
Tierärztliche Hochschule Hannover...neuartige Tierseuchenerreger, Insel Riems-Greifswald 1....
177
Tierärztliche Hochschule Hannover Ausbreitung der klassischen BSE-Prion-Erreger vom Darm über das periphere Nervensystem zum Gehirn INAUGURAL – DISSERTATION zur Erlangung des Grades eines Doktors der Veterinärmedizin - Doctor medicinae veterinariae - (Dr. med. vet.) Vorgelegt von Martin Kaatz Schwedt/Oder Hannover 2014
Transcript of Tierärztliche Hochschule Hannover...neuartige Tierseuchenerreger, Insel Riems-Greifswald 1....

Tierärztliche Hochschule Hannover
Ausbreitung der klassischen BSE-Prion-Erreger vom
Darm über das periphere Nervensystem zum Gehirn
INAUGURAL – DISSERTATION
zur Erlangung des Grades
eines Doktors der Veterinärmedizin
- Doctor medicinae veterinariae -
(Dr. med. vet.)
Vorgelegt von
Martin Kaatz
Schwedt/Oder
Hannover 2014

Wissenschaftliche Betreuung: Prof. Dr. Martin Groschup
Friedrich-Loeffler-Institut, Institut für neue und
neuartige Tierseuchenerreger, Insel Riems-Greifswald
1. Gutachter: Prof. Dr. Martin Groschup
Friedrich-Loeffler-Institut, Institut für neue und
neuartige Tierseuchenerreger, Insel Riems-Greifswald
2. Gutachter: Univ.-Prof. Dr. Wolfgang Baumgärtner
Institut für Pathologie, Tierärztliche Hochschule
Hannover
Tag der mündlichen Prüfung: 17.7.2014

Wer den Blick hebt, sieht keine Grenzen.
aus Japan
Für meine Familie


1 Einleitung .............................................................................................................................. 15
2 Literaturübersicht .................................................................................................................. 16
2.1 Anatomische Grundlagen .................................................................................................. 16
2.1.1 Bauelemente des Nervensystems ................................................................................ 17
2.1.2 Zentrales Nervensystem (ZNS) ................................................................................... 17
2.1.2.1 Rückenmark ......................................................................................................... 17
2.1.2.2 Gehirn/Stammhirn ............................................................................................... 18
2.1.3 Peripheres Nervensystem (PNS) ................................................................................. 20
2.1.3.1 Vegetatives Nervensystem (VNS) ....................................................................... 20
2.1.4 Enterisches Nervensystem (ENS) ............................................................................... 22
2.2 Transmissible Spongiforme Enzephalopathien ................................................................. 23
2.2.1 Das Prion-Protein ........................................................................................................ 24
2.2.1.1 Struktur des zellulären Prion-Proteins (PrPC) ....................................................... 24
2.2.1.2 Eigenschaften und Funktion des PrPC ................................................................. 24
2.2.1.3 Das pathologische Prion-Protein (PrPSc
) ............................................................. 25
2.2.2 Erregerhypothese ........................................................................................................ 25
2.2.3 Transmissible Spongiforme Enzephalopathien der Tiere ........................................... 27
2.2.3.1 Pathologie der Transmissiblen Spongiformen Enzephalopathien ........................ 29
2.2.3.2 Diagnostik ............................................................................................................ 31
2.2.3.3 Scrapie ................................................................................................................. 32
2.3.3.4 Bovine spongiforme Enzephalopathie (BSE) ..................................................... 34
2.3 Pathogenese der TSE-Erkrankungen ................................................................................. 36
2.3.1 Dosis und Inkubationszeiten ....................................................................................... 37
2.3.2 Das lymphatische System ........................................................................................... 38
2.3.3 Das Periphere Nervensystem (PNS) ............................................................................ 41
2.3.3.1 Neuroinvasion ...................................................................................................... 41
2.3.3.2 Ausbreitung von PrPSc
im PNS ............................................................................ 42
2.3.3.3 Ausbreitungsmechanismen im Nervensystem ..................................................... 43
2.3.4 Alternative Ausbreitungswege von PrPSc
im Körper .................................................. 44
2.3.5 Akkumulation von PrPSc
in extraneuronalen Geweben und Ausscheidung ............... 45
2.3.6 Entzündungen und TSE .............................................................................................. 46
3 Tiere, Material und Methoden .............................................................................................. 48

3.1 Tiere .................................................................................................................................. 48
3.1.1 Rinder ......................................................................................................................... 48
3.1.2 Durchführung der Rinder-Pathogenesestudie ............................................................. 48
3.1.2.1 Tierhaltung ........................................................................................................... 48
3.1.2.2 Inokulat und Inokulation ...................................................................................... 48
3.1.2.3 Verhaltensstudie ................................................................................................... 49
3.1.2.4 Probenentnahme ................................................................................................... 49
3.1.2.5 Bestandsbetreuung ............................................................................................... 50
3.1.3 Mäuse .......................................................................................................................... 51
3.1.3.1 Mauslinien ........................................................................................................... 51
3.1.3.2 Inokulate .............................................................................................................. 51
3.1.3.3 Durchführung des Tierversuchs ........................................................................... 52
3.1.3.4 Probenentnahme ................................................................................................... 53
3.1.3.5 Inkubationszeiten ................................................................................................. 53
3.2 Probenmaterial .................................................................................................................. 54
3.3 Histologische Methoden .................................................................................................... 57
3.3.1 Herstellung der histologische Präparate ..................................................................... 57
3.3.2 Hämatoxylin-Eosin-Färbung ...................................................................................... 58
3.3.3 Bewertung der Hämatoxylin-Eosin-Färbung .............................................................. 58
3.3.4 Immunhistologische Untersuchungen ........................................................................ 59
3.3.5 Immunhistologische Kontrollen ................................................................................. 60
3.3.6 Bewertung der immunhistologischen Untersuchungen .............................................. 60
3.4 Proteinbiochemische Methoden ........................................................................................ 61
3.4.1 Homogenatherstellung ................................................................................................ 61
3.4.2 Fällung mit Phosphorwolframsäure (PTA-Fällung) ................................................... 62
3.4.3 Proteinauftrennung mittels SDS-Polyacrylamid-Gel-Elektrophorese ........................ 62
3.4.4 Western Blot ............................................................................................................... 63
3.4.5 Bewertung der Ergebnisse und Kontrollen ................................................................. 64
4 Ergebnisse ............................................................................................................................ 65
4.1 Allgemeine Ergebnisse ...................................................................................................... 65
4.1.1 Klinische Symptomatik .............................................................................................. 65
4.1.2 Ergebnisse der histopathologischen Untersuchung .................................................... 66
4.1.3 Ergebnisse der immunhistochemischen Untersuchung .............................................. 67

4.1.3.1 Quantifizierung der Ergebnisse ........................................................................... 67
4.1.3.2 Lokalisation und Reaktionsmuster von PrPSc
...................................................... 68
4.1.4 Ergebnisse des Maus-Bioassay (Tgbov XV-Mäuse) ................................................... 73
4.2 Zusammenfassende Betrachtung ....................................................................................... 74
4.2.1 Assoziation von Teilergebnissen ................................................................................ 74
4.2.2 Strukturierung der Ergebnisse .................................................................................... 76
4.3 Einzeltierergebnisse .......................................................................................................... 77
4.3.1 Ergebnisse der Tiere ohne Beteiligung der Medulla oblongata ................................. 77
4.3.1.1 IT 28 (16 mpi) ....................................................................................................... 78
4.3.1.2 IT 46 (16 mpi) ....................................................................................................... 79
4.3.1.3 IT 50 (20 mpi) ....................................................................................................... 79
4.3.1.4 IT 60 (20 mpi) ....................................................................................................... 80
4.3.1.5 IT 58 (24 mpi) ....................................................................................................... 81
4.3.1.6 IT 52 (28 mpi) ....................................................................................................... 82
4.3.2 Ergebnisse der Tiere mit geringgradiger Beteiligung der Medulla oblongata ........... 84
4.3.2.1 IT 24 (24 mpi) ....................................................................................................... 84
4.3.2.2 IT 61 (32 mpi) ....................................................................................................... 85
4.3.2.3 IT 25 (40 mpi) ....................................................................................................... 86
4.3.3 Ergebnisse der Tiere mit mittel- bis hochgradiger Beteiligung
der Medulla oblongata ................................................................................................ 89
4.3.3.1 IT 21 (28 mpi) ....................................................................................................... 89
4.3.3.2 Zusammenfassung der weiteren Ergebnisse ........................................................ 91
4.4 Natürlich infiziertes Rind (R09/06).................................................................................... 97
5 Diskussion ............................................................................................................................ 98
5.1 Ziel der Arbeit ................................................................................................................... 98
5.2 Kritische Betrachtung des Untersuchungsmaterials .......................................................... 98
5.3 Allgemeine Betrachtungen .............................................................................................. 100
5.3.1 Ergebnisüberblick ..................................................................................................... 100
5.3.2 Dosis, Inkubationszeiten und klinische Symptomatik .............................................. 101
5.3.3 Zusammenhänge von erhobenen Befunden .............................................................. 102
5.3.4 Vergleich von Feldinfektion vs. experimenteller Studie .......................................... 103
5.4 Pathogenese der BSE ...................................................................................................... 104
5.4.1 Rolle des lymphatischen Systems ............................................................................. 104

5.4.2 Rolle des ENS und Neuroinvasion ........................................................................... 106
5.4.3 Ausbreitung im peripheren Nervensystem ............................................................... 107
5.4.3.1 Ausbreitungsrichtung ......................................................................................... 108
5.4.3.2 Gehirn ................................................................................................................ 108
5.4.3.3 Rückenmark ....................................................................................................... 109
5.4.3.4 Gemischtfaserige Ganglien ................................................................................ 110
5.4.3.5 Sympathikus ....................................................................................................... 112
5.4.3.6 Parasympathikus ................................................................................................ 112
5.4.4 Hypothese zur Ausbreitung im PNS ......................................................................... 113
5.4.5 Transport des PrPSc
................................................................................................... 116
5.5 Pathogenese und Pathologie ............................................................................................ 118
6 Zusammenfassung ............................................................................................................... 120
7 Summary ............................................................................................................................. 122
8 Literaturverzeichnis ............................................................................................................. 124
9 Anhang ................................................................................................................................ 167
9.1 Histologische Methoden .................................................................................................. 167
9.1.1 Entparaffinierungs- und Differenzierungsprotokoll ................................................. 167
9.2 Legende zur Verhaltensbeobachtung der Rinder ............................................................ 167
9.3 Puffer und Lösungen ....................................................................................................... 171
9.3.1 Immunhistochemie ................................................................................................... 171
9.3.2 Hämatoxylin-Eosin-Färbung .................................................................................... 172
9.3.3 Westernblot ............................................................................................................... 173

Abkürzungsverzeichnis
µm Mikrometer
A Alanin
A. bidest. bidestilliertes Wasser
Abb. Abbildung
AG Antigen
AK Antikörper
APS Ammoniumpersulfat
AS Aminosäure
BA Bioassay
BBP12/92 Boviner Brain Pool 12/1992
BHV1 Bovine Herpesvirus-1
bov bovine
BRSV Bovines Respiratorisches Synzytialvirus
BSE Bovine Spongiforme Enzephalopathie
bzw beziehungsweise
CJK, CJD Creutzfeldt-Jakob-Krankheit
CO2 Kohlendioxid
C-terminal Carboxy-terminal
CWD Chronic Wasting Disease
d.h. das heißt
DAB Diaminobenzidintetrahydrochloride
DC Dendritische Zellen
DMNV Dorsaler motorischer Kern des Nervus vagus
DNA Desoxyribonukleinsäure
Dpi days post infection
EDTA Ethylendiamintetraessigsäure
ENS Enterisches Nervensystem
ER Endoplasmatisches Retikulum
EU Europäische Union
exp experimentell
FA Firma
FAE Follikel-assoziiertes Epithel
fCJD familiäre Creutzfeldt-Jakob-Krankheit

FDC Follikulär Dendritische Zellen
FFI Fatale Familiäre Insomnie
FLI Friedrich-Loeffler-Institut
FSE Feline Spongiforme Enzephalopathie
g Gramm
GALT Darm-assoziiertes lymphatisches Gewebe
GG gemischtfaserige Ganglien
Ggl Ganglion
GMGC Ganglion coeliacum-Komplex
GPI-Anker Glykosylphosphatidylinositol-Anker
GSS Gerstmann-Sträussler-Scheinker-Syndrom
GT Ganglion trigeminale
HE Hämatoxylin-Eosin
hu human
i.c. intrazerebral
i.p. intraperitoneal
ID infektiöse Dosis
IgG Immunglobulin G
IHC Immunhistochemie
INNT Institut für neue und neuartige Tierseuchenerreger
IT Infektionstier
Kap Kapitel
kDa Kilo-Dalton
l Liter
Lnn. Lymphonodi
LRS Lymphoretikuläres System
M Molar; Mol/Liter
mab monoklonaler Antikörper
max maximal
min Minute
ml Milliliter
mm Millimeter
mM Millimolar
MO Medulla oblongata

mpi Monate post infectionem
n Anzahl
N Nervus
n.a. nicht auswertbar
n.d. nicht durchgeführt
neg negativ
Nn Nervi
NP 40 Nonidet P 40
N-terminal Amino-terminal
o.g. oben genannt
O.I.E. Office International des Epizooties - Weltorganisation für Tiergesundheit
OHV2 Ovines Herpesvirus Typ 2
ov ovin
PAGE Polyacrylamid-Gel-Elektrophorese
PBS Phosphat-gepufferte Natriumchloridlösung
PCR Polymerasekettenreaktion
PET-Blot Paraffin Embedded Tissue Blot
p.i. post infection
PK Proteinase K
PMCA Protein Misfolding Cyclic Amplification
PNS Peripheres Nervensystem
PP Peyer´sche Platten
Prion Proteinaceous infectious particle
Prnp Prion-Protein-Gen
PrP Prion-Protein
PrP27-30 Proteinase K resistenter Teil des PrPSc
PrPC zelluläres Prion-Protein
PrPSc
Scrapie-Isoform des Prion-Proteins
PS Parasympathikus
PTA Phosphorwolframsäure
PVDF Polyvinylpyrrolidon
Q Glutamin
R Arginin
rER rauhes Endoplasmatisches Retikulum

RM Rückenmark
RNA Ribonukleinsäure
RT Raumtemperatur
S Sympathikus
s. siehe
S.E.M. Standardabweichung des Mittelwertes
SAF Scrapie-assoziierte Fibrillen
sCJD sporadische Creutzfeldt-Jakob-Krankheit
SDS Natriumdodecylsulfat
sog. sogenannt
Tab. Tabelle
TBM Tingible body macrophages
TBS Tris-gepufferte Natriumchloridlösung
TEMED N,N,N’,N’-Tetramethylethylendiamin
TME Transmissible Enzephalopathie der Nerze
TNT tunneling nano tubes
Tris Tris(hydroxymethyl)aminomethan
TSE Transmissible Spongiforme Enzephalopathie
Tween 20 Polyoxyethylen(20)sorbitanmonolaureat
U Unit
u.a. unter anderem
ud undefiniert
UK United Kingdom
UV Ultraviolett
V Valin
vCJD Variante der CJD
VLA Veterinary Laboratories Agency
VNS Vegetatives Nervensystem
WB Western blot
z.B. zum Beispiel
z.T. zum Teil
ZNS Zentrales Nervensystem
ZVO Zirkumventrikuläre Organe
μg Mikrogramm

μl Mikroliter


15 Einleitung
1. EINLEITUNG
Transmissible spongiforme Enzephalopathien (TSE) treten bei unterschiedlichen Spezies auf.
Am bekanntesten sind dabei Scrapie bei Schafen, die chronisch zehrende Krankheit der
Hirsche (CWD) und die Bovine Spongiforme Enzephalopathie (BSE) der Rinder. Bei all
diesen Infektionskrankheiten wird ein pathogenes Prion-Protein als ursächlicher Erreger
vermutet.
Im Jahre 1987 erstmals detailliert beschrieben, geriet BSE durch die sogenannte BSE-Krise in
den Fokus der Öffentlichkeit. Am stärksten waren Rinder im Vereinigten Königreich
betroffen und neben einer Vielzahl von europäischen Ländern wurden Fälle auch in den USA,
Kanada und Japan nachgewiesen. Die Brisanz der Epidemie ergibt sich aus der
Übertragbarkeit der BSE-Erreger auf den Menschen. Die als Variante der Creutzfeldt-Jakob-
Krankheit (vCJD) bekannte Erkrankung wurde vermutlich durch den Verzehr von infiziertem
Rindfleisch ausgelöst.
Als natürlicher Infektionsweg von BSE im Rind wird die orale Aufnahme von verseuchten,
d.h. ungenügend vorbehandelten, Futtermitteln vermutet. Dies führte in den Jahren 1988 bzw.
1996 im Vereinigten Königreich sowie 2001 in der gesamten EU zu einem vollständigen
Fütterungsverbot von Tiermehlen, weshalb es nach ca. 5 Jahren, der durchschnittlichen BSE-
Inkubationszeit bei Rindern, zu rückläufigen Fallzahlen kam. Eine eindeutige Diagnostik
kann bis zum heutigen Zeitpunkt lediglich post mortem durchgeführt werden, da ein
zugelassener Lebendtest trotz interessanter Ansätze nicht zur Verfügung steht. Vor allem aus
Versuchen mit Labortieren (Hamster, Maus) stammen die bisherigen Erkenntnisse über die
Ausbreitung der BSE-Erreger im Tierkörper. Die lange Inkubationszeit und das zoonotische
Potential ließen nur wenige Studien am natürlichen Wirt mit begrenzten Tierzahlen zu. So ist
die Frage nach der Pathogenese von BSE nach oraler Infektion nach wie vor nur
unvollständig beantwortet.
Der am Friedrich-Loeffler-Institut (FLI) durchgeführte Tierversuch zur Pathogenese der BSE
umfasste insgesamt 74 Tiere, von denen 56 Kühe infektiöses Gehirn-Inokulat erhielten. Die
umfassende Probenentnahme an präklinisch und klinisch erkrankten Rindern führte zu einem
einzigartigen Pool von Geweben. Die hier vorgelegte Arbeit basiert auf der Untersuchung von
Gewebeproben aus dem zentralen und peripheren Nervensystem sowie des Darms
ausgewählter Tiere, um die neuronale Ausbreitung des BSE-Agens zum Gehirn darzustellen.

Literaturübersicht 16
2. LITERATURÜBERSICHT
2.1 Anatomische Grundlagen
Für das Verständnis der in dieser Dissertation verwendeten Termini erfolgt zunächst die
Beschreibung der Bestandteile des Nervensystems unter Berücksichtigung der für die
Pathogenese der Transmissiblen Spongiformen Enzephalopathien (TSE) relevanten
Strukturen.
Wenn nicht anders gekennzeichnet, sind die folgenden Angaben dem Lehrbuch der Anatomie
der Haustiere (NICKEL, SCHUMMER, SEIFERLE, 3. Auflage, 1991) entnommen.
Das Nervensystem wird in das Zentrale Nervensystem (ZNS), durch Gehirn und Rückenmark
repräsentiert, und das Periphere Nervensystem (PNS) mit Nerven und Ganglien eingeteilt.
Morphologisch und funktionell ist die Grenze weniger deutlich, da die zum Großteil im
zentralen Anteil liegenden Nervenzellen ihre Axone als periphere Nerven abgeben.
Das Enterische Nervensystem (ENS) ist im Schema der Abbildung 2.1 als eigenständiger
Anteil dargestellt. Anderen Lehrmeinungen zufolge wird es dem vegetativen Anteil
zugeordnet, da seine Steuerung den parasympathischen und sympathischen Nervenbahnen
obliegt.
Abb. 2.1 Hierarchie des Nervensystems

Literaturübersicht 17
2.1.1 Bauelemente des Nervensystems
Das Nervengewebe baut sich aus den funktionell unterschiedlichen Zellelementen, Neuroglia
und den Nervenzellen auf:
Die Neuroglia lassen sich in Astrozyten, Oligodendrozyten, Mikroglia und die
Schwann´schen Zellen unterscheiden, die verschiedene Aufgaben wahrnehmen. Sie
gewährleisten die Funktion des Nervengewebes, indem die unterschiedlichen Gliazellen den
Liquor cerebrospinalis produzieren, den Stoffwechsel ermöglichen, Axone mit Mark- oder
Myelinscheide umhüllen und phagozytierende Eigenschaften besitzen.
Die Nervenzellen bestehen aus dem kernhaltigen Zellkörper, dem Perikaryon, und den
Fortsätzen, das bei jeder Nervenzelle nur einmal vorkommende Axon und die vielfach
auftretenden Dendriten. Die Axone leiten Erregungen vom Zell-Leib weg und können damit
Ausgangspunkt eines peripheren Nervens sein.
Die Gesamtheit der die Perikaryen einbettenden Nerven- und Gliazellfortsätze wird unter dem
Begriff Neuropil zusammengefasst.
Im ZNS lässt sich immer die mehr oder weniger deutlich abgrenzbare graue Substanz
(Substantia grisea) und die weiße Substanz (Substantia alba) unterscheiden. Die graue
Substanz enthält Gliazellen (vor allem Astrozyten) und Neurone, während in der weißen
Substanz die umhüllten Axone verlaufen.
2.1.2 Zentrales Nervensystem (ZNS)
2.1.2.1 Rückenmark
Die Ausdehnung des Rückenmarks erstreckt sich vom ersten Halswirbel bis zum Kreuzbein.
Es wird aufgrund seiner Lage im Wirbelkanal und durch die segmental abgehenden
Rückenmarksnerven (Nn. spinales) in das Halsmark (Pars cervicalis), das Brustmark (Pars
thoracica), das Lendenmark (Pars lumbalis) und das Kreuzmark (Pars sacralis) sowie das
Schwanzmark (Pars caudalis) unterteilt.
Der innere Aufbau des Rückenmarks ist mit der H- oder der Schmetterlingsform der grauen
Substanz und der sie umgebenen weißen Substanz makroskopisch sichtbar (s. Abb. 2.2, S.18).
Im Feinbau weist die graue Substanz bilateral symmetrisch das Dorsalhorn (Cornu dorsale)
und das Ventralhorn (Cornu ventrale) auf. Die jeweiligen Seiten korrespondieren über die

18 Literaturübersicht
Commissura grisea, die ihrerseits in die Substantia intermedia lateralis und die den Canalis
centralis umschließende Substantia intermedia centralis unterteilt werden kann.
Die sensiblen Neurone des Dorsalhorns empfangen Reize über afferente Nervenfasern der
Radix dorsalis und ihrem Spinalganglion (Ganglion spinale). Im Ventralhorn finden sich
große motorische Neuronen, deren efferenten Axone als Radix ventralis gemeinsam mit der
Radix dorsalis als gemischtfaseriger Spinalnerv, Truncus nervi spinalis, den Wirbelkanal
verlassen. Motorische sympathische Wurzelzellen sind in der Substantia intermedia lateralis
lokalisiert.
Die weiße Substanz lässt sich in einen Dorsalstrang (Funiculus dorsalis), einen Lateral- und
Ventralstrang (Funiculus lateralis/ventralis) einteilen. Weiterhin werden Commisura alba,
Fissura mediana ventralis und der Sulcus medianus dorsalis beschrieben. Funktionell besteht
die weiße Substanz aus markhaltigen Nervenfasern, die als Bündel (Tractus) funktionell
unterschiedlich in aufsteigenden und absteigenden Bahnen verlaufen.
Abb. 2.2 Schematische Darstellung des Rückenmarks
Strukturen der grauen Substanz: 1. Cornu ventrale, 2. Cornu dorsale, 3. Commisura grisea;
Strukturen der weißen Substanz: 4. Funiculus anterior, 5. Funiculus lateralis, 6. Funiculus
posterior, 7. Commisura alba, 8. Fissura mediana ventralis; 9. Sulcus medianus dorsalis; 10.
Canalis centralis, 11. Radix ventralis, 12. Radix dorsalis, 13. Ganglion spinale
2.1.2.2 Gehirn/Stammhirn
Als zentrales, übergeordnetes Steuerungsorgan entspringen dem Gehirn 12 paarige Nerven
(Nervi craniales), die ihren Ursprung in den verschiedenen Abschnitten des Gehirns finden.
Je nach Funktion weisen die einzelnen Nerven einen sensiblen, sensorischen oder
motorischen Fasergehalt auf. Neben diesen reinen treten auch gemischte Nerven auf, die

Literaturübersicht 19
durch den Faseraustausch mit Ganglien zusätzlich sympathische und parasympathische
Qualitäten besitzen können.
Ein spezielles Endothel mit tight junctions schützt als Blut-Hirn-Schranke und Blut-Liquor-
Schranke das Gehirn und gewährleistet über aktive und passive Transportvorgänge die
Versorgung. Eine eingeschränkte Blut-Hirn-Schrankenfunktion weisen die
zirkumventrikulären Organe (ZVO) auf, die über ein fenestriertes Endothel Hormone
sezernieren und sensorische Funktionen besitzen.
Abb. 2.3 Schematischer Querschnitt der Medulla oblongata mit den für die TSE Diagnostik
relevanten Kerngebieten, Abb. aus CASALONE et al. 2005
Für die BSE-Routine-Diagnostik wird die Region des Obex im Bereich des verlängerten
Marks (Medulla oblongata) untersucht. Die Medulla oblongata als Teil des Stammhirns
(Myelencephalon) bildet den am weitesten kaudal gelegenen Teil des Gehirns und stellt die
Verbindung zum Rückenmark her. Der Obex selbst bildet mit anderen Strukturen den
kaudalen Abschluss des Daches des IV. Ventrikels.
Die graue Substanz dieses Anteils der Medulla oblongata wird durch mikroskopisch
isolierbarer Nervenzellnester als Kerngebiete (Nuclei) bestimmter Gehirnnerven repräsentiert
(s. Abb. 2.3), die von der weißen Substanz umschlossen und teilweise durchzogen wird.
Funktionell ist die Medulla oblongata damit ein wichtiges Zentrum peripherer Nerven und
kann als Koordinations- und Reflexzentrum charakterisiert werden.

20 Literaturübersicht
2.1.3 Peripheres Nervensystem (PNS)
Das PNS umfasst alle für die Erregungsleitung verantwortlichen Nerven und Ganglien, die
direkt oder indirekt Kontakt zum ZNS haben.
Die Skelettmuskulatur und Sinnesorgane werden durch das somatische System innerviert,
während vegetative oder autonome Nerven und Ganglien für die adäquate Steuerung der
Organe und Organsysteme sorgen.
Im Folgenden werden lediglich die für diese Arbeit relevanten Strukturen beschrieben.
2.1.3.1 Vegetatives Nervensystem (VNS)
Im VNS wird das sympathische vom parasympathischen System differenziert, die sich durch
ihre antagonistische Wirkungsweise und durch die chemische Natur ihrer
Transmittersubstanzen (Sympathikus: Adrenalin, Noradrenalin; Parasympathikus:
Acetylcholin) unterscheiden. Eine schematische Übersicht der für diese Arbeit relevanten
Anteile des vegetativen Nervensystems zeigt die Abb. 2.4 (S.21).
Sympathisches Nervensystem
Der Ursprung des Sympathikus liegt in den Wurzelzellen der Substantia intermedia des
Rückenmarks. Die entsprechenden Axone verlassen den Wirbelkanal über die Ventralwurzel
und finden größtenteils Anschluss an die Ganglienkette des Grenzstranges (Truncus
sympathicus), wo zum Teil die Umschaltung auf das sekundäre Neuron erfolgt. Manche
Fortsätze durchziehen jedoch diese sogenannten Paravertebralganglien und werden erst in den
sogenannten Prävertebralganglien auf das zweite Neuron verschaltet.
Der Halsteil des sympathischen Grenzstranges enthält das Ggl. stellatum und im Kopfbereich
das Ggl. cervicale craniale. Dieses Paravertebralganglion stellt nach erfolgter Umschaltung
größtenteils den sympathischen Anteil der Gehirnnerven. Der Brustteil besteht aus einer Kette
symmetrisch und segmental angeordneter Paravertebralganglien (Ganglia trunci sympathici),
welche die Nervi splanchnici majores abgeben, die als Eingeweidenerven zum Plexus bzw.
Ggl. coeliacum ziehen.
Das Ggl. coeliacum wird zusammen mit dem anatomisch nicht separierbaren Ggl.
mesenteriale craniale als Ganglionkomplex (englisch: Coeliac Mesenteric Ganglion
Complex, CMGC) angesprochen und ist dem Bauchteil des Sympathikus zugeordnet. In enger
Verbindung mit dem Ggl. mesentericum caudale ist es für die sympathische Innervation des

Literaturübersicht 21
Darmsystems verantwortlich. In diesem als Prävertebralganglion fungierende
Nervenzellknoten werden die Fasern des N. splanchnicus auf die sekundären
postganglionären Neurone umgeschaltet und führt durch den Anschluss von Zweigen des N.
vagus der Ganglienkomplex auch parasympathische Fasern.
Parasympathisches System
Der kraniale Teil des Parasympathikus hat seine Ursprung im Mittelhirn und der Medulla
oblongata, während die Wurzelzellen des kaudalen Anteils in der Substantia intermedia des
Kreuzmarks (Pars sacralis) lokalisiert sind. Erstgenannter Teil versorgt die Organe des
Kopfes und der Bauch- und Beckenhöhle, die parasympathische Innervation der
Beckenorgane und des Geschlechtsapparates erfolgt durch den Pars sacralis.
Abb. 2.4 Schematische Darstellung von Anteilen des zentralen und peripheren Nerven-
systems
ZNS: 1-Gehirn, 1a-Ebene des Obex in der Medulla oblongata, 2a-Halsmark, 2b-Brustmark,
2c-Lendenmark, 2d-Kreuzmark des Rückenmarks; sympathisches System: 3-sympathischer
Grenzstrang, 4-Ggl. cervicale craniale, 5-Ggl. stellatum, 6-Nn. splanchnici majores, 7-Nn.
splanchnici lumbales; parasympathisches System: 8-N.vagus, 9-Ggl. nodosum; somatisches
System: 10 Ggl. trigeminale; gemischte Anteile: 11-Ggl. coeliacum, 12-Ggl. mesenteriale
caudale, 13- Plexus aorticus abdominalis (Abb. modifiziert nach (NICKEL, SCHUMMER,
SEIFERLE, 3. Auflage, 1991)

22 Literaturübersicht
Der N. vagus ist der bedeutsamste Nerv des parasympathischen Systems, führt aber auch
somatomotorische, sensible und sympathische Fasern. Im Bereich des Nucleus tractus
solitarii und Nucleus tractus nervi trigemini der Medulla oblongata enden die afferenten
Wurzelfasern, während die parasympathischen Wurzelzellen im Kern des N. vagus (s. Abb.
2.4, S.21) des verlängerten Marks liegen.
Der N. vagus enthält im Nervenstrang selbst z.T. zahlreiche Neuronennester und durchläuft
das Ggl. distale oder nodosum, welches die sekundären Neurone der afferenten Nervenfasern
enthält. Über eine Vielzahl von Ästen ist der N. vagus unter anderem an der Steuerung der
Kehlkopffunktion und des Herzens beteiligt, besitzt aber auch viszerosensible und -
motorische Fasern für die Funktion der meisten Organe im Brust- und Bauchraum. Im
Bauchraum erhält er dabei eine anatomische Verbindung mit den Ggl. coeliacum.
Des Weiteren führt der N. vagus auch einige sympathischen Fasern, die ihm durch das Ggl.
cervicale craniale zugeführt werden.
2.1.4 Enterisches Nervensystem (ENS)
Die folgenden Angaben sind dem Buch „The enteric nervous system“ (FURNESS, 2006)
entnommen und durch weitere Literaturangaben ergänzt.
Das ENS setzt sich aus Neuronen, ihren Axonen und Gliazellen in Form kleiner, aber sehr
zahlreicher Ganglien zusammen. So wird der Darm einerseits autark über Reflexbögen
gesteuert, über ein weitreichend verschaltetes Netzwerk erfolgt aber auch eine intensive
Kommunikation mit dem ZNS.
Die extrinsische Steuerung wird vor allem durch Nerven sympathischer Qualität
gewährleistet (OHMORI et al. 2008), die auch für die direkte Innervation der Peyer`schen
Platten bestimmt werden konnten (DEFAWEUX et al. 2007; CHIOCCHETTI et al. 2008)
und somit eine funktionelle Verbindung zwischen dem Nerven- und dem Immunsystem
darstellen.
Nach der Lokalisation werden zwei Plexus unterschieden. Zu Veranschaulichung der
anatomischen Verhältnisse dient Abb. 2.5 (S.23).
Der Plexus myentericus (Auerbach`scher Plexus) befindet sich zwischen der longitudinalen
und der zirkulären Schicht der äußeren Muskulatur des Verdauungstraktes. Der myenterische
Plexus, dessen Ganglien über sogenannte interganglionäre Stränge vernetzt sind, ist in allen
Abschnitten das Gastrointestinaltraktes zu finden. Dabei wird primär unabhängig vom ZNS

Literaturübersicht 23
die Motilität und Peristaltik des Verdauungssystems gesteuert. Allerdings greifen
Parasympathikus und Sympathikus in die Aktivität des Plexus ein und beeinflussen somit die
Darmmotilität.
Das Auftreten des Plexus submucosus (Meißner`scher Plexus) ist auf die Bereiche des Dünn-
und Dickdarms beschränkt. Die variable Lokalisation der Ganglien und funktionell
verschiedenen Neuronen-Typen konzentrieren sich auf die Submukosa und können dort die
Follikel der Peyer`schen Platten umspannen, während einzelne Neurone aber auch in der
Mukosa zu finden sind. Funktionell ist dieser Plexus für die Innervation der Mukosa, für den
transepithelialen Ionentransport, die Regulation der mukösen Blutversorgung und für
Immunreaktionen verantwortlich (SURPRENANT, 1994).
Abb. 2.5 Schematische Darstellung der Anteile des enterischen Nervensystems
A: Mukosa; B: Muscularis mucosae; C: Submukosa; D: zirkuläre Muskulatur; E:
longitudinale Muskulatur; 1: Peyer´sche Platten; 2: Follikel-assoziiertes Epithel (FAE);
Neurone des 3: internen mukösen Plexus; 4: des externen mukösen Plexus; 5: des
myenterischen Plexus; 6: interganglionäre Stränge
2.2 Transmissible Spongiforme Enzephalopathien
Bei den Transmissiblen Spongiformen Enzephalopathien (TSE) handelt es sich um
übertragbare, progressiv verlaufende neurodegenerative Erkrankungen, die bei verschiedenen
Säugetierspezies auftreten. Als ursächlicher Erreger wird für diese Krankheiten nach
heutigem Wissenstand eine abnormale Form des eigentlich zellulären, d.h. körpereigenen,
Prion-Proteins (PrPC, C von cellular) (PRUSINER 1982; PRUSINER et al. 1990)

24 Literaturübersicht
angenommen. Diese abnormale, fehlgefaltete Isoform akkumuliert bei den bekannten TSE
und wird als PrPSc
(Sc von Scrapie) bezeichnet.
2.2.1 Das Prion-Protein
2.2.1.1 Struktur des zellulären Prion-Proteins (PrPC)
Prion-Protein PrPC ist ein körpereigenes, extrazelluläres und hochkonserviertes Glykoprotein
(RIEK et al. 1996; HORNEMANN et al. 2004; GOSSERT et al. 2005; LYSEK et al. 2005),
welches durch das Prion-Protein-Gen (Prnp) kodiert wird und sich im Genom einer Vielzahl
von Säugetierspezies, Reptilien, Vögeln und Amphibien findet (WOPFNER et al. 1999;
CALZOLAI et al. 2005).
Die posttranslationalen Modifikationen des Proteins im Endoplasmatischen Reticulum und
dem Golgi-Apparat umfassen die Bildung einer Disulfid-Brücke und die Glykosylierung
(PRUSINER 1998; HARRIS 2003), die zur Bildung dreier Isoformen führt: unglykosyliert,
einfach oder zweifach glykosyliert (RUSSELAKIS-CARNEIRO et al. 2002).
Am reifen Protein lässt sich ein N-Terminus mit einer hochkonservierten Oktarepeatregion
und ein globulärer C-terminaler Bereich unterscheiden (RIEK et al. 1996).
2.2.1.2 Eigenschaften und Funktion des PrPC
PrPC ist gut löslich, wird durch Proteinasen vollständig abgebaut und weist zudem im
Gegensatz zu seiner pathologischen Isoform eine höhere Empfindlichkeit gegenüber Hitze,
pH-Wert-Schwankungen und UV-Strahlung auf. Die PrPC-Expression findet bei Säugern vor
allem im ZNS und in geringeren Mengen in nahezu allen Körpergeweben statt (BENDHEIM
et al. 1992; FORD et al. 2002), darunter auch Zellen des Immunsystems (DODELET u.
CASHMAN 1998). Die physiologische Funktion des Proteins wurde von WESTERGARD et
al. (2007) zusammenfassend diskutiert: Neben zytoprotektiven Effekten scheint das Protein
indirekte Auswirkungen auf die neuronale Differenzierung, Reifung und Regeneration zu
haben. Als interaktives Transmembranprotein ist die Beteiligung an Signalkaskaden und bei
der synaptischen Übertragung wahrscheinlich. Darüber hinaus ist PrPC offensichtlich an der
Zelladhäsion beteiligt und bedingt die zelluläre Aufnahme und Bindung von Kupferionen.
Eine aufgrund des hohen Konservierungsgrades vermutete essentielle Rolle des Prion-
ProteinPrion-Proteins wird durch Lebensfähigkeit von PrP- knock-out-Mauslinien (PrP0/0
) in

Literaturübersicht 25
Frage gestellt. Die Herstellung dieser transgenen Mäuse führte zu phenotypisch
unveränderten Tiere (BÜELER et al. 1992).Allerdings berichten andere Studien eine gestörte
neuronale Erregung (COLLINGE et al. 1994; COLLING et al. 1995), eine höhere
Anfälligkeit gegenüber oxidativem Stress (BROWN et al. 1999) und anormale
Schlafrhythmen bei PrP0/0
-Mäusen (TOBLER et al. 1996).
2.2.1.3 Das pathologische Prion-Protein (PrPSc
)
PrPC und das pathologische Prion-Protein (PrP
Sc) besitzen die gleiche Aminosäuresequenz. Es
bestehen jedoch deutliche Unterschiede hinsichtlich ihrer physikalisch-chemischen
Eigenschaften und ihrer Struktur.
So zeigt die pathologische Form des Prion-Proteins eine ausgesprochen hohe
Widerstandsfähigkeit gegenüber chemischen und physikalischen Einflüssen (HUNTER et al.
1964; ALPER et al. 1966, 1967). Neben der Unlöslichkeit in wässrigen Lösungen sind
herkömmliche Desinfektionsmittel gegen PrPSc
kaum oder gar nicht wirksam.
Als wichtigstes Unterscheidungskriterium gilt die partielle Resistenz der pathologischen
Isoform gegen Proteinasen, beispielsweise Proteinase K (PK) (OESCH et al. 1985). Durch die
Inkubation mit PK werden wird ein 27-30 kDa (PrP 27-30) großes Proteinfragment gebildet,
das das Kernstück des PrPSc
darstellt (PRUSINER et al. 1987). Untersuchungen mit
transgenen, PrP27-30
exprimierenden Mäusen zeigten, dass dieses Fragment des Proteins für
die Infektion der Tiere ausreicht (FISCHER et al. 1996). Es aggregiert und bildet
regelmäßige, amyloide Strukturen, die man nach Aufreinigung als Prion-Rods bzw. Scrapie-
assoziierte Fibrillen (SAF) anspricht (PRUSINER et al. 1983). Anhand der charakteristischen
Verschiebung (shift) der molekularen Masse von PrPSc
im Western Blot kann anhand der
Glykosylierungsformen (un-, mono-, di-glykosyliert) der Erreger charakterisiert werden
(KUCZIUS et al. 1998). So dominiert bei BSE anders als bei Scrapie die diglykosylierte
Form (STACK et al. 2002), obwohl eine Diskriminierung von BSE und Scrapie nicht immer
eindeutig möglich ist (BARON et al. 1999).
2.2.2 Erregerhypothese
Aufgrund der Tatsache, dass sich die Infektiosität des Scrapie-Erregers nicht durch
herkömmliche Verfahren und Substanzen zum Abbau von Nukleinsäuren verringern lässt,

26 Literaturübersicht
wurde erstmals durch ALPER et al. (1966) der Verdacht eines infektiösen Proteins geäußert.
Die Anreicherung eines Proteins der Größe von 27-30 kDa aus TSE-infiziertem
Gehirnmaterial (PRUSINER et al. 1980) führte im Folgenden zur Formulierung der protein-
only-hypothesis (OESCH et al. 1985). Diese Theorie postuliert PrPSc
als infektiöses Agens,
welches für die Erkrankung verantwortlich ist und seine Fehlfaltung auf das vorhandene,
körpereigene PrPC überträgt. Die Replikation erfolgt durch die autokatalytische Konversion
von Alpha-helix-reichem PrPC
in die pathologische, Beta-faltblatt-reiche Isoform
(PRUSINER 1998). Diese Konformationsänderung wird als entscheidendes Ereignis in der
Pathogenese der Prion-Erkrankungen diskutiert. Für diesen neuartigen Erreger prägte
PRUSINER aus der Bezeichnung „proteinaceous infectious particle“ den Begriff „Prion“,
dem eine eigenständige Nukleinsäure fehlt (PRUSINER 1998).
Vielfältige Hinweise stützen diese Hypothese So konnte PrPC in vitro durch die Zugabe von
PrPSc
in die fehlgefaltete Form konvertiert werden (GASSET et al. 1992; KOCISKO et al.
1994). Weiterhin gelang mit hoch aufgereinigtem PrPSc
die Infektion von Wildtypmäusen
(PRUSINER 1982). Zudem war in vitro hergestelltes rekombinantes PrPC nach
anschließender Überführung in eine Beta-Faltblatt-reiche Struktur für transgene Mäuse
infektiös (LEGNAME et al. 2004). In einem entgegengesetzten Ansatz erwiesen sich PrPC-
defiziente Mäuse gegen eine Infektion als resistent (BUELER et al. 1993).
Der Nachweis bei Wildtyptieren wäre ein finaler Hinweis für die Gültigkeit der 'protein-only-
hypothesis'. Mittels zyklischer Amplifikation der Protein-Fehlfaltung (engl., protein
misfolding cyclic amplification, PMCA) war in vitro generiertes PrPSc
in der Lage, Wildtyp-
Hamster zu infizieren (CASTILLA et al. 2005). Ein weiteres Experiment generierte ein Prion
aus bakteriell exprimierten PrP, welches nicht nur die typischen Eigenschaften von PrPSc
aufwies, sondern auch ohne Co-Faktoren für Wildtypmäuse infektiös war (WANG et al.
2010). Diese Ergebnisse wurden als Durchbruch für die Gültigkeit der 'protein-only-
hypothesis' gewertet.
Dennoch war die mögliche Beteiligung anderer Faktoren am Infektionsprozess bis dato einer
der Kritikpunkte (ABID u. SOTO 2006). Des Weiteren bestanden Zweifel, dass durch die
unterschiedliche Faltung des Prion-Proteins die Information für verschiedene TSE Stämme
kodiert werden kann. Außerdem konnte in einer Studie mit BSE-infizierten und klinisch
erkrankten Mäusen bei 55% der Tiere kein PrPSc
nachgewiesen werden. Erst nach mehreren
Subpassagen gelang die Detektion von pathologischem Prion-Protein. Aus diesem Grund
wurde ein bisher unbekannter Faktor für die eigentliche, klinische Infektion diskutiert
(LASMEZAS et al. 1997).

Literaturübersicht 27
Eine andere Theorie beschäftigt sich mit der Beteiligung von Viren am Krankheitsgeschehen.
In dieser Virus-Hypothese von DIRINGER et al. (1994) und MANUELIDIS (1994) dient das
Prion-Protein lediglich als Rezeptor für ein bisher nicht bekanntes Virus. Eine Prion-Infektion
hat in vivo wie auch in der Zellkultur einen steigernden Einfluss auf die Expression
bestimmter Retroviren (CARP et al. 1999; STENGEL et al. 2006). Deren Expression förderte
die Freisetzung von Prionen/Infektiosität in den Überstand und die Verbreitung innerhalb der
Kultur (LEBLANC et al. 2006).
Als mögliche Co-Faktoren erleichterten in vitro insbesondere kleine, hoch-strukturierte RNAs
oder unspezifische DNAs die Konversion von Hamster-PrPC
(CORDEIRO et al. 2001;
DELEAULT et al. 2003, 2005). Ergänzend zu vorherigen Studien zur Interaktion von PrPSc
mit RNA (WEISS et al. 1997; DERRINGTON et al. 2002; ADLER et al. 2003;
SUPATTAPONE 2004), wird die Beteiligung eines RNA-haltigen Nukleoprotein-Komplexes
diskutiert (SIMONEAU et al. 2009).
2.2.3 Transmissible Spongiforme Enzephalopathien bei Tieren
Die das ZNS betreffenden Erkrankungen (s. Tab. 2.1, S.28) treten nach einer verhältnismäßig
langen Inkubationszeit von mehreren Monaten bis Jahren auf und enden stets letal (BUDKA
et al. 1995). Sie führen zu neurodegenerativen Schädigungen, die durch spongiforme
Läsionen im Neuropil, Degeneration von Nervenzellen und Astrozytose gekennzeichnet sind
(FIELD u. PEAT 1969). Der progressive Krankheitsverlauf beginnt mit dem Einsetzen von
Symptomen wie Verhaltensänderungen, Bewegungs- und Sensibilitätsstörungen und ist beim
Menschen mit einer fortschreitenden Demenz verbunden.
Ähnlich wie bei anderen neurodegenerativen Erkrankungen wie der Alzheimer´schen oder der
Parkinson´schen Krankheit können auch bei TSEs amyloide Plaques auftreten (SOTO 2003).
Prion-Krankheiten treten nicht nur sporadisch oder hereditär auf, sondern können auch auf
natürliche, akzidentielle oder experimentelle Infektionen zurückgehen (CHANDLER 1961;
CHANDLER u. TURFREY 1972; WELLS et al. 2003). Bedeutsam ist das zoonotische Potentials
der BSE-Erreger (BRUCE et al. 1997) – humane Infektionen sind seit Mitte der neunziger
Jahre als Variante der Creutzfeldt-Jakob-Krankheit (vCJD) bekannt (WILL et al. 1996;
BUDKA 2001).
28 Literaturübersicht
Allen Prion-Erkrankungen gemein ist das Ausbleiben einer detektierbaren humoralen und
zellulären Immunantwort. Wirkungsvolle Therapie-Ansätze gegen TSEs bei Mensch und
Tier gibt es derzeit noch nicht (VANA et al. 2007).
Am lebenden Tier kann anhand der klinischen Symptome lediglich eine Verdachtsdiagnose
gestellt werden. Die Diagnose einer TSE-Erkrankung kann bis dato ausschließlich post
mortem durch biochemische und immunhistochemische Tests gestellt werden.
Tab. 2.1 Liste natürlich auftretender Transmissibler Spongiformer Enzephalopathien
Natürliches
Vorkommen TSE Literatur
Mensch
Kuru ZIGAS u. GAJDUSEK (1957)
Creutzfeldt-Jakob-Krankheit (CJK) (iatrogen, sporadisch, familiär-
genetisch, Variante)
CREUTZFELDT (1920), JAKOB
(1921a; 1921b; 1921c) vCJK: WILL et al. (1996)
Gerstmann-Sträussler-Scheinker-
Syndrom (GSS)
GERSTMANN et al. (1936)
Fatale familiäre Insomnie (FFI) LUGARESI et al. (1986)
Schaf
Scrapie MCGOWAN (1922)
atypische Scrapie BENESTAD et al. 2003
Rind
Bovine spongiforme Enzephalopathie
(BSE) WELLS et al. (1987)
JEFFREY u. WELLS (1988)
atypische BSE CASALONE et al. 2004
BIACABE et al. 2004
Cerviden chronisch zehrende Krankheit der
Hirsche (CWD) WILLIAMS u. YOUNG (1980)
Katzen schwammartige Hirndegeneration der
Katzen (FSE) WYATT et al. (1990)
PEARSON et al. (1992)
Nerz übertragbare Hirndegeneration der
Nerze (TME) BURGER et al. (1965)
HARTSOUGH et al. (1965)

Literaturübersicht 29
2.2.3.1 Pathologie der Transmissiblen Spongiformen Enzephalopathien
Pathologische Veränderungen konnten bisher ausschließlich im ZNS lokalisiert werden. Es
handelt sich dabei um neurodegenerative Prozesse, die nicht mit einem spezifischen
inflammatorischen Geschehen einhergehen.
Histopathologische Veränderungen des Gehirns kommen bei allen TSE´s vor, sie
unterscheiden sich aber hinsichtlich ihrer Ausprägung und Lokalisation. Die im Vordergrund
stehenden spongiformen Veränderungen entstehen bilateral durch das Anschwellen von
Perikarien (neuronale Vakuolen) und der Nervenfortsätze (Auflockerung des Neuropils) bei
gleichzeitigem Verlust intrazellulärer Organellen.
Die histopathologischen Vakuolen bei klassischer BSE sind unabhängig von Rasse, Dosis und
Art der Inokulation (WELLS u. SIMMONS, 1996). Die Läsionen in den Kerngebieten des
Tractus solitarii, Tractus spinalis, N. trigeminus, N. vestibulares und der Substantia grisea
periaquaeductalis der Medulla oblongata (Obex) können für eine BSE-Diagnose
herangezogen werden, die Läsionen erstrecken sich aber auch auf das Cerebellum,
Diencephalon, Mesencephalon und die Pons (WELLS u. WILESMITH, 1995).
Für Scrapie wird das Auftreten von Vakuolen im Mesencephalon, Diencephalon,
Telencephalon, Corpus striatum beschrieben, die aber nicht in den Olivkernen, dem
zerebralen Cortex und dem Hippocampus vorkommen (JEFFREY u. GONZÁLEZ, 2004).
Weiterhin weisen die Ergebnisse verschiedener Studien eine Vielzahl von Variationen
abhängig vom Scrapie-Stamm, Genotyp und anderen individuellen Faktoren auf (ZLOTNIK
1958; WOOD et al. 1997; BEGARA-MCGORUM et al. 2002; LIGIOS et al. 2002).
Die Unterschiede in der neuroanatomischen Verbreitung der Vakuolen nach Adaptation an
definierte Nagetiere können in Form von Läsionsprofilen dargestellt werden, durch die
verschiedene Scrapie-Stämme differenzierbar sind (BEGARA-MCGORUM et al. 2002;
LIGIOS et al. 2002). Auch BSE-Infektionen zeichnen sich durch ein charakteristisches
Läsionsprofil aus, identische Läsionsmuster von BSE und vCJD in Mäusen ließen zudem den
Schluss eines direkten Zusammenhangs zwischen beiden TSE´s zu (BRUCE et al. 1997).
Wie auch die Vakuolisierung sind das Vorkommen einer Gliose und der Verlust von
Nervenzellen keineswegs einheitlich. Bei einer Gliose handelt es sich um eine über das
physiologische Maß hinausgehende erhöhte Anzahl von Gliazellen in einem geschädigten
Bereich des Zentralnervensystems. Der Nachweis eines apoptotischen Neuronenuntergangs in
signifikanten Tierzahlen konnte weder für Scrapie (LYAHYAI et al. 2006) noch für BSE
(THEIL et al. 1999) erbracht werden, obwohl die Steigerung pro-apoptotischer Faktoren bei

30 Literaturübersicht
Scrapie (SERRANO et al. 2009) und der Verlust von Nervenzellen bei BSE-Rindern
(JEFFREY u. HALLIDAY 1994) beschrieben wurden. Für beide Erkrankungen können
allerdings die Veränderungen auch unter einem lichtmikroskopisch detektierbaren Level
liegen (FRASER 1976; WELLS et al. 1989, 1994; SOMERVILLE et al. 1997; CHAPLIN et
al. 1998).
Als weiterer pathologischer Aspekt gilt der Nachweis von PrPSc
-Akkumulationen mittels
Immunhistochemie. Diese treten an den gleichen neuroanatomischen Lokalisationen wie die
spongiformen Veränderungen auf (WELLS u. WILESMITH, 1995).
Tab. 2.2 Pathologische Befunde bei BSE und Scrapie
BSE Scrapie
Immunhistochemie - PrPSc
- Akkumulation (4)
korreliert mit Vakuolisierung Lokalisation korreliert mit Vakuolisierung
Akkumulationen:
perineuronal, linear, extrazellulär Neuropil-assoziiert
fein-punktiert,
(linear, fein-punktiert und grobkörnig-partikulär ,
koaleszieren und perineuronal),
grobkörnig-partikulär
Astrozyten-assoziiert
(sternförmig, subpial, subependymal und
perivaskulär),
Ependymalzellen-assoziiert
(supraependymal)
Endothelzellen-assoziiert
(vaskuläre Plaques)
intraneuronal, intraglial intrazellulär intraneuronal, intraglial
(1) WELLS u. WILESMITH, 1995; (2) JEFFREY et al. 2011; (3) HADLOW et al. 1982,
(4) JEFFREY u. GONZÁLEZ, 2004
Bei der Bewertung der morphologischen und topographischen Muster, dem sogenannten
PrPSc
-profiling, lassen sich für klassische BSE relativ gleichmäßige und einheitliche PrPSc
-
Akkumulationen nachweisen. Dagegen weisen die Gehirne klinischer Scrapie-Fälle eine hohe
Variabilität im PrPSc
-Profil auf (s. Tab. 2.2). Dadurch lassen sich nicht nur Informationen zum
Stamm und Ursprung des Scrapie-Erregers ableiten, es konnte ebenfalls die Abhängigkeit von
der Inokulationsroute und dem Genotyp bei klinisch erkrankten Schafen gezeigt werden
(JEFFREY u. GONZÁLEZ, 2007). Das sogenannte PrPSc
-mapping bedient sich dagegen

Literaturübersicht 31
unterschiedlicher Antikörper, die spezifische Epitope des pathologischen Prion-Proteins
erkennen und erlaubt so die immunhistologische Unterscheidung von BSE und Scrapie.
Die Korrelation zwischen Pathologie, Erregernachweis und Symptomen gestaltet sich
ebenfalls uneinheitlich. So besteht bei einzelnen Individuen die Möglichkeit einer klinischen
TSE-Erkrankung gänzlich ohne nachweisbare pathologische Veränderungen (KONOLD et al.
2008). Während Gliose und Neuronenverlust mit Depositionen des pathologischen Prion-
Proteins assoziiert sind, trifft diese Korrelation nicht auf die spongiformen Veränderungen
und apoptotische Prozesse zu. Als eigentliche Krankheitsursache wird daher der Verlust der
protektiven PrPC-Funktion anstatt einer direkt schädigen Wirkung des PrP
Sc diskutiert
(JEFFREY et al. 2011).
2.2.3.2 Diagnostik
Die Routine-Diagnostik bedient sich zunächst kommerzieller Schnelltests, die im Falle eines
Reagenten weitergehende Untersuchungen in den nationalen Referenzlaboren zur
Diagnosestellung nach sich ziehen. Hier kommen OIE-zertifizierte Testmethoden zur
Anwendung, zu denen die Histopathologie, Immunhistochemie (IHC) und der Scrapie-
assoziierte-Fibrillen (SAF)- Immunoblot gehören.
Abb. 2.6: Vergleich der elektrophoretischen Profile und der Antikörperbindung nach
Proteinase K Verdau, PTA-Präzipitation und Immunoblot bei Verwendung der monoklonalen
Antikörper P4 und L42 (modifiziert nach Gretzschel 2007).

32 Literaturübersicht
Da sich die neuropathologischen Veränderungen aller TSE’s vorrangig in der Obexregion der
Medulla oblongata befinden, wird als Probenmaterial für die Diagnostik das Stammhirn
eingesetzt. Bei Scrapie sollte zur Abklärung möglicher atypischer Fälle auch das Kleinhirn
untersucht werden.
In der histopathologischen Untersuchung auftretenden Vakuolen allein sind jedoch nicht
beweisend. Der direkte Nachweis von PrPSc
-Ablagerungen im Rahmen der
Immunhistochemie unter Verwendung spezifischer Antikörper stellt hingegen einen
wesentlichen Bestandteil der Diagnostik dar.
Als biochemische Methode kommt der Immunoblot zur Anwendung, der nach
elektrophoretischer Auftrennung des mit Proteinase K degradierten Proteins unter
Verwendung spezifischer Antikörper die drei Glykosylierungsformen des pathologischen
Prion-Proteins als einzelne Banden visualisiert. Darüber hinaus kann unter Berücksichtigung
des Antikörperbindungsverhältnisses, des Glykosylierungsverhältnisses und der molekularen
Masse des unglykosylierten PrPSc
eine Charakterisierung des PrPSc
zur Unterscheidung von
BSE und Scrapie vorgenommen werden (s. Abb.2.6, S.31).
2.2.3.3 Scrapie
Scrapie (von engl. scrape „kratzen“, „schaben“) oder Traberkrankheit ist eine übertragbare,
langsam tödlich verlaufende Erkrankung des Gehirns bei Schafen und in geringerem Ausmaß
auch bei Ziegen, deren Erstbeschreibung auf das Jahr 1750 zurückgeht (LEOPOLDT 1750).
Allerdings erwähnt ein 1772 erschienener Aufsatz in England Scrapie-Fälle, die sogar auf das
Jahr 1732 zurückgehen (COMBER 1772). Sie gilt damit als Prototyp der TSE und tritt in
zahlreichen Ländern endemisch bei kleinen Wiederkäuern auf.
Für Scrapie gilt nur die Zusammenfassung der histopathologischen Befunde in Kombination
mit den typischen PrPSc
-Ablagerungen als pathognomisch, da auch bei gesunden Tieren
spongiforme Veränderungen in geringerer Zahl gefunden wurden (ZLOTNIC u. RENNIE
1958, ZIOTNIK 1962).
Das Auftreten klinisch erkrankter und unauffälliger Tiere innerhalb einer Schafherde beruht
auf einer Resistenz, die durch Polymorphismen im Prnp-Genotyp kodiert sind, die vor allem
am Codon 136, 154 und 171 auftreten (GOLDMANN et al. 1990; HUNTER et al. 1994a;
BELT et al. 1995; BOSSERS et al. 1996) (s. Tab. 2.3, S.33). Vor diesem Hintergrund sind
seit 2003 alle EU-Mitgliedsstaaten aufgefordert, Programme zur Resistenzzucht bezüglich
Scrapie aufzustellen.

Literaturübersicht 33
Scrapie- Erreger werden horizontal über die infektiöse Plazenta übertragen (TOUZEAU et al.
2006; BROTHERSTON et al. 1968; PATTISON et al. 1972; DICKINSON et al. 1974).
Allerdings gelingt der Nachweis des PrPSc
nur in der Plazenta von Föten des empfänglichen
Genotyps (ANDREOLETTI et al. 2002; LACROUX et al. 2007) und die Übertragung auf
den Embryo ist nicht obligatorisch (FOSTER et al. 2006).
Schafe infizieren sich direkt an der infektiösen Nachgeburt und indirekt an mit dieser
kontaminiertem Weideland (HOINVILLE, 1996; FOSTER et al. 1996, 2006; VAN KEULEN
et al. 2008). Möglicherweise spielen auch Kot (GONZÁLES et al. 2006) und Urin (SISÓ et
al. 2006) eine Rolle bei der horizontaler Übertragung.
Tab. 2.3 Übersicht Scrapie
Scrapie klassisch atypisch
Symptome Unruhe, Schreckhaftigkeit, Ataxie,
Abmagerung, Juckreiz, Festliegen v.a. Ataxie
(1)
Einfluss des Genotyps PrP
ARR - unempfänglich, PrP
VRQ -
hoch empfänglich (2) (3)
PrPVRQ
- unempfänglich, PrPARR
- hoch empfänglich
(4)
Infektionsquelle Kontamination der Umwelt, direkter
Tierkontakt spontanes Auftreten
(4)
intra-spezies
Übertragung horizontal
Übertragbarkeit auf Schafe
experimentell nachgewiesen (5)
Erregeraufnahme oral
(6), konjunktival
(7), dermal
(8),
gingival (9)
unklar
(1) BENESTAD et al. 2003; (2) VAN KEULEN et al. 2000; (3) ANDREOLETTI et al. 2000;
(4) BENESTAD et al. 2008; (5) SIMMONS et al. 2007; (6) HADLOW et al. 1982; (7)
SCOTT et al. 1993; (8) MOHAN et al. 2004; (9) CARP, 1982
Des Weiteren sprechen PrPSc
-Akkumulationen im Euter (LIGIOS et al. 2005), den
Speicheldrüsen (VACELLARI et al. 2007), der Zunge (CASALONE et al. 2005) und
Infektiosität im Kolostrum und der Milch (LACROUX et al. 2008) für die Möglichkeit einer
Übertragung durch direkten Tierkontakt. Als Erregerreservoir werden zudem Rötelmäuse
aufgrund ihrer hohen Empfänglichkeit gegenüber Scrapie diskutiert (PIENING et al. 2006).
In Norwegen wurde 1998 erstmals eine atypische Variante von Scrapie festgestellt
(BENESTAD 2003, BUSCHMANN et al. 2004). Retrospektive Analysen ergaben aber, dass
atypische Fälle bereits Ende der 80ziger Jahre in Großbritannien auftraten und weltweit mit

34 Literaturübersicht
einer vergleichbaren Prävalenz wie klassischen Scrapie vorkommen (FEDIAEVSKY et al.
2008). Im Gegensatz zur klassischen Form weist sie eine vermehrte Akkumulation des
pathologischen Prion-Proteins vor allem im Kleinhirn und weniger im Stammhirn auf.
Weitere Unterscheidungskriterien sind das fehlende Vorkommen von PrPSc
in
lymphoretikulären Geweben und deutlich unterschiedliche biochemische Eigenschaften
(BENESTAD et al. 2003; BUSCHMANN et al. 2004; GAVIER-WIDEN et al. 2005; LE
DUR et al. 2005).
2.2.3.4 Bovine spongiforme Enzephalopathie (BSE)
Die spongiforme Enzephalopathie der Rinder wurde 1985 in Vereinten Königreich
Großbritannien entdeckt und erstmals 1987 beschrieben (WELLS et al.).
Die Frage nach dem Ursprung konnte noch nicht eindeutig geklärt werden. Diskutiert wurde
zunächst die Übertragung von Scrapie-Erregern aus infizierten Schafen (WILESMITH et al.
1988, WILESMITH et al. 1991). Trotz der erfolgreichen Übertragung von Scrapie auf Rinder
wies die Infektion nach intra-zerebraler Inokulation andere proteinbiochemische und
histopathologische Merkmale auf und scheint daher als Ursache wenig wahrscheinlich
(KONOLD et al. 2006).
Die Hypothese, dass spontan an BSE erkrankte Rinder den Ausgangspunkt für die Epidemie
darstellten (EDDY 1995), fand mit Entdeckung der sporadisch auftretenden, sogenannten
atypischen BSE-Fälle, Unterstützung (BUSCHMANN et al. 2006; CAPOBIANCO et al.
2007).
Die klinischen Symptome (s. Tab. 2.4, S.35) treten erst nach einer sehr langen Inkubationszeit
deutlich in Erscheinung, auf die eine relativ kurze progressive Krankheitsperiode folgt.
Für die Verbreitung unter den Rindern wird die Verfütterung von infektiösem Tiermehl oder
Milchaustauschern angenommen (WILESMITH et al. 1988; PAISLEY u. HOSTRUP-
PEDERSEN 2004). Die Persistenz und Inzidenz des BSE-Erregers bei der
Tiermehlherstellung entstand durch die im Vereinigten Königreich der siebziger Jahre
beschlossenen Reduzierung der Behandlungstemperaturen und -zeiten und durch die
Steigerung der Tiermehlfütterung an Rinder (HÖRNLIMANN 2001). Ein
Verfütterungsverbot von Tiermehl an Wiederkäuer erfolgte Großbritannien bereits 1988, dem
sich die EU 2001 anschloss. So ist die Zahl der aufgetretenen BSE-Fälle in Großbritannien
seit 1993 und in Deutschland seit 2001 rückläufig. Hierzulande wurden bisher insgesamt 413
Fälle registriert (Stand: 10.08.2012).

Literaturübersicht 35
Als natürliche Übertragung gilt die orale Aufnahme infektiösen Materials als sicher, da bisher
weder eine vertikale noch eine horizontale Übertragung durch die Ausscheidung von Sekreten
oder Exkreten beschrieben wurde (CURNOW u. HAU, 1996) und auch im Blut keine
Infektiosität nachgewiesen werden konnte (WELLS et al. 1998; ESPINOSA et al. 2007).
In den letzten Jahren ließ sich das Auftreten zweier atypische BSE-Varianten nachweisen. Es
wird der sogenannte H- Typ (BIACABE et al. 2004) von dem im gleichen Jahr erstmals
detektierten L-Typ (oder Bovine Amyloidotic Spongiform Encephalopathy - BASE)
unterschieden (CASALONE et al. 2004; BARON u. BIACABE, 2006). Beide Formen der
atypischen BSE wurden auch in Deutschland dokumentiert (BUSCHMANN et al. 2006).
Dabei weist der L-Typ durch das etwas geringere Molekulargewicht ein deutlich anderes
Glykosylierungsprofil auf. Die IHC weist PrPSc
-Akkumulationen in Form von amyloiden
Plaques vor allem in Thalamus und Cortex (CASALONE et al. 2004) anstelle der typischen
granulären und linearen Ablagerungen im Hirnstamm auf. Eine an Primaten durchgeführte
Studie zeigte markante klinische und neuropathologische Unterschiede (COMOY et al. 2008).
Die Inokulation humanes PrP exprimierender Mäuse mit dem L-Typ war effizienter als bei
der Verwendung der klassischen Variante (KONG et al. 2008) und Mäuse mit bovinem PrP
wiesen deutlich verkürzte Inkubationszeiten auf (BUSCHMANN et al. 2006).
Tab. 2.4 Übersicht BSE
BSE Klassisch atypisch
Symptome Abmagerung, Verhaltensänderungen,
erhöhte Sensibilität, Ataxie Abmagerung, Verhaltensänderungen,
erhöhte Sensibilität, Ataxie (3)
Einfluss des Genotyps ggf. Polymorphismen oder Depletion
der Promotorregion des Prion-Protein-
Gens (Prnp) und im Intron 1 (1) (2)
keine bekannt
Infektionsquelle infektiöses Tiermehl spontanes Auftreten
intra-spezies
Übertragung nur experimentell oral oder
intra-zerebral intrazerebrale Übertragbarkeit auf
Rinder experimentell nachgewiesen (3)
Erregeraufnahme oral -
(1) HAASE et al. 2007; (2) JULING et al. 2006; (3) BALKEMA-BUSCHMANN et al. 2011
Neben der BSE-Erkrankung beim Hausrind traten auch TSE-Erkrankungen bei exotischen
Wiederkäuern in zoologischen Gärten auf (JEFFREY u. WELLS 1988, KIRKWOOD et al.
1990). Desweiteren wurden in Frankreich und im Vereinten Königreich zwei BSE-
Infektionen bei Ziegen identifiziert (ELOIT et al. 2005; VACCARI et al. 2009).

36 Literaturübersicht
Im Gegensatz dazu lässt sich der H-Typ vor allem durch sein signifikant höheres
Molekulargewicht von klassischer BSE unterscheiden (CASALONE et al. 2004). Die
Infektion von bovines PrP exprimierenden Mäusen ging mit einer Verlängerung der
Inkubationszeit gegenüber der klassischen Form einher (BUSCHMANN et al. 2006).
Grundsätzlich wird angenommen, dass diese atypischen TSE-Formen spontan entstehen
können und angesichts der in der Vergangenheit stattgefundenen Verfütterung von
infektiösem Tiermehl an Wiederkäuer eine mögliche Ursache für die klassische BSE-
Infektion gewesen sein könnten (BIACABE et al. 2004, CASALONE et al. 2004,
BUSCHMANN et al. 2006, CAPOBIANCO et al. 2007). Der im Western-Blot durch einen
Scrapie-spezifischen Antikörper detektierbare H-Typ wirft die Frage auf, ob es sich
tatsächlich um eine neuartigen BSE-Typ handelt oder eher eine natürliche Scrapie-Infektion
beim Rind darstellt (BIACABE et al. 2004).
2.3 Pathogenese der TSE-Erkrankungen
Zur Klärung der TSE-Pathogenese (griechisch, páthos, Leiden und génesis, Entstehung)
wurden verschiedene In vivo-Infektionsstudien mit unterschiedlichen Tierarten durchgeführt.
Die kann die ausgewählte Infektionsroute (oral, intrazerebral oder intraperitoneal) kann dabei
eine wesentlichen Einfluss haben und ergaben mitunter unterschiedliche Ergebnisse.
Experimentelle Studien zu BSE an oral inokulierten Rindern wurden lediglich am damaligen
Veterinary Laboratory Agency, Weybridge, Vereinigtes Königreich Großbritannien und am
Friedrich-Loeffler-Institut, Insel Riems, Deutschland durchgeführt.
Im Folgenden soll die Pathogenese nach Prion-Infektionen bei verschiedenen Tieren erörtert
werden, wobei der Schwerpunkt auf Ergebnissen aus Studien mit dem Scrapie- und BSE-
Erregern liegen soll. In die Verbreitung des PrPSc
ist nach derzeitigem Kenntnisstand neben
lymphoiden Strukturen das PNS über parasympathische und sympathische Bahnen involviert.
Im Zuge einer Scrapie-Infektion weisen Studien auch auf eine mögliche hämatogene
Verbreitung hin. Dagegen wird insbesondere für BSE beim Rind aufgrund strenger Indizien
die neuronale Route favorisiert. Detaillierten Ergebnisse zur anterograden Ausbreitung des
pathologischen Prion-Proteins beim Rind liegen bisher jedoch noch nicht vor (s. Abb. 2.7,
S.37).

Literaturübersicht 37
Abb. 2.7 Möglicher Ausbreitungsweg des PrPSc
bei klassischer Scrapie (modifiziert nach
VAN KEULEN et al. 2002), grau hinterlegt: vermutete Beteiligung bei BSE; 1: Infektion des
GALT; 2: Ausbreitung im lymphatischen System; 3: Neuroinvasion, GC: Ganglion
coeliacum; GN: Ganglion nodosum
2.3.1 Dosis und Inkubationszeiten
Die durch epidemiologische Studien geschätzte Inkubationsperiode einer natürlichen BSE-
Erkrankung liegt bei 5-5,5 Jahren (ARNOLD u. WILESMITH, 2004; WILESMITH et al.
1988). Dies würde im Tierexperiment einer einmaligen Dosis von 0,1g bis 1g eines
Hirnhomogenat-Pools entsprechen (ARNOLD et al. 2007; WELLS et al. 2007).
Unberücksichtigt bleibt bei dieser Betrachtung eine im Feld mögliche mehrfache Aufnahme
infektiösen Materials, die im Nagetiermodell eine erhöhte Infektionswahrscheinlichkeit und
verkürzte Inkubationszeiten zur Folge hatte (DIRINGER et al. 1998; GRAVENOR et al.
2003). Statistischen Schätzungen zufolge liegt das höchste Infektionsrisiko bei Kälbern in den
ersten sechs Lebensmonaten (ARNOLD u. WILESMITH, 2004).

38 Literaturübersicht
Tab. 2.5 Korrelation zwischen Dosis und PrPSc
-Nachweis/Symptomatik
Dosis initialer PrP
Sc- Nachweis
im Obex
definierte klinische Symptome frühestens
nach
1 g 44 mpi (1)
; 42-47 mpi (2)
0,001 g 68 mpi (5)
0,1 g
1 g
53 mpi (5)
45 mpi (5)
100 g 24 mpi (3)
; 27 mpi (4)
; 10 g 44 mpi (1)
41 mpi (5)
30 mpi (1) (2)
100g 31 mpi (5)
; 35 mpi (1)
(1) MASUJIN et al, 2007 (2) ARNOLD et al. 2007 (3) HOFFMANN et al. 2007 (4)
ESPINOSA et al. 2007 (5) WELLS et al. 2007; mpi: Monate post infectionem
Im Maus- und Rindermodell (MCLEAN u. BOSTOCK, 2000; WELLS et al. 2007) ließ sich
zwar keine Schwellendosis für eine Infektion ermitteln, dennoch beeinflusst die Menge des
infektiösen Materials die Zeit bis zum Auftreten definierter klinischer Symptomatik ebenso
wie die erstmalige Detektion von PrPSc
im Hirnstamm (s. Tab. 2.5). Nach der Gabe von hohen
Dosen lässt sich nicht nur PrPSc
im Hirnstamm deutlich früher detektieren, es kommt zudem
zu einer Verringerung der Inkubationszeit und deren Variabilität (WELLS et al. 2007).
Weiterhin konnte nach oraler Verabreichung von 100g Homogenat PrPSc
im ZNS signifikant
früher vor dem Auftreten klinischer Symptome nachgewiesen werden als bei niedriger
dosierten Tieren (ARNOLD et al. 2007).
2.3.2 Das lymphatische System
Der initiale Nachweis nach oraler Scrapie-Inokulation erfolgt in Form von PrPSc
-
Akkumulationen im Darm-assoziierten-lymphatischen Geweben (engl., gut associated
lymphoid tissue, GALT), speziell in den Peyer´schen Platten (MAIGNIEN et al. 1999; VAN
KEULEN et al. 1999; BEEKES u. MCBRIDE, 2000) und der Tonsille (ANDREOLETTI et
al. 2000; VAN KEULEN et al. 2002). Im weiteren Verlauf erfolgt bei Scrapie der Befall des
Retropharyngeal- und der Mesenteriallymphknoten durch die das GALT drainierenden
Lymphgänge (ANDREOLETTI et al. 2000; VAN KEULEN et al. 2002).
Die Mechanismen für die Überwindung der Darmwand können für PrPSc
bisher lediglich
vermutet werden. Sogenannte M-Zellen, die im Follikel-assoziierten Epithel (FAE) der
Peyer´schen Platten lokalisiert sind, können verschieden Pathogene (Viren, Bakterien,
Prionen) transportieren (NEUTRA et al. 1996; HEPPNER et al. 2001). Auch ein

Literaturübersicht 39
enzymatischer Abbau in kleinere PrPSc
-Fragmente und ein vermuteter Transport von
Proteinkomplexen über einen Ferritin-gekoppelten Endozytosemechanismus wurde
beschrieben (MISHIRA et al. 2004). Ebenfalls über Endzytose durch Enterozyten könnte das
pathologische Prion-Protein die Darmwand passieren (JEFFREY et al. 2006). Für Bakterien
konnte eine direkten Aufnahme durch sogenannte dendritische Zellen (DC) gezeigt werden,
die mit ihren Fortsätzen Antigene direkt aus dem Darmlumen aufnehmen können (RESIGNO
et al. 2001).
Für die Beteiligung an der Pathogenese werden DC, sowie Makrophagen (TBM) und
follikulär dendritische Zellen (FDC) diskutiert, da eine direkte Assoziation zwischen diesen
Zellen und PrPSc
-Ablagerungen bei infizierten Hamstern (BEEKES u. MCBRIDE, 2000;
MCBRIDE et al. 2001) und bei Scrapie-inokulierten Schafen (ANDREOLETTI et al. 2000;
JEFFREY u. GONZALES, 2007) hergestellt werden konnte.
Die stationären FDC´s sind in primären B-Zellfollikeln lokalisiert und können, ohne selbst
phagozytotisches Potential zu besitzen, Antigene über einen langen Zeitraum präsentieren
(MANDEL et al. 1980; SHORTMAN et al. 2002). Da das zelluläre Prion-Protein als die
Voraussetzung für eine Infektion (BÜELER et al. 1993) auf den FDC exprimiert wird, besteht
über die Möglichkeit der Konversion von PrPC
zu PrPSc
die Involvierung der FDC in der
Pathogenese (BROWN et al. 1999). Fehlende oder deaktivierte FDC’s verzögern die
Neuroinvasion bei Mäusen (MABBOTT et al. 2000; MONTRASIO et al. 2000) und im
gleichen Tiermodell wird die Akkumulation von PrPSc
im lymphoiden Geweben durch die
Abwesenheit FDC-assoziierter Zytokine verhindert (KLEIN et al. 1997; MABBOTT et al.
2000). Gegen eine essentielle Beteiligung der FDC spricht die unbeeinflusste Pathogenese
transgener Mäuse ohne B-Zellfollikel (PRINZ et al. 2003) und eine Neuroinvasion des Agens
trotz pharmakologischer FDC-Depletion (OLDSTONE et al. 2002).
In den Peyer´schen Platten bilden die Dendritischen Zellen (DC) eine Zellschicht mit engen
Kontakt zu den M-Zellen (KELSALL et al. 1996), die in Versuchen an Ratten PrPSc
aufnahmen und zu den Mesenteriallymphknoten beförderten (HUANG et al. 2002).
Allerdings ist durch die präferierte Migration der DC in die T-Zellregionen der Lymphfollikel
ein Kontakt zu den FDC eingeschränkt (SHORTMAN et al. 2002) und die Fähigkeit zur
Degradation des pathologischen Prion-Proteins in vitro (RYBNER-BARNIER et al. 2006)
lassen Zweifel an einer effektiven Verbreitung von PrPSc
durch die DC aufkommen.
In vitro-Studien konnten eine Degradation des PrPSc
durch zur Phagozytose fähige
Makrophagen zeigen (CARP et al. 1981 und 1982; SASSA et al. 2010), deren
pharmakologische Depletion in vivo mit erhöhten PrPSc
-Ablagerungen einherging

40 Literaturübersicht
(BERINGUE et al. 2000; MAIGNIEN et al. 2005). In Scrapie-infizierten Schafen ließ sich
PrPSc
in Makrophagen detektieren, welches sehr wahrscheinlich von phagozytierten
Fortsätzen der FDC stammt (HERRMANN et al. 2003).
Die Integration mobiler und stationärer Immunzellen in die TSE-Pathogenese ist demnach
noch nicht vollständig geklärt.
Es gibt jedoch auch Studien, die gänzlich gegen eine Beteiligung des lymphatischen Systems
sprechen. Eine reduzierte Empfänglichkeit von immundefizienten Mäusen wurde durch eine
hochdosierte, parenterale Infektion überwunden (LASZEMAS et al. 1996; KLEIN et al.
1998). Die Inokulation verschiedener TSE-Agens in stark innervierte, extraneuronale Gewebe
(z.B. Zunge) von Hamstern und von transgenen Mäusen, die PrPC ausschließlich in
neuronalen Geweben exprimierten, führte zu einer Erkrankung ohne Beteiligung des
Lymphsystems (RACE et al. 2000). Weiterhin sei angemerkt, dass in Fällen atypischer
Scrapie bei Schafen (BENESTAD et al. 2003), bei Schafen mit heterologen VRQ/ARR-
Genotyp (VAN KEULEN et al. 1996) und in experimentell infizierten Schafen (JEFFREY et
al. 2002) die Etablierung einer Erkrankung ohne nachgewiesene Beteiligung lymphatischer
Gewebe verläuft.
Mit dem BSE-Erreger inokulierte Mäuse, Schafe und Primaten zeigen ebenfalls eine
Beteiligung des lymphatischen Systems (FOSTER et al. 1996, 2001; MAIGNIEN et al.
1999; HERZOG et al. 2004) und weisen damit auf eine Wirtsspezifität hin. In experimentell
infizierten Rindern konnte bisher lediglich PrPSc
in vereinzelten Peyer´schen Platten ab dem
4. Monat post infectionem (mpi) detektiert werden (HOFFMANN et al. 2011). Dagegen
wurde für natürlich erkrankte Tiere bisher zwar Infektiosität (BUSCHMANN u.
GROSCHUP, 2005) aber keine PrPSc
-Akkumulationen (TERRY et al. 2003; IWATA et al.
2006) detektiert. Die Ablagerungen waren mit FDC und Makrophagen PrPSc
bereites 24 mpi
assoziiert (HOFFMANN et al. 2007), während in anderen Studien FDC erst in der klinischen
Phase in Erscheinung traten (TERRY et al. 2003). Weitere Studien zeigen Infektiosität in der
Tonsille in einem präklinischen Rind (WELLS et al. 2005) als auch bei Tieren im terminalen
Stadium der Krankheit (ESPINOSA et al. 2007), die aber nicht als Hinweis auf eine
weitergehende lymphatische Ausbreitung verstanden werden wollen.
Die Frage, ob das lymphatische System eine notwendige Voraussetzung, einen optionalen
Mediator oder einen autarken Beteiligten an der Neuroinvasion darstellt, lässt sich mit den
Ergebnissen der bisherigen Studien nicht vollständig beantworten. Für BSE-infizierte Rinder
konnte lediglich das GALT als beteiligte lymphatische Komponente nach experimenteller
Inokulation hoher oraler Dosen nachgewiesen werden.

Literaturübersicht 41
2.3.3 Das periphere Nervensystem (PNS)
2.3.3.1 Neuroinvasion
Für die Neuroinvasion wird die Beteiligung lymphoider Strukturen kontrovers diskutiert.
Studien an verschiedenen Tiermodellen suggerieren (a) die Neuroinvasion über das Lymph-
und Blutsystem, (b) den Aufstieg von PrPSc
/Infektiosität über Strukturen des ENS/PNS nach
initialer Replikation im lymphatischen Gewebe und (c) die direkte Aufnahme des Erregers
über Nervenenden des PNS.
Scrapie und BSE-Infektionen zeigen in verschiedenen Tiermodellen den primären Nachweis
von PrPSc
-Ablagerungen im GALT vor der Einbeziehung des ENS (ANDREOLETTI et al.
2000; VAN KEULEN et al. 2000; MCBRIDE et al. 2001; IWATA et al. 2006; HOFFMANN
et al. 2011). Die Verbindung zwischen immunologischen Zellen insbesondere des GALT´s
und Nervenenden des ENS wird als möglicher Übergang in das neuronale Gewebe
angenommen (HEGGEBØ et al. 2003; PRESS et al. 2004; DEFAWEUX et al. 2005). Dafür
wird neben Zell-Zell-Kontakten auch die Möglichkeit eines aktiven Transports zwischen der
Lymphe und dem ENS in Betracht gezogen (HEGGEBØ et al. 2003). Ebenfalls ist ein
interzellulärer Transport durch von infizierten Zellen (FDC, DC, Makrophagen) abgegebene
Exosomen denkbar (FEVRIER. et al. 2004).
Elektronenmikroskopische Untersuchungen zeigen zwar ein Inokulat-unabhängiges de novo
generiertes PrPSc
in den Peyer´schen Platten, doch der fehlende Nachweis des pathologischen
Prion-Proteins in anderen, möglicherweise als Mediatoren fungierenden Zellen legt die
Möglichkeit einer Neuroinvasion unabhängig vom Befall der lymphoiden Strukturen und des
ENS durch Fasern peripherer Nerven direkt unterhalb der Basallamina nahe (JEFFREY et al.
2006). Bei Rindern konnte ein direkter Kontakt sympathischer Nervenfasern des PNS mit den
FDC’s und mit Makrophagen gezeigt werden (DEFAWEUX et al. 2007). Außerdem wies das
ENS erst im klinischen Stadium von BSE PrPSc
-Ablagerungen sowohl bei Infektionen
natürlichen Ursprungs als auch bei experimentell infizierten Tieren auf (TERRY et al. 2003).
Zudem konnte für DC in vitro eine Verbindung zu Fortsätzen peripherer Neurone gezeigt
werden (DORBAN et al. 2010), ebenso ließ sich im Mausmodell ein direkter Transport von
infektiösem Agens durch DC zu Anteilen des peripheren Nervengewebes beobachten
(AUCOUTURIER et al. 2001).

42 Literaturübersicht
2.3.3.2 Ausbreitung von PrPSc
im PNS
Es sind vor allen Studien bei Schafen und Hamstern, die bisher (I) sympathische Nerven und
alternativ (II) auch den parasympathischen N. vagus als beteiligte Komponenten des PNS bei
der Ausbreitung des TSE-Agens beschreiben.
Der Sympathikus ist dabei in Form der Efferenzen der Nn. splanchnici und der
postganglionären Nervenzellen des Ggl. coeliacum beteiligt, die mit den sympathischen
präganglionären Neuronen in der Substantia intermedia lateralis des Rückenmarks
kommunizieren (MCBRIDE u. BEEKES, 1999; VAN KEULEN et al. 2000, 2002;
MCBRIDE et al. 2001).
Tab. 2.6 Übersicht PrPSc
-Nachweis/Infektiosität von Proben des peripheren Nervensystems
beim Rind
(1) ARNOLD et al. 2007; (2) MASUJIN et al. 2007; (3) KIMURA et al. 2008; (4)
ESPINOSA et al. 2007; # nicht durchgeführt; ud: undefiniert; exp: experimentell; IHC:
Immunhistochemie; WB: Western Blot; BA: Bioassay; mpi: Monate post infectionem; * Bio-
Rad TeSeE
Status
IHC WB BA klinisch Obex mpi Infektion
Ggl.
trigeminale
+ # # - + 32 exp (1)
# + # + + ud natürlich (2)
Ggl. spinale + # # - + 32 exp (1)
Ggl.
stellatum
- - # + + ud natürlich (3)
# + # + + 36 exp (2)
Ggl. cervicale craniale - -* # - + 32 exp (1)
sympathischer
Grenzstrang - # # + + ud natürlich
(3)
N. vagus # + + + + ud natürlich
(2)
- # # + + ud natürlich (3)
Nebenniere # + + + + ud natürlich
(2)
- # # + + ud natürlich (3)
N. ischiadicus # # + - + 30 exp (4)

Literaturübersicht 43
Alternativ könnte der Aufstieg über die efferenten parasympathischen Bahnen des N. vagus
zu den entsprechenden präganglionären Neurone im dorsalen motorischen Kern des N. vagus
(engl. dorsal motor nucleus of the vagus – DMNV) in der Medulla oblongata erfolgen. Dieser
Pfad wird aus der Lokalisation einzelner PrPSc
-positiven Neuronen im DMNV abgeleitet und
als initialer Befall des ZNS interpretiert (BALDAUF et al. 1997; VAN KEULEN et al. 2000,
2002; MCBRIDE et al. 2001).
Indizien sprechen für einen ähnlichen neuronalen Ausbreitungsweg bei BSE. So gelang bei
einem oral infizierten, präklinischen Rind der Nachweis von PrPSc
zum einen im Ggl.
coeliacum, Ggl. mesenteriale caudale und dem ZNS (DMNV, Rückenmark) wie auch im
DMNV des Obex, jedoch ohne weitere Beteiligung des PNS (HOFFMANN et al. 2007). Mit
der deutlichen Beteiligung des ZNS sind weitere Daten zur Ausbreitung im PNS (s. Tab.2.6,
S.42) verfügbar, die allerdings eher auf eine zentrifugale Infektion hindeuten. Indizien für
eine solche sekundäre Ausbreitung nach Replikation des Agens im ZNS ist die nach initialen
Befall des Rückenmarks gleichzeitig auftretende zentrifugale und zentripetale Ausbreitung
mit einer Geschwindigkeit von 0,8-1,0 mm/Tag (BEEKES et al. 1996) und die ebenfalls
zentrifugal erfolgte Infektion des Ischias-Nerven der kontralateralen Seite nach unilateraler
Inokulation in die Fußsohle bei Hamstern (KRATZEL er al., 2007a).
2.3.3.3 Ausbreitungsmechanismen im Nervensystem
Der zugrunde liegende Mechanismus des Transports des infektiösen Agens ist bisher nur
unvollständig geklärt.
Eine der diskutierten Möglichkeiten ist der axonale Transport, der für verschiedene Toxine
wie z.B. das Tetanustoxin (MANNING et al. 1990) und das Choleratoxin (WAN et al. 1982;
AGELUCCI et al. 1996) sowie für Vertreter aus der Familie der Herpesviridae
(TOMISHIMA et al. 2001) gezeigt und für das Tollwutvirus vermutet wird (VON
BARTHELD, 2004). Für das körpereigene PrPC wurde die Möglichkeit einer axonalen
Weiterleitung berichtet (BORCHELT et al. 1994). Dies wird auch aus dem Vergleich der
Inkubationszeiten zu der geschätzten axonalen Ausbreitungsgeschwindigkeit im Maus- und
Hamsterversuch vermutet (KIMBERLIN et al. 1983, 1987; MCBRIDE et al. 2001).
Studien am Nagetiermodell weisen nach Einschränkung des axonalen Transports jedoch keine
Beeinflussung der Inkubationszeit auf (KUNZI et al. 2002; HAFEZPARAST et al. 2005;
KRATZEL et al. 2007a,b). Letztgenannte Autoren beobachteten wie schon GROSCHUP et
al. (1999) adaxonale PrPSc
-Akkumulationen zwischen der Myelinscheide und dem Axon. Für

44 Literaturübersicht
Schwann´sche Zellen konnte in vivo die PrPC-Expression sowie die Konvertierbarkeit und die
Infektiosität dieser Zellen in vitro gezeigt werden. Sie könnten damit als Mediatoren im
Transportprozess für das pathologische Prion-Protein beteiligt sein (FOLLET et al. 2002).
Eine weitere Möglichkeit ist die „Domino“-ähnliche Konversion von PrPC zu PrP
Sc entlang
der Nerven, wie sie von GLATZEL u. AGUZZI (2000) diskutiert wurden.
Verschiedene Zellkulturmodelle zeigen eine Ausschleusung des pathologischen Prion-
Proteins in Exosomen (FEVRIER et al. 2005; VELLA et al. 2007), deren Nachweis in
Scrapie-infizierten Neuronen mittels Elektronenmikroskopie allerdings nicht erbracht werden
konnte (JEFFREY et al. 2009).
Für die Ausbreitung im ZNS kommen verschieden Mechanismen in Frage. So kann PrPSc
von
Membranen der Dendriten freigesetzt werden und sich im extrazellulären Raum zu Fibrillen
zusammenlagern (JEFFREY et al. 1994). Scrapie-infizierte Gehirne von Mäusen zeigten in
Vorläufern der extrazellulären Exosomen eine Anreicherung von PrPSc
(ARNOLD et al.
1995). Sogenannte „Tunneling nanotubes“ (TNTs) sind nanometer-dünne Membran-Kanäle,
die Zellen zu komplexen Netzwerken verknüpfen und für den interzellulären Transfer
unterschiedlicher Biomoleküle verantwortlich sind. Somit können diese Strukturen als weitere
potentielle Kandidaten für den Transfer des infektiösen Agens gehandelt werden (CAUGHEY
et al. 2009; GERDES, 2009).
2.3.4 Alternative Ausbreitungswege von PrPSc
im Körper
Über das eigentliche Vorkommen des Scrapie-Erregers im Blut hinaus (SCHMERR et al.
1999; SAA et al. 2006), konnten Transfusionsexperimente am Schaf die Infektiosität des
Blutes bereits in der präklinischen Phase der Spendertiere zeigen (HUNTER et al. 2002; SISO
et al. 2006, 2009; HOUSTON et al. 2008). Für vCJD wurde von vier bestätigten Fälle einer
Übertragung durch Transfusionen berichtet, bei denen drei Patienten darauffolgend eine
klinische Symptomatik zeigten (LLEWELYN et al. 2004; PEDEN et al. 2004;
http://www.hpa.org.uk).
Der Nachweis von PrPSc
in verschieden peripheren Geweben wird als Folge einer
hämatogenen Infektion bewertet. Beispielsweise sprechen Akkumulationen des
pathologischen Prion-Protein in den Mesenchymzellen der renalen Papille nachdrücklich für
den Befall der Niere nach Filtration des Blutes (SISO et al. 2008).

Literaturübersicht 45
Letztgenannte Studie setzt somit das Vorliegen des infektiösen Agens als zellfreies, lösliches
Molekül im Blutplasma voraus. Unter diesen Voraussetzungen könnte eine hämatogene
Neuroinvasion über die zirkumventrikulären Organe (ZVOs) erfolgen, für die PrPSc
-
Akkumulationen unabhängig von der Inokulationsroute früh und einheitlich bei Schafen
(SISO et al. 2009) ebenso wie in retrospektiven Studien bei oral infizierten, klinisch an BSE
erkrankten Rindern (SISO et al. 2010) gezeigt werden konnte.
2.3.5 Akkumulation von PrPSc
in extraneuronalen Geweben und Ausscheidung
Die in Kap. 2.3.3.2 (s. S. 42) und 2.3.6 (s. S. 46) beschriebenen Ausbreitungswege können
auch zu PrPSc
-Akkumulationen in nicht-neuronalen Geweben führen.
Die experimentelle orale Inokulation von Hamstern und Schafen führte wie auch bei
natürlichen vorkommenden Scrapie-Fällen zum Nachweis von PrPSc
und/oder Infektiosität in
der Muskulatur, einschließlich der Zunge bei präklinischen und klinischen Tieren
(THOMZIG et al. 2003, 2004; ANDREOLETTI et al. 2004; CASALONE et al. 2005,
BALKEMA-BUSCHMANN et al. 2011). Auch bei einer BSE-Infektion konnte die
Beteiligung der Muskulatur gezeigt werden, so wurde bei Hamstern PrPSc
(THOMZIG et al.
2006) und bei Rindern Infektiosität im Musculus semitendinosus detektiert (BUSCHMANN
u. GROSCHUP, 2005).
Mit dem Nachweis infektiösen Fettgewebes bei Mäusen (RACE et al. 2008) wird eine weitere
Möglichkeit von kontaminierten Material in der humane Nahrungskette erfasst. Des Weiteren
konnte Ablagerungen in der Haut experimentell mit Scrapie infizierter Hamster und
klinischen Scrapie-Schafen nachgewiesen werden (THOMZIG et al. 2007). Wie auch bei der
Muskulatur wird ursächlich eine Infektion des Gewebes auf neuronalem Wege nach
anterograder Ausbreitung diskutiert.
Das Scrapie-Agens wurde zudem im Kolostrum und der Milch (LACROUX et al. 2008), in
der Niere (SISO et al. 2006; LIGIOS et al. 2007) sowie in der Speicheldrüse (VASCELLARI
et al. 2007) bei infizierten Schafen detektiert. KRÜGER et al. (2009) unterscheiden
nachgewiesenes PrPSc
im Kot Scrapie-infizierter Hamstern zwischen einer frühen Inokulat-
Ausscheidung (24 und 72 Stunden post infectionem) und marginalen, neu generierten Mengen
bei terminal erkrankten Tieren. In den Experimenten von SAFAR et al. (2005) ließen sich
Hamster infizieren, die Faeces inokulierter Tiere exponiert waren.

46 Literaturübersicht
Die in verschiedenen Studien bei Scrapie gezeigte Präsens von krankheitsassoziiertem Prion-
Protein in Exkreten und Sekreten lässt auf eine horizontalen Übertragung bei Schafen
schließen, während es keine Hinweise für einen solchen Übertragungsweg bei BSE-infizierten
Rindern gibt (CURNOW u. HAU, 1996; WRATHALL et al. 2002).
2.3.6 Entzündungen und TSE
Obwohl das pathologische Prion-Protein auch in verschiedenen Organen ohne entsprechende
Entzündungsprozesse lokalisiert werden konnte (s. Kap. 2.3.5, S.45), scheinen
inflammatorische Prozesse die Darmpassage bzw. die Neuroinvasion und die Ausscheidung
von PrPSc
und/oder Infektiosität zu erleichtern.
So war bei Mäusen die Empfänglichkeit für eine Scrapie-Infektion nach einer Salmonellen-
induzierten Kolitis stark erhöht (SIGURDSON et al. 2009). Es kam zudem zu einer
schnelleren Ausbildung des terminalen Stadiums (THACKRAY et al. 2003).
Entzündungsherde im Abomasum bei Schafen wiesen Ablagerungen von PrPSc
auf
(JEFFREY et al. 2001). Dafür werden vielfältige pathogenetische Mechanismen diskutiert.
Häufig auftretende Darmerkrankungen, hervorgerufen durch Viren, Bakterien oder Parasiten,
beeinflussen die tight junctions des Darmepithels (SAKAGUCHI et al. 2001; MILLING et al.
2007) und induzieren eine gesteigerten Aufnahme von Protein-Antigenen (SODERHOLM et
al. 2004). Durch z.B. gastrointestinale Entzündungen wird die PrPC-Expression erhöht
(PAMMER et al. 2000), was eine effizientere Neuroinvasion zur Folge haben könnte
(THACKRAY et al. 2003). Weiterhin ist über die Aktivierung von DC (BROWN et al. 1999)
als Folgeerscheinung einer Darminfektion eine Beeinflussung der TSE-Pathogenese denkbar.
Der physiologische Status des Verdauungstraktes scheint somit einen Einfluss auf die
Empfänglichkeit für eine TSE-Erkrankung zu haben und könnte sich so auf die Akkumulation
bzw. Replikation des pathologischen Prion-Proteins auswirken.
Außerhalb des Verdauungstraktes wurde bei einem sCJK-Patienten mit sporadischer
Einschlusskörpermyositis beträchtliche PrPSc
-Ablagerungen in der Muskulatur gefunden
(KOVACS et al. 2004). Bei Mäusen führt eine chronische lymphozytäre Entzündung in
verschiedenen Organen zu einer Veränderung des Organtropismus; pathologisches Prion-
Protein wurde in der Niere, dem Pankreas und der Leber entdeckt (HEIKENWALKER et al.
2005). Infolge einer Nephritis kam es zur Ausscheidung von infektiösem Urin (SEEGER et

Literaturübersicht 47
al. 2005) und bei Schafen wurde das Auftreten von PrPSc
im Euter von Tieren mit Mastitis
nachgewiesen (LIGIOS et al. 2005).

Material und Methoden 48
3. TIERE, MATERIAL UND METHODEN
3.1 Tiere
3.1.1 Rinder
Am Friedrich-Loeffler-Institut wurden 2002/2003 insgesamt 74 ausschließlich weibliche
Kälber für die Pathogenesestudie eingestallt. Bei den Rindern handelte es sich um Fleckvieh-
Simmental-Kreuzungen, die aus einem ökologisch geführten Mutterkuh-Betrieb in
Mecklenburg-Vorpommern stammen.
Die Tiere waren vor dem Einstallen gegen das Bovine Parainfluenza 3-Virus, das Bovine
Respiratorische Synzytialvirus (BRSV) und das Bovine Herpesvirus-1 (BHV1) geimpft.
Das Tierversuchsvorhaben wurde unter dem Aktenzeichen LALLF M-V/TSD/7221.3-1.2-
008/07 genehmigt.
Eine Liste der ausgewählten Infektionstiere mit den zugehörigen klinischen Daten findet sich
in Tab. 3.3 (S.56).
3.1.2 Durchführung der BSE-Pathogenesestudie
3.1.2.1 Tierhaltung
Die Haltung der infizierten Rinder erfolgte in einem separaten Stallgebäude der
Sicherheitsstufe L3**. Die anfallenden Abwässer inklusive der Fäkalien aus dem Stall
wurden für vier Stunden bei 136°C autoklaviert und anschließend in der institutseigenen
Kläranlage aufbereitet. Die Tiere der Kontrollgruppe waren in einem Quarantänestall
untergebracht. Die Fütterung der Tiere erfolgte unter Aufsicht mit Heu und Heucobs, die mit
Kraftfutter und Mineralfutter supplementiert wurden. Wasser stand ad libitum zur Verfügung.
3.1.2.2 Inokulat und Inokulation
Insgesamt wurden zwischen Januar und April 2003 56 Rinder peroral mit BSE infiziert, 18
weitere Tiere dienten als negative Kontrolltiere. Am Tag der Inokulation waren die Tiere
zwischen 4 und 6 Monaten alt.

Material und Methoden 49
Die Infektionstiere erhielten hierzu jeweils 100g eines Hirnhomogenates über eine
Schlundsonde oral appliziert, das aus einem Pool von nachweislich BSE-positiven
Rindergehirnen aus dem Vereinigten Königreich hergestellt war. Der über eine Endpunkt-
Titration in transgenen, bovines PrPC-überexprimierenden Tgbov XV-Mäusen ermittelte
Infektionstiter des Homogenates betrug 106.1
ID pro g Gewebe (HOFFMANN et al. 2007).
Die Kontrolltieren erhielten 100g eines negativen Gehirnhomogenates.
3.1.2.3 Verhaltensstudie
Um Verhaltensaufälligkeiten bei den Tieren festzustellen, wurde alle 8 Wochen (beginnend
mit 18 Monaten nach der Inokulation) ein Verhaltenstest durchgeführt. Dabei kamen
unterschiedliche Methoden zum Einsatz, nach deren Bewertung und Auswertung der Status
der Tiere bestimmt wurde:
Status 0 - keine Anzeichen von BSE
Status 1 - möglicherweise BSE, d.h. Verhaltens- oder Sensibilitätsstörungen
treten nur gelegentlich auf
Status 2 - vermutlich BSE, d.h. Störungen treten regelmäßig (bei mindestens
zwei aufeinanderfolgenden Untersuchungen) auf oder mindestens zwei
der Tests auf Übererregbarkeit fallen positiv aus, selbst wenn sie
erstmals beobachtet werden
Status 3 - definitiv BSE, d.h. progressiver Verlauf, auch wenn nicht alle
Symptome/Verhaltens- bzw. Bewegungsstörungen nachweisbar sind
Die Kriterien für die eingesetzten Verhaltenstests befinden sich im Anhang (s. Kap. 9.2,
S.165).
3.1.2.4 Probenentnahme
Vor der Inokulation wurden Blut, Harn- und Liquor-Proben als sogenannte Nullproben
gewonnen und die Tiere danach regelmäßig beprobt (s. Tab 3.1, S.50). Die Liquorentnahme
erfolgte nach Verabreichung von 0,5-1ml Xylazin (Rompun®, Fa. Bayer-Vital, Leverkusen).
Ferner erforderte die Entnahme der Blutproben zeitweise eine Sedation.
Einen Monat nach der Inokulation beginnend, erfolgten die Sektionen in einem 4-monatigen
Rhythmus. Dazu wurden vier oder fünf zufällig ausgewählte Tiere je Sektionszeitpunkt unter
TSE-sterilen Bedingungen seziert. Von einer zufälligen Auswahl wurde abgesehen, wenn

50 Material und Methoden
Tiere eindeutige BSE-Symptomatik aufwiesen oder wenn durch andere tierschutzrelevante
Gründe ein Verbleib im Versuch nicht möglich war.
Tab. 3.1 Beprobungsintervalle für die Entnahme der Körperflüssigkeiten während der
Inkubationszeit
Probe Intervall
Blut 8 Wochen
Harn 3 dpi, 5 dpi,14 dpi*, anschließend 8 Wochen
Liquor 16 Wochen
*dpi: Tage nach der Inokulation
Das Sektionsprotokoll sah eine genau definierte Reihenfolge der Probennahme vor, um eine
Kreuzkontamination auszuschließen. Dazu wurden zunächst die Probenentnahme am Kopf,
Tierkörper und Verdauungstrakt räumlich, personell und instrumentell getrennt. Nach einer
Vorpräparation wurde jede einzelne Probe mit sterilem Einmalbesteck entnommen und in
ebenfalls sterile Einmalbehältnisse verbracht.
Die entnommenen Gewebeproben wurden, abhängig von der geplanten Verwendungs- bzw.
Detektionsmethodik, unterschiedlich behandelt. Bei paarigen Organen, bzw. Geweben
wurden die Proben der rechten Tierkörperseite in 4%ig neutral gepufferten Formalin fixiert,
während das Äquivalent der linken Körperhälfte im Rahmen der BSE-Probenbank bei -70°C
und -20°C gelagert wurde. Lagen die Proben unpaar vor, so wurden sie unter
Berücksichtigung der TSE-Sterilität in zwei Teile präpariert und entsprechend in Formalin
fixiert bzw. eingefroren.
Die weiterführenden Untersuchungen der entnommenen Proben wurden teilweise in L3**-
Laboratorien durchgeführt, die besondere Sicherheitsmaßnahmen wie das Tragen von
Schutzkleidung, separaten Laborschuhen und von Einmal-Handschuhen erforderten.
3.1.2.5 Bestandsbetreuung
Drei Monate nach der Inokulation fand eine prophylaktische Impfung der Tiere gegen
Kälberflechte statt.
Zu Beginn des Jahres 2004 zeigten drei der infizierten Tiere klinische Symptome einer
Infektion mit dem Ovinen Herpesvirus Typ 2 (OHV2)- Infektion. Diese löst bei Rindern das
Bösartige Katarrhalfieber aus, welches sich in einer perakuten Kopf-Augen-Form, einer
intestinalen Form oder einer milden Form äußern kann. Ein Tier verstarb an dieser Krankheit,

Material und Methoden 51
zwei andere wurden euthanasiert. Alle Rinder wurden nach dem Standardprotokoll seziert.
Durch eine umfassende Blutuntersuchung gelang bei sechs Tieren der Nachweis von Antigen,
insgesamt 27 Rinder wiesen mindestens zu einem Untersuchungszeitpunkt (8, 10, 12 14 mpi)
Antikörper gegen das OHV2 auf (s. Tab. 3.3, S.56). Die Kontrolltiere wurden dagegen
negativ getestet.
Angesichts eines Kokzidienbefalls im Darm aller Tiere fand in regelmäßigen Abständen eine
Bekämpfung der Ekto- und Endoparasiten statt.
Weitere gesundheitliche Aspekte stellten temporäre Klauenprobleme, sporadisch auftretende
Infektionen des respiratorischen Systems sowie Hämatome und Abszesse dar. Diese Probleme
konnten durch individuelle Therapien, unter Umständen durch den Einsatz von Antibiotika,
behoben werden.
3.1.3 Mäuse
3.1.3.1 Mauslinien
Mit dem Maus-Bioassay steht eine äußerst sensitive Methode zur Detektion auch geringer
Spuren von Infektiosität in den Proben zur Verfügung. Für die Versuche wurden Tgbov XV-
Mäuse verwendet, die aufgrund einer Überexpression des bovines Prion-Proteins die
Speziesbarriere zwischen Maus und Rind aufheben und sich durch eine sehr hohe
Empfänglichkeit für den BSE-Erreger auszeichnen. Die Vorteile dieser transgenen Mauslinie
im Vergleich zu konventionellen RIII-Mäusen liegen in einer 10.000fach höheren Sensitivität
und deutlich verkürzten Inkubationszeiten (BUSCHMANN u. GROSCHUP 2005). Auch im
Vergleich zum Rinder-Bioassay (HAWKINS et al. 2000) sind die Tgbov XV-Mäuse
bezüglich der Sensitivität deutlich überlegen. Diese am FLI generierten Mäuse wurden dem
Bedarf entsprechend in der Kleintierzucht des Instituts gezüchtet.
Dieser Tierversuch wurde unter dem Aktenzeichen LVL M-V/310-4/7221.3-2.1-012/03
genehmigt.
3.1.3.2 Inokulate
Aus den in der Sektion eingefrorenen und bei -70°C gelagerten Gewebe wurde für jede Probe
circa 1,5 ml steriles Inokulat mit einer Zielkonzentration von 10% hergestellt. Die dafür

52 Material und Methoden
benötigten repräsentativen Anteile wurden frisch präpariert, abgewogen und mit Hilfe von
Skalpell-Klingen zerkleinert. Gemeinsam mit der entsprechenden Menge steriler isotonischer
(0,9%ig) NaCl-Lösung wurde jede Probe in ein Einmal-Schraubdeckelgefäß mit
Keramikkugeln (Precellys Keramik-Set, Fa. Peqlab) gegeben. Je nach Beschaffenheit des
Gewebes waren bis zu acht Durchgänge im Ribolyser bei einer Geschwindigkeit von 6,5 und
einer Laufzeit von 45sek notwendig. Das Inokulat wurde mit einer Kanüle (0,6 x 30mm)
abgehoben und in ein steriles Eppendorf-Reaktionsgefäß überführt. Zur Lagerung wurden die
Proben nochmals bei -20°C eingefroren. Diese wurden vor der Inokulation aufgetaut und für
zwei Minuten mit Ultraschall homogenisiert.
3.1.3.3 Durchführung des Tierversuchs
Für die Inokulation wurden männliche und weibliche Tgbov-XV-Mäuse im Alter von 8 bis 12
Wochen verwendet. Bei einer Gruppengröße von 15 Mäusen pro Gewebeprobe wurden
jeweils 30µl des Inokulates jeder Maus unter Isofluran-Kurznarkose intrazerebral verabreicht.
Von zwei Tieren aus sechs Sektionszeitpunkten zwischen 16 und 36 mpi standen für 11
Proben Inokulate bereit. Aufgrund von unzweifelhaften immunhistologischen Befunden war
für insgesamt 11 ZNS-Proben die Durchführung des Bioassays nicht notwendig. Daraus
ergibt sich die Gesamtzahl inokulierter Mäuse von 2485.
Dreimal wöchentlich wurden alle Mäuse auf das Vorhandensein klinischer Symptome
untersucht. Die folgenden Erscheinungsbilder wurden zur Beurteilung herangezogen:
Apathie
Abmagerung
Ataxie
Verhaltensauffälligkeiten
gestörtes Allgemeinbefinden,
stumpfes und gesträubtes Fell
getrübte Augen
gekrümmter Rücken
Pruritus
Tiere mit spezifischer Symptomatik wurden aus dem Versuch genommen und durch CO2-
Intoxikation euthanasiert. Des Weiteren wurden Tiere mit sonstigen Erkrankungen wie
beispielsweise fehlerhaftem Zahnwuchs oder Tumoren unter Dokumentation der

Material und Methoden 53
Verdachtsdiagnose ebenfalls durch CO2-Intoxikation euthanasiert. Nach maximal 700 Tagen
erfolgte die Euthanasierung aller noch im Versuch befindlichen Mäuse.
3.1.3.4 Probenentnahme
Ausgehend vom Foramen occipitale wurde der Schädel mit einer kleinen Schere an der
rechten Seite aufgeschnitten und das Gehirn heraus präpariert. Mit einem Einmalskalpell
wurde das Gehirn mit einem leicht paramedianen Sagittalschnitt, circa 2mm neben dem
Sulcus longitudinalis, von dorsal nach ventral gerade durchtrennt. Der größere Teil des
Gehirns wurde für die histologischen Untersuchungen in 4%igem, neutral gepufferten
Formalin fixiert. Der kleinere Anteil wurde für die Untersuchungen im Western Blot
verwendet und bei –20°C gelagert.
3.1.3.5 Inkubationszeiten
Die Inkubationszeit ist definiert als die Zeit von der Infektion (Inokulation) bis zum Auftreten
der ersten eindeutigen Krankheitserscheinungen. In den hier durchgeführten Versuchen wurde
jeweils der Todestag der Tiere als Ende der Inkubationszeit definiert und ungeachtet ob eine
Euthanasie wegen eindeutiger klinischer Symptome oder ein natürlicher/pathologischer Tod
eintraf als Parameter zur weiteren Berechnung verwendet. Für jede Tiergruppe einer Probe
wurden hierzu der Mittelwert und die Standardabweichung der Inkubationszeiten aller
positiven Mäuse berechnet.
Wie gezeigt werden konnte (BUSCHMANN u. GROSCHUP, 2005), liegen die
Inkubationszeiten der Tgbov XV-Mäuse nach intrazerebraler Inokulation des hochpositiven
Gehirnmaterials bei über 200 Tagen. Nimmt die Infektiosität ab, kann sich die Inkubationszeit
auf knapp 600 Tage verlängern. So wurden in diesem Versuch alle euthanasierten und tot
aufgefundenen Mäuse mit einer Inkubationszeit von ≥ 100 Tagen im Westernblot auf das
Vorhandensein von PrPSc
untersucht.
Dagegen wurden die Mäuse, die in der Zeit bis zum 100. Tag nach der Inokulation
euthanasiert oder tot aufgefunden wurden, nicht für die weiteren Untersuchungen
berücksichtigt und fanden somit keinen Eingang in die Statistik.

54 Material und Methoden
3.2 Probenmaterial
Die Pathogenese der BSE stand im Fokus dieser Untersuchung. Primäres Ziel war es, neue
Erkenntnisse über den Ausbreitungsweg des PrPSc
vom Darm in das ZNS als Zielorgan zu
erhalten.
Bei der Auswahl der Rinder wurde der Untersuchungszeitraum so bestimmt, dass sowohl
Tiere mit präklinischen, BSE-freiem Status bis hin zu klinischen Fällen erfasst werden
konnten. Die Auswahl fiel auf jeweils zwei Tiere pro Tötungszeitpunkt im Sektionszeitraum
von 16 bis 44 Monate p.i. (also insgesamt 16 Rinder) für die immunhistochemischen
Untersuchung, während Proben der gleichen Tiere im verkürzten Zeitraum von 16 bis 36
Monate p.i. für den Bioassay verwendet wurden.
Der Zielstellung entsprechend kamen Gewebe des zentralen und peripheren Nervensystems
sowie der ilealen Peyer´schen Platten zur Untersuchung. Im Falle der Medulla oblongata
wurde die Ebene des Obex für die Immunhistochemie verwendet, während der kaudale
Medulla-Anteil im Maus-Bioassay zum Einsatz kam.
In Tabelle 3.2 (S.55) sind die Infektionstiere, die Gewebe und die vorgenommenen
Untersuchungen aufgelistet.

Material und Methoden 55
Tab. 3.2 Liste der untersuchten Proben und der durchgeführten Untersuchung
IT
28
IT46
IT50
IT60
IT24
IT58
IT21
IT52
IT09
IT61
IT49
IT11
IT25
IT56
IT22
IT38
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
nd
xn
dn
dn
dn
dn
dn
d
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
nd
xn
dx
nd
nd
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
xx
xx
xx
xx
xx
xx
nd
nd
nd
nd
IHC
xx
xx
xx
xx
xx
xx
xx
xx
BA
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
nd
Darm
ZN
S
So
mati
sch
es
Nerv
en
sy
ste
m
Sy
mp
ath
isch
es
Nerv
en
sy
ste
m
Para
sy
mp
ath
isch
es
Nerv
en
sy
ste
m
Gem
isch
t-fa
seri
ge
An
teile d
es V
NS
Pey
er`
sch
e
Pla
tten
, Il
eu
m
Ob
ex/
cau
d.
Med
ulla
RM
T7/L
3 *
Gg
l. t
rig
em
inale
Nn
. sp
lan
ch
nic
i
Gg
l. c
erv
. cra
n.
Gg
l. s
tellatu
m
N. v
ag
us (
Hals
)
Gg
l. n
od
osu
m
N. v
ag
us
(Bru
st)
Gg
l. c
oeliacu
m
Gg
l. m
esen
t.
cau
d.
Mo
nate
p.i./
Tie
rnu
mm
er
44m
pi
40m
pi
36m
pi
32m
pi
28m
pi
24m
pi
20m
pi
16m
pi
nd: nic
ht durc
hgefü
hrt
, *R
M T
7/L
3: R
ückem
ark
segm
ent lu
mbal 3 u
nd thora
kal 7
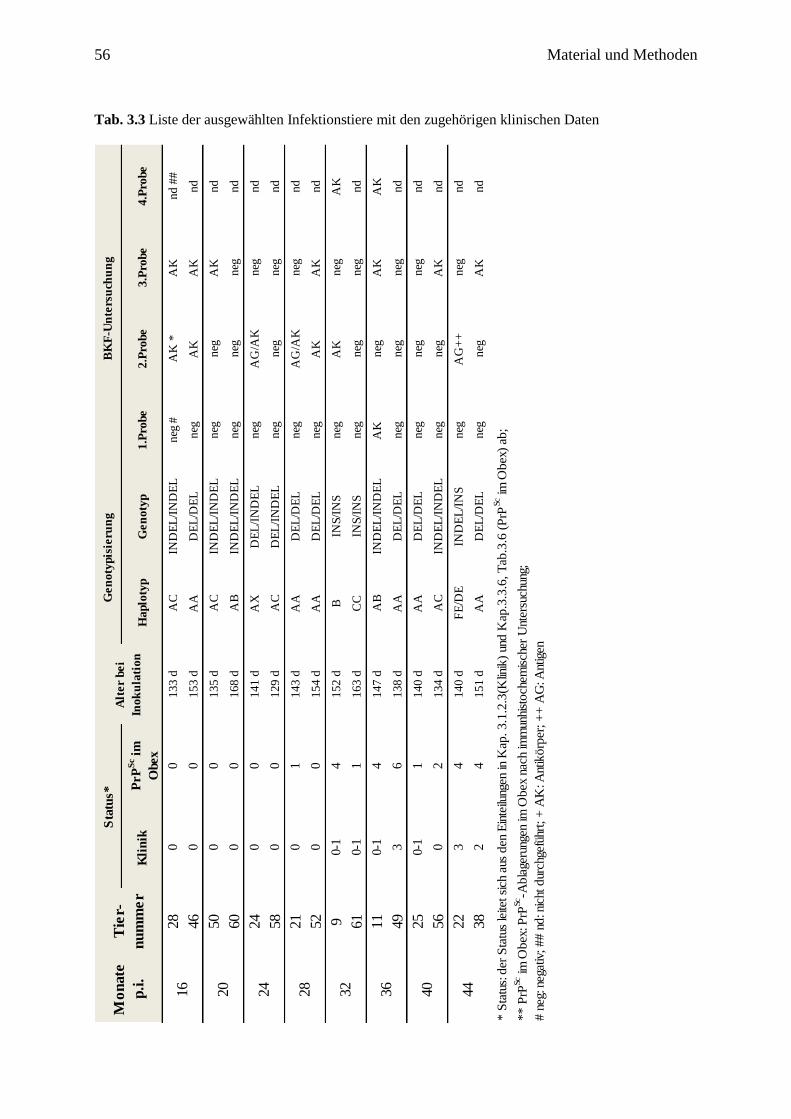
56 Material und Methoden
Tab. 3.3 Liste der ausgewählten Infektionstiere mit den zugehörigen klinischen Daten
K
lin
ikP
rP
Sc i
m
Obex
Haplo
typ
Gen
oty
p1.P
robe
2.P
robe
3.P
robe
4.P
robe
28
00
133 d
AC
IND
EL
/IN
DE
Lneg
#A
K *
AK
nd #
#
46
00
153 d
AA
DE
L/D
EL
neg
AK
AK
nd
50
00
135 d
AC
IND
EL
/IN
DE
Lneg
neg
AK
nd
60
00
168 d
AB
IND
EL
/IN
DE
Lneg
neg
neg
nd
24
00
141 d
AX
DE
L/I
ND
EL
neg
AG
/AK
neg
nd
58
00
129 d
AC
DE
L/I
ND
EL
neg
neg
neg
nd
21
01
143 d
AA
DE
L/D
EL
neg
AG
/AK
neg
nd
52
00
154 d
AA
DE
L/D
EL
neg
AK
AK
nd
90-1
4152 d
BIN
S/I
NS
neg
AK
neg
AK
61
0-1
1163 d
CC
INS/I
NS
neg
neg
neg
nd
11
0-1
4147 d
AB
IND
EL
/IN
DE
LA
Kneg
AK
AK
49
36
138 d
AA
DE
L/D
EL
neg
neg
neg
nd
25
0-1
1140 d
AA
DE
L/D
EL
neg
neg
neg
nd
56
02
134 d
AC
IND
EL
/IN
DE
Lneg
neg
AK
nd
22
34
140 d
FE
/DE
IND
EL
/IN
Sneg
AG
++
neg
nd
38
24
151 d
AA
DE
L/D
EL
neg
neg
AK
nd
BK
F-U
nte
rsu
chu
ng
Mo
nate
p.i
.
Tie
r-
nu
mm
er
Sta
tus*
Alt
er
bei
Inok
ula
tion
Gen
oty
pis
ieru
ng
40
44
16
20
24
28
32
36
* S
tatu
s: d
er S
tatu
s le
itet si
ch a
us d
en E
inte
ilung
en in
Kap
. 3.1
.2.3
(Klin
ik)
und K
ap.3
.3.6
, T
ab.3
.6 (
PrP
Sc im
Obex
) ab
;
** P
rPSc im
Obex
: PrP
Sc-A
bla
geru
ngen
im O
bex
nac
h im
mun
hist
och
emis
cher
Unt
ersu
chun
g;
# n
eg: n
egat
iv; ## n
d: n
icht
dur
chge
führ
t; +
AK
: Ant
ikörp
er; +
+ A
G: A
ntig
en

Material und Methoden 57
3.3. Histologische Methoden
3.3.1 Herstellung der histologischen Präparate
Für die histologischen Untersuchungen wurden in 4%igen, neutral gepufferten Formalin
fixierte Gewebeproben verwendet.
Das zugeschnittene Gewebe wurde zur Verringerung der Oberflächen-Infektiosität für eine
Stunde in Ameisensäure inkubiert und anschließend für 40 Minuten unter fließendem
Leitungswasser gespült. Im Folgenden automatisierten Schritt (ASP 300, Fa. Leica) erfolgte
die Entwässerung des Gewebes in einer aufsteigenden Alkoholreihe, dann wurde der Alkohol
durch das Intermedium Xylol entfernt und mit Paraffin durchtränkt. An der Ausgießstation
(Modell EG1140, Fa. Leica) wurden die Paraffinblöcke gegossen.
Die histologischen Schnitte wurden am Mikrotom (Rotationsmikrotom RM2145, Fa. Leica)
angefertigt.
Abb.3.1 Schematische Darstellung der in der Histologie angewendeten Schnitttechnik
Um eine möglichst hohe Durchdringtiefe zu erreichen, wurden pro Block auf fünf Ebenen
jeweils drei µm dicke Schnitte angefertigt. Zu einer Ebene zählen drei aufeinander folgende
Schnitte, zwei für die immunhistologische Untersuchung und ein Schnitt für den Einsatz bei

58 Material und Methoden
der HE-Färbung. Danach wurden 6 bis 8 Schnitte verworfen bzw. einzelne als Reserve
verwendet und entsprechend gekennzeichnet, d.h. es wurde ein Abstand von circa 18-24µm
zwischen den einzelnen Ebenen erreicht. Demzufolge ermöglichte diese Schnitttechnik, wie
in Abb. 3.1 (S.56) schematisch dargestellt, eine Gesamtdurchdringtiefe von 117 bis 141µm.
Auf Objektträger (IHC: Superfrost plus; HE: Superfrost; Fa. Menzel-Gläser) gezogen,
erfolgte eine Aufbewahrung der Schnitte für mindestens 48h im Wärmeschrank (Incucell, Fa.
MMM Medcenter) bei 60°C für die optimalen Haftung der Präparate.
3.3.2 Hämatoxylin-Eosin-Färbung
Nach der Entparaffinierung der Schnitte in einer absteigenden Alkoholreihe wurden die
Schnitte für 3min in A. bidest. inkubiert. Anschließend erfolgte für 10min die Färbung in
filtrierter Hämatoxylin-Lösung. Die Schnitte wurden dann 15min durch die Inkubation mit
Leitungswasser gebläut.
Nach zweiminütiger Inkubation in 50%igem Ethanol erfolgte die Gegenfärbung der Schnitte
in Eosin für fünf Minuten. In der aufsteigenden Alkoholreihe wurden die Schnitte entwässert
und mit Entellan eingedeckt.
3.3.3 Bewertung der Hämatoxylin-Eosin-Färbung
In der Hämatoxylin-Eosin (HE)-Färbung werden durch das Hämatoxylin alle basophilen
Strukturen blau angefärbt, insbesondere die Desoxyribonukleinsäure (DNA) der Zellkerne
und das raue endoplasmatische Retikulum (rER). Dagegen färbt Eosin als saurer Farbstoff
alle acidophilen (eosinophilen) Strukturen rot, was vor allem die Zellplasmaproteine umfasst.
So wurden die Präparate an einem Lichtmikroskop unter Verwendung von 2,5x, 10x, 20x und
40x Objektiven mit dem Ziel untersucht, morphologische Alterationen zu dokumentieren.
Durch die immunhistochemische und pathologisch-histologische Bewertung von Schnitten
der gleichen Ebene sollten mögliche Zusammenhänge zwischen morphologischen
Alterationen und PrPSc
-Ablagerungen erschlossen werden.

Material und Methoden 59
3.3.4 Immunhistologische Untersuchungen
Vorversuche
Zur Optimierung des Färbeergebnisses wurden Untersuchungen mit verschiedenen
Konjugaten durchgeführt. Dabei wurde die Avidin-Biotin-Komplex (ABC)-Methode
(Vectastain ABC Kit Elite Standard, Fa. Vector Laboratories) dem gebrauchsfertigen
EnVision+TM
(Fa. Dako) gegenübergestellt. Es zeigte sich, dass das EnVision+TM- System
der ABC-Methode hinsichtlich Sensitivität und unspezifischen Hintergrundfärbungen deutlich
überlegen war und daher als Standardkonjugat eingesetzt wurde.
Hauptversuche
In Abhängigkeit vom verwendeten Antikörper wurden verschiedene Vorbehandlungen zur
Demaskierung der Epitope verwendet. Als erster Vorbehandlungsschritt nach der
Entparaffinierung erfolgte eine Inkubation in Ameisensäure für 15min. War eine Färbung mit
dem monoklonalen Antikörper (mab) 6C2 geplant, verlängerte sich diese Inkubationszeit auf
30min. Nach anschließenden Spülen der Objektträger unter fließendem Leitungswasser für
fünf min wurden die Schnitte, zur Blockierung der endogenen Peroxidase, für 30min in
Methanol mit 3% Wasserstoffperoxid inkubiert. Die Verwendung des mab 12F10 erforderte
einen Verdau mit Proteinase K (Fa. Roche). Dazu wurden die Schnitte in PK-Puffer fünf min
äquilibriert und anschließend bei einer Enzymendkonzentration von vier µg/ml für 15min bei
37°C inkubiert. Die Vorbehandlung für den Antikörper 6C2 beinhaltete eine
Hitzebehandlung. Demnach wurden die Objektträger in 0,1M Citratpuffer für 20min bei
121°C in einem Dampfsterilisator autoklaviert (Varioklav, Fa. Thermo Scientific).
Nach der Zusammensetzung des Coverplates® mit den Objektträgern und dem Einbringen in
die Shandon Coverplate® Kassette erfolgte mit einer Spülung mit TBS die Überprüfung des
korrekten Zusammenbaus des Systems.
Für die Standardfärbung wurde der mab 12F10 in der Verdünnung 1:150 in 10% Ziegenserum
in TBS mit 0,03% Natriumazid eingesetzt. Für den mab 6C2 wurde für die Verdünnung 1:50
das gleiche Verdünnungsmedium verwendet. Beide Antikörper wurden über einen Zeitraum
von zwei Stunden bei Raumtemperatur inkubiert. Die Tabelle 3.6 (S.63) gibt eine Übersicht
über die eingesetzten Antikörper und die entsprechenden Konjugate.
Nach der Inkubation wurden die Schnitte dreimalig mit TBS gespült. Als Konjugat wurde
EnVision+TM
(Fa. Dako) mit einer Inkubationszeit von 30min bei Raumtemperatur
verwendet. Nach dreimaligem Spülen mit TBS wurden die Objektträger aus dem
Coverplate®
-Systems entfernt und wieder in eine Küvette verbracht.

60 Material und Methoden
Daraufhin erfolgte für zehn Minuten die Inkubation der Schnitte mit dem Enzymsubstrat
DAB. Nach dreimaligem Waschen in TBS und kurzem Spülen in Aqua bidest. erfolgte die
Gegenfärbung in Hämatoxylin für zehn min, anschließend wurden die Präparate 15 min unter
fließendem Leitungswasser gebläut.
Die Dehydrierung der Schnitte wurde nach der Färbeprozedur in einer aufsteigenden
Alkoholreihe durchgeführt und das Eindecken mit Entellan vorgenommen.
3.3.5 Immunhistologische Kontrollen
Der für die Immunhistochemie eingesetzte zweite Schnitt jeder Ebene wurde mit der
Antikörper-Verdünnungslösung (10% Ziegenserum in TBS mit 0,03% Natriumazid) als
Kontrolle inkubiert.
Positive Kontrollschnitte waren Gehirnschnitte aus der Routinediagnostik, die in
vorangegangenen immunhistochemische Untersuchungen eindeutig das Vorkommen von
PrPSc
zeigten. Zur Kontrolle auf unspezifischen Anfärbungen wurde ebenfalls ein
nachweislich PrPSc
-freies Gehirnpräparat mitgeführt. Für beide Schnitte erfolgte während der
immunhistologischen Untersuchung eine Behandlung sowohl mit dem entsprechenden
Antikörper als auch mit der verwendeten Antikörper- Verdünnungslösung.
3.3.6 Bewertung der immunhistologische Untersuchungen
Tab.3.4 Immunhistologischer Auswertungsindex für Proben des ZNS und VNS
1 vereinzelt positive Zellen (5-10%)
2 einzelne positive Zellen (max. 20%)
3 mehrere positive Zellen (max. 40%)
4 viele positive Zellen (max. 60%)
5 sehr viele positive Zellen (max. 80%)
6 (fast) alle Zellen positiv (80 – 100%)
max.: maximal
Alle fünf Ebenen jedes Blocks wurden mit dem Lichtmikroskop unter Verwendung von 2,5x,
10x, 20x und 40x Objektiven auf das PrPSc
- Akkumulation hin untersucht. Als positives
Bewertungskriterien galten in der Schnittebene liegende, mittel-bis dunkelbraune,
feingranuläre Reaktionsprodukte, die in den Kontrollschnitten nicht nachweisbar waren. Dazu
war ein klarer Strukturbezug notwendig.

Material und Methoden 61
Für die Proben des vegetativen und zentralen Nervensystems wurde die Zahl der positiven
Zellen/Strukturen gezählt bzw. geschätzt und in Prozent zur Gesamtzellzahl/Gesamtstruktur
angegeben (s. Tab. 3.4, S.60).
Für die Schnitte des Ileums wurden Befunde einerseits für die Lymphfollikel der Peyer`schen
Platten und andererseits für Anteile des enterischen Nervensystems erhoben. Tabelle 3.5 zeigt
den Auswertungsindex für die Peyer`schen Platten abhängig vom Reaktionsmuster und
Lokalisation der beobachteten Anfärbung. Das Verhältnis zwischen der Gesamtanzahl und
den positiven Follikeln wurde in der Auswertung ebenfalls berücksichtigt. Für das ENS kam
die der Auswertungsindex des ZNS und VNS (s. Tab. 3.4, S.60) zur Anwendung.
Tab. 3.5 Immunhistologischer Auswertungsindex für die Untersuchung der Peyer`schen Platten des
Ileums
0 negativ
X unvollständig angeschnittener Follikel
1 1-2 Zellen in der Light Zone positiv
2 mehrere Zellen in der Light Zone positiv
3 Makrophagen in Light/Dark Zone positiv
4 Makrophagen in Light/Dark Zone positiv, FDC schwach positiv
5 Makrophagen in Light/Dark Zone positiv, FDC deutlich positiv
FDC: Follikulär-dendritische Zellen
Als eindeutig negativ wurden Anfärbungen labortechnisch bedingter Schnittartefakte
gewertet. Dies trifft auch auf ein in den Schnitten des Ileums auftauchendes granulär
angeordnetes, braunes, oftmals zellassoziiertes Pigment (Hämosiderin) zu, das im
entsprechenden Kontrollschnitt ebenfalls nachweisbar und somit von einem echten, reaktiven
Präzipitat eindeutig abzugrenzen war.
3.4 Proteinbiochemische Methoden
3.4.1 Homogenatherstellung
Zunächst wurde aus den bei -20°C gelagerten Mäusegehirnen 500µl eines 10%igen
Hirnhomogenates hergestellt. Das abgewogene Material wurde zusammen mit dem
neunfachen Volumen (w/v) an Sucrose-DOC-NP40-Lysispuffer versetzt und in Einmal-
Schraubdeckelgefäßen mit Keramik-Kugeln (Precellys Keramik-Set, Fa. Peqlab)
homogenisiert. Dazu waren zwei Durchgängen im Ribolyser (Geschwindigkeit 6500rpm; Zeit

62 Material und Methoden
45sek) ausreichend. Anschließend wurden die Proben bei 6000rpm über fünf min
zentrifugiert, anschließend 200µl des Homogenates für die anschließende Fällungsreaktion in
ein Eppendorf-Reaktionsgefäß aliquotiert und der Rest bei -20°C gelagert.
3.4.2 Fällung mit Phosphorwolframsäure (PTA-Fällung)
Durch die PTA-Fällung kommt es zu einer spezifischen Anreicherung von PrPSc
(WADSWORTH et al. 2001). Phosphorwolframsäure bildet bei neutralem pH-Wert unter der
Anwesenheit von Magnesiumionen spezifische Aggregate mit PrPSc
-Molekülen, aber nicht
mit PrPC-Molekülen, aus. Durch die Zentrifugation dieser Aggregate gelingt eine
Anreicherung von PrPSc
in den entstandenen Pellets.
Für 200µl-Aliqots des Homogenates wurde ein Verdau mit 10µl Proteinase K (Fa. Roche) für
eine Stunde bei 55°C auf einem Thermoschüttler (Fa. Eppendorf) durchgeführt. Der Verdau
wurde mit 4µl PEFA-Block abgestoppt und es folgte ein Denaturierungsschritt über fünf
Minuten bei 95°C. Anschließend wurde 200µl PBS mit 4% Sarkosyl und 32µl 4%iger
Phosphorwolframsäure zugegeben und eine Stunde bei 37°C inkubiert. Durch Zentrifugation
für 30min bei 13.000rpm entstand ein Pellet, das nach Abnahme des Überstandes in 20µl
zweimal Lade-Puffer aufgenommen wurde.
3.4.3 Proteinauftrennung mittels SDS-Polyacrylamid-Gel-Elektrophorese
Die Auftrennung der Proteine erfolgte mittels SDS-PAGE in einer Minigel-Elektrophorese-
Apparatur (BioRad, München) in einem 16%igen, 0,75mm dicken Acrylamidgel. Nach der
Denaturierung der Proben für fünf Minuten bei 95°C wurden 20μl der Ansätze in die
einzelnen Taschen des Sammelgels pipettiert und in einem 4%igen Sammel- oder Taschengel
bei 100 Volt fokussiert. Nach Eintritt in das Trenngel wurde die Spannung auf 200 Volt
erhöht, um die Proteine ihren unterschiedlichen Molekulargewichten entsprechend vollständig
aufzutrennen. Neben einem Proteinmarker (5µl) zur Ermittlung der Molekulargewichte wurde
ebenfalls eine Positivkontrolle mit bovinem PrPSc
(10μl) mitgeführt.

Material und Methoden 63
3.4.4 Western Blot
In einer Semi-Dry-Blotting-Apparatur (BioRad, München) wurden die Proteine auf eine
Immobilon-Transfermembran (Polyvinylpyrrolidon) übertragen, die zuvor fünf min in
Methanol aktiviert und anschließend in Blotting-Puffer äquilibriert wurde. Zwischen
mehreren Lagen ebenfalls mit Blotting-Puffer befeuchteten Whatman®
Chromatographie-
Papier erfolgte der 45 minütige Blot-Vorgang bei einer Spannung von 15 Volt und einer
Stromstärke von 0,3 Ampere pro Gel.
Anschließend wurden die freien Bindungsstellen in 5%iger Magermilch (5%
Magermilchpulver in PBST) eine Stunde abgesättigt. Die Inkubation mit dem primären
Antikörper in der entsprechenden Verdünnung (s.Tab. 3.6) erfolgte eine Stunde bei
Raumtemperatur oder über Nacht bei 4°C. Anschließend wurde die Membran dreimal zehn
min mit PBS-Tween gewaschen und eine Stunde mit dem zweiten Alkalische-Phosphatase-
konjugierten Antikörper (s. Tab. 3.6) versehen. Nach drei weiteren Waschschritten von
jeweils 10min wurde zweimal zwei Minuten mit Assay-Puffer inkubiert und anschließend
mittels CDP-Star entwickelt. Die Intensität der Reaktion wurde im Versa Doc®
-Imager über
einen Zeitraum von 120sek gemessen.
Tab. 3.6 Übersicht über die in der Immunhistologie und in der biochemischen Untersuchung
eingesetzten Antikörper und deren Konjugate
Methode Anti-
körper Epitop Herkunft Verdünnung Konjugat
Verdün
nung Herkunft
IHC
12F10 hu
142-160
Cayman
Chemical,
USA
1 : 150
in TBS-ZS**
EnVisionTM
- Dako,
Hamburg
6C2 bov
117-130
Jan
Langeveld,
CIDC-
Lelystad, NL
1 : 50
in TBS-ZS
Western
Blot L42
ov
145-163 r-biopharm
1:2000
in 5% MM-
PBST
Ziege anti-
Maus IgG
AP*-
gekoppelt
1:2000
in
TBST
Dianova,
Hamburg
hu: human; bov: bovin; ov: ovin; *AP: alkalische Phosphatase; **TBS-ZS: TBS in
Ziegenserum

64 Material und Methoden
3.4.5 Bewertung der Ergebnisse und Kontrollen
Als positives Signal wurde das eindeutige Erkennen der drei Glykosylierungsformen des
PrPSc
als Banden mit der entsprechenden molekularen Masse bewertet. In einigen Fällen war
ein Wiederholungsversuch mit einer erhöhten (bis 500µl) oder verringerten (100µl)
Probenmenge erforderlich.
Als Referenz wurde auf jedem Blot ein definitiv BSE-positives Hirnhomogenat desselben
Mausstammes sowie unbehandeltes PrPC-Hirnhomogenat mitgeführt.

65 Ergebnisse
4. ERGEBNISSE
Für jede Gewebeprobe wurden immunhistochemische und histopathologische
Untersuchungen durchgeführt und für ausgewählte Proben (16 bis 36 mpi) zusätzlich der
Bioassay. Letztere Methode ermöglicht Aussagen zur Infektiosität der Gewebeprobe,
während die Immunhistochemie den direkten Nachweis des pathologischen Prion-Proteins
erbringt.
4.1 Allgemeine Ergebnisse
4.1.1 Klinische Symptomatik
Aus den Resultaten der regelmäßigen klinischen Untersuchungsgänge ergaben sich die
Bewertungen des Krankheitsstatus der einzelnen Rinder. Diese wurden in regelmäßigen
Abständen auf eine erhöhte Sensibilität, abnorme Bewegungsabläufe und Kondition getestet
und in Kategorien von 0 (keine Symptome) bis 3 (definitive BSE-Symptomatik) eingeordnet
(s. Kap.3.1.2.3, S.48).
Abb. 4.1 Numerische Verteilung der in dieser Studie analysierten Rinder nach klinischem
Status
Über die Hälfte der Tiere (n=9) zeigten bis zu ihrer Euthanasie keine klinischen Symptome (s.
Abb. 4.1). Bei erkrankten Tieren zeigte sich in den wiederholten Untersuchungen oft eine
vielgestaltige klinische Symptomatik, die nur schwer als eindeutig für BSE anzusprechen war
(n=5). Diese Tiere reagierten vermehrt sensibel auf Geräusche, Licht und Berührungen am
Kopf und den Gliedmaßen. Weiterhin fielen Veränderungen im Verhalten sowie hinsichtlich

66 Ergebnisse
ihrer Kondition und Bewegung auf. Lediglich bei zwei Tieren war das klinische
Erscheinungsbild so eindeutig, dass sie als BSE (Status 3) eingestuft werden konnten.
4.1.2 Ergebnisse der histopathologischen Untersuchung
Die Auswertung der histopathologischen Untersuchung der Obexregion ergab bei sieben
Tieren spongiforme Veränderungen, von denen zwei als verdächtig, drei als gering- und zwei
als hochgradig bewertet wurden. Eine Verdachtsdiagnose wurde aufgrund marginaler,
spongiformer Auflockerungen des Neuropils auch bei unilateralem Auftreten gestellt.
Hingegen waren bereits geringgradige Degenerationen immer bilateral lokalisiert.
Unabhängig vom Ausprägungsgrad der Erkrankung waren schon in den initialen Stadien der
DMNV, Tractus solitarii, Tractus spinalis N. Trigemini und die Nuclei olivares besonders
häufig betroffen. Beim Auftreten hochgradiger spongiformer Auflockerungen zeigten sich
entsprechende Veränderungen in allen Kerngebieten des Obex (s. Abb. 4.2, S.67).
Neben konstanten Veränderungen im Neuropil aller betroffenen Tiere traten bei drei Rindern
(IT 11, IT 49, beide 36 mpi; IT 22, 44 mpi) zusätzlich vakuoläre Degenerationen in den
Perikaryia des DMNV, der Formatio reticularis und der Nuclei olivares auf.
Beide Tiere mit hochgradigen histopathologisch erkennbaren Alterationen zeigten eine
definitive BSE-Symptomatik. Mit Ausnahme der IT 38 (44 mpi), die den Status 2 zugeordnet
wurde, gingen geringfügige spongiforme Veränderungen mit unspezifischen (IT 09 und IT 61
beide 32 mpi sowie IT 11, 36 mpi) bzw. ohne (IT 56, 40 mpi) klinisch manifestierte
Symptome einher.
Als weiterer pathologischer Befund wurde bei 12 der 16 Tiere eine Gliose diagnostiziert.
Diese trat nicht nur in Form einer diffusen rein numerischen Erhöhung der Zahl der Gliazellen
auf, sondern wies zum Teil auch sogenannte Glia-Knoten auf. Lokalisiert waren diese
Veränderungen hauptsächlich im DMNV, dem Tractus trigeminus und seltener im Nucleus
cuneatus. Die Gliose trat bei Tieren in jedem klinischem Stadium auf, vier Tiere waren dabei
unauffällig, sechs unspezifisch und zwei definitiv erkrankt.

Ergebnisse 67
Abb. 4.2 Hauptlokalisation der spongiformen Enzephalopathie in der Medulla oblongata of
Höhe des Obex; Abb. modifiziert, aus CASALONE et al. 2005
Des Weiteren fielen bei acht Tieren im Zeitraum von 24 bis 44 mpi inflammatorische
Prozesse in verschiedenen Anteilen des PNS auf. Es handelte sich dabei um fokale
geringgradige bis multifokal auftretende mittelgradige lymphohistiozytäre Ganglio-
Neuritiden. Obwohl in allen Anteilen des PNS auftretend, ließ sich eine Tendenz zugunsten
der sympathischen Anteile erkennen, da das Ggl. Stellatum (n=7), das Ggl. Coeliacum (n=6)
und die Nn. Splanchnici (n=5) besonders häufig betroffen waren. Mit einer Ausnahme (IT 58,
24 mpi) zeigten alle betroffenen Tiere sowohl PrPSc
Ablagerungen im PNS und ZNS als auch
eine klinische Symptomatik.
Allen Tieren gemein war das Auftreten einer mittelgradigen eosinophilen Enteritis im distalen
Ileum.
4.1.3 Ergebnisse der immunhistochemischen Untersuchungen
4.1.3.1 Quantifizierung der Ergebnisse
Mittels der Immunhistochemie erfolgte der direkte Nachweis von PrPSc
und ermöglichte die
Beurteilung betroffener zellulärer Strukturen. Für die Proben des Nervensystems wurde die
Zahl der positiven Zellen gezählt bzw. geschätzt. Das Verhältnis zur
Gesamtzellzahl/Gesamtstruktur wird im Auswertungsindex der Tabelle 4.1 (S.71)
wiedergegeben. Die Quantifizierung der positiven Follikel des Ileums erfolgte im Verhältnis

68 Ergebnisse
zur Follikel-Gesamtzahl, während für das stets nur geringgradig betroffene ENS die
Lokalisation der Ablagerung im Plexus submucosus bzw. Plexus muscularis bestimmt wurde.
4.1.3.2 Lokalisation und Reaktionsmuster von PrPSc
Zentrales Nervensystem:
Initial war PrPSc
zunächst auf die Formatio reticularis bzw. den Nucleus tractus solitarii und
den Nucleus motorius nervi trigemini des Obex beschränkt. Im terminalen Stadium hingegen
traten die hochgradigen Akkumulationen von PrPSc
in allen Bereichen der Obexregion auf.
Die initialen Ablagerungen von PrPSc
in der Obexregion waren vorwiegend
zelloberflächenassoziiert diffus feingranulär im Neuropil sowie perineuronal nachweisbar.
Weiterhin waren intragliale Akkumulationen wie auch geringe intraneuronale Ablagerungen
zu beobachten (s. Abb. 4.3A). Mit fortschreitender Akkumulation nahmen alle beschriebenen
Reaktionsmuster an Stärke zu, dabei zeigten insbesondere die extrazellulären Aggregate ein
grobgranuläres bis koaleszierendes Erscheinungsbild und in einigen Arealen dominierte ein
lineares Reaktionsmuster.
Abb. 4.3 PrPSc
-Akkumulation im ZNS
(A) geringe Ablagerungen in der Formatio reticularis der Medulla oblongata (Obex, IT 61,
32 mpi), das einzelnes Neuron zeigt ein intra- und perineuronales Reaktionsmuster. (B)
moderate PrPSc
-Akkumulation sind in der Substantia intermediolateralis des lumbalen
Rückenmarkssegmentes intraneuronal (Asterisk), intraglial (Pfeil) und linear (Pfeilkopf)
nachweisbar. (IT38, 44 mpi).
Immunhistochemie, PrP mAb L42 (A) and PrP mAb 12F10 (B), Nomarski
Interferenzkontrast, Bars 50 µm
Im Rückenmark war zunächst die Substantia intermedia lateralis betroffen (s. Abb. 4.3B),
wobei sich die Ablagerungen im Verlaufe der Erkrankung zunehmend auf das Cornu dorsale,

Ergebnisse 69
das Cornu ventrale sowie auf die Substantia intermedia centralis ausbreiteten. In keinem Fall
konnte pathologisches Prion-Protein in der weißen Substanz des Rückenmarkes detektiert
werden. Die zellulären Reaktionsmuster entsprachen den in der Medulla oblongata
beschriebenen Verhältnissen.
Peripheres Nervensystem:
Abb. 4.4 PrPSc
-Akkumulationen im peripheren Nervensystem
Für das parasympathische Gewebe konnten PrPSc
-Ablagerungen (A) im N. vagus (IT 38, 44
mpi) perineuronal und im geringen Grad auch intraneuronal sowie im (B) Ggl. nodosum (IT
22, 44 mpi) ebenfalls perineuronal nachgewiesen werden. PrPSc
-Akkumulationen im
sympathischen System traten (C) hautsächlich intraneuronal im Ggl. cervicale craniale (IT
11, 36 mpi) und (D) perineuronal im Ggl. stellatum (R09/06) auf.
Die Pfeile zeigen betroffene Neurone.
Immunhistochemie, PrP mAb 12F10; Nomarski Interferenzkontrast, Bars 20µm
Ausschließlich nach ersten detektierbaren Akkumulationen im ZNS traten PrPSc
-positive
Nervenzellen vereinzelt in den Ganglien des PNS auf (s. Abb. 4.4 und Abb. 4.5, S.70). Dabei
war das Ggl. coeliacum bereits nach geringgradiger Beteiligung des ZNS konstant involviert
und wies bei insgesamt sieben Tieren PrPSc
-Akkumulationen auf. Nach hochgradigen

70 Ergebnisse
Ablagerungen im Obex waren im N. vagus von vier Rindern vereinzelte intra- und
perineuronale Reaktionsmuster detektierbar.
Die Reaktionsmuster der in allen untersuchten Anteilen des PNS stets nur vereinzelt
nachweisbaren positiven Neurone zeigten eine deutliche Übereinstimmung. Die dominanten
Ablagerungen befanden sich extrazellulär mit Assoziation zur Zellmembran (perineuronal).
Diffuse intraneuronale Akkumulationen von PrPSc
traten in geringerem Maße auf. Seltener
gelang der intrazelluläre Nachweis von granulärem PrPSc
in den Satellitenzellen, die
eigenständig oder assoziiert mit ebenfalls betroffenen Neuronen sein konnten.
Abb. 4.5 PrPSc
-Akkumulationen im peripheren Nervensystem
Die gemischtfaserigen (A) Ggl. coeliacum (IT 09, 32 mpi) und (B) Ggl. mesenteriale caudale
(IT 09, 32 mpi) weisen deutliche perineuronale als auch geringe intraneuronale
Reaktionsmuster neben einer geringgradigen Beteiligung der Satellitenzellen auf. (C) Eine
dominante Anfärbung von Satellitenzellen konnte im Ggl. trigeminale (IT 49, 36 mpi)
beobachtet werden.
Pfeile zeigen auf betroffene Neurone, Asterisken markieren die Satellitenzellen.
Immunhistochemie, PrP mAb 12F10 (A;B) und PrP mAb 6C2 (C), Nomarski
Interferenzkontrast, Bars 20µm (A;B) und 50 µm (C)

Ergebnisse 71
Eine Sonderstellung nahm das Ggl. trigeminale ein. Die Ergebnisse schienen eng mit denen
der Medulla oblongata assoziiert zu sein, da PrPSc
-Ablagerungen bereits nach geringgradiger
Akkumulation von PrPSc
im Obex nachweisbar waren. Dabei zeigten sich deutliche
Unterschiede zu den anderen untersuchten Anteilen des PNS hinsichtlich der Intensität und
des Reaktionsmusters. Zum einen zeigten bis zu 20% der Zellen im weiteren Verlauf der
Erkrankung eine positive Reaktion. Zum Anderen war ein dominantes intrazellulär-granuläres
Reaktionsmuster der Satellitenzellen dominant.
Peyer`sche Platten und Enterisches Nervensystem :
In den Peyer´schen Platten des Ileums waren in 11 der 16 untersuchten Tiere PrPSc
-
Ablagerungen nachweisbar. Assoziiert mit dem zunehmenden Lebensalter wiesen die Tiere
mit längeren Inkubationszeiten jedoch quantitativ weniger Follikel auf. So war bis 32 mpi mit
einer Ausnahme (IT46, 16 mpi) PrPSc
bei allen Tieren in den Follikeln darstellbar. Dagegen
zeigten lediglich zwei Tiere (IT 49, 36mpi; IT 56, 40 mpi) zum späten
Untersuchungszeitpunkt noch PrPSc
-Ablagerungen in den Follikeln. Die Anzahl positiver
Follikel gegenüber der Gesamtanzahl lag bei den meisten Tieren unter einem Prozent. Bei
drei Tieren fiel hingegen eine deutlich erhöhte Anzahl positiver Follikel auf. So konnte beim
Tier IT 24 (24 mpi) in 5,5% der Follikel (15/272), IT 09 (32 mpi) in 16,7% (2/12) und IT 49
(36 mpi) in 5% der Follikel (3/60) pathologische Prion-Proteine nachgewiesen werden. Die
betroffenen Follikel zeigten vorwiegend multigranuläre bis globuläre, intrazytoplasmatische
Ablagerungen in den mononukleären Zellen des Centrum germinativum, die morphologisch
als Tingible body macrophages (TBM) anzusprechen sind. Des Weiteren waren bei den
Tieren ab 16 mpi in sechs der 11 Rinder mit PrPSc
-Nachweis in den Follikeln fein-granuläre,
netzförmige Akkumulationen zu beobachten, die verstärkt 24, 32 und 36 mpi auftraten und
morphologisch den FDC zugeordnet werden können (s. Abb. 4.6, S.72).
Die Detektion des pathologischen Prion-Proteins in den Plexus des ENS gelang in acht von 16
Rindern. In keinem Fall der untersuchten Peyer’schen Platten trat jedoch eine anatomische
Assoziation zwischen PrPSc
-Ablagerungen in den Follikeln und dem ENS auf. Allerdings ging
die Steigerung der Akkumulationen im ENS mit der bei den älteren Rindern auftretenden
quantitativen Abnahme und der damit verbundenen sinkenden Anzahl positiver Follikel
einher. In der präklinischen Phase war vor allem der Plexus myentericus und erst im späteren
Verlauf der Erkrankung auch der Plexus submucosus betroffen. In den stets vereinzelt
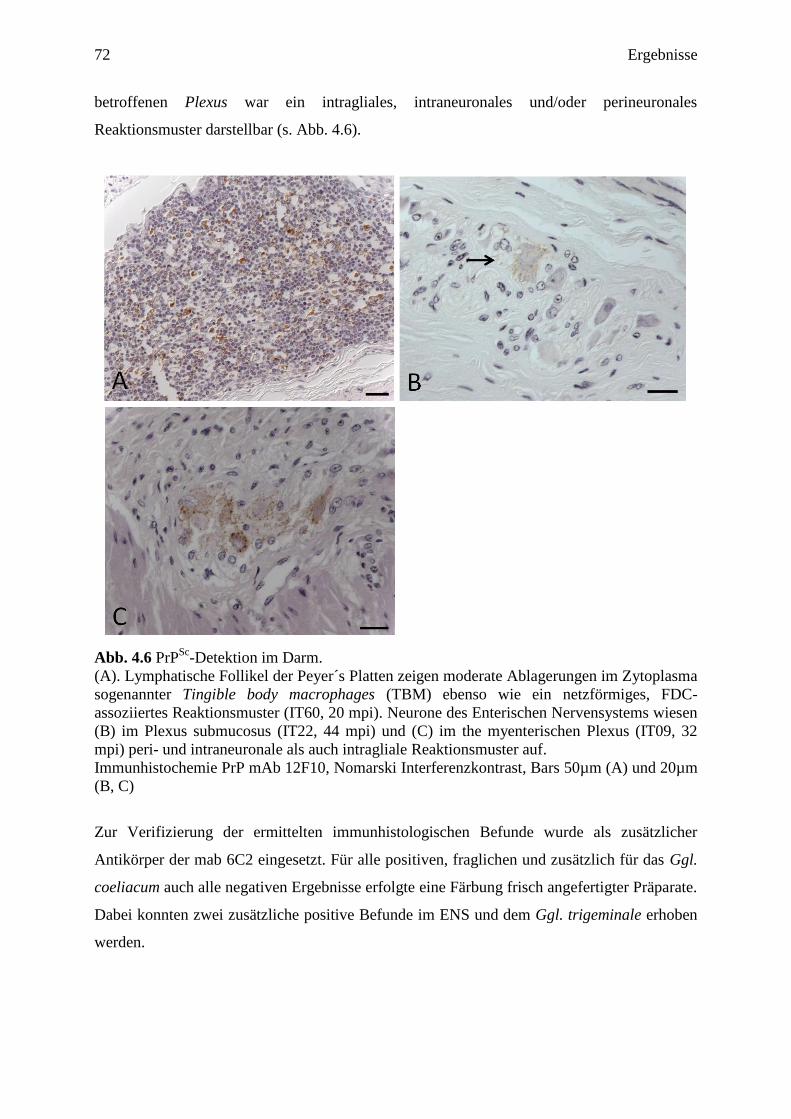
72 Ergebnisse
betroffenen Plexus war ein intragliales, intraneuronales und/oder perineuronales
Reaktionsmuster darstellbar (s. Abb. 4.6).
Abb. 4.6 PrPSc
-Detektion im Darm.
(A). Lymphatische Follikel der Peyer´s Platten zeigen moderate Ablagerungen im Zytoplasma
sogenannter Tingible body macrophages (TBM) ebenso wie ein netzförmiges, FDC-
assoziiertes Reaktionsmuster (IT60, 20 mpi). Neurone des Enterischen Nervensystems wiesen
(B) im Plexus submucosus (IT22, 44 mpi) und (C) im the myenterischen Plexus (IT09, 32
mpi) peri- und intraneuronale als auch intragliale Reaktionsmuster auf.
Immunhistochemie PrP mAb 12F10, Nomarski Interferenzkontrast, Bars 50µm (A) und 20µm
(B, C)
Zur Verifizierung der ermittelten immunhistologischen Befunde wurde als zusätzlicher
Antikörper der mab 6C2 eingesetzt. Für alle positiven, fraglichen und zusätzlich für das Ggl.
coeliacum auch alle negativen Ergebnisse erfolgte eine Färbung frisch angefertigter Präparate.
Dabei konnten zwei zusätzliche positive Befunde im ENS und dem Ggl. trigeminale erhoben
werden.

Ergebnisse 73
4.1.4 Ergebnisse des Maus-Bioassays (Tgbov XV-Mäuse)
Die für diesen Maus-Bioassay verwendeten transgenen Tgbov XV-Mäuse überexprimieren
bovines PrPC um den Faktor 10.000 und sind damit deutlich sensitiver gegenüber einer BSE-
Infektion als andere Mauslinien. Durch diese hohe Empfänglichkeit geben sie das infektiöse
Potential (Infektiosität) einer inokulierten Gewebeprobe wieder, dessen Grad in dieser Studie
über die Transmissionsraten und die Inkubationszeiten bestimmt wurde.
Von den 12 untersuchten Rindern im Zeitraum von 16 bis 36 mpi wiesen 11 Tiere
Infektiosität im PNS und/oder ZNS auf. Die Anzahl erkrankter Mäuse/Gruppe
(Transmissionsrate) zusammen mit dem Zeitraum bis zur Ausprägung klinischer Symptome
(Inkubationszeit) war mit der Infektiosität der jeweiligen Gewebeprobe assoziiert. Bei
niedrigen Transmissionsraten wiesen die Mäuse deutlich längere Inkubationszeiten auf (292
bis 561 Tage) sowie auch eine sehr hohe Variabilität (Standardabweichung von 63 bis 100
Tage) in der Dauer bis zu erkennbaren Krankheitssymptomen. In Gruppen mit hohen
Transmissionsraten ergaben sich bei den Mäusen Inkubationszeiten von im Schnitt 332 Tagen
(mittlere Standardabweichung 39 Tage) bis zur Ausbildung der spezifischen klinischen
Symptomatik und wiesen damit auf eine starke Infektiosität des Ausgangsgewebes hin. Um
die Auswertung der erhobenen Daten zu objektivieren und zu vereinfachen, wurde daher die
Infektiosität der einzelnen Proben lediglich auf Grundlage der Transmissionsraten klassifiziert
(s. Tab.4.1).
Tab. 4.1 Auswertungsindex für Proben des ZNS und PNS
Auswertungs-
index für IHC IHC Intensität Bioassay
1 vereinzelt positive Zellen
(5-10%) geringe Intensität
≤ 20% der Mäuse erkrankt
2 einzelne positive Zellen
(11-20%)
3 mehrere positive Zellen
(21-40%) mittlere Intensität
21%- 60% der Mäuse erkrankt
4 viele positive Zellen
(41-60%)
5 sehr viele positive Zellen
(61-80%) hohe Intensität
> 61% der Mäuse erkrankt
6 (fast) alle Zellen positiv
(81 – 100%)

74 Ergebnisse
4.2 Zusammenfassende Betrachtung
4.2.1 Assoziation von Teilergebnissen
Es wurde untersucht, ob im untersuchten Material eine Assoziation zwischen dem Grad der
PrPSc
-Ablagerungen im ZNS, dem Nachweis pathomorphologischer Veränderungen und dem
Auftreten neurologischer Symptomatik bestand (s. Abb. 4.7).
Abb. 4.7 Korrelation der erhobenen Befunde; bei den Tieren ohne Klinik und spongiformer
Enzephalopathie 40 mpi handelt es sich um zwei unterschiedlich Tiere: *: IT 56, **: IT 25
Tab. 4.2 Zusammenhänge zwischen dem Grad der PrPSc
-Akkumulationen und klinischen
Status
PrPSc
Klinischer Status Anzahl der
betroffenen Tiere
ohne 0 n=6
gering 0 n=1
1 n=1
mittel bis hoch
0 n=2
1 n=1
2 n=1
3 n=2
In Tab. 4.2 wird ersichtlich, dass der Grad der PrPSc
- Ablagerung nicht mit der Ausprägung
der BSE-typischen Symptome assoziiert ist. Bei mittelgradigen PrPSc
-Ablagerungen im
Stammhirn war ein Rind (IT 56, 40 mpi) noch klinisch unauffällig, Weiterhin führten mittel-

Ergebnisse 75
bis hochgradige Akkumulationen bei zwei Rindern ebenfalls nicht immer zu einer sicheren
klinischen Diagnose (IT 09,32 mpi; IT 11, 36 mpi). Auf der anderen Seite traten auch
unspezifische Symptome bereits bei geringen Ablagerungen des pathologischen Prion-
Proteins auf (IT 61, 32 mpi). Klinische Symptome des Status 2 bzw 3 sind allerdings immer
ein Hinweis auf hohe Akkumulationen im Gehirn.
Tab. 4.3 Zusammenhänge zwischen dem Grad der PrPSc
-Akkumulationen und dem Grad der
spongiformen Enzephalopathie
PrPSc
spongiforme Enzephalopathie Anzahl der
betroffenen Tiere
ohne keine n=7
gering keine n=2
Verdacht n=1
mittel bis hoch
Verdacht n=1
geringgradig n=3
hochgradig n=2
Dagegen schien der Grad der Ablagerung des pathologischen Prion-Proteins im ZNS mit der
Ausprägung der spongiformen Enzephalopathie assoziiert zu sein (s. Tab. 4.3). Auch wenn
bei einem Tier (IT 56, 40 mpi) mittelgradige Ablagerungen nur zu einem Verdacht einer
spongiformen Enzephalopathie führen, so gehen hohe PrPSc
-Level stets mit entsprechenden
pathomorphologischen Veränderungen einher.
Tab. 4.4 Zusammenhänge zwischen dem Grad der spongiformen Enzephalopathie und dem
klinischen Status der Tiere
spongiforme Enzephalopathie Klinischer Status Anzahl der
betroffenen Tiere
keine 0 n=8
1 n=1
Verdacht 0 n=1
1 n=1
geringgradig 1 n=2
2 n=1
hochgradig 3 n=2
Es deutete sich zudem eine Assoziation zwischen dem Auftreten klinischer Symptomatik und
dem Grad der spongiformen Enzephalopathie an, da eine klinisch manifestierte BSE-
Erkrankung stets mit hochgradigen pathomorphologischen Veränderungen einherging (s. Tab.
4.4)

76 Ergebnisse
4.2.2 Strukturierung der Ergebnisse
Tab. 4.5 Zusammenfassende Darstellung der Ergebnisse aus der Untersuchung der Medulla
oblongata
Medulla oblongata
Monate p.i. IT-Nummer Klinischer
Status Obex
kaudale
Medulla Grad der
Beteiligung
IHC BA
16
28 0 - 0/12
- 473 (± 104)
46 0 - 0/13
- 448 (± 97)
20
50 0 - 0/10
- 476 (± 108)
60 0 - 0/13
- 464 (± 116)
24
24 0 - 1/7
gering 433
58 0 - 0/9
- 308 (± 36)
28
21 0 + 8/12
hoch (1) 337 (± 37)
52 0 - 0/8
- 420 (± 119)
32
09 0-1 +
n.d. mittel (4)
61 0-1 + 0/9
gering (1) 387 (± 129)
36
11 0-1 +
n.d. mittel (4)
49 3 +
n.d. hoch (6)
40
25 0-1 +
gering (1)
56 0 +
mittel (2-3)
44
22 3 +
mittel (4)
38 2 +
mittel (4)
n.d.: nicht durchgeführt; 8/12: Anzahl positiver Mäuse/Anzahl untersuchter Mäuse; 337±(37):
Inkubationszeit in Tagen (Standardabweichung); Blau: positives Ergebnis mittels IHC; dunkel
grau: positives Ergebnis mittles Bioassay
Für jedes Tier konnte im Verlauf der Untersuchung mindestens ein positives Ergebnis erzielt
werden, sodass grundsätzlich von einer erfolgreichen Infektion ausgegangen werden konnte.
Die mit Hilfe der Tabelle 4.1 (s. S.73) vorgenommene Quantifizierung positiver Ergebnisse
aus dem Maus-Bioassay und der IHC führt mit Bezug auf die Ergebnisse der Medulla
oblongata zur Einteilung der Tiere in drei Gruppen. Entsprechend dem Grad der PrPSc
-

Ergebnisse 77
Akkumulation bzw. der Infektiosität der Probe konnte, ohne Berücksichtigung der
Inkubationszeit, zwischen fehlender, geringer oder mittlerer/hochgradiger Beteiligung der
Medulla oblongata unterschieden werden (s. Tab. 4.5, S.76). Die quantitative Verteilung der
einzelnen Gruppen zeigt Abb. 4.8.
Abb. 4.8 Verteilung nach Zuordnung der Tiere in die Gruppen nach Grad der PrPSc
Akkumulation im Gehirn
4.3. Einzeltierergebnisse
In den folgenden Kapiteln werden die Strukturen beschrieben, bei denen ein positives
Ergebnis erzielt wurde. Untersuchte Proben mit negativen Resultaten bleiben unerwähnt.
Die Zusammenstellung der Ergebnisse aus der histopathologischen Untersuchung erfolgt in
Tabelle 4.25 (s. S.94) im Anschluss an die Besprechung der einzelnen Ergebnisse.
Da bei allen Tieren eine eosinophile Enteritis diagnostiziert wurde, wird dieser Befund bei
den einzelnen Tieren nicht wiederholt aufgeführt.
4.3.1 Ergebnisse der Tiere ohne Beteiligung der Medulla oblongata
Insgesamt sechs Tiere zwischen 16 und 28 mpi wiesen weder Ablagerungen noch Infektiosität
der Medulla oblongata auf, wobei keines der Tiere klinische Symptome zeigte. In den
Peyer´schen Platten des Ileums zeigte nur ein Tier (IT 46, 16 mpi) keine PrPSc
-Ablagerungen.
In den beteiligten Komponenten des PNS wiesen die Tiere mit Ausnahme der IT 58 (24 mpi;
negativ im PNS) geringe Infektiosität entweder im Sympathikus oder Parasympathikus auf.

78 Ergebnisse
Unabhängig davon war das gemischtfaserige Ggl. coeliacum bei vier der sechs Rinder
infektiös. Eine Besonderheit stellt Tier IT 28 dar, das bereits 16 mpi nicht nur pathologisches
Prion-Protein erstmals im ENS sondern auch geringgradige Infektiosität im Rückenmark
aufwies. Eeine Übersicht der Ergebnisse der Rinder ohne Beteiligung der Medulla oblongata
gibt Tab. 4.12 (s. S.81).
4.3.1.1 IT 28 (16 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Von 76 untersuchten Follikeln der Peyer´schen Platten des Ileums konnte in
einem Follikel (≙1,3%) PrPSc
-Ablagerungen bei FDC und TBM nachgewiesen werden. Im
Plexus muscularis des ENS wurden mit Hilfe des Antikörpers 6C2 zudem geringgradige
Akkumulationen an vereinzelten Neuronen und Satellitenzellen festgestellt (s. Tab. 4.6).
Maus-Bioassay: Der thorakale Rückenmarksabschnitt zeigte einen geringen Erregergehalt.
Für das sympathische Nervensystem konnte in den Nn. splanchnici geringe und dem Ggl.
cervicale craniale mittlere Transmissionsraten detektiert werden.
Histopathologische Untersuchung: Die untersuchten Gewebe zeigten keinen besonderen
Befund.
Tab. 4.6 Ergebnisübersicht IT 28, 16 mpi
Klinik 0
Diagnose o.b.B.
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - + + - - - -
BA - + - +(+) -
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
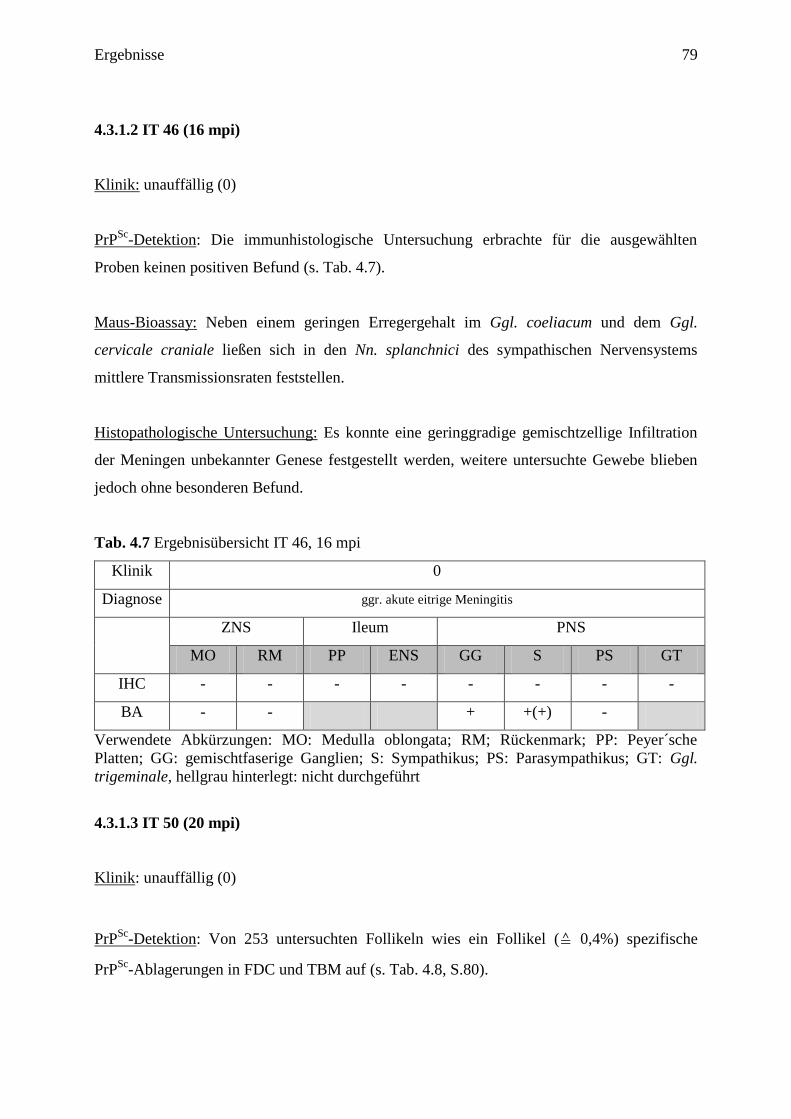
Ergebnisse 79
4.3.1.2 IT 46 (16 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Die immunhistologische Untersuchung erbrachte für die ausgewählten
Proben keinen positiven Befund (s. Tab. 4.7).
Maus-Bioassay: Neben einem geringen Erregergehalt im Ggl. coeliacum und dem Ggl.
cervicale craniale ließen sich in den Nn. splanchnici des sympathischen Nervensystems
mittlere Transmissionsraten feststellen.
Histopathologische Untersuchung: Es konnte eine geringgradige gemischtzellige Infiltration
der Meningen unbekannter Genese festgestellt werden, weitere untersuchte Gewebe blieben
jedoch ohne besonderen Befund.
Tab. 4.7 Ergebnisübersicht IT 46, 16 mpi
Klinik 0
Diagnose ggr. akute eitrige Meningitis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - - - - - - -
BA - - + +(+) -
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
4.3.1.3 IT 50 (20 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Von 253 untersuchten Follikeln wies ein Follikel (≙ 0,4%) spezifische
PrPSc
-Ablagerungen in FDC und TBM auf (s. Tab. 4.8, S.80).
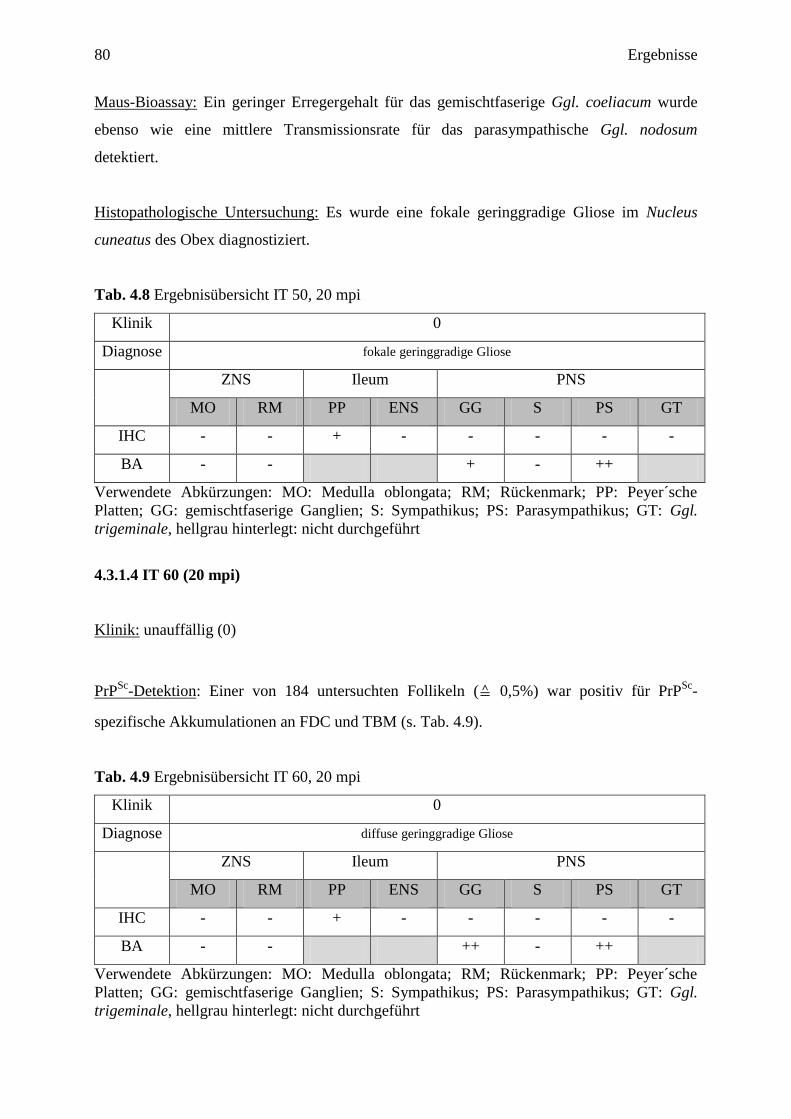
80 Ergebnisse
Maus-Bioassay: Ein geringer Erregergehalt für das gemischtfaserige Ggl. coeliacum wurde
ebenso wie eine mittlere Transmissionsrate für das parasympathische Ggl. nodosum
detektiert.
Histopathologische Untersuchung: Es wurde eine fokale geringgradige Gliose im Nucleus
cuneatus des Obex diagnostiziert.
Tab. 4.8 Ergebnisübersicht IT 50, 20 mpi
Klinik 0
Diagnose fokale geringgradige Gliose
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - + - - - - -
BA - - + - ++
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
4.3.1.4 IT 60 (20 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Einer von 184 untersuchten Follikeln (≙ 0,5%) war positiv für PrPSc
-
spezifische Akkumulationen an FDC und TBM (s. Tab. 4.9).
Tab. 4.9 Ergebnisübersicht IT 60, 20 mpi
Klinik 0
Diagnose diffuse geringgradige Gliose
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - + - - - - -
BA - - ++ - ++
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt

Ergebnisse 81
Maus-Bioassay: Der Bioassay wies sowohl für das gemischtfaserige Ggl. coeliacum als auch
für den parasympathischen N. vagus (Halsteil) mittlere Erregergehalte aus.
Histopathologische Untersuchung: Im Obex dieses Tieres wurde eine diffuse geringgradige
Gliose diagnostiziert.
4.3.1.5 IT 58 (24 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Im Ileum fiel ein von 82 untersuchten Follikeln (≙ 1,2%) mit spezifischen
Ablagerungen an FDC und TBM auf (s. Tab. 4.10).
Maus-Bioassay: Der Bioassay konnte für keine der untersuchten Proben Infektiosität
nachweisen.
Histopathologische Untersuchung: Es fallen inflammatorische Prozesse in mehreren Anteilen
des PNS auf. Im Ggl. coeliacum sind geringgradige lymphohistiozytäre Infiltrationen zu
beobachten, die in den Nn. splanchnici, dem N. vagus und dem Ggl. stellatum eine
mittelgradige Ausprägung erreichen.
Tab. 4.10 Ergebnisübersicht IT 58, 24 mpi
Klinik 0
Diagnose gering- bis mittelgradige, nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - + - - - - -
BA - - - - -
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
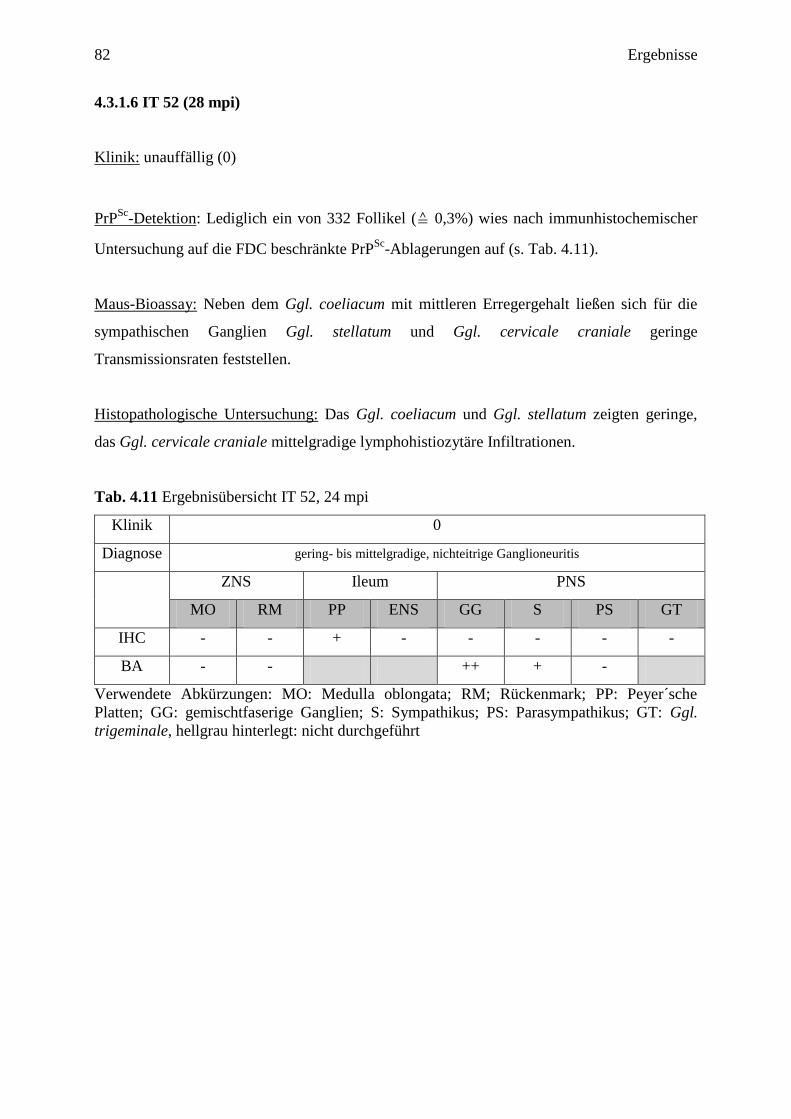
82 Ergebnisse
4.3.1.6 IT 52 (28 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Lediglich ein von 332 Follikel (≙ 0,3%) wies nach immunhistochemischer
Untersuchung auf die FDC beschränkte PrPSc
-Ablagerungen auf (s. Tab. 4.11).
Maus-Bioassay: Neben dem Ggl. coeliacum mit mittleren Erregergehalt ließen sich für die
sympathischen Ganglien Ggl. stellatum und Ggl. cervicale craniale geringe
Transmissionsraten feststellen.
Histopathologische Untersuchung: Das Ggl. coeliacum und Ggl. stellatum zeigten geringe,
das Ggl. cervicale craniale mittelgradige lymphohistiozytäre Infiltrationen.
Tab. 4.11 Ergebnisübersicht IT 52, 24 mpi
Klinik 0
Diagnose gering- bis mittelgradige, nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - + - - - - -
BA - - ++ + -
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt

83 Ergebnisse
Tab. 4.12 Detektion von PrPSc
/Infektiosität ohne Akkumulation in der Medulla oblongata IT
Infe
kti
on
stie
r
Ob
exka
ud
ale
Med
ull
aB
AB
ioas
say
PP
EN
S1
/10
An
zah
l p
osi
tiv
er M
äuse
/An
zah
l un
ters
uch
ter
Mäu
se
0/12
2/13
0/1
3+
+4
28
(±
13
3)
Ink
ubat
ion
szei
t in
Tag
en (
Stan
dard
abw
eich
ung)
47
3 (
± 10
4)
35
44
28
(±
133
)1/
76
- f
/gp
m
0/13
0/1
10
/14
-IH
CIm
mun
his
toch
emie
44
8 (
± 9
7)
43
7 (
± 11
3)
38
1 (±
62
)0
/87
Dar
m
0/10
0/1
20
/12
+3
/60
An
zah
l p
osi
tiv
er F
oll
ikel
/Ges
amta
nza
hl
Fo
llik
el
47
6 (
± 10
8)
45
6 (
± 10
8)
38
9 (
± 8
6)
1/2
53
- f
/gf
fein
-gra
nul
äres
Rea
kti
on
smus
ter
0/13
0/1
40
/13
+g
kug
elfö
rmig
es R
eak
tio
nsm
uste
r
46
4 (
± 11
6)
48
8 (
± 9
2)
45
7 (
± 6
6)
1/18
4 -
f/g
pm
Ple
xus
mus
cula
ris
0/9
0/1
00
/12
+p
sP
lex
us s
ubm
uco
sus
30
8 (
± 3
6)
36
4 (
± 8
7)
34
5 (
± 6
7)
1/8
2 -
f/g
ZN
S
0/8
0/1
10
/10
+(3
)In
dex
für
die
In
ten
sitä
t de
r P
rPS
c -Ak
kum
ulat
ion
42
0 (
± 11
9)
38
1 (±
89
)3
89
(±
107
)1/
33
2 -
f
SN
Sso
mat
isch
es N
erv
ensy
stem
Ggl
.
trig
emin
ale
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
-0
/11
-0
/12
-1/
10-
5/13
-0
/5-
0/9
-0
/12
-0
/11
-
39
8 (
± 7
7)
34
0 (
± 7
7)
37
74
36
(±
77
)5
22
(±
109
)4
51
(± 1
03
)4
39
(±
155
)4
01
(± 1
09
)
-2/
12-
0/1
4-
3/11
-1/
12-
0/9
-0
/11
-0
/9-
0/9
-
40
9 (
± 6
3)
36
5 (
± 8
9)
59
8 (
± 6
7)
47
23
85
(±
124
)4
97
(±
149
)4
95
(±
98
)5
26
(±
113
)
-1/
13-
0/1
1-
0/1
0-
0/1
2-
0/6
-0
/7-
0/9
-5
/13
-
49
53
08
(±
62
)4
33
(±
140
)3
45
(±
64
)4
63
(±
132
)3
76
(±
72
)4
39
(±
132
)5
35
(±
79
)
-7/
15-
0/1
4-
0/1
2-
0/1
1-
0/6
-1/
9-
0/1
2-
0/8
-
32
9 (
± 3
9)
37
0 (
± 7
9)
47
8 (
± 12
6)
36
4 (
± 7
5)
54
9 (
± 9
8)
48
34
04
(±
130
)4
37
(±
88
)
0/5
0/9
0/8
0/1
10
/40
/70
/70
/6
37
2 (
± 12
1)3
35
(±
52
)4
64
(±
155
)3
76
(±
123
)5
30
(±
66
)4
19 (
± 12
4)
36
2 (
± 13
4)
40
7 (
± 10
9)
3/12
0/1
30
/31
/12
1/12
0/9
0/7
0/3
314
(±
25
)3
91
(± 1
35
)4
62
(±
121)
62
44
27
39
4 (
± 10
3)
42
7 (
± 9
8)
37
1 (±
99
)
Par
asy
mp
ath
isch
es N
erv
ensy
stem
Veg
etat
ives
Ner
ven
syst
emSN
S
Per
iph
eres
Ner
ven
syst
em
Kli
nis
cher
Stat
us
IT-
Num
mer
Mo
nat
e
p.i
.
Kli
nis
cher
Stat
us
IT-
Num
mer
Mo
nat
e
p.i
.
Gem
isch
tfas
erig
e A
nte
ile
Sym
pat
his
ches
Ner
ven
syst
em
16
IT 2
80
-
IT
46
0-
--
-
--
Dar
m
dist
ales
Ile
um
IHC
BA
IHC
BA
IHC
BA
Zen
tral
es N
erv
ensy
stem
Rüc
ken
mar
k T
7R
ück
enm
ark
L3
IHC
--
-
20
IT 5
00
-
IT 6
00
-
28
--
--
-
--
24
IT 5
80
--
IT 5
20
--
Ggl
. n
odo
sum
16
IT 2
80
IT
46
0
Ggl
. co
elia
cum
Ggl
. m
esen
tera
le c
aud.
Nn
. sp
lan
chn
ici
N.
vag
us (
Hal
s)
20
IT 5
00
IT 6
00
Ggl
. ce
rvic
ale
cran
.G
gl.
stel
latu
mN
. v
agus
(B
rust
)
--
--
24
IT 5
80
--
--
28
IT 5
20
--
-- -
--
--
-

84 Ergebnisse
4.3.2 Ergebnisse der Tiere mit geringgradiger Beteiligung der Medulla oblongata
Der geringe und damit initiale Befall des Obex konnte bei insgesamt drei Tieren entweder
durch die Detektion von PrPSc
und/oder in Form von Infektiosität nachgewiesen werden und
ging bei zwei dieser Rinder mit unspezifischen klinischen Symptomen einher. Von den drei
betroffenen Tieren war zum frühesten Zeitpunkt der kaudal an den Obex grenzende Anteil der
Medulla oblongata der IT 24 (24 mpi) infektiös, während für zwei weitere Rinder vereinzelte
PrPSc
-positive Neurone mit Beschränkung auf die Formatio reticularis (IT 61, 32 mpi) bzw.
den Nucleus tractus solitarii und den Nucleus motorius nervi trigemini (IT 25, 40 mpi)
auftraten. Für die IT 61 (32 mpi) fiel der Nachweis im Maus-Bioassay negativ aus. Ein
solcher Test wurde aufgrund der Inkubationszeit (40 mpi) beim Tier IT 25 nicht
durchgeführt.
Beim Rind IT 24 (24 mpi) konnte als einzigem Tier dieser Gruppe im Rückenmark weder
Infektiosität noch PrPSc
nachgewiesen werden. Interessanterweise wurden aber mittel- bis
hochgradigen Transmissionsraten für alle untersuchten Anteile des Sympathikus detektiert,
wohingegen die Proben des Parasympathikus in diesem Rind keine positive Befunde
aufwiesen.
Weiterhin fällt auf, dass mit der IT 25 (40 mpi) ein Tier aus der späten Inkubationsperiode
zwar nur geringe PrPSc
-Akkumulationen im Obex aufwies, diese aber bereits im Ggl.
trigeminale zu finden waren.
Bei allen Tieren war der Nachweis des pathologischen Prion-Proteins in den Peyer´schen
Platten und/oder im ENS erfolgreich.
Die Zusammenstellung der Ergebnisse der Rinder mit geringgradiger Beteiligung der Medulla
oblongata erfolgt in Tab.4.16 (s. S.86).
4.3.2.1 IT 24 (24 mpi)
Klinik: unauffällig (0)
PrPSc
-Detektion: Bemerkenswert ist der Nachweis von PrPSc
in den Follikeln der Peyer´schen
Platten. Von 272 Follikeln waren insgesamt 15 positiv (≙ 5.5%) und wiesen ein
Reaktionsmuster spezifisch sowohl für FDC als auch TBM auf. Im Plexus muscularis des
ENS ließen sich an vereinzelten Neuronen und Satellitenzellen geringgradige
Akkumulationen des pathologischen Prion-Proteins detektieren (s. Tab. 4.13, S 85).
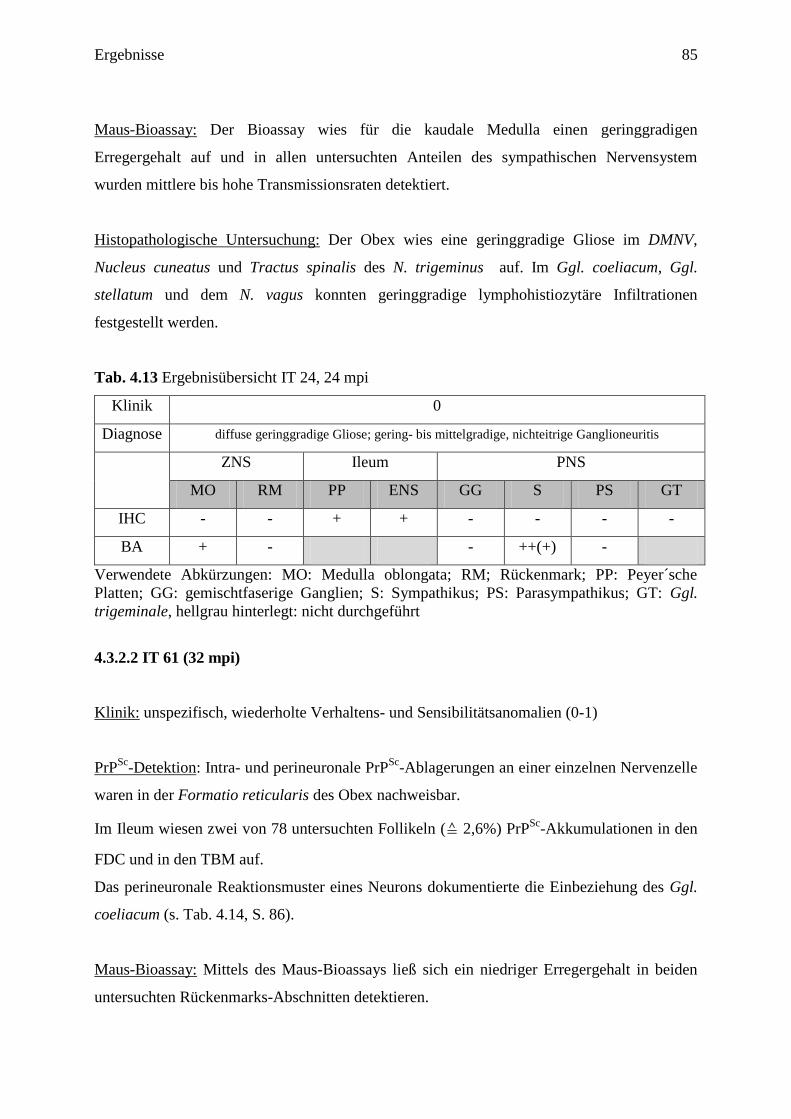
Ergebnisse 85
Maus-Bioassay: Der Bioassay wies für die kaudale Medulla einen geringgradigen
Erregergehalt auf und in allen untersuchten Anteilen des sympathischen Nervensystem
wurden mittlere bis hohe Transmissionsraten detektiert.
Histopathologische Untersuchung: Der Obex wies eine geringgradige Gliose im DMNV,
Nucleus cuneatus und Tractus spinalis des N. trigeminus auf. Im Ggl. coeliacum, Ggl.
stellatum und dem N. vagus konnten geringgradige lymphohistiozytäre Infiltrationen
festgestellt werden.
Tab. 4.13 Ergebnisübersicht IT 24, 24 mpi
Klinik 0
Diagnose diffuse geringgradige Gliose; gering- bis mittelgradige, nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC - - + + - - - -
BA + - - ++(+) -
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
4.3.2.2 IT 61 (32 mpi)
Klinik: unspezifisch, wiederholte Verhaltens- und Sensibilitätsanomalien (0-1)
PrPSc
-Detektion: Intra- und perineuronale PrPSc
-Ablagerungen an einer einzelnen Nervenzelle
waren in der Formatio reticularis des Obex nachweisbar.
Im Ileum wiesen zwei von 78 untersuchten Follikeln (≙ 2,6%) PrPSc
-Akkumulationen in den
FDC und in den TBM auf.
Das perineuronale Reaktionsmuster eines Neurons dokumentierte die Einbeziehung des Ggl.
coeliacum (s. Tab. 4.14, S. 86).
Maus-Bioassay: Mittels des Maus-Bioassays ließ sich ein niedriger Erregergehalt in beiden
untersuchten Rückenmarks-Abschnitten detektieren.
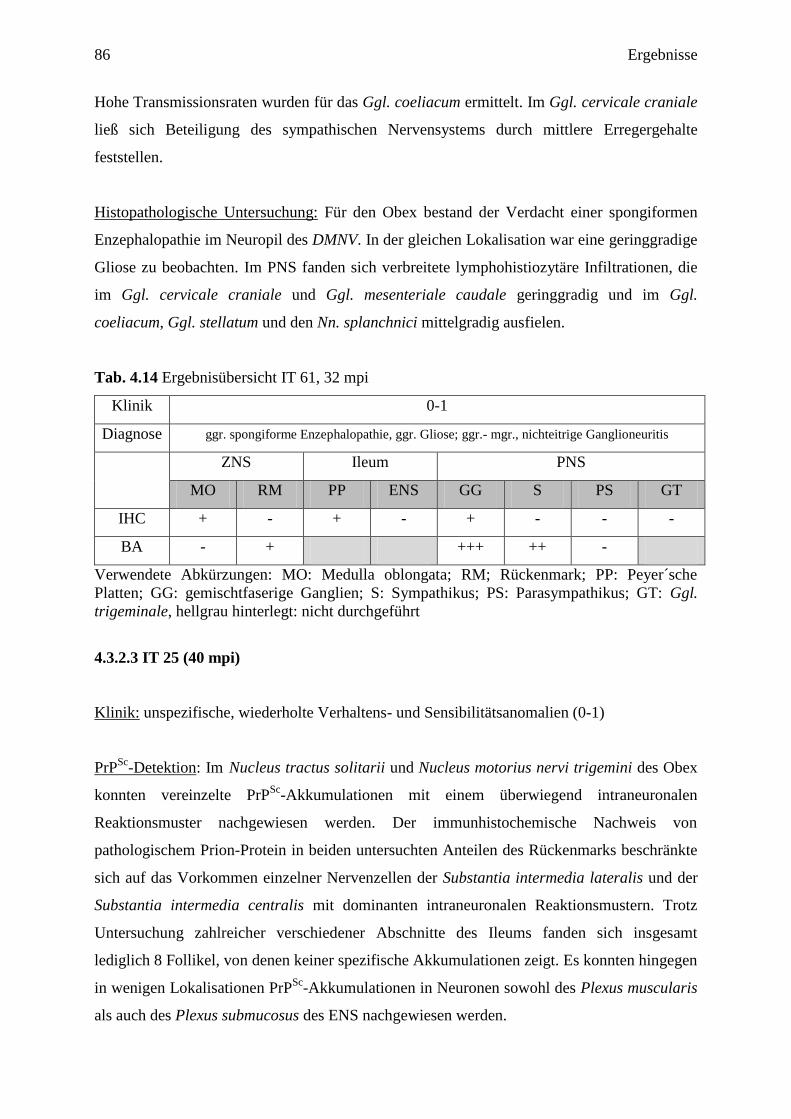
86 Ergebnisse
Hohe Transmissionsraten wurden für das Ggl. coeliacum ermittelt. Im Ggl. cervicale craniale
ließ sich Beteiligung des sympathischen Nervensystems durch mittlere Erregergehalte
feststellen.
Histopathologische Untersuchung: Für den Obex bestand der Verdacht einer spongiformen
Enzephalopathie im Neuropil des DMNV. In der gleichen Lokalisation war eine geringgradige
Gliose zu beobachten. Im PNS fanden sich verbreitete lymphohistiozytäre Infiltrationen, die
im Ggl. cervicale craniale und Ggl. mesenteriale caudale geringgradig und im Ggl.
coeliacum, Ggl. stellatum und den Nn. splanchnici mittelgradig ausfielen.
Tab. 4.14 Ergebnisübersicht IT 61, 32 mpi
Klinik 0-1
Diagnose ggr. spongiforme Enzephalopathie, ggr. Gliose; ggr.- mgr., nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC + - + - + - - -
BA - + +++ ++ -
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
4.3.2.3 IT 25 (40 mpi)
Klinik: unspezifische, wiederholte Verhaltens- und Sensibilitätsanomalien (0-1)
PrPSc
-Detektion: Im Nucleus tractus solitarii und Nucleus motorius nervi trigemini des Obex
konnten vereinzelte PrPSc
-Akkumulationen mit einem überwiegend intraneuronalen
Reaktionsmuster nachgewiesen werden. Der immunhistochemische Nachweis von
pathologischem Prion-Protein in beiden untersuchten Anteilen des Rückenmarks beschränkte
sich auf das Vorkommen einzelner Nervenzellen der Substantia intermedia lateralis und der
Substantia intermedia centralis mit dominanten intraneuronalen Reaktionsmustern. Trotz
Untersuchung zahlreicher verschiedener Abschnitte des Ileums fanden sich insgesamt
lediglich 8 Follikel, von denen keiner spezifische Akkumulationen zeigt. Es konnten hingegen
in wenigen Lokalisationen PrPSc
-Akkumulationen in Neuronen sowohl des Plexus muscularis
als auch des Plexus submucosus des ENS nachgewiesen werden.
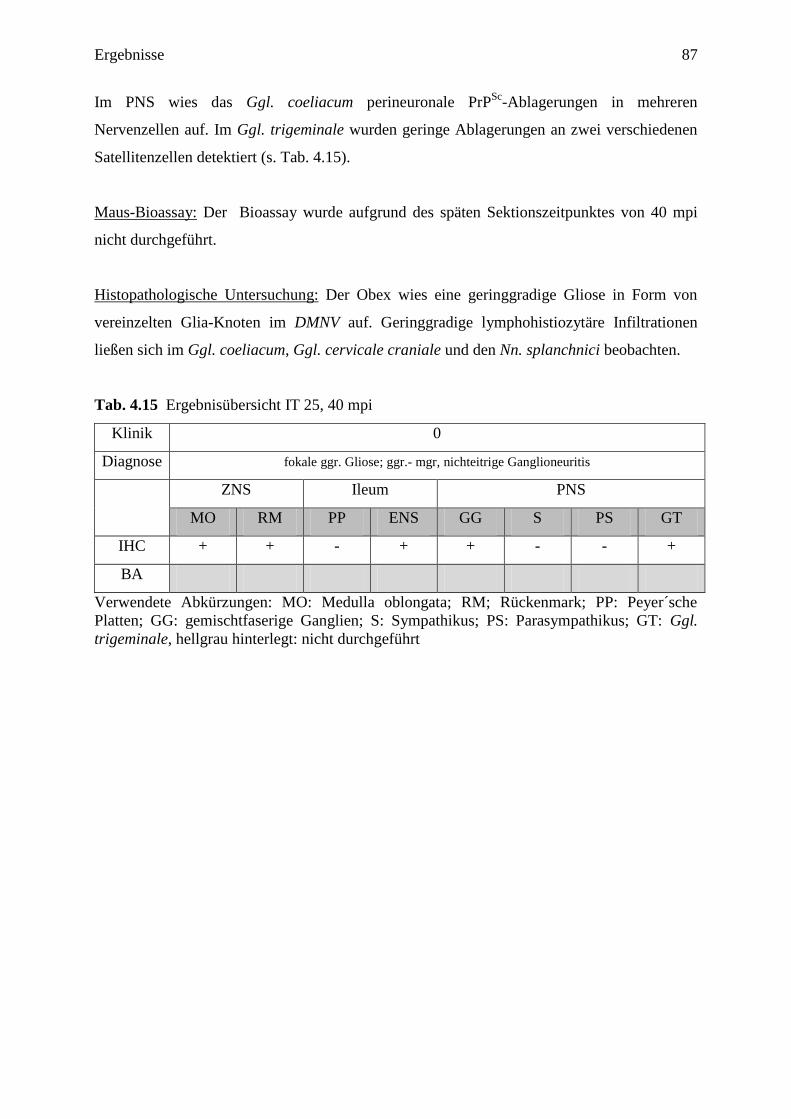
Ergebnisse 87
Im PNS wies das Ggl. coeliacum perineuronale PrPSc
-Ablagerungen in mehreren
Nervenzellen auf. Im Ggl. trigeminale wurden geringe Ablagerungen an zwei verschiedenen
Satellitenzellen detektiert (s. Tab. 4.15).
Maus-Bioassay: Der Bioassay wurde aufgrund des späten Sektionszeitpunktes von 40 mpi
nicht durchgeführt.
Histopathologische Untersuchung: Der Obex wies eine geringgradige Gliose in Form von
vereinzelten Glia-Knoten im DMNV auf. Geringgradige lymphohistiozytäre Infiltrationen
ließen sich im Ggl. coeliacum, Ggl. cervicale craniale und den Nn. splanchnici beobachten.
Tab. 4.15 Ergebnisübersicht IT 25, 40 mpi
Klinik 0
Diagnose fokale ggr. Gliose; ggr.- mgr, nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC + + - + + - - +
BA
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
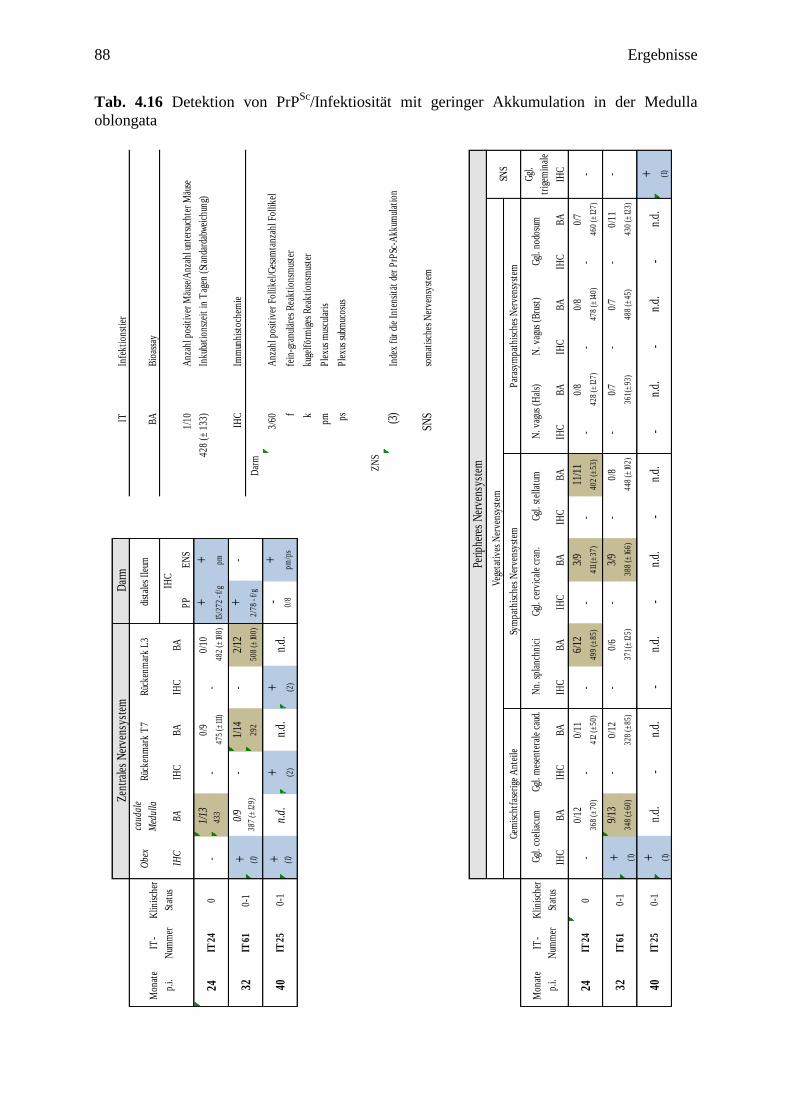
88 Ergebnisse
Tab. 4.16 Detektion von PrPSc
/Infektiosität mit geringer Akkumulation in der Medulla
oblongata IT
Infe
ktio
nsti
er
Obe
xca
udal
e
Med
ulla
BA
Bio
assa
y
PP
EN
S1/
10A
nzah
l pos
itiv
er M
äuse
/Anz
ahl u
nter
such
ter
Mäu
se
1/13
0/9
0/10
++
428
(± 1
33)
Inku
bati
onsz
eit
in T
agen
(St
anda
rdab
wei
chun
g)
433
475
(± 11
1)48
2 (±
108)
15/2
72 -
f/g
pm
+0/
9-
1/14
-2/
12+
-IH
CIm
mun
hist
oche
mie
(1)
387
(± 12
9)29
250
8 (±
100)
2/78
- f/
gD
arm
++
+-
+3/
60A
nzah
l pos
itiv
er F
ollik
el/G
esam
tanz
ahl F
ollik
el
(1)
(2)
(2)
0/8
pm/p
sf
fein
-gra
nulä
res
Rea
ktio
nsm
uste
r
kku
gelf
örm
iges
Rea
ktio
nsm
uste
r
pmP
lexu
s m
uscu
lari
s
psP
lexu
s su
bmuc
osus
ZN
S
(3)
Inde
x fü
r di
e In
tens
ität
der
PrP
Sc-A
kkum
ulat
ion
SNS
som
atis
ches
Ner
vens
yste
m
Ggl
.
trig
emin
ale
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
0/12
0/11
6/12
3/9
11/1
10/
80/
80/
7
368
(± 7
0)41
2 (±
50)
499
(± 8
5)41
1 (±
37)
402
(± 5
3)42
8 (±
127)
478
(± 14
0)46
0 (±
127)
+9/
13-
0/12
-0/
6-
3/9
-0/
8-
0/7
-0/
7-
0/11
-
(1)
348
(± 6
0)32
8 (±
85)
371 (
± 12
5)38
8 (±
166)
448
(± 10
2)36
1 (±
93)
488
(± 4
5)43
0 (±
123)
++
(1)
(1)
IT 2
40
--
IT-
Num
mer
Mon
ate
p.i.
-
Nn.
spl
anch
nici
Ggl
. cer
vica
le c
ran.
40n.
d.
0-1
IT 6
1
n.d.
32
Klin
isch
er
Stat
us
IHC
Klin
isch
er
Stat
us
Perip
here
s N
erve
nsys
tem
24
SNS
Dar
m
dist
ales
Ile
um
BA
IHC
BA
Zen
tral
es N
erve
nsys
tem
Rüc
kenm
ark
T7
Rüc
kenm
ark
L3
IHC
BA
IHC
-n.
d.-
n.d.
-n.
d.-
n.d.
-
--
--
--
N. v
agus
(H
als)
N. v
agus
(B
rust
)G
gl. n
odos
um
--
n.d.
n.d.
-n.
d.-
40IT
25
0-1
n.d.
IT 6
132
IT-
Num
mer
Mon
ate
p.i.
Ggl
. coe
liacu
m
0-1
n.d.
IT 2
5
Gem
isch
tfas
erig
e A
ntei
le
Veg
etat
ives
Ner
vens
yste
m
Par
asym
path
isch
es N
erve
nsys
tem
Ggl
. mes
ente
rale
cau
d.
Sym
path
isch
es N
erve
nsys
tem
Ggl
. ste
llatu
m
24IT
24
0-
0-1

89 Ergebnisse
4.3.3 Ergebnisse der Tiere mit mittel- bis hochgradiger Beteiligung der Medulla
oblongata
Insgesamt zählten sieben Tiere ab dem 28. Monat p.i. zu dieser Gruppe.
Für die IT 21 waren zwar nur geringe PrPSc
-Ablagerungen an einzelnen Neuronen des Obex
nachweisbar, allerdings gab in diesem Fall die hohe Übertragungsrate des Maus-Bioassays
den Ausschlag für die Zuordnung in diese Gruppe.
Dabei waren bei zwei Tieren (IT 21, 28 mpi; IT 56, 40 mpi) keine nachweisbaren klinischen
Symptome feststellbar. Zwei Rinder dieser Gruppe zeigten hingegen definitive Anzeichen für
eine BSE-Erkrankung (IT 49, 36 mpi; IT 22, 44 mpi). Die weiteren Rinder waren entweder
unspezifisch (IT 09, 32 mpi; IT 11, 36 mpi) oder vermutlich (IT 38, 44 mpi) an BSE erkrankt.
Der Maus-Bioassay führte in drei von vier untersuchten Tier zur Detektion von Infektiosität
im gesamten PNS mit hohen, teils maximalen Transmissionsraten. Tier IT 21 wies nur für die
Proben des Sympathikus solch hohe Übertragungsraten auf, die im Parasympathikus lediglich
geringe bis mittlere Intensität erreichten.
Erstmals gelang sporadisch der immunhistochemische Nachweis von PrPSc
in Proben des
Sympathikus und Parasympathikus, wobei vor allem der N. vagus häufiger betroffen war.
Weiterhin konnte bei allen Tieren im Ggl. trigeminale (Ausnahme IT 21) und im Rückenmark
deutliche Ablagerungen des pathologischen Prion-Proteins detektiert werden. Dieser
Nachweis war hinsichtlich des Ileums leicht in Richtung des ENS verschoben, auch weil die
absolute Anzahl der Follikel mit steigender Inkubationsdauer und dem damit einhergehenden
steigenden Alter der Tiere abnahm. Bei Rind IT 38 (44 mpi) blieb die Detektion von PrPSc
im
Ileum ganz aus.
Eine Übersicht zu den Ergebnissen von Tieren mit mittel- bis hochgradiger Beteiligung der
Medulla oblongata gibt Tab. 4.24 s. S.93).
4.3.3.1 IT 21 (28 mpi)
Die im Bioassay nachgewiesene Transmissionsrate führte anstelle des geringen PrPSc
-
Nachweises mittels IHC zur Einbeziehung dieses Tieres in diese Gruppe.
Klinik: Unauffällig (0)
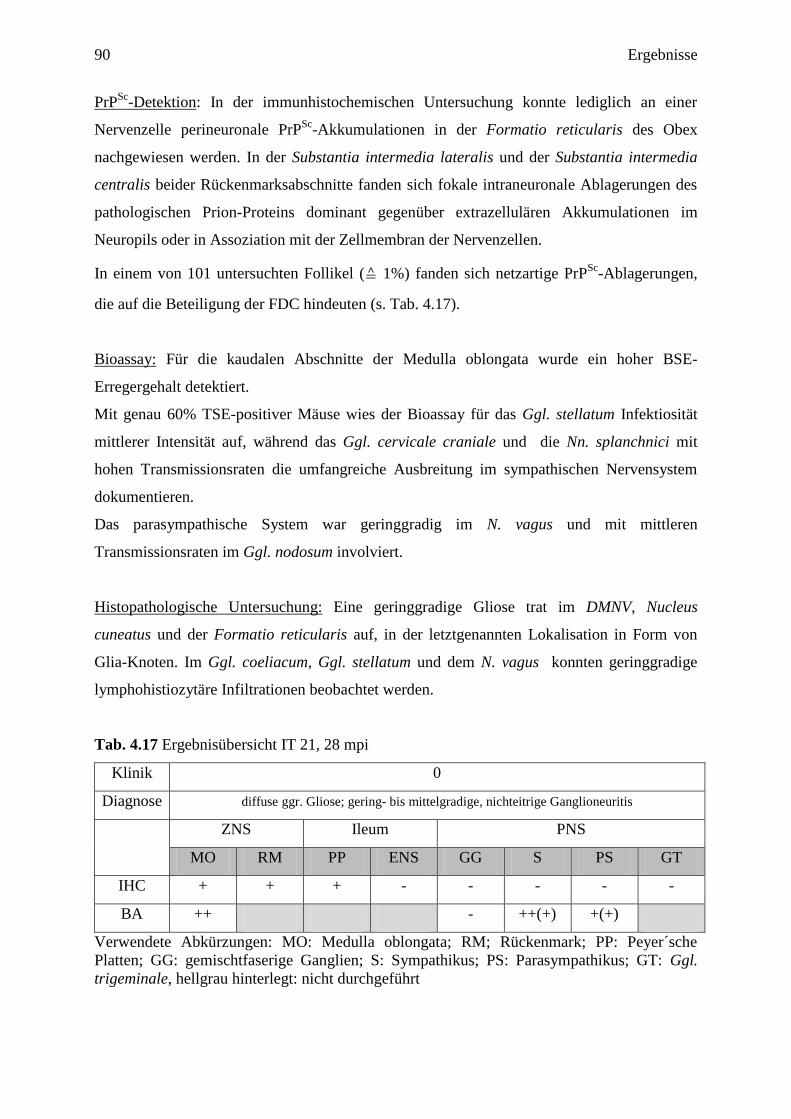
90 Ergebnisse
PrPSc
-Detektion: In der immunhistochemischen Untersuchung konnte lediglich an einer
Nervenzelle perineuronale PrPSc
-Akkumulationen in der Formatio reticularis des Obex
nachgewiesen werden. In der Substantia intermedia lateralis und der Substantia intermedia
centralis beider Rückenmarksabschnitte fanden sich fokale intraneuronale Ablagerungen des
pathologischen Prion-Proteins dominant gegenüber extrazellulären Akkumulationen im
Neuropils oder in Assoziation mit der Zellmembran der Nervenzellen.
In einem von 101 untersuchten Follikel (≙ 1%) fanden sich netzartige PrPSc
-Ablagerungen,
die auf die Beteiligung der FDC hindeuten (s. Tab. 4.17).
Bioassay: Für die kaudalen Abschnitte der Medulla oblongata wurde ein hoher BSE-
Erregergehalt detektiert.
Mit genau 60% TSE-positiver Mäuse wies der Bioassay für das Ggl. stellatum Infektiosität
mittlerer Intensität auf, während das Ggl. cervicale craniale und die Nn. splanchnici mit
hohen Transmissionsraten die umfangreiche Ausbreitung im sympathischen Nervensystem
dokumentieren.
Das parasympathische System war geringgradig im N. vagus und mit mittleren
Transmissionsraten im Ggl. nodosum involviert.
Histopathologische Untersuchung: Eine geringgradige Gliose trat im DMNV, Nucleus
cuneatus und der Formatio reticularis auf, in der letztgenannten Lokalisation in Form von
Glia-Knoten. Im Ggl. coeliacum, Ggl. stellatum und dem N. vagus konnten geringgradige
lymphohistiozytäre Infiltrationen beobachtet werden.
Tab. 4.17 Ergebnisübersicht IT 21, 28 mpi
Klinik 0
Diagnose diffuse ggr. Gliose; gering- bis mittelgradige, nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC + + + - - - - -
BA ++ - ++(+) +(+)
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt

Ergebnisse 91
4.3.3.2 Zusammenfassung der weiteren Ergebnisse
Bei den Tieren mit mittlerer oder starker Beteiligung der Medulla oblongata handelte es sich
um die IT 09, IT 11, IT 49, IT 56, IT 22 und IT 38. Der Sektionszeitpunkt dieser Rinder
variierte zwischen 32 und 44 mpi. Dabei trat bei den einzelnen Rindern eine höchst
unterschiedliche klinische Ausprägung auf. Diese reichte von Symptomfreiheit (IT 56) über
ein unspezifisches Krankheitsbild (IT 09 und IT 11) bis hin zu definitiven BSE-Erkrankungen
(IT 49 und IT 22).
PrPSc
-Detektion: In den untersuchten Anteilen des ZNS waren bei allen Tieren die PrPSc
-
Akkumulationen bzw. Erregergehalte mindestens mittleren Grades nachweisbar. In der
Obexregion fanden sich in sämtlichen Kerngebieten intra- und perineuronale sowie intragliale
Reaktionsmuster. Die extrazelluläre Ablagerungen des Neuropils traten weitverbreitet diffus
fein-bis grobgranulär bis hin zu koaleszierend auf und zeigten sich je nach Kerngebiet auch
deutlich als lineare Akkumulationen. Die Reaktionsmuster der grauen Substanz des
Rückenmarks waren mit denen des Obex vergleichbar, stimmten aber nicht zwangsläufig mit
deren Grad der Intensität überein. Die weiße Substanz des Rückenmarks zeigte keine PrPSc
-
Ablagerungen.
Im Vergleich zu den jüngeren Tieren nahm die Zahl auswertbarer Follikel im Ileum deutlich
ab. In nur drei der sechs Tiere konnten FDC-spezifische PrPSc
-Ablagerungen mit einer
Häufigkeit von 0,8 bis 16,7% detektiert werden. In fünf der sechs Fälle war das ENS
betroffen, wobei bei drei Tieren Akkumulationen parallel in Follikeln und ENS auftraten.
PrPSc
ließ sich multifokal in wenigen vereinzelten Neuronen mit perineuronalen
Reaktionsmustern nachweisen. Obwohl beide Plexus involviert waren, gelang die Detektion
positiver Nervenzellen im muskulären Plexus häufiger.
Im PNS gestaltete sich die Detektion des pathologischen Prion-Proteins uneinheitlich. Konnte
durch die immunhistochemische Untersuchung ein positiver Befund erhoben werden, zeigten
sich stets vereinzelte Neurone mit perineuronalen Reaktionsmuster. Seltener konnte auch eine
intraneuronale und intragliale Akkumulation beobachtet werden. Am häufigsten wurde der
Nachweis für das Ggl. coeliacum geführt (5/6), aber auch der parasympathische N. vagus war
in vier Fällen positiv. Im sympathischen System konnte lediglich einmal PrPSc
im Ggl.
cervicale craniale (IT 11) nachgewiesen werden.
Das Ggl. trigeminale zeigte bei allen Tieren dieser Gruppe hauptsächlich in den
Satellitenzellen spezifische intragliale diffus-granuläre oder verdichtete PrPSc
-Ablagerungen.
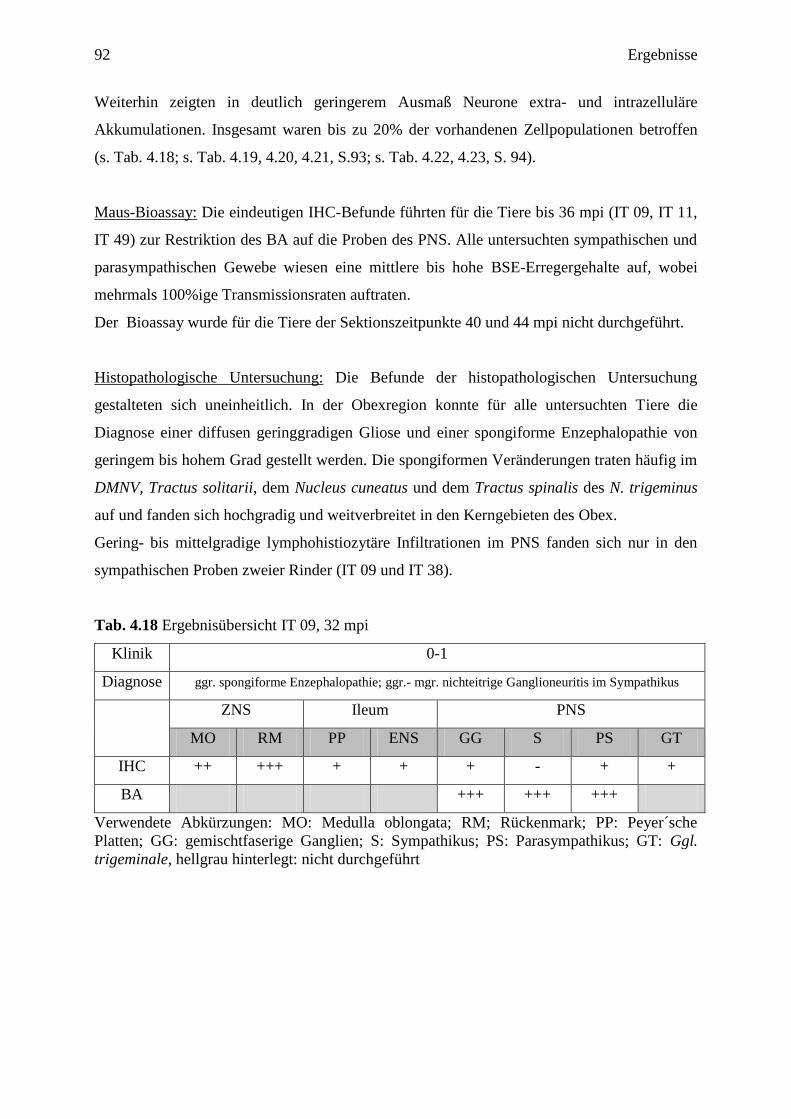
92 Ergebnisse
Weiterhin zeigten in deutlich geringerem Ausmaß Neurone extra- und intrazelluläre
Akkumulationen. Insgesamt waren bis zu 20% der vorhandenen Zellpopulationen betroffen
(s. Tab. 4.18; s. Tab. 4.19, 4.20, 4.21, S.93; s. Tab. 4.22, 4.23, S. 94).
Maus-Bioassay: Die eindeutigen IHC-Befunde führten für die Tiere bis 36 mpi (IT 09, IT 11,
IT 49) zur Restriktion des BA auf die Proben des PNS. Alle untersuchten sympathischen und
parasympathischen Gewebe wiesen eine mittlere bis hohe BSE-Erregergehalte auf, wobei
mehrmals 100%ige Transmissionsraten auftraten.
Der Bioassay wurde für die Tiere der Sektionszeitpunkte 40 und 44 mpi nicht durchgeführt.
Histopathologische Untersuchung: Die Befunde der histopathologischen Untersuchung
gestalteten sich uneinheitlich. In der Obexregion konnte für alle untersuchten Tiere die
Diagnose einer diffusen geringgradigen Gliose und einer spongiforme Enzephalopathie von
geringem bis hohem Grad gestellt werden. Die spongiformen Veränderungen traten häufig im
DMNV, Tractus solitarii, dem Nucleus cuneatus und dem Tractus spinalis des N. trigeminus
auf und fanden sich hochgradig und weitverbreitet in den Kerngebieten des Obex.
Gering- bis mittelgradige lymphohistiozytäre Infiltrationen im PNS fanden sich nur in den
sympathischen Proben zweier Rinder (IT 09 und IT 38).
Tab. 4.18 Ergebnisübersicht IT 09, 32 mpi
Klinik 0-1
Diagnose ggr. spongiforme Enzephalopathie; ggr.- mgr. nichteitrige Ganglioneuritis im Sympathikus
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC ++ +++ + + + - + +
BA +++ +++ +++
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
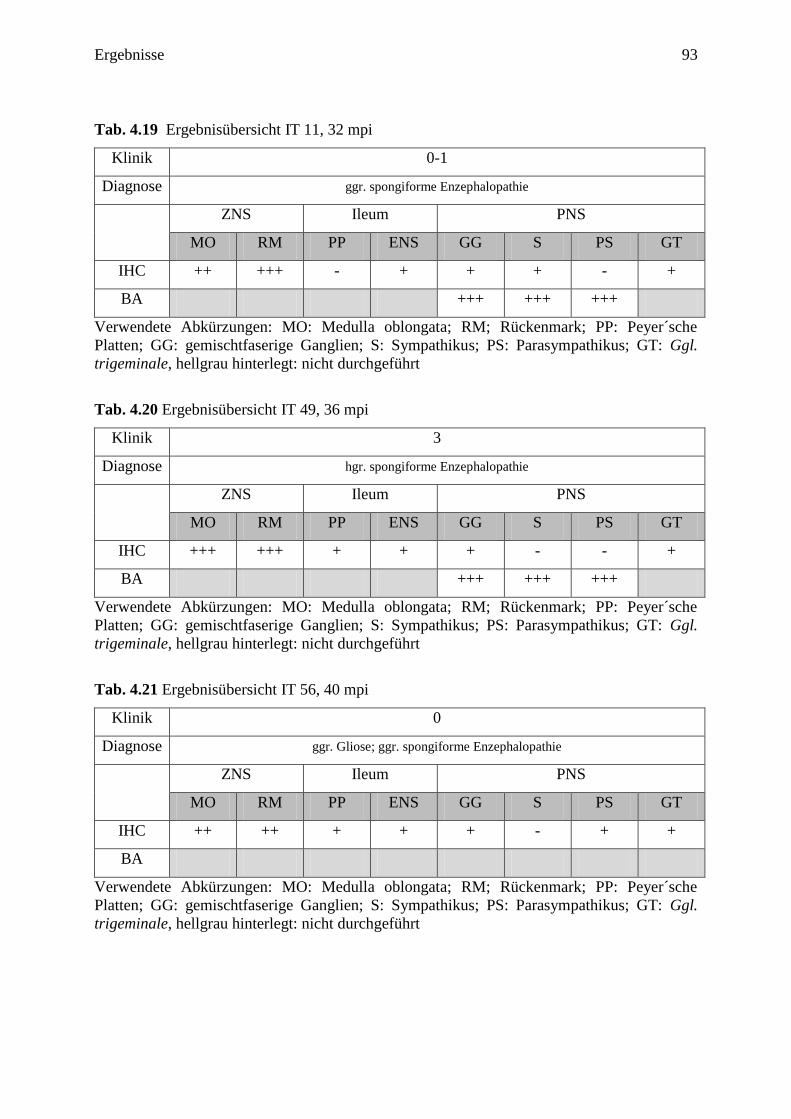
Ergebnisse 93
Tab. 4.19 Ergebnisübersicht IT 11, 32 mpi
Klinik 0-1
Diagnose ggr. spongiforme Enzephalopathie
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC ++ +++ - + + + - +
BA +++ +++ +++
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
Tab. 4.20 Ergebnisübersicht IT 49, 36 mpi
Klinik 3
Diagnose hgr. spongiforme Enzephalopathie
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC +++ +++ + + + - - +
BA +++ +++ +++
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
Tab. 4.21 Ergebnisübersicht IT 56, 40 mpi
Klinik 0
Diagnose ggr. Gliose; ggr. spongiforme Enzephalopathie
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC ++ ++ + + + - + +
BA
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt

94 Ergebnisse
Tab. 4.22 Ergebnisübersicht IT 22, 44 mpi
Klinik 3
Diagnose ggr. Gliose; hgr. spongiforme Enzephalopathie
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC ++ +++ - + + - + +
BA
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt
Tab. 4.23 Ergebnisübersicht IT 38, 44 mpi
Klinik 2
Diagnose fokale ggr. Gliose; ggr. nichteitrige Ganglioneuritis
ZNS Ileum PNS
MO RM PP ENS GG S PS GT
IHC ++ ++ - - + - + +
BA
Verwendete Abkürzungen: MO: Medulla oblongata; RM; Rückenmark; PP: Peyer´sche
Platten; GG: gemischtfaserige Ganglien; S: Sympathikus; PS: Parasympathikus; GT: Ggl.
trigeminale, hellgrau hinterlegt: nicht durchgeführt

95 Ergebnisse
Tab. 4.24 Detektion von PrPSc
/Infektiosität mittlere und hoher Akkumulation in der Medulla
oblongata IT
Infe
kti
on
stie
r
Ob
ex
ka
ud
ale
Med
ull
aB
AB
ioass
ay
PP
EN
S1
/10
An
zah
l p
osi
tiv
er
Mäuse
/An
zah
l un
ters
uch
ter
Mäuse
+8
/13
++
+4
28
(±
13
3)
Ink
ubati
on
szeit
in
Tagen
(Sta
ndard
abw
eic
hun
g)
(1)
33
7 (
± 3
7)
(1)
(1)
1/10
1 -
g
++
++
+IH
CIm
mun
his
toch
em
ie
(4)
(6)
(6)
2/1
2 -
fp
m/p
sD
arm
++
+-
+3
/60
An
zah
l p
osi
tiv
er
Fo
llik
el/
Gesa
mta
nzah
l F
oll
ikel
(4)
(6)
(6)
0/6
9p
mf
fein
-gra
nulä
res
Reak
tio
nsm
ust
er
++
++
+k
kugelf
örm
iges
Reak
tio
nsm
ust
er
(6)
(5)
(5)
3/6
0 -
fp
mp
mP
lex
us
musc
ula
ris
++
++
+p
sP
lex
us
subm
uco
sus
(2-3
)(3
)(3
)1/
124
- f
pm
ZN
S
++
+-
+(3
)In
dex
für
die
In
ten
sitä
t der
PrP
Sc-A
kk
um
ula
tio
n
(4)
(6)
(6)
0/4
1p
m/p
s
++
+-
SN
Sso
mati
sch
es
Nerv
en
syst
em
(4)
(3)
(3)
0/2
3
+
Ggl.
trig
em
inale
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
BA
IHC
0/1
00
/81
0/1
51
1/1
19
/15
2/1
51
/11
4/1
3
35
2 (
± 7
0)
37
2 (
± 1
03
)5
23
(± 9
0)
33
3 (
± 4
1)4
59
(± 5
8)
49
8 (
± 1
00
)5
61
52
5 (
± 5
7)
+1
2/1
5+
15
/15
12
/12
13
/15
11
/11
+1
0/1
21
3/1
51
0/1
4+
(1)
33
1 (±
25
)(1
)2
88
(± 2
6)
34
0 (
± 3
7)
29
8 (
± 2
8)
33
2 (
± 2
3)
(1)
35
9 (
± 5
9)
38
3 (
± 7
0)
34
0 (
± 5
5)
(1)
11
/11
+1
0/1
11
4/1
4+
12
/14
12
/12
8/8
14
/14
9/1
0+
25
7 (
± 3
9)
30
8 (
± 3
5)
311
(± 5
9)
(1)
27
8 (
± 3
9)
32
6 (
± 3
4)
33
9 (
± 5
1)3
31
(± 4
3)
38
9 (
± 8
9)
(1)
+7
/96
/14
14
/14
12
/13
8/8
12
/13
9/1
09
/13
+(1
)2
51
(± 1
7)
46
8 (
± 4
1)3
23
(± 4
0)
28
1 (±
37
)3
37
(± 2
8)
34
8 (
± 2
9)
35
6 (
± 4
1)3
10 (
± 3
6)
(1)
++
+(1
)(1
)(1
)
++
++
(1)
(1)
(1)
(1)
++
+(1
)(1
)(1
)
++
++
+(1
)(1
)(1
)(1
)(1
)
IT 5
6
--
-
28
IT 2
10
n.d
.n
.d.
-
28
IT 2
10
--
--
0
n.d
.
IT 2
23
IT 4
93
N.
vagus
(Bru
st)
n.d
.
Para
sym
path
isch
es
Nerv
en
syst
em
Gem
isch
tfase
rige A
nte
ile
Sy
mp
ath
isch
es
Nerv
en
syst
em
n.d
.
-
Kli
nis
ch
er
Sta
tus
IT-
Num
mer
Mo
nate
p.i
.B
AIH
CB
AIH
C
32
IT 0
90
-1
40
36
44
2IT
38
IT 1
10
-1n
.d.
32
09
0-1
Ggl.
ste
llatu
m
-k
Pk
P
N.
vagus
(Hals
)M
on
ate
p.i
.
R 0
9/0
6n
.d.
--
Ggl.
co
eli
acum
Ggl.
mese
nte
rale
caud.
Nn
. sp
lan
ch
nic
iG
gl.
cerv
icale
cra
n.
kP
Kli
nis
ch
er
Sta
tus
IT-
Num
mer
Peri
ph
ere
s N
erv
en
sy
ste
m
Vegeta
tiv
es
Nerv
en
syst
em
SN
S
Ggl.
no
do
sum
-
--
-
-
--
-
--
-
40
56
0
--
36
11
0-1
-
49
3
44
22
3
38
2
R 0
9/0
6
-
-
-
-
-
--
--
--
-
-
-
Darm
dis
tale
s Il
eum -
kP
n.d
.k
Pn
.d.
kP
kP
--
-
-
Zen
trale
s N
erv
en
sy
ste
m
n.d
.
Rück
en
mark
T7
Rück
en
mark
L3
n.d
.
BA
IHC
n.d
.n
.d.
IHC
n.d
.
kP
kP
-k
P
--
-
kP
kP
kP

96 Ergebnisse
Tab. 4.25 Zusammenfassende Darstellung der histopathologischen Ergebnisse aller
untersuchten Tiere

Ergebnisse 97
4.4 Natürlich infiziertes Rind (R09/06)
Im Obex dieses Rindes ließ sich in hohem Grad das pathologische Prion-Protein in allen
Kerngebieten nachweisen. Intra- und perineuronale Akkumulationen der Nervenzellen sowie
fein- bis grobgranuläre Ablagerungen im Neuropil waren wie bei den experimentell
inokulierten Tieren die häufigsten Reaktionsmuster. Die Proben des Ileums wiesen eine
fortgeschrittene Autolyse auf, die ein Ansprechen der einzelnen Strukturen erschwerte. So
konnte mittels immunhistochemischer Anfärbung weder für die Follikel der Peyer’schen
Platten noch für das ENS ein eindeutig positives Ergebnis erzielt werden. In vereinzelten
Neuronen des Ggl. coeliacum, der sympathischen Ganglien (Ggl. stellatum, Ggl. cervicale
craniale) und des N. vagus konnte PrPSc
in Form des PNS-typischen perineuronalen
Reaktionsmusters detektiert werden. Für das Ggl. trigeminale wurden ebenfalls eindeutige
spezifische PrPSc
-Ablagerungen hauptsächlich in den Satellitenzellen beobachtet. Weitere für
diesen Versuch relevante Proben standen nicht zur Verfügung.

98 Zusammenfassung
5. DISKUSSION
5.1 Ziel der Arbeit
Ziel dieser Arbeit war es, mittels der im Rahmen der deutschen BSE-Pathogenesestudie
gewonnenen Daten und Materialien nach oraler experimenteller den Ausbreitungsweg des
pathologischen Prion-Proteins (PrPSc
) in der präklinischen und klinischen Phase der
Erkrankung aufzuklären. Dafür wurden repräsentative Gewebe vorwiegend aus dem
peripheren Nervensystem auf Infektiosität über den Maus-Bioassay als auch auf PrPSc
-
Ablagerungen in den Proben untersucht.
5.2 Kritische Betrachtung des Untersuchungsmaterials
Das bei der Infektion verwendete infektiöse Hirnhomogenat stammte von englischen an BSE
erkrankten Rindern. Das Homogenat zeigte im SAF-Immunoblot ein für klassische BSE
typisches Glykosylierungs-Profil und wies einen Infektionstiter von 106.1
infektiösen Dosen
(ID) pro g Gewebe auf (HOFFMANN et al, 2007).
Jedes Rind erhielt oral die einmalige Dosis von 100g Inokulat unter der Verwendung einer
Schlundsonde. Die experimentelle Applikation dieser hohen Infektionsdosis soll die
Reproduzierbarkeit und Auswertbarkeit der Infektion verbessern, ist aber wesentlich höher als
die geschätzte natürliche Infektionsdosis von 0,1 bis 1g (WELLS et al. 2007) unter
Feldbedingungen. Deutlich wird dies z.B. durch die wesentlich kürzere Inkubationszeit nach
experimenteller Infektion (ARNOLD et al. 2007). Da jedoch in vorangegangenen Studien
beim Rind äquivalente Dosen eingesetzt wurden (ARNOLD et al. 2007; ESPINOSA et al.
2007; MASUJIN et al. 2007; WELLS et al. 2007), sind die Resultate der vorliegenden Studie
mit denen aus der aktuellen Literatur bekannten Ergebnissen vergleichbar.
Um experimentelle Artefakte z.B. durch Kontaminationen, zu vermeiden, wurde die Sektion
der Rinder nach einem klar definierten Ablauf durchgeführt. So wurden für die
Probenentnahme Einmal-Präparationsbestecke verwendet und die Entnahmen der einzelnen,
unterschiedlich infektiösen Organsysteme, wie beispielsweise Darm und ZNS, räumlich und
personell strikt voneinander getrennt. Ähnlich sorgsame Vorkehrungen wurden später auch
bei der Präparation der Mäusegehirne angewandt.

Diskussion 99
Bisherige Studien wiesen, wenn überhaupt, Ablagerung des PrPSc
nur sehr geringgradig im
PNS BSE- infizierter Tiere nach (ARNOLD et al. 2007; ESPINOSA et al. 2007;
HOFFMANN et al. 2007; MASUJIN et al. 2007). Um eine repräsentative Untersuchung der
Gewebe sicherzustellen, wurden daher bei mindestens 3 Paraffin-Blöcken jeder Probe jeweils
5 Ebenen getestet. Jede Ebene umfasste 5 sukzessive Schnitte und somit eine Tiefe von 15-
25µm. Somit standen Schnitte aus einer Gesamt-Durchdringtiefe von 150-200µm pro Block
für die immunhistochemische und histopathologische Untersuchung zur Verfügung. Der
Anteil der untersuchten Strukturen konnte dadurch deutlich erweitert werden. Frisch
angefertigte Präparate für alle fraglichen Ergebnisse und alle Proben des Ggl. coeliacum und
des Ileums wurden mit einem zweiten Antikörper (6C2) nachuntersucht.
Die verwendete transgene Mauslinie (Tgbov XV) ist hochsensitiv für bereits äußerst geringe
Erregergehalte, da sie das bovine Prion-Protein um den Faktor 10.000 überexprimiert. Eine
spontane, Inokulat-unabhängige Konversion des zellulären Prion-Proteins konnte
ausgeschlossen werden, da diese Mauslinie uninfiziert auch nach über 700 Tagen keine
klinische Erkrankung und kein neu generiertes PrPSc
aufwies (BUSCHMANN u.
GROSCHUP, 2005).
Abhängig von der Untersuchungsmethodik sind Infektiosität und PrPSc
-Ablagerungen als
Parameter für eine BSE-Infektion voneinander abzugrenzen. Der Maus-Bioassay bestimmt
das infektiöse Potential des Ausgangsgewebes über dessen Fähigkeit, in hochempfänglichen
Mäusen BSE zu induzieren. Dagegen weist die IHC direkt die PK-resistente pathologische
Isoform des Prion-Proteins nach. Beobachtungen an Scrapie-infizierten Hamstern (BEEKES
et al. 1996) unterstützten die Prion-Hypothese, in der PrPSc
als alleiniges Pathogen infektiös
ist (PRUSINER et al. 1982, 1987, 1991). Andere Studien wiesen allerdings Infektiosität auch
in Geweben ohne entsprechende Ablagerungen des pathologischen Prion-Proteins nach
(CZUB et al. 1988; BALKEMA-BUSCHMANN et al. 2011).
In der vorliegenden Untersuchung wurden positive Ergebnisse überwiegend mit dem Maus-
Bioassay erzielt, dem lediglich ein einziger Nachweis von PrPSc
ohne Infektiosität (Medulla
oblongata, IT 61, 32 mpi) gegenüberstand. Wies eine Probe gleichzeitig beide BSE-
assoziierte Parameter auf, gingen geringgradige PrPSc
-Akkumulationen in der Regel mit
einem mittel- bis hochgradigen Erregergehalt einher.
Die deutlich höhere Sensitivität des Maus-Bioassays ist hierfür die wahrscheinlichste
Erklärung. Die durchgeführten Untersuchungen basierten jedoch stets auf Stichproben, sodass
negative Befunde auch durch Beprobungsartefakte bedingt sein können.

100 Zusammenfassung
5.3 Allgemeine Betrachtungen
Für das BSE-Agens ist bekannt, dass lediglich die Peyer´schen Platten des Dünndarms und
die Tonsille als lymphatische Komponenten betroffen sind (TERRY et al. 2003; WELLS et
al. 2005; ESPINOSA et al. 2007; HOFFMANN et al. 2011). Darüber hinaus besteht eine
Restriktion auf das Nervensystem (BUSCHMANN u. GROSCHUP, 2005). PrPSc
-
Ablagerungen im Obex und dem Rückenmark lassen eine Ausbreitung des Agens über die
Strukturen des PNS vermuten, wobei wahrscheinlich beim Rind sowohl sympathische als
auch parasympathische Nerven beteiligt sind (HOFFMANN et al. 2007). Welche Strukturen
zu welchem Zeitpunkt involviert sind, konnten die bis dato durchgeführten Studien allerdings
nicht zweifelsfrei bestimmen.
5.3.1 Ergebnisüberblick
Von den 16 ausgewählten mit BSE-Material inokulierten Rindern wiesen alle Tiere PrPSc
und/oder Infektiosität in mindestens einer der 12 untersuchten Gewebe auf. Die ilealen
Peyer`schen Platten waren bei 13 Rindern betroffen und lassen diesen Abschnitt des Darms
als primären Eintrittsstelle des Erregers vermuten. Die Ergebnisse deuten allerdings auf einen
direkten Kontakt des pathologischen Prion-Proteins mit den im Darm lokalisierten Nerven als
Ausgangspunkt für eine weitere Ausbreitung im PNS hin. Die Ausbreitung erfolgt dann vor
allem über autonome Fasern, wobei ein Schwerpunkt auf den sympathischen Strukturen zu
liegen scheint.
Die Nachweise bzw. unterschiedlichen Nachweisintensitäten im ZNS traten bei den einzelnen
Rindern zu mitunter höchst unterschiedlichen Zeitpunkten auf. Daher wurden die Ergebnisse
nicht strikt nach den Inkubationszeiten zusammengefasst, sondern anhand der Ergebnisse im
Obex. Ein besonderes Augenmerk lag dabei auf den sechs Tieren, die negative Befunde in der
Medulla oblongata aufwiesen. Dadurch ließen sich zuverlässige Rückschlüsse auf die
zentripetale Ausbreitung im PNS ziehen, da eine zentrifugale Migration des Agens vom ZNS
aus in diesen Fällen äußerst unwahrscheinlich ist.

Diskussion 101
5.3.2 Dosis, Inkubationszeiten und klinische Symptomatik
Bei der oralen Verabreichung von 1 mg bis 100 g BSE-Homogenat wurde bei Rindern eine
deutliche Dosis-Wirkungsbeziehung hinsichtlich der Inkubationsdauer und dem
Infektionserfolg beobachtet. Schätzungen zufolge liegt die mittlere Inkubationszeit im Feld
bei 5 bis 5,5 Jahren (ARNOLD u. WILESMITH 2004), der vermutlich eine natürliche Dosis
zwischen 0.1g und 1g zugrunde liegt (WELLS et al. 2007). Hohe oral inokulierte Dosen
lassen daher Auswirkungen auf die Transmissionsrate und auf die Länge der Inkubationszeit
erwarten. So beschrieben WELLS et al. (2007) bei einer Dosis von 100g eine
Transmissionsrate von 100%, die auch bei der deutschen BSE-Pathogenesestudie erreicht
wurde. Davon zeugte neben positiven Ergebnissen in mindestens einer der untersuchten
Körperproben auch der Nachweis von Infektiosität bzw. PrPSc
im Darm bei 41 von 46 Tieren
der FLI-Studie (HOFFMANN et al. 2011).
Eine weitere Dosis-Wirkungsbeziehung besteht für den Zeitpunkt der Detektion des
pathologischen Prion-Proteins vor dem Einsetzen klinischer Symptome. Nach Berechnungen
von ARNOLD et al. (2007) war PrPSc
im Obex bei 50% der Tiere, die 100g Inokulat
erhielten, neun Monate vor dem Auftreten einer definitiven Symptomatik nachweisbar.
Wurde lediglich 1g Homogenat verabreicht, verringert sich dieser Zeitraum auf 1,7 Monate.
Diese von Arnold et al. (2007) berechneten Korrelationen waren auch in der FLI-Studie
zutreffend. Die Detektion von PrPSc
bzw. von Infektiosität im Hirnstamm erfolgte acht (28
mpi, IT 21) bzw. 12 Monate (IT 24, 24 mpi) vor den ersten eindeutigen klinischen
Symptomen (36 mpi, IT 49).
Die offensichtlichen Unterschiede sowohl zwischen den einzelnen Pathogenese-Studien als
auch innerhalb der hier untersuchten Tiergruppe sind am ehesten durch eine hohe biologische
Varianz zwischen den einzelnen Tieren erklärbar. Bei einer Dosis von 100g liegt die
Zeitspanne für das Auftreten klinischer Symptome in der Studie von WELLS et al. (2007)
zwischen 31-60 mpi, bei ARNOLD et al. (2007) bei 35-59 mpi und in der FLI-Studie
zwischen 32 und 54 mpi. Diese Varianz verdeutlicht, dass sich die Rinder nicht anhand der
Inkubationszeiten vergleichen ließen. Dieser Tatsache wurde in der eigenen Studie Rechnung
getragen, indem die Ergebnisse der Medulla oblongata als Grundlage für die Einordnung in
eine der drei Gruppen dienten (kein Nachweis, geringer Nachweis, mittel- bis hochgradiger
Nachweis). So konnten die weiteren Resultate aus dem PNS, Rückenmark und Darm von
Tieren des gleichen Inkubationsstatus vergleichend betrachtet werden.

102 Zusammenfassung
Für die immer noch unterschiedlichen Resultate ist neben den individuellen Schwankungen
der einzelnen Rinder die unterschiedliche effektive Dosis der Homogenate bei identisch
verabreichten Mengen von 100g in Betracht zu ziehen. So basierten die oralen Inokulationen
bei den in Großbritannien durchgeführten Experimenten und bei der deutschen
Pathogenesestudie zwar auf ähnlichen Dosen (VLA: 103.10
- 103.5
i.c./i.p. ID50 x g-1
[WELLS
et al. 1996; ARNOLD et al. 2007; MASUJIN et al. 2007]; FLI: 106.1
ID50 x g-1
). Ein direkter
Vergleich der Infektiosität der verwendeten Homogenate ist jedoch aufgrund der
unterschiedlichen für die Titerbestimmung eingesetzten Mauslinien (VLA: RIII; FLI: Tgbov
XV) nur bedingt möglich. Dies wird besonders bei der Gegenüberstellung der Ergebnisse
unter Verwendung desselben Inokulates deutlich (BUSCHMANN u. GROSCHUP 2005). Die
Unterschiede hinsichtlich der Sensitivität des Maus-Bioassays werden sowohl durch höhere
Transmissionsraten (Hirnstamm: 7/15 für RIII; 14/14 für Tgbov XV) als auch verkürzte
Inkubationszeiten (Hirnstamm: 394d für RIII; 208d für Tgbov XV) offensichtlich.
Rasse-und Genotyp-spezifische Effekte bei den Rindern wurden bisher noch nicht
beschrieben (WELLS u. SIMMONS 1996), sind aber als weitere mögliche Co-Faktoren
dennoch nicht vollständig auszuschließen.
5.3.3 Zusammenhänge von erhobenen Befunden
In vorangegangenen Publikationen wurde bereits versucht, Zusammenhänge zwischen der
PrPSc
-Detektion im Gehirn, der vakuolären Degeneration und klinischen Symptomen
herzustellen. Besonders ausführlich diskutierten JEFFREY u. GONZALES (2007) mögliche
Korrelationen für Ergebnisse bei Scrapie, eindeutige Erkenntnisse konnten dabei aber nicht
gewonnen werden.
Eine histopathologisch nachweisbare spongiforme Enzephalopathie konnte eindeutig bei fünf
der 16 hier untersuchten Tiere und bei zwei weiteren Tieren ansatzweise nachgewiesen
werden. Bei all diesen Tieren waren PrPSc
-Akkumulationen nachweisbar, sodass ein
neurotoxischer Einfluss des pathologischen Prion-Proteins möglich erscheint. Morphologisch-
funktionelle, aber auch molekularbiologische Methoden könnten in Zukunft bei der
Ermittlung eines Zusammenhanges zwischen PrPSc
-Ablagerungen und den spongiformen
Auflockerungen hilfreich sein.
Auch im eigenen Untersuchungsgut zeigte das Auftreten klinischer Symptome weder mit
PrPSc
-Ablagerungen noch dem Auftreten spongiformer Veränderungen klare

Diskussion 103
Zusammenhänge. Insbesondere bei der Einschätzung des klinischen Status sind die
subjektiven Einflüsse unterschiedlichen Arbeitsgruppen zu beachten. Obgleich die Parameter
für die Bestimmung der Klinik ähnlich waren, so gestaltete sich die Bestimmung
abweichender Verhaltensweisen insbesondere im frühen Stadium äußerst schwierig.
Die fehlende Korrelation zwischen den verschiedenen Parametern verdeutlicht, dass die
Kenntnisse über die Mechanismen der entscheidenden pathologischen Manifestation einer
BSE-Erkrankung weiterhin fehlen. Die neueste Pathogenesestudie an der VLA (ARNOLD et
al. 2007) scheint aufgrund ihrer hohen Tierzahl geeignet, hierzu weiterführende Erkenntnisse
zu gewinnen.
5.3.4 Vergleich von Feldinfektion vs. experimentelle Studie
Mit der immunhistologischen Untersuchung eines natürlichen BSE-Falls konnte ein Vergleich
zum Reaktionsmuster der experimentell infizierten Rinder gezogen werden. Geboren im März
2000 war das dieses Tier zum Zeitpunkt der Euthanasie 6 Jahre alt und gelangte als klinischer
Verdachtsfall R09/06 zur Untersuchung an das FLI.
Bereits in den Studien von WELLS et al. (2007) wiesen Feldinfektionen und Versuchstiere
nahezu identische PrPSc
-Reaktionsmuster auf. Und so ließen sich auch bei der R09/06 die
gleichartigen Akkumulationen in den vorliegenden Proben des zentralen und peripheren
Nervensystems detektieren. Dabei blieb in Übereinstimmung mit den Daten von TERRY et
al. (2003) ein Nachweis von PrPSc
im Darm aus, möglicherweise bedingt durch die
fortgeschrittenen autolytischen Prozesse. Bei der Verwendung der hohen Dosis in den
experimentellen Studien scheint sich die Ausbreitung des pathologischen Prion-Proteins
allerdings zu beschleunigen (s. Kap 2.3.1, S.37). Vergleichbare Reaktionsmuster legen jedoch
eine enge Beziehung in Bezug auf die in die Pathogenese involvierten Nerven und Ganglien
des PNS zwischen der natürlichen und experimentellen Infektion nahe.

104 Zusammenfassung
5.4. Pathogenese der BSE
5.4.1 Rolle des lymphatischen Systems
Vorherige Studien zeigten, dass von einer weitgehenden Restriktion des BSE-Agens auf das
Nervensystem auszugehen ist (BUSCHMANN u. GROSCHUP, 2005) und dass das
lymphoretikuläre System bis auf wenige Ausnahmen nicht in die Erregerausbreitung
involviert zu sein scheint. So konnte für BSE beim Rind Infektiosität im lymphatischen
System bisher lediglich in der Tonsille (WELLS et al. 2005; ESPINOSA et al. 2007) und den
Peyer‘schen Platten des Jejunums, Ileums und Ileozäkal-Einganges (HOFFMANN et al.
2011) detektiert werden. Entsprechende PrPSc
-Ablagerungen waren zudem in den
lymphatischen Einrichtungen von Jejunum (STACK et al. 2011), Ileum (TERRY et al. 2003;
HOFFMANN et al. 2011) und Kolon (OKADA et al. 2010) nachweisbar. Als die
wahrscheinlichsten Eintrittsorte für den Erreger wurden in der hier vorliegenden Studie somit
nur die Peyer`schen Platten des distalen Ileums berücksichtigt.
Während der PrPSc
-Nachweis bei Scrapie im Schaf bereits nach 30 Tagen p.i. in den
Peyer’schen Platten des distalen Ileums erfolgte (JEFFREY et al. 2006; RYDER et al. 2009),
war diese Zeitspanne beim Rind deutlich länger. Ein erster Nachweis des pathologischen
Prion-Proteins in den darmassoziierten Lymphfollikeln gelang bei experimentell (mit 100g
Inokulat) infizierten Rindern ab dem vierten mpi (HOFFMANN et al. 2011, STACK et al.
2011), gefolgt von beständigen Ablagerungen bis zum 44./45. mpi. Der in der deutschen
BSE-Pathogenese-Studie beobachtete deutliche Peak positiver Follikel 12 und 24 mpi (im
geringeren Ausmaß auch bei 36 mpi) (HOFFMANN et al. 2011), wurde als Verschiebung des
Gleichgewichtes zwischen Akkumulation und Abbau zugunsten der Akkumulation gedeutet
und konnte in der britischen Studie nicht gezeigt werden (STACK et al. 2011). Die eigenen
Ergebnisse waren Teil der erwähnten deutschen Publikation und ergaben bei den meisten
(11/16) Tieren nur in vereinzelten Follikeln ein positives Ergebnis. Lediglich drei Tiere (24
mpi, 32 mpi, 36 mpi) wiesen einen deutlich erhöhten Anteil positiver Follikel zwischen 5%
und 16,7% auf. Die Ursache dafür bleibt unklar, eine lokal begrenzte Aufnahme der Prion-
Erreger aus dem Darmtrakt ist dabei ebenso möglich wie ein bestehendes Gleichgewicht
zwischen Akkumulation und Abbau. Auch eine Akkumulation unterhalb der
Sensitivitätsgrenze der immunhistologischen Untersuchung ist denkbar, obwohl die
Ergebnisse von HOFFMANN et al. (2011) im Bereich des Ileums keine wesentlichen
Unterschiede zwischen Maus-Bioassay und IHC aufwiesen.

Diskussion 105
Wie auch bei den älteren Tieren in der hier vorgelegten Studie ließen auch alle vorherigen
Untersuchungen eine Abnahme der Gesamtzahl untersuchter Follikel im Verlauf des
Versuchs erkennen. Dieses Phänomen ist durch die altersabhängige physiologische Involution
dieser lokalen lymphatischen Komponente erklärbar (DEFAWEUX et al. 2007).
Interessanterweise steht dem bis zu einem gewissen Punkt ein erhöhter Anteil an PrPSc
-
positiven Follikeln gegenüber. Dieser stellte sich in der Studie am FLI als bereits erwähnte
Peaks dar (HOFFMANN et al. 2011). Allerdings war in den Untersuchungen des britischen
VLA-Experiments (STACK et al. 2011) ein eher stetiger Anstieg zu beobachten. In der
späten Inkubationsperiode war beiden Studien (ab ca. 40 mpi) dann der nur noch sporadische
Nachweis von PrPSc
in den Follikeln gemein. Dies konnte auch bei den hier untersuchten
Proben beobachtet werden, hier wiesen vier der fünf negativen Tiere Inkubationszeiten von
36-44 mpi auf. Wahrscheinlich ist die altersabhängige Abnahme der Anzahl der vorhandenen
Follikel für die Reduzierung der Nachweiswahrscheinlichkeit verantwortlich und erklärt
dessen nur noch sporadischen Nachweis zu den späten Inkubationszeitpunkten. Die von
TERRY et al. (2003) diskutierte verringerte Involution von PrPSc
enthaltenen Follikeln ist für
die verbleibenden positiven Nachweise denkbar.
Die von PrPSc
-Akkumulationen betroffenen zellulären Strukturen innerhalb der Follikel
wiesen in allen Studien inklusive der eigenen Ergebnisse stets Reaktionsmuster auf, die eine
Assoziation mit TBM und/oder FDC wahrscheinlich machen. Kontrovers wurden allerdings
die Ergebnisse hinsichtlich des Auftretens der entsprechenden Zelltypen beschrieben.
Während in der frühen Inkubationsperiode zunächst übereinstimmend TBM-assoziierte
Ablagerungen detektiert wurden, waren in der Untersuchung von TERRY et al. (2003) die
FDC erst im klinischen Stadium zusammen mit dem Nachweis von PrPSc
im ZNS involviert.
Im Gegensatz dazu beschrieben HOFFMANN et al. (2011) die Beteiligung von FDC bereits
16 mpi bei einem präklinischen Tier und weiterhin das simultane Auftreten beider Zelltypen
während der gesamten untersuchten Inkubationsperiode. Auch im eigenen Untersuchungsgut
ließ sich keine temporäre Dominanz hinsichtlich der beteiligten Zelltypen feststellen.
Es muss jedoch betont werden, dass die Einbeziehung der FDC zu keinem Zeitpunkt der
BSE-Infektion dem Ausmaß entspricht, wie sie beispielsweise für Scrapie beschrieben ist.
Wie in der Studie von TERRY et al. (2003) so lieferte auch diese Studie keine spezifischen
Ablagerungen in den Follikeln nach einer Feldinfektion. Das hier untersuchte Ileum der
R09/06 wies allerdings eine fortgeschrittene Autolyse auf und ließ deshalb ein eindeutiges

106 Zusammenfassung
Ansprechen der Follikel nicht zu. Die bereits erwähnte altersbedingte Involution der Follikel
erschwerte die Detektion zusätzlich. Des Weiteren ist die Detektionswahrscheinlichkeit
positiver Follikel dosisabhängig. Diese lag nach experimenteller Inokulation von 1g
Homogenat nur bei einem Prozent und stieg erst bei Verabreichung von 100g Inokulat auf
45,5% an (STACK et al. 2011). Somit lässt sich das Ausbleiben einer Detektion auch durch
die natürlich vorkommende geringe Exposition mit infektiösem Material erklären. Weiterhin
könnte die angesprochene Degradation des PrPSc
durch TBM für Akkumulationen unterhalb
der IHC-Nachweisgrenze verantwortlich sein.
Die Frage nach der Rolle des lymphatischen Systems, insbesondere der der Peyer`schen
Platten des Ileums, bei der BSE-Pathogenese lässt sich mit den Ergebnissen dieser Studie
nicht vollständig beantworten. Indizien, wie die des bisher fehlenden Nachweises in natürlich
infizierten Rindern und die frühe Beteiligung des ENS (16 mpi) als möglicher Ausgangspunkt
für eine weitere Ausbreitung, sprechen aber gegen eine essentielle Beteiligung der
Peyer`schen Platten. Vielmehr kann aufgrund der hohen Dosis und dem damit verbundenen
Infektionsdrucks eine lokal begrenzte Akkumulation nach der Aufnahme des BSE-Agens über
den Darm vermutet werden. Warum allerdings bisher noch keine Involvierung von weiteren
lymphatischen Geweben beschrieben wurde, bleibt unklar. Der Einsatz des hochsensitiven
Maus-Bioassay mit der Inokulation verschiedener Lymphgewebe könnte einen möglichen
Ansatz zur weiteren Klärung darstellen.
5.4.2 Rolle des ENS und Neuroinvasion
Resultate aus Studien mit Scrapie zeigten in klinisch erkrankten Schafen PrPSc
-Ablagerungen
im ENS Schafen, die vermutlich auf eine initiale Infektion des lymphatischen Systems
zurückzuführen ist (VAN KEULEN et al. 2000, 2002). Dabei traten eindeutige lokale
Zusammenhänge zwischen positiven Neuronen des ENS und positiven Follikeln auf
(HEGGEBØ et al. 2003). Bei BSE-infizierten Rindern war der Nachweis im ENS zunächst
nur 38 Monate p.i. möglich und auf klinisch erkrankte Tieren beschränkt (TERRY et al.
2003). Nur eines von 98 Rindern wies dabei PrPSc
-Akkumulationen im ENS bei einer Dosis
von 100g Inokulat nach 36 mpi auf, die Reduktion der verabreichten Menge auf 1g führte erst
nach 51 mpi zu einem positiven Ergebnis. Interessanterweise wies das mit der geringen Dosis
infizierte Tier dabei keine Ablagerungen in den Follikeln auf. In der Publikation von

Diskussion 107
HOFFMANN et al. (2011), die die Ergebnisse der vorliegenden Arbeit mit einbezieht, wurde
die Beteiligung des ENS bei 18 von 46 untersuchten Rindern gezeigt. Dabei gelang die
initiale Detektion von PrPSc
im frühen präklinischen Stadium nach 16 und 24 mpi bei zwei
Rindern dieser Untersuchung. Ab dem 32. Monat p.i. waren dann zuverlässig Ablagerungen
nachweisbar. Ausgehend von den Tieren dieser Studie waren beide Plexus mit dem
Schwerpunkt auf dem Plexus myentericus involviert, während im Gegensatz dazu STACK et
al. (2011) ausschließlich die Beteiligung des Plexus submucosus beschreibt. Eine
topographische Assoziation zwischen PrPSc
-Akkumulationen in den Follikeln und im ENS
konnte dabei mit Ausnahme der IT 09 nicht hergestellt werden.
Dieser fehlende Bezug lässt einen frühen pathogenetischen Zusammenhang zwischen beiden
Lokalisationen von PrPSc
eher zweifelhaft erscheinen. Für eine entscheidende Beteiligung des
ENS an der Ausbreitung des BSE-Agens sind die anatomischen Voraussetzungen einerseits
über die Nervenendigungen in der Submukosa nahe dem Darmlumen (FURNESS, 2006) und
andererseits über den direkten Kontakt sympathischer Nervenfasern mit den FDC’s und
Makrophagen der Follikel (DEFAWEUX et al. 2007) am wahrscheinlichsten. DEFAWEUX
et al. (2007) beschrieben weiterhin den direkten Kontakt zwischen intestinal aufgenommenem
PrPSc
und freien Nervenendigungen der Darmschleimhaut für den möglichen Beginn der
Neuroinvasion. Eine Infektionsroute über Nervenendigungen unterhalb der intestinalen
Mukosa wurde auch von JEFFREY et al. (2006) favorisiert. Die geringe Ausprägung der
PrPSc
-Akkumulationen im ENS deutet auf eine kurze Transitphase des Erregers an der Grenze
der immunhistochemischen Detektierbarkeit hin. Diese Nachweiswahrscheinlichkeit steigt
mit der Untersuchung von jeweils fünf Ebenen jedes Paraffinblockes. Diese hier erstmals
angewendete Schnitttechnik kann als Erklärung für die Diskrepanz der Ergebnisse zu früheren
Studien angesehen werden. Die zum späteren Zeitpunkt der Erkrankung vermehrt
beobachteten Anfärbungen könnten ursächlich auf eine vermehrte lokale Akkumulation
zurückzuführen sein. Eine zentrifugale Ausbreitung des PrPSc
nach Replikation im ZNS kann
in diesem Fall jedoch nicht sicher ausgeschlossen werden.
5.4.3 Ausbreitung im peripheren Nervensystem
Basierend auf den Ergebnissen früherer Studien (BUSCHMANN u. GROSCHUP, 2005;
HOFFMANN et al. 2007) wurde der Fokus der vorliegenden Arbeit auf die Ausbreitung des

108 Zusammenfassung
BSE-Agens entlang den Strukturen des PNS während der frühen und späten Phase der
Erkrankung gelegt.
5.4.3.1 Ausbreitungsrichtung
Bei der Betrachtung der Verbreitung des infektiösen BSE-Agens im Körper werden in der
Literatur die Termini zentrifugal und zentripetal unterschieden. Dabei beschreibt zentripetal
die Ausbreitung von PrPSc
zum Gehirn hin. Zu einer zentrifugalen Dissemination ausgehend
vom ZNS in Richtung des PNS kommt es nach der Replikation von PrPSc
im Gehirn und
Rückenmark. (BEEKES u. MCBRIDE 2007; KRATZEL et al. 2007b)
In dieser Studie wurde unter anderem das Ggl. trigeminale untersucht, das über den N.
trigeminus mit einem Kerngebiet im Obex weite Teile des Kopfes sensibel und motorisch
innerviert und damit unabhängig vom PNS des Brust- und Bauchraumes agiert.
Bei der Betrachtung der anatomischen Lage dieses Ganglions war weniger der Zeitpunkt in
der Inkubationsperiode als der Grad der PrPSc
-Akkumulationen im Obex interessant. Von
einem sekundären Befall des Ganglions ausgehend, konnte es als Bestimmung der ersten
zentrifugalen Dissemination herangezogen werden. So lässt sich der Nachweis des
pathologischen Prion-Proteins im Ggl. trigeminale bereits nach äußerst geringer
Akkumulation im Obex (IT 25, 40 mpi) als eine sehr frühzeitige zentrifugale Ausbreitung auf
periphere Strukturen deuten.
Die für die Betrachtung der frühen Pathogenese notwendige Unterscheidung zwischen
zentripetaler und zentrifugaler Ausbreitung lässt sich daher anscheinend nur ohne vorherige
Beteiligung des ZNS feststellen. Diese Voraussetzung konnten die vergleichbaren Studien
allerdings bisher nicht erfüllen (s. Tab.2.6, S.42).
5.4.3.2 Gehirn
Nach den OIE-Statuten ist die Medulla oblongata mit der Obexregion für die BSE-Diagnostik
heranzuziehen. ARNOLD et al. stellten (2007) in ihren Studien zum Nachweis von PrPSc
in
Relation zur klinischen Symptomen nochmals heraus, dass im Obex der früheste Nachweis
von PrPSc
geführt werden kann. Wie für Scrapie (BEEKES et al. 1999; VAN KEULEN et al.
2000, 2002; MCBRIDE et al. 2001) so wird auch für BSE im Rind stets der DMNV als
initiale Eintrittsstelle des pathologischen Prion-Proteins (SCHULZ-SCHAEFFER et al. 2000;
HOFFMANN et al. 2007) angenommen.

Diskussion 109
Im eigenen Untersuchungsgut zeigten sechs Tiere weder PrPSc
-Akkumulation noch
Infektiosität im Gehirn, während drei Tiere eine geringe und sieben Rinder eine mittel- bis
hochgradige Beteiligung der Medulla oblongata aufwiesen. Interessanterweise fiel die
Diskrepanz zwischen den Ergebnissen des Maus-Bioassay und der IHC im Gehirn deutlich
geringer aus als für die Gewebe des PNS, d.h. Infektiosität konnte nicht deutlich vor den
ersten Ablagerungen detektiert werden. Während nahezu alle Tiere sowohl Infektiosität als
auch PrPSc
-Akkumulationen auswiesen, konnte bei einem Tier (IT24, 24 mpi) Spuren von
Infektiosität ohne detektierbare Akkumulationen nachgewiesen werden. Dagegen zeigte ein
weiteres Rind (IT61, 28 mpi) eine lokale, sehr geringgradige neuronale Akkumulation bei
einem negativen Ergebnis im Maus-Bioassay. Neben individuellen Unterschieden zwischen
den Versuchsrindern kann das auf eine schnellere Replikation im ZNS als im PNS hinweisen,
so dass sehr schnell in der IHC detektierbare PrPSc
-Level erreicht werden.
Geringgradige PrPSc
-Ablagerungen traten im Obex erstmals nach 28 mpi und 32 mpi (IT 21,
IT 61) in der Formatio reticularis sowie im Nucleus motorius nervi trigemini und dem
Nucleus tractus solitarii 40 mpi (IT 25) auf. Neben feingranulären Ablagerungen im Neuropil
waren dabei auch extra- und intrazelluläre Akkumulationen einzelner Neurone zu beobachten.
Interessanterweise war der DMNV bei keinem dieser drei Rinder involviert.
Mittel- bis hochgradige PrPSc
-Akkumulationen fanden sich wie in vorangegangenen Studien
zu Scrapie (VAN KEULEN et al. 2000) weitverbreitet in den verschiedenen Regionen des
Obex.
Mögliche Theorien für das Auftreten des PrPSc
in den hier beobachteten Kerngebieten werden
im Kap. 5.4.4 (S.113) diskutiert.
5.4.3.3 Rückenmark
Die zweite Komponente des ZNS stellt das Rückenmark dar. PrPSc
-Ablagerungen waren
zumeist mit Akkumulationen im Obex assoziiert. So ließen sich einzelne PrPSc
-positive
Neurone im ZNS (DMNV) und der Substantia intermedia lateralis des Rückenmarkes bei
präklinischen Schafen mit Scrapie 10 mpi (VAN KEULEN et al. 2000) nachweisen.
Ausgehend vom thorakalen Abschnitt 8-10 (10 mpi) breitete sich das pathologische Prion-
Protein beim Schaf zentrifugal und zentripetal über das gesamte Rückenmark (26 mpi) aus.
Während sich bei einem Rind der deutschen Pathogenesestudie 24 mpi PrPSc
in allen
Abschnitten des Rückenmarks nachweisen ließ (HOFFMANN et al. 2007), konnten WELLS
et al. (1998) Akkumulationen des pathologischen Prion-Proteins 32 mpi bei einem Tier ohne

110 Zusammenfassung
gleichzeitige Ablagerungen im Obex im thorakalen und zervikalen Segment zeigen. Oral
inokulierte Hamster wiesen 89 Tage p.i. PrPSc
Initial in den Thorakalsegmenten 4-9 auf,
wobei den Berechnungen zufolge insbesondere der siebte Abschnitt durch periphere Nerven
infiziert wird (BEEKES et al. 1996). Basierend auf diesen Ergebnissen wurden für die
vorliegende Studie repräsentative Rückenmarksproben aus dem siebten thorakalen und dem
dritten lumbalen Segment untersucht.
Im eigenen Untersuchungsgut stellte die Detektion von Infektiosität im siebten thorakalen
Segment 16 mpi (IT 28) ohne Beteiligung des Obex den bisher frühesten Nachweis im
Rückenmark beim Rind dar. Im Zusammenhang mit geringen Ablagerungen in der Medulla
oblongata ließen sich PrPSc
-Akkumulationen erstmals in beiden untersuchten Abschnitten 28
mpi (IT 21) beobachten. Interessanterweise wies der Obex beim Tier IT 61 (32 mpi) dann ein
positives Neuron auf, während im Thorakal- und Lumbalabschnitt ausschließlich Infektiosität
detektiert wurde. Nach umfangreicher Replikation des pathologischen Prion-Proteins im
Gehirn decken sich die beobachteten Reaktionsmuster sowie die Lokalisation der
Ablagerungen mit den Ergebnissen vorangegangener Studien (VAN KEULEN et al. 2000,
2002; HOFFMANN et al. 2007).
Der Vergleich von Ablagerungen/Infektiosität im Obex und dem Rückenmark zeigte
besonders im präklinischen Stadium keine offensichtliche Assoziation. In keinem Fall war der
Nachweis von Infektiosität in den Rückenmarkssegmenten äquivalent zur Detektion in der
Medulla oblongata. Und auch bei der Betrachtung der Intensität der PrPSc
-Ablagerungen
wichen die Ergebnisse in den untersuchten Anteilen des ZNS voneinander ab. Es lässt sich
somit durchaus vermuten, dass es sich bei der Infektion von Gehirn und Rückenmark um zwei
separate Prozesse handelt. Die Rolle des Rückenmarks in der Pathogenese wird bei der
abschließenden Diskussion möglicher Ausbreitungswege behandelt.
5.4.3.4 Gemischtfaserige Ganglien
Die gemischtfaserigen Ganglien wurden im eigenen Untersuchungsgut durch das Ggl.
coeliacum und das Ggl. mesenteriale caudale repräsentiert.
Das erstgenannte Ganglion fungiert als Prävertebralganglion und schaltet die den Darm
innervierenden Nervenfasern des sympathischen N. splanchnicus auf die sekundären
postganglionären Neurone des Rückenmarkes um. Dabei führt es durch den Anschluss von
Zweigen des N. vagus auch parasympathische Fasern. Im Rahmen von TSE-Erkrankungen
konnte PrPSc
in der Regel bisher nur im Zusammenhang mit Akkumulationen im ZNS im Ggl.

Diskussion 111
coeliacum von Hamstern (MCBRIDE u. BEEKES 1999; MCBRIDE et al. 2001), Schafen
(VAN KEULEN et al. 2000, 2002) und einem Rind (HOFFMANN et al. 2007) nachgewiesen
werden.
In dieser Studie wurde im Ggl. coeliacum ab 16 und 20 mpi Infektiosität in Zusammenhang
mit dem initialen Nachweis von Infektiosität im sympathischen und parasympathischen
Nervensystem detektiert. Der Obex war zu diesem Zeitpunkt nicht betroffen. Die erstmalige
PrPSc
-Akkumulation 32 mpi (IT 09, IT 61) ging wie in der Fallstudie (24 mpi) von
HOFFMANN et al. (2007) mit der Detektion im Obex einher. Mit dem Fortschreiten der
Inkubationszeit waren Ablagerungen/Infektiosität dann konstant nachweisbar. In
Übereinstimmung mit den Thesen vorangegangener Untersuchungen kann das Ggl. coeliacum
als erste Struktur des PNS in der Pathogenese angesehen werden. Der zunächst fehlende
Nachweis im Obex sowie die anatomische Lage dieses Ganglion unterstreichen die
Verlässlichkeit dieser Hypothese. Die funktionelle Rolle wird im Kapitel 5.4.4 (S.113)
ausführlicher diskutiert.
In der o.g. Fallstudie (HOFFMANN et al. 2007) wies zusätzlich auch das Ggl. mesenteriale
caudale des präklinischen Rindes 24 mpi Ablagerungen des pathologischen Prion-Proteins
auf. Dieses Ganglion ist u.a. für die Innervation des Colon descendens und des Rectum
verantwortlich, hat jedoch zusätzlich zahlreiche Verbindungen mit den anderen
Bauchganglien. Sowohl für BSE-infizierte Rinder (OKADA et al. 2010) als auch für Scrapie-
infizierte Schafe (RYDER et al. 2009) konnten bereits PrPSc
-Akkumulationen im Colon
nachgewiesen werden. Diese Akkumulationen könnten eine denkbare Erklärung für die
Einbeziehung des Ggl. mesenteriale caudale sein. Dem gegenüber steht die Fallstudie von
HOFFMANN et al. (2007), bei der sich PrPSc
im Ganglion (bei gleichzeitigen geringgradigen
Befall des Obex), nicht aber im Colon nachweisen ließ. Die Autoren interpretierten dieses
Phänomen als mögliche zentrifugale Ausbreitung, möglicherweise auch über die Verbindung
zu den ebenfalls betroffenen Ggl. coeliacum. Die am eigenen Untersuchungsgut erhobenen
Befunde unterstützten diese Theorie, da sowohl Infektiosität als auch pathologisches Prion-
ProteinPrion-Protein erst nach hoher Akkumulation im ZNS (Obex) detektierbar waren.
Die Beteiligung des Ggl. mesenteriale caudale an der zentripetalen Ausbreitung ließ sich über
die Verbindung zum Ggl. coeliacum oder die angeschlossenen Nn. splanchnici minores zum
lumbalen Anteil des Rückenmarks nicht sicher ausschließen. Letztere Möglichkeit ist
aufgrund der initiale Befalls des thorakalen Abschnittes beim Hamster (BEEKES et al. 1996)
und der IT 28 (16 mpi) dieser Studie aber eher unwahrscheinlich.

112 Zusammenfassung
Da das Ggl. mesenteriale caudale im Gegensatz zum Ggl. coeliacum bei der Probenentnahme
nicht makroskopisch sichtbar war, führte das daraus resultierende Fehlen an auswertbaren
Nervenzellen in einer Vielzahl der Proben zu einer eingeschränkten Aussagekraft und zu
Schwierigkeiten bei der Interpretation der Ergebnisse.
5.4.3.5 Sympathikus
In der vorliegenden Arbeit wurden als sympathische Komponenten der N. splanchnicus, das
Ggl. cervicale craniale (GCC) und das Ggl. stellatum untersucht. Der N. splanchnicus stellt
nicht nur eine funktionelle Verbindung zwischen dem Ggl. coeliacum und dem thorakalen
Rückenmark her, es besteht auch ein direkter Kontakt zum sympathischen Grenzstrang.
Dieser Nervenstrang wiederum enthält sowohl das GCC als auch das Ggl. stellatum.
Über die Beteiligung des sympathischen Nervensystems an der Pathogenese der BSE ist
wenig bekannt, andere Autoren (ARNOLD et al. 2007; KIMURA et al. 2008) berichteten
zudem von negativen Ergebnissen für die Anteile des Sympathikus wie dem GCC, dem
sympathischen Grenzstrang und dem Ggl. stellatum. Der bisher einzige Nachweis von PrPSc
in Anteilen des Sympathikus gelang im Ggl. stellatum bei drei experimentell infizierten
Tieren, die ausnahmslos Ablagerungen des pathologischen Prion-Proteins im Obex aufwiesen
(MASUJIN et al. 2007). In der eigenen Untersuchung konnte hingegen bereits in der sehr
frühen präklinischen Phase eine erste Beteiligung des Sympathikus in Form von Infektiosität
ab 16 mpi ohne gleichzeitige Beteiligung der Medulla oblongata nachgewiesen werden.
Weiterhin erfolgte der Nachweis einer marginalen Infektiosität für die kaudalen Abschnitte
der Medulla oblongata bei gleichzeitig mittleren bis hohen Übertragungsraten für die
untersuchten sympathischen Proben (IT 24, 24 mpi). Ein Nachweis von PrPSc
-
Akkumulationen gelang jedoch lediglich im GCC der IT 11 (36 mpi).
Diese Ergebnisse zeigen den bisher umfangreichsten und frühesten Nachweis von
Infektiosität im sympathischen Teil des PNS von Rindern. Dabei war das GCC besonders
präsent und wies so auf eine entscheidende Rolle des sympathischen Nervensystems in der
Pathogenese der BSE hin (s. Kap. 5.4.4, S.113).
5.4.3.6 Parasympathikus
Aufgrund der initialen Beteiligung des DMNV bei der Akkumulation von PrPSc
in der
Obexregion wurde die zentripetale Ausbreitung des pathologischen Prion-Proteins entlang des

Diskussion 113
N. vagus im Scrapie-Hamster-Modell (BEEKES et al. 1998), bei Scrapie infizierten Schafen
(VAN KEULEN et al. 2002) und auch bei BSE-infizierten Rindern (HOFFMANN et al.
2007) diskutiert. Erst im terminalen Stadium konnte Infektiosität direkt im N. vagus bei
Hamstern (MCBRIDE et al. 2001) und Rindern (MASUJIN et al. 2007) nachgewiesen
werden. PrPSc
-Ablagerungen wurden zudem in Scrapie-infizierten Hamstern nach Beteiligung
des ZNS im Ggl. nodosum detektiert (MCBRIDE et al. 2001). Ohne den direkten Nachweis
im N. vagus in der frühen Inkubationsperiode lässt sich aus den bisherigen Studien daher
lediglich die Beteiligung dieses Nerven an der zentripetalen Ausbreitung vermuten. So
wurden in der vorliegende Studie daher der Hals- und Brustabschnitt des N. vagus sowie
dessen kaudal dem ZNS angeschlossenen Ggl. nodosum untersucht.
Im Gegensatz zu den vorgenannten Autoren wurde im eigenen Untersuchungsgut bereits sehr
früh und ohne Beteiligung des ZNS Infektiosität im N. vagus der IT 60 und im Ggl. nodosum
der IT 50 (beide 20 mpi) detektiert, was eindeutig die frühe zentripetale Beteiligung
parasympathischer Strukturen zeigt. Darüber hinaus gelang es erstmalig bei mehreren Tieren
beginnend 32 mpi (IT 09) PrPSc
-Akkumulationen in eingelagerten Neuronen des N. vagus zu
detektieren. Aufgrund der schwer zu separierenden anatomischen Strukturen des Ggl.
nodosums waren Neurone in den Gewebeproben nur spärlich vorhanden, sodass hier lediglich
ein klinisch erkranktes Tier (IT 22, 44 mpi) PrPSc
-Akkumulationen aufwies. Diese traten
jedoch im Zusammenhang mit mittelgradigen PrPSc
-Ablagerungen im Gehirn und hohen
Transmissionsraten im Maus-Bioassay in allen Anteilen des PNS auf und zeugten bereits von
einer intensiven zentrifugalen Verbreitung des BSE-Agens im Körper.
So bestätigten die Ergebnisse dieser Studie die bereits diskutierte These von der zentripetalen
Ausbreitung entlang des N. vagus. Diese schließt im Gegensatz zu den Vermutungen von
BEEKES et al. (2006) das Ggl. nodosum mit ein. Die Bedeutung des parasympathischen
Systems bei der zentrifugalen Ausbreitung wird im anschließenden Kapitel erläutert.
5.4.4 Hypothese zur Ausbreitung im PNS
Basierend auf den Erkenntnissen der aktuellen Literatur ist das Ileum mit den assoziierten
Peyer’schen Platten der wahrscheinlichste Eintrittsort des BSE-Agens nach oraler Exposition.
Dabei kann anhand der vorliegenden Ergebnisse sowie den Ergebnissen anderer Autoren eine
entscheidende Beteiligung der lymphatischen Komponente bezweifelt werden, vielmehr
gelten die Strukturen des ENS als Ausgangspunkt für die Ausbreitung entlang der peripheren
neuronalen Strukturen auf dem Weg zum Gehirn (s. Kap.5.4.2, S.106)

114 Zusammenfassung
Mit den Ergebnissen aus Scrapie-Studien am Hamster- und Schafmodell (BALDAUF et al.
1997; BEEKES et al. 1998; MCBRIDE & BEEKES, 1999; VAN KEULEN et al. 2000, 2002;
MCBRIDE et al. 2001) wurden zwei Routen des Agens auf dem Weg zum Gehirn diskutiert.
Auch die Fallstudien von HOFFMANN et al. (2007) für BSE am Rind unterstützen diese
Möglichkeiten, die (1) den parasympathischen Weg entlang des N. vagus zum DMNV des
Obex und (2) eine Verbreitung über das Ggl. coeliacum, und sympathische Strukturen (N.
splanchnici) zum Rückenmark als zentrale Komponente des Nervensystems umfassen. Mittels
der in dieser Untersuchung erhobenen Befunde lassen sich die bereits bestehenden Theorien
zur Pathogenese der BSE beim Rind weitgehend bestätigen, darüber hinaus können weitere
Hypothesen formuliert werden. Die Abb. 5.1 (S.116) gibt einen schematischen Überblick zu
den hier vorgestellten Überlegungen.
Die vorliegenden Ergebnisse bestätigen die Vermutung, dass die Ausbreitung des PrPSc
sowohl entlang sympathischer als auch parasympathischer Nervenfasern verläuft. Dabei ist
das Ggl. coeliacum die erste Struktur, in der die den Darm innervierende Nerven
umgeschaltet werden. Im Untersuchungsgut dieser Studie finden sich neben Infektiosität (ab
16 mpi) auch PrPSc
-Ablagerungen an vereinzelten Neuronen bereits in der frühen
Inkubationsphase (ab 32 mpi). Da die Qualität der betroffenen Neurone des
Ganglionkomplexes im Rahmen dieser Untersuchung nicht bestimmt wurde, kann hier
lediglich spekuliert werden, ob der sympathische oder parasympathische Ausbreitungsweg im
ENS oder im Ggl. coeliacum determiniert wird. Die von DEFAWEUX et al. (2007) im ENS
verwendete Markierung für Tyrosin-Hydroxylase (TH) ist spezifisch für sympathische
Nervenfasern und könnte als Doppelfärbung mit PrPSc
angewendet für das Ggl. coeliacum
Aufschluss über die beteiligten Strukturen geben.
Im Darm sind parasympathische Komponenten in Form sensorischer Fasern des N. vagus weit
verbreitet (GOEHLER et al. 1999, 2000). Des Weiteren finden sich efferente Synapsen des N.
vagus in den intrinsischen Ganglien des ENS, die die Darmwand innervierend in der Mukosa
und Submukosa lokalisiert sind (FURNESS et al. 1999). Andererseits wird von einer
intensiven Beteiligung des Sympathikus im ENS (CHIOCCHETTI et al. 2008) und der
umfangreichen sympathischen Innervation lymphatischer Strukturen berichtet (FELTON et
al. 1985, 1988; DEFAWEUX et al. 2007). Diese topographisch-anatomischen
Zusammenhänge lassen die Möglichkeit beider Infektionswege mit einem sympathischen
Schwerpunkt zu.
Das hauptsächlich sympathische Ggl. coeliacum könnte ebenfalls den entscheidenden
Modulator darstellen, da mit der Aufnahme parasympathische Fasern des N. vagus die

Diskussion 115
Voraussetzungen für das Vorkommen beider Routen gegeben sind. Der enge Kontakt beider
qualitativ unterschiedlicher Nervenfasern und Neuronen kann als entscheidender Faktor für
den weiteren Verlauf der Infektion nicht sicher ausgeschlossen werden.
Die Ergebnisse ohne eine simultane Beteiligung der Hirnstammes (IT 28 und 46, 20 mpi; IT
52, 28 mpi) legen den Schluss nahe, dass der Sympathikus eine überragende Rolle bei der
zentripetalen Ausbreitung spielt. Ausgehend vom Ggl. coeliacum verlaufen die Nn.
splanchnici in dorsaler Richtung und nehmen Kontakt zum sympathischen Grenzstrang auf,
um dann im thorakalen Abschnitt des Rückenmarks zu münden. Das im besagten Grenzstrang
eingelagerte GCC stellt durch seine anatomische Lage nahe des Kopfes und der Abgabe
sympathischer Nervenfasern an ein Großteil der Gehirnnerven (unter anderem N. vagus und
den N. trigeminus) möglicherweise den entscheidenden Transmitter für die Ausbreitung des
Erregers in das Gehirn dar. Damit ließen sich die beobachteten initialen PrPSc
-Ablagerungen
in unerwarteten (Formatio reticularis, Nucleus motorius nervi trigemini) oder
parasympathischen (DMNV, Nucleus tractus solitarii) Lokalisationen des Obex erklären. Es
ist herauszustellen, dass es sich dabei entgegen bisheriger Vermutungen um eine Ausbreitung
unter Umgehung des Rückenmarks handeln würde. Trotz des Nachweises von Infektiosität im
Rückenmark eines Tieres 16 mpi, lässt diese Untersuchung Zweifel, dass eine zentripetale
Verbreitung über den N. splanchnicus zwangsläufig den Thorakal-Abschnitt des
Rückenmarks erreicht. Die umfangreiche Einbeziehung sympathische Strukturen neben einer
nur geringgradigen Infektiosität in der Medulla oblongata (IT 24, 24 mpi) veranschaulichen
die Bedeutung des Sympathikus nochmals nachhaltig. Diese Theorie wird ebenfalls durch die
Verzögerung einer Scrapie-Erkrankung bei Mäusen nach Sympathektomie unterstützt
(GLATZEL et al. 2001).
In Einklang mit vorhergehenden Studien zeigen auch die hier vorliegenden
Resultate, dass parasympathische Strukturen auf dem Weg zum Gehirn involviert sind. Ließ
sich bisher aufgrund der Detektion von PrPSc
im DMNV des Obex eine parasympathische
Beteiligung vermuten, kann nun der direkte Nachweis von Infektiosität im N. vagus und im
Ggl. nodosum ohne Beteiligung des ZNS und des sympathischen Systems als entscheidendes
Indiz angesehen werden.
Zusammenfassend lässt sich nicht nur zweifelsfrei die zentripetale Ausbreitung des BSE-
Agens über das PNS nachweisen. Während beide Komponenten des PNS separat in der
Ausbreitung involviert sein können, so überrascht wie deutlich das sympathische
Nervensystem als Schwerpunkt gegenüber einer parasympathischen Ausbreitung determiniert

116 Zusammenfassung
werden konnte. Das Rückenmark ist somit wahrscheinlich als zusätzlicher Infektionsweg zu
bewerten.
Abb. 5.1 Schematische Darstellung zu möglichen Ausbreitung des BSE-Agens im Rind nach
oraler Inokulation
Violett: Zentrales Nervensystem; Rot: Sympathikus; Blau: Parasympathikus
1: Nach Passage der Darmwand wird der Erreger über sympathische oder parasympathische
Fasern aufgenommen und gelangt in das Ggl. coeliacum; 2: Entlang der Fasern des N.
splanchnicus findet die BSE-Erreger Eingang in den sympathischen Grenzstrang; 3: Das Ggl.
cervicale craniale moduliert den Übergang in unterschiedliche Areale des Gehirns; 4: Als
alternative Route kann die Ausbreitung entlang des N. vagus erfolgen; 5. Als zusätzlicher
zentripetaler Infektionsweg könnte über den N. splanchnicus das Rückenmark involviert sein.
5.4.5. Transport des PrPSc
Ein interessanter Aspekt in der Pathogenese von BSE ist der Nachweis des pathologischen
Prion-Proteins im PNS ausschließlich in Assoziation mit Neuronen und Satellitenzellen.
Dabei eröffneten vor allen die Transversalschnitte des N. vagus und des Rückenmarks die
Möglichkeit, PrPSc
auch in den von Markscheiden umhüllten Axonen zu detektieren. Dies
gelang in keiner der Proben dieser Studie und hinterlässt die Frage nach dem eigentlichen
Transportmechanismus des pathologischen Prion-Proteins.

Diskussion 117
Für PrPC wurde ein axonaler Transportmechanismus entlang der Nerven gezeigt
(BORCHELT et al. 1994; BUTOWT et al. 2006), der für die pathologische Isoform bisher
noch nicht erbracht werden konnte. In einer Studie wird eine Translokation von PrPSc
entlang
etablierter axonaler Transportmechanismen (MCBRIDE et al. 2001) vermutet. In den
Untersuchungen von KIMBERLIN et al. (1983) führte eine intraneuronale Inokulation
schneller zum Befall des ZNS als eine extraneuronale Infektion. Die daraus geschlussfolgerte
Ausbreitungsgeschwindigkeit von 1-2 mm/Tag spricht ebenfalls für eine axonale Verbreitung,
wie sie für Makromoleküle (TYTELL et al. 1981) bekannt ist. Einige neurotrope Viren
breiten sich intraneuronal allerdings um das 50fache schneller aus (KIMBERLIN et al. 1983).
Einschränkungen des axonalen Transports am Nagetiermodell weisen jedoch keine
Beeinflussung der Inkubationszeit auf (KUNZI et al. 2002; HAFEZPARAST et al. 2005;
KRATZEL et al. 2007a,b). Als weiteres Indiz für eine adaxonale Ausbreitung werden die
beobachteten PrPSc
-Akkumulationen zwischen der Myelinscheide und dem Axon bewertet
(GROSCHUP et al. 1999; KRATZEL et al. 2007a). Des Weiteren wird eine adaxonale
„Domino“-ähnliche Konversion von PrPC zu PrP
Sc entlang der Nerven diskutiert (GLATZEL
u. AGUZZI, 2000).
Einer Bewertung der Ausbreitungsgeschwindigkeit ausgehend von den Ergebnissen dieser
Studie steht die Problematik der biologischen Varianz entgegen. So handelte es sich bei der
initialen PrPSc
-Detektion im GALT der Rinder nach vier mpi (HOFFMANN et al. 2011) und
der frühesten Detektion im Obex nach 24 mpi um zwei verschiedene Rinder. Die Schätzung
der Länge der für die Ausbreitung relevanten Nervenachse ENS - Ggl. coeliacum - N.
splanchnicus - sympathischer Grenzstrang ist ebenfalls äußerst ungenau. Somit kann die
Ausbreitungsgeschwindigkeit aufgrund der fehlenden fundierten Daten nicht durchgeführt
werden.
Die Limitierung der in dieser Untersuchung nachweisbaren PrPSc
-Akkumulationen auf
Neurone und Gliazellen bedeutet auch, dass der für die Beteiligung der Myelinscheiden
entscheidende Nachweis in der weißen Substanz des Rückenmarks nicht erbracht werden
konnte.
Aufgrund dieser fehlenden Indizien und der langen Inkubationszeit lässt sich daher über eher
langsame intra-axonalen Transportmechanismen spekulieren, wie sie für neurotrope Viren
bereits bekannt sind. So sind zur weiteren Klärung In vivo- und In vitro-Untersuchungen,
möglicherweise unter Verwendung der Elektronenmikroskopie, notwendig.

118 Zusammenfassung
5.5 Pathogenese und Pathologie
In der Literatur werden inflammatorische Prozesse unterschiedlichster Art als mögliche
Einflussfaktoren auf die Pathogenese der TSE-Erkrankungen diskutiert. So führte eine
infektiöse Darmschädigung bei Mäusen zu einer erhöhten Scrapie-Empfänglichkeit
(SIGURDSON et al. 2009) und einer schnelleren Ausbildung des terminalen
Krankheitsstadiums (THACKRAY et al. 2003). Bakterielle, virale oder parasitäre Infektionen
beeinflussten die tight junctions des Darmepithels (SAKAGUCHI et al. 2001; MILLING et
al. 2007) und führten zu einer gesteigerten Aufnahme von Proteinen (SODERHOLM et al.
2004). Speziell für die TSE ist die Erhöhung der PrPC-Expression infolge gastrointestinaler
Entzündungen interessant. Der im Gegensatz zu vergleichbaren Studien in England
(ARNOLD et al. 2007) bis zu 8 Monaten schnellere Krankheitsverlauf in der FLI-Studie ließe
sich durch die bei allen im Versuch befindlichen Rindern beobachtete massive chronisch
aktive oberflächliche eosinophile Enteritis erklären. Der unter Feldbedingungen
unumgängliche Kontakt mit Enteritis-Erregern könnte somit auch bei natürlichen Infektionen
eine nicht unerhebliche Rolle spielen. Ob bei einer vorliegenden Enteritis die Darmpassage
erleichtert wird oder eine erhöhte PrPC-Expression im Darmepithel vorliegt, kann anhand der
hier vorliegenden Ergebnisse jedoch nicht beantwortet werden. Weiterführende Studien mit
Doppelfärbungen für entzündungsassoziierte Marker und für PrPSc
könnten zur Klärung
dieser Fragestellung beitragen.
Bei Mäusen und Schafen wurde unter Einbeziehung des lymphatischen Systems eine
Korrelation von Entzündungen und dem Organtropismus festgestellt. So führen chronische
mononukleäre Entzündungen zum Nachweis des pathologisches Prion-Protein in der Niere,
dem Pankreas und der Leber (HEIKENWALKER et al. 2005). Infektiöser Urin infolge einer
Nephritis (SEEGER et al. 2005) wurde bei Schafen ebenso beobachtet wie das Auftreten des
pathologischen Prion-Proteins im Euter von Tieren mit Mastitis (LIGIOS et al. 2005).
In diesem Zusammenhang muss jedoch betont werden, dass sich in anderen Untersuchungen
PrPSc
in der Niere und der Milch bei Schafen auch ohne entsprechende entzündliche Prozesse
(SISO et al. 2008; LACROUX et al. 2007) detektierten ließ. Die im eigenen
Untersuchungsgut in zahlreichen Proben des PNS festgestellten lymphohistiozytären
Infiltrationen standen ohne kausale Zusammenhänge zu den Befunden aus IHC und BA.
Weiterhin waren altersabhängig deutlich später in der Inkubationsperiode auch die
Kontrolltiere von diesen Entzündungen betroffen (FAST, mündliche Mitteilung). Somit bleibt

Diskussion 119
weiter unklar, inwieweit inflammatorische Prozesse und die BSE-assoziierte Pathologie
verknüpft sind.

120 Zusammenfassung
6. ZUSAMMENFASSUNG
Martin Kaatz
Ausbreitung der klassischen BSE-Prion-Erreger vom Darm über das periphere Nervensystem
zum Gehirn
Wie für die anderen Vertreter der Transmissiblen Spongiformen Enzephalopathien (TSE) gilt
auch für die bovine spongiforme Enzephalopathie (BSE) die Prion-Hypothese, die von einem
pathologischen Prion-Protein (PrPSc
) als Erreger ausgeht. Dabei ist die Pathogenese der BSE
im natürlichen Wirt weitestgehend ungeklärt und die derzeitigen Annahmen basieren zum
Großteil auf Erkenntnissen aus Versuchen an anderen Spezies. Die am Friedrich-Loeffler-
Institut initiierte deutsche BSE-Pathogenesestudie umfasste neben 18 Kontrolltieren 56 oral
inokulierte Rinder. In einem viermonatigen Rhythmus wurden jeweils vier Tiere aus dem
Versuch genommen und seziert. Von zwei Rindern an acht Sektionszeitpunkten (insg. 16
Tiere, 16 bis 44 Monate post infectionem [mpi]) wurden neben Gewebeproben des zentralen
Nervensystems (ZNS) sympathische und parasympathische Proben aus dem peripheren
Nervensystem (PNS) und des Ileums analysiert. Der ausgewählte Untersuchungszeitraum
umfasste mehrheitlich präklinische Tiere, um die zentripetale neuronale Ausbreitung des
pathologischen Prion-Proteins vom Darm zum Gehirn aufzuklären.
Als Nachweisverfahren zur Detektion des BSE-assoziierten Agens kamen mit der
Immunhistochemie (IHC) und dem Maus-Biosassay (Maus-BA) zwei Methoden mit
unterschiedlichen Strategien zum Einsatz. Mittels IHC werden die mikroskopisch
lokalisierbaren PrPSc
-Ablagerungen nachgewiesen. Die Hämatoxylin-Eosin- (HE) Färbung
dient in der TSE-Diagnostik zum Nachweis spongiformer Enzephalopathien als auch zum
Nachweis möglicher pathologischer Veränderungen. Der hochsensitive Maus-Bioassay
ermöglicht Aussagen über den Erregergehalt von intrazerebral in transgene Mäuse
inokulierten Gewebeproben. Die Gehirne klinisch erkrankter Mäuse wurden anschließend
biochemisch auf PrPSc
-Akkumulationen als Nachweis einer BSE-Infektion hin untersucht.
Jedes in dieser Untersuchung analysierte Rind war mindestens in einer der untersuchten
Proben positiv. Klinische Symptome einer BSE-Erkrankung waren bei zwei Rindern (36 und
44 mpi) eindeutig zu beobachten, bei einem Tier (44 mpi) bestand ein Verdacht auf eine ZNS-
Erkrankung und vier Rinder (32 bis 40 mpi) wiesen unspezifische Krankheitsbilder auf. Bei
neun Rindern ließen sich bis hin zu 40 mpi keine klinischen Symptome nachweisen.

Zusammenfassung 121
In der Medulla oblongata des Gehirns waren BSE-Erreger frühestens nach 24 mpi und PrPSc
-
Ablagerungen erstmals ab 28 mpi detektierbar. Der früheste Nachweis im Rückenmark
erfolgte bei einem Tier 16 mpi (Maus-BA) bzw. 28 mpi (IHC). Die histopathologische
Untersuchung des ZNS wies bei sieben Tieren ab 32 mpi eine spongiforme Enzephalopathie
auf. Bei dem Versuch, eine Assoziation zwischen den Ergebnissen der IHC, den
histopathologischen und den klinischen Untersuchungen herzustellen, schien eine Verbindung
zwischen PrPSc
-Ablagerungen und dem Auftreten einer spongiformen Enzephalopathie zu
bestehen.
Für eine zuverlässige Bestimmung des zentripetalen Ausbreitungsweges wurden
ausschließlich Proben von Rindern ausgewertet, bei denen PrPSc
oder der BSE-Erreger im
PNS, aber noch nicht im ZNS nachweisbar waren. So konnte eine terminale zentrifugale
Ausbreitung ausgeschlossen werden. Das eine solche Ausbreitung sehr schnell nach dem
Befall des Gehirns beginn kann, zeigt der erstmalige PrPSc
-Nachweis im Ggl. trigeminale bei
nur geringgradigen PrPSc
-Akkumulation im Obex (IT 25, 40 mpi). In dieser Studie konnte
erstmals bei sechs Tieren (16-28 mpi) BSE-Erreger im PNS deutlich vor dem initialen
Nachweis in der Medulla oblongata detektiert werden. Von diesen Tieren waren vier im
gemischtfaserigen Ggl coeliacum positiv. Interessanterweise ließ sich bei beiden Rindern 16
mpi sowie bei einem weiteren Tier 28 mpi ausschließlich in den sympathischen Geweben des
Ggl. cervicale craniale, Nn. splanchnici bzw. des Ggl. stellatum BSE-Erreger detektieren. Im
Gegensatz dazu waren 20 mpi lediglich parasympathische Anteile (Ggl. nodosum, N. vagus)
betroffen. Unabhängig von diesen Befunden konnte in zahlreichen Ganglien/Nerven eine
geringgradige lymphohistiozytäre Ganglioneuritis festgestellt werden.
Die Vermutungen bisheriger Studien zur zentripetalen Ausbreitung des BSE-Agens anhand
parasympathischer und sympathischer Nerven konnten mit den Ergebnissen dieser Arbeit
zweifelsfrei nachgewiesen werden. Allerdings stellt die dominante und bemerkenswert frühe
Beteiligung des Sympathikus eine neue Erkenntnis dar. Hierbei ist besonders das Ggl.
cervicale craniale herauszustellen, das anatomisch und funktionell Verbindungen zu einer
Vielzahl von Gehirnerven besitzt. Somit könnte diesem Ganglion eine entscheidende
Bedeutung beim Übergang der BSE-Erreger aus dem PNS in die unterschiedlichen
Kerngebiete des Gehirns zukommen. Das Rückenmark, welches bisher neben den N. vagus
als ein primäres zentripetales Ausbreitungsorgan betrachtet wurde, scheint dagegen lediglich
einen zusätzlichen Weg darzustellen. Da alle hier untersuchten Tiere eine Beteiligung des
PNS/ZNS zeigten, liegt die initiale Infektion peripherer Nerven vor dem 16. Monat p.i.. Dies
macht weitere Untersuchungen von Tieren mit einer geringeren Inkubationszeit notwendig.

122 Summary
7. SUMMARY
Martin Kaatz
Spread of classic BSE-Prions from the gut via the peripheral nervous system to the brain
Like other members of the transmissible spongiform encephalopathies (TSE) the prion-
hypothesis is also considered for Bovine Spongiform Encephalopathy (BSE) with a
pathological prion protein (PrPSc
) as the infectious agent. The pathogenesis of BSE is not well
understood and is based mainly on data from other species. Therefore, the Friedrich-Loeffler
institute initiated a trial including 56 orally infected cows and 18 control animals. Every four
month four animals were euthanized and a necropsy was performed. From in total 16 cows at
the time period between 16 and 44 months post infectionem (two cows at each necropsy date)
tissue samples from the central nervous system (CNS), sympathetic and parasympathetic parts
of peripheral nervous system (PNS) and the ileum were examined. The animals (mpi)
included primarily pre-clinical animals to clarify the centripetal spread of the pathological
prion protein from the gut to the brain.
Two different strategies, the immunhistochemistry (IHC) and a mouse bioassay (mouse-BA),
were chosen as detection methods. The IHC detects PrPSc
and enabled the analysis of the
cellular structures involved. The hematoxylin-eosin (HE) staining revealed pathological
diagnostics and a potential association to BSE. The highly sensitive mouse-BA detected the
infectious potential of the examined tissues. These transgenic mice were over-expressing the
physiologic bovine Prion protein which made them highly susceptible to a BSE-infection.
After an intracerebral inoculation a biochemical method (Immunoblot) was used to detect
PrPSc
in the brain of clinical affected mice.
At least one positive result for the examined samples was established for every animal of this
study. Clinical signs were definitively observed for two cows (36 and 44 mpi), one animal
was presumptively affected with BSE and four showed an unspecific disease pattern (32 to 44
mpi).
At the medulla oblongata of the brain we detected infectivity by the mouse-BA initially at 24
mpi and PrPSc
accumulations by IHC at 28 mpi. For the spinal cord the earliest evidence was
found 16 mpi by the mouse bioassay and 28 mpi with the IHC. The histopathologic
examination showed a spongiform encephalopathy for seven cows after 32 mpi. An

Summary 123
association between the revealed results was only established for the PrPSc
accumulation and
the occurrence of a spongiform encephalopathy.
For a reliable determination of the centripetal route it is mandatory to examine animals prior
to an involvement of the brain. The first detection of PrPSc
at the Ggl. trigeminale was
accompanied by a mild PrPSc
accumulation at the Obex (IT 25, 40 mpi). Therefore this
ganglion could indicate a fast centrifugal spread. For the first time this study detected BSE
infectivity in the PNS well before the initial involvement of the Medulla oblongata in six
animals. Four of these cows were positive at the Ggl. coeliacum, which contains both
sympathetic and parasympathetic fibers. Interestingly, both animals at 16 mpi and one animal
28 mpi showed infectivity exclusively in the sympathetic tissues of the Ggl. cervicale
craniale, Nn. splanchnici and Ggl. stellatum respectively. In contrast, in two animals
examined at 20 mpi, only parasympathetic tissues (Ggl. nodosum, N. vagus) revealed positive
results. Irrespective of these findings a mild lymphohistocytic ganglioneuritis was observed in
multiple ganglia and nerves.
In agreement with previous reports, the results of this study showed the centripetal spread of
the BSE-agent along parasympathetic and sympathetic nerves. However, the dominant and
early involvement of the sympathetic nervous system is novel. In this regard, the Ggl.
cervicale craniale has to be emphasized in terms of its anatomical and functional connections
to multiple cranial nerves. Therefore, this ganglion could be considered to play a crucial role
for the spread of the BSE-agent from the PNS to the different nuclei of the brain. Besides the
N. vagus, hitherto the spinal cord was considered to be involved in the primary centripetal
spread of the agent, but our results indicate that this is likely an additional pathway. Because
all animals showed positive findings in the PNS/CNS the definitive determination of the
initial onset of the centripetal spread cannot be confidently ascertained and further
investigations are warranted.

124 Literaturverzeichnis
8. LITERATURVERZEICHNIS
ABID, K., u. C. SOTO (2006):
The intriguing prion disorders.
Cell. Mol. Life Sci. 63, 2342-51
ADLER, V., B. ZEILER, V. KRYUKOV, R. KASCSAK, R. RUBENSTEIN u. A.
GROSSMAN (2003):
Small, highly structured RNAs participate in the conversion of human recombinant PrP(Sen)
to PrP(Res) in vitro.
J. Mol. Biol. 332, 47-57
ALPER, T., D.A. HAIG u. M.C. CLARKE (1966):
The exceptionally small size of the scrapie agent.
Biochem. Biophys. Res. Commun. 22, 278-84
ALPER, T., W.A. CRAMP, D.A. HAIG u. M.C. CLARKE (1967):
Does the agent of scrapie replicate without nucleic acid?
Nature 214, 764-6
ANDREOLETTI, O., P. BERTHON, D. MARC, P. SARRADIN, J. GROSCLAUDE, L.
VAN KEULEN, F. SCHELCHER, J. M. ELSEN u. F. LANTIER (2000):
Early accumulation of PrP(Sc) in gut-associated lymphoid and nervous tissues of susceptible
sheep from a Romanov flock with natural scrapie.
J. Gen. Virol. 81, Pt 12, 3115-26
ANDREOLETTI, O., C. LACROUX, A. CHABERT, L. MONNEREAU, G. TABOURET, F.
LANTIER, P. BERTHON, F. EYCHENNE, S. LAFOND BENESTAD, J.M. ELSEN u.
F. SCHELCHER (2002):
PrP(Sc) accumulation in placentas of ewes exposed to natural scrapie: influence of foetal PrP
genotype and effect on ewe-to-lamb transmission.
J. Gen. Virol. 83, 2607-16

Literaturverzeichnis 125
ANDREOLETTI, O., S. SIMON, C. LACROUX, N. MOREL, G. TABOURET, A.
CHABERT, S. LUGAN, F. CORBIERE, P. FERRE U. G. FOUCRAS, H. LAUDE, F.
EYCHENNE, J. GRASSI u. F. SCHELCHER (2004):
PrP(Sc) accumulation in myocytes from sheep incubating natural scrapie.
Nat. Med. 10, 591-3
ANGELUCCI, A., F. CLASCÁ u. M. SUR (1996):
Anterograde axonal tracing with the subunit B of cholera toxin: a highly sensitive
immunohistochemical protocol for revealing fine axonal morphology in adult and neonatal
brains.
J. Neurosci. Methods 65, 1, 101-12
ARNOLD, J.E., C. TIPLER, L. LASZLO, J. HOPE, M. LANDON u. R.J. MAYER (1995):
The abnormal isoform of the prion protein accumulates in late-endosome-like organelles in
scrapie-infected mouse brain.
J. Pathol. 176, 4, 403-11
ARNOLD, M. E., u. J. W. WILESMITH (2004):
Estimation of the age dependent risk of infection to BSE of dairy cattle in Great Britain.
Prev. Vet. Med. 66, 35-47
ARNOLD, M. E., J.B.M. RYAN, T. KONOLD, M.M. SIMMONS, Y.I. SPENCER, A.
WEAR, M. CHAPLIN, M. STACK, S. CZUB, R. MUELLER, P.R. WEBB, A. DAVIS, J.
SPIROPOULOS, J. HOLDAWAY, S.A.C. HAWKINS, A.R. AUSTIN u. G.A.H. WELLS
(2007):
Estimating the temporal relationship between PrPSc
detection and incubation period in
experimental bovine spongiform encephalopathy of cattle.
J. Gen. Virol. 88, 3198-208
AUCOUTURIER, P., F. GEISSMANN, D. DAMOTTE, G.P. SABORIO, H.C. MEEKER, R.
KASCSAK, R. KASCSAK, R.I. CARP u. T. WISNIEWSKI (2001):
Infected splenic dendritic cells are sufficient for prion transmission to the CNS in mouse
scrapie.
J. Clin. Invest. 108, 703-8

126 Literaturverzeichnis
BALDAUF, E., M. BEEKES u. H. DIRINGER (1997):
Evidence for an alternative direct route of access for the scrapie agent to the brain bypassing
the spinal cord.
J. Gen. Virol. 78, 1187-97
BALKEMA-BUSCHMANN, A., M. EIDEN, C. HOFFMANN, M. KAATZ, U. ZIEGLER,
M. KELLER u. M.H. GROSCHUP (2011):
BSE infectivity in the absence of detectable PrP(Sc) accumulation in the tongue and nasal
mucosa of terminally diseased cattle.
J. Gen. Virol. 92, 467-76
BARON, T. G., J. Y. MADEC u. D. CALAVAS (1999):
Similar signature of the prion protein in natural sheep scrapie and bovine spongiform
encephalopathy-linked diseases.
J. Clin. Microbiol. 37, 11, 3701-4
BARON, T., u. A.G. BIACABE (2006):
Origin of bovine spongiform encephalopathy.
Lancet 367, 9507, 297-8
BEEKES, M., E. BALDAUF u. H. DIRINGER (1996):
Sequential appearance and accumulation of pathognomonic markers in the central nervous
system of hamsters orally infected with scrapie.
J. Gen. Virol. 77, 1925-34
BEEKES, M., u. P.A. MCBRIDE (2000):
Early accumulation of pathological PrP in the enteric nervous system and gut-associated
lymphoid tissue of hamsters orally infected with scrapie.
Neurosci. Lett. 278, 181-4
BEGARA-MCGORUM, I., L. GONZÁLEZ, M. SIMMONS, N. HUNTER, F. HOUSTON u.
M. JEFFREY (2002):
Vacuolar lesion profile in sheep scrapie: Factors influencing its variation and relationship to
disease-specific PrP accumulation.

Literaturverzeichnis 127
J. Comp. Pathol. 127, 59-68
BELT, P.B., I.H. MUILEMAN, B.E. SCHREUDER, J. BOS-DE RUIJTER, A.L. GIELKENS
u. M.A. SMITS (1995):
Identification of five allelic variants of the sheep PrP gene and their association with natural
scrapie.
J. Gen. Virol. 76, 509-17
BENDHEIM, P.E., H.R. BROWN, R.D. RUDELLI, L.J. SCALA, N.L. GOLLER, G.Y.
WEN, R.J. KASCSAK, N.R. CASHMAN u. D.C. BOLTON (1992):
Nearly ubiquitous tissue distribution of the scrapie agent precursor protein.
Neurology 42, 149-56
BENESTAD, S. L., P. SARRADIN, B. THU, J. SCHONHEIT, M. A. TRANULIS u. B.
BRATBERG (2003):
Cases of scrapie with unusual features in Norway and designation of a new type, Nor98.
Vet. Rec. 153, 7, 202-8
BENESTAD, S.L., J.-N. ARSAC, W. GOLDMANN, u. M. NÖREMARK (2008):
Atypical/Nor98 scrapie: properties of the agent, genetics, and epidemiology.
Vet. Res. 39, 4, 19
BERINGUE, V. M. DEMOY, C.I. LASMÉZAS, B. GOURITIN, C. WEINGARTEN, J.P.
DESLYS, J.P. ANDREUX, P. COUVREUR u. D. DORMONT (2000):
Role of spleen macrophages in the clearance of scrapie agent early in pathogenesis.
J. Pathol. 190, 495-502
BIACABE, A.G., J.L. LAPLANCHE, S. RYDER u. T. BARON (2004):
Distinct molecular phenotypes in bovine prion diseases.
EMBO Rep 5, 110-5
BLANCHARD, N., D. LANKAR, F. FAURE, A. REGNAULT, C. DUMONT, G. RAPOSO
u. C. HIVROZ (2002):

128 Literaturverzeichnis
TCR activation of human T cells induces the production of exosomes bearing the
TCR/CD3/zeta complex.
J. Immunol. 168, 7, 3235-41
BORCHELT, D.R., V.E. KOLIATSOS, M. GUARNIERI, C.A. PARDO, S.S. SISODIA u.
D.L. PRICE (1994):
Rapid anterograde axonal transport of the cellular prion glycoprotein in the peripheral and
central nervous systems.
J. Biol. Chem. 269, 14711-4
BOSSERS, A., B.E. SCHREUDER, I.H. MUILEMAN, P.B. BELT u. M.A. SMITS (1996):
PrP genotype contributes to determining survival times of sheep with natural scrapie.
J. Gen. Virol. 77, 2669-73
BROTHERSTON, J.G., C.C. RENWICK, J.T. STAMP, I. ZLOTNIK u. I.H. PATTISON
(1968):
Spread of scrapie by contact to goats and sheep.
J. Comp. Pathol. 78, 9-17
BROWN, D.R., B.S. WONG, F. HAFIZ, C. CLIVE, S.J. HASWELL u. I.M. JONES (1999):
Normal prion protein has an activity like that of superoxide dismutase.
Biochem. J. 344, 1-5
BROWN, K. L., K. STEWART, D.L. RITCHIE, N.A. MABBOTT, A. WILLIAMS, H.
FRASER, I. MORRISONW u. M.E. BRUCE (1999):
Scrapie replication in lymphoid tissues depends on PrP-expressing follicular dendritic cells.
Nature Med. 5, 1308-12
BRUCE, M.E., R.G. WILL, J.W. IRONSIDE, I. MCCONNELL, D. DRUMMOND, A.
SUTTIE, L. MCCARDLE, A. CHREE, J. HOPE, C. BIRKETT, S. COUSENS, H. FRASER
u. C.J. BOSTOCK (1997):
Transmissions to mice indicate that 'new variant' CJD is caused by the BSE agent.
Nature 389, 498-501

Literaturverzeichnis 129
BUDKA, H., A. AGUZZI, P. BROWN, J.M. BRUCHER, O. BUGIANI, F. GULLOTTA, M.
HALTIA, J.J. HAUW, J.W. IRONSIDE u K. JELLINGER, et al. (1995):
Neuropathological diagnostic criteria for Creutzfeldt-Jakob disease (CJD) and other human
spongiform encephalopathies (prion diseases).
Brain Pathol. 5, 459-66
BUDKA, H. (2001):
Portrait der Creutzfeldt-Jakob-Krankheit.
In: HÖRNLIMANN, B., D. RIESNER u. H. A. KRETZSCHMAR (Hrsg.): Prionen und
Prionkrankheiten.
De Gruyter, New York, S. 147-51
BUELER, H., M. FISCHER, Y. LANG, H. BLUETHMANN, H.P. LIPP, S.J. DEARMOND,
S.B. PRUSINER, M. AGUET, C. WEISSMANN (1992):
Normal development and behavior of mice lacking the neuronal cell-surface PrP protein.
Nature 356, 577-82
BUELER, H., A. AGUZZI, A. SAILER, R.A. GREINER, P. AUTENRIED, M. AGUET u. C.
WEISSMANN (1993):
Mice devoid of PrP are resistant to scrapie.
Cell 73, 1339-47
BURGER, D., u. G. R. HARTSOUGH (1965):
Encephalopathy of mink. II. Experimental and natural transmission.
J. Infect. Dis. 115, 4, 393-9
BUSCHMANN, A., A. G. BIACABE, U. ZIEGLER, A. BENCSIK, J. Y. MADEC, G.
ERHARDT, G. LUHKEN, T. BARON u. M. H. GROSCHUP (2004):
Atypical scrapie cases in Germany and France are identified by discrepant reaction patterns in
BSE rapid tests.
J.Virol. Methods 117, 1, 27-36
BUSCHMANN, A., u. M.H. GROSCHUP (2005):

130 Literaturverzeichnis
Highly bovine spongiform encephalopathy-sensitive transgenic mice confirm the essential
restriction of infectivity to the nervous system in clinically diseased cattle.
J. Infect. Dis. 192, 934-42
BUSCHMANN, A., A. GRETZSCHEL, A.G. BIACABE, K. SCHIEBEL, C. CORONA, C.
HOFFMANN, M. EIDEN, T. BARON, C. CASALONE u. M.H. GROSCHUP (2006):
Atypical BSE in Germany-proof of transmissibility and biochemical characterization.
Vet. Microbiol. 117, 103-16
CALZOLAI, L., D.A. LYSEK, D.R. PEREZ, P. GUNTERT u. K. WUTHRICH (2005):
Prion protein NMR structures of chickens, turtles, and frogs.
Proc. Natl. Acad. Sci. USA 102, 651-5
CAPOBIANCO, R., C. CASALONE, S. SUARDI, M. MANGIERI, C. MICCOLO, L.
LIMIDO, M. CATANIA, G. ROSSI, G. DI FEDE, G. GIACCONE, M.G. BRUZZONE, L.
MINATI, C. CORONA, P. ACUTIS, D. GELMETTI, G. LOMBARDI, M.H. GROSCHUP,
A. BUSCHMANN, G. ZANUSSO, S. MONACO, M. CARAMELLI, F. TAGLIAVINI
Conversion of the BASE Prion Strain into the BSE Strain: The Origin of BSE?
PLoS Pathog. 3, 3, e31.doi, 10.1371
CARP, R.I., u. S.M. CALLAHAN (1981):
In vitro interaction of scrapie agent and mouse peritoneal macrophages.
Intervirology 16, 8-13
CARP, R.I. (1982):
Transmission of scrapie by oral route: effect of gingival scarification.
Lancet 1, 170-1
CARP, R.I., u. S.M. CALLAHAN (1982):
Effect of mouse peritoneal macrophages on scrapie infectivity during extended in vitro
incubation.
Intervirology 17, 201-7
CARP, R.I., H.C. MEEKER, V. CARUSO u. E. SERSEN (1999):

Literaturverzeichnis 131
Scrapie strain-specific interactions with endogenous murine leukemia virus.
J. Gen. Virol. 80, 5-10
CASALONE, C., G. ZANUSSO, P. ACUTIS, S. FERRARI, L. CAPUCCI, F. TAGLIAVINI,
S. MONACO u. M. CARAMELLI (2004):
Identification of a second bovine amyloidotic spongiform encephalopathy: molecular
similarities with sporadic Creutzfeldt-Jakob disease.
Proc. Natl. Acad. Sci. USA 101, 3065-70
CASALONE, C., C. CORONA, M.I.CRESCIO, F. MARTUCCI, M. MAZZA, G. RU, E.
BOZZETTA, PL. ACUTIS u. M. CARAMELLI (2005):
Pathological prion protein in the tongues of sheep infected with naturally occurring scrapie.
J. Virol. 79, 9, 5847-9
CASTILLA, J., P. SAA, C. HETZ u. C. SOTO (2005):
In vitro generation of infectious scrapie prions.
Cell 121, 195-206
CAUGHEY, B., G.S. BARON, B. CHESEBRO u. M. JEFFREY (2009):
Getting a grip on prions: oligomers, amyloids, and pathological membrane interactions.
Annu. Rev. Biochem. 78, 177-204
CHANDLER, R.L. (1961):
Encephalopathy in mice produced by inoculation with scrapie brain material.
Lancet 1, 1378-9
CHANDLER, R.L., u. B.A. TURFREY (1972):
Inoculation of voles, Chinese hamsters, gerbils and guinea-pigs with scrapie brain material.
Res. Vet. Sci. 13, 219-24
CHAPLIN, M.J., A.D. ALDRICH u. M.J. STACK (1998)
Scrapie associated fibril detection from formaldehyde fixed brain tissue in natural cases of
ovine scrapie.
Res. Vet. Sci. 64, 41-4

132 Literaturverzeichnis
CHESEBRO, B. (1998):
BSE and prions: uncertainties about the agent.
Science 279, 42-3
CHIOCCHETTI, R., G. MAZZUOLI, V. ALBANESE, M. MAZZONI, P. CLAVENZANI,
G. LALATTA-COSTERBOSA, ML. LUCCHI, G. DI GUARDO, G. MARRUCHELLA u.
J.B. FURNESS (2008):
Anatomical evidence for ileal Peyer's patches innervation by enteric nervous system: a
potential route for prion neuroinvasion
Cell Tissue Res. 332, 2, 185-94
COLCHESTER, A.C., u. N.T. COLCHESTER (2005):
The origin of bovine spongiform encephalopathy: the human prion disease hypothesis.
Lancet 366, 856-61
COLLING, S.B., T.M. KING, J. COLLINGE u. J.G.R. JEFFERYS (1995):
Prion protein null mice: abnormal intrinsic properties of hippocampal CA1 pyramidal cells.
Brain Res. Assic. Abstr. 12 (Abstract)
COLLINGE, J., M.A. WHITTINGTON, K.C.SIDLE, C.J. SMITH, M.S. PALMER, A.R.
CLARKE u. J.G. JEFFERYS (1994):
Prion protein is necessary for normal synaptic function.
Nature 370, 295-7
COMBER, T. (1772):
Real Improvements in Agriculture.
gedruckt für W. Nicoll, London
COMOY, E.E., C. CASALONE, N. LESCOUTRA-ETCHEGARAY, G. ZANUSSO, S.
FREIRE, D. MARCE, F. AUVRE, M.-M. RUCHOUX, S. FERRARI, S. MONACO, N.
SALES, M. CARAMELLI, P. LEBOULCH, P. BROWN, C. I. LASMEZAS u. J.-P. DESLYS
(2008)
Atypical BSE (BASE) Transmitted from Asymptomatic Aging Cattle to a Primate.

Literaturverzeichnis 133
PLoS ONE 3, 8, e3017
CORDEIRO, Y., F. MACHADO, L. JULIANO, M.A. JULIANO, R.R. BRENTANI, D.
FOGUEL u. J.L.SILVA (2001):
DNA converts cellular prion protein into the beta-sheet conformation and inhibits prion
peptide aggregation.
J. Biol. Chem. 276, 49400-9
CREUTZFELDT, H.G. (1920):
Über eine eigenartige herdförmige Erkrankung des Zentralnervensystems.
Z. ges. Neurol. Psychiat. 57, 1-18
CURNOW, R.N., u. C.M. HAU (1996):
The incidence of bovine spongiform encephalopathy in the progeny of affected sires and
dams.
Vet. Rec. 138, 407-8
CZUB, M., H.R. BRAIG u. H. DIRINGER (1988):
Replication of the scrapie agent in hamsters infected intracerebrally confirms the pathogenesis
of an amyloid-inducing virosis.
J. Gen. Virol. 69, 1753-6
DEFAWEUX, V., G. DORBAN, C. DEMONCEAU, J. PIRET, O. JOLOIS, O. THELLIN, C.
THIELEN, E. HEINEN u. N. ANTOINE (2005):
Interfaces between dendritic cells, other immune cells, and nerve fibres in mouse Peyer’s
patches: potential sites for neuroinvasion in prion diseases.
Microsc. Res. Tech. 66, 1-9
DEFAWEUX, V., G. DORBAN, N. ANTOINE, J. PIRET, A. GABRIEL, O. JACQMOT, N.
FALISSE-POIRIER, S. FLANDROY, D. ZORZI u. E. HEINEN (2007):
Neuroimmune connections in jejunal and ileal Peyer's patches at various bovine ages:
potential sites for prion neuroinvasion.
Cell Tissue Res. 329, 1, 35-44

134 Literaturverzeichnis
DELEAULT, N.R., R.W. LUCASSEN u. S. SUPATTAPONE (2003):
RNA molecules stimulate prion protein conversion.
Nature 425, 717-20
DELEAULT, N.R., J.C. GEOGHEGAN, K. NISHINA, R. KASCSAK, R.A. WILLIAMSON
u. S. SUPATTAPONE (2005):
Protease resistant prion protein amplification reconstituted with partially purified substrates
and synthetic polyanions.
J. Biol. Chem. 280, 26873-9
DENZER, K., M. VAN EIJK, M.J. KLEIJMEER, E. JAKOBSON, C. DE GROOT u. H.J.
GEUZE (2000):
Follicular dendritic cells carry MHC class II-expressing microvesicles at their surface.
J. Immunol. 165, 3, 1259-65
DERRINGTON, E., C. GABUS, P. LEBLANC, J. CHNAIDERMANN, L. GRAVE, D.
DORMONT, W. SWIETNICKI, M. MORILLAS, D. MARCK, P. NANDI u. J.L. DARLIX
(2002):
PrP(C) has nucleic acid chaperoning properties similar to the nucleocapsid protein of HIV-1.
C. R. Biol. 325, 17-23
DICKINSON, A.G., J.T. STAMP u. C.C. RENWICK (1974):
Maternal and lateral transmission of scrapie in sheep.
J. Comp. Pathol. 84, 19-25
DIRINGER, H., M. BEEKES u. U. OBERDIECK (1994):
The nature of the scrapie agent: the virus theory.
Ann. NY Acad. Sci. 724, 246-58
DIRINGER, H., J. ROEHMEL u. M. BEEKES (1998):
Effect of repeated oral infection of hamsters with scrapie.
J. Gen. Virol. 79, 609-12
DODELET, V.C., u. N.R. CASHMAN (1998)

Literaturverzeichnis 135
Prion protein expression in human leukocyte differentiation.
Blood 91, 1556-61
DORBAN, G., V. DEFAWEUX, E. HEINEN u. N. ANTOINE (2010):
Spreading of prions from the immune to the peripheral nervous system: a potential
implication of dendritic cells.
Histochem. Cell Biol. 133, 5, 493-504
EDDY, R.G. (1995):
Origin of BSE.
Vet. Rec. 137, 648
EIDEN, M., A. BUSCHMANN, L. KUPFER u. M.H. GROSCHUP (2006):
Synthetic prions.
J. Vet. Med. B. Infect. Dis. Vet. Public Health 53, 251-6
ELOIT, M., K. ADJOU, M. COULPIER, J.J. FONTAINE, R. HAMEL, T. LILIN, S.
MESSIAEN, O. ANDREOLETTI, T. BARON, A. BENCSIK, A.G. BIACABE, V.
BERINGUE, H. LAUDE, A. LE DUR, J.L. VILOTTE, E. COMOY, J.P. DESLYS, J.
GRASSI, S. SIMON, F. LANTIER u. P. SARRADIN (2005):
BSE agent signatures in a goat.
Vet. Rec. 156, 523-4
ESCOLA, J.M., M.J. KLEIJMEER, W. STOORVOGEL, J.M. GRIFFITH, O. YOSHIE u.
H,J. GEUZE (1998):
Selective enrichment of tetraspan proteins on the internal vesicles of multivesicular
endosomes and on exosomes secreted by human B-lymphocytes.
J. Biol. Chem. 273, 32, 20121-7
ESPINOSA, J.C., M. MORALES, J. CASTILLA, M. ROGERS u. J.M. TORRES (2007):
Progression of prion infectivity in asymptomatic cattle after oral bovine spongiform
encephalopathy challenge.
J. Gen. Virol. 88, 1379-83

136 Literaturverzeichnis
FEDIAEVSKY, A., S.C. TONGUE, M. NÖREMARK, D. CALAVAS, G. RU u. P. HOPP
(2008): A descriptive study of the prevalence of atypical and classical scrapie in sheep in 20
European countries.
BMC Vet. Res. 4, 19-43
FEVRIER, B., D. VILETTE, F. ARCHER, D. LOEW, W. FAIGLE, M. VIDAL, H. LAUDE
u. G. RAPOSO (2004):
Cells release prions in association with exosomes.
Proc. Natl. Acad. Sci. USA 101, 9683-8
FEVRIER, B., D. VILETTE, H. LAUDE u. G. RAPOSO (2005):
Exosomes: a bubble ride for prions.
Traffic 6, 10-7
FIELD, E.J., u. A. PEAT (1969):
Structural changes in scrapie affected brain.
Biochem. J. 114, 19P-20P
FISCHER, M., T. RULICKE, A. RAEBER, A. SAILER, M. MOSER, B. OESCH, S.
BRANDNER, A. AGUZZI u. C. WEISSMANN (1996):
Prion protein (PrP) with amino-proximal deletions restoring susceptibility of PrP knockout
mice to scrapie.
Embo J. 15, 6, 1255-64
FOLLET, J., C. LEMAIRE-VIEILLE, F. BLANQUET-GROSSARD, V. PODEVIN-
DIMSTER, S. LEHMANN, J.P. CHAUVIN, J.P. DECAVEL, R. VAREA, J. GRASSI, M.
FONTES u. J.Y. CESBRON (2002):
PrP expression and replication by Schwann cells: implications in prion spreading.
J. Virol. 76, 2434-9
FORD, M.J., L.J. BURTON, R.J. MORRIS u. S.M. HALL (2002):
Selective expression of prion protein in peripheral tissues of the adult mouse.
Neuroscience 113, 177-92

Literaturverzeichnis 137
FOSTER, J.D., M. BRUCE, I. MCCONNELL, A. CHREE u. H. FRASER (1996):
Detection of BSE infectivity in brain and spleen of experimentally infected sheep.
Vet. Rec. 138, 546-8
FOSTER, J.D., N. HUNTER, A.E. WILLIAMS, M.J. MYLNE, W.A. MCKELVEY, J.
HOPE, H. FRASER u. C. BOSTOCK (1996):
Observations on the transmission of scrapie in experiments using embryo transfer.
Vet. Rec. 138, 559-62
FOSTER, J.D., D.W. PARNHAM, N. HUNTER u. M. BRUCE (2001):
Distribution of the prion protein in sheep terminally affected with BSE following
experimental oral transmission.
J. Gen. Virol. 82, 2319-26
FOSTER, J., C. MCKENZIE, D. PARNHAM, D. DRUMMOND, W. GOLDMANN, E.
STEVENSON u. N. HUNTER (2006):
Derivation of a scrapie-free sheep flock from the progeny of a flock affected by scrapie.
Vet. Rec. 159, 42-5
FRASER, H. (1976):
The pathology of natural and experimental scrapie.
In: R.H. Kimberlin (Hrsg.): Slow virus diseases of animals and man.
North-Holland Publishing Company, Amsterdam, S. 267-305
FURNESS, J.B. (2006):
The enteric nervous system
Blackwell Publishing, Massachusetts, Oxfort
GASSET, M., M.A. BALDWIN, D.H. LLOYD, J.M. GABRIEL, D.M. HOLTZMAN, F.
COHEN, R. FLETTERICK u. S.B. PRUSINER (1992):
Predicted alpha-helical regions of the prion protein when synthesized as peptides form
amyloid.
Proc. Natl. Acad. Sci. USA 89, 10940-4

138 Literaturverzeichnis
GAVIER-WIDEN, D., M. NOREMARK, S. BENESTAD, M. SIMMONS, L. RENSTROM,
B. BRATBERG, M. ELVANDER u. C.H. SEGERSTAD (2004):
Recognition of the Nor98 variant of scrapie in the Swedish sheep population.
J. Vet. Diagn. Invest. 16, 562-7
GERDES, HH. (2009):
Prions tunnel between cells.
Nat. Cell. Biol. 11, 3, 235-6
GERSTMANN, J., E. STRÄUSSLER u. I. SCHEINKER (1936):
Über eine eigenartige hereditär-familiäre Erkrankung des Zentralnervensystems; zugleich ein
Beitrag zur Frage des vorzeitigen Alterns.
Z. Ges. Neurol. Psychiat. 154, 736-62
GLATZEL, M., u. A. AGUZZI (2000):
Peripheral pathogenesis of prion diseases.
Microbes. Infect. 2, 613-9
GONZÁLEZ, L., M.P. DAGLEISH, S.J. BELLWORTHY, S. SISÓ, M.J. STACK, M.J.
CHAPLIN, L.A. DAVIS, S.A. HAWKINS, J. HUGHES u. M. JEFFREY (2006):
Postmortem diagnosis of preclinical and clinical scrapie in sheep by the detection of disease-
associated PrP in their rectal mucosa.
Vet. Rec. 158, 10, 325-31
GOSSERT, A.D., S. BONJOUR, D.A. LYSEK, F. FIORITO u. K. WUTHRICH (2005):
Prion protein NMR structures of elk and of mouse/elk hybrids.
Proc. Natl. Acad. Sci. USA, 102, 646-50
GRAVENOR, M.B., N. STALLARD, R. CURNOW u. A.R. MCLEAN (2003):
Repeated challenge with prion disease: the risk of infection and impact on incubation period.
Proc. Natl. Acad. Sci. USA 100, 10960-5
GROSCHUP, M.H., M. BEEKES, P.A. MCBRIDE, M. HARDT, J.A. HAINFELLNER u. H.
BUDKA (1999):

Literaturverzeichnis 139
Deposition of disease-associated prion protein involves the peripheral nervous system in
experimental scrapie.
Acta Neuropathol. (Berl.) 98, 453-7
HAASE, B., M.G. DOHERR, T. SEUBERLICH, C. DROGEMULLER, G. DOLF, P.
NICKEN, K. SCHIEBEL, U. ZIEGLER, M.H. GROSCHUP, A. ZURBRIGGEN u. T. LEEB
(2007):
PRNP promoter polymorphisms are associated with BSE susceptibility in Swiss and German
cattle.
BMC Genet. 8, 15
HADLOW, W.J., R.C. KENNEDY u. R.E. RACE (1982):
Natural infection of Suffolk sheep with scrapie virus.
J. Infect. Dis. 146, 657-64
HAFEZPARAST, M., S. BRANDNER, J. LINEHAN, J.E. MARTIN, J. COLLINGE u. E.M.
FISHER (2005):
Prion disease incubation time is not affected in mice heterozygous for a dynein mutation.
Biochem. Biophys. Res. Commun. 326, 18–22
HARRIS, D.A. (2003):
Trafficking, turnover and membrane topology of PrP.
Br. Med. Bull. 66, 71-85
HARTSOUGH, G.R., u. D. BURGER (1965):
Encephalopathy of mink. I. Epizootiologic and clinical observations.
J. Infect. Dis. 115, 387-92
HEGGEBØ, R., C. M. PRESS, G. GUNNES, M. J. ULVUND, M. A. TRANULIS u. T.
LSVERK (2003):
Detection of PrPSc in lymphoid tissues of lambs experimentally exposed to the scrapie agent.
J. Comp. Pathol. 128, 2-3, 172-81

140 Literaturverzeichnis
HEIKENWALDER, M., N. ZELLER, H. SEEGER, M. PRINZ, P.C. KLÖHN, P.
SCHWARZ, N.H. RUDDLE, C. WEISSMAN u. A. AGUZZI (2005):
Chronic lymphocytic inflammation specifies the organ tropism of prions.
Science 307, 5712, 1107-10
HEPPNER, F.L., A.D. CHRIST, M.A. KLEIN, M. PRINZ, M. FRIED, J.P. RAEHENBUHL
u. A. AGUZZI (2001):
Transepithelial prion transport by M cells.
Nat. Med. 7, 976-7
HERRMANN, L.M., W.P. CHEEVERS, W.C. DAVIS, D.P. KNOWLES u. K.L.
O’ROURKE (2003):
CD21-positive follicular dendritic cells: a possible source of PrP(Sc) in lymph node
macrophages of scrapie-infected sheep.
Am. J. Pathol. 162, 1075-81
HERZOG, C., N. SALES, N. ETCHEGARAY, A. CHARBONNIER, S. FREIRE, D.
DORMONT, J.P. DESLYS u. C.I. LASMEZAS (2004):
Tissue distribution of bovine spongiform encephalopathy agent in primates after intravenous
or oral infection.
Lancet 363, 422-8
HÖRNLIMANN B. (2001):
Prionen und Prionkrankheiten.
In: B. HÖRNLIMANN, D. RIESNER u. H. KRETZSCHMAR (Hrsg.): Prionen und
Prionkrankheiten.
Verlag Walter de Gruyter, Berlin, S. 3-20
HOFFMANN, C., U. ZIEGLER, A. BUSCHMANN, A. WEBER, L. KUPFER, A.
OELSCHLEGEL, B. HAMMERSCHMIDT u. M.H. GROSCHUP (2007):
Prions spread via the autonomic nervous system from the gut to the central nervous system in
cattle incubating bovine spongiform encephalopathy.
J. Gen. Virol. 88, 1048-55

Literaturverzeichnis 141
HOFFMANN, C., M. EIDEN, M. KAATZ, M. KELLER, U. ZIEGLER, R. ROGERS, B.
HILLS, A. BALKEMA-BUSCHMANN, L. VAN KEULEN, J.G. JACOBS u. M.H.
GROSCHUP (2011):
BSE infectivity in jejunum, ileum and ileocaecal junction of incubating cattle.
Vet. Res. 42, 1, 21
HOINVILLE, L.J. (1996):
A review of the epidemiology of scrapie in sheep.
Rev-Off. Int. Epizoot. 15, 827-52
HORNEMANN, S., C. SCHORN u. K. WUTHRICH (2004):
NMR structure of the bovine prion protein isolated from healthy calf brains.
EMBO Rep. 5, 1159-64
HOUSTON, F., S. MCCUTCHEON, W. GOLDMANN, A. CHONG, J. FOSTER, S. SISÓ,
L. GONZÁLEZ, M. JEFFREY u. N. HUNTER (2008):
Prion diseases are efficiently transmitted by blood transfusion in sheep.
Blood 112, 12, 4739-45
HUANG, F.P., C.F. FARQUHAR, N.A. MABBOTT, M.E. BRUCE u. G.G. MACPHERSON
(2002):
Migrating intestinal dendritic cells transport PrP (Sc) from the gut.
J. Gen. Virol. 83, 267-71
HUNTER, G.D., u. G.C. MILLSON (1964):
Studies on the heat stability and chromatographic behaviour of the scrapie agent.
J. Gen. Microbiol. 37, 251-8
HUNTER, N., W. GOLDMANN, G. SMITH u. J. HOPE (1994):
The association of a codon 136 PrP gene variant with the occurrence of natural scrapie.
Arch. Virol. 137, 171-7
HUNTER, N., J. FOSTER, A. CHONG, S. MCCUTCHEON, D. PARNHAM, S. EATON, C.
MACKENZIE u. F. HOUSTON (2002):

142 Literaturverzeichnis
Transmission of prion diseases by blood transfusion.
J. Gen. Virol. 83, 2897-905
IWATA, N., Y. SATO, Y. HIGUCHI, K. NOHTOMI, N. NAGATA, H. HASEGAWA, M.
TOBIUME, Y. NAKAMURA, K. HAGIWARA, H. FURUOKA, M. HORIUCHI, Y.
YAMAKAWA u. T. SATA (2006):
Distribution of PrP(Sc) in cattle with bovine spongiform encephalopathy slaughtered at
abattoirs in Japan.
Jpn. J. Infect. Dis. 59, 100-7
JAKOB, A. 1921:
Über eine der multiplen Sklerose klinisch nahestehende Erkrankung des
Zentralnervensystems (spastische Pseudosklerose) mit bemerkenswertem anatomischem
Befund.
Med. Klin. 17, 372-6
JAKOB, A. (1921):
Über eigenartige Erkrankungen des Zentralnervensystems mit bemerkenswertem
anatomischem Befunde.
Z. ges. Neurol. Psychiat. 64, 147-228.
JAKOB, A. M. (1921):
Über eigenartige Erkrankungen des Zentralnervensystems mit bemerkenswertem
anatomischem Befunde (Spastische Pseudesklerose-Encephalomyelopathie mit disseminierten
Degenerationsherden).
Dtsch. Z. Nervenheilk. 70, 132-46
JEFFREY, M., u. G.A. WELLS (1988):
Spongiform encephalopathy in a nyala (Tragelaphus angasi)
Vet. Pathol. 25, 398-9
JEFFREY, M., C.M. GOODSIR, M.E. BRUCE, P.A. MCBRIDE u. C. FARQUHAR (1994):
Morphogenesis of amyloid plaques in 87V murine scrapie.
Neuropathol. Appl. Neurobiol. 20, 535-42

Literaturverzeichnis 143
JEFFREY, M., u. W. HALLIDAY (1994):
Numbers of neurons in vacuolated and non vacuolated neuroanatomical nuclei in bovine
spongiform encephalopathy affected brains.
J. Comp. Pathol. 110, 287-93
JEFFREY, M., S. MARTIN, J.R. THOMSON, W.S. DINGWALL, I. BEGARA-MCGORUM
u. L. GONZÁLEZ (2001):
Onset and distribution of tissue PrP accumulation in scrapie-affected suffolk sheep as
demonstrated by sequential necropsies and tonsillar biopsies.
J. Comp. Pathol. 125, 1, 48-57
JEFFREY, M., I. BEGARA-MCGORUM, S. CLARK, S. MARTIN, J. CLARK, M.
CHAPLIN u. L. GONZALEZ (2002):
Occurrence and distribution of infection-specific PrP in tissues of clinical scrapie cases and
cull sheep from scrapie-affected farms in Shetland.
J. Comp. Pathol. 127, 4, 264-73
JEFFREY, M., u. L. GONZÁLEZ (2004):
Pathology and pathogenesis of bovine spongiform encephalopathy and scrapie.
Curr. Top. Microbiol. Immunol. 284, 65-97
JEFFREY, M., L. GONZALEZ, A. ESPENES, C.M. PRESS, S. MARTIN, M. CHAPLIN, L.
DAVIS, T. LANDSVERK, C. MACALDOWIE, S. EATON u. G. MCGOVERN (2006):
Transportation of prion protein across the intestinal mucosa of scrapie-susceptible and
scrapie-resistant sheep.
J. Pathol. 209, 4-14
JEFFREY, M., u. L. GONZÁLEZ (2007):
Classical sheep transmissible spongiform encephalopathies: pathogenesis, pathological
phenotypes and clinical disease.
Neuropathol. Appl. Neurobiol. 33, 373-94
JEFFREY, M., G. MCGOVERN, C.M. GOODSIR u. L. GONZALEZ (2009):

144 Literaturverzeichnis
Strain-associated variations in abnormal PrP trafficking of sheep scrapie.
Brain Pathol. 19, 1-11
JEFFREY, M., G. MCGOVERN, S. SISÓ S u. L. GONZÁLEZ (2011):
Cellular and sub-cellular pathology of animal prion diseases: relationship between
morphological changes, accumulation of abnormal prion protein and clinical disease.
Acta Neuropathol. 121, 1, 113-34
JULING, K., H. SCHWARZENBACHER, J.L. WILLIAMS u. R. FRIES (2006):
A major genetic component of BSE susceptibility.
BMC Biol. 4, 33
KELSALL, B.L., u. W. STROBER (1996):
Distinct populations of dendritic cells are present in the subepithelial dome and T cell regions
of the murine Peyer’s patch.
J. Exp. Med. 183, 237-47
KIMBERLIN, R.H., S.M. HALL u. C.A. WALKER (1983):
Pathogenesis of mouse scrapie. Evidence for direct neural spread of infection to the CNS after
injection of sciatic nerve.
J. Neurol. Sci. 61, 315–25
KIMBERLIN, R.H., S. COLE u. C.A. WALKER (1987):
Pathogenesis of scrapie is faster when infection is intraspinal instead of intracerebral.
Microb. Pathog. 2, 6, 405-15
KIMURA, K., u. M. HARITANI (2008):
Distribution of accumulated prion protein in a cow with bovine spongiform encephalopathy.
Vet. Rec. 162, 25, 822-5
KIRKWOOD, J.K., G.A. WELLS, J.W. WILESMITH, A.A. CUNNINGHAM u. S.I.
JACKSON (1990):
Spongiform encephalopathy in an arabian oryx (Oryx leucoryx) and a greater kudu
(Tragelaphus strepsiceros).

Literaturverzeichnis 145
Vet. Rec. 127, 418-20
KLEIN M.A., R. FRIGG, E. FLECHSIG, A.J. RAEBER, U. KALINKE, H.
BLUETHMANN, F. BOOTZ, M. SUTER, R.M. ZINKERNAGEL, A. AGUZZI (1997):
A crucial role for B cells in neuroinvasive scrapie.
Nature 390, 687-91
KLEIN, M.A., R. FRIGG, A.J. RAEBER, E. FLECHSIG, I. HEGYI, R.M. ZINKERNAGEL,
C. WEISSMANN u. A. AGUZZI (1998):
PrP expression in B lymphocytes is not required for prion neuroinvasion.
Nat. Med. 4, 1429-33
KOCISKO, D.A., J.H. COME, S.A. PRIOLA, B. CHESEBRO, G.J. RAYMOND, P.T.
LANSBURY u. B. CAUGHEY (1994):
Cell-free formation of protease-resistant prion protein.
Nature 370, 471-4
KONOLD, T., Y.H. LEE, M.J STACK, C. HORROCKS, R.B. GREEN, M. CHAPLIN, M.M.
SIMMONS, S.A. HAWKINS, R. LOCKEY, J. SPIROPOULOS, J.W. WILESMITH u. G.A.
WELLS (2006):
Different prion disease phenotypes result from inoculation of cattle with two temporally
separated sources of sheep scrapie from Great Britain
BMC Vet. Res. 2, 31
KONOLD, T., G. BONE, A. VIDAL-DIEZ, R. TORTOSA, A. DAVIS, G. DEXTER, P.
HILL, M. JEFFREY, M.M. SIMMONS, M.J. CHAPLIN, S.J. BELLWORTHY u. C.
BERTHELIN-BAKER (2008):
Pruritus is a common feature in sheep infected with the BSE agent.
BMC Vet. Res. 4, 16
KONG, Q., M. ZHENG, C. CASALONE, L. QING, S. HUANG, B. CHAKRABORTY, P.
WANG, F. CHEN, I. CALI, C. CORONA, F. MARTUCCI, B. IULINI, P. ACUTIS, L.
WANG, J. LIANG J, M. WANG, X. LI, S. MONACO, G. ZANUSSO, W.Q. ZOU, M.
CARAMELLI u. P. GAMBETTI (2008):

146 Literaturverzeichnis
Evaluation of the human transmission risk of an atypical bovine spongiform encephalopathy
prion strain.
J. Virol. 82, 7, 3697-701
KOVACS, G.G., E. LINDECK-POZZA, L. CHIMELLI, A.Q. ARAÚJO, A.A. GABBAI, T.
STRÖBEL, M. GLATZEL, A. AGUZZI u. H. BUDKA (2004):
Creutzfeldt-Jakob disease and inclusion body myositis: abundant disease-associated prion
protein in muscle.
Ann. Neurol. 55, 1, 121-5
KRATZEL, C., J. MAI, K. MADELA, M. BEEKES u. D. KRÜGER (2007a):
Propagation of scrapie in peripheral nerves after footpad infection in normal and neurotoxin
exposed hamsters.
Vet. Res. 38, 127-39
KRATZEL, C., D. KRÜGER u. M. BEEKES (2007b):
Prion propagation in a nerve conduit model containing segments devoid of axons.
J. Gen. Virol. 88, 3479-85
KRÜGER D, A. THOMZIG, G. LENZ, K. KAMPF, P. MCBRIDE u. M. BEEKES (2009):
Faecal shedding, alimentary clearance and intestinal spread of prions in hamsters fed with
scrapie.
Vet. Res. 40, 1, 4
KUCZIUS, T., u. M.H. GROSCHUP (1999):
Differences in proteinase K resistance and neuronal deposition of abnormal prion proteins
characterize bovine spongiform encephalopathy (BSE) and scrapie strains.
Mol. Med. 5, 406-18
KUNZI, V., M. GLATZEL, M.Y. NAKANO, U.F. GREBER, F. VAN LEUVEN u. A.
AGUZZI (2002):
Unhampered prion neuroinvasion despite impaired fast axonal transport in transgenic mice
overexpressing four-repeat tau.
J. Neurosci. 22, 7471-77

Literaturverzeichnis 147
LACROUX, C., F. CORBIERE, G. TABOURET, S. LUGAN, P. COSTES, J. MATHEY,
J.M. DELMAS, J.L. WEISBECKER, G. FOUCRAS, H. CASSARD, J.M. ELSEN, F.
SCHELCHER u. O. ANDRÉOLETTI (2007):
Dynamics and genetics of PrP(Sc) placental accumulation in sheep.
J. Gen. Virol. 88, 1056-61
LACROUX, C., S. SIMON, S. L. BENESTAD, S. MAILLET, J. MATHEY, S. LUGAN, F.
CORBIÈRE, H. CASSARD, P. COSTES, D. BERGONIER, J.L. WEISBECKER, T.
MOLDAL, H. SIMMONS, F. LANTIER, C. FERAUDET-TARISSE, N. MOREL, F.
SCHELCHER, J. GRASSI u. O. ANDRÉOLETTI (2008):
Prions in milk from ewes incubating natural scrapie.
PLoS Pathog. 4, 12, e1000238
LASMEZAS, C.I., J.Y. CESBRON, J.P. DESLYS, R. DEMAIMAY, K.T. ADJOU, R.
RIOUX, C. LEMAIRE, C. LOCHT u. D. DORMONT (1996):
Immune system-dependent and -independent replication of the scrapie agent.
J. Virol. 70, 1292-5
LASMEZAS, C.I., J.P. DESLYS, O. ROBAIN, A. JAEGLY, V. BERINGUE, J.M. PEYRIN,
J.G. FOURNIER, J.J. HAUW, J. ROSSIER u. D. DORMONT (1997):
Transmission of the BSE agent to mice in the absence of detectable abnormal prion protein.
Science 275, 402-5
LEBLANC, P., S. ALAIS, I. PORTO-CARREIRO, S. LEHMANN, J. GRASSI, G. RAPOSO
u. J.L. DARLIX (2006):
Retrovirus infection strongly enhances scrapie infectivity release in cell culture.
EMBO J. 25, 2674-85
LE DUR, A., V. BERINGUE, O. ANDREOLETTI, F. REINE, T.L. LAI, T. BARON, B.
BRATBERG, J.L. VILOTTE, P. SARRADIN, S.L. BENESTAD u. H. LAUDE (2005):
A newly identified type of scrapie agent can naturally infect sheep with resistant PrP
genotypes.
Proc. Natl. Acad. Sci. USA 102, 16031-6

148 Literaturverzeichnis
LEGNAME, G., I.V. BASKAKOV, H.O. NGUYEN, D. RIESNER, F.E. COHEN, S.J.
DEARMOND u. S.B. PRUSINER (2004):
Synthetic mammalian prions.
Science 305, 673-6
LEOPOLDT, J.G. (1750):
Nützliche und auf die Erfahrung gegründete Einleitung zu der Land-Wirthschafft.
Verlag Johann Gottlieb Rothen, Sorau, Bd.5
LIGIOS, C., M. JEFFREY, S.J. RYDER, S.J. BELLWORTHY u. M.M. SIMMONS (2002):
Distinction of scrapie phenotypes in sheep by lesion profiling.
J. Comp. Pathol. 127, 1, 45-57
LIGIOS, C., C.J. SIGURDSON, C. SANTUCCIU, G. CARCASSOLA, G. MANCO, M.
BASAGNI, C. MAESTRALE, M.G. CANCEDDA, L. MADAU u. A. AGUZZI (2005):
PrP(Sc) in mammary glands of sheep affected by scrapie and mastitis.
Nat. Med. 11, 1137-8
LIGIOS, C., G.M. CANCEDDA, I. MARGALITH, C. SANTUCCIU, L. MADAU, C.
MAESTRALE, M BASAGNI, M. SABA u. M. HEIKENWALDER (2007):
Intraepithelial and interstitial deposition of pathological prion protein in kidneys of scrapie-
affected sheep.
PLoS One 2, 9, e859
LLEWELYN, C.A., P.E. HEWITT, R.S. KNIGHT, K. AMAR, S. COUSENS, J.
MACKENZIE u. R.G. Will (2004):
Possible transmission of variant Creutzfeldt–Jakob disease by blood transfusion.
Lancet 363, 417-21
LUGARESI, E., R. MEDORI, P. MONTAGNA, A. BARUZZI, P. CORTELLI, A.
LUGARESI, P. TINUPER, M. ZUCCONI u. P. GAMBETTI (1986):
Fatal familial insomnia and dysautonomia with selective degeneration of thalamic nuclei.
N. Engl. J. Med. 315, 16, 997-1003

Literaturverzeichnis 149
LYAHYAI, J., R. BOLEA, C. SERRANO, E. MONLEÓN, C. MORENO, R. ROSTA, P.
ZARAGOZA, J.J. BADIOLA u. I. MARTÍN-BURRIEL (2006):
Correlation between Bax overexpression and prion deposition in medulla oblongata from
natural scrapie without evidence of apoptosis.
Acta Neuropathol. 112, 4, 451-60
LYSEK, D.A., C. SCHORN, L.G. NIVON, V. ESTEVE-MOYA,B. CHRISTEN, L.
CALZOLAI, C. VON SCHROETTER, F. FIORITO, T. HERRMANN, P. GUNTERT u. K.
WUTHRICH (2005):
Prion protein NMR structures of cats, dogs, pigs, and sheep.
Proc. Natl. Acad. Sci. USA, 102, 640-5
MABBOTT, N.A., A. WILLIAMS, C.F. FARQUHAR, M. PASPARAKIS, G. KOLLIAS u.
M.E. BRUCE (2000):
Tumor necrosis factor alpha-deficient, but not interleukin-6-deficient, mice resist peripheral
infection with scrapie.
J. Virol. 74, 3338-44
MABBOTT, N. A., F. MACKAY, F. MINNS u. M.E. BRUCE (2000):
Temporary inactivation of follicular dendritic cells delays neuroinvasion of scrapie.
Nature Med. 6, 719-20
MAIGNIEN, T., C.I. LASMEZAS, V. BERINGUE, D. DORMONT u. J.P. DESLYS (1999):
Pathogenesis of the oral route of infection of mice with scrapie and bovine spongiform
encephalopathy agents.
J. Gen. Virol. 80, 3035-42
MAIGNIEN, T., M. SHAKWEH, P. CALVO, D. MARCE, N. SALES, E. FATTAL, J.P.
DESLYS, P. COUVREUR u. C.I. LASMEZAS (2005):
Role of gut macrophages in mice orally contaminated with scrapie or BSE.
Int. J. Pharm. 298, 293-304
MANDEL, T. E., R.P. PHIPPS, A. ABBOT u. J.G. TEW (1980):

150 Literaturverzeichnis
The follicular dendritic cell: long term antigen retention during immunity.
Immunol. Rev. 53, 29-59
MANNING, K.A., J.T. ERICHSEN u. C. EVINGER (1990):
Retrograde transneuronal transport properties of fragment C of tetanus toxin.
Neuroscience 34, 1, 251-63
MANUELIDIS, L. (1994):
Dementias, neurodegeneration, and viral mechanisms of disease from the perspective of
human transmissible encephalopathies.
Ann. NY Acad. Sci. 724, 259-81
MASUJIN, K., D. MATTHEWS, G.A.H. WELLS, S. MOHRI u. T. YOKOYAMA (2007):
Prions in the peripheral nerves of bovine spongiform encephalopathy-affected cattle.
J. Gen. Virol. 88, 1850-8
MCBRIDE, P.A., u. M. BEEKES (1999):
Pathological PrP is abundant in sympathetic and sensory ganglia of hamsters fed with scrapie.
Neurosci. Lett. 265, 135-8
MCBRIDE, P.A., W.J. SCHULZ-SCHAEFFER, M. DONALDSON, M. BRUCE, H.
DIRINGER, H.A. KRETZSCHMAR u. M. BEEKES (2001):
Early spread of scrapie from the gastrointestinal tract to the central nervous system involves
autonomic fibers of the splanchnic and vagus nerves.
J. Virol. 75, 9320-7
MCGILL, I., u. G.A.H. WELLS (1993):
Neuropathological findings in cattle with clinically suspect but histologically unconfirmed
bovine spongiform encephalopathy (BSE).
J. Comp. Pathol. 108, 241-60
MCGOWAN, J.P. (1922):
Scrapie in sheep.
Scott. J. Agri. 5, 365-75

Literaturverzeichnis 151
MCLEAN, A.R., u. C.J. BOSTOCK (2000):
Scrapie infections initiated at varying doses: an analysis of 117 titration experiments.
Philos. Trans. R. Soc. Lond. B. Biol. Sci. 355, 1043-50
MILLING, S.W., U. YRLID, C. JENKINS, C.M. RICHARDS, N.A. WILLIAMS u. G.
MACPHERSON (2007): Regulation of intestinal immunity: effects of the oral adjuvant
Escherichia coli heat-labile enterotoxin on migrating dendritic cells.
Eur. J. Immunol. 37, 87-99
MISHRA, R.S., S. BASU, Y. GU, X. LUO, W.Q. ZOU, R. MISHRA, R. LI, S.G. CHEN, P.
GAMBETTI, H. FUJIOKA u. N. SINGH (2004):
Protease-resistant human prion protein and ferritin are cotransported across Caco-2 epithelial
cells: implications for species barrier in prion uptake from the intestine.
J. Neurosci. 24, 11280-90
MOHAN, J., K.L. BROWN, C.F. FARQUHAR, M.E. BRUCE u. N.A. MABBOTT (2004):
Scrapie transmission following exposure through the skin is dependent on follicular dendritic
cells in lymphoid tissues.
J. Dermatol. Sci. 35, 101-11
MONTRASIO, F., R. FRIGG, M. GLATZEL, M.A. KLEIN, F. MACKAY, A. AGUZZI u. C.
WEISSMANN (2000):
Impaired prion replication in spleens of mice lacking functional follicular dendritic cells.
Science 288, 1257-9
NEUTRA, M.R., E. PRINGAULT, J.P. KRAEHENBUHL (1996):
Antigen sampling across epithelial barriers and induction of mucosal immune responses.
Annu. Rev. Immunol. 14, 275–300
NICKEL, SCHUMMER, SEIFERLE (1991):
Anatomie der Haustiere
3. Auflage, Bd. 1-3
Verlag Paul Parey, Berlin, Hamburg

152 Literaturverzeichnis
OESCH, B., D. WESTAWAY, M. WÄLCHLI, M.P. MCKINLEY, S.B. KENT, R.
AEBERSOLD, R.A. BARRY, P. TEMPST, D.B. TEPLOW u. L.E. HOOD, et al. (1985):
A cellular gene encodes scrapie PrP 27-30 protein.
Cell 40, 4, 735-46
OHMORI, Y., Y. ATOJI, S. SAITO, H. UENO, Y. INOSHIMA u. N. ISHIGURO (2008):
Localization of sympathetic, parasympathetic and sensory neurons innervating the distal
ileum of the cattle.
J. Vet. Med. Sci. 70, 12, 1289-94
OLDSTONE, M.B., R. RACE, D. THOMAS, H. LEWICKI, D. HOMANN, S. SMELT, A.
HOLZ, P. KONI, D. LO, B. CHESEBRO u. R. FLAVELL (2002):
Lymphotoxin-α- and lymphotoxin-β-deficient mice differ in susceptibility to scrapie:
evidence against dendritic cell involvement.
J. Virol. 76, 4357-63
PAISLEY, L.G., u. J. HOSTRUP-PEDERSEN (2004):
A quantitative assessment of the risk of transmission of bovine spongiform encephalopathy by
tallo-based calf-milk-replacer.
Prev. Vet. Med. 63, 135-49
PAMMER J., H.S. CROSS, Y. FROBERT, E. TSCHACHLER u. G. OBERHUBER (2000):
The pattern of prion-related protein expression in the gastrointestinal tract.
Virchows Arch. 436, 5, 466-72
PATTISON, I.H., M.N. HOARE, J.N. JEBBETT u. W.A. WATSON (1972):
Spread of scrapie to sheep and goats by oral dosing with foetal membranes from scrapie-
affected sheep.
Vet. Rec. 90, 465-8
PEARSON, G.R., J.M. WYATT, T.J. GRUFFYDD-JONES, J. HOPE, A. CHONG, R.J.
HIGGINS, A.C. SCOTT u. G.A. WELLS (1992):
Feline spongiform encephalopathy: fibril and PrP studies.

Literaturverzeichnis 153
Vet. Rec. 131, 14, 307-10
PEDEN, A.H., M.W. HEAD, D.L. RITCHIE, P.J.E. BELL u. P.J.W. IRONSIDE (2004):
Preclinical vCJD after blood transfusion in a PRNP codon 129 heterozygous patient.
Lancet 364, 9433, 527-9
PIENING, N., R. NONNO, M. DI BARI, S. WALTER, O. WINDL, U. AGRIMI, H.A.
KRETZSCHMAR u. U. BERTSCH (2006):
Conversion efficiency of bank vole prion protein in vitro is determined by residues 155 and
170, but does not correlate with the high susceptibility of bank voles to sheep scrapie in vivo.
J. Biol. Chem. 281, 9373-84
PRESS, C. M., R. HEGGEBO u. A. ESPENES (2004):
Involvement of gut-associated lymphoid tissue of ruminants in the spread of transmissible
spongiform encephalopathies.
Adv. Drug Deliv. Rev. 56, 6, 885-99
PRINZ, M., M. HEIKENWALDER, T. JUNT, P. SCHWARZ, M. GLATZEL, F.L.
HEPPNER, Y.X. FU, M. LIPP u. A. AGUZZI (2003):
Positioning of follicular dendritic cells within the spleen controls prion neuroinvasion.
Nature 425, 6961, 957-62
PRUSINER, S.B., D.F. GROTH, S.P. COCHRAN, F.R. MASIARZ, M.P. MCKINLEY u.
H.M. MARTINEZ (1980):
Molecular properties, partial purification, and assay by incubation period measurements of the
hamster scrapie agent.
Biochemistry 19, 4883-91
PRUSINER, S. B. (1982):
Novel proteinaceous infectious particles cause scrapie.
Science 216, 4542, 136-44
PRUSINER, S.B., M.P. MCKINLEY, K.A. BOWMAN, D.C. BOLTON, P.E BENDHEIM,
D.F. GROTH u. G.G. GLENNER (1983):

154 Literaturverzeichnis
Scrapie prions aggregate to form amyloid-like birefringent rods.
Cell 35, 349-58
PRUSINER, S. B., R. GABIZON u. M. P. MCKINLEY (1987):
On the biology of prions.
Acta. Neuropathol. (Berl.) 72, 4, 299-314
PRUSINER, S.B., (1990):
Novel structure and genetics of prions causing neurodegeneration in humans and animals.
Biologicals 18, 247-62
PRUSINER, S. B. (1998):
Prions.
Proc. Natl. Acad. Sci. USA 95, 23, 13363-83
RACE, R., M. OLDSTONE u. B. CHESEBRO (2000):
Entry versus blockade of brain infection following oral or intraperitoneal scrapie
administration: role of prion protein expression in peripheral nerves and spleen.
J. Gen. Virol. 74, 828-33
RACE, B., K. MEADE-WHITE, M.B. OLDSTONE, R. RACE u. B. CHESEBRO (2008):
Detection of prion infectivity in fat tissues of scrapie-infected mice.
PLoS Pathog. 4, 12, e1000232
RESCIGNO, M., M. URBANO, B. VALZASINA, M. FRANCOLINI, G. ROTTA, R.
BONASIO, F. GRANUCCI, J.P. KRAEHENBUHL u. P. RICCIARDI CASTAGNOLI
(2001):
Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to
sample bacteria.
Nat. Immunol. 2, 361-7
RIEK, R., S. HORNEMANN, G. WIDER, M. BILLETER, R. GLOCKSHUBER u. K.
WUTHRICH (1996):
NMR structure of the mouse prion protein domain PrP(121-321).

Literaturverzeichnis 155
Nature 382, 180-2
RUSSELAKIS-CARNEIRO, M., G.P. SABORIO, L. ANDERES u. C. SOTO (2002):
Changes in the glycosylation pattern of prion protein in murine scrapie. Implications for the
mechanism of neurodegeneration in prion diseases.
J. Biol. Chem. 277, 36872-7
RYBNER-BARNIER, C., C. JACQUEMOT, C. CUCHE, G. DORÉ, L. MAJLESSI, M.M.
GABELLEC, A. MORIS, O. SCHWARTZ, J. DI SANTO, A. CUMANO, C. LECLERC u. F.
LAZARINI (2006):
Processing of the bovine spongiform encephalopathy-specific prion protein by dendritic cells.
J. Virol. 80, 10, 4656-63
SAÁ, P., J. CASTILLA u. C. SOTO (2006):
Presymptomatic detection of prions in blood.
Science 313, 5783, 92-4
SAFAR, J.G., S.J. DEARMOND, K. KOCIUBA, C. DEERING, S. DIDORENKO, E.
BOUZAMONDO-BERNSTEIN, S.B. PRUSINER u. P. TREMBLAY (2005):
Prion clearance in bigenic mice.
J. Gen. Virol. 86, 2913-23
SAKAGUCHI, T., S. BRAND u. H.C. REINECKER (2001):
Mucosal barrier and immune mediators.
Curr. Opin. Gastroenterol. 17, 573-7
SASSA, Y., T. YAMASAKI, M. HORIUCHI, Y. INOSHIMA u. N. ISHIGURO (2010):
The effects of lysosomal and proteasomal inhibitors on abnormal forms of prion protein
degradation in murine macrophages.
Microbiol. Immunol. 54, 12, 763-8
SCHMERR, M.J., A.L. JENNY, M.S. BULGIN, J.M. MILLER, A.N. HAMIR, R.C. CUTLIP
u. K.R. GOODWIN (1999):

156 Literaturverzeichnis
Use of capillary electrophoresis and fluorescent labeled peptides to detect the abnormal prion
protein in the blood of animals that are infected with a transmissible spongiform
encephalopathy.
J. Chromatogr. A. 853, 207-14
SCOTT, M., D. GROTH, D. FOSTER, M. TORCHIA, S.L. YANG, S.J. DEARMOND u.
S.B. PRUSINER (1993):
Propagation of prions with artificial properties in transgenic mice expressing chimeric PrP
genes.
Cell 73, 979-88
SEEGER, H., M. HEIKENWALDER, N. ZELLER, J. KRANICH, P. SCHWARZ, A.
GASPERT, B. SEIFERT, G. MIELE u. A. AGUZZI (2005):
Coincident scrapie infection and nephritis lead to urinary prion excretion.
Science 310, 5746, 324-6
SERRANO, C., J. LYAHYAI, R. BOLEA, et al. (2009)
Distinct spatial activation of intrinsic and extrinsic apoptosis pathways in natural scrapie:
association with prion-related lesions.
Vet. Res. 40, 42
SHORTMAN, K., u. Y.-J. LIU (2002):
Mouse and human dendritic cell subtypes.
Nature Rev. Immunol. 2, 151-61
SIGURDSON, C.J., M. HEIKENWALDER, G. MANCO, M. BARTHEL, P. SCHWARZ, B.
STECHER, N.J. KRAUTLER, W.D. HARDT, B. SEIFERT, A.J. MACPHERSON, I.
CORTHESY u. A. AGUZZI (2009):
Bacterial colitis increases susceptibility to oral prion disease.
J. Infect. Dis. 15, 199(2), 243-52
SIMMONS, M.M., T. KONOLD, H.A. SIMMONS, Y.I. SPENCER, R. LOCKEY, J.
SPIROPOULOS, S. EVERITT u. D. CLIFFORD (2007):
Experimental transmission of atypical scrapie to sheep.

Literaturverzeichnis 157
BMC Vet. Res. 3, 20
SIMONEAU, S., M.M. RUCHOUX, N. VIGNIER, P. LEBON, S. FREIRE, E. COMOY, J.
P. DESLYS u. J.G. FOURNIER (2009):
Small critical RNAs in the scrapie agent.
Nature Precedings: hdl:10101/npre.2009.3344.1
SISÓ, S., L. GONZÁLEZ, F. HOUSTON, N. HUNTER, S. MARTIN u. M. JEFFREY
(2006):
The neuropathologic phenotype of experimental ovine BSE is maintained after blood
transfusion.
Blood 108, 2, 745-8
SISÓ, S., L. GONZÁLEZ , M. JEFFREY, S. MARTIN, F. CHIANINI u. P. STEELE (2006):
Prion protein in kidneys of scrapie-infected sheep.
Vet. Rec. 159, 327-8
SISÓ, S., M. JEFFREY, P. STEELE, G. MCGOVERN, S. MARTIN, J. FINLAYSON, F.
CHIANINI u. L. GONZÁLEZ (2008):
Occurrence and cellular localization of PrP(d) in kidneys of scrapie-affected sheep in
the absence of inflammation.
J. Pathol. 215, 2, 126-34
SISÓ, S., M. JEFFREY u. L. GONZÁLEZ (2009):
Neuroinvasion in sheep transmissible spongiform encephalopathies: the role of the
haematogenous route.
Neuropathol. Appl. Neurobiol. 35, 3, 232-46
SISÓ, S., L. GONZÁLEZ u. M. JEFFREY (2010):
Neuroinvasion in Prion Diseases: The Roles of Ascending Neural Infection and Blood
Dissemination.
Interdisciplinary Perspectives on Infectious Diseases, Volume 2010, Art ID 747892,
doi:10.1155/2010/747892

158 Literaturverzeichnis
SÖDERHOLM, J.D., C. STREUTKER, P.C. YANG, C. PATERSON, P.K. SINGH, D.M.
MCKAY, P.M. SHERMAN, K. CROITORU u. M.H. PERDUE (2004):
Increased epithelial uptake of protein antigens in the ileum of Crohn’s disease mediated by
tumour necrosis factor alpha.
Gut 53, 1817-24
SOMERVILLE, R.A., C.R. BIRKETT, C.F. FARQUHAR, N. HUNTER, W. GOLDMANN,
J. DORNAN, D. GROVER, R.M. HENNION, C. PERCY, J. FOSTER u. M. JEFFREY
(1997):
Immunodetection of PrPSc
in spleens of some scrapie-infected sheep but not BSE-infected
cows.
J. Gen. Virol. 78, 2389-96
SOTO, C. (2003):
Unfolding the role of protein misfolding in neurodegenerative diseases.
Nat. Rev. Neurosci. 4, 49-60
SOTO, C., u. J. CASTILLA (2004):
The controversial protein-only hypothesis of prion propagation.
Nat. Med. 10, 63-7
STACK, M. J., M. J. CHAPLIN u. J. CLARK (2002):
Differentiation of prion protein glycoforms from naturally occurring sheep scrapie, sheep-
passaged scrapie strains (CH1641 and SSBP1), bovine spongiform encephalopathy (BSE)
cases and Romney and Cheviot breed sheep experimentally inoculated with BSE using two
monoclonal antibodies.
Acta Neuropathol. 104, 3, 279-86
STENGEL, A., C. BACH, I. VORBERG, O. FRANK, S. GILCH, G. LUTZNY, W.
SEIFARTH, V. ERFLE, E. MAAS, H. SCHATZL, C. LEIB-MOSCH u. A.D.
GREENWOOD (2006):
Prion infection influences murine endogenous retrovirus expression in neuronal cells.
Biochem. Biophys. Res. Commun. 343, 825-31

Literaturverzeichnis 159
SUPATTAPONE, S. (2004):
Prion protein conversion in vitro.
J. Mol. Med. 82, 348-56
SURPRENANT, A. (1994):
Control of the gastrointestinal tract by enteric neurons.
Annu. Rev. Physiol. 56, 117-40
TERRY, L.A., S. MARSH, S.J. RYDER, S.A. HAWKINS, G.A. WELLS u. Y.I. SPENCER
(2003):
Detection of disease-specific PrP in the distal ileum of cattle exposed orally to the agent
of bovine spongiform encephalopathy.
Vet. Rec. 152, 387-92
THACKRAY, A.M., M.A. KLEIN u. R. BUJDOSO (2003):
Subclinical prion disease induced by oral inoculation.
J. Virol. 77, 14, 7991-8
THEIL, D., R. FATZER, R. MEYER, M. SCHOBESBERGER, A. ZURBRIGGEN u. M.
VANDEVELDE (1999):
Nuclear DNA fragmentation and immune reactivity in bovine spongiform encephalopathy.
J. Comp. Pathol. 121, 4, 357-67
THÉRY, C., A. REGNAULT, J. GARIN, J. WOLFERS, L. ZITVOGEL, P. RICCIARDI-
CASTAGNOLI, G. RAPOSO u. S. AMIGORENA (1999):
Molecular characterization of dendritic cell-derived exosomes. Selective accumulation of the
heat shock protein hsc73.
J. Cell. Biol. 147, 3, 599-610
THÉRY, C., M. BOUSSAC, P. VÉRON, P. RICCIARDI-CASTAGNOLI, G. RAPOSO, J.
GARIN u. S. AMIGORENA (2001):
Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment
distinct from apoptotic vesicles.
J. Immunol. 166, 12, 7309-18

160 Literaturverzeichnis
THOMZIG, A., C. KRATZEL, G. LENZ, D. KRÜGER u. M. BEEKES (2003):
Widespread PrP(Sc) accumulation in muscles of hamsters orally infected with scrapie.
EMBO 4, 530-3
THOMZIG A, W. SCHULZ-SCHAEFFER, C. KRATZEL, J. MAI u. M. BEEKES (2004):
Preclinical deposition of pathological prion protein PrP(Sc) in muscles of hamsters orally
exposed to scrapie.
J. Clin. Invest. 113, 1465-72
THOMZIG, A., F. CARDONE, D. KRÜGER, M. POCCHIARI, P. BROWN u. M. BEEKES
(2006):
Pathological prion protein in muscles of hamsters and mice infected with rodent-adapted BSE
or vCJD.
J. Gen. Virol. 87, 251-4
THOMZIG, A., W. SCHULZ-SCHAEFFER, A. WREDE, W. WEMHEUER, B. BRENIG, C.
KRATZEL, K. LEMMER u. M. BEEKES (2007):
Accumulation of pathological prion protein PrPSc in the skin of animals with experimental
and natural scrapie.
PLoS Pathog. 3, 5, e66
TOBLER, I., S.E. GAUS, T. DEBOER, P. ACHERMANN, M. FISCHER, T. RULICKE, M.
MOSER, B. OESCH, P.A. MCBRIDE u. J.C. MANSON (1996):
Altered circadian activity rhythms and sleep in mice devoid of prion protein.
Nature 380, 639-42
TOMISHIMA, M.J., G.A. SMITH U. L.W. ENQUIST (2001):
Sorting and transport of alpha herpesviruses in axons.
Traffic 2, 7, 429-36
TOUZEAU, S., M.E. CHASE-TOPPING, L. MATTHEWS, D. LAJOUS, F. EYCHENNE u.
N. HUNTER, J.D. FOSTER, G. SIMM, J.M. ELSEN u. M.E. WOOLHOUSE (2006):

Literaturverzeichnis 161
Modelling the spread of scrapie in a sheep flock: evidence for increased transmission during
lambing seasons.
Arch. Virol. 151, 735-51
VACCARI, G., C.H. PANAGIOTIDIS, C. ACIN, S. PELETTO, F. BARILLET, P. ACUTIS,
A. BOSSERS, J. LANGEVELD, L. VAN KEULEN, T. SKLAVIADIS, J.J. BADIOLA, O.
ANDREOLETTI, M.H. GROSCHUP, U. AGRIMI, J. FOSTER u. W. GOLDMANN (2009):
State-of-the-art review of goat TSE in the European Union, with special emphasis on PRNP
genetics and epidemiology.
Vet. Res. 40, 48
VANA, K., C. ZUBER, D. NIKLES u. S. WEISS (2007):
Novel aspects of prions, their receptor molecules, and innovative approaches for TSE therapy.
Cell. Mol. Neurobiol. 27, 107-28
VAN KEULEN, L. J., B. E. SCHREUDER, R. H. MELOEN, G. MOOIJ-HARKES, M. E.
VROMANS u. J. P. LANGEVELD (1996):
Immunohistochemical detection of prion protein in lymphoid tissues of sheep with natural
scrapie.
J. Clin. Microbiol. 34, 5, 1228-31
VAN KEULEN, L.J.M., B.E.C. SCHREUDER, M.E.W. VROMANS, J.P.M. LANGEVELD
u. M.A. SMITS (1999)
Scrapie associated prion protein in the gastro-intestinal tract of sheep with natural scrapie.
J. Comp. Pathol. 121, 55-63
VAN KEULEN, L. J., B. E. SCHREUDER, M. E. VROMANS, J. P. LANGEVELD u. M. A.
SMITS (2000):
Pathogenesis of natural scrapie in sheep.
Arch. Virol. Suppl. 16, 57-71
VAN KEULEN, L. J., M. E. VROMANS u. F. G. VAN ZIJDERVELD (2002):
Early and late pathogenesis of natural scrapie infection in sheep.
Apmis 110, 1, 23-32

162 Literaturverzeichnis
VAN KEULEN, L.J.M., A. BOSSERS u. F. VAN ZIJDERVELD (2008):
TSE pathogenesis in cattle and sheep.
Vet. Res. 39, 24
VASCELLARI, M., R. NONNO, F. MUTINELLI, M. BIGOLARO, M.A. DI BARI, F.
MELCHIOTTI, S. MARCON, C. D’AGOSTINO, G. VACCARI, M. CONTE, L. DE
GROSSI, F. ROSONE, F. GIORDANI u. U. AGRIMI (2007):
PrPSc
in salivary glands of scrapie-affected sheep.
J. Virol. 81, 4872-6
VELLA, L.J., R.A. SHARPLES, V.A. LAWSON, C.L. MASTERS, R. CAPPAI u. A.F.
HILL (2007):
Packaging of prions into exosomes is associated with a novel pathway of PrP processing.
J. Pathol. 211, 5, 582-90
VON BARTHELD, C.S. (2004):
Axonal transport and neuronal transcytosis of trophic factors, tracers, and pathogens.
J. Neurobiol. 58, 2, 295-314
WADSWORTH, J.D., S. JOINER, A.F. HILL, T.A. CAMPBELL, M. DESBRUSLAIS, P.J.
LUTHERT u. J. COLLINGE (2001):
Tissue distribution of protease resistant prion protein in variant Creutzfeldt-Jakob disease
using a highly sensitive immunoblotting assay.
Lancet 358, 171-80
WAN, X.C., J.Q. TROJANOWSKI u. J.O. GONATAS (1982):
Cholera toxin and wheat germ agglutinin conjugates as neuroanatomical probes: their uptake
and clearance, transganglionic and retrograde transport and sensitivity.
Brain Res. 243, 2, 215-24
WANG, F., X. WANG, C.-G. YUAN U. J. MA (2010):
Generating a Prion with bacterially expressed recombinant Prion Protein.

Literaturverzeichnis 163
Science 327, 1132, DOI: 10.1126/science.1183748
WEISS, S., D. PROSKE, M. NEUMANN, M.H. GROSCHUP, H.A. KRETZSCHMAR, M.
FAMULOK u. E.L. WINNACKER (1997):
RNA aptamers specifically interact with the prion protein PrP.
J. Virol. 71, 8790-7
WELLS, G. A., A. C. SCOTT, C. T. JOHNSON, R. F. GUNNING, R. D. HANCOCK, M.
JEFFREY, M. DAWSON u. R. BRADLEY (1987):
A novel progressive spongiform encephalopathy in cattle.
Vet. Rec. 121, 18, 419-20
WELLS, G.A.H., R.D. HANCOCK, W.A. COOLEY, M.S. RICHARDS, R.J. HIGGINS u.
G.P. DAVID (1989):
Bovine Spongiform Encephalopathy: Diagnostic Significance of Vacuolar Changes in
Selected Nuclei of the Medulla Oblongata.
Vet. Rec. 125, 521-4
WELLS G.A., M. DAWSON, S.A.C. HAWKINS, R.B. GREEN, I. DEXTER, M.E.
FRANCIS, M.M. SIMMONS, A.R. AUSTIN u. M.W. HORIGAN (1994):
Infectivity in the ileum of cattle challenged orally with bovine spongiform encephalopathy.
Vet. Rec. 135, 40-1
WELLS, G.A.H., u. J.W. WILESMITH (1995)
The neuropathology and epidemiology of bovine spongiform encephalopathy.
Brain Path. 5, 91-103
WELLS, G.A.H., u. M.M. SIMMONS (1996):
The essential lesion profile of bovine spongiform encephalopathy (BSE) in cattle is
unaffected by breed or route of infection.
Neuropathol. Appl. Neurobiol. 22, 453
WELLS, G.A., S.A. HAWKINS, R.B. GREEN, A.R. AUSTIN, I. DEXTER, Y.I. SPENCER,
M.J. CHAPLIN, M.J. STACK u. M. DAWSON (1998):

164 Literaturverzeichnis
Preliminary observations on the pathogenesis of experimental bovine spongiform
encephalopathy (BSE): an update.
Vet. Rec. 142, 103-6
WELLS, G.A., S.A. HAWKINS, A.R. AUSTIN, S.J. RYDER, S.H. DONE, R.B. GREEN, I.
DEXTER, M. DAWSON u. R.H. KIMBERLIN (2003):
Studies of the transmissibility of the agent of bovine spongiform encephalopathy to pigs.
J. Gen. Virol. 84, 1021-31
WELLS, G.A.H., J. SPIROPOULOS, S.A. HAWKINS u. S.J. RYDER (2005):
Pathogenesis of experimental bovine spongiform encephalopathy: preclinical infectivity in
tonsil and observations on the distribution of lingual tonsil in slaughtered cattle
Vet. Rec. 156, 401-7
WELLS, G.A.H., T. KONOLD, M.E. ARNOLD, A.R. AUSTIN, S.A.C. HAWKINS,
M. STACK, M.M. SIMMONS, Y.H. LEE, D. GAVIER-WIDEN, M. DAWSON u. J.W.
WILESMITH (2007):
Bovine spongiform encephalopathy: the effect of oral exposure dose on attack rate and
incubation period in cattle.
J. Gen. Virol. 88, 1363-73
WESTERGARD, L., H.M. CHRISTENSEN u. D.A. HARRIS (2007):
The cellular prion protein (PrP(C)): Its physiological function and role in disease.
Biochim. Biophys. Acta 1772, 629-44
WILESMITH, J. W., G. A. WELLS, M. P. CRANWELL u. J. B. RYAN (1988):
Bovine spongiform encephalopathy: epidemiological studies.
Vet. Rec. 123, 25, 638-44
WILESMITH, J. W., J. B. RYAN u. M. J. ATKINSON (1991):
Bovine spongiform encephalopathy: epidemiological studies on the origin.
Vet. Rec. 128, 9, 199-203

Literaturverzeichnis 165
WILL, R.G., J.W. IRONSIDE, M. ZEIDLER, S.N. COUSENS, K. ESTIBEIRO, A.
ALPEROVITCH, S. POSER, M. POCCHIARI, A. HOFMAN u. P.G. SMITH (1996):
A new variant of Creutzfeldt-Jakob disease in the UK.
Lancet 347, 921-5
WILLIAMS, E.S., u. S. YOUNG (1980):
Chronic wasting disease of captive mule deer: a spongiform encephalopathy.
J. Wildl. Dis. 16, 89-98
WOPFNER, F., G. WEIDENHOFER, R. SCHNEIDER, A. VON BRUNN, S. GILCH, T.F.
SCHWARZ, T. WERNER u. H.M. SCHATZL (1999):
Analysis of 27 mammalian and 9 avian PrPs reveals high conservation of flexible regions of
the prion protein.
J. Mol. Biol. 289, 1163-78
WRATHALL, A.E., K.F.D. BROWN, A.R. SAYERS, G.A.H. WELLS, M.M. SIMMONS,
S.S.J. FARRELLY, P. BELLERBY, J. SQUIRRELL, Y.I. SPENCER, M. WELLS, M.J.
STACK, B. BASTIMAN, D. PULLAR, J. SCATCHERD, L. HEASMAN, J. PARKER,
D.A.R. HANNAM, D.W. HELLIWELL, A. CHREE u. H. FRASER (2002):
Studies of embryo transfer from cattle clinically affected by bovine spongiform
encephalopathy (BSE).
Vet. Rec. 150, 365-78
WUBBOLTS, R., R.S. LECKIE, P.T. VEENHUIZEN, G. SCHWARZMANN, W. MÖBIUS,
J. HOERNSCHEMEYER, J.W. SLOT, H.J. GEUZE u. W. STOORVOGEL (2003):
Proteomic and biochemical analyses of human B cell-derived exosomes. Potential
implications for their function and multivesicular body formation.
J. Biol. Chem. 278, 13, 10963-72
WYATT, J. M. (1990):
Spongiform encephalopathy in a cat.
Vet. Rec. 126, 20, 513
ZIGAS, V., u. D.C. GAJDUSEK (1957):

166 Literaturverzeichnis
Kuru: clinical study of a new syndrome resembling paralysis agitans in natives of the Eastern
Highlands of Australian New Guinea.
Med. J. Aust. 44, 745-54
ZLOTNIK, I., u. J.C. RENNIE (1958):
A comparative study of the incidence of vacuolated neurons in the medulla from apparently
healthy sheep of various breeds.
J. Comp. Path. Therap. 68, 411-15
ZLOTNIK, I. (1962):
The pathology of scrapie: a comparative study of lesions in the brain of sheep and goats.
Acta Neuropathol. Suppl. 1, 61-70

167
9.ANHANG
9.1 Histologische Methoden
9.1.1 Entparaffinierungs- und Differenzierungsprotokoll
Für die Entparaffinierung und Rehydrierung wurden die Schnitte wie folgt behandelt:
5 min Xylol
5 min Xylol
3 min Isopropanol
3 min Isopropanol
3 min 96% Ethanol
3 min 70% Ethanol
3 min 50% Ethanol
In der aufsteigenden Alkoholreihe wurden die Schnitte nach dem folgenden Protokoll
entwässert und am Ende mit Entellan und einem Deckglas auf dem Objektträger fixiert.
2 min 50 % Ethanol
2 min 70 % Ethanol
2 min 96 % Ethanol
2 min Isopropanol I
3 min Isopropanol II
3 min Xylol I
3 min Xylol II
9.2 Legende zur Verhaltensbeobachtung der Rinder
A. Herantreten des Tierpflegers
(1) neugierig, Kopf bleibt nach vorn gestreckt
(2) tritt rückwärts, weicht mit dem Kopf zurück

168
(3) schreckt ruckartig zurück, Kopf kracht heftig in das Fressgatter
B. Droh-Reflex (Hand bewegt sich in Richtung Tierkopf ohne diesen zu berühren)
(1) Tier zuckt nicht zurück, maximal leichtes Blinzeln
(2) Tier bewegt sich rückwärts, Blepharospasmus-Blinzeln
(3) rasches Zurückziehen des Kopfes mit/ohne Blepharospasmus
(4) schwere Überreaktion (Zurückspringen, Kopfschlagen, Schnauben, Brüllen)
C. Sensibilität am Kopf (mittels kleiner Arterienklemme am N. trigeminus/N. facialis)
(Ohr-Reflex, Lidreflex, Berührung der Nasenlöcher und des Flotzmauls)
(1) keine bis leichte Abwehrbewegung
(2) Abwehrbewegungen ohne Kopfschlagen / Schnauben, Kopfregionen können
präzise berührt werden
(3) Abwehrbewegungen mit Kopfschlagen / Schnauben, einzelne Kopfpartien
können nur schwer präzise berührt werden
(4) Schwere Überreaktion (Übererregung des Tieres mit heftigem Kopfschlagen
und extremer Unruhe, so dass Verletzungsgefahr für Tier oder Untersucher
besteht)
D. Blitzlicht-Probe (Test wird wiederholt: 2x bei leichter, 4x bei schwerer Über-Reaktion)
(1) keine Überreaktion beim ersten Test, max. leichtes Zusammenzucken
(2) Tier schreckt nur beim ersten Test zurück
(3) wechselweise keine Reaktion bzw. Überreaktion oder lediglich zwei
Reaktionen
(4) Überreaktion ist wiederholbar (mindestens drei aufeinanderfolgende
Reaktionen)
E. Klatsch-Test oder metallisches Geräusch

169
Ausführung hinter dem Tier stehend um zusätzlichen visuellen Stimulus zu
vermeiden; mind. 3x und weitere 2x, falls das Tier abwechselnd oder bei den ersten
zwei Tests überreagiert – dann jeweils das Tier beruhigen lassen
(1) keine Überreaktion, max. leichtes Zusammenzucken beim ersten Test
(2) Tier schreckt nur beim ersten Test zurück
(3) wechselweise keine Reaktion bzw. Überreaktion oder lediglich zwei
Reaktionen
(4) Überreaktion ist wiederholbar (mindestens drei aufeinanderfolgende
Reaktionen)
F. Besen-Probe (mittels leichter Gerte jedes Bein mindestens ein Mal)
bei Reaktion wiederholen – Tier soll insgesamt ruhig dabei bleiben, ggf. einen
Moment warten)
(1) Tier steht ruhig
(2) Anheben eines oder abwechselnd beider Beine
(3) Leichtes, langsames Ausschlagen eines oder beider Beine
(4) Kräftiges, schnelles Ausschlagen ohne Verzögerung
G. Kondition
Beurteilung von 1-5 siehe Anlage „Body Condition Score“
H. Clipboard-Test
Das Tier steht frei 1-3m vor dem Untersucher, der 5x mit dem Clipboard in dessen
Augenhöhe wedelt – Tier muss vor jedem Wedeln ruhig sein
Achtung: Vorsicht vor aggressiven BSE-Tieren, die ggf. den Untersucher attackieren!
(1) normal (keine Überreaktion, max. leichtes Zusammenzucken beim ersten Test
(2) leichte Reaktion (Tier schreckt nur beim ersten Test zurück)

170
(3) zweifelhaft (wechselweise keine Reaktion bzw. Überreaktion oder lediglich
zwei Reaktionen)
(4) Überreaktion ist wiederholbar (mindestens drei aufeinanderfolgende
Überreaktionen
I. Bewegung
Allgemein – das Tier wird hin- und her getrieben
(1) keine Auffälligkeiten
(2) Lahmheit, steifer Gang - ggf. Wertung von 1-5
(3) Ataxie – ggf. Wertung von 1-5
(4) Hypermetrie – welche Gliedmaßen
Verhalten während der Bewegungs-Beurteilung – freies Laufen
(1) kommt neugierig näher oder bleibt stur stehen, Berührung möglich
(2) leicht kopfscheu – Annährung von vorn möglich, lässt sich nur von hinten
berühren
(3) stark kopfscheu – Annährung nur von hinten möglich, flüchtet aber bei
Berührung
(4) hochgradig furchtsam und unsicher, flieht bei Annährung
Fluchtverhalten – Tier wird getrieben
(1) Berührung möglich, Tier läuft trotz des Treibens ruhig weiter
(2) Annährung möglich, verfällt bei Berührung in schnelleres Tempo
(3) flieht bei Annährung, lässt sich nicht berühren
(4) flüchtet in panischer Angst, heftige Galoppsprünge, Ausschlagen
J Weitere Symptome:

171
Zähneknirschen
Tremor durch Erregung/Nervosität (nicht zur Abwehr von Fliegen!)
Nase lecken
Flotzmaul rümpfen, flehmen ohne normalen Anlass
BSE-typische Verhaltensserien, z.B.:
- Kopfschlagen nach vorn, gefolgt von Kopfschlagen zur Seite mit Lecken der
Flanke
- Flotzmaul rümpfen und Zähneknirschen oder Flehmen gefolgt von Lecken des
Flotzmauls und Gähnen
9.3 Puffer und Lösungen
Soweit nicht anders angegeben, wurden die Puffer und Lösungen in Aqua dest. angesetzt.
9.3.1 Immunhistochemie
Proteinase K-Stammlösung
10 mg PK
in 10 ml Aqua dest.
Gebrauchslösung: 760 µl Proteinase K-Stammlösung in 190 ml PK-Puffer
3,5% gepuffertes Formalin nach Lillie (für 5 l)
500 ml 37% Formol
4500 ml Aqua dest.
20 g NaH2PO4 x H2O
32,5 g Na2HPO4 (wasserfrei) oder
40,75 g Na2HPO4 x 2H2O
10x PK-Puffer pH 8,3 (500ml):
30,28 g Tris
1,46 g EDTA
5 ml Tween

172
Der pH-Wert von 8,3 wird mit 37% HCl eingestellt.
Enzymsubstrat Diaminobenzidintetrahydrochlorid (DAB) pH 7,1
100 mg 3,3′-Diaminobenzidintetrahydrochlorid
Imidazol/HCl-Puffer (0,1 M, pH 7,1) ad 200 ml
70 µl H2O2 (kurz vor Gebrauch zufügen)
10x TBS pH 7,6 (1000 ml)
60,57 g Tris
80,0 g NaCl
Der pH-Wert von 7,6 wird mit 37% HCl eingestellt.
Hämatoxylin
Siehe 3.4.2
9.3.2 Hämatoxylin-Eosin-Färbung
Mayer’s saures Hämatoxilin:
1 g Hämalaun
0,2 g NaJO3
50 g Kalialaun (Kaliumaluminiumsulfat zur Analyse)
50 g Chloralhydrat
1 g Zitronensäure zufügen
in 1000ml Aqua bidest lösen
Eosinlösung
1g Eosin gelblich
1 Tropfen Eisessig
in 100 ml 50% Ethanol lösen

173
9.3.3 Western Blot
APS 10 %
1 g Ammoniumpersulfat
in 10 ml Aqua dest.
Acrylamidgel
Trenngel, 16 %ig (zwei Gele) Sammelgel (zwei Gele)
Aqua dest. 3,0 ml Aqua dest. 6 ml
1,5 M Tris HCl (pH 8,8) 3,75 ml 0,5 M Tris HCl (pH 6,8) 2,5 ml
30 % Bis-Acrylamid 8,0 ml 30 % Bis-Acrylamid 1,3 ml
10 % SDS 750 µl 10 % SDS 500 µl
10 % APS 100 µl 10 % APS 200 µl
TEMED 10 µl TEMED 20 µl
Blottingpuffer für Western-Blot, pH 8,3
25 mM Tris
192 mM Glycin
20% (v/v) Methanol
0,2% (w/v) SDS
Lade-Puffer (10x)
10% (w/v) SDS
0,25 M Tris
25% (v/v) β-Mercaptoethanol
15% (w/v) Saccharose
0,05% (w/v) Bromphenolblau
pH 6,8 mit HCl einstellen, bei -20 °C lagern
Magermilch
5% (w/v) Magermilchpulver in PBS-Tween
Natriumhypochlorid (2,4%ig)
1 L Natriumhypochlorid (12%ig)

174
4 L Aqua dest.
Natronlauge (2 M)
80 g Natriumhydroxid
in 1 L Aqua dest.
PBS (10x)
1,4 M NaCl
27 mM KCl
65 mM Na2HPO4 x 2H2O
15 mM KH2PO4
pH 6,9 überprüfen; autoklavieren (121 °C, 20 min)
PBS mit 4% Sarkosyl (pH 7,4)
Sarkosyl 10 g
PBS ad 250 ml
PBS-Tween
1 L 1x PBS
0,1% (v/v) Tween 20
Pefabloc (0,1 M)
Pefabloc 100 mg
A. bidest. ad 4,17 ml
Powerstrippuffer
0,2 M Glycin
1% (w/v) SDS
pH 2,0 mit konzentrierter Salzsäure einstellen
4% PTA mit 34 mM MgCl2 (pH 7,4)
4 g PTA
691 mg Magnesiumchloridhexahydrat
Aqua. bidest. ad 100 ml

175
Der pH-Wert wurde mit Natronlauge (NaOH) eingestellt.
SDS (10%)
1 g Natriumdodecylsulfat (SDS)
in 10 ml Aqua dest.
Tris HCl (1,5 M; pH 8,8)
90,86 g Tris
in 500 ml Aqua dest.
pH 8,8 mit konzentrierter Salzsäure einstellen
Tris HCl (0,5 M; pH 6,8)
30,29 g Tris
in 500 ml Aqua dest.
pH 6,8 mit konzentrierter Salzsäure einstellen

176
DANKSAGUNG
„Am Ende wird alles gut. Und wenn es nicht gut ist, ist es nicht das Ende.“
Oscar Wilde
Viele Menschen haben mich über die letzten Jahre auf durchaus schwierigen Pfaden begleitet.
Das ich jetzt tatsächlich diese Arbeit zu einem erfolgreichen Abschluss bringen kann,
verdanke ich auch genau diesen Menschen. Und denen möchte ich an dieser Stelle danken.
Auch wenn sie vielleicht nicht namentlich erwähnt sind.
Prof. Dr. Martin H. Groschup, der mir mit der Überlassung des Themas die Tür zur TSE-
Forschung aufgestoßen hat, der viele weitere positive Erfahrungen folgten. Danke für die
Diskussionen und Anregungen, die Korrekturen trotz des vollen Terminkalenders und für Ihre
Geduld.
Prof. Dr. med. vet. Wolfgang Baumgärtner, der als Zweitgutachter ebenfalls entscheidend zur
Fertigstellung der Arbeit beigetragen hat.
Dr. Christine Fast, ohne Ihre Aufmunterungen, Anweisungen, Hilfestellungen und
Korrekturen hätte diese Arbeit niemals fertiggestellt werden können.
Danke an alle Mitarbeiter des INNT des Friedrich-Loeffler-Instituts. Herausstellen möchte ich
dabei Bärbel, Leila, Anja, Jule, Katja, Imke, Anne und Dan, die neben der fachlichen
Unterstützung auch für eine großartige Zeit außerhalb des Instituts gesorgt haben. Besondere
Aufmerksamkeit gilt den Tierpflegern Benn und Volker, die sich trotz aller Strapazen und
Widrigkeiten aufopferungsvoll um die Rinder gekümmert haben.
Und zum Schluss gilt mein Dank meiner Familie. Ihnen ist nicht nur diese Arbeit gewidmet,
es sei Ihnen auch für die bedingungslose Unterstützung in all diesen Jahren von Herzen
gedankt.

177
ERKLÄRUNG
Hiermit erkläre ich, dass ich die Dissertation „Ausbreitung der klassischen BSE-Prion-Erreger
vom Darm über das periphere Nervensystem zum Gehirn“ selbstständig verfasst habe.
Bei der Anfertigung wurden folgende Hilfen Dritter in Anspruch genommen:
• _____________________________________________________________________
• _____________________________________________________________________
Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten
(Promotionsberater oder anderer Personen) in Anspruch genommen. Niemand hat von mir
unmittelbar oder mittelbar entgeltliche Leistungen für Arbeiten erhalten, die im
Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen.
Ich habe die Dissertation an folgenden Institutionen angefertigt:
_________________________________________________________________________
Die Dissertation wurde bisher nicht für eine Prüfung oder Promotion oder für einen ähnlichen
Zweck zur Beurteilung eingereicht.
Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig und der
Wahrheit entsprechend gemacht habe.
___________________________________________
Martin Kaatz


















