
„Kupfer - ein Spurenelement mit vielseitiger biologischer Bedeutung“ ?

Modellierung biologischer und molekularer Systeme Titel
Modellierung biologischer und molekularerSysteme
PD Dr. Ingo R oder
Institut fur Medizinische Informatik, Statistik und Epidemiology
Wintersemester 2009/10
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 1

Modellierung biologischer und molekularer Systeme Einfuhrung
Modell
verschiedene Definitionen nach Meyers Lexion und Duden:
a) in verkleinertem Maßstab vor- oder nachgebildetes Bauwerk/Objekt
b) Studienobjekt des Kunstlers
c) Fotomodell, Mannequin
d) vereinfachtes gegenstandliches od. abstraktes Abbild der Wirklichkeit ...Voraussetzung ist das Bestehen von Analogie (hinsichtlich Strukturen od.Funktionen) zwischen Original und Modell. ...
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 2

Modellierung biologischer und molekularer Systeme Uberblick
Inhaltliche Struktur
• Differenzengleichungen: Losung, Stabilitatsanalyse→ Populationswachstum
• gewohnliche Differentialgleichungen: Stabilitats-/ Phasenraumanalyse→ Populationswachstum, Rauber-Beute Systeme
• Enzymkinetik: Massenwirkungsgesetz, Quasi-Steady-State Approximation→ Enzym-Substrat Reaktion, Enzymhemmung
• Stochastische Prozesse: Markovprozesse, First-Step-Analyse, Simulation→ Populationswachstum
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 3

Modellierung biologischer und molekularer Systeme Uberblick
Literatur
• Edelstein-Keshet L., Mathematical Models in Biology, Birkhauser Mathematics Series,McGraw-Hill Inc., Boston, 1988
• Murray J.D., Mathematical Biology, Springer Verlag, Heidelberg, 1993
• Segel L.A., Modeling dynammic phenomena in molecular and cellular biology, Cam-bridge University Press, Cambridge, 1989
• Alberts B. et al., Molecular biology of the cell. Garland Publishing, Inc. New York, 2002
• Strogatz S.H., Nonlinear Dynamics and Chaos, Westview Press, 2000
• Taylor H.M., Karlin S., An Introduction To Stochastic Modeling, Academic Press, 1998
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 4

Modellierung biologischer und molekularer Systeme Uberblick
Beschreibungsebenen
Molekul: z.B. Stoffwechselprozesse, Genregulationsprozesse
Zelle: z.B. Erregungsleitung durch Nervenzelle
Zellverb ande, Organe: z.B. Tumorwachstum, Gewebsregeneration
Individuum: z.B. Verteilung von Arzneistoffen im Korper
Population: z.B. Konkurrenz in Okosystemen, Epidemieausbreitung
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 5

Modellierung biologischer und molekularer Systeme Uberblick
Methodenklassen (1)
Differenzengleichungen: zeitdiskret, meist keine raumliche Auflosung
Gewohnliche Differentialgleichungen (ODE): zeitkontinuierlich, keine raum-liche Auflosung
Partielle Differentialgleichungen (PDE): zeit-/raumkontinuierlich
Stochastische Differentialgleichungen (SDE): zeitkontinuierlich, meist kei-ne raumliche Auflosung
Zellularautomaten: zeit-/raumdiskret
Individual Particle Systems (IPS) diskret/kontinuierlich in Raum und Zeit
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 6
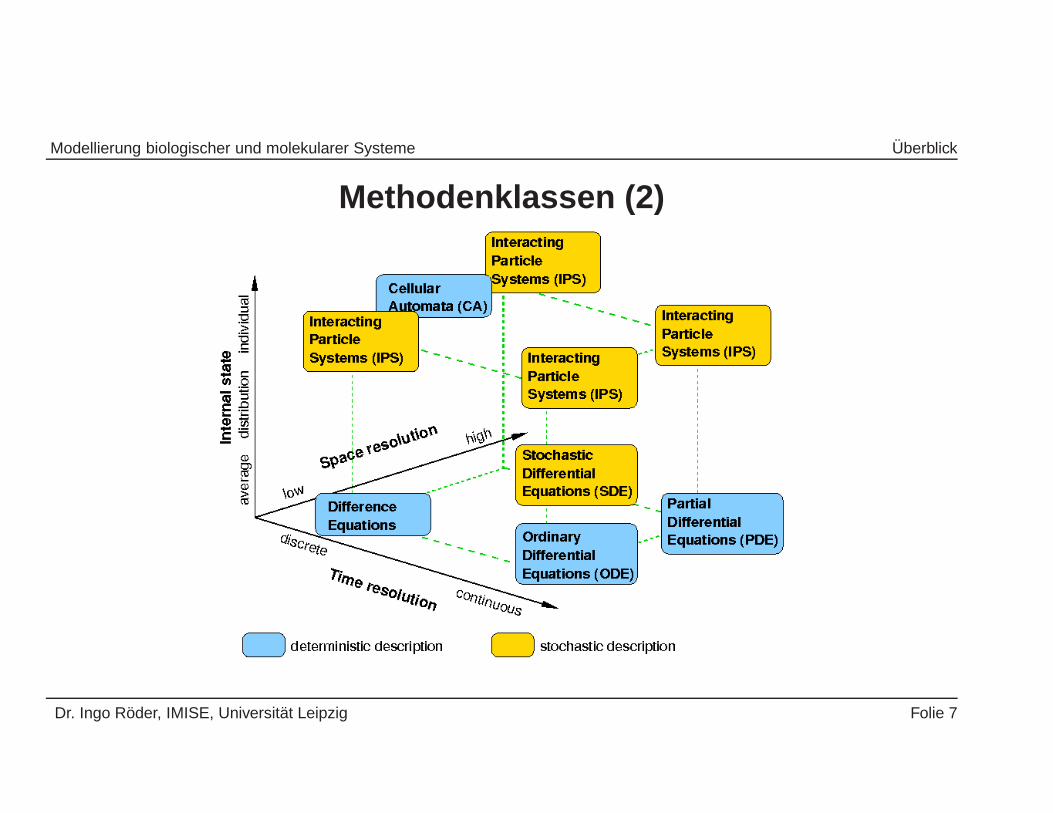
Modellierung biologischer und molekularer Systeme Uberblick
Methodenklassen (2)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 7
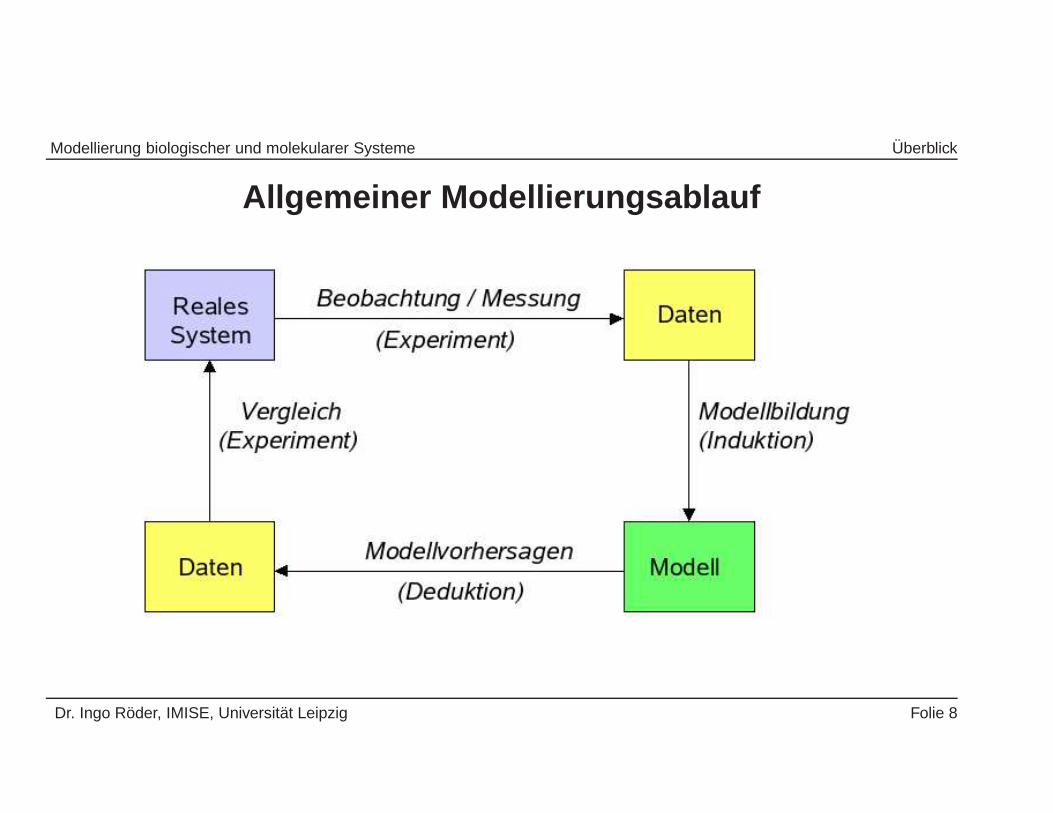
Modellierung biologischer und molekularer Systeme Uberblick
Allgemeiner Modellierungsablauf
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 8

Modellierung biologischer und molekularer Systeme
Populationswachstumsmodelle
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 9

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Populationswachstum
• Diskrete Modelle
– Lineare Differenzengleichungen– Nichtlineare Differenzengleichungen
• Kontinuierliche Modelle
– Gewohnliche Differentialgleichungen– Systeme gewohnlicher Differentialgleichungen
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 10

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Diskrete Modelle
Dieser Abschnitt basiert auf Kapitel 1-3 aus L. Edelstein-Keshet, Mathema-tical Models in Biology, Birkhauser Mathematics Series, McGraw-Hill Inc.,Boston, 1988
• Biologische Systeme:
– Zellwachstum– Insektenpopulation– Jahreszyklus von Pflanzen
• Modellierungsmethoden:
– lineare/nichtlineare Differenzengleichungen
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 11

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Zellteilung (1)
• Betrachte eine Population sich synchron teilender Zellen.
• Die Anzahl der Zellen in Generation i sei mit Mi bezeichnet.
• Die Anzahl von Zellen in aufeinanderfolgenden Generationen ist:
Mn+1 = 2Mn.
• Gegeben eine Anfangszellzahl M0 kann die Populationsgroße nach nGenerationen rekursive bestimmt werden:
Mn+1 = 2Mn = 2(2Mn−1) = 2[2(2Mn−2)] = . . . = 2n+1M0.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 12

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Zellteilung (2)
• Damit erhalt man fur die n-te Generation
Mn = 2nM0.
• Dieses Resultat kann man verallgemeinern, wenn man annimmt, dassin einer Generation jeweils a (a > 0) Nachkommen erzeugt werden. Esfolgt:
Mn = anM0.
• Hierbei wachst die Population fur a > 1, sie schrumpft fur a < 1 und siebleibt konstant fur a = 1.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 13

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Insektenpopulation (1)
• Wir betrachten den Lebenszyklus der Pappelblattlaus (poplar gall aphid).
• Erwachsene Weibchen produzieren Gelege auf Pappelblattern (”Gall”),aus denen Nachwuchs schlupft, von dem ein gewisser Anteil uberlebt.
an = Anzahl erwachsener Weibchen in der n-ten Generation
pn = Anzahl der Nachkommen in der n-ten Generation
m = Mortalitat der Nachkommen
f = Anzahl der Nachkommen pro Weibchen
r = Anteil der Weibchen an den Nachkommen
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 14

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Insektenpopulation (2)
• Unter der vereinfachenden Annahme konstanter Anteile f, r,m folgt
pn+1 = f · an (1)
an+1 = r · (1 − m)pn+1 (2)
⇒ an+1 = f · r(1 − m) · an (3)
• Diese lineare Differenzengleichung 1. Ordnung hat folgende Losung
an = [f · r(1 − m)]n · a0, (4)
wobei f · r(1−m) die pro Kopf Anzahl der erwachsenen Weibchen, welchejedes Mutterweibchen produziert, reprasentiert.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 15

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (1)
Problemstellung:
• Einjahrige Pflanzen produzieren Samen am Enden des Sommers undgehen zugrunde.
• Ein Teil der Samen uberlebt den Winter und wiederrum ein Teil dieserSamen bringt im nachsten Fruhjahr neue Pflanzen hervor.
• Der restliche Teil der Samen kann noch einen weiteren Winter uberste-hen und ggf. im zweiten Fruhjahr Pflanzen hervorbringen.
Aufgabe:
• Formulierung einer Modellbeschreibung;
• Vorhersage der Populationsgroßenentwicklung.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 16

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (2)
Parameter:
γ = Anzahl der produzierten Samen pro Pflanze
α = Anteil der aufgehenden Samen im 1. Jahr
β = Anteil der aufgehenden Samen im 2. Jahr
σ = Anteil der Samen, welche den jeweiligen Winter uberleben
Vereinfachende Annahme:Samen, alter als 2 Jahre konnen nicht mehr keimen.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 17

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (3)
Variablen:
pn = Anzahl der Pflanzen in Generation n.
S1n = Anzahl der 1-Jahres-Samen vor dem Keimen.
S2n = Anzahl der 2-Jahres-Samen vor dem Keimen.
S1n = Anzahl der 1-Jahres-Samen, die nicht gekeimt sind.
S2n = Anzahl der 2-Jahres-Samen, die nicht gekeimt sind.
S0n = Anzahl der jeweils von den Pflanzen produzierten Samen.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 18
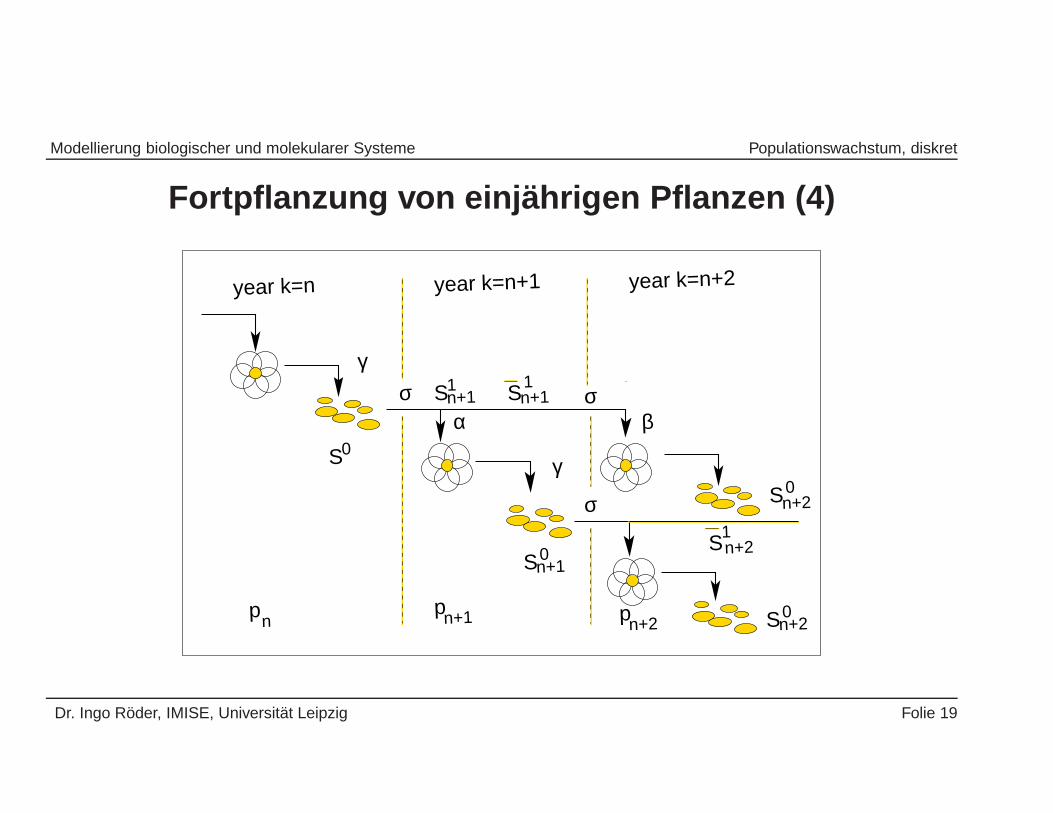
Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (4)
pn+2pn+1pn Sn+2
0
γ
γ
α β
S
S
S
S Sσ σ
σ
S
n+1 n+1
n+1n+2
n+2
0
0
0
1 1
1
year k=n+2year k=n+1year k=n
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 19

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (5)
Modellgleichungen:
pn = αS1n + βS2
n (5)
S1n = (1 − α)S1
n (6)
S2n = (1 − β)S2
n (7)
S0n = γpn (8)
S1n+1 = σS0
n (9)
S2n+1 = σS1
n (10)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 20

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (6)
Einsetzten von Gleichung (8) in (9) liefert
S1n+1 = σ(γpn). (11)
Weiterhin erhalt man in (10) mittels (6)
S2n+1 = σ(1 − α)S1
n. (12)
Betrachte nun (5) fur Generation n + 1:
pn+1 = αS1n+1 + βS2
n+1. (13)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 21

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fortpflanzung von einj ahrigen Pflanzen (7)
Damit erhalt man nun ein System zweier Gleichungen erster Ordnung, wel-che den Zusammenhang von Pflanzen und 1-jahrige Samen beschreibt:
pn+1 = ασγpn + βσ(1 − α)S1n, (14)
S1n+1 = σγpn. (15)
Dieser Zusammenhang kann auch alternativ in Form einer Gleichung zwei-ter Ordnung dargestellt werden:
pn+1 = ασγpn + βσ2(1 − α)γpn−1. (16)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 22

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Einschub: NomenklaturBezeichnung von Differenzengleichungen:
1. Ordnung: pn abhangig nur von pn−1
(z.B. pn = apn−1)
k. Ordnung: pn abhangig von pn−1, pn−2, . . . , pn−k
(z.B. pn = apn−1 + β(1 − a)pn−2)
Linear: pn abhangig nur von linearen Funktionen von pn−i
(z.B.pn = apn−1 + bpn−2)
Nichtlinear: pn abhangig von nichlinearen Funktionen von pn−i
(z.B. pn = rp2n−1 oder pn = spn−1pn−2)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 23

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
System linearer Differenzengleichungen (1)
• Das letzte Beispiel fuhrte auf ein System von zwei linearen Differenzen-gleichungen 1. Ordnung.
• Aquivalent hierzu ist die Reprasentation des Problems in Form einer li-nearen Differenzengleichungen 2. Ordnung moglich.
• Diese beiden Darstellungen konnen immer ineinander uberfuhrt werden.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 24

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
System linearer Differenzengleichungen (2)Betrachte allgemeines System von 2 linearen DGLn 1. Ordnung:
xn+1 = a11xn + a12yn, (17)
yn+1 = a21xn + a22yn. (18)
Einsetzten von (18) in (17) ergibt:
xn+2 = a11xx+1 + a12yn+1
= a11xx+1 + a12(a21xn + a22yn)
= a11xx+1 + a12a21xn + a22(xn+1 − a11xn)
→ xn+2 − (a11 + a22)xn+1 + (a22a11 − a12a21)xn = 0 (19)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 25

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losungsansatz
In den Bspl. Zellteilung und Insektenpopulation hatte die Losung einer linea-ren Differenzengleichung 1. Ordung die Form
xn = Cλn. (20)
Kann die Losung auch fur lineare Differenzengleichungen hoherer Ordung,wie (19), ebenfalls mit einem solchen Ansatz ermittelt werden?
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 26

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Charakteristische Gleichung
Um dies zu testen substituiere den Ansatz xn = λnC in Gleichung (19):
Cλn+2 − (a11 + a22)Cλn+1 + (a22a11 − a12a21)Cλn = 0
Unter der Annahme Cλn 6= 0 (Ausklammerung der trivialen Losung xn = 0)und Division durch den Faktor Cλn erhalt man aus (19)
λ2 − (a11 + a22)λ + (a22a11 − a12a21) = 0 (21)
D.h. der Ansatz xn = Cλn liefert eine Losung fur den Fall, dass λ die qua-dratische Gleichung (21) erfullt.(21) wird als charakteristische Gleichung des Problems (19) bezeichnet.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 27

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losungen der charakteristischen Gleichung
Zur Vereinfachung der Darstellung fuhren wir die folgenden Bezeichnungender Koeffizenten aus Gleichung (21) ein
β = a11 + a22,
γ = a22a11 − a12a21.
Die Losungen der charakteristischen Gleichung (21) lauten damit:
λ1,2 =β ±
√
β2 − 4γ
2(22)
λ1,2 werden als Eigenwerte bezeichnet.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 28

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Allgemeine L osung
Fur lineare Gleichung (wie in Fall (19)) gilt das folgende, sogenannte Super-positionsprinzip. Dieses besagt:
Wenn mehrere, verschiedene Losungen eines linearen Problems be-kannt sind, dann ist auch jede Linearkombination dieser Losungen wiedereine Losung des Problems.
D.h. ausgehend vom Losungsansatz (20) mit den ermittelten Eigenwertenλ1 und λ2 ergibt sich die allgemeine Losung des Problems (19):
xn = A1λn1 + A2λ
n2 (23)
fur λ1 6= λ2.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 29

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losung mittels Matrixschreibweise (1)
Die Losung des Problems (19) bzw. aquivalent dazu (17,18) kann auch mit-tels einer anderen Methodik bestimmt werden. Betrachte hierzu das folgen-de System linearer Gleichungen:
ax + by = 0,
cx + dy = 0. (24)
In Matrixschreibweise kann dies geschrieben werden als:
Mv = 0 mit M =
(
a bc d
)
und v =
(
xy
)
(25)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 30

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losung mittels Matrixschreibweise (2)
• Der Nullvektor v = 0 ist immer eine (triviale) Losung von (24).
• v = 0 ist auch einzige Losung, wenn gilt detM = ad − bc 6= 0
• Im Falle detM = 0 enthalten beide Gleichungen ”dieselbe Information”.D.h. jede Kombination von x und y, die eine der Gleichungen erfullt, istauch Losung des Systems.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 31

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losung mittels Matrixschreibweise (3)Anwendung dieser Schreibweise auf System (17,18):
Vn+1 = MVn mit Vn =
(
xn
yn
)
und M =
(
a11 a12
a21 a22
)
. (26)
Vermittels des schon vorher verwendenten Ansatzes erhalt man eineLosung der Form
Vn =
(
Aλn
Bλn
)
. (27)
Einsetzen dieses Ansatzes in (26) ergibt(
Aλn+1
Bλn+1
)
=
(
a11 a12
a21 a22
)(
Aλn
Bλn
)
. (28)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 32

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losung mittels Matrixschreibweise (4)
Ausmultiplizieren und Division durch den Faktor λn liefert folgendes Glei-chungssystem
0 = A(a11 − λ) + Ba12,
0 = Aa21 + B(a22 − λ). (29)
In Matrixform geschrieben
0 =
(
a11 − λ a12
a21 a22 − λ
)(
AB
)
. (30)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 33

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losung mittels Matrixschreibweise (5)Eine Losung des Systems (29) ist die triviale Losung A = B = 0. Diesesfuhrt allerdings auf V ≡ 0. Um weitere (nichttriviale) Losungen zu erhaltensetzte die Determinate der Koeffizientenmatrix gleich 0:
det
(
a11 − λ a12
a21 a22 − λ
)
= 0 (31)
Dies fuhrt auf(a11 − λ)(a22 − λ) − a12a21 = 0
und damit auf die charakteristische Gleichung der Eigenwerte λ
λ2 − βλ + γ = 0 (32)
wobei β = a11 + a22 und γ = (a11a22 − a12a21) bezeichnet.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 34

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Losung mittels Matrixschreibweise (6)
Die Losungen der characteristischen Gleichung sind bereits bekannt:
λ1,2 =β ±
√
β2 − 4γ
2.
Die Großen β, γ und β2 − 4γ werden wie folgt bezeichnet:
β = a11 + a22 = trM . . . Spur (trace) der Matrix M
γ = a11a22 − a12a21 = detM . . . Determinante von M
β2 − 4γ = disc(M) . . . Diskriminante von M
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 35

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Qualitatives Verhalten linearer Differenzengleichungen (1)
• Eine allgemeine Differenzengleichung m-ter Ordnung hat die Form
a0xn+m + a1xn+m−1 + · · · + amxn = bn.
• Sind die ai(i = 0, . . . ,m) Konstanten und ist bn = 0, so spricht man voneiner homogenen linearen Differenzengleichung mit konstanten Koeffi-zienten. Bisher vorgestellten Losungsverfahren bezogen sich auf diesespezielle Art von Differenzengleichungen. Die Losungen waren in diesemFalle Linearkombinationen von Ausdrucken der Form
xn = Cλn, mit λ . . . Losungen der charakt. Gleichung (Eigenwerte).
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 36

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Qualitatives Verhalten linearer Differenzengleichungen (2)
• Die Anzahl (verschiedener) Basis-Losungen ist durch die Ordnung derD.-Gleichung bestimmt; Im Allgmeinen hat eine D.-Gleichung m-ter Ord-nung (bzw. ein System von m D.-Gleichungen 1. Ordnung) m Basis-Losungen.
• Die allgemeine Losung ist eine lineare Superposition (Linearkombination)der m Basis-Losungen (gegeben, unterschiedliche λ )
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 37
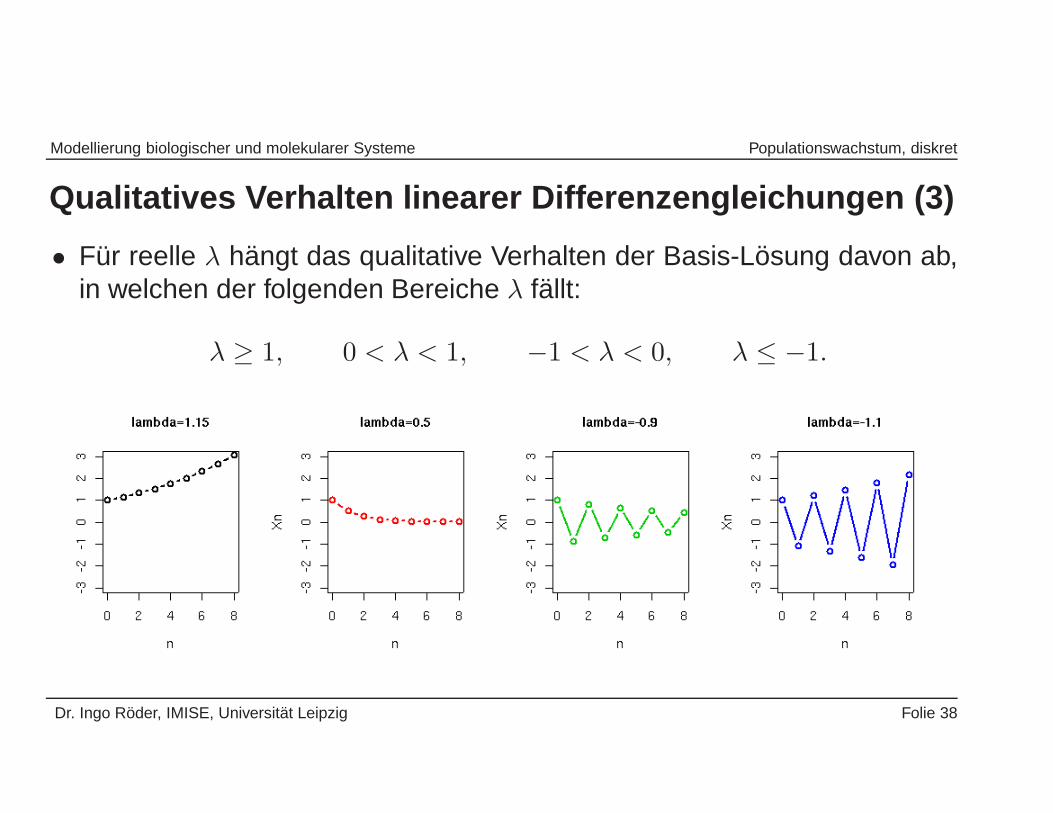
Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Qualitatives Verhalten linearer Differenzengleichungen (3)
• Fur reelle λ hangt das qualitative Verhalten der Basis-Losung davon ab,in welchen der folgenden Bereiche λ fallt:
λ ≥ 1, 0 < λ < 1, −1 < λ < 0, λ ≤ −1.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 38

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Qualitatives Verhalten linearer Differenzengleichungen (4)
• Allgemeine Losungen von linearen Differenzengleichungen der Ordnungm > 1 kombinieren ggf. verschiedene dieser qualitativen Charakteristiken
• Der dominante Eigenwert (das betragsmaßig großte λi) hat dabei denstarksten Einfluß auf das Verhalten; dies bedeutet, dass das Langzeitver-halten (Generationszahl n groß) approximativ beschrieben werden kanndurch
xn+1 ≃ λixn
• Die Dynamik enthalt eine oszillatorische Komponente, wenn einer derEigenwerte negativ (oder komplex) ist.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 39

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Einschub: Komplexe Eigenwerte (1)
Die charakteristische Gleichung λ2−βλ+γ = 0 liefert fur β2 < 4γ komplexeEigenwerte. Die treten als sogenannte konjugierte Paare auf
λ11 = a + bi und λ12 = a − bi
wobei a = β/2 und b = 12|β2 − 4γ|1/2.
Die allgemeine Losung (Linearkombination der Basislosungen) enthalt da-mit Potenzen von komplexen Zahlen
xn = A1(a + bi)n + A2(a − bi)n. (33)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 40

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Einschub: Komplexe Eigenwerte (2)
Um eine solche Losungsstruktur verstehen zu konnen, hier eine kurze Zu-sammenfassung von Rechenregeln komplexer Zahlen.
Es gelten folgende aquivalente Darstellungsformen fur komplexe Zahlen:
a + bi = r(cosΦ + i sin Φ) = reiΦ (34)
a − bi = r(cosΦ − i sin Φ) = re−iΦ (35)
Hierbei gelte:a = r cosΦ , b = r sinΦ. (36)
bzw. aquivalent dazu
r = (a2 + b2)1/2 , Φ = arctan(b/a) (37)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 41

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Einschub: Komplexe Eigenwerte (3)
Damit kann die Potenz einer komplexen Zahl verstanden werden als:(a + bi)n = (reiΦ)n = rneinΦ = c + di, mit c = rn cosnΦ und d = rn sinnΦ.Graphisch interpretiert bedeutet dies eine Rotation des ursprunglichen Vek-tors (a,b) um n · Φ und Streckung auf n-fache Potenz der Originallange.
Imag
inär
e A
chse
Reelle Achse
b
a
r
Φ
(1+i)
(1+i)
(1+i)
(1+i) (1+i)
Reelle Achse
2
(1+i)3
(1+i)4
5
67
Imaginäre Achse
(a) (b)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 42

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Einschub: Komplexe Eigenwerte (4)
Unter Ausnutzung dieser Regeln erhalt man fur die allgemeine Losung mitkomplexen Eigenwerten:
xn = A1(a + bi)n + A2(a − bi)n
= A1rn(cosnΦ + i sinnΦ) + A2r
n(cosnΦ − i sinnΦ)
= B1rn cosnΦ + iB2r
n sinnΦ
mit B1 = A1 + A2 und B2 = A1 − A2. Fur un = rn cosnΦ und vn = rn sinnΦerhalt man
xn = B1un + iB2vn (38)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 43

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Einschub: Komplexe Eigenwerte (5)
Da von einer linearen Differenzengleichung ausgegangen war, kann gezeigtwerden, dass sowohl der reelle als auch der imaginare Anteil von xn =B1un + iB2vn selbst Losungen sind. Auf dieser Basis kann man eine reell-wertige Losung durch Linearkombination von un und vn konstruieren:
xn = C1un + C2vn = rn(C1 cosnΦ + C2 sinnΦ). (39)
Komplexe Eigenwerte (λ = a ± bi) sind somit verbunden mit oszillierenden(reellen) Losungen. Hierbei gilt: wachsende Amplitude fur r =
√
(a2 + b2) >
1, sich verringernde Amplitude fur r =√
(a2 + b2) < 1 oder konstante Am-plitude fur r =
√
(a2 + b2) = 1. Die Frequenz der Oszillation hangt vomVerhaltnis b/a ab.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 44

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Komplexe Eigenwerte - Beispiel
Betrachte:xn+2 − 2xn+1 + 2xn = 0. (40)
Die zugehorige charakteristische Gleichung lautet: λ2 − 2λ + 2 = 0. Diesehat komplexe Losungen λ = 1 ± i. D.h. a = b = 1, so dass
r =√
(a2 + b2) =√
2 und Φ = arctan(b/a) = π/4.
Somit lautet die reellwertige allgemeine Losung von (40)
xn =√
2n[C1 cos(nπ/4) + C2 sin(nπ/4)].
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 45

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Nichtlineare Differenzengleichungen
• Eine nichtlineare Differenzengleichung ist jede Gleichung der Form
xn+1 = f(xn, xn−1, . . .), (41)
wobei f eine nichtlineare Beziehung der Argumente xn, xn−1, . . . dar-stellt.
• Eine analytische Losung einer solchen nichtlinearen Gleichung kann nurwenigen Fallen angegeben werden.
• Ausweg: Numerische Losung, lineare Approximation, qualitative Be-schreibung der Losung.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 46

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Fixpunkte / Steady states
Betrachte zunachst nichtlineare Differenzengleichungen 1. Ordnung:
xn+1 = f(xn). (42)
• Eine stabile Losung (steady state) bezeichnet einen Systemzustand, wel-cher sich uber die Zeit nicht andert.
• Ein solche stabile Losung (auch Fixpunkt genannt) ist definiert durch
xn+1 = xn = x (43)
• Als eine Folgerung aus (42), erfullt x auch
x = f(x) (44)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 47

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit at von Fixpunkten
• Man unterscheidet prinzipiell 2 Arten von Fixpunkten.
– stabil : ”zieht Nachbarzustande an”– instabil : ”stoßt Nachbarzustande ab”
��������
��������
��������
��������
��������
��������
1
2 3
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 48

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Linearisierung (1)
• Um die Stabilitat eines Fixpunktes x zu bestimmen geht man folgendermaßen vor:
1. Betrachte (kleine) Storungen x′n des Fixpunktes x, d.h.
xn = x + x′n (45)
2. Analysiere, ob diese Storung mit der Zeit zu oder abnimmt.
• Diese Vorgehensweise reduziert das Problem auf eine lineare Differen-zengleichung fur die bereits vorgestellte Losungsverfahren angewandtwerden konnen.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 49

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Linearisierung (2)
• Ausgehend von (44) und (45) erhalt man
x′n+1 = xn+1 − x = f(xn) − x = f(x + x′
n) − x. (46)
• Approximation von f mittels Taylorentwicklung unter Ausnutzung der An-nahme: x′
n klein.
f(x + x′n) = f(x) +
(
df
dx
∣
∣
∣
∣
x
)
x′n + O(x′2
n) (47)
• O(x′2n) reprasentiert in der Nahe von x sehr kleine Werte.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 50

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Linearisierung (3)
• Die O(x′2n) werden nun vernachlassigt und man erhalt
x′n+1 ≃ f(x) − x +
(
df
dx
∣
∣
∣
∣
x
)
x′n. (48)
• Unter Beachtung von f(x) = x kann die obige Gleichung geschriebenwerden als
x′n+1 = ax′
n mit a =
(
df
dx
∣
∣
∣
∣
x
)
(49)
• Somit erhalt man aus dem nichtlinearen Problem eine lineare Gleichungfur die Storung x′
n.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 51

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Linearisierung (4)
• Um das Verhalten des nichtlinearen Systems in der Nahe seiner Fix-punkte zu charakterisieren, betrachtet man als das Verhalten der kleinenStorung x′
n:
• Die Losung von (42) wachst fur |a| > 1 und wird kleiner fur |a| < 1.
• Man erhalt hieraus eine Bedingung fur Stabilitat des eines Fixpunktes x:
x is stabiler Fixpunkt von (42) ⇔∣
∣
∣
∣
df
dx|x∣
∣
∣
∣
< 1 (50)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 52

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit atsanalyse - Beispiel (1)
Betrachte das folgende nichtlineare Populationswachstumsmodell
xn+1 =kxn
b + xn, b, k > 0 (51)
Bestimmung der Fixpunkte: Setze x = xn+1 = xn in (51) ein:
x =kx
b + x⇒ x(b + x) = kx ⇒ x(x + b − k) = 0. (52)
Damit erhalt manx = k − b oder x = 0. (53)
Beachte: k > b da negative Populationsgroßen uninteressant sind.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 53

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit atsanalyse - Beispiel (2)
Zur Analyse des Stabilitatsverhaltens bestimme
d
dx
(
kxn
b + xn
)
=k(b + xn) − kxn
(b + xn)2
1. x = 0:
df
dx|x=0 =
kb
b2=
k
b> 1 ⇒ instabil.
2. x = k − b:
df
dx|x=k−b =
kb
(b + k − b)2=
b
k< 1 ⇒ stabil.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 54

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Die logistische Differenzengleichung (1)Gegeben sei folgende Gleichung:
xn+1 = rxn(1 − xn) (54)
Bestimmung der Fixpunkte: x = rx(1 − x) ⇒ rx2 − x(r − 1) = 0.
Damit erhalt man die Fixpunkte: x1 = 0 und x2 = 1 − 1r.
Die Analyse von Storungen um den nichttrivialen Fixpunkt x2 liefert:
x′n+1 = ax′
n, mit a =df
dx
∣
∣
∣
∣
x2
= r(1 − 2x)|x2= 2 − r.
Daraus folgernd gilt: x2 ist stabil wenn gilt |a| < 1, d.h. 1 < r < 3. DieStabilitat hangt also vom Parameter r ab.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 55

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Die logistische Differenzengleichung (2)
• Diese einfache nichtlineare Differenzengleichung (ein Parameter, einfa-che quadratische Nichtlinearitat) wird oft zur Beschreibung einer Popula-tion mit dichteabhangiger Reproduktionsrate verwendet.
• Dabei zu beachten: Außerhalb der Intervalle 0 < x < 1 und 1 < r < 4stirbt die Population aus.
• Desweitern: ”Auffalliges” Verhalten des Systems fur 3 < r < 4 (sieheSimulationsbeispiele)
• Setzt man z.B. r ein klein wenig großer als 3, so scheinen sich stabileOszillationen der Peroide 2 zu ergeben.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 56

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Die logistische Differenzengleichung (3)
• Um die Fixpunkte des zwei-Punkte Zyklus von xn+1 = rxn(1 − xn) (furr > 3) zu bestimmen betrachte die zusammengesetze Abbildung g(x) =f(f(x)).
g(x) = r[rx(1 − x)](1 − [rx(1 − x)])
= r2x(1 − x)[1 − rx(1 − x)] (55)
• Zur Bestimmung des Fixpunktes ¯x von g setze ¯x = g(¯x):
1 = r2(1 − ¯x)[1 − r¯x(1 − ¯x)] (56)
• Dies ist ein kubisches Polynom mit im allgemeinen 3 Losungen.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 57

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Die logistische Differenzengleichung (4)• Beachte: Ein Fixpunkt von xn+1 = f(xn) ist automatisch auch Fixpunkt
von xn+1 = f(f(xn)).
• Damit muss also der bereits erhaltene Fixpunkt 1 − 1/r einer der dreiLosungen von (56) entsprechen und kann somit eliminiert werden.
• Nach Anwendung von ”ein wenig Algebra” verbleibt somit die Bestim-mung der Losungen von x2 −
(
r+1r
)
x +(
r+1r2
)
= 0.
⇒ ¯x1/2 =r + 1 ±
√
(r − 3)(r + 1)
2r. (57)
• Dies ergibt reelle Losungen fur r < −1 oder r > 3. (r > 3 entspricht auchder Bedingung fur welche x = 1 − 1/r instabil wird.)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 58

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Die logistische Differenzengleichung (5)
• Eine Analyse der Stabilitat von ¯x1 und ¯x2 (Bestimmung von df/dx an denStellen ¯x1 und ¯x2) zeigt, dass der 2-Punkt Zyklus nur fur 3 < r < r2 mitr2 ≃ 3.3 stabil ist.
• Was passiert fur r2 < r < 4?
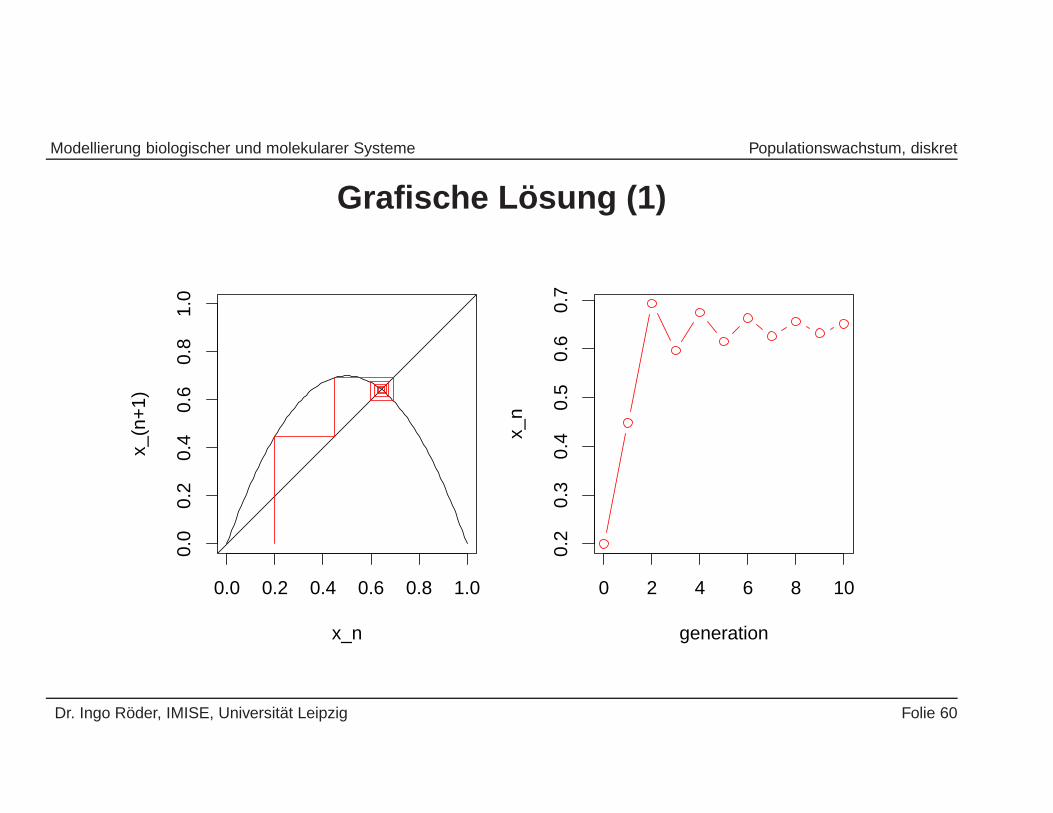
• Hierzu betrachten wir eine grafische Methode der Analyse von Differen-zengleichungen ...
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 59
Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Grafische L osung (1)
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
x_n
x_(n
+1)
0 2 4 6 8 100.
20.
30.
40.
50.
60.
7
generation
x_n
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 60
Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
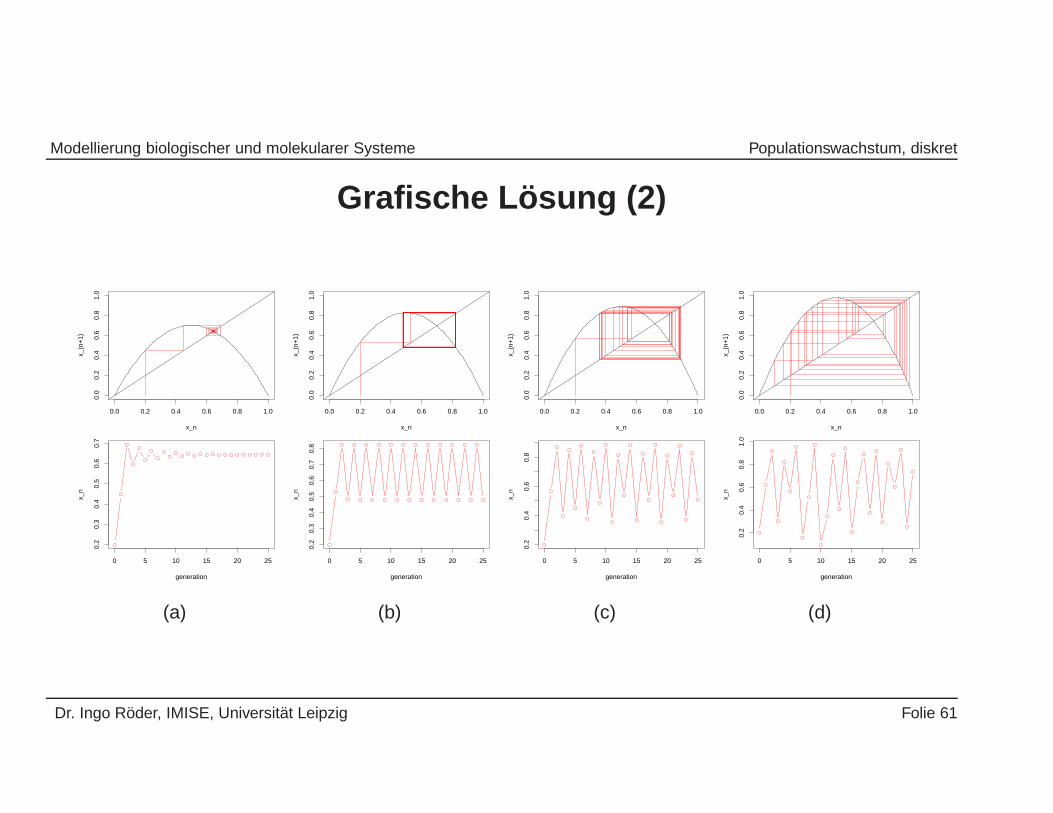
Grafische L osung (2)
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
x_n
x_(n
+1)
0 5 10 15 20 25
0.2
0.3
0.4
0.5
0.6
0.7
generation
x_n
(a)
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
x_n
x_(n
+1)
0 5 10 15 20 25
0.2
0.3
0.4
0.5
0.6
0.7
0.8
generation
x_n
(b)
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
x_n
x_(n
+1)
0 5 10 15 20 25
0.2
0.4
0.6
0.8
generation
x_n
(c)
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
x_n
x_(n
+1)
0 5 10 15 20 25
0.2
0.4
0.6
0.8
1.0
generation
x_n
(d)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 61

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Systeme nichtlinearer Differenzengleichungen
• Verallgemeinerung der vorgestellten Methoden auf Systeme von n nicht-linearen Differenzengleichungen am Beispiel n = 2
xn+1 = f(xn, yn)
yn+1 = g(xn, yn) (58)
• Die stationaren Zustande dieses Systems erfullen die Bedingungen
x = f(x, y)
y = g(x, y) (59)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 62

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit at nichtlinearer Differenzengleichungssysteme (1)• Stabilitatsverhalten wieder anhand des linearisierten Systems analysie-
ren: D.h. betrachte kleine Storung nahe dem Fixpunkt (x, y):
f(x + x′, y + y′) = f(x, y) +∂f
∂x
∣
∣
∣
∣
x,y
x′ +∂f
∂y
∣
∣
∣
∣
x,y
y′ + . . . (60)
• Weiterhin gilt, wie bereits weiter vorn in ahnlicher Weise gezeigt:
x′n+1 = xn+1−x = f(xn, yn)−x = f(x+x′
n, y+y′)−x und f(x, y) = x.
⇒ x′n+1 ≃ ∂f
∂x
∣
∣
∣
∣
x,y
x′n +
∂f
∂y
∣
∣
∣
∣
x,y
y′n
• Eine analoge Darstellung ergibt sich fur g(x, y).
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 63

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit at nichtlinearer Differenzengleichungssysteme (2)
• Damit kann man folgendes lineares Gleichungssystem fur die Storungenx′ und y′ formulieren
x′n+1 = a11x
′n + a12y
′n
y′n+1 = a12x
′n + a22y
′n (61)
wobei
a11 =∂f
∂x
∣
∣
∣
∣
x,y
, a12 =∂f
∂y
∣
∣
∣
∣
x,y
a21 =∂g
∂x
∣
∣
∣
∣
x,y
, a22 =∂g
∂x
∣
∣
∣
∣
x,y
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 64

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit at nichtlinearer Differenzengleichungssysteme (3)
• In Matrixschreibweise erhalt man
x′n+1 = Ax
′n mit A =
(
a11 a12
a21 a22
)
, x′n =
(
x′n
y′n
)
. (62)
• A nennt man Jacobimatrix des Systems (58).
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 65

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit at nichtlinearer Differenzengleichungssysteme (4)Die Ruckfuhrung der Betrachtung auf ein lineares System erlaubt nun dieStabilitatsanalyse in der Nahe eines Fixpuntes (x, y) mit Hilfe der bereitsetablierten Methoden:
1. Bestimme die Losungen (Eigenwerte) der charakteristischen Gleichung
det(A − λI) = 0 bzw. λ2 − βλ + γ = 0
wobei β = a11 + a22 und γ = a11a22 − a12a21.
2. Untersuche, ob die Eigenwerte betragsmaßig kleiner 1 sind.
⇒ Wenn dies der Fall ist, dann handelt es sich um einen stabilen Fixpunkt.
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 66

Modellierung biologischer und molekularer Systeme Populationswachstum, diskret
Stabilit at nichtlinearer Differenzengleichungssysteme (5)
Oft ist es nicht notwending die Eigenwerte explizit zu berechnen, um Sta-bilitat eines Fixpunktes zu bestimmen. Anstatt dessen betrachte folgendeBedingung (fur Gleichungen 2. Ordnung / System zweier Gleichungen 1.Ordnung):
2 > 1 + γ > |β|, d.h. 2 > 1 + detA > |trA| →{
beide Eigenwerte|λi| < 1d.h. Fixpunkt stabil
(63)(Herleitung siehe z.B. Edelstein-Keshet, section 2.8)
Dr. Ingo Roder, IMISE, Universitat Leipzig Folie 67


















