
Neuronal Grammar - Universität Ulm · Neuronal Grammar Phillipp Hehrmann [email protected]...
35
Neuronal Grammar Phillipp Hehrmann [email protected] Stefan Scherer [email protected] Fakult¨ at f¨ ur Informatik Universit¨ at Ulm Zusammenfassung Es wird ein neurowissenschaftlich motiviertes Modell der menschlichen Sprachverar- beitung vorgestellt. Functional Webs, speziell organisierte Verb¨ unde von Neuronen, werden als grundlegende funktionale Einheit der Informationsrepr¨ asentation und -verarbeitung im Gehirn vorgestellt. Aus verschiedenen, spezialisierten Formen von Functional Webs wie etwa Word Webs zu Repr¨ asentation von Worten oder Sequence Sets zur Erkennung von Wortfolgen wird schließlich ein Modell zusammengesetzt, welches – abseits der Pfade g¨ angiger, linguistischer Syntax-Theorien – das Verst¨ and- nis komplexer grammatikalischer Abh¨ angigkeiten erm¨ oglichen k¨ onnte. 8. Februar 2005
Transcript of Neuronal Grammar - Universität Ulm · Neuronal Grammar Phillipp Hehrmann [email protected]...

Neuronal Grammar
Phillipp Hehrmann
Stefan Scherer
Fakultat fur InformatikUniversitat Ulm
Zusammenfassung
Es wird ein neurowissenschaftlich motiviertes Modell der menschlichen Sprachverar-beitung vorgestellt. Functional Webs, speziell organisierte Verbunde von Neuronen,werden als grundlegende funktionale Einheit der Informationsreprasentation und-verarbeitung im Gehirn vorgestellt. Aus verschiedenen, spezialisierten Formen vonFunctional Webs wie etwa Word Webs zu Reprasentation von Worten oder SequenceSets zur Erkennung von Wortfolgen wird schließlich ein Modell zusammengesetzt,welches – abseits der Pfade gangiger, linguistischer Syntax-Theorien – das Verstand-nis komplexer grammatikalischer Abhangigkeiten ermoglichen konnte.
8. Februar 2005

Inhaltsverzeichnis
1 Grundlegende Syntax 4
1.1 Ersetzungsregeln 4
1.2 Zentrale Einbettung 5
1.3 Diskontinuierliche Satzbestandteile und verteilte Worter 6
1.4 Abhangigkeitsgrammatiken 7
1.5 Fragen zu einer neuronalen Grammatik 8
2 Grundlagen zur Sprachverarbeitung im Gehirn 8
2.1 Modellierung von Nervenzellen 9
2.1.1 Naturliche Neurone 9
2.1.2 Kunstliche Neuronenmodelle 11
2.2 Worte im Gehirn 13
2.2.1 Die Topologie des Kortex 13
2.2.2 Functional Webs 16
2.2.3 Word Webs 19
2.3 Neuronale Mechanismen zur Reprasentation serieller Ordnung 21
2.3.1 Grundsatzliche Uberlegungen anhand kunstlicher Neurone 21
2.3.2 Synfire Chains 24
2.3.3 Neuronale Sequenzdetektoren 26
3 Neuronale Grammatik 28
3.1 Erregungszustande von funktionellen Netzen 28
3.2 Neuronale Sets 29
3.3 Schwellwertkontrolle 30
3.4 Sequenzdetektion in Netzwerken von neuronalen Sets 31
3.5 Sequenzerkennung 31
2

3.6 Lexikale Kategorien in neuronalen Sets 33
3.6.1 Warum lexikale Kategorien? 33
3.6.2 Lexikale Kategorien als Mengen von Sequenz-Sets 33
3.6.3 Lexikale Unterscheidungen durch Sequenz-Sets 34
3.6.4 Anforderungen an eine neuronale ’Grammar Machine’ 34
Literatur 35
3

1 Grundlegende Syntax
In diesem Kapitel wird dargestellt, wie man Sprache auf formale Weise defi-nieren kann. Es werden die so genannten Phrasenstruktur-Grammatiken 1 unddie Abhangigkeitsgrammatiken behandelt. Außerdem geht dieses Kapitel dar-auf ein, welche Beschrankungen es bezuglich Sprachen bei neuronalen Netzengibt und was neuronale Netze bieten, was formalen Grammatiken fehlt.
1.1 Ersetzungsregeln
Eine einfache Methode um eine formale Syntax zu definieren sind die Erset-zungsregeln 2 . Eine Ersetzungsregel ist eine Formel, die ausdruckt, dass etwasersetzt werden kann durch etwas anderes. Eine Menge solcher Regeln wirdauch als Phrasenstruktur-Grammatik bezeichnet, wenn auf der linken Seitedes Pfeils immer nur ein Symbol und auf der rechten beliebig viele stehen,wird diese Grammatik auch als kontextfreie Grammatik bezeichnet. Der Pfeilx → y kann so gelesen werden: x wird ersetzt durch y, was auch den Namender Regeln erlautert. Eine Menge solcher Regeln wird nun als Grammatik be-zeichnet. Diese Grammatik hat die Einschrankung, dass sie die Bedeutung vonWortern vollig außer Acht lasst und nur auf die Syntax achtet. Nun folgt einBeispiel einer solchen Grammatik. In diesem Beispiel werden einige Abkurzun-gen bzw. Symbole benutzt, die fur gewisse syntaktische Kategorien stehen. Sbezeichnet das Startsymbol, NP die Nominalphrase, VP die Verbalphrase, Detist der Artikel und V das Verb. Ausdrucke in runden Klammern sind nicht obli-gatorisch und Ausdrucke in geschweiften Klammern sind dann die eigentlichenWorter im Satz.
(1) S → NP VP(2) NP → (Det) N(3) N → V (NP)(4) N → {Betty, machine}(5) V → {laughs, cleans, switches}(6) Det → {the, a}
Mit dieser kleinen Grammatik konnen sowohl sinnvolle als auch unsinnigeSatze gebildet werden, die aber vom syntaktischen Aspekt betrachtet durchausrichtig sind:
(1) Betty cleans the machine.(2) The Betty laughs machine.
1 Phrase structure grammars2 Rewriting rules
4

Zu diesem Zeitpunkt konnte man sich eigentlich fragen, ob ein neuronalesNetz, das eine neuronale Grammatik 3 beinhaltet, nicht gleichermaßen wieeine solche Phrasenstruktur-Grammatik fahig ist, dieses Problem zu losen.Das folgende Teilkapitel wird jedoch zeigen, dass dem nicht so ist.
1.2 Zentrale Einbettung
Der Vorteil, den kontextfreie Grammatiken besitzen, ist, dass die Regeln re-kursiv anwendbar sind und so Satze gebildet werden konnen, die syntaktischkorrekt und sehr komplex sind. Es wurde bewiesen[2], dass neuronale Netzeaquivalent zu regularen Grammatiken sind, die jedoch nicht so machtig sindwie kontextfreie. Kontextfreie Grammatiken ermoglichen es Satze zu bilden, indenen andere Satze eingebettet sind. Diese Einbettung von Satzen wird auchzentrale Einbettung 4 genannt. Nun folgen zwei Beispiele solcher Satze 5 :
(1) The rat (the cat (the dog chased) killed) ate the malt.(2) Anyone (1 who feels (2 that (3 if so many more students (4 whom we
haven’t actually admitted )4 are sitting in on the course (4 than ones wehave )4 (4 that the room had to be changed )4 )3 then probably auditorswill have to be excluded )2 )1 is likely to agree that the curriculum needsrevision.
Was diese Beispiele jedoch auch zeigen, dass es wahrscheinlich gar nicht notigist, dass man unendlich komplexes bzw. tiefes zentrales Einbetten von Satzenin einer neuronalen Grammatik ermoglichen muss, um Sprache sinnvoll ver-stehen zu konnen, da das menschliche Gehirn es auch nicht ohne Hilfe undlanges Nachdenken schafft, vor allem den zweiten Beispielsatz zu verstehen.Es scheint also irgendwo eine Art Grenze der verstandlichen Komplexitat zugeben. Diese Grenze wurde von einigen Wissenschaftlern bei drei eingebette-ten Satzen festgelegt[3]. Um einen Satz wie den ersten Beispielsatz sprechenzu konnen, ist es notig im Speicher bzw. im Gehirn die Information des bereitsGesprochenen zu behalten und zwar Reihenfolge und fehlende Teile. Kontext-freie Grammatiken sind dazu in der Lage, da bewiesen wurde, dass sie aquiva-lent zu einem Kellerautomaten, der unendlich viel Speicherplatz besitzt, sind.Dieser Automat funktioniert nach dem first-in last-out Prinzip und speichertso ein ’Spiegelbild’ des bereits Gesprochenen. Der Satz ist dann fertig, wennder Speicher wieder leer ist.
3 Neuronal grammar4 Center embedding5 Die Klammern wurden zur Verdeutlichung des Einbettens und zur Vereinfachungdes Verstandnisses eingefugt
5

Da es in vielen Sprachen vorkommt, dass zentral eingebettete Satze benutztwerden, muss man neuronale Netze anpassen und verandern, um dieser Bege-benheit gerecht zu werden. Eine Moglichkeit, die Tatsache, dass ein neuronalesNetz mit endlich vielen Neuronen nur endlich viele Zustande besitzt zu umge-hen, ware z.B. ein Wachstum des Netzes zu erlauben[4]. Dieses Wachstum istjedoch nicht unbedingt sehr realistisch, wenn man interne Prozesse des Gehirnsbeschreiben will, da ein unendliches Wachstum des Gehirns alleine durch denSchadelknochen beschrankt ist. Wie schon gesagt, ist dies aber auch nicht un-bedingt notig, da Sprache nur bis zu einer gewissen Komplexitat verstandlichist.
1.3 Diskontinuierliche Satzbestandteile und verteilte Worter
Die vorher definierte Grammatik besteht aus sechs Ersetzungsregeln, die allenicht eine relativ haufig auftretende grammatikalische Besonderheit beschrei-ben. Diese Besonderheit wird auch ein verteiltes Wort 6 genannt. ’Betty swit-ches the machine on.’ ist ein Beispiel fur dies. Es musste mindestens eineRegel verandert werden um dem verteilten Wort ’switches ... on’ gerecht zuwerden.
(1) VP → V (NP) (on)
Diskontinuierliche Satzbestandteile 7 sind ein Uberbegriff fur solche verteiltenWorter, die grammatikalisch gesehen eng miteinander verbunden sind, jedochin geschriebener Sprache durch den Raum bzw. in gesprochener Sprache durchdie Zeit getrennt sind. Die Problematik besteht darin, dass dieser Abstandunterschiedlich lange sein kann, wie folgende Beispiele zeigen:
(1) Betty switched it on.(2) Betty switched the coffee machine on.(3) Betty switched the nice brown coffee machine on.
Um nun solche Satze produzieren bzw. verstehen zu konnen, muss man In-formationen uber die vorherigen Worter speichern und das uber einen unbe-stimmten Zeitraum.
Eine weitere Schwierigkeit besteht darin, dass Worter wie ’switch’ und ’switch... on’ zwar den selben Wortstamm teilen jedoch eine vollig unterschiedlicheBedeutung besitzen, d.h. also, dass die Bedeutung des Wortes moglicherweiseerst am Ende des Satzes eindeutig bestimmt werden kann. Eine Grammatikwie:
6 Distributed word7 Discontinuous constituents
6

(1) a → A b(2) b → switches c(3) c → B d(4) d → C (on),
ermoglicht es zwar, das verteilte Wort ’switches ... on’ zu schreiben, ist jedochnicht korrekt im eigentlichen Sinne, da in (2) schon entschieden werden muss,welches der beiden wortstammgleichen Worter gewahlt wird. Um dies zu losen,wurde vorgeschlagen die Worter genauer zu unterscheiden und mehr lexikaleUntergruppen zu benutzen, wie z.B. ’switch ... on’ in die Gruppe der tran-sitiven Verben mit Partikel zu stecken und das Wort ’switch’ in die Gruppeder transitiven Verben ohne Partikel. Eine Grammatik, die das berucksichtigtware z.B.:
(1) b → switches1 c1(2) c1 → B d1(3) d1 → C on(4) b → switches2 c2(5) c2 → B d2(6) d2 → C
Hierbei wird sichergestellt, dass falls ’switches1’ gewahlt wird, der Partikel’on’ am Ende gesetzt werden muss.
Ein Nachteil dieser Losung fur das Problem der diskontinuierlichen Satzbe-standteile ist jedoch die hohere Komplexitat der Grammatik: Wenn angenom-men wird, dass die Zahl der diskontinuierlichen Satzbestandteile n ist, dannwurde die Zahl der Ersetzungsregeln mit 2n erhoht werden.
1.4 Abhangigkeitsgrammatiken
Ein anderer Ansatz, um Grammatiken zu beschreiben, ist, Worter in ihre lexi-kalen Kategorien einzuteilen und sie in Abhangigkeiten zu ihren Komplemen-ten - die notwendigen Zusatze fur diese Worter - zu stellen. Solche Abhangig-keitsgrammatiken 8 konnen mit Abhangigkeitsregeln beschrieben werden, de-ren Notation folgendermaßen erfolgt: Auf der linken Seite einer runden Klam-mer steht die Kategorie des Wortes, das gerade behandelt wird. In den Klam-mern befindet sich ein Stern-Symbol /*/, welches den Platz des aktuellenWortes besetzt. Links davon stehen eine bzw. mehrere abhangige lexikale Ka-tegorien, die im Satz zeitlich bzw. raumlich vor dem aktuellen Wort stehenund analog hierzu stehen auf der rechten Seite eine bzw. mehrere lexikaleKategorien, die danach auftreten. Eine Beispielgrammatik ware nun:
8 Dependency grammars
7

(1) V14par (N1 /*/ N4 Par)(2) V14 (N1 /*/ N4)(3) V1 (N1 /*/)(4) N1 ((Det) /*/)(5) N4 ((Det) /*/)(6) V14par {switches}(7) V14 {cleans}(8) V1 {laughs}(9) V14par {Betty, machine}
(10) Par {on},
wobei V14par ein transitives Verb mit Partikel, V14 ein transitives Verb, V1ein intransitives Verb, N1 ein Nomen im Nominativ, N4 ein Nomen im Akkusa-tiv und Par einen Partikel bezeichnen. Beispielsatze, die mit dieser Grammatikgebildet werden konnen, sind:
(1) Betty cleans the machine.V14 (N1 /*/ N4 (Det /*/))
(2) Betty switched the machine on.V14par (N1 /*/ N4 (Det /*/) Par)
1.5 Fragen zu einer neuronalen Grammatik
Nachdem nun gezeigt wurde, wie man mit formalen Grammatiken diverseBegebenheiten verschiedener Sprachen losen kann stellen sich nun einige sehrbedeutende Fragen zu neuronalen Grammatiken:
(1) Wie lasst sich die zentrale Einbettung realisieren bzw. wie kann kann mansolche Satze reprasentieren?
(2) Wie konnen diskontinuierliche Satzbestandteile bzw. verteilte Worter rea-lisiert werden?
(3) Wie kann der wiederholte Gebrauch ein und desselben Wortes in einemSatz gespeichert werden?
(4) Wie konnen lexikale Kategorien reprasentiert werden?
Zu all diesen Fragen versuchen wir in den folgenden Kapiteln Antworten bzw.Losungsvorschlage zu finden.
2 Grundlagen zur Sprachverarbeitung im Gehirn
In diesem Abschnitt werden – ausgehend von neurophysiologischen Beobach-tungen – mogliche Modelle zur Reprasentation und Verarbeitung von Infor-
8

mationen und Strukturen vorgestellt, wie beim Verstandnis und bei der Pro-duktion naturlicher Sprache auftreten. Zunachst wird ein vereinfachtes Neuro-nenmodell vorgestellt, das die Grundlage aller weiteren Uberlegungen bildet.Im zweiten Abschnitt wird diskutiert, wie Worter und ihre Semantik im Ge-hirn reprasentiert werden konnten. Zuletzt werden wir einige Mechanismenbetrachten, die darauf aufbauend das Verstandnis grammatikalischer Struktu-ren ermoglichen. Der Schwerpunkt liegt auf einer konzeptuellen Darstellung,die die Entwicklung eines konkreten Modells in Abschnitt 3 vorbereiten undals dessen biologische Rechtfertigung dienen soll.
2.1 Modellierung von Nervenzellen
2.1.1 Naturliche Neurone
Hauptinformationstrager und -prozessor des menschlichen Gehirns sind Neu-rone – grob geschatzt 1011 dieser hochspezialisierten Zellen beinhaltet un-ser Zentralnervensystem. Zwar finden sich in unserem Nervensystem danebenauch noch andere Zelltypen wie etwa Glia-Zellen (Astrozyten, Schwann’scheZellen u.a.). Jedoch dienen diese heutigem Wissen nach vor allem der Auf-rechterhaltung des Stoffwechselhaushalts sowie als mechanisches Stutzgerustder informationsverarbeitenden Nervenzellen.
A.
B.
Abbildung 1. A. Schematische Darstellung eines Neurons (nach [8]). B. SynaptischeKontakte (aus [6])
9

Spezialisiert sind Neurone hinsichtlich der Erzeugung und Ubertragung elek-trischer Impulse in Reaktion auf elektrochemische Reize aus ihrer Umgebung.Abbildung 1 zeigt vereinfacht die Anatomie einer solchen Zelle. Zur Aufnah-me externer Reize dienen dem Neuron die Dendriten, ein oftmals baumartigverzweigter Zellauswuchs bestehend aus Nervenfasern. Die Erzeugung eineseigenen Reizimpulses (Aktionspotenzials) erfolgt – unter bestimmten Bedin-gungen, s.u. – am Axonhugel, dem Ubergang zwischen Zellkorper (Soma) undAxon. Das Axon ist ein weiterer, faserartiger Auswuchs der Zelle, welcher derUbertragung der am Axonhugel erzeugten Aktionspotenziale an andere Neu-rone dient. Die Impulsleitung erfolgt je nach Zelltyp mit Geschwindigkeitenzwischen 1 und 100 m/s. Auch das Axon ist oftmals stark verzweigt. Am Endeeines jeden Astes befindet sich eine Kontaktstelle zu einer dendritischen Fasereines weiteren Neurons. An diesen sog. Synapsen konnen elektrische Impul-se von Zelle zu Zelle ubertragen werden: bei Eintreffen eines prasynaptischen(axonalen) Impulses werden chemische Botenstoffe – sog. Neurotransmitter– freigesetzt. Durch spezifische Rezeptoren kann die postsynaptischen Zellenun auf den Impuls reagieren. Das Eintreffen eines chemischen Reizes beein-flusst die Wahrscheinlichkeit, mit der am postsynaptischen Dendrit wiederumein elektrischer Impuls erzeugt wird, der dann entlang der dendritische Faserbis zum Zellkorper der postsynaptische Zelle wandert. Erhohen die Impulsean einer Synapse die Wahrscheinlichkeit einer postsynaptischen Impulserzeu-gung, so nennt man die Synapse erregend oder exzitatorisch. Verringert siehingegen die Wahrscheinlichkeit, so spricht man von einer hemmenden oderauch inhibitorischen Synapse. Die Bedingung fur die Erzeugung eines Impul-ses am Axonhugel schließlich ist das gleichzeitige Eintreffen ausreichend vielerImpulse aus den Dendriten der Zelle. Wird hierbei eine bestimmt Schwelleuberschritten, so feuert die Zelle und der Impuls wandert entlang des Axonszu den Dendriten weiterer Zellen und so weiter und so fort.
Soweit ist das Funktionsprinzip einer einzelnen Nervenzelle – trotz aller hassli-chen, biochemischen Details, versteht sich – vergleichsweise einfach. Die großeLeistungsfahigkeit unseres Nervensystems entsteht aus dem hohen Vernet-zungsgrad der Neurone untereinander: jede der 1011 Zellen steht mittels synap-tischer Verbindungen in Kontakt mit durchschnittlich 1.000–10.000 weiterenZellen. Dabei entstehen zahllose Ruckkopplungsschleifen, fernerhin sind dieEinflussstarken der Zellen aufeinander variabel. Eine zentrale Frage ist da-her die nach den Mechanismen neuronaler Plastizitat, d.h. nach welchen Re-geln sich die Verbindungsstarken zwischen einzelnen Neuronen im Laufe derZeit verandern. Obwohl die Details dieser Vorgange auch heute noch teilwei-se unklar sind, so scheint sich doch die fruhe Hypothese des amerikanischenPsychologen Donald Hebb prinzipiell bestatigt zu haben:
Wenn ein Axon der Zelle A einer Zelle B nahe genug ist, diese zu erregenund sich wiederholt und dauerhaft an deren Impulserzeugung beteiligt, sogeschieht ein Wachstumsprozess oder eine metabolische Veranderung in ei-
10

ner oder beiden Zellen, wodurch sich die Effizienz von A bezuglich ihrerWirkung auf B erhoht.
D. Hebb, 1949
Besteht also eine hohe, positive Korrelation zwischen der Aktivitat von ZelleA und Zelle B, so wird die Verbindung der beiden langfristig Verstarkt. DieserProzess wird auch als long-term potentiation, kurz: LTP, bezeichnet. Daruberhinaus hat man durch Zellstimulationsexperimente auch noch einen gegentei-ligen Prozess festgestellt: besteht keine oder nur geringe Korrelation zwischender Aktivitat von A und B, so nimmt die Verbindungsstarke ab. Man sprichthierbei analog von LTD, long-term depression, oder auch anti-Hebb´schemLernen. Da die Feuerfrequenz einer Zelle nie negativ ist, gilt die Hebb‘scheRegel in dieser Formulierung allerdings hochstens fur erregende Synapsen.Zwar ließe sich die Regel problemlos fur hemmende Synapsen umformulieren.Die Faktenlage ist jedoch in diesem Fall sparlicher und weit weniger eindeu-tig([11]), so dass eine solche Behauptung aus neurophysiologischer Sicht nochnicht ausreichend gestutzt ware.
2.1.2 Kunstliche Neuronenmodelle
Bei naherer Betrachtung liegen der neuronalen Reizleitung – im vorigen Ab-schnitt weitestgehend ubergangen – komplexe bio- und elektrochemische Pro-zesse zu Grunde. Zwar sind biologisch realistische mathematische Beschrei-bungen dieser Prozesse bekannt. Will man aber das Verhalten eines Neuronsauf diese Weise simulieren, so kommt man nicht umhin, ein verschachteltesSystem von Differentialgleichungen zu berechnen – der Rechenaufwand uber-schreitet bei wachsender Anzahl zu simulierender Neurone schnell die Grenzendes Machbaren. Daher sind vereinfachte Modelle notwendig, die die Simulati-on einer großen Anzahl von Neuronen erlauben. Das alteste dieser abstraktenNeuronenmodelle ist die McCulloch-Pitts-Zelle. Eine solche Zelle hat n binareEingangsleitungen x = x1 . . . xn (entsprechend den Dendriten) sowie einenebenfalls binaren Ausgang (analog zum Axon), dessen Wert sich als Funkti-on f(x) der Eingange berechnet. Des weiteren ist jeder Eingang mit +1 oder−1 gewichtet (entsprechend erregenden und hemmenden Synapsen), zusam-mengefasst als bipolarer Gewichtsvektor w = w1 . . . wn. Zur Berechnung derAusgabe werden die gewichteten Eingangssignale summiert. Uberschreitet dieSumme einen bestimmten Schwellwert θ ∈ R, so ist die Ausgabe 1, andernfalls0. Es ist also
f(x) =
1, falls∑n
i=1 wixi ≥ θ
0, sonst(1)
11

Abbildung 2 zeigt beispielhaft McCulloch-Pitts-Zellen fur die booleschen Funk-tionen UND, ODER und NICHT. Offenbar kann man diese genau wie herkomm-liche logische Gatter parallel und in Reihe schalten und so jede beliebigeboolesche Funktion berechnen – auch wenn ein einzelnes Neuron nur lineartrennbare Funktionen berechnen kann. Lasst man – im Gegensatz zu reinen
Abbildung 2. UND, ODER und NICHT Neurone
Feed-Forward-Architekturen – Ruckkopplungsschleifen zu, so existiert zu jederregularen Sprache ein McCulloch-Pitts-Netz, welches diese erkennt. Genauergesagt sind McCulloch-Pitts-Netze (mit endlicher Neuronenzahl) und endlicheAutomaten als Berechenbarkeitsmodell genau gleich machtig. Die Machtigkeitdes McCulloch–Pitts–Modells andert sich auch dann nicht, wenn man als Ge-wichte wi anstelle von ±1 beliebige reelle Zahlen zulasst. Abbildung 3 demon-striert, wie sich ein Neuron mit reellwertigen Gewichten in ein aquivalentesNeuron mit bipolaren Gewichten umwandeln lasst. In Anbetracht der Dis-
Abbildung 3. Uberfuhrung reeller in bipolare Gewichte
kussion um Hebbs Regel in Abschnitt 2.1.1 ist es jedoch offensichtlich, dassohne eine derartige Erweiterung des Gewichtsvektors eine biologisch plausibleModellierung synaptischer Plastizitat nicht gelingen kann. Auch hat sich ne-ben der hier vorgestellten, binaren Schwellwertfunktion eine Vielzahl andererAusgabefunktionen eingeburgert, darunter beispielsweise die Sigmoide
x 7→ 1
1 + e−x(2)
oder lineare Funktionen mit Schwellwert (siehe Abb. 4):
x 7→
c(x− θ), falls x ≥ θ
0, sonst(3)
Dadurch andert sich auch die Interpretation des Ausgabewertes in Bezug aufden Aktivitatszustand des Neurons. Anstelle einzelner Impulse reprasentiert
12

Abbildung 4. verschiedene neuronale Ausgabefunktionen: A. Binar mit Schwellwert,B. Sigmoid, C. Linear mit Schwellwert (aus [7])
der Ausgabewert bei diesen Funktionen vielmehr die Impulsfrequenz, gemit-telt uber einen gewissen Zeitraum – im Falle der Sigmoiden zusatzlich nor-miert, wobei ein Ausgabewert von 1 der maximalen Feuerfrequenz der Zelleentsprache, unabhangig von deren absolutem Wert.
Der Vollstandigkeit halber sei an dieser Stelle noch angedeutet, wie Hebb’sches(LTP) und anti-Hebb’sches (LTD) Lernen auf kunstliche Neuronen ubertra-gen werden kann (vgl. Anschnitt 2.1.1) . Eine einfache Form von LTP lasstsich folgendermaßen formulieren: Sei wij ∈ R das Gewicht der Verbindungvon Neuron i zu j, und seien ai und aj deren Aktivitat zum Zeitpunkt desLernschrittes. Dann betragt die Veranderung des Gewichtes
∆wij = ηaiaj, (4)
mit Lernrate η. Da ai und aj in natura immer positiv sind (und auch beiallen hier vorgestellten Ausgabefunktionen), kann wij nur standig großer wer-den. Ausgeklugeltere Lernregeln ermoglichen sowohl LTP als auch LTD, auchbei rein positiven Ausgabefunktionen. Beispiele hierfur sind u.a. SejnowskisKovarianz-Regel, Ojas Regel oder die BCM-Regel, siehe [5] fur einen umfas-senden Uberblick.
2.2 Worte im Gehirn
2.2.1 Die Topologie des Kortex
Der fur Sprachproduktion und -verstandnis entscheidende Teil des Gehirnsist elektrophysiologische, bildgebenden sowie Lasionsstudien zufolge die Rin-de des Großhirn, der cerebrale Kortex. Daneben ermoglicht der Kortex nochzahlreiche weitere Formen
”hoheren Denkens“ wie z.B. alle Formen bewusster
sinnlicher Wahrnehmung und bewussten Handelns sowie Lang- und Kurzzeit-gedachtnis. Der Kortex ist in zwei Halften (Hemispharen) geteilt, die nur uberdie Nervenfasern des sog. Balkens (Corpus callosum) miteinander verbundensind. Dabei ist eine der beiden Hemispharen – bei ca. 90% der Menschen die
13
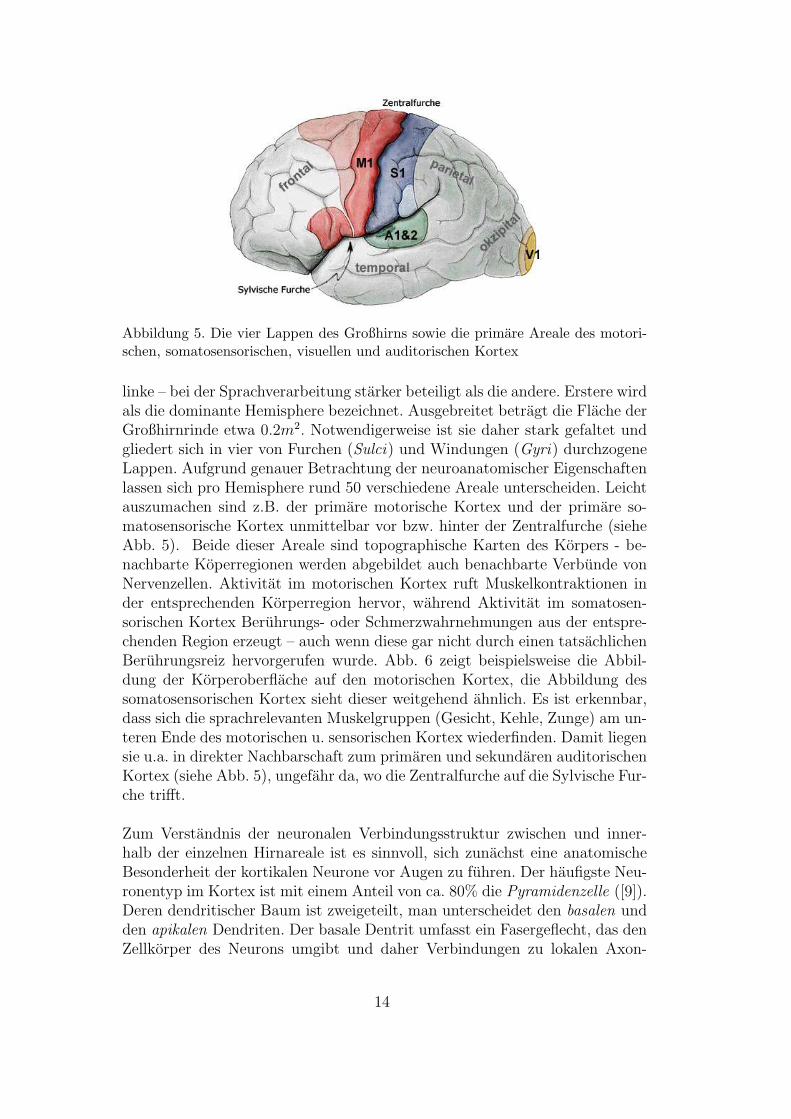
Abbildung 5. Die vier Lappen des Großhirns sowie die primare Areale des motori-schen, somatosensorischen, visuellen und auditorischen Kortex
linke – bei der Sprachverarbeitung starker beteiligt als die andere. Erstere wirdals die dominante Hemisphere bezeichnet. Ausgebreitet betragt die Flache derGroßhirnrinde etwa 0.2m2. Notwendigerweise ist sie daher stark gefaltet undgliedert sich in vier von Furchen (Sulci) und Windungen (Gyri) durchzogeneLappen. Aufgrund genauer Betrachtung der neuroanatomischer Eigenschaftenlassen sich pro Hemisphere rund 50 verschiedene Areale unterscheiden. Leichtauszumachen sind z.B. der primare motorische Kortex und der primare so-matosensorische Kortex unmittelbar vor bzw. hinter der Zentralfurche (sieheAbb. 5). Beide dieser Areale sind topographische Karten des Korpers - be-nachbarte Koperregionen werden abgebildet auch benachbarte Verbunde vonNervenzellen. Aktivitat im motorischen Kortex ruft Muskelkontraktionen inder entsprechenden Korperregion hervor, wahrend Aktivitat im somatosen-sorischen Kortex Beruhrungs- oder Schmerzwahrnehmungen aus der entspre-chenden Region erzeugt – auch wenn diese gar nicht durch einen tatsachlichenBeruhrungsreiz hervorgerufen wurde. Abb. 6 zeigt beispielsweise die Abbil-dung der Korperoberflache auf den motorischen Kortex, die Abbildung dessomatosensorischen Kortex sieht dieser weitgehend ahnlich. Es ist erkennbar,dass sich die sprachrelevanten Muskelgruppen (Gesicht, Kehle, Zunge) am un-teren Ende des motorischen u. sensorischen Kortex wiederfinden. Damit liegensie u.a. in direkter Nachbarschaft zum primaren und sekundaren auditorischenKortex (siehe Abb. 5), ungefahr da, wo die Zentralfurche auf die Sylvische Fur-che trifft.
Zum Verstandnis der neuronalen Verbindungsstruktur zwischen und inner-halb der einzelnen Hirnareale ist es sinnvoll, sich zunachst eine anatomischeBesonderheit der kortikalen Neurone vor Augen zu fuhren. Der haufigste Neu-ronentyp im Kortex ist mit einem Anteil von ca. 80% die Pyramidenzelle ([9]).Deren dendritischer Baum ist zweigeteilt, man unterscheidet den basalen undden apikalen Dendriten. Der basale Dentrit umfasst ein Fasergeflecht, das denZellkorper des Neurons umgibt und daher Verbindungen zu lokalen Axon-
14
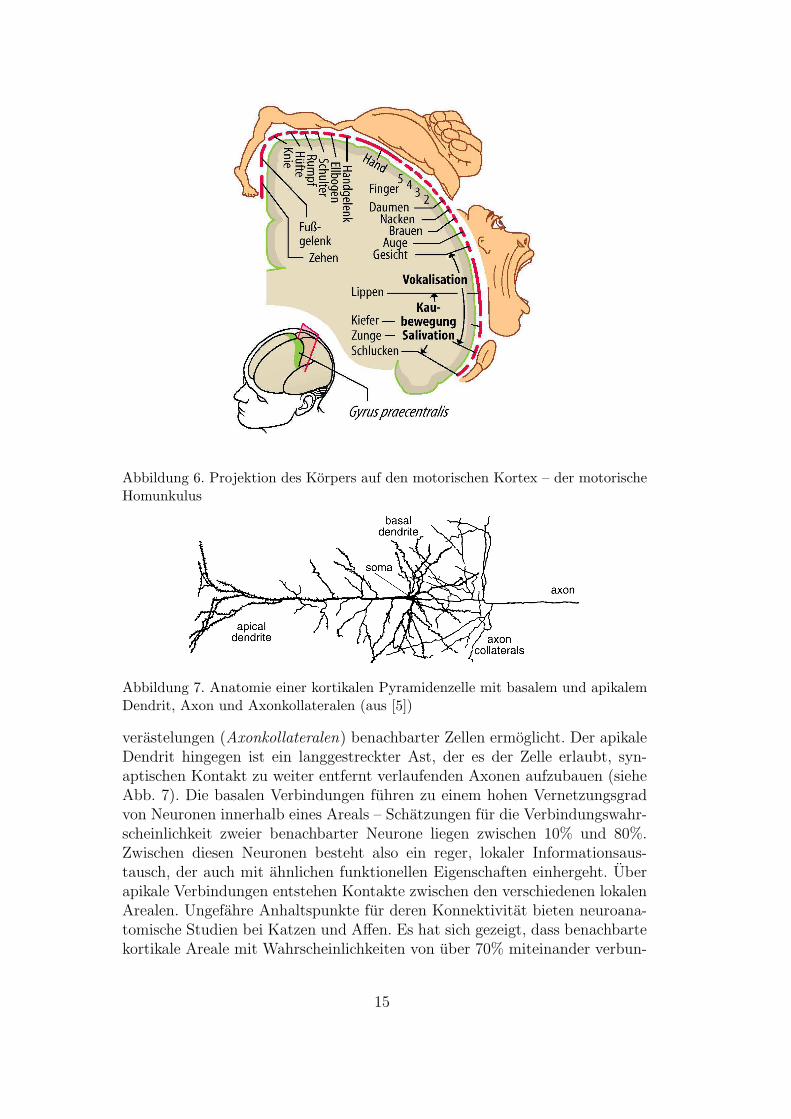
Abbildung 6. Projektion des Korpers auf den motorischen Kortex – der motorischeHomunkulus
Abbildung 7. Anatomie einer kortikalen Pyramidenzelle mit basalem und apikalemDendrit, Axon und Axonkollateralen (aus [5])
verastelungen (Axonkollateralen) benachbarter Zellen ermoglicht. Der apikaleDendrit hingegen ist ein langgestreckter Ast, der es der Zelle erlaubt, syn-aptischen Kontakt zu weiter entfernt verlaufenden Axonen aufzubauen (sieheAbb. 7). Die basalen Verbindungen fuhren zu einem hohen Vernetzungsgradvon Neuronen innerhalb eines Areals – Schatzungen fur die Verbindungswahr-scheinlichkeit zweier benachbarter Neurone liegen zwischen 10% und 80%.Zwischen diesen Neuronen besteht also ein reger, lokaler Informationsaus-tausch, der auch mit ahnlichen funktionellen Eigenschaften einhergeht. Uberapikale Verbindungen entstehen Kontakte zwischen den verschiedenen lokalenArealen. Ungefahre Anhaltspunkte fur deren Konnektivitat bieten neuroana-tomische Studien bei Katzen und Affen. Es hat sich gezeigt, dass benachbartekortikale Areale mit Wahrscheinlichkeiten von uber 70% miteinander verbun-
15

den sind. Fur nichtbenachbarte Areale betragt die Wahrscheinlichkeit nur noch15–30%. Dabei ist zu beachten, dass fast alle Verbindungen zwischen verschie-denen Arealen – sofern sie denn existieren – wechselseitig sind, Informationenalso jeweils sowohl empfangen wie auch gesendet werden. Eine weitere Beob-achtung ist, dass die primaren Areale untereinander nicht direkt verbundensind - der motorische und der somatosensorische Kortex bilden hierbei dieeinzige Ausnahme. Die ubrigen Verbindung bestehen allenfalls indirekt uberUmschaltneurone in nicht-primaren Arealen. Dadurch werden aber gleichzeitigauch komplexere Verschaltungen und damit Informationsverarbeitungsschrittemoglich – ahnlich wie beispielsweise Perzeptrone mit zwei Berechnungsschich-ten komplexere Funktionen berechnen konnen also solche mit nur einer.
2.2.2 Functional Webs
Zieht man nun zu diesen grundlegenden Beobachtungen uber die Verbindungs-strukturen innerhalb des Kortex die in Abschnitt 2.1.1 vorgestellten Mecha-nismen synaptischer Plastizitat – LTP und LTD – in Betracht, so wird manerwarten, dass haufig auftretende Aktivitatsmuster zur Starkung der Gewich-te zwischen den beteiligten Neuronen fuhren. Dies kann sowohl innerhalbeinzelner Areale geschehen, durch die apikalen Verbindungen aber auch inraumlich getrennten Regionen. Sogar die indirekte Verbindung zweier haufigkoaktivierter Neurone aus unterschiedlichen primaren Arealen konnte auf die-se Weise verstarkt werden – durch Verstarkung der jeweiligen Verbindungenzu einem gemeinsamen Umschaltneuron. Korrelation zwischen der Aktivitatbenachbarter Neurone ist aufgrund der allgemein verbreiteten, topographi-schen Form der Reprasentation zu erwarten: benachbarte Neurone innerhalbeines der Wahrnehmungsareale beispielsweise reagieren auf ahnliche Reize.Aber auch selektive Korrelationen zwischen Neuronen verschiedener Arealesind leicht vorstellbar: jedes Mal zum Beispiel, wenn wir durch Aktivitat immotorischen Kortex unsere Stimmbander aktivieren, entstehen im somatosen-sorischen Kortex Empfindungsreize durch die hervorgerufenen Vibrationen.Und mit zeitlicher Verzogerung schlagt sich die Schallwahrnehmung des eige-nen Lautes als Aktivitat im auditorischen Kortex nieder. Vergleichbare Bei-spiele lassen sich auch fur viele weitere Sinnesmodalitaten finden, wie etwagemeinsam auftretenden Geschmacks- und Konsistenzempfindungen beim Es-sen und vieles andere mehr. Auf diese Weise kann es zur Bildung von uberden Kortex verteilten Netzwerken kommen, die haufig zusammen auftretendeAktivitats- oder Reizmuster reprasentieren. Im folgenden werden Netze dieserArt als Functional Web bezeichnet. Zusammengefasst sind Functional Websdefiniert als eine Gruppe von Neuronen, die
(1) untereinander hochgradig vernetzt sind(2) uber mehrere kortikale Hirnareale verteilt sind(3) zusammen als funktionale Einheit wirken
16

(4) voneinander funktional abhangig sind in dem Sinne, dass jedes einzelnefur das optimale Funktionieren des Netzes notwendig ist.
Abgesehen davon, dass sich dieses Modell kortikaler Reprasentation auf naturli-che Weise aus den bereits vorgestellten Prinzipien neuronaler Plastizitat undkortikaler Konnektivitat ergibt, bringt sie auch aus theoretischer Sicht eineReihe von Vorteilen mit sich. Ein Vorteil der Functional Webs liegt in derstochastischen Natur des Feuerverhaltens einzelner Neurone begrundet. DaNeurone unabhangig von externer Stimulation immer wieder spontan Akti-onspotentiale erzeugen, haben sie fur sich genommen ein schlechtes Signal-Rausch-Verhaltnis. Durch Mittelung der Signale mehrerer Neurone lasst sichdieses Verhaltnis entscheidend verbessern. Das Rauschen hebt sich mit wach-sender Anzahl von Neuronen gegenseitig, wahrend das Signal erhalten bleibt.Ebenso ist es eine wunschenswerte Eigenschaft, dass der Verlust eines einzel-nen Neurons nicht den Verlust einer gesamten Reprasentation nach sich zieht.
Wichtig ist es auch, dass zwei Functional Webs nicht aus disjunkten Mengenvon Neuronen bestehen mussen, eine Neuron also Teil mehrerer FunctionalWebs sein. Dies zieht einige Konsequenzen nach sich. Zum einen wird es durchden Uberlapp schwierig zu bestimmen, welche Neurone tatsachlich Teil einesgegebenen Functional Webs sind. Diese Form der Reprasentation beinhaltetalso zwangslaufig eine gewisse Unscharfe (Fuzziness). Zum zweiten gilt es zuvermeiden, dass sich die Functional Webs im Cortex durch ihren wechselseiti-gen Uberlapp nach und nach alle gegenseitig aktivieren. Hierzu bedarf es einesRegulationsprozesses, der sicherstellt, dass der gemittelte kortikale Aktivie-rungsgrad bestimmt Ober- und Untergrenzen nicht uberschreitet (ein volligesAussterben jeglicher Aktivitat ware naturlich ebenso unerwunscht). Prinzipi-ell lasst sich dies durch zusatzliche, inhibitorische Verbindungen der Neuroneuntereinander erreichen, so dass nach dem
”Winner–Takes–All“–Prinzip nur
wenige, maximal aktivierte Functional Webs ihre Aktivitat aufrecht erhaltenkonnen. Ein genaueres Modell fur einen derartigen Mechanismus unter Einbe-ziehung anatomisch bekannter, subkortikaler Ruckkopplungswege findet sichin [1].
Was nun sind die bislang weitgehend ubergangenen, funktionalen Eigenschaf-ten von Functional Webs? Was außer einer etwas stabileren Reprasentationgewinnt man durch sie gegenuber einzelnen Neuronen? Kurz gesagt liegt derentscheidende Vorteil von Functional Webs in ihrer großeren Anzahl qualitativverschiedener Aktivitatszustande. Betrachten wir zunachst ein solches Netz,bei dem ein ausreichend großer Anteil von Neuronen extern stimuliert wird.Durch die die wechselseitigen, erregenden Verbindungen innerhalb des Net-zes kommt es zur Ausbreitung und gegenseitigen Verstarkung der Aktivitatim gesamten Netz, bis schließlich alle enthaltenen Neurone maximal Aktiviertsind. Dieser Vorgang wird als
”Ignition“ (Zundung) des Netzes bezeichnet.
Nutzlich ist dies zur automatischen Erganzung unvollstandiger Reprasentati-
17

Abbildung 8. Verlauf der Aktivitat eines Neurons aus dem temporalen Kortex einesMakaken wahrend eines visuellen Gedachtnistests(aus [12])
onsmuster: man stelle sich beispielsweise ein Functional Web vor, das das se-mantische Konzept
”Mucke“ reprasentiert. Allein die Wahrnehmung des sum-
menden Fluggerausches konnte dann ausreichen, um uns durch Ignition desNetzes sofort alle semantischen Aspekte einer Mucke ins Bewusstsein zu rufen,vom Aussehen eines Facettenauges bis hin zur Abneigung gegen den Juckreiz.Bei der Ignition handelt es sich allerdings nur um eine kurzzeitige vollstandi-ge Aktivierung. Anschließend kommt es durch
”Ermudung“ der Neurone und
moglicherweise auch entgegenwirkenden, regulatorischen Prozessen zu einerReduzierung der Aktivitat im gesamten Web. Dennoch kommt es nicht zueinem plotzlichen Aussterben, da wiederum die gegenseitigen Verbindungenals positive Ruckkopplung wirken und den Abfallsprozess stark abbremsenbzw. die Aktivitat teilweise auf niedrigerem Level aufrechterhalten konnen.Dieser Prozess des allmahlichen Aktivitatsruckgangs wird als
”Reverberation“
(Nachhall) bezeichnet. Im Elektroenzephalogramm (EEG) oder Magnetoen-zephalogramm (MEG) schlagt sich dieser Vorgang in Form hochfrequenterHirnwellen (∼ 30 Hz) nieder. Experimentell konnten derartige Phanomeneu.a. bei Gedachtnistests mit Affen nachgewiesen werden ([12]). Die Aufgabefur den Affen bestand darin, die Form eines nur kurz (0.2s) prasentierten,visuellen Stimulus 16s lang bis zur Prasentation eines Vergleichsstimulus imGedachtnis zu behalten und bei Ubereinstimmung bzw. Verschiedenheit einenentsprechenden Schalter zu betatigen. Abbildung 8 zeigt den Aktivitatsverlaufeines einzelnen Neurons aus dem temporalen Kortex eines Affen. Klar erkenn-bar ist die starke Aktivierung des Neurons gleich nach Prasentation des erstenStimulus, entsprechend der Ignition seines Functional Webs. Darauf folgt einePhase reduzierter, aber nur schwach abfallender Aktivitat bis zur Prasentati-on des Vergleichsstimulus. Diese wurde durch die anschließende Reverberationdes Functional Webs erklart. Im ubrigen gibt dieses Beispiel auch eine Vor-stellung uber den moglichen zeitlichen Umfang des Reverberation-Prozesses.
18

2.2.3 Word Webs
In diesem Abschnitt soll nun vertieft werden, wie Worter mit ihren phone-tischen wie semantischen Aspekten im Gehirn mittels Functional Webs re-prasentiert werden konnten. Die Ausgangsbasis fur eine derartige Reprasen-tation wird bereits im fruhesten Neugeborenenalter geschaffen, besonders mitBeginn der sog. Lallphase im Alter von etwa 6 Monaten. Aktivitat im unterenTeil des primaren motorischen Kortex lost spontane Lautaußerungen aus, dieuber den Umweg des Gehors Aktivitat in den auditorischen Hirnregionen her-vorrufen. Da bei gleicher Stellung des Stimmorgane auch immer ein ahnlicherLaut erzeugt wird, konnen durch Hebb’sches Lernen Netze von Neuronen ent-stehen, die sowohl die akustischen als auch die motorischen Aspekte der haufigerzeugten Phoneme verbinden. Damit ermoglichen sie das imitierende Nach-sprechen wahrgenommener Silben oder Worte – ein Fahigkeit, die Sauglingebereits kurz nach Beginn der Lallphase entwickeln. Durch
”ubergeordnete“
Verbindungen zwischen einzelnen Phonemnetzen konnen dann Reprasentatio-nen fur haufig auftretende Folgen von Phonemen entstehen - der Grundsteinfur neuronale Wortreprasentationen ist gelegt. Eine berechtigte Frage ist, wieder Saugling aus der Wahrnehmung eines kontinuierlichen Sprachflusses ler-nen kann, welche Phoneme uberhaupt zu einem Wort gehoren. Denn nur inden seltensten Fallen werden ihm ja einzelne Worter isoliert vorgesprochen.Auch dies lasst sich jedoch mit Hilfe des Korrelations-Lernprinzips erklaren:Phonemfolgen, die zu ein und dem selben Wort gehoren, tauchen statistischgesehen haufiger auf als die eher zufalligen Lautfolgen, die an Wortgrenzenentstehen. Zur Bildung von Wortreprasentationen scheint es also das Vorspre-chen isolierter Worte nicht notwendig zu sein.
Das bis hierhin vorgeschlagenen Modell wurden auch in elektrophysiologischenStudien experimentell uberpruft. Eine der Vorhersagen des Modells ist die,dass die Wahrnehmung von echten Wortern und Pseudowortern zu unter-schiedlichen Hirnreaktionen fuhrt. Genauer gesagt sollte ein echtes Wort Igni-tion und Reverberation des entsprechenden Wortnetzes hervorrufen, wahrendPseudoworter zu keinerlei solchen Reaktionen fuhren sollten. Tatsachlich wur-den bei EEG- und MEG-Studien bei echten Worten signifikant starkere, hoch-frequente Hirnwellen gemessen als bei Pseudowortern, und zwar nahe der Syl-vischen Furche, wo sich sowohl der auditorische Kortex als auch die Sprachre-levanten motorischen und somatosensorischen Projektionsfelder befinden (vgl.Abschnitt 2.2.1). Somit stimmt das experimentelle Ergebnis also mit der ge-troffenen Vorhersage uberein.
Durch den haufigen Gebrauch von Wortern im Zusammenhang mit bestimm-ten Gegenstanden oder Handlungen konnen die bislang ausschließlich pho-nologischen Wortreprasentationen durch das Prinzip des Korrelationslernensmit semantischen Inhalten verknupft werden. Dies konnten etwa Farbe, Griff-gefuhl und Geruch eines Gegenstandes aus der Umgebung des Sauglings sein,
19

Abbildung 9. Topologie mutmaßlicher Word Webs zur Reprasentation von Worternverschiedener semantischer Kategorien
aber ebenso auch der Bewegungsablauf beim Krabbeln. Functional Webs, dieaus diese Weise sowohl die phonologische Information eines Wortes als auchdessen zugehorige Wahrnehmungs- und Handlungsaspekte beinhalten, werdenim folgenden auch als Word Webs bezeichnet. Word Webs sollten gemaß derbisher angewandten Prinzipien der kortikalen Organisation nicht unspezifischuber den Kortex verteilt sein. Vielmehr wurde man erwarten, dass sich se-mantische Aspekte eines Begriffs in der Topologie seines zugehorigen WordWebs widerspiegelt. Die Bedeutung von Worten wie z.B.
”Hai“ oder
”Wal“
werden den meisten von uns in erster Linie durch visuelle Eindrucke (sprich:Bilder oder Filme) bekannt sein. Im Gegensatz dazu ergibt sich die Bedeutungetwa von Werkzeugnamen viel eher aus der mit ihren Benutzung verbunde-nen Handlungen. Word Webs fur vornehmlich visuell gepragte Begriffe solltendaher uberproportional viele Neurone aus den okzipital gelegenen visuellenArealen enthalten, Word Webs fur handlungsgepragte Begriffe hingegen vieleNeurone im prazentralen, motorischen Kortex. Auch diese Hypothese decktsich mit Schwankungen im Energieverbrauch der betroffenen Areale, die mitHilfe hirnbildgebender Verfahren experimentell beobachtet worden sind. Undes finden sich sogar noch feinere topologische Unterschiede: beim Vergleichzwischen gesichts-, arm- und beinbezogenen Verben (z.B.
”sprechen“,
”grei-
fen“ und”gehen“) zeigte sich unterschiedlich starke Aktivitat entlang des mo-
torischen Kortex, und zwar dergestalt, dass das Zentrum der Aktivitat je nachVerbkategorie im motorischen Projektionsfeld des Gesichts, der Arme bzw. derBeine lag (vgl. Abb. 6). Abbildung 9 zeigt die mutmaßlichen Word Webs furdie hier behandelten Wortkategorien: visuell gepragte und handlungsorientier-te Substantive sowie gesichts-, arm- und beinbezogene Verben.
Ist erstmal ein”Grundstock“ von Wortern im Gehirn reprasentiert, so kann
die Bedeutung neuer, unbekannter Worter auch ihrem sprachlichen Kontexterschlossen werden. Es ist also keinesfalls notwendig, dass jedes neu zu er-
20

lernende Wort in Zusammenhang mit einem konkreten Gegenstand oder einkonkreten Handlung prasentiert werden muss, um dessen phonologische Ei-genschaften mit zusatzlichen, semantischen Aspekten zu verknupfen.
Aus dem hebb’schen Lernprinzip ergibt sich im Ubrigen auch eine Modelldafur, wie sich die Ahnlichkeit verschiedener Wort in ihrer neuronalen Re-prasentation niederschlagen konnte. In Abschnitt 2.2.2 wurde gesagt, dassverschiedene Functional Webs nicht aus disjunkten Gruppen von Neuronenbestehen mussen. Ganz im Gegenteil macht es sogar Sinn, dass sich einzelneWebs teilweise sehr stark uberlappen – dann namlich, wenn sie sich semantischahnlich sind. Word Webs vollstandig synonymer Worter z.B. wurden sich imidealisierten Fall nur in ihrem Phonemteil unterscheiden. Im Gegensatz dazuwurden sich homophone Worter gerade in ihrer Phonemreprasentation uber-lappen und der semantische Teil ihrer Word Webs ware weitgehend disjunkt(siehe Abbildung 10).
Abbildung 10. Topologie mutmaßlicher Word Webs zur Reprasentation von Worternverschiedener semantischer Kategorien
Zusammenfassend lasst sich also festhalten, dass Word Webs ein plausiblesModell der Reprasentation von Worten im Gehirn darstellen, sowohl auf Grundprinzipieller Uberlegungen zu Organisation und Plastizitat des Kortex als auchgestutzt auf experimentelle Ergebnisse aus elektrophysiologischen und funk-tional bildgebenden Studien.
2.3 Neuronale Mechanismen zur Reprasentation serieller Ordnung
2.3.1 Grundsatzliche Uberlegungen anhand kunstlicher Neurone
Nach der vorangegangen Diskussion um die grundlegende Reprasentations-form von Worten im Gehirn wenden wir uns jetzt allmahlich wieder denin Abschnitt 1 aufgeworfenen Fragen nach den Prinzipien einer neuronalenGrammatik zu. Es wird aufgezeigt, mit welchen Mechanismen verschiedenAspekte serieller Ordnung in kunstlichen McCulloch-Pitts-Netzen reprasen-tiert werden konnen. Diese Uberlegungen fuhren zusammen mit dem Konzeptder Functional Webs und Word Webs auf ein konkretes Modell einer neuro-nalen Grammatik hin, das in Abschnitt 3 abschließend vorgestellt wird.
21

Das Neuronenmodell von McCulloch&Pitts wurde in Abschnitt 2.1.2 vorge-stellt. Fur die Abbildungen dieses Abschnitts gelten folgende Konventionen:der Schwellwert eines Neurons ist die Zahl in dessen Inneren, und das Ge-wicht aller Verbindungen betragt jeweils 1, soweit nicht anders angegeben.Wie nun lassen sich Netze von solchen Neuronen nutzen, um Sequenzen vonWortern zu erkennen? Betrachten wir zunachst den einfachsten Fall: gesuchtist ein Netz, das genau einen einzelnen Satz erkennen kann, wobei die einzel-nen Worter dem Netz sequentiell prasentiert werden. In Abbildung 11 sehenwir zwei mogliche solche Netze zur Erkennung des Beispielsatzes
”Betty get
up“. Die Neurone in der Input-Schicht reagieren selektiv auf je eines der dreiWorte. Wird die Wortfolge
”Betty get up“ etwa dem linken der beiden Netze
prasentiert, so lauft folgende Netzreaktion ab (alle Neurone seine zu Beginninaktiv):
(1) Input”Betty“ aktiviert Neuron a.
(2) Input”get“ aktiviert Neuron b, wahrend gleichzeitig Neuron d durch a
aktiviert wird.(3) Input “up“ aktiviert Neuron c, wahrend die Neurone e und f von Neuron
b und d erregt werden(4) Durch ihre gleichzeitige Aktivitat erregen die Neurone c e und f Neuron
g
Man erkennt leicht, dass Neuron g nur dann feuert, wenn der Satz”Betty get
up“ wortlich und ohne Unterbrechung im Input vorkommt. Es konnte damitals interne Reprasentation dieses Satzes genutzt werden. g wird hierbei alsGroßmutter- oder Kardinalzelle bezeichnet, da es alleine ein komplexes, zeit-lich ausgedehntes Ereignis reprasentiert oder vermittelt – man spricht auchvon vermittelter Sequenzerkennung 9 . Im Vergleich dazu kommt das Netz inAbb. 11B mit weniger Neuronen aus. Zwar ist auch hier das letzte Neuron derKette letztendlich eine Kardinalzelle. Zusatzlich dazu geht aber die Erkennungdes Beispielsatzes einher mit Aktivitatswelle, die sich entlang der unteren Rei-he von Neuronen fortpflanzt - ein Phanomen, auf das in Abschnitt ?? nochnaher eingegangen wird.
Einfache Stringdetektoren lassen sich also mit Hilfe kunstlicher NeuronalerNetze implementieren. Eine zentrale Rolle beim Verstandnis von Sprache spieltjedoch aufgrund des Prinzips der zentralen Einbettung Abhangigkeiten zwi-schen Wortern, die im Satz raumlich bzw. zeitlich beliebig weit voneinanderentfernt stehen konnen (vgl. Abschnitt 1.2 und 1.3). Dem sind die bisherigenStringdetektoren jedoch nicht gewachsen – hierzu bedarf der Einfuhrung vonruckgekoppelten Verbindungen. In Abbildung 12 sehen wir zwei Neurone mit
”Gedachtnis“. Nehmen wir an, das linke Neuron zu Beginn einer Wortfolge
voraktiviert. Dann ist es nach Prasentation der Folge nur dann noch aktiv,
9 mediated sequence processing
22

Abbildung 11. Zwei Netze zur Erkennung der Wortfolge ”Betty get up“ A. durcheine Kardinalzelle. B. begleitet von einer Aktivitatswelle in der unteren Zellreihe.
Abbildung 12. Netze zur Erkennung des A. dauerhaften bzw. B. einmaligen Auftre-tens eines Wortes in einer Wortfolge.
wenn kontinuierlich extern stimuliert wurde. Kommt es hingegen zu einer Un-terbrechung der externen Stimulation, so ist das Neuron am Ende inaktiv.Das rechte Neuron dagegen bleibt aktiv, sobald es einmal stimuliert wurde (esbedarf dann freilich noch irgendeiner Art von Reset-Mechanismus). Diese Artvon Verhalten erinnert an den All- bzw. den Existenzquantor aus der Pradika-tenlogik. Speziell das rechte
”Existenz-Neuron“ eignet sich dazu, den Zusam-
menhang zwischen verteilten Satzbestandteilen herzustellen. Dies verdeutlichtAbbildung 13: das abgebildete Netz ist durch seine Ruckkopplungsschleifen inder Lage, alle der folgenden Satze zuerkennen:
(1) Betty switched it on.(2) Betty switched the nice brown coffee machine on.(3) Betty switched the coffee machine she received as a present from her
mother at her 25th birthday on.
Auf diese Weise gelingt es mit neuronalen Netzen, Strukturen zu reprasen-tieren, bei denen das auftreten eines Bestandteils das sichere Auftreten einesanderen Bestandteils bedingt, und zwar mit beliebigem Abstand zwischen denbeiden. Damit sind sie rein stochastische Modelle wie z.B. Markov-Ketten indieser Hinsicht uberlegen. Im Folgenden sollen nun ausgehend von den hierskizzierten Uberlegungen anhand von McCulloch-Pitts-Zellen sollen nun nochzwei auch neurophysiologisch plausible Ansatze zur neuronalen Modellierungserieller Ordnung vorgestellt werden.
23

Abbildung 13. Netz zur Erkennung der Worter ”Betty“, ”switches“ und ”on“ inner-halb einer langeren Wortfolge in genau dieser Reihenfolge und mit beliebig langenUnterbrechungen zwischen den Wortern
2.3.2 Synfire Chains
Der erste Ansatz zielt hierbei nicht primar auf das Erkennen oder die Re-prasentation grammatikalischer Ordnung, sondern auf die Speicherung vonPhonemfolgen. Seien in Analogie zum motorischen Kortex Neurone gegeben,der Aktivierung die Außerung verschiedener Laute hervorruft. Der denkbareinfachste Weg, eine Phonemfolge zu speichern ware es, die entsprechendenNeurone mit erregenden Synapsen zu verbinden. Wird nun das erste Neuronder Folge aktiviert, pflanzt sich die Aktivitat entlang dieser Verbindungen fortund fuhrt dabei zur entsprechenden Lautaußerung. Allerdings hat man festge-stellt, dass die synaptischen Gewichte im Kortex im Regelfall viel zu schwachsind, als dass ein einzelnes Neuron ein anderes erregen konnte. Dazu ist nor-malerweise die annahernd gleichzeitige Aktivitat zahlreicher Input-Neuronenotwendig. Statt dessen ware es also plausibler anzunehmen, dass anstelle ein-zelner Neurone jeweils ganze Neuronengruppen miteinander verbunden sind,wobei alle Neuronen einer Gruppe jeweils synchron von ihrer Vorgangergrup-pe innerhalb der Verbindungskette erregt werden. Diese Form der neuronalenVerschaltung wird als Synfire Chain bezeichnet. Auch experimentell konntenPhanomene beobachtet werden, die sich mit diesem Modell decken. Unter-sucht wurde die Korrelation zwischen dem Feuerverhalten mehrerer Neurone.Dabei fiel auf, dass ein Neuron besonders haufig dann feuerte, wenn zuvorzwei weitere Neurone in einem ganz bestimmten Zeitabstand gefeuert hatten.
Abbildung 14 zeigt eine solche Synfire Chain. Durch Aktivierung aller Neuro-ne der ersten Gruppe entsteht eine Welle synchroner Aktivitat innerhalb derGruppen. Eine nur teilweise Aktivierung hingegen reicht nicht aus, um einesich fortpflanzende Welle zu erzeugen. In Wirklichkeit sind die Gruppen wahr-scheinlich deutlich großer und konnten geschatzt ca. 50–100 Neurone enthal-ten, mit einem Schwellwert jeweils zwischen 5 und 10. Mehrere Synfire Chainskonnen sich teilweise uberlappen und kreuzen, ohne dass sich eine Aktivitats-welle dabei unkontrolliert in alle Richtungen weiter ausbreitet. Die verdeutlichtAbbildung 15. Aktivierung der Neuronen in der Gruppe links oben (
”b“) fuhrt
zu einer Aktivitatswelle, die sich nach rechts unten ausbreitet, nicht aber – ent-
24

Abbildung 14. Ein Synfire Chain bestehend aus 5 Gruppen von je 3 synchron aktivenNeuronen. Der Schwellenwert der Neurone betragt 2.
lang der kreuzende Synfire Chain – nach links unten. Dies wird durch den nurteilweisen Uberlapp der jeweils mittleren Neuronengruppen der beiden Kettengewahrleistet. Die beiden Neurone in Mitte ganz links und ganz rechts dienenalso als Kontextindikator, die bestimmen, in welche Richtung sich die Aktivitatim Uberlapp-Bereich weiter ausbreitet. In der Abbildung ist zudem wieder derBezug zur Speicherung von Phonemfolgen angedeutet. Jedes Wort wurde nachdieser Vorstellung phonetisch durch eine eigene Synfire Chain reprasentiert,die sich mit den Ketten zahlreicher anderer Worter uberschneidet und kreuzt.Freilich bedarf dieses Modell noch der Verfeinerung. Die Geschwindigkeit derWelle ist bisher konstant. Wir aber konnen unsere Sprechgeschwindigkeit nachBelieben variieren. Dies ließe sich aber beispielsweise durch die Regulation derunspezifischen Hintergrundaktivitat erreichen. Durch starkes Hintergrundrau-schen waren die Neuronen schon von vornherein naher an ihrem Schwellwert,so ihre letztendliche Aktivierung schneller von statten ginge als bei schwacherHintergrundaktivitat. Dies konnte auch erklaren, wie es bei hoher Sprechge-schwindigkeit zunehmend zum Nuscheln und Versprechen kommt. Zum einenkonnte es geschehen, dass sich die Aktivitat schneller durch den motorischenKortex ausbreitet, als es die Muskulatur umsetzen kann. Zudem steigt durchdie hohe Voraktivierung der kreuzenden Synfire Chains die Wahrscheinlich-keit, dass sich die Aktivitat entlang eines falschen Pfades, moglicherweise so-gar entlang mehrerer Pfade gleichzeitig ausbreitet. Das Synfire-Chain-Modellzur phonetischen Reprasentation deckt sich also auch mit psycholinguistischenBeobachtungen.
Dennoch erscheint es unwahrscheinlich, dass sich ein Ahnlicher Ansatz – etwaSynfire Chains zwischen Worten anstelle von Phonemen – auf die Verarbei-tung von ganzen Satzen ubertragen laßt. Der zeitliche Abstand zwischen derAktivierung zwei benachbarter Neuronengruppen liegt bei Synfire Chains imMillisekundenbereich, wahrend bei der Verarbeitung von Satzen Verzogerun-gen von einigen Sekunden auftreten konnen, noch dazu von Fall zu Fall varia-bel. Kritisch ist auch die riesige Anzahl moglicher Wortfolgen und daher der
25

Abbildung 15. Kreuzende Synfire Chains zur phonologischen Reprasentation derenglischen Worter ”bat“ und ”tab“
benotigten Synfire Chains. Im Falle von Phonemfolgen liegt die Anzahl dermoglichen Nachfolger zwischen 5 und 20, im Falle von Wort jedoch rund 104.Dies ließe sich Reduzierung durch die Einfuhrung verschiedener Wortkatego-rien. In diesem Falle ist jedoch unklar, wie eine Synfire Chain die Auswahlzwischen verschiedenen Wortern einer Kategorie vornehmen konnte. Und wiekonnte ein solches System – egal ob auf der Ebene von Wortern oder Wort-kategorien – von bereits bekannten auf unbekannte Strukturen generalisieren?Im Folgenden wird ein alternativer Ansatz vorgestellt, der kombiniert mit demModell der Functional Webs diese Probleme uberwinden konnte.
2.3.3 Neuronale Sequenzdetektoren
Kehren wir in Gedanken noch einmal zuruck zu Abbildung 11A. Prasentationdes Beispielsatzes
”Betty get up“ fuhrte zur Aktivierung der Kardinalzelle g.
Die Zellen d, e, f dienen hierbei lediglich der Verzogerung der Aktivitat derInput-Neurone a und b. Aus neurophysiologischer erscheint allerdings ein an-derer Verzogerungsmechanismus wahrscheinlicher. Da die Reizleitung in einerNervenfaser nur mit begrenzter Geschwindigkeit erfolgt, konnte die Verzoge-rung allein durch unterschiedlich Langen der Verbindungsleitungen erfolgen.Solche Verzogerungsstrecken konnten im Kleinhirn bereits nachgewiesen wer-den, die auch dort sicherstellen, dass zeitlich genau abgestimmte Sequenzen
26

Abbildung 16. A. Ein Phrasenstruktur-Baum zur syntaktischen Reprasentation desSatzes ”He comes“. B. Word Webs und Sequenzdetektoren zur Reprasentation syn-taktischer Abhangigkeiten
neuronaler Aktivitat gleichzeitig an einer Kardinalzelle eintreffen und dieseerregen.
Dennoch kann auch diese Art von Sequenzdetektor so kaum als Grundlage zurSprachverarbeitung dienen. Die beobachteten Verzogerungen liegen im Bereichvon deutlich unter 1s, wahrend im Fall von Sprache viele Sekunden zwischenden einzelnen Bestandteilen verteilter Worter liegen konnen – zudem konnendie Verzogerungen von Fall zu Fall stark schwanken. Einen moglichen Aus-weg bietet die Implementierung von Sequenzdetektoren als Funktional Websanstelle einzelner Kardinalzellen. Dieses Modell wird in Abschnitt 3 im De-tail vorgestellt. An dieser Stelle sei nur noch als zusatzliche Motivation nochangedeutet, welche Vorteile eine Grammatik basierend auf Sequenzdetekto-ren gegenuber formalen linguistischen Modellen wie etwa Phrasenstruktur-Grammatiken bieten konnte. Ein in der Linguistik gangiger Ansatz zur Re-prasentation syntaktischer Strukturen sind hierarchische Baume mit syntak-tischen Kategorien als inneren Knoten und den Wortern oder Morphemen 10
eines Satzes als Blattern (siehe Abb. 16A). Ein Problem hierbei ist die Model-lierung von bestimmten Abhangigkeiten zwischen verteilten Satzbestandteilenwie etwa der Kongruenz zwischen Subjekt und Verb-Endung (
”He“ und
”-s“
im abgebildeten Beispiel). Zur Prufung der grammatikalischen Korrektheit ei-nes Satzes musste zusatzlich zum Aufbau eines syntaktisch korrekten Baumes
10 Morphem: kleinste bedeutungstragende Spracheinheit, z.B. ein Wortstamm odereine Verb-Endung
27

gemaß der Phrasenstrukturgrammatik noch derartige Abhangigkeiten in ei-nem weiteren Arbeitsschritt uberpruft werden. Dagegen kommt das Modell inAbbildung 16B basierend auf Sequenzdetektoren mit einem einheitlichen Me-chanismus aus. Die Detektoren sind nicht hierarchisch angeordnet: jeder fursich erkennt ein Wort-/Morphempaar: Pronomen – Verb, Verb – Verb-Endungsowie Pronomen – Verb-Endung. Speziell letzterer gibt Auskunft uber dieKongruenz zwischen Subjekt und Pradikat des Satzes. Zusammengenommenvermittelt – so die Behauptung – der Aktivitatszustand zahlreicher solcherpaarweiser Sequenzdetektoren genugend Information uber die Struktur einesprasentierten Satzes, um seine Bedeutung erschließen zu konnen. Diese Be-hauptung zu Untermauern ist Sinn des folgenden Abschnitts.
3 Neuronale Grammatik
Bisher haben wir neuronale Ensembles und funktionelle Netze betrachtet. Indiesem Kapitel werden zwei neue Begriffe eingefuhrt.
(1) Neuronale Sets, die funktionelle Netze mit speziellen Eigenschaften sind.(2) Sequenz Sets sind, neuronale Sets sind, die Sequenzen von Wortern re-
prasentieren konnen.
3.1 Erregungszustande von funktionellen Netzen
Ein solches Netz agiert aufgrund der starken internen Verbindungen als einefunktionelle Einheit, d.h. wenn ein angemessener Teil des Netzes durch einenInput erregt wird, fuhrt die Konnektivitat dazu, dass alle bzw. ein Großteil derNeuronen aktiviert werden. Diese volle Aktivitat bezeichnet man mit Ignition.Diverse Erschopfungseffekte reduzieren danach kontinuierlich die Aktivitat,die jedoch nicht ganz ausstirbt. Der Rest der Aktivitat, der nicht verloren geht,wird Reverberation genannt. Im Gegensatz zur Ignition ist die Reverberationein lang anhaltender Prozess innerhalb des funktionellen Netzes, jedoch sindbei weitem nicht so viele Neuronen aktiviert wie bei der Ignition. Zusatzlichbesteht die Moglichkeit, dass sich das Netz in einem inaktiven Ruhezustandbefindet.
Im Folgenden wird das neuronale Set eingefuhrt und erlautert weshalb es einSpezialfall dieses funktionellen Netzes ist, das drei Zustande besitzt: Ignition,Reverberation und Ruhe.
28

3.2 Neuronale Sets
Ein neuronales Set zeichnet sich gegenuber einem funktionellen Netz durcheine großere Variabilitat der Zustande aus.
Aktivitat eines neuronalen Sets kann sich in einer Menge solcher Sets uber ex-terne Verbindungen, die schwacher sind als interne Verbindungen, ausbreitenund beeinflusst so benachbarte Sets. Um ein neuronales Set in den Zustandder vollen Aktivitat Ignition zu fuhren, ist immer ein externer Input notwen-dig. Eine solche Aktivierung wirkt sich dann auf andere Sets aus, die dann inden Zustand des Primings gefuhrt werden. Das Priming ist weitaus schwacherals Ignition und Reverberation.
Uber die Zeit verringert sich die Aktivitat aller Zustande mit exponentiellerGeschwindigkeit:
A(Si, t) = A(Si, t0) ∗ e−c∆t, (5)
wobei t0 den Zeitpunkt der externen Aktivierung durch ein anderes Set bzw.durch externen Input, ∆t den Abstand zwischen t0 und t, A(Si, t) die Aktivitatvon Set Si zu Zeitpunkt t0 und c die Steigung des Abfallens der Aktivitatbezeichnet.
Inputs anderer Sets, die sich in den Zustanden Reverberation bzw. Priming be-finden, wirken uber verschiedene Verbindungen auf das neuronale Set. Es wirdangenommen, dass sich die Inputs der anderen im Set summieren. Insofern istdie Aktivitat von Si so zu beschreiben:
A(Si, t) =∑
j
A(Sj, t− 1), (6)
wobei A(Sj, t−1) die Menge der exzitatorischen Stimulation von Set Sj auf Si
zum Zeitpunkt t−1 bezeichnet. Die Stimulation erreicht Set Si zum Zeitpunktt.
Falls sich ein Netz bereits im Zustand Reverberation befindet verandert sichdie Gleichung nur um einen zusatzlichen Term:
A(Si, t) = A(Si, t0) ∗ e−c∆t +∑
j
A(Sj, t− 1) (7)
Der Zustand der Ignition wird dann erreicht, wenn die zeitliche bzw. raumlicheSummation eine Schwelle θ ubersteigt. Nach einer Ignition I wechselt derZustand im nachsten Zeitschritt zur Reverberation R1, wobei R1 der starkste
29

Reverberation Zustand ist. Durch den exponentiellen Abfall gibt es theoretischunendlich viele solche Reverberation Zustande. Kurz gefasst verandern sich dieZustande folgendermaßen:
A(Si, t) > θ ⇒ A(Si, t + 1) = I ⇒ A(Si, t + 2) = R1 (8)
Ein neuronales Set kann sich in vier Zustanden befinden:
AS = {0, P, R, I}, (9)
wobei P und R eigentlich eine Menge von Zustanden bezeichnen. Aufgrund deskontinuierlichen Abfalls der Aktivitat kann es jeweils unendlich viele ZustandeR1, R2, ... und P1, P2, ... geben. Dies ist jedoch unwahrscheinlich in der Rea-litat, weil das Signal-zu-Rausch Verhaltnis die Anzahl beschrankt, weswegenwir auch nur eine endliche Menge von Zustanden betrachten, die ihrer Starkenach sortiert sind, wobei R1, P1 die starksten Zustande bezeichnen. WeitereVereinfachungen sind, dass der zeitliche Abfall der Aktivitat ignoriert wirdund die raumliche und zeitliche Summation der Aktivitat vereinfacht wird.
3.3 Schwellwertkontrolle
Um das Aktivitatslevel in einem gewissen Rahmen zu halten muss eine Re-gulation der Schwellwert eingefuhrt werden. Es wird angenommen, dass einstarker Anstieg der Aktivitat detektiert werden muss und in Folge darauf derSchwellwert angepasst wird. Die Realisierung ist eigentlich relativ einfach. Derextremste Anstieg der Aktivitat ist dann, wenn ein neuronales Set aus demRuhezustand in den Zustand Ignition springt. Dies fuhrt dann zu einer glo-balen Erhohung des Schwellwerts. Diesen Anstieg der Aktivitat mit folgenderSchwellwertkontrolle 11 bezeichnen wir mit dem Symbol I. Die Ignition ohnedarauf folgende Korrektur wird wie bisher mit I bezeichnet. I tritt dann auf,wenn ein Set aktiviert wird, das sich zuvor schon in einem Zustand der Rever-beration bzw. Priming befand. Die globale Inhibition der Aktivitat durch dieSchwellwertregulierung wird so realisiert, dass alle Sets, die sich in Rn bzw.Pn befinden, in die Zustande Rn+1 bzw. Pn+1 herabgestuft werden.
11 Threshold control
30

Abbildung 17. Detektor fur die Sequenz alpha – beta
3.4 Sequenzdetektion in Netzwerken von neuronalen Sets
An dieser Stelle genugt es zu sagen, dass es einen Mechanismus gibt, der zurDetektion von Sequenzen oder seriellen Reihenfolgen 12 herangezogen werdenkann. Dieser Mechanismus konnte beispielsweise uber Verbindungen, die in nureiner Richtung verlaufen, implementiert werden. Da dies jedoch unrealistischist und in der Natur nicht so vorkommt, wird im Folgenden von bidirektionalenVerbindungen ausgegangen, die jedoch asymmetrisch sind, d.h. es gibt in eineRichtung sehr starke und in die andere Richtung eher schwache Verbindungenzwischen zwei neuronalen Sets. Eine Unterscheidung zwischen ’starken’ und’schwachen’ Verbindungen ist selbstverstandlich eine weitere Vereinfachungdes realen Modells. Die starkeren Verbindungen werden in die Richtung desublichen Flusses angenommen, d.h. Sets, die Worter die oft vor gewissen ande-ren gesprochen werden, reprasentieren, haben starke Verbindungen zu einemSequenz-Set und dieses wiederum zu den Sets, die haufig danach gesprocheneWorter reprasentieren. Abbildung 17 zeigt ein Netz, das α vor β akzeptiert:Solche starken Verbindungen, wie zwischen α und γ, ließen sich beispielsweisedurch Hebb’sches-Lernen trainiert, da durch eine solche Regel Synapsen dannverstarkt werden, wenn sowohl pra- als auch postsynaptische Aktivitat vor-handen ist. Im Folgenden wird genauer auf ein Beispiel eines Netzwerks ausneuronalen Sets eingegangen und erlautert, warum das Netz nur dann einenString akzeptiert, wenn dieser in der korrekten Weise prasentiert wird.
3.5 Sequenzerkennung
Es wird im Folgenden angenommen, dass jedes Wort bzw. Morphem im Inputsein Wortnetz 13 bzw. neuronales Set aktiviert und als Konsequenz das Setin den Zustand Ignition bringt und daraufhin in den Zustand Reverberation.Reverberation wiederum bringt andere Sets uber starke Verbindungen in den
12 Serial order13 Word web
31

Zustand Priming. Ignition aktiviert auch uber schwache Verbindungen sichim Ruhezustand befindliche Sets und bringt sie in den Zustand Priming. EinSequenz-Set wird nur dann in den Zustand Ignition gefuhrt, wenn es von einerIgnition eines anderen Sets aktiviert wird und sich zuvor schon in Reverbera-tion bzw. Priming befand. Nun folgt die konkrete Abfolge einer Sequenz (Eswird angenommen, dass die Sequenz AB ein korrekter Satz ist.):
(1) Das Wort A aktiviert die Inputeinheit α.(2) Da α im Ruhezustand war, erfolgt eine volle Aktivierung I und die
Schwellwertregulierung wird aktiviert. Diese hat jedoch keinen weiterenEinfluss, da sich kein Set in einem aktivierten Zustand befand außer α.
(3) α aktiviert und ’primet’ das verbundene Sequenz-Set γ. γ wird jedochnicht vollstandig aktiviert, da es sich nicht in Priming oder Reverberationbefand.
(4) α andert seinen Zustand in R1 und ’primet’ so konstant γ uber die starkeVerbindung zwischen den beiden. Da es auch starke Verbindungen von γzu β gibt, wird auch β von γ ’geprimet’.
(5) Nun erscheint B als Input und β wird vollstandig aktiviert und verandertseinen Zustand in I, da β zuvor schon im Zustand Priming war.
(6) β aktiviert nun mit I den Sequenzdetektor γ, welcher sich nun auch in Ibefindet.
(7) Schlussendlich aktiviert auch γ noch einmal α, was zum Akzeptieren desSatzes fuhrt. Daraufhin ruhen alle Sets wieder.
Damit man nicht glaubt, dass das auch bei einem grammatikalisch falschemSatz passiert, wird der Input in BA geandert:
(1) Der Input B fuhrt zu Ignition I von β. Die Schwellwertregulierung hatkeinen Effekt.
(2) Die Ignition I von β fuhrt zu einem kurzen Priming in γ. Dieses Pri-ming schwindet jedoch schnell wieder, da nur eine schwache Verbindungzwischen β und γ besteht.
(3) β befindet sich in R1 und sowohl α als auch γ sind im Ruhezustand.(4) Es folgt nun A im Input und fuhrt zu I in α.(5) Die Schwellwertregulierung verringert nun die Aktivierung von β zu R2.(6) Am Ende der Sequenz befinden sich also alle Sets in unterschiedlichen
Zustanden: β in R2, γ in P1 und α in R1. Insofern wird die Sequenz BAnicht akzeptiert.
Zusammenfassend sind drei Dinge notwendig, damit ein String akzeptiert wird:
(1) Synchronization - Eine sich durch das Netz fortpflanzende Welle der Igni-tion ’synchronisiert’ alle beteiligten Sets.
(2) Satisfaction - Die Sequenz-Sets, die direkt mit Inputeinheiten verbundensind befinden sich in R1.
32

(3) Visibility - Die Inputeinheiten befinden sich in R1 und sind somit ’sicht-bar’.
3.6 Lexikale Kategorien in neuronalen Sets
Im ersten Kapitel haben wir lexikale Kategorien eingefuhrt um gewisse Abhangig-keiten korrekt zu reprasentieren und um feiner unterscheiden zu konnen. Istdies nun wieder notwendig in Bezug auf das Modell mit neuronalen Sets? Imfolgenden Abschnitt wird dies diskutiert.
3.6.1 Warum lexikale Kategorien?
Nehmen wir nun das Beispielwort aus Kapitel (1) wieder auf: ’switch’ bzw.’switch ... on’. Der Inputdetektor fur den Wortstamm ’switch’ ist wie gehabtein neuronales Set, das reagiert, falls das Wort ’switch’ im Input erscheint. Manbrauchte danach jedoch unterschiedliche Sequenz-Sets, die auf unterschiedlichenachfolgende Worter reagieren. In diesem Beispiel gibt es z.B. einen fur dasWort ’switch’ als Nomen, welches ein Verb erwartet oder fur das Wort ’switch’als transitives Verb, das ein Nomen im Akkusativ erwartet oder auch fur dasWort ’switch ... on’, das zuerst ein Nomen im Akkusativ und dann spater denPartikel ’on’ erwartet. Man sieht also, dass es viele Verzweigungen gibt. Wennman das fur jedes mogliche Wortpaar oder fur jede mogliche Wortsequenzmachen wurde ware die Zahl der Sequenz-Sets astronomisch groß. Ein Weg,dieser Tatsache auszuweichen ware, Worter, so wie schon bei den formalenGrammatiken vorgeschlagen, in lexikale Kategorien einzuteilen. Wenn man dieWorter in etwa 100 lexikale Kategorien einteilt, wie z.B. Nomen im Nominativ,Akkusativ oder Dativ und Verben in transitive, intransitive etc., wurde mandie Zahl der notigen Sequenz-Sets drastisch senken.
3.6.2 Lexikale Kategorien als Mengen von Sequenz-Sets
Wie schon weiter oben beschrieben, wird ein Sequenzmerkmal dadurch re-prasentiert, dass ein Sequenz-Set zuerst im Zustand Ignition ist und dann imZustand Reverberation verweilt und so die Information uber die aufgetreteneSequenz speichert. Eine lexikale Kategorie kann nun als eine Vereinigung vonmehreren Sequenz-Sets gesehen werden. Zwei Beispiele hierfur waren:
(1) Ein Nomen im Nominativ wurde mittels zwei Sequenz-Sets reprasentiertwerden. Das erste erkennt, dass ein Artikel vor dem aktuellen Wort er-schien und das zweite, dass ein Verb dem aktuellen Wort folgt.
(2) Ein Nomen im Akkusativ wurde ebenfalls zwei Sequenz-Sets benotigen.Das erste steht fur ein voranstehendes transitives Verb und das zweite
33

Tabelle 1Ungefahre Anzahl und Großenordnung der neuronalen Sets zur Sprachverarbeitung
Lingusitisch Sets Neuronen
Wort/Morphem 1 Inputeinheit 105 Neuronen
Sequenzmerkmale 1 Sequenz-Set 105 Neuronen
Lexikale Kategorie < 10 Sequenz-Sets < 106 Neuronen
Vokabular 105 Inputeinheiten 1010 Neuronen
Grammatik103 Sequenz-Sets 108 Neuronen
107 Verbindungen zwischen den Sets
fur einen voranstehenden Artikel.
Hierbei ist wichtig zu unterscheiden, dass es Sequenz-Sets gibt, die in die ’Zu-kunft blicken’ und welche, die sich die ’Vergangenheit merken’. Diese Sequenz-Sets konnen direkt aus den Regeln der Abhangigkeitsgrammatiken abgelesenwerden.
3.6.3 Lexikale Unterscheidungen durch Sequenz-Sets
Um Worter, wie ’switch’, eindeutig zu einer lexikalen Kategorie zuordnen zukonnen, ist noch ein zusatzlicher Mechanismus notwendig, der die anderenZuordnungsmoglichkeiten unterdruckt. Es muss gewahrleistet werden, dass daswahrscheinlichste Set, die anderen hemmt und so eine Art ’the-winner-takes-it-all’ Selektion entsteht. Dieser Mechanismus konnte beispielsweise so realisiertwerden, dass erkannt wird, wessen Sequenzmerkmale vollstandig erfullt werdenund so kann entschieden werden, welches Set gewonnen hat gegenuber denen,die nicht alle Merkmale erfullen. Falls die letzten notigen Sequenz-Sets zweierlexikalen Kategorien gleichzeitig im Zustand Ignition sind, ist die Kategorie,die die meisten aktiven Sets besitzt, der Sieger.
3.6.4 Anforderungen an eine neuronale ’Grammar Machine’
In diesem Abschnitt geht es darum mathematisch abzuschatzen, wie vieleSets bzw. Neuronen notwendig sind, um eine Grammatik zu reprasentieren.Wie ein vorhergehender Abschnitt zeigte, kann man die Abhangigkeitsgram-matiken dazu benutzen, fur ein Wort die Anzahl der benotigten Sequenz-Setsabzuschatzen. In einer Grammatik, wie sie im Englischen ublich ist, ist es nahe-zu unmoglich Worter zu finden, die mehr als funf abhangige Worter besitzen.Man kann nun ein mehrdeutiges Wort mit funf moglichen Bedeutungen undjeweils funf abhangigen Wortern als Extremfall fur eine Abschatzung anneh-men. In diesem Fall wurde die Inputeinheit 25 Verbindungen zu Sequenz-Sets
34

benotigen. Nun kann man ein Vokabular von ca. 100000 Wortern annehmen,die durchschnittlich in zwei der 100 lexikalen Kategorien stehen und jeweilsfunf Sequenzmerkmale besitzen. Nun wird fur jedes Wort ein neuronales Setbenotigt. Um jede der lexikalen Kategorien zu reprasentieren, benotigt manjeweils funf Sequenz-Sets, also 500. Man benotigt also nur 500 Sequenzdetek-toren um eine solche Grammatik reprasentieren zu konnen. Man sieht also,dass die Reprasentation der 100000 Worter weitaus aufwandiger ist als dieReprasentation der Sequenzen. Wenn man nun noch etwa 100000 Neuronenfur jedes Set veranschlagt, ergibt das eine maximale Gesamtzahl von 1010
Neuronen, die das Vokabular reprasentieren. Eine ’feinere’ Grammatik, diemehrere lexikale Kategorien reprasentiert, schlagt also gar nicht so stark insGewicht, da auch bei 1000 Kategorien nur 5000 Sequenz-Sets benotigt wer-den. Man sieht also, dass es entscheidend darauf ankommt, wie viele Worterreprasentiert werden sollen.
Literatur
[1] Pulvermuller, P. (2002). The Neuroscience of Language. Cambridge UniversityPress, Cambridge
[2] Chomsky, N. (1963). Formal properties of grammars.
[3] Bach, E., C. Brown, W. Marslen-Wilson (1986). Crossed and nesteddeoendencies in German und Dutch; a psycholinguistic study. Language andCognitive Processes, 1, 249–262.
[4] Petri, C. (1970). Kommunikation mit Automaten. Dessertation: UniversitatBonn.
[5] Dayan, P., L. P. Abbot (2001). Theoretical Neuroscience. MIT Press,Cambridge, MA
[6] Rojas, R. (1993). Theorie der neuronalen Netze. Springer Verlag, Berlin
[7] McLeod, P., K. Plunkett, E. T. Rolls (1998). Introduction to ConnectionistModelling of Cognitive Processes. Oxford University Press, New York
[8] Penrose, R. (1991). Computerdenken. Spektrum Verlag, Heidelberg
[9] Silbernagel, S., A. Depopoulos (2001). Taschenatlas der Physiologie. ThiemeVerlag, Stuttgart
[10] Schmidt, F., G. Thews, F. Lang (1997). Physiologie des Menschen. SpringerVerlag, Berlin
[11] Gaıarsa, J.-L. (2004). Plasticity of GABAergic synapses in the neonatal rathippocampus. J. Cell. Mol. Med. 8(1): 31-37
[12] Miyashita, Y., H. S. Chang (1988). Neural correlate of pictorial short-termmemory in the primate temporal cortey. Nature 331: 307-311
35


















