
Modulation der IL-6-abhängigen Signaltransduktion durch...
108
Modulation der IL-6-abhängigen Signaltransduktion durch die Protein-Tyrosinphosphatase SHP2 Von der Medizinischen Fakultät der Rheinisch-Westfälischen Technischen Hochschule Aachen zur Erlangung des akademischen Grades einer Doktorin der Medizin genehmigte Dissertation vorgelegt von Cornelia Henke-Gendo geb. Gendo aus Bad Soden am Taunus Berichter: Herr Universitätsprofessor Dr. rer. nat. Peter C. Heinrich Herr Universitätsprofessor Dr. med. Hans Merk Tag der mündlichen Prüfung: 13. August 2002 Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfügbar.
Transcript of Modulation der IL-6-abhängigen Signaltransduktion durch...

Modulation derIL-6-abhängigen Signaltransduktion
durch die Protein-Tyrosinphosphatase SHP2
Von der Medizinischen Fakultätder Rheinisch-Westfälischen Technischen Hochschule Aachen
zur Erlangung des akademischen Gradeseiner Doktorin der Medizin genehmigte Dissertation
vorgelegt von
Cornelia Henke-Gendo geb. Gendoaus
Bad Soden am Taunus
Berichter: Herr UniversitätsprofessorDr. rer. nat. Peter C. Heinrich
Herr UniversitätsprofessorDr. med. Hans Merk
Tag der mündlichen Prüfung: 13. August 2002
Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek onlineverfügbar.

.

meinem Großvater gewidmet

.

Wesentliche Teile dieser Arbeit wurden veröffentlicht:
Schaper F., Gendo C., Eck M., Schmitz J., Grimm C., Anhuf D., Kerr I.M. undHeinrich P.C.Activation of the protein tyrosine phosphatase SHP2 via the interleukin-6 signal transducingreceptor gp130 requires tyrosine kinase Jak1 and limits acute-phase protein expressionBiochem. J. (1998) 355: 557-565
Schmitz J., Dahmen H., Grimm C., Gendo C., Müller-Newen G., Heinrich P.C. undSchaper F.The cytoplasmic tyrosine motifs in full-length glycoprotein 130 have different roles in IL-6signal transductionJ. Immunol. (2000) 164:848-854

.

Inhaltsverzeichnis I
Inhaltsverzeichnis
1. Einleitung 1
1.1 Zytokine und Zytokinrezeptoren 1
1.2 Interleukin-6 und der Interleukin-6-Rezeptorkomplex 3
1.3 Die Interleukin-6 induzierte Signaltransduktion 6
1.4 Struktur und Funktion der SHP2 8
1.5 Zielsetzung der Arbeit 11
2. Material und Methoden 13
2.1 Chemikalien und Verbrauchsmaterialien 13
2.1.1 Radiochemikalien 13
2.1.2 Zellkultur 13
2.1.3 Zytokine und Rezeptoren 13
2.1.4 Antikörper 14
2.1.5 Oligonukleotide 15
2.1.6 Plasmide 16
2.2 Allgemeine molekularbiologische Methoden 17
2.2.1 Verwendete Bakterienstämme und deren Kultivierung 17
2.2.2 Herstellung kompetenter Zellen 18
2.2.3 Transformation nach Hanahan 18
2.2.4 Isolierung von Plasmid-DNA aus E. coli 18
2.2.5 Konzentrationsbestimmung von DNA 19
2.2.6 Oligohybridisierung 19
2.2.7 Modifikation von Plasmid-DNA 19
2.2.8 Polymerase-Ketten-Reaktion 21
2.2.9 Analyse der DNA durch Sequenzierungen 23
2.3 Eukaryonte Zellen und deren Kultivierung 24
2.3.1 Kulturmedien 24
2.3.2 Verwendete permanente Zell-Linien 25

II Inhaltsverzeichnis
2.3.3 Kultivierung von Zell-Linien 26
2.3.4 Transiente Transfektion 27
2.3.4 Stabile Transfektion von Ba/F3-Zellen 28
2.4 Biochemische Methoden 28
2.4.1 Herstellung zytosolischer Extrakte 28
2.4.2 Immunpräzipitation 29
2.4.3 SDS-Polyacrylamid Gelelektrophorese 29
2.4.4 Western Blot 30
2.4.5 Detektion von Proteinen mittels verstärkter Lichtemission 30
2.4.6 Herstellung von Kernextrakten 32
2.4.7 Proteinbestimmung nach Bradford 32
2.4.8 Radioaktive Markierung doppelsträngiger DNA 33
2.4.9 Elektrophoretischer Mobilitäts-Shift Assay (EMSA) 33
2.4.10 Immunfluoreszenz-Analysen eukaryonter Zellen 34
2.4.11 FACS-Analysen von eukaryonten Zellen 35
2.4.12 Reportergen-Assays 35
2.5 Konstruktion der ErG-Mutanten 36
2.6 Konstruktion der GrG-Mutanten 38
2.7 Konstruktion der SHP2-Mutanten 39
3. Ergebnisse 41
3.1 Konstruktion und Expression der ErG-Mutanten 41
3.2 Die SHP2-Tyrosinphosphorylierung 42
3.3 Die IL-6-induzierte Tyrosinphosphorylierung der SHP2 benötigt
das Y759 in gp130 47
3.4 Einfluß der SHP2-Tyrosinphosphorylierung auf die STAT-Aktivierung 50
3.5 Rolle der Janus-Kinasen für die SHP2-Phosphorylierung 51
3.6 Assoziation der SHP2 mit den Jak-Kinasen 54
3.7 Rolle der SHP2 in der Akute-Phase-Protein Gen-Induktion 56

Inhaltsverzeichnis III
4. Diskussion 61
4.1 Ausblick 69
5. Zusammenfassung 73
6. Anhang 75
6.1 Literaturverzeichnis 75
6.2 Abkürzungsverzeichnis 85
6.3 Sequenzen 87
Danksagung
Lebenslauf

.

Einleitung 1
1. Einleitung
1.1 Zytokine und Zytokinrezeptoren
Für alle vielzelligen Organismen ist die Koordination ihrer verschiedenen Körperfunktionen
die existentielle Voraussetzung, um in einer Welt mit sich fortlaufend ändernden äußeren Be-
dingungen zu überleben. Wachstum, Differenzierung und Stoffwechsel in den verschiedenen
Organen und Geweben erfordern eine ständige Kommunikation zwischen den Zellen. Im Lau-
fe der Evolution haben sich hierfür zwei Systeme herausgebildet: Einerseits das Nervensy-
stem, welches blitzschnell Sinneseindrücke ins Körperinnere leiten kann, andererseits das
System der Hormone, das eher Minuten bis Stunden braucht und dazu dient, definierte Bedin-
gungen im Körper herzustellen (Homöostase).
Die Hormone ihrerseits sind sowohl bezüglich ihrer chemischen Zusammensetzung als auch
hinsichtlich der Wirkungsweise ihrer Mitglieder sehr inhomogen. Neben den klassischen
Hormonen, die von spezialisierten Organen produziert werden und endokrin über den Blut-
strom auf das/die Zielgewebe einwirken, gibt es die sogenannten Gewebshormone. Sie wer-
den von einer Vielzahl von Zellen nach Stimulation produziert und wirken hauptsächlich pa-
ra- und autokrin. Hierzu gehören die Arachidonsäure-Metabolite sowie Polypeptide und Pro-
teine wie Wachstumsfaktoren und Zytokine.
Als Zytokine bezeichnet man im allgemeinen diejenigen Polypeptid-Gewebshormone, die
regulierend auf die Hämatopoese, die Immunantwort und die Entzündungsreaktion einwirken.
Sie besitzen oft ein pleiotropes Wirkungsspektrum, d.h. dasselbe Zytokin ist in der Lage, auf
unterschiedlichen Zellen verschiedene Effekte auszuüben. Untereinander beeinflussen sie sich
in additiver, z.T. synergistischer aber auch antagonistischer Weise.
Anhand ihrer Struktur lassen sich die Zytokine in verschiedene Familien einteilen. Eine be-
kannte Familie ist die Gruppe der 4-α-helix bundle-Zytokine, so benannt, weil sich ihre
Struktur durch vier antiparallel angeordnete α-Helices auszeichnet. Ein wichtiger Vertreter ist
z.B. das Interleukin-6, aber auch das Erythropoetin oder das G-CSF (Granulocyte Colony-
Stimulating Factor) gehören in diese Familie, ebenso wie die klassischen Hormone Prolaktin
und Wachstumshormon [9].

2 Einleitung
Zytokine vermitteln ihre Wirkung über spezifische Oberflächenrezeptoren, die von ihren Ziel-
zellen exprimiert werden. Genau wie die Zytokine sind auch diese Rezeptoren strukturell und
funktionell miteinander verwandt und werden zu einer Zytokinrezeptor-Superfamilie zusam-
mengefaßt (Tab. 1). Extrazellulär zeichnen sich die Mitglieder dieser Familie neben optional
vorhandenen Immunglobulin-ähnlichen Domänen und/oder Fibronektin Typ III-Domänen
durch den Besitz eines sogenannten Zytokinrezeptor-Homologiemoduls aus. Dieses wird von
zwei je etwa 90 Aminosäuren langen Fibronektin Typ III-Domänen gebildet und ist durch vier
in ihrer Position konservierte Cysteinreste und ein Tryptophan-Serin-X-Tryptophan-Serin-
Motiv (WSXWS; X ist eine nicht konservierte Aminosäure) charakterisiert [10]. Die intra-
zellulären Abschnitte der Zytokinrezeptoren sind wenig homolog und strukturell noch nicht
näher charakterisiert. Sie enthalten jedoch zwei konservierte als box1 und box2 bezeichnete
Regionen. Anders als die sogenannten Rezeptor-Tyrosin-Kinasen konnte bei den Zytokinre-
zeptoren bisher noch keine katalytische Domäne identifiziert werden. Vielmehr sind sie kon-
stitutiv mit Tyrosinkinasen der Familie der Janus-Kinasen (Jak) assoziiert [53, 76], die Ligan-
den-induziert den Rezeptor und verschiedene zytoplasmatische Adapterproteine phosphorylie-
ren und so Signale ins Zellinnere leiten.
Die Zytokinrezeptor-Superfamilie
Klasse I: GH-R, PRL-R, Epo-R, G-CSF-R, Mpl bilden Homodimere
IL-6R, IL-11R, LIF-R, OSM-Rβ, CNTF-R bilden Heterodimere mit gp130
IL-3R, GM-CSF-R, IL-5R bilden Hetero-Oligomere mit βc
IL-2R, IL-4R, IL-7R, IL-9R, IL-13R,
IL-15R
bilden Hetero-Oligomere mit γc
Klasse II: IFN-αR, IFN- βR, TF-R, IL-10R
Tab.1: Die Rezeptoren der Zytokinrezeptor-Superfamilie lassen sich nach strukturellen und funktionellen Ei-genschaften in zwei Klassen einteilen. Die signaltransduzierenden Rezeptorkomplexe der Klasse I bil-den Homodimere aus identischen Rezeptorketten oder Hetero-Oligomere aus Liganden-spezifischen α–Rezeptoren und den gemeinsam genutzten β–Rezeptor-Untereinheiten gp130, βc oder γc (nach Wells,1996 [90]). R = Rezeptor; GH growth hormone, PRL Prolaktin, Epo Erythropoetin, G-CSF granulocytecolony-stimulating factor, Mpl myeloproliferative leukemia receptor, IL Interleukin, LIF leukemia inhi-bitory factor, OSM Oncostatin M, CNTF ciliary neurotrophic factor, GM-CSF granulocyte/ macropha-ge colony-stimulating factor, IFN Interferon, TF tissue factor.

Einleitung 3
1.2 Interleukin-6 und der Interleukin-6-Rezeptorkomplex
Das Zytokin Interleukin-6 (IL-6) wurde ursprünglich als ein von T-Zellen sezerniertes Lym-
phokin identifiziert. Es induziert die Differenzierung von aktivierten B-Zellen zu Antikörper
produzierenden Plasmazellen, daher sein ursprünglicher Name: B-cell stimulating factor 2
[37]. Daneben erfüllt IL-6 noch andere physiologische, aber auch pathophysiologische Funk-
tionen [35, 46]. Es wirkt z.B. als Differenzierungsfaktor für weitere hämatopoetische Vorläu-
ferzellen oder aber als Wachstums- bzw. Aktivierungsstimulans für Plasmozytomzellen und
Osteoklasten. Eine weitere Eigenschaft des IL-6, welches bei Verletzungen oder Infektionen
von Monozyten, Endothelzellen und Fibroblasten ausgeschüttet wird, ist die Auslösung der
Akute-Phase-Reaktion. Letztere ist definiert als eine komplexe Allgemeinantwort des Orga-
nismus auf verschiedene schädigende Einflüsse und umfaßt Reaktionen wie Fieber, Leuko-
zytose und Veränderungen der Expression einer Reihe von in der Leber gebildeten Plasma-
proteinen, den sog. Akute-Phase-Proteinen. Deren bekannteste Vertreter sind das C-reaktive
Protein (CRP) oder das Haptoglobin. Weitere Funktionen des IL-6 sind in Tab. 2 zusammen-
gefaßt.
Um die Signaltransduktion auszulösen, benötigt IL-6 wie andere Zytokine einen Plasmamem-
bran-Rezeptorkomplex. Im Falle des IL-6-Rezeptor-Komplexes besteht dieser aus einer IL-6
bindenden Untereinheit, dem IL-6-Rezeptor (IL-6-R, gp80, CD126), und einer signaltransdu-
zierenden Komponente, dem gp130 (CD130) (Abb. 1). Der humane IL-6-Rezeptor gehört zu
den Klasse I-Rezeptoren der Zytokinrezeptor-Superfamilie. Das transmembranäre Protein,
dessen verhältnismäßig kurzer intrazellulärer Abschnitt ohne Funktion zu sein scheint, hat
eine Größe von 80 kDa. Der extrazelluläre Teil des Rezeptors wird von einer Immunglobulin-
ähnlichen Domäne gebildet, gefolgt von einem Zytokinrezeptor-Homologiemodul [92]. Ne-
ben der membranständigen wurde auch eine lösliche Variante des Rezeptors, sIL-6R, be-
schrieben [39], die entweder durch alternatives Spleißen der gp80-mRNA oder durch limi-
tierte Proteolyse (shedding) des Rezeptors unmittelbar vor der Transmembrandomäne entsteht
[40, 60]. Entscheidend an der Auslösung der Signaltransduktion ist jedoch das Glykoprotein
gp130. Dieses beim Menschen 918 Aminosäuren lange membranständige Protein wird ubi-
quitär exprimiert und vermittelt nicht nur die Signaltransduktion des IL-6, sondern auch die
der sogenannten IL-6-Typ Zytokine. Dazu gehören neben IL-6 auch LIF (leukemia inhibitory
factor), OSM (Oncostatin M), CNTF (ciliary neurotrophic factor), IL-11 und Cardiotrophin-1

4 Einleitung
Die pleiotrope Wirkung von IL-6
Effekte auf B-Zellen: Immunglobulin-ProduktionProliferation von MyelomzellenProliferation von Epstein-Barr-Virus infizierten B-Zellen
Effekte auf T-Zellen: Proliferation und Differenzierung von T-ZellenDifferenzierung von T-Lymphozyten zu zytotoxischen T-ZellenInduktion der Expression des IL-2-RezeptorsInduktion der IL-2-ProduktionVermehrung der Aktivität der natürlichen Killerzellen
Effekt auf hämatopoetischeProgenitorzellen:
Verstärkung pluripotenter hämatopoetischer Koloniebildung
Effekt auf Megakaryozyten: Maturation
Effekte auf Makrophagen: Wachstumsinhibition von myeloiden Leukämie-Zellinien undInduktion ihrer Differenzierung zu Makrophagen
Effekte auf Hepatozyten: Regulation der Akute-Phase-Protein SyntheseProliferation während der Leber-Regeneration
Effekte auf den Knochen-metabolismus:
Stimulation der OsteoklastenentwicklungInduktion der Knochenresorption
Effekte auf Blutgefäße: PDGF-InduktionProliferation der glatten Gefäßmuskulatur
Effekte auf Mesangialzellen Stimulation der Proliferation
Effekte auf Hautzellen Stimulation der Proliferaton von Keratinozyten
Effekt auf das Herz: negativer inotroper Effekt auf Herzmuskelzellen
Effekte auf neuronale Zellen: neuronale Differenzierung von PC12-ZellenErhöhung der Überlebenschancen cholinerger NeuroneInduktion der Synthese adrenokortikotroper Hormone
Effekt auf die Plazenta: Sekretion von Choriogonadotropin aus Trophoblasten
Wirkung auf Tumorzellen: Hemmung des Wachstums von Melanom- und Brustkarzinom-zellen
allgemeine Reaktionen: Erhöhung der Körpertemperatur
Tab. 2: nach Heinrich et al. (1998), Kishimoto et al. (1995) [35, 46]

Einleitung 5
(CT-1) [27, 51, 66, 74, 94]. gp130 besteht im extrazellulären Bereich aus sechs Fibronectin
Typ III-Domänen, von denen die zweite und die dritte ein Zytokinrezeptor-Homologiemodul
mit vier konservierten Cysteinresten und einem WSXWS-Motiv bilden. Es handelt sich hier
also ebenfalls um ein Mitglied der Zytokinrezeptor-Superfamilie. Die Struktur des intrazellu-
lären Teils ist noch nicht bekannt, enthält jedoch die für Zytokinrezeptoren typischen box1-
bzw. box2-Regionen. Desweiteren besitzt gp130 sechs Tyrosinmotive, die eine wichtige Rolle
in der Signaltransduktion spielen.
Die Stöchiometrie des IL-6-Rezeptorkomplexes wird noch diskutiert. Neuere Untersuchungen
weisen darauf hin, daß dieser ein Hexamer aus zwei IL-6-, zwei gp80- und zwei gp130-
Molekülen ist [87]; ein anderes Modell, basierend auf Mutagenese-Studien, schlägt jedoch die
Interaktion von je einem Molekül IL-6 und IL-6R mit zwei gp130-Molekülen vor [31].
Abb.1: Schematische Darstellung des IL-6-Rezeptors und des gp130Der extrazelluläre Teil des IL-6-R besteht aus einer aminoterminalen Immunglobulin (Ig-)-ähnlichenDomäne und aus zwei Fibronektin Typ III-Domänen, die ein Zytokinrezeptor-Homologiemodul mitvier konservierten Cysteinresten (C) und einem konservierten WSXWS-Motiv bilden. Die extrazel-luläre Region des gp130 wird von sechs Fibronektin Typ III-Domänen gebildet, von denen diezweite und die dritte Domäne ein Zytokinrezeptor-Homologiemodul darstellen. Im intrazellulärenTeil enthält gp130 die für Zytokinrezeptoren typischen box1- bzw. box2-Regionen und sechs Tyro-sine bzw. Tyrosinmotive, die z.T. eine wichtige Rolle in der Signaltransduktion spielen.

6 Einleitung
1.3 Die Interleukin-6-induzierte Signaltransduktion
Ihre Wirkung entfalten die verschiedenen Zytokine durch Aktivierung ihrer Zielgene im Zell-
kern. Das Signal dafür wird jedoch an der Zellmembran gesetzt. Es muß von dort in den Kern
weitergeleitet werden. Bei der IL-6-Signaltransduktion kann das Signal über zwei verschiede-
ne Wege in den Zellkern gelangen, zum einem über den Jak/STAT-Signalweg, zum anderen
über die Ras/Raf/MAPK-Kaskade.
Den ersten Schritt dieser Signalübertragung bildet die Bindung von IL-6 an den IL-6-
Rezeptor mit nachfolgender Assoziation und Homodimerisierung von gp130 [61]. Anschlie-
ßend kommt es zur Autophosphorylierung und Aktivierung der konstitutiv an gp130 assozi-
ierten Janus-Kinasen Jak1, Jak2 und Tyk2 [53, 76]. Diese Jak-Kinasen sind durch eine C-
terminale Kinase-Domäne, eine anschließende Kinase-ähnliche Domäne und fünf N-terminale
Jak-Homologie-Domänen (JH1-5) charakterisiert. Aktiviert vermitteln sie die Phosphorylie-
rung von gp130 an spezifischen Tyrosinmotiven, die ihrerseits nun mehreren Proteinen mit
SH2-Domänen (Transkriptionsfaktoren, Adaptermoleküle etc.) als Bindungspartner zu Verfü-
gung stehen [77, 88]. Besondere Bedeutung wird hier den STAT (signal transducer and acti-
vator of transcription)- Faktoren beigemessen. Hierbei handelt es sich um eine Familie von
Transkriptionsfaktoren, die sich durch den Besitz einer DNA-Bindungsdomäne, einer SH2-
sowie einer Tetramerisierungsdomäne auszeichnen [11, 15, 23]. An der IL-6-Signaltrans-
duktion sind zwei dieser Faktoren beteiligt, STAT1 und STAT3, die nach ihrer Phosphorylie-
rung an einem konservierten Tyrosin-Rest homo- bzw. heterodimerisieren, indem sie mit ihrer
SH2-Domäne an das Phosphotyrosin ihres Partners binden [75]. STAT-Faktoren translozieren
über noch unbekannte Mechanismen in den Zellkern, binden dort an die Promotoren ihrer
Zielgene und leiten die Transkription ein [88, 95]. Neben der Tyrosinphosphorylierung wurde
auch eine Serinphosphorylierung der STAT-Faktoren in der C-terminalen Region beobachtet
[54, 96]. Sie erfolgt durch eine noch nicht näher identifizierte Kinase.
Die Ras/Raf/MAPK-Kaskade wird ebenfalls durch IL-6 induziert [48, 62]. Der genaue Me-
chanismus hierfür ist jedoch noch nicht bekannt. Wichtig für die Aktivierung der MAPK
scheint das Tyrosin 759 im gp130 zu sein [72]. Weiterhin sind mehrere Proteine beschrieben
worden, die nach IL-6-Induktion tyrosinphosphoryliert werden und als linker mit Kompo-
nenten der anderer Signalwege assoziieren können, z.B. die Protein-Tyrosinphosphatase
SHP2 [50] oder die Adapterproteine Shc [29] und Vav [52].
Die Signaltransduktion des IL-6 ist schematisch in Abb. 2 wiedergegeben.
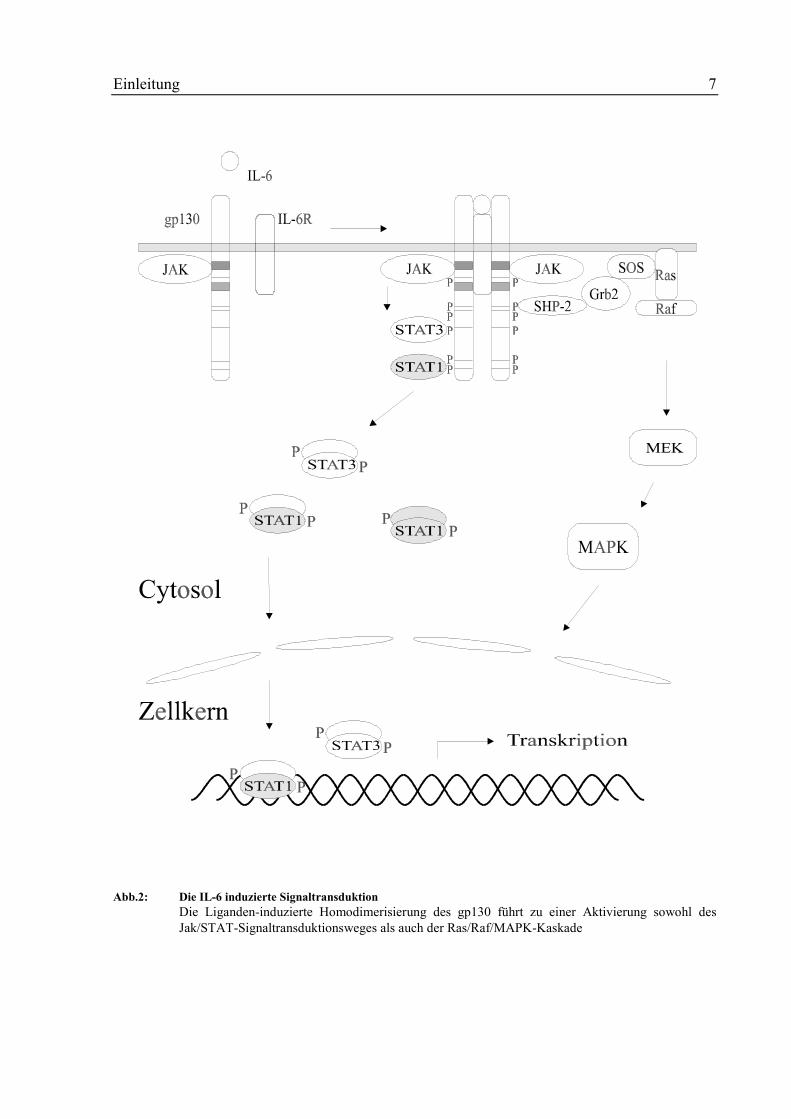
Einleitung 7
Abb.2: Die IL-6 induzierte SignaltransduktionDie Liganden-induzierte Homodimerisierung des gp130 führt zu einer Aktivierung sowohl desJak/STAT-Signaltransduktionsweges als auch der Ras/Raf/MAPK-Kaskade

8 Einleitung
1.4 Struktur und Funktion der SHP2
Im Jahre 1993 entdeckten gleich mehrere Gruppen ein 65 - 68 kDa großes Protein, welches
nach PDGF- und EGF-Stimulation tyrosinphosphoryliert wird [2, 20, 22, 84]. Hierbei handelt
es sich um die Phosphatase SHP2, SH2 domain containing protein-tyrosine-phosphatase 2,
auch beschrieben als PTP1D oder syp. Dieses Protein, dessen humane Variante eine Gesamt-
länge von 585 Aminosäuren hat, wird – im Gegensatz zu ihrer hämatopoetischen Schwester
SHP1 – ubiqitär exprimiert, wobei sich besonders hohe Proteinmengen in Lungen-, Hirn- und
Lebergewebe befinden [2].
Strukturell gehört die SHP2 in die Klasse der nicht-membranständigen Protein-Tyrosin-
phosphatasen (Abb. 3). Sie besteht aus zwei N-terminal gelegenen SH2-Domänen (aa. 6-214)
und einer sich anschließenden Phosphatase-Domäne. Die beiden SH2-Domänen, welche spe-
zifische Phosphotyrosin-abhängige Protein-Protein-Interaktionsstellen darstellen, dienen zur
Rekrutierung der SHP2 an bestimmte Phosphotyrosinmotive, die die Konsensussequenz
pYXX(L/V/I) aufweisen [14]. Diese finden sich z.B. in den die SHP2 aktivierenden Rezepto-
ren. Neben dieser Rekrutierung an die Rezeptoren scheinen die Domänen auch eine suppri-
mierende regulatorische Funktion wahrzunehmen. Pluskey et al. wiesen nach, daß Bindung
zweier Phosphotyrosin-Peptide aus dem IRS-1, die spezifisch mit den SH2-Domänen der
SHP2 interagieren können, zu einer 37-fachen Stimulation der SHP2-Enzymaktivität führt
[67]. Die SHP2 scheint sich also durch Anlagerung ihrer SH2-Domänen an die Phosphatase-
Domäne selbst inhibieren zu können, was vor kurzem durch die Strukturanalyse des kristalli-
sierten Proteins bestätigt werden konnte [38].
Im C-terminalen Bereich besitzt die SHP2 eine Phosphatase-Domäne mit der für nichttrans-
membranäre Phosphatasen typischen Konsensussequenz VHC459XAGXXR [2]. Entscheidend
für die Aktivität dieses Enzyms ist das Cystein 459 innerhalb des katalytischen Zentrums.
Mutation dieser Aminosäure zu einem Serin führt zu einer komplett inaktiven, dominant ne-
gativen Mutante [8, 59]. Eine verringerte enzymatische Aktivität besitzt auch eine andere, in
dem Fall natürliche Variante der SHP2, bei der vier Aminosäuren (ALLQ) an Position 408
innerhalb der Phosphatase-Domäne eingefügt sind. Sie entsteht wahrscheinlich durch alterna-
tives Spleißen der SHP2-Prä-mRNA [3, 58].
Trotz einer Vielzahl von Untersuchungen ist die Funktion der SHP2-Phosphatase-Aktivität in
der Signaltransduktion weiterhin unbekannt. Schon potentielle Substrate werden kontrovers
diskutiert. Hierfür käme neben den aktivierenden Rezeptoren auch SHPS-1, SHP-substrate 1,

Einleitung 9
in Frage [25, 45, 63]. Es handelt sich hierbei um ein kürzlich entdecktes Transmembran-Pro-
tein mit zwei mal zwei potentiellen SHP2-Bindungsstellen, an denen jeweils beide SH2-
Domänen eines Enzyms binden können. Einen Hinweis darauf, daß die SHP2 dieses Protein
tatsächlich dephosphoryliert, bieten diese Veröffentlichungen allerdings nicht. Auch bezüg-
lich der Dephosphorylierung des PDGF-Rezeptors existieren noch offene Fragen. Vogel et al.
[84] waren nicht in der Lage, die Dephosphorylierung dieses Rezeptors durch die SHP2 nach-
zuweisen, obwohl die Dephosphorylierung einer EGF-c-kit-Chimäre in einem Parallelansatz
sehr wohl gezeigt werden konnte. Zwei anderen Gruppen, Kazlauskas et al. und Dechert et al.
[18, 43] gelang eben dieser Nachweis der Dephosphorylierung des PDGF-R durch die SHP2.
Eine Autodephosphorylierung der SHP2 wurde ebenfalls beschrieben [78].
Abb.3: Struktur der SHP2Die SHP2 gehört zu den nicht-transmembranären Protein-Tyrosinphosphatasen. Das 585 Aminosäu-ren lange Protein besitzt im aminoterminalen Bereich zwei SH2-Domänen (SH2). Am C-Terminusbefindet sich eine Protein-Tyrosinphosphatase-Domäne (PTP) mit dem Cystein 459 im katalytischenZentrum. Eine Spleißvariante besitzt vier zusätzliche Aminosäure (ALLQ) an Position 408.
Die SHP2-Tyrosinphosphorylierung wurde jedoch nicht nur im Zusammenhang mit der Akti-
vierung der Rezeptoren PDGF-R oder EGF-R gezeigt. Eine Vielzahl von Rezeptoren ist in der
Lage, die SHP2 zu rekrutieren. Sie beeinflußt sowohl die Signaltransduktion der Tyrosin-
Kinase-Rezeptoren (Insulin-R, FGF-R, c-kit, IGF-R), als auch die der G-Protein-gekoppelten
Rezeptoren (Angiotensin-R) und Zytokinrezeptoren (IL-2R, IL-3R, IL-5R, IL-6R, IL-11R,
GM-CSF-R, IFNα/β-R, Epo-R) [1, 4, 12, 17, 24, 55, 64, 80, 81, 82]. Interessanterweise füh-
ren jedoch nicht nur Rezeptoren im engeren Sinn zu einer Aktivierung der SHP2. So konnte
eine Tyrosinphosphorylierung dieses Proteins in Endothel-Zellen nachgewiesen werden, die
unterschiedlichen physikalischen Rahmenbedingungen wie erhöhter Fließgeschwindigkeit des
umgebenen Mediums oder hypo- bzw. hyperosmolarem Milieu ausgesetzt waren [56]. Diese

10 Einleitung
Phosphorylierung beinhaltet wahrscheinlich die Aktivierung von Mitgliedern der Integrin-
Familie [41, 42].
Bei einer solchen Fülle von aktivierenden und beeinflussenden Faktoren ist es nicht verwun-
derlich, daß die Elimination des SHP2-Genes in der Maus zu einem frühembryonal letalen
Phänotyp führt [70].
Eine weitere Frage ist, wie trotz der Vielzahl der Aktivierungsmöglichkeiten ein spezifisches,
speziell für diesen Signalweg zugeschnittenes Signal weitergeleitet wird. Eine Antwort auf
diese Frage könnte in der Tatsache liegen, daß viele verschiedene Tyrosin-Kinasen in der La-
ge sind, die SHP2 zu phosphorylieren [65, 83, 93]. Auch werden abhängig vom aktivierenden
Zytokin unterschiedliche Tyrosine innerhalb der SHP2 phosphoryliert. So ist z.B. beschrieben
[85], daß nach EGF-R-Stimulation zwei Tyrosine innerhalb der SH2-Domänen und zwei am
C-terminalen Ende der SHP2 (Y546 und Y584) phosphoryliert werden. Bei PDGF-R-
Aktivierung aber wird neben den beiden letzteren Tyrosinen ein weiteres, in der linker-Region
zwischen Phosphatase- und SH2-Domänen gelegenes Tyrosin phosphoryliert. Jak2 soll hin-
gegen für die Phosphorylierung der SHP2 an Tyrosin 304 und 327 verantwortlich sein [93].
Phosphoryliert ist die SHP2 in der Lage, mit vielen SH2-Domänen-haltigen Proteinen, so z.B.
IRS-1, PLC, Grb2 oder Grb7 zu interagieren [7, 44, 47]. Sie ermöglicht so die Erweiterung
des Signals auf andere Signalwege (Adapterfunktion). Ein wichtiger Bindungspartner scheint
in diesem Zusammenhang Grb2 zu sein, da über Grb2 die Ras/Raf/MAPK-Kaskade aktiviert
wird. Wie SHP2 und Grb2 miteinander agieren, hängt interessanterweise von dem aktivieren-
den Rezeptor ab. Nach PDGF-Stimulation bindet Grb2 die SHP2 über seine SH2-Domänen,
im EGF-Signalweg interagieren beide Partner über die SH3-Domänen des Grb2 [85, 91].
Weitere Interaktionspartner bilden die Janus-Kinasen Jak1 und Jak2. Im Jahre 1995 konnte
gezeigt werden, daß SHP2 und Jak2 in 3T3-L1 Maus Präadipozyten stimulationsunabhängig
interagieren können [24]. Die Assoziation beider Proteine konnte durch IL-11-Stimulation
weiter induziert werden. Zwei Jahre später konnte in Co-Transfektionsexperimenten verkürz-
ter SHP2- bzw. Jak2-Fragmente die Interaktionstelle weiter eingegrenzt werden. In der Phos-
phatase scheinen die Aminosäuren 272 bis 322 für die Bindung notwendig zu sein, innerhalb
der Kinase der N-terminale Bereich [93].
Die Notwendigkeit des tyrosinphosphorylierten Y-Motivs YXX(L/V/I) zur Rekrutierung der
SHP2 an einen Rezeptor legt nahe, daß auch im Interleukin-6-Signalweg die SHP2 am
Y759STV des gp130 aktiviert werden kann. In der Tat konnten Stahl et al. bzw. Fukada et al.
dies nachweisen [26, 77]. Sie verwendeten dazu verkürzte bzw. chimäre Rezeptoren. Auch

Einleitung 11
zur Funktion der SHP2 in der IL-6-Signaltransduktion wurden bereits erste Untersuchungen
durchgeführt. Fukada et al. zeigten, daß die IL-6-abhängige Proliferation einer stabil gp130
exprimierenden Ba/F3-Zellinie von der STAT- und der SHP2-Aktivierung beeinflusst wird.
Während die STAT-Aktivierung für die anti-Apoptose verantwortlich ist, löst das Rekrutieren
der SHP2 an den Rezeptor das mitogene Signal aus [26].
1.5 Zielsetzung der Arbeit
Die Protein-Tyrosin-Phosphatase SHP2 wird im Kontext verschiedener Signaltransduktionen
tyrosinphosphoryliert. Auch für die IL-6-abhängige Signaltransduktion konnte diese Phospho-
rylierung nachgewiesen werden. Welche Kinase für diese Reaktion verantwortlich ist, ist bis
jetzt noch nicht bekannt. Als Kandidaten kommen u.a. die ebenfalls an der IL-6 Signaltrans-
duktion beteiligten, konstitutiv an gp130 assoziierten Janus-Kinasen Jak1, Jak2 und Tyk2 in
Frage. Ihre mögliche Beteiligung an der IL-6-abhängigen SHP2-Tyrosinphosphorylierung
sollte hier analysiert werden.
Weiterhin sollte nachgeprüft werden, welchen Einfluss die SHP2-Tyrosinphosphorylierung
auf IL-6-abhängige Reaktionen in der Zelle hat. Es sollte der Frage nachgegangen werden, ob
und wie die Akute-Phase-Gen-Induktion durch die SHP2-Tyrosinphosphorylierung bzw. ihre
enzymatische Aktivität beeinflusst wird.

.

Material und Methoden 13
2. Material und Methoden
2.1 Chemikalien und Verbrauchsmaterialien
Alle Chemikalien wurden in der Qualität pro analysi eingesetzt und von den angegebenen
Quellen bezogen.
2.1.1 Radiochemikalien
[α32P]-dATP Amersham Buchler, Braunschweig
[α35S]-dATP Amersham Buchler, Braunschweig
2.1.2 Zellkultur
DMEM Flüssigmedium mit 4,5 g Glukose und Glutamin Gibco, Eggenstein
DMEM/NUT-Mix (1:1) Flüssigmedium mit F12 und Glutamin Gibco, Eggenstein
Penicillin/Streptomycin Gibco, Eggenstein
Trypsin/EDTA-Lösung (0,05%/ 0,02%) Cytogen, Berlin
Fetales Kälberserum Cytogen, Berlin
2.1.3 Zytokine und Rezeptoren
Interleukin-6 humanes IL-6 wurde nach der von Arcone et al. [6] entwickelten Me-
thode rekombinant hergestellt. Die spezifische Aktivität betrug
1x106 BSF2 U/mg Protein [37].
Erythropoetin rekombinantes humanes Erythropoetin (2,2 x 105 E/mg Protein) wurde
von der Firma Boehringer Mannheim, Penzberg, bezogen.
sIL-6R rekombinanter humaner sIL-6R wurde nach der Methode von
Weiergräber et al. [89] hergestellt und freundlicherweise von A.
Küster, Aachen, zur Verfügung gestellt.

14 Material und Methoden
2.1.4 Antikörper
Antikörper gerichtet gegen:
Phosphotyrosin (4G10) monoklonaler Maus-Antikörper,
Upstate Biotechnology, USA
muriner EpoR polyklonaler Kaninchen-Antikörper,
Upstate Biotechnology, USA
human gp130 cyt polyklonaler Kaninchen-Antikörper gegen den
cytoplasmatischen Teil des gp130,
Upstate Biotechnology, USA
c-myc monoklonaler Maus-Antikörper,
Boehringer Mannheim, Penzberg
Maus-IgG polyklonaler Ziegen-Antikörper
Dako, Dänemark
Maus-IgG HRP HRP gekoppelter polyklonaler Ziegen-Antikörper,
Dako, Dänemark
Kaninchen-IgG HRP HRP gekoppelter polyklonaler Ziegen-Antikörper
Dako, Dänemark
gp130 (BS12) monoklonaler Maus-Antikörper,
Dr. J. Wijdenes, Besançon, Frankreich
gp130 (BP4) monoklonaler Maus-Antikörper,
Dr. J. Wijdenes, Besançon, Frankreich
STAT3-Phosphotyrosin polyklonaler Kaninchen-Antikörper gegen
Y705-phosphoryliertes STAT3,
New England BioLabs, Schwalbach/Ts.
STAT1-Phosphotyrosin polyklonaler Kaninchen-Antikörper gegen
Y701-phosphoryliertes STAT1,
Upstate Biotechnology, USA
Jak1 polyklonaler Kaninchen-Antikörper,
Dr. Ziemiecki, Bern, Schweiz
Jak2 polyklonaler Kaninchen-Antikörper,
Dr. Ziemiecki, Bern, Schweiz

Material und Methoden 15
Tyk2 monoklonaler Maus-Antikörper,
Transduction Laboratories, USA
SHP2 polyklonaler Kaninchen-Antikörper,
Dr. Ullrich, MPI, Martinsried
SHP2 polyklonaler Kaninchen-Antikörper gegen den
N-terminalen Bereich der SHP2,
Santa Cruz, USA
2.1.5 Oligonukleotide
Alle Oligonukleotide wurden von der Firma MWG, Ebersberg, bezogen.
P1 BstEII lr Y 5’- GCCCAGTGGTCACCTCACACTCC -3’
P2 BstEII lr F 5’- GCCCAGTGGTCACCTCACACTCCTCCAAGGCACAATTTT
AATTCAAAAGATCAAATGTTCTCAGATGGCAATTTCAC -3’
P3 Sac rl 5’- GGACAGTGGAGCTCGTGTTTTGTGAAGATTC -3’
P4 SacIAccIII lr YY 5’- CAAAACACGAGCTCCACTGTCCAGTATTCTACCGTGGTA
CACTCCGGATACAGACACCAAGTTCCGTC -3’
P5 SacIAccIII lr FF 5’- CAAAACACGAGCTCCACTGTCCAGTTCTCTACCGTGGTA
CACTCCGGATTCAGACACCAAGTTCCGTC -3’
P6 AspI rl 5’- CCGCCATCGACATGGTCTACTAATTGTAGATC -3’
P7 AspI lr Y 5’- CAATTAGTAGACCATGTCGATGGCGGTGATGGTATTTTG
CC -3’
P8 AspI lr F 5’- CAATTAGTAGACCATGTCGATGGCGGTGATGGTATTTTG
CCCAGGCAACAGTTCTTCAAACAGAACTGCAG -3’
P9 SfuIBsmI rl 5’- CTTTTAGGCATTCCTTCATCAGTCGCAGCCTCCATGCCAA
CTGTTTCGAATCTTTCTACTTGTCCC -3’
P10 BsmIRsRII lr YY 5’- GCGACTGATGAAGGAATGCCTAAAAGTTACTTACCACAG
ACGGTCCGGCAAGGCGGCTACATGCCTC -3’
P11 BsmIRsRII lr FF 5’- GCGACTGATGAAGGAATGCCTAAAAGTTTCTTACCACAG
ACGGTCCGGCAAGGCGGCTTCATGCCTCAGTGAAGG -3’
P12 pSVL 3 rl 5’- GCATTCTAGTTGTGG -3’
P19 Ymyc lr 5’-GTCCGGCAAGGCGGCTACATGCCTCAGGGCGGCGAGCAG
AAGCTGATATCGGAGGAAGATCTGTGAG -3’

16 Material und Methoden
P20 Ymyc rl 5’- GATCCTCACAGATCTTCCTCCGATATCAGCTTCTGCTCG
CCGCCCTGAGGCATGTAGCCGCCTTGCCG -3’
P21 Fmyc lr 5’- GTCCGGCAAGGCGGCTTCATGCCTCAGGGCGGCGAGCA
GAAGCTGATATCGGAGGAAGATCTGTGAG -3’
P22 Fmyc rl 5’- GATCCTCACAGATCTTCCTCCGATATCAGCTTCTGCTCGC
CGCCCTGAGGCATGAAGCCGCCTTGCCG -3’
S1 sypgfp rl 5’- GGCTGGTGCCGGCAAAGCTTTTCTGAAACTTTTCTGCTG
TTGC -3’
S2 sypgfp lr 5’- GGTCCCGGAATTCCCGGGTCTAGAGTCACCATGACATCG
CGGAGATGG -3’
S3 sypmut lr 5’- GGGCCGGTCGTGGTGCAGTCGAGTGCTGGAATTGGCCGG
ACAGGG -3’
S4 sypmut rl 5’- CCCTGTCCGGCCAATTCCAGCACTCGAGTGCACCACGAC
CGGCCC -3’
S8 sypseq lr 345 5’- GGAGGGGAGAAATTTGCC -3’
S17 sypkorr1 lr 5’- GGTTGGACAAGGGAATACGGAGAGAACGG -3’
S18 sypkorr1 rl 5’- CCCTTGTCCAACCTTCGAAAGTTTAAGTTCTCTTAGCG -3’
S20 sypmyc lr 5’- AGCTTTAGAGCAGAAGCTGATATCGGAGGAAGATCT
GGA -3’
S21 sypmyc rl 5’- AGCTTCCAGATCTTCCTCCGATATCAGCTTCTGCTC
TAA -3’
2.1.6 Plasmide
pSVL Expressionvektor für Eukaryonten, Pharmacia, Freiburg
pSVL EG pSVL-Vektor, in den die cDNA eines chimären Rezeptors einkloniert
wurde, der aus dem extrazellulären Anteil des murinen Erythropoetin-Re-
zeptors und der Transmembran- und zytoplasmatischen Domäne des huma-
nen gp130 besteht [28, 36].
pSVL ∆ EcoRI hum gp130 [40‘]
pBS-SK- Klonierungsvektor pBluescript-SK, Stratagene, Heidelberg ohne die Re-
striktionsschnittstellen Eco RV, Acc I, Cla I, Hind III und Sal I
pRC/CMV Expressionsvektor für Eukaryonten, Invitrogen, Breda, Niederlande

Material und Methoden 17
pRK 1D Expressionsvektor für Eukaryonten, der die cDNA der humanen SHP2-
Splice Variante enthält, freundlicherweise zur Verfügung gestellt von A.
Ullrich, MPI, Martinsried
pMBC-1Bgfp bicistronischer Expressionsvektor, der die IRF-1-NLS fusioniert an gfp
enthält [71]
pSBC1 bicistronischer Expressionsvektor mit SV40-Promotor [19]
pCBC1 bicistronischer Expressionsvektor mit CMV-Promotor [19]
pSVL STAT3 pSVL-Vektor, der die murine STAT3-cDNA enthält
pGl3α2M215.luc Expressionvektor, der die cDNA des Luciferase-Gen fusioniert an den α2M-
Promotor enthält [95]
pCR(TM) 3lacZ Expressionsvektor für Eukaryonten, der die cDNA des bakteriellen β-
Galactosidase-Gens enthält, Invitrogen, Breda, Niederlande
pSV2 Neo Expressionsvektor mit Gen, das für eine Neomycinresistenz kodiert
2.2 Allgemeine molekularbiologische Methoden
2.2.1 Verwendete Bakterienstämme und deren Kultivierung
JM83 F’ara ∆(lac-proAB) rspL Φ80dlacZ∆M15
XL-1-Blue supE44, hsdR17, recA1, endA1, gyrA64, relA1, lacF’
[proAB+, lac1q, lac Z∆M15, Tn10 (tetr)]
Die Anzucht rekombinanter Bakterien erfolgte in Luria-Bertani (LB)-Medium bei 37°C auf
einem Schüttler bei 220 UpM in der Regel über Nacht. Alle Bakterien wurden unter Zugabe
der entsprechenden Antibiotika (Ampicillin oder Penicillin, 50 µg/ml Medium) inkubiert.
Die Langzeitaufbewahrung der Bakterien erfolgte in LB-Medium mit 50% Glycerin bei
-80°C.
LB-Medium: 10 g/l Bactotrypton, Difco, USA10 g/l Yeast extract, Difco, USA 5 g/l NaCl
Für die Herstellung von Agarplatten wurden dem LB-Medium 15 g Agar pro Liter Medium
zugesetzt. Ampicillinhaltige Agarplatten mischte man nach Abkühlen des LB-Mediums
1/1000 Volumen Ampicillin eines 50 mg/ml konzentrierten Stocks bei.

18 Material und Methoden
2.2.2 Herstellung kompetenter Zellen
E. coli-Bakterien wurden in 500 ml Tetracyclin-haltigem LB-Medium bis zu einer OD600 von
0,3 bei 37°C geschüttelt. Dann wurde der Kolben rasch im Eiswasserbad abgekühlt und die
Zellen 5 min bei 6000 UpM und 4°C abzentrifugiert. Das Sediment wurde in 50 ml TSB-
Lösung aufgenommen, aliquotiert und sofort in flüssigem Stickstoff schockgefroren. Auf
diese Weise vorbehandelte und bei -80°C gelagerte E. colis konnten über mehrere Jahre zur
Transformation verwendet werden.
TSB-Puffer: Poly-Ethylenglykol (PEG) 4000 10 %DMSO 5 %MgSO4 10 mMMgCl2 10 mM
2.2.3 Transformation von Bakterien nach Hanahan
10 µl Ligationsansatz oder Plasmidlösung (ca. 10-100 ng DNA) wurden mit 100 µl auf Eis
aufgetauter kompetenter Bakteriensuspension für 20 min inkubiert. Der Ansatz wurde für
90 sec auf 42°C erhitzt und anschließend 5 min auf Eis abgekühlt. Nach Zugabe von 1 ml LB-
Medium wurden die Bakterien nun 30 min bei 37°C inkubiert und dann in unterschiedlichen
Konzentrationen mit einem Drigalski-Spatel auf Antibiotika-haltigen Agarplatten ausplattiert.
Die so behandelten Platten wurden für 12-18 h bei 37°C inkubiert und anschließend weiter-
verarbeitet [33].
2.2.4 Isolierung von Plasmid-DNA aus E. coli
2.2.4.1 ...im analytischen Maßstab
Kleine Mengen Plasmid-DNA wurden aus 3 ml-Schüttelkulturen nach den Präparationsvor-
schriften des QIAprep Kits der Firma Qiagen, Hilden, gewonnen.
2.2.4.2 ...im präparativen Maßstab
Plasmid-DNA in präparativem Maßstab wurden aus 250-500 ml Schüttelkulturen ebenfalls
mit Hilfe eines Plasmidpräparationskits der Firma Qiagen, Hilden, gemäß der Anleitung des
Herstellers isoliert.

Material und Methoden 19
2.2.5 Konzentrationsbestimmung von DNA
Die photometrischen Messungen zur Konzentrationsbestimmung der DNA wurden in 1 ml
Quarzküvetten in einem Zweistrahlphotometer durchgeführt. Dazu verdünnte man die DNA
1:100 mit Wasser und maß die Extinktion bei den Wellenlängen 260 und 280 nm. Eine
gemessene Extinktion E260 von 0,1 entspricht einer Konzentration von 5 µg doppelsträngiger
DNA pro ml Lösung in der Küvette, dementsprechend beträgt die Konzentration der
Ausgangslösung 0,5 µg/µl. Über den Quotienten E260/E280 konnte eine Aussage über die
Proteinverunreinigung der DNA getroffen werden, über die Extinktion bei 320 nm eine
Angabe über den Salzgehalt gemacht werden [68].
2.2.6 Oligonukleotid-Hybridisierung
Die zu hybridisierenden Oligonukleotide wurden in einer 150 mM NaCl-Lösung in einem
Heizblock auf 80°C erhitzt und dann nach Ausschalten des Gerätes langsam auf RT
abgekühlt.
Bei Bedarf wurden die Nukleotide im Anschluß an diese Reaktion mit Hilfe der Poly-Nu-
kleotid-Kinase der Firma Boehringer Mannheim, Penzberg, nach Angaben des Herstellers
phosphoryliert werden und standen so nach anschließender Phenol-/Chloroform-Fällung
gereinigt für die Ligation zur Verfügung.
2.2.7 Modifikation von Plasmid-DNA
2.2.7.1 Restriktionsanalyse von Plasmid-DNA
Ein mit Hilfe der analytischen oder präparativen Plasmidpräparation isoliertes Plasmid kann
durch Spaltung mit Restriktionsendonukleasen charakterisiert oder verändert werden.
Für die enzymatischen Spaltungen mit Restriktionsendonukleasen wurden die von den Her-
stellern der einzelnen Enzyme angegebenen Vorschriften angewendet. Zur Spaltung der
Plasmid-DNA wurde für 1 µg Plasmid-DNA die 1- bis 4-fache Menge an Enzymeinheiten
(units) eingesetzt. Die Menge an zugegebenem Enzym sollte 10% des Gesamtvolumens nicht
überschreiten, da das im Kulturpuffer des Enzyms enthaltene Glycerin die Enzymaktivität
inhibiert oder eine Sternaktivität (Spaltung der DNA außerhalb der spezifischen
Restriktionsschnittstellen) verursachen kann.

20 Material und Methoden
Sämtliche Enzyme und die dazugehörigen Reaktionspuffer wurden von den Firmen AGS,
Heidelberg, Boehringer Mannheim, Penzberg, und New England Biolabs, Schwalbach,
bezogen.
2.2.7.2 Elektrophoretische Auftrennung der DNA
Nach einem Verdau mit geeigneten Restriktionsendonukleasen erfolgte die Analyse der DNA
durch Auftrennung der Fragmente nach ihrer Größe im elektrischen Feld. Dazu wurde die
DNA-Lösung mit 1/5 Volumen eines 6x DNA-Probenpuffers versetzt, auf ein Agarosegel
aufgetragen (1-2% Agarose (Biozym, Hameln) in Laufpuffer) und in 1x TAE-Laufpuffer
aufgetrennt. Als Größenstandard diente mit EcoRI/HindIII verdaute Lamba-Phagen-DNA.
Durch Zusatz von Ethidiumbromid zum Gel (1 µg/ml Endkonzentration) war es möglich, die
DNA-Banden unter UV-Licht sichtbar zu machen.
1x TAE-Laufpuffer: 0,4 M Tris-Base1,1 M EDTA0,005 M Natriumacetat0,032 M Essigsäure
6x DNA-Probenpuffer: 15 ml TAE 1x10 ml Glycerin60 mg Bromphenolblau/Xylencyanol
2.2.7.3 Isolierung von DNA aus Agarosegelen
Um bestimmte DNA-Fragmente nach der Gelelektrophorese wieder aus dem Gel zu isolieren,
wurden die entsprechenden Banden unter UV-Licht mit einem Skapell ausgeschnitten und in
ein Eppendorf-Reaktionsgefäß überführt. Die Freisetzung der DNA aus der Gelmatrix erfolgte
nach Herstellerangaben mit Hilfe des QIAquick Kits der Firma Qiagen, Hilden.
2.2.7.4 Dephosphorylierung linearisierter DNA
Um die Wahrscheinlichkeit einer Selbstligation geschnittener Vektoren in einem Ligations-
ansatz zu vermindern und somit den Anteil der durch Insertion geschlossenen Plasmide nach
der Ligation zu erhöhen, wurde die endständige 5’-Phosphatgruppe der geöffneten Vektoren
entfernt. Dies geschah mit Hilfe der alkalischen Phosphatase (calf intestine phosphatase =
CIP) der Firma Boehringer Mannheim, Penzberg, nach Standardvorschrift.

Material und Methoden 21
2.2.7.5 Ligation von DNA-Fragmenten
Bei der Ligation verknüpft die T4-DNA-Ligase DNA-Fragmente, die sowohl kompatible
überstehende, als auch glatte Enden besitzen. Dabei bildet sich eine Phosphodiester-Bindung
zwischen den 3’-OH- und den 5’-P-Enden der DNA aus. Das Enzym wurde von der Firma
Boehringer Mannheim, Penzberg bezogen. Die Reaktion erfolgte unter Standardbedingungen
des Herstellers. In der Regel wurden Vektor und Fragment im Verhältnis 1:1,5 eingesetzt; bei
Ligationen mit synthetischen Oligonukleotiden sollte man den 1000fachen molaren
Überschuß an Fragment verwenden.
2.2.8 Polymerase-Ketten-Reaktion (PCR)
Die PCR ist eine einfache Methode, Nukleinsäuren in einer Lösung zu vervielfältigen. Dabei
wird der natürliche Replikationszyklus der Zelle nachgeahmt, wobei sich die Anzahl der
DNA-Moleküle bei jedem Zyklus verdoppelt. Die Methode basiert auf der Wiederholung
einer Abfolge von drei Schritten mit bestimmten Reaktionsbedigungen:
1. Denaturierung der Template-DNA: Die doppelsträngige Ursprungs-DNA wird bei hohen
Temperaturen inkubiert. Die Doppelstränge dissoziieren und liegen solange in Lösung einzel-
strängig vor, bis die Temperatur abgesenkt wird.
2. Annealing von Primern: Bei Abkühlung der Lösung hybridisieren die Einzelstränge mit
den sogenannten Primern. Dies sind Oligonukleotide, die so gewählt werden, daß sie den zu
amplifizierenden Bereich der DNA flankieren. Sie sind so beschaffen, daß einer der Primer
komplementär zum codierenden Strang diesen bindet, der andere Primer aber den nicht-codie-
renden Strang erkennt. Da die Konzentration der Primer in der Lösung sehr hoch ist, wird die
Ausbildung von Primer-Template-Komplexen gegenüber der Renaturierung der beiden Ein-
zelstränge bevorzugt werden.
3. Primer Extension: Der letzte Schritt ist die von der DNA-Polymerase vermittelte Verlänge-
rung der Primer in den Primer-Template-Komplexen. Diese Extensionsreaktion kann z.B. von
der Klenow Polymerase bewerkstelligt werden. Die Reaktionstemperatur dieses Enzyms liegt
allerdings bei 37°C, so daß es bei jedem nachfolgenden Zyklus zerstört wird und neu
hinzugegeben werden muß. Die Verwendung der Taq-Polymerase erlaubt die Erhöhung der
Temperatur für die Extensionsreaktion auf über 70°C. Die hohe Thermostabilität des Enzyms
bedingt gleichzeitig, daß viele Zyklen der PCR durchlaufen werden können, ohne das neues

22 Material und Methoden
Enzym zugegeben werden muß. Der Vent-Polymerase spricht man zusätzlich eine proof-
reading Eigenschaft zu, welche den Einbau falscher Nukleotide vermehrt verhindern soll [68].
Ein Standardansatz setzte sich wie folgt zusammen:2 x 5 µl Primer (2x 5 pmol/µl)
5 µl Nukleotide (2,5 mM jedes Nukleotides)4 µl MgCl2 (25mM)5 µl Vent-Polymerase-Pufferx µl Template (1ng)
ad 49 µl H2O
Die Proben wurden mit Mineralöl überschichtet, um ein Verdunsten der Flüssigkeit und damit
die Veränderung der Reaktionsbedingungen zu vermeiden. Anschließend wurden sie den
PCR-Prozessor gegeben und die Reaktion gestartet.
Ein Standardprofil für die DNA-Amplifikation mit ungeschnittenen Plasmiden als Template
sah wie folgt aus:
Erste Denaturierung: 94°C 60 secAnnealing (primer-abhängig): 45°C 30 secExtension: 72°C 240 secDenaturierung: 94°C 30 secLetzte Extension: 72°C 600 sec
2.2.8.1 DNA Mutagenese durch PCR
Gezielte Veränderungen der DNA-Sequenz bei der PCR können lediglich an den Enden der
PCR-Produkte, im Bereich der Homologie zu den Primern eingefügt werden. Hierfür plant
man die Oligonukleotide so, daß sie, wenn sie die Mutation aufweisen, nicht vollständig
komplementär zur Template-DNA sind, sondern entweder einzelne missmatches oder sogar
ein längeres nicht komplementäres 5’-Ende aufweisen. Es ist darauf zu achten, daß am 3’-
Ende wenigstens über eine Länge von 10 Basen vollständige Komplementarität besteht. Um
eine sichere Hybridisierung zwischen Primer und Template zu erreichen, sollten die Oligonu-
kleotide möglichst mit einigen Cytosinen oder Guaninen enden, da GC-Bingungen stabiler als
AT-Bindungen sind. Entsprechend lassen sich auch Verkürzungen in die zu exprimierenden
Proteinen einfügen: C-Terminale Deletionen erreicht man durch das Einfügen eines neuen
Stopcodons in die DNA, welches vor dem natürlichen Stop-Codon, also in Richtung 5‘
(upstream) liegen sollte. N-terminale Verkürzungen werden analog dazu durch Entfernen des
ursprünglichen Startcodons und Einführung eines neuen Translationsstartes an beliebiger
Stelle downstream erzeugt. Interne Deletionen oder Punktmutationen erreicht man durch
Ligation über neu eingefügte Restriktionserkennungs-Sequenzen von PCR-Produkten C- und

Material und Methoden 23
N-terminaler Fragmente oder durch die sogenannte Fusions-PCR. Hierbei werden die bei
einer ersten PCR gewonnenen Fragmente zu gleichen Teilen als Matrize einer zweiten PCR
eingesetzt. Als Primer verwendet man entsprechend weiter N- bzw. C-terminal gelegene
Basensequenzen.
2.2.9 Analyse der DNA durch Sequenzierungen
2.2.9.1 Sequenzierung nach Sanger et al. (1977)
Die Sequenzierung erfolgte nach obengenannter Methode mit Hilfe des T7-Sequencing-Kit
der Firma Pharmacia, Freiburg. Zur radioaktiven Markierung der Reaktionsprodukte
verwendete man [α35S]-dATP. Die entstehenden DNA-Fragmente wurden auf einem durch
Zusatz von 7 M Harnstoff denaturierenden 5%-igen Polyacrylamidgel aufgetrennt und durch
Autoradiographie sichtbar gemacht [69].
2.2.9.2 Automatische Sequenzierung mit dem ABI Prism Modell 310 Sequencer
Das Prinzip der automatischen Sequenzierung beruht auf der Anregung und Detektion fluo-
reszenzmarkierter Didesoxynukleotide, die in einer Polymerase-Ketten-Reaktion in ein DNA-
Molekül eingebaut werden.
Zur Präparation der DNA-Proben wurde das PRISM™ Ready Reaction Dye-Desoxy™ Ter-
minator Cycle Sequencing Kit der Firma Perkin Elmer, USA, verwendet.
Der Reaktionsansatz wurde wie folgt angesetzt:
6 µl Terminator Mix1 ng DNA-Matrix5 pmol Oligonukleotid-Primer ad 20 µl H2O
Die DNA-Matrize wurde in 25 Reaktionszyklen in einem PCR-Prozessor unter Einbau der
fluoreszenzmarkierten Didesoxynukleotide amplifiziert. Ein Reaktionszyklus bestand aus:
Erste Denaturierung: 96°C 20 secAnnealing: z.B. 45°C 20 secExtension: 60°C 40 secDenaturierung: 96°C 20 secLetzte Extension: 60°C 240 sec
Nach der Amplifikation wurde die DNA mit 10 µl 3M Na-Acetat, pH 5,2, und 50 µl 100%
Ethanol 20 min lang bei RT gefällt und abzentrifugiert. Das DNA-Sediment wurde in 70%
Ethanol gewaschen und für 10 min bei 37°C getrocknet. Die Proben wurden in TSR-Proben-

24 Material und Methoden
puffer der Firma Applied Biosystems ,Foster City, USA, aufgenommen, für 2 min bei 90°C
denaturiert und anschließend in einem 6%-igen Performance Optimized Polymer (POP6) mit
8% Harnstoff aufgetrennt. Die Elektrophorese erfolgte in einer unbeschichteten Kapillare
(Ø 51 µm) bei 12,5 kV und einer Temperatur von 50°C für ca. 135 min. Eine genaue
Beschreibung der hierfür erforderlichen Computer-Software ist dem User Bulletin ABI Prism
310 Genetic Analyser (PE Applies Biosystems, 1997) zu entnehmen.
Terminator-premix: 1,58 µM A-DyeDesoxy94,74 µM T-DyeDesoxy
0,42 µM G-DyeDesoxy78,95 µM C-DyeDesoxy78,95 µM DITP15,79 µM DDTP15,79 µM DTTP15,79 µM DATP
169,42 mM Tris/HCl pH 9,04,21 mM (NH4)2SO4
42,10 mM MgCl2
0,42 U/µl AmpliTaq-Polymerase
2.3 Eukaryontische Zellen und deren Kultivierung
2.3.1 Kulturmedien
PBS: NaCl 200 mMKCl 2,5 mMNa2HPO4 8 mMKH2PO4 1,5 mM
Hygromycin: 50 mg/ml, bei 4°C gelagert,
Boehringer Mannheim, Penzberg
Neomycin: 125 mg/ml PBS steril, bei -20°C gelagert,
Sigma, Deisenhofen
Puromycin: 0,5 mg/ml in H2O, sterilfiltriert, bei -20°C gelagert,
Sigma, Deisenhofen
s. auch 2.1.2

Material und Methoden 25
2.3.2 Verwendete permanente Zell-Linien
COS-7 adhärent wachsende Affennieren-Zell-Linie (ATCC, CRL1651);
Kultivierung in DMEM
HepG2 adhärent wachsende humane Hepatomzell-Linie (ATCC, HB8058);
Kultivierung in DMEM/NUT-Mix-F12
Ba/F3 nicht adhärent wachsende IL-3-abhängige murine Prä-B-Zell-Linie
(DSM, ACC300)
Kultivierung in DMEM plus 5% BPV-Überstand
X63Ag8-653 BPV-mIL-3
nicht adhärent wachsende murine Myelomzell-Linie, die murines IL-3
sezerniert,
Kultivierung in DMEM
2.3.2.1 Janus-Kinasen defiziente Zell-Linien
Hierbei handelt es sich um diverse Abkömmlinge einer Fibrosarkom-Zell-Linie, die nicht
mehr in der Lage sind, jeweils eine der drei Janus-Kinasen zu exprimieren. Sie wurden
freundlicherweise von Herrn Ian Kerr, Imperial Cancer Research Fund, London, Groß-
britannien, zur Verfügung gestellt.
Ausgangspunkt für die Herstellung dieser Zell-Linien ist eine humane Fibrosarkom-Zell-
Linie, HT1080. Ein Abkömmling dieser Zell-Linie, HPRT–, besitzt einen Defekt im Salvage
Pathway, der zur Bereitstellung von Purinnukleotiden dient. Sie besitzen keine Hypoxanthin-
Phosphoribosyltransferase und können deshalb nicht im sogenanten HAT-(Hypoxanthin-
Aminopterin-Thymidin)-Medium überleben, da der Folsäure-Antagonist Aminopterin die
de novo-Synthese der Purine hemmt, und Hypoxanthin nicht mehr zu Purin umgesetzt werden
kann. Gleichzeitig werden diese Zellen jedoch gegen die toxische Wirkung des Guanin-Ana-
logons 6-Thioguanin (6-TG) resistent.
Die Transfektion eines IFN-α regulierten Gens, das für das bakterielle Enzym Guanosin-
Phosphoribosyltransferase kodiert, stellt IFN-α-abhängig den salvage pathway wieder her.
Derart transfizierte Zellen, 2fTGH-Zellen, überleben in einem IFN-α-haltigen HAT-Medium
und sterben in der Gegenwart von 6-TG. Behandelte man diese Zellen nun mit einem frame-
shift-Mutagen und züchtete sie in einem 6-TG- und IFN-α-haltigen normalen Medium an, so

26 Material und Methoden
überlebten diejenigen Zellen, bei denen der IFN-α-Signalweg zerstört ist. Entstehende Klone
untersuchte man auf ihren Defekt.
Da Jak2 keine Rolle im IFN-α-Signalweg zu spielen scheint, mußte man zur Deletion dieses
Proteins eine andere Strategie wählen. Man transfizierte ein IFN-γ-abhängig exprimiertes
Oberflächen-Protein, CD2, in HT-1080 und wählte die positiven Klone, 2C4, im FACS aus.
Nach Mutagen-Einwirkung wurden hierbei diejenigen Zellen selektiert, die auf IFN-γ nicht
mehr mit Expression des CD2 reagieren. Anschließend suchte man nach dem Defekt [16].
Die so gefundenen Jak-defizienten Zellen wurden im Anschluß durch Transfektion der ent-
sprechenden cDNA rekonstituiert. Es handelt sich dabei im Einzelnen um folgende Zell-
Linien:
2fTGH parentale Zell-Linie
Kultivierung in DMEM-Medium plus Hygromycin
U1A Tyk2-defiziente Zell-Linie
Kultivierung in DMEM-Medium plus Hygromycin
U1A/Tyk2 Tyk2-defiziente Zell-Linie stabil transfiziert mit Tyk2-cDNA
Kultivierung in DMEM-Medium plus Hygromycin und Neomycin
U4A exprimiert eine verkürzte nicht aktive Form von Jak1
Kultivierung in DMEM-Medium plus Hygromycin
U4A/Jak1 U4A stabil transfiziert mit Jak1-cDNA
Kultivierung in DMEM-Medium plus Hygromycin und Neomycin
γ2A Jak2-defiziente Zellen
Kultivierung in DMEM-Medium plus Neomycin
γ2A/Jak2 Jak2-defiziente Zellen stabil transfiziert mit Jak2-cDNA
Kultivierung in DMEM-Medium plus Neomycin und Puromycin
U4A/Jak1(K→E) U4A stabil transfiziert mit katalytisch inaktiver Jak1-cDNA
2.3.3 Kultivierung von Zell-Linien
Die Zellen aller Linien wurden bei 37°C in wasserdampfgesättigter Atmosphäre mit einem
CO2-Gehalt von 5% kultiviert. Die Medien enthielten, sofern nicht anders angegeben, 10%
fetales Kälberserum, 60 mg/ml Penicillin und 100 mg/ml Streptomycin.

Material und Methoden 27
2.3.3.1 Passagierung von Zellen
Zur Weiterkultivierung mußten adhärente Zellen von einer konfluent bewachsenen Petrischale
mit Trypsin/EDTA abgelöst werden. Nach Aufnahme in frischem Kulturmedium entspre-
chender Menge wurden die Zellen auf mehrere neue Schalen in einer Verdünnung 1:2 bis
1:20 verteilt. Nicht adhärente Zellen wurden direkt mit frischem Medium verdünnt und auf
neue Schalen verteilt.
2.3.3.2 Langzeitaufbewahrung
Für die Langzeitaufbewahrung wurden die Zellen einer Platte gegebenenfalls nach Trypsini-
sierung bei 900 UpM abzentrifugiert, in 1 ml Medium mit 10% DMSO aufgenommen und in
Kryoröhrchen aliquotiert. Zur langsamen Abkühlung erfolgte eine Lagerung bei -80°C für ca.
24 Stunden. Anschließend wurden sie in flüssigen Stickstoff überführt. Zum Auftauen mußten
die Kryoampullen möglichst schnell bei 37°C aufgetaut und das DMSO-haltige Medium mit
10 ml kaltem Medium verdünnt werden. Die Zellen wurden dann bei 900 UpM und 4°C ab-
zentrifugiert, in neuem Medium aufgenommen und auf einer Petrischale ausgesät.
2.3.4 Transiente Transfektion
2.3.4.1 Elektroporation
Eine konfluent bewachsene Kulturschale (Ø10cm) wurde zweimal mit 5 ml PBS gewaschen
und 5 min mit 1 ml Trypsin/EDTAösung inkubiert. Die Aktivität des Trypsins stoppte man
durch Zugabe von 4 ml DMEM. Anschließend wurden die Zellen bei 900 UpM
abzentrifugiert und in 1,6 ml DMEM resuspendiert. 800 µl dieser Zellsuspension überführte
man zu der vorgelegten Plasmid-DNA (20-30 µg Plasmid-DNA pro Versuchsansatz) in eine
mit PBS gespülte Elektroporationsküvette. Die Elektroporation erfolgte im Modell T820 der
Firma BTX, USA, mit einem Puls bei einer Spannung von 1,5 kV und einer Pulsdauer von
99 µs. Nach 2-3 Tagen konnten die Zellen weiter verarbeitet werden.
2.3.4.2 Calcium-Phosphat-Präzipitation
Die Zellen einer konfluent bewachsenen Schale wurden am Vortag sorgfältig vereinzelt und
1:8 verdünnt wieder ausgesät. Vor der Transfektion wusch man die Zellen zweimal mit PBS
und inkubierte sie mindestens eine Stunde in DMEM ohne F12. Maximal 30 µg Plasmid-

28 Material und Methoden
DNA wurden in einem sterilen Falcongefäß vorgelegt, mit 125 µl 1M CaCl2 versetzt und ad
500 µl mit Wasser aufgefüllt. Anschließend gab man 500 µl 2xHBS tropfenweise in das vor-
texende Röhrchen. Nach einer Inkubationszeit von mindestens einer halben Stunde wurde die
Lösung auf die Zellen pipettiert und für 5-15 h dort belassen. Nach Ablauf dieser Zeitspanne
wurden die Zellen in geeignetem Kulturmedium weitergezogen.
2x HBS: HEPES 5 g 40 mMNaCl 8 g 274 mMKCl 0,37 g 1 mMNa2HPO4•2H2O 0,125 g 1,4 mMGlukose 1 gH20 ad 500 ml
den pH exakt auf 7,05 mit NaOH eingestellt, sterilfiltriert
2.3.5 Stabile Transfektion von Ba/F3-Zellen
Die stabile Transfektion von Ba/F3-Zellen mit membranständigen gp130-Proteinen erfolgte
durch Elektroporation. Dazu wurden 3,5 x 106 Zellen mit insgesamt 30 µg DNA gemischt und
mit einem 70 msec langen Puls von 200 V elektroporiert. Zur Selektion transfizierter Zellen
wurden 2 µg des Plasmids pSV2 Neo, das für eine Neomycinresistenz kodiert, kotransfiziert.
Nach der Elektroporation wurden die Ba/F3-Zellen zunächst für 24 h ohne Selektionsdruck in
DMEM/IL-3 kultiviert. Anschließend wurden sie in 50 ml Medium mit 1 mg/ml Geneticin
(G418) aufgenommen und in zwei 24-Loch-Zellkulturplatten ausgesät. 16-28 Tage später
wurden erste G418-resistente Klone erkennbar, die in frischen G418-haltigen Medium aufge-
nommen und amplifiziert wurden. Die Expressionskontrolle bzw. -Quantifizierung der gp130-
Proteine erfolgte in der Durchflußzytometrie mit Hilfe eines gegen die extrazelluläre Domäne
des gp130 gerichteten Antiköpers (s. 2.4.11).
2.4 Biochemische Methoden
2.4.1 Herstellung zytosolischer Extrakte
Alle Lösungen wurden eisgekühlt eingesetzt, und alle Arbeitsschritte nach Möglichkeit auf
Eis oder bei 4°C durchgeführt. Die zu lysierenden Zellen wurden zunächst zweimal mit
PBS/Vanadat gewaschen und in 1 ml PBS/Vanadat mit einem Gummischaber von der Schale
gelöst. Danach zentrifugierte man sie ab und resuspendierte sie in 500 µl Lysispuffer. Nach

Material und Methoden 29
einer Inkubation von 30 min auf Eis wurde das Lysat 10 min bei maximaler Rotation
zentrifugiert. Anschließend überführte man der Überstand in ein neues Eppendorf-
Reaktionsgefäß.
Lysis-Puffer: 50 mM HEPES pH 7,5 Vor Gebrauch frisch hinzufügen:150 mM NaCl NaVanadat 1 mM
1 % Triton X-100 NaF 1 mM10 % Glycerol PMSF 0,2 mM
1 mM EGTA Aprotinin 10 ng/ml1,5 mM MgCl2 Leupeptin 10 ng/ml
Pepstatin 10 ng/ml
Für Co-Immunpräzipitationen wurde dem Lysispuffer nur 0,1% Triton X-100 hinzugefügt.
2.4.2 Immunpräzipitation
Zu den Zell-Lysaten wurden je nach Versuch zwischen 1-6 µg Antikörper gegeben. Zeitgleich
wurden 3,5-4 mg Protein A-Sepharose, Pharmacia, Freiburg, in 100 µl Lysispuffer pro Ansatz
aufgenommen, bei Bedarf mit einem Kaninchen-anti-Maus-Antiserum, Dako, Dänemark,
abgesättigt und quellen gelassen. Anschließend erfolgte eine Inkubation in einem Über-Kopf-
Schüttler bei 4°C für mindestens 8 Stunden. Danach gab man je 100 µl gequollene Protein A-
Sepharose zu den Proben und ließ sie für eine weitere Stunde schütteln. Die Sepharose wurde
durch Zentrifugation sedimentiert, dreimal in Lysispuffer gewaschen und schließlich in 30 µl
Lämmli-Probenpuffer mit 5% β-Mercaptoethanol aufgenommen. Den Ansatz kochte man
5 min lang bei 95°C auf, zentrifugierte ihn erneut und trug den Überstand auf ein SDS-
Polyacrylamidgel auf.
2.4.3 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE)
Die Auftrennung von Proteinen erfolgte in einer diskontinuierlichen 7%-igen SDS-Polyacryl-
amid-Gelelektrophorese nach Lämmli [49].
Laufpuffer 5x: 30,25 g Tris-Base144 g Glycin
10 g SDSad 2 l H2O

30 Material und Methoden
Trenngel: 7,34 ml H2O3,36 ml Acrylamidlösung: 29% Acrylamid, 0,8% Bisacrylamid3,80 ml 1,5 M Tris/HCl, pH 8,8
75 µl 10% SDS15 µl TEMED75 µl 20% APS
Sammelgel: 4 ml H2O635 µl Acrylamidlösung: 29% Acrylamid, 0,8% Bisacrylamid313 µl 2 M Tris/HCl pH 6,8
25 µl 10% SDS5 µl TEMED
40 µl 20% APS
2x Lämmli-Probenpuffer 20 % Glycerin4 % SDS
125 mM Tris/HCl0,002 % Bromphenolblau
2.4.4 Western Blot
Durch das Western Blot-semidry-Verfahren [34] können Proteine auf eine Membran über-
tragen und mit Hilfe Enzym markierter Antikörper schnell und spezifisch nachgewiesen wer-
den. Die durch SDS-PAGE (2.4.3) aufgetrennten Proteine wurden durch einen Western Blot
auf eine Polyvinylidenfluorid-Membran (PVDF) übertragen. Hierfür benetzte man die
Membran zuerst 5 sec mit Methanol, anschließend wässerte man sie und legte sie in
Anodenpuffer II. Das Gel wurde 5 min lang im Kathodenpuffer äquilibriert. 15 Whatman-
Papierfilter wurden auf Gelgröße zurechtgeschnitten und mit verschiedenen Puffern getränkt
(siehe Blotaufbau).
Der Blotvorgang dauerte 1 h und wurde bei 0,8 mA/cm2 Membran durchgeführt. Der Western
Blot konnte sofort zur Detektion mit spezifischen Antikörpern verwendet oder bei Bedarf in
TBS-N-Puffer (2.4.5) bei 4°C gelagert werden.
Anodenpuffer I 0,3 M Tris-Base 20 % Methanol
pH 10,4
Anodenpuffer II 0,025 M Tris-Base 20 % Methanol
pH 10,4
Kathodenpuffer 0,04 M ε-Aminocarponsäure 20 % Methanol
pH 9,4

Material und Methoden 31
Abb.4: Western Blot-Aufbau
2.4.5 Detektion von Proteinen mittels verstärkter Lichtemission
Mit der Enhanced Chemiluminescence (ECL) bzw. der Enhanced Chemiluminescence plus
(ECL+) der Firma Amersham, Braunschweig können Proteine, die zuvor mit einem
spezifischen Primärantikörper markiert worden sind, mit Hilfe eines gegen diesen Primäranti-
körper gerichteten Meerrettich-Peroxidase konjungierten Zweitantikörper nachgewiesen
werden. Das Prinzip dieser Methode beruht darauf, daß Peroxidasen in Gegenwart von
Wasserstoffperoxid Diacylhydrazide, z.B. Luminol, oxidieren können. Neben freiem
Stickstoff entsteht dadurch ein aktiviertes Intermediat, das unter Abgabe von Lichtquanten in
seinen Grundzustand zurückfällt. Dieses Licht ist in der Lage, einen Röntgenfilm zu
schwärzen.
Für die Detektion der Proteine wurde die PVDF-Membran (2.4.4) zuerst 30 min in 10%-iger
BSA-Lösung inkubiert, um unspezifische Bindungsstellen abzusättigen. Anschließend wusch
man die Membran 15 min in TBS-N-Puffer und inkubierte dann eine Stunde bei RT mit einer
1:1000 Verdünnung eines Protein-spezifischen Antikörpers in TBS-N. Danach wurde erneut
gewaschen, um ungebundene Antikörper zu entfernen. Es folgte die Inkubation mit dem
Peroxidase-konjugierten spezies-spezifischen Antikörper in einer Verdünnung von 1:2000 für
ca. 30 min. Der so markierte Western Blot wurde mindestens 30 min gewaschen, wobei die
Spüllösung 5-6x gewechselt werden mußte. Die anschließende ECL-Reaktion erfolgt nach
Vorschrift des Herstellers.
Für eine erneute Detektion müssen die Antikörper entfernt werden. Hierzu wurde die Mem-
bran 30 min bei 70°C in einem stripping-Puffer inkubiert und anschließend in TBS-N ge-
waschen.

32 Material und Methoden
TBS-N-Puffer 20 mM Tris/HCl pH 7,6140 mM NaCl
0,01 % Nonidet-P40
stripping-Puffer 100 mM 2-Mercaptoethanol2 % SDS
62,5 mM Tris/HCl, pH6,7
2.4.6 Herstellung von Kernextrakten
Die zu fraktionierenden Zellen wurden nach eventueller Transfektion und Stimulation zwei-
mal mit gekühlten PBS mit 100 µM Na-Vanadat gewaschen und anschließend mit einem
Gummischaber von ihren Schalen gelöst. Die eigentliche Kernextraktion erfolgte wie bei
Andrews und Faller beschrieben [5]: Die Zellen wurden in ein Reaktionsgefäß überführt und
zehn Sekunden lang bei 14000 UpM abzentrifugiert. Das Zellsediment wurde in 400 µl des
hypotonen Puffers A aufgenommen und zehn Minuten auf Eis inkubiert, was zur Schwellung
der Zellen führte. Durch zehn-sekündiges Vortexen erfolgte die Aufschließung der Zellen.
Nach erneutem Zentrifugieren wurde das erhaltene Sediment in 100 µl Puffer C auf-
genommen und 20 min auf Eis inkubiert. Zum Schluß zentrifugierte man die Suspension ein
letztes Mal für 2 min. Der resultierende Überstand, der die Kernproteine der eingesetzten
Zellen enthielt, wurde in ein neues Gefäß überführt und bei -80°C eingefroren. Der Protein-
gehalt konnte mit dem BioRad Protein-Assay (2.4.7) bestimmt werden.
Puffer A: Puffer C:10,0 mM HEPES/KOH, pH 7,8 20,0 mM HEPES/KOH, pH 7,8
1,5 mM MgCl2 1,5 mM MgCl210,0 mM KCl 420,0 mM NaCl
0,5 mM DTT 0,2 mM EDTA0,2 mM PMSF 25 % Glycerin
0,5 mM DTT0,2 mM PMSF1,0 mM Na-Vanadat
2.4.7 Proteinbestimmung nach Bradford
Zum quantitativen Nachweis gelöster Proteine wurde ein Test der Firma BioRad (München)
verwendet. Das Prinzip der Bestimmung basiert auf einer von Bradford [13] beschriebenen
Methode und ermöglicht anhand einer aus 1-12 µg/µl BSA in Wasser erstellten Eichgerade
einen schnelle Quantifizierung der Proteinmenge.

Material und Methoden 33
5 µl der zu untersuchenden Probe wurden mit Wasser auf 800 µl Gesamtvolumen verdünnt
und mit 200 µl BioRad Gebrauchslösung vermischt. Nach fünfminütiger Inkubation erfolgte
die photometrische Bestimmung bei einer Wellenlänge von 595 nm.
2.4.8 Radioaktive Markierung doppelsträngiger DNA
Doppelsträngige DNA mit überhängenden 5’-Enden kann durch den Einbau radioaktiver
Nukleotide (α[32P]-dATP) mit Hilfe des Klenow-Enzyms markiert werden. Die Markierungs-
dauer betrug 30 min bei RT. Die Trennung der radioaktiv markierten DNA von den noch
freien Nukleotiden erfolgte mit dem QIAquick Nucleotide Removal Kit der Firma Qiagen,
Hilden.
Reaktionsansatz: 2,5 pmol DNA(2,5 pmol/µl)6,0 µl 10x Puffer M (Boehringer Mannheim)1,0 µl dCTP (0,5 mM)1,0 µl dGTP (0,5 mM)1,0 µl dTTP (0,5 mM)
45,5 µl H2O2,0 µl [α32P]dATP2,5 µl Klenow-Enzym (5U)
In allen Reaktionen wurde das doppelsträngige „m67SIE“-Oligonukleotid eingesetzt. Diese
Sequenz enthält das sis-induzierbare-Element des humanen c-fos Promotors, welches so ver-
ändert worden ist, daß es die Transkriptionfaktoren STAT1 und STAT3 mit gleich hoher
Affinität bindet [86].
„SIE“ 5’-GATTGACGGGAACTG-3’
„m67SIE“ 5’-GATCCGGGAGGGATTTACGGGAAATGCTG -3’
3’-GCCCTCCCTAAATGCCCTTTACGACTTAA-5’
2.4.9 Elektrophoretischer-Mobilitäts-Shift Assay (EMSA)
Mit Hilfe des EMSA ist es möglich, Wechselwirkungen zwischen Transkriptionsfaktoren und
radioaktiv markierten Oligonukleotiden durch Autoradiographie sichtbar zu machen.
5 µg des Kernextraktes wurden in einem speziellen Reaktionspuffer (eingestellt auf 5000 cpm
pro µl) mit 1 µl einer radioaktiv markierten Sonde (2.4.8) inkubiert. Nach fünfminütiger Inku-
bation bei RT erfolgte der Auftrag des Ansatz auf ein 4%-iges natives Polyacrylamidgel

34 Material und Methoden
aufgetragen mit nachfolgender Elektrophorese (bei einer Gellänge von 20 cm: 4 h bei 200V in
0,25x TBE). Der Laufpuffer wurde dabei solange durch eine Pumpe umgewälzt, bis eine
Laufstrecke von 12 cm erreicht worden ist. Um zu vermeiden, daß freie radioaktive
Oligonukleotide ins obere Pufferreservoir – und damit wieder ins Gel – gelangen, erfolgte die
weitere Auftrennung bis zu einer Laufstrecke von 15,5 cm ohne Umwälzung. Im Anschluß an
die Elektrophorese fixierte man das Gel für 30 min in 10% Methanol /10% Eis-essig, nahm es
dann auf ein 3 mm dickes Whatman-Papier auf, trocknete und autoradiographierte es.
5x Gelshift-Puffer: 50 mM HEPES/KOH, pH 7,85 mM EDTA
25 mM MgCl2
50 % Glycerin
Reaktionsansatz: 5,0 µg Kernextrakt4,0 µl 5x Gelshift-Puffer0,1 µl 1M DTT0,2 µl 200mM PMSF1,0 µl Poly(dI-dC), 1mg/ml2,0 µl BSA, 10 mg/ml
ad 20,0 µl H20Bromphenolblau / Xylencyanol
4-%iges PAA: 6,0 ml Acrylamidlösung 40%4,5 ml Glycerol (Endkonzentration 7,5%)3,0 ml TBE 5x
46,2 ml H2O40 µl TEMED
200 µl APS 20%
2.4.10 Immunfluoreszenz-Analysen eukaryonter Zellen
Transfizierte Zellen wurden auf Deckgläschen, die sich in 24-Loch-Zellkulturplatten befin-
den, ausgesät. Nach drei Tagen, wenn eine Zelldichte von 104-105 Zellen pro Loch erreicht
worden war, wurden die Zellen zweimal mit PBS++ gewaschen und 20 min in 2% Paraformal-
dehyd fixiert. Um die Plasmamembran zu permeabilisieren, enthielten alle folgenden Puffer
0,1% Triton X-100. Zur Unterdrückung einer möglichen Eigenfluoreszenz des Paraformalde-
hyds folgte die Inkubation mit 50 mM NH4Cl in PBS++. Die unspezifischen Antikörperbin-
dungsstellen wurden dann mit 1% BSA in PBS++ blockiert. Anschließend erfolgte die
Inkubation der Zellen mit 1-10 ng spezifischen Antikörper für mindestens 30 min. Nach drei
Waschschritten mit PBS++ + 0,2% BSA erfolgte die Detektion der gebundenen Antikörper mit

Material und Methoden 35
einem FITC-konjugierten speziesspezifischen Immunglobulin in einer 1:200-Verdünnung für
mindestens 30 min. Nach erneuten dreimaligen Waschen wurden die Deckgläschen in
Mowiol, Calbiochem, USA, eingebettet und unter dem Immunfluoreszenzmikroskop analy-
siert [30].
PBS++: 1 mM MgCl2
0,1 mM CaCl 2 in PBS
2.4.11 FACS-Analysen von eukaryonten Zellen
Die Durchflußzytometrie wurde verwendet, um die Oberflächenexpression von gp130-
Mutanten in transfizierten BaF/3-Zellen zu überprüfen.
Dazu erntete man die Zellen, wusch und resuspendierte sie in kaltem FACS-Waschpuffer.
Anschließend erfolgte eine Inkubation mit einem extrazellulären gp130-Antikörper auf Eis für
30 min, Nach Entfernung der überschüssigen Aks erfolgte zur Detektion der gebundenen
spezifischen Immunglobuline eine Inkubation mit 2 µl eines Phycoerythrin-gekoppelten anti-
Maus-IgG-Antikörpers für 20 min im Dunkeln. Die Zellen wurden anschließend erneut
gewaschen und in 400 µl FACS-Waschpuffer resuspendiert. 104 Zellen wurden durch-
flußzytometrisch im FACScalibur der Firma Beckton Dickinson mit einem 488 nm Argonla-
ser analysiert. Die Messung erfolgte nach Herstellerangaben der Firma Beckton Dickinson,
Heidelberg.
FACS-Waschpuffer: 5% FCS in PBS
2.4.12 Reportergen-Assays
Promotoren eines Gens können untersucht werden, indem man sie vor das Gen eines leicht
nachweisbaren Enzyms, z.B die Leuchtkäferluciferase, kloniert. Wird ein solches Konstrukt
in eine eukaryonte Zelle eingebracht, kann die Aktivität dieses Enzyms als ein Maß für die
Menge dieses Enzyms in der Zelle gelten, also als Maß für die Transkriptionsrate seines
Genes. Dies wiederum bedeutet, daß man eine Aussage über die Aktivität des vorgeschalteten
Promotors machen kann. Um die einzelnen Werte einer Versuchsreihe untereinander verglei-
chen zu können, muß man die Transfektionseffizienz kontrollieren. Dies geschieht durch Co-
Transfektion z.B. des β-Galaktosidase-Gens, welches stimulationsunabhängig transkribiert
wird.

36 Material und Methoden
2.4.12.1 Luciferase-Assay
Die Luciferase-Assays wurden mit Hilfe des Luciferase Assay System With Reporter Lysis
Buffer-Kits der Firma Promega, USA, durchgeführt. Zunächst wurden die zu untersuchenden
Zellen einen Tag nach der Transfektion über 20 h stimuliert und dann unter Benutzung des
Reporter Lysis Buffer gemäß den Vorschriften des Herstellers geerntet. 20 µl des erhaltenen
Zell-Lysats wurden in einer lichtundurchlässigen 96-iger Multiwellplatte mit 100 µl Assay
Reagent des Kits versetzt und damit die von der Luciferase katalysierte Licht-emittierende
Reaktion gestartet. Die Lumineszenz maß man über 10 sec im Microlumat 96 P der Firma
EG&G Berthold, Bad Wildbad. Außerdem wurde der Proteingehalt der Zell-Lysate nach
Bradford bestimmt (s. 2.4.7).
2.4.12.2 ββββ-Galaktosidase-Assay
Zum Abgleich der Transfektionseffizienz im β-Galaktosidase-Assay wurden 100 µg Protein
mit 500 µl β-Gal-Mix und 100 µl ONPG, ein chromogenes Substrat der β-Galaktosidase, bei
37°C inkubiert. Sobald eine leichte Gelbfärbung der Proben festzustellen war, wurde die Re-
aktion mit 250 µl 1M Na2CO3, pH 8, gestoppt und die dafür benötigte Zeit festgestellt. Dann
maß man die optische Dichte der Proben bei einer Wellenlänge von 420 nm. Die Aktivität des
Enzyms berechnete sich wie folgt:
Aktivität (U/µg) = (OD420 x 6000) / (Inkubationszeit [min] x 100 µg Protein)
β-Gal-Mix: 60 mM Na2HPO4
40 mM NaH2PO4
1 mM KCl1 mM MgCl2
+ 36,8 µl β-Mercapto-Ethanol / 100 ml Lösung
ONPG (1mg/ml in H2O),
2.4.12.3 Auswertung
X = Luciferase-Wert / β-Gal-Aktivität
2,5 Konstruktion der ErG-Mutanten
Hierbei handelt es sich um gp130 -Mutanten, bei denen die extrazelluläre Domäne durch den
extrazellulären Teil des Epo-R ersetzt worden ist. Im gp130-Anteil bieten neueingefügte Re-

Material und Methoden 37
striktionsschittstellen die Möglichkeit, jeweils eines oder auch mehrere der sechs vorhandenen
Tyrosine gegen Phenylalanine auszutauschen. Diese Phenylalanine erhält man durch gleich-
zeitige Konstruktion einer 6F-Mutante, bei der neben der Einfügung der neuen Schnittstellen
auch alle Tyrosine in Phenylalanine umgewandelt worden sind. So besitzt man zwei
Konstrukte mit identischen Schnittstellen, bei denen man nun einzelne Fragmente beliebig
austauschen kann.
Die ErG-Konstrukte wurden mit Hilfe der PCR bzw. Fusions-PCR-Technik in mehreren
Schritten hergestellt. Die Mutationen wurden über synthetische sense- und antisense-Primer
eingeführt, die die ausgetauschten Basen enthielten. Über den als Matrize dienenden Vektor
pSVL-EG [28, 36] wurden mit den Primerpaaren P1/P3, P4/P6, P7/P9, P10/P12, P2/P3,
P5/P6, P8/P9, P11/P12 acht Fragmente hergestellt. In einem nächsten Schritt wurden über
eine Fusions-PCR jeweils zwei aufeinanderfolgende Fragmente in äquimolaren Verhältnissen
als Matrize eingesetzt. Als Primer verwandte man die äußeren flankierenden Oligonukleotide
(P1/P6, P2/P6, P7/P12, P8/P12). Schließlich wurde in einer zweite Fusion-PCR-Runde die
gesamte Länge des zu verändernden Teiles ligiert (P1/P12, P2/P12). Diese Fragmente wurden
über die Restriktionsschnittstellen EcoRI und BamHI in den Expressionsvektor pSVL-EG
einkloniert (Abb. 5).
Alle so hergestellten Kostrukte wurden durch Sequenzierungen auf ihre Richtigkeit überprüft
und mit Immunfluoreszenzen und EMSA auf ihre Expression und Funktionalität analysiert.
2.6 Konstruktion der GrG-myc-Mutanten
Für die ErG-X-myc-Konstrukte wurde der Bereich zwischen den Restriktionsschnittstellen
Rsr II und BamHI durch synthetische Oligonukleotide (P19/P20 für die Y-Variante, P21/P22
für die F-Variante) ersetzt, die für das c-myc-Epitop fusioniert an das letzte Tyrosinmotiv
kodieren. Als Linker zwischen Rezeptormutante und c-myc-Epitop dienen zwei Glycine.
Für die Reportergen-Assays wurden diese Konstrukte über den Vektor pBS-SK- in den
Expressionvektor pRC/CMV umkloniert.
Mit Hilfe der Restriktionsstellen EcoRI und BamHI kann man den transmembranen bzw. in-
trazellulären Teil der ErG-X-myc-Konstrukte in den Vektor pSVL human gp130 ∆ EcoRI
[40‘] einklonieren. Man erhält so die native cDNA für gp130 einschließlich der eingefügten
Restriktionsstellen und des c-myc-Epitops.
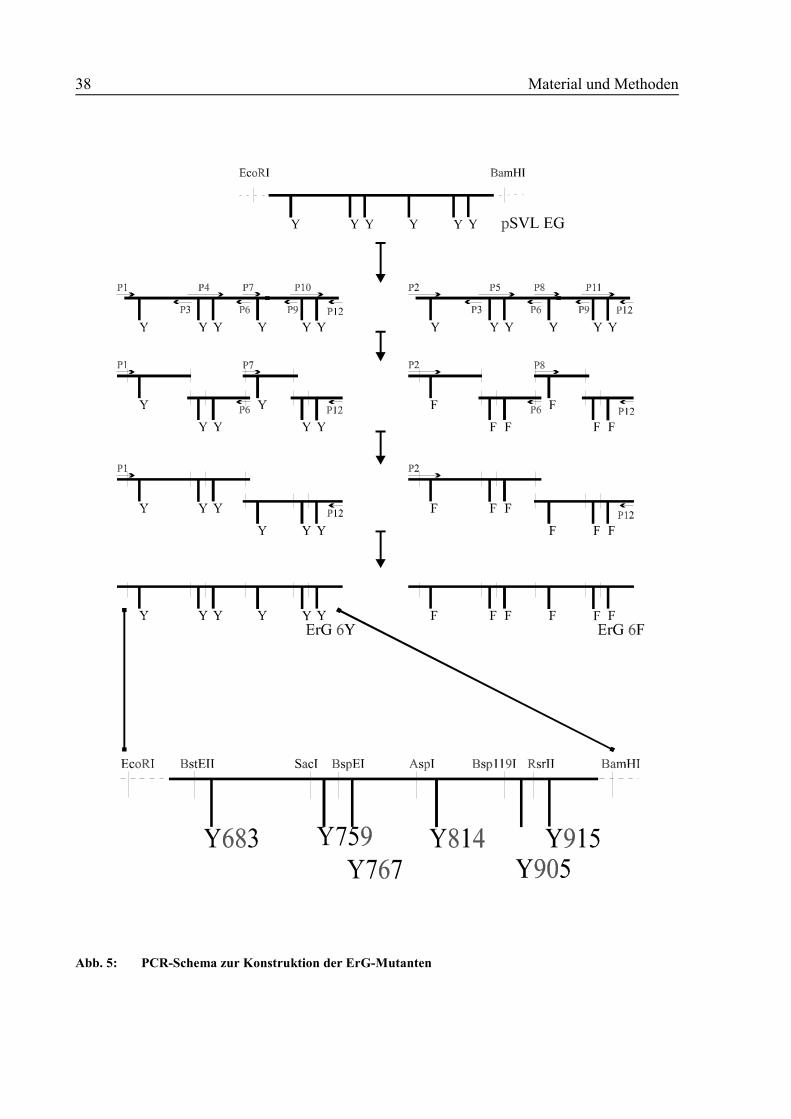
38 Material und Methoden
Abb. 5: PCR-Schema zur Konstruktion der ErG-Mutanten

Material und Methoden 39
Beispielhaft für die rG-Mutanten findet sich die Aminosäuresequenz der cDNA des
GrG YYYYYYmyc-Konstruktes im Anhang.
2.7 Konstruktion der SHP2-Mutanten
Die SHP2-Mutanten wurden über SHP2- Green-Fluorescent Protein (gfp) Fusionsproteine
kloniert, die für die hier erwähnten Versuche allerdings nicht benötigt wurden. Ausgangpunkt
für die Herstellung dieser Fusionsgene war eine PCR über den SHP2-Anteil des Vektors
pRK1D mit den Primern S1/S2 mit anschließenden Restriktionsverdau an den neueingefügten
Schnittstellen Nae I (5’- Ende) und EcoRI (3’-Ende). Das gfp-Gen wurde aus dem Vektor
pMBC1-Bgfp mit Hilfe der Schnittstellen Nae I und Not I isoliert. Beide Fragmente wurden
in den mit Hilfe der Enzyme Not I und EcoRI geöffneten Vektor pSBC1 einkloniert (pSBC1
sypigfp). Hierbei handelt es sich um einen bicistronischen Expressionsvektor mit SV40-Pro-
motor [19]. Dank des Einklonierens einer Hind III-Restriktionsstelle in alle GFP-Fusionsgene
konnte man den gfp-Anteil mittels eines Hind III-Verdaus deletieren. Daraus resultierten
SHP2-Protein-Varianten, die zwei zusätzliche Aminosäuren (Lysin und Alanin) am C-termi-
nalen Ende besaßen.
Es handelt sich im einzelnen um folgende Mutanten:
pSBC1 sypi
Mit obiger Versuchbeschreibung erhielt man ein Plasmid, welches die cDNA für die
Spleißvariante der SHP2 beinhaltet. Diese enthält im Gegensatz zu der erstveröffentlichen
Variante vier zusätzliche Aminosäuren an Position aa 408 (ALLQ) [84].
pSBC1 syp
Bei diesem Konstrukt wurde ausgehend vom obigen pSBC1 sypi das vier Aminosäuren lange
Insert entfernt. Damit entspricht es der erstveröffentlichten Variante der SHP2.
Es wurde mit Hilfe der PCR- und PCR-Fusions-Technik in mehreren Schritten hergestellt.
Als Matrize diente wiederum der Vektor pRK1D. Mit den Primerpaaren S17/S1 wurde ein C-
terminales, mit S8/S18 ein N-terminales Fragment hergestellt und in einer erneuten (jetzt
Fusions-) PCR mit den Primern S1/S8 verbunden. Mit Hilfe der Restriktionsstellen Ban II und
Bgl II konnte das PCR-Produkt in den Vektor pSBC1sypigfp einkloniert werden.
Anschließend erfolgte die die Deletion des gfp-Anteils wie oben beschrieben.

40 Material und Methoden
pSBC1 sypC459S
Durch einen Aminosäurenaustausch an Position 459, wobei das Cystein in ein Serin
gewechselt wurde, erhält man eine katalytisch inaktive Phosphatase [2].
Zur Herstellung dieser Mutante wurde mit den Primer-Paaren S3/S1 und S8/S4 über den Vek-
tor pRK1D eine PCR-Reaktion durchgeführt und die beiden erhaltenen Fragmente in einer
Fusion-PCR mit den Primern S1/S8 verknüpft. Über die Restriktionsschnittstellen Ban II und
Bgl II wurde das letztere PCR-Produkt in den Vektor pSBC1sypigfp einkloniert und der gfp-
Anteil deletiert.
pSBC1 sypmyc, pSBC1 sypimyc, pSBC1 sypC459Smyc
Bei diesen Konstrukten handelt es sich um SHP2-Varianten, an denen ein c-myc-Epitop
(EQKLISEEDL) fusioniert wurde. Man erhielt sie durch Einfügen eines synthetischen, für das
Epitop kodierenden Oligonukleotids (S20, S21) in die C-terminal gelegene Hind III-
Schnittstelle des Proteins.
Um auch in HepG2 arbeiten zu können, wurden die Konstrukte in Folge in den Vektor
pCBC1 umkloniert. Dieser enthält einen CMV-Promotor, entspricht ansonsten aber dem
pSBC1.
Alle Konstrukte wurden mittels Sequenzierungen auf ihre Richtigkeit überprüft. Die Expres-
sion wurde in Western Blots und Immunfluoreszenz-Analysen überprüft.
Beispielhaft für die hier erwähnten Sequenzen ist die cDNA von sypimyc im Anhang wieder-
gegeben.

Ergebnisse 41
3. Ergebnisse
3.1 Konstruktion und Expression der ErG-Mutanten
Die Signaltransduktion des IL-6 wurde bereits vielfältig untersucht. So ist zum Beispiel be-
schrieben worden, daß IL-6-Zugabe zu einer Tyrosinphosphorylierung des gp130 führt [32].
Es ist jedoch nicht bekannt, welche der vorhandenen sechs Tyrosine phosphoryliert werden
können, bzw. welche bei IL-6-Stimulation tatsächlich phosphoryliert werden. Dies und weite-
re Fragestellungen können mit Hilfe verschiedener Rezeptormutanten untersucht werden, bei
denen nur noch jeweils eines der sechs intrazellulären Tyrosine erhalten ist.
Bei diesen sogenannten ErG-Mutanten handelt es sich um gp130-Chimäre, bei denen die ex-
trazelluläre Domäne des gp130 durch den extrazellulären Teil des Erythropoetin (Epo)-R er-
setzt worden ist. Durch diesen Kunstgriff ist es möglich, in einer Zell-Linie, die endogenes
gp130 besitzt, einen gp130-abhängigen Signalweg nachzuempfinden. Der intrazelluläre und
transmembranäre Teil dieser Chimären beinhaltet die entsprechende cDNA des gp 130. Er ist
so konzipiert, daß die einzelnen Mutanten an den Tyrosinmotiven eine beliebige Kombination
an Y→F-Austauschen besitzen.
Abb. 6: Konstruktion der ErG-MutantenBei den ErG-Mutanten handelt es sich um chimäre Rezeptoren, die extrazellulär die extrazelluläreDomäne des Epo-R besitzen. Der transmembranäre und intrazelluläre Teil sind dem gp130 entnom-men. Mittels PCR-Technik sind auf DNA-Ebene zwischen den einzelnen Tyrosinen neue Erken-nungssequenzen für Restriktionsendonukleasen eingefügt worden. Desweiteren ist eine Mutante ge-neriert worden, in der neben diesen neuen Schnittstellen alle Tyrosine in Phenylalanine umgewan-delt worden sind. So besitzt man nun je sechs Tyrosin- bzw. Phenylalanin-Blöcke, die beliebig aus-getauscht werden können.

42 Ergebnisse
Ausgangsvektor für die Klonierarbeiten war das Konstrukt pSVL EG [28, 36]. Mittels PCR-
bzw. Fusions-PCR-Techniken wurden auf DNA-Ebene zwischen den einzelnen Tyrosinen
über stille Mutationen neue Restriktionsstellen eingefügt. Ebenso wurde eine Mutante kon-
struiert, bei der neben diesen neuen Schnittstellen auch alle Tyrosine in Phenylalanine umge-
wandelt worden waren. So erhielt man zwei Konstrukte mit identischen Schnittstellen, bei
denen nun einzelne Fragmente beliebig austauschbar sind (Abb. 6).
Für die hier vorliegende Arbeit war neben dem ErG YYYYYY-Konstrukt, welches dem in-
trazellulären Wildtyp-gp130 entspricht, vor allem das Konstrukt ErG YFYYYY, bei dem das
zweite Tyrosin, Y759, zu Phenylalanin ausgetauscht worden war (= Y759F), von besonderem
Interesse. Die Expression dieser beiden Konstrukte wurde in Western Blots verifiziert. Dazu
wurden COS7-Zellen transient mit der cDNA der beiden Mutanten transfiziert, nach drei Ta-
gen lysiert und die Zell-Lysate in einem SDS-PA-Gel aufgetrennt. Anschließend wurde mit
einem gegen den intrazellulären Bereich des gp130 gerichteten Antikörper in einem Immuno-
blot detektiert (Abb. 7).
Sowohl die Wildtyp- als auch die Y759F-Mutante ließen sich im Western Blot nachweisen.
Abb. 7: Nachweis der Expression der ErG YYYYYY- und ErG YFYYYY-Rezeptorchimären72 h nach Transfektion von COS7-Zellen mit den Vektoren pSVL-Leervektor (Spur 1), pSVL ErGYYYYYY (Spur 2) und pSVL ErG YFYYYY (Spur 3) wurden die Zellen lysiert. Die Zell-Lysatewurden in einer SDS-PAGE aufgetrennt und im Western Blot analysiert. Die Detektion erfolgtenach Inkubation mit einem polyklonalen Antikörper, der gegen den zytoplasmatischen Teil desgp130 gerichtet ist und einem Peroxidase-gekoppelten Zweitantikörper mit der ECL-Reaktion.
3.2 Die SHP2-Tyrosinphosphorylierung
Nachweis der SHP2-Tyrosinphosphorylierung
Die Tyrosinphosphorylierung der SHP2 nach Aktivierung des gp130 durch IL-6, aber auch
IL-11, wurde bereits in einigen Zell-Linien gezeigt, teilweise jedoch unter Verwendung chi-

Ergebnisse 43
märer Rezeptoren [24, 77]. Im ersten Abschnitt dieser Arbeit wurden die Eigenschaften dieser
Aktivierung in den Zell-Systemen HepG2, COS-7 und Ba/F3 genauer untersucht.
Dazu wurden zuerst COS7-Zellen, eine Affennieren-Zell-Linie, mit IL-6/sIL-6R stimuliert
und nach anti-SHP2-Immunopräzipitation im Phosphotyrosin-Western Blot analysiert. Die
Auftragskontrolle erfolgte ebenfalls im Immunoblot mit dem SHP2-Antikörper. Es wurden
mehrere käuflich erwerbliche Phosphotyrosin-Antikörper ausgetestet. Eine sichtbare Reaktion
gab es jedoch nur mit dem 4G10-anti-Phosphotyrosin-Antikörper (Abb. 8).
Abb. 8: Nachweis der SHP2-Tyrosinphosphorylierung in COS7-ZellenCOS7-Zellen wurden 7 min mit 200 U/ml IL-6 und 1 µg/ml sIL-6R stimuliert. Aus den hergestelltenZell-Lysaten aus stimulierten und nicht stimulierten Zellen wurde das SHP2-Protein mittels einesspezifischen anti-SHP2-Antikörpers präzipitiert. Die Auftrennung der präzipitierten Proteine er-folgte in einer SDS-PAGE, ihre anschließende Detektion nach Transfer auf eine PVDF-Membran ineiner ECL-Reaktion. Als spezifische Antikörper wurden der anti-Phosphotyrosin-Antikörper 4G10(Blot: PY) bzw. ein anti-SHP2-Antikörper (Blot: SHP2) verwendet.
Bei etwa 66-70 kDa war im α-PY-Blot eine deutliche Bande zu sehen. Bei der Gegenfärbung
mit einem SHP2-spezifischen Antikörper erhielt man scharfe Banden, die sich jedoch nur
partiell mit der Phosphotyrosin-Bande überdeckten. Je nach Menge an vorhandenem Protein
bzw. je nach Auftrennlänge und Konzentration des SDS-PA-Geles zeigte sich in der anti-
SHP2-Gegenfärbung eine Doppelbande, wobei sich die obere der beiden Banden als dek-
kungsgleich mit der entsprechenden anti-Phosphotyrosin-Bande herausstellte. Weitere Kon-
trollversuche an Ba/F3-Zellen verdeutlichten dies besser. Ba/F3-Zellen stammen von einer
murinen Prä-B-Zell-Linie ab, die kein endogenes gp130 besitzen. Diese Zellen wurden stabil
mit gp130 transfiziert (vgl. Kap. 3.3).
Diese transfizierten Ba/F3-Zellen und nicht transfizierte Kontroll-Zellen wurden mit IL-6/sIL-
6R stimuliert, mit einem SHP2-Antikörper immunpräzipitiert und schließlich im Phosphoty-
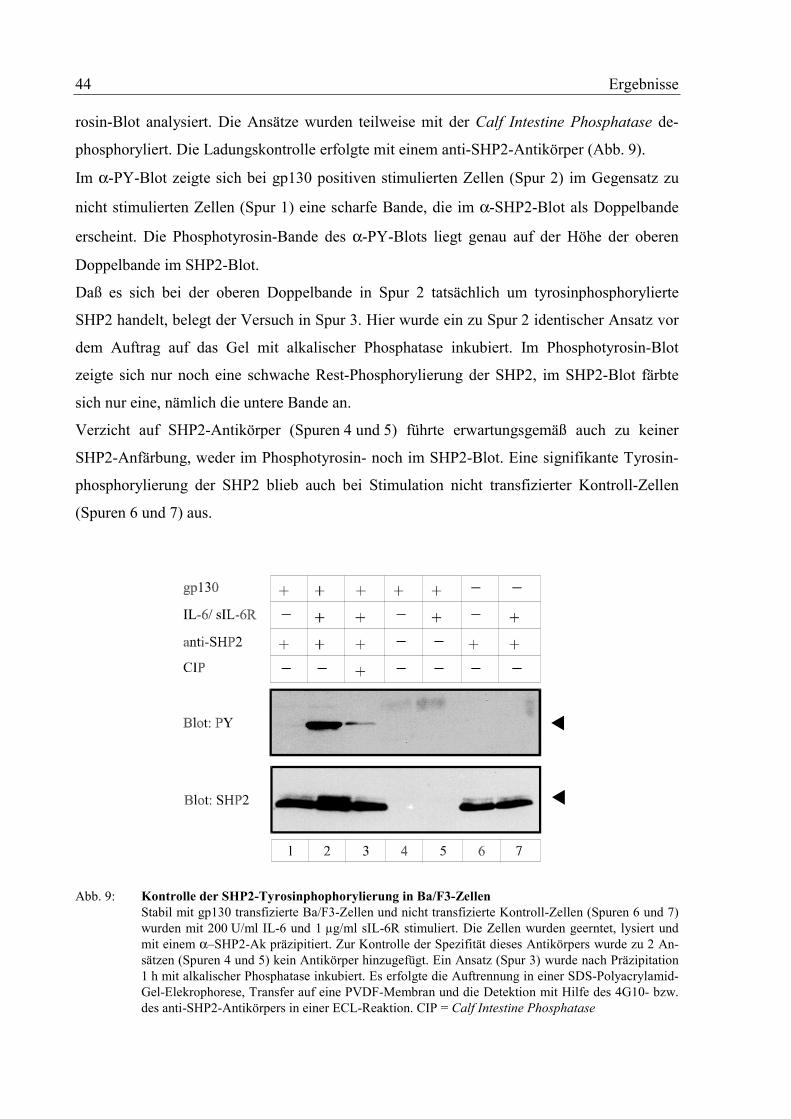
44 Ergebnisse
rosin-Blot analysiert. Die Ansätze wurden teilweise mit der Calf Intestine Phosphatase de-
phosphoryliert. Die Ladungskontrolle erfolgte mit einem anti-SHP2-Antikörper (Abb. 9).
Im α-PY-Blot zeigte sich bei gp130 positiven stimulierten Zellen (Spur 2) im Gegensatz zu
nicht stimulierten Zellen (Spur 1) eine scharfe Bande, die im α-SHP2-Blot als Doppelbande
erscheint. Die Phosphotyrosin-Bande des α-PY-Blots liegt genau auf der Höhe der oberen
Doppelbande im SHP2-Blot.
Daß es sich bei der oberen Doppelbande in Spur 2 tatsächlich um tyrosinphosphorylierte
SHP2 handelt, belegt der Versuch in Spur 3. Hier wurde ein zu Spur 2 identischer Ansatz vor
dem Auftrag auf das Gel mit alkalischer Phosphatase inkubiert. Im Phosphotyrosin-Blot
zeigte sich nur noch eine schwache Rest-Phosphorylierung der SHP2, im SHP2-Blot färbte
sich nur eine, nämlich die untere Bande an.
Verzicht auf SHP2-Antikörper (Spuren 4 und 5) führte erwartungsgemäß auch zu keiner
SHP2-Anfärbung, weder im Phosphotyrosin- noch im SHP2-Blot. Eine signifikante Tyrosin-
phosphorylierung der SHP2 blieb auch bei Stimulation nicht transfizierter Kontroll-Zellen
(Spuren 6 und 7) aus.
Abb. 9: Kontrolle der SHP2-Tyrosinphophorylierung in Ba/F3-ZellenStabil mit gp130 transfizierte Ba/F3-Zellen und nicht transfizierte Kontroll-Zellen (Spuren 6 und 7)wurden mit 200 U/ml IL-6 und 1 µg/ml sIL-6R stimuliert. Die Zellen wurden geerntet, lysiert undmit einem α–SHP2-Ak präzipitiert. Zur Kontrolle der Spezifität dieses Antikörpers wurde zu 2 An-sätzen (Spuren 4 und 5) kein Antikörper hinzugefügt. Ein Ansatz (Spur 3) wurde nach Präzipitation1 h mit alkalischer Phosphatase inkubiert. Es erfolgte die Auftrennung in einer SDS-Polyacrylamid-Gel-Elekrophorese, Transfer auf eine PVDF-Membran und die Detektion mit Hilfe des 4G10- bzw.des anti-SHP2-Antikörpers in einer ECL-Reaktion. CIP = Calf Intestine Phosphatase

Ergebnisse 45
Dosisabhängigkeit der IL-6-induzierten SHP2-Tyrosinphosphorylierung in HepG2-
Zellen
Um eine optimale Stimulationsdosis zu ermitteln, wurden Zellen der humanen Hepatom-Zell-
Linie HepG2 mit rekombinantem IL-6 in steigenden Konzentrationen stimuliert. Das SHP2-
Protein wurde mittels Immunopräzipitation aufgereinigt und im anti-Phosphotyrosin-Western
Blot analysiert (Abb. 10).
Mindestens 100 Einheiten IL-6/ml wurden benötigt, um eine Phosphorylierung der SHP2
sichtbar zu machen. Die maximale SHP2-Phosphorylierung war mit 200 Einheiten IL-6/ml
erreicht. Weitere Dosiserhöhungen führten keiner Steigerung der Antwort.
Abb: 10: Dosisabhängigkeit der SHP2-TyrosinphosphorylierungHepG2-Zellen wurden mit steigenden Konzentrationen IL-6 für 10 min stimuliert. Nach Zell-Ernteund -Lyse erfolgte eine Aufreinigung der SHP2-Proteine in einer SHP2-spezifischen Immunopräzi-pitation. Anschließend wurden die Proteine in einer SDS-PAGE aufgetrennt und in einem WesternBlot mittels des 4G10-Ak auf Tyrosinphosphorylierung untersucht. Die Ladungskontrolle erfolgtemit einem anti-SHP2-Western Blot.
Zeitabhängigkeit der IL-6-induzierten SHP2-Tyrosinphosphorylierung entspricht der-
jenigen des gp130 in HepG2-Zellen
Die Zeitabhängigkeit der SHP2-Tyrosinphosphorylierung wurde ebenfalls in HepG2 unter-
sucht. Die Zellen wurden zu verschiedenen Zeiten nach unterschiedlicher Stimulationsdauer
geerntet (Abb. 11). Nach α-SHP2-Immunopräzipitation zeigte sich im Western Blot eine tran-
siente Tyrosinphosphorylierung der SHP2. Fünf Minuten nach Stimulation war bereits eine
SHP2-Phosphorylierung erkennbar, die nach 40 min nicht mehr nachweisbar war.
In einem Parallelansatz unter gleichen Bedingungen wurde gp130 mit Hilfe eines gegen die
extrazelluläre Domäne dieses Proteins gerichteten Antikörpers präzipitiert und ebenfalls im α-
PY-Blot untersucht (Abb. 12). Die Kinetik der gp130-Phosphorylierung wies große Ähnlich-
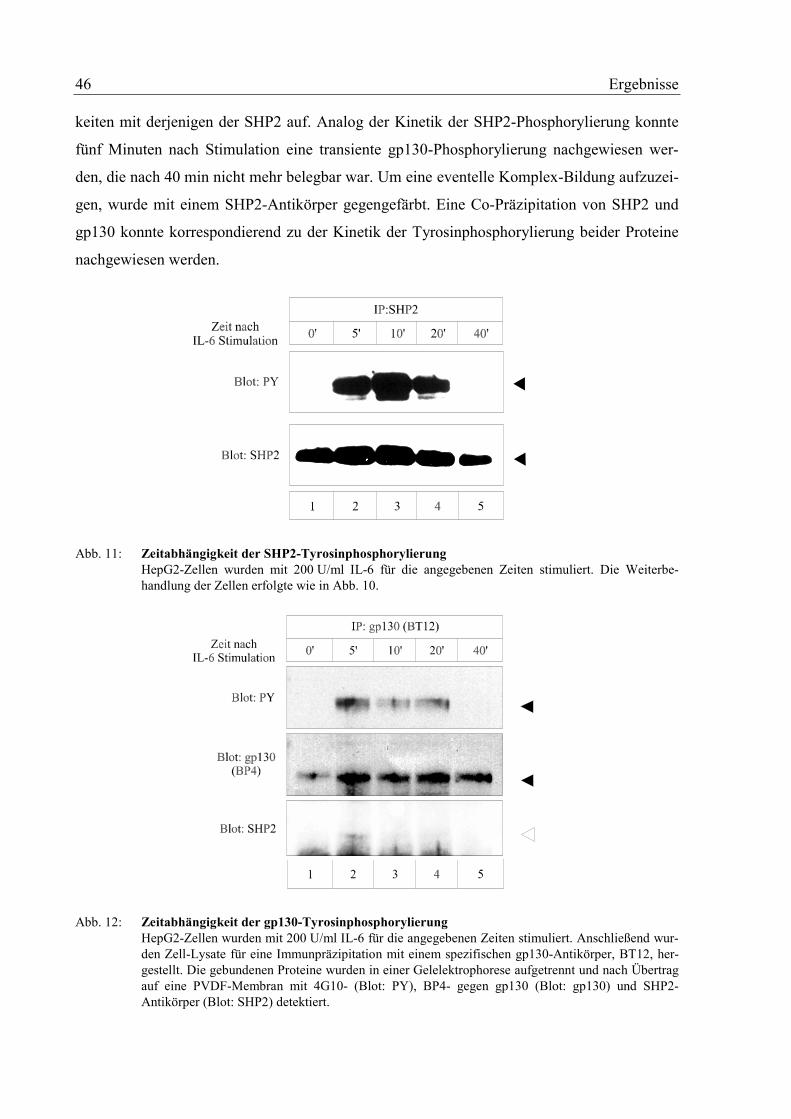
46 Ergebnisse
keiten mit derjenigen der SHP2 auf. Analog der Kinetik der SHP2-Phosphorylierung konnte
fünf Minuten nach Stimulation eine transiente gp130-Phosphorylierung nachgewiesen wer-
den, die nach 40 min nicht mehr belegbar war. Um eine eventelle Komplex-Bildung aufzuzei-
gen, wurde mit einem SHP2-Antikörper gegengefärbt. Eine Co-Präzipitation von SHP2 und
gp130 konnte korrespondierend zu der Kinetik der Tyrosinphosphorylierung beider Proteine
nachgewiesen werden.
Abb. 11: Zeitabhängigkeit der SHP2-TyrosinphosphorylierungHepG2-Zellen wurden mit 200 U/ml IL-6 für die angegebenen Zeiten stimuliert. Die Weiterbe-handlung der Zellen erfolgte wie in Abb. 10.
Abb. 12: Zeitabhängigkeit der gp130-TyrosinphosphorylierungHepG2-Zellen wurden mit 200 U/ml IL-6 für die angegebenen Zeiten stimuliert. Anschließend wur-den Zell-Lysate für eine Immunpräzipitation mit einem spezifischen gp130-Antikörper, BT12, her-gestellt. Die gebundenen Proteine wurden in einer Gelelektrophorese aufgetrennt und nach Übertragauf eine PVDF-Membran mit 4G10- (Blot: PY), BP4- gegen gp130 (Blot: gp130) und SHP2-Antikörper (Blot: SHP2) detektiert.

Ergebnisse 47
3.3 Die IL-6-induzierte Tyrosinphosphorylierung der SHP2 benötigt dasY759 in gp130
Um zu zeigen, daß die SHP2-Tyrosinphosphorylierung nach IL-6-Stimulation tatsächlich über
gp130 induziert wird, wurden COS7-Zellen transient mit dem Expressionskonstrukt der Epo-
R/gp130-Chimäre ErG YYYYYY (s. Kap. 3.1) transfiziert. Gleichzeitig wurde in einem Par-
allelansatz das ErG YFYYYY-Konstrukt exprimiert, um die bereits von mehreren Autoren
beschriebene Abhängigkeit der SHP2-Phosphorylierung vom Tyrosin 759 im COS7-System
nachzuvollziehen [26, 78]. Nach Stimulation mit Epo wurde das SHP2-Protein mittels eines
spezifischen Antikörpers immunpräzipitiert, in einem Gel aufgetrennt und auf eine PVDF-
Membran transferiert. In darauf folgenden Phosphotyrosin-Immun-Blots konnte jedoch weder
bei der „Wildtyp“-Mutante YYYYYY noch bei der Y759F-Mutante, YFYYYY, eine SHP2-
Tyrosinphosphorylierung festgestellt werden, obwohl die transiente Expression der Rezep-
tormutanten – wie in Abb. 7 dargestellt – im Western Blot bereits nachgewiesen werden
konnte. Auch besitzen COS7-Zellen wie oben gezeigt (Abb. 8) prinzipiell das Vermögen, auf
Zytokin-Einwirkung mit einer SHP2-Phosphorylierung zu antworten, was die sichtbare Tyro-
sinphosphorylierung der SHP2 nach IL-6-Stimulation des endogenen gp130 belegte.
Ein Grund für das Mißlingen dieses Experimentes könnte in der transienten Transfektion lie-
gen. Ein Nachteil der transienten Transfektion ist häufig eine geringe Transfektionseffizienz.
Immunfluoreszenzen der ErG-Mutanten in COS7-Zellen zeigten, daß nur wenige Zellen die
transfizierten Rezeptoren exprimierten [73]. Wenn jedoch nur eine geringe Menge von Zellen
die Rezeptoren besitzen, kann auch nur dieser Anteil stimuliert werden. Dadurch wird nur ein
Bruchteil der insgesamt vorhandenen SHP2-Menge tyrosinphosphoryliert: zu wenig, als daß
es der anti-Phosphotyrosin-Antikörper 4G10 nachweisen könnte. Um den prozentualen Anteil
der transfizierten Zellen zu erhöhen, wurden stabil gp130 exprimierende Ba/F3-Zellen herge-
stellt. Ba/F3-Zellen, eine murine Prä-B-Zell-Linie, besitzen kein endogenes gp130, was den
Vorteil bietet, sie nicht mit chimären Rezeptoren transfizieren zu müssen, sondern mit nativer
gp130-cDNA transfizieren zu können. Dazu wurde der extrazelluläre Teil der ErG-Mutanten
durch den entsprechenden nativen Teil des gp130 ersetzt. Man erhält so gp130-Mutanten,
GrG, die im intrazellulären Bereich statt der Tyrosine wahlweise Phenylalanine enthalten
können (vgl. Kap. 2.7). GrG YFYYYY ist also analog der Chimäre ErG YFYYYY eine
gp130-Mutante, bei der das Tyrosin 759 zu Phenylalanin ausgetauscht worden ist (= GrG
Y759F). Die „Mutante“ GrG YYYYYY entspricht dem Wildtyp. Die Expression wurde
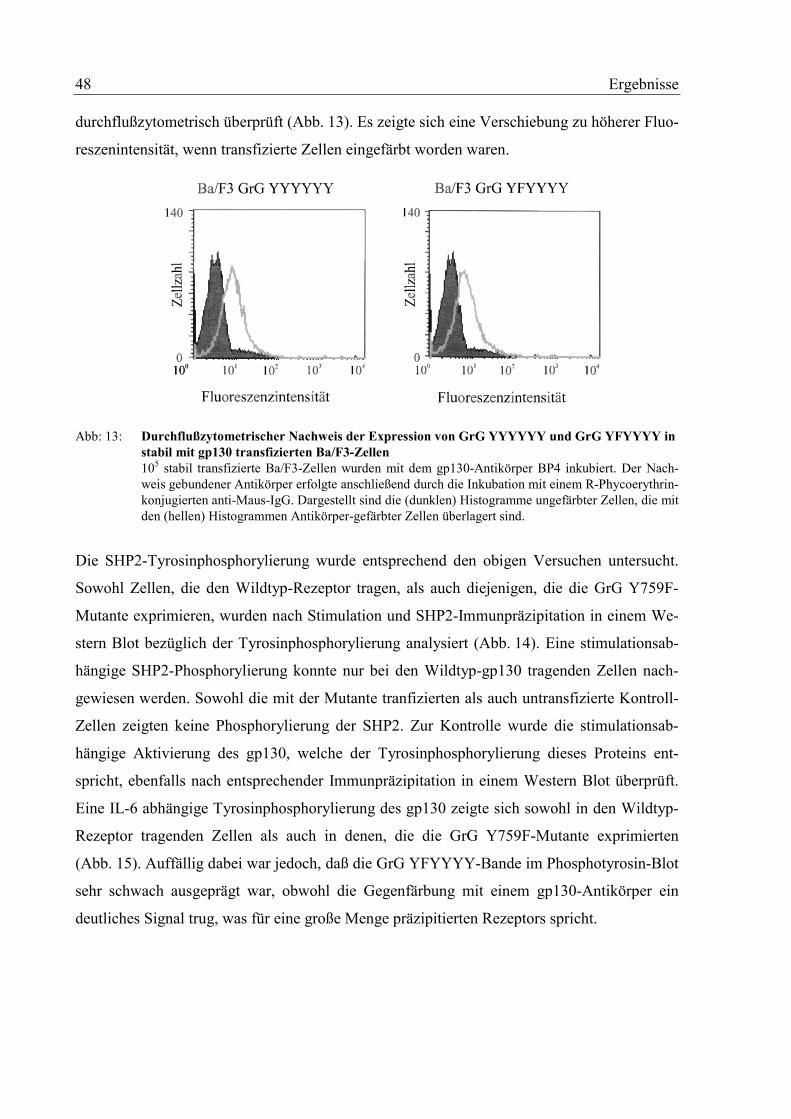
48 Ergebnisse
durchflußzytometrisch überprüft (Abb. 13). Es zeigte sich eine Verschiebung zu höherer Fluo-
reszenintensität, wenn transfizierte Zellen eingefärbt worden waren.
Abb: 13: Durchflußzytometrischer Nachweis der Expression von GrG YYYYYY und GrG YFYYYY instabil mit gp130 transfizierten Ba/F3-Zellen105 stabil transfizierte Ba/F3-Zellen wurden mit dem gp130-Antikörper BP4 inkubiert. Der Nach-weis gebundener Antikörper erfolgte anschließend durch die Inkubation mit einem R-Phycoerythrin-konjugierten anti-Maus-IgG. Dargestellt sind die (dunklen) Histogramme ungefärbter Zellen, die mitden (hellen) Histogrammen Antikörper-gefärbter Zellen überlagert sind.
Die SHP2-Tyrosinphosphorylierung wurde entsprechend den obigen Versuchen untersucht.
Sowohl Zellen, die den Wildtyp-Rezeptor tragen, als auch diejenigen, die die GrG Y759F-
Mutante exprimieren, wurden nach Stimulation und SHP2-Immunpräzipitation in einem We-
stern Blot bezüglich der Tyrosinphosphorylierung analysiert (Abb. 14). Eine stimulationsab-
hängige SHP2-Phosphorylierung konnte nur bei den Wildtyp-gp130 tragenden Zellen nach-
gewiesen werden. Sowohl die mit der Mutante tranfizierten als auch untransfizierte Kontroll-
Zellen zeigten keine Phosphorylierung der SHP2. Zur Kontrolle wurde die stimulationsab-
hängige Aktivierung des gp130, welche der Tyrosinphosphorylierung dieses Proteins ent-
spricht, ebenfalls nach entsprechender Immunpräzipitation in einem Western Blot überprüft.
Eine IL-6 abhängige Tyrosinphosphorylierung des gp130 zeigte sich sowohl in den Wildtyp-
Rezeptor tragenden Zellen als auch in denen, die die GrG Y759F-Mutante exprimierten
(Abb. 15). Auffällig dabei war jedoch, daß die GrG YFYYYY-Bande im Phosphotyrosin-Blot
sehr schwach ausgeprägt war, obwohl die Gegenfärbung mit einem gp130-Antikörper ein
deutliches Signal trug, was für eine große Menge präzipitierten Rezeptors spricht.
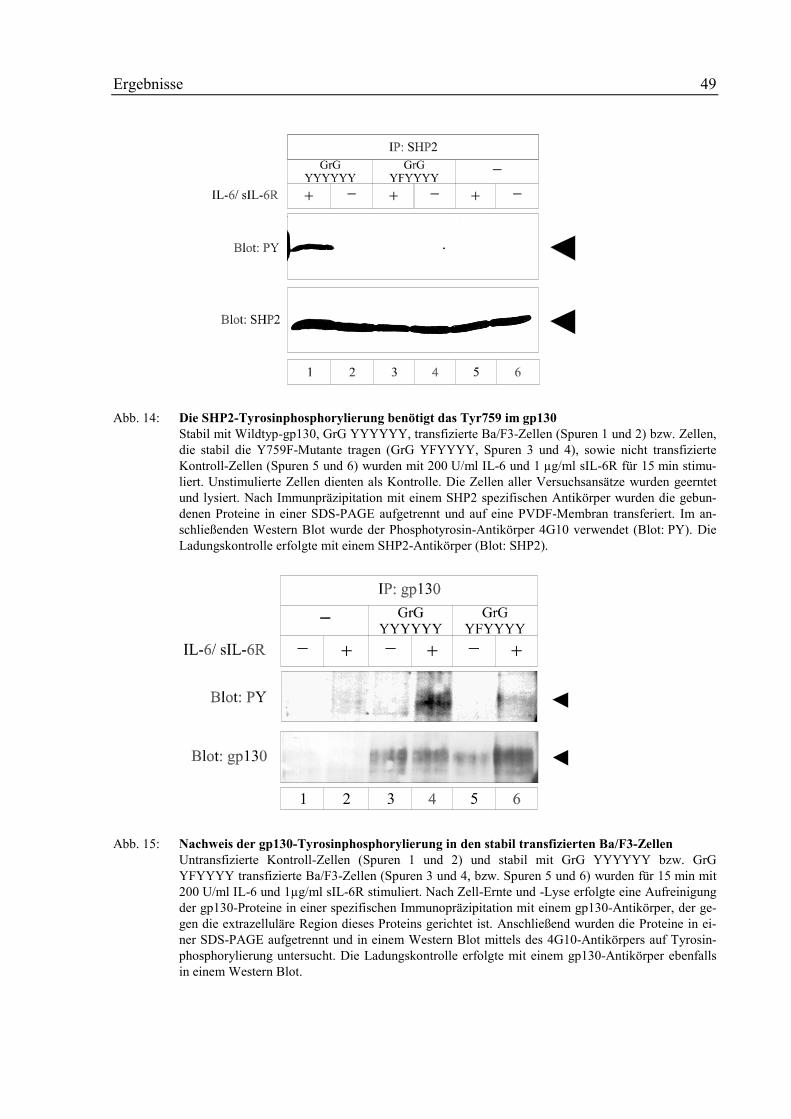
Ergebnisse 49
Abb. 14: Die SHP2-Tyrosinphosphorylierung benötigt das Tyr759 im gp130Stabil mit Wildtyp-gp130, GrG YYYYYY, transfizierte Ba/F3-Zellen (Spuren 1 und 2) bzw. Zellen,die stabil die Y759F-Mutante tragen (GrG YFYYYY, Spuren 3 und 4), sowie nicht transfizierteKontroll-Zellen (Spuren 5 und 6) wurden mit 200 U/ml IL-6 und 1 µg/ml sIL-6R für 15 min stimu-liert. Unstimulierte Zellen dienten als Kontrolle. Die Zellen aller Versuchsansätze wurden geerntetund lysiert. Nach Immunpräzipitation mit einem SHP2 spezifischen Antikörper wurden die gebun-denen Proteine in einer SDS-PAGE aufgetrennt und auf eine PVDF-Membran transferiert. Im an-schließenden Western Blot wurde der Phosphotyrosin-Antikörper 4G10 verwendet (Blot: PY). DieLadungskontrolle erfolgte mit einem SHP2-Antikörper (Blot: SHP2).
Abb. 15: Nachweis der gp130-Tyrosinphosphorylierung in den stabil transfizierten Ba/F3-ZellenUntransfizierte Kontroll-Zellen (Spuren 1 und 2) und stabil mit GrG YYYYYY bzw. GrGYFYYYY transfizierte Ba/F3-Zellen (Spuren 3 und 4, bzw. Spuren 5 und 6) wurden für 15 min mit200 U/ml IL-6 und 1µg/ml sIL-6R stimuliert. Nach Zell-Ernte und -Lyse erfolgte eine Aufreinigungder gp130-Proteine in einer spezifischen Immunopräzipitation mit einem gp130-Antikörper, der ge-gen die extrazelluläre Region dieses Proteins gerichtet ist. Anschließend wurden die Proteine in ei-ner SDS-PAGE aufgetrennt und in einem Western Blot mittels des 4G10-Antikörpers auf Tyrosin-phosphorylierung untersucht. Die Ladungskontrolle erfolgte mit einem gp130-Antikörper ebenfallsin einem Western Blot.

50 Ergebnisse
3.4 Einfluß der SHP2-Phosphorylierung auf die STAT-Aktivierung
Da man durch gp130-Stimulation über die YFYYYY-Mutante selektiv die SHP2-
Phosphorylierung ausschalten kann, eignet sich dieses System zu ersten Untersuchungen der
biologischen Funktion dieser Phosphorylierung. Dazu wurde die Zeitabhängigkeit der
STAT3- bzw. STAT1-Phosphorylierung in den stabil transfizierten Ba/F3-Zellen analysiert
(Abb. 16). Aus Kernextraktion gewonnenes Material wurde in einer SDS-Polyacrylamid-Gel-
Elektrophorese aufgetrennt und sowohl mit einem spezifischen anti-Phospho-STAT3-
Antikörper als auch einem anti-STAT1-Phosphotyrosin-Antikörper im Western Blot unter-
sucht.
vAbb. 16: Einfluß der SHP2-Phosphorylierung auf die STAT-Aktivierung(A) Stabil mit gp130 transfizierte Ba/F3-Zellen (GrG YYYYYY) und mit GrG YFYYYY transfi-zierte sowie untransfizierte Ba/F3-Zellen wurden mit 200 U/ml IL-6 und 1 µg/ml sIL-6R für die an-gegebenen Zeiten stimuliert. 50 µg Gesamtprotein aus anschließend hergestellten Kernextraktenwurden in einer SDS-PAGE aufgetrennt und im Western Blot mittels spezifischer anti-Phospho-STAT3- bzw. anti-Phospho-STAT1-Antikörper untersucht.(B) 5 µg der in (A) vorgestellten Kernextrakte wurden mit einer radioaktiven m67SIE-Sonde inku-biert. Anschließend erfolgte die Analyse der Protein-DNA-Komplexe im EMSA.

Ergebnisse 51
Während nicht transfizierte Kontroll-Zellen und nicht stimulierte Zellen keine STAT3-
Phosphorylierung zeigten, konnte eine transiente STAT3-Phosphorylierung sowohl beim
Wildtyp als auch bei der YFYYYY-Mutante nachgewiesen werden. Interessanterweise führte
die Stimulation der Y759F-Mutante zu einer gesteigerten Antwort. Die Gegenfärbung mit
dem anti-STAT1-Antikörper zeigte dies noch deutlicher. Stimulation der YFYYYY-Mutante
führte zu einer stärkeren und verlängerten STAT1-Phosphorylierung (Abb. 16A).
Diese Beobachtungen konnten sich in anschließenden DNA-Bindungs-Experimenten bestäti-
gen. Kernextrakte aus den Wildtyp bzw. Mutante exprimierenden Ba/F3-Zellen wurden mit
einer STAT1- und STAT3- bindungsspezifischen DNA-Probe vermischt und in einem EMSA
analysiert (Abb. 16B).
In Ba/F3-Zellen, die den Wildtyp gp130-Rezeptor oder die YFYYYY-Mutante exprimierten,
konnte stimulationsabhängig ein DNA/Proteinkomplex aus STAT3-, STAT1-Homodimeren
und STAT1/STAT3-Heterodimeren aufgezeigt werden, nicht transfizierte Kontroll-Zellen
hingegen wiesen dies nicht auf. Die Menge des sichtbaren STAT/DNA-Komplexes war bei
den Zellen, die die Mutante exprimierten, signifikant erhöht und länger nachweisbar.
3.5 Rolle der Janus-Kinasen für die SHP2-Phosphorylierung
Wie oben beschrieben wird die SHP2 in der IL-6-Signaltransduktion tyrosinphosphoryliert.
Auch für die Janus-Kinasen Jak1, Jak2 und Tyk2 wurde eine Tyrosinphosphorylierung und
damit Aktivierung nach IL-6 beschrieben [54, 76]. Im folgenden wurde untersucht, ob und
welche dieser konstitutiv an gp130 assozierten Jaks für die SHP2-Phosphorylierung notwen-
dig sind. Dies wurde mit einer Reihe Jak-defizienter Zell-Linien (vgl. Kap. 2.3.2.1) erarbeitet,
die Abkömmlinge einer humane Fibrosarkom-Zell-Linie sind. Bei diesen ist jeweils eine der
drei Kinasen inaktiviert, bzw. im Anschluß daran retransfiziert worden [16].
Zum Aufbau eines funktionierenden Assays wurde zuerst die parentale Zell-Linie 2fTGH
analysiert. Die Zellen wurden mit IL-6 und sIL-6R stimuliert und anschließend lysiert. Das
SHP2-Protein wurde sodann in einer Immunpräzipitation mit einem SHP2-spezifischen Anti-
körper aufgereinigt und in einem Phosphotyrosin Western Blot analysiert. Als Ladungskon-
trolle diente die Gegenfärbung mit einem SHP2-spezifischen Antikörper (Abb. 17).
Es zeigte sich eine stimulationabhängige SHP2-Phosphorylierung. Interessanterweise war im
Gegensatz zu den anderen untersuchten Zell-Linien bei den Fibrosarkom-Abkömmlingen
zwar ein deutliches, jedoch schwächeres Signal im Western Blot zu verzeichnen.

52 Ergebnisse
Abb. 17: Tyrosinphosphorylierung der SHP2 in 2fTGH-Zellen2fTGH-Zellen wurden mit den angegebenen Mengen IL-6 und 1 µg/ml sIL-6R für 15 min stimuliert.Nach Zell-Ernte und -Lyse wurde mit Hilfe eines spezifischen SHP2-Antikörpers das SHP2-Proteinaus den Zell-Lysaten präzipitiert, und die gebundenen Proteine in einer Gelelektrophorese aufge-trennt. Nach Transfer auf eine PVDF-Membran wurde in einem Western Blot mittels des anti-Phosphotyrosin-Antikörpers 4G10 die SHP2-Tyrosinphosphorylierung analysiert (Blot: PY). DieLadungskontrolle erfolgte mit einem spezifischen SHP2-Antikörper (Blot: SHP2).
Um nun zu erfahren, ob Jak2 eine wesentliche Funktion in der IL-6 induzierten Signaltrans-
duktion spielt, wurden γ2A-Zellen, denen das Jak2-Protein fehlt, stimuliert, mit dem anti-
SHP2-Antikörper immunopäzipitiert und im Western Blot analysiert. Als Kontroll-Zellen
dienten γ2A/Jak2-Transfektanten, die stabil Jak2 exprimieren (Abb. 18A).
In beiden Zell-Linien konnte eine von der IL-6-Stimulation abhängige Tyrosinphosphorylie-
rung der SHP2 beobachtet werden.
Analoge Experimente wurden mit den Zell-Linien U1A bzw. U1A/Tyk2 durchgeführt. U1A-
Zellen sind nicht mehr in der Lage, Tyk2 zu exprimieren, U1A/Tyk2 sind die entsprechenden
mit der Tyk2-cDNA rekonstitutierten Kontroll-Zellen (Abb. 18B).
Wie bei den γ2A bzw. γ2A/Jak2-Zellen zeigte sich eine stimulationsabhängige SHP2-
Tyrosinphosphorylierung sowohl bei den Tyk2-defizienten, als auch bei den retransfizierten
Kontroll-Zellen. Interessanterweise führte eine Überexpression von Jak2 und Tyk2 in den
Kontroll-Zellen nicht zu einer gesteigerten SHP2-Tyrosinphosphorylierung.
Die Funktion der Jak1 wurde in den Zell-Linien U4A und U4A/Jak1 untersucht. Entspre-
chend zu den oben genannten Zell-Linien handelt es sich hierbei um Jak1 defiziente Zellen,
U4A. Zum Vergleich dienten mit Jak1-cDNA stabil retransfizierte Kontroll-Zellen, U4A/Jak1
(Abb. 18C).
Während sich in den U4A-Zellen keine SHP2-Tyrosinphosphorylierung nach IL-6-
Stimulation zeigte, konnte sie in den U4A/Jak1-Transfektanten nachgewiesen werden.
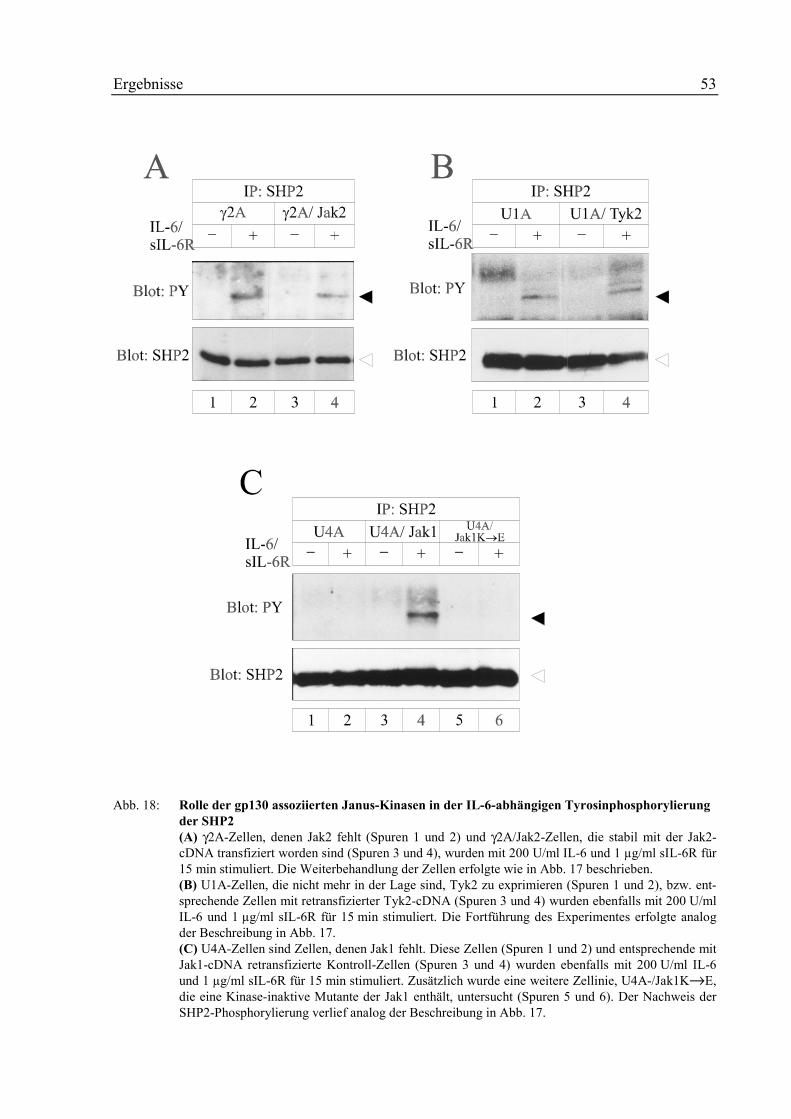
Ergebnisse 53
Abb. 18: Rolle der gp130 assoziierten Janus-Kinasen in der IL-6-abhängigen Tyrosinphosphorylierungder SHP2(A) γ2A-Zellen, denen Jak2 fehlt (Spuren 1 und 2) und γ2A/Jak2-Zellen, die stabil mit der Jak2-cDNA transfiziert worden sind (Spuren 3 und 4), wurden mit 200 U/ml IL-6 und 1 µg/ml sIL-6R für15 min stimuliert. Die Weiterbehandlung der Zellen erfolgte wie in Abb. 17 beschrieben.(B) U1A-Zellen, die nicht mehr in der Lage sind, Tyk2 zu exprimieren (Spuren 1 und 2), bzw. ent-sprechende Zellen mit retransfizierter Tyk2-cDNA (Spuren 3 und 4) wurden ebenfalls mit 200 U/mlIL-6 und 1 µg/ml sIL-6R für 15 min stimuliert. Die Fortführung des Experimentes erfolgte analogder Beschreibung in Abb. 17.(C) U4A-Zellen sind Zellen, denen Jak1 fehlt. Diese Zellen (Spuren 1 und 2) und entsprechende mitJak1-cDNA retransfizierte Kontroll-Zellen (Spuren 3 und 4) wurden ebenfalls mit 200 U/ml IL-6und 1 µg/ml sIL-6R für 15 min stimuliert. Zusätzlich wurde eine weitere Zellinie, U4A-/Jak1K→E,die eine Kinase-inaktive Mutante der Jak1 enthält, untersucht (Spuren 5 und 6). Der Nachweis derSHP2-Phosphorylierung verlief analog der Beschreibung in Abb. 17.

54 Ergebnisse
Dies läßt darauf schließen, daß nur Jak1, nicht aber Jak2 oder Tyk2, essentiell für die SHP2-
Tyrosinphosphorylierung zu sein scheint. Um nun zu überprüfen, ob die Abwesenheit des
Jak1-Proteins an sich oder nur das Fehlen der Kinase-Aktivität dieses Enzyms die defiziente
SHP2-Phosphorylierung bewirkte, wurde die Tyrosinphosphorylierung der SHP2 in U4A-
Zellen untersucht, die stabil mit einer Kinase-inaktiven Jak1-Mutante transfiziert worden wa-
ren (Abb. 18C, Spuren 5 und 6).
Auch die Transfektanten dieser Jak1-Mutante, U4A/Jak1K→E, waren nicht in der Lage, die
SHP2-Phosphorylierung wiederherzustellen, was zeigt, daß nicht nur das Protein Jak1, son-
dern vielmehr seine enzymatische Aktivität für die SHP2-Tyrosinphosphorylierung essentiell
ist.
3.6 Assoziation der SHP2 mit den Janus-Kinasen
Die gesonderte Funktion der Jak1 könnte über eine bevorzugte Komplexbildung der SHP2
mit dieser Kinase erklärt werden. Es wäre möglich, daß SHP2 und Jak1 in vivo eine direkte
Bindung eingehen, wie es für Jak2 bereits in 3T3-L1 Maus-Präadipozyten beschrieben wor-
den ist [24]. Deshalb wurde versucht, eine Co-Immunopräzipitation der einzelnen Kinasen mit
SHP2 nachzuweisen. Dafür wurde die SHP2 nach Stimulation aus den Zellen der verschiede-
nen Fibrosarkom-Zell-Linien mittels eines spezifischen Antikörpers präzipitiert, in einer SDS-
PAGE aufgetrennt und im Western Blot durch Immunoblotting mit den einzelnen Jak-
Antikörpern bezüglich der Co-Präzipitation mit den einzelnen Kinasen untersucht (Abb. 19).
In den U4A-Zellen, die das Jak1-Gen nicht exprimieren, konnte wie erwartet keine Assozia-
tion von SHP2 und Jak1 nachgewiesen werden. Sie dienten lediglich als zusätzliche Kontrolle
für die Spezifität der Jak1-Bande, welche in den U4A/Jak1-Präzipitaten beobachtet wurde. Da
beide Zell-Linien sich nur durch die Existenz dieses einen Proteins unterscheiden, mußte es
sich bei dieser Bande um Jak1 handeln. Interessanterweise zeigte sich die Komplexbildung in
den U4A/Jak1-Zellen sowohl bei stimulierten als auch bei unstimulierten Zellen.
Die Co-Immunopräzipitation zwischen SHP2 und Jak2 bzw. Tyk2 wurde ähnlich getestet. Die
Zell-Linien γ2A und γ2A/Jak2 gaben Aufschluß über die Situation bei Jak2. Genau wie bei
den Jak1-Versuchen zeigte sich bei den Ausgangszellen, γ2A, keine Co-Immunopräzipitation;
bei den γ2A/Jak2-Zellen, die stabil Jak2 exprimieren, konnte eine stimulationsunabhängige
Assoziation von Jak2 und SHP2 nachgewiesen werden. Ähnliches gilt für Tyk2: fehlende
Komplexbildung bei den Tyk-negativen U1A-Zellen, stimulationsunabhängige Co-Immuno-
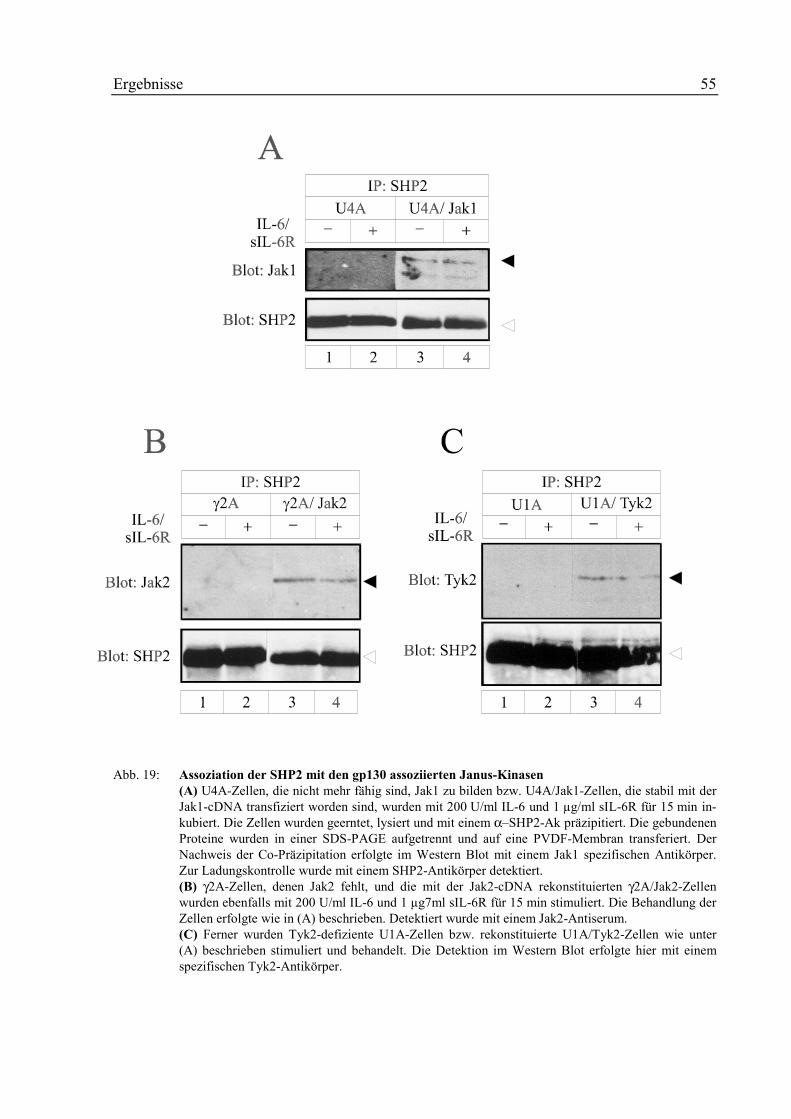
Ergebnisse 55
Abb. 19: Assoziation der SHP2 mit den gp130 assoziierten Janus-Kinasen(A) U4A-Zellen, die nicht mehr fähig sind, Jak1 zu bilden bzw. U4A/Jak1-Zellen, die stabil mit derJak1-cDNA transfiziert worden sind, wurden mit 200 U/ml IL-6 und 1 µg/ml sIL-6R für 15 min in-kubiert. Die Zellen wurden geerntet, lysiert und mit einem α–SHP2-Ak präzipitiert. Die gebundenenProteine wurden in einer SDS-PAGE aufgetrennt und auf eine PVDF-Membran transferiert. DerNachweis der Co-Präzipitation erfolgte im Western Blot mit einem Jak1 spezifischen Antikörper.Zur Ladungskontrolle wurde mit einem SHP2-Antikörper detektiert.(B) γ2A-Zellen, denen Jak2 fehlt, und die mit der Jak2-cDNA rekonstituierten γ2A/Jak2-Zellenwurden ebenfalls mit 200 U/ml IL-6 und 1 µg7ml sIL-6R für 15 min stimuliert. Die Behandlung derZellen erfolgte wie in (A) beschrieben. Detektiert wurde mit einem Jak2-Antiserum.(C) Ferner wurden Tyk2-defiziente U1A-Zellen bzw. rekonstituierte U1A/Tyk2-Zellen wie unter(A) beschrieben stimuliert und behandelt. Die Detektion im Western Blot erfolgte hier mit einemspezifischen Tyk2-Antikörper.

56 Ergebnisse
präzipitation bei mit dem Tyk2-Gen rekonstituierten Zell-Linie, U1A/Tyk2. Auch hier konnte
der negative Befund bei den Kinase-defizienten Zellen als Beleg der Spezifität der Banden bei
den retransfizierten Zellen herangezogen werden.
Die konstitutive Assoziation wirft die Frage nach der Spezifität der Versuche auf. Um nach-
zuweisen, daß die Kinasen nicht unspezfisch mit der Protein A-Sepharose präzipitieren oder
mit sonstigen Ingredentien interagieren, wurde in einem Kontrollversuch ohne SHP2-
Antikörper „präzipitiert“. Hier ließ sich die Co-Immunopräzipitation nicht nachweisen, was
für die Spezifität der Interaktion spricht (Abb. 20).
Zusammenfassend läßt sich sagen, SHP2 und Jak1, Jak2 und Tyk2 bilden konstitutiv einen
Komplex, obwohl nur Jak1 essentiell für die Tyrosinphosphorylierung der SHP2 nach Stimu-
lation mit IL-6 ist.
Abb. 20: Kontrolle der Assoziation der SHP2 mit den Jak-KinasenAls Kontrolle unspezifischer Bindungen der Jak-Kinasen an Protein A-Sepharose oder anderen In-haltsstoffen des Versuchsansatzes wurden die Co-Immunpräzipitationen in Abwesenheit der SHP2-Antikörper durchgeführt. Zell-Lysate der Zell-Linien U4A/ Jak1, 2A/Jak2 und U1A/Tyk2 wurdenmit oder ohne SHP2-Antikörper inkubiert. Nach Präzipitation mit Protein A-Sepharose wurden diegebundenen Proteine in einer SDS-PAGE aufgetrennt, auf eine PVDF-Membran transferiert und mitJak-spezifischen Antikörpern analysiert. Die Präzipitationskontrolle erfolgte mit einem anti-SHP2-Antikörper. (offene Pfeilspitzen: Jak, schwarze Pfeilspitze: SHP2)
3.7 Rolle der SHP2 in der Akute-Phase-Protein-Gen-Induktion
IL-6 ist der Hauptinduktor der Akute-Phase-Protein-Expression in der Leber. Der Mechanis-
mus, der zur Expression dieser Proteine führt, ist gut untersucht und involviert die Aktivie-
rung des Jak-STAT-Signalweges. Interessant ist es nun zu wissen, welche Funktion die SHP2,
deren Nicht-Phosphorylierung mit einer gesteigerten STAT3-Aktivierung einhergeht, in der
Akute-Phase-Reaktion hat.
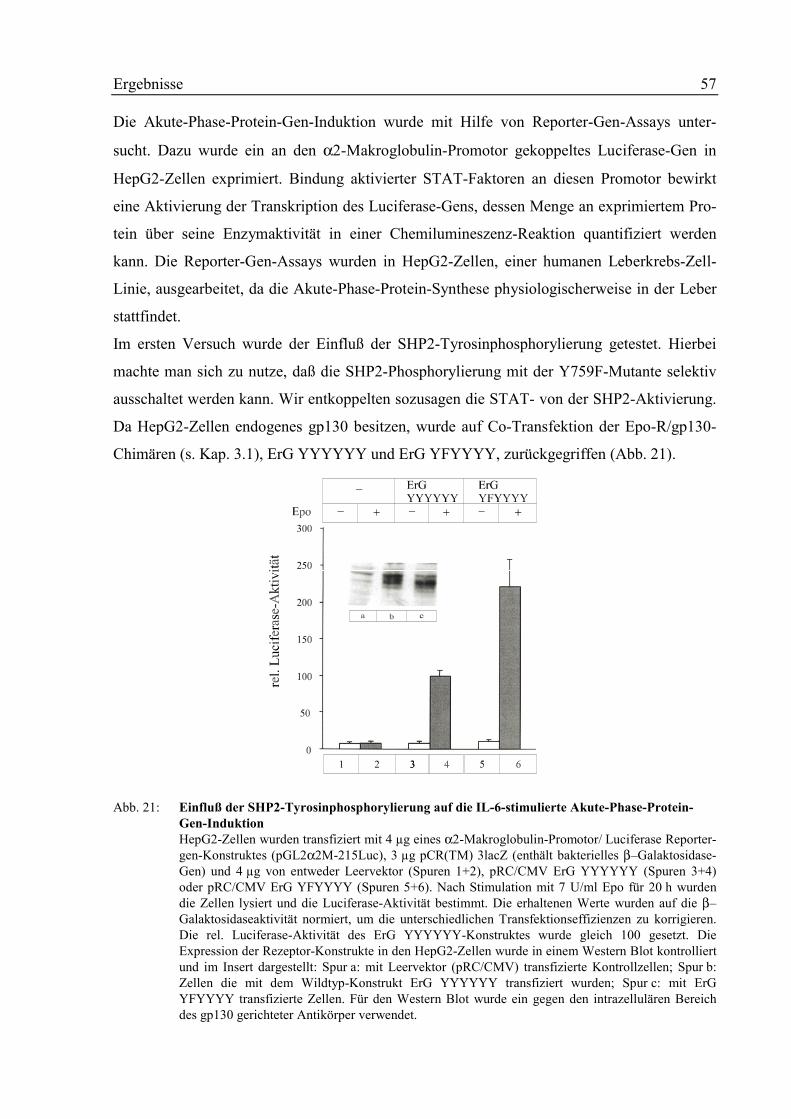
Ergebnisse 57
Die Akute-Phase-Protein-Gen-Induktion wurde mit Hilfe von Reporter-Gen-Assays unter-
sucht. Dazu wurde ein an den α2-Makroglobulin-Promotor gekoppeltes Luciferase-Gen in
HepG2-Zellen exprimiert. Bindung aktivierter STAT-Faktoren an diesen Promotor bewirkt
eine Aktivierung der Transkription des Luciferase-Gens, dessen Menge an exprimiertem Pro-
tein über seine Enzymaktivität in einer Chemilumineszenz-Reaktion quantifiziert werden
kann. Die Reporter-Gen-Assays wurden in HepG2-Zellen, einer humanen Leberkrebs-Zell-
Linie, ausgearbeitet, da die Akute-Phase-Protein-Synthese physiologischerweise in der Leber
stattfindet.
Im ersten Versuch wurde der Einfluß der SHP2-Tyrosinphosphorylierung getestet. Hierbei
machte man sich zu nutze, daß die SHP2-Phosphorylierung mit der Y759F-Mutante selektiv
ausschaltet werden kann. Wir entkoppelten sozusagen die STAT- von der SHP2-Aktivierung.
Da HepG2-Zellen endogenes gp130 besitzen, wurde auf Co-Transfektion der Epo-R/gp130-
Chimären (s. Kap. 3.1), ErG YYYYYY und ErG YFYYYY, zurückgegriffen (Abb. 21).
Abb. 21: Einfluß der SHP2-Tyrosinphosphorylierung auf die IL-6-stimulierte Akute-Phase-Protein-Gen-InduktionHepG2-Zellen wurden transfiziert mit 4 µg eines α2-Makroglobulin-Promotor/ Luciferase Reporter-gen-Konstruktes (pGL2α2M-215Luc), 3 µg pCR(TM) 3lacZ (enthält bakterielles β–Galaktosidase-Gen) und 4 µg von entweder Leervektor (Spuren 1+2), pRC/CMV ErG YYYYYY (Spuren 3+4)oder pRC/CMV ErG YFYYYY (Spuren 5+6). Nach Stimulation mit 7 U/ml Epo für 20 h wurdendie Zellen lysiert und die Luciferase-Aktivität bestimmt. Die erhaltenen Werte wurden auf die β–Galaktosidaseaktivität normiert, um die unterschiedlichen Transfektionseffizienzen zu korrigieren.Die rel. Luciferase-Aktivität des ErG YYYYYY-Konstruktes wurde gleich 100 gesetzt. DieExpression der Rezeptor-Konstrukte in den HepG2-Zellen wurde in einem Western Blot kontrolliertund im Insert dargestellt: Spur a: mit Leervektor (pRC/CMV) transfizierte Kontrollzellen; Spur b:Zellen die mit dem Wildtyp-Konstrukt ErG YYYYYY transfiziert wurden; Spur c: mit ErGYFYYYY transfizierte Zellen. Für den Western Blot wurde ein gegen den intrazellulären Bereichdes gp130 gerichteter Antikörper verwendet.
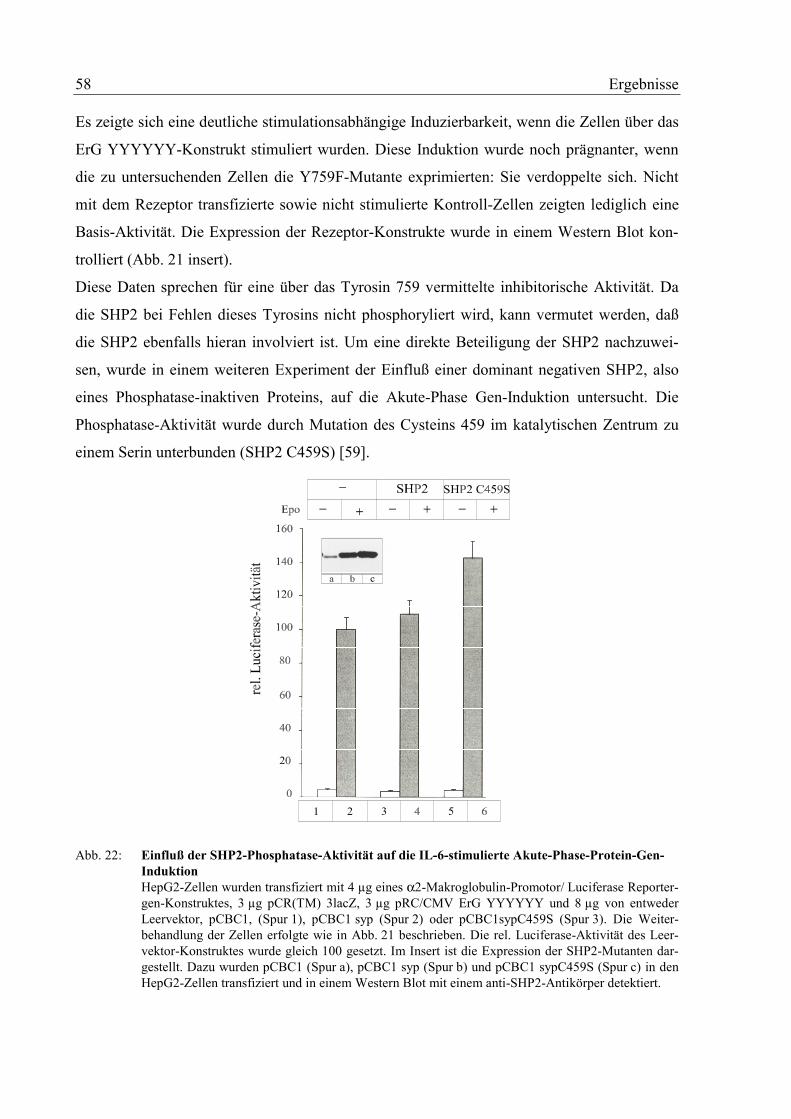
58 Ergebnisse
Es zeigte sich eine deutliche stimulationsabhängige Induzierbarkeit, wenn die Zellen über das
ErG YYYYYY-Konstrukt stimuliert wurden. Diese Induktion wurde noch prägnanter, wenn
die zu untersuchenden Zellen die Y759F-Mutante exprimierten: Sie verdoppelte sich. Nicht
mit dem Rezeptor transfizierte sowie nicht stimulierte Kontroll-Zellen zeigten lediglich eine
Basis-Aktivität. Die Expression der Rezeptor-Konstrukte wurde in einem Western Blot kon-
trolliert (Abb. 21 insert).
Diese Daten sprechen für eine über das Tyrosin 759 vermittelte inhibitorische Aktivität. Da
die SHP2 bei Fehlen dieses Tyrosins nicht phosphoryliert wird, kann vermutet werden, daß
die SHP2 ebenfalls hieran involviert ist. Um eine direkte Beteiligung der SHP2 nachzuwei-
sen, wurde in einem weiteren Experiment der Einfluß einer dominant negativen SHP2, also
eines Phosphatase-inaktiven Proteins, auf die Akute-Phase Gen-Induktion untersucht. Die
Phosphatase-Aktivität wurde durch Mutation des Cysteins 459 im katalytischen Zentrum zu
einem Serin unterbunden (SHP2 C459S) [59].
Abb. 22: Einfluß der SHP2-Phosphatase-Aktivität auf die IL-6-stimulierte Akute-Phase-Protein-Gen-InduktionHepG2-Zellen wurden transfiziert mit 4 µg eines α2-Makroglobulin-Promotor/ Luciferase Reporter-gen-Konstruktes, 3 µg pCR(TM) 3lacZ, 3 µg pRC/CMV ErG YYYYYY und 8 µg von entwederLeervektor, pCBC1, (Spur 1), pCBC1 syp (Spur 2) oder pCBC1sypC459S (Spur 3). Die Weiter-behandlung der Zellen erfolgte wie in Abb. 21 beschrieben. Die rel. Luciferase-Aktivität des Leer-vektor-Konstruktes wurde gleich 100 gesetzt. Im Insert ist die Expression der SHP2-Mutanten dar-gestellt. Dazu wurden pCBC1 (Spur a), pCBC1 syp (Spur b) und pCBC1 sypC459S (Spur c) in denHepG2-Zellen transfiziert und in einem Western Blot mit einem anti-SHP2-Antikörper detektiert.

Ergebnisse 59
Dazu wurde in HepG2, die mit dem Epo-R/gp130-Konstrukt ErG YYYYYY und zusätzlich
entweder mit der Wildtyp-SHP2- oder der SHP2 C459S-cDNA transfiziert wurden, die Akti-
vierung der Transkription des ebenfalls transfizierten Reportergens gemessen (Abb. 22). Die
Expression der SHP2-Varianten wurde in einem Western Blot kontrolliert (Abb. 22 insert).
Kontroll-Zellen, die nur mit den ErG YYYYYY-Konstrukten transfiziert wurden, zeigten die
bereits oben gezeigte Induktion nach Epo-Stimulation. Eine zusätzliche SHP2-Transfektion
erhöhte die Induktion nicht wesentlich, obwohl in einem Kontroll-Western Blot eine größere
Menge an SHP2-Protein nachweisbar war. Wenn jedoch SHP2 C459S- cDNA co-tranfiziert
wurde konnte eine geringe, jedoch signifikant gesteigerte Geninduktion beobachtet werden.
Diese Ergebnisse legen nahe, daß die enzymatische Aktivität der SHP2 und nicht nur das
Protein an sich als Adapter für die Funktion dieses Enzyms in der Akute-Phase Gen-Induktion
wichtig ist.

.

Diskussion 61
4. Diskussion
Über Struktur und Funktion der SHP2 wurde in den letzten Jahren viel diskutiert. Im Gegen-
satz zur hämatopoetischen Variante SHP1, für die sich relativ schnell eine negative regulatori-
sche Funktion herausstellte [21], ist die Rolle der SHP2 in der Signaltransduktion noch
weiterhin umstritten. Die hier vorliegenden Ergebnisse deuten eher auf eine ebenfalls negative
regulatorische Funktion der SHP2 hin. Diese Erkenntnisse und weitere Hypothesen und Ge-
dankenansätze zur Rolle der SHP2 in der IL-6-abhängigen Signaltransduktion sollen im Fol-
genden vorgestellt und erarbeitet werden.
Die IL-6-abhängige SHP2-Tyrosinphosphorylierung
Die Tyrosinphosphorylierung der SHP2 ist nach Stimulation der verschiedensten Zytokin-
Rezeptoren beobachtet worden. Zuerst beschrieben als stimulationsabhängiger Interaktions-
partner der Rezeptor-Tyrosin-Kinasen wie EGF-R, PDGF-R oder Insulin-R [47, 84] stellte
sich schnell heraus, daß die SHP2 auch im Kontext anderer Rezeptor-Aktivierungen phospho-
ryliert werden kann, so z.B. nach Stimulation mit Epo oder zahlreichen Interleukinen
[12, 64, 82]. Aber auch die Aktivierung von 7TM-Rezeptoren wie der Angiotensin-R führen
zu einer Tyrosinphosphorylierung der SHP2 [4]. Interessanterweise wurde eine SHP2-
Phosphorylierung auch in Zellen beobachtet, die veränderten physikalischen Bedingungen
wie Hyperosmolarität des Mediums ausgesetzt waren [56]. Die SHP2 scheint somit ein fast
„universell“ eingesetztes Werkzeug in der Signaltransduktion zu sein. Dies ist vielleicht der
Grund, weshalb dieses Protein in so einer Vielfalt von Zellen in großer Menge exprimiert
wird.
In der vorliegenden Arbeit erfolgte nun die Untersuchung der SHP2-Tyrosinphosphorylierung
in der IL-6-abhängigen Signaltransduktion im Western Blot. Bei genauer Betrachtung eines
anti-Phosphotyrosin Blots und der entsprechenden anti-SHP2-Gegenfärbung wird deutlich,
daß die jeweiligen Banden nicht absolut deckungsgleich übereinander liegen. Durch „un“-
geschickte Wahl der Auftragsmenge oder SDS-PA-Gelkonzentration bzw. -lauflänge lassen
sich im α–SHP2 Blot sogar Doppelbanden sichtbar machen, von denen die obere kongruent
mit der entsprechenden Bande im anti-Phosphotyrosin Blot ist. Dies spricht dafür, daß die
tyrosinphosphorylierte SHP2 in der Elektrophorese langsamer läuft. Ein Grund hierfür könnte

62 Diskussion
in der Ladungsverschiebung der aktivierten SHP2 liegen, da sie dann vermehrt negative La-
dungen besitzt. Ob eine so deutliche Aufwärtsverschiebung der SHP2 bedeutet, daß sie in der
IL-6-abhängigen Signaltransduktion an mutiplen Tyrosinen phosphoryliert wird bzw. an wel-
chen Tyrosinen sie tatsächlich phosphoryliert wird, bleibt weiter zu untersuchen.
Auch die Zeitabhängigkeit der SHP2-Tyrosinphosphorylierung ist untersucht worden. Fünf
Minuten nach Stimulation ist eine deutliche Phosphorylierung im anti-Phosphotyrosin We-
stern Blot nachweisbar. Nach Stimulation über den PDGF-R soll eine entsprechende Reaktion
sogar schon 1 Minute post stimulationem nachweisbar sein [20]. Wenn auch die Funktion der
SHP2 noch nicht geklärt sein mag, die rasche Phosphorylierung dieser Phosphatase spricht für
eine schnelle Aktivierung und damit für ein unverzügliches Eingreifen in die Signaltransduk-
tion.
Die Dauer der SHP2-Tyrosinphosphorylierung scheint ebenfalls gut reguliert zu sein. Bereits
40 min nach Stimulation ist keine Tyrosinphosphorylierung mehr nachweisbar. Interessanter-
weise stimmen diese Zeiten genau mit der Kinetik der Phosphorylierung des gp130 überein,
was für eine gemeinsame Regulation beider Proteine sprechen könnte.
Abhängigkeit der IL-6-induzierten SHP2-Tyrosinphosphorylierung vomTyrosin 759 des gp130
Bindungsstudien mit synthetisch hergestellten Phosphotyrosin-Peptiden ergaben für die N-
terminale SH2-Domäne der SHP2 ein spezifisches Bindungsmotiv, pY-X-X-V [14]. Auch
gp130 besitzt an Position 759-762 mit der Aminosäuresequenz YSTV ein solches Motiv. Dies
legt die Vermutung nahe, daß auch gp130 in der Lage ist, SHP2 zu rekrutieren und zu aktivie-
ren. Mehreren Autoren ist es bereits gelungen, die Abhängigkeit der SHP2-Phosphorylierung
vom Tyrosin 759 des gp130 nachzuweisen. Hierfür wurden jedoch stets verkürzte oder chimä-
re Rezeptoren benutzt [26, 77]. Dank der stabil gp130 exprimierenden Ba/F3-Mutanten
konnte nun erstmals die Abhängigkeit der SHP2-Tyrosinphosphorylierung von Y759 des
gp130 an einem kompletten, nativen gp130-Molekül gezeigt werden. Die vorbeschriebenen
Ergebnisse bestätigten sich. Obwohl Wildtyp-Rezeptoren und Y759F-Mutanten tragende
Zellen etwa die gleiche Anzahl an Rezeptoren exprimierten, konnte nur beim Wild-Typ eine
stimulationsabhängige SHP2-Tyrosinphosphorylierung nachgewiesen werden. Mutation des
Tyrosins 759 zu Phenylalanin führt zu einer kompletten Auslöschung der Phosphorylierung.
Als Kontrolle dieses Experimentes ist es wichtig, neben dem Nachweis einer äquivalenten

Diskussion 63
Expression der Mutanten auch deren erhaltene Reaktionsfähigkeit zu überprüfen, besonders
bei der in ihrer Aminosäuresequenz veränderten GrG YFYYYY-Mutante. Trotz des fehlenden
Tyrosins an Position 759 in der Mutante werden beide Konstrukte weiterhin stimulationsab-
hängig phosphoryliert und scheinen damit prinzipiell fähig zu sein, ihre Funktion in der Si-
gnaltransduktion wahrnehmen zu können.
Interessanterweise ließ sich die SHP2-Tyrosinphosphorylierung nicht in Zellen nachweisen,
die nur transient mit den Epo-R/gp130-Mutanten transfiziert worden sind, obwohl deren Ex-
pression sowohl in Western Blots, in Immunfluoreszenzen [73] als auch nach Abschluß dieser
Arbeit in FACS-Analysen gezeigt werden konnte. Gleichermaßen zeigen EMSA-Studien mit
transient transfizierten gp130-Mutanten in COS7-Zellen die biologische Funktionalität der
Rezeptoren [73]. Das Problem des Nachweises der SHP2-Tyrosinphosphorylierung bei Sti-
mulation über die transient transfizierten Rezeptoren scheint ein ungünstiges Mengenverhält-
nis zwischen phosphorylierter bzw. nicht phosphorylierter SHP2 zu sein. Auf der einen Seite
existiert eine große Menge an SHP2 in den Zellen, auf der anderen wird bei Stimulation über
die heterolog exprimierten Rezeptoren nur eine geringe Menge dieser SHP2 tyrosinphospho-
ryliert, nämlich der Anteil, der aus den Zellen stammt, die die Rezeptoren tragen. Der pro-
zentuale Anteil phosphorylierter SHP2 in einem Zell-Lysat ist also gering. Bei der
nachfolgenden Immunpräzipitation gewinnt man entsprechend zu wenig phosphoryliertes
Protein, als daß man es nachweisen könnte. Bei den stabilen gp130 exprimierenden Ba/F3-
Zellen hingegen sollte jede Zelle die Rezeptoren tragen. Es wird entsprechend ein größerer
Anteil der SHP2 phosphoryliert, der dann nachweisbar ist. Ein weiterer Lösungsansatz dieses
Problems war die zusätzliche transiente Expression einer mit dem c-myc-Epitop markierten
SHP2. In der Annahme, daß jede Zelle, die die cDNA für die Rezeptor-Mutanten erhalten
habe, auch das SHP2-myc-Konstrukt trage, ließe sich mit einer Präzipitation über das c-myc-
Epitop gezielt der Anteil der SHP2 präzipitieren, der aus transfizierten Zellen stammt. Da-
durch würde man den Hintergrund, also SHP2-Moleküle aus nicht transfizierten Zellen, redu-
zieren. Leider war auch hier die Transfektionseffizienz gering und die Präzipitationsfähigkeit
des myc-Antikörpers so schwach, daß zu wenig SHP2-myc präzipitiert wurde, um eine Tyro-
sinphosphorylierung der SHP2 nachzuweisen.

64 Diskussion
Funktion der SHP2-Tyrosinphosphorylierung
Bis jetzt wurden die Eigenschaften und Rahmenbedingungen der SHP2-Tyrosinphosphory-
lierung genauer betrachtet. Wichtiger und interessanter ist jedoch die Funktion der SHP2 in
der IL-6-abhängigen Signaltransduktion.
Bisherige Veröffentlichungen zu diesem Thema sind widersprüchlich. Häufig wurde die
SHP2 nur als Adapterprotein beschrieben, das in der Lage ist, das ankommende Signal auf
mehreren Signalwegen weiterzuleiten. Mit ihren SH2-Domänen interagiert sie mit dem je-
weiligen aktivierenden Rezeptor und leitet über Bindung an Grb2 die Signalkette weiter zur
Ras/Raf/MAPK-Kaskade. Die Enzymaktivität der Phosphatase scheint für diese Funktion der
SHP2 nur von untergeordneter Rolle zu sein. Pazdrak et al. [64] fanden, daß die SHP2 an der
gemeinsamen IL-5-β-Kette aktiviert wird, und daß diese Aktivierung zu einem verlängertem
Überleben von eosinophilen Granulozyten führt. Dies entspricht einer positiven Regulations-
funktion der SHP2. Fukada et al. [26] konnten belegen, daß die stabile Expression von gp130
in Ba/F3-Zellen zu einem IL-6- abhängigen Wachstum dieser Zellen führt. Auch konnten sie
in weiterführenden Untersuchungen an diesem System zeigen, daß SHP2 bzw. ihre Rekrutie-
rung an den Rezeptor entscheidend an der Auslösung des mitogenen Signals in diesen Zellen
beteiligt ist, während die STAT3-Aktivierung das anti-apoptotische Signal auslöst. Auch dies
entspricht einer positiven Regulationsfunktion durch die SHP2.
Die hier vorliegenden Ergebnisse legen eine eher negative Regulation der IL-6-
Signaltransduktion durch die SHP2 nahe. Mutation des Tyrosins 759 zu Phenylalanin führt
zu einer verstärkten und verlängerten STAT-Phosphorylierung bzw. -Aktivierung, die sowohl
in Western Blots als auch in EMSA-Experimenten nachweisbar ist. Da der Austausch des
Tyrosins 759 zu Phenylalanin zudem die Nicht-Phosphorylierung der SHP2 zur Folge hat,
liegt der Schluß nahe, die SHP2 selbst bzw. ihre Tyrosinphosphorylierung beeinflusse die
STAT-Aktivierung, und zwar in einer negativ regulierenden, beschränkenden Weise. Wie
STAT-Faktoren und SHP2 miteinander interagieren können, und ob es vielleicht sogar eine
Enzym-Substrat-Beziehung zwischen SHP2 und STAT-Faktoren gibt, ist bisher noch nicht
erfolgreich untersucht worden. Termination der STAT-Phosphorylierung durch die SHP2-
Phosphatase wäre die schnellste und eine durchaus attraktive Deutung dieser Ergebnisse. Sie
ist jedoch nicht notwendigerweise impliziert. Es wäre ebenfalls denkbar, daß die SHP2 als
Adapterprotein weitere Phosphatasen an den Rezeptorkomplex rekrutieren könnte, oder daß

Diskussion 65
sie durch Dephosphorylierung anderer Signaltransduktionsteilnehmer eine verlängerte STAT-
Aktivierung verhindert. Aber auch andere – noch unbekannte – Y759-abhängige Reaktionen,
die zu einer Verkürzung der STAT-Phosphorylierung führen, sind vorstellbar.
Aussagen über die SHP2-Enzymaktivität und deren Funktion in der Signaltransduktion ge-
stalten sich bisher noch schwierig. Es gibt zu diesem Thema schon etliche Veröffentlichun-
gen, die jedoch noch kein schlüssiges Bild liefern und sich zum Teil widersprechen. Dies liegt
mit Sicherheit auch an der Tatsache, daß bisher noch keine Einigkeit über potentielle Sub-
strate der SHP2 herrscht. Ein Beispiel dafür ist der PDGF-Rezeptor. Vogel et al. [84] meinen
nachweisen zu können, daß die SHP2 zwar durch diesen Rezeptor aktiviert wird, jedoch kei-
nen Einfluß auf seinen Phosphorylierungszustand habe. Dechert et al. [18] schildern hingegen
eben diese Dephosphorylierung des Rezeptors durch die SHP2. Wenn die SHP2 nun wirklich
in der Lage ist, den sie aktivierenden PDGF-R zu dephosphorylieren, käme dies einer negati-
ven regulatorischen Funktion der SHP2 gleich. Die SHP2 würde das Signal terminieren. Eine
weitere Möglichkeit der SHP2, regulierend in die Signaltransduktion einzugreifen, zeigen
Stein-Gerlach et al. [78]. Diese Autoren weisen nach, daß die SHP2 imstande ist, ihren eige-
nen C-Terminus zu dephosphorylieren. Dies bedeutet, daß die SHP2 ihren eigenen Phospho-
rylierungsstatus beeinflußt und so an der Regulation des Signals mitwirkt.
In der hier vorliegenden Arbeit konnte nachgewiesen werden, daß Nicht-Phosphorylierung
der SHP2 mit einer gesteigerten und verlängerten STAT-Aktivierung einhergeht. Im folgen-
den wurde untersucht, ob und welchen Einfluß diese Nicht-Phosphorylierung der SHP2
auf STAT-abhängige Funktionen wie die Akute-Phase-Protein-Gen-Induktion hat.
Es konnte gezeigt werden, daß Stimulation über die Y759F-Mutante und damit die Aus-
schaltung der SHP2-Tyrosinphosphorylierung mit einer erhöhten Aktivierung eines Reporter-
genes einhergeht, welches den STAT1- bzw. STAT3-responsiven α2-Makroglobulin-Pro-
motor besitzt. Dies bedeutet, daß die SHP2 negativ regulierend in die IL-6-abhängige Signal-
transduktion eingreift. Inwiefern an diesem Ergebnis das Protein SHP2 an sich oder seine
Enzymaktivität wichtig sind (und nicht vielleicht eine andere Tyrosin 759-abhängige Reakti-
on), kann durch diesen Versuch noch nicht eindeutig geklärt werden. Um dies zu ergründen,
wurde eine Phosphatase-inaktive, dominant negativ wirkende SHP2-Mutante, SHP2 C459S,
in den Versuchszellen überexprimiert. In den folgenden Reportergen-Assays zeigte sich so-
dann eine gesteigerte Geninduktion. Es liegt also nahe, den beobachteten inhibitorischen Ef-

66 Diskussion
fekt der SHP2-Phosphatase-Aktivität zuzuschreiben und nicht dem einfachen Vorhandensein
eines phosphorylierten Proteins, welches als Adapterprotein bzw. als Andockstelle für weitere
Komponenten der Signaltransduktion dient. Diese Versuche schließen allerdings keineswegs
aus, daß die SHP2 nicht auch als Adapterprotein fungieren kann. Es wird lediglich bewiesen,
daß die Enzymaktivität an der Funktion der SHP2 in der IL-6- abhängigen Signaltransduktion
beteiligt ist.
Diese Ergebnisse decken sich mit denen, die Symes et al. [79] in ähnlichen Versuchen erhal-
ten haben. Diese Autoren finden eine gesteigerte Geninduktion bei Überexpression der SHP2-
SH2-Domänen, welche sie als dominant negativ beschreiben. Diese Bruchstücke, bei denen
fast zwei Drittel des Proteins – inklusive der Phosphatase-Domäne – fehlen, als dominant
negativ, also als nur seiner Enzymaktivität beraubt, zu bezeichnen, ist jedoch sehr gewagt.
Hier ist nicht eindeutig, ob die Gen-Induktion aufgrund fehlender Andockstellen für Adapter-
proteine, die sich ebenfalls in der Phosphatase-Domäne befinden, gesteigert ist oder durch die
fehlende Phosphatase-Aktivität. In den hier vorliegenden Versuchen konzentrierte man sich
vollkommen auf die Enzymaktivität der SHP2. Das Protein wurde in nur einer Aminosäure
verändert. Damit sind alle Bindungsstellen für andockende Proteine erhalten geblieben und
nur die fehlende Phosphatase-Aktivität kann für die Induktion verantwortlich sein.
Zusammenfassend bedeutet dies also, daß die Enzym-Aktivität der SHP2 – im Gegensatz zu
der weitläufigen Meinung, die SHP2 wäre nur eine Adapterprotein – für die IL-6-abhängige
Signaltransduktion durchaus eine wichtige Rolle spielt. Sie wirkt hemmend auf die Akute-
Phase-Protein-Gen-Induktion.
Notwendigkeit der Janus-Kinasen für die SHP2-Tyrosinphosphorylierung
Sowohl IL-6 als auch die anderen IL-6-Typ-Zytokine führen zu einer Aktivierung der Janus-
Kinasen Jak1, Jak2 und Tyk2. Eine besondere Rolle in der IL-6-abhängigen Signaltransduk-
tion kommt dabei Jak1 zu, da die Eliminierung der Jak1-Expression zu einer Ausschaltung
der stimulationsabhängigen Tyrosinphosphorylierung von gp130 und den STAT-Faktoren
führt [30]. Die hier vorliegenden Ergebnisse zur Rolle der Janus-Kinasen in der SHP2-
Phosphorylierung sprechen ebenfalls Jak1 eine entscheidene Rolle zu. In U4A-Zellen, denen
Jak1 fehlt, findet man eine nur noch residuale Tyrosinphosphorylierung der SHP2. Stabile Re-
Transfektion einer Jak1-cDNA in diese Zellen stellt die vollständige Reaktionsfähigkeit der
SHP2 auf IL-6-Stimulation wieder her. Bei Jak2- bzw. Tyk2-defizienten Zellen und den Ki-

Diskussion 67
nase-retransfizierten Kontroll-Zellen ist die stimulationsabhängige SHP2-Tyrosinphospho-
rylierung jedoch nachweisbar.
Die hervorgehobene Rolle der Jak1 muß nicht bedeuten, daß diese Kinase die SHP2 direkt
phosphoryliert, welches die einfachste Erklärung für diese Resultate darstellen würde. Jak1
könnte durch Phosphorylierung von gp130, insbesondere des Tyrosins 759, einfach nur die
Voraussetzungen schaffen, daß die SHP2 stimulationsabhängig an den Rezeptor rekrutiert
werden kann. Eine weitere – noch nicht näher identifizierte – Kinase könnte so die SHP2
phosphorylieren.
Eine andere mögliche Erklärung wäre die direkte Aktivierung einer zweiten Kinase durch
Jak1. Für die Rolle dieser zweiten Kinase kommen verschiedene Kandidaten in Frage. So ist
z.B. nach gp130-Stimulierung in einigen Zell-Linien die Aktivierung der Tyrosinkinasen Hck,
Fes, Btk und Tec beschrieben worden [57]. Aber auch Jak2 bzw. Tyk2 sind in Erwägung zu
ziehen. Die Tatsache, daß weder die Auschaltung der Jak2- noch der Tyk2-Expression die
SHP2-Tyrosinphosphorylierung antastet, muß nicht als Widerspruch zu dieser Interpretation
gelten. Die mangelnde Beeinflußbarkeit der SHP2-Phosphorylierung durch diese Kinasen
könnte mit der Annahme erklärt werden, daß beide Kinasen sich in ihrer Funktion gegenseitig
ersetzen können. Wer die SHP2 letzten Endes phosphoryliert, ist also noch nicht geklärt.
Die Expression einer Kinase-inaktiven Mutante, Jak1K→E, in die Jak1 defizienten U4A-
Zellen ist nicht in der Lage, die insuffiziente SHP2-Tyrosinphosphorylierung in den U4A-
Zellen wiederherzustellen. Dies verdeutlicht, daß nicht das Jak1-Protein an sich, sondern sei-
ne Enzymaktivität für die Signalweiterleitung wichtig ist. Die oben gestellte Frage nach einer
weiteren Kinase bleibt davon jedoch unbeantwortet.
Assoziation der SHP2 mit den Janus-Kinasen
Über die Assoziation von SHP2 mit Kinasen der Jak-Familie ist schon einiges berichtet wor-
den. Fuhrer et al. [24] konnten eine konstitutive Verbindung zwischen SHP2 und Jak2 in
3T3-L1 Maus-Präadipozyten nachweisen, die sich nach IL-11-Stimulation weiter induzieren
läßt. In einer nachfolgenden Veröffentlichung [93] zeigten SHP2/ Jak-Co-Transfektions-
experimente in COS1-Zellen, daß SHP2 mit Jak1 und Jak2, nicht aber mit Jak3 assoziiert.
Nachteil bei diesen Experimenten war allerdings, daß die Überexpression der Janus-Kinasen
dazu führte, daß diese konstitutiv zahlreiche Proteine inklusive sich selbst phosphorylierten.

68 Diskussion
Eine Aussage über die Stimulationsabhängigkeit bzw. den notwendigen Phosphorylierungs-
status der Proteine für diese Assoziation ist somit nicht möglich. Ziel sollte es also sein, in
stimulationsfähigen Zellsystemen zu arbeiten.
Die hier vorgestellten Versuche wurden in den bereits mehrfach erwähnten Fibrosarkom-Zell-
Linien durchgeführt, deren IL-6-abhängige Reaktionsfähigkeit bereits an anderer Stelle nach-
gewiesen wurde [32]. Zudem bieten diese Zellen den Vorteil, mit endogener SHP2 arbeiten zu
können. Es konnte gezeigt werden, daß SHP2 konstitutiv mit Jak1, Jak2 und Tyk2 interagiert.
Stimulation mit IL-6/sIL-6R beeinflusste die Protein-Protein-Interaktion nicht.
Diese Assoziation wird um so vielschichtiger, wenn man berücksichtigt, daß gp130 ebenfalls
als Interaktionspartner von SHP2 und den Janus-Kinasen wirkt. Es stellt sich die Frage, wie,
wann und wo in der Zelle diese drei Faktoren miteinander in Verbindung stehen.
Es ist gezeigt worden, daß gp130 konstitutiv mit den Janus-Kinasen assoziiert ist [53, 76].
Gleichzeitig konnte nachgewiesen werden, daß gp130 und SHP2 ebenfalls in Verbindung
stehen. Diese Assoziation ist jedoch stimulationsabhängig. Kombiniert man diese beiden Be-
funde, kommt man zu dem Schluß, daß SHP2 und die Janus-Kinasen nur im Falle einer Sti-
mulation mit IL-6 im IL-6-Rezeptor-Komplex miteinander interagieren dürften. Als
Verbindungsstück fungiert dann das gp130-Molekül. Dies steht jedoch im Widerspruch zu
den hier vorgestellten Ergebnissen. Hier wurde gezeigt, daß SHP2 und Janus-Kinasen stimu-
lationsunabhängig assoziieren. Beide Daten können nur so in Einklang gebracht werden,
wenn man einen zweiten, nicht an gp130 assoziierten Pool an Janus-Kinasen definiert. Dieser
vermutlich im Cytosol liegende Teil ist in der Lage, die SHP2 direkt, also gp130-unabhängig
zu binden. Gestützt wird diese Interpretation durch die Versuche von Fuhrer et al. [24]. Diese
Autoren zeigten, daß phosphorylierte YSTV-Peptide in der Lage sind, die SHP2-gp130-
Assoziation zu kompetitieren. Die Interaktion von Jak2 und SHP2 blieb davon unbeeinflußt.
Dort sind durch Versuche mit Deletionsmutanten auch mögliche Interaktionsstellen in den
verschiedene Molekülen eingegrenzt worden. In der SHP2 konnte der Bereich zwischen den
Aminosäuren 232-272, der vor der Phosphatase-Domäne liegt, als für die Assoziation mit
Jak2 essentiell herausgestellt werden. Die entsprechende Bindungsstelle im Jak-Molekül liegt
im N-terminalen Bereich. Zusammenfassend bedeutet dies, daß die Janus-Kinasen entweder
mit gp130 an der Zellmembran oder mit der SHP2 im Cytosol assoziiert sind (Abb. 23).
Die Frage nach dem Sinn dieser Komplexbildung, welche eine Assoziation zweier gegensätz-
lich wirkender Enzyme, Kinase ↔ Phosphatase, darstellt, kann nicht befriedigend beantwortet
werden: Mögliche regulatorische Funktion? Rekrutierung der SHP2 an gp130 durch die Ja-

Diskussion 69
nus-Kinasen? Auch eine eventuell bestehende Substrat-Enzym-Beziehung zwischen den Part-
nern ist zur Zeit noch unklar, ebenso wie eine Verbindung beider Proteine im IL-6-Rezeptor-
Komplex. In dieser Arbeit wurde nachgewiesen, daß weder Jak2 noch Tyk2 in der Lage sind,
eine durch Fehlen von Jak1 entstandene mangelhafte SHP2-Phosphorylierung zu rekonstituie-
ren. Auf der anderen Seite sind aber alle drei Kinasen in der Lage, die SHP2 konstitutiv zu
binden, jedoch ohne Auswirkung auf den Phosphorylierungsstatus der SHP2.
Abb. 23: Komplexbildung von gp130, Janus-Kinasen und SHP2 und deren mögliche Lokalisation in derZelleDie Janus-Kinasen sind konstitutiv mit gp130 assoziiert. Ebenso binden sie stimulationsunabhängigan die SHP2. Diese Bindung ist gp130-unabhängig. Im IL-6-Rezeptor-Komplex interagieren alledrei Partner miteinander. Verbindungsglied zwischen SHP2 und Janus-Kinasen ist gp130.
4.1 Ausblick:
Der Rezeptor-Tyrosinmotiv-Baukasten
Durch Einfügen neuer Restriktionsschnittstellen als stille Mutationen in den zytoplasmati-
schen Teil des gp130 bzw. durch gleichzeitigen Austausch aller Tyrosine gegen Phenylalani-
ne bei einer der Mutanten wurde eine Art Baukasten geschaffen. Mit Hilfe dieses Baukastens
ist es relativ leicht möglich, beliebige Kombinationen von Y→F-Mutanten herzustellen und in
weiterführenden Experimenten zu untersuchen. Zum Beispiel ließe sich durch jeweiliges Re-

70 Diskussion
vertieren eines einzelnen Tyrosins in der Phenylalanin-Mutante ermitteln, welche der sechs
Tyrosinmotive bei der Aktivierung des gp130 tatsächlich phosphoryliert werden. Dazu müß-
ten die Rezeptor-Mutanten stabil in Ba/F3-Zellen exprimiert werden und könnten anschlie-
ßend in Phosphotyrosin Blots analysiert werden. Man könnte auch mit Hilfe dieser Mutanten
untersuchen, ob diejenigen Tyrosinmotive im gp130, die teilweise das Potential haben,
STAT-Faktoren zu aktivieren, sich synergistisch auf die STAT-Aktivierung auswirken. Dies
wäre leicht in Reporter-Gen-Assays und EMSAs zu prüfen. Auch der extrazelluläre Teil der
Mutanten ist durch gleiche Restriktionsschnittstellen einfach austauschbar. Es stehen mehrere
extrazelluläre Domänen zur Verfügung: Die des Epo-R, IL-5-R und Wildtyp-gp130. Mit Hilfe
einer unterschiedlichen Kombination von Y→F-Tyrosinmotiven im Zellinneren und ver-
schiedener extrazellulärer Domänen könnte man untersuchen, ob die verschiedenen Signal-
transduktionsschritte (STAT-Aktivierung zum einen, SHP2-Tyrosinphoshorylierung auf der
anderen Seite) an einen einzigen Rezeptorkomplex gebunden sind oder ob es ausreicht, diese
Proteine an unterschiedlichen Chimären aktiviert zur Verfügung zu stellen.
Sind beide Tyrosine 759 im IL-6-Rezeptor-Komplex für die Tyrosinphosphorylierung
der SHP2 notwendig?
In der vorliegenden Arbeit konnte gezeigt werden, daß das Y759 des gp130 essentiell für die
Tyrosinphosphorylierung der SHP2 in der IL-6- abhängigen Signaltransduktion ist. Es ist be-
kannt, daß zwei gp130-Moleküle nach IL-6-Stimulation homodimerisieren. In einem Rezep-
torkomplex befinden sich demnach zwei Tyrosine 759. Interessant wäre nun herauszufinden,
ob wirklich beide Tyrosine für die SHP2-Aktivierung notwendig sind oder ob ein einziges
ausreicht. Dies kann man mit dem hier aufgebauten Assay zur Untersuchung der SHP2-
Tyrosinphosphorylierung gut untersuchen, wenn man den intrazellulär gelegenen Teil der
dargestellten ErG-Mutanten mit einem System mit hetero-oligomeren extrazellulären Anteil
verbindet, z.B. dem IL-5-R-System bestehend aus einem IL-5α-R und einer „common β
chain“.
Weitere Fragen, die sich in diesem Zusammenhang stellen, sind z.B. Fragen nach einer mög-
lichen Regulation der SHP2 durch ihre SH2-Domänen, oder anders formuliert: Bedeutet die
Belegung beider SH2-Domänen eine andere (verstärkte?, verlängerte?) Aktivierung der SHP2
als die Besetzung einer einzelnen SH2-Domäne? Wenn ja, wie muß diese zweite SH2-
Domänen-Bindungsstelle beschaffen sein? Muß sie der Konsensussequenz pY X X V folgen
oder reicht ein anderes vorhandenes Tyrosinmotiv? Zwei Lösungsansätze sind hier vorstell-

Diskussion 71
bar. Entweder brauchen beide Domänen ein Tyrosinmotiv analog des Y759 des gp130. Dann
sind für eine maximal aktivierte SHP2 zwei gp130-Moleküle notwendig. Oder die anderen
Tyrosinmotive im gp130 besitzen ebenfalls das Vermögen, durch Interaktion mit der C-
terminalen SH2-Domäne die Aktivierung der SHP2 im gp130-Kontext zu regulieren. Dies
würde bedeuten, daß für eine maximale Stimulation der SHP2 nur ein gp130 mit mindestens
zwei Tyrosinmotiven nötig ist. Dies könnte man mit Hilfe des hier etablierten Assays durch
den direkten Vergleich der SHP2-Aktivierung an verschiedenen Mutanten im IL-5-R-Sytem
untersuchen, in dem man mit Hilfe des Rezeptor-Tyrosinmotiv-Baukastens entsprechende
Mutanten (z.B. 5α/βrG FYFFFF und 5αrG FYYFFF) herstellt und im Phosphotyrosin Blot
überprüft.
Welche Tyrosine der SHP2 werden im IL-6-Kontext tyrosinphosphoryliert?
Es ist bereits beschrieben, daß die SHP2 je nach stimulierendem Zytokin an unterschiedlichen
Tyrosinen innerhalb des Proteins phosphoryliert werden kann. Durch Mutation sämtlicher
Tyrosine zu Phenylalanin und anschließender Reversion eines einzelnen Tyrosins könnte nach
entsprechender Expression in eukaryonten Zellen eine eventuelle stimulationsabhängige
Phosphorylierung im anti-Phosphotyrosin-Western Blot ausbleiben. Dies wäre ein Beleg da-
für, daß dieses Tyrosin im IL-6-Kontext phosphoryliert würde. Auch könnte man mit diesen
Mutanten untersuchen, ob eventuelle Phosphotyrosin-abhängige Andockstellen für andere
Proteine zerstört werden, was die Adapterfunktion der SHP2 erhellen würde.
Bedeutet Phosphorylierung der SHP2 Aktivierung der Enzymaktivität?
Es ist nicht selbstverständlich anzunehmen, daß die Phosphorylierung der SHP2 gleichzeitig
bedeutet, daß dieses Enzym dadurch tatsächlich aktiviert wird. Man könnte untersuchen, wie
Tyrosinphosphorylierung und Aktivierung des Enzyms zusammenhängen. Dies könnte zum
Beispiel durch Modifikation von bereits beschriebenen Phosphatase-Assays geschehen. Wie
hier gezeigt, kann man mit Hilfe der Stimulation über die Y759F-Mutante selektiv die SHP2-
Tyrosinphosphorylierung ausschalten. Interessant wäre zu untersuchen, ob und wie sich die
Enzymaktivität der SHP2 verändert, wenn sie über die Mutante aktiviert bzw. nicht aktiviert
wird.

.

Zusammenfassung 73
5. Zusammenfassung
Im Jahre 1993 wurde eine Protein-Tyrosinphosphatase, die SH2 domain containing protein-
tyrosine-phosphatase 2 (SHP2), entdeckt, die in einer Vielzahl von verschiedenen Signal-
transduktionen eine Rolle zu spielen scheint. Ziel der hier vorliegenden Arbeit war es, die Be-
deutung der Tyrosinphosphorylierung dieses Enzyms in der IL-6-abhängigen Signaltransduk-
tion genauer zu analysieren.
Nach Stimulation mit IL-6 konnte eine transiente Tyrosinphosphorylierung der SHP2 nach-
gewiesen werden. Zudem zeigte sich eine Komplexbildung dieses Proteins mit dem ebenfalls
tyrosinphosphorylierten Signaltransduktor gp130. Interessanterweise wiesen beide Phospho-
rylierungen den gleichen zeitlichen Verlauf auf, was für eine gemeinsame Regulation der
Proteine spricht.
Um die Abhängigkeit dieser SHP2-Tyrosinphosphorylierung von gp130 genauer zu untersu-
chen, wurde eine Reihe von gp130-Rezeptor-Mutanten hergestellt. Auf DNA-Ebene sind die-
se Mutanten so gestaltet, daß aufgrund neu eingefügter Restriktions-Endonukleasen-
Schnittstellen an jedem der sechs intrazellulär vorhandenen Tyrosinmotive ein Y→F-
Austausch möglich ist. Mittels dieser Mutanten, die stabil in Ba/F3-Zellen transfiziert wurden,
ist erstmals die Abhängigkeit der SHP2-Tyrosinphosphorylierung vom Y759 des gp130 an
einem nativen, ungekürzten Rezeptormolekül nachgewiesen worden.
Mit Hilfe von Fibrosarkom-Zellen, die nicht mehr in der Lage sind, jeweils eine der drei an
gp130 assoziierten Janus-Kinasen, Jak1, Jak2 bzw. Tyk2, zu exprimieren, wurde untersucht,
welche Bedeutung die drei obengenannten Kinasen bei der Tyrosinphosphorylierung der
SHP2 haben. Es stellte sich heraus, daß Jak1 essentiell für die IL-6-abhängige SHP2-
Tyrosinphosphorylierung ist. Die Elimination der Expression dieser Kinase führt zu einer le-
diglich residualen Tyrosinphosphorylierung der SHP2. Die beiden anderen Kinasen, Jak2 und
Tyk2, scheinen eine nur untergeordnete bzw. austauschbare Funktion zu haben, obwohl
nachweislich alle drei Kinasen konstitutiv mit SHP2 assoziieren.
Bei der Herausarbeitung möglicher Wirkungen der SHP2 auf die IL-6-abhängige Signaltrans-
duktion wurde beobachtet, daß der Austausch des Tyrosins 759 zu Phenylalanin, und damit
das Ausbleiben der SHP2-Tyrosinphosphorylierung, zu einer verstärkten und verlängerten
STAT-Aktivierung führt. Zudem geht die Stimulation über die Y759F-Mutante mit einer er-
höhten Aktivität eines IL-6-abhängigen Akute-Phase-Protein-Promotors einher. Die hier vor-

74 Zusammenfassung
liegenden Ergebnisse deuten auf eine negative Regulation der Signaltransduktion durch die
SHP2.
Die SHP2 wird häufig als Adapterprotein beschrieben, dessen Enzymaktivität von unterge-
ordneter Rolle zu sein scheint. Reportergen-Assays mit einer dominant negativ wirkenden
SHP2 zeigen jedoch, daß die Enzymaktivität der SHP2 durchaus eine wichtige Rolle in der
IL-6-abhängigen Signaltransduktion spielt. Ihre Ausschaltung geht mit einer erhöhten Indu-
zierbarkeit der untersuchten Promotoren einher.

Anhang 75
6.1 Literaturverzeichnis
1. Adachi M., Ishino M., Torigoe T., Minami Y., Matozaki T., Miyazaki T.,Taniguchi T., Hinoda Y. und Imai K.Interleukin-2 induces tyrosine phosphorylation of SHP-2 through IL-2 receptor betachainOncogene (1997) 14: 1629-1633
2. Adachi M., Sekiya M., Miyachi T., Matsuno K., Hinoda Y., Imai K. und Yachi A.Molecular cloning of a novel protein-tyrosine phosphatase SH-PTP3 with sequencesimilarity to the src-homology region 2FEBS Let. (1992) 314: 335-339
3. Ahmad S., Banville D., Zhao Z., Fischer E.H. und Shen S.H.A widely expressed human protein-tyrosine phosphatase containing src homology 2domainsProc. Natl. Acad. Sci. USA (1993) 90: 2197-2201
4. Ali M.A., Schieffer B., Delafontaine P., Bernstein K.E., Ling B.N. undMarrero M.B.Angiotensin II stimulates tyrosine phosphorylation and activation of insulin receptorsubstrate 1 and protein-tyrosine phosphatase 1D in vascular smooth muscle cellsJ. Biol. Chem. (1997) 272: 12373-12379
5. Andrews N.C. und Faller D.V.A rapid micropreparation technique of DNA-binding proteins from limiting numbersof mammalian cells.Nucl. Acids Res. (1991) 19: 2499
6. Arcone R., Pucci P., Zappacosta F., Fontaine V., Marloni A., Marino G. undCiliberto G.Single-step purification and structural characterization of human interleukin-6produced in Escherichia coli from a T7 RNA polymerase expression vector.Eur. J. Biochem. (1991) 198: 541-547
7. Arkinstall S., Gillieron C., Vialknecht E. und Maundrell K.A negatively regulatory function for the protein tyrosine phosphatase PTP2C revealedby reconstruction of platelet-derived growth factor receptor signalling inSchizosaccharomyces pombeFEBS Lett. (1998) 422: 321-327
8. Bastien L., Ramachandran C., Liu S. und Adam M.Cloning, expression and mutational analysis of SH-PTP2, human protein-tyrosinephosphataseBiochem. Biophys. Res. Comm. (1993) 196: 124-133
9. Bazan J.F.Hematopoietic receptors and helical cytokinesImmunol. Today (1990) 11: 350-354

76 Anhang
10. Bazan J.F.Structural design and molecular evolution of a cytokine receptor superfamilyProc. Natl. Acad. Sci USA (1990) 87: 6934-6938
11. Becker S., Groner B und Müller C.W.Three-dimensional structure of the Stat3beta homodimer bound to DNANature (1998) 394: 145-151
12. Bone H., Dechert U., Jirik F., Schrader J.W. und Welham M.J.SHP1 and SHP2 protein-tyrosine phosphatases associate with beta c after interleukin-3-induced receptor tyrosine phosphorylation – Identification of potential binding sitesand substratesJ. Biol. Chem. (1997) 272: 14470-14476
13. Bradford M.M.A rapid and sensitive method for the quantitation of microgram quantities of proteinsutilizing the principle of protein-dye bindingAnal. Biochem. (1976) 72: 248-254
14. Case R.D., Piccione E., Wolf G., Bennett A.M., Lechleider R.J., Neel B.G. undShoelson S.E.SH-PTP2/Syp SH2 domain binding specifity is defined by direct interactions withplatelet-derived growth factor beta-receptor, epidermal growth factor receptor andinsulin receptor substrate-1-derived phosphopeptidesJ. Biol. Chem. (1994) 269: 10467-10474
15. Chen X., Vinkemeier U., Zhao Y., Jeruzalmi D., Darnell Jr J.E. und Kuriyan J.Crystal structure of a tyrosine phosphorylated STAT-1 dimer bound to DNACell (1998) 93: 827-839
16. Darnell Jr. J.E., Kerr I.M. und Stark G.R.Jak-STAT pathways and transcriptional activation in response to IFNs and otherextracellular signaling proteins.Science (1994) 264: 1415-1421
17. David M., Zhou G., Pine R., Dixon J.E. und Larner A.C.The SH2 domain-containing tyrosine phosphatase PTP1D is required for interferonα/β-induced gene expressionJ. Biol. Chem. (1996) 271: 15862-15865
18. Dechert U., Adam M., Harder K.W., Clark Lewis I. und Jirik F.Characterization of protein tyrosine phosphatase SH-PTP2. Study of phosphopeptidesubstrates and possible regulatory role of SH2 domains.J. Biol. Chem. (1994) 269: 5602-5611
19. Dirks W., Schaper F., Kirchhoff S., Morelle C. und Hauser H.A multifunctional vector family for gene expression in mammalian cellsGene (1994) 387-388
20. Feng G.S., Hui C.C. und Pawson T.SH2-containing phosphotyrosine phosphatase as a target of protein-tyrosine kinasesScience (1993) 259: 1607-1611

Anhang 77
21. Frearson J.A. und Alexander D.R.The role of phosphotyrosine phosphatases in haematopoietic cell signal transductionBioEssays (1997) 19: 417-427
22. Freeman Jr. R.M., Plutzky J. und Neel B.G.Identification of a human src homology 2-containing protein-tyrosine-phosphatase:A putative homolog of Drosophila corkscrew.Proc. Natl. Acad. Sci. USA (1992) 89: 11239-11243
23. Fu X.Y.A transcription factor with SH2 and SH3 domains is directly activated by an interferonα–induced cytoplasmic protein tyrosin kinase(s)Cell (1992) 790: 323-355
24. Fuhrer D.K., Feng G.S. und Yang Y.C.Syp associates with gp130 and Janus kinase 2 in response to interleukin-11 in 3T3-L1mouse preadipocytesJ. Biol. Chem. (1995) 270: 24826-24830
25. Fujioka Y., Matozaki T., Nogushi T., Iwamatsu A., Yamao T., Takahashi N.,Tsuda T. und Kasuga M.A novel membrane glycoprotein, SHPS-1, that binds the SH2-domain-containingtyrosine phosphatase SHP-2 in response to mitogens and cell adhesionMol. Cell. Biol. (1996) 16: 6887-6899
26. Fukada T., Hibi M., Yamanaka Y., Takahashitezuka M., Fujitani Y.,Yamagushi T., Nakajima K. und Hirano T.Two signals are necessary for cell proliferation induced by a cytokine receptor gp130:Involvement of STAT3 in anti-apoptosisImmunity (1996) 5: 449-460
27. Gearing D.P., Thut C.J., VandeBos T., Gimpel S.D., Delaney P.B., King J.,Price V., Cosman D. und Beckman M.P.Leukemia inhibitory factor receptor is structurally related to the IL-6 signal transducer,gp130EMBO J. (1991) 2839-2848
28. Gerhartz, C., Heesel B., Sasse J., Hemman U., Landgraf C.,Schneider-Mergener J., Horn F., Heinrich P.C. und Graeve L.Differential activation of acute phase response factor/STAT3 and STAT1 via thecytoplasmic domain of the interleukin 6 signal transducer gp130. Definition of a novelphosphotyrosine motif mediating STAT1 activationJ. Biol.Chem. (1996) 271: 12991-12998
29. Giordana V., De Falco G., Chiari R., Quinto I., Pelicci P.G., Bartholomew L.,Delmastro P., Gadina M. Scala G.Shc mediates IL-6 signaling by interacting with gp130 and Jak2J. Immunol. (1997) 158: 4097-4103
30. Graeve L., Drickamer K. und Rodriguez-Boulan E.Polarized endocytosis by Mardin-Darby canine kidney cells transfected withfunctional chicken liver glycoprotein receptorJ. Cell. Biol. (1989) 265: 1216-1224

78 Anhang
31. Grötzinger J., Kurapkat G., Wollmer A., Kalai M. und Rose-John S.The familiy of the IL-6 type cytokines: specificity and promiscuty of the receptorcomplexesProteins (1997) 27: 96-109
32. Guschin D., Rogers N., Briscoe J., Witthuhn B., Watling D., Horn F.,Pellegrini S., Yasukawa K., Heinrich P.C., Stark G.R., Ihle J.N. und Kerr I.M.A major role for the protein tyrosine kinase JAK1 in the JAK/STAT signaltransduction pathway in response to interleukin-6EMBO J. (1995) 14: 1421-1429
33. Hanahan D.Studies on transformation of Escherischia coli with plasmidsJ. Mol. Biol. (1983) 166: 557-580
34. Harlow E. und Lane D.Antibodies. A Laboratory ManualCold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
35. Heinrich P.C., Behrmann I., Müller-Newen G., Schaper F. und Graeve L.Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathwayBiochem. J. (1998) 334: 297-314
36. Hemman U., Gerhartz, C., Heesel B., Sasse J., Kurapkat G., Grötzinger J.,Wollmer A., Zhong Z., Darnell Jr. J.E., Graeve L., Heinrich P.C. und Horn F.Differential activation of acute phase response factor/STAT3 and STAT1 via thecytoplasmic domain of the interleukin 6 signal transducer gp130. Src homology SH2domains define the specificity of Stat factor activationJ. Biol. Chem. (1996) 271: 12999-13007
37. Hirano T., Yasukawa K., Harada H., Taga T., Watanabe Y., Matsuda T.,Kashiwamura S., Nakajima K., Koyoma K., Iwamuta A., Tsunusawa S.,Sakiyama F., Matsui H., Takahara Y., Taniguchi T. und Kishimoto T.Complementary DNA for a novel interleukin (BSF-2) that induces B lymphocytes toproduce immunoglobulin.Nature (1986) 324: 73-76
38. Hof P., Pluskey S., Dhe-Paganon S., Eck M.J. und Shoelson S.E.Crystal Structure of the Tyrosine Phosphatase SHP-2Cell (1998) 92: 441-450
39. Honda M., Yamamoto S., Cheng M., Yasukawa K., Suzuki H., Saito T., Usugi T.,Tokunaga T. und Kishimoto T.Human soluble IL-6-receptor: Its detection and enhanced release by HIV infectionJ. Immunol. (1992) 148: 2175-2180
40. Horiuchi S., Koyanagi Y., Zhou Y., Miyamoto H., Tanaka Y., Waki M.,Matsumoto A., Yamamoto M. und Yamamoto N.Soluble interleukin-6 receptor released from T cells or granulocyte/macrophage celllines and human peripheral blood mononuclear cells is generated through analternative splicing mechanism.Eur. J. Immunol. (1994) 24: 1945-1948

Anhang 79
40‘. Horsten U., Schmitz-Van der Leur H., Müllberg J., Heinrich P.C. andRose-John S.The membran distal half of gp130 is responsible for the formation of a ternarycomplex with IL-6 and Il-6 receptorFEBS Lett. (1995) 360: 43-46
41. Jackson D.E., Kupcho K.R. und Newman P.J.Characterization of phosphotyrosine binding motifs in the cytoplasmic domain ofplatelet/endothelial cell adhesion molecule-1 (PECAM-1) that are required for thecellular association and activation of the protein-tyrosine phosphatase, SHP-2J. Biol. Chem. (1997) 272: 24868-24875
42. Jackson D.E., Ward M.W., Wang R. und Newman P.J.The protein-tyrosine phosphatase SHP-2 binds platelet/endothelial cell adhesionmolecule-1 (PECAM-1) and forms a distinct signaling complex during plateletaggregation. Evidence for a mechanistic link between PECAM-1 and integrin-mediated cellular signalingJ. Biol. Chem. (1997) 272: 6986-6993
43. Kazlauskas A., Feng G.S., Pawson T. und Valius M.The 64-kDa protein that associates with the platelet-derived growth factor receptorBeta subunit via Tyr-1009 is the SH2-containing phosphotyrosine phosphatase SypProc. Natl. Acad. Sci. USA (1993) 90: 6939-6943
44. Keegan K. und Cooper J.A.Use of the two hybrid system to detect the association of the protein-tyrosine-phosphatase, SHPTP2, with another SH2-containing protein Grb7Oncogene (1996) 12: 1537-1544
45. Kharitonenkov A., Chen Z., Sures I., Wang H., Shilling J. und Ullrich A.A family of proteins that inhibit signalling through tyrosine kinase receptorsNature (1997) 386: 181-186
46. Kishimoto T.The biology of interleukin-6Blood (1989) 74: 1-10
47. Kuhne M.R., Pawson T., Lienhard G.E. und Feng G.S.The insulin receptor substrate 1 associates with the SH2-containing phosphotyrosinephosphatase sypJ. Biol. Chem. (1993) 268: 11479-11481
48. Kumar G., Gupta S., Wang S. und Nel A.E.Involvement of Janus kinases, p52shc, Raf-1 and MEK-1 in the IL-6-induced mitogen-activated protein cascade of a growth-responsive B cell lineJ. Immunol. (1994) 153: 4436-4447
49. Lämmli U.K.Cleavage of structural proteins during the assembly of the head of bacteriophage T4Nature (1970) 227: 680-685

80 Anhang
50. Li W., Nishimura R., Kashishian A., Batzer A.G., Kim W.J., Cooper J.A. undSchlessinger J.A new function of a phosphatase:. Linking GRB2-Sos to a receptor tyrosine kinaseMol. Cell. Biol. (1994) 14: 509-517
51. Liu J., Modrell B., Aruffo A., Marken J.S., Taga T., Yasukawa K., Murakami M.,Kishimoto T. und Shoyab M.Interleukin-6 signal transducer gp130 mediates oncostatin M signalingJ. Biol. Chem (1992) 267: 16763-16766
52. Lee I., Narazaki M., Hibi M., Kishimoto T. und Taga T.Vav is associated with signaling molecules gp130, Grb2 and Erk2, and is tyrosinephosphorylated in response to interleukin-6FEBS Lett. (1997) 401: 133-137
53. Lütticken C., Wegenka U.M., Yuan J., Buschmann J., Schindler C., Ziemicki A.,Harpur A.G., Wilks A.F., Yasukawa K., Taga T., Kishimoto T., Barbieri G.,Pellegrini S., Sendtner M., Heinrich P.C. und Horn F.Association of transcription factor APRF and protein kinase JAK1 with the IL-6 signaltransducer gp130Science (1994) 263: 89-92
54. Lütticken C., Coffer P., Yuan J., Schwartz C., Caldenhoven E., Schindler C.,Kruijer W., Heinrich P.C. und Horn F.Interleukin-6-induced serine phosphorylation of transcription factor APRF: evidencefor a role in interleukin-6 target gene inductionFEBS Lett. (1995) 360: 137-143
55. Maegawa H., Ugi S., Ishibashi O., Tachikawa Ide R., Takahara N., Tanaka Y.,Takagi Y., Kikkawa R., Shigeta Y., und Kashiwagi A.Src homology 2 domains of protein tyrosine phosphatase are phosphorylated byinsulin receptor kinase and bind to the COOH-terminus of insulin receptor in vitroBiochem. Biophys. Res. Comm. (1993) 194: 208-214
56. Masuda M., Osawa M., Shigematsu H., Harada N. und Fujiwara K.Platelet endothelial cell adhesion molecule-1 is a major SH-PTP2 binding protein invascular endothelial cells.FEBS Lett. (1997) 408: 331-336
57. Matsuda T., Takahashi-Tezuka M., Fukada T., Okuyama Y., Fujitani Y.,Tsukada S., Mano H., Hirai H., Witte O.N. und Hirano T.Association and activation of Btk and Tec tyrosine kinases by gp130, a signaltransducer of the interleukin-6 family of cytokinesBlood (1995) 85: 627-633
58. Mei L., Doherty C.A. und Huganir R.L.RNA splicing regulates the activity of a SH2 domain-containing protein tyrosinephosphataseJ. Biol. Chem. (1994) 269: 12254-12262
59. Milarski K.L. und Saltiel A.R.Expression of catalytically inactive Syp phosphatase in 3T3 cells blocks stimulation ofmitogen-activated protein kinase by insulin.J. Biol. Chem (1994) 269: 21239-21243

Anhang 81
60. Müllberg J., Oberthür W., Lottspeich F., Mehl E., Dittrich E., Graeve L.,Heinrich P.C. und Rose-John S.The soluble human IL-6 receptor. Mutational characterization of the proteolyticcleavage siteJ. Immunol (1994) 152: 4958-4968
61. Murakami M., Hibi M., Nakagawa N., Nakagawa T., Yasukawa K.,Yamanishi K., Taga T. und Kishimoto T.IL-6-induced homodimerization of gp130 and associated activation of a tyrosinekinaseScience (1993) 260: 1808-1810
62. Nakafuku M., Satoh T. und Kaziro Y.Differentiation factors, including nerve growth factor, fibroblast growth factor andinterleukin-6, induce an accumulation of an acitve Ras*GTP complex in ratpheochromocytoma PC12 cellsJ. Biol. Chem. (1992) 267: 19448-19454
63. Ohnishi H., Kubota M., Ohtake A., Sato K. und Sano S.Activation of protein-tyrosine phosphatase SH-PTP2 by a tyrosine based activationmotif of a novel brain moleculeJ. Biol. Chem. (1996) 271: 25569-25574
64. Pazdrak K., Adachi T. und Alam R.Src homology 2 protein tyrosine phosphatase (SHPTP2)/Src homology 2 phosphatase2 (SHP2) tyrosine phosphatase is a positive regulator of the interleukin 5 receptorsignal transduction pathways leading to the prolongation of eosinophil survivalJ. Exp. Med. (1997) 186: 561-568
65. Peng Z.Y. und Cartwright C.A.Regulation of the Src tyrosine kinase and Syp tyrosine phosphatase by their cellularassociationOncogene (1995) 11: 1955-1962
66. Pennica D., Shaw K.J., Swanson T.A., Moore M., Shelton D.L., Zioncheck K.A.,Rosenthal A., Taga T., Paini N.F. und Wood W.I.Cardiotrophin-1: biological activities and binding to the leukemia inhibitory factorreceptor/gp130 signaling complexJ. Biol. Chem. (1995) 270: 10915-10922
67. Pluskey S., Wandless T.J., Walsh C.T. und Shoelson S.E.Potent stimulation of SH-PTP2 phosphatase activity by simultaneous occupancy ofboth SH2 domainsJ. Biol. Chem. (1995) 270: 2897-2900
68. Sambrook J., Fritsch E.F. und Maniatis T.A laboratory manual.Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
69. Sanger F., Nicklen S. und Coulson A.R.DNA sequencing with chain-terminating inhibitors.Proc. Natl. Acad. USA (1977) 74: 5463-5467

82 Anhang
70. Saxton T.M., Henkemeyer M., Gasca S., Shen R., Rossi D.J., Shalaby F.,Feng G.S. und Pawson T.Abnormal mesoderm patterning in mouse embryos mutant for the SH2 tyrosinephosphatase Shp2EMBO J. (1997) 16: 2352-2364
71. Schaper F., Kirchhoff S., Posern G., Köster M., Oumard A., Sharf R., Levi B.Z.und Hauser H.Functional domains of interferon regulatory protein I (IRF-1)Biochem. J. (1998) 335: 147-157
72. Schiemann W.P., Bartoe J.L. und Nathanson N.M.Box 3-independent signaling mechanisms are involved in leukemia inhibitory factoralpha- and gp130-mediated stimulation of mitogen-activated protein kinase. Evidencefor participation of multiple signaling pathways which converge at Ras.J. Biol. Chem. (1997) 272: 16631-16636
73. Schmitz J.Diplomarbeit an der Westfälischen Wilhelms-Universität Münster1997
74. Schooltink H., Stoyan T., Roeb E., Heinrich P.C. und Rose-John S.Ciliary neurotrophic factor induces acute-phase protein expression in hepatocytes.FEBS Lett. (1992) 314: 280-284
75. Shuai K., Horvath C.M., Tsai Huang L.H., Qureshi S.A., Cowburn D. undDarnell J.E.Interferon activation of the transcription factor STAT91 involves dimerization throughSH2-phosphotyrosyl peptide interactionsCell (1994) 76: 821-828
76. Stahl N., Boulton T.G., Farruggella T., Ip N.Y., Davis S., Witthuhn B.A.,Quelle F.W., Silvennoinen O., Barbieri G., Pellegrini S., Ihle J.N. undYancopoulos G.D.Association and activation of Jak-Tyk kinases by CNTF-LIF-OSM-IL-6 β receptorcomponentsScience (1994) 263: 92-95
77. Stahl N., Farruggella T., Boulton T.G., Zhong Z., Darnell J.E. undYancopoulos G.D.Choice of STATs and other substrates specified by modular tyrosine-based motifs incytokine receptorsScience (1995) 267: 1349-1353
78. Stein Gerlach M., Kharitonenkov A., Vogel W., Ali S. und Ullrich A.Protein-tyrosine phosphatase 1D modulates ist own state of tyrosine phosphorylationJ. Biol. Chem. (1995) 270: 24635-24637
79. Symes A., Stahl N., Reeves S.A., FarruggellaT., Servidei T., Gearan T.,Yancopoulos G. und Fink S.The protein tyrosine phosphatase SHP-2 negatively regulates ciliary neurotrophicfactor induction of gene expressionCurr. Biol. (1997) 7: 697-700

Anhang 83
80. Tang T.L., Freeman Jr. R.M., O’Reilly A.M., Neel B.G. ans Sokol S.L.The SH2-containing protein-tyrosine phosphatase SH-PTP2 is required upstream ofMAP kinase for early Xenopus development.Cell (1995) 80: 473-483
81. Tauchi T., Feng G.S., Marshall M., Shen R., Mantel C., Pawson T. undBroxmeyer H.E.The ubiquitously expressed Syp phosphatase interacts with c-kit and Grb2 inhematopoietic cellsJ.Biol. Chem. (1994) 269: 25206-25211
82. Tauchi T., Feng G.S., Shen R., Hoatlin M., Bagby Jr. G.C., Kabat D., Lu L. undBroxmeyer H.E.Involvement of SH2-containing phosphotyrosine phosphatase Syp in erythropoietinreceptor signal transduction pathwaysJ. Biol. Chem. (1995) 270: 5631-5635
83. Tauchi T., Feng G.S., Shen R., Song H.Y., Pawson T. und Broxmeyer H.E.SH2-containing phosphotyrosine phosphatase Syp is a target of p210bcr-abl tyrosinekinaseJ. Biol. Chem. (1994) 269: 15381-15387
84. Vogel W., Lammers R., Huang J. und Ullrich A.Activation of a phosphotyrosine phosphatase by tyrosine phosphorylationScience (1993) 259: 1611-1614
85. Vogel W. und Ullrich A.Multiple in vivo phosphorylated tyrosine phosphatase SHP-2 engages binding to Grb2via tyrosine 584Cell Growth Differ. (1997) 7: 1589-1597
86. Wagner B.J., Hayes T.E., Hoban C.J. und Cochran B.H.The SIF binding element confers sis/PDGF inducibility onto the c-fos promoterEMBO J. (1990) 9: 4477-4484
87. Ward L.D., Howlett G.J., Discolo G., Yasukawa K., Hamacher A., Moritz R.L.und Simpson R.J.High affinity interleukin-6 receptor is a hexameric complex consisting of twomolecules each of interleukin-6, interleukin-6 receptor, and gp130J. Biol. Chem. (1994) 269: 23286-23289
88. Wegenka U.M., Buschmann J., Lüttiken C., Heinrich P.C. und Horn F.Acute-phase response factor, a nuclear factor binding to acute-phase responseelements, is rapidly activated by interleukin-6 at the posttranslational levelMol. Cell. Biol. (1994) 13: 276-288
89. Weiergraeber O., Hemman U., Kuester A., Mueller-Newen G.,Schneider-Mergener J., Rose-John S., Kurschat P., Brakenhoff J.P.J.,Hart M.H.L., Stabel S. und Heinrich P.C.Soluble human interleukin-6 receptor – Expression in insect cells, purification andcharacterization.Eur. J. Biochem (1995) 324: 661-669

84 Anhang
90. Wells J.A. und de Vos A.M.Hematopoietic receptor complexesAnn. Rev. Biochem. (1996) 65: 609-634
91. Wong L. und Johnson G.R.Epidermal growth factor induces coupling of protein-tyrosine phosphatase 1D toGRB2 via the COOH-terminal SH3 domain of GRB2J. Biol. Chem. (1996) 271: 20981-20984
92. Yamasaki K., Taga T., Hirata Y., Yawata H., Kawanishi Y., Seed B.,Tanigushi T., Hirano T. und Kishimoto T.Cloning and expression of a human interleukin-6 (BSF-2/IFNβ2) receptorScience (1988) 241: 825-828
93. Yin T.G., Shen R., Feng G.S. und Yang Y.C.Molecular characterization of specific interactions between SHP-2 phosphatase andJAK tyrosine kinasesJ. Biol. Chem. (1997) 272: 1032-1037
94. Yin T., Taga T., Tsang M.L., Yasuwaka K., Kishimoto T. und Yang Y.C.Involvement of IL-6 signal transducer gp130 in IL-11-mediated signal transduction.J. Immunol. (1993) 151: 2555-2561
95. Yuan J., Wegenka U.M., Lüttiken C., Buschmann J., Decker T., Schindler C.,Heinrich P.C. und Horn F.The signaling pathways of interleukin-6 and interferon-γ converge by activation ofdifferent transcription factors which bind to common responsive DNA elementsMol. Cell. Biol (1994) 14: 1657-1668
96. Zhang X., Blenis J., Li H.C., Schindler C. und Cheng-Kiang S.Requirement of serine phosphorylation for formation of STAT-promotor complexesScience (1995) 267: 1990-1994

Anhang 85
6.2 Abkürzungsverzeichnis
α anti-A Alaninaa AminosäurenAbb AbbildungAPS AmmoniumperoxidsulfatATP Adenosintriphosphatβ-Gal β-GalaktosidaseBSA bovine serum albuminBSF B-cell stimulating factorC CysteincDNA complementary DNACIP calf intestine phosphatasec-kit receptor of stem cell factorCMV ZytomegalievirusCNTF ciliary neurotrophic factorcpm counts per minuteCRP C-reaktives ProteinCT-1 Cardiotrophin-1DMEM Dulbecco’s modified Eagle MediumDMSO DimethylsulfatDNA DesoxyribonukleinsäureECL enhanced chemiluminiscenceEDTA EthylendiamintetraacetatEGF epidermal growth factorEMSA Elektrophoretischer Mobilitätshift AssayEpo ErythropoetinF PhenylalaninFACS fluorescence activated cell sortingFCS fetal calf serumFGF fibroblast growth factorG-CSF granulocyte colony-stimulating factorgfp green-fluorescent proteinGH growth hormoneGM-CSF granulocyte/macrophage colony-stimulating factorgp Glykoproteingrb growth factor receptor bound proteinh StundeHBS HEPES buffered salineHEPES N-(2-Hydroxyethyl)piperazin-N‘-2-ethanolsulfonsäureHRP horse radish peroxidaseIFN InterferonIg ImmunglobulinIB ImmunblotIL InterleukinIP Immunpräzipitation

86 Anhang
IR Insulin-RezeptorIRF interferon regulatory factorIRS insulin receptor substrateJak Janus-KinaseK LysinkDA KilodaltonL LeucinLB-Medium Luria Bertani-MediumLIF leukemia inhibitory factorM MolarMAPK mitogen activated protein kinaseMEK MAPK-Kinasemin MinuteMpl myeloproliferative leukemia receptorOD optische DichteONPG o-NitrophenylgalaktosidOSM Oncostatin MPBS phosphate buffered salinePDGF platelet derived growth factorPCR Polymerase-KettenreaktionPEG PolyethylenglykolPLC Phospholipase CPMSF PhenylmethylsulfonylfluoridPRL ProlaktinPVDF PolyvinylidenfluoridPY Phospho-TyrosinQ GlutaminR RezeptorRNA RibonukleinsäureRT Raumtemperaturs soluble, löslichS SerinSDS SodiumdodecylsulfatSDS-PAGE SDS-Polyacrylamid-Gel-Elektrophoresesec SekundeSH src-HomologiedomäneSHP2 SH2-domain containing protein-tyrosine-phosphataseSTAT signal transducer and activator of transcriptionSV40 (Affenvirus)Tab TabelleTEMED N, N, N‘, N‘-TetramethylendiaminTF tissue factorTris Tris(hydroxy)aminomethanU unit, EinheitUpM Umdrehungen pro MinuteUV ultraviolettW TryptophanY Tyrosin

Anhang 87
6.3 Sequenzen
6.3.1 Sequenz der GrG YYYYYYmyc-cDNA
M L T L Q T W V V Q A L F I F L T T E ]------------------------------------------------------------- ATGTTGA CGTTGCAGAC TTGGGTAGTG CAAGCCTTGT TTATTTTCCT CACCACTGAA TACAACT GCAACGTCTG AACCCATCAC GTTCGGAACA AATAAAAGGA GTGGTGACTT
20 S T G E L L D P C G Y I S P E S P V V Q ----------------------------------------------------------------- 58 TCTACAGGTG AACTTCTAGA TCCATGTGGT TATATCAGTC CTGAATCTCC AGTTGTACAA AGATGTCCAC TTGAAGATCT AGGTACACCA ATATAGTCAG GACTTAGAGG TCAACATGTT
40 L H S N F T A V C V L K E K C M D Y F H ----------------------------------------------------------------- 118 CTTCATTCTA ATTTCACTGC AGTTTGTGTG CTAAAGGAAA AATGTATGGA TTATTTTCAT GAAGTAAGAT TAAAGTGACG TCAAACACAC GATTTCCTTT TTACATACCT AATAAAAGTA
60 V N A N Y I V W K T N H F T I P K E Q Y ----------------------------------------------------------------- 178 GTAAATGCTA ATTACATTGT CTGGAAAACA AACCATTTTA CTATTCCTAA GGAGCAATAT CATTTACGAT TAATGTAACA GACCTTTTGT TTGGTAAAAT GATAAGGATT CCTCGTTATA
80 T I I N R T A S S V T F T D I A S L N I ----------------------------------------------------------------- 238 ACTATCATAA ACAGAACAGC ATCCAGTGTC ACCTTTACAG ATATAGCTTC ATTAAATATT TGATAGTATT TGTCTTGTCG TAGGTCACAG TGGAAATGTC TATATCGAAG TAATTTATAA
100 Q L T C N I L T F G Q L E Q N V Y G I T ----------------------------------------------------------------- 298 CAGCTCACTT GCAACATTCT TACATTCGGA CAGCTTGAAC AGAATGTTTA TGGAATCACA GTCGAGTGAA CGTTGTAAGA ATGTAAGCCT GTCGAACTTG TCTTACAAAT ACCTTAGTGT
120 I I S G L P P E K P K N L S C I V N E G ----------------------------------------------------------------- 358 ATAATTTCAG GCTTGCCTCC AGAAAAACCT AAAAATTTGA GTTGCATTGT GAACGAGGGG TATTAAAGTC CGAACGGAGG TCTTTTTGGA TTTTTAAACT CAACGTAACA CTTGCTCCCC
140 K K M R C E W D G G R E T H L E T N F T ---------------------------------------------------------------- 418 AAGAAAATGA GGTGTGAGTG GGATGGTGGA AGGGAAACAC ACTTGGAGAC AAACTTCACT TTCTTTTACT CCACACTCAC CCTACCACCT TCCCTTTGTG TGAACCTCTG TTTGAAGTGA
160 L K S E W A T H K F A D C K A K R D T P ----------------------------------------------------------------- 478 TTAAAATCTG AATGGGCAAC ACACAAGTTT GCTGATTGCA AAGCAAAACG TGACACCCCC AATTTTAGAC TTACCCGTTG TGTGTTCAAA CGACTAACGT TTCGTTTTGC ACTGTGGGGG

88 Anhang
180 T S C T V D Y S T V Y F V N I E V W V E ----------------------------------------------------------------- 538 ACCTCATGCA CTGTTGATTA TTCTACTGTG TATTTTGTCA ACATTGAAGT CTGGGTAGAA TGGAGTACGT GACAACTAAT AAGATGACAC ATAAAACAGT TGTAACTTCA GACCCATCTT
200 A E N A L G K V T S D H I N F D P V Y K ----------------------------------------------------------------- 598 GCAGAGAATG CCCTTGGGAA GGTTACATCA GATCATATCA ATTTTGATCC TGTATATAAA CGTCTCTTAC GGGAACCCTT CCAATGTAGT CTAGTATAGT TAAAACTAGG ACATATATTT
220 V K P N P P H N L S V I N S E E L S S I ----------------------------------------------------------------- 658 GTGAAGCCCA ATCCGCCACA TAATTTATCA GTGATCAACT CAGAGGAACT GTCTAGTATC CACTTCGGGT TAGGCGGTGT ATTAAATAGT CACTAGTTGA GTCTCCTTGA CAGATCATAG
240 L K L T W T N P S I K S V I I L K Y N I ----------------------------------------------------------------- 718 TTAAAATTGA CATGGACCAA CCCAAGTATT AAGAGTGTTA TAATACTAAA ATATAACATT AATTTTAACT GTACCTGGTT GGGTTCATAA TTCTCACAAT ATTATGATTT TATATTGTAA
260 Q Y R T K D A S T W S Q I P P E D T A S ----------------------------------------------------------------- 778 CAATATAGGA CCAAAGATGC CTCAACTTGG AGCCAGATTC CTCCTGAAGA CACAGCATCC GTTATATCCT GGTTTCTACG GAGTTGAACC TCGGTCTAAG GAGGACTTCT GTGTCGTAGG
280 T R S S F T V Q D L K P F T E Y V F R I ----------------------------------------------------------------- 838 ACCCGATCTT CATTCACTGT CCAAGACCTT AAACCTTTTA CAGAATATGT GTTTAGGATT TGGGCTAGAA GTAAGTGACA GGTTCTGGAA TTTGGAAAAT GTCTTATACA CAAATCCTAA
300 R C M K E D G K G Y W S D W S E E A S G ----------------------------------------------------------------- 898 CGCTGTATGA AGGAAGATGG TAAGGGATAC TGGAGTGACT GGAGTGAAGA AGCAAGTGGG GCGACATACT TCCTTCTACC ATTCCCTATG ACCTCACTGA CCTCACTTCT TCGTTCACCC
320 I T Y E D R P S K A P S F W Y K I D P S ----------------------------------------------------------------- 958 ATCACCTATG AAGATAGACC ATCTAAAGCA CCAAGTTTCT GGTATAAAAT AGATCCATCC TAGTGGATAC TTCTATCTGG TAGATTTCGT GGTTCAAAGA CCATATTTTA TCTAGGTAGG
340 H T Q G Y R T V Q L V W K T L P P F E A ----------------------------------------------------------------- 1018 CATACTCAAG GCTACAGAAC TGTACAACTC GTGTGGAAGA CATTGCCTCC TTTTGAAGCC GTATGAGTTC CGATGTCTTG ACATGTTGAG CACACCTTCT GTAACGGAGG AAAACTTCGG
360 N G K I L D Y E V T L T R W K S H L Q N ----------------------------------------------------------------- 1078 AATGGAAAAA TCTTGGATTA TGAAGTGACT CTCACAAGAT GGAAATCACA TTTACAAAAT TTACCTTTTT AGAACCTAAT ACTTCACTGA GAGTGTTCTA CCTTTAGTGT AAATGTTTTA
380 Y T V N A T K L T V N L T N D R Y L A T ----------------------------------------------------------------- 1138 TACACAGTTA ATGCCACAAA ACTGACAGTA AATCTCACAA ATGATCGCTA TCTAGCAACC ATGTGTCAAT TACGGTGTTT TGACTGTCAT TTAGAGTGTT TACTAGCGAT AGATCGTTGG

Anhang 89
400 L T V R N L V G K S D A A V L T I P A C ----------------------------------------------------------------- 1198 CTAACAGTAA GAAATCTTGT TGGCAAATCA GATGCAGCTG TTTTAACTAT CCCTGCCTGT GATTGTCATT CTTTAGAACA ACCGTTTAGT CTACGTCGAC AAAATTGATA GGGACGGACA
420 D F Q A T H P V M D L K A F P K D N M L ----------------------------------------------------------------- 1258 GACTTTCAAG CTACTCACCC TGTAATGGAT CTTAAAGCAT TCCCCAAAGA TAACATGCTT CTGAAAGTTC GATGAGTGGG ACATTACCTA GAATTTCGTA AGGGGTTTCT ATTGTACGAA
440 W V E W T T P R E S V K K Y I L E W C V ----------------------------------------------------------------- 1318 TGGGTGGAAT GGACTACTCC AAGGGAATCT GTAAAGAAAT ATATACTTGA GTGGTGTGTG ACCCACCTTA CCTGATGAGG TTCCCTTAGA CATTTCTTTA TATATGAACT CACCACACAC
460 L S D K A P C I T D W Q Q E D G T V H R ----------------------------------------------------------------- 1378 TTATCAGATA AAGCACCCTG TATCACAGAC TGGCAACAAG AAGATGGTAC CGTGCATCGC AATAGTCTAT TTCGTGGGAC ATAGTGTCTG ACCGTTGTTC TTCTACCATG GCACGTAGCG
480 T Y L R G N L A E S K C Y L I T V T P V ----------------------------------------------------------------- 1438 ACCTATTTAA GAGGGAACTT AGCAGAGAGC AAATGCTATT TGATAACAGT TACTCCAGTA TGGATAAATT CTCCCTTGAA TCGTCTCTCG TTTACGATAA ACTATTGTCA ATGAGGTCAT
500 Y A D G P G S P E S I K A Y L K Q A P P ----------------------------------------------------------------- 1498 TATGCTGATG GACCAGGAAG CCCTGAATCC ATAAAGGCAT ACCTTAAACA AGCTCCACCT ATACGACTAC CTGGTCCTTC GGGACTTAGG TATTTCCGTA TGGAATTTGT TCGAGGTGGA
520 S K G P T V R T K K V G K N E A V L E W ----------------------------------------------------------------- 1558 TCCAAAGGAC CTACTGTTCG GACAAAAAAA GTAGGGAAAA ACGAAGCTGT CTTAGAGTGG AGGTTTCCTG GATGACAAGC CTGTTTTTTT CATCCCTTTT TGCTTCGACA GAATCTCACC
540 D Q L P V D V Q N G F I R N Y T I F Y R ----------------------------------------------------------------- 1618 GACCAACTTC CTGTTGATGT TCAGAATGGA TTTATCAGAA ATTATACTAT ATTTTATAGA CTGGTTGAAG GACAACTACA AGTCTTACCT AAATAGTCTT TAATATGATA TAAAATATCT
560 T I I G N E T A V N V D S S H T E Y T L ----------------------------------------------------------------- 1678 ACCATCATTG GAAATGAAAC TGCTGTGAAT GTGGATTCTT CCCACACAGA ATATACATTG TGGTAGTAAC CTTTACTTTG ACGACACTTA CACCTAAGAA GGGTGTGTCT TATATGTAAC
580 S S L T S D T L Y M V R M A A Y T D E G ----------------------------------------------------------------- 1738 TCCTCTTTGA CTAGTGACAC ATTGTACATG GTACGAATGG CAGCATACAC AGATGAAGGT AGGAGAAACT GATCACTGTG TAACATGTAC CATGCTTACC GTCGTATGTG TCTACTTCCA
600 G K D G P E F T F T T P K F A Q G E I E ----------------------------------------------------------------- 1798 GGGAAGGATG GTCCAGAATT CACTTTTACT ACCCCAAAGT TTGCTCAAGG AGAAATTGAA CCCTTCCTAC CAGGTCTTAA GTGAAAATGA TGGGGTTTCA AACGAGTTCC TCTTTAACTT EcoRI

90 Anhang
620 A I V V P V C L A F L L T T L L G V L F ----------------------------------------------------------------- 1858 GCCATAGTCG TGCCTGTTTG CTTAGCATTC CTATTGACAA CTCTTCTGGG AGTGCTGTTC CGGTATCAGC ACGGACAAAC GAATCGTAAG GATAACTGTT GAGAAGACCC TCACGACAAG
640 C F N K R D L I K K H I W P N V P D P S ----------------------------------------------------------------- 1918 TGCTTTAATA AGCGAGACCT AATTAAAAAA CACATCTGGC CTAATGTTCC AGATCCTTCA ACGAAATTAT TCGCTCTGGA TTAATTTTTT GTGTAGACCG GATTACAAGG TCTAGGAAGT
660 K S H I A Q W S P H T P P R H N F N S K ----------------------------------------------------------------- 1978 AAGAGTCATA TTGCCCAGTG GTCACCTCAC ACTCCTCCAA GGCACAATTT TAATTCAAAA TTCTCAGTAT AACGGGTCAC CAGTGGAGTG TGAGGAGGTT CCGTGTTAAA ATTAAGTTTT BstEII 680 D Q M Y S D G N F T D V S V V E I E A N ----------------------------------------------------------------- 2038 GATCAAATGT ATTCAGATGG CAATTTCACT GATGTAAGTG TTGTGGAAAT AGAAGCAAAT CTAGTTTACA TAAGTCTACC GTTAAAGTGA CTACATTCAC AACACCTTTA TCTTCGTTTA
700 D K K P F P E D L K L L D L F K K E K I ----------------------------------------------------------------- 2098 GACAAAAAGC CTTTTCCAGA AGATCTGAAA TTATTGGACC TGTTCAAAAA GGAAAAAATT CTGTTTTTCG GAAAAGGTCT TCTAGACTTT AATAACCTGG ACAAGTTTTT CCTTTTTTAA
720 N T E G H S S G I G G S S C M S S S R P ----------------------------------------------------------------- 2158 AATACTGAAG GACACAGCAG TGGTATTGGG GGGTCTTCAT GCATGTCATC TTCTAGGCCA TTATGACTTC CTGTGTCGTC ACCATAACCC CCCAGAAGTA CGTACAGTAG AAGATCCGGT
740 S I S S S D E N E S S Q N T S S T V Q Y ----------------------------------------------------------------- 2218 AGCATTTCTA GCAGTGATGA AAATGAATCT TCACAAAACA CGAGCTCCAC TGTCCAGTAT TCGTAAAGAT CGTCACTACT TTTACTTAGA AGTGTTTTGT GCTCGAGGTG ACAGGTCATA SacI 760 S T V V H S G Y R H Q V P S V Q V F S R ----------------------------------------------------------------- 2278 TCTACCGTGG TACACTCCGG ATACAGACAC CAAGTTCCGT CAGTCCAAGT CTTCTCAAGA AGATGGCACC ATGTGAGGCC TATGTCTGTG GTTCAAGGCA GTCAGGTTCA GAAGAGTTCT BspEI 780 S E S T Q P L L D S E E R P E D L Q L V ----------------------------------------------------------------- 2338 TCCGAGTCTA CCCAGCCCTT GTTAGATTCA GAGGAGCGGC CAGAAGATCT ACAATTAGTA AGGCTCAGAT GGGTCGGGAA CAATCTAAGT CTCCTCGCCG GTCTTCTAGA TGTTAATCAT
800 D H V D G G D G I L P R Q Q Y F K Q N C ----------------------------------------------------------------- 2398 GACCATGTCG ATGGCGGTGA TGGTATTTTG CCCAGGCAAC AGTACTTCAA ACAGAACTGC CTGGTACAGC TACCGCCACT ACCATAAAAC GGGTCCGTTG TCATGAAGTT TGTCTTGACG AspI 820 S Q H E S S P D I S H F E R S K Q V S S ----------------------------------------------------------------- 2458 AGTCAGCATG AATCCAGTCC AGATATTTCA CATTTTGAAA GGTCAAAGCA AGTTTCATCA TCAGTCGTAC TTAGGTCAGG TCTATAAAGT GTAAAACTTT CCAGTTTCGT TCAAAGTAGT

Anhang 91
840 V N E E D F V R L K Q Q I S D H I S Q S ----------------------------------------------------------------- 2518 GTCAATGAGG AAGATTTTGT TAGACTTAAA CAGCAGATTT CAGATCATAT TTCACAATCC CAGTTACTCC TTCTAAAACA ATCTGAATTT GTCGTCTAAA GTCTAGTATA AAGTGTTAGG
860 C G S G Q M K M F Q E V S A A D A F G P ----------------------------------------------------------------- 2578 TGTGGATCTG GGCAAATGAA AATGTTTCAG GAAGTTTCTG CAGCAGATGC TTTTGGTCCA ACACCTAGAC CCGTTTACTT TTACAAAGTC CTTCAAAGAC GTCGTCTACG AAAACCAGGT
880 G T E G Q V E R F E T V G M E A A T D E ----------------------------------------------------------------- 2638 GGTACTGAGG GACAAGTAGA AAGATTCGAA ACAGTTGGCA TGGAGGCTGC GACTGATGAA CCATGACTCC CTGTTCATCT TTCTAAGCTT TGTCAACCGT ACCTCCGACG CTGACTACTT Bsp119I 900 G M P K S Y L P Q T V R Q G G Y M P Q G ----------------------------------------------------------------- 2698 GGCATGCCTA AAAGTTACTT ACCACAGACG GTCCGGCAAG GCGGCTACAT GCCTCAGGGC CCGTACGGAT TTTCAATGAA TGGTGTCTGC CAGGCCGTTC CGCCGATGTA CGGAGTCCCG RsrII 920 G E Q K L I S E E D L * ---)~~~~~~c-myc-Epitop~~~~~~~~~~~~> 2758 GGCGAGCAGA AGCTGATATC GGAGGAAGAT CTGTGA CCGCTCGTCT TCGACTATAG CCTCCTTCTA GACACT
6.3.2 Sequenz der sypimyc-cDNA
M T S R R W F H P N ]--------------------------------- GTCCCGGAAT TCCCGGGTCT AGAGTCACCA TGACATCGCG GAGATGGTTT CACCCAAATA CAGGGCCTTA AGGGCCCAGA TCTCAGTGGT ACTGTAGCGC CTCTACCAAA GTGGGTTTAT EcoRI 11 I T G V E A E N L L L T R G V D G S F L ----------------------------------------------------------------- 61 TCACTGGTGT GGAGGCAGAA AACCTACTGT TGACAAGAGG AGTTGATGGC AGTTTTTTGG AGTGACCACA CCTCCGTCTT TTGGATGACA ACTGTTCTCC TCAACTACCG TCAAAAAACC
31 A R P S K S N P G D F T L S V R R N G A ----------------------------------------------------------------- 121 CAAGGCCTAG TAAAAGTAAC CCTGGAGACT TCACACTTTC CGTTAGAAGA AATGGAGCTG GTTCCGGATC ATTTTCATTG GGACCTCTGA AGTGTGAAAG GCAATCTTCT TTACCTCGAC
51 V T H I K I Q N T G D Y Y D L Y G G E K ----------------------------------------------------------------- 181 TCACCCACAT CAAGATTCAG AACACTGGTG ATTACTATGA CCTGTATGGA GGGGAGAAAT AGTGGGTGTA GTTCTAAGTC TTGTGACCAC TAATGATACT GGACATACCT CCCCTCTTTA
71 F A T L A E L V Q Y Y M E H H G Q L K E ----------------------------------------------------------------- 241 TTGCCACTTT GGCTGAGTTG GTCCAGTATT ACATGGAACA TCACGGGCAA TTAAAAGAGA AACGGTGAAA CCGACTCAAC CAGGTCATAA TGTACCTTGT AGTGCCCGTT AATTTTCTCT

92 Anhang
91 K N G D V I E L K Y P L N C A D P T S E ----------------------------------------------------------------- 301 AGAATGGAGA TGTCATTGAG CTTAAATATC CTCTGAACTG TGCAGATCCT ACCTCTGAAA TCTTACCTCT ACAGTAACTC GAATTTATAG GAGACTTGAC ACGTCTAGGA TGGAGACTTT
111 R W F H G H L S G K E A E K L L T E K G ----------------------------------------------------------------- 361 GGTGGTTTCA TGGACATCTC TCTGGGAAAG AAGCAGAGAA ATTATTAACT GAAAAAGGAA CCACCAAAGT ACCTGTAGAG AGACCCTTTC TTCGTCTCTT TAATAATTGA CTTTTTCCTT
131 K H G S F L V R E S Q S H P G D F V L S ----------------------------------------------------------------- 421 AACATGGTAG TTTTCTTGTA CGAGAGAGCC AGAGCCACCC TGGAGATTTT GTTCTTTCTG TTGTACCATC AAAAGAACAT GCTCTCTCGG TCTCGGTGGG ACCTCTAAAA CAAGAAAGAC
151 V R T G D D K G E S N D G K S K V T H V ----------------------------------------------------------------- 481 TGCGCACTGG TGATGACAAA GGGGAGAGCA ATGACGGCAA GTCTAAAGTG ACCCATGTTA ACGCGTGACC ACTACTGTTT CCCCTCTCGT TACTGCCGTT CAGATTTCAC TGGGTACAAT
171 M I R C Q E L K Y D V G G G E R F D S L ----------------------------------------------------------------- 541 TGATTCGCTG TCAGGAACTG AAATACGACG TTGGTGGAGG AGAACGGTTT GATTCTTTGA ACTAAGCGAC AGTCCTTGAC TTTATGCTGC AACCACCTCC TCTTGCCAAA CTAAGAAACT
191 T D L V E H Y K K N P M V E T L G T V L ----------------------------------------------------------------- 601 CAGATCTTGT GGAACATTAT AAGAAGAATC CTATGGTGGA AACATTGGGT ACAGTACTAC GTCTAGAACA CCTTGTAATA TTCTTCTTAG GATACCACCT TTGTAACCCA TGTCATGATG
211 Q L K Q P L N T T R I N A A E I E S R V ----------------------------------------------------------------- 661 AACTCAAGCA GCCCCTTAAC ACGACTCGTA TAAATGCTGC TGAAATAGAA AGCAGAGTTC TTGAGTTCGT CGGGGAATTG TGCTGAGCAT ATTTACGACG ACTTTATCTT TCGTCTCAAG
231 R E L S K L A E T T D K V K Q G F W E E ----------------------------------------------------------------- 721 GAGAACTAAG CAAATTAGCT GAGACCACAG ATAAAGTCAA ACAAGGCTTT TGGGAAGAAT CTCTTGATTC GTTTAATCGA CTCTGGTGTC TATTTCAGTT TGTTCCGAAA ACCCTTCTTA
251 F E T L Q Q Q E C K L L Y S R K E G Q R ----------------------------------------------------------------- 781 TTGAGACACT ACAACAACAG GAGTGCAAAC TTCTCTACAG CCGAAAAGAG GGTCAAAGGC AACTCTGTGA TGTTGTTGTC CTCACGTTTG AAGAGATGTC GGCTTTTCTC CCAGTTTCCG
271 Q E N K N K N R Y K N I L P F D H T R V ----------------------------------------------------------------- 841 AAGAAAACAA AAACAAAAAT AGATATAAAA ACATCCTGCC CTTTGATCAT ACCAGGGTTG TTCTTTTGTT TTTGTTTTTA TCTATATTTT TGTAGGACGG GAAACTAGTA TGGTCCCAAC
291 V L H D G D P N E P V S D Y I N A N I I ----------------------------------------------------------------- 901 TCCTACACGA TGGTGATCCC AATGAGCCTG TTTCAGATTA CATCAATGCA AATATCATCA AGGATGTGCT ACCACTAGGG TTACTCGGAC AAAGTCTAAT GTAGTTACGT TTATAGTAGT

Anhang 93
311 M P E F E T K C N N S K P K K S Y I A T ----------------------------------------------------------------- 961 TGCCTGAATT TGAAACCAAG TGCAACAATT CAAAGCCCAA AAAGAGTTAC ATTGCCACAC ACGGACTTAA ACTTTGGTTC ACGTTGTTAA GTTTCGGGTT TTTCTCAATG TAACGGTGTG
331 Q G C L Q N T V N D F W R M V F Q E N S ----------------------------------------------------------------- 1021 AAGGCTGCCT GCAAAACACG GTGAATGACT TTTGGCGGAT GGTGTTCCAA GAAAACTCCC TTCCGACGGA CGTTTTGTGC CACTTACTGA AAACCGCCTA CCACAAGGTT CTTTTGAGGG
351 R V I V M T T K E V E R G K S K C V K Y ----------------------------------------------------------------- 1081 GAGTGATTGT CATGACAACG AAAGAAGTGG AGAGAGGAAA GAGTAAATGT GTCAAATACT CTCACTAACA GTACTGTTGC TTTCTTCACC TCTCTCCTTT CTCATTTACA CAGTTTATGA
371 W P D E Y A L K E Y G V M R V R N V K E ----------------------------------------------------------------- 1141 GGCCTGATGA GTATGCTCTA AAAGAATATG GCGTCATGCG TGTTAGGAAC GTCAAAGAAA CCGGACTACT CATACGAGAT TTTCTTATAC CGCAGTACGC ACAATCCTTG CAGTTTCTTT Insert 391 S A A H D Y T L R E L K L S K V G Q A L ----------------------------------------------------------------- 1201 GCGCCGCTCA TGACTATACG CTAAGAGAAC TTAAACTTTC AAAGGTTGGA CAAGCTCTAC CGCGGCGAGT ACTGATATGC GATTCTCTTG AATTTGAAAG TTTCCAACCT GTTCGAGATG
411 L Q G N T E R T V W Q Y H F R T W P D H ----------------------------------------------------------------- 1261 TCCAGGGGAA TACGGAGAGA ACGGTCTGGC AATACCACTT TCGGACCTGG CCGGACCACG AGGTCCCCTT ATGCCTCTCT TGCCAGACCG TTATGGTGAA AGCCTGGACC GGCCTGGTGC
431 G V P S D P G G V L D F L E E V H H K Q ----------------------------------------------------------------- 1321 GCGTGCCCAG CGACCCTGGG GGCGTGCTGG ACTTCCTGGA GGAGGTGCAC CATAAGCAGG CGCACGGGTC GCTGGGACCC CCGCACGACC TGAAGGACCT CCTCCACGTG GTATTCGTCC Katalytisches Zentrum 451 E S I M D A G P V V V H C S A G I G R T ----------------------------------------------------------------- 1381 AGAGCATCAT GGATGCAGGG CCGGTCGTGG TGCACTGCAG TGCTGGAATT GGCCGGACAG TCTCGTAGTA CCTACGTCCC GGCCAGCACC ACGTGACGTC ACGACCTTAA CCGGCCTGTC
471 G T F I V I D I L I D I I R E K G V D C ----------------------------------------------------------------- 1441 GGACGTTCAT TGTGATTGAT ATTCTTATTG ACATCATCAG AGAGAAAGGT GTTGACTGCG CCTGCAAGTA ACACTAACTA TAAGAATAAC TGTAGTAGTC TCTCTTTCCA CAACTGACGC
491 D I D V P K T I Q M V R S Q R S G M V Q ----------------------------------------------------------------- 1501 ATATTGACGT TCCCAAAACC ATCCAGATGG TGCGGTCTCA GAGGTCAGGG ATGGTCCAGA TATAACTGCA AGGGTTTTGG TAGGTCTACC ACGCCAGAGT CTCCAGTCCC TACCAGGTCT
511 T E A Q Y R F I Y M A V Q H Y I E T L Q ----------------------------------------------------------------- 1561 CAGAAGCACA GTACCGATTT ATCTATATGG CGGTCCAGCA TTATATTGAA ACACTACAGC GTCTTCGTGT CATGGCTAAA TAGATATACC GCCAGGTCGT AATATAACTT TGTGATGTCG

94 Anhang
531 R R I E E E Q K S K R K G H E Y T N I K ----------------------------------------------------------------- 1621 GCAGGATTGA AGAAGAGCAG AAAAGCAAGA GGAAAGGGCA CGAATATACA AATATTAAGT CGTCCTAACT TCTTCTCGTC TTTTCGTTCT CCTTTCCCGT GCTTATATGT TTATAATTCA
551 Y S L A D Q T S G D Q S P L P P C T P T ----------------------------------------------------------------- 1681 ATTCTCTAGC GGACCAGACG AGTGGAGATC AGAGCCCTCT CCCGCCTTGT ACTCCAACGC TAAGAGATCG CCTGGTCTGC TCACCTCTAG TCTCGGGAGA GGGCGGAACA TGAGGTTGCG
571 P P C A E M R E D S A R V Y E N V G L M ----------------------------------------------------------------- 1741 CACCCTGTGC AGAAATGAGA GAAGACAGTG CTAGAGTCTA TGAAAACGTG GGCCTGATGC GTGGGACACG TCTTTACTCT CTTCTGTCAC GATCTCAGAT ACTTTTGCAC CCGGACTACG
591 Q Q Q K S F R K A L E Q K L I S E E D L -------------------------------)~~~~~~~~~c-myc-Epitop~~~~~~~~~~~~ 1801 AACAGCAGAA AAGTTTCAGA AAAGCTTTAG AGCAGAAGCT GATATCGGAG GAAGATCTGG TTGTCGTCTT TTCAAAGTCT TTTCGAAATC TCGTCTTCGA CTATAGCCTC CTTCTAGACC HindIII 611 E A * ~~~~> 1861 AAGCTTAA TTCGAATT HindIII

Danksagung
Ohne die Mithilfe einer ganzen Reihe von Personen wäre die Erstellung dieser Doktorarbeit
nicht möglich gewesen. Ihnen möchte ich an dieser Stelle meine Anerkennung aussprechen.
Ich bedanke mich
bei Herrn Prof. Heinrich für die freundliche Aufnahme in seinen Arbeitskreis, sowie für die
Ermöglichung dieser Arbeit und die darüber hinaus in jeder Hinsicht gewährte Unterstützung,
auch bei der Erstellung von Zeugnissen und Bescheinigungen.
bei Herrn Dr. Fred Schaper, der diese Arbeit maßgeblich betreut hat, für das wissenschaftliche
Arbeiten, welches er mir bei gebracht hat, ganz zu schweigen von den kleinen Tricks, die das
Leben im Labor erleichtern. Davon profitiere ich noch heute.
bei Herrn Jochen Schmitz, Frau Monika Eck, Herrn Carsten Grimm und Herrn Dirk Anhuf für
die erfahrene Unterstützung, besonders in der Endphase dieser Arbeit.
bei Frau Andrea Graf für die Aufarbeitung unendlicher Liter E.coli-Lösungen und die
Bereitstellung ebenso großer Mengen an Puffer und Lösungen; bei Frau Christa Gerlach für
den Nachschub an COS-7-Zellen und deren Bekannten; bei Herrn Marcel Robbertz für die
Photoarbeiten und schließlich bei Frau Hildegard Schmitz-Van de Leur und Frau Andrea
Küster für die Geduld und Mühe, die sie sich mit den Sequenzierungen machten.
bei allen übrigen Mitarbeitern des Institutes für das angenehme Arbeitsklima und die
erfahrene Hilfsbereitschaft, besonders bei Herrn Dr. Elmar Siewert – in Gedanken an alte
Labor 3-Zeiten.
Schließlich möchte ich noch meinen Eltern und Marc danken, die mir nicht nur die
Möglichkeit gegeben haben, Medizin zu studieren, sondern mich immer wieder aufgebaut
haben, wenn es mal wieder eher drunter als drüber ging.

.

Lebenslauf
Cornelia Henke-Gendo geb. Gendo
Geburtsdatum: 21. Oktober 1973 in Bad Soden am Taunus
Schulbildung: 1980-1984: Katholische Grundschule Geilenkirchen
1984-1993: Bischöfliches Gymnasium St. Ursula, Geilenkirchen
Abschluß: Allgemeine Hochschulreife
Studium: ab WS 1993/94: Studium der Medizin an der RWTH Aachen
1995: ärztliche Vorprüfung
1996: 1. Staatsexamen
1996-1997: Promotion am Institut für Biochemie,
Abt. Prof. Heinrich
10.1998-01.1999: Auslandssemester an der
Rijksuniversiteit Limburg in Maastricht (NL)
1999: 2. Staatsexamen
1999-2000: Praktisches Jahr am
Elisabeth-Krankenhaus Mönchengladbach-Rheydt,
Kliniken Maria Hilf GmbH, Wahlfach: Neurologie
2000: 3. Staatsexamen
seit 01.2001: Ärztin im Praktikum in der
Neurologischen Klinik der MH Hannover

.


















