
LA LIMITE MARITIME DE L'EPINETTE BLANCHE (PICEA GLAUCA ... · Guillaume de Lafontaine, Goulwen Dy,...
66
ANN-CATHERINE LALIBERTE LA LIMITE MARITIME DE L'EPINETTE BLANCHE (PICEA GLAUCA (MOENCH) VOSS) SUR LA CÔTE ORIENTALE DE LA BAIE D'HUDSON Mémoire présenté à la Faculté des études supérieures de l'Université Laval dans le cadre du programme de maîtrise en biologie pour l'obtention du grade de maître es sciences (M. Se.) DEPARTEMENT DE BIOLOGIE FACULTÉ DES SCIENCES ET DE GÉNIE UNIVERSITÉ LAVAL QUÉBEC 2006 Ann-Catherine Laliberté, 2006
Transcript of LA LIMITE MARITIME DE L'EPINETTE BLANCHE (PICEA GLAUCA ... · Guillaume de Lafontaine, Goulwen Dy,...

ANN-CATHERINE LALIBERTE
LA LIMITE MARITIME DE L'EPINETTEBLANCHE (PICEA GLAUCA (MOENCH) VOSS)
SUR LA CÔTE ORIENTALE DE LA BAIED'HUDSON
Mémoire présentéà la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en biologiepour l'obtention du grade de maître es sciences (M. Se.)
DEPARTEMENT DE BIOLOGIEFACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVALQUÉBEC
2006
Ann-Catherine Laliberté, 2006

Résumé
Huit sites possédant des peuplements d'épinette blanche (Picea glauca [Moench]
Voss) sur substrat bien drainé le long de la côte en émersion de la baie d'Hudson ont été
étudiés. Les objectifs de la recherche sont de caractériser la limite maritime des arbres et
la limite maritime des forêts en fonction de l'altitude et du climat afin d'estimer le délai
nécessaire entre l'émersion des terres et l'établissement des épinettes dans un contexte
de succession primaire. La végétation évolue le long de la chronoséquence, partant
d'une bande littorale près de la mer jusqu'à un stade de pessière à lichen au niveau de la
limite maritime des forêts. La podzolisation est le processus pédogénétique dominant
dans ce contexte. Parallèlement à la végétation qui change, les sols se développent avec
l'altitude. Ce n'est que 145 à 900 ans suivant l'émersion que les conditions du milieu
permettent l'établissement des espèces arborescentes. Les épinettes de la limite des
arbres sont plus jeunes que les arbres composant la limite des forêts et ces derniers n'ont
pas plus de 120 ans. Aucun macroreste d'épinette blanche n'a été trouvé dans les
paléosols précédant la limite des forêts. Les épinettes blanches les plus âgés sur les sites
d'étude se sont établis à partir de la fin du XIXe siècle.

Avant-propos
J'aimerais tout d'abord remercier mon directeur, Serge Payette, pour ses conseils,
son aide, sa grande disponibilité et son soutien tout au long de ma maîtrise. Il a su me
transmettre sa passion pour l'écologie et les environnements nordiques. Sans lui, cette
expérience n'aurait pas été la même. Je remercie également les membres de mon comité
d'évaluation, Gilles Houle et Michel Caillier, pour leurs conseils.
Merci à Ann Delwaide, Véronique Gravel et Jonathan Régnière qui ont été
vraiment efficaces sur le terrain et avec qui il a été très agréable de travailler. L'aide de
Daniel Marcotte du département des sols et génie agroenvironnemental et de Véronique
Gravel pour les analyses de sol en laboratoire fût grandement appréciée. Les échanges
avec les gens du laboratoire d'écologie végétale ont été d'une grande aide et vont bien
me manquer. Merci à Yann Arlen-Pouliot, Stéphane Boudreau, Sébastien Cyr,
Guillaume de Lafontaine, Goulwen Dy, François Girard, Stefanie Pollock, Amélie Rivet
et Simon Thibault. J'aimerais aussi remercier ma famille, Paul Laliberté, Élizabeth
Vatcher, Laurence Laliberté ainsi que Laurent Dionne, qui m'ont encouragé pendant les
deux dernières années.
Ce travail n'aurait pas pu se réaliser sans l'aide financière du Conseil de
recherche en sciences naturelles et en génie du Canada (CRSNG) et du Ministère des
Affaires indiennes et du Nord (Programme de formation scientifique dans le nord-
PFSN). J'aimerais également remercier le Centre d'études nordiques pour son soutien
logistique.

Table des matières
Résumé ii
Avant-propos iii
Table des matières iv
Liste des figures et des tableaux v
Introduction 7
1. Région d'étude 10
2. Méthodologie 132.1 Choix des sites 132.2 Caractérisation du site et de la végétation 142.3 Échantillonnage des sols 152.4 Analyses de sol 152.5 Âge desépinettes 162.6 Datation au 14C 162.7 Statistiques 17
3. Résultats 183.1 Caractéristiques spécifiques des sites 183.2 Caractéristiques de la végétation 183.3 Limite des arbres '. 293.4 Limite des forêts 343.5 Sols 39
4. Discussion 454.1 Limite maritime de Pépinette blanche 454.2 Influence du climat 464.3 Influence des facteurs locaux 474.4 Expansion récente de l'épinette blanche 494.5 Données historiques 49
Conclusion 52
Bibliographie 53
Annexe 1 Analyses de sol 59

Liste des figures et des tableaux
Figure 1. Sites d'étude 12
Figure 2. Site d'étude typique (site Petite Rivière) 13
Figure 3. Topographie et végétation le long du transect du site Guillaume-Delisle. Lesépinettes représentent la limite des arbres et la limite des forêts 20
Figure 4. Topographie et végétation le long du transect du site Goulet. Les épinettesreprésentent la limite des arbres et la limite des forêts 21
Figure 5. Topographie et végétation le long du transect du site Petite Rivière. Lesépinettes représentent la limite des arbres et la limite des forêts 22
Figure 6. Topographie et végétation le long du transect du site Manitounuk. Lesépinettes représentent la limite des arbres et la limite des forêts 23
Figure 7. Topographie et végétation le long du transect du site Grande Rivière. Lesépinettes représentent la limite des arbres et la limite des forêts 24
Figure 8. Topographie et végétation le long du transect du site Dérive. Les épinettesreprésentent la limite des arbres et la limite des forêts 25
Figure 9. Topographie et végétation le long du transect du site Inuit. Les épinettesreprésentent la limite des arbres et la limite des forêts 26
Figure 10. Topographie et végétation le long du transect du site île Longue. Lesépinettes représentent la limite des arbres et la limite des forêts 27
Figure 11. Altitude des bandes de végétation à partir de la première présence continuedes espèces 28
Figure 12. Structure d'âge des plantules 32
Figure 13. Répartition altitudinale des plantules selon leur âge 33
Figure 14. Âge des arbres à la limite des arbres et à la limite des forêts 36
Figure 15. Altitude de la limite des arbres et de la limite des forêts. Chaque pointreprésente l'altitude moyenne (± 1 écart-type) de tous les arbres échantillonnés...37
Figure 16. Relation entre l'âge des arbres et l'altitude à la limite des arbres et à la limitedes forêts. Etant des valeurs distantes, les épinettes de la limite des arbres du siteGoulet n'ont pas été incluses dans la régression 38
Figure 17. Évolution du pH en fonction de l'altitude. Les points en blanc représentent lesite Manitounuk 43

VI
Figure 18. Teneur en fer-pyro et en aluminium- pyro de l'horizon B des sols de tous lessites étudiés. Les points blancs représentent les échantillons prélevés directementsous les arbres 44
Figure 19. Limite maritime de l'épinette blanche à la localité 18 (54° 50' N, 79° 02' O)en 1948 (tiré de Hustich, 1950; photo E. H. Kranck) 51
Figure 20. Limite maritime de l'épinette blanche au site île Longue(54° 48' N, 79° 06' O) 51
Tableau 1. Pente des sites calculée à partir de la position de la bande à Honckenyapeploides jusqu'à la limite des forêts 18
Tableau 2. Âge (années) estimé du substrat à la limite inférieure de chaque bande devégétation, arrondi à la décennie, en fonction d'un relèvement isostatique de 1,2m/siècle 29
Tableau 3. Arbres échantillonnés à la limite des arbres pour l'ensemble des sites. Lesarbres représentés en caractère gras sont les plus près de la mer et déterminent laposition de la limite des arbres 31
Tableau 4. Arbres échantillonnés à la limite des forêts pour l'ensemble des sites. Lesarbres représentés en caractère gras sont les plus près de la mer et déterminent laposition de la limite des forêts 35
Tableau 5. Dates I4C des paléosols 42
Tableau 6. Teneur en fer et en aluminium extraits au pyrophosphate de sodium, pHCaCb et % C tot de l'horizon B et de l'horizon B enfoui dans les profils renfermantdes paléosols 42
Tableau 7. Age (années) estimé de la surface au moment de la colonisation parl'épinette blanche d'après un taux de relèvement moyen de 1,2 m/siècle 46

Introduction
Les arbres à leur limite de répartition nordique sont sensibles aux changements
climatiques (LaMarche, 1973; Kullman, 1986). Malgré le réchauffement général en
Amérique du Nord depuis la dernière glaciation, certaines périodes ont été moins
favorables que d'autres à la croissance des arbres. Entre 1580 et 1880, les basses
températures du Petit Age Glaciaire ont limité la croissance des arbres (Lamb 1977;
Grove, 1988) et ce n'est qu'à partir du XIXe siècle que le climat s'est réchauffé, que la
densité des peuplements forestiers a augmenté (Morin & Payette, 1984; Payette &
Filion, 1985; Scott et al, 1987; Gajewski et al, 1993; Szeicz & MacDonald, 1995) et
que les formes de croissance des espèces arborescentes à la limite des arbres ont changé
de manière significative (Lavoie & Payette, 1994; Lescop-Sinclair & Payette, 1995;
Lavoie & Payette, 1996).
Lors de la dernière glaciation, l'Inlandsis laurentidien occupait l'ensemble du
Bouclier canadien (Fulton, 1989). Son retrait a débuté vers 15 000 ans avant l'actuel,
mais ce n'est que vers 8000 ans avant l'actuel que la déglaciation de la région de la baie
d'Hudson a débuté. Par la suite, la transgression marine de la mer de Tyrrell a atteint
une altitude de 315 m à Kuujjuarapik et de 270 m dans la partie sud du Lac Guillaume-
Delisle (Hillaire-Marcel & Vincent, 1980). De nos jours, la côte orientale de la baie
d'Hudson est encore en phase de relèvement isostatique post-glaciaire et connaît un taux
de relèvement moyen de 1,1 à 1,3 m/siècle (Hillaire-Marcel, 1976; Allard & Tremblay,
1983; Bégin et al, 1993). Ainsi, connaissant le rythme du relèvement, il est possible de
déterminer l'âge du substrat en mesurant son altitude actuelle par rapport au niveau de la
mer (Svensson & Jeglum, 2003a). L'influence de l'émersion post-glaciaire des côtes
maritimes sur la végétation a fait l'objet de plusieurs travaux le long de la baie
d'Hudson (Audet, 1976; Grégoire & Bégin, 1993; von Mors & Bégin, 1993; Bégin et
al., 1993; Ricard & Bégin, 1999) et du golfe de Bothnie en Europe du Nord qui se
relève à une vitesse de 0,9 m/siècle (Môrner, 1980; Ericson, 1980; Cramer, 1985;
Cramer & Hytteborn, 1987; Ecke & Rydin, 2000; Svensson & Jeglum 2001, 2003a,
2003b).

Au Québec, l'épinette noire (Picea mariana [Mill.] BSP) est la principale espèce
présente à la limite des arbres (Marr, 1948; Hustich, 1950; Payette, 1983). Bien qu'elle
soit parfois présente à l'intérieur du continent (Rousseau, 1968), l'épinette blanche
{Picea glauca [Moench] Voss) est plus concentrée sur la côte de la baie d'Hudson où
elle forme la limite maritime des arbres (Hustich, 1950; Payette, 1983). D'autres
populations d'épinette blanche sont également présentes le long de la côte du Labrador
et le long de la côte sud-est de la baie d'Ungava (Payette, 1993). La limite maritime des
arbres correspond à la frange des arbres (> 2,5 m de hauteur) la plus près de la mer.
L'épinette blanche forme une bande continue large de 5 à 8 kilomètres de la pointe
Louis-XIV (54° 48' N, 79° 06' O) jusqu'au Lac Guillaume-Delisle (54° 11' N, 76° 37' O)
où se trouve la limite latitudinale des forêts maritimes (Payette, 1975). Dans cette
région, l'épinette blanche croît symétriquement et atteint plus de 5 mètres de hauteur, ce
qui suggère des conditions de croissance plutôt favorables. En effet, les épinettes
blanches sont mieux adaptées aux conditions d'humidité importante causées par le
brouillard régulier de la côte, alors que l'épinette noire s'y trouve généralement sous
forme prostrée (Payette, 1975).
L'épinette blanche tolère des conditions climatiques et édaphiques très variables
(Burns & Honkala, 1990). Par contre, dans des conditions extrêmes, la forme de
croissance normale de l'épinette peut être affectée (Payette, 1974). La reproduction de
l'espèce est principalement assurée par les graines qui nécessitent une humidité
suffisante, une température se situant idéalement entre 10 °C et 24 °C et un sol minéral
(Burns & Honkala, 1990). L'épinette blanche est moins adaptée aux feux que l'épinette
noire puisque ses cônes ne sont pas sérotineux. La production de graines est variable
chez l'épinette blanche. Certaines périodes sont plus favorables que d'autres à la
régénération, car la survie des plantules est dépendante du climat hivernal (Hadley &
Smith, 1986) et de la disponibilité en eau au cours de l'été.
Par définition, le substrat initial lors d'une succession primaire, comme les plages
sableuses en voie de soulèvement le long de la baie d'Hudson, ne possède ni banque de
graines ni d'autres banques de diaspores. La dispersion des graines doit s'effectuer de
manière passive (vent, eau, gravité) ou de manière active par les animaux. La distance à
parcourir peut souvent être une barrière importante à la dispersion, d'où l'importance

d'avoir des semenciers à proximité des sites à coloniser (Walker & del Moral, 2003).
Selon la théorie de Connell et Slatyer (1977), la facilitation serait prédominante dans un
contexte de succession primaire, puisque le substrat fraîchement exposé possède
habituellement des propriétés extrêmes (pH, humidité, nutriments, etc.).
Dans les régions où les dépôts sableux marins sont abondants, les phénomènes
d'érosion éolienne sont fréquents. Les plantes jouent un rôle important dans la
stabilisation des dunes littorales (Filion & Morisset, 1983; Imbert & Houle, 2001). Les
parties aériennes favorisent la sédimentation du sable, alors que les racines retiennent le
sable et limitent l'érosion (Forster & Nicholson, 1981). En général, les déplacements de
sable et l'exposition aux embruns diminuent lorsqu'on s'éloigne de la côte. Une fois la
végétation pionnière établie sur le sable littoral, l'épaisseur de l'horizon organique, la
disponibilité en éléments nutritifs, ainsi que l'humidité du sol augmentent et facilitent
l'établissement des arbres lors des étapes plus avancées de la succession (Bégin et al.,
1993; Imbert & Houle, 2000; Svensson & Jeglum, 2003a). Étant donné l'émersion
rapide du continent dans la région de la baie d'Hudson, la végétation forme une
séquence évolutive de la limite des marées, où le substrat affleure, à l'intérieur des
terres. En conséquence, le sol devrait être plus développé avec l'établissement des arbres
et de la forêt. Le développement de la végétation et du sol varie en fonction de la
distance et de l'altitude par rapport à la côte en voie d'émersion.
Dans cette optique, la présente étude vise à décrire les caractéristiques de la limite
maritime des arbres et de la limite maritime des forêts de la côte orientale de la baie
d'Hudson. L'objectif principal est de déterminer le délai entre l'émersion des terres et
l'établissement des arbres et de la forêt et de comparer la position de la limite des arbres
et de la limite des forêts en fonction de la latitude et du climat. Pour ce faire, plusieurs
sites sur la côte orientale de la baie d'Hudson ont été étudiés.

1. Région d'étude
L'aire de recherche se situe dans le domaine maritime de la toundra forestière
(Payette, 1983) entre la Pointe Louis-XIV (54° N, 79° O) et le nord du Lac Guillaume-
Delisle (56° N, 76° O) (Fig. 1). Cette zone occupe une bande large de plusieurs
kilomètres où la limite des arbres est parallèle à la côte. Les forêts occupent
principalement les vallées où les sols sont meubles et humides (Payette et al, 1973). La
principale espèce arborescente présente dans cette région maritime est l'épinette
blanche.
Le climat de la région de Kuujjuarapik-Whapmagoostui est de type subarctique
(Wilson, 1968). La température moyenne annuelle est de -4,4 °C, alors que la
température moyenne mensuelle la plus basse est en janvier (-23,4 °C) et la plus élevée
en août (11,4 °C). Les précipitations moyennes annuelles sont de 650 mm, dont environ
40 % tombent sous forme de neige (Environnement Canada, 2006). Les dépressions
atmosphériques accumulent l'humidité en passant au-dessus de la baie d'Hudson, ce qui
augmente les précipitations sur les côtes adjacentes (Rouse, 1991). Le brouillard est
fréquent le long de la côte; il peut survenir environ 10 % du temps (Audet, 1976).
L'activité éolienne est particulièrement importante le long de la côte orientale de la
baie d'Hudson (Filion & Morisset, 1983). Les vents les plus fréquents à Kuujjuarapik
sont de l'ouest, de l'est sud-ouest et du nord. Les vents les plus forts viennent de l'ouest
et de l'est sud-ouest au début de l'été et en l'automne. L'amplitude moyenne des marées
est de 1,3 m mais peut atteindre jusqu'à 1,9 m (Ruz & Allard, 1994). Alors que le
niveau relatif de la mer baisse, la plage progresse vers la mer en formant de nouvelles
crêtes éoliennes (Ruz & Allard, 1994). Elles sont habituellement formées à partir de
dépôts estuariens ou de dépôts littoraux de la Mer de Tyrrell (Filion & Morisset, 1983).
On estime qu'à Kuujjuarapik-Whapmagoostui un cordon littoral se forme tous les 100
ans (Ruz & Allard, 1994).

I l
Le littoral de la baie d'Hudson présente une alternance de pointes rocheuses et de
baies à plages sableuses. De la Pointe Louis-XIV à la région de Kuujjuarapik-
Whapmagoostui, l'assise rocheuse est un socle archéen de granité et de gneiss, alors que
plus au nord dans la région du Lac Guillaume-Delisle, une couverture protérozoïque de
roches volcano-sédimentaires comme le basalte, le quartzite et la dolomie, repose en
discordance sur le socle archéen et forme des cuestas faisant face à l'ouest. La région
d'étude se trouve dans la zone de pergélisol discontinu (Allard & Séguin, 1987).

12
Figure 1. Sites d'étude.

2. Méthodologie
2.1 Choix des sites
Les cartes de dépôts de surface ainsi que les photographies aériennes de la région
ont été examinées afin de repérer les sites les plus propices pour l'étude de la limite
maritime des épinettes blanches. Les sites sélectionnés devaient avoir un substrat
sableux bien drainé et une séquence de végétation littorale depuis une bande de
végétation pionnière jusqu'à une bande correspondant à une forêt d'épinettes blanches
(Fig. 2). La surface occupée par chaque site sélectionné devait être suffisamment grande
pour faciliter la colonisation des espèces arborescentes et l'établissement d'un transect
d'inventaire. Un survol aérien a permis de sélectionner 8 sites répondant le mieux aux
critères établis. Un transect de 30 m de largeur et d'une longueur variable a été tiré
aléatoirement dans chacun des sites tout en évitant les effets de bordure associés aux
cours d'eau et aux buttes rocheuses.
Figure 2. Site d'étude typique (site Petite Rivière).

14
2.2 Caractérisation du site et de la végétation
Dans chaque site, la microtopographie a été mesurée systématiquement au centre
du transect à l'aide d'un théodolite à infrarouge (Leica-T-1010 précision de 0,0005 m).
Le niveau topographique de référence utilisé dans tous les sites était la position la plus
près de la mer des premières plantes pionnières à coloniser le sable nu, soit Honckenya
peploides. Chaque transect partait de la bande à Honckenya peploides en passant par la
limite des arbres jusqu'à la limite des forêts où les arbres avaient un minimum de 5 m
de hauteur. Toutes les épinettes présentes dans le transect ont été positionnées par
rapport au niveau de référence.
Une analyse linéaire de la végétation a été effectuée systématiquement à tous les
mètres (ou plus si le transect était très long et la végétation uniforme) au centre du
transect par la méthode de la ligne intersecte (Mueller-Dombois & Ellenberg, 1974).
Les espèces présentes étaient identifiées et une classe de recouvrement linéaire le long
de chaque ligne de 1 m de long leur était assignée. Onze classes ont été utilisées : +: < 1
% de recouvrement, 1: 1-10 %, 2: 10-20 %, et ainsi de suite jusqu'à 10 pour un
recouvrement de 90-100 %. L'objectif du relevé de végétation était de caractériser le
site afin de délimiter des bandes de végétation indicatrices de la succession primaire,
comme par exemple la bande littorale, la bande arbustive ou la bande forestière.
Les premières épinettes de plus de 2,5 m de hauteur établis depuis la plage ont été
considérés comme formant la limite des arbres. La limite des forêts comprend les
premiers arbres de plus de 5 m de hauteur, ayant un port symétrique et formant un
peuplement forestier selon la définition de Payette (1975). Toutes les épinettes de moins
de 2,5 m de hauteur (plantules et gaules) et situées avant la limite des arbres dans le
transect ont été échantillonnées afin que leur âge soit déterminé en laboratoire. Tous les
arbres formant la limite des arbres ainsi que 3 à 5 arbres situés à la limite des forêts ont
été coupés au niveau du sol afin de déterminer leur âge. Il en est de même des arbres
morts présents dans chaque site. La hauteur et le diamètre au collet des plantules et des
arbres ont été mesurés.

15
2.3 Échantillonnage des sols
Des échantillons de sol des horizons de surface (FH, A) et des horizons B et C ont
été prélevés dans les bandes de végétation littorale identifiées à l'aide des relevés
linéaires, soit la bande à Honckenya peploides et la bande à Leymus mollis, ainsi que
sous le premier arbre de la limite des arbres et près de celui-ci en milieu ouvert, ainsi
que sous le plus grand arbre de la limite des forêts et près de celui-ci en milieu ouvert.
Les sols échantillonnés correspondent aux principales bandes de végétation littorales de
la chronoséquence primaire. De plus l'échantillonnage des sols sous les arbres donne
une indication directe de l'influence de ceux-ci sur la pédogenèse. Lorsque des
paléosols étaient présents, l'horizon organique enfoui a été échantillonné et daté au
radiocarbone. La datation de charbons de bois et de bois non décomposé permet de
donner l'âge du bois (vivant ou mort) présent à l'époque du feu, alors que la datation de
l'humus donne un âge composite de la matière organique décomposée.
2.4 Analyses de sol
En laboratoire, les sols ont été séchés à 38 °C et tamisés manuellement à 2 mm.
Une fraction de chaque échantillon a également été moulue à 25 \am pour les analyses
chimiques. Puisque les sols étaient constitués principalement de sable, le tamisage de
100 g de sol de chaque horizon C dans une série de tamis de lmm, 500 om, 250 um,
106 um et 53 \im était suffisant pour connaître la granulométrie du matériel d'origine.
Le pH à l'eau (1 : 2) et le pH CaCl2 (1 : 2) (McKeague, 1978) ont été déterminés chez
tous les échantillons. Le pH CaCk est nécessaire pour classifier les sols selon le système
de classification canadienne des sols (Groupe de travail sur la classification des sols,
2002). La conductivité électrique servant à mesurer la salinité relative des sols a été
mesurée pour l'ensemble des échantillons à l'aide d'un conductivimètre Bach-Simpson
(model CDM 2e). La couleur de tous les échantillons secs et humides a été évaluée à
l'aide du code de couleur Munsell (MacBeth, 1994). La présence de carbonates a
également été vérifiée à l'aide de HC1.

16
La concentration en cations échangeables (Ca, Na, Mg, K) extraits à l'acétate
d'ammonium a été mesurée à l'aide d'un spectrophotomètre d'absorption atomique
PerkinElmer (modèle Analyst 200). La teneur en fer et aluminium de l'horizon B
extraits au pyrophosphate de sodium a été mesurée par spectrophotométrie (McKeague,
1978). Enfin, la concentration en carbone et en azote total des horizons de surface et de
l'horizon B a été déterminée par combustion dans un four CNS-2000 de LECO. Les
analyses de sol ont été effectuées au Département des sols et génie agro-
environnemental de la Faculté des sciences de l'agriculture et de l'alimentation de
l'Université Laval.
2.5 Âge des épinettes
L'âge des plantules au collet a été déterminé en tenant compte du dénombrement
des cernes à plusieurs niveaux sur la tige de chaque plantule à cause des problèmes
d'enfouissement reliés à l'éolisation des sables littoraux (Marin & Filion, 1992). L'âge
minimal des arbres a été déterminé par comptage du nombre de cernes annuels sur les
rondelles prélevées à la base des troncs.
2.6 Datation au 14C
Les charbons de bois provenant des horizons organiques ont été prélevés des
paléosols. D'autres paléosols composés uniquement de matière organique décomposée
ont aussi été datés au radiocarbone. Les racines et autres matières organiques fraîches
ont été triées manuellement. Certains échantillons ont été datés au laboratoire de
radiochronologie du Centre d'études nordiques, alors que d'autres ont été envoyés au
laboratoire de l'Université de Californie à Irvine (Keck Carbon Cycle AMS Facility)
lorsque la quantité de carbone recueilli était trop faible pour être datée de façon
conventionnelle. L'étalonnage des dates au radiocarbone conventionnelles a été effectué
à l'aide du programme CALIB REV 5.0.1 (Stuiver & Reimer, 1993) et des données
d'étalonnage intcalO4.14c de Reimer et al. (2004).

17
2.7 Statistiques
En ce qui trait aux analyses portant sur les arbres, tous les sites ont été disposés en
trois sous-groupes selon la latitude afin d'augmenter le nombre d'échantillons à la limite
des arbres. La répartition des sites est la suivante : groupe Nord (Guillaume-Delisle et
Goulet), groupe Centre (Petite Rivière et Manitounuk), groupe Sud (Grande Rivière,
Dérive, Inuit et île Longue). Quant aux analyses portant sur les arbres à la limite des
arbres et à la limite des forêts, des analyses de variance (ANOVA) suivie du test de
Tukey ont permis de vérifier si les sites étaient significativement différents l'un de
l'autre quant à l'âge des arbres, leur altitude et leur distance à la mer. Des régressions
linéaires ont été effectuées pour les données de sol. La normalité et l'homogénéité des
résidus ont été vérifiées (Sokal & Rohlf, 1995). Les analyses statistiques ont été
effectuées avec le logiciel Statistica 7 (StatSoft, 2005). Afin d'estimer l'âge des surfaces
lors de l'établissement des épinettes, un taux de relèvement médian (1,2 m/siècle) entre
les taux de relèvement minimum (1,1 m/siècle) et maximum (1,3 m/siècle) a été utilisé.
L'altitude du matériel d'origine à la base des arbres a servi à déterminer l'âge minimal du
substrat au moment de l'établissement des épinettes une fois l'âge des arbres soustrait.
Cette méthode a déjà été utilisée à plusieurs reprises (von Mors & Bégin, 1993;
Grégoire & Bégin, 1993; Ecke & Rydin, 2000; Svensson & Jeglum, 2003a, 2003b).

3. Résultats
3.1 Caractéristiques spécifiques des sites
De manière générale, les sites étudiés ont tous une pente relativement faible,
inférieure à 6 ° (Tableau 1). Les sites Guillaume-Delisle et Dérive ont le dénivelé le
plus faible, alors que les sites Petite Rivière et Grande Rivière ont la pente la plus
prononcée. En général, les sites présentent la même exposition, mais contrairement aux
autres, le site Guillaume-Delisle est adjacent à une butte rocheuse. La présence évidente
de pergélisol près de la forêt différencie également le site Guillaume-Delisle.
Tableau 1. Pente des sites calculée à partir de la position de la bande à Honckenyapeploides jusqu'à la limite des forêts
SiteGuillaume-Delisle
GouletPetite RivièreManitounuk
Grande RivièreDériveInuit
Ile Longue
Pente (°)1,72,74,32,65,01,82,42,9
3.2 Caractéristiques de la végétation
Les transects d'inventaire de végétation mettent en évidence une chronoséquence
végétale primaire semblable pour l'ensemble des sites. La première plante à coloniser le
sable nu de la plage est toujours Honckenya peploides, plus tolérante à l'érosion. Elle est
rapidement remplacée par Leymus mollis et Lathyrus japonicus, deux plantes
dominantes de la bande littorale qui stabilisent le sable. Une bande de plantes herbacées
relativement homogène et tapissée de bryophytes en sous-étage s'installe une fois le
substrat stabilisé. On y trouve généralement des espèces comme Hierochloe alpina,
Chamerion latifolium, Achillea borealis, Festuca rubra, Trisetum spicatum, Potentilla
spp., plusieurs poacées et des bryophytes comme Drepanocladus uncinatus,
Hylocomium splendens et Dicranum elongatum. Une fois la limite des arbres atteinte, la

19
végétation change rapidement et les lichens accompagnés d'espèces forestières font leur
apparition en synchronie avec les arbres. Cladina rangiferina, Cladina mitis et
éventuellement Cladina stellaris, ainsi que plusieurs espèces de Cladonia sont parmi les
lichens les plus communs. Entre la limite des arbres et la limite des forêts, des arbustes
dont Salix spp. et Betula glandulosa, ainsi que des éricacées comme Empetrum nigrum
et Vaccinium vitis-idaea sont plus fréquents. Ces espèces, accompagnées de Picea
glauca, forment la bande forestière.
La bande littorale, la bande herbacée et la bande forestière sont présentes dans
tous les sites mais leur étendue varie. Une bande dominée par les lichens (lichenaie)
précède parfois la bande forestière. Le site Manitounuk est le seul qui renferme des
espèces distinctes des autres sites à cause du substrat calcaire, mais le patron de
succession primaire demeure le même. Le site Guillaume-Delisle renferme plus
d'arbustes {Salix spp. et Betula glandulosa) tout au long du transect. Les figures 3 à 10
illustrent la répartition générale de la végétation littorale en fonction de la distance à la
mer, de l'altitude et de la topographie dans chaque site. Les valeurs de recouvrement des
histogrammes proviennent du relevé de végétation effectué au centre du transect dans
chaque site. La valeur de recouvrement moyenne à l'intérieur de chaque mètre a été
utilisée et celle-ci a été cumulée lorsque plusieurs espèces ont été regroupées (ex.
herbacées). Étant donné que les arbres de la limite des arbres et des forêts n'ont pas
toujours été rencontrés le long de cette ligne, on constate une certaine discordance entre
Picea glauca et la position réelle de la limite des arbres et des forêts dans chaque figure.
Ces figures permettent de comparer l'altitude et l'étendue des bandes de végétation entre
les sites (Fig. 11) et de déterminer l'âge approximatif des surfaces au moment de
l'apparition des bandes le long de chaque transect (Tableau 2).
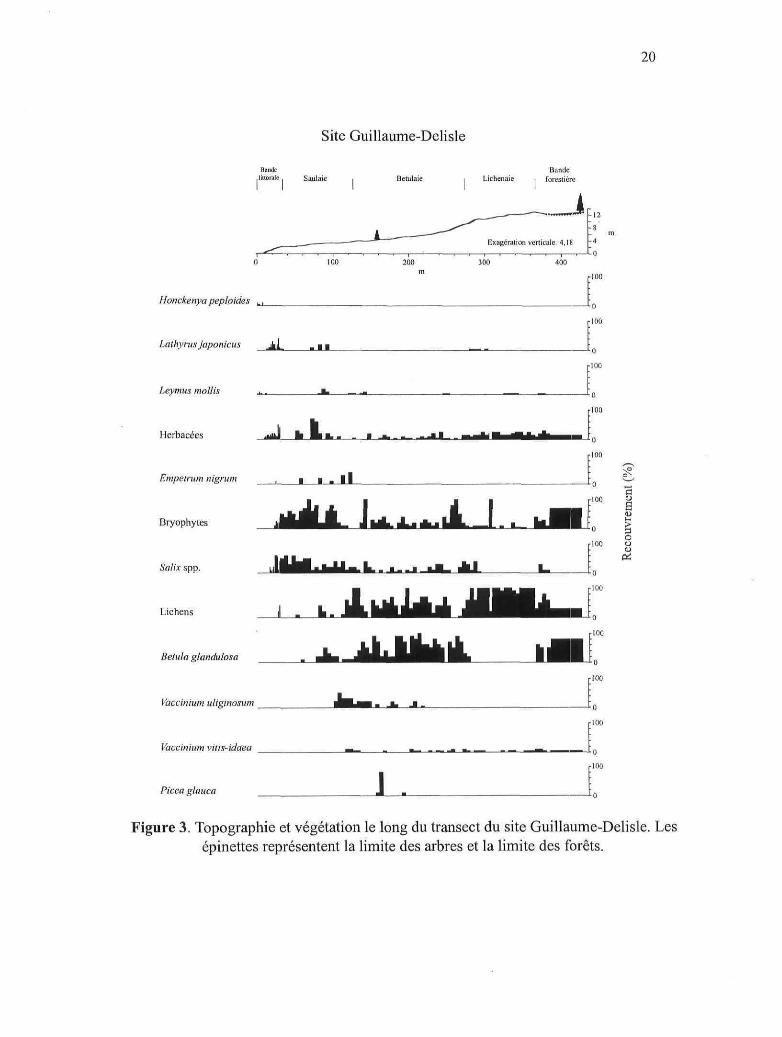
20
Site Guillaume-Delisle
Bandeliuorale Saulaie Lichenaie i forestière
Honckenya peploides k
Lathyrus japonicus
Leymus mollis
Herbacées
Empetrum nigrum
Bryophytes
Salix spp.
Lichens
Betula glandulosa
Vaccinium uliginosum
Vaccinium vilis-idaea
Picea glauca
Figure 3. Topographie et végétation le long du transect du site Guillaume-Delisle. Lesépinettes représentent la limite des arbres et la limite des forêts.

21
Site Goulet
Bande forestière
Honckenya peploides
Lathyrus japonicus
Leymus mollis
Herbacées
Empetrum nigrum
Bryophytes
Salix spp.
Lichens
Vaccinium uliginosum
Vaccinium vilis-idaea
Betula glandulosa
Picea glauca
Figure 4. Topographie et végétation le long du transect du site Goulet. Les épinettesreprésentent la limite des arbres et la limite des forêts.

22
Site Petite Rivière
Bandelittorale i Bande herbacée i Bande forestière
Exagération verticale: 1,42
0 20 40 60 80m
[100
Honckenya peploides lu II ! . . [
100
Lathyrusjaponicus .ilili.li.l... I... II..I I..I..I m.i. . . .0
100
Leymus mollis _ . L0
100 o^
M M I K , , , . . i l lIHIi IHIHIIIHHII.I. l . I I . . . , , i |
Bryophytes
Salix planifolia
••i....i..iiii.li.li.liiiuiiillliiill,iL. ii ilhi.i..
Lichens , . . . . I. . l l l l l
s, Lo
0
10(1
Vaccinium vitis-idaea
Picea glauca - i l . .. IlFigure 5. Topographie et végétation le long du transect du site Petite Rivière. Les
épinettes représentent la limite des arbres et la limite des forêts.

Site Manitounuk
23
Bandelittorale i
Bande herbacéeet saules 1
J
Bande forestière
Exagération vert
Honckenya peploides
Lathyrus japonicus
Leymus mollis
Herbacées
Bryophytes
Shepherdia canadensis
Salix spp.
Lichens
Juniperus communis
Empetrum nigrum
Picea glauca
iso
oL»L , J ..« i.l ^JJÀ, •0
• 100
jj IL, o
100
Iu>3oo'i I
Pi
• J U . I 1 i. iJ .
J • i . lui M ni
^Ji
0
100
0
100
Figure 6. Topographie et végétation le long du transect du site Manitounuk.Les épinettes représentent la limite des arbres et la limite des forêts.

Site Grande Rivière
24
[
Honckenya peploides
Lathyrus japonicus
Leymun mollis
Herbacées
Bryophytes
Lichens
Empetrum nigrum
Vaccinium vilis-idaea
Picea glauca
Bandelittorale
^ — "20
. . . l l l l l l l .H .1 . tf.
H.IHI..|I|.IHI.
Jllllll
Bandeherbacée
.—.—•—-*
40
llllllllllllll
m
. . . . . .
l l H l l , . , .
. . . l l l l l .
...1
J
BandeLichenaie forestière
Exagération verticale: 1,55
60 80
. .. .llillliililllin
Il I1..11.I1..I. . . .h
JiilliiLnlllllil, ni\i
1
- 8- 6- 4- 2- 0
•100
{100
100
100
100
•100
•100
rlO(
rlOO
t(l
aB
; iOo
Figure 7. Topographie et végétation le long du transect du site Grande Rivière. Lesépinettes représentent la limite des arbres et la limite des forêts.
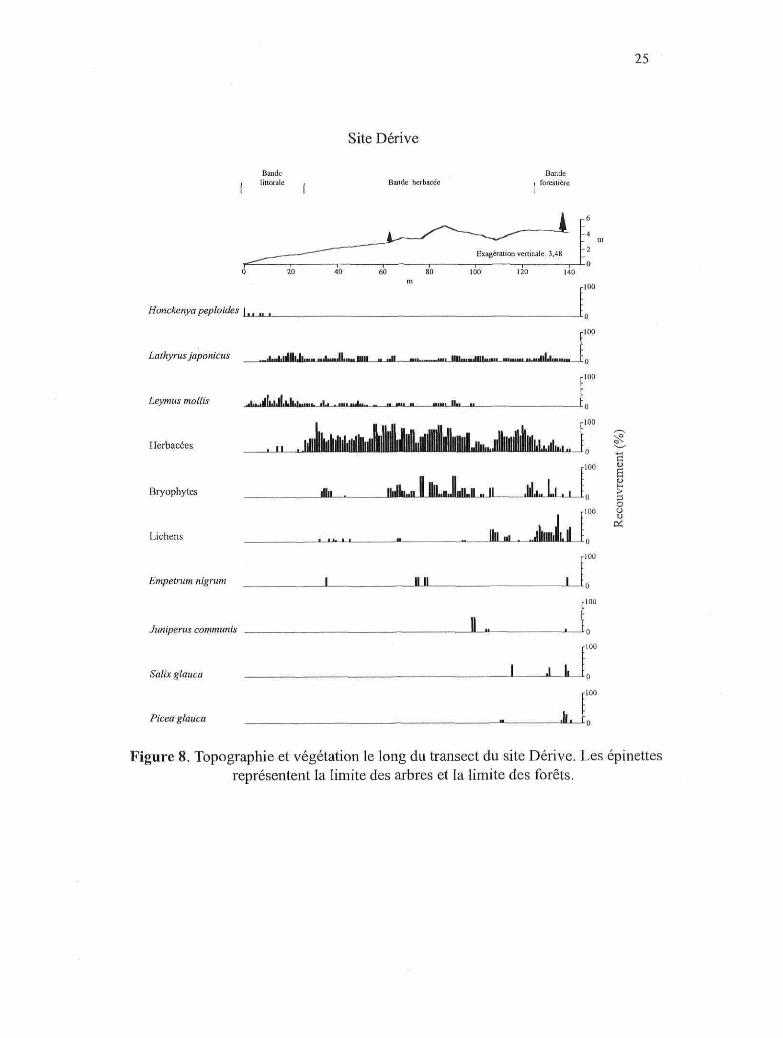
25
Site Dérive
Bandelittorale Bande herbacée
Bandeforestière
Honckenya peploides | . , ,. .
Lathyrus japonicus
Leymus mollis
Herbacées
Bryophytes
Lichens
Empetrum nigrum
Juniperus communis
Salix glauca
Picea glauca
Jbu^JllllilU
; • ; '
>
oo1)
0
100
n
100
r i o o
I .1 I. 0
100
Figure 8. Topographie et végétation le long du transect du site Dérive. Les épinettesreprésentent la limite des arbres et la limite des forêts.

Site Inuit
26
Bandelittorale Bande forestière
Honckenya peploides < III !•!• • •
Lathyrus japonicus
Leymus mollis
Herbacées
Bryophytes
Lichens
Empetrum nigrum
Vaccinium vilis-idaea
Picea glauca
40 60 80 100 120 140
• - . 1 . .I...I...IL. .«.I •• ..I ,l,ll 1.1. J . . . . . . . . . . • ..I.
Figure 9. Topographie et végétation le long du transect du site Inuit. Les epinettesreprésentent la limite des arbres et la limite des forêts.

27
Site île Longue
Bande forestière
Exagération verticale: 2,56
1 1
) 20
. .lllll...!
.. lllll .... ..
••••ini.ni
... .ii
i -40
lllllllllllllilllllllll.Mll
. l . l l . l
m
llllllll
lllllll.
1
• •
l i60 80
i. . i
I l l I l lÉ^^
ill.i. lia . lil.lll uilldl
lh.l,iillnL .ll.illl.
. .lit lll 1
.llllllll., .1....... 1
. Il i
- 2
0
100
0
100
0
100
0
100
3oo
Honckenya peploides
Lathyrus japonicus
Leymus mollis
Herbacées
Bryophytes
Lichens
Juniperus communis
Salix glauca
Picea glauca
Figure 10. Topographie et végétation le long du transect du site île Longue. Lesépinettes représentent la limite des arbres et la limite des forêts.
o
100
o
100
o100
o100

28
25 -,
20 -
15 -
uT3
3io -
M i
• 1czz• •• •
*
Honckenya peploidesLeymus mollis + Lathyrus japonicusHerbacéesAutres (ex. arbustes)Lichens jusqu'à la limite des forêts
Limite des arbres
Limite des forêts
Sites
Figure 11. Altitude des bandes de végétation à partir de la première présence continuedes espèces.

29
Tableau 2. Âge (années) estimé du substrat à la limite inférieure de chaque bande devégétation, arrondi à la décennie, en fonction d'un relèvement isostatique de 1,2m/siècle
Site
Guillaume- DelisleGoulet
Petite RivièreManitounuk
Grande RivièreDériveInuit
île Longue
Honckenyapeploides
00000000
Leymus mollis etLathyrus japonicus
10601080130608040
Herbacées
201603014022012040130
Lichens
730810480360420370240250
3.3 Limite des arbres
Des plantules et gaules situées au-delà de la limite des arbres sont présents dans
tous les sites d'étude (Fig. 12). Les sites nordiques possèdent peu de plantules, soit 3
dans le site Guillaume-Delisle et 13 dans le site Goulet. Avec 168 plantules, le site
Manitounuk possède le plus grand nombre de plantules. Au sud de Kuujjuarapik, les
sites renferment tous entre 70 et 80 plantules, sauf chez le site Dérive qui n'en possède
que 4. Les plantules les plus vieilles et les moins nombreuses se trouvent dans les sites
Guillaume-Delisle et Dérive. Les plantules au-delà de la limite des arbres se situent
rarement à des altitudes inférieures à 2 m (Fig. 13). l'exception étant pour le site
Manitounuk où une quantité importante de plantules se trouvait entre deux crêtes de
plage à une altitude plus basse que 2 m. Il est probable que les plus jeunes et petites
plantules n'aient pas toutes été trouvées sur le terrain. La relation entre l'âge des
plantules au-delà de la limite des arbres et l'altitude n'est pas significative (Fig. 16). Les
arbres du site Goulet ont été exclus de cette relation.
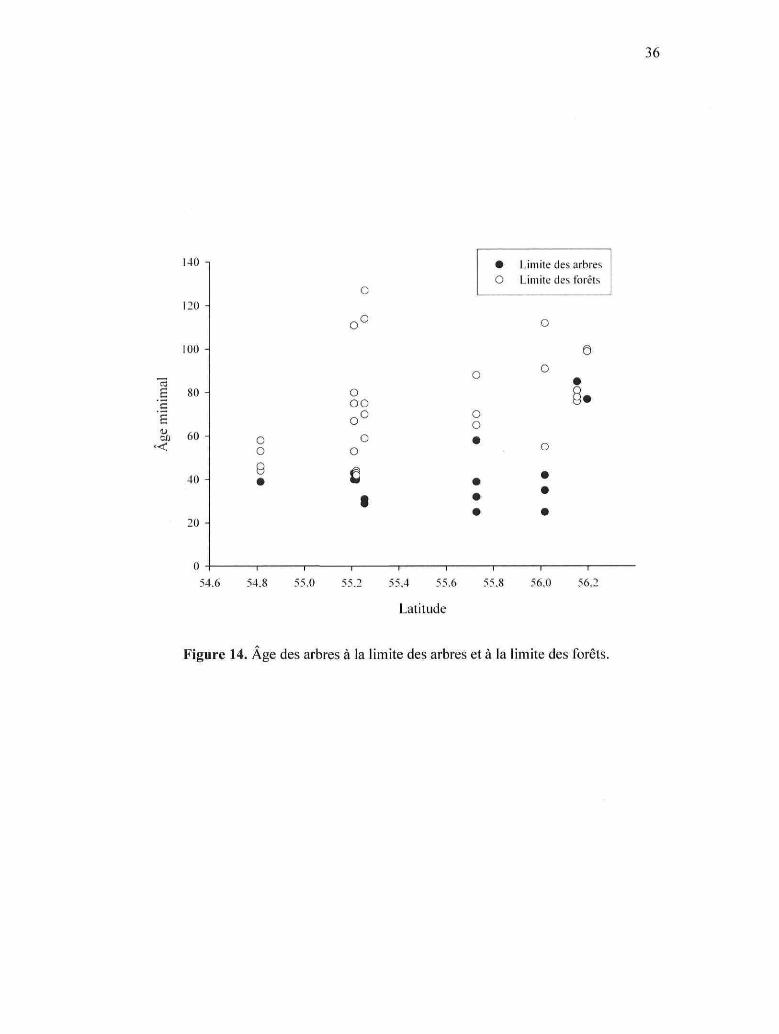
Les arbres du groupe Nord à la limite des arbres sont significativement plus vieux
que ceux des groupes Centre et Sud, alors que l'âge moyen des arbres des groupes
Centre et Sud est semblable (ANOVA, test de Tukey, p < 0,05) (Fig. 14 et 16). La
moyenne d'âge des arbres des groupes Nord, Centre et Sud est respectivement de 79 ± 5
ans (n = 3), 37 ± 11 ans (n = 8) et 38 ± 6 ans (n = 8). Le tableau 3 donne de plus amples
informations sur les arbres échantillonnés à la limite des arbres.

30
La limite des arbres du groupe Nord est significativement plus élevée en altitude
que celle des groupes Centre et Sud (ANOVA, test de Tukey, p < 0,05). L'altitude de la
limite des arbres des groupes Centre et Sud n'est pas significativement différente (p =
0,63). La limite des arbres du site Guillaume-Delisle est à 4,26 m alors que celle du
Goulet est à 10,82 m; c'est donc ce dernier site qui rend la différence significative.
L'altitude moyenne de la limite des arbres chez les sites des groupes Centre et Sud est
de 3,68 ± 1,00 m (» = 6) (Fig. 15 et 16). La relation entre l'âge des épinettes à la limite
des arbres et l'altitude n'est pas significative (Fig. 16).
La distance à la mer de la limite des arbres ne diffère pas significativement entre
les groupes Centre et Sud (ANOVA, test de Tukey, p = 0,99) d'un site à l'autre et se
situe en moyenne autour de 59,19 ± 20,78 m (n = 6) de la mer. Par contre, la limite des
arbres des sites Goulet et Guillaume-Delisle est considérablement plus éloignée de la
mer (groupe Nord : ANOVA, test de Tukey,/» < 0,05), soit respectivement 193,81 m et
158,45 m.

31
Tableau 3. Arbres échantillonnés à la limite des arbres pour l'ensemble des sites. Lesarbres représentés en caractère gras sont les plus près de la mer et déterminent laposition de la limite des arbres
Latitude
56° 1 1 ' 4 7 " N
56° 09' 14" N
56° 09' 14" N
56° 01' 00" N
56° 0 1 ' 0 0 " N
56° 0 1 ' 0 0 " N
55° 43' 36" N
55° 43' 36" N
55° 4 3 ' 3 6 " N
55° 4 3 ' 3 6 " N
55° 43' 36" N
55° 15' 17" N
55° 15' 17" N
55° 1 3 ' 0 8 " N
55° 12' 37" N
55° 12' 37" N
54° 4 8 ' 5 4 " N
54° 48' 54" N
54° 48' 54" N
Site
Guillaume-DelisleGoulet
Goulet
PetiteRivièrePetite
RivièrePetite
RivièreManitounuk
Manitounuk
Manitounuk
Manitounuk
Manitounuk
GrandeRivièreGrandeRivièreDérive
Inuit
Inuit
île Longue
île Longue
île Longue
Ageminimal
77
76
85
35
42
25
39
25
32
58
39
31
29
40
43
40
34
44
39
Diamètre(cm)
24,2
37,5
29,6
26,7
32,7
12,2
9,3
10,4
16,1
15,9
23,3
8,2
10,2
11,9
23,2
17,2
12,1
22,1
32,2
Hauteur(m)
4,1
3,7
4,3
3,43
5,00
2,59
2,5
2,55
3,53
3,6
3,39
4,37
4,05
2,64
3,36
3,22
3,39
3,58
3,38
Altitude(m)
4,26
10,819
14,625
5,881
5,699
4,516
3,679
3,534
3,804
3,84
3,991
5,155
5,124
3,018
2,639
2,989
2,936
2,561
2,81
Distanceà la mer
(m)158,5
193,8
253,7
63,2
61,7
31,7
90,6
94,2
106,4
107,4
114,1
50,5
51,0
63,6
72,0
80,0
38,4
46,8
55,4

20 -
Nord
Guillaume-DelisteC.oulel
Centre
50
I
I •
• • I Petite RivièreW777*. Maniloumik
Sud
11
Grande RivièreDériveI :île Longue
Age
Figure 12. Structure d'âge des plantules.

oo80o°
Nord
• Guillaume-DelisleO Goulet
30
33
12
10
8
6 1
2 -
Centre• Petite RivièreO Manitouiuik
Oo O
3 O o
10 20 30 40
2 •
(1
8 -
6 •
4 -
0 •
Sud
Y
• Grande RivièreO DériveT InuitV île Longue
•
o o o vo
Âge
Figure 13. Répartition altitudinale des plantules selon leur âge.

34
3.4 Limite des forêts
L'âge moyen des arbres échantillonnés à la limite des forêts est de 99 ± 1 ans (n =
3) dans le site Guillaume-Delisle, 78 ± 3 ans (n = 3) au Goulet, 86 ± 30 ans (n = 3) dans
le site Petite Rivière, 74 ± 12 ans (n = 3) dans le site Manitounuk, 89 ± 30 ans (n = 5)
dans le site Grande Rivière, 43 ± 1 ans dans le site Dérive {n = 3), 77 ± 22 ans {n = 5)
dans le site Inuit et 50 ± 6 ans (n = 4) dans le site île Longue. Dans l'ensemble des
arbres échantillonnés, seulement 5 d'entre eux ont plus que 100 ans, le plus âgé ayant au
minimum 127 ans (site Grande Rivière) (Fig. 14 et 16). Seuls les arbres du groupe Sud
sont significativement plus jeunes que ceux des autres groupes (ANOVA, test de Tukey,
p < 0,05). Le tableau 4 donne plus d'information sur les arbres échantillonnés à la limite
des forêts.
Les arbres à la limite des arbres des groupes Centre et Sud sont significativement
plus jeunes que ceux situés à la limite des forêts (ANOVA, test de Tukey, p < 0,05),
alors que l'âge moyen des arbres à la limite des arbres du groupe Nord ne semble pas
être significativement différent de celui des arbres situés à la limite des forêts (p = 0,23)
(Fig. 14 et 16). Dans l'ensemble des sites étudiés, la limite des arbres est
significativement plus basse en altitude que la limite des forêts (ANOVA, test de Tukey,
p < 0,05) (Fig. 15 et 16). La relation entre l'âge des épinettes à la limite des forêts et
l'altitude n'est pas significative (Fig. 16).
De manière générale, l'altitude de la limite des forêts augmente avec la latitude. En
effet, la limite des forêts est à plus de 10 m d'altitude chez le groupe Nord, alors qu'elle
se situe autour de 4 m dans les sites les plus au sud. L'altitude de la limite des forêts
varie significativement d'un site à l'autre (ANOVA, test de Tukey, p < 0,05), sauf chez
les sites Grande Rivière et Petite Rivière où elle est semblable (7,23 ± 0,17 m, n = 8),
ainsi que dans les 3 sites les plus au sud où elle se situe en moyenne à 4,38 ± 0,30 m {n
= 12) (Fig. 15 et 16). La position de la limite des forêts tend également à s'éloigner de la
mer avec la latitude (Tableau 4).
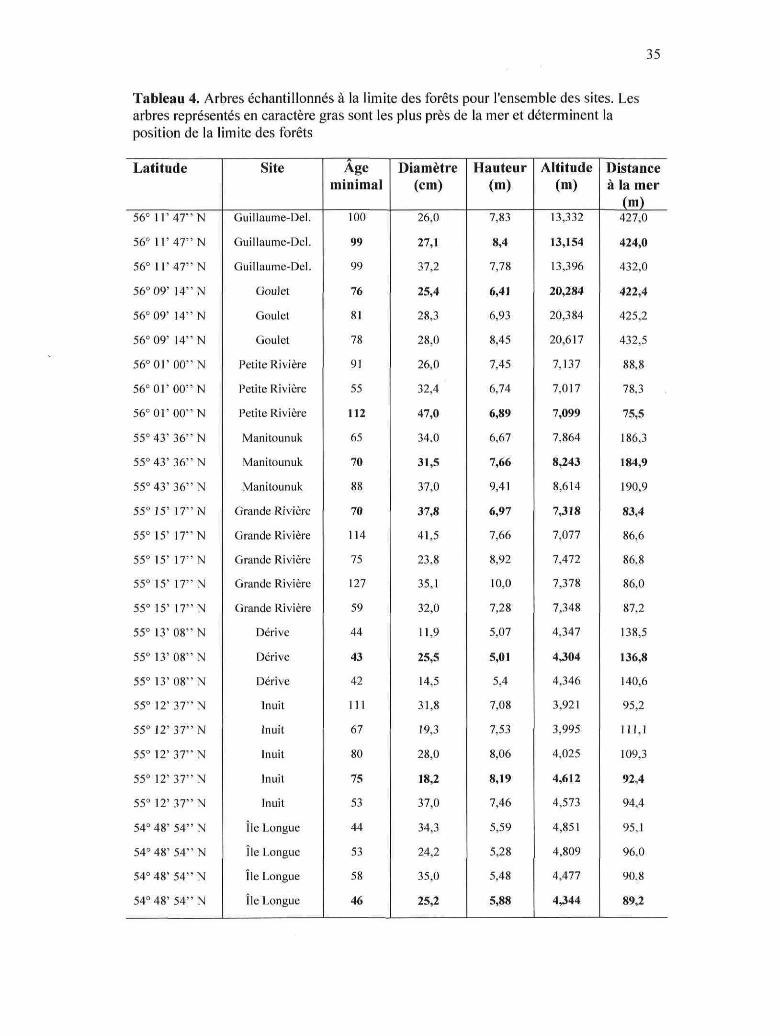
35
Tableau 4. Arbres échantillonnés à la limite des forêts pour l'ensemble des sites. Lesarbres représentés en caractère gras sont les plus près de la mer et déterminent laposition de la limite des forêts
Latitude
56° 1 1 ' 4 7 " N
56° 1 1 ' 4 7 " N
56° 1 1 ' 4 7 " N
56° 09' 14" N
56° 09' 14" N
56° 09' 14" N
56° 0 1 ' 0 0 " N
56° 0 1 ' 0 0 " N
56° 01' 00" N
55° 43' 36" N
55° 4 3 ' 3 6 " N
55° 4 3 ' 3 6 " N
55° 15' 17" N
55° 15' 17" N
55° 15' 17" N
55° 15' 17" N
55° 15' 17" N
55° 13 ' 08"N
55° 1 3 ' 0 8 " N
55° 1 3 ' 0 8 " N
55° 12' 37" N
55° 1 2 ' 3 7 " N
55° 1 2 ' 3 7 " N
55° 1 2 ' 3 7 " N
55° 12' 37" N
54° 4 8 ' 5 4 " N
54° 48' 54" N
54° 48' 54" N
54° 48' 54" N
Site
Guillaume-Del.
Guillaume-Del.
Guillaume-Del.
Goulet
Goulet
Goulet
Petite Rivière
Petite Rivière
Petite Rivière
Manitounuk
Manitounuk
Manitounuk
Grande Rivière
Grande Rivière
Grande Rivière
Grande Rivière
Grande Rivière
Dérive
Dérive
Dérive
Inuit
inuit
Inuit
Inuit
Inuit
île Longue
île Longue
île Longue
île Longue
Ageminimal
100
99
99
76
81
78
91
55
112
65
70
88
70
114
75
127
59
44
43
42
111
67
80
75
53
44
53
58
46
Diamètre(cm)
26,0
27,1
37,2
25,4
28,3
28,0
26,0
32,4
47,0
34,0
31,5
37,0
37,8
41,5
23,8
35,1
32,0
11,9
25,5
14,5
31,8
19,3
28,0
18,2
37,0
34,3
24,2
35,0
25,2
Hauteur(m)
7,83
8,4
7,78
6,41
6,93
8,45
7,45
6,74
6,89
6,67
7,66
9,41
6,97
7,66
8,92
10,0
7,28
5,07
5,01
5,4
7,08
7,53
8,06
8,19
7,46
5,59
5,28
5,48
5,88
Altitude(m)
13,332
13,154
13,396
20,284
20,384
20,617
7,137
7,017
7,099
7,864
8,243
8,614
7,318
7,077
7,472
7,378
7,348
4,347
4,304
4,346
3,921
3,995
4,025
4,612
4,573
4,851
4,809
4,477
4,344
Distanceà la mer
(m)427,0
424,0
432,0
422,4
425,2
432,5
88,8
78,3
75,5
186,3
184,9
190,9
83,4
86,6
86,8
86,0
87,2
138,5
136,8
140,6
95,2
111,1
109,3
92,4
94,4
95,1
96,0
90,8
89,2
140 -,
120 -
100 -
40 -
20 -
0
O
RU1
3S•vM<
80 -
60 - OO
oooo°
oo
• Limite des arbresO Limite des forêts
O
Oo
o
o
o
54.6 54,8 55,0 55,2 55,4 55.6 55.8 56,0 56.2
Latitude
36
Figure 14. Âge des arbres à la limite des arbres et à la limite des forêts.

37
22 -i
20 -
18 -
16 -
14 -
1 ,18 -
6
4
2 -
(I
• Limite des arbresO Limite des forêts
o
O
S?
54,6 54.8 55,0 55.2 55,4 55.6
Latitude
55,8 56.0 56,2
Figure 15. Altitude de la limite des arbres et de la limite des forêts. Chaque pointreprésente l'altitude moyenne (± 1 écart-type) de tous les arbres échantillonnés.

38
sa<
140 -,
120 -
100 -
80 -
60 -
40 -
20 -
• Planlules0 Limite des arbres• Limite des arbres site Goulet• Limite des forêts
y=ll,05-0,069xR2 = 0,069y = 46,76 - 1,735 xR2 = 0,02y = 62,8-1,498 xR2 = 0,098
10 12 14
Altitude (m)
16 18 20 22
Figure 16. Relation entre l'âge des arbres et l'altitude à la limite des arbres et à la limitedes forêts. Étant des valeurs distantes, les épinettes de la limite des arbres du site Goulet
n'ont pas été incluses dans la régression.

39
3.5 Sols
La granulométrie de l'horizon C de tous les sols des sites étudiés est semblable. Le
substrat est une dépôt fluvio-marin constitué de particules ayant un diamètre plus grand
que 50 um (> 95 %) et dont plus de la moitié se situe entre 250 et 500 |im, ce qui le
qualifie de sable moyen. Le degré de podzolisation des sols le long de la
chronoséquence est comparable d'un site à l'autre. Etant dans une région où la
température moyenne annuelle est inférieure à 0 °C, les sols entrent tous dans l'ordre
des cryosols. Cependant, les sols les plus développés se trouvent à la limite des forêts et
correspondent à l'ordre des brunisols dystriques (pH < 5,5) et à celui des brunisols
eutriques (pH > 5,5) (Groupe de travail sur la classification des sols, 2002).
Une relation significative a été trouvée entre l'altitude et le pH CaCb (régression
logarithmique, n = 43, R2 = 0,817, p < 0,05) (Fig. 17). Le pH de l'horizon B diminue
généralement avec l'altitude. Toutefois, les sols du site Manitounuk sont plus alcalins
tout au long du transect, puisqu'ils renferment des carbonates (réaction au HC1)
provenant de coquillages marins. À cause de leur nature différente, ces sols ont donc été
exclus de la régression. Les horizons B des sols du site île Longue possèdent aussi un
pH plus élevé que les sols des autres sites, mais ils n'ont pas réagi au HC1. Il est possible
que le pH plus alcalin du site île Longue soit influencé par des dépôts calcaires sous-
jacents. À partir de 4 à 5 m d'altitude, le pH des sols ne diminue plus de manière notable
(ANOVA, test de Tukey,/? < 0,05) et il varie entre 3,8 et 4,5 (sauf chez les sols des sites
Manitounuk et île Longue où il demeure entre 5 et 7). Quant au pH de l'horizon B,
aucune différence significative entre les sols de la limite des arbres et de la limite des
forêts n'a été trouvée (ANOVA, test de Tukey, p = 0,60). Le pH de l'horizon C tend à
diminuer le long du transect pour tous les sites, mais il reste toujours plus alcalin que
l'horizon B. À part les sites Manitounuk et île Longue dont l'horizon C demeure autour
de 7 tout au long du transect, le pH de cet horizon diminue jusqu'à 4,5 à 4,0 chez les
autres sites aux altitudes les plus élevées.
L'épaisseur de l'horizon B et le % Fe+Al extraits au pyrophosphate de sodium
tendent à augmenter avec l'altitude. Une relation significative entre l'altitude et le %
(Fe+Al)p a été trouvée (régression linéaire, n = 45, R2 = 0,58, p < 0,05) (Fig. 18). La

40
teneur en (Fe+Al)p de l'horizon B des sols de la limite des arbres du groupe Nord est
significativement plus élevée que celle des sols des groupes Centre et Sud (ANOVA,
test de Tukey, p < 0,05). Malgré l'altitude plus élevée de la limite des forêts du groupe
Nord, la teneur en (Fe+Al)p des sols n'est pas significativement différente de celle des
sols des autres groupes (ANOVA, test de Tukey, p = 0,068) et ce, malgré le fait que le
sol du site Guillaume-Delisle ait une teneur en Fe+Al nettement plus grande (0,155 %).
Aucune différence significative n'a été trouvée entre la teneur en fer-pyro et en
aluminium-pyro des sols de la limite des arbres et de la limite des forêts (ANOVA, test
de Tukey, p = 0,076). Les sols situés directement sous les arbres, étant influencés par
l'acidité de la litière d'aiguilles, n'ont pas été traités avec l'ensemble des échantillons,
mais renferment une plus grande teneur en (Fe+Al)p que les sols de milieu ouvert situés
à la même altitude (Fig. 18). Un horizon éluvié (Aej, poivre et sel) ayant moins de 2 cm
d'épaisseur était généralement présent à partir de la bande de végétation herbacée
stabilisée et pouvait atteindre entre 5 et 9 cm (Ae) dans les échantillons directement
sous les arbres.
Le Ctot (%), l'N (%), et les cations échangeables (Ca, Na, Mg, K) de l'horizon
organique de surface sont corrélés positivement les uns avec les autres (corrélation de
Pearson, p < 0,05) sauf pour le sodium avec le calcium (r = 0,30). Il existe également
une relation significative entre les variables décrites précédemment et l'altitude des
horizons de surface (régression linéaire, n = 33, p < 0,05). Par contre, la teneur en Ctot
de l'horizon de surface des sols de la limite des arbres et de la limite des forêts n'est pas
significativement différente (ANOVA, test de Tukey, p = 0,86). Les analyses de
conductivité électrique montrent une diminution rapide du contenu en sels avec la
profondeur dans le sol. Par contre, étant constamment exposé aux embruns et au vent
marin, l'horizon de surface conserve une conductivité élevée tout au long de chaque
transect. L'épaisseur de l'horizon organique de surface varie entre 0 et 4 cm dans les
bandes non forestières; elle est de 3 à 6 cm sous les arbres et atteint jusqu'à 15 cm sous
les arbres à la limite des forêts.
Des paléosols ont été trouvés dans les sites Grande Rivière, Guillaume-Delisle et
Petite Rivière. La couche organique des paléosols des sites Grande Rivière et Petite
Rivière contenait des charbons de très petite taille. Le paléosol du site Grande Rivière,

41
dont l'horizon organique était enfoui à 10 cm de profondeur, s'étend de la limite des
arbres à la limite des forêts. Les charbons du site Grande Rivière n'ont pas pu être
échantillonnés à cause de leur petite taille, de leur petit nombre et de la contamination
par des hyphes de champignons. Les charbons du site Petite Rivière ont été
échantillonnés et datés au radiocarbone (Tableau 5). L'horizon organique de ce paléosol
daté à 80 ± 70 ans étal. BP était enfoui à une quinzaine de centimètres de profondeur et
ce, à partir de la limite des arbres. Le paléosol du site Guillaume-Delisle à la limite des
forêts était constitué de matière organique décomposée d'origine tourbeuse située à une
profondeur de 9 cm. La matière organique des couches supérieure et inférieure du
paléosol a été datée au radiocarbone. Un paléosol a également été échantillonné sur la
terrasse principale du village de Kuujjuarapik. Il renfermait de petits charbons, mais
aucun reste d'aiguilles n'a été trouvé. L'horizon Bmb sous la couche organique des
paléosols des sites Petite Rivière et Grande Rivière est moins développé que l'horizon
Bm (Tableau 6).
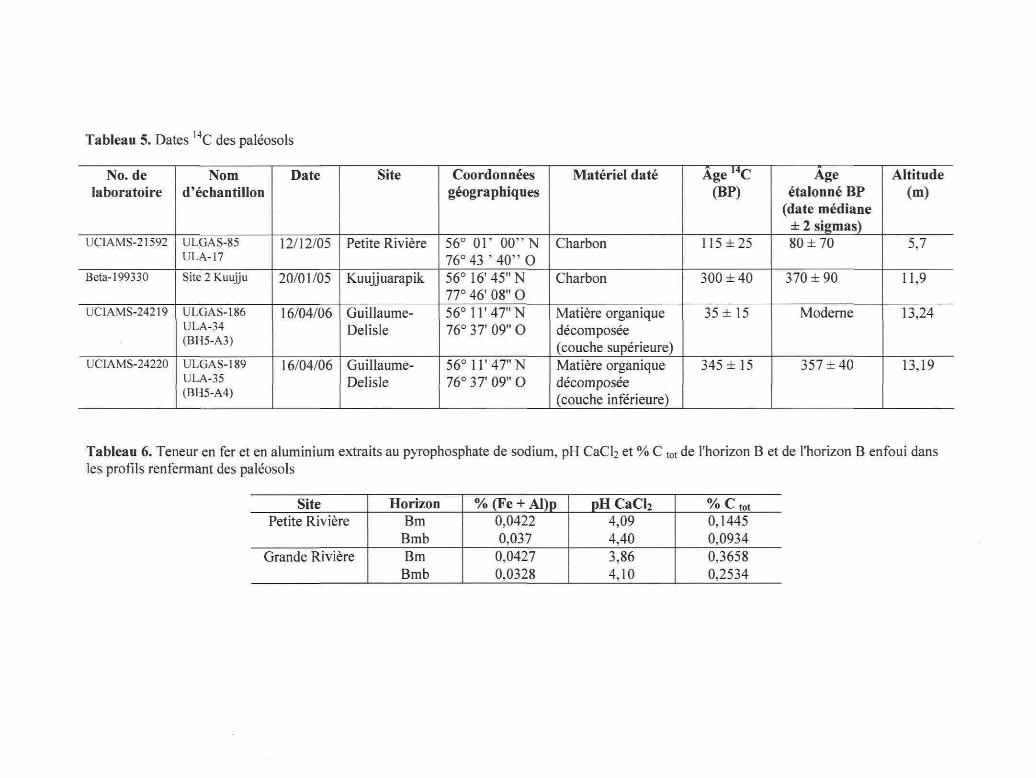
14,Tableau 5. Dates C des paléosols
No. delaboratoire
UCIAMS-21592
Beta-199330
UCIAMS-24219
UCIAMS-24220
Nomd'échantillon
ULGAS-85ULA-17
Site 2 Kuujju
ULGAS-186ULA-34(BH5-A3)
ULGAS-189ULA-35(BH5-A4)
Date
12/12/05
20/01/05
16/04/06
16/04/06
Site
Petite Rivière
Kuujjuarapik
Guillaume-Delisle
Guillaume-Delisle
Coordonnéesgéographiques
56° 01' 00" N76° 43 ' 40"O56° 16'45" N77° 46' 08" O56° 11'47" N76° 37' 09" O
56° 11'47" N76° 37' 09" O
Matériel daté
Charbon
Charbon
Matière organiquedécomposée(couche supérieure)Matière organiquedécomposée(couche inférieure)
Âge14C(BP)
115 ±25
300 ± 40
35 ±15
345 ±15
Ageétalonné BP
(date médiane± 2 sigmas)80 ±70
370 ±90
Moderne
357 ±40
Altitude(m)
5,7
11,9
13,24
13,19
Tableau 6. Teneur en fer et en aluminium extraits au pyrophosphate de sodium, pH CaCb et % C tot de l'horizon B et de l'horizon B enfoui dansles profils renfermant des paléosols
SitePetite Rivière
Grande Rivière
HorizonBmBmbBmBmb
% (Fe + Al)p0,04220,037
0,04270,0328
pH CaCl2
4,094,403,864,10
/O V' tOt
0,14450,09340,36580,2534

43
y = 6,509 -1,106 In xR2 = 0,817p < 0,05
Altitude (m)
Figure 17. Évolution du pH en fonction de l'altitude. Les points en blanc représentent lesite Manitounuk.

44
5 10 15
Altitude (m)
y = 0,0139 +0,005 x
R2 = 0,58
p < 0,05
20 25
Figure 18. Teneur en fer-pyro et en aluminium- pyro de l'horizon B des sols de tous lessites étudiés. Les points blancs représentent les échantillons prélevés directement sous
les arbres.

4. Discussion
4.1 Limite maritime de l'épinette blanche
L'action des vagues sur la côte, surtout lors d'événements de tempête et de
variations de la pression atmosphérique, peut perturber les plantules d'épinette établies à
basse altitude jusqu'à environ 2 m au-dessus du niveau moyen de la mer (Allard &
Tremblay, 1983; Ruz & Allard, 1994). Peu de plantules sont effectivement établies à
une altitude inférieure à 1,9 m. Elles sont généralement trouvées dans les dépressions
humides entre les crêtes de plage; autour le substrat est trop sec pour leur établissement.
Les arbres qui forment la limite des arbres sont relativement jeunes dans tous les
sites étudiés (< 40 ans), sauf ceux des sites les plus nordiques où l'établissement s'est
fait il y a environ 80 ans. Par contre, l'âge des arbres à la limite des forêts est
relativement variable d'un site à l'autre. Dans les sites les plus nordiques, les arbres ne
sont pas plus vieux dans les forêts, mais leur altitude est nettement plus élevée que dans
les autres sites. Le climat étant plus rigoureux vers le nord, les limites maritimes des
arbres et des forêts se trouvent plus en retrait de la mer et à une altitude plus élevée,
donc sur des surfaces plus anciennes. L'altitude ne détermine toutefois pas l'âge des
arbres.
En se basant sur un relèvement de 1,2 m/siècle, un délai de 140 à 470 ans de
succession végétale s'est écoulé avant que les plantules d'épinette blanche aient été en
mesure de s'établir au-delà de la limite actuelle des arbres, à l'exception du site Goulet
où le délai atteint 900 ans (Tableau 7). La formation de la limite des arbres et de la
limite des forêts actuelle s'est faite attendre après un long délai d'émersion allant
respectivement de 180 à 825 ans et de 310 à 1615 ans. Étant donnée le relèvement
isostatique, le sol était donc plus âgé (plus éloigné de la mer et plus élevé en altitude) au
moment de l'établissement des épinettes constituant la limite des forêts qu'au moment de
l'établissement des plantules et des arbres à la limite des arbres. Ceci signifie que les
conditions climatiques des 50 dernières années ont été plus favorables à la colonisation

46
plus près de la mer par l'épinette blanche, particulièrement dans les sites au sud de la
Petite-Rivière-de-la-Baleine.
Des études similaires ont montré que les plantules d'épinette de Norvège (Picea
abies) s'établissent entre 35 et 110 cm (40 à 130 ans) (Ericson, 1972 dans Svensson &
Jeglum, 2003a), 40 cm (45 ans) (Eriscon, 1980), 120 cm (Schwank, 1981 dans
Svensson & Jeglum, 2003a) et 25 et 35 cm (30 à 40 ans) (Svensson & Jeglum, 2003a)
au-dessus du niveau moyen de la mer sur la côte en émersion du Golfe de Bothnie. Les
forêts d'épinette de Norvège prendraient entre 130 et 140 ans avant de se constituer dans
le Golfe de Bothnie (Svensson & Jeglum, 2003b). Cet établissement plus rapide des
épinettes le long de la mer en Scandinavie est probablement dû à des conditions
édaphiques et climatiques plus favorables à l'établissement et à la survie des espèces
arborescentes (ex. humidité du sol, exposition des sites).
Tableau 7. Âge (années) estimé de la surface au moment de la colonisation par lesépinettes blanches actuelles d'après un taux de relèvement moyen de 1,2 m/siècle
Site
Guillaume-DelisleGoulet
Petite RivièreManitounuk
Grande RivièreDériveInuit
île Longue
Plantules
145900470140410195210210
Limitedes
arbres280825350270400210180210
Limitedes
forêts10001615480620540315310315
4.2 Influence du climat
Le nord du Québec a connu des alternances de périodes favorables et défavorables
à la croissance des arbres. Depuis 750 ans, on constate une diminution du nombre de
feux et de l'activité éolienne le long de la baie d'Hudson, ce qui serait une indication de
conditions estivales plus humides (Filion, 1984a, b). Il y aurait également eu une
augmentation des précipitations en hiver au XXe siècle dans le nord du Québec (Bégin,
2001), favorisant ainsi la colonisation des milieux mésiques par les épinettes. Les

47
changements de la forme de croissance de l'épinette noire, passant de la forme arbustive
à la forme arborescente depuis la fin du XIXe siècle sont aussi une conséquence d'étés et
d'hivers plus cléments et humides (Payette et ai, 1985; Lescop-Sinclair & Payette,
1995).
Le phénomène de rétroaction positive enclenché par les peuplements forestiers
assure une certaine protection contre le vent et permet l'accumulation d'un couvert de
neige plus important, permettant aux épinettes d'avoir une croissance normale et un port
symétrique en forêt (Payette et al., 1985). La forme de croissance normale des épinettes
blanches observée jusqu'à la limite latitudinale des forêts maritimes démontre aussi que
cette espèce n'a probablement pas encore atteint son aire de répartition post-glaciaire
maximale en équilibre avec le climat (Payette, 1993). L'abondance de plantules dans la
majorité des sites est aussi probablement reliée à des conditions climatiques favorables.
4.3 Influence des facteurs locaux
Une séquence évidente de colonisation, conséquence du relèvement isostatique,
forme une succession de bandes végétales dans tous les sites étudiés. Les plantes
vasculaires, les bryophytes et les lichens permettent de stabiliser le substrat et
d'augmenter le contenu en nutriments et en matière organique des sols (Kimmins, 1987;
Bâtes, 2000; Imbert & Houle, 2000; Brodo et al, 2001; Svensson & Jeglum, 2003a).
L'humidité du sol étant un facteur limitant à l'établissement de l'épinette blanche, la
matière organique ainsi que la présence de bryophytes et lichens à la surface du sol
contribuent à augmenter la rétention d'eau et à limiter l'évaporation (Bâtes, 2000; Brodo
et al, 2001).
Les plantules d'épinettes s'établissent dans la bande de végétation herbacée (ou
arbustive dans le cas du site Guillaume-Delisle) stabilisée où on observe un important
couvert de bryophytes au sol. Ce n'est que dans le site Goulet que les plantules se sont
établies sur un tapis composé exclusivement de lichens. L'abondance des lichens
explique, du moins en partie, le petit nombre de plantules recensées dans les sites
nordiques (Houle & Filion, 2003). Le couvert lichénique augmente fortement à
proximité de la limite des arbres, ce qui indique que la plupart des lichens se sont établis

48
à la suite de la colonisation des épinettes, sauf près de la limite latitudinale des forêts
maritimes (sites Guillaume-Delisle et Goulet) où ils ont colonisé les sites bien avant
l'établissement de l'épinette blanche. Houle et Filion (2003) ont montré que le couvert
de lichens pouvait être favorable à la croissance des épinettes ayant passé le stade
juvénile.
Les sols sont généralement plus développés en altitude le long de la côte, puisque
dans un contexte de relèvement isostatique, une surface plus élevée en altitude est
exposée depuis plus longtemps à l'influence de la végétation et du climat. Le matériel
d'origine, le climat, le temps, la topographie, les microorganismes, ainsi que l'humidité
du sol, les perturbations et la végétation influencent les processus de pédogenèse
(Jenny, 1941; Chapin et al, 1996). Le mince horizon organique de tous les sols étudiés
le long de la côte de la baie d'Hudson est redevable à une végétation peu productive, à
une litière clairsemée, à un sol acide et à des températures fraîches. La podzolisation est
le processus pédogénétique dominant dans le contexte de succession primaire qui
caractérise la végétation et les sols de la côte de la baie d'Hudson en émersion. Avec
l'établissement des arbres, la litière acide des épinettes ainsi qu'une plus grande
accumulation de neige près de ceux-ci (Payette & Filion, 1985) accélèrent la
podzolisation. Le substrat passe donc d'un non-sol sur la plage, à un régosol dans la
bande herbacée et à un brunisol dans la forêt et sous les arbres. Le travail de Filion
(1984b) a montré que les paléosols enfouis dans les dunes de la toundra forestière sont
le plus souvent des brunisols dystriques ayant une teneur en fer-pyro et en aluminium-
pyro inférieure à 0,15 %. Ces valeurs sont semblables à celles des échantillons pris à la
limite des forêts le long de la côte.
Deux paléosols possédant des charbons de bois datés entre 80 et 370 ans étal. BP,
ainsi qu'un feu datant des années 1950 près de la Pointe Louis-XIV ont été observés.
Les feux sont toutefois peu fréquents et de petite superficie le long de la côte (Payette &
Filion, 1985). Suite au passage des feux, une succession secondaire s'enclenche. La
régénération rapide de l'épinette blanche suivant le passage du feu dans les sites Petite
Rivière et Grande Rivière s'explique par le fait que des semenciers survivants se
trouvaient probablement dans les milieux avoisinants plus humides comme au pied des
buttes rocheuses et le long des cours d'eau (Payette & Filion, 1985). La nature des

49
paléosols avec un horizon B enfoui moins développé que l'actuel et l'absence de
macrorestes d'épinette dans l'horizon organique enfoui suggèrent la présence ancienne
d'une végétation non forestière au-delà de la limite actuelle des forêts.
Bégin et al. (1993) ont montré que l'épinette blanche colonise les sites côtiers
humides environ au même rythme que le taux d'émersion actuel le long de la baie
d'Hudson, avec un délai d'une vingtaine d'années seulement. Cette situation contraste
avec la lente colonisation de l'épinette blanche sur les sols sableux bien drainés de tous
les sites étudiés où l'on constate un asynchronisme marqué avec le taux de relèvement
actuel. La limite de colonisation des épinettes est déterminée par les facteurs de sol et de
végétation qui empêchent la colonisation en dessous d'une certaine altitude, où le
substrat est instable, moins humide et moins développé.
4.4 Expansion récente de l'épinette blanche
Peu d'informations sont disponibles concernant l'arrivée de l'épinette blanche le
long de la côte est de la baie d'Hudson (Payette, 1993). Caccianiga et Payette (sous-
presse) ont signalé la présence de l'épinette blanche seulement à partir du XVIIe siècle
au nord de la région d'étude le long la côte. L'épinette blanche serait en expansion
depuis 400 à 500 ans, avec une densifïcation des populations forestières au cours du
dernier siècle (Payette & Filion, 1985). Le long de la baie d'Hudson, aucun arbre mort
n'a été trouvé entre la plage et la limite des forêts dans l'ensemble des sites, ce qui
confirme la formation très récente de la limite des arbres, ainsi que l'établissement des
forêts primaires au cours du dernier siècle seulement.
4.5 Données historiques
En 1950, Hustich a noté qu'à la limite maritime des arbres près de la Pointe Louis-
XIV (à 4 km au sud du site île Longue) les épinettes blanches étaient relativement
jeunes et que la forêt semblait « avancer vers la mer ». Cette forêt a brûlé quelques
années plus tard, après le passage de l'auteur. Hustich (1950) mentionne que les arbres
endommagés par la neige et le vent sont fréquents le long de la côte et qu'il y a peu de
régénération à la limite maritime (Fig. 19). Il souligne la présence d'un tapis d'épinette

50
blanche rampant. De nos jours, aucune forme de croissance rampante n'a été rencontrée
et les arbres ont un port symétrique (Fig. 20). Selon Hustich (1950), les cônes d'épinette
blanche étaient abondants en 1947 près de la Pointe Louis-XIV, malgré l'absence de
plantules et une croissance lente des arbres. Depuis ce temps, on dénombre maintenant
des arbres âgés entre 34 et 44 ans (âge minimum) à la limite des arbres. Il est possible
que la présence de jeunes arbres à la limite des arbres résulte des années de grande
abondance de cônes notée par l'auteur en 1947.

51
Figure 19. Limite maritime de Pépinette blanche à la localité 18 (54° 50' N, 79° 02' O)en 1948 (tiré de Hustich, 1950; photo E. H. Kranck).
Figure 20. Limite maritime de l'épinette blanche au site île Longue(54° 48' N, 79° 06' O).

Conclusion
La limite maritime des arbres et des forêts le long de la côte orientale de la baie
d'Hudson s'est formée récemment. Aucune preuve macrofossile, comme la présence de
restes d'arbres au sol ou de macrofossiles enfouis dans les paléosols, n'a été trouvée
quant à la présence ancienne d'une limite d'arbres ou d'une limite de forêt plus près de
la mer. Les conditions climatiques depuis un siècle auraient favorisé la progression de
Pépinette blanche vers la mer. Le potentiel de régénération de l'espèce au-delà de la
limite des arbres du côté de la mer est élevé, d'après le grand nombre de plantules qui
s'y sont déjà établies. La limite maritime actuelle des arbres a été formée au cours des
derniers 50 à 80 ans. Les arbres croissant à la limite des forêts sont généralement plus
âgés que ceux que l'on trouve à la limite des arbres, mais ils n'ont pas plus de 130 ans.
Les conditions climatiques plus rigoureuses de la partie nord de la région d'étude sont
probablement responsables de la limite altitudinale plus élevée de la limite des arbres et
de la limite des forêts.
La végétation et les sols de tous les sites d'étude évoluent en conditions de
succession primaire reliées directement au relèvement post-glaciaire de la côte.
L'établissement des épinettes blanches s'est effectué sur une surface littorale dont l'âge
varie entre 145 ans et 900 ans dans le cas des plantules au-delà de la limite des arbres et
des arbres constituant la limite des arbres. Contrairement à ces deux dernières limites,
les arbres de la limite des forêts maritimes se sont établis sur une surface dont l'âge
varie entre 230 et 1275 ans. Ceci signifie que les conditions climatiques récentes ont
permis à l'épinette blanche de s'établir plus près de la mer qu'au moment de
l'établissement des arbres qui forment actuellement la limite des forêts. Si les conditions
climatiques plus douces et humides se maintiennent, les plantules d'épinette blanche
présentes aujourd'hui pourront se développer et assurer la progression de la limite des
arbres vers la mer.

Bibliographie
Allard, M. & G. Tremblay. 1983. La dynamique littorale des îles Manitounuk durantl'Holocène. Zeitschrift fur Geomorpholgie 47: 61-95.
Allard, M. & M. K. Seguin. 1987. Le pergélisol au Québec nordique, bilan etperspectives. Géographie Physique et Quaternaire 41: 141-152.
Audet, R. 1976. Distribution de l'épinette blanche [Picea glauca (Moench) Voss] dansl'estuaire de la Grande Rivière de la Baleine, Nouveau Québec. Mémoire M. Se.Université Laval, Sainte-Foy, Québec.
Bâtes, J. W. 2000. Minerai nutrition, substratum ecology, and pollution. Dans A. J.Shaw et B. Goffînet, éditeurs. Bryophyte Biology, pp. 248-311. CambridgeUniversity Press, Cambridge.
Bégin, Y. 2001. Tree-ring dating of extrême lake levels atthe subarctic-boreal interface.Quaternary Research 55: 133-139.
Bégin, Y., D. Bérubé & M. Grégoire. 1993. Downward migration of coastal conifers asa response to récent land émergence in eastern Hudson Bay. QuaternaryResearch 40: 81-88.
Brodo, I. M., S. D. Sharnoff & S. Sharnoff. 2001. Lichens of North America. YaleUniversity Press, New Haven, Connecticut.
Burns, R. M. & B. H. Honkala. 1990. Silvics of North America. Forest Service, UnitedStates Department of Agriculture, Washington, D.C.
Caccianiga, M. & S. Payette. Sous-presse. Récent advance of white spruce {Piceaglauca (Moench) Voss) in the coastal tundra of the eastern shore of Hudson Bay(Québec, Canada). Journal ofBiogeography.
Chapin, F. S. III, M. S. Torn & M. Tateno. 1996. Principles of ecosystem sustainability.American Naturalist 148: 1016-1037.
Connell, J. H. & R. O. Slatyer. 1977. Mechanisms of succession in natural communitiesand their rôle in community stability and organisation. American Naturalist 111:1119-1144.
Cramer, W. 1985. The effect of sea shore displacement on population âge structure ofcoastal Alnus glutinosa (L.) Gaertn. Holarctic Ecology 8: 265-272.
Cramer W. & H. Hytteborn. 1987. The séparation of fluctuation and long term changein végétation dynamics of a rising sea-shore. Vegetatio 69: 157-167.
Ecke, F. & H. Rydin. 2000. Succession on a land uplift coast in relation to plant strategytheory. Annales Botanici Fennici 37: 163-171.

54
Environnement Canada. 2006. Normales climatiques.http://www.climate.weatheroffice.ec.gc.ca/climate_normals/index_f.html
Ericson, L. 1972. Lëvvegetationen vid norrlandskusten. Norrlandica 6:227-255. DansSvensson, J. S. & J. K. Jeglum. 2003. Primary succession pathway of Norwayspruce communities on land-uplift seashores. Écoscience 10: 96-109.
Ericson, L. 1980. The downward migration of plants on a rising Bothnian sea-shore.Acta Phytogeographica Suecica 68: 61-72.
Filion, L. 1984a. Analyse macrofossile et pollinique de paléosols de dunes en Hudsonie,Québec nordique. Géographie Physique et Quaternaire 38: 113-122.
Filion, L. 1984b. A relationship between dunes, fire and climate recorded in theHolocene deposits of Québec. Nature 309: 543-546.
Filion, L. & P. Morisset. 1983. Eolian landforms along the eastern coast of Hudson Bay,Northern Québec. Dans P. Morisset & S. Payette, éditeurs. Tree-Line Ecology.Proceedings of the Northern Québec Tree-Line Conférence. Nordicana 47: 73-94.
Forster, S. M. & T. H. Nicholson. 1981. Aggregation of sand from a maritime embryodune. SoilBiology andBiochemistry 13: 199-203.
Fulton, R. J. 1989. Avant-propos sur le Quaternaire du Canada et du Groenland. LeQuaternaire du Canada et du Groenland, sous la direction de R. J Fulton,Commission géologique du Canada. Géologie du Canada, vol. 1. Ottawa,Ontario.
Gajewski, K., S. Payette & J. C. Ritchie. 1993. Holocene végétation history at theboréal forest-shrub tundra transition in northern Québec. Journal ofEcology 81:433-443.
Grégoire, M. & Y. Bégin. 1993. The récent development of a mixed shrub conifercommunity on a rapidly emerging coast (Eastern Hudson Bay, subarctic Québec,Canada). Journal of Coastal Research 9: 924-933.
Groupe de travail sur la classification des sols. 2002. Le système canadien declassification des sols (3ieme édition). Agriculture et Agroalimentaire CanadaPublication 1646, Ottawa, Ontario.
Grove, J. M. 1988. The Little IceAge. Methuen and Co. Ltd, Londres.
Hadley, J. L. & W. K. Smith. 1986. Wind effects on needles of timberline conifers:Seasonal influence on mortality. Ecology 67: 12-19.
Hillaire-Marcel, C. 1976. La déglaciation et le relèvement isostatique sur la côte est dela Baie d'Hudson. Cahiers de Géographie de Québec 50: 185-220.

55
Hillaire-Marcel, C. & J.-S. Vincent. 1980. Stratigraphie de l'Holocène et évolution deslignes de rivage au sud-est de la baie d'Hudson, Canada: livret-guide. Paléo-Québec 11: 1-165.
Houle, G. & L. Filion. 2003. The effects of lichens on white spruce seedlingestablishment and juvénile growth in a spruce-lichen woodland of subarcticQuébec. Écoscience 10: 80-84.
Hustich, I. 1950. On the forests on the east coast of Hudson Bay. Acta Geographica 11 :5-83.
Imbert, É. & G. Houle. 2000. Ecophysiological différences among Leymus mollispopulations across a subarctic dune System caused by environmental, notgenetic, factors. New Phytologist 147: 601-608.
Imbert, É. & G. Houle. 2001. Spatio-temporal dynamics of root density in a coastaldune in subarctic Québec, Canada. Journal of Coastal Research 17: 877-884.
Jenny, H. 1941. Factors ofSoil Formation. McGraw-Hill, New York.
Kimmins, J. P. 1987. Forest Ecology. Macmillan Publishing Company, New York.
Kullman, L. 1986. Récent tree-limit history of Picea abies in the southern SwedishScandes. Canadian Journal of Forest Research 16: 761-771.
Lamb, H. H. 1977. Climate- Présent, Past and Future. Methuen and Co. Ltd, Londres.
LaMarche, V. C. Jr. 1973. Holocene climatic variations inferred from treelinefluctuations in the White Mountains, California. Quaternary Research 3: 632-660.
Lavoie, C. & S. Payette. 1994. Récent fluctuations of the lichen-spruce forest limit insubarctic Québec. Journal of Ecology 82: 725-734.
Lavoie C. & S. Payette. 1996. The long-term stability of the boréal forest limit insubarctic Québec. Ecology 77: 1226-1233.
Lescop-Sinclair, K. & S. Payette. 1995. Récent advance of the arctic treeline along theeastern coast of Hudson Bay. Journal of Ecology 83: 929-936.
Marin, P. & L. Filion. 1992. Récent dynamics of subarctic dunes as determined fromtree-ring analysis of white spruces, eastern coast of Hudson Bay, Québec.Quaternary Research 38: 316-330.
Marr, J. W. 1948. Ecology of the forest-tundra ecotone on the east coast of Hudson Bay.Ecological Monographs 18: 117-144.
MacBeth division de Kollmorgen Instruments Corporation. 1994. Munsell Soil ColorCharts. Édition 1994. Baltimore.

56
McKeague, J. A. 1978. Manual ofsoil sampling and methods of analysis. 2ieme édition.Canadian Soil Survey Committee. Canadian Society of Soil Science, Ottawa,Ontario.
Morin, A. & S. Payette. 1984. Expansion récente du mélèze à la limite des forêts(Québec nordique). Canadian Journal ofBotany 62: 1404-1408.
Môrner, N.-A. 1980. The Fennoscandinavian uplift: Geological data and theirgeodynamical implication. Dans N.-A. Morner, éditeur. Earth Rheology,Isostasy and Eustasy. Proceedings of Earth Rheology and Late CenozoicIsostatic Movements, an Interdisciplinary Symposium. Stockholm, Suède.Geodynamics Project: Scientific Report 49: 251 -284.
Mueller-Dombois, D. & H. Ellenberg. 1974. Aims and Methods of Végétation Ecology.John Wiley & Sons, Inc., New York.
Payette, S. 1974. Classification écologique des formes de croissance de Picea glauca(Moench.) Voss et de Picea mariana (Mill.) BSP. en milieux subarctiques etsubalpins. Naturaliste canadien 101: 893-903.
Payette, S. 1975. La limite septentrionale des forêts sur la côte orientale de la baied'Hudson, Nouveau-Québec. Naturaliste canadien 102: 317-329.
Payette, S. 1983. The forest-tundra and présent tree-lines of the northern Québec-Labrador peninsula. Dans P. Morisset & S. Payette, éditeurs. Tree-Line Ecology.Proceedings of the Northern Québec Tree-Line Conférence. Nordicana 47: 3-23.
Payette, S. 1993. The range limit of boréal tree species in Québec-Labrador: anecological and palaeoecological interprétation. Review of Palaeobotany andPalynology 79: 7-30.
Payette, S., L. Filion & J. Ouzilleau. 1973. Relations neige-végétation dans la toundraforestière du Nouveau-Québec, baie d'Hudson. Naturaliste canadien 100: 493-508.
Payette, S. & L. Filion. 1985. White spruce expansion at the tree-line and récentclimatic change. Canadian Journal of Forest Research 15: 241-251.
Payette, S., L. Filion, L. Gauthier & Y. Boutin. 1985. Secular climate change in old-growth tree-line végétation of northern Québec. Nature 31: 135-138.
Reimer, P. J., M. G. L. Baillie, E. Bard, A. Bayliss, J. W. Beck, C. Bertrand, P. G.Blackwell, C. E. Buck, G. Burr, K. B. Cutler, P. E. Damon, R. L. Edwards, R. G.Fairbanks, M. Friedrich, T. P. Guilderson, K. A. Hughen, B. Kromer, F. G.McCormac, S. Manning, C. Bronk Ramsey, R. W. Reimer, S Remmele, J. R.Southon, M. Stuiver, S. Talamo, F. W. Taylor, J. van der Pilcht & C. E.Weyhenmeyer. 2004. IntCalO4 Terrestrial Radiocarbon Age Calibration, 0 - 2 6ka cal BP. Radiocarbon 46: 1029-1058.

57
Ricard, B. & Y. Bégin. 1999. Le développement d'une pessière à épinette blanche et àlichens sur la côte en émersion rapide de la baie d'Hudson au Québecsubarctique. Géographie Physique et Quaternaire 53: 351-364.
Rouse, W. R. 1991. Impacts of Hudson Bay on the terrestrial climate of the Hudson Baylowlands. Arctic and Alpine Research 23: 24-30.
Rousseau, J. 1968. The végétation of the Québec-Labrador Peninsula between 55° and60° N. Naturaliste canadien 95: 469-563.
Ruz, M.-H. & M. Allard. 1994. Foredune development along a subarctic emergingcoastline, eastern Hudson Bay, Canada. Marine Geology 117: 57-74.
Schwank, B. 1981. Landhôjningen och floran. Dans Finska Vetenskaps SocietetensÂrsbok, pp. 157-163. Dans Svensson, J. S. & J. K. Jeglum. 2003. Primarysuccession pathway of Norway spruce communities on land-uplift seashores.Écoscience 10: 96-109.
Scott , P. A., R. I. C. Hansell & D. C. F. Fayle. 1987. Establishment of white sprucepopulations and responses to climatic change at the treeline, Churchill,Manitoba, Canada. Arctic and Alpine Research 19: 45-51.
Sokal, R. R. & F. J. Rohlf. 1995. Biometry. W. H. Freeman and Company, New York.
StatSoft. 2005. Statistica Base et Statistica Advanced Linear/Non-linear Models. Tulsa,Oklahoma.
Stuiver, M. & P. J. Reimer. 1993. Extended 14C database and revised CALIBradiocarbon calibration program. Radiocarbon 35: 215-230.
Svensson, J. S. & J. K. Jeglum. 2001. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline. Forest Ecologyand Management 151: 67-79.
Svensson, J. S. & J. K. Jeglum. 2003a. Primary succession pathway of Norway sprucecommunities on land-uplift seashores. Écoscience 10: 96-109.
Svensson, J. S. & J. K. Jeglum. 2003b. Spatio-temporal properties of tree-species beltsduring primary succession on rising Gulf of Bothnia coastlines. Annales BotaniciFennici 40: 265-282.
Szeicz, J. M. & G. M. MacDonald. 1995. Récent white spruce dynamics at the subarcticalpine treeline of north-western Canada. Journal of Ecology 83: 873-885.
von Mors, I. & Y. Bégin. 1993. Shoreline shrub population extension in response torécent isostatic rebound. Arctic and Alpine Research 25: 15-23.
Walker, L. R. & R. del Moral. 2003. Primary Succession and Ecosystem Rehabilitation.Cambridge University Press, Cambridge.

Wilson, C. 1968. Notes on the climate of Poste-de-la-Baleine, Québec. Nordicana 24:1-93.

Annexe 1
Analyses de sol
Site Guillaume-Dolisle
VégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg(mBq/100g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivrté (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide]
HonckenyjC
BH5-E1
2.23.623.870.40.00.0
0.6700.074
41.44442.5310.0080.0060.0130.0780.121Non
10YR 6,5/110YR 4,5/1
LeymusC
BH5-D1
0.516
25.772.10.00.0
0.4720.08441.4440.0000.006
0.0050.0110.4410.144Non
10YR6/110YR5/1
LathyrusC
BH5-C1
2.010.536.450.80.50.0
0.4090.09717.5940.0000.0120.0070.0190.1180.084Non
10YR6/210YR4/2
^rès de la limite des arbresH (sel poivre)
BH5-B1
-
2.2812.510
446.112528.075
9.4010.203Non
10YR2/210YR 1,7/1
Aej BmBH5-B2
4.312.345.537.30.50.0
0.2080.163
43.79032.64500410.0140.0550.7170.085Non
10YR5/310YR3/3
CSous arbre limite des forêts
LFHBH5-A1
-
7.1665 801
257.945
0.231Non
10YR2/12,5Y 2/1
Ae OhbBH5-A3(haut)BH5-A4(bas)
• Datés au 14C
-
Hb Aeb BmbBH5-A2
0.10.9
37.260.50.80.4
0.0870.1365.865
257.9450.1310.0240.155
0.114Non
10YR5/310YR3/3
Cg

60
Site GouletVégétationHorizon
No.échantillon
Tamisage (%)
sable très grossiersable grossier
sable moyensable fin
sable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)
Mg (meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)
C lot (%)Conductivité (mmhos/cm)
Réaction au HCICouleur (sec)Couleur (humide)
HonckenyaC
BH6-G1
0.128.9
6852.4
0.00.0
6.8100.0929
0.08654.7380.0000.0090.0070.016
0.0350.061
Non
10YR 6,5/110YR4/1
Leymus
CBH6-F1
0.03.9
86.69.50.0
0.0
6.2300.1770 140
48.0910.000
0.0080.0070.016
0.0770.068Non
2,5Y 6/22.5Y 4/2
HerbacéeLF
BH6-E1
-
5.5801.637
0.675310.05011.265
3.1790.292Non
2,5Y 3/3
2.5Y2/1
Sable Mb Bmb
BH6-E2
-
4.5800.2770.179
44.5720.0000.0170.0080.025
0.4050.130Non
2,5Y 5/22,5Y 4/2
C
BH6-E3
0.05.2
79.515.3
0.0
0.0
4.7000.0820.081
21.5040.000
0.061Non
10YR6/210YR4/3
Herbacée
LF
BH6-D1
-
4.6302.4951.810
454.71355.635
6.9460.208Non
2.5Y 3/32.5Y2/1
BmBH6-D2
-
3.9800.318
0.28860.6020.0000.0360.0150.051
0.5530.125
Non10YR4/2
10YR 3/2
Hb BmbBH6-D3
-
4.1300.1520.16818.3760.000
0.0220.0120.0340.176
0.102Non
10YR 5/310YR 4/2
Hb2 Bmb2
BH6-D4
-
4.1100215
0.20615.6390.0000.0260.0140.040
0.2670.075
Non10YR 5/210YR 3/3
CBH6-D5
0.05.9
76.617.40.1
0.0
4.7300.2080.127
21.1130.000
0.070Non
10YR 5/210YR 4/2
Site GouletVégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable linsable très finlimon et argile
Analyses
pH CaCI2Ca (mEq/100 g)Mg(mEq/100g)K (mEq/100 g)Na (mEq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)
Réaction au HCICouleur (sec)Couleur (humide^
Près de la limite des arbresLF Aej Bm1 Bm2
BH6-C1
-
4.2000.2480.1285.8650.0000.0310.0160.0460.1440.096
Non10YR5/210YR4/2
CBH6-C2
0.03.2
78.618.20.00.0
4.6200.2230.08711.3390.000
0.079Non
10YR 6/210YR 4/2
Sous arbre limite des arbreLF
BH6-B1
-
3.360
18.6900.067Non
10YR 2/310YR 1,7/1
Aej BmBH6-B2
-
3.8600.1570.11930.8880.0000.0490.0120.0610.1620.077Non
10YR5/310YR3/2
CBH6-B3
0.07.6
75.616.80.00.0
4.3900.0540.04011.7290.000
0.072Non
10YR6/310YR3/2
Sous arbre limite des forêts
LFHBH6-A1
-
3.790
12i900.371Non
10YR 4/210YR 1,7/1
Aej AheBH6-A2
-
3.6300.1760.180
82.1060.000
1.7790.159Non
10YR4/25Y2/2
BmBH6-A3
-
3.9600.0850.04717.20322.5300.0780.0280.1060.3790.128Non
10YR5/310YR2/3
CBH6-A4
0.01.3
82.615.90.10.2
4.0100.1080.06314.8570.000
_
0.099
Non10YR5/310YR 3/3

61
Végétation
HorizonNo.échantillon
Tamisage (%)
sable très grossier
sable grossiersable moyen
sable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg (meq/10û g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)
Réaction au HCICouleur (sec)
Couleur (humide)
HonckenyaC
BH7-G1
-
6.6300.1370.075
57.0840.0000.0110.0110.0230.0100.093
Non10YR 5,5/210YR4/2
LeymusLF
BH7-F1
-
5.3202.2161.629
486.7743.219
1.9750.266
Non2.5Y 4/32,5Y 3/2
CBH7-F2
0.4
40.049.410.10.10.0
5.6000.0990.089
53.9560.0000.0160.0100.026
0.068Non
2,5Y 5/22,5Y 4/2
Site Pc tito Rivière
Herbacée et bryoLF
BH7-E1
-
4.2802.6851.744
572.79011.265
5.3930.218
Non2.5Y 3/32.5Y 2/1
AehytesBm
BH7-E2
-
4.0000.1770.114
32.8430.0000.0230.0140.0370.1850.092
Non10YR 5/310YR 4/2
CBH7-E3
1.032.058.6
8.20.10.1
4.5000.1780.07910.9480.000
0.072Non
2,5Y 6/32,5Y 4/2
Près de la limiteLF
BH7-D1
-
4.2400.9030.946
276.81617.932
40960.145
Non2,5Y 3/22,5Y2/1
Aeles arbresBm
BH7-D2
-
4.0900.1270.08814.8572.2990.0260.0160.0420.1450.072Non
10YR5/210YR 4/2
Hb BmbBH7-D3
-
4.4000.1390.07618.3760.0000.0220.0150.0370.0930.069Non
10YR5/310YR 3/3
CBH7-D4
1.0
37.048.6
13.20.20.0
4.7100.128
0.0658.211
0.000
0.053Non
2,5Y 5/32,5Y 3/3
Site Petite Rivière
Végétation
HorizonNo.échantillon
Tamisage (%)
sable très grossier
sable grossiersable moyen
sable fin
sable très fin
limon et argile
Analyses
pH CaCI2
Ca (meq/100 g)Mg (meq/100 g)
K (meq/100 g)
Na (meq/100 g)
Fe pyro (%)
Al pyro (%)Fe+AI pyro (%)
C tôt (%)
Conductivité (mmhos/cm)
Réaction au HCI
Couleur (sec)
Couleur (humide)
Sous arbre lim
LFBH7-C1
-
4.2604.371
2.962
830.057
777.054
17.470
0.320
Non
2.5Y4/2
2.5Y2/1
Ae
te des arbres
BmBH7-C2
-
4.050
0.1820.168
24241
26.8980.027
0.015
0.042
0.236
0.196
Non
10YR 5/2
10YR 4/2
Hb
BH7-C3* datation 14C
-
BmbBH7-C4
-
4.310
0.137
0.108
13.293
6.667
0.1690.167
Non
2,5Y 5/4
2,5Y 4/3
CBH7-C5
1.4
34.7
47.815.8
0.2
0.0
4.6300.147
0.085
12.511
18.392
0.111
Non
10YR 5/2
10YR4/2
Près de I
LFH
BH7-B1
-
3.590
0.5780.543
141.536
29.427
2.940
0.128
Non
2,5Y 3/1
2,5Y 2/1
a limite des forêt.Aej Bm
BH7-B2
_
_
-
4.170
0.057
0.072
12.1200.000
0.0320.0180.051
0.251
0.117
Non
10YR5/3
10YR3/2
C
BH7-B3
17.8
67.4
10.83.8
0.1
0.0
5.1100.079
0.065
12.1200.000
_
0.057
Non
10YR 6/3
10YR4/2
Sous arbre limite des forêtsLH
BH7-A1
_
-
3.810
1.0640.131
217.778180.010
_
_
1.9500.148
Non
2,5Y 3/1
2,5Y 2/1
Ae BmBH7-A2
-
4.000
0.0840.098
22.677
0.0000.038
0.020
0.058
0.235
0.094
Non
10YR5/2
10YR3/2
CBH7-A3
44.5
45.47.8
2 3
0.1
0.0
4.2900.047
0.055
21.504
0.000
0.074
Non10YR 5/3
10YR 4/2

62
Site ManitounukVégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg {meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
HonckenyaCBH8-A1
1.523262.912.30.00.0
7.4800.2630.217
58.2560.0000.0110.0060.0160.5360.153Oui
10YR6/110YR4/1
Leymus
cBH8-B1
0.02.568,828.70.00.0
7.3100.2100.153
32.0610.0000.0060.0040.0090.1860.082Non
10YR 6,5/110YR5/2
Salix, bryophytes et plantulesLFBH8-C1
-
7.0007.6852.139
272.5150.000
2.5240.115Non
2,5Y 3/12.5Y 2/1
BmBH8-C2
-
7.2101.0820.420
35.1880.0000.01300110.0250.5150.150Non
10YR 5/210YR 4/2
CBH8-C3
0.17.575.11720.001
7.3500.4430.239
25.0230.000
0.081Non
10YR 6,5/12.5Y 5/2
Près de la limite des arbresLFHBH8-D1
-
6.3907.4652.386
267.04150.577
3.1180.125Non
2.5Y 3/12.5Y 2/1
BmBH8-D2
-
6.9801 0830.379
30.1060.0000.01800100.0280.5110.103Non
2.5Y 5,5/22.5Y 3/2
CBH8-D3
0.825.865.3810.00.0
7.0700.6320.280
26.9780.000
0.106Non
2,5Y 6/22,5Y 4/2
Sous arbre limite des arbresLFHBH8-E1
-
6.3306.4573.168
277.207155 871
3.3370.209Non
2,5Y 3/12,5Y 2/1
Aej BmBH8-E2
-
6.9200.9220.42041.8350.0000.0130.0070.0210.3210.102Non
2,5Y 5/22.5Y 4/2
BH8-E3
1.118 471 98.40.10.1
7.1500.6231.424
27.3690.000
0.092Non
2,5Y 6/22,5Y 4/2
VégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg (meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
Près de la ILFH
BH8-F1
-
5.243.9621.391
277.98969.429
4.8030.147Oui
2,5Y 3/12,5Y 2/1
mite des forêtsAej Bm
BH8-F2
-
7.211.1730.56032.45200000.0180.0110.02804700.135Oui
2.5Y 5/22,5Y 4/2
SI
CBH8-F3
0.315.475.38.80.10 1
7,150.7420.17022.6770.000
0.Ï56Oui
2.5Y 6/22,5Y 4/2
e ManitounukSous arbre limite des forêts
LFHBH8-G1
-
5.079.1776.089
1002.87141054.5407
8.6840.284Non
2,5Y 3/22.5Y 2/1
Sable LFHBH8-G2
-
5.44
7,1570.282Non
2,5Y 3/210YR 1,7/1
Aej BmBH8-G3
-
6.891 252049459.429
00.0160.0080.0230 3860.144Oui
10YR 5/310YR 3/2,5
CBH8-G4
3.438252.25.50.40.3
7.30.8390.38744.181
0
7.8200.111Non
2,5Y 6/22,5Y 4/2

63
Site Grande Rivière
VégétationHorizon
No.échantillon
Tamisage (%)
sable très grossiersable grossier
sable moyensable fin
sable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)
Mg(meq/100g)K (meq/100 g)Na {meq/100 g)
Fe pyro (%)
Al pyro (%)Fe+AI pyro (%)C tôt (%)
Conductivité (mmhos/cm)
Réaction au HCICouleur (sec)Couleur (humide)
Honckenya
C
BH4-A1
15.135.148.0
1.80.00 0
6.950
0.099
0.08132.0610.000
0.0080.006
0.0140.047
0.082Non
2,5Y 6/12.5Y 4/2
Leymus et Lathyms
L Ahj C
BH4-B1
0.421.6
71.36.7
0 00.0
6.070
0.138
0.1694457219.541
0.0060.0050.0111.464
0.097Non
2,5Y 6/2
2,5Y 4/2
Près de la
LFHBH4-C1
-
3.970
1.8811.629
437.510175.642
7.6620.164
Non2,5Y 3/22,5Y 2/1
mile des arbres
Aej BmBH4-C2
-
4.060
0.2610.21418.3760.690
0.0160.010
0.0260.3910.098
Non10YR5/210YR4/2
Hb
BH4-C3•datation 14C
_
_
-
__
BmbBH4-C4
-
4.460
0.2720.1328.9933.908
0.0220.013
0.0350.1380.075
Non10YR 5/310YR 4/3
C
BH4-C5
0.211.480.87.6
0.00.0
4.670
0.0950.075
5.8650.000
0.063Non
2,5Y 6/2
2,5Y 4/3
Site Grande Rivière
VégétationHorizonNo.échantillon
Tamisage (%)
sable très grossier
sable grossiersable moyensable fin
sable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg (meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)
Réaction au HCICouleur (sec)Couleur (humide)
Sous arbreLFH
BH4-D1
-
3.8401.4771.695
374.171142.766
3.2520,147
Non2,5Y 3/2
10YR 1,7/1
mite des arbresAej Bm
BH4-D2
-
3.8600.0580.14322.677
28.50700310.0120.0430.3660.088
Non2,5Y 5/22,5Y 4/2
Hb Sable Hb2 Bm2BH4-D3
_
-
4.1000.1510.14519.5490.0000.021
0.0120.0330.2530.091
Non2,5Y 5/32,5Y 3/3
CBH4-D4
0,0
4,979.016.10.0
0.0
4.6700.1590.09115.6390.000
0.060
Non
2,5Y 6/32,5Y 4/3
Près de la limiteFH
BH4-E1
_
-
3.2301.2191.646
694.386493.590
_
18.0300.184
Non2.5Y2/1
10YR 1,7/1
Aejles forêts
BmBH4-E2
_
-
4.0600.0480.0438.6020.0000.0210.0140.034
0.1860.065
Non2,5Y 5/32.5Y4/2
C
BH4-E3
0.2
31.565.82.50.0
0.0
4.2300.0270.0568.602
45.060
_
0.072
Non2,5Y 5/22,5Y 4/2
Sous arbre limite des forêtsLFH
BH4-F1
-
4.0602.1913.333
489.902577.503
8.1950.265
Non2,5Y 3/3
10YR 1,7/1
Ae Bm
BH4-F2
-
3.8500.0900.22219.549
133.800
0.0310.0150.045
0.5680.087
Non
10YR 4/210YR 5/2
CBH4-F3
0.3
24,972,22.70.00.0
4.2100.0120.07310.55729.657
_
0.065
Non2.5Y 5/32.5Y 4/3

64
Site DériveVégétationHorizonNo.échantillon
Tamisage {%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg (meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
HonckenyaC
BH3-A1
34.048.217.00.80.00.0
7.7700.8080.159
36.75274.9470.0150.0090.0240.1840.140Non
10YR5/110YR 6/1
^eymusL C
BH3-B1
4.359.336.00.30.00.1
7.0000.3780.144
32.45225.2890.0090.0070.0160.2330.117Non
2,5Y 6/22,5Y4/2
Près de la limite des arbresLF
BH3-D1
-
4.900980110.862
891.441202 080
4.9670.255Non
2,5Y 3/210YR 2/1
Aej BmBH3-D2
-
4.8200.5690.21437.1430.0000.0170.0080.0260.3400.100Non
2.5Y 5/22,5Y 4/2
CBH3-D3
0.231.966.41.50.00.0
6.6500.2280.12516.0300.000
0.056Non
2.5Y 6/22,5Y 5/2
Sous arbreLFH
BH3-C1
-
5.49011.77710.862
1429.8252478.987
11.5900.254Non
2,5Y 3/32,5Y 2/1
limite des arbresAej Bm
BH3-C2
-
4.4000.2430.214
40.27111.9550.0220.0090.0310.2240.155Non
2,5Y 5/22,5Y2/1
CBH3-C3
0.330.967.90.90.00.0
67300.2010.12516.4210.000
0.088Non
2,5Y 6/22,5Y 5/2
Site DériveVégétationHorizonNo échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg (meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
Près de laLFH
BH3-E1
-
4.8306.9913.538
483.64672.418
7.3240.194Non
2.5Y3/12,5Y 2/1
imiteAei
des forêtsBm
BH3-E2
-
4.3000.3270.214
39.0980.0000.0220.0090.0310.3590.106Non
2,5Y 5/32,5Y 4/3
CBH3-E3
1.335.862.40.50.00.0
4.6700.2410.15213.2930.000
0.057Non
2.5Y 6/32,5Y 4/2
Sous arbre limite des forêtsLFH
BH3-F1
-
4.7003.6632.222
464.879411.977
4.3280.152Non
2.5Y 3/12,5Y 2/1
Aej BmBH3-F2
-
4.0400.2580.222
26.5870.0000.0260.0120.0390.9110.091Non
2,5Y 5/32,5Y 3/3
CBH3-F3
0.827.670.90.70.00.0
4.7900.1950.14815.6390.000
0.078Non
2.5Y 6/32.5Y 4/3

65
Site InuitVégétationHorizonNo. échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca(meq/100g)Mg(meq/100g)K(meq/100g)Na(meq/100g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
HonckenyaC
BH2-A1
0.846.750.81.7000.0
7.0500.0690.06523.06810.8050.0090.0070.0160.0600.066Non
2,5Y 6/22,5Y 5/2
LeymusL C
BH2-B1
0.433 563.32.90.00.0
6.6000.2800.16828.1510.0000.0100.0070.01700860.077Non
2,5Y 6/22.5Y 4/2
Près de la limite des arbresLFH
BH2-D1
-
4.4602.4651.580
363.61482.073
4.9150.146Non
2,5Y 2/12,5Y 1,7/1
Aej BmBH2-D2
-
4.5800.3150.18937.9250.0000.0230.0160.0390.1310.074Non
2,5Y 5/32.5Y 4/2
CBH2-D3
22.651.924.21.20.00.0
6.3300.5080.137
28.5420.000
0.067Non
10YR5/210YR4/2
Sous arbre limiteLFH
BH2-C1
-
4.2201.4321.810
483.646245.991
4.1430.215Non
2,5Y 3/32,5Y 2/1
Aejles arbres
BmBH2-C2
-
4.8800.2980.222
26.97811.7250.0260.0140.0400.2130.091Non
2:5Y 5/32.5Y 4/2
CBH2-C3
3.848.94551.80.10.0
6.1100.3710.149
26.97847.589
0.096Non
2,5Y5/22,5Y 4/2
Site InuitVégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100g)Mg(meq/100g)K(meq/100g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
Près de laFH
BH2-F1
-
4.4301.5671.366
363.61417.012
0.2340.191Non
2,5Y 3/12,5Y 2/1
imite des forêtsAej Bm
BH2-F2
-
40100.0990133
23.0680.00000550.0360.0910.2010.082Non
10YR 4,5/310YR 3,5/2
CBH2-F3
18.457223.21.20.00.0
6.1500.1450.07614.8577.127
0.061Non
2,5Y 6/22,5Y 5/2
Sous arbre limite des forêtsLFH
BH2-E1
-
3.3700.9911.473
320.606211.506
4.8260.221Non
2,5Y 2/12,5Y 3/1
Aej BmBH2-E2
-
3.8900.1090.12024.63233.3350.0270.0150.0420.1880.094Non
2,5Y 5/32,5Y 1,7/1
CBH2-E3
24.445.326.24.00.00.0
5.2200.1080.089
25.8050.000
0.129Non
2,5Y 5/22.5Y 4/2

66
Site Ile LongueVégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca(meq/100g)Mg(meq/100g)K (meq/100 g)Na(meq/100g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
HonckenyaC
BH1-A1
0.455.942.90.90.00.0
7.2100.7830.272
45.3540.0000.0110.0070.0190.4890.229Non
2,5Y 6/12,5Y 5/1
LeymusC
BH1-B1
0.423.174.22.30.00.0
7.0300.4380.16132 0619.8860.0100.0050.0150.2720.084Non
2,5Y 6/12,5Y 4/1
Près de la limite des arbresLF
BH1-C1
-
4.3700.1871.341
192.75572.878
7.9760.126Non
2,5Y 3/12.5Y 2/1
Aej BmBH1-C2
-
6.6101.2270.51877.0240.0000.0150.0090.0240.5130.142Non
2,5Y 5/22,5Y 4/1
CBH1-C3
10.046.541.61.80.10.1
7.1101.0860.211
21.5040.000
0.103Non
2.5Y6/22,5Y 2/1
Sous arbre limite des arbresLFH
BH1-D1
-
613058044937
395.675769.468
46150.262Non
2,5Y 3/32,5Y 2/1
Aej BmBH1-D2
-
6.9000.9560.59256.69393.1090.0110.0080.0180.4530.140Non
2,5Y 5/22,5Y 3/1
CBH1-D3
0.138.260.21.50.00.0
7.3600.4850.173
28.9330.000
0.114Non
2,5Y 6/12,5Y 5/2
Site île LangueVégétationHorizonNo.échantillon
Tamisage (%)
sable très grossiersable grossiersable moyensable finsable très finlimon et argile
Analyses
pH CaCI2Ca (meq/100 g)Mg (meq/100 g)K (meq/100 g)Na (meq/100 g)Fe pyro (%)Al pyro (%)Fe+AI pyro (%)C tôt (%)Conductivité (mmhos/cm)Réaction au HCICouleur (sec)Couleur (humide)
Près limite des forêtsLF
BH1-E1
-
6.0607.2114.731
654.115159.779
9.6460.301Non
2,5Y 3/22,5Y2/1
Aej BmBH1-E2
-
6.4607.4360.560
37.5340.0000.0250.0110.0360.5060.087Non
2,5Y 4/22,5Y 3/2
CBH1-E3
0.331.365.13.00.20.1
7.1700.5410.58412.9022.759
0.090Non
2,5Y 6/22,5Y 4/2
Sous arbre limite des forêtsLFH
BH1-F1
-
5.4100.0050.288
1435.6902307.253
21.0300.313Non
2,5Y 3/32,5Y 2/1
Ae BmBH1-F2
-
6.9101.2280.535
25.023101.3850.0230.0110.0340.4650.125Non
2,5Y 5/22,5Y 3/1
CBH1-F3
2.679.218.00.20.00.0
7.3800.4660.22214.0750.000
0.104Non
2.5Y 6/12,5Y 4/1
















