TßR-I [Kopien/ng] 606,0 +/- 601,4 923,6 +/- 1669,2 214,9 +/- 135,3
TßR-II [Kopien/ng] 79,3 +/- 61,6 125,1 +/- 289,1 25,1 +/- 50,9
TßR-III [Kopien/ng] 478,4 +/- 657,0 801,0 +/- 1095,6 401,7 +/- 533,1
Tabelle 32: Darstellung des Verhältnisses der Rezeptoren zueinander:
Rezeptor SZ CR TETßR-I/TßR-II 7,67 7,38 8,56
TßR-I/TßR-III 1,27 1,15 0,53
TßR-II/TßR-III 0,17 0,16 0,06
Seite 98

Kapitel 4. Ergebnisse
Alle Korrelationen sind hochsignifikant und der TßR-I korreliert mit dem TßR-II am stärks-
ten. Folgende Korrelationen nach Pearson werden ermittelt:
TßR-I und TßR-II: r=0,945, p<0,01
TßR-I und TßR-III: r=0,606, p<0,01
TßR-II und TßR-III: r=0,587, p<0,01
4.4.2 Einfluß der Steroide auf primäre Knochenzellkulturen
Der Einfluß einer Steroidbehandlung auf die TßR-Expression wird in einer primären Zell-
kultur (ZK 797, siehe Kap. 3.2 Zellkultur) untersucht. Die Zellen weisen die drei Diffe-
renzierungsgrade SZ, CR und TE (Diff.grad: Differenzierungsgrad) auf. Die Steroidbe-
handlung wird mit einer 1 µM Dexamethason-Lösung über verschieden lange Zeiträume
(von 12 bis 96 h) durchgeführt. Diese Versuche werden nachteiligerweise nur mit einer
Zellkultur durchgeführt und somit sind keine sinnvoll berechneten statistischen Angaben
verfügbar. Die Versuche werden auf dem TaqMan7 durchgeführt und absolut quantifi-
ziert. Der Übersichtlichkeit halber sind die Zahlen der Expression der einzelnen Werte re-
lativ zu der jeweiligen Kontrolle und nicht die absoluten Zahlenwerte angegeben. Nach
den vorhandenen Zahlen scheint es eine biphasische Beeinflussung der TßR-Expressi-
on in Abhängigkeit der Behandlungsdauer mit Steroiden zu geben. Meist ist bei der ge-
samten Population ein Abfall der Rezeptorzahlen über die Zeit zu sehen. Eine wesentli-
che Ausnahme stellt der TßR-III in der SZ-Gruppe dar, in der die Expression kontinuier-
lich ansteigt.
Seite 99
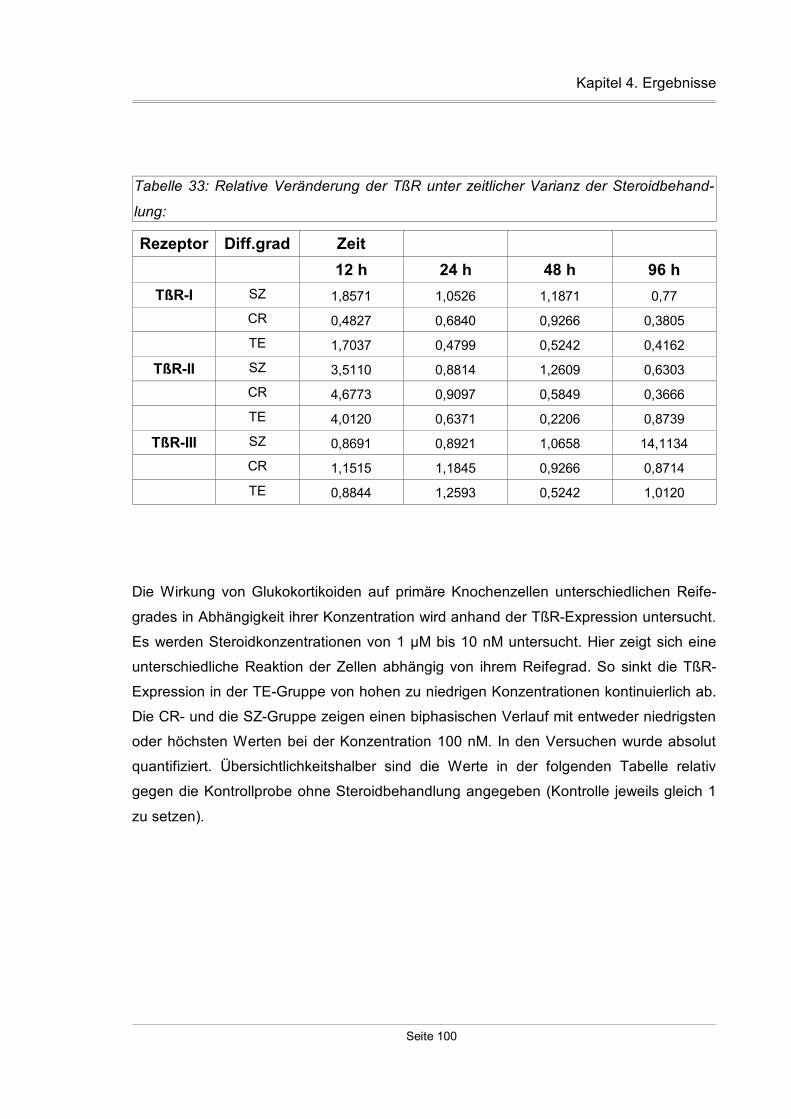
Kapitel 4. Ergebnisse
Tabelle 33: Relative Veränderung der TßR unter zeitlicher Varianz der Steroidbehand-
lung:
Rezeptor Diff.grad Zeit12 h 24 h 48 h 96 h
TßR-I SZ 1,8571 1,0526 1,1871 0,77
CR 0,4827 0,6840 0,9266 0,3805
TE 1,7037 0,4799 0,5242 0,4162
TßR-II SZ 3,5110 0,8814 1,2609 0,6303
CR 4,6773 0,9097 0,5849 0,3666
TE 4,0120 0,6371 0,2206 0,8739
TßR-III SZ 0,8691 0,8921 1,0658 14,1134
CR 1,1515 1,1845 0,9266 0,8714
TE 0,8844 1,2593 0,5242 1,0120
Die Wirkung von Glukokortikoiden auf primäre Knochenzellen unterschiedlichen Reife-
grades in Abhängigkeit ihrer Konzentration wird anhand der TßR-Expression untersucht.
Es werden Steroidkonzentrationen von 1 µM bis 10 nM untersucht. Hier zeigt sich eine
unterschiedliche Reaktion der Zellen abhängig von ihrem Reifegrad. So sinkt die TßR-
Expression in der TE-Gruppe von hohen zu niedrigen Konzentrationen kontinuierlich ab.
Die CR- und die SZ-Gruppe zeigen einen biphasischen Verlauf mit entweder niedrigsten
oder höchsten Werten bei der Konzentration 100 nM. In den Versuchen wurde absolut
quantifiziert. Übersichtlichkeitshalber sind die Werte in der folgenden Tabelle relativ
gegen die Kontrollprobe ohne Steroidbehandlung angegeben (Kontrolle jeweils gleich 1
zu setzen).
Seite 100
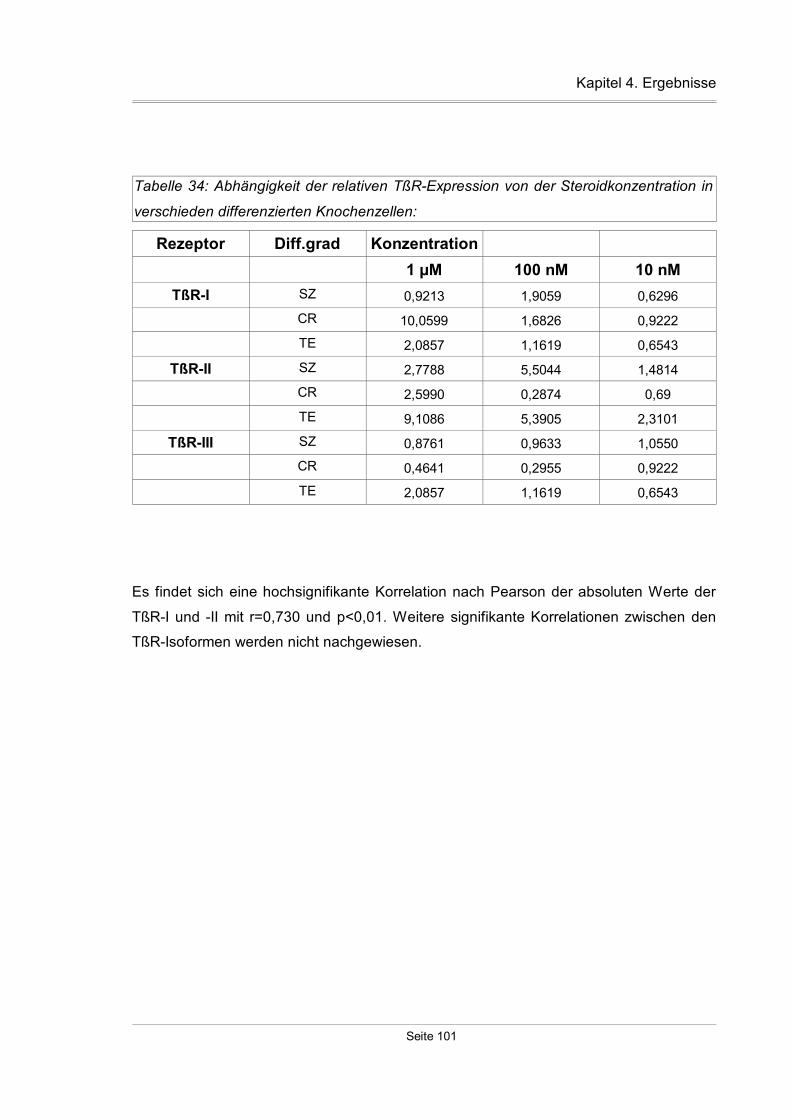
Kapitel 4. Ergebnisse
Tabelle 34: Abhängigkeit der relativen TßR-Expression von der Steroidkonzentration in
verschieden differenzierten Knochenzellen:
Rezeptor Diff.grad Konzentration1 µM 100 nM 10 nM
TßR-I SZ 0,9213 1,9059 0,6296
CR 10,0599 1,6826 0,9222
TE 2,0857 1,1619 0,6543
TßR-II SZ 2,7788 5,5044 1,4814
CR 2,5990 0,2874 0,69
TE 9,1086 5,3905 2,3101
TßR-III SZ 0,8761 0,9633 1,0550
CR 0,4641 0,2955 0,9222
TE 2,0857 1,1619 0,6543
Es findet sich eine hochsignifikante Korrelation nach Pearson der absoluten Werte der
TßR-I und -II mit r=0,730 und p<0,01. Weitere signifikante Korrelationen zwischen den
TßR-Isoformen werden nicht nachgewiesen.
Seite 101

Kapitel 5. Diskussion
5. Diskussion
5.1 Wertigkeit der angewandten Methoden
Das primäre Ziel dieser Arbeit bestand darin, auf einem "real-time"-PCR-Gerät - dem
Light-Cycler7 von Roche - eine verlässliche, valide und sensitive Methode zur cDNA-Be-
stimmung zu etablieren. Die Etablierungsphase wurde mit anderen Proben als den spä-
ter analysierten durchgeführt, da das Ausgangsmaterial in Form von Knochenproben nur
in sehr begrenztem Umfang zur Verfügung stand. Die Untersuchungsmethode mußte
äußerst sensitiv und zuverlässig sein, um mit wenig Ausgangsmaterial reproduzierbare
Ergebnisse zu erhalten und alle Proben analysieren zu können.
Die Durchführung dieser Arbeit zeigte die Aufwendigkeit der Probengewinnung, der Ver-
arbeitung und der Versuche. Probleme ergaben sich einerseits wegen der Instabilität der
RNA gegenüber Auftauen und RNAsen-Aktivierung im Verlauf der Verarbeitung,
andererseits mußte der Knochen gut gereinigt werden, um nur originäre Zellen des
Knochengewebes zu erhalten. Durch ein modifiziertes Extraktionsprotokoll nach Chomc-
zynski und Sacchi konnte reine RNA von hoher Qualität und frei von genomischer DNA
gewonnen und in die Reverse Transkription eingesetzt werden (Chomczynski et al.,
1987). Eine Extraktionskontrolle war durch den Einsatz exonspezifischer Primer in der
PCR möglich, die bei Kontamination eine Doppelbande in der Gelelektrophorese gezeigt
hätte. Bei der Reversen Transkription wurden stets gleiche Reaktionsbedingungen ein-
gehalten, um die Variabilität möglichst gering zu halten. Um Konzentrations-
schwankungen in den Proben zu reduzieren, wurde vor Beginn der Analysen die cDNA
in entsprechenden Mengen mit Wasser verdünnt. Mit den "real-time"-PCR-Geräten ergibt
sich die Möglichkeit der absoluten Quantifizierung. Angaben können in Kopien pro ng
eingesetzte RNA gemacht werden. Dazu werden Verdünnungsreihen aus bereits amplifi-
zierten Proben in einem konstanten Verhältnis von 1:10 als Standardreihen hergestellt
und die Proben gegen diese gemessen. Die anschließende Schmelzkurvenanalyse
ermöglicht die Produktcharakterisierung und darüber werden die zur Berechnung her-
anzuziehenden Proben ermittelt.
Seite 102

Kapitel 5. Diskussion
Die Aussagekraft wird durch das kleine Kollektiv eingeschränkt. Von ursprünglich 55
Proben wurden 48 in die Analyse der TßR-Expression einbezogen. Für diese Kollektiv-
größe ist es schwer, signifikante Aussagen zu treffen. Die Proben zeigen eine große
Schwankungsbreite der Expressionswerte. Ein weiteres Problem stellt die Aufgliederung
des Kollektives in Gruppen von maximal 25 Patienten bei der Analyse nach den Untersu-
chungsvariablen dar. In manchen Fällen besteht das zu untersuchende Kollektiv aus
weniger als zehn Proben. Daher wird meist die tendenzielle Veränderung betrachtet.
Die Versuche haben eine Sensitivität von 99,6 % für den LightCycler7 und 98,4 % für
den TaqMan7 und eine Spezifität von 84,07 % für den TaqMan7 und 81,07 % für den
LightCycler7 ergeben, was deutlich über den Werten der herkömmlichen Gelanalyse-
verfahren liegt. Unterschiede in der Sensitivität und Spezifität bei der PCR beruhen
möglicherweise darauf, daß der TaqMan7 mit halben Ansatzmengen (25 µl anstatt 50 µl)
bestückt worden ist und dies die Analysegenauigkeit schmälern könnte. Die Spezifität
des TaqMan7 ist etwas niedriger, da wegen der Mikrotiterplatten die Kreuzkontamination
möglicherweise etwas größer als bei dem LightCycler7 ist. Nur die HPLC-gestützte
quantitative PCR erreicht ähnlich gute Werte wie die verwendeten „real-time“-PCR-Gerä-
te. Als Nachweisgrenze des HPLC-Verfahrens werden Werte zwischen 0,2 ng und 0,4
ng angegeben. Nachweise zwischen 1,8 und 74,8 fg/µl bei einem Einsatz zwischen 10
und 100 ng cDNA konnten mit der "real-time"-PCR primer- und geräteabhängig regelmä-
ßig durchgeführt werden. Bei der Agarosegelelektrophorese wird die Nachweisgrenze
auf 1-5 ng geschätzt. Konventionelle Nachweisverfahren wie „Northern Blots“ benötigen
10 µg Gesamt-RNA und „RNAse Protection Assays“ (RPA) ca. 5 µg Oligo-dT-RNA, also
ein Mehrfaches an Gesamt-RNA zum sicheren Nachweis einer Probe mit entsprechend
hohen Nachweisgrenzen. Für die Intraassayvarianz der TßR werden primer- und geräte-
abhängig Werte zwischen 0,04 % und 0,7 % und für die Interassayvarianz der TßR
primer- und geräteabhängig Werte zwischen 1,8 % und 2,8 % für die TßR (jeweils be-
rechnet Intraassay- oder Interassayvarianz multipliziert mit 100 und dividiert durch den
Mittelwert) ermittelt. Die Varianzen für PDH liegen wegen eines offenbar nicht-optimalen
Primerdesigns zwischen 10 % und 17 %. Hinweise für ein nicht optimal ausgesuchtes
Primerpaar oder Primingverhalten ergeben sich durch die nicht immer ganz saubere
Schmelzkurvendarstellung und wegen der reduzierten PCR-Effizienz. Die Varianzen für
HPLC-basierte Systeme liegen bei 2 bis 6 % innerhalb und bei 2 bis 9 % zwischen den
Nachweisen (Köhler et al., 1995), respektive zwischen 2 und 3 % nach Vorversuchen
Seite 103

Kapitel 5. Diskussion
(Isken, 2001). Die Varianzen sind bei den „real-time“-PCR-Geräten besser als bei der
HPLC-basierten, quantitativen PCR. Schätzungen der Varianz der Gelelektrophorese-
verfahren liegen zwischen 50 und 200 % (Katz et al., 1990; Henninger et al., 1993).
Die Abhängigkeit der Amplifikationseffektivität von der Effektivität konnte nur für den
LightCycler akzeptabel dargestellt werden. Größten Wert sollte auf die Auswahl des
Primerpaares gelegt und diese nach den Kriterien des Cyclerherstellers mittels deren
Primeranalyseprogrammes synthetisiert werden. Als weiterer „Störfaktor“ können kleine
Fehler in der Verdünnungsreihenherstellung hinzutreten. Die Standardkurve des Light-
Cycler7 kann durch einzelne Wertepaare bestimmt werden, während der TaqMan7 alle
analysierten und vordefinierten Werte zur Berechnung heranzieht. Beides kann zu einer
falschen Lage der Standardkurve führen und die Berechnung der Amplifikationseffektivi-
tät unmöglich machen (Köhler et al., 1995 und PE Biosystems 2001).
Die modernen "real-time"-PCR-Geräte bieten mehr als nur einen Zeitvorteil bei der
Analyse. Eine PCR mit Analyse und Auswertung dauert auf dem LightCycler7 etwa eine
Stunde bei 32, auf dem TaqMan7 zwischen 3 und 4 Stunden bei 96 möglichen Einzel-
untersuchungen. Der Light-Cycler7 bietet die schnellste Detektionszeit. 3 Ansätze
können problemlos an einem Tag durchgeführt und ausgewertet werden. Die Anzahl der
durchführbaren Analysen ist durch die maximal 32 Glaskapillaren im Gerät limitiert. Wird
ein Dreifach-Ansatz gewählt, können maximal 9 Proben analysiert werden. Durch gleich-
zeitige Messung dreier Fluorophoren ist eine Multiplex-PCR möglich. Nachteil dieses
Systems ist der hohe Preis der Verbrauchsmaterialien (etwa 40 Cent pro Glaskapillare
plus Reagenzienkosten). Neuartige Systeme mit Verwendung normaler Reagenzgefäße
sind auf dem Markt und können die Anwendung dieser Technik erleichtern.
Mit dem TaqMan7 können mehr Proben in einem Ansatz analysiert und gegen eine
Standardreihe gemessen werden. Dies führt zu einer größeren Exaktheit der Ergebnisse
und zu einer Reduktion von Reagenzien und Kosten. Das Arbeiten mit Mikrotiterplatten
und die Analyse mittels spezieller Software macht dieses System besonders interessant
zur Untersuchung größerer Kollektive. Die Analyse läuft über vorgegebene Programme
ab. Dies reduziert die Fehlermöglichkeit bei der Auswertung. Eine manuelle Nachbe-
arbeitung der Daten ist über eine Exportfunktion der Software möglich. Zur Auswertung
der Daten der humanen Knochenbiopsien, deren Analyse auf dem LightCycler7 statt-
fand, wurde der einprogrammierte Algorithmus der „second derivative maximum“-Metho-
Seite 104
Jens Regula
Erratum: von der Amplifikatlänge

Kapitel 5. Diskussion
de verwendet. Die humanen Knochenzellkulturen wurden auf dem TaqMan7 analysiert
und die Auswertung fand mittels der implementierten Software statt. Zwischen den TßR-
Expressionswerten beider Versuchsreihen wurden keine Vergleiche durchgeführt. Auf je-
dem Gerät wurde nach einem einheitlichen Schema ausgewertet.
Im Vergleich zu weiteren Analysemethoden ist die "real-time"-PCR zur Zeit das Verfah-
ren, das eine sehr hohe Sensitivität, Spezifität, Intra- und Interassayvarianz bietet. Die
HPLC-gestützte quantitative PCR erreicht nicht ganz so gute Werte. Im Vergleich zu
HPLC und Gelanalyseverfahren sind LightCycler7 und TaqMan7 zu Nachweisen im
Femtogramm-Bereich fähig. Dieses bietet ansonsten nur die Massenspektrometrie, die
noch geringere Mengen nachweisen kann. Der Arbeitsaufwand ist bei der "real-time"-
PCR niedrig, da nur die PCR mit Zugabe der nötigen Reagenzien vorbereitet werden
muß. Als zusätzlicher Arbeitsfaktor kommt die präzise Herstellung der Standardreihen
hinzu. Die Anschaffungskosten für ein "real-time"-PCR-Gerät sind hoch und die Analyse-
kosten pro Probe betragen bis zu 2 Euro, wobei der TaqMan7 in den Verbrauchskosten
günstiger ist. Die vorgefertigten Reagenzien sind für 50 µl Reaktionsansatz berechnet,
bei einer optimierten PCR reichen dagegen 25 µl Reaktionsansatz aus (25 µl wurden in
weiteren Versuchen ohne erkennbare Qualitätseinbußen eingesetzt). Daher können
doppelt so viel Ansätze aus einem Reagenziensatz angefertigt und die Kosten reduziert
werden. Das Arbeiten mit solch kleinen Volumina ist gewöhnungsbedürftig und
beinhaltet die Gefahr der Kreuzkontamination oder Verwechslung. Beide Systeme -
LightCycler7 und Taqman7 - geben ein gleich hohe Sensitivität und Reliabilität an und
sind nach den erhaltenen Ergebnissen durchaus als gleichwertig anzusehen.
Seite 105

Kapitel 5. Diskussion
5.2 Einflüsse auf die PDH-Expression bei relativer Quantifizierung
Für den Vergleich der erhaltenen Daten mit den Daten der RNA-Expression der TGF-ß-
Isoformen wird zusätzlich die PDH-Expression untersucht und in Verhältnis zu der TßR-
Expression gesetzt, also relativ quantifiziert. Unter gewissen Umständen kann es im
Kollektiv zu einer veränderten PDH-Expression gekommen sein, mit dem Nachteil ver-
bunden, daß PDH nicht als ideales „housekeeping gene“ fungiert. Höheres Lebensalter
führt zu einer verminderten Expression der PDH (Stadtman et al., 1992). Eine längerfris-
tige Immobilisation geht mit einer reduzierten Expression der Energiestoffwechselgene
einher (Ward et al., 1986). Fettreiche Kost bei Adipositas (BMI > 25 kg/m^2) bewirkt eine
Erhöhung der PDH-Expression mit einer Insulinresistenz als mögliche Folge (Sugden et
al., 1995). Eine längerfristige Steroidmedikation führt zu erhöhter Transkription des PDH-
Komplexes in der Zelle (Lehninger et al., 2001). Obige Auflistung zeigt, daß die Aus-
sagekraft bei relativer Quantifizierung eingeschränkt ist und es sinnvoll ist, die absolute
Quantifizierung zu präferieren, da die meisten Gene multiplen Einflüssen unterliegen.
Seite 106

Kapitel 5. Diskussion
5.3 Diskussion der Ergebnisse der Knochenstudie
5.3.1 Generelle Aspekte
In dieser Studie wurde die TGF-ß-Rezeptor-Expression der drei Typen untersucht, die
nach bisher bekannten Veröffentlichungen deutlich durch endogene und exogene
Einflüsse reguliert werden. Einfluß auf die Expression haben nachgewiesenermaßen der
Differenzierungsgrad der Knochenzellen, eine Steroidbehandlung, der Hormonstatus
einer Person und einige Krankheiten. Für weitere Parameter sind Änderungen der TGF-
ß-Rezeptor-Expression denkbar, doch in bisherigen Experimenten finden sich diesbe-
züglich nur Hinweise. Außerdem sind Veränderungen der TGF-ß-Expression bei einigen
Erkrankungen außerhalb des Knochens bekannt. Ein Zusammenhang zwischen der
Knochenbelastung und der TGF-ß-Expression stellt sich dar. Alter, Hormonstatus und di-
verse fibrosierende Krankheiten beeinflussen die TGF-ß-Expression. Nur wenig ist dar-
über bekannt, wie sich Änderungen der TGF-ß-Expression auf die der Rezeptoren aus-
wirkt. Da die Wirkung der TGF-ß-Isoformen über die TßR vermittelt wird, ist eine Ver-
änderung der Rezeptorexpression durch TGF-ß nicht unwahrscheinlich (Baylink et al.,
1993). Gebken et al. haben beschrieben, daß die TGF-ß-Rezeptorzahl an der Oberflä-
che von Knochenzellen sich durch eine längere Inkubation mit TGF-ß1 um 95 % redu-
zieren läßt. Dieser Effekt scheint auf posttranslationalen Veränderungen der TGF-ß-Re-
zeptoren zu beruhen, da sich die gemessenen mRNA-Spiegel nur unwesentlich
verändert haben (Gebken et al., 1999). Centrella et al. wiesen auf die nicht unwesentli-
che Bedeutung der posttranskriptionellen und posttranslationalen Modifikationen der
TßR in Knochenzellen hin (Centrella et al., 1996). Eine weitere Möglichkeit ist, daß
mRNA-Transkripte nicht in Proteine translatiert werden. Bei anderen Rezeptorsystemen
ist gezeigt, daß hohe Proteinkonzentrationen über einen „escape“-Mechanismus zu er-
niedrigten Rezeptorexpressionen führen. Umgekehrt führen niedrige Proteinkonzentra-
tionen zu vermehrter Rezeptorexpression. Ob diese klassischen Rezeptorregulationsme-
chanismen im TGF-ß-Rezeptorsystem Gültigkeit haben, ist nicht hinreichend belegt.
Doch diese Mechanismen erklären die gefundenen Ergebnisse gut und werden in der
Diskussion angewandt. In der Literatur finden sich nur sehr wenig Hinweise auf posttran-
Seite 107

Kapitel 5. Diskussion
skriptionelle und posttranslationale Modifikationen der TßR. Da in vielen Untersuchungen
nur geringe Veränderungen der TßR-mRNA gefunden wurden, scheinen posttranskrip-
tionelle Veränderungen wichtig zu sein. Centrella et al. wiesen auf Stabilitätsunterschie-
de zwischen mRNA und Protein der TßR, wahrscheinlich auf intensiver Prozessierung
der mRNA oder der Proteine beruhend, hin (Centrella et al., 1996). Bei der Osteogenesis
imperfecta (OI) sind allgemein erhöhte TßR-Spiegel bei einer nahezu unveränderten
Bindungsaffinität nachweisbar. Die liganden-induzierte Regulationsfähigkeit ist bei OI-
Patienten reduziert (Gebken et al., 2000). Über Verhältnisangaben der Rezeptoren zu-
einander sind in der Literatur wenig Hinweise zu finden. Centrella et al. haben darauf
hingewiesen, daß es mehr auf die relative Expression der Rezeptoren zueinander als auf
die absolute Anzahl ankommt (Centrella et al., 1994). Bisher waren solche Ergebnisse
mit herkömmlichen Verfahren wie „Northern-Blots“ oder PCRs mit Agarosegelelektropho-
rese nicht zu erzielen. Die quantitative PCR ermöglicht mit der absoluten Quantifizierung,
zuverlässige Aussagen über Anzahl, relative und absolute Verhältnisse der Rezeptoren
zueinander zu treffen.
5.3.2 Einfluß des Geschlechts auf die TßR-Expression
Die Daten zur geschlechtsspezifischen Expression liefern keine signifikanten Ergeb-
nisse. Absolut zeigen Männer eine leicht erhöhte Expression der TßR-I und -II, relativ
zeigen Frauen deutlich höhere Expressionswerte als Männer. In der Aufschlüsselung
nach Alter (prä- oder postmenopausal, bzw. älter oder jünger als 45 Jahre) werden deut-
liche Unterschiede zwischen den Geschlechtern in der Hinsicht gesehen, daß bei präme-
nopausalen Frauen niedrigere Werte als bei männlichen Alterskollegen vorliegen. Bei
postmenopausalen Frauen zu Männern über 45 Jahren ist das Verhältnis genau umge-
kehrt. Durchschnittlich zeigen Männer eine höhere Expression.
Literaturangaben zur geschlechtsspezifischen TßR-Expression gibt es nicht. Androgene
erhöhen die TGF-ß-Expression (Kasperk et al., 1990). Bodine et al. zeigten, daß sich die
TGF-ß-mRNA-Expression unter Androgenstimulation nicht verändert, aber die TGF-ß-Ak-
tivität auf ein Mehrfaches steigt (Bodine et al., 1995). Kasperk et al. konnten Unterschie-
de in der Wirkung der Androgene gonadalen und adrenalen Ursprungs zeigen. So stimu-
Seite 108

Kapitel 5. Diskussion
liert Dihydrotestosteron (DHT) die Zellproliferation und die zelldifferenzierende Wirkung
auf den Knochen stärker als Dihydroepiandrostendion (DHEA) oder -sulfat (DHEA-S).
Die Androgenwirkung auf den Knochen wird zum einen über Androgen-Rezeptoren ver-
mittelt, zum anderen fördern Androgene die AP-Aktivität über eine vermehrte TGF-ß-Ak-
tivität (Kasperk et al., 1997). Hofbauer et al. konstruierten eine reife, immortalisierte,
Androgenrezeptor-überexprimierende Osteoblastenzellkultur und untersuchten die
Androgenwirkung auf diese Zellen. Darin wirkten Androgene proliferationshemmend, re-
duzierten die Kollagensynthese und die AP-Aktivität. Diese Effekte konnten durch TGF-
ß-Stimulation aufgehoben werden. Androgene scheinen in reifen Osteoblasten TGF-ß-
vermittelt proliferationshemmend zu wirken (Hofbauer et al., 1998). Diese Studie
verdeutlicht, daß Geschlechtshormone eine von der Zelldifferenzierung im Knochen
abhängige Wirkung entfalten. Östrogene erhöhen die TGF-ß-Expression (Seino et al.,
1994). Yang et al. zeigten in ovarektomierten Ratten wegen einer Abnahme der TGF-ß-
Konzentration eine vermehrte Knochenresorption mit deutlich beschleunigtem
Knochenumbau. Östrogen- oder Raloxifenapplikation konnte den TGF-ß-Abfall und den
Knochenverlust komplett antagonisieren. Östrogene scheinen TGF-ß-vermittelt am Erhalt
der Knochendichte und -struktur beteiligt zu sein (Yang et al., 1996). Einer der Haupt-
gründe für die postmenopausale Osteoporose ist der nach der Menopause sinkende
Östrogenspiegel im Blut. Östrogene wirken meistens, auch in den Osteoblasten und
Osteoklasten, über eine Steuerung der Apoptoseprozesse. Fehlen Östrogene, so leben
die Osteoklasten länger und sorgen für einen vermehrten Knochenabbau. Hughes et al.
fanden Hinweise, daß Östrogene die Apoptose in Osteoklasten TGF-ß-vermittelt indu-
zieren (Hughes et al., 1996). Qu et al. untersuchten in einer Maus-Knochenmarkszell-
kultur die fördernde Wirkung von Östrogen auf die Differenzierung in Richtung reifer
Osteoblast, bewiesen durch einen Anstieg der osteoblastären Marker wie Erhöhung der
AP, des Osteocalcin- und Osteopontin-Gehaltes, des TGF-ß-Spiegels sowie der
Kollagenproduktion. Durch Anti-Östrogene konnten alle Effekte antagonisiert werden.
Dies zeigt eine TGF-ß-vermittelte Förderung der Proliferation der Knochenzellen durch
Östrogene (Qu et al., 1998). Bord et al. untersuchten an Beckenkammbiopsien von Pati-
enten mit oder ohne hochdosierte Östrogentherapie die Expression von lokalen Wachs-
tumsfaktoren und deren Rezeptoren im Knochen mittels semiquantitativer PCR. Für die
mRNA von TGF-ß1 und -2 zeigten sie einen mehrfachen Anstieg bei den behandelten
Patientinnen. Für die Proteinexpression der TGF-ß-Isoformen und Rezeptoren verzeich-
neten sie einen Anstieg bei den behandelten Personen und eine Abnahme der
Osteoklastenpopulation. Sie konnten eine positive Beeinflussung der TGF-ß-Isoformen
Seite 109

Kapitel 5. Diskussion
und der TßR durch Östrogene zeigen (Bord et al., 2001).
Daten über den Einfluß der Geschlechtshormone auf den Knochenstoffwechsel gibt es
viele, doch die TßR-Expression wurde nur selten untersucht. So fördern Östrogene und
Androgene die TGF-ß-Expression, anscheinend jedoch unterschiedlich stark. Die hier
vorgestellten Arbeiten machen eine Beeinflussung der TGF-ß-Rezeptoren durch Sexu-
alhormone wahrscheinlich. Genaue Daten zur TßR-Expression sind schwer zu finden.
Über die Expression der einzelnen Isoformen konnte nichts gefunden werden. Die Da-
ten, daß postmenopausale Frauen eine höhere Expression der TßR zeigen, stehen in
Einklang mit der Tatsache, daß die Östrogenspiegel im Alter sinken, die TGF-ß-Expressi-
on erniedrigt ist und konsekutiv die Rezeptoren hochreguliert werden. Eine Abnahme der
TßR-Expression bei den Männern ließe sich durch ein verändertes Ansprechen auf Si-
gnale erklären. Es scheint Unterschiede in der Wirkungsweise der Geschlechtshormone
auf das TGF-ß-System zu geben. Die Veränderungen der TßR scheinen teilweise
gegenläufig geregelt zu sein bei Vergleich von Männern und Frauen. Ob diese Unter-
schiede IL-6-vermittelt sind, ist nicht auszuschließen. Der Unterschied zwischen relativer
und absoluter Expression zwischen den Gruppen könnte auf eine geschlechts- und hor-
monabhängige PDH-Expression hindeuten. Daten, die diesen Sachverhalt unterstützen,
gibt es derzeit nicht.
5.3.3 Einfluß des Alters auf die TßR-Expression
Die erhaltenen Daten zeigen keine einheitliche und signifikante Änderung der TßR-Ex-
pression. Das Kollektiv der 76- bis 90-Jährigen zeigt eine deutliche Abnahme der re-
lativen Expression, aber bei einer Stichprobe von 3 Personen muß dieses Ergebnis vor-
sichtig interpretiert werden.
Pfeilschifter et al. zeigten, daß die Antwortfähigkeit von Knochenzellkulturen aus
menschlichen Beckenkammbiopsien unterschiedlichen Alters gemessen an der ge-
steigerten DNA-Synthese mit zunehmendem Alter der Spenderzellen deutlich abnimmt
(Pfeilschifter et al., 1993). Nicolas et al. fanden in menschlichen Beckenkammbiopsien
eine altersabhängige Abnahme der TGF-ß- und IGF-1-Konzentration, hinweisend darauf,
Seite 110

Kapitel 5. Diskussion
daß diese Faktoren für den physiologischen Knochenverlust während des Alterungspro-
zesses verantwortlich sein können (Nicolas et al., 1994). Bätge et al. konnten in ihrem
kleinen Patientenkollektiv (n=19, Alter: 2 bis 83 Jahre) eine signifikante Zunahme der
TßR-II-Expression zeigen. Diese Zunahme ging mit einer Abnahme der Rezeptoraffinität
einher. Die TßR-II-Expression wurde mittels semiquantitativer Analyse gegen Glyceralde-
hyd-3-Phosphat-Dehydrogenase (GAPDH) als „housekeeping gene“ untersucht. Die
erhöhte TßR-II-Expression könnte in Zusammenhang mit einer gestörten Balance zwi-
schen Knochenbildung und -resorption stehen (Bätge et al., 2000). Diese Annahme steht
in Einklang mit der altersabhängigen Abnahme der TGF-ß-Expression, da bei redu-
ziertem Proteingehalt die entsprechenden Rezeptoren hochreguliert wird (Pfeilschifter et
al., 1993; Gebken et al., 1999). Eine konstant hohe TGF-ß-Expression unterdrückt den
endgültigen Mineralisationsprozess im Knochen (Harris et al., 1994). Eine veränderte Ex-
pression der TGF-ß-Rezeptoren im Alter ist für andere Gewebe gezeigt. Bei M. Alz-
heimer ist die TßR-Expression in Teilen des Gehirnes, wo reaktiv Gliazellen proliferieren,
erhöht (Lippa et al., 1998). Die belastungsinduzierte TGF-ß-Freisetzung ist bei älteren
Patienten im Vergleich zu jüngeren deutlich reduziert (Neidlinger-Wilke et al., 1995).
TGF-ß1 ist die hauptsächlich von Knochenzellen produzierte TGF-ß-Isoform. In
Knochenzellkulturen von Patienten unterschiedlichen Alters konnte keine Abnahme, son-
dern eine Konstanz der konstitutiven TGF-ß-Sekretion nachgewiesen werden (Bismar et
al., 1999). In Stimulationsexperimenten mit Knochenzellkulturen von jungen und alten
Mäusen konnten Wang et al. eine allgemein verringerte Ansprechbarkeit der alten Popu-
lation auf hormonelle Stimuli zeigen. Die Expression von TGF-ß und Interleukin-1 mRNA
war in der jungen Population deutlich höher (Wang et al., 1999). Untersuchungen von
Gazit et al. zeigten eine Abnahme der TGF-ß-Expression im Alter und eine Zunahme der
TßR-Expression ohne Änderung der Rezeptoraffinität. Die Antwortfähigkeit der Rezepto-
ren auf TGF-ß war im Alter erhöht. Zusätzlich konnten sie zeigen, daß ältere Osteo-
blastenvorläuferzellen weniger TGF-ß produzieren als jüngere und daß älterer Knochen
weniger TGF-ß enthält. Darüber konnten sie die erhöhte Antwortfähigkeit der älteren Po-
pulation erklären. Dies unterstützt die Hypothese, daß in der älteren Population die Ab-
nahme der Synthese und die Verfügbarkeit von in der Knochenmatrix inkorporiertem
TGF-ß für die reduzierte Anzahl und für die Schmälerung des Entwicklungspotentials der
Osteoblastenvorläuferzellen verantwortlich sein kann (Gazit et al., 1998). Mäuse, bei
denen die intrazelluläre Signalweiterleitung der TßR durch ein Fehlen von Smad-3 blo-
ckiert ist, entwickeln eine ausgeprägte Osteopenie. Die Abnahme der Knochendichte
spiegelt die osteoblastäre Unfähigkeit wider, die Osteoklasten adäquat zu kontrollieren.
Seite 111

Kapitel 5. Diskussion
Die Smad-3-induzierte Osteopenie der Mäuse beruht auf einer erniedrigten
Knochenneubildungsrate mit Erhöhung der Osteoblastenzahl und Apoptose. TGF-ß
kann die Differenzierung der Osteoblasten, die in der Anzahl zunehmen und deren
Lebenszeit verkürzt ist, zu Osteozyten in den Smad-3-defizienten Mäusen nicht aufhal-
ten (Borton et al., 2001).
Oben dargelegte Sachverhalte könnten teilweise erklären, warum die Expression der
TßR sich nur unwesentlich verändert. Die bisherigen Studien zeigen keine einheitlichen
Ergebnisse bezüglich der Veränderungen im TGF-ß-System. Die Ergebnisse hängen
deutlich von dem eingesetzten System, ob menschliche oder tierische Zellkultur oder
Tierversuche, ab. Im Alter werden gegenläufige Kompensationsmechanismen aktiviert.
So geht die TGF-ß-Konzentration im Knochen leicht zurück. Dafür zeigen die Rezepto-
ren eine verbesserte Ansprechbarkeit auf TGF-ß-vermittelte Stimuli. Posttranslationale
Modifikationen der Rezeptoren und des Signalweges scheinen mitverantwortlich für die
nur geringen Veränderungen zu sein. Blockierung des Signalweges kann ebenso mitver-
antwortlich sein für eine Osteopenie. Im Alter scheint offensichtlich die Funktionsfähigkeit
der Rezeptoren wichtiger als deren absolute Zahl zu sein. Experimente zur Bindungsaf-
finität konnten nicht durchgeführt werden, da die Knochenproben nicht kultiviert, sondern
direkt der Weiterverarbeitung zugeführt wurden. Das vorhandene, native Material hätte
für derartige Untersuchungen nicht ausgereicht.
5.3.4 TßR-Expression bei pathologischen Veränderungen des Knochens
Die erhaltenen Daten zeigen erkennbare Unterschiede in der TßR-Expression. Die Ex-
pression des TßR-I ist in der Kontrollgruppe am höchsten und in der Traumagruppe am
niedrigsten. Ein signifikanter Unterschied zeigt sich zwischen der Kontrollgruppe mit der
niedrigsten und den Osteoarthrose-Patienten mit der höchsten TßR-II-Expression. Der
TßR-III wird in der Traumagruppe am wenigsten und in der Osteoarthrosegruppe am
meisten nachgewiesen.
Fenwick et al. analysierten Veränderungen im TGF-ß-System in Sehnen bei chronischer,
durch repetitive Reparaturprozesse entstandener Tendinose und in Blutgefäßen als Kon-
Seite 112

Kapitel 5. Diskussion
trolle. In den Blutgefäßen wurden alle TGF-ß-Isoformen und Rezeptoren gefunden, in
pathologisch veränderten Sehnen jedoch nur eine vermehrte TGF-ß2- und TßR-II-Ex-
pression. TßR-I konnte in der Sehnenmatrix nicht detektiert werden. Wegen eines
Mangels an TßR-I ist die Signalkaskade unterbrochen und daher kann die chronische
Läsion nicht heilen (Fenwick et al., 2001). Serra et al. konstruierten einen kinasedefizi-
enten TßR-II, der mit normalen TßR-II um Bindungsmoleküle konkurriert. Dieser Rezep-
tor wurde in Mäusen exprimiert und führte zu einer langsamen, skelettalen Degeneration
und histologisch zu einem Osteoarthrose-ähnlichen Bild. Es wurde vermehrt Kollagen
Typ X im Gelenkknorpel gefunden, der in gesundem Knorpel nicht nachweisbar ist. Ein
Verlust der Antwortfähigkeit auf TGF-ß fördert die terminale chondrozytäre Diffe-
renzierung und geht mit der Entwicklung einer degenerativen Gelenkerkrankung im Sinn
der Osteoarthrose einher (Serra et al., 1997). Steinbrech et al. untersuchten die physio-
logische Knochenheilung nach Defektsetzung an Mäusen und analysierten im Verlauf
die TGF-ß1-, TßR-II-, Osteocalcin- und Kollagen-Expression mittels Immunolokalisation
und „Northern Blot“. Die TGF-ß1-Spiegel stiegen im Verlauf an, sanken am Ende wieder
auf das Ausgangsniveau. Die TßR-II-Expression ging erst zurück, um anschließend weit
über den Ursprungswert anzusteigen. Die Osteocalcin-Produktion wurde herunterge-
regelt. In der Immunolokalisation war der TßR-II anfangs nur in Entzündungszellen nach-
weisbar, später in den Osteoblasten in der Nähe des Frakturspaltes und am Ende der
Heilungsphase in allen Bereichen des neugebildeten Knochens. Dies demonstriert die
temporär differentielle Expression von TGF-ß1 und TßR-II in der Frakturheilung (Stein-
brech et al., 2000). Cillo et al. untersuchten die Zytokinexpression bei der Distraktionsos-
teogenese und fanden, daß die osteoinduktiven Zytokine wie TGF-ß, IGF-1 und IL-6 auf
mRNA-Ebene vermehrt waren, wohin gegen katabole Zytokine nicht verändert waren.
Diese Zytokine scheinen die Hauptmediatoren der mechanischen Zugbelastung auf die
Knochenneubildung zu sein (Cillo et al., 2000). Die Wichtigkeit der osteoinduktiven Zyto-
kine wie TGF-ß, IGF-1 und deren Rezeptoren konnte durch immunhistochemische Fär-
bungen in der Frakturzone durch Eingartner et al. ebenfalls nachgewiesen werden (Ein-
gartner et al., 1999). Ralston et al. konnten keine Unterschiede in der Bildung von Zyto-
kinen (IL-1, TNF-α, TNF-ß, IL-6, FGF, TGF-ß und IGF-1) mittels RT-PCR in Zellen von
gesunden Probanden und von Patienten mit Morbus Paget nachweisen. Für die
Knochenveränderungen bei Morbus Paget könnten entweder posttranslationale Modifi-
kationen dieser Zytokine, damals noch nicht bekannte Faktoren oder eine Störung in
Osteoklasten verantwortlich sein (Ralston et al., 1994). Szekanecz et al. untersuchten
Seite 113

Kapitel 5. Diskussion
Synovialgewebe von Gesunden und von Patienten mit rheumatischer Arthritis (RA) mit-
tels immunohistochemischer Antikörper gegen Endoglin und die TGF-ß-Isoformen. Die
TGF-ß1- und Endoglin-Antikörper reagierten deutlich stärker mit dem pathologischen Ge-
webe und zeigten untereinander eine positive Korrelation. Die Destruktion der Synovia
bei RA scheint teilweise über TGF-ß und Endoglin vermittelt zu sein (Szekanecz et al.,
1995).
Nach oben analysierten Quellen ist eine veränderte TßR-Expression bei Vergleich von
Kontrollen mit Trauma- oder Osteoarthrosepatienten sehr wahrscheinlich, da TGF-ß an
der Pathogenese der Osteoarthrose und an der Frakturheilung wesentlich beteiligt ist. In
den meisten dieser Untersuchungen wurde nur TßR-II analysiert. Eine nicht-signifikante
Erhöhung des TßR-III analog zu Szekanecz et al. konnte bei Osteoarthrosepatienten
nachgewiesen werden. Die Erhöhung des TßR-II bei Osteoarthrose könnte nach Serra et
al. erklärt werden. Die Erniedrigung aller TßR bei Traumapatienten im Vergleich zu den
anderen könnte durch einen Anstieg der TGF-ß-Isoformen und einer konsekutiven Her-
unterregelung der Rezeptoren erklärt werden, da TGF-ß einen wesentlichen Beitrag zu
der Frakturheilung liefert.
5.3.5 Einfluß des Entnahmeortes auf die TßR-Expression
Die gefundenen Daten zeigen Unterschiede in der absoluten und relativen TßR-Ex-
pression. Der TßR-I ist in der Tibia-Gruppe am geringsten und in der Beckenkamm-
Gruppe am stärksten exprimiert. Für den TßR-II verhält es sich genau umgekehrt: nied-
rigste Expression in der Beckenkamm-Gruppe und höchste in der Tibia-Gruppe. Dieses
Ergebnis erreicht ein Signifikanzniveau von p<0,05. Der TßR-III ist in der Tibia-Gruppe
am niedrigsten und in der Hüft-Gruppe am höchsten nachweisbar. Die Gruppe der sons-
tigen Entnahmen ist aufgrund der Größe (n=3) und Heterogenität nicht berücksichtigt.
Tatsächlich ist die Expression der TßR streng örtlich und zeitlich geregelt. Wang et al.
demonstrierten in in-situ-Hybridisierungsexperimenten an Mäusen, daß die TßR-II-Ex-
pression hauptsächlich in mesenchymalem Gewebe und bei Interaktionen zwischen
mesenchymalem und epithelialem Gewebe von außerordentlicher Bedeutung ist (Wang
Seite 114

Kapitel 5. Diskussion
et al., 1995). Lawler et al. wiesen eine Korrelation zwischen der TßR-II und TGF-ß1-Ex-
pression während der Mausentwicklung nach (Lawler et al., 1994). Horner et al. zeigten,
daß durch die TGF-ß-Isoformen unterschiedliche Effekte auf den Knochen mit konseku-
tiven Auswirkungen auf die TßR-Expression ausgeübt werden. Sie konnten alle TGF-ß-
Isoformen und Rezeptoren an Stellen der enchondralen und intramembranären Ossifika-
tion nachweisen. TGF-ß2 konnte in der hypertrophen und mineralisierenden Zone, TGF-
ß3 in der proliferativen und hypertrophen Zone und TGF-ß1 hauptsächlich in der prolife-
rativen Zone in höchsten Konzentrationen nachgewiesen werden. Sie konnten Unter-
schiede bezüglich der lokalen TßR-Expression feststellen. So fand sich ein Verlust der
TßR-I und -II in der hypertrophen Zone. Das Verhältnis der TßR-I und -II zueinander
änderte sich im Verlauf der Wachstumszone des Knochens mit einem Überwiegen des
TßR-II in der Wachstumszone und einem Überwiegen des TßR-I in der Ruhe- und Proli-
ferationszone des Knochens. Diese Ergebnisse sprechen für ein exakt geregeltes und
überlappendes Expressionsmuster der TßR und stützen die These, daß sie in Entwick-
lung und Erhalt des Knochens involviert sind (Horner et al., 1998). Sandberg et al. zeig-
ten, daß TGF-ß1 in wachsendem Schädelknochen an Orten nah der Schädeloberfläche
stärker exprimiert wurde als an tiefer liegenden Orten der Schädelkalotte. Sie sahen dar-
in einen Hinweis darauf, daß TGF-ß1 mitverantwortlich für die Richtung des Knochenan-
baus ist (Sandberg et al., 1988). Es wurde nachgewiesen, daß Veränderungen im TGF-
ß-System die Verknöcherung der Schädelnähte beeinflußt (Oppermann et al., 1995; Lin
et al., 1995). Kabasawa et al. zeigten an Tibiae von Ratten eine örtliche Begrenzung der
TGF-ß- und TßR-Expression. Sie wiesen eine starke Expression des TßR-II in Chondro-
zyten der Proliferationszone, reifenden und hypertrophen Zellschichten, in prolife-
rierenden und reifen Chondrozyten sowie in Fibroblasten des Knochens nach. Sie identi-
fizierten extrazelluläre, matrixproduzierende Chondrozyten und Prä- und Osteoblasten
als die Zielzellen von TGF-ß (Kabasawa et al., 1998). In der veröffentlichten Literatur
finden sich nur wenig Hinweise, wie sich die TßR-Expression unter Berücksichtigung des
Entnahmeortes ändern könnte. Einen großen Einfluß hat mit Sicherheit die auf den
Knochen wirkende Belastung, da TGF-ß an der Transduktion mechanischer Signale auf
den Knochen beteiligt ist (Zhuang et al., 1996). Die örtliche Knochenbildungsrate wird
durch den TGF-ß-Gehalt mitbestimmt (Pfeilschifter et al., 1998).
Demnach müßte ein hoher TGF-ß-Spiegel wegen einer hohen Knochenbildungsrate eine
erniedrigte und umgekehrt eine niedrige Knochenbildungsrate eine hohe TßR-Expressi-
on zur Folge haben. Nach unseren Daten stimmt diese Hypothese für die TßR-I und
Seite 115

Kapitel 5. Diskussion
TßR-III. Der TßR-II ist in proliferierenden Zellen vermehrt und in schon fast ausdiffe-
renzierten Zellen vermindert nachweisbar. Die erhöhte TßR-II-Nachweisbarkeit bei einem
erniedrigten TßR-I-Nachweis spricht für einen nicht abgeschlossenen Knochenumbau,
respektive für einen lebenslang erhöhten Knochenumbau. Unveröffentlichte Daten aus
der Arbeitsgruppe Dr. Hering konnten keine signifikanten Unterschiede in der TGF-ß1-
Expression in Abhängigkeit von der Lokalisation (Femur, Wirbelsäule, Beckenkamm,
Schädelkalotte und Rippen) zeigen. Die Werte der Schädelkalotte lagen signifikant unter
den anderen. Nach weiteren, bisher unveröffentlichten Untersuchungen der Arbeits-
gruppe Dr. Hering konnte ein hochsignifikanter Zusammenhang zwischen der Knochen-
bildungsrate und dem TGF-ß1-Gehalt des Knochens mit Ausnahme des Beckenkammes
hergestellt werden. Das Verhältnis TGF-ß1-Gehalt zu Knochenumbaurate nahm in der
Reihenfolge Wirbelsäule, Rippe, Tibia, Humerus und Hüfte ab. Diese Daten unterstützen
oben Diskutiertes. Das Becken nimmt bezüglich der Belastung und Funktion eine Son-
derstellung ein. Seine Hauptaufgabe ist neben der Stabilisierung und Übertragung der
auf den Körper wirkenden Kräfte mitverantwortlich für die Hämatopoese im
Erwachsenenalter mit entsprechend veränderten Mikroverhältnissen bezüglich der
Wachstumsfaktoren. Daher scheinen die oben angewendeten Diskussionsgrundlagen
nur begrenzt auf das Becken übertragbar zu sein. Da diese Knochen wahrscheinlich
einen unterschiedlichen Diferenzierungsgrad besitzen, sind diesbezüglich schon
Änderungen in der TßR-Expression zu erwarten.
5.3.6 Einfluß der Mobilisation auf die TßR-Expression
Untersucht wurde die TßR-Expression in einem Kollektiv, das nach der Immobilisations-
zeit in teilweise und voll Immobilisierte sowie voll Mobilisierte unterteilt wurde. Da das
Kollektiv der voll Immobilisierten mit 2 Personen sehr klein ist, aber die Unterschiede der
absoluten Expression der TßR-I und -II von den anderen Gruppen sehr groß, jedoch
nicht signifikant, sind, wird dieses Phänomen kurz beleuchtet. Die Unterschiede der voll
Mobilisierten und der teilweise Immobilisierten sind relativ klein, aber zeigen eine Ab-
nahme der absoluten und relativen Expression.
Parfitt et al. wiesen auf den Einfluß der Immobilisation und Schwerelosigkeit auf den
Seite 116

Kapitel 5. Diskussion
Knochenstoffwechsel hin (Parfitt et al., 1981). Die Osteopenie infolge der Schwerelosig-
keit beruht auf reduzierter Knochenneubildung und Abkoppelung der Knochenbildung
von der Knochenresorption. Die Differenzierung des Knochens entspricht einem unreife-
ren Stadium (Bikle et al., 1994 und 1995). Während der Entlastungsphase verlieren die
Knochen am meisten an Knochensubstanz, die unter der Schwerkraft am stärksten be-
lastet werden (Bikle et al., 1999). Gesunde und pathologisch veränderte Knochen zeigen
auf mechanische Belastung eine differente Ansprechbarkeit derart, daß der patholo-
gische Knochen signifikant weniger TGF-ß als Antwort auf die Belastung freisetzt als
gesunder (Neidlinger-Wilke et al., 1995). Eine durch mechanische Beanspruchung indu-
zierte Proliferation der Osteoblasten geht mit einer gesteigerten TGF-ß1-Expression
einher. Dies scheint nicht nur TGF-ß-vermittelt zu sein, da Hemmstoffe des Inositol-Tri-
phosphat-(IP3-) Weges die belastungsinduzierte Proliferation reduzieren (Zhuang et al.,
1996). Drissi et al. zeigten in einem Tiermodell unter Hinterbeinentlastung den Rückgang
der mRNA-Spiegel des TßR-II und der IGF-Rezeptoren. Sie wiesen auf die bisher un-
terschätzte Bedeutung des IGF-Systems bei Entlastung hin (Drissi et al., 1999). Ein
Verlust der Gewichtsbelastung reduziert die Knochenbildung, die Anzahl der Osteo-
blasten und den Differenzierungsgrad in entlastetem Knochen. Die Entlastung führt zu
einer Hemmung der Proliferation und Differenzierung von Osteoprogenitorzellen in vitro
(Kostenuik et al., 1997). Die veränderte Expression der Zytokine bei Gewichtsentlastung
der Knochen ist sehr ortsspezifisch geregelt. Westerlind et al. fanden nach längerer Im-
mobilisation keine veränderte TGF-ß1-Expression in der Wachstumszone des Knochens
junger Tiere, während die Spiegel im Periost gesunken waren (Westerlind et al., 1995).
Eine Entlastung des Skeletts geht mit erniedrigten TGF-ß-Spiegeln einher. Erniedrigte
Spiegel bewirken normalerweise eine Erhöhung der zugehörigen Rezeptoren. Bei kurz-
fristiger Nichtverwendung eines Systems kann analog zu dem Zytokin der entspre-
chende Rezeptor herunterreguliert werden. In den Experimenten von Drissi et al. wurde
unter kurzfristiger Entlastung der TßR-II vermindert exprimiert. Dieses kann als Erklärung
für die durchweg reduzierte Expression der TßR der Teilmobilisierten im Vergleich zu
den Vollmobilisierten dienen. Der erhöhte TßR-I bei Immobilisierten ist nicht auf das
erhöhte Alter der Patienten zurückzuführen, sondern diese Proben stammen aus reifem
Knochen, der eine hohe TßR-I- und eine niedrige TßR-II-Expression zeigt. Eine höhere
relative Expression des TßR-I und -III bei Vollmobilen gegenüber Teilmobilen kann an
einer kurzfristigen Beeinflussung der PDH-Expression unter Ruhebedingungen liegen.
Seite 117

Kapitel 5. Diskussion
5.3.7 TßR-Expression unter längerfristiger Glukokortikoidbehandlung
Die erhaltenen Daten zeigen, daß die absolute Expression aller TßR unter Steroidbe-
handlung leicht abfällt. In der relativen Expression fällt nur der TßR-I ab. TßR-II und -III
zeigen nur geringe Unterschiede in der absoluten und relativen Expression.
Steroide fördern die Ausdifferenzierung der Knochenzellen und Expression der Gene
wie Osteocalcin und Fibronektin, die mit einer fortschreitenden Zelldifferenzierung
einhergehen (Shalhoub et al., 1992). Oursler et al. zeigten, daß Steroide die Expression
lysosomaler Enzyme fördern und daß darüber latentes TGF-ß vermehrt freigesetzt und
aktiviert wird. Dies könnte die Förderung der Differenzierung bei Knochenzellen durch
Steroide mitbewirken (Oursler et al., 1993). Centrella et al. fanden ein verändertes
Bindungsverhalten von TGF-ß1 an die TßR unter Kortisoneinfluß. In „cross-linking“-Expe-
rimenten nahm die Bindung an den TßR-III, der den extrazellulären Gehalt an TGF-ß re-
präsentiert, zu und die Bindung an die TßR-I und -II ab. Eine veränderte TGF-ß-Vertei-
lung in den Speicherorten könnte für die Osteopenie unter Kortisoneinfluß verantwortlich
sein (Centrella et al., 1991). Ein Großteil der Steroidwirkung auf den Knochen, beson-
ders die Effekte auf die differenzierten Osteoblasten, können durch BMP-2 unterdrückt
werden (Centrella et al., 1997). Parrelli et al. konnten ein „glucocorticoid-response-
element“ im TGF-ß1-Gen nachweisen (Parrelli et al., 1998). Iba et al. konnten den posi-
tiven Einfluß der Glukokortikoide auf die Differenzierung der Knochenzellen, gemessen
an der Synthesesteigerung von Osteopontin und an der AP-Aktivitätssteigerung, und
keinen Einfluß auf die Proliferation zeigen (Iba et al., 1995). Iba et al. konnten die stero-
id-induzierte Differenzierung durch TGF-ß dosisabhängig antagonisieren (Iba et al.,
1995). Chang et al. wiesen CBFa1-abhängige Elemente im TßR-I-Gen nach. CBFa1 ist
ein Transkriptionsfaktor, der mit zunehmender Differenzierung der Osteoblasten an-
steigt. Steroide supprimieren die Expression von CBFa1, reduzieren darüber die TßR-I-
Expression und verursachen so einen Teil der negativen Effekte auf den Knochenstoff-
wechsel unter Steroidtherapie (Chang et al., 1998). In einer Osteoblasten-ähnlichen Zell-
kultur konnten Nakayama et al. eine verstärkte Expression von Betaglykan nachweisen.
Diese Erhöhung war zeit- und konzentrationsabhängig. Mittels „cross-linking“-Expe-
rimenten konnten sie eine verstärkte Bindung von TGF-ß1 an den TßR-III, aber nicht an
den TßR-II zeigen. Für die Erhöhung des TßR-III sind transkriptionelle Ursachen mitver-
antwortlich (Nakayama et al., 1994). Der Glukokortikoid-Rezeptor interagiert mit Smad-3
Seite 118

Kapitel 5. Diskussion
und -4 und modifiziert sie so, daß sie die Signale des TGF-ß-Signalweges nicht mehr
korrekt weiterleiten können (Song et al., 1999).
Diese Literaturangaben stützen die gefundenen Daten. Allerdings sind die Effekte der
Steroide auf die Knochenzellen abhängig von der Differenzierung, der Homo- oder Hete-
rogenität des untersuchten Systems, der Einwirkungsdauer, den entsprechend ausge-
bildeten Kompensationsmechanismen und der Wirksamkeit des verwendeten Steroides.
Die eingesetzten humanen Proben zeigen eine höhere Heterogenität als Zellkultursyste-
me bezüglich des Differenzierungsgrades und der Zellreinheit. In den Biopsien konnte
nicht zwischen Osteoblasten, Osteoklasten und Osteozyten unterschieden werden, aber
der Großteil der untersuchten Zellen waren Osteoblasten. Die Patienten wurden über
mehr als 1 Jahr mit Steroiden behandelt. Solche Langzeiteffekte sind in Zellkulturen
schwerlich zu sehen. Unter diesen Voraussetzungen ist es schwer, einen einzelnen
Effekt der Steroide herauszufiltern. Die meisten der bisherigen Untersuchungen wurden
an Zellkulturen durchgeführt und auf TßR-Veränderungen in vivo finden sich kaum Hin-
weise. Unsere Versuche zeigen eine TßR-I/PDH-Abnahme unter Steroiden, die wahr-
scheinlich einer Kontrolle durch CBFa1 unterliegt. Weitere Veränderungen konnten mit-
bedingt durch die Heterogenität nicht nachgewiesen werden. Der Einfluß der Steroide
über die Smads auf den TGF-ß-Signalweg könnte mitverantwortlich sein, daß nur
geringe Unterschiede in der TßR-Expression aufgefallen sind. Untersuchungen an den
Smads und CBFa1 dürften weitere Aufschlüsse darüber geben, inwieweit die Be-
einflussung des Knochenstoffwechsels TGF-ß-vermittelt ist. Eine weitere, zu klärende
Frage ist der Einfluß der posttranslationalen und posttranskriptionellen Veränderungen
auf die Rezeptoren.
5.3.8 Einfluß des BMI auf die TßR-Expression
Die gewonnenen Daten zeigen, daß bei Normalgewichtigen der TßR-I signifikant mehr
und der TßR-III deutlich mehr exprimiert wird wie bei Übergewichtigen. Die Änderungen
des TßR-II sind nur unwesentlich. Die Daten der relativen Expression zeigen ein ähnli-
ches Bild.
Seite 119

Kapitel 5. Diskussion
Geiser et al. konnten an TGF-ß-“knock-out“-Mäusen einen negativen Einfluß auf die
Knochenqualität bei TGF-ß-Verlust zeigen. Der Knochen dieser Tiere zeigte einen redu-
zierten Kalksalzgehalt und eine verringerte Elastizität (Geiser et al., 1998). Nguyen et al.
konnten eine positive Korrelation zwischen der Muskelkraft des Quadricepsmuskels und
dem BMI herstellen (Nguyen et al., 2000). Allerdings ist wahrscheinlich, daß es für diese
Korrelation eine Obergrenze gibt, denn klinisch haben morbid adipöse Leute eine re-
lative Schwäche der Muskulatur. Aus mehreren Arbeiten geht hervor, daß eher die fett-
freie Körpermasse als die gesamte Körpermasse die Knochendichte, die Knochenstabili-
tät und den Kalksalzgehalt des Knochens bestimmt (Witzke et al., 1999; Ravaglia et al.,
2000). Alessi et al. konnten im Fettgewebe eine erhöhte PAI-1- und TGF-ß1-Expression
nachweisen. Da TGF-ß die PAI-1-Expression induziert, ist es wahrscheinlich, daß die
PAI-1-Erhöhung TGF-ß-vermittelt ist. Sie konnten auch eine positive Korrelation sowohl
zwischen der PAI-1-, als auch der TGF-ß1-Expression und dem BMI nachweisen (Alessi
et al., 2000). Für TGF-ß konnte ein Zusammenhang mit dem Knochenumbau und nicht
mit der Knochenmasse gezeigt werden (Pfeilschifter et al., 1998). Leptin scheint nach
Daten von Han et al. den TßR-II hochzuregulieren und in die Aktivierung des TGF-ß-Sys-
tems in fibrosierenden Prozessen bei Adipositas und Diabetes involviert zu sein (Han et
al., 2001). Leptin limitiert die Fähigkeit der Osteoblasten, Matrix zu produzieren. Anderer-
seits stimuliert Leptin direkt das Knochenwachstum via IGF-1 und hemmt die
Osteoklastentätigkeit (Whitfield et al., 2001). Inwieweit Leptin an der Regelung der
Knochendichte und des Kalksalzgehaltes beteiligt ist, ist noch nicht abschließend ge-
klärt. Sato et al. zeigten, daß eine hohe Knochendichte mit einem niedrigen Leptin-
spiegel assoziiert ist (Sato et al., 2001). Im Gegensatz dazu fanden Yamauchi et al., daß
bei Osteoporose-Patienten ein niedriger Leptinspiegel mit einer erhöhten Frakturinzidenz
und einer erniedrigten Knochendichte einhergeht (Yamauchi et al., 2001). Amling et al.
wiesen auf die essentielle Bedeutung der zentralen Regulation des Knochenstoff-
wechsels hin. Leptin scheint einer der Vermittler dieser Regulation zu sein (Amling et al.,
2001).
Diese Daten zeigen, daß Körpermasse, BMI und Leptinspiegel das TGF-ß-System be-
einflussen. Doch über die Regelung ist bisher nichts Genaues bekannt. Die Literatur gibt
Hinweise, daß wegen erhöhter TGF-ß1-Spiegel eine Reduktion des TßR-I möglich ist.
Dies zeigen die gefundenen Werte, die sogar mit p<0,05 signifikant sind. Eine Erhöhung
des TßR-II, wie Han et al. es gefunden haben, ist lediglich in der relativen TßR-II-Ex-
pression zu vermuten. Der erniedrigte TßR-III bei den Adipösen steht nicht in Einklang
Seite 120

Kapitel 5. Diskussion
mit der Tatsache, daß es eine postive Korrelation zwischen TGF-ß1 und dem TßR-III
gibt, da Fettgewebe sehr hohe TGF-ß1-Spiegel aufweist und demnach einen erhöhten
TßR-III nach sich zieht. Welche Regualtionsmechanismen da greifen, ist nicht ganz
schlüssig. Es ist möglich, daß die gefundenen Daten nicht nur durch den BMI direkt, son-
dern indirekt durch einen veränderten Lebensstil, eine Tendenz zu Insulinresistenz und
Diabetes, eine Tendenz zu körperlicher Inaktivität und anderen Veränderungen im
Rahmen des metabolischen Syndroms zustande gekommen sind. Multivariate Korre-
lationen, die diesen Zusammenhang näher beleuchten könnten, sind aufgrund der
Kollektivgröße nicht sinnvoll möglich.
5.3.9 Korrelation zwischen den TßR-Isoformen
Die ermittelten Werte für die Korrelationen sind signifikant. Die Werte der absoluten und
relativen Expression unterscheiden sich in der Form, daß die Werte für die relative Ex-
pression besser miteinander korrelieren. Die TßR-I und TßR-II müssen jeweils zweifach
vorhanden sein, um den heterooligomeren Komplex zu bilden (Derynck et al., 1997).
Diese Tatsache legt den Schluß nahe, daß sich die Expression der TßR-I und -II zuein-
ander in geordneten Verhältnissen bewegen muß. Darüber, warum die beiden TßR je-
weils mit dem TßR-III korrelieren, finden sich in der Literatur nur wenig Hinweise. Es
kann der TßR-III mit dem TßR-II über Ligandenpräsentation in Verbindung treten. TGF-
ß2-Signale werden durch Betaglykan-Interaktion mit dem TßR-II um ein Vielfaches
besser übertragen als ohne Betaglykan. Dies könnte eine Begründung für die Korrelation
zwischen diesen beiden Rezeptoren sein. Die Funktion des TßR-III besteht haupt-
sächlich in einer Art „Reservoir“-Funktion für TGF-ß. Es wäre denkbar, da sich die kon-
stitutive TGF-ß-Expression im Laufe des Lebens nur wenig ändert (Bismar et al., 1999),
daß die Höhe des TßR-III die eigene TGF-ß-Produktion widerspiegelt und darüber mit
den anderen TßR in Verbindung zu bringen ist. Eine positive Korrelation zwischen dem
TGF-ß1-Gehalt im Knochenmehl und der TßR-III-Expression konnte gezeigt werden.
Weitere Korrelationen wurden zwar gefunden, doch da über die Isoform-spezifische
Wirkung von TGF-ß wenig bekannt ist, würden weitere Diskussionen auf spekulativer
Basis geführt. Da eine genaue Zuordnung der Proben zu den histologischen Zonen re-
Seite 121

Kapitel 5. Diskussion
trospektiv nicht möglich ist, kann die isoformspezifische TGF-ß-Expression nicht mit der
TßR-Expression korreliert werden.
5.3.10 Weitere Ergebnisse der Knochenstudie
Weitere erhobene und gemessene Parameter korrelieren miteinander signifikant. Zu-
sammenhänge zwischen diesen Korrelationen sind schon früher beschrieben worden.
Collins et al. konnten zeigen, daß IGF i.S. mit einigen Werten signifikant korreliert: nega-
tiv mit dem Alter und positiv mit dem Osteocalcin-, dem AP- und dem TRAP-Spiegel.
Eine Korrelation von IGF-1 mit der Knochendichte war nicht vorhanden (Collins et al.,
1998). Diese Korrelationen stimmen mit den gefundenen Werten gut überein. Klinisch ist
bekannt, daß Steroide einen sekundären Hyperparathyreodismus induzieren, da Ste-
roide den Kalzium-Spiegel über verschiedene Mechanismen reduzieren. Kompensato-
risch kommt es zu einem Anstieg des Parathormones (sekundärer Hyperparathyreodis-
mus), das bei Erniedrigung des Kalzium-Spiegels ausgeschüttet wird (Schölmerich et al.,
1996). Für die Korrelation zwischen dem Kalziumspiegel im Serum und dem Mobilisa-
tionsstatus fanden mehrere Studien einen erhöhten Kalziumspiegel, wenn die Patienten
entlastet wurden (Zerwekh et al., 1998; Inoue et al., 2000). Wie die erhaltenen Daten be-
stätigen, konnte eine signifikante Korrelation zwischen der AP i.S. und der Mobilisation in
früheren Entlastungsexperimenten festgestellt werden (Kurokouchi et al., 1995; Yamagu-
chi et al., 1991).
Seite 122

Kapitel 5. Diskussion
5.4 Diskussion der Ergebnisse der humanen, primären Knochen-
zellkulturen
5.4.1 Einfluß der TßR auf die Differenzierung
In primären, humanen Knochenzellkulturen wurde untersucht, ob sich die TßR-Expressi-
on im Laufe der Differenzierung verändert. Dazu wurde aus acht Zellkulturen die RNA
extrahiert, revers transkribiert und die erhaltene cDNA mittels TaqMan7 absolut quantifi-
ziert. Bei der Darstellung der Ergebnisse wurde versucht, Unterschiede in den relativen
Verhältnissen der Rezeptoren zueinander darzustellen. Bei dem Vergleich von SZ und
TE sind ein größeres TßR-I/TßR-II-Verhältnis und kleinere TßR-I/TßR-III- und TßR-
II/TßR-III-Verhältnisse zu sehen. Bei dem Vergleich von SZ zu CR fällt eine Reduktion
aller Verhältnisse auf. Bisherige Versuche sind meist mit immortalisierten Zellkulturen
durchgeführt worden. Daher ist zu fragen, ob diese Ergebnisse mit denen der Literatur
übereinstimmen und ob die Verhältnisse übertragbar sind.
Centrella et al. zeigten eine Änderung der TßR-Expression während der Differenzierung
und die Wichtigkeit des Verhältnisses der Rezeptoren zueinander (Centrella et al.,
1995). Retinsäure bewirkt in Knochenzellkulturen eine Stimulierung der Reifung. So sti-
mulierte Zellen exprimieren weniger TGF-ß-Rezeptoren auf der Zelloberfläche. Diffe-
renzierung in den Zellkulturen bewirkt eine Herunterregelung der TßR (Gazit et al.,
1993). Ji et al. wiesen nach der Analyse des TßR-I darauf hin, daß von der Struktur her
eine konstant hohe TßR-I-Expression im Rahmen der Differenzierung möglich ist, die auf
der konstitutiven Expression von CBFa1 und einem CBFa1-abhängigen Element im
TßR-I-Gen beruht (Ji et al., 1998). Nach Takeuchi et al. scheint es, daß die Kombination
der TßR die Antwortfähigkeit von Osteoblasten auf TGF-ß und Änderungen der TßR die
Steuerung der Matrixsynthese und Knochenbildung mitbestimmen (Takeuchi et al.,
1995). Horner et al. zeigten, daß der TßR-II in proliferierenden Osteoblasten vermehrt
nachweisbar ist (Horner et al., 1998). TGF-ß stimuliert die Matrix- und Kollagensynthese.
Im Zuge der Knochenzelldifferenzierung nimmt der Kollagengehalt der Matrix zu. Daher
scheint es, daß durch TGF-ß-vermittelte Kollagenproduktion die TGF-ß-Rezeptorzahl
Seite 123
Jens Regula
Jens Regula
Erratum: von TE zu SZ

Kapitel 5. Diskussion
über eine negative Rückkoppelungsschleife reduziert wird (Takeuchi et al., 1996). Toyo-
no et al. konnten in Odontoblasten ein differenzierungsabhängiges Expressionsmuster
der TßR nachweisen, die TßR-II-Expression war nahezu konstant, während der TßR-I in
seiner Expression anstieg (Toyono et al., 1997).
Die erhaltenen Daten stehen im Einklang mit oben Dargelegtem. Die absolute Expressi-
on aller TßR steigt mit zunehmender Differenzierung, um im ausgereiften Osteozyten
wieder abzufallen. Die Verhältnisse der TßR zueinander zeigen ein anderes Bild. Das
Verhältnis TßR-I/TßR-II wird von SZ zu CR kleiner, beruhend auf dem größeren Gewicht
des TßR-II für die proliferierende Wirkung. Von CR zu TE steigt das Verhältnis stark an,
basierend auf der hohen TßR-I-Expression bei ausdifferenzierten Knochenzellen. Das
Verhältnis TßR-I zu TßR-III ändert sich von SZ zu CR nur wenig. Von CR nach SZ wird
dieses Verhältnis kleiner, sich auf die Bedeutung des TßR-I in differenzierten Zellen
stützend. Das Verhältnis TßR-II/TßR-III ändert sich von SZ zu CR nur unwesentlich. Von
CR zu TE wird das Verhältnis viel kleiner. Da während der Differenzierung der TßR-II an
Bedeutung verliert und Betaglykan auf recht hohem Niveau weiter exprimiert wird, ist der
Abfall dieses Verhältnisses gut zu erklären. Die TGF-ß-Rezeptoren scheinen differentiell
exprimiert zu werden. Das Verhältnis der Rezeptoren zueinander ist von großer Bedeu-
tung.
5.4.2 Einfluß von Glukokortikoiden auf primäre Knochenzellkulturen
Die erhaltenen Daten zeigen, daß Steroide einen dosis-, einwirkungsdauer- und diffe-
renzierungsabhängigen Einfluß auf primäre Knochenzellen haben. Im Zeitverlauf von
einer kurzen zu einer langen Inkubationszeit ist eine klare Tendenz zur Abnahme des
TßR-I in den SZ- und TE-Zellen und bei den TßR-II und -III in CR-Zellen zu sehen. Einen
Anstieg zeigen nur der TßR-III in SZ-Zellen und der TßR-I in CR-Zellen. Die restlichen
Proben zeigen ein heterogenes Bild in der Expression der TßR. Über dem Konzentra-
tionsverlauf von hohen zu niedrigen Konzentrationen des Steroides zeigen TßR-I bei CR
und TE, TßR-II und -III bei TE eine Abnahme der Expression. Einen deutlichen Anstieg
zeigt TßR-III bei SZ. Einen biphasischen Verlauf mit Maximum in der Expression bei der
mittleren Konzentration zeigen TßR-I und -II in der SZ-Gruppe. Einen biphasischen
Seite 124
Jens Regula
Erratum: Von CR nach TE

Kapitel 5. Diskussion
Verlauf mit einem Minimum der Expression zeigen TßR-II und -III in der CR-Gruppe.
Zusätzlich zu oben schon diskutierten Beeinflussungen der TßR-Expression in Knochen-
zellen durch Glukokortikoide werden weitere, wichtige Punkte dargelegt. Die TßR-Ex-
pression scheint differenzierungsabhängig mit einem Überwiegen des TßR-II in undiffe-
renziertem Gewebe, das sich später zu Knochen entwickelt, reguliert zu sein. Iseki et al.
gelang der TßR-Expressionsnachweis immer dann, wenn die Liganden ebenfalls
exprimiert wurden (Iseki et al., 1995). Steinbrech et al. konnte in Knochenheilungsexpe-
rimenten zeigen, daß es im Verlauf der Frakturheilung anfangs zu einer Reduktion und
später zu einer Erhöhung der TGF-ß-Rezeptoren kommt (Steinbrech et al., 2000). Che-
valley et al. konnten einen IGF- und IGFBP-vermittelten Einfluß von Dexamethason mit
Verringerung der Knochenzellproliferation bei höheren Konzentrationen und längerer In-
kubationszeit nachweisen (Chevalley et al., 1996). Centrella et al. wiesen auf die sup-
primierte Bindung von TGF-ß an die TßR unter Glukokortikoideinfluß, die durch BMP-2
antagonisiert werden konnte, hin (Centrella et al., 1997). Nakayama et al. konnten an
Osteoblasten-ähnlichen Zellen eine verstärkte Expression des TßR-III unter Steroidein-
wirkung zeigen (Nakayama et al., 1994). Ji et al. identifizierten ein „glucocorticoid-resp-
onse element“ im TßR-III. Die Promotoraktivität konnte durch Glukokortikoide gesteigert
und durch BMP-2 gehemmt werden (Ji et al., 1999). In Versuchen, in denen die
Knocheninselbildung als Parameter der Differenzierung eingesetzt wurde, gelang Bel-
lows et al. der Nachweis, daß Kortison, Kortisol und Kortikosteron unterschiedliche
Einflüsse auf den Knochenstoffwechsel haben. Dieser Effekt war auch abhängig von
dem verwendeten Zellkultursystem (Bellows et al., 1998). In einer von einem Knochen-
tumor abgeleiteten Zellinie sind die Spiegel des aktiven TGF-ß ohne Aktivierung der
mRNA-Expression durch Glukokortikoide angestiegen. Eine Aktivierung der lysosomalen
Proteasen, die ihrerseits latentes TGF-ß aktivieren, scheint für wesentliche Effekte der
Steroide verantwortlich zu sein (Boulanger et al., 1995; Oursler et al., 1993). Bodine et
al. untersuchten zusätzlich zu TGF-ß die mRNA-Expression von c-fos und c-jun unter
Glukokortikoideinfluß, konnten aber keine wesentlichen Expressionsunterschiede zeigen.
Vielmehr konnten sie das Ergebnis von Boulanger et al. bestätigen, daß Glukokortikoide
die Aktivierung von TGF-ß fördern (Bodine et al., 1995). Die Expression von TGF-ß1
kann durch Glukokortikoide über ein GRE im TGF-ß1-Gen modifiziert werden (Parrelli et
al., 1998). Hulley et al. demonstrierten, daß die Steroidhemmung im MAPK-Signalweg
über die Ebene der mRNA-Expression ausgeübt wird (Hulley et al., 1998). Lian et al.
zeigten Speziesunterschiede bei der Wirkung von Glukokortikoiden auf den Knochen. In
Seite 125

Kapitel 5. Diskussion
der von ihnen verwendeten Zellkultur wurde die Differenzierung durch Steroide ge-
hemmt, wohingegen andere Untersuchungen eine Förderung der Differenzierung durch
Glukokortikoide gezeigt haben (Lian et al., 1997). Unter Glukokortikoideinfluß wird der
TßR-I über einen CBFa1-vermittelten Prozeß auf Knochenzellen herunterreguliert
(Chang et al., 1998).
Die Literatur gibt genügend Hinweise, daß Steroide einen großen Einfluß auf die
Knochenzellen haben und hilft, die gefundenen Ergebnisse zu stützen. Die Effekte der
Steroide auf den TßR-I scheinen wie bei Chang et al. in einer Reduktion der Expression
zu bestehen, wobei es differenzierungs-, zeit- und konzentrationsabhängig kleine Unter-
schiede gibt (Chang et al., 1998). Für die Beeinflussung des TßR-II durch Steroide gibt
es weniger Daten. Auch hier scheint es zeit-, differenzierungs- und konzentrationsabhän-
gige Effekte auf den Knochen zu geben. Im Zeitverlauf sinkt die Expression des TßR-II,
nicht immer linear, ab. Die Expression des TßR-II scheint differenzierungsabhängig auf
steigende Konzentrationen zu reagieren (Iba et al., 1995). Nur die TE-Gruppe zeigt
einen linearen Anstieg bei steigender Steroidkonzentration. Der TßR-III scheint diffe-
renzierungsabhängig die größten Unterschiede aufzuweisen. Dies mag an der Anwesen-
heit eines GRE im TßR-III-Promotor liegen, der differentiell aktiviert werden könnte (Ji et
al., 1999). Der TßR-II scheint in der CR-Gruppe unter längerer Steroidwirkung kontinuier-
lich leicht abzunehmen. Im Konzentrationsverlauf zeigt sich eine konzentrationsabhän-
gige Hemmung der Expression mit einem Minimum bei 100 nM. Eine steigende Expressi-
on bei höheren Steroidkonzentrationen zeigt der TßR-III in der TE-Gruppe, während es
im Zeitverlauf eine polyphasische Expression mit keiner einheitlichen Tendenz gibt. Ste-
roide bewirken bei Stammzellen einen Stopp der Proliferationstätigkeit (Lian et al., 1997).
Im Zeitverlauf zeigt sich das an der Abnahme der TßR-I und -II. Da Stammzellen norma-
lerweise viel TGF-ß produzieren und dementsprechend auch viele TßR besitzen, wirkt
sich eine Abnahme der Rezeptoren auf die Proliferationstätigkeit der Stammzellen aus
(Gazit et al., 1993). Die Signale können nicht in ausreichender Höhe generiert werden,
wie es für die Stammzellen vonnöten wäre. Die höchste Expression der TßR-I und -II
wird bei 100 nM erreicht. Der TßR-III steigt im Zeitverlauf und mit zunehmender Steroid-
konzentration an. Diese Anstiege könnten auf vermehrt verfügbares TGF-ß zurückzufüh-
ren sein, das nicht mehr in ausreichender Menge von den Rezeptoren gebunden werden
kann (Boulanger et al., 1995). In der CR-Gruppe steigt der TßR-I im Zeitverlauf an und
die TßR-II und -III fallen ab, was für eine Förderung der Proliferation spricht. Dies bewir-
ken Steroide in vivo und in vitro. In toto führen steigende Steroidkonzentrationen zu
Seite 126

Kapitel 5. Diskussion
einem Anstieg der TßR-I und -II (Iba et al., 1995). Der TßR-III zeigt ein Expressionstief
bei 100 nM. Im Zeitverlauf sinkt die Expression der TßR-I und -II in der TE-Gruppe. Der
TßR-III zeigt einen uneinheitlichen Verlauf. Die TE-Gruppe besteht hauptsächlich aus
osteozytären Zellen. Diese Reduktion der Rezeptoren scheint das Unvermögen der
Knochenzellen widerzuspiegeln, nicht mehr auf äußere Signale des TGF-ß-Weges ad-
äquat reagieren zu können. Mit zunehmender Steroidkonzentration steigt die Expression
der TßR-I bis -III an. Dieser Effekt könnte promotorvermittelt ablaufen, da zumindest für
den TßR-III ein GRE (Ji et al., 1999) und für den TßR-I eine Beeinflussung über CBFa1
nachgewiesen ist (Chang et al., 1998). Die Glukokortikoide üben ihre Effekte in Abhän-
gigkeit der Differenzierung, der Inkubationszeit und der Konzentration aus. Bezüglich
dieser Zusammenhänge bedarf es weiterer Untersuchungen, da die Auswertung einer
Zellinie nicht repräsentativ ist.
Seite 127

Kapitel 6. Zusammenfassung
6. Zusammenfassung
Der Knochen stellt ein Organsystem dar, das ständig durch Auf-, Um- und Abbau der
Knochensubstanz verändert wird. Transforming Growth Factor beta (TGF-ß) ist ein maß-
geblicher Faktor in der Steuerung dieser Prozesse. Eine Deregulation von TGF-ß kann
pathologische Zustände, unter anderem Arthrose, Fibrose sowie einen verstärkten
Knochenumbau und -abbau, zur Folge haben. Für die Übertragung der TGF-ß-Signale
auf die Zelle werden die TGF-ß-Rezeptoren (TßR) benötigt, die die Signale unter
anderem an die Smads weiterleiten. Die TßR-Expression hängt von diversen Faktoren
ab. Veränderungen der TßR-Wirksamkeit können sowohl auf der mRNA-Ebene als auch
auf der Proteinebene stattfinden.
In dieser Studie wird zum einen die TßR-mRNA-Expression an menschlichen, intraopera-
tiv gewonnenen Knochenproben untersucht, zum anderen die TßR-mRNA-Expression
an humanen, primären Knochenzellkulturen unterschiedlichen Reifegrades mit oder
ohne Einwirkung des hochpotenten Glukokortikoides Dexamethason.
Zur Analyse wurde ein System verwendet, das hochsensitiv und spezifisch ist und es
ermöglicht, aus kleinen Knochenproben zuverlässig RNA zu extrahieren, revers zu trans-
kribieren und zu analysieren. Es mußte zuverlässig arbeiten und kleinste Mengen an
cDNA nachweisen können. Es wurde ein Zwei-Schritt-RT-PCR-Verfahren zur Quantifi-
zierung mit Hilfe der „real-time“-PCR etabliert. Die neuen „real-time“-PCR-Geräte erfüll-
ten alle Bedingungen, die an die Analyseverfahren gestellt wurden. Spezifische Nach-
weise sind damit bis in den Femtogrammbereich bei einem cDNA-Einsatz im Nano-
grammbereich möglich. Außerdem verfügen diese Geräte über eine äußerst hohe Sensi-
tivität und eine hohe Spezifität. Dieses Verfahren ermöglicht es, sowohl absolut als auch
relativ zu quantifizieren und die gewonnenen Proben zuverlässig zu analysieren.
In dieser klinischen Studie ist es erstmals möglich, in einem Kollektiv Korrelationen zwi-
schen den Serumwerten der TGF-ß-Isoformen, der Konzentration der TGF-ß-Isoformen
im Knochenmehl, der mRNA-Expression der TGF-ß-Isoformen und der mRNA-Expressi-
on der TßR zu untersuchen. Es finden sich signifikante Korrelationen zwischen den TGF-
ß-Isoformen im Knochen oder Serum und der TßR-Expression. Es zeigen sich Korre-
lationen zwischen der mRNA-Expression der TGF-ß-Isoformen und der TßR. Das
Kollektiv wird nach verschiedenen Gesichtspunkten gegliedert und innerhalb dieser Kri-
Seite 128

Kapitel 6. Zusammenfassung
terien werden zum Teil signifikante Veränderungen der TßR-Expression sichtbar.
Wesentliche altersspezifische Unterschiede in der TßR-Expression können nicht nachge-
wiesen werden. Geschlechtsspezifische Unterschiede sind nachweisbar. Prämenopau-
sale Frauen zeigen eine niedrigere Expression als postmenopausale. Bei Männern
verhält es sich fast umgekehrt, jüngere Männer zeigen deutlich höhere TßR- Ex-
pressionen als ältere, männliche Probanden. Bei Patienten mit Osteoarthrose kann ein
signifikant erhöhter TßR-II nachgewiesen werden, der durch eine zunehmende Diffe-
renzierung der Knochenzellen erklärt werden kann. Aus der Tibia entnommene Proben
zeigen eine signifikant erhöhte Expression des TßR-II im Vergleich zu Beckenkamm-
proben. Eine Abhängigkeit von der örtlichen Knochenumbaurate, die in der Tibia höher
ist als im Beckenkamm, und erhöhten TGF-ß-Spiegeln könnte für die hohe TßR-II-Ex-
pression verantwortlich sein. Immobilisation scheint eine Veränderung der TßR-Expressi-
on zu bewirken. Kurzfristige Immobilisation erniedrigt alle TßR. Steroide bewirken eine
deutliche Erniedrigung der TßR-I/PDH-Expression, alle weiteren TßR werden ebenfalls
vermindert exprimiert. Diese Änderungen könnten CBFa1-vermittelt sein, da Steroide
CBFa1 hemmen und die Knochenzellen sich CBFa1-vermittelt ausdifferenzieren. Der
Einfluß des BMI auf die TßR-Expression bewirkt eine signifikante Erniedrigung des TßR-I
und eine nicht-signifikante Erniedrigung der TßR-II und -III bei Normalgewichtigen im
Vergleich zu Übergewichtigen. Dies könnte mit dem Einfluß des Leptins auf die Diffe-
renzierung von Knochenzellen zusammenhängen. Es werden hochsignifikante Korre-
lationen zwischen den TßR nachgewiesen, darauf hinweisend, daß das Verhältnis der
Rezeptoren zueinander wichtig ist.
Die Ergebnisse aus den primären Knochenzellkulturen deuten auf eine hohe Diffe-
renzierungsabhängigkeit der TßR-Expression hin. Die höchsten Expressionswerte der
TßR erreichen osteoblastäre Zellen. Ebenfalls scheint es eine differenzierungsabhängige
Wirkung der Glukokortikoide auf Knochenzellen zu geben, die von Einwirkungsdauer
und -konzentration der Glukokortikoide abhängen. In toto scheinen Steroide die TßR-Ex-
pression im Zeitverlauf zu reduzieren. Höhere Konzentrationen der Glukokortikoide be-
wirken im Schnitt eine höhere TßR-Expression.
In dieser Arbeit wird an humanen Knochenproben und -zellkulturen die exogenen und
endogenen Einflüsse auf die TßR-mRNA-Expression mittels quantitativer RT-PCR näher
analysiert. Der Nachweis der differentiellen Expression der TßR-Isoformen liefert neue
Hinweise für das Verständnis des Knochenstoffwechsels.
Seite 129

Kapitel 7. Literatur
7. Literatur
1. Alessi, M.C., Bastelica, D., Morange, P., Berthet, B., Leduc, I., Verdier, M., Geel, O., Juhan-
Vague, I.:
Plasminogen activator inhibitor 1, transforming growth factor-beta1, and BMI are closely associa-
ted in human adipose tissue during morbid obesity.
Diabetes 2000; 49 (8): 1374-1380.
2. Allolio, B., Schulte, H.M.:
Praktische Endokrinologie.
Urban und Schwarzenberg, München, 1996.
3. Amling, M., Schilling, A.F., Haberland, M., Rueger, J.M.:
Leptin: factor in the central nervous system regulation of bone mass. Development of a new un-
derstanding of bone remodeling, skeletal reconstruction, skeletal preservation and skeletal repair.
Orthopade 2001; 30 (7): 418-424.
4. Audi, L., Garcia-Ramirez, M., Carrascosa, A.:
Genetic determinants of bone mass.
Horm Res 1999; 51 (3): 105-123.
5. Batge, B., Feydt, A., Gebken, J., Klein, H., Notbohm, H., Muller, P.K., Brinckmann, J.:
Age-related differences in the expression of receptors for TGF-beta in human osteoblast-like cells
in vitro.
Exp Clin Endocrinol Diabetes 2000; 108 (4): 311-315.
6. Baylink, D.J., Finkelman, R.D., Mohan, S.:
Growth factors to stimulate bone formation.
J Bone Miner Res 1993; 8 Suppl 2 : S565-572.
7. Bellido, T., Jilka, R.L., Boyce, B.F., Girasole, G., Broxmeyer, H., Dalrymple, S.A., Murray, R.,
Manolagas, S.C.:
Regulation of interleukin-6, osteoclastogenesis, and bone mass by androgens. The role of the
androgen receptor.
J Clin Invest 1995; 95 (6): 2886-2895.
Seite 130

Kapitel 7. Literatur
8. Bellows, C.G., Ciaccia, A., Heersche, J.N.:
Osteoprogenitor cells in cell populations derived from mouse and rat calvaria differ in their resp-
onse to corticosterone, cortisol, and cortisone.
Bone 1998; 23 (2): 119-125.
9. Bikle, D.D., Halloran, B.P.:
The response of bone to unloading.
J Bone Miner Metab 1999; 17 (4): 233-244.
10. Bikle, D.D., Harris, J., Halloran, B.P., Currier, P.A., Tanner, S., Morey-Holton, E.:
The molecular response of bone to growth hormone during skeletal unloading: regional diffe-
rences.
Endocrinology 1995; 136 (5): 2099-2109.
11. Bikle, D.D., Harris, J., Halloran, B.P., Morey-Holton, E.:
Altered skeletal pattern of gene expression in response to spaceflight and hindlimb elevation.
Am J Physiol 1994; 267 (6 Pt 1): E822-827.
12. Billiau, A., Edy, V.G., Heremans, H., Van Damme, J., Desmyter, J., Georgiades, J.A., De
Somer, P.:
Human interferon: mass production in a newly established cell line, MG-63.
Antimicrob Agents Chemother 1977; 12 (1): 11-15.
13. Bismar, H., Kloppinger, T., Schuster, E.M., Balbach, S., Diel, I., Ziegler, R., Pfeilschifter, J.:
Transforming growth factor beta (TGF-beta) levels in the conditioned media of human bone cells:
relationship to donor age, bone volume, and concentration of TGF-beta in human bone matrix in
vivo.
Bone 1999; 24 (6): 565-569.
14. Bloom, B.B., Humphries, D.E., Kuang, P.P., Fine, A., Goldstein, R.H.:
Structure and expression of the promoter for the R4/ALK5 human type I transforming growth fac-
tor-beta receptor: regulation by TGF-beta.
Biochim Biophys Acta 1996; 1312 (3): 243-248.
15. Blümcke, S., (Herausgeber):
Pathologie (Lehrbuch).
Walter de Gruyter, Berlin, 1995.
Seite 131

Kapitel 7. Literatur
16. Bodine, P.V., Riggs, B.L., Spelsberg, T.C.:
Regulation of c-fos expression and TGF-beta production by gonadal and adrenal androgens in nor-
mal human osteoblastic cells.
J Steroid Biochem Mol Biol 1995; 52 (2): 149-158.
17. Bonewald, L.F., Dallas, S.L.:
Role of active and latent transforming growth factor beta in bone formation.
J Cell Biochem 1994; 55 (3): 350-357.
18. Bonewald, L.F., Wakefield, L., Oreffo, R.O., Escobedo, A., Twardzik, D.R., Mundy, G.R.:
Latent forms of transforming growth factor-beta (TGF beta) derived from bone cultures: identifica-
tion of a naturally occurring 100-kDa complex with similarity to recombinant latent TGF beta.
Mol Endocrinol 1991; 5 (6): 741-751.
19. Bord, S., Beavan, S., Ireland, D., Horner, A., Compston, J.E.:
Mechanisms by which high-dose estrogen therapy produces anabolic skeletal effects in postmeno-
pausal women: role of locally produced growth factors.
Bone 2001; 29 (3): 216-222.
20. Boulanger, J., Reyes-Moreno, C., Koutsilieris, M.:
Mediation of glucocorticoid receptor function by the activation of latent transforming growth factor
beta 1 in MG-63 human osteosarcoma cells.
Int J Cancer 1995; 61 (5): 692-697.
21. Brand, T., Schneider, M.D.:
Inactive type II and type I receptors for TGF beta are dominant inhibitors of TGF beta-dependent
transcription.
J Biol Chem 1995; 270 (14): 8274-8284.
22. Centrella, M., Casinghino, S., Ignotz, R., McCarthy, T.L.:
Multiple regulatory effects by transforming growth factor-beta on type I collagen levels in osteo-
blast-enriched cultures from fetal rat bone.
Endocrinology 1992; 131 (6): 2863-2872.
23. Centrella, M., Horowitz, M.C., Wozney, J.M., McCarthy, T.L.:
Transforming growth factor-beta gene family members and bone.
Endocr Rev 1994; 15 (1): 27-39.
Seite 132

Kapitel 7. Literatur
24. Centrella, M., Casinghino, S., Kim, J., Pham, T., Rosen, V., Wozney, J., McCarthy, T.L.:
Independent changes in type I and type II receptors for transforming growth factor beta induced by
bone morphogenetic protein 2 parallel expression of the osteoblast phenotype.
Mol Cell Biol 1995; 15 (6): 3273-3281.
25. Centrella, M., Ji, C., Casinghino, S., McCarthy, T.L.:
Rapid flux in transforming growth factor-beta receptors on bone cells.
J Biol Chem 1996; 271 (31): 18616-18622.
26. Centrella, M., McCarthy, T.L., Canalis, E.:
Glucocorticoid regulation of transforming growth factor beta 1 activity and binding in osteoblast-
enriched cultures from fetal rat bone.
Mol Cell Biol 1991; 11 (9): 4490-4496.
27. Centrella, M., Rosen, V., Wozney, J.M., Casinghino, S.R., McCarthy, T.L.:
Opposing effects by glucocorticoid and bone morphogenetic protein-2 in fetal rat bone cell
cultures.
J Cell Biochem 1997; 67 (4): 528-540.
28. Charng, M.J., Kinnunen, P., Hawker, J., Brand, T., Schneider, M.D.:
FKBP-12 recognition is dispensable for signal generation by type I transforming growth factor-beta
receptors.
J Biol Chem 1996; 271 (38): 22941-22944.
29. Chevalley, T., Strong, D.D., Mohan, S., Baylink, D., Linkhart, T.A.:
Evidence for a role for insulin-like growth factor binding proteins in glucocorticoid inhibition of nor-
mal human osteoblast-like cell proliferation.
Eur J Endocrinol 1996; 134 (5): 591-601.
30. Chomczynski, P., Sacchi, N.:
Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extracti-
on.
Anal Biochem 1987; 162 (1): 156-159.
31. Choy, L., Derynck, R.:
The type II transforming growth factor (TGF)-beta receptor-interacting protein TRIP-1 acts as a
modulator of the TGF-beta response.
J Biol Chem 1998; 273 (47): 31455-31462.
Seite 133

Kapitel 7. Literatur
32. Cillo, J.E., Jr., Gassner, R., Koepsel, R.R., Buckley, M.J.:
Growth factor and cytokine gene expression in mechanically strained human osteoblast-like cells:
implications for distraction osteogenesis.
Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2000; 90 (2): 147-154.
33. Clark, D.A., Coker, R.:
Transforming growth factor-beta (TGF-beta).
Int J Biochem Cell Biol 1998; 30 (3): 293-298.
34. Collins, D., Woods, A., Herd, R., Blake, G., Fogelman, I., Wheeler, M., Swaminathan, R.:
Insulin-like growth factor-I and bone mineral density.
Bone 1998; 23 (1): 13-16.
35. Dabovic, B., Chen, Y., Colarossi, C., Obata, H., Zambuto, L., Perle, M.A., Rifkin, D.B.:
Bone abnormalities in latent TGF-{beta} binding protein (Ltbp)-3-null mice indicate a role for Ltbp-
3 in modulating TGF-{beta} bioavailability.
J Cell Biol 2002; 14 : 14.
36. Dallas, S.L., Park-Snyder, S., Miyazono, K., Twardzik, D., Mundy, G.R., Bonewald, L.F.:
Characterization and autoregulation of latent transforming growth factor beta (TGF beta) comple-
xes in osteoblast-like cell lines. Production of a latent complex lacking the latent TGF beta-binding
protein.
J Biol Chem 1994; 269 (9): 6815-6821.
37. Derynck, R.:
TGF-beta-receptor-mediated signaling.
Trends Biochem Sci 1994; 19 (12): 548-553.
38. Derynck, R., Feng, X.H.:
TGF-beta receptor signaling.
Biochim Biophys Acta 1997; 1333 (2): F105-150.
39. Devoto, M., Shimoya, K., Caminis, J., Ott, J., Tenenhouse, A., Whyte, M.P., Sereda, L., Hall,
S., Considine, E., Williams, C.J., Tromp, G., Kuivaniemi, H., Ala-Kokko, L., Prockop, D.J., Spotila,
L.D.:
First-stage autosomal genome screen in extended pedigrees suggests genes predisposing to low
bone mineral density on chromosomes 1p, 2p and 4q.
Eur J Hum Genet 1998; 6 (2): 151-157.
Seite 134

Kapitel 7. Literatur
40. Drissi, H., Lomri, A., Lasmoles, F., Holy, X., Zerath, E., Marie, P.J.:
Skeletal unloading induces biphasic changes in insulin-like growth factor-I mRNA levels and osteo-
blast activity.
Exp Cell Res 1999; 251 (2): 275-284.
41. Dunstan, C.R., Boyce, R., Boyce, B.F., Garrett, I.R., Izbicka, E., Burgess, W.H., Mundy, G.R.:
Systemic administration of acidic fibroblast growth factor (FGF-1) prevents bone loss and increa-
ses new bone formation in ovariectomized rats.
J Bone Miner Res 1999; 14 (6): 953-959.
42. Eingartner, C., Coerper, S., Fritz, J., Gaissmaier, C., Koveker, G., Weise, K.:
Growth factors in distraction osteogenesis. Immuno-histological pattern of TGF-beta1 and IGF-I in
human callus induced by distraction osteogenesis.
Int Orthop 1999; 23 (5): 253-259.
43. Eisman, J.A.:
Genetics of osteoporosis.
Endocr Rev 1999; 20 (6): 788-804.
44. Erlebacher, A., Filvaroff, E.H., Ye, J.Q., Derynck, R.:
Osteoblastic responses to TGF-beta during bone remodeling.
Mol Biol Cell 1998; 9 (7): 1903-1918.
45. Falk, L.A., De Benedetti, F., Lohrey, N., Birchenall-Roberts, M.C., Ellingsworth, L.W., Falty-
nek, C.R., Ruscetti, F.W.:
Induction of transforming growth factor-beta 1 (TGF-beta 1), receptor expression and TGF-beta 1
protein production in retinoic acid-treated HL-60 cells: possible TGF-beta 1-mediated autocrine in-
hibition.
Blood 1991; 77 (6): 1248-1255.
46. Fenwick, S.A., Curry, V., Harrall, R.L., Hazleman, B.L., Hackney, R., Riley, G.P.:
Expression of transforming growth factor-beta isoforms and their receptors in chronic tendinosis.
J Anat 2001; 199 (Pt 3): 231-240.
47. Fernandez-Ruiz, E., St-Jacques, S., Bellon, T., Letarte, M., Bernabeu, C.:
Assignment of the human endoglin gene (END) to 9q34-->qter.
Cytogenet Cell Genet 1993; 64 (3-4): 204-207.
Seite 135

Kapitel 7. Literatur
48. Franzen, P., ten Dijke, P., Ichijo, H., Yamashita, H., Schulz, P., Heldin, C.H., Miyazono, K.:
Cloning of a TGF beta type I receptor that forms a heteromeric complex with the TGF beta type II
receptor.
Cell 1993; 75 (4): 681-692.
49. Garcia-Lozano, J.R., Gonzalez-Escribano, M.F., Valenzuela, A., Garcia, A., Nunez-Roldan, A.:
Association of vitamin D receptor genotypes with early onset rheumatoid arthritis.
Eur J Immunogenet 2001; 28 (1): 89-93.
50. Gazit, D., Ebner, R., Kahn, A.J., Derynck, R.:
Modulation of expression and cell surface binding of members of the transforming growth factor-
beta superfamily during retinoic acid-induced osteoblastic differentiation of multipotential
mesenchymal cells.
Mol Endocrinol 1993; 7 (2): 189-198.
51. Gazit, D., Zilberman, Y., Ebner, R., Kahn, A.:
Bone loss (osteopenia) in old male mice results from diminished activity and availability of TGF-
beta.
J Cell Biochem 1998; 70 (4): 478-488.
52. Gebken, J., Brenner, R., Feydt, A., Notbohm, H., Brinckmann, J., Muller, P.K., Batge, B.:
Increased cell surface expression of receptors for transforming growth factor-beta on osteoblasts
from patients with Osteogenesis imperfecta.
Pathobiology 2000; 68 (3): 106-112.
53. Gebken, J., Feydt, A., Brinckmann, J., Notbohm, H., Muller, P.K., Batge, B.:
Ligand-induced downregulation of receptors for TGF-beta in human osteoblast-like cells from adult
donors.
J Endocrinol 1999; 161 (3): 503-510.
54. Geiser, A.G., Zeng, Q.Q., Sato, M., Helvering, L.M., Hirano, T., Turner, C.H.:
Decreased bone mass and bone elasticity in mice lacking the transforming growth factor-beta1
gene.
Bone 1998; 23 (2): 87-93.
55. Gelbrich, G.:
Statistik für Anwender.
Shaker Verlag, Aachen, 1998.
Seite 136

Kapitel 7. Literatur
56. Ghosh-Choudhury, N., Ghosh-Choudhury, G., Celeste, A., Ghosh, P.M., Moyer, M., Abboud,
S.L., Kreisberg, J.:
Bone morphogenetic protein-2 induces cyclin kinase inhibitor p21 and hypophosphorylation of reti-
noblastoma protein in estradiol-treated MCF-7 human breast cancer cells.
Biochim Biophys Acta 2000; 1497 (2): 186-196.
57. Gori, F., Thomas, T., Hicok, K.C., Spelsberg, T.C., Riggs, B.L.:
Differentiation of human marrow stromal precursor cells: bone morphogenetic protein-2 increases
OSF2/CBFA1, enhances osteoblast commitment, and inhibits late adipocyte maturation.
J Bone Miner Res 1999; 14 (9): 1522-1535.
58. Gortz, B., Fassbender, W.J.:
Genetics of osteoporosis.
Orthopade 2001; 30 (7): 412-417.
59. Gough, A., Sambrook, P., Devlin, J., Lilley, J., Huisoon, A., Betteridge, J., Franklyn, J., Nguy-
en, T., Morrison, N., Eisman, J., Emery, P.:
Effect of vitamin D receptor gene alleles on bone loss in early rheumatoid arthritis.
J Rheumatol 1998; 25 (5): 864-868.
60. Graves, D.T., Valentin-Opran, A., Delgado, R., Valente, A.J., Mundy, G., Piche, J.:
The potential role of platelet-derived growth factor as an autocrine or paracrine factor for human
bone cells.
Connect Tissue Res 1989; 23 (2-3): 209-218.
61. Gross, R., Schölmerich, P., Gross, W., (Herausgeber):
Die Innere Medizin (Lehrbuch).
Schattauer, Stuttgart, 1996.
62. Guise, T.A., Mundy, G.R.:
Cancer and bone.
Endocr Rev 1998; 19 (1): 18-54.
63. Halloran, B.P., Bikle, D.D., Harris, J., Foskett, H.C., Morey-Holton, E.:
Skeletal unloading decreases production of 1,25-dihydroxyvitamin D.
Am J Physiol 1993; 264 (5 Pt 1): E712-716.
Seite 137

Kapitel 7. Literatur
64. Han, D.C., Isono, M., Chen, S., Casaretto, A., Hong, S.W., Wolf, G., Ziyadeh, F.N.:
Leptin stimulates type I collagen production in db/db mesangial cells: glucose uptake and TGF-
beta type II receptor expression.
Kidney Int 2001; 59 (4): 1315-1323.
65. Harris, S.A., Zhang, M., Kidder, L.S., Evans, G.L., Spelsberg, T.C., Turner, R.T.:
Effects of orbital spaceflight on human osteoblastic cell physiology and gene expression.
Bone 2000; 26 (4): 325-331.
66. Harris, S.E., Sabatini, M., Harris, M.A., Feng, J.Q., Wozney, J., Mundy, G.R.:
Expression of bone morphogenetic protein messenger RNA in prolonged cultures of fetal rat cal-
varial cells.
J Bone Miner Res 1994; 9 (3): 389-394.
67. Hata, A., Shi, Y., Massague, J.:
TGF-beta signaling and cancer: structural and functional consequences of mutations in Smads.
Mol Med Today 1998; 4 (6): 257-262.
68. Helvering, L.M., Sharp, R.L., Ou, X., Geiser, A.G.:
Regulation of the promoters for the human bone morphogenetic protein 2 and 4 genes.
Gene 2000; 256 (1-2): 123-138.
69. Henninger, H.P., Hoffmann, R., Grewe, M., Schulze-Specking, A., Decker, K.:
Purification and quantitative analysis of nucleic acids by anion-exchange high-performance liquid
chromatography.
Biol Chem Hoppe Seyler 1993; 374 (8): 625-634.
70. Hering, S.:
Untersuchung zur Regulation von TGFß 1 bis 3 und der spezifischen TGFß-Rezeptor Typen I bis
III in normalem und pathologisch verändertem Knochen.
Habilitation, Medizinische Fakultät Bochum, 2002.
71. Hill, C.S.:
Signalling to the nucleus by members of the transforming growth factor-beta (TGF-beta) superfa-
mily.
Cell Signal 1996; 8 (8): 533-544.
Seite 138

Kapitel 7. Literatur
72. Hill, C.S.:
The Smads.
Int J Biochem Cell Biol 1999; 31 (11): 1249-1254.
73. Hofbauer, L.C., Hicok, K.C., Khosla, S.:
Effects of gonadal and adrenal androgens in a novel androgen-responsive human osteoblastic cell
line.
J Cell Biochem 1998; 71 (1): 96-108.
74. Horie, K., Yamashita, H., Mogi, A., Takenoshita, S., Miyazono, K.:
Lack of transforming growth factor-beta type II receptor expression in human retinoblastoma cells.
J Cell Physiol 1998; 175 (3): 305-313.
75. Horner, A., Kemp, P., Summers, C., Bord, S., Bishop, N.J., Kelsall, A.W., Coleman, N., Comp-
ston, J.E.:
Expression and distribution of transforming growth factor-beta isoforms and their signaling recep-
tors in growing human bone.
Bone 1998; 23 (2): 95-102.
76. Hu, P.P., Datto, M.B., Wang, X.F.:
Molecular mechanisms of transforming growth factor-beta signaling.
Endocr Rev 1998; 19 (3): 349-363.
77. Huang, L., Li, C.:
Leptin: a multifunctional hormone.
Cell Res 2000; 10 (2): 81-92.
78. Hughes, D.E., Dai, A., Tiffee, J.C., Li, H.H., Mundy, G.R., Boyce, B.F.:
Estrogen promotes apoptosis of murine osteoclasts mediated by TGF-beta.
Nat Med 1996; 2 (10): 1132-1136.
79. Hulley, P.A., Gordon, F., Hough, F.S.:
Inhibition of mitogen-activated protein kinase activity and proliferation of an early osteoblast cell
line (MBA 15.4) by dexamethasone: role of protein phosphatases.
Endocrinology 1998; 139 (5): 2423-2431.
Seite 139

Kapitel 7. Literatur
80. Humphries, D.E., Bloom, B.B., Fine, A., Goldstein, R.H.:
Structure and expression of the promoter for the human type II transforming growth factor-beta re-
ceptor.
Biochem Biophys Res Commun 1994; 203 (2): 1020-1027.
81. Iba, K., Chiba, H., Sawada, N., Hirota, S., Ishii, S., Mori, M.:
Glucocorticoids induce mineralization coupled with bone protein expression without influence on
growth of a human osteoblastic cell line.
Cell Struct Funct 1995; 20 (5): 319-330.
82. Iba, K., Sawada, N., Chiba, H., Wewer, U.M., Ishii, S., Mori, M.:
Transforming growth factor-beta 1 downregulates dexamethasone-induced tetranectin gene ex-
pression during the in vitro mineralization of the human osteoblastic cell line SV-HFO.
FEBS Lett 1995; 373 (1): 1-4.
83. Iba, K., Sawada, N., Nuka, S., Chiba, H., Obata, H., Isomura, H., Satoh, M., Ishii, S., Mori, M.:
Phase-Dependent effects of transforming growth factor beta 1 on osteoblastic markers of human
osteoblastic cell line sV-HFO during mineralization.
Bone 1996; 19 (4): 363-369.
84. Inoue, M., Tanaka, H., Moriwake, T., Oka, M., Sekiguchi, C., Seino, Y.:
Altered biochemical markers of bone turnover in humans during 120 days of bed rest.
Bone 2000; 26 (3): 281-286.
85. Iseki, S., Osumi-Yamashita, N., Miyazono, K., Franzen, P., Ichijo, H., Ohtani, H., Hayashi, Y.,
Eto, K.:
Localization of transforming growth factor-beta type I and type II receptors in mouse development.
Exp Cell Res 1995; 219 (2): 339-347.
86. Isken, F.:
Entwicklung und Anwendung einer HPLC gestützten quantitative RNA-Analyse von drei Isoformen
der TGF-ß Familie in humanem Knochen.
Dissertationsarbeit, Medizinische Fakultät Bochum, 2000.
87. Ji, C., Casinghino, S., McCarthy, T.L., Centrella, M.:
Multiple and essential Sp1 binding sites in the promoter for transforming growth factor-beta type I
receptor.
J Biol Chem 1997; 272 (34): 21260-21267.
Seite 140

Kapitel 7. Literatur
88. Ji, C., Casinghino, S., McCarthy, T.L., Centrella, M.:
Cloning, characterization, and expression of the transforming growth factor-beta type I receptor
promoter in fetal rat bone cells.
J Cell Biochem 1996; 63 (4): 478-490.
89. Ji, C., Chen, Y., McCarthy, T.L., Centrella, M.:
Cloning the promoter for transforming growth factor-beta type III receptor. Basal and conditional
expression in fetal rat osteoblasts.
J Biol Chem 1999; 274 (43): 30487-30494.
90. Johnson, D.W., Berg, J.N., Gallione, C.J., McAllister, K.A., Warner, J.P., Helmbold, E.A.,
Markel, D.S., Jackson, C.E., Porteous, M.E., Marchuk, D.A.:
A second locus for hereditary hemorrhagic telangiectasia maps to chromosome 12.
Genome Res 1995; 5 (1): 21-28.
91. Johnson, D.W., Qumsiyeh, M., Benkhalifa, M., Marchuk, D.A.:
Assignment of human transforming growth factor-beta type I and type III receptor genes (TGFBR1
and TGFBR3) to 9q33-q34 and 1p32-p33, respectively.
Genomics 1995; 28 (2): 356-357.
92. Jost, C.F.:
Bedeutung des transforming growth factor beta (TGF-beta) für den Knochenstoffwechsel des Men-
schen in physiologischen und unphysiologischen Situationen.
Dissertationsarbeit, Medizinische Fakultät Bochum, 2000.
93. Junqueira, L.C., Carneiro, J.:
Histologie (Lehrbuch).
Springer, Heidelberg, 1996.
94. Kabasawa, Y., Ejiri, S., Matsuki, Y., Hara, K., Ozawa, H.:
Immunoreactive localization of transforming growth factor-beta type II receptor-positive cells in rat
tibiae.
Bone 1998; 22 (2): 93-98.
95. Kaspar, D., Seidl, W., Ignatius, A., Neidlinger-Wilke, C., Claes, L.:
In vitro cell behavior of human osteoblasts after physiological dynamic stretching.
Orthopade 2000; 29 (2): 85-90.
Seite 141

Kapitel 7. Literatur
96. Kasperk, C., Fitzsimmons, R., Strong, D., Mohan, S., Jennings, J., Wergedal, J., Baylink, D.:
Studies of the mechanism by which androgens enhance mitogenesis and differentiation in bone
cells.
J Clin Endocrinol Metab 1990; 71 (5): 1322-1329.
97. Kasperk, C.H., Wakley, G.K., Hierl, T., Ziegler, R.:
Gonadal and adrenal androgens are potent regulators of human bone cell metabolism in vitro.
J Bone Miner Res 1997; 12 (3): 464-471.
98. Kassem, M., Kveiborg, M., Eriksen, E.F.:
Production and action of transforming growth factor-beta in human osteoblast cultures:
dependence on cell differentiation and modulation by calcitriol.
Eur J Clin Invest 2000; 30 (5): 429-437.
99. Katz, E.D., Dong, M.W.:
Rapid analysis and purification of polymerase chain reaction products by high-performance liquid
chromatography.
Biotechniques 1990; 8 (5): 546-555.
100. Katz, E.D., Haff, L.A., Eksteen, R.:
Rapid separation, quantitation and purification of products of polymerase chain reaction by liquid
chromatography.
J Chromatogr 1990; 512 : 433-444.
101. Khalil, N.:
TGF-beta: from latent to active.
Microbes Infect 1999; 1 (15): 1255-1263.
102. Köhler, T., Laßner, D., Rost, A.K., Thamm, B., Pustowoit, B., H., R.:
Quantitation of mRNA by polymerase chain reaction.
Springer, Heidelberg, 1995.
103. Koike, K., Urata, Y., Matsuo, S., Koike, M.:
Characterization and nucleotide sequence of the gene encoding the human pyruvate dehydrogen-
ase alpha-subunit.
Gene 1990; 93 (2): 307-311.
Seite 142

Kapitel 7. Literatur
104. Koike, K., Urata, Y., Koike, M.:
Molecular cloning and characterization of human pyruvate dehydrogenase beta subunit gene.
Proc Natl Acad Sci U S A 1990; 87 (15): 5594-5597.
105. Kostenuik, P.J., Halloran, B.P., Morey-Holton, E.R., Bikle, D.D.:
Skeletal unloading inhibits the in vitro proliferation and differentiation of rat osteoprogenitor cells.
Am J Physiol 1997; 273 (6 Pt 1): E1133-1139.
106. Kume, K., Satomura, K., Nishisho, S., Kitaoka, E., Yamanouchi, K., Tobiume, S., Nagayama,
M.:
Potential role of leptin in endochondral ossification.
J Histochem Cytochem 2002; 50 (2): 159-170.
107. Kurokouchi, K., Ito, T., Ohmori, S., Kanda, K., Murata, Y., Seo, H.:
Changes in the markers of bone metabolism following skeletal unloading.
Environ Med 1995; 39 (1): 21-24.
108. Langdahl, B.L., Knudsen, J.Y., Jensen, H.K., Gregersen, N., Eriksen, E.F.:
A sequence variation: 713-8delC in the transforming growth factor-beta 1 gene has higher preva-
lence in osteoporotic women than in normal women and is associated with very low bone mass in
osteoporotic women and increased bone turnover in both osteoporotic and normal women.
Bone 1997; 20 (3): 289-294.
109. Lawler, S., Candia, A.F., Ebner, R., Shum, L., Lopez, A.R., Moses, H.L., Wright, C.V., De-
rynck, R.:
The murine type II TGF-beta receptor has a coincident embryonic expression and binding prefe-
rence for TGF-beta 1.
Development 1994; 120 (1): 165-175.
110. Lehninger, A.T., L., N.D.:
Lehninger Biochemie (Lehrbuch).
Springer, Heidelberg, 2001.
Seite 143

Kapitel 7. Literatur
111. Lian, J.B., Shalhoub, V., Aslam, F., Frenkel, B., Green, J., Hamrah, M., Stein, G.S., Stein,
J.L.:
Species-specific glucocorticoid and 1,25-dihydroxyvitamin D responsiveness in mouse MC3T3-E1
osteoblasts: dexamethasone inhibits osteoblast differentiation and vitamin D down-regulates
osteocalcin gene expression.
Endocrinology 1997; 138 (5): 2117-2127.
112. Lin, H.Y., Wang, X.F.:
Expression cloning of TGF-beta receptors.
Mol Reprod Dev 1992; 32 (2): 105-110.
113. Lin, H.Y., Wang, X.F., Ng-Eaton, E., Weinberg, R.A., Lodish, H.F.:
Expression cloning of the TGF-beta type II receptor, a functional transmembrane serine/threonine
kinase.
Cell 1992; 68 (4): 775-785.
114. Lin, K.Y., Nolen, A.A., Gampper, T.J., Jane, J.A., Opperman, L.A., Ogle, R.C.:
Elevated levels of transforming growth factors beta 2 and beta 3 in lambdoid sutures from children
with persistent plagiocephaly.
Cleft Palate Craniofac J 1997; 34 (4): 331-337.
115. Linkhart, T.A., Mohan, S., Baylink, D.J.:
Growth factors for bone growth and repair: IGF, TGF beta and BMP.
Bone 1996; 19 (1 Suppl): 1S-12S.
116. Lippa, C.F., Flanders, K.C., Kim, E.S., Croul, S.:
TGF-beta receptors-I and -II immunoexpression in Alzheimer's disease: a comparison with aging
and progressive supranuclear palsy.
Neurobiol Aging 1998; 19 (6): 527-533.
117. Longo, M.C., Berninger, M.S., Hartley, J.L.:
Use of uracil DNA glycosylase to control carry-over contamination in polymerase chain reactions.
Gene 1990; 93 (1): 125-128.
118. Lopez-Casillas, F., Cheifetz, S., Doody, J., Andres, J.L., Lane, W.S., Massague, J.:
Structure and expression of the membrane proteoglycan betaglycan, a component of the TGF-
beta receptor system.
Cell 1991; 67 (4): 785-795.
Seite 144

Kapitel 7. Literatur
119. Lopez-Casillas, F., Wrana, J.L., Massague, J.:
Betaglycan presents ligand to the TGF beta signaling receptor.
Cell 1993; 73 (7): 1435-1444.
120. Mann, V., Hobson, E.E., Li, B., Stewart, T.L., Grant, S.F., Robins, S.P., Aspden, R.M., Ral-
ston, S.H.:
A COL1A1 Sp1 binding site polymorphism predisposes to osteoporotic fracture by affecting bone
density and quality.
J Clin Invest 2001; 107 (7): 899-907.
121. Marcelli, C., Yates, A.J., Mundy, G.R.:
In vivo effects of human recombinant transforming growth factor beta on bone turnover in normal
mice.
J Bone Miner Res 1990; 5 (10): 1087-1096.
122. Massague, J.:
Receptors for the TGF-beta family.
Cell 1992; 69 (7): 1067-1070.
123. Massague, J.:
TGF-beta signal transduction.
Annu Rev Biochem 1998; 67 : 753-791.
124. Massague, J., Andres, J., Attisano, L., Cheifetz, S., Lopez-Casillas, F., Ohtsuki, M., Wrana,
J.L.:
TGF-beta receptors.
Mol Reprod Dev 1992; 32 (2): 99-104.
125. Massague, J., Blain, S.W., Lo, R.S.:
TGF-beta signaling in growth control, cancer, and heritable disorders.
Cell 2000; 103 (2): 295-309.
126. Mathew, S., Murty, V.V., Cheifetz, S., George, D., Massague, J., Chaganti, R.S.:
Transforming growth factor receptor gene TGFBR2 maps to human chromosome band 3p22.
Genomics 1994; 20 (1): 114-115.
Seite 145

Kapitel 7. Literatur
127. McAllister, K.A., Grogg, K.M., Johnson, D.W., Gallione, C.J., Baldwin, M.A., Jackson, C.E.,
Helmbold, E.A., Markel, D.S., McKinnon, W.C., Murrell, J., et al.:
Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic
telangiectasia type 1.
Nat Genet 1994; 8 (4): 345-351.
128. Miyazono, K.:
TGF-beta receptors and signal transduction.
Int J Hematol 1997; 65 (2): 97-104.
129. Miyazono, K., ten Dijke, P., Heldin, C.H.:
TGF-beta signaling by Smad proteins.
Adv Immunol 2000; 75 (1): 115-157.
130. Miyazono, K., ten Dijke, P., Yamashita, H., Heldin, C.H.:
Signal transduction via serine/threonine kinase receptors.
Semin Cell Biol 1994; 5 (6): 389-398.
131. Mundy, G.R.:
Regulation of bone formation by bone morphogenetic proteins and other growth factors.
Clin Orthop 1996; (324): 24-28.
132. Mundy, G.R., Boyce, B., Hughes, D., Wright, K., Bonewald, L., Dallas, S., Harris, S., Ghosh-
Choudhury, N., Chen, D., Dunstan, C., et al.:
The effects of cytokines and growth factors on osteoblastic cells.
Bone 1995; 17 (2 Suppl): 71S-75S.
133. Murakami, T., Yamamoto, M., Ono, K., Nishikawa, M., Nagata, N., Motoyoshi, K., Akatsu, T.:
Transforming growth factor-beta1 increases mRNA levels of osteoclastogenesis inhibitory factor in
osteoblastic/stromal cells and inhibits the survival of murine osteoclast-like cells.
Biochem Biophys Res Commun 1998; 252 (3): 747-752.
134. Murata, H., Zhou, L., Ochoa, S., Hasan, A., Badiavas, E., Falanga, V.:
TGF-beta3 stimulates and regulates collagen synthesis through TGF-beta1-dependent and in-
dependent mechanisms.
J Invest Dermatol 1997; 108 (3): 258-262.
Seite 146

Kapitel 7. Literatur
135. Nakashima, T., Sasaki, H., Tsuboi, M., Kawakami, A., Fujiyama, K., Kiriyama, T., Eguchi, K.,
Ichikawa, M., Nagataki, S.:
Inhibitory effect of glucocorticoid for osteoblast apoptosis induced by activated peripheral blood
mononuclear cells.
Endocrinology 1998; 139 (4): 2032-2040.
136. Nakayama, H., Ichikawa, F., Andres, J.L., Massague, J., Noda, M.:
Dexamethasone enhancement of betaglycan (TGF-beta type III receptor) gene expression in
osteoblast-like cells.
Exp Cell Res 1994; 211 (2): 301-306.
137. Neidlinger-Wilke, C., Stalla, I., Claes, L., Brand, R., Hoellen, I., Rubenacker, S., Arand, M.,
Kinzl, L.:
Human osteoblasts from younger normal and osteoporotic donors show differences in proliferation
and TGF beta-release in response to cyclic strain.
J Biomech 1995; 28 (12): 1411-1418.
138. Neidlinger-Wilke, C., Wilke, H.J., Claes, L.:
Cyclic stretching of human osteoblasts affects proliferation and metabolism: a new experimental
method and its application.
J Orthop Res 1994; 12 (1): 70-78.
139. Nguyen, T.V., Center, J.R., Eisman, J.A.:
Osteoporosis in elderly men and women: effects of dietary calcium, physical activity, and body
mass index.
J Bone Miner Res 2000; 15 (2): 322-331.
140. Nicolas, V., Prewett, A., Bettica, P., Mohan, S., Finkelman, R.D., Baylink, D.J., Farley, J.R.:
Age-related decreases in insulin-like growth factor-I and transforming growth factor-beta in femoral
cortical bone from both men and women: implications for bone loss with aging.
J Clin Endocrinol Metab 1994; 78 (5): 1011-1016.
141. Ogawa, S., Hosoi, T., Shiraki, M., Orimo, H., Emi, M., Muramatsu, M., Ouchi, Y., Inoue, S.:
Association of estrogen receptor beta gene polymorphism with bone mineral density.
Biochem Biophys Res Commun 2000; 269 (2): 537-541.
Seite 147

Kapitel 7. Literatur
142. Opperman, L.A., Nolen, A.A., Ogle, R.C.:
TGF-beta 1, TGF-beta 2, and TGF-beta 3 exhibit distinct patterns of expression during cranial su-
ture formation and obliteration in vivo and in vitro.
J Bone Miner Res 1997; 12 (3): 301-310.
143. Oursler, M.J., Riggs, B.L., Spelsberg, T.C.:
Glucocorticoid-induced activation of latent transforming growth factor-beta by normal human
osteoblast-like cells.
Endocrinology 1993; 133 (5): 2187-2196.
144. Padgett, R.W.:
Intracellular signaling: Fleshing out the TGFbeta pathway.
Curr Biol 1999; 9 (11): R408-411.
145. Padgett, R.W., Cho, S.H., Evangelista, C.:
Smads are the central component in transforming growth factor-beta signaling.
Pharmacol Ther 1998; 78 (1): 47-52.
146. Padgett, R.W., Das, P., Krishna, S.:
TGF-beta signaling, Smads, and tumor suppressors.
Bioessays 1998; 20 (5): 382-390.
147. Parfitt, A.M.:
Quantum concept of bone remodeling and turnover: implications for the pathogenesis of osteopo-
rosis.
Calcif Tissue Int 1979; 28 (1): 1-5.
148. Parfitt, A.M.:
Bone effects of space flight: analysis by quantum concept of bone remodelling.
Acta Astronaut 1981; 8 (9-10): 1083-1090.
149. Parrelli, J.M., Meisler, N., Cutroneo, K.R.:
Identification of a glucocorticoid response element in the human transforming growth factor beta 1
gene promoter.
Int J Biochem Cell Biol 1998; 30 (5): 623-627.
Seite 148

Kapitel 7. Literatur
150. Pearce, G., Tabensky, D.A., Delmas, P.D., Baker, H.W., Seeman, E.:
Corticosteroid-induced bone loss in men.
J Clin Endocrinol Metab 1998; 83 (3): 801-806.
151. PerkinElmerBiosystems:
„DNA/RNA Real-Time Quantitative PCR“ (P/N: 4304965 Rev. B) und „SYBR Green PCR and RT-
PCR Reagents“.
Applied Biosytems, Foster City, USA, 2001.
152. Pettersson, U., Nordstrom, P., Alfredson, H., Henriksson-Larsen, K., Lorentzon, R.:
Effect of high impact activity on bone mass and size in adolescent females: A comparative study
between two different types of sports.
Calcif Tissue Int 2000; 67 (3): 207-214.
153. Pfeilschifter, J., Bonewald, L., Mundy, G.R.:
Characterization of the latent transforming growth factor beta complex in bone.
J Bone Miner Res 1990; 5 (1): 49-58.
154. Pfeilschifter, J., Diel, I., Pilz, U., Brunotte, K., Naumann, A., Ziegler, R.:
Mitogenic responsiveness of human bone cells in vitro to hormones and growth factors decreases
with age.
J Bone Miner Res 1993; 8 (6): 707-717.
155. Pfeilschifter, J., Wolf, O., Naumann, A., Minne, H.W., Mundy, G.R., Ziegler, R.:
Chemotactic response of osteoblastlike cells to transforming growth factor beta.
J Bone Miner Res 1990; 5 (8): 825-830.
156. Pfeilschifter, J., Diel, I., Scheppach, B., Bretz, A., Krempien, R., Erdmann, J., Schmid, G.,
Reske, N., Bismar, H., Seck, T., Krempien, B., Ziegler, R.:
Concentration of transforming growth factor beta in human bone tissue: relationship to age, meno-
pause, bone turnover, and bone volume.
J Bone Miner Res 1998; 13 (4): 716-730.
157. Pfeilschifter, J., Pignat, W., Leighton, J., Marki, F., Vosbeck, K., Alkan, S.:
Transforming growth factor beta 2 differentially modulates interleukin-1 beta- and tumour-necrosis-
factor-alpha-stimulated phospholipase A2 and prostaglandin E2 synthesis in rat renal mesangial
cells.
Biochem J 1990; 270 (1): 269-271.
Seite 149

Kapitel 7. Literatur
158. Pouponnot, C., Jayaraman, L., Massague, J.:
Physical and functional interaction of SMADs and p300/CBP.
J Biol Chem 1998; 273 (36): 22865-22868.
159. Qu, Q., Perala-Heape, M., Kapanen, A., Dahllund, J., Salo, J., Vaananen, H.K., Harkonen,
P.:
Estrogen enhances differentiation of osteoblasts in mouse bone marrow culture.
Bone 1998; 22 (3): 201-209.
160. Ralston, S.H., Hoey, S.A., Gallacher, S.J., Adamson, B.B., Boyle, I.T.:
Cytokine and growth factor expression in Paget's disease: analysis by reverse-transcription/poly-
merase chain reaction.
Br J Rheumatol 1994; 33 (7): 620-625.
161. Ravaglia, G., Forti, P., Maioli, F., Nesi, B., Pratelli, L., Cucinotta, D., Bastagli, L., Cavalli, G.:
Body composition, sex steroids, IGF-1, and bone mineral status in aging men.
J Gerontol A Biol Sci Med Sci 2000; 55 (9): M516-521.
162. Reiss, M.:
TGF-beta and cancer.
Microbes Infect 1999; 1 (15): 1327-1347.
163. Roberts, A.B.:
Molecular and cell biology of TGF-beta.
Miner Electrolyte Metab 1998; 24 (2-3): 111-119.
164. Robey, P.G., Termine, J.D.:
Human bone cells in vitro.
Calcif Tissue Int 1985; 37 (5): 453-460.
165. Robinson, J.A., Harris, S.A., Riggs, B.L., Spelsberg, T.C.:
Estrogen regulation of human osteoblastic cell proliferation and differentiation.
Endocrinology 1997; 138 (7): 2919-2927.
166. Robinson, J.A., Riggs, B.L., Spelsberg, T.C., Oursler, M.J.:
Osteoclasts and transforming growth factor-beta: estrogen-mediated isoform-specific regulation of
production.
Endocrinology 1996; 137 (2): 615-621.
Seite 150

Kapitel 7. Literatur
167. Rodriguez, C., Chen, F., Weinberg, R.A., Lodish, H.F.:
Cooperative binding of transforming growth factor (TGF)-beta 2 to the types I and II TGF-beta re-
ceptors.
J Biol Chem 1995; 270 (27): 15919-15922.
168. Russell-Aulet, M., Shapiro, B., Jaffe, C.A., Gross, M.D., Barkan, A.L.:
Peak bone mass in young healthy men is correlated with the magnitude of endogenous growth
hormone secretion.
J Clin Endocrinol Metab 1998; 83 (10): 3463-3468.
169. Sandberg, M., Autio-Harmainen, H., Vuorio, E.:
Localization of the expression of types I, III, and IV collagen, TGF-beta 1 and c-fos genes in deve-
loping human calvarial bones.
Dev Biol 1988; 130 (1): 324-334.
170. SantaLucia, J., Jr.:
A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics.
Proc Natl Acad Sci U S A 1998; 95 (4): 1460-1465.
171. Sato, M., Takeda, N., Sarui, H., Takami, R., Takami, K., Hayashi, M., Sasaki, A., Kawachi,
S., Yoshino, K., Yasuda, K.:
Association between serum leptin concentrations and bone mineral density, and biochemical
markers of bone turnover in adult men.
J Clin Endocrinol Metab 2001; 86 (11): 5273-5276.
172. Schiebler, T., Schmidt, W., Zilles, K.:
Anatomie: Zytologie, Histologie, Entwicklungsgeschichte, makroskopische und mikroskopische
Anatomie des Menschen (Lehrbuch).
Springer, Heidelberg, 1995.
173. Seino, Y.:
Cytokines and growth factors which regulate bone cell function.
Acta Astronaut 1994; 33 : 131-136.
174. Seitzer, U., Batge, B., Acil, Y., Muller, P.K.:
Transforming growth factor beta 1 influences lysyl hydroxylation of collagen I and reduces steady-
state levels of lysyl hydroxylase mRNA in human osteoblast-like cells.
Eur J Clin Invest 1995; 25 (12): 959-966.
Seite 151

Kapitel 7. Literatur
175. Serra, R., Johnson, M., Filvaroff, E.H., LaBorde, J., Sheehan, D.M., Derynck, R., Moses,
H.L.:
Expression of a truncated, kinase-defective TGF-beta type II receptor in mouse skeletal tissue pro-
motes terminal chondrocyte differentiation and osteoarthritis.
J Cell Biol 1997; 139 (2): 541-552.
176. Shalhoub, V., Conlon, D., Tassinari, M., Quinn, C., Partridge, N., Stein, G.S., Lian, J.B.:
Glucocorticoids promote development of the osteoblast phenotype by selectively modulating ex-
pression of cell growth and differentiation associated genes.
J Cell Biochem 1992; 50 (4): 425-440.
177. Siggelkow, H., Niedhart, C., Kurre, W., Ihbe, A., Schulz, A., Atkinson, M.J., Hufner, M.:
In vitro differentiation potential of a new human osteosarcoma cell line (HOS 58).
Differentiation 1998; 63 (2): 81-91.
178. Song, C.Z., Tian, X., Gelehrter, T.D.:
Glucocorticoid receptor inhibits transforming growth factor-beta signaling by directly targeting the
transcriptional activation function of Smad3.
Proc Natl Acad Sci U S A 1999; 96 (21): 11776-11781.
179. Sporn, M.B.:
TGF-beta: 20 years and counting.
Microbes Infect 1999; 1 (15): 1251-1253.
180. Stadtman, E.R., Starke-Reed, P.E., Oliver, C.N., Carney, J.M., Floyd, R.A.:
Protein modification in aging.
Exs 1992; 62 : 64-72.
181. Steinbrech, D.S., Mehrara, B.J., Rowe, N.M., Dudziak, M.E., Luchs, J.S., Saadeh, P.B.,
Gittes, G.K., Longaker, M.T.:
Gene expression of TGF-beta, TGF-beta receptor, and extracellular matrix proteins during mem-
branous bone healing in rats.
Plast Reconstr Surg 2000; 105 (6): 2028-2038.
182. Stroschein, S.L., Wang, W., Zhou, S., Zhou, Q., Luo, K.:
Negative feedback regulation of TGF-beta signaling by the SnoN oncoprotein.
Science 1999; 286 (5440): 771-774.
Seite 152

Kapitel 7. Literatur
183. Sugden, M.C., Orfali, K.A., Holness, M.J.:
The pyruvate dehydrogenase complex: nutrient control and the pathogenesis of insulin resistance.
J Nutr 1995; 125 (6 Suppl): 1746S-1752S.
184. Szekanecz, Z., Haines, G.K., Harlow, L.A., Shah, M.R., Fong, T.W., Fu, R., Lin, S.J., Rayan,
G., Koch, A.E.:
Increased synovial expression of transforming growth factor (TGF)-beta receptor endoglin and
TGF-beta 1 in rheumatoid arthritis: possible interactions in the pathogenesis of the disease.
Clin Immunol Immunopathol 1995; 76 (2): 187-194.
185. Takenoshita, S., Hagiwara, K., Nagashima, M., Gemma, A., Bennett, W.P., Harris, C.C.:
The genomic structure of the gene encoding the human transforming growth factor beta type II re-
ceptor (TGF-beta RII).
Genomics 1996; 36 (2): 341-344.
186. Takeuchi, Y., Fukumoto, S., Matsumoto, T.:
Relationship between actions of transforming growth factor (TGF)-beta and cell surface expression
of its receptors in clonal osteoblastic cells.
J Cell Physiol 1995; 162 (3): 315-321.
187. Takeuchi, Y., Nakayama, K., Matsumoto, T.:
Differentiation and cell surface expression of transforming growth factor-beta receptors are regula-
ted by interaction with matrix collagen in murine osteoblastic cells.
J Biol Chem 1996; 271 (7): 3938-3944.
188. Toyono, T., Nakashima, M., Kuhara, S., Akamine, A.:
Temporal changes in expression of transforming growth factor-beta superfamily members and
their receptors during bovine preodontoblast differentiation in vitro.
Arch Oral Biol 1997; 42 (7): 481-488.
189. Tsuji, S., Munkhbat, B., Hagihara, M., Tsuritani, I., Abe, H., Tsuji, K.:
HLA-A*24-B*07-DRB1*01 haplotype implicated with genetic disposition of peak bone mass in heal-
thy young Japanese women.
Hum Immunol 1998; 59 (4): 243-249.
Seite 153

Kapitel 7. Literatur
190. Verschueren, K., Huylebroeck, D.:
Remarkable versatility of Smad proteins in the nucleus of transforming growth factor-beta ac-
tivated cells.
Cytokine Growth Factor Rev 1999; 10 (3-4): 187-199.
191. von Ahsen, N., Wittwer, C.T., Schutz, E.:
Oligonucleotide Melting Temperatures under PCR Conditions: Nearest-Neighbor Corrections for
Mg2+, Deoxynucleotide Triphosphate, and Dimethyl Sulfoxide Concentrations with Comparison to
Alternative Empirical Formulas.
Clin Chem 2001; 47 (11): 1956-1961.
192. Vostriukhina, O.A., Nikiforova, I.F., Shtam, T.A., Kantorov, S.L., Kovalev, V.K., Vasil'ev,
S.V., Pozharisskii, K.M., Lantsov, V.A.:
Damage to the transforming growth factor TGF-beta type II receptor gene and microsatellite in-
stability in carcinoma cells of the gastrointestinal tract.
Vopr Onkol 1998; 44 (6): 667-671.
193. Wang, T., Donahoe, P.K., Zervos, A.S.:
Specific interaction of type I receptors of the TGF-beta family with the immunophilin FKBP-12.
Science 1994; 265 (5172): 674-676.
194. Wang, X., Schwartz, Z., Yaffe, P., Ornoy, A.:
The expression of transforming growth factor-beta and interleukin-1beta mRNA and the response
to 1,25(OH)2D3' 17 beta-estradiol, and testosterone is age dependent in primary cultures of
mouse-derived osteoblasts in vitro.
Endocrine 1999; 11 (1): 13-22.
195. Wang, X.F., Lin, H.Y., Ng-Eaton, E., Downward, J., Lodish, H.F., Weinberg, R.A.:
Expression cloning and characterization of the TGF-beta type III receptor.
Cell 1991; 67 (4): 797-805.
196. Wang, Y.Q., Sizeland, A., Wang, X.F., Sassoon, D.:
Restricted expression of type-II TGF beta receptor in murine embryonic development suggests a
central role in tissue modeling and CNS patterning.
Mech Dev 1995; 52 (2-3): 275-289.
Seite 154

Kapitel 7. Literatur
197. Ward, G.R., MacDougall, J.D., Sutton, J.R., Toews, C.J., Jones, N.L.:
Activation of human muscle pyruvate dehydrogenase with activity and immobilization.
Clin Sci (Lond) 1986; 70 (2): 207-210.
198. Whitfield, J.F.:
Leptin: brains and bones.
Expert Opin Investig Drugs 2001; 10 (9): 1617-1622.
199. Witzke, K.A., Snow, C.M.:
Lean body mass and leg power best predict bone mineral density in adolescent girls.
Med Sci Sports Exerc 1999; 31 (11): 1558-1563.
200. Wrana, J.L., Attisano, L.:
The Smad pathway.
Cytokine Growth Factor Rev 2000; 11 (1-2): 5-13.
201. Yamaguchi, M., Ozaki, K., Hoshi, T.:
Simulated weightlessness and bone metabolism: impairment of insulin effect on alkaline phospha-
tase activity in bone tissue.
Res Exp Med 1991; 191 (2): 113-119.
202. Yamauchi, M., Sugimoto, T., Yamaguchi, T., Nakaoka, D., Kanzawa, M., Yano, S., Ozuru,
R., Sugishita, T., Chihara, K.:
Plasma leptin concentrations are associated with bone mineral density and the presence of verte-
bral fractures in postmenopausal women.
Clin Endocrinol (Oxf) 2001; 55 (3): 341-347.
203. Yang, N.N., Bryant, H.U., Hardikar, S., Sato, M., Galvin, R.J., Glasebrook, A.L., Termine,
J.D.:
Estrogen and raloxifene stimulate transforming growth factor-beta 3 gene expression in rat bone: a
potential mechanism for estrogen- or raloxifene-mediated bone maintenance.
Endocrinology 1996; 137 (5): 2075-2084.
204. Yu, Y.S., Suzuki, Y., Yoshitomo, K., Muramatsu, M., Yamaguchi, N., Sugano, S.:
The promoter structure of TGF-beta type II receptor revealed by "oligo-capping" me-
thod and deletion analysis.
Biochem Biophys Res Commun 1996; 225 (1): 302-306.
Seite 155

Kapitel 7. Literatur
205. Zerwekh, J.E., Ruml, L.A., Gottschalk, F., Pak, C.Y.:
The effects of twelve weeks of bed rest on bone histology, biochemical markers of bone turnover,
and calcium homeostasis in eleven normal subjects.
J Bone Miner Res 1998; 13 (10): 1594-1601.
206. Zhang, R., Supowit, S.C., Hou, X., Simmons, D.J.:
Transforming growth factor-beta2 mRNA level in unloaded bone analyzed by quantitative in situ
hybridization.
Calcif Tissue Int 1999; 64 (6): 522-526.
207. Zhang, Y., Derynck, R.:
Regulation of Smad signalling by protein associations and signalling crosstalk.
Trends Cell Biol 1999; 9 (7): 274-279.
208. Zhou, H., Choong, P.C., Chou, S.T., Kartsogiannis, V., Martin, T.J., Ng, K.W.:
Transforming growth factor beta 1 stimulates bone formation and resorption in an in-vivo model in
rabbits.
Bone 1995; 17 (4 Suppl): 443S-448S.
209. Zhuang, H., Wang, W., Tahernia, A.D., Levitz, C.L., Luchetti, W.T., Brighton, C.T.:
Mechanical strain-induced proliferation of osteoblastic cells parallels increased TGF-beta 1 mRNA.
Biochem Biophys Res Commun 1996; 229 (2): 449-453.
Seite 156

Kapitel 8. Danksagung
8. Danksagung
Mein Dank gilt:
Prof. Dr. H. Schatz für die Bereitstellung der hervorragenden Laborarbeitsplätze
Prof. Dr. A.F.H. Pfeiffer für die Bereitstellung und den Unterhalt der von mir genutztenArbeitsplätze sowie für das LightCycler-Testgerät;
PD Dr. S. Hering für die sehr gute Betreuung, die zahlreichen und produktiven Diskussionen sowie für die Übernahme der Doktorvaterschaft;
Dr. R. Assert und Dr. M. Osterhoff in besonderem Maße sowie einigen weiteren Mitarbei-tern des endokrinologischen Labors für die gute Betreuung und Zusammen-arbeit im Labor und für die zahlreichen Diskussionen;
Prof. Dr. A. Bufe und Dipl. Biol. S. Löseke für die Bereitstellung und Einarbeitung am„TaqMan“;
Dr. Frank Isken für die hervorragend durchgeführte Extraktion und Konservierung derKnochenproben;
Prof. Dr. G. Muhr und der chirurgischen Abteilung des Bergmannsheils Bochum für dieAsservierung der Knochenproben;
Dr. A. Battmann für die Bereitstellung der primären Knochenzellkulturen und fürKorrekturen am Manuskript;
Dr. D. Schultzki und der Firma Roche, Mannheim, für die freundliche Bereitstellung des„LightCycler“ und der Erlaubnis, die eingebundenen Grafiken zu verwenden;
Kerstin Löschmann und der BioCarta Europe GmbH, Hamburg, für die Genehmigung,die eingebundene Grafik zu verwenden;
Katja Serova (Lehrstuhl für Sozialwissenschaftliche Methodenlehre und Statistik) für dieBesprechung der statistischen Ergebnisauswertung;
Der Firma Perkin-Elmer, Darmstadt, für die freundliche Bereitstellung der Grafiken unddie technische Unterstützung am „TaqMan“;
Meinen Eltern für die dauerhafte Unterstützung und für die Korrekturen am Manuskript;
Meinen Brüdern Jörg und Jan für die Hilfe bei diversen Computer- und Internetfragenund für die Korrekturen am Manuskript.
Seite 157

Kapitel 9. Lebenslauf
9. Lebenslauf
Persönliche Daten:
Name: Jens Ulrich RegulaAnschrift Handschuhsheimer Landstrasse 87, 69121 HeidelbergGeburtsdatum: 18. Mai 1976Geburtsort: HeidelbergKonfession: EvangelischFamilienstand: Ledig
Schulbildung:
1982 - 1986 Hans-Geiger-Schule, Neustadt/Weinstraße1986 - 1995 Leibniz-Gymnasium, Neustadt/Weinstraße1995 Abitur (Note: 1,6)
Hochschulbildung:
1995 - 2001 Studium der Humanmedizin an der Ruhr-Universität Bochum1997 Ärztliche Vorprüfung (Note: 2,66)1998 1. Abschnitt der ärztlichen Prüfung (Note: 2,0)2000 2. Abschnitt der ärztlichen Prüfung (Note: 1,66)2000 - 2001 Praktisches Jahr im Bergmannsheil Bochum (Innere Medizin und
Neurologie) und im Regionalspital Interlaken, Schweiz (Chirurgie)2001 3. Abschnitt der ärztlichen Prüfung (Note: 2,0, Gesamtnote: 1,83)
Seit 1. Mai 2002 Arzt im Praktikum an der neurologischen Universitätsklinik
Heidelberg (Direktor: Prof. Dr. W. Hacke) und in der neurologisch-
psychiatrische Gemeinschaftspraxis Dr. Reifschneider, Frau Un-
sorg und Dr. Ries, Erbach im Odenwald
Heidelberg, im Mai 2003
Seite 158