
Thrombopoietinproduktion in Wildtyp- und Interleukin-6 ... · Aus dem Institut für Physiologie der...
56
Aus dem Institut für Physiologie der Universität zu Lübeck Direktor: Prof. Dr. med. W. Jelkmann Thrombopoietinproduktion in Wildtyp- und Interleukin-6-"knock-out"-Mäusen mit akuter Entzündung Inaugural-Dissertation zur Erlangung der Doktorwürde der Universität zu Lübeck - Aus der Medizinischen Fakultät - vorgelegt von Helen Burmester aus Hamburg Lübeck 2008
-
Upload
duonghuong -
Category
Documents
-
view
217 -
download
0
Transcript of Thrombopoietinproduktion in Wildtyp- und Interleukin-6 ... · Aus dem Institut für Physiologie der...

Aus dem Institut für Physiologie
der Universität zu Lübeck
Direktor: Prof. Dr. med. W. Jelkmann
Thrombopoietinproduktion
in Wildtyp- und Interleukin-6-"knock-out"-Mäusen mit akuter Entzündung
Inaugural-Dissertation
zur
Erlangung der Doktorwürde
der Universität zu Lübeck
- Aus der Medizinischen Fakultät -
vorgelegt von
Helen Burmester
aus Hamburg
Lübeck 2008

2
1. Berichterstatter: Prof. Dr. med. Wolfgang Jelkmann
2. Berichterstatter: Prof. Dr. med. Peter Maria Rob

3
Inhaltsverzeichnis
Inhaltsverzeichnis ............................................................................. 3
Abkürzungsverzeichnis .................................................................... 4
1 Einleitung ............................................................................... 6 1.1 Physiologie der Thrombozyten ............................................................................ 6 1.2 Biochemische Eigenschaften von Thrombopoietin .............................................. 8 1.3 Orte der Thrombopoietinproduktion .................................................................. 10 1.4 Regulation der Megakaryopoiese und Thrombozytenproduktion ....................... 11 1.5 Thrombopoietinregulation bei Entzündungen und die Bedeutung von
Zytokinen .......................................................................................................... 14 1.6 Fragestellung .................................................................................................... 15
2 Material und Methoden ........................................................ 16 2.1 Versuchstiere .................................................................................................... 16 2.2 In vitro Inkubation von Vollblut .......................................................................... 17 2.3 IL-6 Bestimmung ............................................................................................... 18 2.4 Thrombopoietinbestimmung im Plasma ............................................................ 19 2.5 RNA-Extraktion ................................................................................................. 19 2.6 Reverse Transkription ....................................................................................... 20 2.7 Polymerase Kettenreaktion ............................................................................... 20 2.8 Thrombopoietin mRNA Quantifizierung mittels kompetitiver PCR ..................... 21 2.9 Thrombopoietin mRNA Bestimmung mit "Real-Time" PCR ............................... 23 2.10 Statistik ............................................................................................................. 24 2.11 Puffer und Lösungen ......................................................................................... 25 2.12 Chemikalien und Reagenzien ........................................................................... 27
3 Ergebnisse ........................................................................... 28 3.1 IL-6 Produktion in ex-vivo Vollblutkulturen ........................................................ 28 3.2 In-vivo IL-6 Produktion nach der Applikation von LPS oder Terpentin ............... 29 3.3 Plasma IL-6 Konzentration in Terpentin behandelten Wildtyp- und IL-6-
"knockout"-Mäusen ........................................................................................... 30 3.4 Plasma Thrombopoietin Konzentration in Terpentin behandelten Wildtyp- und IL-
6-"knockout"-Mäusen ........................................................................................ 31 3.5 Thrombopoietin mRNA Quantifizierung mittels kompetitiver PCR ..................... 32 3.6 Thrombopoietin mRNA Quantifizierung mittels "Real-Time" PCR ..................... 36 3.7 Einfluss der Terpentinapplikation auf das Blutbild der Tiere .............................. 37
4 Diskussion ........................................................................... 41
5 Zusammenfassung .............................................................. 45
Referenzen ....................................................................................... 46
Veröffentlichung .............................................................................. 54
Danksagung ..................................................................................... 55
Lebenslauf ....................................................................................... 56

4
Abkürzungsverzeichnis
% (v/v) Volumenanteil
% (w/w) Gewichtsanteil
Abb. Abbildung
bp Basenpaare
ca. circa
cDNA komplementäre DNA
CP Crossing Point
CSF Kolonien-stimulierender Faktor
c-mpl Gen für MPL
DNA Desoxyribonukleinsäure
DNase Desoxyribonuklease
dNTP Desoxyribonukleosidtriphosphat
EDTA Ethylendiamintetraacetat
EPO Erythropoietin
fg Femtogramm
g Gramm
GAPDH Glycerinaldehyd-3-phosphat-Dehydrogenase
GTC Guanidiniumthiocyanat
h Stunden
HCl Salzsäure
IFN Interferon
i.m. intramuskulär
IL Interleukin
i.p. intraperitoneal
JAK Janus-Kinase
kb Kilobasenpaare
kDa Kilodalton
LPS Lipopolysaccharid
min Minuten
ml Milliliter
MOPS 3-Morpholinopropansulfonsäure
MPL TPO-Rezeptor

5
mRNA Boten-RNA
NaCl Kochsalz
nM nanomolar
nl Nanoliter
PBS Phosphat-gepufferte Salzlösung
PCR Polymerasekettenreaktion
pg Pikogramm
rh IL-6 rekombinantes Interleukin 6
RNA Ribonukleinsäure
RNase Ribonuklease
RT reverse Transkription
s.c. subkutan
SCF Stammzellfaktor
SD Standardabweichung
sek Sekunden
sog. sogenannte
TAE-Puffer Tris-Acetat-EDTA-Puffer
Tg transgen
TNF Tumornekrosefaktor
TPO Thrombopoietin
Tris Tris-(hydroxymethyl)-aminomethan
U Enzym-Aktivitätseinheit
Wt wildtyp
µl Mikroliter
z.B. zum Beispiel

Einleitung
6
1 Einleitung
1.1 Physiologie der Thrombozyten
Thrombozyten spielen bei der primären Hämostase eine wichtige Rolle. Nach einer
Gefäßverletzung heften sie sich an Bindegewebsfasern des Subendothels,
verformen sich und bilden stachelartige Fortsätze. Sie setzen gefäßverengende und
aggregationsfördernde Stoffe frei. Es bildet sich der sog. weiße
Abscheidungsthrombus, der die Blutung zum Stillstand bringt. Bei
Thrombozytopenien oder –pathien ist die primäre Hämostase gestört, und es kommt
zu verstärkten Blutungen nach Schnittverletzungen, zu oberflächlichen Hämatomen
und petechialen Blutungen (6). Normalerweise ist die Thrombozytenkonzentration im
Blut relativ konstant. Sie zeigt allerdings beim Menschen interindividuelle
Schwankungen zwischen 150 und 400 pro nl (78). Ein Thrombozytenverlust, z.B.
während operativer Eingriffe, führt innerhalb von 1-2 Wochen zu einer reaktiven
sekundären Thrombozytose. Aus dieser Beobachtung wurde schon frühzeitig
gefolgert, dass die Thrombozytenbildung humoral geregelt sei (59).
Thrombozyten stammen ebenso wie die Erythrozyten, Monozyten und Granulozyten
von pluripotenten Stammzellen des Knochenmarks ab (CFU-GEMM, s. Abb.1-1). Die
Differenzierungsreihe von der pluripotenten Stammzelle bis zu den Megakaryozyten
umfasst ca. 10 Teilungsschritte. Die Megakaryopoiese wird durch verschiedene
Zytokine gefördert wie z.B. den Stammzellfaktor, den Granulozyten/Monozyten-
Kolonien stimulierenden Faktor (GM-CSF), Erythropoietin und die Interleukine (IL) 1,
3, 4, 6, 7 und 11 (3; 10). Seit 1994 ist gesichert, dass die Megakaryopoiese
außerdem durch ein spezifisches Hormon gefördert wird, das allgemein
Thrombopoietin genannt wird (5; 70; 55; 74; 83). Der Entdeckung des Hormons ging
die Charakterisierung seines Rezeptors voraus. Dieser wurde ursprünglich MPL
genannt. Sein Gen c-mpl ist das zelluläre Homolog des Onkogens v-mpl, eines
murinen Retrovirus, welches eine myeloproliferative Leukämie verursacht
("Myeloproliferative leukemia oncogene-encoded receptor" (80; 60; 75)).
Thrombopoietin hemmt die Apoptose megakaryozytärer Vorläufer und fördert deren

Einleitung
7
Proliferation und Differenzierung (9; 66). Außerdem beschleunigt es die
Megakaryozytensequestrierung und steigert die Aggregationsfähigkeit zirkulierender
Thrombozyten (48). Thrombopoietin wirkt dabei nicht direkt auf die
Plättchenaggregation, sondern fördert die Aktivierung verschiedener
Gerinnungsfaktoren. Es erhöht die Thrombozyten-Fibrinogen Bindung und deren
Adhäsion an Kollagen, fördert die Freisetzung von Adenosin-Diphosphat und
sensibilisiert die Thrombozyten gegenüber der Wirkung von Thrombin (14).
TPOSCFIL-3
IL-6IL-11TPO
Megakaryozyt
ThrombozytenCFU-Mega
CFU-Bas
CFU-Eo
CFU-GM
BFU-E
CFU-GEMM
IL-3, IL-4, CSF
GM-CSF, IL-3, IL-5
GM-CSF, G-CSF
GM-CSF, M-CSF
EPO
Basophiler Granulozyt
Eosinophiler Granulozyt
Neutrophiler Granulozyt
Monozyt
Erythrozyt
Abb. 1-1: (Verändert nach Wolber und Jelkmann, News Physiol Sci 17: 6-10,
2002) Darstellung eines vereinfachten Schemas der Hämatopoiese. Die
Proliferation und Differenzierung der Vorläuferzellen (CFU, colony-forming unit)
von Megakaryozyten (M, Meg), Granulozyten (G), Monozyten (M) und
Erythrozyten (E) wird durch Wachstumsfaktoren (TPO, Thrombopoietin; IL,
Interleukin; SCF, Stammzellfaktor; CSF, Kolonien-stimulierende Faktoren; EPO,
Erythropoietin) gesteuert.

Einleitung
8
1.2 Biochemische Eigenschaften von Thrombopoietin
Das menschliche Thrombopoietin-Gen wurde auf Chromosom 3 q27-28 lokalisiert.
Es umfasst 6 Kilobasenpaare und enthält 5 kodierende Exons (26; 30). Die
molekulare Masse beträgt 70 kDa. Zusätzlich sind weitere Isoformen identifiziert
worden, die durch alternatives „splicing“ entstehen. Die verschiedenen Isoformen
sind in unterschiedlicher Menge über die verschiedenen Gewebe verteilt, ihre
physiologische Rolle ist jedoch noch unklar (89).
Das primäre Translationsprodukt des menschlichen Thrombopoietingens ist aus 353
Aminosäuren zusammengesetzt. Diese schließen ein Starterpeptid aus 21
Aminosäuren ein. Zirkulierendes Thrombopoietin (TPO) besteht aus 332
Aminosäuren. Die biologische Halbwertszeit des Hormons ist mit 20-30 h beim
Menschen länger als die anderer hämopoietischer Wachstumsfaktoren. Das reife
TPO-Molekül besteht aus zwei Domänen: Die ersten 153 Aminosäuren bilden den
NH2-terminalen Abschnitt und sind für die Bindung an den Thrombopoietinrezeptor
notwendig. Die zweite Domäne bildet der COO-terminale Abschnitt, der 177
Aminosäuren umfasst.
Die NH2-terminale Domäne zeigt eine 23%ige Sequenzhomologie zum
menschlichen Erythropoietin (EPO), weitere 27% der Aminosäuren sind dem EPO
auch funktionell ähnlich. Die letzten 40 Aminosäuren der EPO-ähnlichen Domäne
beinhalten vier Cystein-Reste, die wahrscheinlich essentiell für die Vermittlung der
biologischen Aktivität des TPO sind (19).
Der COO-terminale Abschnitt weist 6 N- und ca. 8 O-gekoppelte
Kohlenhydratseitenketten sowie 2 Bisulfidbrücken auf. Er gewährleistet die lange
Lebenszeit des Moleküls dadurch, dass er wahrscheinlich die schnelle Entfernung
von Thrombopoietin aus der Zirkulation verhindert (37). Außerdem spielt diese
Domäne eine Rolle bei der effizienten Sekretion von TPO (53).
Große Hoffnung wurde in die klinische Anwendung von Thrombopoietin bei
verschiedenen, mit Thrombozytopenien einhergehenden, Krankheitsbildern gesetzt
(53). Gegenstand der intensiven klinischen Forschung waren zwei rekombinante
humane Thrombopoietinpräparate:
Ein glykosyliertes, vollständiges rekombinantes humanes Thrombopoietinpräparat
(rHu-TPO) und ein mit Polyethylenglykol verbundenes, verkürztes rekombinantes

Einleitung
9
Thrombopoietin-Analogon (PEG-rHu-MGDF: rekombinanter humaner
„megakaryocyte growth and development factor“) (s. Abb.1-2).
1.Thrombopoietin cDNA
SC CT C T C TTSN NT N STSTSSSTSS NNN
ss
ss
ss
ss
2. rHu-TPO
3. PEG-rHu-MGDF
EPO-gleiche Domäne Kohlenhydratreiche Domäne
PEG
Kohlenhydratseitenketten
Abb. 1-2: (Verändert nach Kaushansky, N Engl J Med 339: 746-754, 1998)
(1) Modell der Thrombopoietin-cDNA bzw. des kodierten Polypeptids mit einer
Sequenzhomologie der NH2-terminalen Domäne zum Erythropoietin. Die
Aminosäuren mit wichtigen Eigenschaften beinhalten Serin (S), Cystein (C),
Threonin (T) und Asparagin (N). (2) Das glykosylierte, vollständige rekombinante
humane Thrombopoietinpräparat (rHu-TPO) und (3) das mit Polyethylenglykol
verbundene, verkürzte rekombinante Thrombopoietin-Analogon (PEG-rHu-MGDF).

Einleitung
10
1.3 Orte der Thrombopoietinproduktion
Untersuchungen zur Lokalisation der Thrombopoietin produzierenden Organe
wurden zunächst an Versuchstieren durchgeführt. Thrombopoietin-mRNA wurde in
Leberextrakten fetaler sowie in Leber- und Nierenextrakten adulter Tiere
nachgewiesen. Andere Gewebe bzw. Zellen, die nach Literaturangaben
Thrombopoietin-mRNA exprimieren können, sind in vivo glatte Muskelzellen, Milz,
Gehirn, Lunge, Darm und Knochenmark sowie in der Zellkultur Endothelzellen,
Fibroblasten und Hepatozyten (Übersichten in (20; 45; 77; 30)). Untersuchungen an
den Organen verstorbener menschlicher Feten und Neugeborener haben gezeigt,
dass über 95% der gesamten Thrombopoietin-mRNA des Menschen in der Leber
exprimiert wird (84). Tierexperimentelle Studien deuten an, dass – anders als bei der
Synthese des verwandten Hormons Erythropoietin – die Leber zeitlebens das
dominierende Organ der Thrombopoietinsynthese bleibt (65). Erythropoietin wird im
fetalen und neonatalen Lebensabschnitt hauptsächlich in der Leber gebildet,
während die mRNA-Expression in der Niere ab der 30. Schwangerschaftswoche
stark zunimmt (17) und im Erwachsenalter Hauptbildungsort für Erythropoietin ist
(43).
In-situ-Hybridisierungsstudien ergaben, dass die Thrombopoietin-mRNA
exprimierenden Zellen der Leber Hepatozyten und die der Niere proximale
Tubuluszellen sind (77). Klinische Untersuchungen stehen im Einklang mit der
Vorstellung, dass die Leber das wichtigste Organ der Thrombopoietinproduktion
beim Menschen ist. Patienten mit Leberzirrhose leiden an einer Thrombozytopenie,
die zumindest partiell auf einer Einschränkung der Thrombopoietingenexpression
beruht (57; 40; 86). Es wurden stark erniedrigte Thrombopoietinkonzentrationen im
Plasma dieser Patienten gefunden (69). Nach erfolgreicher Lebertransplantation
steigt die Thrombopoietinkonzentration im Plasma der Transplantierten an, und die
Plättchenzahl im Blut normalisiert sich. Dagegen ist nie berichtet worden, dass es bei
einer chronischen Niereninsuffizienz zu einer Thrombozytopenie aufgrund eines
Thrombopoietinmangels kommt.

Einleitung
11
1.4 Regulation der Megakaryopoiese und Thrombozytenproduktion
Thrombopoietin ist der wichtigste physiologische Faktor, der die Megakaryopoiese
und die Thrombozytenproduktion reguliert (19; 51). Es induziert die Proliferation der
„colony forming units“ der Megakaryozyten (CFU-Meg) und die Entwicklung der
reifen polyploiden Megakaryozyten, welche zu der Bildung von Thrombozyten führt
(55).
Zusätzlich wirkt Thrombopoietin zusammen mit anderen Zytokinen auf die
Proliferation von Vorläuferzellen der Erythrozyten (47) und auf primitive
hämatopoietische Vorläuferzellen (73).
In der Thrombopoiese spielen nach den bisherigen Studien drei Regelmechanismen
eine Rolle.
Der als erstes beschriebene Mechanismus ist die „feed back“-Regulation (52) (Abb.
1-3). Thrombopoietin wird in einer konstanten Menge hauptsächlich in der Leber,
aber auch in der Niere produziert. Es wird angenommen, dass die
Thrombopoietinkonzentration im Blutplasma vor allem von der Internalisierung des
Hormons durch seine Zielzellen abhängt (23; 76; 21; 63; 71). Der MPL-Rezeptor der
Thrombozyten kann Thrombopoietin binden und prozessieren. Dadurch wird in
normalen menschlichen Thrombozyten die Tyrosin Phosphorylierung verschiedener
Signalproteine aktiviert, wie unter anderem die der Janus Kinase 2 (JAK 2) und von
Shc , Stat3 und Stat5 (61). Somit wird um so mehr Thrombopoietin aus dem
Blutplasma entfernt, desto mehr Plättchen und Megakaryozyten vorhanden sind.
Folglich ist die Thrombopoietinkonzentration im Plasma bei Thrombozytopenie hoch
und bei normaler oder vergrößerter Megakaryozyten- und Thrombozytenmasse
niedrig. Tierexperimentelle und klinische Untersuchungen unterstützen dieses
Konzept. In Mäusen, denen der Thrombopoietinrezeptor MPL fehlt (MPL-“knockout“-
Mäuse), sind durch eine sehr hohe Plasmathrombopoietinkonzentration
gekennzeichnet (23). Nach Transfusion normaler Thrombozyten fällt die
Plasmathrombopoietinkonzentration sehr rasch ab (23). Umgekehrt kommt es bei
Tumorpatienten nach Chemotherapie zu einer Thrombozytopenie, die einen inversen
Anstieg der Thrombopoietinkonzentration im Blut verursacht (33).
Als weiterer Regelmechanismus wurde die Rolle der Thrombozyten als Speicherort
für das Thrombopoietin beschrieben, wobei das Thrombopoietin nach Aktivierung der
Thrombozyten freigesetzt wird (24).

Einleitung
12
Der dritte Mechanismus ist die Steigerung der Thrombopoietinproduktion durch
erhöhte mRNA-Expression im Knochenmarkstroma bei Thrombozytopenie.
Verschiedene Arbeitsgruppen haben tierexperimentelle Untersuchungen
durchgeführt, um zu prüfen, ob die Thrombopoietingenexpression in der Leber und in
anderen Thrombopoietin produzierenden Organen von der
Thrombozytenkonzentration im Blut abhängt. Die meisten Arbeiten haben gezeigt,
dass eine solche Beziehung nicht besteht. Der Thrombopoietin-mRNA Gehalt der
Leber war in Tieren mit einer experimentell erzeugten Thrombozytopenie oder
Thrombozytose gleich groß wie in Normaltieren (58; 23; 16; 76). Abweichend wurde
in verschiedenen Spezies – und auch beim Menschen - gefunden, dass es bei einer
Thrombozytopenie zu einer zusätzlichen Thrombopoietin-mRNA Expression in
Stromazellen des Knochenmarks kommt (77; 36).
Darauf aufbauend wurden in neueren Studien an Ratten mit systemischen
entzündlichen Erkrankungen, die mit einer reaktiven Thrombozytose einhergehen,
eine gesteigerte Thrombopoietin-mRNA Expression in der Leber gefunden. Diese
Reaktion könnte einen Mechanismus der Infektionsabwehr darstellen und liefert neue
Erkenntnisse über die Regulation der Thrombopoietin-mRNA Expression bei
Entzündungen (85).

Einleitung
13
LeberNiere
Thrombopoietin
Thrombozyten
MegakaryozytenHypoplasie:Thrombozytopenie
Hohe PlasmaThrombopoietinKonzentrationen
MegakaryozytenHyperplasie:Thrombozytose
Niedrige PlasmaThrombopoietinKonzentrationen
Knochenmark
“Steady-state” ThrombozytenProduktion
Abb. 1-3: Modell der Thrombopoietinregulation. Leber und Niere produzieren
Thrombopoietin (TPO) und geben es ans Blut ab.
Bei einer Thrombozytopenie (linke Seite der Abbildung) wird nur wenig des
freigesetzten TPOs metabolisiert, woraus eine hohe TPO-Konzentration im Plasma
resultiert, die das hypoplastische Knochenmark stimuliert. Zusätzlich produziert das
Knochenmarkstroma TPO (gebogener, dicker Pfeil). Die hohen TPO-
Konzentrationen helfen, die Thrombozyten- und Megakaryozytenproduktion wieder
anzuregen.
Während einer Thrombozytose (rechte Seite der Abbildung) entfernen die
zahlreichen Thrombozyten einen Grossteil des TPOs aus dem Blut und die
Produktion durch die Stromazellen im Knochenmark ist stark reduziert. Folglich steht

Einleitung
14
wenig TPO zur Verfügung, um auf die Megakaryozyten zu wirken (gebogener,
dünner Pfeil). Es kommt zu einer “steady-state“ Thrombozytenproduktion.
1.5 Thrombopoietinregulation bei Entzündungen und die Bedeutung von
Zytokinen
Abweichend von den normalen Regelmechanismen ist bei entzündlichen Prozessen
sowohl die Thrombozyten- als auch die Thrombopoietinkonzentration im Blut
abnormal hoch (12; 42), es kommt zur sog. reaktiven Thrombozytose. Während der
akuten Entzündungsphase steigt die Thrombopoietinkonzentration stark an und
erreicht in der ersten Woche den höchsten Wert. Die Thrombozytenzahl hingegen ist
in der ersten Woche unverändert und erreicht die Höchstwerte erst zwei bis drei
Wochen nach Entzündungsbeginn (39). Aufgrund dieser Ergebnisse und aufgrund
von Versuchen an gesunden Menschen, deren Thrombozytenzahlen 12 Stunden
nach Thrombopoietin Injektion ihre Höchstwerte erreichten (31), lässt sich vermuten,
dass der Thrombopoietinanstieg die Thrombozytose in akuten Infektionen induziert
und Teil einer Akut-Phase-Reaktion ist (85). Diese Reaktion findet sich
beispielsweise bei Autoimmunerkrankungen wie Morbus Crohn, bei Infektionen, nach
einem Trauma, nach chirurgischen Eingriffen und bei malignen Erkrankungen vor
Beginn der Chemotherapie. Prinzipiell könnte der Verlust der inversen Beziehung
zwischen der Thrombozytenmasse und der Thrombopoietinkonzentration auf einer
verlängerten Lebenszeit des Hormons oder auf einer abnormal gesteigerten
Stimulation der Thrombopoietinproduktion beruhen. Experimentelle Hinweise auf
eine verlängerte Thrombopoietinlebenszeit bei entzündlichen Erkrankungen sind in
der Literatur nicht zu finden. Die zweite Erklärungsmöglichkeit ist wahrscheinlicher.
Das Zytokin Interleukin-6 (IL-6) stimuliert die Reifung von Megakaryozyten und die
Thrombozytenbildung im Knochenmark (4; 64), außerdem steigert es die Synthese
verschiedener Akut-Phase-Proteine in der Leber (32). In Zellkulturversuchen mit
humanen Hepatomzellen wurde gefunden, dass das proentzündliche Zytokin IL-6 die
Thrombopoietin-mRNA Expression steigert. Die Promoter Sequenz des
Thrombopoietin-Gens enthält außerdem 13 mutmaßliche IL-6-Antwort-Sequenzen.
Andere Zytokine wie IL-1, IL-11 oder der Tumornekrosefaktor α (TNF α) zeigen keine
solche Wirkung (88). Bei entzündlichen Erkrankungen ist neben Thrombopoietin also

Einleitung
15
auch die IL-6 Konzentration im Blut erhöht und diese Werte korrelieren stark
miteinander (39; 38). Aufgrund dieser Untersuchungen entstand die Hypothese, dass
die hohen IL-6 Konzentrationen und eine Thrombozytose bei entzündlichen
Prozessen und malignen Erkrankungen durch einen IL-6-induzierten
Thrombopoietinanstieg hervorgerufen werden könnten (44). Es wurde außerdem
gezeigt, dass die Verabreichung von rekombinantem IL-6 (rhIL-6) im Versuchstier
eine Thrombozytose hervorruft. In Tumorpatienten bewirkt die Gabe von rhIL-6 eine
Zunahme der Plättchenkonzentration im Blut (82). Durch die wiederholte Applikation
hoher Dosen von IL-6 lässt sich im Versuchstier (Maus) ein Anstieg der
Plasmathrombopoietinkonzentration erwirken (44). Inwieweit endogen gebildetes IL-6
bei entzündlichen Prozessen die Thrombopoietin-mRNA Expression und
Thrombopoietinproduktion steigert, ist bislang unbekannt.
1.6 Fragestellung
Die Untersuchungen, über die hier berichtet wird, sollten einem besseren
Verständnis der Vermittlerrolle von IL-6 bei der gesteigerten
Thrombopoietinproduktion bei entzündlichen Prozessen dienen. Hierzu wurden
tierexperimentelle Studien an Wildtyp-Mäusen (Bl/6) und an transgenen IL-6
defizienten Mäusen (IL-6-"knockout") durchgeführt. In den Tieren wurde durch
subkutane Applikation von Terpentin ein steriler Abszess gesetzt. Bestimmungen
der IL-6- und Thrombopoietinkonzentration im Plasma sowie des Thrombopoietin-
mRNA Gehaltes im Lebergewebe der Tiere erbrachten Ergebnisse, die die
Hypothese stützen, dass Thrombopoietin zur Gruppe derjenigen Akut-Phase-
Proteine gehört, deren Syntheserate durch IL-6 gesteigert wird.

Material und Methoden
16
2 Material und Methoden
2.1 Versuchstiere
Für die Untersuchungen wurden Mäuse des Stammes C57 Bl/6 verwendet. IL-6-
defiziente (IL-6-"knockout") Mäuse nach Rückkreuzung auf den genetischen
Hintergrund des Stammes C57Bl/6 wurden freundlicherweise von Herrn Prof.
Manfred Kopf (Institut für Molekulare Biomedizin, ETH Zürich, Schweiz) zur
Verfügung gestellt. In den IL-6 defizienten Tieren ist das IL-6-Gen im zweiten Exon
(erstes kodierendes Exon) durch eine Neomycin-resistente Kassette zerrissen, so
dass sie kein IL-6 produzieren (49). Die Mäuse wurden in der Tierhaltung der
Vorklinischen Institute unter konventionellen Bedingungen bei freiem Zugang zu
Standardfutter und Wasser und einem 12 h Hell-Dunkel-Zyklus gehalten und
vermehrt. Experimente wurden mit männlichen Tieren mit einem Körpergewicht
von 25-32 g durchgeführt. Um akute Entzündungsreaktionen zu erzeugen, wurde
den Tieren subkutan (s.c.) 150 µl Terpentin (Karl Roth GmbH, Karlsruhe) unter die
Rückenhaut appliziert. Kontrolltiere erhielten 150 µl sterile physiologische
Kochsalzlösung. Die Injektion von Terpentinöl führt zu einem sterilen Abszess
(27). Nach Literaturangaben (8) bewirkt die Terpentinölinjektion in Mäusen einen
ausgeprägteren IL-6 Anstieg im Plasma als die Gabe von bakteriellem
Lipopolysaccharid (LPS). Zum Vergleich wurde in dieser Arbeit einigen Tieren
intraperitoneal LPS (0,1 µg/g Körpergewicht, Stammlösung 10 µg/ml in
physiologischer Kochsalzlösung; Escherichia coli, Serotyp 0:III:B4; Fa. Sigma,
Taufkirchen) verabreicht.
6 h, 24 h oder 72 h später wurden die Tiere intramuskulär (i.m.) mit Ketamin (25
mg/ml) und Pentobarbital (30 mg/ml) narkotisiert und aus der Vena cava caudalis
Blut entnommen. Zur Erstellung des Blutbildes wurden EDTA beschichtete
Spritzen (1,35 mg Na2-EDTA/ml Blut) und Blutröhrchen benutzt. Das Plasma
wurde durch 15 min Zentrifugation bei 2900 rpm und 20°C gewonnen (Eppendorf
Zentrifuge 5402; Eppendorf, Hamburg). Die Leber und die Nieren wurden
entnommen, sofort in flüssigem Stickstoff tiefgefroren und bis zur Aufarbeitung bei
–80°C gelagert. Der sterile Abszess der mit Terpentinöl behandelten Mäuse wurde

Material und Methoden
17
durch Eröffnen der Rückenhaut sichtbar gemacht, bei den Kontrolltieren war
dieser nicht nachweisbar. Untersuchungen des Blutbildes wurden mit einem
veterinärmedizinischen Blut-Analysegerät durchgeführt (ABC Vet, Scil animal care
company GmbH, Viernheim). Mess- und Normalwerte-Kennung waren auf die
Spezies "Maus" eingestellt.
Die Versuche waren durch das zuständige Ministerium am 4. September 2000
genehmigt worden (Nr. 21-1c/00).
2.2 In vitro Inkubation von Vollblut
Um abzusichern, dass die IL-6 "knockout"-Mäuse kein IL-6 produzieren, wurden in
vitro Studien durchgeführt. Dabei wurden Vollblutproben der Wildtyp- und IL-6
"knockout"-Tiere nach Stimulation mit LPS im ex vivo Modell nach der für
menschliches Blut beschriebenen Methode (18; 56) auf IL-6 Bildung untersucht. Den
unbehandelten Tieren wurde in Äthernarkose durch Herzpunktion Blut in Lithium-
Heparin-Monovetten (Sarstedt, Nürmbrecht) entnommen. Je 50 µl heparinisiertes
Vollblut wurde im zweifach Ansatz pro Blutspender unverdünnt unter sterilen
Bedingungen in die – mit Inkubationslösung versetzten – Zellkulturplatten überführt
(24er Platten, Nunc, Wiesbaden). Die Inkubationslösungen bestanden aus 450 µl
Zellkulturmedium RPMI 1640 (Invitrogen, Karlsruhe) mit 100 ng/ml LPS und
Antibiotika (100 µg/ml Streptomycin, 100 U/ml Penicillin). Die Suspensionskulturen
wurden 6 h bzw. 24 h im Brutschrank inkubiert (37°C, Inkubationsgas 5% CO2 in Luft,
Wasserdampfsättigung). Anschließend wurde die Suspension 10 min bei 1000 rpm
und 4°C zentrifugiert und der Überstand zur enzymimmunologischen IL-6
Bestimmung bei -40°C eingefroren.

Material und Methoden
18
2.3 IL-6 Bestimmung
Zur Bestimmung der IL-6 Konzentration im Mausplasma bzw. in den
Vollblutkulturüberständen wurde zunächst das kommerzielle
enzymimmunologische (ELISA) Testbesteck Quantikine M Maus-IL-6 der Fa. R&D
Systems (Minneapolis, MN, USA, Katalog-Nr. M6000) verwendet. Dabei stellte
sich heraus, dass entgegen den Literaturangaben (67) das Testbesteck, das
polyklonale Antikörper gegen Maus IL-6 enthielt, für die Plasmaproben keine
validen Ergebnisse lieferte. Dies war erkenntlich an den hohen IL-6 Werten (ca.
150 pg/ml) im Plasma normaler Wildtyp- und IL-6 "knockout" -Tiere und der
Beobachtung, dass Verdünnungsreihen mit Mausplasma keinen parallelen Verlauf
gegenüber dem rekombinanten Maus IL-6 Standard lieferten. Nach Schriftwechsel
mit der Herstellerfirma wurde im September 2002 ein neues Testsystem auf den
Markt gebracht (M6000B), das sich für die Messung von IL-6 im Mausplasma als
geeignet erwies. Im Ergebnisteil sind nur die mit dem neuen Testbesteck
(M6000B) bestimmten Plasma IL-6 Werte gezeigt. Dabei wurden 50 µl der zu
untersuchenden Probe bzw. des rekombinanten IL-6 Standards mit
Inkubationspuffer 2 h bei Raumtemperatur in den mit monoklonalem Anti-IL-6
Antikörper beschichteten Mikrotiterplatten auf einer Schüttelplattform inkubiert,
anschließend fünfmal gewaschen, mit Meerrettichperoxidase gekoppeltem
polyklonalem Anti-IL-6 Antikörper erneut 2 h bei Raumtemperatur inkubiert,
wiederum fünfmal gewaschen und mit dem Substratpuffer mit Chromogen (H2O2
und Tetramethylbenzidin) 30 min inkubiert. Die Reaktion wurde mit Salzsäure
(HCl) gestoppt und die Absorption mittels eines Mikrotiterplatten-Photometers
(Rainbow; SLT, Kreilsheim) bei einer Wellenlänge von 450 nm mit einem
Referenzfilter von 570 nm ermittelt. Es wurden Doppelbestimmungen
durchgeführt. Das untere Detektionslimit betrug 3 pg/ml. Die Inter- und Intra-Test-
Variationskoeffizienten betrugen < 10% bzw. < 5%.

Material und Methoden
19
2.4 Thrombopoietinbestimmung im Plasma
Zur Bestimmung der Thrombopoietinkonzentration im Mausplasma wurde das
kommerzielle enzymimmunologische Testbesteck Quantikine M Maus-
Thrombopoietin der Fa. R & D Systems (Katalog-Nr. MTP00) verwendet. Nach
Herstellerangaben beträgt die untere Detektionsgrenze 20 pg/ml Thrombopoietin,
die Koeffizienten für die Intra- und Interassay-Variabilität ≤ 8%.
2.5 RNA-Extraktion
Die tiefgefrorenen Organe wurden in 4 mol/l Guanidiniumthiocyanat (GTC; ca. 10
ml pro g Gewebe) auf Eis homogenisiert (Ultra Turrax T25; IKA-Labortechnik,
Stauffen im Breisgau). Die Gesamt-RNA wurde mittels saurer Phenolextraktion
aus dem Gewebehomogenat nach (15) isoliert. Die Methode nutzt die
unterschiedliche Verteilung von DNA, RNA und Proteinen zwischen der
organischen und der wässrigen Phase, in welcher die RNA verbleibt. Das
Originalprotokoll (15) wurde für ein Zellhomogenatvolumen von 700 µl adaptiert,
welches mit 560 µl Phenol (pH 4,5) und 280 µl Phenol: Chloroform: Isoamylalkohol
im Verhältnis 25:24:1 (Lösungen von Sigma) sowie 70 µl 2 mol/l Natriumacetat-
Puffer (pH 4,0) in einem 1,5 ml Reaktionsgefäß gemischt wurde. Die Ansätze
wurden 20 min auf Eis inkubiert und anschließend bei 12.000 rpm und 4°C 20 min
zentrifugiert (Eppendorf Zentrifuge 5402; Eppendorf, Hamburg). Nach der
Zentrifugation wurde die obere wässrige Phase abpipettiert und mit einem
gleichen Volumen Isopropanol (Merck, Darmstadt) versetzt. Nach einer 30
minütigen Präzipitationsphase bei –80°C wurde die RNA-Lösung wie o.a.
zentrifugiert. Das Präzipitat wurde in 300 µl GTC resuspendiert und erneut mit
Isopropanol gefällt. Anschließend wurde die RNA mit 75% Ethanol gewaschen,
getrocknet und in 20-40 µl mit Diäthylpyrocarbonat behandeltem Wasser
aufgenommen.
Die Konzentration der RNA-Lösungen wurde anhand der Extinktion bei 260 nm
ermittelt. Zudem wurde der Quotient der Absorptionen bei 260 und 280 nm zur
Prüfung auf Verunreinigungen der RNA durch Proteine oder Phenol bestimmt. Nur

Material und Methoden
20
RNA mit einem Quotienten zwischen 1,8 und 2,0 wurde verwendet. Alle RNA-
Lösungen wurden bei –80°C aufbewahrt.
2.6 Reverse Transkription
Vor der reversen Transkription wurden 5 µg Gesamt-RNA auf einem
denaturierenden Agarose-Gel visualisiert, um Degradationen und ungenaue
Konzentrationsbestimmungen ausschließen zu können. Die RNA wurde mit 6 µl
Ladepuffer gemischt und 10 min bei 68° C im Wasserbad denaturiert. Auf einem
1,1%igen Agarose-Gel mit 2% Formaldehyd wurde die RNA in MOPS-Puffer
elektrophoretisch aufgetrennt. Anschließend wurden 5 µg Gesamt-RNA
transkribiert (84). Der vollständige Reaktionsansatz von 25 µl enthielt 1 µg
oligo(dT)15 als Startermolekül für die Polymerase, je 500 nM dNTPs und 100 U der
reversen Transkriptase (Maus- Moloney Leukämievirus reverse
Transkriptase/MMLV TR). Die RNA und die Oligonukleotide wurden in 14,5 µl
Wasser gemischt und für 15 min bei 68°C denaturiert. Nach 15 minütigem
Abkühlen bei 4°C wurden die restlichen Reagenzien hinzugefügt und der Ansatz
45 min bei 42°C und 45 min bei 52°C inkubiert. Durch 10 minütiges Erhitzen auf
100°C wurde das Enzym inaktiviert und der Ansatz auf 4°C heruntergekühlt. Die
so gewonnene cDNA wurde bis zur weiteren Verwendung bei -20°C aufbewahrt.
2.7 Polymerase Kettenreaktion
Vor der Quantifizierung wurden qualitative PCR-Analysen für Thrombopoietin und
Glycerinaldehyd-3-Phosphat-Dehydrogenase (GAPDH) cDNAs durchgeführt.
Das PCR-Signal für GAPDH diente dabei als Kontrolle für die Integrität und die
gleichmäßige Konzentration der cDNA.
Es wurde 1 µl cDNA in einem 50 µl-Ansatz benutzt, der 0,75 U Taq-Polymerase
(Gibco BRL Life Technologies, Karlsruhe), je 200 nM dNTPs und je 400 nM
Oligonukleotide im vom Enzymhersteller mitgelieferten Reaktionspuffer enthielt.
Zur Denaturierung wurden die Reaktionsansätze 3 min auf 94°C erhitzt.
Anschließend wurden 27 - 35 PCR-Zyklen von 1 min Denaturierung bei 94°C, 90 s

Material und Methoden
21
Anheftung bei 55°C für die GAPDH- und 60°C für m-TPO-Startermoleküle (s.Tab
2-1) und 3 min Elongation bei 72°C durchgeführt. Der abschließende
Elongationsschritt dauerte 10 min.
Danach wurden 20 µl des Reaktionsansatzes mit 2 µl DNA-Ladepuffer versetzt
und zur Agarose-Gelelektrophorese eingesetzt. Es wurden Gele mit 2% (w/w)
Agarose und 0,5 µg/ml Ethidiumbromid in TAE-Puffer, welcher auch als
Elektrophoresepuffer diente, benutzt. Die Länge der PCR-Produkte wurde durch
Vergleich mit einem Molekulargewichtsstandard ermittelt (100 bp DNA Standard).
Name Position Sequenz TAn
5'-GAPDH 4296 - 4315 5'-ATC ATC CCT GCC TCT ACT GG-3'
55°C 3'-GAPDH 4533 - 4552 5'-TGG GTG TCG CTG TTG AAG TC-3'
5'-mTPO 229 – 247 5'-CTC TGT CCA GCC CCG TAG C-3'
60°C
3'-mTPO 525 - 542 5'-CCC CAA GAG GAG GCG AAC-3'
Tab. 2-1: Als Startermoleküle verwendete Oligonukleotide. 5'-GAPDH und 3'-
GAPDH dienten zur Amplifizierung von GAPDH-cDNA (PCR-Produkt 256 bp) und
5'-mTPO und 3'-mTPO von TPO-cDNA (314 bp). Die angegebenen Positionen
bezeichnen das erste und das letzte Nukleotid, an das das Oligonukleotid bindet.
TAn, Anheftungstemperatur in der PCR.
2.8 Thrombopoietin mRNA Quantifizierung mittels kompetitiver PCR
Zur Quantifizierung der Thrombopoietin-mRNA wurde zunächst die klassische
Methode nach Gilliland et al. (28) in der Modifikation von Siebert und Larrick (72)
verwendet. Dabei wird eine unbekannte Menge cDNA mit der bekannten Menge
einer Kompetitor-DNA in derselben PCR koamplifiziert und konkurriert dabei mit

Material und Methoden
22
der TPO-cDNA um die Bindung der Oligonukleotide. Das PCR-Produkt dieses
Kompetitors besitzt eine Größe von 252 bp.
Die kompetitive PCR wurde ebenso durchgeführt wie die qualitative PCR für TPO
(siehe Abschnitt 2.7), zusätzlich zu 1 µl cDNA wurden dem Ansatz auch 1 µl
Kompetitorlösung mit bekannter Konzentration als interner Standard zugesetzt.
Für jede cDNA-Probe wurden vier PCR-Reaktionen mit unterschiedlichen
Konzentrationen an Kompetitor angesetzt. Von der cDNA und den
Kompetitorlösungen wurden 1:10 Vorverdünnungen hergestellt, um Fehler beim
Pipettieren zu reduzieren. Die Lösungen wurden mit den übrigen Reagenzien
vermischt.
Nach der PCR-Reaktion und Agarose-Gel Elektrophorese sind die aus der cDNA
und dem Kompetitor gebildeten PCR-Produkte anhand ihrer Größe unterscheidbar
(314 bp für m-TPO und 252 bp für den Kompetitor), so dass aus
Äquivalenzmengen die Ausgangs-mRNA-Menge abgeschätzt werden kann.
Aus der Äquivalenzkonzentration wurde die TPO-mRNA Konzentrationen pro
Mikrogramm Gesamt-RNA errechnet. Dabei wurden die molekulare Masse des
Kompetitors, die in der RT eingesetzte Menge Gesamt-RNA, die RT-Effizienz und
die Tatsache, dass der Kompetitor doppelsträngig ist, wohingegen die cDNA
einzelsträngig vorliegt, in folgender Formel berücksichtigt:
cTPO-mRNA = 2000 * q / (MKomp * cRNA * fRT)
cTPO-mRNA TPO-mRNA-Konzentration [amol/µg Gesamt-RNA]
q Äquvalenzkonzentration [fg Kompetitor/50 µL PCR-Ansatz]
MKomp molekulare Masse des Kompetitors [g/mol]
cRNA Gesamt-RNA-Konzentration [µg/µL RT-Ansatz]
fRT RT-Effizienz
Die so gewonnenen Ergebnisse bestätigten jedoch den bekannten Nachteil einer
großen Intra- und Inter-Assay Variation. Daher wurden die Proben zusätzlich den
neueren Light-Cycler Verfahren der “Real-Time“ PCR unterzogen. Da zum
Zeitpunkt der hier berichteten Untersuchungen ein solches Gerät am Institut für
Physiologie der Universität zu Lübeck nicht verfügbar war, wurde die Möglichkeit
genutzt, die Messungen am Institut für Physiologie der Universität Duisburg-Essen
durchzuführen (Direktor Prof. Dr. Joachim Fandrey). Dabei wurde davon

Material und Methoden
23
ausgegangen, dass die Präzision (geringe Intra- und Inter-Assay Variation) der
“Real-Time“ PCR Bestimmung größer als bei der bislang praktizierten
kompetitiven PCR ist.
2.9 Thrombopoietin mRNA Bestimmung mit "Real-Time" PCR
Für die Quantifizierung der TPO cDNA wurden der 5´-Primer 5´-CCT GGG AGA
ATG GAA AAC CC-3´ und der reverse 3´-Primer 5´-ACT GTC CTC GTG CTG
CCA TC-3´ verwendet, der ein Amplifikationsprodukt von 108 Basenpaaren ergibt.
Bei dieser Methode wird ein Farbstoff in das PCR-Produkt eingebaut und zum
Fluoreszieren gebracht. Die Fluoreszenz wird einmal pro Zyklus am Ende der
Elongationsphase gemessen und zeitgleich auf dem Bildschirm dargestellt.
Dadurch kann der Anstieg des PCR–Produktes Zyklus für Zyklus verfolgt werden.
In Abhängigkeit von der Ausgangskonzentration wird ab einem bestimmten Zyklus
ein spezifisches Signal messbar. Dieser Wert, an dem die Proben erstmals eine
ausreichende Konzentration aufweisen, um aus dem Hintergrund hervorzutreten,
wird auch als Crossing-Point (CP) bezeichnet und lieferte die Grundlage für die
Quantifizierung.
Es gibt mehrere Fluoreszenzformate, von denen SYBR Green gewählt wurde
(SYBR-Green, GeneAmp 5700 Sequence Detection System, Applied Biosystem,
Weiterstadt, Germany). Dabei handelt es sich um einen Fluoreszenzfarbstoff, der
in doppelsträngige DNA eingebaut wird. Am Beginn der Reaktion enthält der
Reaktionsansatz die denaturierte DNA, die Primer und den Farbstoff SYBR Green.
Die ungebundenen Moleküle des Farbstoffs strahlen nur eine schwache
Fluoreszenz aus. Nach dem Binden der Primer an die DNA können Moleküle des
Farbstoffs an die doppelsträngig vorliegende DNA binden. Dadurch kommt es zu
einem sichtbaren Anstieg der Fluoreszenz. Während der Elongationsphase binden
immer mehr Farbstoffmoleküle an die neusynthetisierte DNA. Wenn die Reaktion
kontinuierlich am Monitor aufgezeigt wird, ist es möglich, den Anstieg der
Fluoreszenz und damit des Amplifikationsproduktes zeitgleich (“Real-Time“) zu
verfolgen. Während der Denaturierungsphase, die sich anschließt, erlischt das
Signal, da die DNA dann wieder einzelsträngig vorliegt (11).

Material und Methoden
24
Für die Quantifizierung wurde dann eine 2-Schritt-“Real-Time“ PCR mit einem
Denaturierungsschritt bei 95°C für 10 min und anschließend 40 Zyklen bei 95°C
für 15 s und 60°C für 1 min durchgeführt.
Als interner Standard diente das TPO-cDNA Amplifikationsprodukt, das nach
Gelelektrophorese aufgereinigt worden war. Die Konzentration dieses
aufgereinigten DNA-Fragments wurde durch Messung der OD bei 260 nm
bestimmt und in Konzentrationen von 1 pg/µl bis 0,1 fg/µl als Standard in der PCR
eingesetzt. Zusätzlich wurde mit einer “Real-Time“ PCR für β-Aktin mit dem 5´-
Primer 5´-CCC TCT GAA CCC TAA GGC CA-3´ und dem reversen 3´-Primer 5´-
GGG ACA ACA CAG CCT GGA TG-3´ für 25 Zyklen bei 95°C für 15 s und 57°C
für 1 min die β-Actin cDNA quantifiziert. Die Ergebnisse der Quantifizierung
werden als Quotient der TPO-cDNA zur β-Aktin-cDNA dargestellt.
2.10 Statistik
Normalverteilte Daten sind mit Mittelwert ± Standardabweichung angegeben. Die
Bestimmung der Signifikanz der Unterschiede erfolgte bei gleicher
Standardverteilung bei 2 Gruppen mit dem ungepaarten Student´s T-Test.
Nicht-normalverteilte Ergebnisse sind als Box-Plot mit Median aufgetragen. Zur
Abschätzung der Signifikanz der Unterschiede zwischen einer Kontrollgruppe
(unbehandelte oder NaCl-behandelte Kontrolltiere) und Tieren mit entzündlicher
Reaktion (Terpentin- oder LPS- Applikation) wurden bei 2 Gruppen die nicht-
parametrischen Verfahren nach Mann-Whitney bzw. bei mehr als 2 Gruppen der
Dunn`s Test angewendet (GraphPad InStat Version 3,05; GraphPad Software,
San Diego, Ca, USA). P < 0,05 wurde als signifikant angesehen.
Der Dixon Test wurde als Ausreißertest benutzt.

Material und Methoden
25
2.11 Puffer und Lösungen
DEPC-Wasser:
0,1% (v/v) DEPC in Wasser
über Nacht rühren, autoklavieren
DNA-Ladepuffer (1:10 mit Probe zu mischen):
10 mM Tris-Acetat, pH 8,0
1 mM EDTA
50% (v/v) Glycerin
0,25% (w/w) Bromphenolblau
75% (v/v) Ethanol:
75% (v/v) Ethanol
in DEPC-Wasser
GTC-Lösung:
4 M GTC
25 mM Natriumacetat
filtrieren mit Whatman-Filter Nr. 1
0,1 M β-Mercaptoethanol
MOPS-Puffer:
40 mM MOPS, pH 7,0
10 mM Natriumacetat
1 mM EDTA
autoklavieren
Natriumacetat-Puffer:
2 M Natriumacetat, pH 4,0
autoklavieren

Material und Methoden
26
PBS :
8,5 mM Na2HPO4
1,5 mM KH2HPO4
137 mM NaCl
3 mM KCl
mit 85% (v/v) H3PO4 auf pH 7,3 einstellen
autoklavieren oder steril filtrieren
Phenol: (Fertiglösung von Sigma)
Phenol
äquilibriert mit 0,1 M Natriumcitrat, pH 4,3
Phenol:Chloroform:Isoamylalkohol: (Fertiglösung von Sigma)
50% (v/v) Phenol
48% (v/v) Chloroform
2% (v/v) Isoamylalkohol
äquilibriert mit 10 mM Tris-HCl, pH 8,0, 1 mM EDTA
RNA-Ladepuffer (1:1 mit Probe mischen):
40 mM MOPS, pH 7,0
10 mM Natriumacetat
1 mM EDTA
7% (v/v) Formaldehyd
50% (v/v) Formamid
10% (v/v) Glycerin
0,05% (w/w) Bromphenolblau
40 µg/ml Ethidiumbromid
TAE-Puffer:
10 mM Tris-Acetat, pH 8,0
1 mM EDTA

Material und Methoden
27
2.12 Chemikalien und Reagenzien
Soweit nicht anders erwähnt, wurden alle Chemikalien oder Reagenzien bei
Sigma (Deisenhofen), Roth (Karlsruhe) oder Merck (Darmstadt) im höchst-
möglichen Reinheitsgrad erworben. Ausnahmen waren:
100 bp DNA Standard Gibco BRL Life Technologies, Karlsruhe
Agarose Gibco BRL Life Technologies, Karlsruhe
dNTPs Takara Biomedicals, Shiga, Japan
Ethidiumbromid Gibco BRL Life Technologies, Karlsruhe
FKS Sigma, Deisenhofen; Seromed Biochrom KG, Berlin
oligo(dT)15 MWG Biotech, Ebersberg
PCR-Startermoleküle MWG Biotech, Ebersberg
Restriktionsenzyme MBI Fermentas, St. Leon-Rot
reverse Transkriptase Promega, Mannheim
rNTPs Promega, Mannheim
Taq-DNA-Polymerase Gibco BRL Life Technologies, Karlsruhe
Zellkulturmedien RPMI 1640, Invitrogen, Karlsruhe
Ergebnisse
28
3 Ergebnisse
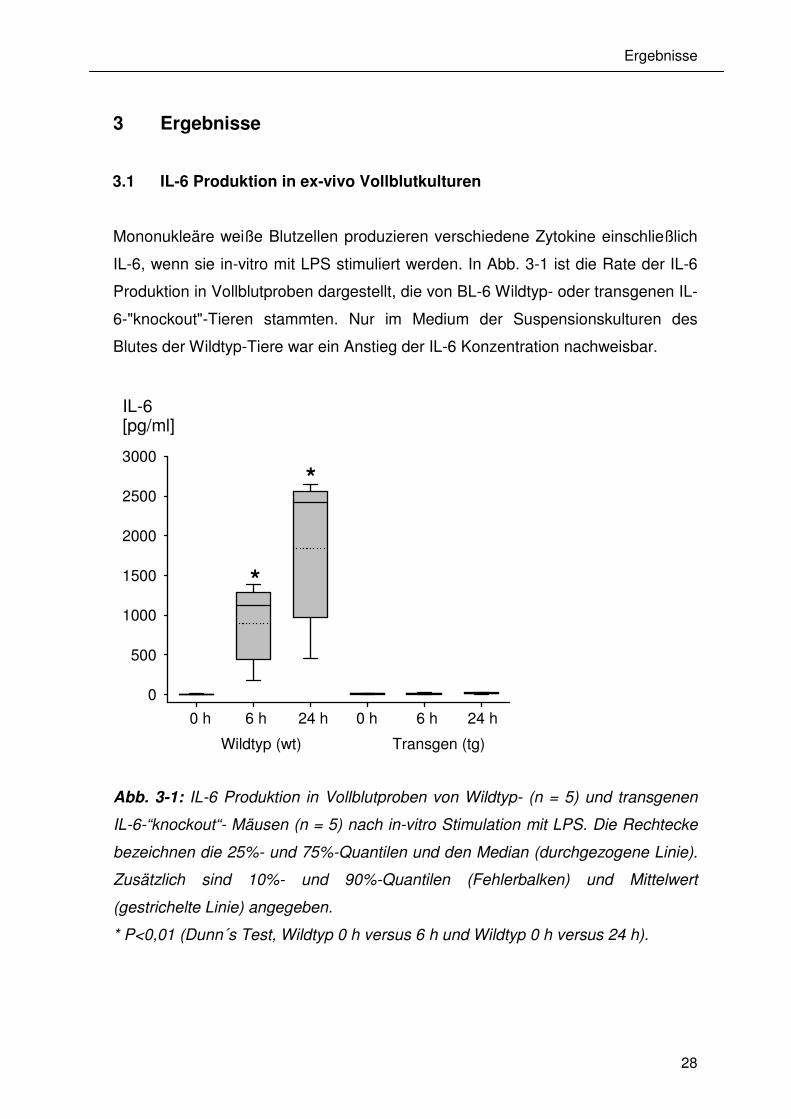
3.1 IL-6 Produktion in ex-vivo Vollblutkulturen
Mononukleäre weiße Blutzellen produzieren verschiedene Zytokine einschließlich
IL-6, wenn sie in-vitro mit LPS stimuliert werden. In Abb. 3-1 ist die Rate der IL-6
Produktion in Vollblutproben dargestellt, die von BL-6 Wildtyp- oder transgenen IL-
6-"knockout"-Tieren stammten. Nur im Medium der Suspensionskulturen des
Blutes der Wildtyp-Tiere war ein Anstieg der IL-6 Konzentration nachweisbar.
0
500
1000
1500
2000
2500
3000
IL-6[pg/ml]
0 h 6 h 24 h 0 h 6 h 24 h
Wildtyp (wt) Transgen (tg)
*
*
Abb. 3-1: IL-6 Produktion in Vollblutproben von Wildtyp- (n = 5) und transgenen
IL-6-“knockout“- Mäusen (n = 5) nach in-vitro Stimulation mit LPS. Die Rechtecke
bezeichnen die 25%- und 75%-Quantilen und den Median (durchgezogene Linie).
Zusätzlich sind 10%- und 90%-Quantilen (Fehlerbalken) und Mittelwert
(gestrichelte Linie) angegeben.
* P<0,01 (Dunn´s Test, Wildtyp 0 h versus 6 h und Wildtyp 0 h versus 24 h).

Ergebnisse
29
3.2 In-vivo IL-6 Produktion nach der Applikation von LPS oder Terpentin
In Abb. 3-2 ist die IL-6 Konzentration im Plasma von Wildtyp-Mäusen, denen
entweder 300 µl LPS (0,1 µg/g Körpergewicht; i.p.) oder Terpentinöl (s.c.)
appliziert worden war, gegenübergestellt. In den LPS-behandelten Tieren war die
IL-6 Konzentration 6 h nach der Behandlung gegenüber der IL-6 Konzentration im
Plasma von unbehandelten Wildtyp-Tieren etwa verdoppelt (13,7 pg/ml versus
23,1 pg/ml, jeweils Median). Demgegenüber war die IL-6 Konzentration im Plasma
der mit Terpentinöl behandelten Tieren auf 169 pg/ml angestiegen.
0
100
200
300
400
500
600
700
Unbehandelt LPS Terpentin
IL-6 [pg/ml]
*
Abb. 3-2: IL-6 Produktion in Plasmaproben von Wildtyp- Mäusen nach Applikation
von LPS (n = 5) oder Terpentin (n = 6). Die Rechtecke bezeichnen die 25%- und
75%-Quantilen und den Median (durchgezogene Linie). Zusätzlich sind 10%- und
90%-Quantilen (Fehlerbalken), Ausreißer (Kreise) und Mittelwert (gestrichelte
Linie) angegeben.
* P<0,001 (Dunn´s Test, Terpentin versus unbehandelt).
Ergebnisse
30
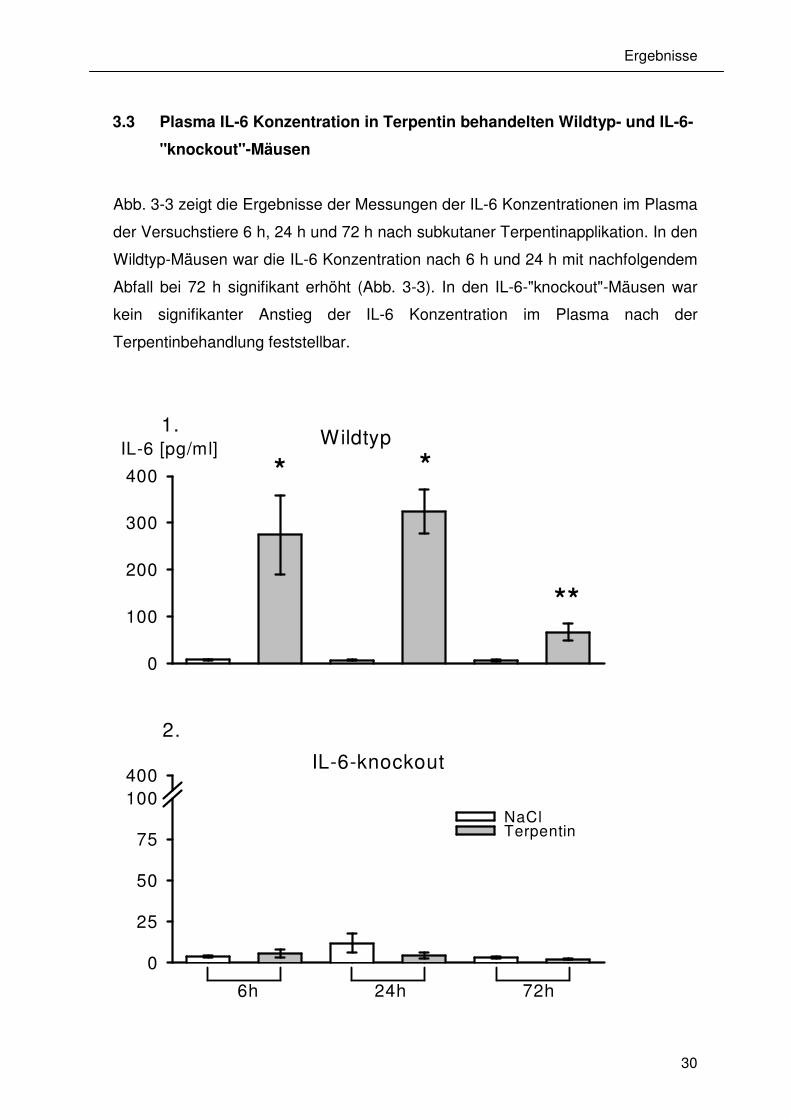
3.3 Plasma IL-6 Konzentration in Terpentin behandelten Wildtyp- und IL-6-
"knockout"-Mäusen
Abb. 3-3 zeigt die Ergebnisse der Messungen der IL-6 Konzentrationen im Plasma
der Versuchstiere 6 h, 24 h und 72 h nach subkutaner Terpentinapplikation. In den
Wildtyp-Mäusen war die IL-6 Konzentration nach 6 h und 24 h mit nachfolgendem
Abfall bei 72 h signifikant erhöht (Abb. 3-3). In den IL-6-"knockout"-Mäusen war
kein signifikanter Anstieg der IL-6 Konzentration im Plasma nach der
Terpentinbehandlung feststellbar.
0
100
200
300
400
0
25
50
75
100400
NaClTerpentin
6h 24h 72h
IL-6-knockout
IL-6 [pg/ml]
* *
**
Wildtyp
2.
1.

Ergebnisse
31
Abb. 3-3: IL-6 Konzentration in Plasmaproben von (1) Wildtyp- (n = 6) und (2) IL-
6-“knockout“- Mäusen (n = 6) nach Applikation von Kochsalzlösung (weiße
Säulen) oder Terpentin (graue Säulen). Es sind die Standardabweichungen als
Balken angegeben.
* P<0,01, * * P<0,05 ( Mann-WhitneyTest).
3.4 Plasma Thrombopoietin Konzentration in Terpentin behandelten
Wildtyp- und IL-6-"knockout"-Mäusen
Die Thrombopoietinkonzentration im Plasma NaCl behandelter Wildtyp-Mäuse
betrug ca. 8 pg/ml (Abb. 3-4). Die interindividuelle Streuung war relativ breit. Zu
allen untersuchten Zeitpunkten war die Plasmathrombopoietinkonzentration in den
Terpentin behandelten Wildtyp-Mäusen gegenüber der in den NaCl behandelten
Kontrolltieren erhöht, wobei der Unterschied nur nach 6 h und 24 h das
Signifikanzniveau erreichte. In den IL-6-“knockout“-Tieren war zu keinem Zeitpunkt
ein signifikanter Unterschied der Plasmathrombopoietinkonzentration zwischen
den NaCl behandelten Kontrolltieren und den Terpentin behandelten Mäusen
feststellbar (Abb. 3-4).

Ergebnisse
32
0
600
1200
1800
Wildtyp
IL-6-knockout
6h 24h 72h
0
600
1200
1800
TPO [pg/ml]NaClTerpentin
1.
2.
*
*
Abb. 3-4: Thrombopoietin Konzentration in Plasmaproben von (1) Wildtyp- (n = 6)
und (2) IL-6-“knockout“- Mäusen (n = 6) nach Applikation von Kochsalzlösung
(weiße Säulen) oder Terpentin (graue Säulen). Es sind die
Standardabweichungen als Balken angegeben.
* P<0,05 (Student´s T-Test, Terpentinbehandlung versus NaCl-Behandlung ).
3.5 Thrombopoietin mRNA Quantifizierung mittels kompetitiver PCR
TPO-mRNA konnte in Leber und Niere von allen Mäusen nachgewiesen werden.
Die TPO-PCR zeigte leichte Unterschiede zwischen den verschiedenen Organen
der selben Maus und Unterschiede zwischen den unterschiedlich behandelten
Mäusen, wie in Abbildung 3-5 ersichtlich.

Ergebnisse
33
TPO
GAPDH
Wt 6
h N
aCl
Wt 2
4 h
NaC
l
Wt 7
2 h
NaC
l
Wt 6
h T
erpe
ntin
Wt 2
4 h
Terp
entin
Wt 7
2 h
Terp
entin
L L L L L LN N N N N N
Neg
ativ
kont
rolle
Kom
petit
or
Abb. 3-5: Ergebnisse der qualitativen Analyse zur TPO-mRNA-Bestimmung in
Lebern (L) und Nieren (N) von Wildtyp- (Wt) Mäusen nach Behandlung mit
steriler Kochsalzlösung (NaCl) oder Terpentin für 6, 24 oder 72 Stunden (h).
Abbildung 3-6 zeigt repräsentative Agarose-Gele der kompetitiven PCR für
TPO in Lebern und Nieren der Wildtyp- und IL6-“knockout“-Mäuse nach
Injektion von steriler Kochsalzlösung oder Terpentin. In den Leberproben der
Wildtyp-Mäuse ist schon nach 6 Stunden ein Anstieg der TPO mRNA
ersichtlich (hier von 2,5 fg auf 12,2 fg), nicht hingegen bei den IL6-“knockout“-
Tieren.
TPO mRNA Leber 6h
Wt NaCl Tg NaClWt Terpentin Tg Terpentin
cDNAKomp.
20 fg
48,8
fg
40 fg
40 fg
10 fg
24,4
fg
20 fg
20 fg5 fg
12,2
fg
10 fg
10 fg
2,5
fg
6,1
fg
5 fg
5 fg
1.

Ergebnisse
34
TPO mRNA Niere 6h
Wt NaCl Tg NaClWt Terpentin Tg Terpentin
cDNAKomp.
40 fg
40 fg
40 fg
40 fg
20 fg
20 fg
20 fg
20 fg
10 fg
10 fg
10 fg
10 fg5 fg
5 fg
5 fg
5 fg
2.
Abb. 3-6: Exemplarische Ergebnisse der kompetitiven PCR zur TPO mRNA-
Bestimmung in (1) Lebern und (2) Nieren von Wildtyp- (Wt) und IL-6- “knockout“-
(Tg) Mäusen nach Applikation von Kochsalzlösung (NaCl) oder Terpentin nach 6
Stunden.
Die obere Bande besteht aus dem PCR-Produkt der TPO-cDNA, die untere Bande
aus dem PCR-Produkt des TPO-Kompetitors (Komp.). Die PCR-Ansätze
enthielten die gleiche Menge cDNA. Die eingesetzte Menge an Kompetitor ist
unter den Abbildungen angegeben. Die Pfeile zeigen den Äquivalenzpunkt der
Kompetitor- und der TPO-cDNA-Konzentration.
Die Quantifizierung der Leberproben ergab zwischen 0,382 und 7,460 amol TPO-
mRNA/µg Gesamt-RNA, die der Nierenproben zwischen 0,573 und 4,586 amol
TPO-mRNA/µg Gesamt-RNA (Abb. 3-7).
Die Unterschiede der Wildtyp-Mäuse, die mit Terpentin gegenüber steriler
Kochsalzlösung behandelt wurden, waren statistisch signifikant (ungepaarter T-
Test). Die Expression 6 h nach Terpentin- (Median 1,86) versus 6 h nach
Kochsalzlösungbehandlung (Median 0,68) ergab einen p-Wert von <0,01,
24 h nach Terpentin- (Median 2,01) versus 24 h nach Kochsalzlösungbehandlung
(Median 0,80) ergab einen p-Wert von <0,01 und die Expression 72 h nach
Terpentin- (Median 3,45) versus 72 h nach Kochsalzlösungbehandlung (Median
1,08) zeigten einen p-Wert von <0,02.

Ergebnisse
35
0
1
2
3
4
5
0
1
2
3
4
5
TP O [am ol/µg R N A ]
N aC lTerpentin
6h 24h 72h
Leber
N iere
* **
***
1.
2 .
Abb 3-7: Vergleich der Thrombopoietin-mRNA Expression (kompetitive RT-PCR)
in (1) Lebern und (2) Nieren von Wildtyp-Mäusen nach Applikation von
Kochsalzlösung (weiße Säulen) oder Terpentin (graue Säulen), nach 6, 24 und 72
Stunden (jeweils n = 6). Es sind die Standardabweichungen als Balken
angegeben.
* P<0,01, * * P<0,01, * * * P<0,05 ( ungepaarter T-Test vs. NaCl-Kontrolle).

Ergebnisse
36
3.6 Thrombopoietin mRNA Quantifizierung mittels "Real-Time" PCR
Die Messungen mittels "Real-Time“ PCR bestätigen die Ergebnisse der
kompetitiven PCR.
Abb. 3-8 zeigt, dass die Terpentinapplikation nach 6 h in den Wildtypmäusen eine
Zunahme der Thrombopoietin mRNA in der Leber bewirkte, der Gehalt an
Thrombopoietin mRNA in den IL-6-“knockout“-Tieren war niedriger als in den
Wildtyp-Tieren. Alle Werte für die Thrombopoietin mRNA wurden auf den Gehalt
an β-Aktin mRNA bezogen. Die Expression 6 h nach Terpentin- versus 6 h nach
Kochsalzlösungbehandlung ergab einen p-Wert von <0,05.
Abb 3-8: In der “Real-Time“ PCR gemessene Thrombopoietin-mRNA Expression
in der Leber von Wildtypmäusen (Wt) (n = 4) und IL-6-“knockout“-Mäusen (n = 4) 6
Stunden nach Applikation von Terpentin. Als mRNA Referenz wurde β-Aktin
benutzt. Es sind die Standardabweichungen als Balken angegeben. * P<0,05 (
ungepaarter T-Test vs. NaCl-Kontrolle ).

Ergebnisse
37
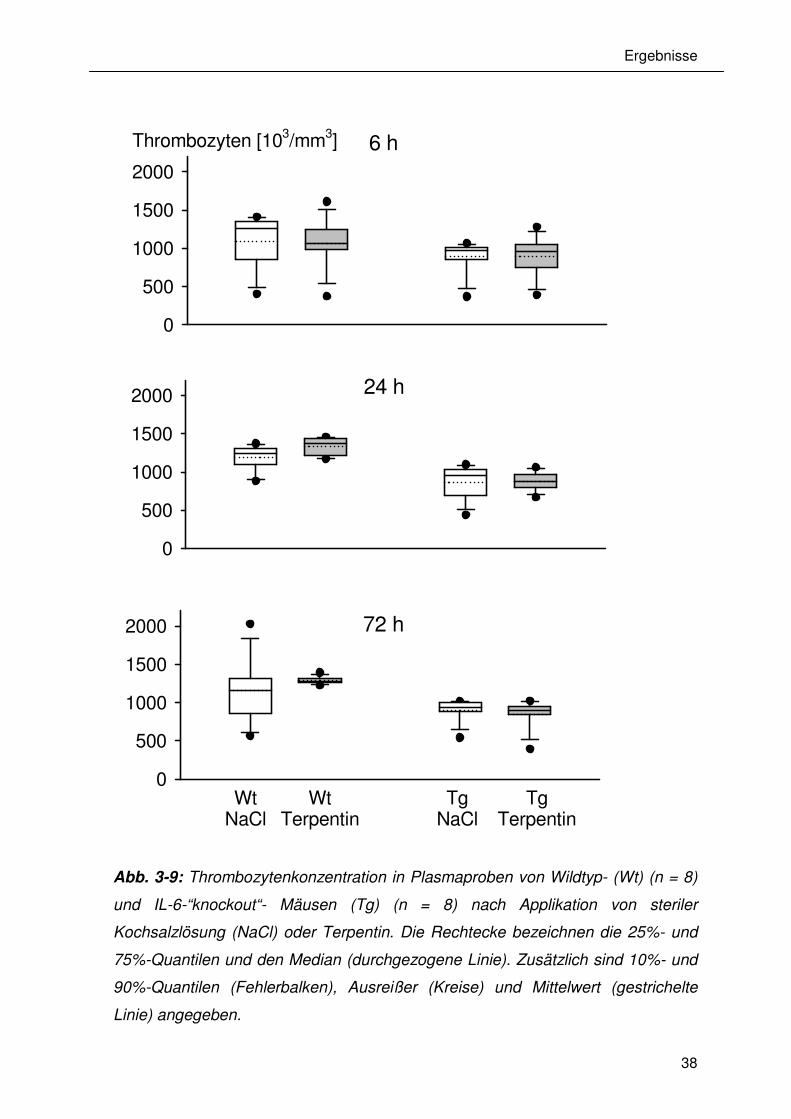
3.7 Einfluss der Terpentinapplikation auf das Blutbild der Tiere
In Abb. 3-9, 3-10 und 3-11 sind die Konzentrationen der Thrombozyten,
Erythrozyten und Leukozyten 6 h, 24 h und 72 h nach der Erzeugung eines
sterilen Abszesses durch die Applikation von Terpentinöl gezeigt. Weder in den
Wildtyp-Tieren noch in den IL-6-"knockout"-Tieren bewirkte die Entzündung eine
signifikante Veränderung der jeweiligen Zellzahl. Als Nebenbefund wurde
festgestellt, dass die Plazebo (NaCl) behandelten IL-6-"knockout"-Tiere zu allen
untersuchten Zeitpunkten signifikant niedrigere Erythrozytenkonzentrationen als
die NaCl behandelten Wildtyp-Tiere aufwiesen.
Ergebnisse
38
0
500
1000
1500
2000
6 hThrombozyten [103/mm3]
0
500
1000
1500
2000 24 h
0
500
1000
1500
2000 72 h
WtNaCl
TgNaCl
WtTerpentin
TgTerpentin
Abb. 3-9: Thrombozytenkonzentration in Plasmaproben von Wildtyp- (Wt) (n = 8)
und IL-6-“knockout“- Mäusen (Tg) (n = 8) nach Applikation von steriler
Kochsalzlösung (NaCl) oder Terpentin. Die Rechtecke bezeichnen die 25%- und
75%-Quantilen und den Median (durchgezogene Linie). Zusätzlich sind 10%- und
90%-Quantilen (Fehlerbalken), Ausreißer (Kreise) und Mittelwert (gestrichelte
Linie) angegeben.

Ergebnisse
39
0
3
6
9
12
15 6 h
Erythrozyten [106/mm3]
0
3
6
9
12
15 24 h
0
3
6
9
12
15 72 h
WtNaCl
TgNaCl
WtTerpentin
TgTerpentin
Abb. 3-10: Erythrozytenkonzentration in Plasmaproben von Wildtyp- (Wt) (n = 8)
und IL-6-“knockout“- Mäusen (Tg) (n = 8) nach Applikation von steriler
Kochsalzlösung (NaCl) oder Terpentin. Die Rechtecke bezeichnen die 25%- und
75%-Quantilen und den Median (durchgezogene Linie). Zusätzlich sind 10%- und
90%-Quantilen (Fehlerbalken), Ausreißer (Kreise) und Mittelwert (gestrichelte
Linie) angegeben.

Ergebnisse
40
0
2
4
6
8
106 hLeukozyten [103/mm3]
0
2
4
6
8
10 24 h
0
2
4
6
8
1072 h
WtNaCl
TgNaCl
WtTerpentin
TgTerpentin
Abb. 3-11: Leukozytenkonzentration in Plasmaproben von Wildtyp- (Wt) (n = 8)
und IL-6- “knockout“- Mäusen (Tg) (n = 6-8) nach Applikation von steriler
Kochsalzlösung (NaCl) oder Terpentin. Die Rechtecke bezeichnen die 25%- und
75%-Quantilen und den Median (durchgezogene Linie). Zusätzlich sind 10%- und
90%-Quantilen (Fehlerbalken), Ausreißer (Kreise) und Mittelwert (gestrichelte
Linie) angegeben.

Diskussion
41
4 Diskussion
In der vorliegenden Arbeit wurde die TPO-Bildung, die TPO-Konzentration im Blut
und die Thrombozytenzahl unter pathophysiologischen Aspekten in
Wildtypmäusen und IL-6-“knockout“-Mäusen untersucht.
Die Leber wurde als das Organ mit dem größten Anteil an der TPO-Synthese
identifiziert (65). Die TPO-Produktion in der Leber ist aber nicht unter allen
Umständen konstant, sondern kann durch pathophysiologische Einflüsse
verändert werden. Daher wurde hier an Wildtyp- und IL-6-“knockout“-Mäusen der
Einfluss von IL-6 auf die TPO-Bildung bei einer akuten Entzündung untersucht.
Von dem Zytokin IL-6 ist bekannt, dass es die Reifung von Megakaryozyten und
die Bildung von Blutplättchen stimulieren kann (64; 4). Andererseits ist IL-6 auch in
der Lage, die Genexpression und Proteinproduktion in der Leber anzuregen, z.B.
die Bildung von EPO (22).
IL-6 ist der Hauptvermittler bei der Bildung von Akut-Phase-Proteinen (32).
Tatsächlich ist die Konzentration von Plasma TPO im Blut von Patienten mit
reaktiver Thrombozytose durch maligne, infektiöse oder autoimmune
Erkrankungen oft abnormal hoch (13; 81; 34; 7).
Auch nach großen Operationen steigt der Plasma TPO Spiegel früher an als die
Thrombozytenzahl. Das bedeutet, dass TPO auch verantwortlich ist für das
Entstehen der postoperativen Thrombozytose (25).
Daher ist es berechtigt anzunehmen, dass die TPO Produktion Teil der Akut-
Phase-Reaktion bei Entzündungen, neoplastischem Wachstum oder
immunologischen Prozessen ist.
Die Fähigkeit von IL-6, die TPO Produktion in menschlichen Hepatomzellkulturen
zu stimulieren wurde durch verschiedene Arbeitsgruppen gezeigt (25; 44; 88),
auch wenn ein gegenteiliger Artikel vorliegt (35).
Die in vitro Studien haben IL-6 als das bedeutendste immunmodulatorische Peptid
identifiziert, das die hepatische TPO-Synthese bei entzündlichen Prozessen
stimuliert. IL-1 (88), IL-11 (88), Tumor Nekrose Faktor α (TNF-α) (88) und

Diskussion
42
Interferone (68; 87) erhöhen die TPO mRNA Expression und die TPO-Synthese in
menschlichen Hepatomzelllinien jedoch nicht. Unter den verschiedenen getesteten
Zytokinen an Kulturen primärer Rattenhepatozyten wurde herausgefunden, dass
der growth factor/scatter factor in einer Studie die TPO mRNA Expression
stimuliert (90), in einer anderen hingegen nicht (41).
Die wichtigste neue Erkenntnis unserer Untersuchungen ist, dass die TPO mRNA
der Leber und die TPO Blutspiegel der Wildtypmäuse nach Verursachen eines
sterilen Abszesses durch die Injektion von Terpentin erhöht sind, während dieser
Effekt bei den IL-6-“knockout“-Mäusen nicht auftrat.
Unsere Ergebnisse zeigen eine wichtige Verbindung zu anderen Studien, in denen
durch die Behandlung mit LPS eine Entzündung in Ratten erzeugt wurde und
daraufhin erhöhte TPO-mRNA Spiegel in der Leber (85) nachgewiesen werden
konnten und im Mausversuch sowohl TPO-mRNA und TPO im Plasma nach der
Injektion von IL-6 angestiegen sind (44).
Aufgrund dieser Ergebnisse liegt es nahe anzunehmen, dass die Stimulation der
TPO-mRNA Expression und die darausfolgende erhöhte TPO-Synthese in der
Leber im Rahmen einer entzündlichen Reaktion auf die Wirkung von IL-6
zurückzuführen sind.
Die Beobachtung, dass IL-6 die TPO-Produktion in Hepatomzellen und in
Wildtypmäusen steigern kann, liefert eine Erklärung für die relativ hohen TPO-
Spiegel bei den Patienten mit reaktiver Thrombozytose durch eine entzündliche
Erkrankung. Das IL-6 kann direkt durch Anregung der Thrombopoiese eine
Erhöhung der Blutplättchen verursachen (64; 4).
Hierbei müsste es aber eigentlich zu einem Absinken des TPO-Spiegels kommen,
da durch die vermehrte Bindung von TPO an den MPL Rezeptor mehr TPO aus
dem Blut entfernt werden müsste. Wenn aber das IL-6 die Hepatozyten zur
Bildung von mehr TPO anregt, dann führt eine gesteigerte TPO-Bildung zu einem
höheren TPO-Spiegel.
Während die Leber als TPO-Syntheseort feststeht, ist wenig über die Beteiligung
der Nieren am zirkulierenden TPO bekannt. Die TPO-Konzentration ist bekanntlich
erhöht bei Patienten mit terminalen Nierenerkrankungen aufgrund der reduzierten
Thrombozytenzahl (1; 54).

Diskussion
43
In dieser Arbeit war die Entzündung nicht mit einem erhöhten TPO-mRNA Gehalt
der Nieren in den Wildtyp-Mäusen assoziiert. Die konstante Produktion von TPO
in den Nieren könnte eine Erklärung liefern für die Beobachtung, dass die Plasma-
TPO-Konzentration nur mäßig ansteigt trotz der deutlich erhöhten TPO-mRNA
Menge in der Leber der Wildtypmäuse mit Entzündung. Tatsächlich war die
relative zur gesamten Konzentration von TPO-mRNA von Leber- und
Nierengewebeproben fast gleich. Die selbe Beobachtung wurde in Bezug auf die
TPO-mRNA Expression in Leber, Nieren, Milz und Knochenmark von
menschlichen Feten und Neugeborenen gemacht (84). In dieser Studie wurde aus
den Ergebnissen geschlossen, dass die Leber aufgrund des viel größeren
Organgewichtes Hauptsyntheseort von TPO ist (84).
Folglich ist der mäßige Anstieg der Plasma-TPO-Konzentration in den
Wildytpmäusen mit Entzündung eher durch den Mechanismus der
Wiederaufnahme von zirkulierendem TPO durch die Zielzellen (52; 23; 16; 21), als
durch die in Leber und Niere synthetisierte Menge zu erklären.
Neben dem Effekten auf die hepatische TPO-Genexpression stimuliert IL-6 die
Proliferation weniger differenzierter hämatopoetischer Vorläuferzellen, Dieser
Effekt könnte eine Erklärung für die beobachtete erniedrigte basale
Erythrozytenzahl in IL-6-“knockout“-Mäusen sein. Einzelne IL-6 Injektionen an
Ratten bewirkten eine vorübergehende Stimulation der Erythropoiese und eine
Retikulozytose (79). Der erniedrigte Hämatokrit bei Krebspatienten unter einer IL-6
Therapie wurde auf ein durch IL-6 erhöhtes Plasmavolumen zurückgeführt, wobei
der Mechanismus bisher nicht geklärt ist (2). Zusätzlich scheint die Anzahl der
Thrombozyten im Blut in den IL-6-“knockout“-Mäusen reduziert zu sein.
In Synergie mit TPO, IL-3 und IL-1 stimuliert IL-6 die Bildung und Reifung der
Megakaryozyten und die Produktion der Pro-Plättchen. Klinische Versuche an
thrombozytopenischen Patienten mit soliden Tumoren (82) oder
myelodysplastischen Syndromen (29) haben gezeigt, dass die Therapie mit IL-6
die Thrombopoiese bei Menschen fördert.
IL-6 wirkt synergistisch mit IL-1 und TNF-α bei der Induktion der Akut-Phase-
Antwort. Akut-Phase-Proteine, die bei Infektionen oder Entzündungen zunehmen,
dienen wahrscheinlich protektive Funktionen. Das beinhaltet auch hämostatische

Diskussion
44
Funktionen (z.B. Fibrinogen), antithrombotischen Funktionen und mikrobizide und
phagozytische Funktionen (z.B. Komplementfaktoren, C-reaktives Protein) (62).
Daraus ergibt sich die Frage nach der Rolle von TPO in der Familie der Akut-
Phase Proteine. Zielgerichtet schützt eine große Menge von Thrombozyten den
Organismus vor Blutverlusten durch krankheitsbedingten Schaden der
Gefäßwand. Außerdem spielen die Thrombozyten eine wichtige Rolle bei der
Hämostase. Sie zeigen Merkmale immunkompetenter Zellen und sind an der
antimikrobiellen Abwehr beteiligt (91; 46). Thrombozyten sind die ersten zellulären
Bestandteile, die durch Gewebsverletzung oder das Eindringen von Pathogenen
am Ort des Schadens der Gefäßwand akkumulieren. Stimulierte Blutplättchen
setzen eine Vielzahl von Zytokinen und anderen Mediatoren der Immunantwort
frei. Zusätzlich unterstützen sie die Leukozyten und Endothelzellen bei der
Kontrolle der Entzündungsreaktion, z.B. dadurch, dass sie die transendotheliale
Migration der Leukozyten in das umliegende Gewebe unterstützen.
Elektronenmikroskopische Studien haben gezeigt, dass Thrombozyten Bakterien
und Viren internalisieren durch das offene kanalikuläre System und durch
Vakuolen, welche mit α-Granula fusionieren (92). Diese α-Granula der
Thrombozyten enthalten antibakterielle Proteine, sog. Thrombozidine, die das
Abtöten der Bakterien unterstützen (50). Die Blutplättchen werden generell
unterschätzt für ihren Beitrag an der antimikrobiellen Abwehr, obwohl Ergebnisse
vorliegen, dass thrombozytopenische Patienten anfälliger für Infektionskrankheiten
sind (91) .

Zusammenfassung
45
5 Zusammenfassung
Klinische und experimentelle Studien zeigen, dass die Thrombopoietin-
Genexpression bei akuten Entzündungen gesteigert ist. Um die Rolle von IL-6 in
diesem Prozess zu klären, wurden Blutbilder, Plasma TPO-Konzentrationen und
die hepatische und renale TPO-mRNA-Konzentrationen in Wildtyp- und IL-6-
“knockout“- Mäusen mit sterilen Abszessen durch subkutane Injektion von
Terpentinöl untersucht. Diese Behandlung hat nach 72 h keine Veränderungen im
Blutbild gezeigt. Die Anzahl der Thrombozyten und Erythrozyten waren nach allen
Behandlungen in den IL-6-“knockout“-Tieren geringfügig niedriger als in den
Wildtypmäusen.
Die IL-6- und TPO-Konzentrationen im Plasma nach Terpentininjektion waren nur
in den Wildtypmäusen erhöht. Außerdem zeigte die Behandlung mit Terpentin in
diesen Tieren einen Anstieg der hepatischen TPO-mRNA-Konzentration, welche
mittels kompetitiver PCR (RT-PCR) und “Real-Time“ PCR bestimmt wurde. Die
renale TPO-mRNA-Konzentration blieb unverändert. Die TPO-mRNA-
Konzentration in den Lebern der IL-6-“knockout“-Mäuse nach Terpentininjektion
war nicht erhöht.
Diese Ergebnisse unterstützen das Konzept, dass sich TPO wie ein Akut-Phase-
Protein verhält, dadurch, dass seine Synthese durch IL-6 in der Leber induziert
wird.

Referenzen
46
Referenzen
(1) Ando M, Iwamoto Y, Suda A, Tsuchiya K, Nihei H. New insights into the
thrombopoietic status of patients on dialysis through the evaluation of megakaryocytopoiesis in bone marrow and of endogenous thrombopoietin levels. Blood 2001; 97(4):915-921.
(2) Atkins MB, Kappler K, Mier JW, Isaacs RE, Berkman EM. Interleukin-6-associated anemia: determination of the underlying mechanism. Blood 1995; 86(4):1288-1291.
(3) Avraham H. Regulation of megakaryocytopoiesis. Stem Cells Dayt 1993; 11(6):499-510.
(4) Baatout S. Interleukin-6 and megakaryocytopoiesis: an update. Annals of Hematology 1996; 73(4):157-162.
(5) Bartley TD, Bogenberger J, Hunt P, Li YS, Lu HS, Martin F et al. Identification and cloning of a megakaryocyte growth and development factor that is a ligand for the cytokine receptor Mpl. Cell 1994; 77(7):1117-1124.
(6) Bick RL. Platelet function defects: a clinical review. Semin Thromb Hemost 1992; 18(2):167-185.
(7) Bjerre A, Ovstebo R, Kierulf P, Halvorsen S, Brandtzaeg P. Fulminant meningococcal septicemia: dissociation between plasma thrombopoietin levels and platelet counts. Clin Infect Dis 2000; 30(4):643-647.
(8) Boelen A, Maas MA, Lowik CW, Platvoet MC, Wiersinga WM. Induced illness in interleukin-6 (IL-6) knock-out mice: a causal role of IL-6 in the development of the low 3,5,3'-triiodothyronine syndrome. Endocrinology 1996; 137(12):5250-5254.
(9) Borge OJ, Ramsfjell V, Cui L, Jacobsen SE. Ability of early acting cytokines to directly promote survival and suppress apoptosis of human primitive CD34+CD38- bone marrow cells with multilineage potential at the single-cell level: key role of thrombopoietin. Blood 1997; 90(6):2282-2292.
(10) Burstein SA. Cytokines, platelet production and hemostasis. Platelets 1997; 8:93-104.
(11) Bustin SA. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol 2000; 25(2):169-193.
(12) Cerutti A, Custodi P, Duranti M, Cazzola M, Balduini CL. Circulating thrombopoietin in reactive conditions behaves like an acute phase reactant. Clin Lab Haematol 1999; 21(4):271-275.

Referenzen
47
(13) Cerutti A, Custodi P, Duranti M, Noris P, Balduini CL. Thrombopoietin levels in patients with primary and reactive thrombocytosis. Br J Haematol 1997; 99(2):281-284.
(14) Chen J, Herceg-Harjacek L, Groopman JE, Grabarek J. Regulation of platelet activation in vitro by the c-Mpl ligand, thrombopoietin. Blood 1995; 86(11):4054-4062.
(15) Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 1987; 162(1):156-159.
(16) Cohen-Solal K, Villeval JL, Titeux M, Lok S, Vainchenker W, Wendling F. Constitutive expression of Mpl ligand transcripts during thrombocytopenia or thrombocytosis. Blood 1996; 88(7):2578-2584.
(17) Dame C, Fahnenstich H, Freitag P, Hofmann D, Abdul NT, Bartmann P et al. Erythropoietin mRNA expression in human fetal and neonatal tissue. Blood 1998; 92(9):3218-3225.
(18) De Groote D, Zangerle PF, Gevaert Y, Fassotte MF, Beguin Y, Noizat-Pirenne F et al. Direct stimulation of cytokines (IL-1 beta, TNF-alpha, IL-6, IL-2, IFN-gamma and GM-CSF) in whole blood. I. Comparison with isolated PBMC stimulation. Cytokine 1992; 4(3):239-248.
(19) de Sauvage FJ, Hass PE, Spencer SD, Malloy BE, Gurney AL, Spencer SA et al. Stimulation of megakaryocytopoiesis and thrombopoiesis by the c-Mpl ligand. Nature 1994; 369(6481):533-538.
(20) Eaton DL, de Sauvage FJ. Thrombopoietin: the primary regulator of megakaryocytopoiesis and thrombopoiesis. Exp Hematol 1997; 25(1):1-7.
(21) Emmons RV, Reid DM, Cohen RL, Meng G, Young NS, Dunbar CE et al. Human thrombopoietin levels are high when thrombocytopenia is due to megakaryocyte deficiency and low when due to increased platelet destruction. Blood 1996; 87(10):4068-4071.
(22) Faquin WC, Schneider TJ, Goldberg MA. Effect of inflammatory cytokines on hypoxia-induced erythropoietin production. Blood 1992; 79(8):1987-1994.
(23) Fielder PJ, Gurney AL, Stefanich E, Marian M, Moore MW, Carver-Moore K et al. Regulation of thrombopoietin levels by c-mpl-mediated binding to platelets. Blood 1996; 87(6):2154-2161.
(24) Folman CC, Linthorst GE, Van Mourik J, Van Willigen G, De Jonge E, Levi M et al. Platelets release thrombopoietin (TPO) upon activation: another regulatory loop in thrombocytopoiesis? Thromb Haemost 2000; 83:923-930.

Referenzen
48
(25) Folman CC, Ooms M, Kuenen B, de Jong SM, Vet RJWM, De Haas M et al. The role of thrombopoietin in post-operative thrombocytosis. Br J Haematol 2001; 114(1):126-133.
(26) Foster DC, Sprecher CA, Grant FJ, Kramer JM, Kuijper JL, Holly RD et al. Human thrombopoietin: gene structure, cDNA sequence, expression, and chromosomal localization. Proc Natl Acad Sci U S A 1994; 91(26):13023-13027.
(27) Gershenwald JE, Fong YM, Fahey TJ, Calvano SE, Chizzonite R, Kilian PL et al. Interleukin 1 receptor blockade attenuates the host inflammatory response. Proc Natl Acad Sci U S A 1990; 87(13):4966-4970.
(28) Gilliland G, Perrin S, Blanchard K, Bunn HF. Analysis of cytokine mRNA and DNA: detection and quantitation by competitive polymerase chain reaction. Proc Natl Acad Sci U S A 1990; 87(7):2725-2729.
(29) Gordon MS, Nemunaitis J, Hoffman R, Paquette RL, Rosenfeld C, Manfreda S et al. A phase I trial of recombinant human interleukin-6 in patients with myelodysplastic syndromes and thrombocytopenia. Blood 1995; 85(11):3066-3076.
(30) Gurney AL, Kuang WJ, Xie MH, Malloy BE, Eaton DL, de Sauvage FJ. Genomic structure, chromosomal localization, and conserved alternative splice forms of thrombopoietin. Blood 1995; 85(4):981-988.
(31) Harker LA, Roskos L, Marzec UM, Carter RA, Cherry JA, Sundell B et al. Effects of megakaryocyte growth and development factor on platelet production, platelet life span, and platelet function in healthy human volunteers. Blood 2000; 95(8):2514-2522.
(32) Heinrich PC, Castell JV, Andus T. Interleukin-6 and the acute phase response. Biochem J 1990; 265(3):621-636.
(33) Heits F, Katschinski DM, Wilmsen U, Wiedemann GJ, Jelkmann W. Serum thrombopoietin and interleukin 6 concentrations in tumour patients and response to chemotherapy-induced thrombocytopenia. Eur J Haematol 1997; 59(1):53-58.
(34) Heits F, Stahl M, Ludwig D, Stange EF, Jelkmann W. Elevated serum thrombopoietin and interleukin-6 concentrations in thrombocytosis associated with inflammatory bowel disease. J Interferon Cytokine Res 1999; 19:757-760.
(35) Hino M, Nishizawa Y, Tagawa S, Yamane T, Morii H, Tatsumi N. Constitutive expression of the thrombopoietin gene in a human hepatoma cell line. Biochem Biophys Res Commun 1995; 217(2):475-481.
(36) Hirayama Y, Sakamaki S, Matsunaga T, Kuga T, Kuroda H, Kusakabe T et al. Concentrations of thrombopoietin in bone marrow in normal subjects and in patients with idiopathic thrombocytopenic purpura, aplastic anemia,

Referenzen
49
and essential thrombocythemia correlate with its mRNA expression of bone marrow stromal cells. Blood 1998; 92(1):46-52.
(37) Hokom MM, Lacey D, Kinstler OB, Choi E, Kaufman S, Faust J et al. Pegylated megakaryocyte growth and development factor abrogates the lethal thrombocytopenia associated with carboplatin and irradiation in mice. Blood 1995; 86(12):4486-4492.
(38) Hsu HC, Tsai WH, Jiang ML, Ho CH, Hsu ML, Ho CK et al. Circulating levels of thrombopoietic and inflammatory cytokines in patients with clonal and reactive thrombocytosis. J Lab Clin Med 1999; 134(4):392-397.
(39) Ishiguro A, Suzuki Y, Mito M, Shimbo T, Matsubara K, Kato T et al. Elevation of serum thrombopoietin precedes thrombocytosis in acute infections. Br J Haematol 2002; 116(3):612-618.
(40) Ishikawa T, Ichida T, Matsuda Y, Sugitani S, Sugiyama M, Kato T et al. Reduced expression of thrombopoietin is involved in thrombocytopenia in human and rat liver cirrhosis. J Gastroenterol Hepatol 1998; 13(9):907-913.
(41) Ishikawa T, Ichida T, Matsuda Y, Sugitani S, Sugiyama M, Kato T et al. Expression of hepatic thrombopoietin mRNA in primary cultured hepatocytes and in rats with acute liver injury or bone marrow suppression with or without cirhosis. J Gastroenterol Hepatol 2000; 15:647-653.
(42) Jelkmann W. The role of the liver in the production of thrombopoietin compared to erythropoietin. Eur J Gastroenterol Hepatol 2001; 13:791-801.
(43) Jelkmann W. Erythropoietin: structure, control of production, and function. Physiol Rev 1992; 72(2):449-489.
(44) Kaser A, Brandacher G, Steurer W, Kaser S, Offner FA, Zoller H et al. Interleukin-6 stimulates thrombopoiesis through thrombopoietin: role in inflammatory thrombocytosis. Blood 2001; 98(9):2720-2725.
(45) Kaushansky K. Thrombopoietin. N Engl J Med 1998; 339(11):746-754.
(46) Klinger MH, Jelkmann W. Role of blood platelets in infection and inflammation. J Interferon Cytokine Res 2002; 22(9):913-922.
(47) Kobayashi M, Laver JH, Kato T, Miyazaki H, Ogawa M. Recombinant human thrombopoietin (Mpl ligand) enhances proliferation of erythroid progenitors. Blood 1995; 86(7):2494-2499.
(48) Kojima H, Hamazaki Y, Nagata Y, Todokoro K, Nagasawa T, Abe T. Modulation of platelet activation in vitro by thrombopoietin. Thromb Haemost 1995; 74(6):1541-1545.

Referenzen
50
(49) Kopf M, Baumann H, Freer G, Freudenberg M, Lamers M, Kishimoto T et al. Impaired immune and acute-phase responses in interleukin-6-deficient mice. Nature 1994; 368(6469):339-342.
(50) Krijgsveld J, Zaat SA, Meeldijk J, van Veelen PA, Fang G, Poolman B et al. Thrombocidins, microbicidal proteins from human blood platelets, are C-terminal deletion products of CXC chemokines. J Biol Chem 2000; 275(27):20374-20381.
(51) Kuter DJ, Beeler DL, Rosenberg RD. The purification of megapoietin: a physiological regulator of megakaryocyte growth and platelet production. Proc Natl Acad Sci U S A 1994; 91(23):11104-11108.
(52) Kuter DJ, Rosenberg RD. The reciprocal relationship of thrombopoietin (c-Mpl ligand) to changes in the platelet mass during busulfan-induced thrombocytopenia in the rabbit. Blood 1995; 85(10):2720-2730.
(53) Linden HM, Kaushansky K. The glycan domain of thrombopoietin enhances its secretion. Biochemistry 2000; 39:3044-3051.
(54) Linthorst GE, Folman CC, van Olden RW, dem Borne AE. Plasma thrombopoietin levels in patients with chronic renal failure. Hematol J 2002; 3(1):38-42.
(55) Lok S, Kaushansky K, Holly RD, Kuijper JL, Lofton-Day CE, Oort PJ et al. Cloning and expression of murine thrombopoietin cDNA and stimulation of platelet production in vivo. Nature 1994; 369(6481):565-568.
(56) Louis E, Franchimont D, Piron A, Gevaert Y, Schaaf-Lafontaine N, Roland S et al. Tumour necrosis factor (TNF) gene polymorphism influences TNF-alpha production in lipopolysaccharide (LPS)-stimulated whole blood cell culture in healthy humans. Clin Exp Immunol 1998; 113(3):401-406.
(57) Martin TG, Somberg KA, Meng YG, Cohen RL, Heid CA, de Sauvage FJ et al. Thrombopoietin levels in patients with cirrhosis before and after orthotopic liver transplantation. Ann Intern Med 1997; 127(4):285-288.
(58) McCarty JM, Sprugel KH, Fox NE, Sabath DE, Kaushansky K. Murine thrombopoietin mRNA levels are modulated by platelet count. Blood 1995; 86(10):3668-3675.
(59) McDonald TP. Thrombopoietin: its biology, purification, and characterization. Exp Hematol 1988; 16(3):201-205.
(60) Methia N, Louache F, Vainchenker W, Wendling F. Oligodeoxynucleotides antisense to the proto-oncogene c-mpl specifically inhibit in vitro megakaryocytopoiesis. Blood 1993; 82(5):1395-1401.
(61) Miyakawa Y, Oda A, Druker BJ, Kato T, Miyazaki H, Handa M et al. Recombinant thrombopoietin induces rapid protein tyrosine phosphorylation of Janus kinase 2 and Shc in human blood platelets. Blood 1995; 86(1):23-27.

Referenzen
51
(62) Moshage H. Cytokines and the hepatic acute phase response. J Pathol 1997; 181(3):257-266.
(63) Nagata Y, Shozaki Y, Nagahisa H, Nagasawa T, Abe T, Todokoro K. Serum thrombopoietin level is not regulated by transcription but by the total counts of both megakaryocytes and platelets during thrombocytopenia and thrombocytosis. Thromb Haemost 1997; 77(5):808-814.
(64) Navarro S, Debili N, Le Couedic JP, Klein B, Breton-Gorius J, Doly J et al. Interleukin-6 and its receptor are expressed by human megakaryocytes: in vitro effects on proliferation and endoreplication. Blood 1991; 77(3):461-471.
(65) Nomura S, Ogami K, Kawamura K, Tsukamoto I, Kudo Y, Kanakura Y et al. Cellular localization of thrombopoietin mRNA in the liver by in situ hybridization. Exp Hematol 1997; 25(7):565-572.
(66) Osada M, Komeno T, Todokoro K, Takizawa M, Kojima H, Suzukawa K et al. Immature megakaryocytes undergo apoptosis in the absence of thrombopoietin. Exp Hematol 1999; 27(1):131-138.
(67) Panoskaltsis-Mortari A, Ingbar DH, Jung P, Haddad IY, Bitterman PB, Wangensteen OD et al. KGF pretreatment decreases B7 and granzyme B expression and hastens repair in lungs of mice after allogeneic BMT. Am J Physiol Lung Cell Mol Physiol 2000; 278(5):L988-L999.
(68) Peck-Radosavljevic M, Wichlas M, Pidlich J, Sims P, Meng G, Zacherl J et al. Blunted thrombopoietin response to interferon alfa-induced thrombocytopenia during treatment for hepatitis C. Hepatology 1998; 28(5):1424-1429.
(69) Peck-Radosavljevic M, Zacherl J, Meng YG, Pidlich J, Lipinski E, Langle F et al. Is inadequate thrombopoietin production a major cause of thrombocytopenia in cirrhosis of the liver? J Hepatol 1997; 27(1):127-131.
(70) Sandor G. Über die blutbildende Wirkung des Serums von Tieren, die in verdünnter Luft gehalten wurden. Z Gesamte Exp Med 1932; 82:633-646.
(71) Sato T, Fuse A, Niimi H, Fielder PJ, Avraham H. Binding and regulation of thrombopoietin to human megakaryocytes. Br J Haematol 1998; 100(4):704-711.
(72) Siebert PD, Larrick JW. Competitive PCR. Nature 1992; 359(6395):557-558.
(73) Sitnicka E, Lin N, Priestley GV, Fox N, Broudy VC, Wolf NS et al. The effect of thrombopoietin on the proliferation and differentiation of murine hematopoietic stem cells. Blood 1996; 87(12):4998-5005.

Referenzen
52
(74) Sohma Y, Akahori H, Seki N, Hori T, Ogami K, Kato T et al. Molecular cloning and chromosomal localization of the human thrombopoietin gene. FEBS Lett 1994; 353(1):57-61.
(75) Souyri M. Mpl: from an acute myeloproliferative virus to the isolation of the long sought thrombopoietin. Semin Hematol 1998; 35(3):222-231.
(76) Stoffel R, Wiestner A, Skoda RC. Thrombopoietin in thrombocytopenic mice: evidence against regulation at the mRNA level and for a direct regulatory role of platelets. Blood 1996; 87(2):567-573.
(77) Sungaran R, Markovic B, Chong BH. Localization and regulation of thrombopoietin mRNA expression in human kidney, liver, bone marrow, and spleen using in situ hybridization. Blood 1997; 89(1):101-107.
(78) Trowbridge EA, Martin JF, Slater DN, Kishk YT, Warren CW, Harley PJ et al. The origin of platelet count and volume. Clin Phys Physiol Meas 1984; 5(3):145-170.
(79) Ulich TR, del Castillo J, Yin SM, Egrie JC. The erythropoietic effects of interleukin 6 and erythropoietin in vivo. Exp Hematol 1991; 19(1):29-34.
(80) Vigon I, Mornon JP, Cocault L, Mitjavila MT, Tambourin P, Gisselbrecht S et al. Molecular cloning and characterization of MPL, the human homolog of the v-mpl oncogene: identification of a member of the hematopoietic growth factor receptor superfamily. Proc Natl Acad Sci U S A 1992; 89(12):5640-5644.
(81) Wang JC, Chen C, Lou LH, Mora M. Blood thrombopoietin, IL-6 and IL-11 levels in patients with agnogenic myeloid metaplasia. Leukemia 1997; 11(11):1827-1832.
(82) Weber J, Yang JC, Topalian SL, Parkinson DR, Schwartzentruber DS, Ettinghausen SE et al. Phase I trial of subcutaneous interleukin-6 in patients with advanced malignancies. J Clin Oncol 1993; 11(3):499-506.
(83) Wendling F, Maraskovsky E, Debili N, Florindo C, Teepe M, Titeux M et al. cMpl ligand is a humoral regulator of megakaryocytopoiesis. Nature 1994; 369(6481):571-574.
(84) Wolber E-M, Dame C, Fahnenstich H, Hofmann D, Bartmann P, Jelkmann W et al. Expression of the thrombopoietin gene in human fetal and neonatal tissues. Blood 1999; 94(1):97-105.
(85) Wolber E-M, Fandrey J, Frackowski U, Jelkmann W. Hepatic thrombopoietin mRNA is increased in acute inflammation. Thromb Haemost 2001; 86(6):1421-1424.
(86) Wolber E-M, Ganschow R, Burdelski M, Jelkmann W. Hepatic thrombopoietin mRNA levels in acute and chronic liver failure of childhood. Hepatology 1999; 29(6):1739-1742.

Referenzen
53
(87) Wolber E-M, Haase B, Jelkmann W. Thrombopoietin production in human hepatic cell cultures (HepG2) is resistant to interferon-α, -β and -γ treatment. J Interferon Cytokine Res 2002; 22:1185-1189.
(88) Wolber E-M, Jelkmann W. Interleukin-6 increases thrombopoietin production in human hepatoma cells HepG2 and Hep3B. J Interferon Cytokine Res 2000; 20(5):499-506.
(89) Wu X, Nakayama M, Adamson JW. Identification of five new isoforms of murine thrombopoietin mRNA. Biochem Biophys Res Commun 2000; 276:137-143.
(90) Yamashita K, Matsuoka H, Ochiai T, Matsushita R, Kubuki Y, Suzuki M et al. Hepatocyte growth factor/scatter factor enhances the thrombopoietin mRNA expression in rat hepatocytes and cirrhotic rat livers. J Gastroenterol Hepatol 2000; 15(1):83-90.
(91) Yeaman MR. The role of platelets in antimicrobial host defense. Clin Infect Dis 1997; 25(5):951-968.
(92) Youssefian T, Drouin A, Masse JM, Guichard J, Cramer EM. Host defense role of platelets: engulfment of HIV and Staphylococcus aureus occurs in a specific subcellular compartment and is enhanced by platelet activation. Blood 2002; 99:4021-4029.

Veröffentlichungen
54
Veröffentlichung
Burmester H, Wolber E-M, Freitag P, Fandrey J, Jelkmann W, Thrombopoietin
production in wild-type and interleukin-6 knockout mice with acute inflammation.
Journal of Interferon & Cytokine Research 25:407-413, 2005.

Danksagung
55
Danksagung
Ich danke Herrn Professor Jelkmann und Frau Doktor Wolber für die Bereitstellung
des Themas und die fachkundige Betreuung.
Bei Herrn Professor Jelkmann möchte ich mich außerdem für seine Geduld und die
Hilfe bei meiner Arbeit und der Publikation herzlich bedanken.
Herrn Professor Fandrey danke ich für die Durchführung der “Real-Time“ PCR.
Herrn Professor Knopf bin ich für die Bereitstellung der adulten IL-6- “knockout“-
Mäuse sehr dankbar.
Frau Dorothea Brennecke danke ich für die administrative Hilfe.
Weiterhin danke ich allen Mitarbeiterinnen und Mitarbeitern des Institutes für
Physiologie für die Unterstützung und freundliche Hilfe bei meiner Arbeit.
Ganz besonders möchte ich mich bei meiner Familie bedanken, die niemals den
Glauben an mich aufgibt und mich immer unterstützt.

Lebenslauf
56
Lebenslauf
Persönliche Daten Name Helen Burmester
Geburtsdatum und – ort 10.11.1975 in Hamburg
Religion evangelisch-lutherisch Beruflicher Werdegang Seit Feb 2008 Assistenzärztin für Pädiatrie
in der Fachklinik Gaißach Okt 2004 – Jan 2008 Assistenzärztin für Pädiatrie
in der Kinderklinik Dritter Orden, München
Juli – Sep 2004 Ärztin im Praktikum für Pädiatrie in der Kinderklinik Dritter Orden, München
Hochschulausbildung Okt 1996 - Mai 2004 Studium der Humanmedizin an der
Universität zu Lübeck Sep 1999 - Apr 2000 Studium der Humanmedizin an der
Rijksuniversiteit Groningen, Niederlande im Rahmen des ERASMUS-Programms der EU Juli 1999 United States Medical Licensing Examination
(USMLE) Step 1 Forschung Seit Feb 2001 Dissertation am Institut für Physiologie der
Universität zu Lübeck (Prof. Jelkmann)
April - Nov 2000 Forschungsprojekt an der Klinik für Anästhesiologie und operative Intensivmedizin
der Humboldt-Universität zu Berlin, Charité (Prof. Petzelt) Thema: „Hypoxie-induzierte Neurotransmitterfreisetzung, die Regulation durch Calciumionen“
Schulausbildung 1982-1995 Grundschule und Gymnasium in Hamburg Abschluss: Allgemeine Hochschulreife


















