Untersuchung GABAerger Neurone im Piriformen Cortex und ... · Aus dem Institut für Pharmakologie,...
239
Aus dem Institut für Pharmakologie, Toxikologie und Pharmazie der Tierärztlichen Hochschule Hannover Untersuchung GABAerger Neurone im Piriformen Cortex und anderen Epilepsie- relevanten Gehirnregionen der Ratte in verschiedenen Modellen der Temporallappenepilepsie THESE zur Erlangung des Grades eines PHILOSOPHICAL DOCTOR - Ph.D. - im Fachgebiet Pharmakologie durch die Tierärztliche Hochschule Hannover vorgelegt von Christian Freichel aus Hannover Hannover 2001
Transcript of Untersuchung GABAerger Neurone im Piriformen Cortex und ... · Aus dem Institut für Pharmakologie,...

Aus dem Institut für Pharmakologie, Toxikologie und Pharmazie der Tierärztlichen Hochschule Hannover
Untersuchung GABAerger Neurone im Piriformen Cortex und anderen Epilepsie-relevanten Gehirnregionen der Ratte in
verschiedenen Modellen der Temporallappenepilepsie
THESE
zur Erlangung des Grades eines
PHILOSOPHICAL DOCTOR - Ph.D. -
im Fachgebiet Pharmakologie
durch die Tierärztliche Hochschule Hannover
vorgelegt von Christian Freichel
aus Hannover
Hannover 2001

Supervisor: Prof. Dr. W. Löscher Betreuungsgruppe: Univ.-Prof. Dr. W. Löscher Univ.-Prof. Dr. E. Zimmermann Univ.-Prof. Dr. H. Bigalke 1. Gutachten: Prof. W. Löscher (Institut für Pharmakologie,
Toxikologie und Pharmazie der Tierärztlichen Hochschule Hannover) Prof. E. Zimmermann (Institut für Zoologie der Tierärztlichen Hochschule Hannover) Prof. H. Bigalke (Zentrum für Pharmakologie und Toxikologie der Medizinischen Hochschule Hannover)
2. Gutachten: Prof. G. F. Walter (Institut für Neuropathologie der Medizinischen Hochschule Hannover) Datum der mündlichen Prüfung: 22.11.2001 gefördert durch eine Sachbeihilfe der Deutschen Forschungsgemeinschaft und ein Promotionsstipendium der BAYER AG, Leverkusen

Für meine Familie


1 Einführung..........................................................................................1
2 Übersicht ............................................................................................2
2.1 Epilepsie...................................................................................................... 2
2.1.1 Definition und Bedeutung ........................................................................ 2
2.1.2 Tiermodelle der Temporallappenepilepsie............................................... 4
2.1.2.1 Das Kainat-Modell ............................................................................. 4
2.1.2.2 Das Lithium-Pilocarpin-Modell........................................................... 5
2.1.2.3 Das Amygdala-Kindling Modell ......................................................... 5
2.1.2.4 Neuropathologische Veränderungen in Tiermodellen der Temporallappenepilepsie .................................................................. 7
2.2 Das GABAerge System .............................................................................. 9
2.3 Gehirnregionen......................................................................................... 13
2.3.1 Piriformer Cortex ................................................................................... 13
2.3.2 Bedeutung des Piriformen Cortex.......................................................... 15
2.3.3 Basolaterale Amygdala.......................................................................... 18
2.3.4 Hippocampusformation.......................................................................... 18
2.3.5 Substantia nigra pars reticulata ............................................................. 19
2.4 Untersuchung GABAerger Neurone ....................................................... 20
2.4.1 Methoden zur Untersuchung GABAerger Neurone ............................... 20
2.4.2 GABAerge Neurone unter Kontrollbedingungen.................................... 26
2.4.3 GABAerge Neurone in Tiermodellen der Temporallappenepilepsie ...... 30
2.5 Fragestellung............................................................................................ 34
3 Material und Methoden....................................................................36
3.1 Versuchstiere............................................................................................ 36
3.2 Histologie .................................................................................................. 37
3.2.1 Gehirnpräparation und Anfertigung von Gewebeschnitten .................... 37
3.2.1.1 Nicht-radioaktive in-situ-Hybridisierung, Immunhistologie und Elektrodenlokalisation ..................................................................... 37
3.2.1.2 Radioaktive in-situ-Hybridisierung................................................... 38
3.2.2 Thioninfärbung....................................................................................... 39
3.3 Vergleichende Darstellung GABAerger Neurone mittels in-situ-Hybridisierung und Immunhistochemie ................................................. 40
3.3.1 Prinzip der in-situ-Hybridisierung........................................................... 40
3.3.2 Etablierung der nicht-radioaktiven in-situ-Hybridisierung....................... 42

3.3.2.1 Herstellung Digoxigenin-markierter RNA-Sonden ........................... 42
3.3.2.1.1 Transformation von Bakterien................................................... 43
3.3.2.1.2 Anzucht von Bakterien .............................................................. 43
3.3.2.1.3 Präparation von Plasmid-DNA .................................................. 44
3.3.2.1.4 Restriktion von Plasmid-DNA.................................................... 45
3.3.2.1.5 In-vitro-Transkription und alkalische Hydrolyse ........................ 46
3.3.2.1.6 Überprüfung der Markierungseffizienz von RNA-Sonden ......... 48
3.3.2.2 Protokoll der nicht-radioaktiven in-situ-Hybridisierung..................... 50
3.3.3 Etablierung der radioaktiven in-situ-Hybridisierung ............................... 52
3.3.3.1 Markierung von Oligonukleotid-Sonden .......................................... 52
3.3.3.2 Protokoll der radioaktiven in-situ-Hybridisierung ............................. 53
3.3.4 Validierung der etablierten Protokolle.................................................... 56
3.3.4.1 Validierung der nicht-radioaktiven in-situ-Hybridisierung................. 56
3.3.4.1.1 Einsatz von Kontrollsonden ...................................................... 56
3.3.4.1.2 Neutralrotgegenfärbung ............................................................ 57
3.3.4.1.3 Modifikationen am Protokoll zur nicht-radioaktiven in-situ-Hybridisierung ....................................................................... 58
3.3.4.2 Validierung der radioaktiven in-situ-Hybridisierung ......................... 58
3.3.5 Vorversuche zur parallelen Darstellung GABAerger Neurone an Gewebeschnitten eines Tieres .............................................................. 59
3.3.5.1 Vorversuche zur Auswahl geeigneter Antikörperkonzentrationen ... 59
3.3.5.2 Fixierung ......................................................................................... 60
3.3.6 Immunhistochemie für GABA und GAD................................................. 60
3.4 Tiermodelle der Temporallappenepilepsie............................................. 62
3.4.1 Das Lithium-Pilocarpin-Modell und das Kainsäure-Modell..................... 62
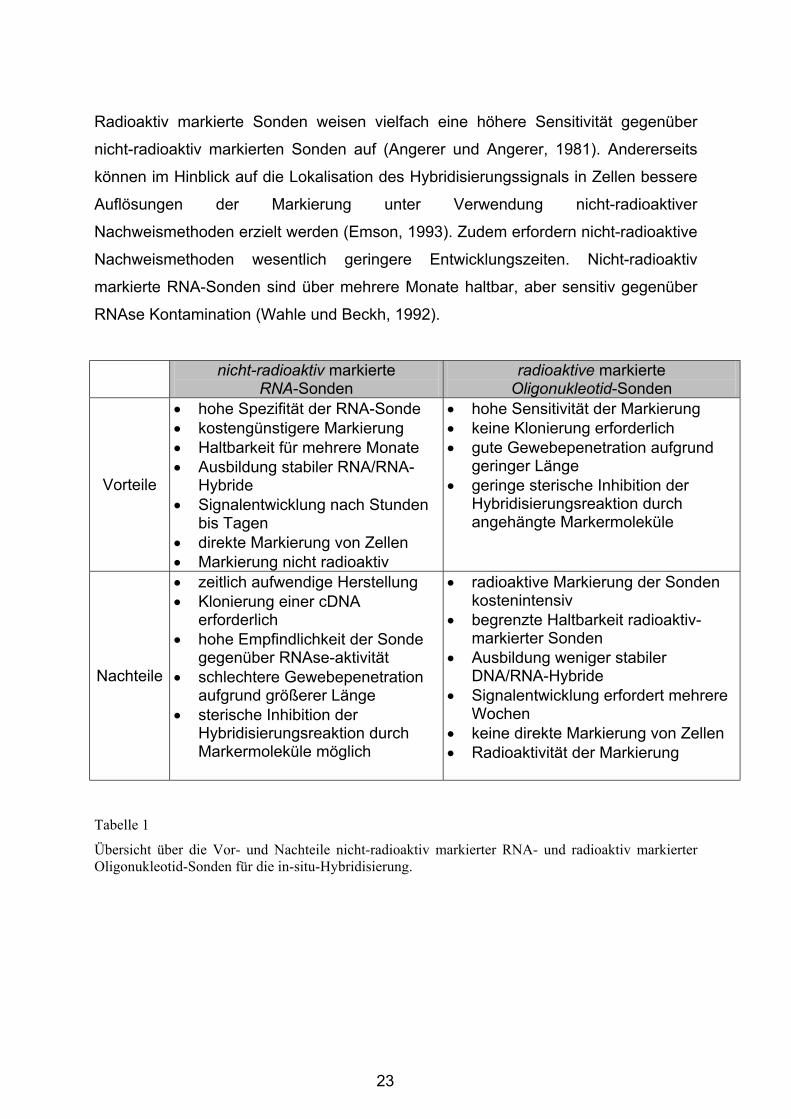
3.4.2 Das Amygdala-Kindling-Modell.............................................................. 64
3.4.2.1 Stereotaktische Implantation von Stimulationselektroden ............... 64
3.4.2.2 Kindling-Protokoll ............................................................................ 66
3.5 Untersuchungen an Ratten nach Status epileptikus induziert durch Kainat/Lithium-Pilocarpin ........................................................................ 70
3.5.1 Versuchsaufbau und -durchführung ...................................................... 70
3.5.2 Bestimmung der Dichte GABAerger Neurone ....................................... 71
3.5.3 Untersuchung der Expression von GAD-mRNA im Gyrus dentatus ...... 75
3.5.4 Untersuchung neuropathologischer Veränderungen ............................. 75
3.5.5 Versuchsauswertung und Statistik......................................................... 76

3.6 Untersuchungen an voll gekindelten Ratten.......................................... 78 3.6.1 Versuchsaufbau und -durchführung ...................................................... 78
3.6.2 Elektrodenlokalisation............................................................................ 80
3.6.3 Bestimmung der Dichte GABAerger Neurone ....................................... 80
3.6.4 Versuchsauswertung und Statistik......................................................... 80
4 Ergebnisse........................................................................................81
4.1 Etablierung und Validierung der in-situ-Hybridisierung ....................... 81
4.1.1 Vergleich methodischer Aspekte ........................................................... 81
4.1.2 Darstellung GABAerger Neurone mit in-situ-Hybridisierung .................. 83
4.1.2.1 Generelles Erscheinungsbild........................................................... 83
4.1.2.2 Regionsspezifisches Erscheinungsbild .......................................... 85
4.2 Untersuchung GABAerger Neurone unter Kontrollbedingungen ........ 88
4.2.1 Generelles Erscheinungsbild der Immunhistochemie............................ 88
4.2.2 Vergleich des regionsspezifischen Erscheinungsbildes ........................ 88
4.3 Untersuchungen an Ratten mit Status epileptikus induziert durch Kainat/ Lithium-Pilocarpin ....................................................................... 94
4.3.1 Ergebnisse der Behandlung mit Kainat/ Lithium-Pilocarpin ................... 94
4.3.2 Bestimmung der Dichte GABAerger Neurone ....................................... 95
4.3.2.1 Anteriorer Piriformer Cortex ............................................................ 96
4.3.2.2 Zentraler Piriformer Cortex............................................................ 101
4.3.2.3 Posteriorer Piriformer Cortex......................................................... 106
4.3.2.4 Basolaterale Amygdala ................................................................. 111
4.3.2.5 Hippocampusformation ................................................................. 113
4.3.2.6 Substantia nigra pars reticulata..................................................... 120
4.3.3 Untersuchung der Expression von GAD-mRNA im Gyrus dentatus .... 124
4.3.4 Neuropathologische Veränderungen ................................................... 128
4.3.4.1 Neurodegeneration im limbischen System.................................... 128
4.3.4.2 Läsionen in der Substantia nigra pars reticulata ........................... 132
4.4 Untersuchungen an voll gekindelten Ratten........................................ 135
4.4.1 Ergebnisse des Kindling ...................................................................... 135
4.4.2 Bestimmung der Dichte GABAerger Neurone ..................................... 138
4.4.2.1 Anteriorer Piriformer Cortex .......................................................... 139
4.4.2.2 Zentraler Piriformer Cortex............................................................ 143
4.4.2.3 Posteriorer Piriformer Cortex......................................................... 147
4.4.2.4 Basolaterale Amygdala ................................................................. 151

4.4.2.5 Hippocampusformation ................................................................. 154
4.4.2.6 Substantia nigra pars reticulata..................................................... 158
5 Diskussion......................................................................................163
5.1 Methodik.................................................................................................. 163
5.2 Diskussion der Untersuchungsergebnisse.......................................... 167
5.2.1 Untersuchung GABAerger Neurone unter Kontrollbedingungen ......... 167
5.2.2 Untersuchungen an Ratten mit Status epileptikus induziert durch Kainat/ Lithium-Pilocarpin .................................................................... 173
5.2.3 Untersuchungen an voll gekindelten Ratten ........................................ 179
5.2.4 Bedeutung der Untersuchungsergebnisse .......................................... 187
6 Zusammenfassung/Summary.......................................................190
7 Literatur ..........................................................................................194
8 Anhang............................................................................................209
8.1 Substanzen ............................................................................................. 209
8.1.1 Substanzen für die in-situ-Hybridisierung ............................................ 209
8.1.2 Substanzen für die Histologie und Immunhistochemie ........................ 210
8.1.3 Sonstige Substanzen........................................................................... 211
8.2 Enzyme und Substrate........................................................................... 211
8.3 Antikörper ............................................................................................... 212
8.4 Puffer und Lösungen ............................................................................. 212
8.4.1 Allgemeine Histologie .......................................................................... 212
8.4.1.1 Nicht-radioaktive in-situ-Hybridisierung, Immunhistologie, Elektrodenlokalisation ................................................................... 212
8.4.1.2 Radioaktive in-situ-Hybridisierung................................................. 213
8.4.2 Herstellung Digoxigenin-markierter RNA-Sonden ............................... 214
8.4.3 Nicht-radioaktive in-situ-Hybridisierung ............................................... 216
8.4.4 Radioaktive in-situ-Hybridisierung ....................................................... 221
8.4.5 Immunhistochemie .............................................................................. 225

ABKÜRZUNGSVERZEICHNIS
Abkürzung Bedeutung BCIP Brom-chlor-indol-phosphat
DAB 3,3`-Diaminobenzidin
DEN dorsaler Endopiriformer Nukleus
DEPC Diethylpyrocarbonat
DIG Digoxigenin
DNA Desoxyribonukleinsäure
E. coli Escherichia coli
2-DG 2-Deoxyglucose
EDTA Ethylendiamintetraessigsäure
EN Endopiriformer Nukleus
Fa. Firma
g Gramm
GABA γ-Aminobuttersäure
GAD Glutamatdecarboxylase
i.p. intraperitoneal
Kap. Kapitel
kg Kilogramm
KGW Körpergewicht
M Molar
LB Medium Luria Bertani Medium
mg Milligramm
ml Milliliter
mm Millimeter
mM Millimolar
mRNA messenger-Ribonukleinsäure
ms Millisekunden
NBT Nitroblau-Tetrazoliumsalz
µCi Mikrocurie
µg Mikrogramm
µl Mikroliter

µm Mikrometer
n Anzahl von Tieren in einer Versuchsgruppe
NBT Nitroblau-Tetrazoliumsalz
NaCl Natriumchlorid
ng Nanogramm
PC Piriformer Cortex
S.E./S.E.M. Standardfehler (engl.:standard error/standard error mean)
VEN ventraler Endopiriformer Nukleus
SE Status epileptikus
TBS Tris-gepufferte, 0,9%ige Natriumchlorid-Lösung
ZNS Zentralnervensystem



1
1 Einführung
Epilepsien zählen zu den häufigsten chronischen Erkrankungen des zentralen
Nervensystems des Menschen. Eine verbreitete Form ist die
Temporallappenepilepsie (Engel et al., 1997), welche durch komplex-fokale Anfälle
gekennzeichnet ist, die sich vielfach als besonders therapieresistent erweisen
(Leppik, 1992).
Die Ätiologie und die Pathomechanismen der Epilepsien werden bisher nicht oder
nur in Ansätzen verstanden. Als eine mögliche Ursache der Epileptogenese werden
Störungen im Gleichgewicht der Mechanismen neuronaler Erregung und Hemmung
diskutiert. Die „GABA-Hypothese“ der Epilepsie (Burnham 1989, Löscher 1989)
postuliert einen Verlust GABAerger Inhibition als Vorraussetzung für die
Epileptogenese, welcher infolge wiederholter paroxysmaler neuronaler Aktivität im
Bereich des Fokus und assoziierter Strukturen letztlich in einem Überwiegen
exzitatorischer Einflüsse resultiert. McNamara (1988) und Dingledine et al. (1990)
vertreten dagegen in ihrer „Glutamat−Hypothese“ die Auffassung, dass ein initiales
Überwiegen der exzitatorischen glutamatergen Transmission die epileptische
Aktivität im Fokus verstärkt und propagiert.
Der Piriforme Cortex (PC) ist eine Gehirnregion, der zunehmende Bedeutung in der
Generierung und Propagierung epileptischer Aktivität zugesprochen wird. Der Verlust
GABAerger Neurone im zentralen PC nach Amygdala-Kindling (Lehmann et al.,
1998), einem Tiermodell der Temporallappenepilepsie, deutet auf eine Beteiligung
GABAerger Mechanismen in diesem Zusammenhang hin.
Ziel dieser Arbeit war die Charakterisierung GABAerger Neurone im PC und anderen
für die Epileptogenese bedeutenden Hirnregionen anhand des Nachweises
verschiedener Syntheseabschnitte im GABA-Metabolismus. Hierfür wurde die
Methode der in-situ-Hybridisierung zur Detektion der mRNA des GABA-
synthetisierenden Enzyms Glutamatdecarboxylase (GAD) etabliert und mit
immuncytochemischen Nachweisen für GAD und GABA verglichen. Mit den
Untersuchungen im Rahmen dieser Arbeit war weiterhin das Ziel verbunden, die
eingesetzten Nachweismethoden hinsichtlich ihrer Aussagekraft und
Einsatzmöglichkeiten unter basalen und pathophysiologischen Bedingungen zu
beurteilen.

2
2 Übersicht
2.1 Epilepsie
2.1.1 Definition und Bedeutung
Der Begriff Epilepsie bzw. Epilepsien beschreibt alle Zustände und Krankheiten, die
wiederholt zu epileptischen Anfällen führen. Diese Anfälle sind dabei Ausdruck
plötzlicher exzessiver Entladungen von Neuronen in Teilen oder im gesamten
zentralen Nervensystem. Das Entstehen unkontrollierter Erregung kann durch
pathologische Veränderungen oder genetische Ursachen bedingt sein, die zu einer
Störung des Gleichgewichtes zwischen neuronaler Erregung und Hemmung führen.
Vielfach treten auch kryptogene Formen von Epilepsien auf, die eine unbekannte
Genese haben (Matthes und Schneble, 1992).
Die klinischen Phänomene epileptischer Anfälle entstehen in Abhängigkeit von der
Lokalisation und der Zahl der vorübergehend epileptischen Neurone und zeigen sich
in Veränderungen des Verhaltens, des Bewusstseins und der Wahrnehmung.
Basierend auf den enzephalographischen Befunden und den klinischen Symptomen
während eines Anfalls erfolgte eine Klassifizierung durch die INTERNATIONAL
LEAGUE AGAINST EPILEPSY (1981, 1989) in fokale, generalisierte und
unklassifizierbare Anfälle sowie Status epileptikus (SE).
Fokale oder partielle Anfälle haben einen lokalen Ursprung, den epileptischen Fokus,
auf den sich die epileptische Aktivität begrenzt. Einfach−fokale Anfälle verlaufen
ohne Beeinträchtigung des Bewusstseins, während komplex−fokale Anfälle mit
Bewusstseinsverlust einhergehen. Komplex-fokale Anfälle können sekundär
generalisieren und zu klonischen oder tonisch/klonischen Anfällen führen.
Primär generalisierte Anfälle zeichnen sich von Beginn an durch gleichzeitige
neuronale Entladungen in beiden Hirnhemisphären aus. Diese zeigen sich in Form
von Absencen, sowie myoklonischen, klonischen, tonischen, tonisch-klonischen oder
atonischen Anfällen. Ein SE liegt vor, wenn ein epileptischer Anfall länger als 10-15
min dauert oder der Patient zwischen den Anfällen das vollständige Bewusstsein
nicht wieder erlangt.

3
Unter der Bevölkerung westlicher Industrienationen treten Epilepsien mit einer
Prävalenz von 0,5-1 % auf und zählen somit zu den häufigsten ZNS-Erkrankungen
(US Department of Health, Education and Welfare, 1977; Janz, 1979; Keränen und
Riekkingen, 1988). Die verbreitetste Anfallsform stellen komplex−fokale Anfälle mit
oder ohne sekundäre Generalisation dar, von denen etwa 40−50% aller Patienten
betroffen sind (Löscher und Schmidt, 1993, 1994). Bei 70−85 % dieser Fälle befindet
sich der Focus epileptischer Aktivität in den Temporallappen des Gehirns, besonders
im Hippocampus und der Amygdala. Dementsprechend wird für diese Form der
Erkrankung die Bezeichnung „Temporallappenepilepsie“ verwendet (Wolf, 1994).
Für Tiere wird ein gehäuftes Auftreten von Epilepsien bei Hunden, seltener bei
Katzen beschrieben. Diese zeigen meist einen fokalen Beginn, und generalisieren
sekundär zu tonisch-klonischen Anfällen (Löscher, 1994).
Der Therapieerfolg bei Epilepsien ist zumeist unbefriedigend. So sind etwa 70 % der
komplex−fokalen Epilepsien bei Erwachsenen therapieresistent. Zudem besitzen
einige der eingesetzten Medikamente ein relativ hohes Nebenwirkungspotential.
Die Untersuchung der pathophysiologischen Grundlagen der
Temporallappenepilepsie sowie die Entwicklung verbesserter Behandlungsstrategien
erfordert den Einsatz geeigneter Tiermodelle (Löscher und Schmidt, 1988). Als
solche haben sich das Kainsäure-Modell (Ben−Ari et al., 1981) und das Lithium-
Pilocarpin-Modell (Honchar et al., 1983) erwiesen. Zu den am besten untersuchten
Tiermodellen zählt nach wie vor das Kindling−Modell (Goddard et al., 1969) der
Temporallappenepilepsie.

4
2.1.2 Tiermodelle der Temporallappenepilepsie
2.1.2.1 Das Kainat-Modell
Durch die systemische oder intracerebrale Applikation von Kainsäure bzw. Kainat
können bei Ratten Symptome ausgelöst werden, die neuropathologisch,
elektrographisch und im Verhalten denen der Temporallappenepilepsie des
Menschen sehr ähnlich sind (Ben-Ari et al., 1980; Nadler, 1981). Kainsäure entfaltet
seine Wirkung als starkes Excitotoxin durch Bindung an Kainatrezeptoren, die einen
Subtyp des Glutamatrezeptors darstellen. Glutamat ist der quantitativ wichtigste
excitatorische Neurotransmitter im ZNS. Die Bindung an den Rezeptor öffnet einen
Kanal zum Einstrom von Na+- und K+-Ionen, wodurch eine Membrandepolarisation
eintritt. Im Hippocampus aktiviert Kainat sowohl inhibitorische Interneurone als auch
excitatorische Pyramidenzellen (Ben-Ari und Cossart, 2000). Die Kainatrezeptoren in
der CA3 Region des Hippocampus besitzen eine hohe Affinität für Kainat, während
für die Rezeptoren der CA1 Region eine niedrige Affinität beschrieben wurde (Berger
et al., 1983; Foster et al., 1981). Eine hohe Konzentration an Kainat-Rezeptoren
besitzt auch der Bulbus olfaktorius, der eine starke Verbindung zum PC unterhält
(Sperk, 1994).
Häufig zeigen Ratten im Anschluss an einen Kainat-induzierten SE spontan
auftretende epileptische Anfälle, die noch Monate nach der Behandlung beobachtet
werden können (Cavalheiro et al., 1982). Im Kainat-Modell sind Parallelen in der
Anfallssymptomatik zum Kindling-Modell zu beschreiben (Sperk, 1994)
(Kap. 2.1.2.3), was vermuten lässt, das in beiden Modellen zum Teil ähnliche
anatomische Wege in der Krampfausbreitung beschritten werden (Löscher und
Ebert, 1996a). Analog zum Kindling-Modell kommt es nach der Applikation von
Kainat ebenfalls zu einer permanenten Absenkung der Krampfschwelle (Feldblum
und Ackermann, 1987; Marksteiner et al., 1990).
Als Nachteil gegenüber dem Kindling-Modell ist das Auftreten massiver
neurodegenerativer Veränderungen nach einem induzierten SE zu werten. Zudem
stellt die Behandlung mit Kainat eine starke Belastung für die Tiere dar, die in einer
relativ hohen Mortalität resultiert.

5
2.1.2.2 Das Lithium-Pilocarpin-Modell
Durch die systemische Applikation des Muskarinrezeptor-Agonisten Pilocarpin kann
bei der Ratte analog zum Kainat-Modell ein SE induziert werden. Im Anschluss
entwickeln diese Tiere vielfach wiederholt spontane komplex-fokale Anfälle in
Verbindung mit klinischen Symptomen und neuropathologischen Alterationen, die
denen der Temporallappenepilepsie des Menschen ähneln (Turski, 1989). Im
Gegensatz zu Kainsäure löst Pilocarpin starke periphere Nebenwirkungen aus,
welche die Applikation von peripher wirksamen Muskarinrezeptor-Antagonisten im
Vorfeld der Behandlung erforderlich machen. Die Applikation hoher Dosen von
Pilocarpin führt nicht immer zur Auslösung eines SE und ist zudem mit einer hohen
Mortalität verbunden. Die Vorbehandlung der Versuchstiere mit Lithium erlaubt den
Einsatz geringerer Dosierungen von Pilocarpin (Clifford et al., 1987), wodurch eine
Verringerung der Mortalität erreicht werden kann. Hinsichtlich des
Wirkungsmechanismus von Pilocarpin wird angenommen, dass die Induzierung des
SE auf der Aktivierung des cholinergen Systems beruht, während die weiteren
Veränderungen auf die Wirkung von Glutamat zurückzuführen sind (Goodman,
1998).
2.1.2.3 Das Amygdala-Kindling Modell
Goddard et al. (1969) beschrieben bei Versuchstieren, denen täglich zunächst
subkonvulsive elektrische Ströme in bestimmten Bereichen des limbischen Systems
appliziert worden waren, das Auftreten von Krämpfen. Diese nahmen in Dauer und
Schweregrad rasch zu und endeten schließlich in sekundär generalisierten tonisch-
klonischen Anfällen. Sie bezeichneten dieses Phänomen als „Kindling“ (engl.: „to
kindle“, etwas entfachen/anzünden).
Elektrisches Kindling bei der Ratte ist eines der wichtigsten Modelle für
komplex−fokale Anfälle, dem klinischen Bild der Temporallappenepilepsie des
Menschen. Dementsprechend oft findet dieses Modell Verwendung bei der
Aufklärung der Pathophysiologie dieser Anfallsform, ihrer fokalen Epileptogenese
und der Wirkung von Arzneimitteln auf fokale und sekundär generalisierte Anfälle
(Sato et al., 1990).

6
„Kindling“ beschreibt den Prozess der Epileptogenese, welcher durch die Auslösung
paroxysmaler Nachentladungen über Elektroden im Gehirn in Gang gesetzt wird.
Aufgrund klinischer Befunde bei Epileptikern ist der Hippocampus als ein wichtiger
Ausgangspunkt paroxysmaler Aktivität beim Menschen identifiziert worden. Im
Kindling-Modell läuft die Epileptogenese dagegen schneller ab, wenn die Stimulation
in limbischen Hirnregionen außerhalb des Hippocampus, etwa in der Amygdala
erfolgt (Goddard et al., 1969). Im Amygdala-Kindling-Modell wird bei den
Versuchstieren die Reiz- und Ableitelektrode in die basolaterale Amygdala, dem
größten Kerngebiet der Amygdala, implantiert. Durch regelmäßige, zunächst
subkonvulsive Stimulation („kindeln“) entwickeln die Versuchstiere zunehmend
schwerere Krampfanfälle. Die im Elektroenzephalogramm aufgezeichneten
paroxysmalen Nachentladungen nehmen parallel in Dauer und Amplitude zu.
Die fünf Anfallsstadien, die nach Racine (1972) an gekindelten Ratten differenziert
werden können, werden folgendermaßen definiert:
Stadium I: Immobilität, schwacher Fazialklonus (Schließen eines oder beider
Augen, Zittern der Tasthaare, stereotypes Schnüffeln)
Stadium II: Zunahme der Schwere der Fazialklonien (klonische Kaubewegungen
und Kopfnicken)
Stadium III: Klonus der ipsilateral zur Stimulationselektrode befindlichen
Vorderextremität (zusätzlich zu Symptomen der Stadien I und II)
Stadium IV: Klonus beider Vorderextremitäten mit Aufrichten
Stadium V: Vollbild des sekundär generalisierten Anfalls (Aufrichten der Tiere mit
anschließendem Verlust der Stellreflexe, nach hinten Überfallen,
generalisierte klonische Krämpfe)
Die Stadien I und II ähneln dem Erscheinungsbild beim Patienten mit
komplex−fokalen Anfällen, Stadium III ist ebenfalls ein fokaler Anfall. Das Bild
generalisierter Krämpfe stellt sich in den Stadien IV und V dar (Löscher und Schmidt,
1988). Der Prozess der Epileptogenese ist nach mehreren Anfällen des Stadiums V
beendet. Nach Abschluss der Kindlingprozesses gelten die Tiere als „voll gekindelt“
und behalten ihre Empfindlichkeit gegenüber elektrischer Stimulation auch über
längere stimulationsfreie Zeiträume bei.

7
Ein entscheidender Vorteil des Kindling-Modells liegt in der experimentell
steuerbaren stufenweise fortschreitenden Epileptogenese, was die Untersuchung
dieses Geschehens zu beliebigen Zeitpunkten des Kindlings erlaubt. Andererseits
setzt die Auslösung von Krampfaktivität eine elektrische Stimulation voraus, das
Auftreten spontaner Anfälle wird nach Kindling im Gegensatz zu den anderen
Modellen seltener beobachtet. Weiterhin erfordert Kindling den Einsatz unstimulierter
Kontrolltiere mit implantierten Elektroden, um zwischen Einflüssen der Elektroden auf
biochemische und funktionelle Prozesse und Kindling-induzierten Veränderungen
differenzieren zu können (Löscher et al., 1999).
2.1.2.4 Neuropathologische Veränderungen in Tiermodellen der Temporallappenepilepsie
Geeignete Tiermodelle zur Untersuchung der Temporallappenepilepsie des
Menschen müssen neben den klinischen Symptomen und
elektroenzephalographischen Befunden auch die neuropathologischen
Veränderungen widerspiegeln können. Im Hippocampus, der vielfach als
Ausgangspunkt paroxysmaler Aktivität ausgemacht werden kann, treten bei vielen
Patienten mit komplex-fokalen Epilepsien chronisch degenerative Veränderungen
auf. Diese sogenannten „Hippocampus-Sklerosen“ wurden schon 1880 von Sommer
beschrieben. Bei etwa 2/3 aller Patienten mit therapieresistenter
Temporallappenepilepsie ist eine Hippocampus-Sklerose nachweisbar, die sich
durch Zellverluste und Gliosen im Bereich der CA-Regionen sowie im Hilus des
Gyrus dentatus auszeichnet (Babb und Brown, 1987; Pfeifer, 1992; Babb und
Pretorius, 1993). Ein weiteres neurologisches Phänomen im Zusammenhang mit
komplex-fokalen Anfällen des Menschen ist das „mossy-fiber-sprouting“ im Bereich
des Gyrus dentatus. Dieser Begriff beschreibt Umgestaltungen in der Ausbildung und
der synaptischen Verschaltung der Axone von Körnerzellen, den Moosfasern, als
Reaktion auf Neuronenverluste im Hilus des Gyrus dentatus (Cascino et al., 1988;
Houser et al., 1990). Analoge Prozesse neuronaler Reorganisation werden auch bei
Tieren mit spontanen Anfällen nach einem Kainat- oder Pilocarpin-induziertem SE
(Cronin und Dudek, 1988; Mello et al., 1993) sowie im Kindling-Modell gesehen
(Sutula, 1991).

8
In Tiermodellen mit chemisch-induziertem SE treten ausgeprägte neurodegenerative
Veränderungen auf. So konnte 24-36 h nach einem Kainat-induzierten SE bei Ratten
in bestimmten Hirnregionen ein erheblicher Neuronenverlust festgestellt werden, der
vielfach von Ödembildung begleitet wurde (Sperk et al., 1983). Zu den am stärksten
von neurodegenerativen Alterationen betroffenen Hirnregionen zählten der Piriforme
und der Entorhinale Cortex sowie die Amygdala. Im Hippocampus waren Neurone in
der Region CA3 und dem Hilus des Gyrus dentatus betroffen, in der CA1 Region
waren Neurone dagegen zu diesem Zeitpunkt noch nicht sehr stark geschädigt
(Sperk, 1994). Ein ähnliches Bild ergab sich in den beschriebenen Hirnregionen auch
im Pilocarpin-Modell (Turski et al., 1984, 1986; Mello und Covolan, 1996).
Neuere Untersuchungen zeigten, dass es bereits wenige Stunden nach Induzierung
eines selbsterhaltenden SE zur Schädigung von Neuronen in verschiedenen
Hirnregionen kommt. Im Vergleich beider Modelle in Hinblick auf das Ausmaß
regionsspezifischer Neuronenschädigung wurde deutlich, dass ein durch Pilocarpin
induzierter SE innerhalb kürzerer Zeit zu ausgeprägteren Neuronenschäden führt
(Covolan und Mello, 2000). In beiden Modellen waren bereits nach 2,5 h veränderte
Neurone in den Schichten II und III des PC sowie im Hilus des Gyrus dentatus
festzustellen. In der Amygdala sowie im Hippocampus traten geschädigte Neurone
dagegen erst nach 8-48 h auf. Für die Substantia nigra pars reticulata wurden neben
schnell einsetzenden ausgeprägten Neuronenschäden eine intensive Schwellung
des Neuropils und Gliosen infolge des SE beschrieben (Covolan und Mello, 2000).
Im Kindling-Modell fallen neurodegenerative Veränderungen im limbischen System
wesentlich diskreter aus. Frühere Untersuchungen ergaben keine Hinweise auf
Gewebedegeneration durch Kindling (Goddard, 1972; Goddard und McIntyre, 1974),
später mehrten sich Hinweise auf Zellverluste nach elektrischer Stimulation. Cavazos
und Sutula (1990) wiesen den Verlust von Neuronen im Hilus des Gyrus dentatus
infolge von Tractus perforans-Stimulation nach, welcher mit der Zahl der
generalisierten Anfälle zunahm. Diese Arbeitsgruppe zeigte in einer weiteren
Untersuchung Zellverluste im Hippocampus und anderen limbischen Regionen als
Resultate des Kindlings verschiedener Lokalisationen (Cavazos et al., 1994).

9
2.2 Das GABAerge System
γ−Aminobuttersäure (GABA) nimmt unter den inhibitorischen Neurotransmittern des
zentralen Nervensystems eine herausragende Rolle bei der Kontrolle neuronaler
Schaltkreise ein. 30−40 % aller Synapsen des ZNS sind GABAerg (Fonnum, 1987).
GABA entfaltet seine inhibitorische Wirkung durch Hyperpolarisation der Membran
nachgeschalteter Neurone (postsynaptische Inhibition). Störungen der GABAergen
Transmission spielen eine wesentliche Rolle bei verschiedenen neurologischen
Erkrankungen. Dies wird neben den Epilepsien u. a. für Dystonien/Dyskinesien,
Morbus Parkinson, Morbus Alzheimer und Chorea Huntington angenommen (Bowery
und Nistico, 1989).
GABA kommt ubiquitär im Gehirn vor, wobei die Konzentrationen regionsabhängig
variieren (Löscher et al., 1989). Dies zeigte sich auf zellulärer Ebene in einer
unterschiedlichen Dichte GABA-immunreaktiver Neurone in verschiedenen
Gehirnregionen (Hendry et al., 1987; Najlerahim et al., 1990). Neben ihrer Funktion
als Interneurone treten GABAerge Neurone auch als Projektionsneurone im ZNS auf,
etwa in den Basalganglien (Parent und Hazrati, 1995), als striato-nigrale Neurone
(Ribak et al., 1980) und in Form von Purkinjezellen des Cerebellums (Oertel und
Mugnaini, 1984).
GABA entsteht durch Decarboxylierung von Glutamat durch das Enzym
Glutamatdecarboxylase (GAD), wobei Glutamat über den sogenannten GABA−Shunt
aus dem Zitratzyklus bereitgestellt wird. Die GAD tritt im zentralen Nervensystem in
zwei bisher bekannten Isoformen mit einem Molekulargewicht von 65.000 Da
(GAD65) bzw. 67.000 Da (GAD67) auf (Kaufman et al., 1986), welche beide GABA
synthetisieren können. GAD65 und GAD67 werden von zwei unterschiedlichen
Genen codiert, die in Ihrer Sequenz zu 65 % übereinstimmen (Erlander et al., 1991;
Kaufman et al., 1991; Martin et al., 1991 a, b). Die Gensequenz für eine Isoform
weist im Vergleich zwischen Maus, Ratte, Katze und Mensch eine Übereinstimmung
von 95 % auf (Kaufman et al, 1986; Julien et al., 1990; Erlander et al., 1991).
10
Beide Formen des Enzyms sind Pyridoxalphospat−abhängig. GAD67 tritt stets als
Holoenzym auf, während GAD65 zu 50 % als Apoenzym vorliegt (Erlander et al.,
1991, Kaufman et al., 1991; Martin et al., 1991a).
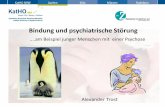
Abbildung 1
Vereinfachte Darstellung des Stoffwechsels und der synaptischen Übertragung von GABA. GABA wird durch die GAD aus Glutamat (Glu) gebildet und in Vesikeln von Neuronen gespeichert (3). Der Release von GABA in den synaptischen Spalt führt postsynaptisch über den GABAA- Rezeptor zu Aktivierung eines Chlorid-Ionophors bzw. über den GABAB-Rezeptor zu einer G-Protein vermittelten Aktivierung von Calcium- (Ca²+) und Kaliumkanälen (K+). GABA kann seine Freisetzung über präsynaptische Autorezeptoren (A) modulieren. GABA wird zu Succinatsemialdehyd (SSA) abgebaut und nach Oxidation zu Bernsteinsäure (SC) in den Zitratzyklus eingeschleust. Die Pfeile auf der Abbildung stellen Stoffbewegungen und Stoffumwandlungen dar. Beeinflussungen dieser Prozesse sind durch ein + (Aktivierung) oder ein - (Hemmung) gekennzeichnet (Nach Starke, 1996).

11
GABA wird in Vesikeln der Terminalen von Neuronen als sogenannter
„Transmitterpool“ gespeichert und durch Aktionspotentiale unter dem Einfluss von
Calcium in den synaptischen Spalt freigesetzt. Die Wirkung von GABA kann über die
Aktivierung von GABAA-, GABAB- oder GABAC-Rezeptoren erfolgen. Postsynaptisch
vermittelt die Aktivierung des GABAA-Rezeptors die Öffnung eines assoziierten
Chlorid-Kanals, der über einen Influx von Chlorid zu einer schnellen
Hyperpolarisation des Neurons führt. Die Aktivierung des GABAB-Rezeptors induziert
post- wie auch präsynaptisch über die Kopplung an Kalium- oder Calciumkanäle eine
langsame Hyperpolarisation. GABAC-Rezeptoren steuern Chlorid-Kanäle. Teilweise
wird diese Gruppe der GABA-Rezeptoren auch als eine spezielle Form des GABAA-
Rezeptors angesehen (Barnard et al., 1998).
Die Inaktivierung GABAerger Wirkung wird im wesentlichen über eine Aufnahme des
Transmitters aus dem synaptischen Spalt präsynaptisch in Neurone aber auch in
Gliazellen bewirkt. In beiden Zellentypen wird GABA durch das Pyridoxalphosphat-
abhängige Enzym GABA-Transaminase vornehmlich zu Succinylsemialdehyd
desaminiert und nach Oxidation zu Bernsteinsäure in den Zitratzyklus eingeschleust
(Salganikoff und De Robertis, 1965). Ein geringer Anteil des von Neuronen
aufgenommenen GABA wird in Vesikeln des „metabolischen Pools“ gespeichert.
Die Mehrheit GABAerger Neurone im ZNS exprimiert die Gene für beide Isoformen
der GAD (Erlander et al., 1991; Mercugliano et al., 1992; Esclapez et al., 1993;
Feldblum et al., 1993; Esclapez et al., 1994; Hendrickson et al., 1994; Houser und
Esclapez, 1994). Frühere Untersuchungen in Neuronen des Cerebellums ergaben
eine Verteilung von GAD67 über weite Bereiche der Nervenzelle inklusive des
Somas, während GAD65 hauptsächlich in den Terminalen lokalisiert war (Kaufman
et al., 1991). Martin und Rimval (1993) ordneten daraufhin GAD65 dem
Transmitterpool von GABA zu, in welchem GABA hauptsächlich als Neurotransmitter
synthetisiert wird, während GAD67 dem metabolischen Pool zugerechnet wurde.
Neuere Untersuchungen wiesen jedoch GAD67 auch in Synaptosomen und
Nervenendigungen nach (Erlander et al, 1991; Esclapez et al., 1994; Sloviter et al.,
1996), was gegen eine strikte Zuordnung der Isoformen zu den beiden Pools spricht.

12
Hinsichtlich der Existenz zweier Isoformen der GAD für die Synthese von GABA
besteht weiterhin Klärungsbedarf (Soghomonian und Martin, 1998). Untersuchungen
an GAD-knock-out Mäusen legen den Schluss nahe, dass die GAD67 für den
überwiegenden Teil der GABA-Synthese in Neuronen verantwortlich ist. Ein Fehlen
von GAD67 erwies sich bei solchen Tieren als Letalfaktor, da nur 7 % der GABA-
Konzentration naiver Tiere erreicht wurden und eine um 80 % reduzierte GAD-
Aktivität auftrat. An GAD65-knock-out Mäusen konnten keine Unterschiede in der
intracerebralen GABA-Konzentration festgestellt werden. Diese Tiere zeigten
allerdings eine erhöhte Anfälligkeit für Krämpfe (Asada et al., 1996; Asada et al.,
1997). Weiterhin wird GAD67-mRNA in der Mehrzahl der Regionen des ZNS stärker
exprimiert und das vor allem in Neuronen, die tonisch feuern oder über längere Zeit
feuern können (Feldblum et al., 1993; Esclapez et al., 1993, 1994; Hendrickson et
al., 1994). Zudem treten GAD67 und GAD67-mRNA früher in der Entwicklung des
Gehirns auf (Greif et al., 1991; Greif et al., 1992) und lassen sich durch Läsionen
stärker induzieren (Feldblum et al., 1990, Soghomonian und Martin, 1998).
Die Regulation der GABA-Synthese kann kurzfristig über eine Beeinflussung der
GAD-Aktivität oder langfristig über die Konzentration des Enzyms in GABAergen
Zellen erfolgen. Die Mechanismen, die der Regulation von GAD zugrunde liegen,
sind dabei bisher nur ansatzweise geklärt (Martin und Rimval, 1993; Soghomonian
und Martin, 1998). Es wird vermutet, dass GAD65 hauptsächlich über den Cofaktor
Pyridoxalphosphat reguliert wird (Rimvall et al., 1993; Rimvall und Martin, 1994). Die
Aktivität der GAD67 unterliegt zum überwiegenden Teil der Kontrolle der
Genexpression, wobei auch posttranskriptionale Faktoren eine regulierende Funktion
haben. Die Tatsache, das GAD67 aber nicht GAD67-mRNA durch einen
intrazellulären Anstieg von GABA supprimiert wird, weist auf eine Bedeutung von
Regulationsmechanismen auf der Ebene der Translation und des Proteinabbaus hin.
Die Expression von GAD65 wird dagegen durch die intrazelluläre GABA
Konzentration nicht beeinflusst (Rimvall et al., 1992; Rimvall et al., 1993; Rimvall und
Martin, 1994). Einige Autoren sehen die Funktion von GAD65 als synaptisches
Reservoir von Enzymaktivität an Terminalen, welches unter Bedingungen erhöhter
GABAerger Transmission aktiviert werden kann (Erlander und Tobin, 1991; Martin,
1993; Esclapez et al., 1994).

13
2.3 Gehirnregionen
Paroxysmale Aktivität findet ihren Ursprung bei Epileptikern vielfach in
Gehirnregionen, die zum limbischen System gerechnet werden. Neben dem
Hippocampus und der Amygdala stellt der primäre olfaktorische oder PC eine
„limbische Struktur“ dar. Das limbische System bildet mit dem Corpus striatum und
dem Neocortex die Hauptanteile des Telenzephalons. Eine weitere Hirnregion mit
Bedeutung für die Epilepsieforschung ist die Substantia nigra, der eine maßgebliche
Funktion im Hinblick auf die Generalisation von Krampfaktivität limbischen Ursprungs
zukommt (Depaulis et al., 1994).
2.3.1 Piriformer Cortex
Die Bezeichnung „Piriformer Cortex“ (PC) wurde aufgrund der birnenartigen
(piriformen) Form dieser Struktur bei einigen Säugetieren, so etwa bei der Katze,
gewählt. Der PC stellt das größte Gebiet des olfactorischen Cortex der Säugetiere
dar (Shipley et al., 1995), beim Menschen ist diese Struktur im Verhältnis relativ klein
(Paxinos, 1990). Der PC der Ratte ist bei einer Länge von 5 mm sowie einer Breite
von 3 mm an der rostro−lateralen Oberfläche des Vorderhirns unterhalb der rhinalen
Fissur gelegen (Paxinos und Watson, 1986). Die Literatur beschreibt vielfach einen
anterioren und einen posterioren Teil des PC. Der Übergang zwischen beiden
Anteilen wird durch das Verschwinden des lateralen olfaktorischen Traktes (LOT) an
der Oberfläche des PC bei gleichzeitiger Zunahme der Schicht III sowie des
anhängenden ventralen endopiriformen Nukleus (EN) gekennzeichnet (Haberly und
Price, 1978; Hönack et al., 1991). Diese umschriebene Region im Übergang
zwischen anteriorem und posteriorem PC wird im weiteren als zentraler PC
bezeichnet.
Obwohl phylogenetisch relativ alt und nur aus drei Schichten bestehend, ist der PC in
Zellmorphologie und Physiologie sowie lokaler Vernetzung dem sechsschichtigen
Neocortex sehr ähnlich. Die oberflächlich gelegene Schicht I zeichnet sich durch eine
geringe Zahl GABAerger Horizontalzellen aus, hier dominieren die Ausläufer
glutamaterger Zellen tieferer Schichten.

14
Schicht I kann unterteilt werden in eine obere Schicht, welche afferente Fasern vom
Bulbus olfactorius enthält, und eine untere oder tiefe Schicht mit den afferenten
Axonen anderer Teile des PC sowie benachbarter Nuclei (Price, 1973). Die Schicht II
des PC ist charakterisiert durch eine große Zahl an Zellkörpern von Semilunarzellen
in Schicht IIa und von Pyramidenzellen in Schicht IIb. Letztere entsenden apikale
Dendriten sowohl in Schicht I aber auch basale Dendriten in Schicht III (Haberly,
1983; Haberly und Bower, 1984; Martinez et al., 1987). Beide Zelltypen sind
glutamaterg (Watanake und Kawana, 1984), wohingegen die Interneurone in dieser
Schicht GABAerg sind. In der Schicht III treten große GABAerge multipolare Zellen
gleichmäßig verteilt auf, während Pyramidenzellen fast ausschließlich in der oberen
Hälfte der Schicht zu finden sind (Haberly und Feig, 1983; Martinez et al., 1987).
Medial der Schicht III findet sich ein Gebiet mit starker Anhäufung multipolarer
Zellen. Dieser endopiriforme Nucleus (EN) gliedert sich in einen zelldichteren
dorsalen Anteil (dorsaler EN/DEN) und einen weniger zelldichten ventralen Anteil
(ventraler EN/VEN). Der EN wird aufgrund seiner räumlichen Nähe und der
ausgeprägten Verbindung zum PC häufig als dessen vierte Schicht bezeichnet
(Tseng und Haberly, 1989).
Innerhalb des PC bilden die Zellen der einzelnen Schichten ein komplexes Netzwerk.
Erregung aus dem Bulbus olfactorius führt zur Stimulation exzitatorischer
Pyramidenzellen der Schichten II und III als auch der in der Schicht I gelegenen
GABAergen Horizontalzellen (Haberly und Bower, 1984). Die Pyramidenzellen leiten
die Erregung zum einen an nahe gelegene multipolare Zellen weiter (Haberly und
Presto, 1986), treten jedoch auch mit weiter entfernt gelegenen Pyramidenzellen der
Schichten II und III in Verbindung (Haberly und Behan, 1983). Horizontalzellen und
multipolaren Zellen fällt eine inhibitorische Funktion gegenüber den Pyramidalzellen
zu. Während die Horizontalzellen dabei in Form eines „feedforwards“ eine direkte
Hemmung auf die Pyramidalzellen ausüben, tritt dieser Effekt durch multipolare
Zellen erst infolge der Erregung durch Pyramidenzellen in Form eines „feedbacks“
ein (Haberly und Feig, 1983).

15
Die Verarbeitung olfaktorischer Informationen ist die Hauptaufgabe des PC.
Dementsprechend erhält diese Struktur Informationen aus dem Bulbus olfactorius
(Price, 1973) aber auch aus dem basalen Vorderhirn, dem Thalamus und
Hypothalamus, sowie aus dem Hirnstamm (Löscher und Ebert, 1996b).
Der PC projiziert zum ipsilateralen Bulbus olfactorius (Luskin und Price, 1983) sowie
zu Inselgebieten des Neocortex (Haberly, 1990) und zu verschiedenen Kernen der
Amygdala (Veening, 1978; Wakefield, 1980). Weiterhin steht der PC mit dem
Entorhinalen Cortex, dem Subiculum (Krettek und Price, 1977c, 1987b), dem
mediodorsalen Thalamus (Krettek und Price, 1977b), dem Hypothalamus sowie dem
ventralen Putamen (Krettek und Price, 1987a, b) in Verbindung. Durch die
Verbindung zum lateralem enthorinalen Cortex und ventralem Subiculum (Krettek
und Price, 1977c) unterhält der PC Anschluss an die wichtigsten ein− und
ausgehenden Strukturen des Hippocampus (Amaral und Witter, 1995). Für die
Ausbreitung epileptischer Aktivität im limbischen System ist neben dieser Verbindung
der Kontakt des PC zur Amygdala von entscheidender Bedeutung.
Dementsprechend kann sich Krampfaktivität sowohl im Kreislauf zwischen PC und
Amygdala als auch über den enthorinalem Cortex zwischen Hippocampus und PC
ausbreiten (Löscher und Ebert, 1996b).
2.3.2 Bedeutung des Piriformen Cortex
Die Untersuchungen zur Entstehung von Epilepsien mit komplex-fokalen Anfällen
konzentrierten sich vielfach auf den Hippocampus und die Substantia nigra (Stark
1992, Löscher und Ebert, 1996b). Es mehren sich jedoch zunehmend Hinweise
darauf, dass der PC bezüglich der Krampfentstehung und –ausbreitung eine
Schlüsselpositition im limbischen System einnimmt. Eine mögliche Ursache in dieser
Hinsicht liegt im dreischichtigen Aufbau des PC begründet, in denen inhibitorische
Prozesse im Gegensatz zum Neocortex wahrscheinlich schwächer ausgeprägt sind.

16
Die Empfindlichkeit des PC gegenüber elektrischer Stimulation konnte in
vergleichenden Untersuchungen im limbischen System gezeigt werden, in denen der
PC sowie der Bulbus olfactorius mit der geringsten Zahl an Stimulationen gekindelt
werden konnten. Dieser Prozess erforderte im Hippocampus dagegen eine viel
höhere Zahl an Stimulationen (Racine et al., 1989). Als weiterer Hinweis für die
besondere Stellung des PC ist das frühe Auftreten interiktaler epileptiformer
EEG−Potentiale im Kindling zu werten. Diese interiktale Spikes (IIS) werden vom PC
schon sehr früh generiert, unabhängig davon, welcher Teil des limbischen Systems
stimuliert wurde (Racine et al., 1988). Mit Hilfe der 14C−2−Deoxyglukose−Technik
(2−DG−Technik) als Marker für erhöhte Stoffwechselaktivität konnte gezeigt werden,
dass Anteile der Amygdala und des PC schon in der Anfangsphase des Kindlings
aktiviert werden. Andere Regionen werden dagegen erst nach Erreichen des
Vollbildes generalisierter Krämpfe aktiviert (Engel et al., 1978; Ackermann et al.,
1986). Weiterhin wurde das Protoncogen c−fos, ein Indikator für den
Aktivitätszustand von Neuronen, in frühen Stadien des Amygdala-Kindlings
besonders im posterioren PC exprimiert (Ebert und Löscher, 1995).
Es mehren sich zudem Hinweise darauf, dass Anteile des PC als funktionell
anatomische Einheiten aufzufassen sind (Löscher und Ebert, 1996b). Piredda und
Gale (1985) identifizierten ein umschriebenes Gebiet im anterioren PC (Präpiriformer
PC), in dem sich durch Applikation geringer Mengen des GABAA-
Rezeptorantagonisten Bicucullin generalisierte Krämpfe bei Ratten auslösen ließen.
Diese Struktur wurde von den Autoren aufgrund dieser Eigenschaft als „Area
tempestas“ (AT) bezeichnet (Piredda und Gale, 1986). In neueren Studien konnte
jedoch auch für den posterioren PC eine Krampfsensitivität gegenüber Bicucullin
nachgewiesen werden (Ebert et al., 2000). Hönack et al. (1991) und Löscher et al.
(1995) untersuchten in ihren Studien die Empfindlichkeit des PC gegenüber
elektrischer Stimulation. Die meisten Regionen zeigten eine hohe Empfindlichkeit
besonders in Hinblick auf die Höhe der initialen Nachentladungsschwelle
unbehandelter Ratten und die Herabsetzung der Nachentladungsschwelle voll
gekindelter Ratten (Löscher et al., 1995). Der zentrale PC erwies sich in diesen
Untersuchungen als besonders sensitiv gegenüber elektrischer Stimulation auch im
Vergleich zur Area tempestas (Hönack et al., 1991; Löscher et al., 1995).

17
Für die sekundäre Generalisation fokaler Krampfaktivität ausgehend von der
Amygdala oder dem Hippocampus ist die Ausbreitung diese Aktivität über
Zwischenstrukturen auf den Hirnstamm Vorraussetzung. Der PC kann als eine
solche Struktur fungieren und zur Verstärkung und Ausbreitung von Krampfaktivität
limbischen Ursprungs beitragen. Dies wird zusätzlich durch die vielfältigen
Verbindungen des PC zu anderen Gebieten auch außerhalb des limbischen Systems
begünstigt (Löscher und Ebert, 1996b). Die besondere Bedeutung des zentralen PC
bei der Ausbreitung von Krampfaktivität wurde in Läsionsstudien deutlich. Durch
große unilaterale Läsionen im posterioren PC verlangsamte sich die initiale
Kindlingsentwicklung vor allem bei Tieren, bei denen die Läsionen auch den
zentralen PC mit einschlossen (Wahnschaffe et al., 1993). Diese Erkenntnisse
fanden Bestätigung im Zusammenhang mit der bilateralen selektiven Läsionierung
des zentralen PC, die im Gegensatz zu Läsionen in der Area tempestas und im
posterioren PC den Prozess des Amygdala-Kindlings verlangsamen konnten
(Schwabe et al., 2000a). In beiden Versuchsansätzen konnte der Kindling-Prozess
jedoch nicht unterdrückt werden (Wahnschaffe et al., 1993; Schwabe et al., 2000a).
GABAerge Mechanismen scheinen an der Krampfausbreitung im Amygdala-Kindling
beteiligt zu sein. Für den zentralen PC konnte eine Abnahme GABA-immunreaktiver
Neurone durch Kindling nachgewiesen werden, die in den anterioren und posterioren
Anteilen dieser Struktur nicht auftrat (Lehmann et al., 1998). Eine Erhöhung der
GABA-Konzentration durch bilaterale Mikroinjektionen des GABA-
Transaminaseinhibitors Vigabatrin hatte im zentralen PC die stärksten
antikonvulsiven Effekte, führte aber auch im anterioren und posterioren PC zu einer
Erhöhung der Nachentladungsschwelle bei gekindelten Ratten. Durch die bilaterale
Erhöhung der GABA-Konzentration im zentralen PC konnte auch die Entwicklung
des Kindlings verzögert werden. Mikroinjektion des GABAA-Rezeptor-Agonisten
Muscimol hatten ebenfalls eine antikonvulsive Wirkung sowie eine Erhöhung der
Nachentladungsschwelle bei voll gekindelten Ratten zur Folge (Schwabe et al., 1999;
Schwabe et al., 2000b).

18
2.3.3 Basolaterale Amygdala
Das Kerngebiet der Amygdala (Mandelkern) der Ratte wird unterteilt in die
olfaktorische, die mediale, die zentrale, sowie die basolaterale Amygdala. Dieses
Kerngebiet besitzt weitreichende Aufgaben hinsichtlich der Modulation emotionaler
und endokriner Prozesse und spielt eine zentrale Rolle beim Lernen. Die
basolaterale Amygdala setzt sich aus dem lateralen, dem basolateralen, dem
ventralen basolateralen und dem basomedialen Nucleus zusammen (Alheid et al.,
1995) und stellt den größten Anteil der Gesamtstruktur dar. Anhand morphologischer
Kriterien können drei Klassen von Neuronen unterschieden werden. Die Neurone der
Klasse I besitzen wahrscheinlich die Funktion von Projektionsneuronen. McDonald
(1982) definierte kleine Stellatzellen als Klasse II-Neurone und neurogliaforme Zellen
als Klasse III-Neurone, welche als lokale inhibitorische GABAerge Interneurone
fungieren.
2.3.4 Hippocampusformation
Die Hippocampusformation setzt sich aus dem Entorhinalen Cortex, dem Para- und
Presubiculum, dem Subiculum proper sowie dem Ammonshorn und dem Gyrus
dentatus zusammen. Der Hippocampus im engeren Sinne umfasst das Ammonshorn
und den Gyrus dentatus. Das Ammonshorn unterteilt sich in die Regionen CA1, CA2
und CA3, die im Querschnitt vier Schichten aufweisen (Stratum oriens, -pyramidale, -
radiatum, lacunosum moleculare). Das Stratum pyramidale ist durch eine sehr hohe
Dichte an Zellsomata glutamaterger Pyramidenzellen gekennzeichnet (Switzer et al.,
1985). GABAerge Neurone besitzen einen Anteil von 10-15 % an der gesamten
Neuronenpopulation des Hippocampus (Woodson et al., 1989). Der Gyrus dentatus
gliedert sich in den Hilus, welcher auch als polymorphe Schicht bezeichnet wird,
sowie die Körnerzellschicht und die Molekularschicht. Die prinzipalen Zellen der
Körnerzellschicht stellen Körnerzellen dar, welche wie die Mooszellen des Hilus,
Glutamat als exzitatorischen Neurotransmitter benutzen. In beiden Schichten sind
GABAerge Neurone vorhanden, so in Form von Korbzellen am Übergang von der
Körnerzellschicht zum Hilus (Amaral und Witter, 1995).

19
Die wichtigsten Fasereingänge der Hippocampusformation bei der Ratte kommen
von der Amygdala, dem Raphe Kern, dem Claustrum, dem medialen Septum sowie
dem Nucleus supramammillaris des Hypothalamus (Amaral und Witter, 1995). Die
intrahippocampale Innervation verläuft hauptsächlich unidirektional. Der Gyrus
dentatus erhält seinen Input zu einem gewissen Teil über den Perforant Pathway aus
dem Entorhinalen Cortex. Die Körnerzellen des Gyrus dentatus innervieren über
Moosfasern („mossy fibers“) zum einen die CA3 Region, treten aber auch mit
Mooszellen im Hilus in Verbindung. Die Axone der Pyramidenzellen aus der Region
CA3 ziehen als sogenannte Schaffer-Collateralen zu Pyramidenzellen der CA1
Region (Switzer et al., 1985).
2.3.5 Substantia nigra pars reticulata
Die Substantia nigra wird wie das Corpus Striatum (Striatum), der Globus pallidus
und der Nucleus subthalamicus (STN) zu den Basalganglien gezählt. Damit ist sie im
Verschaltungssystem derjenigen Hirnzentren, die die Motorik beeinflussen,
eingebunden und liefert einen wesentlichen Anteil an der Kontrolle und der
Modulation von Bewegungsabläufen und -impulsen. Die Substantia nigra ist im
Mesencephalon lokalisiert und unterteilt sich in eine dopaminreiche Pars compacta
und eine Pars reticulata, welche von GABAergen Neuronen dominiert wird (Flaherty
und Graybiel, 1994). Die Bezeichnung als „schwarze Substanz“ leitet sich von der
dunklen Pigmentierung der dopaminergen Neurone in der Substantia nigra pars
compacta ab, die besonders beim erwachsenen Menschen ausgeprägt ist, bei der
Ratte jedoch nicht auftritt. Das Striatum, die Eingangsstruktur der Basalganglien,
entsendet GABAerge Projektionsneurone zur Substantia nigra pars reticulata und
dem Globus pallidus internus, den Ausgangsstrukturen der Basalganglien. Durch
diese Projektionen werden GABAerge Efferenzen der Substantia nigra pars reticulata
gehemmt, so zum Thalamus (nigro-thalamischer Pathway), zum Superioren
Colliculus des Tectums, zur reticularen Formation sowie zum Tecmentum (Fallon und
Loughlin, 1995).

20
2.4 Untersuchung GABAerger Neurone
2.4.1 Methoden zur Untersuchung GABAerger Neurone
Die Untersuchung GABAerger Neurone kann grundsätzlich durch den Nachweis
verschiedener Syntheseabschnitte im GABA-Metabolismus erfolgen. Neben der
Detektion der mRNA für die Isoformen des GABA synthetisierenden Enzyms GAD
erlaubt der direkte immunhistochemische Nachweis dieses Enzyms sowie seines
Reaktionsproduktes GABA die Darstellung GABAerger Neurone.
Die in-situ-Hybridisierung stellt eine Methode zur Lokalisation von DNA− und
mRNA−Sequenzen in Geweben, Zellen und intrazellulären Strukturen mittels
spezifischer Sonden dar. Die Methode basiert auf der Fähigkeit von
Nukleinsäuremolekülen, sich bei Komplementarität ihrer Basensequenz zu
Hybridmolekülen zusammenzulagern. Der Vorteil dieser Technik gegenüber der
Genanalyse mittels Southern- oder Northern-blot besteht darin, dass das
Hybridisierungssignal direkt im biologischen Präparat sichtbar wird. Dadurch lassen
sich auch die räumlichen und zeitlichen Expressionsmuster von Genen bzw. durch
diese kodierte Neurotransmitter bestimmen. Die Methode erlaubt außerdem eine
quantitative Abschätzung der mRNA−Konzentration in Zellen und dadurch
Rückschlüsse über das Verhältnis zwischen mRNA−Synthese einerseits und
Produktion, Morphogenese und Feinstruktur von Proteinen andererseits (Leitch et al.,
1994). Harrison et al. (1974) konnten unter Verwendung einer tritiummarkierter cDNA
die Expression des Globingens in fetalen Leberzellen mit in-situ-Hybridisierung
nachweisen. In vitro−transkribierte einzelsträngige RNA−Sonden wurden erstmals
zur Hybridisierung mit Histon−mRNA in Seeigelembryonen von der Arbeitsgruppe um
Cox (1984) verwendet.

21
Bei der in-situ-Hybridisierung finden heutzutage mehrheitlich einzelsträngige RNA-
oder DNA-Sequenzen als Sonden Verwendung (Emson, 1993). Die Herstellung von
RNA-Sonden erfordert den Einbau der nachzuweisenden Gensequenz als cDNA in
einen geeigneten Transkriptionsvektor, der Transkriptionsstartpunkte für RNA-
Polymerasen enthält. Die Synthese erfolgt mittels in-vitro-Transkription entlang des
linearisierten Transkriptionsvektors in Gegenwart von Nukleotiden (Abbildung 2A).
Einzelsträngige Oligonukleotid-Sonden bestehen aus kurzen DNA-
Nukleotidsequenzen, die gewöhnlich mit einer Länge von 10 bis 50 Basenpaaren im
DNA-Syntheseautomaten hergestellt werden.
Abbildung 2 A/B
Vereinfachte Darstellung der Herstellung nicht-radioaktiv (A) und radioaktiv markierter Sonden (B) für die in-situ-Hybridisierung. A. Die Synthese Digoxigenin (DIG)-markierter RNA-Sonden setzt die Klonierung einer cDNA in einen geeigneten Transkriptionsvektor voraus. Die Markierung der RNA-Sonden erfolgt mittels in-vitro-Transkription am linearisierten Vektor durch Inkorporation DIG-markierter Nukleotide. B. Mittels DNA-Syntheseautomaten hergestellte Oligonukleotid-Sonden können durch das Anhängen 35Schwefel-gekoppelter Nukleotide markiert werden. Die Detektion des Hybridisierungssignals erfordert die Markierung der im
Hybridisierungsversuch eingesetzten Sonden, welche durch Inkorporation (Abbildung
2A) oder das Anhängen markierter Nukleotide an die Sonde erfolgen kann
(Abbildung 2B). Neben der Verwendung radioaktiver Markermoleküle werden
zunehmend nicht-radioaktive Markierungstechniken angewandet (Bloch et al., 1990;
Kiyama et al., 1990a, b). Radioaktiv markierte Nukleotide enthalten 32Phosphor, 35Schwefel oder Tritium, als nicht radioaktive Markermoleküle kommen Digoxigenin
(DIG) und Biotin zum Einsatz.
Bakterium
cDNA
3`
Oligonukleotid -Sonde
35S-ATP
35S-ATP markierte Oligonukleotid-Sonde
DIG-markierte RNA-Sonde
Transkriptionsvekor
In vitro-Transkription
A
linearisierter Vektor
Markierungsreaktion
B

22
Für die in-situ-Hybridisierung werden die markierten Sonden mit dem Gewebe
inkubiert, welches die nachzuweisende Gensequenzen enthält. Die Detektion der
Hybride erfolgt in Abhängigkeit von der Markierung der Sonde. Sonden, die nicht-
radioaktive Marker tragen, können unter Verwendung von Antikörpern
immunhistochemisch nachgewiesen werden (Abbildung 3A). Radioaktiv markierte
Sonden erfordern die autoradiographische Detektion mittels Röntgenfilm oder einer
photosensitiven Emulsion (Abbildung 3B).
Abbildung 3 A/B
In-situ-Hybridisierung unter Verwendung nicht-radioaktiv markierter RNA-Sonden (A) bzw. radioaktiv markierter Oligonukleotid-Sonden (B). Im Anschluss an die Hybridisierung erfolgt die immunhistologische Detektion Digoxigenin-(DIG)-markierter RNA-Sonden mittels eines Antikörpers und anschließender Farbreaktion (A). Das Signal radioaktiv markierter Oligonukleotid-Sonden wird autoradiographisch unter Verwendung eines Röntgenfilms oder einer fotosensiblen Emulsion nachweisbar (B). Hinsichtlich der verwendeten Sonden ergeben sich Vor- und Nachteile in
Abhängigkeit von der Sondenstruktur, der Art der Markierung sowie der
nachzuweisenden Gensequenzen (Tabelle 1). Radioaktiv markierte Oligonukleotid-
Sonden gelangen aufgrund ihrer geringen Länge leicht in das Gewebe, bilden aber
weniger stabile Hybride beim Nachweis von mRNA aus. DIG-markierte RNA-Sonden
führen zu stabilen RNA/RNA-Hybride. Augrund ihrer Länge und der inkorporierter
Nachweismoleküle können sich Nachteile für die in-situ-Hybridisierung ergeben.
Röntgenfilm
Fotosensible Emulsion Anti-DIG Antikörper
DIG-markierte RNA-Sonde Gewebeschnitte 35S-ATP markierte
Oligonukloetid-Sonde
Gewebeschnitte mit hybridisierter Sonde
Gewebeschnitte
Gewebeschnitte mit hybridisierter Sonde
Nicht-radioaktive in situ Hybridisierung Radioaktive in situ Hybridisierung
Immunhistologische Detektion Autoradiographische Detektion
B A
23
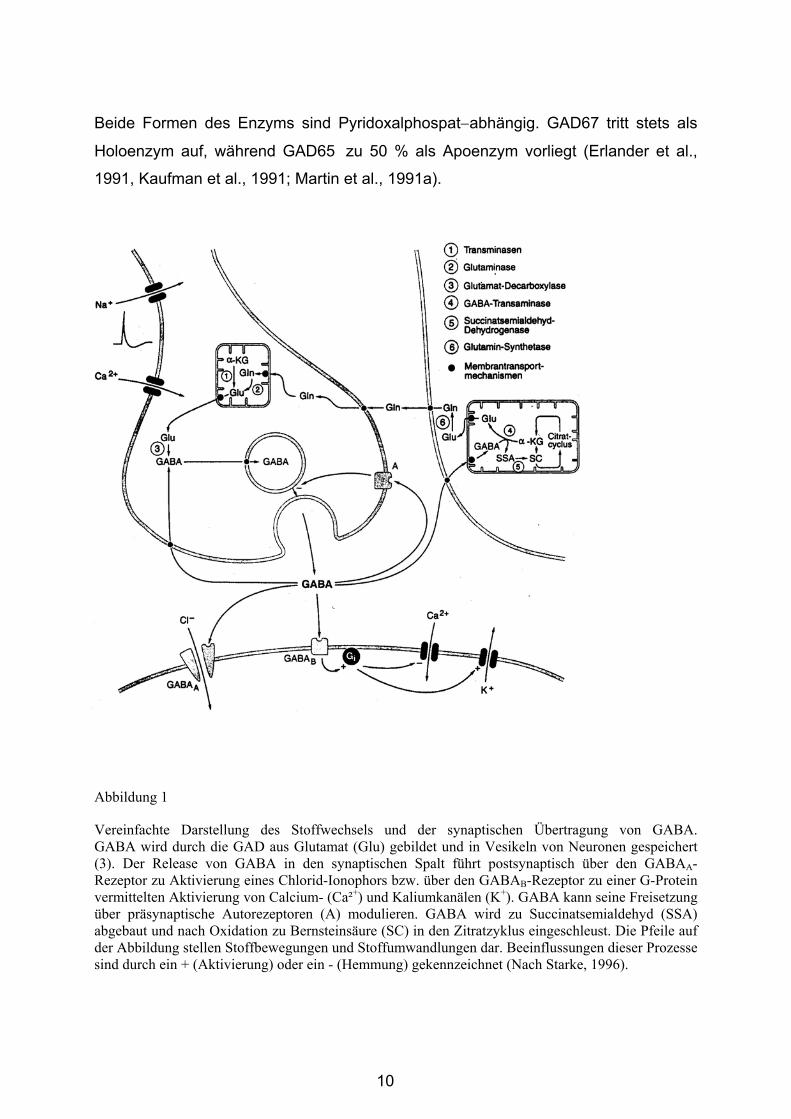
Radioaktiv markierte Sonden weisen vielfach eine höhere Sensitivität gegenüber
nicht-radioaktiv markierten Sonden auf (Angerer und Angerer, 1981). Andererseits
können im Hinblick auf die Lokalisation des Hybridisierungssignals in Zellen bessere
Auflösungen der Markierung unter Verwendung nicht-radioaktiver
Nachweismethoden erzielt werden (Emson, 1993). Zudem erfordern nicht-radioaktive
Nachweismethoden wesentlich geringere Entwicklungszeiten. Nicht-radioaktiv
markierte RNA-Sonden sind über mehrere Monate haltbar, aber sensitiv gegenüber
RNAse Kontamination (Wahle und Beckh, 1992).
Tabelle 1
Übersicht über die Vor- und Nachteile nicht-radioaktiv markierter RNA- und radioaktiv markierter Oligonukleotid-Sonden für die in-situ-Hybridisierung.
nicht-radioaktiv markierte RNA-Sonden
radioaktive markierte Oligonukleotid-Sonden
Vorteile
• hohe Spezifität der RNA-Sonde • kostengünstigere Markierung • Haltbarkeit für mehrere Monate • Ausbildung stabiler RNA/RNA-
Hybride • Signalentwicklung nach Stunden
bis Tagen • direkte Markierung von Zellen • Markierung nicht radioaktiv
• hohe Sensitivität der Markierung • keine Klonierung erforderlich • gute Gewebepenetration aufgrund
geringer Länge • geringe sterische Inhibition der
Hybridisierungsreaktion durch angehängte Markermoleküle
Nachteile
• zeitlich aufwendige Herstellung • Klonierung einer cDNA
erforderlich • hohe Empfindlichkeit der Sonde
gegenüber RNAse-aktivität • schlechtere Gewebepenetration
aufgrund größerer Länge • sterische Inhibition der
Hybridisierungsreaktion durch Markermoleküle möglich
• radioaktive Markierung der Sonden kostenintensiv
• begrenzte Haltbarkeit radioaktiv-markierter Sonden
• Ausbildung weniger stabiler DNA/RNA-Hybride
• Signalentwicklung erfordert mehrere Wochen
• keine direkte Markierung von Zellen • Radioaktivität der Markierung

24
Die Immunhistochemie erlaubt die licht- oder elektronenmikroskopische
Untersuchung von Strukturen wie Neuronen anhand der von ihnen synthetisierten
Neurotransmitter. Neben der Charakterisierung der Morphologie einzelner Zelltypen
und neuroanatomischer Bestimmungen erlaubt diese Methode auch die
Quantifizierung markierter Strukturen. Gibb et al. (1979) legten mit der Entwicklung
spezifischer Antikörper gegen Dopamin-β-Hydroxylase den Grundstein für
immunhistologische Untersuchungen im Bereich der Neurowissenschaften.
Die Immunhistochemie ist eine histologische Technik, die immunologische und
histochemische Methoden miteinander verbindet (Harlow und Lande, 1988; Cuello,
1980). Hierbei werden die nachzuweisenden Strukturen durch spezifische Antikörper
gegen bestimmte Antigene, die in den nachzuweisenden Strukturen vorliegen,
detektiert und durch gekoppelte Farbreaktionen sichtbar gemacht. Die Antikörper
werden entweder als polyklonaler Antikörper von einem dem Antigen exponierten
Tier oder als monoklonaler Antikörper in Hybridomzellen gewonnen. Das Prinzip der
Immunhistochemie basiert heutzutage auf der von Coons und Kaplan (1950)
entwickelten Methode. Das zu untersuchende Gewebe wird mit dem Antikörper
inkubiert, der gegen ein Antigen gerichtet ist, welches in der zu untersuchenden
Struktur lokalisiert ist. Hierdurch entsteht ein Antigen-Antikörper-Komplex an der
Zielstruktur, der direkt oder indirekt nachgewiesen werden kann.
Bei der direkten Methode trägt der primäre Antikörper einen Farbstoff oder ein
Enzym. Der indirekte Nachweis läuft über ein sekundäres Antikörper-System, bei
dem ein zweiter Antikörper den am Antigen haftenden Erstantikörper erkennt. Dies
bietet zum einen den Vorteil, dass nur der sekundäre Antikörper markiert werden
muss, welcher gegen viele verschiedene primäre Antikörper aus einer Spezies
gerichtet sein kann. Zudem lässt sich durch die indirekte Markierung die Sensitivität
des Primärantikörpers erhöhen und gleichzeitig die Hintergrundfärbung reduzieren.
Die am weitesten verbreitete Methode ist die sogenannte Strept-ABC-Technik
(Strept-Avidin-Biotin-Complex), welche von HSU et al. (1981) eingeführt wurde.
Hierbei finden Sekundärantikörper Verwendung, welche an ihren Fc-terminalen
Endstücken Biotin-Moleküle tragen. Da jedes diese Biotin-Moleküle wiederum
mehrere Streptavidin-Moleküle binden können, wird eine Amplifikation erreicht.

25
Mittels an Streptavidin konjugierter Farbstoffe oder Enzyme kann das Antigen
schließlich sichtbar gemacht werden. Häufig gebrauchte Enzyme in der
Immunhistochemie sind die Meerrettichperoxidase (HRP) und die Alkalische
Phosphatase (AP) (Abbildung 4).
Abbildung 4
Prinzip der Immunhistochemie unter Verwendung des Streptavidin-Biotin-Komplexes.
Der primäre (1.) Antikörper bindet an das Antigen im histologischen Schnitt. Der 2. Antikörper bindet an spezifische Erkennungsregionen des ersten Antikörpers. Über Biotin-Moleküle (B), die an den 2. Antikörper gekoppelt sind, werden mehrere Streptavidin-Moleküle (S) gebunden, die mit einem Enzym (E) konjugiert sind, welches eine Farbreaktion katalysiert. Durch dieses Verfahren kann eine Amplifikation des Signals erreicht werden (modifiziert nach Wolf, 1997).
Biotin
B
B E
E
Antigen
Strept- avidin
S
S

26
2.4.2 GABAerge Neurone unter Kontrollbedingungen
Mit der immunhistochemischen Untersuchung des GABAergen Systems wurde
bereits in den 70er Jahren des letzten Jahrhunderts begonnen (Saito et al., 1974;
Roberts 1976). Die frühe immunhistochemische Darstellung GABAerge Neurone
unter Einsatz polyklonaler anti-GABA-Antikörper erforderte Vorbehandlungen des
Gewebes mit Agenzien wie Kolchizin, um eine ausreichende GABA-Konzentration im
Perikaryon durch Hemmung des GAD-Transports zu erreichen (Ribak et al., 1978;
McDonald, 1985; McDonald und Pearson, 1989). Die heute erhältlichen GABA-
Antikörper weisen eine höhere Sensitivität auf, sodass entsprechende
Vorbehandlungen nicht mehr erforderlich sind. Grundsätzlich ist bei der
immunhistochemischen Darstellung GABAerger Neurone der Einsatz von GAD-
Antikörpern vorzuziehen, da diese ausschließlich in Neuronen exprimiert wird.
Subpopulationen GABAerger Neurone werden vermehrt anhand des Nachweises
Calcium-bindender Proteine charakterisiert (Hendry et al., 1989, Sloviter, 1989),
weiterhin können GABA-Rezeptoren sowie ihre Subtypen und GABA-Transporter-
Systeme immunhistochemisch nachgewiesen werden.
Untersuchungen GABAerger Neurone wurden vielfach unter Einsatz der
beschriebenen immunhistochemischen Nachweise durchgeführt (Babb et al., 1988;
Soriano et al., 1989; Woodson et al., 1989). Die Bestimmung der Zellmorphologie
und intracerebralen Verteilung dieser Zellpopulation erfordert die Markierung von
Zellsomata, die vielfach mit immunhistologischen Methoden nicht erreicht werden
konnte. Unter Einsatz der in situ−Hybridisierung gegen GAD-mRNA ist dagegen die
Markierung von Zellkörpern möglich, da die mRNA ausschließlich im Zytoplasma
exprimiert wird (Esclapez et al., 1993, 1994; Houser und Esclapez, 1994). Die in-situ-
Hybridisierung zur Untersuchung der Expression von mRNA für das GABA
synthetisierende Enzym GAD wurde in früheren Studien hauptsächlich mit radioaktiv
markierten Sonden durchgeführt, die gegen die mRNA für GAD67 gerichtet waren
(Wuenschell et al., 1986; Chesselet et al., 1987; Ferraguti et al., 1990; Najlerahim et
al., 1990).

27
Die Identifizierung der für die GAD-Isoformen codierenden Gensequenzen erlaubte
die differenzierte Darstellung (Erlander et al, 1991; Mercugliano et al., 1992;
Feldblum et al., 1993, Esclapez et al., 1993, 1994; Esclapez und Houser, 1994),
wobei zunehmend nicht-radioaktiv markierte Sonden zum Einsatz kamen (Esclapez
et al., 1993, 1994; Esclapez und Houser, 1994). Sowohl mit der radioaktiven als auch
mit der nicht-radioaktiven in-situ-Hybridisierung gegen GAD-mRNA ließen sich in
diesen Studien Zellpopulationen darstellen, die bereits immunhistochemisch als
GABAerge Neurone identifiziert worden waren. Dabei ergaben sich Hinweise auf
regionsabhängige Unterschiede in der Expression des Enzyms (Ferraguti et al.,
1990), welche sich in weiterführende Studien unter Verwendung von Sonden gegen
die mRNA für beide Isoformen der GAD bestätigten (Erlander et al., 1991;
Mercugliano et al., 1992; Feldblum et al., 1993). Diese Arbeitsgruppen wiesen nach,
dass die mRNA für GAD67 in den meisten Hirnregionen stärker exprimiert wird als
die GAD65-mRNA, so etwa in den Basalganglien und im Cortex. Weiterhin stellten
sie Unterschiede zwischen der Dichte und der Anzahl markierter Neurone in
einzelnen Hirnregionen fest.
Die basale Expression von GAD-mRNA ist in den genannten Studien für die
Substantia nigra pars reticulata, den Hippocampus sowie die Amygdala beschrieben
worden. Der PC, dem zunehmende Bedeutung in der Generierung und Propagierung
epileptischer Aktivität im limbischen System zugesprochen wird, ist dagegen bis
heute in dieser Hinsicht nur unzureichend charakterisiert. Die Informationen über die
Expression von GAD-mRNA in dieser Hirnregion beschränkten sich auf
Übersichtsarbeiten unter Verwendung radioaktiv markierter Sonden zum Nachweis
von GAD67-mRNA (Ferraguti et al., 1990; Najlerahim et al., 1990). Zudem wurde in
einem Teil der Arbeiten die durchschnittliche Expression der GAD-mRNA für
bestimmte Hirnregionen nur mittels Röntgenfilm und nicht auf zellulärer Ebene
beurteilt. Ferraguti et al. (1990) beschrieben in diesem Zusammenhang für die tiefen
Schichten des PC eine mittlere Intensität und den EN eine schwache Intensität der
optischen Dichte des Expressionssignals im Röntgenfilm.

28
Die Populationen GABAerger Neurone des PC wurden in der Vergangenheit
immunhistologisch zunehmend detaillierter charakterisiert (Haberly et al., 1987;
Lehmann et al., 1996; Löscher et al., 1998; Ekstrand et al., 2001), wobei sich
teilweise widersprüchliche Ergebnisse hinsichtlich der Zahl markierter Zellen in den
einzelnen Schichten herausstellten. Löscher et al. (1998) beschrieben die höchste
Zahl und Dichte GABA-immunreaktiver Neurone für die sehr zellreiche Schicht II des
PC weiblicher Wistar-Ratten. Im zentralen PC ergab sich zudem eine signifikant
höhere Dichte GABAerger Zellen im Vergleich zu den anterioren und posterioren
Anteilen der Struktur. Der Nachweis von GAD in Neuronen des PC mittels
Immunhistochemie erbrachte die höchste Zahl immunreaktiver Zellen ebenfalls in der
Schicht II des PC, die Zahl GAD-immunreaktiver Zellen fiel im Vergleich zum GABA-
Nachweis aber niedriger aus (Löscher et al., 1998). Im Gegensatz hierzu stehen die
Ergebnisse der Arbeitsgruppe um Haberly (1987), von denen die höchste Dichte
GABA- und GAD-immunreaktiver Zellen in der Schicht III des PC sowie im
angrenzenden EN des Opposums quantifiziert wurde. Auch Mugnaini und Oertel
(1985) beschrieben für die Schicht II des PC der Ratte einen Anteil GAD-
immunreaktiver Zellen von unter 5 %, während diese Neurone in der Schicht III
weniger als 15 % der gesamten Neuronenpopulation ausmachten.
Auch für andere Regionen des limbischen Systems sind unterschiedliche Ergebnisse
hinsichtlich der Darstellung GABAerger Neurone in Abhängigkeit der gewählten
Nachweismethode beschrieben worden. In den glutamatergen Körnerzellen des
Stratum granulosum des Gyrus dentatus wurde unter Kontrollbedingungen eine
GABA-Immunreaktivität in Zellsomata (Lehmann et al., 1996; Sloviter et al., 1996)
sowie den Ausläufern dieser Zellen, den Moosfasern, festgestellt (Sloviter et al.,
1996). Anderer Arbeitgruppen zeigten dagegen keine Immunreaktivität für GABA in
diesen Strukturen (Ding et al., 1998). Mit Immunhistochemie für GAD konnten in
einigen Untersuchungen weder Körnerzellen noch deren Ausläufer markiert werden
(Mugnaini und Oertel, 1985; Lehmann et al., 1996; Esclapez und Houser, 1999).
Sloviter et al. (1996) beschrieben in diesem Zusammenhang eine Immunreaktivität
für GAD67 aber nicht für GAD65 in Moosfasern. In den Somata von Körnerzellen ließ
sich unter Kontrollbedingungen keine der beiden Isoformen immunhistochemisch
darstellen.

29
Die Literatur beschreibt bezüglich der Dichte und Zahl GABAerger Neurone nach
Untersuchungen mittels in-situ-Hybridisierung für GAD-mRNA und
Immunhistochemie für GAD oder GABA unterschiedliche Ergebnisse. Ferraguti et al.
(1990) stellten eine schlechte Korrelation zwischen den Ergebnissen ihrer Studie zur
Expression der mRNA für GAD67 und den Angaben von Mugnaini und Oertel (1985)
hinsichtlich der regionalen Verteilung GAD-immunreaktiver Neurone im ZNS fest.
Diese Arbeitsgruppe konnte im Striatum, dessen Neurone zum überwiegenden Teil
GAD-immunreaktiv sind, nur eine geringe Expression von GAD67-mRNA
nachweisen. Esclapez und Mitarbeiter (1994) beschrieben in den meisten der von
ihnen untersuchten Hirnregionen dagegen eine Übereinstimmung der Expression
von GAD-mRNA und dem immunhistochemischen Nachweis der GAD-Isoformen.
Eine Ausnahme bildete in diesem Zusammenhang der Globus pallidus. In diesem
Teil der Basalganglien waren Zellkörper für GAD65-mRNA markiert, während sich
nur die Terminalen für das zugehörige Protein anfärbten. In den Körnerzellen des
Gyrus dentatus ließ sich mit der in-situ-Hybridisierung keine GAD-mRNA
nachweisen, es wurde jedoch eine verstärkte Hintergrundfärbung für GAD67-mRNA
in dieser Zellschicht beschrieben (Houser und Esclapez, 1994).

30
2.4.3 GABAerge Neurone in Tiermodellen der Temporallappenepilepsie
Die GABA-Hypothese der Epilepsie (Burnham, 1989; Löscher, 1989) postuliert einen
Verlust GABA-vermittelter Inhibition im Bereich des Fokus und assoziierter
Strukturen infolge wiederholter paroxysmaler neuronaler Aktivität. Ein daraus
resultierendes Übergewicht exzitatorischer Einflüsse wird als Vorraussetzung für die
Epileptogenese angenommen.
Ein Verlust GABAerger Interneurone konnte im hippocampalen Gewebe von
Epileptikern (Houser et al., 1990) nachgewiesen werden. Basierend auf diesen
Erkenntnissen wurden Untersuchungen in Tiermodellen der Epilepsie hinsichtlich
entsprechender Veränderungen in der Anzahl GABAerger Neurone durchgeführt.
Obenaus et al. (1993) beschrieben einen Verlust GABAerger Neurone, welche
GAD65-mRNA exprimierten, im Hilus des Gyrus dentatus nach Pilocarpin-
induziertem SE. Die Expression von GAD67-mRNA wurde in dieser Studie nicht
untersucht. Eine Woche nach Applikation von Pilocarpin waren GAD65-mRNA-
positive Neurone im Hilus um 44 % und nach 2 Wochen um 39 % reduziert. In der
Körnerzell- sowie der Molekularschicht des Gyrus dentatus zeigten sich GABAerge
Neurone in ihrer Zahl dagegen nicht vermindert.
Unter Verwendung des Kainat-Modells mit spontanen Anfällen konnte gezeigt
werden, dass Somatostatin-immunreaktive Neurone mit 83 % den größten Anteil
untergehender GABAerger Neurone im Hilus des Gyrus dentatus darstellen, welche
GAD67-mRNA exprimieren (Buckmaster und Jongen-Relo, 1999).
Im Amygdala-Kindling-Modell zeigte sich bei voll gekindelten Ratten, die einer
40tägigen Stimulationspause unterzogen wurden, eine signifikante Reduktion GABA-
immunreaktiver Neurone im Bereich des zentralen PC (Lehmann et al., 1998). Diese
war auf die ipsilaterale Seite beschränkt und betrug abhängig von der untersuchten
Schnittebene zwischen 17 % und 26 %. In der basolateralen Amygdala erfolgte
dagegen eine beidseitige Abnahme GABA-immunreaktiver Neurone infolge des
Kindlings, die mit durchschnittlich 36 % höher ausfiel als im PC. Für den
Hippocampus und den Gyrus dentatus wurden dagegen keine signifikanten
Veränderungen der Zahl GABA-positiver Zellen festgestellt (Lehmann et al., 1996).

31
Der Einfluss des Amygdala-Kindlings auf die Dichte GABAerge Neurone wurde auch
in anderen Studien deutlich. 2-6 Monate nach Abschluss des Kindlingprozesses war
eine Abnahme GABA-immunreaktiver Neurone in der contralateralen lateralen und
basolateralen Amygdala von bis zu 62 % festzustellen (Callahan et al., 1991).
Tuunanen et al. (1997) untersuchten die Auswirkung des Amygdala-Kindlings auf die
Anzahl GABA- und Somatostatin-immunreaktiver Neurone in den Kernen der
Amygdala und im Hilus des Gyrus dentatus. Die Untersuchungen wurden 6 Monate
nach 5 gekindelten generalisierten Anfällen durchgeführt. Während die Zahl
Somatostatin-immunreaktiver Neurone in der Amygdala durch Kindling verringert
wurde, hatten die Stimulationen keine Auswirkungen auf die Dichte GABA-
immunreaktiver Neurone. Im Hilus des Gyrus dentatus zeigten sich ebenfalls keine
Unterschiede in der Dichte GABAerger Neurone.
Durch hippocampales Kindling mittels Stimulation der Schaffer-Collateralen konnte
im Anschluss an eine Stimulationspause von 24 Tagen eine 35 %ige Reduktion
GABA-immunreaktiver Neurone in der Region CA1 ipsilateral zur
Stimulationselektrode nachgewiesen werden (Kamphuis et al., 1986). In einer
weiterführenden Untersuchung (Kamphuis et al., 1987) zeigten Tiere 22-24 h nach 6
stimulierten Nachentladungen ipsilateral eine 10 %ige Zunahme GABA-
immunreaktiver Zellen, während contralateral eine 17 %ige Abnahme zu verzeichnen
war. Nach 14 stimulierten Nachentladungen zeigte sich bei Ratten ebenfalls nach
22-24 h eine 38 %ige Zunahme GABA-positiver Neurone, voll gekindelte Tiere
wiesen dagegen nur contralateral eine Zunahme auf. Die Quantifizierung
Parvalbumin-positver Zellen, einer weiteren Subpopulation GABAerger Interneurone,
ergab höhere Zellzahlen bei stimulierten und voll gekindelten Tieren, die sofort nach
Abschluss der Versuche perfundiert wurden. Nach einer Stimulationspause von 31
Tagen waren keine Veränderungen für Parvalbumin-positive Zellen nachzuweisen,
während sich GABA-positive aber Parvalbumin-negative Zellen in der Region CA1
um 50 % reduziert zeigten (Kamphuis et al., 1989).

32
Anhand der aufgeführten Studien wurde deutlich, dass Populationen GABAerger
Neurone in verschiedenen Gehirnregionen unterschiedlich empfindlich auf
Krampfaktivität reagieren (Houser und Esclapez, 1996). Aus diesen Erkenntnissen
resultierten Untersuchungen, die sich zunehmend mit Alterationen des GABA-
Metabolismus überlebender Zellen in Tiermodellen der Temporallappenepilepsie
beschäftigten.
Schwarzer und Sperk (1995) beschrieben im Kainat-Modell eine um mehr als 300 %
gesteigerte Expression von GAD67-mRNA in Körnerzellen des Gyrus dentatus 6 h
nach Applikation von Kainat. Dieser Effekt erwies sich als transient und reduzierte
sich nach 24 h auf etwa 150 % im Vergleich zur Kontrolle, war aber mit einer
verstärkten GAD-Immunreaktivität in Moosfasern korreliert. Hingegen ließ sich zu
keinem Zeitpunkt der Untersuchung die mRNA für GAD65 in Körnerzellen
nachweisen. In GABAergen Zellen des Gyrus dentatus wurde nach 7 Tagen eine
ansteigende Expression von GAD-mRNA für beide Isoformen beobachtet, welche
besonders stark in Typ 1 Korbzellen am Übergang der Körnerzellschicht zum Hilus
des Gyrus dentatus ausfiel. Diese Beobachtung wurde von den Autoren als Ausdruck
einer vermehrten GABAergen Transmission in Reaktion auf die Krampfaktivität
gewertet.
Analoge Ergebnisse lieferte eine Studie der Arbeitsgruppe um Ding (1998) im Kainat-
Modell. Sie beschrieben ebenfalls die transiente Expression von GAD67-mRNA in
Körnerzellen des Gyrus dentatus 4 und 6 h nach Applikation, während die mRNA für
GAD65 nicht nachgewiesen werden konnte. Eine verstärkte GABA-Immunreaktivität
in Moosfasern konnte im Hilus und in der Region CA3 des Hippocampus nach 6-8 h
beobachtet werden. Diese war zeitlich verbunden mit einem Anstieg von
extrazellulärem GABA in der Region CA3 und wurde von den Autoren als Folge der
Freisetzung von GABA aus den Moosfasern gedeutet. Ein Unterschied in der Anzahl
und Intensität der Anfärbung GABAerger Interneurone im Hippocampus und im
Gyrus dentatus konnte im Untersuchungszeitraum nicht festgestellt werden.

33
Esclapez und Houser (1999) beschrieben eine verstärkte Expression von GAD65-
und GAD67-mRNA in GABAergen Neuronen der Hippocampusformation von Ratten
mit spontanen Anfällen infolge eines Pilocarpin-induzierten SE. Die Expression der
mRNA für GAD65 war um 76 % im Vergleich zur Kontrolle erhöht, für die GAD67-
mRNA ergab sich eine Steigerung von 52 %. Parallel durchgeführte
immunhistologische Nachweise erbrachten eine verstärkte Immunreaktivität für beide
Isoformen in Terminalen, welche besonders intensiv für GAD65 ausfiel.
Die Aktivierung des lateralen Enthorinalen Cortex durch Applikation von Quisqualat
erhöhte die Anzahl GAD67-mRNA-positiver Neurone im Stratum oriens, Stratum
radiatum sowie Stratum pyramidale in der Region CA1 des Hippocampus
(Falkenberg et al., 1997). Die Induktion von GAD-mRNA wurde von einem Anstieg
extrazellulärer GABA in CA1 begleitet, wobei die höchste Dosis Quisqualat zu einem
6fach höheren Gehalt an GABA in dieser Region führte. Die Expression von GAD65
wurde in dieser Studie nicht untersucht.
Sloviter et al. (1996) konnten durch Stimulation des Perforant Path epileptiforme
Entladungen in Körnerzellen des Gyrus dentatus und Pyramidenzellen des
Hippocampus auslösen. Dies führte zu einer Induktion von Immunreaktivität für
GAD65 und GAD67 sowie GABA selektiv in Körnerzellen. Dagegen war die Anzahl
GAD- bzw. GABA-immunreaktiver Pyramidenzellen und Neurone im Hilus des Gyrus
dentatus nicht verändert. Der Nachweis von GAD65- und GAD67-mRNA in
Körnerzellen erwies sich als abhängig von der Dauer der Stimulation. Während
GAD67-mRNA bereits nach sechsstündiger Stimulation nachgewiesen werden
konnte, erforderte die Induktion der GAD65-mRNA Stimulationen über 24 h. Bei
solchen Tieren war GAD67-mRNA noch bis zu 48 h nachzuweisen, GAD65-mRNA
wurde dagegen nicht mehr in detektierbarer Menge exprimiert. Immunhistochemisch
waren beide Enzyme jedoch noch bis zu 4 Tage nachzuweisen.
Babb et al. (1989) beschrieben eine reduzierte Immunreaktivität in GAD-
immunreaktiven Neuronen in der Körnerzellschicht des Gyrus dentatus als Reaktion
auf Kindling im Entorhinalen Cortex. Dieser Effekt erwies sich als transient und war
nach 3 und 7 Tagen nicht mehr nachweisbar. In der CA3-Region sowie der
Substantia nigra trat kein vergleichbares Geschehen auf.

34
2.5 Fragestellung
Veränderungen der GABAergen Inhibition werden als eine Ursache für die
Entstehung und Ausbreitung epileptischer Aktivität im ZNS angesehen (Houser und
Esclapez, 1996). Verluste GABAerger Neurone sowie Alterationen des GABA-
Metabolismus überlebender Neurone in Tiermodellen der Temporallappenepilepsie
unterstützen diese Hypothese. Allerdings sind die Ergebnisse aufgrund der
unterschiedlichen Aussagekraft der angewendeten histologischen
Nachweismethoden vielfach nicht direkt vergleichbar.
Der PC ist zunehmend von Interesse im Hinblick auf die Entstehung und Ausbreitung
epileptischer Aktivität limbischen Ursprungs (Löscher und Ebert, 1996b). GABAerge
Neurone sind hinsichtlich einer Beteiligung inhibitorischer Mechanismen in diesem
Zusammenhang immunhistochemisch für GABA und GAD charakterisiert worden
(Löscher et al., 1998), die Verhältnisse von GAD auf der Ebene der mRNA-
Expression sind dagegen noch unzureichend dokumentiert.
Im Rahmen dieser Arbeit soll daher die Expression der mRNA für die beiden
Isoformen der GAD mittels in-situ-Hybridisierung unter Kontrollbedingungen
untersucht und mit dem immunhistochemischen Nachweisen von GAD und GABA
verglichen werden. Dazu wurden zwei unterschiedliche Protokolle zur Durchführung
der situ Hybridisierung etabliert.
Im Kainat-Modell der Temporallappenepilepsie konnte bereits eine verstärkte
Expression von GAD67-mRNA in Körnerzellen des Gyrus dentatus nachgewiesen
werden (Schwarzer und Sperk, 1995; Ding et al., 1998). Die Auswirkungen akuter
Krampfaktivität auf den Metabolismus GABAerger Neurone des PC werden im
Kainat- und im Pilocarpin-Modell nach Induktion eines selbsterhaltenden SE mit in-
situ-Hybridisierung und Immunhistochemie untersucht.
Im Amygdala-Kindling-Modell ist eine beständige Abnahme GABA-immunreaktiver
Neurone im PC ipsilateral zur Stimulationselektrode beschrieben worden (Lehmann
et al., 1998). In diesem Modell sollen mögliche chronische Veränderungen der
Expression von GAD-mRNA in GABAergen Neuronen untersucht und die
Reproduzierbarkeit der beschriebenen Neuronenverluste mittels in-situ-
Hybridisierung überprüft werden.

35
Neben dem PC erstrecken sich die Untersuchungen mit der Amygdala und der
Hippocampusformation auf weitere bedeutende Schlüsselstrukturen im
Zusammenhang mit der Entstehung von Temporallappenepilepsie. Die Substantia
nigra pars reticulata, der eine besonderer Rolle bei der Generalisation von
epileptischer Aktivität limbischen Ursprungs zukommt, bildet einen weiteren
Schwerpunkt der Untersuchungen.

36
3 Material und Methoden
3.1 Versuchstiere
Für alle Versuche fanden ausschließlich weibliche Wistar-Ratten (Versuchstierzucht
Harlan-Winkelmann, Borchen) Verwendung. Die Tiere hatten bei Ihrer Ankunft ein
Gewicht von 190-220 g. Ratten für Dauerimplantationsexperimente wurden sofort
nach der Ankunft einzeln in Makrolonkäfige vom Typ III gesetzt, für sonstige
Versuche erfolgte die Unterbringung in Gruppen zu 5-6 Tieren in Gruppenkäfigen
vom Typ IV. Als Einstreu dienten Sägespäne (Altromin; Fa. Altrogge, Lage).
Im betreffenden Stall wurden ausschließlich weibliche Ratten gehalten. Während der
gesamten Versuchszeit erfolgte die Haltung unter kontrollierten Umweltbedingungen
bei einer Umgebungstemperatur von 22-24°C und einer Luftfeuchtigkeit von
50-60 %. Der Hell-Dunkel-Zyklus von 12 Stunden wurde von einem Lichtprogramm
gesteuert; die Hellphase dauerte von 6 bis 18 Uhr MEZ. Es bestand freier Zugang zu
Futterpellets (Altromin 1324-Standarddiät, Altromin, Lage) und Leitungswasser in
Makrolonflaschen.
Einmal wöchentlich wurden die Tiere in Käfige mit frischer Einstreu umgesetzt sowie
Trinkwasser und Futterpellets erneuert. Das Umsetzen erfolgte jeweils in
genügendem zeitlichen Abstand zu den Versuchen, um eine Beeinflussung der Tiere
zu vermeiden. Die Ratten wurden nach Anlieferung mindestens 7 Tage an die neue
Umgebung gewöhnt. Zugang zum Tierstall hatten nur Experimentatoren und
zuständige Tierpfleger.

37
3.2 Histologie
Im folgenden wird die in-situ-Hybridisierung mit Digoxigenin (DIG)-markierten RNA-
Sonden als nicht-radioaktive, die Hybridisierung mittels 35Schwefel-radioaktiv-
markierter Oligonukleotidsonden als radioaktive in-situ-Hybridisierung bezeichnet.
Sämtliche für die Histologie verwendeten Lösungen sind im Anhang aufgeführt.
3.2.1 Gehirnpräparation und Anfertigung von Gewebeschnitten
3.2.1.1 Nicht-radioaktive in-situ-Hybridisierung, Immunhistologie und Elektrodenlokalisation
Zur Gewinnung der Gehirne nach Abschluss der Versuche wurden die Ratten
intraperitoneal mit 540 mg/kg Chloralhydrat tief narkotisiert, bis keine Reflexe mehr
auftraten. Für die transcardiale Perfusion mit einer Fixanslösung erfolgte zunächst
die Eröffnung der Bauch- und Brusthöhle. Nach Freilegen des Herzens und
Eröffnung des Perikards wurde eine Knopfkanüle durch die linke Herzkammer bis in
die Aorta vorgeschoben und das rechte Herzohr eröffnet, um einen Abfluss von Blut
und Perfusat aus dem Körperkreislauf zu ermöglichen. Durch einen Dreiwegehahn
konnte nun über die angeschlossene Knopfkanüle wahlweise eine Spülflüssigkeit
oder einer Fixanzlösung eingeleitet werden.
Mittels Schwerkraftperfusion wurde zunächst für etwa zwei Minuten das Blut des
Körperkreislaufes durch Spülung mit 0,01 M phosphatgepufferte 0,9 % NaCl-Lösung
(PBS, pH 7,4) entfernt. Anschließend wurde die Perfusion mit 250 ml einer 4 %igen
0,1 M phosphatgepufferten Formaldehydlösung (pH 7,4) für 15 min fortgesetzt.
Die fixierten Gehirne verblieben für weitere 30 min in situ im Tierkörper bei +4 oC. Bei
implantierten Tieren konnte der Elektrodenaufbau inklusive Elektrode entfernt, die
Schädelhöhle vorsichtig eröffnet und das Gehirn nach Entfernen der Hirnhäute
entnommen werden. Das so gewonnene Untersuchungsmaterial wurde nochmals für
30 min in 30 ml einer 4 %igen 0,1 M phosphatgepufferten Formaldehydlösung (pH
7,4) bei Raumtemperatur nachfixiert und anschließend dreimal in jeweils 50 ml 0,1 M
Phosphatpuffer gespült.

38
Zur Kryoprotektion wurde das Gehirn in 30 ml einer 0,1 M phosphatgepufferten 30
%igen Saccharoselösung (pH 7,4) für 48-72 h bei +4 oC gelagert. Nachdem die
Gehirne in der Saccharoselösung abgesunken waren, erfolgte die Aufarbeitung. Mit
Hilfe eines Gefriermikrotoms (Frigomobil Modell 1205, Fa. Jung, Heidelberg) konnten
30 µm dicke Transversalschnitte der zu untersuchenden Hirnregionen angefertigt
werden. Der Umfang der Schnittserien beinhaltete Strukturen zwischen +2,2 mm bis
-6,8 mm von Bregma, womit der größte Teil des limbischen Systems sowie die
Substantia nigra erfasst wurde. Die so gewonnenen Schnitte wurden in acht Serien
in 0,1 M Phosphatpuffer aufgefangen, so dass sich in jeder Serie Schnitte mit einem
Abstand von ca. 240 µm befanden.
Die erste Serie jedes Tieres diente generell zur Beurteilung der zu untersuchenden
Hirnregionen, bei Ratten mit implantierten Elektroden wurden sie zusätzlich zur
Überprüfung der Elektrodenlokalisation herangezogen. Von den weiteren Serien
wurden solche für die Immunhistologie sofort im Anschluss an das Schneiden dem
Nachweisprotokoll unterzogen, während die Schnittserien für die nicht-radioaktive in-
situ-Hybridisierung bis zur Weiterverarbeitung in einer kryoprotektiven Lösung (siehe
8.4.1.1) bei -20 °C gelagert wurden.
3.2.1.2 Radioaktive in-situ-Hybridisierung
Für die radioaktive in-situ-Hybridisierung mit Oligonukleotidsonden wurden die
Ratten mit einer Guillotine (Eigenbau, Institut für Pharmakologie) dekapitiert, der
Schädel mittels einer Zange geöffnet und die unfixierten Gehirne vorsichtig aus der
Schädelkalotte präpariert. Nach Entfernung des Kleinhirns konnte das Vorderhirn mit
der kaudalen Seite auf eine runde Metallscheibe aufgebracht und 1-2 min direkt in
Isopentan als Gefriermedium fixiert werden. Das Isopentan war zuvor mittels
flüssigem Stickstoff auf eine Temperatur von etwa -80 °C gekühlt worden.
Nach Abschluss der Fixation wurde das Gefriermedium für 2 min bei
Raumtemperatur und für weitere 60 min bei -80 °C aus dem Gehirngewebe
abgedampft, da Rückstände des Isopentans die Struktur des Hirngewebes
schädigen konnten. Bis zur Weiterverarbeitung wurden die Gehirne bei -80 °C
gelagert.

39
Die Anfertigung von 20 µm dicken Gewebeschnitten erfolgte mit Hilfe eines
Kryostaten (Leica, Wezlar). Die angefertigten Schnitte wurden direkt auf doppelt mit
Gelatine beschichtete Objektträger aufgenommen, was die Aufarbeitung der
einzelnen Gehirne in einer einzigen Serie ermöglichte. Bis zur weiteren Verarbeitung
wurden die Objektträger mit den Gewebeserien bei -20 °C verschlossen gelagert.
3.2.2 Thioninfärbung
Zur Beurteilung von Neurodegeneration und Überprüfung der korrekten
Elektrodenlokalisation in der basolateralen Amygdala wurde eine Schnittserie jedes
Tieres durch eine Nissl-Färbung mit Thionin zur Darstellung von Neuronen gefärbt.
Die in anterior-posteriorer Reihenfolge sortierten Schnitte wurden mit Hilfe von
flüssiger Gelatine-Chromalaun-Lösung auf entfettete Objektträger aufgezogen. Nach
Abtrocknen erfolgte die Rehydratisierung über eine absteigende Alkoholreihe (jeweils
drei min in 96, 70 und 50 %igem Alkohol sowie destilliertem Wasser) und die
Färbung für 70-90 s in einer wässrigen Thionin-Lösung.
Über eine aufsteigende Alkoholreihe wurden die Schnitte wieder entwässert. Die
ausreichende Konservierung der Schnitte erfolgte im Anschluss für jeweils drei min in
einer Terpineol-Xylolersatzmedium-Mischung und zweimal in Xylolersatzmedium-
Lösung. Direkt im Anschluss wurden die Schnitte auf dem Objektträger mit einem
Deckgläschen und Entellan (Firma Merck AG, Darmstadt) eingedeckt.

40
3.3 Vergleichende Darstellung GABAerger Neurone mittels in-situ-Hybridisierung und Immunhistochemie
Die Vorversuche zur vergleichenden Untersuchung GABAerger Neurone mittels in-
situ-Hybridisierung und Immunhistochemie umfassten folgende Schritte:
1. Etablierung eines Protokolls für die nicht-radioaktive sowie die radioaktive in-situ-
Hybridisierung zum Nachweis der mRNA von Isoformen der GAD in GABAergen
Neuronen (Kap. 3.3.2, 3.3.3)
2. Versuche zur Validierung und Auswahl des geeigneten in-situ-Hybridisierungs-
Protokolls (Kap. 3.3.4)
3. Versuche zur Kombination von in-situ-Hybridisierung mit Immunhistochemie zum
Nachweis von GAD und GABA in Schnittserien eines Tieres (Kap. 3.3.5, 3.3.6)
3.3.1 Prinzip der in-situ-Hybridisierung
Die in-situ-Hybridisierung dient zur Darstellung von Nukleinsäuren am Ort ihrer
Expression in Geweben („in situ“) mittels markierter Nukleinsäurensonden. Zum
besseren Verständnis der folgenden Kapitel sind die einzelnen Schritte der
Protokolle eingangs dargestellt.
A. Die Durchführung der Protokolle erforderte unterschiedliche Sonden:
Für die nicht-radioaktive in-situ-Hybridisierung wurden Digoxigenin (DIG)-markierte
RNA-Sonden durch in-vitro-Transkription an cDNA von GAD65 und GAD67
synthetisiert. Die radioaktive in-situ-Hybridisierung erfolgte unter Einsatz kommerziell
hergestellter DNA-Sonden, welche radioaktiv markiert wurden.

41
B. Die Protokolle zur in-situ-Hybridisierung umfassten 4 grundsätzliche Schritte:
1. Vorbehandlung der Gewebeschnitte
Durch die Schritte der Vorbehandlung wurde eine bessere Penetration der Sonde
an den Ort der Hybridisierung erreicht. Zu diesem Zweck wurde das zu
untersuchenden Gewebes enzymatisch mit Proteasen bzw. Agenzien zur
Perforation von Zellenmembranen behandelt.
2. Hybridisierung
Die Hybridisierung ermöglichte die Ausbildung von Hybriden aus Sonden und
nachzuweisenden Nukleotidsequenzen in Zellen. Die für den Vorgang
erforderlichen Bedingungen waren von der Art und der Länge der Sonde
abhängig und bestimmten die Zusammensetzung des Hybridisierungspuffer
sowie die Hybridisierungstemperatur.
3. Nachbehandlung
Die Nachbehandlung diente der Erhöhung der Spezifität des Signals in Zellen.
Sie beinhaltete die Schaffung von stringenten Bedingungen, die dem Anhaften
der Sonde besonders an Nukleotidmolekülen falscher Sequenz entgegenwirkten.
Die Stringenz ließ sich bei der nicht-radioaktiven in-situ-Hybridisierung durch den
Einsatz von Kontrollsonden in Parallelexperimenten prüfen. Ein Entfernen
unspezifisch gebundener Sonden und Kontrollsonden konnte bei der Verwendung
von RNA-Sonden durch enzymatischen Abbau erreicht werden. Weiterhin kamen
bei den Versuchen Reagenzien (z.B. Formamid) oder Lösungsreihen mit
abgestuften Salzkonzentrationen bei von der Art und Länge der Nachweissonde
abhängigen Temperaturen zum Einsatz.
4. Detektion der Hybride in situ
Der Nachweis spezifisch im Gewebe gebundener Sonden erfolgte bei der nicht-
radioaktiven in-situ-Hybridisierung mittels eines Antikörper-Alkalische-
Phosphatase-Konjugates mit anschließender Farbreaktion zur Darstellung
positiver Zellen. Bei der radioaktiven in-situ-Hybridisierung wurde das Signal in
Form reduzierter Silberkörner im Röntgenfilm oder in einer Fotoemulsion auf den
Objektträgern sichtbar.

42
3.3.2 Etablierung der nicht-radioaktiven in-situ-Hybridisierung
Die Etablierung der nicht-radioaktiven in-situ-Hybridisierung erfolgte anhand von
Protokollen der Arbeitsgruppe von Prof. C. Houser, Department of Biology, University
of California, Los Angeles, USA, die ebenfalls die Plasmidvektoren (Bluescript®, Fa.
Stratagene, San Diego, USA) mit der cDNA zur Synthese der RNA-Sonden zur
Verfügung stellte.
3.3.2.1 Herstellung Digoxigenin-markierter RNA-Sonden
Die Durchführung der Versuche zur nicht-radioaktiven in-situ-Hybridisierung setzte
die Synthese von RNA-Sonden zur Detektion der Zielsequenzen sowie
Kontrollsonden voraus. Der Herstellungsprozess gliederte sich dabei in folgende
Arbeitsschritte:
1. Transformation von Bakterienzellen und Vermehrung von Plasmidvektoren
(Kap. 3.3.2.1.1 und 3.3.2.1.2)
2. Präparation der Plasmidvektoren aus den Trägerorganismen (Kap. 3.3.2.1.3)
3. Restriktion der Plasmidvektoren (Kap. 3.3.2.1.4)
4. Herstellung DIG-markierter RNA-Sonden mittels in-vitro-Transkription und
Alkalischer Hydrolyse (Kap. 3.3.2.1.5)
5. Bestimmung der Konzentration von RNA-Sonden im Dot-Blot (Kap. 3.3.2.1.6)
Das gesamte Aqua bidest. für die Versuche im Zusammenhang mit der nicht-
radioaktiven in-situ-Hybridisierung wurde mit dem RNAse-inhibitor
Diethylpyrocarbonat (DEPC) behandelt. Im folgenden wird für dieses Wasser der
Begriff Aqua bidest./DEPC verwendet.

43
3.3.2.1.1 Transformation von Bakterien
Für die Transformation kamen apathogene E. coli vom Stamm dh5α (Fa. Invitrogen,
San Diego, USA) zum Einsatz. Als Vektoren für die cDNA für GAD Isoformen wurden
Trägerplasmide (Bluescript®, Fa. Stratagene, San Diego, USA) eingesetzt, welche
den Empfängerbakterien eine Ampicillinresistenz verliehen, die eine Selektion positiv
transformierter Zellen mittels dieses Antibiotikums ermöglichte.
Für die Transformation wurden ca. 500 ng Plasmid mit 50 µl Bakterienzellen
suspendiert und 30 min auf Eis inkubiert. Darauf wurde das Gemisch 1 min bei 42 °C
erwärmt, um die Aufnahme der Vektoren durch die Bakterien zu erreichen und erneut
5 min gekühlt. Nach Zugabe von 1 ml Luria-Bertani (LB)-Medium (Fa. Sigma,
Deisenhofen) erfolgte die Inkubation bei 37 °C für 45 min. Das durch die
Zentrifugation entstandene Bakterienpellet wurde resuspendiert, mittels einer Pipette
auf einer Selektivagarplatte mit 100 mg/ml Ampicillin ausgestrichen und für 12 h bei
37 °C inkubiert. Durch diese Vorgehensweise wurde sichergestellt, dass sich nur
Bakterien vermehren konnten, die das Plasmid mit der cDNA für GAD aufgenommen
hatten. Die gewachsenen Kolonien konnten auf dem Nährboden ohne Verlust der
Vermehrungsfähigkeit bis zu 4 Wochen bei 4 °C gelagert werden. Für längere
Zeiträume erfolgte die Herstellung von Bakteriendauerkulturen, welche bei -80 °C
gelagert wurden (Kap. 3.3.2.1.2).
3.3.2.1.2 Anzucht von Bakterien
Die Gewinnung von cDNA in einer ausreichenden Menge machte die Vermehrung
der Plasmid-tragenden Bakterien erforderlich. Hierfür wurde eine einzelne
Bakterienkolonie von einer Ampicillin-Selektivagarplatte in 3 ml LB-Medium
überimpft, wobei das Wachstum anderer unerwünschter Keime durch Zugabe von
6 µl einer Ampicillin-Lösung (Fa. Sigma, Deisenhofen) verhindert wurde. Diese
„Startkultur“ wurde 8 h bei 37° C unter starkem Rütteln inkubiert. Nach Ende der
Inkubationszeit konnten 50 µl der Startkultur in 12,5 ml LB-Medium mit 25 µl
Ampicillin-Lösung für mindestens 12 h bei 37° C auf dem Rüttler inkubiert werden.
Zur Herstellung von Bakteriendauerkulturen wurde 1 ml einer Startkultur mit 375 µl
Glycerin (Fa. Merck, Darmstadt) gemischt und bei -80° C gelagert.

44
3.3.2.1.3 Präparation von Plasmid-DNA
Zur Gewinnung der Plasmid-DNA aus den Bakterien wurde die Bakterienkultur 15
min bei 6000 rpm in einer Ultrazentrifuge (Centrikon T2000, Fa. Kontron Instruments,
München) bei 4°C zentrifugiert. Die Plasmide wurden unter Verwendung eines
Qiagen-High-Speed-MIDI-Kits (Fa. Qiagen, Hilden) präpariert.
Die pelletierten Bakterien wurden in 6 ml Puffer 1 resuspendiert und darauf mit 6 ml
Puffer 2 gemischt und 5 min bei Raumtemperatur inkubiert. Das Gemisch wurde mit
6 ml Puffer 3 versetzt, um die Lyse der Bakterien und damit das Freisetzen der
Plasmide herbeizuführen, der Ansatz unmittelbar in einen Qiagenfilter gegeben und
10 min bei Raumtemperatur inkubiert. Nach Ablauf der Inkubationszeit wurde der
Inhalt des Qiagenfilters in die Qiagentip-Säule filtriert, wodurch die Bakterien-DNA
von Bestandteilen der lysierten Bakterienzellen getrennt wurde.
Das gereinigte Filtrat passierte die Matrix der Qiagentip-Säule, wobei ausschließlich
Plasmid-DNA an das Substrat gebunden wurde und damit eine Trennung von der
übrigen Bakterien-DNA erzielt werden konnte. Nach Waschen der Säule mit 20 ml
Puffer QC konnte die gebundene Plasmid-DNA durch Spülen der Säule mit 5 ml des
Elutionspuffers QF gewonnen werden. Das Eluat wurde zum Ausfällen der Plasmid-
DNA mit 3,5 ml Isopropanol gemischt und mittels eines High-Speed-Midi-Filters
gewonnen. Die erhaltene Plasmid-DNA wurde in 20 µl Aqua bidest./DEPC gelöst.
Zur Abschätzung der Menge an präparierter Plasmid-DNA wurde eine
photometrische Bestimmung der Konzentration durchgeführt (Photometer Varion
DMS 70, Fa. Varion, Australien) durchgeführt. Hierfür wurden 1 µl der gelösten
Plasmid-DNA mit 499 µl Aqua bidest. gemischt und bei einer Wellenlänge von
260 nm gemessen.
Anhand des ermittelten Wert für die Absorption der Probe konnte mit folgender
Formel die Konzentration der DNA in ng/µl bestimmt werden:
Konzentration der DNA in ng/µl = Absorption bei 260 nm x 500 x 50

45
3.3.2.1.4 Restriktion von Plasmid-DNA
Die Herstellung von RNA-Sonden für die nicht-radioaktive in-situ-Hybridisierung
mittels in-vitro-Transkription erforderte die Linearisierung des die cDNA enthaltenden
Plasmids. Durch die Restriktion wurde das ringförmige Plasmid in die offene,
linearisierte Form überführt, um das Anheften der RNA-Polymerase an die Plasmid-
DNA in der in-vitro-Transkription zu ermöglichen.
Die Restriktion der Plasmid-DNA erfolgte mit Restriktionsenzymen (Fa. Roche
Diagnostics, Mannheim), die spezifische Schnittstellen in der Plasmid-DNA erkennen
konnten. Für die Restriktion der Klone 90 und 92 zur Herstellung der Nachweis- und
Kontrollsonde gegen GAD65-mRNA wurde das Enzym Hind III aus Hämophilus
influenzae verwendet, für die Restriktion der Klone 14 und 18 zur Herstellung der
Sonde gegen GAD67-mRNA kam das Enzym Sal I aus Streptomyces albus zum
Einsatz.
1 µl Restriktionsenzym und 2 µl 10fach Restriktionspuffer wurden mit der
entsprechenden Menge an gelöster Plasmid-DNA mit Aqua bidest./DEPC ad 20 µl
komplettiert. Dieser Ansatz wurde 3-4 h bei 37° C inkubiert. Mit dem beschriebenen
Ansatz für die Restriktionsreaktion konnten bis zu 20 µg Plasmid-DNA linearisiert
werden. Da jedoch im Verlauf der Versuche festgestellt wurde, dass der Verlust an
Plasmid-DNA durch die Restriktionsreaktion mit der Menge an eingesetztem Plasmid
zunahm, wurden maximal 10 µg Plasmid-DNA in einer Restriktionsreaktion
eingesetzt.
Die linearisierte Plasmid-DNA wurde unter Verwendung von ElutipR-Säulen (Fa.
Schleicher und Schüll, Dassel) von Proteinen und Salzen gereinigt, da diese die
folgenden Reaktionsschritte stören konnten.
Vor der Aufreinigung wurde die Matrix einer ElutipR-Säule mit 5 ml eines Low Salt
Puffers gewaschen und 2 h bei Raumtemperatur inkubiert. Der aufzureinigende
Restriktionsansatz wurde in 1 ml eines Low Salt Puffer suspendiert und über die
Säule gegeben, wodurch das Anheften der Plasmid-DNA an die Matrix ermöglicht
wurde. Es folgte das Spülen der Säule mit 2 ml Low Salt Puffer. Die Elution der
Plasmid-DNA aus der Matrix geschah mit 0.5 ml eines High Salt Puffers.

46
1 2 3 4 5
Der Erfolg der Restriktionsreaktion wurde durch Analyse der aufgereinigten Plasmid-
DNA in einem 1,5 %igen Agarosegel überprüft, da für die Durchführung der in-vitro-
Transkription die vollständige Linearisierung der gesamten eingesetzten Plasmid-
DNA Vorraussetzung war (Abbildung 5).
Abbildung 5
Überprüfung der Restriktionsreaktion mittels Agarosegel-elektrophorese. Die linearisierten Plasmide mit der cDNA für GAD65 und GAD67 sind in den Spuren 3 und 5 als einzelne Banden sichtbar. In Spur 2 und 4 sind die ungeschnittenen Plasmide mit der cDNA für GAD65 bzw. GAD67 zum Vergleich aufgetragen. (Spur 1: DNA-Längenmarker)
3.3.2.1.5 In-vitro-Transkription und alkalische Hydrolyse
Die Herstellung Digoxigenin (DIG)-markierter RNA-Sonden für die nicht-radioaktive
in-situ-Hybridisierung erfolgte mittels in-vitro-Transkription. Bei dieser Reaktion
wurde unter Einsatz einer RNA-Polymerase (Fa. Roche Diagnostics, Mannheim) eine
RNA-Sonde spezifischer Sequenz und Länge entlang einer linearisierten cDNA
transkribiert. Die Markierung der Sonde konnte dadurch erreicht werden, dass ein
Teil der in die RNA eingebauten Nukleotide mit einem DIG-Molekül konjugiert waren.
Der Standardansatz gemäß Herstellerangaben (Fa. Roche Diagnostics, Mannheim)
für die Reaktion bestand aus 2 µl Nukleotiden, 2 µl Transkriptionspuffer für die RNA-
Polymerase, 1 µl RNAse-Inhibitor und 2 µl RNA-Polymerase. Ca. 1 µg linearisierte
Plasmid-DNA wurde als Synthesevorlage für die RNA-Polymerase hinzugegeben
und der Ansatz mit Aqua bidest./DEPC komplettiert.

47
1 2 3 4 5 6
Nach Inkubation bei 37 °C für 2 h wurde die Reaktion mit 2 µl einer 0.2 M EDTA-
Lösung pH 8.0 abgestoppt und mit 2,5 µl 4 M Lithiumchloridlösung und 79 µl 100%
Ethanol für mindestens 1 h bei -70°C oder bei -20°C über Nacht präzipitiert. Der
Ansatz wurde in 50 µl Aqua bidest./DEPC gelöst.
Da nur RNA-Sonden mit korrekter Basenlänge von 2,3 kb für den weiteren
Herstellungsprozess verwendet werden konnten, wurden die synthetisierten RNA-
Sonden auf vollständigen Länge in einem 1,5 % Agarosegel geprüft. Dabei erfolgte
ein Vergleich mit einem RNA-Längenmarker (Fa. Ferments, St. Leon-Roth) und einer
RNA-Sonde (Fa. Roche Diagnostics, Mannheim) bekannter Länge (0,7 kb) und
Konzentration (100 ng/µl).
Für die Penetration der synthetisierten RNA-Sonden in das Untersuchungsgewebe
während der in-situ-Hybridisierung war eine Verkürzung der Sonden Vorraussetzung,
da die RNA-Sonden mit einer Länge von 2,3 kb nicht in die zu untersuchenden
Zellen gelangten. Durch den Vorgang der alkalischen Hydrolyse wurden die
einzelnen RNA-Stränge in eine Vielzahl kleinerer Fragmente von 0,16 kb gespalten.
Abbildung 6
Gelelektrophoretische Überprüfung von RNA-Sonden nach in-vitro-Transkription und alkalischer Hydrolyse. Darstellt sind die Nachweis- und die Kontrollsonde für GAD65-mRNA in mit einer Länge von 2,3 kb in Spur 3 und 5. Die Spuren 4 und 6 enthalten die gleichen Sonden nach alkalischer Hydrolyse mit einer Länge von 0,16 kb. Zum Vergleich ist in Spur 2 eine RNA-Sonde von 0,7 kb aufgetragen. Spur 1 enthält einen RNA-Längenmarker.
Für die alkalische Hydrolyse wurden 50 µl RNA-Sonde mit 30 µl 0,2 M
Natriumcarbonat (Fa. Merck, Darmstadt) und 20 µl 0,2 M Natriumhydrogebcarbonat
(Fa. Merck, Darmstadt) 53 min bei 60 °C inkubiert.

48
Die Hydrolyse wurde durch 3 µl 3 M Natrium-Acetat pH 5,5 und 5 µl
10% Essigsäure abgestoppt und durch Zugabe von 10,8 µl 4 M Lithiumchlorid und
324 µl 100 % Ethanol gefällt. Nach Entsalzen mit 70 % Ethanol wurde der Ansatz in
50 µl Aqua bidest./DEPC gelöst und mit 1 µl RNAse-Inhibitor versetzt.
Zum Abschluss wurde wiederum in einem 1,5 %igen Agarosegel überprüft, ob durch
die alkalische Hydrolyse Sonden der gewünschten Länge entstanden waren. Im
Gegensatz zur Agarosegelelektrophorese der Transkriptionsreaktion war hier bei
erfolgreicher Reaktion keine scharfe Bande definierter Länge sichtbar (Abbildung 6).
3.3.2.1.6 Überprüfung der Markierungseffizienz von RNA-Sonden
Bevor die durch in-vitro-Transkription hergestellten RNA-Sonden in den
Hybridisierungsversuchen zum Einsatz kamen, war es notwendig, die Menge
synthetisierter RNA-Sonden festzustellen. So konnte die in der in-situ-Hybridisierung
einzusetzende Konzentration für die jeweilige Sonde ermittelt und gleichzeitig
sichergestellt werden, dass nur Nachweis- und Kontrollsonden mit vergleichbarer
Markierungseffizienz für die Versuche zum Einsatz kamen.
Im sogenannten „Dot-Blot“ wurden die hergestellten Sonden mit einer DIG-
markierten RNA-Sonde von bekannter Länge und Konzentration (Fa. Roche
Bioscence, Mannheim) verglichen. Das Prinzip dieser Methode beruht auf der
Übertragung und Fixierung von Nukleinsäuren auf eine Membran („blotten“) und
deren farbigen Nachweis auf dem Punkt des Auftragens („dot“).
Die zum Vergleich eingesetzte RNA-Sonde lag in eine definierten Länge von 0,76 kb
in einer Konzentration von 100 ng/µl vor. Von dieser Sonde wurden Verdünnungen
von 20 ng, 10 ng und 1 ng hergestellt. Die zu überprüfenden Sonden wurden jeweils
1:25, 1:50 und 1:100 in Aqua bidest./DEPC verdünnt. Die Verdünnungsansätze
wurden auf eine Nylonmembran (Fa. Roche Diagnostics, Mannheim) aufpipettiert
(„geblottet“), und 3 min auf einem Transiluminator (Modell Ti 1, Fa. Biometra,
Göttingen) fixiert. Alle Protokollschritte mit Ausnahme der Farbreaktion wurden bei
Raumtemperatur unter Schwenken durchgeführt.

49
A
B
C 1:1 1:25 1:50 1:100
100 ng 20 ng 10 ng 1 ng
1:1 1:25 1:50 1:100
Die Membran wurde im Anschluss 1 min mit 20 ml Puffer 1 mit 0,3 %igem Tween
20 und daran anschließend 30 min mit 20 ml Puffer 2 gewaschen. Die Detektion der
Sonden auf der Membran erfolgte mit einem Antikörper gegen Digoxigenin, der mit
alkalischer Phosphatase konjugiert war (DIG Markierungskit, Fa. Roche Diagnostics,
Mannheim). Der Antikörper wurde Puffer 2 in einer Verdünnung von 1:5000
zugegeben und für 30 min inkubiert. Nach zweimaligem Waschen mit Puffer 1 für
jeweils 15 min wurde 2 min mit Puffer 3 äquilibriert. Für die Farbreaktion wurde die
Membran 30 min in einer Nitroblau-Tetrazoliumsalz (NBT)/Brom-chlor-indol-phosphat
(BCIP)-Lösung (DIG Markierungskit®, Fa. Roche Diagnostics, Mannheim) unbewegt
im Dunkeln inkubiert. Das Abstoppen der Farbreaktion erfolgte durch Waschen der
Membran mit Puffer 4 für 5 min.
Abbildung 7
Dot Blot zur Überprüfung der Markierungseffizienz der Sonde für GAD67-mRNA . Dargestellt sind die Verdünnungen der Nachweis- (B) und der Kontroll-Sonden (C) im Vergleich zu einer RNA-Sonde bekannter Konzentration (A). Die aufgetragenen Sonden lagen in vergleichbarer Konzentration von etwa 100 ng/µl vor. Zwischen Nachweis- und Kontrollsonden bestand kein Unterschied in der Markierungseffizienz (3B und C).
Die Konzentration der hergestellten Sonden konnte anhand der Intensität der
Farbreaktionen abgeschätzt werden. Für die nicht-radioaktive in-situ-Hybridisierung
wurden Nachweis- und Kontrollsonden in einer Konzentration von ca. 200 ng/µl
eingesetzt.

50
3.3.2.2 Protokoll der nicht-radioaktiven in-situ-Hybridisierung
Die histologischen Schnitte für die nicht-radioaktive in-situ-Hybridisierung wurden wie
unter 3.2.1.1 beschrieben angefertigt und bis zur Verwendung in den Versuchen bei
-20 °C gelagert. Sämtliche Protokollschritte mit Ausnahme der Hybridisierung und
der Farbreaktion der Alkalischen Phosphatase wurden unter Schwenken auf einem
Rüttler durchgeführt.
Zu Beginn des Protokolls wurden die Gewebeschnitte 15 min an die
Raumtemperatur äquilibriert und zweimal für jeweils 10 min in 5 ml PBS gespült, um
anhaftende kryoprotektive Lösung zu entfernen. Die folgenden Inkubationsschritte
dienten zur Erhöhung der Permeabilität des Gewebes und um eine Penetration der
Sonden in die Zellen zu ermöglichen. Hierfür wurde zu Beginn mit 3 ml
0,02 N Salzsäure für 10 min gewaschen. Nach zweimaligem Spülen in PBS erfolgte
eine Inkubation in 3 ml 0,01 % Triton in PBS für 3 min und ein erneutes zweimaliges
Waschen mit PBS. Des weiteren wurde 10 min mit 3 ml einer Lösung inkubiert,
welche 0,25 µg/ml des Enzyms Proteinase K in 50 mM Tris, pH 7.4, und 5 mM EDTA
enthielt. Die Enzymaktivität beendet, indem für 10 min in 3 ml 2 mg Glycin/ml PBS
gewaschen wurde.
Vor der Inkubation mit RNA-Sonden wurde das Gewebe für 60 min mittels eines
Prehybridisierungspuffers (Kap. 8.4.3) an die Hybridisierungsbedingungen
äquilibriert. Die für die Hybridisierung verwendeten Sonden wurden in der in
Vorversuchen ermittelten Konzentration von etwa 200 ng/µl dem
Hybridisierungspuffer (Kap. 8.4.3) zugesetzt, 3 min bei 100° C erhitzt, und im
Anschluss 10 min auf Eis gekühlt. Die Hybridisierung erfolgte in 3 ml
Hybridisierungspuffer für 20 h bei 50 °C.
Nach Abschluss der Reaktion wurde das Reaktionsgefäß auf Raumtemperatur
abgekühlt und der Hybridisierungspuffer und nicht gebundene RNA-Sonde durch
Spülung mit 3 ml einer 4x Natriumcitrat (sodium saturated Saline, SSC) Lösung mit
10 mM Natriumthiosulfat aus dem Gewebe entfernt. Anschließend erfolgte eine
Inkubation in 3 ml RNAse-Puffer mit 0,05 mg/ml des Enzyms RNAse für 30 min bei
37 °C, um unspezifisch im Gewebe gebundene Sonden abzubauen.

51
Der Puffer für diesen Reaktionsschritt war vor Beginn der Inkubation auf 37° C
erwärmt worden. Danach wurde für 30 min mit RNAse-Puffer bei Raumtemperatur
gespült. Im Folgenden konnte durch Inkubationen in 2 x SSC/10 mM
Natriumthiosulfat bei Raumtemperatur und für 30 min in 0,1x SSC/10 mM Thiosulfat
bei 55 °C ein Anstieg der Stringenz bewirkt werden.
Vor der Zugabe des Antikörpers wurde mit Immundetektionspuffer 1 pH 7,5 für
10 min sowie mit einem Blockierungspuffer für 1 h gewaschen, wodurch
unspezifische Anheftung des Antikörpers verhindert wurde. Zur Detektion der
hybridisierten Sonden in den Neuronen wurde ein Schaf-anti-DIG-Antikörper mit
gekoppelter alkalischer Phosphatase (Fa. Roche Diagnostics, Mannheim) verwendet.
Die Inkubation erfolgte mit einer Verdünnung des Antikörpers von 1:1000 in
Immundetektionspuffer 1 unter Zusatz von 0,3 % Triton bei +4 °C für 20 h.
Ungebundener Antikörper wurde nach Ende der Reaktion durch dreimaliges
Waschen mit Immundetektionspuffer 1 für 30 min entfernt und anschließend 10 min
mit Immundetektionspuffer 2, pH 9,5, äquilibriert. Für die Farbreaktion der
alkalischen Phosphatase zur Sichtbarmachung des gebundenen Antikörpers wurden
die Gewebeschnitte in Immundetektionspuffers 2 überführt, dem 17 µl NBT und
17,5 µl BCIP pro 5 ml zugesetzt worden waren. Die Reaktionsgefäße wurden mit
Aluminiumfolie lichtdicht verschlossen und während der Entwicklung nicht bewegt.
Die Länge der Inkubation richtete sich nach dem jeweiligen Isoenzym der GAD,
welches es nachzuweisen galt. Für den Nachweis von GAD67-mRNA wurde nach
3-4 h eine optimale Anfärbung erreicht, während die Detektion der GAD65-mRNA 5-
6 h erforderte. Der Abschluss der Reaktion wurde durch Spülen der Gewebeschnitte
mit Immundetektionspuffer 3, pH 8,0 erreicht.
Nach Beendigung des Protokolls wurden die entwickelten Schnitte in einer
Petrischale von anterior nach posterior sortiert, mit Hilfe des Immundetektionspuffers
3 auf entfettete Objektträger aufgezogen und nach kurzem Trocknen mit Kaisers
Chromgelatine® (Fa. Merck, Darmstadt) eingedeckt.

52
3.3.3 Etablierung der radioaktiven in-situ-Hybridisierung
Für die Etablierung wurde das Protokoll zur in-situ-Hybridisierung mit radioaktiv
markierten Oligonukleotiden der Arbeitsgruppe von Prof. G. Sperk aus dem Institut
für Pharmakologie der Universität Innsbruck, Österreich zu Grunde gelegt.
3.3.3.1 Markierung von Oligonukleotid-Sonden
Für die Vorversuche zur Etablierung der in-situ-Hybridisierung mit radioaktiv
markierten Oligonukleotidsonden wurde eine Sonde gegen GAD67-mRNA gewählt.
Die Sonde besaß eine Länge von 32 Nukleotiden mit der Sequenz ttc-ttg-gag-gcc-
tgc-ctc-tcc-ctg-aag-gca-ct (Schwarzer und Sperk, 1995) und war damit komplementär
zu den Basen 318-349 der GAD67-mRNA der Ratte (Julien et al., 1990). Die
Synthese erfolgte durch die Fa. Microsynth, Balgach, CH.
Die Detektion der gebundenen Sonde im Gewebe machte eine Markierung
erforderlich. Diese wurde durch Inkubation mit dem Enzym Terminale Transferase
(Roche Diagnostics, Mannheim) erreicht, das 35Schwefel-markiertes ATP (35S-dATP,
New England Nuclear, Köln) an das Ende der Sonde koppelte.
Für die Hybridisierung von bis zu 50 Gewebeschnitten wurde folgender Ansatz
pipettiert:
35S-dATP 50 µCi in 4 µl
Oligonukleotidsonde 5 pMol in 2 µl (2.5 pMol/µl)
Reaktionspuffer 2 µl
CoCl2 1 µl
Terminale Transferase 1 µl
Der Ansatz wurde 30 min bei 37° C inkubiert und die Reaktion des Enzyms durch
Zugabe von 180 µl Tris-EDTA-(TE) Puffer gestoppt. Zum Fällen der Nukleotidsonden
wurden 4 µl t-RNA, 15 µl 4 M NaCl-Lösung und 660 µl 96 % Ethanol zugegeben und
für mindestens 1 h bei -80 °C gekühlt.

53
Nach Zentrifugation für 30 min bei 13.000 rpm konnte der Überstand mit nicht
gebundenem 35
S-dATP abpipettiert und die pelletierten Sonden in 50 µl Tris-
EDTA-Puffer mit 0.5 M Dithiothreitol (DTT) gelöst werden. Der gewonnene
Überstand wurde für die Überprüfung der Effizienz der Markierungsreaktion
aufbewahrt.
Für die Überprüfung der Markierungsreaktion wurden die Aktivitäten von 1 µl der
gelösten Sonde und 10 µl des Überstand in jeweils 5 ml Szintillatorflüssigkeit (Fa.
Baker, Holland) in einem Beta-counter (LS 5000, Fa. Beckmann, München)
bestimmt. Aus den beiden ermittelten Werten konnte die gesamte Aktivität des
Ansatzes in counts per minute (cpm) berechnet und der jeweilige Prozentanteil der
Sonde und des Überstandes an der Gesamtaktivität berechnet werden. Dabei wurde
ein Anteil der an die Sonde gebundenen Aktivität von mindestens 80 % der
Gesamtaktivität als ausreichend angesehen.
3.3.3.2 Protokoll der radioaktiven in-situ-Hybridisierung
Die histologischen Schnitte für die radioaktive in-situ-Hybridisierung wurden wie unter
3.2.1.2 beschrieben angefertigt und bis zur Verwendung bei -20 °C gelagert.
Die Fixierung der Gewebeschnitte erfolgte für 10 min in eiskalter 2 %iger
Paraformaldehydlösung in PBS. Alle weiteren Schritte der Vorbehandlung wurden
bei Raumtemperatur durchgeführt. Nach einer zweimaligen Spülung in PBS wurde
für 10 min in 200 ml Triethanolamin-Puffer pH 8,2 unter Zusatz von 500 µl
Essigsäureanhydrid inkubiert, um unspezifische Anheftungen der Sonde im Gewebe
zu verringern. Im Anschluss passierten die Schnitte eine aufsteigende Alkoholreihe,
an die sich eine Behandlung mit Chloroform für 5 min anschloss, die der Entfettung
der Gewebeschnitte diente. Das Chloroform wurde durch eine absteigende
Alkoholreihe entfernt und die histologischen Schnitte getrocknet.

54
Für die Hybridisierung eines Gewebeschnittes wurden 50 µl Hybridisierungspuffer
(Kap. 8.4.3) mit einer Aktivität von 35S-dATP von 1-2 Mio. cpm eingesetzt. Die
Schnittpräparate wurden gleichmäßig mit Hybridisierungspuffer beschichtet, mit
einem Deckglas versehen und 18 h bei 43° C in feuchten Kammer inkubiert.
Nach Abschluss der Hybridisierung wurden die Schnitte für 10 min auf
Raumtemperatur heruntergekühlt. Das Nachwaschen zum Entfernen ungebundener
Nachweissonden begann mit dreimaligem Spülen in einer 2x SSC/50 % Formamid-
Lösung unter Zusatz von 100 µl 1M DTT und 0,01% Triton bei Raumtemperatur.
Dann wurde zweimal für 15 min bei 40 °C in 2x SSC/50 % Formamid mit
100 µl 1 M DTT und anschließend zweimal in 2x SSC/50 % Formamid ohne weitere
Zusätze gewaschen. Nach zweimaligem Spülen in 1x SSC-Lösung und
zweimaligem Spülen in Aqua bidest. wurden die Gewebeschnitte durch
70 %igen Alkohol passiert und getrocknet.
Die Detektion der Hybride aus markierter Sonde und der nachzuweisenden mRNA
erfolgte zum einen mit Hilfe eines Röntgenfilms und zum anderen durch direktes
Beschichten der Objektträger mit einer fotosensitiven Emulsion. Der Nachweis
mittels Röntgenfilm kam dabei zur Überprüfung des Erfolges der Hybridisierung zum
Einsatz, da nur bei Auftreten von Signal im Film die Detektion auf zellulärer Ebene
mit Hilfe der photosensitiven Emulsion durchgeführt wurde.
Für die Detektion mittels Röntgenfilm (Hyperfilm ß-max ; Fa. Amersham, Freiburg)
wurde das histologische Material nach Abschluss des Hybridisierungsprotokolls in
der Dunkelkammer lichtdicht in eine Filmkassette (Fa. Eickemeyer, München)
verbracht und der Film für 14 Tage bei Zimmertemperatur exponiert. Die Entwicklung
des Films erfolgte mit Illford Phenisol® (Fa. Illford, Mobberley, England) für 2-3 min in
Abhängigkeit von der Stärke des sich entwickelten Signals. Nach kurzem
Zwischenwässern in Aqua bidest. wurde das Signal mit Illford Hypam® (Fa. Illford,
Mobberley, England) für 7 min fixiert und nach abschließendem Wässern getrocknet.

55
Das Aufbringen der fotosensitiven Emulsion (Hypercoat LM-1®, Amersham, Freiburg)
für den Nachweis des Hybridisierungssignals in Neuronen wurde in der
Dunkelkammer durchgeführt. Die Emulsionslösung wurde für 15 min bei 43 °C
erwärmt, in ein Gefäß (Hypercoat Dipping vessel®, Fa. Amersham, Freiburg)
überführt und die Objektträger durch Eintauchen beschichtet. Die Träger wurden für
12 h in völliger Dunkelheit getrocknet und für 4 Wochen bei +4 °C exponiert.
Die Entwicklung wurde mit Illford Phenisol® für 3 min durchgeführt. 4 min erforderte
die Fixierung des Signals mit Illford Hypam®. Nach 10 min unter fließendem
Leitungswasser und Nachspülen mit Aqua bidest. wurden die entwickelten
Objektträger getrocknet.
Die Zuordnung des Hybridisierungssignals in Form reduzierter Silberkörner in der
Photoemulsion zu darunter liegenden Zellen im Gewebe machte eine
Zellgegenfärbung erforderlich. Dabei war für die Auswertung von entscheidender
Bedeutung, das nur eine schwache Anfärbung der Zellen erreicht wurde, da eine zu
starke Anfärbung die Detektion der Silberkörner verhindert hätte.
Für die Gegenfärbung wurden die Gewebeschnitte für jeweils 5 min in 100 %, 95 %
und 70 % Alkohol hydratisiert und mit doppelt destilliertem Wasser abgespült. Es
folgte die Färbung der Zellen in einer 0,5 %igen Cresylviolettacetat-Lösung (Kap.
8.4.4) für maximal 20 s. Einer Inkubation in Aqua bidest. für 1 min folgte das
Dehydrieren in 70%, 95% und 2x 100 % Alkohol für 5 min und das Eindecken mit
EnteIlan® (Fa. Merck, Darmstadt).

56
3.3.4 Validierung der etablierten Protokolle
Zur Etablierung und Validierung der oben beschriebenen Protokolle zur in-situ-
Hybridisierung wurden umfangreiche Vorversuche durchgeführt. Während der
Etablierung der Ausgangsprotokolle wurden die erzielten Ergebnisse in der
Markierung GABAerger Neurone mit der Literatur verglichen (Esclapez et al., 1993;
Schwarzer und Sperk, 1995).
Im Anschluss folgten Experimente zur Validierung mit Modifikationen der Protokollen
mit der Absicht, mögliche Veränderungen in der Anzahl und Intensität spezifisch
markierter Neurone in untersuchten Hirnregionen wie etwa dem PC zu erzielen. Am
Schluss dieser Versuche stand die Auswahl des in-situ-Hybridisierungsprotokolls für
die weiteren Versuche.
3.3.4.1 Validierung der nicht-radioaktiven in-situ-Hybridisierung
3.3.4.1.1 Einsatz von Kontrollsonden
Die Spezifität der RNA-Sonden gegen GAD65- und GAD67-mRNA wurde durch den
Einsatz von Kontrollsonden für beide Isoformen der GAD überprüft. Die
Kontrollsonden wurden wie die Nachweissonden mittels in-vitro-Transkription (Kap.
3.3.2.1.5) hergestellt, besaßen jedoch eine zur nachzuweisenden mRNA identische
Nukleotidsequenz.
Bei der Durchführung der in-situ-Hybridisierung erfolgte der Einsatz von Nachweis-
und Kontrollsonden unter gleichen Versuchsbedingungen in einer Konzentration von
200 ng/ml Hybridisierungspuffer in parallelen Versuchen. Zudem wurden eine
Gewebeserie ohne den Zusatz von Sonde dem Protokoll unterzogen.
Zunächst galt es sicherzustellen, dass Nachweis- und Kontrollsonden während der
Inkubation in vergleichbarer Menge im Gewebe binden. Zu diesem Zweck wurde ein
Teil der Gewebeschnitte nach der Hybridisierung nicht den Nachwaschschritten des
Protokolls unterzogen, sondern sofort für die Detektion prozessiert, während für die
übrigen Schnitte das vollständigen Protokoll durchgeführt wurde.

57
Nach Abschluss der Farbreaktion zeigte sich den Gewebeschnitten, bei denen auf
ein Nachwaschen verzichtet worden war, für Nachweis- und Kontrollsonden ein
vergleichbar starkes unspezifisches Signal in den Somata von Neuronen in
unterschiedlichen Hirnregionen.
In Versuchen mit vollständig durchgeführtem Protokoll waren nur in Gewebeserien
nach Inkubation mit der Nachweissonde Neurone markiert, während in den mit den
Kontrollsonden behandelten Geweben kein Signal in den Zellen zu erkennen war
(Abbildung 8). In Gewebeserien, die im Hybridisierungspuffer ohne Zusatz von
Nachweis- oder Kontrollsonden behandelt worden waren, entstand kein Signal in den
Zellen.
Abbildung 8
Ergebnisse der Validierung der nicht-radioaktiven in-situ-Hybridisierung für GAD67-mRNA. Unter Einsatz von Nachweissonden wurden in den Schichten I- III (Pfeile) des zentralen PC Neurone spezifisch markiert (A), während sich mit Kontrollsonden kein Signal in Zellen ergab (B) Kalibrierbalken für A und B: 100 µm.
3.3.4.1.2 Neutralrotgegenfärbung
Im Zuge der Vorversuche zur Entwicklung der nicht-radioaktiven in-situ-
Hybridisierung wurde versucht, eine Gegenfärbung der entwickelten Schnitte mit
Neutralrot zu erzielen. Hierzu wurden die auf Objektträger aufgezogenen
Gewebeschnitte für 4 min in Neutralrot gefärbt, in Aqua bidest. gespült und über eine
aufsteigende Alkoholreihe entwässert und eingedeckt. Die Färbung von Neuronen
erwies sich als unzureichend und erschwerte zudem die Detektion des
Hybridisierungssignals in den Somata markierter Zellen, so dass auf eine weitere
Verbesserung der Methode verzichtet wurde.
A B
I II
III I
II III

58
3.3.4.1.3 Modifikationen am Protokoll zur nicht-radioaktiven in-situ-Hybridisierung
Im weiteren Verlauf der Vorversuche wurde der Einfluss von Modifikationen am
Protokoll zur nicht-radioaktiven in-situ-Hybridisierung untersucht.
Eine Erhöhung der Sondenkonzentration auf 300 bzw. 400 ng/ml
Hybridisierungspuffer führte wie eine Herabsetzung der Stringenz in den
Nachwaschschritten nicht zu einer Erhöhung der Zahl spezifisch markierter Zellen
etwa im PC. Vielmehr war nur eine Zunahme an unspezifischer Hintergrundfärbung
insbesondere in zelldichteren Regionen wie der Schicht II des PC und in den
Pyramidenzellschichten des Hippocampus zu bemerken.
Weiterhin wurden Experimente mit verlängerten Inkubationszeiten für die
Farbreaktion während der Detektion durchgeführt, um eine mögliche Darstellung von
Neuronen mit geringerem Gehalt an mRNA für GAD zu überprüfen. Hierfür wurden
die Entwicklungszeiten für die Farbreaktion für die GAD67 auf 5 und 6 h und für die
GAD65 auf 7 und 8 h erhöht. Diese Modifikationen führten jedoch ebenfalls nur zur
Verstärkung des unspezifischen Hintergrundsignals.
3.3.4.2 Validierung der radioaktiven in-situ-Hybridisierung
Die Vorversuche zur Etablierung der radioaktiven in-situ-Hybridisierung gemäß des
oben beschriebenen Protokolls (Kap. 3.3.3) wurden mit der Sonde gegen GAD67-
mRNA durchgeführt. Zur Validierung des Protokolls erfolgten nach Absprache mit der
Arbeitsgruppe von Prof. G. Sperk (Institut für Pharmakologie, Universität Innsbruck,
Österreich) Experimente mit Modifikationen an der Stringenz der Nachwaschschritte.
Zur Herabsetzung der Stringenz wurden in einer ersten Versuchsreihe Lösungen mit
einer erhöhten Konzentration an SSC bei gleichzeitiger Herabsetzung des Gehaltes
an Formamid getestet. Die Nachwaschtemperatur von 40 °C wurde zunächst nicht
verändert. In weiteren Versuchen wurde mit herabgesetzten
Nachwaschtemperaturen von 30, 35 und 37 °C experimentiert.

59
Die Zusammensetzung der Nachwaschlösung (2x SSC/50 % Formamid) war dabei
nicht verändert. Bei jeder Versuchsreihe wurden zur Kontrolle Schnitte nach den
Bedingungen des Ausgangsprotokolls behandelt.
Mit keiner der beschriebenen Modifikationen konnte eine Veränderung in der Anzahl
spezifisch positiv markierter Zellen erreicht werden; es kam abhängig von der
Herabsetzung der Stringenz ausschließlich zu einer Verstärkung von unspezifischem
Signal in den untersuchten Hirngebieten.
Auf eine Überprüfung der Hybridisierungsbedingungen durch Versuche mit
Kontrollsonden wurde verzichtet, da für die weiteren Versuche ausschließlich die
nicht-radioaktive in-situ-Hybridisierung eingesetzt wurde.
3.3.5 Vorversuche zur parallelen Darstellung GABAerger Neurone an Gewebeschnitten eines Tieres
Für die weiteren Experimente wurde die nicht-radioaktive in-situ-Hybridisierung
gemäß dem beschriebenen Ausgangsprotokoll (3.3.2.2) durchgeführt. Die
methodischen Aspekte, welche zur Auswahl des Protokolls führten, sind im
Ergebnisteil unter 4.1.1 dargestellt.
3.3.5.1 Vorversuche zur Auswahl geeigneter Antikörperkonzentrationen
Um für die Immunhistochemie im Free-Floating-Verfahren eine intensive
Immunreaktivität ohne den Einsatz einer unnötig hohen Konzentration an Antikörpern
zu erreichen, wurden die eingesetzten Antikörper in verschiedenen Verdünnungen
getestet. Für die primären Antikörper gegen GABA und GAD erwiesen sich
Verdünnungen von 1:1000 als ausreichend; für das sekundäre Antikörpersystem aus
dem Kaninchen wurden Verdünnungen von 1:500 gewählt. Die Streptavidin-
Meerrettich-Peroxidase wurde in einer Verdünnung von 1:400 eingesetzt. In
Experimenten, in denen keine primären Antikörper gegen GABA und GAD eingesetzt
wurden, konnte keine spezifische Anfärbung von Zellen beobachtet werden.

60
3.3.5.2 Fixierung
Die Durchführung von in-situ-Hybridisierung und Immunhistochemie an
Gewebeserien eines Tieres erforderte die Auswahl einer geeigneten Fixierung.
Die für die Immunhistochemie zum Nachweis von GABA und GAD am Institut
etablierte Perfusionsfixation mit einer Lösung aus 2 % Paraformaldehyd und 2 %
Glutaraldehyd führte bei der nicht-radioaktiven in-situ-Hybridisierung zu starker
unspezifischen Hintergrundfärbung und war damit ungeeignet. Das
Ausgangsprotokoll für die nicht-radioaktive in-situ-Hybridisierung war mit einer
Fixierung durch 4 %igem Paraformaldehyd ohne Zusatz von Glutaraldehyd
eingeführt worden. Nach Immunhistochemie an derart fixierten Gewebeschnitten
zeigte sich jedoch eine ungenügende Markierung von Neuronen.
Im weiteren Verlauf wurde mit 4 %igen Paraformaldehydlösungen unter Zusatz von
0,5, 1 sowie 1,5 % Glutaraldehyd experimentiert. Hierbei zeigte die Verwendung
eines Anteils von 0,5 % Glutaraldehyd das beste Ergebnis. Es kam zu einer guten
Fixierung des Hirngewebes bei optimaler Darstellung von Neuronen mit den
unterschiedlich eingesetzten histologischen Untersuchungsmethoden.
3.3.6 Immunhistochemie für GABA und GAD
Zu Beginn wurden die Gewebeschnitt dreimal in jeweils 5 ml Tris-gepufferter, 0,9 %
Natriumchloridlösung (Tris-buffered-Saline, TBS, pH 7,6) gewaschen und
anschließend 30 min in 5 ml TBS unter Zusatz von 75 µl H2O2 vorinkubiert, um
endogene Peroxidasen im Gewebe zu inaktivieren. Nach erneutem dreimaligem
Spülen mit TBS erfolgte die 60minütige Inkubation in 2 ml „Block-Lösung“ zur
„Absättigung“ von unspezifischen Proteinbindungsvalenzen in den Schnitten mit
bovinem Serumalbumin (BSA) und Kaninchenserum, der Spendertierart, aus der der
sekundäre Antikörper gewonnen wurde (Kap. Anhang). Für weitere 10 min wurde in
3 ml „Carrier-Lösung“ inkubiert, um die Gewebeschnitte an die Bedingungen für den
Primärantikörper gegen GABA bzw. GAD zu äquilibrieren. Die Behandlung mit
jeweils einem primären Antikörper (anti-GABA Maus Ig, 1:1000 bzw. anti-GAD Schaf
Ig) erfolgte für 20 Stunden bei +4 °C im Free-Floating-Verfahren auf einem Rüttler.

61
Nach einer dreimaligen TBS-Spülung und erneuter Inkubation in „Carrier-Lösung“ für
10 min wurden die Schnitte für 90 min in dem sekundären Antikörper (Kaninchen-
Anti-Maus bzw. Kaninchen-Anti-Ziege Ig, biotinyliert, 1:500) inkubiert. Ungebundener
sekundärer Antikörper wurde durch dreimaliges Waschen mit TBS entfernt. Nach
einer abschließenden 90minütigen Behandlung in Streptavidin-Meerrettich-
Peroxidase (HRP, 1:400 in 3 ml TBS) war die Inkubation beendet. Das Streptavidin
bindet hierbei an den biotinylierten Sekundärantikörper. Überschüssige Streptavidin-
Meerrettich-Peroxidase konnte durch viermaliges Spülen in TBS beseitigt werden.
Zur Detektion der Antikörper-Konjugate mit 3,3`-Diaminobenzidin (DAB) wurden die
Schnitte für 15 min mit einer 0,0062 %igen DAB-Lösung (4 ml Tris/Nickel-Lösung
und 30 µl DAB) vorinkubiert, um eine gleichmäßige Anfärbung der histologischen
Schnitte zu gewährleisten, danach mit 2 µl H2O2 pro 4 ml DAB-Lösung versetzt und
für weitere 15 min inkubiert. Nach Abschluss der Farbreaktion wurde nochmals
dreimal mit TBS gespült. Die Schnitte wurden anschließend nach anterior-posterior-
Ebene sortiert und mit Gelatine-Lösung auf entfettete Objektträger aufgezogen. Nach
dem Trocknen wurden sie über eine aufsteigende Alkoholreihe (50, 70, 80 und 96 %
für jeweils 1 min) passiert, in Xylolersatzmedium konserviert und mittels
Deckgläschen und Entellan® eingedeckt.

62
3.4 Tiermodelle der Temporallappenepilepsie
3.4.1 Das Lithium-Pilocarpin-Modell und das Kainsäure-Modell
Das Lithium-Pilocarpin-Modell sowie das Kainsäure-Modell stellen vielfach
verwendete Modelle für Temporallappenepilepsie dar. Da beide Modelle im Versuch
parallel verwendet wurden, werden die durchgeführten Protokolle zusammen
besprochen.
24 h vor Applikation von Pilocarpin zur Induktion eines selbsterhaltenden SE wurde
weiblichen Wistar-Ratten Lithiumchlorid (Fa. Sigma-Aldrich-Chemie, Steinheim) in
einer Dosierung von 127 mg/kg in 3 ml/kg physiologischer NaCl-Lösung i.p.
verabreicht. Weiterhin erhielten diese Tiere 30 min vor dem Applikationszeitpunkt
von Pilocarpin 1 mg/kg Methylscopolaminbromid (Fa. Sigma-Aldrich-Chemie,
Steinheim) in 2 ml/kg physiologischer Kochsalzlösung i.p. zur Abschwächung
peripherer Nebenwirkungen. Das Pilocarpin (Fa. Sigma-Aldrich-Chemie, Steinheim)
wurde in einer Dosierung von 30 mg/kg in 3 ml/kg 0,9 %iger NaCl-Lösung i.p.
verabreicht.
Kainsäure wurde bei den Tieren in einer Dosierung von 10 mg/kg in 3 ml/kg
physiologischer NaCl-Lösung i.p. injiziert.
In der folgenden Beobachtungszeit wurden das Auftreten fokaler und generalisierter
Anfälle, sowie der Beginn und das Ende eines SE protokolliert. Weitere
Verhaltensauffälligkeiten wie Ataxie oder Hypermotorik wurden ebenfalls vermerkt.

63
Die Bewertung der Anfallsschwere erfolgte nach Baran et al. (1995):
Skala der Krampfschwere
Anfallscharakteristika (klinische Symptomatik)
Wet dog shakes (WDS)
keine motorischen Zwangsbewegungen
Stadium I Immobilität, schwacher Fazialklonus (Schließen eines oder beider Augen, Zittern der Tasthaare, stereotypes Schnüffeln)
Stadium II Schwerer Fazialklonus in Form von Kaubewegungen und Kopfnicken
Stadium III unilateraler/bilateraler Vorderextremitätenklonus
Stadium IV Aufrichten und bilateraler Vorderextremitätenklonus
Stadium V bilateraler Vorderextremitätenklonus mit Verlust der Stellreflexe (nach hinten fallen)
Stadium VI Status epileptikus
Tabelle 2
Klassifikation der Anfallsstadien nach Baran et al. (1995)
Die Stadien I-III bezeichnen fokale Anfallsaktivität, während die Stadien IV-V
generalisierte Krämpfe darstellen.
Die Ratten, die einen selbsterhaltenden SE entwickelten, erhielten 90 min nach
Beginn des SE 10 mg/kg Diazepam (Fa. Ratiopharm, Ulm) i.p. zur Unterbrechung
der Krampfaktivität. Diese Tiere wurden zusammen mit den Tieren der
Kontrollgruppe 8 h nach Applikation von Kainat, Pilocarpin bzw. physiologischer
Kochsalzlösung gemäß 3.2.1.1 perfundiert. Der Zeitpunkt der Perfusion wurde in
Anlehnung an die Studien von Ding et al. (1989) gewählt, die 8 h nach Applikation
von Kainat zur Auslösung eines selbsterhaltenden SE eine Expression von GAD67-
mRNA in Körnerzellen des Hilus des Gyrus dentatus nachwiesen. Alle Tiere aus den
behandelten Gruppen, bei denen das Kriterium eines 90 minütigen selbsterhaltenden
SE nicht erfüllt wurde, wurden nicht für die weiteren Untersuchungen verwendet.

64
3.4.2 Das Amygdala-Kindling-Modell
3.4.2.1 Stereotaktische Implantation von Stimulationselektroden
Die Implantation der Elektroden für die Stimulation der basolateralen Amygdala
erfolgte unter Zuhilfenahme eines Stereotakten (Fa. Kopf, Tujunga, USA). Für die
Operation wurden die Ratten mit Chloralhydrat (Fa. Merck, Darmstadt; 360 mg/kg
Körpergewicht in 10 ml/kg destilliertem Wasser i.p.) narkotisiert.
Die Oberkiefer-Halterung des stereotaktischen Apparates wurde auf -3,3 mm ventral
der Interauralinie eingestellt, wodurch die Kreuzungspunkte der
Schädelknochennähte und Bezugspunkte des stereotaktischen Atlas (Paxinos und
Watson 1998), Lambda und Bregma, auf gleicher Höhe zu liegen kamen. Die
Kopfbehaarung wurde im Operationsbereich rasiert, die Haut mit 70 %igem Alkohol
mehrfach desinfiziert und Haut und Unterhaut in der Medianebene rostrocaudal
inzisiert. Das Periost der Schädeloberfläche wurde bis zur angrenzenden Muskulatur
entfernt und die Schädeloberfläche gesäubert, um Bregma, den Schnittpunkt der
Sutura sagittalis und der Sutura coronalis, sichtbar zu machen.
Die dreidimensionalen Koordinaten für die Implantation der Elektrode ergaben sich
in Anlehnung an den stereotaktischen Atlas von Paxinos und Watson (1998), der die
Lage der Hirnstrukturen in Bezug zu Bregma wiedergibt. Die aus dem Atlas
ablesbaren Koordinaten bestimmen die rostrocaudale, laterale und ventrale
Entfernung von Bregma. Die Koordinaten für die Elektrodenimplantation in die
rechte basolaterale Amygdala betrugen -2,2 mm rostrocaudal, -4,8 mm lateral und
rechts sowie-8,5 mm ventral.
Die stereotaktischen Koordinaten von Bregma wurden mit einer Nadel bestimmt.
Anhand dieser Werte erfolgte die Berechnung und Markierung der Koordinaten für
die Trepanationsstelle der Schädeloberfläche in rostrocaudaler und lateraler
Richtung. Weiterhin wurden drei Trepanationsstellen für die Aufnahme der
Erdungsschraube gegenüber dem Bohrloch für die bipolare Elektrode und für zwei
weitere Bohrlöcher rostral und caudal der Elektrode im Os frontale dexter und im Os
parietale dexter für Fixationsschrauben markiert. Die Trepanation der Schädeldecke
geschah unter Verwendung eines 0,9 mm starken Dentalbohrers (Fa. Dremel,
Wisconsin, USA) unter Schonung der Dura mater.
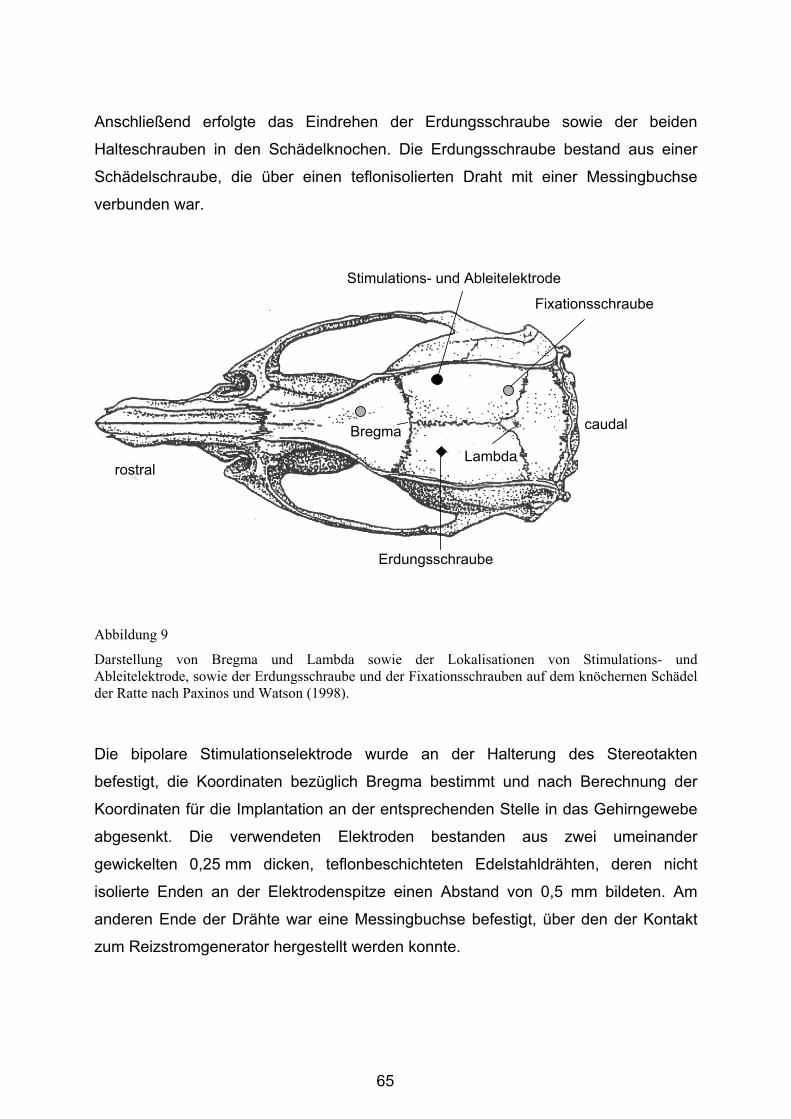
65
Anschließend erfolgte das Eindrehen der Erdungsschraube sowie der beiden
Halteschrauben in den Schädelknochen. Die Erdungsschraube bestand aus einer
Schädelschraube, die über einen teflonisolierten Draht mit einer Messingbuchse
verbunden war.
Abbildung 9
Darstellung von Bregma und Lambda sowie der Lokalisationen von Stimulations- und Ableitelektrode, sowie der Erdungsschraube und der Fixationsschrauben auf dem knöchernen Schädel der Ratte nach Paxinos und Watson (1998).
Die bipolare Stimulationselektrode wurde an der Halterung des Stereotakten
befestigt, die Koordinaten bezüglich Bregma bestimmt und nach Berechnung der
Koordinaten für die Implantation an der entsprechenden Stelle in das Gehirngewebe
abgesenkt. Die verwendeten Elektroden bestanden aus zwei umeinander
gewickelten 0,25 mm dicken, teflonbeschichteten Edelstahldrähten, deren nicht
isolierte Enden an der Elektrodenspitze einen Abstand von 0,5 mm bildeten. Am
anderen Ende der Drähte war eine Messingbuchse befestigt, über den der Kontakt
zum Reizstromgenerator hergestellt werden konnte.
rostral caudal
Lambda Bregma
Stimulations- und Ableitelektrode Fixationsschraube
Erdungsschraube

66
Die Elektrode, die Erdungsschraube sowie die beiden Halteschrauben wurden mit
einem kaltpolymerisierenden Kunststoff (Paladur; Fa. Kutzer, Weinheim) an der
Schädeloberfläche fixiert, wobei dem Paladur 2,1 % Gentamicinsulfat (Sigma
Aldrich, München) beigemischt war, um Wundinfektionen vorzubeugen. Nach
Trocknung der Schicht konnten die Drähte der Elektrode auseinandergewickelt und
ausgerichtet werden. Die Messingbuchsen der Elektrode und der Erdungsschraube
wurden in einen dreipoligen Steckerkontakt eingeführt und aus Paladur ein ca. 2 cm
hoher Aufbau geformt, der alle Drahtteile umhüllte und nach Entfernen des Steckers
den Steckkontakt an der Spitze freiließ.
Der Operationsbereich wurde mit Ethacridinlactat (Rivanol; Fa. Chinosol, Hannover)
desinfiziert und die Hautränder mit Knopfheften aus nichtresorbierbaren Nahtmaterial
adaptiert. Zur Wundinfektionsprophylaxe wurden den Ratten im Anschluss an die
Operation 20 mg/kg KGW Gentamicin (Frieso-Gent; Fa. Essex Tierarznei,
München) i. m. sowie an den nächsten zwei Tagen dieselbe Dosis s.c. injiziert. An
den darauffolgenden fünf Tagen wurden zweimal täglich 100 mg/kg
Chloramphenicolsuccinat (Fa. Sigma Aldrich, München) i. m. injiziert.
3.4.2.2 Kindling-Protokoll
Nach einer postoperativen Rekonvaleszenz von 14 Tagen wurde mit dem Kindling-
Protokoll begonnen. Hierfür wurden die Ratten über ein dreipoliges, abgeschirmtes
und flexibles Kabel mit einer elektrischen Stimulationseinheit (Fa. Sachs Elektronik,
Hugstetten) verbunden. Hierdurch konnte über die implantierte Elektrode in der
basolateralen Amygdala stimuliert und gleichzeitig über einen angeschlossenen
Thermoschreiber (Picker, Schwarzer-Uniskript UD 210) kontinuierlich das EEG der
Hirnregion auf Endlosmillimeterpapier aufgezeichnet werden. Bei dem
Stimulationsstrom handelte es sich um monophasische Rechteckimpulse von einer
Millisekunde Dauer und einer Frequenz von 50 Hz für die Dauer von einer Sekunde.

67
Am ersten Versuchstag wurde bei jeder Ratte der Kindling-Gruppe die initiale Nachentladungsschwelle (afterdischarge threshold, ADT) bestimmt.
Dieser Parameter beschreibt die Reizstromstärke in µA, die nötig ist, um mit dem
beschriebenen Stimulationsstrom im EEG ein Nachentladungsmuster von
mindestens drei Sekunden auszulösen. Hierzu wurde ausgehend von 15 µA die
Stromstärke in einem Abstand von je einer Minute jeweils um etwa 20 % erhöht.
Die fixen Stufen für die Stimulationen betrugen 15, 18, 21, 25, 30, 36, 42, 50, 60,
75, 90, 110, 130, 160, 200, 240, 280, 330, 400, 480, 590, 710, 840, 1000 µA.
Am Tag 2 des Protokolls begann der eigentliche Kindlingprozess. Hierfür wurden die
Ratten der Kindling-Gruppe einmal täglich in der Zeit zwischen 13 und 15 Uhr mit
einer Stromstärke von 500 µA stimuliert.
Zur Bewertung der motorischen und elektroenzephalographischen Krampfaktivität
wurden bei jeder Stimulation die folgenden Krampfparameter bestimmt:
1. Die Krampfschwere (seizure severity) gekindelter Anfälle wird auf einer
sechsstufigen Skala nach Racine (1972), modifiziert nach Löscher und Schmidt
(1988), eingeteilt:
Im Stadium 0 zeigen die Tiere trotz Nachentladungen im EEG keine sichtbaren
Verhaltensänderungen. Das Stadium I ist durch Nachentladungen in Spike-Wave-
Formation verbunden mit einer Immobilität des Tieres und zumeist dem Schließen
des Auges ipsilateral zur Stimulationselektrode gekennzeichnet. Weiterhin können
teilweise schwache Fazialklonien, Zittern der Tasthaare oder stereotypes
Schnüffeln hinzukommen. Im Stadium II treten zusätzlich zu den Symptomen aus
Stadium I schwere Fazialklonien in Form klonischer Kaubewegungen und
Kopfnicken auf, die in Stadium III durch einen ipsilateral zur Stimulation
auftretenden Vorderextremitätenklonus ergänzt werden. Stadium IV bedeutet ein
Aufrichten der Tiere in Verbindung mit einem bilateralen
Vorderextremitätenklonus, wobei im Stadium V zusätzlich zu diesem Verhalten ein
Verlust der Stellreflexe auftritt, infolge dessen die Tiere nach hinten überfallen.
Die Stadien I-III repräsentieren fokale Krampfaktivität, wohingegen die Stadien IV
und V eine Generalisierung des Krampfgeschehens bedeuten (Löscher und
Schmidt, 1988).

68
Rangskala der Krampfschwere
Anfallscharakteristika (klinische Symptomatik)
Stadium 0 keine motorischen Zwangsbewegungen
Stadium I Immobilität, schwacher Fazialklonus (Zittern der Tasthaare, stereotypes Schnüffeln, Schließen des ipsilateralen Auges)
Stadium II verstärkter Fazialklonus in Form von Kaubewegungen und Kopfnicken
Stadium III unilateraler Vorderextremitätenklonus
Stadium IV Aufrichten und bilateraler Vorderextremitätenklonus
Stadium V Aufrichten mit nachfolgendem Gleichgewichtsverlust, Hintenüberfallen und generalisierten Klonien
Tabelle 3
Klassifikation der klinischen Anfallsstadien nach Racine (1972)
2. Die Anfallsdauer (seizure duration, SD1 und SD2) beschreibt die Zeit vom
Beginn der Stimulation bis zum Ende der sichtbaren Verhaltensänderungen.
SD1 ist definiert als die Zeitspanne fokaler bzw. generalisierter motorischer
Krampfaktivität, wobei die nach generalisierten Krämpfen auftretende fokale
Krampfaktivität nicht berücksichtigt wird. SD2 beinhaltet die Zeit vom Beginn der
Stimulation bis zum Ende der Immobilitätsphase nach der das Tier wieder normale
Reaktionsbereitschaft zeigt. Dementsprechend ist bei Ratten nach ausschließlich
fokaler Krampfaktivität SD1 identisch mit SD2, da diese Tiere direkt nach dem
Anfall normal reaktionsfähig sind.
3. Die Nachentladungsdauer (afterdischarge duration, ADD) wird definiert als die
Zeit, in der im EEG Spikes mit einer mindestens dreifachen Amplitude zur Grundlinie
und einer Frequenz von mindestens einem Hertz auftreten. Kurze
Unterbrechungen im Spikemuster bis maximal drei Sekunden werden vernachlässigt.
In einigen seltenen Fällen treten im Verlauf der Nachentladung im EEG
Veränderungen in Amplitude, Frequenz oder Form der Spikes auf, welche sich bis
zum Ende der Entladung fortsetzen.

69
Dementsprechend wird die Zeit ab der Stimulation bis zum Ende der ersten
Spikeperiode als ADD1 beschrieben, während die Zeit der gesamten
Nachentladung als ADD2 angegeben wird. Das Auftreten einer zweiten
Spikeperiode nach einer Ruhephase wird als sekundäre Nachentladung
bezeichnet.
Abbildung 10 Beispiel für die elektroenzephalographische Entwicklung von Nachentladungen abgeleitet aus der basolateralen Amygdala einer gekindelten Ratte. Die Dauer der motorischen Krämpfe (Seizure duration, SD1) sowie die Dauer der Nachentladungen (Afterdischarge duration, ADD 1 und ADD 2) sind jeweils in Bezug auf den Zeitpunkt der Stimulation gemessen.
Im Verlauf des Kindlingprotokolls nahm die Krampfsymptomatik in Schweregrad und
Dauer mit der Zahl der Stimulationen schrittweise zu, bis die Ratten zuletzt
generalisierte tonisch-klonische Anfälle (Stadium V) zeigten. Nach zehn Krämpfen
des Stadium V wurden die Tiere als voll gekindelt bezeichnet. Zum Abschluss des
Protokolls erfolgte nach einer Stimulationspause von sieben Tagen die Bestimmung
der „post-Kindling“ ADT basierend auf dem Schema der initialen ADT.
Krampf (generalisierter motorischer Anfall)
Stimulation
physiol. EEG
SD 1
500 µA10 s
sekundäre Nachentladungen
Nachentladungsdauer(ADD1+ ADD2)
Nachentladungen

70
3.5 Untersuchungen an Ratten nach Status epileptikus induziert durch Kainat/Lithium-Pilocarpin
3.5.1 Versuchsaufbau und -durchführung
Für die Untersuchungen wurden 42 Ratten verwendet, von denen 8 Tiere als
Kontrollgruppe eingesetzt wurden. Mit den Versuchen wurde frühestens 7 Tage nach
Anlieferung der Tiere begonnen, damit eine ausreichende Adaptation der Tiere
gewährleistet war. Zusätzlich wurden die Tiere vor der Durchführung der eigentlichen
Versuche durch Verbringen in den Versuchsraum zur geplanten Versuchszeit
zwischen 7.00 und 9.00 Uhr und Applikation von 3 ml/kg physiologischer
Kochsalzlösung i.p. an die Untersuchungsbedingungen gewöhnt.
Versuchsplan 1. Ankunft der Tiere 7 Tage vor Beginn der Versuche
2. • Applikation von 10 mg/kg Kainat bzw. 30 mg/kg Pilocarpin i.p. zur
Auslösung eines selbsterhaltenden SE
• Unterbrechung des SE nach 90 min durch 10 mg/kg Diazepam i.p.
• Perfusion 8 h nach Applikation von Kainat, Pilocarpin bzw. 0,9%iger NaCl-
Lösung
3. Histologische Aufarbeitung der Tiere
4. Immunhistochemie für GABA und GAD
5. Nicht-radioaktive situ Hybridisierung für GAD65- und GAD67-mRNA
6. Codierung der Gewebeschnitte und Auszählung GABAerger Neurone in
festgelegten Hirnregionen
7. Untersuchung der Expression von GAD65- bzw. GAD67-mRNA in Körner-
und Korbzellen des Gyrus dentatus
8. Untersuchung neuropathologischer Veränderungen
9. Auswertung der Ergebnisse und Statistik
Tabelle 4
Übersicht über den Versuchsplan zur Untersuchung GABAerger Neurone und Neurodegeneration in Ratten mit selbsterhaltendem SE 8 h nach Applikation von Kainat bzw. Lithium-Pilocarpin.

71
Die Tiere der Kontrollgruppe erhielten 3 ml/kg 0,9 % ige NaCL-Lösung i.p. appliziert.
Bei 18 Tieren wurde Kainat und bei 16 Tieren Pilocarpin appliziert (Kap. 3.4.1). Die
Ratten, die einen SE entwickelten, erhielten 90 min nach Beginn des SE Diazepam
zur Unterbrechung der Krampfaktivität. 8 h Stunden nach Applikation von Kainat,
Pilocarpin bzw. physiologischer Kochsalzlösung erfolgte die Perfusion der Tiere.
Die Aufarbeitung der Tiere erfolgte in den folgenden 48 bis 72 Stunden, wobei
jeweils Tiere der einzelnen Gruppen parallel behandelt wurden. Nach Durchführung
der Immunhistochemie und der nicht-radioaktiven in-situ-Hybridisierung erfolgte die
Auszählung von Neuronen, die Auswertung von Neurodegeneration und die
Messung von Anfärbung im Gyrus dentatus.
3.5.2 Bestimmung der Dichte GABAerger Neurone
Die Zählung markierter Zellen erfolgte mit Hilfe eines Bildanalysesystems (Kontron
Elektronik, KS 300). Über eine Kamera (Zeiss Axioskop, Jena) wurde das
Mikroskopbild auf einen Bildschirm übertragen.
Vor Beginn der Auszählung wurden definierte Schnittebenen für die Auswertung in
den unterschiedlichen Hirnregionen unter zu Hilfenahme des Atlas nach Paxinos und
Watson (1998) festgelegt. Das Auffinden der Schnittebenen geschah durch
Orientierung an anatomischen Strukturen im Gewebe, denen exakte Koordinaten im
Bezug auf Bregma zugeordnet werden konnten. Da die Abstände zwischen den
einzelnen Schnitten bekannt waren, konnten nach Auffinden der Bezugsstruktur die
Untersuchungsschnittebenen rechnerisch ermittelt werden.
Im anterioren PC wurde im Bereich der Area tempestas +2,2 mm von Bregma
ausgewertet. Dabei diente das Corpus callosum, welches sich bei +1,6 mm mit dem
Corpus callosum der contralateralen Gehirnseite verbindet, als Anhaltspunkt für das
Auffinden. Der zentrale PC bei -0,8 mm konnte durch das Verschwinden der
vorderen Kommissur bei -0,6 mm und dem Beginn des Kerns des lateralen
olfaktorischen Traktes bei -1,3 mm eingegrenzt werden. Für den posterioren PC
diente das Auftreten des ventralen Hippocampus als Anhaltspunkt für das Auffinden
der Koordinaten -3,8 mm.

72
Die basolaterale Amygdala wurde -2,1 mm caudal von Bregma ausgewertet, wo sie
anhand des Nucleus intercallaris, der direkt an diese Hirnregion angrenzt, auffindbar
war. Der Hippocampus wurde in der Schnittebene -3,0 mm caudal von Bregma
untersucht, ausgehend vom Auftreten des ventralen Hippocampus bei -3,8 mm.
Die Zählbereiche in der anterioren und posterioren Substantia nigra pars reticulata
lagen bei -4,8 mm bzw. -5,8 mm in Bezug auf Bregma. Diese konnten rechnerisch
nach Festlegung der Schnittebenen mit den Anfangs- (-4,3 mm) bzw.
Endkoordinaten (-6,8 mm) der Struktur aufgefunden werden.
Im Bereich des anterioren PC wurde in den Schichten I und II, sowie im VEN und im
DEN ausgewertet. Im zentralen und posterioren PC wurde die Dichte GABAerger
Neurone zusätzlich in der Schicht III bestimmt.
In der basolateralen Amygdala und der Substantia nigra pars reticulata erfolgte die
Auszählung nach Zentrierung der Region im Mikroskop. Innerhalb der
Hippocampusformation wurden positive Neurone in der Regionen CA1, CA3, CA3c,
sowie im Hilus des Gyrus dentatus und im Bereich der Körnerzellschicht des Gyrus
dentatus ausgewertet (Abbildung 11).
Abbildung 11
Darstellung der Regionen zur Erfassung der Dichte GABAerger Neurone im PC (A-C), der basolateralen Amygdala (D), in der Hippocampusformation (E) sowie der anterioren (F) und posterioren Substantia nigra pars reticulata (G). Die Zählbereiche innerhalb der Regionen sind als schwarze Flächen markiert. Die angegebenen Zahlen stellen die anterior-posteriore Schnittebene in mm zu Bregma dar.
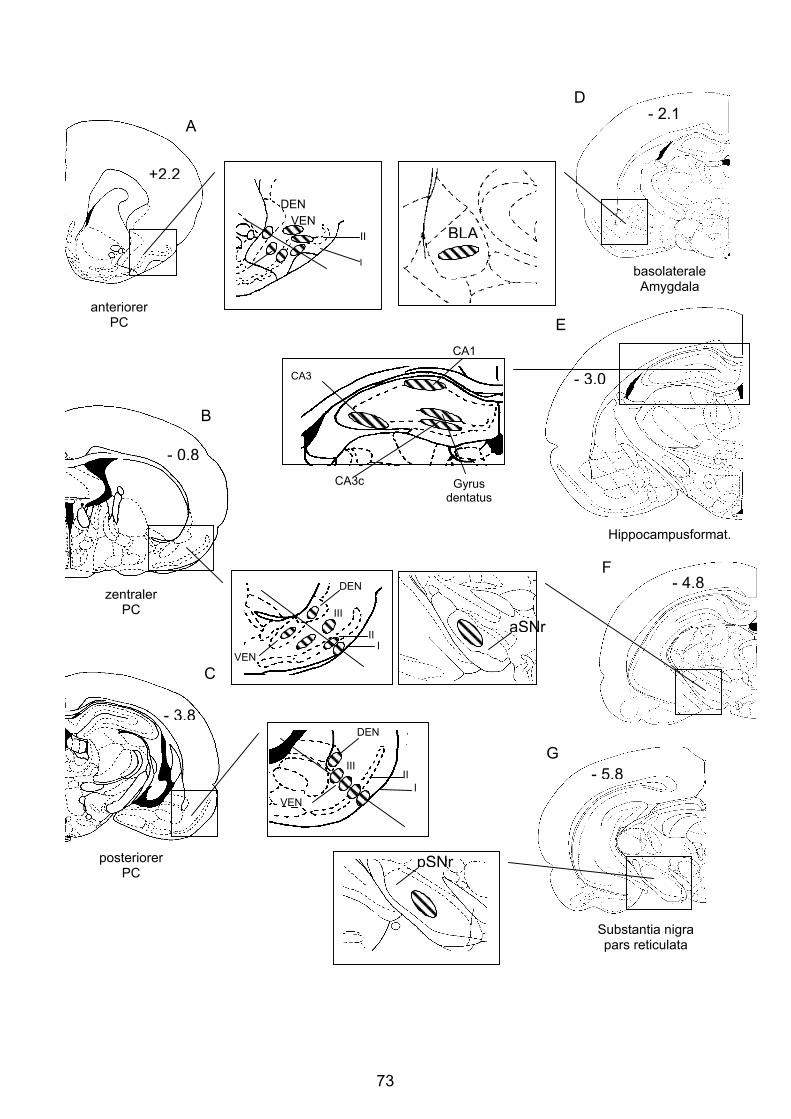
73
DEN
VEN
III
II I
- 3,8
- 0,8
+2,2
DEN
VEN
III II
I
DEN
II
I
VEN
- 2,1
- 3,0
- 4,8
- 5,8
pSNr
aSNr
CA1
CA3
CA3c Gyrus dentatus
BLA
A
F
G
C
B
D
E
posteriorer PC
zentraler PC
anteriorer PC
basolaterale Amygdala
Hippocampusformat.
Substantia nigra pars reticulata

74
Im Vorfeld der Zählungen wurden die Schnitte von Personen, die nicht in der
Versuchsdurchführung involviert waren, verschlüsselt. Damit war dem Untersucher
die Unterscheidung zwischen Kontrollen und behandelten Tieren erst nach
Abschluss der Zählungen möglich.
In jeder der festgelegten Schnittebenen konnte beidseitig mit Hilfe einer auf dem
Bildschirm des Bildanalysesystems befestigten Folie mit 25 Quadraten definierter
Seitenlänge ausgewertet werden. Dafür wurden die zu untersuchenden Regionen bei
400facher Vergrößerung auf dem Bildschirm so eingestellt, dass ein Auszählen von
Neuronen innerhalb von 10 Quadraten möglich war. Im Bereich der Schicht III des
PC, des EN, der basolateralen Amygdala und der Substantia nigra pars reticulata
wurden die auszuzählenden 10 Quadrate zusätzlich per Zufallszahlen aus der
Gesamtzahl von 25 Quadraten ermittelt.
In jeder Region bzw. Subregion war das Zählgebiet durch anatomische
Anhaltspunkte genau definiert (Abbildung 11). Nach dieser Vorgehensweise konnten
dreimal 10 Quadrate gezählt werden, wobei das Gesichtsfeld für jede Zählung
innerhalb des umschriebenen Zählbereiches willkürlich verschoben wurde, um
repräsentative Ergebnisse zu erhalten.
Die Zählungen erfassten nur solche Neurone, deren deutlich markierte Somata
während des Durchmusterns des Schnittes komplett innerhalb der Quadrate lagen
bzw. von der dorsalen und/oder rechten Seite eines Quadrates geschnitten wurden.
Nicht berücksichtigt wurden Neurone, welche sich in der ersten Focusebene des
Schnittes scharf stellen ließen, um Mehrfachzählungen zu vermeiden (Methode des
optischen Disektors nach Saper (1996) und West (1999).

75
3.5.3 Untersuchung der Expression von GAD-mRNA im Gyrus dentatus
Im Bereich der Körnerzellschicht des Gyrus dentatus wurden Unterschiede in der
Expression für GAD mRNA in Körner- und Korbzellen untersucht. Dies erfolgte mit
Hilfe des Bildanalysesystems durch Bestimmung der diffusen Dichte als Maß für die
Intensität der Anfärbung von Zellen. Für diese Vorgehensweise musste initial die
Lichtstufe am Mikroskop für die Messungen festgelegt werden, in der eine möglichst
lineare Verteilung der ermittelten Grauwerte gegeben war. Dazu erfolgte die
Bestimmung der Grauwerte von 20 Feldern eines Kalibrierungsstreifens (Fa.
Eastman Kodak, Rochester, New York, USA) mit definierten diffusen Dichten.
Für die Untersuchungen wurden die Schnitte kodiert. Die Messungen wurden für
GAD65- und GAD67-mRNA im Hippocampus -3,0 mm caudal von Bregma bei
400facher Vergrößerung beidseitig durchgeführt. Nach Einlesen eines digitalen
Bildes wurde die zu untersuchende Region mit dem Zeigegerät umfahren.
Zur Bestimmung der Hintergrundfärbung wurde in jedem Schnitt zu Beginn der
mittlere Grauwert einer Fläche von 35000 µm2 im Bereich der internen Kapsel
bestimmt. Zur Erhebung der Werte im Gyrus dentatus wurde das Gesichtsfeld derart
eingestellt, das die dorsale Flexur der Körnerzellschicht durch das Zentrum des
Gesichtsfeldes lief. Im Anschluss wurde die Intensität der Anfärbung der
Körnerzellschicht bestimmt. Zusätzlich wurden die Anfärbungen von drei Neuronen
ermittelt, welche anhand ihrer Morphologie und Lage am Übergang der
Körnerzellschicht zum Hilus des Gyrus dentatus als Korbzellen identifiziert wurden.
3.5.4 Untersuchung neuropathologischer Veränderungen
Die Beurteilung von Neurodegeneration und die Auswertung von Läsionen erfolgte
mit dem Bildanalysesystem KS 300 in Thionin gefärbten Schnittserien. Als Läsion
wurden solche Gebiete definiert, die keine morphologisch unveränderten Zellen mehr
aufwiesen.

76
Im Bereich der Substantia nigra pars reticulata wurden zunächst die Flächen der
Läsionen in den einzelnen Schnittebenen nach dem Einlesen eines digitalen Bildes
bei 25facher Vergrößerung durch Umfahren bestimmt. Als Grenze der Läsionen
wurde der Übergang zu Gewebe mit Neuronen unveränderter Morphologie gewählt.
Anhand der ermittelten Fläche konnte unter Einbeziehung des Abstandes zwischen
den Schnittebenen (240 µm) das Volumen der Läsionen mittels folgender Formel
berechnet werden:
Läsionsvolumen [mm3 ] = (Fläche 1 +Fläche 2 +Fläche n) x 240 µm
In den untersuchten Hirnregionen des limbischen Systems wurde das Ausmaß der
Neurodegeneration anhand eines Scoresystems beurteilt (Tabelle 5). Dabei wurden
solche Neurone als geschädigt beurteilt, die deutlich erkennbare Abweichungen von
der normaler Morphologie zeigten.
Score Ausmaß der Neurodegeneration
0 Keine sichtbare Schädigung von Neuronen
1 <50 % der Neurone geschädigt
2 >50 % der Neurone geschädigt
3 100 % der Neurone geschädigt
Tabelle 5
Scoresystem für die Beurteilung von Neurodegeneration im limbischen System von Tieren nach SE induziert durch Kainat bzw. Lithium-Pilocarpin.
3.5.5 Versuchsauswertung und Statistik
Die Statistik erfolgte unter zu Hilfenahme des EDV-Programms Sigma Stat (1.0) für
Windows, wobei ein p<0,05 für alle Tests als signifikant betrachtet wurde.
Für die statistische Auswertung wurden die codierten Daten im Anschluss an die
Auszählung GABAerger Neurone den jeweiligen Tieren in den Versuchsgruppen
abhängig von den Nachweisen zugeordnet. Für den deskriptiven Teil der Statistik
wurde zunächst für jede Region bzw. Schicht der Mittelwert aus den beidseitig
gezählten Werten für jedes Tier ermittelt.

77
Der Umrechnung der Anzahl von Neuronen einer Region bezogen auf das Volumen
ging die Bestimmung der Fläche im Präparat voraus, die der ausgezählten Fläche
von 10 Quadraten am Bildschirm unter Berücksichtigung der Vergrößerung des
Mikroskops entsprach. Bei einer bekannten Schnittdicke von 30 µm konnte die
Neuronenanzahl/mm3 mit folgender Formel berechnet werden:
Anzahl Neurone/mm3 = Anzahl markierter Neurone in 10 Feldern x 2292,53
Anhand der Einzeldaten ergaben sich die Gruppenmittelwerte und deren
Standardfehler. Die Gruppenmittelwerte der drei Versuchsgruppen für die 4
verschiedenen Nachweise aus den einzelnen Regionen wurden mittels einer
einfaktoriellen Varianzanalyse (analysis of variance, ANOVA) untersucht und
anschließend ein zweiseitiger t-Test zwischen den einzelnen Gruppen durchgeführt.
Das Datenmaterial zur Untersuchung der Expression von GAD mRNA im Gyrus
dentatus wurde nach der Erhebung den Versuchsgruppen zugeordnet. Es folgte die
Berechnung der Mittelwerte der Grauwerte für die untersuchten Regionen und die
Umrechung in diffuse Dichte. Des weiteren wurde bei jedem Tier der Mittelwert für
die diffuse Dichte des Hintergrundes vom Wert für die Dichte der Körnerzellschicht
und der Korbzellen subtrahiert, um den Einfluss eines möglichen Unterschiedes in
der Intensität der Gesamtfärbung des Gewebeschnittes ausschließen zu können.
Abschließend folgte die Berechnung der Gruppenmittelwerte sowie des zugehörigen
Standardfehlers. Um Aussagen über die Signifikanz der Ergebnisse treffen zu
können, wurde eine einfaktorielle Varianzanalyse (Analysis of Variance, ANOVA) der
drei Versuchsgruppen und anschließend ein zweiseitiger t-Test zum Vergleich der
diffusen Dichten der Färbung von Körner- und Korbzellen durchgeführt.
Für die Werte der Ausmessung von Läsionen erfolgte eine deskriptive Statistik, in der
Gruppenmittelwerte und der zugehörige Standardfehler berechnet wurden.

78
3.6 Untersuchungen an voll gekindelten Ratten
3.6.1 Versuchsaufbau und -durchführung
Für die Versuche wurden 34 Ratten verwendet, von denen 14 Tieren eine
Stimulationselektrode in die basolaterale Amygdala implantiert wurde. Danach
erfolgte die Aufteilung in drei Gruppen:
Gruppe 1: naive, weibliche Wistar-Ratten (n=10)
Gruppe 2 : Tiere, denen eine Elektrode in die basolaterale Amygdala implantiert
wurde (implantierte bzw. schein-gekindelte Tiere) (n=12)
Gruppe 3: Tiere, die nach Implantation einer Stimulationselektrode dem oben
beschriebenen Kindling-Protokoll unterzogen wurden (n=12)
Im Anschluss einer vierzehntägigen Pause nach der Operation wurde bei den Tieren
der Gruppe 3 die initiale ADT bestimmt. Daran anschließend erfolgte einmal täglich
nachmittags in der Zeit von 13 bis 16 Uhr die Stimulation der Tiere mit 500 µA.
Während des Kindlingprozesses der Gruppe 3 wurden jeweils Tiere der anderen
beiden Gruppen in den Stimulationsraum verbracht, wobei bei Tieren der Gruppe 2
zusätzlich das Anstecken an den Stimulator durchgeführt wurde, ohne jedoch eine
Stimulation auszulösen. Diese Maßnahmen dienten der Schaffung einheitlicher
Versuchsbedingungen für die Tiergruppen.
Nachdem die Tiere der Gruppe 3 zehn Krämpfe des Stadiums V gezeigt hatten,
galten sie als „voll-gekindelt“. Nach sieben Tagen wurde einmalig die post-kindling
ADT bestimmt, um den Erfolg des Kindlings zu überprüfen.
42 Tage nach Erreichen des Stadiums „voll-gekindelt“ wurden die Tiere perfundiert.
Hierfür wurden in der Zeit zwischen 13 und 16 Uhr jeweils eine gleich große Anzahl
von Tieren jeder Versuchsgruppe ausgewählt, um die Einflussfaktoren für alle
Gruppen einheitlich zu halten.
Die Gehirne der Tiere wurden histologisch aufgearbeitet und die
Elektrodenlokalisation im Thioninschnitt bestimmt.
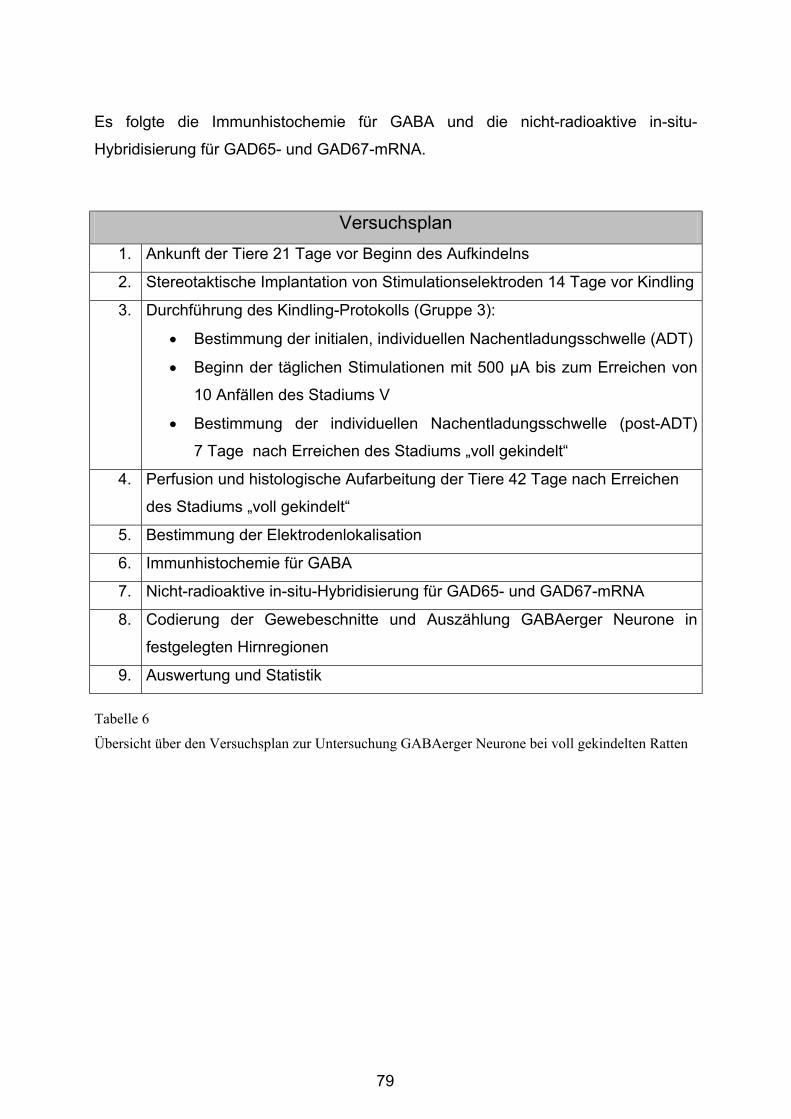
79
Es folgte die Immunhistochemie für GABA und die nicht-radioaktive in-situ-
Hybridisierung für GAD65- und GAD67-mRNA.
Versuchsplan 1. Ankunft der Tiere 21 Tage vor Beginn des Aufkindelns
2. Stereotaktische Implantation von Stimulationselektroden 14 Tage vor Kindling
3. Durchführung des Kindling-Protokolls (Gruppe 3):
• Bestimmung der initialen, individuellen Nachentladungsschwelle (ADT)
• Beginn der täglichen Stimulationen mit 500 µA bis zum Erreichen von
10 Anfällen des Stadiums V
• Bestimmung der individuellen Nachentladungsschwelle (post-ADT)
7 Tage nach Erreichen des Stadiums „voll gekindelt“
4. Perfusion und histologische Aufarbeitung der Tiere 42 Tage nach Erreichen
des Stadiums „voll gekindelt“
5. Bestimmung der Elektrodenlokalisation
6. Immunhistochemie für GABA
7. Nicht-radioaktive in-situ-Hybridisierung für GAD65- und GAD67-mRNA
8. Codierung der Gewebeschnitte und Auszählung GABAerger Neurone in
festgelegten Hirnregionen
9. Auswertung und Statistik
Tabelle 6
Übersicht über den Versuchsplan zur Untersuchung GABAerger Neurone bei voll gekindelten Ratten

80
3.6.2 Elektrodenlokalisation
Zur Auffindung der Elektrodenlokalisation wurden die Thionin-gefärbten Schnittserien
unter einem Auflichtmikroskop bei 25- und 100-facher Vergrößerung durchgemustert,
der tiefste Punkt des Elektrodenstichkanals bestimmt und die Koordinaten in
Millimetern von Bregma anhand umliegender Strukturen zugeordnet.
Als Anhaltspunkte konnten dabei Strukturen herangezogen werden, die an die
basolaterale Amygdala angrenzten und von anterior nach posterior scharf
abgrenzbar waren. Zu diesen Strukturen zählte der Nucleus intercallaris (IN), der in
Höhe von -2,1 mm im Querschnitt zungenförmig erscheint und sich bei -2,3 mm
zweiteilt. Da die Abstände zwischen den einzelnen Schnitten (240 µm) bekannt
waren, wurde der erste Schnitt bevor sich der IN zweiteilte mit -2,1 mm festgelegt.
Dementsprechend konnte über die Multiplikation der Schnittzahl mit 240 µm die
exakte rostrocaudale Lokalisation der Elektrode bestimmt werden.
Die ermittelten Koordinaten wurden mit den Atlasbildern und Koordinatenangaben
von Paxinos und Watson (1998) verglichen und in entsprechenden
Auswertungsbögen festgehalten. Die Tiere der implantierten Gruppen, bei denen
sich die Spitze des Stichkanals außerhalb der basolateralen Amygdala befand,
wurden von den weiteren Untersuchungen ausgeschlossen.
3.6.3 Bestimmung der Dichte GABAerger Neurone
Die Auszählung GABAerger Neurone wurde wie in Kap. 3.5.2. beschrieben
durchgeführt. Im Bereich der basolateralen Amygdala musste ipsilateral zur
Stimulationsseite aufgrund des Elektrodenstichkanals bei einigen Tieren die
Schnittebene -1,8 bzw. -2,3 alternativ ausgewertet werden.
3.6.4 Versuchsauswertung und Statistik
Die Auswertung der erhobenen Daten zur Dichte GABAerger Neurone erfolgte wie
unter 3.5.5 beschrieben nach contra- und ipsilateral getrennt.

81
4 Ergebnisse
4.1 Etablierung und Validierung der in-situ-Hybridisierung
Ein Ziel dieser Arbeit bestand in der Untersuchung der Expression der mRNA für die
Isoformen des GABA synthetisierenden Enzyms GAD in GABAergen Neuronen des
PC sowie anderer für die Epileptogenese relevanten Hirnregionen. Zu diesem
Zweck wurden ein Protokoll für die nicht-radioaktive in-situ-Hybridisierung gegen
GAD65- und GAD67-mRNA sowie ein Protokoll für die radioaktive in-situ-
Hybridisierung zum Nachweis der GAD67-mRNA etabliert. Die Ergebnisse der
Vorversuche zur Etablierung der Protokolle sind im Kap. 3.3.4 im Zusammenhang
dargestellt. Aufgrund der erzielten Ergebnisse und der Erfahrungen aus den
Vorversuchen wurde die nicht-radioaktive in-situ-Hybridisierung als geeignetes
Protokoll für die Untersuchungen ausgewählt.
4.1.1 Vergleich methodischer Aspekte
Für beide Methoden zur in-situ-Hybridisierung war eingangs ein hoher Zeitaufwand
für die Schaffung RNAse freier Arbeitsbedingungen verbunden. Dieser erwies sich im
Verlauf der Etablierung als gerechtfertigt, da keine Störungen der Versuche durch
RNAse Aktivität auftraten.
Während der Durchführung der Vorversuche traten bei der Gewinnung und Lagerung
des Untersuchungsmaterials sowie bei der Durchführung der Protokolle zur in-situ-
Hybridisierung keine Probleme auf. Dagegen erwies sich die Herstellung der
Nachweissonden als störanfälliger. Vielfach wiesen die mittels in-vitro-Transkription
synthetisierten RNA-Sonden nicht die erforderliche Länge auf. Nach wenigen
gelungenen Ansätzen konnten jedoch Sonden in ausreichender Menge für die
Untersuchungen hergestellt werden. In Vorversuchen konnte weiterhin durch den
Einsatz ungeschnittener Sonden in der nicht-radioaktiven in-situ-Hybridisierung durch
die zu große Länge der Sonden keine Markierung von Zellen erreicht werden, dies
war erst nach Durchführung der alkalischen Hydrolyse der RNA-Sonden möglich.

82
Die Markierungsreaktion der Oligonukleotid-Sonden für die radioaktive in-situ-
Hybridisierung erwies sich als weniger störungsanfällig, allerdings konnten die
markierten Sonden aufgrund der begrenzten Halbwertszeit des radioaktiven 35Schwefels nicht gelagert werden. Dagegen zeigten die RNA-Sonden auch nach
mehrmonatiger Lagerung noch eine ausreichende Markierungseffizienz in den
Versuchen.
Mit der nicht-radioaktiven und der radioaktiven in-situ-Hybridisierung konnten in der
überwiegenden Zahl der Vorversuche Neurone für die mRNA der GAD markiert
werden. Durch den Einsatz von Kontrollsonden in parallel durchgeführten
Experimenten zur nicht-radioaktiven in-situ-Hybridisierung erwies sich das Signal für
beide Isoformen als spezifisch. Als ein entscheidender Vorteil der Verwendung nicht-
radioaktiv markierter RNA-Sonden zeigte sich in diesem Zusammenhang der deutlich
geringere Zeitaufwand für die Entwicklung des Signals. In den Versuchen zur nicht-
radioaktiven in-situ-Hybridisierung war diese bereits nach wenigen Stunden
abgeschlossen, während für die Exposition bei der radioaktiven in-situ-Hybridisierung
vier Wochen erforderlich waren.
Bei der Quantifizierung GABAerger Zellen waren die mittels nicht-radioaktiver in-situ-
Hybridisierung markierten Neurone eindeutig an gefärbten Zellsomata zu
identifizieren. Das Signal der radioaktiven in-situ-Hybridisierung war dagegen in
einigen der untersuchten Regionen nicht immer eindeutig zuzuordnen. Dieses
Problem trat in zelldichten Strukturen wie der Körnerzellschicht des Gyrus dentatus
auf, da sich die Silberkörnung teilweise über die Grenzen des Zytoplasmas der
GAD67-mRNA-exprimierenden Zelle hinaus auch auf benachbarte Zellen erstreckte
(Abbildung 12 A/B).
Durch die in Kap. 3.3.4 beschrieben Modifikationen an den etablierten Protokollen
zur in-situ-Hybridisierung konnten keine Veränderungen in der Anzahl und Intensität
spezifisch markierter Zellen im Vergleich zu den Ausgangsprotokollen erreicht
werden. Eine höhere Sensitivität der radioaktiven in-situ-Hybridisierung konnte
zudem unter den beschriebenen Versuchsbedingungen in den untersuchten
Hirnregionen nicht nachgewiesen werden (Kap. 4.1.2).

83
Für die weiteren Untersuchungen im Rahmen der Arbeit wurde die Methode der
nicht-radioaktiven in-situ-Hybridisierung ausgewählt. Hierfür sprachen neben den
oben aufgeführten Vorteilen auch die geringeren Kosten für die Durchführung sowie
die bessere Kombinierbarkeit mit den Protokollen zur Immunhistochemie.
4.1.2 Darstellung GABAerger Neurone mit in-situ-Hybridisierung
4.1.2.1 Generelles Erscheinungsbild
Die nicht-radioaktive in-situ-Hybridisierung wurde mit RNA-Sonden gegen die mRNA
für GAD65 und GAD67 durchgeführt. Positiv markierte Zellen zeigten eine bläuliche
Anfärbung, welche sich auf das Zytoplasma und kurze Abschnitte der Zellausläufer
der Neurone beschränkte (Abbildung 12 A). Der Zellkern war in den meisten Fällen
nicht markiert. Die Art der Anfärbung erlaubte demnach eine Charakterisierung der
Zellen anhand der Größe, Form der Somata und des Größenverhältnisses von
Zellkern und Zytoplasma. Eine Gegenfärbung der histologischen Schnitte zur
Darstellung von Neuronen mit Neutralrot konnte nicht etabliert werden, eine
Anfärbung anderer Zelltypen wie Asytrozyten wurde jedoch nicht beobachtet.
Abbildung 12 A/B
Darstellung der Signalentwicklung mit der nicht-radioaktiven (A) sowie radioaktiven in-situ-Hybridisierung (B) am Beispiel positiv für GAD67-mRNA markierter Korbzellen im Gyrus dentatus einer naiven Ratte. Nach Durchführung der nicht-radioaktiven in-situ-Hybridisierung entstand das Farbsignal direkt im Zytoplasma (A, Pfeile), während positive Zellen nach der radioaktiven in-situ-Hybridisierung durch schwarze Silberkörner gekennzeichnet waren (B, Pfeile). Die zelluläre Auflösung des Signals der nicht-radioaktiven in-situ-Hybridisierung ist deutlich besser im Vergleich zur radioaktiven in-situ-Hybridisierung, bei der die Markierung teilweise nicht eindeutig zuzuordnen war. Kalibrierbalken für A und B: 50 µm
A B

84
Mit der radioaktiven in-situ-Hybridisierung unter Verwendung einer Oligonukleotid-
Sonde gegen GAD67-mRNA erfolgte keine direkte Markierung in Zellen. Vielmehr
entstand das Signal in Form schwarzer Silberkörnern in einer fotosensitiven
Emulsion über dem Gewebe, wodurch eine Zuordnung des Signals zu Zellen nach
Gegenfärbung der Gewebeschnitte mit Cresylviolettacetat möglich war. Positiv
markierte Zellen waren durch eine massive Ansammlung von Silberkörnern
gekennzeichnet (Abbildung 12 B).
Mit beiden Protokollen zur in-situ-Hybridisierung wurden GABAerge Neurone in
Regionen des limbischen Systems und der Substantia nigra pars reticulata markiert.
Weiterhin fanden sich GAD-mRNA-positive Neurone im retikulären Nukleus des
Thalamus, im Striatum und Globus pallidus sowie im Neocortex. Im Gegensatz
hierzu waren in der von dopaminergen Neuronen dominierten Substantia nigra pars
compacta nur vereinzelte Somata markiert. In zellfreien Arealen wie der internen
Kapsel ergab sich keine spezifische Anfärbung.
Mit nicht-radioaktiver in-situ-Hybridisierung wurden die meisten Neurone für die
mRNA beider Isoformen der GAD markiert, es waren jedoch Unterschiede in der
Intensität der Anfärbung zwischen den beiden Isoformen in einzelnen Regionen zu
bemerken. Zudem ließen sich mit der Sonde gegen GAD67-mRNA unter gleichen
Versuchsbedingungen bei gleicher Entwicklungszeit der Farbreaktion eine höhere
Anzahl an Neurone markieren als mit der Sonde gegen GAD65-mRNA.
Dementsprechend stieg nach längerer Inkubationszeit nur noch die Zahl der GAD65-
mRNA-positiven Zellen. Zudem schien das Signal bei Versuchen zur Darstellung von
GAD 67-mRNA dunkler auszufallen (Abbildung 14 A-C). Da in den Vorversuchen im
Zusammenhang mit der radioaktiven in-situ-Hybridisierung nur die Sonde gegen
GAD67-mRNA verwendet wurde, konnten hierbei keine entsprechenden Vergleiche
angestellt werden.

85
4.1.2.2 Regionsspezifisches Erscheinungsbild
Unter Kontrollbedingungen wurden mit der nicht-radioaktiven und der radioaktiven in-
situ-Hybridisierung für GAD67-mRNA in den untersuchten Hirnregionen eine
vergleichbare Anzahl von Neuronen dargestellt. Dementsprechend war subjektiv
unter den festgelegten Versuchsbedingungen kein Unterschied in der Sensitivität in
Bezug auf die Zahl markierter Zellen zwischen den Protokollen festzustellen.
Im retikulären Nukleus des Thalamus waren nahezu alle Neurone gleichstark für
GAD65- und GAD67-mRNA markiert. Mit der radioaktiven in-situ-Hybridisierung
wurde über allen Neuronen eine starke Anhäufung von Silberkörnern gesehen.
Mit der Sonde gegen GAD67-mRNA ließen sich im Striatum die Mehrzahl der
Neurone schwach bis mittelstark anfärben. Ein geringer Anteil der
Neuronenpopulation war dagegen stark für diese Isoform der GAD markiert. Diese
Neurone waren gleichmäßig im Striatum verteilt und unterschieden sich in Größe und
Form nicht von den schwächer gefärbten Neuronen. Ein vergleichbares Bild ergab
sich mit der radioaktiven in-situ-Hybridisierung. Die Anfärbung der Neurone für
GAD65-mRNA ließ sich als schwach bis mittelstark beschreiben, stark markierte
Zellen waren in dieser Region nicht zu bemerken.
Im allen Schichten des PC sowie des darunter liegenden EN wurden Neurone für die
mRNA beider Isoformen der GAD markiert. Die größte Dichte an Zellen fand sich
dabei in der Schicht III und im DEN und VEN, während in der Schicht II und Schicht I
des PC eine geringere Zahl an Zellen dargestellt wurde. Innerhalb der einzelnen
Schichten waren die Zellen gleichmäßig verteilt. Ein Unterschied zwischen den
Zellen bezüglich der Intensität der Anfärbung für eine Isoform war subjektiv nicht zu
beobachten, GAD67-mRNA-positive Neurone wiesen eine mittlere bis starke,
GAD65-mRNA- markierte Neurone eine mittlere Intensität in der Färbung auf. Das
Ergebnis des radioaktiven Nachweises erbrachte vergleichbare Ergebnisse
(Abbildung 13 A-C).

86
Abbildung 13 A-C
Darstellung GABAerger Neurone im posterioren PC markiert für GAD65- (A) und GAD67-mRNA (B/C). In der Schicht III des posterioren PC war eine höhere Dichte positiver Zellen festzustellen (Pfeile) als in Schicht II (Pfeilköpfe). Mit nicht-radioaktiver (B) und radioaktiver in-situ-Hybridisierung (C) wurden vergleichbare Anzahlen an Neuronen positiv für GAD67-mRNA markiert. Kalibrierbalken für A-C: 50 µm
Die verschiedenen Kerngebiete der Amygdala wiesen Unterschiede in der Dichte
markierter Neurone auf. Dabei enthielten der zentrale Kern die größte Dichte, der
mediale Nukleus enthielt eine mittlere Dichte und die basolaterale Amygdala die
geringste Dichte an markierten Neuronen. In der basolateralen Amygdala waren
keine Unterscheide zwischen den Isoformen und den beiden angewendeten
Protokollen auffällig.
In allen Regionen des Hippocampus fanden sich nach der Durchführung beider
Protokolle markierte Neurone. Dabei erschien die Anzahl und das Verteilungsbild
grundsätzlich ähnlich für die beiden GAD Isoformen. In der CA1 Region fanden sich
die meisten markierten Neurone im Stratum pyramidale. In der Region CA3 waren
die markierten Neurone gleichmäßig in allen Schichten verteilt, CA3c zeigte eine
geringere Dichte in der Pyramidenzellschicht und der angrenzenden dendritischen
Schicht.
A B
C
II
III
II III
II III

87
Im Gyrus dentatus war die größte Dichte markierter Zellen an der ventralen Seite der
Körnerzellschicht und im Hilus zu sehen. Unter Kontrollbedingungen ergab sich keine
spezifische Anfärbung von Körnerzellen. Für GAD67-mRNA war eine verstärkte
Hintergrundfärbung in dieser Region auffällig, welche auch in der Literatur
beschrieben wurde (Esclapez et al., 1994).
In der Substantia nigra (SN) ergab sich in der pars reticulata eine starke Markierung
für GAD67-mRNA mit beiden in-situ-Hybridisierungsprotokollen, während die
Anfärbung für GAD65-mRNA von mittlerer Intensität war (Abbildung 14).
Abbildung 14 A-C
Darstellung GABAerger Neurone (Pfeile) in der anterioren Substantia nigra pars reticulata einer naiven Ratte markiert für GAD65- (A) und GAD67-mRNA (B/C). Die GABAergen Neurone dieser Region wiesen eine hohe Expressionsrate für GAD67-mRNA auf, was anhand der intensiven Markierung der Zellen nach nicht-radioaktiver (B) und radioaktiver in-situ-Hybridisierung (C) deutlich wurde. Die Anfärbung für GAD65-mRNA war dagegen von mittlerer Intensität (A). Kalibrierbalken für A-C: 50 µm
A B
C

88
4.2 Untersuchung GABAerger Neurone unter Kontrollbedingungen
Im folgenden Abschnitt der Arbeit wurde die Expression der mRNA für die Isoformen
der GAD mit dem immunhistochemischen Nachweis von GAD und GABA in
GABAergen Neuronen unter Kontrollbedingungen verglichen.
4.2.1 Generelles Erscheinungsbild der Immunhistochemie
Mit der Immunhistochemie gegen GAD und GABA wurden die Somata von Neuronen
tief schwarz markiert, während der Nukleolus meistens nicht gefärbt wurde. Zum Teil
waren die proximalen Ausläufer der Zellen zu erkennen, wodurch sich diese als
Neurone charakterisieren ließen. Eine Anfärbung von Astrozyten, die auch einen
Uptake-Mechanismus für GABA besitzen, wurde nicht beobachtet. Im Gegensatz zur
in-situ-Hybridisierung erfolgte mit Immunhistochemie eine Anfärbung von
Terminalen, welche besonders ausgeprägt für GAD in der Substantia nigra pars
reticulata beobachtet wurde. Unterschiede in der Intensität der Anfärbung zwischen
den beiden Nachweisen konnten nicht festgestellt werden, es ergaben sich jedoch in
den untersuchten Regionen Differenzen in der Dichte dargestellter Neuronen .
4.2.2 Vergleich des regionsspezifischen Erscheinungsbildes
Mit der Immunhistochemie zum Nachweis von GABA und GAD in Neuronen konnten
in den untersuchten Regionen des limbischen Systems sowie in der Substantia nigra
pars reticulata Zellen in unterschiedlicher Dichte dargestellt werden.
Im PC waren GABA- und GAD-immunreaktive Zellen in allen Schichten zu finden.
Die höchste Dichte markierter Zellen trat für beide Nachweise in Schicht II auf, wobei
ein höherer Anteil an Neuronen positiv für GABA war. Für die Schicht III sowie den
EN ergaben sich vergleichbare Zelldichten. Diese Verteilung wurde bereits in der
Literatur beschrieben (Löscher et al., 1998).
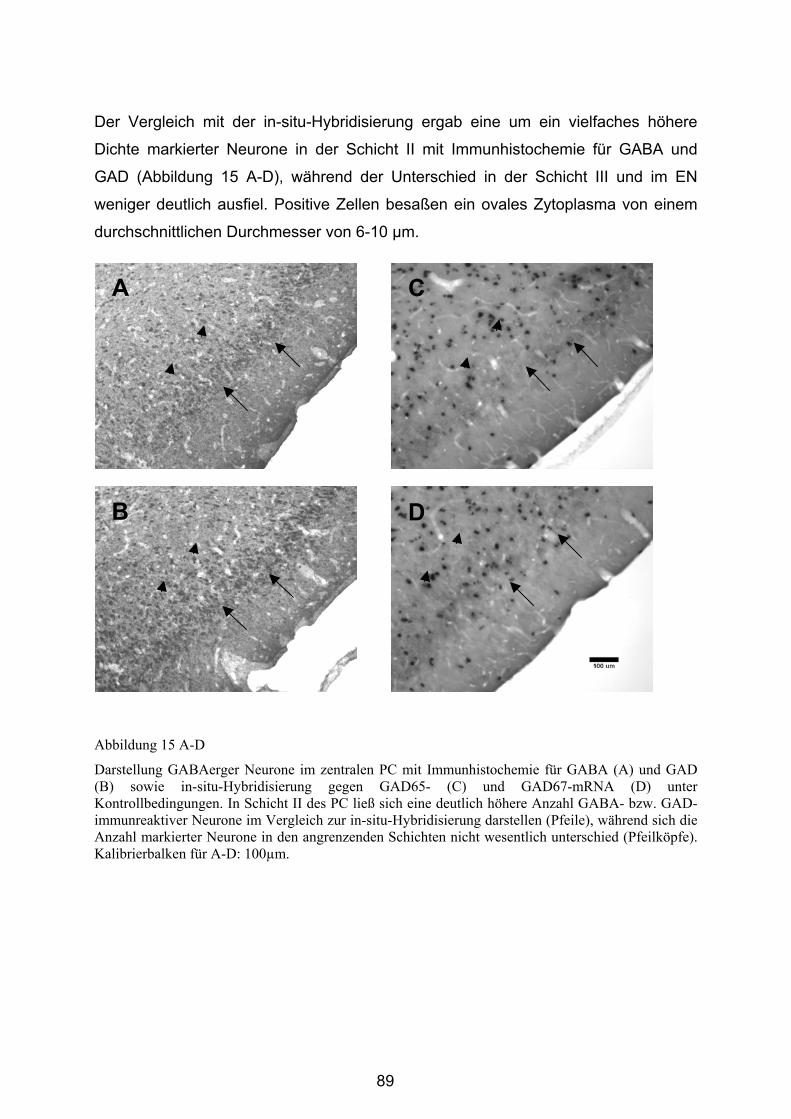
89
Der Vergleich mit der in-situ-Hybridisierung ergab eine um ein vielfaches höhere
Dichte markierter Neurone in der Schicht II mit Immunhistochemie für GABA und
GAD (Abbildung 15 A-D), während der Unterschied in der Schicht III und im EN
weniger deutlich ausfiel. Positive Zellen besaßen ein ovales Zytoplasma von einem
durchschnittlichen Durchmesser von 6-10 µm.
Abbildung 15 A-D
Darstellung GABAerger Neurone im zentralen PC mit Immunhistochemie für GABA (A) und GAD (B) sowie in-situ-Hybridisierung gegen GAD65- (C) und GAD67-mRNA (D) unter Kontrollbedingungen. In Schicht II des PC ließ sich eine deutlich höhere Anzahl GABA- bzw. GAD-immunreaktiver Neurone im Vergleich zur in-situ-Hybridisierung darstellen (Pfeile), während sich die Anzahl markierter Neurone in den angrenzenden Schichten nicht wesentlich unterschied (Pfeilköpfe). Kalibrierbalken für A-D: 100µm.
A C
D B
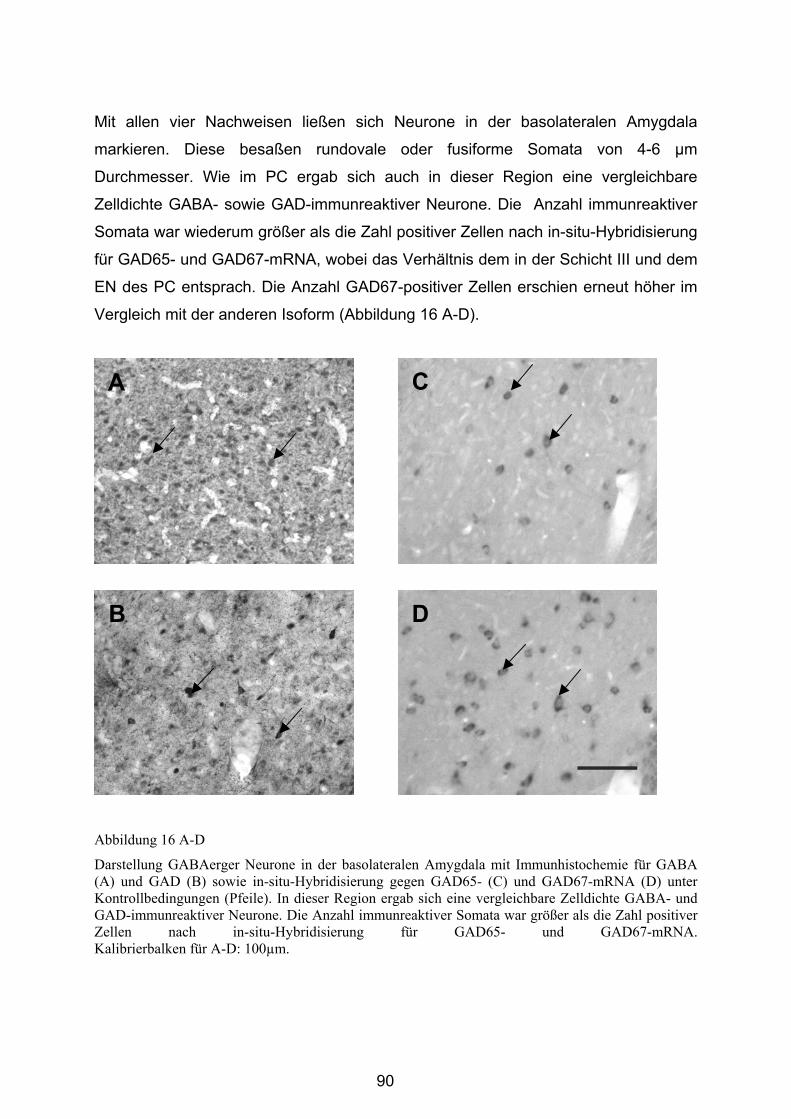
90
Mit allen vier Nachweisen ließen sich Neurone in der basolateralen Amygdala
markieren. Diese besaßen rundovale oder fusiforme Somata von 4-6 µm
Durchmesser. Wie im PC ergab sich auch in dieser Region eine vergleichbare
Zelldichte GABA- sowie GAD-immunreaktiver Neurone. Die Anzahl immunreaktiver
Somata war wiederum größer als die Zahl positiver Zellen nach in-situ-Hybridisierung
für GAD65- und GAD67-mRNA, wobei das Verhältnis dem in der Schicht III und dem
EN des PC entsprach. Die Anzahl GAD67-positiver Zellen erschien erneut höher im
Vergleich mit der anderen Isoform (Abbildung 16 A-D).
Abbildung 16 A-D
Darstellung GABAerger Neurone in der basolateralen Amygdala mit Immunhistochemie für GABA (A) und GAD (B) sowie in-situ-Hybridisierung gegen GAD65- (C) und GAD67-mRNA (D) unter Kontrollbedingungen (Pfeile). In dieser Region ergab sich eine vergleichbare Zelldichte GABA- und GAD-immunreaktiver Neurone. Die Anzahl immunreaktiver Somata war größer als die Zahl positiver Zellen nach in-situ-Hybridisierung für GAD65- und GAD67-mRNA. Kalibrierbalken für A-D: 100µm.
A C
D B
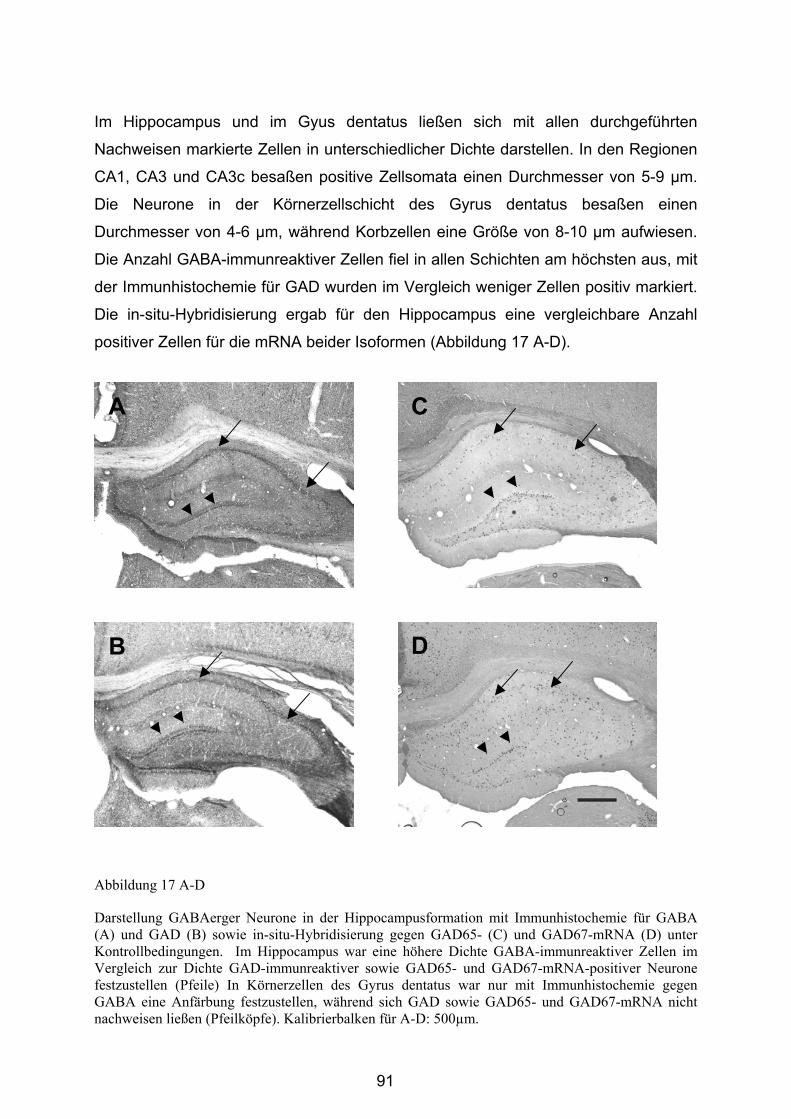
91
Im Hippocampus und im Gyus dentatus ließen sich mit allen durchgeführten
Nachweisen markierte Zellen in unterschiedlicher Dichte darstellen. In den Regionen
CA1, CA3 und CA3c besaßen positive Zellsomata einen Durchmesser von 5-9 µm.
Die Neurone in der Körnerzellschicht des Gyrus dentatus besaßen einen
Durchmesser von 4-6 µm, während Korbzellen eine Größe von 8-10 µm aufwiesen.
Die Anzahl GABA-immunreaktiver Zellen fiel in allen Schichten am höchsten aus, mit
der Immunhistochemie für GAD wurden im Vergleich weniger Zellen positiv markiert.
Die in-situ-Hybridisierung ergab für den Hippocampus eine vergleichbare Anzahl
positiver Zellen für die mRNA beider Isoformen (Abbildung 17 A-D).
Abbildung 17 A-D
Darstellung GABAerger Neurone in der Hippocampusformation mit Immunhistochemie für GABA (A) und GAD (B) sowie in-situ-Hybridisierung gegen GAD65- (C) und GAD67-mRNA (D) unter Kontrollbedingungen. Im Hippocampus war eine höhere Dichte GABA-immunreaktiver Zellen im Vergleich zur Dichte GAD-immunreaktiver sowie GAD65- und GAD67-mRNA-positiver Neurone festzustellen (Pfeile) In Körnerzellen des Gyrus dentatus war nur mit Immunhistochemie gegen GABA eine Anfärbung festzustellen, während sich GAD sowie GAD65- und GAD67-mRNA nicht nachweisen ließen (Pfeilköpfe). Kalibrierbalken für A-D: 500µm.
A C
D B

92
Im Gyrus dentatus ließen sich unter Kontrollbedingungen nur mit Immunhistochemie
für GABA Neurone in der Körnerzellschicht markieren. Mit Immunhistochemie für
GAD erfolgte die Färbung von Korbzellen in vergleichbarer Anzahl zur in-situ-
Hybridisierung. Im Hilus des Gyrus dentatus wurden wiederum die meisten positiven
Zellsomata nach Immunhistochemie für GABA gesehen, die Anzahl GAD-
immunreaktiver Neurone erschien etwas geringer im Vergleich zur in-situ-
Hybridisierung (Abbildung 17 A-D).
In der Substantia nigra pars reticulata besaßen GABAerge Neurone mit 15-20 µm
den größten Durchmesser aller untersuchten Hirnregionen. Hier wurde mit allen
Nachweisen die Mehrzahl der Zellen dargestellt. Die Dichte markierter Neurone fiel
im posterioren Teil der Region höher aus als im anterioren. Zudem erschien die
Anzahl positiver Neurone nach in-situ-Hybridisierung höher als nach den
immunhistochemischen Nachweisen. Insgesamt zeigten die Neurone in der
Substantia nigra pars reticulata für allen Nachweise eine höhere Intensität in der
Anfärbung im Vergleich zu markierten Zellen im limbischen System. Auffällig war
weiterhin die hohe Dichte GAD- und GABA-immunreaktiver Terminalen
(Abbildung 18 A-D).

93
Abbildung 18 A-D
Darstellung GABAerger Neurone in der posterioren Substantia nigra pars reticulata mit Immunhistochemie für GABA (A) und GAD (B) sowie in-situ-Hybridisierung gegen GAD65- (C) und GAD67-mRNA (D). Im Gegensatz zum PC ergab sich unter Kontrollbedingungen mit allen Nachweisen eine vergleichbare Dichte markierter Zellen (Pfeile). Die Anzahl GAD- und GABA-immunreaktiver Terminalen war in der Substantia nigra pars reticulata im Vergleich zu den anderen untersuchten Hirnregionen auffällig (Pfeilköpfe). Kalibrierbalken für A-D: 50 µm.
A
B
C
D
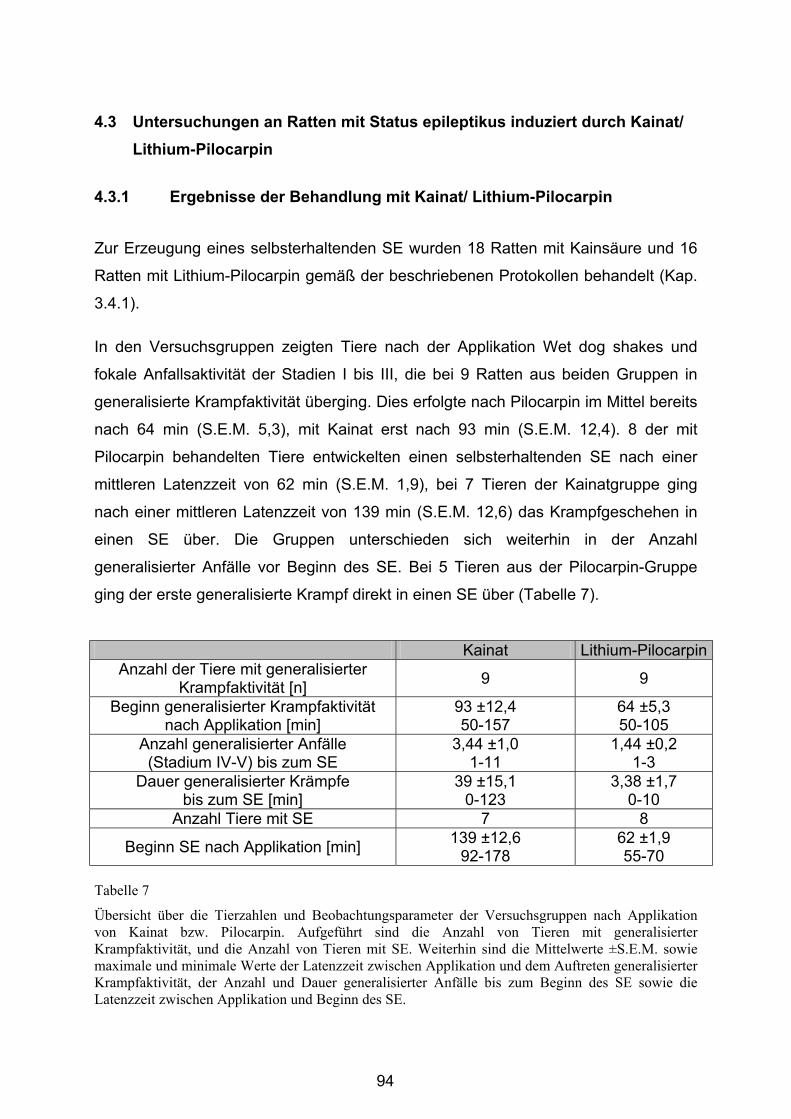
94
4.3 Untersuchungen an Ratten mit Status epileptikus induziert durch Kainat/ Lithium-Pilocarpin
4.3.1 Ergebnisse der Behandlung mit Kainat/ Lithium-Pilocarpin
Zur Erzeugung eines selbsterhaltenden SE wurden 18 Ratten mit Kainsäure und 16
Ratten mit Lithium-Pilocarpin gemäß der beschriebenen Protokollen behandelt (Kap.
3.4.1).
In den Versuchsgruppen zeigten Tiere nach der Applikation Wet dog shakes und
fokale Anfallsaktivität der Stadien I bis III, die bei 9 Ratten aus beiden Gruppen in
generalisierte Krampfaktivität überging. Dies erfolgte nach Pilocarpin im Mittel bereits
nach 64 min (S.E.M. 5,3), mit Kainat erst nach 93 min (S.E.M. 12,4). 8 der mit
Pilocarpin behandelten Tiere entwickelten einen selbsterhaltenden SE nach einer
mittleren Latenzzeit von 62 min (S.E.M. 1,9), bei 7 Tieren der Kainatgruppe ging
nach einer mittleren Latenzzeit von 139 min (S.E.M. 12,6) das Krampfgeschehen in
einen SE über. Die Gruppen unterschieden sich weiterhin in der Anzahl
generalisierter Anfälle vor Beginn des SE. Bei 5 Tieren aus der Pilocarpin-Gruppe
ging der erste generalisierte Krampf direkt in einen SE über (Tabelle 7).
Kainat Lithium-Pilocarpin Anzahl der Tiere mit generalisierter
Krampfaktivität [n] 9 9
Beginn generalisierter Krampfaktivität nach Applikation [min]
93 ±12,4 50-157
64 ±5,3 50-105
Anzahl generalisierter Anfälle (Stadium IV-V) bis zum SE
3,44 ±1,0 1-11
1,44 ±0,2 1-3
Dauer generalisierter Krämpfe bis zum SE [min]
39 ±15,1 0-123
3,38 ±1,7 0-10
Anzahl Tiere mit SE 7 8
Beginn SE nach Applikation [min] 139 ±12,6 92-178
62 ±1,9 55-70
Tabelle 7
Übersicht über die Tierzahlen und Beobachtungsparameter der Versuchsgruppen nach Applikation von Kainat bzw. Pilocarpin. Aufgeführt sind die Anzahl von Tieren mit generalisierter Krampfaktivität, und die Anzahl von Tieren mit SE. Weiterhin sind die Mittelwerte ±S.E.M. sowie maximale und minimale Werte der Latenzzeit zwischen Applikation und dem Auftreten generalisierter Krampfaktivität, der Anzahl und Dauer generalisierter Anfälle bis zum Beginn des SE sowie die Latenzzeit zwischen Applikation und Beginn des SE.

95
Die Unterbrechung des SE erfolgte nach 90 min mit Diazepam. 8 h nach Applikation
von Kainat bzw. Pilocarpin wurden die Tiere perfundiert. Eine Ratte verendete im
weiteren Verlauf des Versuches. Dementsprechend standen für die weiteren
Untersuchungen 6 bzw. 8 Tiere nach SE induziert durch Kainsäure bzw. Lithium-
Pilocarpin sowie 8 Kontrolltiere zur Verfügung.
4.3.2 Bestimmung der Dichte GABAerger Neurone
Für die Zählung GABAerger Neurone in den Versuchsgruppen wurde nach der
Methode des sog. Optischen Disektors (Saper, 1996 und West, 1999) vorgegangen,
wie sie in Kap. 3.5.2 erläutert wurde. Dafür wurden die festgelegten Zählbereiche
innerhalb der zu untersuchenden Regionen anhand oben beschriebener
Vorgehensweise aufgesucht. Die Auswertung erfolgte an verschlüsselten
Objektträgern, um subjektive Einflüsse durch den Untersucher auf das Ergebnis der
Zählungen ausschließen zu können. Bei der Zählung wurden nur Zellen
berücksichtigt, die eine unveränderte Morphologie aufwiesen. Nach der Zuordnung
der Daten zu den einzelnen Versuchsgruppen und Nachweisen erfolgte die
Berechnung der Neuronendichte auf die Anzahl von Neuronen/mm3 (Kap. 3.5.5). Die
Gruppenmittelwerte und die zugehörigen Standardfehler für die verschiedenen
Regionen und Schichten wurden ermittelt und verglichen. Dazu erfolgte für jeden
Nachweis eine einfaktorielle analysis of variance (ANOVA) der Neuronendichte
zwischen den Versuchsgruppen in den einzelnen Schichten der untersuchten
Gehirnregionen.
Für die Auswertung standen 8 Kontrolltiere, 6 Ratten mit SE induziert durch Kainat
sowie 8 Tiere mit SE induziert durch Lithium-Pilocarpin zur Verfügung. Eine
Verringerung der Anzahl auswertbarer Serien aufgrund der Beschädigung von
Gewebeschnitten bei der Herstellung bzw. durch mangelhafte Anfärbung ist bei der
Darstellung der untersuchten Hirnregionen aufgeführt.

96
4.3.2.1 Anteriorer Piriformer Cortex
Im anterioren PC wurden die Neuronendichten in den Schichten I und II sowie im EN
bestimmt. Da sich Schicht III erst im weiteren Verlauf des PC nach caudal deutlich
abgrenzen lässt, wurde die zwischen Schicht II und dem DEN gelegene Schicht VEN
definiert.
Für die Kontrollgruppe verringerte sich die Anzahl auswertbarer Tiere auf 7 für jeden
Nachweis, in der Kainat-Gruppe konnten für die GAD-Immunhistochemie nur 5 Tiere
berücksichtigt werden. In der Pilocarpin-Gruppe konnten für die in-situ-Hybridisierung
gegen GAD67-mRNA 6 Tiere, für die anderen Nachweise jeweils 7 Tiere
berücksichtigt werden.
Unter Kontrollbedingungen wurde mit in-situ-Hybridisierung für GAD65- und GAD67-
mRNA die höchste Dichte markierter Neurone im VEN bestimmt. Dabei waren im
Mittel 8645 Neurone/mm3 (S.E.M. 687,8) positiv für GAD65-mRNA und 8760
Neurone/mm3 (S.E.M. 896,6) positiv für GAD67-mRNA. Die zweithöchste Dichte
besaß der DEN, gefolgt von Schicht II und Schicht I. Die Anzahl GAD67-mRNA-
positiver Neurone lag dabei im Mittel für jede Schicht über den Werten für GAD65-
mRNA (Tabelle 8).
Anteriorer PC GAD65
in-situ-Hybridisierung GAD67
in-situ-Hybridisierung
Neurone/mm3 Neurone/mm3 Gruppe n I II VEN DEN n I II VEN DEN
491 4583 8645 6557 818 6687 8760 7107 Kontrolle 7 ±284,3 ±699,2 ±687,8 ±481,4 7 ±128,8 ±1037,8 ±896,6 ±527,3 1398 5415 5857 4901 635 6334 6082 4202 SE nach
Kainat 6 ±660,2 ±302,6 ±492,9 ±621,3 6 ±212,7 ±464,7 ±749,7 ±574,0 1527 4725 4640 3874 509 6373 5766 4207 SE nach
Pilocarpin 7 ±323,2 ±442,5 ±382,9 ±568,5 6 ±305,4 ±742,3 ±774,2 ±623,8
Tabelle 8
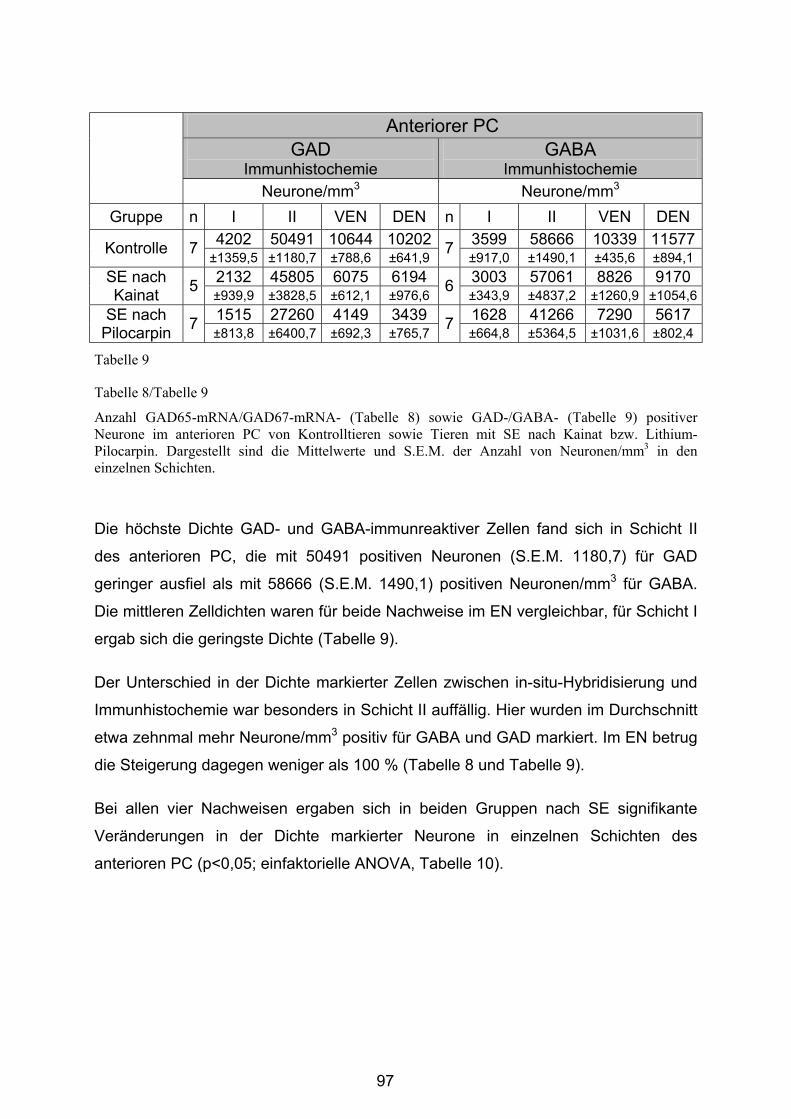
97
Anteriorer PC GAD
Immunhistochemie GABA
Immunhistochemie
Neurone/mm3 Neurone/mm3 Gruppe n I II VEN DEN n I II VEN DEN
4202 50491 10644 10202 3599 58666 10339 11577 Kontrolle 7 ±1359,5 ±1180,7 ±788,6 ±641,9 7 ±917,0 ±1490,1 ±435,6 ±894,1 2132 45805 6075 6194 3003 57061 8826 9170 SE nach
Kainat 5 ±939,9 ±3828,5 ±612,1 ±976,6 6 ±343,9 ±4837,2 ±1260,9 ±1054,6 1515 27260 4149 3439 1628 41266 7290 5617 SE nach
Pilocarpin 7 ±813,8 ±6400,7 ±692,3 ±765,7 7 ±664,8 ±5364,5 ±1031,6 ±802,4
Tabelle 9
Tabelle 8/Tabelle 9
Anzahl GAD65-mRNA/GAD67-mRNA- (Tabelle 8) sowie GAD-/GABA- (Tabelle 9) positiver Neurone im anterioren PC von Kontrolltieren sowie Tieren mit SE nach Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die Mittelwerte und S.E.M. der Anzahl von Neuronen/mm3 in den einzelnen Schichten.
Die höchste Dichte GAD- und GABA-immunreaktiver Zellen fand sich in Schicht II
des anterioren PC, die mit 50491 positiven Neuronen (S.E.M. 1180,7) für GAD
geringer ausfiel als mit 58666 (S.E.M. 1490,1) positiven Neuronen/mm3 für GABA.
Die mittleren Zelldichten waren für beide Nachweise im EN vergleichbar, für Schicht I
ergab sich die geringste Dichte (Tabelle 9).
Der Unterschied in der Dichte markierter Zellen zwischen in-situ-Hybridisierung und
Immunhistochemie war besonders in Schicht II auffällig. Hier wurden im Durchschnitt
etwa zehnmal mehr Neurone/mm3 positiv für GABA und GAD markiert. Im EN betrug
die Steigerung dagegen weniger als 100 % (Tabelle 8 und Tabelle 9).
Bei allen vier Nachweisen ergaben sich in beiden Gruppen nach SE signifikante
Veränderungen in der Dichte markierter Neurone in einzelnen Schichten des
anterioren PC (p<0,05; einfaktorielle ANOVA, Tabelle 10).
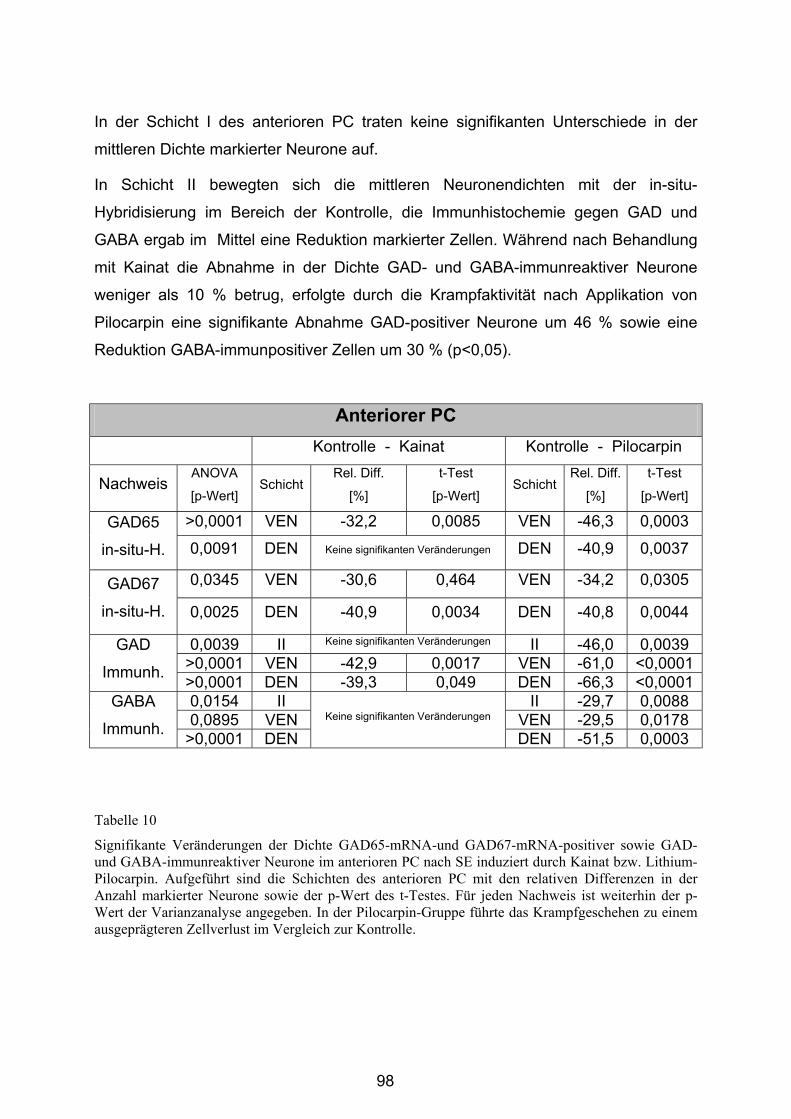
98
In der Schicht I des anterioren PC traten keine signifikanten Unterschiede in der
mittleren Dichte markierter Neurone auf.
In Schicht II bewegten sich die mittleren Neuronendichten mit der in-situ-
Hybridisierung im Bereich der Kontrolle, die Immunhistochemie gegen GAD und
GABA ergab im Mittel eine Reduktion markierter Zellen. Während nach Behandlung
mit Kainat die Abnahme in der Dichte GAD- und GABA-immunreaktiver Neurone
weniger als 10 % betrug, erfolgte durch die Krampfaktivität nach Applikation von
Pilocarpin eine signifikante Abnahme GAD-positiver Neurone um 46 % sowie eine
Reduktion GABA-immunpositiver Zellen um 30 % (p<0,05).
Anteriorer PC Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA
[p-Wert] Schicht
Rel. Diff.
[%]
t-Test
[p-Wert] Schicht
Rel. Diff.
[%]
t-Test
[p-Wert]
>0,0001 VEN -32,2 0,0085 VEN -46,3 0,0003 GAD65
in-situ-H. 0,0091 DEN Keine signifikanten Veränderungen DEN -40,9 0,0037
0,0345 VEN -30,6 0,464 VEN -34,2 0,0305 GAD67
in-situ-H. 0,0025 DEN -40,9 0,0034 DEN -40,8 0,0044
0,0039 II Keine signifikanten Veränderungen II -46,0 0,0039 >0,0001 VEN -42,9 0,0017 VEN -61,0 <0,0001
GAD
Immunh. >0,0001 DEN -39,3 0,049 DEN -66,3 <0,0001 0,0154 II II -29,7 0,0088 0,0895 VEN VEN -29,5 0,0178
GABA
Immunh. >0,0001 DEN Keine signifikanten Veränderungen
DEN -51,5 0,0003
Tabelle 10
Signifikante Veränderungen der Dichte GAD65-mRNA-und GAD67-mRNA-positiver sowie GAD- und GABA-immunreaktiver Neurone im anterioren PC nach SE induziert durch Kainat bzw. Lithium-Pilocarpin. Aufgeführt sind die Schichten des anterioren PC mit den relativen Differenzen in der Anzahl markierter Neurone sowie der p-Wert des t-Testes. Für jeden Nachweis ist weiterhin der p-Wert der Varianzanalyse angegeben. In der Pilocarpin-Gruppe führte das Krampfgeschehen zu einem ausgeprägteren Zellverlust im Vergleich zur Kontrolle.

99
In den Versuchsgruppen lagen die Mittelwerte im VEN für alle Nachweise unterhalb
der Kontrollwerte. Die Behandlung mit Kainat und Lithium-Pilocarpin führte zur
signifikanten Reduktion von GAD65- und GAD67-mRNA sowie GAD-positiven
Neuronen, die Anzahl GABA-immunreaktiver Zellen wurde nur in der Pilocarpin-
Gruppe statistisch signifikant verringert (Abbildung 19 A-D). Während die Zellverluste
durch Kainat zwischen 31-42 % betrugen, reduzierte sich die Dichte nach Pilocarpin
um 34-61 %, wobei GAD-markierte Zellen im Mittel am stärksten betroffen waren.
Im DEN waren mit allen Methoden in der Pilocarpin-Gruppe signifikant weniger
Neurone nachzuweisen (Abbildung 19 A-D). Positive Neurone für GAD65- bzw.
GAD67-mRNA reduzierten sich um durchschnittlich 41 %, die Verminderung GAD-
und GABA-immunpositiver Zellen betrug dagegen 66 % sowie 52 %. Durch Kainat
verringerte sich die Dichte für GAD67-mRNA- und GAD-markierter Zellen signifikant
um 41 bzw. 39 %. Die mittlere Abnahme der Neuronendichte, dargestellt mit den
übrigen Nachweisen, war mit 21-25 % nicht signifikant.
Abbildung 19 A-D
Anzahl GAD65-mRNA- (A), GAD67-mRNA- (B) sowie GAD- (C) und GABA- (D) positiver Neurone/mm3 in den unterschiedlichen Schichten des anterioren PC. Dargestellt sind Mittelwerte ±S.E.M. von Tieren mit SE induziert durch Kainat bzw. Lithium-Pilocarpin im Vergleich zur Kontrolle. Unter Kontrollbedingungen wurden in allen Schichten höhere Dichten immunreaktiver Neurone festgestellt. In der Pilocarpin-Gruppe waren immunreaktive Zellen signifikant reduziert (C/D), während die Dichten GAD65- und GAD67-mRNA-positiver Neurone näherungsweise Kontrollniveau besaßen. Signifikante Unterschiede zwischen Kontrollen und behandelten Tieren sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit anschließendem t-Test]

100
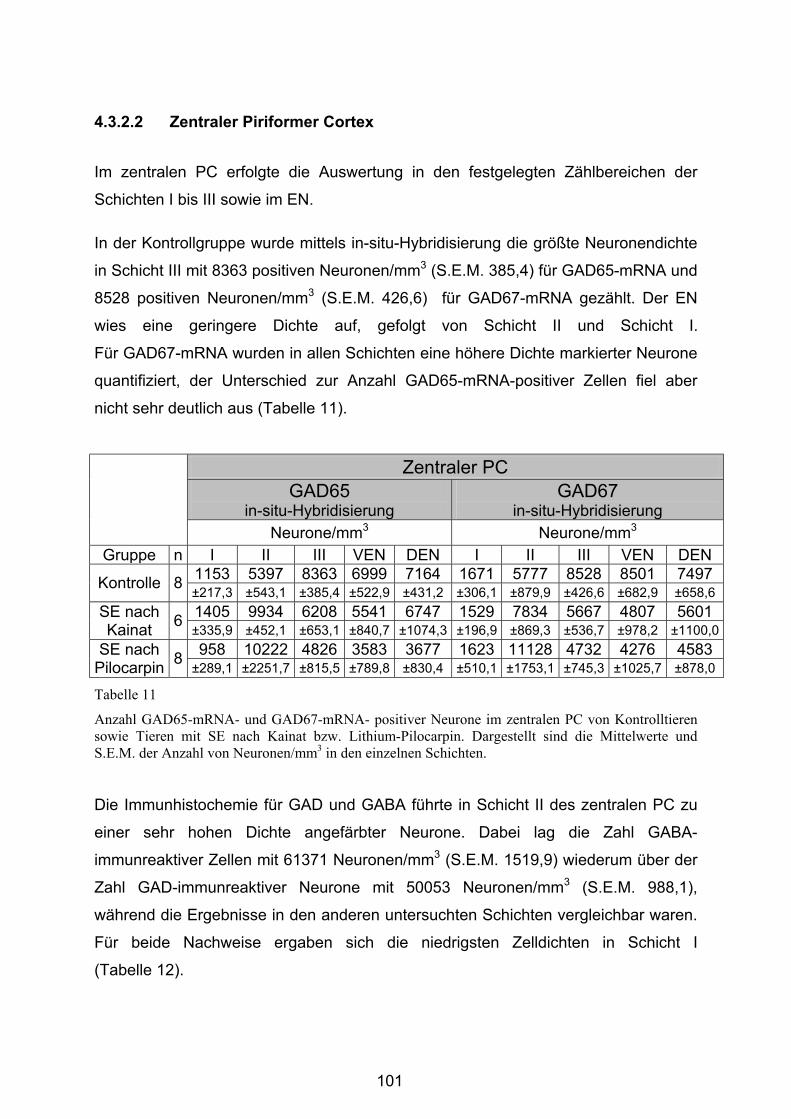
101
4.3.2.2 Zentraler Piriformer Cortex
Im zentralen PC erfolgte die Auswertung in den festgelegten Zählbereichen der
Schichten I bis III sowie im EN.
In der Kontrollgruppe wurde mittels in-situ-Hybridisierung die größte Neuronendichte
in Schicht III mit 8363 positiven Neuronen/mm3 (S.E.M. 385,4) für GAD65-mRNA und
8528 positiven Neuronen/mm3 (S.E.M. 426,6) für GAD67-mRNA gezählt. Der EN
wies eine geringere Dichte auf, gefolgt von Schicht II und Schicht I.
Für GAD67-mRNA wurden in allen Schichten eine höhere Dichte markierter Neurone
quantifiziert, der Unterschied zur Anzahl GAD65-mRNA-positiver Zellen fiel aber
nicht sehr deutlich aus (Tabelle 11).
Zentraler PC GAD65
in-situ-Hybridisierung GAD67
in-situ-Hybridisierung
Neurone/mm3 Neurone/mm3 Gruppe n I II III VEN DEN I II III VEN DEN
1153 5397 8363 6999 7164 1671 5777 8528 8501 7497 Kontrolle 8 ±217,3 ±543,1 ±385,4 ±522,9 ±431,2 ±306,1 ±879,9 ±426,6 ±682,9 ±658,6 1405 9934 6208 5541 6747 1529 7834 5667 4807 5601 SE nach
Kainat 6 ±335,9 ±452,1 ±653,1 ±840,7 ±1074,3 ±196,9 ±869,3 ±536,7 ±978,2 ±1100,0 958 10222 4826 3583 3677 1623 11128 4732 4276 4583 SE nach
Pilocarpin 8 ±289,1 ±2251,7 ±815,5 ±789,8 ±830,4 ±510,1 ±1753,1 ±745,3 ±1025,7 ±878,0
Tabelle 11
Anzahl GAD65-mRNA- und GAD67-mRNA- positiver Neurone im zentralen PC von Kontrolltieren sowie Tieren mit SE nach Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die Mittelwerte und S.E.M. der Anzahl von Neuronen/mm3 in den einzelnen Schichten.
Die Immunhistochemie für GAD und GABA führte in Schicht II des zentralen PC zu
einer sehr hohen Dichte angefärbter Neurone. Dabei lag die Zahl GABA-
immunreaktiver Zellen mit 61371 Neuronen/mm3 (S.E.M. 1519,9) wiederum über der
Zahl GAD-immunreaktiver Neurone mit 50053 Neuronen/mm3 (S.E.M. 988,1),
während die Ergebnisse in den anderen untersuchten Schichten vergleichbar waren.
Für beide Nachweise ergaben sich die niedrigsten Zelldichten in Schicht I
(Tabelle 12).
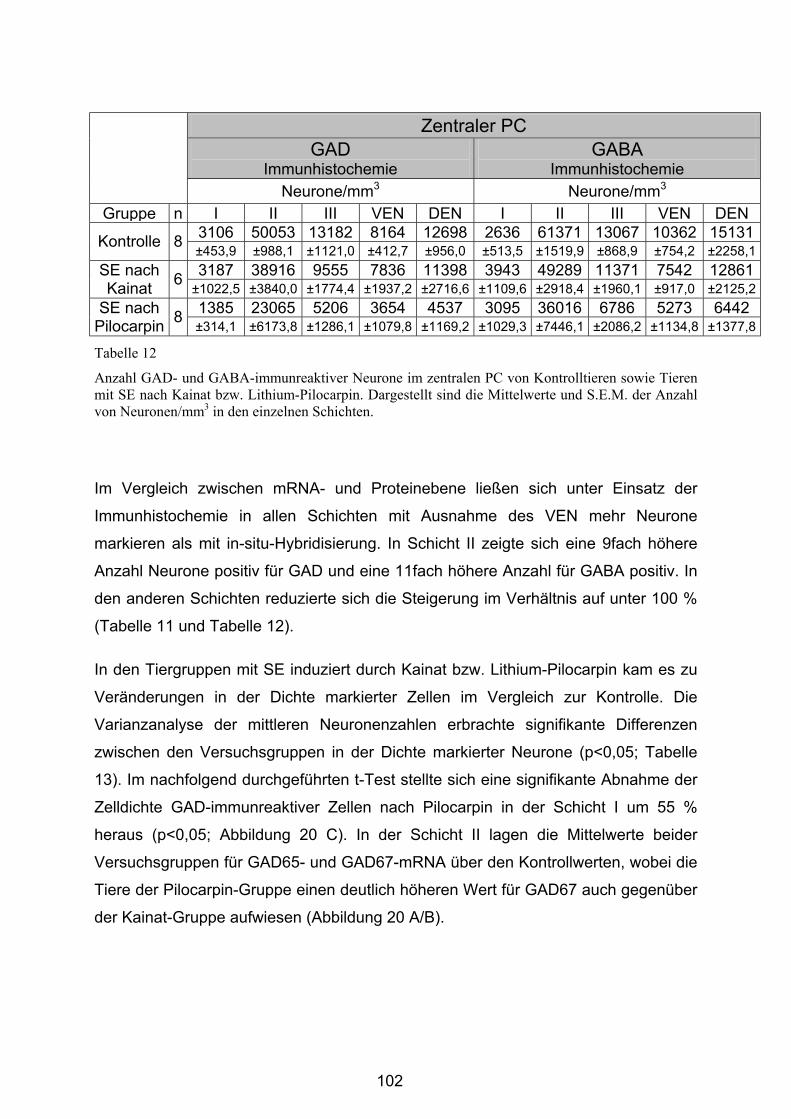
102
Zentraler PC GAD
Immunhistochemie GABA
Immunhistochemie
Neurone/mm3 Neurone/mm3 Gruppe n I II III VEN DEN I II III VEN DEN
3106 50053 13182 8164 12698 2636 61371 13067 10362 15131 Kontrolle 8 ±453,9 ±988,1 ±1121,0 ±412,7 ±956,0 ±513,5 ±1519,9 ±868,9 ±754,2 ±2258,1 3187 38916 9555 7836 11398 3943 49289 11371 7542 12861 SE nach
Kainat 6 ±1022,5 ±3840,0 ±1774,4 ±1937,2 ±2716,6 ±1109,6 ±2918,4 ±1960,1 ±917,0 ±2125,2 1385 23065 5206 3654 4537 3095 36016 6786 5273 6442 SE nach
Pilocarpin 8 ±314,1 ±6173,8 ±1286,1 ±1079,8 ±1169,2 ±1029,3 ±7446,1 ±2086,2 ±1134,8 ±1377,8
Tabelle 12
Anzahl GAD- und GABA-immunreaktiver Neurone im zentralen PC von Kontrolltieren sowie Tieren mit SE nach Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die Mittelwerte und S.E.M. der Anzahl von Neuronen/mm3 in den einzelnen Schichten.
Im Vergleich zwischen mRNA- und Proteinebene ließen sich unter Einsatz der
Immunhistochemie in allen Schichten mit Ausnahme des VEN mehr Neurone
markieren als mit in-situ-Hybridisierung. In Schicht II zeigte sich eine 9fach höhere
Anzahl Neurone positiv für GAD und eine 11fach höhere Anzahl für GABA positiv. In
den anderen Schichten reduzierte sich die Steigerung im Verhältnis auf unter 100 %
(Tabelle 11 und Tabelle 12).
In den Tiergruppen mit SE induziert durch Kainat bzw. Lithium-Pilocarpin kam es zu
Veränderungen in der Dichte markierter Zellen im Vergleich zur Kontrolle. Die
Varianzanalyse der mittleren Neuronenzahlen erbrachte signifikante Differenzen
zwischen den Versuchsgruppen in der Dichte markierter Neurone (p<0,05; Tabelle
13). Im nachfolgend durchgeführten t-Test stellte sich eine signifikante Abnahme der
Zelldichte GAD-immunreaktiver Zellen nach Pilocarpin in der Schicht I um 55 %
heraus (p<0,05; Abbildung 20 C). In der Schicht II lagen die Mittelwerte beider
Versuchsgruppen für GAD65- und GAD67-mRNA über den Kontrollwerten, wobei die
Tiere der Pilocarpin-Gruppe einen deutlich höheren Wert für GAD67 auch gegenüber
der Kainat-Gruppe aufwiesen (Abbildung 20 A/B).
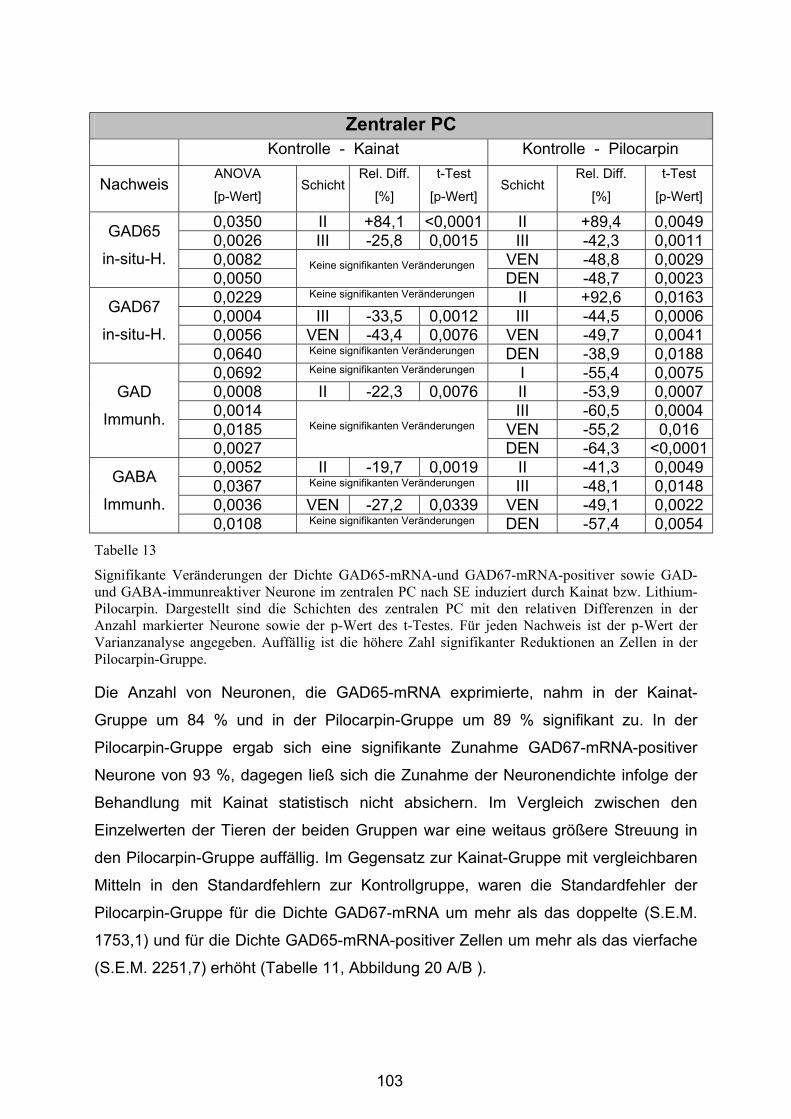
103
Zentraler PC Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA
[p-Wert] Schicht
Rel. Diff.
[%]
t-Test
[p-Wert] Schicht
Rel. Diff.
[%]
t-Test
[p-Wert]
0,0350 II +84,1 <0,0001 II +89,4 0,0049 0,0026 III -25,8 0,0015 III -42,3 0,0011 0,0082 VEN -48,8 0,0029
GAD65
in-situ-H. 0,0050
Keine signifikanten Veränderungen DEN -48,7 0,0023
0,0229 Keine signifikanten Veränderungen II +92,6 0,0163 0,0004 III -33,5 0,0012 III -44,5 0,0006 0,0056 VEN -43,4 0,0076 VEN -49,7 0,0041
GAD67
in-situ-H. 0,0640 Keine signifikanten Veränderungen DEN -38,9 0,0188 0,0692 Keine signifikanten Veränderungen I -55,4 0,0075 0,0008 II -22,3 0,0076 II -53,9 0,0007 0,0014 III -60,5 0,0004 0,0185 VEN -55,2 0,016
GAD
Immunh.
0,0027 Keine signifikanten Veränderungen
DEN -64,3 <0,0001 0,0052 II -19,7 0,0019 II -41,3 0,0049 0,0367 Keine signifikanten Veränderungen III -48,1 0,0148 0,0036 VEN -27,2 0,0339 VEN -49,1 0,0022
GABA
Immunh. 0,0108 Keine signifikanten Veränderungen DEN -57,4 0,0054
Tabelle 13
Signifikante Veränderungen der Dichte GAD65-mRNA-und GAD67-mRNA-positiver sowie GAD- und GABA-immunreaktiver Neurone im zentralen PC nach SE induziert durch Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die Schichten des zentralen PC mit den relativen Differenzen in der Anzahl markierter Neurone sowie der p-Wert des t-Testes. Für jeden Nachweis ist der p-Wert der Varianzanalyse angegeben. Auffällig ist die höhere Zahl signifikanter Reduktionen an Zellen in der Pilocarpin-Gruppe.
Die Anzahl von Neuronen, die GAD65-mRNA exprimierte, nahm in der Kainat-
Gruppe um 84 % und in der Pilocarpin-Gruppe um 89 % signifikant zu. In der
Pilocarpin-Gruppe ergab sich eine signifikante Zunahme GAD67-mRNA-positiver
Neurone von 93 %, dagegen ließ sich die Zunahme der Neuronendichte infolge der
Behandlung mit Kainat statistisch nicht absichern. Im Vergleich zwischen den
Einzelwerten der Tieren der beiden Gruppen war eine weitaus größere Streuung in
den Pilocarpin-Gruppe auffällig. Im Gegensatz zur Kainat-Gruppe mit vergleichbaren
Mitteln in den Standardfehlern zur Kontrollgruppe, waren die Standardfehler der
Pilocarpin-Gruppe für die Dichte GAD67-mRNA um mehr als das doppelte (S.E.M.
1753,1) und für die Dichte GAD65-mRNA-positiver Zellen um mehr als das vierfache
(S.E.M. 2251,7) erhöht (Tabelle 11, Abbildung 20 A/B ).

104
Im Gegensatz zur gesteigerten Expression von GAD-mRNA verringerte sich die
Dichte GAD- und GABA-immunpositiver Neurone in der Schicht II signifikant, wobei
die Mittelwerte der Pilocarpin-Gruppe unter denen der Kainat-Gruppe lagen
(Abbildung 20 C/D). Die Mittelwerte der Kainat-Gruppe waren mit 22 % für GAD und
20 % für GABA auf gleichem Niveau zu finden, mit Pilocarpin wurden GABA-positive
Neurone um 41 % und für GAD markierte Zellen um 54 % verringert. In Schicht III
und dem angrenzenden EN wiesen die Mittelwerte aller Nachweise geringere Werte
als die Kontrolle auf, für Pilocarpin ergaben sich zudem niedrigere Mittel als für die
Gruppe mit SE nach Kainat (Abbildung 20 A-D). Ein Lithium-Pilocarpin-induzierter SE
führte zu einer signifikanten Reduktion in der Neuronendichte unter Anwendung aller
Nachweise in Schichten III und EN, die relativen Differenzen der Mittelwerte zur
Kontrollgruppe fielen dabei für die GAD- Immunhistochemie mit bis zu 65 % am
deutlichsten aus (Tabelle 13). Für die Abnahmen in der Neuronenzahl der Kainat-
Gruppe erwiesen sich die Mittelwerte der in-situ-Hybridisierung für GAD65-mRNA in
Schicht III und für GAD67-mRNA auch im VEN als statistisch signifikant. GABA-
immunreaktive Neurone waren in dieser Versuchsgruppe im VEN ebenfalls
signifikant vermindert.
Abbildung 20 A-D
Anzahl GAD65-mRNA- (A), GAD67-mRNA- (B) sowie GAD- (C) und GABA- (D) positiver Neurone/mm3 in den unterschiedlichen Schichten des zentralen PC. Dargestellt sind Mittelwerte ±S.E.M. von Tieren mit SE nach Kainat bzw. Lithium-Pilocarpin im Vergleich zur Kontrolle. Während es in Schicht II zur Zunahme in der Dichte GAD65- und GAD67-mRNA-positiver Neurone kam (A/B), war in dieser Schicht eine Reduktion in der Dichte immunreaktiver Zellen festzustellen (C/D). In der Schicht III sowie im EN ergaben sich mit allen Nachweisen geringere Neuronendichten. Signifikante Unterschiede zwischen Kontrollen und behandelten Tieren sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit anschließendem t-Test].
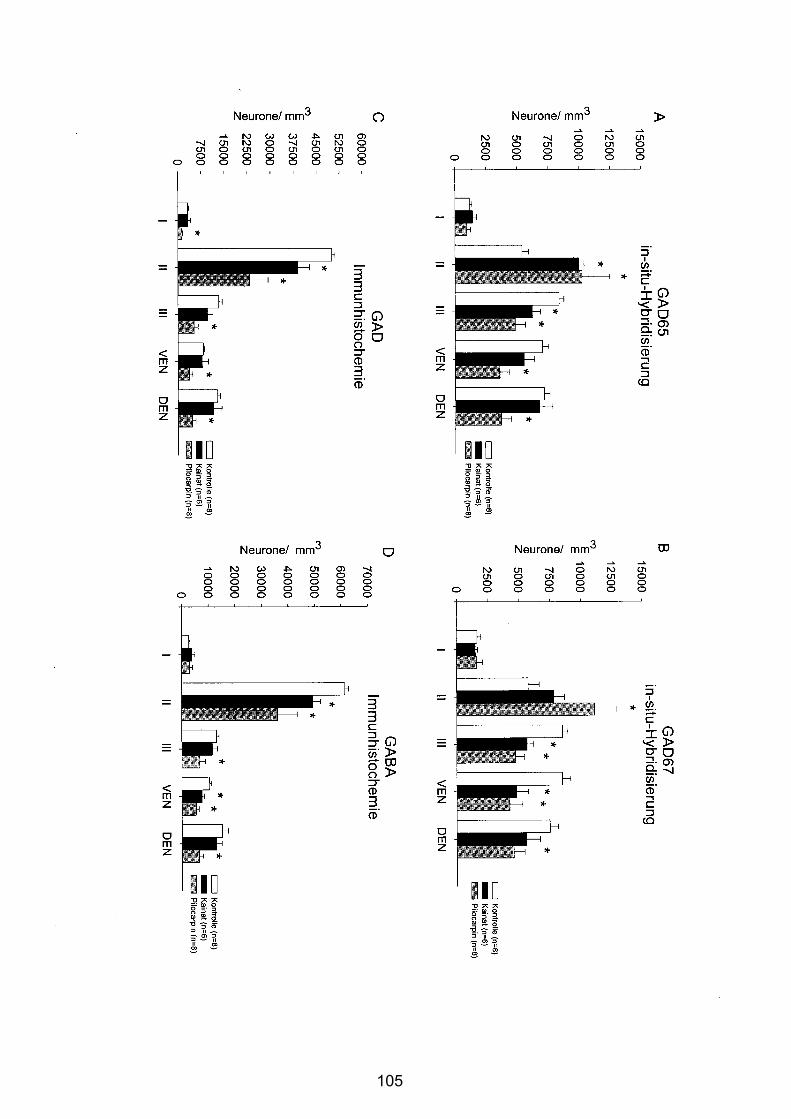
105

106
4.3.2.3 Posteriorer Piriformer Cortex
Im posterioren PC wurde die Auswertung in den definierten Zählbereichen der
Schichten I bis III sowie im EN durchgeführt.
In der Kontrollgruppe fanden sich nach in-situ-Hybridisierung die höchsten Dichten
GAD65- und GAD67-mRNA-positiver Neurone in der Schicht III, die Anzahl für
GAD67-mRNA gefärbter Zellen war in allen Schichten größer als für GAD65-mRNA.
Auffällig war eine gesteigerte Dichte markierter Zellen in Schicht II im Vergleich zum
anterioren und zentralen PC. Im posterioren PC wurden mit 8505 positiven
Neuronen/mm3 für GAD67-mRNA (S.E.M. 1072,9) eine höhere Dichte an Zellen
bestimmt als im angrenzenden DEN und VEN (Tabelle 14).
Im posterioren PC ergaben sich die höchsten Zelldichten mit der Immunhistochemie
wiederum in Schicht II. 59033 GABA-immunreaktive Zellen (S.E.M. 1730,9) wurden
gegenüber 49427 GAD-immunreaktiven Neuronen/mm3 (S.E.M. 1939,5) in dieser
Schicht quantifiziert. In Schicht III und dem EN ergaben sich vergleichbare Werte für
beide Nachweise (Tabelle 15).
Posteriorer PC GAD65
in-situ-Hybridisierung GAD67
in-situ-Hybridisierung
Neurone/mm3 Neurone/mm3 Gruppe n I II III VEN DEN I II III VEN DEN
2820 6121 9124 6969 7795 3118 8505 9422 7886 8253 Kontrolle 8 ±644,2 ±564,0 ±708,4 ±520,4 ±632,7 ±742,8 ±1072,9 ±641,9 ±499,8 ±582,3 3691 9629 7703 6946 7061 3118 10500 7703 7061 7336 SE nach
Kainat 6 ±749,7 ±894,1 ±1199,0 ±937,6 ±928,5 ±334,7 ±917,0 ±866,6 ±848,2 ±816,1 3003 9147 4310 3576 3439 2728 9789 5112 4402 6006 SE nach
Pilocarpin 8 ±619,0 ±1501,6 ±988,1 ±921,6 ±816,1 ±683,2 ±1558,9 ±1001,8 ±859,7 ±1091,2
Tabelle 14
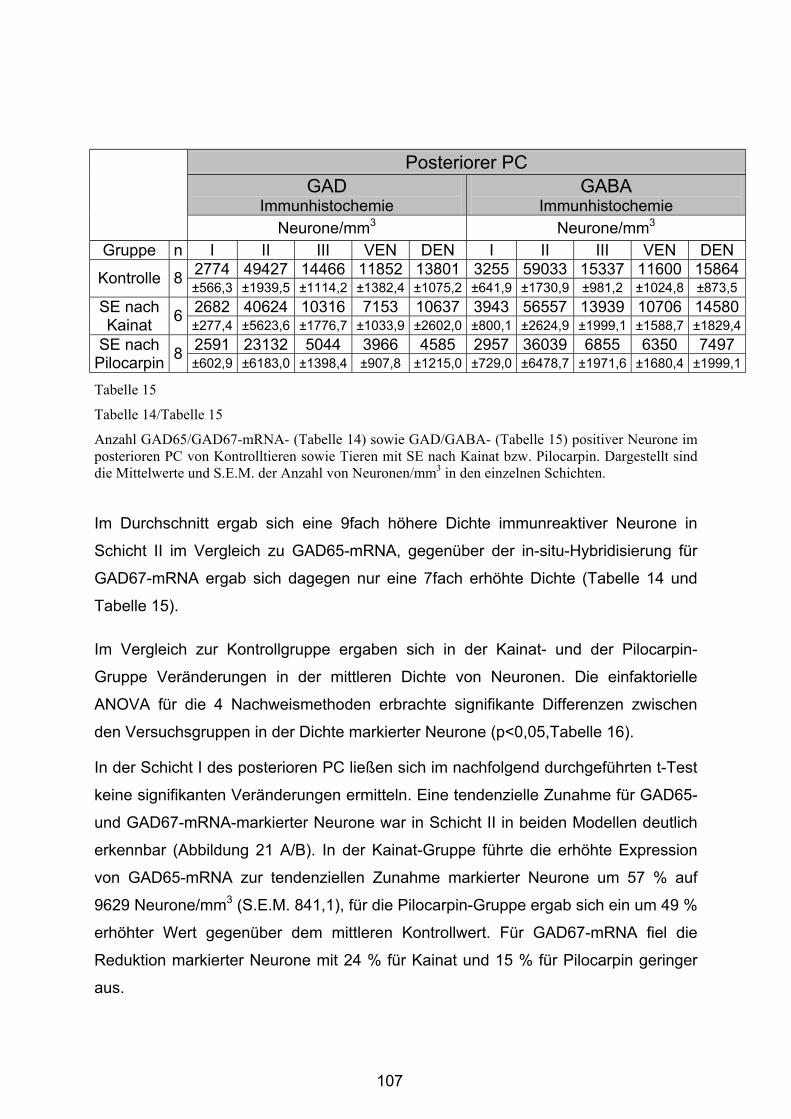
107
Posteriorer PC GAD
Immunhistochemie GABA
Immunhistochemie
Neurone/mm3 Neurone/mm3 Gruppe n I II III VEN DEN I II III VEN DEN
2774 49427 14466 11852 13801 3255 59033 15337 11600 15864 Kontrolle 8 ±566,3 ±1939,5 ±1114,2 ±1382,4 ±1075,2 ±641,9 ±1730,9 ±981,2 ±1024,8 ±873,5 2682 40624 10316 7153 10637 3943 56557 13939 10706 14580 SE nach
Kainat 6 ±277,4 ±5623,6 ±1776,7 ±1033,9 ±2602,0 ±800,1 ±2624,9 ±1999,1 ±1588,7 ±1829,4 2591 23132 5044 3966 4585 2957 36039 6855 6350 7497 SE nach
Pilocarpin 8 ±602,9 ±6183,0 ±1398,4 ±907,8 ±1215,0 ±729,0 ±6478,7 ±1971,6 ±1680,4 ±1999,1
Tabelle 15
Tabelle 14/Tabelle 15
Anzahl GAD65/GAD67-mRNA- (Tabelle 14) sowie GAD/GABA- (Tabelle 15) positiver Neurone im posterioren PC von Kontrolltieren sowie Tieren mit SE nach Kainat bzw. Pilocarpin. Dargestellt sind die Mittelwerte und S.E.M. der Anzahl von Neuronen/mm3 in den einzelnen Schichten.
Im Durchschnitt ergab sich eine 9fach höhere Dichte immunreaktiver Neurone in
Schicht II im Vergleich zu GAD65-mRNA, gegenüber der in-situ-Hybridisierung für
GAD67-mRNA ergab sich dagegen nur eine 7fach erhöhte Dichte (Tabelle 14 und
Tabelle 15).
Im Vergleich zur Kontrollgruppe ergaben sich in der Kainat- und der Pilocarpin-
Gruppe Veränderungen in der mittleren Dichte von Neuronen. Die einfaktorielle
ANOVA für die 4 Nachweismethoden erbrachte signifikante Differenzen zwischen
den Versuchsgruppen in der Dichte markierter Neurone (p<0,05,Tabelle 16).
In der Schicht I des posterioren PC ließen sich im nachfolgend durchgeführten t-Test
keine signifikanten Veränderungen ermitteln. Eine tendenzielle Zunahme für GAD65-
und GAD67-mRNA-markierter Neurone war in Schicht II in beiden Modellen deutlich
erkennbar (Abbildung 21 A/B). In der Kainat-Gruppe führte die erhöhte Expression
von GAD65-mRNA zur tendenziellen Zunahme markierter Neurone um 57 % auf
9629 Neurone/mm3 (S.E.M. 841,1), für die Pilocarpin-Gruppe ergab sich ein um 49 %
erhöhter Wert gegenüber dem mittleren Kontrollwert. Für GAD67-mRNA fiel die
Reduktion markierter Neurone mit 24 % für Kainat und 15 % für Pilocarpin geringer
aus.
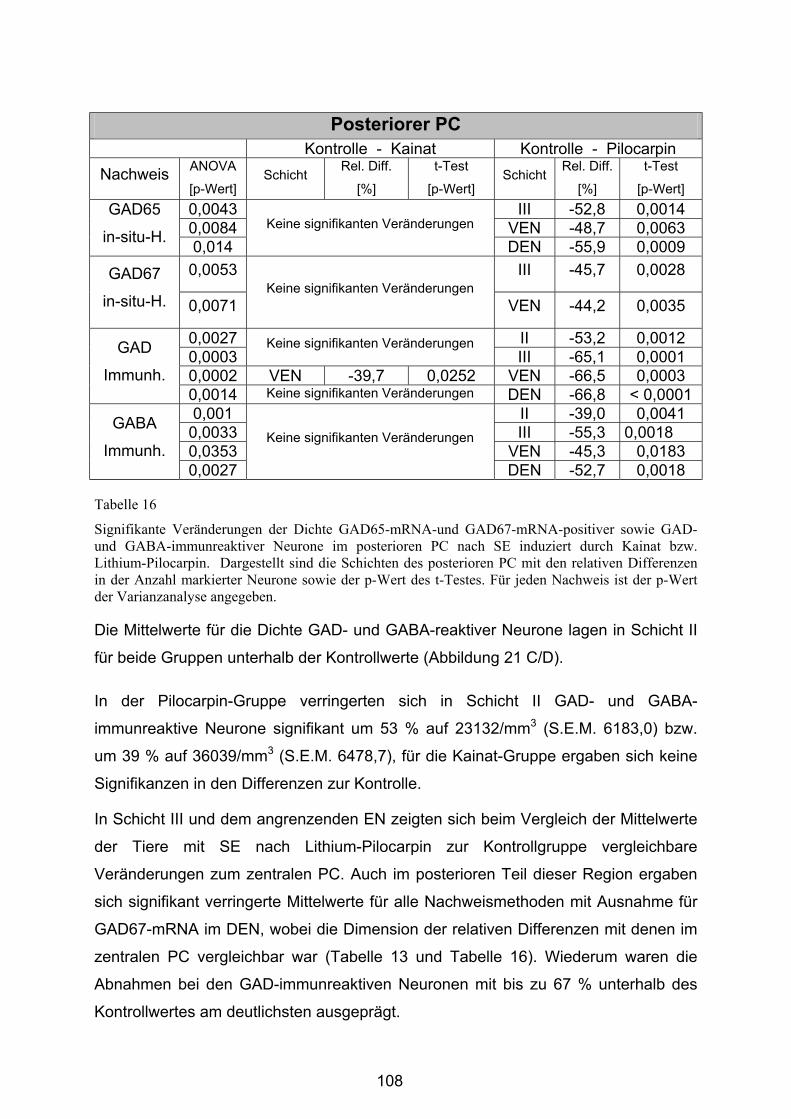
108
Posteriorer PC Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA
[p-Wert] Schicht Rel. Diff.
[%]
t-Test
[p-Wert] Schicht Rel. Diff.
[%]
t-Test
[p-Wert] 0,0043 III -52,8 0,0014 0,0084 VEN -48,7 0,0063
GAD65
in-situ-H. 0,014 Keine signifikanten Veränderungen
DEN -55,9 0,0009 0,0053 III -45,7 0,0028 GAD67
in-situ-H. 0,0071 Keine signifikanten Veränderungen
VEN -44,2 0,0035
0,0027 II -53,2 0,0012 0,0003
Keine signifikanten Veränderungen III -65,1 0,0001
0,0002 VEN -39,7 0,0252 VEN -66,5 0,0003
GAD
Immunh. 0,0014 Keine signifikanten Veränderungen DEN -66,8 < 0,0001 0,001 II -39,0 0,0041
0,0033 III -55,3 0,0018 0,0353 VEN -45,3 0,0183
GABA
Immunh. 0,0027
Keine signifikanten Veränderungen
DEN -52,7 0,0018
Tabelle 16
Signifikante Veränderungen der Dichte GAD65-mRNA-und GAD67-mRNA-positiver sowie GAD- und GABA-immunreaktiver Neurone im posterioren PC nach SE induziert durch Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die Schichten des posterioren PC mit den relativen Differenzen in der Anzahl markierter Neurone sowie der p-Wert des t-Testes. Für jeden Nachweis ist der p-Wert der Varianzanalyse angegeben.
Die Mittelwerte für die Dichte GAD- und GABA-reaktiver Neurone lagen in Schicht II
für beide Gruppen unterhalb der Kontrollwerte (Abbildung 21 C/D).
In der Pilocarpin-Gruppe verringerten sich in Schicht II GAD- und GABA-
immunreaktive Neurone signifikant um 53 % auf 23132/mm3 (S.E.M. 6183,0) bzw.
um 39 % auf 36039/mm3 (S.E.M. 6478,7), für die Kainat-Gruppe ergaben sich keine
Signifikanzen in den Differenzen zur Kontrolle.
In Schicht III und dem angrenzenden EN zeigten sich beim Vergleich der Mittelwerte
der Tiere mit SE nach Lithium-Pilocarpin zur Kontrollgruppe vergleichbare
Veränderungen zum zentralen PC. Auch im posterioren Teil dieser Region ergaben
sich signifikant verringerte Mittelwerte für alle Nachweismethoden mit Ausnahme für
GAD67-mRNA im DEN, wobei die Dimension der relativen Differenzen mit denen im
zentralen PC vergleichbar war (Tabelle 13 und Tabelle 16). Wiederum waren die
Abnahmen bei den GAD-immunreaktiven Neuronen mit bis zu 67 % unterhalb des
Kontrollwertes am deutlichsten ausgeprägt.

109
In der Kainat-Gruppe zeigten sich die Abnahmen in der Neuronendichte im Vergleich
zum zentralen PC weniger deutlich ausgeprägt. Zwar waren auch in dieser Gruppe
die mittleren Neuronendichten mit allen Nachweisen unterhalb der Kontrolle
angesiedelt, es ergab sich jedoch nur mit der GAD-Immunhistochemie eine
signifikante Reduktion von Zellen im VEN.
Abbildung 21 A-D
Anzahl GAD65-mRNA- (A), GAD67-mRNA- (B) sowie GAD- (C) und GABA- (D) positiver Neurone/mm3 in den unterschiedlichen Schichten des posterioren PC von Tieren mit SE induziert durch Kainat bzw. Lithium-Pilocarpin im Vergleich zur Kontrolle. In Schicht II lagen die mittleren Dichten GAD65- und GAD67-mRNA-positiver Neurone wie im zentralen PC über den Kontrollwerten. In der Schicht III sowie im EN ergaben sich geringere Neuronendichten mit allen Nachweisen nach SE induziert durch Pilocarpin. Dargestellt sind die Mittelwerte ±S.E.M. für jede Gruppe. Signifikante Unterschiede zwischen Kontrollen und behandelten Tieren sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit anschließendem t-Test].
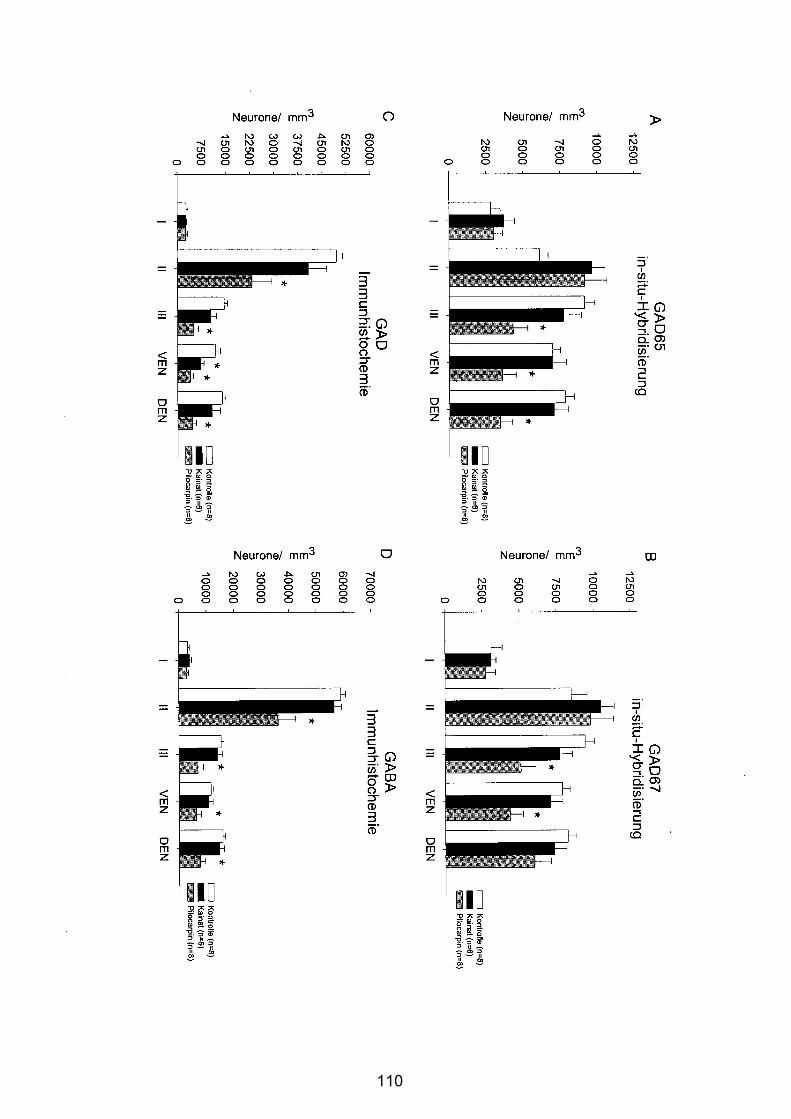
110

111
4.3.2.4 Basolaterale Amygdala
In der basolateralen Amygdala wurden markierte Neurone in der Schnittebene -2,1
mm caudal von Bregma erfasst. In der Versuchsgruppe mit SE induziert durch
Lithium-Pilocarpin musste die Anzahl auswertbarer Serien für die Nachweise von
GAD67-mRNA und GAD Immunhistochemie aufgrund mangelhafter Anfärbung auf 7
Serien reduziert werden.
Die höchste Dichte an Neuronen wurde in den jeweiligen Kontrollgruppen für GAD-
und GABA-immunreaktive Zellen bestimmt (11119 Neurone/mm3; S.E.M. 1137,1
bzw. 11417 Neurone/mm3; S.E.M. 1100,4), die Anzahl GAD65- und GAD67-mRNA-
positiver Neurone/mm3 war im Mittel um 27 % bzw. 22 % geringer (Tabelle 17).
Basolaterale Amygdala in-situ-Hybridisierung Immunhistochemie n GAD65 n GAD67 n GAD n GABA
Gruppe Neurone/mm3 8207 8780 11119 11417 Kontrolle 8 ±832,2 8 ±781,8 8 ±1137,1 8 ±1100,4 5227 7015 8528 12105 SE nach
Kainat 6 ±772,6 6 ±437,9 6 ±1187,5 6 ±1448,9 5021 5456 5685 8689 SE nach
Pilocarpin 8 ±609,8 7 ±593,8 7 ±614,4 8 ±1155,4
Tabelle 17
Anzahl positiver Neurone/mm3 nach in-situ-Hybridisierung für GAD65- und GAD67-mRNA sowie Immunhistochemie für GAD und GABA.. Dargestellt sind die Mittelwerte und S.E.M. der Anzahl von Neuronen/mm3 in der basolateralen Amygdala von Kontrolltieren sowie Tieren mit SE induziert durch Kainat bzw. Lithium-Pilocarpin.
Der Vergleich der mittleren Neuronenzahlen nach den verschiedenen Nachweisen
auf mRNA- bzw. Proteinebene erbrachte signifikante Differenzen zwischen den
Versuchsgruppen in der Dichte GAD65-mRNA-, GAD67-mRNA- und GAD-markierter
Neurone (p<0,05, einfaktorielle ANOVA, Tabelle 18) jedoch nicht für GABA-
immunreaktive Zellen.
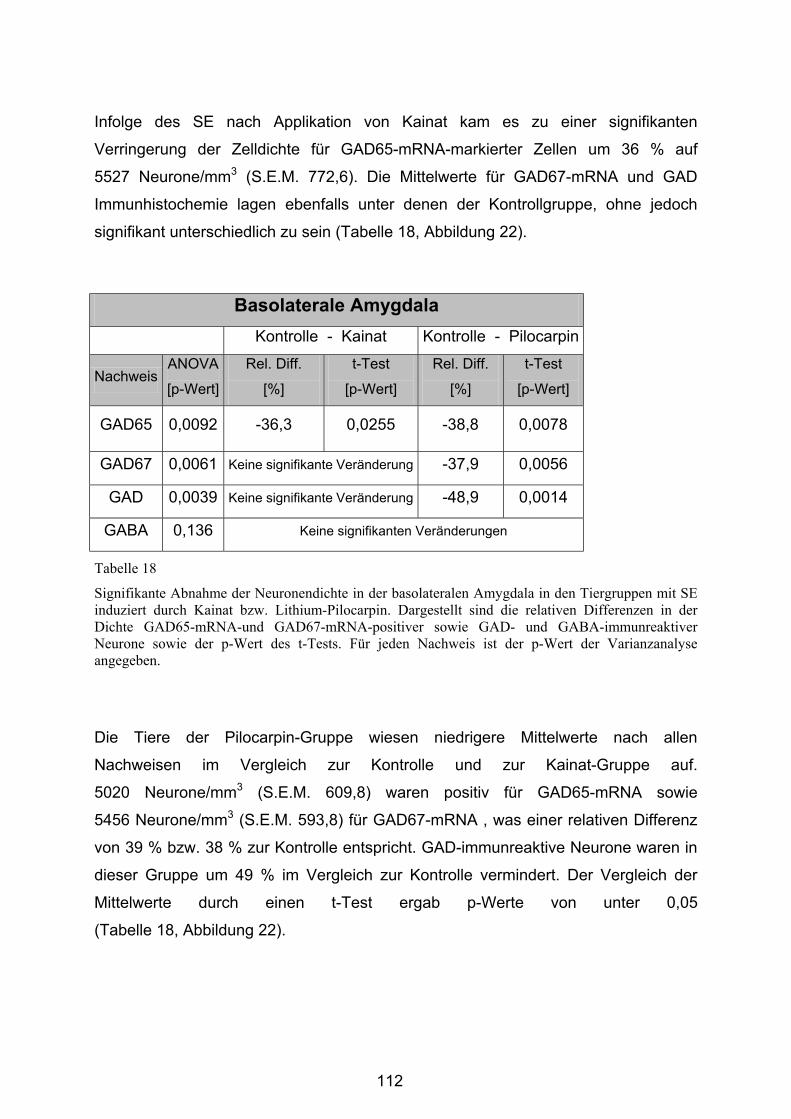
112
Infolge des SE nach Applikation von Kainat kam es zu einer signifikanten
Verringerung der Zelldichte für GAD65-mRNA-markierter Zellen um 36 % auf
5527 Neurone/mm3 (S.E.M. 772,6). Die Mittelwerte für GAD67-mRNA und GAD
Immunhistochemie lagen ebenfalls unter denen der Kontrollgruppe, ohne jedoch
signifikant unterschiedlich zu sein (Tabelle 18, Abbildung 22).
Basolaterale Amygdala Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert]
GAD65 0,0092 -36,3 0,0255 -38,8 0,0078
GAD67 0,0061 Keine signifikante Veränderung -37,9 0,0056
GAD 0,0039 Keine signifikante Veränderung -48,9 0,0014
GABA 0,136 Keine signifikanten Veränderungen
Tabelle 18
Signifikante Abnahme der Neuronendichte in der basolateralen Amygdala in den Tiergruppen mit SE induziert durch Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die relativen Differenzen in der Dichte GAD65-mRNA-und GAD67-mRNA-positiver sowie GAD- und GABA-immunreaktiver Neurone sowie der p-Wert des t-Tests. Für jeden Nachweis ist der p-Wert der Varianzanalyse angegeben.
Die Tiere der Pilocarpin-Gruppe wiesen niedrigere Mittelwerte nach allen
Nachweisen im Vergleich zur Kontrolle und zur Kainat-Gruppe auf.
5020 Neurone/mm3 (S.E.M. 609,8) waren positiv für GAD65-mRNA sowie
5456 Neurone/mm3 (S.E.M. 593,8) für GAD67-mRNA , was einer relativen Differenz
von 39 % bzw. 38 % zur Kontrolle entspricht. GAD-immunreaktive Neurone waren in
dieser Gruppe um 49 % im Vergleich zur Kontrolle vermindert. Der Vergleich der
Mittelwerte durch einen t-Test ergab p-Werte von unter 0,05
(Tabelle 18, Abbildung 22).

113
GAD65 in-situ-H
GAD67 in-situ-H
GAD Immunh.
GABA Immunh.
Neu
rone
/ m
m3
0
2500
5000
7500
10000
12500
15000
Kontrolle Kainat Pilocarpin
* * **
Abbildung 22
Anzahl GAD65- und GAD67-mRNA sowie GAD und GABA positiver Neurone/mm3 in der basolateralen Amygdala. Dargestellt sind die Mittelwerte ± S.E.M. von jeweils 8 Kontrolltieren, 6 Ratten mit SE nach Kainat und 8 (7 GAD67 und GAD) Ratten mit SE nach Lithium-Pilocarpin. Signifikante Unterschiede zwischen Kontrollen und behandelten Tieren sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit anschließendem t-Test].
4.3.2.5 Hippocampusformation
In der Hippocampusformation wurde innerhalb der Regionen CA1, CA3, CA3c sowie
im Hilus und der dorsalen Flexur des Gyrus dentatus ausgewertet.
Unter Kontrollbedingungen wurde mit allen Nachweisen in der Region CA1 die
höchste Dichte positiver Neurone/mm3 in den drei untersuchten Regionen des
Hippocampus ermittelt, gefolgt von CA3 und CA3c. Im Gegensatz zu den
Ergebnissen im PC und der basolateralen Amygdala war die Dichte GAD65-mRNA-
positiver Neurone hier höher als für GAD67-mRNA (Tabelle 19 und Tabelle 20). Mit
Immunhistochemie für GAD ließen sich mehr Zellen als mit in-situ-Hybridisierung
nachweisen, die Dichte GABA-immunreaktiver Zellen erwies sich in den CA
Regionen jedoch mehr als doppelt so hoch wie die Dichte GAD-immunreaktiver
Neurone (Tabelle 21 und Tabelle 22).
Im Hilus des Gyrus dentatus zeigten sich unter bei Kontrolltieren keine deutlichen
Unterschiede in der Dichte mittels in-situ-Hybridisierung und Immunhistochemie
dargestellter Neurone. In dieser Region wurden für GAD-Immunhistochemie mit 5961
Neuronen/mm3 (S.E.M. 527,3) der geringste Wert ermittelt.
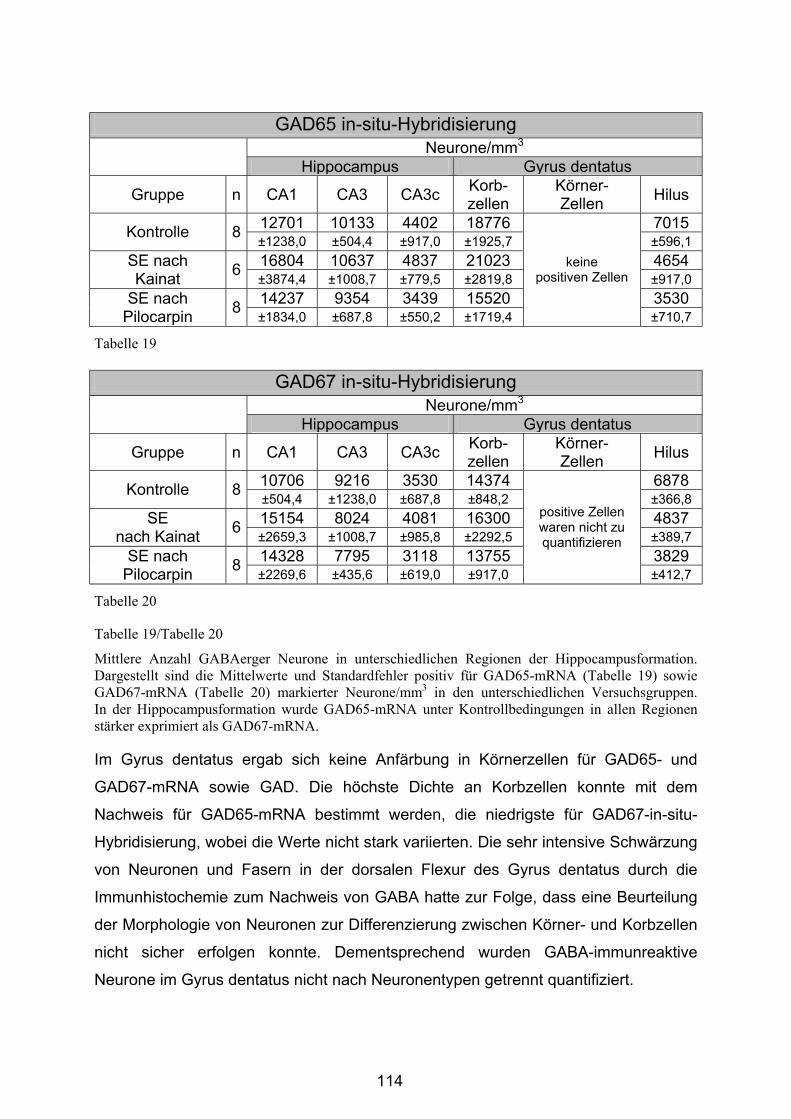
114
GAD65 in-situ-Hybridisierung Neurone/mm3
Hippocampus Gyrus dentatus
Gruppe n CA1 CA3 CA3c Korb- zellen
Körner- Zellen Hilus
12701 10133 4402 18776 7015 Kontrolle 8 ±1238,0 ±504,4 ±917,0 ±1925,7 ±596,1 16804 10637 4837 21023 4654 SE nach
Kainat 6 ±3874,4 ±1008,7 ±779,5 ±2819,8 ±917,0 14237 9354 3439 15520 3530 SE nach
Pilocarpin 8 ±1834,0 ±687,8 ±550,2 ±1719,4
keine positiven Zellen
±710,7
Tabelle 19
GAD67 in-situ-Hybridisierung Neurone/mm3
Hippocampus Gyrus dentatus
Gruppe n CA1 CA3 CA3c Korb- zellen
Körner- Zellen Hilus
10706 9216 3530 14374 6878 Kontrolle 8 ±504,4 ±1238,0 ±687,8 ±848,2 ±366,8 15154 8024 4081 16300 4837 SE
nach Kainat 6 ±2659,3 ±1008,7 ±985,8 ±2292,5 ±389,7 14328 7795 3118 13755 3829 SE nach
Pilocarpin 8 ±2269,6 ±435,6 ±619,0 ±917,0
positive Zellen waren nicht zu quantifizieren
±412,7
Tabelle 20
Tabelle 19/Tabelle 20
Mittlere Anzahl GABAerger Neurone in unterschiedlichen Regionen der Hippocampusformation. Dargestellt sind die Mittelwerte und Standardfehler positiv für GAD65-mRNA (Tabelle 19) sowie GAD67-mRNA (Tabelle 20) markierter Neurone/mm3 in den unterschiedlichen Versuchsgruppen. In der Hippocampusformation wurde GAD65-mRNA unter Kontrollbedingungen in allen Regionen stärker exprimiert als GAD67-mRNA.
Im Gyrus dentatus ergab sich keine Anfärbung in Körnerzellen für GAD65- und
GAD67-mRNA sowie GAD. Die höchste Dichte an Korbzellen konnte mit dem
Nachweis für GAD65-mRNA bestimmt werden, die niedrigste für GAD67-in-situ-
Hybridisierung, wobei die Werte nicht stark variierten. Die sehr intensive Schwärzung
von Neuronen und Fasern in der dorsalen Flexur des Gyrus dentatus durch die
Immunhistochemie zum Nachweis von GABA hatte zur Folge, dass eine Beurteilung
der Morphologie von Neuronen zur Differenzierung zwischen Körner- und Korbzellen
nicht sicher erfolgen konnte. Dementsprechend wurden GABA-immunreaktive
Neurone im Gyrus dentatus nicht nach Neuronentypen getrennt quantifiziert.
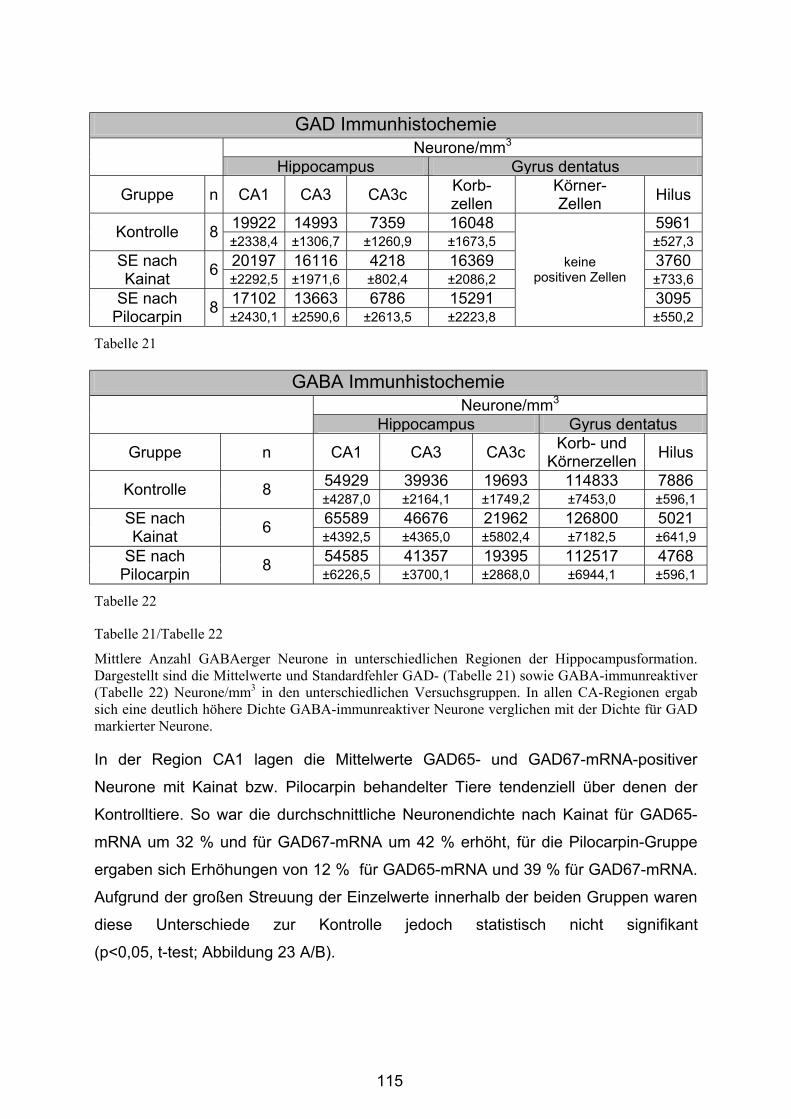
115
GAD Immunhistochemie Neurone/mm3
Hippocampus Gyrus dentatus
Gruppe n CA1 CA3 CA3c Korb- zellen
Körner- Zellen Hilus
19922 14993 7359 16048 5961 Kontrolle 8 ±2338,4 ±1306,7 ±1260,9 ±1673,5 ±527,3 20197 16116 4218 16369 3760 SE nach
Kainat 6 ±2292,5 ±1971,6 ±802,4 ±2086,2 ±733,6 17102 13663 6786 15291 3095 SE nach
Pilocarpin 8 ±2430,1 ±2590,6 ±2613,5 ±2223,8
keine positiven Zellen
±550,2
Tabelle 21
GABA Immunhistochemie Neurone/mm3
Hippocampus Gyrus dentatus
Gruppe n CA1 CA3 CA3c Korb- und Körnerzellen Hilus
54929 39936 19693 114833 7886 Kontrolle 8 ±4287,0 ±2164,1 ±1749,2 ±7453,0 ±596,1 65589 46676 21962 126800 5021 SE nach
Kainat 6 ±4392,5 ±4365,0 ±5802,4 ±7182,5 ±641,9 54585 41357 19395 112517 4768 SE nach
Pilocarpin 8 ±6226,5 ±3700,1 ±2868,0 ±6944,1 ±596,1
Tabelle 22
Tabelle 21/Tabelle 22
Mittlere Anzahl GABAerger Neurone in unterschiedlichen Regionen der Hippocampusformation. Dargestellt sind die Mittelwerte und Standardfehler GAD- (Tabelle 21) sowie GABA-immunreaktiver (Tabelle 22) Neurone/mm3 in den unterschiedlichen Versuchsgruppen. In allen CA-Regionen ergab sich eine deutlich höhere Dichte GABA-immunreaktiver Neurone verglichen mit der Dichte für GAD markierter Neurone.
In der Region CA1 lagen die Mittelwerte GAD65- und GAD67-mRNA-positiver
Neurone mit Kainat bzw. Pilocarpin behandelter Tiere tendenziell über denen der
Kontrolltiere. So war die durchschnittliche Neuronendichte nach Kainat für GAD65-
mRNA um 32 % und für GAD67-mRNA um 42 % erhöht, für die Pilocarpin-Gruppe
ergaben sich Erhöhungen von 12 % für GAD65-mRNA und 39 % für GAD67-mRNA.
Aufgrund der großen Streuung der Einzelwerte innerhalb der beiden Gruppen waren
diese Unterschiede zur Kontrolle jedoch statistisch nicht signifikant
(p<0,05, t-test; Abbildung 23 A/B).

116
Die Dichte GAD-immunreaktiver Neurone war in der Gruppe Kainat-behandelter
Tiere in CA1 um 19 % erhöht, während die Mittelwerte der GABA-immunreaktiven
Neurone auf einem vergleichbaren Niveau zur Kontrollgruppe lagen. In der
Pilocarpin-Gruppe ergaben sich umgekehrte Verhältnisse; hier verringerte sich die
Anzahl positiver Neurone für GAD um durchschnittlich 14 %, während die Dichte
GABA-positiver Zellen zur Kontrolle vergleichbar war ( Abbildung 23 C/D).
Beim Vergleich der Zählergebnisse aus den Regionen CA3 und CA3c zeigten sich in
den mit Kainat bzw. Lithium-Pilocarpin behandelten Gruppen mit keiner der
durchgeführten Nachweismethoden einheitliche Veränderungen in Form einer Zu-
oder Abnahme der Neuronendichte. Die Unterschiede in den Mittelwerten bewegten
sich im Bereich von 5 bis 22 % im Vergleich zu den Werten der Kontrollgruppen.
Auffällig war ein um 43 % verringerter Mittelwert GAD-immunreaktiver Neurone in
CA3c, für den sich aber keine statistische Absicherung ergab.
Abbildung 23 A-D
Anzahl GAD65- (A) und GAD67-mRNA (B) sowie GAD- (C) und GABA- (D) positiver Neurone/mm3 in den verschiedenen Regionen der Hippocampusformation. Dargestellt sind die Mittelwerte ±S.E.M. von Tieren mit SE induziert durch Kainat bzw. Lithium-Pilocarpin im Vergleich zur Kontrolle. Körnerzellen positiv für GAD67-mRNA konnten aufgrund der Intensität der Anfärbung nicht quantifiziert werden. GAD65-mRNA-positive sowie GAD-immunreaktive Körnerzellen traten nicht auf. Mit der Immunhistochemie gegen GABA konnte aufgrund der sehr intensiven Anfärbung im Gyrus dentatus nicht zwischen Körner- und Korbzellen differenziert werden. Deshalb wurden beide Zelltypen zusammen quantifiziert (D). Signifikante Unterschiede ergaben sind nur im Hilus des Gyrus dentatus (G. dent.) und sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit anschließendem t-Test].
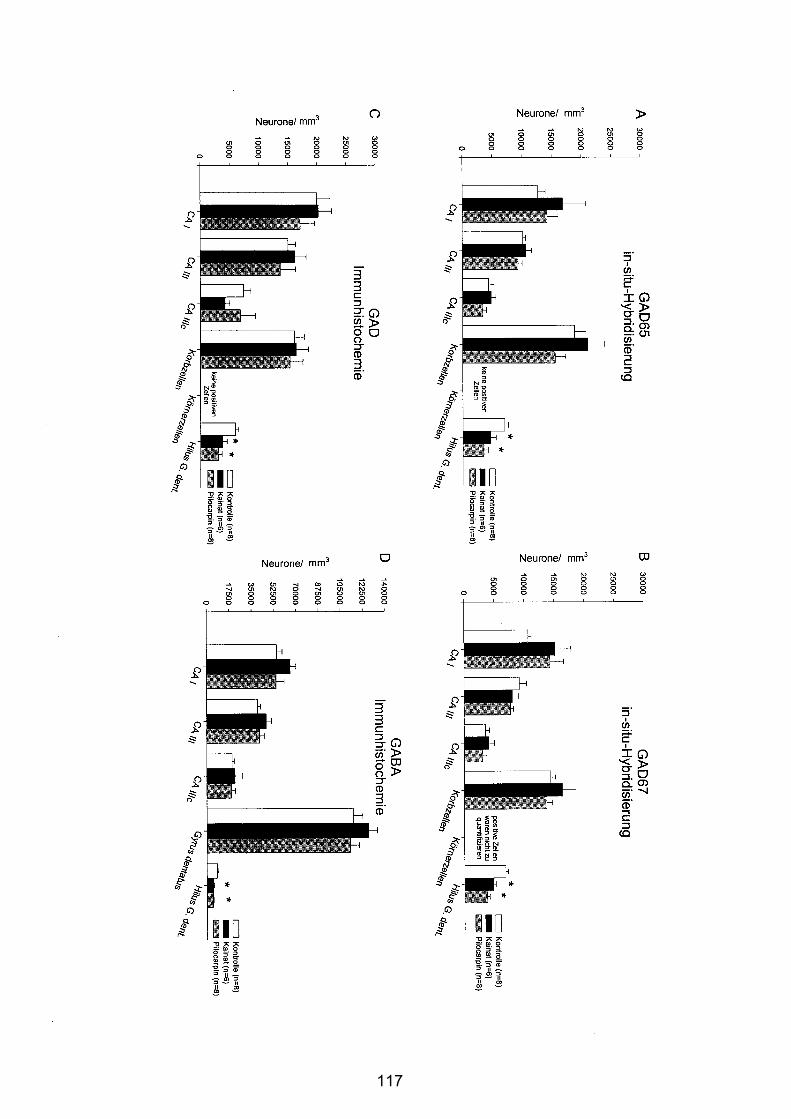
117

118
Im Kainat-Modell der Temporallappenepilepsie wurde die verstärkte Expression von
GAD67-mRNA in Körnerzellen im Gyrus dentatus 6 h nach Applikation von Kainat
mit radioaktiver in-situ-Hybridisierung beschrieben, während sich für GAD65 mRNA
in diesem Zeitintervall keine signifikanten Veränderungen zur Kontrolle ergaben. Die
Expression beider Isoformen in Korbzellen war zu diesem Zeitpunkt ebenfalls nicht
verändert (Schwarzer und Sperk, 1995).
In Anlehnung an diese Untersuchungsergebnisse sollte die Quantifizierung von
Neuronen im Gyrus dentatus getrennt nach Körner- und Korbzellen durchgeführt
werden. Im Zuge der Auswertungen zeigte sich eine Anfärbung von Körnerzellen für
GAD67-mRNA in einigen der Tiere der Gruppen mit induziertem SE. Diese konnte
jedoch nicht anhand von Zellzählungen quantifiziert werden, da durch die Intensität
der Anfärbung keine eindeutigen Identifikation von Zellen mehr möglich war. Um
dennoch Veränderungen in der Expression untersuchen zu können, wurde die
diffuse Dichte der Anfärbung in der Körnerzellschicht bestimmt und verglichen. Die
Ergebnisse dieser Untersuchungen sind in Kap. 4.3.3 dargestellt. In keiner der drei
Versuchsgruppen traten GAD65-mRNA-positive oder GAD-immunreaktive
Körnerzellen auf.
Im Gyrus dentatus wurden bei Tieren mit SE nach Kainsäure mit der in-situ-
Hybridisierung für GAD65- und GAD67-mRNA im Mittel 12-13 % mehr Korbzellen
markiert, die GAD-Immunhistochemie erbrachte für diese Gruppe keine
Abweichungen vom Niveau der Kontrollgruppe. In der Pilocarpin-Gruppe wich die
mittlere Anzahl positiv für GAD67-mRNA- und GAD-markierter Korbzellen nicht
auffallend vom Mittelwert der Kontrolle ab, während sich GAD65-mRNA-markierte
Zellen im Mittel um 17 % verringerten (Abbildung 23 A-C).
Mit der Immunhistochemie gegen GABA konnte aufgrund der sehr intensiven
Anfärbung im Gyrus dentatus nicht zwischen Körner- und Korbzellen differenziert
werden. Deshalb wurden beide Zelltypen zusammen quantifiziert. Die Pilocarpin-
Gruppe wies einen der Kontrollgruppe vergleichbaren Mittelwert GABA-
immunreaktiver Neurone auf, während der Durchschnitt in der Gruppe mit Kainsäure
behandelter Tiere um 10 % höher lag (Abbildung 23 D).

119
Im Hilus des Gyrus dentatus führte die Anfallsaktivität in beiden Gruppen zu einer
signifikanten Reduktion der Zelldichte mit allen Nachweisen. Durch Kainat verringerte
sich die Zahl GAD65- und GAD67-mRNA-positiver Neurone um 30-34 %, für GAD
und GABA im Mittel um 36-37 %. Bei den mit Pilocarpin behandelten Tieren konnten
40-50 % weniger Neurone im Vergleich zur Kontrolle nachgewiesen werden
(Tabelle 23).
Hilus des Gyrus dentatus Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA
[p-Wert] Rel. Diff. [%] t-Test [p-Wert] Rel. Diff. [%] t-Test [p-Wert]
GAD65 in-situ-H. 0,0066 -33,7 0,0408 -49,7 0,0020
GAD67 in-situ-H. >0,0001 -29,7 0,0030 -44,3 <0,0001
GAD Immunh. 0,0047 -36,9 0,025 -48,1 0,0018
GABA Immunh. 0,0024 -36,3 0,0072 -39,5 0,0024
Tabelle 23
Signifikante Abnahme der Dichte GAD65-mRNA-und GAD67-mRNA-positiver sowie GAD- und GABA-immunreaktiver Neurone im Hilus des Gyrus dentatus. Dargestellt sind die unterschiedlichen Nachweismethoden mit den relativen Differenzen der Versuchsgruppen zu den Kontrollgruppen sowie die Ergebnissen des nachfolgenden t-Tests. In beiden Tiergruppen mit SE induziert durch Kainat bzw. Lithium-Pilocarpin ließen sich mit allen durchgeführten Nachweisen eine signifikante Reduktion GABAerger Neurone nachweisen.
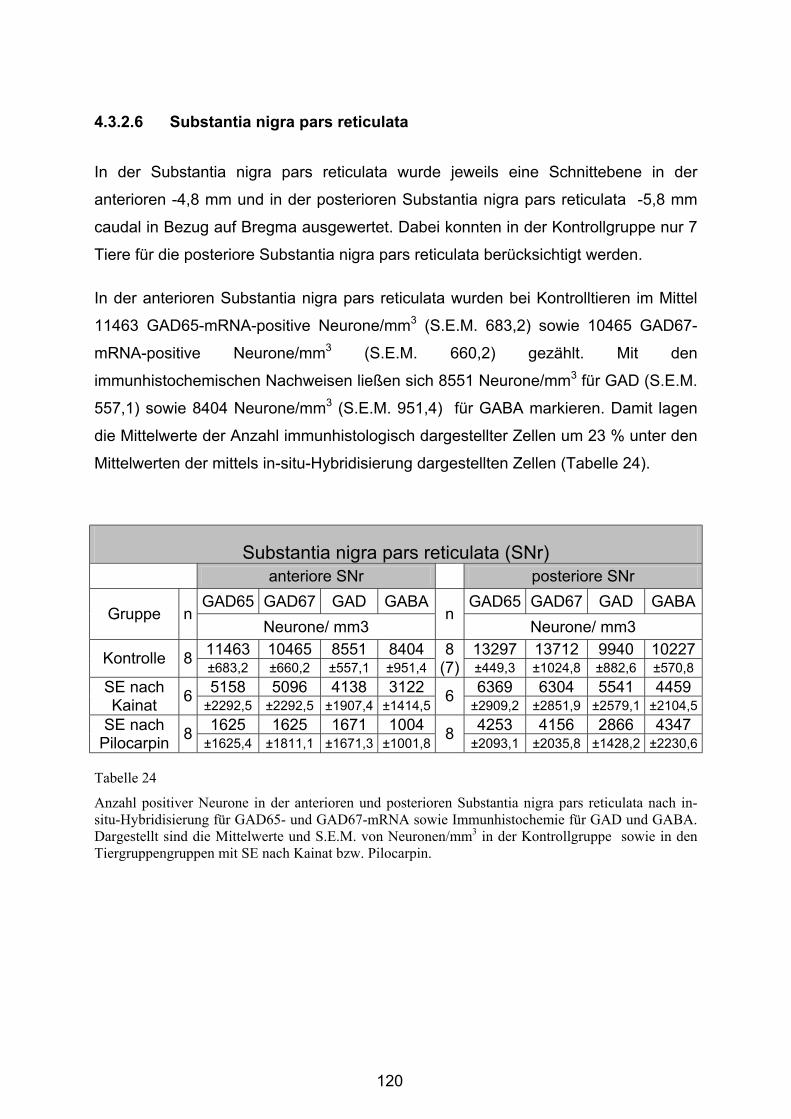
120
4.3.2.6 Substantia nigra pars reticulata
In der Substantia nigra pars reticulata wurde jeweils eine Schnittebene in der
anterioren -4,8 mm und in der posterioren Substantia nigra pars reticulata -5,8 mm
caudal in Bezug auf Bregma ausgewertet. Dabei konnten in der Kontrollgruppe nur 7
Tiere für die posteriore Substantia nigra pars reticulata berücksichtigt werden.
In der anterioren Substantia nigra pars reticulata wurden bei Kontrolltieren im Mittel
11463 GAD65-mRNA-positive Neurone/mm3 (S.E.M. 683,2) sowie 10465 GAD67-
mRNA-positive Neurone/mm3 (S.E.M. 660,2) gezählt. Mit den
immunhistochemischen Nachweisen ließen sich 8551 Neurone/mm3 für GAD (S.E.M.
557,1) sowie 8404 Neurone/mm3 (S.E.M. 951,4) für GABA markieren. Damit lagen
die Mittelwerte der Anzahl immunhistologisch dargestellter Zellen um 23 % unter den
Mittelwerten der mittels in-situ-Hybridisierung dargestellten Zellen (Tabelle 24).
Substantia nigra pars reticulata (SNr) anteriore SNr posteriore SNr
GAD65 GAD67 GAD GABA GAD65 GAD67 GAD GABA Gruppe n
Neurone/ mm3 n
Neurone/ mm3 11463 10465 8551 8404 13297 13712 9940 10227 Kontrolle 8 ±683,2 ±660,2 ±557,1 ±951,4
8 (7) ±449,3 ±1024,8 ±882,6 ±570,8
5158 5096 4138 3122 6369 6304 5541 4459 SE nach Kainat 6 ±2292,5 ±2292,5 ±1907,4 ±1414,5 6 ±2909,2 ±2851,9 ±2579,1 ±2104,5
1625 1625 1671 1004 4253 4156 2866 4347 SE nach Pilocarpin 8 ±1625,4 ±1811,1 ±1671,3 ±1001,8 8 ±2093,1 ±2035,8 ±1428,2 ±2230,6
Tabelle 24
Anzahl positiver Neurone in der anterioren und posterioren Substantia nigra pars reticulata nach in-situ-Hybridisierung für GAD65- und GAD67-mRNA sowie Immunhistochemie für GAD und GABA. Dargestellt sind die Mittelwerte und S.E.M. von Neuronen/mm3 in der Kontrollgruppe sowie in den Tiergruppengruppen mit SE nach Kainat bzw. Pilocarpin.

121
Im posterioren Anteil der Substantia nigra pars reticulata wurden für alle Nachweise
höhere Dichten markierter Neurone in den Kontrollgruppen ermittelt. Für GAD65- und
GAD67-mRNA-positive Neurone ergaben sich wiederum Mittelwerte, die über denen
der immunhistochemisch dargestellten Zellen lagen. Wurden mit 13297
Neuronen/mm3 für GAD65-mRNA (S.E.M. 449,3) und 13712 Neuronen/mm3 für
GAD67-mRNA vergleichbare Zahlen mit der in-situ-Hybridisierung dargestellt, so
fielen die Mittelwerte der GAD- (9940 Neurone/mm3; S.E.M. 882,6) bzw. GABA-
positiven Zellen (10227 Neurone/mm3; S.E.M. 570,8) wie in der anterioren Substantia
nigra pars reticulata geringer aus (25 % relative Differenz zu den Mittelwerten der in-
situ-Hybridisierung, Tabelle 24).
In der Kainat- und der Pilocarpin-Gruppe kam es zur Ausbildung von Läsionen in der
Substantia nigra pars reticulata unterschiedlichen Ausmaßes (Kap. 4.3.4.2). In der
Gruppe mit Kainat behandelter Tiere waren bei 3 von 6 Ratten keine Neurone
erhalten, durch Pilocarpin wurden aus der Versuchsgruppe von 8 Tieren die
Substantia nigra pars reticulata in ihrer gesamten Ausdehnung bei 5 Ratten zerstört,
bei 2 Tieren dieser Gruppe waren keine Neurone im anterioren Teil der Substantia
nigra pars reticulata mehr erhalten. Die Mittelwerte der Pilocarpin-Gruppe waren
dementsprechend in allen Nachweisen unterhalb derer der Kainat-Gruppe
angesiedelt, wobei die mittleren Werte in der anterioren Substantia nigra pars
reticulata in dieser Gruppe am weitesten unter denen der Kontrolle lagen (Abbildung
24 A/B).
Die Abnahme in der Dichte markierter Neurone in den Gruppen mit Tieren nach SE
erwies sich in der Varianzanalyse als statistisch signifikant zur Kontrolle (p<0,05;
einfaktorielle ANOVA). Durch die neurodegenerativen Veränderungen reduzierte sich
in der Kainat-Gruppe in der anterioren Substantia nigra pars reticulata die mittlere
Dichte GAD65, GAD67 und GAD positiver Neurone zwischen 51 und 55 %, bei
GABA um 63 %. In der Pilocarpin-Gruppe waren die mittleren Werte aller Nachweise
zwischen 81 und 88 % geringer (p<0,05, t-Test, Tabelle 25).

122
In der posterioren Substantia nigra pars reticulata nahm die Anzahl markierter
Neurone/mm3 in der Kainat-Gruppe für GAD65, GAD67 und GABA zwischen 52 und
56 % signifikant ab. Der Nachweis von GAD erbrachte eine Reduktion um 44 %,
welche sich statistisch nicht absichern lies (p>0,05, t-Test). Durch Pilocarpin
verringerten sich GABA-immunreaktive Neurone um 58 % , die Anzahl der durch in-
situ-Hybridisierung und GAD-Immunhistochemie gefärbten Zellen war zwischen 68
und 71 % signifikant vermindert (p<0,05, t-Test, Tabelle 25).
Anteriore Substantia nigra pars reticulata Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA [p-Wert] Rel. Diff. [%] t-Test [p-Wert] Rel. Diff. [%] t-Test [p-Wert]
GAD65 in-situ-H. 0,0005 -55,0 0,0117 -85,8 0,0186
GAD67 in-situ-H. 0,0016 -51,3 0,0256 -84,5 0,0002
GAD Immunh. 0,0070 -51,6 0,0271 -80,5 0,0016
GABA Immunh. 0,0002 -62,8 0,0074 -88,1 0,0001
Posteriore Substantia nigra pars reticulata
Kontrolle - Kainat Kontrolle - Pilocarpin
Nachweis ANOVA [p-Wert] Rel. Diff. [%] t-Test [p-Wert] Rel. Diff. [%] t-Test [p-Wert]
GAD65 in-situ-H. 0,0069 -52,1 0,0186 -68,0 0,0009
GAD67 in-situ-H. 0,0052 -54,0 0,0183 -69,7 0,0009
GAD Immunh. 0,0200 keine signifikanten
Veränderungen -71,2 0,013
GABA Immunh. 0,0400 -56,4 0,0109 -57,5 0,0229
Tabelle 25
Signifikante Veränderungen der Dichte GAD65-mRNA-und GAD67-mRNA-positiver sowie GAD- und GABA-immunreaktiver Neurone in der anterioren und posterioren Substantia nigra pars reticulata infolge eines SE induziert durch Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die relativen Differenzen in der Anzahl markierter Neurone sowie der p-Wert der einfaktoriellen Varianzanalyse und des anschließend durchgeführten t-Tests.
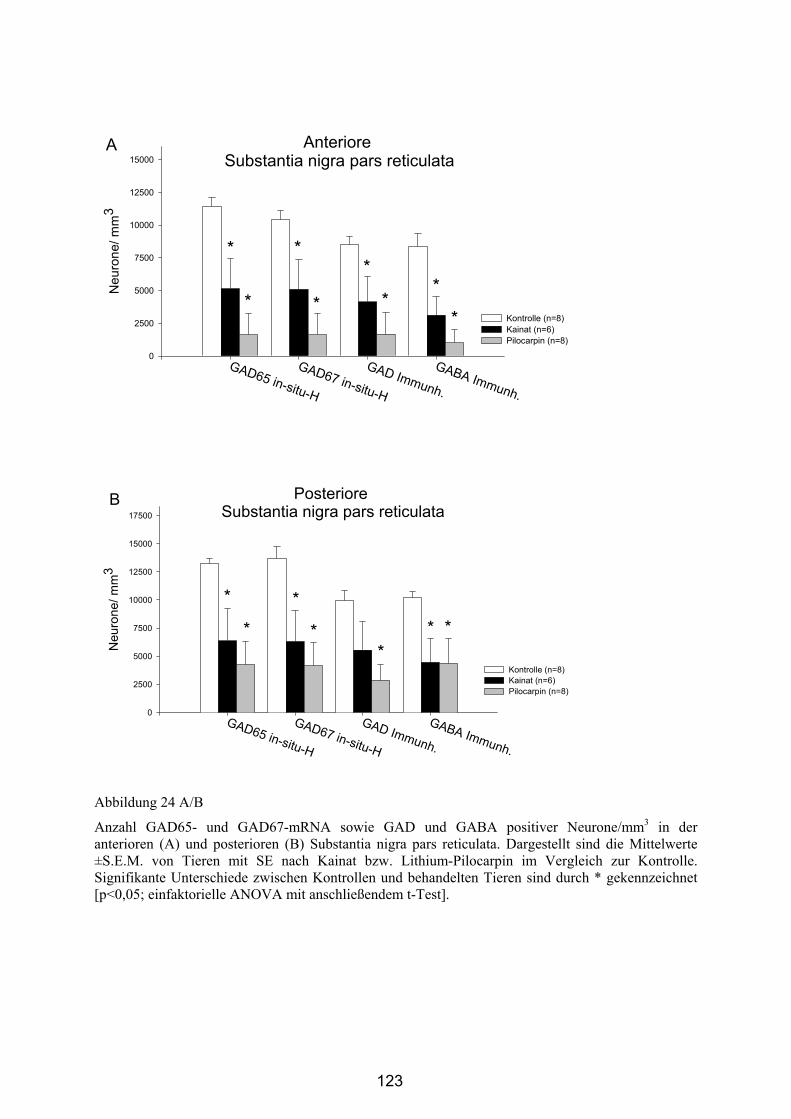
123
Anteriore Substantia nigra pars reticulata
GAD65 in-situ-H
GAD67 in-situ-H
GAD Immunh.
GABA Immunh.
Neu
rone
/ mm
3
0
2500
5000
7500
10000
12500
15000
Kontrolle (n=8)Kainat (n=6)Pilocarpin (n=8)
* *
* **
**
*
A
Posteriore Substantia nigra pars reticulata
GAD65 in-situ-H
GAD67 in-situ-H
GAD Immunh.
GABA Immunh.
Neu
rone
/ mm
3
0
2500
5000
7500
10000
12500
15000
17500
**
* * **
*
B
Kontrolle (n=8)Kainat (n=6)Pilocarpin (n=8)
Abbildung 24 A/B
Anzahl GAD65- und GAD67-mRNA sowie GAD und GABA positiver Neurone/mm3 in der anterioren (A) und posterioren (B) Substantia nigra pars reticulata. Dargestellt sind die Mittelwerte ±S.E.M. von Tieren mit SE nach Kainat bzw. Lithium-Pilocarpin im Vergleich zur Kontrolle. Signifikante Unterschiede zwischen Kontrollen und behandelten Tieren sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit anschließendem t-Test].

124
4.3.3 Untersuchung der Expression von GAD-mRNA im Gyrus dentatus
Schwarzer und Sperk (1995) beschrieben eine vermehrte Expression von GAD67-
mRNA in Körnerzellen des Gyrus dentatus im Kainsäure-Modell der
Temporallappenepilepsie mit radioaktiver in-situ-Hybridisierung. Die Expressionsrate
für GAD65-mRNA war zu diesem Zeitpunkt nicht verändert. Im Rahmen der eigenen
Untersuchung sollte überprüft werden, ob sich entsprechende Veränderung auch mit
Hilfe der nicht-radioaktiven in-situ-Hybridisierung im Kainat- und im Lithium-
Pilocarpin-Modell in Körnerzellen nachweisen lassen.
In der Kontrollgruppe war weder für GAD65- noch für GAD67-mRNA eine
spezifischen Anfärbung von Körnerzellen zu erkennen. GABAerge Neurone an der
ventralen Seite der dorsalen Flexur des Stratum granulosum exprimierten die mRNA
für beide GAD-Isoformen. Anhand der Größe und Morphologie der Somata wurden
diese Zellen als Korbzellen identifiziert, wobei eine eindeutige Beschreibung
aufgrund der fehlenden Anfärbung der Zellausläufer nicht zweifelsfrei erfolgen
konnte. Im Bereich der Körnerzellschicht war in beiden Gruppen nach SE induziert
durch Kainat bzw. Lithium-Pilocarpin im Gyrus dentatus einiger Tiere eine deutliche
Anfärbung der Körnerzellen für GAD67-mRNA zu bemerken (Abbildung 25 B/C). Für
GAD65-mRNA und GAD- Protein konnte dagegen keine Anfärbung festgestellt
werden (Abbildung 23 A/C).
Abbildung 25 A-C
Darstellung der Anfärbung von Körnerzellen (Pfeile) im Gyrus dentatus (GD) infolge gesteigerter Expression von GAD67-mRNA nach SE induziert durch Kainat (B) bzw. Lithium- Pilocarpin (C). Aufgrund der intensiven Anfärbung konnten markierter Körnerzellen in einigen Serien nicht quantifiziert werden. In der Kontrollgruppe (A) war keine Anfärbung in dieser Zellschicht festzustellen. In allen Gruppen waren GABAerge Neurone am Übergang an der Innenseite der Körnerzellschicht und im Hilus des Gyrus dentatus für GAD67-mRNA markiert (Pfeilköpfe). Kalibrierbalken für A-C: 100 µm.
GD A B C

125
Durch die starke Intensität der Anfärbung ließen sich bei einem Teil der Tiere die
Somata der gefärbten Körnerzellen nicht mehr eindeutig voneinander unterscheiden,
so dass eine Zählung der markierten Zellen nicht durchgeführt werden konnte.
Zudem waren die Zellen in dieser Schicht sehr kompakt angeordnet, wodurch die
Genauigkeit der Zählung zusätzlich beeinträchtigt worden wäre.
Um eine Aussage über Unterschiede in der Expression für GAD67-mRNA zwischen
den Versuchgruppen machen zu können, wurde die Messung der diffusen Dichte der
Anfärbung als Parameter ausgewählt. Die Messungen wurden mit einem
Bildanalysesystem gemäß Kap. 3.5.3 in der Schnittebene -3,0 mm caudal von
Bregma durchgeführt. Dabei erfolgte eingangs für jeden Gewebeschnitt die
Bestimmung der diffusen Dichte der Färbung des Hintergrundes in der internen
Kapsel. Zusätzlich wurde die diffuse Dichte für GAD65-mRNA in Körnerzellen sowie
für beide Isoformen in Korbzellen bestimmt. Die Messungen wurden an
verschlüsselten Objektträgern durchgeführt.
In der Kontrollgruppe betrug der Mittelwert der diffusen Dichte des Hintergrundes für
die GAD65-mRNA 0,20 (S.E.M. 0,03) und besaß damit ein zu den beiden anderen
Versuchsgruppen vergleichbares Niveau. Für die GAD67-mRNA fiel die Dichte des
Hintergrundes im Mittel mit 0,26 (S.E.M. 0,03) in der Kontrollgruppe dunkler aus
(Tabelle 26). Der Vergleich der Mittelwerte der Versuchsgruppen ergab keine
statistisch signifikanten Unterschiede zwischen den diffusen Dichten der
Hintergrundfärbung in den Gruppen (p>0,05; einfaktorielle ANOVA).
Für die Untersuchung einer vermehrten Expression von GAD-mRNA im Gyrus
dentatus erfolgte die Berechnung der relativen Zunahme der diffusen Dichte der
Anfärbung für die Körner- und Korbzellen im Vergleich zum Hintergrund in jeder der
drei Versuchsgruppen. Hierfür wurde die Differenz der diffusen Dichte der Anfärbung
von Körner- und Korbzellen und der diffusen Dichte des Hintergrundes für jedes Tier
berechnet. Die Zunahmen in der diffusen Dichte sind in Tabelle 26 aufgeführt.

126
GAD65 in-situ-Hybridisierung
GAD67 in-situ-Hybridisierung
Hinter-grund
Körnerzellen (rel. Zunahme zum Hintergr.)
Korbzellen (rel. Zunahme zum Hintergr.)
Hinter-grund
Körnerzellen (rel. Zunahme zum Hintergr.)
Korbzellen (rel. Zunahme zum Hintergr.)
Gruppe n diffuse Dichte diffuse Dichte 0,20 +0,02 +0,29 0,26 +0,02 +0,31 Kontrolle 5 ±0,03 ±0,02 ±0,04 ±0,03 ±0,02 ±0,05 0,21 +0,02 +0,33 0,23 +0,13 +0,33 SE nach
Kainat 5 ±0,03 ±0,05 ±0,07 ±0,04 ±0,03 ±0,05 0,18 +0,05 +0,33 0,21 +0,17 +0,36 SE nach
Pilocarpin 5 ±0,03 ±0,03 ±0,03 ±0,02 ±0,05 ±0,05
Tabelle 26
Darstellung der diffusen Dichte der Anfärbung des Hintergrundes in der internen Kapsel sowie von Körner- und Korbzellen im Gyrus dentatus nach in-situ-Hybridisierung für GAD65- und GAD67-mRNA. Aufgeführt sind die Mittelwerte und Standardfehler der diffusen Dichte des Hintergrundes sowie der relativen Zunahmen der diffusen Dichte der Anfärbung von Körner- und Korbzellen für die drei Versuchsgruppen.
Mit in-situ-Hybridisierung für GAD65-mRNA lagen die Mittelwerte der Zunahme an
diffuser Dichte der Anfärbung in Körnerzellen für alle Versuchsgruppen auf
vergleichbarem Niveau, gleiches galt für die Anfärbung der Korbzellen (Tabelle 26,
Abbildung 26 A/B). Die Intensität der Anfärbung von Korbzellen lag dabei im Mittel
der drei Versuchsgruppen um 162 % über dem Hintergrundwert. Die in-situ-
Hybridisierung für GAD67-mRNA ergab vergleichbare Zunahmen der diffusen
Dichten in der Anfärbung von Korbzellen mit durchschnittlich 144 % im Vergleich zur
Internen Kapsel.
Expression von GAD mRNA im Gyrus dentatus Kontrolle - Kainat Kontrolle - Pilocarpin Körnerzellen Korbzellen Körnerzellen Korbzellen
Nachweis Rel. Diff.
[%]
t-Test
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert]
GAD65 keine signifikanten Veränderungen GAD67 +550 0,0144 keine sign. Veränd. +750 0,0151 keine sign. Veränd.
Tabelle 27
Signifikante Zunahme der Expression von GAD67-mRNA in Körnerzellen des Gyrus dentatus von Tieren mit SE induziert durch Kainat bzw. Lithium-Pilocarpin. Dargestellt sind die relativen Differenzen in der Zunahme der diffusen Dichte zur Kontrolle und der Wert des nachfolgend durchgeführten t-Tests. Die Expression von GAD65-mRNA war infolge des SE nicht verändert. Zudem ergaben sich keine Veränderungen in der Expression der mRNA für beide Isoformen der GAD in Korbzellen.
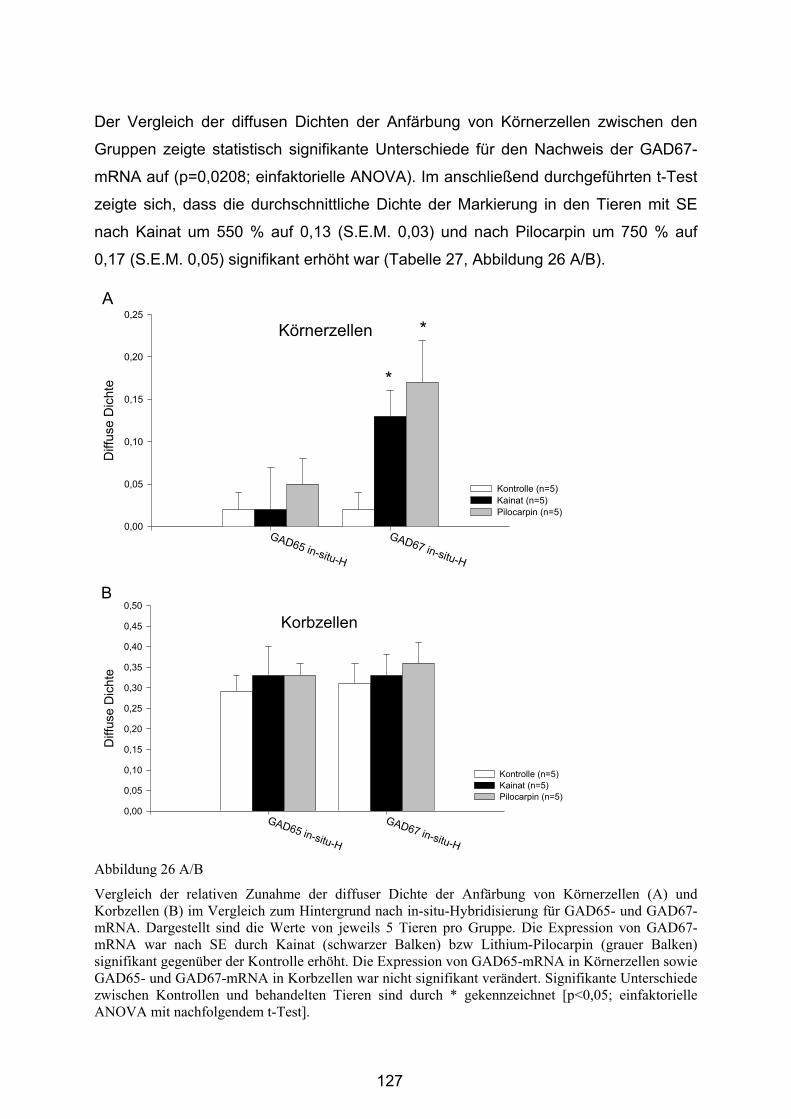
127
Der Vergleich der diffusen Dichten der Anfärbung von Körnerzellen zwischen den
Gruppen zeigte statistisch signifikante Unterschiede für den Nachweis der GAD67-
mRNA auf (p=0,0208; einfaktorielle ANOVA). Im anschließend durchgeführten t-Test
zeigte sich, dass die durchschnittliche Dichte der Markierung in den Tieren mit SE
nach Kainat um 550 % auf 0,13 (S.E.M. 0,03) und nach Pilocarpin um 750 % auf
0,17 (S.E.M. 0,05) signifikant erhöht war (Tabelle 27, Abbildung 26 A/B).
Körnerzellen
GAD65 in-situ-H
GAD67 in-situ-H
Diff
use
Dic
hte
0,00
0,05
0,10
0,15
0,20
0,25
*
*A
Kontrolle (n=5)Kainat (n=5)Pilocarpin (n=5)
Korbzellen
GAD65 in-situ-H
GAD67 in-situ-H
Diff
use
Dic
hte
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0,45
0,50
Kontrolle (n=5)Kainat (n=5)Pilocarpin (n=5)
B
Abbildung 26 A/B
Vergleich der relativen Zunahme der diffuser Dichte der Anfärbung von Körnerzellen (A) und Korbzellen (B) im Vergleich zum Hintergrund nach in-situ-Hybridisierung für GAD65- und GAD67-mRNA. Dargestellt sind die Werte von jeweils 5 Tieren pro Gruppe. Die Expression von GAD67-mRNA war nach SE durch Kainat (schwarzer Balken) bzw Lithium-Pilocarpin (grauer Balken) signifikant gegenüber der Kontrolle erhöht. Die Expression von GAD65-mRNA in Körnerzellen sowie GAD65- und GAD67-mRNA in Korbzellen war nicht signifikant verändert. Signifikante Unterschiede zwischen Kontrollen und behandelten Tieren sind durch * gekennzeichnet [p<0,05; einfaktorielle ANOVA mit nachfolgendem t-Test].

128
4.3.4 Neuropathologische Veränderungen
Neuropathologische Veränderungen sind für das Kainat- und das Lithium-Pilocarpin-
Modell der Temporallappenepilepsie in der Literatur beschrieben worden (Sperk,
1994; Covolan und Mello, 2000). Die auftretende Neurodegeneration war dabei
abhängig von der jeweiligen Hirnregion und dem Zeitabstand nach Applikation von
Kainat bzw. Pilocarpin.
In den beiden Versuchsgruppen wurde der induzierte SE nach 90 min mit Diazepam
unterbrochen und die Tiere 8 h nach Applikation perfundiert, um ausgedehnte
neurodegenerative Veränderungen zu verhindern. Trotzdem war in beiden
Tiergruppen eine Neurodegeneration in unterschiedlichem Ausmaß zu beschreiben.
Die Veränderungen betrafen hauptsächlich den PC (Kap. 4.3.4.1) und die Substantia
nigra pars reticulata (Kap. 4.3.4.2).
4.3.4.1 Neurodegeneration im limbischen System
Im PC, der BLA sowie im Hippocampus erfolgte die Beurteilung des Ausmaßes der
Neurodegeneration in diesen Regionen anhand eines Scoresystems, welches den
Anteil von Neuronen mit abweichender Morphologie prozentual zu der Zahl nicht
veränderter Neurone angab (Kap. 3.5.4). Die Stufe 0 beschrieb dabei keine
sichtbarer Neuronenschädigung, die Stufe 1 gab eine Schädigung von unter, Stufe 2
eine Schädigung von über 50 % alle Neurone an. Waren keine morphologisch
unveränderten Neurone mehr erhalten, wurde mit Stufe 3 bewertet.
In der Amygdala sowie im Hilus des Gyrus dentatus hatte die Untersuchung
GABAerger Neurone mit in-situ-Hybridisierung und Immunhistochemie eine
signifikante Reduktion GABAerger Neurone nach Kainat- bzw. Lithium-Pilocarpin
induziertem SE ergeben (Kap. 4.3.2.4 und 4.3.2.5). Im Gegensatz dazu waren im
Nissl-gefärbten Gewebeschnitt im Bereich der basolateralen Amygdala sowie im
Hippocampus keine sichtbaren Zellveränderungen oder Zellverluste festzustellen.

129
In diesen Regionen hätte nur durch Auszählung der Neurone eine Aussage über das
Ausmaß des Zellunterganges gemacht werden können.
Im PC wurde der Umfang der Zellschädigung im anterioren, im zentralen und im
posterioren PC in jeweils einer Schnittebene +2,2 mm, -0,8 mm und -3,8 mm in
Bezug auf Bregma bestimmt. Die Beurteilung erfolgte beidseitig in der Schicht I bis III
sowie im EN. Bei 4 von 6 Tieren der Gruppe mit Kainat-induziertem SE traten
neurodegenerative Veränderungen im PC auf, in der Gruppe mit SE nach Pilocarpin
bei allen 8 Ratten. In beiden Versuchsgruppen nahmen die Schädigungen von
anterior nach posterior zu, wobei Schicht III sowie der VEN am stärksten betroffen
waren (Abbildung 27 A/B).
Abbildung 27 A/B
Histologische Aufnahmen von Nissl-gefärbten transversalen Gehirnschnitten des zentralen PC eines Kontrolltieres (A) und eines Tieres mit SE induziert durch Lithium-Pilocarpin (B). In der Schicht II (Pfeile) sowie den angrenzenden Schichten des zentralen PC des Tieres mit SE induziert durch Lithium-Pilocarpin waren ausgeprägte neurodegenerative Veränderungen festzustellen, die das Gewebe stark aufgelockert erscheinen ließen. Kalibrierbalken für A und B: 500 µm
A
PC
DEN
B DEN

130
Im anterioren PC traten in der Kainat-Gruppe keine sichtbaren morphologischen
Veränderungen auf, die Behandlung mit Lithium-Pilocarpin führte bei 2 Tieren zu
Veränderungen in der Schicht III und im angrenzenden VEN (Abbildung 28 A).
Bei der Betrachtung des zentralen PC war die Schicht III am stärksten von
Neurodegeneration betroffen. Zwei Tiere der Pilocarpin-Gruppe wiesen in dieser
Schicht keine morphologisch unveränderten Neurone auf, bei den übrigen Tieren der
Gruppe betraf die Schädigung mehr als 50 % der Zellen. Die Veränderungen in der
Kainat-Gruppe betrafen nur 2 Tiere. Diese zeigten einen Anteil von weniger als 50 %
geschädigter Zellen in der Schicht III und dem EN (Abbildung 28 B).
In beiden Tiergruppen war der posteriore PC am stärksten neurodegenerativ
verändert. Von den 6 Ratten der Kainat-Gruppe zeigten zwei Tiere bis zu 50 % und
weitere 2 Tiere über 50 % geschädigte Neurone in Schicht III. In der Gruppe mit SE
nach Lithium-Pilocarpin waren die Neurone in den Schichten II und III sowie im EN
prozentual stärker verändert als im zentralen PC (Abbildung 28 C).
I II VEN DEN
Scor
e de
r Neu
rone
nsch
ädig
ung
0
1
2
3
KainatPilocarpin
anteriorer PC A
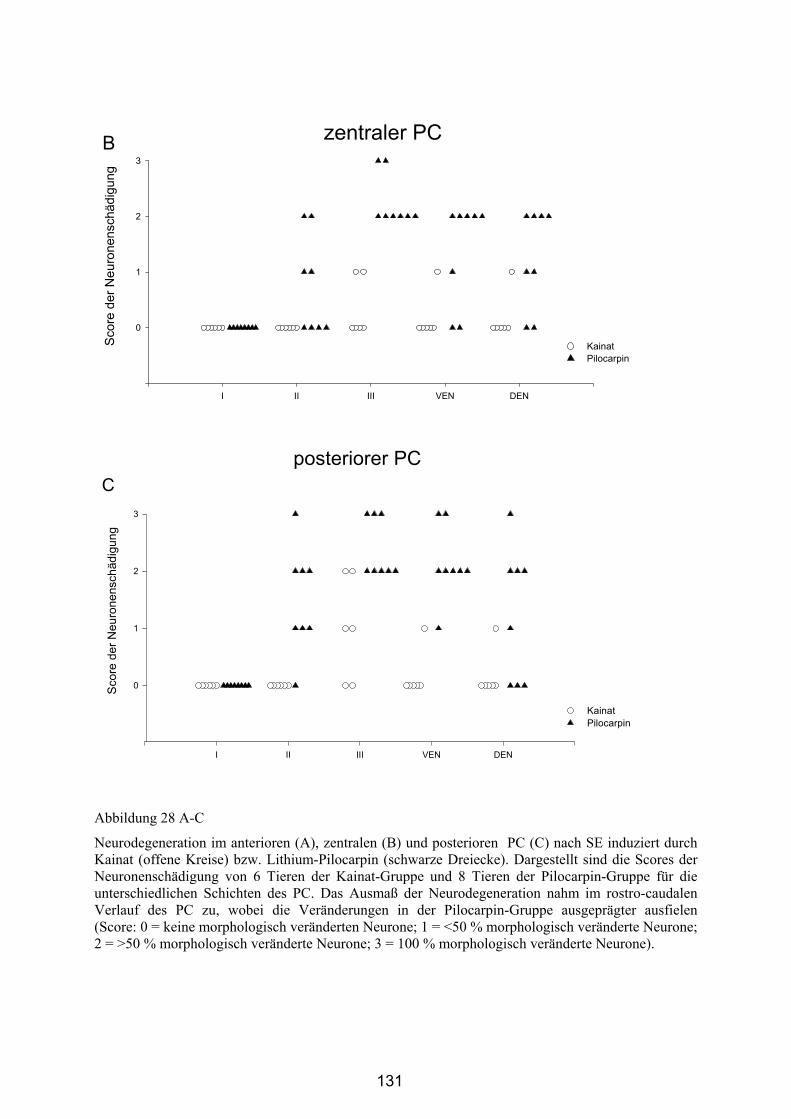
131
I II III VEN DEN
Scor
e de
r Neu
rone
nsch
ädig
ung
0
1
2
3
zentraler PC
KainatPilocarpin
B
I II III VEN DEN
Scor
e de
r Neu
rone
nsch
ädig
ung
0
1
2
3
KainatPilocarpin
posteriorer PC C
Abbildung 28 A-C
Neurodegeneration im anterioren (A), zentralen (B) und posterioren PC (C) nach SE induziert durch Kainat (offene Kreise) bzw. Lithium-Pilocarpin (schwarze Dreiecke). Dargestellt sind die Scores der Neuronenschädigung von 6 Tieren der Kainat-Gruppe und 8 Tieren der Pilocarpin-Gruppe für die unterschiedlichen Schichten des PC. Das Ausmaß der Neurodegeneration nahm im rostro-caudalen Verlauf des PC zu, wobei die Veränderungen in der Pilocarpin-Gruppe ausgeprägter ausfielen (Score: 0 = keine morphologisch veränderten Neurone; 1 = <50 % morphologisch veränderte Neurone; 2 = >50 % morphologisch veränderte Neurone; 3 = 100 % morphologisch veränderte Neurone).

132
4.3.4.2 Läsionen in der Substantia nigra pars reticulata
In der Tiergruppe mit Kainat-induziertem SE traten bei 3 von 6 Ratten in der
anterioren und posterioren Substantia nigra pars reticulata Läsionen auf. Als Läsion
wurden solche Gebiete definiert, die keine morphologisch unveränderten Zellen mehr
aufwiesen. Nach Lithium-Pilocarpin waren bei 7 Tieren der Versuchsgruppe
Läsionen in der Substantia nigra pars reticulata unterschiedlichen Ausmaßes
festzustellen. Bei 5 Ratten dieser Gruppe war der anteriore und posteriore Abschnitt
der Region betroffen, bei 2 Tieren beschränkten sich die sichtbaren Veränderungen
auf den anterioren Teil der Struktur.
Die Substantia nigra pars reticulata erstreckt sich in einem Bereich von -4,3 mm bis -
6,3 mm caudal von Bregma. Zur Dokumentation der Läsionen wurden die Flächen in
den betroffenen Schnittebenen mit dem Bildanalysesystems vermessen und die
individuellen Läsionsvolumina anhand des bekannten Abstandes zwischen den
Schnitten berechnet.
Abbildung 29 A/B
Histologische Aufnahmen von Nissl-gefärbten transversalen Gehirnschnitten der anterioren Substantia nigra pars reticulata (aSNr) eines Kontrolltieres (A) und eines Tieres mit SE induziert durch Lithium-Pilocarpin (B). Die Substantia nigra pars reticulata des Tiere mit SE war in ihrer gesamten Ausdehnung läsioniert (Pfeilköpfe), während in der angrenzenden Substantia nigra pars compacta (SNc) keine neuropathologischen Veränderungen auffällig waren. Kalibrierbalken für A und B: 500 µm
A B
aSNr
SNc
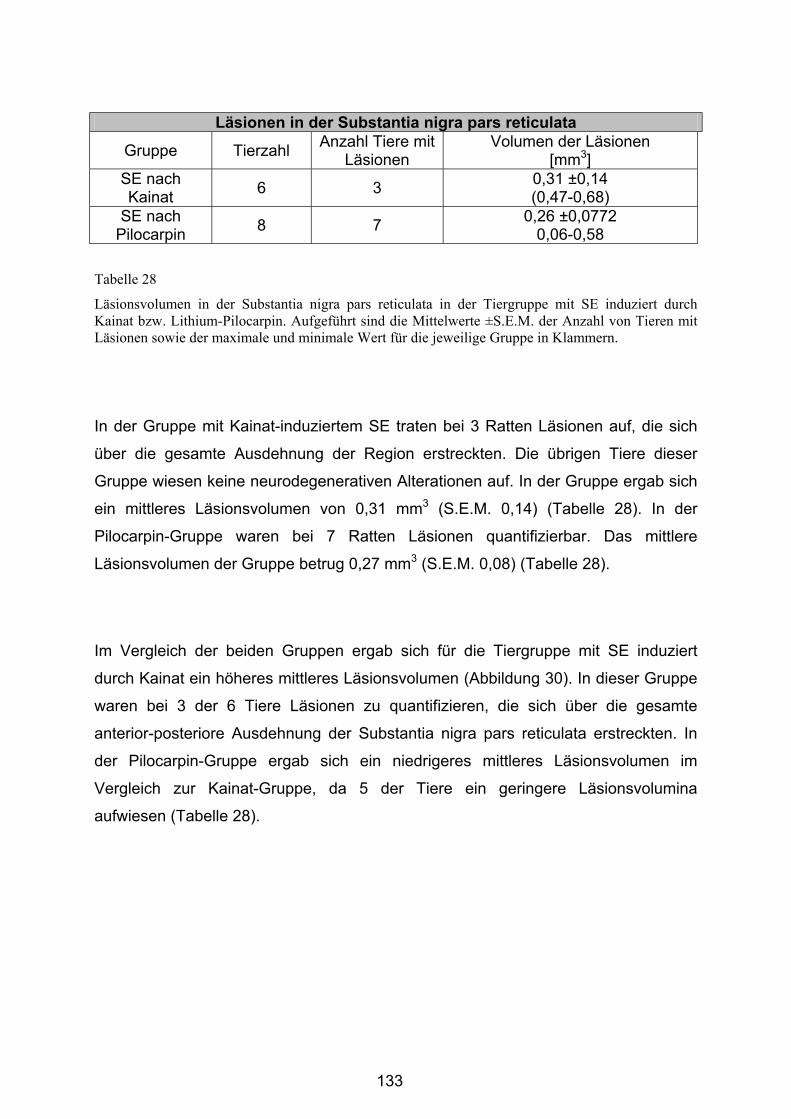
133
Läsionen in der Substantia nigra pars reticulata Gruppe Tierzahl Anzahl Tiere mit
Läsionen Volumen der Läsionen
[mm3] SE nach Kainat 6 3 0,31 ±0,14
(0,47-0,68) SE nach
Pilocarpin 8 7 0,26 ±0,0772 0,06-0,58
Tabelle 28
Läsionsvolumen in der Substantia nigra pars reticulata in der Tiergruppe mit SE induziert durch Kainat bzw. Lithium-Pilocarpin. Aufgeführt sind die Mittelwerte ±S.E.M. der Anzahl von Tieren mit Läsionen sowie der maximale und minimale Wert für die jeweilige Gruppe in Klammern.
In der Gruppe mit Kainat-induziertem SE traten bei 3 Ratten Läsionen auf, die sich
über die gesamte Ausdehnung der Region erstreckten. Die übrigen Tiere dieser
Gruppe wiesen keine neurodegenerativen Alterationen auf. In der Gruppe ergab sich
ein mittleres Läsionsvolumen von 0,31 mm3 (S.E.M. 0,14) (Tabelle 28). In der
Pilocarpin-Gruppe waren bei 7 Ratten Läsionen quantifizierbar. Das mittlere
Läsionsvolumen der Gruppe betrug 0,27 mm3 (S.E.M. 0,08) (Tabelle 28).
Im Vergleich der beiden Gruppen ergab sich für die Tiergruppe mit SE induziert
durch Kainat ein höheres mittleres Läsionsvolumen (Abbildung 30). In dieser Gruppe
waren bei 3 der 6 Tiere Läsionen zu quantifizieren, die sich über die gesamte
anterior-posteriore Ausdehnung der Substantia nigra pars reticulata erstreckten. In
der Pilocarpin-Gruppe ergab sich ein niedrigeres mittleres Läsionsvolumen im
Vergleich zur Kainat-Gruppe, da 5 der Tiere ein geringere Läsionsvolumina
aufwiesen (Tabelle 28).
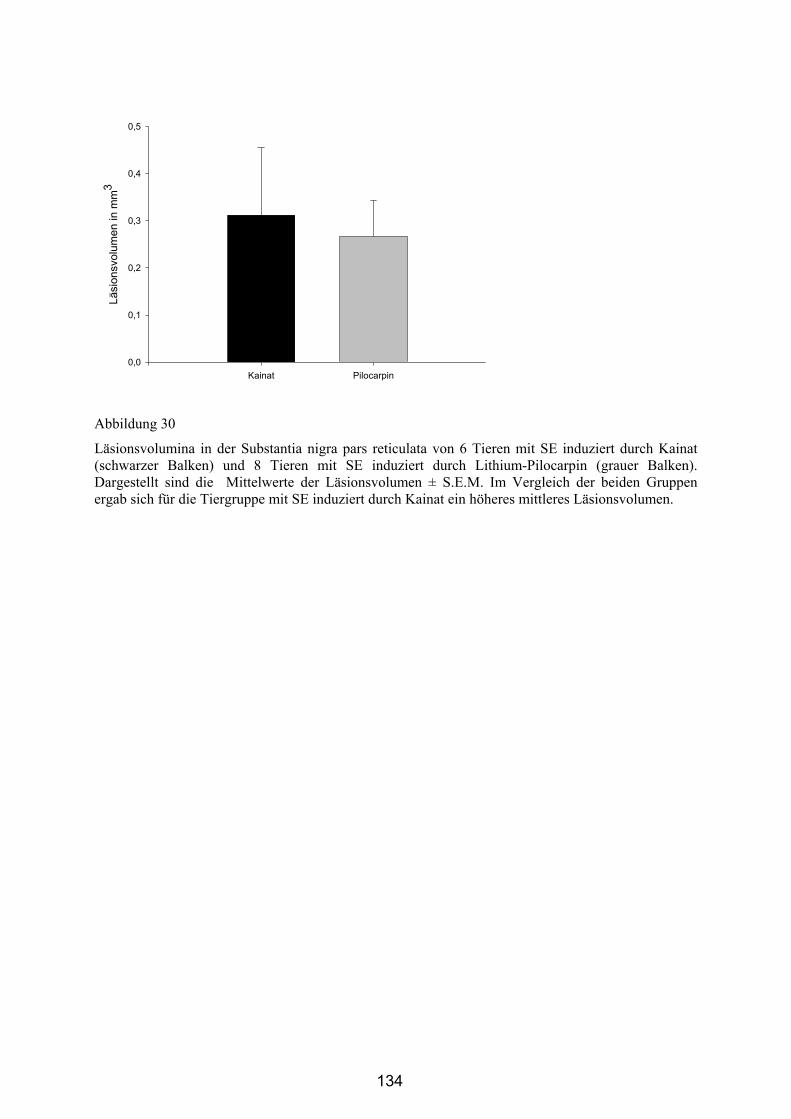
134
Kainat Pilocarpin
Läsi
onsv
olum
en in
mm
3
0,0
0,1
0,2
0,3
0,4
0,5
Abbildung 30
Läsionsvolumina in der Substantia nigra pars reticulata von 6 Tieren mit SE induziert durch Kainat (schwarzer Balken) und 8 Tieren mit SE induziert durch Lithium-Pilocarpin (grauer Balken). Dargestellt sind die Mittelwerte der Läsionsvolumen ± S.E.M. Im Vergleich der beiden Gruppen ergab sich für die Tiergruppe mit SE induziert durch Kainat ein höheres mittleres Läsionsvolumen.

135
4.4 Untersuchungen an voll gekindelten Ratten
Im Kindling-Modell der Temporallappenepilepsie wurden in verschiedenen
Untersuchungen Veränderungen des GABAergen Systems beschrieben. Bei Wistar-
Ratten konnte 40 Tage nach Erreichen des Stadiums „voll gekindelt“ eine Abnahme
GABA-immunreaktiver Neurone im zentralen PC ipsilateral zur Stimulationselektrode
in der rechten basolateralen Amygdala nachgewiesen werden
(Lehmann et al., 1998). In der gleichen Studie wurde zudem eine beidseitige
Reduktion in der Dichte GABA-immunreaktiver Neurone für die basolaterale
Amygdala beschrieben. Sonnenberg et al. (1991) zeigten in der Substantia nigra voll
gekindelter Ratten eine verminderte Expression von GAD-mRNA mit radioaktiver in-
situ-Hybridisierung 30 min nach dem letzten gekindelten Anfall.
Die Untersuchung der Expression von mRNA für die beiden Isoformen der GAD
durch Amygdala-Kindling 42 Tage nach Erreichen des Stadiums „voll gekindelt“ war
Gegenstand des folgenden Teils der Arbeit. Zusätzlich wurden in den
Versuchsgruppen GABAerger Neurone mit Immunhistochemie gegen GABA
untersucht.
4.4.1 Ergebnisse des Kindling
Zu Beginn der auf dem Kindling basierenden Untersuchungen erfolgt zunächst eine
Darstellung der während des Kindling-Prozesses der Gruppe gekindelter Tiere
erfassten Parameter (Tabelle 29) sowie die Ergebnisse der Elektrodenlokalisationen
(Abbildung 31).
Innerhalb der Gruppe der gekindelten Tiere variierten die initialen
Nachentladungsschwellen (initiale ADT) zwischen 75 und 240 µA, der Mittelwert
betrug 136 µA. Bereits am ersten Tag der Stimulation zeigten alle Tiere die
Symptomatik des Stadiums I, die ersten Anfälle des Stadiums V wurden nach 2 bis
10 weiteren Stimulationen gezeigt (Mittelwert 5,3 Stimulationen). Das Erreichen des
voll gekindelten Zustandes nach 10 Anfällen des Stadiums V erforderte mindestens
12 bis maximal 21 Stimulationen.
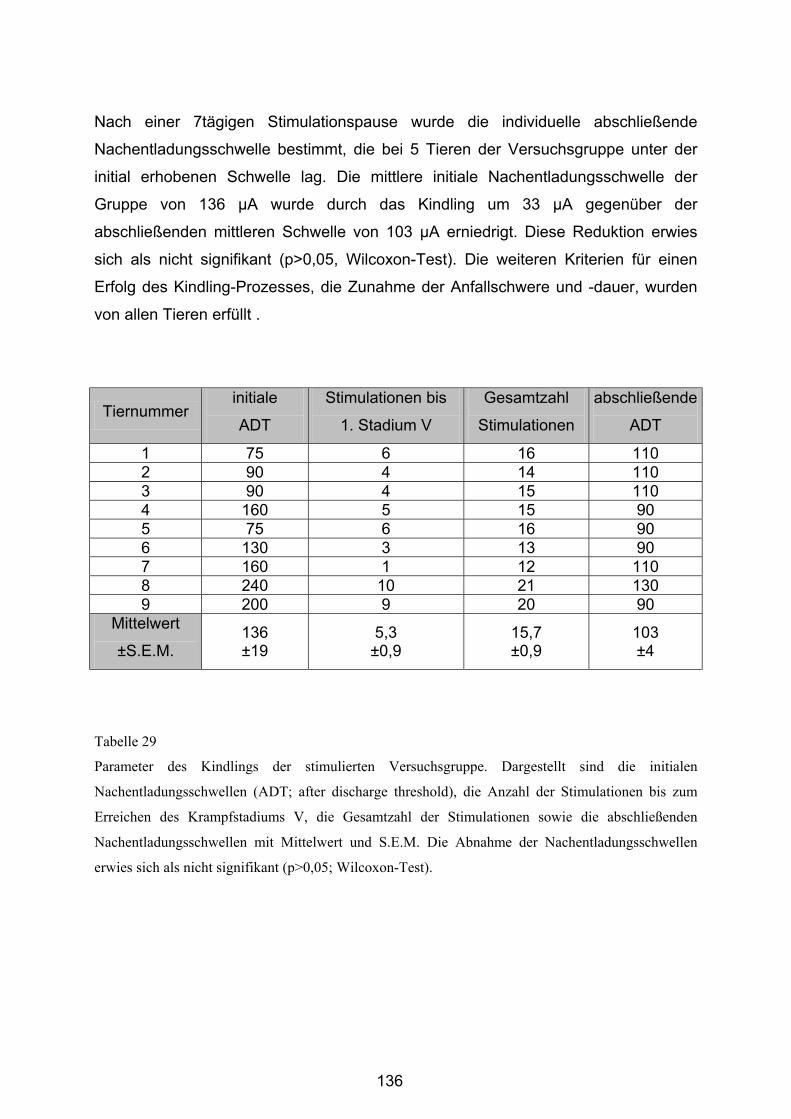
136
Nach einer 7tägigen Stimulationspause wurde die individuelle abschließende
Nachentladungsschwelle bestimmt, die bei 5 Tieren der Versuchsgruppe unter der
initial erhobenen Schwelle lag. Die mittlere initiale Nachentladungsschwelle der
Gruppe von 136 µA wurde durch das Kindling um 33 µA gegenüber der
abschließenden mittleren Schwelle von 103 µA erniedrigt. Diese Reduktion erwies
sich als nicht signifikant (p>0,05, Wilcoxon-Test). Die weiteren Kriterien für einen
Erfolg des Kindling-Prozesses, die Zunahme der Anfallschwere und -dauer, wurden
von allen Tieren erfüllt .
Tiernummer initiale
ADT
Stimulationen bis
1. Stadium V
Gesamtzahl
Stimulationen
abschließende
ADT
1 75 6 16 110 2 90 4 14 110 3 90 4 15 110 4 160 5 15 90 5 75 6 16 90 6 130 3 13 90 7 160 1 12 110 8 240 10 21 130 9 200 9 20 90
Mittelwert
±S.E.M. 136 ±19
5,3 ±0,9
15,7 ±0,9
103 ±4
Tabelle 29
Parameter des Kindlings der stimulierten Versuchsgruppe. Dargestellt sind die initialen
Nachentladungsschwellen (ADT; after discharge threshold), die Anzahl der Stimulationen bis zum
Erreichen des Krampfstadiums V, die Gesamtzahl der Stimulationen sowie die abschließenden
Nachentladungsschwellen mit Mittelwert und S.E.M. Die Abnahme der Nachentladungsschwellen
erwies sich als nicht signifikant (p>0,05; Wilcoxon-Test).

137
BLA
BLA
BLA
- 2,1
- 1,8
- 2,3
Durch Verlust des Steckeraufbaus reduzierte sich die Tierzahl in der Gruppe der
implantierten Tiere auf 8 und in der Kindling-Gruppe auf 11 Ratten. Die
Elektrodenlokalisationen der implantierten und gekindelten Tiere sind in Abbildung 31
dargestellt. Die Elektrodenspitzen beider Tiergruppen lagen rostro-caudal zwischen
-1,8 bis -2,3 posterior von Bregma.
Abbildung 31
Lokalisationen der Stimulationselektrode in der rechten basolateralen Amygdala (BLA) von implantierten (n=8) und gekindelten Tieren (n=11). Die Lokalisationen der Elektrodenspitzen der implantierten Tiere sind als graue Kreise, die der gekindelten Tiere als schwarze Dreiecke in entsprechenden Transversalschnitten nach Paxinos und Watson (1998) markiert, deren Lage jeweils in mm zu Bregma angegeben ist.
Bei einer Ratte aus der Gruppe der implantieren Tiere lag der tiefste Punkt des
Elektrodenstichkanals dorsal, bei zwei Tieren der Kindling-Gruppe ventral der
basolateralen Amygdala. Diese Tiere wurden von der Auswertung ausgeschlossen.
Dadurch standen in der implantierten Gruppe 7 Tiere und in der Kindling-Gruppe 9
Tiere für die weiteren Untersuchungen zur Verfügung.

138
4.4.2 Bestimmung der Dichte GABAerger Neurone
Zur Untersuchung von Kindling-induzierten Veränderungen in der Expression von
GAD-mRNA in GABAergen Neuronen wurden Gewebeserien von voll gekindelten
Ratten, welche 6 Wochen nach dem 10. generalisierten Anfallsstadium V perfundiert
worden waren, der nicht-radioaktiven in-situ-Hybridisierung für GAD65- und GAD67-
mRNA unterzogen. An einer weiteren Gewebeserie jedes Tieres wurde die
Immunhistochemie für GABA durchgeführt. Nach Abschluss der Versuche erfolgte
die Quantifizierung positiv gefärbter Neurone nach der Methode des optischen
Disektors in den festgelegten Hirnregionen (Kap. 3.6.3) an verschlüsselten
Objektträgern. Die Zählung erfolgte zunächst für GAD65-mRNA und GAD67-mRNA
im zentralen PC, der basolateralen Amygdala sowie in der anterioren und posterioren
Substantia nigra. Im anterioren und posterioren PC sowie im Hippocampus wurden
nicht alle Tiere der Versuchsgruppen für die in-situ-Hybridisierung ausgewertet, da in
diesen Regionen aufgrund der Literatur keine ausgeprägten Veränderungen erwartet
wurden. Die mit GABA- Immunhistochemie behandelten Serien wurden im Bereich
der Substantia nigra pars reticulata ausgewertet, da für diese Region in früheren
Untersuchungen keine Daten erhoben wurden. Nach Abschluss der Auszählung
wurden die Daten decodiert und den Tieren der drei Versuchsgruppen den
Nachweisen entsprechend zugeordnet. Die Berechnung der Neuronendichten wurde
für die beiden Hirnhemisphären getrennt durchgeführt.
Die Darstellung GABAerger Neurone mittels nicht-radioaktiver in-situ-Hybridisierung
entsprach dem im Kap. 4.1.2 beschriebenen Erscheinungsbild. Ein Unterschied in
der Intensität der Anfärbung für GAD65- bzw GAD67-mRNA markierter Neurone
zwischen den drei Versuchsgruppen konnte nicht festgestellt werden (Abbildung 32).
Beim Vergleich der beiden Isoformen erschien die Anfärbung für die GAD67-mRNA
wiederum etwas dunkler.
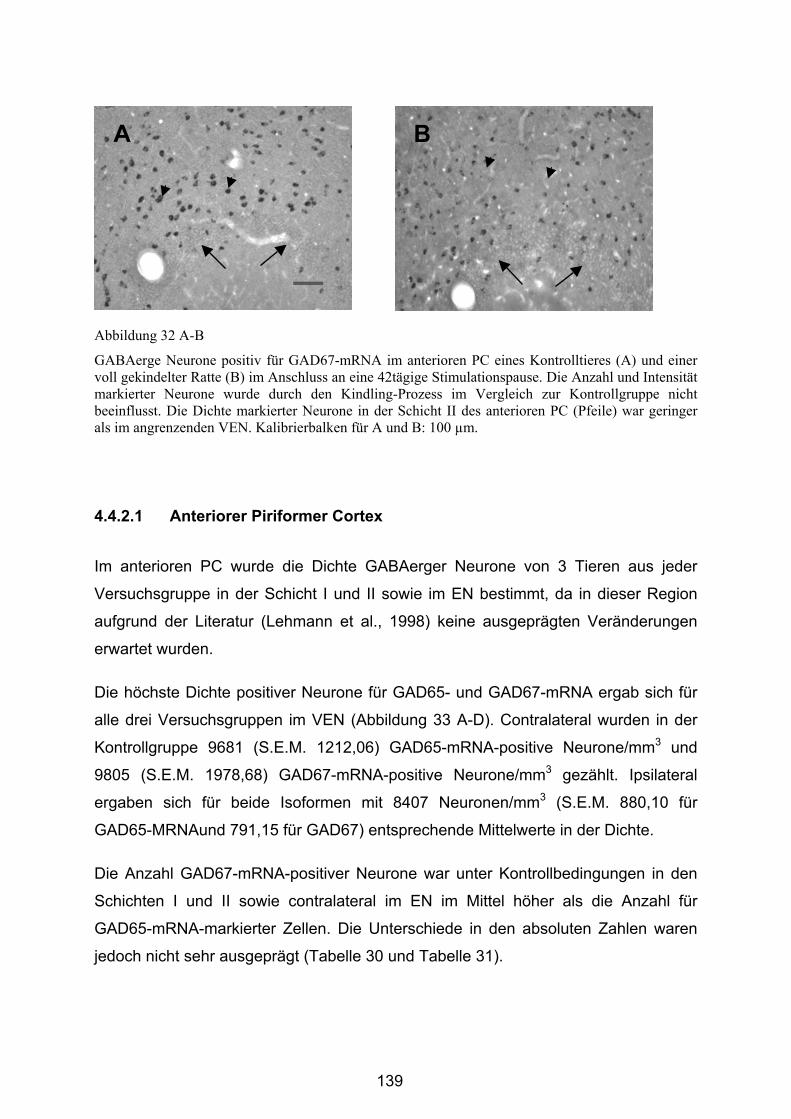
139
Abbildung 32 A-B
GABAerge Neurone positiv für GAD67-mRNA im anterioren PC eines Kontrolltieres (A) und einer voll gekindelter Ratte (B) im Anschluss an eine 42tägige Stimulationspause. Die Anzahl und Intensität markierter Neurone wurde durch den Kindling-Prozess im Vergleich zur Kontrollgruppe nicht beeinflusst. Die Dichte markierter Neurone in der Schicht II des anterioren PC (Pfeile) war geringer als im angrenzenden VEN. Kalibrierbalken für A und B: 100 µm.
4.4.2.1 Anteriorer Piriformer Cortex
Im anterioren PC wurde die Dichte GABAerger Neurone von 3 Tieren aus jeder
Versuchsgruppe in der Schicht I und II sowie im EN bestimmt, da in dieser Region
aufgrund der Literatur (Lehmann et al., 1998) keine ausgeprägten Veränderungen
erwartet wurden.
Die höchste Dichte positiver Neurone für GAD65- und GAD67-mRNA ergab sich für
alle drei Versuchsgruppen im VEN (Abbildung 33 A-D). Contralateral wurden in der
Kontrollgruppe 9681 (S.E.M. 1212,06) GAD65-mRNA-positive Neurone/mm3 und
9805 (S.E.M. 1978,68) GAD67-mRNA-positive Neurone/mm3 gezählt. Ipsilateral
ergaben sich für beide Isoformen mit 8407 Neuronen/mm3 (S.E.M. 880,10 für
GAD65-MRNAund 791,15 für GAD67) entsprechende Mittelwerte in der Dichte.
Die Anzahl GAD67-mRNA-positiver Neurone war unter Kontrollbedingungen in den
Schichten I und II sowie contralateral im EN im Mittel höher als die Anzahl für
GAD65-mRNA-markierter Zellen. Die Unterschiede in den absoluten Zahlen waren
jedoch nicht sehr ausgeprägt (Tabelle 30 und Tabelle 31).
A B
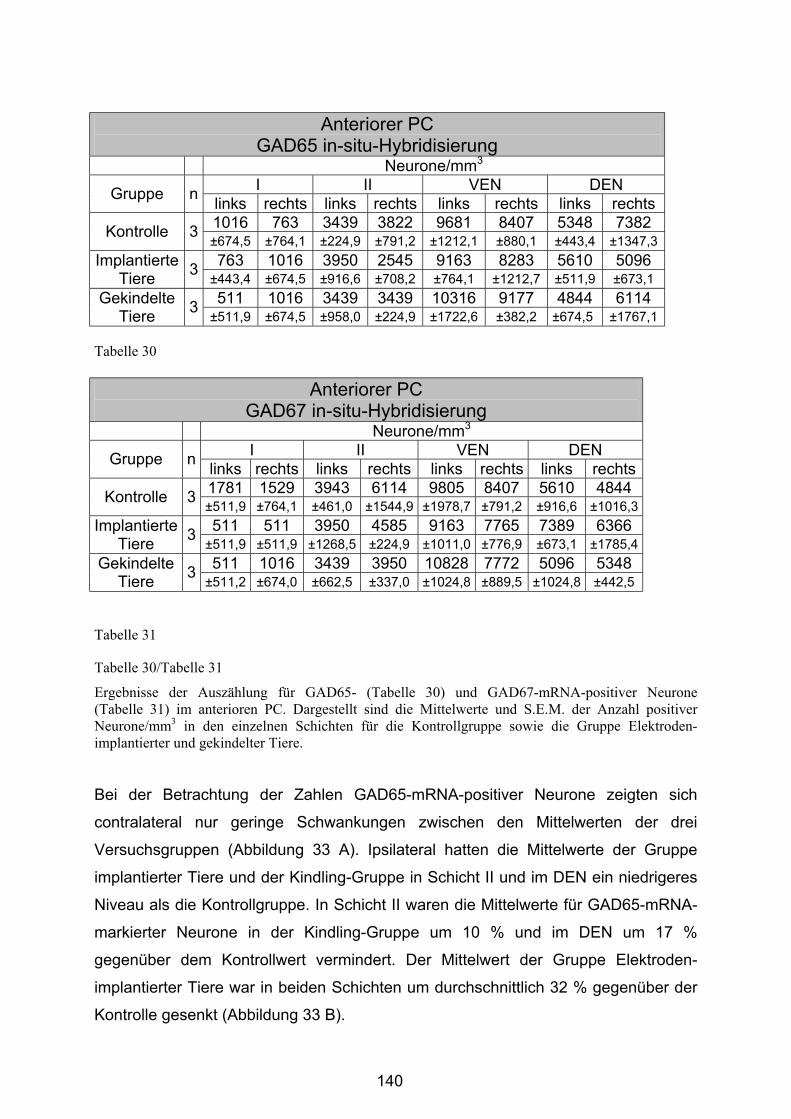
140
Anteriorer PC GAD65 in-situ-Hybridisierung
Neurone/mm3 I II VEN DEN Gruppe n links rechts links rechts links rechts links rechts
1016 763 3439 3822 9681 8407 5348 7382 Kontrolle 3 ±674,5 ±764,1 ±224,9 ±791,2 ±1212,1 ±880,1 ±443,4 ±1347,3 763 1016 3950 2545 9163 8283 5610 5096 Implantierte
Tiere 3 ±443,4 ±674,5 ±916,6 ±708,2 ±764,1 ±1212,7 ±511,9 ±673,1 511 1016 3439 3439 10316 9177 4844 6114 Gekindelte
Tiere 3 ±511,9 ±674,5 ±958,0 ±224,9 ±1722,6 ±382,2 ±674,5 ±1767,1
Tabelle 30
Anteriorer PC GAD67 in-situ-Hybridisierung
Neurone/mm3 I II VEN DEN Gruppe n links rechts links rechts links rechts links rechts
1781 1529 3943 6114 9805 8407 5610 4844 Kontrolle 3 ±511,9 ±764,1 ±461,0 ±1544,9 ±1978,7 ±791,2 ±916,6 ±1016,3 511 511 3950 4585 9163 7765 7389 6366 Implantierte
Tiere 3 ±511,9 ±511,9 ±1268,5 ±224,9 ±1011,0 ±776,9 ±673,1 ±1785,4 511 1016 3439 3950 10828 7772 5096 5348 Gekindelte
Tiere 3 ±511,2 ±674,0 ±662,5 ±337,0 ±1024,8 ±889,5 ±1024,8 ±442,5
Tabelle 31
Tabelle 30/Tabelle 31
Ergebnisse der Auszählung für GAD65- (Tabelle 30) und GAD67-mRNA-positiver Neurone (Tabelle 31) im anterioren PC. Dargestellt sind die Mittelwerte und S.E.M. der Anzahl positiver Neurone/mm3 in den einzelnen Schichten für die Kontrollgruppe sowie die Gruppe Elektroden-implantierter und gekindelter Tiere.
Bei der Betrachtung der Zahlen GAD65-mRNA-positiver Neurone zeigten sich
contralateral nur geringe Schwankungen zwischen den Mittelwerten der drei
Versuchsgruppen (Abbildung 33 A). Ipsilateral hatten die Mittelwerte der Gruppe
implantierter Tiere und der Kindling-Gruppe in Schicht II und im DEN ein niedrigeres
Niveau als die Kontrollgruppe. In Schicht II waren die Mittelwerte für GAD65-mRNA-
markierter Neurone in der Kindling-Gruppe um 10 % und im DEN um 17 %
gegenüber dem Kontrollwert vermindert. Der Mittelwert der Gruppe Elektroden-
implantierter Tiere war in beiden Schichten um durchschnittlich 32 % gegenüber der
Kontrolle gesenkt (Abbildung 33 B).

141
Der Nachweis von GAD67-mRNA in Neuronen zeigte beidseitig deutlich geringere
Mittelwerte in Schicht I des anterioren PC. Waren contralateral 1781 Neurone/mm3
(S.E.M. 511,9) in der Kontrollgruppe zu zählen, ergaben sich für die beiden anderen
Gruppe relative Differenzen von 71 % zu diesem Wert ( Tabelle 31, Abbildung 33 C).
Ipsilateral besaßen Kontrolltiere in der Schicht I im Mittel 1529 Neurone/mm3, der
Mittelwerte der Kindling-Gruppe lag um 34 % darunter, die Abnahme für die Gruppe
implantierter Tiere entsprach der anderen Hemisphäre (Tabelle 31, Abbildung 33 D).
In Schicht II lagen die Mittelwerte GAD67-mRNA-positiver Neurone der Gruppen
implantierter und gekindelter Tiere ipsilateral um 25 % bzw. 35 % unter den basalen
Werten, wobei jedoch eine große Streuung innerhalb der Kontrollgruppe festgestellt
wurde. Contralateral traten in Schicht II keine deutlichen Veränderungen auf,
gleiches galt für den VEN (Tabelle 31, Abbildung 33 C/ D).
Im DEN besaßen implantierte Tiere contralateral mit 7389 Neuronen/mm3 (S.E.M.
673,1) und ipsilateral mit 6366 Neuronen/mm3 (S.E.M. 1785,4) die höchsten
Mittelwerte, die mittleren Neuronendichten der anderen Gruppen lagen auf
vergleichbarem Niveau darunter (Abbildung 33 C/D).
Eine Verringerung in der Neuronendichte der gekindelten Tiere in einigen Schichten
des anterioren PC ging mit einer Reduktion von vergleichbarem oder größerem
Ausmaß in der Gruppe der implantierten Tiere einher. Die Varianzanalyse der
Untersuchungsergebnisse erbrachte keine statistische Signifikanz für die
dargestellten Veränderungen zwischen den Versuchsgruppen (p>0,05; einfaktorielle
ANOVA).
Abbildung 33 A-D
Darstellung der Dichte GAD65- (A/B) bzw. GAD67-mRNA-positiver Neurone (C/D) in den unterschiedlichen Schichten des anterioren PC contralateral und ipsilateral zur Stimulationselektrode. Aufgeführt sind die Anzahl markierter Neurone/mm3 sowie der zugehörige Standardfehler. Signifikante Unterschiede in der Anzahl markierter Neurone/mm3 zwischen den Versuchsgruppen bestanden nicht (p>0,05; einfaktorielle ANOVA).

142

143
4.4.2.2 Zentraler Piriformer Cortex
Für die Auswertung im Bereich des zentralen PC reduzierte sich die Anzahl der
Serien aufgrund mangelhafter Anfärbung in jeder Gruppe um jeweils ein Tier. In der
Kontrollgruppe mussten zwei Serien der nach der in-situ-Hybridisierung für GAD65-
mRNA ausgeschlossen werden. Dementsprechend wurden 8 bzw. 9 Kontrolltiere für
GAD65 bzw. GAD67 und 6 implantierte und 8 gekindelte Ratten ausgewertet.
In der Kontrollgruppe fand sich die niedrigste Dichte markierter Neurone in Schicht I
des zentralen PC.
Nach in-situ-Hybridisierung für GAD65-mRNA war die Dichte positiver Zellen mit
7451 Neuronen/mm3 (S.E.M. 746,9) in Schicht III contralateral zur
Stimulationselektrode im Mittel am höchsten, gefolgt vom DEN, dem VEN und
Schicht II mit jeweils abnehmender Dichte. Ipsilateral ergaben sich für die Schicht III
sowie den EN Mittelwerte auf vergleichbarem Niveau, während Schicht II wiederum
eine geringere Dichte aufwies (Tabelle 32).
Die in-situ-Hybridisierung für GAD67-mRNA ergab für alle 3 Versuchsgruppen
höhere Werte in der mittleren Neuronendichte in Schicht II im Vergleich zur GAD65-
mRNA (Tabelle 33, Abbildung 34 C-D). Contralateral lag die Anzahl markierter
Neurone/mm3 in der Kontrollgruppe mit 6282 Neuronen/mm3 (S.E.M. 278,8) um 57
%, ipsilateral mit 6458 Neuronen/mm3 um 83 % über den Werten der Kontrolle für
GAD65. Damit besaß die Schicht II des PC beidseitig die zweitgrößte Dichte
GAD67-mRNA- markierter Neurone nach Schicht III (Tabelle 33). Im Vergleich
ergaben sich für die Schicht III sowie den EN keine maßgeblichen Unterschiede in
der Dichte für GAD65- und GAD67-mRNA-markierter Zellen.
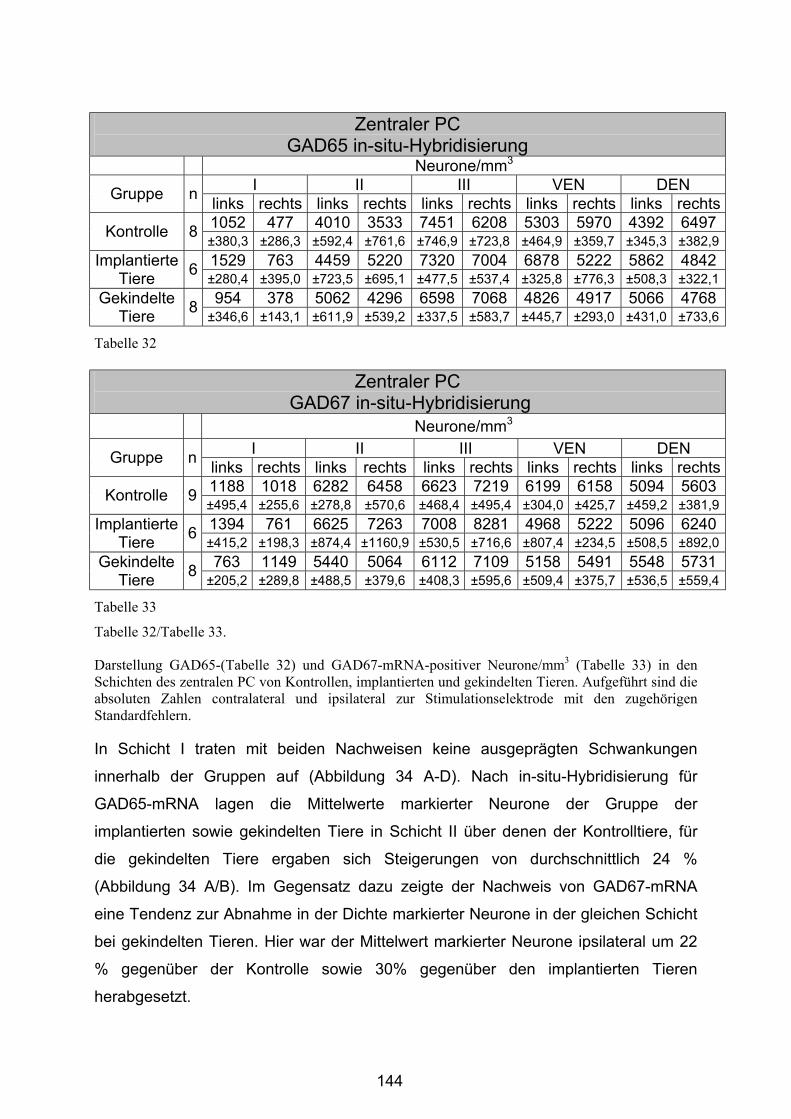
144
Zentraler PC GAD65 in-situ-Hybridisierung
Neurone/mm3 I II III VEN DEN Gruppe n links rechts links rechts links rechts links rechts links rechts
1052 477 4010 3533 7451 6208 5303 5970 4392 6497 Kontrolle 8 ±380,3 ±286,3 ±592,4 ±761,6 ±746,9 ±723,8 ±464,9 ±359,7 ±345,3 ±382,9 1529 763 4459 5220 7320 7004 6878 5222 5862 4842 Implantierte
Tiere 6 ±280,4 ±395,0 ±723,5 ±695,1 ±477,5 ±537,4 ±325,8 ±776,3 ±508,3 ±322,1 954 378 5062 4296 6598 7068 4826 4917 5066 4768 Gekindelte
Tiere 8 ±346,6 ±143,1 ±611,9 ±539,2 ±337,5 ±583,7 ±445,7 ±293,0 ±431,0 ±733,6
Tabelle 32
Zentraler PC GAD67 in-situ-Hybridisierung
Neurone/mm3 I II III VEN DEN Gruppe n links rechts links rechts links rechts links rechts links rechts
1188 1018 6282 6458 6623 7219 6199 6158 5094 5603 Kontrolle 9 ±495,4 ±255,6 ±278,8 ±570,6 ±468,4 ±495,4 ±304,0 ±425,7 ±459,2 ±381,9 1394 761 6625 7263 7008 8281 4968 5222 5096 6240 Implantierte
Tiere 6 ±415,2 ±198,3 ±874,4 ±1160,9 ±530,5 ±716,6 ±807,4 ±234,5 ±508,5 ±892,0 763 1149 5440 5064 6112 7109 5158 5491 5548 5731 Gekindelte
Tiere 8 ±205,2 ±289,8 ±488,5 ±379,6 ±408,3 ±595,6 ±509,4 ±375,7 ±536,5 ±559,4
Tabelle 33
Tabelle 32/Tabelle 33.
Darstellung GAD65-(Tabelle 32) und GAD67-mRNA-positiver Neurone/mm3 (Tabelle 33) in den Schichten des zentralen PC von Kontrollen, implantierten und gekindelten Tieren. Aufgeführt sind die absoluten Zahlen contralateral und ipsilateral zur Stimulationselektrode mit den zugehörigen Standardfehlern.
In Schicht I traten mit beiden Nachweisen keine ausgeprägten Schwankungen
innerhalb der Gruppen auf (Abbildung 34 A-D). Nach in-situ-Hybridisierung für
GAD65-mRNA lagen die Mittelwerte markierter Neurone der Gruppe der
implantierten sowie gekindelten Tiere in Schicht II über denen der Kontrolltiere, für
die gekindelten Tiere ergaben sich Steigerungen von durchschnittlich 24 %
(Abbildung 34 A/B). Im Gegensatz dazu zeigte der Nachweis von GAD67-mRNA
eine Tendenz zur Abnahme in der Dichte markierter Neurone in der gleichen Schicht
bei gekindelten Tieren. Hier war der Mittelwert markierter Neurone ipsilateral um 22
% gegenüber der Kontrolle sowie 30% gegenüber den implantierten Tieren
herabgesetzt.

145
Contralateral zur Stimulationselektrode erwies sich der Unterschied mit 13 % zur
Kontrolle und 18 % gegenüber implantierten Tieren geringer (Abbildung 34 C/D).
In Schicht III des zentralen PC ergaben sich keine ausgeprägten Veränderungen mit
beiden Nachweisen. Im VEN lagen die Mittelwerte GAD65-mRNA-positiver Neurone
gekindelter Tiere unterhalb der Werte der beiden anderen Gruppen, dabei betrug die
größte relative Differenz contralateral 30 % zur Gruppe der implantierten Tiere
(Abbildung 34 A/B). Der Nachweis von GAD67-mRNA erbrachte Mittelwerte für
gekindelte Tiere, die contralateral um 17 % und ipsilateral um 11 % unterhalb derer
der Kontrolle lagen, während die mittleren Werte der implantierten Tiere sich
beidseitig nur als geringgradig niedriger erwiesen (Abbildung 34 C/D).
Die mittlere Dichte von GAD65-mRNA-positiven Neuronen/mm3 wich im DEN
ipsilateral um -27 % vom basalen Wert ab, unterschied sich aber gleichzeitig nur um
2 % vom Wert implantierter Tiere. Contralateral lag der Wert der Kindling-Gruppe um
15 % über der Kontrolle, nach dem Nachweis für GAD67-mRNA war auf der
gleichen Seite ein Anstieg von 9 % gegenüber beiden anderen Gruppen zu
verzeichnen (Abbildung 34 C/D).
Die dargestellten Veränderungen in der Dichte GAD65- bzw. GAD67-mRNA-positiver
Neurone wurden mit einer zweifaktoriellen Varianzanalyse für wiederholte
Messungen verglichen. Dabei ergaben sich keine signifikanten Unterschiede in
Abhängigkeit von der Schicht und Behandlung zwischen den Versuchsgruppen
(p>0,05; einfaktorielle ANOVA).
Abbildung 34 A-D
Anzahl GAD65- (A/B) und GAD67-mRNA-positiver Neurone (C/D) in den Schichten des zentralen PC contralateral und ipsilateral zur Stimulationselektrode. Neben den Gruppenmittelwerten sind die zugehörigen Standardfehler dargestellt. Die Dichte positiver Neurone für GAD67-mRNA war in Schicht II durch Kindling beidseitig tendenziell erniedrigt. Der Nachweis von GAD65-mRNA zeigte keine entsprechende Tendenz. Die dargestellten Unterschiede erwiesen sich als nicht signifikant [p>0,05, einfaktorielle ANOVA].
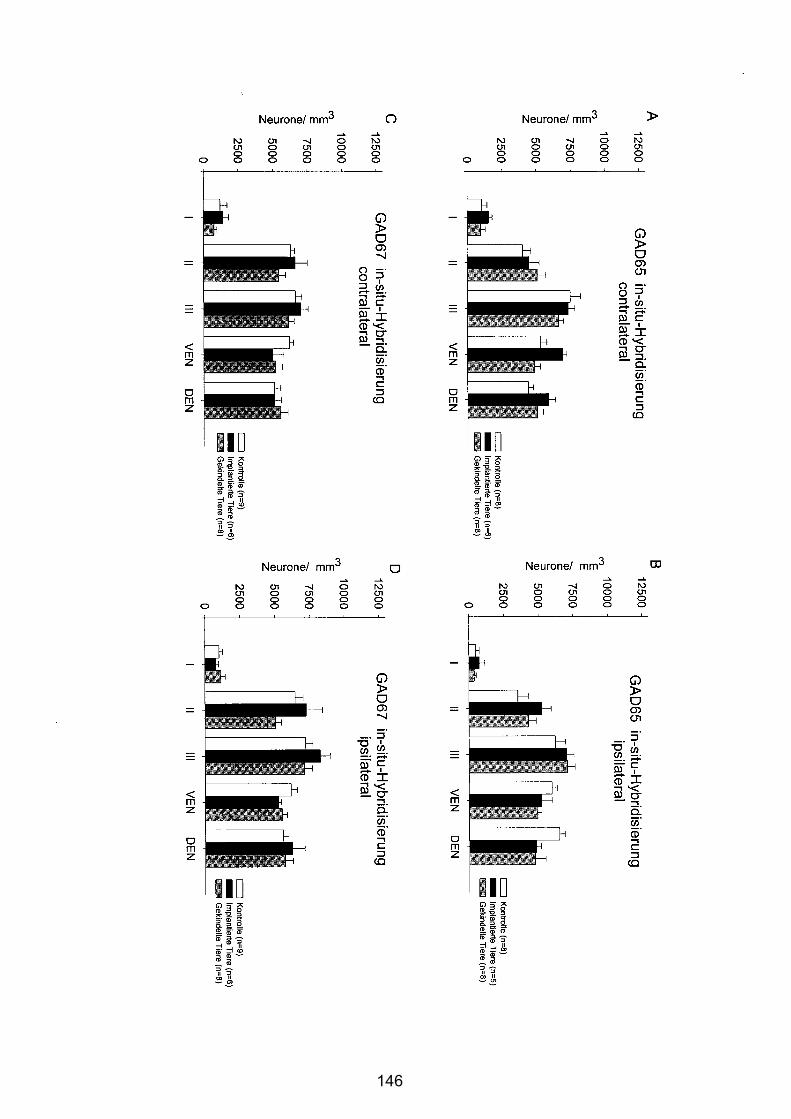
146

147
4.4.2.3 Posteriorer Piriformer Cortex
Im posterioren PC wurden 3 Tiere aus jeder Versuchsgruppe in der Auswertung
berücksichtigt, da für diese Region aufgrund der Literatur (Lehmann et al., 1998)
keine ausgeprägten Veränderungen erwartet wurden. Statistisch signifikanten
Veränderungen in der Dichte GAD65- bzw. GAD67-mRNA-positiver Neurone
konnten zwischen den Versuchsgruppen nicht festgestellt werden (p>0,05;
einfaktorielle ANOVA).
Nach in-situ-Hybridisierung für GAD65-mRNA wurden in der Schicht III mit 9934
Neuronen/mm3 (S.E.M. 1162,31) contralateral und 9170 Neuronen/mm3 (S.E.M.
437,87) ipsilateral die größte Anzahl an Zellen quantifiziert. Im DEN war die Dichte
dieser Neurone im Mittel geringer, gefolgt vom VEN und der Schicht II. Die
niedrigste Zelldichte ergab sich für GAD65-mRNA in Schicht I mit contralateral 1277
Neuronen/mm3 (S.E.M. 259,06) und ipsilateral 511 (S.E.M. 511,23) Neuronen/mm3
(Tabelle 34, Abbildung 35 A/B).
Die in-situ-Hybridisierung zur Detektion von GAD67-mRNA ergab im posterioren PC
eine gleiche Abstufung der mittleren Zelldichte in den untersuchten Schichten wie der
Nachweis der GAD65-mRNA (Abbildung 35 C/D). Die mittleren Neuronendichte aller
Schichten lagen beidseitig über denen für GAD65-mRNA. Besonders deutlich zeigte
sich der Unterschied in der Schicht II, in der contralateral mit 6878 positiven
Neuronen/mm3 (S.E.M. 2338,4) ein Unterschied von 80 % zur gleichseitigen
Kontrolle bei GAD65-mRNA bestand. Ipsilateral wurden mit 6114 Neuronen/mm3 bei
einem vergleichbaren Standardfehler (S.E.M. 1169,19) doppelt so viele Zellen für
GAD67-mRNA markiert (Tabelle 34 und Tabelle 35).

148
Posteriorer PC GAD65 in-situ-Hybridisierung
Neurone/mm3 I II III VEN DEN Gruppe n links rechts links rechts links rechts links rechts links rechts
1277 511 3822 3056 9934 9170 5601 5089 8407 8918 Kontrolle 3 ±259,1 ±511,2 ±442,5 ±1171,5 ±1162,3 ±437,9 ±259,1 ±1348,0 ±1162,3 ±1348,0 2293 763 2033 3056 9422 7641 7130 4844 8155 9170 Implantierte
Tiere 3 ±1322,8 ±442,5 ±674,0 ±442,5 ±1111,9 ±880,3 ±1111,9 ±674,0 ±259,1 ±887,2 1781 1529 3569 2804 8918 8414 5610 6366 7641 7389 Gekindelte
Tiere 3 ±674,0 ±442,5 ±1554,3 ±674,0 ±504,4 ±1322,8 ±917,0 ±917,0 ±880,3 ±1785,9
Tabelle 34
Posteriorer PC GAD67 in-situ-Hybridisierung
Neurone/mm3 I II III VEN DEN Gruppe n links rechts links rechts links rechts links rechts links rechts
2286 2040 6878 6114 10706 15032 7641 8407 9943 10699 Kontrolle 3 ±756,5 ±252,2 ±2338,4 ±1169,2 ±0,0 ±1100,4 ±756,5 ±1536,0 ±756,5 ±756,5 3049 1536 6619 6878 11974 10699 7130 5862 11722 7902 Implantierte
Tiere 3 ±1329,7 ±0,0 ±1673,5 ±1925,7 ±1421,4 ±1169,2 ±1123,3 ±1100,4 ±2934,4 ±504,4 1268 2040 5601 5096 10699 9170 7130 7641 8659 8911 Gekindelte
Tiere 3 ±917,0 ±252,2 ±674,0 ±511,2 ±1162,3 ±437,9 ±1111,9 ±1162,3 ±1024,8 ±1669,0
Tabelle 35
Tabelle 34/Tabelle 35
Mittelwerte und S.E.M. der Anzahl GAD65- (Tabelle 34) und GAD67-mRNA-positiver Neurone/mm3 (Tabelle 35) im posterioren PC. Signifikante Unterschiede in der Dichte zwischen den drei Versuchsgruppen bestanden nicht (p>0,05, einfaktorielle ANOVA). In Schicht II des posterioren PC waren die Mittelwerte GAD65-mRNA-positiver
Neurone in der Gruppe der gekindelten Tiere beidseitig geringgradig gegenüber
denen der Kontrollgruppe vermindert. Nach der in-situ-Hybridisierung für GAD67-
mRNA lagen mittleren Neuronendichten der gekindelten Tiere unterhalb derer der
anderen Versuchsgruppen. Die Differenz betrug zur Kontrolle contralateral 19 % und
ipsilateral 17 %. Im Vergleich zur Gruppe implantierter Tiere waren contralateral
15 % und ipsilateral 26 % weniger Neurone vorhanden (Abbildung 35 A-D).
Bei der Betrachtung von Schicht III war nur die sehr hohe Zelldichte in der
Kontrollgruppe mit 15032 GAD67-mRNA-positiven Neuronen/mm3 (S.E.M. 1100,41)
ipsilateral zur Stimulationselektrode auffällig. Diese führte zu einer 39 %igen
Differenz zur Gruppe der gekindelten Tiere (Abbildung 35 D).

149
Im VEN ergaben sich durch den Kindling-Prozeß keine eindeutigen Veränderungen
in der Zelldichte. Während mit beiden Nachweisen contralateral keine oder nur eine
geringe Abnahme des Mittelwertes im Vergleich zur Kontrolle auftrat, war nach in-
situ-Hybridisierung für GAD65-mRNA die Anzahl positiver Neurone/mm3 in der
Kindling-Gruppe um 25 % erhöht, wohingegen eine 9 %ige Reduktion GAD67-mRNA
positiver Neurone zu verzeichnen war (Abbildung 35 B und D).
Im DEN ergaben sich für beide Nachweise geringere Werte in der Gruppe
gekindelter Tiere im Vergleich zur Kontrolle, wobei die Reduzierung GAD67-mRNA-
positiver Neurone beidseitig 17 % betrug. Die Mittelwerte der Elektroden-
implantierten Gruppe waren mit Ausnahme des ipsilateralen Wertes für GAD67-
mRNA ebenfalls höher angesiedelt.
Abbildung 35 A-D
Darstellung der Mittelwerte GAD65- (A/B) und GAD67-mRNA-positiver Neurone (C/D) im posterioren PC mit zugehörigen Standardfehlern. Unter Kontrollbedingungen führte die Expression von GAD67-mRNA zu einer höheren Anzahl markierter Neurone. Eine Tendenz zur Abnahme GAD67-mRNA-markierter Neurone durch Kindling waren in Schicht II zu beobachten. Die Veränderungen zwischen den Gruppen erwiesen sich in der Varianzanalyse als nicht signifikant (p>0,05; einfaktorielle ANOVA).
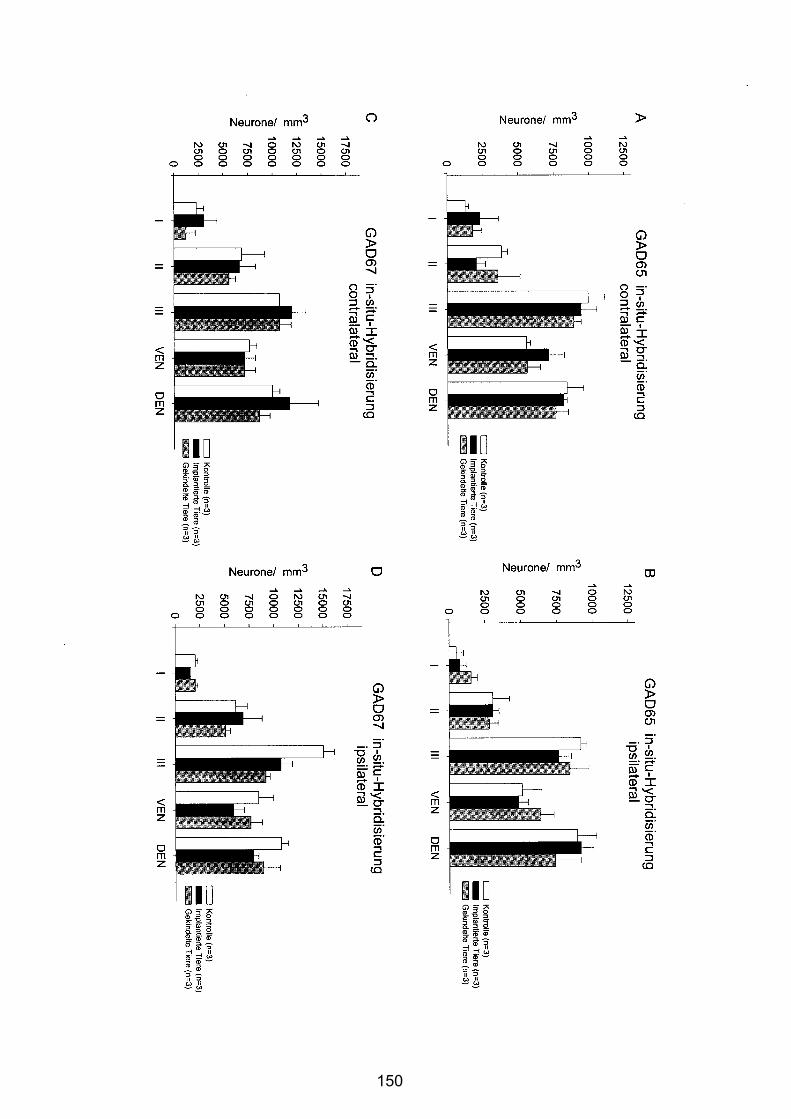
150
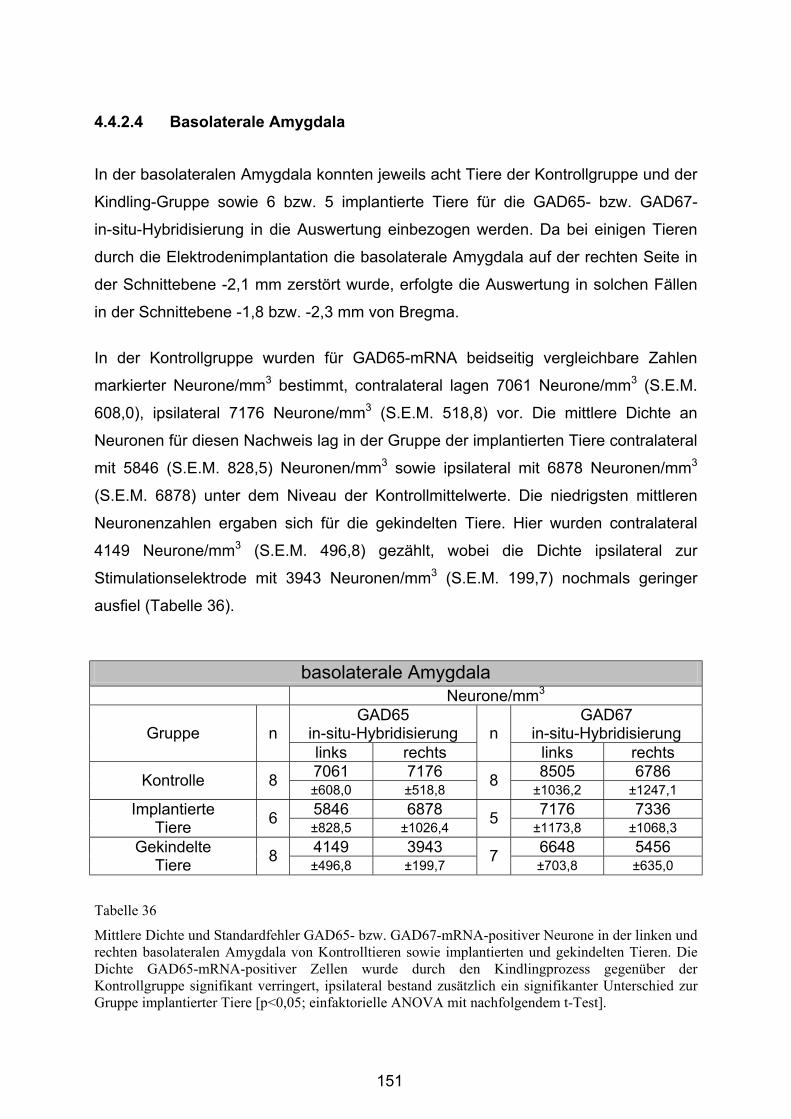
151
4.4.2.4 Basolaterale Amygdala
In der basolateralen Amygdala konnten jeweils acht Tiere der Kontrollgruppe und der
Kindling-Gruppe sowie 6 bzw. 5 implantierte Tiere für die GAD65- bzw. GAD67-
in-situ-Hybridisierung in die Auswertung einbezogen werden. Da bei einigen Tieren
durch die Elektrodenimplantation die basolaterale Amygdala auf der rechten Seite in
der Schnittebene -2,1 mm zerstört wurde, erfolgte die Auswertung in solchen Fällen
in der Schnittebene -1,8 bzw. -2,3 mm von Bregma.
In der Kontrollgruppe wurden für GAD65-mRNA beidseitig vergleichbare Zahlen
markierter Neurone/mm3 bestimmt, contralateral lagen 7061 Neurone/mm3 (S.E.M.
608,0), ipsilateral 7176 Neurone/mm3 (S.E.M. 518,8) vor. Die mittlere Dichte an
Neuronen für diesen Nachweis lag in der Gruppe der implantierten Tiere contralateral
mit 5846 (S.E.M. 828,5) Neuronen/mm3 sowie ipsilateral mit 6878 Neuronen/mm3
(S.E.M. 6878) unter dem Niveau der Kontrollmittelwerte. Die niedrigsten mittleren
Neuronenzahlen ergaben sich für die gekindelten Tiere. Hier wurden contralateral
4149 Neurone/mm3 (S.E.M. 496,8) gezählt, wobei die Dichte ipsilateral zur
Stimulationselektrode mit 3943 Neuronen/mm3 (S.E.M. 199,7) nochmals geringer
ausfiel (Tabelle 36).
basolaterale Amygdala Neurone/mm3
GAD65 in-situ-Hybridisierung
GAD67 in-situ-Hybridisierung Gruppe n
links rechts n
links rechts 7061 7176 8505 6786 Kontrolle 8 ±608,0 ±518,8 8 ±1036,2 ±1247,1 5846 6878 7176 7336 Implantierte
Tiere 6 ±828,5 ±1026,4 5 ±1173,8 ±1068,3 4149 3943 6648 5456 Gekindelte
Tiere 8 ±496,8 ±199,7 7 ±703,8 ±635,0 Tabelle 36
Mittlere Dichte und Standardfehler GAD65- bzw. GAD67-mRNA-positiver Neurone in der linken und rechten basolateralen Amygdala von Kontrolltieren sowie implantierten und gekindelten Tieren. Die Dichte GAD65-mRNA-positiver Zellen wurde durch den Kindlingprozess gegenüber der Kontrollgruppe signifikant verringert, ipsilateral bestand zusätzlich ein signifikanter Unterschied zur Gruppe implantierter Tiere [p<0,05; einfaktorielle ANOVA mit nachfolgendem t-Test].

152
Die Auszählung GAD67-mRNA-positiver Neurone ergab in allen drei Gruppen mit
Ausnahme der contralateralen Seite in der Kontrollgruppe höhere Mittelwerte an
Neuronen/mm3 im Vergleich zur GAD65-in-situ-Hybridisierung. Analog zur in-situ-
Hybridisierung für GAD65-mRNA wies die Gruppe der gekindelten Ratten auch für
GAD67-mRNA die geringsten Mittelwerte in der Anzahl gefärbter Neurone der drei
Versuchsgruppen auf. Contralateral wurde eine Dichte von 6648 Neuronen/mm3
(S.E.M. 703,8) und ipsilateral von 5456 Neuronen/mm3 (S.E.M. 635,0) bestimmt
(Tabelle 36, Abbildung 36).
contralateral ipsilateral
Neu
rone
/ m
m3
0
2500
5000
7500
10000
12500
KontrolleImplantierte TiereGekindelte Tiere
GAD65 in-situ-Hybridisierung
* *°
A
contralateral ipsilateral
Neu
rone
/ m
m3
0
2500
5000
7500
10000
12500GAD67
in-situ-Hybridisierung
B
KontrolleImplantierte TiereGekindelte Tiere
Abbildung 36 A/B
Vergleich der mittleren Dichte GAD65- (A) und GAD67-mRNA- (B) positiver Neurone pro mm3 in der basolateralen Amygdala. Dargestellt sind die Mittelwerte ±S.E.M. von 8 Kontrolltieren (weiße Balken), 6 (5 GAD67) implantierten Tieren (schwarze Balken) und 7 gekindelten Tieren (graue Balken). Die Dichte GAD65-mRNA-positiver Neurone wurde durch Kindling beidseitig signifikant gegenüber der Kontrolle (*) sowie ipsilateral auch gegenüber der Gruppe implantierter Tiere verringert (°). Signifikante Unterschiede in der Dichte GAD67-mRNA-positiver Neurone zwischen Kontrollen und gekindelten Tieren bzw. implantierten Tieren bestanden nicht [p<0,05; einfaktorielle ANOVA mit nachfolgendem t-Test].

153
Die Varianzanalyse der mittleren Dichten GAD65-mRNA-markierter Neurone in der
basolateralen Amygdala erbrachte signifikante Differenzen zwischen den
Versuchsgruppen (p<0,05; einfaktorielle ANOVA). Im nachfolgend durchgeführten
t-Test stellte sich beidseitig eine signifikante Abnahme der Zelldichte GAD65-mRNA-
positiver Neurone in der Kindling-Gruppe gegenüber der Kontrollgruppe heraus
(p<0,05, Tabelle 37), ipsilateral war die Abnahme auch gegenüber der Gruppe der
implantieren Tiere signifikant. Die Differenz entsprach contralateral durchschnittlich
41 % gegenüber der Kontrollgruppe und ipsilateral 45 % gegenüber der
Kontrollgruppe sowie 43 % gegenüber der Gruppe implantierter Tiere. Die
Verringerung der Dichte GAD67-mRNA-positiver Neurone in der Kindling-Gruppe
war dagegen nicht signifikant (p>0,05; einfaktorielle ANOVA).
Basolaterale Amygdala
GAD65
in-situ-Hybridisierung
GAD67
in-situ-Hybridisierung
Gruppe ANOVA
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert]
Rel. Diff.
[%]
t-Test
[p-Wert] contralateral 0,0132 -41,2 0,0005 Kontrolle-
Gekindelt ipsilateral 0,0259 -45,0 0,0001 contralateral keine signifikanten Veränderungen Implantiert-
Gekindelt ipsilateral 0,0132 -42,7 0,0112
keine signifikanten
Veränderungen
Tabelle 37
Darstellung der signifikanten Abnahme der Zelldichte durch Kindling in der basolateralen Amygdala. Mit in-situ-Hybridisierung für GAD65-mRNA konnte eine signifikante Abnahme der Zelldichte beidseitig zur Kontrolle nachgewiesen werden, im Vergleich zur Gruppe implantierter Tiere lag nur auf der ipsilateralen Seite eine signifikante Reduktion vor. Die in-situ-Hybridisierung für GAD67-mRNA ergab beidseitig nur tendenziell eine Abnahme markierter Neurone durch den Kindlingprozess.

154
4.4.2.5 Hippocampusformation
In der Hippocampusformation wurden 4 Tiere bzw. 3 Tiere aus jeder Versuchsgruppe
nach in-situ-Hybridisierung für GAD65-mRNA bzw. GAD67-mRNA ausgewertet, da
aufgrund der Literatur (Lehmann et al., 1998) keine ausgeprägten Veränderungen
erwartet wurden. Es konnten keine statistisch signifikanten Veränderungen in der
Dichte GAD65- bzw. GAD67-mRNA-positiver Neurone zwischen den
Versuchsgruppen festgestellt werden (p>0,05, einfaktorielle ANOVA).
Die höchste Dichte GAD65- und GAD67-mRNA-positiver Neurone im Hippocampus
wurde für Korbzellen im Gyrus dentatus ermittelt. Contralateral konnten in der
Kontrollgruppe 12609 Neurone/mm3 (S.E.M. 733,61) für GAD65-mRNA sowie 14512
Neurone/mm3 (S.E.M. 1925,73) für GAD67-mRNA markiert werden. Ipsilateral
wurden für beide Nachweise im Mittel höhere Werte erhoben, so waren 15658
Neurone/mm3 (S.E.M. 1604,77) positiv für GAD65-mRNA und 15291 Neurone/mm3
(S.E.M. 435,58) positiv für GAD67-mRNA (Tabelle 38, Tabelle 39).
Mit beiden Nachweismethoden ließ sich in der Kontrollgruppe eine Abnahme in der
Dichte markierter Zellen von der Region CA1 hin zur Region CA3c beschreiben, im
Hilus des Gyrus dentatus wurden für GAD67-in-situ-Hybridisierung contralateral eine
höhere Anzahl an Neuronen/mm3 beziffert als in CA3. Ansonsten lagen die mittleren
Kontrollwerte in der Hilusregion unterhalb der Werte der Kontrollen im Bereich von
CA3 (Abbildung 37 A-D).
In Körnerzellen des Gyrus dentatus konnte nach Durchführung der in-situ-
Hybridisierung in keiner der drei Versuchsgruppen eine Expression von GAD65- und
GAD67-mRNA nachgewiesen werden. Demnach war 42 Tage nach Abschluss des
Kindlingprozesses im Gegensatz zu den Versuchgruppen mit einem SE induziert
durch Kainat bzw. Lithium-Pilocarpin keine gesteigerte Expression von GAD67-
mRNA in Körnerzellen festzustellen.
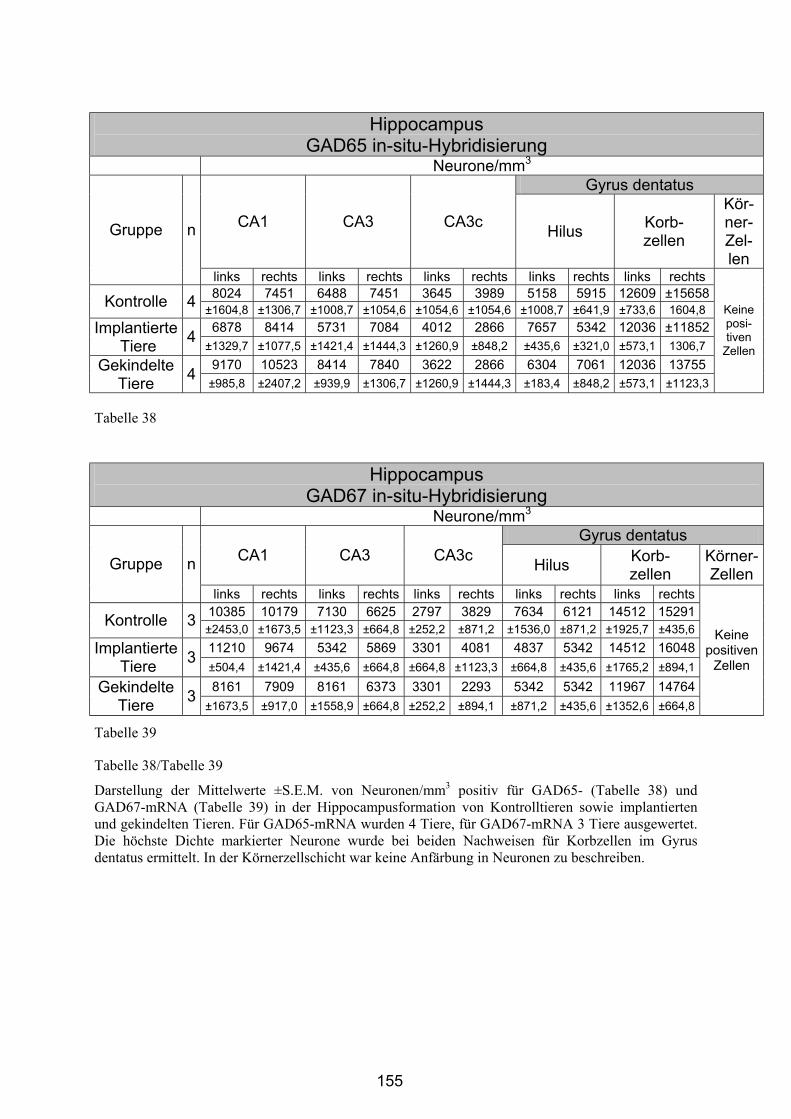
155
Hippocampus GAD65 in-situ-Hybridisierung
Neurone/mm3 Gyrus dentatus
CA1 CA3 CA3c Hilus Korb- zellen
Kör- ner- Zel-len
Gruppe n
links rechts links rechts links rechts links rechts links rechts 8024 7451 6488 7451 3645 3989 5158 5915 12609 ±15658 Kontrolle 4 ±1604,8 ±1306,7 ±1008,7 ±1054,6 ±1054,6 ±1054,6 ±1008,7 ±641,9 ±733,6 1604,8 6878 8414 5731 7084 4012 2866 7657 5342 12036 ±11852 Implantierte
Tiere 4 ±1329,7 ±1077,5 ±1421,4 ±1444,3 ±1260,9 ±848,2 ±435,6 ±321,0 ±573,1 1306,7 9170 10523 8414 7840 3622 2866 6304 7061 12036 13755 Gekindelte
Tiere 4 ±985,8 ±2407,2 ±939,9 ±1306,7 ±1260,9 ±1444,3 ±183,4 ±848,2 ±573,1 ±1123,3
Keine posi- tiven
Zellen
Tabelle 38
Hippocampus GAD67 in-situ-Hybridisierung
Neurone/mm3 Gyrus dentatus
CA1 CA3 CA3c Hilus Korb- zellen
Körner- Zellen Gruppe n
links rechts links rechts links rechts links rechts links rechts 10385 10179 7130 6625 2797 3829 7634 6121 14512 15291 Kontrolle 3 ±2453,0 ±1673,5 ±1123,3 ±664,8 ±252,2 ±871,2 ±1536,0 ±871,2 ±1925,7 ±435,6 11210 9674 5342 5869 3301 4081 4837 5342 14512 16048 Implantierte
Tiere 3 ±504,4 ±1421,4 ±435,6 ±664,8 ±664,8 ±1123,3 ±664,8 ±435,6 ±1765,2 ±894,1 8161 7909 8161 6373 3301 2293 5342 5342 11967 14764 Gekindelte
Tiere 3 ±1673,5 ±917,0 ±1558,9 ±664,8 ±252,2 ±894,1 ±871,2 ±435,6 ±1352,6 ±664,8
Keine positiven
Zellen
Tabelle 39
Tabelle 38/Tabelle 39
Darstellung der Mittelwerte ±S.E.M. von Neuronen/mm3 positiv für GAD65- (Tabelle 38) und GAD67-mRNA (Tabelle 39) in der Hippocampusformation von Kontrolltieren sowie implantierten und gekindelten Tieren. Für GAD65-mRNA wurden 4 Tiere, für GAD67-mRNA 3 Tiere ausgewertet. Die höchste Dichte markierter Neurone wurde bei beiden Nachweisen für Korbzellen im Gyrus dentatus ermittelt. In der Körnerzellschicht war keine Anfärbung in Neuronen zu beschreiben.

156
In der Region CA1 des Hippocampus lagen die mittleren Werte GAD65-mRNA-
positiver Neurone/mm3 gekindelter Tiere beidseitig tendenziell über denen der beiden
anderen Versuchsgruppen. Contralateral ergab sich eine relative Differenz zur
Kontrollgruppe von 14 %, zur Elektroden-implantierten Gruppe von 33 %. Ipsilateral
belief sich der Unterschied zur Kontrolle auf 41 %, allerdings wies der Mittelwert von
10523 Neuronen/mm3 mit ±2407,16 einen vergleichsweise hohen Standardfehler auf
(Abbildung 37 A/B). Die in-situ-Hybridisierung für GAD67-mRNA ergab in der
Kindling-Gruppe beidseitig Mittelwerte, die im Durchschnitt 22-23 % unterhalb derer
der anderen Gruppen angesiedelt waren (Abbildung 37 C/D).
In der Region CA3 wurden contralateral im Vergleich zur Kontrolle für GAD65- und
GAD67-mRNA tendenziell höhere Mittelwerte positiver Neurone durch Kindling
gesehen, wobei die Dichte in der Gruppe implantierter Tiere für beide Isoformen im
Vergleich dazu durchschnittlich 50 % niedriger war (Abbildung 37 A/C). Ipsilateral
bestanden keine deutlichen Unterschiede zwischen den Versuchsgruppen. In der
Region CA3c war ein geringerer Mittelwert GAD67-mRNA-markierter Zellen
ipsilateral zur Stimulationselektrode in der Gruppe der gekindelten Tiere auffällig. Die
Mittelwerte der beiden anderen Gruppen zeigten im Durchschnitt 40 % höhere
Dichten an. Contralateral sowie nach in-situ-Hybridisierung für GAD65-mRNA
ergaben sich keine deutlichen Schwankungen.
Die ermittelten Werte GAD65-mRNA-markierter Neurone für die Kindling-Gruppe im
Hilus des Gyrus dentatus lagen contralateral um 22 % und ipsilateral um 19 % über
den Basalwerten, nach in-situ-Hybridisierung für GAD67-mRNA wurden Werte
contralateral um 30 % und ipsilateral um 13 % unterhalb der Kontrolle ermittelt. Für
die Korbzellen fiel in der Kindling-Gruppe contralateral ein um 18 % reduzierter
Mittelwert GAD67-mRNA-markierter Neurone/mm3 gegenüber beiden anderen
Gruppen auf (Abbildung 37 C).
Abbildung 37 A-D
Darstellung der Anzahl von Neuronen/mm3 positiv markiert für GAD65- (A/B) und GAD67-mRNA (C/D) in der Hippocampusformation von Kontrolltieren, implantierten und gekindelten Tieren. Dargestellt sind die Mittelwerte mit zugehörigen Standardfehler (S.E.M.). Während sich für GAD65-mRNA in CA1 und im Hilus des Gyrus dentatus Tendenzen einer erhöhten Neuronendichte in der Kindling-Gruppe zeigten, wiesen diese Tiere nach in-situ-Hybridisierung für GAD67-mRNA in diesen Regionen im Mittel geringere Dichten auf. Signifikante Veränderungen wurden in der Varianzanalyse nicht ermittelt (p>0,05, einfaktorielle ANOVA).

157
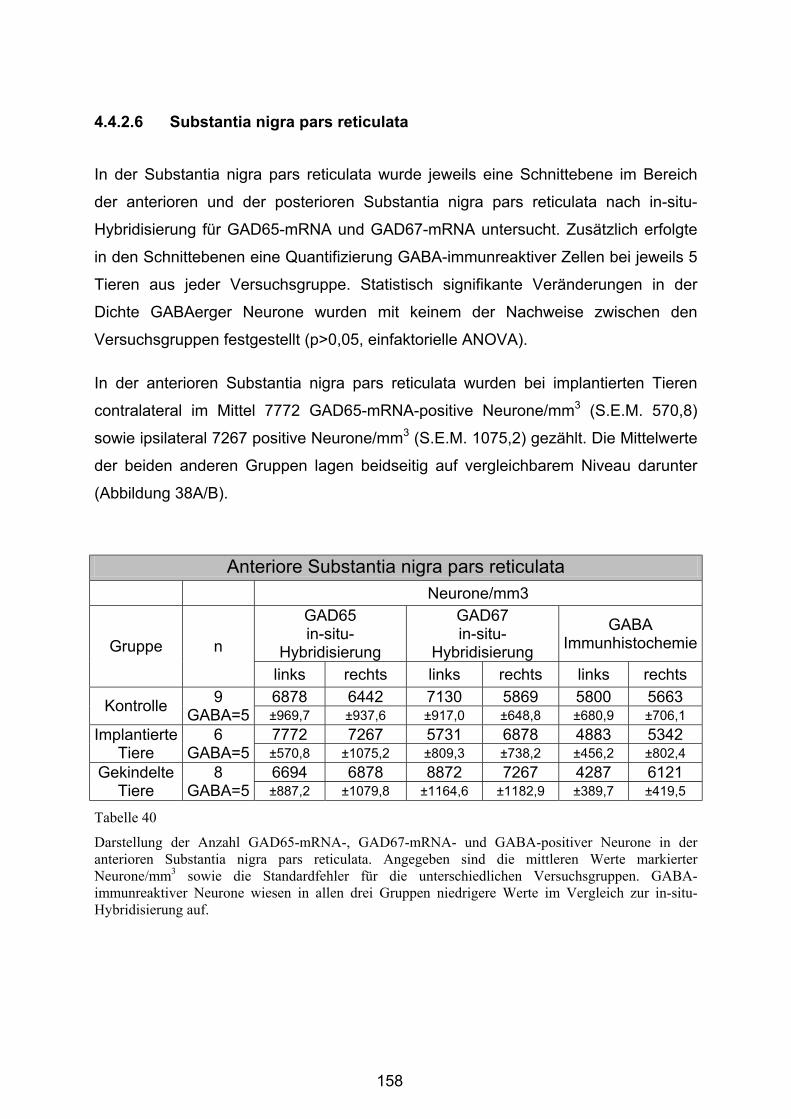
158
4.4.2.6 Substantia nigra pars reticulata
In der Substantia nigra pars reticulata wurde jeweils eine Schnittebene im Bereich
der anterioren und der posterioren Substantia nigra pars reticulata nach in-situ-
Hybridisierung für GAD65-mRNA und GAD67-mRNA untersucht. Zusätzlich erfolgte
in den Schnittebenen eine Quantifizierung GABA-immunreaktiver Zellen bei jeweils 5
Tieren aus jeder Versuchsgruppe. Statistisch signifikante Veränderungen in der
Dichte GABAerger Neurone wurden mit keinem der Nachweise zwischen den
Versuchsgruppen festgestellt (p>0,05, einfaktorielle ANOVA).
In der anterioren Substantia nigra pars reticulata wurden bei implantierten Tieren
contralateral im Mittel 7772 GAD65-mRNA-positive Neurone/mm3 (S.E.M. 570,8)
sowie ipsilateral 7267 positive Neurone/mm3 (S.E.M. 1075,2) gezählt. Die Mittelwerte
der beiden anderen Gruppen lagen beidseitig auf vergleichbarem Niveau darunter
(Abbildung 38A/B).
Anteriore Substantia nigra pars reticulata Neurone/mm3
GAD65 in-situ-
Hybridisierung
GAD67 in-situ-
Hybridisierung
GABA Immunhistochemie Gruppe n
links rechts links rechts links rechts 6878 6442 7130 5869 5800 5663 Kontrolle 9
GABA=5 ±969,7 ±937,6 ±917,0 ±648,8 ±680,9 ±706,1 7772 7267 5731 6878 4883 5342 Implantierte
Tiere 6
GABA=5 ±570,8 ±1075,2 ±809,3 ±738,2 ±456,2 ±802,4 6694 6878 8872 7267 4287 6121 Gekindelte
Tiere 8
GABA=5 ±887,2 ±1079,8 ±1164,6 ±1182,9 ±389,7 ±419,5
Tabelle 40
Darstellung der Anzahl GAD65-mRNA-, GAD67-mRNA- und GABA-positiver Neurone in der anterioren Substantia nigra pars reticulata. Angegeben sind die mittleren Werte markierter Neurone/mm3 sowie die Standardfehler für die unterschiedlichen Versuchsgruppen. GABA-immunreaktiver Neurone wiesen in allen drei Gruppen niedrigere Werte im Vergleich zur in-situ-Hybridisierung auf.

159
Mit in-situ-Hybridisierung gegen GAD67-mRNA wurden bei gekindelten Tiere auf
beiden Seiten die höchsten Mittelwerte berechnet, die jedoch gleichzeitig die
höchsten Standardfehler aufwiesen. Contralateral waren im Mittel 8872 GAD67-
mRNA-positive Neurone/mm3 (S.E.M. 1164,4) und ipsilateral 7267 Neurone/mm3
(S.E.M. 1182,9) vorhanden. Damit lagen die mittleren Werte contralateral um 24 %
über den Werten der Kontrollgruppe und 54 % über denen der implantierten Tiere.
Die relative Differenz fiel ipsilateral mit 24 % zur Kontrolle und 6 % zur Gruppe der
implantierten Tiere geringer aus (Abbildung 38 A/B).
Mit Immunhistochemie zum Nachweis von GABA ergaben sich im Vergleich zur in-
situ-Hybridisierung beidseitig niedrigere Mittelwerte für alle drei Versuchsgruppen in
der anterioren Substantia nigra pars reticulata (Abb. 17 A/B). Contralateral wurde für
die Kontrollgruppe die höchste Dichte GABA-immunreaktiver Neurone mit 5800
Neuronen/mm3 (S.E.M. 680,9) ermittelt, die Elektroden-implantierte Gruppe wies
einen 16 % geringeren Wert, die Gruppe gekindelter Tiere lag im Durchschnitt um 26
% unter dem Kontrollniveau. Mit 6121 Neurone/mm3 (S.E.M. 419,5) besaßen die
gekindelten Tiere contralateral den höchsten Mittelwert der drei Gruppen
(Tabelle 40, Abbildung 38 A/B)
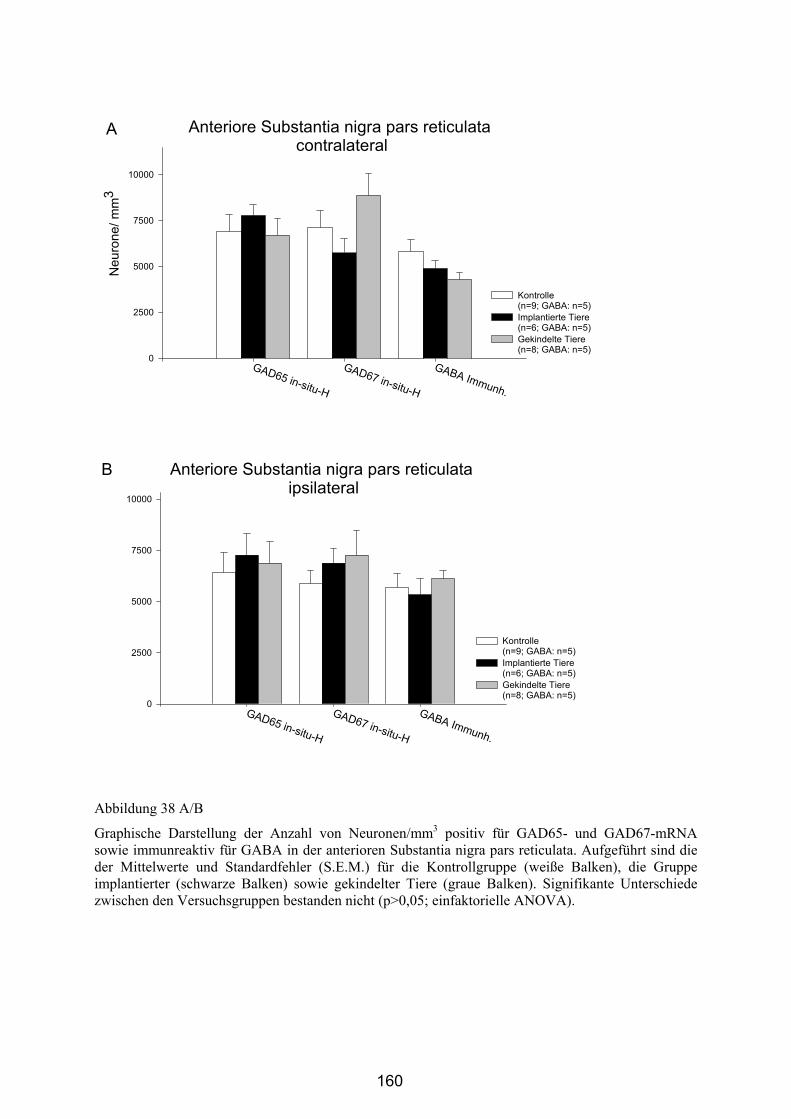
160
GAD65 in-situ-H
GAD67 in-situ-H
GABA Immunh.
Neu
rone
/ mm
3
0
2500
5000
7500
10000
Kontrolle(n=9; GABA: n=5)Implantierte Tiere(n=6; GABA: n=5)Gekindelte Tiere(n=8; GABA: n=5)
Anteriore Substantia nigra pars reticulata contralateral
A
GAD65 in-situ-H
GAD67 in-situ-H
GABA Immunh.
0
2500
5000
7500
10000
B Anteriore Substantia nigra pars reticulata ipsilateral
Kontrolle(n=9; GABA: n=5)Implantierte Tiere(n=6; GABA: n=5)Gekindelte Tiere(n=8; GABA: n=5)
Abbildung 38 A/B
Graphische Darstellung der Anzahl von Neuronen/mm3 positiv für GAD65- und GAD67-mRNA sowie immunreaktiv für GABA in der anterioren Substantia nigra pars reticulata. Aufgeführt sind die der Mittelwerte und Standardfehler (S.E.M.) für die Kontrollgruppe (weiße Balken), die Gruppe implantierter (schwarze Balken) sowie gekindelter Tiere (graue Balken). Signifikante Unterschiede zwischen den Versuchsgruppen bestanden nicht (p>0,05; einfaktorielle ANOVA).

161
Im posterioren Anteil der Substantia nigra pars reticulata wurden mit allen
Nachweisen höhere Dichten markierter Neurone in den Untersuchgruppen im
Vergleich zum anterioren Anteil der Struktur ermittelt, wobei die Dichte GABA-
immunreaktiver Zellen wiederum im Vergleich zur in-situ-Hybridisierung im Mittel
niedriger angesiedelt war (Tabelle 41, Abbildung 39A/B).
Posteriore Substantia nigra pars reticulata Neurone/mm3
GAD65 in-situ-
Hybridisierung
GAD67 in-situ-
Hybridisierung
GABA Immunhistochemie Gruppe n
links rechts links rechts links rechts 12563 11554 12059 11623 10569 8414 Kontrolle 9
GABA=5 ±1141,7 ±1114,2 ±1011,0 ±1215,0 ±834,5 ±538,7 12999 11210 11715 12357 8551 7634 Implantierte
Tiere 6
GABA=5 ±862,0 ±1178,4 ±1056,9 ±974,3 ±919,3 ±965,2 11944 11279 9560 10316 8551 7634 Gekindelte
Tiere 8
GABA=5 ±946,8 ±1153,1 ±1217,3 ±967,4 ±287,8 ±724,4
Tabelle 41
Darstellung der Anzahl GAD65-, GAD67- und GABA-positiver Neurone in der posterioren Substantia nigra pars reticulata. Angegeben sind die mittleren Werte markierter Neurone/mm3 sowie die Standardfehler für die unterschiedlichen Versuchsgruppen. Analog zur anterioren Substantia nigra pars reticulata ergaben sich mit GABA Immunhistochemie geringere Mittelwerte in allen Gruppen.
Für GAD65-mRNA ergaben sich beidseitig nur geringe Schwankungen in den
Mittelwerten positiver Neurone der drei Gruppen. Die Dichte GAD67-mRNA-positiver
Neurone der implantierten Tiere fiel im Vergleich zur Kontrolle nur gering verändert
aus, während die Werte der Kindling-Gruppe tendenziell contralateral um 21 % und
ipsilateral um 11 % unter den basalen Werten lagen (Abbildung 39 A/B).
Die Dichte GABA-immunreaktiver Neuronen war in der Gruppe der Kontrolltiere
contralateral mit 10569 Neuronen/mm3 (S.E.M. 10569) und contralateral mit 8414
Neuronen/mm3 (S.E.M. 538,7) tendenziell am höchsten. Die Mittelwerte der beiden
anderen Tiergruppen lagen contralateral um 19 % und contralateral um 9 % auf
vergleichbarem Niveau darunter.

162
GAD65 in-situ-H
GAD67 in-situ-H
GABA Immunh.
Neu
rone
/ mm
3
0
2500
5000
7500
10000
12500
15000
A Posteriore Substantia nigra pars reticulata contralateral
Kontrolle(n=9; GABA: n=5)Implantierte Tiere(n=6; GABA: n=5)Gekindelte Tiere(n=8; GABA: n=5)
GAD65 in-situ-H
GAD67 in-situ-H
GABA Immunh.
Neu
rone
/ mm
3
0
2500
5000
7500
10000
12500
15000
Posteriore Substantia nigra pars reticulata ipsilateral
B
Kontrolle(n=9; GABA: n=5)Implantierte Tiere(n=6; GABA: n=5)Gekindelte Tiere(n=8; GABA: n=5)
Abbildung 39 A/B
Graphische Darstellung der Anzahl von Neuronen/mm3 positiv für GAD65- und GAD67-mRNA sowie immunreaktiv für GABA in der posterioren Substantia nigra pars reticulata. Aufgeführt sind die der Mittelwerte und Standardfehler (S.E.M.) für die Kontrollgruppe (weiße Balken), die Gruppe implantierter (schwarze Balken) sowie gekindelter Tiere (graue Balken). Signifikante Unterschiede zwischen den Versuchsgruppen bestanden nicht (p>0,05; einfaktorielle ANOVA).

163
5 Diskussion
5.1 Methodik
In der vorliegenden Arbeit wurde die Methode der in-situ-Hybridisierung zum
Nachweis der mRNA für GAD-Isoformen in GABAergen Neuronen verwendet.
Dabei kamen nicht-radioaktiv markierte RNA-Sonden und radioaktiv markierte
Oligonukleotid-Sonden zum Einsatz. Zwischen den beiden etablierten in-situ-
Hybridisierungsprotokollen konnten hinsichtlich der Zahl markierter Zellen unter den
gegebenen Versuchsbedingungen keine Unterschiede in der Sensitivität festgestellt
werden. Beide Methoden erlaubten die Darstellung von Zellen, die sich durch geringe
mRNA-Gehalte auszeichnen. Zu diesen gehörte die Mehrzahl der GABAergen
Neurone des Striatums, welche auch in früheren Untersuchungen unter Verwendung
radioaktiver in-situ-Hybridisierung zum Nachweis der mRNA für GAD67 durch eine
geringe Anzahl von Silberkörnern markiert waren (Chesselet et al., 1987). Dieses
Bild zeigte sich mit der nicht-radioaktiven in-situ-Hybridisierung in Form einer
schwachen Intensität der Anfärbung dieser Neurone (Esclapez et al., 1993).
Mit beiden Methoden konnte dagegen der Nachweis von GAD-mRNA in der
überwiegenden Mehrzahl GABA- und GAD-immunreaktiver Interneurone in der
Schicht II des PC nicht erfolgen, woraus sich ebenfalls eine vergleichbare Sensitivität
ableiten lässt. Diese Schlussfolgerung wird durch Studien untermauert, in denen die
Nachweiseffizienz von RNA-Sonden gegen GAD-mRNA mit radioaktiver und mit
nicht-radioaktiver Markierung verglichen wurde. Unabhängig von der Art der
Markierung konnte GAD-mRNA im Slot-blot in gleicher Verdünnung nachgewiesen
werden (Esclapez et al., 1993). Dementsprechend ist zu vermuten, dass die
radioaktive Markierung der in den eigenen Versuchen eingesetzten RNA-Sonden
nicht zu einer entscheidenden Verbesserung der Sensitivität geführt hätte. Da sich
die radioaktive in-situ-Hybridisierung bei vergleichbarer Sensitivität als ungleich zeit-
und kostenintensiver gegenüber der nicht-radioaktiven Variante der Methode
herausstellte, wurde das Protokoll der nicht-radioaktiven in-situ-Hybridisierung für
die weiteren Untersuchungen verwendet.

164
Unter Einsatz der nicht radioaktiven in-situ-Hybridisierung war in der Mehrzahl der
untersuchten Hirnregionen eine höhere Dichte GAD67- als GAD65-exprimierender
Zellen festzustellen. Weiterhin wurde infolge induzierter Krampfaktivität
ausschließlich die Expression der GAD67-Isoform in Körnerzellen nachgewiesen,
während die mRNA für die GAD65 nicht detektiert werden konnte. Methodische
Aspekte können als Ursache für die beschriebenen Unterschiede zwischen den
Isoformen weitestgehend vernachlässigt werden. Die in den eigenen Experimenten
eingesetzten RNA-Sonden wurden hinsichtlich ihrer Spezifität getestet und wiesen
keine Kreuzreaktivität auf (Erlander et al., 1991; Esclapez et al., 1993). Die
verwendeten Sonden wurden bei identischen Bedingungen unter Verwendung der
gleichen Reagenzien hergestellt und besaßen die gleiche Länge. Dadurch kann
davon ausgegangen werden, dass während der Synthese der Sonden eine
vergleichbare Anzahl DIG-markierter Nukleotide inkorporiert wurde. Weiterhin
erfolgte die Überprüfung der Sonden mittels Dot-Blot, wodurch gewährleistet wurde,
dass nur RNA-Sonden vergleichbarer Markierungseffizienz für die Versuche zum
Einsatz kamen. Die Spezifität der Zellmarkierung wurde durch Kontrollsonden
überprüft, die ebenfalls unter den beschriebenen Bedingungen hergestellt wurden.
Unter Einsatz der Kontrollsonden konnte unter gleichen Versuchsbedingungen kein
spezifisches Signal in Zellen erzielt werden.
Weiterhin ist hinsichtlich der Differenzen in der Zahl GAD65- und GAD67-mRNA-
exprimierender Zellen zu berücksichtigen, dass unterschiedliche Inkubationszeiten
für die Farbreaktion zur Detektion der beiden Isoformen durchgeführt wurden.
GAD67-mRNA wird in den meisten Gehirnregionen in höherem Maße exprimiert als
GAD65-mRNA (Feldblum et al. 1993; Esclapez et al., 1993, 1994; Hendrickson et
al., 1994). Mehrere Studien belegen, dass mit radioaktiver und nicht-radioaktiver in-
situ-Hybridisierung längere Inkubationszeiten erforderlich sind, um mRNA
darzustellen, die in niedrigeren Konzentrationen in Neuronen vorliegt (Kessler et al.,
1990; Esclapez et al., 1993). Eine identische Entwicklungszeit für beide Isoformen
hätte dementsprechend entweder zu einer nicht ausreichenden Darstellung von
GAD65-mRNA-markierten Neuronen geführt oder die unspezifische Färbung von
Zellen bei der in-situ-Hybridisierung für GAD67-mRNA verstärkt.

165
Eine direkte farbliche Markierung von Zellsomata erwies sich in den Vorversuchen zu
dieser Arbeit als ein entscheidender Vorteil der nicht-radioaktiven in-situ-
Hybridisierung gegenüber der radioaktiven Variante der Methode. Allerdings führte
die Induktion von GAD67-mRNA in Körnerzellen des Hippocampus bei einigen
Schnittserien zu einer intensiven Färbung, wodurch die Anzahl markierter Zellen
aufgrund undeutlicher Zellgrenzen nicht mehr zweifelsfrei zu bestimmen war. Die
Quantifizierung des Ausmaßes der mRNA-Expression erfolgte deshalb anhand der
optischen Dichte der Anfärbung. Der Einsatz der nicht-radioaktiven in-situ-
Hybridisierung zur Bestimmung unterschiedlicher Expressionsraten von mRNA ist
mehrfach beschrieben worden (Larsson et al., 1991; Larrson und Hougaard, 1993,
1994). Dies basierte auf den Erkenntnissen von Studien, dass eine verstärkte
Anfärbung von Neuronen als Ausdruck eines erhöhten Gehaltes an GAD-mRNA
anzusehen ist (Kessler et al., 1990; Esclapez et al., 1993). Dabei ist zu
berücksichtigen, dass die Farbintensität nicht unbedingt in direktem Bezug zu der
absoluten mRNA-Menge in Zellen stehen muss, da enzymatischen Farbreaktionen
keine linearen Reaktionen darstellen. Demnach kommt der Dauer der
Entwicklungsreaktion entscheidende Bedeutung zu. Dieses zeigte sich auch in
Studien von Esclapez und Houser (1999), die eine vermehrte Expression von GAD-
mRNA im Pilocarpin-Modell anhand einer gesteigerten optischen Dichte der
Anfärbung hippocampaler Neurone nachweisen konnten. In diesen Versuchen
resultierte eine Inkubationszeit von 2 h in einer deutlich intensiveren Färbung von
Neuronen der Tieren mit spontanen Anfällen, wohingegen dieser Unterschied nach
vierstündiger Entwicklungszeit im Vergleich zur Kontrolle deutlich reduziert war. Da
die Autoren ein derartiges Geschehen nur für die Hippocampusformation
beschrieben, wurden für die eigenen Untersuchungen die etablierten
Inkubationszeiten angewendet, um auch in anderen Regionen des limbischen
Systems wie dem PC und der Amygdala eine optimale Markierungen von Zellen zu
erreichen. Zudem war eine vermehrte Expression von GAD-mRNA in GABAergen
Neuronen im Hippocampus von anderen Arbeitsgruppen nicht beschrieben worden
(Schwarzer und Sperk, 1995; Ding et al., 1998).

166
Der in den eigenen Experimenten verwendete monoklonale anti-GABA-Antikörper
zeichnet sich durch eine sehr hohe Sensitivität im Vergleich zu polyklonalen GABA-
Antiseren aus. Ein Problem bei der Detektion von GABA besteht darin, dass diese
Aminosäure aufgrund ihrer niedermolekularen Größe nur nach Konjugation an ein
Aldehyd mit Antikörpern detektiert werden kann. Beim Nachweis GABAerger
Neurone ist demnach grundsätzlich die immunhistochemische Detektion von GAD
vorzuziehen, da dieses Enzym zum einen nur in Neuronen exprimiert wird und im
Gegensatz zu GABA ein Vollantigen darstellt, das für die Detektion nicht konjugiert
werden muss. Allerdings erwiesen sich GAD-Antikörper in der Mehrzahl von Studien
als weniger sensitiv (Somogyi et al., 1984; Spreafico et al., 1988), was auch in den
eigenen Experimenten deutlich wurde. Unter Berücksichtigung der Literatur
(Lehmann et al., 1996; Lehmann et al., 1998) und eigener Vorversuche hinsichtlich
der Spezifität der Antikörper (3.3.5.1) sind die mit der Immunhistologie erzielten
Ergebnisse als spezifisch anzusehen.

167
5.2 Diskussion der Untersuchungsergebnisse
5.2.1 Untersuchung GABAerger Neurone unter Kontrollbedingungen
Ein Ziel dieser Arbeit beinhaltete die Charakterisierung der Expression von mRNA für
die Isoformen der GAD im PC, dem zunehmende Bedeutung im Zusammenhang mit
der Entstehung und Ausbreitung epileptischer Aktivität im limbischen System
beigemessen wird (Löscher und Ebert, 1996b). Während GABAerge Neurone im PC
in der Vergangenheit immunhistologisch zunehmend besser charakterisiert wurden
(Haberly et al., 1987; Lehmann et al., 1998; Löscher et al., 1998; Ekstrand et al.,
2001), beschränken sich die Informationen über die Expression von GAD-mRNA auf
wenige Übersichtsarbeiten unter Verwendung radioaktiver in-situ-Hybridisierung
(Ferraguti et al., 1990; Najlerahim et al., 1990). Meines Wissens ist die vorliegende
Arbeit die erste Studie, in der die Expression von beiden Isoformen der GAD mittels
nicht-radioaktiver in-situ-Hybridisierung im Vergleich zum immunhistologischen
Nachweis von GAD und GABA an Gewebeserien eines Tieres untersucht wurde.
Die eigenen Ergebnisse der Darstellung GABAerger Neurone mit nicht-radioaktiver
in-situ-Hybridisierung gegen die mRNA von Isoformen der GAD sowie
Immunhistochemie gegen GAD und GABA unter Kontrollbedingungen entsprechen
weitestgehend den Erkenntnisse in der Literatur (Esclapez et al., 1993, 1994; Houser
und Esclapez, 1994; Lehmann et al., 1996, Löscher et al., 1998). Unter
Kontrollbedingungen stellten sich einige grundsätzliche Tendenzen heraus. So
ergaben sich neben allen untersuchten Regionen besonders in der
Hippocampusformation nach Durchführung der Immunhistochemie höhere Dichten
GABA- als GAD-immunreaktiver Neurone. Diese Beobachtung entspricht Hinweisen,
dass sich unter Verwendung von Antikörpern gegen GABA in den meisten Fällen
höhere Neuronenzahlen erfassen lassen als dies unter Einsatz gegen GAD
gerichteter Immunseren der Fall ist (Somogyi et al., 1984; Spreafico et al., 1988).

168
In den untersuchten Hirnregionen war mit Ausnahme der Hippocampusformation
eine stärkere Expression von GAD67-mRNA festzustellen. Auch diese Ergebnisse
decken sich mit den Angaben, dass GAD67-mRNA in den meisten Hirnregionen
stärker exprimiert wird als GAD65-mRNA, so in den Basalganglien, im Neocortex
und im Cerebellum (Feldblum et al., 1993; Esclapez et al., 1993, 1994; Hendrickson
et al., 1994). Für die Hippocampusformation wurden dagegen vergleichbare
Expressionsraten der beiden Isoenzyme dokumentiert (Houser und Esclapez, 1994).
Im Vergleich der Immunhistochemie mit der in-situ-Hybridisierung war im Retikulären
Nukleus des Thalamus die überwiegende Mehrzahl von Neuronen intensiv für GAD
und GABA sowie GAD-mRNA markiert. Ein vergleichbares Bild hinsichtlich der Zahl
erfasster GABAerger Zellen ergab sich in der Substantia nigra pars reticulata sowie
der basolateralen Amygdala. Im PC zeigte sich unter Kontrollbedingungen indes ein
deutliche Diskrepanz hinsichtlich der Anzahl markierter Zellen in Abhängigkeit von
der eingesetzten Nachweismethode. In der Schicht II war die Dichte GABA- und
GAD-immunreaktiver Neurone in der gesamten Region bis zu zehnmal größer als die
Dichte mRNA-positiver Neurone nach in-situ-Hybridisierung. Eine Anfärbung von
Pyramidenzellen wurde nicht beobachtet. In den anderen Schichten des PC sowie im
angrenzenden EN ergaben sich dagegen Unterschiede in der Dichte von unter
100 %.
Aufgrund der in Kap. 5.1 diskutierten Maßnahmen zur Überprüfung der Spezifität der
verwendeten Nachweise können methodische Aspekte im Hinblick auf diese
Ergebnisse vernachlässigt werden. Zudem ist die Immunreaktivität einer großen Zahl
von Neuronen unter Anwendung der in dieser Studie verwendeten Antikörper gegen
GAD und GABA für die Schicht II des PC bereits belegt worden (Lehmann et al.,
1998; Löscher et al., 1998). Grundsätzlich ist davon auszugehen, dass GABA- bzw.
GAD-immunreaktive Neurone die mRNA für GAD exprimieren. In diesem
Zusammenhang lässt sich spekulieren, dass die mRNA für die Isoformen der GAD
in solchen Zellen unter Kontrollbedingungen nur in geringem Maße exprimiert wird
und mit der in-situ-Hybridisierung unter den gegebenen Versuchbedingungen nicht
detektiert werden kann.

169
Cao und Mitarbeiter (1998) beschrieben ein ähnliches Phänomen bei der
Untersuchung hippocampaler Neurone in Zellkulturen. Obwohl in diesen Zellen die
mRNA für beide Isoformen der GAD mittels PCR nachgewiesen werden konnte,
ließen sich mit Immunhistochemie für GABA und GAD nur Teile der Zellpopulation
markieren. Mit der nicht-radioaktiven in-situ-Hybridisierung für GAD65- und GAD67-
mRNA konnte wiederum nur ein geringerer Prozentsatz der immunhistologisch
positiven Neurone erfasst werden.
Weiterhin ließe sich als Ursache für die Unterschiede in Betracht ziehen, dass die
mRNA in den meisten GABAergen Neuronen der Schicht II des PC in höheren
Konzentrationen vorliegt, aber nicht oder nur in unzureichendem Ausmaß detektiert
werden kann. Da die Steuerung der GAD-Synthese auch posttranskriptional reguliert
wird, könnte ein Anheften von Proteinen, welche die Translation der mRNA
inhibieren, auch die Hybridisierung zumindest eines Teils der eingesetzten Sonde
verhindern (Bailey, 1991). Dies würde bedeuten, dass mittels in-situ-Hybridisierung
mRNA nur in bestimmten Funktionszuständen detektieren werden könnte.
Mehrere Indizien sprechen für die Hypothese, dass die mRNA für GAD-Isoformen in
der Mehrzahl GAD- bzw. GABA-immunreaktiver Neurone im PC unterhalb des
Detektionslevels der in-situ-Hybridisierung exprimiert wird. Unter
Kontrollbedingungen konnten in den eigenen Untersuchungen Differenzen bezüglich
der Expression für GAD-mRNA auch in anderen Hirnregionen festgestellt werden. Im
Striatum zeigte sich die Mehrzahl der Neurone immunreaktiv für GABA und GAD.
Eine vergleichbare Anzahl von Zellen war für die mRNA beider Isoformen der GAD
markiert, die eine schwache Anfärbung für GAD65-mRNA zeigten. Die Intensität der
Markierung für GAD67-mRNA war in der Mehrzahl der Neurone ebenfalls als
schwach bis mittelstark einzustufen, während ein geringer Prozentsatz der
Neuronenpopulation diese Isoform stark exprimierte (vgl. Esclapez et al., 1993).
In früheren Studien wurde eine starke Expression von GAD67-mRNA für Neurone
beschrieben, die sich durch eine tonische Feuerrate oder die Fähigkeit, über längere
Zeitabstände tonisch feuern zu können, auszeichnen. Zu diesen gehören
GABAergen Neurone in der Substantia nigra pars reticulata (Delong et al., 1983;
Hikosaka und Wurtz, 1983) und im Globus pallidus (Deniau und Chevalier, 1985).

170
Die Projektionsneurone des Striatums, welche die Mehrheit der Neurone in dieser
Region bilden, sind durch eine geringere Feuerrate charakterisiert (Wilson, 1990)
und zeigten nur geringe Anfärbung für GAD67-mRNA (Esclapez et al., 1993).
In elektrophysiologischen Untersuchungen konnte festgestellt werden, dass
GABAerge Interneurone im PC Aktionspotentiale aufwiesen, die denen der
Substantia nigra pars reticulata sehr ähnlich waren, aber eine weitaus geringere
Feuerrate als die nigralen Neurone besaßen (Bloms-Funke et al., 1999). Bei der
Untersuchung an Slicepräparaten des PC zeigten zudem nur wenige Neurone eine
spontane Feueraktivität ganz im Gegensatz zu den Neuronen anderen kortikale oder
subkorticaler Regionen (Löscher und Ebert, 1996b). In den eigenen Experimenten
wiesen die GABAergen Neurone der Substantia nigra pars reticulata eine deutliche
Expression für GAD-mRNA und intensive Immunreaktivität für GAD und GABA als
Hinweise einer hohen GABAergen Transmission unter Kontrollbedingungen auf. Für
die Mehrzahl der GABAergen Neurone in der Schicht II des PC ist zu vermuten, dass
diese aufgrund ihrer niedrigen basalen Feuerrate keiner hohen Enzymaktivität der
GAD bedürfen und dementsprechend nur niedrige Expressionsraten für GAD-mRNA
aufweisen. Erhöhte Expressionsraten wären für Bedingungen zu erwarten, die eine
vermehrte GABAerge Inhibition als Reaktion auf gesteigerte Aktivität im Netzwerk
des PC erforderlich machen. Eine solche Induktion der Expression von GAD-mRNA
konnte in GABAergen Neuronen des PC nach Auslösung eines SE erreicht werden
(Kap. 5.2.2).
Unter Kontrollbedingungen wurde die höchste Dichte GAD-mRNA-positiver Neurone
in der Schicht III des PC festgestellt. Dieses Ergebnis deckt sich mit den Angaben
von Haberly et al. (1987), welche die höchste Dichte GABA- und GAD-
immunreaktiver Zellen für diese Schicht des PC an Präparaten des Opposums unter
Verwendung polyklonaler Antikörper beschrieben. Die Übereinstimmung mit den
Ergebnissen dieser Arbeitsgruppe legt den Schluss nahe, dass GABAerge Neurone
in der Schicht III des PC höhere Gehalte an GABA und GAD in Verbindung mit einer
gesteigerten Syntheserate für GAD-mRNA aufweisen. Dies ermöglicht die Detektion
der Mehrzahl diese Neurone sowohl mit in-situ-Hybridisierung als auch mit
polyklonalen Antikörpern von geringerer Sensitivität.

171
In der Hippocampusformation wurden unter Kontrollbedingungen ebenfalls
Unterschiede in der Dichte markierter Zellen in Abhängigkeit von den angewendeten
Nachweisen festgestellt. In den Körnerzellen des Gyrus dentatus ließ sich nur für
GABA eine Anfärbung erzielen, nicht jedoch für die GAD und die mRNA des
Enzyms. Eine GABA-Immunreaktivität dieser glutamatergen Neurone wurde bereits
von verschiedenen Arbeitsgruppen beschrieben (Lehmann et al., 1996; Sloviter et
al., 1996). Sloviter et al. (1996) wiesen zudem GABA-Immunreaktivität in den Axonen
dieser Zellen, den Moosfasern, nach. Unter Kontrollbedingungen wurde für GAD
meines Wissens bisher keine Immunreaktivität in Körnerzellsomata beschrieben
(Lehmann et al., 1996; Sloviter et al., 1996; Esclapez und Houser, 1999). Eine
Anfärbung für GAD67 beschränkte sich in früheren Untersuchungen auf die
Ausläufer dieser Zellen, während sich für GAD65 in unbehandelten Tieren keine
immunpositiven Strukturen ergaben (Sloviter et al., 1996). Houser und Esclapez
(1994) beschrieben für die Körnerzellen des Gyrus dentatus eine vermehrte
Hintergrundfärbung nach Durchführung der in-situ-Hybridisierung für GAD67-mRNA,
welche sie als unspezifisch beurteilten. Vergleichbare Beobachtungen waren auch in
den eigenen Untersuchungen zu machen, eine spezifische Markierung dieser Zellen
für GAD67-mRNA konnte unter Kontrollbedingungen nicht nachgewiesen werden.
Hinsichtlich des Nachweises von GABA in glutamatergen Körnerzellen wurde
spekuliert, ob diese Neurone zur Aufnahme dieses Neurotransmitters befähigt sind.
Allerdings konnten GABA-Transporter bisher nur in GABAergen Neuronen
nachgewiesen werden (Augood et al., 1995) und Glutamat-Transporter besitzen
aufgrund ihrer Spezifität offensichtlich nicht die Fähigkeit zur Aufnahme von GABA
(Fykse et al., 1992). Eine mögliche Funktion der GAD in Neuronen, die GABA nicht
als Neurotransmitter verwenden, wurde im Bezug auf die Beteiligung am GABA-
Shunt postuliert. Solche Zellen würden ausschließlich einen „metabolischen GABA-
Pool“ aufweisen, was bisher aber nicht belegt werden konnte. Von Interesse sind
Überlegungen unter Berücksichtigung der Fähigkeit von Neurone, ihren
Neurotransmitterphänotyp in Reaktion auf verschiedene Einflüsse kurzfristig ändern
können (Rao et al., 1993) . Eine solche „standby neurotransmitter“ Hypothese würde
einem exzitatorischen Neuron erlauben, auf paroxysmale Aktivität in seiner
Umgebung durch einen Release von GABA zu reagieren (Cao et al., 1996).

172
Im Hippocampus stellte sich im Stratum pyramidale entlang der Regionen CA1-CA3
eine hohe Dichte GABA-immunreaktiver Neurone dar, die bereits von Lehmann et al.
(1996) sowie Meldrum et al. (1987) dokumentiert wurde. Der immunhistochemische
Nachweis der GAD führte zu deutlich weniger markierten Zellen in diesen Regionen.
Diese Ergebnisse stehen in Übereinstimmung mit den Angaben von Mugnaini und
Oertel (1985), die unter Verwendung des gleichen Antikörpers im Stratum pyramidale
des Hippocampus zwischen 5-15 % positive Neurone für GAD beschrieben. Eine
Anfärbung von Pyramidenzellen wurde bei der Verwendung beider Antikörper nicht
beobachtet. Die Isoformen der GAD wurden im Hippocampus in vergleichbarem
Maße exprimiert, die Anzahl GAD-mRNA-positiver Neurone zeigte sich wiederum
niedriger im Vergleich zur Dichte immunpositiver Neurone. Diese Beobachtung war
besonders für Pyramidenzellschicht der CA1 Region auffällig, in der Houser und
Esclapez (1994) analog zu ihren Beobachtungen im Gyrus dentatus in den
Schnittserien einiger Tiere eine unspezifische Färbung für GAD67-mRNA
beschrieben. Hinweise auf die Synthese von GABA in hippocampalen
Pyramidenzellen erbrachte der Nachweis von mRNA für die GAD-Isoformen in
diesen Zellen (Cao et al.,1996). Der immunhistochemische Detektion von GABA
gelang jedoch nicht in allen Zellen, so dass die Arbeitsgruppe postulierte, dass in der
Mehrzahl dieser glutamatergen Neurone keine nachweisbaren Konzentrationen an
GABA synthetisiert werden.

173
5.2.2 Untersuchungen an Ratten mit Status epileptikus induziert durch Kainat/ Lithium-Pilocarpin
Die Auswirkung akuter Krampfaktivität auf GABAerge Neurone sowie die Expression
von GAD-mRNA im PC bildete einen Untersuchungsschwerpunkt im folgenden
Abschnitt der Studie.
In beiden Tiergruppen konnten infolge des Kainat- bzw. Lithium-Pilocarpin
induzierten SE neurodegenerative Veränderungen in den untersuchten Hirnregionen
festgestellt werden. Von diesen Alterationen waren vor allem der PC und die
Substantia nigra pars reticulata betroffen, während in der Amygdala und der
Hippocampusformation keine Schädigungen an Neuronen beurteilt werden konnten.
Im Vergleich zwischen den Gruppen traten zudem bei den mit Lithium-Pilocarpin
behandelten Tieren stärkere Schäden der Neuronenpopulationen auf. In diesem
Zusammenhang wäre zu vermuten, dass das Ausmaß der Neuronenschädigung
größtenteils auf die Länge des Status zurückzuführen ist (Lemos und Cavalheiro,
1995). Da bei allen Tieren der SE nach einer identischen Zeitspanne unterbrochen
wurde, erscheint dies als Erklärung der Unterschiede nicht wahrscheinlich. Zudem
sind Unterschiede im Ausmaß der Neurodegeneration zwischen dem Kainat- und
dem Pilocarpin-Modell beschrieben worden. Ein Pilocarpin-induzierter SE führte nach
kürzerer Zeit zu ausgeprägteren Gewebeveränderungen im Vergleich zur
Behandlung mit Kainat (Covolan und Mello, 2000). Die Autoren beschrieben zudem
Differenzen in der Latenzzeit zwischen dem Beginn des SE und dem Auftreten
geschädigter Zellen in einzelnen Hirnregionen. Im PC, in der Substantia nigra pars
reticulata sowie im Hilus des Gyrus dentatus waren geschädigte Neurone bereits
nach 2,5 h nachzuweisen, während Zellen in der Amygdala und den CA-Regionen
des Hippocampus erst nach 8-48 h verstärkt betroffen waren.
Krampfaktivität infolge des Kainat bzw. Lithium-Pilocarpin induzierten SE führte bei
einigen Tieren zu ausgeprägter Neurodegeneration im PC. Diese betraf vornehmlich
die Schicht III und den VEN im zentralen und posterioren PC, während sich der
anteriore PC durch weniger ausgeprägte Neuronenschäden auszeichnete.

174
Für die Pilocarpin-Gruppe ergaben sich signifikante Reduktionen in der Dichte von
Neuronen positiv für die mRNA von GAD-Isoformen sowie GAD- und GABA-
immunreaktiver Zellen im gesamten PC. Nach Kainat-induziertem SE fielen die
Abnahmen in diesen Schichten im Vergleich zur Kontrolle geringer aus.
In der Schicht II des zentralen PC mit Lithium-Pilocarpin behandelter Tiere trat eine
verstärkte Expression von mRNA für beide Isoformen der GAD auf, während die Zahl
immunreaktiver Zellen für GABA und GAD reduziert war. In der Kainat-Gruppe war
die Expression von GAD65-mRNA verstärkt, GAD67-mRNA war dagegen nur
tendenziell erhöht. Im posterioren PC ergab sich für beide Gruppen ein
vergleichbares Bild mit einer tendenziell höheren Dichte positiver Zellen nach in-situ-
Hybridisierung für beide Isoformen der GAD.
Eine verstärkte Expression von GAD-mRNA in GABAergen Neuronen der
Hippocampusformation ist für das Kainat-Modell und das Pilocarpin-Modell der
Temporallappenepilepsie beschrieben worden (Schwarzer und Sperk, 1995;
Esclapez und Houser, 1999). In Bezug auf die eigenen Untersuchungsergebnisse ist
zu vermuten, dass die feed-forward- und feed-back-Mechanismen GABAerger
Inhibition infolge induzierter akuter Krampfaktivität mit überschießender Erregung im
neuronalen Netzwerk des PC verstärkt aktiviert wurden. Eine erhöhte GABAerge
Transmission würde gesteigerte Syntheseraten für GABA voraussetzen, welche die
vermehrte Expression von GAD erfordern. Die Zunahme der intrazellulären mRNA-
Konzentration ist demzufolge als Erklärung für eine erhöhte Dichte GAD-mRNA-
positiver Neurone im PC anzunehmen. Zudem wird die eigene Hypothese
untermauert, dass der überwiegende Teil der GABAerger Interneurone im PC die
mRNA für GAD-Isoformen unter Kontrollbedingungen in einer Menge exprimiert, die
mittels in-situ-Hybridisierung nicht detektiert werden kann (Kap. 5.2.1).
Im Gegensatz zu der gesteigerten Dichte GAD-mRNA-positiver Neurone wurde eine
Abnahme GABA- und GAD-immunreaktiver Neurone in der Schicht II des zentralen
PC festgestellt. Da die überwiegende Mehrzahl GABAerger Neurone unter
Kontrollbedingungen immunhistologisch erfasst wird, ist anzunehmen, dass mittels
diese Nachweise nur eine Reduktion immunreaktiver Zellen festgestellt werden kann.

175
Zudem ist beschrieben worden, dass eine vermehrte Expression von GAD-RNA mit
einem Anstieg der Immunreaktivität für GAD und GABA in Terminalen von Neuronen
einherging und nicht zu einer vermehrten Anfärbung von Zellsomata führte.
Dementsprechend war eine Induktion von GAD67-mRNA in Körnerzellen des Gyrus
dentatus von einer verstärkten Immunreaktivität in Moosfasern begleitet, die Somata
der Zellen zeigten dagegen keine Immunreaktivität für GAD und GABA (Schwarzer
und Sperk, 1995; Ding et al., 1998). Parallel zur gesteigerten Immunreaktivität konnte
ein Anstieg des extrazellulärem GABA Gehaltes im Innervationsgebiet dieser Fasern
nachgewiesen werden, was darauf hindeutet, dass eine verstärkte Expression von
GAD-mRNA mit der Freisetzung von GABA aus Terminalen einhergeht (Ding et al.,
1998).
Die Expression von GAD-mRNA wurde in den eigenen Untersuchungen für beide
Isoformen des Enzyms in der Schicht II des PC induziert. Eine aktivitätsabhängige
Regulation ist dagegen bisher hauptsächlich für GAD67 beschrieben worden (Litwak
et al., 1990; Drengler und Oltmans, 1993; Benson et al., 1994; Drengler et al., 1996).
Zudem konnte in der Mehrzahl von Studien durch Läsionen und andere Insulte ein
stärkere Expression von GAD67 und GAD67-mRNA induziert werden (Feldblum et
al., 1990; Soghomonian und Martin, 1998). Hierfür sprechen auch Erkenntnisse aus
dem Kainat-Modell, für das eine schnelle aber transiente Hochregulation von
GAD67-mRNA in Körnerzellen des Hippocampus beschrieben wurde, wogegen die
Induktion von GAD65-mRNA in diesen Zellen nicht nachgewiesen werden konnte
(Schwarzer und Sperk, 1995; Ding et al., 1998). Neuere Untersuchungen ergaben
Hinweise auf eine Regulation von GAD65 auf der Ebene der Transkription in
Verbindung mit epileptischer Aktivität. Esclapez und Houser (1999) belegten bei
Ratten mit spontanen epileptischen Anfällen infolge der Behandlung mit Pilocarpin
eine verstärkte Expression beider GAD-Isoformen in GABAergen Neuronen der
Hippocampusformation. Im Kainat-Modell kam es zu einer verzögerten aber
dauerhaften Expression von mRNA für beide Isoformen in GABAergen Korbzellen
(Schwarzer und Sperk, 1995). Die Ergebnisse dieser Studien lassen jedoch eher
vermuten, dass die GAD65 Expression hauptsächlich infolge chronischer Einflüsse
verändert wird.

176
Weitere Hinweise in diese Richtung stammen von Bowers und Mitarbeitern (1998),
die eine verstärkte Expression von GAD65-mRNA ausschließlich durch chronische
Stressstimuli auslösen konnten, während GAD67-mRNA bereits durch akute Stimuli
induziert wurde.
In den eigenen Untersuchungen wurde die Expression der mRNA für beide
Isoformen infolge akuter Krampfaktivität induziert. Eine mögliche Erklärung lässt sich
aus den funktionellen Unterschieden zwischen den beiden GAD-Isoformen ableiten.
Die GAD67 repräsentiert den überwiegenden Teil enzymatischer Aktivität in
GABAergen Neuronen und ist maßgeblich für die Synthese von GABA unter basalen
Bedingungen verantwortlich. Für GAD65 wird dagegen die Funktion als Reservoir an
induzierbarer Enzymaktivität im Bereich der Axonterminalen von Neuronen
angenommen, der eine besondere Bedeutung für die GABA-Synthese im Bereich der
Synapsen zukommt (Kaufman et al., 1991; Martin und Rimvall, 1993; Esclapez et al.,
1994). Die überschießende neuronale Aktivität im PC lässt eine gesteigerte
GABAerge Inhibition vermuten, die eine hohe GABA-Syntheserate in Interneuronen
erfordert. Diese scheint nicht durch die Aktivität der GAD67 sowie die Aktivierung von
apoGAD65 an den Synapsen allein ausgeglichen werden zu können, wodurch neben
der gesteigerten Expression von GAD67-mRNA zusätzlich die vermehrte Expression
von GAD65-mRNA erforderlich wird.
Die starke Aktivierung von Neuronen durch Kainat und Lithium-Pilocarpin führt im PC
zu Neurodegeneration, die im Gegensatz zu anderen Regionen des limbischen
Systems wie dem Hippocampus schon nach kurzer Zeit zum Untergang der
überwiegenden Mehrzahl der Neurone im PC führt (Sperk, 1994; Turski et al., 1986).
Es wäre denkbar, dass die Mechanismen, die auch im Hippocampus zu einer
verstärkten Expression von mRNA beider GAD-Isoformen in GABAergen
Interneuronen führen, im PC in einem viel engeren Zeitrahmen ablaufen und aus
diesem Grund die Induktion der GAD65-mRNA infolge akuter Anfallsaktivität
nachgewiesen werden konnte.

177
Im Gyrus dentatus wurde infolge Kainat- und Lithium-Pilocarpin-induziertem SE
GAD67-mRNA in Körnerzellen exprimiert, während GAD65-mRNA nicht
nachgewiesen werden konnte. Die schnelle aber transiente Expression von GAD67-
mRNA in glutamatergen Körnerzellen war für das Kainat-Modell im Vorfeld
beschrieben worden (Schwarzer und Sperk, 1995; Ding et al., 1998). Entsprechende
Angaben hinsichtlich des Pilocarpin-Modells sind mir aus der Literatur nicht bekannt.
In den Untersuchungen der genannten Arbeitsgruppen konnte die Expression von
GAD65-mRNA in Körnerzellen ebenfalls nicht induziert werden.
In der Hippocampusformation waren GABAerge Neurone im Hilus des Gyrus
dentatus reduziert, während für die Zahl GABAerger Korbzellen keine
Veränderungen auftraten. Die Neurone im Hilus des Gyrus dentatus sind als sehr
empfindlich gegenüber Krampfaktivität beschrieben worden (Turski et al, 1986;
Obenaus et al., 1993). Dies belegen auch die Untersuchungen von Covolan und
Mello (2000), die nach Kainat- und Pilocarpin-induziertem SE analog zum PC bereits
nach 2,5 h geschädigte Neurone in dieser Region nachweisen konnten. Aufgrund der
eigenen Ergebnisse kann jedoch nicht zweifelsfrei bewiesen werden, dass die
reduzierte Dichte GABAerger Neurone bereits Ausdruck eines Untergang dieser
Neurone ist. Ein reduzierte intrazelluläre Konzentration der nachzuweisenden mRNA
und Proteine in Neuronen muss ebenfalls als Erklärung in Betracht gezogen werden.
Den Verlust GABAerger Neurone im Hilus beschrieben Obenaus et al. (1993) infolge
Pilocarpin-induzierter Krampfaktivität. Dieser wurde begleitet von einer Reduktion
Nissl-gefärbter Neurone und Gliaproliferation, wohingegen GABAerge Korbzellen von
der Krampfaktivität nicht beeinflusst wurden. Für das Kainat-Modell konnte der
Nachweis des Verlustes GABAerger Neurone im Hilus ebenfalls erbracht werden
(Buckmaster und Jongen-Relo, 1999). Diese Erkenntnisse lassen vermuten, dass
GABAerge Neurone im Hilus des Gyrus dentatus bereits wenige Stunden nach
Auslösung eines SE untergehen oder zumindest irreversibel geschädigt sind.

178
Im Hippocampus ergaben sich keine signifikanten Veränderungen hinsichtlich der
Dichte GABAerger Neurone. In diesem Zusammenhang wurde berichtet, dass in den
Regionen CA1 und CA3 erst nach 24 h bzw. 48 h geschädigte Zellen in Folge eines
induzierten SE auftraten (Covolan und Mello, 2000). Dementsprechend sind
ausgeprägte Zellschäden in dem Zeitrahmen der eigenen Untersuchungen nicht
wahrscheinlich. Hierfür sprechen auch die Erfahrungen, dass die Unterbrechung
eines induzierten SE mit Diazepam ausgeprägte Zellschäden besonders in der
Region CA3 verhindert (Ben-Ari und Cossart, 2000).
Die Auslösung eines SE durch Applikation von Kainat bzw. Lithium-Pilocarpin führte
in der Amygdala und der Substantia nigra zu ausgeprägter Neurodegeneration
(Turski et al., 1986; Schmidt-Kastner et al., 1991; Sperk, 1994). In der Substantia
nigra pars reticulata sind die ausgeprägten neurodegenerativen Veränderungen
vermutlich das Resultat von Übererregung nigraler Neurone mit nachfolgenden
metabolischen Entgleisungen (Schmidt-Kastner et al., 1991). Im Rahmen der
eigenen Untersuchungen ließen sich in der basolateralen Amygdala sowie in der
anterioren und posterioren Substantia nigra pars reticulata verringerte Dichten
GABAerger Neurone sowohl mit in-situ-Hybridisierung als auch mit
Immunhistochemie darstellen. In der Amygdala waren in der Pilocarpin-Gruppe mit
allen Nachweisen Verluste GABAerger Neurone nachzuweisen, während die
Neuronendichten in der Kainat-Gruppe im Vergleich zur Kontrolle tendenziell weniger
stark reduziert waren. Diese Ergebnisse stehen im Einklang mit Covolan und Mello
(2000) die bei Tieren mit SE induziert durch Pilocarpin bereits nach 2,5 h
geschädigte Zellen in der basolateralen Amygdala fanden, während diese in der
Gruppe Kainat-behandelter Tiere erst nach 8 h auftraten. In der Substantia nigra pars
reticulata wurden in der Kainat-Gruppe weniger Tiere mit Läsionen gesehen als in
der Pilocarpin-Gruppe. Covolan und Mello (2000) sahen in dieser Region
ausgeprägte Zellschäden sowie eine intensive Schwellung des Neuropils 8-48 h
nach Beginn des SE in mit Kainat und Pilocarpin behandelten Tieren.

179
5.2.3 Untersuchungen an voll gekindelten Ratten
Im Gegensatz zu Veränderungen aufgrund akuter Krampfaktivität nach Kainat- und
Lithium-Pilocarpin-induziertem SE war die Untersuchung möglicher chronischer
Veränderungen hinsichtlich der GAD-Expression Ziel dieses Teils der Studie. Zu
diesem Zweck wurden GABAerge Neuronen im PC sowie anderen limbischen
Hirnregionen und der Substantia nigra pars reticulata an voll gekindelten Ratten
untersucht. Die Tiergruppe war im Anschluss an den letzten gekindelten
generalisierten Anfall einer 42tägigen Stimulationspause unterzogen worden, um
anfallsbedingte Einflüsse auf die Versuchsergebnisse ausschließen zu können.
In der basolateralen Amygdala, dem Ausgangspunkt der elektrischen Stimulation des
angewendeten Protokolls, war die Dichte GAD65-mRNA-postiver Neurone in der
Gruppe gekindelter Tiere beidseitig gegenüber der Kontrollgruppe und ipsilateral zur
Gruppe implantierter Tiere vermindert. Nach der in-situ-Hybridisierung zum Nachweis
von GAD67-mRNA ließ sich ein solcher Effekt nur tendenziell und hauptsächlich auf
der ipsilateralen Seite nachweisen.
In vorausgegangenen immunhistochemischen Untersuchungen konnten Verluste
GABAerger Neurone in den Kerngebieten der Amygdala nach Kindling dokumentiert
werden. Callahan et al. (1991) beschrieben eine Abnahme GABA-immunreaktiver
Neurone von bis zu 62 % für die contralaterale Amygdala, Lehmann et al. (1998)
konnten den beidseitigen Verlust dieser Neurone in geringerem Ausmaß nach
Kindling nachweisen. Hinsichtlich der selektiven Degeneration bestimmter
Subpopulationen GABAerger Neurone fand die Arbeitsgruppe um Tuunanen (1997)
6 Monate nach 5 gekindelten generalisierten Anfällen keine Veränderungen in der
Zahl GABA-immunreaktiver Neurone in der contralateralen Amygdala. Die Dichte
Somatostatin-immunreaktiver Neurone war in den verschiedenen Kernen der
Amygdala dagegen um durchschnittlich 68 % gesenkt.

180
Bezüglich des eigenen Ergebnisses einer verringerten Dichte von Neuronen in der
Amygdala müssen Veränderungen im Volumen der Struktur bei gekindelten Ratten
aber auch methodische Aspekte als mögliche Ursachen in Betracht gezogen werden.
Eine reduzierte Zahl GAD-mRNA-positiver Neurone könnte ebenfalls aus einer
kindling-induzierten verringerten Expression von GAD-mRNA unterhalb der
Detektionsgrenze der in-situ-Hybridisierung resultieren. Dementsprechend kann mit
der angewendeten Nachweismethode für GAD-mRNA eine tatsächliche
Degeneration GABAerger Neurone nicht zweifelsfrei bewiesen werden. Zudem
ergeben sich aus der Literatur widersprüchliche Erkenntnisse hinsichtlich
neurodegenerativer Effekte gekindelter sekundär generalisierter Anfälle auf die
Neuronenpopulation der Amygdala. 6 Monate nach fünf generalisierten Anfällen
konnten keine Unterschiede in der Gesamtzahl von Neuronen in den verschiedenen
Kernen der Amygdala gekindelter Ratten festgestellt werden. Dies lässt vermuten,
dass neurodegenerative Veränderungen bezüglich der Neuronensubpopulationen in
der Amygdala bei der Bestimmung von Neuronengesamtzahlen mimikriert werden
können (Tuunanen und Pitkänen, 2000).
Zahlreiche Studien weisen jedoch darauf hin, dass eine Reduktion GABAerger
Neurone in der Amygdala nach Kindling auf den Untergang dieser Neurone und nicht
auf das Unterschreiten von Nachweisgrenzen zurückzuführen ist. Löscher und
Schwark (1987) sahen neben einer Abnahme der GABA-Konzentration und der
GAD-Aktivität von etwa 30 % auch eine Reduktion der GABA-Rezeptorbindung von
63 % nach Amygdala-Kindling. Zudem konnte von einer anderen Arbeitsgruppe eine
Reduktion an extrazellulären GABA-Gehalten durch Amygdala-Kindling von bis zu
60 % für beide Seiten der Struktur nachgewiesen werden (Kaura et al., 1995).
Itagaki und Kimura (1986) belegten die Abnahme GABA-Transaminase-positiver
Neurone in der Amygdala für das gleiche Modell. Hinweise auf eine Reduktion
GABAerger Inhibition ergaben sich auch aus der Untersuchung von Slices
Amygdala-gekindelter Ratten. In solchen Präparaten war die GABA-Rezeptor-
vermittelte Inhibition verringert, was auf den Untergang inhibitorischer GABAerger
Interneurone zurückgeführt wurde (Rainnie et al., 1992). Grundsätzliche
Anhaltspunkte für die Auslösung neurodegenerativer Veränderungen in der
Amygdala durch Kindling lieferte weiterhin der Nachweis einer Aktivierung von
Astrozyten in diesem Kerngebiet (Racine et al., 1989).

181
Amygdala-Kindling induzierte eine Aktivierung von Mikroglia im PC und im
angrenzenden EN (Ebert et al., 1996). Vorrausgegangene Untersuchungen hatten
bereits Belege für Neuronenverluste im EN nach Amygdala-Kindling geliefert
(Cavazos et al., 1994). Für den PC wurde später eine chronische Reduktion GABA-
immunreaktiver Neurone durch Amygdala-Kindling beschrieben (Lehmann et al,
1998), welche sich auf einen eng umschriebenen Bereich im ipsilateralen zentralen
PC beschränkte.
Dem PC wird eine Schlüsselrolle bei der Ausbreitung und Verstärkung epileptischer
Aktivität, welche von der Amygdala ausgeht, zugesprochen (Löscher und Ebert,
1995). Diese Annahme wurde auch von Burchfield und Applegate (1989) unterstützt,
die aufgrund der stufenweisen Entwicklung des Kindling-Phänomens eine Einteilung
in drei Phasen postulierten. Die erste Phase geht mit fokalen Veränderungen im
Bereich der stimulierten Region einher und äußert sich nur in geringer
Krampfaktivität. In der mittleren Phase breitet sich die epileptische Aktivität aus,
verbleibt aber in krampfsensiblen neuronalen Netzwerken des Vorderhirns, vor allem
in der Amygdala-PC-Region. Dies äußert sich in partiell konvulsivem Verhalten der
Ratten mit starken Fazialklonien und einseitigem Vorderextremitätenklonus. Die
letzte Phase manifestiert sich in generalisierter Krampfaktivität unter Einbeziehung
des Hirnstammes.
Neben dem Verlust GABAerger Neurone im zentralen PC (Lehmann et al., 1998)
belegten auch andere Studien die Beteiligung GABAerger Mechanismen an der
Krampfausbreitung in dieser Region. Durch die bilaterale Erhöhung der lokalen
GABA-Konzentration infolge von Mikroinjektionen des GABA-Transaminaseinhibitors
Vigabatrin im zentralen PC konnte die Kindling-Entwicklung bei Ratten verzögert
werden. Dies äußerte sich besonders in einer größeren Anzahl von Stimulationen,
die für den Übergang vom fokalen Krampfstadium III zum ersten generalisierten
Krampfanfall erforderlich war (Schwabe et al., 1999). Dementsprechend wurden
durch die Anstieg der GABAergen Inhibition im zentralen PC die Mechanismen, die
einen Übergang von lokal begrenzter in generalisierte Krampfaktivität verhindern,
verstärkt.

182
Eine Erhöhung der GABA-Konzentration hatte im zentralen PC gekindelter Ratten
die stärksten antikonvulsiven Effekte, führte aber auch im anterioren und posterioren
PC zu einer Erhöhung der Nachentladungsschwelle. Durch die direkte Beeinflussung
von GABAA-Rezeptoren im PC mittels Mikroinjektion von Muscimol konnten ebenfalls
eine antikonvulsive Wirkung sowie eine Erhöhung der Nachentladungsschwelle bei
voll gekindelten Ratten ausgelöst werden, die Effekte waren aber von kürzerer Dauer
als nach Vigabatrin (Schwabe et al., 1999; Schwabe et al., 2000).
In den eigenen Untersuchungen konnten keine signifikante Effekte des Amygdala-
Kindlings auf die Dichte GAD65- bzw. GAD67-mRNA-positiver Neurone im PC
gezeigt werden. In Schicht II des zentralen PC war die Dichte der Neurone, welche
die GAD67-mRNA exprimierten, in der Gruppe gekindelter Tiere jedoch tendenziell
gegenüber den beiden anderen Versuchsgruppen erniedrigt. Zudem fiel diese
Reduktion ipsilateral zur Stimulationselektrode stärker aus als contralateral. Der
Nachweis der GAD65-mRNA erbrachte keine entsprechenden Tendenzen, hier lagen
die mittleren Werte dagegen beidseitig über denen der Kontrollgruppen.
Eine mögliche Erklärung dafür, dass die beschriebene Reduktion GABA-
immunreaktiver Zellen mittels in-situ-Hybridisierung für GAD-mRNA nicht
reproduziert werden konnte, lässt sich aus der unterschiedlichen Zahl markierter
GABAerger Neurone ableiten. Im Vergleich der Nachweise unter
Kontrollbedingungen konnten immunhistochemisch bis zu 10fach höhere Zahlen
GABAerger Zellen im PC erfasst werden. Da bei der Untersuchung GABA-
immunreaktiver Neurone nach Kindling eine Verringerung von maximal 26 %
beobachtet wurde (Lehmann et al., 1998), wäre vorstellbar, dass eine diskrete
Abnahme GABAerger Neurone mittels in-situ-Hybridisierung für GAD67-mRNA nur
tendenziell nachzuweisen war. Außerdem wurde diese Isoform besonders in Schicht
II des zentralen und des posterioren PC in höherem Maße als GAD65 exprimiert.
Dementsprechend wäre zu vermuten, dass sich eine Verringerung in der Anzahl
GABAerger Neurone eher auf die Dichte GAD67- als GAD65-mRNA-positiver
Neurone ausgewirkt haben könnte.

183
Eine Abnahme GABAerger Zellen hätte andererseits infolge einer verstärkten
Expression besonders von GAD65-mRNA maskiert werden können.
Elektrophysiologische Untersuchungen von Neuronen des zentralen PC gekindelter
Ratten lieferten dahingehende Hinweise. Gernert et al. (2000) beschrieben einen
Anstieg der Aktivität von Neuronen im zentralen PC voll gekindelter Ratten im
Anschluss an eine 35tägige Stimulationsunterbrechung. Diese vermutlich
GABAergen Interneurone am Übergang zwischen Schicht II und Schicht III wiesen
eine um ca. 90 % erhöhte Feuerrate auf, die mit einer um 70 % verringerten
Empfindlichkeit gegenüber Glutamat einherging. Aufgrund dieser herabgesetzten
Sensitivität wurde von der Arbeitsgruppe postuliert, dass solche Interneurone eine
geringere Feedback-inhibition auf Pyramidenzellen ausüben könnten. Durch den
Untergang GABAerger Neurone infolge des Kindling-Prozesses (Lehmann et al.,
1998) wäre die GABAerge Inhibition im zentralen PC zusätzlich vermindert und
erhöhte Aktivität GABAerger Interneurone als kompensatorische Reaktion auf die
verminderte GABAerge Inhibition zu werten (Gernert et al., 2000).
In Anbetracht dieser Erkenntnisse könnte eine vermehrte Expression von GAD-
mRNA in GABAerge Interneuronen des zentralen PC gekindelter Ratten als Reaktion
auf eine gesteigerte GABAerger Transmission auftreten. Im Rahmen meiner Arbeit
konnte gezeigt werden, dass in GABAergen Neurone des zentralen PC die
Expression von GAD-mRNA unter dem Einfluss akuter paroxysmaler Aktivität
induziert wird. Eine chronisch erhöhte GAD-Expression ist für Interneurone in der
Hippocampusformation im Kainat- und im Pilocarpin-Modell beschrieben worden
(Schwarzer und Sperk, 1995; Houser und Esclapez, 1999). In diesen
Untersuchungen wurde zudem eine verstärkte Expression von GAD65-mRNA
beobachtet, für die eine Funktion in Form eines Reservoirs an enzymatischer
Aktivität hinsichtlich der GABA-Synthese im Bereich von Synapsen postuliert wird
(Erlander und Tobin, 1991; Martin, 1993; Esclapez et al., 1994). Die vermehrt aktiven
GABAergen Interneuronen im zentralen PC gekindelter Ratten könnten aufgrund
ihrer verstärkte Expression von GAD-mRNA mit der in-situ-Hybridisierung detektiert
werden, aber gleichzeitig hinsichtlich des Untergangs anderer GABAerger Neurone
zu einem falsch negativem Ergebnis führen.

184
Da die chronischen Veränderungen hinsichtlich der Abnahme GABA-immunreaktiver
Neurone und der Aktivitätszunahme überlebender Interneurone nicht sehr deutlich
ausgeprägt sind (Gernert et al., 2000), wäre eine solche Erklärung für die eigenen
Untersuchungsergebnisse durchaus wahrscheinlich.
Der Substantia nigra pars reticulata wird analog zum PC eine entscheidende Rolle
für die Ausbreitung von Krampfaktivität, die im limbischen System generiert wird,
zugesprochen (Löscher und Ebert, 1996a). Dieser Struktur kontrolliert das
Übergreifen paroxysmaler Aktivität auf den Hirnstamm und damit die Generalisation
von Anfällen (Depaulis et al., 1994). Mikroinjektionen von GABA oder GABAerger
Substanzen in Substantia nigra pars reticulata konnten das Auftreten von Krämpfen
in verschiedenen Epilepsiemodellen abschwächen oder ganz verhindern (Gale 1988,
1992; Löscher und Ebert, 1996a) sowie die Kindlingentwicklung Amygdala-
gekindelter Ratten verzögern (Löscher et al., 1987).
Die eigenen Untersuchungen des von GABAergen Neuronen dominierten retikulären
Teils der SN erbrachten keine chronischen Veränderungen durch Kindling
hinsichtlich der Expression von GAD65- und GAD67-mRNA sowie der Dichte GABA-
immunreaktiver Neurone. Dieses Ergebnis war zunächst überraschend, da sich in
der Literatur Hinweise auf eine Beeinflussung der GABAergen Inhibition in der
Substantia nigra pars reticulata finden. 7 Tage nach Kindling war die Aktivität der
GAD in Synaptosomen nigraler Neurone verringert (Löscher und Schwark, 1985,
1987), während die GABA-Gehalte in den Nervenendigungen vermindert aber nicht
signifikant gegenüber den Kontrollwerten reduziert waren (Löscher und Schwark,
1987). Im Zusammenhang mit Kindling wurde weiterhin eine geringere Expression
von GAD-mRNA in der Substantia nigra pars reticulata beschrieben (Sonnenberg et
al., 1991). Das Zeitfenster der Untersuchung lag zwischen 30 min und 24 h nach
dem letzten gekindelten Anfall. Die verringerte Expression von GAD-mRNA fanden
sich allein in der Substantia nigra pars reticulata von Tieren, die 30 min nach dem
letzten Anfall untersucht wurden und beschränkten sich zudem auf einzelne Neurone
in dieser Region. Die Veränderungen fielen so diskret aus, dass verringerte GAD-
mRNA in Blot-experimenten oder im Röntgenfilm nicht nachgewiesen werden
konnten.

185
Die eigenen Untersuchungen zielten auf den Nachweis chronischer Veränderungen
der Expression von GAD-mRNA durch Amygdala-Kindling ab. Dementsprechend
wurden sie an Tieren durchgeführt, die nach dem letzten gekindelten Anfall einer
42tägigen Stimulationspause unterzogen wurden. Grundsätzlich muss davon
ausgegangen werden, dass Modifikationen, die auf akuter Krampfaktivität infolge
eines gekindelten Anfalls beruhen, bis zu 2 Wochen existent bleiben können,
wohingegen alle Veränderungen nach 4 Wochen als permanent und kindling-
assoziiert anzusehen sind (Burnham und Cottrell, 1990). Entsprechende
Erkenntnisse lieferten elektrophysiologische Untersuchungen, die eine deutlich
veränderte Aktivität von Neuronen in der Substantia nigra pars reticulata während
gekindelter Anfälle beschrieben (Bonhaus et al., 1986), wohingegen sich nach zwei
bis drei Wochen keine Unterschiede mehr in hinsichtlich der Aktivität und der
Empfindlichkeit gegenüber verschiedenen Transmittern nachweisen ließen
(Waszczak et al., 1988). Folglich sind die Ergebnisse der obigen Studien bezüglich
einer Beeinflussung der GABA-Synthese im Vergleich mit den eigenen Ergebnisse
eher als anfalls- denn als kindling-assoziiert zu werten.
Kindling-assoziierte Veränderungen, die immunhistologisch (Lehmann et al., 1998)
und elektrophysiologisch nachgewiesen wurden (Gernert et al., 2000), waren meist
nicht sehr ausgeprägt. Hinsichtlich der eigenen Untersuchungen in der Substantia
nigra pars reticulata kann nicht zweifelsfrei ausgeschlossen werden, dass mögliche
diskrete Unterschiede in der Expression von GAD-mRNA auftraten, die anhand der
verwendeten Nachweisprotokolle nicht detektiert werden konnten. Esclapez und
Houser (1999) belegten, dass methodischen Aspekten eine entscheidende Rolle bei
der Bestimmung von Differenzen der Expression von GAD-mRNA mittels nicht-
radioaktiver in-situ-Hybridisierung zukommt. So fielen Unterschiede in der
Expression von GAD-mRNA zwischen den Versuchsgruppen nach einer
Entwicklungszeit von 2 h wesentlich deutlicher aus und reduzierten sich nach einer
Reaktionszeit von 4 h (Esclapez und Houser, 1999). Demnach hätte ein Unterschied
in der Expression von GAD-mRNA in den eigenen Versuchen durch die gewählten
Inkubationszeiten von 3-4 h für GAD67-mRNA und 5-6 h für GAD65-mRNA maskiert
werden können.

186
Neben den eigenen Untersuchungsergebnissen lassen auch die Ergebnisse anderer
Studien an einer entscheidenden Rolle der Substantia nigra pars reticulata im
Kindling zweifeln (Löscher und Ebert, 1996a). Eine Läsionierung dieser Struktur blieb
ohne Effekt auf die Kindlingentwicklung nach Stimulationen der basolateralen
Amygdala und des PC (Wahnschaffe und Löscher, 1990a). Läsionen in der SN
hatten weiterhin keinen Einfluss auf die Wirkung GABA-mimetischer Antiepileptika
(Wahnschaffe und Löscher, 1990b), was wiederum dagegen spricht, dass die SN die
antikonvulsiven Effekte erzeugt, die durch Verstärkung der GABAergen Transmission
entstehen (Löscher und Ebert, 1996a).
Dem Hippocampus wurde oftmals eine Funktion als primärer oder sekundärer Focus
im Zusammenhang mit Epilepsien zugesprochen. Dies resultiert aus der Fähigkeit
der Struktur, über Rückkopplungsschleifen mit extrahippocampalem Gewebe lange
und wiederkehrende Nachentladungen zu bilden (Lothman et al., 1991). Die Rolle
dieser Struktur im Kindling ist jedoch in dieser Hinsicht nicht klar bewiesen, da
Läsionsstudien keinen eindeutigen Einfluss auf den Kindlingsprozess zeigten
(Löscher und Ebert, 1996a). Ebenso widersprüchlich sind die Ergebnisse bezüglich
der Verluste GABAerger Neurone im Hippocampus infolge von Kindling. Konnte nach
hippocampalem Kindling eine Abnahme GABAerger Neurone erreicht werden
(Kamphuis et al., 1986, 1989), so beschrieben Studien unter Verwendung des
Amygdala-Kindling-Modells keine derartigen Veränderungen (Lehmann et al., 1996;
Sonnenberg et al., 1991, Tuunanen et al., 1997). Dies ist dahingehend
verwunderlich, da sich diese Modelle in der Kindlingsentwicklung und in der
Ausprägung der Krampfparameter sehr ähnlich sind (Löscher et al., 1998).
Die eigenen Untersuchungsergebnisse ergaben keine Hinweise auf eine Wirkung
des Amygdala-Kindlings auf die GAD-Expression in der Hippocampusformation und
stehen damit in guter Übereinstimmung mit den oben angeführten Arbeiten. Auch
Babb et al. (1989) sahen nach Kindling im Entorhinalen Cortex eine reduzierte GAD-
Immunreaktivität in Neuronen des Gyrus dentatus, die nach einer Woche wieder
Kontrollniveau erreicht hatte.

187
5.2.4 Bedeutung der Untersuchungsergebnisse
Ein Verlust GABA-vermittelter Inhibition wird als eine mögliche Ursache für die
Entstehung und Verbreitung von epileptischer Aktivität in Tiermodellen und in Bezug
auf die Epilepsien des Menschen angenommen (Bradford, 1995; Glass und
Dragunow, 1995; Houser, 1991; Löscher, 1989, 1993; Lothman, 1992). Hierfür
sprechen die antikonvulsive Wirkung GABA-mimetischer Wirkstoffe und die
konvulsive Wirkung von GABA-Antagonisten sowie Inhibitoren der GAD, des GABA
synthetisierenden Enzyms. In der vorliegen Arbeit wurden GABAerge Neurone
mittels in-situ-Hybridisierung für die beiden Isoformen der GAD sowie
immunhistochemisch für GAD und GABA untersucht. Dabei wurden neben
Kontrollbedingungen auch die Auswirkungen akuter und chronischer
pathophysiologischer Veränderungen untersucht.
Die Ergebnisse meiner Arbeit unterstreichen die große Bedeutung der Auswahl
geeigneter Nachweismethoden im Hinblick der Fragestellung der durchzuführenden
Experimente. Unter Kontrollbedingungen konnte im PC eine ausgeprägte Diskrepanz
in der Anzahl markierter Neurone nach in-situ-Hybridisierung und Immunhistochemie
festgestellt werden. Demnach ist eine quantitative Erfassung GABAerger Neurone in
dieser Region ausschließlich unter Einsatz des immunhistochemischen Nachweises
von GAD oder besser GABA möglich, da mittels in-situ-Hybridisierung für GAD-
mRNA die Mehrzahl der Neurone nicht erfasst werden kann. In anderen von mir
untersuchten Hirnregionen wie der Amygdala, der Substantia nigra und dem Gyrus
dentatus konnte die Mehrzahl GABAerger Neurone mit in-situ-Hybridisierung und
Immunhistochemie markiert werden. Dennoch ist der immunhistochemische
Nachweis aufgrund des geringeren Zeit- und Arbeitsaufwandes grundsätzlich für die
Darstellung vorzuziehen, wenn eine Quantifizierung dieser Neuronenpopulation
durchgeführt werden soll.
Bei der Auswahl histologischer Nachweismethoden ist zu berücksichtigen, dass
mittels in-situ-Hybridisierung und der Immunhistochemie für GAD und GABA
unterschiedliche Substrate und Syntheseabschnitte im GABA-Metabolismus
detektiert werden, die zudem verschiedenen Regulationsprozessen unterliegen und
zeitlich versetzt ablaufen.

188
Im PC war eine vermehrte Expression von GAD-mRNA infolge eines induzierten SE
festzustellen, wohingegen sich mittels Immunhistochemie keine Hinweise auf eine
verstärkte GABAerge Transmission ergaben. Aus diesen Ergebnissen lässt sich
ableiten, dass bei histologischen Untersuchungen eine Kombination von in-situ-
Hybridisierung und Immunhistochemie anzustreben ist, da beide Methoden
verschiedene Informationen in Bezug auf den GABA-Metabolismus liefern, die für die
Gesamtbeurteilung der Ergebnisse erforderlich sein können. Aus den eigenen
Erfahrungen ist dabei der wesentlich höhere Aufwand der radioaktiven in-situ-
Hybridisierung nicht gerechtfertigt, da sie keine Vorteile gegenüber der nicht-
radioaktiven Variante der Methode in Hinblick auf die Sensitivität bietet.
Unter Berücksichtigung des Zeitaufwandes für die Methodik und spätere Auswertung
muss allerdings bei der Versuchplanung geklärt werden, ob in den Studien
Veränderungen zu erwarten sind, die einen hohen methodischen Aufwand
rechtfertigen. Im Rahmen der eigenen Studien wäre die in-situ-Hybridisierung für
GAD67-mRNA und der Einsatz der Immunhistochemie zum Nachweis von GABA für
die Ermittlung der Ergebnisse unter akuten Krampfbedingungen ausreichend
gewesen. Der Einsatz der in-situ-Hybridisierung bei der Untersuchung diskreter
Veränderungen, die etwa nach Amygdala-Kindling auftreten, ist aufgrund der
eigenen Erfahrungen nicht sinnvoll.
Neben der beschriebenen Kombination histologischer Nachweisverfahren erscheint
die Verwendung der Mikrodialyse-Technik als eine mögliche sinnvolle Erweiterung
bei der Untersuchung von Veränderungen GABAerger Inhibition in Tiermodellen zur
Temporallappenepilepsie. Diese Technik kann zur Bestimmung des extrazellulären
GABA-Gehaltes in bestimmten Hirnregionen eingesetzt werden, um z. B. zu belegen,
dass eine erhöhte Expression von GAD-mRNA in Verbindung mit einer gesteigerten
GABA-Immunreaktivität Ausdruck vermehrter GABAergen Transmission ist.

189
Die Bedeutung GABAerger Mechanismen hinsichtlich der Rolle des PC in der
Pathophysiologie der Temporallappenepilepsie erfordert weitere Untersuchungen.
Einen Ansatzpunkt bildet die Klärung einer spezifischen Beteiligung von
Subpopulationen GABAerger Zellen an den Verlusten GABAerger Neurone nach
Amygdala-Kindling (vgl. Lehmann et al., 1998). GABAerge Neurone lassen sich
anhand colokalisierender Substanzen in verschiedene Untergruppen unterteilen
(Kosaka et al., 1987). Hierfür haben sich besonders die Kalzium-bindenden Proteine
Parvalbumin und Calbindin-D (28K) bewährt (Celio, 1990). In vorausgegangenen
Untersuchungen unserer Arbeitsgruppe trat nach Kindling eine reduzierte
Immunreaktivität für Calbindin-D (28K) in Neuronen der tiefen Schichten des
anterioren und posterioren PC auf. Dieser Effekt ging verloren, wenn voll gekindelte
Tiere einen Tag vor der Tötung nochmals stimuliert wurden. Einen
entgegengesetzten Effekt ergab die Untersuchung im Bezug auf Parvalbumin, hier
zeigte sich erst eine Abnahme der Immunreaktivität nach einmaliger Stimulation voll
gekindelter Ratten. Diesen Ergebnissen kann mittels immunhistochemischer
Doppelmarkierungsstudien für GABA und die Kalzium-bindenden Proteine
nachgegangen werden, ein Einsatz der in-situ-Hybridisierung erscheint in diesem
Zusammenhang nicht angebracht.
Veränderungen von GABAA−Rezeptoren und damit verbundene
Funktionsänderungen im PC erscheinen als ein weiteres interessantes
Untersuchungsvorhaben. In Untersuchungen des Hippocampus konnte gezeigt
werden, dass Krampfaktivität die Zusammensetzung der Untereinheiten dieses
Rezeptortyps beeinflusst (Brooks-Kayal et al., 1998; Nusser et al., 1998).
Die Klärung einer solchen Fragestellung im PC könnte unter Einsatz der in-situ-
Hybridisierung zum Nachweis der mRNA für die Subunits von GABAA-Rezeptoren
durchgeführt werden.

190
6 Zusammenfassung
Eine reduzierte GABAerge Inhibition wird als eine mögliche Ursache der Entstehung
von Temporallappenepilepsien des Menschen vermutet. Diese Erkrankung resultiert
in komplex-fokalen Anfällen, die sich häufig durch Therapieresistenz auszeichnen.
Der PC stellt eine Hirnregion dar, der im Zusammenhang mit der Entstehung und
Ausbreitung von Krampfaktivität eine bedeutende Rolle beigemessen wird. Ein früher
beschriebener dauerhafter Verlust GABA-immunreaktiver Neurone im zentralen PC
nach Amygdala-Kindling, einem Modell der Temporallappenepilepsie, deutet auf
eine Beteiligung GABAerger Mechanismen an diesem Geschehen hin. Die
GABAergen Neurone des PC waren bereits mehrfach Ziel von Untersuchungen unter
Einsatz immunhistochemischer Methoden, die Verhältnisse der Expression des
GABA synthetisierenden Enzyms GAD sind jedoch in dieser Region nicht
ausreichend charakterisiert. GAD tritt im ZNS in zwei bisher bekannten Isoformen als
GAD65 und GAD67 auf, welche sich hinsichtlich der Expression in einzelnen
Hirnregionen unterscheiden.
Für die Untersuchung der Expression von mRNA für die GAD-Isoformen im PC und
anderen für die Epileptogenese relevanten Gehirnregionen wurden zwei Protokolle
zur in-situ-Hybridisierung etabliert. Unter Einsatz nicht-radioaktiv und radioaktiv
markierter Sonden trat kein Unterschied in der Sensitivität hinsichtlich der Anzahl
markierter Zellen auf, weshalb die weiteren Studien unter Einsatz der nicht-
radioaktiven in-situ-Hybridisierung durchgeführt wurden.
Unter Kontrollbedingungen konnten in der Mehrzahl der untersuchten
Gehirnregionen eine höhere Dichte GAD- und GABA-immunreaktiver Neurone im
Vergleich zur Anzahl GAD65- und GAD67-mRNA-positiver Neurone bestimmt
werden. Besonders ausgeprägt zeigten sich die Unterschiede in der Schicht II des
PC, die eine bis zu 10fach höhere Dichte immunreaktiver Neurone aufwies.
Zur Untersuchung der Auswirkungen von Krampfaktivität auf die Expression von
GAD-Isoformen wurde bei weiblichen Wistar Ratten ein selbsterhaltender SE durch
Applikation von Kainat- bzw. Lithium-Pilocarpin ausgelöst. Dieser wurde nach 90 min
mit Diazepam abgebrochen und die Tiere 8 h nach Applikation von Kainat bzw.
Lithium-Pilocarpin perfundiert.

191
In Körnerzellen des Gyrus dentatus wurde die Expression von GAD67-mRNA, aber
nicht von GAD65-mRNA induziert. In der Schicht II des zentralen PC wurde eine
verstärkte Expression von GAD65- und GAD67-mRNA festgestellt, während die
immunhistochemischen Nachweise nur eine Reduktion GABAerger Zellen ergaben.
In der Amygdala, dem Hilus des Gyrus dentatus sowie der Substantia nigra pars
reticulata war unter Einsatz der in-situ-Hybridisierung und der Immunhistochemie
eine reduzierte Dichte GABAerger Neurone nachzuweisen. In den tieferen Schichten
des PC und der Substantia nigra waren zudem ausgeprägte neuropathologische
Veränderungen zu beschreiben.
Im Amygdala-Kindling Modell der Temporallappenepilepsie wurde chronischen
Veränderungen der Expression von GAD-Isoformen in GABAergen Neuronen
nachgegangen. Dazu wurden weibliche Wistar Ratten stimuliert, bis 10 sekundär
generalisierte Anfälle auftraten, und nach einer anschließenden Stimulationspause
von 42 Tagen perfundiert, um transiente, anfallsbedingte Effekte ausschließen zu
können. Im Bereich der basolateralen Amygdala gekindelter Tiere traten beidseitig
Verluste GAD65-mRNA positiver Neurone auf, die Dichte GAD67-mRNA
exprimierender Zellen war dagegen nur tendenziell erniedrigt. Die Quantifizierung
GAD-mRNA positiver Neurone des PC, der Hippocampusformation sowie der
Substantia nigra pars reticulata erbrachte keine signifikanten Veränderungen.
Die Ergebnisse der durchgeführten Studien zeigen, dass die verschiedenen
Syntheseabschnitte im GABA-Metabolismus unter basalen und
pathophysiologischen Bedingungen unterschiedlich beeinflusst werden. Dies betont
die Notwendigkeit der Kombination geeigneter histologischer Nachweismethoden für
die Untersuchung des GABAergen Systems in Tiermodellen der
Temporallappenepilepsie.

192
Summary
Christian Freichel
Investigation of GABAergic neurons in the piriform cortex and other epilepsy-related brain regions of the rat in different models of temporal lobe epilepsy
A deficit in GABAergic inhibition is hypothesized to underlie human temporal lobe
epilepsy. This disease results in complex-focal seizures, which commonly display
resistance to pharmacological treatment. The piriform cortex (PC) is thought to be
critically involved in the generation and propagation of seizure activity. Recently, a
chronic loss of GABA-immunoreactive neurons in the central PC has been described
in the amygdala-kindling model of temporal lobe epilepsy, indicating that GABA
mediated mechanisms might be involved in this regard. Although GABAergic neurons
in the PC have been investigated immunhistochemically for GABA and its
synthesizing enzyme glutamic acid decarboxylase (GAD), the expression patterns of
GAD in the PC need to be further investigated. The brain contains at least two
isoforms of GAD, termed GAD65 and GAD67, which differ in the level of expression
among brain regions.
In order to investigate the expression of mRNA encoding for GAD-Isoforms in the PC
and other brain regions thought to be involved in epileptogenisis, two different
protocols for in situ hybridization were established. Using non-radioactively or
radioactively labelled probes, no differences in the number of cells marked for GAD-
mRNA were obvious. Therefore, non-radioactive in situ hybridization was utilized for
further studies.
Basal conditions revealed a higher number of GAD- and GABA-immunoreactive
neurons in the majority of the investigated brain regions compared to cell counts
obtained by in situ hybridization. This result was most striking in layer II of the PC,
with immunoreactive neurons displaying a 10fold higher density of GABAergic
neurons compared to in situ hybridization.
Furthermore, possible alterations in the expression of GAD isoforms provoked by
seizure activity were investigated. For this purpose a self-sustained status epilepticus
was induced in female Wistar rats by administration of kainate and lithium-
pilocarpine, respectively, which was terminated after 90 min with diazepam.

193
All animals were perfused 8 h post application. An upregulation of GAD67 but not
GAD65-mRNA was determined in granule cells of the hippocampus in both models.
In the PC, seizure activity resulted in an upregulation of GAD-mRNA in layer II of the
central PC, while the number of GAD and GABA positive neurons was decreased.
In the amygdala, the hilus of the gyrus dentatus, and the substantia nigra pars
reticulata a decrease of GABAergic neurons was detected using in situ hybridization
and immunhistochemistry. Finally, in parts of the substantia nigra pars reticulata and
the central and posterior parts of the PC a marked neurodegeneration occurred.
Possible chronic alterations in the expression of GAD-mRNA were investigated in the
amygdala-kindling model. Following 10 stimulated secondary generalized seizures,
kindled animals were perfused 42 days after the last elicited convulsion to avoid
transient effects related to the stimulation process. Neurons expressing GAD65-
mRNA were decreased bilaterally in the basolateral amygdala, while neurons
containing GAD67-mRNA were not significantly reduced. Cell counts in the PC, the
hippocampal formation, and the substantia nigra pars reticulata revealed no
significant changes compared to controls.
The present data indicate that individual stages in the metabolism of GABA are
differently regulated under basal and pathophysiological conditions. The results
strongly suggest a combination of different histological techniques if alterations in the
GABA system are investigated in animal models of temporal lobe epilepsy.

194
7 Literatur
Ackermann, R. F., Chungani, H. T., Handforth, S., Moshe, S., Caldecott−Hatazard and Engel, J. (1986) Autoradiographic studies of the cerebral metabolism and blood flow in rat amygdala kindling. In: J. A. Wada (Hrsg.): Kindling 3. Raven Press, New York, S. 73−90.
Alheid, G. F., de Olmos, J. S. und Beltramino, C. A. (1995) Amygdala and extended Amygdala. In: G. Paxinos, G. (Hrsg): The Rat Nervous System, second eds., Academic Press, San Diego, S. 495-478.
Amaral, D. G. und Witter, M. P. (1995) Hippocampal formation. In: G. Paxinos (Hrsg): The Rat Nervous System, second ed. Academic Press, San Diego, S. 443-494.
Angerer, L. M. und Angerer, R. C. (1981) Detection of poly A+ RNA in sea urchin eggs and embryos by quantitative in situ hybridization. Nucleic Acids Res. 9, 2819-2840.
Asada, H., Kawamura, Y., Maruyama, K., Kume, H., Ding, R. G., Kanbara, N., Kuzume, H., Sanbo, M., Yagi, T. und Obata, K. (1997) Cleft palate and decreased brain gamma-aminobutyric acid in mice lacking the 67-kDa isoform of glutamic acid decarboxylase. Proc. Natl. Acad. Sci. U.S.A. 94, 6496-6499.
Asada, H., Kawamura, Y., Maruyama, K., Kume, H., Ding, R., Ji, F. Y., Kanbara, N., Kuzume, H., Sanbo, M., Yagi, T. und Obata, K. (1996) Mice lacking the 65 kDa isoform of glutamic acid decarboxylase (GAD65) maintain normal levels of GAD67 and GABA in their brains but are susceptible to seizures. Biochem. Biophys. Res. Commun. 229, 891-895.
Augood, S. J., Herbison, A. E. und Emson, P. C. (1995) Localization of GAT-1 GABA transporter mRNA in rat striatum: cellular coexpression with GAD67 mRNA, GAD67 immunoreactivity, and parvalbumin mRNA. J. Neurosci. 15, 865-874.
Babb, T. L. und Brown, W. J. (1987) Pathological findings in epilepsy. In: J. Engel (Hrsg.): Surgical treatment of the epilepsies. Raven Press, New York, S. 511-540.
Babb, T. L. und Pretorius, J. K. (1993) Pathological substrates of epilepsy. In: E. Wyllie (Hrsg.): The treatment of epilepsy: Principles and Practice. Lea and Febiger, Philadelphia, S. 55-70.
Babb, T. L., Pretorius, J. K., Kupfer, W. R. und Brown, W. J. (1988) Distribution of glutamate−immunoreactive neurons and synapses in the rat and monkey hippocampus: light and electron microscopy. J. Comp. Neurol. 278, 121−138.
Babb, T. L., Pretorius, J. K., Kupfer, W. R. und Feldblum, S. (1989) Recovery of decreased glutamate decarboxylase immunoreactivity after rat hippocampal kindling. Epilepsy Res. 3, 18-30.
Bailey, J. M. und Verma, M. (1991) Analytical procedures for a cryptic messenger RNA that mediates translational control of prostaglandin synthase by glucocorticoids. Anal. Biochem. 196, 11-18.
Baran, H., Gramer, M., Hönack, D. und Löscher, W. (1995) Systemic administration of kainate induces marked increases of endogenous kynurenic acid in various brain regions and plasma of rats. Eur. J. Pharmacol. 286, 167-175.
Barnard, E. A., Skolnick, P., Olsen, R. W., Mohler, H., Sieghart, W., Biggio, G., Braestrup, C., Bateson, A. N. und Langer, S. Z. (1998) International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function. Pharmacol. Rev. 50, 291-313.

195
Ben Ari, Y., Tremblay, E., Riche, D., Ghilini, G. und Naquet, R. (1981) Electrographic, clinical and pathological alterations following systemic administration of kainic acid, bicuculline or pentetrazole: metabolic mapping using the deoxyglucose method with special reference to the pathology of epilepsy. Neuroscience 6, 1361-1391.
Ben-Ari, Y. und Cossart, R. (2000) Kainate, a double agent that generates seizures: two decades of progress. Trends Neurosci. 23, 580-587.
Ben-Ari, Y.,Tremblay, E., und Ottersen, O. P. (1980) Injections of kainic acid into the amygdaloid complex of the rat: an electrographic, clinical and histological study in relation to the pathology of epilepsy. Neuroscience 5, 515-528.
Benson, D. L., Huntsman, M. M. und Jones, E. G. (1994) Activity-dependent changes in GAD and preprotachykinin mRNAs in visual cortex of adult monkeys. Cereb. Cortex. 4, 40-51.
Berger, M. und Ben-Ari, Y. (1983) Autoradiographic visualisation of [3H]kainic acid receptor subtypes in the rat hippocampus. Neurosci. Lett. 39, 237-242.
Bloch, B., Guitteny, A. F., Chouham, S., Mougin, C., Roget, A. und Teoule, R. (1990) Topography and ontogeny of the neurons expressing vasopressin, oxytocin, and somatostatin genes in the rat brain: an analysis using radioactive and biotinylated oligonucleotides. Cell Mol. Neurobiol. 10, 99-112.
Bloms-Funke, P., Gernert, M., Ebert, U. und Löscher, W. (1999) Extracellular single-unit recordings of piriform cortex neurons in rats: influence of different types of anesthesia and characterization of neurons by pharmacological manipulation of serotonin receptors. J. Neurosci. Res. 55, 608-619.
Bonhaus, D. W., Walters, J. R. und McNamara, J. O. (1986) Activation of substantia nigra neurons: role in the propagation of seizures in kindled rats. J. Neurosci. 6, 3024-3030.
Bowers, G., Cullinan, W. E. und Herman, J. P. (1998) Region-specific regulation of glutamic acid decarboxylase (GAD) mRNA expression in central stress circuits. J. Neurosci. 18, 5938-5947.
Bowery, N. G. u. G. Nistico (1989) GABA: Basic aspects and clinical applications. Pythargora Press, Rom, Mailand.
Bradford, H. F. (1995) Glutamate, GABA and epilepsy. Prog.Neurobiol. 47, 477-511. Brooks-Kayal, A. R., Shumate, M. D., Jin, H., Rikhter, T. Y. und Coulter, D. A. (1998)
Selective changes in single cell GABA(A) receptor subunit expression and function in temporal lobe epilepsy. Nat. Med. 4, 1166-1172.
Buckmaster, P. S. und Jongen-Relo, A. L. (1999) Highly specific neuron loss preserves lateral inhibitory circuits in the dentate gyrus of kainate-induced epileptic rats. J. Neurosci. 19, 9519-9529.
Burchfield, J. L. und Applegate, C. D. (1989). Stepwise progression of kindling: perspectives from the kindling antagonism model. Neurosci. Biobehav. Rev. 13, 289-299.
Burnham, W. M. und Cottrell, G. A. (1990) The GABA hypothesis of kindling. In: J. A. Wada (Hrsg.): Kindling 4. Raven Press, New York.
Callahan, P. M., Paris, J. M., Cunningham, K. A. und Shinnick-Gallagher, P. (1991) Decrease of GABA-immunoreactive neurons in the amygdala after electrical kindling in the rat. Brain Res. 555, 335-339.
Cao, Y., Wilcox, K. S., Martin, C. E., Rachinsky, T. L., Eberwine, J. und Dichter, M. A. (1996) Presence of mRNA for glutamic acid decarboxylase in both excitatory and inhibitory neurons. Proc. Natl. Acad. Sci. U. S. A. 93, 9844-9849.
Cascino, G. D., Ring, S. R., King, P. J., Brown, R. H. und Chiappa, K. H. (1988) Evoked potentials in motor system diseases. Neurology 38, 231-238.

196
Cavalheiro, E. A., Riche, D. A. und Le Gal, La Salle (1982) Long-term effects of intrahippocampal kainic acid injection in rats: a method for inducing spontaneous recurrent seizures. Electroencephalogr. Clin. Neurophysiol. 53, 581-589.
Cavazos, J. E. und Sutula, T. P. (1990) Progressive neuronal loss induced by kindling: a possible mechanism for mossy fiber synaptic reorganization and hippocampal sclerosis. Brain Res. 527, 1-6.
Cavazos, J. E., Das, I. und Sutula, T. P. (1994) Neuronal loss induced in limbic pathways by kindling: evidence for induction of hippocampal sclerosis by repeated brief seizures. J. Neurosci. 14, 3106-3121.
Celio, M. R. (1990) Calbindin D-28k and parvalbumin in the rat nervous system. Neuroscience 35, 375-475.
Chesselet, M. F., Weiss, L., Wuenschell, C., Tobin, A. J. und Affolter, H. U. (1987) Comparative distribution of mRNAs for glutamic acid decarboxylase, tyrosine hydroxylase, and tachykinins in the basal ganglia: an in situ hybridization study in the rodent brain. J. Comp. Neurol. 262, 125-140.
Clifford, D. B., Olney, J. W., Maniotis, A., Collins, R. C. und Zorumski, C. F. (1987) The functional anatomy and pathology of lithium-pilocarpine and high- dose pilocarpine seizures. Neuroscience 23, 953-968.
Coons, A. H. und Kaplan, M. H. (1950) Localization of antigen in tissue cells. II. Improvements in a method for the detection of antigen by means of fluorescent antibody. J. Exp. Med. 91, 1-14.
Covolan, L. und Mello, L. E. (2000) Temporal profile of neuronal injury following pilocarpine or kainic acid-induced status epilepticus. Epilepsy Res. 39, 133-152.
Cox, K. H., Deleon, D. V., Angerer L. M. und Angerer, R. C. (1984) Detection of mRNA in sea urchin embryos by in situ−hybridization using asymmetric RNA probes. Dev. Biol. 101, 485−502.
Cronin, J. und Dudek, F. E. (1988) Chronic seizures and collateral sprouting of dentate mossy fibers after kainic acid treatment in rats. Brain Res. 474, 181-184.
Cuello, A. C., Milstein, C. und Priestley, J. V. (1980) Use of monoclonal antibodies in immunocytochemistry with special reference to the central nervous system. Brain Res. Bull. 5, 575-587.
DeLong, M. R. (1983) The neurophysiologic basis of abnormal movements in basal ganglia disorders. Neurobehav. Toxicol. Teratol. 5, 611-616.
Deniau, J. M. und Chevalier, G. (1985) Disinhibition as a basic process in the expression of striatal functions. II. The striato-nigral influence on thalamocortical cells of the ventromedial thalamic nucleus. Brain Res. 334, 227-233.
Depaulis, A., Vergnes, M. und Marescaux, C. (1994) Endogenous control of epilepsy: the nigral inhibitory system. Prog. Neurobiol. 42, 33-52.
Ding, R., Asada, H. und Obata, K. (1998) Changes in extracellular glutamate and GABA levels in the hippocampal CA3 and CA1 areas and the induction of glutamic acid decarboxylase-67 in dentate granule cells of rats treated with kainic acid. Brain Res. 800, 105-113.
Drengler, S. M. und Oltmans, G. A. (1993) Rapid increases in cerebellar Purkinje cell glutamic acid decarboxylase (GAD67) mRNA after lesion-induced increases in cell firing. Brain Res.615, 175-179.

197
Drengler, S. M., Lorden, J. F., Billitz, M. S. und Oltmans, G. A. (1996) Adrenergic agents inhibit rapid increases in cerebellar Purkinje cell glutamic acid decarboxylase (GAD67) mRNA levels after climbing fiber lesions or reserpine treatment. J. Neurosci.16, 1844-1851.
Ebert, U. und Löscher, W. (1995) Strong induction of c-fos in the piriform cortex during focal seizures evoked from different limbic brain sites. Brain Res. 671, 338-344.
Ebert, U. Schoen, S. W. und Löscher, W. (1996) Remote activation of microglia in the ventral posterior thalamic neuclei by kindling electrodes. Neuroforum, 2 (suppl.) H:155.
Ebert, U. und Löscher, W. (1995b) Strong induction of c-fos in the piriform cortex during focal seizures evoked from different limbic brain sites. Brain Res. 671, 338-344.
Ebert, U., Wlaz, P. und Löscher, W. (2000) High susceptibility of the anterior and posterior piriform cortex to induction of convulsions by bicuculline. Eur. J. Neurosci. 12, 4195-4205.
Ekstrand, J. J., Domroese, M. E., Feig, S. L., Illig, K. R. und Haberly, L. B. (2001) Immunocytochemical analysis of basket cells in rat piriform cortex. J. Comp. Neurol. 434, 308-328.
Emson, P. C. (1993) In-situ hybridization as a methodological tool for the neuroscientist. Trends Neurosci. 16, 9-16.
Engel, J. Jr., Wolfson, L. und Brown, L. (1978) Anatomical correlates of electrical and behavioral events related to amygdaloid kindling. Am. Neurol. 3, 538-544.
Erlander, M. G. und Tobin, A. J. (1991) The structural and functional heterogeneity of glutamic acid decarboxylase: a review. Neurochem. Res. 16, 215-226.
Erlander, M. G. und Tobin, A. J. (1991) The structural and functional heterogeneity of glutamic acid decarboxylase: a review. Neurochem. Res. 16, 215-226.
Erlander, M. G., Tillakaratne, N. J., Feldblum, S., Patel, N. und Tobin, A. J. (1991) Two genes encode distinct glutamate decarboxylases. Neuron 7, 91-100.
Esclapez, M. und Houser, C. R. (1999) Up-regulation of GAD65 and GAD67 in remaining hippocampal GABA neurons in a model of temporal lobe epilepsy. J. Comp. Neurol. 412, 488-505.
Esclapez, M., Tillakaratne, N. J. K.; Tobin, A.J.; Houser, C. R. (1993) Comparative localization of mRNAs encoding two forms of glutamic acid decarboxylase with nonradioactive in situ hybridization methods. J. Comp. Neurol. 331, 339−362.
Esclapez, M., Tillakaratne, N. J., Kaufman, D. L., Tobin, A. J. und Houser, C. R. (1994) Comparative localization of two forms of glutamic acid decarboxylase and their mRNAs in rat brain supports the concept of functional differences between the forms. J. Neurosci. 14, 1834-1855.
Falkenberg, T., Lindefors, N., O‘Connor, W. T., Zachrisson, O., Camilli, F. und Ungerstedt, U. (1997) GABA release and GAD67 mRNA expression in rat hippocampus following entorhinal cortex activation. Brain Res. Mol. Brain Res. 48, 413-416.
Fallon, J. H. und Loughlin, S. E. (1995) Substantia nigra. In: G. Paxinos (Hrsg): The Rat Nervous System, second eds., Academic Press, San Diego, S. 443-494.
Feldblum, S. und Ackermann, R. F. (1987) Increased susceptibility to hippocampal and amygdala kindling following intrahippocampal kainic acid. Exp. Neurol. 97, 255-269.
Feldblum, S., Ackermann, R. F. und Tobin, A. J. (1990) Long-term increase of glutamate decarboxylase mRNA in a rat model of temporal lobe epilepsy. Neuron 5, 361-371.

198
Feldblum, S., Erlander, M. G. und Tobin, A. J. (1993) Different distributions of GAD65 and GAD67 mRNAs suggest that the two glutamate decarboxylases play distinctive functional roles. J. Neurosci. Res. 34, 689-706.
Ferraguti, F., Zoli, M., Aronsson, M., Agnati, L. F., Goldstein, M., Filer, D. und Fuxe, K. (1990) Distribution of glutamic acid decarboxylase messenger RNA-containing nerve cell populations of the male rat brain. J. Chem. Neuroanat. 3, 377-396.
Flaherty, A. W. und Graybiel, A. M. (1994) Anatomy of the basal ganglia. In: Fahn, S. u. Marsden C. C. (Hrsg.): Movement disorders 3. Butterworth-Heinemann, Oxford, 3-27.
Fonnum, F. (1987) Biochemistry, anatomy, and pharmacology of GABA neurons. In: H. Y. Meltzer (Hrsg.): Psychopharmacology. The third generation of progress. Raven Press, New York, S. 173−182.
Foster, A. C., Mena, E. E., Monaghan, D. T. und Cotman, C. W. (1981) Synaptic localization of kainic acid binding sites. Nature 289, 73-75.
Fykse, E. M., Iversen, E. G. und Fonnum, F. (1992) Inhibition of L-glutamate uptake into synaptic vesicles. Neurosci. Lett. 135, 125-128.
Gale K und Piredda S (1986) Identification of a discrete forebrain site responsible for the genesis of chemically induced seizures. In: G. Nistico, P. L. Morselli, K. G. Lloyd, R. G. Fariello, J. J. Engel (Hrsg.):Neurotransmitters, Seizures, and Epilepsy III. Elsevier, Amsterdam, S. 229-234.
Gale, K. (1988) Progression and generalization of seizure discharge: anatomical and neurochemical substrates. Epilepsia 29 Suppl 2, S15-S34.
Gale, K. (1992) GABA and epilepsy: basic concepts from preclinical research. Epilepsia 33 Suppl 5, S3-12.
Gernert, M., Bloms-Funke, P., Ebert, U. und Löscher, W. (2000) Kindling causes persistent in vivo changes in firing rates and glutamate sensitivity of central piriform cortex neurons in rats. Neuroscience 99, 217-227.
Gibb, J. W. und Kogan, F. J. (1979) Influence of dopamine synthesis on methamphetamine-induced changes in striatal and adrenal tyrosine hydroxylase activity. Naunyn Schmiedebergs Arch. Pharmacol. 310, 185-187.
Glass, M. und Dragunow, M. (1995) Neurochemical and morphological changes associated with human epilepsy. Brain Res. Brain Res. Rev. 21, 29-41.
Goddard, G. V. (1972): Long term alteration following amygdaloid stimulation. in : B. Eleftheriou (Hrsg.): The neurobiology of the amygdala. Plenum Press, New York, S. 581-596.
Goddard, G. V. und McIntyre, D. (1974) Some properties of a lasting epileptogenic trace kindled by repeated electrical stimulation of the amygdala in mammals. In: L. V. Laitinent und K. E. Livingstone (Hrsg): Surgical approaches in psychiatry. University Park Press, Baltimore, S. 109-115.
Goddard, G. V., McIntyre, D. und Leech, E. (1969) A permanent change in brain funktion resulting from daily electrical stimulation. Exp. Neurol. 25, 295−330.
Goodman, J. H. (1998) Experimental models of status epilepticus. In: S. L. Peterson und T. E. Albertson (Hrsg.) Neuropharmacology methods in epilepsy research. CRC Press, Boca Raton, S. 95-125.
Greif, K. F., Erlander, M. G., Tillakaratne, N. J. und Tobin, A. J. (1991) Postnatal expression of glutamate decarboxylases in developing rat cerebellum. Neurochem. Res. 16, 235-242.

199
Greif, K. F., Tillakaratne, N. J., Erlander, M. G., Feldblum, S. und Tobin, A. J. (1992) Transient increase in expression of a glutamate decarboxylase (GAD) mRNA during the postnatal development of the rat striatum. Dev. Biol. 153, 158-164.
Haberly, L. B. (1983) Structure of the piriform cortex of the opossum. I. Description of neuron types with golgi methods. J. Comp. Neurol. 213, 163-187.
Haberly, L. B. (1990) Olfactory cortex. In: G. M. Shepherd (Hrsg.): The Synaptic Organization of the Brain. Oxford University Press, New York, S. 317-345.
Haberly, L. B. und Price, J. L. (1978) Association and commissural fiber system of the olfactory cortex of the rat. I. Systems originating in the piriform cortex and adjacent areas. J. Comp. Neurol. 178, 711-740.
Haberly, L. B. und Behan, M. (1983) Structure of the piriform cortex of the opossum. III. Ultrastructural characterization of synaptic terminals of association and olfactory bulb afferent fibers. J. Comp. Neurol., 219, 448−460.
Haberly, L. B. und Bower, J. M. (1984) Analysis of association fiber system in piriform cortex with intracellular recording and staining techniques. J. Neurophysiol. 51, 90-112.
Haberly, L. B. und Feig, S. L. (1983) Structure of the piriform cortex of the opossum. II. Fine structure of cell bodies and neuropil. J. Comp. Neurol. 216, 69-88.
Haberly, L. B. und Presto, S. (1986) Ultrastructural analysis of synaptic relationships of intracellulärly stained pyramidal cell axons in piriform cortex. J. Comp. Neurol. 248, 464−474.
Haberly, L. B., Hansen, D. J., Feig, S. L. und Presto, S. (1987) Distribution and ultrastructure of neurons in opossum piriform cortex displaying immunoreactivity to GABA and GAD and high-affinity tritiated GABA uptake. J. Comp. Neurol. 266, 269-290.
Harlow, E. und Lane, D. P. (1988) Antibodies-a laboratory manual. 13. Aufl. C. Spring Habor Lab., New York.
Harrison, P. R., Conkie, D., Affara, N. und Paul, J. (1974) In situ localization of globin messenger RNAformation I. during mouse fetal liver development. J. Cell Biol. 63, 401−413.
Hendrickson, A. E., Tillakaratne, N. J., Mehra, R. D., Esclapez, M., Erickson, A., Vician, L. und Tobin, A. J. (1994) Differential localization of two glutamic acid decarboxylases (GAD65 and GAD67) in adult monkey visual cortex. J. Comp. Neurol. 343, 566-581.
Hendry, S. H., Jones, E. G., Emson, P. C., Lawson, D. E., Heizmann, C. W. und Streit, P. (1989) Two classes of cortical GABA neurons defined by differential calcium binding protein immunoreactivities. Exp. Brain. Res 76, 467-472.
Hendry, S. H., Schwark, H. D., Jones, E. G. und Yan, J. (1987) Numbers and proportions of GABA-immunoreactive neurons in different areas of monkey cerebral cortex. J. Neurosci. 7, 1503-1519.
Hikosaka, O. und Wurtz, R. H. (1983) Effects on eye movements of a GABA agonist and antagonist injected into monkey superior colliculus. Brain Res. 272, 368-372.
Hönack, D., Wahnschaffe, U. und Löscher, W. (1991) Kindling from stimulation of a highly sensitive locus in the posterior part of the piriform cortex. Comparison with amygdala kindling and effects of antiepileptic drugs. Brain Res. 538, 196-202.
Honchar, M. P., Olney, J. W. und Sherman, W. R. (1983) Systemic cholinergic agents induce seizures and brain damage in lithium-treated rats. Science 220, 323-325.
Houser, C. R. (1991) GABA neurons in seizure disorders: a review of immunocytochemical studies. Neurochem. Res. 16, 295-308.

200
Houser, C. R. und Esclapez, M. (1994) Localization of mRNAs encoding two forms of glutamic acid decarboxylase in the rat hippocampal formation. Hippocampus. 4, 530-545.
Houser, C. R. und Esclapez, M. (1996) Vulnerability and plasticity of the GABA system in the pilocarpine model of spontaneous recurrent seizures. Epilepsy Res. 26, 207-218.
Houser, C. R., Miyashiro, J. E., Swartz, B. E., Walsh, G. O., Rich, J. R. und Delgado-Escueta, A. V. (1990) Altered patterns of dynorphin immunoreactivity suggest mossy fiber reorganization in human hippocampal epilepsy. J. Neurosci. 10, 267-282.
Hsu, S.-M., Raine, L. und fanger, H. (1981) Use of avidin-biotin-peroxidase complex (ABC) immunoperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. 29, 577-580.
INTERNATIONAL LEAGUE AGAINST EPILEPSY/ COMISSION ON CLASSIFICATION AND TERMINOLOGY (1989): Proposal for revised classification of epilepsies and epileptic syndroms. Epilepsia 30, 389-399.
INTERNATIONAL LEAGUE AGAINST EPILEPSY/ COMISSION ON CLASSIFICATION AND TERMINOLOGY (1981):Proposal for revised clinical and electroencephalographic classification of epileptic seizures. Epilepsia 22, 489-501.
Itagaki, S. und Kimura, H. (1986) Retardation of resynthesis of GABA-transaminase in some brain regions of amygdala-kindled rats. Brain Res. 381, 77-84.
Janz, D. (1979) Epidemiologie und Klassifikation von Epilepsien und epileptischen Anfällen. Akt. Neurol. 6, 189-196.
Julien, J. F., Samama, P. und Mallet, J. (1990) Rat brain glutamic acid decarboxylasesequence deduced from a cloned cDNA. J. Neurochem. 54, 703-705.
Kamphuis, W., Huisman, E., Wadman, W. J., Heizmann, C. W. und Lopes, da Silva FH (1989) Kindling induced changes in parvalbumin immunoreactivity in rat hippocampus and its relation to long-term decrease in GABA-immunoreactivity. Brain Res. 479, 23-34.
Kamphuis, W., Wadman, W. J., Buijs, R. M. und Lopes, da Silva FH (1987) The development of changes in hippocampal GABA immunoreactivity in the rat kindling model of epilepsy: a light microscopic study with GABA antibodies. Neuroscience 23, 433-446.
Kamphuis, W., Wadman, W. J., Buijs, R. M. und Lopes, da Silva FH (1986) Decrease in number of hippocampal gamma-aminobutyric acid (GABA) immunoreactive cells in the rat kindling model of epilepsy. Exp. Brain Res. 64, 491-495.
Kaufman, D. L., Houser, C. R. und Tobin, A. J. (1991) Two forms of the gamma-aminobutyric acid synthetic enzyme glutamate decarboxylase have distinct intraneuronal distributions and cofactor interactions. J. Neurochem. 56, 720-723.
Kaufman, D. L., McGinnis, J. F., Krieger, N. R. und Tobin A. J. (1986) Brain glutamate decarboxylase cloned in λgt−11: fusion protein produce γ−aminobutyric acid. Science 232, 1138−1140.
Kaura, S., Bradford, H. F., Young, A. M., Croucher, M. J. und Hughes, P. D. (1995) Effect of amygdaloid kindling on the content and release of amino acids from the amygdaloid complex: in vivo and in vitro studies. J. Neurochem. 65, 1240-1249.
Keränen, T. und Riekkinen, P. (1988): Severe Epilepsy: diagnostic and epidemiological aspects. Acta. Neurol. Scand. 57, 7-14.
Kessler, C., Holtke, H. J., Seibl, R., Burg, J. und Muhlegger, K. (1990) Non-radioactive labeling and detection of nucleic acids. I. A novel DNA labeling and detection system based on digoxigenin: anti-digoxigenin ELISA principle (digoxigenin system). Biol. Chem. Hoppe Seyler 371, 917-927.

201
Kiyama, H., Emson, P. C. und Tohyama, M. (1990) Recent progress in the use of the technique of non-radioactive in situ hybridization histochemistry: new tools for molecular neurobiology. Neurosci. Res. 9, 1-21.
Kiyama, H., Emson, P. C., Ruth, J. und Morgan, C. (1990) Sensitive non-radioisotopic in situ hybridization histochemistry: demonstration of tyrosine hydroxylase gene expression in rat brain and adrenal. Brain Res. Mol. Brain Res. 7, 213-219.
Kosaka, T., Kosaka, K., Heizmann, C. W., Nagatsu, I, Wu, J. Y., Yanaihara, N. und Hama, K. (1987) An aspect of the organization of the GABAergic system in the rat main olfactory bulb: laminar distribution of immunohistochemically defined subpopulations of GABAergic neurons. Brain Res. 411, 373-378.
Krettek, J. E. und Price, J. L. (1977b) Projections from the amygdaloid complex to the cerebral cortex and thalamus in the rat and cat. J. Comp. Neurol. 172, 687-722.
Krettek, J. E. und Price, J. L. (1977c) Projections from the amygdaloid complex and adjacent olfactory structures to the entorhinal cortex and to the subiculum in the rat and cat. J. Comp. Neurol. 172, 687-722.
Krettek, J. E. und Price, J. L. (1978a) Amygdaloid projections to subcortical structures within the basal forebrain and brainstem in the rat and cat. J. Comp. Neurol. 178, 225-254.
Krettek, J. E. und Price, J. L. (1978b) A description of the amygdaloid complex in the rat and cat with observations on intra-amygdaloid axonal connections. J. Comp. Neurol. 178, 255-280.
Larsson, L. I. und Hougaard, D. M. (1993) Sensitive detection of rat gastrin mRNA by in situ hybridization with chemically biotinylated oligodeoxynucleotides: validation, quantitation, and double-staining studies. J. Histochem. Cytochem. 41, 157-163.
Larsson, L. I. und Hougaard, D. M. (1994) Glass slide models for immunocytochemistry and in situ hybridization. Histochemistry 101, 325-331.
Larsson, L. I., Traasdahl, B. und Hougaard, D. M. (1991) Quantitative non-radioactive in situ hybridization. Model studies and studies on pituitary proopiomelanocortin cells after adrenalectomy. Histochemistry 95, 209-215.
Lehmann, H., Ebert, U. und Löscher, W. (1998) Amygdala-kindling induces a lasting reduction of GABA-immunoreactive neurons in a discrete area of the ipsilateral piriform cortex. Synapse 29, 299−309.
Lehmann, H., Ebert, U. und Löscher, W. (1996) Immunocytochemical localization of GABA immunoreactivity in dentate granule cells of normal and kindled rats. Neurosci. Lett. 212, 41-44.
Leitch, A. R., Schwarzacher, T., Jackson, D. und Leitch, I. J.(1994) In situ−hybridisation. Spektrum Akademischer Verlag: Heidelberg.
Lemos, T. und Cavalheiro, E. A. (1995) Suppression of pilocarpine-induced status epilepticus and the late development of epilepsy in rats. Exp. Brain Res. 102, 423-428.
Leppik, I. E. (1992) Intractable epilepsy in adults. Epilepsy Res. Suppl. 5, 7-11. Litwak, J., Mercugliano, M., Chesselet, M. F. und Oltmans, G. A. (1990) Increased glutamic
acid decarboxylase (GAD) mRNA and GAD activity in cerebellar Purkinje cells following lesion-induced increases in cell firing. Neurosci. Lett. 116, 179-183.
Löscher, W. (1994) Neue Antiepileptica - ein Fortschritt für die Behandlung epileptischer Tiere ? Kleintierpraxis 39, 325-323.
Löscher, W. (1989) GABA and the epilepsies. Experimental and clinical considerations. In: N. G. Bowery und G. Nistico (Hrsg.): GABA. Basic aspects and clinical applications. Phythagora Press, Rom. S. 260-300.

202
Löscher, W. (1993) Basic aspects of epilepsy. Curr. Opin. Neurol. Neurosurg. 6, 223-232. Löscher, W. und Ebert, U. (1996a) Basic mechanisms of seizure propagation: targets for
rational drug design and rational polypharmacy. Epilepsy Res. Suppl. 11, 17-43. Löscher, W. und Ebert, U. (1996b) The role of the piriform cortex in kindling. Prog. Neurobiol.
50, 427-481. Löscher, W. und Schmidt, D. (1988) Which animal models should be used in the search for
new antiepileptic drugs ? A proposal based on the experimental and clinical considerations. Epilepsy Res. 2, 145-181.
Löscher, W. und Schmidt, D. (1993) New drugs for the treatment of epilepsy. Curr. Opin. Invest. Drugs 2, 1067-1095.
Löscher, W. und Schmidt, D. (1994) Strategies in antiepileptic drug development: is rational drug design superior to random screening and structural variations?. Epilepsy Res. 17, 95-134.
Löscher, W. und Schwark, W. S. (1985) Evaluation of different GABA receptor agonists in the kindled amygdala seizure model in rats. Exp. Neurol. 89, 454-460.
Löscher, W. und Schwark, W. S. (1987) Further evidence for abnormal GABAergic circuits in amygdala-kindled rats. Brain Res. 420, 385-390.
Löscher, W., Czuczwar, S. J., Jackel, R. und Schwarz, M. (1987) Effect of microinjections of gamma-vinyl GABA or isoniazid into substantia nigra on the development of amygdala kindling in rats. Exp. Neurol. 95, 622-638.
Löscher, W., Hönack, D. und Gramer, M. (1989) Use of inhibitors of gamma-aminobutyric acid (GABA) transaminase for the estimation of GABA turnover in various brain regions of rats: a reevaluation of aminooxyacetic acid. J. Neurochem. 53, 1737-1750.
Löscher, W., Hönack, D. und Gramer, M. (1999) Effect of depth electrode implantation with or without subsequent kindling on GABA turnover in various rat brain regions. Epilepsy Res. 37, 95-108.
Löscher, W., Lehmann, H. und Ebert, U. (1998) Differences in the distribution of GABA- and GAD-immunoreactive neurons in the anterior and posterior piriform cortex of rats. Brain Res. 800, 21-31.
Löscher, W., Wahnschaffe, U. Hönack, D. und Rundfeldt, C. (1995) Susceptibility of different cell layers of the anterior and posterior part of the piriform cortex to electrical stimulation and kindling: comparison with the basolateral amygdala and „area tempestas“. Neuroscience 66, 265-276.
Lothman, E. W. (1992) Basic mechanisms of the epilepsies. Curr. Opin. Neurol. Neurosurg. 5, 216-223.
Lothman, E. W., Bertram, E. H., III and Stringer, J. L. (1991) Functional anatomy of hippocampal seizures. Prog. Neurobiol. 37, 1-82.
Luskin, M. B. und Price, J. L. (1983) The topographic organization of associational fibers of the olfactory system in the rat, including centrifugal fibers to the olfactory bulb. J. Comp. Neurol. 216, 264-291.
Marksteiner, J., Prommegger, R. und Sperk, G. (1990) Effect of anticonvulsant treatment on kainic acid-induced increases in peptide levels. Eur. J. Pharmacol. 181, 241-246.
Martin, D. L. (1993) Short-term control of GABA synthesis in brain. Prog. Biophys. Mol. Biol. 60, 17-28.
Martin, D. L. und Rimvall, K. (1993) Regulation of gamma-aminobutyric acid synthesis in the brain. J. Neurochem. 60, 395-407.

203
Martin, D. L., Martin, S. B., Wu, S. J. und Espina, N. (1991b) Regulatory properties of brain glutamate decarboxylase (GAD): the apoenzyme of GAD is present principally as the smaller of two molecular forms of GAD in brain. J. Neurosci. 11, 2725-2731.
Martin, D. L., Martin, S. B., Wu, S. J. und Espina, N. (1991a) Cofactor interactions and the regulation of glutamate decarboxylase activity. Neurochem. Res. 16, 243-249.
Martinez, M. C., Blanco, J., Bullon, M. M. und Aguido, F. J. (1987) Structure of the piriform cortex of the adult rat. A Golgi study. J. Hirnforsch. 28, 341-348.
Matthes, A. u. Schneble, H. (1992) Epilepsien: Diagnostik und Therapie für Klinik und Praxis. 5. Auflage, Thieme, Stuttgart.
McDonald, A. J. (1982) Neurons of the lateral and basolateral amygdaloid nuclei: a Golgi study in the rat. J. Comp Neurol. 212, 293-312.
McDonald, A. J. (1985) Immunohistochemical identification of gamma-aminobutyric acid- containing neurons in the rat basolateral amygdala. Neurosci. Lett. 53, 203-207.
McDonald, A. J. und Pearson, J. C. (1989) Coexistence of GABA and peptide immunoreactivity in non-pyramidal neurons of the basolateral amygdala. Neurosci. Lett. 100, 53-58.
Meldrum, B. S., Swan, J. H., Ottersen, O. P. und Storm-Mathisen, J. (1987) Redistribution of transmitter amino acids in rat hippocampus and cerebellum during seizures induced by L-allylglycine and bicuculline: an immunocytochemical study with antisera against conjugated GABA, glutamate and aspartate. Neuroscience 22, 17-27.
Mello, L. E. und Covolan, L. (1996) Spontaneous seizures preferentially injure interneurons in the pilocarpine model of chronic spontaneous seizures. Epilepsy Res. 26, 123-129.
Mello, L. E., Cavalheiro, E. A., Tan, A. M., Kupfer, W. R., Pretorius, J. K., Babb, T. L. und Finch, D. M. (1993) Circuit mechanisms of seizures in the pilocarpine model of chronic epilepsy: cell loss and mossy fiber sprouting. Epilepsia 34, 985-995.
Mercugliano, M., Soghomonian, J. J., Qin, Y., Nguyen, H. Q., Feldblum, S., Erlander, M. G., Tobin, A. J. und Chesselet, M. F. (1992) Comparative distribution of messenger RNAs encoding glutamic acid decarboxylases (Mr 65,000 and Mr 67,000) in the basal ganglia of the rat. J. Comp Neurol. 318, 245-254.
Mugnaini, E. und Oertel, W. H. (1985) An atlas of the distribution of GABAergic neurons and terminals in the rat CNS as revealed by GAD immunhisto-chemistry. In: A. Björklund und T. Hökfeld (Hrsg.):Handbook of Chemical Neuroanatomy, Vol. 4: GABA and neuropeptides in the CNS Part I. Elsevier, Amsterdam, S. 436-608.
Nadler, J. V. (1981) Minireview. Kainic acid as a tool for the study of temporal lobe epilepsy. Life Sci. 29, 2031-2042.
Najlerahim, A., Harrison, P. J., Barton, A. J., Heffernan, J. und Pearson, R. C. (1990) Distribution of messenger RNAs encoding the enzymes glutaminase, aspartate aminotransferase and glutamic acid decarboxylase in rat brain. Brain Res. Mol. Brain Res. 7, 317-333.
Nusser, Z., Hajos, N., Somogyi, P. und Mody, I. (1998) Increased number of synaptic GABA(A) receptors underlies potentiation at hippocampal inhibitory synapses. Nature 395, 172-177.
Obenaus, A., Esclapez, M. und Houser, C. R. (1993) Loss of glutamate decarboxylase mRNA-containing neurons in the rat dentate gyrus following pilocarpine-induced seizures. J. Neurosci. 13, 4470-4485.

204
Oertel, W. H. und Mugnaini, E. (1984) Immunocytochemical studies of GABAergic neurons in rat basal ganglia and their relations to other neuronal systems. Neurosci. Lett. 47, 233-238.
Parent, A. und Hazrati, L. ( 1995) Functional anatomy of the basal ganglia. I.The cortico-basal-ganglia-thalamo-cortical loop. Brain Res. Rev. 20, 91-127.
Paxinos G und Watson C (1998) The Rat Brain in Stereotaxic Coordinates. Academic Press: Sydney.West, M. J. (1999) Stereological methods for estimating the total number of neurons and synapses: issues of precision and bias. Trends Neurosci. 22, 51-61.
Paxinos, G. (1990) The Human Nervous System. Academic Press, New York. Paxinos, G. und Watson, C. (1986) The Rat Brain in Stereotaxic Coordinates. Academic
Press, Sydney. Pfeifer, J. (1993) Neuronale Schäden durch Epilepsien. Thieme, Stuttgart. Piredda, S. und Gale, K. (1985) A crucial epileptogenic site in the deep prepiriform cortex.
Nature 317, 623-625. Price, J. L. (1973) An autoradiographic study of complementary laminar patterns of
termination of afferent fibers to the olfactory cortex. J. Comp. Neurol. 150, 87-108. Racine, R. J. (1972) Modification of seizure activity by electrical stimulation: Motor seizure.
Electrenceph. Clin. Neurophysiol. 32, 281-294. Racine, R. J., Ivy, G. O. und Milgram, N. W. (1989) Kindling: clinical relevance and
anatomical substrate. In: T. G. Bolwig und M. R. Trimble (Hrsg.): The Clinical Relevance of Kindling. John Wiley and Sons, Chichester, S. 15-34.
Racine, R. J., Ivy, G. O. und Milgram (1989). Kindling: clinical relevance and anatomical substrate. in: Bowling, Y. G. und Trimble, M. E. (Hrsg.): The clinical relevance of kindling. Wiley & sons, Chichester, S. 15-34.
Racine, R. J., Mosher, J. M. und Kairiss, E. W. (1988) The role of the piriform cortex in the generation of interiktal spikes in the kindled preparation. Brain Res. 454, 264−274.
Rao, M. S., Sun, Y., Vaidyanathan, U., Landis, S. C. und Zigmond, R. E. (1993) Regulation of substance P is similar to that of vasoactive intestinal peptide after axotomy or explantation of the rat superior cervical ganglion. J. Neurobiol. 24, 571-580.
Ribak, C. E., Vaughn, J. E. und Roberts, E. (1980) GABAergic nerve terminals decrease in the substantia nigra following hemitransections of the striatonigral and pallidonigral pathways. Brain Res. 192, 413-420.
Ribak, C. E., Vaughn, J. E. und Saito, K. (1978) Immunocytochemical localization of glutamic acid decarboxylase in neuronal somata following colchicine inhibition of axonal transport. Brain Res. 140, 315-332.
Rimvall, K. und Martin, D. L. (1992) Increased intracellular gamma-aminobutyric acid selectively lowers the level of the larger of two glutamate decarboxylase proteins in cultured GABAergic neurons from rat cerebral cortex. J. Neurochem. 58, 158-166.
Rimvall, K. und Martin, D. L. (1994) The level of GAD67 protein is highly sensitive to small increases in intraneuronal gamma-aminobutyric acid levels. J. Neurochem. 62, 1375-1381.
Rimvall, K., Sheikh, S. N. und Martin, D. L. (1993) Effects of increased gamma-aminobutyric acid levels on GAD67 protein and mRNA levels in rat cerebral cortex. J. Neurochem. 60, 714-720.

205
Roberts, E. (1976) Imuncytochemistry of the GABA system- a novel approach to an old neurotransmitter. In: J. A. Ferrendelli, B. S. Mc Ewen und S. H. Snyder (Hrsg.): Neurotransmitters, hormones andreceptors, novel approaches. Neuroscience Symposia 1, Soc. Neurosci., Bethesda, S. 123-138.
Saito, K., Barber, R., Wu, J., Matsuda, T., Roberts, E. und Vaughn, J. E. (1974) Immunohistochemical localization of glutamate decarboxylase in rat cerebellum. Proc. Natl. Acad. Sci. U. S. A. 71, 269-273.
Salganikoff, K. und DeRobertis, E. (1965) Subcellular ditribution of the enzyme of the glutamic acid, glutamine and gammaaminobutyric acid cycle in the rat brains. J. Neurochem. 12, 287−309.
Saper, C. B. (1996) Any way you cut it: a new journal policy for the use of unbiased counting methods. J. Comp. Neurol. 364, 5.
Sato, M., Racine, R. J. und McIntyre, D. C. (1990) Kindling: basic mechanisms and clinical validity. Electroenceph. Clin. Neurophysiol. 76, 459-472.
Schmidt-Kastner, R., Heim, C. und Sontag, K. H. (1991) Damage of substantia nigra pars reticulata during pilocarpine-induced status epilepticus in the rat: immunohistochemical study of neurons, astrocytes and serum-protein extravasation. Exp. Brain Res. 86, 125-140.
Schwabe, K., Ebert, U. und Löscher, W. (1999) Microinjections of vigabatrin in the central piriform cortex retard amygdala kindling in rats. Epilepsia 40 Suppl. 7 9-10.
Schwabe, K., Ebert, U. und Löscher, W. (2000a) Bilateral lesions of the central but not anterior or posterior parts of the piriform cortex retard amygdala kindling in rats. Neuroscience 101, 513-521.
Schwabe, K., Ebert, U. und Löscher, W. (2000b) Bilateral microinjection of vigabatrin into the central piriform cortex is antiepileptogenic and anticonvulsant in amygdala kindling of rats. Europ. J. Neurosci. 12 Suppl. 11, 210.
Schwarzer, C. und Sperk, G. (1995) Hippocampal granule cells express glutamic acid decarboxylase-67 after limbic seizures in the rat. Neuroscience 69, 705-709.
Shipley, M. T., McLean, J. H. und Ennis, M. (1995) The olfactory system. In: G. Paxinos (Hrsg.) The Rat Nervous system, Vol. 2. Academic Press, San Diego, S. 899-928.
Sloviter, R. S. (1987) Decreased hippocampal inhibition and selective loss of interneurons in experimental epilepsy. Science 235, 73-76.
Sloviter, R. S., Dichter, M. A., Rachinsky, T. L., Dean, E., Goodman, J. H., Sollas, A. L. und Martin, D. L. (1996) Basal expression and induction of glutamate decarboxylase and GABA in excitatory granule cells of the rat and monkey hippocampal dentate gyrus. J. Comp. Neurol. 373, 593-618.
Soghomonian, J. J. und Martin, D. L. (1998) Two isoforms of glutamate decarboxylase: why? Trends Pharmacol. Sci. 19, 500-505.
Sommer, W. (1880) Erkrankung des Ammonshornes als ätiologisches Moment in der Epilepsie. Arch. Psychiatrie Nervenkrankh. 10, 613-675.
Somogyi, P., Hodgson, A. J., Smith, A. D., Nunzi, M. G., Gorio, A. und Wu, J. Y. (1984) Different populations of GABAergic neurons in the visual cortex and hippocampus of cat contain somatost. J. Neurosci. 4, 2590-2603.
Sonnenberg, J. L., Frantz, G. D., Lee, S., Heick, A., Chu, C., Tobin, A. J. und Christakos, S. (1991) Calcium binding protein (calbindin-D28k) and glutamate decarboxylase gene expression after kindling induced seizures. Brain Res. Mol. Brain Res 9, 179-190.

206
Soriano, E., Cobas, A., Fairen, A. (1989) Neurogenesis of glutamic acid decarboxylase immunoreaktive cells in the hippocampus of the mouse. I: regio superior und inferior. J. Comp. Neurol. 281, 586−602.
Sperk, G. (1994) Kainic acid seizures in the rat. Prog. Neurobiol. 42, 1-32. Sperk, G., Lassmann, H., Baran, H., Kish, S. J., Seitelberger, F. und Hornykiewicz, O. (1983)
Kainic acid induced seizures: neurochemical and histopathological changes. Neuroscience 10, 1301-1315.
Spreafico, R., De Biasi, S., Frassoni, C. und Battaglia, G. (1988) A comparison of GAD- and GABA-immunoreactive neurons in the first somatosensory area (SI) of the rat cortex. Brain Res. 474, 192-196.
Stark, L. G. (1992) Kindling: amygdaloid neuronal networks and actions of convulsant and antiepileptic drugs. In: C. L. Faingold und G. H. Fromm (Hrsg.): Drugs for control of Epilepsy: Actions on Neuronal Networks Involved in Seizure Disorders. CRC Press, Boca Raton, S. 195-212.
Starke, K. (1996) Grundlagen der Pharmakologie des Nervensystems. In: W. Forth, D. Henschler, W. Rummel und K. Starke (Hrsg.) Allgemeine und spezielle Pharmakologie und Toxikologie. Spektrum Akademischer Verlag,Mannheim.
Sutula, T. P. (1991) Reactive changes in epilepsy: cell death and axon sprouting induced by kindling. Epilepsy Res. 10, 62-70.
Switzer, R. C., DeOlmos, J. und Heimer (1985) Olfactory system. In: G. Paxinos (Hrsg):The Rat Nervous System. Academic Press, San Diego, S. 1-36.
Tseng, G. F. und Haberly, L. B. (1989) Characterization of synaptically mediated fast and slow inhibitory processes in piriform cortex in an in vitro slice preparation. J. Neurophysiol. 59, 1352-1376.
Turski, L. (1989) Die konvulsive Wirkung von Pilocarpin bei Ratten: Pharmakologische, elektroenzephalographische und morphologische Untersuchung zur Bedeutung zentraler cholinerger Mechanismen für die Krampfgenese. Hrissanthi Ikonomidou, Berlin.
Turski, L., Cavalheiro, E. A., Sieklucka-Dziuba, M., Ikonomidou-Turski, C., Czuczwar, S. J. und Turski, W. A. (1986) Seizures produced by pilocarpine: neuropathological sequelae and activity of glutamate decarboxylase in the rat forebrain. Brain Res. 398, 37-48.
Turski, W. A., Cavalheiro, E. A., Bortolotto, Z. A., Mello, L. M., Schwarz, M. und Turski, L. (1984) Seizures produced by pilocarpine in mice: a behavioral, electroencephalographic and morphological analysis. Brain Res. 321, 237-253.
Tuunanen, J. und Pitkänen, A. (2000) Do seizures cause neuronal damage in rat Amygdala kindling? Epilepsy Res. 39, 171-176.
Tuunanen, J., Halonen, T. und Pitkänen, A. (1997) Decrease in somatostatin-immunoreactive neurons in the rat amygdaloid complex in a kindling model of temporal lobe epilepsy. Epilepsy Res. 26, 315-327.
US Department of Health, Education and Welfare (1977) Plan for nationwide action on epilepsy. D.H.E.W. Publ.No.(NIH). 1 und 4, 78-276.
Veening, J. G. (1978) Cortical afferents of the amygdaloid complex in the rat. Neurosci. Lett. 8, 191-195.
W. A. (1986) Seizures produced by pilocarpine: neuropathological sequelae and activity of glutamate decarboxylase in the rat forebrain. Brain Res. 398, 37-48.

207
Wahle, P. und Beckh, S. (1992) A method of in situ hybridization combined with immunocytochemistry, histochemistry, and tract tracing to characterize the mRNA expressing cell types in heterogeneous neuronal populations. J. Neurosci. Methods 41, 153-166.
Wahnschaffe, U. und Löscher, W. (1990a) Selective bilateral destruction of substantia nigra has no effect on kindled seizures induced from stimulation of amygdala or piriform cortex in rats. Neurosci. Lett. 113, 205-210.
Wahnschaffe, U. und Löscher, W. (1990b)Effect of selective bilateral destruction of the substantia nigra on antiepileptic drug actions in kindled rats. Eur. J. Pharmacol. 186, 157-167.
Wahnschaffe, U., Ebert, U. und Löscher, W. (1993) The effect of lesions of the posterior piriform cortex on amygdala kindling in the rat. Brain Res. 615, 295-303.
Wakefield, C. (1980) The topographical organization and laminar origin of some cortico-amygdaloid connections. Neurosci. Lett. 20, 21-24.
Waszczak, J. V., Applegate, C. D. und Burchfield, J. L. (1988) Kindling does not cause persistent changes in firing rates or transmitter sensitivities of substantia nigra pars reticulata neurons. Brain Res. 455, 115-122.
Watanake, K. und Kawana, E. (1984) Selective retrograde transport of striated D−aspartate from the olfactory to the anterior olfactory nulceus, piriform cortex and nuclei of the lateral olfactory tract in the rat. Brain Res., 296, 148−151.
Wilson, C. J. (1990) Basal ganglia. In: G. M. Sheperd (Hrsg.): The synaptic Organization of the brain. Oxford University Press, New York ,S. 279-316.
Wolf, P. (Hrsg.) (1994) Epileptic Seizures and Syndromes. John Libbey, London.
Woodson, W., Nitecka, L. und Ben−Ari, Y. (1989) Organization of the GABAergic system in the rat hippocampal formation: a quantitative immunocytochemical study. J. Comp. Neurol. 280, 254−271.
Yagi, T. und Obata, K. (1997) Cleft palate and decreased brain gamma-aminobutyric acid in mice lacking the 67-kDa isoform of glutamic acid decarboxylase. Proc. Natl. Acad. Sci. U.S.A. 94, 6496-6499.

208
Publikationsliste
Potschka, H., C. Freichel, and W. Löscher (2000)
Study of ATP-dependent transport pf phenytoin at the blood brain barrier.
Nauny-Schmiedeberg´s Arch. of Pharmacology. 361 (Suppl. 4), 368.
Freichel, C., H. Potschka, and W. Löscher (2000)
Basal expression and seizure-induced alteration of glutamate decarboxylae isoforms
in limbic brain regions of the rat.
Nauny-Schmiedeberg´s Arch. of Pharmacology. 362 (Suppl. 4-5 ), 7.
Freichel, C., H. Potschka, and W. Löscher (2000)
Seizure-induced alteration of glutamate decarboxylae isoforms in different animal
models of temporal lobe epilepsy.
Nauny-Schmiedeberg´s Arch. of Pharmacology. 363 (Suppl. 4), 359.

209
8 Anhang
8.1 Substanzen
8.1.1 Substanzen für die in-situ-Hybridisierung
Substanz Bezugsquelle (Fa.)
Bovines Serum Albumin Sigma, Deisenhofen
Chloroform Fluka, Buchs, CH
Cresylviolettacetat Fluka, Buchs, CH
DEPC Roth, Karlsruhe
Dextransulfat Sigma, Deisenhofen
DTT Sigma, Deisenhofen
EDTA Sigma, Deisenhofen
Essigsäureanhydrid Fluka, Buchs, CH
Ficoll Pharmacia-Biotech, Uppsala, Schweden
Formamid Sigma, Deisenhofen
Glycine Sigma, Deisenhofen
KH2PO4 Merck, Darmstadt
Levamisol Sigma, Deisenhofen
Maleinsäure Sigma, Deisenhofen
Na2HPO4 Merck, Darmstadt
NaCl Merck, Darmstadt
NaH2PO4 Merck, Darmstadt

210
Substanz Bezugsquelle (Fa.)
Natriumcitrat Merck, Darmstadt
Natrium-Thiosulfat Sigma, Deisenhofen
Paraform Fluka, Buchs, CH
Pipes Sigma, Deisenhofen
Poly A Roche Diagnostics, Mannheim
Polyvinylpyrrolidon Sigma, Deisenhofen
RNAse A Sigma, Deisenhofen
Salm Sperm DNA Sigma, Deisenhofen
SDS Sigma, Deisenhofen
Triethanolamin Merck, Darmstadt
Tris Merck, Darmstadt
Tris-HCl Sigma, Deisenhofen
Tris-Base Sigma, Deisenhofen
Triton X−100 Sigma, Deisenhofen
t-RNA Sigma, Deisenhofen
8.1.2 Substanzen für die Histologie und Immunhistochemie
Substanz Bezugsquelle (Fa.)
Bovines Serum Albumin (BSA) Sigma, Deisenhofen
Diaminobenzidin Sigma, Deisenhofen
Kaninchenserum DAKO, Hamburg
Trisaminomethan Fluka, Buchs, CH
Wasserstoffperoxid Merck, Darmstadt

211
8.1.3 Sonstige Substanzen
Substanz Bezugsquelle (Fa.)
Chloralhydrat Merck, Darmstadt
Chromalaun Merck, Darmstadt
Kaisers Chromgelatine Merck, Darmstadt
Entellan Merck, Darmstadt
Gelatine Merck, Darmstadt
Gentamycin (Friso-Gent) Pitman-Moore, Burgwedel
Gentamycinsulfat Sigma, Deisenhofen
8.2 Enzyme und Substrate
Enzym Verwendung Bezugsquelle (Fa.)
SAl I Restriktion Plasmid DNA Roche Diagnostics, Mannheim
Hind III Restriktion Plasmid DNA Roche Diagnostics, Mannheim
RNA-Polymerase In-vitro-Transkription Roche Diagnostics, Mannheim
NBT-Markierungs-Mix In-vitro-Transkription Roche Diagnostics, Mannheim
Proteinase K Nicht-radioaktive ISH Sigma, Deisenhofen
Ribonuklease A Nicht-radioaktive ISH Sigma, Deisenhofen
Terminale Transferase
Nr.220 582 Radioaktive ISH Roche Diagnostics, Mannheim

212
8.3 Antikörper
Antikörper Verwendung Bezugsquelle
Monoklonal anti-GABA,
Aszites Maus, klone GB-69 Immunhistochemie Fa. Sigma, Deisenhofen
Polyklonal anti-GAD,
Schaf Immunhistochemie
National Institute of Health,
Bethesda, MD, USA
Anti-Maus Ig
Antiserum, Kaninchen Immunhistochemie Fa. DAKO, Hamburg
Anti-Ziege Ig
Antiserum, Kaninchen Immunhistochemie Fa. DAKO, Hamburg
Anti-DIG-Alkalische Phoshatase
Polyklonal
Antiserum, Schaf
Nicht-radioaktive in-
situ-Hybridisierung
Fa. Roche Diagnostics,
Mannheim
8.4 Puffer und Lösungen
8.4.1 Allgemeine Histologie
8.4.1.1 Nicht-radioaktive in-situ-Hybridisierung, Immunhistologie, Elektrodenlokalisation
Thionin-Lösung
1 M Essigsäure 100 ml
1 M Natronlauge 6 ml
Aqua bidest. 864 ml
Thionin 1,25 g
Essigsäure, Natronlauge und destilliertes Wasser wurden gemischt und zusammen
auf 60 °C erhitzt. Thionin wurde hinzugefügt , eine Stunde lang unter Rühren gelöst
und die Lösung anschließend filtriert. Diese Lösung war bei 60 °C einen Monat lang
haltbar.

213
7 % ige Gelatine-Chromalaun-Lösung:
Gelatine 3,5 g
Chromalaun 0,35 g
Aqua bidest. 500 ml
Der Ansatz wurde bis zum vollständigen Lösen der Gelatine langsam auf 60 °C
erwärmt. Die Zugabe von Chromalaun diente der Konservierung.
Gefrierlösung zur Lagerung der Serien für die nicht-radioaktive in-situ-Hybridisierung
10 x PBS 125 ml
Ethylenglykol 75 ml
Glucose 75 g
Aqua bidest. ad 250 ml
DEPC 125 µl
8.4.1.2 Radioaktive in-situ-Hybridisierung
Gelatine zum Beschichten von Objektträgern:
1,12 g Gelatine
240 ml Aqua bidest.
Das Gemisch aus Gelatine und Aqua bidest. wurde auf dem Rührer erwärmt bis die
Gelatine gelöst war und anschließend ohne Erwärmen weitergerührt. Nach Abkühlen
wurde der Lösung 9,5 ml Chromalaun zur Konservierung zugefügt. Die Objektträger
wurden zweimal in der Gelatinelösung gedippt und senkrecht über Nacht getrocknet.
Die beschichteten Objektträger wurden bis zur Verwendung staubfrei bei
Raumtemperatur gelagert.
Chromalaun-Lösung:
CrKSO 41 g
Aqua bidest 25 ml
Das Gemisch wurde nach vollständiger Lösung des Chromalaun steril gefiltert und
bei 4 °C.

214
8.4.2 Herstellung Digoxigenin-markierter RNA-Sonden
Allgemeine Hinweise:
Alle Lösungen, welche RNAse frei sein mussten, wurden 0,05 %ig mit DEPC versetzt
und über Nacht bei 37° C inkubiert. Die Lösungen wurden 21 min bei 121° C
autoklaviert, um das DEPC zu zerstören.
Low Salt Puffer für ElutipR-Säulen pH 7,4
NaCl 7,3 g
Tris HCl 0,788 g
EDTA 0,093 g
Aqua bidest. 250 ml
DEPC 125 µl
High Salt Puffer für ElutipR-Säulen pH 7,4
NaCl 29,2 g
Tris HCl 0,788 g
EDTA 0,093 g
Aqua bidest. 250 ml
DEPC 125 µl
0,2 M EDTA pH 8,0
EDTA 7,4 g
Aqua bidest. 100 ml
DEPC 50 µl
3 M Na-acetat pH 5,5
Nahacetat 24,6 g
Aqua bidest. 100 ml
DEPC 50 µl

215
Lösung für Alkalische Hydrolyse
NaHCO3 0,672 g
Na2CO3 1,27 g
Aqua bidest. 100 ml
DEPC 50 µl
Puffer 1 für Dot-Blot pH 7,5
Maleinsäure 11,61 g
NaCl 8,76 g
Aqua bidest. 1000 ml
Puffer 2 für Dot-Blot
Blocking-Reagenz 1 g
Puffer 1 100 ml
Puffer 3 für Dot-Blot pH 9,5
Tris−HCl 15,76 g
NaCl 5,84 g
MgCl2−6 H2O 10,16 g
Aqua bidest. 1000 ml
Puffer 4 für Dot-Blot pH 8,0
Tris−HCl 1,57 g
EDTA 0,37 g
Aqua bidest. 1000 ml

216
8.4.3 Nicht-radioaktive in-situ-Hybridisierung
Allgemeine Hinweise:
8.4.2
1x PBS pH 7,4
NaCl 8 g
KCl 0,2 g
Na2PO4 1,44g
KH2PO4 0,24 g
Aqua bidest. 1000 ml
DEPC 500 µl
Proteinase K Stocklösung
Proteinase K 1 mg/ml 20 µl
1 M Tris pH 7,4 5 ml
250mM EDTA 2 ml
Aqua bidest./DEPC ad 100 ml
Lagerung bei-20° C
1 M Tris pH 7,4
Tris HCl 66,1 g
Tris Base 9,7 g
Aqua bidest. 500 ml
DEPC 250 µl
250mM EDTA
EDTA 46,5 g
Aqua bidest. 500 ml
DEPC 250 µl

217
5M NaCl
NaCl 73,05 g
Aqua bidest. 500 ml
DEPC 250 µl
20 x Hybridisierungssalzlösung pH 7,2
5M NaCl 60 ml
PIPES 3, 46 g
EDTA 3,72 g
Aqua bidest. ad 250 ml
Zum Lösen auf 50° C erwärmen
DEPC 125 µl
10 % SDS
SDS 10 g
Aqua bidest./DEPC ad 100 ml
100 x Denhardts Lösung
Polyvinylpyrrolidon 2 g
Ficoll 2 g
BSA 2 g
Aqua bidest./DEPC ad 100 ml
Lagerung bei-20° C
5 mg/ml SSDNA
SSDNA 100 mg
Aqua bidest./DEPC 5 ml
Das Gemisch wurde 15 min bei 121° C autoklaviert, um die DNA zu
denaturieren und bei-20° C gelagert.
10 mg/ml Poly A
Poly A 100 mg
Aqua bidest./DEPC 10 ml
218
50 % Dextransulfat
Dextransulfat 5 g
Aqua bidest./DEPC 10 ml
Lagerung auf-20° C
Prehybridisierungspuffer: Lösung Endkonzentration für 100 ml
100 % Formamid 50 % 50 ml 20 x Hybridisierungssalzlösung 5x 25 ml 10 % SDS 0,2 % 2 ml 100 x Denhardts Lösung 1 x 1 ml 5 mg/ml SSDNA 250 µg/µl 5 ml 10 mg/ml Poly A 250 µg/µl 2,5 ml Aqua bidest./DEPC 2,5 ml Aqua bidest./DEPC ad 100 ml SSDNA, Poly A und Aqua bidest./DEPC wurden gemischt, 3 min bei 100° C erhitzt und auf Eis gekühlt. Dieser Ansatz wurde dem Hybridisierungspuffer zugegeben, auf 100 ml Aqua bidest./DEPC aufgefüllt und bei-20° C gelagert.
Hybridisierungspuffer: Lösung Endkonzentration für 100 ml
100 % Formamid 50 % 50 ml 20 x Hybridisierungssalzlösung 5x 25 ml 10 % SDS 0,2 % 2 ml 100 x Denhardts Lösung 1 x 1 ml DTT 100 mM 0,77 g 50 % Dextransulfat 5 % 5 ml 5 mg/ml SSDNA 250 µg/µl 5 ml 10 mg/ml Poly A 250 µg/µl 2,5 ml Aqua bidest./DEPC 2,5 ml Aqua bidest./DEPC ad 100 ml SSDNA, Poly A und Aqua bidest./DEPC wurden gemischt, 3 min bei 100° C erhitzt und im Anschluss auf Eis gekühlt. Dieser Ansatz wurde dem Puffer zugegeben, auf 100 ml Aqua bidest./DEPC aufgefüllt und bei-20° C gelagert.

219
RNAse Puffer
5M NaCl 100 ml
1M Tris pH 8,0 10 ml
250 mM EDTA 4 ml
1M NaThiosulfat 10 ml
Aqua bidest. ad 1000ml
5 M NaCl
NaCl 73,05 g
Aqua bidest. 500 ml
1 M Tris pH 8,0
Tris HCl 66,1 g
Tris Base 9,7 g
Aqua bidest. 500 ml
250 mM EDTA
EDTA 46,5 g
Aqua bidest. 500 ml
1 M NaThiosulfat
NaThiosulfat 158,11 g
Aqua bidest. 1000 ml
20x SSC
NaCl 43,83 g
Natriumcitrat 88,23 g
Aqua bidest. 250 ml
4x SSC mit 10mM NaThiosulfat pH 7,0
20x SSC 100 ml
1M NaThiosulfat 5 ml
Aqua bidest. ad 500 ml

220
2x SSC mit 10mM NaThiosulfat pH 7,0
20x SSC 50 ml
1M NaThiosulfat 5 ml
Aqua bidest. ad 500 ml
0,1x SSC mit 10mM NaThiosulfat pH 7,0
20x SSC 2,5 ml
1M NaThiosulfat 5 ml
Aqua bidest. ad 500 ml
Immunodetektionspuffer 1 pH 7,5
Tris−HCl 15,76 g
NaCl 8,76 g
Aqua bidest. 1000 ml
Immunodetektionspuffer 2 pH 9,5
Tris−HCl 15,76 g
NaCl 5,84 g
MgCl2−6 H2O 10,16 g
Aqua bidest. 1000 ml
Immunodetektionspuffer 3 pH 8,0
Tris−HCl 1,57 g
EDTA 0,37 g
Aqua bidest. 1000 ml

221
8.4.4 Radioaktive in-situ-Hybridisierung
Allgemeine Hinweise
Die Lösungen für die radioaktive in-situ-Hybridisierung wurden nicht mit DEPC
behandelt. Die Reagenzien wurden jedoch ausschließlich für den Ansatz dieser
Lösungen verwendet, um eine Kontamination mit RNAsen zu verhindern. Die für das
Ansetzen der Lösungen eingesetzten Glasgefäße wurden durch Erhitzen (8 h bei
180° C) RNAse frei gemacht. Für das Einstellen des pH-Wertes wurden HCl bzw.
NaCl Lösungen mit Aqua bidest./ DEPC angesetzt.
Tris-EDTA-Puffer pH 8,0
Tris 60 mg
EDTA 18,5 mg
Aqua bidest. 50 ml
4 M NaCl
NaCl 11,68 g
Aqua bidest. 50 ml
TE Puffer mit 0,5 M DTT
DTT 3,85 g
TE-Puffer pH 8,0 ad 50 ul
20x SSC pH 7,2
NaCI 8,77 g
Natriumcitrat 4,41 g
Aqua bidest. 50 ml
Lagerung bei -20°C.
1 M Na2HPO4
Na2HPO4 6,85 g
Aqua bidest. 50 ml
Lagerung bei -20° C

222
1 M NaH2PO4
NaH2PO4 7,1 g
Aqua bidest. 50 ml
Lagerung bei -20° C
1M Natriumphosphatpuffer pH 6,5
Na2HPO4 25 ml
Die Lösung wurde durch von Zugabe von 1 M NaH2PO4 Lösung auf einen ph-
Wert von 6,5 eingestellt und bei-20° C gelagert.
20x Denhardts Lösung
Ficoll 1 g
BSA 1 g
Polyvinylpyrrolidon 1 g
Aqua bidest. 50 ml
Der Ansatz wurde mit einem Steril-Filter 0,45 mm filtriert, aliquotiert und bei-
20° C gelagert.
t-RNA 25mg/ml
t-RNA 25mg
steriles Wasser 1 ml
Die Lagerung erfolgte bei -20° C.
Salm Sperm DNA Der Ansatz wurde durch kurzes Aufkochen denaturiert, auf Eis gekühlt und bei
-20° C gelagert.
0.1M EDTA pH 8,0
EDTA 1,86g
Aqua bidest. 50 ml
Lagerung bei -20° C

223
10% SDS pH 7,0
SDS 3g
Aqua bidest. 30 ml
10x PBS
NaCl 9 g
Na2PO4 0,815 g
KH2PO4 0,122 g
Aqua bidest. 100 ml
10x Triethanolamin
Triethanolamin 14,9 g
NaCl 9 g
Aqua bidest. 100 ml
Für die in-situ-Hybridisierung wurden 40 ml der Stocklösung in 300 ml Aqua
bidest. gegeben, der pH-Wert auf 8,2 eingestellt und der Ansatz auf 400 ml
komplettiert.
20x SSC Puffer-Stocklösung pH 7,2
NaCl 87,7g
Natriumcitrat 44,1 g
Aqua bidest. 500 ml
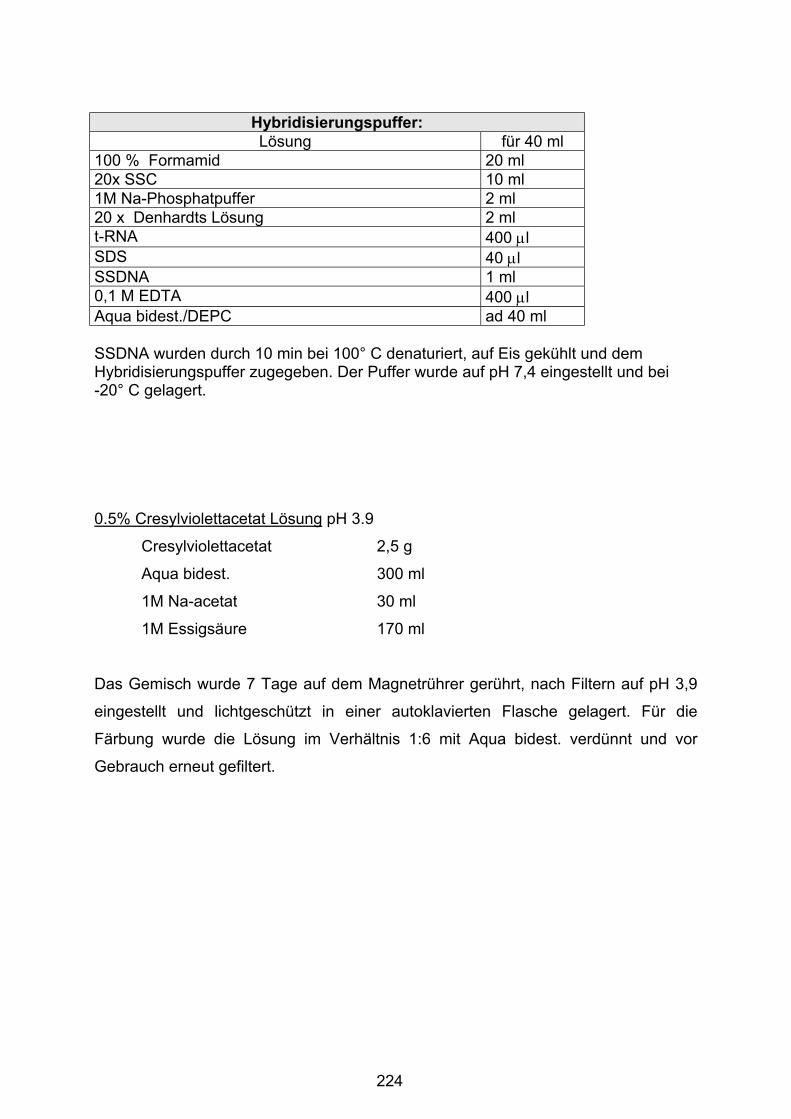
224
Hybridisierungspuffer: Lösung für 40 ml
100 % Formamid 20 ml 20x SSC 10 ml 1M Na-Phosphatpuffer 2 ml 20 x Denhardts Lösung 2 ml t-RNA 400 µl SDS 40 µl SSDNA 1 ml 0,1 M EDTA 400 µl Aqua bidest./DEPC ad 40 ml SSDNA wurden durch 10 min bei 100° C denaturiert, auf Eis gekühlt und dem Hybridisierungspuffer zugegeben. Der Puffer wurde auf pH 7,4 eingestellt und bei -20° C gelagert.
0.5% Cresylviolettacetat Lösung pH 3.9
Cresylviolettacetat 2,5 g
Aqua bidest. 300 ml
1M Na-acetat 30 ml
1M Essigsäure 170 ml
Das Gemisch wurde 7 Tage auf dem Magnetrührer gerührt, nach Filtern auf pH 3,9
eingestellt und lichtgeschützt in einer autoklavierten Flasche gelagert. Für die
Färbung wurde die Lösung im Verhältnis 1:6 mit Aqua bidest. verdünnt und vor
Gebrauch erneut gefiltert.

225
8.4.5 Immunhistochemie
„Carrier-Lösung“:
Tris-gepufferte, 0,9 % Natriumchlorid-Lösung (pH 7,6) 100 ml
Rinderserumalbumin 1 g
Kaninchenserum 1 ml
„Block-Lösung“:
„Carrier-Lösung“ 4,5 ml
Rinderserumalbumin 100 mg
Kaninchenserum 0,5 ml
Tris/Nickel-Lösung:
Nickelammoniumsulfat 0,3 g
TBS (pH 7,6) 50 ml
Diaminobenzidin-Lösung:
Ansatz (entspricht 3,3 mg 3,3`-Diaminobenzidin) 30 µl
Tris/Nickel-Lösung 4 ml
TBS:
50 mM Tris-Lösung (pH 7,6) 1l
Natriumchlorid 9 g

226

227
DANKSAGUNG Herrn Prof. W. Löscher gilt mein großer Dank für die Überlassung des Themas, die wissenschaftliche Anleitung bei der Durchführung der Versuche und der Abfassung dieser Dissertation sowie für die jederzeit gewährte freundliche Unterstützung. Weiterhin bedanke ich mich bei Frau Dr. H. Potschka und Herrn PD. Dr. U. Ebert für die Geduld bei der Beantwortung und Diskussion aller Fragen, die Anleitung zur praktischen Durchführung der Experimente sowie zur statistischen Auswertung der Versuche; Frau Prof. E. Zimmermann und Herrn Prof. H. Bigalke für die Betreuung während des Ph.D.-Studiums; Herrn M. Weißing, Frau C. Bartling, Frau D. Pieper für die hilfreiche Unterstützung bei der Anfertigung der histologischen Präparate; der Arbeitsgruppe von Herrn Prof. G. Sperk (Institut für Pharmakologie, Universität Innsbruck, Österreich) für die Anleitung zur Etablierung der radioaktiven in-situ-Hybridisierung; der Arbeitsgruppe von Frau Prof. C. Houser (Department of Biology, University of California, Los Angeles, USA) für die Überlassung von Plasmiden mit der cDNA für GAD65 und GAD67; den Arbeitsgruppen von Herrn Prof. G. Gerlach (Institut für Mikrobiologie, Tierärztliche Hochschule, Hannover) und Herrn Prof. H. Naim (Institut für physikalische Chemie, Tierärztliche Hochschule, Hannover) für die Einweisung in die Transformation kompetenter Zellen; Herrn Briese für die Anfertigung der bipolaren Elektroden; Frau Zivkovic und den übrigen Tierpflegern für die sorgsame Betreuung meiner Versuchstiere; allen ehemaligen und derzeitigen Mitarbeitern des Institutes für die freundliche Aufnahme und die stets gewährte Hilfsbereitschaft; der Bayer AG, Leverkusen, für die Bereitstellung eines Promotionsstipendiums; der Deutschen Forschungsgemeinschaft für die Bereitstellung von Sachmitteln im Rahmen des Antrags Lo 274/5. Mein besonderer Dank gilt meiner Familie, die mir durch ihre finanzielle und moralische Unterstützung die Anfertigung dieser Dissertation ermöglich hat.













![Cortex vinatura vinyl_fertigparkett[1]](https://static.fdokument.com/doc/165x107/55844c2ed8b42adf748b493c/cortex-vinatura-vinylfertigparkett1.jpg)