
LERNEN UND ADAPTATION IM VISUELLEN …elib.suub.uni-bremen.de/diss/docs/00010953.pdf · genannten...
105
LERNEN UND ADAPTATION IM VISUELLEN SYSTEM DES MENSCHEN Sven Wischhusen vorgelegt dem Fachbereich 2 (Biologie/Chemie) der Universität Bremen als DISSERTATION zur Erlangung des akademischen Grades DOKTOR DER NATURWISSENSCHAFTEN (Dr. rer. nat.)
Transcript of LERNEN UND ADAPTATION IM VISUELLEN …elib.suub.uni-bremen.de/diss/docs/00010953.pdf · genannten...

LERNEN UND ADAPTATION IM VISUELLEN SYSTEM DES MENSCHEN
Sven Wischhusen
vorgelegt dem Fachbereich 2 (Biologie/Chemie) der Universität Bremen
als
DISSERTATION
zur Erlangung des akademischen Grades
DOKTOR DER NATURWISSENSCHAFTEN (Dr. rer. nat.)

i
Inhaltsverzeichnis
Gutachter .................................................................................................................................. iiEigenanteil und Erklärung...................................................................................................... iiÜbersicht über Publikationen und Manuskripte .................................................................iiiDanksagung.............................................................................................................................. iv
1. Einleitung .............................................................................................................................. 11.1 Ein erster Überblick ......................................................................................................... 1 1.2 Plastizität, Lernen und Adaptation: Begriffsabgrenzung und Beispiele .......................... 2 1.3 Neuronale Korrelate der Sensomotorik............................................................................ 8 1.4 Sensomotorische Plastizität am Beispiel der Prismenadaptation ................................... 11
2. Allgemeine Methodik ......................................................................................................... 152.1 Verhaltensuntersuchungen und Psychophysik ............................................................... 15 2.2 Sensomotorische Aufgaben: Zeige- und Wurfbewegungen........................................... 16 2.3 Datenauswertung............................................................................................................ 19
3. Motivation und Zusammenfassung der Studien ............................................................. 203.1 Motivation der Studien................................................................................................... 20 3.2 Zusammenfassung von Studie 1..................................................................................... 22 3.3 Zusammenfassung von Studie 2..................................................................................... 23 3.4 Zusammenfassung von Studie 3..................................................................................... 25
4. Fazit ..................................................................................................................................... 27
5. Studie 1 Incomplete visuomotor adaptation despite extensive training..................... 28
6. Studie 2 Task-specificity of prism adaptation: no transfer from pointing to throwing.................................................................................................................................................. 50
7. Studie 3 Effects of training conditions on spatial generalization of prism adaptation.................................................................................................................................................. 71
Abkürzungsverzeichnis.......................................................................................................... 92
Literaturverzeichnis............................................................................................................... 93

ii
Gutachter
1. Gutachter: Professor Dr. Manfred Fahle, Universität Bremen
2. Gutachter: Professor Dr. Ben Godde, Jacobs University Bremen
Tag des Kolloquiums: 17.04.2008
Eigenanteil und Erklärung
Die wissenschaftlichen Untersuchungen, auf denen diese Dissertation beruht, habe ich
selbständig durchgeführt und ausgewertet. Die vorliegenden Manuskripte habe ich
eigenständig verfasst und lediglich die endgültige Fassung mit meinem Betreuer und
Mitautoren, Herrn Professor Manfred Fahle, überarbeitet.
Ich erkläre hiermit, dass ich die Arbeit ohne unerlaubte fremde Hilfe angefertigt und keinerlei
anderen Quellen und Hilfsmittel als die angegebenen benutzt habe. Ferner sind alle wörtlich
oder inhaltlich anderen Werken entnommenen Stellen im Text als solche kenntlich gemacht.
Sven Wischhusen
Bremen, 29.02.2008

iii
Übersicht über Publikationen und Manuskripte
Die vorliegende Dissertation beruht kumulativ auf folgenden drei Manuskripten, die als
Fachartikel zur Veröffentlichung in internationalen neurowissenschaftlichen Fachzeitschriften
eingereicht wurden:
Fachartikel
Wischhusen, S. & Fahle, M. (2008). Incomplete visuomotor adaptation despite extensive training. Vision Research (eingereicht).
Wischhusen, S. & Fahle, M. (2008). Task-specificity of prism adaptation: no transfer from pointing to throwing. Experimental Brain Research (eingereicht).
Wischhusen, S. & Fahle, M. (2008). Effects of training conditions on spatial generalization of prism adaptation. Journal of Motor Behavior (eingereicht).
Konferenzbeiträge
Fahle, M., Wischhusen, S., & Spang, K. (2005). Prism adaptation by gain control. Perception, 34 (Suppl.), 28th European Conference on Visual Perception, A Coruña, Spain.
Wischhusen, S. & Fahle, M. (2005). Prism adaptation: no generalization from one visuomotor task to another. Workshop “Preemptive Perception”, Hanse Institute for Advanced Study, Delmenhorst, Germany.
Wischhusen, S. & Fahle, M. (2006). Is prism adaptation complete? 5th Forum of European Neuroscience, Vienna, Austria.
Fahle, M., Eggert, T., & Wischhusen, S. (2006). Perceptual learning in a visual masking task. Workshop “Visual Masking and the Dynamics of Vision and Consciousness”, Hanse Institute for Advanced Study, Delmenhorst, Germany.
Fahle, M., Wischhusen, S., & Spang, K. (2006). Prism adaptation and normalization of eye-hand coordination. Perception, 35 (Suppl.), 29th European Conference on Visual Perception, St. Petersburg, Russia.
Fahle, M., Eggert, T., Wischhusen, S., & Spang, K. (2006). Perceptual learning with masked stimuli. Perception, 35 (Suppl.), 29th European Conference on Visual Perception, St. Petersburg, Russia.
Wischhusen, S., Schütze, C., & Fahle, M. (2007). Prism adaptation in a patient with damage to the right parietal cortex – a case study. 31st Göttingen Neurobiology Conference, Göttingen, Germany.

iv
Danksagung
Zuallererst gilt mein ganz herzlicher Dank meinem Betreuer, Herrn Professor Manfred Fahle,
für die interessante und lehrreiche Zeit in seiner Arbeitsgruppe und dafür, dass er mir die
Möglichkeit gegeben hat, meine Doktorarbeit zu schreiben und dabei einen tiefen Einblick in
die naturwissenschaftliche Forschung zu bekommen. Für die Unterstützung und die
konstruktive Kritik während der gesamten Zeit bedanke ich mich sehr!
Darüber hinaus danke ich Herrn Professor Ben Godde von der Jacobs University Bremen
dafür, dass er sich bereit erklärt hat, seinen Sachverstand als „zweiter“ Gutachter für diese
Arbeit zur Verfügung zu stellen.
Mein besonderer Dank gilt den Kollegen der Arbeitsgruppe „Human-Neurobiologie“ am
Institut für Hirnforschung, die im Laufe der Zeit zu guten Freunden geworden sind: Dani
Högl, Sirko Straube, Cathleen Grimsen und Tina Friederich.
Ein herzliches Dankeschön geht auch an die Mitarbeiter und Studenten aus der Arbeitsgruppe,
die mich unterstützt und mir bei allen kleinen und großen Problemen stets geholfen haben.
Des Weiteren möchte ich mich bei allen freiwilligen Versuchspersonen, die mit vollem
Einsatz und viel Geduld an den Experimenten teilgenommen haben, bedanken. Diese Arbeit
hat mir immer sehr viel Freude bereitet!
Ein großer Dank geht aber auch an meine Eltern und meine Familie in Worpswede und umzu,
die mich einerseits immer wieder auf den „Boden der Tatsachen“ zurückgebracht haben,
deren echter Unterstützung ich mir jedoch während der ganzen Zeit immer sicher sein konnte.
Außerdem danke ich meinen Freunden Alex, Jan-Dirk, Jan Christoph, Jörg und Niels einfach
dafür, dass sie immer da und für allerlei lustige Aktionen zu haben waren!

Einleitung 1
1. Einleitung
1.1 Ein erster Überblick
Nach einem Gegenstand – beispielsweise einer Kaffeetasse auf einem Tisch – zielgerichtet zu
greifen ist eigentlich keine komplizierte Angelegenheit. Oder doch? An der Ausführung einer
solchen Aufgabe sind ganz verschiedene Gehirnsysteme beteiligt: Zunächst muss das Objekt
visuell lokalisiert werden. Die visuelle Information über die Position des Objekts wird in
einem nächsten Schritt mit der sensorisch-propriozeptiven Information über die aktuelle
Körperposition in Bezug zum Objekt abgeglichen und für die Bewegungsplanung
umgewandelt („sensomotorische Transformation“). Auf der Basis des Abgleichens
sensorischer und motorischer Koordinaten kann die Bewegung schließlich geplant und
ausgeführt werden (Krakauer & Ghez, 2000). Dass es im Normalfall keine große Mühe
bereitet, nach einer Kaffeetasse zu greifen, begründet sich aus der funktionellen Kopplung
von Sensorik und Motorik (kurz: Sensomotorik1), einer Kopplung, die so zuverlässig und
automatisiert ist, dass wir uns über die komplexe Leistung, die das Gehirn für die erfolgreiche
Ausführung einer Bewegung vollbringt, in der Regel nicht bewusst sind.
Diese funktionelle Kopplung von Sensorik und Motorik kann bei gesunden menschlichen
Versuchspersonen gezielt experimentell manipuliert werden, beispielsweise mit Hilfe einer
Prismenbrille, durch welche die Versuchsperson die visuelle Welt als seitlich versetzt
wahrnimmt. Aufgrund der Prismenbrille kommt es zu einer Nicht-Übereinstimmung zwischen
sensorischen und motorischen Koordinaten mit der Folge, dass Bewegungen zunächst in
Richtung der prismatischen Versetzung abweichen (vgl. Redding et al., 2005). Um in dem
genannten Beispiel zu bleiben: Versuchspersonen würden es mit Prismenbrille zunächst nicht
schaffen, die Kaffeetasse zielgerichtet zu greifen.
Eine weitere wichtige Eigenschaft des Gehirns ist die Fähigkeit, seine Organisation kurzfristig
und flexibel neuen Gegebenheiten anzupassen: Diese Neuroplastizität ermöglicht somit, dass
die sensomotorischen Koordinaten während des Tragens einer Prismenbrille neu miteinander
abgeglichen und gekoppelt werden. Im Verlauf dieses Prozesses werden die Bewegungen an
die neuen Bedingungen angepasst, d.h. es kommt zu einer sensomotorischen Adaptation.
1 Kurzer Ausflug in die Nomenklatur: Neben dem Begriff der „Sensomotorik“, der die funktionelle Verknüpfung von sensorischer Information (visuell, auditiv, somatosensorisch, propriozeptiv) und Motorik beschreibt, findet sich auch der Begriff der „Visuomotorik“, bei dem die Verknüpfung speziell von visueller Information und Motorik hervorgehoben wird (vgl. Birbaumer & Schmidt, 2003).

Einleitung 2
Plastische Prozesse bilden damit die Grundlage der sensomotorischen Adaptation, in deren
Verlauf die Bewegungen trotz der (experimentellen) Manipulation eine immer größere
Genauigkeit aufweisen. Die Kaffeetasse könnte folglich nach einigen Versuchen trotz der
Prismenbrille zielgerichtet ergriffen werden.
Prismenadaptation ist ein Beispiel für einen Adaptationsprozess im sensomotorischen
System. Die Untersuchung der Mechanismen und Charakteristika dieses
Adaptationsprozesses kann wichtige Antworten auf die Frage liefern, in welcher Art Sensorik
und Motorik funktionell miteinander verknüpft sind und welches die Merkmale und Faktoren
der Neuroplastizität im sensomotorischen System sind. Es geht folglich darum, wie und mit
welchen Mechanismen es das Gehirn schafft, neuronale Repräsentationen schnell neuen
Wahrnehmungsbedingungen anzupassen.
In der vorliegenden Arbeit wurden verschiedene Ausprägungsformen von Neuroplastizität im
visuellen System – Lernen und insbesondere Adaptation – unter Verwendung des
Prismenadaptations-Paradigmas2 bei gesunden menschlichen Versuchspersonen
psychophysikalisch untersucht. Zunächst sollen jedoch wichtige Begriffe wie Plastizität,
Lernen und Adaptation sprachlich voneinander abgegrenzt und anhand von Beispielen in
einen konzeptuellen Zusammenhang gestellt werden.
1.2 Plastizität, Lernen und Adaptation: Begriffsabgrenzung und Beispiele
Plastizität
In einer sich fortlaufend ändernden Umwelt muss das zentrale Nervensystem (ZNS) die
Fähigkeit aufweisen, seine strukturelle und funktionelle Organisation stets diesen neuen
Umweltbedingungen anzugleichen, um dem Organismus angepasstes Verhalten zu
ermöglichen. Diese Eigenschaft des Gehirns, sich neuen Bedingungen anzupassen, bezeichnet
man als neuronale Plastizität, kurz Neuroplastizität (Sterr, 2008). Neuroplastische Prozesse
sind insbesondere in Arealen der Großhirnrinde (Cortex) untersucht und nachgewiesen
worden; eine Übersicht zur corticalen Neuroplastizität geben Buonomano & Merzenich
(1998). Dabei treten Veränderungen sowohl auf der Ebene von Synapsen und Neuronen als
auch von größeren Neuronenverbänden auf. Sterr (2008) unterscheidet Ebenen molekularer, 2 Schon an dieser Stelle sei darauf hingewiesen, dass die plastischen Veränderungen im Gehirn, die im Zusammenhang mit der Prismenadaptation auftreten, nicht auf das visuelle System im engeren Sinne, d.h. auf die frühen visuellen Areale (V1, V2 etc.), beschränkt sondern vielmehr auch in „höheren“ multimodalen sowie motorischen Arealen zu finden sind.

Einleitung 3
struktureller und funktioneller Plastizität. Neuronale Veränderungen, die im Zusammenhang
mit Entwicklung und Reifung (Entwicklungsplastizität), nach Läsionen des ZNS sowie durch
Erfahrung und Übung auftreten, sind folglich nur aufgrund plastischer Prozesse im Gehirn
möglich.
Lernen – allgemein
Die Anpassungsfähigkeit des Gehirns äußert sich u.a. durch Lernen, worunter man ganz
allgemein den Erwerb von Fähigkeiten und Fertigkeiten durch Übung versteht. Menzel (2001,
S. 504) definiert Lernen als „[…] die Fähigkeit, Verhalten aufgrund individueller Erfahrung
so zu ändern, daß es veränderten Situationen besser angepaßt ist“ und unterstreicht damit den
biologischen Anpassungswert von Lernen für den Organismus. Lernen umfasst eine
längerfristige Verhaltensanpassung aufgrund von Erfahrung, wobei die funktionelle
Grundlage von Lernprozessen in der Plastizität der beteiligten neuronalen Strukturen
begründet ist. Je nach untersuchter neurobiologischer Domäne unterscheidet man zwischen
kognitivem, perzeptuellem und motorischem Lernen, wobei die beiden letzteren für die
vorliegende Arbeit von größerer Bedeutung sind.
Perzeptuelles Lernen
Perzeptuelles Lernen, eine andauernde Verbesserung der Wahrnehmungsleistung durch
Übung bzw. Erfahrung, verdeutlicht, dass auch (frühe) sensorische Areale des Gehirns beim
erwachsenen Menschen durch Übung modifizierbar und damit plastisch sind (Fahle, 2002).
Perzeptuelle Lernprozesse bei Erwachsenen, die mit neuronalen Veränderungen auf frühen
Ebenen der sensorischen Informationsverarbeitung einhergehen müssen, waren insofern eine
Überraschung, weil lange Zeit die Auffassung vertreten wurde, dass plastische
Veränderungen der sensorischen Cortices auf ein relativ kleines Zeitfenster während der
frühen Individualentwicklung – die sog. „kritische Periode“ – beschränkt seien (Fahle, 2002).
Perzeptuelles Lernen liefert damit den experimentellen Nachweis dafür, dass auch bei der
sensorischen Informationsverarbeitung im adulten Gehirn plastische Prozesse auftreten
können, die zu dauerhaften Veränderungen der Wahrnehmungsleistung führen und damit der
o.g. Definition von Lernen entsprechen.
Perzeptuelles Lernen wurde beim Menschen insbesondere im visuellen (siehe Fahle, 2004;
Gilbert et al., 2001; Karmarkar & Dan, 2006; Seitz & Watanabe, 2005), somatosensorischen
(z.B. Godde et al., 2000; Hodzic et al., 2004; Pleger et al., 2003) sowie im auditorischen
System (siehe Dahmen & King, 2007) mit psychophysikalischen und neurophysiologischen

Einleitung 4
Methoden untersucht; hierbei konnten durch Lernen induzierte neuronale Veränderungen in
den frühen bzw. primären corticalen sensorischen Projektionsarealen (V1, S1 und A1) bei
entsprechenden sensorischen Aufgaben nachgewiesen werden.
Die durch perzeptuelles Lernen hervorgerufenen Verbesserungen der Wahrnehmungsleistung
weisen häufig eine sehr hohe Spezifität für die trainierten Reiz-Eigenschaften auf (z.B.
Orientierung bzw. Position des Reizes). Diese Spezifität ist ein starker Hinweis darauf, dass
die neuronalen Korrelate des perzeptuellen Lernens in frühen sensorischen Arealen zu finden
sind, da hier einfache Reiz-Merkmale in räumlich geordneter Form repräsentiert werden und
diese frühen Stufen der sensorischen Informationsverarbeitung ein günstiges Signal-zu-
Rausch-Verhältnis aufweisen (Fahle, 2004; 2005; für eine alternative Erklärung siehe Mollon
& Danilova, 1996). Neuronale Korrelate perzeptuellen Lernens finden sich aber auch in
„späteren“ sensorischen Arealen, wobei die Selektion der entsprechenden Areale durch Top-
Down-Modulationen entsprechend den Erfordernissen der sensorischen Aufgabe vermittelt
wird (Fahle, 2004).
Motorisches Lernen
Auch im motorischen System, das die Ausführung zielgerichteter Bewegungen plant und
steuert, finden Funktionsanpassungen durch Lernen statt: Motorisches Lernen ist, ähnlich wie
perzeptuelles Lernen, implizit, d.h. der Lernprozess läuft beim Lernenden unbewusst ab
(Konczak, 2003). Dabei kann motorisches Lernen allerdings nicht isoliert betrachtet werden,
da immer eine enge Verknüpfung zwischen sensorischer Informationsverarbeitung und
Bewegungsausführung besteht. Die Unterscheidung zwischen perzeptuellem und
motorischem Lernen ist im Prinzip konzeptuell und zeigt an, auf welchem Gehirnsystem
(sensorisch vs. motorisch) der Untersuchungsschwerpunkt liegt; im praktisch-experimentellen
Zusammenhang kommt es dagegen immer zum sensomotorischen Lernen (vgl. Konczak,
2003). Hierzu passt auch die Beobachtung, dass sowohl beim perzeptuellen als auch beim
motorischen Lernen auf der physiologischen Ebene ganz ähnliche Mechanismen wirksam
sind, was darauf hindeutet, dass beide Lernprozesse auf gleichen neuronalen
Funktionsprinzipen beruhen (Paz et al., 2004).
Welches sind die Charakteristika motorischen Lernens? Während motorische Reflexe und
einfache Bewegungsprogramme3 im Genom eines Organismus encodiert sind, setzen
komplexere motorische Verhaltensweisen in einer wechselnden Umwelt die fortlaufende 3 Ein Bewegungsprogramm, die zeitliche Abfolge von motorischen Kommandos, führt letztlich zur Bewegung eines bestimmten motorischen Effektors, z.B. des Arms (Shadmehr & Wise, 2005).

Einleitung 5
Anpassung und Erweiterung von bestehenden Bewegungsprogrammen voraus. Laut
Shadmehr & Wise (2005), die als Modell Reich- und Zeigebewegungen des Arms verwenden,
umfasst motorisches Lernen neben a) der Erweiterung des Bewegungsrepertoires um neue
Fertigkeiten (engl. „skill acquisition“) auch b) die Modifikation bzw. Anpassung eines bereits
bestehenden Bewegungsprogramms an neue Bedingungen (engl. „motor adaptation“), wobei
beide Prozesse zur Stabilität und Kontrolle der Bewegung beitragen.
Während beim perzeptuellen Lernen Wahrnehmungsleistungen durch Übung verbessert
werden und damit zum „adaptiven“ Verhalten des Organismus beitragen, stehen beim
motorischen Lernen neue bzw. angepasste Bewegungsprogramme im Vordergrund, die ein
effizienteres Verhalten ermöglichen. Anders ausgedrückt: Wird beim perzeptuellen Lernen
die Informationsverarbeitung des sensorischen Eingangs optimiert, kommt es beim
motorischen Lernen zu Verbesserungen des motorischen Ausgangs bzw. der Ausführung
einer Bewegung.
Die von Shadmehr & Wise (2005) vorgenommene Unterscheidung zwischen dem Erwerb
neuer motorischer Fertigkeiten einerseits und motorischer Adaptation andererseits ist insofern
sinnvoll, als es bei der motorischen Adaptation zur Anpassung eines vorhandenen
Bewegungsprogramms an neue externe Bedingungen kommt, d.h. die Leistungsfähigkeit des
motorischen Systems wird bei veränderten Bedingungen durch adaptive Prozesse
wiederhergestellt. Shadmehr & Wise (2005, S. 47): „Adaptation involves changes in motor
performance that allow the motor system to regain its former capabilities in altered
circumstances“. Der Prozess der motorischen Adaptation gewährleistet folglich eine
erfolgreiche Bewegungsausführung unter veränderten Bedingungen.
Welcher Art können diese veränderten „externen“ Bedingungen sein? Experimentell kann die
erfolgreiche Bewegungsausführung durch Prismen, durch welche die visuelle Welt
beispielsweise seitlich versetzt wahrgenommen wird, gestört werden. Infolge der durch die
Prismen verursachten Manipulation des sensomotorischen Systems weichen zielgerichtete
Armbewegungen zunächst ab, aber im Verlauf der Adaptation wird die Genauigkeit der
Armbewegungen wiederhergestellt (siehe Abschnitt 1.4). Während dieses Prozesses wird
keine neue motorische Fertigkeit hervorgebracht, sondern ein bestehendes
Bewegungsprogramm wird an neue (sensomotorische) Bedingungen angepasst, um
zielgerichtete und präzise Armbewegungen trotz der Prismen zu ermöglichen. Darüber hinaus
können Adaptationsprozesse unter Verwendung externer Kraftfelder (engl. „force field
adaptation“) oder mit Hilfe von rotierter visueller Rückmeldung (engl. „rotation adaptation“)
untersucht werden (Shadmehr & Wise, 2005).

Einleitung 6
Adaptation: Motorisch, perzeptuell, sensomotorisch
Ähnlich wie der Begriff „Lernen“ wird auch der Begriff „Adaptation“ in verschiedenen
Kontexten und auf ganz verschiedene neurobiologische Phänomene angewandt, weshalb eine
einheitliche Definition schwierig ist. Ganz allgemein versteht man unter Adaptation eine
kurzfristige Anpassung an neue Bedingungen (Wade & Verstraten, 2005), was gut mit der
o.g. Begriffsabgrenzung von motorischer Adaptation zu vereinen ist. Eine sensorische
Adaptation (z.B. Licht-Adaptation) tritt dagegen durch eine längere sensorischer Stimulation
auf und verschiebt den Arbeitsbereich eines biologischen Sensors (Palmer, 1999). Eine
perzeptuelle Adaptation betrifft eine Veränderung des Perzepts, d.h. des subjektiven
Wahrnehmungsinhalts (z.B. Bewegungsnacheffekt, siehe Huk et al., 2001; Ibbotson, 2005)
oder beeinflusst die sensomotorische Integration, wie z.B. bei der Prismenadaptation (Wade
& Verstraten, 2005).
Aus dieser Einleitung ergibt sich in Bezug auf die Prismenadaptation ein konzeptuelles
Problem: Autoren, die die perzeptuellen Veränderungen infolge der Prismenadaptation
betonen, sehen in dem Prozess ein Beispiel für eine perzeptuelle Adaptation (z.B. Palmer,
1999; Wade & Verstraten, 2005), während Autoren, die mit adaptiven motorischem
Veränderungen (z.B. Shadmehr & Wise, 2005) argumentieren, den Prozess eher in den
Bereich der motorischen Adaptation einordnen. Für beide Standpunkte gibt es experimentelle
Belege, wobei eine rein perzeptuelle Perspektive die starke motorische Komponente der
Adaptation außer Acht lässt, während eine rein motorische Betrachtungsweise vernachlässigt,
dass während der Prismenadaptation auch Veränderungen im perzeptuellen System auftreten
können.
Prismenadaptation ist folglich ein gutes Beispiel für einen integrativen biologischen
Adaptationsprozess, der sich genau an der Schnittstelle von Wahrnehmungssystem und
Motorik befindet und sich daher weder ausschließlich in die perzeptuelle noch in die
motorische Domäne einordnen lässt – vielmehr sind beide Systeme an diesem
Adaptationsprozess beteiligt. Um der Tatsache gerecht zu werden, dass sowohl im sensorisch-
perzeptuellen – insbesondere im visuellen – System als auch im motorischen System
plastische Veränderungen wirksam werden können, wird Prismenadaptation in der
vorliegenden Arbeit als sensomotorischer Adaptationsprozess verstanden, der eine durch
Prismen induzierte Störung der Sensomotorik kurzfristig und schnell kompensiert.
Was unterscheidet sensomotorische Adaptation von sensomotorischem Lernen? Ein
wesentlicher Unterschied betrifft die Zeitskalen, auf denen beide Prozesse operieren:

Einleitung 7
Während es durch Lernen zu länger andauernden Veränderungen in der neuronalen
Verarbeitung kommt, wird das sensomotorische System durch Adaptation kurzfristig (im
Zeitbereich von wenigen Sekunden bis Minuten) neuen Bedingungen angepasst. Der
Adaptationsmechanismus operiert flexibel innerhalb eines kurzen Zeitfensters, um die
ursprüngliche Leistungsfähigkeit des Systems wiederherzustellen.
Ein wesentliches Charakteristikum von Adaptationsprozessen – seien sie sensorisch,
perzeptuell oder motorisch – ist das Auftreten sog. Nacheffekte infolge der Adaptation
(Palmer, 1999; Wade & Verstraten, 2005). Während das Lernen neuer perzeptueller oder
motorischer Fertigkeiten nicht dazu führt, dass nach „Abschluss“ des Lernens Nacheffekte
auftreten, ist eine Adaptation an neue Bedingungen immer mit einer De-Adaptation
verbunden (Nacheffekt) sobald die alten Bedingungen wiederhergestellt sind (Shadmehr &
Wise, 2005). Nacheffekte zeigen somit an, dass der Adaptationsprozess auf einen aktuellen
Kontext ausgerichtet ist.
Auch bei der Prismenadaptation tritt ein negativer Nacheffekt auf, d.h. zielgerichtete
Armbewegungen weichen zunächst in der dem Prismeneffekt entgegen gesetzten Richtung ab
(Fernandez-Ruiz & Diaz, 1999; Harris, 1965; Redding et al., 2005). Der durch die
Prismenadaptation hervorgerufene Nacheffekt weist ferner darauf hin, dass der
Adaptationsprozess nicht ausschließlich auf kognitiven Mechanismen, einer „bewussten“
Korrektur, beruht (Palmer, 1999): Obwohl nach dem Entfernen der Prismen die
ursprünglichen sensomotorischen Bedingungen wiederhergestellt sind, weichen die
Bewegungen in der Gegenrichtung ab, was darauf hinweist, dass durch die Prismenadaptation
ein neuer funktioneller Abgleich („Verrechnung“) von sensorischen und motorischen
Koordinaten stattgefunden hat (s.u.).
Diese kurzfristige und schnelle Anpassung an neue sensomotorische Bedingungen
charakterisiert den Prozess der Prismenadaptation. Dass durch längerfristiges
sensomotorisches Training unter Verwendung von Prismen neben Adaptations- auch
„klassische“ Lernprozesse aktiviert werden, die zu einer neuen Fertigkeit führen, machen die
Ergebnisse von Martin et al. (1996b) deutlich. In dieser psychophysikalischen Studie wurde
gezeigt, dass menschliche Versuchspersonen nach mehreren Wochen Training zwischen
Bedingungen mit und ohne Prismenbrille unmittelbar wechseln konnten, d.h. es war ihnen
möglich, je nach Kontext das passende Bewegungsprogramm auszuwählen. Nach dem
Training waren weder Prismen- noch Nacheffekt zu beobachten, was den Schluss nahe legt,
dass die Versuchspersonen durch sensomotorisches Lernen zwischen den Bedingungen mit
und ohne Prismen hin- und herwechseln konnten.

Einleitung 8
Diese Ergebnisse verdeutlichen, dass der Übergang zwischen Adaptation und Lernen im
sensomotorischen System fließend ist und sich beide Prozesse gegenseitig bedingen. In den
Untersuchungen der vorliegenden Arbeit spielen durch Lernen bedingte Veränderungen des
sensomotorischen Systems eine untergeordnete Rolle; vielmehr wurden die kurzfristigen
Anpassungsmechanismen untersucht, was als Prismenadaptation bezeichnet werden soll.
1.3 Neuronale Korrelate der Sensomotorik
Verschiedene sensorische (insbesondere visuelle), sensomotorische und motorische Areale
des Gehirns tragen zur Prismenadaptation bei. In diesem Abschnitt werden die neuronalen
Korrelate des sensomotorischen Systems daher kurz vorgestellt.
Analyse durch das visuelle System
Die Ausführung einer zielgerichteten visuell-geführten Bewegung zu einem Objekt ist
unmittelbar mit der sensorischen Analyse und Lokalisation des Objekts im Raum verknüpft
(Sensomotorik, s.o.). Die räumliche Lokalisation ist im Wesentlichen eine Funktion des
visuellen Systems, da dieses Sinnessystem eine räumlich hochauflösende dreidimensionale
Analyse der Außenwelt ermöglicht (Wurtz & Kandel, 2000b).
Nach dem peripheren visuellen System (Auge und Netzhaut) sowie dem visuellen Thalamus
als subcorticale Zwischenstation stellt der primäre visuelle Cortex (V1) am hinteren
Okzipitalpol das erste corticale Analysemodul dar, in dem visuelle Basismerkmale (z.B.
Orientierung) in räumlich (retinotop) geordneter Weise repräsentiert sind (Wurtz & Kandel,
2000a). In den hierarchisch nachgeschalteten visuellen Arealen (V2, V3, V5/MT etc.) erfolgt
eine zunehmend spezialisierte neuronale Verarbeitung von Muster-, Farb- und
Bewegungsinformation, wobei funktionell zwei getrennte visuelle Verarbeitungspfade
unterschieden werden: a) Ein sog. ventraler „Was-Pfad“, der vom okzipitalen in den
temporalen Cortex zieht und mit Objektwahrnehmung in Zusammenhang steht und b) ein sog.
dorsaler „Wo-Pfad“, der sich in den parietalen Cortex erstreckt und visuell-räumliche
Funktionen aufweist (Grill-Spector & Malach, 2004; Wurtz & Kandel, 2000b). Um die
visuelle Verarbeitung handlungsrelevanter räumlicher Information im dorsalen Pfad zu
betonen wurde dieser von Goodale & Milner (1992) als „vision-for-action“-Pfad bezeichnet –
im Gegensatz zum ventralen Pfad, der auf die Verarbeitung visueller Information für die
Wahrnehmung („vision-for-perception“) spezialisiert ist.

Einleitung 9
Neuere Untersuchungen deuten auf dynamische Interaktionen zwischen beiden Pfaden hin
(Goodale & Westwood, 2004), wobei kürzlich in einer Human-fMRT-Studie von Konen &
Kastner (2008) gezeigt werden konnte, dass es auch im dorsalen Pfad zu Objekt-spezifischen
neuronalen Aktivierungen kommt. Ein wesentlicher Unterschied in der neuronalen
Verarbeitung der beiden Pfade könnte in der Codierung der visuellen Information, d.h. in der
Art der Referenzsysteme, begründet sein (vgl. Schenk, 2006): Während der ventrale Pfad
visuelle Information in einem Objekt-bezogenen (allozentrischen) Referenzsystem
verarbeitet, encodiert der dorsale Pfad visuelle Information Subjekt-bezogen, also in einem
egozentrischen Koordinatensystem.
Sensomotorische Transformation im posterioren parietalen Cortex
Der erste Schritt bei der Planung einer Bewegung ist die visuell-räumliche Analyse des
Objekts durch die corticalen Areale des dorsalen Pfades. Da das visuelle System und das
motorische System unterschiedliche räumliche Bezugssysteme aufweisen, kommt es in einem
intermediären Schritt zur Umwandlung der visuellen (retinalen) Koordinaten in motorische
(Arm-zentrierte) Koordinaten für die Bewegungsplanung, ein Prozess, der als
sensomotorische Transformation bezeichnet wird und eine wesentliche Funktion des
posterioren parietalen Cortex (PPC) darstellt (Andersen et al., 2004). In verschiedenen
neuronalen Populationen im PPC können sowohl sensorische als auch motorische
Aktivierungen nachgewiesen werden, so dass der PPC als sensomotorische Schnittstelle für
die Planung von Augen- und Armbewegungen angesehen werden kann (Buneo & Andersen,
2006). Neben dem Areal LIP (engl. „lateral intraparietal“), das bei der Planung von
Augenbewegungen aktiviert wird, ist für zielgerichtete Armbewegungen das Areal PRR (engl.
„parietal reach region“) von Bedeutung, das zunächst beim Affen und später in einer fMRT-
Studie von Connolly et al. (2003) auch beim Menschen nachgewiesen werden konnte.
Für die Planung einer visuell-geführten Bewegung wird im PPC eine Umwandlung der
retinalen in motorische Koordinaten vollzogen, wobei die retinalen Koordinaten mit
sensorischen Informationen über die aktuelle Augen-, Kopf- und Körperposition
(propriozeptive Signale) in Bezug zum Objekt kombiniert werden; dabei existieren
verschiedene Modelle darüber, ob diese Informationen auf neuronaler Ebene seriell,
kombinatorisch oder direkt miteinander abgeglichen werden (Andersen et al., 2004). Die
Transformation in motorische Koordinaten, die auf das Effektororgan, z.B. den Arm, zentriert
sind, umfasst folglich auch die Integration verschiedener sensorischer Modalitäten im PPC
(Andersen et al., 1997). Eine PET-Studie von Clower et al. (1996) konnte zeigen, dass es

Einleitung 10
während der Prismenadaptation zu neuronalen Aktivierungen im PPC – in der Hemisphäre
kontralateral zum sich bewegenden Arm – kommt, was darauf hindeutet, dass die
sensomotorischen Repräsentationen im PPC plastischen Veränderungen unterliegen können
(s.u.).
Kontrolle von Bewegung durch corticale und subcorticale motorische Areale
Das periphere motorische System steuert letztlich die Ausführung einer Bewegung durch ein
motorisches Effektororgan auf Basis des komplexen räumlich-zeitlichen Zusammenspiels der
beteiligten Muskeln. Die zentralnervöse Vorbereitung der Bewegung erfolgt jedoch durch
corticale und subcortiale motorische Areale, die spezifisch und Kontext-bezogen an der
Planung und Kontrolle der Bewegung beteiligt sind bzw. diese modulieren: Der motorische
Cortex untergliedert sich in den primären motorischen Cortex (M1), das supplementär-
motorische Areal (SMA) sowie den prämotorischen Cortex (PMC). Körperteile werden in M1
somatotop repräsentiert, wobei die Größe der Repräsentation eine Funktion der Feinheit der
motorischen Steuerung ist (Blickhan, 2001). Eingänge erhält der motorische Cortex von den
Basalganglien, dem Kleinhirn (über den Thalamus) und aus den sensorischen Cortices; den
Hauptausgang bildet die sog. Pyramidenbahn, die „absteigend“ in das Rückenmark projiziert
(ebd.). Es besteht eine enge Verknüpfung zwischen dem motorischen und dem posterior
anliegenden somatosensorischen Cortex, in dem die Körperteile ebenfalls somatotop
repräsentiert sind. Das SMA und der PMC spielen insbesondere bei der Vorbereitung einer
Handlung eine Rolle; neuronale Aktivität im PMC liefert kontextuelle und räumliche
Information für die Bewegungsplanung, wobei enge wechselseitige Verbindungen zum PPC
bestehen, also dem Bereich des Gehirns, in dem die sensomotorische Transformation der
visuellen in motorische Koordinaten erfolgt (Shadmehr & Wise, 2005).
Ein wesentliches neuronales Substrat motorischer Kontrolle ist das Kleinhirn (Cerebellum),
das insbesondere bei der Feinabstimmung von Bewegungen und bei der Bewegungskorrektur
durch die Generierung eines neuronalen Fehlersignals eine wichtige Rolle spielt (Ghez &
Thach, 2000). Neben diesen eher allgemeinen motorischen Funktionen wird das Kleinhirn
auch mit motorischem Lernen und Adaptationsprozessen in Zusammenhang gebracht, eine
Vorstellung, die auf theoretische Arbeiten von Marr (1969) sowie Albus (1971) zurückgeht
und für die es eine Reihe von experimentellen Belegen gibt (eine Übersicht dazu liefern z.B.
Glickstein, 2007; Halsband et al., 2006; Thach et al., 1992). Auch für die Prismenadaptation
spielt das Kleinhirn eine entscheidende Rolle – neben dem posterioren parietalen Cortex wird
es als das wesentliche neuronale Substrat der plastischen Anpassungsprozesse, die während

Einleitung 11
der Prismenadaptation wirksam sind, diskutiert; die entsprechenden experimentellen Belege
werden im weiteren Verlauf bzw. in den Manuskripten näher vorgestellt.
1.4 Sensomotorische Plastizität am Beispiel der Prismenadaptation
Prozesse und Mechanismen
Wie im vorigen Abschnitt beschrieben, sind an der Planung und Ausführung einer
zielgerichteten Armbewegung ganz verschiedene Teile des Gehirns mit unterschiedlichen
funktionellen Eigenschaften beteiligt, wobei die hochgradig automatisierte sensomotorische
Bewegungsplanung in der Regel die präzise und erfolgreiche Ausführung einer Bewegung
gewährleistet.
Setzt man dagegen einer Versuchsperson eine geeignete Prismenbrille auf, durch welche sie
die visuelle Welt um einen bestimmten Winkelbetrag beispielsweise in der horizontalen
Ebene versetzt wahrnimmt, weichen zielgerichtete visuell-geführte Armbewegungen zunächst
in der Richtung der prismatischen Versetzung seitlich ab. Wodurch wird dieser Prismeneffekt
ausgelöst? Eintreffendes Licht wird durch die Prismen gebrochen und dabei zur Prismenbasis
abgelenkt, was zur Folge hat, dass die Strahlen auch im Augenhintergrund seitlich abgelenkt
ankommen. Damit das Ziel weiterhin visuell fixiert werden kann, wird eine gleichsinnige
kompensatorische Augenbewegung in die Gegenrichtung der Prismenbasis ausgeführt, d.h.
die Augenstellung im Kopf wird unter dem Einfluss der Prismen verändert. Die manipulierte
Augenstellung führt dazu, dass der Arm zu dem „virtuellen“ – als seitlich versetzt
wahrgenommen – Ziel geleitet wird und nicht die tatsächliche physikalische Zielposition
erreicht. Der Prismeneffekt ergibt sich folglich primär aus der Planung einer Armbewegung
zu einem als versetzt wahrgenommenen Ziel, was eine direkte Folge der durch die Prismen
verursachten veränderten Augenstellung im Kopf ist (vgl. Ghez & Thach, 2000; Martin et al.,
1996b). Die Größe des Prismeneffekts ist dabei proportional zur Stärke der optischen
Versetzung durch die Prismen (Fernandez-Ruiz & Diaz, 1999). Der initiale Prismeneffekt
fällt jedoch meist geringer aus als aufgrund der optischen Eigenschaften der Prismen
rechnerisch zu erwarten wäre (dazu siehe Redding & Wallace, 2004).
Der Prozess der Prismenadaptation, der bereits von Helmholtz (1867) beschrieben wurde,
umfasst die zunehmende Verringerung des seitlichen motorischen Fehlers aufgrund visueller
Rückmeldung über die räumliche Genauigkeit der Armbewegung. Im Verlauf der Adaptation
werden die Armbewegungen durch Training – aktives Ausüben und Wiederholen der
motorischen Aufgabe – korrigiert und an die veränderten sensomotorischen Bedingungen

Einleitung 12
angepasst bis das Ziel trotz der Prismenbrille wieder getroffen wird (Harris, 1965; Redding et
al., 2005).
Dass es sich dabei um einen „klassischen“ Adaptationsprozess handelt, zeigt der negative
Nacheffekt, der auftritt, wenn die Versuchsperson nach erfolgter Adaptation die
sensomotorische Aufgabe ohne Prismenbrille ausführt: In diesem Fall weichen die
Armbewegungen zunächst in der dem Prismeneffekt entgegengesetzten Richtung seitlich ab,
d.h. es tritt erneut ein motorischer Fehler auf, der aber im weiteren Verlauf des
sensomotorischen Trainings vollständig abgebaut wird (Harris, 1965; Redding et al., 2005).
Der Nacheffekt lässt darauf schließen, dass es im Verlauf des Adaptationsprozesses durch
Training zu einer neuen funktionellen Kopplung innerhalb des sensomotorischen Systems
gekommen ist; durch diese wird sichergestellt, dass die intendierten Armbewegungen trotz
der neuen sensomotorischen Bedingungen zielgerichtet und präzise ausgeführt werden
können. Der Prozess des Neu-Abgleichs sensomotorischer Koordinaten setzt funktionelle
Plastizität der beteiligten neuronalen Strukturen voraus (s.o.). Mit dem Verschwinden des
Nacheffekts ist die De-Adaptation abgeschlossen und die ursprüngliche sensomotorische
Kopplung wieder hergestellt.
Ohne an dieser Stelle auf die Details des entsprechenden Modells eingehen zu können, sei
festgehalten, dass Redding & Wallace (2002; 2003a) bzw. Redding et al. (2005) die
Prismenadaptation im wesentlichen auf zwei adaptive Mechanismen im Gehirn zurückführen:
a) einen Mechanismus strategischer (Re-)Kalibrierung (engl. „strategic re-/calibration“) und
b) einen Mechanismus räumlicher (Neu-)Ausrichtung (engl. „spatial re-/alignment“). Diese
beiden Mechanismen interagieren dynamisch und tragen gemeinsam zum adaptiven Verhalten
bei. Dabei führt der („kognitive“) Kalibrierungs-Mechanismus zu einer schnellen Reduktion
des motorischen Fehlers während des Tragens der Prismenbrille, d.h. die Genauigkeit des
motorischen Ausgangs steigt mit der Anzahl der ausgeführten Armbewegungen an. Parallel
dazu operiert der („sensomotorische“) Neuausrichtungs-Mechanismus und gleicht die
Koordinatensysteme (visuell, propriozeptiv, motorisch) räumlich-funktionell neu miteinander
ab, wodurch die räumlichen Bezugssysteme der Sensomotorik unter dem Einfluss der
Prismenbrille adaptiv in Übereinstimmung gebracht werden.
In einer neueren Untersuchung dissoziieren Redding & Wallace (2006a) beide Mechanismen,
indem sie das Generalisierungsmuster des Nacheffekts auf neue Aufgaben miteinander
vergleichen; leider tragen sie weiter zur sprachlichen Unschärfe im Zusammenhang mit der
Prismenadaptation bei, indem sie „recalibration“ als ein Beispiel für „kognitives Lernen“
anführen, während „realignment“ ein Beispiel für „perzeptuelles Lernen“ sei (S. 1006f.).

Einleitung 13
Eine Studie von Michel et al. (2007) deutet darauf hin, dass sich beide Mechanismen
bedingen, wobei der kognitive Mechanismus jedoch zu einer schwächeren Adaptation im
Vergleich zum sensomotorisch-automatischen Mechanismus führt: Erfolgte die Adaptation an
eine gegebene prismatische Versetzung abrupt, also mit einer starken bewusst-kognitiven
Komponente, war der Nacheffekt schwächer als bei einer progressiven Adaptation während
der sich die Versuchsperson über die prismatische Versetzung nicht „bewusst“ war. Das
bedeutet, dass der „kognitive“ Mechanismus zwar schnell die prismatische Versetzung
kompensieren kann, dieser jedoch nicht zu einem so präzisen funktionellen Abgleich
sensomotorischer Koordinaten führt wie der „automatische“ Mechanismus.
Im Prinzip sind mindestens drei Subsysteme der Sensomotorik als Locus der Prismen-
induzierten Veränderungen im Verlauf des Adaptationsprozesses denkbar: a) die räumliche
Codierung im visuellen System, b) die Lokalisation der Körperteile (insbesondere des Arms)
im Raum durch das propriozeptive System und c) die Bewegungsausführung durch das
motorische System. Folglich könnte die Prismenadaptation visuell, propriozeptiv oder
motorisch realisiert werden, wobei es für alle drei Erklärungsmöglichkeiten experimentelle
Hinweise gibt (siehe z.B. Harris, 1963; 1965; Martin et al., 1996b; Redding et al., 2005;
Uhlarik & Canon, 1971). Wahrscheinlich ist, dass es in allen drei Subsystemen zu schnellen
adaptiven Veränderungen kommen kann, wobei die relativen Beiträge von der
sensomotorischen Aufgabe, dem experimentellen Kontext sowie den spezifischen
experimentellen Bedingungen wie beispielsweise der Art der sensorischen Rückmeldung etc.
abhängen.
Neuronale Substrate der Verarbeitung
Neben einer Vielzahl psychophysikalischer Studien über funktionelle Aspekte der
Prismenadaptation liegen auch einige Studien über die beteiligten neuronalen Substrate vor.
So konnten bereits Weiner et al. (1983) zeigen, dass menschliche Patienten mit einer Läsion
des Kleinhirns Probleme bei der Prismenadaptation aufweisen und dementsprechend auch der
Nacheffekt erheblich reduziert ist, was auf eine Rolle des Kleinhirns beim funktionellen
Abgleich sensomotorischer Koordinaten hinweist. Ähnliche Ergebnisse lieferte die
Patientenstudie von Martin et al. (1996a), die den Vorteil hatte, dass aufgrund des Einsatzes
bildgebender neuroradiologischer Verfahren (Magnetresonanz- bzw. Computertomographie)
eine genauere Analyse über Art und Ausmaß der cerebellären Schädigung möglich war. Auch
die Ergebnisse der Studien von Morton & Bastian (2004) sowie Pisella et al. (2005) lassen auf

Einleitung 14
eine wesentliche Rolle des Kleinhirns bei der Prismenadaptation schließen. In
Übereinstimmung mit den Befunden beim Menschen konnte beim Rhesusaffen (Macaca
mulatta) nachgewiesen werden, dass focale Läsionen des Kleinhirns die Fähigkeit zur
Prismenadaptation massiv beeinträchtigen (Baizer et al., 1999).
Als corticales neuronales Substrat der Prismenadaptation wird der posteriore parietale Cortex
diskutiert, was sowohl in der bereits genannten PET-Studie von Clower et al. (1996) bei
Normalpersonen als auch in Patientenstudien u.a. von Newport & Jackson (2006) sowie
Newport et al. (2006) nachgewiesen werden konnte. Diese Ergebnisse weisen darauf hin, dass
die sensomotorischen Repräsentationen im PPC schnellen plastischen Veränderungen
unterzogen werden können, wobei dieses möglicherweise durch die Aktivität eines parieto-
cerebellären Netzwerks vermittelt wird.
Dass die Prismenadaptation auch einen Einfluss auf höhere Ebenen der neuronalen
Informationsverarbeitung hat (insbesondere auf die Raumrepräsentation), legen Studien zur
Prismenadaptation bei Neglect-Patienten nahe. Neglect ist eine Störung der räumlichen
Repräsentation der Außenwelt, die auf eine meist rechtsseitige Läsion im parietalen Cortex
zurückgeht und sich bei Patienten in Form eines Defizits bei der Orientierung auf Reize in der
kontraläsionalen Seite des Raumes äußert (Redding & Wallace, 2006b; Rode et al., 2003).
Rossetti et al. (1998) konnten nachweisen, dass es bei Patienten mit linksseitigem Neglect
nach der Adaptation an rechtsversetzende Prismen (Nacheffekt nach links) bei
neuropsychologischen Tests von räumlichen Aufgaben zu länger anhaltenden Verbesserungen
kommt. Dies zeigt, dass die plastischen Veränderungen im Verlauf der Prismenadaptation
räumliche Repräsentationen spezifisch beeinflussen.

Allgemeine Methodik 15
2. Allgemeine Methodik
2.1 Verhaltensuntersuchungen und Psychophysik
Methodisch lässt sich diese Arbeit in den Bereich der Psychophysik einordnen, in der die
quantitativen Beziehungen zwischen Reizgröße und subjektiver Empfindungsgröße erfasst
werden (Birbaumer & Schmidt, 2003). Durch das systematische Variieren einer bestimmten
(physikalisch definierten) Reizgröße und das Messen der darauf folgenden subjektiven
Empfindung bzw. des Verhaltens einer Versuchsperson wird auf die zugrunde liegenden
Mechanismen und Prozesse geschlossen. Psychophysikalische Methoden eignen sich folglich
besonders gut, um die funktionellen Merkmale eines Prozesses zu charakterisieren, wobei der
Vorteil dieses methodischen Ansatz auch darin liegt, dass für die Untersuchungen keine
experimentellen Eingriffe bei der Versuchsperson vorgenommen werden müssen, d.h. es
handelt sich um eine nicht-invasive Methode.
Der psychophysikalische Ansatz soll am Beispiel der Prismenadaptation näher erläutert
werden: Der Versuchsperson wird eine geeignete experimentelle Aufgabe gestellt,
beispielsweise die Ausführung einer visuell-geführten Armbewegung zu einem Ziel, wobei
die Genauigkeit der Armbewegungen unter verschiedenen Bedingungen (z.B. mit vs. ohne
Prismenbrille) kontinuierlich aufgezeichnet und gemessen wird. Mit Hilfe dieses Verfahrens
wird eine quantitative Beziehung zwischen den physikalischen Reizeigenschaften (in diesem
Fall z.B. Stärke und Richtung der prismatischen Versetzung) und dem beobachtbaren
Verhalten (Genauigkeit der Armbewegung) unter verschiedenen Bedingungen hergestellt, um
auf die zugrunde liegenden Prozesse und Mechanismen zu schließen. Die
psychophysikalische Untersuchung der Prismenadaptation charakterisiert den Prozess folglich
auf der Verhaltensebene und liefert über die geeignete Variation der experimentellen
Bedingungen Erkenntnisse über die funktionellen Aspekte der Adaptation. Beispielsweise
kann man durch das Messen der Größe des Nacheffekts unter verschiedenen Bedingungen die
Stabilität bzw. die Generalisierbarkeit des Adaptationsprozesses untersuchen (siehe Studie 3).
Bei der Interpretation psychophysikalischer Daten muss jedoch berücksichtigt werden, dass
Aussagen über die zugrunde liegenden neuronalen Prozesse und Substrate nur indirekt
möglich sind, da keine neurophysiologischen Signale, sondern Verhaltensantworten gemessen
werden. Um sowohl funktionelle als auch neuronal-strukturelle Aspekte der

Allgemeine Methodik 16
Prismenadaptation zu untersuchen wäre folglich ein Paradigma geeignet, das (indirekte)
psychophysikalische und (direkte) neurophysiologische bzw. bildgebende Methoden
miteinander kombiniert. Gerade bei menschlichen Versuchspersonen sind
neurophysiologischen Untersuchungen jedoch enge ethische und methodische Grenzen
gesetzt. Allerdings können psychophysikalische Untersuchungen zur Adaptationsfähigkeit bei
Patienten mit anatomisch umschriebenen Läsionen des ZNS ebenfalls wertvolle
experimentelle Belege für die Beteiligung bestimmter neuronaler Strukturen liefern (s.o.).
2.2 Sensomotorische Aufgaben: Zeige- und Wurfbewegungen
In der vorliegenden Arbeit wurde die Prismenadaptation bei zwei sensomotorischen Aufgaben
– Zeige- bzw. Wurfbewegungen – psychophysikalisch untersucht.
Zeigebewegungen im peripersonalen Raum
Bei der Zeige-Aufgabe (Fig. 2.1a) bestand die Aufgabe der Versuchsperson darin, an einem
Tisch sitzend zielgerichtete visuell-geführte Zeigebewegungen zu einem bestimmten Ziel
auszuführen und dabei das Ziel unter visueller Fixation immer so genau wie möglich zu
treffen. Die Versuchsperson konnte ihren Arm zu Beginn der Bewegung nicht sehen, so dass
die Arm-Trajektorie im Wesentlichen nur auf Basis der visuellen Information über die
räumliche Position des Ziels in Bezug zur Versuchsperson geplant werden konnte. Auch
während der Trajektorie konnte keine visuelle Korrektur erfolgen, da der Bewegungspfad für
die Versuchsperson aufgrund einer undurchsichtigen Tischplatte unsichtbar war (Ausnahme:
Studie 2); es handelte es sich im Übrigen um eine ballistische Bewegung. Am Ende der
Bewegung konnte die Versuchsperson die Spitze des Zeigefingers in Bezug zum Ziel sehen
und erhielt somit stets visuelle Rückmeldung über die räumliche Genauigkeit der Bewegung
(Fehlersignal). Die einzelnen Bewegungen wurden in rascher Folge ausgeführt und die
Versuchsperson konnte aufgrund der visuellen Rückmeldung die Bewegungen schrittweise
korrigieren.
Da sich das Ziel innerhalb des räumlichen Arbeitsbereichs befand, der mit dem Arm direkt
erreicht und damit aktiv exploriert werden kann, handelte sich um Bewegungen im Nahraum,
d.h. im peripersonalen Raum, in dem die sensomotorische Kontrolle von Armbewegungen
aufgrund des dreidimensionalen Sehens sehr präzise ist (siehe auch Previc, 1998). Der
peripersonale Raum wird auch als „action space“ bezeichnet, wodurch betont werden soll,
dass in diesem räumlichen Bereich bewegungsrelevante Aktivität eine große Rolle spielt. Die

Allgemeine Methodik 17
Mehrzahl der Bewegungen erfolgt im Nahraum unter visueller Kontrolle – hieraus ergibt sich,
dass Zeigebewegungen im Nahraum bei Versuchspersonen in der Regel hochgradig trainiert
und abgestimmt sind. Um die generelle Genauigkeit von Zeigebewegungen auf ein visuelles
Ziel im Nahraum zu messen, wurde in den Experimenten jeweils auch eine sog. Baseline-
Messung (ohne Prismenbrille; s.u.) vorgenommen.
Die Genauigkeit der Zeigebewegungen wurde mit Hilfe eines PC-kontrollierten Messsystems
der Firma Zebris Medical (Isny, Deutschland) erfasst, das den räumlichen Abstand zwischen
Ziel und Zeigefinger in allen drei Raumdimensionen durch Analyse der
Laufzeitverzögerungen von Ultraschallsignalen mit einer Genauigkeit von ca. 1 mm
berechnen konnte. Da die prismatische Versetzung jeweils in der horizontalen Ebene erfolgte,
wurde entsprechend der horizontale Abstand zwischen Ziel und Zeigefinger am Ende der
Trajektorie ermittelt.
Fig. 2.1 Skizze des experimentellen Aufbaus der (A) Zeige- und (B) Wurf-Aufgaben.
Wurfbewegungen im extrapersonalen Raum
Neben Zeigebewegungen wurden auch Wurfbewegungen untersucht, wobei die Aufgabe der
Versuchsperson darin bestand, einen Ball so genau wie möglich zu einem Ziel an einer 2 m
entfernten Leinwand zu werfen. Dabei sollte das Ziel immer visuell fixiert werden; bei den
Würfen handelte es sich um „seitliche“ Oberhand-Würfe wie sie beispielsweise auch von
Martin et al. (1996a; 1996b; 2002) sowie Fernandez-Ruiz & Diaz (1999) untersucht wurden.
Die Versuchsperson erhielt visuelle Rückmeldung über die Genauigkeit des aktuellen Wurfes,
da der Ball an der mit Klettmaterial beschichteten Leinwand hängen blieb; somit konnten die
Würfe – analog zu den Zeigebewegungen – schrittweise korrigiert werden.

Allgemeine Methodik 18
Anders als bei der Zeige-Aufgabe befand sich das Ziel bei der Wurf-Aufgabe jenseits des
räumlichen Bereichs, der mit den Armen direkt erreicht werden kann, d.h. es handelte sich
hier um eine motorische Aufgabe im Fernraum (extrapersonaler Raum), in dem
wahrnehmungsbezogene räumliche Information ein wichtige Rolle spielt (vgl. Previc, 1998).
Wurfbewegungen weisen gegenüber Zeigebewegungen einen höheren Grad an motorischer
Komplexität auf; hinzu kommt, dass das Werfen nicht zum „klassischen“ Alltags-
Bewegungsrepertoire einer Versuchsperson gehört und dieses Bewegungsmuster damit auch
nicht so hochgradig trainiert ist wie das der Zeigebewegungen; aufgrund dessen begannen die
Wurf-Experimente jeweils mit einer kurzen Trainings- bzw. Gewöhnungsphase. Im
Anschluss hieran erfolgte die Messung der Wurf-Genauigkeit ohne Prismenbrille (Baseline-
Messung). Die räumliche Genauigkeit der Würfe wurde mit einem PC-kontrollierten
optischen System ermittelt, das die horizontale Entfernung zwischen dem Ziel und der
Position des Balls mit einer räumlichen Auflösung von ca. 1 cm feststellen konnte4.
Ablauf
Der Ablauf sowohl der Zeige- als auch der Wurf-Experimente gliederte sich standardmäßig in
drei Phasen bzw. Bedingungen (siehe Redding et al., 2005), wobei je nach Fragestellung ein
hiervon leicht abgewandelter Ablauf realisiert wurde (das gilt insbesondere für Studie 2): a)
eine Prä-Prismenbedingung, in der die Genauigkeit der Bewegungen ohne Prismenbrille
aufgezeichnet wurde (Baseline-Messung), b) eine Prismenbedingung, während der die
Adaptation an die prismatische Versetzung erfolgte und c) eine Post-Prismenbedingung, in
welcher der Nacheffekt gemessen und die De-Adaptation vollzogen wurde. Dabei wurden pro
Bedingung mindestens 30 zielgerichtete Bewegungen ausgeführt, wobei diese Anzahl
insbesondere in der Prismenbedingung der Wurf-Experimente nicht unterschritten werden
sollte, um eine ausreichende Trainings-bedingte Adaptation sicherzustellen. Eine detaillierte
Beschreibung des experimentellen Ablaufs befindet sich im Methoden-Teil der Manuskripte.
4 Nach jedem Wurf wurde ein Digitalphoto von der Leinwand mit der Position des Balls aufgenommen. Die Auswertung der elektronischen Bilddaten erfolgte mit Hilfe einer von Herrn Dipl.-Phys. D. Trenner entwickelten Matlab-basierten Software.

Allgemeine Methodik 19
2.3 Datenauswertung
Für jede einzelne Bewegung wurde der horizontale Abstand („horizontaler Fehler“) zwischen
dem Ziel und dem Endpunkt der Bewegung in metrischen Einheiten bestimmt und als
Funktion der Anzahl der Bewegungen in der jeweiligen Bedingung graphisch aufgetragen.
Bei den Zeigebewegungen wurde der horizontale Abstand zwischen Ziel und Zeigefinger auf
dem höchsten Punkt der Trajektorie ermittelt, da sich Ziel und Zeigefinger in diesem Fall auf
einer Höhe befanden (Tischkante als gemeinsame Begrenzung) und die Bewegung somit
nicht mehr fortgesetzt werden konnte. Bei den Wurfbewegungen war der Endpunkt der
Bewegung durch die Position des Balls auf der Leinwand definiert; auch hier wurde
ausschließlich der Abstand vom Ziel zum Zentrum des Balls in der horizontalen Ebene
bestimmt.
Die Rohdaten einer Versuchsperson wurden darüber hinaus individuell einer sog. „Baseline-
Korrektur“ unterzogen, um eine Normalisierung der Daten zu erreichen. Dafür wurde das
arithmetische Mittel der horizontalen Fehler der Bewegungen in der Prä-Prismenbedingung
gebildet und von jedem einzelnen Datenpunkt subtrahiert. Dieses Verfahren wurde auch von
Clower & Boussaoud (2000) angewandt und hat den Vorteil, dass der Datensatz individuell
um eine eventuelle Vorzugsrichtung („Bias“) der Bewegung bei einer bestimmten Aufgabe
korrigiert wird, was eine bessere Vergleichbarkeit der Effekte zwischen Versuchspersonen
ermöglicht. Gerade bei der Wurf-Aufgabe war dieser „Bias“ von Versuchsperson zu
Versuchsperson unterschiedlich stark, wodurch eine Normalisierung der Daten notwendig
erscheint. Alle weiteren Analysen wurden auf Basis der Baseline-korrigierten Daten
durchgeführt.
In der Datenanalyse lag neben dem zeitlichen Verlauf von Adaptation und De-Adaptation ein
besonderer Focus auf der Größe von Prismen- bzw. Nacheffekt. Für die Ermittlung der Größe
des Prismeneffekts wurde der horizontale Fehler als Funktion der ersten vier Bewegungen der
Prismenbedingung aufgetragen und es wurde eine lineare Regressionsgerade angelegt; der
Schnittpunkt der Regressionsgeraden mit der y-Achse wurde als Maß für die Größe des
initialen Prismeneffekts verwendet. Entsprechend wurde bei der Ermittlung des Nacheffekts
vorgegangen. Das verwendete mathematische Verfahren berücksichtigt, dass Prismen- bzw.
Nacheffekt nicht bereits nach der ersten Bewegung vollständig verschwunden sind, sondern
eine schnelle schrittweise Reduktion des horizontalen Fehlers während der ersten
Bewegungen erfolgt.

Motivation und Zusammenfassung der Studien 20
3. Motivation und Zusammenfassung der Studien
3.1 Motivation der Studien
Das Ziel der vorliegenden Arbeit bestand darin, bestimmte funktionelle Aspekte der
Prismenadaptation bei gesunden menschlichen Versuchspersonen psychophysikalisch zu
untersuchen um Antworten auf die Frage zu liefern, mit welchen plastischen Mechanismen es
das Gehirn erreicht, seine funktionelle Organisation schnell und reversibel neuen
Bedingungen anzupassen. Dafür wurde das Paradigma der Prismenadaptation gewählt, da es
sich besonders gut eignet, schnelle sensomotorische Adaptationsprozesse beim Menschen mit
nicht-invasiven psychophysikalischen Methoden zu charakterisieren.
Studie 1: Vollständigkeit des Adaptationsprozesses
In Studie 1 wurde die Vollständigkeit des Adaptationsprozesses bei zwei verschiedenen
Aufgaben (Zeigen vs. Werfen) und zwei verschiedenen Richtungen der prismatischen
Versetzung (links vs. rechts) nach ausgedehntem sensomotorischen Training untersucht. Die
Motivation für diese Studie geht auf Beobachtungen aus Pilot-Experimenten zu
Wurfbewegungen zurück, bei denen die Versuchspersonen am Ende der Prismenadaptation
(nach 30 Bewegungen) einen deutlichen horizontalen Restfehler in Richtung der
prismatischen Versetzung aufwiesen. Ein ähnlich hoher Restfehler von Wurfbewegungen ist
beispielsweise auch in den Ergebnissen von Fernandez-Ruiz & Diaz (1999, Fig. 1)
auszumachen, wobei dieser Effekt in der genannten Studie jedoch nicht diskutiert wird.
Ausgehend von dieser Beobachtung wurde eine Studie konzipiert, die die Vollständigkeit des
Adaptationsprozesses nach ausgedehntem Training systematisch prüfen sollte. Die Anzahl der
Bewegungen während der Prismenbedingung wurde von 30 auf 120 bzw. 240 (Kontroll-
Experiment) erhöht, um ein ausgedehntes Training der entsprechenden Aufgabe zu
gewährleisten. Für den Fall, dass die Vollständigkeit der Adaptation und damit die
Genauigkeit, mit der sensomotorische Koordinaten im Verlauf der Adaptation neu
miteinander abgeglichen werden, nur vom Ausmaß des Trainings abhängt, sollten sich die
Bewegungen im Verlauf des Adaptationsprozesses einem mittleren Fehler von „Null“
angleichen. Die Vollständigkeit der Prismenadaptation wurde bei zwei Aufgaben untersucht,
um hochgradig trainierte Bewegungen im Nahraum (Zeigen) mit weniger stark trainierten und
komplexeren Bewegungen im Fernraum (Werfen) miteinander zu vergleichen.

Motivation und Zusammenfassung der Studien 21
Studie 2: Aufgaben-Spezifität der Prismenadaptation
Die Konzeption von Studie 2 war durch die Frage motiviert, inwieweit es zu einer
Generalisierung der Prismenadaptation von einer trainierten auf eine untrainierte
sensomotorische Aufgabe kommt. Das Ausmaß der Generalisierung spiegelt die Spezifität des
Adaptationsprozesses für den aktuellen Aufgabenkontext wider. Das Generalisierungsmuster
liefert somit wichtige Erkenntnisse über die Art der Informationsverarbeitung (siehe Poggio
& Bizzi, 2004). In Studie 2 wurde die Generalisierung des Prismen- bzw. Nacheffekts von
einer Zeige-Aufgabe auf eine Wurf-Aufgabe gemessen. Eine vollständige Generalisierung auf
die Wurf-Aufgabe würde auf einen „globalen“ Adaptationsprozess unabhängig vom
Aufgabenkontext hindeuten. Umgekehrt würde eine schwache bzw. ausbleibende
Generalisierung zeigen, dass die adaptiven Veränderungen im Gehirn jeweils spezifisch für
die aktuell ausgeführte Aufgabe vorgenommen werden. Einen Hinweis hierauf lieferte bereits
die Studie von Martin et al. (1996b). Parallel hierzu wurde in Studie 2 untersucht, welchen
Einfluss die Art der sensorischen Rückmeldung während der Ausführung der Zeige-Aufgabe
auf die Generalisierung der Adaptation zur Wurf-Aufgabe hat. Die Versuchsperson erhielt im
ersten Experiment nur (terminale) visuelle Rückmeldung über die Zeigebewegung am Ende
der Bewegung, während im zweiten Experiment der komplette Bewegungspfad von der
Versuchsperson visuell verfolgt werden konnte (kontinuierliche Rückmeldung). Bereits von
Uhlarik & Canon (1971) wurde vorgeschlagen, dass die Adaptation je nach Art der
Rückmeldung in unterschiedlichen sensorischen Subsystemen vollzogen werde; dieser
Mechanismus würde folglich ein unterschiedliches Generalisierungsmuster in Abhängigkeit
von der Art der sensorischen Rückmeldung vorhersagen.
Studie 3: Räumliche Generalisierung des Nacheffekts
Die Frage nach dem räumlichen Generalisierungsmuster des Nacheffekts stand in Studie 3 im
Mittelpunkt. Der Nacheffekt wurde als kritische Messvariable gewählt, da dieser weniger
stark von kognitiven Einflüssen überformt ist; es wurden ausschließlich Zeigebewegungen
untersucht. Anders als in Studie 2 kam es nach erfolgter Prismenadaptation nicht zu einem
Wechsel der sensomotorischen Aufgabe, sondern es wurde getestet, inwieweit der Nacheffekt
auf nicht-trainierte Zielpositionen im Raum generalisiert. Dabei wurde in zwei Experimenten
die Art des sensomotorischen Trainings während der Prismenadaptation systematisch variiert:
Entweder wurden die Zeigebewegungen während der Adaptation nur zu einem Ziel in
geblockter Form durchgeführt (Experiment 1) oder das Training erfolgte abwechselnd zu zwei

Motivation und Zusammenfassung der Studien 22
verschiedenen Zielen im Raum (Experiment 2). In Experiment 2 wurde insbesondere
untersucht, wie die Generalisierung des Nacheffekts auf Ziele bei räumlichen Interpolations-
bzw. Extrapolationsaufgaben ausfällt. Studie 3 war folglich durch die Frage motiviert, ob die
Form des sensomotorischen Trainings während der Prismenadaptation einen Einfluss auf das
räumliche Generalisierungsmuster ausübt. Darüber hinaus liefert das Generalisierungsmuster
des Nacheffekts Erkenntnisse über die Art der adaptiven Veränderungen der räumlichen
Repräsentationen. Die Untersuchung orientierte sich dabei inhaltlich an den Studien von
Bedford (1993) sowie Redding & Wallace (2006a).
3.2 Zusammenfassung von Studie 1
In Studie 1 wurde die Vollständigkeit des Adaptationsprozesses bei Zeige- bzw.
Wurfbewegungen unter Verwendung links- bzw. rechtsversetzender Prismen bei vier
unabhängigen Gruppen rechtshändiger gesunder Versuchspersonen untersucht. Jedes
Experiment bestand dabei aus drei Bedingungen (Prä-, Prismen- und Post-Bedingung), in
denen jeweils 120 zielgerichtete Bewegungen ausgeführt wurden, um ein ausgedehntes
Training der sensomotorischen Aufgabe zu gewährleisten.
Erwartungsgemäß trat in allen vier Gruppen in der Prismenbedingung ein ausgeprägter
horizontaler Fehler in Richtung der prismatischen Versetzung auf, der durch Training deutlich
reduziert wurde. Nach dem Entfernen der Prismen trat ein Nacheffekt in der Gegenrichtung
auf, der in allen Gruppen durch Training im weiteren Verlauf der Post-Prismenbedingung
vollständig verschwand. Die Vollständigkeit des Adaptationsprozesses (Prismenbedingung)
hing dabei nicht von der Richtung der prismatischen Versetzung, sondern von der Art der
sensomotorischen Aufgabe ab. Während bei der Zeige-Aufgabe am Ende der
Prismenbedingung ein horizontaler Restfehler statistisch nicht nachgewiesen werden konnte,
trat bei der Wurf-Aufgabe nach 120 Bewegungen ein signifikanter horizontaler Restfehler
auf. Bei beiden Aufgaben nahm die Reduktion des horizontalen Fehlers während der
Prismenbedingung einen asymptotischen Verlauf, wobei allerdings bei der Wurf-Aufgabe ein
Restfehler zu beobachten war, der durch das Anlegen einer Exponentialfunktion an die Daten
quantifiziert wurde. Während folglich bei Zeigebewegungen ein nahezu vollständiger
Adaptationsprozess mit einer kompletten Kompensation der prismatischen Versetzung zu
beobachten war, deuten die Ergebnisse für die Wurfbewegungen auf einen unvollständigen
Adaptationsprozess mit einem inhärenten Restfehler hin. Dies konnte durch ein Kontroll-
Experiment, bei dem die Anzahl der Wurfbewegungen während der Prismenbedingung auf

Motivation und Zusammenfassung der Studien 23
240 verdoppelt wurde, bestätigt werden. Selbst unter diesen nochmals verlängerten
Adaptationsbedingungen war ein Restfehler statistisch nachweisbar, was stark darauf
hindeutet, dass bei Wurfbewegungen neben der reinen Anzahl von Bewegungen noch andere
Faktoren eine Rolle für die Adaptation spielen. Einer dieser Faktoren ist die allgemeine
Variabilität bei der Ausführung der Aufgabe: Für die Wurfbewegungen fand sich eine
signifikante positive Korrelation zwischen der allgemeinen Variabilität der Wurfbewegungen
und der Größe des Restfehlers am Ende der Prismenbedingung. Je höher die Variabilität,
desto größer der verbleibende Restfehler, d.h. umso unvollständiger die Adaptation. Diese
lineare Beziehung konnte nur bei der Wurf-Aufgabe nachgewiesen werden.
Die Ergebnisse von Studie 1 zeigen, dass das Gehirn bei der Ausführung von
Zeigebewegungen im Nahraum die sensomotorischen Koordinaten im Verlauf der
Prismenadaptation sehr präzise funktionell miteinander abgleichen kann, wodurch kein
Restfehler am Ende der Adaptation nachzuweisen ist. Die genaue adaptive Abstimmung der
Sensomotorik setzt ein hohes Maß an funktioneller Plastizität voraus, was wiederum in der
Verhaltensrelevanz von präzisen Zeigebewegungen im Nahraum begründet liegen könnte.
Dagegen scheint bei Wurfbewegungen im Fernraum die allgemeine Variabilität die
Vollständigkeit des Adaptationsprozesses zu begrenzen. Dieses könnte damit
zusammenhängen, dass das (neuronale) Fehlersignal, welches durch die Verrechnung von
motorischen Fehlern innerhalb eines gewissen Zeitfensters generiert wird, um die
Bewegungen zu korrigieren, aufgrund der großen Variabilität ein ungünstiges Signal-zu-
Rausch-Verhältnis aufweist und es somit zu einer unvollständigen Bewegungskorrektur mit
einem Restfehler kommt.
3.3 Zusammenfassung von Studie 2
In Studie 2 wurde getestet, ob Prismen- bzw. Nacheffekt von einer Zeige-Aufgabe auf eine
Wurf-Aufgabe generalisieren. Das Ausmaß der Generalisierung wurde bei zwei unabhängigen
Gruppen rechtshändiger gesunder Versuchspersonen ermittelt, die während der Zeige-
Adaptation jeweils unterschiedliche sensorische Rückmeldungen erhielten. Im ersten
Experiment war visuelle Information über die Genauigkeit der Zeigebewegung nur am Ende
der Trajektorie verfügbar; dagegen war im zweiten Experiment der gesamte Pfad der
Zeigebewegung während der Adaptation sichtbar. Im Anschluss an die Adaptation an
rechtsversetzende Prismen durch Ausführung der Zeige-Aufgabe wurde die Generalisierung
a) des Prismen- bzw. b) des Nacheffekts auf die Wurf-Aufgabe getestet.

Motivation und Zusammenfassung der Studien 24
Die Ergebnisse beider Experimente zeigen ein einheitliches Bild, das auf eine hohe Aufgaben-
Spezifität der Prismenadaptation hinweist (vgl. Martin et al., 1996b). Behielten die
Versuchspersonen nach erfolgter Zeige-Adaptation die Prismen aufgesetzt und sollten direkt
im Anschluss Wurfbewegungen zum Ziel ausführen, trat bei dieser untrainierten Aufgabe ein
ausgeprägter horizontaler Fehler in Richtung der prismatischen Versetzung auf. Die
Reduktion dieses seitlichen Fehlers durch das Trainieren der Wurf-Aufgabe erfolgte
asymptotisch und war von einem konventionellen Adaptationsprozess nicht zu unterscheiden.
Die Größe des „zweiten“ Prismeneffekts nach dem Aufgaben-Wechsel glich dabei der Größe
des Standard-Prismeneffekts für die Wurf-Aufgabe. Dabei trat der Prismeneffekt nach dem
Aufgaben-Wechsel unabhängig von der Art der sensorischen Rückmeldung während der
vorangegangenen Zeige-Adaptation auf, folglich führte keine der beiden
Rückmeldungsformen zu einer stärkeren Generalisierung auf die neue Aufgabe. Auch beim
Nacheffekt trat keine Generalisierung von der Zeige- zur Wurf-Aufgabe auf: Setzten die
Versuchspersonen nach der Zeige-Adaptation die Prismen ab und führten dann
Wurfbewegungen aus, war bei der neuen Aufgabe kein Nacheffekt zu beobachten, was darauf
hindeutet, dass auch der Nacheffekt unter den gegebenen Bedingungen nicht generalisiert.
Die Ergebnisse von Studie 2 legen den Schluss nahe, dass die plastischen Veränderungen, die
im Zusammenhang mit der Prismenadaptation auftreten, hochgradig spezifisch für den
Kontext bzw. die aktuelle sensomotorische Aufgabe vorgenommen und nicht ohne weiteres
von einem Bewegungskontext auf einen neuen übertragen werden können. Darüber hinaus
führte keine den beiden Formen von sensorischer Rückmeldung (terminal vs. kontinuierlich)
zu einer stärkeren Generalisierung auf die neue Aufgabe. Andererseits gibt es experimentelle
Belege dafür (vgl. Redding & Wallace, 2006a; Uhlarik & Canon, 1971), dass bei terminaler
Rückmeldung die adaptiven Veränderungen insbesondere die visuelle Codierung betreffen,
was eine stärkere Generalisierung der Adaptation von einer Aufgabe auf die andere unter
diesen Bedingungen vorhersagen würde. Die Ergebnisse des Experiments mit terminaler
Rückmeldung sind mit dieser Sichtweise nicht in Einklang zu bringen, da auch in diesem Fall
keine Generalisierung auf die neue Aufgabe nachweisbar war. Die ausgeprägte Aufgaben-
Spezifität der Prismenadaptation, die sich in Studie 2 zeigt, ist möglicherweise auch darauf
zurückzuführen, dass es sich bei Zeige- bzw. Wurfbewegungen um sehr unterschiedliche
Bewegungsfamilien handelt, was eine Generalisierung erschwert.

Motivation und Zusammenfassung der Studien 25
3.4 Zusammenfassung von Studie 3
In Studie 3 wurde die räumliche Generalisierung des Nacheffekts auf verschiedene
Zielpositionen bei zwei unterschiedlichen Formen von sensomotorischem Training
untersucht. Hierfür wurden zwei unabhängige Gruppen rechtshändiger gesunder
Versuchspersonen gebildet. In Experiment 1 erfolgte das sensomotorische Training während
der Prismenadaptation immer in geblockter Form zu einer von drei möglichen Zielpositionen
und im Anschluss wurde die Generalisierung des Nacheffekts auf eine nicht-trainierte
Zielposition getestet. In Experiment 2 führten die Versuchspersonen dagegen während der
Prismenadaptation Zeigebewegungen zu zwei verschiedenen Zielpositionen im Raum aus,
wodurch ein spezifischer Arbeitsbereich adaptiert wurde. In direktem Anschluss an dieses
gemischte Training wurde die Generalisierung des Nacheffekts entweder auf ein Ziel jenseits
des adaptierten räumlichen Bereichs (Extrapolationsaufgabe) oder auf ein Ziel innerhalb des
adaptierten Bereichs (Interpolationsaufgabe) gemessen.
Das Generalisierungsmuster des Nacheffekts nach Training in geblockter Form zeigt
unabhängig von der adaptierten Zielposition eine nahezu vollständige Generalisierung des
Nacheffekts auf nicht-trainierte Ziele im Raum. Wurde nach der Prismenadaptation zu einem
bestimmten Ziel die Größe des Nacheffekts an einem anderen Ziel getestet, so trat auch hier
ein Nacheffekt auf, was auf eine räumliche Generalisierung des Nacheffekts nach geblocktem
sensomotorischen Training hinweist. Dieses Generalisierungsmuster zeigt an, dass die
Adaptation an die prismatische Versetzung auch räumliche Positionen betrifft, die nicht direkt
während der Prismenadaptation trainiert wurden. Darüber hinaus deutet die lineare
Generalisierung des Nacheffekts zu untrainierten Zielen darauf hin, dass die räumlichen
Repräsentationen im Verlauf der Prismenadaptation in ihrer Gesamtheit verschoben bzw.
angeglichen werden, was gut mit den Ergebnissen von Bedford (1993) vereinbar ist. Nach
gemischtem sensomotorischen Training zeigt sich ein komplexeres Generalisierungsmuster
des Nacheffekts, wobei das Ausmaß der Generalisierung von der Position des getesteten Ziels
abhängt: Während der Extrapolations-Nacheffekt für das linke Ziel größer als der
entsprechende Standard-Nacheffekt war, verhielt es sich beim Extrapolations-Nacheffekt für
das rechte Ziel umgekehrt. Bei einer Interpolationsaufgabe zum zentralen Ziel fiel der
Generalisierungs-Nacheffekt ebenfalls geringer als der Standard-Nacheffekt aus. Die
Ergebnisse von Experiment 2 lassen darauf schließen, dass es im sensomotorischen System
infolge des gemischten Trainings nach der Prismenadaptation zu einer leichten aber
systematischen Unterschätzung der räumlichem Distanz zum Ziel kommt, durch die je nach

Motivation und Zusammenfassung der Studien 26
Richtung, in der das Ziel liegt, die Größe des Generalisierungs-Nacheffekts erhöht bzw.
erniedrigt wird. Dabei scheint die Richtungsinformation einen stärkeren Einfluss auf die
Größe des Nacheffekts zu haben als die Art der räumlichen Aufgabe (Extrapolation vs.
Interpolation). Studie 3 bekräftigt damit, dass die Prismenadaptation räumliche
Repräsentationen im Gehirn verändert, wobei auch die Form des sensomotorischen Trainings
einen spezifischen Einfluss auf die Art dieser Veränderungen hat.

Fazit 27
4. Fazit
Menschliche Versuchspersonen weisen eine bemerkenswert schnelle Fähigkeit auf,
zielgerichtete Zeige- oder Wurfbewegungen anzupassen, wenn es durch Prismen zu einer
Manipulation der sensomotorischen Kopplung kommt. Die Fähigkeit zur Prismenadaptation
setzt die funktionelle Plastizität des sensomotorischen Systems voraus und das experimentelle
Paradigma der Prismenadaptation eignet sich gut dazu, verhaltensdynamische Prozesse
psychophysikalisch zu untersuchen.
Die funktionellen Charakteristika des Adaptationsprozesses hängen von Faktoren wie der
Variabilität bei der Ausführung einer zielgerichteten Bewegung (Studie 1), der
sensomotorischen Aufgabe, die während der Adaptation ausgeführt wird (Studie 2) und dem
räumlichen Kontext der Aufgabe sowie der Art des sensomotorischen Trainings ab (Studie 3).
Der Adaptationsprozess ist dabei spezifisch für die aktuell ausgeführte Aufgabe und
generalisiert nicht auf eine Bewegung aus einer anderen Bewegungsfamilie; dagegen kommt
es zu einer räumlichen Generalisierung der Adaptation auf neue Ziele, sofern dieselbe
sensomotorische Aufgabe (Zeigen) ausgeführt wird.
Der Prozess der Prismenadaptation weist bereits auf Verhaltensebene ein hohes Maß an
Komplexität auf; um sowohl die funktionellen als auch die strukturellen Grundlagen der
Prismenadaptation besser zu verstehen, wäre die Kombination von psychophysikalischen und
neurophysiologischen bzw. bildgebenden Methoden ein viel versprechender Ansatz. Bei der
experimentellen Untersuchung darf in keinem Fall die Komplexität des Adaptationsprozesses
unterschätzt werden, was sich auch in einem abschließenden Zitat von Redding et al. (2005,
S. 440) widerspiegelt: „Prism adaptation is not a simple process“. Wie wahr.

Studie 1: Vollständigkeit des Adaptationsprozesses 28
5. Studie 1 Incomplete visuomotor adaptation despite extensive training
Sven Wischhusen & Manfred Fahle
Abstract
Prism adaptation demonstrates that visuomotor representations in the brain are rapidly
adjusted when prisms induce a displacement of the visual world. We investigated the speed
and extent of prism adaptation after extensive visuomotor training. Human subjects had to
perform visually-guided throwing or pointing movements towards a target before, during and
after wearing prism goggles that shifted the visual world laterally either to the right or left. In
each condition of each task, 120 movements were performed to ensure extensive training. Our
results indicate that prism adaptation was not complete when subjects performed throwing
movements since throws still deviated laterally at the end of the adaptation period; even after
an extended training of 240 throws, the remaining lateral deviation amounted to 29 mm. That
is, training did not yield complete adaptation for throwing movements. When subjects
performed pointing movements the remaining lateral deviation at the end of the adaptation
period failed to reach significance. In both tasks, removal of the prisms led to an aftereffect
which completely disappeared in the course of further training. The findings of incomplete
prism adaptation are discussed in terms of movement variability in combination with an
adaptive neural control system exhibiting a finite capacity for evaluating movement errors.
Introduction
Prism adaptation served as an experimental paradigm to study the mechanisms of adaptation
in neural representations of the physical world as a response to changes in sensory signals.
This adaptation demonstrates plasticity in the adult human brain, as do priming, perceptual,
and sensorimotor learning (Ahissar & Hochstein, 2004; Fahle, 2002).
Phenomenology of prism adaptation
Suitable prisms deflect the ray paths laterally and shift the retinal images of the visual world
causing a deviation between the felt direction of gaze versus position of arm: Subjects
wearing prism goggles initially fail to execute accurate goal-directed movements (e.g.
pointing, reaching, or throwing) towards a visual target because their sensory and motor

Studie 1: Vollständigkeit des Adaptationsprozesses 29
coordinate systems do not correspond properly (Martin et al., 1996b; 2001; 2002; Norris et
al., 2001). The movements’ lateral error is proportional to the magnitude of the prismatic
deflection applied, although it is a common observation that initial prism effects are smaller
than expected according to the optics of the prisms (e.g. Redding & Wallace, 2003b; Redding
& Wallace, 2004). Within a few trials under visual feedback, the movements’ lateral error is
reduced and trajectories recover high spatial accuracy. That is, subjects adapt to the optical
displacement of the visual world, a process of sensorimotor adaptation termed “prism
adaptation” (e.g. Bedford, 1999; Fernandez-Ruiz & Diaz, 1999; Harris, 1963; 1965; 1980;
Held & Freedman, 1963; Redding et al., 2005; Redding & Wallace, 2003a).
After removal of the prisms, on the other hand, movements initially deviate in the direction
opposite to the prismatic displacement, indicating a negative aftereffect which completely
vanishes in the course of further visuomotor practice if feedback is provided (e.g. Fernandez-
Ruiz & Diaz, 1999; Harris, 1965; 1980; Newport et al., 2006; Redding et al., 2005; Weiner et
al., 1983; Welch et al., 1974). The occurrence of an aftereffect is considered to prove “true”
adaptation because it indicates a spatial realignment of sensorimotor representations through
adaptation rather than a purely cognitive adaptation (Newport & Jackson, 2006). According to
the model of Redding & Wallace (2002) two closely related processes operate during prism
adaptation: first, a strategic recalibration mechanism based on cognitive control leading to
rapid error reduction during prism exposure and second a rather slow spatial realignment
mechanism yielding correspondence between different sensorimotor reference frames. During
prism adaptation, sensorimotor representations are rapidly adjusted in an experience-
dependent way to ensure high precision of eye-hand coordination which is essential to interact
with objects in space, followed by a slower process of spatial realignment.
The process of prism adaptation demonstrates that even in the adult brain sensorimotor
representations can maintain a rather high degree of plasticity in order to adjust neural
representations to altered environmental conditions. In this respect, similarities exist with the
effects of perceptual learning (see Fahle, 2008, in press; Gilbert et al., 2001).
Neural substrates of prism adaptation
Although a large number of studies on prism adaptation have been conducted in both humans
and non-human primates, the neural mechanisms and underlying neural substrates remain to
be elucidated. Converging experimental evidence from both monkeys (e.g. Baizer et al., 1999;
Kitazawa & Yin, 2002) and humans (e.g. Luaute et al., 2006; Martin et al., 1996a; 2002;
Pisella et al., 2005; Weiner et al., 1983) suggests a crucial role of the cerebellum during the

Studie 1: Vollständigkeit des Adaptationsprozesses 30
adaptation process, which is consistent with the view that the cerebellum is a major
subcortical structure for motor learning and movement adaptation (for reviews see Doyon &
Benali, 2005; Marr, 1969; Robinson, 1995; Shadmehr & Wise, 2005).
Furthermore, two studies on a patient with bilateral damage to the posterior parietal cortex
(PPC) found specific deficits in the ability to adapt to the optical displacement induced by
prisms, suggesting that the PPC is involved in the adaptive spatial updating of sensorimotor
representations (Newport et al., 2006; Newport & Jackson, 2006). In line with these findings,
a PET study on prism adaptation by Clower et al. (1996) reported adaptation-related
activation in a focal region of the PPC contralateral to the reaching limb confirming the
functional role of the PPC in the integration and alignment of different sensory (e.g. visual
and proprioceptive) input signals for motor output (Scherberger & Andersen, 2003; Shadmehr
& Wise, 2005; but cf. Redding et al., 2005). In an animal study, Kurata & Hoshi (1999)
demonstrated that monkeys with temporary inactivation of the ventral premotor cortex (PMv)
had deficits in prism adaptation. All the brain areas mentioned are known to be involved in
motor control.
In addition, a study on long-term adaptation (over several months) to a left-right reversal of
the visual field in monkeys found functional changes of neurons’ properties in primary visual
cortex (V1) indicating that long-term adaptation to reversal of the visual field may involve
large-scale reorganization – probably entirely through recalibration – even on a very early
level of visual cortical processing (Sugita, 1996).
In summary, prism adaptation involves a functional realignment of different sensory input
signals which is likely accomplished by plasticity of an extended cortico-cerebellar network
in the brain, with recalibration possibly achieved primarily in cerebral cortex and realignment
realized primarily in cerebellum (Redding et al., 2005).
Speed and extent of prism adaptation
In most studies examining prism adaptation in humans, the number of trials during the
adaptation period varied between 20 and 40 movements (e.g. Fernandez-Ruiz et al., 2000;
2003; 2004; Fernandez-Ruiz & Diaz 1999; Kitazawa et al., 1995; Martin et al., 1996a; 1996b;
Roller et al., 2001). Typically, this period of visuomotor training was sufficient for adaptation
to take place since error reduction occurred and aftereffects were observed upon removal of
the prisms. However, when we conducted pilot experiments in which subjects had to throw a
ball towards a visual target while wearing prism goggles, movements deviated laterally in the
direction of the prismatic displacement even at the end of the adaptation period comprising 30

Studie 1: Vollständigkeit des Adaptationsprozesses 31
trials. That is, although subjects were able to adjust their throws on a trial by trial basis and
movements gradually approached the target, throwing accuracy at the end of the prism
condition was clearly worse than in the baseline condition due to an incomplete compensation
of the prism effect.
We wondered whether this incompleteness of adaptation in the throwing task was due to an
insufficient number of trials during the adaptation period and therefore increased the number
of trials substantially to ensure extensive visuomotor training. A remaining lateral deviation of
movements in the direction of the prismatic displacement following an extensive period of
training would indicate that the remaining error was not caused primarily by an insufficient
number of trials. Rather, different amounts of adaptation could result from different extents of
neural plasticity in the task-related sensorimotor representations.
In order to control for effects of task-specificity, completeness of prism adaptation was
examined in two different visuomotor tasks, namely pointing and throwing. These two tasks
test movements of different skill level, and of different distances. Hence, investigating both
tasks allows to compare the completeness of prism adaptation to near space, a high level skill,
with a low level skill to far space. The results indeed highlight important differences between
adaptation under these different conditions.
Our results demonstrate a clearly incomplete adaptation process. Spatial deviation remained
significant in the Throwing Experiment even after extensive adaptation. On the other hand, the
remaining lateral deviation in the Pointing Experiment failed to reach significance indicating
complete adaptation. To the best of our knowledge, this incompleteness of adaptation has so
far not received any attempt at explanation. We here conclude that sensorimotor
representations for pointing exhibit more experience-dependent plasticity in order to ensure
high precision of visually-guided movements in the peripersonal workspace. More
importantly, the prismatic displacement is not completely compensated even after extensive
training for throwing movements. This remaining error indicates a limited integration capacity
of the neural processes underlying this type of plasticity.
Methods
Experimental setup and procedure
In the Throwing Experiment, subjects’ task was to throw softballs (24 g, 5.0 cm diameter) as
accurately as possible towards a visual target on a 1.5 m x 1.5 m wide wall. The target was a
blue spot (2.0 cm diameter) attached to the wall at a height of 156 cm. Subjects stood upright

Studie 1: Vollständigkeit des Adaptationsprozesses 32
with their mid-sagittal plane aligned with the target at a viewing distance of 2.0 m (Fig. 5.1a).
With their head being unrestrained, subjects always had an unobstructed binocular view of the
target. The experiment was conducted under daylight illumination.
The wall was layered with Velcro material and the softball adhered to the wall after each
throw. In order to assess spatial accuracy of the throws, a photo was taken after each throw
using a digital camera which was kept at a fixed location during the entire experiment. A
Matlab-based program (Matlab release 12.1) was used to readout the jpeg-files generated by
the camera and to compute the horizontal and vertical distances between the target and the
position of the ball in a metric system (cm) by means of trigonometry. Spatial accuracy was
around 1 cm.
The wedge prisms of high optical quality (Carl Zeiss, Oberkochen/Germany) used shifted the
visual image laterally by 17 deg. either to the right (base-left prisms, 30 diopters) or left
(base-right prisms). Subjects had to perform overhand throws always using the right arm
while fixating the visual target and were instructed not to look at their arm when throwing in
order to avoid on-line corrections of the pre-planned movement. After a brief instruction,
subjects were asked to perform about 10 test throws to get used to the task.
The Throwing Experiment consisted of three conditions in the following order: i) pre-prism
condition (PRE; baseline measurement, prisms off), ii) prism condition (PRISM; adaptation,
prisms on) and iii) post-prism condition (POST; aftereffect, prisms off). In each condition,
120 throws had to be performed to ensure extensive training of the visuomotor task.
Throwing movements were moreover studied in an additional Control Experiment in which
the number of trials per condition was modified. Specifically, after a PRE condition of 60
throws, subjects had to execute 240 throws while wearing a prism goggle (PRISM condition);
the POST condition thereafter included 90 throws. This experiment was conducted only with
rightward shifting prisms and aimed to evaluate the effects of a further extended adaptation
period on the completeness of prism adaptation.
The spatial offset between the target and the impact of the ball was clearly visible after each
throw, hence immediate visual feedback about the precision of the throw was provided.
Between conditions, subjects had to keep the eyes closed in order to prevent habituation and
to minimize spatial reorientation which could decrease both the magnitude of the prism effect
and the aftereffect.

Studie 1: Vollständigkeit des Adaptationsprozesses 33
Fig. 5.1 A) Experimental setup of the Throwing Experiment in which subjects stood upright in front of a Velcro-layered wall at a distance of 2 m. Subjects’ body midline was centered with the target. The task was to throw a ball to the target. After each throw, a photo was taken using a digital camera. B) In the Pointing Experiment, subjects were sitting at a table and performed pointing movements towards a visual target which was fixed at the table’s edge. Subjects could not correct their movement on-line once it had started because of the table top being opaque but received direct visual feedback about their index finger’s position at the end of the trajectory. A PC-driven ultrasound measuring unit was placed in front of the table for assessing the movements’ spatial accuracy. Target and subject’s index finger were equipped with an ultrasound marker (microphone). C) Experimental groups tested.
In the Pointing Experiment, subjects had to perform speedy but accurate ballistic pointing
movements towards a visual target always using the right arm (Fig. 5.1b). During the
experiment, subjects were sitting at a table with their mid-sagittal plane aligned with the target
which was a small white disk fixed at the table’s edge at a distance of 65 cm from the subject.
Since the table-top was opaque, subjects could not see and hence not correct their movements
on-line once the movement had started. Consequently, the trajectory of the arm had to be
planned purely feed-forward on the basis of visual information about the target location in
space. Subjects viewed the target binocularly and had visual feedback about the position of
the index finger in relation to the target only at the end of the arm movement and could
therefore adjust the trajectory on a trial by trial basis, based on this trial to trial knowledge of
their results.
The movements’ accuracy was assessed using a PC-driven ultrasound measuring system
operating with high spatial resolution based on the travelling time of ultrasound pulses (Zebris
Medical, Isny/Germany). Ultrasound signals were sent out by an ultrasound measuring unit
and recorded by ultrasound microphones being attached to both the target and the subject’s

Studie 1: Vollständigkeit des Adaptationsprozesses 34
index finger. By means of a computer program, the horizontal distance between the index
finger and the target position was quantified for the endpoint of each individual movement
(measuring rate: 100 Hz).
Similar to the Throwing Experiment, the Pointing Experiment consisted of a PRE, PRISM,
and POST condition, each with 120 movements. Subjects we instructed to fixate the target
and to keep the eyes closed between conditions in order to prevent visual reorientation in
space. The wedge prisms used shifted the visual image laterally by 17 deg. either to the right
or left.
Experimental groups
Five experimental groups of right-handed human subjects were formed (Fig. 5.1c). For the
Throwing Experiment, two groups of 8 subjects each were studied: Group 1 (1 male, 7 female;
mean age 23.5 years) was exposed to rightward shifting prisms while group 2 (3 male, 5
female; mean age 23.4 years) was exposed to leftward shifting prisms. For the Pointing
Experiment, another two groups of 10 subjects each were formed. Members of group 3 (3
male, 7 female; mean age 23.9 years) were exposed to rightward shifting prisms while
members of group 4 (1 male, 9 female; mean age 24.1 years) wore leftward shifting ones in
the PRISM condition. In the Control Experiment, another 10 subjects (5 male, 5 female; mean
age 24.5 years) were exposed to rightward shifting prisms.
Subjects reported no history of brain or eye disorders and had normal or corrected-to-normal
visual acuity. Prior to participation, subjects gave informed consent in accordance with the
“Declaration of Helsinki” without knowledge of the exact purpose of the study. This type of
experiments was approved by the ethics committee of Bremen University.
Data analysis
We calculated the horizontal distance between the endpoint of the movement (Throwing
Experiment: position of the ball on the wall; Pointing Experiment: top of index finger at the
end of the trajectory) and the target for each movement in each condition and subject.
For further analysis, the data were baseline corrected: that is, we subtracted the mean of
deviations in the PRE condition from each data point (see also Clower & Boussaoud, 2000).
Baseline correction was conducted for each condition of each subject individually. The mean
of the PRE condition mirrors a subject’s general task-related visuomotor accuracy and
indicates a potential spatial bias of the movements. This bias averaged -0.7 cm (± 0.2 SEM) in
the Throwing Experiment, -0.8 cm (± 0.1 SEM) in the Pointing Experiment, and -0.3 cm (±

Studie 1: Vollständigkeit des Adaptationsprozesses 35
0.3 SEM) in the Control Experiment. The baseline correction normalized the data by
removing any spatial bias which could mask the effects of the prismatic displacement on the
movements’ accuracy and also allowed a better comparison between different subjects since
each set of data was normalized individually. A third advantage was the possibility of
conventional statistical analysis, i.e. to test the data against the value zero corresponding to
the individually normalized target of the task. Consequently, all data shown are baseline
corrected.
In order to analyze the data in terms of completeness of the adaptation process, we divided
each condition (PRE, PRISM, POST) into blocks of 30 subsequent throws each and calculated
the mean amount of target deviation for each block and every subject individually. Blocks
were averaged across subjects and tested against the value of zero (i.e. no deviation from
normalized target) using a two-sided t-test. In the analysis, we focused on the last block of the
PRISM and POST conditions since this value reflects the remaining lateral deviation.
Specifically, a last block of the PRISM condition differing significantly from zero indicates a
remaining lateral deviation at the end of the adaptation period hence suggesting incomplete
adaptation.
Results
Time course of prism adaptation in the Throwing and Pointing Experiment
Averaged data of experimental groups 1-4 are presented in Fig. 5.2 and Fig. 5.3. The lateral
deviation of movements’ endpoints from the target is plotted as a function of trial number in
the PRE, PRISM, and POST conditions, where negative values represent a leftward deviation
and positive values indicate a rightward deviation.
In the PRE condition, the lateral deviation from the target was close to zero in all four
experimental groups suggesting accurate motor performance. At the beginning of the PRISM
condition, movements deviated according to the direction of the prismatic deflection
introduced. Group 1 (throwing task, rightward shifting prisms) showed an initial prism effect
of +42.8 cm (± 4.2) decreasing to +4.3 cm (± 2.6) (Fig. 5.3a, upper panel). The negative
aftereffect averaged -38.8 cm (± 2.0) and declined to +3.5 cm (± 1.2). The decrease of
throwing movements’ lateral deviation in the course of both the PRISM and the POST
condition was statistically highly significant as revealed by a paired two-sided t-test
comparing trial 1 and trial 120 (p < .001).

Studie 1: Vollständigkeit des Adaptationsprozesses 36
Fig. 5.2 Lateral deviation of movements from the target in the PRE, PRISM and POST condition as a function of movement number in the (A) Throwing Experiment and the (B) Pointing Experiment for the rightward shifting prisms groups (upper panel) and the leftward shifting prisms groups (lower panel). Positive values on the ordinate indicate a rightward deviation from the target whereas negative values denote a leftward one (units in Throwing Experiment: cm; Pointing Experiment: mm). All data shown are baseline-corrected (means and SEM).
Group 2 (throwing task, leftward shifting prisms) exhibited an initial prism effect (throw 1) of
-41.1 cm (± 2.2 SEM) which was reduced to -3.2 cm (± 2.4) at the end of the PRISM
condition (throw 120) (Fig. 5.3a, lower panel). Upon removal of the prisms (POST condition,
throw 1), throwing movements initially deviated by +36.7 cm (± 3.7) to the right indicating a
negative aftereffect which was reduced to -0.8 cm (± 2.4) at the end of the POST condition
(throw 120). Again, the reduction of lateral deviations was highly significant in both
conditions (p < .001).
In the group of subjects wearing the rightward shifting prisms in the Pointing Experiment
(group 3), the prism effect declined from +166.5 mm (± 12.6) to -13.3 mm (± 7.9) in the
PRISM condition and the aftereffect diminished from -58.1 mm (± 11.0) to -14.2 mm (± 6.7)

Studie 1: Vollständigkeit des Adaptationsprozesses 37
(Fig. 5.3b, upper panel). Error reduction was significant in both the PRISM condition (p <
.001) and the POST condition (p < .01).
In group 4 (pointing task, leftward shifting prisms), the prism effect amounted to -78.8 mm (±
22.4) decreasing to -12.4 mm (± 8.4) (Fig. 5.3b, lower panel). Upon removal of the prisms,
the aftereffect reached +117.3 mm (± 11.1) and declined to -5.2 mm (± 6.1) at the end of the
POST condition. In both conditions, the decrease of the pointing movements’ lateral deviation
in the course of training was statistically significant (PRISM condition: p < .01; POST
condition: p < .001).
Fig. 5.3 Mean lateral deviation from the target of the first movement (black bars) and the last movement (white bars) in the PRISM and POST condition of the (A) Throwing Experiment and the (B) Pointing Experimentfor the rightward shifting prisms groups (upper panel) and the leftward shifting prisms groups (lower panel) (means and SEM). Asterisks indicate a statistically significant difference of the lateral deviation between the first movement and the last movement as revealed by a paired t-test.
To sum up, each experimental group showed a marked lateral deviation of movements in the
direction of the prismatic displacement once the prisms were introduced. The lateral error was
significantly reduced (Throwing Experiment) or even removed (Pointing Experiment) in the
course of visuomotor training. Conversely, upon removal of the prisms, an aftereffect in the
opposite direction emerged which likewise disappeared in the course of the POST condition.

Studie 1: Vollständigkeit des Adaptationsprozesses 38
Completeness of prism adaptation: block analysis
To assess whether complete adaptation was accomplished through practice in the Throwing
and Pointing Experiments, we divided each condition into blocks of 30 subsequent
movements and calculated the mean lateral deviation from the target for each block
individually. Averaged data are shown in Fig. 5.4.
Fig. 5.4 Block analysis of movements’ mean lateral deviation from the target in the PRE, PRISM and POST conditions in the (A) Throwing Experiment and the (B) Pointing Experiment for the rightward shifting prisms groups (upper panel) and the leftward shifting prisms groups (lower panel). For each subject, each condition was divided into four blocks of 30 subsequent movements and block means were calculated. Data shown represent group averages (means and SEM). Asterisks indicate blocks differing significantly from zero as revealed by a t-test.
In group 1 (throwing task, rightward shifting prisms; Fig. 5.4a, upper panel) no significant
lateral deviation was found for any block of the PRE condition (p > .05). However, in the
PRISM condition that followed, each block differed highly significantly from zero (p < .001)
and the block-averaged lateral deviation was explicit even in block 4 where it reached +5.0
cm (± 0.9). In the POST condition, the lateral deviation amounted to -10.8 cm (± 1.0) in block

Studie 1: Vollständigkeit des Adaptationsprozesses 39
1 and -2.6 cm (± 0.5) in block 2 yielding a significant difference from zero (block 1: p < .001;
block 2: p < .01). In contrast, the mean lateral deviation of blocks 3 and 4 was very close to
zero (+0.1 cm in both blocks) and statistical analysis did not show a significant difference
from zero (p > .05).
In group 2 (throwing task, leftward shifting prisms; Fig. 5.4a, lower panel), the first block of
the PRE condition differed significantly from zero (p < .05) which was presumably due to a
larger movement variability at the beginning of the condition. In the remaining three blocks of
the PRE condition, movements’ mean lateral deviation was negligible. In the PRISM
condition, the mean of block 1 reached -12.6 cm (± 1.1 SEM), sharply decreasing to -3.6 cm
(± 1.0) in block 2, further decreasing to -1.8 cm (± 0.8) in block 3 and increasing to -2.3 cm (±
0.7) in block 4. All blocks were found to differ significantly from zero (block 1: p < .001;
block 2: p < .01; block 3: p < .05; block 4: p < .05). In the POST condition, the average
aftereffect was +11.5 cm (± 1.2) in block 1 and decreased to +2.6 cm (± 0.8) in block 2,
reached +0.4 cm (± 0.6) in block 3 and +0.2 cm (± 0.4) in block 4. While block 1 (p < .001)
and block 2 (p < .05) differed significantly from zero, this was not the case for the last two
blocks of the POST condition (p > .05).
Both groups of the Throwing Experiment showed an identical pattern of results: in the PRISM
condition, the mean lateral deviation decreased across blocks but a significant lateral
deviation was still present in the last block suggesting that subjects did not regain baseline-
like motor accuracy in the course of training. Conversely, in the POST condition only the first
two blocks differed significantly from zero and throwing movements did not deviate laterally
in the last two blocks. Here, subjects’ visuomotor performance returned to the pre-prism
accuracy.
In group 3 (pointing task, rightward shifting prisms; Fig. 5.4b, upper panel), no block of the
PRE condition differed significantly from zero (p > .05) and the mean lateral deviation was
marginal. In the PRISM condition, block 1 reached +20.6 mm (± 6.9) and showed a
significant difference from zero (p < .05) whereas in the other blocks the averaged lateral
deviation was small and did not differ significantly from zero (p > .05). In the POST
condition, only the first block differed significantly from zero (-18.3 mm ± 5.3; p < .01) while
the remaining blocks did not (p > .05)
Group 4 (pointing task, leftward shifting prisms; Fig. 5.4b, lower panel) showed a similar
pattern of results as the former group. The results of the four blocks of the PRE condition
were close to the normalized target position – although blocks 1 and 4 yielded a significant

Studie 1: Vollständigkeit des Adaptationsprozesses 40
difference from zero (block 1: p < .05; block 4: p < .01). In the PRISM condition, the
averaged lateral deviation of pointing movements decreased stepwise from block 1 (-30.6 mm
± 9.4) to block 4 (-4.9 mm ± 3.3). While the results of the first three blocks differed
significantly from zero (block 1: p < .01; block 2: p < .001; block 3: p < .05), this did not
apply for block 4 (p > .05). In the POST condition, only block 1 with a mean lateral deviation
of +13.3 mm (± 4.3) differed significantly from zero (p < .05), whereas the remaining blocks
were close to zero and without a significant deviation (p > .05).
In the PRISM condition, both groups of the Pointing Experiment showed a similar pattern of
results: while the first blocks exhibited a considerable lateral deviation due to the prismatic
deflection introduced, the last block of 30 pointing movements did not differ significantly
from zero. That is, subjects’ pointing accuracy returned to a pre-prism exposure level in the
course of the adaptation period. This result represents the most important difference to the
Throwing Experiment in which a significant lateral deviation remained even at the end of the
PRISM condition. Furthermore, within the course of the POST condition, pointing
movements rapidly reached baseline-like accuracy since the lateral deviation was no longer
significantly different from zero in blocks 2-4. This is essentially identical to the results of the
Throwing Experiment.
Control Experiment: extended visuomotor training in the PRISM condition
Similar to the Throwing Experiment, subjects had to perform throwing movements in the
Control Experiment (Fig. 5.5). In the PRISM condition, the number of throws was doubled to
240 trials in order to evaluate the effect of an extended training period on the completeness of
the adaptation process. In the PRE condition (60 throws), the lateral deviation of throws was
close to zero (Fig. 5.5a). When the prisms were introduced (PRISM condition), the first throw
deviated strongly by +49.1 cm (± 1.7) to the right but subjects decreased their movements’
lateral errors significantly (p < .001; paired two-sided t-test comparing throw 1 and 240) on a
trial by trial basis during the PRISM condition, i.e. they adapted. Removal of the prisms
(POST condition) led to an aftereffect of -43.7 cm in the first throw but the lateral deviation
of throws was significantly (p < .001) reduced by throw 90. An exponential decay function
fitted to the averaged data of the PRISM condition showed an offset of +2.9 cm (Tab. 5.1).
The adaptation process observed in the Control Experiment was slightly slower than in the
conventional Throwing Experiment.
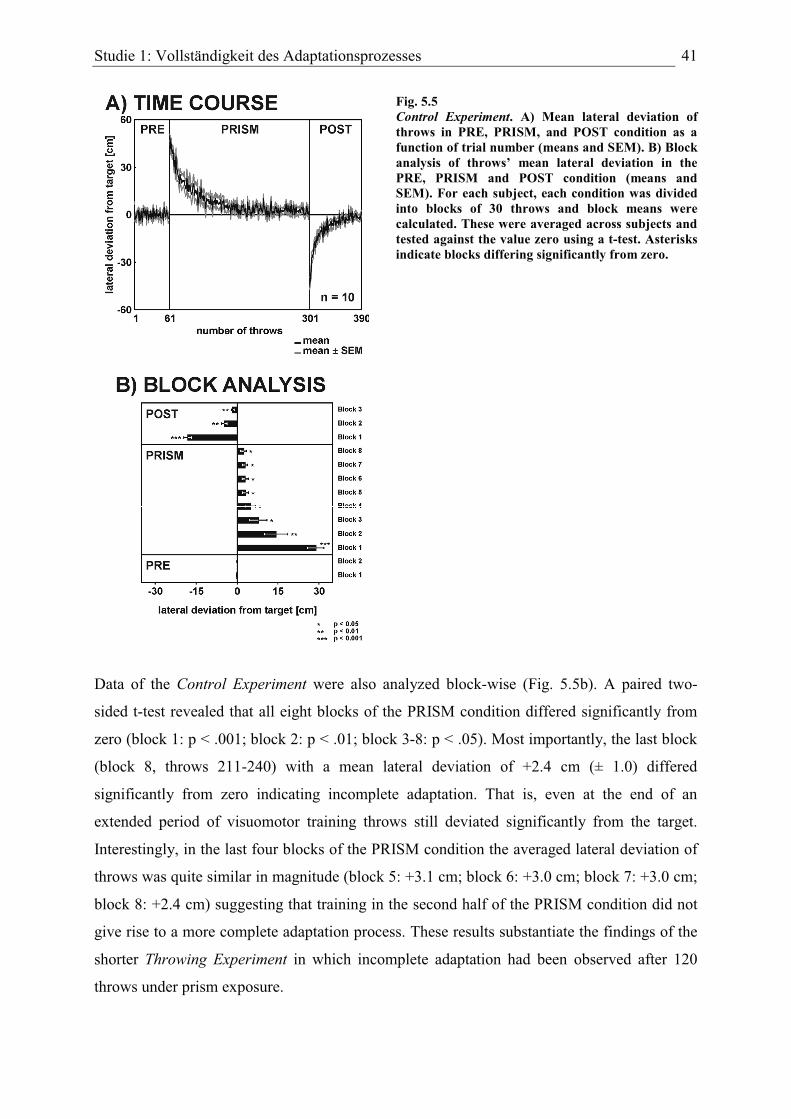
Studie 1: Vollständigkeit des Adaptationsprozesses 41
Fig. 5.5 Control Experiment. A) Mean lateral deviation of throws in PRE, PRISM, and POST condition as a function of trial number (means and SEM). B) Block analysis of throws’ mean lateral deviation in the PRE, PRISM and POST condition (means and SEM). For each subject, each condition was divided into blocks of 30 throws and block means were calculated. These were averaged across subjects and tested against the value zero using a t-test. Asterisks indicate blocks differing significantly from zero.
Data of the Control Experiment were also analyzed block-wise (Fig. 5.5b). A paired two-
sided t-test revealed that all eight blocks of the PRISM condition differed significantly from
zero (block 1: p < .001; block 2: p < .01; block 3-8: p < .05). Most importantly, the last block
(block 8, throws 211-240) with a mean lateral deviation of +2.4 cm (± 1.0) differed
significantly from zero indicating incomplete adaptation. That is, even at the end of an
extended period of visuomotor training throws still deviated significantly from the target.
Interestingly, in the last four blocks of the PRISM condition the averaged lateral deviation of
throws was quite similar in magnitude (block 5: +3.1 cm; block 6: +3.0 cm; block 7: +3.0 cm;
block 8: +2.4 cm) suggesting that training in the second half of the PRISM condition did not
give rise to a more complete adaptation process. These results substantiate the findings of the
shorter Throwing Experiment in which incomplete adaptation had been observed after 120
throws under prism exposure.
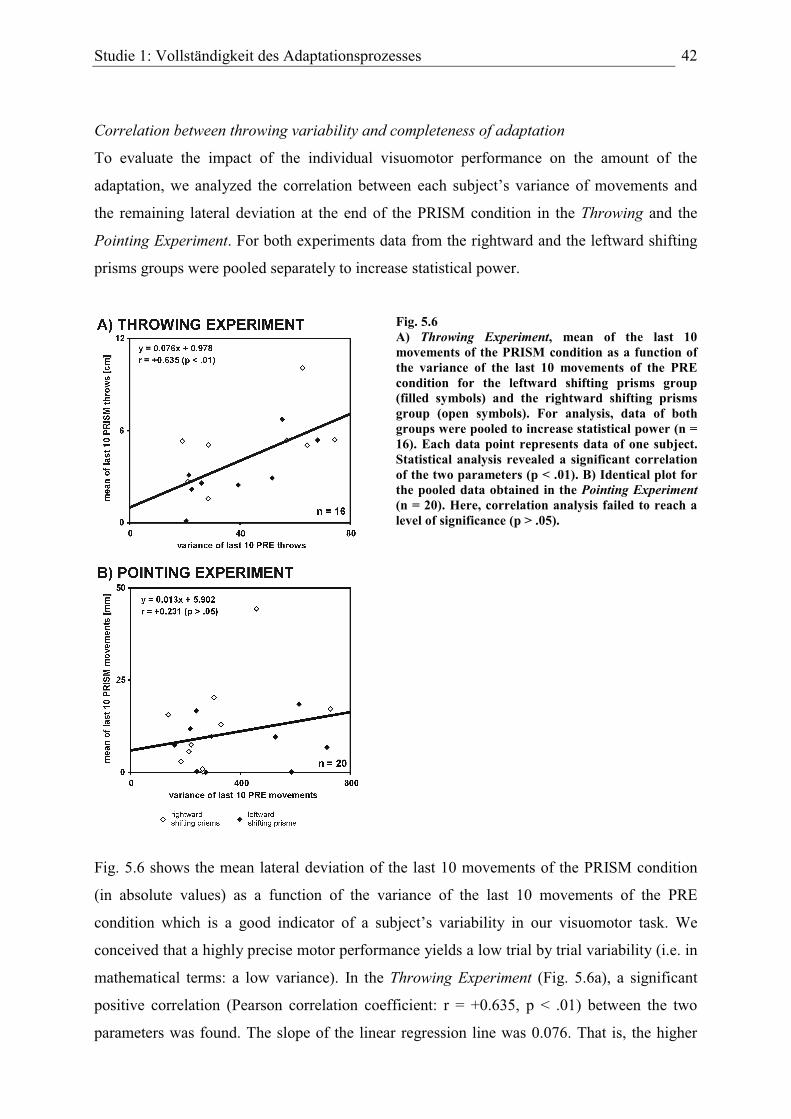
Studie 1: Vollständigkeit des Adaptationsprozesses 42
Correlation between throwing variability and completeness of adaptation
To evaluate the impact of the individual visuomotor performance on the amount of the
adaptation, we analyzed the correlation between each subject’s variance of movements and
the remaining lateral deviation at the end of the PRISM condition in the Throwing and the
Pointing Experiment. For both experiments data from the rightward and the leftward shifting
prisms groups were pooled separately to increase statistical power.
Fig. 5.6 A) Throwing Experiment, mean of the last 10 movements of the PRISM condition as a function of the variance of the last 10 movements of the PRE condition for the leftward shifting prisms group (filled symbols) and the rightward shifting prisms group (open symbols). For analysis, data of both groups were pooled to increase statistical power (n = 16). Each data point represents data of one subject. Statistical analysis revealed a significant correlation of the two parameters (p < .01). B) Identical plot for the pooled data obtained in the Pointing Experiment(n = 20). Here, correlation analysis failed to reach a level of significance (p > .05).
Fig. 5.6 shows the mean lateral deviation of the last 10 movements of the PRISM condition
(in absolute values) as a function of the variance of the last 10 movements of the PRE
condition which is a good indicator of a subject’s variability in our visuomotor task. We
conceived that a highly precise motor performance yields a low trial by trial variability (i.e. in
mathematical terms: a low variance). In the Throwing Experiment (Fig. 5.6a), a significant
positive correlation (Pearson correlation coefficient: r = +0.635, p < .01) between the two
parameters was found. The slope of the linear regression line was 0.076. That is, the higher

Studie 1: Vollständigkeit des Adaptationsprozesses 43
the general variability of throws in the PRE condition, the higher was the remaining lateral
deviation of throws in the PRISM condition, and vice versa. Thus, the amount of remaining
error of an individual seems to be related to this individual’s throwing variability. In contrast,
no significant correlation existed in the Pointing Experiment (p > .05; Fig. 5.6b). Here, the
movements’ variance was only weakly correlated with the amount of adaptation (r = +0.231)
and the slope of the corresponding linear regression line was shallow (0.013).
Tab. 5.1 Averaged data of the PRISM and POST condition from each experimental group were fitted with an exponential decay function of the form . Whereas a1 represents an estimate of the spatial error of the initial movement, a2 indicates the speed of the adaptation (PRISM) and de-adaptation (POST) process and a3 gives an estimate of the remaining spatial offset of the movements at the end of the condition.
Discussion
In the present study, we examined the effects of extensive training of a visuomotor task on the
completeness of prism adaptation in human subjects and found that adaptation was
incomplete in the Throwing Experiment. Adaptation in the Pointing Experiment was more
complete, with no significant error remaining at the end of the (much faster) adaptation
process. The amount of prism adaptation therefore depended on the precise visuomotor task
performed but was unrelated to the direction of the visual world’s lateral displacement
induced by the prisms. That is, both the rightward and the leftward shifting prisms produced
incomplete adaptation in the Throwing Experiment. The results of the Control Experiment in
which the number of throws during prism exposure was doubled to 240 are consistent with the
findings of the Throwing Experiment: Even after 240 throws a significant lateral deviation
was present strongly indicating that after a rather fast period of improvement, even a very
large number of throws does not produce a perfect adjustment of sensory and motor
coordinate systems. Even after this extensive practice of 240 throws the mean remaining error
was as large as almost 30 mm, a deviation that can be easily perceived – and should be
corrected.
312)( aeaxf xa +=

Studie 1: Vollständigkeit des Adaptationsprozesses 44
Accurate pre-exposure visuomotor performance was evident in all groups since the average
lateral deviation of both throwing and pointing movements was small in the PRE condition.
Introducing the prisms initially shifted movements away from the target but subjects rapidly
recovered visuomotor accuracy in the course of training. During exposure, the movements’
lateral deviation decayed exponentially and this process of error reduction is supposed to rely
on a rapidly operating strategic recalibration mechanism and a slower realignment mechanism
as suggested by the model of Redding & Wallace (2002). In all groups studied, the reduction
of the movements’ lateral deviation within the PRISM condition was pronounced and
statistically reliable (see Fig. 5.3) suggesting that the strategic recalibration mechanism
worked well. As expected and well documented by earlier studies, a distinct negative
aftereffect in the direction opposite to the prismatic displacement occurred as soon as the
prisms were removed. In all groups, the negative aftereffect completely disappeared in the
course of training, consistent with the view that newly acquired visuospatial coordinations are
more labile (Fernandez-Ruiz & Diaz, 1999).
Speed and extent of prism adaptation depend on the visuomotor task
Precise pre-exposure visuomotor performance, a pronounced prism effect and a significant
error reduction during exposure as well as the appearance of a negative aftereffect are well
documented results in the prism adaptation literature – so what is the main novel finding of
our study?
As to be expected, (near) complete prism adaptation took place in the Pointing Experiment.
The most surprising result is that even extensive training did not yield complete adaptation for
throwing movements. Although subjects were able to reduce their movements’ lateral error in
the course of the PRISM condition, they did not reach pre-exposure accuracy levels even
through extensive throwing training. Rather, throwing movements deviated systematically in
the direction of the prismatic displacement throughout the adaptation period – irrespective of
whether this period covered 120 or even 240 throws. Hence, prism adaptation was clearly
incomplete for throwing movements.
An exponential decay function fitted to the averaged data (Tab. 5.1) of the Throwing and
Control Experiment shows that the results of the PRISM condition asymptotically approach
not zero deviation but maintain a certain “persisting deviation” that is related to a subject’s
variability of results. The offset of the decay function amounted to +58 mm (r² = 0.93) in the
rightward shifting prisms group and to -25 mm (r² = 0.92) in the leftward shifting prisms

Studie 1: Vollständigkeit des Adaptationsprozesses 45
groups. In the Control Experiment, after 240 throws the decay function still exhibited an
offset of +29 mm (r² = 0.96). At first glance, this does not seem to be a large effect but note
that +29 mm is the value the function is approaching to for an infinite number of throws – a
value which is clearly offset relative to the target.
In contrast, the same mathematical procedure applied to the PRISM condition data of the
Pointing Experiment yielded an offset of only -3 mm (r² = 0.92) for the rightward shifting
prisms group and -7 mm (r² = 0.90) for the leftward shifting prisms group. These small
deviations failed to reach significance.
Which other factors are conceived to affect the adaptation process? Kitazawa et al. (1995)
have shown that both the rate and the amount of prism adaptation (measured as the magnitude
of the aftereffect) critically depend on the availability of visual feedback information after the
completion of the movement. In humans, a delay of > 50 ms between the end of a reaching
movement and the onset of visual feedback significantly reduced both the rate and the amount
of prism adaptation. Therefore, the adaptation mechanism primarily accesses visual feedback
information within a temporal window of ~ 50 ms after movement completion. If visual
feedback is delayed by more than 50 ms, the adaptation process becomes slower and less
complete, i.e. aftereffects decrease (but note that aftereffects are still observed). These results
were confirmed in a monkey study by Kitazawa & Yin (2002). Apart from the fact that the
Kitazawa et al. study measured the amount of prism adaptation indirectly by taking the
magnitude of the aftereffect, the task-related difference in the completeness of the adaptation
process observed in our data cannot easily be explained by means of delayed visual feedback
information about the outcome of the movement since immediate visual feedback was always
provided in both tasks. Subjects perceived the spatial offset between the endpoint of the
movement and the target as soon as the movement was completed, hence there was no delay
of visual information apart from the “flight time” of the balls in the Throwing Experiment.
This lack of feedback information during the flight time is unlikely to explain incomplete
prism adaptation in the Throwing Experiment.
In a recent study, Michel et al. (2007) compared the magnitude of the negative aftereffect in
two groups of human subjects: The first group was adapted to prisms in a stepwise manner
from 2 deg. to 10 deg. optical displacement while the second group was adapted in a single
step to a 10 deg. optical displacement. While the first group was not aware of the progressive
prismatic shift during exposure to the prisms, the second group experienced the abrupt shift of
the visual world when exposed to the prisms. Most importantly, the negative aftereffect was

Studie 1: Vollständigkeit des Adaptationsprozesses 46
significantly larger in the “unaware” (multiple step) group as compared to the “aware” (single
step) group. This led to the conclusion that unawareness of prismatic exposure enhances
adaptation while awareness is suggested to play a detrimental role on the adaptation process.
Following this line of argumentation, complete adaptation might be accomplished by active
extensive training of a visuomotor task while leaving the subjects unaware of the prismatic
shift induced. In our experiments, we employed a horizontal shift of the visual world by 17
deg. either to the right or to the left and this large and easily detectable amount of visual
displacement made subjects aware of the prismatic shift. This applies for the Throwing
Experiment as well as for the Pointing Experiment and hence a difference in the “state of
awareness” of the prismatic shift cannot explain why complete adaptation was only observed
in the Pointing Experiment where error reduction during the PRISM condition led to
visuomotor accuracy similar to the PRE condition. That is, although subjects viewed the
target as laterally displaced through the prisms, their pointing movements became as precise
as without prisms during the course of training. In contrast, error reduction during prism
exposure in the Throwing Experiment did not reach pre-exposure levels of visuomotor
accuracy.
Plasticity of visuomotor representations
One possible explanation for the task-related difference in the completeness of prism
adaptation could be different extents of neural plasticity in the task-related visuomotor
representations. The amount of practice-induced reorganization of visuomotor representations
may critically depend upon the spatial requirements in the task-related workspace.
Generally, the near (peripersonal) space of a person within arms’ reach is primarily concerned
with highly skilled motor activities (that is why peripersonal space is also termed “action
space”) whereas the (extrapersonal) space beyond arms’ reach is mainly devoted to perception
(Previc, 1998). It has been shown that prism adaptation can affect neural representations of
both peripersonal and extrapersonal space (Berberovic & Mattingley, 2003).
In our experiments, subjects’ workspace differed according to the task performed: while
throwing movements were executed within extrapersonal space, the pointing movements
covered peripersonal space. A precise eye-hand coordination in peripersonal space is of high
behavioral relevance and requires the continuous and spatially fine-tuned adjustment of the
underlying visuomotor representations.
When the prisms are introduced, precise eye-hand coordination is suddenly disrupted and
pointing movements initially exhibit a distinct lateral deviation. An “internal” error signal (see

Studie 1: Vollständigkeit des Adaptationsprozesses 47
next section) triggers the updating of visuomotor representations for pointing, a process
requiring neural plasticity. The experience-dependent adjustment of visuomotor
representations progresses until visuomotor performance is as accurate as it was before the
displacement of the visual world.
The incompleteness of prism adaptation observed especially for the throwing movements in
extrapersonal space may be attributed to a lesser extent of plasticity in the corresponding
visuomotor representation possibly due to a less direct coupling between visual feedback
regarding external objects (the position of the ball on the wall) versus body parts (position of
the hand) or else to the difference between a high level skill (pointing) and a low level one
(throwing). To be clear: there was substantial adaptation to the prismatic deflection during the
Throwing Experiment since both error reduction and aftereffects were observed. The main
difference to the Pointing Experiment is that the completeness of spatial compensation during
prism exposure was clearly less pronounced in the Throwing Experiment. Therefore, the
visuomotor representations for throwing movements were not as fast and precisely adjusted
by visual feedback as it was the case for the pointing representations.
We cannot completely exclude the possibility that a further increase in the number of
throwing trials during exposure could in principle lead to complete prism adaptation, but as
demonstrated in the Control Experiment, even doubling the number of training throws did not
lead to complete adaptation and deviations did not even converge towards zero remaining
error. Instead, the asymptotic adaptation curves are clearly offset from zero (Tab. 5.1).
Throwing variability limits complete prism adaptation if the neural integration capacity is
finite
In general, even skilled or well-practiced movements vary from trial to trial, yielding a certain
degree of movement variability which may result from variability during the execution of a
specific movement (e.g. van Beers et al., 2004) or from variability during motor preparation
or from both. Churchland et al. (2006) demonstrated in a monkey single-cell study that a
substantial amount (> 50 %) of movement variability resulted from neural variability during
movement preparation in cortical motor areas.
We found a significant positive correlation between the individual’s (general) throwing
variability and the remaining lateral deviation of throws at the end of the PRISM condition
(see Fig. 5.6). With increasing movement variability, the remaining lateral error increased in a
linear way. Conversely, the lower the variability of movements the lower was the remaining

Studie 1: Vollständigkeit des Adaptationsprozesses 48
lateral error at the end of the adaptation period. What may be the functional relevance of this
observation?
We hypothesize that the size of the remaining error critically depends upon a neural error
signal that subjects have to generate by integrating throwing errors over a number of
sequential trials. In individuals exhibiting a high degree of throwing variability, the
“averaged” error signal is corrupted by a large proportion of noise and thus the remaining
lateral deviation at the end of the adaptation period is large. On the other hand, individuals
with a lower variability generate an error signal with a better signal-to-noise ratio and as a
consequence, the adjustment of throwing movements is spatially more fine-tuned resulting in
a smaller remaining lateral deviation at the end of the adaptation period.
However, this variability as such would only slow down adaptation but cannot by itself
explain the large residual error. It is important in this context that each individual’s
(sensorimotor) system has a finite integration capacity to integrate throwing errors within a
temporal interval in order to form an error signal. The results are compatible with a neural
system with a limited “memory” that tries to optimize behavior by adapting specific
parameters. If we adopt the reasonable assumption that the system has a limited integration
capacity over time, given a chosen confidence level and limited number of throws (n), the
variance of throws ( ²) determines the point in time at which adaptation stops since the system
is no longer “sure” that additional adaptation is required. This is the case if the confidence
interval calculated on the basis of ² and n includes zero deviation. On the basis of a given
confidence level (i.e. .05), “n” can be calculated by the following formula where x is the
mean lateral deviation over n throws:
If the individual’s integration capacity is small, only few trials can be averaged to form the
error signal and the – in mathematical terms – “standard error” of this signal is quite large.
Conversely, individuals with a larger integration capacity integrate the error signals over a
larger number of trials. Since the proportion of noise within an average decreases as a
function of the square root of the number of trials, the internal error signal is less corrupted by
noise and the adjustment of throwing movements is more accurate. Therefore, the remaining
lateral deviation at the end of the adaptation period would be less pronounced in subjects with
a large integration capacity.
This model fits very well with recent experiments on eye-hand-coordination in balancing a
(virtual) stick. Patzelt et al. (2007) were able to model human control dynamics in much detail
2²2���
����
�=
xn δ

Studie 1: Vollständigkeit des Adaptationsprozesses 49
for this complex system by assuming that the memory of humans for on-line adaptation is
rather short (in their case in the second range). They conclude that the human nervous system
employs adaptive motor control using only a very limited memory of past observations for
parameter adaptation – well in line with our results in the Throwing Experiment.
On the basis of our experiments, we can only speculate about the neural generators of a
movement-related error signal. However, Kitazawa et al. (1998) reported that in the
cerebellum complex spikes of Purkinje cells convey information about the relative error of
reaching movements. Besides that, Greger & Norris (2005) have shown that even the simple
spike firing of cerebellar neurons was modulated by the spatial accuracy of a goal-directed
movement. The error-related neural signal originating in the cerebellum might trigger the
functional adjustment of visuomotor representations (both realignment and recalibration) in
the course of prism adaptation.
In the present study, we demonstrate that the completeness of prism adaptation depends on the
visuomotor task tested. Whereas extensive training of pointing movements led to a near
complete compensation of the prismatic shift, prism adaptation was incomplete when subjects
executed goal-directed throwing movements and a significant lateral deviation was observed
even at the end of the adaptation period. The incompleteness of adaptation during throwing
movements may be attributed to visuomotor variability coupled with a limited capacity to
integrate errors over time and thus to evaluate the remaining mean error. Failing to realize the
remaining error obviously limits the extent of compensation through training. This
phenomenon may well be present in other types of adaptation processes.
Acknowledgments
Parts of this study were presented on a poster at the “5th Forum of European Neuroscience”
(FENS), Vienna/Austria, July 2006. The authors would like to thank D.C. Högl for the
assistance in collecting the data and D. Trenner for supplying the computer program for
assessing the data. We are also grateful to C. Grimsen for the valuable comments and
suggestions on the interpretation of the data. The authors declare that they have no competing
financial interests. Supported by grant 01GQ0705 (Bernstein programme) of the German
Federal Ministry of Education and Research (BMBF).

Studie 2: Aufgaben-Spezifität der Prismenadaptation 50
6. Studie 2 Task-specificity of prism adaptation: no transfer from pointing
to throwing
Sven Wischhusen & Manfred Fahle
Abstract
Adaptation to a lateral shift of the visual world induced by prisms involves the experience-
dependent adjustment of neural representations according to task demands. The present study
examined the task-specificity of prism adaptation under different visual feedback conditions
in humans. First, subjects adapted to the prisms by performing pointing movements;
thereafter, the transfer of prism adaptation and aftereffect to an untrained throwing task was
tested. During adaptation in the pointing task, visual feedback was provided either at the
terminus of the movement (Exp. 1) or during its entire trajectory (Exp. 2). In both feedback
conditions, neither prism adaptation nor the aftereffect transferred to the untrained throwing
task. Switching to the throwing task evoked a second adaptation process when subjects kept
on wearing the prisms; removing the prisms after adaptation in the pointing task did not
produce an aftereffect in the throwing task. The same pattern of results was observed when
both task and arm trained were switched after prism adaptation indicating a lack of
intermanual transfer. The acquired visuomotor remapping was not accessible after the switch-
over to the novel task suggesting a strong task-specificity of prism adaptation irrespective of
the type of visual feedback.
Introduction
In a changing environment, the brain’s functional organization has to be continuously
adjusted to match the requirements of the physical world. The retention of neural plasticity in
the adult human brain becomes manifest in priming, perceptual learning, and adaptation
(Ahissar & Hochstein, 2004; Fahle, 2002). In the present study, prism adaptation was used as
an experimental approach to study i) the mechanisms adjusting visuomotor representations
according to prior experience and ii) the factors governing the generalization of visuomotor
adaptation.

Studie 2: Aufgaben-Spezifität der Prismenadaptation 51
Prism adaptation as an example of visuomotor adaptation
Suitable optical wedge prisms lead to a lateral deflection of the ray paths on the retina causing
a perceived horizontal shift of the visual world. As a consequence of the prism-induced
disparity between the direction of gaze and the felt arm position subjects wearing prism
goggles initially execute inaccurate visually guided movements which deviate in the direction
of the prismatic displacement. Under normal conditions, however, human subjects rapidly
adapt to the lateral displacement of the visual world: within a limited number of movements
under visual feedback, the lateral error is gradually reduced and movements recover high
spatial accuracy in the course of further practice. This process of re-establishing a functional
correspondence between sensory and motor coordinate systems is designated “prism
adaptation” (e.g. Bedford, 1999; Fernandez-Ruiz & Diaz, 1999; Harris, 1963; 1965; Martin et
al., 1996b; Welch, 1978).
When the prisms are removed subsequent to adaptation, movements initially deviate in the
direction opposite to the prismatic displacement, indicating a negative aftereffect which
completely disappears in the course of further practice (e.g. Fernandez-Ruiz & Diaz, 1999;
Fernandez-Ruiz et al., 2004; Harris, 1965; Newport et al., 2006; Norris et al., 2001; Redding
et al., 2005; Weiner et al., 1983; Welch et al., 1974). The occurrence of an aftereffect
indicates the experience-dependent adjustment of visuomotor representations in the course of
prism adaptation and therefore proves “true” visuomotor adaptation rather than an adaptation
process primarily based on cognitive control (Newport & Jackson, 2006).
During prism adaptation, visuomotor representations are adjusted according to task demands
in order to ensure a high precision of eye-hand coordination which is essential to manually
interact with objects in space. The adjustment of visuomotor representations is functionally
based on two closely related but separable mechanisms operating during adaptation: i) a
strategic control mechanism leading to a rapid sequential reduction of movement errors
during prism exposure and ii) a spatial realignment mechanism functionally re-linking sensory
and motor coordinate systems (for a theoretical framework see Redding et al., 2005; Redding
& Wallace, 2003a).
Activity of a cortico-cerebellar network during prism adaptation
What is known about the underlying neural processes during prism adaptation? Converging
experimental evidence suggests a contribution of different brain areas to the adaptation
process. Studies conducted in both monkeys (e.g. Baizer et al., 1999; Kitazawa & Yin, 2002)
and human neurological patients (e.g. Luaute et al., 2006; Martin et al., 1996a; 2002; Morton

Studie 2: Aufgaben-Spezifität der Prismenadaptation 52
& Bastian 2004; Pisella et al., 2005; Weiner et al., 1983) point to a specific role of the
cerebellum during prism adaptation. It is well established that the cerebellum in general plays
an important role in motor control and adaptation (Marr, 1969; Robinson, 1995; Thach et al.,
1992). Specifically, the spatial realignment mechanism yielding correspondence between
sensory and motor coordinate systems in the course of prism adaptation is mainly a cerebellar
function (Redding et al., 2005). Consistent with this fact, a reduction of aftereffects in human
cerebellar patients has been reported (Weiner et al., 1983) – but reduced aftereffects were also
observed in human patients suffering from basal ganglia disorders (Fernandez-Ruiz et al.,
2003).
Furthermore, the ability to adapt to a prismatic displacement of the visual world is explicitly
impaired in patients with damage to the posterior parietal cortex (PPC) (Newport et al., 2006;
Newport & Jackson, 2006), a cortical module being important for the planning and on-line
control of goal-directed visuomotor behavior (Buneo & Andersen, 2006). Neuroimaging
evidence for the involvement of the PPC during prism adaptation is supplied by a PET study
conducted by Clower et al. (1996) who found adaptation-related activation of the PPC
contralateral to the reaching limb. This suggests that the PPC is involved in the updating of
visuomotor representations in the course of adaptation to laterally displaced vision. Kurata &
Hoshi (1999) demonstrated that monkeys with a pharmacological inactivation of the ventral
premotor cortex (PMv) were impaired in adapting to the prismatic displacement suggesting a
contribution of frontal motor areas during visuomotor adaptation.
A prism-induced left-right reversal of the visual field, which is a highly artificial manipulation
of the visuomotor input-output relationship, has been shown to trigger neural modifications
even in early sensory cortical areas, such as primary visual cortex (V1): in monkeys wearing
left-right reversing prism goggles, a portion of V1 neurons began to respond to visual stimuli
presented in the ipsilateral visual field after a few months suggesting that this type of
adaptation is mediated by plastic processes in early visual processing (Sugita, 1996).
Psychophysical and functional MRI data from human subjects provide additional evidence for
neural modifications in early visual cortex in response to a reversed visual world (Miyauchi et
al., 2004; Tanaka et al., 2007).
Specificity of prism adaptation
The rapid adaptive remapping of visuomotor representations during prism exposure requires
the execution of a particular visuomotor task, for example goal-directed pointing or throwing
movements. In this respect, an important issue is whether a newly acquired visuomotor

Studie 2: Aufgaben-Spezifität der Prismenadaptation 53
mapping – the “result” of prism adaptation – transfers (generalizes) to another task which
differs from the exposure task. Generalization means to apply a function from previous
experience to a new situation. The understanding of the neural mechanisms underlying
generalization is essential for studying adaptive processes both in the sensory and motor
domains (Poggio & Bizzi, 2004).
Using the prism adaptation paradigm, a number of psychophysical studies have addressed the
task-specificity of the adaptation process and the conditions under which generalization from
one task to another occurs. For example, Martin et al. (1996b) demonstrated the specificity of
the negative aftereffect for the trained arm with no intermanual transfer to the untrained arm.
In a second experiment, the transfer of the aftereffect from overhand throws (exposure task) to
underhand throws (post-exposure task) was tested. Here, no aftereffect occurred when
subjects performed underhand throws after having adapted to the prismatic displacement by
executing overhand throws. These results suggest an adaptation process in which movement-
related parameters are adjusted in a task-specific way.
Moreover, Baily (1972) has shown incomplete transfer of prism adaptation from slow to fast
pointing movements. Consistent with this finding, Kitazawa et al. (1997) reported specificity
of prism adaptation for the velocity of reaching movements.
The amount of generalization from a trained task to an untrained one critically depends upon
the similarity between these tasks regarding, for example, movement kinematics, arm trained,
arm posture, velocity of movement, limb weighting etc. With increasing dissimilarity between
these two tasks, generalization decreases, a phenomenon that can be described in terms of a
generalization gradient (Baraduc & Wolpert, 2002; Redding & Wallace, 2006a).
Experimental questions
In the present study, we addressed the task-specificity of prism adaptation by testing the
generalization of a prism-induced visuomotor remapping from a pointing task to a throwing
task. More specifically, human subjects were adapted to a prism-induced rightward shift of
the visual world by executing visually-guided pointing movements. Thereafter, the direct
transfer of i) prism adaptation (prism goggles remain on after adaptation in the pointing task)
and ii) the aftereffect (removing prism goggles after adaptation) to the untrained throwing task
was tested. If the newly acquired visuomotor mapping would completely transfer, a switch-
over to the throwing task should not result in a substantial lateral deviation of throws (i.e.
small/absent prism effect) when keeping on prism goggles. Accordingly, a task-unspecific
adaptation process should yield an aftereffect after switching to the throwing task. On the

Studie 2: Aufgaben-Spezifität der Prismenadaptation 54
other hand, task-specific prism adaptation should produce a large prism effect and an absent
aftereffect upon switch-over to the throwing task. This pattern of results would indicate that
the new visuomotor mapping is not accessible when switching to the untrained task.
In order to investigate the influence of visual feedback availability on the generalization of
prism adaptation to the untrained task, two experiments with different visual feedback
conditions during pointing adaptation were carried out: in the first experiment (Terminal
Vision Experiment) subjects could use the tip of their index finger as a visual feedback signal
only at the terminus of the movement whereas the whole arm trajectory was visible in the
second experiment (Full Vision Experiment). In this context, we asked whether a specific type
of feedback information during adaptation would give rise to different amounts of
generalization from the trained to the untrained visuomotor task.
Methods
Experimental setup and procedure
In two experiments (Terminal Vision Experiment, Exp. 1; Full Vision Experiment, Exp. 2), we
tested the transfer of prism adaptation from a pointing task to a throwing task (Fig. 6.1). First,
subjects underwent an adaptation procedure while performing speedy but accurate ballistic
pointing movements to a visual target always using the right arm. In this pointing task,
subjects were sitting at a table with their body midline centered with the target which was a
small white mark at the table’s edge at a distance of 65 cm. Subjects viewed the target
binocularly.
Before adaptation, subjects always performed 30 pointing movements without wearing prism
goggles in order to familiarize with the task. Thereafter, subjects executed the same number
of pointing movements while wearing prism goggles which shifted the visual word laterally
by 17 deg. to the right (adaptation procedure, pointing task). Thirty pointing movements
during exposure to the prisms had to be performed in order to make sure that subjects adapted
sufficiently to the prismatic displacement (e.g. Newport & Jackson, 2006). The results of pilot
experiments confirmed that a total of 30 pointing movements were adequate for adaptation to
take place since proper aftereffects occurred upon removal of the prisms.
The only difference between the Terminal Vision Experiment and the Full Vision Experiment
was the kind of visual feedback information that subjects received during the pointing task. In
the Terminal Vision Experiment, the table top was opaque and subjects could not see and
hence not correct their trajectory before the movement had finished. In this case, the pointing

Studie 2: Aufgaben-Spezifität der Prismenadaptation 55
movement was planned purely feed-forward on the basis of the visual information about the
target location in space. In this experiment, subjects had visual information about the position
of the index finger in relation to the target only at the end of the arm movement. Because both
visual and proprioceptive feedback is available concurrently only at the terminal part of the
movement, this condition has also been termed “terminal exposure” (Redding et al., 2005; see
also Uhlarik & Canon, 1971).
Fig. 6.1 Experimental setup: In the pointing task, subjects adapted to the lateral displacement of the visual world by executing 30 goal-directed pointing movements while wearing prism goggles. Subjects were sitting at a table and had to point with their right arm as accurate as possible to a target at the table’s edge. Immediately after this adaptation procedure, the transfer of prism adaptation from the pointing task to the untrained throwing task was tested. Subjects stood upright in front of a Velcro-layered wall and had to perform throwing movements to a visual target. The transfer of prism adaptation (prisms on) and the aftereffect (prisms off) was tested either for the ipsilateral (adapted, right) arm and the contralateral (unadapted, left) arm. A digital photo was taken from a fixed position after each throw and stored on a PC for assessing spatial accuracy of the movements.
In contrast, the table top was removed in the Full Vision Experiment and hence subjects had
visual information about the ongoing arm movement during the entire trajectory. This
condition of “concurrent exposure” in which visual and proprioceptive feedback is available
concurrently during the whole movement causes a faster adaptation process (Redding et al.,
2005). Nevertheless, on-line corrections of movements during the Full Vision Experiment
were rather small since subjects fixated the target and did not attend to the arm trajectory.
Immediately after the adaptation procedure in the pointing task, the transfer of prism
adaptation to and the aftereffect in the throwing task were tested. In the throwing task,
subjects had to throw softballs (24 g, 5 cm diameter) as accurately as possible towards a
visual target on a 1.5 m x 1.5 m wide wall. The target was a blue spot (2 cm diameter)
attached to the wall at a height of 156 cm. Subjects stood upright at the same position as in the

Studie 2: Aufgaben-Spezifität der Prismenadaptation 56
pointing task with their mid-sagittal plane aligned with the target wall at a viewing distance of
2 m. The head being unrestrained, subjects always had an unobstructed binocular view of the
target. Subjects were instructed to “Throw, where you see the target” and had to perform
overhand throws while fixating the target without looking at their arm when throwing in order
to prevent on-line corrections of the planned movement. The spatial offset between the impact
of the ball and the target was clearly visible after each throw providing direct visual feedback
about the precision of the throw.
The wall was layered with Velcro material and the softball adhered to the wall after each
throw. In order to assess spatial accuracy of the throws, a photo was taken after each throw
using a digital camera which was kept at a fixed location during the experiments. A Matlab-
based computer program (Matlab release 12.1) was used to readout the electronic images
(.jpeg format) and to compute the horizontal distances between the position of the ball on the
wall and the target in a metric system by means of trigonometry with a spatial accuracy of ~ 1
cm (corresponding to ~ 0.3 deg. of visual angle at the throwing distance of 2 m).
Generalization of prism adaptation and the aftereffect from the pointing task to the throwing
task was tested in four transfer conditions: i) transfer of prism adaptation to ipsilateral arm
(pointing task: right arm, throwing task: adapted right arm), ii) transfer of aftereffect to
ipsilateral arm, iii) transfer of prism adaptation to contralateral arm (pointing task: right arm,
throwing task: unadapted left arm) and iv) transfer of aftereffect to contralateral arm. Note
that these denotations do not necessarily imply that any effects did actually transfer but
simply represent an unambiguous labeling of conditions.
Before each transfer condition, subjects adapted to the prismatic displacement in the pointing
task (adaptation procedure, see above); to test transfer of prism adaptation to the throwing
task, subjects had to throw balls while wearing prism goggles. Conversely, to test transfer of
the aftereffect, subjects adapted during the pointing task and removed the prism goggles
directly before the throwing task. Each transfer condition consisted of 30 trials and was
followed by the same number of throws without prism goggles in order to de-adapt subjects.
The order of transfer conditions was pseudo-randomized across subjects. To minimize spatial
reorientation which may decrease the magnitude of both effects, subjects kept the eyes closed
between conditions.
The “contralateral” conditions served as controls: after adaptation in the pointing task using
the right arm, the transfer to the untrained throwing task was tested with the unadapted left
arm. Neither a transfer of prism adaptation nor of the aftereffect to the throwing task was

Studie 2: Aufgaben-Spezifität der Prismenadaptation 57
expected due to the lack of intermanual transfer of prism adaptation under controlled
conditions (e.g. Baizer et al., 1999; Kitazawa et al., 1997; Martin et al., 1996b).
At the end of the experiment, separate measurements of right and left arm’s throwing
performance were conducted (standard conditions). For these measurements, subjects had to
perform 30 throws before (PRE), during (PRISM) and after (POST) wearing prisms goggles.
These recordings of conventional prism adaptation and aftereffect (without switching tasks)
served to compare the time course and magnitude of effects in the standard conditions with
the time course and magnitude of effects in the transfer conditions.
Experimental groups
For the Terminal Vision Experiment, eight healthy right-handed human subjects (3 male, 5
female) aged 22-26 years (mean 23.6 years) were recruited (Terminal Vision Group). Another
eight subjects (3 male, 5 female) aged 21-26 years (mean 23.6 years) participated in the Full
Vision Experiment (Full Vision Group). None of the subjects reported a history of
neurological or neuropsychological disorders and all had normal or corrected-to-normal visual
acuity; subjects were naïve to the exact purpose of the study.
The experiments were conducted in accordance with the “Declaration of Helsinki” and
subjects gave informed consent prior to participation. This type of experiments was approved
by the ethics committee of Bremen University.
Data analysis
The horizontal distance between the position of the ball on the wall and the target was
calculated for each individual throw. For further analysis, the data were baseline corrected;
that is, we subtracted the mean of all throws of the PRE condition from each data point of
each condition (see also Clower & Boussaoud, 2000). This was done for each condition with
regard to the corresponding arm and each subject individually. The mean of the PRE
condition mirrors a subject’s general arm-specific throwing performance and indicates a
potential spatial bias of the movements. In the Terminal Vision Experiment, this bias averaged
0.0 cm (± 0.5 SEM) for the right arm and -2.9 cm (± 0.7) for the left arm. A bias of -0.4 cm (±
0.4) in the right arm and -2.6 cm (± 0.6) in the left arm was recorded in the Full Vision
Experiment. The baseline correction normalized the data by removing any spatial bias which
could have masked the effects of prismatic displacement on the movements’ accuracy and
also allowed a better comparison between subjects and arms since each set of data was
normalized individually. Consequently, all data shown are baseline corrected.

Studie 2: Aufgaben-Spezifität der Prismenadaptation 58
In order to assess the magnitude of effects in the standard conditions and the transfer
conditions, we calculated the magnitude of the effects by fitting a linear regression line to the
first four data points of each condition and took the intersection of this regression line with
the ordinate as measure of the (initial) prism effect and aftereffect, respectively. This method
was applied to all data instead of using the result of the first movement because the regression
takes into account that i) prism-induced movement errors decrease over the first trials and do
not completely disappear after the first movement and ii) the decrease of movement errors
over the first few trials is mathematically best modeled by a linear function.
Results
The logic of both experiments was as follows: subjects were prism-adapted by executing
right-arm pointing movements, and immediately thereafter the transfer of prism adaptation or
the aftereffect to the throwing task was tested. In order to evaluate the amount of
generalization from one task to the other, we compared the size of the effects in the “transfer
conditions” with the size of the effects in the “standard conditions”, i.e. with conventional
prism adaptation and aftereffect while performing throwing movements without switching
between tasks.
Time course of effects in the standard conditions
Averaged data of both experimental groups for the right and left arm are presented in Fig. 6.2.
The lateral deviation (in cm) of the endpoints of the throwing movements from the target is
plotted as a function of trial number before (PRE), during (PRISM), and after (POST)
wearing prism goggles. Negative values represent a leftward deviation, positive a rightward
deviation.
Before wearing prism goggles (PRE condition), subjects’ throwing movements only exhibited
a very small lateral deviation in both arms suggesting accurate visuomotor performance. Note
that the variability of throws was somewhat higher in the left arm which is likely due to the
right-handedness of all subjects studied.
Right Arm To wear the prism goggles (PRISM condition) initially led to a marked lateral
deviation of throwing movements in the direction of the prismatic displacement. In the right
arm (Fig. 6.2a) of the Terminal Vision Group the prism effect (throw 1) amounted +41.4 cm
(± 6.2 SEM) and decreased to +10.0 cm (± 1.6) at the end of this condition (throw 30). Upon

Studie 2: Aufgaben-Spezifität der Prismenadaptation 59
removal of the prisms (POST condition), a negative aftereffect of -32.2 cm (± 4.8) occurred
(throw 1) decreasing to -2.1 cm (± 1.7) at throw 30. The reduction of lateral throwing errors
was statistically significant in both conditions (PRSIM: p < .01; POST: p < .001) as revealed
by a paired two-sided student’s t-test comparing throws 1 and 30. The movements’ lateral
deviation recorded in the right arm of the Full Vision Group at the beginning of the PRISM
condition averaged +41.0 cm (± 5.4) and was reduced to +8.6 cm (± 3.0) by throw 30.
Throwing without prisms (POST condition) produced an aftereffect of -24.3 cm (± 4.0) which
almost completely disappeared at the end of the condition (mean -0.7 cm, SEM ± 2.6). The
reduction of movement errors was statistically significant in both conditions (p < .01).
Fig. 6.2 Time course of effects in the standard conditions: Mean lateral deviation (in cm) of throwing movements from the target in the PRE, PRISM and POST condition in the Terminal Vision Group (n = 8; filled diamonds) and the Full Vision Group (n = 8; open circles) for the (A) right arm and the (B) left arm. Negative values indicate a leftward deviation, positive a rightward. All data are baseline corrected (means and SEM).
Left Arm In the left arm (Fig. 6.2b), the Terminal Vision Group showed an initial prism
effect of +31.9 cm (± 4.7) which was reduced to +2.8 cm (± 3.4) at the end of the PRISM
condition. In the first throw of the POST condition, the aftereffect averaged -36.5 cm (± 5.8)
and declined to -0.6 cm (± 3.7) at throw 30. As for the right arm, the gradual reduction of

Studie 2: Aufgaben-Spezifität der Prismenadaptation 60
movement errors was statistically significant in both conditions (PRISM: p < .001; POST: p <
.01). In the left arm of the Full Vision Group, the prism effect reached +41.5 cm (± 3.2),
declining to +10.3 cm (± 4.6) at the end of the PRISM condition. Upon removal of the prism
goggles (POST), an aftereffect of -34.5 cm (± 5.5) occurred which was reduced to -0.4 cm (±
3.7). Again, error reduction within the conditions was statistically significant (p < .01).
Taken together, it is clear that introducing the prisms led to a marked lateral deviation of
throwing movements in the direction of the prismatic displacement (i.e. rightward) but the
movements recovered high spatial accuracy within the PRISM condition, hence subjects
showed prism adaptation. Removal of the prisms evoked a leftward aftereffect, vanishing over
the course of visuomotor practice. These results are in good agreement with the literature.
Time course of effects in the transfer conditions
After prism adaptation in the pointing task using the right arm, subjects kept on wearing the
prism goggles but had to execute throwing movements with the ipsilateral (adapted, right)
arm and the transfer of prism adaptation was tested (Fig. 6.3a, upper panel).
In the Terminal Vision Group, the initial throw after switching between tasks showed a
marked rightward deviation of +35.9 cm (± 6.8) indicating a “second” prism effect; with
practice, the lateral deviation of throws declined to +9.4 cm (± 3.2) at the end of the condition
(throw 30). The Full Vision Group showed the same pattern of results; here, the initial
deviation of +50.1 cm (± 5.2) was reduced to +14.8 cm (± 3.4). In both groups, subjects
passed a “second” adaptation process (resembling conventional prism adaptation, see standard
condition) and the error reduction with practice was statistically significant (Terminal Vision
Group: p < .05; Full Vision Group: p < .001).
To test the transfer of the aftereffect from the pointing to the throwing task in the ipsilateral
(right, adapted) arm, prism goggles were removed after adaptation (Fig. 6.3a, lower panel). In
the Terminal Vision Group, the first throw deviated slightly leftwards by -5.9 cm (± 2.8) and
the subsequent throws were close to the target. In the other group tested, the first throw
deviated by -12.0 cm (± 5.7) and all other throws were close to zero. Although there was no
apparent aftereffect and the throwing movements were quite accurate throughout the
condition, the results of the first and the last throw were statistically different (both groups: p
< .05). Nevertheless, there was no explicit transfer of the aftereffect from pointing to throwing
in the right (ipsilateral) arm which is in line with the finding that prism adaptation did not
transfer to the throwing task at all.
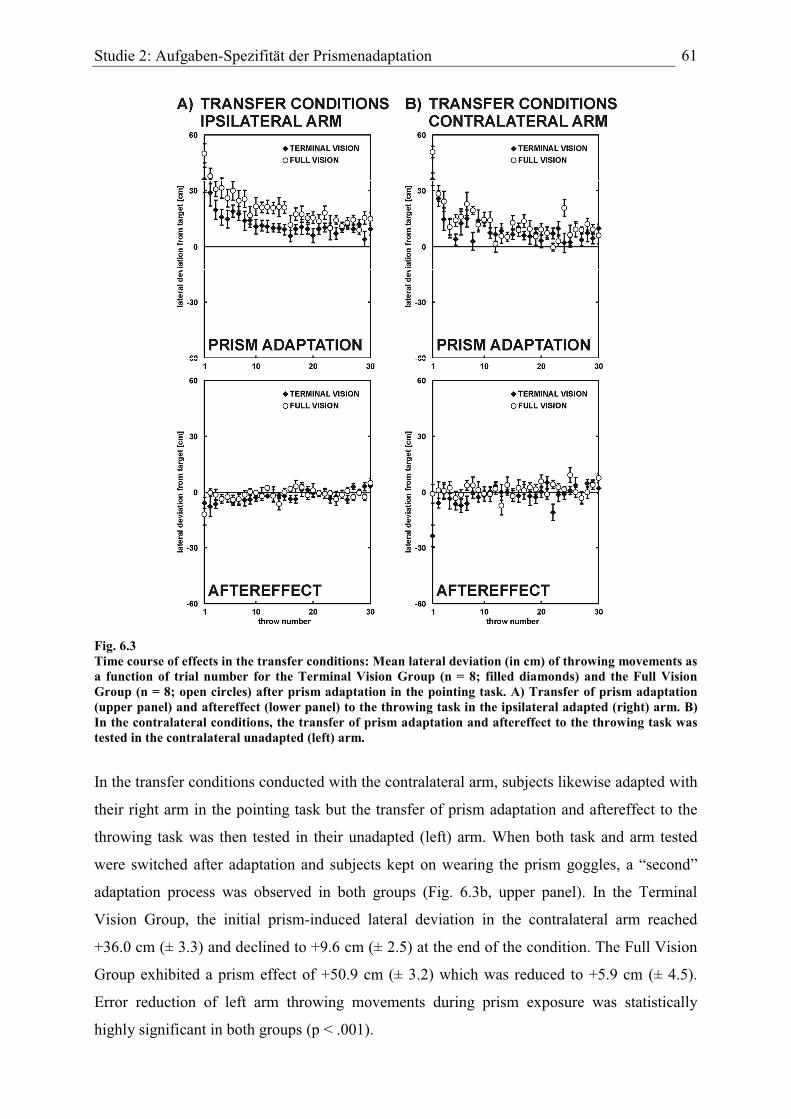
Studie 2: Aufgaben-Spezifität der Prismenadaptation 61
Fig. 6.3 Time course of effects in the transfer conditions: Mean lateral deviation (in cm) of throwing movements as a function of trial number for the Terminal Vision Group (n = 8; filled diamonds) and the Full Vision Group (n = 8; open circles) after prism adaptation in the pointing task. A) Transfer of prism adaptation (upper panel) and aftereffect (lower panel) to the throwing task in the ipsilateral adapted (right) arm. B) In the contralateral conditions, the transfer of prism adaptation and aftereffect to the throwing task was tested in the contralateral unadapted (left) arm.
In the transfer conditions conducted with the contralateral arm, subjects likewise adapted with
their right arm in the pointing task but the transfer of prism adaptation and aftereffect to the
throwing task was then tested in their unadapted (left) arm. When both task and arm tested
were switched after adaptation and subjects kept on wearing the prism goggles, a “second”
adaptation process was observed in both groups (Fig. 6.3b, upper panel). In the Terminal
Vision Group, the initial prism-induced lateral deviation in the contralateral arm reached
+36.0 cm (± 3.3) and declined to +9.6 cm (± 2.5) at the end of the condition. The Full Vision
Group exhibited a prism effect of +50.9 cm (± 3.2) which was reduced to +5.9 cm (± 4.5).
Error reduction of left arm throwing movements during prism exposure was statistically
highly significant in both groups (p < .001).

Studie 2: Aufgaben-Spezifität der Prismenadaptation 62
The aftereffect did not transfer from pointing to throwing in the contralateral arm (Fig. 6.3b,
lower panel), but the first throw deviated by -23.7 cm (± 5.8) in the Terminal Vision Group.
All subsequent throws within this condition were close to zero (throw 30: mean +2.1, SEM ±
2.9). A paired two-sided t-test comparing the first and the last throw of this condition revealed
a significant difference (p < .01). In the Full Vision Group, no transfer of the aftereffect
across task and arm occurred: all throws showed a small lateral deviation (throw 1: mean -0.9
cm, SEM ± 4.9). A direct comparison of throw 1 and throw 30 did not show any significant
difference (p > .05).
Direct comparison between standard conditions and transfer conditions within groups
In a next step, we directly compared the magnitude of the effects in the transfer conditions
with the magnitude of the effects assessed in the standard conditions in the corresponding
arm. Remember that these effects were assessed on the basis of a linear regression line
through the first four data points (see methods).
In the Terminal Vision Group, the prism effect in the standard condition (standard prism
effect) reached +40.1 cm (± 5.6) when throws were executed with the right arm (Fig. 6.4a,
upper panel); when the tasks were switched and subjects threw the ball with the right arm
while wearing prism goggles (transfer condition, ipsilateral arm), another prism effect of
+35.6 cm (± 7.3) occurred. That is, switching the task led to another prism effect although
subjects had already adapted to the optical displacement by executing pointing movements.
The difference between the standard and the transfer prism effect was statistically not
significant (p > .05) as tested by a paired two-sided t-test. The same pattern of results applies
for the Full Vision Group: the right arm’s standard prism effect of +38.4 cm (± 5.0) and the
slightly higher transfer prism effect of +47.0 cm (± 5.2) in the ipsilateral arm did not differ
significantly. These results indicate that switching to the throwing task evoked a prism effect
similar to the conventional prism effect, hence prism adaptation in both groups did not
transfer to the novel task.
In the Terminal Vision Group, right arm throwing after removal of the prisms produced a
standard aftereffect of -32.0 cm (± 4.6) (Fig. 6.4a, lower panel). When the prisms were
removed and the task was switched after adaptation (transfer condition), throwing movements
in the ipsilateral (right) arm did not deviate distinctly and a transfer aftereffect of -7.2 cm (±
3.3) was recorded. In the Full Vision Group, the standard right arm aftereffect averaged -23.9
cm (± 3.0) and the transfer aftereffect in the ipsilateral arm reached -8.0 cm (± 3.7). Whereas
proper right arm aftereffects occurred in the standard conditions in both groups, almost no
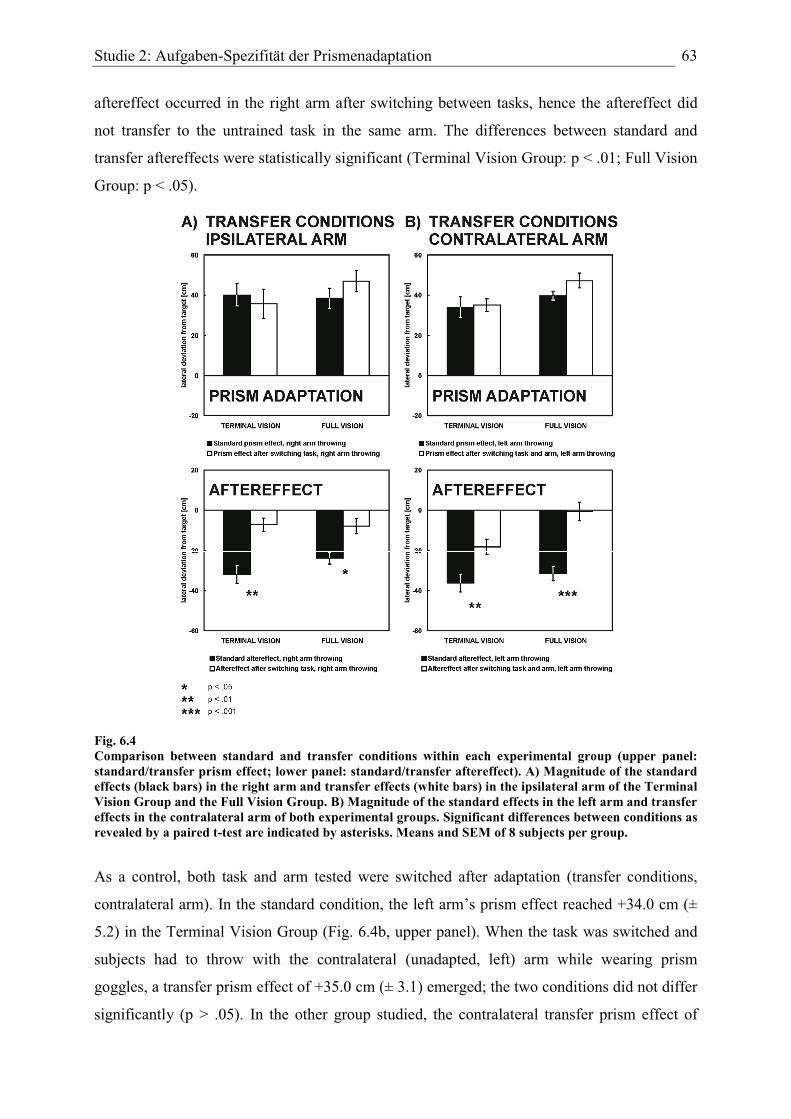
Studie 2: Aufgaben-Spezifität der Prismenadaptation 63
aftereffect occurred in the right arm after switching between tasks, hence the aftereffect did
not transfer to the untrained task in the same arm. The differences between standard and
transfer aftereffects were statistically significant (Terminal Vision Group: p < .01; Full Vision
Group: p < .05).
Fig. 6.4 Comparison between standard and transfer conditions within each experimental group (upper panel: standard/transfer prism effect; lower panel: standard/transfer aftereffect). A) Magnitude of the standard effects (black bars) in the right arm and transfer effects (white bars) in the ipsilateral arm of the Terminal Vision Group and the Full Vision Group. B) Magnitude of the standard effects in the left arm and transfer effects in the contralateral arm of both experimental groups. Significant differences between conditions as revealed by a paired t-test are indicated by asterisks. Means and SEM of 8 subjects per group.
As a control, both task and arm tested were switched after adaptation (transfer conditions,
contralateral arm). In the standard condition, the left arm’s prism effect reached +34.0 cm (±
5.2) in the Terminal Vision Group (Fig. 6.4b, upper panel). When the task was switched and
subjects had to throw with the contralateral (unadapted, left) arm while wearing prism
goggles, a transfer prism effect of +35.0 cm (± 3.1) emerged; the two conditions did not differ
significantly (p > .05). In the other group studied, the contralateral transfer prism effect of

Studie 2: Aufgaben-Spezifität der Prismenadaptation 64
+47.3 cm (± 3.8) was higher than the left arm’s prism effect as measured in the standard
condition (mean +39.7 cm, SEM ± 2.3) but this difference did not reach statistical
significance (p > .05). In both groups, switching task and arm tested evoked a “second” prism
effect in almost the same manner as in the standard condition, hence the adaptation did not
generalize across tasks and arms and subjects had to undergo another adaptation process.
The standard aftereffect in the left arm of the Terminal Vision Group (Fig. 6.4b, lower panel)
reached -36.3 cm (± 4.4); after removal of the prisms and switching both task and arm tested,
contralateral throwing movements initially showed a leftward deviation of -18.3 cm (± 3.7)
which is about 50 % of the left arm’s standard aftereffect. Yet, the difference between the two
conditions was statistically significant (p < .01), indicating no more than a partial transfer of
the aftereffect across tasks and arms. In the Full Vision Group, the left arm’s standard
aftereffect was -31.6 cm (± 3.8). When both task and arm tested were switched (transfer
aftereffect, contralateral arm), throwing movements showed hardly any deviation from the
target (mean -0.6 cm, SEM ± 4.6). The difference between these conditions was statistically
highly significant (p < .001). To sum up, neither prism adaptation nor the aftereffect
transferred to the untrained task in the unadapted arm.
Comparison between Terminal Vision Group and Full Vision Group
A direct comparison of the standard effects and transfer effects between experimental groups
is presented in Fig. 6.5. The standard effects for both groups were comparable: an unpaired
two-sided student’s t-test did not reveal any significant differences – neither for the prism
effect nor for the aftereffect in the right or left arm (p > .05). The same applies for the transfer
effects in the ipsilateral arm: the magnitude of the prism effect after switching tasks did not
differ significantly between both groups (p > .05). In line with these results, the transfer
aftereffects were equally small in both groups, hence there was no difference in the amount of
transfer between the groups studied. Prism adaptation and the aftereffect simply did not
transfer across tasks in the ipsilateral arm.
However, when comparing the magnitude of the transferred prism effect in the contralateral
arm, a significant difference between groups appeared (p < .05). Whereas the transferred
prism effect amounted to +35.0 cm (± 3.1) in the Terminal Vision Group, the Full Vision
Group exhibited a significantly higher effect of +47.3 cm (± 3.8). In the Terminal Vision
Group, the transferred aftereffect in the contralateral arm reached +18.3 cm (± 3.7) whereas
the same condition yielded a marginal effect of -0.6 cm (± 4.6) in the Full Vision Group. The
difference between groups was statistically significant (p < .01).

Studie 2: Aufgaben-Spezifität der Prismenadaptation 65
Fig. 6.5 Direct between-group comparison of the (A) standard effects and the (B) transfer effects. Black bars represent the Terminal Vision Group and white bars stand for the Full Vision Group. Significant differences between groups as revealed by an unpaired t-test are indicated by asterisks. Means and SEM of 8 subjects per group.
Discussion
Adaptation to a lateral displacement of the visual world induced by prisms requires the
execution of a particular task in order to trigger the experience-dependent remapping of
visuomotor representations. In the present study, we examined whether a prism-induced
visuomotor remapping acquired in one task is transferred to another task. Subjects adapted in
a pointing task and the transfer effects of both prism adaptation and the aftereffect to an
untrained throwing task were tested. Our results show a strong task-specificity of the
adaptation process since neither prism adaptation nor the aftereffect transferred across tasks.
Furthermore, the type of visual feedback provided during adaptation in the pointing task had
no differential impact on the pronounced task-specificity. The experiment with visual
feedback on the movement’s precision at the terminal part of the movement (Exp. 1) as well
as the experiment with full visibility of the trajectory during adaptation (Exp. 2) lacked
generalization of prism adaptation from the trained to the untrained task.
Specifying visual feedback conditions
In our study, the availability of visual and proprioceptive feedback (i.e. sensory information
from receptors in the muscles, and joints) during movement execution varied according to
experiments: in the Terminal Vision Experiment, proprioceptive information about the moving
limb was available during the entire movement whereas visual information was available only

Studie 2: Aufgaben-Spezifität der Prismenadaptation 66
at the terminal part of the movement (terminal exposure). Hence, both visual and
proprioceptive feedback information coincided only at the end of the movement representing
the most important difference to the Full Vision Experiment where both sensory modalities
were concurrently available during the entire movement (concurrent exposure).
It has been suggested (e.g. Redding & Wallace, 1988; Uhlarik & Canon, 1971) that the extent
of adaptive remapping of each modality during the adaptation process depends on the specific
prism-exposure conditions: under terminal exposure, spatial localization of the arm relies
more on proprioception of the arm since visual information is only available at end of the
movement; hence, visual coordinates are remapped (“adapted”) whereas proprioceptive
coordinates remain unchanged. Conversely, during concurrent exposure vision forms the
dominant sensory input and proprioceptive coordinates are remapped in the course of
adaptation (of course, in the end, also the visual “spatial” information relies partly on
proprioception of the eye position).
This view has important implications for the generalization of prism adaptation from one task
to another as examined in our study. An adaptive remapping of the visual system taking place
under terminal exposure would predict a transfer of prism adaptation across tasks since the
remapping of visual coordinates would – once completed – affect motor planning irrespective
of the task. On the contrary, adaptation of the proprioceptive system of the arms (especially
with concurrent exposure) would be arm- and task-specific since only the current task-specific
configuration of the moving limb is adapted. Whereas the model described above predicts
lack of generalization to the untrained task only under concurrent exposure, our results show
task-specific prism adaptation irrespective of the type of visual feedback. What may be the
reason for this deviation from the model’s predictions?
Task-specificity and direction-dependent weighting of sensory inputs
For the planning and control of goal-directed limb movements, both visual and proprioceptive
input signals about arm position are combined and weighted to generate an estimate of limb
localization in space (Bays & Wolpert, 2007). However, in the horizontal plane the precision
of visual and proprioceptive localization varies with the direction being considered: whereas
proprioception provides more precise spatial information in depth (i.e. in a radial direction
from the observer) than in azimuth (i.e. in a direction orthogonal to that), the opposite applies
for vision which is more precise in azimuth than in depth (van Beers et al., 2002) and this is
supposed to result in a direction-dependent weighting of sensory inputs for optimal
integration (Bays & Wolpert, 2007).

Studie 2: Aufgaben-Spezifität der Prismenadaptation 67
It has been shown experimentally that spatial localization in depth relies more on
proprioception whereas spatial judgments in azimuth are computed on the basis of visual
information. Thus, for judgments involving the azimuth vision is weighted stronger than
proprioception leading to adaptation of arm-muscle proprioception (van Beers et al., 2002).
According to this model, the direction of an experimental manipulation is the essential factor
for the type of adaptation. In the case of laterally displacing prisms, a conflict of visual and
proprioceptive sensory inputs is introduced in azimuth which causes adaptation of the
modality with the lower spatial resolution in that direction, i.e. proprioception. This type of
adaptation, affecting the felt arm position is specific for the current configuration of the
moving limb, in other words, it is task- and trajectory-dependent. Since the type of arm
trajectory during a pointing movement clearly differs from a throwing movement,
proprioceptive adaptation of the pointing trajectory does not transfer to a throwing trajectory.
A simple way of explaining the pronounced task-specificity irrespective of the type of visual
feedback observed in our study is to attribute a large portion of the adaptation process to a
remapping of the proprioceptive system as a consequence of the flexible direction-dependent
weighting of visual and proprioceptive input signals. This would imply that it is not the type
of exposure condition (concurrent vs. terminal) which determines the type of adaptation
(proprioceptive vs. visual) but instead the direction of the visual-proprioceptive conflict
(azimuth vs. depth) is the crucial factor for the type of adaptation.
Task- and trajectory-specific prism adaptation
In a related study, Marotta et al. (2005) reported task-specific prism adaptation to a left-right
reversal of the visual field; this severe artificial manipulation of the relationship between
visual input and motor output is apparently compensated for on the basis of task-specific
transformations. In our study, however, even a moderate lateral shift of the visual world by 17
deg. resulted in a task-specific adaptation process agreeing well with previous studies
addressing the specificity of prism adaptation. In particular, Martin et al. (1996b) reported that
prism adaptation did not transfer from overhand throws to underhand throws in the same arm.
In their study, subjects executed overhand throws during prism adaptation. After removal of
the prisms, they executed underhand throws and no aftereffect occurred. However, the authors
attribute this task-specificity mainly to motor adaptation and an adaptive change in the
relationship between the direction of gaze and the direction of throw. Other studies as well
have shown that adaptation is very specific for the critical task-related motor parameters like

Studie 2: Aufgaben-Spezifität der Prismenadaptation 68
arm posture (Baraduc & Wolpert, 2002), velocity of arm movements (Kitazawa et al., 1997),
or the weight of the limb moving (Fernandez-Ruiz et al., 2000).
In this context, it is important to keep in mind that switching from a trained to an untrained
task always involves a complex alteration of motor parameters, hence task-specific adaptation
can also be interpreted as a trajectory-specific adaptation; switching the task is closely related
to an alteration of movement parameters, hence a novel trajectory has to be planned.
Therefore, it is indisputable that the adaptation process must involve a strong motor
component as suggested by the studies mentioned (e.g. Baraduc & Wolpert, 2002; Martin et
al., 1996b). It is very likely that the sensorimotor system employs both proprioceptive and
motor adaptation mechanisms in order to rapidly restore a correspondence between sensory
and motor coordinate systems. The amount of adaptation within the sensory (i.e. visual, and
proprioceptive) and motor system may vary according to the exact spatial and temporal
requirements of a given task.
Further aspects: speed of adaptation and lack of intermanual transfer
Subjects adapting to the prism-induced displacement of the visual world by executing
pointing movements could not rely on their acquired visuomotor remapping when they had to
perform throwing movements. In the untrained task, subjects had to adapt anew when wearing
prisms and passed a second adaptation process although they had already adapted to the
prisms immediately before. The time course and speed of the second adaptation process after
switching between tasks was very similar to the conventional adaptation process (see Fig. 6.2
and Fig. 6.3) suggesting that the visuomotor remapping formed during pointing adaptation
could not be retrieved for the throwing task – it was functionally not accessible for the
untrained task. This conclusion is derived from the experimental finding that no aftereffect
occurred when the prisms were removed and the tasks were switched.
Moreover, we fitted an exponential decay function of the form
312)( aeaxf xa +=
to the individual data of both the standard and transfer prism conditions in order to assess the
speed of the adaptation process which is reflected by the parameter a2. A paired two-sided t-
test did not reveal significant differences between conditions suggesting that the visuomotor
remapping acquired during pointing did not even facilitate (i.e. speed up) the adaptation
process after switching between tasks. Rather, the speed of adaptation was the same as that of
the conventional adaptation process. This holds true for both experimental groups and
underlines the pronounced task-specificity irrespective of the type of visual feedback.

Studie 2: Aufgaben-Spezifität der Prismenadaptation 69
After switching both task and arm tested (contralateral conditions), subjects showed a second
adaptation process when they kept on wearing the prisms which was expected since most
studies find that the adaptation process is restricted to the exposed arm (e.g. Baizer et al.,
1999; Baraduc & Wolpert, 2002; Kitazawa et al., 1997; Martin et al., 1996b). In both
experimental groups, the initial prism effect after switching both task and arm tested was as
large as the conventional left arm prism effect and subjects reduced the movements’ lateral
errors gradually with comparable speed. Due to the lack of intermanual transfer and the strong
task-specificity of prism adaptation it is not surprising that there was no transfer when both
variables were changed (i.e. task and arm tested). Again, these results highlight the strong
specificity of the acquired visuomotor remapping and confirm that neural adjustments
underlying the adaptation process do not occur within the visual system since this would yield
a transfer across both task and arm.
In this context, we do not want to keep secret odd results observed in five of eight subjects of
the Terminal Vision Group: after adaptation in the pointing task using the right arm, these
subjects exhibited an aftereffect in the throwing task in the unadapted arm (see Fig. 6.3b,
lower panel, first black diamond). It was, however, only the first throw which deviated
laterally whereas the other throws were close to the target. These results would imply that the
aftereffect transferred across both task and arm but not across tasks using the same arm! Since
this can hardly be the case we infer that this effect may rather reflect cognitive strategies
employed by some of our subjects, and indeed, one should never forget that in these types of
adaptation experiments cognitive strategies can, in principle, interact with the automatic and
pre-conscious adaptation mechanisms that constantly update the relationship between
sensorimotor representations of the physical world.
Conclusions
The central goal of this study was to assess whether prism adaptation during pointing
movements transfers to untrained throwing movements. By systematically varying the
exposure conditions during adaptation (terminal vs. concurrent visual feedback) we wanted to
investigate whether specific types of exposure conditions would lead to higher or lower
amounts of generalization across tasks. Our results strongly suggest task-specific prism
adaptation irrespective of the type of visual feedback during exposure. The specificity of the
adaptation process can hardly be explained in terms of an adaptive remapping of visual
coordinates in the course of practice. Rather, we propose that the task-specificity can be

Studie 2: Aufgaben-Spezifität der Prismenadaptation 70
explained as a result of an adaptive proprioceptive remapping. Since the prism-induced
manipulation occurs in azimuth where vision has a higher spatial resolution, visual inputs are
weighted more heavily and proprioceptive inputs are adapted in a task-dependent way in order
to rapidly restore accurate eye-hand coordination. However, it is clear that adaptive changes
may also occur within the motor system; both sensory and motor adaptation mechanisms
might operate in a dynamic and mutual manner. From a biological perspective, “local” task-
specific adaptation seems to be the optimal (and more saving) strategy for the brain to adjust
neural representations in response to short-term changes of the physical world or the own
body.
Acknowledgments
Parts of this study were presented on a poster at the workshop on “Preemptive Perception” at
the Hanse Institute for Advanced Study, Delmenhorst/Germany, October 2005. The authors
would like to thank D.C. Högl for assistance in collecting the data and D. Trenner for the
programming work. The authors declare that they have no competing financial interests.
Supported by grant 01GQ0705 (Bernstein programme) of the German Federal Ministry of
Education and Research (BMBF).

Studie 3: Räumliche Generalisierung des Nacheffekts 71
7. Studie 3 Effects of training conditions on spatial generalization of prism
adaptation
Sven Wischhusen & Manfred Fahle
Abstract
Prism adaptation is often very specific to the exact experimental context of the particular
visuomotor task. Here, we investigated whether the type of visuomotor training (i.e. blocked
vs. mixed) during prism adaptation affects the pattern of spatial generalization. In Experiment
1, subjects adapted to a lateral shift of the visual world by executing pointing movements to a
single target in rapid succession (blocked training) and generalization of the aftereffect to an
untrained target was tested. Aftereffects mostly transferred to other target positions indicating
pronounced generalization across space. In Experiment 2, subjects adapted by pointing
alternately to two target locations, a form of mixed training. Generalization of aftereffects to
targets located beyond the adapted visuomotor workspace depended upon the direction tested;
pointing to a target within the adapted workspace yielded smaller aftereffects. Results are
discussed in terms of a scaled spatial representation as a consequence of prism adaptation
through mixed training.
Introduction
Neural representations of the physical world are continuously shaped through experience to
assure adequate behavior in a dynamic environment. Updating the brain’s spatial
representations in response to altered external conditions requires neural plasticity of the
structures involved (Steven & Blakemore, 2004). Perceptual learning (e.g. Ahissar &
Hochstein, 2004; Fahle, 2002; 2004) and visuomotor adaptation demonstrate the preservation
of human brain plasticity during adulthood even on rather elementary levels of processing.
Here, we tested whether the form of visuomotor training during prism adaptation would
influence the pattern of spatial generalization of the adaptive behavior.
Prism adaptation demonstrates visuomotor plasticity
Prism adaptation is a form of visuomotor adaptation in which subjects adapt to a prism-
induced perturbation of the visual world – usually a lateral shift – while executing goal-

Studie 3: Räumliche Generalisierung des Nacheffekts 72
directed visually guided movements to a specific target (for reviews see e.g. Bedford, 1999;
Harris, 1965; Redding et al., 2005). The (wedge) prisms in front of the subjects’ eyes
introduce deviations between the perceived direction of the target and the position of the arm
(Martin et al., 1996b; 2002). As a result, movements initially deviate markedly from the target
in the direction of the prism-induced optical shift (prism effect). Typically, visual feedback
regarding the arm movement is provided signaling the difference between the actual and the
desired endpoint of the movement. The resultant error signal serves to adjust the movements
on a trial by trial basis (“error reduction phase”) leading to increasingly accurate arm
movements under prism exposure. As a result, movements recover high spatial accuracy in
the course of the adaptation process (Redding et al., 2005).
When the prisms are removed subsequent to adaptation, movements initially deviate laterally
in the direction opposite to the prismatic shift. This negative aftereffect decreases rapidly
during the course of continuing practice with feedback (e.g. Fernandez-Ruiz & Diaz, 1999;
Fernandez-Ruiz et al., 2000; 2004; Redding et al., 2005). Functionally, the aftereffect
demonstrates the adaptive re-mapping of sensory (i.e. visual and proprioceptive) and motor
coordinates in the course of prism adaptation suggesting short-term plasticity of the
underlying neural structures. The course of de-adaptation following removal of the prisms is
somewhat faster than the prism adaptation process because the visuomotor system recovers
rapid access to the stored original representation for eye-hand coordination (Fernandez-Ruiz
& Diaz, 1999). Furthermore, the aftereffect suggests that the adaptation process is not
primarily of cognitive origin because if it were, removal of the prisms should almost
immediately lead to unbiased pre-exposure like visuomotor behavior.
Neuroanatomical findings
Mirroring the complexity of prism adaptation, a variety of brain areas – cortical as well as
subcortical – have been identified as contributing to the adaptation process. Consistent with
the view that the cerebellum is an important neural substrate for motor skills (e.g. Hikosaka et
al., 2002; Thach et al., 1992), several studies conducted with humans (Martin et al., 1996a;
2002; Morton & Bastian, 2004; Pisella et al., 2005; Weiner et al., 1983) and non-human
primates (Baizer et al., 1999; Kitazawa & Yin, 2002) indicate that cerebellar structures are
involved in the adaptation process. In addition, contributions of frontal motor areas like
ventral premotor cortex (Kurata & Hoshi, 1999) and “classical” motor structures like the basal
ganglia (Fernandez-Ruiz et al., 2003) have been demonstrated experimentally.

Studie 3: Räumliche Generalisierung des Nacheffekts 73
In addition, strong evidence indicates that parietal cortical areas also contribute to prism
adaptation (Clower et al., 1996; Newport et al., 2006; Newport & Jackson, 2006) compatible
with the role of parietal cortex in sensorimotor integration (Andersen & Buneo, 2003; Buneo
& Andersen, 2006). A number of studies even report ameliorating effects of prism adaptation
on hemispatial neglect, a neuropsychological disorder following unilateral damage of parietal
cortex (e.g. Luaute et al., 2006; Redding & Wallace, 2006b; Rode et al., 2003; Rossetti et al.,
1998; Serino et al., 2006). Obviously, spatial representations in the parietal lobe undergo
plastic changes during prism adaptation. Moreover, left-right reversing prisms strongly
manipulating visuomotor coordination change representations in early visual areas suggesting
that neural plastic processes may occur even in early sensory (e.g. visual) cortices (Miyauchi
et al., 2004; Sugita, 1996; Tanaka et al., 2007). To sum up, prism adaptation seems to be
achieved by an extended network of motor, sensorimotor, and sensory brain areas
contributing to sensorimotor plasticity to different extents depending on the requirements of a
given visuomotor task.
Psychophysical findings: Specificity versus generalization
The prism adaptation paradigm is well suited to study the short-term brain mechanisms
adjusting sensorimotor representations according to specific task demands. An important issue
concerns the specificity of the adaptation process: Is the acquired visuomotor mapping
specific to the precise training conditions as is usually the case in perceptual learning (cf.
Fahle, 2005) or does it generalize to untrained conditions? This pattern of generalization can
provide important information about the underlying processing system (Poggio & Bizzi,
2004).
Several studies on prism adaptation highlight a specific adaptation process where the amount
of generalization critically depends on the similarity between training (adaptation) and testing
conditions. For example, prism adaptation during overhand throwing movements did not
generalize when underhand throws were subsequently tested (Martin et al., 1996b). Similarly,
adaptation was found to be specific for movement velocity during prism exposure.
Generalization decreased as the difference in velocities between trained and tested movement
increased (Kitazawa et al., 1997). Furthermore, Fernandez-Ruiz et al. (2000) showed that
prism adaptation only generalized when limb weight was identical for training and testing
conditions. In an experiment using a virtual-reality setup closely related to the “classical”
prism adaptation paradigm, Baraduc & Wolpert (2002) demonstrated the specificity of the
adaptation process for starting limb posture. As the tested posture increasingly differed from

Studie 3: Räumliche Generalisierung des Nacheffekts 74
the initially trained posture generalization decreased constituting a generalization gradient. In
these examples the adaptation mechanism precisely incorporated the specific task-related
parameters during prism exposure. The more the conditions differed between training and
testing, the less did the system employ the “adaptive strategy” for the novel conditions.
Besides this type of local adaptation yielding an associative generalization gradient along a
given dimension, experimental evidence exists for a more global type of adaptation. For
example, Bedford (1989; 1993) and Redding & Wallace (2006a) both demonstrated by means
of a simple pointing task that adaptation generalized to untrained target locations in space
suggesting a rigid shift in the mapping of all points in the spatial maps involved in the task, a
form of linear generalization. That is, adaptation to a specific target transferred to other
targets in space although these had not been trained. Similarly, a study in which subjects were
adapted to hemiprisms (i.e. prisms covering only the upper or lower visual hemifield) found
that the system initially modified visuomotor transformations globally in the whole spatial
representation, i.e. it performed a rigid linear shift; yet, with differential feedback the system
was also able to locally adjust the visuomotor map (Fernandez-Ruiz et al., 2006). These two
processes (global vs. local adjustment) seem to operate on different time scales
Experimental questions
The present psychophysical study on prism adaptation was motivated by the question whether
or not the pattern of spatial generalization in a visually guided pointing task is influenced by
the type of visuomotor training. Since the aftereffect is less affected by cognitive strategies, it
was taken as the critical measure and spatial generalization was tested at one of three possible
target locations.
In the first experiment, subjects adapted to the prism-induced lateral shift by pointing thirty
times to a visual target in space. Thereafter the prisms were removed and the generalization of
the aftereffect to an untrained target was tested. During adaptation, subjects pointed
sequentially to a single visual target in rapid succession, a form of “blocked training”. In case
of local adaptation an associative generalization gradient (cf. Bedford, 1993; Redding &
Wallace, 2006a) is expected to emerge, decreasing with spatial distance between targets
trained and tested. In contrast, extensive generalization across targets in space would indicate
a rigid linear shift of the entire visuomotor map as the underlying cause of this adaptation
process.
The second experiment investigated whether a “mixed training” procedure during prism
adaptation would give rise to a different pattern of spatial generalization. To test this question,

Studie 3: Räumliche Generalisierung des Nacheffekts 75
subjects pointed alternately to two targets in space during prism adaptation, a form of mixed
training. After adaptation, generalization of the aftereffect to the untrained target was
measured. Due to the mixed training, different parts of the visuomotor workspace were
adapted since the pointing movements to two separate targets covered a relatively wide range
of directions. This type of experimental design made it therefore possible to differentiate
between conditions in which generalization was tested in a target located beyond (i.e.
extrapolation) versus within (i.e. interpolation) the adapted workspace. In this respect, we
asked whether a specific type of workspace adaptation would lead to different amounts of
generalization.
Methods
Task, setup, and procedure
The visuomotor task required subjects to perform accurate visually-guided pointing
movements towards a distant target using the right arm. Subjects were sitting at a table and
executed ballistic movements to one of three targets located at the table’s edge at a viewing
distance of 65 cm. The three targets (left, central, and right) were spatially separated by a
distance of 25 cm from each other (Fig. 7.1a). To ensure that subjects could reach each of the
three targets equally well, the distal part of the right shoulder faced the central target.
Subjects were instructed to point as accurately as possible to the perceived target location.
They could not visually track their movement after its start due to a nontransparent table-top.
Consequently, the trajectories were largely planned feed-forward on the basis of the perceived
target position relative to the body. At the end of the movement, however, subjects saw the tip
of their index finger and so received terminal visual feedback about the precision of the
movement. This type of feedback condition has been termed “terminal exposure” (cf. Redding
et al., 2005). The movements’ spatial accuracy was assessed using a PC-controlled measuring
system (Zebris Medical, Isny/Germany) based on the travelling time of ultrasound pulses
which were emitted by three senders and picked up by microphones attached to the index
finger and the targets. A computer program calculated the distance between the finger and the
target in all three spatial directions. The spatial resolution of the measuring system was < 1
mm. The high quality wedge prisms (Carl Zeiss, Oberkochen/Germany) used were mounted
into conventional spectacle frames and produced a perceived lateral shift of the visual world
by ~ 17 deg. to the left (i.e. base-right prisms; 30 diopters); subjects always viewed the target
binocularly.

Studie 3: Räumliche Generalisierung des Nacheffekts 76
Fig. 7.1 A) Experimental setup: Subjects were sitting at a table and had to perform pointing movements to one of three targets at the table’s edge. Terminal visual feedback about the precision of the executed movement was provided. The spatial accuracy of the movements was assessed using a PC-controlled ultrasound measuring device. Conditions tested in (B) Experiment 1 and (C) Experiment 2; L = left target, C = central target, R = right target; for details see text.
Experiment 1 In Experiment 1, spatial generalization of the aftereffect from a trained
target (prism adaptation) to an untrained target was examined. Each experimental block (Fig.
7.1b) consisted of two conditions with 30 pointing movements each. At first, subjects adapted
to the prism-induced optical displacement by executing pointing movements to one of the
three targets (“target trained”) in rapid succession. After adaptation, the prism goggles were
removed and generalization of the aftereffect to another target was tested (“target tested”).
This experiment covered a “blocked training” procedure: During prism adaptation, subjects
pointed to a single target in space and the transfer of the aftereffect was assessed for another
single target location.
The generalization of the aftereffect was probed systematically for each target (including
trained = tested target) leading to a total of 9 (3x3) discrete experimental blocks ([trained: left,
central, right] x [tested: left, central, right]). Each block started with an adaptation procedure
and in the following condition, spatial generalization of the aftereffect was tested. In order to
minimize effects of habituation, subjects had to keep their eyes closed between conditions.
Order effects were minimized by counter-balancing the order of blocks tested across subjects.
At the beginning of the experiment, spatial accuracy of the movements without prism goggles
(PRE, baseline condition) was recorded for each target individually. This served to control for
possible effects of target position on the movements’ accuracy and to correct the data for
directional biases (baseline correction; see Data analysis).

Studie 3: Räumliche Generalisierung des Nacheffekts 77
Experiment 2 Similar to Experiment 1, spatial generalization of the aftereffect to an
untrained visual target was tested in Experiment 2. However, the basic difference to the first
experiment was that during prism adaptation subjects had to point alternately to two targets
(i.e. mixed training during prism exposure). Each experimental block consisted of two
conditions (Fig. 7.1c); in the first condition, subjects pointed alternately to two targets (30
movements per target trained) while wearing prism goggles. In the following condition (30
pointing movements), the prism goggles were removed and generalization of the aftereffect to
the untrained target was measured (“target tested”).
The spatial configuration of the three targets allowed both an extrapolation task and an
interpolation task. In the extrapolation task, the left (resp. right) and the central target were
trained during adaptation and generalization of the aftereffect to the right (resp. left) target
was tested. This probes the extrapolation of the aftereffect to a visual target located beyond
the adapted visuomotor workspace. Testing extrapolation to the left versus the right target
allowed to compare the effects of an extrapolation in the direction of the aftereffect (i.e.
extrapolation to right target) and in the direction opposite to the aftereffect (i.e. extrapolation
to left target).
The extrapolation task was the counterpart of the interpolation task. In this case, the left and
the right targets were trained alternately during prism adaptation, and generalization of the
aftereffect to the central target was examined. This test of generalization formed an
interpolation task since the central target was located within the adapted visuomotor
workspace. Interpolation of the aftereffect to the central target was tested twice. In one block,
adaptation started with pointing to the left target while in the other block adaptation started
with the right target. To sum up, generalization of the aftereffect to an untrained target was
tested in two extrapolation (left and right target) and two interpolation (2x central target)
blocks. The order of these four blocks was counter-balanced across subjects.
The movements’ spatial accuracy was recorded for each target before (PRE), during
(PRISM), and after (POST) wearing prism goggles. In each condition, 30 pointing movements
had to be performed. These measurements were carried out i) to reveal potential target-related
spatial biases, ii) to trace the temporal courses of prism adaptation (PRISM) and de-adaptation
(POST), and iii) to evaluate the size of the standard aftereffect in each target. In the analysis,
we compared the size of the standard aftereffect with the size of the extrapolation and
interpolation aftereffects for the corresponding target.

Studie 3: Räumliche Generalisierung des Nacheffekts 78
Data analysis
For each individual trial, the horizontal distance between the endpoint of the movement and
the target was assessed (in mm) and analyzed as a function of trial number. For further
analysis, a baseline correction was performed. That means, we subtracted the average lateral
deviation of pointing movements of the baseline condition (PRE, no prisms) of a specific
target from each data point of the corresponding target. This normalization procedure,
inspired by Clower & Boussaoud (2000), was performed on each subject’s data individually.
Why is normalization desirable? The mean of the baseline condition reflects a subject’s
general visuomotor performance with regard to a specific target and reveals a potential target-
related directional bias. Removing this target-related bias leads to a normalized set of data and
therefore allows a better comparison of effect sizes between targets and subjects.
Furthermore, baseline correction removes effects which are due to finger-target geometry (i.e.
an inherent rightward deviation of movements when pointing to the right target). In
Experiment 1, the spatial bias averaged -7.4 mm (± 0.7 SEM) in the left target, +16.1 mm (±
0.9) in the central target and +29.4 mm (± 1.0) in the right target. Spatial biases of -4.6 mm (±
0.8) in the left target, +11.0 mm (± 0.6) in the central target and +28.4 mm (± 0.7) in the right
target were recorded in Experiment 2.
To assess the size of aftereffects, a linear regression line was fitted to the first four data points
of each condition and the intersection of the regression line with the y-axis was taken as a
measure of the aftereffect. Why not simply taking the result of the first movement?
Aftereffects do not completely vanish after the first movement but decrease stepwise over the
first few trials. Our method incorporates this early and rapid decrement of lateral errors, and
reduces noise in the data by using four data points rather than a single one. Moreover, the
slope of the regression line indicates the speed of de-adaptation (aftereffect).
Subjects
A total of 20 right-handed human subjects with no reported history of neurological or
ophthalmologial disorders was recruited. In Experiment 1, 10 subjects (5 female, 5 male) aged
23-29 years (mean 25.4 years) participated. Another 10 subjects (5 female, 5 male) aged 21-
29 years (mean 25.1 years) took part in Experiment 2. All subjects had normal or corrected-
to-normal (contact lenses) vision. Subjects gave written informed consent prior to
participation and were naïve to the exact experimental question of the study. All experiments
were carried out in accordance with the “Declaration of Helsinki” and this type of
psychophysical experiments was approved by the ethics committee of Bremen University.
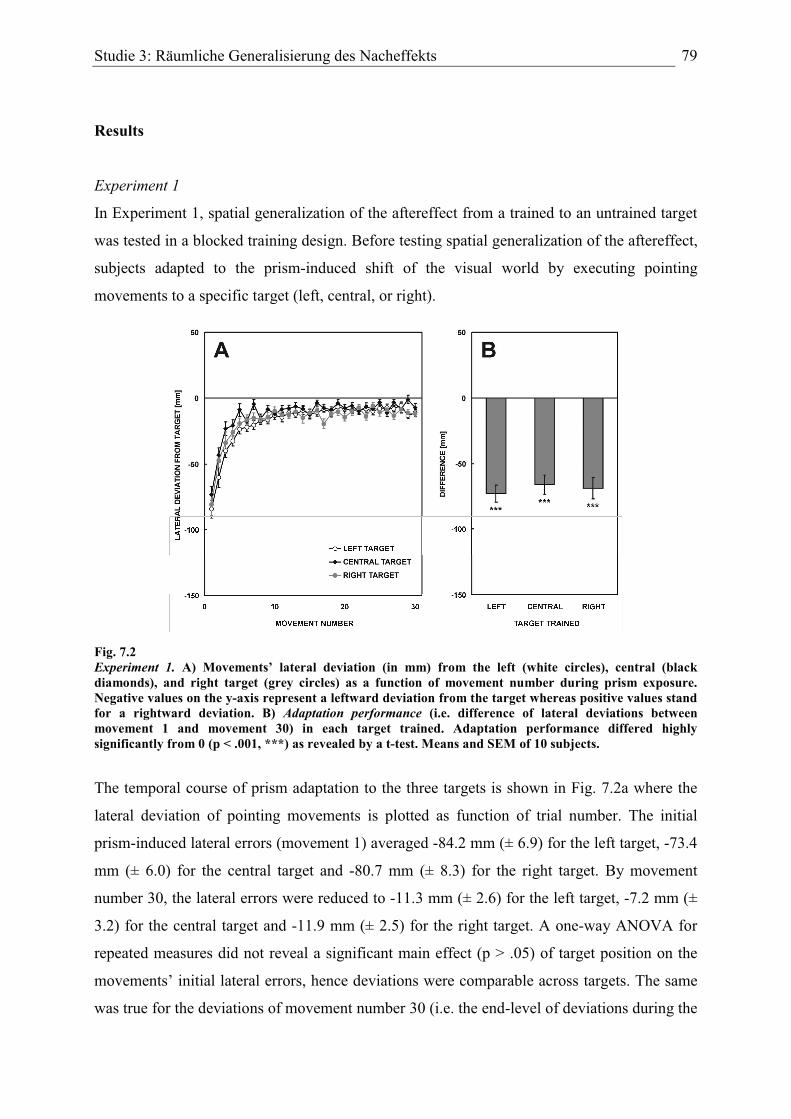
Studie 3: Räumliche Generalisierung des Nacheffekts 79
Results
Experiment 1
In Experiment 1, spatial generalization of the aftereffect from a trained to an untrained target
was tested in a blocked training design. Before testing spatial generalization of the aftereffect,
subjects adapted to the prism-induced shift of the visual world by executing pointing
movements to a specific target (left, central, or right).
Fig. 7.2 Experiment 1. A) Movements’ lateral deviation (in mm) from the left (white circles), central (black diamonds), and right target (grey circles) as a function of movement number during prism exposure. Negative values on the y-axis represent a leftward deviation from the target whereas positive values stand for a rightward deviation. B) Adaptation performance (i.e. difference of lateral deviations between movement 1 and movement 30) in each target trained. Adaptation performance differed highly significantly from 0 (p < .001, ***) as revealed by a t-test. Means and SEM of 10 subjects.
The temporal course of prism adaptation to the three targets is shown in Fig. 7.2a where the
lateral deviation of pointing movements is plotted as function of trial number. The initial
prism-induced lateral errors (movement 1) averaged -84.2 mm (± 6.9) for the left target, -73.4
mm (± 6.0) for the central target and -80.7 mm (± 8.3) for the right target. By movement
number 30, the lateral errors were reduced to -11.3 mm (± 2.6) for the left target, -7.2 mm (±
3.2) for the central target and -11.9 mm (± 2.5) for the right target. A one-way ANOVA for
repeated measures did not reveal a significant main effect (p > .05) of target position on the
movements’ initial lateral errors, hence deviations were comparable across targets. The same
was true for the deviations of movement number 30 (i.e. the end-level of deviations during the
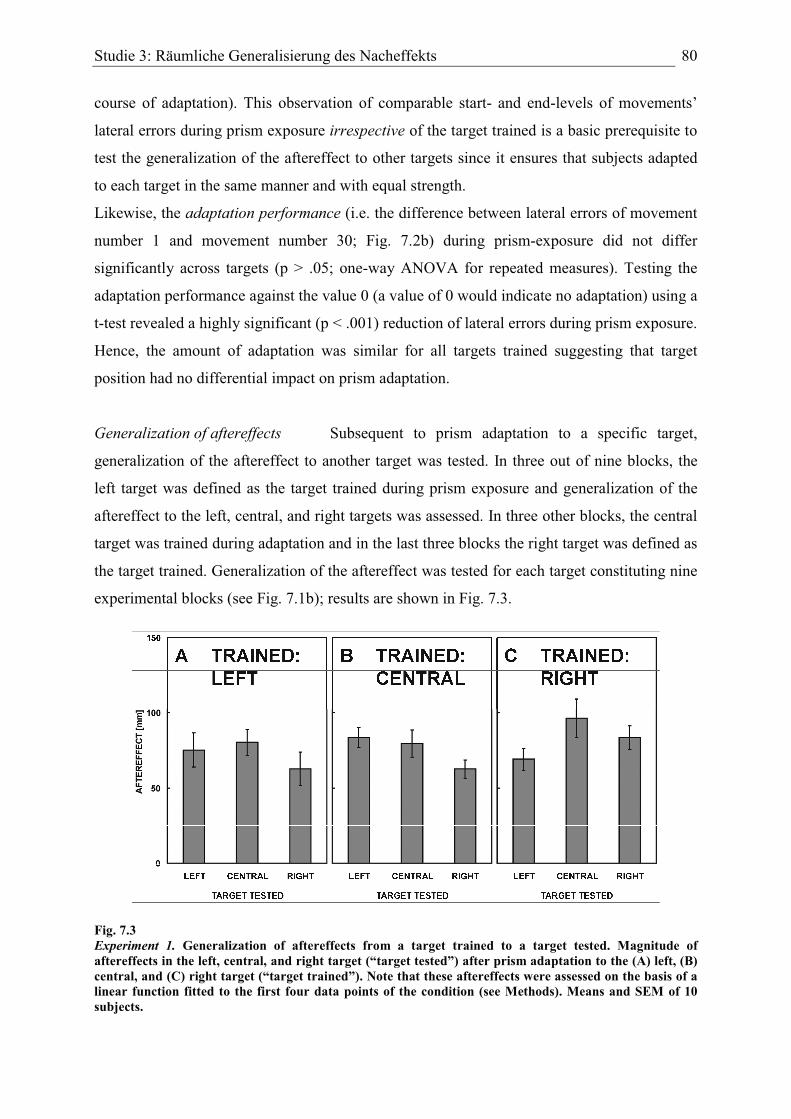
Studie 3: Räumliche Generalisierung des Nacheffekts 80
course of adaptation). This observation of comparable start- and end-levels of movements’
lateral errors during prism exposure irrespective of the target trained is a basic prerequisite to
test the generalization of the aftereffect to other targets since it ensures that subjects adapted
to each target in the same manner and with equal strength.
Likewise, the adaptation performance (i.e. the difference between lateral errors of movement
number 1 and movement number 30; Fig. 7.2b) during prism-exposure did not differ
significantly across targets (p > .05; one-way ANOVA for repeated measures). Testing the
adaptation performance against the value 0 (a value of 0 would indicate no adaptation) using a
t-test revealed a highly significant (p < .001) reduction of lateral errors during prism exposure.
Hence, the amount of adaptation was similar for all targets trained suggesting that target
position had no differential impact on prism adaptation.
Generalization of aftereffects Subsequent to prism adaptation to a specific target,
generalization of the aftereffect to another target was tested. In three out of nine blocks, the
left target was defined as the target trained during prism exposure and generalization of the
aftereffect to the left, central, and right targets was assessed. In three other blocks, the central
target was trained during adaptation and in the last three blocks the right target was defined as
the target trained. Generalization of the aftereffect was tested for each target constituting nine
experimental blocks (see Fig. 7.1b); results are shown in Fig. 7.3.
Fig. 7.3 Experiment 1. Generalization of aftereffects from a target trained to a target tested. Magnitude of aftereffects in the left, central, and right target (“target tested”) after prism adaptation to the (A) left, (B) central, and (C) right target (“target trained”). Note that these aftereffects were assessed on the basis of a linear function fitted to the first four data points of the condition (see Methods). Means and SEM of 10 subjects.

Studie 3: Räumliche Generalisierung des Nacheffekts 81
When the left target was defined as the target trained (Fig. 7.3a) and the aftereffect was tested
for the same target (trained = tested), the aftereffect reached +75.3 mm (± 11.4). Testing the
aftereffect for the central target after training the left target resulted in an aftereffect of +80.3
mm (± 8.5). A smaller aftereffect of +62.8 mm (± 11.0) was recorded for the right target after
left target prism adaptation. A one-way ANOVA for repeated measures yielded no significant
main effect (p > .05) of target position on the magnitude of the aftereffects suggesting
complete generalization of aftereffects to all targets tested following blocked visuomotor
training to the left target.
For prism adaptation to the central target (Fig. 7.3b), pointing to the left target after removal
of the prisms yielded an aftereffect of +83.6 mm (± 6.5); the aftereffect for the central target
after adapting with the central target (trained = tested) was slightly smaller (+79.6 mm, ± 9.0).
The aftereffect decreased to +62.5 mm (± 6.1) for testing the right target after adaptation to
the central target. Here, a one-way ANOVA revealed a significant main effect (p < .05) of
target position on the magnitude of the aftereffect indicating a partial generalization of
aftereffects after prism adaptation to the central target.
When the right target was defined as the target trained during prism adaptation (Fig. 7.3c), an
aftereffect of +69.1 mm (± 7.3) was recorded when testing the left target. The aftereffect
increased to +96.3 mm (± 12.8) when it was tested for the central target after right target
prism adaptation. When generalization was assessed for the right target after adapting with the
right target (trained = tested), the aftereffect reached +83.5 mm (± 7.9). A one-way ANOVA
for repeated measures did not show a significant effect (p > .05) of target position on the
magnitude of the aftereffects indicating generalization across target locations tested.
In addition, the size of aftereffects was analyzed as a function of the spatial distance between
the target trained during prism adaptation and the target tested (Fig. 7.4). Data of conditions
were pooled according to the spatial distance between targets trained and those tested. When
the targets trained were identical to those tested (i.e. trained: left, tested: left etc.) the
aftereffect reached on average +79.4 mm (± 7.2). Proceeding one “target step” to the right
after prism adaptation (i.e. +250 mm; trained: left, tested: central; or else trained: central,
tested: right) decreased the aftereffect to +71.4 mm (± 5.6). Further increasing the spatial
distance between targets trained and tested by two “target steps” to the right (i.e. +500 mm;
trained: left, tested: right) reduced the aftereffect even more to +62.8 mm (± 11.0).
In the leftward direction, increasing the spatial distance between the target trained and the
target tested by -250 mm (i.e. one “target step” to the left; trained: right, tested: central; or

Studie 3: Räumliche Generalisierung des Nacheffekts 82
else trained: central, tested: left) increased the aftereffect to +89.9 mm (± 8.4). However,
when the spatial difference between targets trained and tested was as large as -500 mm (i.e.
two “target steps” to the left; trained: right, tested: left), the aftereffect dropped markedly to
+69.1 mm (± 7.3). Although there was an apparent stepwise trend within this set of data, a
one-way ANOVA for repeated measures did not show a significant main effect (p > .05) of
the spatial distance between targets trained and those tested on the magnitude of the
aftereffect, confirming (almost) complete generalization across target locations. To sum up,
under blocked visuomotor training conditions (i.e. a single target is trained during prism
adaptation) the aftereffect generalized to a first approximation linearly to untrained target
locations.
Fig. 7.4 Experiment 1. Magnitude of generalization aftereffects as a function of the spatial distance between the target trained and the target tested. Negative values on x-axis indicate that the target tested (generalization aftereffect) lies leftward from the trained target during prism adaptation whereas positive values denote that the target tested is located rightward from the target trained. Means and SEM of 10 subjects.
Experiment 2
In Experiment 2, subjects adapted by executing pointing movements alternately to two visual
targets in space and after this “mixed training” procedure generalization of the aftereffect to a
target located beyond (i.e. extrapolation task) or within (i.e. interpolation task) the adapted
visuomotor workspace was tested. In order to compare extrapolation and interpolation
aftereffects with the standard aftereffect in each target, the experiment began with recordings
of conventional prism adaptation and de-adaptation (aftereffect) for each target (single-target
pointing).
The lateral deviation of pointing movements as a function of trial number is displayed in Fig.
7.5a (adaptation) and Fig. 7.5b (de-adaptation) for each target. The movements’ initial prism-
induced lateral error (movement number 1) reached -97.0 mm (± 8.6) for the left target, -86.7
mm (± 6.9) for the central target, and -91.7 mm (± 10.9) for the right target. A one-way

Studie 3: Räumliche Generalisierung des Nacheffekts 83
ANOVA for repeated measures did not reveal any significant differences between target
positions (p > .05). In the course of prism adaptation, lateral errors were reduced to -12.2 mm
(± 4.3) for the left target, -5.4 mm (± 12.0) for the central target and -6.4 mm (± 6.1) for the
right target (movement number 30). As in Experiment 1, end-levels of prism adaptation were
comparable across targets (ANOVA: no significant main effect; p > .05) indicating that
subjects adapted in the same manner irrespective of target position in space.
Fig. 7.5 Experiment 2. Movements’ lateral deviation (in mm) from the left (white circles), central (black diamonds), and right target (grey circles) as a function of movement number during (A) prism adaptation and (B) de-adaptation (single-target pointing). Negative values on the y-axis represent a leftward deviation from the target whereas positive values stand for a rightward deviation. C) Adaptation performance (i.e. difference of lateral deviations between movement 1 and movement 30) and de-adaptation performance(i.e. difference of lateral deviations between movement 31 and movement 60) in the left (white bars), central (black bars) and right target (grey bars). Adaptation and de-adaptation performances differed highly significantly from 0 (p < .001, ***) as revealed by a t-test. Means and SEM of 10 subjects.
Removing the prisms (de-adaptation, Fig. 7.5b) led to pronounced initial lateral errors to the
right (aftereffect; movement number 31) amounting to +70.1 mm (± 4.2) for the left target,
+99.7 mm (± 10.2) for the central target and +76.7 mm (± 6.8) for the right target. A one-way
ANOVA for repeated measures revealed a significant main effect (p < .01) of target position
on the initial lateral deviation which is likely due to the increased aftereffect measured for the
central target. The end-levels of de-adaptation (movement number 60) on the other hand were
comparable across targets (ANOVA: no significant main effect; p > .05); lateral errors
reached -2.9 mm (± 1.9) for the left target, +5.5 mm (± 5.5) for the central target and -7.1 mm
(± 4.5) for the right target.

Studie 3: Räumliche Generalisierung des Nacheffekts 84
Adaptation performance (i.e. the difference of lateral errors between movement number 1 and
movement number 30) and de-adaptation performance (i.e. the difference of lateral errors
between movement number 31 and movement number 60) did not differ significantly
(ANOVA; p > .05) across targets (Fig. 7.5c) confirming that the improvement through
practice was largely the same irrespective of target position. Yet, adaptation and de-adaptation
performances differed highly significantly from 0 (t-test; p < .001). Together, these findings
confirm that adaptive behaviors were comparable across targets during single-target pointing.
Extrapolation aftereffects During prism adaptation in Experiment 2, subjects pointed
alternately to two target locations. After adapting for a specific range of the visuomotor
workspace generalization of the aftereffect to a target located beyond the adapted workspace
(i.e. extrapolation) was tested. The end-levels of mixed training adaptation for each target (i.e.
the mean lateral deviation of the last 5 movements) were assessed in order to compare the
completeness of the adaptation process. For the right target, the lateral error at the end-level
reached -5.5 mm (± 3.6) and was not significantly different from 0 (t-test; p > .05) whereas
for the central target the end-level of -8.7 mm (± 3.4) differed significantly from 0 (p < .05)
suggesting that adaptation to the central target was slightly incomplete. After adapting to the
right and the central target the extrapolation aftereffect (Fig. 7.6a, left panel) for the left target
reached +95.0 mm (± 4.1) whereas the standard aftereffect for the left target only amounted to
+68.9 mm (± 4.6) and this difference was statistically clearly significant (p < .01) as revealed
by a paired two-sided t-test. Thus, in comparison with the standard aftereffect for the left
target, the extrapolation aftereffect was significantly increased.
An extrapolation aftereffect for the right target was measured after adapting to both the left
and central targets (Fig. 7.6a, right panel). End-levels of mixed training adaptation for both
targets did not differ significantly from 0 (p > .05) indicating complete adaptation. The
extrapolation aftereffect for the right target reached +64.2 mm (± 6.8) whereas the standard
aftereffect for the right target was larger and averaged +78.8 mm (± 6.7). However, statistical
analysis did not show any significant difference between these two conditions (p > .05).
Interpolation aftereffects To measure an interpolation aftereffect, i.e. a possible
generalization of the aftereffect to a target located within the adapted visuomotor workspace,
the left and right targets were trained during prism adaptation and the aftereffect was assessed
for the central target. For the left target, the end-level of mixed training adaptation reached
+3.2 mm (± 2.6) and did not differ significantly from 0 (p > .05). However, the end-level for

Studie 3: Räumliche Generalisierung des Nacheffekts 85
the right target amounted to -19.1 mm (± 6.3) and was statistically different from 0 (p < .05)
indicating incomplete adaptation for the right target during mixed training.
The interpolation aftereffect for the central target amounted to +78.2 mm (± 4.5) and was
significantly (p < .05) smaller that the standard aftereffect for the central target (+93.0 mm, ±
8.4) as shown by a paired two-sided t-test (Fig. 7.6a, central panel). That is, the interpolation
task yielded a significantly smaller aftereffect than the standard aftereffect for the central
target.
Fig. 7.6 Experiment 2. A) Magnitude of standard aftereffects (black bars) and extrapolation (white bars) and interpolation (grey bar) aftereffects in the left, central, and right target. Note that these aftereffects were assessed on the basis of a linear function fitted to the first four data points of the condition (see Methods). B) Difference between the extrapolation aftereffect and the standard aftereffect (white bars) and between the interpolation aftereffect and the standard aftereffect (grey bar) in the corresponding target. Means and SEM of 10 subjects.
The standard aftereffect for specific targets was subtracted from the extrapolation and
interpolation aftereffects, respectively for each subject individually to evaluate the differences
between standard and extrapolation/interpolation aftereffects (Fig. 7.6b). The left target
extrapolation aftereffect was +26.1 mm (± 6.3) larger than the left target standard aftereffect

Studie 3: Räumliche Generalisierung des Nacheffekts 86
(p < .01; t-test against the value 0). The extrapolation aftereffect for the right target was -14.6
mm (± 10.2) smaller than the standard aftereffect for the right target but this difference was
not statistically significant (p > .05). In case of an interpolation, the aftereffect for the central
target was -14.8 mm (± 5.6) smaller than the central target standard aftereffect and this
difference was statistically significant (p < .05).
Discussion
Prism adaptation demonstrates the rapid experience-dependent updating of sensorimotor
representations when a lateral shift of the visual world is introduced by prisms. In the present
study, we addressed the spatial specificity of the adaptation process under different training
conditions.
Experiment 1: Linear generalization across target locations
In Experiment 1, visuomotor training during prism adaptation occurred as the usual blocked
training. Subjects adapted by pointing to a single visual target and immediately subsequent to
adaptation we tested generalization of the aftereffect to another visual target. The overall
pattern of results was as follows: Aftereffects largely transferred to other target locations
irrespective of whether the left, central, or right target was trained in a blocked fashion during
adaptation. Pointing to a target different from that trained during adaptation did not decrease
the size of the aftereffect. This linear generalization suggests a global type of adaptation
causing a relatively rigid shift of the entire visuomotor mapping which is in line with a
number of previous studies (e.g. Bedford, 1989; 1993; Fernandez-Ruiz et al., 2006; Redding
& Wallace, 2006a).
In the experimental setting used, each of the three visual targets had a distance of 25 cm to its
“neighbor”. Subjects were oriented such that the distal part of the right shoulder faced the
central target in order to ensure that subjects could reach out to each target in a convenient
way. This reduced the risk to obtain asymmetries in pointing behaviors simply due to the
mechanics of the body. Furthermore, our baseline correction method removed directional
biases related to target location. As is obvious from Fig. 7.2, adaptive behaviors did not
depend upon the spatial location of the target hence subjects adapted with equal strength to
each of the three targets. In particular, equal adaptation performances allowed a fair
comparison of the pattern of generalization between targets.

Studie 3: Räumliche Generalisierung des Nacheffekts 87
In case of nonlinear generalization, increasing the spatial distance between the targets trained
and tested should lead to a decrease of the aftereffect (i.e. a generalization gradient) indicating
that the adaptation mechanism is specific to the exact spatial context of the task executed. A
decreased or even absent aftereffect in an untrained target therefore would indicate that the
system resorts to the original visuomotor mapping as conditions between training and testing
differ sufficiently. However, since considerable and often not significantly smaller aftereffects
occurred even in targets spatially remote from the target trained generalization seems to be
linear.
It is important to keep in mind that the angular distances between visual targets amounted to
approx. 21 deg. (left to central target, or central to right target). That is, switching to another
target after adaptation necessarily included the execution of a new motor trajectory with
different angular arm posture parameters. Although our study did not record target-related
differences according to arm posture the linear generalization of aftereffects clearly shows
that adaptation was not specific to arm geometry (but see also Baraduc & Wolpert, 2002).
Therefore, it is unlikely that a type of motor adaptation specific to movement-related
parameters during task execution occurred. In other words: The aftereffect generalized across
quite different motor trajectories.
Surprisingly, a small but significant (p = .048) effect of target position on the size of the
aftereffect was revealed when the central target was defined as the target trained during
adaptation suggesting a partial generalization under these conditions (see Fig. 7.3b). A
possible explanation of this effect is that adaptation to the central target was somewhat easier
due to body position and that there was an effect of symmetric target positions with respect to
the body on the size of the aftereffect. However, given the fact that differences between these
aftereffects were quite small the overall pattern of linear generalization nevertheless remains
valid.
Linear generalization across locations due to internal constraints
What does the pattern of linear generalization of aftereffects across different visual locations
tell us about the characteristics of the adaptation process? Prism adaptation by pointing to a
discrete visual target in rapid succession produced a global type of spatial remapping affecting
untrained visual locations in the corresponding spatial map. This rigid transformational shift
suggests that task-related spatial representations in the visuomotor brain were modified as a
whole which is consistent with the results of both Bedford (1989; 1993) and Redding &
Wallace (2006a). A conceivable explanation of this finding is that the brain relies on internal

Studie 3: Räumliche Generalisierung des Nacheffekts 88
constraints about the physical world (cf. Bedford, 1989; 1993). Although experience is limited
to a narrow range of the workspace (i.e. pointing to a single target) spatial representations are
updated and shifted as a unit because the brain “assumes” that the represented space is regular
and uniform (a fundamental physical constraint). The prism-induced shift of the visual world
is experienced and compensated for in a local part of the spatial map but since internally
represented physical constraints predict that space is always uniform the spatial remapping
affects all points in space. This means that the rigid linear shift (i.e. global adaptation) results
from adaptation in a local part of the workspace in combination with an internal prediction on
the characteristics of the physical world, i.e. the linearity of space. In another conceptual
framework, this would correspond to a shift in the perceived position of the eyes for the
purpose of arm movements to all types of targets. This result is far from trivial given the lack
of generalization of prism adaptation between different types of movements to the same target
(cf. Baraduc & Wolpert, 2002; Martin et al., 1996b).
A study by Fernandez-Ruiz et al. (2006) in which hemiprisms were used has shown, however,
that topographical modifications in a regional part of the spatial map – as opposed to global
generalization – are also possible under certain conditions. These authors used a different task
than we did (i.e. throwing movements in far space) and a more important difference to our
study is that their subjects received 250 training trials during adaptation. Since the higher
number of training trials allowed more time for interactions, the system was capable of
performing specific local modifications in the spatial map. In our study, visuomotor
experience was restricted to 30 pointing movements in a local part of the workspace; yet
adaptation was global presumably reflecting a tendency to use linear functions as “default” for
the rapid updating of spatial representations (see also Bedford, 1993). Moreover, subjects had
no indication or benefit from using a regional modification of their visuomotor
representations.
Another aspect concerns the localization of adaptive shifts subject to the type of sensory
feedback information during adaptation. There is experimental evidence that under terminal
exposure (i.e. the finger tip is visible only at the end of the trajectory) shifts are larger in the
so-called visual eye-head system than in the proprioceptive hand-head system (see e.g.
Redding & Wallace, 2006a; Uhlarik & Canon, 1971). Our subjects received terminal visual
feedback during prism adaptation. Consequently, an adaptive shift predominantly localized in
visual coding necessarily affects all other points in the spatial map – this mechanism would
also explain the pattern of linear generalization of aftereffects observed in Experiment 1. Both
processes – linear generalization based upon internal physical constraints and the adaptive

Studie 3: Räumliche Generalisierung des Nacheffekts 89
visual shift under terminal exposure – may dynamically add to the rapid adaptive
performance.
Experiment 2: Generalization depends upon the direction tested
Experiment 2 addressed the pattern of spatial generalization of the aftereffect to an untrained
target location under another aspect. A fundamental difference to the first experiment was that
subjects did not perform pointing movements to a single target but alternately pointed to two
different target locations during prism adaptation. In particular, this form of mixed training
adaptation sampled a specific range of the subject’s visuomotor workspace. Following
“workspace adaptation” by mixed training the prisms were removed and generalization of the
aftereffect to a target located beyond (i.e. extrapolation) versus within (i.e. interpolation) the
adapted workspace was tested.
The pattern of generalization varied with the type of task (extrapolation vs. interpolation) and
the direction tested. Whereas the extrapolation aftereffect in the left target was larger than the
standard aftereffect in the left target, the extrapolation aftereffect in the right target was
smaller than the standard aftereffect. For interpolation, the aftereffect in the central target was
likewise reduced compared to the standard aftereffect (see Fig. 7.6). Hence, under these
conditions the pattern of spatial generalization of aftereffects was more complex than that
obtained after a blocked training adaptation procedure (Experiment 1). These results indicate
that the size of the aftereffect decreases for targets in the direction of the aftereffect and
increases for targets in the direction opposite to the aftereffect. This phenomenon of varying
sizes of aftereffects according to direction was investigated in detail in another study (cf.
Fahle et al., 2005). It is noteworthy that the effect of prism adaptation seems not to depend on
inter- vs. extrapolation as in the case for example of object recognition (cf. Bulthoff &
Edelman, 1992).
Underestimation of spatial distances following mixed training adaptation
The amount of generalization of the aftereffect to an untrained target following mixed training
adaptation depended upon the direction tested – how can this pattern of results be explained
with regard to the underlying processing? A possible way to interpret the data is to argue that
the visuomotor system underestimated spatial distances as a consequence of the mixed
training adaptation procedure.
Testing generalization of the aftereffect in the left target (i.e. extrapolation) included prism
adaptation by pointing to the central and right target. In order to point to the left target after

Studie 3: Räumliche Generalisierung des Nacheffekts 90
adaptation, the visuomotor system has to assess the actual spatial distance and to program a
compensatory trajectory in the leftward direction. However, if the spatial distance to the left
target is underestimated as a consequence of mixed training adaptation, the planned trajectory
to the left will become too short and therefore exhibits a certain rightward offset from the
target. This rightward offset and the standard rightward aftereffect sum up and so the
aftereffect recorded in the left target after mixed training adaptation is larger than the
corresponding standard aftereffect (see Fig. 7.6a, left panel). Underestimating the spatial
distance also accounts for the decreased extrapolation aftereffect in the right target compared
with the standard aftereffect. In this case, underestimation leads to a rightward trajectory
which under-compensates the actual distance to the right target and this effect subtracts from
the standard aftereffect. Hence the extrapolation aftereffect is smaller than the standard
aftereffect (see Fig. 7.6a, right panel).
This difference between extrapolation aftereffect and standard aftereffect was more
pronounced (and statistically significant) in the left than in the right target (see Fig. 7.6b).
This may be a consequence of the subject’s body position in space: Since the right shoulder
faced the central target the whole body was slightly shifted in the direction of the left target
and it is possible that this asymmetry of body position with respect to the targets enforced
underestimation of the distance to the left target.
The interpolation aftereffect in the central target was also smaller than the standard aftereffect
(see Fig. 7.6a, central panel) and this difference can also be explained in terms of an
underestimated spatial distance: The compensatory trajectory to the right was too short for the
actual distance and this effect was subtracted from the standard aftereffect. Interestingly, the
mean of both extrapolation aftereffects (+79.6 mm; ± 5.2) virtually equals the interpolation
aftereffect for the central target of +78.2 mm (± 4.5) suggesting a linear relationship for
underestimating spatial distances.
In visuomotor rotation learning (e.g. Krakauer et al., 2000) the extent of generalization
increases as training involves larger amounts of the workspace. In a psychophysical study on
dynamics learning, however, Mattar & Ostry (2007) have shown that the generalization after
training with two targets flanking the tested target location was as large as after training with
multiple targets suggesting that generalization depended upon interpolation between localized
learning. In our study, the interpolation aftereffect was significantly smaller than the standard
aftereffect in the central target hence adaptation by pointing to the flanking targets (i.e. left,
and right) did not entail full generalization.

Studie 3: Räumliche Generalisierung des Nacheffekts 91
The results of Experiment 2 are compatible with the notion that the visuomotor system
underestimates spatial distances as a consequence of the mixed training prism adaptation
procedure and employs a scaled linear function for generalization. In combination with a
rightward workspace shift obtained through mixed training, the adaptation mechanism might
involve a downscaling of spatial representations in order to restore accurate visuomotor
behavior. After adaptation, the downscaled spatial representation is applied to untrained
locations leading to a systematical underestimation of spatial distances.
These results are in line with previous studies on the performance of normals in cognitive
spatial tasks (e.g. line bisection, or number line bisection) after prism adaptation reporting that
especially leftward-shifting prisms affect higher-level spatial representations in the brain due
to hemispheric asymmetry (e.g. Berberovic & Mattingley, 2003; Colent et al., 2000; Loftus et
al., 2008). The role of the mixed training seems to involve an enhancement of the spatial re-
scaling by providing additional visuomotor exploration of the workspace; in contrast, if
training during adaptation is restricted to a single location in the workspace as it was in
Experiment 1, there is no enhancement effect hence generalization is linear and not scaled.
Acknowledgments
The authors declare that they have no competing financial interests. The authors would like to
thank C. Grimsen and S. Straube for the valuable comments on the manuscript.

Abkürzungsverzeichnis 92
Abkürzungsverzeichnis
A1 Primärer auditorischer Cortex
cm Zentimeter
fMRT Funktionelle Magnetresonanztomographie
LIP Laterales intraparietales Areal (engl. lateral intraparietal area)
M1 Primärer motorischer Cortex
mm Millimeter
PET Positronen-Emissions-Tomographie
PMC Prämotorischer Cortex
PMv Ventraler prämotorischer Cortex
PPC Posteriorer parietaler Cortex
PRR Parietale Reich-Region (engl. parietal reach region)
SEM Standardfehler (engl. standard error of the mean)
SMA Supplementär-motorisches Areal
V1 Primärer visueller Cortex
V2 Sekundärer visueller Cortex
VP Versuchsperson
ZNS Zentrales Nervensystem (Gehirn und Rückenmark)

Literaturverzeichnis 93
Literaturverzeichnis
Ahissar, M. & Hochstein, S. (2004). The reverse hierarchy theory of visual perceptual learning. Trends in Cognitive Sciences, 8, 457-464.
Albus, J. S. (1971). A theory of cerebellar function. Mathematical Biosciences, 10, 25-61.
Andersen, R. A. & Buneo, C. A. (2003). Sensorimotor integration in posterior parietal cortex. In A.M.Siegel, R. A. Andersen, H.-J. Freund, & D. Spencer (Hrsg.), Advances in Neurology, Vol.93 The parietal lobes (S. 159-177). Philadelphia, PA: Lippincott Williams & Wilkins.
Andersen, R. A., Meeker, D., Pesaran, B., Breznen, B., Buneo, C., & Scherberger, H. (2004). Sensorimotor transformations in the posterior parietal cortex. In M.S.Gazzaniga (Hrsg.), The cognitive neurosciences (3. Aufl., S. 463-474). Cambridge, MA: MIT Press.
Andersen, R. A., Snyder, L. H., Bradley, D. C., & Xing, J. (1997). Multimodal representation of space in the posterior parietal cortex and its use in planning movements. Annual Review of Neuroscience, 20, 303-330.
Baily, J. S. (1972). Adaptation to prisms: do proprioceptive changes mediate adapted behaviour with ballistic arm movements? Quarterly Journal of Experimental Psychology, 24,8-20.
Baizer, J. S., Kralj-Hans, I., & Glickstein, M. (1999). Cerebellar lesions and prism adaptation in macaque monkeys. Journal of Neurophysiology, 81, 1960-1965.
Baraduc, P. & Wolpert, D. M. (2002). Adaptation to a visuomotor shift depends on the starting posture. Journal of Neurophysiology, 88, 973-981.
Bays, P. M. & Wolpert, D. M. (2007). Computational principles of sensorimotor control that minimize uncertainty and variability. Journal of Physiology, 578, 387-396.
Bedford, F. L. (1989). Constraints on learning new mappings between perceptual dimensions. Journal of Experimental Psychology.Human Perception and Performance, 15, 232-248.
Bedford, F. L. (1993). Perceptual and cognitive spatial learning. Journal of Experimental Psychology.Human Perception and Performance, 19, 517-530.
Bedford, F. L. (1999). Keeping perception accurate. Trends in Cognitive Sciences, 3, 4-11.
Berberovic, N. & Mattingley, J. B. (2003). Effects of prismatic adaptation on judgements of spatial extent in peripersonal and extrapersonal space. Neuropsychologia, 41, 493-503.
Birbaumer, N. & Schmidt, R. F. (2003). Biologische Psychologie. (5. Aufl.) Berlin: Springer.
Blickhan, R. (2001). Motorische Systeme bei Vertebraten. In J.Dudel, R. Menzel, & R. F. Schmidt (Hrsg.), Neurowissenschaft. Vom Molekül zur Kognition (2. Aufl., S. 191-213). Berlin: Springer.

Literaturverzeichnis 94
Bulthoff, H. H. & Edelman, S. (1992). Psychophysical support for a two-dimensional view interpolation theory of object recognition. Proceedings of the National Academy of Sciences of the United States of America, 89, 60-64.
Buneo, C. A. & Andersen, R. A. (2006). The posterior parietal cortex: sensorimotor interface for the planning and online control of visually guided movements. Neuropsychologia, 44,2594-2606.
Buonomano, D. V. & Merzenich, M. M. (1998). Cortical plasticity: from synapses to maps. Annual Review of Neuroscience, 21, 149-186.
Churchland, M. M., Afshar, A., & Shenoy, K. V. (2006). A central source of movement variability. Neuron, 52, 1085-1096.
Clower, D. M. & Boussaoud, D. (2000). Selective use of perceptual recalibration versus visuomotor skill acquisition. Journal of Neurophysiology, 84, 2703-2708.
Clower, D. M., Hoffman, J. M., Votaw, J. R., Faber, T. L., Woods, R. P., & Alexander, G. E. (1996). Role of posterior parietal cortex in the recalibration of visually guided reaching. Nature, 383, 618-621.
Colent, C., Pisella, L., Bernieri, C., Rode, G., & Rossetti, Y. (2000). Cognitive bias induced by visuo-motor adaptation to prisms: a simulation of unilateral neglect in normal individuals? Neuroreport, 11, 1899-1902.
Connolly, J. D., Andersen, R. A., & Goodale, M. A. (2003). FMRI evidence for a 'parietal reach region' in the human brain. Experimental Brain Research, 153, 140-145.
Dahmen, J. C. & King, A. J. (2007). Learning to hear: plasticity of auditory cortical processing. Current Opinion in Neurobiology, 17, 456-464.
Doyon, J. & Benali, H. (2005). Reorganization and plasticity in the adult brain during learning of motor skills. Current Opinion in Neurobiology, 15, 161-167.
Fahle, M. (2002). Introduction. In M.Fahle & T. Poggio (Hrsg.), Perceptual learning (S. IX-XX). Cambridge, MA: MIT Press.
Fahle, M. (2004). Perceptual learning: a case for early selection. Journal of Vision, 4, 879-890.
Fahle, M. (2005). Perceptual learning: specificity versus generalization. Current Opinion in Neurobiology, 15, 154-160.
Fahle, M. (2008). Perceptual learning and sensory plasticity. In L.R.Squire (Hrsg.), New encyclopedia of neuroscience (S. ?). Amsterdam: Elsevier (im Druck).
Fahle, M., Wischhusen, S., & Spang, K. M. (2005). Prism adaptation by gain control. Perception, 34 (Supplement), 79.
Fernandez-Ruiz, J. & Diaz, R. (1999). Prism adaptation and aftereffect: specifying the properties of a procedural memory system. Learning & Memory, 6, 47-53.

Literaturverzeichnis 95
Fernandez-Ruiz, J., Diaz, R., Aguilar, C., & Hall-Haro, C. (2004). Decay of prism aftereffects under passive and active conditions. Cognitive Brain Research, 20, 92-97.
Fernandez-Ruiz, J., Diaz, R., Hall-Haro, C., Vergara, P., Mischner, J., Nunez, L. et al. (2003). Normal prism adaptation but reduced after-effect in basal ganglia disorders using a throwing task. European Journal of Neuroscience, 18, 689-694.
Fernandez-Ruiz, J., Diaz, R., Moreno-Briseno, P., Campos-Romo, A., & Ojeda, R. (2006). Rapid topographical plasticity of the visuomotor spatial transformation. Journal of Neuroscience, 26, 1986-1990.
Fernandez-Ruiz, J., Hall-Haro, C., Diaz, R., Mischner, J., Vergara, P., & Lopez-Garcia, J. C. (2000). Learning motor synergies makes use of information on muscular load. Learning & Memory, 7, 193-198.
Ghez, C. & Thach, W. T. (2000). The cerebellum. In E.R.Kandel, J. H. Schwartz, & T. M. Jessell (Hrsg.), Principles of neural sciences (4. Aufl., S. 832-852). New York: McGraw-Hill.
Gilbert, C. D., Sigman, M., & Crist, R. E. (2001). The neural basis of perceptual learning. Neuron, 31, 681-697.
Glickstein, M. (2007). What does the cerebellum really do? Current Biology, 17, R824-R827.
Godde, B., Stauffenberg, B., Spengler, F., & Dinse, H. R. (2000). Tactile coactivation-induced changes in spatial discrimination performance. Journal of Neuroscience, 20, 1597-1604.
Goodale, M. A. & Milner, A. D. (1992). Separate visual pathways for perception and action. Trends in Neurosciences, 15, 20-25.
Goodale, M. A. & Westwood, D. A. (2004). An evolving view of duplex vision: separate but interacting cortical pathways for perception and action. Current Opinion in Neurobiology, 14,203-211.
Greger, B. & Norris, S. (2005). Simple spike firing in the posterior lateral cerebellar cortex of Macaque Mulatta was correlated with success-failure during a visually guided reaching task. Experimental Brain Research, 167, 660-665.
Grill-Spector, K. & Malach, R. (2004). The human visual cortex. Annual Review of Neuroscience, 27, 649-677.
Halsband, U. & Lange, R. K. (2006). Motor learning in man: a review of functional and clinical studies. Journal of Physiology (Paris), 99, 414-424.
Harris, C. S. (1963). Adaptation to displaced vision: visual, motor, or proprioceptive change? Science, 140, 812-813.
Harris, C. S. (1965). Perceptual adaptation to inverted, reversed, and displaced vision. Psychological Review, 72, 419-444.
Harris, C. S. (1980). Insight or out of sight?: Two examples of perceptual plasticity in the human adult. In C.S.Harris (Hrsg.), Visual coding and adaptability (S. 95-149). Hillsdale, NJ: Lawrence Erlbaum.

Literaturverzeichnis 96
Held, R. & Freedman, S. J. (1963). Plasticity in human sensorimotor control. Science, 142,455-462.
Helmholtz, H. (1867). Handbuch der physiologischen Optik. Leipzig: Leopold Voss.
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H. (2002). Central mechanisms of motor skill learning. Current Opinion in Neurobiology, 12, 217-222.
Hodzic, A., Veit, R., Karim, A. A., Erb, M., & Godde, B. (2004). Improvement and decline in tactile discrimination behavior after cortical plasticity induced by passive tactile coactivation. Journal of Neuroscience, 24, 442-446.
Huk, A. C., Ress, D., & Heeger, D. J. (2001). Neuronal basis of the motion aftereffect reconsidered. Neuron, 32, 161-172.
Ibbotson, M. R. (2005). Physiological mechanisms of adaptation in the visual system. In C.W.G.Clifford & G. Rhodes (Hrsg.), Fitting the mind to the world. Adaptation and after-effects in high-level vision (S. 17-45). Oxford: Oxford University Press.
Karmarkar, U. R. & Dan, Y. (2006). Experience-dependent plasticity in adult visual cortex. Neuron, 52, 577-585.
Kitazawa, S., Kimura, T., & Uka, T. (1997). Prism adaptation of reaching movements: specificity for the velocity of reaching. Journal of Neuroscience, 17, 1481-1492.
Kitazawa, S., Kimura, T., & Yin, P. B. (1998). Cerebellar complex spikes encode both destinations and errors in arm movements. Nature, 392, 494-497.
Kitazawa, S., Kohno, T., & Uka, T. (1995). Effects of delayed visual information on the rate and amount of prism adaptation in the human. Journal of Neuroscience, 15, 7644-7652.
Kitazawa, S. & Yin, P. B. (2002). Prism adaptation with delayed visual error signals in the monkey. Experimental Brain Research, 144, 258-261.
Konczak, J. (2003). Motorisches Lernen. In H.-O.Karnath & P. Thier (Hrsg.), Neuropsychologie (S. 669-676). Berlin: Springer.
Konen, C. S. & Kastner, S. (2008). Two hierarchically organized neural systems for object information in human visual cortex. Nature Neuroscience, 11, 224-231.
Krakauer, J. & Ghez, C. (2000). Voluntary movement. In E.R.Kandel, J. H. Schwartz, & T. M. Jessell (Hrsg.), Principles of neural sciences (4. Aufl., S. 756-781). New York: McGraw-Hill.
Krakauer, J. W., Pine, Z. M., Ghilardi, M. F., & Ghez, C. (2000). Learning of visuomotor transformations for vectorial planning of reaching trajectories. Journal of Neuroscience, 20,8916-8924.
Kurata, K. & Hoshi, E. (1999). Reacquisition deficits in prism adaptation after muscimol microinjection into the ventral premotor cortex of monkeys. Journal of Neurophysiology, 81,1927-1938.

Literaturverzeichnis 97
Loftus, A. M., Nicholls, M. E. R., Mattingley, J. B., & Bradshaw, J. L. (2008). Left to right: Representational biases for numbers and the effect of visuomotor adaptation. Cognition (im Druck).
Luaute, J., Michel, C., Rode, G., Pisella, L., Jacquin-Court, Costes, N. et al. (2006). Functional anatomy of the therapeutic effects of prism adaptation on left neglect. Neurology, 66, 1859-1867.
Marotta, J. J., Keith, G. P., & Crawford, J. D. (2005). Task-specific sensorimotor adaptation to reversing prisms. Journal of Neurophysiology, 93, 1104-1110.
Marr, D. (1969). A theory of cerebellar cortex. Journal of Physiology, 202, 437-470.
Martin, T. A., Greger, B. E., Norris, S. A., & Thach, W. T. (2001). Throwing accuracy in the vertical direction during prism adaptation: not simply timing of ball release. Journal of Neurophysiology, 85, 2298-2302.
Martin, T. A., Keating, J. G., Goodkin, H. P., Bastian, A. J., & Thach, W. T. (1996a). Throwing while looking through prisms. I. Focal olivocerebellar lesions impair adaptation. Brain, 119 ( Pt 4), 1183-1198.
Martin, T. A., Keating, J. G., Goodkin, H. P., Bastian, A. J., & Thach, W. T. (1996b). Throwing while looking through prisms. II. Specificity and storage of multiple gaze-throw calibrations. Brain, 119 ( Pt 4), 1199-1211.
Martin, T. A., Norris, S. A., Greger, B. E., & Thach, W. T. (2002). Dynamic coordination of body parts during prism adaptation. Journal of Neurophysiology, 88, 1685-1694.
Mattar, A. A. & Ostry, D. J. (2007). Modifiability of generalization in dynamics learning. Journal of Neurophysiology, 98, 3321-3329.
Menzel, R. (2001). Neuronale Plastizität, Lernen und Gedächtnis. In J.Dudel, R. Menzel, & R. F. Schmidt (Hrsg.), Neurowissenschaft. Vom Molekül zur Kognition (2. Aufl., S. 478-526). Berlin: Springer.
Michel, C., Pisella, L., Prablanc, C., Rode, G., & Rossetti, Y. (2007). Enhancing visuomotor adaptation by reducing error signals: single-step (aware) versus multiple-step (unaware) exposure to wedge prisms. Journal of Cognitive Neuroscience, 19, 341-350.
Miyauchi, S., Egusa, H., Amagase, M., Sekiyama, K., Imaruoka, T., & Tashiro, T. (2004). Adaptation to left-right reversed vision rapidly activates ipsilateral visual cortex in humans. Journal of Physiology (Paris), 98, 207-219.
Mollon, J. D. & Danilova, M. V. (1996). Three remarks on perceptual learning. Spatial Vision, 10, 51-58.
Morton, S. M. & Bastian, A. J. (2004). Prism adaptation during walking generalizes to reaching and requires the cerebellum. Journal of Neurophysiology, 92, 2497-2509.
Newport, R., Brown, L., Husain, M., Mort, D., & Jackson, S. R. (2006). The role of the posterior parietal lobe in prism adaptation: Failure to adapt to optical prisms in a patient with bilateral damage to posterior parietal cortex. Cortex, 42, 720-729.

Literaturverzeichnis 98
Newport, R. & Jackson, S. R. (2006). Posterior parietal cortex and the dissociable components of prism adaptation. Neuropsychologia, 44, 2757-2765.
Norris, S. A., Greger, B. E., Martin, T. A., & Thach, W. T. (2001). Prism adaptation of reaching is dependent on the type of visual feedback of hand and target position. Brain Research, 905, 207-219.
Palmer, S. E. (1999). Vision science. Photons to phenomenology. Cambridge, MA: MIT Press.
Patzelt, F., Riegel, M., Ernst, U., & Pawelzik, K. (2007). Self-organised critical noise amplification in human closed loop control. Eingereicht.
Paz, R., Wise, S. P., & Vaadia, E. (2004). Viewing and doing: similar cortical mechanisms for perceptual and motor learning. Trends in Neurosciences, 27, 496-503.
Pisella, L., Rossetti, Y., Michel, C., Rode, G., Boisson, D., Pelisson, D. et al. (2005). Ipsidirectional impairment of prism adaptation after unilateral lesion of anterior cerebellum. Neurology, 65, 150-152.
Pleger, B., Foerster, A. F., Ragert, P., Dinse, H. R., Schwenkreis, P., Malin, J. P. et al. (2003). Functional imaging of perceptual learning in human primary and secondary somatosensory cortex. Neuron, 40, 643-653.
Poggio, T. & Bizzi, E. (2004). Generalization in vision and motor control. Nature, 431, 768-774.
Previc, F. H. (1998). The neuropsychology of 3-D space. Psychological Bulletin, 124, 123-164.
Redding, G. M., Rossetti, Y., & Wallace, B. (2005). Applications of prism adaptation: a tutorial in theory and method. Neuroscience and Biobehavioral Reviews, 29, 431-444.
Redding, G. M. & Wallace, B. (1988). Components of prism adaptation in terminal and concurrent exposure: organization of the eye-hand coordination loop. Perception & Psychophysics, 44, 59-68.
Redding, G. M. & Wallace, B. (2002). Strategic calibration and spatial alignment: a model from prism adaptation. Journal of Motor Behavior, 34, 126-138.
Redding, G. M. & Wallace, B. (2003a). Dual prism adaptation: calibration or alignment? Journal of Motor Behavior, 35, 399-408.
Redding, G. M. & Wallace, B. (2003b). First-trial adaptation to prism exposure. Journal of Motor Behavior, 35, 229-245.
Redding, G. M. & Wallace, B. (2004). First-trial adaptation to prism exposure: artifact of visual capture. Journal of Motor Behavior, 36, 291-304.
Redding, G. M. & Wallace, B. (2006a). Generalization of prism adaptation. Journal of Experimental Psychology.Human Perception and Performance, 32, 1006-1022.

Literaturverzeichnis 99
Redding, G. M. & Wallace, B. (2006b). Prism adaptation and unilateral neglect: review and analysis. Neuropsychologia, 44, 1-20.
Robinson, F. R. (1995). Role of the cerebellum in movement control and adaptation. Current Opinion in Neurobiology, 5, 755-762.
Rode, G., Pisella, L., Rossetti, Y., Farne, A., & Boisson, D. (2003). Bottom-up transfer of sensory-motor plasticity to recovery of spatial cognition: visuomotor adaptation and spatial neglect. Progress in Brain Research, 142, 273-287.
Roller, C. A., Cohen, H. S., Kimball, K. T., & Bloomberg, J. J. (2001). Variable practice with lenses improves visuo-motor plasticity. Cognitive Brain Research, 12, 341-352.
Rossetti, Y., Rode, G., Pisella, L., Farne, A., Li, L., Boisson, D. et al. (1998). Prism adaptation to a rightward optical deviation rehabilitates left hemispatial neglect. Nature, 395,166-169.
Schenk, T. (2006). An allocentric rather than perceptual deficit in patient D.F. Nature Neuroscience, 9, 1369-1370.
Scherberger, H. & Andersen, R. A. (2003). Sensorimotor transformation in the posterior parietal cortex. In L.M.Chalupa & J. S. Werner (Hrsg.), The visual neurosciences (S. 1324-1336). Cambridge, MA: MIT Press.
Seitz, A. & Watanabe, T. (2005). A unified model for perceptual learning. Trends in Cognitive Sciences, 9, 329-334.
Serino, A., Angeli, V., Frassinetti, F., & Ladavas, E. (2006). Mechanisms underlying neglect recovery after prism adaptation. Neuropsychologia, 44, 1068-1078.
Shadmehr, R. & Wise, S. P. (2005). The computational neurobiology of reaching and pointing. A foundation for motor learning. Cambridge, MA: MIT Press.
Sterr, A. (2008). Neuronale Plastizität. In S.Gauggel & M. Herrmann (Hrsg.), Handbuch der Neuro- und Biopsychologie (S. 44-53). Göttingen: Hogrefe.
Steven, M. S. & Blakemore, C. (2004). Cortical plasticity in the adult human brain. In M.S.Gazzaniga (Hrsg.), The cognitive neurosciences (3. Aufl., S. 1243-1254). Cambridge, MA: MIT Press.
Sugita, Y. (1996). Global plasticity in adult visual cortex following reversal of visual input. Nature, 380, 523-526.
Tanaka, Y., Miyauchi, S., Misaki, M., & Tashiro, T. (2007). Mirror symmetrical transfer of perceptual learning by prism adaptation. Vision Research, 47, 1350-1361.
Thach, W. T., Goodkin, H. P., & Keating, J. G. (1992). The cerebellum and the adaptive coordination of movement. Annual Review of Neuroscience, 15, 403-442.
Uhlarik, J. J. & Canon, L. K. (1971). Influence of concurrent and terminal exposure conditions on the nature of perceptual adaptation. Journal of Experimental Psychology, 91,233-239.

Literaturverzeichnis 100
van Beers, R. J., Haggard, P., & Wolpert, D. M. (2004). The role of execution noise in movement variability. Journal of Neurophysiology, 91, 1050-1063.
van Beers, R. J., Wolpert, D. M., & Haggard, P. (2002). When feeling is more important than seeing in sensorimotor adaptation. Current Biology, 12, 834-837.
Wade, N. J. & Verstraten, F. A. J. (2005). Accommodating the past: a selective history of adaptation. In C.W.G.Clifford & G. Rhodes (Hrsg.), Fitting the mind to the world. Adaptation and after-effects in high-level vision (S. 83-101). Oxford: Oxford University Press.
Weiner, M. J., Hallett, M., & Funkenstein, H. H. (1983). Adaptation to lateral displacement of vision in patients with lesions of the central nervous system. Neurology, 33, 766-772.
Welch, R. B. (1978). Perceptual modification. Adapting to altered sensory environments. New York: Academic Press.
Welch, R. B., Choe, C. S., & Heinrich, D. R. (1974). Evidence for a three-component model of prism adaptation. Journal of Experimental Psychology, 103, 700-705.
Wurtz, R. H. & Kandel, E. R. (2000a). Central visual pathways. In E.R.Kandel, J. H. Schwartz, & T. M. Jessell (Hrsg.), Principles of neural sciences (4. Aufl., S. 523-547). New York: McGraw-Hill.
Wurtz, R. H. & Kandel, E. R. (2000b). Perception of motion, depth, and form. In E.R.Kandel, J. H. Schwartz, & T. M. Jessell (Hrsg.), Principles of neural sciences (4. Aufl., S. 548-571). New York: McGraw-Hill.












![Ereignis-korrelierte fMRT-Studie zu differentiellen ... · perzeptuelles Lernen, sowie die klassische Konditionierung [127]. Das nicht-deklarative Gedächtnis ist entscheidend zur](https://static.fdokument.com/doc/165x107/5b9f5d3d09d3f2d0208d0f85/ereignis-korrelierte-fmrt-studie-zu-differentiellen-perzeptuelles-lernen.jpg)




![Diplomarbeit Leyrer 3.0 upload - CORE · Aktivität im ACC auch durch rein perzeptuelles Rezipieren von ... [15]. Wenn Patienten nun lernen würden, die Aktivität ... Sie wissen](https://static.fdokument.com/doc/165x107/5b9f16e309d3f25b318c8743/diplomarbeit-leyrer-30-upload-core-aktivitaet-im-acc-auch-durch-rein-perzeptuelles.jpg)
