
Universität Ulm Protokoll zu den Grundübungen ... · 1.3 Einteilung und Anwendungsbereiche der...
27
1 Universität Ulm Protokoll zu den Grundübungen Pflanzenphysiologie und Molekulare Botanik Wintersemester 2011/2012 Versuch H2: Reportergenaktivität in transgenen Pflanzen Tutorin: Vorgelegt von: Abgegeben am: Version:
Transcript of Universität Ulm Protokoll zu den Grundübungen ... · 1.3 Einteilung und Anwendungsbereiche der...

1
Universität Ulm
Protokoll zu den Grundübungen Pflanzenphysiologie und Molekulare Botanik
Wintersemester 2011/2012
Versuch H2: Reportergenaktivität in transgenen Pflanzen
Tutorin:
Vorgelegt von:
Abgegeben am:
Version:

2
Inhaltsverzeichnis:
1. Theorieteil 2
1.1 Historischer Abriss zur Biotechnologie 2
1.2 Gentechnologie 2
1.3 Einteilung und Anwendungsbereiche der Bio- und Gentechnologie 2
1.4 Diskussion: Vor-und Nachteile der Gentechnologie gegenüber der Biotechnologie 6
1.5 Der Einsatz von Tier- und Pflanzenzellen muss in der Gentechnik sinnvoll 7
gewählt werden
1.6 Gentransfer in Natur und Labor 8
1.6.1 Gentransfer in der Natur 8
1.6.2 Gentransfer im Labor: Methoden 10
1.7 Regulierende Einflüsse der Genaktivität 14
1.8 Nachweis und Regulation von Genaktivität 15
2. Durchführung 18
2.1 Qualtitativer Nachweis der Promotoraktivität im Gewebe durch in-situ-Färbung 18
2.2 Fluorimetrische Bestimmung der GUS-Aktivität (quantitativ) 18
3. Ergebnisse 20
3.1 Qualtitativer Nachweis der Promotoraktivität im Gewebe durch in-situ-Färbung 20
3.2 Fluorimetrische Bestimmung der GUS-Aktivität (quantitativ) 20
4. Diskussion 23
4.1 Qualtitativer Nachweis der Promotoraktivität im Gewebe durch in-situ-Färbung 23
4.2 Fluorimetrische Bestimmung der GUS-Aktivität (quantitativ) 24
5. Quellen 25

3
1. Theorieteil
1.1 Historischer Abriss zur Biotechnologie
Bei der Biotechnologie handelt es sich um eine interdisziplinäre Wissenschaft, auf die
Menschen bereits seit Jahrtausenden zurückgreifen. Kennzeichnend hierfür ist die
Benutzung lebender Organismen oder biologisch synthetisierten Materials für
technische Anwendungen. Etablierte Einsatzfelder der Biotechnolgie sind
beispielsweise die Bier-oder Weinherstellung und die Verarbeitung von
Milchprodukten. Dabei wird auf Mikroorganismen wie z.B. der Bäckerhefe
(Saccharomyces cervisiae) oder Milchsäurebakterien zurückgegriffen.
1.2 Gentechnologie
Es bedarf einer strikten Abtrennung zwischen der Gentechnologie von der
Biotechnologie. Der Begriff Gentechnologie bezeichnet bestimmte Verfahren der
Biotechnologie, die Kentnisse aus der Molekularbiologie und/oder der Genetik
erfordern und mit dem Einbau in-vitro veränderter DNA in Organismen Hand in Hand
gehen. Gentechnologie beinhaltet also immer eine synthetische Veränderung oder
Neuzusammensetzung der DNA. Das bedeutet in der Praxis, dass bestimmte Gene
von Organismen, die eine bestimmte Stoffwechseleigenschaft, also ein Produkt oder
eine Fähigkeit aufweisen, entnommen werden und in andere Zellen eingefügt
werden. Ziel ist es, dass diese Eigenschaft dann von einem anderen
Wunschorganismus exprimiert wird. Es werden beispielsweise Enzyme wie
Chymosin oder Pepsin, die früher aus den Mägen junger Wiederkäuer entnommen
wurden, heute aber mithilfe von Mikroorganismen herstellen kann, verwendet.
1.3 Einteilung und Anwendungsbereiche der Bio- und
Gentechnologie
Beide Disziplinien werden nach Farben in ihre jeweiligen Anwendungsgebiete
separiert. Aufgrund der Ganzheit der Forschung, werden diese Grenzen mittlerweile
immer wieder durchbrochen. Im Folgenden wird die klassische Trennung betrachtet.

4
Bei der Roten Bio-und Gentechnologie handelt es sich um diejenigen Bereiche, die
in der Medizin zur Anwendung kommen und die Gesundheit des Menschen zum Ziel
haben. Dazu gehört die Entwicklung verschiedener Impfstoffe, Therapeutika,
Modellorganismen, die zur Entwicklung neuer Therapeutika benötigt werden, die
Produktion von Wirkstoffen durch genetisch veränderte Organismen und die dazu
erforderlichen Plattformtechnologien. Ein Beispiel hierfür ist die Herstellung von
Arzneimitteln wie etwa Insulin, das heutzutage mithilfe von Mikroorganismen
hergestellt wird. Die dafür notwendigen Gene wurden einer menschlichen Pankreas
entnommen.
Braune Bio-und Gentechnologie befasst sich mit der Biotechnologie der Umwelt und
damit nicht zuletzt der Aufrechterhaltung von bestimmten Lebensräumen und der
Grauen Bio-und Gentechnologie werden Verfahren zur Beseitigung von anfallenden
Abfällen diskutiert.
Blaue Bio-und Gentechnologie sind bio-und gentechnologische
Anwendungsmethoden auf Lebewesen aus dem Meer. Dabei wird große Hoffnung in
tieflebende Bakterien, welchen man wirtschaftlich nutzbare Stoffwechselwege
zutraut, gesetzt.
Die Grüne Bio-und Gentechnologie befasst sich speziell mit Pflanzen um deren
Eigenschaften als Nutzpflanzen zu verbessern. Ferner werden pflanzliche Enzyme
und deren Wirkprinzipien genauer durchleuchtet.
In asiatischen Entwicklungsländern ist Vitamin A-Mangel weit verbreitet. Um dem
entgegen zu wirken hat ein Freiburger Forschungsteam Wildtyp-Reis dahingehend
transformiert, dass diese nun im Endosperm ausreichend Provitamin A (= �-Carotin)
synthetisieren kann, welches dann vom Menschen zu seinen aktiven Formen
umgewandelt werden kann. Es wurden hierzu zwei Gene fremder Arten übertragen.
Zum einen waren dies das Phytoensynthase-Gen der Narzisse (Narcissus
pseudonarcissus) und zum anderen das Caratindesaturase-Gen des Bakteriums
Erwinia uredovora. Abbildung 2 zeigt die erzielten Farbveränderungen.

5
Abbildung 1: Golden Rice und Wildtyp-Reis
Quelle: http://www.gatesfoundation.org/agriculturaldevelopment/PublishingImages/golden-rice-hero.jpg. Stand: 17.12.11
Weiße Bio-und Gentechnologie befasst sich mit dem industriellen Bereich dieser
Technologien. Hierbei wird nach weiteren industriell nutzbaren Enzymen und
Stoffwechselwegen gesucht. Ferner werden optimierte Bioverfahrenstechniken
angestrebt.
Die Käseherstellung dient als Beispiel einer seit Jahrtausenden betriebenen
biotechnischen Anwendung, die im Zuge der Prozessoptimierung mithilfe von
gentechnologischen Verfahrensweisen verbessert wurde. Milcheiweiß wird mithilfe
der proteaseaktiven Enzyme Chymosin und Pepsin die die Milch eindicken, zum
Ausfallen gebracht. Früher wurden diese Enzyme aus den Mägen von Kälbern
entnommen. Heutzutage wird jedoch die Stoffwechselfähigkeit von besonders
modifiziertern Schimmelpilzen oder Hefen genutzt. Die Vorteile liegen auf der Hand.
Die Enzymgewinnung erfolgt unkomplizierter und die Verunreinigungen, mit denen
bei der Enzymentnahme aus den Kälbermägen gerechnet werden müssen, entfallen.
Abbildung 1 dient zur Veranschaulichung des gentechnischen Verfahrens.

6
Abbildung 2: Chymosingewinnung. Quelle: http://www.gm.rt.schule-
bw.de/lehrer/schmid/gentechnik/bilder/folie24.gif. Stand: 17.12.11
Ein Beispiel aus der Pflanzenoptimierung ist die sogenannte Flavr-Savr-Tomate, die
in den 90er Jahren auf den Markt kam. Bei ihr wurde das Enzym Polygalacturonase
mittels RNA-Interferenz blockiert, wodurch der Abbau von Pektin, einem Stützprotein
der primären Zellwand, verhindert wurde und die Pflanze dadurch selbst nach langen
Transportwegen nicht matschig wurde. Aufgrund des faden Geschmacks ist der
Verkauf kurz darauf wieder eingestellt worden.
Die Firma Monsanto veröffentliche Ende des letzten Jahrhunderts die mit dem
Bodenbakterium Agrobacterium tumefaciens transformierte Pflanze Roundup Ready
Soja. Diese ist mit herbizidresistenten Genen ausgestattet. „Praktischerweise“
verkauft die Firma das passende Herbizid, nämlich Glyphosat, gleich mit. Es wurde
ein geringerer Schädlingsbefall der Pflanzen, um eine höhere Produktivität zu
erzielen, angestrebt.
Die gentechnisch veränderte Kartoffelsorte Amflora revolutionierte die Papier-, Textil-
und Klebstoffindustrie, indem sie die aufwendigen Trennungsverfahren der beiden
Stärkearten Amylose und Amylopektin weitgehend überflüssig machte. Der Grund
hierfür ist, dass mithilfe einer RNA-Interferenz die die Translation von Genen, die an
dem Aufbau der Amylose beteiligt sind, behindert werden und es nur noch zu einer
Translation von Amylopektin kommt.

7
Zuletzt sei nicht außer Acht gelassen, dass es sich hier nur um Beispiele handelt. Die
Vielzahl moderner Verfahren ist kaum noch zu überschauen.
1.4 Diskussion: Vor-und Nachteile der Gentechnologi e gegenüber
der Biotechnologie
Die Gentechnik liefert praktische Verfahrensmethoden, die dem Wohle der
Menschheit dienen. Die Wissenschaft ist heutzutage soweit, dass Erbkrankheiten
gentherapeutisch behandelt werden können. Ferner können durch die
Pränataldiganostik werdende Mütter über den Zustand ihres Kindes aufgeklärt
werden und so frühzeitig über mögliche Krankheiten oder Behinderungen des Kindes
unterrichtet werden.
Durch die Gentechnik werden die genetischen Grenzen zwischen den Arten
weitgehend aufgehoben. Beispiele hierzu wurden in 1.3 bereits genannt. Als Folge
lässt sich eine immense Fülle neuer Organismen kreieren. All dies dient oft der
Optimierung von Bioprozessen und spart damit Kosten ein. Ein weiterer großer
Vorteil besteht darin, dass die gewünschten Produkte durch gezielte Expression von
Mikroorganismen viel weniger verunreinigt sind. Es kann oft auf aufwendige und
teure Reinigungsverfahren verzichtet werden. Letzten Endes kann durch
gentechnischen Einsatz also die Qualität vieler Lebensmittel und anderen Produkten
verbessert werden.
Gentechnologie ist oft forschungseffizienter und billiger als die dementsprechende
Biotechnologie. Durch in – vitro Rekombination gewünschter Merkmale entfällt das
Rückkreuzen, wie es nach den Mendel’schen Regeln notwendig wäre. Dadurch
entfallen das Heranreifen bestimmter Filialgenerationen und die erst dann mögliche
Überprüfung der Ergebnisse. Eine große Zeitersparnis wird möglich. Außerdem
können die gewünschten Merkmale gezielt in den Organismus geschleust werden,
wodurch sich die Spezifität erhöht und ungewollte Merkmalsexpression
unwahrscheinlich wird.
Wenn eine gentechnisch veränderte Pflanze allerdings nicht isoliert wird, so wird sie
unweigerlich ihr genetisches Material mit der Umwelt austauschen. Dies entspricht
einem vertikalen Gentransfer. Oft haben diese Pflanzen einen Selektionsvorteil. Sie

8
sind resistent gegen Schädlinge oder vertragen schlechtes Wetter besser. Es wird
befürchtet, dass dies zu einer Einschränkung der genetischen Vielfalt führen kann.
Die physiologischen und phänotypischen Folgen eines transgenen Organismus
können nicht vorhergesagt werden. Es ist einfach noch viel zu wenig über die
komplizierten Wechselwirkungen innerhalb eines Organismus bekannt, um die
Folgen überblicken zu können. Schwerwiegende Wechselwirkungen innerhalb eines
Organismus und mit dessen Umwelt sind nicht ausgeschlossen. Es bleibt ferner die
Frage bestehen, welche Langzeitfolgen genetisch veränderte Lebewesen bewirken.
Transgene Pflanzen können ferner auch Nützlinge schädigen. Ein damit
zusammenhängender horizontaler Gentransfer lässt sich nicht ausschließen. Daher
sind die Fähigkeiten und Fertigkeiten der Gentechnik nach wie vor mit Vorsicht zu
genießen.
Ethnische Bedenken werden von den „Würdeträgern“ der Kirche und
Umweltschützern ausgesprochen, vor allem wenn es um Genanalyse und Forschung
beim Menschen geht. Es wird befürchtet, den Mensch irgendwann katalogisieren zu
können und dadurch in die Schöpfung, aus welcher Sicht auch immer, erheblich
einzugreifen. Es ist nicht auszuschließen, dass viele Menschen aus dem Instinkt
heraus Angst vor den neuen Techniken haben.
1.5 Der Einsatz von Tier-und Pflanzenzellen muss in der Gentechnik
sinnvoll gewählt werden
Wird mit Pflanzen gearbeitet, so kann aus deren Eigenschaft, dass alle Zellen
totipotent sind ein großer Nutzen gezogen werden. Totipotentz bedeutet, dass eine
Zelle auf alles Erbgut eines Organismus zugreifen kann und diesen prinzipiell
komplett regenerieren könnte. Bestimmte Pflanzen, denken wir an den
Modellorganismus Arabidopsis thaliana, haben eine kurze Generationszeit und
können viele Nachkommen zeugen. Außerdem sind sie billiger in der Haltung und
pflegeleichter wie z.B. ein Säugetier.
Ein Nachteil bei der gentechnischen Arbeit mit Pflanzen ist sicherlich die Zellwand,
die es zu durchdringen gilt. Pflanzen sind weniger zur Synthese bestimmter
Stoffwechselprodukte wie beispielweise Bakterien oder tierische Zellen geeignet, da

9
sie oft nur geringe Produktmengen erzeugen. Es bleibt das ungeklärte Problem mit
Monokotyledonen, die mit dem häufig für transformierende Vorgänge verwendeten
Bodenbakterium Agrobacterium tumefaciens keine Transformation erfahren.
Nutzpflanzen wie Weizen, Reis und Mais müssen daher mithilfe anderer, weniger
effektiver Methoden behandelt werden. Aufgrund der Polyploidie der Pflanzenzellen
ist es nur schwer diese vollständig zu transformieren, da alle Chromosomensätze
gentechnisch verändert werden müssen. Besonders Kallusgewebe ist anfällig für
somaklonale Variation. Das bedeutet, dass sich der Genotyp einer Pflanze durch
häufige Fehler während der Replikation von selbst spontan abwandelt (Mutation).
1.6 Gentransfer in Natur und Labor
Hierbei handelt es sich um jegliche Übertragung von Genen zwischen Organismen.
Es wird eine Einteilung zwischen horizontalem und vertikalem Gentransfer
vorgeschlagen. Als horizontalen (= lateraler) Gentransfer wird eine Genübertragung
außerhalb der geschlechtlichen Fortpflanzung und über Artgrenzen hinweg
bezeichnet. Beim vertikalen Gentransfer wird von Genübertragung oder Weitergabe
an einen Nachkommen gesprochen.
1.6.1 Gentransfer in der Natur
Bei Prokaryoten sind uns 3 mögliche Arten des Gentransfers bekannt.
Als Konjugation wird die Übertragung von Genomteilen (DNA -Übertragung) einer
Spenderzelle (Donor mit �� − Faktor) auf eine Empfängerzelle (Rezipient mit
�� − ������) durch einen direkten Zellkontakt bezeichnet. Konjugation ist über die
Artgrenzen hinweg möglich. Diese kommt meist mithilfe des F-Pilus (F für Fertilität)
zustande. Die Stelle, von der aus der Transfer startet, heißt oriT-Squenz (origin of
replication Transfer). Abbildung 3 zeigt diesen Vorgang bei gram-negativen E. coli
Bakterien.

Abbildung 3: Konjugation bei
http://upload.wikimedia.org/wikipedia/commons/f/f3/Konjugation
Als Transformation wird die
Organismus bezeichnet. Dabei wird
wurde das Genom des Wirtsorganismus
mithilfe von Rezeptoren selbst bestimmte DNA ins Cytosol aufnehmen.
ist der Begriff für eine Transformation
Als Transduktion wird der Gentransfer zwischen Bakterien und Viren, also auch de
indirekte, kontaktlose Austausch zweier Bakterien miteinander, sowie die Infektion
mit einem viralen Vektor
unterschieden. Virulente Phagen tun dies nicht. Da sie ihre eigene DNA nicht ins
Wirtsgenom einbauen, kann beim Ausschneiden selbiger keine Wirts
Phagenkopf gelangen.
Bei der allgemeinen Transduktion
Wirts-DNA im Phagenkopf.
homologer Stelle eingebaut.
Bei der speziellen Transduktion
Bakteriophage spezifische
Abbildung 3: Konjugation bei E. coli. Quelle:
http://upload.wikimedia.org/wikipedia/commons/f/f3/Konjugation-ecoli.svg. Stand: 17.12.11
die nicht-virale Übertragung von freiliegender
Dabei wird Fremd-DNA ins Cytoplasma übertragen
wurde das Genom des Wirtsorganismus transformiert. Kompetente Zellen können
mithilfe von Rezeptoren selbst bestimmte DNA ins Cytosol aufnehmen.
Transformation bei Eukaryoten.
Gentransfer zwischen Bakterien und Viren, also auch de
indirekte, kontaktlose Austausch zweier Bakterien miteinander, sowie die Infektion
mit einem viralen Vektor von Genen bezeichnet. Es wird zwischen 2 Arten
Virulente Phagen tun dies nicht. Da sie ihre eigene DNA nicht ins
Wirtsgenom einbauen, kann beim Ausschneiden selbiger keine Wirts
Transduktion befindet sich im lytischen Zyklus
DNA im Phagenkopf. Diese wird dann beim nächsten befallen
homologer Stelle eingebaut.
Transduktion schneidet im lytischen Zyklus ein temperenter
e Teile des Wirtsgenoms aus, die die Phagen
10
. Stand: 17.12.11
virale Übertragung von freiliegender DNA auf einen
DNA ins Cytoplasma übertragen. Damit
. Kompetente Zellen können
mithilfe von Rezeptoren selbst bestimmte DNA ins Cytosol aufnehmen. Transfektion
Gentransfer zwischen Bakterien und Viren, also auch der
indirekte, kontaktlose Austausch zweier Bakterien miteinander, sowie die Infektion
Es wird zwischen 2 Arten
Virulente Phagen tun dies nicht. Da sie ihre eigene DNA nicht ins
Wirtsgenom einbauen, kann beim Ausschneiden selbiger keine Wirts-DNA in den
m lytischen Zyklus ein Stück der
Diese wird dann beim nächsten befallenen Bakterium an
m lytischen Zyklus ein temperenter
, die die Phagen-DNA

11
flankieren und baut diese zusammen mit seinem Erbgut in dessen Phagenkopf ein.
Bei erneutem Befall werden diese DNA-Segmente mit transferiert.
1.6.2 Gentransfer im Labor: Methoden
Im Folgenden werden gezielte gentechnische Methoden vorgestellt, die der
Übertragung spezifischer Genen dienen.
Bei den Techniken zum direkten Gentransfer handelt es sich um Verfahrensweisen,
bei denen das gewünschte Genmaterial unmittelbar in die Zelle eingeführt wird. Im
Folgenden werden einige Verfahren vorgestellt.
Bei der Mikroinjektion wird mit feinsten Glaskanülen das Genmaterial (freie DNA)
vorwiegend in tierische Zellen eingebracht (meist direkt in den Zellkern). Bei
pflanzlichen Zellen ist meist der Druck zu hoch, beim Einstechen würde der
Innendruck der Zelle zusammenbrechen und die Zelle dadurch sterben. Außerdem
ist dieses Verfahren sehr langwierig.
Abbildung 4: Mikroinjektion. Quelle: http://www.medizin.uni-
halle.de/zra/media/Bilder/Labor/mikroinjektion2_250.jpg. Stand: 17.12.11
Um eine Protoplastentransformation durchführen zu können, werden die Zellwände
durch Enzyme (Zellulasen und Pektinasen) entfernt. Als Folge liegt nur noch ein
membranumhüllter Protoplast vor. Um diese Zelle nun transformieren zu können,
muss dessen Membran dahingehend manipulieren werden, dass sie für Fremd-DNA
kompetent wird. Dies geschieht entweder im PEG-Bad (Polyethylenglycol) oder
mittels einer Calciumbehandlung (z.B. mit CaC��).

12
Nach Entfernung der Zellwände zweier zu transformierender Zellen werden diese
entweder mit Calcium oder PEG zur Verschmelzung gebracht, es entsteht ein Cybrid.
DNA-Material aus den Zellkernen wird ausgetauscht. Das Verschmelzungsprodukt
der Kerne bezeichnet man als Hybrid. Dieses Verfahren wird als Protoplastenfusion
bezeichnet.
Aus einer sogenannten Particle Gun werden aus Wolfram oder Gold bestehende
Mikroprojektile, auf deren Oberfläche das Genmaterial aufgetragen wird, in Richtung
eines Zellverbands (Blätter, Kallus, embryonales Gewebe, etc.) mithilfe eines
Druckgradienten beschleunigt. Beim Aufprall auf die Zellwand wird diese
durchdrungen und beschädigt, aber die Zelle ist oft in der Lage, sich zu regenerieren.
Aus so transformierten Zellen kann eine ganze transgene Pflanze entstehen.
Bei der Elektroporation wird ein elektrisches Feld an die Zellen angelegt. Es sollte
darauf geachtet werden, dass die Zelllösung möglichst salzarm ist, da es sonst zu
Kurzschlüssen kommen kann. Nun werden sehr kurze, jedoch kräftige
Elektroschocks ausgelöst, die die Zellmembran kurzzeitig permeabel, oder auch
löchrig machen. Während dieser Zeit kann ein Makromolekül, wie etwa die DNA
durch die Membran eintreten.
Bei der Lipofektion verwendet man Vesikel, die aus einer unpolaren
Lipiddoppelschicht bestehen (sog. Liposomen, allg. Micellen). Diese können mit der
Zellmembran aufgrund ihrer elektrischen Beschaffenheit fusionieren und deren Inhalt
ins Cytosol absondern. Wird nun in so ein Vesikel DNA eingebracht, so kann diese in
die gewünschte Zelle wandern. Das technisch am Einfachsten umzusetzende
Verfahren funktioniert mithilfe von kationischen Lipidvesikeln, deren Eigenladung die
der negativ geladenen DNA aufhebt. Als Folge entsteht ein unpolarer Komplex, der
von der Zelle aufgenommen werden kann.
Der indirekte Gentransfer geschieht mithilfe sog. Vektoren (= DNA-Vehikel). Vektoren
sind Träger des gewünschten Genabschnitts, der zusammen mit dem Vektor in einen
Wirt eingebracht werden kann. Sie sind in der Lage, ihr Erbgut zu replizieren. Es
folgen Beispiele für Vektoren.
Bei den Plasmiden und Cosmiden handelt es sich um außerhalb des Zellkern
vorliegendes, relative kleines, ringförmiges Erbgut mit der Größe von 1 – 25 kBp. Oft
werden darauf Resistenzgene codiert. Aufgrund der Replikationsunabhängigkeit vom
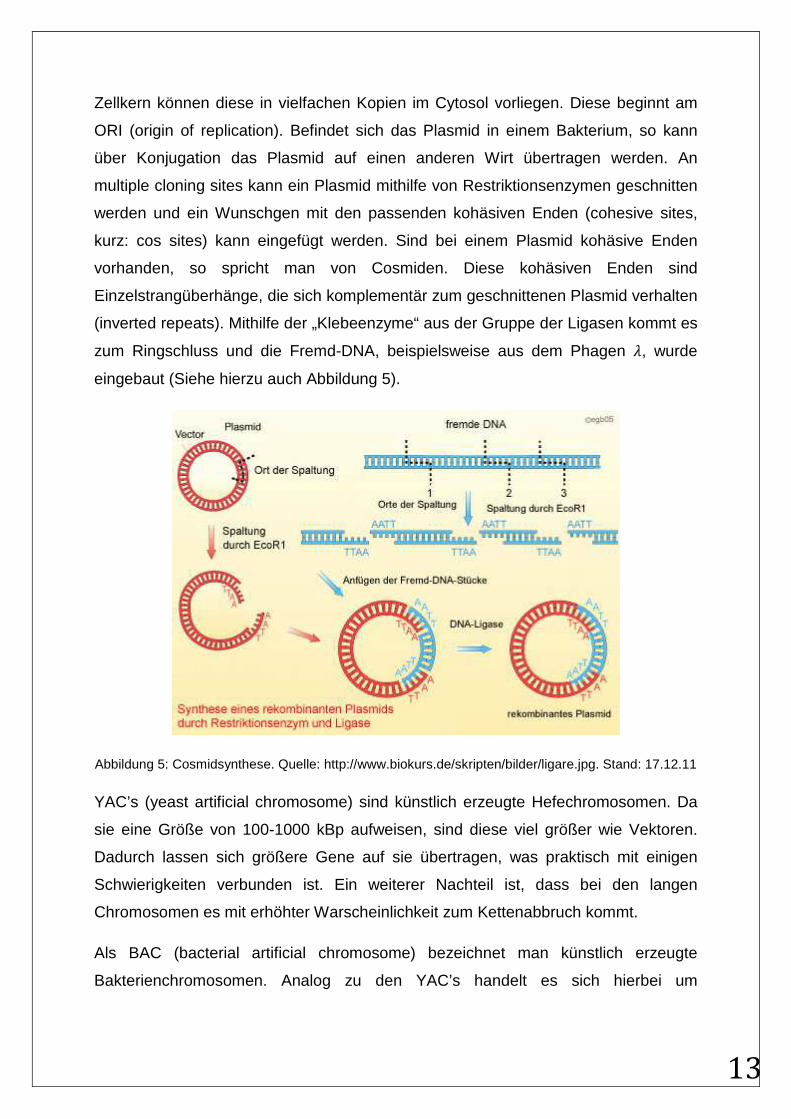
13
Zellkern können diese in vielfachen Kopien im Cytosol vorliegen. Diese beginnt am
ORI (origin of replication). Befindet sich das Plasmid in einem Bakterium, so kann
über Konjugation das Plasmid auf einen anderen Wirt übertragen werden. An
multiple cloning sites kann ein Plasmid mithilfe von Restriktionsenzymen geschnitten
werden und ein Wunschgen mit den passenden kohäsiven Enden (cohesive sites,
kurz: cos sites) kann eingefügt werden. Sind bei einem Plasmid kohäsive Enden
vorhanden, so spricht man von Cosmiden. Diese kohäsiven Enden sind
Einzelstrangüberhänge, die sich komplementär zum geschnittenen Plasmid verhalten
(inverted repeats). Mithilfe der „Klebeenzyme“ aus der Gruppe der Ligasen kommt es
zum Ringschluss und die Fremd-DNA, beispielsweise aus dem Phagen , wurde
eingebaut (Siehe hierzu auch Abbildung 5).
Abbildung 5: Cosmidsynthese. Quelle: http://www.biokurs.de/skripten/bilder/ligare.jpg. Stand: 17.12.11
YAC’s (yeast artificial chromosome) sind künstlich erzeugte Hefechromosomen. Da
sie eine Größe von 100-1000 kBp aufweisen, sind diese viel größer wie Vektoren.
Dadurch lassen sich größere Gene auf sie übertragen, was praktisch mit einigen
Schwierigkeiten verbunden ist. Ein weiterer Nachteil ist, dass bei den langen
Chromosomen es mit erhöhter Warscheinlichkeit zum Kettenabbruch kommt.
Als BAC (bacterial artificial chromosome) bezeichnet man künstlich erzeugte
Bakterienchromosomen. Analog zu den YAC’s handelt es sich hierbei um

14
synthetische Bakterienplasmide, die aus dem single-copy F-Plasmid des Bakteriums
E. coli bestehen und bis zu 300 kBp’s groß sein können.
Auch Viren können als Vektoren für bestimmte Gene dienen und diese in Bakterien
einschleußen. Man spricht in diesem Zusammenhang von Phagenvektoren. Beim
Phagen liegt die maximale Vektorkapazität bei 25 kbp und ist damit deutlich
geringer wie bei den BAC’s und YAC’s.
Mithilfe des Bodenbakteriums Agrobakterium tumefaciens als Vektor kann DNA in
eine Zelle eingeführt werden. Dieses Bakterium befällt in der Natur oft verwundete
Wurzeln und wird von Phenolen angelockt, die von der Wunde abgesondert werden.
Als Folge des Befalls bilden sich an der Pflanze Wucherungen, sogenannte
Wurzelhalsgallen aus. Der Grund für die Bildung eines solchen Tumors liegt auf dem
Genom des Bakteriums. Agrobakterium tumefaciens besitzt ein Ti-Plasmid
(tumorinduzierendes Plasmid), welches die Information für tumorinduzierende Gene
enthält. Nach der Transformation bestimmter Abschnitte des Plasmids, der
sogenannten T-DNA (Transfer-DNA) über einen F-Pilus in die DNA einer Wirtszelle,
beginnt die Pflanze mit der Synthese von Bakterienproteinen mithilfe des Bauplans
aus deren Genen. Dazu zählen vir-Gene (Virulenzgene, die die phytopathogene
Virulenz festlegen), Gene für die Auxin-und Cytokininsynthese (Hormone, die die
Pflanze wachsen lassen und damit dem Bakterium mehr Opin bescheren) und Gene
für die Opinsynthese, von denen sich das Bakterium ernähren kann, indem es
zusätzlich Gene synthetisiert, die dem Opinkatabolismus dienen. Left Border und
Right Border grenzen die Gene für Auxin-, Cytokinin- und Opinsynthese ab. Teile des
Ti-Plasmids werden herausgeschnitten und zur Übertragung freigegeben. An
sogenannten ori-Stellen (origin of replication) wird die DNA-Replikation eingeleitet
(Abbildung 6).

15
Abbildung 6: Ti-Plasmid. Quelle: http://upload.wikimedia.org/wikipedia/commons/8/89/Ti_Plasmid.jpg.
Stand: 17.12.11
Manipulativ werden Teile der T-DNA, wie etwa die Gene für die Hormonsynthese
herausgeschnitten und andere, gewünschte Gene eingebaut. Um diese DNA später
von anderen selektieren zu können, werden Selektionsmarker wie z.B.
Antibiotikaresistenzen hinter einem prokaryotischen Promotor mit eingebaut. Das nun
erhaltene Ti-Plasmid wird in Escherichia coli eingeführt und durch dessen Replikation
vermehrt. Nun wird das Antibiotikum der Resistenz hinzugegeben, wodurch nur
Zellen mit der Resistenz überleben. Nach Aufreinigung und Isolierung des Plasmids
wird dieses durch Transformation wieder in Agrobakterium tumefaciens eingeführt.
Wurde in diesem Bakterium bereits ein zweites Plasmid durch das eben
beschriebene Verfahren eingebracht, so spricht man von einem binären
Vektorsystem. Das erste beschriebene Plasmid ist für die Kodierung der T-DNA
verantwortlich und das Zweite Plasmid (Helferplasmid) enthält eine vir-Stelle, welche
für die Überführung der T-DNA essentiell ist. Binäre Vektoren sind einfacher in der
Handhabung. Zum einen sind sie stabiler, weil das gewünschte Gen auf einem
Vektor ohne vir-Region liegt und der Vektor damit um die vir-Region kürzer ist und
zum anderen weil bei dieser Technik weniger unerwünschtes Erbgut (Gene für die
Opinsynthese, sowie für die Wachstumshormone Cytokinin und Auxin) übertragen
wird. Nun kann eine Pflanze mit dem Bakterium infiziert werden. Damit es zur
gewünschten Genexpression kommt, muss in der Wunschsequenz ein
eukaryotischer Promotor eingebaut sein. Der Arbeitsfortschritt ist wieder mit einem
Antibiotikum, dessen Resistenz auf der T-DNA kodiert wurde, zu überprüfen. Hierbei

16
handelt es sich um ein Antibiotikum, welches auch für Pflanzen toxisch ist (z.B. ein
Kanamcin)
1.7 Regulierende Einflüsse der Genaktivität
Promotoren sind Nukleotid-Sequenzen auf der DNA, die als essentieller
Genbestandteil deren Expression regulieren. Das am 5‘-Ende liegende Element
wechselwirkt mit Proteinen, die die Transkription starten (Transkriptionsfaktoren), da
sie RNA Polymerasen binden können. Auch weiter entfernte Basensequenzen
können am Promotor beteiligt sein. Diese werden unterschieden in Enhancer, die die
Genexpression fördern und Silencer, die diese hemmen (sog. cis-Elemente, weil sie
auf der DNA liegen).
Bei Prokaryoten findet man meistens die Abfolge TATA, die sogenannte TATA-Box
(Pribnow-Box), welche bei der -10-Region liegt (Upstream, Zahl gibt an, wie viele
Nukleotide vor dem Transkriptionsstart lokalisiert) und eine TTGACA-Box, welche bei
der -35-Region liegt. Je geringer die Sequenz von der Box-Sequenz abweicht desto
wahrscheinlicher ist eine Transkription, in der die -Untereinheit
(Transkriptionsfaktor) an die -10 Region und die -35 Region bindet.
Promotoren bei Eukaryoten sind wesentlich komplizierter und es kann sich ihnen nur
mit bioinformatischen Methoden angenähert werden. Der Kernpromotor (am
Transkriptionsstart liegend) erstreckt sich meist von -37 bis +32 und besteht oft aus
einer TATA-Box und weiteren Elementen. Der Proximale Promotor ist zwischen -50
und -200 lokalisiert und besteht aus weiteren Boxen wie etwa einer CAAT-Box, oder
eine GC-Box. Der Transkriptionsfaktor TFIID bindet direkt an die TATA-Box und
komplexiert bei Transkriptionsstart mit weiteren Faktoren.
Promotoren könne mithilfe bestimmter bindender Stoffe manipuliert werden, wodurch
die Polymerase nicht andocken kann und es zu keiner Transkription kommen kann.
1.8 Nachweis und Regulation von Genaktivität
Mithilfe von Reporter-oder Indikatorgenen kann die Expression eines anderen Gens
indirekt nachgewiesen werden. Meist handelt es sich bei diesen Genen um solche,

17
deren Genprodukte sich leicht nachweisen lassen, sei es durch Fluorimetrie oder
einfache enzymatische Tests. Wird beispielsweise ein Fusionsprotein mit GFP (green
fluorescent protein) verwendet, so wird dieses hinter demselben Promotor postiert,
wird ein Ti-Plasmid genutzt, so werden Gene innerhalb derselben Border-Sequenz
der T-DNA eingebracht, wie das eigentlich eingebrachte Gen. Zur Kontrolle wird ein
Selektionsmarker außerhalb der Border-Sequenz angebracht, um den
Transformationserfolg überprüfen zu können. Reportergene eigenen sich auch oft
zur quantitativen Bestimmung. In Eukaryoten verwendet man oft das GUS-Gen (� –
Glucuronidase, v.a. bei Pflanzenzellen) oder das GFP-Gen (green fluorescent
protein, eines der wichtigsten Reportergene), da diese normalerweise in deren
Genom nicht vorkommen.
Das transkribierte � –Glucuronidase-Enzym hydrolysiert das Substrat X-Glucuronid
nach dessen Zugabe, welches daraufhin blaues Licht reflektiert, dessen Intensität
gemessen werden kann. Das X-Gluc (5-Brom-4-chlor-3indolyl-�-D-Glucuronid), fällt
nach Reaktion mit dem Enzym (Spaltung einer glykosidischen Bindung) zu einem
schwer löslichen Indigofarbstoff aus. Um die Intensität messen zu können, muss die
Zelle aufgebrochen und damit getötet werden. Weitere in-vitro-Analysen sind
beispielsweise mit dem Substrat 4-Methylumbelliferyl-beta-D-Glucuronid möglich,
dass mit dem Enzym zum fluoreszierenden 4-Methylumbelliferon umgesetzt wird.
Biolumineszenz kann durch die Oxidation von Luciferinen mithilfe von Luciferasen,
welche oft als Reporterproteine eingesetzt werden, erzeugt werden. Der Vorteil liegt
darin, dass die Zelle je nach Toxizität des Luciferins hierbei nicht getötet werden
muss.
Das aus dem CAT-Gen transkribierten Enzym Chloramphenicol Acetyltransferase
dient zur Acetylierung eines radioaktiv markierten Substrats (Chloramphenicol). Der
quantitative Nachweis erfolgt mithilfe eins CAT-Assays (spezielle
Dünnschichtchromatographie).
Oft bringen Reportergene auch Selektionsmarker mit sich. Eine quantitative Analyse
entfällt, jedoch überleben nur die Zellen auf einer Nährlösung, die mit den
bestimmten Antibiotika angereichert wurde, die die Antibiotikaresistenzen aufweisen.
Die RNA-Interferenz (RNA-Silencing) dient der zielgerichteten Ausschaltung von
Genen. Trifft in einer Zelle mRNA auf eine passende antisense-mRNA, so lagern sich

18
diese zu einer doppelsträngigen dsRNA zusammen. Diese ist dann funktionslos und
kann in dieser Gestalt nicht vom Ribosom translatiert werden. Das unerwünschte
Gen wurde stillgelegt. dsRNA wird von einem Dicer-Enzym (Hächsler-) zu kurzer (21-
26 bp langer) doppelsträngiger siRNA verdaut, welche nun mit einem
Enzymkomplex, den sogenannten RISC-Komplex (RNA induced silencing complex)
ausbilden. Hier wird die DNA zu einem Einzelstrang zersetzt und die im Komplex
verbleibende siRNA kann mit einer mRNA interagieren und diese schneiden. Gene
lassen sich also gezielt ausschalten, indem man eine künstlich hergestellte
komplementäre Nucleotidsequenz hinter einen geeigneten Promotor in das
Zellgenom einbringt. In der Natur dient dieses Verfahren zur Erkennung
doppelsträngiger Viren RNA. Der Wirkmechanismus wurde in Abbildung 6
dargestellt.
Abbildung 7: RNA- Interferenz. Quelle: http://www.biosicherheit.de/basisinfo/466.rna-interferenz-rnai-
junge-bahnbrechende-entdeckung.html. Stand 17.12.11
Wird von einem Gen zu viel RNA transkribiert, so kommt es zu keiner
Proteinsynthese. Es wird angenommen, dass eine Überrepräsentation eines Gens
eine unabhängige RNA-Polymerase-Aktivität (RNA-dependant RNA-polymerase,
kurz RdRP) startet, die die mRNA zu einer dsRNA synthetisiert. Als Folge wird die
dsRNA wieder in den Prozess der RNA-Interferenz eingeschleust. Dieser Prozess
wird als Co-Suppression bezeichnet. Er wurde zufällig entdeckt, als Forscher
versuchten in Petunien ein weiteres Gen für den Syntheseweg der Anthocyane
einzubauen, um diese noch intensiverer blau zu färben. Als Ergebnis erhielten sie
jedoch teilweise weiße Blüten.

19
2. Durchführung
Grundsätzlich kann zur Durchführung das Praktikumsskript zu Rate gezogen werden.
Einige Details wurden jedoch in der Praxis verändert. Die Versuchsabwandlungen
werden in diesem Abschnitt wiedergegeben.
2.1 Qualtitativer Nachweis der Promotoraktivität im Gewebe durch
in-situ-Färbung
Es wurden die Farbmarkierungen mittels der Hydrolyse von MUG (4-Methyl-�-D-
umbelliferylglucuonid) durch das Enzym �-Galactosidase in Arabidopsis thaliana 4
verschiedener Genotypen miteinander verglichen.
Linie 1: Wildtyp (Negativkontrolle)
Linie 2: Promotor CaMV35S vor GUS-Leseraster (Positivkontrolle)
Linie 3: Promotor AtBCAT-1 vor GUS-Leseraster (zu bestimmende Linie)
Linie 4: Promotor AtBCAT-4 vor GUS-Leseraster (zu bestimmende Linie)
Von jeder der oben genannten Linien wurden Blüten, junge Blätter, Stengel und
Wurzel entnommen und auf eine 24-Well-Platte aufgetragen und zu jedem Loch
wurde 400 µl Reaktionslösung C (X- Gluc) gegeben. Aufgrund der Hydrolyse von X-
Glc (5-Brom-4-chlor-3indolyl-�-D-Glucuronid) mithlfe des Enzyms �-Galactosidase
beginnt dieses sich blau zu färben. Aufgrund der Tatsache, dass das Enzym bei
37°C sein Wirkoptimum entfaltet, wird die Well-Plat te in ein 37°C warmes Wasserbad
gelegt. Danach wurde Lösung C durch 400 µl 70%igen Ethanol ersetzt, wodurch sich
das Präparat entfärbte. Die Well-Platte wurde der nächsten Gruppe zur Verfügung
gestellt. Hier wurde die Well-Platte der vorherigen Gruppe betrachtet.
2.2 Fluorimetrische Bestimmung der GUS-Aktivität (q uantitativ)
Es wurden von jeder Linie ca. 100 mg Blattmaterial entnommen und in ein
Eppendorfgefäß gegeben und in flüssigen Stickstoff eingefroren, wodurch die Zellen

20
aufbrachen. Anschließend wurde das Blattmaterial zermörsert. In jedes
Eppendorfgefäß wurden 100 µl des Extraktionspuffers zugegeben und großzügig
gevortext. Danach wurde bei 13000 rpm 15 Minuten lang zentrifugiert, um grobe
Zellbestandteile, wie etwa Zellwandfragmente vom Zentrifugat abzutrennen. 12
weitere Eppendorfgefäßge wurden mit 1900 µL Stopplösung befüllt und wie folgt
beschriftet.
Abbildung 8: Beschriftung der Eppendorfgefäße
Vier weitere Eppendorfgefäße wurden mit je 400 µL der Reaktionslösung F (MUG)
befüllt und der Überstand der Zentrifugation zugegeben und bei 37 °C inkubiert.
Während der Inkubation wird das MUG in der Reaktion von den Enzymen aus dem
Blattmaterialzentrifugationsüberstand in Methylumbelliferon umgesetzt. Nach den
jeweiligen Inkubationszeiten wurden je 100 µL aus dem inkubierten Pflanzenmaterial
entnommen und zu den vorgelegten 1900 µL Stopplösung entsprechend der
Zeitreihe zugegeben. Diese Lösung sorgt für ein basisches Milieu, welches die
Enzymaktivität, dessen Optimum bei einem pH-Wert von 4-5 liegt, hemmt. Dann
wurde das Fluorimeter mit 10 mM Kalibrierlösung (enthält das blau fluoreszierende 4-
Methylumbelliferon) auf 100 % kalibriert und die jeweiligen Messungen in der
Quarzküvette auf Fluoreszenz untersucht.
+
-
P1
P4
P1
P4
P1
P4
P1
P4
P1
P4
21
3. Ergebnisse
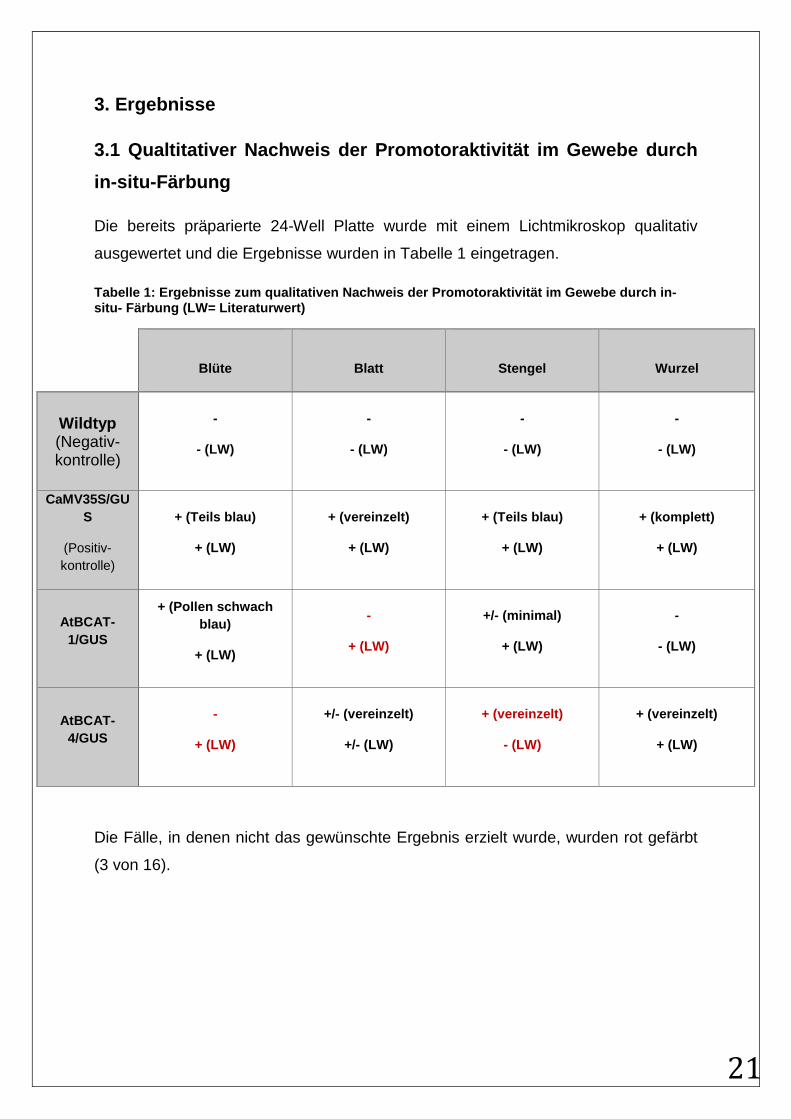
3.1 Qualtitativer Nachweis der Promotoraktivität im Gewebe durch
in-situ-Färbung
Die bereits präparierte 24-Well Platte wurde mit einem Lichtmikroskop qualitativ
ausgewertet und die Ergebnisse wurden in Tabelle 1 eingetragen.
Tabelle 1: Ergebnisse zum qualitativen Nachweis der Promotoraktivität im Gewebe durch in- situ- Färbung (LW= Literaturwert)
Blüte
Blatt
Stengel
Wurzel
Wildtyp (Negativ-kontrolle)
-
- (LW)
-
- (LW)
-
- (LW)
-
- (LW)
CaMV35S/GUS
(Positiv-kontrolle)
+ (Teils blau)
+ (LW)
+ (vereinzelt)
+ (LW)
+ (Teils blau)
+ (LW)
+ (komplett)
+ (LW)
AtBCAT-1/GUS
+ (Pollen schwach blau)
+ (LW)
-
+ (LW)
+/- (minimal)
+ (LW)
-
- (LW)
AtBCAT-4/GUS
-
+ (LW)
+/- (vereinzelt)
+/- (LW)
+ (vereinzelt)
- (LW)
+ (vereinzelt)
+ (LW)
Die Fälle, in denen nicht das gewünschte Ergebnis erzielt wurde, wurden rot gefärbt
(3 von 16).
22
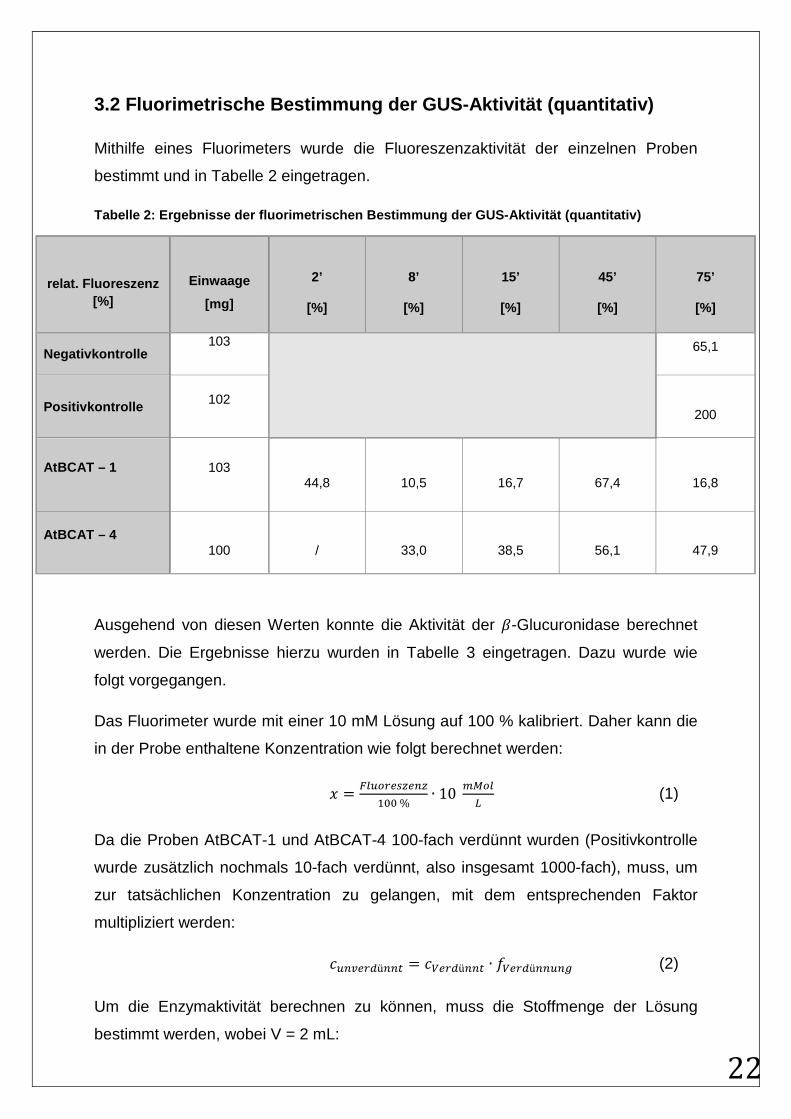
3.2 Fluorimetrische Bestimmung der GUS-Aktivität (q uantitativ)
Mithilfe eines Fluorimeters wurde die Fluoreszenzaktivität der einzelnen Proben
bestimmt und in Tabelle 2 eingetragen.
Tabelle 2: Ergebnisse der fluorimetrischen Bestimmu ng der GUS-Aktivität (quantitativ)
relat. Fluoreszenz [%]
Einwaage
[mg]
2’
[%]
8’
[%]
15’
[%]
45’
[%]
75’
[%]
Negativkontrolle 103
65,1
Positivkontrolle 102
200
AtBCAT – 1 103
44,8
10,5
16,7
67,4
16,8
AtBCAT – 4
100
/
33,0
38,5
56,1
47,9
Ausgehend von diesen Werten konnte die Aktivität der �-Glucuronidase berechnet
werden. Die Ergebnisse hierzu wurden in Tabelle 3 eingetragen. Dazu wurde wie
folgt vorgegangen.
Das Fluorimeter wurde mit einer 10 mM Lösung auf 100 % kalibriert. Daher kann die
in der Probe enthaltene Konzentration wie folgt berechnet werden:
� =��������
��%∙ 10
����
� (1)
Da die Proben AtBCAT-1 und AtBCAT-4 100-fach verdünnt wurden (Positivkontrolle
wurde zusätzlich nochmals 10-fach verdünnt, also insgesamt 1000-fach), muss, um
zur tatsächlichen Konzentration zu gelangen, mit dem entsprechenden Faktor
multipliziert werden:
�����ü��� = ���ü��� ∙ ����ü����� (2)
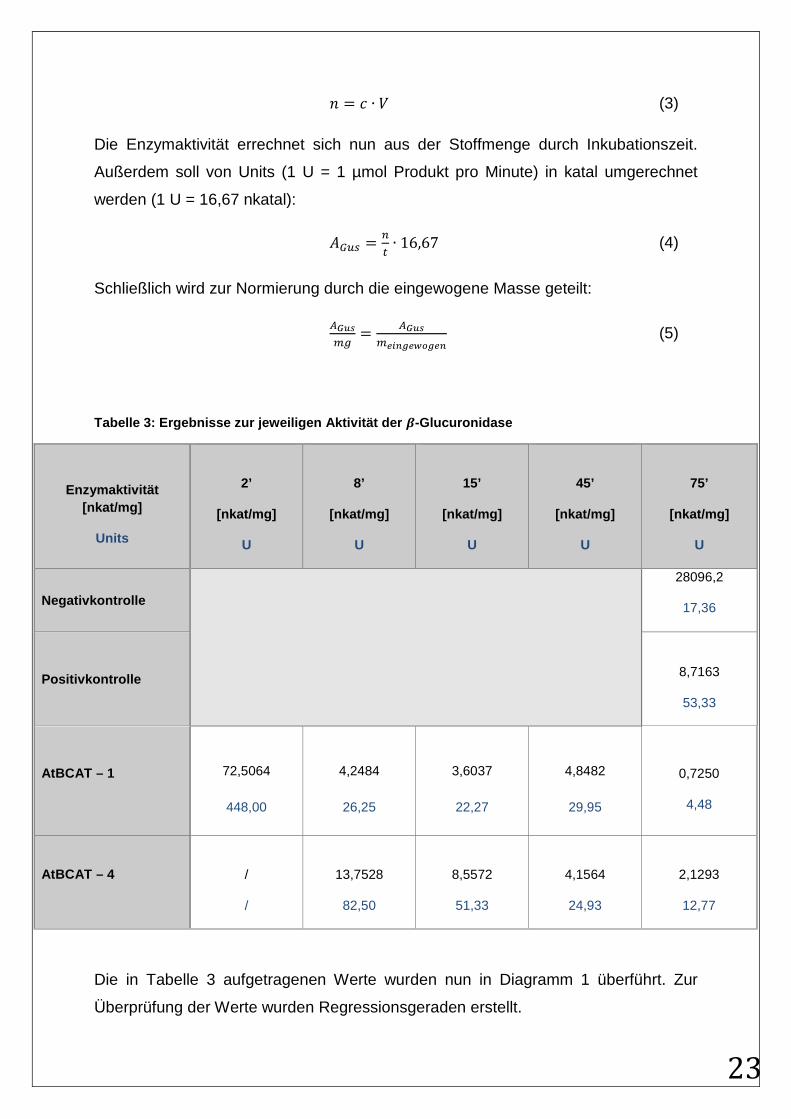
Um die Enzymaktivität berechnen zu können, muss die Stoffmenge der Lösung
bestimmt werden, wobei V = 2 mL:
23
� = ∙ � (3)
Die Enzymaktivität errechnet sich nun aus der Stoffmenge durch Inkubationszeit.
Außerdem soll von Units (1 U = 1 µmol Produkt pro Minute) in katal umgerechnet
werden (1 U = 16,67 nkatal):
��� =�
�∙ 16,67 (4)
Schließlich wird zur Normierung durch die eingewogene Masse geteilt:
����
��=
����
���������� (5)
Tabelle 3: Ergebnisse zur jeweiligen Aktivität der �-Glucuronidase
Enzymaktivität [nkat/mg]
Units
2’
[nkat/mg]
U
8’
[nkat/mg]
U
15’
[nkat/mg]
U
45’
[nkat/mg]
U
75’
[nkat/mg]
U
Negativkontrolle
28096,2
17,36
Positivkontrolle
8,7163
53,33
AtBCAT – 1
72,5064
448,00
4,2484
26,25
3,6037
22,27
4,8482
29,95
0,7250
4,48
AtBCAT – 4
/
/
13,7528
82,50
8,5572
51,33
4,1564
24,93
2,1293
12,77
Die in Tabelle 3 aufgetragenen Werte wurden nun in Diagramm 1 überführt. Zur
Überprüfung der Werte wurden Regressionsgeraden erstellt.

24
Diagramm 1: Grafische Auswertung der Enzymaktivität
Es wird sichtbar, dass beide eingezeichneten Regressionsgeraden streng monoton
fallen. Allerdings fällt die Gerade von AtBCAT–1 viel steiler wie die von AtBCAT–4.
4. Diskussion
4.1 Qualtitativer Nachweis der Promotoraktivität im Gewebe durch
in-situ-Färbung
Aus Tabelle 1 wurden die Ergebnisse ersichtlich. Positiv-und Negativkontrolle
hingegen entsprechen gänzlich den Erwartungen. Die Expression der Positivkontrolle
kam durch einen konstitutiven Promotor zustande, der gewebsunspezifische Aktivität
aufwies. Da die Negativkontrolle das zu exprimierende Gen �-Glucuronidase nicht
besitzt, konnte es zu keinem dementsprechenden Phänotyp (Blaufärbung) kommen.
Bei AtBCAT-1/GUS und AtBCAT-4/GUS wurde das Gen hinter 2 gewebespezifische
Promotoren kloniert, wodurch sich nur manche Pflanzenorgane färbten. AtBCAT-
1/GUS weißt keine Genexpression in den Blättern auf. Dies kann daran liegen, dass
zu alte Blätter getestet wurden, bei denen keine Genaktivität mehr stattfand. Die
Vorgänge in den Pflanzenzellen werden mit zunehmendem Alter langsamer. Die nur
minimale Genexpression im Stengel kann ein Hinweis darauf sein, dass es sich um
eine bereits ältere Pflanze gehandelt haben könnte. Es kam also zu einer
-10000
0
10000
20000
30000
40000
50000
60000
70000
80000
0 10 20 30 40 50 60 70 80
En
zym
ak
tiv
itä
t p
ro M
ass
e [
nk
at/
g]
Zeit [min]
AtBCAT - 1
AtBCAT - 4
Linear (AtBCAT - 1)
Linear (AtBCAT - 4)

25
schwächeren Enzymaktivität in Blättern und Stängeln dieser Linie, wie eigentlich
erwartet wurde. Wie erwartet färbte sich das Blütengewebe blau ein. Die Wurzeln
färbten sich nicht. AtBCAT-4/GUS weißt Enzymaktivät im Stengel auf, wo eigentlich
keine sein sollte, wohingegen in der Blüte keine Enzymaktivität festgestellt werden
konnte. Dies kann darauf hin deuten, dass das Pflanzengewebe mit. einer anderen
Linie vertauscht worden ist. Teilweise erschien farbiges Gewebe unter dem
Mikroskop zu klein und wurde einfach übersehen. Bei Blättern und Wurzeln hingegen
kam es zu der erwarteten Blaufärbung.
4.2 Fluorimetrische Bestimmung der GUS-Aktivität (q uantitativ)
Durch den Vergleich der Enzymaktivitäten konnte die Aktivität der Promotoren
miteinander verglichen werden. Der konstitutive Promotor der Positivkontrolle stellte
den Oberwert der Enzymaktivität dar. In der Negativkontrolle sollte eigentlich keine
Enzymaktivität zu beobachten gewesen sein, jedoch wurde ein relativ hoher Wert für
die Fluoreszenz gemessen. Das kann zwei Gründe haben.
Die Quarzküvette, die für alle Messungen verwendet wurde, war von den vorherigen
Lösungen verunreinigt. Wäre dies der Fall, so sind beinahe alle Messungen davon
betroffen.
Aufgrund des Alters des Fluorimeters ist es durchaus denkbar, dass es hier zu
erheblichen Messabweichungen aus technischen Gründen kam.
AtBCAT–1 und AtBCAT–4 zeigen geringere Enzymaktivität wie die Positivkontrolle,
wobei AtBCAT–1 der aktivere ist. Werden die beiden Regressionsgeraden
miteinander verglichen, so fällt auf, dass die Gerade von AtBCAT–1 viel steiler
verläuft, da das Substrat schneller mit dem Enzym interagiert, dadurch die
Substratkonzentration abnimmt. Bei Substratüberschuss bleibt das Enzym immer
besetzt. Bei AtBCAT–4 hingegen bleibt die Substratkonzentration und damit die
Enzymaktivität relativ gleich. Es wäre zu erwarten gewesen, dass die beiden
normierten Regressionsgeraden waagrecht verlaufen. Leider konnten die
Verdünnungsfaktoren der einzelnen Proben nicht mehr nachvollzogen werden.
Daher ist die Gesamtenzymaktivität der unverdünnten Proben unbekannt. Es wurde

26
immer wieder ein Teil aus den Proben entnommen. Dadurch kam es zu einer
Abnahme der Enzymmenge. Als Folge wurde weniger Umsatz erzielt.
5. Quellen
1) Stryer; Biochemie 6.Auflage 2007 Spektrum akademischer Verlag
2) Heß, Dieter; Pflanzenphysiologie, 10. Auflage, 1999, Ulmer Verlag
3) Skript zum Grundpraktikum Pflanzenphysiologie und molekulare Botanik WS
2011/12
4) Campbell, Neil A.; Biologie, 2. Korrigierter Nachdruck 2000, Spektrum Verlag
5) http://de.wikipedia.org/wiki/Transfektion
6) http://de.wikipedia.org/wiki/Biotechnologie
7) http://de.wikipedia.org/wiki/Gentechnologie
8) http://de.wikipedia.org/wiki/Bt-Mais#Bt-Mais
9) http://de.wikipedia.org/wiki/Flavr-Savr-Tomate
10) http://de.wikipedia.org/wiki/Golden_Rice
11) http://de.wikipedia.org/wiki/Konjugation_(Biologie)
12) http://de.wikipedia.org/wiki/Transduktion_(Genetik)
13) http://de.wikipedia.org/wiki/Transformation_(Genetik)
14) http://wiki.zum.de/images/a/ad/GAtechnischeEntwicklung.pdf
15) http://de.wikipedia.org/wiki/Transfektion
16) http://de.wikipedia.org/wiki/Protoplastenfusion
17) http://de.wikipedia.org/wiki/Elektroporation
18) http://de.wikipedia.org/wiki/Vektor_(Biologie)
19) http://de.wikipedia.org/wiki/Plasmid

27
20) http://de.wikipedia.org/wiki/Cosmid
21) http://de.wikipedia.org/wiki/YAC
22)http://de.wikipedia.org/wiki/Gr%C3%BCne_Gentechnik#Transformation_durch_Ag
robacterium_tumefaciens
23) http://de.wikipedia.org/wiki/Promotor_(Genetik)
24) http://de.wikipedia.org/wiki/Enhancer_(Genetik)
25) http://de.wikipedia.org/wiki/Gr%C3%BCn_fluoreszierendes_Protein
26) http://de.wikipedia.org/wiki/Cosuppression
27) http://en.wikipedia.org/wiki/RNA-dependent_RNA_polymerase










![[GENDOPING DOPING DER ZUKUNFT] - gentechnologie · PDF file1 Kurzfassung In meiner Facharbeit setze ich mich mit der Thematik des Gendopings auseinander, vor allem in Bezug auf die](https://static.fdokument.com/doc/165x107/5a78c7fa7f8b9a83238c575c/gendoping-doping-der-zukunft-gentechnologie-kurzfassung-in-meiner-facharbeit.jpg)







