
PATHOGENITÄTSVERGLEICH VON SALMONELLA … Kultureller und molekularbiologischer Nachweis von S....
117
Aus dem Institut für Tierhygiene und Öffentliches Veterinärwesen der Veterinärmedizinischen Fakultät der Universität Leipzig PATHOGENITÄTSVERGLEICH VON SALMONELLA TYPHIMURIUM DT104 - WILDTYP UND SALMONELLA TYPHIMURIUM - DELETIONSMUTANTEN (sseD::aphT & invC::aphT) IN PERSISTENT INFIZIERTEN SCHWEINEN Inaugural-Dissertation zur Erlangung des Grades eines Doctor medicinae veterinariae (Dr. med. vet.) durch die Veterinärmedizinische Fakultät der Universität Leipzig eingereicht von Haukur Lindberg Sigmarsson aus Reykjavík Leipzig, 2012
Transcript of PATHOGENITÄTSVERGLEICH VON SALMONELLA … Kultureller und molekularbiologischer Nachweis von S....

Aus dem
Institut für Tierhygiene und Öffentliches Veterinärwesen
der Veterinärmedizinischen Fakultät der Universität Leipzig
PATHOGENITÄTSVERGLEICH VON SALMONELLA
TYPHIMURIUM DT104 - WILDTYP UND SALMONELLA
TYPHIMURIUM - DELETIONSMUTANTEN (sseD::aphT &
invC::aphT) IN PERSISTENT INFIZIERTEN SCHWEINEN
Inaugural-Dissertation
zur Erlangung des Grades eines
Doctor medicinae veterinariae (Dr. med. vet.)
durch die Veterinärmedizinische Fakultät
der Universität Leipzig
eingereicht von
Haukur Lindberg Sigmarsson
aus Reykjavík
Leipzig, 2012

Mit Genehmingung der Veterinärmedizinischen Fakultät der Universität Leipzig Dekan: Prof. Dr. Uwe Truyen Betreuer: Prof. Dr. Dr. Andreas Hensel Gutachter: Prof. Dr. Dr. Andreas Hensel Bundesinstitut für Risikobewertung des Bundesministeriums für Ernährung,
Landwirtschaft und Verbraucherschutz (BMELV), Berlin Gutachter: Prof. Dr. Michael Wendt Klinik für kleine Klauentiere und Forensische Medizin / Ambulatorische Klinik,
Stiftung Tierärztliche Hochschule Hannover Tag der Verteidigung: 10.07.2012

Meiner Familie

Inhaltsverzeichnis
INHALTSVERZEICHNIS
ABKÜRZUNGSVERZEICHNIS
TABELLENVERZEICHNIS
ABBILDUNGSVERZEICHNIS
1 EINLEITUNG ....................................................................................................... 1
2 LITERATURÜBERSICHT.................................................................................. 4
2.1 Salmonella ............................................................................................................... 4
2.1.1 Taxonomie ............................................................................................................... 4
2.1.2 Epidemiologie ......................................................................................................... 6
2.1.2.1 Salmonellose Mensch .............................................................................................. 7
2.1.2.2 Salmonellose Schwein ............................................................................................. 9
2.1.3 Pathogenitätsfaktoren ............................................................................................ 12
2.1.3.1 Chromosomal kodierte Pathogenitätsfaktoren ...................................................... 13
2.1.3.2 Plasmid-kodierte Pathogenitätsfaktoren ................................................................ 18
2.1.4 Mutationen ............................................................................................................ 20
2.1.5 Diagnostik von Salmonellosen .............................................................................. 23
2.1.5.1 Kultureller Nachweis ............................................................................................. 23
2.1.5.2 Voranreicherung .................................................................................................... 24
2.1.5.3 Selektivanreicherung ............................................................................................. 25
2.1.5.4 Selektivnährböden ................................................................................................. 26
2.1.5.5 PCR-Nachweis ...................................................................................................... 29
3 MATERIAL UND METHODEN ...................................................................... 32
3.1 Tiere und Aufstallung ............................................................................................ 32
3.2 Versuchsdesign ...................................................................................................... 33
3.2.1 Orale Infektion der Tiere per Schlundsonde ......................................................... 36
3.2.2 Klinische Untersuchung der Versuchstiere ........................................................... 38
3.2.3 Blutentnahme und hämatologische Untersuchung ................................................ 39
3.2.4 Untersuchung auf Salmonellen ............................................................................. 39
3.2.4.1 Kulturelle Diagnostik ............................................................................................ 39

Inhaltsverzeichnis
3.2.4.2 PCR ....................................................................................................................... 40
3.3 Statistische Auswertung ........................................................................................ 42
4 ERGEBNISSE ..................................................................................................... 43
4.1 Ergebnisse der klinischen Untersuchung .............................................................. 43
4.2 Hämatologische Befunde ...................................................................................... 47
4.3 Kultureller und molekularbiologischer Nachweis von S. Typhimurium DT104 .. 56
4.3.1 Ausscheidung mit den Faeces ............................................................................... 56
4.3.2 Besiedlung von Organen und Geweben ................................................................ 57
4.3.3 Qualitative PCR zur Identifizierung und Nachweis von S. Typhimurium DT104 ........................................................................................ 60
5 DISKUSSION ...................................................................................................... 62
5.1 Methodische Überlegungen ................................................................................... 63
5.2 Klinische Symptomatik ......................................................................................... 63
5.3 Weißes Blutbild ..................................................................................................... 64
5.4 Ausscheidung und Organbesiedlung ..................................................................... 67
5.5 Schlussbetrachtung ................................................................................................ 70
6 ZUSAMMENFASSUNG .................................................................................... 71
7 SUMMARY .......................................................................................................... 73
8 LITERATURVERZEICHNIS ........................................................................... 75
9 ANHANG ..................................................................................................... I - XIV

Abkürzungsverzeichnis
ABKÜRZUNGSVERZEICHNIS
A Adenin Abb. Abbildung aphT Aminoglykosid 3`-Phosphotransferase BfR Bundesinstitut für Risikobewertung BPL Brilliantgrün-Phenolrot-Lactose BPLS-Medium Brilliantgrün-Phenolrot-Sucrose-Agarmedium ca circa cAMP zyklisches Adenosin-3,5-monophosphat °C Grad Celsius C Cytosin CT Choleratoxin d. h das heißt DM Deletionsmutante DNA Desoxyribonukleinsäure ECDC European Centre for Disease Prevention and Control EDTA Ethylendiamintetraessigsäure bzw. Ethylendiamintetraacetat EFSA European Food Safety Authority ELISA Enzyme Linked Immunosorbent Assay E. coli Escherichia coli EHEC Enterohämorrhagische Eschericia coli g Erdbeschleunigung G Guanin GKZ Gesamtkeimzahl ID Infektionsdosis IfSG Infektionsschutzgesetz INFOSAN International Food Safety Authorities Network invC Invasin C i.p. intra peritoneal ISO International Organisation for Standardization, Genf, Schweiz i.v. intravenös kb Kilobasen KBE Kolonie bildende Einheiten kDa Kilodalton kg Kilogramm LB-Boullion Luria-Bertani-Boullion LD Lethaldosis LD50 Lethaldosis, bei der 50 % der Tiere sterben LPS Lipopolysaccharid LT hitzelabile Form MHC Major Histokompatibilitäts-Komplexe Mg Milligramm min Minute ml Milliliter mm3 Kubikmillimeter mRNA messenger Ribonukleinsäure nm Nanometer PBS Phosphate-buffered saline

Abkürzungsverzeichnis
PCR Polymerase-Kettenreaktion RKI Robert Koch-Institut RNA Ribonukleinsäure rpm Rotation per minute RV-Medium Rappaport-Vassiliadis-Medium S. Salmonella S. Tm. Salmonella Typhimurium s Standardabweichung sec Sekunde ser. Serovar SGI Salmonella Genomische Insel sp. Spezies SPI Salmonella Pathogenitätsinsel SPIs Salmonella Patogenitätsinseln Spp. Spezies spv Salmonella plasmid virulence ssa Secretion System Apparatus ssc Secretion System Chaperone sse Secretion System Effector ssr Secretion System Regulator ST hitzestabile Form stn Salmonella enterotoxin Gen subsp. Subspezies s. S. siehe Seite T Thymin Taq Thermus aquatus TBE TRIS-Borat-EDTA TLR Toll-like-Rezeptor TNF Tumor Nekrose Faktor TRIS Tris-(hydroxymethyl)-aminomethan TTSS Type Three Secretion System U Umdrehungen u.ä. und ähnliches US United States UV-Licht Ultraviolettes Licht V. Vena (v/v) Volumenprozent WHO World Health Organization XLD-Agarmedium Xylose-Lysin-Desoxycholat-Agarmedium z.B. zum Beispiel ZKBS Zentrale Kommission für die Biologische Sicherheit µl Mikroliter

Tabellenverzeichnis
TABELLENVERZEICHNIS
Tabelle 1. Wichtige Anreicherungs- und Differentialnährmedien für Salmonellendiagnostik .................................................................................... 27
Tabelle 2. Übersicht über die in den Infektionsversuchen eingesetzten Bakterienstämme ............................................................................................. 35
Tabelle 3. Gewebeproben zur Untersuchung auf Salmonella Typhimurium DT104-Wildtyp und auf die S. Typhimurium DT104 Deletionsmutanten invC::aphT und sseD::aphT aus persistent infizierten Schweinen ...................................................................................... 36
Tabelle 4. Score für die Beurteilung der Befunde der klinischen Examination im Rahmen der Untersuchung zur Pathogenität von Salmonella Typhimurium DT104 Wildtyp und Salmonella Typhimurium Deletionsmutanten (sseD::aphT & invC::aphT) in persistent infizierten Schweinen. ..................................................................................... 38
Tabelle 5. Primersequenzen zum Nachweis des Wildtyps von S. Typhimurium DT104 und dessen invC::aphT- und sseD::aphT-Deletionsmutanten in Faeces und Gewebeproben von experimentell infizierten Schweinen ............ 42
Tabelle 6. Körpergewichte (Mittelwert ± s) der mit dem Wildtyp DT 104 und mit den Deletionsmutanten invC::aphT bzw. sseD::aphT infizierten Schweine in kg ................................................................................................ 47
Tabelle 7. Mittelwerte der Körpertemperatur (°C) der, mit dem S. Typhimurium DT104-Wildtyp, der Deletionsmutante-invC::aphT, der Deletionsmutante-sseD::aphT infizierten Schweine und die dazugehörige Standardabweichung .................................................................... I
Tabelle 8. Mittelwerte des Körpergewichts der mit dem S. Typhimurium DT104-Wildtyp, der Deletionsmutante-invC::aphT, der Deletionsmutante-sseD::aphT infizierten Schweine mit der dazugehörigen Standardabweichung und Signifikanz ................................................................ I
Tabelle 9. Klinischer Score des Allgemeinbefindens der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, Deletionsmutante invC::aphT und Deletionsmutante sseD::aphT .......................................................................... II
Tabelle 10. Klinischer Score des Durchfallerscheinens der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, Deletionsmutante invC::aphT und Deletionsmutante sseD::aphT ......................................................................... III
Tabelle 11. Die Mittelwerte der Gesamtleukozytenzahl (103/mm3) der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten ........................................................................ V
Tabelle 12. Die Mittelwerte der relativen Lymphozytenzahlen der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent ...................................................... V
Tabelle 13. Die Mittelwerte der relativen Zahlen der neutrophilen stabkernigen Granulozyten der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent .......... VI

Tabellenverzeichnis
Tabelle 14. Die Mittelwerte der relativen Zahlen der neutrophilen segmentkernigen Granulozyten der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent ......................................................................... VI
Tabelle 15. Die Mittelwerte der relativen Zahlen der eosinophilen Granulozyten der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent ............................................. VII
Tabelle 16. Die Mittelwerte der relativen Monozytenzahlen der Infektionsgruppen, S Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent .................................................... VII
Tabelle 17. Qualitativer Nachweis von S. Typhimurium DT104 in Faeces ................... VIII
Tabelle 18. Qualitativer Nachweis von S. Typhimurium DT104 in untersuchten Organ- und Gewebeproben .......................................................................... VIII
Tabelle 19. Quantitativer Nachweis von S. Typhimurium DT104-Infektionsstämme in untersuchten Organ- und Gewebeproben ...................... IX
Tabelle 20. Quantitativer Nachweis der S. Typhimurium DT104-Infektionsstämme in Organ- und Gewebeproben mit der dazugehörigen deskriptiven Statistik und Signifikanzprüfung ........................ X
Tabelle 21. Positiver Nachweis von S. Typhimurium DT104-Wildtyp in untersuchten Organ- und Gewebeproben ....................................................... XII
Tabelle 22. Positiver Nachweis von S. Typhimurium DT104 invC::aphT-Mutante in untersuchten Organ- und Gewebeproben ................................................. XIII
Tabelle 23. Positiver Nachweis von S. Typhimurium DT104 sseD::aphT-Mutante in untersuchten Organ- und Gewebeproben ................................................. XIV

Abbildungsverzeichnis
ABBILDUNGSVERZEICHNIS
Abbildung 1. Schematische Darstellung des Versuchablaufes zur Pathogenität von Salmonella Typhimurium DT104 Wildtyp und Salmonella Typhimurium Deletionsmutanten (sseD::aphT & invC::aphT) in persistent infizierten Schweinen. ...................................................................................................... 34
Abbildung 2. Infektion von Schweinen mit Salmonella-Symposium mittels Magenschlundsonde ........................................................................................ 37
Abbildung 3. Allgemeinbefinden der mit S. Typhimurium DT104 infizierten Schweine .... 43
Abbildung 4. Durchfall der mit S. Typhimurium DT104 infizierten Schweine .................... 44
Abbildung 5. Innere Körpertemperatur (Mittelwert ± s) der mit S. Typhimurium DT104 infizierten Schweine. ....................................................................................... 45
Abbildung 6. Gesamtleukozytenzahl (Mittelwert ± s) der infizierten Schweine. ................. 48
Abbildung 7. Gesamtleukozytenzahl (Mittelwert ± s) der infizierten Schweine. ................. 48
Abbildung 8. Prozentualer Anteil an Lymphozyten (Mittelwert ± s) bei infizierten Schweinen. ...................................................................................................... 49
Abbildung 9. Prozentualer Anteil an Lymphozyten (Mittelwert ± s) bei infizierten Schweinen. ...................................................................................................... 50
Abbildung 10. Prozentualer Anteil neutrophiler stabkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. ...................................................... 51
Abbildung 11. Prozentualer Anteil neutrophiler segmentkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. ...................................................... 51
Abbildung 12. Prozentualer Anteil neutrophiler stabkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. ...................................................... 52
Abbildung 13. Prozentualer Anteil neutrophiler segmentkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. ...................................................... 53
Abbildung 14. Prozentualer Anteil eosinophiler Granulozyten (Mittelwert ± s) der infizierten Schweine. ....................................................................................... 54
Abbildung 15. Prozentualer Anteil eosinophiler Granulozyten (Mittelwert ± s) der infizierten Schweine. ....................................................................................... 54
Abbildung 16. Prozentualer Anteil an Monozyten (Mittelwert ± s) bei infizierten Schweinen. ...................................................................................................... 55
Abbildung 17. Prozentualer Anteil an Monozyten (Mittelwert ± s) bei infizierten Schweinen. ...................................................................................................... 56
Abbildung 18. Kultureller Nachweis von S. Typhimurium DT104 (in % der infizierten Tiere) in Faeces infizierter Schweine. ............................................................. 57
Abbildung 19. Kultureller Nachweis von S. Typhimurium (in % der infizierten Schweine) in Gewebeproben. ............................................................................................ 59
Abbildung 20. Quantitativer Nachweis des Wildtyps (WT) DT 104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT in Gewebeproben. ........ 60

Abbildungsverzeichnis
Abbildung 21. PCR-Produkte zum Nachweis von S. Typhimurium DT104-Wildtyp sowie der invC::aphT- und sseD::aphT-Deletionsmutanten in ausgewählten Gewebeproben experimentell infizierter Schweine. ....................................... 61
Abbildung 22. Nachweis von S. Typhimurium DT104-Wildtyp, Deletionsmutanten in Faeces von ausgewählten Schweinen, die entweder mit der sseD::aphT (I)- oder der invC::aphT-Mutante (II) infiziert wurden. ................................. 61

Einleitung
1
1 EINLEITUNG
Salmonella (S.) Typhimurium ist einer der bedeutendsten Erreger humaner Gastroenteritiden.
In der Europäischen Union (EU) wurden im Jahr 2009 108 614 bestätigte humane
Salmonellose-Fälle gemeldet. Die Anzahl der gemeldeten Salmonellosefälle in der EU zeigt
im Vergleich zum Jahr 2008 einen Rückgang von 17,4 % (EFSA und ECDC2011). Allein im
Jahr 2006 wurden dem Robert Koch-Institut (RKI) 52 575 Salmonellosen bei Menschen in
Deutschland gemeldet (RKI 2007). S. Typhimurium DT 104 wurde als zweithäufigster der
wichtigen Salmonellose-Erreger genannt und hat durch seine chromosomal fixierte
Multiresistenz gegenüber Chemotherapeutika epidemiologisch sehr an Bedeutung gewonnen
(PRAGER et al. 1999; THRELFALL 2000; THRELFALL et al. 2003; HELMS et al. 2005;
ZHAO et al. 2005; MULVEY et al. 2006; ZHAO et al. 2007; LAN et al. 2009). Im Jahr 2009
wurden deutlich weniger Fälle gemeldet (31 408). Mit einem Rückgang von 27 % zum
Vorjahr (42 921) trug S. Typhimurium erstmalig einen signifikanten Beitrag zu sinkenden
Zahlen bei bakteriellen Erkrankungen bei (RKI 2010). Der Trend setzte sich 2010 fort, mit
19 % geringeren Erkrankungszahlen (25 307). Die Salmonellose liegt jedoch an der zweiten
Stelle der durch Bakterien verursachten Erkrankungen in Deutschland (RKI 2011c).
Allein in Sachsen erfasste man 2010 insgesamt 1954 infizierte Menschen (RKI 2011a). Nach
Studien aus anderen Ländern wird jedoch von einer 50-fach höheren Inzidenz der Fälle
ausgegangen. Hieraus ergibt sich für Deutschland eine volkswirtschaftliche und
gesundheitspolitische Bedeutung, die sich in hohen Aufwendungen für Behandlungen und
Kontrollverfahren widerspiegelt.
In Dänemark wurden die Kosten für Salmonellosen bei Menschen im Jahr 2001 auf 15,5
Millionen US-Dollar geschätzt. Die jährlichen Aufwendungen für die Kontrollprogramme für
Menschen, Tiere und Lebensmittel betrugen 14,1 Millionen US-Dollar. Durch die Kontrollen
wurden schätzungsweise 25,5 Millionen US-Dollar öffentlicher Ausgaben eingespart
(WEGENER et al. 2003; KORSGAARD et al. 2005; INFOSAN 2005).
Der Hauptübertragungsweg von S. Typhimurium vom Tier auf den Menschen ist
kontaminiertes Schweinefleisch (RKI 2006; BOYEN et al. 2008; EFSA und ECDC2011).
Somit stellt das Schlachtschwein ein bedeutendes Reservoir für diesen Zoonoseerreger dar.
Eine besondere Rolle nimmt dabei das persistent infizierte Schwein ein. Da Infektionen mit S.

Einleitung
2
Typhimurium beim Schwein überwiegend symptomlos verlaufen, bleibt diese
Infektionsquelle trotz aller Maßnahmen meist unentdeckt.
Die Bekämpfung von Salmonellen in Nutztierbeständen durch eine aktive Immunisierung
gewinnt immer mehr an Bedeutung. Steigende Kosten in Hygiene und Management, sowie
die Zunahme von Resistenzen stellen ein Problem dar. Die bisherigen Maßnahmen, wie
Kontrollen, Einsatz von Antibiotika und Konkurrenzausschluss oder deren Kombinationen,
gelangen zunehmend an ihre Grenzen. Insbesondere der Einsatz von Antibiotika bei
Lebensmittel liefernden Tieren, löst zudem bei immer mehr Menschen Angst vor Resistenzen
aus (ZHANG-BARBER et al. 1999). Bei den heutigen Impfstrategien handelt es sich
vorwiegend um eine Anwendung von Lebendimpfstoff aus vermehrungsfähigen Bakterien
von bestimmten, stabil attenuierten Salmonella-Stämmen. Die Überlegenheit der
Lebendvakzine gegenüber Inaktivat- und Subunitvakzinen konnte in mehreren
tierexperimentellen Untersuchungen bestätigt werden (ROBERTSSON et al. 1983; MARTIN
1992; KARASOVA et al. 2009). Die Wirksamkeit von Salmonella-Lebendimpfstoffen setzt
die Anheftung (Adhäsion), das Eindringen (Invasion) und das Überleben und Vermehren in
der Wirtszelle und lymphoretikulärem Gewebe voraus (MITOV et al. 1992). Dies sind die
drei wichtigen pathogenetischen Schritte von Salmonelleninfektionen. Jedoch ist
entscheidend, dass die in den Impfstoffen enthaltenen Stämme eine verminderte Virulenz
aufweisen und phänotypisch von Salmonella-Feldstämmen zu unterscheiden sind, z.B. durch
Auxotrophien und Resistenzen gegen Antibiotika.
Um Mechanismen der Infektion und der Persistenz des Erregers in den verschiedenen
Geweben des Schweins besser zu verstehen, werden im Rahmen dieser Arbeit die Bedeutung
zweier Virulenzfaktoren mittels des Einsatzes zweier Deletionsmutanten in experimentellen
Infektionensversuchen untersucht. Die hierfür eingesetzten Mutanten wurden durch
Gendeletionen erzeugt, bei denen dadurch von einer verminderten Virulenz auszugehen ist.
Die Mutante I ist durch P22-Transduktion von sseD::aphT aus dem Stamm MvP101
(MEDINA et al. 1999) in den DT104-Stamm erzeugt worden. Das sseD-Gen wird auf der
Salmonella pathogenecity island 2 kodiert, die für die intrazelluläre Überlebensfähigkeit der
Salmonellen in Makrophagen verantwortlich gemacht wird. OCHMAN und Mitarbeiter
(1996) sowie HENSEL und Mitarbeiter (1998) konnten zeigen, dass derartige Mutanten eine
kürzere Überlebenszeit in Makrophagen aufweisen.
Die Mutante II ist durch P22- Transduktion der invC::aphT (Kanamycin-Resistenz)-Kassette
von SB566 (EICHELBERG et al. 1994) in den Wildtyp DT104-Stamm konstruiert worden.

Einleitung
3
Die Deletionsmutanten wurden, wie bereits von EICHELBERG und Mitarbeiter (1994)
beschrieben, konstruiert. Die Deletion des invC-Gens, das sich auf der Salmonella
Patogenicity Island 1 befindet, verursacht in vitro eine verminderte Fähigkeit zur epithelialen
Invasion (GALLOIS et al. 2001).
Ein Wildtyp-Stamm der Salmonella Typhimurium DT104 (MARG et al. 2001; ROESLER et
al. 2004; ARNOLD et al. 2004), vom Schwein isoliert, wurde für den Vergleich zu den vorher
genannten Deletionsmutanten eingesetzt. Dieser in experimentellen Infektionen durchgeführte
Vergleich sollte Erkenntnisse zur unterschiedlichen Organbesiedlung, Veränderungen in
Blutparametern und klinischen Symptomen bringen. Die so erlangten Befunde wiederum
sollten Rückschlüsse auf die Mechanismen der Infektion und durch Persistenz ermöglichen,
um damit die Rolle der Virulenzfaktoren in diesem Infektiongeschehen zu deuten.

Literaturübersicht
4
2 LITERATURÜBERSICHT
2.1 Salmonella
2.1.1 Taxonomie
Die Gattung Salmonella (S.) gehört zur Familie Enterobacteriaceae. Salmonellen sind
gramnegative, fakultativ anaerobe, fast ausnahmslos bewegliche, gerade Stäbchenbakterien
mit einer Größe von 0,7-1,5 x 2,0-5,0 µm (SELBITZ 1992). Die Gattung Salmonella gehört
zu den wichtigsten Vertretern dieser Familie, welche zur Gamma-Untergruppe der
Proteobakterien und somit zu den Eubakterien gezählt werden. Die taxonomischen Einteilung
der einzelnen Salmonella-Isolate basierte über Jahrzehnte auf deren Reaktionen mit Seren
(STACKBRANDT et al. 1988).
So wurden beispielsweise neue epidemiologisch bedeutsame Salmonellen als eigenständige
Spezies definiert. Die übrigen Vertreter sollten hierbei nur als Serovare bezeichnet werden
(DARWIN und MILLER 1999; BRENNER et al. 2000).
KAUFFMANN (1961) beschrieb alle bis dahin bekannten Isolate als eine einzige eigene
Spezies. Später teilte KAUFFMANN (1966) das Genus Salmonella auf der Basis
biochemischer Charakteristika in vier Subgenera ein, wobei jedes Serovar als Spezies
ausgewiesen wurde. Einen entscheidenden Ansatz in der Entwicklung der taxonomischen
Einteilung setzten CROSA et al., (1973) mit dem Beweis der genetischen Verwandtschaft der
Salmonella Subgenera I (enterica), II (salamae) und IV (houtenae) und deren Serovaren,
sowie aller Serovaren von S. arizonae, die einer Spezies zuzuordnen sind (BRENNER et al.
2000).
REEVES et al., (1989) schlugen vor, die Spezies Salmonella anhand von genetischen
Merkmalen, in zwei Subspezies Salmonella enterica und Salmonella bongori einzuteilen.
Gegenwärtig werden die beiden Salmonella-Spezies S. Choleraesuis und S. bongori
anerkannt (POPOFF et al. 1994). Die Spezies S. Choleraesuis beinhaltet die Subspezies: S.
Choleraesuis, S. salamae, S. arizonae, S. diarizonae und S. houtenae (POPOFF et al. 1994).
Der Name „Choleraesuis“ stand gleichzeitig für eine der Spezies als auch für ein Serovar,
was für Verwirrung sorgte. Von LE MINOR und POPOFF (1987) wurde deshalb der

Literaturübersicht
5
präzisierende Vorschlag gemacht, sowohl die Spezies als auch die Subspezies S. Choleraesuis
unter der Speziesbezeichnung Salmonella enterica zusammenzufassen (EUZEBY 1999).
Diese Umbenennung wurde im Jahr 2010 vom „International Commitee of Systematic
Bacteriology“ offiziell anerkannt.
Innerhalb der einzelnen Subspezies der Spezies Salmonella werden verschiedene Serovare mit
diversen Stämmen unterschieden. Ursprünglich wurden fast alle neuen Serovare, welche im
White-Kauffmann-Le Minor-Schema aufgeführt sind, mit Eigennamen belegt. Aus Gründen
der Übersichtlichkeit wurde dieses Prinzip letztlich jedoch nur bei den Eigennamen der zur
Subspezies S. enterica zählenden Serovare beibehalten. Demnach müsste beispielsweise die
korrekte Schreibweise für S. Typhimurium lauten: S. enterica subsp. enterica ser.
Typhimurium. Zur praktischen Vereinfachung der Schreibweise wird von vielen Autoren nun
einfach die Bezeichnung S. typhimurium verwendet. Um den Serovarcharakter deutlich zu
machen, wird der Serovarname trotzdem groß geschrieben, z. B. S. Typhimurium. Die
Vertreter der anderen Subspezies werden durch die Angaben der Antigenformel benannt
(SELBITZ et al. 1995; TINDALL et al. 2005; GRIMONT und WEILL 2007).
Die serologische Typisierung innerhalb der Familie Enterobacteriaceae basiert auf dem
Vorkommen von Antigendeterminanten. Salmonellen haben wie alle Vertreter dieser Familie
Oberflächen- (somatische oder O) und Geißel- (H, Phase I und II) Antigene. Das somatische
„O“-Antigen ist ein hitzestabiles Lipopolysaccharid und ein Bestandteil des in der Zellwand
lokalisierten Lipopolysaccharid-Protein-Komplexes. Die Polysaccharidketten des Antigens
bestimmen die jeweilige serologische Spezifität. Das Geißel- („H“) Antigen ist dagegen ein
hitzelabiles, in den Geißeln sitzendes Protein.
Außerdem können zusätzliche Hüllenantigene (K-Antigene) vorkommen. Heute werden diese
Kapselantigene als „Vi-Antigene“, „Vi“ welches von Virulenz abgeleitet wird, bezeichnet.
Diese sind als Bauelemente der Mikrokapsel der eigentlichen Zellwand aufgelagert und
besitzen Polysaccharidstruktur. Die „Vi-Antigene“ sind nur bei den Serovaren S. Typhi, S.
Paratyphi C und S. Dublin vorhanden.
Die unterschiedliche Zusammensetzung der Antigene ist anhand einer Antigenformel aus dem
White-Kauffmann-Le Minor-Schema, welches bis heute insgesamt 2610 Serovare definiert
(GUIBOURDENCHE et al. 2010), zu entnehmen. Die O-Antigene werden mit arabischen
Ziffern bezeichnet. Stämme mit gemeinsamen Haupt-O-Antigenen werden in O-
Antigengruppen, die mit Buchstaben gekennzeichnet sind, zusammengefasst. Die Serovare
der gleichen Hauptgruppe können mit Hilfe der H-Antigene weiter differenziert werden. Die

Literaturübersicht
6
Isolate, welche identische O- und H-Antigene ausweisen, sollten dem gleichen Serovar
zugeordnet werden.
Eine weitere Methode zur Differenzierung ist die Lysotypie von serologisch definierten
Serovaren. Basierend auf der unterschiedlichen Empfänglichkeit der einzelnen Stämme
innerhalb des selben Serovars gegenüber einem selektierten Set von Bakteriophagen erlauben
Bakteriophagen eine weitere Einteilung der Salmonellen (GRIMONT et al. 2000). Anders als
bei S. Typhi und S. Paratyphi, bei denen Internationale Lysotypiesysteme vorhanden sind,
werden für die Untersuchung von S. Typhimurium verschiedene Phagensätze genutzt. FELIX
und CALLOW (1943) etablierten erstmalig ein derartiges System. Dieses wurde später von
CALLOW (1959) erweitert. ANDERSON et al. (1977) modifizierten es zu der Form, die
heute noch weltweit am häufigsten eingesetzt wird (SCHMIEGER 1999).
Der Phagentyp DT104 hat, begründet durch seine chromosomal fixierte Multiresistenz
gegenüber Chemotherapeutika, seit den neunziger Jahren stark an Bedeutung gewonnen und
wird inzwischen mit am häufigsten isoliert (SCHMIEGER und SCHICKLMAIER 1999;
THRELFALL 2000; RABSCH et al. 2001; THRELFALL et al. 2003; HELMS et al. 2005;
ZHAO et al. 2005; MULVEY et al. 2006; ZHAO et al. 2007; LAN et al. 2009; RKI 2011b).
2.1.2 Epidemiologie
Salmonellen gehören weltweit zu den bedeutendsten bakteriellen Infektionserregern bei
Menschen und Tieren. Nach DINJUS (1998) und für die European Food Safety Authority
(EFSA) stellen sämtliche Salmonellen potentielle Krankheitserreger dar. Dabei stellt sie ihre
Rolle als Zoonoseerreger besonders in den Mittelpunkt des Interesses (EFSA 2006). Das
natürliche Habitat von Salmonellen ist der Darm von Tieren und Menschen. Durch spezielle
Resistenzmechanismen können sie jedoch auch monatelang in einer geeigneten Umwelt
überleben (GUTHIRE 1992).
In den letzten Jahren wurde ein fortlaufend steigender Anteil von Salmonellosen beim
Menschen verzeichnet, die durch kontaminierte Lebensmittel tierischer Herkunft ausgelöst
werden. Hier treten besonders Rohfleischprodukte und Konsumeier als Verursacher der
Infektionen in den Vordergrund. Ihre Kontamination erfolgt entweder primär beim Tier oder
sekundär im Verlauf der Gewinnung, Be- und Verarbeitung sowie bei der Abgabe an den
Verbraucher. Die Gefahr einer primären Kontamination besteht immer dann, wenn das
lebende Tier selbst infiziert ist und zum Ausgangspunkt für den Eintrag von Salmonellen in

Literaturübersicht
7
die Lebensmittelkette werden kann (SELBITZ 1992; STEINBACH und KROELL 1999;
BOTTELDOORN et al. 2003; VAN PARYS et al. 2011).
Die Gefahr für die menschliche Gesundheit erfordert, dass Strategien zur Verhinderung der
Übertragung von Salmonellen durch Lebensmittel entwickelt werden mussten. Diese bestehen
aus einer zielgerechten Kombination von antiinfektiösen, veterinärhygienischen und
produktionsorganisatorischen Maßnahmen (MEYER 1999; MAJOWICZ et al. 2010). Durch
dieses Vorgehen ist eine vollständige Elimination der Kolonisierung zwar nicht möglich,
jedoch eine Verminderung der Gesamtkeimzahlen im Tier (WILCOCK und SCHWARTZ
1992).
Einzelne Serovare weisen dabei ein unterschiedliches epidemiologisches Verhalten auf
(MEYER et al. 1993; SELBITZ 1993), welche auf Unterschieden in ihrer Wirtsanpassung der
einzelnen Vertreter des Genus Salmonella basieren. So existieren Serovare, die ausschließlich
an den Menschen angepasst sind, wie S. Typhi und S. Paratyphi. An bestimmte Tierarten
angepasste Vertreter sind zum Beispiel S. Gallinarum beim Huhn, S. Abortusovis beim Schaf
und S. Choleraesuis beim Schwein. Die Serovare Enteritidis und Typhimurium weisen keine
Wirtsanpassung auf und stellen die Haupterreger von Zoonosen dar (HOLLINGER 2000;
EFSA und ECDC 2011). Die Ursachen, die eine Wirtsspezifität bewirken, sind bisher noch
nicht geklärt (PASCOPELLA et al. 1995). Durch die Unterschiede zwischen adaptierten und
nicht adaptierten Serovaren ergeben sich Konsequenzen hinsichtlich der
Bekämpfungsstrategien und Infektionsvermeidung, da bei adaptierten Salmonellen im
Gegensatz zu nicht adaptierten Salmonellen eine Senkung des Infektionsdrucks im Bestand
möglich ist (SELBITZ 1991). Die Problematik besteht darin, dass diese nicht adaptierten
Serovare nach erfolgter Infizierung, über längeren Zeitraum ohne klinische Symptomatik
persistieren können (WOOD et al. 1989; SCHERER et al. 2008) und somit ohne geeignete
Kontrollmaßnahmen bzw. Bekämpfungsstrategien unerkannt bleiben.
2.1.2.1 Salmonellose Mensch
Die Salmonellose wird als die wichtigste Infektionskrankheit des Gastrointestinaltraktes des
Menschen in Industrieländern beschrieben (WHO 2005; MAJOWICZ et al. 2010). Klinisch
ist sie geprägt von gastroenteritischen Geschehen mit lokalem, sich selbst begrenzenden
Charakter und Durchfall (Enteritis), der das auffälligste Krankheitssymptom darstellt. Auf
Grund dessen wurden die Erreger auch als Enteritissalmonellen bezeichnet. Die

Literaturübersicht
8
Hauptvertreter stammen dabei aus der Subspezies 1 von Salmonella enterica. Jedoch gelten
alle Salmonellen als totipotent für den Menschen und sind als klinisch gleich potent pathogen
anzusehen.
Die durch eine Salmonelleninfektion hervorgerufenen Krankheitsbilder beim Menschen
lassen sich in zwei Gruppen gliedern. Zum einen in die typhöse Form, welche durch
humanadaptierte Serovare ausgelöst wird und zum anderen in die enteritische Form.
Verursacher bei dieser Gruppe sind tieradaptierte und nichtadaptierte Serovare. Demnach
wird die enteritische Salmonellose durch alle Salmonella-Stämme ausgelöst, außer durch S.
Typhi und S. Paratyphi (TSCHÄPE und BOCKEMÜHL 2002; GERLACH und HENSEL
2007; COBURN et al. 2007).
Infektionen werden vorwiegend durch Umweltkontaminationen oder Nahrungsmittel
verursacht (SANDER 1993; MAJOWICZ et al. 2010). Weitere Infektionswege können die
Übertragung von Mensch zu Mensch oder von Tier zu Mensch sein. Bei letzteren handelt es
sich jedoch um Infektketten mit einer sehr geringen Bedeutung (RKI 1997).
Bei Enteritissalmonellen handelt es sich um einen gesundheitspolitisch relevante Erreger, da
das Erkennen einer Erkrankung beim Tier aufgrund des vorwiegend symptomlosen oder
persistenten und subklinischen Verlaufs nicht möglich ist. Beim Menschen jedoch bildet sich
ein deutlich klinisches Bild einer Infektion aus.
Die Infektionsdosis (ID50) für den Menschen wird auf ≥ 105 Keime geschätzt. Die Klinik ist
gekennzeichnet durch Brechdurchfall, kolikartige Bauchschmerzen oder auch Diarrhö, häufig
im Zusammenhang mit hohem Fieber. Diese Symptome können bereits wenigen Stunden
nach Infektion in Erscheinung treten. Die Infektion bleibt meist lokal beschränkt
(Magendarmtrakt) und ist üblicherweise selbstlimitierend. Das heißt, bei milden Verläufen ist
mit einer Heilung nach einer Woche zu rechnen. Eine besondere Gefahr stellt die
Salmonellose für Kleinkinder, Abwehrgeschwächte und alte Menschen dar, da sie hier zu
Organbesiedelung, Septikämie und Arthritis führen kann, dem so genannten typhoiden
Verlauf (HOF et al. 2000; COBURN et al. 2007). Bei dem Serovar S. Typhimurium DT 104
ergeben sich aufgrund seiner breiten Antibiotikamehrfachresistenz, die auf der genomischen
Pathogenitätsinsel SGI 1 kodiert ist (MULVEY et al. 2006) Komplikationen bei der Therapie,
wenn es, wie oben erwähnt, zu einem typhoiden Krankheitsverlauf kommt.
Salmonellosen können epidemisch vorkommen oder sporadisch auftreten. Etwa 85 % aller
Enteritissalmonellen in Deutschland kommen sporadisch vor. Es ist noch nicht geklärt, was
die jeweils dominanten Stämme, welche verstärkt auftreten oder dann aber sogar

Literaturübersicht
9
verschwinden können, befähigt, sogenannte Epidemietypen zu sein. Ein typischer Vertreter
der epidemisch auftretenden Salmonellen ist S. Typhimurium.
Die Epidemiologie von Salmonellosen ist charakerisiert durch die Abhängigkeit von der
Prävalenz in den Tierbeständen, den Überlebens- und Vermehrungsbedingungen im Schlacht-
und Lebensmittelbetrieb, der Virulenz des Erregerklons und dem jeweiligen Immunstatus der
Bevölkerung. Somit ist auch der Anstieg der Erkrankungszahlen in den wärmeren Monaten zu
begründen (KRAUSE 2001).
Seit dem Jahr 2005 sind die Zahlen der Salmonellose-Erkrankungen rückläufig die
wahrscheinlich auf zahlreiche antiepidemische und antiinfektiöse Strategien sowie staatliche
Kontrollmaßnahmen zurückzuführen sind. Bei etwa 65 % davon gelten S. Enteritidis und S.
Typhimurium als Verursacher (EFSA und ECDC 2011). Es besteht die Annahme, dass die
Dunkelziffer bei weitem höher ist. Begründet ist dies mit einem Studienvergleich von anderen
Industrieländern. So vermutet man, dass in Großbritannien und den USA für jede bekannte
Salmonellose 3 bis 50 unerkannt bleiben (WHEELER et al. 1999; VOETSCH et al. 2004).
Im Jahr 2010 wurden dem RKI 25 307 Fälle einer Salmonellose gemeldet, die klinisch-
epidemiologisch und klinisch-labordiagnostisch bestätigt waren (RKI 2011a).
Wie bereits erwähnt, stellt das kontaminierte Lebensmittel die Hauptinfektionsquelle für den
Menschen dar. Aus diesem Grund ist die Verbraucheraufklärung vor allem im Hinblick auf
die Küchenhygiene eine der wichtigsten Präventivmassnahmen (TSCHÄPE und
BOCKEMÜHL 2002). Dies gilt umso mehr, da Salmonellen auf und in Lebensmitteln in
einen Ruhezustand treten oder durch chemische bzw. thermische Noxen subletal geschädigt
werden können, dann nicht mehr kultivierbar und nachweisbar aber immer noch infektiös
sind.
2.1.2.2 Salmonellose Schwein
Die Salmonelleninfektion beim Schwein ist gekennzeichnet durch zwei
Problemschwerpunkte. Zum einen handelt es sich um die im eigentlichen Sinn bezeichnete
Salmonellose, die zur klinischen Erkrankung der Tiere führt. Ausgelöst wird die Erkrankung
durch die an das Schwein adaptierten Serovare S. Choleraesuis und S. Typhisuis. Der zweite
Schwerpunkt ist die fleischhygienische Problematik, welche durch klinisch unauffällige
Ausscheider im Schlachtprozess entsteht und mit einer Kontaminierung der Schlachtprodukte
einhergeht. Verursacher hierfür können eine Vielzahl von Serovaren sein.

Literaturübersicht
10
Die beiden speziesadaptierten Erreger (S. Choleraesuis, S. Typhisuis) sind inzwischen in
Deutschland bedeutungslos. Die durch sie ausgelösten primären Salmonellosen gehen in
Schweinebeständen mit einer hohen Verlustrate einher (STEINBACH und KROELL 1999).
Das in den letzten Jahren von dem Nationalen Referenzlabor für Salmonellen am häufigsten
nachgewiesene Salmonella-Serovar ist S. Typhimurium. Betrachtet man die Jahre 1998 bis
2002 ist eine Zunahme bei diesem Serovar zu verzeichnen. Im Jahr 2002 war der Anteil der S.
Typhimurium Infektionen 88 %. An zweiter Stelle befindet sich S. Derby mit 6 %. Weitere
beim Schwein bedeutsame Serovare sind S. Infantis (0,9 %), S. Livingstone (3,7 %), S.
Enteritidis und S. London (0,9 %). Die in den Schweinebeständen dominierenden Serovare
werden ebenfalls bei Lebensmittelkontaminationen nachgewiesen (DORN et al. 2003).
In den Jahren 2004 – 2008 fiel der Anteil von S. Typhimurium auf 61,1 % zurück
(FRIEDRICH et al. 2010). Im Jahr 2009 wurden 17 % weniger S. Typhimurium-Isolate
(43,8 %) diagnostiziert. Dafür konnte ein Anstieg von 23 % der S. enterica subsp. Enterica
serovar 1,4,[5],12:i:-. (35,6 %) registriert werden. Damit war die monophasische Variante der
S. ser. Typhimurium, die S. enterica subsp. Enterica serovar 1,4,[5],12:i:-. im Jahr 2009 an
zweiter Stelle und die S. ser. Derby (5,7 %) auf Rang drei (FRIEDRICH et al. 2010;
FRIEDRICH et al. 2011).
Bei 61,9 % der S. Typhimurium-Isolate handelte es sich 2001 um den multiresistenten
Lysotyp DT104, der inzwischen gegen bis zu 17 verschiedene Antibiotika Resistenzen
aufweist. Von den im Jahr 2009 eingesandten Proben, in denen S. Typhimurium festgestellt
wurde, konnten 33,9 % als Phagentyp DT 104 identifiziert werden. Generell weisen
Salmonella-Isolate aus Schweinebeständen ein sehr hohes Resistenzniveau auf (POPPE et al.
2002; DORN et al. 2003; PERRON et al. 2007; FRIEDRICH et al. 2010; FRIEDRICH et al.
2011).
Basierend auf Beobachtungen aus dem Feld und intensiver experimenteller Arbeiten am
Tiermodell Schwein kann die Salmonelloseerkrankung des Schweines in verschiedene
Verlaufsformen unterschieden werden, die allerdings fließend ineinander übergehen können:
1. Latente Form
2. Enteritische Form
3. Lokal begrenzte und abszedierende Form
4. Systemische Verlaufsform.

Literaturübersicht
11
Die verschiedenen Formen treten abhängig vom Serovar, der Abwehrlage des Tieres, von
äußeren Stressoren und dem Manifestationsort auf (MARG et al. 2001; SCHERER et al.
2008).
Nach einer Inkubationszeit von zwei Tagen bis mehreren Wochen kann sich je nach den
erwähnten Bedingungen eine akute, subakute oder chronische Salmonellose entwickeln.
Bei der akuten Form steht die Septikämie im Vordergrund. Symptome sind
Lungenerkrankungen, Blauverfärbungen an Ohren, Bauch und Schenkelinnenflächen. Nicht
selten treten Todesfälle auf.
Entwickelt sich eine subakute und chronische Salmonellose, kann man zu Beginn
Verstopfungen beobachten, die in profusen Durchfall übergehen. Überlebende Tiere bleiben
zumeist Kümmerer.
Die latente Form der Salmonellose erscheint symptomlos. Die latent infizierten Schweine sind
jedoch üblicherweise Salmonellenausscheider (GAREIS 1995; BOYEN et al. 2008). Diese
Tiere gelten, als persistente Salmonellenträger, die über Wochen bis Monate Salmonellen
ausscheiden können (WOOD et al. 1989; SCHERER et al. 2008). Eine der häufigsten
Ursachen für eine Infektion ist der fäkal-orale Übertragungsweg durch Einschleppung latent
infizierter Tiere in den Bestand. Diese latent infizierten Ausscheider bleiben aufgrund der
fehlenden klinischen Symptome als eine ständige Infektionsquelle unerkannt (SELBITZ
2002; PERRON et al. 2008; CLOTHIER et al. 2010). Eine ebenso große Rolle spielt die
indirekte Erregerübertragung durch belebte oder unbelebte Vektoren. Die bedeutendsten
belebten Vektoren sind Nager, deren Kot mitunter 105 KBE/g Salmonellen enthalten kann
(HENZLER und OPITZ 1992). Zu den unbelebten Vektoren zählen Abwässer und
Futtermittel. Der Eintrag durch Futter hat in den letzten Jahren aufgrund der vorgeschriebenen
Hitzebehandlung deutlich abgenommen. Im Jahr 2009 waren lediglich 1,8 % der untersuchten
Schweinefutterproben kontaminiert (HARTUNG und KÄSBOHRER 2011). Nicht zu
unterschätzen ist die Möglichkeit der Infektion im Stall selbst, denn durch die hohe Tenazität
der Salmonellen gestaltet sich die Bekämpfung problematisch (BAGGESEN et al. 1997).
Dadurch ergibt sich die höhere Inzidenz in Mastbetrieben (BERENDS et al. 1996). Laut der
2006/2007 durchgeführten EU-Grundlagenstudie zur Erhebung der Prävalenz von
Salmonellen bei Mastschweinen gelten 10,3 % der Mastschweine in der EU als infiziert.
Dagegen betrifft es Zucht- und Ferkelanlagen zu 31,8 %. In Deutschland betrug der Anteil
positiver Mastschweine 12,7 %, während Zucht- und Ferkelerzeugerbetriebe zu 22,4 %
positiv waren (EFSA 2008; BfR 2008; EFSA 2009). Interessant ist, dass die Seroprävalenz

Literaturübersicht
12
bei Mastschweinen in Deutschland mit 32,3 % deutlich höher lag als der Anteil
bakteriologisch positiver Tiere (BfR 2008; EFSA 2009; EFSA und ECDC 2011).
2.1.3 Pathogenitätsfaktoren
Die beiden Salmonellenspezies, S. enterica und S. bongori sind als obligate Pathogene für
Mensch und Tier einzustufen. Im Einzelfall und nach der entsprechenden Prüfung können
Serovare und Stämme als avirulent für betroffene Menschen oder das Tier angesehen werden
(SELBITZ 2002).
Die Salmonella-Pathogenese ist ein komplexes und multifaktorielles Phänomen (WALLIS
und GALYOV 2000; MALIK-KALE et al. 2011). Die Pathogenität von Salmonella sowie die
Interaktionen zwischen Salmonellen und der Wirtszelle auf molekularer Ebene sind bereits
gut untersucht (SANTOS et al. 2003; MCGHIE et al. 2009; SABBAGH et al. 2010;
HALLSTROM und MCCORMICK 2011). Gleichwohl ist die Vielzahl und das
Ineinandergreifen von Virulenzmechanismen der Salmonellen weitgehend unbekannt, was auf
die enormen Unterschiede der Pathogenität in der jeweiligen Zielspezies zurückzuführen ist
(RAHMAN et al. 2004). Nur wenige Isolate wurden auf ihre Pathogenität in der jeweiligen
Zielspezies hin getestet. Die zwei erfolgreichsten Tiermodelle, die bis heute eingesetzt werden
und zur Aufklärung der Virulenzmechanismen von Salmonella dienen, sind Kälber- und
Mäusemodelle. Studien am Schwein sind vergleichsweise selten (FEDORKA-CRAY et al.
2000; SANTOS et al. 2001; NIEWOLD et al. 2007; MEURENS et al. 2009).
Die meisten Erkenntnisse über Virulenzfaktoren der Salmonellen wurden in In vitro-
Versuchen gewonnen. Diese zeigen aber, dass Rückschlüsse auf die medizinische Relevanz
und auf die klinische Bedeutung für den betroffenen Patienten nicht immer möglich sind.
Erschwerend kommt eine geringe Wirtsadaptation und vielfältige klinische Manifestation der
Salmonella-Serovare hinzu, die aus der Kombination des Serovars und dem involvierten Wirt
resultiert (WALLIS und GALYOV 2000; WATSON et al. 2000; SANTOS et al. 2003;
PERRON et al. 2008).

Literaturübersicht
13
2.1.3.1 Chromosomal kodierte Pathogenitätsfaktoren
Die Virulenz der meisten infektiösen Bakterien setzt potente Pathogenitätsfaktoren voraus.
Schätzungsweise werden etwa 4 % des Genoms von Salmonella Typhimurium, welches über
200 Gene kodiert, für eine letale Infektion bei Mäusen benötigt. Ein großer Teil dieser Gene
befindet sich als eine Reihe von Genen, welche eine funktionelle Einheit bilden, in
spezifischen Regionen des Genoms, den so genannten Salmonella Patogenicity Islands (SPIs).
Diese können auch als Pathogenicity Islets vorkommen, die lediglich aus einem oder wenigen
Genen bestehen (MARCUS et al. 2000). Insgesamt sind bereits 21 dieser Pathogenitätsinseln
(SPIs) identifiziert. Einzelne Isolate der verschiedenen Serovare können unterschiedlich viele
Pathogenitätsinseln aufweisen. Die genaue Funktion der einzelnen Pathogenitätsinseln und
ihre Interaktion und Regulation sind jedoch nur teilweise bekannt (SABBAGH et al. 2010;
HALLSTROM und MCCORMICK 2011).
Eines der wichtigsten Pathogenitätsmerkmale der Salmonellen ist die Fähigkeit, im Darm zu
kolonisieren und sowohl M-Zellen als auch Enterozyten zu invadieren. Diese Eigenschaft von
Salmonellen in Wirtszellen einzudringen, wird als „Invasion“ bezeichnet. Untersuchungen
zeigen, dass eine Invasion von Salmonellen in den Wirt nur dann möglich ist, wenn die
Wirtszelle aktiv an diesem Prozess beteiligt ist (GALAN et al. 1993).
Die Salmonella-Pathogenitätsinsel 1 (SPI 1) ist zur Zeit die am besten charakterisierte von
allen und ist in allen Linien der beiden Salmonella-Spezies (S. bongori und S. enterica) zu
finden (MARCUS et al. 2000; LOSTROH und LEE 2001). Die SPI 1 ist 40 kb groß und in
der Centisom-63-Region des Salmonella-Cromosoms lokalisiert. Sie kodiert für insgesamt 33
Proteine. Darunter befinden sich alle Komponenten des Typ III-Sekretionssystems und einige
seiner Regulator- und Effektor-Proteine. Das Typ III-Sekretionssystem wird zusammen von
der Salmonella-Pathogenitätsinsel 1 und der Salmonella-Pathogenitätsinsel 2 kodiert und
transportiert Virulenzfaktoren und Effektor-Proteine in Richtung oder direkt in die
Wirtszellen. Das von der SPI 2 kodierte Typ III-Sekretionssystem ist jedoch strukturell und
funktionell von dem, welches auf der Salmonella-Pathogenitätsinsel 1 kodiert ist,
verschieden. Die auf der SPI 1 kodierten Proteine sind für die Invasion in die nicht
professionell phagozytierenden Zellen erforderlich (GROISMAN und OCHMAN 1993;
MARCUS et al. 2000; HANEDA et al. 2009; AGBOR und MCCORMICK 2011). Die
Invasion in nicht professionell phagozytierende Zellen wird für die Besiedlung innerer
lymphatischer Organe als sehr wichtig angesehen. Die nicht-invasiven Mutanten zeigten in

Literaturübersicht
14
vivo eine verminderte Fähigkeit, die Milz und Leber zu kolonisieren und somit in oral
infizierten Mäusen ein attenuiertes klinisches Erscheinungsbild zu erzeugen. Viele
Beobachtungen haben gezeigt, dass für S. Typhimurium die Invasion als solche nicht immer
erforderlich ist, um gezielt Organe des Wirtsorganismus zu erreichen. So ist z.B. S.
Typhimurium, der die gesamte SPI 1 fehlt, nicht in der Lage, kulturelle Gewebezellen zu
invadieren. Es besitzt jedoch die Fähigkeit, in vivo Organe systemisch zu besiedeln.
Salmonellen haben ausserdem die Möglichkeit einen alternativen Weg zu gehen, um die Milz
zu erreichen. Sie dringen in die CD 18+-Makrophagen ein und benutzen sie als Vehikel. Diese
Erkenntnisse wurden im Rahmen einer intragastrischen Infektion in BALB/c Mäusen erlangt
(LOSTROH und LEE 2001). Eines der auf der Salmonella Pathogenicity Island 1
lokalisierten Gene ist das bereits 1989 entdeckte invC-Gen (GALAN und CURTISS 1989). Es
kodiert für ein 47 kDa-Polypeptid, das in seiner Sequenz homolog zu Vertretern der F0F1-
ATPase-Proteinfamilie verschiedener Mikroorganismen ist. Das Protein weist zudem eine
signifikante ATPase-Aktivität auf (EICHELBERG et al. 1994). Trotz der zentralen Rolle
dieser ATPasen in der Sekretion des Typ III-Sekretionsapparates, ist wenig über deren
Mechanismen bekannt. Bewiesen ist hingegen, dass gereinigtes rekombinantes invC sowohl
in monomerer sowie in multimerer Form vorkommt (wahrscheinlich als Hexamer). Weitere
Untersuchungen lassen vermuten, dass das invC in mindestens drei diskrete Domänen
unterteilt ist. Die erste dieser Domänen ist die Amino-terminale-Domäne, die wahrscheinlich
die Verbindung von invC mit der Membran vermittelt. Die zweite ist die zentrale katalytische
und die dritte eine Carboxyl-terminale Domäne, die möglicherweise an Protein-Protein-
Interaktionen, welche für die Funktion des Typ III-Sekretionssystems wichtig sind, beteiligt
ist (AKEDA und GALAN 2004).
Weitere Pathogenitätsmerkmale der Salmonellen sind das intrazelluläre Überleben und die
Fähigkeit sich intrazellulär zu replizieren. Die dafür verantwortlichen Gene werden von der
Salmonella-Pathogenicity 2 (SPI 2) kodiert (ZURAWSKI und STEIN 2004; GERLACH und
HENSEL 2007).
Die molekularen Funktionen der SPI 2 sind vergleichsweise noch nicht so gut charakterisiert
wie die von der SPI 1. Jedoch konnte in Mäuse-Infektionsversuchen gezeigt werden, dass SPI
2-Mutanten eine erheblich schwächere Virulenz entfalteten und damit die Fähigkeit, in
infizierten Wirtsorganen wie Milz und Leber zu proliferieren, verloren haben. Zwar können
solche Mutanten bei einer oralen Infektion die Peyerschen Platten kolonisieren, jedoch sind

Literaturübersicht
15
sie nicht in der Lage, sich darüber hinaus weiter zu verbreiten. Es konnte auch bereits in In
vitro-Versuchen gezeigt werden, dass SPI 2-Deletionsmutanten eine kürzere Überlebenszeit
und verminderte intrazelluläre Replikation aufwiesen (HANSEN-WESTER und HENSEL
2001).
Die Salmonella-Pathogenitätsinsel 2 bildet eine Insertion im tRNAval-Gen und ist in zwei
Segmente geteilt, die wahrscheinlich durch verschiedene horizontale Gentransfers erworben
wurden. Die 40 kb große SPI 2 liegt bei Centisom 31 und kodiert für mehr als 40 Gene. Die
vier verschiedenen Arten von Genen werden in Operone eingeteilt, welchen verschiedene
Funktionen zuzuordnen sind. Diese werden als Gene für Strukturproteine des
Sekretionsapparates (ssa = secretion system apparatus), Gene für sezernierte Effektor-Proteine
(sse = secretion system effector), Gene die als Regulatoren fungieren (ssr = secretion system
regulator) und Gene die für Chaperone kodieren (ssc = secretion system chaperone), definiert.
Die SPI2 kodiert ein Zweikomponentensystem, das für die Expression aller Gene die für das
Sekretionssystem Typ III der Pathogenitätsinsel 2 notwendig sind. Am Ende der Insel,
beginnend bei Centisom 30, befindet sich ein etwa 26 kb großer Bereich, in dem die Gene für
das Sekretionssystem Typ III lokalisiert sind (MARCUS et al. 2000; WATERMAN und
HOLDEN 2003).
Der von der SPI 2 kodierte Sekretionsapparat dient dem Transfer einer Reihe von Effektor-
Proteinen, die für das intrazelluläre Überleben und die systemische Infektion notwendig sind.
Ein Teil dieses Systems ist eine auf der Bakterienoberfläche lokalisierte Translokatorstruktur,
die aus den drei Proteinen, SseB, SseC und SseD besteht (ZURAWSKI und STEIN 2004). Im
Gegensatz zu den anderen Sekretionsapparaten, die die Proteine durch die Wirtszellmembran
translozieren, erfolgt hier der Transport durch die Vakuolenmembran (HOLDEN 2002). Die
SseCD bilden eine vermeintliche Translokator-Poren-Untereinheit, die in der Membran der
Wirtszelle lokalisiert ist. Bei SseB handelt es sich vermutlich um eine scheidenartige
Untereinheit, deren Funktion darin besteht, den Translokator in der Bakterienmembran mit
der Porenuntereinheit zu verbinden. Zusätzlich wurde eine zentrale Domäne innerhalb des
SseD-Proteins, welches Bindungen beeinflusst, beschrieben (ZURAWSKI und STEIN 2004).
Das von der Salmonella-Pathogenicity-Island kodierte Sekretionssystem ist somit ein
multifunktionelles Virulenzsystem, das nach dem Eintritt in die eukariontische Zelle aktiviert
wird. Auch wenn viele Effektorproteine bereits identifiziert sind, können Effektor-Funktionen
nur wenigen davon zugeschrieben werden. Daher ist vermutlich eine große Anzahl weiterer
Effektoren, die sich an diesem Prozess beteiligen, bisher unentdeckt (WATERMAN und
HOLDEN 2003).

Literaturübersicht
16
Abgesehen von den verschiedenen, eng kontrollierten Pathogenitätsinseln, die in
unterschiedlichen Stadien der Infektion wirksam werden, scheint eine hoch entwickelte
Regulation des Bakterienstoffwechsels zu existieren. Salmonellen verfügen über eine Reihe
von adaptierten Mechanismen, die es ihnen ermöglichen, den verschiedensten
Umgebungseinflüssen entgegen zu wirken. Diese Herausforderungen sind unter anderem
solche wie oxidativer Stress, Nährstoffmangel und Verdauungsenzyme, die in
unterschiedlichen Stadien der Infektion auftreten. Eine Gruppe von Genen, die in der
metabolischen Adaption eine Rolle spielen, sind die so genannten „starvation-stress response
Gene“ (SPECTOR 1998). Diese Gene kodieren spezifische metabolische Funktionen, die für
das Überleben der Salmonellen in bestimmten Stadien der Infektion notwendig sind und
damit als Virulenz-Determinanten betrachtet werden müssen.
Die Fähigkeit der Salmonellen, die Ausscheidung von Darmsekret zu induzieren, wird zum
Teil der Produktion von Enterotoxinen zugeschrieben. Bislang wurde bei einigen Salmonella-
Isolaten ein Choleratoxin-ähnliches Enterotoxin beschrieben, das in der Struktur, Funktion
und Antigenität, dem Choleratoxin- (CT) sowie dem von Escherichia coli hitzelabilen- (LT-
I)-Enterotoxin ähnelt (CHOPRA et al. 1987; PRASAD et al. 1990; 1992). CHOPRA et al.
(1999) konnten zeigen, dass Mutationen des Salmonella enterotoxin-Gens (stn) von S.
Typhimurium im Mäuseinfektionsmodel zu einer signifikanten Verminderung der gesamten
Virulenz führt. Jedoch ist die genaue Funktion dieses Toxins nicht vollständig geklärt.
Weiterhin konnten drei verschiedene Zytotoxine bei Salmonellen beschrieben werden. Zwei
dieser Toxine sind in ihrer Funktion weitestgehend unbekannt. Ein membranassoziiertes
Zytotoxin mit geringem Molekulargewicht wurde von REITMEYER et al. (1986)
beschrieben. Die von ASHKENAZI et al.(1988) untersuchten Isolate produzierten alle ein
hitzelabiles-Zytotoxin und waren empfindlich für eine Trypsinbehandlung, was darauf
hinweist, dass der toxisch aktive Teil ein Protein ist. Ein drittes Toxin ist das so genannte
Salmolysin, ein zellassoziiertes Hämolysin (LIBBY et al. 1990). Das Salmolysin wird von
dem slyA-Gen kodiert und hat keine signifikanten Homologien zu anderen beschriebenen
Genen. Das slyA-Gen kommt dennoch auch bei Shigella und enteroinvasiven E. coli-Isolaten
vor. Salmolysin ist ein essentieller Pathogenitätsfaktor in Salmonella, der für das
intrazelluläre Überleben in Peritoneal-Makrophagen und für die Virulenz im
Mäuseinfektions-Modell verantwortlich ist (LIBBY et al. 1994).
Neuere Untersuchungen zeigen, dass das slyA-Gen eine komplexere Funktion aufweist, als
zuerst vermutet werden konnte. Das SlyA-Protein ist ein Mitglied der MarR-Familie von
Transskriptionsfaktoren, die eine wichtige Rolle bei der bakteriellen Virulenz spielen. Es

Literaturübersicht
17
konnte gezeigt werden, dass SlyA eine selbstregulierende Funktion hat. Hierzu konnte eine
Verbindung zu einem anderen regulierenden Faktor, dem PhoP-PhoQ-
Zweikomponentensystem, nachgewiesen werden. Dieser kontrolliert die
Kapselkomponentenmodifikation der Salmonellen. Dies beruht auf der Mg(II)-
Ionenkonzentration, die im Phagosom niedriger ist als im extrazellulären Raum. Das PhoP-
PhoQ-System erkennt die unterschiedlichen Ionenkonzentrationen von Ca(II) und Mg(II),
wodurch die Lokalisation des Bakteriums bestimmt wird und sich das System den jeweiligen
Oberflächenstrukturen anpassen kann. Das SlyA selbst wird dabei nicht direkt von Mg(II)-
Ionen beeinflusst, sondern über das PhoP-PhoQ-Zweikomponentensystem (NORTE et al.
2003).
Unterschiede in der Lipopolysaccharid- (LPS-) und Flagellen-Struktur der Salmonellen
generieren die antigenetischen Variationen, die sich in den über 2500 Serovaren
widerspiegeln. Desweiteren haben Variationen der Lipopolysaccharide Einfluss auf die
Virulenz des jeweiligen Stammes.
Die Gene der rfb-Region des Salmonellen-Chromosoms kodieren die Enzyme, welche für die
Synthese von Zuckern sowie deren Organisation in den Polysaccharid-Seitenketten des
Lipopolysaccharids verantwortlich sind (LINDQVIST et al. 1994; FITZGERALD et al.
2003). Die zweite weniger variable Komponente des LPS der Salmonellen stellt das Lipid A
dar. Da das Lipid A in den meisten In vivo- und In vitro-Testsystemen die biologisch aktive
Komponente des Lipopolysaccharids ist, wird es als „endotoxic principle“ (Endotoxische
Prinzip) des Endotoxins bezeichnet (SEYDEL et al. 2005). Die rfb-Region ist polymorph und
führt somit zu einem hohen Polymorphismus der Polysaccharidketten (FIERER und GUINEY
2001). Die dritte Komponente des Lipopolysaccharides bildet das Kern-Oligosaccharid (core
oligosaccharide). Diese weitaus weniger variable Kernregion wird in eine äußere und innere
unterteilt und verbindet die O-spezifischen Seitenketten mit dem Lipid A (RIETSCHEL und
BRADE 1992; RIETSCHEL et al. 1993; RYCROFT 2000). Bei den meisten Serotypen bildet
das intakte LPS zum Schutz vor Umwelteinflüssen eine Schicht um das Bakterium und
schützt es somit auch vor Phagozytose durch Makrophagen bzw. vor der
Komplementkaskaden vermittelten Abtötung (SAXEN et al. 1987; ROBBINS et al. 1992).
Hinsichtlich der Koloniemorphologie lassen sich bei den Salmonellen S- (smooth) und R-
(rough) Formen unterscheiden, wobei sich der Begriff auf die Ausbildung der Kolonieränder
bezieht. In der Ultrastruktur erklären sich diese phänotypischen Differenzen mit der
unterschiedlichen Exprimierung des O-Antigens. Ursache hierfür sind Defekte an Genen,

Literaturübersicht
18
welche speziell Phosphoryl- und Glykosyltransferasen kodieren, was eine Synthese
unvollständiger LPS-Moleküle bedingt (KLENA et al. 1992). Zum besseren Verständnis
infektiöser Mechanismen ist anzumerken, dass die S-Formen bei Salmonellen hinsichtlich der
Länge ihrer Polysaccharide stark differieren und neben vollständigen (S) Seitenketten auch
gleichseitig R-Formen exprimiert werden (SEYDEL et al. 1993). Gelangt das
Lipopolysaccharid in den Herzkreislauf des Wirtes führt es zu Immunzellstimulation, aus der
die Freisetzung von Zytokinen wie TNF-α (Tumor Nekrose Faktor α) und Interleukinen
resultiert. Die systemische Produktion dieser Entzündungsmediatoren zeigt sich klinisch
charakteristisch in Symptomen einer Sepsis durch Hypotonie, Fieber, respiratorischer
Dysfunktion, die zu multiplen Organversagen und Tod führen kann (MUELLER et al. 2005).
In geringen Mengen wirken die Lipopolysaccharide stimulierend auf das Immunsystem des
Wirtes, höhere Mengen dagegen führen zu dramatischen pathophysiologischen Reaktionen
(SCHLETTER et al. 1995).
Flagellen der Salmonellen sind auch potente Antigene, welche bei der Serotypisierung anhand
des Kauffmann-White-Schemas Anwendung finden. Die Geißel-Antigene (Flagellen, H-
Antigene) werden in zwei Phasen exprimiert, jedoch exprimiert jedes einzelne Bakterium nur
eine von beiden Phasen. Es wird vermutet, dass dieser Wechsel der Antigene ein Weg für das
Bakterium ist, zeitweise der humoralen Abwehr des Wirtes zu entkommen (FIERER und
GUINEY 2001). Flagellen spielen eine entscheidende Rolle bei der Adhäsion und der
Invasion in verschiedenen kultivierten Zellen. Neuere Untersuchungen zeigen, dass Flagellen
auch bei Entzündungen eine spezifische Rolle übernehmen können. Sie aktivieren die
Sekretion von Entzündungsmediatoren wie NF-κB und Interleukin 8 über den TLR5-Rezeptor
(Toll-Like-Receptor 5) in humanen epithelialen Zellmodellen. Eine verminderte Virulenz von
nicht beweglichen und nicht chemotaktischen Salmonellen-Mutanten kann hauptsächlich von
der weniger effizienten Interaktion mit dem zäkalen Epithelium herrühren (STECHER et al.
2004).
2.1.3.2 Plasmid-kodierte Pathogenitätsfaktoren
Die für die Pathogenität von Enterobakterien verantwortlichen Virulenzfaktoren werden
überwiegend von Plasmiden kodiert. Die Salmonella-Virulenzplasmide (spv) wurden erst in
den 80-iger Jahren entdeckt. Diese Entdeckung bezieht sich auf ein relativ großes, 90 kbp
umfassendes Plasmid, welches bei fast allen S. Typhimurium-Isolaten beobachtet wurde

Literaturübersicht
19
(ROTGER und CASADESUS 1999). Auch einige der Salmonella enterica subspezies
enterica tragen diese sogenannten „Low-Copy“-Plasmide. Insbesondere die wirtsadaptierten
Salmonella-Serovare besitzen vermehrt Virulenzplasmide.
Das Vorhandensein dieser Virulenzplasmide korreliert positiv mit der Virulenz der
betreffenden Stämme, weshalb sie bereits kurz nach ihrer Entdeckung den Namen
„Virulenzplasmide“ erhielten. Es konnte im Rahmen experimenteller Infektionen von Mäusen
gezeigt werden, dass das Vorhandensein dieses Plasmids die LD50 um den Faktor 10 auf 106
in Abhängigkeit von Serovar und Art der Inokulation verringert. Dieser als „spv-Effekt“
bezeichnete Vorgang wurde auch in anderen Tierarten beobachtet (HELMUTH et al. 1985;
TINGE und CURTISS 1990; FIERER et al. 1992; LIBBY et al. 1997; ROTGER und
CASADESUS 1999; MAZURKIEWICZ et al. 2008; GUINEY und FIERER 2011). Die
Größe der Virulenzplasmide kann in Abhängigkeit von Serovar zwischen 50 und 140 kb
variieren. Die Virulenzplasmide der Subspezies I von S. enterica, haben einen Abschnitt, der
stark homolog ist und als „Salmonella plasmid virulence“-Locus (spv-Locus) bezeichnet wird.
Dieser 7,8 kb große spv-Locus besteht aus vier Strukturgenen (spvABCD), die als Operon
organisiert sind und vom spvR-Gen reguliert werden (GUINEY et al. 1994; LESNICK et al.
2001). Das sogenannte SpvR-Protein ist der Hauptregulator dieser Strukturgene. Es bindet an
bestimmte Regionen zweier Promotoren (den eigenen und an spvA) und initiiert die
Transkription. Dafür ist zusätzlich der Sigma-Faktor-RpoS notwenig, der vorwiegend in
Stresssituationen vorliegt. Als Folge dieses Prozesses werden vor allem solche Proteine
gebildet, die Bakterienzellen vor schädigenden Umwelteinflüssen schützen (LESNICK et al.
2001; DONG und SCHELLHORN 2010; GUINEY und FIERER 2011). Genetische Analysen
haben ergeben, dass die spvR- und spvBC-Gene für die Virulenz essentiell sind, während spvA
und spvD dafür irrelevant sind (GUINEY und FIERER 2011).

Literaturübersicht
20
2.1.4 Mutationen
Bakterien weisen eine Genomstabilität auf, die im Normalfall durch Reparaturmechanismen
und Restriktionsenzyme gegen fremde DNA sowie Transferbarrieren gewährleistet wird. Im
Verlauf der evolutionären Anpassung bildeten sich jedoch kontinuierlich Mutationen auf
Grund horizontaler Gentransfers und Umlagerungen genetischer Varianten heraus.
Genrearrangements, also die Neuordnung der sich im Genom befindlichen einzelnen Gene
und Gencluster, sind die Folge von Selektionen.
Spricht man vom horizontalen Gentransfer, handelt es sich um eine Genübertragung durch
Konjugation, Transformation oder Transduktion. Diese drei Transfermechanismen
ermöglichen Bakterien eine erfolgreiche Verteilung und schnelle Resistenzentwicklung.
Bei der Konjugation handelt es sich um einen Plasmid-Transfer. Eine plasmidhaltige Zelle,
der so genannte Donor, gibt über einen Sex-Pilus Kopien von Plasmiden an eine andere Zelle,
den Rezipienten, weiter (KRACZKIEWICZ-DOWJAT et al. 1984). Diesen
extrachromosomalen Informationsaustausch beschrieben bereits Lederberg und Tatum (1946).
Bei der Transduktion erfolgt die Plasmidübertragung durch Bakteriophagen (FALKOW
1975). Da sich diese Vektoren jedoch nur an bestimmte Rezeptoren der Zelloberfläche
anheften können, bleibt der jeweilige Austausch auf eine Bakterienspezies, bzw. gram-
verwandte Spezies, beschränkt (SAUNDERS 1984).
Bei der Transformation werden freie, fremde DNA-Stücke in die Bakterienzelle
aufgenommen und können nun durch homologe Rekombination identischer Sequenzen in das
Genom integriert werden (LEVY 1992). Solch ein Austausch von Genen ist unter
verschiedenen Genera möglich (SAUNDERS 1984). Die Transformation hat jedoch im
Vergleich zur Transduktion und Konjugation kaum Bedeutung, da der Verlust von
Zellwandanteilen des Erregers, als eine Bedingung für die Transformation, diesen
Mikroorganismus empfindlicher gegenüber Umwelteinflüssen macht (WERCKENTHIN und
SCHWARZ 1997).
Eine Mutation führt zu einer Veränderung der DNA-Basen-Sequenz. Dies verschafft der
Mutante gegenüber ihrem Wildtyp oftmals einen entscheidenden Überlebensvorteil. Man
unterscheidet bei Mutationen zwei Formen, die Punkt- und die Rastermutation. Bei einer
Punktmutation werden einzelne Nukleotidbasen ausgetauscht. Sie stellt somit die einfachste

Literaturübersicht
21
Form der Mutation dar. Ihr Vorkommen liegt bei einer Häufigkeit von 1 Mutante pro 106
Keimen. Bei Rastermutationen finden größere Veränderungen im Bereich des Erbgutes statt.
Ursachen dafür sind Insertionen (Hinzufügen von Basenpaaren), Deletionen (Verlust von
Basenpaaren) und Inversionen (Herausschneiden und Einsetzen eines DNA-Moleküls in
umgekehrter Reihenfolge) (HUTCHINSON und DONNELLAN 1996).
Von besonderem Interesse ist die Mutation unter dem Gesichtspunkt der Weitergabe von
Resistenzgenen. Hierbei erfolgt eine Unterteilung in spontane und adaptive Formen der
Mutation (MORRIS et al. 1998). Spontanmutationen unterliegen dem Zufallsprinzip. Sie
finden während der Zellteilung bei 0 bis 5 Mutanten pro 108 Mikroorganismen statt
(LECLERC et al. 1996). Im Gegensatz dazu sind adaptive Mutationen die Folge eines
Selektionsdruckes, sie ereignen sich in Zellen, die sich nicht in der Vermehrungsphase
befinden. Allerdings bleibt diese Veränderung auf eine Spezies beschränkt, führt aber zu
räumlich begrenzten Resistenzen. Bei pathogenen S. enteric- und E. col-Stämmen wurden
vergleichsweise hohe Mutationsraten beschrieben. Die Mutanten dieser Stämme weisen eine
1000-fach höhere Rifampicin-Resistenz im Vergleich zum Wildtyp auf. Auch fanden DNA-
Rekombinationen zwischen verschiedenen Bakteriengenera statt, wodurch es zu einer
rasanten Ausbreitung der Resistenzen kam (LECLERC et al. 1996).
Das Ausschalten bestimmter Virulenzgene in Form von Deletionsmutanten, führt zu einer
Attenuierung, welche genauer untersucht und beurteilt werden kann. So ermöglichen
künstlich herbeigeführte Mutationen einen genauen Einblick in das Zusammenspiel zwischen
Erreger und Organismus.
Bei dem Wildtypstamm von S. Typhimurium DT104 wurde durch eine P22-Transduktion der
invC::aphT-Kassette von SB566 eine SPI-1 Mutante konstruiert (EICHELBERG et al. 1994).
Eine SPI-2 Mutante konnte ebenfalls mit Hilfe der P22-Transduktion von sseD::aphT-
Kassette geschaffen werden. Bei der Bezeichnung aphT handelt es sich um eine Kanamycin-
Resistenz-Kassette, mit deren Hilfe ein Erkennen der S. Typhimurium-Klone, bei welchen
durch homologe Rekombination das intakte Gen gegen das mutierte Gen ausgetauscht wurde,
realisierbar wird. Bei solchen Genkassetten handelt es sich um kleine mobile genetische
Einheiten. Sie enthalten üblicherweise nur ein Resistenzgen und eine spezifische
Rekombinationsstelle (RECCHIA und HALL 1995). Sie sind in ein Integron eingefügt, was
durch ihre Mobilität deutlich wird (PARTRIDGE et al. 2001). Ihre Expression steht in
Abhängigkeit von der Lage des Promotor zum Integron (TENOVER und SCHABERG 1998).

Literaturübersicht
22
Ein essentieller Schritt in der Pathogenese von S. Typhimurium ist das Eindringen des
Erregers in seine Wirtszelle, um von dort tiefere Gewebe zu erreichen. Der genetische Locus
Inv, der sich auf SPI1 befindet, ermöglicht dies nicht nur den Salmonella spp., sondern auch
anderen Enterobakterien. Mutationen in der Inv-Genregion führten zu einer 50 % höheren LD
in einem Infektionsversuch an BALB/c Mäusen nach per oraler Infektion (GALAN und
CURTISS 1989).
InvC gehört zum invABC-Operon. Es kodiert Proteine mit einem Molekulargewicht von 15
bis 47 kDa. Mit Hilfe der Gensequenzierung können deutliche Homologien mit Proteinen
anderer Mikroorganismen aufgezeigt werden, z.B. Spa47 der Shigella spp. (VENKATESAN
et al. 1992) und YscN der Yersinia spp. (WOESTYN et al. 1994). Dies spricht für eine hohe
Übereinstimmung der Gene. Bei einer „nonpolaren Mutation“ von invC konnte an Henle-407-
Zellen ein Anheften des Erregers an kultivierte Zellen nachgewiesen werden, ein Eindringen
in die Zellen fand jedoch nicht statt. Die Ursache hierfür liegt in der Unfähigkeit des Erregers,
die typischen und auch notwendigen Zytoskelettveränderungen in der Wirtszelle
herbeizuführen, was als Indikator für das Unvermögen der Bakterien angesehen wird, Signale
an die zu infizierende Zelle zu übermitteln (GINOCCHIO et al. 1994). InvC weist eine hohe
ATPase-Aktivität auf. Die Hydrolyse von ATP dient Salmonellen zur Energiegewinnung für
den Proteintransport in die Wirtszelle, wodurch erst das Eindringen in diese nicht
phagozytierenden Zellen ermöglicht wird. Die SPI-1 TTSS-assoziierte ATPase invC ist aus
drei funktionellen Domänen zusammengesetzt, welche jeweils mit verschiedenen essentiellen
Aktivitäten des Proteins verbunden sind. Das als Domäne 1 bezeichnete Polypeptid besteht
aus ca. 120 Aminosäuren, die den Kontakt mit der Bakterienmembran vermittelt. Domäne 2
stellt den zentral gelegenen katalytischen Teil dar. Domäne 3 bezeichnet den Bestandteil des
Proteins, der unabdingbar für die Funktion des TTSS-Apparates ist. Dieser vermittelt die
Protein- Protein- Interaktionen.
InvC existiert als Monomer oder Hexamer mit einem Molekulargewicht von 47 kDa (AKEDA
und GALAN 2004).
Ein weiteres Kennzeichen der Salmonella-Pathogenese ist die Fähigkeit, intrazellulär zu
überleben und sich zu vermehren. Ermöglicht wird dieser Phänotyp durch die Virulenzgene
auf der SPI 2. Bei SseD handelt es sich um ein Effektorprotein, dessen Aufgabe darin besteht,
die Zellphysiologie zum Vorteil des Bakteriums zu modulieren. Auch werden durch sie
verschiedene Immunitätsmechanismen des Wirtes beeinflusst (WATERMAN und HOLDEN
2003).

Literaturübersicht
23
SseD zeigt eine deutliche Sequenzähnlichkeit zu Sekretionsapparat-Proteinen EspB und EspD
von E. coli und YopB und YopD von Yersinien. In In vitro-Versuchen konnte gezeigt
werden, dass dieses Translokatorprotein im sauren Medium sezerniert wird und dann mit
SseB, ebenfalls einem Effektorprotein der SPI 2, auf der Bakterienoberfläche eine komplexe
Struktur bildet (KLEIN und JONES 2001).
Es besteht ein obligates Zusammenspiel der drei Translokatorproteine SseA, B und D.
Aufgrund der Sequenzhomologien bereits entschlüsselter Enterobakterien wird vermutet, dass
SseD zusammen mit SseC eine Translokator-Poren-Untereinheit in der Vakuole der
Wirtszelle ausbildet. SseB dient zur Verbindung zwischen Translokator der
Bakterienmembran und der Pore. Sie sind damit für den Transport verschiedener Proteine aus
der Bakterienzelle in die Wirtszelle unabdingbar (ZURAWSKI und STEIN 2004).
2.1.5 Diagnostik von Salmonellosen
2.1.5.1 Kultureller Nachweis
Salmonellen stellen keine hohen Ansprüche an Nährmedien, jedoch sollte man beachten, dass
ihre Isolierung abhängig vom physiologischen Zustand der Salmonellen im Probenmaterial,
ihrer Anzahl in der Probe und der Begleitflora ist. Generell gestaltet sich der Nachweis aus
klinischen Proben (Fäzesproben, Organproben u.ä.) leichter als aus Umwelt-, Lebensmittel-
oder Futtermittelproben.
Bei der Entnahme klinischer Proben müssen ebenfalls erhebliche Einflussfaktoren
berücksichtigt werden. Handelt es sich bei dem Probanden beispielsweise um einen latenten
Keimträger, so liegen die Salmonellen in einer weitaus geringeren Anzahl als bei einer akuten
Salmonellose vor. Somit besteht die Notwendigkeit einer Erreger-Anreicherung, die einer
kontinuierlichen Weiterentwicklung im Bezug zur Lebensmittelsicherheit unterliegt. Bis
heute sind zahlreiche unterschiedliche Verfahren beschrieben, die jedoch erhebliche
Abweichungen bezüglich ihrer maximalen Nachweisrate aufweisen. Nicht nur aus diesem
Grund ist der amtliche Nachweis von Salmonellen im Lebensmittel Gegenstand des
Lebensmittel- und Futtermittelgesetzbuches (LFGB, § 64) geworden, welches sich dabei eng
an die Norm 6579:2002 der International Organization for Standardization (ISO) anlehnt
(ANONYMUS 2007).

Literaturübersicht
24
Der Direktausstrich als Nachweisverfahren eignet sich nur für den Fall einer akuten
Salmonellose.
Bei anderen Formen der Erkrankung ist das Verhältnis von Salmonellen zur Darmflora zu
gering, was eine vorangehende Anreicherung nötig macht (WALTMAN 2000).
Es konnte durch verschiedene Untersuchungen belegt werden, dass eine Voranreicherung der
Salmonellen einen positiven Effekt auf die Salmonellen-Isolierungsrate hat (SCHLUNDT und
MUNCH 1993).
Der Salmonellennachweis in Umgebungsproben oder Futter- und Lebensmitteln ist deshalb
schwieriger, weil hier die Begleitflora zahlenmäßig deutlich überlegen ist. Es sollte bevorzugt
mit selektiven Medien gearbeitet werden. Da jedoch die Salmonellen durch verschiedene
Herstellungsverfahren oder Umgebungseinflüsse subletal geschädigt vorliegen können, kann
es zu deren Abtötung durch die Selektivmedien kommen (EIGNER et al. 2001).
Zu berücksichtigen ist die enge Verwandtschaft innerhalb der Enterobacteriaceae, sodass
durch Kreuzreaktionen mit ähnlichen Spezies falsch positive Ergebnisse auftreten können.
Aus diesem Grund wurden zahlreiche Selektivnährmedien erprobt, die genau auf die
physiologischen Eigenschaften der Salmonellen zugeschnitten sind, und die andere
Mikroorganismen, die im natürlichen Milieu der Enterobakterien vorkommen, in ihrem
Wachstum hemmen (WALTMAN 2000).
Daraus ergibt sich für den kulturellen Nachweis der Salmonellen eine festgelegte Reihenfolge
bestimmter Schritte: 1. nicht-selektive Voranreicherung, 2. selektive Anreicherung,
3. Ausstrich auf Selektivnährböden.
Insgesamt werden für den kulturellen Nachweis von Salmonellen vier Tage benötigt, da nach
jedem Schritt eine Inkubationszeit von 18 bis 24 Stunden notwendig ist (ANONYMUS 2007).
2.1.5.2 Voranreicherung
Bei der Voranreicherung erfolgt die Stabilisierung und Aktivierung subletal geschädigter
Salmonellen durch die Gewährleistung optimaler Nährstoffbedingungen.
Die Voranreicherung schützt die Organismen vor den toxischen Zusätzen, die sich in den
selektiven Anreicherungsmedien befinden. Daraus ergibt sich der Vorteil, dass man eine
größere Menge Probenmaterial und auch getrocknete oder tiefgefrorene Proben untersucht
werden können (MAYR und ROLLE 1993; KÜHN und TSCHÄPE 1995).

Literaturübersicht
25
Es existieren verschiedene, international anerkannte Voranreicherungsmedien, die
beispielsweise eine Ringerlösung mit Brilliantgrün, die Laktose- oder die Tryptikase-
Bouillon nützen (D´AOUST 1981). In der ISO 6579, die amtliche Verfahren zur
Untersuchung auf Salmonellen in Deutschland regelt, wird gepuffertes Pepton-Wasser zur
Anreicherung gefordert. Bei Pepton-Wasser handelt es sich um eine nährstoffreiche und
hemmstofffreie Bouillon, die ein intensives Wachstum und eine hohe Wiederbelebungsrate
von subletal geschädigten Salmonellen ermöglicht. Gleichzeitig werden die sich ungünstig
auswirkenden pH-Wert-Schwankungen durch den Phosphat-Puffer ausgeglichen (PIETZSCH
1981).
Von DAVIES und WRAY (1997) wurde eine Methode zur erhöhten Salmonellen-Ausbeute
über eine Voranreicherung entwickelt, bei welcher mit Antikörpern beschichtete
Zelluloseschwämme, in gepuffertem Pepton-Wasser zum Einsatz kommen. Bei 670
Umgebungs- und Schweinekotproben stieg die Anzahl der positiven Isolate um 40 % im
Vergleich zur Standard-Kulturmethode.
Kein einziges eingesetztes Anreicherungsmedium führt allerdings zu einer Verkürzung der
Inkubationszeit, ihre Dauer ist von größerer Bedeutung als die des verwendeten Mediums
(D´AOUST 1981).
2.1.5.3 Selektivanreicherung
Der Schritt der Selektivanreicherung dient der Hemmung der Begleitflora, was durch
verschiedene Zusätze, z.B. Antibiotika, Gallensalze oder Farbstoffe gewährleistet wird.
Bei gramnegativen Begleitkeimen gestaltet sich dies schwieriger, da eine zeitgleiche
Hemmung des Salmonellenwachstums vermieden werden soll. Daher wählt man für eine
zusätzliche Hemmung der Begleitflora und Unterdrückung des Proteus-Schwärmens eine
Bebrütungstemperatur von 42 °C (MAYR und ROLLE 1993).
Das Rappaport-Vassiliadis-Medium (RV-Medium) eignet sich gut zur Anreicherung von
Salmonellen. Dieses Medium weist durch den Zusatz von Malachitgrün eine hohe Selektivität
für Salmonellen auf, da Salmonellen gegen diesen Stoff resistent sind (BAGER und
PETERSEN 1991). Außerdem wird die Fähigkeit der Salmonellen sich bei hohem
osmotischem Druck und niedrigem pH-Wert vermehren zu können, genutzt. Um den Vorteil
der hohen Selektivität und Spezifität dieses Mediums nicht herabzusetzen, kann man

Literaturübersicht
26
allerdings nur geringe Volumina inokulieren. Dieses Verhältnis sollte idealerweise 1:100
betragen.
Die Erstbeschreiber dieses Selektivmediums sind RAPPAPORT et al. (1956). Eine
Modifizierung erfolgte durch VASSILIADIS und Mitarbeiter, indem der Anteil des
Malachitgrün gesenkt und eine Bebrütungstemperatur von 42 °C eingeführt wurde
(VASSILIADIS et al. 1976). Die Nutzung des RV-Mediums setzt eine vorangegangene
Anreicherung voraus.
2.1.5.4 Selektivnährböden
Durch die selektive Voranreicherung erfolgt eine Erhöhung der Keimzahl an Salmonellen in
der Probe wodurch dann eine Kultivierung auf festen Selektivnährböden möglich wird.
Deren Funktionen gleichen denen der selektiven Anreicherungsmedien, wie Hemmung der
Begleitflora und Schaffung einer optimalen Grundlage für das Salmonellenwachstum.
Aufgrund von Farbumschlägen durch Indikatoren oder einer Schwärzung durch
Sulfidausfällungen in den Nährmedien kann man auf das Vorhandensein von Salmonellen
schließen (DEDIE K et al. 1993). Die Funktionsweise der Indikatoren beruht auf
verschiedenen biochemischen Eigenschaften der isolierten Keime. So verstoffwechseln
Bakterien Saccharose (Salmonella nicht), Xylose, Laktose (S. Arizonae und S. Diarizonae)
und andere Substrate, was zu pH-Wert-Änderungen und somit zu einen Farbumschlag führt.
Jedoch wird mittels dieser Methoden eine serologische und biochemische Untersuchung nicht
ersetzt, da auch andere Keime vergleichbare Farbreaktionen auslösen können.
Eine reichhaltige Auswahl solcher Selektivnährböden ist kommerziell erhältlich. Ein kurzer
Überblick wird durch die Tabelle 1 gegeben (SELBITZ 1992).

Literaturübersicht
27
Tabelle 1. Wichtige Anreicherungs- und Differentialnährmedien für Salmonellendiagnostik
Anreicherungsmedien Differentialnährmedien
Voranreicherungen
Tetrathionat-Medien
• Kaliumtetrathionat-Kristallviolett-
Anreicherung nach Preuss
Selenit-Medien
• Natriumselenit-Lactose-Bouillon
nach Leifson
• Selenit-Brilliantgrün-Mannit-
Anreicherungsbouillon
nach Osborne und Stokes
Magnesiumchlorid-Malachitgrün-Medium
nach Rappaport
Lactosespaltung
• Wasserblau-Metachromgelb-Agar nach
Gassner
• Brilliantgrün-Phenolrot-Agar nach
Kauffmann
• MacConkey-Agar
• Fuchsin-Lactose-Agar nach Endo
• Lackmus-Lactose-Agar nach Drigalski
und Conradi
• Salmonella-Shigella-Agar
• Desoxycholat-Citrat-Agar nach Leifson
• Eosin-Methylenblau-Agar nach Levine
• Hektoen-Agar
• Salmonella-Agar nach Önoz
Sulfidbildung
• Xylose-Lysin-Desoxycholat-Agar
• Wismut-Sulfit-Agar
Weitere Medien
• Galle-Chryosidin-Glycerol-Nährboden
nach Ziesché
Durch die ISO-Norm 6579:2002 wird der Ausstrich auf XLD-Agar und einem weiteren
selektiven Nährboden gefordert (ANONYMUS 2007).
Die verschiedenen Nährmedien werden nach ihrem Selektivitätsgrad (gering, mäßig, hoch)
eingeordnet. Das BPLS-Agar gilt nach der Einordnung als hoch selektiv und das XLD-Agar
als mäßig selektiv (LIEBESWAR und ALLERBERGER 2001).

Literaturübersicht
28
Der von Taylor (1965) entwickelte Xylose-Lysine–Deoxycholate-Agar-(XLD-Agar) dient der
Isolierung und Differenzierung pathogener Enterobacteriaceae, insbesondere der Spezies
Salmonella und Shigella.
Nach geeigneter Anreicherung ist seine Überlegenheit gegenüber anderen Selektivnährböden
durch eine weit höhere kultivierbare Anzahl an Salmonellen gekennzeichnet (DUNN und
MARTIN 1971; BHAT und RAJAN 1975). Bei dem verwendeten Indikator handelt es sich
um Phenolrot, welches nach Gelb umschlägt, wenn Laktose, Xylose und Saccharose
verstoffwechselt werden, was jeweils mit einer Säurebildung einhergeht. Die Inhaltsstoffe
Thiosulfat und Eisen(III)-salz dienen zum Nachweis der Schwefelwasserstoffbildung, die man
an einer Schwarzverfärbung der Kolonien infolge der Ausfällung von Eisensulfid erkennt. Die
Hemmwirkung dieses Agarmediums ist allerding relativ schwach. Er nutzt dazu
Natriumdesoxycholat gegen coliforme Keime.
Der Selektivnährboden Brilliant-Grün-Phenol-Rot-Laktose-Agarmedium (BPL-Agar) ist zur
Isolierung aller Salmonellen geeignet, mit Ausnahme von S. Typhi aus Kot, Urin,
pathologischem Material oder Lebensmitteln. Dieser im Jahr 1925 von KRISTENSEN et al.
hergestellte BPL-Agar wurde von KAUFFMANN modifiziert (1935).
Der Nährboden enthält ebenfalls den Indikator Phenolrot, der bei Wachstum unter
Säurebildung zu einem gelben Farbumschlag führt. Das Prinzip ist das gleiche, wie das des
XLD-Agarmediums.
Bei Salmonella erfolgt kein Abbau von Laktose und Saccharose. Dadurch entwickelt sich ein
basisches Milieu und die Kolonien stellen sich in diesem Medium rosarot mit einem roten Hof
dar. Durch den Zusatz von Brilliantgrün wird das Wachstum grampositiver Keime, Shigellen
und Salmonella Typhi gehemmt.
Der Unterschied zwischen dem 1935 entwickelten BPL-Agar und dem in dieser Arbeit
verwendeten BPLS-Agar (Brilliant-Grün-Phenol-Rot-Laktose-Sucrose-Agarmedium) liegt in
der Gewährleistung einer optimierten Nährstoffgrundlage, die zu einem besseren Wachstum
von Salmonellen führt. Gleichzeitig wurde zur Hemmung der Begleitflora der Gehalt an
Brilliantgrün erhöht. Durch die enthaltene Saccharose wird eine zusätzliche Differenzierung
zwischen den Laktose- und Saccharose-negativen Mikroorganismen (Salmonella) und allen
Laktose-negativen oder Saccharose-positiven Keimen erreicht.

Literaturübersicht
29
2.1.5.5 PCR-Nachweis
Um auch geringste Mengen DNA nachzuweisen zu können, wurde eine spezifische Technik
zur Vervielfältigung von DNA entwickelt (MULLIS und FALOONA 1987). Diese Technik
der Polymerase-Ketten-Reaktion (polymerase chain reaction) wurde seitdem intensiv
weiterentwickelt und stellt heute ein unverzichtbares Verfahren mit Anwendungen auf sehr
zahlreichen Gebieten der Mikrobiologie dar. Charakteristisch für diese Methode der DNA-
Analytik sind ihre Schnelligkeit in der Durchführung und ihre hohe Sensitivität.
Das Prinzip beruht auf einem sich ständig wiederholenden DNA-Vermehrungszyklus, wobei
als direkte Replikationsmatrize genomische oder rekombinante DNA, auch Template genannt,
verwendet wird.
Es werden zunächst die Doppelstränge der DNA durch Erhitzen aufgetrennt. Dadurch können
sich kurze, einzelsträngige DNA-Sequenzen, die sogenannten Primer, an die jeweils
komplementären Regionen der DNA anheften. Die Primer stellen den Startbereich für eine
hitzestabile Polymerase dar, die so den einzelnen DNA-Strang komplementär nachbilden
kann. Auf diese Weise werden nur gesuchte DNA-Abschnitte vermehrt, da die vorgelegten
Primer in ihrer Sequenz definiert und synthetisiert sind. Durch sie werden die Sensitivität,
Spezifität und Reproduzierbarkeit dieser Methode geprägt (RYCHLIK 1995).
Theoretisch findet bei jedem Zyklus eine Verdopplung der gesuchten Sequenzen statt, es
handelt sich um ein exponentielles Geschehen, das mit der Formel N=N02n beschrieben
werden kann. Dabei ist N die Anzahl der vervielfältigten Abschnitte, N0 die
Ausgangsmolekülanzahl und n die Anzahl der Zyklen. Unter idealen Bedingungen könnte
man also von einem exponentiellen Zuwachs der Ziel-DNA ausgehen, praktisch jedoch ist die
Effektivität der PCR niemals 100 %. Es gilt die Formel N= N0(1+E) n. Dies ergibt sich durch
den sich ausbildenden Plateaueffekt, der sich an die exponentielle Phase anschließt.
Ursachen dafür können eine Akkumulation des Produktes, ein Verbrauch der
Reaktionskomponenten oder das Absinken des Polymerase- Template- Verhältnis sein.
Für das Amplifikat stehen verschiedene Nachweissysteme zur Verfügung, z. B. der Southern-
Blot oder die Agarose- Gel- Elektrophorese. Jeder dieser Nachweise ist mit einem relativ
hohen Zeitaufwand verbunden.
Mit der PCR-Methode kann lediglich eine qualitative Aussage getroffen werden.
Im Jahr 1991 wurde die wohl bedeutendste PCR-Methode zum Salmonellennachweis
veröffentlicht (WIDJOJOATMODJO et al. 1991). Dabei werden Abschnitte des
Replikationsgens oriC amplifiziert. Im nachfolgenden Jahr entwickelten die gleichen Autoren

Literaturübersicht
30
ein Verfahren zur Extraktion von Salmonellen-DNA aus klinischen Proben, bei der
spezifische Antikörper mit magnetischen Partikeln beladen werden (WIDJOJOATMODJO et
al. 1992). In Kombination mit ihrer 1991 entwickelten PCR nannten sie dieses Verfahren
„Magnetic Immuno-Polymerase-Chain-Reaction“-(MIPA). Mit Hilfe dieses Verfahrens waren
(WIDJOJOATMODJO et al. 1992) und später andere Autoren (WIDJOJOATMODJO et al.
1992; FLUIT et al. 1995; FAVRIN et al. 2003) in der Lage, Salmonellen sowohl aus Stuhl-
als auch Lebensmittelproben mit hoher Detektionsrate zu isolieren (105 KBE/g Stuhl bzw. 1 -
3 KBE/25 g bzw. ml Lebensmittel).
Eine von RAHN und Mitarbeiter (1992) entwickelte Salmonellen-PCR detektiert das von
GALAN und CURTISS entdeckte invA-Gen (GALAN und CURTISS 1989; GALAN und
CURTISS 1991; GALAN et al. 1992). Bei dieser PCR wurde eine Sensivität von 99,4 % und
eine Spezifität von 100 % erzielt. Das Rahn’sche Verfahren wurde später von ARNOLD et al.
(2004) modifiziert und damit optimiert. Dadurch gelang es, sowohl Sensivität als auch
Spezifität zu erhöhen, da keine Kreuzreaktionen mit Genomabschnitten zeitgleich präsenter E.
coli mehr auftraten.
Neben oben genannten eigenen sich andere Genabschnitte zum Nachweis von Salmonellen.
So verwendeten BRUMME (2006) und RÖSLER (2007) Abschnitte des invC- sowie sseD-
Gens, um Mutanten vom Wildtyp von S. Typhimurium nachzuweisen.
Im Jahr 1991 gelang es OLSEN et al., einen 2,3 kb großen DNA-Bereich zu identifizieren,
der bei 20 verschiedener Salmonella-Serovaren identisch war und sich zum Nachweis von
Salmonellen mittels PCR eignete. Diesen Bereich nutzen auch AABO und Mitarbeiter (1993)
die zeigen konnten, dass sich ein 429 bp großes Fragment dieses Bereiches noch besser zum
Nachweis von Salmonellen eignet, da es offensichtlich einen stark konservierten
Genomabschnitt repräsentiert und für mindestens 213 Serovare spezifisch ist. Die zum
Nachweis dieses 429 bp großes Fragments notwendigen Oligonukleotidprimer sind in der
DIN 10135 (ANONYMUS 1999) als solche gelistet, die sich zum Nachweis von Salmonellen
mittels PCR eignen. Diese DIN einhält auch die Primer, die RAHN et al. (1992) nutzte. Diese
DIN sagt jedoch auch, dass auch andere Oligonukleotide zum Nachweis von Salmonellen
eingesetzt werden können, wenn deren Eignung fachlich geprüft wurde.
Neben klassischen PCR Protokollen sind solche erwähnenswert, die auf der sogenannten
nested PCR basieren. Gängige nested PCR sind in den unlängst erschienenen Artikeln von
WAAGE et al. (1999) und RATHNAYAKA (2011) resümiert. Dabei werden in der Regel
folgende Genabschnitte zur Amplifikation genutzt: das bereits oben beschriebene 2,3 kb

Literaturübersicht
31
große Salmonella-DNA-Fragment (AABO et al. 1993; WAAGE et al. 1999) bzw. das fliC-
Gen (TOURON et al. 2005; RATHNAYAKA 2011).
Bei einer nested PCR erfolgt ein initialer PCR-Schritt (externe PCR), dem ein zweiter folgt.
Bei Letzterem werden Abschnitte des initial amplifizierten Genomabschnittes nochmals
vermehrt. Derartige Verfahren sind generell sensitiver als konventionelle PCR. Nachteil ist,
dass die Gefahr von Kontaminationen wesentlich höher ist. Jüngere Verfahren basieren auf
der Real-time PCR. Dabei werden Zielgene bzw. Genomabschnitte amplifiziert, an die sich
spezifische Farbstoffe binden. Das Ausmass der Farbstoffbindung wird gemessen; je mehr
Farbstoff gebunden ist, desto höher ist die Konzentration des Amplifikates und damit des zu
testenden Agens wie Salmonellen in der Testsubstanz. Durch eine Real-time PCR ist neben
dem schlichten Nachweis („Ja/Nein“) auch eine Quantifizierung möglich (KUBISTA et al.
2006).

Material und Methoden
32
3 MATERIAL UND METHODEN
3.1 Tiere und Aufstallung
Insgesamt wurden 25 männliche Hybrid-Schweine (Landrasse x Edelschwein) in die
Untersuchungen einbezogen. Das Tierversuchsvorhaben wurde vom Regierungspräsidium
Leipzig genehmigt (Tierversuchsvorhaben Nr. 02/03). Die Tiere entstammen aus einem
Herdbuch-Zuchtbetrieb einer Erzeugergemeinschaft aus Niedersachsen, der in der
Vergangenheit stets negative serologische und bakteriologische Salmonellenbefunde zu
verzeichnen hatte. Die Tiere waren zu Versuchsbeginn 6 Wochen alt, frei von klinischen
Symptomen und durchschnittlich 10,3 kg (s +- 1,2) schwer. Die Schweine, die vor
Versuchsbeginn serologisch mittels eines kommerziellen ELISA-Tests (Salmotype LPS-
ELISA; LDL, Leipzig, Deutschland) und mikrobiologisch mittels einer Kotprobe (in
Anlehnung an ISO 6579 zur Untersuchung auf Salmonellen) auf Salmonellen untersucht und
als frei befundet worden waren, wurden in die Versuche aufgenommen. Die Muttertiere
wurden auf gleiche Weise getestet und waren ebenso frei von Salmonellen befundet.
Dieselben Schweine fanden in der Dissertation von BRUMME (2006) und in der Habilitation
von RÖSLER (2007) Verwendung. In genannten Arbeiten wurde sich unter anderem
serologischen und immunologischen Aspekten der Infektion gewidmet.
Sämtliche Versuche fanden im voll klimatisierten Versuchstierstall des Institutes für
Tierhygiene und Öffentliches Veterinärwesen an der Veterinärmedizinischen Fakultät der
Universität Leipzig statt. Die Tiere wurden paarweise auf Vollspaltenboden gehalten und
erhielten Ferkelaufzuchtsfutter (FZ I; Agrargenossenschaft Laas e.G.) und Wasser ad libitum.
Das Futter wurde den Tieren 20 Stunden vor Versuchsbeginn (Infektion) entzogen.

Material und Methoden
33
3.2 Versuchsdesign
Das Versuchsdesign ist in Abbildung 1 schematisch dargestellt. Innerhalb der ersten Woche
nach Einstallung wurden die Schweine an die neue Umgebung adaptiert. Danach erfolgte die
Infektion mit entsprechenden Salmonellen (Tabelle 2) mittels Magensonde (Fa. Rüsch,
Kernen, Deutschland; Abbildung 2). Bei acht Tieren erfolgte die Infektion mit einem
Wildtyp-Stamm von S. Typhimurium DT104, der aus dem Schwein isoliert und bereits in
einem vorangegangenen Infektionsversuch eingesetzt worden war (MARG et al. 2001;
ROESLER et al. 2004; ARNOLD et al. 2004). Weitere neun Tiere wurden mit der
invC::aphT-Deletionsmutante infiziert, während bei den verbleibenden acht Tieren die
Deletionsmutante sseD::aphT Verwendung fand (Tabelle 2). Die in den Infektionsversuchen
eingesetzten Deletionsmutanten wurden von der Arbeitsgruppe um Herrn Professor Wolf-
Dietrich Hardt (Institut für Mikrobiologie der Eidgenössischen Technischen Hochschule
Zurich), aus dem S. Typhimurium DT104-Wildstamm konstruiert und freundlicherweise zur
Verfügung gestellt.

Material und Methoden
34
Abbildung 1. Schematische Darstellung des Versuchablaufes zur Pathogenität von Salmonella Typhimurium DT104 Wildtyp und Salmonella Typhimurium Deletionsmutanten (sseD::aphT & invC::aphT) in persistent infizierten Schweinen.
Schweine n = 25
≈ 1 Woche Adaptionszeit
Orale Infektion per Magenschlundsonde
1 x 1011 KBE
S. Typhimurium DT104 Wildtyp
n = 8
Mutante II invC::aphT
n = 9
Mutante I sseD::aphT
n = 8
Beobachtungszeitraum 20 Tage
Täglich: Klinische Untersuchung
Tag 1. bis 7. p.i. Kotprobenentnahme zur Bestätigung der Persistenz
2 x pro Woche: Gewichtskontrolle,
Blutentnahme, Kotprobenentnahme zur
Bestätigung der Persistenz
Tötung am 21. Tag nach
Infektion
Organentnahmen
Aufarbeitung der Organproben für weiterführende Untersuchungen

Material und Methoden
35
Tabelle 2. Übersicht über die in den Infektionsversuchen eingesetzten Bakterienstämme
Bezeichnung Herkunft Spezies/Charakteristika
Wildtyp S. Typhimurium
Stammsammlung des Instituts für Bakteriologie und Mykologie der Veterinärmedizinischen Fakultät
der Universität Leipzig
S. Typhimurium DT104
(Isolat vom Schwein) (SCHOLZ et al. 2001;
MARG et al. 2001; ARNOLD et al. 2004)
SPI-2 Mutante M809
(Mutante I)
Institut für Mikrobiologie der Eidgenössischen Technischen
Hochschule Zurich Prof. Wolf-Dietrich Hardt
Die Mutante ist durch P22-
Transduktion von sseD::aphT aus dem Stamm MvP101
(MEDINA et al. 1999) in den DT104-Stamm erzeugt
worden.
SPI-1 Mutante M810
(Mutante II)
Institut für Mikrobiologie der Eidgenössischen Technischen
Hochschule Zurich Prof. Wolf-Dietrich Hardt
Die Mutante ist durch P22-
Transduktion der invC::aphT (Km-resistenz)-Kassette von SB566 (EICHELBERG et al. 1994) in den Wildtyp DT104-
Stamm konstruiert worden .
Der Infektion schloß sich eine Beobachtungsphase von 20 Tagen an. Während dieser Zeit
wurden die Tiere täglich klinisch untersucht, zweimal pro Woche gewogen (digitale Waage
der Fa. Soehnle, Backnang, Deutschland) und dabei Blut für die hämatologische
Untersuchung gewonnen. Der kulturelle Nachweis von Salmonellen erfolgte im Kot. Dazu
wurde Rektalkot täglich in der ersten Woche post inocculationem und dann an den
Wiegeterminen entnommen. Somit sollten die persistent infizierten Tiere identifiziert und die
Ausscheidung von Salmonellen über den Versuchszeitraum, auch in Abwesenheit klinischer
Symptome, festgehalten werden. Am 21. Tag nach der Infektion wurden die Tiere unter
(Ketamin-Xylazin)-Anästhesie durch Bolzenschuss und Blutentzug getötet und die in Tabelle
3 gelisteten Organe bzw. Organteile für den Nachweis von Salmonellen gewonnen.

Material und Methoden
36
Tabelle 3. Gewebeproben zur Untersuchung auf Salmonella Typhimurium DT104-Wildtyp und auf die S. Typhimurium DT104 Deletionsmutanten invC::aphT und sseD::aphT aus persistent infizierten Schweinen
Organ- und Gewebeproben Abkürzung
Tonsille To
Mandibularlympknoten MaLy
Lunge (Mittellappen) Lu
Lungenlymphknoten LuLy
Leber Le
Milz Mi
Jejunum (kaudaler Abschnitt) Je
Ileum (distaler Abschnitt) Il
Colon + Inhalt Co
Caecum + Inhalt Ca
Jejunal-Lymphknoten Jely
Ileocaecal-Lymphknoten Icly
Colon-Lymphknoten Coly
Muskel Mu
Gallenflüssigkeit Gf
3.2.1 Orale Infektion der Tiere per Schlundsonde
Die Schweine wurden über eine Magensonde (Fa. Rüsch, Kernen, Deutschland; Abbildung 2)
mit den entsprechenden Salmonellen (Tabelle 2) infiziert (MARG et al. 2001; ROESLER et
al. 2004; ARNOLD et al. 2004). Von jedem der genutzten Salmonellenstämme wurden
insgesamt 1 x 1011 KBE in 50 ml PBS aufgenommen und je Tier verabreicht. Die Tiere
wurden vor der Verabreichung des jeweiligen Salmonella Stammes mit 0,4 mg/kg Azaperon
(Stresnil) sediert.

Material und Methoden
37
Abbildung 2. Infektion von Schweinen mit Salmonella-Symposium mittels Magenschlundsonde (MARG et al. 2001; ROESLER et al. 2004; ARNOLD et al. 2004).
Zur Herstellung der Infektionsdosen wurden Übernachtkulturen in einem 50 ml-Kolben mit
Luria-Bertani-Boullion (LB)-Bouillon im Schüttelinkubator aerob bei 37 °C angesetzt. Am
darauffolgenden Tag wurden davon jeweils 2,5 ml entnommen. Damit wurden 12 ml frische
LB-Boullion, die sich in einem 300 ml fassenden Erlenmeyer-Kolben befanden, überimpft.
Insgesamt wurden 16 dieser Kolben angesetzt, im Schüttelinkubator bis zur mittleren
logarithmischen Phase inkubiert und anschließend bei 2000 x g für 10 min zentrifugiert. Der
Überstand wurde verworfen und das erhaltene Pellet einmal in PBS (Phosphat Buffered
Saline; pH 7,2) resuspendiert und nochmals zentrifugiert.
Vor Bestimmung der Gesamtkeimzahl pro Milliliter Infektionskultur wurde jeder Ansatz
1:100 mit PBS verdünnt. Die Bestimmung erfolgte in einer Thoma-Zählkammer
(Kammervolumen 0,00025 mm3). Diese verfügt über 16 Großquadrate, die jeweils 16
Kleinquadrate aufweisen. In dieser Studie wurden insgesamt fünf Großquadrate gewählt und
in diesen jeweils vier Kleinquadrate ausgezählt. Aus den Ergebnissen der Auszählung der
fünf Großquadrate wurde der Mittelwert ermittelt und mit 4 x 106 multipliziert, um die
Gesamtkeimzahl in der Verdünnung zu erhalten. Durch erneute Multiplikation mit 100 wurde
die Gesamtkeimzahl in der Ausgangskultur bestimmt und diese dann mit PBS verdünnt, um
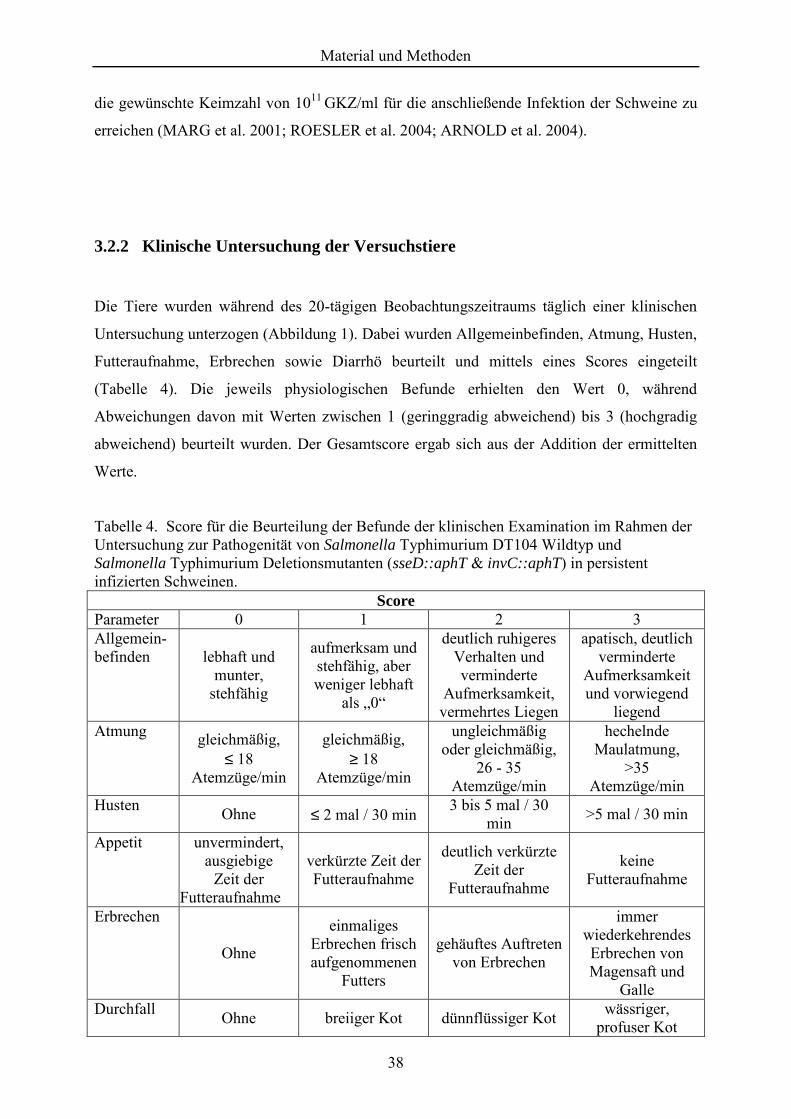
Material und Methoden
38
die gewünschte Keimzahl von 1011 GKZ/ml für die anschließende Infektion der Schweine zu
erreichen (MARG et al. 2001; ROESLER et al. 2004; ARNOLD et al. 2004).
3.2.2 Klinische Untersuchung der Versuchstiere
Die Tiere wurden während des 20-tägigen Beobachtungszeitraums täglich einer klinischen
Untersuchung unterzogen (Abbildung 1). Dabei wurden Allgemeinbefinden, Atmung, Husten,
Futteraufnahme, Erbrechen sowie Diarrhö beurteilt und mittels eines Scores eingeteilt
(Tabelle 4). Die jeweils physiologischen Befunde erhielten den Wert 0, während
Abweichungen davon mit Werten zwischen 1 (geringgradig abweichend) bis 3 (hochgradig
abweichend) beurteilt wurden. Der Gesamtscore ergab sich aus der Addition der ermittelten
Werte.
Tabelle 4. Score für die Beurteilung der Befunde der klinischen Examination im Rahmen der Untersuchung zur Pathogenität von Salmonella Typhimurium DT104 Wildtyp und Salmonella Typhimurium Deletionsmutanten (sseD::aphT & invC::aphT) in persistent infizierten Schweinen.
Score Parameter 0 1 2 3 Allgemein-befinden lebhaft und
munter, stehfähig
aufmerksam und stehfähig, aber weniger lebhaft
als „0“
deutlich ruhigeres Verhalten und verminderte
Aufmerksamkeit, vermehrtes Liegen
apatisch, deutlich verminderte
Aufmerksamkeit und vorwiegend
liegend Atmung gleichmäßig,
≤ 18 Atemzüge/min
gleichmäßig, ≥ 18
Atemzüge/min
ungleichmäßig oder gleichmäßig,
26 - 35 Atemzüge/min
hechelnde Maulatmung,
>35 Atemzüge/min
Husten Ohne ≤ 2 mal / 30 min 3 bis 5 mal / 30 min >5 mal / 30 min
Appetit unvermindert, ausgiebige
Zeit der Futteraufnahme
verkürzte Zeit der Futteraufnahme
deutlich verkürzte Zeit der
Futteraufnahme
keine Futteraufnahme
Erbrechen
Ohne
einmaliges Erbrechen frisch aufgenommenen
Futters
gehäuftes Auftreten von Erbrechen
immer wiederkehrendes Erbrechen von Magensaft und
Galle Durchfall Ohne breiiger Kot dünnflüssiger Kot wässriger,
profuser Kot

Material und Methoden
39
3.2.3 Blutentnahme und hämatologische Untersuchung
Die Entnahme von Blut erfolgte aus der V. cava cranialis. Hierfür wurde das Entnahmesystem
Vacuette® als Probegefäße und Holdex® (Fa. Greiner bio-one, Essen) als Adapter für die
Einmalkanülen (Ø 0,90 x 40 mm, Sterican® B. Braun Melsungen AG, Melsungen)
verwendet. Das Entnahmesystem nutzt EDTA als Antikoagulanz. Das Blut wurde noch am
selben Tag gekühlt an das Labor Fa. Invitro (Wien, Österreich) geschickt. Am
darauffolgenden Tag erfolgte dort die Bestimmung des weißen Blutbildes.
3.2.4 Untersuchung auf Salmonellen
3.2.4.1 Kulturelle Diagnostik
Zur Untersuchung auf Salmonella wurden Medien verwendet, wie sie vor der Modifikation
im Jahre 2008 laut ISO 6579 vorgeschrieben waren. Dabei handelte es sich um gepufferte
Pepton-Bouillon (Fa. Merck, Darmstadt) zur nicht selektiven Voranreicherung und
Rappaport-Vassiliadis (RV)-Medium (Fa. Merck) zur Selektivanreicherung.
Als Selektivnährboden kam Xylose-Lysine-Desoxycholat (XLD)-Agarmedium (Fa. Merck),
XLD-Agarmedium mit 100 µl/ml Kanamycinsulfat (Fa. AppliChem, Darmstadt) und
Brilliantgrün-Phenolrot-Saccharose (BPLS)-Agarmedium (Fa. Merck) zur Anwendung.
Die bakteriologische Untersuchung der Proben erfolgte im Labor in Anlehnung an die ISO
6579. Steril entnommenes Organmaterial wurde in Spiritus getaucht, äußerlich über einem
Bunsenbrenner abgeflammt und jeweils 5 g in 45 ml Pepton-Bouillon eingewogen. Das
Gewebe-Pepton-Bouillon-Gemisch wurde in einen Stomacherbeutel (Fa. Seward, London,
UK) gegeben und anschließend bei höchster Stufe im Stomacher 400 (Fa. Seward) für 1 min
homogenisiert. Die Stomacherbeutel wurden entnommen und anschließend bei 37 °C für 24
Stunden aerob bebrütet. Für die Untersuchungen des Kots wurde jeweils 1 g in 9 ml Pepton-
Boullion gelöst, durch einen Exzenterschüttler gemischt und wie oben für die
Stomacherbeutel beschrieben inkubiert. Aus allen Pepton-Bouillon-Voranreicherungen
wurden zweimal 100 µl entnommen und in jeweils ein neues Reagenzröhrchen überführt, das

Material und Methoden
40
9,9 ml Rappaport-Vassiliadis (RV)–Medium enthielt. Die Reagenzröhrchen wurden für 24
Stunden bei 42 °C inkubiert. Die Inhalte der zwei Reagenzröhrchen einer Voranreicherung
wurden anschließend parallel jeweils auf XLD-Agarmedium und auf BPLS-Agarmedium mit
Hilfe eines Glasstabes ausgestrichen und bei 37 °C für 24 Stunden aerob inkubiert.
Anschließend wurden die Agarmedien ausgewertet und verdächtig erscheinende Kolonien
einer Objektträgerschnellagglutination unterzogen (es wurden spezifische Agglutinationsseren
der Firma Sifin, Berlin, verwendet). XLD-Agarmedium (mit 100 µl/ml Kanamycinsulfat; Fa.
AppliChem, Darmstadt) wurde genutzt, um Salmonella-Mutanten vom Wildstamm zu
unterscheiden.
Eine Quantifizierung von Salmonellen erfolgte nur bei den Organproben. Dazu wurde dem
Stomacherbeutel Homogenisat nach der 1-minütigen Behandlung im Stomacher entnommen
und logarithmische Verdünnungsreihen (bis 10-9) hergestellt. Von der erhaltenen Suspension
wurden 25 µl im Dreifachansatz auf XLD-Agarmedium ausgetropft. Weitere 25 µl wurden im
Mikrotitiersystem in 10er-Schritten verdünnt und ebenfalls auf XLD-Agarmedium
ausgetropft. Die XLD-Agarmedien wurden für 24 Stunden aerob bebrütet und anschließend
ausgezählt, um die Keimzahlen zu bestimmen.
3.2.4.2 PCR
Alle durch Objektträgerschnellagglutination als Salmonella-positiv bewerteten Kolonien aus
Gewebe- und Kotproben wurden mittels PCR untersucht, um die beiden Mutanten von
einander und vom Wildstamm zu unterscheiden.
Vor der DNA-Extraktion wurden die Kolonien auf Nähragarmedium (Fa. Merck)
ausgestrichen und für 24 Stunden bei 37 °C aerob inkubiert, vorsichtig mehrere Kolonien mit
einem Glasstab abgehoben, in 500 µl Lysis-Puffer mit Proteinase K gesetzt und mit einem
Exzenterschüttler vermischt. Danach erfolgte eine einstündige Inkubation bei 56 °C.
Anschließend wurden die Proben für 10 Min in ein 94 °C heißes Wasserbad platziert, um die
Proteinase zu inaktivieren. Als letzter Schritt im Rahmen der Extraktion erfolgte eine
Zentrifugation bei 13.000 rpm (Biofuge Fa Heraeus Instruments, Osterode) für 5 min. Der
Überstand wurde gewonnen und die darin enthaltende DNA als Templete für die PCR
verwendet. Die PCR erfolgte wie bei BRUMME (2006) und RÖSLER (2007) beschrieben.

Material und Methoden
41
Der PCR-Ansatz betrug 50 µl. Fünf µl des Überstandes wurden zu 43 µl Readymix (Fa.
Peqlab, Biotechnologie GmbH, Erlangen) gegeben und durch 1 µl des jeweiligen Forward-
und Reverse-Primers (1:10) ergänzt. Als Primer fanden die in Tabelle 5 dokumentierten
Oligonukleotide (Fa. VBC-Genomics GmbH Wien, Österreich) Verwendung. Dabei handelte
es sich um Primer, die entweder an die invC- oder sseD-Region des Genoms von
S. Typhimurium binden. Beide Primerpaare binden sowohl beim Wildstamm als auch bei
beiden in dieser Arbeit verwendeten Mutanten. Allerdings unterscheiden sich die PCR-
Produkte in ihrer Größe. Werden inv-C Primer verwendet, ist das PCR-Produkt bei der
invC::aphT-Mutante größer als das des Wildtyps und der sseD::aphT-Mutante; jedoch sind
die PCR-Produkte von Wildtyp und sseD::aphT-Mutante dann gleichgroß (2.5 kb versus
jeweils 1.5 kb). Ähnlich verhält es sich, wenn die sseD::aphT-Mutante nachzuweisen ist und
entsprechende Primer genutzt werden. Dann ist das PCR-Produkt der sseD::aphT-Mutante
größer als das des Wildtyps und der invC::aphT-Mutante, die beide gleichgroß sind (1.8 kb
versus jeweils 1.0 kb). Jede Probe wurde im Doppelansatz mit beiden Primerpaaren
untersucht. Das sollte eine klare Differenzierung des Salmonellentyps ermöglichen und dabei
helfen, Kontaminationen auszuschließen. Die PCR wurde wie folgt durchgeführt: Nach einer
initialen Denaturierung bei 95 °C und finalen Elongation bei 72 °C für jeweils 5 min wurden
insgesamt 30 Zyklen durchgeführt. Sie bestanden jeweils aus einer Denaturierung bei 94 °C
für 40 sec, einem Annealing bei 60 °C für 30 sec und einer Elongation bei 72 °C für 70 sec.
Als Thermocycler diente ein Gerät vom Typ „Tpersonal“ (Biometra-biomedizinische
Analytik GmbH, Göttingen).
Die PCR-Produkte wurden anschließend durch Gelelektrophorese aufgetrennt. Dazu wurden
zweiprozentige (v/v) Agarosegele (Agarose der Firma Fa. Peqlab, Biotechnologie GmbH
Erlangen; gelöst in TRIS-Borat-EDTA-(TBE)-Puffer), eine horizontale
Gelelektrophoresekammer (Fa. Biorad, München) und TBE als Laufpuffer verwendet. In jede
Geltasche wurden 10 µl des Produkts sowie eine Positiv- und eine Negativkontrolle
aufgetragen. Zur Größenbestimmung wurden 4 µl eines Längenmarkers (peqGOLD Leiter-
Mix, Fa. Peqlab, Biotechnologie GmbH, Erlangen) mitgeführt. Die Auftrennung der Produkte
erfolgte bei 140 Volt über 30 min. Zur Anfärbung der Produkte wurde das Gel 20 min in
Ethidiumbromid (Fa. Amresco, Solon, Ohio, USA) inkubiert und kurz in Aquabidest
gewaschen. Die Detektion und die fotographische Dokumentation der PCR-Produkte erfolgte
mit dem Geldokumentationssystem Gel DocTM 2000 (Fa. Biorad, München) unter UV-Licht
der Wellenlänge von 254 nm.

Material und Methoden
42
Tabelle 5. Primersequenzen zum Nachweis des Wildtyps von S. Typhimurium DT104 und dessen invC::aphT- und sseD::aphT-Deletionsmutanten in Faeces und Gewebeproben von experimentell infizierten Schweinen S. Typhimurium-
Stamm Primer* Oligonukleotid-Sequenz
invC::aphT-Mutante II
invC-f invC-r
5´-GCC GGC GAA CAA TAG ACT GCT T-3´ 5´-TCT CAT CTG CCT TGA ATG GGT TTT-3´
sseD::aphT-Mutante I
sseD-f sseD-r
5´-CAA CCG AGG GCG CCA GTC AGA-3´ 5´-TAT CCC GCG TAA AAG TCC CAG T-3´
*Fa. VBC-Genomics GmbH Wien, Österreich; (BRUMME 2006; ROESLER 2007)
3.3 Statistische Auswertung
Die statistische Auswertung wurde mit dem Statistikprogramm SPSS® Version 18 (Software
GmbH München) durchgeführt. Die Daten wurden mit dem Shapiro-Wilks-Test auf
Normalverteilung überprüft. Die Scores für die klinischen Parameter sowie für die Werte der
quantitativen bakteriologischen Untersuchungen waren nicht normalverteilt. Für die Scores
wurden Mittelwert und Standardabweichung angegeben. Die Daten wurden mit Hilfe der
ANOVA mit dem Bonferroni-Test ausgewertet. Waren die Varianzen inhomogen, wurde mit
dem Dunett-T3-Test gearbeitet. Veränderungen im Beobachtungszeitraum wurden mit der
Varianzanalyse für Messwiederholung, dem gepaarten t-Test und einer Bonferroni-
Adjustierung durchgeführt. Bei Unterschieden in den Ergebnissen wurde anschließend mit
dem Levene-Test auf Homogenität der Varianzen überprüft.
Bei den nicht normalverteilten Scores für die klinischen Parameter sowie für die Werte der
quantitativen bakteriologischen Untersuchungen erfolgte die statistische Auswertung mit dem
Kruskal-Wallis-Test und dem U-Test nach Mann und Whitney. Häufigkeitsvergleiche
qualitativer Merkmale wurden mit dem mit Chi²- und dem Exaktem Test nach Fisher
überprüft. Unterschiede zwischen Ergebnissen mit P≤0,05 wurden als signifikant angesehen.

Ergebnisse
43
4 ERGEBNISSE
4.1 Ergebnisse der klinischen Untersuchung
Alle acht mit Salmonella Typhimurium DT104-Wildtyp infizierten Schweine wurden krank,
d.h. sie wiesen bei einem oder mehren der beurteilten Parameter von der Norm abweichende
Befunde auf. Generell ist festzustellen, dass Tiere, die mit einer der Mutanten infiziert
wurden, wesentlich seltener krank und selten schwerwiegend krank waren.
Das Allgemeinbefinden war unabhängig vom Infektionsstamm generell nur geringgradig
gestört (drei Tiere in der mit dem Wildstamm infizierten Gruppe und ein Tier in der Gruppe
der mit Mutante I infizierten Schweine; kein Tier in der Gruppe der mit Mutante II infizierten
Tiere; Abbildung 3).
Abbildung 3. Allgemeinbefinden der mit S. Typhimurium DT104 infizierten Schweine (durchschnittlicher Score; 0 – 3 für nicht bis hochgradig gestört). Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104 , der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT. * = Signifikanter Unterschied zwischen Tieren, die mit dem Wildtyp bzw. der invC::aphT-Mutante infiziert wurden (P≤0,05).

Ergebnisse
44
Alle der mit dem Wildstamm infizierten Tiere entwickelten Durchfall (Abbildung 4). Am
ersten Tag nach der Infektion hatten 37,5 % (n = 3) der Schweine dünnflüssigen bis wässrigen
Durchfall, 87,5 % (n = 7) am zweiten und alle 8 Schweine am dritten Tag. Dabei variierte die
Dauer des Durchfalls. Vier (50 %) der Schweine hatten über sieben Tage durchgehend
Durchfall. Ein (12,5 %) Schwein litt 14 Tage unter profusem Durchfall (Abbildung 4). Die
verbleibenden drei wiesen nur 1 bis 2 Tage mittelgradigen Durchfall auf.
Eine Diarrhö trat wesentlich seltener auf, wenn Tiere mit Mutanten infiziert wurden
(Abbildung 4). In der Gruppe der mit der invC::aphT-Mutante infizierten Schweine zeigten
nur drei der Tiere eine geringgradig verminderte Kotkonsistenz (Score1), die auch nur bis 3
Tage anhielt. In der Gruppe der mit sseD::aphT-Mutante infizierten Tiere war nur ein
Schwein für 1 Tag an Diarrhö erkrankt. Dessen Kot war von breiiger Konsistenz.
Abbildung 4. Durchfall der mit S. Typhimurium DT104 infizierten Schweine (durchschnittlicher Score; 0 – 3 für kein bis wässriger/profuser Kot). Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT. * = Signifikante Unterschiede (P≤0,05) zwischen Tieren der Wildtypgruppe und der beiden Mutantengruppen.

Ergebnisse
45
In allen drei Gruppen wurden Tiere beobachtet, die eine Körpertemperatur ≥ 40 °C aufwiesen
(Abbildung 5). Dies betraf drei (37,5 %) der mit dem Wildstamm infizierten und fünf
(55,6 %) der mit der invC::aphT-Mutante infizierten Tiere, während 4 der Tiere, die mit
Mutante sseD::aphT infiziert wurden, eine derartige Körpertemperatur entwickelten. Bei
Tieren, die mit dem Wildstamm oder der Mutante sseD::aphT infiziert wurden, stieg die
Körpertemperatur ab Tag 2 post infectionem, bei Tieren, die mit der invC::aphT-Mutante
infiziert wurden einen Tag später. Während sich die Körpertemperatur Wildstamm-infizierter
Tiere sukzessive bis Tag 5 auf durchschnittlich 39 °C normalisierte, wiesen einzelne Tiere,
die mit Mutanten infiziert wurden, intermittierend auch zu späteren Zeitpunkten (z.B. Tage 8,
12 oder 14) erhöhte Körpertemperaturen bis zu 40,5 °C auf. Nicht immer traten zeitgleich
zum Temperaturanstieg andere klinische Symptome wie verminderte Fresslust oder Diarrhö
auf. Bei 3 Tieren der invC::aphT-Mutantengruppe und 4 Tieren der sseD::aphT-
Mutantengruppe war lediglich erhöhte Körpertemperatur zu beobachten.
Abbildung 5. Innere Körpertemperatur (Mittelwert ± s) der mit S. Typhimurium DT104 infizierten Schweine. Tag.0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT.

Ergebnisse
46
Ein Schwein, das mit dem Wildstamm infiziert wurde, fraß weniger als vor der Infektion. Die
Verminderung der Futteraufnahme begann am Tag 2 und hielt 2 Tage an. Erbrechen wurde
bei einem Schwein zwischen den Tagen 2 und 3 beobachtet. Es war ein gehäuftes Erbrechen
(Score 2) in diesem Zeitraum zu beobachten. Tiere der zwei Mutanten-Infektionsgruppen
zeigten weder eine Verminderung der Nahrungsaufnahme noch Erbrechen.
Bei allen drei Infektionsgruppen war die Atmung während des Beobachtungszeitraums
unbeeinträchtigt.
Detailierte Daten der klinischen Untersuchung sind den Tabellen 9 und 10 im Anhang zu
entnehmen.
Aus technischen Gründen konnten die Schweine der Gruppe „sseD::aphT“ am dritten Tag
und die der Gruppen „sseD::aphT“ sowie „invC::aphT“ am 21 Tag nach der Infektion nicht
gewogen werden, so dass entsprechende Körpergewichte fehlen. So wird das Körpergewicht
vom Tag 17 nach der Infektion als Endgewicht genutzt, um eine Vergleichbarkeit zu
gewährleisten (Tabelle 6).
Das durchschnittliche Körpergewicht der Tiere der drei Versuchsgruppen unterschied sich am
Tag der Infektion nicht signifikant. Alle Tiere nahmen bis zum Tag 16 post infectionem zu.
Die Gewichtzunahme variierte jedoch im Vergleich zwischen den Gruppen. Sie war bei mit
der Mutante invC::aphT infizierten Tieren größer als bei Tieren, die mit dem Wildstamm
infiziert wurden, und zwar durchschnittlich 9 kg versus 6 kg; (P≤0,05). sseD::aphT infizierte
Tiere nahmen ca. 7 kg zu (nicht signifikant im Vergleich aller Gruppen).

Ergebnisse
47
Tabelle 6. Körpergewichte (Mittelwert ± s) der mit dem Wildtyp DT 104 und mit den Deletionsmutanten invC::aphT bzw. sseD::aphT infizierten Schweine in kg
Tag p.i. WT DT 104 n = 8
DM-invC::aphT n = 9
DM-sseD::aphT n = 8
0 8,9 ± 1,3d 10,3 ± 1,2d 9,9 ± 1,1d
3 9,0 ± 1,2B,d 11,6 ± 1,5A,c,d nicht verfügbar1 7 9,6 ± 1,4B,d 13,6 ± 1,8A,b,c 10,9 ± 1,0B,c,d
10 11,2 ± 1,5B,c,d 14,7 ± 1,5A,b 12,5 ± 1,9B,b,c 14 13,0 ± 1,9B,b,c 17,7 ± 1,8A,a 14,0 ± 2,6B,b 17 14,8 ± 2,1B,a,b 19,2 ± 2,6A,a 16,5 ± 1,8A,B,a 21 16,8 ± 2,5a nicht verfügbar1 nicht verfügbar1
1 Tiere konnten aus technischen Gründen nicht gewogen werden. p.i. = post infectionem. WT DT104 = Wildtyp, DM = Deletionsmutante
(A, B) Werte unterscheiden sich signifikant in Zeilen (P≤0,05). (a–d) Werte unterscheiden sich signifikant in Spalten (P≤0,05).
4.2 Hämatologische Befunde
Tiere aller drei Gruppen wiesen ein weißes Blutbild auf, welches sich im Anschluss an die
Infektion veränderte. Dabei unterschieden sich die Gruppen in der Qualität der
Veränderungen.
Nach Infektion mit dem Wildtyp stieg bei allen Schweinen die Gesamtleukozytenzahl bis zum
Tag 10, um dann langsam bis zum Tag 21 wieder abzufallen, ohne das die Ausgangswerte
erreicht werden konnten (P≤0,05; Abbildung 6). Auch bei Tieren, die mit der Mutante
sseD::aphT infiziert wurden, kam es zu einem Anstieg der Gesamtleukozytenzahl. Dieser war
allerdings kurz und nur auf den Tag 3 post infectionem beschränkt. Die
Gesamtleukozytenzahl der Schweine, welche die Mutante invC::aphT erhielten, fluktuierte
und wies am Tag 14 nach der Infektion maximale Werte auf, ohne das die Unterschiede
statistisch signifikant waren.
Die Gesamtleukozytenzahl ist in Abbildung 8 zusätzlich grafisch als Boxplots dargestellt, um
den Vergleich zwischen den Gruppen an den jeweiligen Beprobungstagen zu erleichtern. Die
Gruppen unterschieden sich an einem oder mehreren der Beobachtungstage signifikant

Ergebnisse
48
voneinander (P≤0,05), ohne das ein klares, dem Infektionsstamm zuzuordnendes Muster zu
erkennen war.
Abbildung 6. Gesamtleukozytenzahl (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT. (a–c) Werte unterscheiden sich signifikant zwischen den Tagen für eine und dieselbe Infektionsgruppe (P≤0,05).
Abbildung 7. Gesamtleukozytenzahl (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante bzw. der Deletionsmutante sseD::aphT, (A–C) Werte unterscheiden sich signifikant zwischen den Infektionsgruppen für ein und denselben Tag (P≤0,05).

Ergebnisse
49
Ähnlich zu Veränderungen in der Gesamtleukozytenzahl veränderten sich einzelne
Zellpopulationen des Differentialblutbildes. Nach der Infektion sank der prozentuale Anteil
der Lymphozyten bei den Tieren, die mit dem Wildtyp bzw. der Mutante invC::aphT infiziert
wurden (Abbildung 8). Während sich invC::aphT infizierte Tiere davon schon am Tag 7
erholt hatten bzw. höhere Werte als zum Zeitpunkt vor der Infektion aufwiesen, fiel der Anteil
der Lymphozyten der mit dem Wildstamm infizierten Tiere nochmals (wenn auch nicht
signifikant), um auf diesem Level bis zum Ende des Beobachtungszeitraums zu verbleiben.
Im Vergleich dazu blieb der prozentuale Anteil der Lymphozyten bei Tieren, die mit der
Mutante sseD::aphT infiziert wurden, relativ konstant (Abbildung 8).
Abbildung 8. Prozentualer Anteil an Lymphozyten (Mittelwert ± s) bei infizierten Schweinen. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT. (a–c) Werte unterscheiden sich signifikant zwischen den Tagen für eine und dieselbe Infektionsgruppe (P≤0,05).
Zwischen den drei Gruppen gab es zum Teil erhebliche Unterschiede (Abbildung 9). So
wiesen Tiere der mit dem Wildstamm infizierten Gruppe am Tag 0 (d.h. vor der Infektion)
mehr Lymphozyten als Tiere auf, die Mutante sseD::aphT erhielten, während Letztere an den
Tagen 17 und 21 post infectionem den höchsten Anteil hatten (P≤0,05). Tiere, die Mutante
invC::aphT erhielten, dominierten dahingehend am Tag 7 nach der Infektion (P≤0,05).

Ergebnisse
50
Abbildung 9. Prozentualer Anteil an Lymphozyten (Mittelwert ± s) bei infizierten Schweinen. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (A,B) Werte unterscheiden sich signifikant zwischen den Infektionsgruppen für ein und denselben Tag (P≤0,05).
Während die Tiere, die den Wildtyp erhielten, nach der Infektion mit einem deutlichen
Anstieg des prozentualen Anteils mit neutrophilen Granulozyten (nachfolgend „Neutrophile“
genannt) reagierten, blieb eine derartige Reaktion bei mit Mutanten infizierten Tieren
weitestgehend aus (ohne Abbildung).
Der Anstieg des Anteils neutrophiler Granulozyten mit dem Wildstamm infizierter Tiere war
auf eine Zunahme des Anteils sowohl stab- wie auch segmentkernige Granulozyten
zurückzuführen (Abbildungen 10 und 11). Bei Tieren, welche die invC::aphT-Mutante
erhielten, blieb dieser Anteil jeweils relativ konstant. Im Vergleich dazu sank der Anteil an
stab- wie auch segmentkernige Granulozyten sseD::aphT-Mutante infizierter Tiere sukzessive
während des Beobachtungszeitraums (P≤0,05).

Ergebnisse
51
Abbildung 10. Prozentualer Anteil neutrophiler stabkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (a,b) Werte unterscheiden sich signifikant zwischen den Tagen für eine und dieselbe Infektionsgruppe (P≤0,05).
Abbildung 11. Prozentualer Anteil neutrophiler segmentkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (a,b) Werte unterscheiden sich signifikant zwischen den Tagen für eine und dieselbe Infektionsgruppe (P≤0,05).

Ergebnisse
52
Im Gruppenvergleich für die jeweiligen Untersuchungstage fluktuieren die segmentkernigen
wesentlich stärker als die stabkernigen Neutrophilen. Die Stabkernigen unterschieden sich
lediglich am Tag 3 nach der Infektion; mit dem Wildstamm infizierte Tiere wiesen einen
höheren Prozentsatz dieser Zellen auf als Tiere, die mit der Mutante sseD::aphT infiziert
wurden (P≤0,05; Abbildung 12). Die Anzahl Segmentkerniger war mal bei der einen, mal bei
der anderen Gruppe höher bzw. niedriger (P≤0,05; Abbildung 13). Ein klares Muster war
nicht zu erkennen.
Abbildung 12. Prozentualer Anteil neutrophiler stabkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (A,B) Werte unterscheiden sich signifikant zwischen zwischen den Infektionsgruppen für ein und denselben Tag (P≤0,05).

Ergebnisse
53
Abbildung 13. Prozentualer Anteil neutrophiler segmentkerniger Granulozyten (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (A,B) Werte unterscheiden sich signifikant zwischen zwischen den Infektionsgruppen für ein und denselben Tag (P≤0,05).
Die relative Anzahl der eosinophilen Granulozyten zeigte bei allen Gruppen geringe
Schwankungen. Die Gruppe der mit dem Wildstamm infizierten Tiere zeigte dabei die
stärksten Fluktuationen, die scheinbar einer gewissen Regelmäßigkeit unterlagen (niedrig-
hoch-niedrig; Abbildung 14). Unterschiede zwischen den Gruppen gab es an den Tagen 7, 14
und 17 nach der Infektion. Mit dem Wildstamm infizierte Tiere wiesen an diesen Tagen
relativ mehr eosinophile Granulozyten als Tiere der Vergleichsgruppen auf (Abbildung 15).

Ergebnisse
54
Abbildung 14. Prozentualer Anteil eosinophiler Granulozyten (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT. (a,b) Werte unterscheiden sich zwischen den Tagen für eine und dieselbe Infektionsgruppe (P≤0,05).
Abbildung 15. Prozentualer Anteil eosinophiler Granulozyten (Mittelwert ± s) der infizierten Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (A,B) Werte unterscheiden sich zwischen den Infektionsgruppen für ein und denselben Tag (P≤0,05).

Ergebnisse
55
Ähnlich wie die eosinophilen Granoluzyten verhielt sich auch der relative Gehalt an
Monozyten relativ stabil (konstant in der Gruppe der mit dem Wildtyp infizierten Tiere;
geringe Veränderungen in beiden Mutantengruppen; Abbildung 16). Im Vergleich der
Gruppen waren Unterschiede an den Tagen 10, 17 und 21 deutlich; auch hier hatten Wildtyp-
infizierte Tiere relativ mehr Monozyten (Abbildung 17).
Abbildung 16. Prozentualer Anteil an Monozyten (Mittelwert ± s) bei infizierten Schweinen. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (a,b) Werte unterscheiden sich zwischen den Tagen für eine und dieselbe Infektionsgruppe (P≤0,05).

Ergebnisse
56
Abbildung 17. Prozentualer Anteil an Monozyten (Mittelwert ± s) bei infizierten Schweinen. Tag 0 = Tag der Infektion mit dem Wildtyp(WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (A,B) Werte unterscheiden sich zwischen den Infektionsgruppen für ein und denselben Tag (P≤0,05).
4.3 Kultureller und molekularbiologischer Nachweis von S. Typhimurium DT104
4.3.1 Ausscheidung mit den Faeces
Die Untersuchung der Faeces auf S. Typhimurium DT104 erfolgte in Anlehnung an die
ISO 6579. Vor der Infektion waren alle Schweine frei vom Stamm Salmonella Typhimurium.
Unabhängig vom Infektionsstamm (d.h. Wildtyp oder Mutante) schieden 100 % der
infizierten Schweine bis Tag 5 nach der Infektion den jeweiligen Stamm mit den Faeces aus
(Abbildung 18).
An nachfolgenden Untersuchungstagen war dieser Prozentsatz variabel, ohne dass sich die
Gruppen im Prozentsatz positiv befundeter Tiere unterschieden (Abbildung 18). Er vaiierte
zwischen 62,5 bis 100 % bei Tieren, die den Wildtyp erhielten, betrug 55,6 % bis 100 % bei

Ergebnisse
57
Tieren, die mit der invC::aphT-Mutante infiziert wurden und 75 % bis 100 % bei Schweinen,
welche die sseD::aphT-Mutante bekamen.
Abbildung 18. Kultureller Nachweis von S. Typhimurium DT104 (in % der infizierten Tiere) in Faeces infizierter Schweine. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT.
4.3.2 Besiedlung von Organen und Geweben
Insgesamt wurden 375 Gewebeproben kulturell auf S. Typhimurium DT104 untersucht.
Jeweils 120 stammten von Tieren, die mit dem S. Typhimurium DT104-Wildtypstamm bzw.
der sseD::aphT-Mutante infiziert wurden, während 135 Proben von Schweinen zur
Untersuchung gelangten, welche die invC::aphT-Mutante erhielten. Die Untersuchung
erfolgte in Anlehnung an die ISO 6579.
Von den untersuchten Gewebeproben waren insgesamt 218 (58,1 %) positiv. Davon entfielen
79 (65,8 %) auf Gewebeproben der mit der sseD::aphT-Mutante infizierten Schweine, 77
(64,2 %) auf Tiere, die den S. Typhimurium DT104-Wildtypstamm erhielten, während nur 62

Ergebnisse
58
(45,9 %) Proben der mit der der invC::aphT-Deletionsmutante infizierten Schweine positiv
waren (P≤0.05).
Insgesamt stammten 120 Gewebeproben von Schweinen (n = 8), welche mit dem Wildstamm
von S. Typhimurium DT104 infiziert wurden. Alle Tiere dieser Gruppe waren positiv in den
nachfolgend genannten Organen: Tonsille, Ileum, Caecum Colon, Mandibularlymphknoten
und Ileo-Caecal-Lymphknoten (Abbildung 19). Bei fünf Tieren konnte S. Typhimurium
DT104 im Lungengewebe und bei 4 Tieren im Lungenlymphknoten nachgewiesen werden.
Bei zwei Tieren gelang der Nachweis in der Leber und Muskulatur. Alle Tiere waren negativ
in Milz und Galle.
Von den Tieren (n = 9), die mit der invC::aphT-Mutante infiziert wurden, entstammten 135
Gewebeproben. Alle Tiere waren in Mandibularlymphknoten und im Colon positiv
(Abbildung 19). Häufig wurde die invC::aphT-Mutante auch aus Proben des Caecums, den
Ileo-Cecal- sowie Colon- und Jejunal-Lymphknoten isoliert. Jejunum, Lunge und
dazugehörige Lymphknoten waren jeweils nur bei einem Tier positiv, während Leber, Milz
und Gallenflüssigkeit negativ blieben.
Die 120 Gewebeproben der Tiere (n = 8), die mit der sseD::aphT-Mutante infiziert wurden,
waren wie folgt für genannte Mutante positiv (Abbildung 19): bei allen Tieren Tonsille,
Ileum, Caecum, Colon, Mandibular-, Ileo-Cecal-, und Colon-Lymphknoten; bei sieben Tieren
Jejunallympknoten; bei sechs Tieren Jejunum; bei vier Tieren Lunge und
Lungenlymphknoten; bei zwei Tieren Leber; bei jeweils einem Tier Milz und Muskel. Die
Gallenflüssigkeit war bei allen Probanden negativ.

Ergebnisse
59
Abbildung 19. Kultureller Nachweis von S. Typhimurium (in % der infizierten Schweine) in Gewebeproben. Tag 0 = Tag der Infektion mit dem Wildtyp (WT) DT104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT, (A–B) Werte unterscheiden sich signifikant für ein und dasselbe Gewebe (P≤0,05). # = kein Nachweis. To = Tonsille; Maly = Mandibular-Lymphknoten; Lu = Lunge; LuLy = Lungen-Lymphknoten; Le = Leber; Mi = Milz; Je = Jejunum; Jely = Jejunal-Lymphknoten; Il = Ileum; Icly = Ileocaecal-Lymphknoten; Ca = Caecum; Co = Colon; Coly = Colon-Lymphknoten; Mu = Muskel; Gf = Gallenflüssigkeit.
Die Tonsillen waren gruppenunabhängig am stärksten besiedelt (Abbildung 20). Im Vergleich
der Gruppen war die Deletionsmutante-sseD::aphT dabei offensichtlich effektiver als die
Deletionsmutante-invC::aphT. Auch bei anderen Organen gab es Unterschiede. Wie bei den
Tonsillen wurde die Deletionsmutante-sseD::aphT auch häufiger in mandibularen, jejunalen
Lymphknoten und Ileo-Caecal-Lymphknoten gefunden (Abbildung 20). In anderen Organen
wie z.B. Illeum, Caecum und Jejunum gab es dagegen keine Unterschiede zwischen den
Stämmen.

Ergebnisse
60
Abbildung 20. Quantitativer Nachweis des Wildtyps (WT) DT 104, der Deletionsmutante invC::aphT bzw. der Deletionsmutante sseD::aphT in Gewebeproben. Die y-Achse zeichnet die Werte als Exponent zur Basis 10. To = Tonsille; Maly = Mandibular-Lymphknoten; Je = Jejunum; Il = Ileum; Co = Colon; Jely = Jejunal-Lymphknoten; Icly = Ileocaecal-Lymphknoten; Coly = Colon-Lymphknoten. (A,B) Werte unterscheiden sich signifikant zwischen den Gruppen für ein und denselben Tag (P≤0,05).
4.3.3 Qualitative PCR zur Identifizierung und Nachweis von S. Typhimurium DT104
Insgesamt wurden 252 positive Kotproben und alle 218 kulturell positiven Gewebeproben mit
Hilfe der PCR untersucht. Alle Proben waren S. Typhimurium positiv. Es wurde
ausschließlich der jeweils zur Infektion verwendete Stamm gefunden. Die Ergebnisse der
PCR sind in den Abbildungen 21 und 22 veranschaulicht.

Ergebnisse
61
Abbildung 21. PCR-Produkte zum Nachweis von S. Typhimurium DT104-Wildtyp sowie der invC::aphT- und sseD::aphT-Deletionsmutanten in ausgewählten Gewebeproben experimentell infizierter Schweine. M = Längenmarker; a = Positivkontrolle invC::aphT-Mutante; b = Positivkontrolle sseD::aphT-Mutante; c = Positivkontrolle DT104 Wildtyp mit invC-Primer; d = Positivkontrolle DT104 Wildtyp mit sseD::aphT-Primer, ÷ = Negativkontrolle, To = Tonsille; Maly = Mandibular-Lymphknoten; Icly = Ileocaecal-Lymphknoten.
Abbildung 22. Nachweis von S. Typhimurium DT104-Wildtyp, Deletionsmutanten in Faeces von ausgewählten Schweinen, die entweder mit der sseD::aphT (I)- oder der invC::aphT-Mutante (II) infiziert wurden. M = Marker, a = invC::aphT-Mutante-Positivkontrolle, ÷ = Negativkontrolle, I = Kotproben der sseD::aphT-Mutantengruppe, II = Kotproben der invC::aphT-Mutantengruppe, b = sseD::aphT-Mutante Positivkontrolle.

Diskussion
62
5 DISKUSSION
Ziel dieser Arbeit war der Pathogenitätsvergleich des Wildstamms von Salmonella
Typhimurium DT104 zu seinen Deletionsmutanten sseD::aphT und invC::aphT. Dieses
Serovar ist ubiquitär und weist keine Wirtsspezifität auf. Alle landwirtschaftlichen Nutztiere
können sich infizieren und mit zum Teil schwerwiegender klinischer Symptomatik erkranken.
Gleiches gilt für den Menschen, der sich durch Umgang mit infizierten Tieren und von Tieren
stammenden Produkten anstecken kann. Dem Schwein, vor allem aber
Schweinefleischprodukten, kommt dabei herausragende Bedeutung zu. Wenn auch dies nicht
der öffentlichen Wahrnehmung entspricht, wo eher die unlängst in 2011 aufgetretene
Epidemie mit EHEC (Enterohämorrhagische Escherichia coli) in Deutschland diskutiert wird,
ist die Situation bezüglich S. Typhimurium DT104 nicht weniger dramatisch, da sich jährlich
immer noch viele Tausend Menschen mit S. Typhimurium DT104 infizieren und erkranken.
Ähnlich wie bei EHEC ist besonders brisant, dass S. Typhimurium DT104 über eine
Multiresistenz gegenüber Antibiotika verfügt und somit Erkrankungen schwer therapierbar
sind. Insofern ist das hier beschriebene Forschungsvorhaben von großer Relevanz, da es dazu
beiträgt, die Pathogenitätsmechanismen von S. Typhimurium DT104 besser zu verstehen,
wirksam zu bekämpfen bzw. Infektionen mit diesem Bakterium zu verhindern.
Wie schon vormals bei Hühnern und Mäusen gezeigt (PORTER et al. 1993; MAHAN et al.
1993; FINLAY 1994; HENSEL et al. 1995) demonstrierten die in dieser Arbeit generierten
Ergebnisse eindrucksvoll, dass Infektionen mit dem Wildtyp von S. Typhimurium zu einer
schweren Erkrankung beim Schwein führen können. Es wurde ferner gezeigt, dass beide in
dieser Arbeit verwendeten Mutanten weniger krankmachend sind. Basierend auf den
Ergebnissen dieser Arbeit ist davon auszugehen, dass die Deletionen in den sseD::aphT bzw.
invC-Bereichen tatsächlich zu Veränderungen bestimmter Eigenschaften geführt haben, die
Teil der Pathogenitätsmechanismen für das Schwein sind. Es ist zu vermuten, dass invC in die
Steuerung des Invasionsvermögens eingebunden ist.

Diskussion
63
5.1 Methodische Überlegungen
In dieser Arbeit wurden zur Untersuchung auf Salmonella Medien verwendet, wie sie vor der
Modifikation im Jahre 2008 laut ISO 6579 vorgeschrieben waren. Grund dafür ist, dass die
hier beschriebenen Untersuchungen vor oben genannter Modifikation stattfanden. Die
Modifikation betrifft die Selektivanreicherung mittels Rappaport-Vassiliadis (RV)-Medium.
Während vor der Modifikation flüssiges Medium verwendet wurde, schreibt die modifizierte
ISO 6579 im Anhang D vor, halbfestes Rappaport-Vassiliadis-Medium (MRSV-Medium)
zum Nachweis von Salmonellen in Kot- und Umweltproben zu nutzen. Dadurch sollten
sowohl Sensitivität und Spezifität der Salmonellen-Diagnostik erhöht als auch Arbeits- und
Material-Aufwand reduziert werden (ANONYMUS 2005; ANONYMUS 2007). Ob einzelne
Salmonellen-positive Proben deshalb in dieser Arbeit unerkannt blieben, bleibt offen. Wenn
dieser Fehler tatsächlich systematisch bestand, war er gruppenübergreifend ohne
Konsequenzen für den Gruppenvergleich. Kulturell als positiv befundete Proben waren
hingegen tatsächlich positiv, da sie alle mittels PCR untersucht und bestätigt wurden.
5.2 Klinische Symptomatik
Die unterschiedliche Virulenz der in dieser Arbeit verwendeten S. Typhimurium DT104
Stämme konnte eindeutig demonstriert werden. Je nach Stamm variierte die klinische
Symptomatik zum Teil erheblich. Schwere klinischeVerläufe wurden nach Infektion mit dem
Wildtyp von S. Typhimurium DT104 beobachtet, während nach Infektion mit den Mutanten
vergleichsweise milde klinische Erscheinungen auftraten. Das betraf sowohl Morbidität,
Krankheitsintensität als auch die Dauer der Erkrankung. Während alle mit dem Wildtyp
infizierten Tiere mit unterschiedlichen Symptomatiken klinisch krank waren, wurden bei
Verwendung der Mutanten nur einzelne Tiere krank. Fast alle der in dieser Studie beurteilten
klinischen Parameter waren bei einem oder mehreren mit dem Wildtyp infizierten Tieren zum
Teil zeitgleich verändert, während bei Verwendung der Mutanten mal der eine, mal der
andere Paramter verändert war und äußerst selten dieselben Tiere betraf oder Veränderungen
zeitglich zu beobachten waren. Traten klinische Veränderungen auf, waren die Symptome
stark ausgeprägt, wenn mit dem Wildtyp infiziert wurde. Mutanten verursachten dagegen nur
milde klinische Veränderungen. Waren Schweine mit dem Wildtyp infiziert und erst einmal

Diskussion
64
krank, hielt die Erkrankung über meherer Tage an. Nach Verwendung der Mutanten waren
Krankheitssymptome eher sporadischer Natur und verschwanden schon am darauffolgenden
Tag. Es ist sicher berechtigt zu konstatieren, dass sich nach Infektion mit dem Wildtyp von S.
Typhimurium DT104 eine Erkrankung per definitionem (WIESNER und RIBBECK 1991)
entwickelte, während die Deletionsmutanten wohl eher untypisch milde klinische
Verlaufsformen auslösten. Das zeigt auch die Gewichtsentwicklung, die bei den mit dem
Wildtyp infizierten Tieren geringer als bei den jenigen Schweinen war, die mit Mutanten oral
inokkuliert wurden. Bekanntlich fressen kranke Tiere weniger und nehmen während der Zeit
der Erkrankung weniger zu (BALAJI et al. 2000). So ist allein anhand der klinischen
Symptomatik zu vermuten, dass Deletionen im sseD-Gen oder invC-Gen die
Pathogenitätsseigenschaften der Salmonellen abschwächen. Das gilt wie in dieser Arbeit
gezeigt offensichtlich für das Schwein genauso wie dies für die Maus gezeigt werden konnte
(HEESEMANN und HENSEL 2000) und das Huhn (PORTER et al. 1993). Die in dieser
Arbeit beobachtete klinische Symptomatik nach Infektion mit dem Wildtyp ist bekannt und
wurde wiederholt beschrieben (WOOD et al. 1989; WOOD und ROSE 1992; FEDORKA-
CRAY et al. 1994; NIELSEN et al. 1995; SCHERER et al. 2008). Die Beobachtung milderer
Verläufe durch die in dieser Arbeit verwendeten Deletionsmutanten ist neu. Da sich die
Mutanten nicht wesentlich in den durch sie hervorgerufenen klinischen Erscheinungen
unterschieden, ist davon auszugehen, dass die durch die Deletion hervorgerufenen
Funktionsänderungen oder gar –verluste in den deletierten Genomsabschnitten ähnlich
effektiv in der Abmilderung klinisch relevanter Pathogenitätsmechanismen waren. Wie
eingangs erwähnt, steuert sseD das Überleben der Salmonellen, während invC für die
Invasivität verantwortlich ist. Die vorhandenen klinischen Symptomatiken lassen darauf
schließen, dass invC-deletierte Mutanten im Unterschied zum Wildtyp vergleichsweise selten
in den Organismus gelangten, während Mutanten mit Deletion im sseD-Bereich zwar in den
Organismus gelangten, jedoch effektiver als der Wildtyp und die invC::aphT-Mutante aus
dem Organismus des Schweins eleminiert wurden.
5.3 Weißes Blutbild
Anhand der Beschreibung des weißen Blutbildes sollte geprüft werden, ob und wie effektiv
sich die einzelnen Salmonellenstämme der Immunabwehr des Wirtsorganismus widersetzen

Diskussion
65
können. Wie schon vormals beschrieben (COE et al. 1992; VAN DIEMEN et al. 2002), stieg
die Gesamtleukozytenzahl nach Infektion mit dem Wildtyp bis zum Tag 10 nach der Infektion
an, um danach langsam bis zum Versuchsende abzufallen. Diese offensichtlich für das
Schwein typische Reaktion (COE et al. 1992; VAN DIEMEN et al. 2002) blieb aber bei
denjenigen Tieren, die mit den Deletionsmutanten infiziert wurden, aus. SseD::aphT-
Infektionen führten zwar zu einem Anstieg in der Anzahl der Leukozyten am Tag nach der
Infektion. Dieser Anstieg war jedoch transient und schon am darauffolgenden Tag wieder
abgeklungen. Klassisch und wie von Schilling (zitiert aus KRAFT und DÜRR 2005)
beschrieben, verhielten sich die Lymphozyten, wenn mit dem Wildstamm infiziert wurde.
Nach einem initialen protrahierten und moderaten Abfall bis zum Tag 7 nach der Infektion,
erfolgte eine ebenso protrahierte Erholung. Der initiale Abfall in der Anzahl an Lymphozyten
war möglicherweise das Resultat eines eingangs gestiegenen Verbrauchs vermutlich vor allem
zytotoxischer T-Lymphozyten, die bei der inaten d.h. sofortigen Immunantwort auf die
Infektion beteiligt waren. Dieses klassische Muster fehlte nach Infektion mit den
Deletionsmutanten. Lymphozyten sind Bestandteil der humoralen Immunabwehr.
Offensichtlich wurde dieses durch den Wildstamm effektiver stimuliert als nach den
Infektionen mit den Mutanten. Dieser postulierte Effekt kann schwerlich allein nummerisch
begründet sein, da nach o.g. Theorie sseD::aphT-Mutanten in gleicher Anzahl wie der
Wildtyp in den Organismus gelangt sein müssten. Die Lymphozytenkurve des Wildtyps
unterschied sich jedoch von beiden Mutanten in gleicher Weise. Möglicherweise besitzen
solchermaßen spezifisch deletierte Salmonellen und solche des Wildtypes unterschiedliche
immunogene Eigenschaften. Da es lediglich die Deletion ist, die Mutanten vom Wildtyp
unterscheiden, ist logisch anzunehmen, dass ein Zusammenhang zur Deletion besteht.
Wodurch auch immer letztlich verursacht, empfehlen sich beide Deletionsmutanten aufgrund
der verminderten Immunantwort nicht als Kandidaten für Impfstoffe. Auch wenn, wie
gewünscht, kaum klinische Symptome auftraten, ist mit einer eher schwachen humoralen
Immunantwort zu rechnen. Unterschiede in der Immunantwort im Vergleich von Wildtyp und
Mutanten sind auch im Vergleich der neutrophilen Granulozyten ersichtlich. Wie zu erwarten,
verursachte der Wildtyp eine eindeutige Reaktion (Anstieg) im Anschluss an die Infektion.
Dieses Phänomen wurde schon vormals beschrieben (VAN DIEMEN et al. 2002) und
bestätigt, dass neutrophile Granulozyten bei einer Infektion mit Salmonellen wie auch bei
anderen bakteriellen Infektionen Teil der unspezifischen Immunabwehr sind. Dieser Effekt
blieb nach den Infektionen mit den Deletionsmutanten weitestgehend aus.

Diskussion
66
Offensichtlich unterscheiden sich Wildtyp und Mutante auch in Eigenschaften, welche die
zelluläre Immunantwort des Wirtsorganismus beeinflussen. In diesem Zusammenhang sind
nochmals Überlegungen zur Klinik angebracht. Von vielen anderen bakteriellen Infektionen
beim Schwein einschließlich des vor allem durch coliforme Bakterien verursachte MMA-
Syndroms ist bekannt, dass klinische Verlaufsformen (inklusive Fieber, Inappetenz und
Erbrechen) durch chemotaktische Substanzen wie Interleukine und Zerfallsprodukte der
Bakterien (Endotoxine) verursacht werden (REINER et al. 2007). Bakterielle
Zerfallsprodukte entstehen auch als Resultat der zellulären Abwehr. Neutrophile Granulozten
sind zur Phagozytose befähigt (WIESNER und RIBBECK 1991). Wo Abwehrmechanismen
mit neutrophilen Granulozyten versagen oder nur vermindert funktionieren, wie in dieser
Studie bei der Infektion mit den Deletionsmutanten möglicherweise erfolgt, werden auch
wenige chemotaktische Substanzen vorhanden sein und bakterielle Zerfallsprodukte entstehen
und deshalb möglicherweise auch weniger/keine klinische Symptome auftreten. Die hier
skizzierten „pathways“ finden durch die Ergebnisse der Arbeit von Brumme Bestätigung
(BRUMME 2006; BRUMME et al. 2007). In diesen Arbeiten wurden dieselben Schweine
und gleiches Versuchsdesign genutzt, sich jedoch vornehmlich auf die Untersuchung von
Entzündungsmediatoren (Interleukin 2 und 4 sowie Interferon γ) und Antikörpern (IgA, G und
M) konzentriert. Die Expression aller drei Entzündungsmediatoren war bei den jenigen
Schweinen, die mit dem Wildtyp infiziert wurden, höher als bei den Tieren, welche die
Deletionsmutante erhielten. Ähnliches war bei den untersuchten spezifischen Antikörpern
festzustellen. Es wurde unter anderem geschlussfolgert, dass sowohl die zelluläre wie auch
die humorale Immunantwort bei mit dem Wildstamm infizierten Tieren ausgeprägter als bei
Schweinen ist, die mit einer der beiden Deletionsmutanten infiziert wurden. Interssant ist
auch, dass sich beide Mutanten in der Immunantwort unterschieden. Die durch sseD::aphT-
ausgelöste Reaktion (gemessen an der Expression von Interleukin 4 und Interferon γ) war
heftiger als bei Verwendung der invC::aphT- Deletionsmutante (BRUMME 2006; BRUMME
et al. 2007). Ähnliches wurde in der vorliegenden Arbeit bei der klinischen und
labordiagnostischen Beurteilung sowie bei der Bewertung der Invasivität beobachtet.
Die Ergebnisse der Bestimmung eosinophiler Granulozyten und von Monozyten waren
indifferent und erlauben keine substantielle Diskussion im Hinblick auf die Pathogenität des
jeweiligen Infektionsstammes. Eosinohile Granulozyten sind mononukleäre Zellen, die

Diskussion
67
vermehrt bei Allergien auftreten und bei der Abwehr von Parasitosen und an der
Rekonvaleszenz des gestörten Organs/Gewebes beteiligt sind (EDFORS-LILJA et al. 1994;
KRAFT und DÜRR 2005). Monozyten sind phagozytierende Zellen und wesentlicher
Bestandteil der unspezifischen zellulären Abwehr (EDER 1987). Die Anzahl von Monozyten
im Blut unterliegt generell großen Schwankungen (EDER 1987). Folgt man der klassischen
Infektiologie, wäre zu erwarten gewesen, dass die eosinophilen Granulozyten in der Phase der
Rekonvaleszenz einige Zeit nach der vorangegangenen Infektion mit S. Typhimurium
ansteigen (KRAFT und DÜRR 2005). Dieses klare Muster war, wie oben angedeutet, bei
keinem der verwendeten Infektionsstämme zu beobachten.
5.4 Ausscheidung und Organbesiedlung
Die Untersuchungen der Kotproben zum kulturellen Nachweis von S. Typhimurium DT104-
Wildtyp und Deletionsmutanten erfolgten wie in der ISO 6579 vorgeschrieben. Altrock
(2000) empfahl, diese Untersuchungen kontinuierlich und in engen Intervallen durchzuführen,
um eine sichere Aussage über die Dauer der Ausscheidung treffen zu können. Dies wurde
befolgt und der Beprobungsplan analog engmaschig konzipiert. Prinzipiell lassen die
Ergebnisse der Kultur folgende Schlussfolgerungen zu: Nach oraler Infektion von
Salmonellen werden die Schweine Ausscheider. In diesem Verhalten unterschieden sich
Wildtyp nicht von Mutanten. Die Salmonellen werden gleichwohl intermittierend
ausgeschieden. Auch in dieser generellen Eigenschaft verhielten sich die in dieser Arbeit
verwendeten Stämme gleich. Zwar wurden in der Anzahl „Ausscheider je Untersuchungstag“
vereinzelt Unterschiede beobachtet, jedoch weitab von jeglicher Signifikanz. Das Schweine S.
Typhimurium intermittierend ausscheiden können, ist bekannt (WOOD et al. 1989; WOOD
und ROSE 1992; FEDORKA-CRAY et al. 1994; NIELSEN et al. 1995; SCHERER et al.
2008). Es wird vermutet, dass latent infizierte Tiere dann ausscheiden, wenn sie gestresst
werden oder andere die Ausscheidung begünstigende Konstellationen eintreten (ISAACSON
et al. 1999; LARSEN et al. 2003; LOYNACHAN et al. 2004; BOUGHTON et al. 2007;
WILLIAMS et al. 2008). Es konnte zudem beobachtet werden, dass Salmonellen
ausgeschieden werden, ohne dass die Tiere zwangsläufig klinisch erkranken. Obwohl dies
schon seit langem bekannt ist (WOOD et al. 1989; FEDORKA-CRAY et al. 2000; BOYEN et
al. 2008; SCHERER et al. 2008; VAN PARYS et al. 2011), zeigen auch diese Versuche, dass

Diskussion
68
latente Infektionen typisch sind. Latente Träger wurden vor allem in der Gruppe der mit den
Deletionmutanten infizierten Tiere beobachtet. Unsere Befunde bestätigen, dass die Fähigkeit
zur Persistenz offensichtlich nicht an die Virulenz gekoppelt ist (WALES et al. 2011). Der
Fakt, dass einzelne Tiere (vor allem die mit Deltionsmutanten infizierten) viele Tage nach der
vorangegangenen Infektion, wenn auch nur milde Krankheitsymtome aufwiesen, mag als
zusätzliches Indiz einer faktoriellen Genese der Salmonellose des Schweines gelten.
Möglicherweise erfuhren diese Schweine eine für sie unbeherrschbare Belastung (zum
Beispiel als Folge der Versuchsdurchführung), die zu kurzzeitigen milden klinischen
Verlaufsformen führten.
Zum Nachweis der Invasiviät der verwendeten Salmonellenstämme wurden verschiedenste
Gewebe- und Organproben sowohl kulturell als auch mittels PCR auf Salmonellen untersucht.
Alle in der Kultur positiven Ergebnisse konnten in der PCR bestätigt werden. Die kulturellen
Anzucht- Ergebnisse dieser Arbeit sind damit zuverlässig validiert. Generell ist festzuhalten:
Alle Stämme, d.h. sowohl Wildtyp als auch die Mutanten sind invasiv, da sie sich prinzipell
in fast allen der beprobten Organe nachweisen ließen. Damit bestätigt sich, dass sich
Salmonella Typhimurium über das lymphatische und blutführende System in Organismen
ausbreiten kann und verschiedene Zelltypen invadiert (SANTOS et al. 2003; HAPFELMEIER
et al. 2005; HARAGA et al. 2008). Makrophagen dienen dabei als Vehikel. S. Typhimurium
kann in ihnen überleben und sich dort sogar vermehren (ALTHOUSE et al. 2003). Allerdings
gab es nicht unerhebliche Unterschiede zwischen den in dieser Arbeit verwendeten Stämmen,
vor allem im Hinblick auf das Muster der Organbesiedlung, aber auch mit Blick auf die
Häufigkeit des Befalls einzelner Organe. Interessant war, dass sich die sseD::aphT-Mutante
als gleichermaßen invasiv wie der Wildtyp herausstellte. Obwohl die Invasivität des Wildtyps
bekannt ist (MEYERHOLZ et al. 2002; NIEWOLD et al. 2007), erstaunt dieses Ergebnis
deshalb, da bisher angenommen wurde, dass die sseD::aphT-Mutante über eine verminderte
Überlebensfähigkeit in Makrophagen verfügt und darum recht effektiv durch den Organismus
eliminiert wird. Basierend auf den Ergebnissen dieser Arbeit – vor allem unter
Berücksichtigung der ausbleibenden zellulären Immunantwort – kann vermutet werden, dass
sich diese Mutante womöglich (auch) recht effektiv vor der zellulären Immunabwehr des
Wirtsorganismus maskieren konnte. Etwaige dafür zugrunde liegende Mechanismen müssen
noch untersucht werden. Fakt ist, dass sich die pathogenen Eigenschaften von sseD-Gen beim
Schwein vor allem im Hinblick auf dessen Fähigkeit zur Invasion deutlich von denen bei der

Diskussion
69
Maus unterscheiden, bei der sich sseD-Gen von Salmonella als wenig invasiv herausstellte
(HENSEL 2000; HEESEMANN und HENSEL 2000; GALLOIS et al. 2001; HOLZER und
HENSEL 2010). Dieses Ergebnis verdeutlicht auch, wie wichtig vergleichende
Untersuchungen an bzw. mit der jeweiligen Zielspezies sind.
Im Gegensatz zu sseD bewies invC eine geringere Invasivität bzw. Fähigkeit, sich im
Wirtsorganismus zu verbreiten und Organe zu kolonisieren. Dieses Ergebnis ist nicht
unerwartet, da invC, wie schon mehrfach erwähnt, über ein deletiertes und damit in seiner
Funktion verändertes invC Gen verfügt. Sämtliche Schweine waren unabhängig vom
verwendeten Infektionsstamm in Tonsillen, Mandibularlymphknoten und im Colon mit
Salmonellen besiedelt. Diese Besiedlungsorte sind typisch und gehören zu denjenigen, in
denen Salmonellen bevorzugt persistieren (WOOD et al. 1989; WOOD und ROSE 1992;
HURD et al. 2001; MARG et al. 2001; ROESLER et al. 2004; SCHERER et al. 2008;
SZABO et al. 2008; VAN PARYS et al. 2011). Andere Organe, wie Leber, Muskulatur und
Milz waren wesentlich seltener besiedelt oder kulturell negativ. Dieses Ergebnis ist fleisch-
und lebensmittelhygienisch von Bedeutung, da es sich bei Leber und Muskulatur um vom
Schwein stammende Vorprodukte handelt, die verarbeitet oder unverarbeitet in die
menschliche Nahrungskette gelangen.
Die Ergebnisse dieser Arbeit in ihrer Gesamtheit zeigen, dass der Wildtyp von S.
Typhimurum erwartungsgemäß pathogener ist als die beiden Deletionsmutanten sseD::aphT
und invC::aphT. Diese Interpretation bezieht sich vor allem auf die klinische Symptomatik
und auf Veränderungen der Blutbilder, die sich vor allem beim Wildtyp manifestierten. In der
Fähigkeit zur Ausscheidung (auch intermittierend) bestanden bei den Versuchsschweinen
keine Unterschiede. Anders in der Invasivität (d.h. Fähigkeit der Verbreitung im Organsimus
und Besiedlung von Organen); invC::aphT invadierte seltener als sseD::aphT, die ein ähnlich
starkes Invasionsvermögen wie der Wildtyp aufwies. Damit korrespondieren die Ergebnisse
dieser Arbeit gut mit denen von BRUMME et al. (2007) und BRUMME (2006), die, wie oben
erwähnt, vornehmlich basierend auf immunologischen Parametern ebenso fand, dass die
sseD::aphT-Deletionsmutante pathogener als die invC::aphT-Mutante ist.

Diskussion
70
5.5 Schlussbetrachtung
Angesichts des zoonotischen Potentials von S. Typhimurium und vor allem auch durch die
Gesetzgebung legitimierten Anstrengungen zur Elimination dieses Bakteriums aus der
menschlichen Nahrungskette durch stringente Monitoring- und Eradikationsprogramme sowie
durch die Einhaltung einwandfreier Hygienestandards mag sich die Bedeutung der
Salmonellen eher durch ihre Fähigkeit zur Invasivität, Persistenz und dauerhaften
Ausscheidung als denn allein durch die klinischen Symptome beim Wirt definieren. Wird
dieses als Bewertungsgrundlage genutzt, ist zumindest die Infektion mit der sseD::aphT-
Deletionsmutante nicht weniger klinisch auffällig (und in diesem Sinne „gefährlich“) als die
Infektion mit dem Wildtyp von S. Typhimurium. Ohne erkennbare klinische Erscheinungen
bei den Versuchsschweinen auszulösen, invadierte dieser Stamm den Wirtsorganismus in
einer Weise, wie sie nur vom Wildtyp bekannt ist. Da für die Verbreitung der Salmonellen im
Organismus Makrophagen als Vehikel dienen, muss zukünftig die Frage beantwortet werden,
ob seeD prinzipiell für die Überlebensfähigkeit von S. Typhimurium in Makrophagen kodiert,
oder ob es nicht auch eine spezies-spezifische Komponente gibt. Anders hingegen
invC::aphT, die sich bei Betrachtung aller genannten Kriterien wie klinische Symptomatik
und Invasivität als weniger pathogen erwies. Diese Ergebnisse lassen vermuten, dass die
Deletion im invC-Locus des Bakteriengenoms tatsächlich zu einer Veränderung in dessen
Funktion führte, die dann im Infektionsgeschehen beim Schwein mit einer Beinträchtigung
der Invasivität der invC::aphT-Mutante einherging.

Zusammenfassung
71
6 ZUSAMMENFASSUNG
Haukur Lindberg Sigmarsson
PATHOGENITÄTSVERGLEICH VON SALMONELLA
TYPHIMURIUM DT104 WILDTYP UND SALMONELLA
TYPHIMURIUM DELETIONSMUTANTEN (sseD::aphT & invC::aphT) IN PERSISTENT INFIZIERTEN SCHWEINEN Institut für Tierhygiene und Öffentliches Veterinärwesen der Veterinärmedizinischen Fakultät der Universität Leipzig Eingereicht im Februar 2012 74 Seiten, 22 Abbildungen, 23 Tabellen, 241 Literaturstellen, 14 Seiten Anhang Schlüsselworte: Salmonella Typhimurium DT104, Deletionsmutante, invC-Gen, sseD-Gen, Schwein, experimentelle Infektion
Salmonella (S.) Typhimurium DT104 ist ein gram-negatives Bakterium. Es weist keine Wirtsspezifität auf und gilt als Zoonoseerreger. Jährlich erkranken daran allein in Deutschland mehrere Tausend Menschen unter dem Bild einer schwerwiegenden Diarrhö mit zum Teil tödlichem Ausgang. Das Schwein gilt als eines der Reservoire für S. Typhimurium DT104 des Menschen. S. Typhimurium DT104 gelangt über vom Schwein stammende Produkte in den menschlichen Verzehr. Die Kontrolle von S. Typhimurium DT104 einschließlich effektiver Eradikationsmassnahmen in unseren Schweinebeständen ist deshalb von entscheidender Bedeutung, um den Eintrag dieses Bakteriums in die menschliche Nahrungskette wenn möglich zu eliminieren. Dafür ist das Verständnis über S. Typhimurium DT104 einschließlich der Kenntnis seine Pathogenitätseigenschaften notwendig. Ziel dieser Arbeit waren Untersuchungen zur Pathogenität von S. Typhimurium DT104. Dabei wurden der Wildstamm mit zwei seiner Deletionsmutanten (sseD::aphT und invC::aphT) verglichen.
Die Untersuchungen erfolgten im Infektionsversuch an insgesamt 25 sechs Wochen alten männlichen Schweinen, die in einem vollklimatisierten Versuchsstall gehalten wurden. Den Tieren wurde im Anschluss an eine einwöchige Akklimatisierungsphase eines der nachfolgenden Stämme von S. Typhimurium DT104 oral in einer Konzentration von 1 x 1011 KBE verabreicht: Wildtyp (n = 8 Schweine), Deletionsmutante seeD::aphT (n = 8) und Deletionsmutante invC::aphT (n = 9). Bei den Mutanten handelt es sich um Varianten von S. Typhimurium DT104, die an den entsprechenden Abschnitten des Bakteriumgenoms (d.h.

Zusammenfassung
72
sseD-Gen bzw. invC-Gen) deletiert wurden. SseD regelt die Überlebensfähigkeit von S. Typhimurium in Makrophagen, invC dessen Invasionsvermögen. Im Mäusemodel war die Pathogenität beider Mutanten deutlich vermindert. Nach der Infektion schloss sich ein 20 tägiger Beobachtungszeitraum an, während dessen nachfolgend genannte Parameter erfasst bzw. Proben genommen wurden: klinische Symptome (Allgemeinbefinden, Erbrechen, Durchfall, Futteraufnahme, Atmung, Temperatur); Blutentnahme für Erstellung des weißen Blutbildes; Kotentnahme zum Nachweis der Ausscheidung von S. Typhimurium. Einen Tag nach Ende der Beobachtung wurden die Tiere getötet und Proben von insgesamt 15 Organen (unter anderem Tonsille; Colon und Caecum sowie dazugehörige Lymphknoten; Leber; Milz; Muskulatur) genommen. Kot sowie Gewebeproben wurden kulturell und, wenn positiv, auch mittels PCR untersucht.
Alle mit dem Wildtyp infizierten Schweine wurden mehr oder weniger stark krank. Häufig zeigten erkrankte Schweine zeitgleich mehrere Krankheitssymptome (z. B. Erbrechen und Durchfall). Die Erkrankung hielt über mehrere Tage an. Im Vergleich dazu waren die Krankheitssymptome der Tiere, die mit Mutanten infiziert wurden, mild. Nur wenige Tiere erkrankten und dann auch nur kurzzeitig. Gewöhnlich war nur einer der erfassten Parameter verändert. Typische Veränderungen im weißen Blutbild waren nur bei Wildtyp-infizierten Tieren zu beobachten, während Tiere beider Mutanten kaum auf die Infektion reagierten. Alle 25 infizierten Tiere schieden S. Typhimurium mit dem Kot während der ersten 5 Tage post inocculationem aus. Danach wurden in allen drei Gruppen etwa gleichviel intermittierende Ausscheider beobachtet. Zwischen 65 und 67 % der Gewebeproben der mit dem Wildtyp und mit der sseD::aphT-Mutante infizierten Tiere waren sowohl in der Kultur als auch mittels PCR S. Typhimurium positiv, während dieser Anteil nach Infektion mit invC::aphT nur 49 % betrug. Alle Tiere waren in Mandibularlymphknoten und im Colon positiv, während S. Typhimurium nur selten in Muskulatur und Leber nachzuweisen war.
Die Ergebnisse dieser Arbeit bestätigen, dass Infektionen mit dem Wildtyp von S. Typhimurium zu einer schweren Erkrankung führen können. Gleichzeitig konnte gezeigt werden, dass beide in dieser Arbeit verwendeten Mutanten weniger krankmachend sind. Es muss davon ausgegangen werden, dass die Deletionen in den sseD- bzw. invC-Bereichen tatsächlich zu Veränderungen bestimmter Eigenschaften geführt haben, die Teil der Pathogenitätsmechanismen für das Schwein sind. Im Unterschied zur Maus war sseD beim Schwein allerdings invasiv. Es kann vermutet werden, dass die durch sseD kodierten Pathogenitätseigenschaften von S. Typhimurium bei der Maus anders als beim Schwein wirken und somit unterschiedliche Bedeutung haben. Da die invC::aphT-Mutante jedoch, und wie erwartet, wesentlich schwächer als Wildtyp und sseD::aphT invadierte, ist davon auszugehen, dass die Deletion im invC Bereich das Invasionsvermögen der Mutante beim Schwein ähnlich wie bei der Maus verringerte.

Summary
73
7 SUMMARY
Haukur Lindberg Sigmarsson
COMPARISON OF THE PATHOGENICITY OF SALMONELLA
TYPHIMURIUM DT104 WILD TYPE AND SALMONELLA
TYPHIMURIUM DELETIONSMUTANTS (sseD::aphT &
invC::aphT) IN PERSISTENT INFECTED PIGS
Institute of Animal Hygiene and Veterinary Public Health Faculty of Veterinary Medicine, University of Leipzig Submitted February 2012 74 pages, 22 figures, 23 tables, 241 references, 14 pages appendices Keywords: Salmonella Typhimurium DT104, deletion mutant, invC gene, sseD gene, swine,
experimental infection
Salmonella (S.) Typhimurium DT104 is a gram-negative bacteria. It does not have a strong host specifity and is considered a zoonotic pathogen. In Germany, several thousand people get sick annually with the clinical picture of a severe diarrhea which occasionally can also lead to death. The pig is considered a reservoir for infections of humans. S. Typhimurium DT104 is transmitted to human through pig products that are offered for food consumption. The control of S. Typhimurium DT104 including effective measures for eradication in our pig industry is therefore crucial in the attempt to eliminate the entry of this bacteria into the human food chain. To achieve this goal it is necessary to understand S. Typhimurium DT104 including the mechanisms that regulates its pathogenicity. The aim of the present study was to investigate into pathogenic properties of S. Typhimurium DT104. For this reason the wild type was compared to two of its mutants (i.e. sseD::aphT und invC::aphT). In this investigation, a total of 25 six weeks old male pigs were used for an infection trail. Animals were confined in a fully climatized pig barn. After being moved into the barn the pig were allowed to acclimatize for one week, and were then infected with S. Typhimurium DT104 orally in a concentration of 1 x 1011 cfu as follows: wild type (n = 8 pigs), mutant seeD::aphT (n = 8) as well as mutant invC::aphT (n = 9). The mutants are variants of

Summary
74
S. Typhimurium DT104 that have been modified at the respective loci of the bacterial genome (i.e. at the sseD-gene and invC-gene, respectively). While sseD is involved in regulating of the survival ability of S. Typhimurium in macrophages, invC regulates its ability to invade. Using the mouse model a reduced patogenicity has been demonstrated for both mutants. The day after infection a 20-days observation period commenced and the following parameters were determined, or samples taken during that period: clinical symptoms (general health, vomiting, diarrhea, feed consumption, breathing, body temperature); white blood cell count; collection of feaces for the detection of S. Typhimurium. One day after the last day of the observation period all animals were sacrificed and specimens taken from a total of 15 organs that includes tonsils; colon and caecum as well as lymph nodes, respectively; liver; spleen; muscle). All faecal samples and tissue specimens were subjected to cultural analysis as well as PCR. All animals that received the wild type of S. Typhimurium got more or less sick. Sick animals often had more than one clinical symptom at the same time (e.g. vomiting and diarrhea). The disease often lasted over several days. In comparison, animals that received either of the mutants were only slightly sick. Moreover, only a few animals were sick and the sickness then disappeared quickly. There was also usually only one clinical parameter that was affected. Typical changes of the white blood count was only observed in animals that were infected with the wild type, while animals that received either of the mutants did not show much as the result of infection. All 25 infected animals shed S. Typhimurium via feaces within the first week of infection. After this time animals of all three groups started to shed intermittently. Between 65 and 67 % of the animals that were infected with either the wild type of S. Typhimurium or the sseD::aphT-mutant were found positive for S. Typhimurium with both culture and PCR. In comparison, the percentage of specimens that were positive after infection with invC::aphT was 49 %. In all pigs S. Typhimurium was found in lymph nodes of the mandibular gland and in colon, while it was rarely observed in liver and muscle. The results of this study confirm that, infections with the wild type of S. Typhimurium can lead to severe sickness. At the same time it could be demonstrated that, the mutants that were used in this study are less “sick-making”. It is assumes that, the modifications that occurred to the sseD and invC regions resulted in changes of certain properties that are part of the mechanisms that regulate the pathogenicity if the bacteria in pigs. In contrast to mice sseD has been shown to be invasive in the pig. It is assumed that, sseD codes for pathogenicity properties that act in a different manner in pigs than in mice and thus may have a different significance for the pig. However, the fact that the invC::aphT mutant did invade to a much slighter extend than sseD::aphT suggest that the modification that occurred at the invC region did reduce the invasion capability of that mutant.

Literaturverzeichnis
75
8 LITERATURVERZEICHNIS
Aabo S, Rasmussen OF, Rossen L, Sorensen PD, Olsen JE. Salmonella identification by the polymerase chain reaction. Mol Cell Probes. 1993;7(3):171-8.
Agbor TA, McCormick BA. Salmonella Effectors: Important players modulating host cell function during infection. Cell Microbiol. 2011.
Akeda Y, Galan JE. Genetic analysis of the Salmonella enterica type III secretion-associated ATPase InvC defines discrete functional domains. J Bacteriol. 2004;186(8):2402-12.
Althouse C, Patterson S, Fedorka-Cray P, Isaacson RE. Type 1 fimbriae of Salmonella enterica serovar Typhimurium bind to enterocytes and contribute to colonization of swine in vivo. Infect Immun. 2003;71(11):6446-52.
Anderson ES, Ward LR, Saxe MJ, de Sa JD. Bacteriophage-typing designations of Salmonela typhimurium. J Hyg (Lond). 1977;78(2):297-300.
Anonymus. Verfahren zum Nachweis von Salmonellen mit der Polymerase-Kettenreaktion (PCR). DIN 10135. 1999. Berlin, Deutsches Institut für Normung, e.V.
Anonymus. Mikrobiologie von Lebensmitteln und Futtermitteln - Horizontales Verfahren zum Nachweis von Salmonella spp. - Änderung 1 Anhang D: Nachweis von Salmonella spp. in Tierkot und in Proben aus Primärproduktion (ISO6579:2002/DAmd 1:2005). 2005. International Organization for Standardization.
Anonymus. Mikrobiologie von Lebensmitteln und Futtermitteln - Horizontales Verfahren zum Nachweis von Salmonella spp. (ISO 6579:2002+Amd 1:2007); Deutsche Fassung EN ISO 6579:2002+A1:2007. 2007. International Organization for Standadization.
Arnold T, Scholz HC, Marg H, Rosler U, Hensel A. Impact of invA-PCR and culture detection methods on occurrence and survival of salmonella in the flesh, internal organs and lymphoid tissues of experimentally infected pigs. J Vet Med B Infect Dis Vet Public Health. 2004;51(10):459-63.
Ashkenazi S, Cleary TG, Murray BE, Wanger A, Pickering LK. Quantitative analysis and partial characterization of cytotoxin production by Salmonella strains. Infect Immun. 1988;56(12):3089-94.
Bager F, Petersen J. Sensitivity and specificity of different methods for the isolation of Salmonella from pigs. Acta Vet Scand. 1991;32(4):473-81.
Baggesen DL, Skov MN, Brown DJ, Bisgaard M. Separation of Salmonella typhimurium DT2 and DT135: molecular characterization of isolates of avian origin. Eur J Epidemiol. 1997;13(3):347-52.
Balaji R, Wright KJ, Hill CM, Dritz SS, Knoppel EL, Minton JE. Acute phase responses of pigs challenged orally with Salmonella typhimurium. J Anim Sci. 2000;78(7):1885-91.

Literaturverzeichnis
76
Berends BR, Urlings HA, Snijders JM, van KF. Identification and quantification of risk factors in animal management and transport regarding Salmonella spp. in pigs. Int J Food Microbiol. 1996;30(1-2):37-53.
BfR (Bundesinstitut für Risikobewertung). Grundlagenstudie zur Erhebung der Prävalenz von Salmonellen in Mastschweinen. Bericht des BfR vom 20. Februar 2008. Endbericht gemäß Entscheidung 2006/668/EG. 2008. Bundesinstitut für Risikobewertung (BfR). 20-2-2008.
Bhat P, Rajan D. Comparative evaluation of desoxycholate citrate medium and xylose lysine desoxycholate medium in the isolation of shigellae. Am J Clin Pathol. 1975;64(3):399-404.
Botteldoorn N, Heyndrickx M, Rijpens N, Grijspeerdt K, Herman L. Salmonella on pig carcasses: positive pigs and cross contamination in the slaughterhouse. J Appl Microbiol. 2003;95(5):891-903.
Boughton C, Egan J, Kelly G, Markey B, Leonard N. Rapid infection of pigs following exposure to environments contaminated with different levels of Salmonella typhimurium. Foodborne Pathog Dis. 2007;4(1):33-40.
Boyen F, Haesebrouck F, Maes D, Van IF, Ducatelle R, Pasmans F. Non-typhoidal Salmonella infections in pigs: a closer look at epidemiology, pathogenesis and control. Vet Microbiol. 2008;130(1-2):1-19.
Brenner FW, Villar RG, Angulo FJ, Tauxe R, Swaminathan B. Salmonella nomenclature. J Clin Microbiol. 2000;38(7):2465-7.
Brumme S, Arnold T, Sigmarsson H, Lehmann J, Scholz HC, Hardt WD et al. Impact of Salmonella Typhimurium DT104 virulence factors invC and sseD on the onset, clinical course, colonization patterns and immune response of porcine salmonellosis. Vet Microbiol. 2007.
Brumme S. Charakterisierung der Virulenzfaktoren invC und sseD des Typ III-Sekretionssystems von Salmonella Typhimurium DT104 mittels ex vivo Zytokin-Expression im experimentell infizierten Schwein [Dissertation med. vet.] Univ. Leipzig; 2006.
CALLOW BR. A new phage-typing scheme for Salmonella typhi-murium. J Hyg (Lond). 1959;57:346-59.
Chopra AK, Houston CW, Peterson JW, Prasad R, Mekalanos JJ. Cloning and expression of the Salmonella enterotoxin gene. J Bacteriol. 1987;169(11):5095-100.
Chopra AK, Huang JH, Xu X, Burden K, Niesel DW, Rosenbaum MW et al. Role of Salmonella enterotoxin in overall virulence of the organism. Microb Pathog. 1999;27(3):155-71.
Clothier KA, Kinyon JM, Frana TS. Comparison of Salmonella serovar isolation and antimicrobial resistance patterns from porcine samples between 2003 and 2008. J Vet Diagn Invest. 2010;22(4):578-82.
Coburn B, Grassl GA, Finlay BB. Salmonella, the host and disease: a brief review. Immunol Cell Biol. 2007;85(2):112-8.

Literaturverzeichnis
77
Coe NE, Frank DE, Wood RL, Roth JA. Alteration of neutrophil function in BCG-treated and non-treated swine after exposure to Salmonella typhimurium. Vet Immunol Immunopathol. 1992;33(1-2):37-50.
Crosa JH, Brenner DJ, Ewing WH, Falkow S. Molecular relationships among the Salmonelleae. J Bacteriol. 1973;115(1):307-15.
Darwin KH, Miller VL. Molecular basis of the interaction of Salmonella with the intestinal mucosa. Clin Microbiol Rev. 1999;12(3):405-28.
Davies RH, Wray C. Use of antibody-coated cellulose sponges for enhanced isolation of salmonella. Lett Appl Microbiol. 1997;25(4):246-8.
Dedie K, Bockemühl J, Kühn H, Volkmer K-J, Weinke T. Bakterielle Zoonosen bei Tier und Mensch. Enke Verlag, Stuttgart; 1993.
Dinjus U, Hanel I, Rabsch W, Helmuth R. Studies of the presence of the virulence factors, adhesion, invasion, intracellular multiplication and toxin formation in salmonellas of different origin. Zentralbl Bakteriol. 1998;287(4):387-98.
Dong T, Schellhorn HE. Role of RpoS in virulence of pathogens. Infect Immun. 2010;78(3):887-97.
Dorn C, Schroeter A, Helmuth R. Salmonellenvorkommen beim Schwein-epidemiologische Situation und Bewertung des Verbraucherschutzrisikos. Prakt Tierarzt. 2003;84(12):930-6.
Dunn C, Martin WJ. Comparison of media for isolation of salmonellae and shigellae from fecal specimens. Appl Microbiol. 1971;22(1):17-22.
D´Aoust JY. Update on Penrichment and Selective Enrichment Conditions for Detection of Salmonella in Foods. J Food Prot. 1981;44:369-74.
Eder H. Blut und Lymphe. In: Wittke G, (Hrsg.). Lehrbuch der Veterinär-Physiologie. Berlin und Hamburg: Verlag Paul Parey; 1987: 160-207.
Edfors-Lilja I, Wattrang E, Magnusson U, Fossum C. Genetic variation in parameters reflecting immune competence of swine. Vet Immunol Immunopathol. 1994;40(1):1-16.
EFSA (European Food Safety Authority). Opinion of the Scientific Panel on Biological Hazards on the request from the Commission related to "Risk assessment and mitigation options of Salmonella in pigproduction"1. The EFSA Journal. 2006;341(2090):1-131.
EFSA (European Food Safety Authority). Report of the Task Force on Zoonoses Data Collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, in the EU, 2006-2007, Part A: Salmonella prevalence estimates. The EFSA Journal. 2008;135:1-111.
EFSA (European Food Safety Authority). SIENTIFIC REPORT OF EFSA; Analysis of the baseline survey on the prevalence of Salmonella in holdings with breeding pigs in the EU, 2008 Part A: Salmonella prevalence estimates. The EFSA Journal. 2009;7(12)(1377).

Literaturverzeichnis
78
EFSA (European Food Safety Authority), ECDC (European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2009. The EFSA Journal. 2011;9(2090).
Eichelberg K, Ginocchio CC, Galan JE. Molecular and functional characterization of the Salmonella typhimurium invasion genes invB and invC: homology of InvC to the F0F1 ATPase family of proteins. J Bacteriol. 1994;176(15):4501-10.
Eigner U, Reissbrodt R, Hammann R, Fahr AM. Evaluation of a new chromogenic medium for the isolation and presumptive identification of Salmonella species from stool specimens. Eur J Clin Microbiol Infect Dis. 2001;20(8):558-65.
Euzeby JP. Revised Salmonella nomenclature: designation of Salmonella enterica (ex Kauffmann and Edwards 1952) Le Minor and Popoff 1987 sp. nov., nom. rev. as the neotype species of the genus Salmonella Lignieres 1900 (approved lists 1980), rejection of the name Salmonella choleraesuis (Smith 1894) Weldin 1927 (approved lists 1980), and conservation of the name Salmonella typhi (Schroeter 1886) Warren and Scott 1930 (approved lists 1980). Request for an opinion. Int J Syst Bacteriol. 1999;49 Pt 2:927-30.
Falkow S. Infectious multiple drug resistance. London: Pion Limited; 1975.
Favrin SJ, Jassim SA, Griffiths MW. Application of a novel immunomagnetic separation-bacteriophage assay for the detection of Salmonella enteritidis and Escherichia coli O157:H7 in food. Int J Food Microbiol. 2003;85(1-2):63-71.
Fedorka-Cray PJ, Gray JT, Wray C. Salmonella Infection in Pigs. In: Wray C, Wray A, (Hrsg.). Salmonella in Domestic Animals. New York: CABI Publishing; 2000: 191-208.
Fedorka-Cray PJ, Whipp SC, Isaacson RE, Nord N, Lager K. Transmission of Salmonella typhimurium to swine. Vet Microbiol. 1994;41(4):333-44.
Felix A, Callow BR. Typing of paratyphoid B bacilli by means of Vi bacteriophage. BMJ. 1943;127-30.
Fierer J, Guiney DG. Diverse virulence traits underlying different clinical outcomes of Salmonella infection. J Clin Invest. 2001;107(7):775-80.
Fierer J, Krause M, Tauxe R, Guiney D. Salmonella typhimurium bacteremia: association with the virulence plasmid. J Infect Dis. 1992;166(3):639-42.
Finlay BB. Molecular and cellular mechanisms of Salmonella pathogenesis. Curr Top Microbiol Immunol. 1994;192:163-85.
Fitzgerald C, Sherwood R, Gheesling LL, Brenner FW, Fields PI. Molecular analysis of the rfb O antigen gene cluster of Salmonella enterica serogroup O:6,14 and development of a serogroup-specific PCR assay. Appl Environ Microbiol. 2003;69(10):6099-105.
Fluit AC, Widjojoatmodjo MN, Verhoef J. Detection of Salmonella species in fecal samples by immunomagnetic separation and PCR. J Clin Microbiol. 1995;33(4):1046-7.
Friedrich A, Dorn C, Schroeter A, Szabo I, Jaber M, Berendonk G et al. [Report on Salmonella isolates in livestock, food and feed, received at the German national reference

Literaturverzeichnis
79
laboratory for Salmonella during 2004-2008]. Berl Munch Tierarztl Wochenschr. 2010;123(7-8):265-77.
Friedrich A, Szabo I, Dorn C, Schroeter A, Jaber M, Berendonk G et al. [Report on Salmonella isolates received at the German National Reference Laboratory for Salmonella in the year 2009]. Berl Munch Tierarztl Wochenschr. 2011;124(9-10):401-10.
Galan JE, Curtiss RI. Cloning and molecular characterization of genes whose products allow Salmonella typhimurium to penetrate tissue culture cells. Proc Natl Acad Sci U S A. 1989;86(16):6383-7.
Galan JE, Curtiss RI. Distribution of the invA, -B, -C, and -D genes of Salmonella typhimurium among other Salmonella serovars: invA mutants of Salmonella typhi are deficient for entry into mammalian cells. Infect Immun. 1991;59(9):2901-8.
Galan JE, Ginocchio C, Costeas P. Molecular and functional characterization of the Salmonella invasion gene invA: homology of InvA to members of a new protein family. J Bacteriol. 1992;174(13):4338-49.
Galan JE, Miller VL, Portnoy D. Discussion of in vitro and in vivo assays for studying bacterial entry into and survival within eukaryotic cells. Infect Agents Dis. 1993;2(4):288-90.
Gallois A, Klein JR, Allen LA, Jones BD, Nauseef WM. Salmonella pathogenicity island 2-encoded type III secretion system mediates exclusion of NADPH oxidase assembly from the phagosomal membrane. J Immunol. 2001;166(9):5741-8.
Gareis M. Salmonellae - A Survey. Fleischwirtsch. 1995;75(8):954-7.
Gerlach RG, Hensel M. Salmonella pathogenicity islands in host specificity, host pathogen-interactions and antibiotics resistance of Salmonella enterica. Berl Munch Tierarztl Wochenschr. 2007;120(7-8):317-27.
Ginocchio CC, Olmsted SB, Wells CL, Galan JE. Contact with epithelial cells induces the formation of surface appendages on Salmonella typhimurium. Cell. 1994;76(4):717-24.
Grimont PAD, Weill FX. Antigenic Formulae of the Salmonella Serovars. WHO Collaborating Center for Reference and Research on Salmonella, Institut Pasteur, (Hrsg.). 9. 2007. Paris.
Grimont P, Grimont F, BP. Taxonomy of the Genus Salmonella. In: Wray C, Wray A, (Hrsg.). Salmonella in Domestic Animals. New York: CABI Publishing; 2000: 1-17.
Groisman EA, Ochman H. Cognate gene clusters govern invasion of host epithelial cells by Salmonella typhimurium and Shigella flexneri. EMBO J. 1993;12(10):3779-87.
Guibourdenche M, Roggentin P, Mikoleit M, Fields PI, Bockemuhl J, Grimont PA et al. Supplement 2003-2007 (No. 47) to the White-Kauffmann-Le Minor scheme. Res Microbiol. 2010;161(1):26-9.
Guiney DG, Fang FC, Krause M, Libby S. Plasmid-mediated virulence genes in non-typhoid Salmonella serovars. FEMS Microbiol Lett. 1994;124(1):1-9.

Literaturverzeichnis
80
Guiney DG, Fierer J. The Role of the spv Genes in Salmonella Pathogenesis. Front Microbiol. 2011;2:129.
Guthire RK. Control of Salmonella spread. Salmonella. Boca Raton, Ann Abor, London: CRC Press; 1992: 63-80.
Hallstrom K, McCormick BA. Salmonella Interaction with and Passage through the Intestinal Mucosa: Through the Lens of the Organism. Front Microbiol. 2011;2:88.
Haneda T, Ishii Y, Danbara H, Okada N. Genome-wide identification of novel genomic islands that contribute to Salmonella virulence in mouse systemic infection. FEMS Microbiol Lett. 2009;297(2):241-9.
Hansen-Wester I, Hensel M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001;3(7):549-59.
Hapfelmeier S, Stecher B, Barthel M, Kremer M, Muller AJ, Heikenwalder M et al. The Salmonella pathogenicity island (SPI)-2 and SPI-1 type III secretion systems allow Salmonella serovar typhimurium to trigger colitis via MyD88-dependent and MyD88-independent mechanisms. J Immunol. 2005;174(3):1675-85.
Haraga A, Ohlson MB, Miller SI. Salmonellae interplay with host cells. Nat Rev Microbiol. 2008;6(1):53-66.
Hartung M, Käsbohrer A. Erreger von Zoonosen in Deutschland im Jahr 2009. 17-56. 2011. Bundesinstitut für Risikobewertung (BfR).
Heesemann J, Hensel M. Infektionsmodelle, Yersinien, Salmonellen, Shigellen und Listerien. In: Heesemann J, Hacker J, (Hrsg.). Molekulare Infektionsbiologie. Heidelberg, Berlin: Spektrum Akademischer Verlag; 2000: 220-222.
Helms M, Ethelberg S, Molbak K. International Salmonella Typhimurium DT104 infections, 1992-2001. Emerg Infect Dis. 2005;11(6):859-67.
Helmuth R, Stephan R, Bunge C, Hoog B, Steinbeck A, Bulling E. Epidemiology of virulence-associated plasmids and outer membrane protein patterns within seven common Salmonella serotypes. Infect Immun. 1985;48(1):175-82.
Hensel M. Salmonella pathogenicity island 2. Mol Microbiol. 2000;36(5):1015-23.
Hensel M, Shea JE, Gleeson C, Jones MD, Dalton E, Holden DW. Simultaneous identification of bacterial virulence genes by negative selection. Science. 1995;269(5222):400-3.
Hensel M, Shea JE, Waterman SR, Mundy R, Nikolaus T, Banks G et al. Genes encoding putative effector proteins of the type III secretion system of Salmonella pathogenicity island 2 are required for bacterial virulence and proliferation in macrophages. Mol Microbiol. 1998;30(1):163-74.
Henzler DJ, Opitz HM. The role of mice in the epizootiology of Salmonella enteritidis infection on chicken layer farms. Avian Dis. 1992;36(3):625-31.

Literaturverzeichnis
81
Hof H, Dörries R, Müller RL. Mikrobiologie. Stuttgart, Germany: George Thieme Verlag; 2000.
Holden DW. Trafficking of the Salmonella vacuole in macrophages. Traffic. 2002;3(3):161-9.
Hollinger K. Epidemiology and Salmonellosis. In: Wray C, Wray A, (Hrsg.). Salmonella in domestic animals. New York: CABI Publishing; 2000: 341-353.
Holzer SU, Hensel M. Functional dissection of translocon proteins of the Salmonella pathogenicity island 2-encoded type III secretion system. BMC Microbiol. 2010;10:104.
Hurd HS, Gailey JK, McKean JD, Rostagno MH. Rapid infection in market-weight swine following exposure to a Salmonella typhimurium-contaminated environment. Am J Vet Res. 2001;62(8):1194-7.
Hutchinson F, Donnellan JEJr. A general database for DNA sequence changes induced by mutagenesis of several bacterial and mammalian genes. Nucleic Acids Res. 1996;24(1):172-6.
INFOSAN (The International Foodsafety Authoritis Network). INFOSAN Information Note No 3 / 2005 - Salmonella - Antimicrobial-resitant Salmonella. 3. 2005.
Isaacson RE, Firkins LD, Weigel RM, Zuckermann FA, DiPietro JA. Effect of transportation and feed withdrawal on shedding of Salmonella typhimurium among experimentally infected pigs. Am J Vet Res. 1999;60(9):1155-8.
Karasova D, Sebkova A, Vrbas V, Havlickova H, Sisak F, Rychlik I. Comparative analysis of Salmonella enterica serovar Enteritidis mutants with a vaccine potential. Vaccine. 2009;27(38):5265-70.
Kauffmann F. Weitere Erfahrungen mit dem kombinierten Anreicherungsverfahren für Salmonella Bazillen. Zeitschrift Hyg Infektionskrnk. 1935;117:26-32.
Kauffmann F. The species definition in the Enterobacteriaceae. Int Bull Bacteriol Nomenclat Taxon. 1961;11:5-6.
Kauffmann F. Das Salmonella subgenus IV. Ann Immunol Hungar. 1966;9:77-80.
Klein JR, Jones BD. Salmonella pathogenicity island 2-encoded proteins SseC and SseD are essential for virulence and are substrates of the type III secretion system. Infect Immun. 2001;69(2):737-43.
Klena JD, Pradel E, Schnaitman CA. Comparison of lipopolysaccharide biosynthesis genes rfaK, rfaL, rfaY, and rfaZ of Escherichia coli K-12 and Salmonella typhimurium. J Bacteriol. 1992;174(14):4746-52.
Korsgaard H, Wegener HC, Helms M. [The societal cost of zoonotic Salmonella infections and other foodborne bacterial infections in Denmark]. Ugeskr Laeger. 2005;167(7):760-3.
Kraczkiewicz-Dowjat A, Wolska KI, Kunicki-Goldfinger WJ. Mechanism of conjugation and recombination in bacteria. XXI. Role of F factor genes in post-conjugational recombination in Escherichia coli K-12. Acta Microbiol Pol. 1984;33(1):11-24.

Literaturverzeichnis
82
Kraft W, Dürr UM. Klinische Labordiagnostik in der Tiermedizin. 6. Auflage. Stuttgart; New York: Schattauer; 2005.
Krause G. Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten. Berlin: Mercedes Druck; 2001.
Kristensen M., Lester V., Jurgens A. Use of trypsinized casein, brom-thymol blue, brom-cresol-purple, phenol-red and brilliant green for bacteriological nutrient media. Br J Exp Pathol. 1925;6:291-9.
Kubista M, Andrade JM, Bengtsson M, Forootan A, Jonak J, Lind K et al. The real-time polymerase chain reaction. Mol Aspects Med. 2006;27(2-3):95-125.
Kühn H, Tschäpe H. Salmonellosen des Menschen - Epidemiologische und ätiologische Aspekte. 3/95. Auflage. MMV Medizin Verlag München; 1995.
Lan R, Reeves PR, Octavia S. Population structure, origins and evolution of major Salmonella enterica clones. Infect Genet Evol. 2009;9(5):996-1005.
Larsen ST, McKean JD, Hurd HS, Rostagno MH, Griffith RW, Wesley IV. Impact of commercial preharvest transportation and holding on the prevalence of Salmonella enterica in cull sows. J Food Prot. 2003;66(7):1134-8.
Le Minor L, Popoff MY. Request for an opinion. Designation of Salmonella enterica sp. nov., nom. Rev., as the type and only species of the Genus Salmonella. Int J Syst Bacteriol. 1987;37:465-8.
LeClerc JE, Li B, Payne WL, Cebula TA. High mutation frequencies among Escherichia coli and Salmonella pathogens. Science. 1996;274(5290):1208-11.
Lederberg J, Tatum EL. Novel genotypes in mixed cultures of bichemical mutants of bacteria. Cold Spring Harbour Symp Quant Biol. 1946;11:113.
Lesnick ML, Reiner NE, Fierer J, Guiney DG. The Salmonella spvB virulence gene encodes an enzyme that ADP-ribosylates actin and destabilizes the cytoskeleton of eukaryotic cells. Mol Microbiol. 2001;39(6):1464-70.
Levy SB. The Antibiotic Paradox: How Miracle Drugs are Destroying the Miracle. New York: Plenium Press; 1992.
Libby SJ, Adams LG, Ficht TA, Allen C, Whitford HA, Buchmeier NA et al. The spv genes on the Salmonella dublin virulence plasmid are required for severe enteritis and systemic infection in the natural host. Infect Immun. 1997;65(5):1786-92.
Libby SJ, Goebel W, Ludwig A, Buchmeier N, Bowe F, Fang FC et al. A cytolysin encoded by Salmonella is required for survival within macrophages. Proc Natl Acad Sci U S A. 1994;91(2):489-93.
Libby SJ, Goebel W, Muir S, Songer G, Heffron F. Cloning and characterization of a cytotoxin gene from Salmonella typhimurium. Res Microbiol. 1990;141(7-8):775-83.

Literaturverzeichnis
83
Liebeswar G, Allerberger F. Standardisierung und Qualitätssicherung in der mikrobiologischen Diagnostik-Richtlinien. Wien: Bundesministerium für Soziale Sicherheit und Generationen; 2001.
Lindqvist L, Schweda KH, Reeves PR, Lindberg AA. In vitro synthesis of CDP-d-abequose using Salmonella enzymes of cloned rfb genes. Production of CDP-6-deoxy-D-xylo-4-hexulose, CDP-3,6-dideoxy-D-xylo-4-hexulose and CDP-3,6-dideoxy-D-galactose, and isolation by HPLC. Eur J Biochem. 1994;225(3):863-72.
Lostroh CP, Lee CA. The Salmonella pathogenicity island-1 type III secretion system. Microbes Infect. 2001;3(14-15):1281-91.
Loynachan AT, Nugent JM, Erdman MM, Harris DL. Acute infection of swine by various Salmonella serovars. J Food Prot. 2004;67(7):1484-8.
Mahan MJ, Slauch JM, Mekalanos JJ. Selection of bacterial virulence genes that are specifically induced in host tissues. Science. 1993;259(5095):686-8.
Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O'Brien SJ et al. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis. 2010;50(6):882-9.
Malik-Kale P, Jolly CE, Lathrop S, Winfree S, Luterbach C, Steele-Mortimer O. Salmonella - at home in the host cell. Front Microbiol. 2011;2:125.
Marcus SL, Brumell JH, Pfeifer CG, Finlay BB. Salmonella pathogenicity islands: big virulence in small packages. Microbes Infect. 2000;2(2):145-56.
Marg H, Scholz HC, Arnold T, Rosler U, Hensel A. Influence of long-time transportation stress on re-activation of Salmonella typhimurium DT104 in experimentally infected pigs. Berl Munch Tierarztl Wochenschr. 2001;114(9-10):385-8.
Martin G. Eigenschaften von Salmonellen und ihre Beziehungen zu Stimulierung von Immunreaktionen. Mh Vet Med. 1992;47:632-6.
Mayr A, Rolle M. Medizinische Mikrobiologie, Infektions-und Seuchenlehre für Tierärzte, Biologen, Agrarwissenschaftler und Interessierte aus benachbarten Fachgebieten. Stuttgart: Enke Verlag; 1993.
Mazurkiewicz P, Thomas J, Thompson JA, Liu M, Arbibe L, Sansonetti P et al. SpvC is a Salmonella effector with phosphothreonine lyase activity on host mitogen-activated protein kinases. Mol Microbiol. 2008;67(6):1371-83.
McGhie EJ, Brawn LC, Hume PJ, Humphreys D, Koronakis V. Salmonella takes control: effector-driven manipulation of the host. Curr Opin Microbiol. 2009;12(1):117-24.
Medina E, Paglia P, Nikolaus T, Muller A, Hensel M, Guzman CA. Pathogenicity island 2 mutants of Salmonella typhimurium are efficient carriers for heterologous antigens and enable modulation of immune responses. Infect Immun. 1999;67(3):1093-9.
Meurens F, Berri M, Auray G, Melo S, Levast B, Virlogeux-Payant I et al. Early immune response following Salmonella enterica subspecies enterica serovar Typhimurium infection in porcine jejunal gut loops. Vet Res. 2009;40(1):5.

Literaturverzeichnis
84
Meyer H. [Animals as sources of infections in humans--salmonellosis]. Dtsch Tierarztl Wochenschr. 1999;106(8):344-51.
Meyer H, Steinbach G, Methner U. [Control of salmonella infections in animal herds--basis for a reduction of salmonella entries into food]. Dtsch Tierarztl Wochenschr. 1993;100(7):292-5.
Meyerholz DK, Stabel TJ, Ackermann MR, Carlson SA, Jones BD, Pohlenz J. Early epithelial invasion by Salmonella enterica serovar Typhimurium DT104 in the swine ileum. Vet Pathol. 2002;39(6):712-20.
Mitov I, Denchev V, Linde K. Humoral and cell-mediated immunity in mice after immunization with live oral vaccines of Salmonella typhimurium: auxotrophic mutants with two attenuating markers. Vaccine. 1992;10(1):61-6.
Morris A, Kellner JD, Low DE. The superbugs: evolution, dissemination and fitness. Curr Opin Microbiol. 1998;1(5):524-9.
Mueller M, Brandenburg K, Dedrick R, Schromm AB, Seydel U. Phospholipids inhibit lipopolysaccharide (LPS)-induced cell activation: a role for LPS-binding protein. J Immunol. 2005;174(2):1091-6.
Mullis KB, Faloona FA. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987;155:335-50.
Mulvey MR, Boyd DA, Olson AB, Doublet B, Cloeckaert A. The genetics of Salmonella genomic island 1. Microbes Infect. 2006;8(7):1915-22.
Nielsen B, Baggesen D, Bager F, Haugegaard J, Lind P. The serological response to Salmonella serovars typhimurium and infantis in experimentally infected pigs. The time course followed with an indirect anti-LPS ELISA and bacteriological examinations. Vet Microbiol. 1995;47(3-4):205-18.
Niewold TA, Veldhuizen EJ, van der MJ, Haagsman HP, de Wit AA, Smits MA et al. The early transcriptional response of pig small intestinal mucosa to invasion by Salmonella enterica serovar typhimurium DT104. Mol Immunol. 2007;44(6):1316-22.
Norte VA, Stapleton MR, Green J. PhoP-responsive expression of the Salmonella enterica serovar typhimurium slyA gene. J Bacteriol. 2003;185(12):3508-14.
Ochman H, Soncini FC, Solomon F, Groisman EA. Identification of a pathogenicity island required for Salmonella survival in host cells. Proc Natl Acad Sci U S A. 1996;93(15):7800-4.
Olsen JE, Aabo S, Nielsen EO, Nielsen BB. Isolation of a Salmonella-specific DNA hybridization probe. APMIS. 1991;99(2):114-20.
Partridge SR, Recchia GD, Stokes HW, Hall RM. Family of class 1 integrons related to In4 from Tn1696. Antimicrob Agents Chemother. 2001;45(11):3014-20.
Pascopella L, Raupach B, Ghori N, Monack D, Falkow S, Small PL. Host restriction phenotypes of Salmonella typhi and Salmonella gallinarum. Infect Immun. 1995;63(11):4329-35.

Literaturverzeichnis
85
Perron GG, Quessy S, Bell G. A reservoir of drug-resistant pathogenic bacteria in asymptomatic hosts. PLoS One. 2008;3(11):e3749.
Perron GG, Quessy S, Letellier A, Bell G. Genotypic diversity and antimicrobial resistance in asymptomatic Salmonella enterica serotype Typhimurium DT104. Infect Genet Evol. 2007;7(2):223-8.
Pietzsch O. Salmonella. In: Blobel H, Schliesser T, (Hrsg.). Handbuch der bakteriellen Infektionen bei Tieren. Jena: VEB Fischer Verlag; 1981.
Popoff MY, Bockemuhl J, McWhorter-Murlin A. Supplement 1993 (no. 37) to the Kauffmann-White scheme. Res Microbiol. 1994;145(9):711-6.
Poppe C, Ziebell K, Martin L, Allen K. Diversity in antimicrobial resistance and other characteristics among Salmonella typhimurium DT104 isolates. Microb Drug Resist. 2002;8(2):107-22.
Porter SB, Tinge SA, Curtiss R, III. Virulence of Salmonella typhimurium mutants for White Leghorn chicks. Avian Dis. 1993;37(2):265-73.
Prager R, Liesegang A, Rabsch W, Gericke B, Thiel W, Voigt W et al. Clonal relationship of Salmonella enterica serovar typhimurium phage type DT104 in Germany and Austria. Zentralbl Bakteriol. 1999;289(4):399-414.
Prasad R, Chopra AK, Chary P, Peterson JW. Expression and characterization of the cloned Salmonella typhimurium enterotoxin. Microb Pathog. 1992;13(2):109-21.
Prasad R, Chopra AK, Peterson JW, Pericas R, Houston CW. Biological and immunological characterization of a cloned cholera toxin-like enterotoxin from Salmonella typhimurium. Microb Pathog. 1990;9(5):315-29.
Rabsch W, Tschape H, Baumler AJ. Non-typhoidal salmonellosis: emerging problems. Microbes Infect. 2001;3(3):237-47.
Rahman H, Streckel W, Prager R, Tschape H. Presence of sopE gene & its phenotypic expression among different serovars of Salmonella isolated from man & animals. Indian J Med Res. 2004;120(1):35-8.
Rahn K, De Grandis SA, Clarke RC, McEwen SA, Galan JE, Ginocchio C et al. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol Cell Probes. 1992;6(4):271-9.
RAPPAPORT F, KONFORTI N, NAVON B. A new enrichment medium for certain Salmonellae. J Clin Pathol. 1956;9(3):261-6.
Rathnayaka RMUS. Evaluation of five DNA extraction methods in the detection of Salmonella enterica from meat using nested PCR. JAS; Vol 6, No 1 (2011). 2011.
Recchia GD, Hall RM. Gene cassettes: a new class of mobile element. Microbiology. 1995;141 ( Pt 12):3015-27.
Reeves MW, Evins GM, Heiba AA, Plikaytis BD, Farmer JJ, III. Clonal nature of Salmonella typhi and its genetic relatedness to other salmonellae as shown by multilocus enzyme

Literaturverzeichnis
86
electrophoresis, and proposal of Salmonella bongori comb. nov. J Clin Microbiol. 1989;27(2):313-20.
Reiner G, Hepp S, Holweger K. [Phenotypic and genetic variation in hematological traits of Pietrain-Meishan-F2 crossbred pigs]. Dtsch Tierarztl Wochenschr. 2007;114(11):429-36.
Reitmeyer JC, Peterson JW, Wilson KJ. Salmonella cytotoxin: a component of the bacterial outer membrane. Microb Pathog. 1986;1(5):503-10.
Rietschel ET, Brade H. Bacterial endotoxins. Sci Am. 1992;267(2):54-61.
Rietschel ET, Kirikae T, Schade FU, Ulmer AJ, Holst O, Brade H et al. The chemical structure of bacterial endotoxin in relation to bioactivity. Immunobiology. 1993;187(3-5):169-90.
RKI (Robert Koch-Institut). Epidemiologisches Bulletin; Zur Situation bei ausgewählten meldepflichtigen Infektionskrankheiten im Jahr 1996: Teil 1: Darminfektionen (Gastroenteritiden). 8, 49-53. 1997.
RKI (Robert Koch-Institut). Epidemiologisches Bulletin; Zur Situation bei wichtigen Infektionskrankheiten in Deutschland: Ausgewählte Zoonosen im Jahr 2005: Durch Lebensmittel übertragbare bakterielle gastrointestinale Infektionen. 41, 351-362. 2006.
RKI (Robert Koch-Institut). Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2006. Berlin: Robert Koch-Institut (RKI); 2007.
RKI (Robert Koch-Institut). Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2009. Berlin: Robert Koch-Institut (RKI); 2010.
RKI (Robert Koch-Institut). Epidemiologisches Bulletin; Meldepflichtige Infektionskrankheiten; Jahresstatistik 2010. 14, 105-116. 2011a.
RKI (Robert Koch-Institut). Epidemiologisches Bulletin; Salmonellose; Aus der Arbeit des Nationalen Referenzzentrums für Salmonellen und andere bakterielle Enteritiserreger. 15, 125-136. 2011b.
RKI (Robert Koch-Institut). Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2010. Berlin: Robert Koch-Institut (RKI); 2011c.
Robbins JB, Chu C, Schneerson R. Hypothesis for vaccine development: protective immunity to enteric diseases caused by nontyphoidal salmonellae and shigellae may be conferred by serum IgG antibodies to the O-specific polysaccharide of their lipopolysaccharides. Clin Infect Dis. 1992;15(2):346-61.
Robertsson JA, Lindberg AA, Hoiseth S, Stocker BA. Salmonella typhimurium infection in calves: protection and survival of virulent challenge bacteria after immunization with live or inactivated vaccines. Infect Immun. 1983;41(2):742-50.
Roesler U. Charakterisierung der Porzinen Salmonella Typhimurium DT104-Infektion und Maßnahmen zur Salmonellen-Reduktion in Schweinemast- und Schweinezuchtbetrieben [Dissertation med. vet.] Leipzig: Univ. Leipzig; 2007.

Literaturverzeichnis
87
Roesler U, Marg H, Schroder I, Mauer S, Arnold T, Lehmann J et al. Oral vaccination of pigs with an invasive gyrA-cpxA-rpoB Salmonella Typhimurium mutant. Vaccine. 2004;23(5):595-603.
Rotger R, Casadesus J. The virulence plasmids of Salmonella. Int Microbiol. 1999;2(3):177-84.
Rychlik W. Selection of primers for polymerase chain reaction. Mol Biotechnol. 1995;3(2):129-34.
Rycroft AN. Structure, Funktion and Synthesis of Surface Polysaccharides in Salmonella. In: Wray C, Wray A, (Hrsg.). Salmonella in Domestic Animals. New York: CABI Publishing; 2000: 19-34.
Sabbagh SC, Forest CG, Lepage C, Leclerc JM, Daigle F. So similar, yet so different: uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiol Lett. 2010;305(1):1-13.
Sander J. [Pathogenesis of salmonella infections in humans]. Dtsch Tierarztl Wochenschr. 1993;100(7):283-5.
Santos RL, Tsolis RM, Baumler AJ, Adams LG. Pathogenesis of Salmonella-induced enteritis. Braz J Med Biol Res. 2003;36(1):3-12.
Santos RL, Zhang S, Tsolis RM, Kingsley RA, Adams LG, Baumler AJ. Animal models of Salmonella infections: enteritis versus typhoid fever. Microbes Infect. 2001;3(14-15):1335-44.
Saunders JR. Genetics and evolution of antibiotic resistance. Br Med Bull. 1984;40(1):54-60.
Saxen H, Reima I, Makela PH. Alternative complement pathway activation by Salmonella O polysaccharide as a virulence determinant in the mouse. Microb Pathog. 1987;2(1):15-28.
Scherer K, Szabo I, Rosler U, Appel B, Hensel A, Nockler K. Time course of infection with Salmonella typhimurium and its influence on fecal shedding, distribution in inner organs, and antibody response in fattening pigs. J Food Prot. 2008;71(4):699-705.
Schletter J, Heine H, Ulmer AJ, Rietschel ET. Molecular mechanisms of endotoxin activity. Arch Microbiol. 1995;164(6):383-9.
Schlundt J, Munch B. A comparison of the efficiency of Rappaport-Vassiliadis, tetrathionate and selenite broths with and without pre-enrichment for the isolation of Salmonella in animal waste biogas plants. Zentralbl Bakteriol. 1993;279(3):336-43.
Schmieger H. Molecular survey of the Salmonella phage typing system of Anderson. J Bacteriol. 1999;181(5):1630-5.
Schmieger H, Schicklmaier P. Transduction of multiple drug resistance of Salmonella enterica serovar typhimurium DT104. FEMS Microbiol Lett. 1999;170(1):251-6.
Scholz HC, Arnold T, Marg H, Rosler U, Hensel A. Improvement of an invA-based PCR for the specific detection of Salmonella typhimurium in organs of pigs. Berl Munch Tierarztl Wochenschr. 2001;114(9-10):401-3.

Literaturverzeichnis
88
Selbitz HJ. Lehrbuch der Veterinärmedizinischen Bakteriologie. Stuttgart: Gustav Fischer Verlag Jena; 1992.
Selbitz HJ. Adaption of Salmonella Strains to Their Host and Its Significans for Zoonosis. Prakt Tierarzt. 1993;74:1102.
Selbitz HJ. Bakterielle Krankheiten der Tiere. In: Mayr A, (Hrsg.). Medizinische Mikrobiologie, Infektions- und Seuchenlehre. Suttgart: Enke Verlag; 2002: 417-588.
Selbitz H-J. Die Virulenz der Salmonellen-Faktoren und Mechanismen. Prakt Tierarzt. 1991;7:589-92.
Selbitz H-J, Sinell H-J, Sziegoleit A. Taxonomie und Nomenklatur. Das Salmonellen-Problem. Jena, Stuttgart: Gustav Fischer Verlag; 1995: 17-19.
Seydel U, Labischinski H, Kastowsky M, Brandenburg K. Phase behavior, supramolecular structure, and molecular conformation of lipopolysaccharide. Immunobiology. 1993;187(3-5):191-211.
Seydel U, Schromm AB, Brade L, Gronow S, Andra J, Muller M et al. Physicochemical characterization of carboxymethyl lipid A derivatives in relation to biological activity. FEBS J. 2005;272(2):327-40.
Spector MP. The starvation-stress response (SSR) of Salmonella. Adv Microb Physiol. 1998;40:233-79.
Stackbrandt E, R.G.E.Murray, H.Truper. Proteobacteria classis nov., a name for the phylogenetic taxon that includes the purple bacteria and their relatives. Int J Syst Bacteriol. 1988;38:321-5.
Stecher B, Hapfelmeier S, Muller C, Kremer M, Stallmach T, Hardt WD. Flagella and chemotaxis are required for efficient induction of Salmonella enterica serovar Typhimurium colitis in streptomycin-pretreated mice. Infect Immun. 2004;72(7):4138-50.
Steinbach G, Kroell U. [Salmonella infections in swine herds--epidemiology and importance for human diseases]. Dtsch Tierarztl Wochenschr. 1999;106(7):282-8.
Szabo I, Scherer K, Roesler U, Appel B, Nockler K, Hensel A. Comparative examination and validation of ELISA test systems for Salmonella typhimurium diagnosis of slaughtering pigs. Int J Food Microbiol. 2008;124(1):65-9.
Taylor WI. Isolation of shigellae. I. Xylose lysine agars; new media for isolation of enteric pathogens. Am J Clin Pathol. 1965;44(4):471-5.
Tenover FC, Schaberg DR. Molecular biology of resistance. In: Benett JV, Brachman PS, (Hrsg.). Hospital Infections. Philadelphia, USA: Lippincott-Raven Publishers; 1998: 237-247.
Threlfall EJ. Epidemic salmonella typhimurium DT 104--a truly international multiresistant clone. J Antimicrob Chemother. 2000;46(1):7-10.
Threlfall EJ, Fisher IS, Berghold C, Gerner-Smidt P, Tschape H, Cormican M et al. Antimicrobial drug resistance in isolates of Salmonella enterica from cases of salmonellosis in

Literaturverzeichnis
89
humans in Europe in 2000: results of international multi-centre surveillance. Euro Surveill. 2003;8(2):41-5.
Tindall BJ, Grimont PA, Garrity GM, Euzeby JP. Nomenclature and taxonomy of the genus Salmonella. Int J Syst Evol Microbiol. 2005;55(Pt 1):521-4.
Tinge SA, Curtiss RI. Isolation of the replication and partitioning regions of the Salmonella typhimurium virulence plasmid and stabilization of heterologous replicons. J Bacteriol. 1990;172(9):5266-77.
Touron A, Berthe T, Pawlak B, Petit F. Detection of Salmonella in environmental water and sediment by a nested-multiplex polymerase chain reaction assay. Res Microbiol. 2005;156(4):541-53.
Tschäpe H, Bockemühl J. Lebensmittelübertragene Salmonellose in Deutschland. Bundesgesundheitsbl. 2002;45:491-6.
van Diemen PM, Kreukniet MB, Galina L, Bumstead N, Wallis TS. Characterisation of a resource population of pigs screened for resistance to salmonellosis. Vet Immunol Immunopathol. 2002;88(3-4):183-96.
Van Parys A, Boyen F, Leyman B, Verbrugghe E, Haesebrouck F, Pasmans F. Tissue-Specific Salmonella Typhimurium Gene Expression during Persistence in Pigs. PLoS One. 2011;6(8):e24120.
Vassiliadis P, Pateraki E, Papaiconomou N, Papadakis JA, Trichopoulos D. New Procedure of Salmonella Enrichment. Ann Microb. 1976;B127(2):195-200.
Venkatesan MM, Buysse JM, Oaks EV. Surface presentation of Shigella flexneri invasion plasmid antigens requires the products of the spa locus. J Bacteriol. 1992;174(6):1990-2001.
Voetsch AC, Van Gilder TJ, Angulo FJ, Farley MM, Shallow S, Marcus R et al. FoodNet estimate of the burden of illness caused by nontyphoidal Salmonella infections in the United States. Clin Infect Dis. 2004;38 Suppl 3:S127-S134.
von Altrock A, Schütte A, Hildebrandt G. Untersuchungen aus Deutschland zu dem EU-Projekt "Salmonella in pork (Salinpork)-1.Mitteilung:Untersuchungen in den Beständen. Berl Munch Tierarztl Wochenschr. 2000;113:191-201.
Waage AS, Vardund T, Lund V, Kapperud G. Detection of low numbers of Salmonella in environmental water, sewage and food samples by a nested polymerase chain reaction assay. J Appl Microbiol. 1999;87(3):418-28.
Wales AD, Cook AJ, Davies RH. Producing Salmonella-free pigs: a review focusing on interventions at weaning. Vet Rec. 2011;168(10):267-76.
Wallis TS, Galyov EE. Molecular basis of Salmonella-induced enteritis. Mol Microbiol. 2000;36(5):997-1005.
Waltman WD. Methods for the Cultural Isolation of Salmonella. In: Wray C, Wray A, (Hrsg.). Salmonella in Domestic Animals. New York: CABI Publishing; 2000: 355-372.

Literaturverzeichnis
90
Waterman SR, Holden DW. Functions and effectors of the Salmonella pathogenicity island 2 type III secretion system. Cell Microbiol. 2003;5(8):501-11.
Watson PR, Paulin SM, Jones PW, Wallis TS. Interaction of Salmonella serotypes with porcine macrophages in vitro does not correlate with virulence. Microbiology. 2000;146 ( Pt 7):1639-49.
Wegener HC, Hald T, Lo Fo WD, Madsen M, Korsgaard H, Bager F et al. Salmonella control programs in Denmark. Emerg Infect Dis. 2003;9(7):774-80.
Werckenthin C, Schwarz S. [Resistance to protein biosynthesis inhibitors in Staphylococci: resistance genes and their spread--a review]. Berl Munch Tierarztl Wochenschr. 1997;110(9):324-32.
Wheeler JG, Sethi D, Cowden JM, Wall PG, Rodrigues LC, Tompkins DS et al. Study of infectious intestinal disease in England: rates in the community, presenting to general practice, and reported to national surveillance. The Infectious Intestinal Disease Study Executive. BMJ. 1999;318(7190):1046-50.
WHO (World Health Organization). Drug-resistant Salmonella. WHO, (Hrsg.). Fact Sheet N°139, 1-5. 2005. Geneva, Food Safety Department WHO/Geneva. Media centre; Fact Sheets.
Widjojoatmodjo MN, Fluit AC, Torensma R, Keller BHI, Verhoef J. Evaluation of the Magnetic Immuno Pcr Assay for Rapid Detection of Salmonella. Eur J Clin Microbiol Infect Dis. 1991;10(11):935-8.
Widjojoatmodjo MN, Fluit AC, Torensma R, Verdonk GP, Verhoef J. The magnetic immuno polymerase chain reaction assay for direct detection of salmonellae in fecal samples. J Clin Microbiol. 1992;30(12):3195-9.
Wiesner E, Ribbeck R. Wörterbuch der Veterinärmedizin. 3. Auflage. Jena; Stuttgart: Gustav Fischer Verlag; 1991.
Wilcock BP, Schwartz K. Salmonellosis. In: Leman AD, Straw BE, Mengeling WE, D´Allaire S, Taylor DJ, (Hrsg.). Diseases of Swine. Iowa: Iowa State University Press; 1992: 570-583.
Williams JL, Minton JE, Patterson JA, Marchant FJ, Eicher SD. Lairage during transport of eighteen-kilogram pigs has an impact on innate immunity and commensal bacteria diversity in the intestines. J Anim Sci. 2008;86(5):1232-44.
Woestyn S, Allaoui A, Wattiau P, Cornelis GR. YscN, the putative energizer of the Yersinia Yop secretion machinery. J Bacteriol. 1994;176(6):1561-9.
Wood RL, Pospischil A, Rose R. Distribution of persistent Salmonella typhimurium infection in internal organs of swine. Am J Vet Res. 1989;50(7):1015-21.
Wood RL, Rose R. Populations of Salmonella typhimurium in internal organs of experimentally infected carrier swine. Am J Vet Res. 1992;53(5):653-8.
Zhang-Barber L, Turner AK, Barrow PA. Vaccination for control of Salmonella in poultry. Vaccine. 1999;17(20-21):2538-45.

Literaturverzeichnis
91
Zhao S, Fedorka-Cray PJ, Friedman S, McDermott PF, Walker RD, Qaiyumi S et al. Characterization of Salmonella Typhimurium of animal origin obtained from the National Antimicrobial Resistance Monitoring System. Foodborne Pathog Dis. 2005;2(2):169-81.
Zhao S, McDermott PF, White DG, Qaiyumi S, Friedman SL, Abbott JW et al. Characterization of multidrug resistant Salmonella recovered from diseased animals. Vet Microbiol. 2007;123(1-3):122-32.
Zurawski DV, Stein MA. The SPI2-encoded SseA chaperone has discrete domains required for SseB stabilization and export, and binds within the C-terminus of SseB and SseD. Microbiology. 2004;150(Pt 7):2055-68.

Anhang
I
9 ANHANG
Tabelle 7. Mittelwerte der Körpertemperatur (°C) der, mit dem S. Typhimurium DT104-Wildtyp, der Deletionsmutante-invC::aphT, der Deletionsmutante-sseD::aphT infizierten Schweine und die dazugehörige Standardabweichung
Mittelwerte der Körpertemperatur der infizierten Versuchsschweine in °C
Tag nach der Infektion
DM-invC::aphT DM-sseD::aphT Wildtyp DT104
Mittelwert Standard-
abweichung Mittelwert Standard-
abweichung Mittelwert Standard-
abweichung 0 39,7 0,5 39,4 0,2 38,8 0,3 1 38,9 0,3 39,4 0,4 39,2 0,5 2 39,1 0,4 39,8 0,4 40 0,5 3 39,6 0,3 39,6 0,4 39,3 0,3 4 39,6 0,4 39,3 0,4 39,4 0,6 5 39,4 0,3 39,7 0,2 39,1 0,6 6 39,2 0,3 39,5 0,2 39,2 0,6 7 39,8 0,3 39,4 0,2 39,2 0,4 8 39,9 0,3 39,4 0,2 39,2 0,6 9 39,5 0,3 39,4 0,3 39,2 0,4
10 39,4 0,2 39,5 0,1 39,3 0,3 DM = Deletionsmutante
Tabelle 8. Mittelwerte des Körpergewichts der mit dem S. Typhimurium DT104-Wildtyp, der Deletionsmutante-invC::aphT, der Deletionsmutante-sseD::aphT infizierten Schweine mit der dazugehörigen Standardabweichung und Signifikanz
Mittelwerte für das Körpergewicht der infizierten Schweine in kg
Tag WT DT 104
n = 8 Mittelwert ± s
DM-invC::aphT n = 9
Mittelwert ± s
DM-sseD::aphT n = 8
Mittelwert ± s
0 8,9 ± 1,3d 10,3 ± 1,2d 9,9 ± 1,1d
3 9,0 ± 1,2B,d 11,6 ± 1,5A,c,d nicht gemessen 7 9,6 ± 1,4B,d 13,6 ± 1,8A,b,c 10,9 ± 1,0B,c,d
10 11,2 ± 1,5B,c,d 14,7 ± 1,5A,b 12,5 ± 1,9B,b,c 14 13,0 ± 1,9B,b,c 17,7 ± 1,8A,a 14,0 ± 2,6B,b 17 14,8 ± 2,1B,a,b 19,2 ± 2,6A,a 16,5 ± 1,8A,B,a 21 16,8 ± 2,5a nicht gemessen nicht gemessen
Am Tag 3 konnten die Gruppe DM-sseD::aphT und am Tag 21. die Gruppen DM-sseD::aphT und DM-invC::aphT aus technischen Gründen nicht gemessen werden.
WT DT104 = Wildtyp, DM = Deletionsmutante A – B = Werte unterscheiden sich signifikant (P≤0,05) in Zeilen. a – d = Werte unterscheiden sich signifikant (P≤0,05) in Spalten.

Anhang
II
Tabelle 9. Klinischer Score des Allgemeinbefindens der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, Deletionsmutante invC::aphT und Deletionsmutante sseD::aphT
Klinischer Score für das Allgemeinbefinden
Tag nach der Infektion
Deskriptive Statistik
WT DT104 (A)
DM-invC::aphT
(B)
DM-sseD::aphT
(C)
Signifikanzprüfung zwischen den Gruppen
0
n 8 9 8 A:B p= 1,0 Mittelwert (±
SD) 0a 0a 0a
Standardabw. 0 0 0 A:C p=1,0 min 0 0 0 max 0 0 0 B:C p= 1,0
1
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 0,125
Standardabw. 0 0 0,354 A:C p= 0,317 min 0 0 0 max 0 0 1 B:C p= 0,289
2
n 8 9 8 A:B p= 0,289 Mittelwert 0,250 0 0
Standardabw. 0,707 0 0 A:C p= , 317 min 0 0 0 max 2 0 0 B:C p= 1,0
3
n 8 9 8 A:B p= 0,289 Mittelwert 0,125 0 0
Standardabw. 0,354 0 0 A:C p= 0,317 min 0 0 0 max 1 0 0 B:C p= 1,0
4
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 0
Standardabw. 0 0 0 A:C p= 1,0 min 0 0 0 max 0 0 0 B:C p= 1,0
5
n 8 9 8 A:B p= 0,05 Mittelwert 0,250a 0b 0b
Standardabw. 0,378 0 0 A:C p= 0,06 min 0 0 0 max 1 0 0 B:C p= 1,0
6
n 8 9 8 A:B p= 0,121 Mittelwert 0,125 0 0
Standardabw. 0,232 0 0 A:C p= 0,143 min 0 0 0 max 0,5 0 0 B:C p= 1,0
7
n 8 9 8 A:B p= 0,289 Mittelwert 0,063 0 0
Standardabw. 0,177 0 0 A:C p= 0,317 min 0 0 0 max 0,5 0 0 B:C p= 1,0
8 - 21
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 0
Standardabw. 0 0 0 A:C p= 1,0 min 0 0 0 max 0 0 0 B:C p= 1,0
WT DT104 = Wildtyp, DM = Deletionsmutante
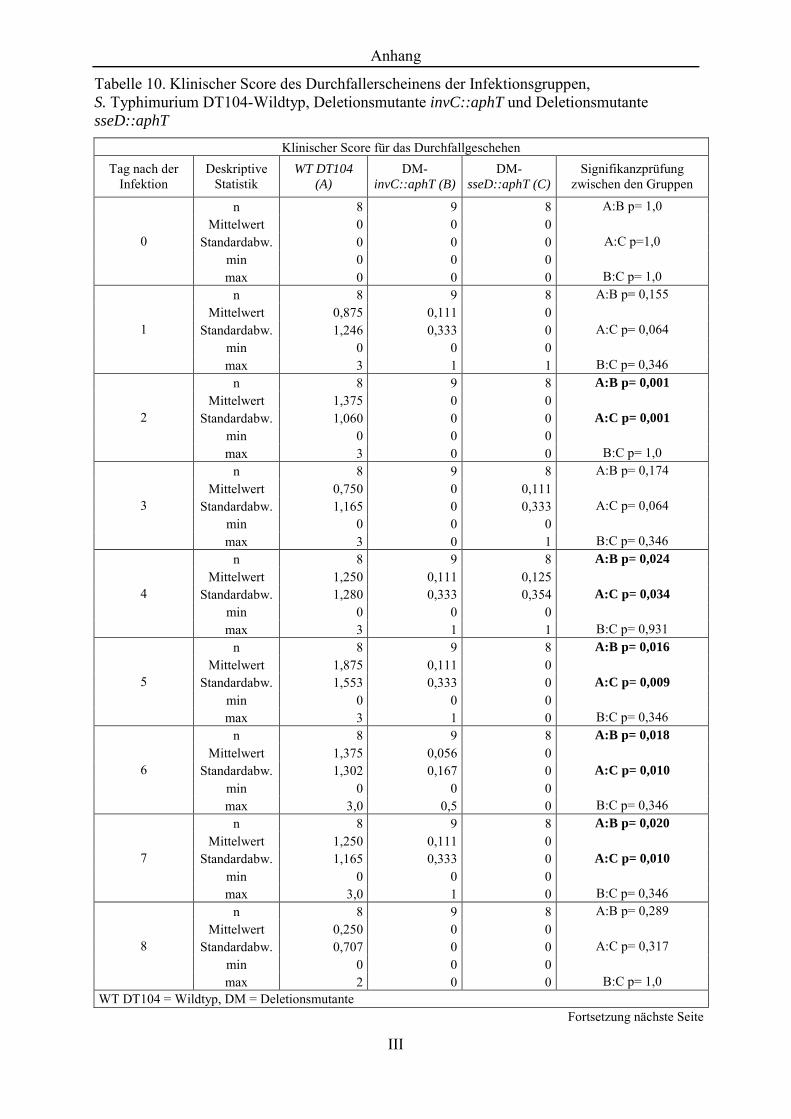
Anhang
III
Tabelle 10. Klinischer Score des Durchfallerscheinens der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, Deletionsmutante invC::aphT und Deletionsmutante sseD::aphT
Klinischer Score für das Durchfallgeschehen Tag nach der
Infektion Deskriptive
Statistik WT DT104
(A) DM-
invC::aphT (B) DM-
sseD::aphT (C) Signifikanzprüfung
zwischen den Gruppen
0
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 0
Standardabw. 0 0 0 A:C p=1,0 min 0 0 0 max 0 0 0 B:C p= 1,0
1
n 8 9 8 A:B p= 0,155 Mittelwert 0,875 0,111 0
Standardabw. 1,246 0,333 0 A:C p= 0,064 min 0 0 0 max 3 1 1 B:C p= 0,346
2
n 8 9 8 A:B p= 0,001 Mittelwert 1,375 0 0
Standardabw. 1,060 0 0 A:C p= 0,001 min 0 0 0 max 3 0 0 B:C p= 1,0
3
n 8 9 8 A:B p= 0,174 Mittelwert 0,750 0 0,111
Standardabw. 1,165 0 0,333 A:C p= 0,064 min 0 0 0 max 3 0 1 B:C p= 0,346
4
n 8 9 8 A:B p= 0,024 Mittelwert 1,250 0,111 0,125
Standardabw. 1,280 0,333 0,354 A:C p= 0,034 min 0 0 0 max 3 1 1 B:C p= 0,931
5
n 8 9 8 A:B p= 0,016 Mittelwert 1,875 0,111 0
Standardabw. 1,553 0,333 0 A:C p= 0,009 min 0 0 0 max 3 1 0 B:C p= 0,346
6
n 8 9 8 A:B p= 0,018 Mittelwert 1,375 0,056 0
Standardabw. 1,302 0,167 0 A:C p= 0,010 min 0 0 0 max 3,0 0,5 0 B:C p= 0,346
7
n 8 9 8 A:B p= 0,020 Mittelwert 1,250 0,111 0
Standardabw. 1,165 0,333 0 A:C p= 0,010 min 0 0 0 max 3,0 1 0 B:C p= 0,346
8
n 8 9 8 A:B p= 0,289 Mittelwert 0,250 0 0
Standardabw. 0,707 0 0 A:C p= 0,317 min 0 0 0 max 2 0 0 B:C p= 1,0
WT DT104 = Wildtyp, DM = Deletionsmutante Fortsetzung nächste Seite
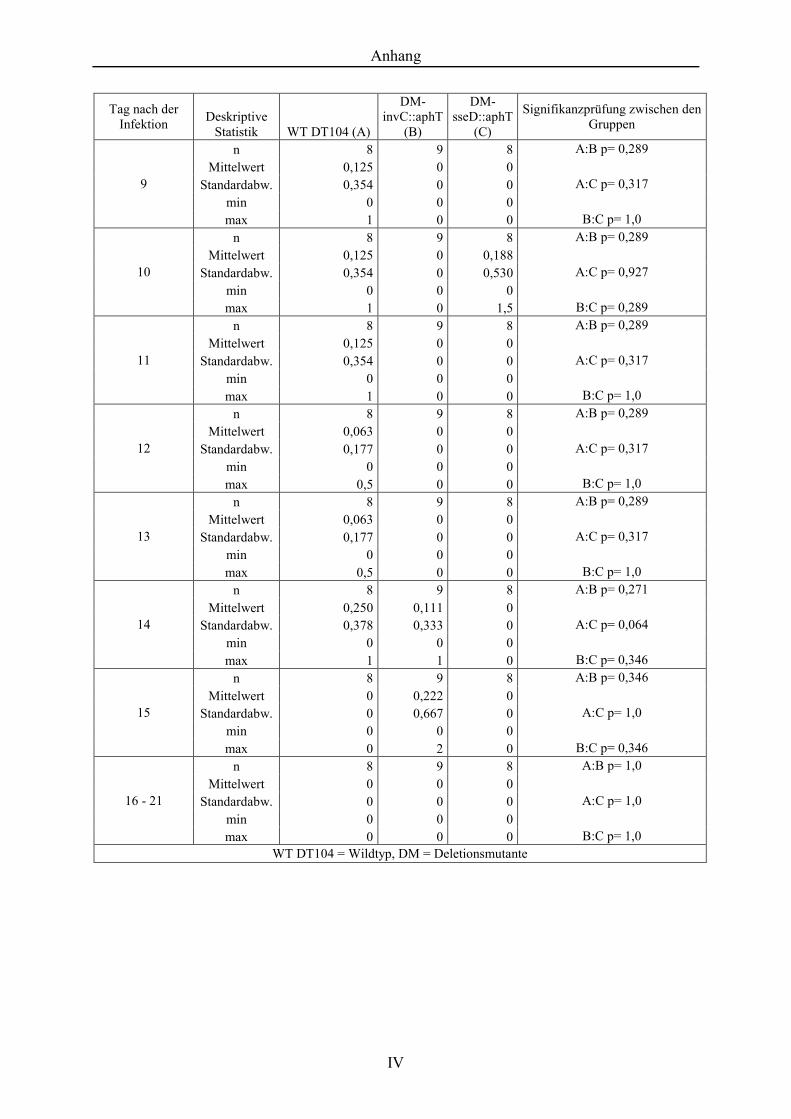
Anhang
IV
Tag nach der
Infektion Deskriptive Statistik WT DT104 (A)
DM-invC::aphT
(B)
DM-sseD::aphT
(C)
Signifikanzprüfung zwischen den Gruppen
9
n 8 9 8 A:B p= 0,289 Mittelwert 0,125 0 0
Standardabw. 0,354 0 0 A:C p= 0,317 min 0 0 0 max 1 0 0 B:C p= 1,0
10
n 8 9 8 A:B p= 0,289 Mittelwert 0,125 0 0,188
Standardabw. 0,354 0 0,530 A:C p= 0,927 min 0 0 0 max 1 0 1,5 B:C p= 0,289
11
n 8 9 8 A:B p= 0,289 Mittelwert 0,125 0 0
Standardabw. 0,354 0 0 A:C p= 0,317 min 0 0 0 max 1 0 0 B:C p= 1,0
12
n 8 9 8 A:B p= 0,289 Mittelwert 0,063 0 0
Standardabw. 0,177 0 0 A:C p= 0,317 min 0 0 0 max 0,5 0 0 B:C p= 1,0
13
n 8 9 8 A:B p= 0,289 Mittelwert 0,063 0 0
Standardabw. 0,177 0 0 A:C p= 0,317 min 0 0 0 max 0,5 0 0 B:C p= 1,0
14
n 8 9 8 A:B p= 0,271 Mittelwert 0,250 0,111 0
Standardabw. 0,378 0,333 0 A:C p= 0,064 min 0 0 0 max 1 1 0 B:C p= 0,346
15
n 8 9 8 A:B p= 0,346 Mittelwert 0 0,222 0
Standardabw. 0 0,667 0 A:C p= 1,0 min 0 0 0 max 0 2 0 B:C p= 0,346
16 - 21
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 0
Standardabw. 0 0 0 A:C p= 1,0 min 0 0 0 max 0 0 0 B:C p= 1,0
WT DT104 = Wildtyp, DM = Deletionsmutante

Anhang
V
Tabelle 11. Die Mittelwerte der Gesamtleukozytenzahl (103/mm3) der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten
Tabelle 12. Die Mittelwerte der relativen Lymphozytenzahlen der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent
Tag nach der
Infektion
WT DT 104 n
Mittelwert ± s
DM-invC::aphT n
Mittelwert ± s
DM-sseD::aphT n
Mittelwert ± s
0 8 66,8 ± 15,7A,a
9 47,4 ± 15,3A,B,a,b
8 43,0 ± 14,5B,b
3 8 47,3 ± 14,7b
8 36,9 ± 12,7b
8 45,4 ± 12,2b
7 8 35,0 ± 8,9B,b
9 60,8 ± 16,6A,a
8 42,6 ± 10,4B,b
10 8 36,8 ± 10,1b
9 54,0 ± 14,1a,b
8 48,1 ± 17,5b
14 8 37,8 ± 8,0b
9 50,1 ± 19,0b
8 48,4 ± 17,8a,b
17 8 47,5 ± 9,0B,b
9 56,1 ± 11,9B,a,b
8 71,9 ± 7,5A,a
21 8 48,4 ± 5,7B,b
6 46,2 ± 15,3B,a,b
7 71,7 ± 13,5A,a
A –C = Werte unterscheiden sich signifikant (P≤0,05) in Zeilen a – c = Werte unterscheiden sich Signifikant (P≤0,05) in Spalten
WT DT104 = Wildtyp, DM = Deletionsmutante
Tag nach der
Infektion
WT DT 104 n
Mittelwert ± s
DM-invC::aphT n
Mittelwert ± s
DM-sseD::aphT n
Mittelwert ± s
0 8
8,0 ± 2,1C,c 9
16,1 ± 3,1A 8
13,9 ± 2,7A,B,a,b,c
3 8
15,3 ± 3,5a,b 8
19,7 ± 5,8 8
19,5 ± 4,0a
7
8 18,1 ±4,0a,b
9 20,7 ± 5,5
8 15,2 ± 3,9a,b
10
8 20,1 ± 5,1a
9 20,3 ± 7,4
8 15,3 ± 2,8a,b
14
8 17,6 ± 3,9A,B,a,b
9 25,6 ± 10,4A
8 12,0 ± 2,3C,b,c
17
8 15,7 ± 0,8A,B,a,b
9 22,8 ± 8,2A
8 12,9 ± 3,0B,b,c
21
8 14,6 ± 1,3b
6 14,4 ± 7,4A,B
7 9,9 ± 2,6B,c
A –C = Werte unterscheiden sich signifikant (P≤0,05) in Zeilen a – c = Werte unterscheiden sich Signifikant (P≤0,05) in Spalten
WT DT104 = Wildtyp, DM = Deletionsmutante

Anhang
VI
Tabelle 13. Die Mittelwerte der relativen Zahlen der neutrophilen stabkernigen Granulozyten der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent
Tag nach der
Infektion
WT DT 104 n
Mittelwert ± s
DM-invC::aphT n
Mittelwert ± s
DM-sseD::aphT n
Mittelwert ± s
0 8 0,9 ± 1,4b
9 1,8 ± 1,3
8 1,8 ± 1,3a
3 8 4,6 ± 3,0A,a
8 1,9 ± 1,9A,B,
8 1,0 ± 0,9B,a,b
7 8 1,1 ± 0,6b
9 1,7 ± 2,5
8 0,5 ± 0,8a,b
10 8 1,1 ± 0,6b
9 2,2 ± 2,7
8 0,5 ± 1,1a,b
14 8 1,5 ± 1,3b
9 0,8 ± 0,8
8 1,3 ± 1,1a,b
17 8 1,5 ± 1,2b
9 1,8 ± 1,8
8 1,0 ± 0,9a,b
21 8 2,3 ± 2,1a,b
6 1,2 ± 1,2
7 0,1 ± 0,4b
A – C = Werte unterscheiden sich Signifikant (P≤0,05) in Zeilen a – c = Werte unterscheiden sich Signifikant (P≤0,05) in Spalten
WT DT104 = Wildtyp, DM = Deletionsmutante
Tabelle 14. Die Mittelwerte der relativen Zahlen der neutrophilen segmentkernigen Granulozyten der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent
Tag nach der
Infektion
WT DT 104 n
Mittelwert ± s
DM-invC::aphT n
Mittelwert ± s
DM-sseD::aphT n
Mittelwert ± s
0 8 22,6 ± 13,8B,b
9 45,0 ± 15,3A,
8 46,4 ± 12,5A,a
3 8 40,5 ± 16,9a,b
8 55,5 ± 9,7
8 49,1 ± 11,5a
7 8 54,9 ± 11,7A,a
9 33,8 ± 16,3B
8 52,1 ± 8,8A,a
10 8 54,0 ± 10,1a
9 37,1 ± 15,5
8 46,8 ± 17,1a
14 8 53,4 ± 7,8a
9 45,4 ± 17,9
8 41,0 ± 14,2a,b
17 8 41,4 ± 8,7A,a
9 34,2 ± 11,4A,B
8 23,6 ± 7,4B,b
21 8 39,3 ± 7,0A,B,a,b
6 47,7 ± 14,7A
7 24,1 ± 12,4B,b
A – C = Werte unterscheiden sich Signifikant (P≤0,05) in Zeilen a – c = Werte unterscheiden sich Signifikant (P≤0,05) in Spalten
WT DT104 = Wildtyp, DM = Deletionsmutante

Anhang
VII
Tabelle 15. Die Mittelwerte der relativen Zahlen der eosinophilen Granulozyten der Infektionsgruppen, S. Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent
Tag nach der
Infektion
WT DT 104 n
Mittelwert ± s
DM-invC::aphT n
Mittelwert ± s
DM-sseD::aphT n
Mittelwert ± s
0 8
2,8 ± 2,3a,b 9
1,2 ± 1,4 8
1,1 ± 1,0 3 8
0,75 ± 0,7b 8
2,3 ± 1,5 8
1,3 ± 1,3
7 8
2,9 ± 2,0A,a,b 9
0,6 ± 0,7B 8
1,4 ± 1,1A,B
10 8
1,6 ± 1,6a,b 9
1,0 ± 0,9 8
1,5 ± 1,3
14 8
3,5 ± 1,4A,a 9
0,9 ± 0,9B 8
1,0 ± 1,2B
17
8 2,3 ± 1,7A,a,b
9 0,9 ± 1,6A,B
8 0,4 ± 0,5B
21
8 2,8 ± 1,8a,b
6 2,3 ± 2,1
7 1,1 ± 1,4
A – C = Werte unterscheiden sich Signifikant (P≤0,05) in Zeilen a – c = Werte unterscheiden sich Signifikant (P≤0,05) in Spalten
WT DT104 = Wildtyp, DM = Deletionsmutante
Tabelle 16. Die Mittelwerte der relativen Monozytenzahlen der Infektionsgruppen, S Typhimurium DT104-Wildtyp, invC::aphT- und sseD::aphT-Deletionsmutanten in Prozent
Tag nach der
Infektion
WT DT 104 n
Mittelwert ± s
DM-invC::aphT n
Mittelwert ± s
DM-sseD::aphT n
Mittelwert ± s
0 8 6,75 ± 4,1
9 4,6 ± 1,3a,b
8 7,5 ± 3,2a
3 8 6,8 ± 3,6
8 3,5 ± 1,3b
8 3,3 ± 1,4b
7 8 6,0 ± 3,0
9 3,2 ± 1,8b
8 3,4 ± 1,6b
10 8 6,5 ± 3,2A
9 5,4 ± 2,5A,B,a,b
8 3,1 ± 1,5B,b
14 8 3,5 ± 1,9
9 2,8 ± 1,9b
8 5,3 ± 2,9a,b
17 8 6,5 ± 3,6A,B
9 6,9 ± 1,5A,a
8 3,1 ± 1,5B,b
21 8 6,75 ± 3,4A
6 2,7 ± 2,7B,b
7 2,7 ± 1,6B,b
A – C = Werte unterscheiden sich Signifikant (P≤0,05) in Zeilen a – c = Werte unterscheiden sich Signifikant (P≤0,05) in Spalten
WT DT104 = Wildtyp, DM = Deletionsmutante
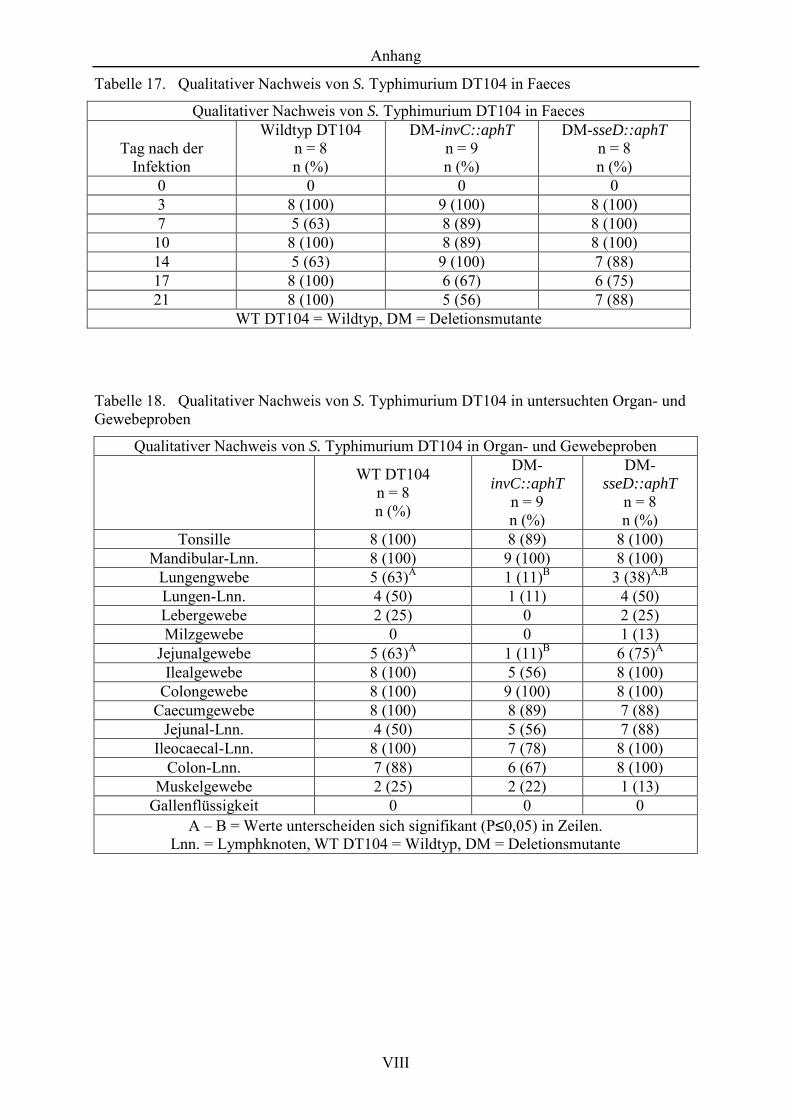
Anhang
VIII
Tabelle 17. Qualitativer Nachweis von S. Typhimurium DT104 in Faeces
Qualitativer Nachweis von S. Typhimurium DT104 in Faeces
Tag nach der Infektion
Wildtyp DT104 n = 8 n (%)
DM-invC::aphT n = 9 n (%)
DM-sseD::aphT n = 8 n (%)
0 0 0 0 3 8 (100) 9 (100) 8 (100) 7 5 (63) 8 (89) 8 (100)
10 8 (100) 8 (89) 8 (100) 14 5 (63) 9 (100) 7 (88) 17 8 (100) 6 (67) 6 (75) 21 8 (100) 5 (56) 7 (88)
WT DT104 = Wildtyp, DM = Deletionsmutante
Tabelle 18. Qualitativer Nachweis von S. Typhimurium DT104 in untersuchten Organ- und Gewebeproben
Qualitativer Nachweis von S. Typhimurium DT104 in Organ- und Gewebeproben
WT DT104
n = 8 n (%)
DM-invC::aphT
n = 9 n (%)
DM-sseD::aphT
n = 8 n (%)
Tonsille 8 (100) 8 (89) 8 (100) Mandibular-Lnn. 8 (100) 9 (100) 8 (100)
Lungengwebe 5 (63)A 1 (11)B 3 (38)A,B
Lungen-Lnn. 4 (50) 1 (11) 4 (50) Lebergewebe 2 (25) 0 2 (25) Milzgewebe 0 0 1 (13)
Jejunalgewebe 5 (63)A 1 (11)B 6 (75)A
Ilealgewebe 8 (100) 5 (56) 8 (100) Colongewebe 8 (100) 9 (100) 8 (100)
Caecumgewebe 8 (100) 8 (89) 7 (88) Jejunal-Lnn. 4 (50) 5 (56) 7 (88)
Ileocaecal-Lnn. 8 (100) 7 (78) 8 (100) Colon-Lnn. 7 (88) 6 (67) 8 (100)
Muskelgewebe 2 (25) 2 (22) 1 (13) Gallenflüssigkeit 0 0 0
A – B = Werte unterscheiden sich signifikant (P≤0,05) in Zeilen. Lnn. = Lymphknoten, WT DT104 = Wildtyp, DM = Deletionsmutante
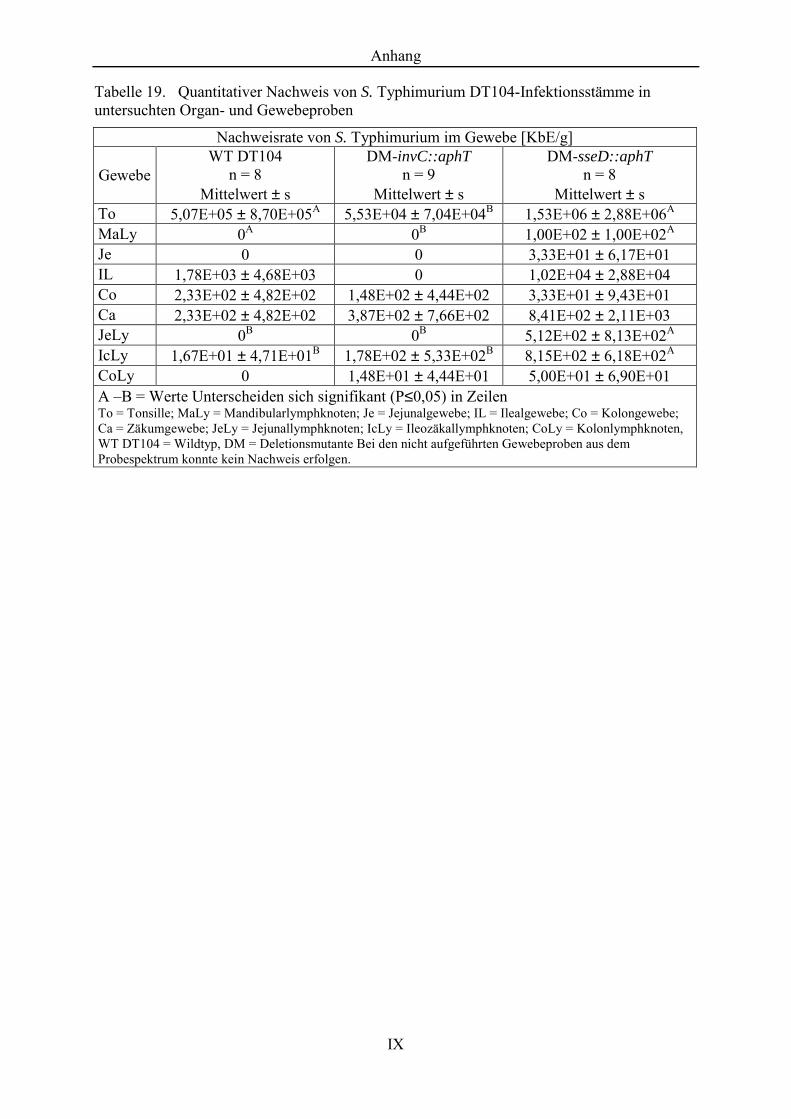
Anhang
IX
Tabelle 19. Quantitativer Nachweis von S. Typhimurium DT104-Infektionsstämme in untersuchten Organ- und Gewebeproben
Nachweisrate von S. Typhimurium im Gewebe [KbE/g]
Gewebe WT DT104
n = 8 Mittelwert ± s
DM-invC::aphT n = 9
Mittelwert ± s
DM-sseD::aphT n = 8
Mittelwert ± s To 5,07E+05 ± 8,70E+05A 5,53E+04 ± 7,04E+04B 1,53E+06 ± 2,88E+06A MaLy 0A 0B 1,00E+02 ± 1,00E+02A
Je 0 0 3,33E+01 ± 6,17E+01 IL 1,78E+03 ± 4,68E+03 0 1,02E+04 ± 2,88E+04 Co 2,33E+02 ± 4,82E+02 1,48E+02 ± 4,44E+02 3,33E+01 ± 9,43E+01 Ca 2,33E+02 ± 4,82E+02 3,87E+02 ± 7,66E+02 8,41E+02 ± 2,11E+03 JeLy 0B 0B 5,12E+02 ± 8,13E+02A
IcLy 1,67E+01 ± 4,71E+01B 1,78E+02 ± 5,33E+02B 8,15E+02 ± 6,18E+02A
CoLy 0 1,48E+01 ± 4,44E+01 5,00E+01 ± 6,90E+01 A –B = Werte Unterscheiden sich signifikant (P≤0,05) in Zeilen To = Tonsille; MaLy = Mandibularlymphknoten; Je = Jejunalgewebe; IL = Ilealgewebe; Co = Kolongewebe; Ca = Zäkumgewebe; JeLy = Jejunallymphknoten; IcLy = Ileozäkallymphknoten; CoLy = Kolonlymphknoten, WT DT104 = Wildtyp, DM = Deletionsmutante Bei den nicht aufgeführten Gewebeproben aus dem Probespektrum konnte kein Nachweis erfolgen.

Anhang
X
Tabelle 20. Quantitativer Nachweis der S. Typhimurium DT104-Infektionsstämme in Organ- und Gewebeproben mit der dazugehörigen deskriptiven Statistik und Signifikanzprüfung
Nachweisrate von S. Typhimurium im Gewebe [KbE/g]
Gewebe Deskriptive Statistik WT DT104 (A)
DM-invC::aphT
(B)
DM-sseD::aphT
(C)
Signifikanzprüfung zwischen den
Gruppen
To
n 8 9 8 A:B p= 0,136 Mittelwert 5,07E+05 5,53E+04 1,53E+06
Median 1,86E+05 2,79E+04 3,81E+05 Standardabw. 8,70E+05 7,04E+04 2,88E+06
A:C p= 0,529 min 0 0 3,64E+02 max 2,56E+06 1,81E+05 8,48E+06
25 1,65E+04 2,00E+02 4,48E+04 B:C p= 0,034 Perzentile 50 1,86E+05 2,79E+04 3,81E+05
75 6,77E+05 1,12E+05 1,51E+06
MaLy
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 1,00E+02
Median 0 0 0 Standardabw. 0 0 1,85E+02
A:C p= 0,064 min 0 0 5,33E+02 max 0 0 0
25 0 0 0 B:C p= 0,050 Perzentile 50 0 0 0
75 0 0 1,33E+02
Je
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 3,33E+01
Median 0 0 0 Standardabw. 0 0 6,17E+01
A:C p= 0,143 min 0 0 0 max 0 0 1,33E+02
25 0 0 0 B:C p= 0,121 Perzentile 50 0 0 0
75 0 0 1,00E+02
IL
n 8 9 8 A:B p= 0,122 Mittelwert 1,78E+03 0 1,02E+04
Median 0 0 0 Standardabw. 4,68E+03 0 2,88E+04
A:C p= 0,644 min 0 0 0 max 1,33E+04 0 8,13E+04
25 0 0 0 B:C p= 0,289 Perzentile 50 0 0 0
75 7,00E+02 0 0
Co
n 8 9 8 A:B p= 0,514 Mittelwert 2,33E+02 1,48E+02 3,33E+01
Median 0 0 0 Standardabw. 4,82E+02 4,44E+02 9,43E+01
A:C p= 0,441 min 0 0 0 max 1,33E+03 1,33E+03 2,67E+02
25 0 0 0 B:C p= 1,0 Perzentile 50 0 0 0
75 4,00E+02 0 0 Fortsezung nächste Seite

Anhang
XI
Gewebe Deskriptive Statistik
WT DT104 (A)
DM-invC::aphT (B)
DM-sseD::aphT
(C)
Signifikanzprüfung zwischen den Gruppen
Ca
n 8 9 8 A:B p= 0,720 Mittelwert 2,33E+02 3,87E+02 8,41E+02
Median 0 0 0 Standardabw. 4,82E+02 7,66E+02 2,11E+03
A:C p= 0,469 min 0 0 0 max 1,33E+03 2,13E+03 6,06E+03
25 0 0 0 B:C p= 0,589 Perzentile 50 0 0 6,67E+01
75 4,00E+02 6,73E+02 3,33E+02
JeLy
n 8 9 8 A:B p= 1,0 Mittelwert 0 0 5,12E+02
Median 0 0 6,67E+01 Standardabw. 0 0 8,13E+02
A:C p= 0,027 min 0 0 0 max 0 0 2,30E+03
25 0 0 0 B:C p= 0,020 Perzentile 50 0 0 6,67E+01
75 0 0 8,82E+02
IcLy
n 8 9 8 A:B p= 1,0 Mittelwert 1,67E+01 1,78E+02 8,15E+02
Median 0,00 0 5,64E+02 Standardabw. 4,71E+01 5,33E+02 6,81E+02
A:C p= 0,001 min 0 0 1,33E+02 max 1,33E+02 1,60E+03 1,94E+03
25 0 0 2,67E+02 B:C p= 0,003 Perzentile 50 0 0 5,64E+02
75 0 0 1,48E+03
CoLy
n 8 9 8 A:B p= 0,346 Mittelwert 0 1,48E+01 5,00E+01
Median 0 0 0 Standardabw. 0 4,44E+01 6,90E+01
A:C p= 0,063 min 0 0 0 max 0 1,33E+02 1,33E+02
25 0 0 0 B:C p= 1,0 Perzentile 50 0 0 0
75 0 0 1,33E+02 To = Tonsille; MaLy = Mandibularlymphknoten; Je = Jejunalgewebe; IL = Ilealgewebe; Co = Kolongewebe; Ca = Zäkumgewebe; JeLy = Jejunallymphknoten; IcLy = Ileozäkallymphknoten; CoLy = Kolonlymphknoten, WT DT104 = Wildtyp, DM = Deletionsmutante

Anhang
XII
Tabelle 21. Positiver Nachweis von S. Typhimurium DT104-Wildtyp in untersuchten Organ- und Gewebeproben
Nachweis von S. Typhimurium DT104-Wildtyp in Organ- und Gewebeproben
Kulturelle Untersuchungen PCR-Untersuchungen
n (%) n (%)
Tonsille 8 (100) 8 (100) Mandibular-Lnn. 8 (100) 8 (100)
Lungengwebe 5 (63) 5 (63) Lungen-Lnn. 4 (50) 4 (50) Lebergewebe 2 (25) 2 (25) Milzgewebe 0 0
Jejunalgewebe 5 (63) 5 (63) Ilealgewebe 8 (100) 8 (100)
Colongewebe 8 (100) 8 (100) Caecumgewebe 8 (100) 8 (100)
Jejunal-Lnn. 4 (50) 4 (50) Ileocaecal-Lnn. 8 (100) 8 (100)
Colon-Lnn. 7 (88) 7 (88) Muskelgewebe 2 (25) 2 (25)
Gallenflüssigkeit 0 0 Positiver Befund für jeweiliges Verfahren. Lnn. = Lymphknoten

Anhang
XIII
Tabelle 22. Positiver Nachweis von S. Typhimurium DT104 invC::aphT-Mutante in untersuchten Organ- und Gewebeproben
Nachweis von S. Typhimurium DT104 invC::aphT-Mutante inOrgan- und Gewebeproben
Kulturelle Untersuchung PCR-Untersuchung
n (%) n (%)
Tonsille 8 (89) 8 (89) Mandibular-Lnn. 9 (100) 9 (100)
Lungengwebe 1 (11) 1 (11) Lungen-Lnn. 1 (11) 1 (11) Lebergewebe 0 0 Milzgewebe 0 0
Jejunalgewebe 1 (11) 1 (11) Ilealgewebe 5 (56) 5 (56)
Colongewebe 9 (100) 9 (100) Caecumgewebe 8 (89) 8 (89)
Jejunal-Lnn. 5 (56) 5 (56) Ileocaecal-Lnn. 7 (78) 7 (78)
Colon-Lnn. 6 (67) 6 (67) Muskelgewebe 2 (22) 2 (22)
Gallenflüssigkeit 0 0 Positiver Befund für jeweiliges Verfahren. Lnn. = Lymphknoten

Anhang
XIV
Tabelle 23. Positiver Nachweis von S. Typhimurium DT104 sseD::aphT-Mutante in untersuchten Organ- und Gewebeproben
Nachweis von S. Typhimurium DT104 sseD::aphT-Mutante in Organ- und Gewebeproben
Kulturelle Untersuchung PCR-Untersuchung
n (%) n (%)
Tonsille 8 (100) 8 (100) Mandibular-Lnn. 8 (100) 8 (100)
Lungengwebe 3 (38) 3 (38) Lungen-Lnn. 4 (50) 4 (50) Lebergewebe 2 (25) 2 (25) Milzgewebe 1 (13) 1 (13)
Jejunalgewebe 6 (75) 6 (75) Ilealgewebe 8 (100) 8 (100)
Colongewebe 8 (100) 8 (100) Caecumgewebe 7 (88) 7 (88)
Jejunal-Lnn. 7 (88) 7 (88) Ileocaecal-Lnn. 8 (100) 8 (100)
Colon-Lnn. 8 (100) 8 (100) Muskelgewebe 1 (13) 1 (13)
Gallenflüssigkeit 0 0 Positiver Befund für jeweiliges Verfahren. Lnn. = Lymphknoten

Danksagung
10 DANKSAGUNG Im Anschluss an meine Arbeit möchte ich all jenen, die mit Worten und Taten zum gelingen dieser Arbeit beigetragen haben, ganz herzlich danken. Insbesondere danke ich:
• Herrn Prof. Dr. Dr. A. Hensel für die Überlassung dieses sehr interessanten Themas, für seine Betreuung und für die fachliche Unterstützung bei der Anfertigung dieser Arbeit.
• Herrn. Prof. Dr. U. Rösler möchte ich mich ebenfalls für die Unterstützung bei der
Durchführung der Versuche und für die fachliche Hilfe jeglicher kritischer Phasen der Laborarbeit bzw. Auswertung, sowie für die freundliche Begleitung der Arbeit bedanken.
• Weiterhin danke ich Herrn Prof. Dr. U. Truyen für die Unterstützung während meiner
Zeit am Institut.
• Herrn Prof. Dr. A. Sobiraj für die Unterstützung und für das entgegengebrachte Vertrauen.
• Dr. T. Arnold für die Beantwortung aller offenen Fragen rund um die PCR und die
stets gewährte freundschaftliche Unterstützung.
• Dr. T. Herold für die stets aufmunternde Worte und die ständige Unterstützung in allen Bereichen.
• Allen an den Infektionsversuchen beteiligten Doktoranden des Instituts für
Tierhygiene und Öffentliches Veterinärwesen, bestehend aus: Peter Burger, Steffi Brumme, Eugénie Zeeuw, Steffi Mauer, Melanie Hassel und Katharina Monig.
• Evelin Brumme und Nadja Leinecker für die tatkräftige und stets gewährte Hilfe im
Labor.
• Allen Angehörigen des Instituts für Tierhygiene und Öffentliches Veterinärwesen für die Ständige Unterstützung jeglicher Art.
• Familie Kauffold für die Freundschaft, die großartige Unterstützung, das
entgegengebrachte Vertrauen und die stets gewährte Hilfe.
• Reinhard Willig und Lydia Hildebrandt für die Freundschaft und die Unterstützung jeglicher Art.
• Meinen Eltern und meiner Freundin Almut Prkno für ihre unermüdliche Geduld und
allseits gewährte Unterstützung.
• Allen anderen ungenannten Personen, die mit Worten und Taten zum gelingen dieser Arbeit geholfen haben.





![A quantitative analysis of Salmonella Typhimurium ... · persons per year), but typhoid fever cases have a higher mortality rate (1%) [11]. In both diseases, infection with Salmonella](https://static.fdokument.com/doc/165x107/5d46f55288c993bd0a8b66fe/a-quantitative-analysis-of-salmonella-typhimurium-persons-per-year-but.jpg)




