
Marine Nahrungsnetze und Trophiestufen · groesste-tier-der-welt-im-portraet.html Ein Extrem:...
86
Marine Nahrungsnetze und Trophiestufen Biologie für die Geowissenschaften Stefanie Ismar WS 2017-2018 worldoceanreview.com oceanworld.tamu.edu
Transcript of Marine Nahrungsnetze und Trophiestufen · groesste-tier-der-welt-im-portraet.html Ein Extrem:...

Marine Nahrungsnetze und Trophiestufen
Biologie für die Geowissenschaften
Stefanie Ismar
WS 2017-2018
worldoceanreview.com
oceanworld.tamu.edu

Einführung - Marine Nahrungsnetze
• Pelagische Systeme
Gesellschaften des Freiwasserbereichs
• Benthische Systeme
Gesellschaften der Organismen, die in oder auf dem
Seeboden leben

• Im Gegensatz zu terrestrischen Systemen (z.B.
Ungulaten – Gras) fast keine reinen Herbivoren
• Biochemische Unterschiede zwischen pflanzlicher und
tierischer Biomasse sind weniger ausgeprägt als an
Land (C:N Verhältnis, Proteingehalt, strukturelle
Polymere)
• Größe und Beweglichkeit sind wichtiger für die
Nahrungswahl
Herbivorie – Omnivorie - Carnivorie

• Plankton: “driftende“ Organismen der pelagischen Zone; • ihre Bewegung und Transport sind größtenteils der Strömung unterworfen
• Funktionelle Gruppen
– Phytoplankton: pflanzliche (O2-abspaltende) Photosynthese – Zooplankton: tierische Ernährungsform – Ernährung durch partikuläre organische
Substanz (POC) – Bacterioplankton: typischerweise heterotroph – Ernährung durch gelöste
organische Substanz (DOC), an Redox-Oberflächen auch chemoautotrophe Bacteria (häufiger im Sediment)
• Phytoplankton-Zooplankton Unterscheidung nicht immer klar gegeben:
einige mixotrophe Gruppen
Plankton

• Nekton: aktiv schwimmende Organismen; Fische, Cephalopoden, marine Säuger
• Übergang zum Plankton fließend; Krill, Fischlarven
Nekton
Was bestimmt, ob ein Organismus zum Plankton oder zum Nekton zählt?

Re = a v ρ η-1
• a: charakteristische Länge [m]
• v: Fließgeschwindigkeit [m s-1]
• ρ: Dichte der Flüssigkeit [kg m-3]
• η: dynamische Viskosität [kg m-1 s-1]; water at 20°C ca. 10-3 kg m-1 s-
1
typische Reynoldszahlen: schwimmender Wal 108
schwimmender Hering 105
flüchtender Mesozooplankter (1mm) 102
schwimmender Ciliat (100 µm) 10-1
sinkende, große Diatomee 10-2
Wasserstrom durch einen Zooplankton Filter (1 µm) 10-3
flagelliertes Bacterium 10-4
Reynoldszahl
nach Sommer 1998

laminarer Strom um einen Partikel
diffuse Grenzschicht
Wasser “klebt” an der Zelle
die Sinkgeschwindigkeit folgt Stokessche Gesetz
Konsequenzen einer geringen Reynoldszahl

v = 2g r2 (ρ'- ρ) (9 ηΦ)-1
• v: Sinkgeschwindigkeit (<0: Auftriebsgeschwindigkeit) [m s-1]
• g: Erdbeschleunigung [9.8 m s-2]
• r: Radius einer volumengleichen Kugel [m]
• ρ': Dichte des sinkenden Partikels [kg m-3]
• ρ: Dichte des umgebenden Mediums
• η: dynamische Viskosität des umgebenden Mediums [kg m-1 s-1]
• Φ: Formwiderstand, dimensionslos, 1 für eine Kugel, Maximum ca. 4
Stokessches Gesetz

große Partikel sinken schneller (quadratische Abhängigkeit)
dichtere Partikel sinken schneller (Dichte-Differenz zu Meerwasser entscheidend!)
Form-Widerstand weniger entscheidend als Größe oder Dichte
Typische Sinkgeschwindigkeiten:
• Coscinodiscus wailesii (große Diatomee; ca. 75 µm): ca. 9 m d-1
• mittelgroße Diatomee (102 to 103 µm3 cell volume): ca. 1 m d-1
• große nicht-Diatomeen: 10-1 m d-1
• kleine Diatomeen (< 100 µm3): 10-2 m d-1
Konsequenzen des Stokesschen Gesetz für Phytoplankton

Größenkl. Von - bis typische Vertreter
Nano- 2-20 µm Flagellaten, Ciliaten
Mikro- 20-200 µm Flagellaten, Ciliaten, Rotatorien, Nauplius-Larven
Meso- 0.2 - 2 mm Ciliaten, Copepoden, Cladoceren, Appendicularien
Makro- 2 mm - 2 cm Copepoden, Krill, Appendicularien
Mega- >2 cm Ctenophora, Cnidaria, Krill, Chaetognathen, Salpen
Plankton-Größenklassen

• Wofür ist Körpergröße auschlaggebend?

Größe entscheidendster Faktor beim ‚Wer frisst Wen‘
‚Große Fische fressen kleine Fische‘ Pieter Bruegel the Elder (1526–1569)
XQE024hSXOrhwQ at Google Cultural Institute maximum zoom level, Gemeinfrei, https://commons.wikimedia.org/w/index.php?curid=21791927

Wichtigstes Kriterium zur Bestimmung des Nahrungsspektrums
Verhältnis Nahrungsgröße : Körpergröße
• ca. 1:103 bei Filtrierern (mm - µm)
• ca. 1:102 bei Partikel-/ Beutegreifern
• fast 1:1 bei einigen Protozoen
Größe und Nahrungswahl
nach Sommer 1998

Blauwal Balaenoptera musculus 30 m
ernährt sich von Krill Euphausia superba
3 - 5 cm
1000 : 1
https://en.wikipedia.org/wiki/Antarctic_krill http://www.t-online.de/leben/familie/id_75214698/blauwal-das-groesste-tier-der-welt-im-portraet.html
Ein Extrem: Filtrierer

Salpa maxima 10 cm filtert Bacterien, Algen,
Protozoen 1 - 1000 µm
100 – 100 000 : 1
http://smrt.ccel.ca/2014/01/28/salpa-maxima/
Ein Extrem: Filtrierer

Größe
wichtig für • metabolische Raten und Wachstumsraten
• Austausch von Materie mit der Umgebung (O:V-Verhältnis)
• Hydromechanik (Reynoldszahl, Stokessches Gesetz)
• Prädatoren
• Nahrungsspektrum
• Methodik der Probennahme
nach Sommer 1998
Plankton: Form und Funktion

Beutegröße und Clearance Rate
Copepoden: macrophag
Tunicaten, Muschel-Veliger:
microphag
Sommer 2005 aus:
A. Katechakis et al (2004) J Plankton Res 26:589-603
F. Sommer et al (2000) Mar Ecol Progr Ser 199:43-63
U. Sommer et al (2002) Hydrobiologia 484:11-20
U. Sommer et al (2002) Mar Ecol Progr Ser 239:251-261

Trophische Ebene
• Ernährungsstufe (oder Trophiestufe): Gesamtheit der Organismen mit gleicher Position in der Nahrungskette

Marine pelagische Nahrungskette
Sekundärkonsument trophische Ebene 3 P3 = 3.4 g m-2 y-1…0.09 P2 ,0.013 P1
B3 = 2 g m-2
Primärkonsument trophische Ebene 2 P2 = 38.4 g m-2 y-1.......0.14 P1
B2 = 1.5 g m-2
Primärproduzent trophische Ebene 1 P1 = 276 g m-2 y-1
B1 = 4 g m-2
Tait 1981, Br. Channel
Größenverhältnis: 10-100 : 1
Größenverhältnis: 10-100 : 1

Energiebilanz der Heterotrophen
Ingestion: I
Assimliation: A = I – D D: Defacation
Produktion: P = A – R – E R: Respiration
E: Exkretion
Assimilations-Effizienz: AQ = A/I
Brutto-Wachstumseffizienz: K1 = P/I
Netto-Wachstumseffizienz: K2 = P/A
Ökologische Effizienz: E = Ppredator/Pprey
Effizienz der Nahrungsumsetzung
nach Sommer 2002
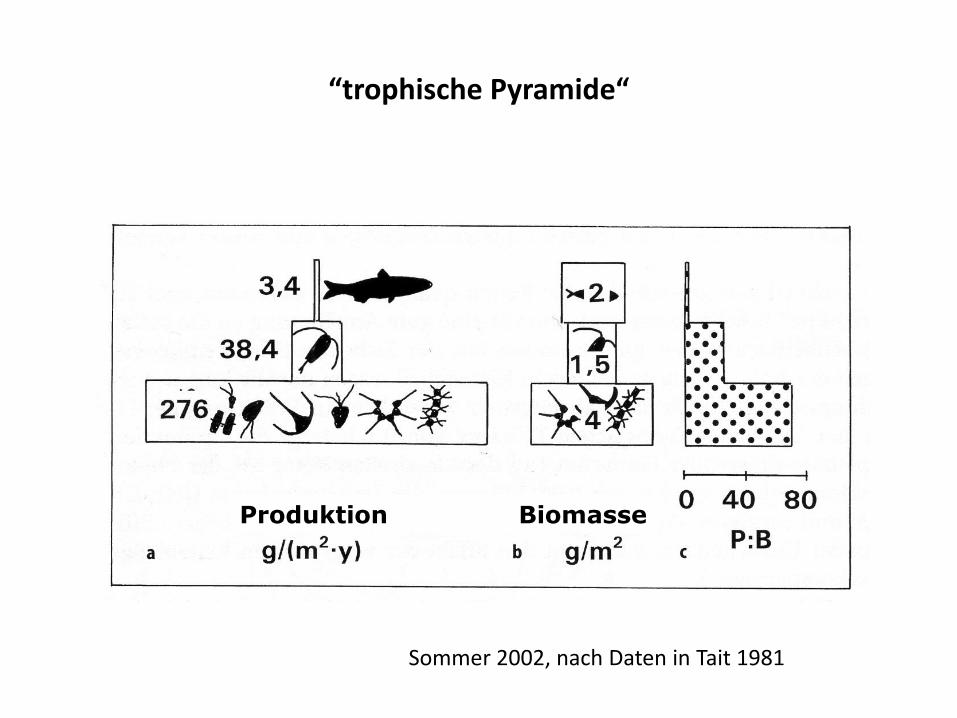
“trophische Pyramide“
Produktion Biomasse
Sommer 2002, nach Daten in Tait 1981
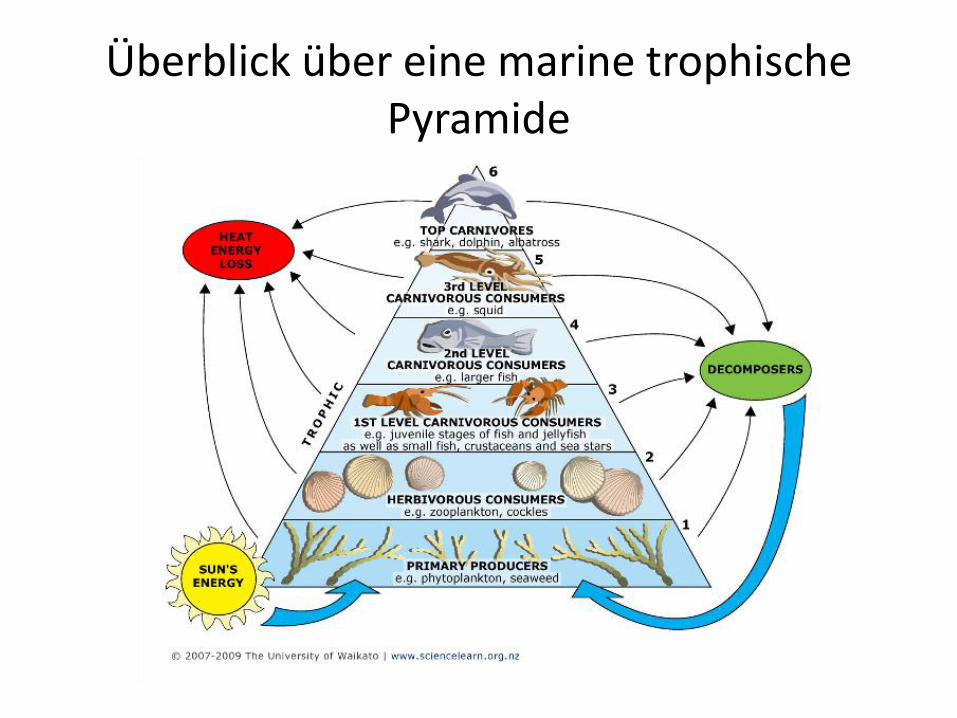
Überblick über eine marine trophische Pyramide
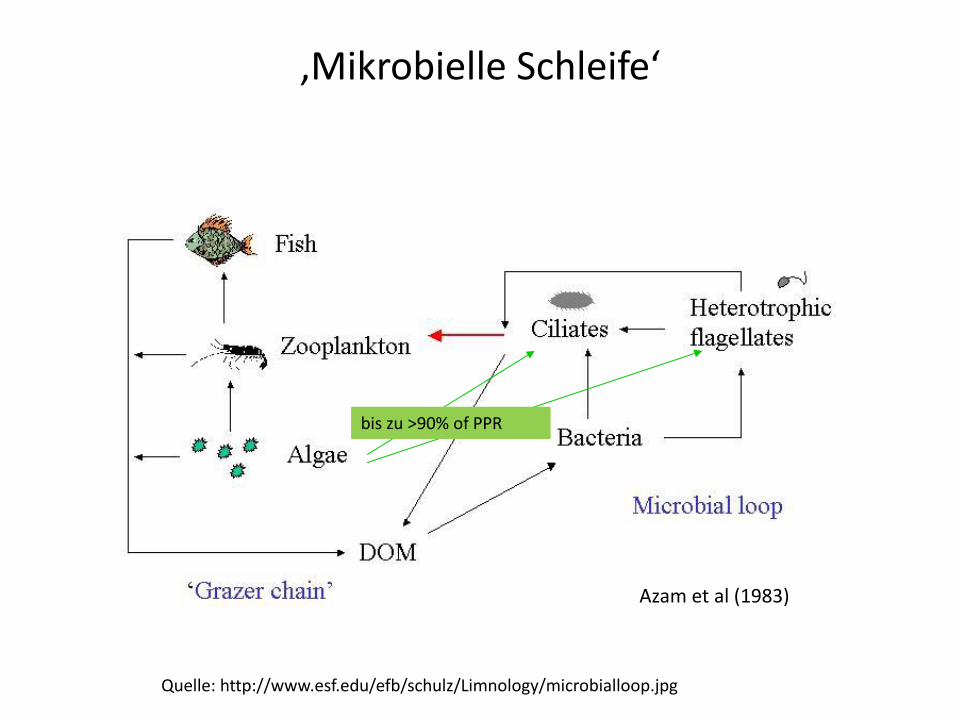
Quelle: http://www.esf.edu/efb/schulz/Limnology/microbialloop.jpg
bis zu >90% of PPR
Azam et al (1983)
‚Mikrobielle Schleife‘

Pelagisches Nahrungsnetz
Sommer et al. (2002)
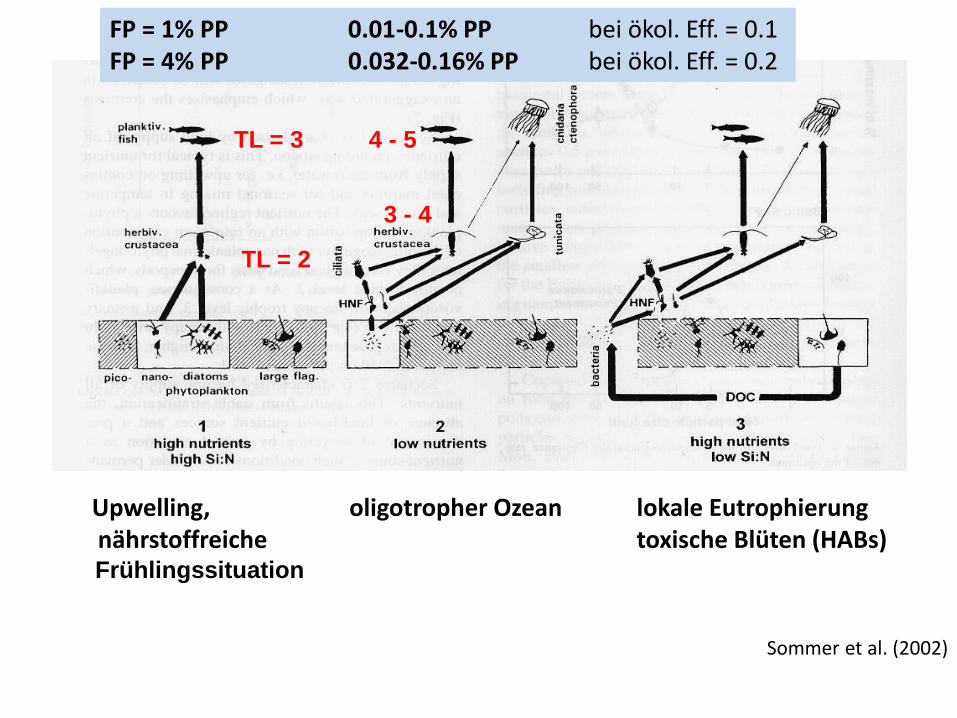
Upwelling, oligotropher Ozean lokale Eutrophierung nährstoffreiche toxische Blüten (HABs) Frühlingssituation
Sommer et al. (2002)
TL = 2
TL = 3
3 - 4
4 - 5
FP = 1% PP 0.01-0.1% PP bei ökol. Eff. = 0.1 FP = 4% PP 0.032-0.16% PP bei ökol. Eff. = 0.2

Die klassische Rolle: Apexprädator und Omnivor:
Paraphysomonas frisst Quadricilia frisst
Bacterien Cryothecomonas und
Chrysochromulina
Qu
Photos: M. Moustaka-Guoni
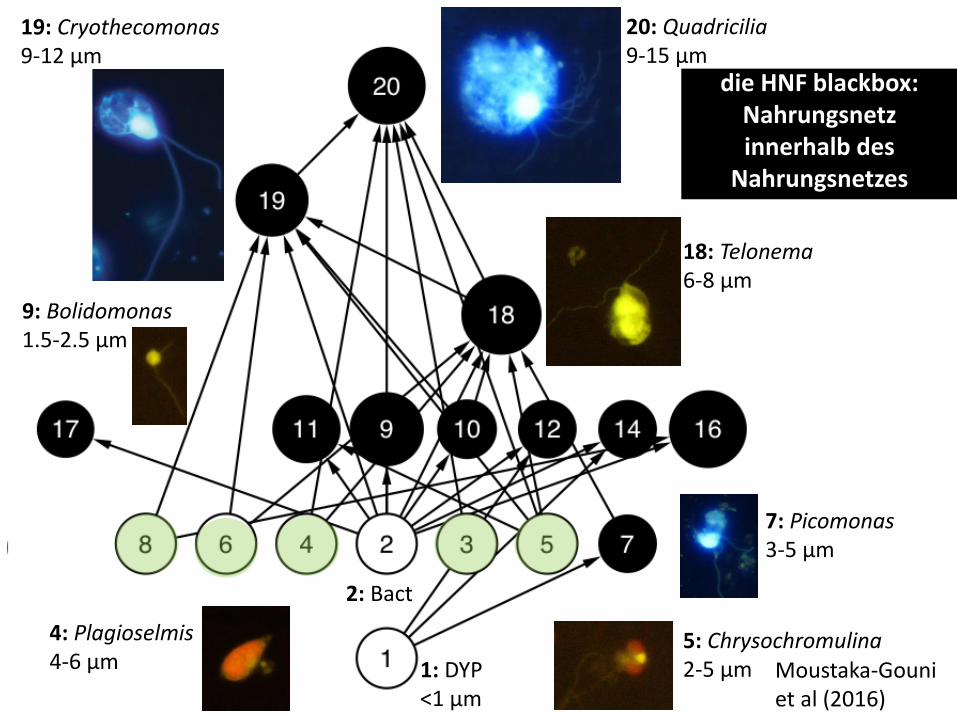
20: Quadricilia 9-15 µm
18: Telonema 6-8 µm
7: Picomonas 3-5 µm
19: Cryothecomonas 9-12 µm
9: Bolidomonas 1.5-2.5 µm
4: Plagioselmis 4-6 µm
5: Chrysochromulina 2-5 µm 1: DYP
<1 µm
2: Bact
die HNF blackbox:
Nahrungsnetz innerhalb des
Nahrungsnetzes
Moustaka-Gouni et al (2016)
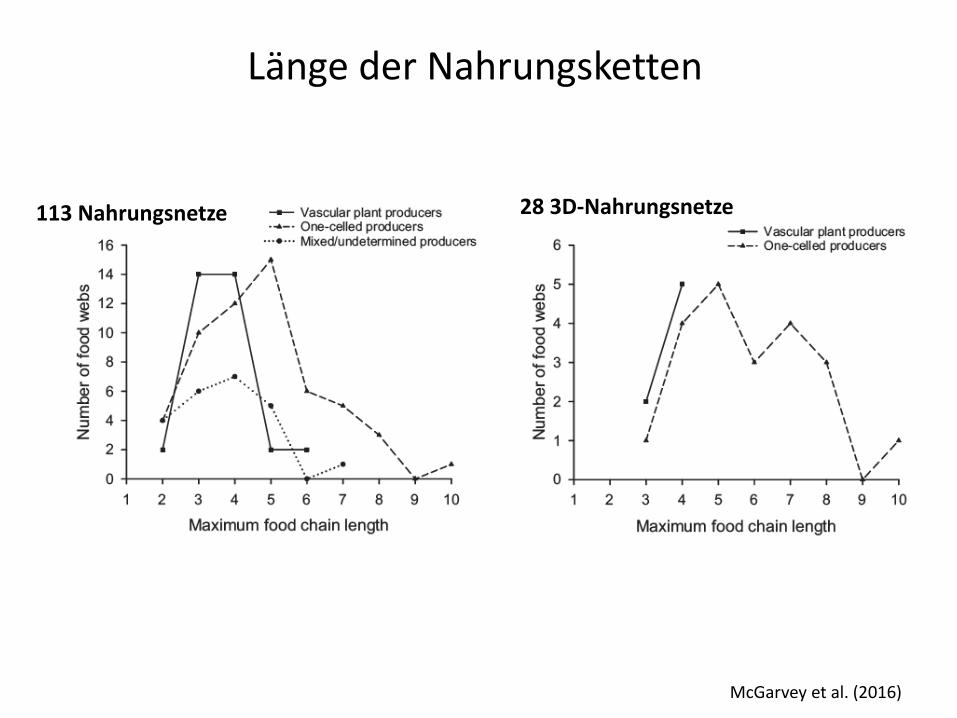
McGarvey et al. (2016)
all 113 food webs
28 3D-Nahrungsnetze 113 Nahrungsnetze
Länge der Nahrungsketten

Hussey NE et al (2014) Rescaling the trophic structure of marine food webs Ecology Letters 17: 239–250
δ15N
2 TL Protisten addiert, 8 TL im Pelagial möglich
Trophieskalierung mariner Nahrungsnetze

Umwelteinflüsse auf marine pelagische Produzenten und ihre Konsumenten
Marine Nahrungsnetze und Trophiestufen
WS 2017-18

Stellt ungefähr 50 % der globalen Primärproduktion an Biomasse (Field et al. 1998)
Globale annuelle NPP in Gramm pro Quadratmeter und Jahr (Field et al. 1998)
Marines Phytoplankton

coolantarctica.com
Ökologische Rollen von Phytoplankton und Zooplankton
Basis der marinen pelagischen Nahrungsnetze
Wichtigstes trophisches Bindeglied zu höheren Nahrungsnetz-Ebenen
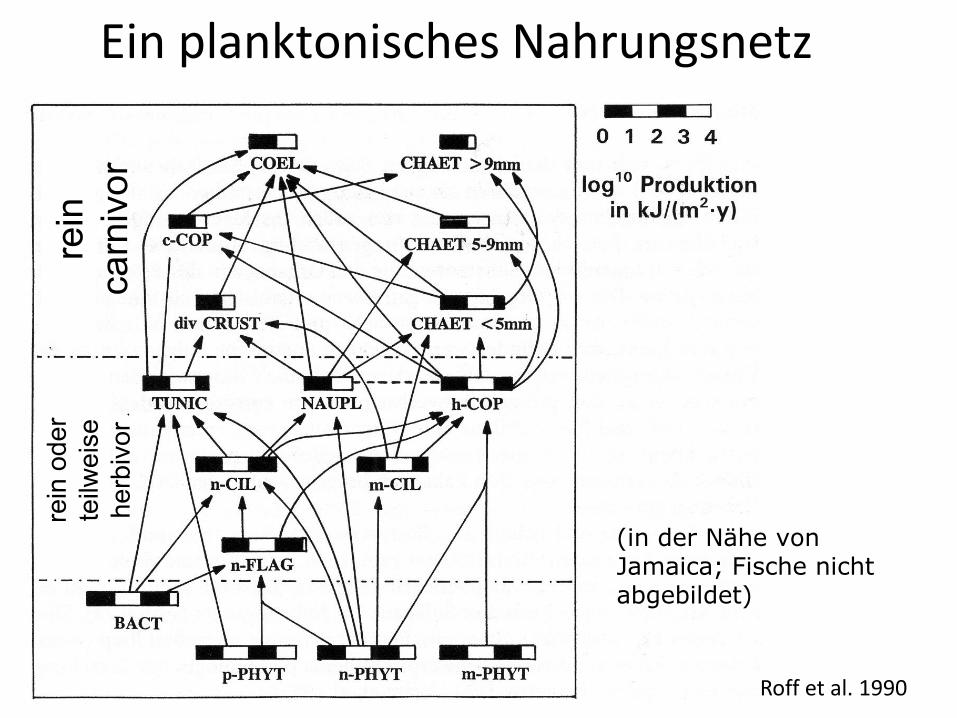
Ein planktonisches Nahrungsnetz
Roff et al. 1990
(in der Nähe von Jamaica; Fische nicht abgebildet)

Ökologischer Kontext der Phytoplankton-Zooplankton Interaktionen
oceanworld.tamu.edu

globale Erwärmung
Eutrophierung
Ozeanversauerung
globale Erwärmung
Eutrophierung
Ozeanversauerung
Ökologischer Kontext der Phytoplankton-Zooplankton Interaktionen

• nimmt in acht von zehn Ozeanregionen ab
• geschätzte Abnahmerate von ~1% des globalen Medians pro Jahr
Langzeitentwicklung der Phytoplankton-Biomasse
Boyce et al. 2010

Boyce et al. 2010
Langzeitentwicklung der Phytoplankton-Biomasse

Mackas et al. 2012; SAHFOS Zeitseriendaten
Zeitverschiebung in Phytoplanktonblüten

Lewandowska & Sommer 2010
Erwärmung und Lichtverfügbarkeit wirken sich auf den Phytoplankton-Biomassepeak
aus

Sommer et al. 2015
Erwärmungs- und Versauerungsauswirkungen auf Phytoplanktonblüten

Peter & Sommer 2015
Interaktive Effekte von Temperatur und Nährstofflimitation auf Phytoplanktongröße

• Verbreitung
• Phänologie
• Größe
Drei wesentliche “universal responses” auf Erwärmung
in der Essenz Verteilungsänderungen in Zeit und/oder Raum, die Abundanzänderungen bewirken
können auch mit Änderungen der Peak-Ausprägung einhergehen
Gardner et al. 2011

Größe – die dritte beschriebene universelle Reaktion auf Erwärmung
• Größenänderungen passieren auf Gesellschafts-, Populations-, und/oder auf Individuum Ebene
Daufresne et al. 2009

Beaugrand et al. 2002
- 10° latitudinale Ausdehnung Warmwasser-adaptierter Arten - verbunden mit Abnahme kaltadaptierter Arten
- großskalige NA Reorganisation von Zooplanktongemeinschaften
Verbreitungsänderungen von Zooplankton
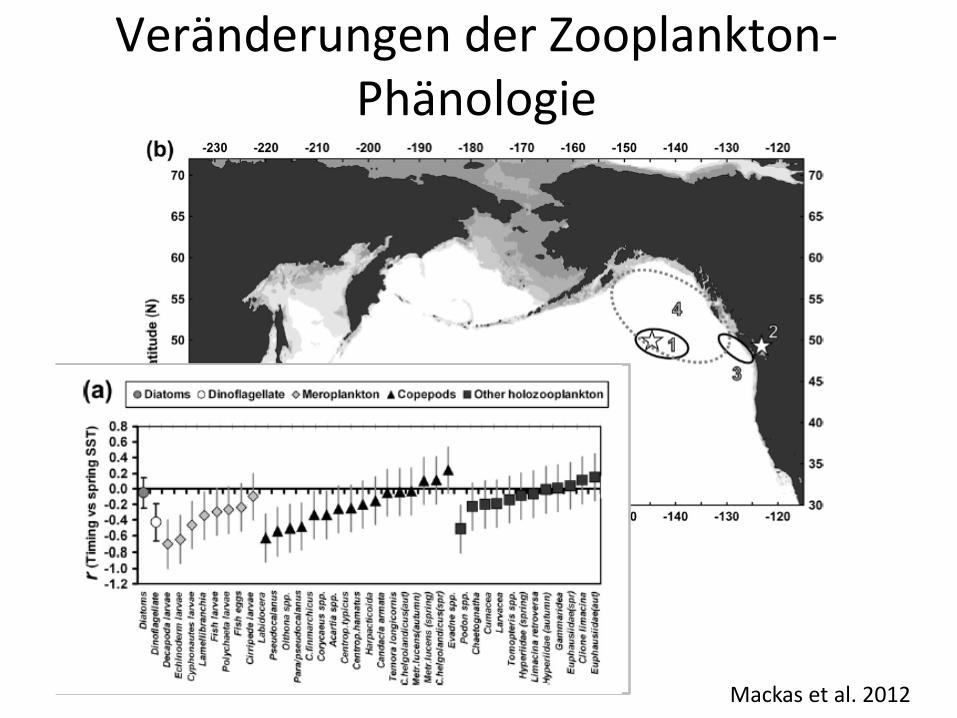
Mackas et al. 2012
Veränderungen der Zooplankton-Phänologie

Mackas et al. 2012
Veränderungen der Zooplankton-Phänologie

Edwards & Richardson 2004
Unterschiedliche Phänologie-Änderungen auf verschiedenen trophischen Ebenen

‘Trophic mismatch’
Thackeray et al. 2012

Räumliche Entkopplung von Biomasse-Peaks – spatial ‘trophic mismatch’
McGinty et al. 2014

Die Energie fließt bottom-up NO RECYCLING
Materie fließt bottom-up RECYCLING
Wie ist das System kontrolliert?

Klarwasserphase: Periode geringer Phytoplankton-Biomasse trotz guter Wachstumsbedingungen (Licht, Nutrienten), typischerweise nach der Fruehjahrsbluete. Induktion durch grazing in limnischen Systemen gut etabliert, in der Ozeanographie noch immer kontrovers (zwei mögliche Ursachen):
• grazing
• Sedimentation aufgrund von Aggregation nach ausgedehnter Blüte (Nutrienten verbraucht)
Top-down Kontrolle
nach Sommer 2005

British Channel
Klarwasserphase
Bautista et al 1992

• bottom-up: positive Korrelation zwischen angrenzenden Trophieebenen
mehr Nährstoffe – mehr Phytoplankton – mehr Zooplankton – mehr Fisch
typischerweise bei großskaligen Vergleichen verschiedener Ökosysteme
• top-down: negative Korrelation zwischen angrenzenden Trophieebenen
mehr Fisch – weniger Zooplankton – mehr Phytoplankton
Typischerweise bei Kurzzeit-Experimenten und Vergleichen zwischen Ökosystemen ähnlicher Produktivität
Trophische Kaskade
Transmission von top-down Effekten über >2 Trophieebenen
Top-down versus bottom-up Kontrolle
nach Sommer 2005

Lachs Biomasse (g m-2)
3 Standorte
• über Standorte hinweg: mehr
Zooplankton – mehr Lachs
• innerhalb Standorten: mehr Lachs –
weniger Zooplankton
Top-down versus bottom-up Kontrolle – abhängig vom Maßstab
aus Sommer 2005

Trophische Kaskade
Ostsee
Aurelia
herbivores Mesozooplankton
Phytoflagellaten
Behrends & Schneider 1995

Volle Kaskade: Biomasse einer ganzen trophischen Ebene betroffen
Partielle Kaskade: nur funktionale Gruppen innerhalb der trophischen Ebenen betroffen – Kompensation durch andere funktionale Gruppen.
Volle versus partielle Kaskaden

Partielle Kaskade: Hopavågn summer 2001 Mesokosmen mit unterschiedlichen Copepoden-Dichten besetzt
nach Tag 7: Gesamtbiomasse Phytoplankton kein Effekt Nanophytoplankton erhöht Microphytoplankton unterdrückt
Sommer et al. 2004

Wachstum von Nanoflagellaten kompensiert Microphytoplankton-Verluste partielle Kaskade: Copepoden Ciliaten Nanoflagellaten
Sommer et al. 2004
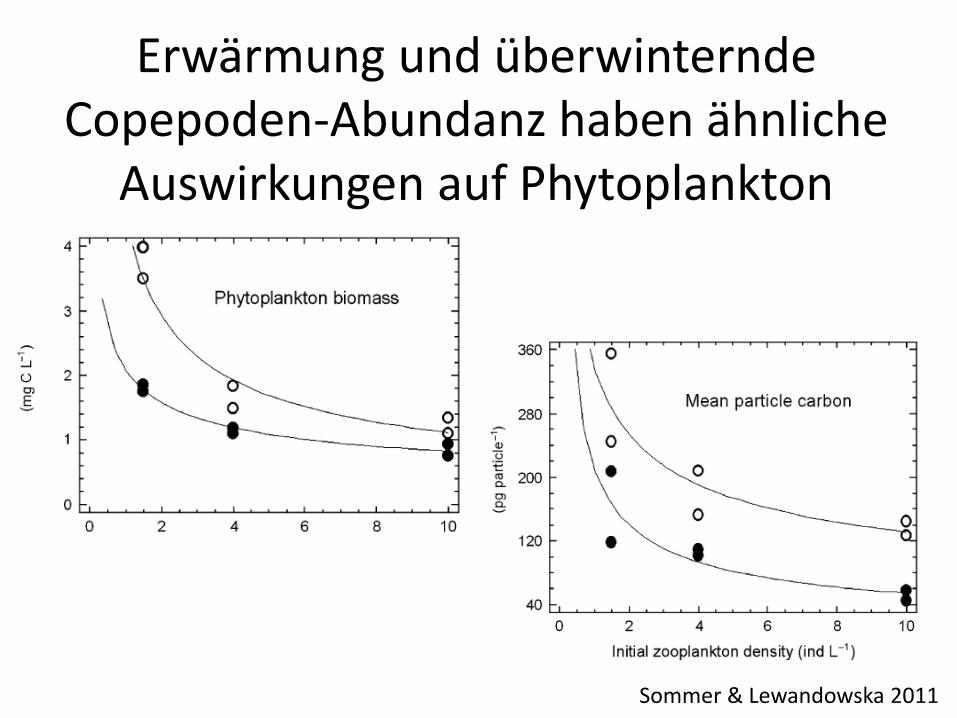
Sommer & Lewandowska 2011
Erwärmung und überwinternde Copepoden-Abundanz haben ähnliche
Auswirkungen auf Phytoplankton
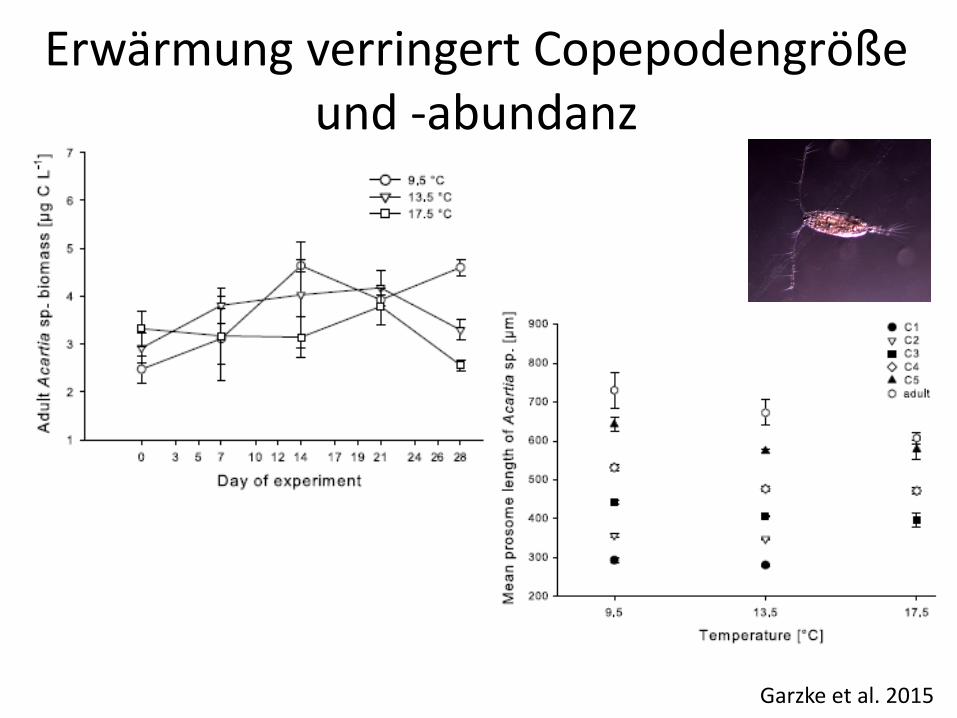
Garzke et al. 2015
Erwärmung verringert Copepodengröße und -abundanz

Versauerung wirkt antagonistisch, und teil-kompensierend auf Erwärmundseffekte
Garzke et al 2016

Garzke et al 2017
Versauerungs- und Erwärmungseffekte auf Zooplankton Resource-Use-Efficiency

Lewandowska et al. 2014
Auswirkungen von Erwärmung auf Plankton hängen mit
Nährstoffverfügbarkeit zusammen

Änderung der Dinoflagellatenabundanz
Hinder et al. 2012
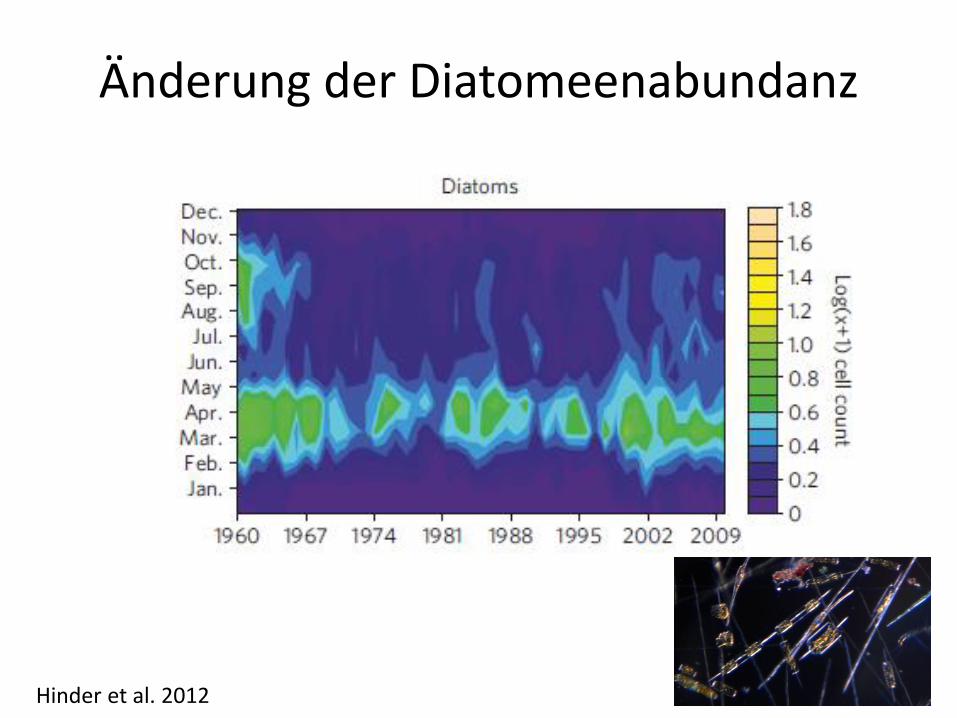
Änderung der Diatomeenabundanz
Hinder et al. 2012
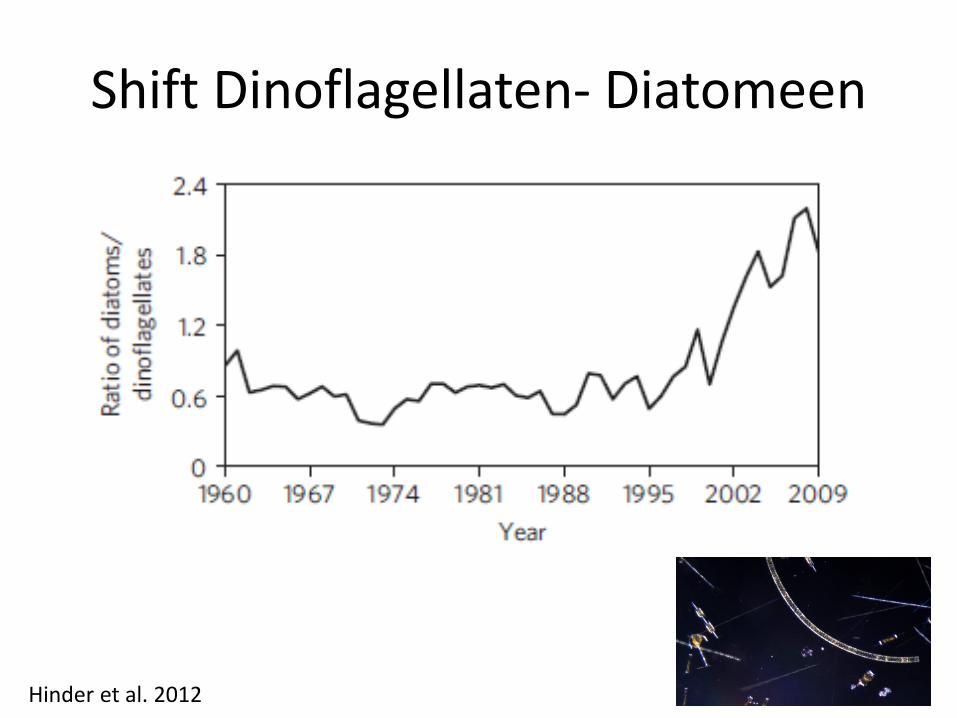
Shift Dinoflagellaten- Diatomeen
Hinder et al. 2012

Shift Dinoflagellaten - Diatomeen
• Mögliche Auswirkungen auf Zooplankton?
– Nahrungsverfügbarkeit
– Fressbarkeit
– Nahrungsqualität
– Toxizität

Edwards et al. 2006
Zunahme toxischer Phytoplanktonblüten
Harmful Algal Blooms (HABs)

Edwards et al. 2006
Räumliche Muster der toxischen Blütenanomalien unter Klimawandel

Bi et al. 2014
Fettsäurezusammensetzung verschiedener Phytoplankter unter verschiedenem Nährstoffangebot
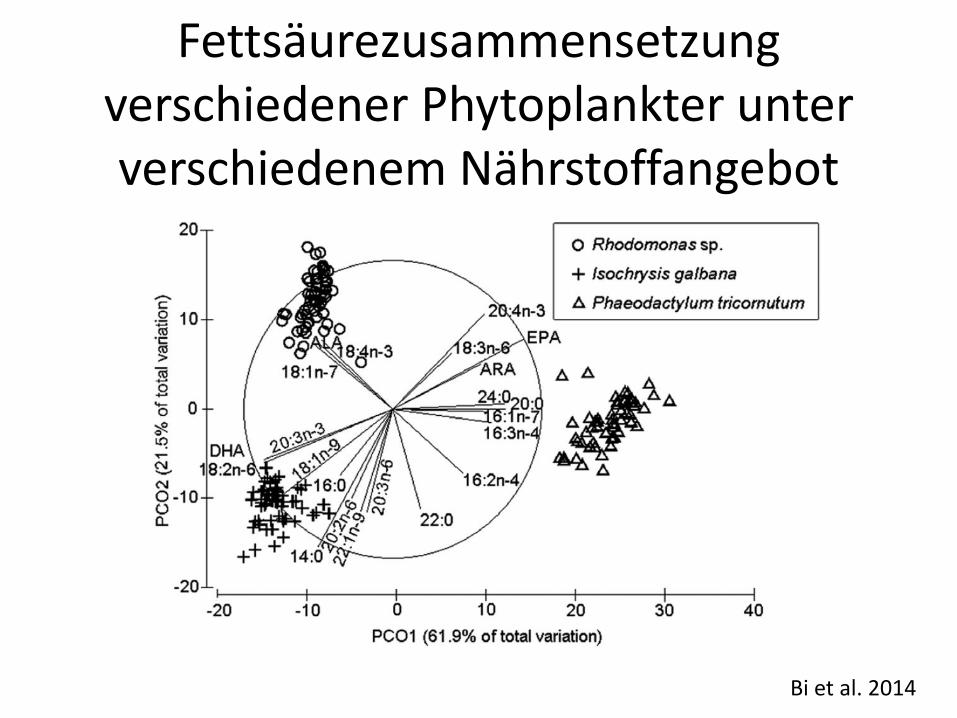
Bi et al. 2014
Fettsäurezusammensetzung verschiedener Phytoplankter unter verschiedenem Nährstoffangebot

Bi et al. 2017
Simultane Auswirkungen von Erwärmung, Nährstofflimitation und Versauerung

Bi et al. Biogeosci Disc
… und auf Emiliania
huxleyi, einen Coccolithophor

Eignung verschiedener Nahrungsquellen zur Acartia Entwicklung
• Entwicklungsgeschwindigkeiten hängen von der Nahrungsabundanz ab
Ismar et al. 2008
Photo S. Ismar
Michaelis-Menten equation

Nahrungsabhängigkeit der Eiproduktion bei Acartia
Teixera et al. 2010 Photo S. Ismar

Rossol et al. 2012
CO2-induzierte Beeinträchtigung der Nahrungsqualität
Erhöhtes CO2 änderte singifikant den Fettsäure-Gehalt und –zusammensetzung der Diatomee Thalassiosira pseudonana
und wirkte sich auf Copepoden-Wachstum und –Fortpflanzung aus.

Hansson et al. 2013
Nahrungskettenlänge kann Effekte des Klimawandels beeinflussen

• Insgesamt verstärkte Beweislage für abnehmende Abundanzen – Wissensstand für Phytoplankton besser als für Zooplankton
• Ausmaß der Abnahme wird debattiert
• Erwärmung zeigt stärkste Auswirkungen auf globaler Ebene; ist aber auch weiterhin der best-untersuchte Faktor des Klimawandels
• Artspezifische Effekte spielen eine große Rolle
• Eigenschaften der Nahrungsnetze können Effekte modifizieren
Umwelteinflüsse auf die Phytoplankton-Zooplankton Schnittstelle
Erwärmung
Eutrophizierung
Versauerung
Erwärmung
Eutrophizierung
Versauerung

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
SAHFOS Zeitserie Phytoplankton
Mackas et al. 2012

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
Edwards et al. 2006
Anstieg HABs

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
Edwards et al. 2006
Anstieg HABs

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
Puelles & Molinero 2013
Shift des pelagischen Ökosystems des westlichen Mittelmeers in den späten 1990igern.

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
Molinero et al. 2013
Interannuelle Varianzverteilung der Zooplanktongesellschaft im englischen Kanal

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
Condon et al. 2012
Zunahme an gelatinösen Planktonblüten.

Warum ist Quantifizierung mariner Nahrungsnetze aktuell wichtig?
McInnes et al. 2017
www.tonykeenebirds.co.uk
Schwarzbrauenalbatross
Campbell Albatross

Literatur • Alsterberg et al (2013) PNAS 110(21):8603-8608.
• Azam et al (1983) Mar Ecol Progr Ser 10:257-26.
• Barofsky et al (2010) J Plankton Res 32(3):263-272.
• Bautista B, Harris RP, Tranter PRG, Harbour D (1992) J Plankton Res 14:691-703.
• Behrends G, Schneider G (1995) Mar Ecol Progr Ser 127:39-45
• Bi R, Arndt C, Sommer U (2014) J Phyc 50:117:130.
• Bi R, Ismar SMH, Sommer U, Zhao M (2017) Limnol Oceanogr 62:334-347.
• Bi R, Ismar SMH, Sommer U, Zhao M (in rev) Biogeosciences Disc.
• Bodini et al (in press) Limnol Oceangogr.
• Boyce et al (2010) Nature 466:591-595.
• Condon et al (2013) PNAS 110:1000-1005.
• Dutkiewicz et al (2015) Nat Clim Change 5:1002.
• Edwards & Richardson (2004) Nature 430:881-883.
• Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998) Science 281:237-240.
• Gardner JL Peters A, Kearney MR, Joseph L, Heinsohn R (2011)Trends Ecol Evol 26:285–291.
• Garzke J, Ismar SMH, Sommer U (2015) Oecologia 177:849-860.
• Garzke J, Hansen T, Ismar SMH, Sommer U (2016) PloS ONE 11(5): e0155952.
• Garzke J, Sommer U, Ismar SMH (2017) Aq Sci. 79:733–748.
• Hansen T, Burmeister A, Sommer U (2009) Rapid Comm Mass Spectrom 23:3387-3393.
• Hansson et al (2013) Nat Clim Change 3:228-233.
• Hillebrand H, Sommer U (1996) J Plankton Res 18(2):295-301.
• Hinder et al (2012) Nat Clim Change 2:271-275.
• Hussey NE et al (2014) Ecology Letters 17: 239–250.
• Ismar SMH, Hansen T, Sommer U (2008) Mar Bio 154:335-343.
• Jordan et al (2014) MEPS 512:89-98.
• Jungbluth et al (2013) Mar Biol 160:3125-3141.
• Katechakis et al (2004) J Plankton Res 26:589-603.
• Kuerten et al (2016) Progr Oceanogr 140:69-90.
• Lewandowska A & Sommer U (2010) MEPS 405:101-111.
• Lewandowska et al (2014) Ecol Lett 18:614-623.
• Loh et al (2014) Fishery Biology 85:307-328.
• Mackas et al (2012) Progr Oceanogr 97-100:31-62.
• McGarvey, Dowling, Cohen (2016) Am Nat 188:76-86
• McGinty et al (2014) J Plankton Res 36(6):1446-1460.
• McInnes et al (2017) Mol Ecol 26:4831-4835.
• Mittermayr A, Hansen T, Sommer U (2014a) MEPS 497:93-103.
• Mittermayr A, Fox SE, Sommer U (2014b) MEPS 505:95-105.
• Molinero JC, Reygiondeau, Bonnet D (2013) Marine Environmental Research 89:91-96.
• Moustaka-Gouni et al (2016) Protist 167:389-410
• Nejstgaard et al (2003) Limnol Oceanogr: Methods 1:29-38.
• Nejstgaard et al (2008) Mar Biol 153:565–577.
• Peters (1983)
• Peter KH & Sommer U (2013) PLoS ONE 8(9):e71528.
• Peter KH & Sommer U (2015) Ecology & Evolution 5(5):1011-1024.
• Renkawitz et al (2015) MEPS 538: 197–211.
• Roff et al (1990)
• Rossol et al (2012) PLoS One 7(4):e34737.
• Sommer F et al (2000) Mar Ecol Progr Ser 199:43-63.
• Sommer U (2002) Biologische Meereskunde. 2nd Edition. Springer Verlag pp 475.
• Sommer U et al (2002) Hydrobiologia 484: 11–20.
• Sommer U et al (2002) Mar Ecol Progr Ser 239:251-261.
• Sommer U (1988) Hydrobiologia 161:125-131
• Sommer U (2005) Biologische Meereskunde. 2 nd Ed. Springer Verlag pp 475.
• Sommer U & Lewandowska A (2011)
• Sommer U, Sommer F, Feuchtmayr H, Hansen T (2004) Protist 154: 91-98.
• Tait (1981)
• Teixera et al (2010) Ann Bras Acad Sci 82(2):333-339.
• Thackeray SJ (2012) 34(12):1001-1010.
• Treml et al (2012) Int Comp 52(4): 525-537.















![Present lec osz2009.ppt [Kompatibilit tsmodus]) · 2018. 5. 15. · OpAmp spielt Limiter, Parallelkreis mit Güte Q filtert Grundwelle aus ... Pierce Colpitts Clapp Gate Modified](https://static.fdokument.com/doc/165x107/60c2c60fe0ecca136a5f41fa/present-lec-kompatibilit-tsmodus-2018-5-15-opamp-spielt-limiter-parallelkreis.jpg)


