
Pioneer woody species in southern Brazilian grasslands ... · TECHNISCHE UNIVERSITÄT MÜNCHEN...
205
TECHNISCHE UNIVERSITÄT MÜNCHEN Lehrstuhl für Vegetationsökologie Pioneer woody species in southern Brazilian grasslands: Life history traits and population dynamics Julia-Maria Hermann Vollständiger Abdruck der von der Fakultät Wissenschaftszentrum Weihenstephan für Ernährung, Landnutzung und Umwelt der Technischen Universität München zur Erlangung des akademischen Grades eines Doktors der Naturwissenschaften genehmigten Dissertation. Vorsitzender: Univ.-Prof. Dr. L. Trepl Prüfer der Dissertation: 1. Univ.-Prof. Dr. J. Pfadenhauer 2. Univ.-Prof. Dr. A. Fischer Die Dissertation wurde am 08.09.2008 bei der Technischen Universität München eingereicht und durch die Fakultät Wissenschaftszentrum Weihenstephan für Ernährung, Landnutzung und Umwelt am 11.12.2008 angenommen.
Transcript of Pioneer woody species in southern Brazilian grasslands ... · TECHNISCHE UNIVERSITÄT MÜNCHEN...

TECHNISCHE UNIVERSITÄT MÜNCHEN
Lehrstuhl für Vegetationsökologie
Pioneer woody species in southern Brazilian grasslands:
Life history traits and population dynamics
Julia-Maria Hermann
Vollständiger Abdruck der von der Fakultät Wissenschaftszentrum Weihenstephan für Ernährung, Landnutzung und Umwelt der Technischen Universität München zur Erlangung des akademischen Grades eines
Doktors der Naturwissenschaften genehmigten Dissertation. Vorsitzender: Univ.-Prof. Dr. L. Trepl Prüfer der Dissertation:
1. Univ.-Prof. Dr. J. Pfadenhauer 2. Univ.-Prof. Dr. A. Fischer
Die Dissertation wurde am 08.09.2008 bei der Technischen Universität München eingereicht und durch die Fakultät Wissenschaftszentrum Weihenstephan für Ernährung, Landnutzung und Umwelt am 11.12.2008 angenommen.

Acknowledgements
For their generosity, advice and support in many ways I would like to express my sincere gratitude to – meus agradecimentos sinceros para:
My supervisors, professors Dr. Jörg Pfadenhauer and Dr. Valério De Patta Pillar.
Friends and colleagues from Universidade Federal do Rio Grande do Sul, Technische Universität München, Fachhochschule Rottenburg and Eberhard-Karls-Universität Tübingen: Wolfram Adelmann; Carolina Blanco; Eric Burgert; Josi Cerveira; Alex Dümig; Leandro Duarte; Wolf Engels; Alessandra Fidelis; Eduardo Forneck; Birgit Harter-Marques; Heinrich Hasenack e o LABGEO; Roland Irslinger and his diplomands; Rafael Machado; Melina Marchesini; Nelson Matzenbacher; Juliano Morales; Gerhard Overbeck; Maria Luiza Porto; Rainer Radtke; Peter Schad; Simone Schromm; Glauco Schüssler; Mirco Solé; Enio Sosinski.
As secretárias fantásticas – Silvana, Andreia, Charlotte Tork, Michaela Bücherl – e o equipe famoso do Pró-Mata – Dona Ero, Cristiane, Cleudia, Cleonir, Jonas, Leandro e Ricardo.
Um abraço grande especial pela ajuda enorme no campo para: Jonas Homem; Felipe Lohmann e amigos, Ricardo Mello, Glauco Schüssler; a segurança da UFRGS; Marcos Westphalen; Laura Prates da Silva.
Um abraço grande especial também para sua gentileza e generosidade excepcional a:Sandra Cristina Müller e Fernando; Martin e Tati; Dona Ero com toda família Homem; Nelcy, Jorge e Jorge Alberto.
My sincere apologies to anyone who was unintentionally forgotten in this list.
Working permission in the CPCN Pró-Mata was given by the Instituto do Meio Ambiente, Pontífica Universidade do Rio Grande do Sul; estou muito obrigada á Ricardo Mello, Betina Blochtein e Jorge Villwock.
Working permission on Fazenda “Trés Estrelas” was given by Sr. Escobar, São Francisco de Paula; work was made possible by Sebastião Avila de Lima – estou muito obrigada.
Funding was provided by Deutsche Forschungsgemeinschaft (DFG), within the framework of projects PF 120/10-1 and PF 120/10-2: „Einfluss von Feuer und Beweidung auf das Wald-Grasland-Mosaik im südlichen Brasilien (Rio Grande do Sul)“ [Influence of fire and grazing on the forest-grassland mosaic in southern Brazil (Rio Grande do Sul)].
Dedicated to - dedicado para
Barbara, Konrad & Matthias HermannDona Eronita Homem
Dona Nelcy Martins da Silva

1
Table of contents
1. Introduction.....................................................................................................................7 1.1. Objective and framework of the study ......................................................................7 1.2. Geography of the study regions..............................................................................11 1.3. Origin of the forest-grassland mosaic .....................................................................15 1.4. The main study area and study species in detail....................................................18
1.4.1. Vegetation.......................................................................................................18 1.4.2. Study species .................................................................................................21 1.4.3. Soils ................................................................................................................24 1.4.4. Microclimate....................................................................................................27
1.5. Terminology............................................................................................................29 1.6. Statistics .................................................................................................................31
2. Seedling emergence of Woody Species in Grassland: Comparison of a Grassland
and a Forest Species ...........................................................................................................32 2.1. Summary ................................................................................................................32 2.2. Introduction.............................................................................................................33 2.3. Study setup.............................................................................................................34
2.3.1. Study sites and species ..................................................................................34 2.3.2. Seed collection, treatment and storage ..........................................................35 2.3.3. Experiment setup............................................................................................35 2.3.4. Evaluation .......................................................................................................37
2.4. Results....................................................................................................................39 2.4.1. Overview.........................................................................................................39 2.4.2. Baccharis uncinella.........................................................................................40 2.4.3. Myrsine parvula ..............................................................................................41
2.5. Discussion ..............................................................................................................42 2.5.1. Possible drawbacks of the experiment design................................................42 2.5.2. Some evidence of safe site characteristics.....................................................43 2.5.3. Seed dispersal in space and time...................................................................45 2.5.4. Consequences for forest-grassland dynamics................................................48
2.6. Appendix.................................................................................................................50
3. Survival and Growth of Woody Seedlings in Grassland: Comparison of a
Grassland and a Forest Species.........................................................................................51 3.1. Summary ................................................................................................................51 3.2. Introduction.............................................................................................................52 3.3. Study setup.............................................................................................................53
3.3.1. Study sites and species ..................................................................................53 3.3.2. Experiment preparation ..................................................................................53 3.3.3. Monitoring .......................................................................................................55 3.3.4. Evaluation .......................................................................................................55
3.4. Results....................................................................................................................57 3.4.1. Mortality ..........................................................................................................57 3.4.2. Stem and leaf development ............................................................................60 3.4.3. Seedling performance.....................................................................................68
3.5. Discussion ..............................................................................................................69 3.5.1. Escarpment compared to inland sites.............................................................69 3.5.2. Inherent species traits: Baccharis compared to Myrsine in nurseries.............70 3.5.3. Impact of management: Pasture compared to abandoned grassland ............72

2
3.5.4. Impact of established vegetation: Station nursery compared to tall shrub abandoned grassland.....................................................................................................75 3.5.5. Inhibition versus facilitation.............................................................................76
3.6. Appendix.................................................................................................................79
4. Recruitment of a Pioneer Forest Tree in Grassland: Root suckers vs. Seedlings.80 4.1. Summary ................................................................................................................80 4.2. Introduction.............................................................................................................81 4.3. Study area and species ..........................................................................................82 4.4. Material and methods .............................................................................................82
4.4.1. “New” root sucker and seedling stem allometry and growth rate....................82 4.4.2. Recruitment in naturally established populations............................................84 4.4.3. Short-distance versus long-distance dispersal ...............................................85 4.4.4. Growth rate of naturally established new and elder recruits ...........................86 4.4.5. Growth rate of naturally established ramets in different grassland types .......87
4.5. Results I: “New” root sucker and seedling allometry and growth rate ....................88 4.5.1. Stem allometry................................................................................................88 4.5.2. Growth rate .....................................................................................................89
4.6. Results II: New recruitment in naturally established populations............................90 4.6.1. Recruit numbers and origin in different grassland types.................................90 4.6.2. Preferred recruitment microsites of root suckers and seedlings .....................91 4.6.3. Short-distance versus long-distance dispersal ...............................................91
4.7. Results III: Individual growth rates in naturally established populations.................94 4.7.1. New and elder recruits in tall shrub grassland................................................94 4.7.2. Mixed root sucker-seedling populations in different grassland types..............94
4.8. Discussion ..............................................................................................................96 4.8.1. What are root suckers, and what do they serve for? ......................................96 4.8.2. Differences between root suckers and seedlings ...........................................99 4.8.3. A forest species as grassland coloniser: Root sucker versus seedling recruitment....................................................................................................................101
4.9. Appendix...............................................................................................................106
5. Population Structure and Dynamics of Woody Species in Grassland: Comparison
of a Grassland and a Forest Species ...............................................................................107 5.1. Summary ..............................................................................................................107 5.2. Introduction...........................................................................................................108 5.3. Study setup...........................................................................................................109
5.3.1. Study area and species ................................................................................109 5.3.2. Study setup...................................................................................................109 5.3.3. Evaluation .....................................................................................................110
5.4. Results..................................................................................................................114 5.4.1. Population structure 2005.............................................................................115 5.4.2. Population dynamics 2005-2006 ..................................................................116 5.4.3. Individual growth rate 2005-2006 .................................................................117 5.4.4. Projection of individual and population growth .............................................119 5.4.5. Influence of Baccharis dieback on Myrsine population and individual development .................................................................................................................121
5.5. Discussion ............................................................................................................122 5.5.1. Origin of Baccharis and Myrsine...................................................................122 5.5.2. Population development in pasture ..............................................................125 5.5.3. Population development in abandoned grassland........................................127 5.5.4. Predicting successional sequences..............................................................131

3
5.6. Appendix...............................................................................................................135
6. Post-disturbance Survival and Growth of Woody Species in Grassland: A
Comparison of Grassland and Forest Species ...............................................................143 6.1. Summary ..............................................................................................................143 6.2. Introduction...........................................................................................................144 6.3. Study setup...........................................................................................................145
6.3.1. Study areas and species ..............................................................................145 6.3.2. Sampling design and treatments ..................................................................146 6.3.3. Evaluation .....................................................................................................147
6.4. Results..................................................................................................................149 6.4.1. Overview of results .......................................................................................149 6.4.2. Resprouting success and timing...................................................................150 6.4.3. Resprouting vigour........................................................................................151 6.4.4. Size dependence of resprouting success and vigour ...................................156 6.4.5. Complementary processes of recovery ........................................................157
6.5. Discussion ............................................................................................................160 6.5.1. Dissimilarities between study areas..............................................................160 6.5.2. Grassland shrub vs. forest tree resprouting..................................................165
7. Final discussion .........................................................................................................170 7.1. Seed arrival at the site..........................................................................................170 7.2. A final comment on facilitation and inhibition........................................................171 7.3. Changes in community invasibility with time.........................................................172 7.4. Origin of pioneer forest species in grassland........................................................176 7.5. Outlook .................................................................................................................184
8. References ..................................................................................................................186
Table of figures
Fig. 1.1: Topographic map of Brazil. ........................................................................................7 Fig. 1.2: A stretch of Araucaria forest surrounded by grasslands ............................................8 Fig. 1.3: Topographical units of Rio Grande do Sul. ..............................................................12 Fig. 1.4: View of the lowland study site Morro Santana in Porto Alegre.................................14 Fig. 1.5: Section from a topographic map scale 1:100.000, sheet „Cambará do Sul“. ...........16 Fig. 1.6: View from observation tower, southwest border of the Pró-Mata grasslands ..........17 Fig. 1.7: Grasslands of the fazenda „Trés Estrelas“, cattle-grazed and burnt. .......................19 Fig. 1.8: Tall tussock grassland of the “Pró-Mata” area where cattle and fire were excluded in 1994. ......................................................................................................................................20 Fig. 1.9: Tall shrub grassland of the “Pró-Mata” area.............................................................20 Fig. 1.10: Branch of Baccharis uncinella de Candolle, flowering. ..........................................22 Fig. 1.11: Myrsine parvula (Mez) Otegui. Left: From Otegui (1998), slightly modified. Right: Specimen growing by the roadside in a Pró-Mata grassland. ................................................23 Fig. 1.12: A highland cambisol on acidic effusive bedrock underneath grassland .................24 Fig. 2.1: Left: Baccharis uncinella seedlings emerging in soil profile. Right: Detail................50 Fig. 3.1: Left: Myrsine seedling transplanted into abandoned grassland. Right: Identification tags.........................................................................................................................................54 Fig. 3.2: Seedlings of grassland shrub Baccharis uncinella and forest tree Myrsine parvula retrieved from various types of grassland after planting-out for 1 year. .................................79 Fig. 4.1: Comparison of stem allometry between new root suckers and seedlings from tall shrub-grassland and open grassland .....................................................................................88

4
Fig. 4.2: Adolescent Myrsine parvula excavated in abandoned grassland ............................92 Fig. 4.3: More or less even-aged root suckers compared side by side with seedlings harvested in a seedling transplant experiment.....................................................................106 Fig. 5.1: Below: Relationship between gap size and the relative contribution of various types of vegetative and generative growth from Bazzaz (1984). Above: Suggested correlation of Myrsine parvula life history traits with disturbance intensity in her original forest habitat. 124 Fig. 5.2: Only a question of time: Dieback of canopy-forming Baccharis in tall shrub abandoned grassland; photograph taken June 2006. ..........................................................133 Fig. 5.3: Above: Symptoms of severe infection on a branch of Baccharis uncinella in a declining tall shrub grassland stand. Below: Results of phytopathological analysis of an individual of Baccharis trimera. ............................................................................................142 Fig. 6.1: Resprouted basal area one year after disturbance as linear function of original basal area ......................................................................................................................................157 Fig. 6.2: Contrasts in resprouted shoot number and allometry: Forest tree species Myrsine parvula (Myrsinaceae), Siphoneugenia reitzii (Myrtaceae) and grassland shrub species Baccharis ochracea (Asteraceae). .......................................................................................167 Fig. 7.1: Principal pathways of grassland-Araucaria forest succession from Klein (1960), slightly modified....................................................................................................................176 Fig. 7.2: Profile of a moderately to lightly disturbed Araucaria forest on typical highland cambisol from Seger et al. (2005), modified.........................................................................179 Fig. 7.3: Profile of a moderately to lightly disturbed Araucaria forest on gleysol from Seger et al. (2005), modified...............................................................................................................180
Index of tables
Tab. 1.1: Excerpts from Jungblut & Pinto (1997a): Soil parameters from three profiles investigated in Pró-Mata within the first year after exclosure from management ..................25 Tab. 1.2: Excerpt from Holl (1999): Comparison of soil parametres in recently abandoned pasture and adjacent forest at a study site in southern Costa Rica .......................................26 Tab. 1.3: Comparison of microclimate between different types of grassland and nurseries in the “Pró-Mata” area.. ..............................................................................................................28 Tab. 1.4: Excerpt from Holl (1999): Comparison of microclimate in cleared grassland areas, recently abandoned pasture and adjacent forest at a study site in southern Costa Rica.......29 Tab. 2.1: Seedling emergence statistics of grassland shrub species Baccharis uncinella and forest tree species Myrsine parvula........................................................................................39 Tab. 2.2: SE% calculated as median and average +/- 1SD of replicate plots/containers ......40 Tab. 3.1: Seasonal variation in mortality of Baccharis uncinella (BU) grassland shrub seedlings and of Myrsine parvula (MP) forest tree seedlings and principal death causes and syndromes in pasture, abandoned grassland and nurseries..................................................57 Tab. 3.2: Overview of per cent survival and size development of seedlings of grassland shrub Baccharis uncinella and forest tree Myrsine parvula transplanted into nursery, pasture and abandoned grassland plots and monitored for 1 year. ...........................................................61 Tab. 3.3: Significance of differences in stem and leaf growth a) between pairs of grassland and nursery types per species; b) between species per grassland and nursery type............62 Tab. 3.4: Overview of final sizes of seedlings of grassland shrub Baccharis uncinella and forest tree Myrsine parvula transplanted into nursery and grassland plots and monitored for 1 year. .......................................................................................................................................63 Tab. 3.5: Significance of differences in final stem and leaf size a) between pairs of grassland and nursery types per species; b) between species per grassland and nursery type............63

5
Tab. 3.6: Overview of stem and leaf allometry of seedlings of grassland shrub Baccharis uncinella and forest tree Myrsine parvula transplanted into nursery and grassland plots and monitored for 1 year. ..............................................................................................................66 Tab. 3.7: Significance of differences in final stem and leaf size a) between pairs of grassland and nursery types per species; b) between species per grassland and nursery type (T-Test for independent samples; bottom table). ................................................................................67 Tab. 3.8: Seedling performance indices calculated from percentage data on seedling height and basal area growth, survival and emergence....................................................................68 Tab. 4.1: Size-age classes and some of their characteristics relevant to population dynamics...............................................................................................................................................86 Tab. 4.2: Comparison of absolute height and per cent height increase after one year in the field of new root suckers and seedlings in different types of grassland. ...............................89 Tab. 4.3: Average proportions (individual no.s/100m²) of Myrsine root sucker and seedling recruitment in naturally established populations in different types of abandoned grassland .90 Tab. 4.4: Numbers of new root sucker recruits in grassland plots 2006 related to numbers of potential feeder individuals in the same plots with and without potential feeders in adjacent forest border plots. .................................................................................................................91 Tab. 4.5: Spatial dispersal potential of populations of generatively reproducing species vs. vegetatively spreading (i.e. clonal) species: Annual rates of spread and pollen travel distances set in relation to maximum height individuals may achieve. ..................................93 Tab. 4.6: Comparison of absolute height and per cent height increase after one year between Myrsine root suckers and seedlings of different age-size classes, naturally established in tall shrub abandoned grassland...................................................................................................94 Tab. 4.7: Comparison of absolute height and per cent height increase after one year between Myrsine juveniles of different size-age classes, naturally established in tall tussock and tall shrub abandoned grassland...................................................................................................95 Tab. 4.8: What height can be expected in new root suckers and seedlings of a certain basal diameter?. ............................................................................................................................106 Tab. 5.1: Size-age classes and some of their characteristics relevant to population dynamics.. ............................................................................................................................111 Tab. 5.2: Size-age structure, spatial structure and (Myrsine only) individual morphology of naturally established populations in different types of grassland. Top: Myrsine population structure. Bottom: Baccharis population structure................................................................114 Tab. 5.3: 1-year natality vs. mortality in mixed stands of Myrsine parvula and Baccharis uncinella. ..............................................................................................................................116 Tab. 5.4: Percent of surviving individuals of Myrsine and Baccharis in naturally established populations in different types of grassland that increased in size or suffered a decrease in size during the one-year observation period. .......................................................................118 Tab. 5.5: Parameters of growth of intact individuals of Myrsine and Baccharis during the one-year observation period (abandoned grassland only) ..........................................................118 Tab. 5.6: Influence of Baccharis tall shrub canopy density on individual density and growth rates of Myrsine juveniles.....................................................................................................121 Tab. 5.7: Myrsine parvula (Mez) Otegui population size-age structure, spatial structure and individual morphology: Variation among plots......................................................................135 Tab. 5.8: Baccharis uncinella A.P. de Candolle live population size-age structure, spatial structure and individual morphology: Variation among plots................................................136 Tab. 5.9: Baccharis uncinella A.P. de Candolle size-age structure of dead: Variation among plots......................................................................................................................................137 Tab. 5.10: Myrsine parvula (Mez) Otegui and Baccharis uncinella A.P. de Candolle 1 year population dynamics. Variation among plots........................................................................138

6
Tab. 5.11: Myrsine parvula (Mez) Otegui and Baccharis uncinella A.P. de Candolle: Projection of individual growth in different types of grassland..............................................139 Tab. 6.1: Comparison of resprouting success, timing and vigour between grassland low shrubs and forest trees from highland sites (cut) and lowland sites (burnt). ........................149 Tab. 6.2 (Tab. 6.1 cont.): Further characteristics of resprouting in grassland low shrubs and forest trees from highland sites (cut) and lowland sites (burnt)............................................150 Tab. 6.3: Top: Comparison of resprouted shoot numbers height relative to pre-disturbance shoot numbers between pooled datasets of highland and lowland grassland shrubs and forest trees. ..........................................................................................................................151 Tab. 6.4: Top: Comparison of resprouted maximum shoot height relative to pre-disturbance maximum shoot height between pooled datasets of highland and lowland grassland shrubs and forest trees. ...................................................................................................................152 Tab. 6.5: Comparisons of resprouted shoot height growth rate between pairs of highland and lowland grassland shrub and forest tree species during three seasons following disturbance.............................................................................................................................................153 Tab. 6.6: Top: Comparison of resprouted individual basal area (=sum of shoot basal area) relative to pre-disturbance individual basal area between pooled datasets of highland and lowland grassland shrubs and forest trees...........................................................................154 Tab. 6.7: Comparison of resprouted individual basal area (=sum of shoot basal area) relative to pre-disturbance individual basal area between pooled datasets of grassland shrubs and forest trees. ..........................................................................................................................155 Tab. 6.8: Comparison of absolute resprouted shoot numbers, maximum shoot basal diameter and shoot allometry (ratio stem basal diameter:height) between species............................155 Tab. 6.9: Complementary to Fig. 4: Values of relative basal area to be expected in grassland shrubs and forest trees one year after disturbance..............................................................157 Tab. 7.1: Phytosociological studies reviewed for statistics in Tab. 7.2.................................177 Tab. 7.2: Ten pioneer tree species in forest-grassland succession and their roles/positions in moderately to lightly disturbed Araucaria forest ...................................................................178

Hermann, J.-M. PhD thesis 1: Introduction
7
1. Introduction
1.1. Objective and framework of the study „Suppose that you had to design a plant to do well in a particular environment. Where would you start? ...Under conditions of moderate temperatures, on well-irrigated, neutral, non-toxic soils, away from severe exposure, the other plant species would almost certainly be trees and, f you want your plant to do well, then it had better be a tree as well. But what kind of tree?...How long will it live, and how long will it wait before flowering for the first time? How tall will it grow and how much will it invest in shoot versus root, and in support structures like trunk and branches versus productive tissues like leaves and flowers? What kind of seeds will it produce, and how are they to be dispersed? Will they germinate at once, or will they exhibit some form of protracted dormancy? These are questions of plant life history, and each of the 250 000 living vascular plant species possesses a more or less unique combination of traits such as these, reflecting both their phylogeny and their recent ecological circumstances.“ M.J. Crawley (1997a, p.73)
Rio Grande do Sul and its two neighbour federal states, Santa Catarína and Paraná, share a fascinating landscape: Hills and plains are covered by a mosaic of grasslands and forests. The native grasslands merge with the Pampa of Argentina and Uruguay in the south and may be broadly viewed as an extension of this biome (Walter 1984; Leite & Klein 1990). Native forests – subtropical in the lowland, humid temperate on the highland – gradually spread over these grasslands (e.g. Behling 2004, 2007), although at a slower rate than large-scale humid climate would suggest (Pillar & Quadros 1997).
This phenomenon is currently being investigated by a German-Brazilian research team coordinated by professors Dr. Jörg Pfadenhauer, Technische Universität München, and Dr. Valério de Patta Pillar, Federal University of Rio Grande do Sul. However, the scientific debate among European and Brazilian scientists on the origin and maintenance of the South Brazilian grasslands may be traced back as far as the onset of the 20th century.
Fig. 1.1: Topographic map of Brazil (Felipe 2006, slightly modified). The black square indicates my
lowland study area, located in Porto Alegre, the capital of the federal state of Rio Grande do Sul; the white
square indicates the main study area Pró-Mata on the High Plateau of Rio Grande do Sul.

Hermann, J.-M. PhD thesis 1: Introduction
8
Fig. 1.2: A stretch of Araucaria forest surrounded by grasslands. Highland of Rio Grande do Sul,
November 2003. Photographer: J.-M. Hermann.
Earlier scientists broadly divide into three schools, each seeking to explain grassland resistance to forest expansion by climate, soil or fire (Alvim 1954, excerpts reprinted in Leite & Klein 1990; see 1.3). Nowadays, the leading dogma is this: If forest establishment is not limited by precipitation (and this is not the case, at least not in the highland region), then fire (and grazing) limit forest establishment (Pillar & Quadros 1997).
This hypothesis is strongly corroborated by the following phenomena: Grasslands have traditionally been used as pasture and burnt at 1-2 year intervals, and the transition between pasture and forest is frequently as sharp as if cut with a razor blade (see Fig. 1.2). Pasture abandonment, on the other hand, results in a pronounced increase in biomass and abundance of both grasses and woody species – small grassland shrubs, tall grassland shrubs and forest species (e.g. Oliveira & Pillar 2004, Waechter at al. 1984). The influence of fire is considered so predominant that characteristics of colonising species such as life history traits, population dynamics and performance of juveniles and adults in different grassland environments have only recently received increased attention.
Such attention is warranted by the following observations: Firstly, shrubs and trees, do establish in grazed and burnt grassland. Some shrubs must be considered an integral part of the native grasslands but others are genuine forest species. As Pillar & Quadros (1997) point out, fires may be ignited by lightning – even without human interference – in the Araucaria-broadleaf forests of the highland region. However, fire frequencies naturally differ between forest and grassland by at least one order of magnitude (see e.g. Peet 1992). So how do forest species cope with the grassland habitat?
Secondly, what happens when fire and grazing are excluded? It is tempting, at first sight of a large area of abandoned pasture, to assume a simple chronological sequence of successional stages dominated by small grassland shrubs, tall grassland shrubs and forest species. A closer look, however, reveals that pioneer forest species establish within various types of managed and abandoned grassland matrix, although to different extent. Klein (1960) reports a broad set of pioneer communities in grassland-forest-succession, characterised by at minimum a dozen forest tree species. Oliveira & Pillar (2004) approach the question of successional sequences with great caution, outlining several pathways of succession evident from evaluation of aerial photographs of the highland study area Pró-Mata, although they do not doubt that all grasslands will eventually develop into forest.

Hermann, J.-M. PhD thesis 1: Introduction
9
This leads to several major questions: Which traits enable pioneer woody species – of both grassland and of forest origin – to establish successfully in a grassland matrix? Which characteristics of grassland itself – or of abiotic and biotic conditions associated with grassland predominance, including anthropogenic impacts such as fire and grazing – may prevent establishment of either type of woody species or, in other words, account for the slow speed of forest expansion?
The objective of my study is to give first answers to these questions by investigating the performance of juvenile to adult life stages of selected pioneer woody species in selected types of grassland under both experimentally manipulated conditions (sowing, transplant, and cutting experiment) as well as under non-manipulated conditions (monitoring structure and dynamics of naturally established populations). These investigations provide the following basic data:
a) Quantitative data on colonisation success and successional pathways. Although previous studies (Oliveira 2003, Machado 2004) have documented a general increase in woody species cover and abundance, have demonstrated the existence of multiple successional pathways and highlighted a number of species as pioneers in grassland, this study is one of the first to quantitatively describe recruitment, mortality, population size-age structure, individual density and individual growth rate of pioneer woody species by monitoring tagged individuals in permanently marked plots in naturally established populations in grassland.
b) Traits that are presumably linked to a species’ ability to colonise grassland, and may help to explain the observed patterns of succession in space and time. Many of these characteristics - such as rapidity of germination/seedling emergence, longevity of seeds in soil, stem and leaf growth rates, regenerative ability and the ability to spread clonally - may be summed up under the term “life-history traits” (Crawley 1997a). In recent decades, such traits have been increasingly considered an important factor driving succession, even more so where communities are dominated by relatively few species (e.g. Peet & Christensen 1980). Peet (1992) traces a general trend towards mechanistic and reductionist approaches – „a process is broken into its component parts so as to determine how the workings of the larger system are consequences of lower-order processes“ (p.103-104) – in successional theory from the early 1970’s to the late 1980’s. He writes: „Collectively, these papers suggest that successional change at the community or ecosystem level can be understood, at least in part, as a consequence of population processes of the component species, which in turn might be understood as consequences of species attributes, such as life history and physiological characteristics“ (p.104). Pickett (1982) concludes, from a long-term study of oldfield succession in New Jersey, U.S.A., that „in those instances where the ecophysiology, life history characteristics, and population properties are known, they do indeed correlate with the population pattern in oldfield succession. However, very few species have been studied from this perspective [...]“ (p.55).
In order to explain observed patterns of seedling performance in different grassland types (chapter 3) and in contrast to root sucker performance (chapter 4), I also evaluate traits such as stem and leaf allometry that are more appropriately termed morphological traits, although the ability to modify these parameters can also be included among life-history traits in a broad sense.
The major part of the work is concerned with species performance at the juvenile stage, presumably the most vulnerable stage: High early mortality is a characteristic of relatively long-lived species, particularly trees and shrubs (Fenner 1987b). It is therefore assumed that the quantification of seed and juvenile performance of principal pioneers in different grassland types, and the detection of causes for possibly differential performance, are of major importance for the explanation of patterns and processes in grassland-forest-succession.

Hermann, J.-M. PhD thesis 1: Introduction
10
Furthermore, focus is on the following: The principal study area is located in the highland of Rio Grande do Sul. Within this
study area, focus is on the contrast between pasture – i.e. native grassland subjected to traditional management methods – and abandoned grassland, i.e. former pasture excluded from fire and cattle-grazing for ca. one decade. Within abandoned grassland, two contrasting structural types are compared: Tall tussock grassland, which accounts for the larger proportion of the area (Oliveira & Pillar 2004; pers. obs.), and tall shrub grassland, which due to its tall shrub canopy corresponds most closely to the notion of an initial stage of forest (“Vorwald”). Furthermore, focus is on two study species that establish in particular abundance in both pasture and abandoned grassland of the region: Forest tree Myrsine parvula (Mez) Otegui and grassland tall shrub Baccharis uncinella de Candolle, principal canopy former in tall shrub grassland.
This is the “red thread” that may be followed through chapters 2-5, in which study setups diverge only in some detail. The part study in chapter 6 is set up on a broader scale: Two species groups, grassland shrubs and forest trees, are studied as to their reaction to aboveground biomass loss, integrating data from two study areas in the lowland and highland of Rio Grande do Sul1.
At the end of each chapter, I discuss how life-history and morphological traits of my selected species contribute to the observed patterns and processes in forest-grassland-succession. Furthermore, I discuss what they reveal about environmental conditions in the original habitat of these species, i.e. their habitat prior to joining in forest expansion over grassland on the Southern Brazilian highland. I have included this topic because I believe it is critical to understanding the process of grassland-forest succession, although there is some danger of introducing circularity here; such would be the case if I concluded from a given species’ behaviour observed in grassland on the conditions under which this behaviour evolved in the species’ original habitat, and finally discussed, on the basis of presumed original habitat conditions, why this species is successful at colonising grassland or not. It is clearly necessary to perform actual observations of the species in her original habitat before that final step, and I hope that my conjectures are an incentive for doing so. – I further discuss – in a broader regional context – to what extent both species traits and the resulting interactions with established grassland vegetation account for the slow speed of forest expansion.
Ideally, any study should begin with non-experimental observations in space and time. Based on these results and through inductive reasoning – i.e. comparison with available data from similar systems - , the scientist forms first models, then hypotheses that may be tested experimentally. On rejection of the tested hypotheses, the process is repeated (see e.g. Gibson 2002). The antagonism between grass-dominated and shrub- or tree-dominated vegetation has received considerable attention worldwide and a large body of literature deals with mechanisms by which either grasses or woody species achieve predominance (e.g. reviews by Belsky 1991, Scholes & Walker 1993, Wilson 1998). On this basis, a number of detailed hypotheses might by formed a priori and experiments designed to test them. However, for a number of reasons – e.g. the frequent lack of basic data on species and populations of the study area –, instead of implementing few elaborate experiments, I opted for exploratory and standard statistical analyses of a larger number of 1 The different part studies presented in chapters 2-6 were not initiated synchronously and logistic constraints only gradually became apparent; resultant overthrows of experiment designs account in part for the incongruities in study setups. The regeneration study was initiated first, on a relatively broad scope. Poor seed set in Myrtaceae prevented, among other factors, usage of the same set of species in sowing experiments initiated later. Sowing experiments contained mowed plots, intended to simulate a post-fire environment. In transplant experiments initiated following sowing experiments, this treatment was omitted due to low number of available seedlings.

Hermann, J.-M. PhD thesis 1: Introduction
11
relatively simple experiments and monitoring studies. The questions or objectives underlying these setups are given at the beginning of each chapter.
This strategy has proved relatively safe insofar as most part studies could be completed successfully, but undoubtedly safety here trades off against accuracy. The interpretability and value of results depend entirely on careful sampling at sufficiently short intervals, a large sample size and on the availability of studies conducted in similar context from which plausible explanations for observed patterns can be obtained. Even if all of these requirements are fulfilled, proximate causes for the observed phenomena remain unknown. No matter how plausible the explanations given, in a strict sense they remain hypothetical until proven or rejected by more elaborate experimental setups. Nevertheless, some results obtained during this study support general observations made worldwide, or add a new perspective to topics of more global interest. For example,
1. the discrepancy between structure and density of naturally established juvenile populations, and performance of seeds and seedlings under experimental conditions especially in the two structural types of abandoned grassland, may be regarded as a “test” for the Connell & Slatyer (1977) models;
2. the contrasts observed in modes of origin of Myrsine parvula juveniles in grassland illustrate differences between root suckers and seedlings in general, and link to the question of costs and benefits of clonal growth in woody species, particularly when a new habitat is colonised.
1.2. Geography of the study regions This paragraph is a brief introduction to the geology, soils, climate and vegetation of
the lowland and highland areas of Rio Grande do Sul where parts of this study were conducted. The principal study area – encompassing the research centre “Pró-Mata” and the adjacent fazenda “Trés Estrelas” - is located at 29°48’ S, 50°21’W at 900-920 m a.s.l. in the municipality of São Francisco de Paula, and is described in more detail in the following paragraph. The other study area, “Morro Santana” – “Santana hill”, is located at 30°03’ S, 51°07’ W, 311 m a.s.l. in a suburban area of the state capital Porto Alegre.
Porto Alegre is considered part of the topographical unit (equivalent to the German term „Landschaftsraum“) „Southriograndean High Plateau“ – „Planalto Sul-Rio-Grandense“ (Herrmann & Rosa 1990). Large parts of this rapidly expanding city are located at sea level, along the shore of Guaíba lagoon, but the municipality also encompasses 20 hills - „morros“ – that are outcrops of acidic Precambrian bedrock (Kaul 1990).
The research station Pró-Mata and the nearest town, São Francisco de Paula, are part of the topographical unit „Araucaria High Plateau“ – „Planalto das Araucárias“ (Herrmann & Rosa 1990) – underlain by acidic effusive bedrock of Tertiary origin (Kaul 1990) and named after Araucaria angustifolia, the characteristic forest tree species in this region. The Araucaria High Plateau rises to elevations of ca. 1300 m a.s.l. towards the northeast border of the state, and to 1600-1800 m a.s.l. in the neighbour states Santa Catarina and Paraná (ibid.).

Hermann, J.-M. PhD thesis 1: Introduction
12
Fig. 1.3: Topographical units of Rio Grande do Sul, from Herrmann & Rosa (1990, slightly modified). Dark
green: „Araucaria High Plateau“ („Planalto das Araucárias“), encompassing the highland study area.
Red: „Southriograndean High Plateau“ („Planalto Sul-Rio-Grandense“), encompassing the lowland study
area.
In the following, these areas are referred to as „lowland“ and „highland“ (or Porto Alegre and Pró-Mata), respectively. The transition between highland and lowland is gradual at the southwest border of the high plateau but very abrupt towards the litoral region in the east; this steep transitional region is referred to as the „escarpment“ (see also Fig. 1.5).
The climate is categorised as subtropical humid in the lowland and humid temperate in the highland area (Cfa and Cfb, respectively, according to the Koeppen classification). Both areas receive their maximum of precipitation towards the middle and end of the southern winter, i.e. from July to August; there is no dry season (Nimer 1990). Annual precipitation is ca. 1.5 times higher, annual average temperature ca. four degrees lower in Pró-Mata than in Porto Alegre (ca. 2000mm and 1400mm, and 15°C and 19°C, respectively; ibid.).
Nocturnal frosts with temperatures <0°C occur on average 10-15 times per year on Pró-Mata, 0-1 times per year in Porto Alegre (Nimer 1990)2. According to data compiled by Hennerich et al. (2005), temperatures <2°C occur on 10 days per year and the period of possible frost occurrence lasts 75 days in the highland; frosts thus occur ca. twice as often and the period of possible frost occurrence lasts ca. twice as long in the highland as in the lowland area (highland data from Passo Fundo, RS; lowland data from Pelotas, RS). Frosts in the highland are usually registered first in the beginning of June, last at the end of August (ibid.); according to the authors, there have been no significant changes in this frost regime between 1931 and 2000 (ibid.), although their data suggest a trend towards onset of the frost period later in the year, and consequently a decrease in length of the frost period. The period between approximately the middle of March and the middle of May is considered climatic autumn; climatic winter lasts until the end of October and the month of November is considered climatic spring (DCRS 2004).
2 Brazilian climatologists distinguish between „black frost“ – „geada preta“, referring to leaf necroses appearing at temperatures <0°C – and „white frost“ – „geada branca“, equivalent to „hoarfrost“ in English and „Rauhreif“ in German. „Cold nights“ – „noites frías“ – are nights with temperatures <0°C and occurrence of „black frost“. „White frost“ may occur even at temperatures a few degrees above zero. Nimer (1990) provides separate data on frequency of black and white frost, Hennerich et al. (2005) sum up both types of events per year.
Research Center „Pró-Mata“ and Fazenda „Trés Estrelas“
Morro Santana, Porto Alegre

Hermann, J.-M. PhD thesis 1: Introduction
13
In the year 2005, in which the majority of experiments and the population monitoring presented in this thesis were initiated, climate diverged from long-year averages twice: The state suffered an unusual late summer drought until the middle of March that ended with torrential rainfalls on March 13th, 2005. Precipitation on that day exceeded total precipitation of the preceding month (Journal „Zero Hora“, March 14th 2005). This dry period may have influenced resprout experiments on Morro Santana and in Pró-Mata, and Baccharis uncinella seedling emergence experimentally tested in Pró-Mata. Occurrence of the first frost night (<0°C) in the Pró-Mata region on May 26th, 2005, was not too unusual in comparison with long-term data of Hennerich et al. (2005), but the frost events on November 1st and 8th, 2005, were unusually late so that damage to transplanted seedlings in Pró-Mata and the adjacent fazenda were probably greater than they would have been in an average year.
The soil ontogeny in Southern Brazil has generally been influenced by long, undisturbed development, relatively base-poor bedrock and subtropical to tropical climate (Watzlawick et al. undated). Nevertheless, soils differ considerably in texture and colour between the highland and lowland study sites: Soils on Morro Santana are coarse-grained, litholic, reddish-yellow soils (categorised as „podzolic“, Moser 1990), whereas soils under grasslands of the Pró-Mata region are fine-grained, rich in organic matter and characterised by a blackish-brown A-horizon. Brazilian soil scientists usually categorise the latter as „humic cambisols“ (e.g. Moser 1990, Jungblut & Pinto 1997a). Recently, however, Dümig et al. 2008b concluded - based on analyses of soil chemistry and physical properties such as Al and Fe contents, phosphate retention and bulk density -, that a considerable proportion of soils underneath grassland in the study area Pró-Mata are more correctly termed Andisols, whereas they classify the remainder of grassland soils as Umbrisols according to the classification system of the World Reference Base for Soil Resources 2006; for detail, please see Dümig et al. 2008b.
Both types of soil are highly acidic and - as a consequence of low pH (see e.g. Scheffer & Schachtschabel 2002) – have excess contents of Al3+ (<50% in the lowland, >50% in the highland; Moser 1990), low base saturation and fertility (ibid.). They are easily eroded on removal of the vegetation cover (lowland: Moser 1990; highland: pers. obs.). Slight water deficiency is possible in the lowland region but not considered relevant in the highland region (Moser 1990). The reddish colour of lowland soil is due to a Fe2O3 –content of 5-10% (ibid.).
Both areas were, prior to the arrival of European settlers, covered by a mosaic of native grasslands and forests. Grasslands owe their outstanding species diversity - almost 800 grass and 200 legume species - to the coexistence of summer C4 and winter C3 species; seasonal climatic variation determines shifts in relative predominance of C3 and C4 species (Nabinger et al. 2000). A common distinction – based on agronomic considerations – is between „pure“ and „dirty“ grasslands: The first are dominated by tussocky and low-growing hemicrytophytes, interspersed with rhizomatous geophytes and chamaephytes; the latter contain larger percentages of chamaephytes (Leite & Klein 1990). The predominant native grass genera are of the C4-type: Aristida, Elyonurus, Erianthus, Panicum, Paspalum, Schizachyrum, Trachypogon and Andropogon, with Andropogon lateralis („capim-caninha“) accouting for 50-60% of cover in highland grasslands (ibid.). The predominant shrub and forb genera are Baccharis ssp. (Rambo 1952 names 90 species for the south of Brazil) and Vernonia ssp. of the family Asteraceae, and Eryngium ssp. of the family Apiaceae (Leite & Klein 1990); these are of the C3-type (A. Dümig, pers. comm.).
Forests of the lowland study area contain elements of the Atlantic rainforest („Floresta Ombrofila Densa“) and the Seasonal Semideciduous Forest („Floresta Estacional Semidecidual“) (Leite & Klein 1990). The Atlantic rainforest is marked by an outstanding biodiversity – approximately 700 tree species, half of which are known to occur only in this forest type (ibid.); originally, this forest type extended all along the Atlantic coast from the

Hermann, J.-M. PhD thesis 1: Introduction
14
Amazon region to southern Brazil; only fragments remain today (Watzlawick et al. undated; Leite & Klein 1990). Partial deciduousness of the Seasonal Forest is related to a period of four months with average temperatures below 15°C (Teixeira & Coura Neto 1986 in Leite & Klein 1990).
At elevations above 600 m a.s.l. the Atlantic rainforest merges into Mixed Araucaria broadleaf forest („Floresta Ombrofila Mista“). This forest type extends up to approximately 1200 m a.s.l. (above this level, grasslands predominate; Watzlawick et al. undated) and is characterised by a mixture of angiosperm and gymnosperm species as well as a mixture of elements of austral-antarctic and american-andean floras (Leite & Klein 1990). Rambo (1956) collected 239 species in an Araucaria forest fragment on the highland of Rio Grande do Sul. In more or less undisturbed forests, Araucaria angustifolia is emergent from, or dominant in the canopy while evergreen trees and shrubs dominate the lower tree strata, among them many species of the families Lauraceae and Myrtaceae (Walter 1984; Leite & Klein 1990). Not least because of the economic value of Araucaria, these forests, too, were severely exploited: Leite & Klein (1990) estimate that the mixed Araucaria-broadleaf forest was reduced to ca. 10% of its original extension; even this figure may be too high (see Watzlawick et al. undated and references therein).
In the highland study area, grasslands of the fazenda „Trés Estrelas“ are cattle-grazed – intensity is approximately 0.5 head of cattle per hectare - and used to be burned on an annual or biennial basis until prohibition of pasture burning (S. Avila de Lima, pers. comm.). Fires were ignited especially towards the end of winter when much standing dead grass biomass had accumulated (see also next paragraph). Grasslands of the Research Center „Pró-Mata“ used to be managed in the same way until closure of the area in 1994; since then, the major part of the grasslands have undergone undisturbed development.
Fig. 1.4: View of the lowland study site (chapter 6) on Morro Santana in the state capital, Porto Alegre, in
January (midsummer) 2005. Relatively high fire frequency at this site is indicated by the standing dead
tree in the background. Small tree foreground left: Myrsine umbellata Mart.; multi-stemmed individual
foreground center: Myrciaria cuspidata O.Berg, which occurs as a tree in adjacent forests. Photographer:
J.-M. Hermann.
Grasslands on Morro Santana in the lowland study area have not been used as pasture in recent decades, and consequently are not burnt for management. However, fires recurrently spread from the surrounding suburbs. Estimation of return intervals is not easy as fires vary greatly in intensity and extension and illegal access of the area has increased during the past years (the area belongs to the State University UFRGS and access was

Hermann, J.-M. PhD thesis 1: Introduction
15
originally intended to be restricted to students and scientists). Müller et al. (2007) give a figure of three to five year intervals but it is safe to assume shorter intervals - of two to three years - locally (pers. obs.; G. Overbeck, pers. comm).
1.3. Origin of the forest-grassland mosaic Rambo (1953, 1958) concluded, based on floristic composition and prevalence of
species with xerophytic character, that the southern Brazilian grasslands are relicts from a cooler and drier postglacial period and would be replaced by forest under present-day climate in absence of human interference. Recently, palynological studies have confirmed that these grasslands had their major extension during the last glacial and post-glacial period, „reflecting a cold and dry climate” (Behling 1998, p.153). Herbivores played only a minor role in their maintenance through most of this period: Large savannah vertebrates of the tribes Equidae and Proboscidae, immigrant from North America, were extinct by the end of the Pleistocene (Webb 1978) so that until the arrival of European settlers, grasslands were not subject to grazing by ungulates.
This is a feature that South Brazilian grasslands share with the intermountain prairie of North America: Enclosed between the Coastal Mountain Range and the Sierra Nevada to the west and the Rocky Mountains to the east, these grasslands are characterised by an arid climate and cold winters even today and were largely avoided by ungulate grazers (bison) in pre-colonisation times (Mack & Thompson 1982)3. The climate of Southern Brazil, however, gradually turned more humid (e.g. Behling 1998, 2001) – and still grasslands resisted forest invasion.
How slowly forest has expanded in the study area can be illustrated by a simple exercise: Fig. 1.5 shows a section from a topographical map scale 1:100.000 which encompasses the main study area „Pró-Mata“ and the adjacent fazenda „Trés Estrelas“ (sheet „Cambará do Sul“). This map is based on aerial photographs from 1965 and gives a fairly good impression of the natural distribution of forest, as large-scale clearcutting did not occur in this relatively remote region and silviculture of non-native species had not yet been extensively practiced (T. de Couto, pers. comm.). (Note, however, that the lower regions of the escarpment, formerly covered in Atlantic rainforest, were deforested by earlier settlers.)
Using this map and a standard wheel map measurer, I determined the total length of forest-grassland boundary running within three kilometres distance from high plateau escarpment versus total length of forest-grassland boundary located further away from the escarpment. A trench of very ragged topography of ca. 40km length in the surroundings of Itaimbezinho canyon was omitted; in another area near the National Forest of São Francisco de Paula, forest on fairly level terrain was so fragmented as to suggest influence of logging, and was also omitted. Of the remainder of more or less continuous forest-grassland-border (ca. 170 km), ca. 80% run within three kilometres from the high plateau escarpment and only 20% have expanded further inland.
3 Originally, the North American intermountain grasslands were characterised by tussock grasses growing within a matrix of mosses, lichens and liverworts, contrary to this, grasslands of the Great Plains and the Great Lakes region east of the Rocky mountains (under a more humid climate and grazed by Bison) featured a mixture of native prostrate and tussock grasses (Mack & Thompson 1982). On arrival of European settlers and introduction of domestic livestock, native and non-native rhizomatous species spread in the intermountain grasslands, the latter more aggressively (ibid.). Such processes have also taken place in Southern Brazil; all managed grasslands are characterised by prostrate and tussock grasses and non-native grasses (e.g. Eragrostis plana – „capim-annoni“) spread aggressively in the occasionally drier subtropical lowland areas of Rio Grande do Sul (A. Jacques, pers. comm.). Similarities between North American prairie and Southern Brazilian grasslands are explored in some more detail in chapter 6.

Hermann, J.-M. PhD thesis 1: Introduction
16
Fig. 1.5: Section from a topographic map scale 1:100.000, sheet „Cambará do Sul“ (DSG 1981; kindly
forwarded by Ministério do Exército, Departamento de Engenharía e Comunicações, Porto Alegre, in
2006). Grasslands of the highland study area Pró-Mata and part of the adjacent fazenda „Trés Estrelas“
are enclosed in the white rectangle. Map quadrats are 2x2 km, contour lines are at 40 m equidistance; the
sharp decline of the escarpment and the belt of forest along the High Plateau Border are well
recognisable.
Behling et al. (2004) date the beginning of Araucaria gallery forest migration along streams in Rio Grande do Sul at around 4300 yr BP, and the expansion of Araucaria forest over highland grasslands at 1100 years BP. This mirrors results from the neighbour states Santa Catarina and Paraná where Behling (1998) dated increased expansion of Araucaria forest at ca. 1000 years BP and 1500 years BP, respectively. Behling (2007) gives a figure of 2000 years. Assuming a period of 1000 years, it must be concluded that forest in the major part of my study region has advanced no more than on average three metres per year onto the High Plateau. If a period of 2000 years is assumed, the rate of expansion approaches 1m/year. This matches short-term observations in the main study area Pró-Mata, where forest species expansion into grassland during ca. one decade of abandonment remained largely confined to a 10 m wide belt along forest borders (see 1.3 and Fig. 1.6; Oliveira 2003).
With all caution that such a crude evaluation warrants, the rate of forest expansion must be considered slow indeed. In the forest-tundra transition zone of Alaska, during the past 50 years, forest has expanded on average 2.55 km over tundra in lowlands and 0.1 km at the treeline (Chapin et al. 2005). Davis (1987) reports that during the Postglacial, in Michigan, U.S.A., pioneer species such as birch (Betula spec.) spread into deglaciated regions at average rates of 100-400m/year and even late-successional species – shade-tolerant, long-lived and slow-growing such as beech (Fagus spec.) – advanced into pioneer forest at rates of up to 200m/year. Similar data are reported from Europe (see chapter 4).

Hermann, J.-M. PhD thesis 1: Introduction
17
Fig. 1.6: View from an observation tower at the southwest border of the Pró-Mata grasslands, where the
transition from escarpment to High Plateau coincides with the transition from forest to grasslands. Half-
spherical shrubs with grey-green foliage are individuals of approximately 3m height and 10-12 years of
age of tall grassland shrub Baccharis uncinella de Candolle. Photographer: J.-M. Hermann.
Prior to the establishment of the Pró-Mata research centre, there was some debate on whether grasslands in the region might be man-made, i.e. created by logging or slash-and-burn culture of native forests. However, this possibility can now be largely ruled out, as δ13C-analyses (Dümig et al. 2008a) and palynological analyses (M. Ebner, pers.comm.) of grassland soil confirm the existence of grasslands long before the advent of European settlers in this particular region; according to Dümig et al. 2008a, they date back to the early and mid Holocene period (6000–8000 yr BP) at minimum. Native people inhabited Rio Grande do Sul earlier but did not fell trees extensively, and agriculture was confined to naturally established forest clearances (Koch & Corrêa 2002). Earlier in the 20th century, Hueck (1952) also expressed some doubts as to this theory, as to his knowledge neither forest soil horizons nor charcoal had been found underneath grasslands.
It is understandable that human influence should be suspected, bearing in view the profound changes in the Riograndean landscape and land use brought about by European settlers within only a few centuries. Forests were severely exploited, as mentioned in the previous paragraph. Vast areas of native grasslands encountered by the first settlers provided the basis for extensive cattle husbandry: The first 1500 head of cattle were introduced in Rio Grande do Sul by the Jesuit father 1634 and by the end of the 20th century their number was estimated at 13 million (Nabinger et al. 2000). European colonisation and, consequently, livestock breeding spread from the southern lowland up the High Plateau. São Francisco de Paula and Canela, the two highland towns located at closest aerial distance to the Pró-Mata study area, acquired city rights in 1903 and 1944, respectively (Municipality São Francisco de Paula 2008; Municipality Canela 2008).
In spite of the economic importance of livestock breeding, the techniques of cattle husbandry have always remained on a very basic level in the Southriograndean highland. Supplementary feeding during winter is not the custom (A. Jacques, pers. comm.). Instead, cattle numbers are adjusted to fodder availability during winter – on average one head of cattle per 2-3 ha in the highland, Mohrdieck (1993) -, resulting in an accumulation of excess live, then standing dead biomass from the end of summer through winter. Pastures were

Hermann, J.-M. PhD thesis 1: Introduction
18
traditionally burnt at the end of winter in order to remove this dead biomass and “encourage” resprouting (Evangelista et al. 1993).
This practice is a continuous source of conflict between fazendeiros and environmentalists. The situation is aggravated by the fact that a Federal Law of 1998 permits “controlled” pasture burning in confined areas but a stricter law applies in the Federal State of Rio Grande do Sul: The State Forestry Code of 1992 (Código Florestal Estadual, law nº 9.519/92, art. 28) expressly prohibits fire in forests and all other types of vegetation unless it serves for control of dangerous weeds and pests, and has been previously authorised (see e.g. MPRS 2008). Pasture fires still occur accidentally.
Although the nature of pre-colonisation fire regimes - i.e. frequency and intensity of grassland fires and sources of ignition - is not well known, it is save to assume that for the past 300 years the Southriograndean grasslands have been subjected to disturbance of unprecedented severity. Nevertheless, before palynological studies were available, and in the light of grassland existence even before the arrival of settlers, their origin and maintenance was controversially debated. Alvim (1954, excerpts reprinted in Leite & Klein 1990) distinguished three main streams, or schools, each regarding climate, soil or fire as the principal or sole explanation.
In a synthesis, Pillar & Quadros (1997) propose that water budget, fire and grazing are principally responsible for the currently observed distribution of grassland and forest, although water shortage is considered irrelevant on the High Plateau. Behling and collaborators claim that Araucaria forest expansion on the highland may be linked to a “very humid climate without a significant dry period” (Behling 1998, p.154), but also point out a decrease of fire evidence in pollen cores coinciding with evidence of forest expansion (e.g. Behling 1997, Behling et al. 2004).
This suggests that fire slowed down forest expansion but did not cause regression of forests to grassland. The topic will be further discussed in chapters 3, 5 and 6 of this study.
1.4. The main study area and study species in detail
1.4.1. Vegetation
The “Nature Research and Conservation Centre” – “Centro das Pesquisas e da Conservação da Natureza” - Pró-Mata, established in cooperation of the Pontífica Universidade Católica of Rio Grande do Sul, of Eberhard-Karls-Universität Tübingen and Fachhochschule Rottenburg (both Germany), is located in the southeast corner of the Araucaria High Plateau. It is a zone of convergence of three major ecosystems - grasslands, mixed Araucaria-broadleaf forest and Atlantic rain forest. These ecosystems form, roughly, a sequence from the high regions of the plateau down the escarpment to the coast (e.g. Leite & Klein 1990). Protection and study of these ecosystems as well as recovery of degraded areas were among the principal objectives on establishment of Pró-Mata in 1994 (Bertoletti & Teixeira 1995), with a focus on forest ecosystems – hence the name (“Pró-Mata”: “for the forest”).
In 2001, Baaske et al. classified, of a total area of ca. 4900 ha, 64% of the vegetation cover as exploited primary forest, ca. 20% as secondary forest, ca. 3% as “capoeira” - a shrub formation developing on deforested areas - and 3% as grassland. Total grassland area thus amounts to ca. 150 ha; this figure was confirmed by Oliveira & Pillar (2004). These native grasslands were excluded from cattle grazing and fire around 1994, i.e. 11 to 12 years prior to this study. Since then tussock grasses, principally Andropogon lateralis Nees, have formed tall densely entangled mats of live and dead biomass and woody species abundance,

Hermann, J.-M. PhD thesis 1: Introduction
19
cover and height – although principally of grassland shrubs - have increased significantly (Oliveira & Pillar 2004; pers. obs.).
This development is concomitant with observations in other areas of Rio Grande do Sul: Pott (1974) coined the term „Andropogonisation“ for the phenomenon of tussock grasses predominating as early as two to three years after cessation of fire and grazing, even if prostrate species characterised the grassland community before. Tall grass tussock and shrub cover, principally of the genus Baccharis, also developed in the ecological station Aracurí within two years after abandonment (Waechter et al. 1984) and in abandoned grasslands of the National Park Aparados da Serra (Monteiro & Couto in prep.; pers. obs.).
Ca. one decade after abandonment, the Pró-Mata grasslands represented a continuum of structural types, composed of different proportions of tussock grasses, low shrubs and tall shrubs. The two end points of this continuum are represented by „tall tussock grassland“ - formed by a dense, up to 1m high matrix of live and dead grass biomass, interspersed with grassland shrubs of about the same height and erect to straggling growth - and „tall shrub grassland“, characterised by a canopy of 3-4m high Baccharis uncinella shrubs over a patchy tussock grass layer (Oliveira & Pillar 2004; pers. obs.). Oliveira & Pillar (2004) concluded from aerial photographs that in 1994 ca. 5% of grasslands were of the tall shrub type and ca. 8.5% of the tall tussock type („low shrub grassland“ in their terminology) while the remainder of the area was more or less shrub-free. In the meantime, low grassland shrubs have spread through most of the grassland area (pers. obs.). The relative influence of grassland sites with and without predominance of tall shrubs on establishment of woody species is investigated in chapters 2-5.
The private fazenda „Trés Estrelas“ adjacent to Pró-Mata continued to be managed in the traditional way: Cattle density can be estimated at ca. 0.5 head of cattle per hectare (S. Avila de Lima, pers. comm.) and fires are still frequent especially in the grasslands close to the settlement. In chapter 5 I discuss a more relaxed disturbance regime apparent in population structure of a pioneer woody species community in a more remote area of the fazenda: These observations were made in two grassland islands at ca. one hour riding distance from the settlement.
Fig. 1.7: Grasslands of the fazenda „Trés Estrelas“, cattle-grazed and burnt on a regular basis until
prohibition of pasture fires. Photographer: J.-M. Hermann.

Hermann, J.-M. PhD thesis 1: Introduction
20
Fig. 1.8: Grassland of the neighbour area “Pró-Mata” where cattle and fire were excluded in 1994; this
area, however, suffered another fire accidentally in 1999, ca. 4 years before this picture was taken.
Foreground: So-called tall tussock grassland, composed principally of C4-grass tussocks and
interspersed with small C3 grassland shrubs of straggling growth. Background, adjacent to forest border:
Ca. 3 year old Baccharis uncinella thicket, which will eventually develop into tall shrub grassland.
Photographer: J.-M. Hermann.
Fig. 1.9: So-called tall shrub grassland of the “Pró-Mata” area, characterised by C3-grassland shrubs –
principally Baccharis uncinella – of up to 4.5 m height. These shrubs are of similar age as those in Fig.
1.6, but formed a more or less closed canopy at least temporarily (chapter 5). Photographer: J.-M.
Hermann.
On establishment of my population monitoring plots in late summer 2005 (chapter 5), the average height of the grass matrix in pasture, tall tussock and tall shrub abandoned grassland was determined by „direct measurement“, as described by Stewart et al. (2001, p.1148): One hand is placed „lightly on the vegetation at the level below which about 80% of

Hermann, J.-M. PhD thesis 1: Introduction
21
vegetation is estimated by eye to be growing (thus ignoring occasional tall stalks)“ and this height is read on a ruler. In relation to its simplicity, this method yields fairly consistent results when small, medium and tall swards have to be compared (see Stewart et al. 2001). Usually one such measurement per m² was made and median values were then calculated for larger plots.
Median height of the pasture matrix was 10cm and ca. 20% of height values were between 1 and 3 cm, i.e. may be considered representative of gaps extending more or less to the soil surface. Median height of the tall-tussock abandoned grassland matrix was 40 cm with gaps making up only a minor proportion (2%) statistically, although human and animal tracks and trails account for larger percentages of open ground locally (pers. obs.). Tall-shrub abandoned grassland offered the largest percentages of gaps in the grass matrix, with median vegetation height only 5cm, 20% of values between 1 and 3 cm and another 15% of values at 0 cm; in these cases the ground was covered by shrub and grass litter (rarely fully exposed).
1.4.2. Study species
1.4.2.1. Baccharis uncinella de Candolle
Baccharis uncinella de Candolle– referred to as “Baccharis” in chapters 1 to 4 -, a shrub species of the family Asteraceae, never occurs inside closed-canopy forest but thrives abundantly along roadsides, in logged pine plantations and in the abandoned Pró-Mata grasslands. In some areas, solitary shrubs are scattered in the grass matrix, in others - especially adjacent to forest borders - Baccharis shrubs form belts or carpets up to 4.5 m height and with a loosely closed canopy (hereafter tall shrub grassland) (Morales 2003, Rafael 2004; pers. obs.).
Short description (data from Barroso & Bueno 2002 unless otherwise stated): Synonyms: Baccharis discolor Baker Popular name: Vassoura, vassoura-lageana. Description: [Young] branches tomentose to glabrescent. Leaves sessile, obovate to
elliptic, 6-15 mm long, 4-6 mm wide, undersurface greyish tomentose, margins inrolled, apex obtuse, mucronate, one-nerved.
Baccharis stems of individuals more than one year old become increasingly twisted and irregular in diameter, with deep lengthwise fissures in the bark especially at the base (pers. obs.). The root system is shallow and wind-throw can sometimes be observed in solitary individuals on shallow soils (pers. obs.).
Phenology: Flowering principally in September and October, occasionally until March, fruiting soon afterwards. Dioecious (male and female flowers on separate shrubs). Fruit is an achaene, cylindrical, ca. 1.5 mm long.
In the highland area studied here – the southeast corner of the Araucaria High Plateau of Rio Grande do Sul - fruit maturation and wind-dispersal occurs in midsummer from the middle to December until the middle of January (pers. obs.).
Geographical distribution and habitat: “Common shrub” in the southeast and south of Brazil. In the federal state Santa Catarina, occurrence almost exclusively on the High Plateau, in so-called “campos sujos” (“dirty grasslands”) at the borders of forest islands, secondary forests, or even in clearances in Araucaria forests. Also found in the litoral regions of Santa Catarina, demonstrating the floristic link between this and Serra region.
Rambo (1952) classified Baccharis uncinella (there under the synonym of B. discolour) as ‘espécie setentrional’, i.e. belonging to a group of systematic units that have their centre of distribution in the hot regions of South America. The genus Baccharis constitutes the major part of this group in Southern Brazil (90 species of a total of 376

Hermann, J.-M. PhD thesis 1: Introduction
22
species). The geographical range of B. uncinella extends between Minas Gerais (Central Brazil) and Rio Grande do Sul. The Asteraceae family in general, being little developed in rainforest, finds its distributional limit in the grasslands and savannahs (“Cerrados”) of Central Brazil.
Fig. 1.10: Branch of Baccharis uncinella de Candolle, flowering December (midsummer) 2005. The spirally
leaf insertion is characteristic of this species. Photographer: J.-M. Hermann.
1.4.2.2. Myrsine parvula (Mez) Otegui
Myrsine parvula (Mez) Otegui – referred to as “Myrsine” in chapters 1 to 4 -, of the family Myrsinaceae, is the most successful pioneer forest species in terms of both frequency and cover in the abandoned Pró-Mata grasslands and in parts of the neighbour fazendas (Morales 2003, Rafael 2004; pers. obs.).
Short description (data from Otegui 1998 unless otherwise stated): Synonyms: Rapenea parvula Mez in Engl., Rapanea megapotamica Mez. in Engl.,
Rapanea lorentziana Mez in Engl., Myrsine lorentziana (Mez) Arechav. Popular name: Capororoca Description: Small tree of 2-5 m height and grey, smooth bark. Leaves glabrous,
cartaceous, elliptic to obovate, 6.5-12 cm long, 2-3.5 cm wide, apex pointed, base decurrent, leaf margins entire, pedicels 10-15 mm long. Inflorescences consist of 4-10 flowers, either staminate or pistillate [arising directly from branches; see Fig. 1.11].
Phenology: Flowering from May to July. Myrsine fruits are near-spherical drupes, 3-4mm in diameter in M. parvula. In the highland area studied here fruit maturation and dispersal occurs in late summer, approximately from the middle of February until the end of March (pers. obs.).
Myrsine fruits are surrounded by a thin, black-purplish pericarp and consumed and dispersed by birds (pers. obs.; Pineschi 1990, Pascotto 2007).
Geographical distribution and habitat: According to Otegui (1998), the species is typical of coastal forests and gallery forests of Brazil, Paraguay and Argentina.
However, it has also been recorded in several studies of forest fragments on the Araucária High Plateau (see chapter 7). Duarte et al. (2006b) classify the species as “generalist”, as both reproductive adults and recruits occur across a range of pioneer stages

Hermann, J.-M. PhD thesis 1: Introduction
23
of grassland-forest-succession, from solitary perch trees to small and large forest patches (capões), in the Pró-Mata area.
It should not be ignored here that Myrsine species are notoriously difficult to identify. Important identification traits such as leaf size, leaf morphology, flowering and fruiting phenology vary according to vegetative or generative origin, site conditions and geographical position to far greater extent than available keys suggest (chapters 3 and 4; see also Pineschi 1990 who reports e.g. two flowering periods per year in certain species).
It is not clear whether the ability to grow clonally by production of root suckers is a locally restricted phenomenon, considered unimportant or largely unknown. I found a single reference to the ability of native species Myrsine ferruginea Spr. to form „horizontal subterraneous stems“ on sand dunes (Backes & Irgang 2002, p.220). In my study area, root suckering is one of the key characteristics of pioneer Myrsine populations (chapter 4) but was not noticed until my own pilot surveys in 2003/2004.
Identification of Myrsine parvula as principal pioneer in grasslands of my study area was based on a variety of sources: Previous studies by Morales (2003) and Machado (2004), an identification key by Otegui (1998) – the most elaborate available to date – and, last but not least, consultation of UFRGS botanists and a preliminary Pró-Mata species list (Matzenbacher, N.I. in prep., manuscript kindly forwarded by Prof. Nelson Matzenbacher, UFRGS). Herbarised plant material of all principal study sites was deposited at Pró-Mata Research Station.
Fig. 1.11: Myrsine parvula (Mez) Otegui. Left: From Otegui (1998), slightly modified. Right: Specimen
growing by the roadside in a Pró-Mata grassland, flowering September (end of winter) 2005. The brown
spots on the leaves are not species characteristics; they indicate infection with a rust fungus.
Photographer: J.-M. Hermann.

Hermann, J.-M. PhD thesis 1: Introduction
24
1.4.3. Soils
Tab. 1.1 contain excerpts of data provided by Jungblut & Pinto (1997a) who investigated soil profiles underneath grassland and surrounding forest early after establishment of the Pró-Mata research area, i.e. when grassland soil still bore traces of cattle husbandry from the previous decades. The first two soil profiles represent typical highland cambisols developed on acidic effusive bedrock. Cambisols derive their name from the „cambic“ B-horizon that distinctly differs from the horizons above and the bedrock below in colour or texture (Watzlawick et al. undated; see Fig. 1.12). They characterise undulating to more or less level ground at highest elevations of the Pró-Mata area and support mostly grassland, but also stretches of Araucaria forest surrounding grasslands close to the escarpment (Jungblut & Pinto 1997a).
The soil type of the third profile, classified by Jungblut & Pinto as „Terra Bruna“, typically develops on diabasic or basaltic bedrock at elevations above 800 m (Watzlawick et al. undated). B-horizons of Terra Bruna contain high proportions of clay and iron oxides, hence the name-giving brown colour (ibid.). Terra Bruna characterises the forested upper regions of the escarpment in the Pró-Mata area, surrounding the highland cambisol zone like a belt (Jungblut & Pinto 1997 a, b).
Fig. 1.12: A highland cambisol on acidic effusive
bedrock underneath grassland, as described in
Tab. 1.1 (profile 1). The characteristically black A-
horizon with its high organic matter content,
reminiscent of Eurasian steppe soils, stands out
clearly against a yellowish brown B-horizon.
Picture taken along service road in the Pró-Mata
research area (hence the rocks and gravel at the
bottom of the picture). Photographer: J.-M.
Hermann.
Of the various minerals and nutrients analysed by Jungblut & Pinto (1997a), I chose specifically potassium as it is important for stomatal control processes (Mohr & Schopfer 1992) and thus presumably plays a key role for forest species colonising more open grassland (see also microclimate descriptions, next paragraph). Grasses in tension zones between grassland and forest are typically are less dependent on stomatal control as they
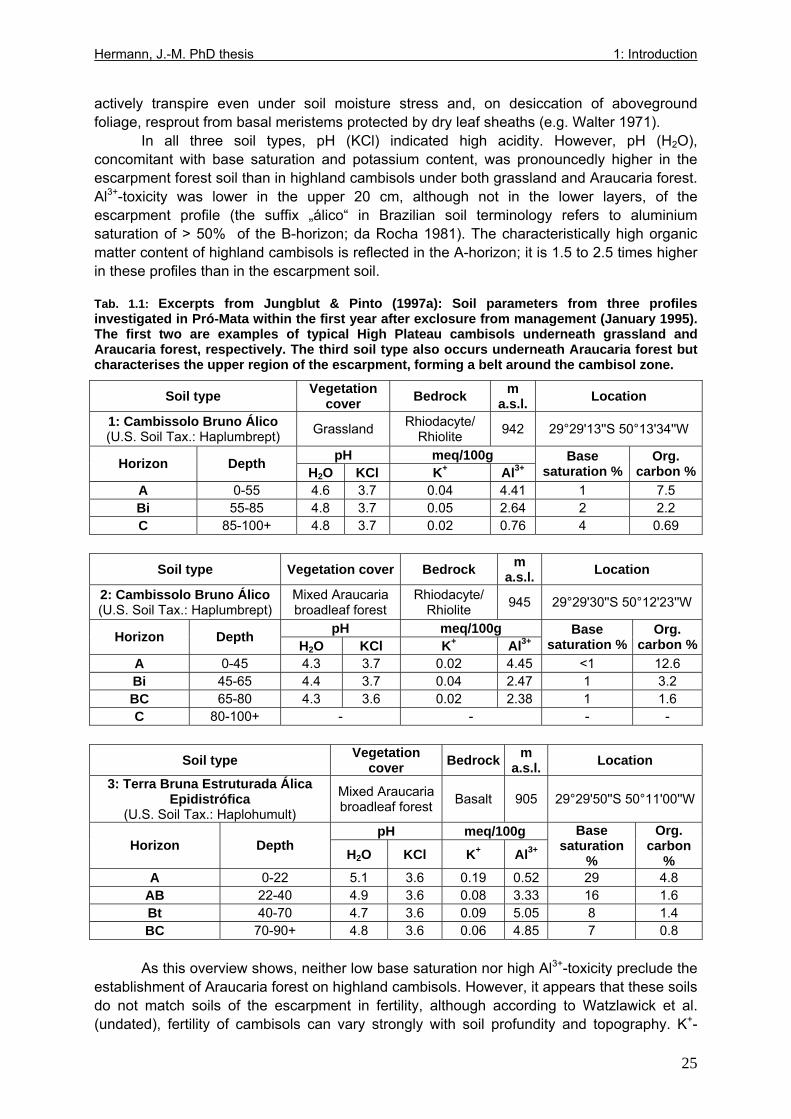
Hermann, J.-M. PhD thesis 1: Introduction
25
actively transpire even under soil moisture stress and, on desiccation of aboveground foliage, resprout from basal meristems protected by dry leaf sheaths (e.g. Walter 1971).
In all three soil types, pH (KCl) indicated high acidity. However, pH (H2O), concomitant with base saturation and potassium content, was pronouncedly higher in the escarpment forest soil than in highland cambisols under both grassland and Araucaria forest. Al3+-toxicity was lower in the upper 20 cm, although not in the lower layers, of the escarpment profile (the suffix „álico“ in Brazilian soil terminology refers to aluminium saturation of > 50% of the B-horizon; da Rocha 1981). The characteristically high organic matter content of highland cambisols is reflected in the A-horizon; it is 1.5 to 2.5 times higher in these profiles than in the escarpment soil.
Tab. 1.1: Excerpts from Jungblut & Pinto (1997a): Soil parameters from three profiles
investigated in Pró-Mata within the first year after exclosure from management (January 1995).
The first two are examples of typical High Plateau cambisols underneath grassland and
Araucaria forest, respectively. The third soil type also occurs underneath Araucaria forest but
characterises the upper region of the escarpment, forming a belt around the cambisol zone.
Soil type
Vegetation
cover
Bedrock
m
a.s.l.
Location
1: Cambissolo Bruno Álico (U.S. Soil Tax.: Haplumbrept) Grassland Rhiodacyte/
Rhiolite 942 29°29'13''S 50°13'34''W
pH meq/100g
Horizon Depth
H2O KCl K
+
Al3+
Base
saturation %
Org.
carbon %
A 0-55 4.6 3.7 0.04 4.41 1 7.5 Bi 55-85 4.8 3.7 0.05 2.64 2 2.2 C 85-100+ 4.8 3.7 0.02 0.76 4 0.69
Soil type Vegetation cover Bedrock
m
a.s.l.
Location
2: Cambissolo Bruno Álico (U.S. Soil Tax.: Haplumbrept)
Mixed Araucaria broadleaf forest
Rhiodacyte/ Rhiolite 945 29°29'30''S 50°12'23''W
pH meq/100g
Horizon Depth
H2O KCl K
+
Al3+
Base
saturation %
Org.
carbon %
A 0-45 4.3 3.7 0.02 4.45 <1 12.6 Bi 45-65 4.4 3.7 0.04 2.47 1 3.2 BC 65-80 4.3 3.6 0.02 2.38 1 1.6 C 80-100+ - - - -
Soil type
Vegetation
cover
Bedrock
m
a.s.l.
Location
3: Terra Bruna Estruturada Álica
Epidistrófica (U.S. Soil Tax.: Haplohumult)
Mixed Araucaria broadleaf forest Basalt 905 29°29'50''S 50°11'00''W
pH meq/100g
Horizon Depth
H2O KCl K
+
Al3+
Base
saturation
%
Org.
carbon
%
A 0-22 5.1 3.6 0.19 0.52 29 4.8 AB 22-40 4.9 3.6 0.08 3.33 16 1.6 Bt 40-70 4.7 3.6 0.09 5.05 8 1.4 BC 70-90+ 4.8 3.6 0.06 4.85 7 0.8
As this overview shows, neither low base saturation nor high Al3+-toxicity preclude the establishment of Araucaria forest on highland cambisols. However, it appears that these soils do not match soils of the escarpment in fertility, although according to Watzlawick et al. (undated), fertility of cambisols can vary strongly with soil profundity and topography. K+-
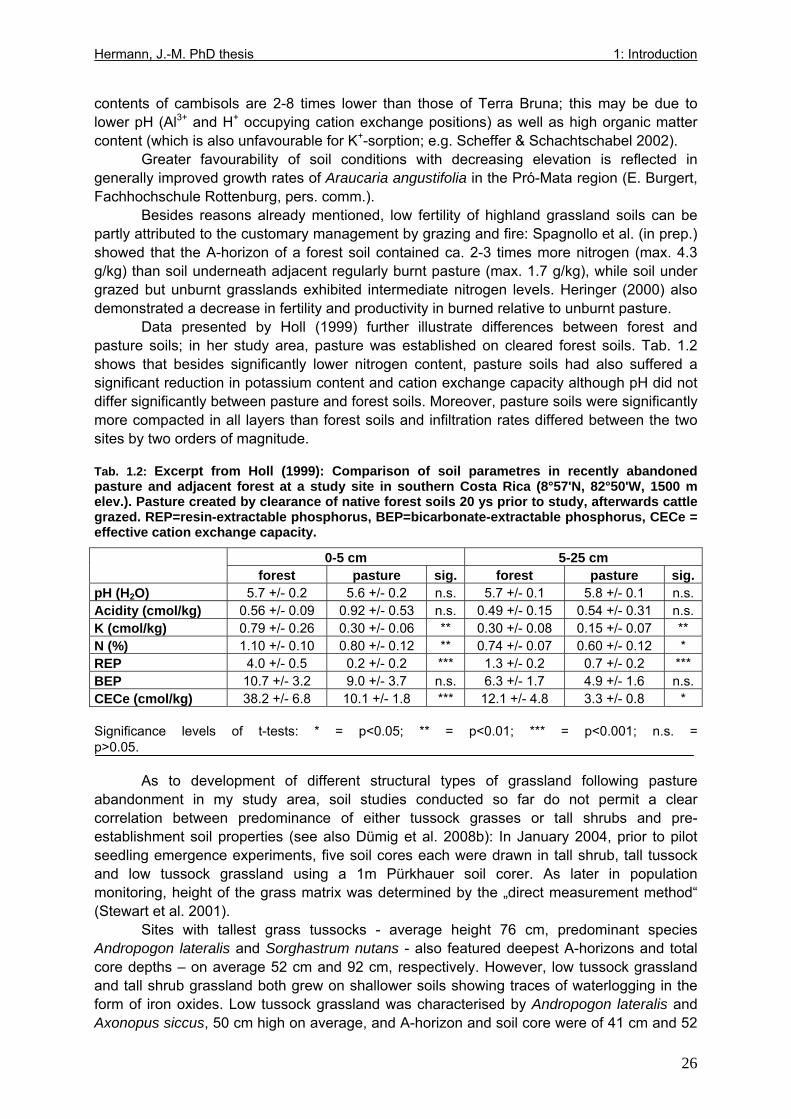
Hermann, J.-M. PhD thesis 1: Introduction
26
contents of cambisols are 2-8 times lower than those of Terra Bruna; this may be due to lower pH (Al3+ and H+ occupying cation exchange positions) as well as high organic matter content (which is also unfavourable for K+-sorption; e.g. Scheffer & Schachtschabel 2002).
Greater favourability of soil conditions with decreasing elevation is reflected in generally improved growth rates of Araucaria angustifolia in the Pró-Mata region (E. Burgert, Fachhochschule Rottenburg, pers. comm.).
Besides reasons already mentioned, low fertility of highland grassland soils can be partly attributed to the customary management by grazing and fire: Spagnollo et al. (in prep.) showed that the A-horizon of a forest soil contained ca. 2-3 times more nitrogen (max. 4.3 g/kg) than soil underneath adjacent regularly burnt pasture (max. 1.7 g/kg), while soil under grazed but unburnt grasslands exhibited intermediate nitrogen levels. Heringer (2000) also demonstrated a decrease in fertility and productivity in burned relative to unburnt pasture.
Data presented by Holl (1999) further illustrate differences between forest and pasture soils; in her study area, pasture was established on cleared forest soils. Tab. 1.2 shows that besides significantly lower nitrogen content, pasture soils had also suffered a significant reduction in potassium content and cation exchange capacity although pH did not differ significantly between pasture and forest soils. Moreover, pasture soils were significantly more compacted in all layers than forest soils and infiltration rates differed between the two sites by two orders of magnitude.
Tab. 1.2: Excerpt from Holl (1999): Comparison of soil parametres in recently abandoned
pasture and adjacent forest at a study site in southern Costa Rica (8°57'N, 82°50'W, 1500 m
elev.). Pasture created by clearance of native forest soils 20 ys prior to study, afterwards cattle
grazed. REP=resin-extractable phosphorus, BEP=bicarbonate-extractable phosphorus, CECe =
effective cation exchange capacity.
0-5 cm 5-25 cm
forest pasture sig. forest pasture sig.
pH (H2O) 5.7 +/- 0.2 5.6 +/- 0.2 n.s. 5.7 +/- 0.1 5.8 +/- 0.1 n.s.
Acidity (cmol/kg) 0.56 +/- 0.09 0.92 +/- 0.53 n.s. 0.49 +/- 0.15 0.54 +/- 0.31 n.s. K (cmol/kg) 0.79 +/- 0.26 0.30 +/- 0.06 ** 0.30 +/- 0.08 0.15 +/- 0.07 ** N (%) 1.10 +/- 0.10 0.80 +/- 0.12 ** 0.74 +/- 0.07 0.60 +/- 0.12 * REP 4.0 +/- 0.5 0.2 +/- 0.2 *** 1.3 +/- 0.2 0.7 +/- 0.2 *** BEP 10.7 +/- 3.2 9.0 +/- 3.7 n.s. 6.3 +/- 1.7 4.9 +/- 1.6 n.s. CECe (cmol/kg) 38.2 +/- 6.8 10.1 +/- 1.8 *** 12.1 +/- 4.8 3.3 +/- 0.8 * Significance levels of t-tests: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
As to development of different structural types of grassland following pasture
abandonment in my study area, soil studies conducted so far do not permit a clear correlation between predominance of either tussock grasses or tall shrubs and pre-establishment soil properties (see also Dümig et al. 2008b): In January 2004, prior to pilot seedling emergence experiments, five soil cores each were drawn in tall shrub, tall tussock and low tussock grassland using a 1m Pürkhauer soil corer. As later in population monitoring, height of the grass matrix was determined by the „direct measurement method“ (Stewart et al. 2001).
Sites with tallest grass tussocks - average height 76 cm, predominant species Andropogon lateralis and Sorghastrum nutans - also featured deepest A-horizons and total core depths – on average 52 cm and 92 cm, respectively. However, low tussock grassland and tall shrub grassland both grew on shallower soils showing traces of waterlogging in the form of iron oxides. Low tussock grassland was characterised by Andropogon lateralis and Axonopus siccus, 50 cm high on average, and A-horizon and soil core were of 41 cm and 52

Hermann, J.-M. PhD thesis 1: Introduction
27
cm average depth, respectively. Under tall shrub grassland, average depths of A-horizon and soil core were 42 cm and 57 cm, respectively.
While this survey indicates a correlation between height of tussock grasses and soil profundity, a similar correlation exists between soil depth and growth vigour of Baccharis uncinella canopies (pers. obs.), so that it cannot be generally concluded that tall shrubs achieve dominance on shallower soils than tussock grasses. However, site conditions favourable to tall shrub establishment can partly be inferred from seedling emergence and growth (chapters 2, 3 and 5).
1.4.4. Microclimate
Accompanying seedling emergence and transplant experiments, microclimate was recorded using Tinytag® probes and data loggers on four days in mid southern summer 2006. PAR, air humidity and air temperature 2 cm above soil surface were recorded on one clear day and one foggy day; values represented here are averages of those two days. Foggy days were January 31st 2006 and February 2nd 2006, clear days February 3rd – 4th 2006. Two measurement days were required as four sets of loggers had to be divided among eight types of microsite.
Seaward highland areas are characterised by dense fogs („serração“) especially between the end of winter and midsummer (October to January), ascending, often within minutes, up the escarpment and through canyons. Likewise, convective rains and thunderstorms are frequent. During this period, the number of days with fine weather more or less matches that of days with either rain or fog (pers. obs. 2004-2005).
Loggers were placed in four types of microsite representative of abandoned grassland and pasture: Tall tussock grassland of ca. 75 cm height, medium height tussock grassland of ca. 50 cm height, tall shrub grassland with a very sparse grass matrix and a mown grass sward of few centimetres height. Experiments were also conducted in two “nurseries” – fenced, shadecloth-covered exclosures - , one of them situated near Pró-Mata station (“Station nursery”), ca. 0.5 km from the escarpment, and the other near Pró-Mata entrance (“field nursery”), at ca. 5 km aerial distance further inland. In order to determine the effect of shadecloth cover, one set of loggers was placed underneath shadecloth and the other in a short grass sward in the immediate surroundings. Measurements in grassland and in nurseries were made on alternate days so that results also indicate the effect of distance from the escarpment on climate. Results from the mown grass sward and uncovered field nursery sward were pooled to represent climatic conditions in low-growing pasture. Pasture climate measurements in situ were considered too hazardous – in spite of low cattle density, experimental constructions had been repeatedly destroyed in the past.
For evaluation, “night time" values recorded between 7:50 p.m. - 7:10 a.m. (defined by lowest level PAR in tall shrub grassland) were subtracted and daytime averages of PAR, temperature, temperature amplitude and humidity were calculated. The following parameters were chosen to best describe relative microclimatic conditions: Average PAR, temperature amplitude, and minimum humidity (loggers frequently failed at maximum values of humidity).
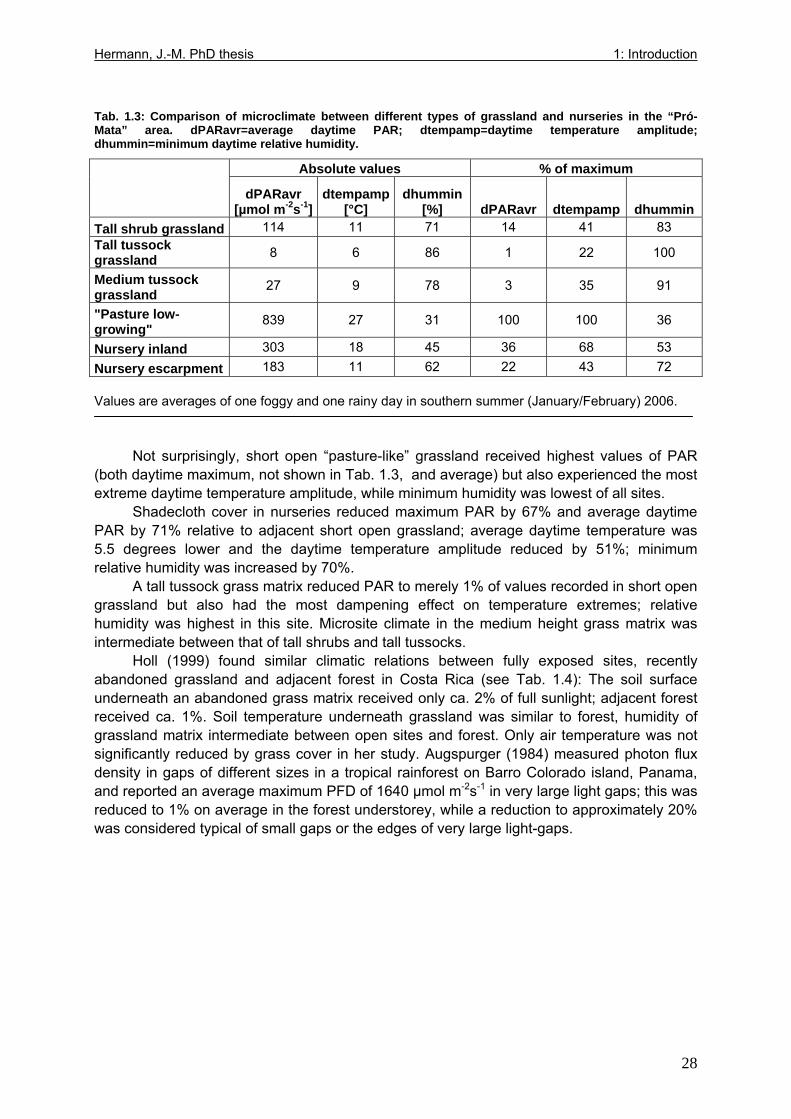
Hermann, J.-M. PhD thesis 1: Introduction
28
Tab. 1.3: Comparison of microclimate between different types of grassland and nurseries in the “Pró-
Mata” area. dPARavr=average daytime PAR; dtempamp=daytime temperature amplitude;
dhummin=minimum daytime relative humidity.
Absolute values % of maximum
dPARavr
[µmol m-2
s-1
]
dtempamp
[°C]
dhummin
[%] dPARavr dtempamp dhummin
Tall shrub grassland 114 11 71 14 41 83 Tall tussock
grassland
8 6 86 1 22 100
Medium tussock
grassland
27 9 78 3 35 91
"Pasture low-
growing"
839 27 31 100 100 36
Nursery inland 303 18 45 36 68 53 Nursery escarpment 183 11 62 22 43 72 Values are averages of one foggy and one rainy day in southern summer (January/February) 2006.
Not surprisingly, short open “pasture-like” grassland received highest values of PAR
(both daytime maximum, not shown in Tab. 1.3, and average) but also experienced the most extreme daytime temperature amplitude, while minimum humidity was lowest of all sites.
Shadecloth cover in nurseries reduced maximum PAR by 67% and average daytime PAR by 71% relative to adjacent short open grassland; average daytime temperature was 5.5 degrees lower and the daytime temperature amplitude reduced by 51%; minimum relative humidity was increased by 70%.
A tall tussock grass matrix reduced PAR to merely 1% of values recorded in short open grassland but also had the most dampening effect on temperature extremes; relative humidity was highest in this site. Microsite climate in the medium height grass matrix was intermediate between that of tall shrubs and tall tussocks.
Holl (1999) found similar climatic relations between fully exposed sites, recently abandoned grassland and adjacent forest in Costa Rica (see Tab. 1.4): The soil surface underneath an abandoned grass matrix received only ca. 2% of full sunlight; adjacent forest received ca. 1%. Soil temperature underneath grassland was similar to forest, humidity of grassland matrix intermediate between open sites and forest. Only air temperature was not significantly reduced by grass cover in her study. Augspurger (1984) measured photon flux density in gaps of different sizes in a tropical rainforest on Barro Colorado island, Panama, and reported an average maximum PFD of 1640 µmol m-2s-1 in very large light gaps; this was reduced to 1% on average in the forest understorey, while a reduction to approximately 20% was considered typical of small gaps or the edges of very large light-gaps.
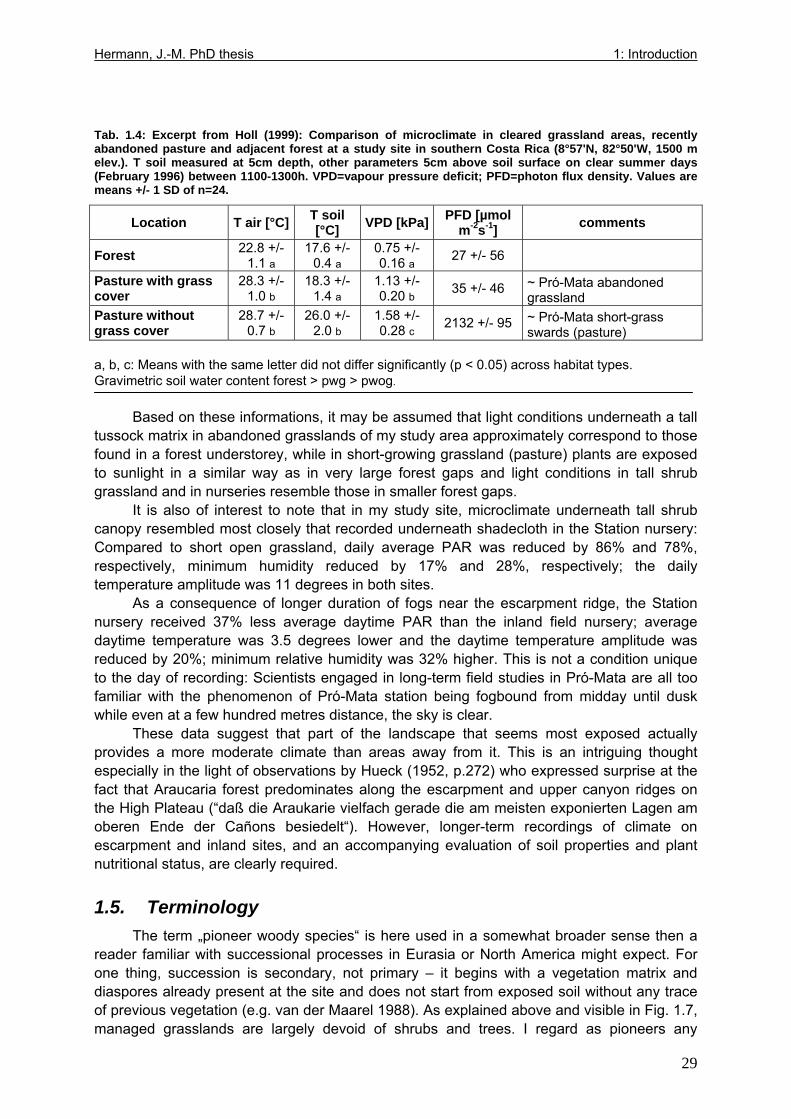
Hermann, J.-M. PhD thesis 1: Introduction
29
Tab. 1.4: Excerpt from Holl (1999): Comparison of microclimate in cleared grassland areas, recently
abandoned pasture and adjacent forest at a study site in southern Costa Rica (8°57'N, 82°50'W, 1500 m
elev.). T soil measured at 5cm depth, other parameters 5cm above soil surface on clear summer days
(February 1996) between 1100-1300h. VPD=vapour pressure deficit; PFD=photon flux density. Values are
means +/- 1 SD of n=24.
Location T air [°C]
T soil
[°C]
VPD [kPa]
PFD [µmol
m-2
s-1
]
comments
Forest
22.8 +/- 1.1 a
17.6 +/- 0.4 a
0.75 +/- 0.16 a 27 +/- 56
Pasture with grass
cover
28.3 +/- 1.0 b
18.3 +/- 1.4 a
1.13 +/- 0.20 b 35 +/- 46 ~ Pró-Mata abandoned
grassland Pasture without
grass cover
28.7 +/- 0.7 b
26.0 +/- 2.0 b
1.58 +/- 0.28 c 2132 +/- 95 ~ Pró-Mata short-grass
swards (pasture) a, b, c: Means with the same letter did not differ significantly (p < 0.05) across habitat types. Gravimetric soil water content forest > pwg > pwog.
Based on these informations, it may be assumed that light conditions underneath a tall
tussock matrix in abandoned grasslands of my study area approximately correspond to those found in a forest understorey, while in short-growing grassland (pasture) plants are exposed to sunlight in a similar way as in very large forest gaps and light conditions in tall shrub grassland and in nurseries resemble those in smaller forest gaps.
It is also of interest to note that in my study site, microclimate underneath tall shrub canopy resembled most closely that recorded underneath shadecloth in the Station nursery: Compared to short open grassland, daily average PAR was reduced by 86% and 78%, respectively, minimum humidity reduced by 17% and 28%, respectively; the daily temperature amplitude was 11 degrees in both sites.
As a consequence of longer duration of fogs near the escarpment ridge, the Station nursery received 37% less average daytime PAR than the inland field nursery; average daytime temperature was 3.5 degrees lower and the daytime temperature amplitude was reduced by 20%; minimum relative humidity was 32% higher. This is not a condition unique to the day of recording: Scientists engaged in long-term field studies in Pró-Mata are all too familiar with the phenomenon of Pró-Mata station being fogbound from midday until dusk while even at a few hundred metres distance, the sky is clear.
These data suggest that part of the landscape that seems most exposed actually provides a more moderate climate than areas away from it. This is an intriguing thought especially in the light of observations by Hueck (1952, p.272) who expressed surprise at the fact that Araucaria forest predominates along the escarpment and upper canyon ridges on the High Plateau (“daß die Araukarie vielfach gerade die am meisten exponierten Lagen am oberen Ende der Cañons besiedelt“). However, longer-term recordings of climate on escarpment and inland sites, and an accompanying evaluation of soil properties and plant nutritional status, are clearly required.
1.5. Terminology The term „pioneer woody species“ is here used in a somewhat broader sense then a
reader familiar with successional processes in Eurasia or North America might expect. For one thing, succession is secondary, not primary – it begins with a vegetation matrix and diaspores already present at the site and does not start from exposed soil without any trace of previous vegetation (e.g. van der Maarel 1988). As explained above and visible in Fig. 1.7, managed grasslands are largely devoid of shrubs and trees. I regard as pioneers any

Hermann, J.-M. PhD thesis 1: Introduction
30
species that establish, as soon as conditions become less inhibitory, in sufficient individual density as to influence establishment of later-successional species.
Adjacent to forest borders, such pioneer woody species communities are composed of both grassland shrub species and forest woody species. The implications of this are discussed in chapters 5 and 7.
The term “seedling” is applied to any juvenile originated from seed, from youngest plants in which only two cotyledons are present (chapter 2, seedling emergence) to plants that possess at least two true leaves in addition to cotyledons (chapter 3, transplant experiment) and individuals up to 1m height naturally established from seed (chapters 4, 5). Fenner (1987b) pointed out that, strictly, this term is appropriate only as long as the plant mobilises any of its seed reserves, but regonised the difficulty of finding a uniformly applicable definition that serves for use in the field.
A “population” of plants has been defined as a collection of individuals belonging to the same species, living in the same area (Silvertown & Lovett Doust 1993). I have pragmatically restricted the “area” of each of my study populations to the interior of 10x10m monitoring plots (chapters 4, 5).
The term „disturbance“ is here used for „any relatively discrete event in time that disrupts ecosystem, community, or population structure and changes resources, substrate availability, or the physical environment“ (White & Pickett 1985). The term „disruption“ indicates vegetation removal and gap creation. Disturbance in this sense encompasses fire and cattle impacts (herbivory, trampling) in grassland as well as anthropogenic and natural processes in forest, such as logging, fire, treefall. It is also implicit that either the survivors of the disturbance or new colonists respond to an increased availability of resources following disturbance (Glenn-Lewin & van der Maarel 1992). Not implicit in this concept of disturbance are changes over evolutionary time spans, such as climate change, or events that occur so infrequently that there is little chance of plants adapting to them (for these, Harper 1977 proposed the term „catastrophe“).
Some years after Pickett & White’s (1985) monograph, van Andel & collaborators proposed the following definition: „Disturbance is a change in conditions which interferes with the normal functioning of a given biological system“ (van Andel & van den Bergh 1987). The intriguing notion behind this definition is that certain species or even ecosystems are so well adapted to fire, grazing etc. that cessation of these impacts may interfere more negatively with progress and completion of their life cycle than their continuation. This thought is worth keeping in mind for discussion in chapter 5. However, the van Andel-definition does not seem to have become widely popular. One major reason would be the difficulty of defining, semantically and practically, the „normal state“ of an organism or community4 (see White & Pickett 1985; Glenn-Lewin & van der Maarel 1992).
I substituted the term „grassland“ for the Brazilian term „campo“ in order to facilitate discussion in the context of other grass-dominated ecosystems worldwide. However, the Brazilian term „campo“ strictly refers not only to grassland, but to any type of vegetation composed of grasses and shrubs (although not forest species) (Pillar & Quadros 1997).
Brazilian terms in local names have not been changed. „Morro Santana“ translates as „Santana hill“. The Research Center „Pró-Mata“ was established „for the forest“, and its neighbour settlement is the „Fazenda Trés Estrelas“ which may be translated as „Three Stars cattle ranch“ or „farm“.
4 In one publication, van Andel and collaborators themselves yield to the temptation of simply proclaiming a desired state as „normal“ state („In such cases, a change in abundance of species should not a priori be considered disturbance of the community. It depends on the criteria for development, chosen before the start of management“ - van Andel et al. 1991, p.144, on grassland succession following abandonment).

Hermann, J.-M. PhD thesis 1: Introduction
31
1.6. Statistics As experiment setups and sampling designs were kept as simple as possible, so
were statistical analyses. Complex multifactorial analyses were precluded by small sample size even on pooling of data. Results are mostly presented in the form of tabular data. In these tables, I usually give median values - which are more strictly appropriate in description of trends in non-normally distributed data - but also average values plus/minus one standard deviation. The latter is the more customary form of representation of results in scientific studies, and this is also the information that ANOVA and T-Tests operate on. In the accompanying texts, for the sake of brevity, only median values are repeated.
Statistical significance of differences in means between species, individual size-age classes and sites was tested by One-Way ANOVA and Tamhane’s T2-Test when more than two alternative treatments were involved (e.g. comparison of a species’ growth rate across more than two grassland and nursery types), and by a T-Test for independent samples when only two alternative treatments were compared (e.g. comparison of performance of two species within one grassland type). This was done using the program package SPSS 14.0.1. In two cases (chapters 4 and 6) linear regression was employed to explore differences between species groups along a range of individual sizes. This was performed using Microsoft Excel (Windows XP).
Although my data were usually non-normally distributed, priority was given to parametrical tests. This is a topic that divides scientists (see e.g. Green 1979, Waite 2000): Non-parametric methods are more conservative and there is a low probability of type I errors, i.e. concluding on treatment effects when in fact there were none. Parametric methods make more assumptions on underlying properties of data that are rarely met in ecological studies, but they are often more powerful statistical tools and there is a low probability of type II errors, i.e. concluding there were no biological effects in a treatment when in fact there were.
The procedures employed here have proved so robust against certain abnormalities of ecological data (see e.g. Harris 1975, Green 1979 and references therein) that part of the scientific community tend towards a pragmatic approach, and accept the use of parametrical methods if the intention of a study is descriptive and exploratory and the results are cautiously interpreted (e.g. Green 1979, Jager & Looman 1987, McGarigal et al. 2000).
Harris (1975, p.31) provides a guideline on which properties data should minimally have for error levels in parametric ANOVA, or two-tailed tests generally, not to be seriously affected: The number of observations per treatment should be greater than „about 10“. The ratio of largest to smallest sample size (n per treatment) should not exceed 4. The ratio of largest to smallest within-sample variance should not exceed 20. I have followed these guidelines, accepting 10 as the minimum sample size per treatment (see also Holl 1999; De Steven 1991 a, b).
Samples were also usually transformed in order to improve homogeneity of variances which is, strictly, a prerequisite for ANOVA. As typical in population studies, size distributions within age or treatment groups were often skewed and variance was greater than the mean so that ln-transformation was considered appropriate (Gibson 2002, Waite 2000). As the only exception, seedling emergence percentage data were arcsin-transformed (Waite 2000).

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
32
2. Seedling emergence of Woody Species in Grassland:
Comparison of a Grassland and a Forest Species
2.1. Summary • Pilot surveys 2004/2005 revealed low seedling numbers of forest tree Myrsine
parvula in naturally established populations, especially in pasture and tall-tussock abandoned grassland, and of grassland shrub Baccharis uncinella in any type of grassland. During a population monitoring 2005-2006, recruitment of Baccharis seedlings was near zero throughout, whereas in Myrsine populations of abandoned grassland, root suckers accounted for the majority of new recruits (chapters 4, 5). Are seedling populations seed- or mcirosite-limited? I checked for availability of microsites by experimental seed input, and for natural seedling emergence in unsown control plots, in four types of grassland: Pasture, tall tussock and tall shrub abandoned grassland, and grassland temporarily cleared of aboveground vegetation and litter by mowing.
• Seeds were collected in local populations, Myrsine seeds scarified and fridge-stored. 100 seeds per species were sown in field plots randomly arranged in a block design, leaving one unsown plot per species per replicate block for quantification of background seedling emergence. For estimation of germinability, replicates of 100 seeds per species were sown in containers with sterilised grassland soil in nurseries, protected from ant and rodent herbivory. Experiments started in late summer 2005: Baccharis was sown in 2 cohorts at the beginning and end of February 2005, Myrsine at the end of March 2005. Monitoring at 2-6 week intervals was conducted during 1-1.25 years. Newly emerged seedlings were censused and removed.
• Of Baccharis only few seedlings were recorded in the field – maximum relative seedling emergence was 2% of sown seed in cleared grassland. Anecdotic evidence also points to bare ground a prerequisite for abundant natural seedling emergence while my pilot experiments demonstrated that full exposure to sunlight is not required.
• In nurseries, both Baccharis cohorts experienced a flush of seedling emergence 2-3 weeks after sowing, clearly coinciding with a rainfall event following a prolonged summer draught in the 2nd cohort. Relative seedling emergence rate was 24% of sown seed, only half of that observed in pilot experiments in 2004. Seedling emergence largely came to an end only 1 month after sowing and there was no indication for formation of a seed bank.
• Contrary to this, Myrsine maximum seedling emergence in the field was delayed until 10 to 13 months after sowing and peaked in mid to late southern summer 2006. Seedling emergence during 1.25 years was highest in tall-tussock abandoned grassland (average 17% of sown), followed by tall-shrub abandoned grassland (11% of sown) and pasture (6% of sown) within 15 months of observation. Seedlings eventually emerged in every field plot except two in cleared grassland. In nurseries, seeds germinated earlier (southern spring 2005) and at slightly higher rates (average 40% of sown within 1 year).
• I conclude that for Baccharis seedling establishment in large numbers, availability of bare ground and favourable climatic conditions must largely coincide with seed maturation. Myrsine populations in grassland are in part seed limited, as a comparison of experimental and background seedling emergence indicates. Seed input requires closer investigation.
• Viability of Myrsine seeds in soil for more than one year following dispersal allows the species to emerge over a wider range of structural types of grassland and largely

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
33
avoids exposure of young seedlings to harsher winter climate. Closer investigation of the relationship between seed morphology, dormancy and longevity of seeds of co-occurring forest species might help to understand why some assume pioneer roles in grassland and others do not.
2.2. Introduction My own pilot surveys of natural seedling emergence of two grassland and one forest
woody species in abandoned grassland in late summer 2004 as well as in pasture plots in late winter 2004 revealed only low seedling density per site – no more than a few seedlings per m² in either managed or abandoned grassland.
According to Harper (1977) and Eriksson & Ehrlén (1992), presence or absence as well as density of a seedling population depend on frequency of seeds as well as frequency of „safe sites“. Eriksson & Ehrlén’s (1992) review – although exclusively of woodland species - indicates that only in a minority of cases, species are exclusively seed limited.
In a crude sense, the number of seedlings appearing is equivalent to the number of “safe sites” sensu Harper (1977) in a given site. Safe sites provide the precise germination and emergence requirements of a particular seed, i.e. dormancy breaking stimuli, abiotic conditions and resource requirements for germination to be initiated, the absence of hazards such as competition, predators, toxicity, pre-emergence pathogens etc. (Harper 1977). Factors such as dormancy breaking stimuli and resource requirements depend, in turn, on a variety of factors such as seed size and shape.
Here, by introducing equal amounts of seeds of two pioneer woody species into different structural types of grassland, and by contrasting seedling emergence in sown plots with “background” seedling emergence in unsown plots, I assess both relative emergence success of the two species and relative limitation of natural seedling emergence by seeds or safe-sites. The questions underlying my experiment design are: How do natural seedling emergence and emergence percentages of experimentally sown seed differ between different structural types of managed and abandoned grassland and between the grassland shrub species Baccharis uncinella L. (hereafter Baccharis) and pioneer forest tree Myrsine parvula Mez. (Otegui), hereafter Myrsine? What conclusions can be drawn on relative safe site availability and natural seed input? How does this affect grassland-forest dynamics in the region? The latter question is addressed in more detail in chapter 3 where seedling emergence values are combined with data on seedling survival and growth obtained in a seedling transplant experiment. In doing so, I assess to what extent experimentally observed performance at this early life stage corresponds to patterns of natural recruitment of each species in the studied grassland types.
Crawley (1990, p. 129) remarked that „it is curious that the simple experiment of sowing extra seeds and recording the number of recruits has been carried out so seldom”. Reasons for this will be discussed in more detail at the end of this chapter. Here, it shall be briefly pointed out that such simple experiments do not allow to distinguish, for example, between relative impact of seed predation and abiotic conditions. However, relative limitation by seed availability, microsite availability and predation is at the centre of many recent works on seedling emergence (e.g. Eriksson & Ehrlén 1992, Turnbull et al. 2000), although there is still some confusion of terminology. Whereas e.g. Hulme (1996) investigates the relative impact of abiotic and biotic factors, Eriksson & Ehrlén (1992), when speaking of “microsite limitation”, actually refer to “safe sites” sensu Harper as their sources do not allow to distinguish between predation and the impact of abiotic, i.e. microsite, conditions.
Furthermore, my experiments yielded no information on length of primary dormancy of seeds in the soil (i.e. length of the period following dispersal during which a seed remains

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
34
viable but unable to germinate; e.g. Baskin & Baskin 1998). They strictly indicate only the length of the period over which seeds may remain viable, or “persistent” in the soil. During this period they may enter a state that Baskin & Baskin (1998) refer to as “secondary dormancy” but that according to Thompson et al. (2003) is not “genuinely dormancy”. This distinction between primary dormancy and persistence is considered increasingly important, as seeds may persist in the soil for a longer period than conventional dormancy-breaking experiments indicate (Thompson et al. 2003; Ooi et al. 2007), but it is rarely made in earlier studies and literature reviews (Thompson et al. 2003). Thus, in discussing my results in the context of these texts, it was not possible to avoid the term “dormancy” when truly either primary dormancy or mere persistence or both conditions may apply.
2.3. Study setup
2.3.1. Study sites and species
This study was conducted in two adjacent areas on the High Plateau of Rio Grande do Sul: 1. Native grasslands in the Nature Conservation and Research Centre “Pró-Mata” (hereafter: Pró-Mata) that were excluded from cattle grazing and fire in 1994, 11 years prior to the experiment. 2. The neighbour fazenda “Trés Estrelas” where grasslands are cattle-grazed (approximately 0.5 head of cattle per hectare) and some of the experimental plots experienced burning – i.e. a complete scorch of aboveground biomass - in August 2005, ca. 6 months after experiment initiation.
In the abandoned grassland, two major structural types have developed: Tall-shrub grassland, characterised by a 3-4m high canopy of Baccharis uncinella shrubs above a patchy tussock grass layer, and tall-tussock grassland where tussock grasses form a dense, up to 1m high matrix of live and dead biomass, interspersed with grassland shrubs of about the same height (Morales & Pillar 2004, pers. obs.). Experimental plots were evenly divided between these two types.
Variations in height and density of pasture grassland are introduced by intensity of cattle grazing and trampling, soil profundity/depth, shading by forest borders and time since last fire. Plots were divided between swards of prostrate and short-grazed tussock grasses of few cm height (hereafter referred to as low-growing pasture) and areas where grass tussocks and forbs of 10-30 cm height predominated (hereafter referred to as high-growing pasture). “Nurseries” were areas protected from rodent herbivory by fences of ca. 0.8 m aboveground height and 0.2 m belowground depth, protected from leaf-cutting ants by regular application of formicide and protected from climate extremes by shade-cloth. One of these areas was located in a man-made, formerly forested grassland area near Pró-Mata research station, close to the escarpment (hereafter referred to as Station nursery), the other at ca. 5 km aerial distance inland, in an abandoned grassland area near Pró-Mata entrance (hereafter referred to as field nursery).
The two species studied were grassland tall shrub Baccharis uncinella de Candolle and pioneer forest tree Myrsine parvula (Mez) Otegui.
Baccharis seeds are cylindrical with ca. 1mm maximum diameter and wind-dispersed by pappuses. Myrsine seeds are spherical, 3-4mm in diameter and consumed and dispersed by birds; there have been no observations of other animal groups (e.g. small mammals) contributing to seed dispersal.
In my own samples, collected in the study area in southern summer 2004/2005 1000-seed-weight was 0.0777 g in Baccharis and 14.7749 g in Myrsine.
For further details on study area and species please see chapter 1.

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
35
2.3.2. Seed collection, treatment and storage
Baccharis seeds were collected in midsummer 2004 (middle to end of December) from three populations, two of them located in abandoned grassland inside the Pró-Mata area, another located at ca. 10 km aerial distance along the roadside. Seeds were hand-selected, discoloured and misshapen seeds discarded, and stored in paper bags at room temperature until use. In order to facilitate seed counting, I removed pappuses by gently rubbing the seeds through a kitchen sieve, although Baskin & Baskin (1998) advise against treatments that do not reflect conditions in nature.
Myrsine fruit, which gradually mature over a period of several weeks, were collected during late summer 2005 (end of February until middle of March) from three populations in the transition zone from forest to abandoned grassland in the Pró-Mata area. Fruit were soaked in water for some days prior to removal of fruit skins by rubbing, then fridge-stored in plastic bags until use (following instruction by Lorenzi 1992 and C. Costella, pers. comm.). Seeds floating at the water surface were discarded. Removing fruit skins served for imitation of the “treatment” Myrsine seeds usually receive on passage through the bird gut: Wenny (2000) states that dispersal by birds is important for removal of pulp - without which germination was reduced or zero in his study - but does not otherwise affect germination. Pineschi (1990) confirms better germination of Myrsine seeds after gut passage than in intact fruit, in which germination was zero in his study.
By chance, shortly prior to the beginning of the experiment, a sample of naturally dispersed Myrsine seeds were obtained from bird faeces along service roads in the Pró-Mata region. Seeds were selected by soaking and removal of floating seeds; storage until experiment beginning was at outside temperature. On examination of seed coats of manually scarified specimens and those obtained from bird faeces with a stereomicroscope, I found that neither treatment had inflicted apparent mechanical damage to the seed coats.
Myrsine seeds treated and stored in the described fashion for pilot experiments in 2004 retained germinability for more than one year. Baccharis seeds collected in early 2004 and stored in paper bags did no longer germinate successfully after the same period (pers. obs.). However, I observed no obvious differences in seedling emergence between the two 2005 Baccharis cohorts (see below) in spite of longer storage of seeds in the latter.
2.3.3. Experiment setup
Sowing experiments were combined with experimental transplants of Baccharis and Myrsine seedlings in grazed and burnt pasture grassland as well as tall tussock and tall shrub abandoned grassland (see following chapter). I selected grassland patches on more or less level ground to minimise lateral seed transport in heavy rain, and at no more than 20 m distance from the forest border; within patches that fulfilled these requirements, experiment plots were randomly placed. This procedure may be referred to as “stratified random sampling” (Müller-Dombois & Ellenberg 1974).
For the sowing experiment only, 10 areas in tall tussock grassland were temporarily cleared of aboveground vegetation by mowing tussocks at the base, and litter was superficially removed. By this treatment, relatively large patches of bare soil were exposed, such as might be the case after a grassland fire.
Field plots were set up in a block design of 0.5 x 0.5m plots lined up adjacent to one another. Each block contained one sown and one unsown plot per species and, in the case of Baccharis, per cohort, the positions of which were randomly assigned. Unsown plots served for quantification of “background” seedling emergence by naturally dispersed seeds. Sown plots received 100 seeds each, placed underneath existing vegetation onto the surface of litter or bare soil as according to Harper (1977) „many (perhaps most?) seeds germinate

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
36
on the soil surface“ (p.137) and it was not the objective of this study to test how many seeds, if dropped from above, get caught in the vegetation matrix. Naturally emerged seedlings were removed from all plots prior to the experiment.
As several studies (e.g. De Steven 1991a, Everham et al. 1996) indicate an inhibitory effect of litter layers on emergence of smaller seeds, I also removed the litter layer in half of the abandoned grassland plots prior to the experiment. However, the effect was of short duration – litter was quickly replaced from senescent Baccharis shrub canopies and standing dead biomass of grass tussocks. By the time Myrsine seedlings emerged, all traces of this treatment had disappeared and plots were pooled into one sample each of tall shrub and tall tussock abandoned grassland.
Control sowings were conducted in two nurseries - one located in grassland near the Pró-Mata area entrance, the other next to the research station – which were fenced and treated with formicide to exclude rodent and ant herbivory. This procedure did not allow for accurate quantification of viability and germinability of seeds but was necessary to demonstrate that complete failure of seedling emergence in the field could not have been due to sterile seed. Germination chambers, which would have allowed for more accurate tests of germinability, were not available.
In nurseries, 100 seeds per replicate were placed in 5–inch diameter flower containers filled with grassland soil previously sterilized at 100°C for 24 hours. Pots were placed in shallow 1x1m basins in the ground so that rims were more or less level with the soil surface, and were shuffled within and between basins at each survey. Basins were uniformly covered with a single layer of shade-cloth, as the influence of different degrees of exposure to sunlight had already been tested in pilot surveys 2004. Weeds, especially mosses, developed abundantly in the pots towards the end of the 1-year survey period and were manually removed to avoid damage to remaining seeds. Three pots per nursery were left unsown to check for background germination, undesired in this case; however, none was observed.
In the case of Baccharis, the experiment was conducted twice during late summer 2005: The 1st cohort was sown February 2nd 2005 (after seed maturity and dispersal had largely ceased in naturally established Baccharis populations) in 20 blocks each in pasture and abandoned grassland, evenly divided between high- and low-growing swards in the first, and tall tussock and tall shrub grassland in the latter. Sowing was also in 10 blocks in cleared grassland; however, five of these plots were found inundated to a level several inches above the soil surface after heavy rainfalls in March 2005, and discarded from further survey and analyses. 10 sown containers were divided between nurseries. The 2nd cohort was sown February 26th 2005 (after peak seedling emergence in the 1st cohort), with the number of blocks in pasture reduced to ten for logistic reasons.
Basically the same setup was used for Myrsine, sown March 31st 2005. 10 sown containers were placed in the field nursery, five in the Station nursery. Seeds obtained from bird faeces were placed in five containers with 53 seeds each in the Station nursery.
At each survey afterwards, I censused and removed numbers of seedlings emerged, i.e. raised above the soil surface and, in the case of Baccharis, usually with cotyledons already unfolded. Myrsine cotyledons require several weeks to discard seed coats. Germination in a strict sense (i.e. emergence of the radicle) occurred largely unobserved in litter or soil layers, and possibly at higher rates than seedling emergence. Baccharis seedling emergence of both cohorts was surveyed at intervals of 10-14 days until the end of March. Afterwards, surveys of both Baccharis 2nd cohort and Myrsine seedling emergence were combined with surveys of transplanted seedlings, i.e. occurred at 2-3 week intervals until spring 2005, and at 6-week intervals afterwards until closure of the experiment. Baccharis plots and pots were only checked cursorily during this latter period as pilot experiments 2004

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
37
had shown that seedling emergence would continue at very low rates, if at all, after the first early flush.
Myrsine seedlings can be clearly identified in the field whereas Baccharis seedlings are no more than a few millimetres high and relatively nondescript. Several times, seedlings of doubtful identity were transplanted into containers and taken to the laboratory for further observation but usually did not survive this treatment. However, trends of seedling emergence would not be altered even if it were assumed that incognito seedlings were indeed of Baccharis uncinella.
At the end of April 2006, as conversion of the pasture area into arable land seemed imminent, both sowing and transplant experiment were closed and sown containers removed from the nursery. However, rumours proved wrong; the pasture plots remained intact and Myrsine seedling emergence in all grassland field plots could be surveyed once more in June 2006, totalling an observation period of 14.5 months.
Some reference is made to the pilot experiments conducted one year prior to this study. The setup of these experiments differed in the some respects: Plots were smaller (0.25 x 0.25 m), the observation period was shorter (approximately six months) and emerged seedlings were not removed but marked with toothpicks (in order to trace seedling survival), a procedure that quickly proved inefficient especially due to the small size of Baccharis seedlings. However, these experiments did reveal some effects of different degrees of exposure (fully exposed to sunlight versus cover by one or two layers of shadecloth) on seedling emergence of the two species in nurseries.
2.3.4. Evaluation
Seedling emergence success or safe site availability, respectively, are expressed by the following parameters: Total emergence percentages and the proportion of plots or containers in which emergence was observed.
Seedling emergence percentages (SE%) given in table Tab. 2.1 were obtained by summing seedling numbers emerged in sown plots, subtracting seedling numbers emerged in unsown control plots (“background seedling emergence”) and relating the remainder to the sum of sown seeds per species and site type (grassland/nursery). This procedure may be criticised as seeds and seedlings are treated as replicates, and differences among plots within a given site type are eliminated (see Green 1979). However, there was no other way of obtaining comparative values across all site types and both species as seedling emergence in many cases was low or zero. For the same reason, I decided not to separately evaluate seedling emergence of the two Baccharis cohorts.
Seedling emergence was considered “negligible” if amounting to less than one per cent of sown seeds over the entire observation period, or if observed in only one of 15 or 20 control plots (Baccharis).
Seasonal change is fairly distinct within the temperate humid highland area (see chapter 1), and I have tried to trace coincidence of peak seedling emergence with certain seasons although sampling dates did not always exactly coincide with duration of seasons meteorologically or according to calendar. “Autumn” samples were obtained in March and April, “winter” samples from May to the middle of October, “spring” sampling occurred from the middle of October to the end of November and “summer” sampling from December through February.
Background seedling emergence served not only for correction of experimentally derived values but is also one indicator of species performance under natural conditions. Based on seedling emergence percentages and quantity of background emergence, I give an estimate of actual quantity of natural seed input in tall shrub grassland, and of the

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
38
quantities of seed input that would be necessary for natural seedling emergence to be observed in the other site types.
Myrsine seedling emergence in untreated grassland plots and nurseries was sufficiently high and frequent for computerized statistical evaluation, using plots as replicates. In this case, background seedling emergence in a given plot was subtracted from seedling emergence in the sown plot of the same block. In order to maintain the minimum n=10, I did not divide between high-growing or low-growing pasture, nor field and Station nursery. Variances within site type samples did not exceed bounds named by Harris (1975; see chapter 1) but in order to improve equality of variances, data were arcsin transformed prior to ANOVA, as appropriate for percentage data (Waite 2000). I performed one-way ANOVA and Post-Hoc Tamhane’s T2-Test using the package SPSS 14.0.1.
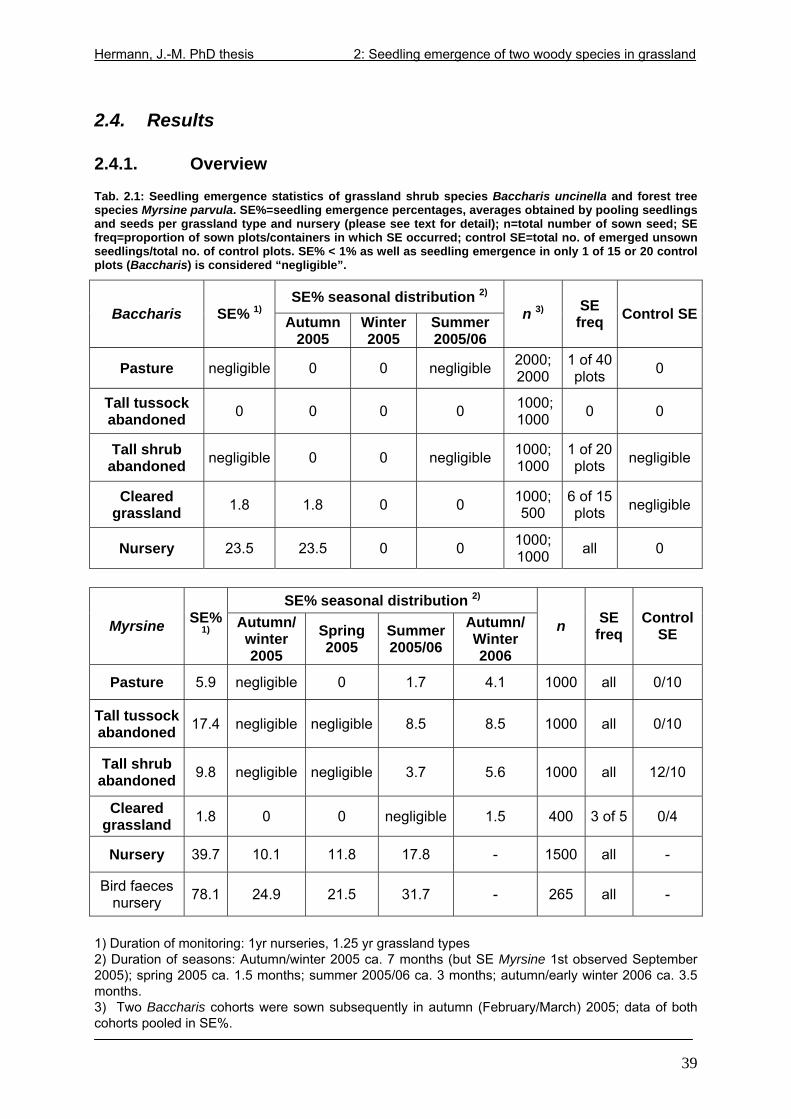
Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
39
2.4. Results
2.4.1. Overview
Tab. 2.1: Seedling emergence statistics of grassland shrub species Baccharis uncinella and forest tree
species Myrsine parvula. SE%=seedling emergence percentages, averages obtained by pooling seedlings
and seeds per grassland type and nursery (please see text for detail); n=total number of sown seed; SE
freq=proportion of sown plots/containers in which SE occurred; control SE=total no. of emerged unsown
seedlings/total no. of control plots. SE% < 1% as well as seedling emergence in only 1 of 15 or 20 control
plots (Baccharis) is considered “negligible”.
SE% seasonal distribution 2)
Baccharis SE% 1)
Autumn
2005
Winter
2005
Summer
2005/06
n 3)
SE
freq
Control SE
Pasture negligible 0 0 negligible 2000; 2000
1 of 40 plots 0
Tall tussock
abandoned
0 0 0 0 1000; 1000 0 0
Tall shrub
abandoned
negligible 0 0 negligible 1000; 1000
1 of 20 plots negligible
Cleared
grassland
1.8 1.8 0 0 1000; 500
6 of 15 plots negligible
Nursery 23.5 23.5 0 0 1000; 1000 all 0
SE% seasonal distribution
2)
Myrsine SE%
1)
Autumn/
winter
2005
Spring
2005
Summer
2005/06
Autumn/
Winter
2006
n SE
freq
Control
SE
Pasture 5.9 negligible 0 1.7 4.1 1000 all 0/10
Tall tussock
abandoned
17.4 negligible negligible 8.5 8.5 1000 all 0/10
Tall shrub
abandoned
9.8 negligible negligible 3.7 5.6 1000 all 12/10
Cleared
grassland
1.8 0 0 negligible 1.5 400 3 of 5 0/4
Nursery 39.7 10.1 11.8 17.8 - 1500 all -
Bird faeces nursery 78.1 24.9 21.5 31.7 - 265 all -
1) Duration of monitoring: 1yr nurseries, 1.25 yr grassland types 2) Duration of seasons: Autumn/winter 2005 ca. 7 months (but SE Myrsine 1st observed September 2005); spring 2005 ca. 1.5 months; summer 2005/06 ca. 3 months; autumn/early winter 2006 ca. 3.5 months. 3) Two Baccharis cohorts were sown subsequently in autumn (February/March) 2005; data of both cohorts pooled in SE%.
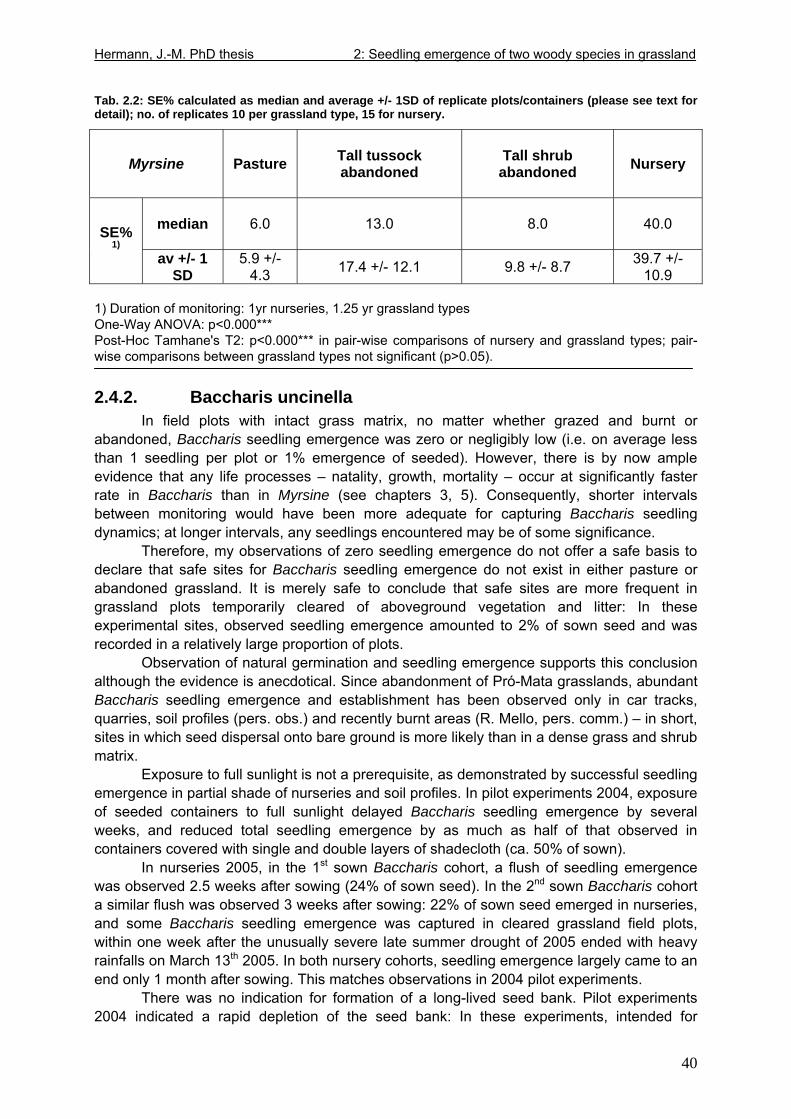
Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
40
Tab. 2.2: SE% calculated as median and average +/- 1SD of replicate plots/containers (please see text for
detail); no. of replicates 10 per grassland type, 15 for nursery.
Myrsine Pasture
Tall tussock
abandoned
Tall shrub
abandoned
Nursery
median 6.0 13.0 8.0 40.0 SE%
1)
av +/- 1
SD
5.9 +/- 4.3 17.4 +/- 12.1 9.8 +/- 8.7 39.7 +/-
10.9 1) Duration of monitoring: 1yr nurseries, 1.25 yr grassland types One-Way ANOVA: p<0.000*** Post-Hoc Tamhane's T2: p<0.000*** in pair-wise comparisons of nursery and grassland types; pair-wise comparisons between grassland types not significant (p>0.05).
2.4.2. Baccharis uncinella
In field plots with intact grass matrix, no matter whether grazed and burnt or abandoned, Baccharis seedling emergence was zero or negligibly low (i.e. on average less than 1 seedling per plot or 1% emergence of seeded). However, there is by now ample evidence that any life processes – natality, growth, mortality – occur at significantly faster rate in Baccharis than in Myrsine (see chapters 3, 5). Consequently, shorter intervals between monitoring would have been more adequate for capturing Baccharis seedling dynamics; at longer intervals, any seedlings encountered may be of some significance.
Therefore, my observations of zero seedling emergence do not offer a safe basis to declare that safe sites for Baccharis seedling emergence do not exist in either pasture or abandoned grassland. It is merely safe to conclude that safe sites are more frequent in grassland plots temporarily cleared of aboveground vegetation and litter: In these experimental sites, observed seedling emergence amounted to 2% of sown seed and was recorded in a relatively large proportion of plots.
Observation of natural germination and seedling emergence supports this conclusion although the evidence is anecdotical. Since abandonment of Pró-Mata grasslands, abundant Baccharis seedling emergence and establishment has been observed only in car tracks, quarries, soil profiles (pers. obs.) and recently burnt areas (R. Mello, pers. comm.) – in short, sites in which seed dispersal onto bare ground is more likely than in a dense grass and shrub matrix.
Exposure to full sunlight is not a prerequisite, as demonstrated by successful seedling emergence in partial shade of nurseries and soil profiles. In pilot experiments 2004, exposure of seeded containers to full sunlight delayed Baccharis seedling emergence by several weeks, and reduced total seedling emergence by as much as half of that observed in containers covered with single and double layers of shadecloth (ca. 50% of sown).
In nurseries 2005, in the 1st sown Baccharis cohort, a flush of seedling emergence was observed 2.5 weeks after sowing (24% of sown seed). In the 2nd sown Baccharis cohort a similar flush was observed 3 weeks after sowing: 22% of sown seed emerged in nurseries, and some Baccharis seedling emergence was captured in cleared grassland field plots, within one week after the unusually severe late summer drought of 2005 ended with heavy rainfalls on March 13th 2005. In both nursery cohorts, seedling emergence largely came to an end only 1 month after sowing. This matches observations in 2004 pilot experiments.
There was no indication for formation of a long-lived seed bank. Pilot experiments 2004 indicated a rapid depletion of the seed bank: In these experiments, intended for

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
41
quantification of predation, Baccharis seeds had been placed in basins in the ground and left covered only by original vegetation, or covered additionally with shade-cloth with ca. 1mm spacing of threads which would admit only smallest insects and microorganisms. Within 1.5 weeks in late summer 2004, only ca. 25% of intact seeds remained in open basins in cleared grassland and tall tussock grassland and ca. 20% in tall shrub grassland. Cloth-cover reduced seed disappearance, but still substantial proportions of seeds disappeared: Ca. 25% in cleared grassland, ca. 50% in tall tussock grassland and ca. 60% in tall shrub grassland. Losses were lower in Myrsine in all sites.
2.4.3. Myrsine parvula
For Myrsine, seedling emergence safe sites are most frequent in tall tussock abandoned grassland, where emergence percentage in 1.25 years was on average 17% of sown seed, and seedling emergence was observed in all sown plots. The latter was also the case in the other untreated grassland types, but emergence percentage amounted to an intermediate 10% in tall shrub abandoned grassland, and to only 6% in pasture.
In the nurseries, seedling emergence not only set in earliest - from southern winter 2005 onwards – but seedling emergence percentage was also higher (on average 40% of sown in only 1 year) than in the field. In seeds collected from bird faeces and placed in nurseries near Pró-Mata station, seedling emergence amounted to 78% of sown and was thus 1.7 times as high as in manually scarified seeds at the same site (46% of sown).
In initially cleared grassland plots, Myrsine seedling emergence was very low (1%) and restricted to grass tussocks resprouted by autumn 2006. In pilot experiments 2004, no seedling emergence had been recorded in seeded containers exposed to full sunlight; in containers placed under single and double layers of shade-cloth, seedling emergence had set in at around the same time (midwinter 2004) but was ca. four times higher in the latter than in the first.
Myrsine seedling emergence was first observed in nurseries in the beginning of September 2005, i.e. the actual period of winter seedling emergence was 1.5-2 months, of similar length as the spring observation period. The summer observation period was three months. Taking this into account, emergence from hand-picked seeds in the nurseries appears fairly evenly distributed across seasons while in seeds collected from bird faeces, seedling emergence during winter and summer was slightly higher than during summer. In contrast to this, Myrsine seedling emergence in field plots was zero or negligibly low until summer. In tall shrub grassland, pasture and cleared plots more than half of seedling emergence was delayed until autumn and early winter, i.e. occurred one year or more after sowing.
Background seedling emergence was observed only in tall shrub abandoned grassland. Ca. 10% of sown seeds emerged and background seedling emergence amounted to one seedling per 0.25m², which indicates a natural seed input of 40 seeds per m² - supposing that germination and emergence behaviour of naturally dispersed seeds matches that of hand-picked and artificially scarified seeds in my experiment. However, emergence percentages of seeds retrieved from bird-faeces in the study area were approximately twice as high as those of artificially treated seeds so that estimates of natural seed input had better be adjusted to 20-40 seeds/m² in tall shrub grassland. Note that Myrsine seeds remained viable in the soil or litter of field plots for at least one year, so that background seedling emergence may have originated from seed crops of more than one year (2005, 2004 and perhaps further back).
Ca. 17% (roughly the fifth part) of sown seeds emerged in tall tussock grassland; as background germination was zero, natural seed input at soil surface must have been less than 10-20 per m². In pasture, only 6% (roughly the twentieth part) of sown seeds emerged;

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
42
natural seed input would have to be at minimum 40-80 seeds per m², i.e. twice to four times as high as in abandoned grassland, to record only one naturally emerged seedling per plot. In cleared areas, with only 1% seedling emergence observed, 200-400 seeds per m² would be required to record only one naturally emerged seedling per plot.
Evaluation of replicate plots (Tab. 2.1Tab. 2.2) revealed considerable variation between plots and containers within sites. The trends in seedling emergence percentages were of course the same as in pooled data, but differences between grassland types were not significant at the p<0.05 level. Seedling emergence was significantly higher (p<0.000) only in nurseries compared to grassland types.
2.5. Discussion
2.5.1. Possible drawbacks of the experiment design
Six-week monitoring intervals were sufficient for capturing Myrsine seedling emergence percentage and rates per season. Baccharis seedlings, however, differ by at least one order of magnitude in initial size from Myrsine seedlings, not surprisingly given their relative seed sizes. De Steven (1991a) found three-week intervals sufficient for monitoring of seedling emergence of a wind-dispersed pioneer tree (Pinus taeda) in the field. In this study, logistic constraints imposed monitoring at six-week intervals (parallel to transplant monitoring) from the second month after sowing. This means that if any flushes of Baccharis seedling emergence – similar to those observed in the nurseries – occurred in the field with a time lag of one month or more, these could have been missed. In other words, although I made a point of having observed seedling emergence instead of seed germination, even this may have been strictly the case only in Myrsine seedlings.
Another bias in favour of Myrsine was introduced by sowing equal amounts of seeds per cohort, although it is a common procedure to standardise seed input (e.g. Baskin & Baskin 1998, De Steven 1991a). However, high seed production is typical in many small-seeded, wind-dispersed species (e.g. Fenner 1987a, Ehrlén & van Groenendael 1998 and references therein). A review by Moles & Westoby (2004) reveals a general tendency for seedlings of large-seeded species to perform better than those of small-seeded species under conditions of drought, shade etc., which small-seeded species may compensate to some degree by such high seed input. Under natural establishment conditions, low availability of safe sites in grassland might thus be compensated by high Baccharis seed input to greater extent than my experiment revealed, even though two sown cohorts of Baccharis were compared with only one sown cohort of Myrsine.
This provides one answer to Crawley’s question on why simple sowing experiments are not more often performed: They may reveal relative density of safe sites in different types of vegetation under a given seed input, but in order to truly understand populations processes seed input ought to be based on previous census or at least realistic estimates of seed rain, which automatically renders the experiment less simple. Moreover, the fate of seeds is difficult or impossible to trace, and interpreting results may actually provide more questions than answers. It is not possible to distinguish further between microsite limitation and herbivore limitation, and while such information may yet be considered dispensable in the initial stages of basic research, they may be of considerable practical importance e.g. in forestry studies. (Of course, Crawley himself already gives this answer, hinting at the complex factorial experiments necessary to determine limiting factors of seedling establishment.)
Emergence percentages were considerably higher in seeds collected from bird faeces and stored at outside temperature than in hand-picked, artificially scarified and fridge-stored

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
43
seeds, although in all seeds, no matter whether untreated or treated, seedling emergence was delayed for at minimum a quarter of a year. As such, good germination of seeds after bird gut passage is not surprising: E.g. Pineschi (1990) noted zero germination of Myrsine seeds when fruits were hand-picked and untreated, whereas passage through the bird gut resulted in some germination (although only in three of seven Myrsine species, and at percentages of ca. 10 to maximum 40%). Wenny (2000) found zero germination of seeds in intact fruit of bird-dispersed Ocotea endresiana, whereas pulp removal either by bird gut passage or by hand resulted in 98% germination. Ellison et al. (1993) reported enhanced germination in high-light demanding Melastome species when seeds were collected from bird droppings. However, as according to Wenny (2000) scarification by gut passage and soaking-and-rubbing treatments do not seem to differ essentially in their effects on germination, there must be other factors contributing to differential seedling emergence in my experiments:
Firstly, according to some authors, seeds may enter the dormant state more than once, and dormancy may then be broken only under a successively more specific range of conditions (e.g. Harper 1977, Baskin & Baskin 1998; but see the current debate on terminology mentioned in 2.2). Only artificially treated seeds were exposed first to fridge-temperatures, then again to outside winter temperatures. Cool temperatures are known to break dormancy in many forest species (Kozlowski et al. 1991); in tropical pioneer species, however, dormancy is broken or germination is triggered, respectively, by increasing light levels or temperature fluctuations (Vázquez-Yánez & Segovia 1984). It is possible that artificially treated seeds entered dormancy more than once and had more specific requirements for dormancy-breaking than bird-dispersed seeds, but this clearly requires research under laboratory conditions.
Another possibility is that birds are more efficient at selecting good quality seeds. As described in 2.3.2, I submitted seeds to a “floating test” to separate empty seeds, but nevertheless birds may be yet more efficient. Johnson & Adkisson (1985) found that jays feeding on beech nuts chose exclusively viable beech nuts although the crop contained substantial proportions (almost 90%) of empty nuts. However, Pascotto (2007) observed birds feeding on both mature and immature Myrsine fruit, although she did not quantify the relative proportions of immature and mature seeds consumed.
Note also that seedling emergence behaviour of seeds collected at the forest-grassland boundary may differ from that of seeds that are dispersed into grassland from source trees located further inside the forest. Growth conditions experienced by the parent plant may strongly influence proportions of dormant seeds and germination requirements of a seed crop (e.g. Grime et al. 1986). In the following chapter, experimentally determined seedling emergence and performance of transplanted seedlings will be joined in order to discuss whether results obtained in this fashion accurately reflect chances of seedling establishment in grassland under natural conditions.
2.5.2. Some evidence of safe site characteristics
Baccharis emerged successfully on bare mineral soil in containers in the nurseries, on the walls of a soil profile underneath adult Baccharis canopy (see appendix), and what little seedling emergence was observed in field experiments also occurred almost exclusively on bare soil surfaces of mown areas. Seedling emergence was apparently cued in at least one case by a heavy rainfall event after a prolonged period of drought; Bock & Bock (1997) report similar behaviour – i.e. precipitation as a major cue for seedling emergence - in a seeder-sprouter Baccharis species (B. pteronoides) in Arizona grassland. Due to their small size Baccharis seeds may fall into cracks and crevices in the soil surface which provide for a

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
44
hideaway during unfavourable climatic conditions, and for better surface contact once soil is moistened, as has been proposed by De Steven (1991b) for Pinus taeda seeds in oldfields.
Vegetation cover and litter layer may both help to retain sufficient moisture for germination (De Steven 1991a, Everham et al. 1996, Holl 1999), but the benefit depends on a variety of factors such as litter quality, thickness of the litter layer and seed size. Facelli (1994) discovered that oak litter harboured large amounts of arthropod predators of cotyledons of Ailanthus altissima, thus indirectly increasing seedling mortality in his study. Only when predators were removed, litter had an indirect positive effect on seedling emergence, survival and growth by suppressing competing herbs, although seedling emergence was delayed (ibid.). If litter layer thickness exceeds the length of the radicles and access to mineral soil water and nutrients is thus denied to seedlings, seedlings will probably succumb to drought and starvation. Emergence of small-seeded early successional forest species may be inhibited by litter layers in this way, as demonstrated e.g. by De Steven (1991a) for Pinus taeda and by Everham et al. (1996) for Cecropia. In my study area, litter layers generally accumulate to greater extent in abandoned grassland than in pasture, and are thickest under adult Baccharis canopy; Baccharis litter has higher tannin contents than grass litter and is thus decomposed at a slower rate (A. Dümig, pers. comm.). This offers one possible reason for little to no observed Baccharis seedling emergence in abandoned grassland – although is does not explain why seedling emergence was similarly low in pasture.
Sunlight filtered through a leaf canopy has a lower red-far red ratio than sunlight above canopy, and this is known to inhibit germination of many species of open habitats (e.g. Fenner 1987a, Kozlowski 1991 and references therein). Sala (1988) reports that for an ungrazed grassland of the flooding pampa in Argentina, which intercepted 80% of the incoming radiation, the red:far red ratio had a daily average of 1.2 above the canopy and 0.4 below the canopy. Similar conditions must be assumed for both of my abandoned grassland types as well as taller-growing patches of pasture. My pilot experiments in nurseries 2004 showed, however, that although Baccharis seedling emergence was possible in full exposure to sunlight, it was clearly higher underneath shadecloth. Although this does not rule out the possibility that grass tussocks and tall shrubs may alter light quality in such a way as to inhibit Baccharis germination, it demonstrates that full sunlight is not required.
Myrsine seeds, contrary to Baccharis seeds, appear to depend critically on vegetation or litter cover to help them retain sufficient moisture for germination. Emergence was never observed in seedlings in which not at least part of the pericarp was moist, and seeds do not survive prolonged exposure to drought; prior to pilot experiments 2004, seeds were stored in paper bags and lost viability after a few weeks, evident from brown discolouration of the embryo (C. Costella, pers. comm.). As my microclimate data (see 1.4.3) show, exposure to irradiation trades off against microsite moisture levels, and it may be for this reason that sites exposed to full sunlight are relatively less suitable for Myrsine seedling emergence than for Baccharis seedling emergence. Moreover, on bare mineral soil, their chances of hiding and benefiting from increased surface contact in cracks must be considerably lower than for Baccharis; noticeably, what little Myrsine seedling emergence was observed in mown areas occurred inside resprouting grass tussocks. Containers in nurseries, although filled with sterile mineral soil initially, were infested with moss by the time seedling emergence peaked and although mosses were removed by hand repeatedly, they probably helped seeds to retain moisture, along with reduced irradiation and increased air humidity underneath shadecloth (see chapter 1).
There was some indication that – besides sufficient moisture – temperature rather than light regime cues germination in Myrsine. Seedling emergence was highest in tall tussock grassland with lowest PAR, and in those sites peaked during midsummer. Similarly, Holl (1999) noted that early successional Costarican forest species did not germinate better

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
45
in cleared pasture plots than in intact pasture grassland, although grass cover reduced photon flux density 5 cm above soil surface to merely 2% of that above grass canopy, thus providing similar conditions as tall tussock abandoned grassland in my study area. Cersósimo (1993) recorded highest seedling density of a Myrsine species in forests of São Paulo in midsummer, followed by another shorter peak in late summer after heavy rainfall. Lorenzi (1998) recommends “half shade” for commercial propagation of Myrsine umbellata.
Myrsine seedling emergence was higher in the deep shade of tall tussock grassland than in the lighter shade of tall shrub grassland. As already mentioned, the experiment design does not allow for a clear distinction between microsite limitation and predator limitation of total seedling emergence. There is, however, some indication from other studies that seed predation is relatively low both in open sites - since predators avoid exposure, as suggested by Gill & Marks (1991) - and in very dense vegetation - due to reduced seed visibility, suggested by De Steven (1991a); both studies were conducted in U.S. east coast oldfields. DeSimone & Zedler (1999), comparing seed predation in a mosaic of sage scrub and grass prairie in California, found seed predation higher underneath shrubs than in grass. This leads to the hypothesis that in my study area, seed predation has a significant impact only in tall shrub grassland.
It is interesting that the observed trends in Myrsine seedling emergence percentages – highest in tall tussock grassland, lowest in pasture and intermediate in tall shrub grassland – were not statistically significant when regarding plots as replicates, due to high within-sample variation. It is possible that statistical significance could be achieved by increasing sample size; on the other hand, sample size is not critically low and there are plausible explanations for similarities between grassland types. Pasture that is not intensely grazed and escapes burning for one year or more features high and dense grass tussocks, similar to tall tussock abandoned grassland. The same applies to tall shrub abandoned grassland under canopy dieback (see chapter 5).
2.5.3. Seed dispersal in space and time
After one flush of seedling emergence within one month after sowing in nurseries in each cohort, Baccharis seedling emergence practically ceased; nor was a delay in seedling emergence of several months observed in pilot experiments 2004. Furthermore, Baccharis seedling emergence after experimental mowing of grassland plots in 2005 was either zero, or low compared to seedling recruitment observed in small grassland shrub Calea phyllolepis, although biomass removal occurred within one month after Baccharis seed maturation and dispersal and adult reproductive individuals were established within dispersal distance from mowed plots (see chapter 6). Seeds stored in paper bags at room temperature lost viability within one year.
Rapid germination has been reported in two other Brazilian Baccharis species (Ferreira et al. 2001, Gomes & Fernandes 2002): B. trimera (which co-occurs with B. uncinella on the Southriograndean High Plateau) germinated within less than five days after sowing at 20°C to 30°C under laboratory conditions (Ferreira et al. 2001). At only 20%, laboratory germination percentage of B. trimera (ibid.) was similarly low as nursery seedling emergence percentage in B. uncinella. Gomes & Fernandes (2002) noted that B. dracunculifolia germinated more successfully in light at temperatures of 20-30°C, but at 15°C germinated equally well in light and shade. They proposed that this behaviour contributes to the species’ particular success as coloniser of open areas but also ensures fast germination as temperatures decrease towards the end of the rainy season, which coincides with seed maturity of B. dracunculifolia in their study area.
Summing up, there is some indication that Baccharis uncinella, and perhaps the genus Baccharis as a whole, does not form a long-lived seed bank, although seed bank

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
46
studies are clearly required to confirm this assumption. This seems unusual as small-seeded species tend to have longer dormancy than large-seeded species (e.g. Crawley 1997a); moreover, pioneer species in secondary succession clearly benefit from being already on site, in the form of dormant seeds, when disturbance opens the vegetation canopy (Fenner 1987a).
On the other hand, lack of dormancy and a short lifespan of seeds (only a few days e.g. in Salix) is known in several genera and species commonly successful in initial stages of succession (e.g. in Populus, Canham & Marks 1985; Salix, Schreiber 1997). Alvarez-Byulla & Martinez-Ramos (1990), in a study of seed bank and seed rain of the tropical pioneer tree Cecropia obtusifolia, found that due to high predation and pathogen attack, seeds persisted for no more than ca. one year in the soil and a seed bank could be maintained only by continuous and copious seed production and seed rain. If Baccharis – whether due to inherent traits or external factors – cannot efficiently disperse a given seed cohort in time, if furthermore seedling emergence is apparently restricted to a relatively narrow range of microhabitats – always bearing in mind that my experimental seed input may have been unrealistically low – and only a minor proportion of seeds germinate, yet the species establishes in patches of high individual density across a landscape, the key to such success must lie in one or more of the following:
Firstly, this may be achieved by high seed production as previously mentioned. Secondly, dispersal in space may be particularly effective. Dispersal of Baccharis
seeds is progressively enhanced by individual growth of reproductive adults to around four metres height, which must seem an astonishing feature to a Middle European scientist familiar only with smaller-growing pioneer Asteraceae, e.g. Senecio in clear-cuts of beech forests (Dierschke 1988). Linked to the latter trait is a relatively long lifespan (ca. one decade at least – see chapter 5): Although individuals reach reproductive maturity only after a few years, they may produce several cohorts of seeds in successive years. This is crucial as Baccharis seedling emergence exhibits considerable year-to-year variation – in 2004 pilot experiments, seedling emergence percentage in relatively sheltered nurseries amounted to ca. 50%, as opposed to only 25% in the following year.
Thirdly, seeds are dispersed fairly synchronously during dry spells in midsummer (pers. obs.) so that if suitable germination conditions soon follow, seedlings may yet achieve appreciable height growth during the remainder of the warm season (see also chapter 3).
Eriksson (1996) proposed that species with efficient dispersal in space but not time build up metapopulations: These may be understood as a network of population patches spread across a landscape in which, continually, some patches become extinct due to deteriorating conditions and other patches recruit in unoccupied, favourable sites at seed dispersal distance. This, in turn, would imply that my study species originated, and may persist, in a landscape and under a disturbance regime in which bare ground becomes available at regular intervals but disturbance-free intervals are long enough for individuals to reach reproductive maturity, and to reproduce several times.
Although animal-dispersed seeds generally do not match wind-dispersed seeds in travelling distance and speed (e.g. van der Valk 1992), fruit morphology in the genus Myrsine encourages relatively far dispersal in space by offering relatively little fruit pulp compared to seed mass. According to Stiles & Rosselli (1993), birds tend to swallow such fruit whole (“gulpers”) and fly away whereas fleshy fruit tend to be consumed by “mashers” in situ so that seeds are dispersed more or less underneath the parent plant. Moreover, abundant production of fruit and easy accessibility of many fruits on leafless sections of branches encourage avid consumption by birds (Pineschi 1991, Pascotto 2007; see also Fig. 1.11). Actual predation, i.e. consumption of the seed endosperm, is rare; this is indicated by the fact that Myrsine seeds collected from bird faeces in my study did not exhibit major mechanical

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
47
damage, and confirmed by Pineschi (1991) who observed only four out of 104 bird species crushing seeds of Myrsine fruit they fed on (those four were of the family of true parrots).
Successful dispersal away from parent plants shows e.g. in the study by Cubiña & Aide (2001) who report one Myrsine species among the few (three out of 35 species) dispersed from secondary forest into pasture at more than four metres distance from the forest border (see also below). Naturally, however, fruits with little pulp not only encourage gulping but are also rapidly digested. This, again, reduces the probability of seed transport over distances of several hundreds of metres. Both the actual nutritional value of Myrsine fruit pulp and the duration of gut passage of fruits remain to be investigated. Seed dispersal distances of several kilometres as reported elsewhere (e.g. 4 km for beech nut dispersal by blue jays in North America; Johnson & Adkisson 1985) are most likely achieved by dispersors that cache undigested fruits. The presence of such hoarding animal dispersors, too, remains to be verified.
A hard seed coat (pericarp) presumably offers protection to the embryo during gut passage and during dormancy. In this study, seedling emergence was observed over a period of 15 months after sowing, and some more may have occurred unobserved after closure of the experiment. Moreover, and contrary to Baccharis, Myrsine fruit maturation is extended over a period of several weeks to months. These two traits, in combination with bird dispersal as described above, should account for effective dispersal of Myrsine seeds in both time and space, allowing a given cohort to attempt establishment over more than one vegetation period and across a much wider range of microsites than was observed in Baccharis – although, other than in Baccharis, seedling emergence would rarely occur under full exposure to sunlight.
Sarukhán (1980) provides one of the few references on dormancy in tropical and subtropical forest species. According to this author, dormancy is rare at least in tropical species of mature forest, and he interprets fast germination and seedling emergence as a means to escape predators. This makes it seem unusual that Myrsine seeds remained viable in soil for more than one year following dispersal. By “US standards”, however, seed dormancy – or viability in soil, respectively - of 3-7 years is regarded as “short-term storage”: According to Canham & Marks (1985) this is a typical trait of several temperate zone gap trees (e.g. Fraxinus, Liriodendron) and species such as Prunus serotina in their study region, in which some seeds germinate every year, irrespective of disturbance, building up a pool of buried seeds and saplings that enables the population to exploit occasional canopy openings, even if seed production and dispersal in the particular year of disturbance are low. Indeed Baaske (1999), on investigation of size distributions of a Myrsine population in an exploited forest of the Pró-Mata area, suggested that the species behaves in just this fashion, with seeds germinating in the shade but seedlings benefiting from competitive release through canopy openings.
In the broader context of migration from the original subtropical forest habitat to the temperate grassland habitat, prolonged viability in soil is probably a key to Myrsine’s greater success as a pioneer relative to most other forest species co-occurring at the forest border, although microsite conditions may ultimately have greater impact. Other than in Baccharis, seed maturation occurs more towards the end of summer than during midsummer, and seedling growth rate is inherently lower especially in winter (chapter 3), so that on immediate emergence Myrsine seedlings could not benefit to a similar extent from the warm season and would run a greater risk of mortality in winter. This can be offset by delaying seedling emergence until spring (as observed in nurseries) or summer (as observed in tall tussock abandoned grassland).
However, in pasture, seedling emergence peaked only during autumn, one year after sowing, i.e. it was most delayed in those microsites were seedlings are in greatest need of a boost in size increase prior to winter (see chapter 3). In tall shrub abandoned grassland,

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
48
seedling emergence was also higher in autumn than in summer, but in those sites the delay is probably less critical for seedling development. Microclimatic conditions are clearly more extreme especially in short-growing pasture than in abandoned grassland. It may be assumed that especially in pasture and cleared areas, insufficient moisture prevented Myrsine emergence. This would signify that microsite conditions influence not only the length of seed viability in soil, but also determine whether such prolonged viability actually serves to improve chances of seedling survival and growth, concomitant with Ehrlén & van Groenendael’s (1998) conclusion that habitat traits overrule seed traits. Seed storage naturally also comes at the cost of prolonged exposure to predators, pathogens etc., which may partly account for the fact that even under most favourable field conditions, in tall tussock grassland, seedling emergence was only ca. half as high as in nurseries.
2.5.4. Consequences for forest-grassland dynamics
Summing up, seed traits and seedling emergence behaviour give a first indication that Baccharis uncinella is better adapted to a landscape in which periodic disturbances – perhaps at intervals of one decade – open areas with bare mineral soil or only a very thin litter layer. Myrsine parvula, on the other hand, seems better equipped for seedling emergence in a forest understorey or in relative small gaps, e.g. caused by tree fall. This is concluded from the fact that seedling emergence was high under tall tussock and talls shrub layers, which correspond approximately to a forest understorey or a small gap in light intensity, but not in pasture, which receives similarly high irradiation as the interior of a large forest gap (see 1.4.3).
These observations also comply with the statement that “in general, the larger the seed, the less the disturbance necessary for successful establishment” (Crawley 1997a, p.110) or, alternatively, that the larger the seed mass – and the lower, consequently, the rate of exhaustion of seed reserves –, the higher the probability of a seedling to survive under deep shade (Thompson 1987 and references therein; Saverimuttu & Westoby 1996).
Several questions outside the scope of this study remain to be answered in order to correctly interpret the meaning of my results for natural population dynamics of the two species in grassland. How high is seed rain into grassland, and how strong is variation in seed rain between types of grassland and between years? Only in tall shrub grassland, both experimental and natural seedling emergence were high enough to give an estimate of natural seed input. The experiment revealed that safe site availability is more limiting in this structural type than in tall tussock grassland; yet natural seedling emergence was considerable in the first and zero in the latter, which suggests that seed limitation at the soil surface is lower in tall shrub than in tall tussock abandoned grassland.
The relevance of tall shrub grassland for grassland-forest succession is discussed in more detail in chapter 5. Here, it shall merely be pointed out that tall tussock grassland accounts for the far majority of the abandoned grassland area in Pró-Mata, so that the focus of investigation may have to be on this structural type and on seed rain both above and below the vegetation layer.
Whereas some authors suggest that a dense matrix of live and dead biomass may obstruct seed dispersal to the soil surface (e.g. Noack 1999 for Pró-Mata grasslands five years after abandonment; Ellenberg 1996 for abandoned pastures in Middle Europe), Schreiber (1997) notes that the grass matrix of a relatively rich abandoned pasture site had apparently no such effect on wind-dispersed Fraxinus seed dispersal and successful seedling establishment in his study. Either way, Hutchings’ (1986) statement that seed interception by standing vegetation has not been studied appears to be still valid today.
Moreover, in my study site, the density of reproductive Myrsine individuals has been estimated to be twice as high in forest border plots adjacent to tall shrub than adjacent to tall

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
49
tussock grassland (chapter 5), which may offer the most straightforward explanation for natural seed input being, apparently, twice as high in first as in the latter.
As to pasture, this study clearly showed that grazed and burnt grassland offers the lowest density of safe sites for Myrsine seedling emergence next to bare ground, but there is evidence from various studies in the neotropics that under natural conditions seed limitation may act prior to safe site limitation. No seed rain of forest species into pasture was detected by Holl (1999) beyond five metres distance, and by Cubiña & Aide (2001) beyond eight metres distance from the forest border.
Alcantará et al. (2000) report that seed dispersing birds avoid open areas – similar to seed predators, see before – but shrub patches slightly increase the attraction of grassland areas. In their study, seed shadows never extended further than six metres from source trees in otherwise open grassland; where shrub patches were scattered across the grassland, seed shadows still decreased dramatically (by 90%) within three metres from the source tree but were at least twice as long (twelve metres from source tree) as in open areas. The study by Alcantará and collaborators offers another possible explanation for relatively high natural seedling emergence of Myrsine in tall shrub grassland. Avoidance of open areas by forest-dwelling birds was confirmed by M. Marchesini (pers. comm.) in the Pró-Mata area.
Moreover, it cannot be ruled out that some of my pasture plots, even within 10 metres distance from the forest border, may have been simply too far off to detect background seedling emergence. Besides behaviour of disperser birds, the low number of reproductive individuals in the adjacent forest border must impose constraints on seed input.
Needless to say, it may be of interest to extend the study of seed rain, dormancy and seedling emergence requirements to other forest species that establish occasionally in grassland. So far, such evidence is anecdotical. Psidium cattleyanum seed collected from mammal faeces, and sown in nurseries in autumn 2005, experienced a peak of seedling emergence in the following spring (L. Prates and I, unpubl. data), similar to Myrsine seeds obtained from bird faeces. Fruit of both species mature in late summer to autumn, seeds have hard coats, and both species establish successfully from seed in grassland, although Psidium is less abundant than Myrsine. Contrary to this, several forest species with soft-coated seeds (in which the endosperm can be squeezed out by applying only light pressure between thumb and forefinger) and mid- to late summer frutification, such as Myrceugenia myrcioides, Myrcia retorta and Casearia decandra, were found to germinate few weeks or months afterwards, in autumn and early winter. This was observed both in seeds sown in nurseries, and in seeds naturally dispersed in faeces and underneath fruiting trees in the field (pers.obs.; L. Prates and I, unpubl. data). While Myrceugenia occurs in abundance similar to Psidium in grasslands, the latter two species are practically never found outside the forest border.
Only a few forest species (Myrsine parvula, Siphoneugenia reitzii) have, to my current knowledge, the ability to reproduce vegetatively, and thus prolong a genetic individual’s time span and the period over which it may produce seeds (see chapter 4); the majority of forest species rely exclusively on generative reproduction for population persistence. Closer investigation of the relationship between forest species seed morphology, dormancy and longevity of seeds, and seedling survival in grassland may thus be crucial to understanding mechanisms of succession from grassland to forest.

Hermann, J.-M. PhD thesis 2: Seedling emergence of two woody species in grassland
50
2.6. Appendix
Fig. 2.1: Left: Baccharis uncinella seedlings emerge abundantly on the walls and floor of a soil profile by
A. Dümig underneath a canopy of adult Baccharis uncinella in late summer 2005. Right: Detail.
Photographer: J.-M. Hermann.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
51
3. Survival and Growth of Woody Seedlings in Grassland:
Comparison of a Grassland and a Forest Species
3.1. Summary • Grassland shrub Baccharis uncinella and forest tree Myrsine parvula are among the
few pioneer woody species that occur with high frequency and abundance at the forest-grassland transition zone of the highland of Rio Grande do Sul, Southern Brazil. Pilot surveys, however, indicate relatively low forest tree seedling recruitment in both pasture and abandoned grassland. Grassland shrub seedlings recruit in low numbers or not at all in either type of grassland. How do mortality and growth differ between Myrsine forest tree and Baccharis grassland shrub seedlings in managed and abandoned grassland?
• Myrsine and Baccharis seedlings were raised in nurseries and in southern autumn 2005, at <10cm initial height, transplanted into the following sites: Cattle-grazed and burnt pasture (low-growing and high-growing type), abandoned grassland (tall- tussock and tall-shrub-dominated type) and nurseries (one located near the escarpment, one 5km inland near grassland plots). Mortality and growth were monitored for one year after transplant.
• More Myrsine seedlings survived in abandoned grassland (85% of planted) than in pasture (27%). Of Baccharis 54% survived in pasture, 30% in abandoned grassland. In pasture, Myrsine seedlings succumbed principally to winter climate (22% of dead), cattle influence (24%) and nutrient/water deficiency (17%). In Baccharis seedlings, which were not physically damaged by winter climate, fire accounted for 21% and cattle influence for 26% of dead. In abandoned grassland, 71% of Baccharis dead apparently starved at low light levels and/or were suffocated by dense vegetation and litter, both conditions that forest species Myrsine could cope better with.
• Myrsine seedlings resprouted even after severe biomass loss from stem stumps of few centimeters height. However, resprouters in pasture barely recovered their original height in 11 months whereas resprouters in nurseries attained median 2.5-5 times original size. Of field plots, tall tussock abandoned grassland was most favourable for Myrsine growth; physical damage from climate extremes was low and final size was median 3.5 times original height. Baccharis seedlings grew more vigorously in both pasture and nurseries than Myrsine seedlings, attaining median 5-6 times original height in 11 months in the first and 8-10 times original height in the latter. Presumably in reaction to climate extremes in pasture and water and nutrient stress in tall-shrub abandoned grassland, Baccharis modified principally stem allometry and leaf morphology, Myrsine stem and leaf allometry.
• These results help to explain why the two species are among the most successful woody pioneers in certain types of grassland. On the other hand, they also indicate – together with results from chapter 2 – the crucial role of seed dispersal, and they suggest that besides management influence (cattle, fire), climate and soil conditions must be incorporated into models of forest-grassland-dynamics for the highlands of Southern Brazil.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
52
3.2. Introduction In pilot surveys 2004 and a population monitoring from 2005-2006 in grassland
adjacent to forest border, numbers of naturally established seedlings of forest tree Myrsine parvula Mez. (Otegui), hereafter Myrsine, were found to be low especially in pasture and tall tussock abandoned grassland (chapter 5).
In tall shrub abandoned grassland, natural Myrsine seed input has been estimated to be 2000-4000 per 100 m² (chapter 2). In population monitoring plots, the average density of Myrsine seedlings <10cm high was 147 seedlings per 100 m² in the year 2005 (my own data), while based on experimental seedling emergence percentages, at minimum 200-400 seedlings per m² may be expected. The difference is not drastic, yet suggests constraints on successful seedling establishment in addition to seedling emergence safe site limitation. Moreover, only ca. 20% of Myrsine seedlings <10cm observed in 2005 recruited into the >10cm size class within one year (my own data).
Seedlings of grassland shrub species Baccharis uncinella L. (hereafter Baccharis) did not recruit into the >10cm size class at all in either pasture or abandoned grassland plots (chapter 5), although this species forms dense stands in grassland locally.
Why is this so? How do seedling mortality and growth differ between Myrsine and Baccharis seedlings, and between pasture and abandoned grassland dominated by either tussock grasses or tall shrubs? In the part study presented in the previous chapter, I introduced seeds of both species into these grassland types in order to assess their relative suitability for colonisation independent of variations in seed rain, and to obtain a coarse estimate of natural seed input. In this part study, I introduce nursery-grown seedlings in order to assess relative seedling establishment success of the two species, or, put in a different way, relative limitation of seedling establishment in different grassland types. In order to trace proximate causes for the observed responses, I attempt to quantify causes of mortality based on circumstantial evidence, and I investigate stem and leaf size and build (allometry and morphology) as these factors are known to be related to species success or failure under different environmental conditions (e.g. Cain 1950, Givnish 1987).
I then integrate data on seedling emergence, survival and growth in order to assess to what extent experimentally observed performance at this early life stage corresponds to patterns of natural recruitment of each species in the studied grassland types. This comparison also serves as a “test” of the Connell & Slatyer (1977) models of inhibition, facilitation and tolerance, which address the potential competitive effects of established, early-successional species on invading, later-successional species. Connell & Slatyer’s publication has been an incentive to various high-quality studies of mechanisms in secondary succession, although the models as such have often proved not altogether satisfactory as the same studies revealed ambiguous effects of a given vegetation type and/or or found that the effect of established vegetation alone did not account for patterns of natural seedling establishment (e.g. Holl 1998, DeSteven 1991 a,b, Gill & Marks 1991).
Forest proceeds from the escarpment onto the High Plateau of Rio Grande do Sul (see 1.2). As a by-product of practical considerations in the experimental setup (establishment of one nursery in natural grassland at some kilometres distance from the escarpment, one on the escarpment adjacent to the research station), I can demonstrate some differences in seedling performance between these two types of environment, and discuss possible underlying causes. I further discuss to what extent relative seedling performance is related to species characteristics, to direct and indirect influence of grassland management and abandonment, and to competition by full-grown tall shrub canopies. In the final paragraph, my results are put into the context of the Connell & Slatyer models as I ask whether any of the three studied types of grassland may conclusively be regarded as

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
53
inhibitory or facilitative to establishment of either of the two study species, or if not, what additional factors may regulate grassland colonisation by these species.
3.3. Study setup
3.3.1. Study sites and species
This study was conducted in two adjacent areas on the High Plateau of Rio Grande do Sul: 1. Native grasslands in the Nature Conservation and Research Centre “Pró-Mata” (hereafter: Pró-Mata) that were excluded from cattle grazing and fire in 1994, 11 years prior to the experiment. 2. The neighbour fazenda “Trés Estrelas” where grasslands are cattle-grazed (approximately 0.5 head of cattle per hectare) and some of the experimental plots experienced burning – i.e. a complete scorch of aboveground biomass - in August 2005, ca. 4 months after experiment initiation.
Study sites were distributed across two structural types of abandoned grassland in the Pró-Mata area, two structural types of pasture grassland of the neighbour fazenda, and nurseries at two locations in the Pró-Mata area.
In the abandoned grassland, two major structural types have developed: Tall-shrub grassland, characterised by a canopy of 3-4m high Baccharis uncinella shrubs above a patchy tussock grass layer, and tall tussock grassland where tussock grasses form a dense, up to 1m high matrix of live and dead biomass, interspersed with grassland shrubs of about the same height (Morales & Pillar 2004; pers. obs.). Transplant plots were evenly divided between these two types.
Height and density of pasture grassland varies according to intensity of cattle grazing and trampling, soil profundity/depth, shading from forest borders and time since last fire. Plots were divided between swards of prostrate and short-grazed tussock grasses of few cm height (hereafter referred to as low-growing pasture) and areas where grass tussocks and forbs of 10-30 cm height predominated (hereafter referred to as high-growing pasture).
“Nurseries” were areas protected from rodent herbivory by fences of ca. 0.8 m aboveground height and 0.2 m belowground depth, protected from leaf-cutting ants by regular application of formicide and protected from climate extremes by shade-cloth. One of these areas was located in a man-made, formerly forested grassland area near Pró-Mata research station, close to the escarpment (hereafter referred to as Station nursery), the other at ca. 5 km aerial distance inland, in an abandoned grassland area near Pró-Mata entrance (hereafter referred to as field nursery).
3.3.2. Experiment preparation
In southern autumn 2004, Myrsine and Baccharis seeds had been sown into pots filled with sieved grassland soil and left to germinate beneath single layers of shade-cloth in the Station nursery. Around New Year 2004/2005, emerged seedlings were transplanted into 50 ml PVC coffee (“cafezinho”) cups filled with a 2:1 mixture of sieved grassland soil and commercial humus-rich potting soil. This was to allow seedlings to develop a reasonably firm root ball before transplant. In order to harden plants and further reduce the risk of death from transplant shock, shade-cloths were removed, gradually increasing duration of exposure to daylight until seedlings remained permanently exposed for some weeks prior to transplant.
A series of misfortunes made it necessary to pot new cohorts of seedlings several times5. As a consequence, by the end of April 2006, ca. 300 potted individuals per species
5 An rust fungus infection spread among seedlings in January 2006; survivors recovered well after application of a copper sulphate solution. Many seedlings, especially of Baccharis, succumbed - due

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
54
were available but differed considerably in initial size. In order to control for this variation, height and largest leaf length of each individual were measured while still potted, individuals were then divided into size classes by help of PC-ORD 10.0 cluster analysis, and equal numbers of individuals per size class were transplanted into each field and nursery plot. Under field conditions, seedling emergence of Myrsine occurs only in midsummer and late summer (chapter 2), so that initial sizes of transplanted seedlings are probably fairly representative of the size of naturally germinated seedlings at the beginning of winter. Average initial seedling size also differed significantly between Baccharis and Myrsine seedlings (41mm in Baccharis, 37mm in Myrsine), which mirrored, even at the onset of the experiment, different growth rates between the two species.
Seedlings were transplanted on April 24th 2005, i.e. in southern autumn. Five lines of 10 individuals each, with distances of 40-60 cm between individuals, were placed in each grassland type. For logistic reasons, transplant lines were placed adjacent to experimental plots of seedling emergence (chapter 2), previously established at maximum 20 m distance from forest borders and on more or less level ground. Within sites conforming to these requirements, lines were randomly placed; this type of sampling may be described as “stratified random sampling” (Müller-Dombois & Ellenberg 1974).
Each individual received an identification tag (rings of plastic-coated wire pinned down with iron cramps) which greatly facilitated localisation of the plants afterwards, and allowed to trace back individual history prior to transplant and its possible effects on fate of seedlings. In plots with high initial, numbers of transplants were supplemented by one to three at first survey in May 2005, as it was not yet clear to what extent transplant shock might have accounted for high mortality.
Fig. 3.1: Left: Myrsine seedling transplanted into abandoned grassland. Right: Identification tags greatly
facilitated recovery of transplant positions even after pasture burning. Photographer: J.-M. Hermann.
In the fenced nurseries, seedlings were planted into beds one metre square filled with sieved grassland soil and covered with a single layer of shade-cloth. The intention was to exclude as many factors as possible that were assumed to impede optimal growth in grassland, i.e. fire, herbivory, competition, climate extremes. However, as Baccharis growth exceeded all expectations, crowding effects in the beds could no longer be ruled out from midsummer onwards. Asymmetric competition between species presumably occurred as well as intraspecific competition among Baccharis seedlings. Results are therefore not representative of species’ growth at physiological optimum (sensu Ellenberg) but – as also pointed out by De Steven (1991b) who faced the same problem towards the end of her
to handling errors - to the exceptional summer drought in February 2006. In March 2006, more plants of both species were trampled beyond recovery by stray horses, a circumstance not foreseen on construction of the nursery fences.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
55
transplant study – they nevertheless give an valuable idea of growth potential in comparison with plant performance in field plots.
Two field nursery plots were invaded by guinea pigs three months after transplant and these individuals are excluded from further analysis.6
The design of the experiment may be described as 2-factor factorial design (grassland/nursery types x species) but the arrangement is not in completely randomized blocks as would be the ideal case (e.g. Gibson 2002).
3.3.3. Monitoring
Surveys of seedling survival, symptoms of decreasing vitality and death causes were conducted at intervals of 2 weeks until 2 months after transplant while mortality was highest, then at 3-4 week intervals until spring (November 2005) and at 6-week intervals afterwards.
Seedling height and basal diameter were measured for the first time one month after transplant (May 2005), and monitored at 6-week intervals afterwards. After midwinter (September 2005) digital calliper rules permitted precise measurement of basal diameter, which increased in importance as a substitute measure of biomass gain as more and more individuals branched. Stem allometry is described by the stem height:diameter ratio.
The intention was to accompany seedling performance in the field for one year after first survey but imminent conversion of the pasture area into arable land led to closure of the experiment at the end of April 2006, 12 months after transplant and 11 months after first field survey. At the final field survey, largest leaf length was measured again and plants were harvested, photographed and fridge-stored. Leaf allometry values were obtained some weeks afterwards from a random sample of leaves stripped off stored plants, using 50 leaves per plant per grassland and nursery type.
3.3.4. Evaluation
Individuals are treated as replicates in analyses. Ideally, averages of transplant lines and plots should be treated as replicates but this was precluded by the initially low number of lines and plots, and mortality of all but one or two individuals in some lines. Efforts were made to balance the sampling design as best possible initially; however, random small-scale heterogeneity of planting sites (especially grass cover) is so high and individual differences in history and traits are so variable as to lend individuals the status of true replicates.
ANOVA (analysis of variance between grassland and nursery types per species) was combined with post-hoc pairwise comparisons using Tamhane´s T2-Test, which does not assume variance equality. Contrasts between the two species per nursery and grassland type were tested for significance by a T-test for independent samples. All data (except leaf allometry) were ln-transformed prior to analysis in order to improve equality of variances. Analyses were performed using SPSS 14.1.1.
Surrounding vegetation may have ambiguous effects on seedling establishment – for example, germination may be favoured while seedling growth is impeded (see 3.5.5). Following the example of De Steven (1991b), in order to estimate the net effects of certain vegetation matrices, I calculated some simple indices from percentages of seedling emergence (chapter 2), mortality and stem growth. My “seedling growth index“ is 1-year height increase multiplied by 1-year basal area increase. The „seedling performance index“ is the former multiplied by 1-year survivorship (per cent of planted seedlings. This index, multiplied by seedling emergence (per cent of sown seed) gives the “seed-seedling
6 The guinea pig incident did, however, draw attention to an interesting fact: The rodents did not predate any forest tree seedlings within their reach so may not have developed a taste for non-grassland species yet.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
56
performance index”. These indices demonstrate by which order of magnitude establishment site favourability differs between species and grassland types, although they must be interpreted with great caution. They can certainly not replace observations in natural populations but they can supplement them, i.e. help to help to explain high numbers of natural seedling establishment or, otherwise, rule out inhibitory effects if natural seedling establishment is low.
Size development is described by the following parametres: Absolute length of stems and largest leaves at the end of the experiment (after ca. 1year in the field), size of stem height and basal area after 11 months in the field relative to initial size, and size of leaves after 1year in the field relative to initial size. The latter three were obtained by dividing stem height, or basal diameter, or leaf length after a given observation period by values measured at the beginning of that period, and subtracting 100. These parameters, especially if summarised, are considered representative of the relative growth rate of each species in different grassland types, although they are not equivalent to the classical “relative growth rate” (RGR), defined as the product of unit leaf rate and leaf area ratio in time (see e.g. Hutchings 1997a for detail). Hence they are also referred to as “1-year relative height increase” etc.
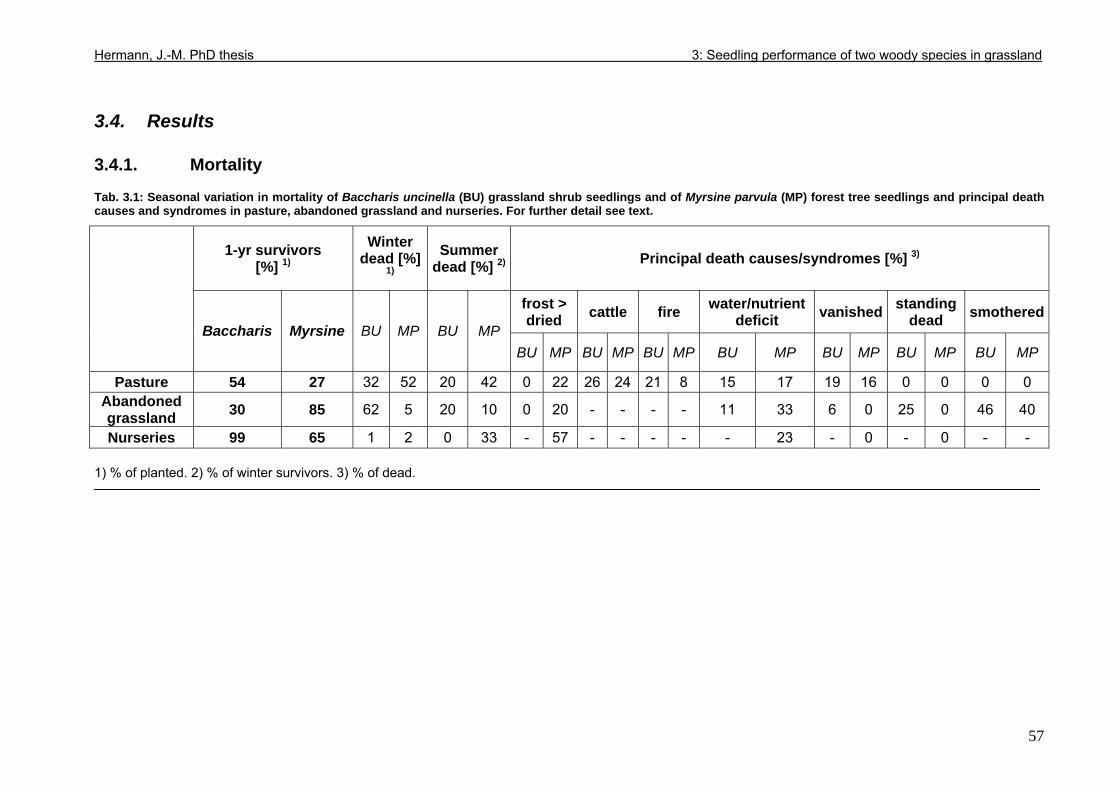
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
57
3.4. Results
3.4.1. Mortality
Tab. 3.1: Seasonal variation in mortality of Baccharis uncinella (BU) grassland shrub seedlings and of Myrsine parvula (MP) forest tree seedlings and principal death
causes and syndromes in pasture, abandoned grassland and nurseries. For further detail see text.
1-yr survivors
[%] 1)
Winter
dead [%]
1)
Summer
dead [%] 2) Principal death causes/syndromes [%]
3)
frost >
dried
cattle fire
water/nutrient
deficit
vanished
standing
dead
smothered
Baccharis Myrsine BU MP BU MP BU MP BU MP BU MP BU MP BU MP BU MP BU MP
Pasture 54 27 32 52 20 42 0 22 26 24 21 8 15 17 19 16 0 0 0 0 Abandoned
grassland
30 85 62 5 20 10 0 20 - - - - 11 33 6 0 25 0 46 40
Nurseries 99 65 1 2 0 33 - 57 - - - - - 23 - 0 - 0 - - 1) % of planted. 2) % of winter survivors. 3) % of dead.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
58
3.4.1.1. Mortality rates Contrasting mortality rates of grassland shrub and forest tree seedlings in managed
and abandoned grassland were among the most striking results of the experiment: The pasture environment proved as inhospitable for forest tree seedlings as abandoned grassland was for grassland shrub seedlings.
One year after transplant, on average 30% of planted grassland shrub seedlings were still alive in abandoned grassland and 54% survived in pasture grassland. Of planted forest tree seedlings, on average 85% were still alive in abandoned grassland but only 27% survived in pasture. In the nurseries practically all (99%) grassland shrub seedlings survived whereas a substantial proportion of forest tree seedlings – 35% – had succumbed in these sites.
The above table contains data on winter mortality of seedlings, expressed as percent of planted seedlings dead 0-6 months after transplant, and summer mortality of seedlings, expressed as percent of winter survivors dead 6-12 months after transplant. In pasture, the winter season turned out more fatal than the summer season for both grassland shrub and forest tree seedlings. In high-growing pasture, mortality of both species peaked in midwinter (4.5 months after transplant); so did mortality of forest tree seedlings in low-growing pasture (not shown in Tab. 3.1).
In abandoned grassland, numbers of grassland shrub survivors decreased rapidly within two months after transplant (also not shown in Tab. 3.1) and winter mortality was clearly higher than summer mortality. In forest tree seedlings transplanted into these sites, however, summer mortality was slightly higher, and this trend was even more pronounced in forest tree seedlings in nurseries.
3.4.1.2. Death causes/syndromes As Fenner (1987b, p.39) writes, “in practice, determining the causes of death of
seedlings presents formidable problems”. Here, some conclusions may be drawn with confidence from circumstantial evidence but other cases involve much guesswork. In no case death of a plant was witnessed at the hour, and the remains especially of grassland shrub seedlings decayed rapidly. It is more appropriate to speak of death “syndromes” rather than causes, and any percentages given must be understood as approximate figures.
“Cattle” influence, i.e. grazing and trampling, was assumed whenever plants were found pulled up next to the plot, or disappeared along with their identification tag. Such cases were restricted to pasture plots, and were more frequent in low-growing than in high-growing pasture. Some plants were found broken or sites were covered in dung. Summed up, these cases accounted for ca. 1/4 of mortality in both Baccharis and Myrsine seedlings in pasture. Cattle were probably also involved in some cases of “vanished” individuals, i.e. plants disappeared while tags remained fixed to the ground, which occurred more frequently in pasture than in other sites, and accounted for 19% of grassland shrub and 16% of forest tree mortality there.
Frosts first occurred at the end of May 2005 (1 month after transplant) and last in the beginning of spring (1st week of November 2005). Frost and winter cold damaged foliage and apical meristems of major proportions of Myrsine seedlings. In some cases this caused immediate death of the plant but more frequently, plants were reduced to stumps that dried out after several weeks or even months; this delay partly explains the peak in Myrsine mortality in nurseries in summer (see also 3.4.1.3). Death after previous frost damage accounted for 57% of dead in nurseries, ca. 1/5 of dead in pasture, most of them in low-growing sites, and even affected some seedlings in gaps of abandoned grassland. Closed

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
59
grass matrices of abandoned sites sheltered forest tree seedlings from climate extremes (see also growth rates, 3.4.2.1).
In contrast, grassland shrub seedlings suffered no physical damage from frost. The midwinter peak in shrub seedling mortality especially in high-growing pasture may be due to more frequent visitation of these sites by cattle as fodder availability decreased.
Fire occurred in pasture towards the end of winter (August 2005) and accounted for ca. 1/5 of winter mortality of grassland shrubs in pasture. In forest tree seedlings it was a death cause of relatively small importance, and there may be two reasons for this: Firstly, the patch-wise spread of the fire (only a minor proportion of transplant sites, located exclusively in high-growing pasture, were burnt) and secondly, the fact that winter climate affected this species sooner.
“Standing dead” Baccharis seedlings were found exclusively in abandoned grassland, greyish dry leaves still attached to the stem. Similar symptoms were observed in plants deliberately “starved” (watered but enclosed in brown paper bags; pers. obs.). A more or less closed high shrub canopy reduces daily average PAR to ca. 10% of that received by open short grassland (chapter 1), possibly below the level required for grassland shrubs to reach light compensation point - although in nurseries, where PAR amounted to 20 to 30% of open area values, grassland shrub seedlings achieved considerably higher growth rates than forest tree seedlings. Severe competition for soil resources by surrounding grass tussocks and adult shrubs may be another cause of starvation. Furthermore, secondary causes of starvation, e.g. deficiencies of root systems, cannot be ruled out, even more so as there is evidence of fungal infections (genus Pythium) in tall-shrub stands of the study region (see chapter 5).
“Smothered” plants disappeared entirely in the dense grass matrix and grass litter of abandoned grassland (a condition that was aggravated by handling the vegetation during monitoring7). Plausible death causes are starvation - dense grass cover reduces average PAR to merely 1% of open area PAR -, suffocation or breaking of stems under cover. During the first months after transplant, almost half of grassland shrub dead (46%) succumbed under these conditions. Together with “standing dead”, 71% of mortality in abandoned grassland must probably be attributed to detrimental influence of the surrounding grass, shrub and litter matrix. Generally, grass and litter cover were better tolerated by forest tree seedlings - note that although 40% of forest tree dead died “smothered”, the absolute numbers behind this figure were lower than in grassland shrubs.
Impaired uptake of water and nutrients was assumed when leaves turned yellow and were shed, beginning with eldest leaves8. This was the second most important death syndrome of Myrsine seedlings in nurseries and abandoned grassland and also one of the principal death syndromes of either species in pasture. However, primary causes of such deficits could not be determined (distinguishing, by visual plant symptoms, between primary and secondary causes of drought is generally problematic; Kozlowski et al. 1991, Larcher 2005), and they may have been as variable as types of transplant sites - conditions of substrate, climate, infection of the root systems etc. may all have contributed to some degree.
7 In tall tussock abandoned grassland especially, standing vegetation consists of entangled live and dead biomass, and invariably, on opening and closing, dead biomass would become detached and accumulate on the soil surface and the seedling plant. 8 A minor proportion of plants were affected by yellowing and shedding of leaves from the apex downwards which can be attributed with more confidence to an infection by fungi e.g. of the genera Verticillium or Pythium. Both types thrive particularly well under warm temperatures in excessively humid soils (M. Dalbosco pers. comm.; see also e.g. Schütt et al. 2002).

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
60
3.4.1.3. “Ned”s and Resprouters From the beginning of winter onwards, substantial proportions of forest tree seedlings
lost all foliage and the upper part of the stem, in some cases several times. Such individuals were initially considered dead, until the discovery that they may resprout, as long as the remaining stem stump does not wilt or dry out, even if that stump is no more than a few millimetres high. These seedlings were nicknamed “Ned”s - Not Entirely Dead.
“Ned”s were principally a winter phenomenon in pasture and nurseries. During the entire one-year observation period, proportions of plants recorded as “Ned” at least once amounted to 43% of planted seedlings in pasture, 31% in nurseries and 9% in abandoned grassland. Ca. half of all “Ned’s” (48%) died, the other half resprouted. In survivor statistics (Tab. 3.1), “Ned’s” entered as fully alive individuals until terminal damage.
By far the majority of all “Ned” cases (78%) were observed in winter (between June and October 2005) and were victims of frost. 12% of cases were probably caused by root or soil resource deficiency (see previous paragraph), which also occurred sporadically in springtime and in abandoned grassland.
In pastures, the average period during which an individual would be recorded as “Ned” – and then either resprout or die – was 6.5 weeks. Nursery “Ned”s would resprout a little faster (after 5 weeks on average) or last a little longer before dying (7.5 weeks on average). By the end of the experiment, all “Ned’s” except one had either resprouted or died.
“Ned” numbers may have been even higher, as a further 26% of planted Myrsine seedlings were recorded resprouting after biomass loss, which in almost all cases could also be attributed to frost. The majority of these resprouters were recorded during winter in the nurseries (73% to 76%).
Summing up, 43% of all planted Myrsine seedlings suffered more or less severe biomass loss and resprouted. In nurseries and pasture, proportions of affected plants were so high that for these sites, non-resprouters are excluded from size development statistics. Although overall resprouting success of Myrsine seedlings was considerable, resprouting vigour differed greatly between grassland and nursery types, as growth rates and final sizes show (see following paragraph).
3.4.2. Stem and leaf development
3.4.2.1. Stem and leaf size development In the pair-wise comparisons described in the following, differences are termed
significant at p=0.05 level unless otherwise stated. While absolute height development of stems is of interest with regard to the
surrounding grass matrix, basal area development gives a more accurate impression of biomass gain as most of grassland shrub individuals branched during the observation period, and forest tree resprouters especially in Station nursery developed two or more basal resprouts.
Size development in pasture grassland, abandoned grassland and nurseries mirrored opposing preferences of the two species in a similar way as mortality did (3.4.1). Final height of Baccharis seedlings was generally significantly higher than that of forest trees and growth rates of height and basal area significantly surpassed those of Myrsine seedlings in all sites except abandoned grassland; more vigorous stem growth had occurred in this species even prior to transplant (see 3.3.2). However, forest tree seedlings excelled grassland shrubs significantly in final largest leaf length in all sites, and in leaf length increase in all sites except pasture.
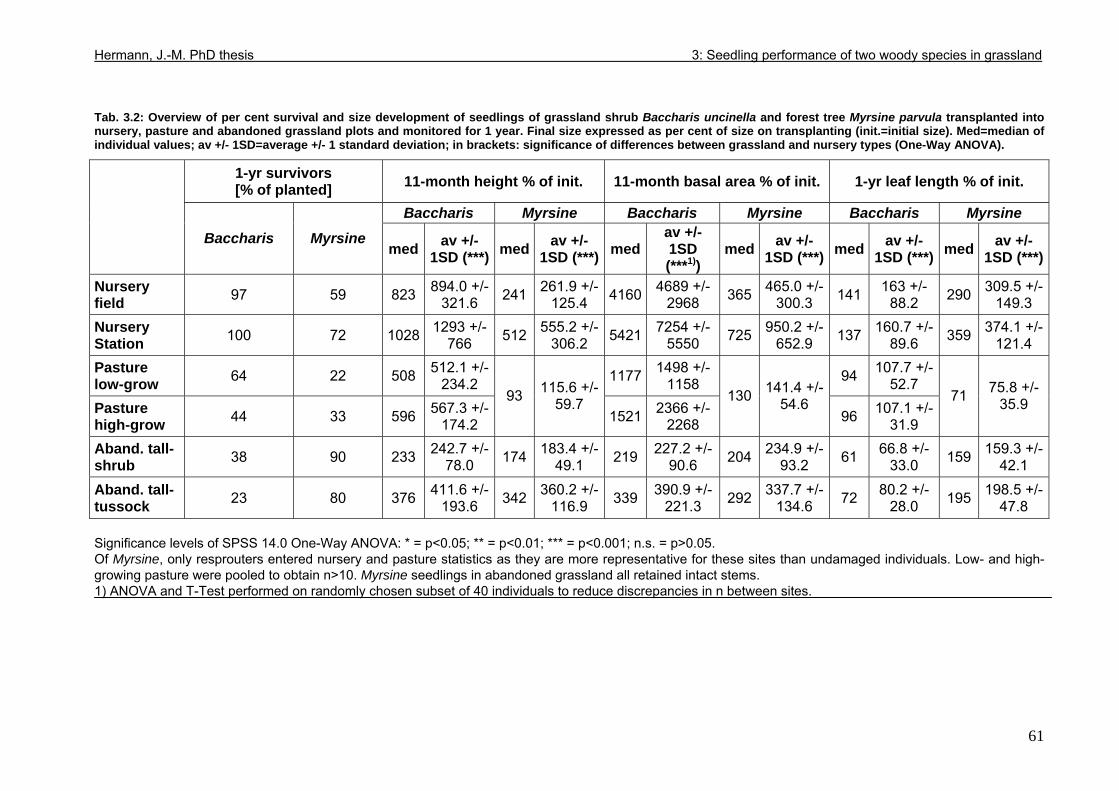
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
61
Tab. 3.2: Overview of per cent survival and size development of seedlings of grassland shrub Baccharis uncinella and forest tree Myrsine parvula transplanted into
nursery, pasture and abandoned grassland plots and monitored for 1 year. Final size expressed as per cent of size on transplanting (init.=initial size). Med=median of
individual values; av +/- 1SD=average +/- 1 standard deviation; in brackets: significance of differences between grassland and nursery types (One-Way ANOVA).
1-yr survivors
[% of planted]
11-month height % of init. 11-month basal area % of init. 1-yr leaf length % of init.
Baccharis Myrsine Baccharis Myrsine Baccharis Myrsine Baccharis Myrsine
med
av +/-
1SD (***)
med
av +/-
1SD (***)
med
av +/-
1SD
(***1)
)
med
av +/-
1SD (***)
med
av +/-
1SD (***)
med
av +/-
1SD (***)
Nursery
field
97 59 823 894.0 +/- 321.6 241 261.9 +/-
125.4 4160 4689 +/- 2968 365 465.0 +/-
300.3 141 163 +/- 88.2 290 309.5 +/-
149.3 Nursery
Station
100 72 1028 1293 +/- 766 512 555.2 +/-
306.2 5421 7254 +/- 5550 725 950.2 +/-
652.9 137 160.7 +/- 89.6 359 374.1 +/-
121.4 Pasture
low-grow
64 22 508 512.1 +/- 234.2 1177 1498 +/-
1158 94 107.7 +/- 52.7
Pasture
high-grow
44 33 596 567.3 +/- 174.2
93 115.6 +/- 59.7
1521 2366 +/- 2268
130 141.4 +/- 54.6
96 107.1 +/- 31.9
71 75.8 +/- 35.9
Aband. tall-
shrub
38 90 233 242.7 +/- 78.0 174 183.4 +/-
49.1 219 227.2 +/- 90.6 204 234.9 +/-
93.2 61 66.8 +/- 33.0 159 159.3 +/-
42.1 Aband. tall-
tussock
23 80 376 411.6 +/- 193.6 342 360.2 +/-
116.9 339 390.9 +/- 221.3 292 337.7 +/-
134.6 72 80.2 +/- 28.0 195 198.5 +/-
47.8 Significance levels of SPSS 14.0 One-Way ANOVA: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05. Of Myrsine, only resprouters entered nursery and pasture statistics as they are more representative for these sites than undamaged individuals. Low- and high-growing pasture were pooled to obtain n>10. Myrsine seedlings in abandoned grassland all retained intact stems. 1) ANOVA and T-Test performed on randomly chosen subset of 40 individuals to reduce discrepancies in n between sites.
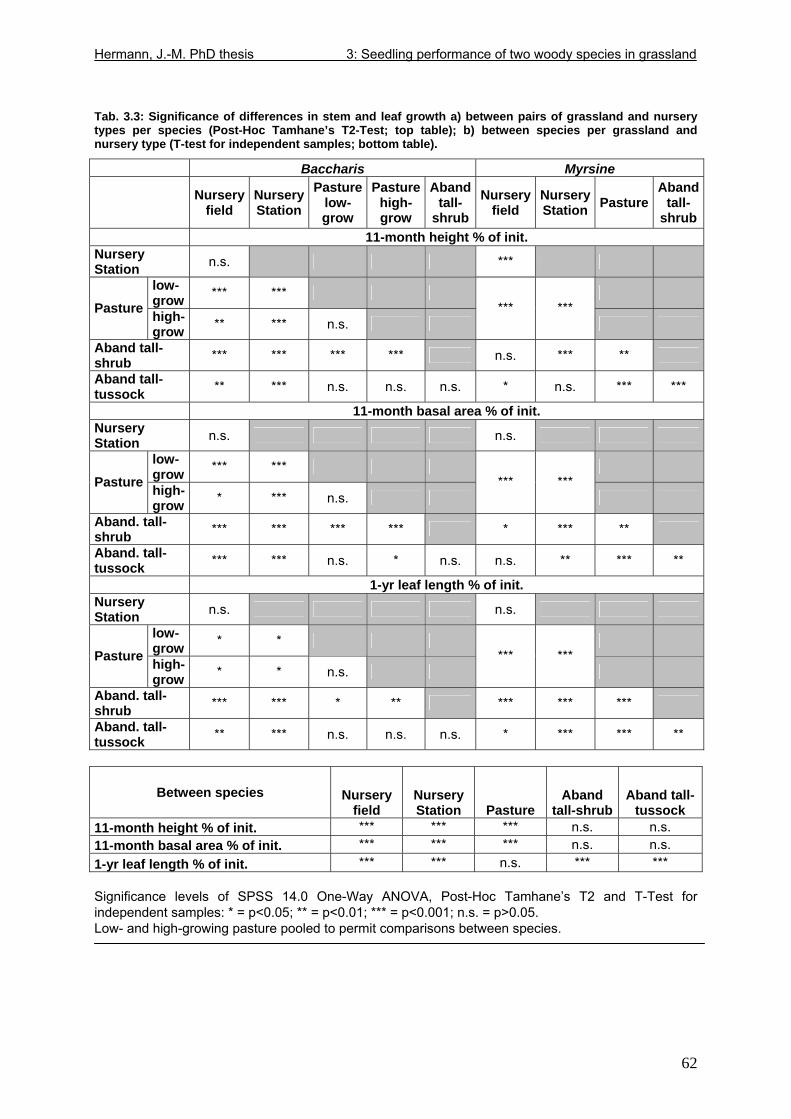
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
62
Tab. 3.3: Significance of differences in stem and leaf growth a) between pairs of grassland and nursery
types per species (Post-Hoc Tamhane’s T2-Test; top table); b) between species per grassland and
nursery type (T-test for independent samples; bottom table).
Baccharis Myrsine
Nursery
field
Nursery
Station
Pasture
low-
grow
Pasture
high-
grow
Aband
tall-
shrub
Nursery
field
Nursery
Station
Pasture
Aband
tall-
shrub
11-month height % of init.
Nursery
Station
n.s. ***
low-
grow
*** *** Pasture
high-
grow
** *** n.s. *** ***
Aband tall-
shrub
*** *** *** *** n.s. *** **
Aband tall-
tussock
** *** n.s. n.s. n.s. * n.s. *** ***
11-month basal area % of init.
Nursery
Station
n.s. n.s.
low-
grow
*** *** Pasture
high-
grow
* *** n.s. *** ***
Aband. tall-
shrub
*** *** *** *** * *** **
Aband. tall-
tussock
*** *** n.s. * n.s. n.s. ** *** **
1-yr leaf length % of init.
Nursery
Station
n.s. n.s.
low-
grow
* * Pasture
high-
grow
* * n.s. *** ***
Aband. tall-
shrub
*** *** * ** *** *** ***
Aband. tall-
tussock
** *** n.s. n.s. n.s. * *** *** **
Between species Nursery
field
Nursery
Station Pasture
Aband
tall-shrub
Aband tall-
tussock
11-month height % of init. *** *** *** n.s. n.s. 11-month basal area % of init. *** *** *** n.s. n.s. 1-yr leaf length % of init. *** *** n.s. *** *** Significance levels of SPSS 14.0 One-Way ANOVA, Post-Hoc Tamhane’s T2 and T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05. Low- and high-growing pasture pooled to permit comparisons between species.
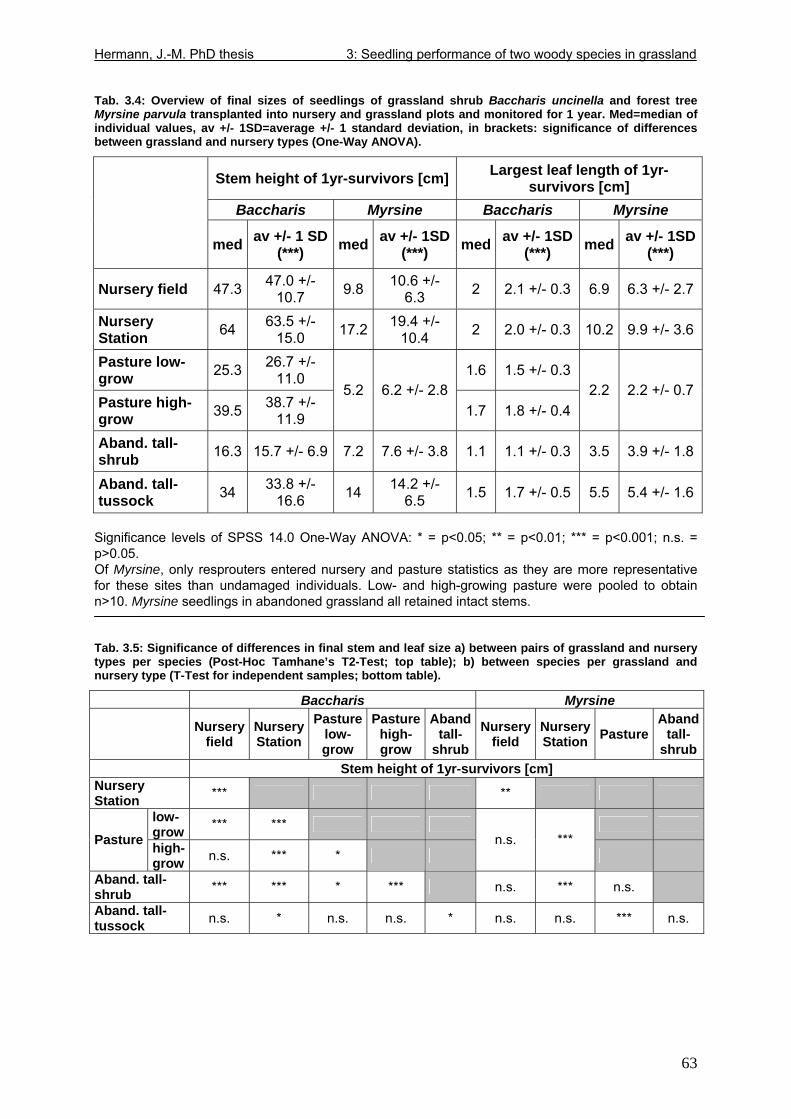
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
63
Tab. 3.4: Overview of final sizes of seedlings of grassland shrub Baccharis uncinella and forest tree
Myrsine parvula transplanted into nursery and grassland plots and monitored for 1 year. Med=median of
individual values, av +/- 1SD=average +/- 1 standard deviation, in brackets: significance of differences
between grassland and nursery types (One-Way ANOVA).
Stem height of 1yr-survivors [cm]
Largest leaf length of 1yr-
survivors [cm]
Baccharis Myrsine Baccharis Myrsine
med
av +/- 1 SD
(***)
med
av +/- 1SD
(***)
med
av +/- 1SD
(***)
med
av +/- 1SD
(***)
Nursery field 47.3 47.0 +/- 10.7 9.8 10.6 +/-
6.3 2 2.1 +/- 0.3 6.9 6.3 +/- 2.7
Nursery
Station
64 63.5 +/- 15.0 17.2 19.4 +/-
10.4 2 2.0 +/- 0.3 10.2 9.9 +/- 3.6
Pasture low-
grow
25.3 26.7 +/- 11.0 1.6 1.5 +/- 0.3
Pasture high-
grow
39.5 38.7 +/- 11.9
5.2 6.2 +/- 2.81.7 1.8 +/- 0.4
2.2 2.2 +/- 0.7
Aband. tall-
shrub
16.3 15.7 +/- 6.9 7.2 7.6 +/- 3.8 1.1 1.1 +/- 0.3 3.5 3.9 +/- 1.8
Aband. tall-
tussock
34 33.8 +/- 16.6 14 14.2 +/-
6.5 1.5 1.7 +/- 0.5 5.5 5.4 +/- 1.6
Significance levels of SPSS 14.0 One-Way ANOVA: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05. Of Myrsine, only resprouters entered nursery and pasture statistics as they are more representative for these sites than undamaged individuals. Low- and high-growing pasture were pooled to obtain n>10. Myrsine seedlings in abandoned grassland all retained intact stems.
Tab. 3.5: Significance of differences in final stem and leaf size a) between pairs of grassland and nursery
types per species (Post-Hoc Tamhane’s T2-Test; top table); b) between species per grassland and
nursery type (T-Test for independent samples; bottom table).
Baccharis Myrsine
Nursery
field
Nursery
Station
Pasture
low-
grow
Pasture
high-
grow
Aband
tall-
shrub
Nursery
field
Nursery
Station
Pasture
Aband
tall-
shrub
Stem height of 1yr-survivors [cm]
Nursery
Station
*** **
low-
grow
*** *** Pasture
high-
grow
n.s. *** * n.s. ***
Aband. tall-
shrub
*** *** * *** n.s. *** n.s.
Aband. tall-
tussock
n.s. * n.s. n.s. * n.s. n.s. *** n.s.
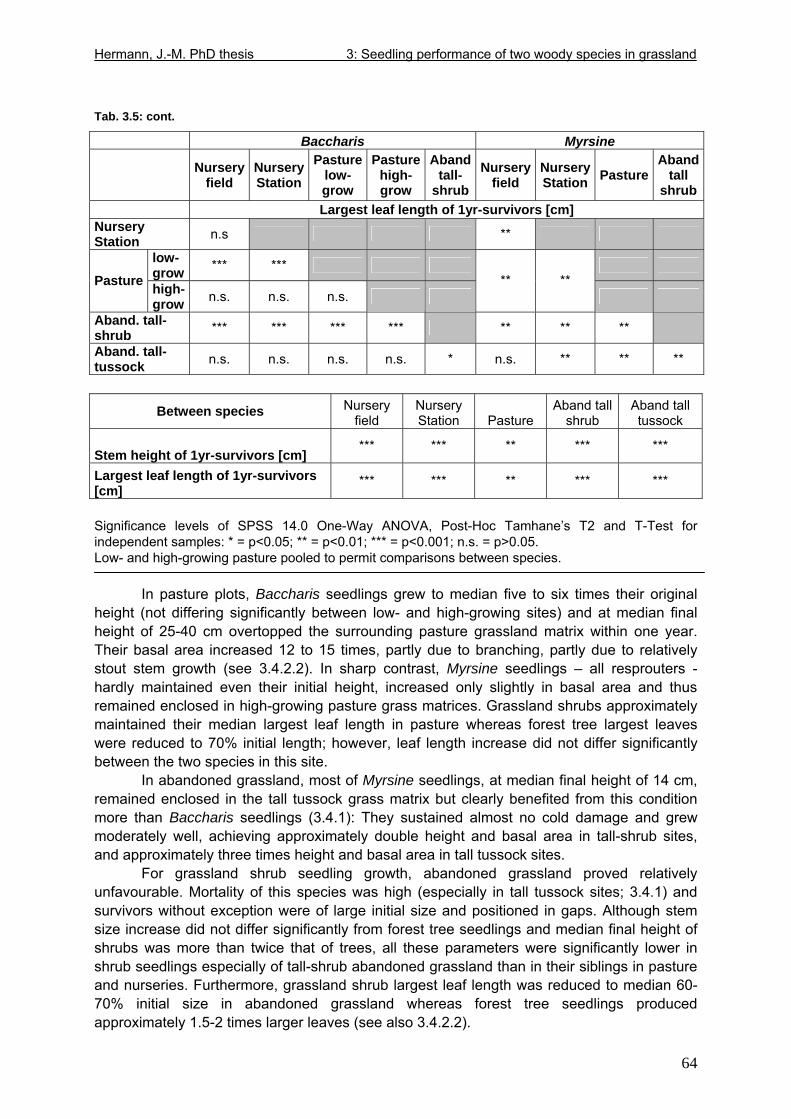
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
64
Tab. 3.5: cont.
Baccharis Myrsine
Nursery
field
Nursery
Station
Pasture
low-
grow
Pasture
high-
grow
Aband
tall-
shrub
Nursery
field
Nursery
Station
Pasture
Aband
tall
shrub
Largest leaf length of 1yr-survivors [cm]
Nursery
Station
n.s **
low-
grow
*** *** Pasture
high-
grow
n.s. n.s. n.s. ** **
Aband. tall-
shrub
*** *** *** *** ** ** **
Aband. tall-
tussock
n.s. n.s. n.s. n.s. * n.s. ** ** **
Between species Nursery
field Nursery Station Pasture
Aband tall shrub
Aband tall tussock
Stem height of 1yr-survivors [cm]
*** *** ** *** ***
Largest leaf length of 1yr-survivors
[cm]
*** *** ** *** ***
Significance levels of SPSS 14.0 One-Way ANOVA, Post-Hoc Tamhane’s T2 and T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05. Low- and high-growing pasture pooled to permit comparisons between species.
In pasture plots, Baccharis seedlings grew to median five to six times their original
height (not differing significantly between low- and high-growing sites) and at median final height of 25-40 cm overtopped the surrounding pasture grassland matrix within one year. Their basal area increased 12 to 15 times, partly due to branching, partly due to relatively stout stem growth (see 3.4.2.2). In sharp contrast, Myrsine seedlings – all resprouters - hardly maintained even their initial height, increased only slightly in basal area and thus remained enclosed in high-growing pasture grass matrices. Grassland shrubs approximately maintained their median largest leaf length in pasture whereas forest tree largest leaves were reduced to 70% initial length; however, leaf length increase did not differ significantly between the two species in this site.
In abandoned grassland, most of Myrsine seedlings, at median final height of 14 cm, remained enclosed in the tall tussock grass matrix but clearly benefited from this condition more than Baccharis seedlings (3.4.1): They sustained almost no cold damage and grew moderately well, achieving approximately double height and basal area in tall-shrub sites, and approximately three times height and basal area in tall tussock sites.
For grassland shrub seedling growth, abandoned grassland proved relatively unfavourable. Mortality of this species was high (especially in tall tussock sites; 3.4.1) and survivors without exception were of large initial size and positioned in gaps. Although stem size increase did not differ significantly from forest tree seedlings and median final height of shrubs was more than twice that of trees, all these parameters were significantly lower in shrub seedlings especially of tall-shrub abandoned grassland than in their siblings in pasture and nurseries. Furthermore, grassland shrub largest leaf length was reduced to median 60-70% initial size in abandoned grassland whereas forest tree seedlings produced approximately 1.5-2 times larger leaves (see also 3.4.2.2).

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
65
Even in most favourable grassland sites, either species remained below their potential growth rate, as evident from comparison with individual development in nurseries, especially the Station nursery. In the nurseries, Baccharis seedlings grew to approximately 8-10 times their initial height and 40-50 times their initial basal area. In Myrsine seedlings, there was also a trend towards maximum growth of stems and leaves in the Station nursery, although height increase did not differ significantly between this site and tall tussock abandoned grassland. Notably, Station nursery individuals suffered frost damage to considerably extent – as in pasture, only resprouters entered statistics - whereas individuals remained undamaged in tall tussock grassland. Largest leaf length increase of both species was also highest in the nurseries, although differences between nursery and grassland sites were more pronounced (more often highly significant) in forest tree seedlings than grassland shrub seedlings.
In spite of large within-site differences, grassland shrub seedlings generally grew to significantly taller stature than forest tree seedlings whereas the latter developed significantly larger leaves.
3.4.2.2. Stem and leaf allometry and morphology In stem and leaf allometry (stem height:basal diameter ratio), the following phenomena
were striking: Myrsine seedlings generally developed stouter stems than Baccharis seedlings (the
difference was significant at p=0.00 level in all sites and in both seasons). Comparison of grassland shrub seedlings from high-growing pasture with forest tree seedlings from tall tussock abandoned grassland (the respective grassland sites in which the two species grew best) reveals that shrub seedlings invested 2.5-3 times more in height growth, relative to diameter growth, than forest tree seedlings.
In all grassland types, stems were stouter at the end of winter than at the end of summer. Furthermore, at the end of winter, stems were stouter in pasture than in abandoned grassland in both species. In forest trees this was enforced by frost and cold damage to stems but in grassland shrub seedlings, which did not suffer damage to apical meristems, it was obviously an acquired trait.
During summer, damaged forest tree seedlings in nurseries and pasture resprouted and by the end of summer they achieved about the same stem height:diameter ratio that undamaged forest tree seedlings in abandoned grassland had featured by the end of winter. Only resprouters in pasture continued with significantly stouter stems than the rest. Most slender stems were produced by undamaged seedlings in tall tussock abandoned grassland: End-of-summer stem allometry differed significantly between this and all other sites (including tall-shrub abandoned grassland) at p=0.00 level.
Baccharis seedlings in nursery developed a densely branched habit during summer while basal area increased; thus the stem height:diameter ratio remained approximately the same as in winter in the Station nursery, and even decreased in the field nursery. Individuals in field plots stretched so that end-of-summer stem allometry did not differ significantly between any of these sites, although grassland shrub seedlings in low-growing pasture retained the stoutest growth of all grassland sites.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
66
Tab. 3.6: Overview of stem and leaf allometry of seedlings of grassland shrub Baccharis uncinella and forest tree Myrsine parvula transplanted into nursery and
grassland plots and monitored for 1 year. End-of-winter survey performed October 2005, five months after transplant. End-of summer survey performed April 2006, at
closure of the experiment. Med=median of individual values, av +/- 1SD=average +/- 1 standard deviation, in brackets: significance of differences between grassland
and nursery types (One-Way ANOVA).
End-of-winter stem allometry
[height:dbas]
End-of-summer stem allometry
[height:dbas]
End-of-summer leaf allometry
[length:width]
Baccharis Myrsine Baccharis Myrsine Baccharis Myrsine
med
av +/- 1SD
(***1)
)
med
av +/- 1SD
(***)
med
av +/- 1SD
(***1)
)
med
av +/- 1SD
(***)
med
av +/- 1SD
(n.s.)
med
av +/- 1SD
(***)
Nursery field 100 98.1 +/- 23.4 14 15.4 +/- 8.0 85 90.0 +/- 15.9 26 27.6 +/- 9.0 3.1 3.2 +/- 0.4 2.7 2.7 +/- 0.4
Nursery Station 102 103.0 +/- 20.0 19 17.6 +/- 7.7 101 104.8 +/- 19.9 36 34.9 +/- 10.4 3.2 3.2 +/- 0.4 3.3 3.3 +/- 0.3
Pasture low-
grow
65 63.6 +/- 22.2 91 103.3 +/- 38.0 3.2 3.2 +/- 0.4
Pasture high-
grow
84 84.5 +/- 35.8 8 11.2 +/- 8.1
135 132.5 +/- 35.0 21 20.1 +/- 4.9
3 3 +/- 0.4 2.2 2.3 +/- 0.4
Aband. tall-
shrub
90 86.7 +/- 26.8 26 27.2 +/- 7.2 127 124.9 +/- 37.8 32 35.7 +/- 13.5 3.2 3.3 +/- 0.8 2.6 2.7 +/- 0.4
Aband. tall-
tussock
107 104.1 +/- 18.1 35 33.4 +/- 11.2 127 145.4 +/- 60.2 54 52.7 +/- 14.9 3.1 3.1 +/- 0.4 2.8 2.8 +/- 0.4
Significance levels of SPSS 14.0 One-Way ANOVA: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05. Of Myrsine, only resprouters entered nursery and pasture statistics as they are more representative for these sites than undamaged individuals. Low- and high-growing pasture were pooled to obtain n>10. Myrsine seedlings in abandoned grassland all retained intact stems. Leaf allometry measured on randomly chosen sample of 50 harvested leaves per species.
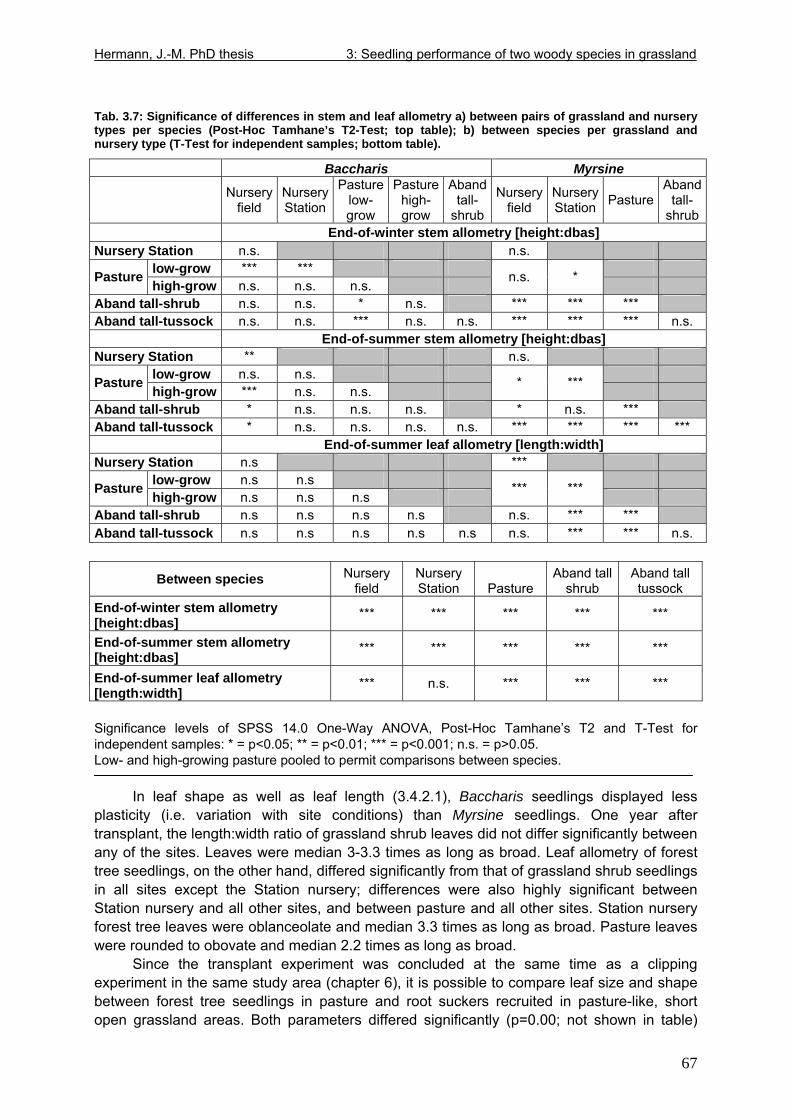
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
67
Tab. 3.7: Significance of differences in stem and leaf allometry a) between pairs of grassland and nursery
types per species (Post-Hoc Tamhane’s T2-Test; top table); b) between species per grassland and
nursery type (T-Test for independent samples; bottom table).
Baccharis Myrsine
Nursery field
Nursery Station
Pasture low-grow
Pasture high-grow
Aband tall-
shrub
Nursery field
Nursery Station Pasture
Aband tall-
shrub End-of-winter stem allometry [height:dbas]
Nursery Station n.s. n.s. low-grow *** ***
Pasture
high-grow n.s. n.s. n.s. n.s. *
Aband tall-shrub n.s. n.s. * n.s. *** *** *** Aband tall-tussock n.s. n.s. *** n.s. n.s. *** *** *** n.s.
End-of-summer stem allometry [height:dbas]
Nursery Station ** n.s. low-grow n.s. n.s.
Pasture
high-grow *** n.s. n.s. * ***
Aband tall-shrub * n.s. n.s. n.s. * n.s. *** Aband tall-tussock * n.s. n.s. n.s. n.s. *** *** *** ***
End-of-summer leaf allometry [length:width]
Nursery Station n.s *** low-grow n.s n.s
Pasture
high-grow n.s n.s n.s *** ***
Aband tall-shrub n.s n.s n.s n.s n.s. *** *** Aband tall-tussock n.s n.s n.s n.s n.s n.s. *** *** n.s.
Between species Nursery
field Nursery Station Pasture
Aband tall shrub
Aband tall tussock
End-of-winter stem allometry
[height:dbas]
*** *** *** *** ***
End-of-summer stem allometry
[height:dbas]
*** *** *** *** ***
End-of-summer leaf allometry
[length:width]
*** n.s. *** *** ***
Significance levels of SPSS 14.0 One-Way ANOVA, Post-Hoc Tamhane’s T2 and T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05. Low- and high-growing pasture pooled to permit comparisons between species.
In leaf shape as well as leaf length (3.4.2.1), Baccharis seedlings displayed less plasticity (i.e. variation with site conditions) than Myrsine seedlings. One year after transplant, the length:width ratio of grassland shrub leaves did not differ significantly between any of the sites. Leaves were median 3-3.3 times as long as broad. Leaf allometry of forest tree seedlings, on the other hand, differed significantly from that of grassland shrub seedlings in all sites except the Station nursery; differences were also highly significant between Station nursery and all other sites, and between pasture and all other sites. Station nursery forest tree leaves were oblanceolate and median 3.3 times as long as broad. Pasture leaves were rounded to obovate and median 2.2 times as long as broad.
Since the transplant experiment was concluded at the same time as a clipping experiment in the same study area (chapter 6), it is possible to compare leaf size and shape between forest tree seedlings in pasture and root suckers recruited in pasture-like, short open grassland areas. Both parameters differed significantly (p=0.00; not shown in table)
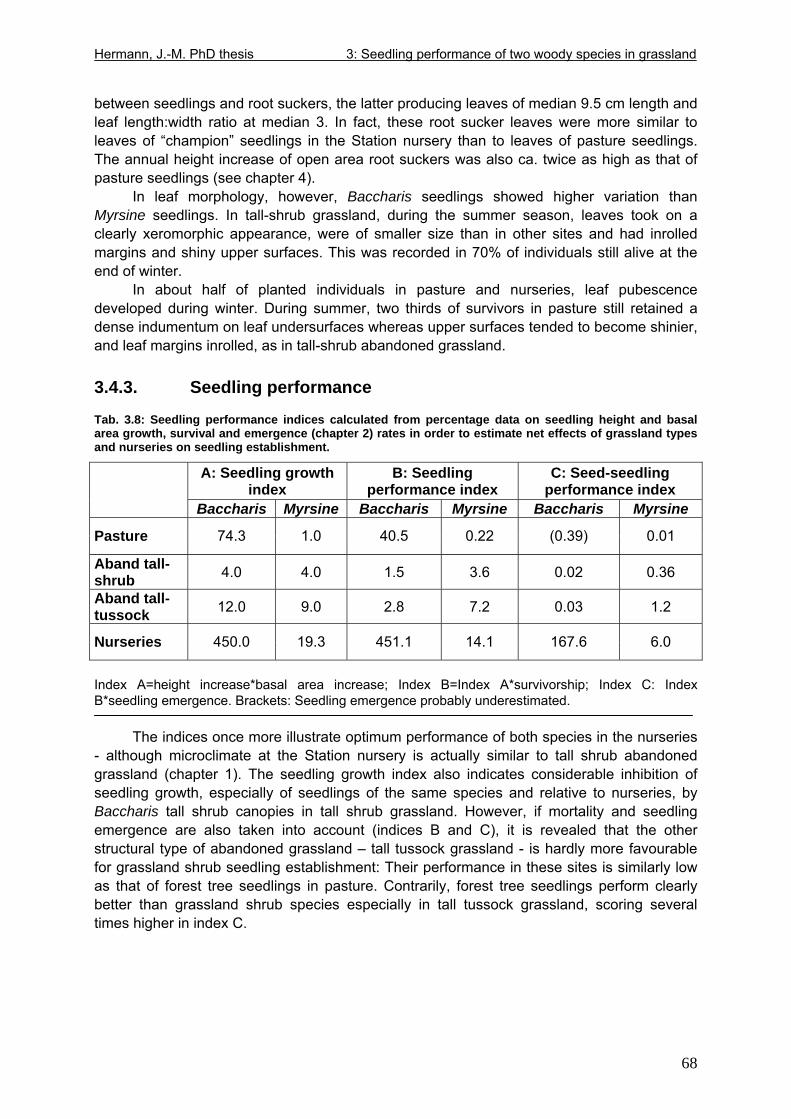
Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
68
between seedlings and root suckers, the latter producing leaves of median 9.5 cm length and leaf length:width ratio at median 3. In fact, these root sucker leaves were more similar to leaves of “champion” seedlings in the Station nursery than to leaves of pasture seedlings. The annual height increase of open area root suckers was also ca. twice as high as that of pasture seedlings (see chapter 4).
In leaf morphology, however, Baccharis seedlings showed higher variation than Myrsine seedlings. In tall-shrub grassland, during the summer season, leaves took on a clearly xeromorphic appearance, were of smaller size than in other sites and had inrolled margins and shiny upper surfaces. This was recorded in 70% of individuals still alive at the end of winter.
In about half of planted individuals in pasture and nurseries, leaf pubescence developed during winter. During summer, two thirds of survivors in pasture still retained a dense indumentum on leaf undersurfaces whereas upper surfaces tended to become shinier, and leaf margins inrolled, as in tall-shrub abandoned grassland.
3.4.3. Seedling performance
Tab. 3.8: Seedling performance indices calculated from percentage data on seedling height and basal
area growth, survival and emergence (chapter 2) rates in order to estimate net effects of grassland types
and nurseries on seedling establishment.
A: Seedling growth
index
B: Seedling
performance index
C: Seed-seedling
performance index Baccharis Myrsine Baccharis Myrsine Baccharis Myrsine
Pasture 74.3 1.0 40.5 0.22 (0.39) 0.01
Aband tall-
shrub
4.0 4.0 1.5 3.6 0.02 0.36
Aband tall-
tussock
12.0 9.0 2.8 7.2 0.03 1.2
Nurseries 450.0 19.3 451.1 14.1 167.6 6.0
Index A=height increase*basal area increase; Index B=Index A*survivorship; Index C: Index B*seedling emergence. Brackets: Seedling emergence probably underestimated.
The indices once more illustrate optimum performance of both species in the nurseries - although microclimate at the Station nursery is actually similar to tall shrub abandoned grassland (chapter 1). The seedling growth index also indicates considerable inhibition of seedling growth, especially of seedlings of the same species and relative to nurseries, by Baccharis tall shrub canopies in tall shrub grassland. However, if mortality and seedling emergence are also taken into account (indices B and C), it is revealed that the other structural type of abandoned grassland – tall tussock grassland - is hardly more favourable for grassland shrub seedling establishment: Their performance in these sites is similarly low as that of forest tree seedlings in pasture. Contrarily, forest tree seedlings perform clearly better than grassland shrub species especially in tall tussock grassland, scoring several times higher in index C.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
69
3.5. Discussion
3.5.1. Escarpment compared to inland sites
Since Station and field nursery were constructed in similar ways - both offering herbivore exclusion and flat beds filled with grassland soil - differences in mortality and growth between the two sites reflect a slight but perceptible change in macroclimate (chapter 1) and perhaps other site conditions. Both species achieved maximum growth near the escarpment, Myrsine even although handicapped by cold and frost damage in winter.
Notably, Myrsine leaf size and allometry achieved in the Station nursery was not matched by seedlings anywhere in the field, but by Myrsine root suckers (harvested after a cutting experiment, see chapter 4). Root suckers can achieve high initial growth rates across a broad range of environmental conditions by regular supply of assimilates either stored in roots or provided by linked individuals (see chapter 4). It may be hypothesized that one or several factors at the Station nursery allow for similarly constant rates of resource supply. For example, microclimate in the Station nursery is characterised by relatively narrow amplitudes in temperature and PAR as well as higher air humidity than in the field nursery, probably as a consequence of longer duration of fogs (chapter 1). This may allow for continuous stomatal aperture and consequently, more constant rates of photosynthesis than in sites exposed to full sunlight; constant assimilation may even compensate low nutrient availability of soils to some degree, as in so-called “peinomorphous” bog plants (Ellenberg 1996). The hypothesis is intriguing but there is, possibly, a considerable flaw in the argument: Namely, that fog may as well cause a decrease photosynthetic rate as light intensity is reduced and gas exchange is decreased or inhibited by excess humidity around leaves (e.g. Mohr & Schopfer 1992). On the other hand, neither drastically reduced PAR nor high humidity in tall tussock grassland (chapter 1) prevented Myrsine from achieving highest growth rates in this grassland type.
Although a plausible explanation cannot yet be given, the contrast between Station and field nursery calls to mind in how many aspects forest expansion over grassland in this area differs from studies of secondary succession e.g. in Middle European abandoned pastures and North American oldfields: Here, a mixture of temperate, subtropical and tropical species are invading a habitat that is (yet) classified as temperate. How relevant may these climatic differences be? According to Larcher (e.g. Larcher 2005), some adaptations to cold and frost may be expected in subtropical species as a group, such as protection of freezing-sensitive parts by supercooling; but he also points out that episodic frosts, such as occur in the Riograndean highland, are more potentially harmful than periodic frosts against which plants may be gradually hardened off.
Recently, Brando & Durigan (2004) reported that half of 57 species in a Cerrado community had all of their aerial parts killed by frost. The majority of individuals, however, belonged to frost-resistant species and suffered no visible damage. The authors therefore suggest that frost does exert selective power on community composition and causes gap formation even where soil resource supply may allow for closed vegetation cover, but does not preclude the establishment of a Cerrado community as such. However, their study is concerned largely with subtropical species in a subtropical climate. Can the impact of frost and of cooler temperatures on species growth rates be sufficient to preclude subtropical species in temperate grassland?
In a study of similar context, Cunningham & Read (2003) exposed seedlings of temperate and tropical Australian rainforest species to a range of day/night temperature regimes typical of temperate to tropical climates. They found that although subtropical tree seedlings achieved highest relative growth rate (hereafter RGR) at high temperatures, and a

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
70
“temperate” regime reduced their RGR by 40-50%, RGR at these lower temperatures was not significantly lower than that of temperate tree species. Although these results help to explain relative proportions of the two groups of species in a given location, the authors judge that their results do not offer a conclusive explanation for distributions of ecosystems and species along natural climatic gradients in their study area; among other reasons, they did not test for low temperature tolerance and minimum temperature under which seedlings may survive. My study has shown that severe cold and frost damage to tree seedlings may occur, but the severity of impact – whether seedlings can recover and eventually outgrow the zone of frost danger – depends, among other things, on management and structure of surrounding vegetation, as will be discussed in more detail below.
Both studies viewed together indicate that subtropical tree seedlings generally should have best chances of establishment by achieving as much initial growth as possible during the summer, when their growth rates match at least those of co-occurring subtropical and temperate tree seedlings, even if not those of co-occurring grassland species. As has been proposed at the end of chapter 1, species that can delay seedling emergence until spring, or disperse only towards the end of winter, should be at an advantage over species that germinate in autumn or at the beginning of winter. This remains to be verified across a broader range of species and study sites. However, it is safe to assume that although forest tree growth rates and forest expansion may be slowed down in highland relative to lowland areas due to climate, this is not sufficiently severe to preclude establishment of forest generally. Note also that due to the occurrence of unusually late frosts in spring, frost damage may have been overestimated in my experiment, as already pointed out in 1.2
As described in 1.4.3, there also exist considerable differences between soils of the escarpment and High Plateau inland. I have excluded this factor from the discussion so far as transplanted individuals remained largely rooted within the grassland soil spread in the nurseries (this became apparent upon harvesting at the end of the experiment). Moreover, soils in the vicinity of the research station were much altered during construction of the building. However, in comparisons of natural seedling establishment along the gradient from High Plateau rim to interior, soil conditions would clearly have to be taken into account, along with extensive studies of climate and ecophysiology of species.
3.5.2. Inherent species traits: Baccharis compared to Myrsine in
nurseries
Concomitant with results from the seedling emergence experiment described in chapter 2, both species achieved considerably better survival and growth in nurseries than under field conditions. There is no reason to assume that conditions in nurseries were representative of the physiological optimum for seedling growth; however, herbivory could be largely avoided, shade-cloth provided for a moderate microclimate and 1st – order neighbour competitors between species were omitted in evaluation.
Considering that incoming radiation was only between 22-36% of fully exposed sites, the excellent performance of grassland shrub Baccharis may be surprising: Not only did stems and leaves grow to larger size than in any grassland site, final stem size and 11-month size increase also surpassed those of forest tree Myrsine. However, Augspurger (1984) studied seedling survival and growth of 18 wind-dispersed rainforest trees in a screened exclosure under two different light intensities: A “sun” treatment – in which radiation was, however, reduced to approximately 20% of full sunlight, i.e. similar to my nursery environments – and a deep shade environment where conditions were similar to a forest understorey, or to tall tussock grassland in my study. She found widely different light requirements among her study species, but generally seedling growth was arrested or slowed down to almost zero only in the deep shade environment, presumably after

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
71
exhaustion of seed reserves. Seedling leaves, too, were not larger in deep shade but at higher light intensity.
Likewise, Everham et al. (1996) demonstrated for five early successional forest species that “gap light” improved seedling emergence relative to a closed forest canopy; gap light in their study was approximately 93 µmol m-2s-1, i.e. similar to conditions under tall shrub canopy in my study. – It follows that even very small-seeded species or species exhibiting apparent “pioneer” behaviour cannot a priori be assumed to require full sunlight for growth. On the contrary, photosynthesis saturates at a fraction of full sunlight (25% of full sunlight according to Crawley 1991a) so that maximum light intensities cannot be exploited even by a species adapted to open habitats.
Although the ability of Myrsine to resprout in grassland even from smallest seedling stumps – “Neds” – did come as a surprise in this study, it is plausible that a forest species should evolve such a trait. By “planting” plastic seedling models into neotropical rainforest and temperate old-growth hardwood forest, respectively, Clark & Clark (1989) and McCarthy & Facelli (1990) demonstrated to what extent seedlings may suffer mechanical damage by litterfall, vertebrate activity etc.: 82% of seedlings were damaged during one year in subtropical forest, 55% during 10 months in temperate forest. Clark & Clark (1985) reported that loss of apical meristems was common in seedlings of the Costarican tropical wet forest tree Dipteryx panamensis but did not significantly affect seedling survivorship; of seedlings surviving to the age of one year in the field in their study, 81% had lost at least one apical meristem and 28% had lost two or more meristems. Resprouting occurred in the majority of surviving seedlings, as in pasture and nursery Myrsine seedlings in my study.
Instead, Clark & Clark (1985) proposed herbivory on the first seedling leaves as a factor of major importance in limitation of tree regeneration. In my study, herbivory was apparently of minor importance but meristem damage was in many cases accompanied by loss of all foliage. Resprouting thus had to be achieved without any initial support by photosynthetically active tissue, which indicates investment from a very early age in assimilate storage and buds, and such a capacity may be extraordinary in a forest species seedling after all. Hoffmann (2000), comparing seedling performance of forest tree species and Cerrado woody species subjected to fire, found that forest species in his study were unable to survive fire until three years old. Eight of nine Cerrado species survived fire when younger, but this was achieved, as Hoffmann suggests, by an extraordinarily deep taproot system suitable for both nutrient storage and uptake. Myrsine seedlings, as clearly apparent on harvesting seedlings at the end of the experiment, do not possess such a taproot.
Moreover, the investment in resprouting can only pay off if growth conditions eventually get beneficial enough for seedlings not only to compensate biomass loss, but to increase further in size. In pasture, Myrsine seedlings did not achieve this during the 11-month observation period. It would have been desirable to continue observations for at least one more year in this case.
Baccharis also showed some seedling resprouting ability – ca. half of the individuals involuntarily predated by guinea pigs resprouted along stem stumps in the field nurseries. But these individuals retained at least part of their foliage, and adolescent and adult individuals have been shown not to survive complete aboveground biomass removal (chapter 6). It may therefore be concluded that in this species, overall priority is on fast resource acquisition and growth to reproductive stage and tall stature (which enhances seed dispersal) rather than preparation for biomass loss.
As to higher growth rates in Baccharis relative to Myrsine, it must be pointed out that some bias in favour of Baccharis uncinella may have been introduced by the experiment setup, namely, by using pure grassland soil in nursery flat beds. Grassland soils are inherently nutrient-poor, so that mycorrhization may be a key to successful establishment of woody species (on the frequency of the phenomenon of mycorrhization see e.g. Ellenberg

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
72
1996) The possibly high relevance of mycorrhization for seedling survival in nutrient-poor soils was pointed out by Fenner (1987b). Breuninger et al. (2000) investigated diversity of AM (arbuscular mycorrhiza)-forming fungi in grassland and forest of the Pró-Mata area as well as mycorrhization of Araucaria seedlings transplanted into the same sites. Both diversity of AM-forming fungi and degrees of mycorrhization of Araucaria seedlings were lower in grassland than in forest, which may have contributed to relatively poor performance of Araucaria seedlings in grassland in their study. Therefore, it cannot be ruled out that poor mycorrhization contributed to relatively little size gain of Myrsine seedlings, compared to Baccharis seedlings, even under protection in nurseries in my study. Baccharis is more likely to be adapted to genera of AM-forming fungi found in grassland.
However, DeSteven (1991b) observed striking contrasts in comparing seedling performance of early successional Pinus taeda with five later-successional hardwood forest species in oldfields of North Carolina. All species in her study are wind-dispersed but Pinus achieves dominance earliest, as Baccharis does relative to Myrsine and other forest species in my study area. During 36 months, Pinus seedlings acquired several times taller height than any of the hardwoods in both presence and absence of oldfield vegetation. DeSteven concludes that the species may overcome any competitive suppression by young oldfield vegetation within one or two years, independent of year-to-year climatic variation. Her results underline the physiological differences that exist between early-successional and later-successional woody species, independent of whether the first are relatively more favoured by particular types of AM-forming fungi at a given site (but see also discussion in chapter 5).
3.5.3. Impact of management: Pasture compared to abandoned
grassland
Ongoing pasture management had a significant impact on seedling mortality of both Baccharis and Myrsine. At least one quarter of death cases in each species could be attributed by circumstantial evidence to cattle grazing, trampling and dung deposition. The actual impact of cattle was probably even higher, as cattle activities also offer a plausible explanation for seedlings “vanishing” without any trace. End-of-winter fire (in August 2005) accounted for one fifth of dead in Baccharis, a substantial proportion although lower than expected; fire influence turned out to be very patchy and only a minor part of transects were almost completely burnt. Fire was a less important cause of seedling mortality in Myrsine because frosts (the first occurring on May 26 2005, one month after transplant) affected plants earlier. Episodic frost and cold spells continued over a rather unusually long period of five months, until the first week of November 2005, in spring.
As explained before, it could not be determined whether winter damage was caused by freezing or by physiological drought; both may occur when temperatures drop below zero, but physiological drought in particular may be aggravated by some microsite conditions typical of pasture, such as soil compaction by trampling and greater exposure to wind in short swards; the latter condition is also pointed out by Leite & Klein (1990). Myrsine showed remarkable resistance by resprouting even from defoliated seedling stem stumps. As discussed in the first paragraph, this may be a pre-adaptation evolved under recurrent mechanical damage in Myrsine’s original forest habitat but nevertheless appears an astonishing feature in seedlings not even one year old.
Possibly, the very removal of part of the stem, which resulted in a reduced surface:volume ratio of the stem, aided in efficient use of what little resources could have been stored in stem base and roots. Facilitation of resprouting by a flush in resource availability following pasture fire was not apparent: Nursery seedlings did not experience burning but achieved greater size gain on resprouting than pasture seedlings. Mortality

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
73
following biomass loss was greater in pasture, and resprouting was delayed for a longer time relative to nurseries.
Strikingly, Baccharis seedlings did not suffer apparent physical damage or even mortality in relation to winter climate. In a beautiful demonstration of the differences between physiological and ecological optimum sensu Ellenberg (e.g. Ellenberg 1996), Baccharis performed very well under part shade in the relatively competition-free nursery environment, but in competition with grassland vegetation performed best under most extreme microclimatic conditions in pasture. While seedling emergence experiments failed to detect any differences in “emergence safe site” availability between pasture and abandoned grassland, perhaps because seed input was unrealistically low (see previous chapter), better chances for natural seedling establishment in relatively short-growing grassland showed clearly in seedling survival and growth, with stem growth alone being 6-18 times higher than in either type of abandoned grassland.
How was this achieved? Firstly, Baccharis stems were significantly stouter – relatively short and thick – in pasture, a common phenomenon under climate extremes, as a reduced surface:volume ratio assists in energy conservation (e.g. Cain 1950). Secondly, they developed a dense indumentum which may serve to reduce water loss through leaves especially in wind-swept areas (e.g. Crawley 1997a). Note that seedlings in tall shrub grassland developed small leathery leaves with tightly inrolled margins – which may indicate water stress – but no indumentum. In this grassland type, as discussed before, competition is probably high but climate is moderated (chapter 1).
Thirdly, according to Kozloswki et al. (1991), biomass production is more strongly linked to leaf traits (such as surface area, duration and exposure) than to other morphological an physiological parameters. Leaf traits are usually very plastic (Crawley 1991a) whereas in Baccharis, neither small size nor the intermediate 3:1 length-width ratio of leaves changed significantly over a range of microsites. This indicates a strong pre-adaptation of this species to a habitat characterised by the following:
1. Recurrent fires, which may be fuelled by plants with similar morphology (Bond & van Wilgen 1996).
2. Extremes in microclimate or soil nutrient status. According to Givnish (1987), leaf size tends to increase with humidity and decrease with irradiance and low fertility, in the latter case even if soil moisture is sufficient. The decrease in sun-exposed sites is due to a trade-off between photosynthetic rate and energetic costs of transpiration; in broad leaves, temperature and transpiration increase relatively more than in small leaves when sun-exposed (Givnish 1987; see also Cain 1950). An adaptation of Baccharis to sunny environments is further indicated by the spirally insertion of leaves on erect twigs (Leigh 1972 in Givnish 1987).
According to Rambo (1952) the genus Baccharis has its centre of distribution in the tropical regions of Brazil, which makes both explanations – adaptation to sun-exposed sites or to fire – seem possible. In fact, however, these two explanations overlap, as post-burn areas also display climate extremes.
Notably, Myrsine leaf size and allometry in pasture also shifted towards relatively small, short, rounded leaves. But Myrsine is unable to invest stem and leaves with a clothing of hairs, which Baccharis did especially in pasture and which may help to prevent water loss through evapotranspiration (Crawley 1991a) under high irradiation and in wind-exposed sites.
Finally, although photosynthetic rate does not alone account for biomass gain, early successional woody species do tend to have higher maximum rates of photosynthesis in high-light environments than later successional species; high light may in fact decrease photosynthetic rates of the latter (Kozlowski et al. 1991).

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
74
In heavily shaded environments, however, especially tall tussock grassland in this study, early-successional species are more easily starved as the decrease in photosynthesis exceeds the decrease in dark respiration (Kozlowski et al. 1991). Initial seed size is also likely to play a role: The larger the seed mass – and the lower, consequently, the rate of exhaustion of seed reserves –, the higher the probability of a seedling to survive under deep shade (Thompson 1987 and references therein; Saverimuttu & Westoby 1996). This offers one explanation for why even smallest Myrsine seedlings, unless smothered by litter deposition, performed better than Baccharis seedlings in tall tussock grassland. Furthermore, Augspurger & Kelly (1984), in a study of wind-dispersed tropical tree species, found that shade tolerance was more closely correlated with resistance to pathogens than with seed size. Fungal infections currently observed in adult Baccharis populations (chapter 5) cannot be ruled out as a death cause in Baccharis seedlings in both tall shrub and tall tussock grassland, although clear symptoms of such infection were observed only in a minor proportion (not shown in Tab. 3.1).
All in all, the performance index indicates that Baccharis seedlings were inhibited to similar extent in tall shrub and tall tussock grassland – survival tended to be better in the first but growth tended to be better in the second; seedling emergence was practically zero in both. This reflects the current situation in naturally established populations of this species in abandoned grassland in which seedling recruitment is rare or zero (chapter 5).
Forest species Myrsine, on the other hand, is more shade-tolerant than Baccharis in the seedling stage and establishes better in abandoned grassland whereas in a high-light environment such as pasture, her performance is greatly reduced. Everham et al. (1996) concluded that light in gaps is a “universally good cue” for germination and seedling survival of forest species, but of course forest gaps come in different sizes and it must be assumed that only the largest (such as following clear-cutting) are characterised by similar extremes in abiotic parameters as pasture; in fact, “gap light” in Everham et al.’s study was of similar intensity as light underneath tall shrub canopies in my study.
It turns out that pasture poses a series of obstacles to successful establishment of Myrsine seedlings: Seed rain is probably low initially, as may be inferred from a number of studies (see previous chapter), seedling emergence is relatively low and delayed until the end of the summer season, mortality is high (although it might be even higher if the species were not able to resprout at a very early age) and growth is poor. It might be hypothesized that pasture patches with a higher-growing grass matrix offer some shelter to seedlings relative to areas with very short-grazed and prostrate grasses, but that such shelter comes at an increased risk of being burnt. Bond & van Wilgen (1996) coined the term “gullivers” for such individuals that might dominate a community when adults, but are held down by lower-growing but flammable surrounding vegetation. However, Myrsine survivor numbers in pasture were not high enough and the observation period was too short to test for this hypothesis.
As discussed in the preceding paragraph, Myrsine seedlings suffer less severe growth reduction under presumably high competition from adult Bacchars shrubs than Baccharis seedlings, and they perform considerably better in tall shrub grassland than in pasture. However, seedling performance is clearly reduced in tall shrub relative to tall tussock grassland especially if seedling emergence percentages are integrated in the performance index. This confirms the suspicion that low numbers of naturally established seedlings in tall tussock grassland must be related to seed limitation above or underneath the tussock grass matrix or both; i.e. seed rain may be reduced relative to tall shrub grassland, or seed dispersal to the soil surface may be obstructed by the grass layer, as discussed before.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
75
3.5.4. Impact of established vegetation: Station nursery compared
to tall shrub abandoned grassland
As shown in chapter 1, within-stand climate of tall shrub abandoned grassland is, of all grassland types investigated, most similar to microclimate underneath shade-cloth in the Station nursery, and these stands established on the same type of soil that was used to fill flat beds in the nurseries (although that soil was collected from underneath tussock grassland, not tall shrub grassland). So if Baccharis and Myrsine achieved maximum growth in the Station nursery but only comparatively poor growth in tall shrub grassland, this may be attributed to further direct and indirect modifications of that environment by an adult Baccharis canopy – such as alteration of light quality, soil water and nutrient availability, pathogens etc. This is of special interest because, paradoxically, higher densities of naturally established Myrsine seedlings were recorded in this than in any other type of grassland (chapter 5).
Growth reduction relative to the Station nursery was significant, and leaf morphology and size also differed conspicuously between sites especially in Baccharis, with Baccharis seedlings under shrub canopy producing small leathery leaves with inrolled margins. Such morphology may be observed in plants of water-limited habitats as well as in habitats where nutrients are more limiting (Crawley 1991a; Ellenberg 1996). In the particular case of this experiment, water limitation may probably be ruled out, as precipitation is generally high, transplants were made only after the 2005 summer drought and seedlings were transplanted directly into mineral soil, not into the litter layer that hypothetically may expose naturally emerged seedlings to drought.
Ellenberg (1996) relates how in Middle European bog plants, peinomorphous build enables plants to maintain stomatal aperture: This saves potassium (an element otherwise required for opening and closing of stomata, but particularly scarce in such environments) and allows for relatively continuous nutrient uptake, which may offset low nutrient availability to some degree. It is plausible that nutrients should be more limiting in tall shrub grassland than in other types of grassland: For example, nitrogen contents are initially low in pasture (chapter 1), legumes that might replenish the nitrogen pool are eventually outcompeted on abandonment of grassland, considerably amounts are bound up in the tall shrub biomass and litter remineralisation is relatively slow.
There are several further possible explanations for greater limitation of woody seedlings by shrubs than by grasses in secondary succession. Holl (1998) suggests, firstly, allelopathy (however, Ellenberg 1996 presumes that this is a relatively rare phenomenon), and, secondly, different rooting depths of seedlings, shrubs and grasses: In Holl’s study area grass roots concentrate in the first 5 cm of soil while shrubs and seedlings both explore a wider range of depths. However, this seems to contradict Wilson (1993) who found that belowground competition was more severe for tree seedlings in grassland than in forest.
Some studies find forest seedling establishment and growth enhanced by shrub cover, but apparently, for this to be possible, both microclimate and soil conditions must be improved by the shrubs. In the case of Cordia multispicata, a shrub colonising Amazonian pasture, the beneficial effect is in acquisition of soil resources and production of easily decomposable litter; in addition, the bright fruits of this species make it a preferred bird perch, enhancing seed dispersal from forest to grassland (Vieira et al. 1994). In the case of Prosopsis glandulosa in subtropical grasslands of Texas, nitrogen fixation accounts for the beneficial effect on later-successional species (e.g. Archer et al. 1988, Brown & Archer 1989, Archer 1991).
Summing up, the possibility that high natural seedling establishment of Myrsine in tall shrub grassland is due to optimal conditions for seedling growth may be ruled out. Notably, in a direct comparison of growth rates between naturally established Myrsine suckers and

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
76
seedlings in tall shrub grassland, only root suckers achieved a growth rate similar to that of seedlings in the Station nursery (chapter 4). This leaves two possible explanations, as already indicated in chapter 1: Higher seed rain due to greater attraction of dispersors, and greater seed input at the soil surface due to higher availability of gaps. However, it is also of interest to note that growth reduction relative to the Station nursery was considerably more severe in Baccharis seedlings than in Myrsine seedlings. As evident from the growth performance index, size increase of Myrsine seedlings was five times lower in tall shrub grassland than in nurseries combined whereas size increase of Baccharis seedlings remained more than a hundred times lower. It appears that Myrsine, as a species achieving dominance later in succession than Baccharis, copes better with reduced resource levels under an established vegetation canopy.
3.5.5. Inhibition versus facilitation
The succession models of inhibition, facilitation and tolerance proposed by Connell & Slatyer (1977) (hereafter referred to as CS-models) address the potential competitive effects of established, early-successional species on invading, later-successional species. Since their publication, various studies have been devoted to detecting whether a given pioneer species or early-successional plant community is inhibitory, facilitative, or tolerant towards later-successional species, i.e. whether performance of the latter is reduced, enhanced or unaffected by presence of the first.
It appears that when researchers examine in detail the effects of certain secondary succession communities on introduced forest species in different stages of development - by seed addition, transplant and vegetation removal experiments – they are ultimately led to conclude that consistent trends and patterns do not exist, i.e. that in fact several of the CS-models apply in a given area at a given point or at successive points in time, depending on species and developmental stage of introduced forest tree specimens. Holl (1998), DeSteven 1991 (a, b) and Gill & Marks (1991), on studying abandoned subtropical pasture and young oldfield vegetation in the northeast and east of the USA, respectively, concluded that with few exceptions, vegetation cover tends to enhance forest tree seedling emergence and survival but reduces seedling growth relative to areas in which vegetation is removed. There are some parallels here to the performance of Myrsine seedlings in abandoned grassland relative to pasture and nurseries in my study – a direct comparison with these studies is not possible as surrounding vegetation was only experimentally removed in seedling emergence experiments.
Connell et al. (1987) agree that “the possibility of simultaneous contrasting mechanisms was not foreseen” but also point out that “the net effect of such interactions must be either positive, negative or zero” (p.136).
Another difficulty becomes apparent whenever two or more species and vegetation types are compared: Inhibition and facilitation are relative terms. For example, in this study, both types of abandoned grassland may be judged to have some inhibitory effect on transplanted seedlings, since seedling growth was nowhere in the field as high as in the nurseries; but relative to tall shrub grassland, tall tussock grassland must clearly be considered the less inhibitory, or more facilitative community at least for Myrsine seedlings.
This problem is not resolved by Connell et al.’s proposal to focus on net effects – validation of the models is still achieved only in comparison with one or more other vegetation types (not only controls in nurseries: for compared to these, all types of vegetation reduce growth).
Is there a way of expressing, in quantitative terms, how much more inhibitory or facilitative a certain community is towards a certain species than another community? De Steven’s performance index, adapted by me, is a summary of experimentally derived

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
77
quantitative data on seedling emergence, mortality and transplant; one might say it represents a bottom-to-top approach towards a net effect.
However, as De Steven (1991b) points out, even this index does not fully explain patterns of natural seedling establishment in her study, and she postulates that such patterns seem to depend strongly on “differential seed rain, which tends to be viewed as a relatively stochastic factor in succession” (De Steven 1991b, p. 1088). Gill & Marks (1991), on summarising their experimental data, conclude on the importance of seed crops of species adjacent to the field at the time of field abandonment.
Two parallels to my study can be drawn. The apparent paradox of abundant natural seedling establishment in tall shrub grassland has already been discussed above; another phenomenon that appears a paradox in the light of my experiments is the fairly high density of naturally seed-established Myrsine individuals in population monitoring plots in pasture (see also chapter 5). These patterns can – probably - only be explained by seed rain counts and population monitoring, both over a period of several years.
Holl (1998, 1999) arrives at a similar conclusion, by a more precise route: She supplemented seedling performance experiments in abandoned pasture with experiments in adjacent forest and with seed rain counts along a transect from forest into pasture, and found that even in recently abandoned pasture, grasses modified microclimatic conditions so that photon flux density, vapour pressure deficit and soil temperature at ground surface did not differ significantly between recently abandoned pasture and forest. Germination of most of her study species was similar in forest and recently abandoned pasture; nor did aboveground biomass and root biomass differ significantly between seedlings grown in grassland and forest. Seed rain of animal-dispersed species, on the other hand, decreased “dramatically” in pasture >5 m from the forest edge.
Thus, while in an earlier paper the author suggests a direct inhibitory effect of pasture vegetation on forest seedling establishment (Holl 1998), her final conclusion is that “although a number of factors impede forest recovery in abandoned pastures, these results suggest that the most important limitation is lack of seed dispersal“ (Holl 1999, p.229).
In the light of observations by Alcantará et al. (2000) and M.Marchesini (pers.comm.), pasture may still be considered to have an indirect inhibitory effect in not offering attractions to seed-dispersing birds. In the case of Cordia multispicata cited above (Vieira et al. 1996) attraction of seed-dispersing birds was included among facilitative traits. However, since the CS-models have already been subjected to much semantic debate, it is perhaps wiser not to stretch the concept further, and to distinguish instead between the process of seed dispersal, and the conditions a seed experiences once dispersed into a given plant community. This also facilitates comparisons between studies, as relatively few studies encompass all stages of colonisation, from seed dispersal to seedling emergence and establishment.
In another oldfield study in the northern USA, Berkowitz et al. (1995) focussed on “net effects” of established vegetation on transplanted forest tree seedlings as follows: They skipped the seedling emergence stage and performed transplants across a broad range of early successional vegetation types and study sites. These authors confirm a tendency for established vegetation to slow down seedling growth but also point out that where this was not the case, physical stresses were higher: “Competition and facilitation traded off along a site-quality gradient, with the result that tree seedling growth was uniformly low” (p.1156). Thus, their findings still do not quite explain observed differences in natural establishment of tree seedlings in different types of oldfield vegetation in their study region.
Berkowitz et al. then suggest that “duration of competition” should be taken into account in investigation of patterns of natural seedling establishment: “Duration of competition” referring to the time span necessary for a seedling to outgrow competition by surrounding vegetation. It can be deduced that this parameter integrates species traits, site conditions and post-dispersal effects of established vegetation. The approach is interesting

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
78
for two reasons: It may provide the desired quantitative measure for comparison of seedling performance between various species and types of established vegetation. At the same time, it demonstrates that a focus on “net effects”, and an attempt to avoid the contradictory terminology of the CS-models, leads back to competition as the one phenomenon that an ecologist must reckon with “anywhere and at any time” (Ellenberg 1996, p.118).
Would it be wise, then, to altogether abandon the CS-terminology? Probably not; after all, there remain several phenomena – such as improvement of soil quality, alleviation of climate extremes – that can not be satisfactorily included in the term “competition”. However, caution is warranted in the use of CS-terminology9.
It follows from Connell et al.’s statement that inhibition, facilitation and tolerance in their sense can only be diagnosed with confidence if seedling emergence and establishment are followed in population studies, preferably over a period long enough to allow species B to reach reproductive maturity in presence of species A. Likewise, diagnosis of Berkowitz et al.’s “duration of competition” requires long-term observations. It is either not possible, or unusually good luck is required, for a scientist to unravel mechanisms of vegetation succession from a single snapshot-in-time study.
None of my findings so far provide an answer to the question whether grassland is a “favourable” habitat for forest tree seedlings at all, i.e. whether seedling establishment is facilitated relative to forest, or at least not more inhibited, as Holl (1999) demonstrated. If future studies prove that forest species’ establishment in grassland is not in the first place slowed down by low seed rain, as discussed above, then a direct comparison of growth conditions for seedlings in grassland and forest may prove helpful in determining why forest expansion over grassland is so slow.
9 Pickett et al. (1987) point out that use of the “tolerance” may cause confusion, as it refers to a physiological trait of either species A or B as well as to the effect of species A on performance of species B in the CS-models. This, too, can be illustrated by an example from my study - adult Baccharis canopy reduces seedling growth, i.e. may be considered inhibitory in this respect; but Myrsine in the seedling stage is apparently more tolerant of such resource limitation than Baccharis.

Hermann, J.-M. PhD thesis 3: Seedling performance of two woody species in grassland
79
3.6. Appendix
Fig. 3.2: A summer’s harvest: Seedlings of grassland shrub Baccharis uncinella (example circled black)
and forest tree Myrsine parvula (circled white) retrieved from various types of grassland after planting-out
for 1 year. In the background: Seedlings from abandoned grassland – tall shrub grassland on left table,
tall tussock grassland on right table. Foreground: Seedlings from pasture. Photographer: J.-M. Hermann.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
80
4. Recruitment of a Pioneer Forest Tree in Grassland: Root
suckers vs. Seedlings
4.1. Summary • The Southern Brazilian highland is characterised by a mosaic of native forest and
grasslands. Both managed and abandoned grassland pose various obstacles to forest species seedling emergence and establishment. Yet in abandoned grassland pioneer forest tree Myrsine has recruited stands of several hundreds of individuals/100m². How is this achieved?
• Joining data from experiments and naturally established populations, I compare a) stem morphology and growth rate of newly recruited root suckers and seedlings in tall shrub and tall tussock abandoned grassland as well as in pasture and short open grassland, b) relative importance of vegetative spread and generative reproduction in colonisation of these sites by Myrsine and c) growth rates of elder root suckers and seedlings in naturally established mixed root sucker-seedling populations.
• New Myrsine seedling recruits developed significantly stouter stems than new root sucker recruits in all types of grassland, presumably as root suckers, supplied continuously with water and nutrients through their feeder root, can invest principally in height growth. In abandoned grassland, new root sucker recruits grew ca. 1.3 times higher than seedlings of the same basal diameter, in pasture and short open grassland ca. 1.2 times higher.
• One-year growth rate of new root sucker recruits was significantly higher, and significantly taller height was attained than in new seedling recruits in all types of grassland. Root suckers increased in height at similar rates in all types of grassland, attaining 4-5 times their original height within one year, whereas seedling size and 1-yr height gain differed significantly between grassland types. Within one year, new root sucker recruits attained twice the height of seedlings in abandoned grassland and ca. three times the height of seedlings in pasture and short open grassland.
• In Myrsine populations of tall tussock and tall shrub abandoned grassland, root sucker production has been the primary mode of recruitment in recent years, accounting for on average 91% of elder recruits (10-100cm height 2005) and 95% of new recruits (>=10cm 2006) in the first, and 67% of elder recruits and 78% of new recruits in the latter grassland type.
• Root suckers accounted for only a minor proportion of recruitment in pasture 2005. Root sucker production was observed 2006 following pasture burning but recruitment was generally very low at 2 new recruits/100m², as opposed to 39 and 83 new recruits/100m² in tall tussock and tall shrub abandoned grassland, respectively. This is attributed to scarcity of potential feeder individuals (>1m tall) in grassland and adjacent pasture.
• Root suckers lose advantage in growth rate of seedlings with increasing size and age in mixed root sucker-seedling populations of abandoned grassland. Height and one-year growth rate differed significantly between root suckers and seedlings in new recruits (>=10 cm height 2006) and in small elder recruits (10-25 cm height 2005) but not in tall elder recruits (25-100 cm height 2005). Juveniles in this size-age class also show differentiated responses to different structural types of grassland: In mixed root sucker-seedling samples, tall elder recruits from tall shrub abandoned grassland show significantly higher one-year height growth rates than individuals from tall tussock abandoned grassland. The same trend was observed in comparison of pure root sucker samples between these two grassland types.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
81
• I propose that this is because suckers grow too large for feeder roots to give adequate supply and/or depend largely on their newly formed adventive root system; they are physiologically no longer different from seedlings. I also suggest that in spite of the species’ pioneer status, recruitment by root suckers may be less efficient in grassland than in the species’ original forest habitat.
• Due to relatively short “dispersal” distances of root suckers, 75-80% of individuals per 100m² plot are concentrated in a 0-5m strip along the forest border both in pasture and in grassland abandoned for 12 years. Recruitment from seed is thus indispensable for long-range colonisation of grassland (formation of forest islands several dozens or hundreds of metres away from forest border) within historical timespans, and where potential feeder individuals are scarce at the forest border (pasture).
• Investigation of genetic diversity and performance of juveniles in forest relative to grassland is required in order to better understand population dynamics, and predict future development of Myrsine populations in grassland.
4.2. Introduction As shown in chapters 2 and 3, both managed and abandoned grassland pose various
obstacles to seedling emergence and establishment of forest tree Myrsine parvula. Yet in abandoned grassland pioneer Myrsine stands of several hundreds of individuals/100m² have developed. How is this achieved?
My own pilot surveys in 2003 revealed, in several woody species of the region - both grassland shrubs and forest species, among them Myrsine - the ability to spread by clonal growth in grassland. Parallels to “polycormon succession” e.g. by Prunus spinosa in European fallow areas and abandoned grasslands (e.g. Ellenberg 1996, Schreiber 1997), to Aspen stands (Populus tremuloides) in North American Prairie (e.g. Anderson 1982, Peltzer 2002) and to post-fire woody species recruitment in the Brazilian Cerrado (Hoffmann 1998) suggest themselves, yet the phenomenon has not been included in Southern Brazilian forest-grassland models so far.
This must be partly attributed to a lack of comparative data on morphology and growth of root suckers and seedlings that might allow for rapid surveys of origin of juveniles in the field. Hoffmann (1998) stated that in his post-burn study sites, seedlings and root suckers could be easily distinguished by their morphology. Is this generally true? What exactly are the differences? In order to answer this question, I first investigate how relatively young and even-aged Myrsine root suckers and seedlings differ in stem allometry and growth rate.
Secondly, I explore how these differences affect recruitment in Myrsine populations naturally established in managed and abandoned grassland. The relative importance of vegetative spread vs. generative reproduction in recruitment of a forest woody species in a Southern Brazilian grassland is quantified for the first time. I hypothesized, when initiating this part study, that recruitment by root suckering would be more frequent than recruitment from seed in tall tussock grassland, and vice versa in pasture and tall shrub grassland, on the assumption that seed dispersal and germination would be inhibited to greater extent by accumulation of grass biomass in tall tussock grassland than in the other two types of grassland.
Other than clonal growth in herbaceous species, clonal growth in woody species has only since the beginning of the 1990’s received increased attention (Jenik 1994) and many open questions on mechanisms, costs and benefits remain. For example, Peterson & Jones (1997) point out that while the superior colonising ability of clonal species is amply

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
82
documented, there is surprisingly little direct evidence that clonal integration improves competitive ability in these situations. Moreover, there is as yet no consensus on the question whether root sucker production essentially serves for genet perpetuation (Peterson & Jones 1997) or foraging (Klimešová & Martínková 2004). I argue that clarity on this point is essential for judging the efficiency of Myrsine as a grassland coloniser, and point out some evidence for each of these hypotheses found in this and other studies.
Furthermore, I demonstrate how growth rate differs between root suckers and seedlings in new recruits and elder recruit classes. I ask whether similarity and dissimilarity between recruits of these two types of origin may be related to a change in physiological integration of root suckers with feeder individuals with age, discussing possible costs and benefits of physiological integration in the particular context of forest species invading grassland habitats.
As to possible long-term consequences of the phenomena observed here, I argue that root sucker recruitment is a process complementary but not alternative to seedling recruitment, and I briefly discuss possible costs and benefits of predominantly clonal recruitment for the progress of grassland-forest-succession in the longer term, including rate of spatial expansion of populations.
4.3. Study area and species The data presented here were compiled from various part studies in two adjacent
sites on the High Plateau of Rio Grande do Sul: 1. Native grasslands in the Nature Conservation and Research Centre Pró-Mata (hereafter: Pró-Mata) that were excluded from cattle grazing and fire in 1994, i.e. 11 years prior to first survey. 2. The neighbour fazenda “Trés Estrelas” where grasslands are grazed by cattle (approximately 0.5 head of cattle per hectare) and the majority of plots experienced burning – i.e. a complete scorch of aboveground biomass – once during the monitoring period, in August 2005. The focus is on pioneer forest tree Myrsine parvula (Mez) Otegui. For details on study area and species please see chapter 1.
4.4. Material and methods
4.4.1. “New” root sucker and seedling stem allometry and growth
rate
Quantities and proportions of root suckers and seedlings differ greatly in naturally established Myrsine populations (see 4.6.1). In order to obtain sufficiently large sample sizes for comparison of stem allometry and growth rate between root suckers and seedlings, data collected in naturally established populations were supplemented with experimentally obtained data.
For methodology of population monitoring please see 4.4.2. Supplementary seedling data were obtained from an experiment in which nursery-
grown seedlings were transplanted into field plots (chapter 3). The following data are included in analyses: Stem height and diameter recorded April 2006, i.e. after one year in the field, and 11-month relative growth rate in the field, observed from May 2005 to April 2006.
Supplementary root sucker data were obtained from a cutting experiment conducted in the Pró-Mata area (chapter 6). Myrsine individuals were cut and surrounding grassland mowed to soil surface level in January 2005. Root suckers emerged ca. three months after mowing and were harvested in April 2006 after ca. one year of growth. Mowed areas

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
83
retained a relatively short, open grass layer during this period and can thus be considered structurally equivalent to pasture plots (rendering a pooled “short grassland” dataset).
I refer to individuals in these pooled datasets as “new” as for most of them, individual ages between 9 months and 1.5 years are known or may be safely assumed; only naturally established seedlings in abandoned grassland may have persisted in populations at small sizes (“Oscars”) for a longer period10. I did not include elder individuals in this analysis, as I reasoned that with increasing age, branching and damage to apical meristems would introduce greater variation in stem allometry; also, sucker or seedling origin could be identified in the field with greatest confidence in youngest individuals.
Stem allometry is investigated by exploring the relationship between stem height and basal diameter. Stem height (from root crown to bottom of apical inrolled leaf) was measured with a standard folding ruler. Basal diameter 1cm above root crown was measured with non-digital calliper rules during initiation of population monitoring and transplant experiments; these instruments were sometimes of poor quality and introduced greater errors in measured values than the digital calliper rules used from September 2005 onwards.
Even in pooled datasets, the range of sampled sizes differed greatly especially between abandoned grassland and pasture. Regression curves facilitate the comparison of root suckers and seedlings within the range of overlapping sampled sizes (see Fig. 4.1). However, only 2006 data on stem height and basal diameter were considered sufficiently precise for this kind of analysis.
In comparison of stem allometry between different types of origin in different types of grassland (4.5.1), basal diameter was chosen as independent variable, height as dependent variable (although in fact the two parameters are interdependent). Multi-stemmed individuals were excluded but analysis includes individuals that resprouted after loss of apical meristems; the majority of individuals had intact stems.
Linear regression graphs were created from ln-transformed data using Microsoft Excel (Windows XP) and regression was tested for significance using SPSS 14.0.1. These analyses were performed with separate tall tussock and tall shrub abandoned grassland datasets as well as with a pooled abandoned grassland dataset, for comparison with the pooled “short grassland” dataset (including pasture and mowed sites). The latter analysis (see Fig. 4.1) includes a sample of 150 root suckers and seedlings each randomly drawn from the pooled abandoned grassland dataset, and 50 root suckers and 25 seedlings each randomly drawn from the pooled short grassland dataset.
By random choice of data in this as well as the following analysis, I attempt to reduce the risk of comparing 1st-order neighbour root suckers and seedlings from naturally established populations. Samples are thus brought closer to the ideal condition of independence of one another, which is – strictly – a prerequisite for the statistical analyses performed.
A measure of relative growth rate was obtained dividing stem height after a given observation period by stem height at the beginning of that period, and subtracting 100 (Relative growth rate = (ht2*ht1
-1)-100). This parameter should not be confused with classical “relative growth rate” - RGR -, which is defined as the product of unit leaf rate and leaf area ratio in time - see e.g. Hutchings 1997a for detail; therefore it is also referred to as “1-year
10 There may be some confusion with so-called new recruits, a term I coined for a certain size-age class of naturally established individuals in population monitoring (see following paragraph). In order to facilitate distinction, I have left the term “new” in quotation marks, and avoided the term “recruit” in this joint analysis of population monitoring and experimental data. Throughout the other parts of my thesis, when reference is made to new recruits, that term applies exclusively to naturally established individuals of smallest size.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
84
relative height increase” etc. With the exception of seedling transplant data, these values are approximations (see 4.4.4).
Pair-wise comparisons of new root sucker and seedling allometry were performed by SPSS 14.0.1 T-Tests for independent samples. For these analyses I used sets of values obtained from regression but restricted the range of calculated values to overlap with the truly sampled range of both partners (see Tab. 4.8).
For comparison of growth rate and stem height between new root suckers and seedlings, median values were calculated from a sample of 50 root suckers and seedlings randomly drawn from tall tussock and tall shrub abandoned grassland datasets each, and 50 root suckers and 25 seedlings randomly drawn from the pooled “short grassland” dataset. Pair-wise comparisons (T-Test for independent samples) were performed on the same samples, ln-transformed in order to improve homogeneity of variances.
4.4.2. Recruitment in naturally established populations
Proportions of root sucker and seedling recruitment were determined in naturally established Myrsine populations. For one year, between 2005 and 2006, recruitment, mortality and individual growth rates of Myrsine individuals >=10cm high were tracked in four 10x10m plots each in tall tussock and tall shrub abandoned grassland, and in eight plots of same size in grazed and burnt pasture. These plots are adjacent to the forest border and opposite forests are located at no more than 200 metres distance so that Myrsine seed input by birds crossing between forests may occur (pers. obs.; see chapter 5). Surveys took place between late autumn and early winter (April to June) of each year. Study plots in abandoned grassland remained undisturbed during the study period whereas pasture plots were cattle-grazed and most of them were burnt in August 2005.
Individual origin from root suckering or seed was determined for so-called elder recruits at 1st survey 2005 and for so-called new recruits at 2nd survey 2006 (size-age classes are explained in the following paragraph). In most cases, and most easily in new recruits, the mode of origin could be confirmed by careful exposure of the root crown, as lateral feeder roots usually run close to the soil surface. Doubtful identities in elder recruits were either back-checked during 2nd survey or, alternatively, the allometry regression function (see previous paragraph) provided a helpful guideline: Taking basal diameter of a certain individual of doubtful origin as a basis, I calculated what height this individual might be expected to have if it originated from either roots or seeds, and then checked whether actual (measured) height was closer to one or the other calculated value.
Nevertheless, a minor proportion of misidentifications cannot be ruled out, firstly, as root suckers develop adventive root systems that may have been confounded with root systems of seedlings, and secondly, as the regression function was calculated from youngest individual data and stem diameter-height relation may change with age.
Origin was not recorded in adolescent and adult individuals (>1m and >2.5m height, respectively), but as recruits predominate over elder individuals at present, I could account for the origin of ca. 83% of pasture populations, 86% of tall shrub grassland populations and ca. 90% of tall tussock grassland populations.
During census of new recruits in 2006, I also made notes on structure of the surrounding vegetation matrix: Open/closed grass layer, presence/absence of grassland shrubs or forbs, moss and litter cover. Based on a frequently cited definition by Brokaw (1982), I define gaps as holes in the matrix of tussock grasses and grassland shrubs that extend all the way to ground level. They may be understood as sites in which seeds can reach the soil surface more easily than in a closed grass layer, and light competition is less severe, but they do not necessarily receive full sunlight as a more or less closed tall shrub

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
85
canopy may yet extend above the grass layer11. How Myrsine juvenile growth is influenced by density of this tall shrub layer is treated in more detail in chapter 5.
In 2005, I censused the number of individuals >1.5m height in 10x10m plots in the adjacent forest border. During the course of the resprouting experiment conducted parallel to population monitoring (see chapter 6), I found that root suckering was zero to low in individuals <1 m height but occurred in the majority of individuals >1 m height. Density of individuals >1.5 m height may thus be considered fairly representative of density of potential feeder individuals. In grassland plots, for which a complete dataset of individual numbers and sizes was available for both 2005 and 2006 survey, all individuals >1m were included in the group of potential root sucker producers.
Furthermore, in 2006, a cursory survey of relative proportions of root suckers and seedlings among recruits was conducted in the forest border adjacent to abandoned grassland plots. The forest border was walked in - approximately - a 1x1m grid and the origin of one juvenile <1m high per grid point was noted. In forest border adjacent to tall shrub abandoned grassland, a total of 200 juveniles in an area of ca. 200 m² was sampled in this fashion; in forest border adjacent to tall tussock abandoned grassland, I sampled a total of 120 juveniles in an area of approximately twice that size.
Size of Myrsine clones was not delimited, nor was genetic diversity of the populations investigated. Such information may be helpful in interpreting long-term behaviour of pioneer populations in grassland (see 4.8), but may be considered dispensable if the focus is on interactions of Myrsine juveniles and surrounding grass and shrub matrices; in such community studies, it is the behaviour of the total population of ramets that matters (van der Valk 1992).
Values of root sucker, seedling and feeder individual density given in the following (individual numbers per 100 m²) are averages of the four monitoring plots per abandoned grassland type, and the eight pasture monitoring plots. Due to small sample size, variation within plots was not explored and statistical significance of differences in means was not tested.
For further details of the study setup please see chapter 5.
4.4.3. Short-distance versus long-distance dispersal
Naturally established seedling and sucker population densities in relation to distance from the forest border were calculated from individual x/y-coordinates recorded during population monitoring.
These data, especially if viewed together with absolute individual numbers, indicate the relative potential of root suckers and seedlings for dispersal in space. However, in order to better interpret these data, it is helpful to know at approximately what distance from a feeder individual root suckers may emerge.
Evidence for this was collected on two occasions: Root suckers were harvested together with portions of their feeder roots on closure of the cutting experiment, at ca. one year of age; and five adolescent Myrsine individuals, all between 1.1 m and 1.6 m high, were excavated with as much of their root systems and root suckers intact as possible in the same populations in 2004. The first allowed an estimate of the minimum diameter a root must have in order to support root suckers; the second indicated at what distance from an individual roots of sufficient thickness may be found. 11 Brokaw (1982) proposed his definition for gaps in forest so the reference is not straightforward. It is helpful to imagine an abandoned grassland in cross-section: Light competition eventually reduces the number of grass tussocks per unit area relative to pasture while tilllers grow taller, and live and dead biomass become entangled at some distance above ground, resembling the stems and crown canopy of a forest.
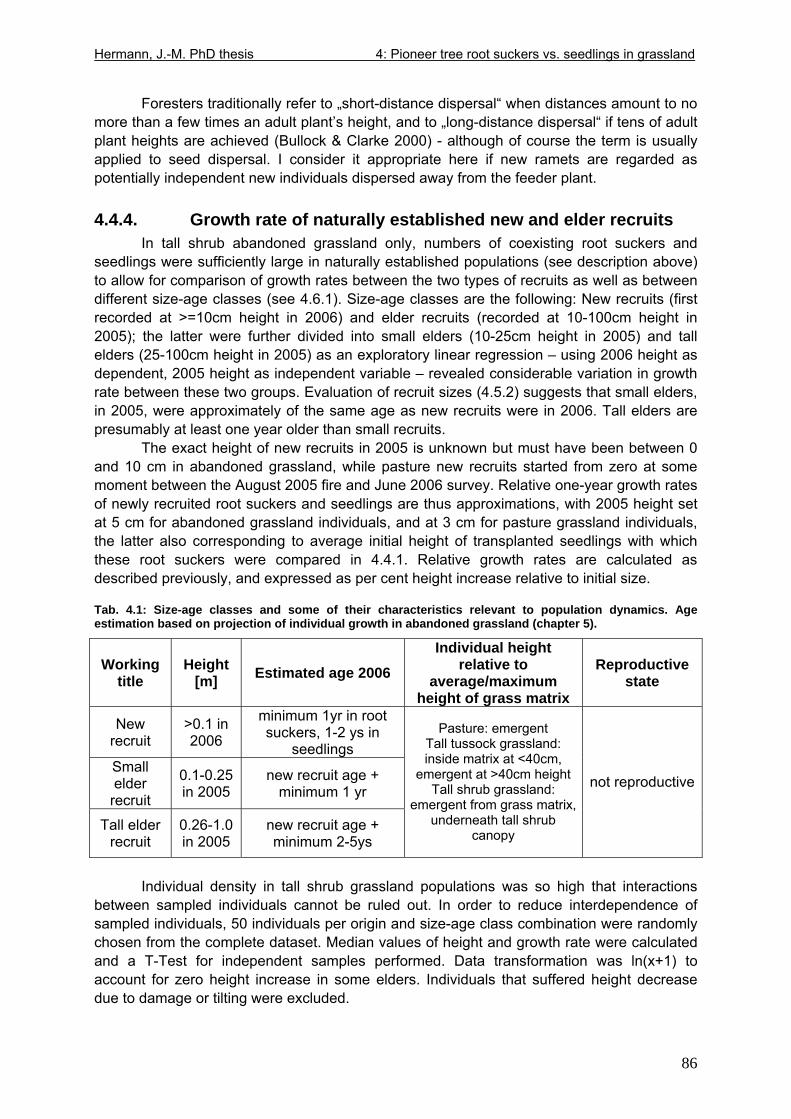
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
86
Foresters traditionally refer to „short-distance dispersal“ when distances amount to no more than a few times an adult plant’s height, and to „long-distance dispersal“ if tens of adult plant heights are achieved (Bullock & Clarke 2000) - although of course the term is usually applied to seed dispersal. I consider it appropriate here if new ramets are regarded as potentially independent new individuals dispersed away from the feeder plant.
4.4.4. Growth rate of naturally established new and elder recruits
In tall shrub abandoned grassland only, numbers of coexisting root suckers and seedlings were sufficiently large in naturally established populations (see description above) to allow for comparison of growth rates between the two types of recruits as well as between different size-age classes (see 4.6.1). Size-age classes are the following: New recruits (first recorded at >=10cm height in 2006) and elder recruits (recorded at 10-100cm height in 2005); the latter were further divided into small elders (10-25cm height in 2005) and tall elders (25-100cm height in 2005) as an exploratory linear regression – using 2006 height as dependent, 2005 height as independent variable – revealed considerable variation in growth rate between these two groups. Evaluation of recruit sizes (4.5.2) suggests that small elders, in 2005, were approximately of the same age as new recruits were in 2006. Tall elders are presumably at least one year older than small recruits.
The exact height of new recruits in 2005 is unknown but must have been between 0 and 10 cm in abandoned grassland, while pasture new recruits started from zero at some moment between the August 2005 fire and June 2006 survey. Relative one-year growth rates of newly recruited root suckers and seedlings are thus approximations, with 2005 height set at 5 cm for abandoned grassland individuals, and at 3 cm for pasture grassland individuals, the latter also corresponding to average initial height of transplanted seedlings with which these root suckers were compared in 4.4.1. Relative growth rates are calculated as described previously, and expressed as per cent height increase relative to initial size.
Tab. 4.1: Size-age classes and some of their characteristics relevant to population dynamics. Age
estimation based on projection of individual growth in abandoned grassland (chapter 5).
Working
title
Height
[m]
Estimated age 2006
Individual height
relative to
average/maximum
height of grass matrix
Reproductive
state
New recruit
>0.1 in 2006
minimum 1yr in root suckers, 1-2 ys in
seedlings Small elder
recruit
0.1-0.25 in 2005
new recruit age + minimum 1 yr
Tall elder recruit
0.26-1.0 in 2005
new recruit age + minimum 2-5ys
Pasture: emergent Tall tussock grassland: inside matrix at <40cm,
emergent at >40cm height Tall shrub grassland:
emergent from grass matrix, underneath tall shrub
canopy
not reproductive
Individual density in tall shrub grassland populations was so high that interactions
between sampled individuals cannot be ruled out. In order to reduce interdependence of sampled individuals, 50 individuals per origin and size-age class combination were randomly chosen from the complete dataset. Median values of height and growth rate were calculated and a T-Test for independent samples performed. Data transformation was ln(x+1) to account for zero height increase in some elders. Individuals that suffered height decrease due to damage or tilting were excluded.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
87
4.4.5. Growth rate of naturally established ramets in different
grassland types
A comparison of growth rates between individuals of different size-age classes – as described above - was also performed on mixed root-sucker seedling datasets from tall tussock and tall shrub abandoned grassland. Samples of 100 individuals were randomly chosen per age-size class and grassland type; they thus contain root suckers und seedlings in the same proportions as the entire sampled populations. Median values of height and growth rate were calculated and a T-Test for independent samples was performed as described in 4.4.4.
In order to find out how the obtained results may have been influenced by root sucker predominance in populations, I also compared height and growth rate of pure root sucker samples between the two types of abandoned grassland. A sample of 50 individuals per grassland type and age class was randomly drawn and compared by T-Test as described before.
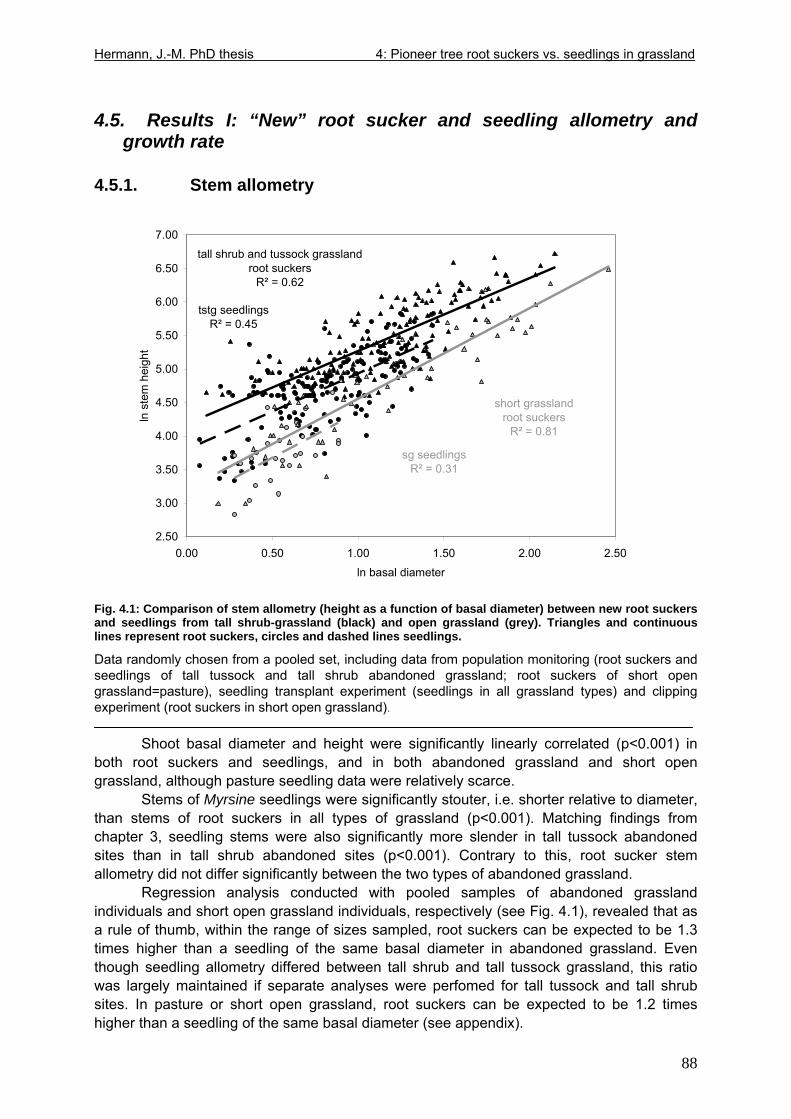
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
88
4.5. Results I: “New” root sucker and seedling allometry and growth rate
4.5.1. Stem allometry
tall shrub and tussock grassland root suckers
R² = 0.62
tstg seedlingsR² = 0.45
sg seedlings R² = 0.31
short grasslandroot suckers
R² = 0.81
2.50
3.00
3.50
4.00
4.50
5.00
5.50
6.00
6.50
7.00
0.00 0.50 1.00 1.50 2.00 2.50
ln basal diameter
ln s
tem
hei
ght
Fig. 4.1: Comparison of stem allometry (height as a function of basal diameter) between new root suckers
and seedlings from tall shrub-grassland (black) and open grassland (grey). Triangles and continuous
lines represent root suckers, circles and dashed lines seedlings.
Data randomly chosen from a pooled set, including data from population monitoring (root suckers and seedlings of tall tussock and tall shrub abandoned grassland; root suckers of short open grassland=pasture), seedling transplant experiment (seedlings in all grassland types) and clipping experiment (root suckers in short open grassland).
Shoot basal diameter and height were significantly linearly correlated (p<0.001) in both root suckers and seedlings, and in both abandoned grassland and short open grassland, although pasture seedling data were relatively scarce.
Stems of Myrsine seedlings were significantly stouter, i.e. shorter relative to diameter, than stems of root suckers in all types of grassland (p<0.001). Matching findings from chapter 3, seedling stems were also significantly more slender in tall tussock abandoned sites than in tall shrub abandoned sites (p<0.001). Contrary to this, root sucker stem allometry did not differ significantly between the two types of abandoned grassland.
Regression analysis conducted with pooled samples of abandoned grassland individuals and short open grassland individuals, respectively (see Fig. 4.1), revealed that as a rule of thumb, within the range of sizes sampled, root suckers can be expected to be 1.3 times higher than a seedling of the same basal diameter in abandoned grassland. Even though seedling allometry differed between tall shrub and tall tussock grassland, this ratio was largely maintained if separate analyses were perfomed for tall tussock and tall shrub sites. In pasture or short open grassland, root suckers can be expected to be 1.2 times higher than a seedling of the same basal diameter (see appendix).
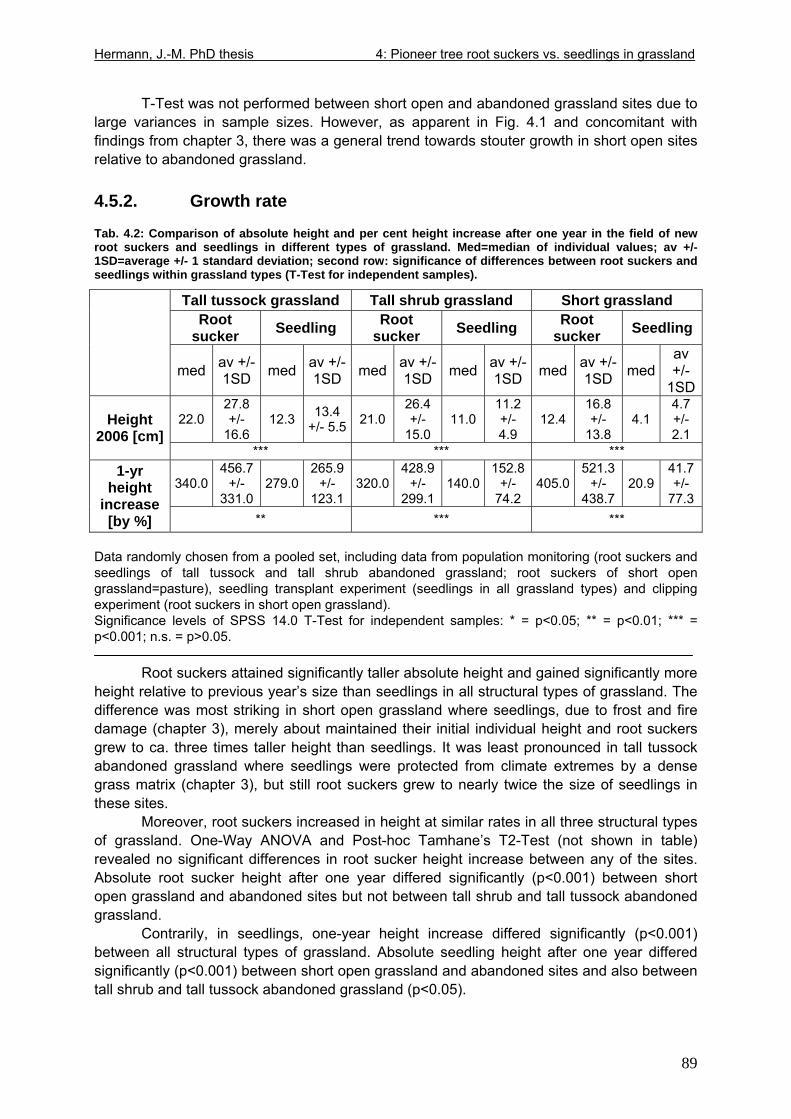
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
89
T-Test was not performed between short open and abandoned grassland sites due to large variances in sample sizes. However, as apparent in Fig. 4.1 and concomitant with findings from chapter 3, there was a general trend towards stouter growth in short open sites relative to abandoned grassland.
4.5.2. Growth rate
Tab. 4.2: Comparison of absolute height and per cent height increase after one year in the field of new
root suckers and seedlings in different types of grassland. Med=median of individual values; av +/-
1SD=average +/- 1 standard deviation; second row: significance of differences between root suckers and
seedlings within grassland types (T-Test for independent samples).
Tall tussock grassland Tall shrub grassland Short grassland
Root
sucker
Seedling
Root
sucker
Seedling
Root
sucker
Seedling
med av +/- 1SD med av +/-
1SD med av +/- 1SD med av +/-
1SD med av +/- 1SD med
av +/-
1SD
22.0 27.8 +/-
16.6 12.3 13.4
+/- 5.5 21.0 26.4 +/-
15.0 11.0
11.2 +/- 4.9
12.4 16.8 +/-
13.8 4.1
4.7 +/- 2.1
Height
2006 [cm]
*** *** ***
340.0 456.7
+/- 331.0
279.0 265.9
+/- 123.1
320.0428.9
+/- 299.1
140.0152.8
+/- 74.2
405.0 521.3
+/- 438.7
20.941.7 +/-
77.3
1-yr
height
increase
[by %] ** *** *** Data randomly chosen from a pooled set, including data from population monitoring (root suckers and seedlings of tall tussock and tall shrub abandoned grassland; root suckers of short open grassland=pasture), seedling transplant experiment (seedlings in all grassland types) and clipping experiment (root suckers in short open grassland). Significance levels of SPSS 14.0 T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
Root suckers attained significantly taller absolute height and gained significantly more
height relative to previous year’s size than seedlings in all structural types of grassland. The difference was most striking in short open grassland where seedlings, due to frost and fire damage (chapter 3), merely about maintained their initial individual height and root suckers grew to ca. three times taller height than seedlings. It was least pronounced in tall tussock abandoned grassland where seedlings were protected from climate extremes by a dense grass matrix (chapter 3), but still root suckers grew to nearly twice the size of seedlings in these sites.
Moreover, root suckers increased in height at similar rates in all three structural types of grassland. One-Way ANOVA and Post-hoc Tamhane’s T2-Test (not shown in table) revealed no significant differences in root sucker height increase between any of the sites. Absolute root sucker height after one year differed significantly (p<0.001) between short open grassland and abandoned sites but not between tall shrub and tall tussock abandoned grassland.
Contrarily, in seedlings, one-year height increase differed significantly (p<0.001) between all structural types of grassland. Absolute seedling height after one year differed significantly (p<0.001) between short open grassland and abandoned sites and also between tall shrub and tall tussock abandoned grassland (p<0.05).
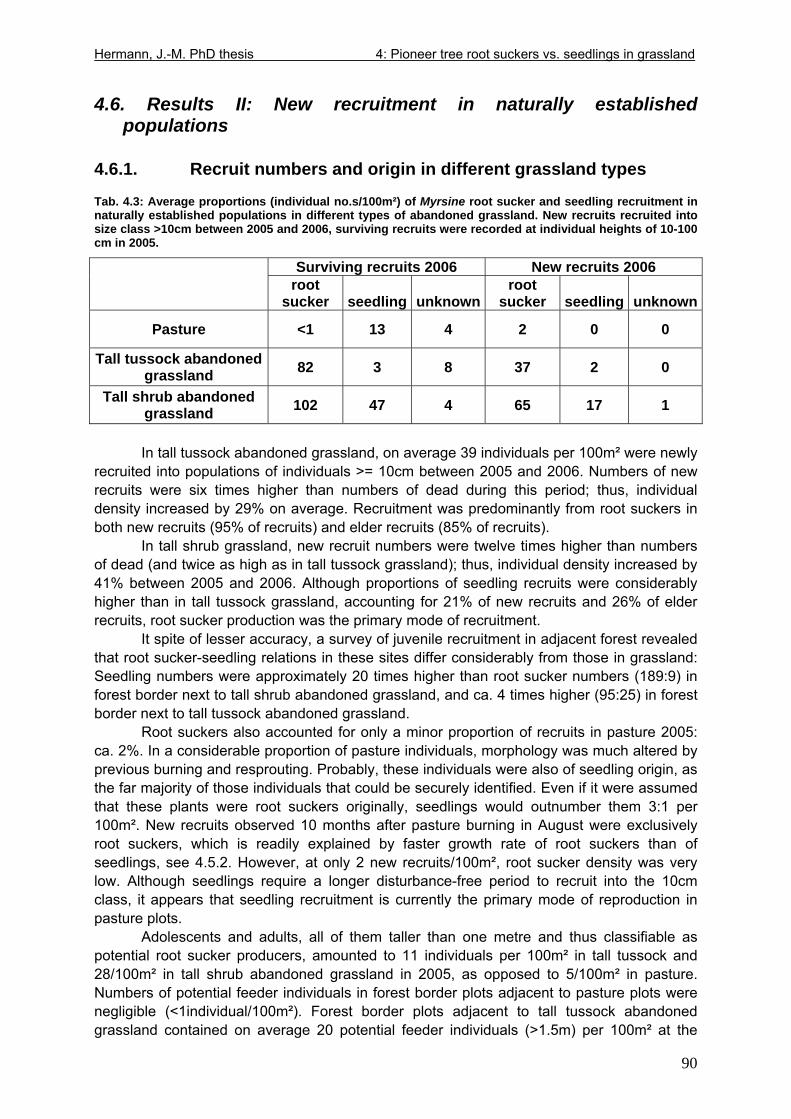
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
90
4.6. Results II: New recruitment in naturally established populations
4.6.1. Recruit numbers and origin in different grassland types
Tab. 4.3: Average proportions (individual no.s/100m²) of Myrsine root sucker and seedling recruitment in
naturally established populations in different types of abandoned grassland. New recruits recruited into
size class >10cm between 2005 and 2006, surviving recruits were recorded at individual heights of 10-100
cm in 2005.
Surviving recruits 2006 New recruits 2006
root
sucker seedling unknown
root
sucker seedling unknown
Pasture <1 13 4 2 0 0
Tall tussock abandoned
grassland
82 3 8 37 2 0
Tall shrub abandoned
grassland
102 47 4 65 17 1
In tall tussock abandoned grassland, on average 39 individuals per 100m² were newly
recruited into populations of individuals >= 10cm between 2005 and 2006. Numbers of new recruits were six times higher than numbers of dead during this period; thus, individual density increased by 29% on average. Recruitment was predominantly from root suckers in both new recruits (95% of recruits) and elder recruits (85% of recruits).
In tall shrub grassland, new recruit numbers were twelve times higher than numbers of dead (and twice as high as in tall tussock grassland); thus, individual density increased by 41% between 2005 and 2006. Although proportions of seedling recruits were considerably higher than in tall tussock grassland, accounting for 21% of new recruits and 26% of elder recruits, root sucker production was the primary mode of recruitment.
It spite of lesser accuracy, a survey of juvenile recruitment in adjacent forest revealed that root sucker-seedling relations in these sites differ considerably from those in grassland: Seedling numbers were approximately 20 times higher than root sucker numbers (189:9) in forest border next to tall shrub abandoned grassland, and ca. 4 times higher (95:25) in forest border next to tall tussock abandoned grassland.
Root suckers also accounted for only a minor proportion of recruits in pasture 2005: ca. 2%. In a considerable proportion of pasture individuals, morphology was much altered by previous burning and resprouting. Probably, these individuals were also of seedling origin, as the far majority of those individuals that could be securely identified. Even if it were assumed that these plants were root suckers originally, seedlings would outnumber them 3:1 per 100m². New recruits observed 10 months after pasture burning in August were exclusively root suckers, which is readily explained by faster growth rate of root suckers than of seedlings, see 4.5.2. However, at only 2 new recruits/100m², root sucker density was very low. Although seedlings require a longer disturbance-free period to recruit into the 10cm class, it appears that seedling recruitment is currently the primary mode of reproduction in pasture plots.
Adolescents and adults, all of them taller than one metre and thus classifiable as potential root sucker producers, amounted to 11 individuals per 100m² in tall tussock and 28/100m² in tall shrub abandoned grassland in 2005, as opposed to 5/100m² in pasture. Numbers of potential feeder individuals in forest border plots adjacent to pasture plots were negligible (<1individual/100m²). Forest border plots adjacent to tall tussock abandoned grassland contained on average 20 potential feeder individuals (>1.5m) per 100m² at the
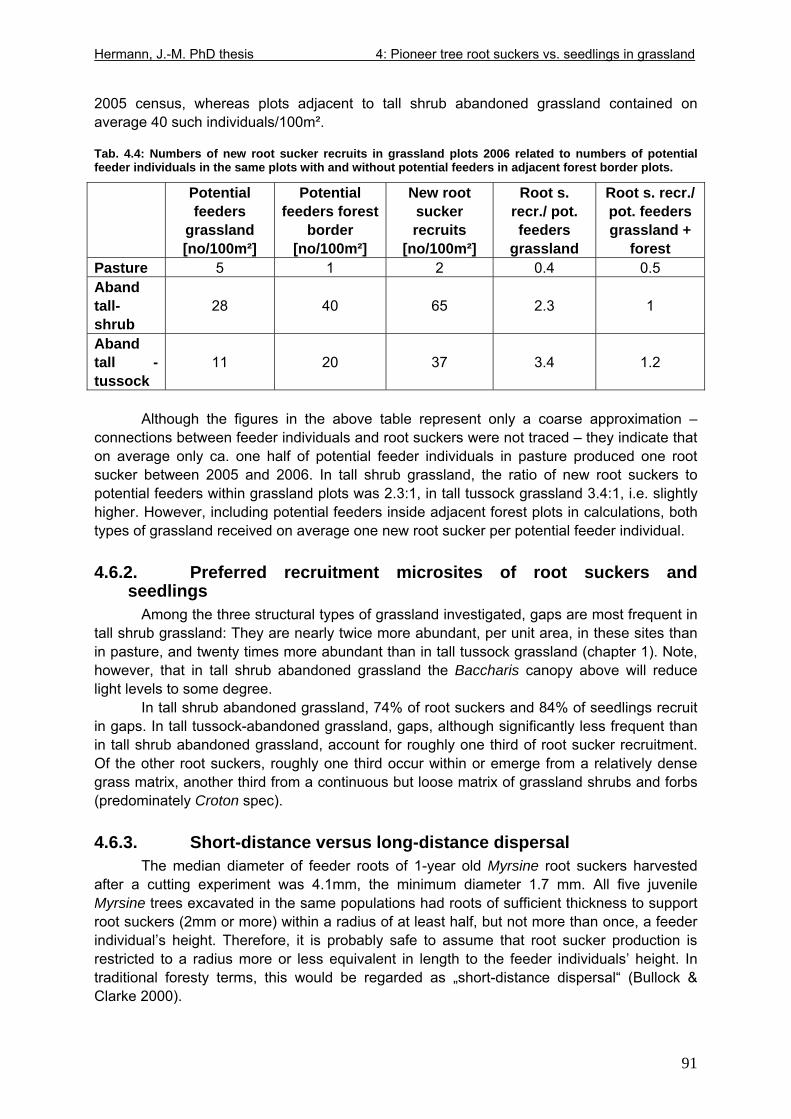
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
91
2005 census, whereas plots adjacent to tall shrub abandoned grassland contained on average 40 such individuals/100m².
Tab. 4.4: Numbers of new root sucker recruits in grassland plots 2006 related to numbers of potential
feeder individuals in the same plots with and without potential feeders in adjacent forest border plots.
Potential
feeders
grassland
[no/100m²]
Potential
feeders forest
border
[no/100m²]
New root
sucker
recruits
[no/100m²]
Root s.
recr./ pot.
feeders
grassland
Root s. recr./
pot. feeders
grassland +
forest
Pasture 5 1 2 0.4 0.5 Aband
tall-
shrub
28 40 65 2.3 1
Aband
tall -
tussock
11 20 37 3.4 1.2
Although the figures in the above table represent only a coarse approximation –
connections between feeder individuals and root suckers were not traced – they indicate that on average only ca. one half of potential feeder individuals in pasture produced one root sucker between 2005 and 2006. In tall shrub grassland, the ratio of new root suckers to potential feeders within grassland plots was 2.3:1, in tall tussock grassland 3.4:1, i.e. slightly higher. However, including potential feeders inside adjacent forest plots in calculations, both types of grassland received on average one new root sucker per potential feeder individual.
4.6.2. Preferred recruitment microsites of root suckers and
seedlings
Among the three structural types of grassland investigated, gaps are most frequent in tall shrub grassland: They are nearly twice more abundant, per unit area, in these sites than in pasture, and twenty times more abundant than in tall tussock grassland (chapter 1). Note, however, that in tall shrub abandoned grassland the Baccharis canopy above will reduce light levels to some degree.
In tall shrub abandoned grassland, 74% of root suckers and 84% of seedlings recruit in gaps. In tall tussock-abandoned grassland, gaps, although significantly less frequent than in tall shrub abandoned grassland, account for roughly one third of root sucker recruitment. Of the other root suckers, roughly one third occur within or emerge from a relatively dense grass matrix, another third from a continuous but loose matrix of grassland shrubs and forbs (predominately Croton spec).
4.6.3. Short-distance versus long-distance dispersal
The median diameter of feeder roots of 1-year old Myrsine root suckers harvested after a cutting experiment was 4.1mm, the minimum diameter 1.7 mm. All five juvenile Myrsine trees excavated in the same populations had roots of sufficient thickness to support root suckers (2mm or more) within a radius of at least half, but not more than once, a feeder individual’s height. Therefore, it is probably safe to assume that root sucker production is restricted to a radius more or less equivalent in length to the feeder individuals’ height. In traditional foresty terms, this would be regarded as „short-distance dispersal“ (Bullock & Clarke 2000).

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
92
Fig. 4.2: Adolescent Myrsine parvula (ca. 1.4 m height) excavated in abandoned grassland in the Pró-Mata
area in 2004. Origin of root suckers pointed out by red arrows. This individual may originally have been a
root sucker itself, judging by the relatively thick lateral root (bearing small root sucker) extending to the
right. However, the adventive root system is sufficiently well developed to support two more root suckers
(small shoot adjacent to individual, medium-sized shoot on the left). Photographer: J.-M. Hermann.
A similar trend is apparent in Tab. 4.5: Here, I compiled data on annual rates of dispersal of generatively reproducing species, and annual rates of vegetative spread in clonal species, and related them to maximum plant height in order to facilitate comparison across plant growth forms.
Note that the North American tree species Populus grandidentata takes the lead among clonal plants, achieving dispersal distances equivalent to nearly times six times adult individual height in one year. Nevertheless, since distances do not exceed more than a few times the height of an adult plant, they would also be classified as short-distance dispersers. Moreover, the other clonal species listed do not spread annually at distances of more, or even as much as their own adult height. Among generatively reproducing genera and species, similarly short dispersal distances are reported e.g. in Fagus and Tsuga canadensis, both of them late-successional – i.e. shade-tolerant, long-lived and slow-growing (see also Davis 1987). Note that the values given here are pollen travel distances extrapolated from palynological data, from which actual seed dispersal distances may differ: E.g. Johnson & Adkisson (1985) report dispersal of Fagus nuts by birds (blue jays) across 4 km.
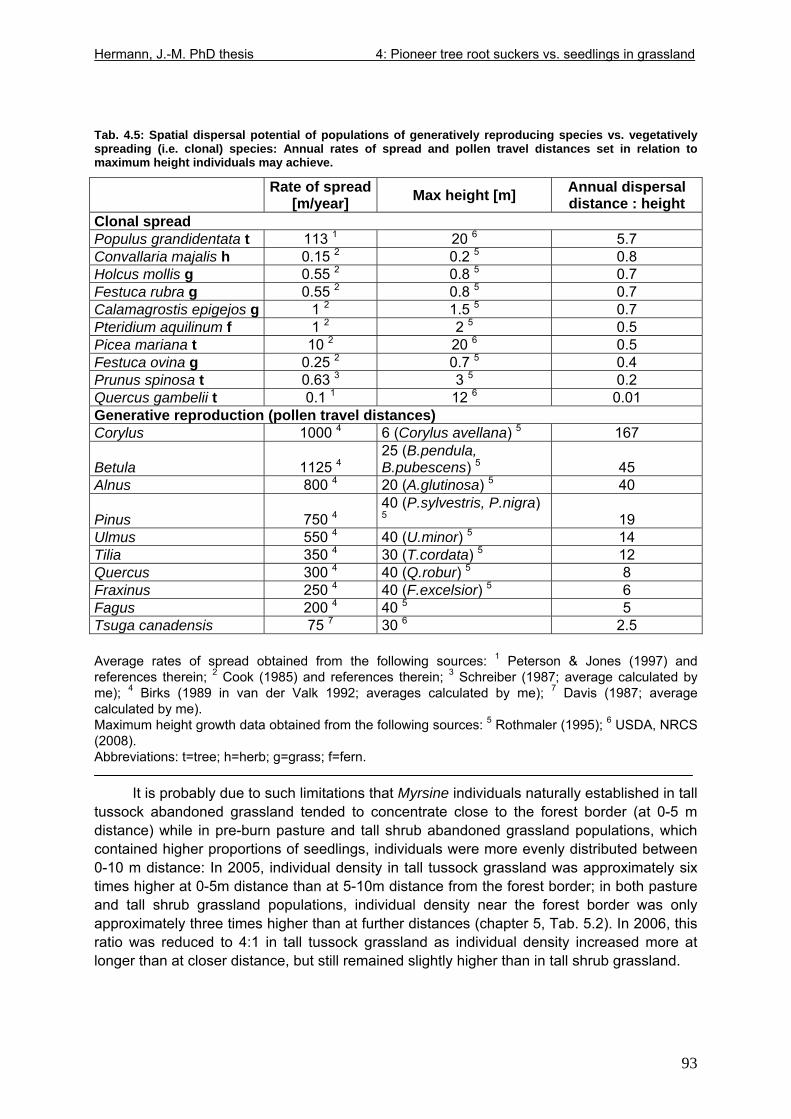
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
93
Tab. 4.5: Spatial dispersal potential of populations of generatively reproducing species vs. vegetatively
spreading (i.e. clonal) species: Annual rates of spread and pollen travel distances set in relation to
maximum height individuals may achieve.
Rate of spread
[m/year]
Max height [m]
Annual dispersal
distance : height
Clonal spread
Populus grandidentata t 113 1 20 6 5.7 Convallaria majalis h 0.15 2 0.2 5 0.8 Holcus mollis g 0.55 2 0.8 5 0.7 Festuca rubra g 0.55 2 0.8 5 0.7 Calamagrostis epigejos g 1 2 1.5 5 0.7 Pteridium aquilinum f 1 2 2 5 0.5 Picea mariana t 10 2 20 6 0.5 Festuca ovina g 0.25 2 0.7 5 0.4 Prunus spinosa t 0.63 3 3 5 0.2 Quercus gambelii t 0.1 1 12 6 0.01 Generative reproduction (pollen travel distances)
Corylus 1000 4 6 (Corylus avellana) 5 167
Betula 1125 4 25 (B.pendula, B.pubescens) 5 45
Alnus 800 4 20 (A.glutinosa) 5 40
Pinus 750 4 40 (P.sylvestris, P.nigra) 5 19
Ulmus 550 4 40 (U.minor) 5 14 Tilia 350 4 30 (T.cordata) 5 12 Quercus 300 4 40 (Q.robur) 5 8 Fraxinus 250 4 40 (F.excelsior) 5 6 Fagus 200 4 40 5 5 Tsuga canadensis 75 7 30 6 2.5
Average rates of spread obtained from the following sources: 1 Peterson & Jones (1997) and references therein; 2 Cook (1985) and references therein; 3 Schreiber (1987; average calculated by me); 4 Birks (1989 in van der Valk 1992; averages calculated by me); 7 Davis (1987; average calculated by me). Maximum height growth data obtained from the following sources: 5 Rothmaler (1995); 6 USDA, NRCS (2008). Abbreviations: t=tree; h=herb; g=grass; f=fern.
It is probably due to such limitations that Myrsine individuals naturally established in tall
tussock abandoned grassland tended to concentrate close to the forest border (at 0-5 m distance) while in pre-burn pasture and tall shrub abandoned grassland populations, which contained higher proportions of seedlings, individuals were more evenly distributed between 0-10 m distance: In 2005, individual density in tall tussock grassland was approximately six times higher at 0-5m distance than at 5-10m distance from the forest border; in both pasture and tall shrub grassland populations, individual density near the forest border was only approximately three times higher than at further distances (chapter 5, Tab. 5.2). In 2006, this ratio was reduced to 4:1 in tall tussock grassland as individual density increased more at longer than at closer distance, but still remained slightly higher than in tall shrub grassland.
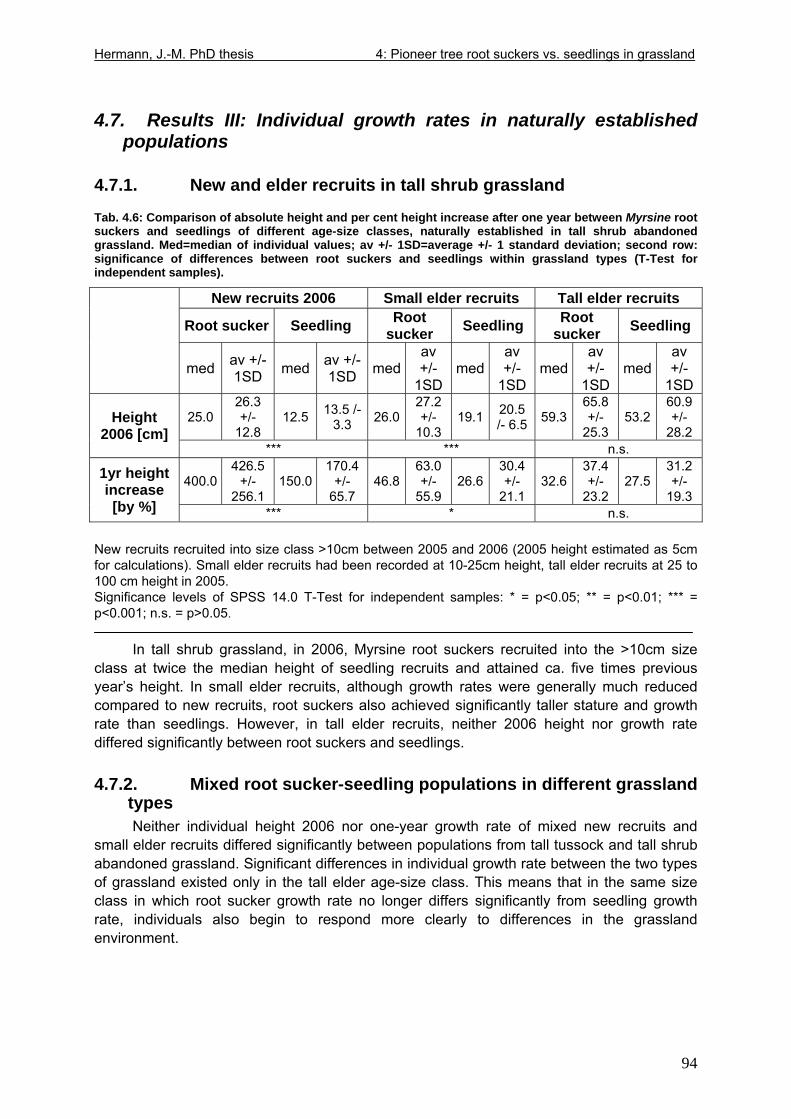
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
94
4.7. Results III: Individual growth rates in naturally established populations
4.7.1. New and elder recruits in tall shrub grassland
Tab. 4.6: Comparison of absolute height and per cent height increase after one year between Myrsine root
suckers and seedlings of different age-size classes, naturally established in tall shrub abandoned
grassland. Med=median of individual values; av +/- 1SD=average +/- 1 standard deviation; second row:
significance of differences between root suckers and seedlings within grassland types (T-Test for
independent samples).
New recruits 2006 Small elder recruits Tall elder recruits
Root sucker Seedling
Root
sucker
Seedling
Root
sucker
Seedling
med av +/- 1SD med av +/-
1SD medav +/-
1SD med
av +/-
1SD med
av +/-
1SD med
av +/-
1SD
25.0 26.3 +/-
12.8 12.5 13.5 /-
3.3 26.027.2 +/-
10.3 19.1 20.5
/- 6.5 59.3 65.8 +/-
25.3 53.2
60.9 +/-
28.2 Height
2006 [cm]
*** *** n.s.
400.0 426.5
+/- 256.1
150.0 170.4
+/- 65.7
46.863.0 +/-
55.9 26.6
30.4 +/-
21.1 32.6
37.4 +/-
23.2 27.5
31.2 +/-
19.3
1yr height
increase
[by %] *** * n.s. New recruits recruited into size class >10cm between 2005 and 2006 (2005 height estimated as 5cm for calculations). Small elder recruits had been recorded at 10-25cm height, tall elder recruits at 25 to 100 cm height in 2005. Significance levels of SPSS 14.0 T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
In tall shrub grassland, in 2006, Myrsine root suckers recruited into the >10cm size class at twice the median height of seedling recruits and attained ca. five times previous year’s height. In small elder recruits, although growth rates were generally much reduced compared to new recruits, root suckers also achieved significantly taller stature and growth rate than seedlings. However, in tall elder recruits, neither 2006 height nor growth rate differed significantly between root suckers and seedlings.
4.7.2. Mixed root sucker-seedling populations in different grassland
types
Neither individual height 2006 nor one-year growth rate of mixed new recruits and small elder recruits differed significantly between populations from tall tussock and tall shrub abandoned grassland. Significant differences in individual growth rate between the two types of grassland existed only in the tall elder age-size class. This means that in the same size class in which root sucker growth rate no longer differs significantly from seedling growth rate, individuals also begin to respond more clearly to differences in the grassland environment.
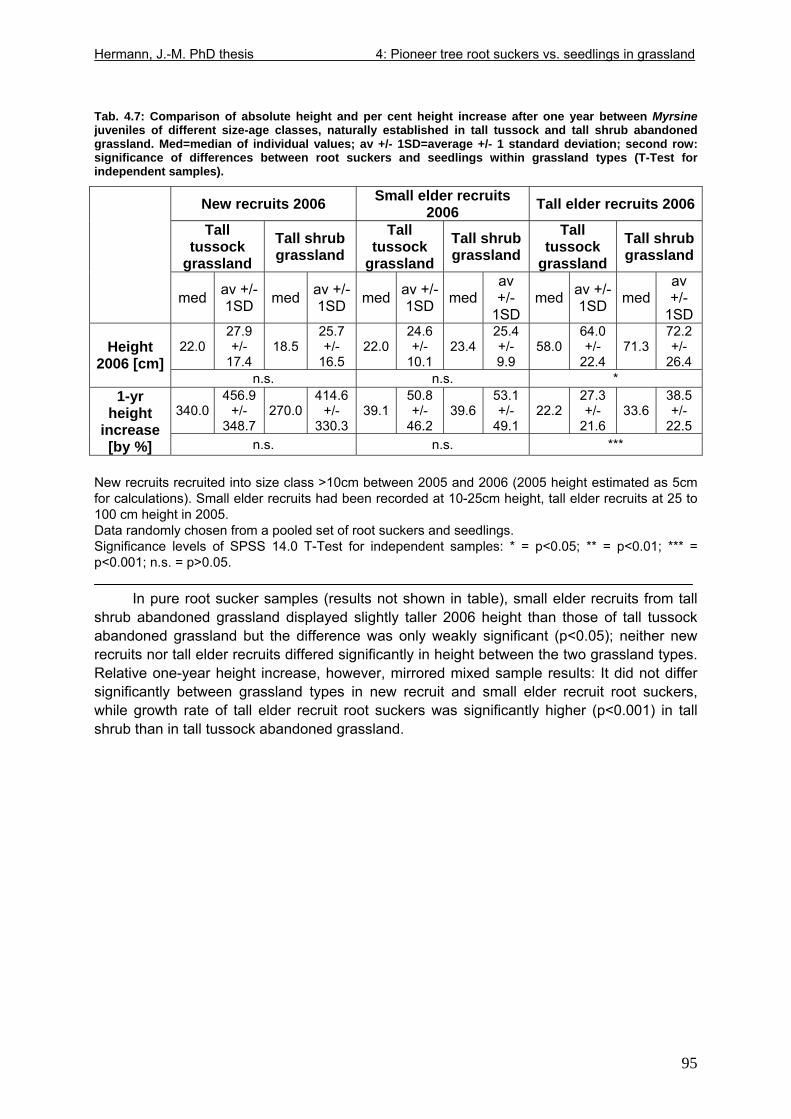
Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
95
Tab. 4.7: Comparison of absolute height and per cent height increase after one year between Myrsine
juveniles of different size-age classes, naturally established in tall tussock and tall shrub abandoned
grassland. Med=median of individual values; av +/- 1SD=average +/- 1 standard deviation; second row:
significance of differences between root suckers and seedlings within grassland types (T-Test for
independent samples).
New recruits 2006
Small elder recruits
2006
Tall elder recruits 2006
Tall
tussock
grassland
Tall shrub
grassland
Tall
tussock
grassland
Tall shrub
grassland
Tall
tussock
grassland
Tall shrub
grassland
med av +/- 1SD med av +/-
1SD med av +/- 1SD med
av +/-
1SD med av +/-
1SD med av +/-
1SD
22.0 27.9 +/-
17.4 18.5
25.7 +/-
16.5 22.0
24.6 +/-
10.1 23.4
25.4 +/- 9.9
58.0 64.0 +/-
22.4 71.3
72.2 +/-
26.4 Height
2006 [cm]
n.s. n.s. *
340.0 456.9
+/- 348.7
270.0 414.6
+/- 330.3
39.150.8 +/-
46.2 39.6
53.1 +/-
49.1 22.2
27.3 +/-
21.6 33.6
38.5 +/-
22.5
1-yr
height
increase
[by %] n.s. n.s. *** New recruits recruited into size class >10cm between 2005 and 2006 (2005 height estimated as 5cm for calculations). Small elder recruits had been recorded at 10-25cm height, tall elder recruits at 25 to 100 cm height in 2005. Data randomly chosen from a pooled set of root suckers and seedlings. Significance levels of SPSS 14.0 T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
In pure root sucker samples (results not shown in table), small elder recruits from tall shrub abandoned grassland displayed slightly taller 2006 height than those of tall tussock abandoned grassland but the difference was only weakly significant (p<0.05); neither new recruits nor tall elder recruits differed significantly in height between the two grassland types. Relative one-year height increase, however, mirrored mixed sample results: It did not differ significantly between grassland types in new recruit and small elder recruit root suckers, while growth rate of tall elder recruit root suckers was significantly higher (p<0.001) in tall shrub than in tall tussock abandoned grassland.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
96
4.8. Discussion
4.8.1. What are root suckers, and what do they serve for?
Root suckers represent a type of clonal growth as opposed to clonal reproduction: The first term relates to the capacity to form potentially independent ramets (i.e. usually a shoot with associated roots) by vegetative means, the second term relates to the ability to form genetically identical units from a single somatic cell (de Kroon & van Groenendael 1997). The elder individual from whose root system a new sucker arises is usually referred to as the parent or mother plant, more seldom as the „ortet“ (e.g. Yeh et al 1995). Here I prefer to use the terms „feeder“ plant or root; this is unusual but intuitively appealing, and helps to avoid confusion of the population processes of generative and vegetative reproduction.
Root suckers develop on lateral meristems of roots usually spreading horizontally close to the soil surface (DeByle 1964) by either programmed reiteration - in which new individuals arise from existing meristems as a normal part of development - or traumatic reiteration, in which new individuals are produced under the stimulus of stress (Tiffney & Niklas 1985). Suckers are linked to the feeder plants initially but may become physiologically independent on growth of adventive roots and interruption of the connection to the feeder individual.
According to reviews by Romberger et al. (1993), Jenik (1994) and Peterson & Jones (1997), the ability to sprout from roots or rhizomes is shared by some of the most widespread North American and European tree and shrub genera, among them pioneers in primary and secondary succession such as Populus, Salix and Robinia, small trees of forest borders and hedgerows such as Pyrus, Malus and Prunus as well as later-successional genera such as Quercus and Fagus.
Records of root suckering in South American species are comparatively scarce in the reviews cited; this may be due to a relative scarcity of studies of this phenomenon, rather than absence of it on the South American continent. Hoffmann (1998) reports root suckering in one shrub and two tree species – Rourea induta, Myrsine guianensis and Roupala montana – of the Brazilian Cerrado, Cirne & Scarano (2001) in the Brazilian restinga tree species Andira legalis, Veblen et al. (2003) in the northern Patagonian tree species Araucaria araucana and Fitzroya cupressoides.
Clonally growing woody species have not received similar attention in scientific studies as clonal herbaceous species - in a 1985 paper, Harper briefly touches the subject but adds that „most tree do not behave like this“ (p. 4). One exception are root suckering Populus species, especially Trembling Aspen (Populus tremuloides Michx.), in the United States and Canada. There, in the semiarid range between Sierra Nevada and Rocky Mountains as well as the Great Lakes Region, Aspen clones form stands of considerable commercial value to both timber industry and tourism; hence comprehensive studies have been carried out especially by the US Department of Agriculture and Forestry.
About one decade following Harper’s statement, Jenik (1994) wrote that „only recently has clonal growth in botany and ecology been clearly and broadly interpreted in order to satisfy the dendrologists“ (p.292). However, so far there seems to be no standardised definition and physiology and mechanisms of root sucker initiation remain rather vague. In fact, there is no agreement on whether sprouting from underground parts in woody plants is primarily a means of ensuring genet survival, or foraging in a patchy environment. Klimešová & Martínková (2004) propose that the latter is more often the case, as the element of foraging is inherent in root growth. Peterson & Jones (1997) propose that the first is more often the case but acknowledge the need of more detailed studies of e.g. clonal integration.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
97
Apparently, there is also some confusion or disagreement on what type of underground structure woody plants sprout from, namely, whether sprouting occurs from roots or rhizomes. For example, Peterson & Jones (1997) list Aspen as a rhizome sprouter, while authors of all Aspen studies cited here refer to Aspen sprouts as root suckers. This confusion may arise, initially, from the definition of the term „rhizome“. According to Tiffney & Niklas (1985, p. 39-40), rhizomes „are horizontal extensions of a single plant that possess the ability to form adventitious roots“; in a „proper morphological sense“ (they later add), rhizomes represent „a horizontal stem bearing roots“. Klimešová & Martínková (2004) declare that biennial plants and trees „do not possess clonal growth organs of stem origin“ (p.678) and even suggest that „the evolution of clonality via root spacers was probably not only independent of, but also alternative to that mediated by stem spacers“ (p.679). Jenik (1994) states that rhizomes are principally found in woody monocot species.
This is not merely a taxonomic debate. If rhizomes are horizontal stems, they represent an extra construction on the plant body that only pays off by subsequent risk-spreading, resource acquisition etc. A root system is constructed for soil resource acquisition in the first place. Sprouting from roots may thus be a highly efficient means of foraging for both light and soil resources – see Klimešová & Martínková (2004). In comparison of my results with studies from the USA, I will follow the lead of the majority and refer to Aspen sprouts as root suckers. In the case of Myrsine, origin of suckers from roots was never in doubt; the structures from which suckers arose clearly resembled other roots in the root system, not the stems (for example, bark of Myrsine stems is always smooth; that of roots is finely fissured).
Authors also differ in their conception of how far away from the feeder individual stem a sprout must be located to be regarded as a root sucker. Kammesheidt (1999), for example, does not distinguish between sprouts from roots and root crown. Peterson & Jones (1997) distinguish between „coppice shoots“ arising from the root crown and „root suckers“ arising from roots. Tiffney & Niklas (1985) include basal sprouting and root sprouting in one category of „clumped“ clonal growth, although they suggest that the first allows more to explore for resources, and the second for efficient exploitation of resources when encountered (so that species capable of both basal and root suckering have an advantage e.g. over rhizomatous species). In practice, especially in my resprouting experiments (chapter 6), basal sprouts and root sprouts often formed a continuum from stem stumps to roots. Nevertheless, I attempted to restrict sampling to ramets arising at such a distance from the feeder individual that they might be considered an extension of the genet in space, even if this distance was no more than a few centimetres; in the population study described in this chapter, even in burnt pasture, this distinction was easier than in the resprout study.
Root sprouting is generally thought to be triggered by a release from apical dominance, i.e. interruption of the flow of inhibitory auxin from the feeder individual’s apical meristem to the roots (e.g. Cook 1985). An important question is whether release from apical dominance necessarily involves physical damage to feeder plant or roots. Forestry studies in the USA have demonstrated that in Aspen, apical dominance is so strong as to be broken only by substantial damage such as cutting of feeder individuals or severing of roots: Farmer (1962) reports that undamaged individuals in his study did not produce any suckers; Fraser et al. (2004) report a twofold increase of root sucker numbers on injured roots relative to uninjured roots. However, Romberger et al. (1993) point out that meristems of wound-callus origin are often ephemeral due to inadequate vascular connections (see also Bosela & Ewers 1997), contrary to root-shoot meristems in intact roots.
Furthermore, strength of apical dominance apparently varies among species: Peterson & Jones (1997) report that clones of some woody species have a distinctly „domed“ appearance, with new recruitment principally occurring on the periphery, whereas others display a mixed distribution of younger and older ramets, and they suggest that the first type

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
98
of spatial distribution indicates strong apical dominance, the latter weak apical dominance. However, Cook (1985) states that apical dominance generally decreases with distance from the feeder individual. Either way, while it is not possible to distinguish between clones in a study area it is also not possible to tell whether a mixed size distribution indicates weak apical dominance, or is simply due to intermingling of several clones.
Generally, similar to clonal herbaceous species (Cook 1985), clonality among trees and shrubs is more frequently observed in harsh climates, resource-poor or highly disturbed areas than in „benign“ habitats (Peterson & Jones 1997). Koop (1987) highlights root suckering as „almost the only method of forest regeneration near the limits of tree growth on the Wadden islands“ (p.103) as well as its importance in colonisation of forest gaps and riverine forests. Whittaker (1993), on investigation of shrub populations in glacier foreland succession in south-central Norway, demonstrated that these populations first recruit from seed (in a previously established matrix of pioneer herbs) and clonal growth predominates only in mature heath, more than two centuries after initial establishment. Nevetheless, it might be worthwhile to check whether auxin flow is in some way linked to reproductive status of the plant; in other words, whether vegetative spread is induced directly by failure to reproduce generatively, or because reproductive individuals detect unfavourable conditions for seedling growth. Such behaviour would support the hypothesis that root sprouting serves for perpetuation of the genet.
High irradiation in gaps may have a dual effect: Firstly, reducing the levels of inhibitory auxin on the upper side of surface-near or exposed roots (on influence of light on auxin levels see e.g. Mohr & Schopfer 1992); secondly, raising soil temperature to a critical level required for root sprouting (Maini & Horton 1966). In my study, root sucker recruitment was highest in gaps in tall shrub grassland; more importantly, in tall tussock grassland one third of root suckers recruited in gaps in the grass matrix, although systematic sampling of vegetation heights demonstrated that the far majority of the area is covered in a more or less closed grass and shrub matrix. Root suckers seem to „find“ gaps where they concentrate e.g. along trails; this indicates foraging. Considering that in both tropical and temperate forests, gaps account for only minor proportions of the total area (ca. 1% according to Denslow 1987, Lorimer 1989) and light is considered the principal limiting factor (Denslow 1987), the ability to forage for light by both vertical and horizontal growth clearly seems advantageous for a forest species. However, the major proportion of root suckers did not recruit in gaps but within or even underneath grass and low shrub cover. Likewise, Koop (1987) and Kammesheidt (1999) propose that root suckering is frequently employed in heavily shaded forest areas where generative reproduction is inhibited. This, again, would favour the genet perpetuation hypothesis.
Although the above does not represent a comprehensive review, it indicates that genet perpetuation and resource foraging are not strictly alternative strategies. Some of the confusion may be resolved simply by re-phrasing terms and concepts. For example, genet perpetuation may be viewed as a type of foraging in time as opposed to foraging in space. At this point, the importance of distinguishing between basal (or coppice) shoots and root shoots becomes obvious. Basal shoots „may help to perpetuate the organism through periods of ecological instability and/or the senescence of the parent individual“ (Tiffney & Niklas 1985 p.41), i.e. may serve for foraging in the future. Of course root suckers may also fulfil this purpose, but initial costs are higher; basal shoots usually have a rootstock at their immediate disposal, whereas sucker production involves costs for transport and construction of new adventive root systems. Consequently, there should be an additional premium – in the form of increased safety, increased resource availability etc. This is discussed in some more detail in chapter 6.
Position of new shoots relative to the original individual must be taken more into account, but it is also necessary to investigate possible triggers of root sucker production.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
99
Regarding Myrsine root sucker recruitment in grassland, the questions are: Is root suckering a means of pre-empting sites in which light, space or perhaps other resources are available in abundance? Is it a reaction to growth limitation in a habitat that differs considerably from the one the species involved in? Or are both processes involved?
4.8.2. Differences between root suckers and seedlings
It has been shown that seedling stems exhibit stouter growth (lower height at a given basal diameter) than root sucker stems. This can be interpreted as a consequence of seedlings’ need to compete for soil resources, and to invest in storage of assimilates, whereas root suckers, supplied continuously with water and nutrients through their feeder root, need to compete principally for light, and thus invest relatively more in height growth.
Better fit of the root sucker regression curves than of the seedling regression curve (i.e. lesser within-sample variance) might indicate more regular growth under regular resource supply, but may also be an artefact of the evaluation, as a wider range of sizes was sampled in root suckers than in seedlings. Other results obtained here underline more clearly the importance of physiological integration for root suckers in early life.
For example, greater extremes e.g. in microclimate of pasture and mowed areas are powerful enough to cause further reduction of the height-diameter ratio in both root suckers and seedlings relative to abandoned grassland, concomitant with findings from the seedling transplant experiment (chapter 3). Stouter growth in this case may be a consequence of ‚pruning’ by cold or frost damage or herbivory (probably by deer, observed in mowed areas); it may also be a trait acquired for the purpose of energy conservation, as a similar trend was observed in stem allometry of undamaged Baccharis shrub seedlings in pasture. Nevertheless, on resprouting, root suckers still benefit from external supply to such extent as to achieve significantly taller height relative to diameter, and higher growth rate than seedlings: 1yr-growth rates of new root sucker recruits did not differ significantly between either type of abandoned grassland and pasture or mowed areas.
Differences in leaf size and allometry between pasture seedlings and root suckers from mowed areas were also remarkable (see previous chapter): Pasture seedlings produced small round leaves whereas mowed area root sucker leaves maintained the species-specific obovate shape, and their size several times surpassed that of seedling leaves. Mowed area root sucker leaves were, in fact, most similar in size and shape to leaves of seedlings grown under moderate mciroclimate and reduced competition in the nurseries.
These results demonstrate that in stands of recently recruited, fairly even-aged individuals stem allometry and growth rate may indeed be sufficient to distinguish with confidence between root suckers and seedlings, as Hoffmann (1998) did. On the other hand, such stands would only develop after large-scale disturbance such as fire, in which case root suckers probably intermingle with basal resprouts that look virtually alike. It may then be indispensable to consider root systems and position of sprouts to remaining stem stumps. However, in my study of Myrsine population structure in pasture, recurrent disturbances had so altered morphology of root systems that origin of a considerable proportion of individuals remained obscure (chapter 5). Only genetic analysis could tell with certainty whether those individuals originated from seeds or represent fragments of clones.
Differences between root suckers and seedlings also become less pronounced with age, as demonstrated by comparison of new and elder recruit growth rates. Root suckers eventually develop their own adventive root system; moreover, supply from feeder individuals may decrease relative to ramet size with age for a number of reasons discussed below. Whatever the underlying cause, it appears that root suckers in the tall elder recruit stage (i.e. from an estimated age of three years onwards) do not greatly differ from seedlings in

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
100
physiology. Unlike younger size-age classes, tall elder recruits as a group – although still containing more root suckers than seedlings – show a clearly differentiated response to the surrounding type of grass matrix: Height growth was significantly higher in tall shrub grassland than in tall tussock grassland.
As discussed above, it is not known what exactly triggers root sucker production in Myrsine in my study area. However, there is little reason to assume that substantial mechanical damage to feeder individuals or roots was the cause, in which case root sucker support would be limited by the amount photosynthetates stored in roots, and consequently by length of the severed piece of root (Perala 1978). When root connections remain largely intact, feeder individuals may support ramets to considerable extent and for a long time. Zahner & DeByle (1965) demonstrated, by severing connections in US Populus forest stands of different ages established after clear-cutting and fire, that new adventive root systems contribute little to growth for at least the first six years following establishment, and they still account for only approximately half of total growth by the 25th year. De Byle (1964) concluded that root connections between feeder individual and ramet remain alive and functioning for at least twice that time span.
These findings contrast strongly with those of Peltzer (2002), who performed severing experiments on Aspen root suckers newly established in US Prairie adjacent to forest (after hailstorm damage to feeder trees at the forest border). She found that „clonal integration tended to improve ramet survival and growth but these trends were often not significant“ (p.194); moreover, „clonal integration did not alter the effect of competition from neighbouring vegetation (...), suggesting that interspecific interactions may be more important than physiological integration in this system“ (ibid.) – i.e. presence or absence of a surrounding grass matrix had a more pronounced effect than presence or absence of connections to feeders.
In comparison of these two studies, the following attracts attention: Zahner and DeByle worked inside Populus forest stands that were occasionally disturbed but nevertheless had been in place for several decades; in Peltzer’s study, Aspen newly colonised grassland from an adjacent forest border, as Myrsine does in my study area. Does it follow that Aspen distinguishes between forest and non-forest habitats, and adjusts allocation to ramets accordingly? Apparently this basic question yet remains to be answered not just for clonal trees and shrubs, but clonal plants in general, as Fischer & van Kleunen (2001) stated that plasticity in clonal integration had not been studied to date of their publication. Similarly, Peterson & Jones (1997) point out that while the superior colonising ability of clonal species is amply documented, there is surprisingly little direct evidence that clonal integration improves competitive ability in these situations (see also Pennings & Callaway 2000).
In theory, a number of reasons may account for relatively early physiological independence of forest tree root suckers in grassland habitats, and I will briefly outline them below. The costs and benefits of physiological integration in general have been reviewed e.g. by Pitelka & Ashmun (1985). Here, I focus on the case that clonal growth occurs on colonisation of a new habitat type - always bearing in mind that such relatively early physiological independence has merely been inferred in both my and Peltzer’s (2002) study, whereas DeByle (1964) confirmed the existence of long-term functioning connections by tracers.
1. The feeder individual cannot supply root suckers any further, e.g. due to damage to connections or to resource limitation.
2. The feeder individual no longer needs to supply the ramet, e.g. because the ramet has outgrown surrounding vegetation, or because resources are so plentiful that even reciprocal support of the feeder individual by the new ramet may occur. The latter suggestion is offered by Peltzer (2002) but seems rather unlikely in the case of my

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
101
study area, given the relative nutrient poverty of grassland soil and intense belowground competition by grasses. Wilson (1993) found tree seedlings more limited by belowground competition in grassland than in forest, and the same probably applies to the adventive root systems of suckers.
3. The feeder individual does not benefit from further supply – revenue of nutrients, photosynthetates etc. is too low compared to investment. In any case, but especially in the latter, it is important to recall what habitat a given
root suckering species evolved in, or at least which habitat was inhabited successfully prior to colonisation of the new habitat. Myrsine is a forest tree, and inside forest, root suckering of trees generally serves to exploit medium gaps (Bazzaz 1984). In such a habitat, at least a transient increase in nutrients may be expected e.g. due to increased litter remineralisation rates. Remember also the discrepancy in nutrient status between highland and escarpment forest soils (chapter 1): Although base saturation of highland cambisols is not as low as to preclude Araucaria forest establishment, it is clearly higher on the escarpment from where these forests have expanded onto the grasslands.
Canham & Marks (1985), on reviewing gap species’ architecture, write: „[These studies] suggest that a high efficiency of growth following a given disturbance [...] will often involve a pattern of branching and leaf display that is less efficient in exploiting other disturbances“ (p.212). They refer to crown architecture, but the point is worth considering for root suckering which, after all, can be viewed as a type of branching at or near the soil surface - Harper 1977 uses the descriptive image of a tree laid on its side.
What follows from the above considerations is that although root suckering does make Myrsine a more successful grassland coloniser than other forest species that lack the ability of clonal growth, she should not a priori be considered a particularly efficient coloniser. It is currently not possible to tell whether root suckering Myrsine individuals forage efficiently for resources not available in forest and forest border, to what extent this is triggered by reaction to environmental cues outside forest or constraints on their vertical growth or generative reproduction inside forest, and whether the evolutionary time span over which this occurred was long enough to select for fittest grassland colonisers, or whether the phenomenon is so young that species in fact „mistakes“ grassland for a forest environment.
Only a cursory survey was made in the forest border adjacent to grassland plots, but this survey nevertheless revealed that extensive root sucker recruitment (i.e. ramet density exceeding seedling density) is a phenomenon restricted to grassland. Secondly, the ratio of new root sucker recruits to potential feeders (if the latter are summed over grassland and forest) is not essentially greater in tall tussock than in tall shrub grassland; i.e. there is no clear indication of „favourability“ of either one or the other grassland type for root sucker recruitment.
At the end of the previous paragraph, it has been asked whether root suckering of Myrsine serves for the perpetuation of genets or for foraging, or both. At the end of this paragraph, the question may be extended to whether the strategy works as efficiently – or more or less efficiently – for Myrsine in grassland as in her original forest habitat. Principal questions that remain to be resolved are: What exactly triggers root sucker growth, and to what extent may feeder individuals coordinate placement and support of ramets?
4.8.3. A forest species as grassland coloniser: Root sucker versus
seedling recruitment
Some confusion in clonal plant population studies arises from the ambiguous use of the term „individual“. The term always refers to a (potentially) physiologically independent plant, but this may be either a discrete genetic individual derived from a single zygote or a ramet derived by lateral growth of a genet that has, or may, become structurally independent

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
102
on severing or decay of connections (e.g. Crawley 1997b). For one thing, this is a consequence of the fact that ramets are often more easily identifiable in the field than genets (e.g. Eriksson 1992; but see Zahner & Crawford 1965).
If the extent of a genet remains hidden unless genetic analyses are performed, if dynamics of physiologically rather than genetically distinct individuals are monitored and if, moreover, it is observed that ramets establish more successfully than seedlings, this may lead to regarding ramet and seedling recruitment as alternative strategies. Ecologists, who tend to focus on costs of dispersal and establishment rather than costs and benefits of generative reproduction, are quite liable to do so (Cook 1985); e.g. Hoffmann & Moreira (2002) write that „rather than producing large cohorts of seedlings following burning, some Cerrado species have evolved the strategy of producing large cohorts of suckers“ (p.164). However, this approach has been criticised early (e.g. Abrahamson 1980) because ecological and genetic differences are confounded: It is assumed that easier establishment (a short-term benefit for the population) incurs greater fitness (a long-term benefit).
Strictly speaking, or at least until otherwise proven12, clonal growth must be regarded as a process complementary, but not alternative, to generative reproduction. It extends the space and the time span over which a genet may attempt generative reproduction, but also trades off against generative reproduction as resources are limited. The implications of this for southern Brazilian grassland-forest dynamics - in which Myrsine takes a leading role - are briefly discussed in the following.
Given the clear „head start“ that newly emerged root suckers receive relative to seedlings, it is not surprising that more root suckers than seedlings recruited during my one year population study (remember that only individuals entering the size class of >10cm between 2005 and 2006 were counted as new recruits). However, the root sucker-seedling ratio observed in new recruits is not contradictory to that observed in elder juveniles in the same grassland plots. It is slightly higher in new recruits, but such a trend is common in clonal plant populations, and attributed to expansion of clones after initial seedling recruitment (e.g. Eriksson 1992, Kreher et al. 2000, Barsoum et al. 2004)
My initial hypothesis that seedling recruitment should predominate over root sucker recruitment in tall shrub grassland was inspired by Hoffmann’s (1998) Cerrado study, and admittedly phrased under the assumption that seedlings and root suckers are more or less equivalent units of population expansion. It was also not confirmed. More root suckers than seedlings recruit in tall shrub grassland, and relative height increase averaged over pooled samples of new recruits does not differ significantly between tall tussock or tall shrub
12 There has been some debate on whether evolutionarily relevant genetic variation may occur within clones – particularly, whether beneficial somatic mutations in ramets may ultimately lead to reproduction under conditions in which the original genetic outfit would not have allowed a genet to do do. Recently, Poore and collaborators (see e.g. Monro & Poore 2004) have confirmed such processes in macroalgae. However, Harper (1988, p.439) strongly cautioned against over-excitement on this subject, pointing out: „If the phenomenon were at all frequent it would have served as a powerful tool for plant breeders! This has not happened.“ Indeed, Klekowski (1995) demonstrated that very long timespans would be neccessary for somatic mutations with advantageous phenotypic effects to occur in higher plants; this could not be offset by the potentially limitless lifespan of perennial plants as deleterious somatic mutations accumulating at the same time irrevocably reduce the genet’s capacity of generative reproduction. The current mood is best described in terms of guarded expectancy. On detection of (surprisingly) high genetic variation within clonal species’ populations, authors tend to „bar“ somatic mutation, and conclude that seedling recruitment must be more frequent than previously thought (e.g. Yeh et al. 1995, Kreher et al. 2000, Suvanto & Latva-Karjanmaa 2005; but see Tuskan et al. 1996). Yet Lamont & Wiens (2003) and Fischer & van Kleunen (2004) propose that the subject deserves further research. In my discussion and references cited therein, however, it is implicit that evolution occurs via selection of traits obtained by generative reproduction.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
103
grassland. However, both proportion and individual density of seedling recruits are higher than in tall tussock grassland. Density of reproductive individuals was approximately twice as high in forest border plots adjacent to tall shrub grassland than in forest border plots adjacent to tall tussock grassland, but this alone cannot account for the discrepancy in seedling numbers. As already proposed at the end of the previous chapter, a major benefit of tall shrub over tall tussock grassland seems to lie in the availability of gaps in the grass matrix, which allow seeds to reach the soil surface more easily.
Relatively high densities of naturally established seedlings in pasture plots – although lower than in abandoned grassland - indicate that conditions for seedling establishment are at least occasionally more beneficial than my seedling emergence and transplant experiments (chapters 2 and 3) suggested. Contrary to the experimental plots, most of which were flanked only on one side by continuous forest and by grassland extending over hundreds of metres on the other side, population monitoring plots were placed between closely opposed forest patches. This might provide for both higher seed rain and shelter than was given in experimental plots. Further peculiarities of these sites are discussed in chapter 5. Generally, the experimental plots must be considered as more representative of most of the study area’s grassland-forest mosaic.
Low absolute numbers of root sucker recruits in pasture relative to abandoned grassland must be attributed at least in part to low absolute numbers of potential feeder individuals. However, the ratio of potential feeders to root suckers was also lower in pasture than in abandoned grassland: I.e. the latter supports at least twice as many root suckers per potential feeder individual than pasture. Possible reasons for this are discussed in chapter 6. Here, I recapitulate that abundant root sucker recruitment of Myrsine represents not only a deviation from population patterns inside forest but is obviously a relatively recent phenomenon in the study area, increasing in importance relative to seedling recruitment since abandonment of former pastures in the Pró-Mata area.
The necessity to distinguish between genets (single-stem seedlings and clones originally established from seed) as evolutionarily relevant units, and ramets as „merely“ physiologically independent units becomes obvious as the question of future prospects of Myrsine populations in their new grassland habitat is raised. What may be the consequences for a population if clonal spread is more frequent than seedling establishment in a new habitat, i.e. genetic diversity apparently decreases relative to the original habitat?
At a given point in time, investment in generative reproduction trades off against investment in vegetative growth, and vice versa (e.g. Lamont & Barrett 1988, Lovett Doust & Lovett Doust 1988, Paté et al. 1990, 1991). However, a delay in generative reproduction due to clonal spread may eventually be compensated in terms of genet fitness, as ramets themselves develop to generatively reproductive individuals; the seed-producing genet may thus exploit a wider range of resources in time and space than one individual in a fixed location might have done. It has been proposed that ramet production should be integrated in fitness measures (Pan & Price 2002), and models such as that of Winkler & Fischer (1999) aim at predicting which proportions of clonal spread and generative reproduction should result in maximum fitness under diverse site conditions.
Winkler & Fischer predict, for example, that if spatial extension of a single disturbance is much larger than the potential distance of vegetative spread of a plant, then equal investment in clonal growth and generative reproduction should result in maximum fitness. They also predict that in undisturbed areas, or where disturbed patches are no larger than the plant may spread vegetatively, clonal growth should predominate to yield maximum fitness. While the latter prediction is apparently reflected in Myrsine populations in tall tussock grassland, pasture plots do not behave according to the model. This may be so simply because current disturbance frequencies neither allow Myrsine individuals to reach reproductive maturity, nor, in the majority of cases, to reach a size at which they may support

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
104
root suckers. In other words, besides spatial extension of disturbance and clones, it may be necessary to include disturbance frequencies and species’ life histories into such models.
This discussion links to the discussion begun in the previous paragraph. Obviously, if root sucker recruitment in grassland should turn out to be both a means of perpetuating Myrsine genets, and foraging efficiently for resources not available inside forest and forest border, this should increase genet fitness. If grassland proves unprofitable ground for colonisation, or a habitat in which Myrsine forages less efficiently than in forest, this should decrease genet fitness.
As in the discussion on physiological integration, however, it must be remembered that these considerations are yet theoretical. They may serve as guidelines for future research but it would be premature to forecast Myrsine population development on the basis of currently available data. Nothing or only little is known about the extent of genets - although a review by Ellstrand & Roose 1987 shows that intermediate genetic diversity and evenness tend to be the rule in multiclonal plant populations -, about performance of seedlings and root suckers over more than one decade, and temporal and spatial dynamics of the surrounding grass and shrub matrices. Tall shrub grassland, the grassland formation that was relative favourable to seedling establishment during the first decade following abandonment, is currently in decline (chapter 5). In tall tussock grassland, root sucker recruits predominate at present; but it remains to be seen – maybe another decade hence - whether conditions do not become more favourable for seedling establishment as grassland community structure changes with age (see also chapter 6).
As in the previous chapter’s discussion on inhibition or facilitation of Myrsine establishment in different types of grassland, a conclusive answer requires not only longer-term observations of population development in grassland but also in forest and forest border. Only in this way it may be possible to tell whether Myrsine individuals increase their fitness by extending root suckers into grassland.
It is of interest to note, however, that although the ability of clonal growth was recorded only in a few forest species in the course of this study (in the Pró-Mata area, only in Myrtacea Siphoneugenia reitzii), it was observed in a greater number of herbaceous and woody grassland species, some of them among the most abundant species in pasture and abandoned grassland: Grassland low shrubs Calea phyllolepis and Croton tenuisissimus both spread by layering; the latter species locally forms thickets of similar density as e.g. Rubus agg. does along roadsides of Middle Europe. Sprouting from subterranean organs is observed in grassland shrubs Agarista nummularia and Baccharis trimera as well as the rosette forb Eryngium horridum and the fern Pteridium aquilinum, both noxious pasture weeds. Needless to say, grasses themselves employ vegetative spread to great success. Of two of these species, it is known that seedling recruitment in grassland is rare (Fidelis 2004 on Eryngium horridum; Calea phyllolepis, my own unpubl. data). So, independent of whether these species maximise fitness in the long run, possessing the ability of clonal growth seems advantageous at least in the short term for species in these grasslands.
In the United States, Aspen establishes from root suckers both in semiarid intermountain prairie of Utah (e.g. Mitton & Grant 1996) and in the Great Lakes Prairie of Wisconsin (Anderson 1982) and Michigan (e.g. DeByle 1964), while establishment from seeds only appears to be common in the moist, humid forests of New England (Mitton & Grant 1996). These clones have been found to cover tens of hectares, include hundreds of ramet trees, and estimated to be thousands of years in age (e.g. Kemperman & Barnes 1976).
Another consequence of predominately clonal spread, more obvious to the eye than genet fitness, is relative slow expansion of Myrsine populations into grassland. In both pasture and tall shrub grassland (where considerable proportions of recruits were seedlings) Myrsine individual density in a 5 m broad strip of grassland adjacent to the forest border was

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
105
ca. three times higher than at 5-10m distance. In tall tussock grassland, Myrsine populations composed principally of root suckers were more concentrated close to the forest border, where individual density was six times higher near the forest border than further away, and individual density over entire plots was only approximately half as high as in tall shrub abandoned grassland.
Tab. 4.5 indicates that as a rule, „dispersal“ distances of vegetatively spreading species are more restricted than those of generatively reproducing species (see also Cook 1985). They clearly contrast with long-distance dispersal on the scale of tens of adult plant heights that wind-dispersed seeds (e.g. genera Corylus, Betula, Alnus) may achieve; usually, such species are most frequent among the first plants to arrive at a site (van der Valk 1992). Indeed, Egler (1954) treats root suckers as a „peripheral phenomenon“ and does not include them in his model of initial floristic composition.
This, in turn, signifies that while root suckers establish more readily in grassland than seedlings generally, expansion of the entire forest community over grassland will occur only at a slow rate, as new recruitment of most forest tree and shrub species remains largely restricted to the forest border and – perhaps - the relatively narrow stretch of grassland eventually dominated by Myrsine clones in their canopy-forming stage. Tall shrub communities may provide for increased seedling recruitment, but perhaps only temporarily. It is curious that Myrsine root suckers, rather than wind-dispersed forest species, are first to arrive at the site of colonisation. This will be further discussed in chapters 6 and 7.

Hermann, J.-M. PhD thesis 4: Pioneer tree root suckers vs. seedlings in grassland
106
4.9. Appendix
Fig. 4.3: More or less even-aged root suckers, harvested in a clipping experiment, compared side by side
with seedlings harvested in a seedling transplant experiment; exposure is similar at both sites (mown and
regrown grass matrix in resprouting experiment, short- to medium height pasture in transplant
experiment). Photographer: J.-M. Hermann.
Tab. 4.8: What height can be expected in new root suckers and seedlings of a certain basal diameter?
Calculations based on regression curves shown in Fig. 4.1. Bold: Overlap of calculated and sampled
values. In brackets: No or little overlap.
Abandoned grassland Short open grassland
dbas [mm]
Expected root sucker height [mm]
Expected seedling
height [mm]
Expected ratio height root
sucker:seedling
Expected root sucker height [mm]
Expected seedling
height [mm]
Expected ratio height root
sucker:seedling
1.5 (101) (75) (1.4) (43) 35 (1.2) 2 139 104 1.4 63 52 1.2
2.5 178 133 1.3 85 (70) (1.2) 3 219 164 1.3 109 (89) (1.2)
3.5 259 196 1.3 134 (110) (1.2) 4 301 (226) (1.3) 161 (132) (1.2)
4.5 343 (258) (1.3) 188 (155) (1.2) 5 385 (291) (1.3) 217 (179) (1.2)
5.5 428 (324) (1.3) 246 (204) (1.2) 6 471 (357) (1.3) 277 (229) (1.2)

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
107
5. Population Structure and Dynamics of Woody Species in
Grassland: Comparison of a Grassland and a Forest
Species
5.1. Summary • The highland area of Rio Grande do Sul, Southern Brazil, is characterised by a
mosaic of grasslands of postglacial origin, and mixed Araucaria-broadleaf forests. Grasslands have traditionally been cattle-grazed and burnt at 1-2 year intervals. Forests expand over grassland, although only at a slow rate.
• On observation of grassland areas prior to and following abandonment, the hypothesis of a successional sequence suggests itself: More or less pure grass swards tall-tussock abandoned grassland tall-shrub abandoned grassland forest. However, evaluation of aerial photographs by Morales & Pillar (2004) as well as my pilot surveys 2004 suggested that more than one successional sequence is possible. Each structural type of abandoned grassland and pasture may develop into stands that can be considered initial stages of forest floristically, if not structurally.
• How do population structure and short-term dynamics differ between tall grassland shrub Baccharis uncinella and pioneer forest tree Myrsine parvula in different structural types of grassland? How are they affected by ongoing management in pasture as opposed to ca. one decade of abandonment? What development can be predicted for the following decade(s)?
• In autumn/early winter 2005, eight 10x10m study plots were established in pasture and abandoned grassland each; the latter were divided four by four between tall-tussock and tall-shrub abandoned grassland. All are adjacent to the forest border, contain mixed populations of Myrsine and Baccharis, and opposite forests are at no more than 200 metres distance so that seed input by birds crossing between forests may occur. All Myrsine and Baccharis individuals were tagged, measured and their position in the plot (x/y coordinates) recorded. Areas were revisited one year afterwards for census of 1yr mortality, growth and new recruitment into >10cm size class.
• In pasture, fire and cattle grazing limit individual and population growth of both species. Parts of the studied areas apparently experienced longer than usual periods without burning (3-4 years instead of 1-2 years) in the past. If this disturbance regime continues, populations of either species may persist in the area but not increase, and will depend largely on external refill of seed and bud bank.
• Baccharis recruitment has come largely to a standstill in both types of abandoned grassland ca. one decade after abandonment. Predominately adult populations of tall shrub grassland experienced a decline in individual density of ca. 50% during 3-4 years prior to last survey. If mortality continues at this rate, individual density will be similarly low as in tall tussock grassland within ca. two decades after abandonment. There is no indication that dieback is triggered by Myrsine populations recruiting under Baccharis canopy. Rather, density-dependent processes seem to be operating.
• Myrsine individual density increased by 30-40% within one year in both types of abandoned grassland. It is to be expected that in the following decade, density will increase at progressively slower rates due to density-dependent mortality and, in tall shrub grassland, due to the decline in Baccharis canopy cover.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
108
• Already, Baccharis dieback seems to account for lower Myrsine juvenile recruitment under lighter than under denser canopy. The impact on dieback on Myrsine individual growth rate, however, was not significant in most cases. Both phenomena may be attributed to a variety of factors, e.g. a simultaneous competitive release of tussock grasses previously suppressed by the Baccharis canopy.
• I suggest that the two species mimic a “classical” successional sequence from wind-dispersed to animal-dispersed species in pioneer forests, but in fact represent a final stage of pyric succession and an initial stage of forest succession in grassland, respectively.
• Myrsine populations in abandoned grassland probably represent the next generation of canopy formers. The majority of forest border species is not capable of compensating limitations of seedling establishment in grassland by root sucker recruitment, and may depend largely on canopy-building ability of Myrsine for successful establishment. However, limited knowledge and (too) short-term observations still preclude exact predictions of sequences and duration of succession from grassland to forest.
5.2. Introduction Cattle grazing and fire have been largely excluded from the area of the Nature
Conservation and Research Center Pró-Mata since 1994. In absence of management, live and standing dead grass biomass have accumulated and woody species have established abundantly. The majority of these are grassland shrubs, ranging from small species with ca. 1m maximum height and erect or straggling growth – characteristic of so-called tall-tussock grassland - to tall species with several metres height. Predominant among the latter is Baccharis uncinella A.P. de Candolle, characteristic of so-called tall shrub grassland in which it may temporarily form more or less closed canopies. Among forest species, Myrsine parvula (Mez) Otegui stands out as most successful pioneer tree species in grassland, spreading from forest borders into both abandoned and managed grasslands of the study area.
Based on an evaluation of aerial photographs of the area, Oliveira (2003) proposed a general sequence of secondary succession from pasture swards via tall tussock grassland (low-shrub grassland in his terminology) and tall shrub grassland to forest. However, Oliveira & Pillar (2004) conclude – and my own preliminary field surveys in 2004 suggest - that this is only one of several sequences. Apparently each structural type of pasture and abandoned grassland may develop into stands that represent initial stages of forest floristically, if not structurally.
In this study, I observe and analyse structure and short-term population dynamics of grassland tall shrub Baccharis uncinella A.P. de Candolle and pioneer forest tree Myrsine parvula (Mez) Otegui in mixed species stands in pasture, tall tussock and tall shrub abandoned grassland. I summarise data collected in this study and presented in the previous chapters to discuss what they reveal about the original habitat of the two study species, i.e. the habitat these species evolved in before joining in grassland-forest-succession on the Southriograndean highland 1000-2000 years BP. I discuss how population structure and dynamics are influenced both by species traits and by ongoing management as opposed to ca. one decade of abandonment, and I attempt to predict the development of these stands during the following decade.
A surprise element was introduced in this study by rapid and extensive dieback of Baccharis stands. Although abandoned grassland plots were chosen so as to best represent the contrast between tall tussock and tall shrub grassland, tall shrub dieback opened gaps or areas of minimum cover of several metres diameter in the high shrub canopy during only one

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
109
year. This gave rise to the question whether and how Baccharis dieback already affects population structure and growth of Myrsine juveniles under their canopy. Studies in another fire-prone ecosystem, Californian chaparral, which may experience fire-free intervals of several decades, have not yielded conclusive evidence of competitive release of seedlings in the understorey even after such long timespans (e.g. Keeley 1992). Openings in a forest canopy, on the other hand, “drive the forest cycle” by release of immature saplings (Whitmore 1989, p.536). It may therefore be hypothesized that gaps opening in the Baccharis canopy release juveniles of forest tree Myrsine recruited under canopy from competition.
„The idea of predictable sequences of species replacements is still central to much ecological thinking“ (Bond & van Wilgen (1996, p.149). Nevertheless, interpretation of patterns of secondary succession in space and time is so difficult that a unifying theory or model of successional pathways and mechanisms has not yet been found (e.g. Kahmen & Poschlod 2004). At the end of this chapter, I briefly discuss this subject and some factors that seem to specifically introduce impredictability in patterns of grassland-forest succession in Southern Brazil.
5.3. Study setup
5.3.1. Study area and species
This study was conducted in two adjacent sites on the High Plateau of Rio Grande do Sul: 1. Native grasslands in the Nature Conservation and Research Centre Pró-Mata (hereafter: Pró-Mata) that were excluded from cattle grazing and fire in 1994, i.e. 11 years prior to first survey. 2. The neighbour fazenda “Trés Estrelas” where grasslands are grazed by cattle (approximately 0.5 head of cattle per hectare) and the majority of plots experienced burning – i.e. a complete scorch of aboveground biomass - in August 2005, ca. 3 months after 1st and 9 months before 2nd survey. Principal objects of research are grassland tall shrub Baccharis uncinella de Candolle and pioneer forest tree Myrsine parvula (Mez) Otegui. For further details on study area and species please see chapter 1.
5.3.2. Study setup
For population monitoring, eight 10x10m plots were established in pasture and abandoned grassland each; the latter were divided half and half between tall-tussock and tall-shrub abandoned grassland. All plots are adjacent to the forest border, contain mixed populations of Baccharis and Myrsine, and opposite forests are located at no more than 200 metres distance so that seed input by birds crossing between forests may occur13.
From southern autumn to early winter 2005 (April to June), in abandoned grassland, all live Myrsine and Baccharis individuals >10cm height were marked with aluminium tags fixed to plastic-coated wire, and the following parameters were recorded: X/y-coordinates within plots, stem number and stem basal diameters, maximum stem height and individual
13 According to M. Marchesini (pers. comm.), frugivore forest birds avoid grassland areas unless attracted e.g. by a fruit-bearing tree at a certain distance, and by chance, I obtained an estimate of what distances may be acceptable to these birds. As reported in chapter 2, a sample of bird faeces containing Myrsine seeds was collected in the Pró-Mata area in southern autumn 2005. Faeces was dropped along a stretch of service road (between main gate and „casinha dos pesquisadores“) lined by grassland and forest on either side. These forest borders are at a distance of approximately 200 metres opposite one another and Myrsine populations are fairly dense in this area, extending from forest borders into grasslands; moreover, there were no perches above those spots where faeces was found, so presumably it was dropped during flight by birds crossing between forests.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
110
vitality on a 3-point scale (0=low, 1=medium, 2=good, i.e. with majority of foliage intact and vigorously green), as well as evidence of predation or pathogen attack. Of Myrsine individuals of 10-100cm height, the origin - seedling or root sucker - was determined by careful exposure of root and root crown; this subject has been treated in detail in the previous chapter. Practical considerations (time limitation and difficulty of tagging) dictated the omission of individuals < 10cm height from this part study. Evidence on survival and growth of seedlings at this stage has been provided by the seedling emergence and transplant experiments described in chapter 2 and 3.
Coordinates and basal diameter were also recorded of dead individuals’ remains (standing dead) in 2005. The far majority of standing dead were Baccharis uncinella; as woody parts and bark of this species decay rapidly, by my estimation these individuals succumbed 1-3 years prior to the 2005 survey.
I refrained from tagging in pasture as losses by cattle or fire influence could be foreseen. Identification of individuals in pasture plots was exclusively by coordinates.
Plots were revisited one year afterwards (maintaining the chronological order of survey of 2005) for censuses of mortality and new recruitment into >10cm size class as well as measurement of height and basal diameter growth of tagged individuals. Further monitoring was not possible within the time scope of this study but individuals remained tagged after the 2006 survey.
5.3.3. Evaluation
5.3.3.1. Population structure
Population structure is described by the following parameters: Individual distribution in size-age classes, in groups of individual morphology and relative to distance from the forest border. All values described and discussed are averages of plots. Structure per plot is listed in the appendix.
Abandoned grassland populations that consist predominately of one-stemmed individuals are subdivided into size-age classes as described in Tab. 5.1. Multiple-stemmed Myrsine individuals were sorted by size of the tallest basal shoot, tilted Myrsine individuals by maximum height of vertical shoots above ground. Baccharis standing dead with known basal diameter were sorted into these size-age classes by way of regression. The formula was obtained using basal diameter of a sample of live individuals as independent variable and height as dependent variable. Data were ln-transformed prior to analysis and r² of the linear regression was 0.477.
Assignment of generatively reproductive state in Myrsine adults is based on personal observations; vegetatively reproductive state became apparent during the resprout study described in chapter 6. Population monitoring plots were not accessible during the period of Baccharis seed maturity (December 2005), so alternatively proportions of reproducing adolescents and adults were recorded in a sample of 50 individuals total in another population growing under similar conditions in the Pró-Mata area. Additionally, in surveys May-June 2006, monitored abandoned grassland plots were checked for remains of inflorescences in the litter layer; litter of Baccharis inflorescences was found in all stands. Harter (1999) determined an approximate ratio 1:1 of males and females in Baccharis stands similar to those of my tall shrub grassland plots. Thus, although sex was not identified in my studied individuals, I can assume that seed set and input occurred in all stands.
For description of size-age distribution of pasture populations, a different approach had to be used: Only negligible proportions of individuals attained more than 1m height in 2005. However, root sucker production observed in 2006 indicates presence of adolescent individuals. In fact, total basal area may be more representative of size as individuals in
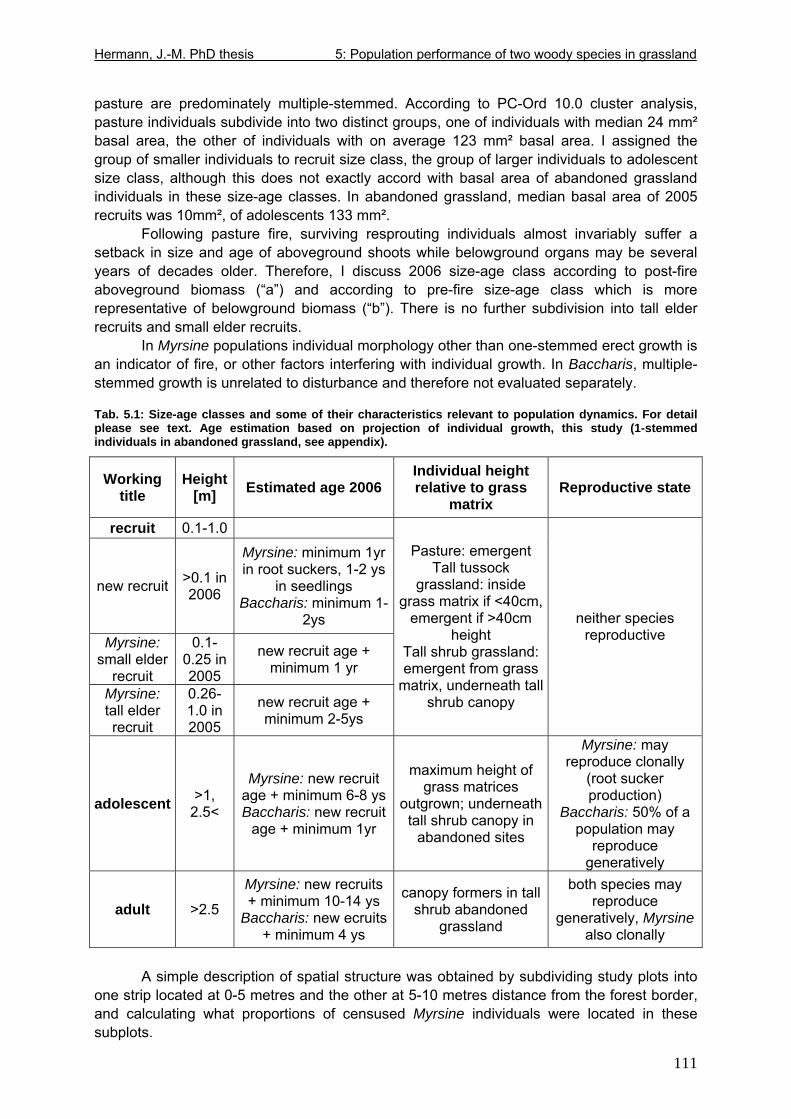
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
111
pasture are predominately multiple-stemmed. According to PC-Ord 10.0 cluster analysis, pasture individuals subdivide into two distinct groups, one of individuals with median 24 mm² basal area, the other of individuals with on average 123 mm² basal area. I assigned the group of smaller individuals to recruit size class, the group of larger individuals to adolescent size class, although this does not exactly accord with basal area of abandoned grassland individuals in these size-age classes. In abandoned grassland, median basal area of 2005 recruits was 10mm², of adolescents 133 mm².
Following pasture fire, surviving resprouting individuals almost invariably suffer a setback in size and age of aboveground shoots while belowground organs may be several years of decades older. Therefore, I discuss 2006 size-age class according to post-fire aboveground biomass (“a”) and according to pre-fire size-age class which is more representative of belowground biomass (“b”). There is no further subdivision into tall elder recruits and small elder recruits.
In Myrsine populations individual morphology other than one-stemmed erect growth is an indicator of fire, or other factors interfering with individual growth. In Baccharis, multiple-stemmed growth is unrelated to disturbance and therefore not evaluated separately.
Tab. 5.1: Size-age classes and some of their characteristics relevant to population dynamics. For detail
please see text. Age estimation based on projection of individual growth, this study (1-stemmed
individuals in abandoned grassland, see appendix).
Working
title
Height
[m]
Estimated age 2006
Individual height
relative to grass
matrix
Reproductive state
recruit 0.1-1.0
new recruit >0.1 in 2006
Myrsine: minimum 1yr in root suckers, 1-2 ys
in seedlings Baccharis: minimum 1-
2ys Myrsine:
small elder recruit
0.1-0.25 in 2005
new recruit age + minimum 1 yr
Myrsine: tall elder recruit
0.26-1.0 in 2005
new recruit age + minimum 2-5ys
Pasture: emergent Tall tussock
grassland: inside grass matrix if <40cm,
emergent if >40cm height
Tall shrub grassland: emergent from grass
matrix, underneath tall shrub canopy
neither species reproductive
adolescent
>1, 2.5<
Myrsine: new recruit age + minimum 6-8 ys Baccharis: new recruit
age + minimum 1yr
maximum height of grass matrices
outgrown; underneath tall shrub canopy in
abandoned sites
Myrsine: may reproduce clonally
(root sucker production)
Baccharis: 50% of a population may
reproduce generatively
adult >2.5
Myrsine: new recruits + minimum 10-14 ys
Baccharis: new ecruits + minimum 4 ys
canopy formers in tall shrub abandoned
grassland
both species may reproduce
generatively, Myrsine also clonally
A simple description of spatial structure was obtained by subdividing study plots into one strip located at 0-5 metres and the other at 5-10 metres distance from the forest border, and calculating what proportions of censused Myrsine individuals were located in these subplots.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
112
5.3.3.2. Population dynamics
Individuals were considered dead both when standing dead were recorded, or individuals not retrieved on 2nd survey. This could be confidently done in abandoned grassland, as tagging and x/y-coordinates offered double security of rediscovery and identification. In pasture misidentification - although in a minority of cases - cannot be ruled out. Recruitment and mortality values described and discussed are averages of the four monitoring plots per abandoned grassland type, and the eight pasture monitoring plots. Due to small sample size, variation within plots was not explored and statistical significance of differences in means was not tested. Structure per plot is, however, listed in the appendix in order to illustrate variance between sampled plots.
5.3.3.3. Individual growth
Size increase is here represented as the percentage by which individuals increased relative to their initial height (between 1st and 2nd survey); e.g. a relative increase by 100% means that the individual doubled its size during the observation period. As in the transplant experiment (chapter 3) and in the previous chapter, values thus represent a relative growth rate, although they are not equivalent to the classical “relative growth rate” (RGR), defined as the product of unit leaf rate and leaf area ratio in time (see Hutchings 1997a for detail).
Basal area is considered more representative for biomass gain than height in multiple-stemmed and much branched individuals. 1-yr relative size increase is therefore calculated from basal area data in Baccharis individuals, Myrsine individuals in pasture and Myrsine adolescents in abandoned grassland, and from height data in Myrsine recruits and adolescents of abandoned grassland. In abandoned grassland, only individuals that did not suffer size losses enter in calculations; in pasture, resprouters with decreased basal area at 2nd survey are more representative and relative size increase is given of both these and intact individuals. Myrsine adult and Baccharis recruit numbers were generally very low and thus enter in description of size-age-structure but not analyses of relative 1-yr individual growth rate.
5.3.3.4. Projection of individual and population growth Although Baccharis uncinella and Myrsine parvula coexist in naturally established
populations, direct comparison of the two species within size-age classes was largely precluded by opposing size-age structure in these populations – stands contain practically no Baccharis recruits but many adults, and vice versa in Myrsine. However, based on seedling growth rates known from transplant experiments (chapter 3) and growth rates of elder size classes recorded in this study, I projected individual growth of either species after disturbance in pasture, and over more or less one decade of undisturbed growth in abandoned grassland (see appendix for detail). Based on these very simple models I draw some conclusions on development of mixed populations of the two species since abandonment, and discuss possible trajectories of development during the following years.
Needless to say, as only short-term data on recruitment and growth are available, these projections must be viewed with great caution (lack of realism in such data has e.g. been criticised by Crawley 1990) – and they clearly cannot replace further monitoring. They should best be viewed as a basis for planning future studies, i.e. as a basis for estimating when to conduct another survey of the marked plots, and for hypothesising on their developmental stage at that time.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
113
5.3.3.5. Influence of Baccharis dieback Based on cover-abundance estimations at 2nd survey 2006, I compiled subsamples of
100m² total area each of lighter and denser Baccharis canopy, composed of patches with minimum size 2.5x2.5m14, and containing equal numbers of patches at 0-5 and 5-10 m distance from the forest. Average Baccharis canopy cover was ca. 10% in the “lighter” subsample and ca. 30% in the “denser” subsample.
A similar subsample of plots at 0-5 and 5-10m distance was compiled from tall tussock grassland plots; hence individual growth values given here differ slightly from those in the previous chapter.
I then calculated individual density and and median one-year relative height increase of individuals of different size-age classes for these subsamples. In order to minimize effects of pseudoreplication, height increase was calculated not from all individuals but from a randomly chosen sample of 30-50 individuals (depending on total individual number) per size-age class. Significance of differences between samples was tested by SPSS 10.0 One-Way ANOVA of ln-transformed data.
In doing so, I substitute a spatial for a temporal series - denser tall shrub lighter tall shrub tall tussock grassland – so that all interpretations must be made with extreme caution.
14 This subdivision of monitoring plots into 2.5x2.5m subplots had been employed during both 1st and 2nd survey in order to facilitate fieldwork; cover-abundance values of the Baccharis canopy were available for each subplot, and median growth rates of Myrsine juveniles per subplot could be tracked by individual x/y-coordinates.
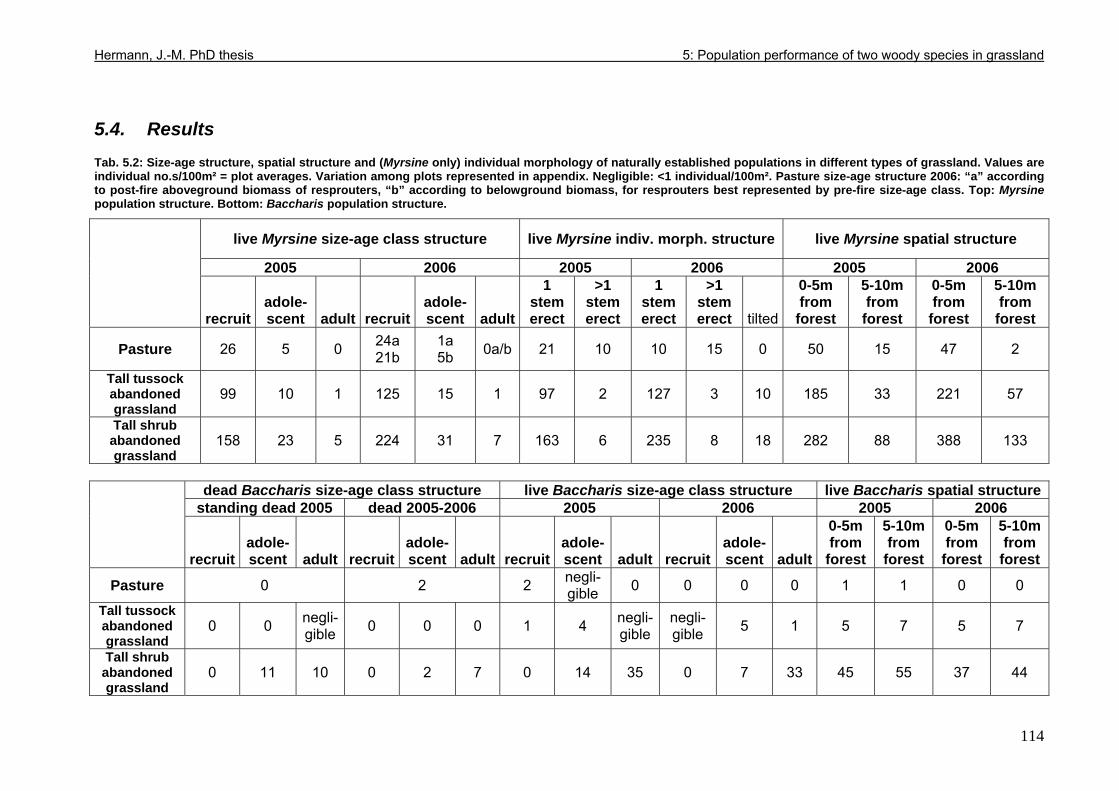
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
114
5.4. Results Tab. 5.2: Size-age structure, spatial structure and (Myrsine only) individual morphology of naturally established populations in different types of grassland. Values are
individual no.s/100m² = plot averages. Variation among plots represented in appendix. Negligible: <1 individual/100m². Pasture size-age structure 2006: “a” according
to post-fire aboveground biomass of resprouters, “b” according to belowground biomass, for resprouters best represented by pre-fire size-age class. Top: Myrsine
population structure. Bottom: Baccharis population structure.
live Myrsine size-age class structure live Myrsine indiv. morph. structure live Myrsine spatial structure
2005 2006 2005 2006 2005 2006
recruit
adole-
scent adult recruit
adole-
scent adult
1
stem
erect
>1
stem
erect
1
stem
erect
>1
stem
erect tilted
0-5m
from
forest
5-10m
from
forest
0-5m
from
forest
5-10m
from
forest
Pasture 26 5 0 24a 21b
1a 5b 0a/b 21 10 10 15 0 50 15 47 2
Tall tussock
abandoned
grassland
99 10 1 125 15 1 97 2 127 3 10 185 33 221 57
Tall shrub
abandoned
grassland
158 23 5 224 31 7 163 6 235 8 18 282 88 388 133
dead Baccharis size-age class structure live Baccharis size-age class structure live Baccharis spatial structure
standing dead 2005 dead 2005-2006 2005 2006 2005 2006
recruit
adole-
scent adult recruit
adole-
scent adult recruit
adole-
scent adult recruit
adole-
scent adult
0-5m
from
forest
5-10m
from
forest
0-5m
from
forest
5-10m
from
forest
Pasture 0 2 2 negli-gible 0 0 0 0 1 1 0 0
Tall tussock
abandoned
grassland
0 0 negli-gible 0 0 0 1 4 negli-
gible negli-gible 5 1 5 7 5 7
Tall shrub
abandoned
grassland
0 11 10 0 2 7 0 14 35 0 7 33 45 55 37 44

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
115
5.4.1. Population structure 2005
5.4.1.1. Myrsine Myrsine populations in both pasture and abandoned grassland had a size structure
typical of growing populations. Pasture populations contained no adult individuals and on average 84% of live individuals were recruits. In abandoned grassland, populations of tall tussock sites contained on average 90% of recruits and one adult individual per 100m², populations of tall shrub sites contained on average 85% of recruits and 5 adult individuals per 100m².
Likewise, in all grassland types the majority of individuals were concentrated within 0-5m distance from the forest border: 77% of individuals in pasture, 85% in tall tussock abandoned grassland and 76% in tall shrub abandoned grassland. With exception of only one individual each in tall shrub and tall tussock sites, all adolescent and adult individuals were contained within this range.
Individual numbers per 100m² were on average 31 in pasture, 109 in tall tussock abandoned grassland and 186 in tall shrub abandoned grassland. True individual density differs slightly less between the two types of abandoned grassland due to the higher concentration of individuals near the forest border in tall tussock grassland previously mentioned.
One third of Myrsine individuals in pasture had more than one stem in 2005, which in this species is indicative of resprouting from base after aboveground biomass loss, e.g. by complete scorch. In contrast, the far majority of individuals in abandoned grassland were single-stemmed: 98% in tall tussock sites, 96% in tall shrub sites.
5.4.1.2. Baccharis All Baccharis populations were characterised by a relatively uniform size-age structure. Populations in pasture consisted almost exclusively of recruits. In tall tussock abandoned grassland populations, adolescent individuals predominated. Populations in tall shrub abandoned sites contained three times more adults than adolescents and no recruits, and individuals were evenly distributed within 0-10 m from the forest border. As in Myrsine, population density differed greatly between grassland types and was lowest in pasture (2 individuals/100m²), intermediate in tall tussock abandoned grassland (6 individuals/100m²) and highest in tall shrub abandoned grassland (49 individuals/100m²). Baccharis populations of tall shrub abandoned grassland also contained on average 21 standing dead individuals/100m². Judging by basal area, almost equal proportions of these had belonged to the adolescent and adult size class.
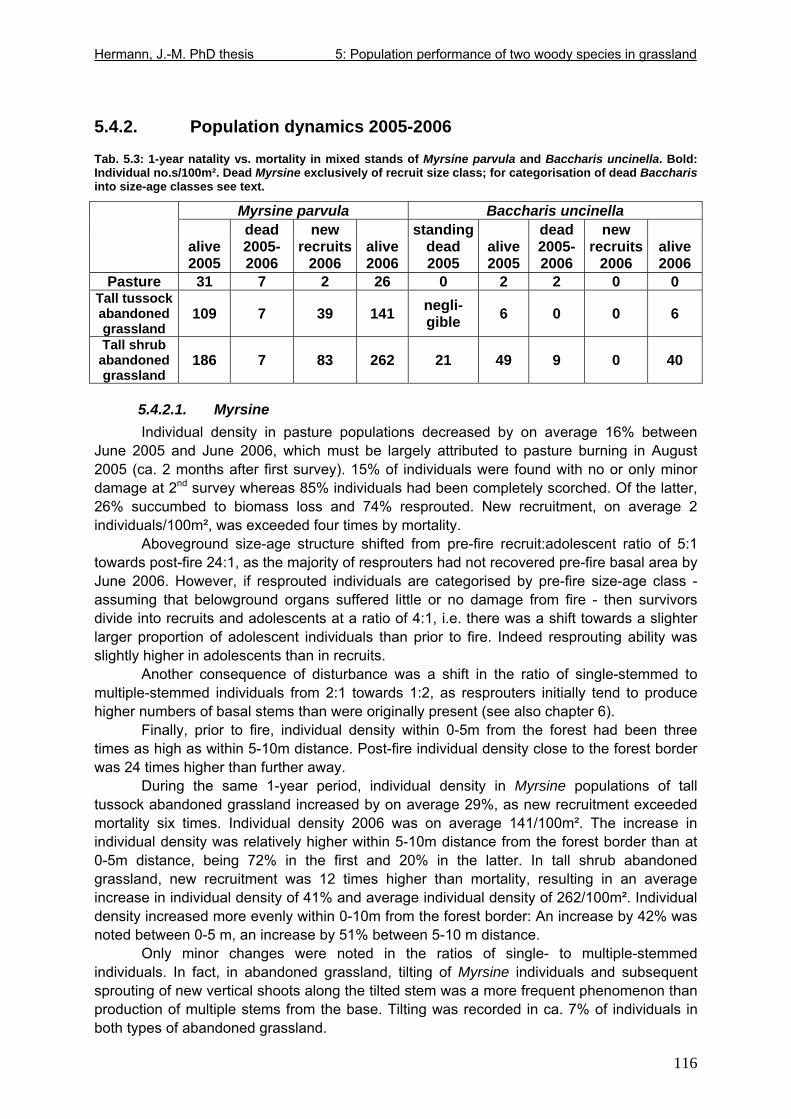
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
116
5.4.2. Population dynamics 2005-2006
Tab. 5.3: 1-year natality vs. mortality in mixed stands of Myrsine parvula and Baccharis uncinella. Bold:
Individual no.s/100m². Dead Myrsine exclusively of recruit size class; for categorisation of dead Baccharis
into size-age classes see text.
Myrsine parvula Baccharis uncinella
alive
2005
dead
2005-
2006
new
recruits
2006
alive
2006
standing
dead
2005
alive
2005
dead
2005-
2006
new
recruits
2006
alive
2006
Pasture 31 7 2 26 0 2 2 0 0
Tall tussock
abandoned
grassland
109 7 39 141
negli-
gible
6 0 0 6
Tall shrub
abandoned
grassland
186 7 83 262 21 49 9 0 40
5.4.2.1. Myrsine Individual density in pasture populations decreased by on average 16% between
June 2005 and June 2006, which must be largely attributed to pasture burning in August 2005 (ca. 2 months after first survey). 15% of individuals were found with no or only minor damage at 2nd survey whereas 85% individuals had been completely scorched. Of the latter, 26% succumbed to biomass loss and 74% resprouted. New recruitment, on average 2 individuals/100m², was exceeded four times by mortality.
Aboveground size-age structure shifted from pre-fire recruit:adolescent ratio of 5:1 towards post-fire 24:1, as the majority of resprouters had not recovered pre-fire basal area by June 2006. However, if resprouted individuals are categorised by pre-fire size-age class - assuming that belowground organs suffered little or no damage from fire - then survivors divide into recruits and adolescents at a ratio of 4:1, i.e. there was a shift towards a slighter larger proportion of adolescent individuals than prior to fire. Indeed resprouting ability was slightly higher in adolescents than in recruits.
Another consequence of disturbance was a shift in the ratio of single-stemmed to multiple-stemmed individuals from 2:1 towards 1:2, as resprouters initially tend to produce higher numbers of basal stems than were originally present (see also chapter 6).
Finally, prior to fire, individual density within 0-5m from the forest had been three times as high as within 5-10m distance. Post-fire individual density close to the forest border was 24 times higher than further away.
During the same 1-year period, individual density in Myrsine populations of tall tussock abandoned grassland increased by on average 29%, as new recruitment exceeded mortality six times. Individual density 2006 was on average 141/100m². The increase in individual density was relatively higher within 5-10m distance from the forest border than at 0-5m distance, being 72% in the first and 20% in the latter. In tall shrub abandoned grassland, new recruitment was 12 times higher than mortality, resulting in an average increase in individual density of 41% and average individual density of 262/100m². Individual density increased more evenly within 0-10m from the forest border: An increase by 42% was noted between 0-5 m, an increase by 51% between 5-10 m distance.
Only minor changes were noted in the ratios of single- to multiple-stemmed individuals. In fact, in abandoned grassland, tilting of Myrsine individuals and subsequent sprouting of new vertical shoots along the tilted stem was a more frequent phenomenon than production of multiple stems from the base. Tilting was recorded in ca. 7% of individuals in both types of abandoned grassland.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
117
5.4.2.2. Baccharis Baccharis populations in pasture were eradicated by fire: No individual survived
complete scorch in August 2005 and no new recruitment occurred until the 2006 survey. Since none of the individuals had reached the reproductive stage prior to fire and there is no indication so far that the species forms a long-lived seed bank, new colonisation of the pasture sites depends entirely on external seed input.
During the same period, Baccharis populations in tall tussock abandoned grassland maintained their 2005 status. Neither mortality nor new recruitment were recorded.
Baccharis populations in tall shrub abandoned grassland suffered a decrease in individual density by 18% on average. Although in all plots, individuals of reproductive size-age class predominate and reproductive activity was confirmed by inflorescence parts found in the litter layer, no new recruitment occurred. Mortality amounted to 9 individuals/100m² on average. Relative to 2005 individual numbers per size class, mortality affected ca. 15% of the adolescent subpopulation and ca. 20% of the adult subpopulation.
Shading-out is one possible reason for high Baccharis mortality in adolescents but less likely in canopy-formers. Phytopathological analysis has revealed presence of fungi of the genus Pythium in Baccharis populations of the area (see appendix). This fungus attacks the root systems of plants of a variety of and thrives particularly well under warm climate in soils with high to excessive humidity, as typical of the region (M. Dalbosco pers. comm.; see also e.g. Schütt et al. 2002). It is also responsible for “damping off-disease”, personally observed in Myrtaceae tree seedlings raised on grassland soil in the Pró-Mata Station nursery.
5.4.3. Individual growth rate 2005-2006
5.4.3.1. Myrsine 70% of surviving Myrsine individuals in pasture suffered a decrease in basal area.
Median relative basal area increase was -63%, i.e. these individuals had recovered median 27% of pre-fire basal area ca. 10 months after fire. In 30% of individuals - with few exceptions undamaged by fire -, basal area increased. Median relative growth rate of these individuals was 63% in 1 year. Size increase was calculated from pooled samples of pre-fire recruits and adolescents as they did not differ significantly in this trait.
In tall tussock grassland, 14% of survivors suffered a height decrease in both 2005 recruit and adolescent size class. In tall shrub grassland, height decrease affected 9% of recruit survivors and 10% of adolescent survivors. Tilting frequently causes a temporary height decrease - until new vertical shoots have attained the same height above ground as the tilted original stem formerly had. Further reasons for height decrease are damage to apical meristems by chafing (especially in Croton spec. matrices in tall tussock grassland and in Baccharis spec. canopies in tall shrub grassland), fungus and frost. Damage to leaves and apical meristems by mammalian herbivory (probably deer) was a rare phenomenon and was, in fact, noted more frequently in pasture plots prior to fire than in any of the abandoned grassland plots.
Growth of recruits with positive size increase differed significantly between tall tussock and tall shrub grassland only in the size-age class of tall elder recruits (chapter 4). In adolescents, relative increase of neither height nor basal area differed significantly between the two types of grassland, although there was a trend towards relatively higher basal area increase in tall tussock grassland, and towards relatively higher height increase in tall shrub grassland.
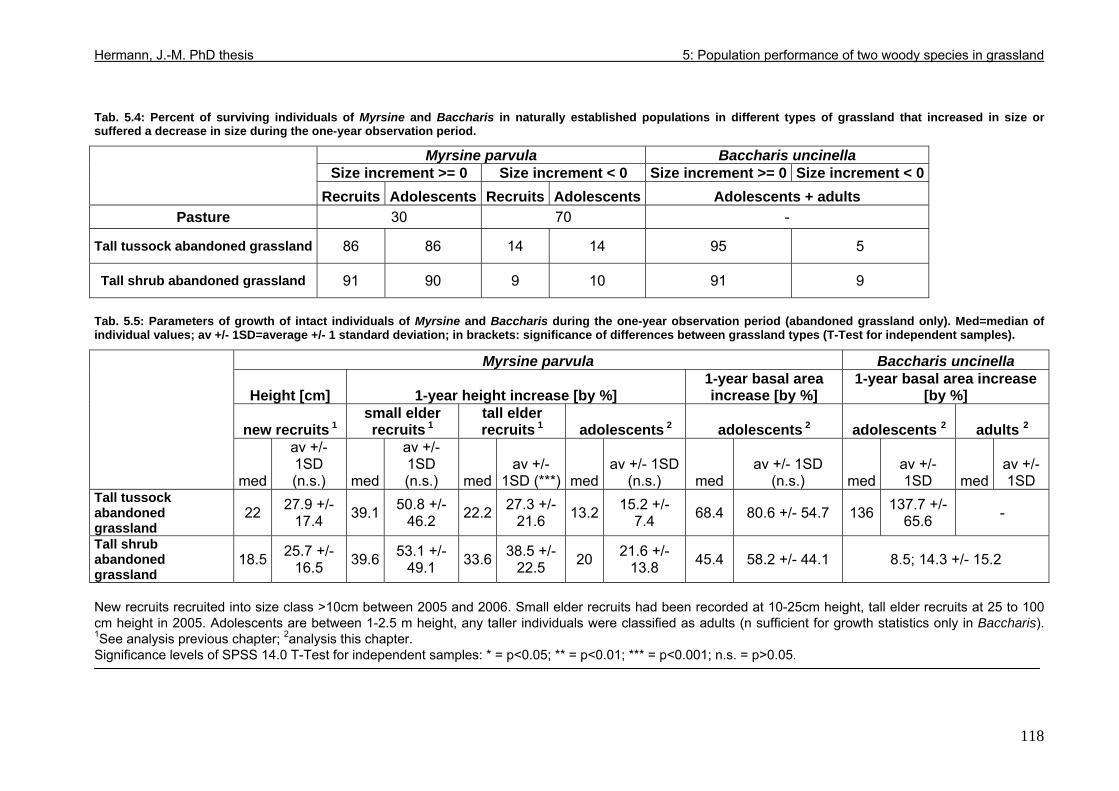
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
118
Tab. 5.4: Percent of surviving individuals of Myrsine and Baccharis in naturally established populations in different types of grassland that increased in size or
suffered a decrease in size during the one-year observation period.
Myrsine parvula Baccharis uncinella Size increment >= 0 Size increment < 0 Size increment >= 0 Size increment < 0
Recruits Adolescents Recruits Adolescents Adolescents + adults
Pasture 30 70 -
Tall tussock abandoned grassland 86 86 14 14 95 5
Tall shrub abandoned grassland 91 90 9 10 91 9
Tab. 5.5: Parameters of growth of intact individuals of Myrsine and Baccharis during the one-year observation period (abandoned grassland only). Med=median of
individual values; av +/- 1SD=average +/- 1 standard deviation; in brackets: significance of differences between grassland types (T-Test for independent samples).
Myrsine parvula Baccharis uncinella
Height [cm] 1-year height increase [by %]
1-year basal area
increase [by %]
1-year basal area increase
[by %]
new recruits 1
small elder
recruits 1
tall elder
recruits 1
adolescents 2
adolescents 2
adolescents 2
adults 2
med
av +/- 1SD (n.s.) med
av +/- 1SD (n.s.) med
av +/- 1SD (***) med
av +/- 1SD (n.s.) med
av +/- 1SD (n.s.) med
av +/- 1SD med
av +/- 1SD
Tall tussock
abandoned
grassland
22 27.9 +/- 17.4 39.1 50.8 +/-
46.2 22.2 27.3 +/- 21.6 13.2 15.2 +/-
7.4 68.4 80.6 +/- 54.7 136 137.7 +/- 65.6 -
Tall shrub
abandoned
grassland
18.5 25.7 +/- 16.5 39.6 53.1 +/-
49.1 33.6 38.5 +/- 22.5 20 21.6 +/-
13.8 45.4 58.2 +/- 44.1 8.5; 14.3 +/- 15.2
New recruits recruited into size class >10cm between 2005 and 2006. Small elder recruits had been recorded at 10-25cm height, tall elder recruits at 25 to 100 cm height in 2005. Adolescents are between 1-2.5 m height, any taller individuals were classified as adults (n sufficient for growth statistics only in Baccharis). 1See analysis previous chapter; 2analysis this chapter. Significance levels of SPSS 14.0 T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
119
5.4.3.2. Baccharis With the exception of only one adult individual, relative basal area increase of
surviving Baccharis in tall shrub abandoned grassland was positive and vitality was good. Relative increase of both height and basal area of Baccharis adolescents were higher than in Myrsine adolescents in the same plots: 22% and 134%, respectively, in Baccharis; 15% and 76%, respectively, in Myrsine. Due to relatively low individual density, 1st-order neighbourhood – and thus high competition - between adolescents of the two species may be largely ruled out. Generally higher growth rates in Baccharis than in Myrsine were also observed in seedlings experimentally transplanted into pasture, tall tussock grassland and nurseries. Only in tall shrub grassland, seedlings of the two species matched one another in relative growth rate (chapter 3).
Contrary to this, Baccharis individual growth in tall shrub grassland illustrates maturity and decline of these populations. 9% of surviving Baccharis adolescents and adults suffered a decrease in basal area due to gradual dieback of basal stems and in 24% of survivors, basal area remained stagnant throughout the year. Median relative basal area increase, which did not differ significantly between adolescents and adults, was 8%. Medium vitality was recorded in 52% of surviving adolescents and 30% of surviving adults; in another 4% of adolescents and 17% of adults, vitality was low.
5.4.4. Projection of individual and population growth
5.4.4.1. Pasture Population growth rate of Myrsine in pasture is difficult to estimate as neither annual
mortality in absence of pasture fire nor annual seedling recruitment rate in pasture are known. Observed post-fire recruitment was exclusively from root suckers, and was very low compared to recruitment rates in abandoned grassland, presumably due to relatively low numbers of potential feeder individuals (see chapter 4). If new recruitment occurs at the same rate during the following years but mortality in absence of disturbance remains as low as in abandoned grassland, then populations may attain pre-fire individual density four years after fire at the earliest.
Median basal area of adolescent Myrsine individuals prior to fire was 123mm². 10 months after fire, resprouters had recovered median 27% of original basal area. Assuming 60% annual relative basal area increase (63% were recorded in undamaged individuals), these individuals may be expected to fully recover original height within four years after fire.
As to Baccharis individual growth in pasture, I assume the following based on results from chapter 3: New Baccharis recruits may attain ca. 30 cm height and 5 mm² basal area after 1year in pasture. Relative height increase will be ca. 100% and relative basal area increase ca. 200% during the following 6 months. Median height of Baccharis individuals in pre-fire pasture populations was 55 cm, median basal area 35 mm². New recruits may achieve that same size within as little as two years. However, I have no basis for estimation of when and at what rates new Baccharis recruitment – which depends entirely on external seed input - will take place. No new recruitment was recorded 10 months after fire, so that populations will attain pre-fire status three years after fire at the earliest, and only if areas remain unburnt.
From the third year, in the adolescent stage, part of the population may begin to produce seed but height growth rate decreases. In Baccharis adolescents of tall tussock abandoned grassland, it was median 22%. Assuming similar growth rates of adolescents in pasture, individuals may be expected to reach the fully reproductive and canopy-forming adult stage only after five to six disturbance-free years.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
120
At least once prior to the 2005 fire, pasture populations were subject to a similar disturbance, as evident from the large proportion of multiple-stemmed Myrsine individuals. Multiple-stemmed growth occurs in both recruits – the predominating size-age class – and adolescents. Multiple-stemmed recruits may have suffered disturbance even in the year before 1st survey. Patches with adolescent Myrsine individuals and Baccharis individuals, however, must have remained undisturbed for slightly longer periods than the customary 1-2 year intervals and will recover pre-fire status at the earliest three to four years after fire. The patchiness of pasture fire, as already apparent in chapter 3, is further illustrated here.
Adult Baccharis are scarce in the surroundings of the pasture plots and low-growing grassland shrubs that characterise tall-tussock abandoned grassland in Pró-Mata are almost absent. If cattle and fire were excluded from the area, the development would most likely be towards a type of tall-tussock grassland structurally similar but floristically different from those monitored in this study.
5.4.4.2. Tall tussock abandoned grassland On the basis of observed Myrsine recruit and adolescent growth rates, it can be
estimated that in tall tussock abandoned grassland, individuals of this species may enter adolescent size-age class (and become potential root sucker feeder individuals) at ca. 8 years age, and the adult size-age class (genetically reproductive) within ca. 15 years. As 1st and 2nd survey took place 11 and 12 years after abandonment, it is not surprising that adult numbers were negligibly low. At survey 2006, erect adolescent individuals were of median 1.35 m height and may attain adult size-age in another five years.
One crucial question that cannot be answered by my short-term observations is how mortality rates in both types of abandoned grassland will develop during the following decade. Apparently density-dependent mortality has not yet set in, as mortality is exceeded by far by new recruitment. According to a report by Shepperd (2000), Populus tremula clones stimulated by severing of feeder roots recruited 111 suckers in a 24 m² patch of meadow adjacent to Aspen forest (444/100 m²). Myrsine individual density even in tall shrub grassland, at on average 262/100 m², is still considerably below this value.
Baccharis individuals were predominately of adolescent size class with a median height of 1.75 m at survey 2006. At annual relative height increase of 22%, these individuals may enter the adult size-age class within two years after 2nd survey. However, at individual densities of only 6/100m² and in absence of further recruitment, there is no evidence that these stands may form the more or less closed canopies that temporarily characterise tall shrub grassland. This also implies that – as in pasture – Baccharis and Myrsine populations develop largely independent of one another.
5.4.4.3. Tall shrub abandoned grassland Myrsine individuals may be expected to grow up to adolescent and adult size-age
class within a shorter period - ca. 7 years and 11 years, respectively - in tall shrub abandoned grassland than in tall tussock abandoned grassland. This helps to explain the relatively high density of adolescent and adult individuals in these populations 11 to 12 years after abandonment. At survey 2006, erect adolescent individuals were of median 1.65 m height and may attain adult size-age in another two years.
Tall-shrub abandoned grassland stands are temporarily characterised by more or less closed Baccharis canopies. These Baccharis populations declined in density and vitality not only between 1st and 2nd survey but also during ca. 1-3 years prior to 1st survey. Summing up standing dead and dead between 1st and 2nd survey, individual density decreased by 50 % in those 2-3 years. Since roughly half of adolescents and adults were recorded with medium to low vitality at 2nd survey, a further decline seems imminent. If mortality continues
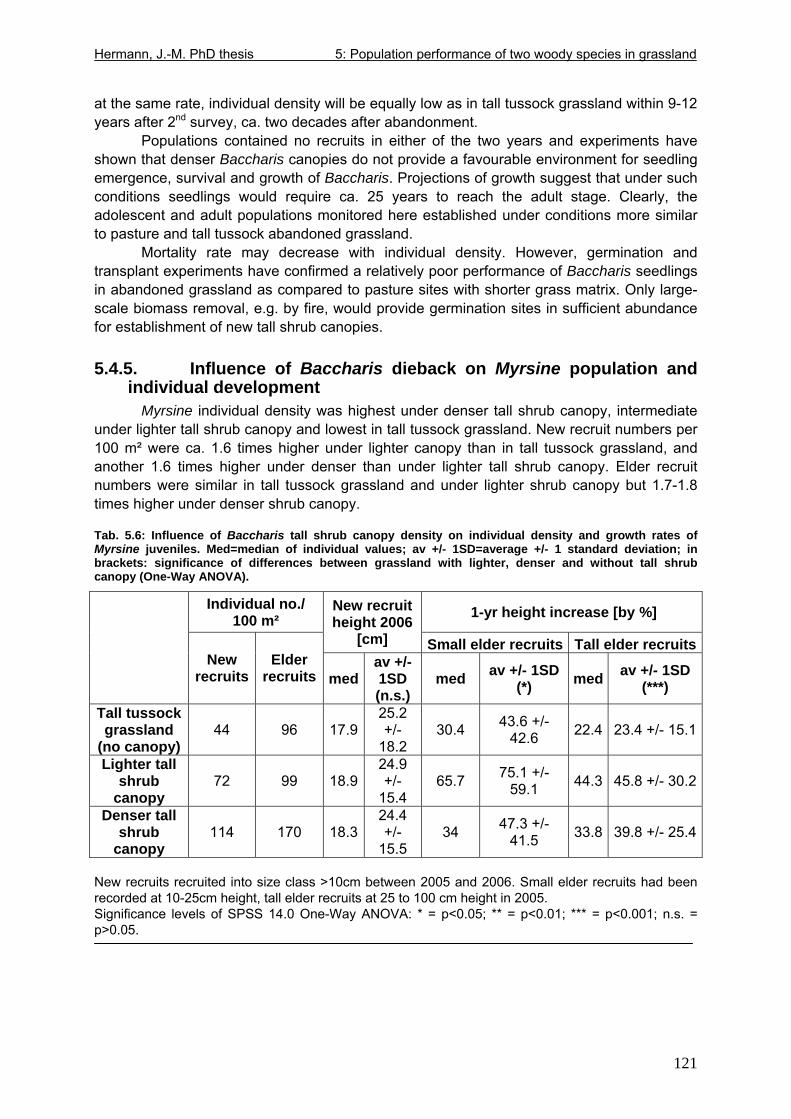
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
121
at the same rate, individual density will be equally low as in tall tussock grassland within 9-12 years after 2nd survey, ca. two decades after abandonment.
Populations contained no recruits in either of the two years and experiments have shown that denser Baccharis canopies do not provide a favourable environment for seedling emergence, survival and growth of Baccharis. Projections of growth suggest that under such conditions seedlings would require ca. 25 years to reach the adult stage. Clearly, the adolescent and adult populations monitored here established under conditions more similar to pasture and tall tussock abandoned grassland.
Mortality rate may decrease with individual density. However, germination and transplant experiments have confirmed a relatively poor performance of Baccharis seedlings in abandoned grassland as compared to pasture sites with shorter grass matrix. Only large-scale biomass removal, e.g. by fire, would provide germination sites in sufficient abundance for establishment of new tall shrub canopies.
5.4.5. Influence of Baccharis dieback on Myrsine population and
individual development
Myrsine individual density was highest under denser tall shrub canopy, intermediate under lighter tall shrub canopy and lowest in tall tussock grassland. New recruit numbers per 100 m² were ca. 1.6 times higher under lighter canopy than in tall tussock grassland, and another 1.6 times higher under denser than under lighter tall shrub canopy. Elder recruit numbers were similar in tall tussock grassland and under lighter shrub canopy but 1.7-1.8 times higher under denser shrub canopy.
Tab. 5.6: Influence of Baccharis tall shrub canopy density on individual density and growth rates of
Myrsine juveniles. Med=median of individual values; av +/- 1SD=average +/- 1 standard deviation; in
brackets: significance of differences between grassland with lighter, denser and without tall shrub
canopy (One-Way ANOVA).
Individual no./
100 m²
1-yr height increase [by %] New recruit
height 2006
[cm] Small elder recruits Tall elder recruits
New
recruits
Elder
recruits med
av +/-
1SD
(n.s.)
med
av +/- 1SD
(*)
med
av +/- 1SD
(***)
Tall tussock
grassland
(no canopy)
44 96 17.925.2 +/-
18.2 30.4 43.6 +/-
42.6 22.4 23.4 +/- 15.1
Lighter tall
shrub
canopy
72 99 18.924.9 +/-
15.4 65.7 75.1 +/-
59.1 44.3 45.8 +/- 30.2
Denser tall
shrub
canopy
114 170 18.324.4 +/-
15.5 34 47.3 +/-
41.5 33.8 39.8 +/- 25.4
New recruits recruited into size class >10cm between 2005 and 2006. Small elder recruits had been recorded at 10-25cm height, tall elder recruits at 25 to 100 cm height in 2005. Significance levels of SPSS 14.0 One-Way ANOVA: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
122
The influence of shrub canopy density on individual height and 1-year relative height increase was less pronounced:
Neither ANOVA nor pairwise comparison by Tamhane’s T2-Test revealed any significant differences in new recruit height between tall tussock grassland and either lighter or denser tall shrub grassland.
One-year height increase of small elder recruits tended to be highest under lighter shrub canopy, contrasting especially with tall tussock grassland, which accounts for the weak significance of within-sample variance revealed by ANOVA. However, this trend was not pronounced enough to be confirmed by Tamhane’s T2-Test.
One-year height increase of tall elder recruits was significantly higher under both lighter and denser tall shrub canopy (both p<0.01) than in tall tussock grassland.
I also performed a T-Test for independent samples (not shown in table) on a dataset including sites with lighter and denser tall shrub canopy but not tall tussock grassland. However, this yielded only one weakly significant trend (p<0.05) towards higher 1-year height increase of small elder recruits under lighter than under denser shrub canopy. Canopy cover had no perceptible influence on either new recruit height or elder recruit height increase.
5.5. Discussion
5.5.1. Origin of Baccharis and Myrsine
During the course of my study, there has been some debate on the origin of Baccharis uncinella. The species does not perform well under customary grassland management. She occurs in large areas of abandoned grassland and along roadsides, but also in islands of abandoned pasture inside forest as well as in logged pine plantations. Must Baccharis perhaps be viewed as a species of forest clearances rather than a grassland species, a oversized tropic equivalent to Senecio or Epilobium species that colonise European forest clearances (e.g. Dierschke 1988, Ellenberg 1996)? The hypothesis is intriguing but so far there is rather more evidence against it than in favour of it.
Firstly, it is in grasslands that the genus Baccharis predominates (Rambo 1952; Leite & Klein 1990). Major surges are recorded on relaxation or cessation of fire and livestock grazing (see e.g. Waechter et al. 1984 for Baccharis articulata and B. uncinella in the Ecological Station of Aracuri, Rio Grande do Sul; furthermore, B. uncinella spreads in abandoned grasslands of the National Park Aparados da Serra, pers. obs.). This phenomenon was also reported in seeder-sprouter Baccharis pteronoides in pastures of Southeastern Arizona (Kenney et al. 1986, Bock & Bock 1997) and in Baccharis pilularis ssp. consanguinea, a common shrub in Californian chaparral (e.g. Hobbs & Mooney 1985). There is also a Brazilian vernacular name for Baccharis-dominated formations: “vassoural”, which may be freely translated as “broomland”, and thus reminds the reader of fire-prone ecosystems of the U.S.A. west coast with their various “broom” and “brush” species.
Although it appears that cessation of fire promotes Baccharis, and populations clearly cannot maintain themselves in one place if fires recur at intervals of only a few years – as in most pastures - there are several indices that the species functions best in a landscape where large-scale disturbances recur at greater frequency than inside forests.
As discussed in chapter 2, Baccharis seeds apparently disperse well in space but not in time, so for the metapopulation to persist, there must be chances of vegetation disturbance within seed dispersal distance of a reproductive population before that population enters the senescent stage; population monitoring showed that this stage may be reached approximately one decade after establishment. Baccharis adults usually do not survive fire, but fire may clearly have a beneficial effect in breaking down litter and opening

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
123
gaps, in short, providing for just such conditions in which Baccharis seedlings establish abundantly. This occurred in part of the Pró-Mata grasslands in vicinity of the neighbour fazenda that accidentally caught fire in 1999, five years after exclosure (R. Mello pers. comm.).
Moreover, not only is Baccharis foliage flammable (pers. obs.) but leaf size and morphology (small and sclerophyllous) and branch architecture are not untypical of so-called fire fuel plants (Bond & van Wilgen 1996). An adult population with overlapping crowns may thus encourage burning and help to provide sites for establishment of a new population (see also Crawley 1997a).
Myrsine, on the other hand, is still essentially a forest species. It may be regarded as certain that Myrsine has not developed on highland soil for more than 2000 years BP when the genus, together with Araucaria, first moved to higher elevations of the High Plateau (Behling 2007). Unless the genus has some extraordinarily fast mechanisms of speciation at its disposal (such as hybridisation coupled with polyploidy), this time span must probably be considered too short for evolution of a subspecies particularly well adapted to forest border-grassland transitions, so that adaptations to a forest habitat are still of major importance for successful colonisation of such sites.
Forests feature a wide range of site conditions – several strata exist, smaller gaps are opened by tree- and limbfall, larger gaps e.g. by wind-throw, fire or infection. There are some indications in which parts of the mosaic Myrsine might have figured originally:
The species is efficiently dispersed by birds (chapter 2) and recruits seedlings in the shade rather than in full exposure to sunlight (chapters 2 and 3); by root suckering, genets may be extended in time and space, although only at a relatively short distance (chapter 4); seedlings can resprout (chapter 3), and so can the majority of adolescents and adults in a population (this chapter, chapter 6). Individuals also readily sprout new vertical stems when tilted, observed in ca. 7% of individuals in both tall tussock and tall shrub grassland.
Bazzaz (1984) correlated gap sizes in tropical rainforest to principal modes of growth and regeneration by which these gaps are subsequently filled. As Fig. 5.1 illustrates, small disturbances or canopy openings, respectively, benefit mostly suppressed seedlings, saplings and sprouts, including root suckers. Gaps of intermediate size are exploited by species with dormant seeds as well as species whose seeds are dispersed to the site following the disturbance, both of which may apply to Myrsine.
An elaborate model of seedfall, juvenile and adult recruitment in tropical tree fall gaps – in this case, openings created by fall of several trees - was presented by Schupp et al. (1989). These authors suggested that although seed and seedling survival of small-seeded, animal-dispersed species should be best in recent treefall gaps – due to lower incidence of diseases -, the probability of survival and growth until the adult stage should be highest at the interface of maturing gaps and the understorey of surrounding old-growth stands, where small frugivore disperser species tend to take refuge.
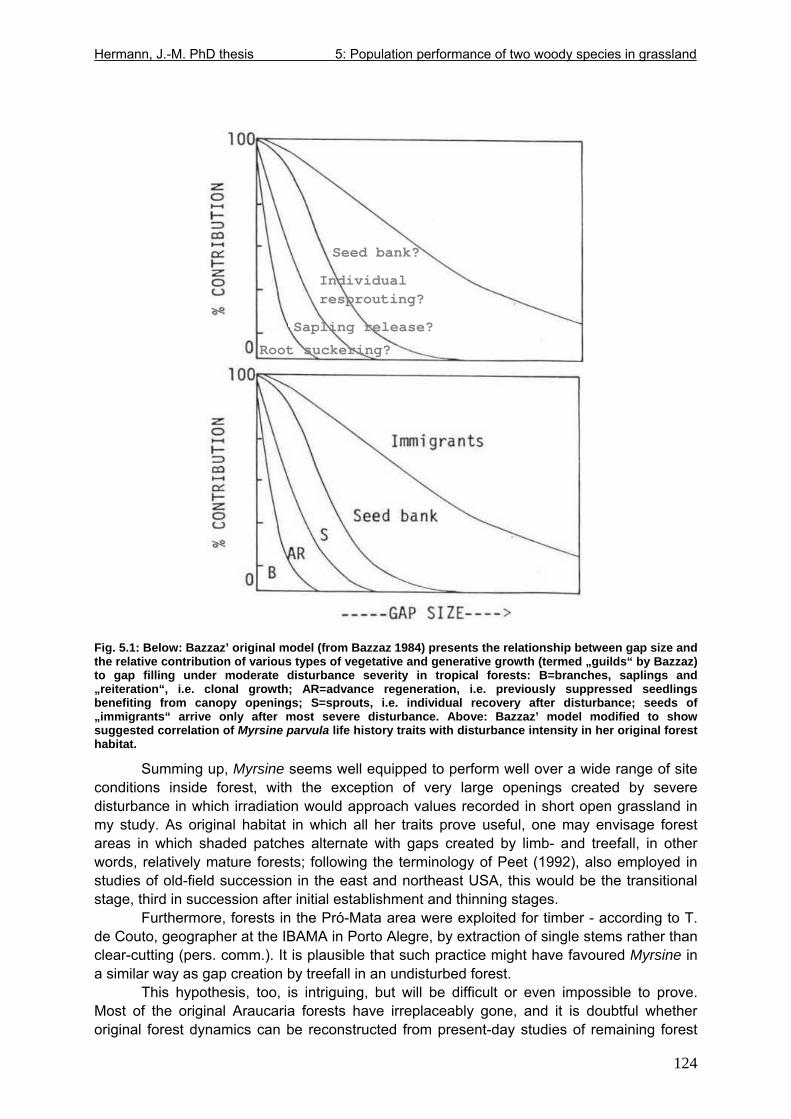
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
124
Fig. 5.1: Below: Bazzaz’ original model (from Bazzaz 1984) presents the relationship between gap size and
the relative contribution of various types of vegetative and generative growth (termed „guilds“ by Bazzaz)
to gap filling under moderate disturbance severity in tropical forests: B=branches, saplings and
„reiteration“, i.e. clonal growth; AR=advance regeneration, i.e. previously suppressed seedlings
benefiting from canopy openings; S=sprouts, i.e. individual recovery after disturbance; seeds of
„immigrants“ arrive only after most severe disturbance. Above: Bazzaz’ model modified to show
suggested correlation of Myrsine parvula life history traits with disturbance intensity in her original forest
habitat.
Summing up, Myrsine seems well equipped to perform well over a wide range of site conditions inside forest, with the exception of very large openings created by severe disturbance in which irradiation would approach values recorded in short open grassland in my study. As original habitat in which all her traits prove useful, one may envisage forest areas in which shaded patches alternate with gaps created by limb- and treefall, in other words, relatively mature forests; following the terminology of Peet (1992), also employed in studies of old-field succession in the east and northeast USA, this would be the transitional stage, third in succession after initial establishment and thinning stages.
Furthermore, forests in the Pró-Mata area were exploited for timber - according to T. de Couto, geographer at the IBAMA in Porto Alegre, by extraction of single stems rather than clear-cutting (pers. comm.). It is plausible that such practice might have favoured Myrsine in a similar way as gap creation by treefall in an undisturbed forest.
This hypothesis, too, is intriguing, but will be difficult or even impossible to prove. Most of the original Araucaria forests have irreplaceably gone, and it is doubtful whether original forest dynamics can be reconstructed from present-day studies of remaining forest
Root suckering?
Sapling release?
Individual resprouting?
Seed bank?

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
125
fragments (Leite & Klein 1990), although an increasing number of detailed plant sociological studies are available and several authors attempt to categorise species by successional groups (e.g. Rosário 2004, Ramos & Boldo 2007).
Seger et al. (2005) recorded the occurrence of Myrsine parvula in the understorey of two relatively undisturbed Araucaria forests – one on typical highland cambisol and one on gleysol –, but with relatively low importance values. The species was predominant in an area of exploited forest studied by Baaske (1999) and also in the understorey of forest nuclei in Pró-Mata grassland ca. 10 years after abandonment (Machado 2004).
The categorisation of forest species as early-, mid- and late-successional is frequent (see also 7.4), but many authors prefer to classify forest species by their ability to exploit gaps of different sizes inside forest, based primarily on rapidity of response to increased light levels in the form of either seeds or (suppressed) seedlings. In a 1987 review, Denslow related life-history attributes of tropical rainforest trees and shrubs to the gap-understorey mosaic as follows: Large open areas of relatively permanence - such as multiple treefall gaps or abandoned agricultural land - are colonised by “tropical ruderal species”, which correspond to “classical pioneer” species e.g. in good dispersal in time and space and in high-light saturated photosynthetic and respiration rates; in smaller gaps, seedlings may germinate but not survive until reproductive age. However, these species account for only a small proportion of the tree flora. “Large gap species” tolerate periodically lower light levels than ruderals, but still require high light for germination, and gaps large enough to allow them to reach reproductive age before canopy closure. “Small gap species” may survive in understorey light conditions but depend on some canopy opening for completion of their life cycle (Denslow 1987 and references therein).
In order to simplify terminology, Swaine & Whitmore (1988) strongly advocated the simple division of species into “pioneer” and “non-pioneer/climax” species, according to whether they germinate only in gaps that receive some full sunlight, or may germinate in the shade and are released in small gaps. According to Whitmore (1989), this division seems applicable to many forests at many latitudes, although he recognises the broad continuum of light responses in either category; e.g. Brokaw (1985) noted that three tree species commonly regarded as pioneers on Barro Colorado island, Panama, greatly differed in shade tolerance, with one one species depending entirely on large gaps for recruitment and another, slow growing species able to exploit even small treefall gaps.
Irrespective of this debate, it appears that no matter which classification and terminology is used, Myrsine does not fit the description of a “classical pioneer” or “large gap-coloniser” but seems best equipped to colonise small to intermediate gap. This topic will be discussed in some more detail in chapter 7, where I investigate whether any such forest species participate in grassland-forest-succession on the Southern Brazilian highland.
5.5.2. Population development in pasture
As concluded in chapters 2 and 3, the origin of so-called tall shrub grasslands probably lies in areas much influenced by cattle grazing, trampling and fire, i.e. covered by a mosaic of prostrate and short grasses and bare ground. In such areas Baccharis seedlings may establish abundantly, although only subsequent abandonment of grazed and trampled areas can have allowed individuals to grow to the adult stage in such densities that crowns overlap. A tall and dense grass and shrub matrix greatly inhibits Baccharis seedling performance.
Under present conditions both Baccharis and Myrsine populations in pastures of my study area are seed sink populations, i.e. their persistence depends entirely on external seed sources, as individuals of both species are impeded from reaching the generatively

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
126
reproductive stage15. Nevertheless, it is unusal for woody plant individuals to occur in such density in pastures of the study region at all, and there is some indication that my population monitoring plots have been subjected to longer fire-free intervals than the customary one or two years. I conjectured that, following the 2005 fire, three to four years at minimum would be necessary for populations to recover pre-fire individual density and sizes. A similarly long time span would have elapsed between the 2005 fire and at least one other fire that - judging by predominance of multiple-stemmed Myrsine individuals - these populations must have experienced previously (see also chapter 6).
There are numerous examples of woody plant cover and abundance increasing on relaxation of fire regimes cited throughout this work, and I will give only one more example here. Following monitoring of an area of the Southern African Nylsvley savannah subjected to controlled burning, it was suggested that the observed increase in woody plant biomass (e.g. Burkea spec., Terminalia spec. in so-called broad-leaved savannah) could be largely attributed to a shift from an estimated historical 3-year fire regime towards burning at ca. 4-5 and, during the final 20 years, 5-7 year intervals. Apparently, as little as half a decade’s rest between fires greatly improves chances of tree survival in such a habitat. However, comparison with a neighbour area unburnt for several decades clearly demonstrated that woody plant biomass was kept at about half the quantity it might achieve if limited only by competition (see Scholes & Walker 1993 and references therein).
In short, the occasional occurrence of adult forest trees and shrubs in pastures of my study region is possible only due to spatial heterogeneity of the grassland fires, i.e. grassland patches escaping biomass loss for more than 1-2 years, and perhaps even longer intervals than indicated here. Note that patchiness of fire was apparent not only in population monitoring, but also in the seedling emergence and transplant experiments (chapters 2 and 3).
Such a relaxed fire regime may be explained, in the case of the population monitoring plots, by the relatively remote location at several kilometres distance from the fazenda settlement, and surrounded by forest. However, overgrazing is another possible cause: If the grass cover is temporarily so much reduced that neither cattle fodder nor fire fuels are available, then seedling survival chances must clearly increase (see chapter 3), even more so on coincidence with favourable climatic conditions such as a mild winter. Overgrazing is possible in spite of the low number of head of cattle per hectare, as cattle graze selectively. Boldrini (1993), who investigated the impact of three levels of cattle grazing intensity on pasture structure in the Central Depression of Rio Grande do Sul, detected overgrazing not only at highest but also at lowest grazing intensity; in the latter case, patches of preferred forage plants were overgrazed and others left practically untouched.
Moreover, population monitoring revealed that pasture fire affected individuals within few metres distance from the forest border less than individuals further away. Apparently, proximity to the forest border reduces fire intensity and promotes individual survival, perhaps due to greater humidity in the shade. A considerable proportion of Myrsine individuals benefited from this, remaining unburnt, but Baccharis individuals were so scarce that none escaped the fire.
The majority of burnt Myrsine individuals resprouted after fire – in similar proportion as observed in the resprout experiment described in the following chapter. It would be interesting to know for how long established individuals may persist under repeated biomass
15 The terms “source” and “sink” are here based on Eriksson’s (1996) review; Eriksson gives an example of sink populations in the clonal woodland herb Rubus saxatilis, in which occasionally seedlings establish at individual densities too low to allow for genetic reproduction (partner-limited fruit-set).

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
127
loss. This question, however, can be resolved only by long-term monitoring or perhaps year-ring counts on root systems.
Whether Myrsine population dynamics under these conditions may be termed “succession” at all, or whether the term fluctuation is more appropriate, depends largely on the time scale of observation: The term “fluctuation”, according to van der Maarel (1988), is applied if a certain quantitative community composition reappears over time, i.e. there is no direction of change towards either pasture or forest. This may be the case if fire recurs at intermediate frequencies, as inferred from structure of my monitored populations, and only some such fire cycles are observed. One year with particularly favourable conditions for seedling establishment, a further delay in fire occurrence or, conversely, increased fire frequency might all induce such directional changes.
5.5.3. Population development in abandoned grassland
5.5.3.1. Baccharis Considering the scarcity of Baccharis individuals in managed pasture, there is little
doubt that the dense Baccharis stands with temporarily more or less closed canopy - that characterise so-called tall-shrub abandoned grassland - developed largely following pasture abandonment and exclusion of fire in the Pró-Mata area. Populations exhibit a relatively even size-age structure, consisting mostly of adults, with individuals evenly distributed across 10 m distance from the forest border. Even-aged cohorts in which age is correlated closely to time since last disturbance are typical of species recruiting in large light-gaps (e.g. Keeley 1992 for obligate seeding shrubs in Californian chaparral; Alvarez-Byulla 1986 in Martínez-Ramos et al. 1989 on shade-intolerant pioneer tree Cecropia obtusifolia; Veblen 1989 on wind-dispersed Nothofagus ssp. dominating pioneer forests on landslides ).
The precipitated decline of tall shrub canopies that became apparent even during my very short observation period, was somewhat unexpected. In the year or two prior to my study, judging by stump sizes of dead individuals, dieback affected considerable proportions of intermediate-size individuals, so may have been largely a consequence of intraspecific competition (especially shading-out). During the year of my study, however, there was an increased proportion of canopy-formers among the dead, and symptoms of fungal infection are present throughout these populations. Supposing that in pasture prior to abandonment, Baccharis individuals could merely have grown to small sizes of one or a few years old, these canopy-formers at the time of my survey were no more than 12-15 years old.
Unfortunately, Baccharis wood is unsuitable for year-ring counts (E.Burgert and J.Morales, pers. comm.) so that maximum age of healthy individuals cannot be determined. It is possible that senescence sets in naturally around one decade after establishment, and makes individuals more susceptible to infection. However, the presence of Pythium fungi indicates that adverse soil conditions also play a role.
In either case, Baccharis dieback is due to age-dependent and density-dependent physiological processes. Myrsine juveniles established under their canopy possibly exert some competitive pressure on Baccharis, as Barnes & Archer (1999) demonstrated for woody species juveniles established underneath pioneer Prosopsis shrubs in subtropical US grasslands. However, in this case one might expect higher density of Myrsine recruits under lighter than under denser Baccharis canopy; the opposite is the case. More importantly, only a couple of Myrsine individuals had reached a size of 5 metres or taller and would count as a tree by common conception (Ellenberg 1996). Thus, shading out by Myrsine can be clearly ruled out.
In the surroundings of declining tall shrub populations – in so-called tall tussock grassland - conditions are similarly inhibitory to Baccharis seedling establishment as under

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
128
their own canopy. There is no indication that under current conditions, tall tussock stands develop towards tall shrub stands. Rather, judging by recruit density, the first woody canopy above the tall tussock grass layer will be established by Myrsine herself. As a consequence, the presumed network of Baccharis population patches that continuously establish and become extinct (metapopulation) is reduced to occasionally recruiting individuals scattered across the landscape.
While such a distribution might have some advantages, e.g. escape from resource limitation by neighbours, and decreased spread of pathogens, there is also at least one known drawback. Harter (1999) found that Baccharis species do not set seed unless insect-pollinated. Moreover, the species is dioecious, and female flowers are less attractive to pollinators than male flowers. Harter therefore proposed that the formation of dense clusters with intertwining branches of male and female individuals (sex ratios are similar) greatly improves chances of pollination. This theory could be tested by seed counts on solitary individuals in tall tussock grassland versus individuals in dense stands.
Keeley and Zedler (1978), assessing the relative importance of resprouters and seeders in California chaparral, proposed that obligate seeders should eventually be favoured as fire-free intervals increased. Their model was based on the assumption that ca. 100 years following a fire, self-thinning between sprouting shrubs would create gaps for seeders or, alternatively, new fires would be so intense that high mortality among resprouters would create gaps for seeders. A similar trend emerges from a review by Recher & Christensen (1981) of fire-prone ecosystems in Australia.
However, Keeley (1992) modified this statement: Canopy gaps created by thinning do indeed appear in chaparral after long disturbance-free intervals, but these benefit “perhaps” saplings, and only saplings of “fire-resistant” species that resprout after fire and newly recruit from seed under shade and in litter layers decades afterwards. “Obligate seeders” (that do not resprout – like Baccharis) recruit only in large gaps within the first years after fire. Keeley also pointed out that chaparral sites rarely remain undisturbed for more than about 50 years.
Summing up, it seems unlikely that closed-canopy Baccharis stands may newly develop in the Pró-Mata area in absence of a large-scale disturbance such as fire. It is to be expected that eventually, former tall shrub stands will either be replaced by shrubs and trees - as happens in Australian heathlands and in certain Eucalyptus species that degenerate in absence of fire (Recher & Christensen 1981) – or the area will be retrieved by grasses that dominated the area prior to establishment of tall shrub stands, and persisted suppressed in their shade, a process described by Walter (1971) for Acacia detinens thickets excluded from grazing in African savannah.
On the other hand, the probability of a new fire increases with every year that dry grass biomass accumulates in the Pró-Mata grasslands. Already two parts of the area have burnt accidentally since abandonment: In 1999, five years after abandonment, flames spread from the adjacent fazenda and consumed several hectares of grassland. This area is now partly covered by dense Baccharis stands that established following this fire (R. Mello pers. comm.), i.e. about eight years old. Another area was ignited in 2003 by a faulty electricity line in spite of rainfall some hours previously, although this fire did not consume more than a few hundreds of square metres (pers. obs.)

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
129
5.5.3.2. Myrsine Contrary to the (perhaps temporary) population decline in post-burn pasture, Myrsine
populations in both structural types of abandoned grassland are clearly expanding, although to different extent.
The previous chapters have shown that Myrsine seedlings recruiting under Baccharis canopy are subjected to various inhibitory factors, e.g. considerable competition. But climate extremes are buffered and, due to shading, gaps in the grass matrix are more abundant, so that seedling recruit numbers far exceed those of tall tussock grassland. Since Myrsine also recruits by root suckers in both types of abandoned grassland, individual density in tall shrub grassland was approximately twice as high as in tall tussock grassland. Relative to the latter, tall shrub grassland may be regarded as having a net facilitative effect on Myrsine.
Tilting of individuals is a phenomenon that may be due to root suckering but was not discussed in the previous chapter. I suspect that root suckers are more prone to tilting than seedlings while their adventive root systems are inadequately developed. However, this condition affects only a minority of individuals, and these readily sprout new vertical shoots and thus do not suffer more than a temporary setback in height growth. I mention it principally because it gives another hint of Myrsine’s good pre-adaptation to relatively mature forest. Individuals already established in tall shrub grassland may be expected to cope fairly well with the observed decline of the tall shrub canopy. A decrease in recruitment appears to be the most pronounced reaction of Myrsine populations in these areas. Both new and elder Myrsine recruits occur in greater density underneath denser than underneath lighter Baccharis canopy; this may be attributed to higher density of gaps in the grass matrix, which are principal recruitment sites for both root suckers and seedlings, in heavier shade.
However, in the light of Myrsine population development across tall shrub grassland sites, reduced recruitment is not necessarily negative. On the contrary: during the year of observation, Myrsine individual density increased by 30% in tall tussock grassland and 40% in tall shrub grassland; if individual numbers continued to increase at this rate, density-dependent thinning within the following decade would seem inevitable. In other words, Myrsine recruit numbers may be expected to decrease whether Baccharis dieback occurs or not.
The impact of Baccharis dieback on Myrsine individual growth during the observation period was weak – in other words, there was little evidence for competitive release. Only small elder recruits (with an estimated age of at least two years at 2nd survey) showed significantly higher growth rates underneath lighter than underneath denser Baccharis canopy, possibly indicative of competitive release. In tall elder recruits (with an estimated age of at least three years at 2nd survey), growth rates were significantly different only between tall tussock grassland and tall shrub grassland, independent of density of the tall shrub canopy within the latter. Growth rate of new recruits (with a minimum age of one year at 2nd survey) did not differ significantly between lighter and denser Baccharis canopy, nor between tall tussock grassland and averaged samples of tall shrub grassland.
The sample of adolescent Myrsine individuals was comparatively small, and although these individuals tended towards smaller height growth rate but higher basal area growth rate in tall tussock grassland relative to tall shrub grassland, this trend was not significant.
In another fire-prone ecosystem, Californian chaparral, woody species also benefit little from competitive release. A flush of recruitment of obligate seeders follows immediately after burning but afterwards, for periods as long as 60 years, seedlings recruit exclusively in the shade even if gaps in the canopy are present (Keeley 1992); this phenomenon led Keeley to coin the term “gap-avoiders”. The gaps noted by Keeley were not formed by shrub dieback and had no litter layer, which may explain their inhospitability for seedlings.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
130
However, Callaway (1992) studied the effects of artificial canopy openings in chaparral and reported that although shrub removal enhanced seedling emergence of two Quercus species – both occurring in chaparral and woodland – subsequent seedling survival was equally low in gaps as in pure grassland. In Tyler’s (1995) study, shrub removal did not even enhance seedling emergence of woody species (shrubs and subshrubs), perhaps due to low rainfall. Callaway (1992) suggested that instead of being released by shrub dieback, Quercus species eventually outcompete their nurse shrubs.
The importance of gaps for release of juveniles in forests has, on the other hand, been amply documented worldwide (e.g. Bazzaz 1984, Denslow and references therein 1987, Whitmore 1989, Veblen 1989, Canham 1989). I have argued above that Myrsine is apparently well adapted to a habitat characterised by occasional canopy openings, with the exception of very large gaps. What may explain her unpronounced response to Baccharis canopy dieback?
Firstly, clonal origin of recruits may have dampened reactions to canopy opening. Since the absolute sampled area of populations was low, it was not possible to distinguish, within these patches, between different origin (root sucker/seedling) of recruits. A general predominance of root sucker over seedling recruitment in both tall tussock and tall shrub grassland was demonstrated in the previous chapter. There, I proposed that root sucker support by feeder individuals decreases with age, and influence of site conditions on growth rate increases accordingly. It is not surprising that especially new recruit samples did not show a perceptible reaction to height and density of the surrounding grass and shrub matrices.
The subsample of small elder recruits under lighter tall shrub canopy, on the other hand, had a relatively high proportion of seedlings (the ratio of root suckers to seedlings was 1:1.7), other than the recruit subsample under denser canopy (ratio 1:2.3). This may explain why individuals of this size class displayed a weak trend towards of competitive release, i.e. higher growth rate under lighter canopy. In mixed samples obtained from entire populations with root sucker predominance, individual growth rate of this size-age class did not differ significantly between grassland types, i.e. seemed still largely governed by feeder root supply (chapter 4).
Secondly, according to Canham (1989), whether a species can successfully exploit increased light levels or not depends largely on her ability to modify physiological and morphological traits, such as photosynthetic rate and leaf angle. Reduced competition by Baccharis comes at the cost of exposure to high irradiation. I have pointed out before (1.4.4) that light intensity under a more or less closed Baccharis canopy (outside grass tussocks) is approximately the same as in a small forest gap; on canopy dieback, light intensity would then approach those in very large forest gaps.
Thirdly, release from competition by Baccharis would not only influence Myrsine individuals, but also grasses and grassland shrubs still present in the same plots. Tussock grasses almost certainly benefit to greater extent than any forest juveniles present, as they are of the C4 type and consequently display increased and more efficient photosynthesis at high light (e.g. Mohr & Schopfer 1992). Moreover, soils underneath Baccharis canopies still essentially resemble grassland rather than forest soils (see Dümig et al. 2008 a,b). Such a process has been described by Walter (1971) for Acacia detinens thickets excluded from grazing in African savannah.
The ratio of root sucker to seedling recruits might be expected to increase with Baccharis dieback as root suckers are more likely to recruit inside a more or less closed grass matrix than seedlings. So far, however, such a trend is not perceptible; on the contrary, Myrsine samples from underneath denser Baccharis canopy contained slightly higher proportions of root suckers than samples from underneath lighter canopy. It follows from these considerations that a longer monitoring period would be necessary in order to assess

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
131
possible competitive release of both grass and tree species established under the formerly intact Baccharis canopy.
How would a large-scale disturbance such as fire – presumably the prerequisite for re-establishment of Baccharis-dominated stands – affect Myrsine populations in their current state? There is no doubt that population growth would suffer a setback of several years (as observed in pasture and in the resprout experiment of chapter 6) but not to zero, as the majority of individuals would probably survive the fire.
5.5.4. Predicting successional sequences
Bond & van Wilgen (1996) write: „The idea of predictable sequences of species replacements is still central to much ecological thinking“ (ibid. p.149). The reasons for this are manifold. In the case of the southern Brazilian grasslands, the interest is not just in understanding past patterns of vegetation development, but also – perhaps more importantly – unraveling mechanisms of seed dispersal, emergence, establishment and persistence, and the influence predominant species exert on one another, may facilitate predictions of future forest expansion under ongoing changes in land use. Pine plantations planted today will be clear-cut 30-40 years hence; grassland converted to arable land today may be abandoned sooner, as soil is not everywhere suitable for this type of cultivation – first fallows were noted even at the end of my fieldwork period in 2006.
Successional theory has been extensively reviewed during the late 1980’s and 1990’s (e.g. Pickett et al. 1987, Kozlowski et al. 1991, Glenn-Lewin & van der Maarel 1992, Bond & van Wilgen 1996). One common feature of models developed since the 1970’s is an emphasis on „life history and competitive relations of component species rather than the emergent properties of communities“ (Peet & Christensen 1980, p. 132); the „view of succession as primarily a species replacement process driven by reaction to plant-controlled (i.e. autogenic) modification“ (Glenn-Lewin & Van der Maarel 1992, p.5) is rejected.
However, partly due to ambiguous terminology, a synthesis of these models has not yet been achieved (e.g. Kozlowski et al. 1991). On the contrary, patterns of secondary succession in space and time, especially following grassland abandonment, are apparently notoriously difficult to interpret. Schreiber (1997), on finding that none of the secondary successional models available at the time of his study served to explain his observations of grassland development following abandonment, attributed this – among other factors – to the fact that many of these models are based on observations in fallow arable land, where succession was not initiated with a more or less closed vegetation layer already present, a fact that Glenn-Lewin & van der Maarel (1992) also noted with concern. Kahmen & Poschlod (2004), in continuation of Schreiber’s study, write that „so far, ecologists have not been able to establish one general, unifying theory for successionally induced changes in species composition following abandonment. In fact, the course of succession seems to be unique for each site and year“ (p.21).
In Schreiber’s study, 14 grassland sites in Baden-Württemberg, south Germany, have been monitored since abandonment in 1975. Schreiber (1997) described colonisation of some of these grassland sites by long-distance wind-dispersed species (e.g. Fraxinus excelsior) or by root suckering species (Prunus spinosa), yet a clear development towards forest could be recognised only in fertile sites in which woody species seedlings established as early as one year after abandonment, and achieved dominance early. As a general trend, Schreiber noted a retrogression of sites towards initial floristic composition, often with grasses increasing in cover. Similarly, Pickett (1982), on studying species curves in time in oldfields abandoned for 20 years on the U.S.A. east coast, reported that characteristic forest species (many of them animal-dispersed and shade-tolerant) had established by the 20th

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
132
year, but various species displayed bimodal and multimodal peaks that he could not explain at the time.
Kahmen & Poschlod’s study (2004) represents a continuation and re-evaluation of Schreiber’s data after 25 years of abandonment. The authors tried to reduce the observed diversity in species composition by focussing on frequency of certain plant functional traits at six sampling dates between 1975 and 2000. However, this approach did not yield many consistent trends either. For example, small and large seed size were traits of similar importance (while large-seeded species might be expected to predominate at later stages of succession). The relative importance of short-distance vs. long-distance vegetative spread (less or more than 1m, respectively), fluctuated strongly, although during the last decade there was a trend towards longer-distance vegetative spread. An ordination of traits and sites at sampling dates 1975 and 2000 yielded only one strong positive correlation with the main axis, namely, with woody species cover per site and time.
The plant functional type approach, i.e. grouping taxonomically different units into functionally similar groups, has been judged useful for comparisons across ecosystems and regions (e.g. Duckworth et al. 2000), but it seems to follow from this attempt that at a given time and in a given site and region, other factors than functionally similar groups may have to be taken into account in order to explain an observed successional sequence – such as seed rain, vegetation structure, and perhaps behaviour of predominant species that obviously represent particularly successful conglomerates of traits. This has emerged from discussions in the previous chapters and will be recapitulated in chapter 7.
There are some peculiarities to my study area that are not likely to facilitate interpretation and prediction of successional sequences. One of them is that neither Baccharis nor Myrsine fit the image of a “classical” pioneer in forest succession. Only at first glance do these two species follow a successional sequence from wind-dispersed, light-demanding, early-colonising woody species to animal-dispersed and more shade-tolerant later-successional species, such as ecologists familiar with secondary succession especially in Middle European or North American forests would expect. Textbook examples of such sequences are described e.g. by Dierschke (1988) for logged Middle European beech forests - earliest colonizers are light-loving herbs; these are eventually shaded out by Rubus thickets, and these in turn are shaded out by early-successional wind-dispersed trees such as Betula pendula – and by Peet (1992) for North Carolina Piedmont oldfields, in which pines shade out herbs and grasses and hardwoods later shade out the pines.
The two species Baccharis and Myrsine have originated from two types of ecosystem – fire-prone grassland-type ecosystems and forest – that are generally believed to obey different assembly rules. In interpretation of vegetation development in fire-prone ecosystems, since Noble & Slatyer’s (1980) publication, the emphasis is on „the interplay between disturbance and the timing of key life-history events“ while „competition is virtually ignored“ (Bond & van Wilgen p. 150). This is based on the assumption that „competitive interactions of adult plants do not have time to have an impact on species composition before another disturbance occurs“ (Kozlowski et al 1991, p. 407). In models of forest succession or primary succession, on the other hand, species interactions play a crucial role (e.g. Connell & Slatyer 1977, Tilman 1990).
Here is one possible answer as to why succession proceeds somewhat haphazardly in Pró-Mata. There is some ambiguity in the effects of Baccharis canopies on Myrsine recruits – protection from climate extremes apparently comes at the cost of intense resource competition (chapter 3). Nutrient status of highland grassland soil is initially lower than forest soil at lower elevations (chapter 1), and it is not difficult to envisage how this situation may be aggravated by a species that, adapted to recurrent fires and flushes of remineralised nutrients, invests large amounts of nutrients in rapid growth. However, such “wasteful” behaviour of Baccharis yet remains to be proved by investigations of physiology.

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
133
In spite of competitive effects, Baccharis canopies offer the most favourable conditions for Myrsine recruitment; but they undergo a precipitated decline little more than one decade after establishment while Myrsine recruits are still far from attaining the canopy stage, whereas often species that arrived on site and established early in succession following a large-scale disturbance are overtopped and outcompeted by species that established in their shade (e.g. Dierschke 1988, Veblen 1989; but see Whitmore 1989). Moreover, such tall shrub grassland accounts for only a minor proportion of the abandoned grassland area in the first place (ca. 5% according to Morales & Pillar 2004).
Co-occurrence of Baccharis and Myrsine individuals in relatively high density in tall-shrub grassland may thus reflect shared preferences for areas strongly altered by human activity rather than directionality of succession (cf Wilson 1998). However, some positive synergism between forest border and grassland populations has also been suggested, namely, shrubs encouraging bird transit across grassland, which in turn would increase seed input in both grassland and forest border (see chapter 2).
Judging by Myrsine individual densities and sizes in grassland relative to other forest species, there is little reason to doubt that this species will form the first pioneer forest canopy. Further spatial expansion of established populations into grassland may also be anticipated, as during my observation period individual density increased more at 5-10 m distance than at 0-5 m distance from the forest border, especially in tall tussock grassland. It remains to be seen what effect Myrsine canopies have on later-successional species.
Fig. 5.2: Only a question of time: Dieback of canopy-forming Baccharis in tall shrub abandoned
grassland; photograph taken June 2006. Although recruit numbers of forest tree Myrsine (bottom left) are
lower under lighter than under denser Baccharis canopy and Myrsine individual growth rate was not
consistently higher in lighter areas, there is little reason to doubt that the species will eventually follow
Baccharis as a canopy-former. Photographer: J.-M. Hermann.
Already, seedlings and juveniles of tree and shrub species of the family Myrtaceae (Myrcia, Siphoneugenia, Psidium), Melastomataceae (Miconia) and Aquifoliaceae (Ilex spec.) have emerged to notable extent underneath tall shrub canopies in my monitoring plots. However, Myrsine populations in grassland consist largely of root suckers, mirroring e.g. colonisation of North American Prairie by Aspen (Populus tremuloides, chapter 4). Ellison & Houston (1958) found that belowground competition in Aspen forest stands recruited from

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
134
root suckers was considerable - dry matter production of herbaceous species was reduced by up to one order of magnitude relative to plots in which Aspen roots had been trenched.
Of the other species named, only Siphoneugenia reitzii possesses this ability of spreading vegetatively, and Siphoneugenia root suckers are much less frequent than Myrsine root suckers (chapter 6). Thus, recruitment of these later-successional species into taller strata or even the canopy may be largely delayed until breakdown of the Myrsine populations – and since the species can rejuvenate by root sucker recruitment, stands may remain stable for a long time. However, this can only be found out by long-term monitoring.
Crawley (1997a, p.74) writes: „Plants have life histories that ‚work’ in the sense that they are capable of persistence (...), but they are bundles of traits cobbled together from countless compromises. They do not represent multidimensional optima.“ He refers to species, of course, but the metaphor applies just as well to community composition. My study area is a zone in which three major ecosystems and two climatic zones converge. Added to this, for perhaps 100-150 years forest areas were subjected to logging and grassland areas were subject to cattle grazing and fire, i.e. more severe biomass removal than species could have experienced previously, until those impacts ceased abruptly little more than a decade prior to this study.
It is not reasonable to suppose that communities could instantaneously revert to more or less directional pre-colonisation successional sequences, if such sequences ever existed – as according to Behling and collaborators (see chapter 1) forest expansion on the highland must also be regarded as a relatively recent phenomenon.
A factor not mentioned so far is randomness. Although the importance of randomness in shaping successional pathways is nowadays acknowledged (Hutchings 1997b), the diagnosis should be made with great caution. With so many variables yet unknown, there is some danger of proclaiming randomness when actually an underlying pattern - e.g. of topography, management influences, major wind-directions etc. – has not been recognised; even more so as with increasing time since abandonment, management impacts are increasingly difficult to trace.
The impression that most phenomena described here are relatively unique grows stronger as the view is extended beyond Pró-Mata. Baccharis does play a prominent role in secondary succession in some other areas excluded from grazing and fire, e.g. the Ecological Station of Aracuri and the National Park Aparados da Serra, both located on the High Plateau of Rio Grande do Sul (pers. obs.). Myrsine, however, is mentioned as a prominent forest pioneer species only by a couple of authors (Oliveira 2003, Machado 2004; Behling et al. 2007). This phenomenon will be discussed in more detail in chapter 7.
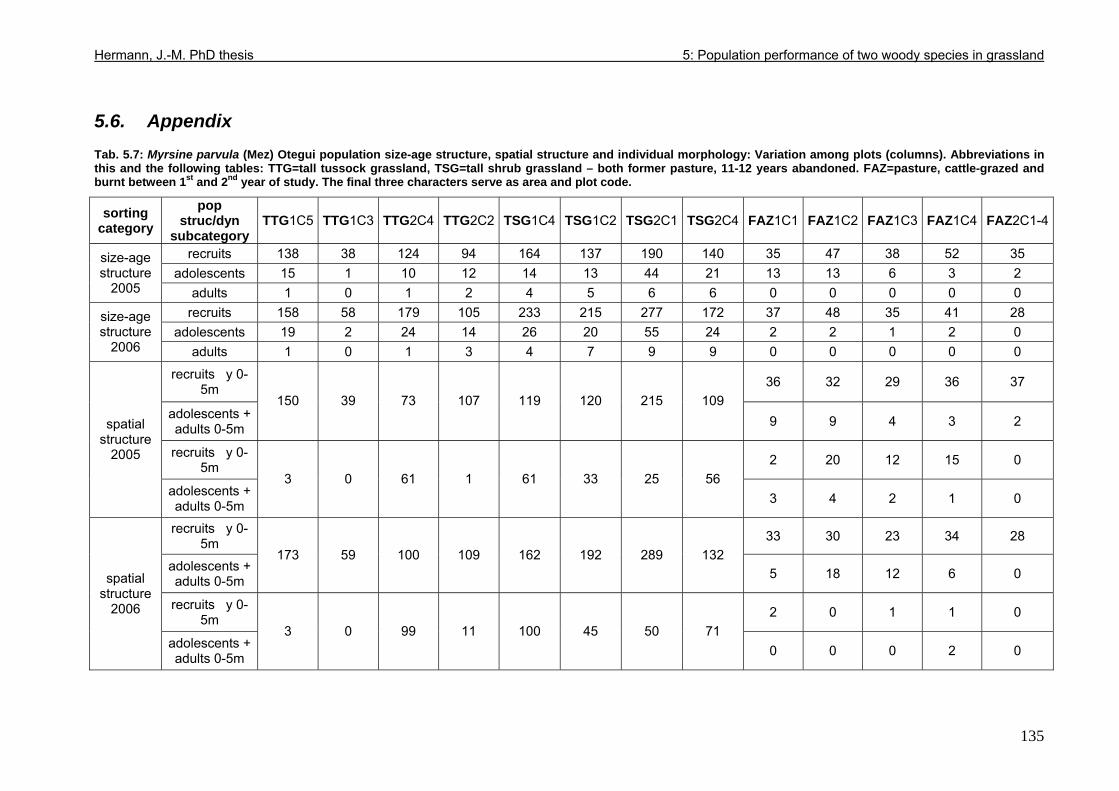
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
135
5.6. Appendix Tab. 5.7: Myrsine parvula (Mez) Otegui population size-age structure, spatial structure and individual morphology: Variation among plots (columns). Abbreviations in
this and the following tables: TTG=tall tussock grassland, TSG=tall shrub grassland – both former pasture, 11-12 years abandoned. FAZ=pasture, cattle-grazed and
burnt between 1st
and 2nd
year of study. The final three characters serve as area and plot code.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
recruits 138 38 124 94 164 137 190 140 35 47 38 52 35 adolescents 15 1 10 12 14 13 44 21 13 13 6 3 2
size-age structure
2005 adults 1 0 1 2 4 5 6 6 0 0 0 0 0 recruits 158 58 179 105 233 215 277 172 37 48 35 41 28
adolescents 19 2 24 14 26 20 55 24 2 2 1 2 0 size-age structure
2006 adults 1 0 1 3 4 7 9 9 0 0 0 0 0
recruits y 0-5m 36 32 29 36 37
adolescents + adults 0-5m
150 39 73 107 119 120 215 109 9 9 4 3 2
recruits y 0-5m 2 20 12 15 0
spatial structure
2005
adolescents + adults 0-5m
3 0 61 1 61 33 25 56 3 4 2 1 0
recruits y 0-5m 33 30 23 34 28
adolescents + adults 0-5m
173 59 100 109 162 192 289 132 5 18 12 6 0
recruits y 0-5m 2 0 1 1 0
spatial structure
2006
adolescents + adults 0-5m
3 0 99 11 100 45 50 71 0 0 0 2 0
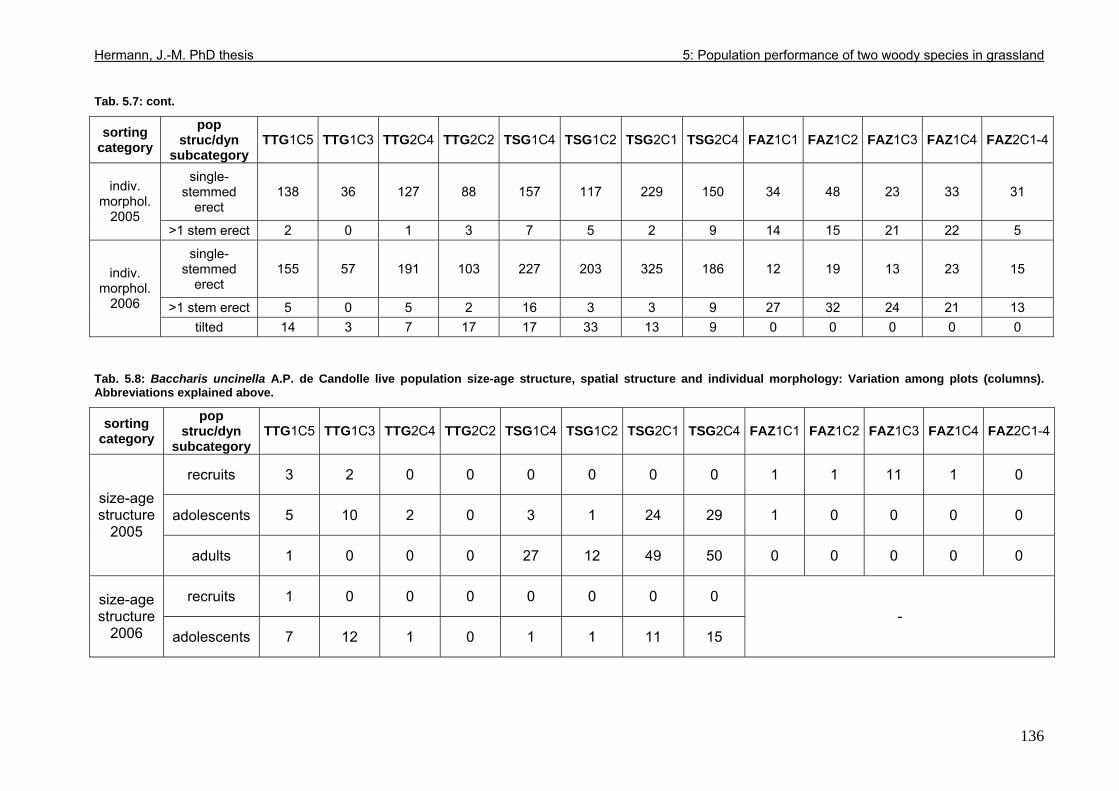
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
136
Tab. 5.7: cont.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
single-stemmed
erect 138 36 127 88 157 117 229 150 34 48 23 33 31 indiv.
morphol. 2005
>1 stem erect 2 0 1 3 7 5 2 9 14 15 21 22 5
single-stemmed
erect 155 57 191 103 227 203 325 186 12 19 13 23 15
>1 stem erect 5 0 5 2 16 3 3 9 27 32 24 21 13
indiv. morphol.
2006
tilted 14 3 7 17 17 33 13 9 0 0 0 0 0
Tab. 5.8: Baccharis uncinella A.P. de Candolle live population size-age structure, spatial structure and individual morphology: Variation among plots (columns).
Abbreviations explained above.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
recruits 3 2 0 0 0 0 0 0 1 1 11 1 0
adolescents 5 10 2 0 3 1 24 29 1 0 0 0 0 size-age structure
2005 adults 1 0 0 0 27 12 49 50 0 0 0 0 0
recruits 1 0 0 0 0 0 0 0 size-age structure
2006 adolescents 7 12 1 0 1 1 11 15 -
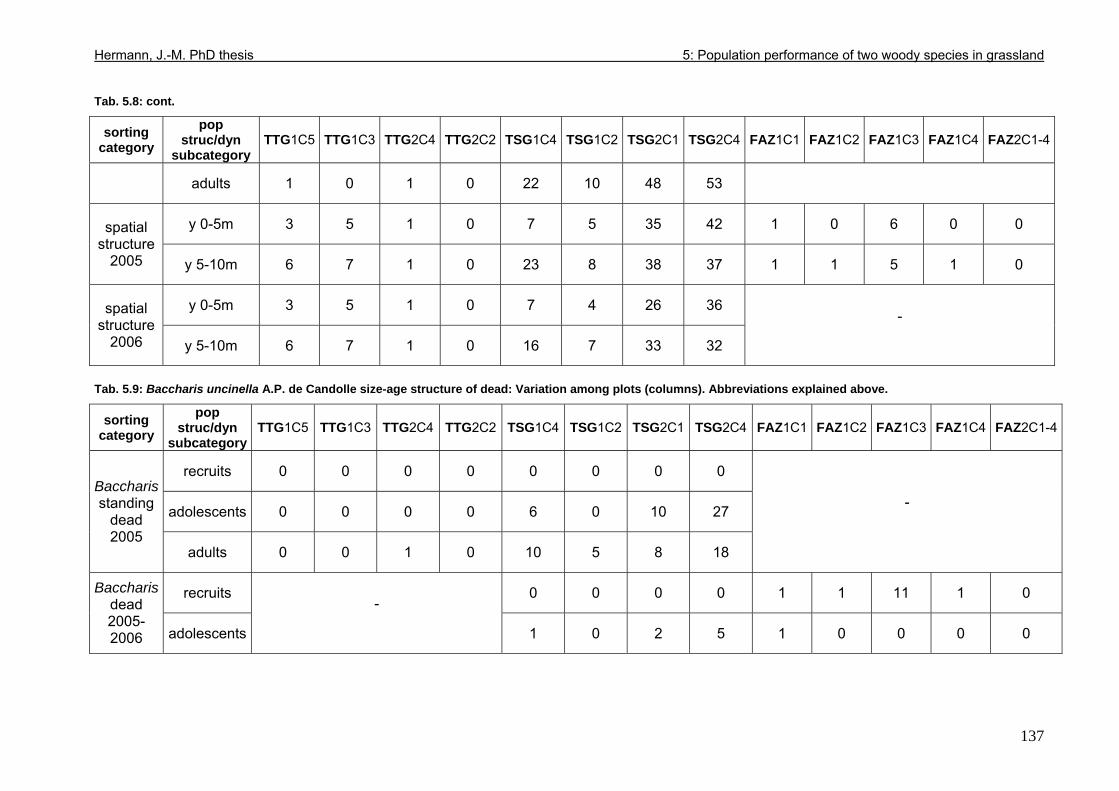
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
137
Tab. 5.8: cont.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
adults 1 0 1 0 22 10 48 53
y 0-5m 3 5 1 0 7 5 35 42 1 0 6 0 0 spatial structure
2005 y 5-10m 6 7 1 0 23 8 38 37 1 1 5 1 0
y 0-5m 3 5 1 0 7 4 26 36 spatial structure
2006 y 5-10m 6 7 1 0 16 7 33 32
-
Tab. 5.9: Baccharis uncinella A.P. de Candolle size-age structure of dead: Variation among plots (columns). Abbreviations explained above.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
recruits 0 0 0 0 0 0 0 0
adolescents 0 0 0 0 6 0 10 27
Baccharis standing
dead 2005
adults 0 0 1 0 10 5 8 18
-
recruits 0 0 0 0 1 1 11 1 0 Baccharis dead 2005-2006 adolescents
-
1 0 2 5 1 0 0 0 0
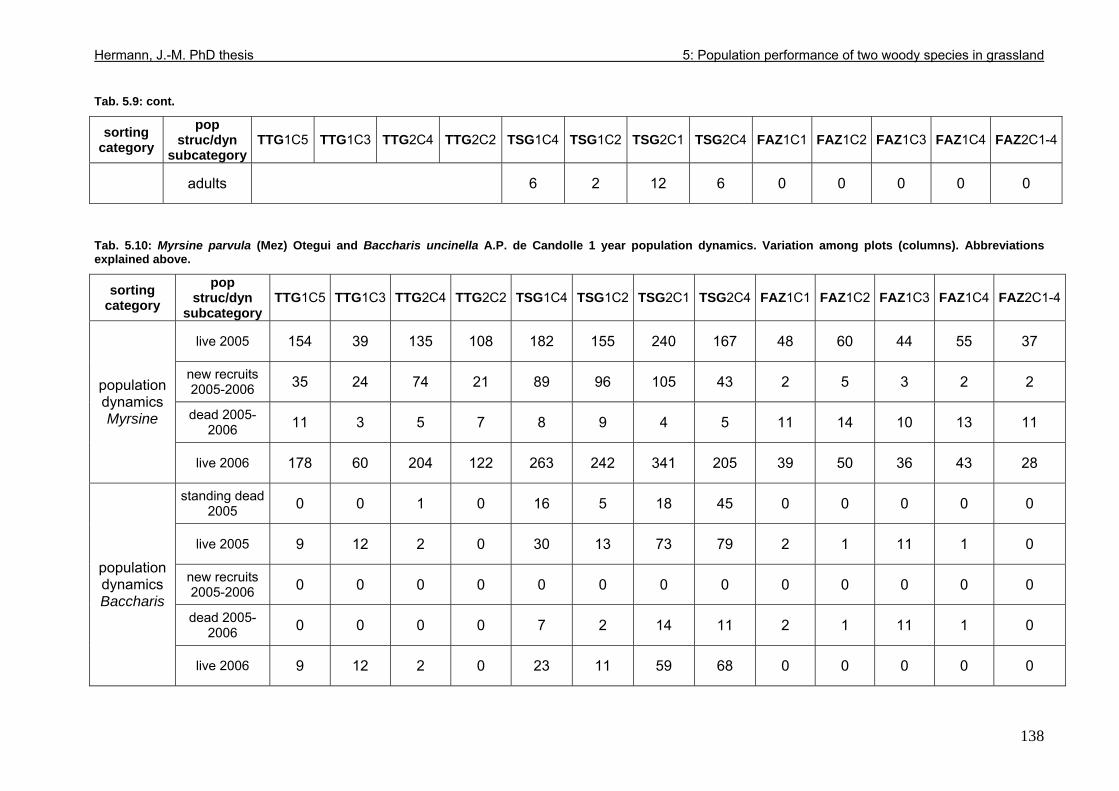
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
138
Tab. 5.9: cont.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
adults 6 2 12 6 0 0 0 0 0
Tab. 5.10: Myrsine parvula (Mez) Otegui and Baccharis uncinella A.P. de Candolle 1 year population dynamics. Variation among plots (columns). Abbreviations
explained above.
sorting
category
pop
struc/dyn
subcategory
TTG1C5 TTG1C3 TTG2C4 TTG2C2 TSG1C4 TSG1C2 TSG2C1 TSG2C4 FAZ1C1 FAZ1C2 FAZ1C3 FAZ1C4 FAZ2C1-4
live 2005 154 39 135 108 182 155 240 167 48 60 44 55 37
new recruits 2005-2006 35 24 74 21 89 96 105 43 2 5 3 2 2
dead 2005-2006 11 3 5 7 8 9 4 5 11 14 10 13 11
population dynamics Myrsine
live 2006 178 60 204 122 263 242 341 205 39 50 36 43 28
standing dead 2005 0 0 1 0 16 5 18 45 0 0 0 0 0
live 2005 9 12 2 0 30 13 73 79 2 1 11 1 0
new recruits 2005-2006 0 0 0 0 0 0 0 0 0 0 0 0 0
dead 2005-2006 0 0 0 0 7 2 14 11 2 1 11 1 0
population dynamics Baccharis
live 2006 9 12 2 0 23 11 59 68 0 0 0 0 0
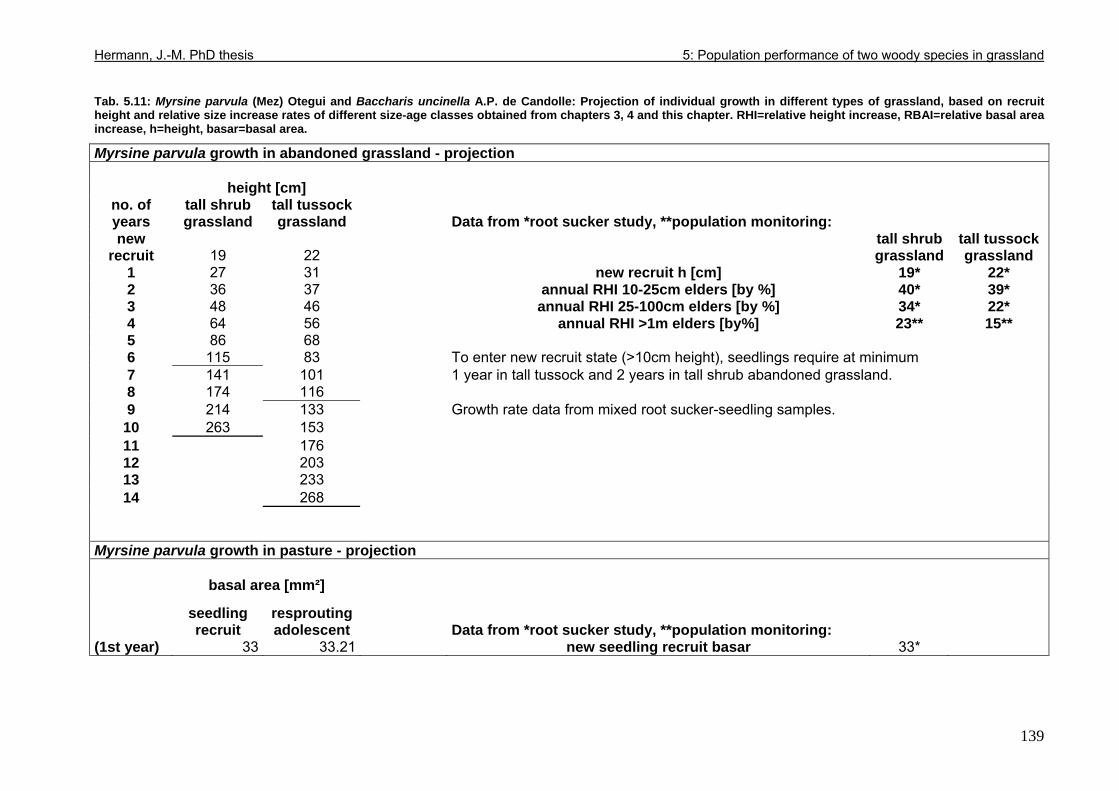
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
139
Tab. 5.11: Myrsine parvula (Mez) Otegui and Baccharis uncinella A.P. de Candolle: Projection of individual growth in different types of grassland, based on recruit
height and relative size increase rates of different size-age classes obtained from chapters 3, 4 and this chapter. RHI=relative height increase, RBAI=relative basal area
increase, h=height, basar=basal area.
Myrsine parvula growth in abandoned grassland - projection
height [cm] no. of
years
tall shrub
grassland
tall tussock
grassland Data from *root sucker study, **population monitoring: new
recruit 19 22 tall shrub
grassland
tall tussock
grassland
1 27 31 new recruit h [cm] 19* 22*
2 36 37 annual RHI 10-25cm elders [by %] 40* 39*
3 48 46 annual RHI 25-100cm elders [by %] 34* 22*
4 64 56 annual RHI >1m elders [by%] 23** 15**
5 86 68 6 115 83 To enter new recruit state (>10cm height), seedlings require at minimum 7 141 101 1 year in tall tussock and 2 years in tall shrub abandoned grassland. 8 174 116 9 214 133 Growth rate data from mixed root sucker-seedling samples.
10 263 153 11 176 12 203 13 233 14 268
Myrsine parvula growth in pasture - projection basal area [mm²]
seedling
recruit
resprouting
adolescent Data from *root sucker study, **population monitoring: (1st year) 33 33.21 new seedling recruit basar 33*
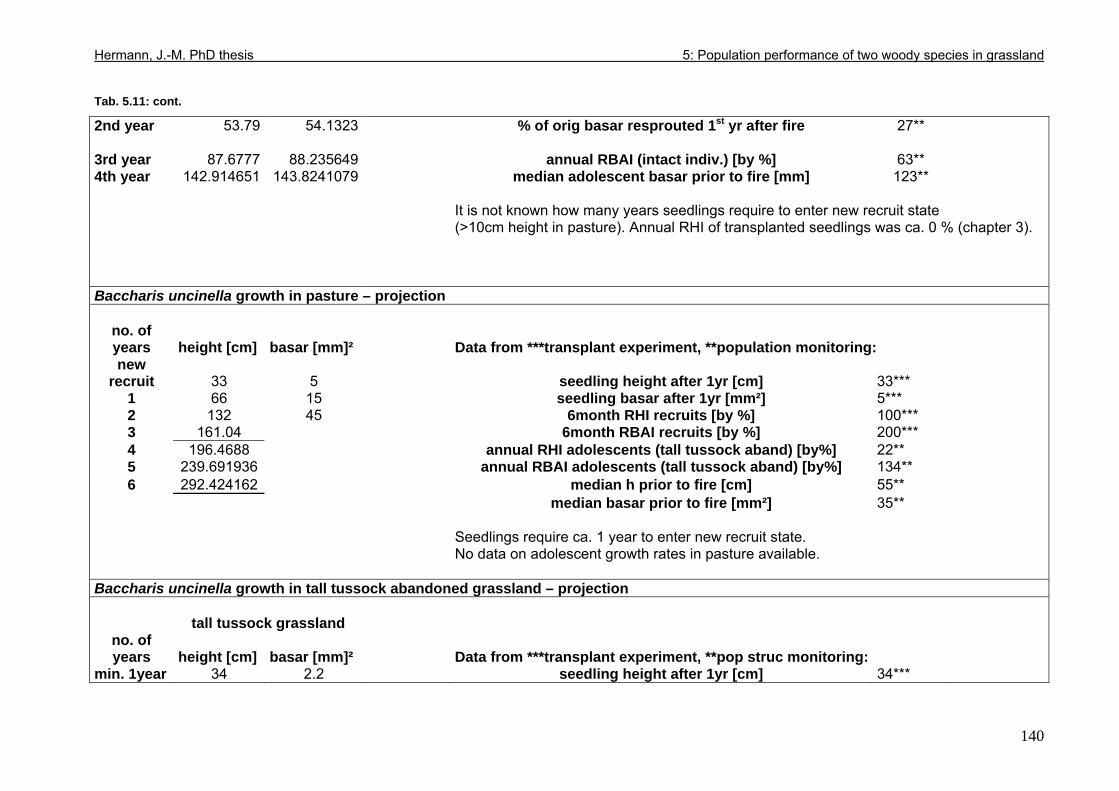
Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
140
Tab. 5.11: cont.
2nd year 53.79 54.1323 % of orig basar resprouted 1st
yr after fire 27** 3rd year 87.6777 88.235649 annual RBAI (intact indiv.) [by %] 63** 4th year 142.914651 143.8241079 median adolescent basar prior to fire [mm] 123** It is not known how many years seedlings require to enter new recruit state (>10cm height in pasture). Annual RHI of transplanted seedlings was ca. 0 % (chapter 3). Baccharis uncinella growth in pasture – projection
no. of
years height [cm] basar [mm]² Data from ***transplant experiment, **population monitoring: new
recruit 33 5 seedling height after 1yr [cm] 33*** 1 66 15 seedling basar after 1yr [mm²] 5*** 2 132 45 6month RHI recruits [by %] 100*** 3 161.04 6month RBAI recruits [by %] 200*** 4 196.4688 annual RHI adolescents (tall tussock aband) [by%] 22** 5 239.691936 annual RBAI adolescents (tall tussock aband) [by%] 134** 6 292.424162 median h prior to fire [cm] 55**
median basar prior to fire [mm²] 35** Seedlings require ca. 1 year to enter new recruit state. No data on adolescent growth rates in pasture available. Baccharis uncinella growth in tall tussock abandoned grassland – projection
tall tussock grassland no. of
years height [cm] basar [mm]² Data from ***transplant experiment, **pop struc monitoring: min. 1year 34 2.2 seedling height after 1yr [cm] 34***

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
141
Tab. 5.11: cont.
+6months 68 4.4 seedling basar after 1yr [mm²] 2.2*** +6months 136 8.8 6month RHI recruits [by %] 100***
17.6 6month RBAI recruits [by %] 100*** 3rd year 165.92 35.2 annual RHI adolescents [by%] 22**
70.4 annual RBAI adolescents [by%] 134** 4th year 202.4224 140.8 min basar adolescents [mm²] 300**
281.6 min basar adults (tall shrub aband) [mm²] 1250** 5th year 246.955328 563.2 6th year 301.2855 1317.888 Seedlings require ca. 1 year to enter new recruit state. 7th year 3083.85792

Hermann, J.-M. PhD thesis 5: Population performance of two woody species in grassland
142
Fig. 5.3: Above: Symptoms of severe infection on a branch of Baccharis uncinella (foreground) in a
declining tall shrub grassland stand (background): In the initial stages, leaves at the apices of branches
turn yellow and brown; in the final stages shown here, leaves are shed or appear dried out along the
entire length of branches, with branch apices hanging down. Photographer: J.-M. Hermann. Below:
Results of phytopathological analysis of an individual of Baccharis trimera exhibiting the same
symptoms in a Pró-Mata grassland, conducted by the UFRGS laboratory of phytopathology and
communicated by L. Prates January 4, 2007, via e-mail. The diagnosis is root rot caused by the oomycete
Pythium spec., which attacks a range of host species and thrives particularly well in humid and badly
drained soils at warm temperatures.
Análise: Exame direto em microscópios estereoscópio e óptico; inoculação em meio de cultura BDA (batat-dextrose-ágar); isolamento com iscas (discos de folhas de laranjeiras); comparação das estruturas encontradas com as ilustradas na bibliografia. Resultado: O resultado das análises do material descrito acima indicou tratar-se Podridão das raízes causada pelo oomiceto Pythium sp., estruturas características, oosporos e esporângios, foram detectados na amostra. Diagnosticado por: Biológa M. Sc. Marisa Dalbosco. Informações complementares: Esse patôgeno sobrevive no solo na forma de oosporos, apresenta elevado número de espécies vegetais hospedeiras e é favorecido por excessiva umidade, drenagem inadequada do solo e temperaturas altas. A doença é mais severa em solos úmidos, sendo comum em áreas com excesso de umidade. A disseminação ocorre principalmente pela água e por equipamentos que tranportam solo infetado.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
143
6. Post-disturbance Survival and Growth of Woody Species
in Grassland: A Comparison of Grassland and Forest
Species
6.1. Summary • The grasslands of Southern Brazil are relicts from the last postglacial and nowadays
are usually grazed and burnt. Native forests, favoured by present-day climate, are expanding over grasslands, although at a slow rate. Regular destruction of aboveground biomass is generally held responsible for low abundance of woody species, especially forest species, in grasslands.
• Many species of forest border shrubs and trees in the study region have been found to resprout after fire. However, better adaptation of grassland shrub species to high-frequency disturbance (at intervals of few years) can be expected. How do resprouting ability, size dependence and vigour differ between grassland shrubs and pioneer forest species in grassland?
• This study integrates data from two regions of Rio Grande do Sul, Brazil: Study area Morro Santana is located in the subtropical humid lowland and was burnt on New Year’s Eve 2004/2005. Study area Pró-Mata is located in the temperate humid highland and was cut and mowed at the beginning of January 2005. Per area, studied species included two grassland shrub species from the family Asteraceae and two forest tree species, one each of the families Myrtaceae and Myrsinaceae. Ca. 20 individuals, roughly divided between small and large size (<1m and >1m original height, respectively) were tagged and post-disturbance survival and growth were monitored for one year at 2-4 month intervals.
• Only one tall-growing grassland shrub species failed to resprout entirely. Of the other grassland shrub species (all low-growing) every individual resprouted, independent of area and size, as early as one month after disturbance, and by the end of the third month onset of flowering was observed. Of forest trees, all Myrtaceae individuals resprouted. Ca. 1/5 of highland Myrsine and 2/3 of lowland Myrsine failed to resprout.
• Independent of study area, size and treatment, resprout growth was more rapid and more vigorous in grassland shrub species than in forest tree species. Grassland shrub height growth was boosted during the first five months after resprouting and shoots were significantly closer to pre-disturbance maximum height in grassland shrubs than in forest trees in both highland and lowland after one year. One year after biomass loss, total basal area of grassland shrub resprouts was median 77% of pre-disturbance basal area, as opposed to median 8-11% in forest trees.
• Low grassland shrubs are of smaller stature than forest trees, but linear regression of original vs. resprouted basal area confirmed higher resprouting vigour of grassland shrubs relative to forest trees within the range in which sampled sizes overlap, particularly in small-size individuals. While a small grassland shrub individual would recover pre-disturbance basal area entirely within one year, a forest tree of the same original size would recover merely 12%. This trait, together with early onset of resprouting and flowering, is interpreted as a better adaptation of small grassland shrubs to high-frequency disturbances.
• Large individuals of highland forest tree species also resprouted from roots. Root suckers are transitional between individual recovery and recruitment of new individuals, and may allow population growth even under disturbance regimes that retard reproductive maturity of individuals. They were rarely observed in the lowland area.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
144
• The subtropical lowland environment, on the other hand, proved more favourable for growth. Lowland species approached pre-disturbance maximum shoot height approximately twice as fast as highland species.
• Successfully resprouting forest trees display two extreme types of resprouted shoot morphology and abundance (many slender vs. few stout shoots) that might assist in energy conservation in the microclimatic extremes of a post-fire environment. (More) successfully resprouting grassland shrubs tend to be intermediate between these extremes, i.e. share the best of both strategies.
• Present-day grassland management (fire at intervals of 1 or 2 years plus influence of cattle grazing/browsing and trampling) impose upon forest woody species higher taxes of biomass loss than they could have adapted to in their native ecosystem. If any, only grassland subshrubs and shrubs could survive under such continuous biomass loss. The occasional persistence of forest shrub and tree individuals in grassland must be attributed to high versatility of some species as well as to the patchiness of grassland fire.
6.2. Introduction In the High Plateau region of Rio Grande do Sul, precipitation and soil water
availability are considered particularly favourable for forest species growth; in accordance with Rambo’s conclusions, cattle grazing and fire are held largely responsible for present-day scarcity of forest trees and shrubs in these grasslands (Pillar & Quadros 1997). Indeed, grassland fires usually do not spread more than a few metres inside forest borders (pers. obs.).
There are exceptions to the rule: Fire destroyed e.g. 165.000 ha of high plateau native forest in the neighbour state Paraná in 1963 (Koch & Corrêa 2002). Moreover, resprouting ability is not an unusual trait in forest species. Many forest border shrub and tree species of both highland and lowland regions have been found to resprout after more or less complete scorch in pasture fires: Among them are members of the Myrtaceae family such as Siphoneugenia, Myrceugenia, Myrciaria and Myrcia, the genera Miconia and Tibouchina of the Melastomataceae family, various Myrsine species (Myrsinaceae), Sebastiana spec. and, last but not least, Araucaria angustifolia, the characteristic species of the Planalto forests (pers. obs.). Kammesheidt (1999) studied a variety of forests under different disturbance regimes in Paraguay and Venezuela and noted, for example, sprouting from aboveground plant parts in 58% of species in forest fallows (following slash-and-burn agriculture) and in 34% of species in logged stands.
Fire frequencies naturally differ between forest and grassland by at least one order of magnitude (see e.g. Peet 1992). However, some evidence has been given in the previous chapter that although customary fire frequencies (1-2 years in the course of pasture management) reduce populations of forest tree Myrsine parvula in grassland, these populations may persist or even expand if patches escape disturbance for as little as half a decade. Bond & van Wilgen (1996), on reviewing models of tree growth in African savannahs, found that in spite of limitations to seedling establishment, each model predicts eventual dominance of trees over grasses; they propose that this is so because adult savannah trees are fire-resistant and models usually ignore other causes of adult mortality. Therefore, in order to give a conclusive answer to the question why grassland-forest-succession is so slow, it is necessary to quantify not only growth and mortality of juveniles but also of elder size classes in grassland. The focus in this part study is on forest tree individuals of sizes that correspond to recruit and adolescent age classes in pop studies.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
145
Grassland vegetation in Rio Grande do Sul also consists, besides grasses and herbs, of numerous shrub and subshrub species e.g. of the genus Baccharis (Asteraceae) that never occur inside forest. They must either have co-evolved with their coexisting herbaceous species, or spread from ecosystems with similar disturbance regimes. This phenomenon is not unique to Southern Brazil but has only recently received increased attention. Prairies of the North American Great Plains are composed not only of C4 grasses but also of native C3 shrub species e.g. of the genus Cornus and Rhus that were considered „invasive“ by McCarron & Knapp (2001). These authors found, on comparison of ecophysiological traits of C3 shrubs and a dominant C4 grass, that the two components of Prairie were neither functional equivalents nor functionally distinct. More recently, such shrub species have been acknowledged as integral parts of the prairie ecosystem (Heisler et al. 2003).
It may be expected that Southern Brazilian grassland shrub species resprout more successfully – faster and more vigorously – than even the most successful pioneer forest species. Comparing disturbance responses of these species with those of forest trees may improve our understanding of success or failure of the latter in grassland colonisation.
How do survival and resprouting vigour differ quantitatively and qualitatively between grassland shrub and forest tree species established in grassland, and subject to total or almost total loss of aboveground biomass? How relevant, compared to resprouting, are complementary processes of individual or population survival, e.g. seedling and root sucker recruitment? How can diffential resprouting success of the two woody species groups be explained? As modifications of shoot allometry apparently enhanced Baccharis uncinella seedling performance in short-growing grassland (pasture; chapter 3), I hypothesize that morphology and clustering of resprouts account at least in part for differences for resprouting sucess.
In order to approach general applicability of the answers given, this study integrates data from two areas in Rio Grande do Sul subjected to two different kinds of disturbance, namely, clipping and burning. It is therefore necessary to extract carefully any irregularities possibly introduced by the experiment design prior to discussing differential performance of the two species groups in the context of studies in North American Prairie and, finally, in the context of Southern Brazilian grassland-forest-dynamics.
6.3. Study setup
6.3.1. Study areas and species
By coincidence, it was possible to integrate data from a clipping experiment in Araucaria High Plateau grassland (hereafter referred to as highland grassland) and from a burnt grassland site on one of the granitic hills in the municipality of Porto Alegre, Rio Grande do Sul (hereafter referred to as lowland grassland). For a general characterisation of the two study areas please see chapter 1.
In each site, post-disturbance surveys included two grassland shrub species and two forest tree species predominant in the transition zone from grassland to forest border.
Species studied in the highland site are: Baccharis uncinella D.C. (Asteraceae), a one- to few-stemmed grassland shrub that may attain around 4m height; Calea phyllolepis Baker (Asteraceae), a few-stemmed grassland shrub or subshrub growing largely within the grass matrix; Myrsine parvula (Mez) Otegui (Myrsinacea), a one- to few-stemmed forest tree and Siphoneugenia reitzii D.Legr. (Myrtaceae), a forest tree species occurring in both single-stemmed tree and multi-stemmed form in the grassland.
Species studied in the lowland site are: Baccharis ochracea Spreng. and Eupatorium ligulaefolium Hook. et Arn. (Asteraceae), two small-growing, one- to multi-stemmed

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
146
grassland shrubs; tree species Myrsine umbellata Mart. (Myrsinaceae) and Myrciaria cuspidata O.Berg (Myrtaceae) corresponding in growth form and shoot morphology to their highland counterparts. As Müller at al. (2005) report, multiple-stemmed woody individuals tend to predominate in recently burnt grassland patches (3 months to one year following fire) whereas the majority of individuals in late post-fire recovery sites (3–4 years following fire) are single-stemmed.
6.3.2. Sampling design and treatments
In both highland and lowland site, a minimum of 20 individuals per species, growing inside a grass-dominated matrix and outside the closed forest canopy, were randomly chosen. In order to assess a species’ or species group’s chances of survival in grassland under different disturbance regimes, it should be known how resprouting ability changes with size (and age); therefore samples were divided between “small” individuals (up to 1m tall and within maximum height of the grass matrix) and “large” individuals (above 1m tall). Besides maximum shoot height, number and basal diameter of basal shoots per individual were recorded.
In the highland area, root sucker production accounts for a major proportion of forest species’ establishment in grasslands (see root sucker chapter). This was taken into account by choosing “isolated individuals” (growing at a distance of at least twice their own height from equal-sized individuals of the same species) whenever possible, and additional sampling of shoot number and size within a circle of 1m radius around large principal individuals.
In the first week of January 2005, all highland individuals were cut down, leaving stumps of no more than a few centimetres height. The sampled surrounding circles were mown and litter was superficially removed. The intention was to imitate a fire of intermediate intensity, since grassland burning has been prohibited in recent years, but in essence the treatment was as in woodland coppicing.
In the highland area, individuals were first sampled prior to cutting whereas in the lowland area, surveys started only one month after a fire had swept the grasslands on New Year’s Eve 2004. All sampled individuals but one had been completely scorched. On account of the precarious location of this study area (field workers are escorted by armed guards), the field protocol had to allow for rapid sampling. Original basal diameters were measured only in small individuals and partly measured, partly estimated in larger ones. In sampling original stem height, errors may have been higher and sampling was less well balanced between small and large individuals than in the highland grassland. In order to sample a sufficient number of individuals (n=20) of the lowland Myrsine tree species, several individuals growing within the forest border had to be included.
Both study sites were surveyed 1, 3, 5, 9 and 12 months after cutting and fire. Individuals were monitored for resprouting success, vigour, onset of flowering, appearance of seedlings or root suckers16 in surrounding circles and evidence of herbivory or pathogens.
16 Resprouting after cutting and fire has been known for a long time in horticulture and forestry and consequently there is an abundance of synonymous or similar terms for the product, such as “epicormic shoot”, “sucker”, “water sprout”, “water shoot” and “sap shoot”. Some terms seem to be applied more specifically to certain plant parts (e.g. shoots on main branches: “water sprout”; shoots at or near the base of a plant: “basal shoot”; shoots from lateral, near-surface roots: “suckers”). In this study, the term “sucker” is used for new sprouts from roots, and the more general term “shoot” for new sprouts on any other plant part. In essence they all refer to a fast-growing shoot newly arising from adventitious (dormant) buds on any aerial or near-surface part of a plant, often after canopy removal or thinning, which destroys apical dominance and increases light levels.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
147
Parameters of resprouting vigour are: Resprouted number of shoots at the stem base and – in cut individuals only - on stem stumps, maximum shoot height and total basal area of shoots. Since measuring all shoots was considered too time-consuming, I restricted sampling to basal diameter of largest, smallest and a random selection (approximately one in five to ten shoots) of shoots per individual; resprouted basal area was then calculated from an average of these values. In Myrtaceae individuals, shoots were too numerous and crowded to reach all bases so diameters were partly estimated.
In mowed circles, at each survey, seedlings and root suckers were censused and an approximate value of total root sucker basal area was obtained as described before.
6.3.3. Evaluation
Although it is a fortunate coincidence that a „real“ burnt area should have become available at the same time that cutting was performed in the highland area, it has already become apparent from the description of the sampling design that pseudoreplication is considerable in this study. Highland and lowland sites differ not only in treatment but also in climate, soils and flora (chapter 1). Congruity is given only in the co-occurrence of two distinct groups of woody species – grassland shrubs and forest species – in grassland, and in loss of aboveground biomass.
Similarities and especially dissimilarities in observed responses between the two sites must therefore be interpreted with great caution, and in consideration of any available circumstantial evidence.
Besides considerable variation in size among sampled individuals, the study species differ considerably in architecture and shoot morphology, so in order to facilitate comparison and pooling, individual parameters of resprouting vigour were set in reference to values prior to disturbance. Resprouted shoot number, height and basal area per individual are expressed as per cent of pre-disturbance values and referred to as “relative shoot number”, “relative maximum height” and “relative basal area”.
Root sucker basal area is set in reference to summed-up basal area of potential feeder individuals. Besides the cut principal, I here considered as potential feeder individuals all neighbour individuals that possessed >10mm basal diameter (roughly equivalent to > 1m height), and were located at such proximity to the cut individual that root systems can be assumed to overlap with the mown circle (assuming that root system radius is more or less equivalent to individual height). After estimating what proportion of a neighbour’s root systems overlapped with the circle, an adequate proportion of its basal area was included in calculations.
In spite of efforts to balance the sampling, individual size distributions were skewed and an overlap of grassland shrub and forest tree samples over the entire range of sampled sizes could not be achieved. Therefore resprouting vigour per species and species group were characterised by the median of resprouted individuals and data were ln-transformed for comparative analyses.
Pair-wise comparisons between species by Tamhane’s T-Test were always performed first. However, in order to trace general patterns as far as possible, data of grassland shrub species and forest tree species were also pooled per site and pair-wise comparisons were made between these groups.
In order to determine the influence of size (age) on resprouting vigour within that range where sampled original sizes of grassland shrubs and forest trees overlap, data of the two species groups were submitted to a curve fitting procedure, using pre-disturbance individual basal area as independent variable and post-disturbance basal area as dependent variable. In this particular case, grassland shrub and forest tree data were pooled across

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
148
areas and treatments, resulting in individual numbers of around 50 per species group after outlier exclusion.
In spite of small sample sizes, curve fitting was also performed in order to obtain an estimate of how much individuals of a given size invest in root sucker production, using root sucker basal area as dependent variable and and feeder individual basal area as independent variable.
The linear regression presented here was calculated using Microsoft Excel (Windows XP). All other analyses, including the curve-fitting procedure employed prior to calculation of linear regression, were performed using SPSS 14.1.1.
Highland grassland shrub Baccharis uncinella failed to resprout throughout the experiment, and in lowland tree Myrsine umbellata only a minor proportion of individuals resprouted. Both species were omitted from statistical analyses.
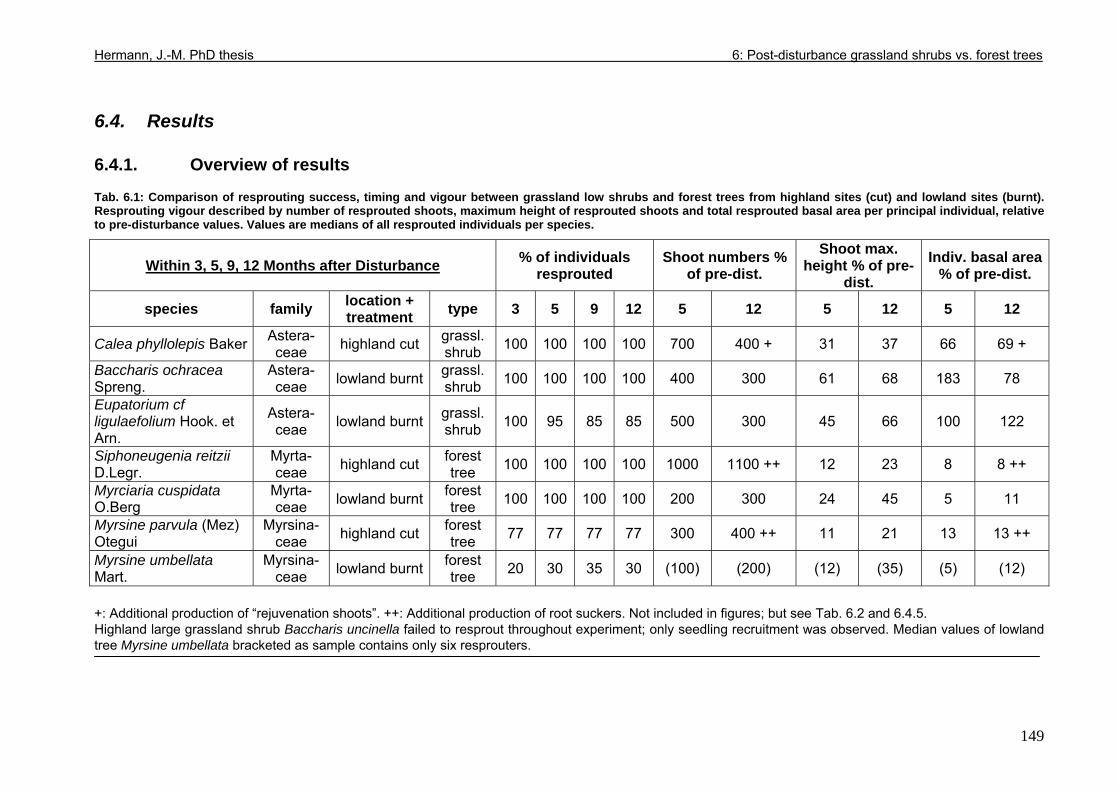
Hermann, J.-M. PhD thesis 6: Post-disturbance grassland shrubs vs. forest trees
149
6.4. Results
6.4.1. Overview of results
Tab. 6.1: Comparison of resprouting success, timing and vigour between grassland low shrubs and forest trees from highland sites (cut) and lowland sites (burnt).
Resprouting vigour described by number of resprouted shoots, maximum height of resprouted shoots and total resprouted basal area per principal individual, relative
to pre-disturbance values. Values are medians of all resprouted individuals per species.
Within 3, 5, 9, 12 Months after Disturbance
% of individuals
resprouted
Shoot numbers %
of pre-dist.
Shoot max.
height % of pre-
dist.
Indiv. basal area
% of pre-dist.
species family
location +
treatment
type 3 5 9 12 5 12 5 12 5 12
Calea phyllolepis Baker Astera-ceae highland cut grassl.
shrub 100 100 100 100 700 400 + 31 37 66 69 +
Baccharis ochracea Spreng.
Astera-ceae lowland burnt grassl.
shrub 100 100 100 100 400 300 61 68 183 78
Eupatorium cf ligulaefolium Hook. et Arn.
Astera-ceae lowland burnt grassl.
shrub 100 95 85 85 500 300 45 66 100 122
Siphoneugenia reitzii D.Legr.
Myrta-ceae highland cut forest
tree 100 100 100 100 1000 1100 ++ 12 23 8 8 ++
Myrciaria cuspidata O.Berg
Myrta-ceae lowland burnt forest
tree 100 100 100 100 200 300 24 45 5 11
Myrsine parvula (Mez) Otegui
Myrsina-ceae highland cut forest
tree 77 77 77 77 300 400 ++ 11 21 13 13 ++
Myrsine umbellata Mart.
Myrsina-ceae lowland burnt forest
tree 20 30 35 30 (100) (200) (12) (35) (5) (12)
+: Additional production of “rejuvenation shoots”. ++: Additional production of root suckers. Not included in figures; but see Tab. 6.2 and 6.4.5. Highland large grassland shrub Baccharis uncinella failed to resprout throughout experiment; only seedling recruitment was observed. Median values of lowland tree Myrsine umbellata bracketed as sample contains only six resprouters.
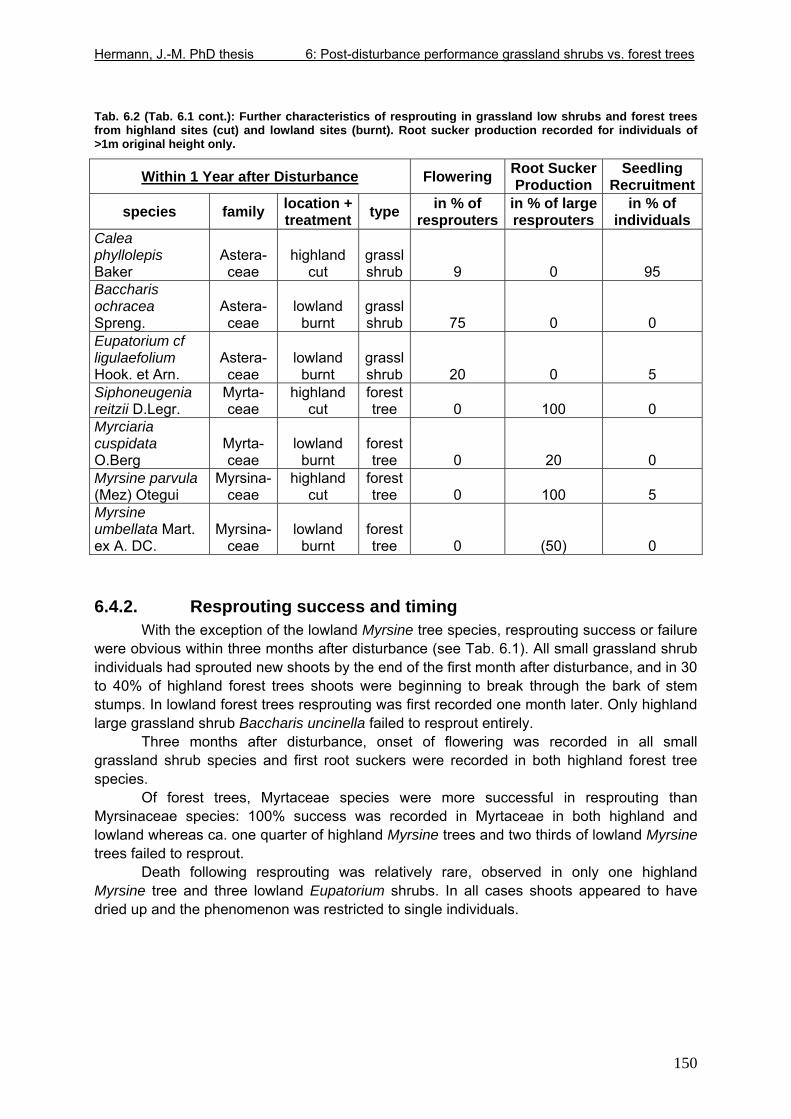
Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
150
Tab. 6.2 (Tab. 6.1 cont.): Further characteristics of resprouting in grassland low shrubs and forest trees
from highland sites (cut) and lowland sites (burnt). Root sucker production recorded for individuals of
>1m original height only.
Within 1 Year after Disturbance Flowering
Root Sucker
Production
Seedling
Recruitment
species family
location +
treatment
type
in % of
resprouters
in % of large
resprouters
in % of
individuals
Calea phyllolepis Baker
Astera-ceae
highland cut
grassl shrub 9 0 95
Baccharis ochracea Spreng.
Astera-ceae
lowland burnt
grassl shrub 75 0 0
Eupatorium cf ligulaefolium Hook. et Arn.
Astera-ceae
lowland burnt
grassl shrub 20 0 5
Siphoneugenia reitzii D.Legr.
Myrta-ceae
highland cut
forest tree 0 100 0
Myrciaria cuspidata O.Berg
Myrta-ceae
lowland burnt
forest tree 0 20 0
Myrsine parvula (Mez) Otegui
Myrsina-ceae
highland cut
forest tree 0 100 5
Myrsine umbellata Mart. ex A. DC.
Myrsina-ceae
lowland burnt
forest tree 0 (50) 0
6.4.2. Resprouting success and timing
With the exception of the lowland Myrsine tree species, resprouting success or failure were obvious within three months after disturbance (see Tab. 6.1). All small grassland shrub individuals had sprouted new shoots by the end of the first month after disturbance, and in 30 to 40% of highland forest trees shoots were beginning to break through the bark of stem stumps. In lowland forest trees resprouting was first recorded one month later. Only highland large grassland shrub Baccharis uncinella failed to resprout entirely.
Three months after disturbance, onset of flowering was recorded in all small grassland shrub species and first root suckers were recorded in both highland forest tree species.
Of forest trees, Myrtaceae species were more successful in resprouting than Myrsinaceae species: 100% success was recorded in Myrtaceae in both highland and lowland whereas ca. one quarter of highland Myrsine trees and two thirds of lowland Myrsine trees failed to resprout.
Death following resprouting was relatively rare, observed in only one highland Myrsine tree and three lowland Eupatorium shrubs. In all cases shoots appeared to have dried up and the phenomenon was restricted to single individuals.
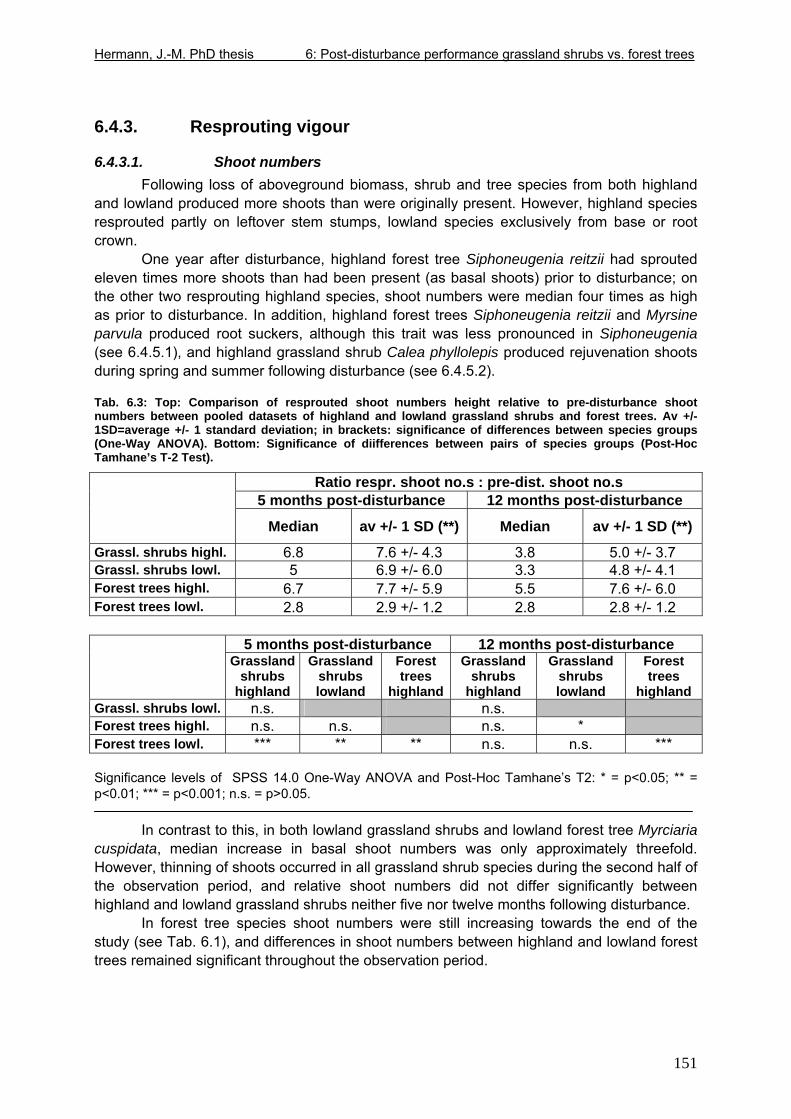
Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
151
6.4.3. Resprouting vigour
6.4.3.1. Shoot numbers Following loss of aboveground biomass, shrub and tree species from both highland
and lowland produced more shoots than were originally present. However, highland species resprouted partly on leftover stem stumps, lowland species exclusively from base or root crown.
One year after disturbance, highland forest tree Siphoneugenia reitzii had sprouted eleven times more shoots than had been present (as basal shoots) prior to disturbance; on the other two resprouting highland species, shoot numbers were median four times as high as prior to disturbance. In addition, highland forest trees Siphoneugenia reitzii and Myrsine parvula produced root suckers, although this trait was less pronounced in Siphoneugenia (see 6.4.5.1), and highland grassland shrub Calea phyllolepis produced rejuvenation shoots during spring and summer following disturbance (see 6.4.5.2).
Tab. 6.3: Top: Comparison of resprouted shoot numbers height relative to pre-disturbance shoot
numbers between pooled datasets of highland and lowland grassland shrubs and forest trees. Av +/-
1SD=average +/- 1 standard deviation; in brackets: significance of differences between species groups
(One-Way ANOVA). Bottom: Significance of diifferences between pairs of species groups (Post-Hoc
Tamhane’s T-2 Test).
Ratio respr. shoot no.s : pre-dist. shoot no.s
5 months post-disturbance 12 months post-disturbance Median av +/- 1 SD (**) Median av +/- 1 SD (**)
Grassl. shrubs highl. 6.8 7.6 +/- 4.3 3.8 5.0 +/- 3.7 Grassl. shrubs lowl. 5 6.9 +/- 6.0 3.3 4.8 +/- 4.1 Forest trees highl. 6.7 7.7 +/- 5.9 5.5 7.6 +/- 6.0 Forest trees lowl. 2.8 2.9 +/- 1.2 2.8 2.8 +/- 1.2
5 months post-disturbance 12 months post-disturbance
Grassland
shrubs
highland
Grassland
shrubs
lowland
Forest
trees
highland
Grassland
shrubs
highland
Grassland
shrubs
lowland
Forest
trees
highland
Grassl. shrubs lowl. n.s. n.s. Forest trees highl. n.s. n.s. n.s. * Forest trees lowl. *** ** ** n.s. n.s. *** Significance levels of SPSS 14.0 One-Way ANOVA and Post-Hoc Tamhane’s T2: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
In contrast to this, in both lowland grassland shrubs and lowland forest tree Myrciaria cuspidata, median increase in basal shoot numbers was only approximately threefold. However, thinning of shoots occurred in all grassland shrub species during the second half of the observation period, and relative shoot numbers did not differ significantly between highland and lowland grassland shrubs neither five nor twelve months following disturbance.
In forest tree species shoot numbers were still increasing towards the end of the study (see Tab. 6.1), and differences in shoot numbers between highland and lowland forest trees remained significant throughout the observation period.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
152
6.4.3.2. Maximum shoot height
Tab. 6.4: Top: Comparison of resprouted maximum shoot height relative to pre-disturbance maximum
shoot height between pooled datasets of highland and lowland grassland shrubs and forest trees. Av +/-
1SD=average +/- 1 standard deviation; in brackets: significance of differences between species groups
(One-Way ANOVA). Bottom: Significance of diifferences between pairs of species groups (Post-Hoc
Tamhane’s T-2 Test).
Shoot max. height % of pre-disturbance height
5 months post-disturbance 12 months post-disturbance
Median av +/- 1 SD (***) Median av +/- 1 SD (***)
Grassl. shrubs highl. 30.8 30.6 +/- 6.3 36.8 37.3 +/- 10.4 Grassl. shrubs lowl. 49.4 52.8 +/- 20.8 67 69.1 +/- 25.4 Forest trees highl. 10.6 10.8 +/- 5.1 22.1 21.5 +/- 8.4 Forest trees lowl. 24.3 24.9 +/- 7.9 45 46.3 +/- 11.6
5 months post-disturbance 12 months post-disturbance
Grassland
shrubs
highland
Grassland
shrubs
lowland
Grassland
shrubs
highland
Grassland
shrubs
lowland
Grassland
shrubs
highland
Grassland
shrubs
lowland
Grassl. shrubs lowl. *** ***
Forest trees highl. *** *** *** *** Forest trees lowl. * *** *** n.s. * *** Significance levels of SPSS 14.0 One-Way ANOVA and Post-Hoc Tamhane’s T2: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
In grassland shrub species, the major part of height growth occurred within five months after disturbance (Tab. 6.1). In both areas, grassland shrub species thus attained significantly taller relative height, i.e. approached pre-disturbance maximum height significantly faster, than forest trees.
Differences between highland and lowland environment were most pronounced in height growth of new shoots: In both lowland grassland shrubs and forest trees, relative maximum shoot height was significantly larger than in their highland counterparts five months as well as one year after disturbance.
It may be said that, approximately, lowland forest trees approached pre-disturbance maximum height twice as fast and lowland grassland shrubs nearly twice as fast as the same species groups did in the highland area.
Height increase was relatively low in the highland region especially during the winter months. This is demonstrated in Tab. 6.5 by comparison of seasonal shoot height increase among a subset of species. The subset contains highland and lowland shrub species Calea phyllolepis and Eupatorium ligulaefolium (both relatively little affected by herbivory) and highland and lowland tree species Siphoneugenia reitzii and Myrciaria cuspidata; these species pairs were chosen for similarity in morphology, and for possessing relatively intact resprouts.
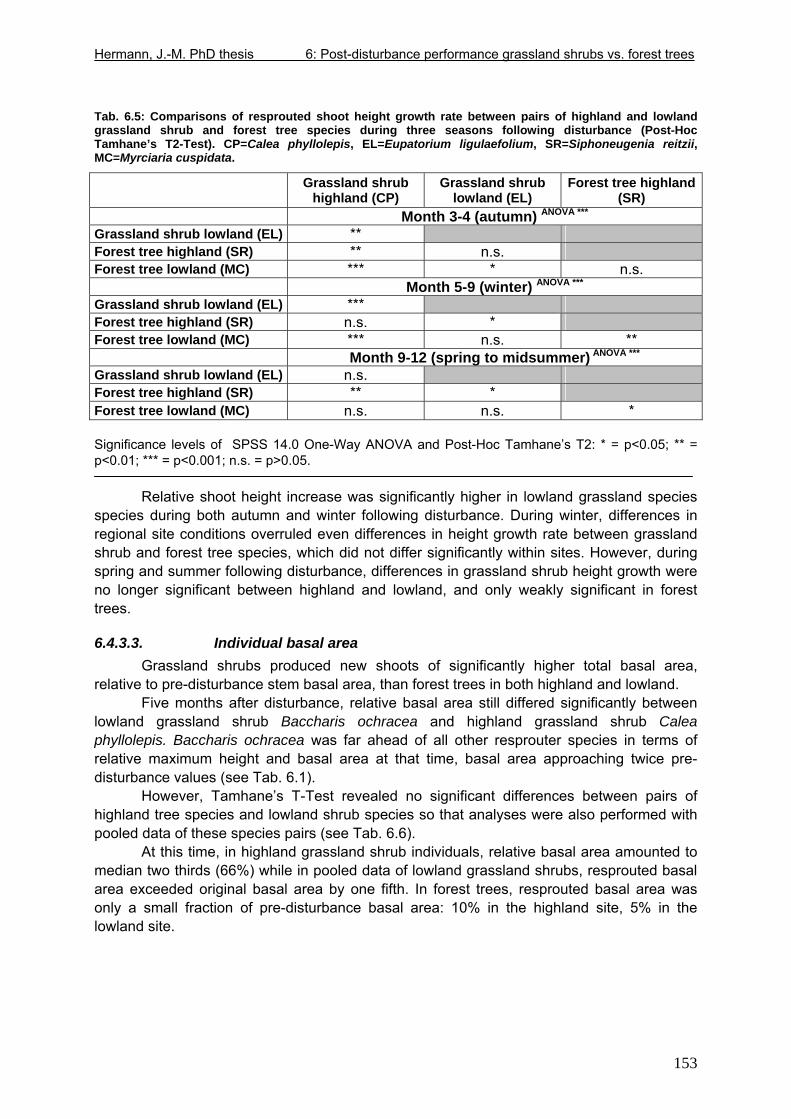
Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
153
Tab. 6.5: Comparisons of resprouted shoot height growth rate between pairs of highland and lowland
grassland shrub and forest tree species during three seasons following disturbance (Post-Hoc
Tamhane’s T2-Test). CP=Calea phyllolepis, EL=Eupatorium ligulaefolium, SR=Siphoneugenia reitzii, MC=Myrciaria cuspidata.
Grassland shrub
highland (CP)
Grassland shrub
lowland (EL)
Forest tree highland
(SR)
Month 3-4 (autumn) ANOVA ***
Grassland shrub lowland (EL) ** Forest tree highland (SR) ** n.s. Forest tree lowland (MC) *** * n.s. Month 5-9 (winter)
ANOVA ***
Grassland shrub lowland (EL) *** Forest tree highland (SR) n.s. * Forest tree lowland (MC) *** n.s. ** Month 9-12 (spring to midsummer)
ANOVA ***
Grassland shrub lowland (EL) n.s. Forest tree highland (SR) ** * Forest tree lowland (MC) n.s. n.s. * Significance levels of SPSS 14.0 One-Way ANOVA and Post-Hoc Tamhane’s T2: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
Relative shoot height increase was significantly higher in lowland grassland species species during both autumn and winter following disturbance. During winter, differences in regional site conditions overruled even differences in height growth rate between grassland shrub and forest tree species, which did not differ significantly within sites. However, during spring and summer following disturbance, differences in grassland shrub height growth were no longer significant between highland and lowland, and only weakly significant in forest trees.
6.4.3.3. Individual basal area Grassland shrubs produced new shoots of significantly higher total basal area,
relative to pre-disturbance stem basal area, than forest trees in both highland and lowland. Five months after disturbance, relative basal area still differed significantly between
lowland grassland shrub Baccharis ochracea and highland grassland shrub Calea phyllolepis. Baccharis ochracea was far ahead of all other resprouter species in terms of relative maximum height and basal area at that time, basal area approaching twice pre-disturbance values (see Tab. 6.1).
However, Tamhane’s T-Test revealed no significant differences between pairs of highland tree species and lowland shrub species so that analyses were also performed with pooled data of these species pairs (see Tab. 6.6).
At this time, in highland grassland shrub individuals, relative basal area amounted to median two thirds (66%) while in pooled data of lowland grassland shrubs, resprouted basal area exceeded original basal area by one fifth. In forest trees, resprouted basal area was only a small fraction of pre-disturbance basal area: 10% in the highland site, 5% in the lowland site.
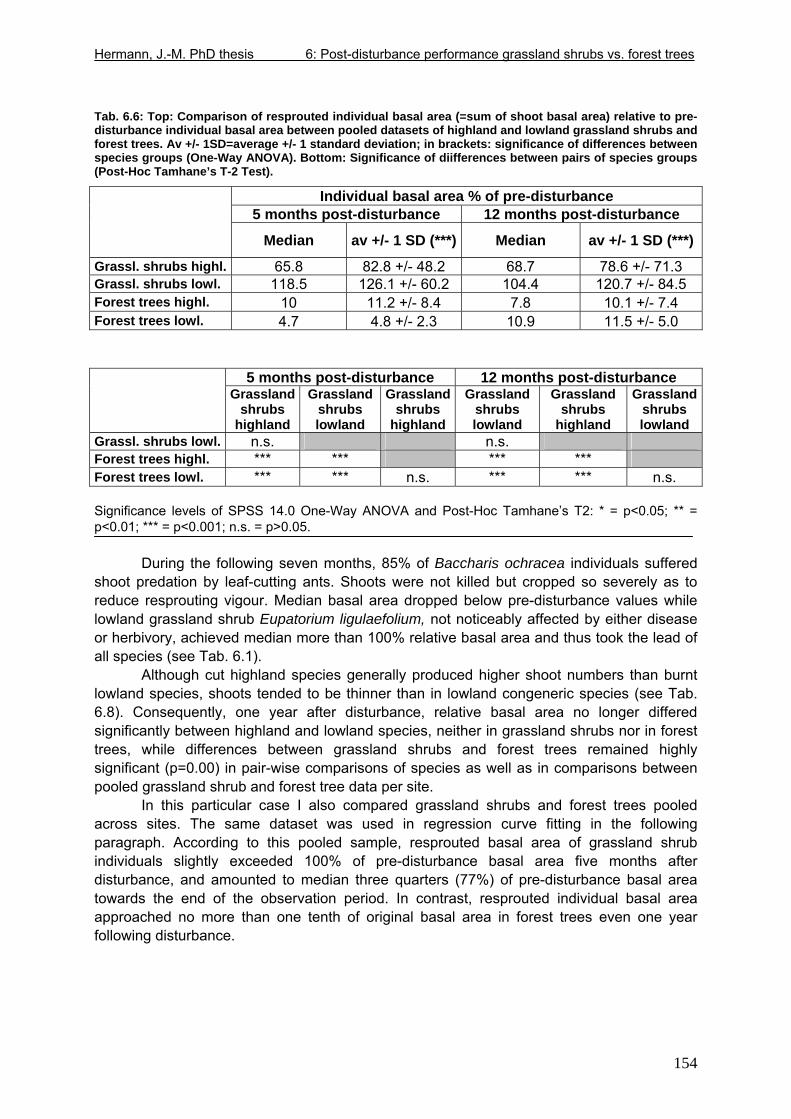
Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
154
Tab. 6.6: Top: Comparison of resprouted individual basal area (=sum of shoot basal area) relative to pre-
disturbance individual basal area between pooled datasets of highland and lowland grassland shrubs and
forest trees. Av +/- 1SD=average +/- 1 standard deviation; in brackets: significance of differences between
species groups (One-Way ANOVA). Bottom: Significance of diifferences between pairs of species groups
(Post-Hoc Tamhane’s T-2 Test).
Individual basal area % of pre-disturbance
5 months post-disturbance 12 months post-disturbance Median av +/- 1 SD (***) Median av +/- 1 SD (***)
Grassl. shrubs highl. 65.8 82.8 +/- 48.2 68.7 78.6 +/- 71.3 Grassl. shrubs lowl. 118.5 126.1 +/- 60.2 104.4 120.7 +/- 84.5 Forest trees highl. 10 11.2 +/- 8.4 7.8 10.1 +/- 7.4 Forest trees lowl. 4.7 4.8 +/- 2.3 10.9 11.5 +/- 5.0
5 months post-disturbance 12 months post-disturbance
Grassland
shrubs
highland
Grassland
shrubs
lowland
Grassland
shrubs
highland
Grassland
shrubs
lowland
Grassland
shrubs
highland
Grassland
shrubs
lowland
Grassl. shrubs lowl. n.s. n.s. Forest trees highl. *** *** *** *** Forest trees lowl. *** *** n.s. *** *** n.s. Significance levels of SPSS 14.0 One-Way ANOVA and Post-Hoc Tamhane’s T2: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
During the following seven months, 85% of Baccharis ochracea individuals suffered shoot predation by leaf-cutting ants. Shoots were not killed but cropped so severely as to reduce resprouting vigour. Median basal area dropped below pre-disturbance values while lowland grassland shrub Eupatorium ligulaefolium, not noticeably affected by either disease or herbivory, achieved median more than 100% relative basal area and thus took the lead of all species (see Tab. 6.1).
Although cut highland species generally produced higher shoot numbers than burnt lowland species, shoots tended to be thinner than in lowland congeneric species (see Tab. 6.8). Consequently, one year after disturbance, relative basal area no longer differed significantly between highland and lowland species, neither in grassland shrubs nor in forest trees, while differences between grassland shrubs and forest trees remained highly significant (p=0.00) in pair-wise comparisons of species as well as in comparisons between pooled grassland shrub and forest tree data per site.
In this particular case I also compared grassland shrubs and forest trees pooled across sites. The same dataset was used in regression curve fitting in the following paragraph. According to this pooled sample, resprouted basal area of grassland shrub individuals slightly exceeded 100% of pre-disturbance basal area five months after disturbance, and amounted to median three quarters (77%) of pre-disturbance basal area towards the end of the observation period. In contrast, resprouted individual basal area approached no more than one tenth of original basal area in forest trees even one year following disturbance.
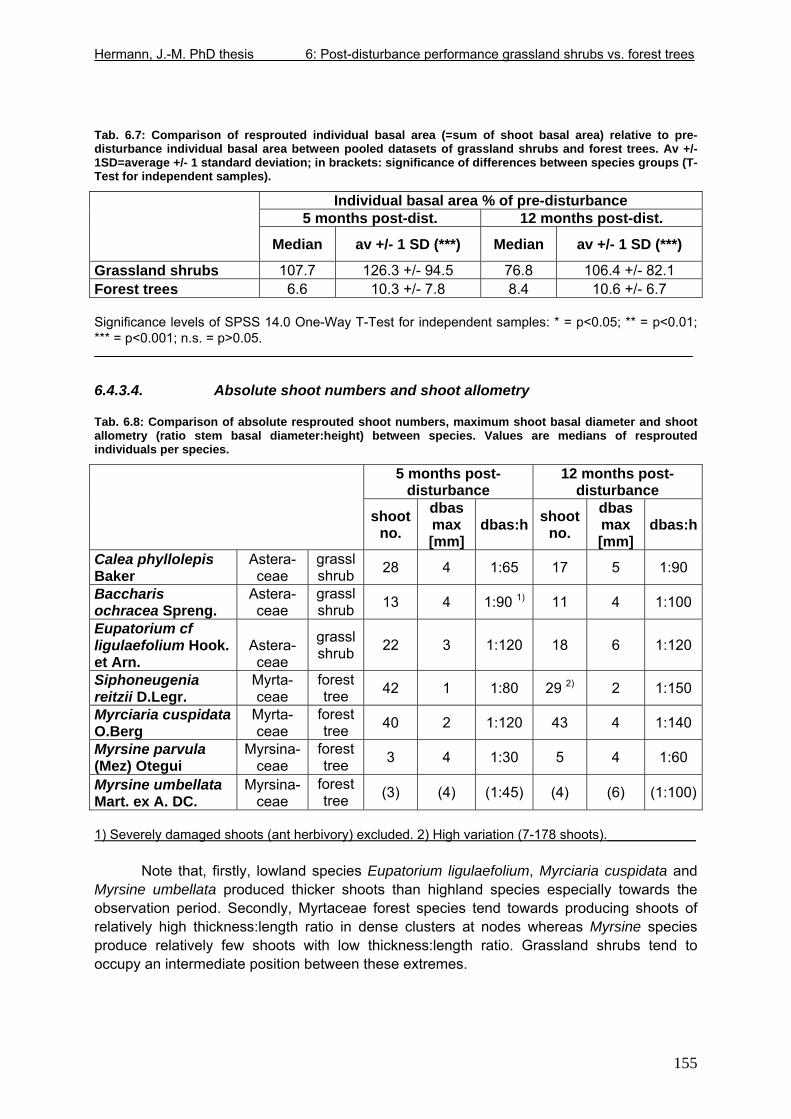
Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
155
Tab. 6.7: Comparison of resprouted individual basal area (=sum of shoot basal area) relative to pre-
disturbance individual basal area between pooled datasets of grassland shrubs and forest trees. Av +/-
1SD=average +/- 1 standard deviation; in brackets: significance of differences between species groups (T-
Test for independent samples).
Individual basal area % of pre-disturbance
5 months post-dist. 12 months post-dist. Median av +/- 1 SD (***) Median av +/- 1 SD (***)
Grassland shrubs 107.7 126.3 +/- 94.5 76.8 106.4 +/- 82.1 Forest trees 6.6 10.3 +/- 7.8 8.4 10.6 +/- 6.7 Significance levels of SPSS 14.0 One-Way T-Test for independent samples: * = p<0.05; ** = p<0.01; *** = p<0.001; n.s. = p>0.05.
6.4.3.4. Absolute shoot numbers and shoot allometry
Tab. 6.8: Comparison of absolute resprouted shoot numbers, maximum shoot basal diameter and shoot
allometry (ratio stem basal diameter:height) between species. Values are medians of resprouted
individuals per species.
5 months post-
disturbance
12 months post-
disturbance
shoot
no.
dbas
max
[mm]
dbas:h
shoot
no.
dbas
max
[mm]
dbas:h
Calea phyllolepis Baker
Astera-ceae
grassl shrub 28 4 1:65 17 5 1:90
Baccharis ochracea Spreng.
Astera-ceae
grassl shrub 13 4 1:90 1) 11 4 1:100
Eupatorium cf ligulaefolium Hook.
et Arn. Astera-ceae
grassl shrub 22 3 1:120 18 6 1:120
Siphoneugenia reitzii D.Legr.
Myrta-ceae
forest tree 42 1 1:80 29 2) 2 1:150
Myrciaria cuspidata O.Berg
Myrta-ceae
forest tree 40 2 1:120 43 4 1:140
Myrsine parvula (Mez) Otegui
Myrsina-ceae
forest tree 3 4 1:30 5 4 1:60
Myrsine umbellata Mart. ex A. DC.
Myrsina-ceae
forest tree (3) (4) (1:45) (4) (6) (1:100)
1) Severely damaged shoots (ant herbivory) excluded. 2) High variation (7-178 shoots)._______________
Note that, firstly, lowland species Eupatorium ligulaefolium, Myrciaria cuspidata and
Myrsine umbellata produced thicker shoots than highland species especially towards the observation period. Secondly, Myrtaceae forest species tend towards producing shoots of relatively high thickness:length ratio in dense clusters at nodes whereas Myrsine species produce relatively few shoots with low thickness:length ratio. Grassland shrubs tend to occupy an intermediate position between these extremes.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
156
6.4.4. Size dependence of resprouting success and vigour
In both highland and lowland grassland shrubs as well Myrtaceae forest trees, resprouting success was size-independent within the sampled range of sizes, as every individual resprouted successfully. In Myrsine forest trees, results are less clear: Non-resprouted highland Myrsine individuals are all of small size. Non-resprouted lowland Myrsine individuals, however, are evenly divided between small and large size class.
Resprouting vigour depends clearly on size, as revealed by regression curve fitting. For regression, total shoot basal area one year after disturbance was chosen as the dependent variable and original stem basal area as the independent variable, since in relative basal area differences between grassland shrubs and forest trees were not overruled by differences between highland and lowland sites (see 6.4.3.3).
For both species groups, the linear function was highly significant (p=0.00, SPSS), although curve fit was clearly better in forest trees than in grassland shrubs (grassland shrub linear function r²=0.41, forest tree linear function r²=0.77). Slightly better curve fit was achieved for grassland shrubs by a power function (r²=0.45); this signifies that grassland shrubs of small basal area resprout slightly less vigorously than the linear function suggests.
The regression shows that, firstly, grassland shrubs produce resprouts of significantly higher total basal area than forest trees. This already became apparent by comparison of sample medians (see 6.4.3.3), but is confirmed here specifically for that range in which sampled individual sizes of the two species groups overlap. Secondly, resprouting vigour is higher in small individuals than in large individuals. This trend is even more pronounced in grassland shrubs than in forest trees, even if the power function is considered instead of the linear function.
Tab. 3 contains some values of relative basal area to be expected in grassland shrubs and forest trees of a given basal diameter one year after disturbance, based on linear and power regression formulas. This is to give an idea of how much resprouting vigour differs between the two species groups differ and decreases with size. In a resprouting grassland shrub of 55 mm² original basal area (ln 4), one year after disturbance, relative basal area amounts to 90% to 100% according to power regression and linear regression, respectively. A grassland shrub individual of 403 mm² original basal area (ln 6) achieves merely 45% to 47% relative basal area according to power regression and linear regression, respectively. In forest tree individuals of same original sizes, calculated relative basal area amounts to 12% in the smaller and 9% in the larger individual. I.e. in grassland shrubs and forest tree individuals of small size – growing within average height of the grassland matrix – resprouted basal area differs by approximately one order of magnitude. In individuals of tall size – growing outside the grassland matrix – there is a tendency towards decreased resprouting vigour, and this is more pronounced in grassland shrubs than in forest trees of the same size.
Similar trends were observed and linear regression remained significant for both grassland shrubs and forest trees when lowland and highland site were evaluated separately.
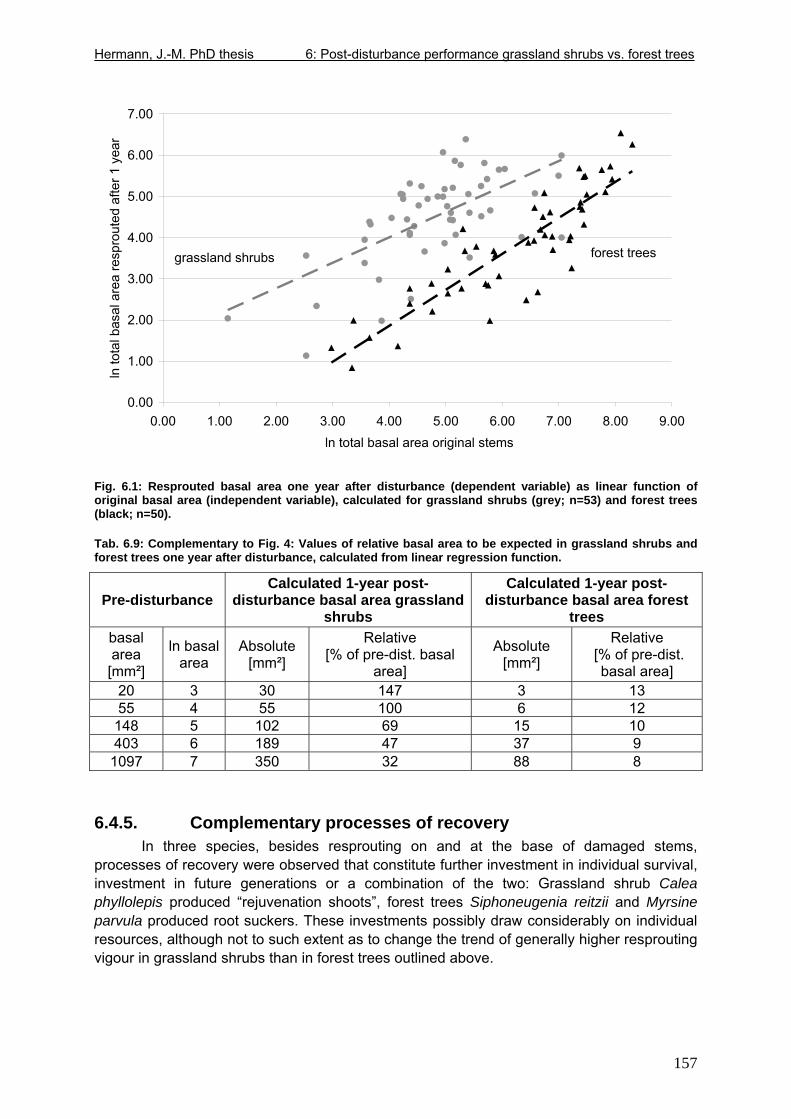
Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
157
grassland shrubs forest trees
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
0.00 1.00 2.00 3.00 4.00 5.00 6.00 7.00 8.00 9.00 ln total basal area original stems
ln to
tal b
asal
are
a re
spro
uted
afte
r 1 y
ear
Fig. 6.1: Resprouted basal area one year after disturbance (dependent variable) as linear function of
original basal area (independent variable), calculated for grassland shrubs (grey; n=53) and forest trees
(black; n=50).
Tab. 6.9: Complementary to Fig. 4: Values of relative basal area to be expected in grassland shrubs and
forest trees one year after disturbance, calculated from linear regression function.
Pre-disturbance Calculated 1-year post-
disturbance basal area grassland
shrubs
Calculated 1-year post-
disturbance basal area forest
trees basal area [mm²]
ln basal area
Absolute [mm²]
Relative [% of pre-dist. basal
area]
Absolute [mm²]
Relative [% of pre-dist.
basal area] 20 3 30 147 3 13 55 4 55 100 6 12
148 5 102 69 15 10 403 6 189 47 37 9
1097 7 350 32 88 8
6.4.5. Complementary processes of recovery
In three species, besides resprouting on and at the base of damaged stems, processes of recovery were observed that constitute further investment in individual survival, investment in future generations or a combination of the two: Grassland shrub Calea phyllolepis produced “rejuvenation shoots”, forest trees Siphoneugenia reitzii and Myrsine parvula produced root suckers. These investments possibly draw considerably on individual resources, although not to such extent as to change the trend of generally higher resprouting vigour in grassland shrubs than in forest trees outlined above.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
158
6.4.5.1. Root sucker recruitment Root sucker production was observed in all tree species, although in the lowland area
the proportion of root resprouters was too low to be statistically relevant (see Tab. 6.2) and average numbers of root suckers per potential feeder individual were negligible, i.e. even lower than in post-burn pasture in the highland region (see chapter 4). Contrary to the highland area where root suckers account for a major proportion of recruitment in abandoned grassland, pre-disturbance root sucker recruitment is also very rare in tree populations in grasslands of the lowland site, Morro Santana, (S.Müller, pers. comm.) although one of the two Myrsine species occurring on Morro S. – M. guianensis – is known to abundantly recruit root suckers following fire in the Brazilian Cerrado (Hoffmann 1998).
Knowledge of correlations between root sucker production and feeder individual size would greatly improve interpretation of population dynamics and reactions to disturbance. However, other than resprouts on stem stumps and at stem bases, root suckers develop an adventive root system; this root system initially supplements resources supplied by the principal individual and eventually make root suckers physiologically independent from the feeder individual. They represent a transition between recovery of the principal individual and recruitment of new individuals into the population (see also chapter 4). It was not possible to trace links between sampled root suckers and feeder individuals by excavation, so the following data represent merely a coarse quantitative estimate of root sucker production.
Pre-disturbance total basal area of root suckers correlated best with summed-up basal area of all potential feeder individuals (i.e. including those individuals that were not located inside the sampled circle, but whose root systems overlapped with this circle), although for Siphoneugenia, the correlation was not significant at p=0.05 level (Siphoneugenia: p=0.08; Myrsine p=0.01). Post-disturbance basal area of root suckers (one year after disturbance) correlated best with summed-up pre-disturbance basal area of the cut principal and all potential feeder individuals within the mown circle (Siphoneugenia: p=0.05; Myrsine p=0.01). This suggests that loss of apical dominance by cutting and mowing, and perhaps increased light influx at ground level, were powerful triggers of root sucker production.
In both cases, across the entire sampled range of sizes, Siphoneugenia seems to invest relatively little in root sucker production. Post-disturbance total basal area of root suckers amounted to 2-3% of original total basal area of feeder individuals. In Myrsine, relative root sucker basal area was ca. 10 times higher than in Siphoneugenia. It thus accounts for, and at the same time compensates, reduced stem resprouting in larger individuals (6.4.4). My data are too scanty to calculate exact quantities of root sucker production per cut principal individual, but it is not unreasonable to assume that root sucker production doubled relative resprouted basal area of the larger individuals, i.e. that a total of 25 to 30% of original basal area was recovered within one year by both stem and root sprouting. In burnt pasture on a neighbour ranch, root sucker production was very low but resprouting Myrsine recovered median 27% of pre-fire basal area within 10 months (chapter 5).
Entire Myrsine populations in grasslands of the study area have higher individual numbers and density than populations of Siphoneugenia or indeed any other forest species of the highland study area, to which root sucker recruitment largely contributes (see also chapter 3). However, while it is a predominant strategy in population growth in areas undisturbed for ca. one decade, root suckers appear to be second in line after stem resprouts in the hierarchy of resprouting, as they emerged later than stem resprouts.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
159
6.4.5.2. Rejuvenation shoots „Rejuvenation shoots“, in this study, were unique to small grassland shrub Calea
phyllolepis. These shoots emerge exclusively from the rootstock and consist initially of green, densely pubescent stems with leaves reduced almost to size of scales. Normal-sized leaves, arranged in whorls of three per node as in resprouted shoots, appear at the tip only after a few weeks. Both types of shoots eventually develop decussate leaf insertion in their adult and senescent stages. Rejuvenation shoots are frequently observed in naturally established populations in abandoned grasslands of the study area, i.e. serve prolongation of individual life spans even in absence of disturbance.
First rejuvenation shoots were observed five months after disturbance. Nine months after disturbance they occurred in 73% of individuals and the median ratio of total basal area of resprouted shoots versus total basal area of rejuvenation shoots was ca. 3:1. No data are available from the survey one year after disturbance. On the assumption that this ratio did not change, basal area of resprouted shoots (see Tab. 6.1) and rejuvenation shoots would have summed up to median 90% of pre-disturbance basal area by that time.
6.4.5.3. Seedling recruitment Highland tall grassland shrub Baccharis uncinella, the only non-resprouter, recruited
seedlings in surrounding circles in 35% of individuals, with maximum seedling density 5/m² five months after disturbance. Seedling recruitment was considerably higher in highland small grassland shrub Calea phyllolepis, with seedlings in surrounding circles of 90% of the individuals and maximum seedling density 25/m² five months after disturbance. Early emergence of first seedlings (recorded three months after disturbance) suggests that in Calea, seedlings recruited from a seed bank rather than from post-disturbance flowering with which it coincided temporally.
With only two exceptions, none of the highland forest trees and neither grassland nor forest lowland species recruited from seed in surrounding circles throughout the observation period.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
160
6.5. Discussion Due to the lack of true replication in my study design, and the difficulties in
interpretation arising from this condition, I have divided the following discussion into two parts: In the first, I will try to explore why certain trends differ between my two study sites. The second part is devoted to discussion of similar trends across lowland and highland sites, focussing on the contrast between grassland shrubs and forest trees, and on possible causes for differential performance of these two groups of woody species.
6.5.1. Dissimilarities between study areas
6.5.1.1. Resprouted shoot numbers Several authors have pointed out that comparisons of clipping and fire treatments are
problematic since resprouted shoot numbers tend to be higher in clipped than in burnt individuals (e.g. Vesk & Westoby 2004a, Lloret et al. 1999, Lloret & López-Soria 1993). Lloret & López-Soria (1993) found that mortality and biomass of resprouts did not differ significantly between clipping treatments and fires of low to medium heat, but significant differences appeared in comparison to high temperature fire. Since the fire on Morro Santana was judged to be exceptionally intense (M. Ebner, pers. comm.), results must be cautiously explored for differences probably introduced by treatment. Such differences are most conspicuous in highland and lowland resprout numbers.
As a rule, priority is on resprouting from the topmost portion of any surviving aboveground parts, and resprouting is restricted to the plant base only if trees and stems have been completely scorched (e.g. Bellingham & Sparrow 2000, Cirne & Scarano 2001). Obviously, resprouting from stem or crown conveys an advantage in canopy recovery and competition for light. Storage of nutrients in the stem, as documented for Australian Myrtaceae by Paté et al. (1990), would facilitate stem resprouting but remains to be verified in Myrtaceae of my study area.
Another reason why clipped individuals tend to sprout more shoots than plants scorched to the base is that the number of available buds increases with size and surface area of leftover stem stumps (see e.g. Shackleton 2001). It is therefore not surprising that in my study, higher numbers of resprouted shoots were recorded in cut highland species than in burnt lowland species.
Another consequence that suggests itself only at second thought is the similarity in resprouted basal area of highland and lowland species. Taking advantage of these similarities, data were pooled across areas and served for a general comparison of basal area recovery in grassland shrubs and forest trees.
Shackleton (2001) examined the effect of height of coppice cuts on number of resprouted shoots and on cumulative resprouted shoot length per stump. His results show that cumulative shoot length depended more on original individual size than stump size; in other words, an individual of a given size left with a small or tall stump would produce few long or many small shoots, respectively. However, these results are for a single African savannah tree (Terminalia sericea). In my case, given the significant between-site differences in maximum resprouted shoot height, the possibility can not be ruled out that if lowland species had retained part of their aboveground stems (as highland species did) they would have resprouted higher numbers of shoots, and as they produce thicker shoots, total resprouted basal area might then have been significantly higher in lowland than in highland species.
Nutrient status following clipping and burning must also be taken into account. Causes of increased productivity following disturbance were investigated by clipping and

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
161
burning e.g. in the Nylsvley savannah (see Scholes & Walker 1993 and references therein): It appears that about 2/3 of increased productivity following fire may be attributed to necromass removal (which accounts for high pre-disturbance light competition) and about 1/3 are due to a nutrient flush caused by increased remineralisation rates. In my study, cut highland individuals were largely deprived of this nutrient flush, even more so as litter was removed in order to expose soil. On the other hand, these individuals had grown undisturbed – and thus been allowed to store photosynthetates to greater extent – for a considerably longer period than individuals in the lowland area.
6.5.1.2. Resprouted shoot height Highland and lowland site differ considerably in climate, inherent soil properties (chapter 1) and, as mentioned in the previous paragraph, post-disturbance soil nutrient status. Among the investigated parameters of resprouting vigour, relative resprout height is the one in which these differences in abiotic factors seem to have the most pronounced effect, as demonstrated by winter and summer growth rates.
In calculating height growth, some bias in favour of lowland species may have been introduced. Firstly, original height was measured in live individuals in the highland area and in standing dead in the lowland area, so that original height of the latter may have been underestimated by around 10% in my estimation. Highland species also originate a few centimetres above ground on stem stumps, although this should not provide for an additional gain of more than 1-3% of original height. Although these factors certainly reduce statistical significance of differences in height growth between the two sites, these differences remain considerable.
More favourable growth conditions in the lowland area are also indicated, indirectly, by recovered basal area: This parameter did not differ significantly between highland and lowland species - although presumably less buds were available in lowland species, highland species had a head start of ca. one month in resprouting and, moreover, highland species had grown undisturbed for at least 10 years prior to the experiment, whereas lowland grassland is subjected to fire at intervals of as little as two years.
Resprouting of Myrsinaceae, however, was more favoured in the highland area. Lowland resprouters recovered more of their original height but highland individuals recovered more of their basal area – partly because of resprouting along stem stumps, but also because of additional root sucker production.
Differences between forest trees and grassland shrubs were so pronounced as to overrule differences in site conditions (see below).
6.5.1.3. Resprout failure In Myrsine parvula, the ability to resprout has been observed even in the seedling
stage, although damaged seedlings require a benign environment, e.g. moderate microclimate and absence of further disturbance, in order to gain more biomass than was lost (see chapter 3). However, a considerable proportion of highland individuals and most of lowland individuals failed to resprout, contrary to Myrtaceae individuals all of which resprouted successfully. Individual numbers were too small for statistical evaluation, but as a trend, mortality occurred independent of size in burnt individuals; according to Lloret & Lopez-Soria (1993) this is typical of high-intensity fires.
It is possible that bark and buds of the genus Myrsine are more heat-sensitive than in Myrtaceae. This is corroborated by observations of cracking and drying bark on highland Myrsine stem stumps during the February 2005 drought; however, contrary to scorched individuals, most of these were still able to resprout from the root crown. Drought damage was not observed in Myrtaceae individuals.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
162
Death following resprouting was a relatively rare and localised event and apparently caused by physiological drought rather than real drought (as, after all, individuals had established successfully in those locations and resprouted successfully in spite of the late summer drought). Pathogens introduced by injuries are one possible cause; so is excess fertilization by urine of territorial canines (stray dogs and campo fox, Dusycyon and Cerdocyon spec.) which are fairly frequent in both study sites. Anthills were frequently erected on newly bared ground and enclosed one Myrsine tree and one lowland Eupatorium shrub; these individuals were omitted from monitoring.
6.5.1.4. Seedling recruitment Seedling recruitment in the surroundings of sampled individuals in the highland area
was higher in resprouter Calea phyllolepis than in non-resprouter Baccharis uncinella, and in the lowland area was practically zero. Overbeck et al. (2005b) noted an increase in species diversity following fire on Morro Santana – species diversity in their experimental plots was highest ca. one year after burning and decreased in the following years – and they suggested that this was due to a surge in recruitment opportunities while competition by resprouting caespitose grasses was still low. The possibility cannot be ruled out that conditions for seedling emergence were atypically unfavourable during the year of my study – not only was the New Year’s Eve fire very intense, but post-disturbance environmental stresses were certainly aggravated by the late summer drought 2005. However, a more elaborate recent study by A.Fidelis and collaborators confirms resprouting as the predominant mode of regeneration in the lowland area: Post-disturbance seedling emergence is low whereas most species possess underground storage organs from which they can resprout, such as rhizomes in grasses and xylopodia in shrubs, which leads the authors to conclude that the bud bank is more important than the seed bank for post-fire regeneration (Fidelis et al. in prep a,b).
Morgan (2001) followed seedling establishment of herbaceous and grass species over three years in Australian grasslands with different fire histories and stated that seedlings of natives were absent or rare although adults were abundant in the standing flora. Carrington & Keeley (1999), on comparison of post-fire seedling recruitment between mediterranean climate and subtropical scrub communities, stated that seedling recruitment tends to be higher in mediterranean climate sites, irrespective of nutrient status; climate therefore seems to play a major role than soil characteristics. Exact reasons for low seedling recruitment in the Southern Brazilian lowland are only partly known so far; Overbeck et al. (2005a) found that other than e.g. in fire-prone Mediterranean ecosystems, exposure to fire did not stimulate germination in any of 9 species tested in their study, although seeds of several herbaceous and grass species of the area retained the ability to germinate even after exposure to temperatures above 100°C.
It is surprising that highland tall shrub Baccharis uncinella, as the only non-resprouter in the study, should not recruit more successfully. There was no indication of dormancy of Baccharis uncinella seeds in the seedling emergence experiment. Baccharis uncinella and the two lowland shrub species Baccharis ochracea and Eupatorium ligulaefolium produce seeds with pappuses that may be wind-dispersed over relatively long distances, and perhaps they also share lack of dormancy and restriction of recruitment to particularly favourable conditions occurring occasionally in time and space. However, in lowland grassland shrubs the apparent scarcity of seedling recruitment safe sites is offset to some degree by a prolonged lifespan of genets, as most individuals of a population resprout successfully, and some of them even flower and seed within few months afterwards. Possible consequences of developmental constraints on Baccharis uncinella populations have been discussed in chapter 4.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
163
Calea phyllolepis seeds bear a crown of small teeth but no pappus. Possibly, there is some animal dispersal – with teeth allowing seeds to clinging to fur and plumage -, but obviously seeds are also shed in the immediate neighbourhood of seed-bearing adults. Timing of Calea seedling emergence prior to the peak of seed set in surrounding populations – observed in this experiment as well as in areas cleared for the seedling emergence experiment – suggests the ability to remain viable in soil for at least three quarters of a year. 1000-seed-weight of Calea phyllolepis is ca. 10 times as high as that of Baccharis (pers. obs.), conferring greater robustness to seedlings. If the species is able to survive not only cutting but also fire – which seems likely, as excavations of individuals revealed swollen rootstocks presumably serving for assimilate storage – populations have a dual advantage following fire, as most individuals resprout and new seedlings are recruited at the same time. Considering this ability, it is all the more curios that the species is confined to the abandoned grasslands of Pró-Mata; it was never observed outside the Station area.
6.5.1.5. Root sucker and rejuvenation shoot production Root sucker recruitment was generally higher in the highland cutting experiment and
in naturally established populations in abandoned grassland than in highland pasture and in lowland populations. What may induce individuals of a species to spread horizontally by sprouting from roots in addition to sprouting at the stem base?
In chapter 4, the discussion focussed on why a species should produce root suckers at all. Here, the discussion is about why an individual should sprout at shorter or longer distances from the stem. Once more, it becomes apparent why a clear distinction between root suckers and coppice shoots (located at the stem base or root crown) may be useful. There is some conceptual overlap between the question raised here and the question of costs and benefits of producing long or short spacers in rhizomes and stolons (discussed e.g. by Lovett Doust 1981, Harper 1985).
Spreading laterally instead of vertically may confer an advantage in the form of risk-spreading (Cook 1985), but in frequently burnt grassland, this does not seem to be the case. Ramets are just as likely to be burnt as mother plants, and more likely to succumb to fire damage, unless alternating burnt and unburnt patches are smaller than the root system of a vegetatively reproductive individual, and patches may escape burning for several years – so that root suckers may grow both physiologically independent from the feeder individual, and large enough to survive the fire.
There is also no obvious advantage in energy-conservation. On the contrary: My results show a tendency towards densely clustered growth especially in relatively long and thin shoots and I suggested that this confers an advantage under exposure to high irradiation and temperature extremes. Scarce root sucker recruitment on Morro Santana may therefore be a consequence of frequent disturbances in the present and past, although this suggestion is dissatisfactory considering high root sucker recruitment in the Cerrado, where fires are similarly frequent. Hoffmann (1998) predicts maximum root sucker recruitment in Myrsine guianensis – the species also occurring on Morro Santana – at 2-3 year fire return intervals.
If individuals spread laterally in spite of a high risk of losing shoots, there may be a premium in the form of resource acquisition. I will briefly discuss this possibility for light, nutrients and water. In the following, the discussion overlaps with that of chapter 4, in which it became apparent that one principal question - whether sprouting from roots is more a survival mechanism or more a foraging mechanisms - is not yet resolved.
Light may be a limiting resource for individuals inside the forest border, and may induce them to sprout from root systems extending into grassland. Inside burnt grassland, however, light competition appears relatively relaxed. There was no indication, even one year after burning and mowing, of resprouting solitary trees in grassland being overtopped by

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
164
neighbour grasses. Thus, for the majority of individuals sampled in this experiment, root suckering does not seem to offer a premium in the form of light acquisition.
Transient high nutrient availability may be expected following fire (e.g. McCarron & Knapp 2003, Scholes & Walker 1993 and references therein, Bond & van Wilgen 1996 and references therein). Exploiting this nutrient flush is certainly of particular importance in nutrient-poor grassland soils. Do root suckers allow the genet to exploit resources more rapidly? Obviously, this depends on whether or not suckers develop an adventive root system so soon that the investment in their initial growth is compensated by resources taken up through the adventive roots, and by assimilates produced. If this is not the case and the investment in root sucker production pays off only in the following years or decades, they are perhaps best seen as a bonus product - produced only if high nutrient supply is available to the genet without their contribution, or if the principal individual has remained undamaged.
This is indicated by three observations: Firstly, various individuals resprouted only from the base or stem stumps, but none resprouted exclusively from roots. Secondly, there was a time lag between stem resprouting and root resprouting. Thirdly, root sucker recruitment was relatively high in abandoned highland grassland undisturbed for ca. eleven years prior to the experiment, relative to adjacent pasture that experienced burning (see chapter 5). Fire should have provided for a nutrient flush in pasture; on the other hand, during the relatively long undisturbed period, individuals in the neighbour area may have stored greater amounts of photosynthetates.
Anderson (1982) reported several unpublished results of burns conducted by the University of Wisconsin in prairie of the Great Lakes region. Their control burns, like fire ad cutting in my experiment, were conducted during summer and probably had a more severe impact on woody species than a dormant season burn. Grassland herbaceous and shrub species resprouted as early as 1.5-2 weeks after burning; other woody resprouted with a greater time lag. Aspen root suckers, however, did not reappear even in the following growing season. This seems concomitant with my hypothesis. On the other hand, Hoffmann’s (1998) models of relative importance of vegetative and generative reproduction in Brazilian Cerrado shrub and tree species under different fire frequencies, predict that in two of three root-suckering species vegetative reproduction should be highest at shortest fire return intervals of 1-3 years.
Water shortage is more likely in soils of the lowland region than in the highland region (chapter 1), and this may indirectly account for low root sucker recruitment: In the coarse-grained soils of Morro Santana, water acquisition is probably improved by deep root systems rather than the horizontally spreading, near-surface roots from which suckers typically emerge (DeByle 1964, Tiffney & Niklas 1985). Excavations of some root systems may be sufficient to determine whether laterally spreading roots are at all present or not.
However, discussions in the previous chapters have shown that it is helpful also to consider conditions of the original forest habitat. It cannot be ruled out that the tendency to produce few root suckers is inherited from conditions encountered there.
The rejuvenation shoots observed in highland grassland shrub Calea phyllolepis differed morphologically from basal resprouts and were also observed in uncut individuals. It is interesting that the species thus possesses the ability to react to biomass loss by resprouting from basal buds (traumatic reiteration) and to perpetuate reproductive individuals in absence of disturbance (programmed reiteration). Keeley (1992) describes the same behaviour in a group of Californian chaparral shrubs that survive fire by regeneration from the root crown, and in the absence of fire continuously regenerate their canopy with basal sprouts. This phenomenon is apparently similarly rare in the lowland site, relative to the highland site, as root suckering. This seems to make lowland shrub species even more critically dependent on seedling recruitment, and the absence of such recruitment all the more curious.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
165
6.5.2. Grassland shrub vs. forest tree resprouting
Across a range of abiotic conditions, treatments and sample sizes, grassland shrubs resprout faster and more vigorously than forest species. In addition to this, they possess the ability to flower and produce seed within few months after disturbance. In one species (Calea phyllolepis), seedling emergence (presumably from a seed bank) was observed at the same time as resprouting, and within one year after disturbance the species produced sucker-like basal shoots that are a regular mode of perennation in absence of disturbance. Grassland shrubs may thus largely proceed in the annual growth cycle as if no interruption in growth had occurred.
Rapid post-disturbance flowering and seedling emergence in addition to resprouting does not match the resprouter-seeder dichotomy often postulated (see e.g. Vesk & Westoby 2004a and references therein). Bellingham & Sparrow (2001) propose a linear trade-off between resprouting and seeding in woody species in fire-prone ecosystems. More recently, however, Vesk & Westoby (2004a) reviewed the literature and pointed out that the dichotomy is only pronounced in cases of (nearly) complete biomass removal, and that in most cases – as in this study - a continuum between sprouting and non-sprouting behaviour can be observed.
There was a tendency towards decreasing resprouting ability with size and age in both forest trees and grassland shrubs. This tendency was more pronounced in grassland shrubs, especially if taking additional root suckering of large forest trees into account (which was not included in linear regression); Cirne & Scarano (2001) found that in the tree species Andira legalis, vigour of resprouting by coppice shoots and root suckers (following fire in coastal sand dunes) was proportional to plant size. However, many grassland shrub individuals were already reproductive whereas forest trees were not, so that this trend does not necessarily reflect decreased vigour of growth but simply a trade-off between vegetative growth and generative reproduction (see e.g. Hodgkinson 1998, Pate et al. 1990 on Australian tree species).
Typically, studies in savannahs and grasslands invaded by trees focus on the antagonism between grasses and trees, trying to link their relative occurrence to soil texture, soil-water relations and plant-water relations (see reviews by Belsky 1991, Wilson 1998). However, the low shrubs investigated in my study are an integral part of grassland ecosystem and at first glance, differences between these shrubs and forest trees appear as pronounced as if grasses and trees were compared.
This phenomenon is not unique to Southern Brazil but has only recently received increased attention. As mentioned in chapter 1, some profound compositional and structural similarities exist between the South American „Pampa“ biome and the North American „prairies“ – not surprisingly, as migrations between the two landmasses occurred during the Pleistocene (Webb 1978). Of particular interest is a comparison between my study sites and the prairies of the Great Plains and Great Lakes region (east of the Rocky Mountains). Grasslands of both areas feature a mixture of C3 and C4 grasses (winter and summer aestivation, respectively) (Mack & Thompson 1982). Great Plains grasslands share predominant grass genera such as Andropogon and Sorghastrum (Heisler et al. 2003, McCarron & Knapp 2001) with highland grassland in my study, and they contain native C3 shrub species e.g. of the genus Cornus and Rhus that were considered „invasive“ e.g. by McCarron & Knapp (2001) but more recently have been acknowledged as integral parts of the prairie ecosystem (Heisler et al. 2003).
In experimental burns in Great Lakes region prairie, shrub species resprouted as early as 1.5-2 weeks after burning (Anderson 1982). Heisler et al. (2003) investigated performance of four species of native shrubs under different fire frequencies in Kansas prairie (Great Plains) and found that these shrubs persisted even under annual burning, and

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
166
expanded under burning at four year (intermediate) intervals. Such observations lead McCarron & Knapp (2001) to ask whether grasses and shrubs are „functionally distinct“, and they compared ecophysiological traits of a dominant C4 grass (Andropogon gerardii) in Kansas prairie with traits of three C3 shrub species. They detected better water use efficiency in Andropogon than in the shrub species, but in each of the other traits investigated (e.g. photosynthetic rate, xylem pressure potential), Andropogon was matched by either one of more of the shrub species. Moreover, relative physiological superiority of Andropogon was reduced in unburnt grassland – this phenomenon seems mirrored in low shrub expansion (especially of Calea phyllolepis) in abandoned Pró-Mata grasslands.
Summing up, dominant C3 shrubs were neither functionally distinct nor functional equivalents to the dominant C4 grass in their study area. Similarly, my above statement - that differences between grassland shrubs and forest trees mirror differences between grasses and forest trees – will, on examination, probably prove partly incorrect and partly correct. Of course, if the statement is ever tested it should be supplemented with measurements of ecophysiology of resprouting forest trees. For the time being, I can only judge on resource use efficiency in a broad sense from data on resprouting vigour and morphology. It appears that grassland shrubs achieve faster recovery at least in part by better energy conservation in a post-burn (or post-mowing) environment. This conclusion follows, apparently paradoxically, from the observation that numbers of resprouted shoots exceed those of original (pre-burn) basal shoots.
Irrespective of treatment differences, grassland and forest woody species shared the trait of resprouting several times more shoots than were originally present. This phenomenon has been reported from fire-prone ecosystems worldwide (e.g. Lloret & López-Soria 1993, Cirne & Scarano 2001, McCarron & Knapp 2003) but is relatively little discussed. It appears that species generally give priority to recovery of shoot basal area over recovery of shoot height (see also Bellingham & Sparrow 2000). This can be viewed as economical resource use for several reasons: Firstly, transport costs between roots and new foliage are reduced; secondly, light competition in a post-burn environment is relatively relaxed. In my study, one year after disturbance, even resprouting forest species seemed in little danger of being overtopped by neighbour grasses and shrubs.
However, in grassland species in my study, losses in shoot basal area were in fact temporarily overcompensated. Shoots were visibly crowded in Myrtaceae forest trees and grassland shrubs; in the latter, self-thinning was confirmed by a decline in resprouted shoot numbers within eleven months after resprouting in this study. A general decrease in shoot numbers of resprouters with increasing time span since last fire emerged from a previous study by Müller et al. (2005). So the question is raised if and when it might be more economical for a resprouter to invest in fewer shoots from the start17.
Clustered growth as such confers several advantages: In pasture, it may protect inner shoots from damage as the tussock growth of grasses does (e.g. Walter 1984). In areas subjected to recurrent fire, inner shoots may be protected from burning (see e.g. Bond & van Wilgen 1996, Walter 1984). Rutherford (1981, in Scholes & Walker 1993) monitored woody plant survival following so-called “slow” and “fast” burns in savannah of the South African
17 The possibility that species do display some wasteful behaviour cannot be ruled out. Harper (1978, p. 6) wrote: „In fact, selfish and physiologically inefficient behaviour may often pay in increased fitness“. He supplied an example of an annual grass that might benefit from wasteful resource use „provided that it completed its life cycle before it exhausted its needed resources“, and would put his perennial competitors at disadvantage into the bargain. It is implicit in his choice that he does not expect such behaviour in perennials that remain in the same site for a longer period, although clonal species may move about to some extent. In any case, wasteful behaviour seems similarly difficult to prove as randomness in community assembly (chapter 5), as the investigator must ascertain not to have overlooked any underlying logical patterns.

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
167
Nylsvley. In slow burns, the heat is concentrated closer to the ground than in fast burns, and this tended to increase mortality of woody plants less than 25 cm tall, with the exception of multi-stemmed shrubs in which Rutherford found zero mortality.
As a third possible advantage, clustered growth helps in energy conservation. Mack & Thompson (1982) propose this as a principal cause for tussock grass predominance in the North American Intermountain grasslands under arid and winter-cold conditions. Similarly, post-burn (or post-mowing) environments are characterised by microclimatic extremes such as high irradiation and temperature amplitudes, as may be inferred from microclimate comparisons between grassland types (chapter 1; see also Bond & van Wilgen 1996).
On viewing absolute numbers of produced resprouts side by side with shoot allometry (see Tab. 6.8), it appears that Myrtaceae forest species tend towards producing shoots of relatively high thickness:length ratio in dense clusters at nodes. The opposite extreme is displayed by Myrsine, who produces relatively few shoots with low thickness:length ratio.
It has been shown in chapters 3 and 4 that stout shoot growth is more pronounced in open areas than under cover of grasses or shrubs, and I hypothesized in those chapters that energy conservation is aided by a reduction in surface:volume ratio. In grassland shrubs as a group, there is a tendency towards combining the benefits of both strategies, i.e. towards production of clusters of relative stout shoots. Lowland shrub Eupatorium ligulaefolium most closely approaches shoot arrangement and morphology of such a “super-resprouter”. Post-burn size gain was highest in this species; however, the species was also unattractive to leaf-cutting ants, which heavily predated resprouted shoots of co-occurring Baccharis ochracea shrubs.
Fig. 6.2: Contrasts in resprouted shoot number and allometry: Forest tree species Myrsine parvula
(Myrsinaceae; left) with few stout shoots and Siphoneugenia reitzii (Myrtaceae; right) with many slender
shoots. Center: Grassland shrub species Baccharis ochracea (Asteraceae), intermediate in resprout
number and allometry. Photographer: J.-M. Hermann.
An important question remaining to be answered is for how long woody species populations may persist under repeated biomass loss in grasslands. Coppicing is an ancient craft in Europe and is currently being promoted as sustainable method of woodland management; supporters of this practice claim that individual life spans of coppiced trees may exceed those of undamaged specimens by centuries (see e.g. Living Countryside Ltd. 2003). However, it stands to reason that recovery of coppiced individuals depends on sufficient length of disturbance-free intervals plus continuous resource supply or efficient nutrient recycling (or both). Occasionally authors refer to “senescence” of rootstocks but this topic has remained rather vague. DeByle (1964) discovered, on excavation of an Aspen clone root system, that rootstocks underneath dead stems and stumps were maintained alive

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
168
and functioning by neighbour ramets. Consequently, it should be possible for new basal shoots to maintain rootstocks alive at least at the periphery – provided that they are not limited by one or more of the factors named.
Paté et al. (1990) found, in a wide range of fire-resprouter species, high starch concentrations and swollen organs belowground where they are protected from aboveground disturbances. Such storage organs were discovered in individuals of highland grassland shrub Calea phyllolepis excavated by me and, more recently, a detailed study has revealed storage organs such as xylopodia in grassland shrubs of the lowland study area (Fidelis et al. in prep. b). This is another trait that enables these species to recover rapidly from aboveground biomass loss, and persist in lowland grassland at fire return intervals of 2-3 years although Overbeck (2004) concludes that only fire frequencies of more than three years favour establishment of shrubs over grasses and herbaceous species.
Could South Brazilian grassland shrubs also persist at increased fire intervals? Heisler et al. (2003) found that annual fires in Great Plains Prairie “effectively prevented the recruitment of new woody species, but even with this high fire frequency, shrub cover increased slightly” (p.423). It is not impossible that some grassland shrubs in my lowland study area might achieve the same: Although Eupatorium ligulaefolium individuals had recovered only approximately two thirds of original maximum height one year after burning, resprouted basal area exceeded original basal area by ca. 20%.
In highland pastures, grassland shrub individuals - with the exception of Baccharis trimera, a subshrub - are as scarce as forest tree individuals. Since some seed input at least of wind-dispersed species should occur, shrub species apparently do not withstand the combined pressure of ranging cattle and fires at intervals of one to two years. Calea phyllolepis individuals returned to the annual cycle of flowering and rejuvenation within one year but recovered neither original height nor basal area, even including basal area of rejuvenation shoots.
It is not likely that forest tree species could withstand fire return intervals of one to two years even in lowland where recovery is faster than in highland. Structure and composition of naturally established forest species populations in a relatively remote highland pasture suggested that the area had not been burnt for at minimum 3-5 years prior to a 2005 fire, and that the same time span would be required after fire for Myrsine populations to recover individual numbers alone (see chapter 5). Perala (1979) found that Aspen clones could not withstand coppicing at rotations of 4 years or less. If it were possible to perform dendrochronology on roots of shrub and tree individuals, this could provide information on both the age of individuals, and disturbance return intervals they experienced in the past (J. Morales, pers. comm.)
Summing up, there remain several interesting fields of research that may help to better understand the observed patterns: For example, a comparison of ecophysiology of grassland shrubs vs. forest trees following the example of McCarron & Knapp (2003); an investigation of whether tree species store substantial proportions of nutrients in roots or, as in Australian Myrtaceae species (Pate et al. (1990), in the stem, which would put them at a clear disadvantage under recurrent loss of aboveground biomass; lastly, type and concentration of buds, as proposed by Vesk & Westoby (2004b). Possibly, grassland shrubs invest in a type of bud or meristem that can be activated more rapidly after disturbance than those of forest trees, and also concentrate these buds and meristems close to or below ground level, well protected from severe disturbance and in close proximity to stored assimilates.
It must be assumed that if forest species manage to colonise grasslands at all this is, for one thing, due to high versatility of some species. For another, forest trees clearly benefit from the patchiness of grassland fires as well as from shelter of river valleys and rocky outcrops (pers. obs.). Unless the fire is exceptionally intense or spreading of flames is

Hermann, J.-M. PhD thesis 6: Post-disturbance performance grassland shrubs vs. forest trees
169
supervised and directed by man, its impact varies with fuel load, climate, time of day, proximity of forest borders etc., so that some patches may escape burning for several years. If this were not the case, even the most flexible and fast-growing of forest species could not master individual survival or even population growth in grassland.

Hermann, J.-M. PhD thesis Final discussion
170
7. Final discussion
The objective of this chapter is to offer an incentive to future, more detailed research of patterns and mechanisms of woody species establishment, especially of forest trees, in Southern Brazilian grassland. In the first three paragraphs, some of my results and conclusions are embedded in a general framework of theories related to secondary succession that may help to direct future research, and interpret results (although this framework is not concise; a concise framework for successional studies in general was presented by Pickett et al. 1987). The fourth paragraph links to the discussion on the phenomenon of multiple successional pathways, and on possible origins of pioneer species, that was begun in chapter 5. I collect some evidence that composition of pioneer communities in grassland-forest-succession in Southern Brazil differs fundamentally from composition of pioneer communities in secondary successional series reported e.g. from Europe and North America, and I discuss some possible causes and consequences of these differences.
7.1. Seed arrival at the site All successional models make some implicit or explicit assumptions about the timing
and quantity of diaspores of colonisers arriving at a site (see e.g. Peet 1992) but few authors offer alternative hypotheses. One of these authors, Egler (1954), suggested that most of the species eventually dominant enter a community in the early stages of development (and that successional sequences, consequently, are merely an expression of differential longevity of these species).
This model of initial floristic composition does not seem to apply in my study area, although seed bank studies have not been performed and the conclusion is based on observations of forest species seedling emergence in population monitoring plots. Maximum species diversity of forest tree and shrub seedlings was noted in high shrub abandoned grassland, but clearly did not represent a full complement of forest species. The sample contained some of the most abundant Myrtaceae species at the forest border, one or two species of Melastomataceae and Ilex and – mostly – Myrsine parvula seedlings. Across the entire sampled grassland area of 1600 m², only two juveniles of Araucaria angustifolia occurred. Seed input of forest species is therefore better described by Egler’s alternative model, “relay floristics”, in which species arrive successively at the site, which is rendered suitable for their establishment by preceding colonisers. Animal behaviour is probably a crucial factor limiting seed dispersal from forest into grassland (see chapters 2 and 3).
Attention should also be payed to the bud bank from which grassland species and some forest species may recruit new shoots. Root suckering is excluded from Egler’s models as a „peripheral phenomenon“, but for the time being it is the principal mode of expansion of Myrsine individuals especially in tall tussock grassland where seed dispersal is apparently inhibited (see chapters 2 and 3).
Eriksson’s (1989) models of seedling recruitment, although developed as a supplement to clonal life histories, may provide a useful framework for classification of seedling emergence behaviour of forest species in forest, which in turn should relate to their chances of establishment in grassland. E.g. species in which seedlings emerge within few days or weeks after dispersal in forest (repeated seedling recruitment, or RSR) would be critically dependent on encountering beneficial, „forest-like“ conditions as soon as dispersed into grassland. Species that are not adapted to immediate beneficial conditions in forest, and are thus well dispersed in space and time (recruitment in windows of opportunity, or RWO), should establish in grassland with greater facility.

Hermann, J.-M. PhD thesis Final discussion
171
Once a vegetation type of several strata has established – such as tall shrub abandoned grassland in Pró-Mata –, disturbances that affect principally the canopy will benefit previously suppressed plants, but not necessarily provoke an increase in seedling emergence. Seedling establishment of later-successional species – in my case, forest species in grassland – will be noticeably favoured only if the lower strata of vegetation are disturbed.
This has been indicated by seedling numbers in tall shrub stands affected to greater or lesser extent by canopy dieback (chapter 5) as well as by Keeley’s (1992) study of long-time undisturbed chaparral and by an experimental study in early secondary successional Pine forests of the North Carolina Piedmont, in which Peet & Christensen (1980) experimentally created gaps in either understorey or canopy (further explained below). Berkowitz et. al. (1995) point out that where vegetation contains many gaps due to reduced soil resources in oldfield sites, seedling growth of forest species is also poor. But as a general trend they suggest that „disturbance of the canopy of low-growing species will reduce resistance to tree invasion in all but the extremely stressful environments“ (p.1167). Myrsine root suckers also recruited predominately in such gaps in the grass matrix (chapter 4).
Large-scale disturbances such as fire, however, that leave bare soil exposed for several months, are more likely to favour establishment of Baccharis uncinella and other grassland species whose seedlings perform comparatively well in such sites (chapters 2 and 3).
7.2. A final comment on facilitation and inhibition Once a seed is dispersed through the layer of established vegetation, the expected
effects of any type of established grass or shrub vegetation that is of larger initial height than a seedling may be summed up as follows:
1. Relative to a competition-free environment (such as in nurseries), seedling emergence will be reduced for both grassland and forest species. Relative to bare ground or short-growing vegetation, seedling emergence will be reduced for grassland species and increased for forest species. There is one exception: Smallest seeds (of only one to a few mm diametre) may be inhibited from emergence by a litter layer if it is thicker than the extension of their radicle. Several forest-border species (Miconia spec., Ilex spec., Psidium cattleyanum) have such small seeds.
2. Relative to a competition-free environment, seedling growth will be reduced for both grassland and forest species. Relative to bare ground or short-growing vegetation, seedling growth will be reduced for grassland species and increased for forest species.
3. Forest seedling survival will be equal or higher than in a competition-free environment while grass seedling survival will be equal or lower. There are two exceptions: In pasture, surrounding vegetation may increase the risk of being burnt (although sample size in the seedling transplant experiment was too small to demonstrate this). Under shrub cover, mammal herbivory may be high (DeSimone & Zedler; however, such an effect was not observed in my study).
4. The net effect of abandoned grassland should tend to be positive for forest species and negative for grassland species.
This summary is partly inferred from my own results, partly deduced from similar studies on inhibition and facilitation in secondary succession (see also chapter 3) – further investigations in order to confirm these trends would be desirable. However, unravelling seedling emergence and establishment conditions separately is an interesting but time-consuming occupation, and complexity is added – as the list indicates – by relative validity of

Hermann, J.-M. PhD thesis Final discussion
172
the terms inhibition and faciliation. A certain vegetation type cannot be declared inhibitory or facilitative in general, but only with reference to certain life stages of certain invading species.
In avoidance of such contradictions, Connell et al. (1987) proposed to focus on the description of „net effects“. As apparent from the oldfield study by Berkowitz et al. (1995), this approach leads back to the expression of plant-plant or plant-community relations in terms of „competition“. Since their transplant experiments failed to detect consistent community-specific trends (but old-field communities do resist to natural tree seedling establishment in „apparently different“ ways), these authors suggest that „duration of competition“ may be more helpful in explaining observed patterns of natural seedling establishment than intensity of competition at one given point in time; „duration of competition“ being expressed by the number of years until a seedling outgrows the competitive effects of surrounding vegetation. Moreover, according to Berkowitz et al. (ibid.), competition trades off against site conditions to some extent. It can be easily envisaged that a seedling establishing in short-growing vegetation – such as a pasture sward – may outgrow competing vegetation faster than a seedling establishing in tall shrubland, but as a drawback will be exposed to greater extent e.g. to trampling and microclimatic extremes.
In other words, while detailed examination of developmental stages of seedlings in different types of vegetation may be avoided by focussing on net effects, longer-term observations across various sites are still required in order to confirm relative favourability of a site. Interpretation will also be facilitated if there is agreement upon a set of growth “standards” – such as a species’ average lifespan, height growth, age and duration of generative reproduction at physiological or ecological optimum – that parameters of growth measured in a given site can be compared to. Holl’s (1999) approach – comparison of performance of tree seedlings invading pasture to performance of seedlings in that species’ original forest habitat – may be regarded as a step in that direction.
A number of population parameters are implicit in „duration of competition“: It depends not only on species-specific traits and the way these traits change through time but also on effects of established vegetation and the way these change through time. Further above, gap availability has been linked to changes in community structure in time and space that involved a decrease in competition. Is it possible to simplify monitoring studies by identifying general stages of community development in which competition is reduced?
7.3. Changes in community invasibility with time It has long been known that successional sequences are „composed of broadly
overlapping population curves through time“ (Pickett 1982, p.45). This concept is more straightforward in a study area such as mine in which relative few species dominate the communities, at least in abandoned areas. If developmental stages of individuals and populations as well as average duration of these stages can be clearly identified, then these stages may be classified according to their accessibility for seed-dispersed forest species.
An example for such an approach is given by Christensen & Peet (1984). These authors studied old-field communities in North Carolina, USA, with the objective of determining whether species composition converges monotonically towards a site-specific climax; in other words, whether compositional differences between sites become smaller with time since old-field abandonment, and whether species composition becomes more easily predictable on the basis of environmental parameters – represented by soil properties in their study.
They concluded that convergence does not necessarily increase in a simple, monotonic fashion; predictability of stand composition, based on soil pH, was best in stands of intermediate and oldest age. One possible explanation they offer is non-monotonic

Hermann, J.-M. PhD thesis Final discussion
173
variation in the intensity of competition: Competition is low during the establishment phase early in succession, increases during the thinning phase, and relaxes again during the transition phase as first gaps appear. The steady-state phase is once more characterised by intense competition but often precluded by disturbance (see also Pickett et al. 1987).
I will briefly explore which of these stages can be recognised in the current stage of grassland-forest succession in Pró-Mata, allthough Peet (1992) cautions that there is no reason to assume general applicability of Christensen’s and his results. Note also that one author – Klein (1960; see following paragraph) – reports, in succession from grassland to Araucaria forest, the eventual convergence of a broad set of pioneer communities to intermediate-late successional communities uniformly dominated by one or two crown species, which indicates the operation of similar processes.
„Establishment phase“: This phase follows upon a disturbance that temporarily removes most of aboveground vegetation. Such disturbance can occur by fire (and grazing, as in my study area), logging or clear-cutting or agricultural practices (as in Christensen & Peet’s study area). Most pastures of the study area are burned at 1-2 year intervals, or used to be burnt that frequently before the practice became illegal. In the relatively remote area of the fazenda where my population monitoring plots were located, intervals are at least twice longer than customary, as inferred from population structure (chapter 5). Fire and cattle were largely excluded from Pró-Mata in 1994; one area adjacent to the fazenda burnt accidentally in 1999, five years after abandonment. In the meantime, a firebreak has been established in order to rule out such accidents.
In the establishment phase, competition is relatively relaxed and correlation with soil parameters may be weak if many species attempt establishment (as proposed by Christensen & Peet). Whether forest species can exploit this stage depends upon a variety of factors: seed dispersal, seed availability and degree of exposure of the seed, i.e. gap size. Seed availability is related to developmental stage and density of individuals in the adjacent forest border, and this, in turn, depends on whether timber is extracted or forest are included in cattle rangeland. Density of Myrsine trees was clearly higher in Pró-Mata forest borders 11-12 years after abandonment than in forest borders of the adjacent fazenda.
Development at this stage clearly differs between grasslands and oldfields: In grassland, much of the belowground biomass remains intact and resprouting individuals rapidly pre-empt the space; in oldfields relatively nutrient-rich bare ground accounts for most of the area, and first recruitment is from diaspores, although perennials and grasses soon follow (e.g. Pickett 1982; Bornkamm 1988). Glenn-Lewin & van der Maarel (1992) voice concern at the fact that so much of secondary successional theory is derived from oldfields, and Schreiber (1997) suspects that for the same reason, existing models cannot satisfactory explain abandoned grassland development in his study.
Considering the triple constraint of site pre-emption by grasses and forbs, animal dispersers avoiding grassland (chapter 2) and initially relatively low numbers of fruit-bearing trees in forest borders, it is not surprising that grassland seeder species, especially Baccharis spec., should have been principal colonisers of the newly abandoned Pró-Mata grasslands. If part of these areas were burnt now, establishment of forest trees from seed would probably be higher, on the basis that higher density of fruit-bearing trees increases chances of seed dispersal into grassland even if dispersers still largely avoid open areas. Also, Myrsine populations already established in grasslands would not have to start from zero: adolescent Myrsine individuals in grassland have fairly good chances of resprouting, although they are still at a disadvantage compared to grasses and grassland shrubs (chapter 6). This seems to have been the case in the Ecological Station of Aracurí, where grassland-forest succession initially proceeded at similarly slow speed as in Pró-Mata – including establishment of Baccharis shrub canopies – and the experimental burning of one such area resulted in increased recruitment of forest species (A.Teixeira, pers. comm.).

Hermann, J.-M. PhD thesis Final discussion
174
„Thinning phase“: This phase is characterised by strong competition between established individuals, with the result that those species best suited to given site conditions achieve dominance, and community composition correlates better with abiotic parameters than before (as proposed by Christensen & Peet). Recruitment should be favoured in species whose seeds can emerge in the shade (chapter 2), provided that the canopy is not so dense as to preclude dispersal, but seedlings remain suppressed. In pasture, thinning among individuals can be relevant in patches that have remained unburnt for several years, and where abiotic conditions are not so stressful as to preclude sward closure. In Pró-Mata, tall shrub grassland stands established following accidental burning in 1999 were in this stage in 2005-2006, i.e. five to six years after establishment. Tall tussock grassland stands still seemed characterised by strong competition among grasses and grassland shrubs, judging by the fact that natural Baccharis seedling emergence was low, and ramet recruitment predominated over seedling recruitment not only in Myrsine but also in the characteristic small grassland shrub Calea phyllolepis (my own data).
„Transition phase“: This stage is characterised by gap creation through senescence and death of canopy individuals. Competition is once more relaxed and correlations of community composition with abiotic parameters become weaker (as proposed by Christensen & Peet). Accessibility of such stands to seeds and new seedling recruits depends on whether more than one discrete vegetation layer is present, and whether openings appear in one or all of these, as already pointed out above. Peet & Christensen (1980) demonstrated that seedling recruitment is favoured if both canopy and understorey are opened, or gaps are created in the understorey. Canopy openings, however will probably result primarily in competitive release of saplings already present in the understorey; this would be followed by another thinning phase in which recruitment from seed is largely precluded.
In tall shrub grassland stands of Pró-Mata, this process was captured 11-12 years after abandonment; Myrsine recruitment was lower in areas with very sparse Baccharis canopy than in areas with denser canopy, and competitive release was not obvious in all size-age stages of Myrsine, which indicates that tussock grasses and grassland shrubs still present under canopy benefited from Baccharis dieback as well. Chances for new seedling recruitment at this stage seem best for species that according to Jelinski & Cheliak’s (1992) and Eriksson & Fröborg’s (1998) terminology exploit „windows of opportunity“, i.e. are well dispersed in both space and time. Otherwise, species that recruit at earlier stages and persist under dense canopy are at an advantage.
One of the most interesting questions remaining is whether gap creation eventually occurs in the other major structural type of abandoned grassland - tall tussock grassland - by local dieback of tussocks or shrubs, or whether seedling establishment of forest species remains largely precluded until the few vegetatively spreading forest species have formed a canopy. On the other hand, much dead grass biomass has accumulated and fire risk increases with every year. Peet (1992) names the following fire return intervals for the Great Lakes region of the USA prior to European settlement: 2-5 years in grassland, 50 years in stands of pioneer species Aspen and Pine, 20 years in the ecotone between such stands and grassland. For the same reason, any „steady-state phase“ of abandoned grassland can probably be ruled out.
Summing up, it is possible to recognise, across my study area, vegetation types in the developmental stages of establishment, thinning and transition. This is due to communities being dominated by few species, so that community dynamics (so far) largely reflect life history and population dynamics of these species. Transitions from one stage into another do not occur simultaneously across the landscape, and thinning and transition stages will probably occur repeatedly in a given site before a proper forest is established - e.g. in the case of a tall shrub grassland site first among Baccharis individuals, then among grasses

Hermann, J.-M. PhD thesis Final discussion
175
and Myrsine individuals, then among individuals of species predominately establishing under canopy of Myrsine, whose identity is not yet known. Judging by species composition of seedlings, this role may be assumed by Myrsine herself, by Myrtaceae of the forest border or a mixture of both.
It follows that in any attempt to correlate vegetation composition to abiotic parameters – no matter whether the units of description are species or plant functional types –, attention should also be paid to whether competition in a given site is currently intense or relaxed. Also, the development towards a „zonal climax community” should not be expected, not only because of the probability of recurrent disturbances but because such communities by definition establish only on more or less level terrain (see e.g. Ellenberg 1996), and this condition is rarely met in the study area. Rather, the topography is undulating and frequently intersected by canyons so that a mosaic of azonal, zonal and perhaps extrazonal communities to develop.
E. Burgert (Irslinger et al. 2001) and A. Dümig (pers. comm.) both measured decreased soil water contents under forest vegetation and Baccharis canopies relative to grassland. Grassland soil, in fact, takes up water like a sponge due to its high content of organic matter; Walter (1984) suggest this as a major factor limiting forest tree species in the south Brazilian grasslands (“Unter dem Grasland findet man tief humose, leicht vernässende Böden, die den Baumarten auf sauren Böden wahrscheinlich wenig zusagen.”- p.259). A. Dümig proposed that soil drainage by woody pioneer vegetation is an essential prerequisite for development of a complete forest community (pers. comm.).
There is some supporting evidence for such limitations, although this is anecdotical. One example: During preparations of my seedling emergence experiment, ten areas were mowed, and to facilitate this task I chose areas with minimum proportion of woody species in the vegetation cover. Five of these areas had to be omitted after been found inundated 20 cm high after heavy rainfalls in the middle of March 2005, which indicates that waterlogging limited woody species’ establishment - even though predominance in this area was clearly of grassland shrubs that may be expected to cope somewhat better with such conditions than forest trees. Another example: Myrtaceae seedlings raised in grassland soil in nurseries were regularly extinguished by damping-off disease, an infection with fungi of the type Pythium (also diagnosed in dying Baccharis communities) that thrives under warm climates in soils of excess humidity. Fortunately for this study, Myrsine seedlings were unaffected by the disease.
Another possibly limiting factor that has only been investigated for Araucaria angustifolia so far is decreased mycorrhization due to scarcity of AM-fungi in grassland relative to forest (Breuninger et al. 2000); a highly interesting question is whether Myrsine root suckers may act as vectors for transport of such fungi into grassland soil, although on arrival those fungi would still be confronted with unfavourable substrate conditions.
This leads back to the discussion begun in chapter 5: It is still not or little known how pioneer tree and shrub species perform in grassland relative to forest. Does grassland offer substantially higher amounts of certain resources? Do pre-adaptations of forest species allow them to exploit these resources efficiently?

Hermann, J.-M. PhD thesis Final discussion
176
7.4. Origin of pioneer forest species in grassland A concise report about successional pathways from grassland to Araucaria forest on
the southern Brazilian High Plateau was published by Klein (1960): According to this author, the initial stages of succession are characterised by a multitude of pioneer associations, each of which is, however, dominated by a relatively low number – usually only two or three - of pioneer species or genera. For example, Klein found that in pioneer forests of the association Siphoneugenietum-Myrceugenieteum along the eastern border of the Serra Geral (including my study region), the two name-giving species Myrceugenia euosma and Siphoneugenia reitzii comprised as much as 80-90% of the vegetation in terms of abundance (ibid.). It seems that even if successional pathways and patterns in the Pró-Mata study area are unique to this particular section of the High Plateau (see chapter 5), their very uniqueness and simple species composition make them fit into a general pattern.
Fig. 7.1: Principal pathways of grassland-Araucaria forest succession described by Klein (1960), slightly
modified. „Campo“, as explained in chapter 1, is the common name for grassland interspersed to greater
or lesser extent with shrubs. The terms „xerossera“ and „hidrossera“ refer to well drained and more
humid sites, respectively. The “Myrsinetum” typical of the Pró-Mata study area was added by me.
Klein (1960) presents eight pioneer associations characterised by the following species (see Fig. 7.1, Tab. 7.2): Araucaria angustifolia, Campomanesia xanthocarpa, Myrceugenia euosma, Capsicodendron dinisii, Siphoneugenia reitzii, Schinus terebinthifolius, S. molle, S. lentiscifolius, Clethra scabra and Ateleia glazioviana. Added to these may be the „Myrsinetum“ dominated by Myrsine parvula typical of the Pró-Mata study area, plus pioneer communities characterised e.g. by Myrcia bombycina (Klein 1963) and Xanthoxylum
Myrsinetum
?
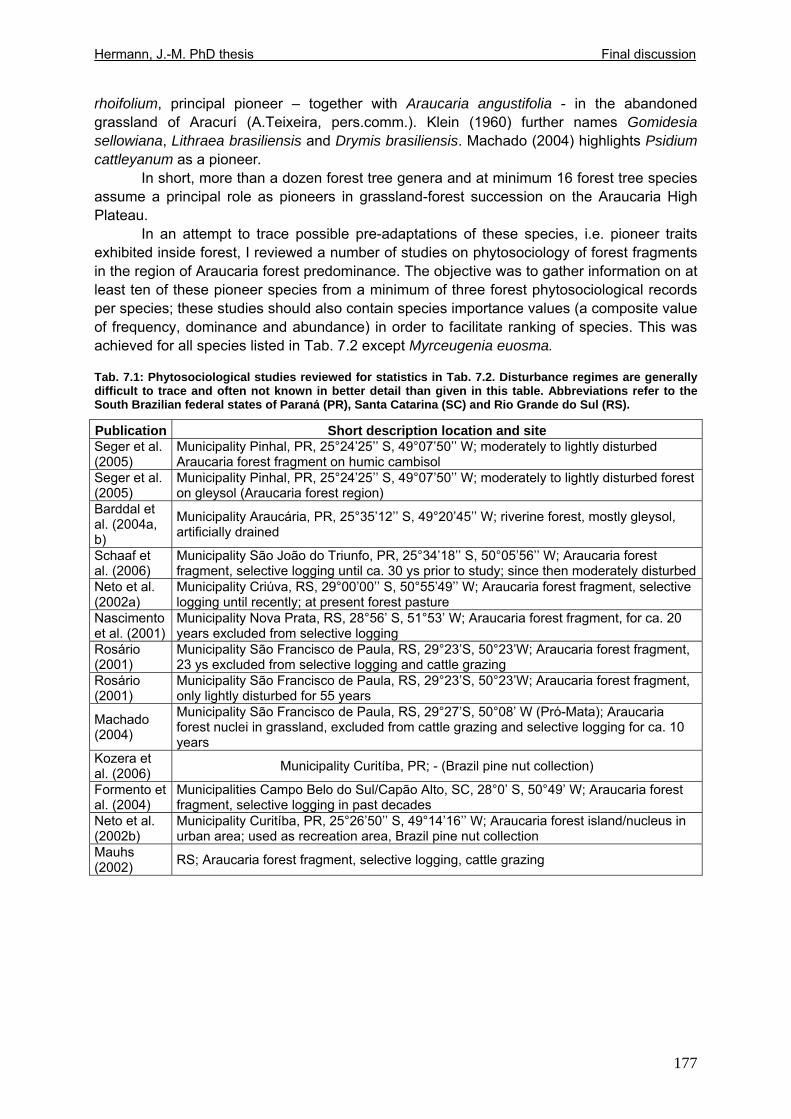
Hermann, J.-M. PhD thesis Final discussion
177
rhoifolium, principal pioneer – together with Araucaria angustifolia - in the abandoned grassland of Aracurí (A.Teixeira, pers.comm.). Klein (1960) further names Gomidesia sellowiana, Lithraea brasiliensis and Drymis brasiliensis. Machado (2004) highlights Psidium cattleyanum as a pioneer.
In short, more than a dozen forest tree genera and at minimum 16 forest tree species assume a principal role as pioneers in grassland-forest succession on the Araucaria High Plateau.
In an attempt to trace possible pre-adaptations of these species, i.e. pioneer traits exhibited inside forest, I reviewed a number of studies on phytosociology of forest fragments in the region of Araucaria forest predominance. The objective was to gather information on at least ten of these pioneer species from a minimum of three forest phytosociological records per species; these studies should also contain species importance values (a composite value of frequency, dominance and abundance) in order to facilitate ranking of species. This was achieved for all species listed in Tab. 7.2 except Myrceugenia euosma.
Tab. 7.1: Phytosociological studies reviewed for statistics in Tab. 7.2. Disturbance regimes are generally
difficult to trace and often not known in better detail than given in this table. Abbreviations refer to the
South Brazilian federal states of Paraná (PR), Santa Catarina (SC) and Rio Grande do Sul (RS).
Publication Short description location and site
Seger et al. (2005)
Municipality Pinhal, PR, 25°24’25’’ S, 49°07’50’’ W; moderately to lightly disturbed Araucaria forest fragment on humic cambisol
Seger et al. (2005)
Municipality Pinhal, PR, 25°24’25’’ S, 49°07’50’’ W; moderately to lightly disturbed forest on gleysol (Araucaria forest region)
Barddal et al. (2004a, b)
Municipality Araucária, PR, 25°35’12’’ S, 49°20’45’’ W; riverine forest, mostly gleysol, artificially drained
Schaaf et al. (2006)
Municipality São João do Triunfo, PR, 25°34’18’’ S, 50°05’56’’ W; Araucaria forest fragment, selective logging until ca. 30 ys prior to study; since then moderately disturbed
Neto et al. (2002a)
Municipality Criúva, RS, 29°00’00’’ S, 50°55’49’’ W; Araucaria forest fragment, selective logging until recently; at present forest pasture
Nascimento et al. (2001)
Municipality Nova Prata, RS, 28°56’ S, 51°53’ W; Araucaria forest fragment, for ca. 20 years excluded from selective logging
Rosário (2001)
Municipality São Francisco de Paula, RS, 29°23’S, 50°23’W; Araucaria forest fragment, 23 ys excluded from selective logging and cattle grazing
Rosário (2001)
Municipality São Francisco de Paula, RS, 29°23’S, 50°23’W; Araucaria forest fragment, only lightly disturbed for 55 years
Machado (2004)
Municipality São Francisco de Paula, RS, 29°27’S, 50°08’ W (Pró-Mata); Araucaria forest nuclei in grassland, excluded from cattle grazing and selective logging for ca. 10 years
Kozera et al. (2006) Municipality Curitíba, PR; - (Brazil pine nut collection)
Formento et al. (2004)
Municipalities Campo Belo do Sul/Capão Alto, SC, 28°0’ S, 50°49’ W; Araucaria forest fragment, selective logging in past decades
Neto et al. (2002b)
Municipality Curitíba, PR, 25°26’50’’ S, 49°14’16’’ W; Araucaria forest island/nucleus in urban area; used as recreation area, Brazil pine nut collection
Mauhs (2002) RS; Araucaria forest fragment, selective logging, cattle grazing
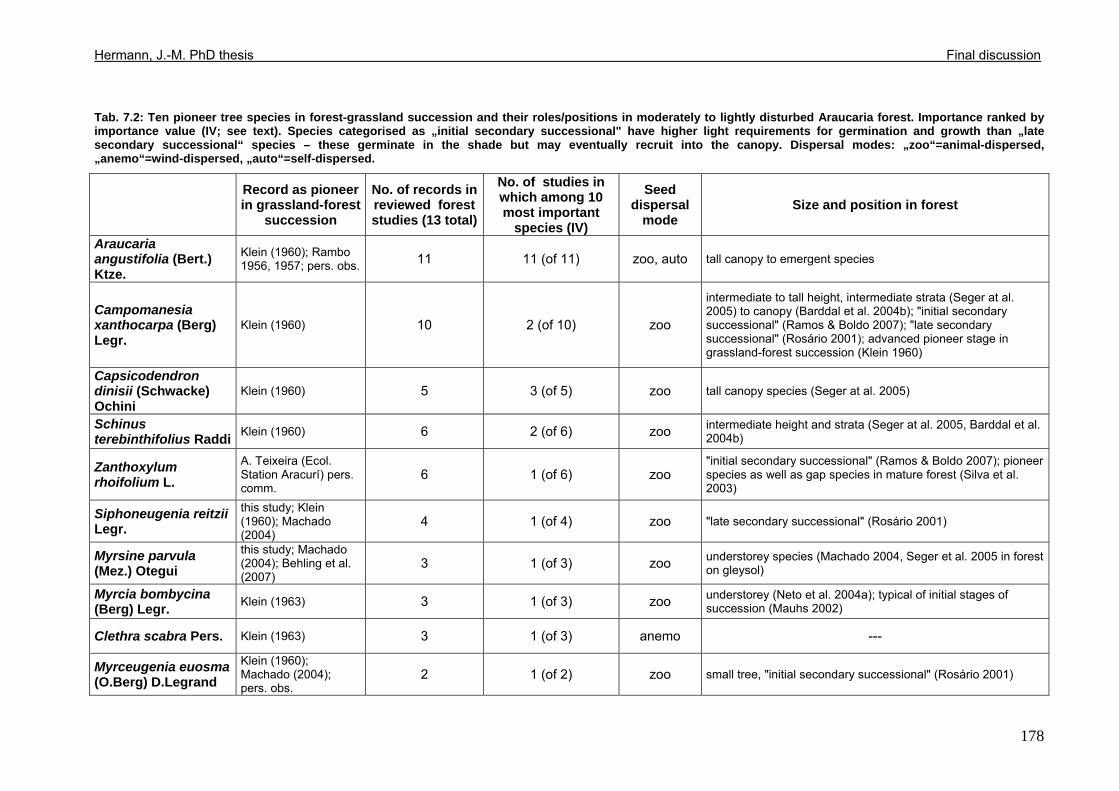
Hermann, J.-M. PhD thesis Final discussion
178
Tab. 7.2: Ten pioneer tree species in forest-grassland succession and their roles/positions in moderately to lightly disturbed Araucaria forest. Importance ranked by
importance value (IV; see text). Species categorised as „initial secondary successional" have higher light requirements for germination and growth than „late
secondary successional“ species – these germinate in the shade but may eventually recruit into the canopy. Dispersal modes: „zoo“=animal-dispersed,
„anemo“=wind-dispersed, „auto“=self-dispersed.
Record as pioneer
in grassland-forest
succession
No. of records in
reviewed forest
studies (13 total)
No. of studies in
which among 10
most important
species (IV)
Seed
dispersal
mode
Size and position in forest
Araucaria angustifolia (Bert.)
Ktze. Klein (1960); Rambo 1956, 1957; pers. obs. 11 11 (of 11) zoo, auto tall canopy to emergent species
Campomanesia xanthocarpa (Berg)
Legr. Klein (1960) 10 2 (of 10) zoo
intermediate to tall height, intermediate strata (Seger at al. 2005) to canopy (Barddal et al. 2004b); "initial secondary successional" (Ramos & Boldo 2007); "late secondary successional" (Rosário 2001); advanced pioneer stage in grassland-forest succession (Klein 1960)
Capsicodendron dinisii (Schwacke)
Ochini Klein (1960) 5 3 (of 5) zoo tall canopy species (Seger at al. 2005)
Schinus terebinthifolius Raddi
Klein (1960) 6 2 (of 6) zoo intermediate height and strata (Seger at al. 2005, Barddal et al. 2004b)
Zanthoxylum rhoifolium L.
A. Teixeira (Ecol. Station Aracurí) pers. comm.
6 1 (of 6) zoo "initial secondary successional" (Ramos & Boldo 2007); pioneer species as well as gap species in mature forest (Silva et al. 2003)
Siphoneugenia reitzii Legr.
this study; Klein (1960); Machado (2004)
4 1 (of 4) zoo "late secondary successional" (Rosário 2001)
Myrsine parvula (Mez.) Otegui
this study; Machado (2004); Behling et al. (2007)
3 1 (of 3) zoo understorey species (Machado 2004, Seger et al. 2005 in forest on gleysol)
Myrcia bombycina
(Berg) Legr. Klein (1963) 3 1 (of 3) zoo understorey (Neto et al. 2004a); typical of initial stages of succession (Mauhs 2002)
Clethra scabra Pers. Klein (1963) 3 1 (of 3) anemo ---
Myrceugenia euosma (O.Berg) D.Legrand
Klein (1960); Machado (2004); pers. obs.
2 1 (of 2) zoo small tree, "initial secondary successional" (Rosário 2001)

Hermann, J.-M. PhD thesis Final discussion
179
The difficulties of tracing more or less natural Araucaria forest dynamics have already been mentioned (chapters 1 and 5). Nevertheless, the following trends emerge:
1. Nearly all pioneer species explicitly named here are animal-dispersed (zoochorous). The only exception is wind-dispersed (anemochorous) Clethra scabra, and this species ranks bottom of the list as far as species importance in forest fragments is concerned. This high proportion of zoochory among pioneers (15 out of 16 species, or 94%) is reflected in the arboreal component of a Podocarp forest with Araucaria studied by Giongo & Waechter (2007): Of 43 woody species, 88% were zoochorous species, 7% anemochorous, the remainder autochorous species.
2. Pioneer species in grassland recruit from virtually every type of habitat found in a forest. They include tall-growing, dominant canopy species of widespread occurrence such as Araucaria angustifolia and Campomanesia xanthocarpa; species of intermediate height (Schinus terebinthifolius) as well as subcanopy and small gap species that appear or achieve predominance in only a few locations across the Planalto - e.g. Myrsine parvula, the principal pioneers of my study area, and Siphoneugenia reitzii, which was infrequently recorded in the reviewed forest studies, although the Siphoneugenietum was highlighted by Klein (1960) as one of the most widely distributed pioneer associations. Of Xanthoxylum rhoifolium, Silva et al. (2003) report the occurrence as pioneer species in initial stages of succession as well as in gaps in mature forest – the same dual role that I proposed, based on observed traits, for Myrsine parvula (chapter 5).
Fig. 7.2: Profile of a moderately to lightly disturbed Araucaria forest on typical highland cambisol from
Seger et al. (2005), modified. Painted grey: The emergent, name-giving species Araucaria angustifolia,
canopy species Capsicodendron dinisii and Lithraea brasiliensis – here recorded in the subcanopy – all
participate as pioneers in grassland-forest succession (Klein 1960).

Hermann, J.-M. PhD thesis Final discussion
180
Fig. 7.3: Profile of a moderately to lightly disturbed Araucaria forest on gleysol from Seger et al. (2005),
modified. Painted grey: Small-growing gap or understorey species Myrsine parvula (center) and Psidium cattleyanum (left) occur as pioneers in grassland as well as intermediate-height Schinus terebinthifolius
and Lithraea brasiliensis (Klein 1960; Machado 2004; this study).
Grassland-forest succession on the Araucaria High Plateau thus differs in some fundamental aspects from secondary succession on grassland and arable land in Europe and North America, where forest stands are often initiated by a relatively small set of wind-dispersed, fast-growing and often short-lived tree genera such as Salix, Betula and Pinus. This phenomenon has not remained unobserved: Heringer & Jacques (2001) deny Araucaria the status as pioneer species for the very reason that in terms of dispersal, height and longevity, she represents the exact opposite of a classical type. However, I think that it may be more rewarding not to question the phenomenon as such but to investigate the underlying causes.
Why have these species emerged as pioneers in grassland? Which traits enable them to assume the role of pioneers in a habitat so significantly different from their original one - even subdominant species of only local importance that are apparently neither excellent dispersers nor excellent competitors? And why is the first position in grassland-forest succession, apparently in absence of forest species of similar behaviour, occasionally occupied by wind-dispersed, fast-growing grassland shrub species such as Baccharis uncinella (chapter 5; see also Waechter et al. 1984)?
The phenomenon is not restricted to the Araucaria High Plateau: Myrciaria cuspidata is one of the principal forest pioneer species on Morro Santana (Müller & Forneck 2004) but occurs as a small understorey tree in forests of the same region (Brack et al. 1998 in Müller & Forneck 2004). Andira legalis, a Fabacea investigated by Cirne & Scarano (2001), forms monospecific thickets of root suckers of ca. 2.5 m height in full exposure to sunlight in the restinga of São Paulo; in adjacent forest, she occurs as small tree of more or less 5 m height.
One answer may lie in regimes of large-scale disturbance in recent millennia. The number of tree genera especially in Europe has been depleted, relative to subtropical and tropical areas, during the ice ages (e.g. Ellenberg 1996); at the same time, environmental conditions during glacier advance and retreat exerted strong selective pressure for wind-dispersed, fast-growing pioneer species. In South America, only the southernmost tip of Argentina – today Patagonia – was glaciated (Ab’Sáber 1977 in Leite & Klein 1990). Montane forests of this region are nowadays characterised by large-scale disturbance such as landslides and fires, and wind-dispersed arboreal species e.g. of the genera Nothofagus

Hermann, J.-M. PhD thesis Final discussion
181
and Austrocedrus predominate as pioneers on exposed soil in this region (Veblen 1989, Veblen et al. 2003).
Araucaria angustifolia probably assumes a similarly unique position among pioneers as she does in the forest community. The species produces large seeds – similar in size to Pará nuts – with a carbohydrate-rich endosperm. Seeds are consumed by at least two species of the family of Jays – Cyanocorax caeruleus and C. chrysops – as beechnuts are in Europe and the USA, and the latter species sometimes caches seeds in soil, thus contributing to dispersal and germination (Koch & Corrêa 2002). Araucaria also possesses broad, leathery to cartaceous needles and coarse bark and can resprout after fire unless very young (pers.obs.). All of these traits may be advantageous in seedling establishment in resource-poor and fire-prone habitats. However, Araucaria also used to be the most important timber species of the highland forests (Koch & Corrêa 2002), and if she is still largely absent from grasslands of Pró-Mata and the adjacent fazenda today, this may be partly due to the fact that seed-bearing adults are yet scarce. Moreover, seed longevity is short: Seed germinability decreases by 50% within four months following maturation, and entirely within one year (Zimmermann-Inui in Walter 1984).
As to the other pioneer species, I may hypothesize with Pickett (1982) on two possible keys to their success: Firstly, genetic differentiation between individuals establishing in lighter and darker habitats; secondly, plasticity. Pickett (1982) proposed these mechanisms on observing that several tree species occurred not only in oldfields of New Jersey, USA, as early as 20 years after abandonment, but also in old-growth forests of that region.
Plasticity of a species is expressed by the degree to which morphological or physiological traits of individuals in a population may be adjusted – more or less reversibly – to environmental conditions. In a strict sense, this excludes genetically introduced, i.e. inheritable, variation in these traits as well as constraints on development imposed directly by the environment, e.g. by low nutrient availability (see e.g. Grime et al. 1986). Phenotypic plasticity is displayed by Myrsine parvula in leaf morphology (chapter 3) as well as plant architecture – referring to prolific root suckering in this species (chapter 4). However, rapid genetic differentiation in this species seems also possible due to efficient means of seed dispersal (chapter 2). Of course, a combined strategy is also imaginable.
Certainly, both strategies may be useful under such conditions as forest species in my study region have experienced in the distant and recent past: Natural forest gap dynamics, timber exploitation and other types of human influence, and, last but not least, an extraordinarily high number of co-occurring species, as the convergence of different climates (among other factors) has made Southern Brazil one of the biodiversity hotspots of this world (Leite & Klein 1990).
Upon reflection, one may ask why the number of pioneer forest species in grassland is not even higher than the round dozen named so far. Why has such an extraordinarily large pool of forest species not yielded more successful pioneers in grassland? I might spontaneously list several species occurring in the Pró-Mata region – sometimes in immediate vicinity of the grasslands – that seem to possess some ideal pioneer traits. What constrains these species?
For example, two shrub species of the genus Miconia (Melastomataceae) occur at the forest border and occasionally in grassland. They have bright fruit attractive to birds and mesophyllous to leathery leaves. Why do they not assume the role of pioneer shrubs as Cordia multispicata does in Amazonian grassland (Viera et al. 1994)?
An arborescent Fabacea, Mimosa scabrella, occurs as a pioneer tree in the Southriograndean lowland (S. Müller, pers. comm.) and in experimental plantations in Pró-Mata near the escarpment attains tree stature within 5 years (R. Mello, pers. comm.). Why does this species not figure in grassland-forest succession, replenishing soil nitrogen as

Hermann, J.-M. PhD thesis Final discussion
182
Prosopsis glandulosa does in subtropical rangelands in Texas, USA? High moisture content and low pH of the grassland soils may act restrictively – as a rule, Fabaceae prefer dry, nitrogen-poor and calcareous soils (Strasburger et al. 1991) and notably, in European wetlands, nitrogen fixation is by Alnaceae and not by Fabaceae – but this is another hypothesis that remains to be investigated.
These two examples shall suffice here; what they serve for is to introduce a final thought: There are constraints on either of the two strategies named; versatility comes at a cost. This is best expressed in another quotation of Crawley (1997a, p. 101): „Some species may be ‚Jacks of all trades’ but no single species can be ‚master of all’.“
According to palynological data, forests and grasslands have existed side by side on the Araucaria High Plateau for no more than one or two millennia (Behling et al. 2004, Behling 2007). Thus, colonisation of the new habitat by genetic differentiation would require fairly rapid evolution, i.e. would seem possible only if species’ life spans are relatively short and during this period, many seeds are produced. This is even more important in colonisation of a habitat in which seed dispersal, due to disperser behaviour, is more impeded than inside the original habitat.
Contrary to this, high phenotypic plasticity comes at the cost of evolution slowed down. Although the ability to modify traits in response to environmental cues is inherited, the modifications themselves are not heritable. Plasticity is thus “interpreted as a mechanism replacing or supplementing genetic variation, as a buffer against spatial or temporal variability in habitat conditions” (Grime et al. 1986, p.8). In other words, plasticity in a species reduces the urgency of adapting to changing environmental conditions via natural selection.
Moreover, there are immediate costs involved e.g. in maintenance of the complex apparatus that is necessary for detection of, and reaction to, a variety of environmental cues (Givnish 2002). These costs may trade off against other life processes such as vertical growth and reproductive growth. In the longer run, plasticity may increase the probability of population persistence but reduce the likelihood of genetic change; evolution may be slowed down18. There is some conceptual overlap with the discussion on costs and benefits of clonal growth that was briefly touched in chapter 4. In fact, clonal growth may be viewed as an expression of phenotypic plasticity in a broad sense, and it has already been pointed out that vegetative growth in this fashion also trades off against generative reproduction.
Considering these options, it seems little likely that a forest species may cope with drastically altered conditions of microclimate, soil and vegetation structure in grassland, and at the same time may disperse seeds for hundreds or even thousands of metres, and grow up to reproductive size and age within a decade or less. This may help to explain why there is a relatively large total number of pioneer species across the High Plateau and at the same time, as in my study area Pró-Mata, only one single species predominates in a given area.
Some interesting evidence emerges from a recent study by Rozendaal et al. (2006). On examining leaf characteristics of 36 tropical forest species exposed to high light and shade, the authors found that short-lived pioneers that experience consistently high light levels exhibited low plasticity – this is concomitant with my observations of Baccharis seedling size and allometry in chapter 3. Plasticity was also low in tall-growing species, although these are believed to „experience large ontogenetic changes in irradiance during 18 The trade-off between costs and benefits of plasticity at the individual and population level has received increased attention recently (e.g van Kleunen et al. 2000, Hulme 2008) and first models have been developed (e.g. Price et al. 2003): According to these authors, high levels of plasticity increase the probability of population persistence but reduce the likelihood of genetic change whereas intermediate levels should be optimal for both population persistence and adaptive evolution. The cost of reduced fitness may be greater in habitats species are not accustomed to (demonstrated e.g. by Steinger et al. (2003) for Sinapis arvensis grown in sun and in shade).

Hermann, J.-M. PhD thesis Final discussion
183
their life cycle“ (p.207; see also Bazzaz 1984). It was highest in shade-tolerant subcanopy species that however, according to the authors, „experience consistently low light levels“ (ibid.).
Summing up, three hypotheses may be formulated: Forest species may be pioneers in grassland if a) genetic differentiation is rapid, or b) phenotypic plasticity is high; and c) these traits are more frequent in subcanopy species than dominants.
The topic is of particular interest as it may link investigations of grassland-forest dynamics to a new field of research that scientists will have to deal with within one or a few decades, namely, the behaviour of grassland and forest species in fallow areas of arable land as well as in logged pine plantations. The landscape changes currently brought about by agriculture and forestry are so sudden, drastic and occur on such a large spatial scale – as Overbeck et al. (2007) point out, 25% of the southern Brazilian grassland biome have already been lost during the past three decades – that only rapid evolution or high plasticity would seem to enable native species to persist in these areas.

Hermann, J.-M. PhD thesis Final discussion
184
7.5. Outlook ...But Christopher Robin wasn’t listening. He was looking at Pooh. ‚Pooh,’ he said, ‚where did you find that pole?’ Pooh looked at the pole in his hands. ‚I just found it,’ he said. ‚I thought it ought to be useful. I just picked it up.’ ‚Pooh,’ said Christopher Robin solemnly, ‚the Expedition is over. You have found the North Pole!’
A.A. Milne, Winnie-The-Pooh
Garrad: “Nonetheless, it is Science. And what is Science, Anson?” Anson: “Oh, I always forget that one, don’t I?” [Pause] “Science is dispassionate.”
Christopher Monger, The Englishman who went up a hill but came down a mountain
In the excitement and relief that comes with completion of a study and the gain of new insights, it is all the more important to resist the temptation of declaring „the North Pole discovered“. Instead, the best I can hope for is that this work will provide a useful basis for future scientists who intend to investigate establishment strategies of forest species outside forest in more detail. Existing hypotheses may be fine-tuned, new hypotheses may be developed. Here are some proposals:
1. Many species of the High Plateau Araucaria forest are of Austral-Antarctic origin.
Nevertheless, a gradient of favourability for forest species establishment and growth exists between grasslands of subtropical lowland and temperate highland as well as between escarpment and inland areas on the High Plateau. Better growth conditions near the escarpment are partly due to higher frequency and longer duration of fogs during the summer which dampen irradiation and temperature extremes, partly to nutrient status of soils.
2. Root sucker recruitment ensures Myrsine population spread into grassland even under poor conditions of seed dispersal and seedling establishment. However, root suckers originally developed as a means of individual spread into forest gaps and they are relatively less efficient in grassland. Abundant sucker recruitment is cued by high light availability in grasslands but soil nutrient availability is low, resulting in early cut-off of supply from the feeder plant. Within three years after recruitment, root suckers lose their relative advantage in resource acquisition over co-occurring seedlings. Relatively slow individual growth further slows down spatial expansion of the population.
3. Clonal growth is an expression of high plasticity. Forest species establishing in grassland are mostly subdominants from adjacent forest; they owe their superior colonising ability to rapid and intense genotypic selection (fast and abundant generative reproduction) or to high plasticity (as expressed e.g. by clonal growth) which enables them to offset selection pressure.

Hermann, J.-M. PhD thesis Final discussion
185
Consequently, future studies should include: 1. Transects from the Escarpment ridge to the interior of the High Plateau, in order to
check for influences of climate and soil on forest-grassland dynamics. 2. Comparison of the performance of seedlings and root suckers in grassland and the
interior of forest. 3. Functional similarities between the various forest species acting as pioneers across
the High Plateau. Genetic studies might be of interest in order to delimit clone size and to trace possible
hybridisation between Myrsine species. Last but not least, as soon as such areas become available, logged plantations and
fallows on arable land should also be included.

Hermann, J.-M. PhD thesis References
186
8. References
Alcántara, J.M., Rey, P.J., Valera, F. & Sánchez-Lafuente, A.M. 2000: Factors shaping the seedfall pattern of a bird-dispersed plant. Ecology 81(7): 1937-1950.
Alvarez-Byulla, E.R. & Martinez-Ramos, M. 1990: Seed bank versus seed rain in the regeneration of a tropical pioneer tree. Oecologia 84(3): 314-325.
Anderson, R.C. 1982: An evolutionary model summarizing the roles of fire, climate, and grazing animals in the origin and maintenance of grasslands. In: Estes, J.R., Tyrl, R.J. & Brunken, J.N. (eds.): Grasses and grasslands: Systematics and ecology. University of Oklahoma Press, Norman.
Archer, S. 1991: Development and stability of grass/woody mosaics in a subtropical savanna parkland, Texas, U.S.A. In: Werner, P.A. (ed.): Savanna ecology and management. Australian perspectives and intercontinental comparisons. Blackwell Scientific Publications, Oxford.
Archer, S., Scifres, C. & Bassham, C.R. 1988: Autogenic succession in a subtropical savanna: Conversion of grassland to thorn woodland. Ecological monographs 58(2): 111-127.
Augspurger, C.K. 1984: Light requirements of neotropical tree seedlings: A comparative study of growth and survival. Journal of Ecology 72: 777-795.
Augspurger, C.K. & Kelly, C.K. 1984: Pathogen mortality of tropical tree seedlings: experimental studies of the effects of dispersal distance, seedling density, and light conditions. Oecologia 61(2): 211-217.
Baaske, R. 1999: Artenzusammensetzung und Struktur eines stark exploitierten Araukarienwaldbestandes an der östlichen Randstufe des südbrasilianischen Planaltos. Diploma Thesis Fachhochschule Rottenburg, Germany.
Baaske, R., Matzenbacher, N.I. & Sobral, M. 2001: Vegetationskartierung des Projektgebietes [Pró-Mata, Rio Grande do Sul, Brasilien] unter Verwendung von Farbinfrarot-Luftbildern. In: Engels, W., Hampp, R. & Tzschupke, W. (eds.): Zwischenbericht zum BMBF-Projekt 01LT0011/7 „Araukarienwald“. Forschungsschwerpunkt Region 4: Mata Atlântica. URL: http://www.uni-tuebingen.de/entw-phys/CD/2000.pdf#page=60 [retrieved 2008-08-17].
Backes, P. & Irgang, B. 2002: Arvores do Sul: guia de identificação e interesse ecológico. Editora Pallotti, Porto Alegre.
Barddal, M. Lacerda, Roderjan, C. Vellozo, Galvão, F. & Curcio, G. Ribas 2004a: Fitossociologia do sub-bosque de uma floresta ombrófila mista aluvial, no município de Araucária, PR. (Phytosociology of the understory of a floodplain forest, at Araucária, PR.) Ciência Florestal (Santa Maria) 14(1): 35-45.
Barddal, M. L., Roderjan, C. Vellozo, Galvão, F.& Curcio, G. Ribas 2004b: Caracterização florística e fitossociológica de um trecho sazonalmente inundável de floresta alluvial, em Araucária, PR. (Floristic and phytosociological characterization of a floodplain forest, at Araucaria, PR.) Ciência Florestal (Santa Maria) 14(2): 37-50
Barnes, P.W. & Archer, S. 1999: Tree-shrub interactions in a subtropical savanna parkland: competition or facilitation? Journal of Vegetation Science 10: 525-536.
Barroso, G. Maciel & Bueno, O. Leites 2002: Compostas. 5. Subtribo: Baccharidinae. (Flora Ilustrada Catarinense.) Itajai, Santa Catarina, Brasil.

Hermann, J.-M. PhD thesis References
187
Barsoum, N., Muller, E. & Skot, L. 2004: Variations in levels of clonality among Populus nigra L. stands of different ages. Evolutionary ecology 18:601-624.
Baskin, C.C. & Baskin, J.M. 1998: Seeds. Ecology, biogeography and evolution of dormancy and germination. Academic Press, San Diego.
Bazzaz, F.A. 1984: Dynamics of wet tropical forests and their species strategies. In: Medina, E., Mooney, H.A. & Vázquez-Yánez, C. (eds.): Physiological ecology of plants of the wet tropics. (Tasks for vegetation science 12.) Dr W. Junk Publishers, The Hague.
Behling, H. 1997: Late Quaternary vegetation, climate and fire history of the Araucaria forest and campos region from Serra Campos Gerais, Paraná state (South Brazil). Review of Paleobotany and Palynology 97 (1997): 109-121.
Behling, H. 1998: Late Quaternary vegetational and climatic changes in Brazil. Review of Paleobotany and Palynology 99: 143-156
Behling, H. 2001: South and southeast Brazilian grasslands during late Quaternary times: a synthesis. Palaeogeography, Palaeoclimatology, Palaeoecology 2710: 1-9.
Behling, H. 2007: Late Quaternary vegetation, fire and climate dynamics of Serra do Araçatuba in the Atlantic coastal mountains of Paraná State, southern Brazil. Vegetation History and Archaeobotany 16(2-3): 77-85.
Behling, H., Pillar, V. de Patta, Orlóci, L. & Bauermann, S.G. 2004: Late Quaternary Araucaria forest, grassland (Campos), fire and climate dynamics, studied by high-resolution pollen, charcoal and multivariate analysis of the Cambará do Sul core in southern Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology 203: 277-297.
Bellingham, P.J. & Sparrow, A.D. 2000: Resprouting as a life history strategy in woody plant communities. Oikos 89(2): 409-416.
Belsky, A.J. 1991: Tree/grass ratios in East African savannas: a comparison of existing models. In: Werner, P.A. (ed.): Savanna Ecology and Management. Australian Perspectives and intercontinental comparisons. Blackwell Scientific, Oxford.
Berkowitz, A. R., Canham, C.D. & Kelly, V.R. 1995: Competition vs. Facilitation of tree seedling growth and survival in early successional communities. Ecology 76(4): 1156-1168.
Bertoletti, J.J. & Teixeira, M.B. 1995: Centro de Pesquisas e Conservação da Natureza Pró-Mata: Termo de Referência. Divul. Mus. Ciênc. Tecnol. – UBEA/PUCRS 2: 1-47.
Bock, C.E. & Bock, J.H. 1997: Shrub densities in relation to fire, livestock grazing, and precipitation in an Arizona desert grassland. Southwestern Naturalist 42(2): 188-193.
Boldrini, I. Iob 1993: Dinâmica de vegetaçao de uma pastagem natural sob diferentes níveis de oferta de forragem e típos de solos, Depressao Central, RS. PhD thesis, Universidade Federal do Rio Grande do Sul - UFRGS, Porto Alegre, RS.
Bond, W.J. & van Wilgen, B.W. 1996: Fire and plants. (Population and Community Biology Series 14.) Chapman & Hall, New York.
Bornkamm, R. 1988: Mechanisms of succession on fallow lands. Plant Ecology 77(1-3): 95-101.
Bosela, M.J. & Ewers, F.W. 1997: The mode of origin of root buds and root sprouts in the clonal tree Sassafras albidum (Lauraceae). American Journal of Botany 84(11): 1466-1481.
Brandão, P. Monteiro & Durigan, G. 2004: Changes in cerrado vegetation after disturbance by frost (São Paulo State, Brazil). Plant Ecology 175(2): 202-215.

Hermann, J.-M. PhD thesis References
188
Bredenkamp, G.J., Spada, F. & Kazmierczak, E. 2002: On the origin of northern and southern hemisphere grasslands. Plant Ecology 163: 209-229.
Breuninger, M., Einig, W., Magel, E., Cardoso, E. & Hampp, R. 2000: Mycorrhiza of Brazil Pine (Araucaria angustifolia [Bert. O. Ktze.]). Plant Biology 2(1): 4–10.
Brokaw, N.V.L. 1982: The definition of a treefall gap and its effect on measures of forest dynamics. Biotropica 14: 158-160.
Brokaw, N.V.L. 1987: Gap phase regeneration of three pioneer tree species in a tropical forest. Journal of Ecology 75: 9-19.
Brown, J.R. & Archer, S. 1989: Woody plant invasion of grasslands: establishment of honey mesquite (Prosopis glandulosa var. glandulosa) on sites differing in herbaceous biomass and grazing history. Oecologia 80: 19-26.
Bullock, J.M. & Clarke, R.T. 2000: Long distance seed dispersal by wind: measuring and modelling the tail of the curve. Oecologia 124(4): 506-521.
Cain, S.A. 1950: Life-forms and phytoclimate. The Botanical Review 16(1): 1-32.
Callaway, R.M. 1992: Effect of shrubs on recruitment of Quercus douglasii and Quercus lobata in California. Ecology 73(6): 2118-2128.
Canham, C.D. 1989: Different responses to gaps among shade-tolerant species. Ecology 70(3): 548-550.
Canham, C.D. & Marks, P.L. 1985: The response of woody plants to disturbance: patterns of establishment and growth. In: Pickett, S.T.A. & White, P.S. (eds.): The ecology of natural disturbance and patch dynamics. Academic Press, Inc., Orlando.
Carrington, M.E. & Keeley, J.E. 1999: Comparison of post-fire seedling establishment between scrub communities in Mediterranean and non-mediterranean climate ecosystems. Journal of Ecology 87: 1025-1036.
Cersósimo, L. Fonseca 1993: Variações espaciais e temporais no estabelecimento de plântulas em trecho de floresta secundária em São Paulo, SP. Master Thesis Universidade de São Paulo – USP, São Paulo, SP, Brazil.
Chapin, F.S. III, Sturm, M. Serreze, M.C., McFadden, J.P., Key, J.R., Lloyd, A.H., McGuire, A.D., Rupp, T.S., Lynch, A.H., Schimel, J.P., Beringer, J., Chapman, W.L., Epstein, H.E., Euskirchen, E.S., Hinzman, L.D., Jia, G., Ping, C.-L., Tape, K.D., Thompson, C.D.C., Walker, D.A. & Welker, J.M. 2005: Role of land-surface changes in arctic summer warming. Science 310(5748): 657 – 660.
Christensen, N.L. & Peet, R.K. 1984: Convergence during secondary forest succession. Journal of Ecology 72: 25-36.
Christensen, N.L. 1985: Shrubland fire regimes and their evolutionary consequences. In: Pickett, S.T.A. & White, P.S. (eds.): The ecology of natural disturbance and patch dynamics. Academic Press, Inc., Orlando.
Cirne, P.& Scarano, F. 2001: Resprouting and growth dynamics after fire of the clonal shrub Andira legalis (Leguminosae) in a sandy coastal plain in south-eastern Brazil. Journal of Ecology 89: 351-357.
Clark, D.B. & Clark, D.A. 1985: Seedlings dynamics of a tropical tree: Impacts of herbivory and meristem damage. Ecology 66(6): 1884-1892.

Hermann, J.-M. PhD thesis References
189
Clark, D.B. & Clark, D.A. 1989: The role of physical damage in the seedling mortality regime of a neotropical rain forest. Oikos 55(2): 225-230.
Connell, J.H. & Slatyer, R.O. 1977: Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist 111: 1119-1144.
Connell, J.H., Noble, I.R. & Slatyer, R.O. 1987: On the mechanisms producing successional change. Oikos 50(1): 136-137.
Cook, R.E. 1985: Growth and development in clonal plant populations. In: Jackson, J.B.C., Buss, L.W. and Cook, R.E. (eds.): Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Crawley, M.J. 1990: The population dynamics of plants. Philosophical transactions of the Royal Society of London, B Series 330: 125-140.
Crawley, M.J. 1997a: Life History and Environment. In: Crawley, M.J. (ed.): Plant Ecology. (2nd. edition.). John Wiley, New York.
Crawley, M.J. 1997b: Sex. In: Crawley, M.J. (ed.): Plant Ecology. (2nd. edition.). John Wiley, New York.
Cubiña, A. & Aide, T. M.2001: The effect of distance from forest edge on seed rain and soil seed bank in a tropical pasture. Biotropica 33(2). 260-267.
Cunningham, S. & Read, J. 2003: Comparison of temperate and tropical rainforest tree species: growth responses to temperature. Journal of Biogeography 30:143-153.
da Rocha, H.O. 1981: Die Böden und geomorphologischen Einheiten der Region von Curitiba (Paraná-Brasilien). Freiburger Bodenkundliche Abhandlungen 10. (Schriftenreihe des Instituts für Bodenkunde und Waldernährungslehre der Albert-Ludwigs-Universität Freiburg i.Br.). URL: http://www.bodenkunde.uni-freiburg.de/veroeffentlichungen/fba/inhalt_10 [retrieved 2008-08-17].
Davis, M.B. 1987: Invasions of forest communities during the holocene: Beech and hemlock in the Great Lakes Region. In: Gray, J., Crawley, M.J. & Edwards, P.J. (eds.): Colonization, succession and stability. Blackwell Scientific Publications, London.
DCRS (Defesa Civil do Rio Grande do Sul) 2004: Tendência climática para o outono de 2004. (Boletim da Rede de Estação de Climatologia Urbana de São Leopoldo, divulgado 2004-03-19.) URL: http://www.defesacivil.rs.gov.br/ [retrieved 2008-08-17].
de Kroon, H. & van Groenendael, J. (eds.) 1997: The ecology and evolution of clonal plants: Glossary. Backhuys Publishers, Leiden, The Netherlands.
De Steven, D. 1991a: Experiments on mechanisms of tree establishment in old-field succession: Seedling emergence. Ecology 72(3): 1066-1075.
De Steven, D. 1991b: Experiments on mechanisms of tree establishment in old-field succession: Seedling survival and growth. Ecology 72(3): 1076-1088.
DeByle, N.V. 1964: Detection of functional intra-aspen root connections by tracers and excavation. Forest Science 10: 386-396.
Denslow, J. Sloane 1987: Tropical rainforest gaps and tree species diversity. Annual Review of Ecology and Systematics 18: 431-451.
DeSimone, S. A. & Zedler, P.H. 1999: Shrub seedling establishment in unburned Californian coastal sage scrub and adjacent grassland. Ecology 80(6): 2018-2032.

Hermann, J.-M. PhD thesis References
190
Dierschke, H. 1988: Pflanzensoziologische und ökologische Untersuchungen in Wäldern Süd-Niedersachsens. IV. Vegetationsentwicklung auf langfristigen Dauerflächen von Buchenwald-Kahlschlägen. Tuexenia 8: 307-326.
DSG (Diretoria de Serviço Geográfico-Brasil) 1981: Mapa de topografía da região sul do Brasil-1:100000, folha Cambará do Sul. (Baseada em fotografías aéreas de 1965 do AST – 10/USAF). Ministério do Exército, Departamento de Engenharía e Comunicações, Brasil.
Duarte, L. da Silva, Marchesini, M. dos Santos, Hartz, S.M. & Pillar, V. de Patta 2006a: Role of nurse plants in Araucaria Forest expansion over grassland in south Brazil. Austral Ecology 31(4): 520–528.
Duarte, L. da Silva, Machado, R.E., Hartz, S.M. & Pillar, V. de Patta 2006b: What saplings can tell us about forest expansion over natural grasslands. Journal of Vegetation Science 17: 799-808.
Duckworth, J.C., Kent, M. & Ramsay, P.M. 2000: Plant functional types: an alternative to taxonomy in plant community description in biogeography? Progress in Physical Geography 24: 515-542.
Dümig, A., Schad, P., Rumpel, C., Dignac, M.-F. & Kögel-Knabner, I. 2008a: Araucaria forest expansion on grassland in the southern Brazilian highlands as revealed by 14C and δ13C studies. Geoderma 145(1-2): 143-157.
Dümig, A., Schad, P., Kohoka, M., Beyerlein, P., Schwimmer, W. & Kögel-Knabner, I. 2008b: A mosaic of nonallophanic Andosols, Umbrisols and Cambisols on rhyodacite in the southern Brazilian highlands. Geoderma 145(1-2): 158-173.
Ehrlén, J. & van Groenendael, J.M. 1998: The trade-off between dispersability and longevity: An important aspect of plant species diversity. Applied Vegetation Science 1(1): 29-36.
Ellenberg, H. 1996: Vegetation Mitteleuropas mit den Alpen. (5th edition.) Ulmer, Stuttgart.
Ellison, L. & Houston, W.R. 1958: Production of herbaceous vegetation in openings and under canopies of Western Aspen. Ecology 39(2): 337-345.
Ellison, A.M., Denslow, J. Sloan, Loiselle, B.A. & Brenés, D.M. 1993: Seed and seedling ecology of neotropical Melastomataceae. Ecology 74(6): 1733-1749.
Ellstrand, N.C. & Roose, M.L. 1987: Patterns of genotypic diversity in clonal plant species. American Journal of Botany 74(1): 123-131.
Eriksson, O. & Ehrlén, J. 1992: Seed and microsite limitation of recruitment in plant populations. Oecologia 91: 360-364.
Eriksson, O. & Fröborg, H. 1996: „Windows of opportunity“ for recruitment in long-lived clonal plants: experimental studies of seedling establishment in Vaccinium shrubs. Can. J. Bot. 74: 1369-1374.
Eriksson, O. 1989: Seedling dynamics and life Histories in clonal plants. Oikos 55(2): 231-238.
Eriksson, O. 1992: Population structure and dynamics of the clonal dwarf shrub Linnaea borealis. Journal of Vegetation Science 3(1): 61-68.
Eriksson, O. 1996: Regional Dynamics of Plants: A Review of Evidence for Remnant, Source-Sink and Metapopulations. Oikos, Vol. 77(2): 248-258.
Evangelista, A.R., de Carvalho, M. Mesquita & Curi, N. 1993: Uso de fogo em pastagens. In: Favoretto, V., Rodrigues, L.R. de Andrade & Reis, R. Andrade (eds.): Simpósio sobre Ecossistema de Pastagens II, Jaboticaba, RS, Brazil.

Hermann, J.-M. PhD thesis References
191
Everham, E.M. III, Myster, R.W. & VanDeGenachte, E. 1996: Effects of light, moisture, temperature, and litter on the regeneration of five tree species in the tropical montane wet forest of Puerto Rico. American Journal of Botany 83(8): 1063-1068.
Facelli, J. M. 1994: Multiple indirect effects of plant litter affect the establishment of woody seedlings in old fields. Ecology 75(6): 1727-1735.
Farmer, R.E., Jr. 1962: Aspen root sucker formation and apical dominance. Forest Science 8: 403-410.
Fenner, M. 1987a: Seed characteristics in relation to succession. In: Gray, J., Crawley, M.J. & Edwards, P.J. (eds.): Colonization, succession and stability. Blackwell Scientific Publications, London.
Fenner, M. 1987b: Seedlings. New Phytologist 106 (Suppl.): 35-47.
Felipe, J. 2006: Topografia do Brasil (map). (Published online in Internet on Wikipedia Brasil.) URL: http://www.melhordawiki.com.br/wiki/Imagem:Brazil_topo_en2.PNG#file [retrieved 2008-08-17].
Ferreira, A.G., Cassol, B., da Rosa, S. Galli Taylor, da Silveira, T. Sales, Stival, A.L. & Silva, A.A. 2001: Germinação de sementes de Asteraceae nativas no Rio Grande do Sul, Brasil. Acta Botanica Brasilica 15(2): 231-242.
Fidelis, A. 2004: Effects of disturbance on population ecology of Eryngium horridum Malme in grasslands in Southern Brazil. Master Thesis Technische Universität München – TUM, Freising-Weihenstephan, Germany.
Fidelis, A., Blanco, C., Müller, S.C., Pillar, V. De Patta & Pfadenhauer, J. (in prep. a): Vegetation regeneration in subtropical grasslands in Brazil: Effects of fire and mowing.
Fidelis, A., Appezzato-da-Glória, B., Müller, S.C., Pillar, V. De Patta & Pfadenhauer, J. (in prep. b): Fire and bud bank: Their importance for vegetation regeneration in subtropical grasslands in Brazil.
Fischer, M. & van Kleunen, M. 2001: On the evolution of clonal plant life histories. Evolutionary Ecology 15(4-6): 565-582.
Formento, S., Schorn, L. A. & Ramos, R. A. Bley 2004: Dinâmica estrutural arbórea de uma floresta ombrófila mista em Campo Belo do Sul, SC. (Tree structural dynamics of an area of mixed rain forest in Campo Belo do Sul, Brazil.) Cerne (Lavras) 10(2): 196-212.
Fraser, E.C., Lieffers, V. J. & Landhäusser, S.M. 2004: Wounding of aspen roots promotes suckering. Can. J. Bot. 82(3): 310–315
Gibson, D.J. 2002: Methods in comparative plant population ecology. Oxford University Press, Oxford.
Gill, D. S. & Marks, P. L. 1991: Tree and shrub seedling colonization of old fields in central New York. Ecological Monographs 61(2): 183-205.
Giongo, C. & Waechter, J. L. 2007: Composição florística e espectro de dispersão das espécies arbóreas de uma floresta mista com Podocarpus, Rio Grande do Sul. Revista Brasileira de Biosciências (Porto Alegre) 5(2): 333-335.
Givnish, T.J. 1987: Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints. New Phytologist 106 (Suppl.): 131-160.
Givnish, T.J. 2002: Ecological constraints on the evolution of plasticity in plants. Evolutionary Ecology 16: 213-242.

Hermann, J.-M. PhD thesis References
192
Glenn-Lewin, D.C. & van der Maarel, E. 1992: Patterns and processes of vegetation dynamics. In: Glenn-Lewin, D.C. & Veblen, T.T. (eds.): Plant succession: theory and prediction. (Population and community biology series 11.) Chapman & Hall, London.
Gomes, V. & Fernandes, G. Wilson 2002: Germinação de aquênios de Baccharis dracunculifolia D.C. (Asteraceae). Acta Botanica Brasilca 16(4): 421-427.
Green, R.H. 1979: Sampling design and statistical methods for environmental biologists. John Wiley & Sons, New York.
Grime, J.P., Crick, J.C. & Rincon, J.E. 1986: The ecological significance of plasticity. In: Jennings, D.H. & Trewavas, A.J. (eds.): Plasticity in plants. (Symposia of the Society for Experimental Biology XXXX.) The Society for Experimental Biology, Cambridge.
Harper, J.L. 1977: Population Biology of Plants. Academic Press Ltd. (Harcourt Brace Jovanovich Publishers), London.
Harper, J.L. 1978: Plant relations in pastures. In: Wilson, J.R. (ed.): Plant Relations in Pastures. CSIRO Publishing, Collingwood, Victoria.
Harper, J.L. 1985: Modules, branches and the capture of resources. In: Jackson, J.B.C., Buss, L.W. and Cook, R.E. (eds.): Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Harper, J.L. 1988: An apophasis of plant population biology. In: Davy, A.J., Hutchings, M.J. & Watkinson, A.R. (eds.): Plant Population Ecology. (The 28th Symposium of the British Ecological Society, Sussex 1988.) Blackwell Scientific Publications, Oxford.
Harris, R. J. 1975: A primer of multivariate statistics. Academic Press (Harcourt Brace Jovanovich Publishers), New York.
Harter, B. 1999: Bienen und ihre Trachtpflanzen im Araukarien-Hochland von Rio Grande do Sul, mit Fallstudien zur Bestäubung von Pionierpflanzen. PhD Thesis Eberhard-Karls-Universität Tübingen, Germany.
Heisler, J.L., Briggs, J.M. & Knapp, A.K. 2003: Long-term pattern of shrub expansion in a C4-dominated grassland: fire frequency and the dynamics of shrub cover and abundance. American Journal of Botany 90: 423-428.
Hennerich, J., Ughini, S., Nicknich, F., da Cunha, G.R., Pires, J.L., Luiz, A.R.M. & Pasinato, A. 2005: Identificação de sinais de mudanças no regime de geadas no Rio Grande do Sul. (Identifying signals of change in the frost regime of Rio Grande do Sul.) Proceedings of the XIV. Brazilian Congress of Agrometeorology in Campinas, SP, Brasil. UNICAMP, EMBRAPA. URL: www.cnpt.embrapa.br/biblio/do/p_do71_tc31-1.PDF [retrieved 2007-12-06].
Heringer, I. & Jacques, A. V. Àvila 2001: Adaptação das plantas ao fogo: Enfoque na transição floresta-campo. (Plants adaptation to burning: Forest-grassland transition.) Ciência Rural 31(6): 1085-1090.
Heringer, I. 2000: Efeitos do fogo por longo periodo e de alernativas de manejo sobre o solo e a vegetação de uma pastagem natural. PhD thesis, Universidade Federal do Rio Grande do Sul - UFRGS, Porto Alegre, RS.
Herrmann, M.L. de Paula & Rosa, R. de Oliveira 1990: Relevo. In: IBGE (Instituto Brasilerio de Geografía e Estatística) (ed.): Geografía do Brasil, Região Sul, Rio de Janeiro.

Hermann, J.-M. PhD thesis References
193
Hobbs, R.J. & Mooney, H.A. 1985: Vegetative regrowth following cutting in the shrub Baccharis pilularis ssp. consanguinea (DC) C. B. Wolf. American Journal of Botany 72(4): 514-519.
Hodgkinson, K.C. 1998: Sprouting success of shrubs after fire: height-dependent relationships for different strategies. Oecologia 115: 64-72.
Hoffmann, W.A. & Moreira, A.G. 2002: The role of fire in population dynamics of woody plants. In: Oliveira, P.S. & Marquis, R.J. (eds.): The cerrados of Brazil. Ecology and natural history of a Neotropical Savanna. Columbia University Press, New York.
Hoffmann, W.A. 1996: The effects of fire and cover on seedling establishment in a neotropical savanna. Journal of Ecology 84: 383-393.
Hoffmann, W.A. 1998: Post-burn reproduction of woody plants in a neotropical savanna: the relative importance of sexual and vegetative reproduction. Journal of Applied Ecology 35: 422-433.
Hoffmann, W.A. 2000: Post-establishment seedling success in the Brazilian Cerrado: A comparison of savanna and forest species. Biotropica 32(1): 62-69
Holl, K.D. 1998: Effects of above-and below-ground competition of shrubs and grass on Calophyllum brasiliense (Camb.) seedling growth in abandoned tropical pasture. Forest Ecology and Management 109: 187-195.
Holl, K.D. 1999: Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate and soil. Biotropica 31(2): 229-242.
Hueck, K. 1952: Verbreitung und Standortansprüche der brasilianischen Araukarie (Araucaria angustifolia). Forstwissenschaftliches Centralblatt 71(9/10): 272-289.
Hulme, P.E. 1996: Natural regeneration of yew (Taxus baccata L.): microsite, seed or herbivore limitation? Journal of Ecology 84: 853-861.
Hulme, P.E. 2008: Phenotypic plasticity and plant invasions: is it all Jack? Functional Ecology 22(1): 3–7.
Hutchings, M.J. 1986: Plant population biology. In: Moore, P.D. & Chapman, S.B. (eds.): Methods in plant ecology. Blackwell Scientific Publications, Oxford.
Hutchings, M.J. 1997: Resource allocation patterns in clonal herbs and their consequences for growth. In: Bazzaz, F.A. & Grace, J. (eds.): Plant resource allocation. Academic Press, San Diego.
Hutchings, M.J. 1997: The structure of plant populations. In: Crawley, M.J. (ed.): Plant Ecology. (2nd. edition.). John Wiley, New York.
Irslinger, R., Burgert, E. & Schumacher, M.V. 2001: Bodenuntersuchungen – Dynamik der Wasserspannung [Pró-Mata, Rio Grande do Sul, Brasilien]. In: Engels, W., Hampp, R. & Tzschupke, W. (eds.): Zwischenbericht zum BMBF-Projekt 01LT0011/7 „Araukarienwald“. Forschungsschwerpunkt Region 4: Mata Atlântica. URL: http://www.uni-tuebingen.de/entw-phys/CD/2000.pdf#page=60 [retrieved 2008-08-17].
Jager, J.C. & Looman, C.W.N. 1987: Data collection. In: Jongman, R.H.G., ter Braak, C.J.F. & van Tongeren, O.F.R. (eds.): Data analysis in community and landscape ecology. Pudoc, Wageningen]
Jelinski & Cheliak 1992: Genetic diversity and spatial subdivision of Populus tremuloides in a heterogenous landscape. American Journal of Botany 79:728-736.

Hermann, J.-M. PhD thesis References
194
Jenik, J. 1994: Clonal growth in woody plants. A review. Folia Geobotanica et Phytotaxonomica (Praha) 29: 291-306.
Johnson, W.C. & Adkisson, C.S. 1985: Dispersal of beech nuts by blue jays in fragmented landscapes. American Midland Naturalist 113 (2): 319-324.
Jungblut, M. & Pinto, L.F.S. 1997a: Levantamento de solos do Centro de Pesquisas e Conservação da Natureza Pró-Mata. Divul. Mus. Ciênc. Tecnolo. - UBEA/PUCRS 3: 29-94 (print).
Jungblut, M. & Pinto, L.F.S. 1997b: Mapa de solos do Centro de Pesquisas e Conservação da Natureza Pró-Mata, escala 1:90.000. (Excerpt from Divul. Mus. Ciênc. Tecnolo. - UBEA/PUCRS 3: 29-94, published online in Internet.) URL: http://www.pucrs.br/ima/promata/diag.htm [retrieved 2008-08-17].
Kahmen, S. & Poschlod, P. 2004: Plant functional trait responses to grassland succession over 25 years. Journal of Vegetation Science 15: 21-32.
Kammesheidt, L. 1999: Forest recovery by root suckers and above-ground sprouts after slash-and-burn agriculture, fire and logging in Paraguay and Venezuela. Journal of Tropical Ecology 15: 143-157.
Kaul, P.F. Teixeira 1990: Geología. In: IBGE (Instituto Brasilerio de Geografía e Estatística) (ed.): Geografía do Brasil, Região Sul, Rio de Janeiro.
Keeley, J.E. & Zedler, P.H. 1978: Reproduction of chaparral shrubs after fire: A comparison of sprouting and seeding strategies. The American Midland Naturalist 99(1): 142-161.
Keeley, J.E. 1992: Recruitment of seedlings and vegetative sprouts in unburned chaparral. Ecology 73(4): 1194-1208.
Kemperman, J.A. & Barnes, B.V. 1976: Clone size in American aspens. Canadian Journal of Botany 54: 2603-2607.
Kenney, W.R., Bock, J.H. & Bock, C.E. 1986: Responses of the shrub Baccharis pteronoides to livestock exclosure in southeastern Arizona. American Midland Naturalist 116(2): 429-431.
Klein, R.M. 1960: O aspeto dinâmico do pinheiro brasilieiro. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 12: 17-44.
Klein, R.M. 1963: Observações e considerações sôbre a vegetação do Planalto Nordeste Catarinense. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 15: 39-56.
Klekowski, E., Jr. 1995: Somatic mutation theory of clonality. In: de Kroon, H. & van Groenendael, J. (eds.): The ecology and evolution of clonal plants. Backhuys Publishers, Leiden, The Netherlands.
Klimešová, J. & Martínková, J. 2004: Intermediate growth forms as a model for the study of plant clonality functioning: An example with root sprouters. Evolutionary Ecology 18: 669-681.
Koch, Z. & Corrêa, M.C. 2002: Araucária: a floresta do Brasil meridional. (Araucária: The Meridional Brazil Forest.) Olhar Brasileiro, Curítiba, Brazil.
Koop, H. 1987: Vegetative reproduction of trees in some European natural forests. Plant Ecology 72(2): 103-110.
Kozera, C., Dittrich, V. A. de Oliveira & Silva, S. Menezes 2006: Fitossociologia do componente arbóreo de um fragmento de floresta ombrófila mista montana, Curitiba, PR, Brasil. Floresta (Curitiba) 36 (2): 225-237.

Hermann, J.-M. PhD thesis References
195
Kozlowski, T.T., Kramer, P.J.& Pallardy, S.G. (eds.) 1991: The physiological ecology of woody plants. Academic Press, Inc., San Diego.
Kreher, S.A., Foré, S.A. & Collins, B.S. 2000: Genetic variation within and among patches of the clonal species, Vaccinium stamineum L. Molecular Ecology 9 (9): 1247–1252.
Lamont, B.B. & Barrett, G.J. 1988: Constraints on seed production and storage in a root-suckering Banksia. Journal of Ecology 76: 1069-1082.
Lamont, B.B. & Wiens, D. 2003: Are seed set and speciation rates always low among species that resprout after fire, and why? Evolutionary Ecology 17: 277-292.
Larcher, W. 2005: Climatic constraints drive the evolution of low temperature resistance in woody plants. J. Agric. Meteorol. 61(4): 189-202.
Leite, P. Furtado & Klein, R.M. 1990: Vegetação. In: IBGE (Instituto Brasilerio de Geografía e Estatística) (ed.): Geografía do Brasil, Região Sul, Rio de Janeiro.
Living Countryside Ltd 2003: The ancient art of coppicing. URL: http://www.coppicing.com/index.html [retrieved 2008-08-17]. Living Countryside Ltd, Petersfield, Hampshire GU31 4AD, U.K.
Lloret, F. & López-Soria, L. 1993: Resprouting of Erica multiflora after experimental fire treatments. Journal of Vegetation Science 4: 367-374.
Lloret, F., Verdú, M., Flores-Hernández, N. & Valiente-Banuet, A. 1999: Fire and resprouting in Mediterranean Ecosystems: Insights from an external biogeographical region, the mexical shrubland. American Journal of Botany 86(12): 1655-1661.
Lorenzi, H. 1992: Árvores brasileiras. Manual de identificação e cultivo de plantas arbóreas do Brasil. Editora Plantarum, Nova Odessa, SP, Brazil.
Lorimer, C.G. 1989: Relative effects of small and large disturbances on temperate hardwood forest structure. Ecology 70(3): 565-567.
Lovett Doust, J. & Lovett Doust, L. 1988: Modules of production and reproduction in a dioecious clonal shrub, Rhus typhina. Ecology 69(3): 741-750.
Lovett Doust, L. 1981: Population dynamics and local specialization in a clonal perennial (Ranunculus repens). I. The dynamics of ramets in contrasting habitats. Journal of Ecology 69: 743-755.
Machado, R. Engelman 2004: Padrões vegetacionais em capões de floresta com Araucária no Planalto Nordeste do Rio Grande do Sul, Brasil. Master Thesis Universidade Federal do Rio Grande do Sul – UFRGS, Porto Alegre, RS, Brazil.
Mack, R.N. & Thompson, J.N. 1982: Evolution in steppe with few large, hooved mammals. The American Naturalist 119(6): 757-773.
Maini, J.S. & Horton, K.W. 1966: Vegetative propagation of Populus ssp. 1. Influence of temperature on formation and initial growth of Aspen suckers. Can. J. Bot. 44: 1183-1189.
Martínez-Ramos, M., Alvarez-Byulla, E. & Sarukhán, J. 1989: Tree demography and gap dynamics in a tropical rain forest. Ecology 70(3): 555-558.
Matzenbacher, N.I. in prep.: Contribuição ao conhecimento da flórula do Pró-Mata II (Levantamento preliminar – listagem ampliada). Updated October 31st 2003. Manuscript in preparation kindly forwarded by Prof. N Matzenbacher, UFRGS, in 2004.

Hermann, J.-M. PhD thesis References
196
Mauhs, J. 2002: Fitossociologia e regeneração natural de um fragmento de floresta ombrófila mista exposto a perturbaçõs antrópicas. Master Thesis Universidade do Vale do Rio dos Sinos – UNISINOS, São Leopoldo, RS, Brazil.
McCarron, J.K. & Knapp, A.K. 2001: C3 woody plant expansion in a C4 grassland: Are grasses and shrubs functionally distinct? American Journal of Botany 88(10): 1818-1823.
McCarron, J.K. & Knapp, A.K. 2003: C3 shrub expansion in a C4 grassland: Positive post-fire responses in resources and shoot growth. American Journal of Botany 90(10): 1496–1501.
McCarthy, B.C. & Facelli, J.M. 1990: Microdisturbances in old fields and forests: implications for woody seedling establishment. Oikos 58: 55-60.
McGarigal, K., Cushman, S. & Stafford, S. 2000: Multivariate Statistics for wildlife and ecology research. Springer, New York.
Mohr, H. & Schopfer, P. 1992: Pflanzenphysiologie. (4. Auflage.) Springer Verlag, Berlin.
Mohrdieck, K.H. 1993: Formações campestres do Rio Grande do Sul. In: Campo nativo: Melhoramento e Manejo. Proceedings of the Ivth symposium of Federacite, Esteio, Rio Grande so Sul, Brazil.
Moles, A.T. & Westoby, M. 2004: Seedling survival and seed size: a synthesis of the literature. Journal of Ecology 92: 372-383.
Monro, K. & Poore, A.G.B. 2004: Selection in Modular Organisms: Is Intraclonal Variation in Macroalgae Evolutionarily Important? American Naturalist 163: 564–578.
Monteiro, S.A. & Couto, T. de P. in prep.: Registro aerofotográfico do avanço das florestas subtropicais sobre os campos do Planalto Meridional. Manuscript in preparation kindly forwarded by T. Couto, IBAMA Porto Alegre, in 2006.
Morgan, J.W. 2001: Seedling recruitment patterns over 4 years in an Australian perennial grassland community with different fire histories. Journal of Ecology 89: 908-919.
Moser, J.M. 1990: Solos. In: IBGE (Instituto Brasilerio de Geografía e Estatística) (ed.): Geografía do Brasil, Região Sul, Rio de Janeiro.
MPRS (Ministério Público do Rio Grande do Sul) 2008: O Combate as Queimadas na Legislação Federal e Estadual. URL: http://www.mp.rs.gov.br/ambiente/doutrina/id18.htm [retrieved 2008-08-17]. Ministério Público do Rio Grande do Sul, 90050-190 Porto Alegre, RS, Brasil.
Müller, S.C. & Forneck, E. Dias 2004: Forest-grassland mosaics in the hills of Porto Alegre city: a study case of forest expansion patterns in Santana hill, Rio Grande do Sul, Brazil. In: Porto, M.L. (ed.): Proceedings of the Workshop „Proteção e Manejo da Vegetação Natural de Porto Alegre com base em pesquisa de Padrões e dinâmica de vegetação“ (Porto Alegre, Nov 5-8 2004). Editora UFRGS, Porto Alegre.
Müller, S.C., Overbeck, G.E., Pfadenhauer, J. & Pillar, V. De Patta 2007: Plant Functional Types of Woody Species Related to Fire Disturbance in Forest–Grassland Ecotones. Plant Ecology 189(1): 1-14.
Müller-Dombois, D. & Ellenberg, H. 1974: Aims and methods of vegetation ecology. John Wiley, New York.
Municipality São Francisco de Paula 2008: A cidade – história. (Official Web page). Last updated 2006-06-30. URL: http://www.saochico.com.br/acidade.htm [retrieved 2008-08-17]. Prefeitura Municipal de São Francisco de Paula, 95400-000 RS, Brasil.

Hermann, J.-M. PhD thesis References
197
Municipality Canela 2008: Histórico. (Official Web page). URL: http://www.canela.rs.gov.br/?opcao=historia [retrieved 2008-08-17]. Prefeitura Municipal de Canela, 95680-000 RS, Brasil.
Nabinger, C., de Moraes, A. & Maraschin, G.E. 2000: Campos in Southern Brazil. In: Lemaire, G., Hodgson, J., de Moraes, A., Carvalho, P.C. de F., Nabinger, C. (eds.): Grassland Ecophysiology and Grazing Ecology. CAB International Publishing.
Nascimento, A. R. Terra, Longhi, S. J. & Brena, D. A. 2001: Estrutura e padrões de distribuição espacial de espécies arbóreas em uma amostra de floresta ombrófila mista em Nova Prata, RS. (Structure and spatial distribution patterns of tree species in a Mixed Ombrophyllous Forest sample in Nova Prata, RS.) Ciência Florestal (Santa Maria) 11(1): 105-119.
Neto, R. Marques Rondon, Kozera, C., Andrade, R. do Rocio de, Cecy, A. T., Hummes, A. P., Fritzsons, E., Caldeira, M. V. Winckler, Maciel, M. de Nazaré & de Souza, M. Koch Fernandes 2002 b: Caracterização florística e estrutural de um fragmento de floresta ombrófila mista, em Curitiba, PR – Brasil. (Floristic and structural characterisation of a fragment of mixed mbrophylous forest in Curitiba, PR- Brazil.) Floresta (Curitiba) 32 (1): 3-16.
Neto, R. Marques Rondon, Watzlawick, L. Farinha, Caldeira, M. V. Winchler & Schoeninger, E. R. 2002 a: Análise florística e estrutural de um fragmento de floresta ombrófila mista montana, situado em Criúva, RS - Brasil. (Floristic and strcutural analysis of a montane mixed ombrophylous forest fragment in Criúva, RS – Brazil.) Ciência Florestal (Santa Maria) 12(1): 29-37.
Nimer, E.1990: Clima. In: IBGE (Instituto Brasilerio de Geografía e Estatística) (ed.): Geografía do Brasil, Região Sul, Rio de Janeiro.
Noack, W. 1999: Synökologische Untersuchung der Sekundärvegetation auf ehemaligem Weideland im Pró-Mata Projekt-Gebiet, Rio Grande do Sul, Brasilien. Diploma thesis Eberhard-Karls-Universität Tübingen, Germany.
Noble, I.R. & Slatyer, R.O. 1980: The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 43: 5-21.
Oliveira, J. Morales & Pillar, V. de Patta 2004: Vegetation dynamics on mosaics of Campos and Araucaria forest between 1974 and 1999 in Southern Brazil. Community Ecology 5(2): 197-202.
Oliveira, J. Morales 2003: Padrões e processos espaço-temporais em ecotonos de campos e floresta com Araucária, em São Francisco de Paula, RS. Master Thesis, Universidade Federal do Rio Grande do Sul – UFRGS, Porto Alegre, Brazil.
Oliveira, M. de L.A.A. de, Balbueno, R.A. & Senna, R.M. 2005: Levantamento florístico de fragmentos florestais na bacia hidrográfica do rio Gravataí, Rio Grande do Sul, Brasil. (Floristic survey of forest fragments in the Gravataí river basin, Rio Grande do Sul, Brazil.) Iherengia (Sér. Bot.) 60(2): 269-284.
Ooi, M. K.J., Auld, T.D. & Whelan, R.J. 2007: Distinguishing between persistence and dormancy in soil seed banks of three shrub species from fire-prone southeastern Australia. Journal of Vegetation Science 18(3): 405-412.
Otegui, M. 1998: Sinopsis del género Myrsine L. (Myrsinaceae) en el Cono Sur de América del Sur. Candollea 53: 133-157.

Hermann, J.-M. PhD thesis References
198
Overbeck, G. 2004: Fire in subtropical grasslands: Factor of environmental impact or management tool? In: Porto, M.L. (ed.): Proceedings of the Workshop „Proteção e Manejo da Vegetação Natural de Porto Alegre com base em pesquisa de Padrões e dinâmica de vegetação“ (Porto Alegre, Nov 5-8 2004). UFRGS, Porto Alegre.
Overbeck, G.E., Müller, S.C., Pillar, V. De Patta & Pfadenhauer, J. 2005a: No heat-stimulated germination found in herbaceous species from burned subtropical grassland. Plant Ecology 184(2): 237-243.
Overbeck, G.E., Müller, S.C., Pillar, V. De Patta & Pfadenhauer, J. 2005b: Fine-scale post-fire dynamics in southern Brazilian subtropical grassland. Journal of Vegetation Science 16(6): 655-664.
Overbeck, G.E., Müller, S.C., Fidelis, A., Pfadenhauer, J., Pillar, V. De Patta, Blanco, C.C., Boldrini, I.I., Both, R. & Forneck, E.D. 2007: Brazil's neglected biome: The South Brazilian Campos. Perspectives in Plant Ecology, Evolution and Systematics 9(2): 101-116.
Pan, J.J. & Price, J.S. 2002: Fitness and evolution in clonal plants: The impact of clonal growth. Evolutionary Ecology 15: 583-600.
Pascotto, M.C. 2007: Rapanea ferruginea (Ruiz & Pav.) Mez. (Myrsinaceae) como uma importante fonte alimentar para as aves em uma mata de galeria no interior do estado de São Paulo. Revista Brasilieira de Zoologia 24 (3): 735-741.
Pate, J.S., Froend, R.H., Bowen, B.J., Hansen, A. & Kuo, J. 1990: Seedling growth and storage characteristics of seeder and resprouter species of Mediterranean-type ecosystems of S. W. Australia. Annals of Botany 65: 585-601.
Pate, J.S., Meney, K.A. & Dixon, K.W. 1991: Contrasting growth and morphological characteristics of fire-sensitive (obligate seeder) and fire-resistant (resprouter) species of Restionaceae (S. Hemisphere Restiads) from South-Western Western Australia. Australian Journal of Botany 39(6): 505-525.
Peet, R.K. & Christensen, N.L. 1980: Succession: A population process. Vegetatio 43: 131-140.
Peet, R.K. 1992: Community structure and ecosystem function. In: Glenn-Lewin, D.C. & Veblen, T.T. (eds.): Plant succession: theory and prediction. (Population and community biology series 11.) Chapman & Hall, London.
Peltzer, D.A. 2002: Does clonal integration improve competitive ability? A test using Aspen (Populus tremuloides [Salicaceae]) invasion into prairie. American Journal of Botany 89(3): 494-499.
Pennings, S.C. & Callaway, R.M. 2000: The advantage of clonal integration under different ecological conditions: A community-wide test. Ecology 81(3): 709-716.
Perala, D. 1978: Aspen Sucker Production and Growth from Outplanted Root Cuttings. USDA For. Serv. Res. Note NC-24, Washington. URL: http://ncrs.fs.fed.us/pubs/rn/rn_nc241.pdf [retrieved 2008-08-17].
Peterson, C.J. & Jones, R.H. 1997: Clonality in woody plants: A review and comparison with clonal herbs. In: De Kroon, H. & van Groenendael, J. (eds.) 1997: The ecology and evolution of clonal plants. Backhuys Publishers, Leiden, The Netherlands.
Pickett, S.T.A. 1982: Population patterns through twenty years of oldfield succession. Vegetatio 49: 45-59.

Hermann, J.-M. PhD thesis References
199
Pickett, S.T.A., Collins, S.L. & Armesto, J.J. 1987: Models, mechanisms and pathways of succession. The Botanical Review 53(3): 335-371.
Pillar, V. De Patta & de Quadros, L.F. 1997: Grassland-forest boundaries in Southern Brazil. Coenoses 12 (2-3): 119-126.
Pineschi, R.B. 1990: Aves como dispersores de sete espécies de Rapanea (Myrsinaceae) no maciço do Itatiaia, estados do Rio de Janeiro e Minas Gerais. Ararajuba 1: 73-78.
Pitelka, L.F. & Ashmun, J.M. 1985: Physiology and integration of ramets in clonal plants. In: Jackson, J.B.C., Buss, L.W. and Cook, R.E. (eds.): Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Pott, A. 1974: Leventamento ecológico da vegetação de um campo natural sob três condições: pastejado, excluido e melhorado. Master thesis Universidade Federal do Rio Grande do Sul - UFRGS, Porto Alegre.
Price, T.D., Qvarnström, A. & Irwin, D.E. 2003: The role of phenotypic plasticity in driving genetic evolution. Proceedings of the Royal Society of London, B Series 270: 1433-1440.
Rambo, B. 1952: Análise geográfica das Compostas Sulbrasileiras. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 4: 87-159
Rambo, B.S.J. 1953: História da flora do planalto riograndense. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 5(5): 185-232.
Rambo, B.S.J. 1954: História da flora do Litoral Riograndense. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 6: 113-172.
Rambo, B.S.J. 1956: A flora fanerogamica dos aparados riograndenses. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 7: 235-298.
Rambo, B.S.J. 1957: Regenwald und Kamp in Rio Grande do Sul. Sellowia (Anais Botânicos do Herbário “Barbosa Rodrigues”; Itajaí) 9(8): 257-298.
Rambo, B.S.J. 1958: Die alte Südflora in Brasilien. Pesquisas, sér. Bot 2: 177-198.
Ramos, A. J. Kegler & Boldo, E. 2007: Diversidade florística e aspectos fitossociológicos de formações florestais em estágio sucessional secundário na floresta ombrófila mista, município de Caxias do Sul-RS. (Resumos do II Congresso Brasileiro de Agroecologia.) Rev. Bras. Agroecologia 2(1): 111-116.
Recher, H.F. & Christensen, P.E. 1981: Fire and the evolution of the Australian biota. In Keast, A. (ed.): Ecological Biogeography of Australia. Dr. W. Junk Publishers, The Hague.
Romberger, J.A., Hejnowicz, Z. & Hill, J.F. 1993: Plant Structure: Function and development. A treatise on anatomy and vegetative development, with special reference to woody plants. Springer-Verlag, Berlin.
Rosário, D. de Almeida Pires do 2001: Padrões florísticos e tipos funcionais em floresta com Araucária e suas relações com o solo. Master Thesis Universidade Federal do Rio Grande do Sul – UFRGS, Porto Alegre, RS, Brazil.
Rothmaler, W. 2005: Gefäßpflanzen: Atlasband. (Exkursionsflora von Deutschland Band 3.) 9. Auflage (Hrsg. Jäger, E.E. & Werner, K.). Gustav Fischer Verlag, Jena.
Sala, O.E. 1988: The effect of herbivory on vegetation structure. In: Werger, M.G.A., van der Aart, P.J.M., During, H.J. & Verhoeven, J.T.A.: Plant form and vegetation structure. SPB Academic Publishing, The Hague.

Hermann, J.-M. PhD thesis References
200
Sarukhán, J. 1980: Demographic problems in tropical systems. In: Solbrig, O. (ed.): Demography and evolution in plants populations. (Botanical Monographs 15.) Blackwell Scientific Publications, Oxford.
Saverimuttu, T. & Westoby, M. 1996: Seedling longevity under deep shade in relation to seed size. Journal of Ecology 84:681-689.
Schaaf, L. Budant, Filha, A. Figueiredo, Galvão, F., Sanquetta, C. R. & Longhi, S. J. 2006: Modificações florístico-estruturais de um remanescente de floresta ombrófila mista montana no período entre 1979 e 2000. (Floristic and structural changes in an Ombrophyllous Mixed Forest between the years 1979 and 2000.) Ciência Florestal (Santa Maria) 16(3): 271-291.
Scheffer, F. & Schachtschabel, P. 2002: Lehrbuch der Bodenkunde. 15. Auflage. Spektrum Akademischer Verlag, Heidelberg.
Schreiber, K.-F. 1997: Grundzüge der Sukzession in 20-jährigen Grünland-Bracheversuchen in Baden-Württemberg. (Characteristics of succession in 20 years’ abandoned grassland studies in Baden-Württemberg.) Forstwissenschaftliches Centralblatt 116: 243-258.
Schütt, P., Schuck, H.J. & Stimm, B. (eds.) 2002: Lexikon der Baum- und Straucharten. Morphologie, Pathologie, Ökologie und Systematik wichtiger Baum- und Straucharten. Nikol Verlagsgesellschaft mbH & Co. KG, Hamburg.
Schupp, E.W., Howe, H.F. & Levey, D.J. 1989: Arrival and survival in tropical treefall gaps. Ecology 70(3): 562-564.
Sebens, K.P. & Thorne, B.L. 1985: Coexistence of clones, clonal diversity, and the effects of disturbance. In: Jackson, J.B.C., Buss, L.W. and Cook, R.E. (eds.): Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Seger, C. D., Dlugosz, F. L., Kurasz, G., Martinez, D. Tyszka, Ronconi, E., de Melo, L. A. Nunes, de Bittencourt, S. M., Brand, M. A., Carniattto, I., Galvão, F. & Roderjan, C. Vellozo 2005: Levantamento florístico e análise fitosociológica de um remanescente de floresta ombrófila mista localizado no município de pinhais, Paraná-Brasil. (Floristic composition and phytosciological analysis of a Mixed Araucaria Forest remainder in Pinhais county, Paraná state – Brazil.) Floresta (Curitiba) 35(2): 291-302.
Shackleton, C.M. 2001: Managing regrowth of an indigenous savanna tree species (Terminalia sericea) for fuelwood: the influence of stump dimensions and post-harvest coppice pruning. Biomass and Bioenergy 20(4): 261-270.
Shepperd, E. 2000: Manipulations to regenerate Aspen ecosystems. In: USDA Forest Service RMRS (eds.): Sustaining Aspen in Western Landscapes. Proceedings of the Symposium 13-15 June 2000, Grand Junction, CO. URL: http://www.fs.fed.us/rm/pubs/rmrs_p018/rmrs_p018_355_366.pdf [retrieved 2008-08-17].
Scholes, R.J. & Walker, B.H. 1993: An African Savanna. Synthesis of the Nylsvley study. Cambridge University Press, Cambridge.
Silva, A.F. da, Oliveira, R.V. de, Santos, N.R. Loureiro & Paula, A. de 2003: Composição florística e grupos ecológicos das espécies de um trecho e floresta semidecídua submontana de Fazenda São Geraldo, Viçosa-MG. Revista Árvore (Viçosa-MG) 27(3): 311-319.
Silvertown, J.W. & Lovett Doust, J. 1993: Introduction to plant population biology. Blackwell Scientific Publications, Oxford.

Hermann, J.-M. PhD thesis References
201
Spagnollo, E., Amado, T.J. Carneiro, Jacques, A.V. Avila, Conceição, P.C., Lemainski, C.L. & Dellamea, R.B. Cereser in prep.: Carbono e nitrogênio nas frações leve e estável da matéria orgânica em campo natural submetido a queima. Manuscript in preparation kindly forwarded by A. Jacques, UFRGS, in 2005.
Steinger, T., Roy, B.A. & Stanton, M.L. 2003: Evolution in stressful environments II: adaptive value and costs of plasticity in response to low light in Sinapis arvensis. Journal of Evolutionary Biology 16(2): 313-323.
Stewart, K.E.J., Bourn, N.A.D. & Thomas, J.A. 2001: An evaluation of three quick methods commonly used to assess sward height in ecology. Journal of Applied Ecology 38: 1148-1154.
Stiles, F.G. & Rosselli, L. 1993: Consumption of fruits of the Melastomataceae by birds: how diffuse is coevolution? In: Fleming, T.H. & Estrada, A. (eds.): Frugicory and seed dispersal: ecological and evolutionary aspects. Kluwer Academic Publishers, Dordrecht.
Strasburger, E., Noll, F. Schenck, H. & Schimper, A.F.W. (eds.) 1991: Lehrbuch der Botanik für Hochschulen (33rd edition). Gustav Fischer Verlag, Stuttgart.
Suvanto, L.I. & Latva-Karjanmaa, T.B. 2005: Clone identification and clonal structure of the European aspen (Populus tremula). Molecular Ecology 14 (9): 2851–2860.
Swaine, M.D. & Whitmore, T.C. 1988: On the definition of ecological species groups in tropical rain forests. Vegetatio 75: 81-86.
Teixeira, M. B., Coura-Neto, A. B., Pastore, U. & Filho, A. L. R. Ranger 1986: Vegetação; as regiões fitoecológicas, sua natureza, seus recursos econômicos; estudo fitogeográfico. In: IBGE (Ed.). Levantamento de recursos naturais. Vol. 33, pp. 541-632. Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro.
Thompson, K. 1987: Seeds and seed banks. New Phytologist 106 (Suppl.): 23-34.
Thompson, K., Ceriani, R.M., Bakker, J.P. & Bekker, R.M. 2003: Are seed dormancy and persistence in soil related? Seed Science Research 13: 97-100
Tiffney, B.H. & Niklas, K.J. 1985: Clonal growth in land plants: A paleobotanical perspective. In: Jackson, J.B.C., Buss, L.W. and Cook, R.E. (eds.): Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Tilman, D. 1990: Constraints and Tradeoffs: Toward a Predictive Theory of Competition and Succession. Oikos 58(1): 3-15.
Tuskan, Francis, K.E., Russ, S.L., Romme, W.H. & Turner, M.G. 1996: RAPD markers reveal diversity within and among clonal and seedling stands of aspen in Yellowstone National Park, U.S.A. Canadian Journal of Forest Research 26(12): 2088–2098.
Tyler, C.M. 1995: Factors contributing to postfire seedling establishment in chaparral: direct and indirect effects of fire. Journal of Ecology 83: 1009-1020.
USDA, NRCS (United States Department of Agriculture, Natural Resources Conservation Center) 2008: The PLANTS Database. URL: http://plants.usda.gov [retrieved 2008-08-17]. National Plant Data Center, Baton Rouge, LA 70874-4490 USA.
van Andel, J. & van den Bergh, J.P. 1987: Disturbance in grasslands: Outline of the theme. In: van Andel, J., Bakker, J.P. & Snaydon, R.W. (eds.): Disturbance in grasslands: Causes effects and processes. (Geobotany 10). Dr W. Junk Publishers, Dordrecht.

Hermann, J.-M. PhD thesis References
202
van Andel, J., van Baalen, J. & Rozijn, N.A.M.G. 1991: Population ecology of plant species in disturbed forest and grassland habitats. In: Rozema, J. & Verkleij, J.A.C. (eds.): Ecological Responses to Environmental Stresses. Kluwer Academic Publishers, Dordrecht, The Netherlands.
van der Maarel, E. 1988: Vegetation dynamics: patterns in time and space. Vegetatio 77: 7-19.
van der Valk, A. G. 1992: Establishment, colonization and persistence. In: Glenn-Lewin, D.C. & Veblen, T.T. (eds.): Plant succession: theory and prediction. (Population and community biology series 11.) Chapman & Hall, London.
van Kleunen, M., Fischer, M. & Schmid, B. 2000: Costs of plasticity in foraging characteristics of the clonal plant Ranunculus reptans. Evolution 54(6): 1947-1955.
Vázquez-Yanes, C. & Segovia, A. Orozco 1984: Ecophysiology of seed germination in the tropical humid forests of the world: a review. In: Medina, E., Mooney, H.A. & Vázquez-Yánez, C. (eds.): Physiological ecology of plants of the wet tropics. (Tasks for vegetation science 12.) Dr W. Junk Publishers, The Hague.
Veblen, T.T. 1989: Tree regeneration responses to gaps along a transandean gradient. Ecology 70(3): 541-543.
Veblen, T.T., Kitzberger, T., Raffaele, E. & Lorenz, D.C. 2003: Fire history and vegetation changes in Northern Patagonia, Argentina. In: Veblen, T.T., Baker, W.L., Montenegro, G. & Swetnam, T.W. (eds.): Fire and climatic change in temperate ecosystems of the Western Americas. (Ecological studies 160.) Springer, Berlin.
Vesk, P.A. & Westoby, M. 2004a: Sprouting ability across diverse disturbances and vegetation types worldwide. Journal of Ecology 92: 310-320.
Vesk, P.A. & Westoby, M. 2004b: Funding the bud bank: A review of the costs of buds. Oikos 106(1): 200-208.
Vieira, I.C.G., Uhl, C. & Nepstad, D. 1994: The role of the shrub Cordia multispicata Cham. as a ‘succession facilitator’ in an abandoned pasture, Paragominas, Amazonia. Vegetatio 115(2): 91-99.
Waechter, J.L., Cestaro, L.A. & Miotto, S.T.S. 1984: Vegetation types in the Ecological Station of Aracuri, Esmeralda, Rio Grande do Sul, Brazil. Phytocoenologia 12 (2/3): 261-269.
Waite, S. 2000: Statistical ecology in practice. A guide to analysing environmental and ecological field data. Prentice Hall, London.
Walter, H. 1971: Ecology of tropical and subtropical vegetation. Oliver & Boyd, Edinburgh.
Walter, H. 1984: Spezielle Ökologie der tropischen und subtropischen Zonen. In: Walter, H. & Breckle, S.-W. (eds.): Ökologie der Erde 2. (UTB für Wissenschaft: Grosse Reihe.) Gustav Fischer Verlag, Stuttgart.
Watzlawick, L. Farinha, Nutto, L., Spathelf, P., Reif, A., Caldeira, M.V. Winckler & Sanquetta, C.R. undated: Die phytogeografischen Einheiten von Paraná, Brasilien. (Published online in Internet on Freiburger Dokumentenserver FreiDok, University of Freiburg, Germany.) URL: http://www.freidok.uni-freiburg.de/volltexte/1036/pdf/Distiller_Livro%20Fitogeografia_24.05.pdf [retrieved 2008-08-17].
Webb, S.D. 1978: A history of savanna vertebrates in the New World. Part II: South America and the Great Interchange. Ann. Rev. Ecol. Syst. 9: 393-426.

Hermann, J.-M. PhD thesis References
203
Wenny, D.G. 2000: Seed dispersal, seed predation, and seedling recruitment of a neotropical montane tree. Ecological Monographs 70(2): 331-351.
White, P.S. & Pickett, S.T.A. 1985: Natural disturbance and patch dynamics: an introduction. In: Pickett, S.T.A. & White, P.S. (eds.): The ecology of natural disturbance and patch dynamics. Academic Press, Inc., Orlando.
Whitmore, T.C. 1989: Canopy gaps and the two major groups of forest trees. Ecology 70(3): 536-538.
Whittaker, R.J. 1993: Plant population patterns in a glacier foreland succession: pioneer herbs and later-colonizing shrubs. Ecography 16: 117-136.
Wilson, S.D. 1993: Belowground competition in forest and prairie. Oikos 68: 146-150.
Wilson, S.D. 1998: Competition between grasses and woody plants. In: Cheplick, G.P. (ed.): Population biology of grasses. Cambridge University Press, Cambridge.
Winkler, E. & Fischer, M. 1999: Two fitness measures for clonal plants and the importance of spatial aspects. Plant Ecology 141: 191–199.
Yeh, F.C., Chong, D. K.X. & Yang, R.-C. 1995: RAPD variation within and among natural populations of Trembling Aspen (Populus tremuloides Michx.) from Alberta. The Journal of Heredity 86(6): 454-460.
Zahner, R. & Crawford, N.A. 1965: The clonal concept in Aspen site relations. In: Youngberg, C.T. (ed.): Forest-soil relationships in North America. Oregon State University Press, Corvallis, Oregon.
Zahner, R. & DeByle, N.V. 1965: Effect of pruning the parent root on growth of Aspen suckers. Ecology 46(3): 373-375.


















