

Prof. Dr. Silvio O. Rizzoli 1. Stunde: Wiederholung der...
101
1 Prof. Dr. Silvio O. Rizzoli 1. Stunde: Wiederholung der Eigenschaften von Neuron und Axon. Elektrische Synapsen. Wie bereits in früheren Vorlesungen erklärt: • Membranen, ihre Eigenschaften und Permeabilität für verschiedene Elemente • Organisation des Neurons: Zellkörper, Dendriten, Axonhügel, Axon • Myelin und seine Funktion • Generierung von Aktionspotentialen • spannungsabhängige Na + - und K + -Kanäle • Diese Elemente leiten Informationen in Form elektrischer Signale in der Zelle weiter. • Um Informationen zwischen Zellen weiterzuleiten, ist eine spezialisierte Kontaktstruktur zwischen den beiden Zellen notwendig: die Synapse. Es gibt im Wesentlichen zwei Formen von Synapsen: • chemische Synapsen, an denen das elektrische Signal in ein chemisches Signal umgewandelt wird, um anschließend wieder in ein elektrisches übersetzt zu werden • elektrische Synapsen, an denen das elektrische Signal über direkte Membrankontakte zwischen zwei Neuronen übertragen wird Elektrische Synapsen: Vorteile ggü. chemischen Synapsen: • Geschwindigkeit, Einfachheit Nachteile ggü. chemischen Synapsen: • begrenzte Modulierbarkeit • Umkehrung des Signals praktisch unmöglich: Depolarisation kann nicht in Hyperpolarisation umgesetzt werden • elektrische Synapsen sind bidirektional, was wiederum die Modulation von Signalen erschwert Organisation elektrischer Synapsen: • Gap Junctions • 6 Connexine in jeder der beiden verbundenen Membranen formen je 1 Connexon, auch Halbkanal (Hemichannel) genannt • 2 Connexone gegenüberliegender Membranen bilden eine Gap Junction • hohe Permeabilität: kleine Moleküle (von Ionen bis zu Aminosäuren) können von einer Zelle in die andere übertreten • zum Beispiel: Na + , K + , Ca 2+ , cAMP, IP3, Glucose. Welche Wirkung haben diese Moleküle in der Nachbarzelle? • Nachbarzellen sind, in etwa, isopotential • Porengröße kann moduliert werden, z.B. durch pH oder elektrische Spannung • besonders wichtig in Herzmuskelzellen und in Glia • In Neuronen findet man sie nicht so häufig wie chemische Synapsen, aber sie könnten für die Synchronisation bestimmter Neuronengruppen essentiell sein und können oft in oszillierenden oder rhythmuserzeugenden Systemen gefunden werden. Unterschiede zwischen Gap Junctions und Tight Junctions: Tight Junctions sind Verbindungen zwischen Membranen benachbarter Zellen, deren Funktion es ist, die Passage von Material durch das Gewebe zu verhindern. Sie schirmen das Gewebe gegen den Ein- oder Austritt von verschiedenen Elementen ab.
Transcript of Prof. Dr. Silvio O. Rizzoli 1. Stunde: Wiederholung der...

1
Prof. Dr. Silvio O. Rizzoli 1. Stunde: Wiederholung der Eigenschaften von Neuron und Axon. Elektrische Synapsen. Wie bereits in früheren Vorlesungen erklärt:
• Membranen, ihre Eigenschaften und Permeabilität für verschiedene Elemente • Organisation des Neurons: Zellkörper, Dendriten, Axonhügel, Axon • Myelin und seine Funktion • Generierung von Aktionspotentialen • spannungsabhängige Na+- und K+-Kanäle • Diese Elemente leiten Informationen in Form elektrischer Signale in der Zelle weiter. • Um Informationen zwischen Zellen weiterzuleiten, ist eine spezialisierte
Kontaktstruktur zwischen den beiden Zellen notwendig: die Synapse. Es gibt im Wesentlichen zwei Formen von Synapsen:
• chemische Synapsen, an denen das elektrische Signal in ein chemisches Signal umgewandelt wird, um anschließend wieder in ein elektrisches übersetzt zu werden
• elektrische Synapsen, an denen das elektrische Signal über direkte Membrankontakte zwischen zwei Neuronen übertragen wird
Elektrische Synapsen: Vorteile ggü. chemischen Synapsen:
• Geschwindigkeit, Einfachheit Nachteile ggü. chemischen Synapsen:
• begrenzte Modulierbarkeit • Umkehrung des Signals praktisch unmöglich: Depolarisation kann nicht in
Hyperpolarisation umgesetzt werden • elektrische Synapsen sind bidirektional, was wiederum die Modulation von Signalen
erschwert Organisation elektrischer Synapsen:
• Gap Junctions • 6 Connexine in jeder der beiden verbundenen Membranen formen je 1 Connexon,
auch Halbkanal (Hemichannel) genannt • 2 Connexone gegenüberliegender Membranen bilden eine Gap Junction • hohe Permeabilität: kleine Moleküle (von Ionen bis zu Aminosäuren) können von
einer Zelle in die andere übertreten • zum Beispiel: Na+, K+, Ca2+, cAMP, IP3, Glucose. Welche Wirkung haben diese
Moleküle in der Nachbarzelle? • Nachbarzellen sind, in etwa, isopotential • Porengröße kann moduliert werden, z.B. durch pH oder elektrische Spannung • besonders wichtig in Herzmuskelzellen und in Glia • In Neuronen findet man sie nicht so häufig wie chemische Synapsen, aber sie könnten
für die Synchronisation bestimmter Neuronengruppen essentiell sein und können oft in oszillierenden oder rhythmuserzeugenden Systemen gefunden werden.
Unterschiede zwischen Gap Junctions und Tight Junctions: Tight Junctions sind Verbindungen zwischen Membranen benachbarter Zellen, deren Funktion es ist, die Passage von Material durch das Gewebe zu verhindern. Sie schirmen das Gewebe gegen den Ein- oder Austritt von verschiedenen Elementen ab.

2
2. Stunde: Die chemische Synapse. Am Beispiel der neuromuskulären Endplatte Skelettmuskelfasern werden durch die neuromuskuläre Endplatte erregt (Neuromuscular Junction, NMJ). Muskeln sind efferent innerviert von Motoneuronen. Ein Motoneuron innerviert mehrere Muskelfasern = eine motorische Einheit. Überblick über die Struktur verschiedener Elemente prä- und postsynaptischer Zellen:
• Präsynapse: Vesikel, Mitochondrien, Aktive Zone, Mikrotubuli, Actin • Postsynapse : Aktive Zone, Rezeptoren, Endosomen, Mitochondrien, Mikrotubuli,
Ribosomen, raues ER, Actin • wichtiger Unterschied zwischen Prä- und Postsynapse = Proteinsynthese
Überblick über die chemische Signalübertragung an der neuromuskulären Endplatte:
• Aktionspotenziale • über myelinisierte Nervenfasern zu NMJ geleitet • hier sind präsynaptische Endknöpfchen, in Kontakt mit Muskelfasern • Ca2+ steigt in der NMJ, durch spannungsabhängige Ca2+ Kanäle • Exozytose von präsynaptischen Vesikeln [Mechanismen in 3. Stunde] • Freisetzung von Acetylcholin • ~200 Vesikel freigesetzt pro Endplatte, ~2-7-fach mehr als benötigt für ein
Aktionspotenzial im Muskel • Diffusion von Acetylcholin durch den synaptischen Spalt • Bindung an nikotnische Acetlycholinrezeptoren, AChR (im Sarkolemma); Oberfläche
ist an dieser Stelle vergrößert durch subsynaptische Einfaltungen, hohe Rezeptordichte • Acetlycholinrezeptoren = Kationenkanäle [Mechanismen in 5. Stunde] • Depolarisation => Endplattenpotential (exzitatorisches postsynaptisches Potential,
EPSP, ~40-70 mV), überschwellig • Aktionspotenzial im Muskel: Depolarisation (Na+), später Repolarisation (K+) • Acetylcholin wird durch ACh-Esterase in Acetat und Cholin gespalten; Cholin wird in
die Präsynapse gepumpt (Na+/Cholin-Kotransporter) • AChRs können durch Tubocurarin blockiert werden. Tubocurarin ist ein kompetetiver,
reversibler Inhibitor (was bedeutet das?). Bungarotoxin, ein nicht-kompetetiver, irreversibler Inhibitor (was bedeutet das?) kann die Rezeptoren ebenfalls blockieren.
• medizinische Anwendung von Tubocurarin-Analoga • medizinische Anwendung von ACh-Esterase-Inhibitoren
Die entscheidenden Schritte in der Entdeckung chemischer synaptischer Signalübertragung:
• Entdeckung von mEPPs (“miniature postsynaptic potentials”) durch Sir Bernard Katz (1940-1950)
• Summierung solcher mEPPs, resultierend in größeren Potentialen, entdeckt durch Del Castillo und Katz (1952) und später durch Boyd und Martin (1955). Ergebnis dieser Studien: Quanta – einzelne Einheiten, die spontanen Potentialen entsprechen; größere Potentiale sind die Summe einzelner dieser Einheiten
• Entdeckung von Fusion synaptischer Vesikel mit der Plasmamembran durch Elektronenmikroskopie: Couteaux and Pecot-Dechavassine, 1970; Heuser and Reese, 1973, Heuser et al., 1979
• verschiedene Experimente, die Fusion zu zeigen: Doppel-Patch-Clamp-Experimente, durchgeführt um 2000, die Voltage-Clamp- und Kapazitätsmessungen nutzten. Was sind diese beiden Arten von Messungen?
• fluoreszensbasierte Messungen von synaptischer Vesikelfreisetzung: pH-sensitive GFPs und lipophile Farbstoffe

3
3. Stunde: Exozytosemechanismen Prinzipien der Membranfusion: Gründe warum zwei Membranen nicht auf einfache Art und Weise miteinander fusionieren können:
• Struktur (Doppelmembran) • Präsenz von polaren (geladenen) Gruppen auf den Außenseiten von beiden
Membranen • Folglich wird Energie benötigt um die beiden Membranen zu destabilisieren • Zuerst wird die Membran destabilisiert • Dann fusionieren die beiden äußeren Lipidschichten (Hemifusion) • Drittens bildet sich die Fusionspore • Viertens expandiert die Fusionspore
Moleküle die an der Fusion synaptischer Vesikel beteiligt sind:
• SNAREs: Synaptobrevin auf dem synaptischen Vesikel und Syntaxin/SNAP-25 auf der Plasmamembran
• Interaktionsmechanismus: gegenseitiges Erkennen und Bildung von trans-Komplexen • SNARE Interaktionen: umeinander Winden, Zippering, Energiefreisetzung • Die immense Widerstandskraft des SNARE Komplexes: SDS-Resistenz,
Temperaturresistenz (70-80°C) • Das Konzept der Hysterese bei SNARE Interaktionen • Die Modulation der Fusion durch andere Proteine, wie z.B. Munc Proteine. Woher
kommt die unc-Namensgebung historisch? • Was passiert mit SNARE Komplexen nach der Fusion: sie werden cis-Komplexe • Trennung der cis-Komplexe durch ATP-ase Aktivität
Medizinisches Interesse an den SNARE Mechanismen:
• Verschiedene Clostridumtoxine spalten verschiedene SNARE Moleküle • Medizinische Beteiligung: z.B. kosmetische Eingriffe mit Botulinumtoxin • Tetanus: Infektionen mit Clostridium tetani, die größtenteils inhibitorische Synapsen
betreffen • Symptome von Wundstarrkrampf: Steifheit, krampfartige Spasmen der
Skelettmuskulatur; Risus sardonicus, Opisthotonus Weitere wichtige Elemente für die Kontrolle der Vesikelfusion:
• Auslösung durch Calcium erfordert eine gute Organisation der aktiven Zonen. Aktive Zonen bestehen aus dichtem Material das sowohl Calciumkanäle als auch synaptische Vesikel vereint. Die Vesikel werden also sehr nahe an die Quelle des Calciumeintritts in die Synapse gebracht.
• Die Struktur der aktiven Zone ist immer noch unbekannt • Was jedoch klar ist, ist dass die Vesikel als erstes an den aktiven Zonen docken
müssen • Sie werden somit gut positioniert und können sowohl den Calciumeintritt detektieren
als auch fusionieren • Der Calciumsensor der Vesikel ist Synaptotagmin, ein Transmembranprotein des
synaptischen Vesikels • Der Mechanismus der Calciumdetektion: in Abwesenheit von Calcium bindet
Synaptotagmin an die Membran der synaptischen Vesikel. Es blockiert

4
möglicherweise auch die SNARE Interaktionen. In Gegenwart von Calcium bindet Synaptotagmin an Calcium Ionen und die Plasmamembran wodurch die Vesikel etwas näher an die Membran gebracht werden und die SNAREs sich um sich winden und die Fusion einleiten können
4. Stunde: Exozytosemechanismen Was passiert mit den Proteinen der synaptischen Vesikel nach der Fusion mit der Plasmamembran? Mehrere Möglichkeiten:
• Erste Möglichkeit: Die Fusionspore öffnet sich nur ein bisschen und schließt sich wieder.
• Dieser Mechanismus heißt Kiss-and-Run. Es ist nicht offenkundig, wie das von einem molekularen Standpunkt her passieren könnte.
• Zweite Möglichkeit: Der Vesikel kollabier in die Plasmamembran und die Moleküle des Vesikels verbleiben als eine Gruppe, verschieden vom Rest der Membran
• Zahlreiche Experimente wurden durchgeführt um dies zu testen: z.B. Immunfluoreszenzversuche die Synaptotagmin Antikörper verwenden und die Moleküle mit Fluoreszenzmikroskopen sichtbar machen
• Ergebnis: Die Moleküle scheinen zusammen zu bleiben – obwohl manche Wissenschaftler diesen Standpunkt anzweifeln und hinweisen auf:
• Die dritte Möglichkeit: Der Vesikel kollabiert in die Plasmamembran und die Moleküle diffundieren überall in der Plasmamembran.
Egal ob die Vesikelmoleküle zusammenbleiben oder nicht müssen die folgenden Ereignisse stattfinden um die Moleküle von der Plasmamembran zurückzuholen:
• Irgendein molekularer Mechanismus muss die Vesikelmoleküle anpeilen und sie vom Rest der Plasmamembran unterscheiden
• Dies wird durch Moleküle, die Synaptotagmin erkennen bewerkstelligt • Durch “Coincidence detection” binden sie Synaptotagmin nur wenn es sich auf der
Plasmamembran befindet: Sie binden sowohl Plasmamembranlipide und Synaptotagmin gleichzeitig
• Die Synaptotagmin erkennenden Moleküle rekrutieren Clathrin • Clathrin bildet eine dreidimensionale Struktur um die Moleküle des synaptischen
Vesikels • Die Struktur wird zu einem fußballförmigen „Ball“, der “gecoatetes” Vesikel genannt
wird • Schließlich wird der Vesikel durch andere Moleküle von der Plasmamembran getrennt • Dieser Mechanismus ist derselbe, der in der Biochemie für die Endozytose von
Membranrezeptoren gelehrt wird (Transferrinrezeptor, LDL Rezeptor, EGF Rezeptor etc.)
Ereignisse, die nach der Trennung des gecoateten Vesikels von der Plasmamembran stattfinden:
• Der Clathrin “Coat” wird verloren. Un-coating Moleküle greifen das eine Loch im Coat an und „schälen“die Clathrinmoleküle ab. Wo könnte sich dieses Loch befinden?
• Der Vesikel ist nun frei vom Coat. Aber ist es ein perfekter Vesikel? Er hat noch keinen Neurotransmitter.

5
• Die meisten Neurotransmitter sind kleine Moleküle: ACh, Glutamat, GABA. Sie müssen zurück in den Vesikel gepumpt werden.
• Dies passiert oft indem ein Antiport System verwendet wird. • Als erstes wird der Vesikel durch eine Protonenpumpe azidifiziert • Dann werden die Protonen gegen Neurotransmitter ausgetauscht • Nun ist das Vesikel bereit, neu zu fusionieren.
5. Stunde: postsynaptische Rezeptoren Die Neurotransmitter-Moleküle binden an Rezeptoren der postsynaptischen Membran und können eine Depolarisation oder eine Hyperpolarisation der postsynaptischen Zelle verursachen. Es gibt zwei Hauptkategorien von postsynaptischen Rezeptoren: Ionotrope Rezeptoren:
• Die Rezeptoren bilden Kanäle in postsynaptischen Plasmamembranen. Diese sind ligandengesteuerte Ionenkanäle.
• Beispiele: nikotinische Acetylcholin-Rezeptoren, in der Plasmamembran der Muskelzellen (Muskelfaser) Struktur: pentamerische Proteine, die den Strom von Ionen erlauben (permeabel für Ionen)
• Die Bindung von ACh an den AChR verursacht eine Depolarisation. Wie kommt es dazu, wenn gleichzeitig Na+-Ionen in die Zellmembran hineinströmen und K+-Ionen aus der Zellmembran herausströmen?
• Ein Aktionspotential wird dann in der Muskelzellen generiert. Von großer Bedeutung ist die Lokalisation der Na+ -Kanäle für die Generierung von Aktionspotentialen: sie sind unmittelbar unterhalb der AChR in den postsynaptischen Einfaltungen.
• Im ZNS verwenden die meisten Synapsen Glutamat als Neurotransmitter. • Zwei Haupkategorien von Glutamat-Rezeptoren sind: AMPA- und NMDA-
Rezeptoren. Eine dritte Kategorie, die Kainat-Rezeptoren, sind den AMPA-Rezeptoren ähnlich.
• Beim normalen Membranpotential werden die NMDA-Rezeptoren durch Magnesium-Ionen blockiert.
• Die NMDA-Rezeptoren werden mehr permeabel für Ca2+ als die AMPA-Rezeptoren. • Wie würde sich diesen Einfluss auf die Depolarisation in der Zelle auswirken? • Viele der hemmenden Transmissionen sind durch GABA oder Glycin vermittelt.
GABAA sind ionotrope Rezeptoren. Sie sind Chlorid-selektive Poren. Welche Folge hat das für das Zellmembranpotential?
• Der Glycin-Rezeptor ist auch ein ionotroper Rezeptor und für Chlorid-Ionen permeabel. Diese Rezeptoren werden von Strychnin, ein pflanzliches Alkaloid, blockiert. Welche Konsequenz folgt daraus: eine Hyper- oder eine Hypoerregbarkeit?
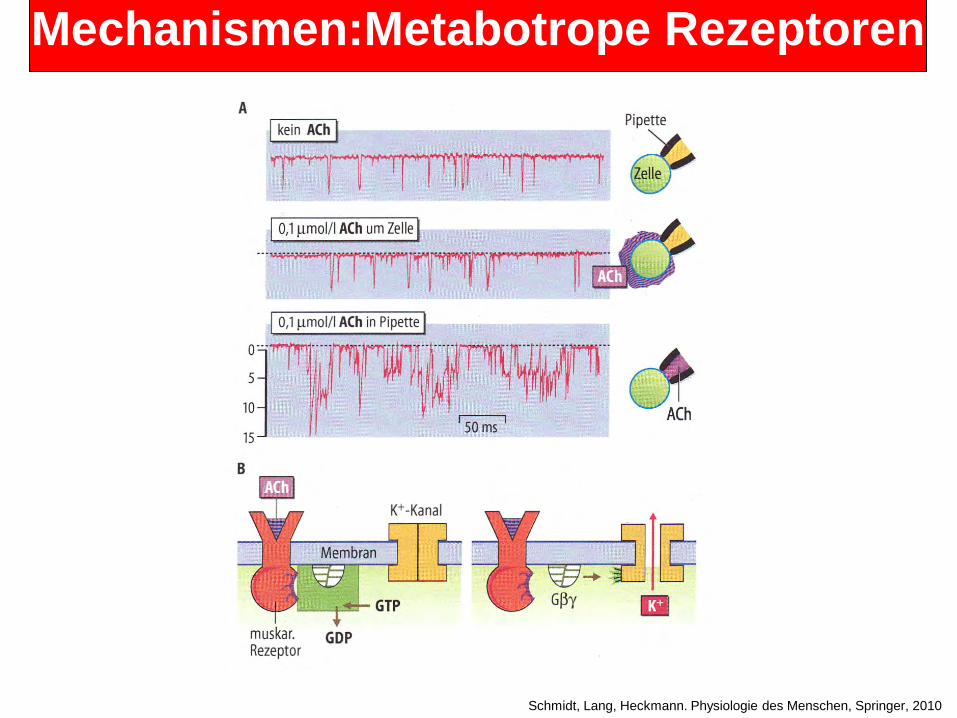
Metabotrope Rezeptoren: • Typischerweise sind sie 7-Transmembrandomänen-Proteine.

6
• Sie werden zu einer Depolarisation oder einer Hyperpolarisation der Membran führen, aber langsamer als die ionotrope Rezeptoren. Viele interagieren mit G-Proteinen in der Membran.
• G-Proteine bestehen aus einem Komplex von alpha-, beta- und gamma Untereinheiten. Nach Aktivierung bindet der metabotrope Rezeptor an das G-Protein. Das G-Protein wird in alpha -und beta/gamma-Untereinheiten dissoziiert durch den Austausch von GDP gegen GTP (in der alpha-Untereinheit des G-Proteins). Sowohl die alpha- und die beta-/gamma-Untereinheiten können danach mit spezifischen Effektoren interagieren. Schließlich hydrolisiert die alpha-Untereinheit GTP zu GDP, und bindet wieder an der beta /gamma Untereinheit solange bis zur nächsten Aktivierung. Beispiele für G-Proteine:
• Gs-gekoppelte-Rezeptoren: exzitatorisch: aktiviert das cAMP-System, indem die Produktion cAMP von ATP stimuliert wird. Die cAMP-abhängige Proteinkinase A (PKA) phosphoriliert viele andere Proteine.
• Gi-gekoppelte-Rezeptoren: inhibitorisch, da es die Produktion von cAMP hemmt. • Gq-gekoppelte-Rezeptoren: exzitatorisch, unter Aktivierung von Phospholipase C
(PLC) wird Phosphatidylinositol-4,5-biphosphat (PIP2) in Inositoltriphosphat (IP3) und Diacylglycerol (DAG) hydrolisiert. IP3 stimuliert die Freisetzung von Ca2+ aus dem endoplasmatischen Reticulum.
• Beispiele für metabotrope Rezeptoren • Muskarinische Acetylcholin-Rezeptoren (Agonist: Muskarin). Wird durch Atropin
blockiert. • Einige interagieren mit Gi, vor allem in Herzzellen und bewirken eine Erhöhung der
K+-Leitfähingkeit und führen somit zu einer Hyperpolarisation. • Andere interagieren mit Gq, zum Beispiel in Muskeln, die in der Akkomodation des
Auges beteiligt sind. Durch die Hemmung der K+-Leitfähigkeit bewirken sie eine längere Depolarisation.
• Metabotrope Glutamat-Rezeptoren. Typischerweise werden sie mit Gq assoziiert, aber auch mit Gi. Sie führen zu einer Depolarisation durch die Hemmung der K+-Leitfähigkeit und der Erhöhung der Na+-Leitfähigkeit. Eine Hemmung ist möglich.
• GABAB-Rezeptoren. Sie interagieren mit verschiedenen G-Proteine (i, o). In dem ZNS, führt die Aktivierung von K+-Kanäle zu einer Hyperpolarisation.
6. Stunde: Synaptische Integration: Summation, Inhibition Synaptische Integration= Summe der Information von mehreren Neuronen. Beide exzitatorische und inhibitorische Signale können summiert werden. Arten der Summation:
• Divergenz: die Information von einem Neuron wird zu mehreren Neuronen übertragen.
• Konvergenz: Die Information von mehreren Neuronen wird auf ein einziges Neuron übertragen.
• Zeitliche Summation: eine Synapse feuert mehrmals nacheinander und depolarisiert (oder hyperpolarisiert) damit stark die postsynaptische Zelle.
• Räumliche Summation: zwei oder mehrere Synapsen aus verschiedenen Neuronen feuern gleichzeitig auf die gleiche postsynaptische Zelle.
Beispiele für Summation und synaptische Integrationsprozesse:

7
Reziproke/antagonistische Hemmung in Muskeln:
• Ein Muskel wird ausgedehnt. • Muskelspindelnafferenzen werden dadurch aktiviert und bilden erregende Synapsen
(Transmitter: Glutamat) mit alpha-Motoneuronen der Agonisten und der glycinergischen Interneuronen.
• Die Interneuronen inaktivieren über Glycin die Alpha-Motoneuronen der Antagonisten
• Bedeutung: die Aktivierung des Agonisten führt in wenigen Zehnteln von Millisekunden zu einer Hemmung der ipsilateralen Antagonisten
Solche Effekte werden für die Lokomotion benutzt:
• Viel komplexer, mehr als nur einfache Aktivierung und Hemmung von einzelnen Synapsen.
• Die Bewegung erfolgt durch komplexe Abfolgen von Kontraktionen der Beinmuskulatur.
• Die rhythmische Aktivität wurde 1911 von Graham Brown entdeckt. • Spinale Rhythmusgeneratoren: für das rhythmisches Gehen, Laufen oder Schwimmen. • Zentraler Muskelgenerator (Central pattern generator, CPG) genannt • Wird in die Vorlesungen über „Muskel und Motorik“ weiter diskutiert
Laterale Hemmung/Inhibition in Sensorik:
• Neurone in visuellen und somatosensorischen Systemen werden z.B. in das Zentrum ihres rezeptiven Feldes erregt und von einem mehr oder minder großen und ebensolchen regelmäßig geformten Umfeld gehemmt.
• Die primären Afferenzen von diesem Bereich sind mit Interneuronen verbunden, die an den betreffenden zentralen Neuronen hemmende Synapsen bilden.
• Kontrastverschärfung. • Wichtig z.B für die Augen: Information über die Helligkeitsunterschiede im Bild wird
geliefert => also über die Begrenzungen einzelner Bildelemente (viel wichtiger als die Information über die absolute Helligkeit).
• Die laterale Hemmung ermöglicht eine präzise Somatotopie im sensorischen Kortex. 7. Stunde: Synaptische Plastizität Lernen: durch Wachstum und Verbindungen von Nervenzellen (Plastizität) Hebb-Regel: „Wenn ein Axon des Neurons X nahe genug an einem Neuron Y liegt, so dass Zelle Y wiederholt von Neuron X erregt wird, so wird die Effizienz von Neuron X für die Erregung von Neuron Y durch einen Wachstumprozess oder eine Stoffwechseländerung in beiden (oder einem der beiden) Neuronen erhöht“. Möglichkeiten von Plastizität:
• Veränderung synaptischer Transmission • Veränderungen der Modulation von Interneuronen • Bildung neuer Synapsen
Bekannte Mechanismen:

8
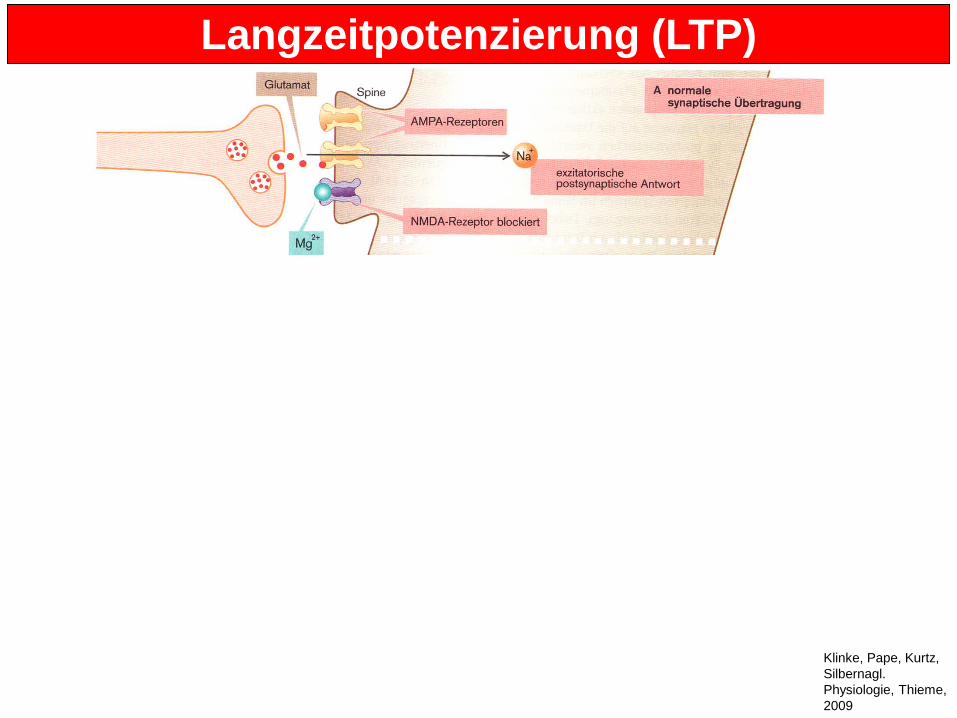
Langzeitpotenzierung (LTP) • Am Anfang: Normale synaptische Übertragung • NMDA-Rezeptoren sind durch Mg2+ blockiert • Nur AMPA-Rezeptoren sind aktiv => Na+-Einstrom => Erregung • Durch starke, wiederholte Erregung werden NMDA-Rezeptoren de-blockiert • Ca2+-Einstrom • CaM-Kinase wird über Calmodulin aktiviert • AMPA-Rezeptoren werden phosphoryliert (durch CaM-Kinase) • => Steigerung der postsynaptischen Antwort • Auch Einbau von Rezeptoren in post-synaptische aktive Zonen, die vorher
internalisiert in Endosomen vorlagen • Später auch de novo Proteinsynthese, z.B. von Rezeptoren. [Prä- oder post-
synaptisch?] • Die dendritischen Spines wachsen dadurch und werden stärker • Ergebnis: Verstärkung von Synapsen und Mobilisierung ruhender Synapsen durch
Einbau zusätzlicher Rezeptoren. • Homosynaptische Langzeitpotenzierung: nur die stimulierte Synapse wird verstärkt • Heterosynaptischen Langzeitpotenzierung: Verstärkung von nicht stimulierten
Synapsen an demselben Neuron Langzeitdepression (LTD)
• Am Anfang: Normale synaptische Übertragung. • Durch Glutamat werden metabotrope Rezeptoren aktiviert • G-Proteine aktivieren Phospholipase C • PIP2 wird in IP3 und DAG gespalten • IP3 öffnet Kanäle im ER (endoplasmatischen Retikulum) und setzt Ca2+ frei • DAG aktiviert Protein Kinase C • => Erhöhung der intrazellulären Ca2+-Konzentration • synaptische Depression.
Andere Arten synaptischer Änderungen: Beispiel von Autoimmunität:
• Myasthenia gravis. Symptome: Ptosis, Muskelermüdung • Autoimmunreaktion, Lebenszeit der AChRs im Muskel wird stark verkürzt.

9
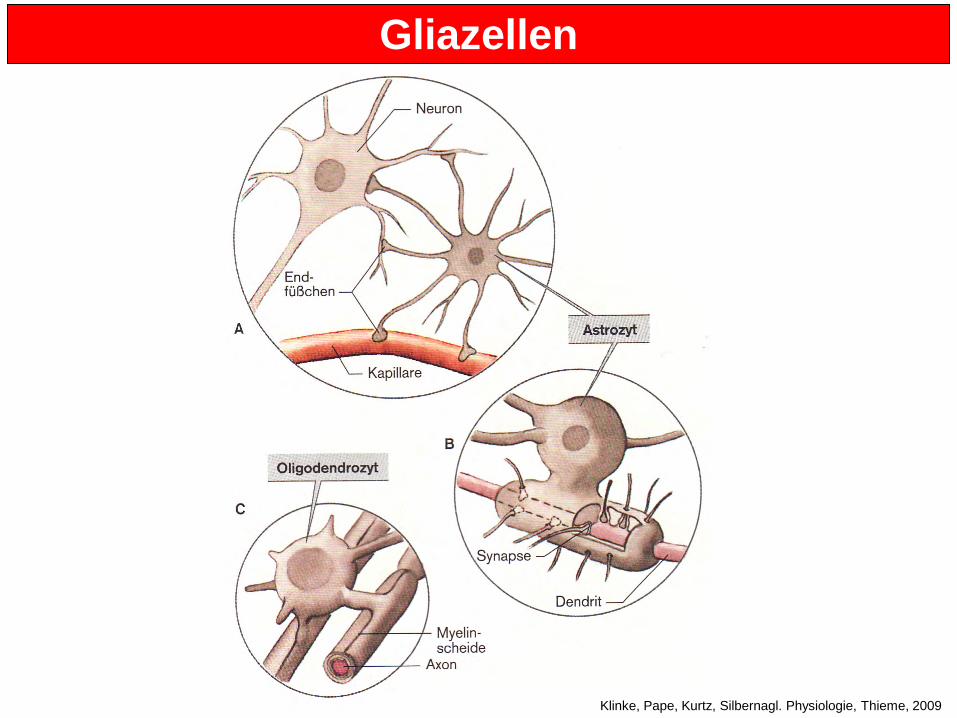
8. Stunde: Gliazellen Die Zahl der Gliazellen im Gehirn ist fünf- bis zehnmal höher als die Zahl der Neuronen. Es gibt verscheiedene Typen von Gliazellen (Astrozyten, Oligodendrozyten and Mikroglia), die aber alle einige Eigenschaften teilen:
• Ihr Membranpotential ist stark negativ (zwischen -80 und -90 mV) • Sie sind nicht erregbar (können keine Aktionspotentiale ausbilden) • Sie haben keine spannungs-abhängigen Na+-Kanäle • Sie haben metabolische Funktionen und helfen Neuronen, ihre Aufgaben zu
bewältigen Astrozyten:
• Groß, sternförmig, mit vielen Endfüßchen (Fortsätze) • Sie kontaktieren sowohl Neurone als auch Blutgefäße • Sind wirken beim Austausch von Subtraten wie Aminosäuren und Zuckern mit und
auch von Molekülen, die bei der Neurotransmitterproduktion eine Rolle spielen • Astrozyten haben Neurotransmitter-Rezeptoren und werden daher bei synaptischer
Aktivität stimuliert. Die Auswirkungen dieser Stimulation sind noch nicht bekannt. • Ihre Fortsätze umschließen die Synapsen, bis hin zum synaptischen Spalt • Einige Wissenschaftler betrachten Astrozyten als die “dritte Zell” der Synapse • Sie seien an Prozessen wie der Regulierung der Diffusion von Neurotransmittern aus
dem synaptischen Spalt beteiligt. Sie können Neurotransmitter nach der Exozytose mit Hilfe von Neurotransmitter-Transportern aufnehmen um sicherzustellen, dass diese nicht im synaptischen Spalt verbleiben
• Mutationen in Glutamat-Transportern von Gliazellen führt zum Tod. Warum? Was passiert, wenn Glutamat nicht aus den Regionen um Synapsen herum entfernt wird? Excitotoxizität.
• Können auch K+ aufnehmen, das von Neuronen freigesetzt wird = räumlicher K+-Puffer. Wie könnte die K+-Aufnahme funtionieren?
Oligodendrozyten
• Sie bilden die Myelinscheide aus, die die Axone im ZNS ummantelt • Ein Oligodendrozyt can mehrere Axone myelinisieren • Störungen der Oligodendrozyten gehören zu den Hauptauslösern von Multipler
Sklerose • Es handelt sich dabei um eine Autoimmunerkrankung, die durch die Zerstörung der
Myelinscheide durch Makrophagen charakterisiert ist • Dies führt zu einer Blockade der Neurotransmission. Wie?
Symptome: Empfindungsstörungen, beispielsweise Sehstörungen, Parese spastische Störungen Mikroglia
• Sie sind die Makrophagen des Gehirns • Wie alle Makrophagen können sie zu verschiedenen Stellen wandern • Nach Aktivierung können sie sich schnell vermehren • Sie könne phagozytieren und unerwünschte Zellen (Bakterien) zerstören • Sie können sich anhäufen, beispielsweise in Amyloid-Plaques bei Alzheimer. Wie?

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen

Figure 10-1 Molecular Biology of the Cell (© Garland Science 2008)
Membranen

Plasmamembran aus Phospholipiden
Hydrophob
Hydrophil
Lipophob
Lipophil
Kopfgruppe +
Phosphat +
Glycerol +
2 Fettsäurereste

Hydratation: Anlagerung von Wassermolekülen an Ionen oder polare moleküle
Plasmamembran aus Phospholipiden

Ionenkanälen: Proteinkomplexe

Na-Kanälen
Eijkelkamp et al., 2012

Das Aktionspotential (AP)
Detlev Schild

Leitwerte
Was konnte
Alles-oder-Nichts
hier bedeuten?
Detlev Schild
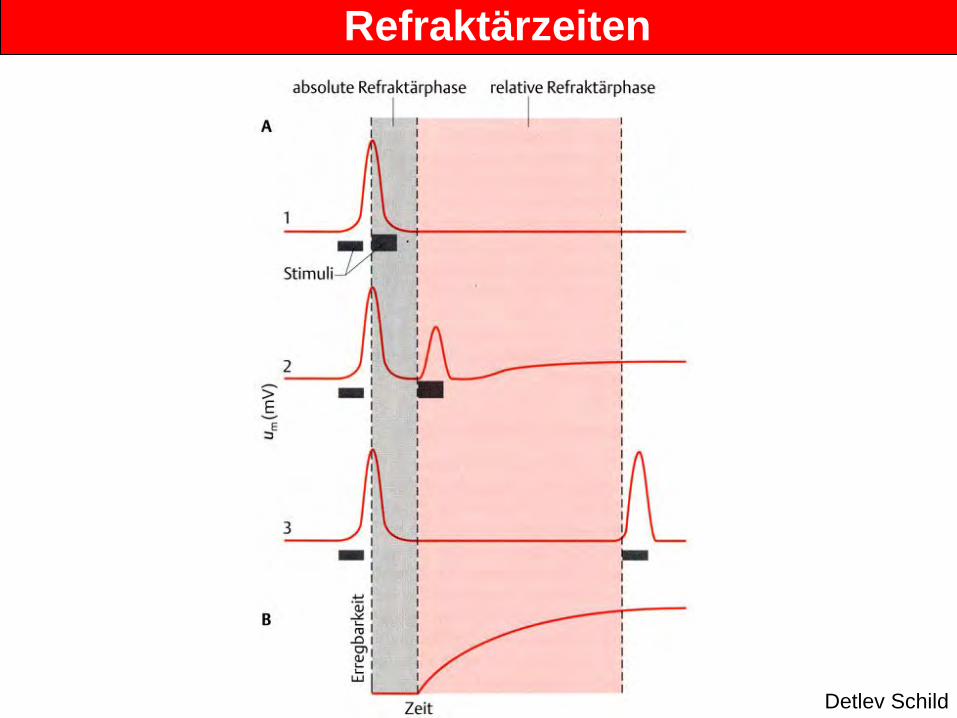
Refraktärzeiten
Detlev Schild
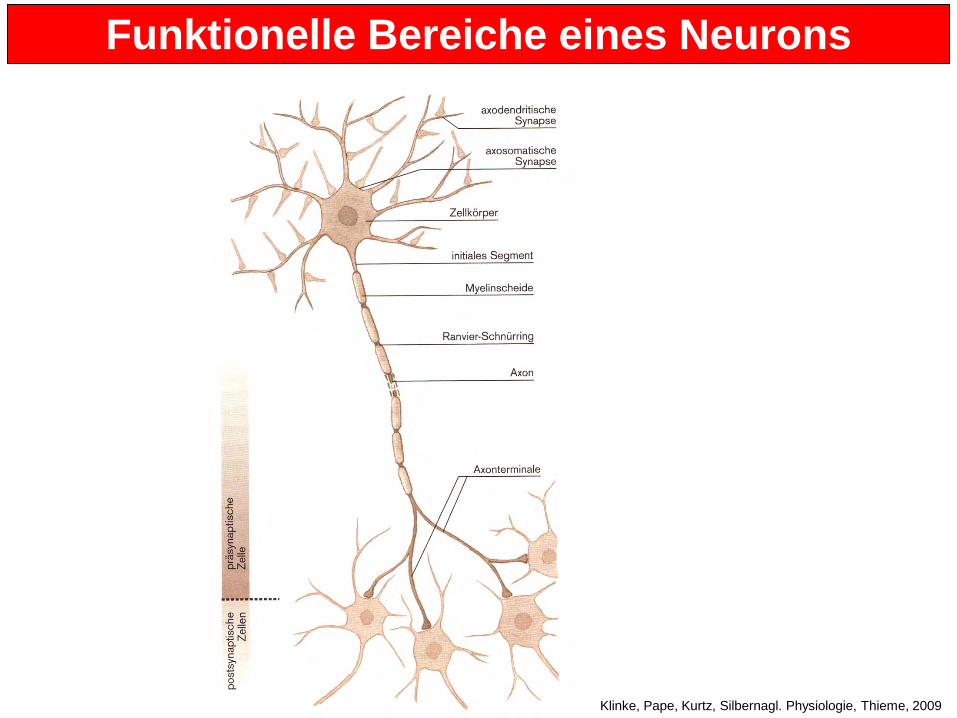
Funktionelle Bereiche eines Neurons
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009
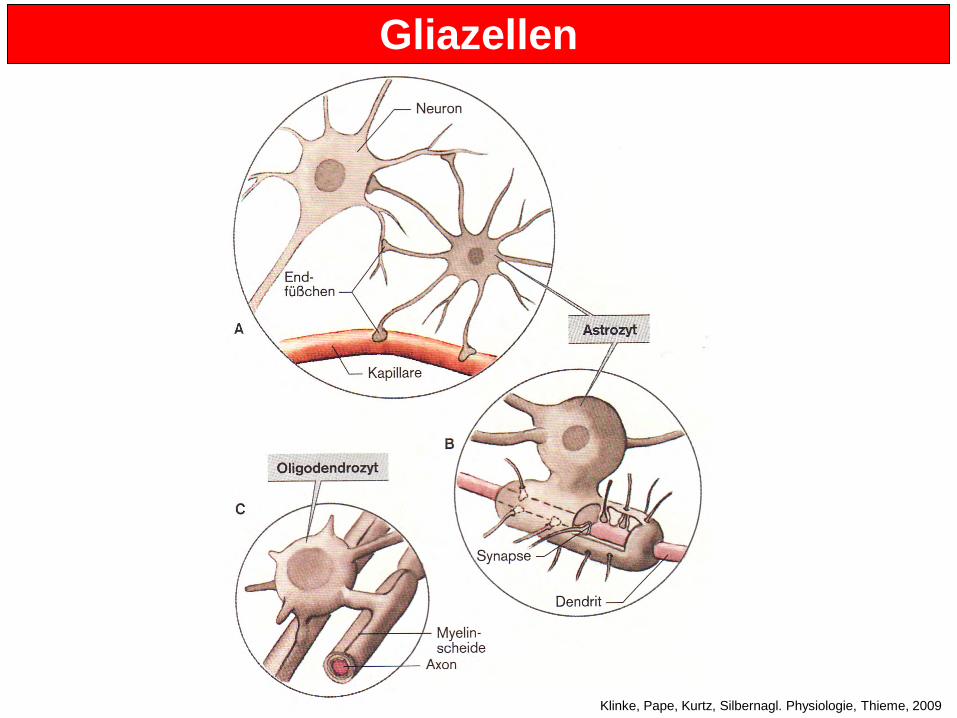
Gliazellen
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Informationsleitung
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Synapsen

Gap Junctions sind Poren von eine Zelle in eine benachbarte Zelle
Detlev Schild

Struktur von Gap Junctions
Atomkraftmikroskopie
http://www.asu.edu/news/research/research_images/afm.jpg
Sosinsky et al., 2005

Detlev Schild
Gap junctions sind Poren von eine Zelle in eine benachbarte Zelle

Gap junctions sind nicht Tight Junctions!
Epithelia
Permeabilität
Warum?
Detlev Schild

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen
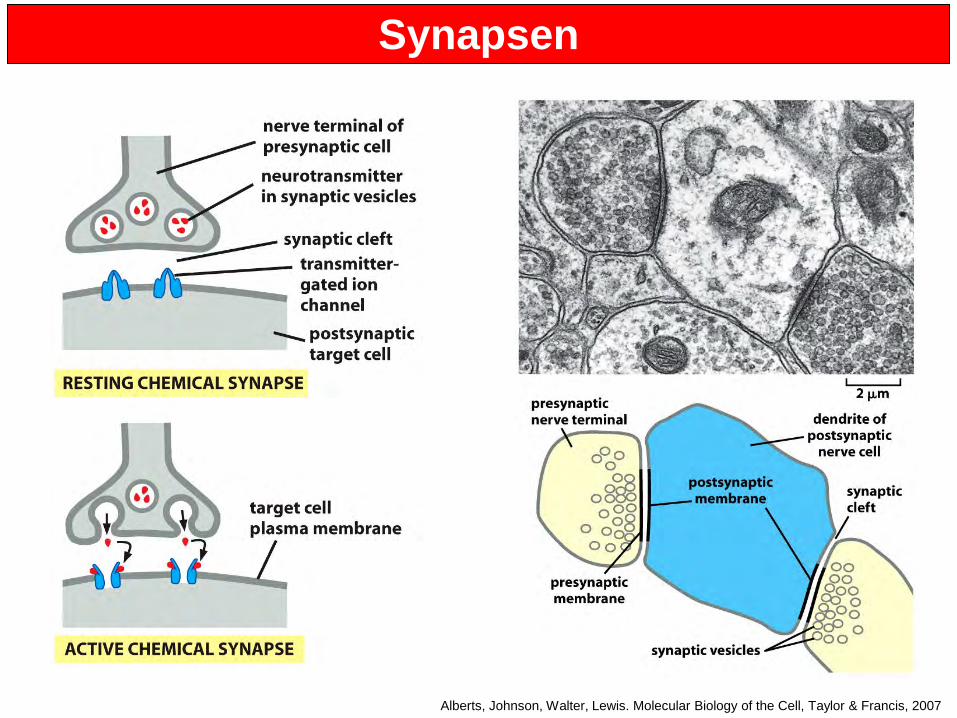
Synapsen
Alberts, Johnson, Walter, Lewis. Molecular Biology of the Cell, Taylor & Francis, 2007

Synapsen
Birks, 1960

Minis vs. EPSC (EPP)
EPSCs
Katz, J Neurocytol, 1993; Rizzoli and Betz, J Neurosci, 2002

Elektrophysiologie
Del Castillo and Katz, J Physiol, 1955; Rizzoli and Betz, Science, 2004
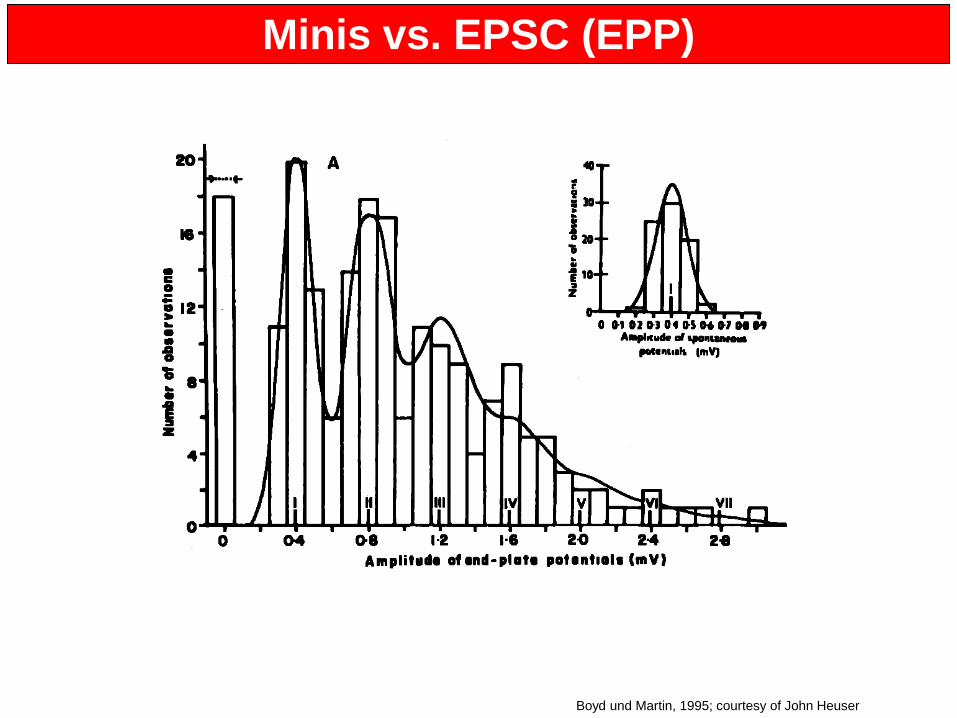
Minis vs. EPSC (EPP)
Boyd und Martin, 1995; courtesy of John Heuser

Minis vs. EPSC (EPP)
Boyd und Martin, 1995; courtesy of John Heuser

Minis vs. EPSC (EPP)
Boyd und Martin, 1995; courtesy of John Heuser

Heuser and Reese, 1973
Nicht-stimulierte Synapse

Heuser and Reese, 1973
Stimulierte Synapse

Heuser and Reese, 1973
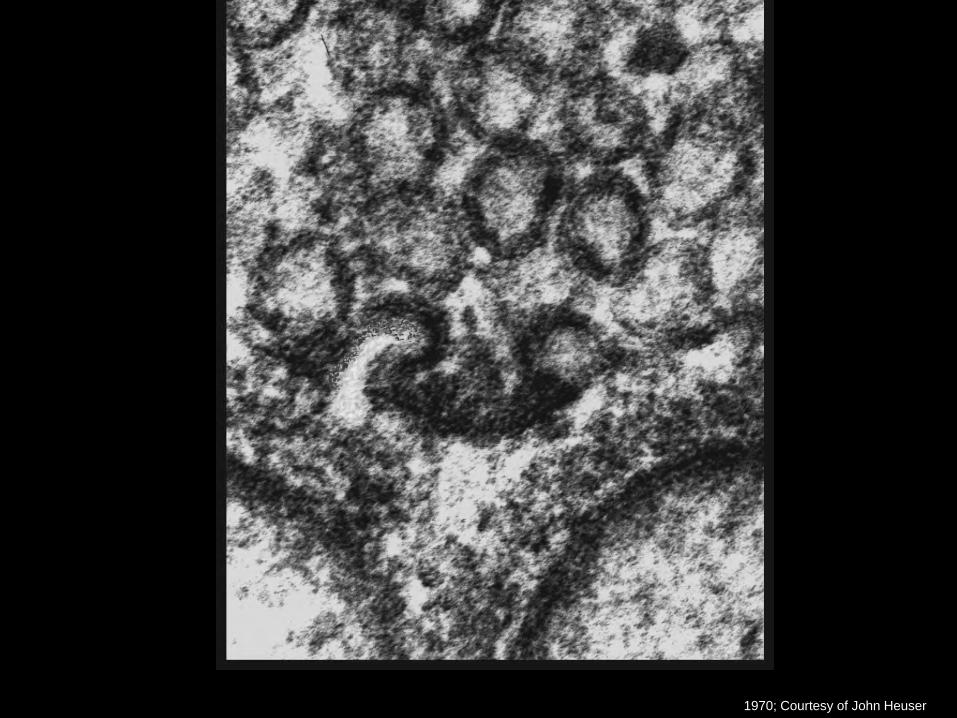
1970; Courtesy of John Heuser

1970; Courtesy of John Heuser

1970; Courtesy of John Heuser

1970; Courtesy of John Heuser

1970; Courtesy of John Heuser
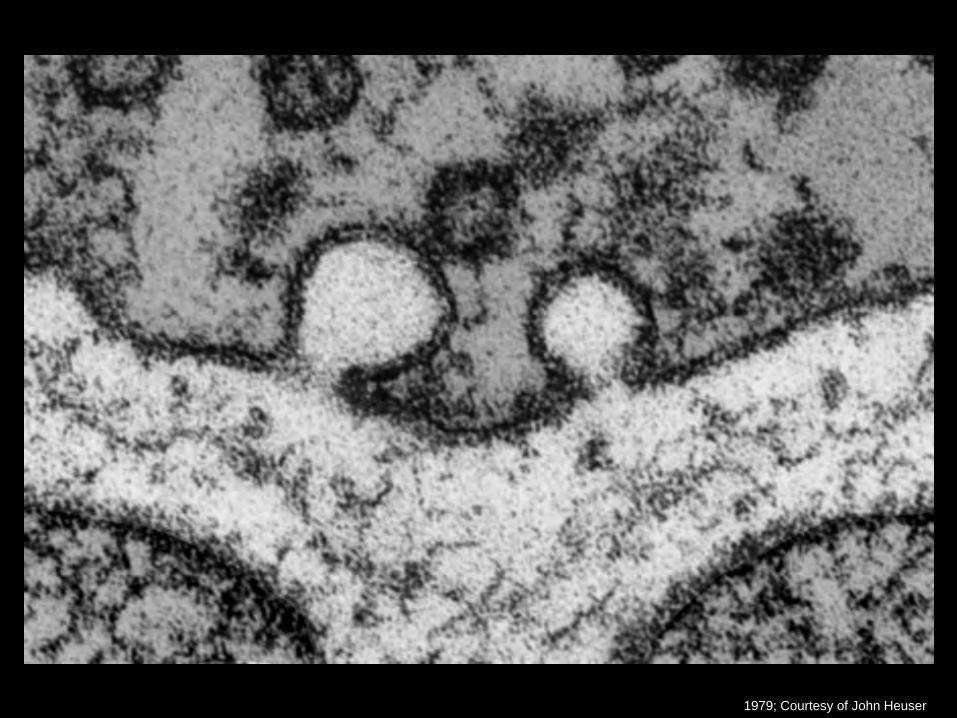
1979; Courtesy of John Heuser

Endplatte
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Subsynaptische Einfaltungen
Slater, J Neurocytol, 2003

Pharmakologie:
Courtesy of Detlev Schild

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen
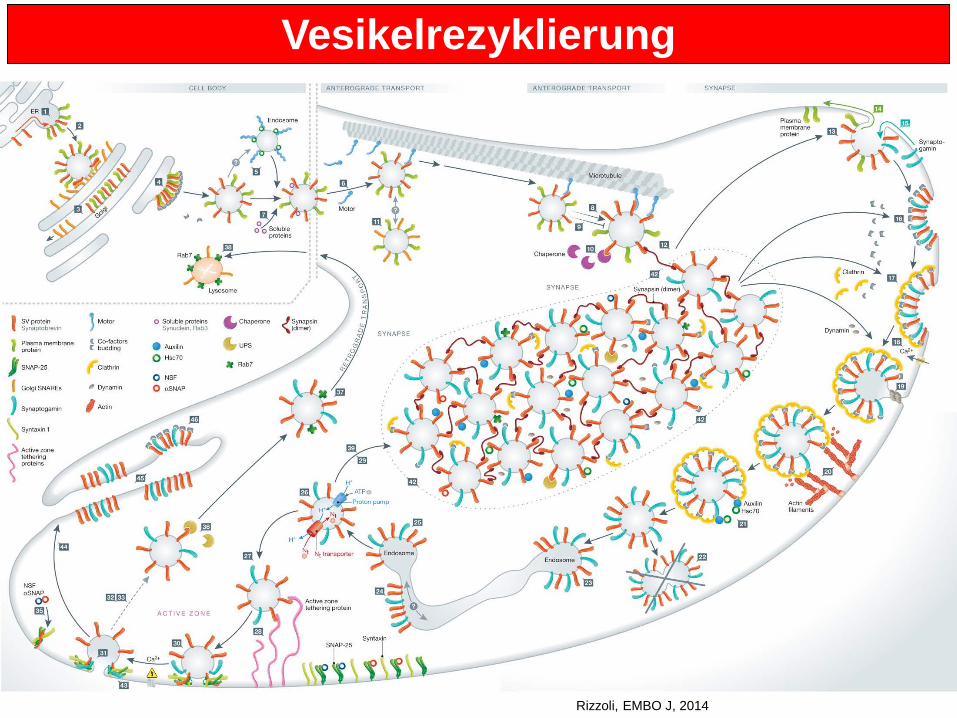
Vesikelrezyklierung
Rizzoli, EMBO J, 2014

Vesikelfusion: docking, SNAREs, synaptotagmin
Rizzoli, EMBO J, 2014

Freisetzung (SNAREs)
SNARES
Jahn and Scheller, Nat Rev Molec Cell Biol, 2006

Post-Vesikelfusion: SNARE freisetzung
Rizzoli, EMBO J, 2014

Pharmakologie: SNAREs

Tetanus
Sir Charles Bell's portrait of a man with generalized tetanus and evident opisthotonus and risus sardonicus.(Original in the Royal College of Surgeons of Edinburgh, Scotland)
Courtesy of Diethelm Richter

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen

Vesikelrezyklierung
Rizzoli, EMBO J, 2014

Endozytosis
McMahon and Boucrot, 2012

Endozytosis
McMahon and Boucrot, 2012

Un-coating und re-filling
Rizzoli, EMBO J, 2014

Sekundär aktiver Transport: Anti – und Symporter
Detlev Schild

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen

Ionenkanälen: Proteinkomplexe
Electronmikroskopie

Tollkirsche = Atropa belladonna (Parasympatholytikum)

Tollkirsche = Atropa belladonna (Parasympatholytikum)
Atropin ↑ Bindet an den muskarinischen Rezeptoren des Parasympathikus (ACh) Wirkung ACh ↓ Lähmung des parasympathisch innervierten sphincter pupillae (Nervus oculomotorius) Mydriasis

Mechanismen: Ionotrope Rezeptoren
Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010
Mechanismen:Metabotrope Rezeptoren
Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010

Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010
Öffnungszeiten

AMPA vs. NMDA Rezeptoren
Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen

Reizverarbeitung: Konvergenz, Divergenz
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Summation – räumliche und zeitliche

Reizverarbeitung: Hemmung
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Reziproke antagonistische Hemmung
Schmidt pg 137. abb 7.7
Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010

Reziproke antagonistische Hemmung
Klinke pg 768. abb 23.6 A
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Autogene Hemmung
Klinke, pg 768, abb 23.6 B
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Verschiedene Afferenzen
Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010

Lokomotion
Kandel, Schwarts, Jessel, Principles of Neural Science, McGraw-Hill, 2012

Komplexität von Beugereflex/Streckreflex
Klinke, pg 770, abb 23.7
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Rhytmische Aktivität, G. Brown um 1911
Kandel, Schwarts, Jessel, Principles of Neural Science, McGraw-Hill, 2012

Neunauge
Kandel, pg 745, 37-5
Kandel, Schwarts, Jessel, Principles of Neural Science, McGraw-Hill, 2012

Lateralen Inhibition (Hemmung)
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen


Cajal, ~1880-1890: Spines


Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010
Zelluläre Mechanismen des Lernens

Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010
Zelluläre Mechanismen des Lernens

Zelluläre Mechanismen des Lernens

Zelluläre Mechanismen des Lernens

Schmidt, Lang, Heckmann. Physiologie des Menschen, Springer, 2010
Langzeitpotenzierung (LTP)
Langzeitpotenzierung (LTP)
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009
Langzeitpotenzierung (LTP)

Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009
Langzeitpotenzierung (LTP)

Langzeitdepression (LTD)

Klinke, pg 836, fig 25.13b
Langzeitdepression (LTD)

Myasthenia gravis

1. Stunde: Übersicht über Eigenschaften des Neurons und des Axons. Elektrische Synapsen.
2. Stunde: Die chemische Synapse. Die neuromuskuläre Endplatte
als Beispiel. 3. Stunde: Die Mechanismen der Exozytose. 4. Stunde: Die Mechanismen der Endozytose. 5. Stunde: Postsynaptische Rezeptoren. 6. Stunde: Synaptische Integration: Summierung, Inhibition. 7. Stunde: Synaptische Plastizität. 8. Stunde: Gliazellen.
Neuronen und Synapsen
Gliazellen
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Ionale Homeostase
Na+
Energie- Versorgung
GLUT2
GLUT1
GLUT3
NT Aufnahme
NT-Versorgung
Astrozyten
Courtesy of Diethelm Richter

Oligodendrozyten
Klinke, Pape, Kurtz, Silbernagl. Physiologie, Thieme, 2009

Migration
Pathologie
Proliferation
C. Boucsein 2001
Amöboide Mikroglia
Makrophagen
Courtesy of Diethelm Richter


















