
Vergleichende Untersuchung der Zellapoptose mononukleärer ... · cantly higher in leukapheresis...
96
Vergleichende Untersuchung der Zellapoptose mononukleärer Zellen aus Leukozytapheresen und Leukozyten-Reduktion-System-Kammern Der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg zur Erlangung des Doktorgrades Dr. med. vorgelegt von Bettina Ziehe aus Ingolstadt
Transcript of Vergleichende Untersuchung der Zellapoptose mononukleärer ... · cantly higher in leukapheresis...

Vergleichende Untersuchung der Zellapoptose mononukleärer Zellen
aus Leukozytapheresen und Leukozyten-Reduktion-System-Kammern
Der Medizinischen Fakultät
der Friedrich-Alexander-Universität
Erlangen-Nürnberg
zur
Erlangung des Doktorgrades Dr. med.
vorgelegt von
Bettina Ziehe
aus
Ingolstadt

Als Dissertation genehmigt von der
Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Vorsitzender des Promotionsorgans: Prof. Dr. med. Dr. h. c. J. Schüttler
Gutachter: Prof. Dr. Erwin Strasser
Gutachter: Prof. Dr. Reinhold Eckstein
Tag der mündlichen Prüfung: 12. Januar 2015

In Liebe und Dankbarkeit für meine Familie
und in Gedenken an meinen Vater

Inhaltsverzeichnis Seite
1. Zusammenfassung und Abstract 1
1.1 Zusammenfassung 1
1.1.1 Hintergrund und Ziele 1
1.1.2 Methoden 1
1.1.3 Ergebnisse und Beobachtungen 1
1.1.4 Praktische Schlussfolgerung 2
1.2 Summary 3
1.2.1 Background and Objectives 3
1.2.2 Materials and Methods 3
1.2.3 Results 3
1.2.4 Conclusion 4
2. Einleitung 5
2.1 Tumorvakzinen aus mononukleären Zellen 5
2.2 Die Rolle mononukleärer Zellen im Immunsystem 5
2.2.1 Die Bedeutung des Immunsystems 5
2.2.2 Funktion der Immunzellen 6
2.2.3 Die Entwicklung der Immunzellen 7
2.2.4 Mononukleäre Zellen und ihre Funktion 8
2.2.4.1 Lymphozyten 8
2.2.4.2 Monozyten und antigenpräsentierende Zellen 9
2.3 Die Apoptose 9
2.3.1 Die Bedeutung der Apoptose 9
2.3.2 Die Stadien der Apoptose 10
2.3.3 Nachweismethoden der Frühapoptose 11
2.3.3.1 Veränderung des mitochondrialen Potentials 11
2.3.3.2 Veränderung an der Plasmamembran 11
2.3.4 Nachweismethoden der Spätapoptose 11
3. Fragestellung 13
4. Material und Methoden 14
4.1 Materialien 14
4.1.1 Geräte 14

4.1.2 Einmalmaterialien 14
4.1.3 Lösungen und Reagenzien 14
4.1.4 Antikörper 15
4.2 Studienbeschreibung 15
4.2.1 Gewinnung der Leukozytenkonzentrate 16
4.2.1.1 Leukozytapherese mittels COM.TEC Zellseparator 16
4.2.1.2 Thrombozytapherese mittels Trima Accel Zellseparator 17
4.2.2 Aufbereitung der Aphereseprodukte 18
4.3 Methoden 18
4.3.1 Grundlagen der FACS Messung 18
4.3.2 Apoptosedetektion mittels Annexin V und 7AAD 21
4.3.2.1 Messeinstellung für die Annexin V-7AAD Methode 22
4.3.3 Apoptosedetektion mittels JC 1 Markierung 24
4.3.3.1 Messeinstellung für die JC 1 Methode 25
4.3.4 Apoptosedetektion mittels Apo-BrdU Markierung 27
4.3.4.1 Messeinstellung für die Apo-BrdU Methode 29
5. Statistische Auswertung 32
6. Ergebnisse 34
6.1 Spenderdaten 34
6.2 Daten des Aphereselaufs 34
6.3 Produktspezifische Daten 35
6.4 Blutzellen vor der Spende 35
6.5 Blutzellen in den Aphereseprodukten 36
6.6 Vergleich von Spenderdaten und Produktdaten 37
6.6.1 Blutzellen vor der Spende und in LRS-Kammern 37
6.6.2 Blutzellen vor der Spende und in Leukozytenkonzentraten 38
6.6.3 Annexin V-7AAD-Messung von Tregs 39
6.6.4 JC 1 Messung von Lymphozyten und Monozyten 42
6.6.5 BrdU Messung von mononukleären Zellen 43
6.7 Darstellung mittels unterschiedlicher Analysen 44
6.7.1 Annexin V-7AAD positive Tregs 44
6.7.2 JC 1 detektierte Lymphozyten und Monozyten 51
6.7.3 BrdU detektierte mononukleäre Zellen 54
6.8 Einfluss der Kurzzeitlagerung 57
6.8.1 Annexin V-7AAD positive Zellen im Aphereseprodukt 57
6.8.2 JC 1 markierte Zellen im Aphereseprodukt 62
6.8.3 BrdU markierte Zellen im Aphereseprodukt 65

6.9 Biochemische Parameter in den Aphereseprodukten 66
6.9.1 Glukoseverbrauch 66
6.9.2 Laktatanstieg 69
7. Diskussion 73
8. Literaturverzeichnis 82
9. Anlage 89

1
1. Zusammenfassung und Abstract
1.1 Zusammenfassung
1.1.1 Hintergrund und Ziele
Der Einsatz mononukleärer Zellen in Immuntherapien zur Behandlung von Krebs-
und Autoimmunerkrankungen hat dazu geführt, diese Zellen in hoher Ausbeute und
maximaler Reinheit gewinnen zu wollen. Als Mittel der Wahl hat sich die Leukozyt-
apherese erwiesen, doch auch den Leukozyten-Reduktion-System-Kammern, in
denen Leukozyten während der Thrombozytapherese angereichert werden, kommt
immer größere Bedeutung zu. Ziel dieser Arbeit war es, die Zellqualität mono-
nukleärer Zellen aus Leukozytapheresaten und Leukozyten-Reduktion-System-
Kammern mittels drei verschiedener Apoptose-Methoden während einer Kurzzeit-
lagerung von 48 Stunden vergleichend zu untersuchen.
1.1.2 Methoden
Von zehn Spendern wurden jeweils 10 ml Blut, das bei der Leukozytapherese und
aus Leukozyten-Reduktion-System-Kammern gewonnen wurde, in Beutelsysteme
aus CO2 und O2 durchlässigem Fluorethylenpropylen Teflon überführt und gelagert.
Die Messungen erfolgten innerhalb von drei Stunden nach Gewinnung, sowie nach
sechs Stunden, 24 Stunden und 48 Stunden Lagerung. Das Apoptoseverhalten der
mononukleären Zellen wurde mittels Annexin V und 7-Aminoactinomycin D (7AAD),
5,5',6,6'-tetra-chloro-1,1',3,3'-tetraethylbenzimidazolcarbocyanin Jodid (JC 1) und 5-
Bromo-deoxyuridin (BrdU) analysiert. Zusätzlich wurden die biochemischen Para-
meter Glukose und Laktat bestimmt.
1.1.3 Ergebnisse und Beobachtungen
Die Leukozytenkonzentration liegt in den LRS-Kammern signifikant höher als in den
Leukozytapheresaten. Bedingt durch das höhere Produktvolumen der Leukozyt-
apheresate weisen diese aber insgesamt mehr Leukozyten auf als die LRS-
Kammern. Während in den Leukozytaphereseprodukten die Verunreinigung mit re-
sidualen Thrombozyten sehr hoch war, zeigten die LRS-Kammern eine signifikant
höhere Verunreinigung mit Erythrozyten. Der Anteil apoptotischer und nekrotischer
mononukleärer Zellen lag in den Leukozytapheresaten signifikant höher als in den

2
LRS-Kammern. In beiden Leukozytenprodukten konnte ein starker Apoptoseanstieg
nach 24 Stunden nachgewiesen werden. Ebenfalls wurde eine starke Korrelation
zwischen dem Anstieg an apoptotischen mononukleären Zellen und der Zunahme
der Laktatkonzentration nachgewiesen.
1.1.4 Praktische Schlussfolgerungen
Die Apoptose-Methoden zeigten für die LRS-Kammern eine höhere Zellviabilität als
die Leukozytapheresate. Es wird vermutet, dass bei der Gewinnung mononukleärer
Zellen der oxidative Stress bei der Zellanreicherung in den LRS-Kammern deutlich
geringer ist als bei der Leukozytapherese. Beide Leukozytenprodukte weisen teil-
weise noch eine hohe Verunreinigung mit unerwünschten Blutzellen auf. Sollte eine
weitere Reduktion dieser Zellen bei gleichzeitig hoher Leukozytenausbeute möglich
sein, könnte dies die Zellqualität weiterhin verbessern. Ebenso spielt die metaboli-
sche Aktivität der Zellen für die Qualität eine große Rolle. So konnte ein deutlicher
Zusammenhang zwischen Laktatkonzentration und der Apoptoserate nachgewiesen
werden.

3
1.2 Summary
1.2.1 Background and Objectives
The use of mononuclear cells in immunotherapy for the treatment of cancer and
autoimmune diseases has led to want to gain these cells in high yield and maximum
purity. As method of choice, the leukapheresis has proven, but also the leukocyte
reduction system chambers in which leukocytes are accumulated during platelet-
pheresis becomes more and more important. The aim of this study was to compare
the quality of mononuclear cells of leukapheresis and leukocyte reduction system
chambers using three different apoptosis methods during a short-term storage of 48
hours.
1.2.2 Materials and Methods
10 ml of blood of ten blood donors, which was obtained from leukapheresis products
and leukocyte reduction system chambers was transferred and stored into bag sys-
tems of CO2 and O2 permeable Fluorethylenpropylen Teflon. Measurements were
made within three hours after collection, and after six hours, 24 hours and 48 hours
of storage. The apoptosis of the mononuclear cells was analyzed by Annexin V and
7-Aminoactinomycin D (7AAD), 5,5´,6,6'-tetra-chloro-1,1',3,3'tetraethylbenzimidazol-
carbocyanine iodide (JC 1) and 5-Bromo-deoxyuridine (BrdU). In addition, the bio-
chemical parameters Glukose and lactate were measured.
1.2.3 Results
The leucocyte concentration in the LRS chambers was significantly higher than in
the leukapheresis products. Due to the higher product volume of leukocyte aphere-
sis products have leukocytes were obtained from it compared to the LRS chambers.
While in leukapheresis products the contamination with residual platelets was very
high, LRS chambers showed a significantly higher contamination with residual red
blood cells. The proportion of apoptotic and necrotic mononuclear cells was signifi-
cantly higher in leukapheresis products than in the LRS chambers. In both leukocyte
products a strong increase of apoptosis could be detected after 24 hours. In addi-
tion, a strong correlation between the increase of apoptotic mononuclear cells and
the increase in lactate concentration could be detected.

4
1.2.4 Conclusion
By use of three apoptosis methods leucocytes obtained from LRS chambers
showed higher cell viability than for the leukapheresis products. It is assumed that in
the recovery of mononuclear cells the oxidative stress in cell accumulation is in the
LRS chambers significantly lower than in the leukapheresis. Both leukocyte products
contain residual blood cells. If a further reduction of the residual cells should be pos-
sible without leucocyte loss, this could still improve cell quality. The metabolic activi-
ties of cells also play a major role for the quality. Thus, a significant correlation be-
tween the lactate concentration and the rate of apoptosis could be detected.

5
2. Einleitung
2.1 Tumorvakzinen aus mononukleären Zellen
Die intensive Forschung zur Krebsbehandlung in den letzten Jahren führte dazu,
dass den mononukleären Zellen heute eine bedeutende Rolle zukommt. Es wurden
bereits einige Vakzinen für die Tumortherapie hergestellt, deren immunologische
Grundlage auf mononukleären Zellen basieren. Nach Applikation autologer Tumor-
zellen oder daraus gewonnener Oligopeptide zur spezifischen Immunisierung, soll
es nach Bindung an potente antigenpräsentierende Zellen wie den dendritischen
Zellen zu einer T-Zell Antwort kommen [9; 60]. Im Jahr 2010 wurde in der USA Sipu-
leucel-T zugelassen, eine autologe therapeutische Tumorvakzine aus aktivierten
mononukleären Zellen des peripheren Blutes, welche zur Behandlung des hormon-
resistenen Prostatakarzinoms im metastasierten Stadiums eingesetzt wird [9]. Wei-
tere Tumorvakzinen aus mononukleären Zellen, die zur Immuntherapie bei Malig-
nem Melanom und Nierenzellkarzinom eingesetzt werden sollen, werden derzeit in
klinischen Studien erprobt. Man verspricht sich davon, dem Patienten Abwehrzellen,
die in vitro mit Tumorantigenen beladen wurden zu retransfundieren, um damit das
Immunsystem auf die noch nicht erkannten Krebszellen aufmerksam zu machen.
Auch monoklonale Antikörper, die aus einem B-Lymphozytenklon generiert werden,
werden heute in der Behandlung von Krebs und Autoimmunerkrankungen einge-
setzt [9]. Daher ist es von großer Bedeutung, eine maximale Ausbeute dieser Zellen
mit hoher Reinheit zu erhalten. Es gibt daher eine Reihe von Untersuchungen zu
verschiedenen Zellseparatoren, die sich mit der Optimierung der Geräteeinstellung
zur Verbesserung des Leukozytaphereseproduktes beschäftigen [34; 77]. Es zeigte
sich, dass nicht nur im Rahmen von Leukozytapheresen eine hohe Zellausbeute an
mononukleären Zellen erzielt werden kann, sondern dass eine hohe Zellkonzentra-
tion an mononukleären Zellen bei Thrombozytapheresen in den Leukozyten-
Reduktion-System-Kammern (LRSC, leucoreduction system chamber) angereichert
werden und somit als Leukozytenquelle zur Verfügung stehen [54].
2.2 Die Rolle mononukleärer Zellen im Immunsystem
2.2.1 Die Bedeutung des Immunsystems
Das Immunsystem spielt eine bedeutende Rolle in der Entwicklung der Lebewesen.
Die Homöostase zwischen Zellerneuerung und Zellapoptose ist dabei von entschei-
dender Bedeutung. Ohne Immunsystem gäbe es keine Schutzfunktion zum Erhalt

6
unseres körperlichen Wohlergehens. Menschen, mit schweren angeborenen oder
erworbenen Immundefekten, zeigen eine hohe Anfälligkeit gegenüber pathogenen
Keimen. Da das Immunsystem hier nicht in der Lage ist adäquat zu reagieren, er-
kranken diese Menschen an opportunistischen Infektionen, die häufig auch zum
Tode führen. Der Schutz des Immunsystems beschränkt sich aber nicht nur auf kör-
perfremde pathogene Keime, es bewahrt uns auch vor körpereigenen Prozessen,
die unter Umständen tödlich verlaufen könnten. So entstehen tagtäglich im gesun-
den Organismus Tumorzellen, die allerdings in der Regel sofort erkannt und elimi-
niert werden. Ebenso besitzt unser Immunsystem Mechanismen, um uns vor auto-
immunen Prozessen zu bewahren. Dies lässt darauf schließen, dass Menschen, die
an Tumorerkrankungen oder Autoimmunerkrankungen leiden, eine Fehlfunktion im
Immunsystem aufweisen [33].
2.2.2 Funktion der Immunzellen
Im Immunsystem besteht eine enge Beziehung zwischen zellulären und humoralen
Bestandteilen sowie zu den lymphatischen Organen. Es lassen sich sowohl eine
angeborene als auch eine erworbene Immunabwehr unterscheiden. Die angeborene
Immunabwehr agiert innerhalb von Stunden nach Aktivierung. Die Zellen der ange-
borenen Immunabwehr, die Granulozyten, Natürlichen Killerzellen, Monozyten und
Makrophagen erkennen mit speziellen Rezeptoren für pathogene Strukturen die
krankheitsverursachende molekulare Struktur, das sogenannte PAMP der Erreger
[68]. Zusätzlich werden humorale Abwehrmechanismen, wie das Komplementsys-
tem, Lysozyme und Interferone aktiviert. Im Gegensatz zur angeborenen Abwehr
muss sich die erworbene Immunabwehr erst entwickeln. Sie wird erst nach Tagen
aktiv. Hierbei spielen zwei Zelltypen eine große Rolle. Die T-Lymphozyten für die
zelluläre Immunantwort und die B-Lymphozyten für die humorale Immunantwort.
Beide Zelltypen erkennen spezielle Proteinstrukturen, die Antigene auf Krankheits-
erregern. Aktivierte B-Lymphozyten produzieren nach Antigenkontakt Antikörper,
während die T-Lymphozyten mittels einer Subpopulation selbst in der Lage sind die
infizierte Zelle abzutöten [55].

7
Abb.1: Übersicht über die Abwehrmechanismen
Abwehrmechanismus
Angeborene Abwehr Adaptive Abwehr
Unspezifische Erkennung Selektive Erkennung von
von Krankheitserreger Krankheitserregern
↓ ↓ ↓ ↓
humoral zellulär humoral zellulär
↓ ↓ ↓ ↓
Komplement Granulozyten B-Lymphozyten T-Lymphozyten
Interferone Makrophagen
Lysozyme Natürliche Killerzellen
Modifiziert nach Abb. 1.14 aus [55]
2.2.3 Die Entwicklung der Immunzellen
Die spezialisierten Zellen des Immunsystems entstehen im Knochenmark aus pluri-
potenten Stammzellen.
Abb.2: Hämatopoetischer Stammbaum
B-Lymphozyt
Lymphoide T-Lymphozyt
Vorläuferzelle
Pluripotente NK-Zellen
Stammzelle
Megakaryozyten Thrombozyten
Myeloische
Vorläuferzelle Erythropoetische Erythrozyten
Vorläuferzelle
Eosinophile
Vorläuferzelle Basophile Granulozytäre und Neutrophile Granulozyten
Vorläuferzelle Monozytäre Vorläuferzelle
Monozyten
Eosinophile Granulozyten Basophile Granulozyten

8
Aus den lymphoiden Vorläuferzellen entstehen die für die erworbene Immunabwehr
wichtigen T- und B-Lymphozyten sowie die Natürlichen Killerzellen (NK-Zellen). Aus
den myeloischen Vorläuferzellen differenzieren sich über weitere Vorläuferzellen die
Granulozyten, Monozyten, Thrombozyten und Erythrozyten. Lymphozyten und Mo-
nozyten zählen zu den mononukleären Zellen, deren Bedeutung in der Tumorthera-
pie bereits erläutert wurde. Die Hämatopoese wird über unterschiedliche Wachs-
tumsfaktoren wie die Interleukine reguliert. Die Zufuhr dieser Faktoren ermöglicht
es, die Entwicklung der differenzierten Zellen, Vorläuferzellen und Stammzellen in
der Peripherie zu induzieren, um sie mittels Leukozytapherese zu gewinnen [62].
2.2.4 Mononukleäre Zellen und ihre Funktion
2.2.4.1 Lymphozyten
Die Lymphozyten entstehen im Knochenmark aus Lymphoblasten. Während B-
Lymphozyten im Knochenmark ausreifen, wandern T-Vorläuferzellen in den Thymus
aus, wo sie durch Zytokine proliferieren. Der Proliferation der Vorläuferzellen folgt
die Entwicklung von Antigenrezeptoren und anschließend die Selektion, damit kör-
pereigene Proteine nicht angegriffen werden. Durch die Aktivierung durch antigen-
präsentierende Zellen differenzieren sich aus den naiven T-Lymphozyten CD 4 posi-
tive T-Helferzellen, zytotoxische CD 8 positive T-Lymphozyten und regulatorische T-
Lymphozyten. Die Helferzellen aktivieren bevorzugt zytotoxische T-Zellen und Mak-
rophagen. Zytotoxische T-Zellen phagozytieren Krankheitserreger. Die B-Lympho-
zyten produzieren nach Aktivierung Antikörper und Gedächtniszellen [64]. Etwa 5-
10% der CD4 positiven T-Zellen beinhalten einen Teil regulatorischer T-Zellen
(Tregs), die sich entweder im Thymus ausbilden oder in der Peripherie aus naiven
CD4 positiven CD25 positiven T-Zellen durch den Transkriptionsfaktor Foxp3 indu-
ziert werden [14]. Regulatorische T-Zellen (Treg) schützen durch ihren supprimie-
rende Eigenschaft auf das Immunsystem vor autoimmunen Prozessen. Eine gestör-
te Immunabwehr könnte so ausufern, dass auf pathogene Keime mit einer verstärk-
ten Entzündungsreaktion reagiert wird, woraus sich weitere Erkrankungen entwi-
ckeln könnten [44]. Deshalb verhindern die CD4 positiven CD25 positiven Tregs
durch die Sekretion von Interleukin 10 und TGF β die Zytokinausschüttung von CD4
positiven und CD8 positiven T-Zellen und unterdrücken deren Proliferation [10]. Sie
wirken aber nicht nur auf Zellen, die sich gegen den eigenen Körper richten. Auf
natürliche Killerzellen und zytotoxische T-Zellen können sie ebenfalls einen suppri-
mierenden Effekt haben, wodurch ein Tumorwachstum gefördert werden kann. Da-

9
her haben sie einen hohen Stellenwert als Verlaufsmarker in der immunmodulieren-
den Tumortherapie, wobei eine hohe Anzahl an regulatorischen T-Zellen eine
schlechtere Prognose bedeuten kann [26]. Diverse Studien beschäftigen sich auch
im Rahmen der bei Transplantationen auftretenden Graft-versus-Host-Disease mit
dem Einsatz von regulatorischen T-Zellen. Die Tregs sollen die Immunantwort ge-
genüber den fremden Spenderantigenen unterdrücken, so dass eine lebenslange
medikamentöse Immunsuppression nicht mehr erforderlich ist [31; 42; 47].
2.2.4.2 Monozyten und antigenpräsentierende Zellen
Monozyten sind die größten Immunzellen im Blut. Sie entstehen im Knochenmark
aus Monoblasten und zirkulieren nur wenige Tage im Blut, bevor sie ins Gewebe
auswandern. Ungefähr zwei bis acht Prozent der Blutzellen stellen Monozyten dar.
Sie fungieren als antigenpräsentierende Zellen, betreiben Phagozytose und sezer-
nieren Zytokine. Die in ihrer Granula enthaltenen sauren Hydrolasen und Peroxi-
dasen sind für die intrazelluläre Fragmentierung von pathogenen Keimen notwen-
dig. Die Erkennung der antigenen Eigenschaften erfolgt dabei über spezifische Re-
zeptoren. Nach Auswanderung ins Gewebe differenzieren sie sich zu Makrophagen.
Auch von den Monozyten existieren mehrere Subpopulationen, deren gemeinsamer
Oberflächenmarker CD 14 ist. Zu dem Monozyten-Makrophagen-System zählen
auch noch andere antigenpräsentierende Zellen wie die Dendritischen Zellen, die
aus Monozyten generiert werden können. Wie die Monozyten und Makrophagen
sind sie auf die Aktivierung durch Zytokine aus infizierten Zellen angewiesen. Ihre
Aufgabe besteht darin, körperfremde Moleküle auf zu nehmen, zu phagozytieren
und anschließend die auf ihrer Oberfläche exprimierten Antigenfragmente T-Zellen
zu präsentieren. Dadurch kommt es zur Proliferation der T-Zellen und letztlich zur
spezifischen zellulären Immunantwort [37].
2.3 Die Apoptose
2.3.1 Die Bedeutung der Apoptose
Die Apoptose, auch programmierter Zelltod genannt, stellt einen komplexen Mecha-
nismus der Zelle dar kontrolliert abzusterben, um in ihrem Erbgut geschädigte Zel-
len gezielt zu eliminieren. Dieser Vorgang ist physiologisch und für die Gewebeho-
möostase von großer Bedeutung, da sich Zellproliferation und Zelltod im Gleichge-
wicht halten [84]. Im Gegensatz zur Nekrose, bei der die Zelle eine Schädigung

10
aufweist und daraufhin zugrunde geht, findet hier keine Entzündungsreaktion statt.
Die Apoptose ist für die Ausbildung von Körperstrukturen wichtig und sorgt dafür,
dass Zellzahl und Gewebegröße ausbalanciert bleiben [30]. Bedeutsam ist sie auch
für die Eliminierung von kanzerogenen Zellen und von T-Zellen mit fehlerhafter Dif-
ferenzierung oder solchen, die sich gegen körpereigene Strukturen richten [40]. Stö-
rungen dieses Prozesses begünstigen die Entstehung maligner Neoplasien, da
fehlgesteuerte Zellen überleben können.
2.3.2 Die Stadien der Apoptose
Eine apoptotische Zelle durchläuft verschiedene Phasen während ihres Absterbens.
Zunächst kommt es durch Dehydrierung zur Schrumpfung der Zelle (2).
Abb. 3: Stadien der Apoptose [64]
1 2 3 4
Durch Veränderungen im Kalzium- und pH-Haushalt der Zelle kommt es zur Verän-
derung des mitochondrialen Potentials. Die daraus resultierende Abflachung
des negativen Gradienten des mitochondrialen Potentials kann durchflußzytomet-
risch bestimmt werden. Auf molekularer Ebene kommt es zur Chromatinkondensie-
rung (2) mit folgender Fragmentierung des Chromatins (3). Durch die Zelldehydrie-
rung kommt es auch zur Kondensation des Zytoplasmas und einer Umverteilung der
Phospholipide an der Plasmamembran. Beendet wird der apoptotische Prozess mit
der Bildung sogenannter apoptotic bodies (4), membranumhüllte kleine Restteilstü-
cke, die letztendlich durch Phagozytose abgebaut werden [19; 33]. Sowohl die Ver-
änderung der Plasmamembran als auch die DNA-Fragmentierung sind durchflußzy-
tometrisch analysierbar.

11
2.3.3. Nachweismethoden der Frühapoptose
2.3.3.1 Veränderungen der mitochondrialen Funktion
Wie bereits erwähnt durchlaufen apoptotische Zellen verschiedene Stadien mit cha-
rakteristischen Veränderungen. Ein sehr früher Prozess während der Apoptose ist
die Depolarisation des mitochondrialen Membranpotentials. Poren in der Außenseite
der Mitochondrienmembran ermöglichen mitochondrialen Proteinen den Austritt ins
Zytosol. Während der Apoptose kommt es zur Öffnung dieser Poren, so dass Proto-
nen einem negativen Gradienten folgend ausströmen und somit eine Aufhebung der
asymmetrischen Verteilung bedingen. Sichtbar wird dieser Prozess durch die Ver-
änderung des mitochondrialen Membranpotentials. In nicht apoptotischen Zellen
können durch den negativen Gradienten kationische lipophile Fluorochrome wie JC1
oder Rhodamin 123 im Mitochondrium akkumulieren. Durch die Depolarisation der
Mitochondrienmembran ist die Zelle dazu allerdings nicht mehr in der Lage. Der
Verlust der Akkumulationsfähigkeit ist mittels Durchflußzytometrie durch Abnahme
der Fluorochromintensität nachweisbar [19; 85].
2.3.3.2 Veränderungen an der Plasmamembran
Relativ früh im apoptotischen Prozess transloziert Phosphatidylserin von der Innen-
seite der Plasmamembran auf die Außenseite. Zu Beginn bleibt die Membranintegri-
tät noch erhalten während der Verlust der Integrität ein Zeichen für Nekrose dar-
stellt. Proteine wie Annexin V, die eine hohe Affinität zu Phospholipiden und insbe-
sonder Phosphatidylserin haben, eignen sich daher besonders zum Nachweis. Da
der alleinige Nachweis mit Annexin V nicht beweisend für eine apoptotische Zelle
ist, wird mit 7AAD (7-Amino-Actinomycin D) oder anderen Kernfärbungen gegenge-
färbt. 7AAD ist nur bei Verlust der Membranintegrität in der Lage zwischen Guanin
und Cytosin in der DNA zu interkalieren und beweist somit einen nekrotischen Pro-
zess. Durch Markierung von Annexin V mit Phycoerythrin (PE) oder Fluoresceiniso-
thiocyanat (FITC) lässt sich der apoptotische Vorgang durchflußzytometrisch dar-
stellen [19; 86].
2.3.4 Nachweismethoden der Spätapoptose
Am Ende der Apoptose kommt es durch die Endonukleaseaktivität von Effektor-
caspasen zur Spaltung der DNA. Hierbei entstehen sowohl doppelsträngige, nie-

12
dermolekulare Fragmente als auch einzelsträngige, hochmolekulare DNA-Frag-
mente. Das entstehende 3`-Hydroxy-Ende dieser Fragmente kann mittels modifizier-
ter markierter Nukleotide in einer enzymatisch katalysierten Reaktion detektiert wer-
den. Als Enzym eignet sich TdT (terminal desoxynucleotidyl-transferase), das zu
einer matrizenunabhängigen Bindung von bromierten Deoxyuridintriphosphat
(BrdUTP) an das 3´-Hydroxy-Ende der Strangbrüche führt. Durch Zellfärbung mit
markiertem Antikörper gegen das bromierte dUTP erfolgt der Apoptosenachweis
mittels Durchflußzytometer [19; 30].

13
3. Fragestellung
Ziel der Arbeit war es, mononukleäre Zellen, die entweder durch Leukozytapherese
gewonnen, oder durch Thrombozytapherese in LRS-Kammern angereichert wurden,
hinsichtlich verschiedener Apoptose-Methoden, mit Annexin V/7-AAD, JC1 und
BrdUTP, während einer Kurzzeitlagerung von 48 Stunden zu analysieren.
Diese analytischen Methoden wurden am Durchflusszytometer durchgeführt.
Zusätzlich sollte die Qualität der Zellen und biochemische Parameter miteinander
vergleichen werden. Ebenso sollte der Zellverlust durch die Kurzzeitlagerung von 48
Stunden im Lagerbeutel ermittelt werden.
Auf Grundlage dieser Zielstellungen ergab sich folgende Fragestellung:
1. Wie unterscheiden sich die Spenderdaten bei Leukozytapherese und
Thrombozytapherese?
2. Welche Unterschiede zeigen die Leukozytenprodukte aus den jeweiligen
Spendearten, Leukozyt- und Thrombozytapherese?
3. Welche Unterschiede zeigen sich in der Zellqualität hinsichtlich lebender,
apoptotischer und nekrotischer Zellen, die konkret mit folgenden Methoden
ermittelt wurden:
a. Annexin V- und 7AAD- positive regulatorische T-Zellen
b. JC1 Nachweis bei der Lymphozyten- und Monozytenpopulation
c. Strangbrüche bei mononukleären Zellen mit der BrdU Methode
4. Welchen Einfluss hat die Kurzzeitlagerung auf die Zellqualität im jeweiligen
Aphereseprodukt, die mit den oben genannten Methoden ermittelt wurden?
5. Welchen Aussagewert haben die einzelnen Analysemethoden?
6. Wie unterscheiden sich die beiden Aphereseprodukte im Glukoseverbrauch
und in der Laktatkonzentration (biochemische Parameter) voneinander? Be-
steht ein Zusammenhang zwischen Apoptoserate und Laktatanstieg?
Ist es vorstellbar, dass die Massenspektrometrie als zusätzliche Analyseme-
thode für eine weitergehende Analyse spezifischer Moleküle (z.B.: Amino-
säuren) während der Zelllagerung sinnvoll ist?

14
4. Material und Methoden
4.1 Materialien
4.1.1 Geräte
Zur Probenmessung wurde ein automatisiertes Multicolor Durchflußzytometer
(FACS Calibur, BD Biosciences, Heidelberg) verwendet, welches sowohl zur Zel-
lanalyse als auch zur Zelldifferenzierung fähig ist. Für die Datenerfassung und Da-
tenanalyse wurde als Software Cellquest pro (BD Biosciences, Heidelberg) verwen-
det. Diese Software speichert formell alle Parametermessungen und Instrument
Settings automatisch in einem FCS (Flow Cytometry Standard) data file auf einem
Macintosh Computer ab. Die Blutbildbestimmung wurde an einem automatischen
Analysiergerät (Sysmex KX-21 N, Sysmex, Norderstedt) durchgeführt, welches 17
verschiedene hämatologische Parameter aus Vollblut mit einer Durchsatzrate von
60 Proben pro Stunde bestimmen kann. Reagenzien wurden im Kühlschrank bei
4°C (Kirsch, Offenburg) oder im Tiefkühlschrank bei -70°C (Heraeus, Hanau) auf-
bewahrt. Die Proben für den APO-BrdU Test lagerten bis zur Messung im -30°C
Tiefkühlschrank (Kirsch, Offenburg). Weiterhin wurden für die Probenaufbereitungen
ein 37°C warmes Wasserbad (GFL, Burgwedel), eine Zentrifuge (Hettich, Tuttlingen)
sowie eine Biofuge Fresco (Heraeus, Hanau) verwendet.
4.1.2 Einmalmaterialien
Die Blutprodukte wurden in 25 ml und 7 ml Beutelsystemen (CellGenix, Gaithers-
burg, USA), welche aus CO2 und O2 durchlässigem Fluorethylenpropylen (FEP)
Teflon bestehen und Gasaustausch im geschlossenen Beutel durch die FEP-
Teflonfolie vollziehen, gelagert. Zur Bestimmung der Glukose- und Laktatwerte wur-
den 1,3 ml fassende Natrium-Fluorid Röhrchen (Sarstedt, Nümbrecht) verwendet.
Für die durchflußzytometrische Messung waren 5 ml Polystyrene Round-Bottom
Gefäße und Falcon Röhrchen (BD Biosciences, Heidelberg) notwendig. Ebenso
kamen Eppendorf Gefäße (Eppendorf, Hamburg) zum Einsatz.
4.1.3 Lösungen und Reagenzien
Zur Zelllyse wurde zehnfach konzentrierte Pharm Lyse (BD Biosciences, Heidel-
berg), ein Lysereagenz auf Ammoniumchlorid Basis, verwendet, die vor Gebrauch

15
mit Aqua dest. verdünnt wurde. Als Waschpuffer wurde PBS verwendet, der 140
mM NaCl, 2,7 mM KCl, 10 mM Na2HPO4 und 1,8 mM KH2PO4 enthält. Der JC1
Antikörper (Lyophilisat) wurde in 100% DMSO-Lösung aufgelöst und aliquotiert bei
-30°C bis zu sechs Monate gelagert. Zur Zellfixierung wird 10% Formaldehyd, das in
der Apotheke bezogen und verdünnt wurde. Zur Aufbewahrung der Zellen wurden
diese mit 70% Ethanol fixiert. Zur Messung und Reinigung des Durchflußzytometers
wurden die von BD Biosciences empfohlenen FACS Flow, FACS Rinse und FACS
Clean Reagenzien verwendet.
Die verwendeten Kits Annexin V Apoptosis Detection Kit I, BD Mitoscreen Kit,
sowie APO-BRDU Kit wurden alle von BD Biosciences, Heidelberg bezogen [Anlage
1].
4.1.4 Antikörper
Gemäß Herstellerangaben stammt der CD 4 APC Antikörper Klon SK3 (BD
Biosciences, Heidelberg) aus der Hybridisierung von Maus NS-1 Myelomzellen mit
Zellen von der Milz von BALB/c Mäusen immunisiert mit menschlichen T-
Lymphozyten aus dem peripheren Blut. Der CD 25 PE Antikörper Klon CE (BD
Biosciences, Heidelberg) stammt ebenfalls aus der Hybridisierung von Maus NS-1
Myelomzellen mit Zellen von der Milz von BALB/c Mäusen immunisiert jedoch mit
PHA-aktivierten menschlichen T-Lymphozyten. Der 7-AAD (7-Amino-Actinomycin D)
Antikörper (BD Biosciences, Heidelberg) dient der DNS Färbung.
4.2 Studienbeschreibung
Es wurden zehn Patienten (zwei Frauen, acht Männer) untersucht, die sowohl Mo-
nozytenspender als auch Thrombozytenspender sind. Der Zeitraum der Messungen
erstreckte sich von Januar 2011 bis Juli 2011. Für die Studie wurden mononukleäre
Zellen aus Leukozytenkonzentraten analysiert, welche durch Leukapherese gewon-
nen wurden. Als Zellseparator wurde COM.TEC (Fresenius, Bad Homburg) einge-
setzt. Jeweils 10 ml des Konzentrates wurden steril in 25 ml Beutelsysteme abge-
füllt. Von denselben Spendern wurden auch LRS-Kammern, die bei der Separation
von Thrombozytenkonzentraten als Nebenprodukt entstanden sind, hinsichtlich der
mononukleären Zellen untersucht. Für die Thrombozytapherese wurde der Zellsepa-
rator Trima Accel verwendet. Die Abfüllung des Inhalts der LRS-Kammern von 10 ml
erfolgte in jeweils zwei 7 ml Beutelsysteme. Die Proben wurden von den Blutspen-
dern der Transfusionsmedizinischen und Hämostaseologischen Abteilung der Uni-

16
versität Erlangen zur Verfügung gestellt. Die Probanden waren über die Studie auf-
geklärt und mit der Untersuchung der Blutproben einverstanden. Die Probandenauf-
klärung ist Bestandteil des Ethikvotums Nr. 3489 vom 9.Mai 2006 mit dem Titel
„Evaluierung von Testsystemen für die Apoptosemessung von mononukleären Zel-
len in gelagerten Leukozytenkonzentraten“.
4.2.1 Gewinnung der Leukozytenkonzentrate
4.2.1.1 Leukozytapherese mittels COM.TEC Zellseparator
Die Leukozytapherese wurde mit dem COM.TEC Zellseparator (Fresenius Hemo-
care GmbH) mit dem Programm autoMNC durchgeführt. Ein Zweinadel-Verfahren
ermöglicht die kontinuierliche Blutentnahme aus dem arteriellen Schenkel und Blut-
rückführung über den venösen Schenkel des Apheresesets. Nachdem das ACD-A
zu Vollblut (Acid citrate dextrose Formula A) im Verhältnis eins zu zehn eingestellt
wurde, erfolgt die Auftrennung in thrombozytenreiches Plasma und Blutzellen in der
PL1-Separationskammer mittels Differentialzentrifugation bei einer Zentrifugendreh-
zahl von 1500 Umdrehungen pro Minute. Die Auftrennung der Blutzellen ist durch
unterschiedliche Größen-, Dichte- und Sedimentationseigenschaften der Zellen
möglich [17]. In der sogenannten Sammelphase und in der Verdichtungsphase ist
eine automatische Trenngrenzenregelung aktiv. Als Trenngrenzenregler fungieren
acht optische Ports, die die Phasengrenze zwischen thrombozytenreichem Plasma
und Zellphase der Leukozyten und Erythrozyten konstant bei einem Wert von sie-
ben zu eins halten. Dadurch wird die Stabilität der abzusammelnden Zellschicht in
der sogenannten Sammelphase gewährleistet. Aufrechterhalten wird dieser Wert
durch eine Software-kontrollierte Plasmapumpe. Sieben optische Ports registrieren
dabei die Zellphase, während der achte Port das thrombozytenreiche Plasma er-
fasst. In den nachfolgenden Phasen (Spillover-Phase und Buffy-Coat-Phase) ist die
Trenngrenzenregelung deaktiviert. Der Blutfluss beträgt hier 15 ml pro Minute im
Gegensatz zur Sammelphase mit 50 ml pro Minute. Die Plasmapumpe ermöglicht
die Retransfusion des Plasmas zusammen mit dem Restblut an den Spender. Wäh-
renddessen entsteht in der Separationskammer zwischen der Erythrozyten- und
Plasmaschicht der sogenannte Buffy Coat, der die Leukozyten beinhaltet. Unter
einer reduzierten Blutflussrate von 15 ml pro min wird der Buffy Coat in der Separa-
tionskammer verdichtet. Anschließend erfolgt der Transport des Buffy Coats mittels
Plasma- und Zellenpumpe in einen Produktbeutel. Nach Abschluss der Zellaphere-
se werden aus diesem Buffy Coat mononukleäre Zellen isoliert. Beim autoMNC

17
Programm arbeitet die Plasmapumpe mit einem optischen Sensor, dem sogenann-
ten Spillover-Sensor, der die durch das thrombozytenreiche Plasma entstehende
Trübung, misst. Durch diesen Sensor, der eine Spillover-Klemme kontrolliert, wird
automatisch das Spillover-Volumen, das Volumen, das von der Plasmapumpe benö-
tigt wird, um den Buffy Coat aus der Separationskammer zu befördern, kontrolliert.
Nach Erreichen der voreingestellten 15 Sammelzyklen wird die Zellapherese nach
Rückführung des Restblutes an den Spender automatisch beendet [77; 79].
4.2.1.2 Thrombozytapherese mittels Trima Accel Zellseparator
Die Gewinnung von Leukozyten aus LRS-Kammern erfolgte im Rahmen der Throm-
bozytenherstellung mit dem Trima Accel Zellseparator (Gambro BCT, Lakewood,
USA). Diese Leukozytengewinnung aus LRS-Kammern gelingt nur mit Hilfe des
Trima Accel Zellseparators, da gezeigt werden konnte, dass die LRS-Kammern der
Cobe Spectra nur einen verschwindend geringen Anteil an Leukozyten enthalten
[78]. Bei dem verwendeten Zellseparator erfolgt die Blutentnahme und Blutrückgabe
mittels Ein-Nadel-System. Die Separation der Erythrozyten, Leukozyten, Throm-
bozyten und des Plasmas nach spezifischem Gewicht erfolgt ebenfalls durch Zentri-
fugation. Die Erythrozyten werden durch den kontinuierlichen Blutfluss wieder aus
der Separationskammer befördert. Innerhalb der Zentrifuge befindet sich die konisch
geformte ungefähr 10 ml fassende LRS-Kammer, in der sich die Trennung von
Thrombozyten und Leukozyten vollzieht. Die LRS-Kammer dient hierbei der Abrei-
cherung der Leukozyten im Thrombozytenprodukt. Die Leukozyten verbleiben in der
Kammer, da sie über eine höhere Dichte als die Thrombozyten verfügen, während
die Thrombozyten in den Beutel für das Thrombozytenkonzentrat abgesammelt
werden. Das technische Prinzip hierbei beruht darauf, dass die Restleukozyten bei
der Thrombozytenseparation in dieser Kammer mittels Flußbettfiltration abgefangen
werden. Der überwiegende Anteil der in der Kammer verbleibenden Zellen sind mo-
nonukleäre Zellen [54]. Im Vergleich zur Leukozytapherese verbleibt in den LRS-
Kammern nur ein kleiner Teil der Leukozyten, während der Großteil dieser Zellen
dem Spender wieder zugeführt wird. Wie hoch die Ausbeute an mononukleären
Zellen ausfällt, hängt von den Separatoreinstellungen wie Zitratbeimengung zur An-
tikoagulation und Blutflussgeschwindigkeit und der Verfahrensdauer ab [13; 54]. Da
die Zellen der LRS-Kammer als Nebenprodukt bei der Thrombozytapherese gewon-
nen werden, unterscheidet sich vor allem ihr Volumenanteil von dem der Leukozy-
tenkonzentrate, während die Zellkonzentration pro Volumeneinheit teilweise sogar
höher liegt [78].

18
4.2.2 Aufbereitung der Aphereseprodukte
Zur Bestimmung der Leukozytenzahl wurde das Blutbild an einem Sysmex Analy-
sengerät bestimmt. Vor der durchflußzytometrischen Messung wurden sowohl die
Leukozytenkonzentrate als auch das LRS-Kammern Material auf 10000 Leukozyten
pro µl Blut eingestellt, wobei die Verdünnung mit PBS erfolgte. Die Lagerung der
abgefüllten Beutelsysteme erfolgte bei Raumtemperatur zwischen 22°C und 26°C.
Jedes Konzentrat und LRS-Kammern Material wurde in einem Zeitraum von drei,
sechs, 24 und 48 Stunden nach Gewinnung am FACS gemessen. Zur jeweils ersten
Messung aus dem Leukozytenkonzentrat wurde parallel ein vom Spender stam-
mendes EDTA-Blutröhrchen, welches vor der Leukozytenspende gewonnen wurde,
untersucht. Zusätzlich wurde das EDTA-Blut zusammen mit dem Konzentrat hin-
sichtlich des Gehalts an Leukozyten, CD14 positiven Monozyten, Erythrozyten,
Thrombozyten und des Hämatokrits analysiert. Der Leukozytenanteil sowohl in den
Leukozytenkonzentraten als auch im EDTA-Blut wurde auf 100% gesetzt. Da die
CD14 positiven Zellen eine Subpopulation der Leukozyten sind, wurden sie als pro-
zentualer Anteil bezogen auf die Leukozyten angegeben. Die Leukozytenzahlen vor
der Spende aller Probanden lagen im für die Spende erlaubten Bereich zwischen
3000 und 10000 Leukozyten pro µl, so dass eine Verdünnung des EDTA-Blutes
nicht notwendig war. Jedes verwendete Untersuchungsmaterial wurde nach dem-
selben Protokoll aufbereitet und analysiert. Zur Bestimmung der biochemischen
Parameter Glukose und Laktat wurden zu Beginn einer jeden Messung aus den
unverdünnten Untersuchungsmaterialien 500 µl Blut in ein Natriumfluorid Röhrchen
pipettiert. Die Fluoridröhrchen wurden bei 3800xg für 10 min zentrifugiert und das
Plasma bis zum Ende einer Messreihe, längstens bis zu zwei Wochen bei minus
70°C eingefroren. Anschließend erfolgte der Transport zur Kinderklinik der Universi-
tät Erlangen, die die Bestimmung der Glukose- und Laktatwerte durchführten.
4.3 Methoden
4.3.1 Grundlagen der FACS Messung
Die Abkürzung FACS steht für Fluorescence Analysing Cell Sorting. Das Durchfluß-
zytometer besteht aus einem Flüssigkeitssystem zum Transport und Fokussierung
der Zellen, sowie einer Optik zur Detektierung der Lichtsignale und Elektronik, wobei
die optischen in elektronische Signale umgewandelt werden. Die Durchflußzytomet-
rie am FACS Gerät ermöglicht es Zellen quantitativ anhand ihrer Streulicht- und
Fluoreszenzeigenschaften zu analysieren. Die Methode beruht auf der Messung

19
und Analyse von Signalen, die dadurch zustande kommen, dass Zellen in einer
Suspension, die eine Glaskapillare durchströmen, einen Laser passieren. Das Vor-
wärtsstreulicht (Forward Scatter FSC) steht in Relation zur Beugung des Lichts,
wodurch die Zellen nach ihrer relativen Größe differenziert werden können. Das
Vorwärtsstreulicht wird entlang der Achse des einfallenden Lichtes gemessen.
Abb. 4: Darstellung der Vorwärtsstreulichtmessung
Das Seitwärtsstreulicht (Sidewards Scatter SSC) korreliert mit der Brechung und
Reflexion des Lichts und differenziert dadurch die Zellen anhand ihrer relativen Gra-
nularität oder internen Komplexität. Es wird in einem 90°-Winkel zum einfallenden
Licht gemessen. Dabei gilt je größer eine Zelle ist, desto stärker ist das Signal des
Vorwärtsstreulichts bzw. je granulierter eine Zelle, desto größer das Seitwärtsstreu-
lichtsignal [8].
Laserstrahl Vorwärtsstreu- lichtdetektor
Große Zellen erzeugen ein großes Vorwärts- streulichtsignal
Probenfluß
20
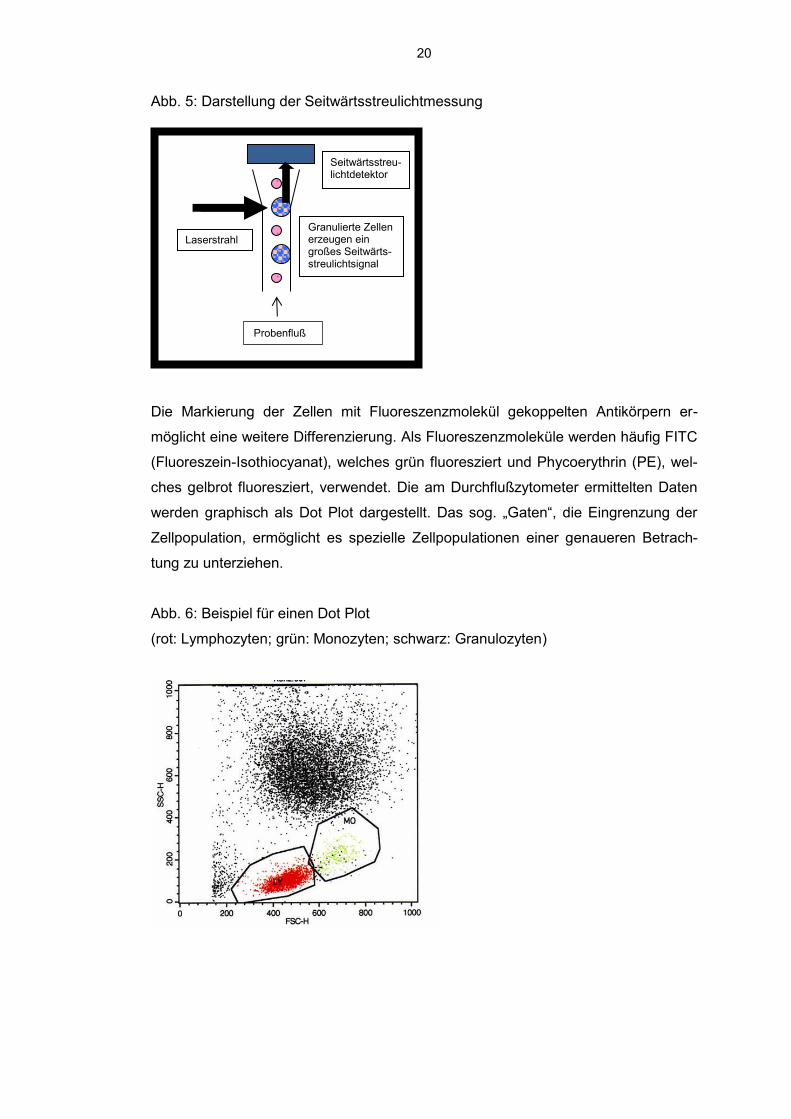
Abb. 5: Darstellung der Seitwärtsstreulichtmessung
Die Markierung der Zellen mit Fluoreszenzmolekül gekoppelten Antikörpern er-
möglicht eine weitere Differenzierung. Als Fluoreszenzmoleküle werden häufig FITC
(Fluoreszein-Isothiocyanat), welches grün fluoresziert und Phycoerythrin (PE), wel-
ches gelbrot fluoresziert, verwendet. Die am Durchflußzytometer ermittelten Daten
werden graphisch als Dot Plot dargestellt. Das sog. „Gaten“, die Eingrenzung der
Zellpopulation, ermöglicht es spezielle Zellpopulationen einer genaueren Betrach-
tung zu unterziehen.
Abb. 6: Beispiel für einen Dot Plot
(rot: Lymphozyten; grün: Monozyten; schwarz: Granulozyten)
Laserstrahl
Seitwärtsstreu- lichtdetektor
Granulierte Zellen erzeugen ein großes Seitwärts- streulichtsignal
Probenfluß

21
4.3.2 Apoptosedetektion mittels Annexin V und 7-AAD
Mit der Annexin V Färbung, welches bevorzugt an negativ geladene Phospholipide
wie Phosphatidylserin bindet, kann eine Frühphase der Apoptose aufgezeigt werden
[24; 40]. Konjugiert mit dem Fluoreszenzmolekül PE oder FITC kann es als sensiti-
ver Apoptosemarker durchflußzytometrisch nachgewiesen werden. Die Plasma-
membran in lebenden Zellen zeigt eine asymmetrische Phospholipid Verteilung.
Phosphatidylcholin und Sphingomyelin befinden sich auf der Außenseite während
Phosphatidylserin auf der Innenseite lokalisiert ist. Durch Verlust dieser Asymmetrie
wird das Phosphytidylserin nach außen transloziert, so dass das Annexin V daran
binden kann. Eine Differenzierung zu nekrotischen Zellen ist durch die Verknüpfung
der Annexin V Färbung mit dem kationischen Farbstoff 7-Amino-Actinomycin-D
(7AAD) möglich, da 7AAD nur in die Zelle dringen kann, wenn die Zellintegrität ver-
loren geht [19]. 7-AAD weist spezifisch DNA nach, so dass es in Zellen mit noch
intakter Plasmamembran nicht eindringen kann. Demnach zeigen sich apoptotische
Zellen Annexin V positiv und 7AAD negativ, während nekrotische Zellen Annexin V
positiv und 7AAD positiv sind [84].
Gegenstand der vorliegenden Untersuchungsreihe war der Apoptosenachweis
in regulatorischen T-Zellen (Tregs), weshalb initial die Leukozytenkonzentrate mit
CD4 und CD25 Antikörpern versetzt wurden, um zwischen CD4 positiven CD25
positiven Tregs und CD4 positiven CD25 negativen Tregs differenzieren zu können.
Die prozentualen Anteile an CD25 positiven und negativen Zellen in den beiden
unterschiedlichen Leukozytenprodukten beziehen sich auf den Anteil an T-Helfer-
zellen. Das heißt der T-Helferzellenanteil wurde 100% gleichgesetzt. Davon wird der
prozentuale Anteil an CD4 und CD25 positiven T-Zellen angegeben, während der
prozentuale Anteil an CD4 positiven und CD25 negativen T-Zellen sich aus der Dif-
ferenz zu 100% errechnet.
Zunächst werden 50 µl des Leukozytenkonzentrats mit 10 µl CD25 PE und 2,5
µl CD4 APC versetzt und eine Viertelstunde bei Raumtemperatur im Dunkeln inku-
biert. Das Leukozytenkonzentrat wurde zuvor mit PBS auf 10000 Leukozyten pro µl
Blut verdünnt. Anschließend erfolgt die zehnminütige Zelllyse mit 2 ml Pharm Lyse
(BD Biosciences) im Dunkeln. Vor Verwendung wird eine einfachkonzentrierte
Pharm Lyse mit Aqua dest. hergestellt. Nach Zentrifugation (5 min, 500 x g, 20°C)
wird der Überstand verworfen, das Zellpellet mit 2 ml PBS gewaschen und wie-
derum zentrifugiert (5 min, 500 x g, 20°C), mit anschließendem Dekantieren des
Überstandes. Es folgt die Apoptosefärbung mit 2,5 µl FITC Annexin V, 10 µl 7-AAD
und 100 µl Annexin Puffer. Der zehnfach konzentrierte Puffer muss vor Gebrauch
mit Aqua dest. zu einem einfach konzentrierten Puffer verdünnt werden. Es wird

22
eine Viertelstunde bei Raumtemperatur unter Lichtabschluss inkubiert. Nach an-
schließendem Zufügen von 400 µl einfachkonzentrierten Annexin Puffer wird die
Färbereaktion gestoppt, so dass die Probe durchflußzytometrisch gemessen werden
kann.
4.3.2.1 Messeinstellung für die Annexin V/ 7-AAD Methode
Zur optimalen Darstellung der einzelnen Zellpopulationen waren einige Einstellun-
gen am FACS Gerät notwendig. AMPS definiert die Spannungs- und Verstärkerein-
stellungen, welche sowohl vom FACS Typ als auch von der Art der Zellen und Pro-
benvorbereitung abhängig sind. Um die Lymphozyten optimal zwischen 400 und
600 im Dot Plot darzustellen, so dass auch die Monozyten- und Granulozytenpopu-
lation noch gut sichtbar sind, wurde AMPS im Vorwärtsscatter (FSC) auf 1,0 gestellt.
Im Seitwärtsscatter (SSC) wurden die Lymphozyten ebenfalls zur optimalen Darstel-
lung so eingestellt, dass sie im Bereich zwischen 100 und 200 liegen. Dies wurde
durch eine Voltageeinstellung im SSC Parameter auf 389 erreicht. Die Fluoreszenz-
intensitäten von Annexin V (FL-1) wurden in der Spannungseinstellung auf 638, von
CD25 PE (FL-2) auf 692, von 7AAD (FL-3) auf 813 und CD4 APC (FL-4) auf 675
eingestellt (siehe Tab. 1).
Tab.1: Darstellung der Instrumenteinstellungen („Instrument-Settings“)
Parameter Voltage AMPS Mode
FSC E00 1.00 Lin
SSC 389 1.00 Lin
FL-1 638 1.00 Log
FL-2 692 1.00 Log
FL-3 813 1.00 Log
FL1-A 1.00 Lin
FL-4 675 Log
Da mehr als eine Fluoreszenz simultan gemessen wird, ist auch eine Einstellung der
Kompensation erforderlich, um eine spektrale Überlappung auszugleichen. Die
Fluoreszenzintensität wird mit FL abgekürzt.
23
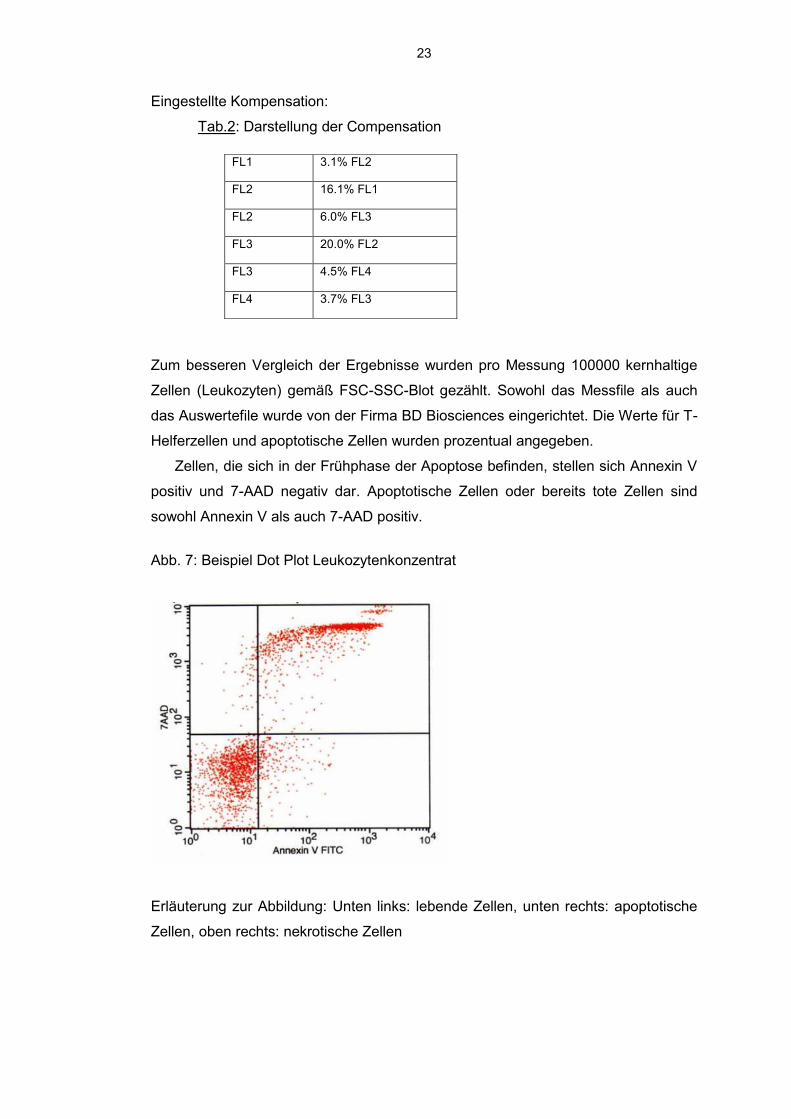
Eingestellte Kompensation:
Tab.2: Darstellung der Compensation
Zum besseren Vergleich der Ergebnisse wurden pro Messung 100000 kernhaltige
Zellen (Leukozyten) gemäß FSC-SSC-Blot gezählt. Sowohl das Messfile als auch
das Auswertefile wurde von der Firma BD Biosciences eingerichtet. Die Werte für T-
Helferzellen und apoptotische Zellen wurden prozentual angegeben.
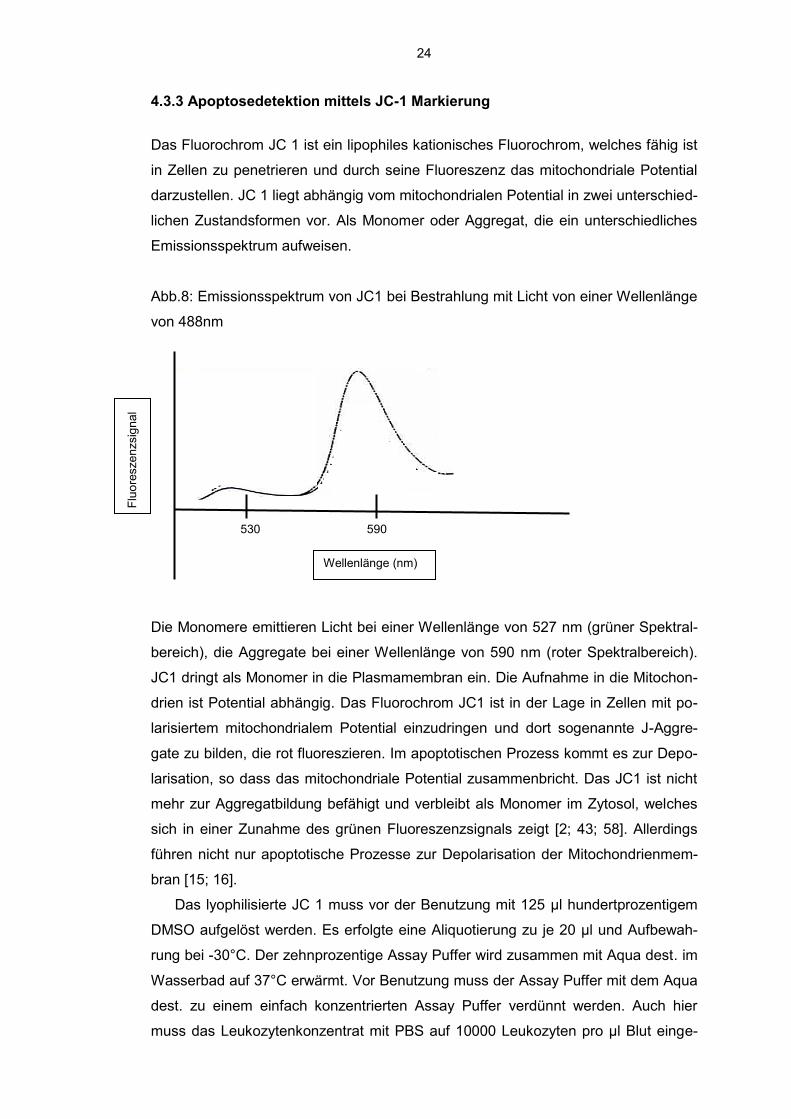
Zellen, die sich in der Frühphase der Apoptose befinden, stellen sich Annexin V
positiv und 7-AAD negativ dar. Apoptotische Zellen oder bereits tote Zellen sind
sowohl Annexin V als auch 7-AAD positiv.
Abb. 7: Beispiel Dot Plot Leukozytenkonzentrat
Erläuterung zur Abbildung: Unten links: lebende Zellen, unten rechts: apoptotische
Zellen, oben rechts: nekrotische Zellen
FL1 3.1% FL2
FL2 16.1% FL1
FL2 6.0% FL3
FL3 20.0% FL2
FL3 4.5% FL4
FL4 3.7% FL3
24
4.3.3 Apoptosedetektion mittels JC-1 Markierung
Das Fluorochrom JC 1 ist ein lipophiles kationisches Fluorochrom, welches fähig ist
in Zellen zu penetrieren und durch seine Fluoreszenz das mitochondriale Potential
darzustellen. JC 1 liegt abhängig vom mitochondrialen Potential in zwei unterschied-
lichen Zustandsformen vor. Als Monomer oder Aggregat, die ein unterschiedliches
Emissionsspektrum aufweisen.
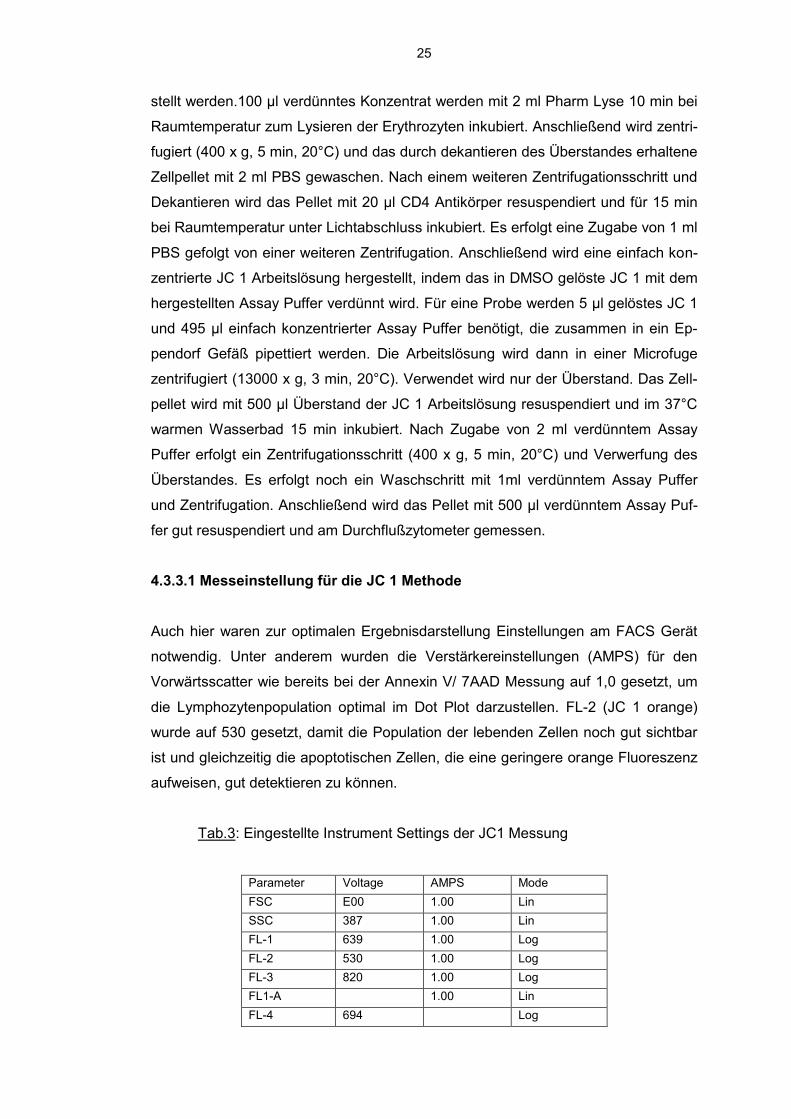
Abb.8: Emissionsspektrum von JC1 bei Bestrahlung mit Licht von einer Wellenlänge
von 488nm
530 590
Die Monomere emittieren Licht bei einer Wellenlänge von 527 nm (grüner Spektral-
bereich), die Aggregate bei einer Wellenlänge von 590 nm (roter Spektralbereich).
JC1 dringt als Monomer in die Plasmamembran ein. Die Aufnahme in die Mitochon-
drien ist Potential abhängig. Das Fluorochrom JC1 ist in der Lage in Zellen mit po-
larisiertem mitochondrialem Potential einzudringen und dort sogenannte J-Aggre-
gate zu bilden, die rot fluoreszieren. Im apoptotischen Prozess kommt es zur Depo-
larisation, so dass das mitochondriale Potential zusammenbricht. Das JC1 ist nicht
mehr zur Aggregatbildung befähigt und verbleibt als Monomer im Zytosol, welches
sich in einer Zunahme des grünen Fluoreszenzsignals zeigt [2; 43; 58]. Allerdings
führen nicht nur apoptotische Prozesse zur Depolarisation der Mitochondrienmem-
bran [15; 16].
Das lyophilisierte JC 1 muss vor der Benutzung mit 125 µl hundertprozentigem
DMSO aufgelöst werden. Es erfolgte eine Aliquotierung zu je 20 µl und Aufbewah-
rung bei -30°C. Der zehnprozentige Assay Puffer wird zusammen mit Aqua dest. im
Wasserbad auf 37°C erwärmt. Vor Benutzung muss der Assay Puffer mit dem Aqua
dest. zu einem einfach konzentrierten Assay Puffer verdünnt werden. Auch hier
muss das Leukozytenkonzentrat mit PBS auf 10000 Leukozyten pro µl Blut einge-
Flu
ore
sze
nzsig
na
l
Wellenlänge (nm)
25
stellt werden.100 µl verdünntes Konzentrat werden mit 2 ml Pharm Lyse 10 min bei
Raumtemperatur zum Lysieren der Erythrozyten inkubiert. Anschließend wird zentri-
fugiert (400 x g, 5 min, 20°C) und das durch dekantieren des Überstandes erhaltene
Zellpellet mit 2 ml PBS gewaschen. Nach einem weiteren Zentrifugationsschritt und
Dekantieren wird das Pellet mit 20 µl CD4 Antikörper resuspendiert und für 15 min
bei Raumtemperatur unter Lichtabschluss inkubiert. Es erfolgt eine Zugabe von 1 ml
PBS gefolgt von einer weiteren Zentrifugation. Anschließend wird eine einfach kon-
zentrierte JC 1 Arbeitslösung hergestellt, indem das in DMSO gelöste JC 1 mit dem
hergestellten Assay Puffer verdünnt wird. Für eine Probe werden 5 µl gelöstes JC 1
und 495 µl einfach konzentrierter Assay Puffer benötigt, die zusammen in ein Ep-
pendorf Gefäß pipettiert werden. Die Arbeitslösung wird dann in einer Microfuge
zentrifugiert (13000 x g, 3 min, 20°C). Verwendet wird nur der Überstand. Das Zell-
pellet wird mit 500 µl Überstand der JC 1 Arbeitslösung resuspendiert und im 37°C
warmen Wasserbad 15 min inkubiert. Nach Zugabe von 2 ml verdünntem Assay
Puffer erfolgt ein Zentrifugationsschritt (400 x g, 5 min, 20°C) und Verwerfung des
Überstandes. Es erfolgt noch ein Waschschritt mit 1ml verdünntem Assay Puffer
und Zentrifugation. Anschließend wird das Pellet mit 500 µl verdünntem Assay Puf-
fer gut resuspendiert und am Durchflußzytometer gemessen.
4.3.3.1 Messeinstellung für die JC 1 Methode
Auch hier waren zur optimalen Ergebnisdarstellung Einstellungen am FACS Gerät
notwendig. Unter anderem wurden die Verstärkereinstellungen (AMPS) für den
Vorwärtsscatter wie bereits bei der Annexin V/ 7AAD Messung auf 1,0 gesetzt, um
die Lymphozytenpopulation optimal im Dot Plot darzustellen. FL-2 (JC 1 orange)
wurde auf 530 gesetzt, damit die Population der lebenden Zellen noch gut sichtbar
ist und gleichzeitig die apoptotischen Zellen, die eine geringere orange Fluoreszenz
aufweisen, gut detektieren zu können.
Tab.3: Eingestellte Instrument Settings der JC1 Messung
Parameter Voltage AMPS Mode
FSC E00 1.00 Lin
SSC 387 1.00 Lin
FL-1 639 1.00 Log
FL-2 530 1.00 Log
FL-3 820 1.00 Log
FL1-A 1.00 Lin
FL-4 694 Log

26
Die JC 1 Messung benötigte auch eine Änderung der Treshold Einstellungen im
FSC-H Parameter auf 150. Treshold definiert den Schwellenwert, eine untere Sig-
nalgrenze, die vom entsprechenden Signal der Zelle überschritten werden muss,
damit die Zelldaten dargestellt werden können.
Tab.4: Treshold Einstellungen der JC1 Messung
Prim.Param. Sec.Param.
150 FSC-H FSC-H
52 SSC-H SSC-H
52 FL 1-H FL 1-H
52 FL 2-H FL 2-H
52 FL 3-H FL 3-H
52 FL 4-H
JC 1 Monomer emittiert bei 527 nm, nach Anregung bei 490 nm. Die Aggregate
emittieren bei 590 nm. Das Fluoreszenzspektrum beider JC 1 Zustandsformen liegt
im grünen Farbskalenbereich, der im FL-1 Kanal des Durchflußzytometers gemes-
sen wird. J-Aggregate bewirken eine reversible Verschiebung des Farbspektrums
von grün nach orange, die im FL-2 Kanal gemessen wird. Monomere sind nicht in
der Lage diese orange Fluoreszenz zu zeigen. Das Fluoreszenzsignal im FL-2 Ka-
nal ist deshalb sehr gering. Da sich die Aggregate nur bilden, wenn die Mitochon-
drienmembran polarisiert ist, kommt es bei Depolarisation zur Umwandlung in Mo-
nomere. Da die Depolarisation mit dem Apoptoseprozess in Verbindung steht,
nimmt in apoptotischen Zellen die orange Fluoreszenz ab.
Zum Vergleich der Leukozytenkonzentrate wurden pro Messung 10000 Ereig-
nisse gezählt.
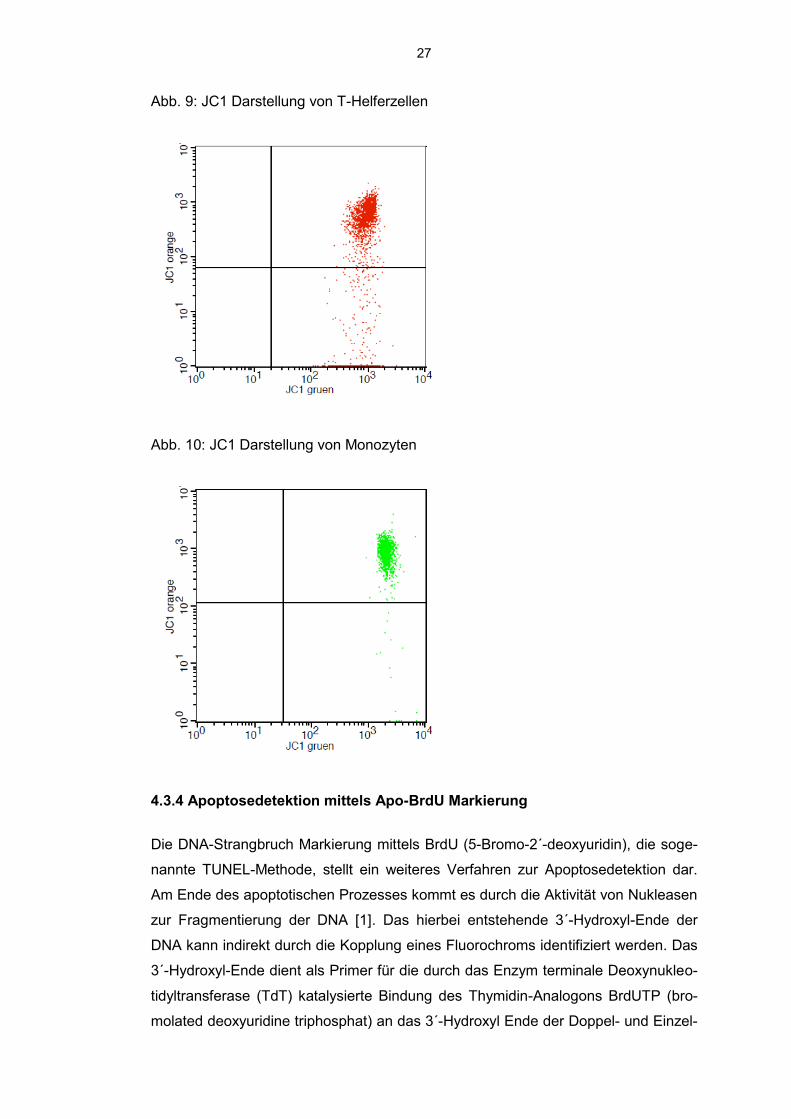
27
Abb. 9: JC1 Darstellung von T-Helferzellen
Abb. 10: JC1 Darstellung von Monozyten
4.3.4 Apoptosedetektion mittels Apo-BrdU Markierung
Die DNA-Strangbruch Markierung mittels BrdU (5-Bromo-2´-deoxyuridin), die soge-
nannte TUNEL-Methode, stellt ein weiteres Verfahren zur Apoptosedetektion dar.
Am Ende des apoptotischen Prozesses kommt es durch die Aktivität von Nukleasen
zur Fragmentierung der DNA [1]. Das hierbei entstehende 3´-Hydroxyl-Ende der
DNA kann indirekt durch die Kopplung eines Fluorochroms identifiziert werden. Das
3´-Hydroxyl-Ende dient als Primer für die durch das Enzym terminale Deoxynukleo-
tidyltransferase (TdT) katalysierte Bindung des Thymidin-Analogons BrdUTP (bro-
molated deoxyuridine triphosphat) an das 3´-Hydroxyl Ende der Doppel- und Einzel-

28
strang-DNA. Diese Bindung kann durch einen Anti-BrdU Antikörper, der mit dem
Fluorochrom FITC (Fluoreszein Isothiocyanat) gekoppelt ist, kenntlich gemacht wer-
den [18]. Voraussetzung ist eine Fixierung der fragmentierten DNA mit Formaldehyd
und die Erhöhung der Zellpermeabilität mittels Ethanol.
Wieder wird zu Beginn das Leukozytenkonzentrat auf 10000 Leukozyten pro µl
Blut verdünnt. In einem ersten Schritt erfolgt die Fixierung der Zellen. Zu 300 µl Leu-
kozytenkonzentrat wird 3 ml Pharm Lyse gegeben, um die Erythrozyten zu lysieren.
Nach einer zehnminütigen Inkubation bei Raumtemperatur wird zentrifugiert (500 x
g, 5 min, 20°C) und der Überstand verworfen. Es erfolgt ein Waschschritt mit 5 ml
PBS und anschließender Zentrifugation. Die zehnprozentige Formaldehydlösung
wird mit PBS verdünnt, um eine einprozentige Lösung zu erhalten. Vor Zugabe von
1 ml Formaldehydlösung (1%) muss das durch Zentrifugation erhaltene Zellpellet
gut gevortext werden. Inkubiert wird für 30 min auf einem Eisblock im 4°C Kühl-
schrank. Anschließend wird zentrifugiert (500 x g, 5 min, 20°C) und der Überstand
verworfen. Das Zellpellet wird mit 5 ml PBS gewaschen und noch einmal zentrifu-
giert. Dann erfolgt das Aufträufeln von 1 ml kalten 70% Ethanol, wobei bereits vor
und während des Aufträufelns das Pellet gut gevortext werden muss. Die Proben
werden so bei -20°C aufbewahrt und gesammelt.
Im zweiten Schritt erfolgt die Markierung und Färbung der Zellen. Die gesam-
melten Proben werden zusammen mit jeweils 1 ml der positiven und negativen Kon-
trollzellen zentrifugiert (300 x g, 5 min, 20°C) und der Überstand zur Ethanolentfer-
nung abpipettiert. Das Pellet wird anschließend mit 1 ml Wash Buffer resuspendiert
und wieder zentrifugiert. Dieser Waschschritt wird ein zweites Mal durchgeführt.
Dabei kann der Überstand durch Dekantieren entfernt werden. Jedes Zellpellet wird
mit 50 µl DNA Labeling Solution resuspendiert und für 60 min im 37°C warmen
Wasserbad inkubiert. Die DNA Labeling Solution für eine Probe enthält 10 µl Re-
action Buffer, 0,75 µl TdT Enzyme, 8 µl Brd-UTP und 32,25 µl Aqua dest. Alle 15
min werden die Zellen im Wasserbad kurz aufgeschüttelt. Am Ende der Inkuba-
tionszeit wird jeder Probe 1 ml Rinse Buffer zugefügt und zentrifugiert. Der Über-
stand wird abpipettiert und das Pellet wiederum mit 1 ml Rinse Buffer resuspendiert.
Anschließend muss noch einmal zentrifugiert und der Überstand abpipettiert wer-
den. Das Zellpellet wird jetzt in 0,1 ml Antibody Staining Solution aufgenommen und
30 min im Dunkeln bei Raumtemperatur inkubiert. Die Antibody Staining Solution
besteht pro Probe aus 5 µl FITC-labeled Anti-BrdU und 95 µl Rinsing Buffer. An-
schließend wird jeder Probe 500 µl PI/RNase Staining Buffer zugefügt, 30 min im
Dunkeln bei Raumtemperatur inkubiert und gemessen.
29
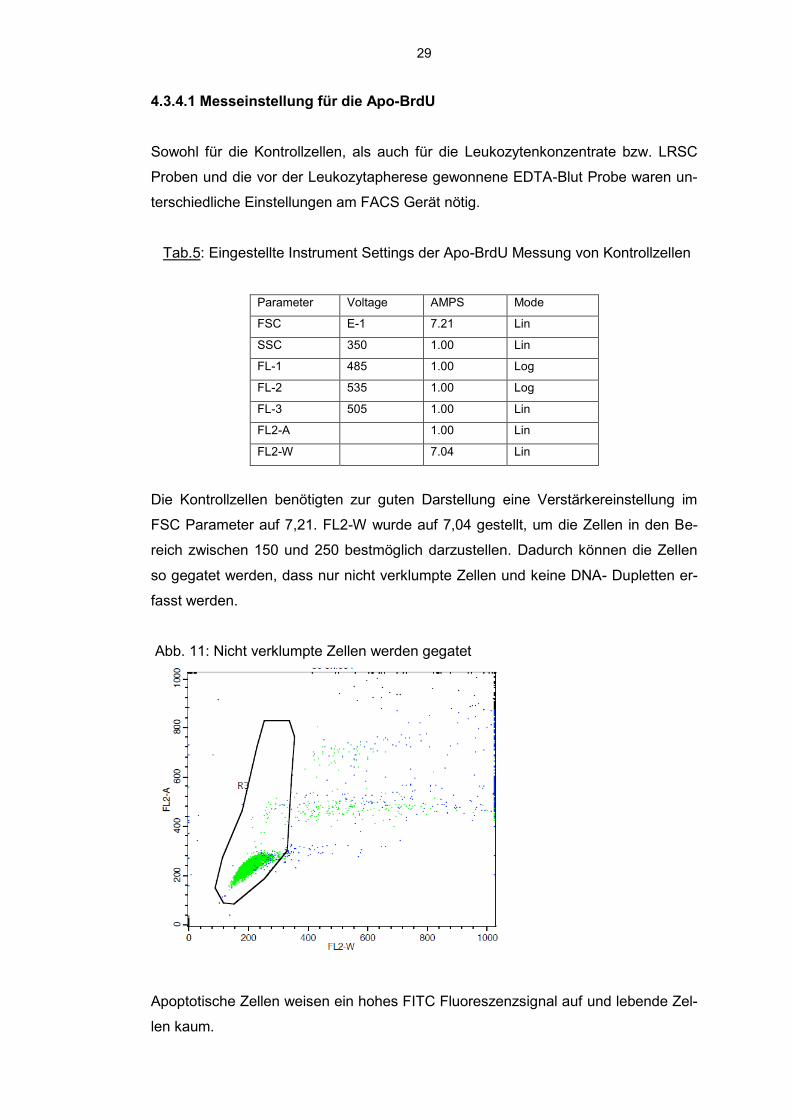
4.3.4.1 Messeinstellung für die Apo-BrdU
Sowohl für die Kontrollzellen, als auch für die Leukozytenkonzentrate bzw. LRSC
Proben und die vor der Leukozytapherese gewonnene EDTA-Blut Probe waren un-
terschiedliche Einstellungen am FACS Gerät nötig.
Tab.5: Eingestellte Instrument Settings der Apo-BrdU Messung von Kontrollzellen
Parameter Voltage AMPS Mode
FSC E-1 7.21 Lin
SSC 350 1.00 Lin
FL-1 485 1.00 Log
FL-2 535 1.00 Log
FL-3 505 1.00 Lin
FL2-A 1.00 Lin
FL2-W 7.04 Lin
Die Kontrollzellen benötigten zur guten Darstellung eine Verstärkereinstellung im
FSC Parameter auf 7,21. FL2-W wurde auf 7,04 gestellt, um die Zellen in den Be-
reich zwischen 150 und 250 bestmöglich darzustellen. Dadurch können die Zellen
so gegatet werden, dass nur nicht verklumpte Zellen und keine DNA- Dupletten er-
fasst werden.
Abb. 11: Nicht verklumpte Zellen werden gegatet
Apoptotische Zellen weisen ein hohes FITC Fluoreszenzsignal auf und lebende Zel-
len kaum.
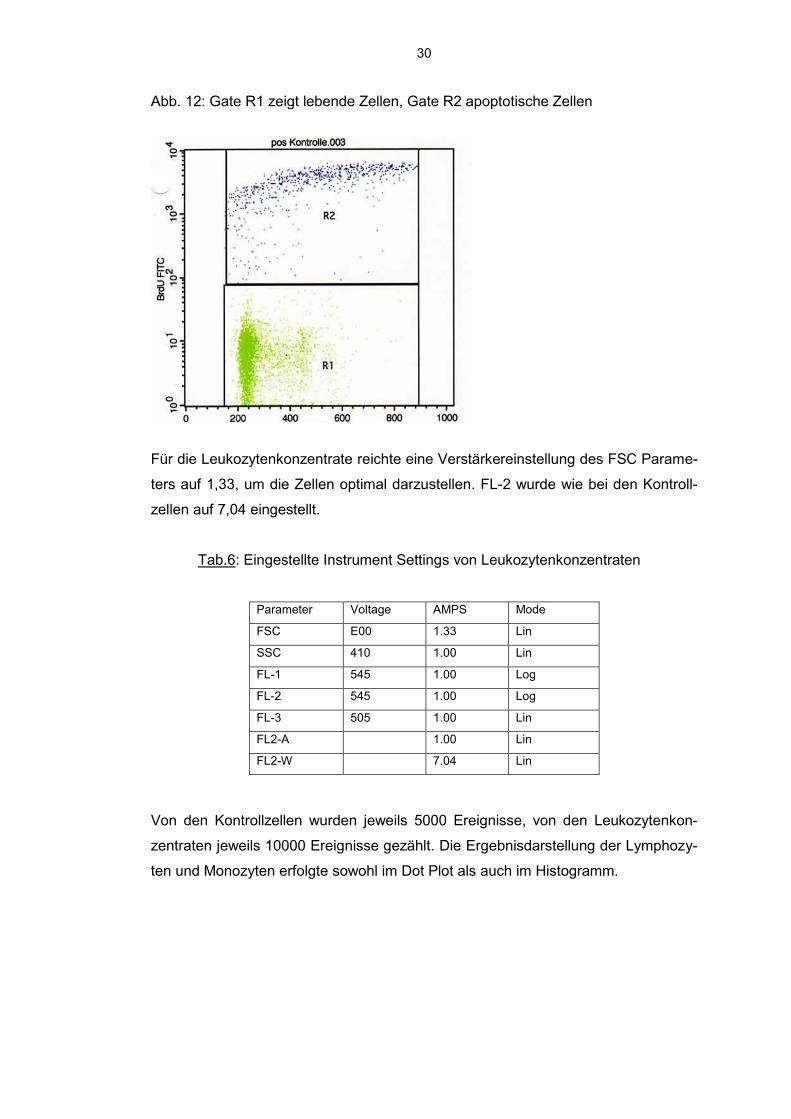
30
Abb. 12: Gate R1 zeigt lebende Zellen, Gate R2 apoptotische Zellen
Für die Leukozytenkonzentrate reichte eine Verstärkereinstellung des FSC Parame-
ters auf 1,33, um die Zellen optimal darzustellen. FL-2 wurde wie bei den Kontroll-
zellen auf 7,04 eingestellt.
Tab.6: Eingestellte Instrument Settings von Leukozytenkonzentraten
Parameter Voltage AMPS Mode
FSC E00 1.33 Lin
SSC 410 1.00 Lin
FL-1 545 1.00 Log
FL-2 545 1.00 Log
FL-3 505 1.00 Lin
FL2-A 1.00 Lin
FL2-W 7.04 Lin
Von den Kontrollzellen wurden jeweils 5000 Ereignisse, von den Leukozytenkon-
zentraten jeweils 10000 Ereignisse gezählt. Die Ergebnisdarstellung der Lymphozy-
ten und Monozyten erfolgte sowohl im Dot Plot als auch im Histogramm.
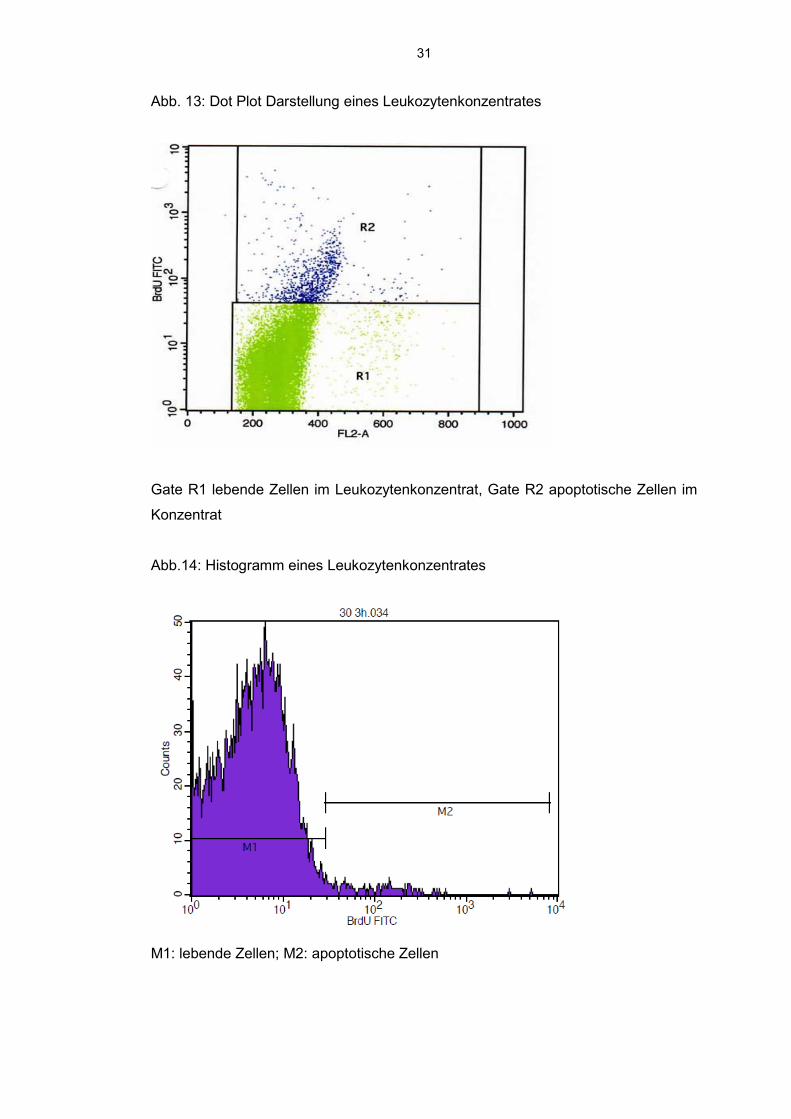
31
Abb. 13: Dot Plot Darstellung eines Leukozytenkonzentrates
Gate R1 lebende Zellen im Leukozytenkonzentrat, Gate R2 apoptotische Zellen im
Konzentrat
Abb.14: Histogramm eines Leukozytenkonzentrates
M1: lebende Zellen; M2: apoptotische Zellen

32
5. Statistische Auswertung Zur Auswertung der Daten wurde das Statistikprogramm SPSS Version 19 für
Windows benutzt. Zunächst wurden mittels deskriptiver Analyse die Mittelwerte und
Standardabweichungen der einzelnen Datensätze errechnet. Anschließend folgte
die Überprüfung der Daten auf Normalverteilung. Da eine kleine Spenderfallzahl (n
kleiner 50) vorlag, wurde zur Prüfung auf Normalverteilung der Shapiro-Wilks-Test
angewandt. Kam es zu einer Abweichung von der Normalverteilung mit einem Signi-
fikanzniveau von über 5%, d.h. der p-Wert ist kleiner als 0,05, lag keine Normalver-
teilung der Werte vor. Anschließend wurden die Daten, je nachdem ob Normalver-
teilung vorlag oder nicht, mit unterschiedlichen nicht-parametrischen Tests statist-
isch ausgewertet. Um die Mittelwerte zweier unterschiedlicher Stichproben zu ver-
gleichen, kam bei Normalverteilung der T-Test für zwei unabhängige Stichproben
zur Anwendung. Lag keine Normalverteilung vor, wurde der U-Test nach Wilcoxon-
Mann-Whitney für zwei unabhängige Stichproben verwendet. Signifikante Unter-
schiede wurden bei p-Werten kleiner als 0,05 und hochsignifikante Unterschiede bei
p-Werten kleiner als 0,01 angenommen. Die statistische Auswertung erforderte
auch den Vergleich mehrerer Messzeitpunkte bei denselben Probanden. Auch hier
wurde zuerst auf Normalverteilung mit dem Shapiro-Wilks-Test geprüft. Bei Normal-
verteilung wurde zum Mittelwertvergleich die einfaktorielle Varianzanalyse mit Mess-
wiederholung für abhängige Werte verwendet. Dieser Test zeigt auf, ob sich Mittel-
werte einer Gruppe im Laufe der Zeit verändern. Die Verwendung der Varianzana-
lyse mit Messwiederholung setzt Sphärizität voraus. Unter Spärizität versteht man
die Homogenität der Varianzen der einzelnen Messzeitpunkte und Homogenität der
Kovarianzen der Zusammenhänge zwischen den Messzeitpunkten. Dies wird mit
dem Mauchly Test überprüft. Der Mauchly Test wird unter der Annahme durchge-
führt, dass die Varianz der Messzeitpunkte gleich ist und die Kovarianz einen Wert
von 0 aufweist. Bei einem nichtsignifikanten p-Wert von ›0,05, kann diese Annahme
beibehalten werden. Der Mauchly Test auf Sphärizität hat bei einer kleinen Fallzahl
nur eine begrenzte Aussagekraft, da die Sphärizität verletzt werden kann ohne sig-
nifikante Werte anzuzeigen. Im Gegensatz dazu zeigt er bei großen Fallzahlen oft
signifikante Werte auf, obwohl die Sphärizität nicht verletzt wurde. Um eine mög-
lichst genaue Auswertung zu erhalten und da es sich um eine kleine Fallzahl han-
delte, wurde deshalb der Signifikanzwert im Test auf Innersubjekteffekte immer un-
ter Annahme der Greenhouse-Geisser Korrektur angegeben. Unter Test auf In-
nersubjekteffekte versteht man einen Test für Messwiederholungsfaktoren und ihre
Interaktionen, d.h. beispielsweise der Einfluss der Lagerungszeit auf die Apoptose
der Zellen. Lag keine Normalverteilung vor, kam die Friedman Zweifach-Rang-

33
varianzenanalyse für abhängige Stichproben zur Anwendung. Auch hier wurde ein
signifikanter Unterschied bei einem p-Wert kleiner als 0,05 und ein hochsignifikanter
Unterschied bei einem p-Wert kleiner als 0,01 angenommen. Zur Überprüfung eines
Zusammenhangs zwischen zwei Variablen, wurde der Korrelationskoeffizient be-
stimmt. Bei Normalverteilung wurde die Korrelation nach Pearson berechnet, wäh-
rend bei nicht-normalverteilten Werten die Korrelation nach Kendall-Tau und Spear-
man-Rho verwendet wurde. Bei Werten nahe 0 bestand kein Zusammenhang zwi-
schen den Variablen, nahe ± 1 bestand ein starker Zusammenhang [11; 23; 57; 61;
65].
34
6.Ergebnisse
6.1 Spenderdaten
In die statistische Auswertung ging eine Spenderfallzahl von zehn ein, wovon acht
Spender männlich und zwei Spender weiblich waren. Das durchschnittliche Alter lag
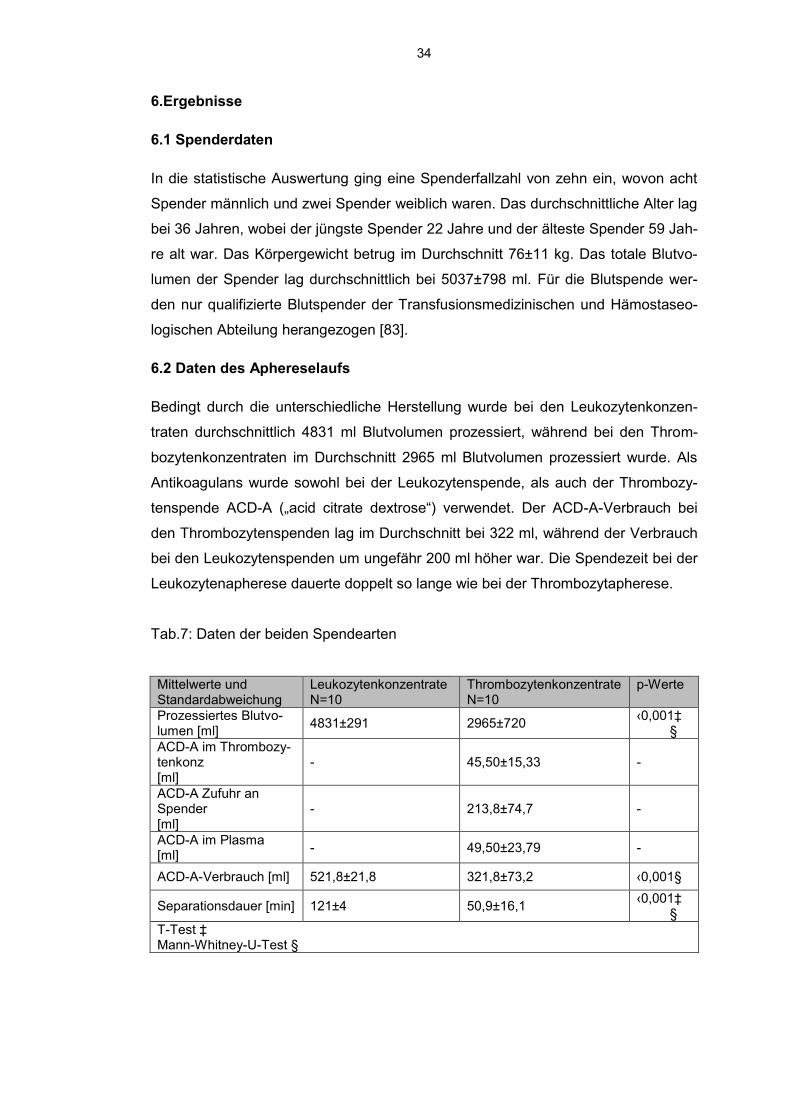
bei 36 Jahren, wobei der jüngste Spender 22 Jahre und der älteste Spender 59 Jah-
re alt war. Das Körpergewicht betrug im Durchschnitt 76±11 kg. Das totale Blutvo-
lumen der Spender lag durchschnittlich bei 5037±798 ml. Für die Blutspende wer-
den nur qualifizierte Blutspender der Transfusionsmedizinischen und Hämostaseo-
logischen Abteilung herangezogen [83].
6.2 Daten des Aphereselaufs
Bedingt durch die unterschiedliche Herstellung wurde bei den Leukozytenkonzen-
traten durchschnittlich 4831 ml Blutvolumen prozessiert, während bei den Throm-
bozytenkonzentraten im Durchschnitt 2965 ml Blutvolumen prozessiert wurde. Als
Antikoagulans wurde sowohl bei der Leukozytenspende, als auch der Thrombozy-
tenspende ACD-A („acid citrate dextrose“) verwendet. Der ACD-A-Verbrauch bei
den Thrombozytenspenden lag im Durchschnitt bei 322 ml, während der Verbrauch
bei den Leukozytenspenden um ungefähr 200 ml höher war. Die Spendezeit bei der
Leukozytenapherese dauerte doppelt so lange wie bei der Thrombozytapherese.
Tab.7: Daten der beiden Spendearten
Mittelwerte und Standardabweichung
Leukozytenkonzentrate N=10
Thrombozytenkonzentrate N=10
p-Werte
Prozessiertes Blutvo-lumen [ml]
4831±291 2965±720 ‹0,001‡ §
ACD-A im Thrombozy-tenkonz [ml]
- 45,50±15,33 -
ACD-A Zufuhr an Spender [ml]
- 213,8±74,7 -
ACD-A im Plasma [ml]
- 49,50±23,79 -
ACD-A-Verbrauch [ml] 521,8±21,8 321,8±73,2 ‹0,001§
Separationsdauer [min] 121±4 50,9±16,1 ‹0,001‡ §
T-Test ‡ Mann-Whitney-U-Test §
35
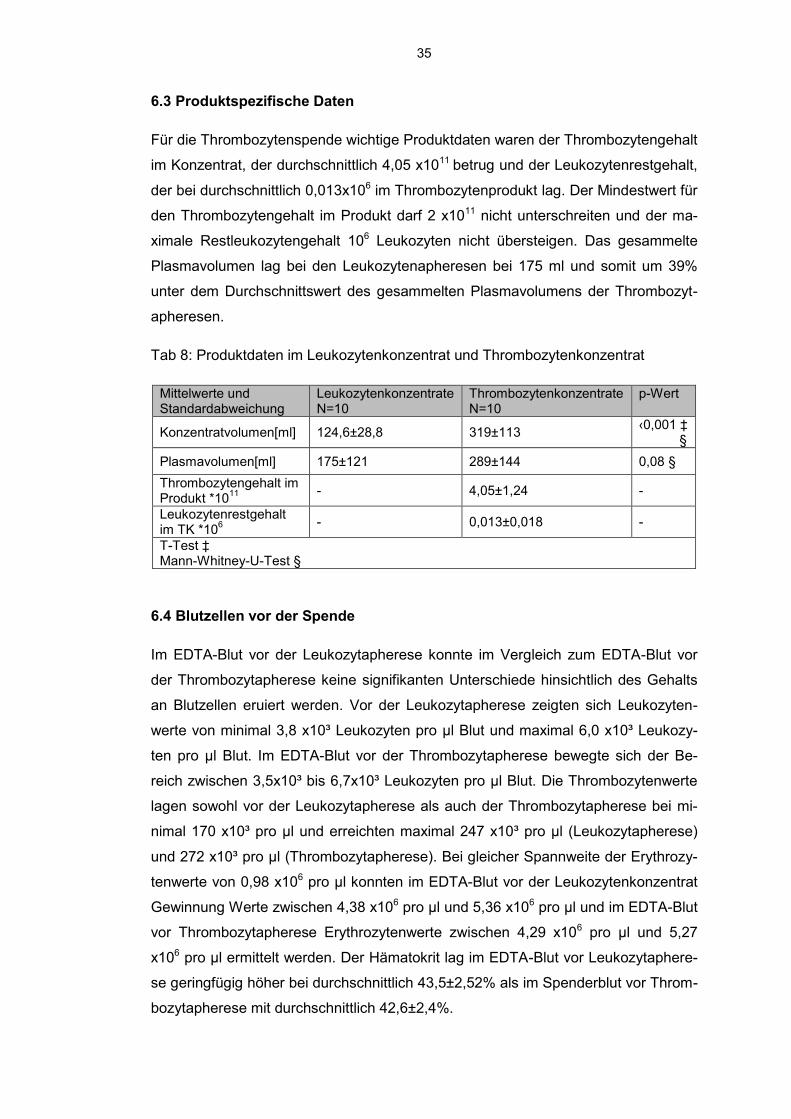
6.3 Produktspezifische Daten
Für die Thrombozytenspende wichtige Produktdaten waren der Thrombozytengehalt
im Konzentrat, der durchschnittlich 4,05 x1011 betrug und der Leukozytenrestgehalt,
der bei durchschnittlich 0,013x106 im Thrombozytenprodukt lag. Der Mindestwert für
den Thrombozytengehalt im Produkt darf 2 x1011 nicht unterschreiten und der ma-
ximale Restleukozytengehalt 106 Leukozyten nicht übersteigen. Das gesammelte
Plasmavolumen lag bei den Leukozytenapheresen bei 175 ml und somit um 39%
unter dem Durchschnittswert des gesammelten Plasmavolumens der Thrombozyt-
apheresen.
Tab 8: Produktdaten im Leukozytenkonzentrat und Thrombozytenkonzentrat
Mittelwerte und Standardabweichung
Leukozytenkonzentrate N=10
Thrombozytenkonzentrate N=10
p-Wert
Konzentratvolumen[ml] 124,6±28,8 319±113 ‹0,001 ‡ §
Plasmavolumen[ml] 175±121 289±144 0,08 §
Thrombozytengehalt im Produkt *10
11
- 4,05±1,24 -
Leukozytenrestgehalt im TK *10
6 - 0,013±0,018 -
T-Test ‡ Mann-Whitney-U-Test §
6.4 Blutzellen vor der Spende
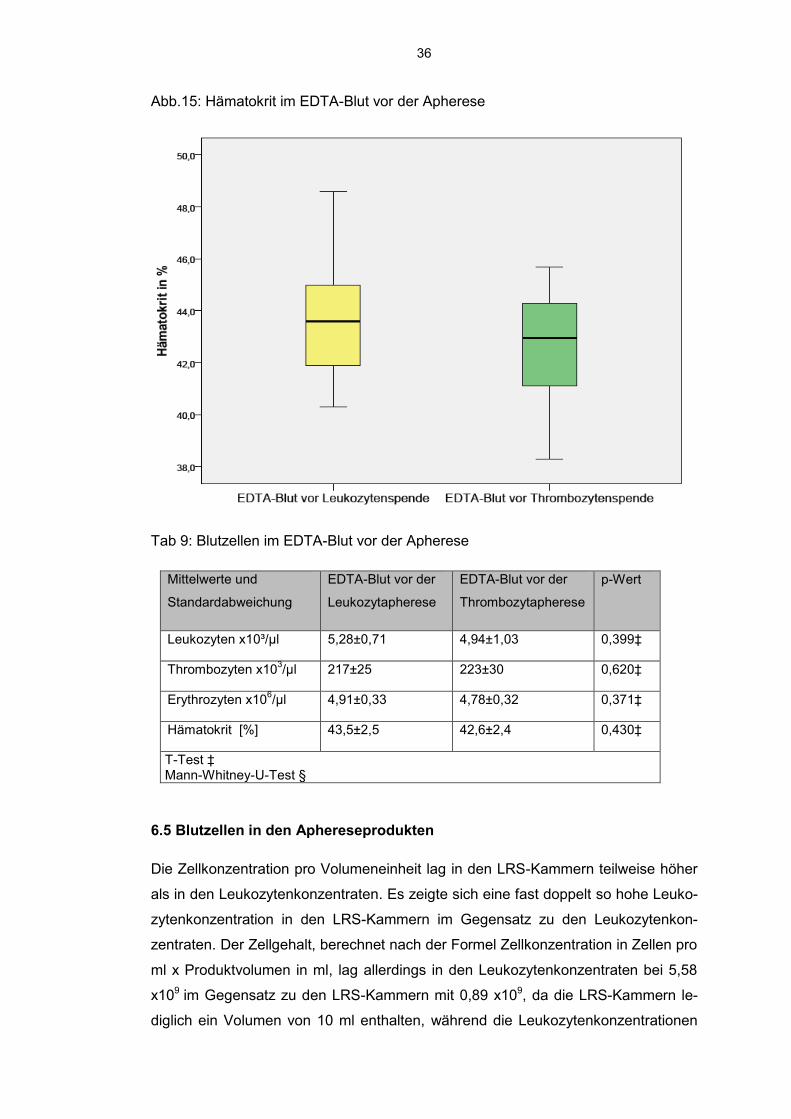
Im EDTA-Blut vor der Leukozytapherese konnte im Vergleich zum EDTA-Blut vor
der Thrombozytapherese keine signifikanten Unterschiede hinsichtlich des Gehalts
an Blutzellen eruiert werden. Vor der Leukozytapherese zeigten sich Leukozyten-
werte von minimal 3,8 x10³ Leukozyten pro µl Blut und maximal 6,0 x10³ Leukozy-
ten pro µl Blut. Im EDTA-Blut vor der Thrombozytapherese bewegte sich der Be-
reich zwischen 3,5x10³ bis 6,7x10³ Leukozyten pro µl Blut. Die Thrombozytenwerte
lagen sowohl vor der Leukozytapherese als auch der Thrombozytapherese bei mi-
nimal 170 x10³ pro µl und erreichten maximal 247 x10³ pro µl (Leukozytapherese)
und 272 x10³ pro µl (Thrombozytapherese). Bei gleicher Spannweite der Erythrozy-
tenwerte von 0,98 x106 pro µl konnten im EDTA-Blut vor der Leukozytenkonzentrat
Gewinnung Werte zwischen 4,38 x106 pro µl und 5,36 x106 pro µl und im EDTA-Blut
vor Thrombozytapherese Erythrozytenwerte zwischen 4,29 x106 pro µl und 5,27
x106 pro µl ermittelt werden. Der Hämatokrit lag im EDTA-Blut vor Leukozytaphere-
se geringfügig höher bei durchschnittlich 43,5±2,52% als im Spenderblut vor Throm-
bozytapherese mit durchschnittlich 42,6±2,4%.
36
Abb.15: Hämatokrit im EDTA-Blut vor der Apherese
Tab 9: Blutzellen im EDTA-Blut vor der Apherese
Mittelwerte und
Standardabweichung
EDTA-Blut vor der
Leukozytapherese
EDTA-Blut vor der
Thrombozytapherese
p-Wert
Leukozyten x10³/µl 5,28±0,71 4,94±1,03 0,399‡
Thrombozyten x103/µl 217±25 223±30 0,620‡
Erythrozyten x106/µl 4,91±0,33 4,78±0,32 0,371‡
Hämatokrit [%] 43,5±2,5 42,6±2,4 0,430‡
T-Test ‡ Mann-Whitney-U-Test §
6.5 Blutzellen in den Aphereseprodukten
Die Zellkonzentration pro Volumeneinheit lag in den LRS-Kammern teilweise höher
als in den Leukozytenkonzentraten. Es zeigte sich eine fast doppelt so hohe Leuko-
zytenkonzentration in den LRS-Kammern im Gegensatz zu den Leukozytenkon-
zentraten. Der Zellgehalt, berechnet nach der Formel Zellkonzentration in Zellen pro
ml x Produktvolumen in ml, lag allerdings in den Leukozytenkonzentraten bei 5,58
x109 im Gegensatz zu den LRS-Kammern mit 0,89 x109, da die LRS-Kammern le-
diglich ein Volumen von 10 ml enthalten, während die Leukozytenkonzentrationen
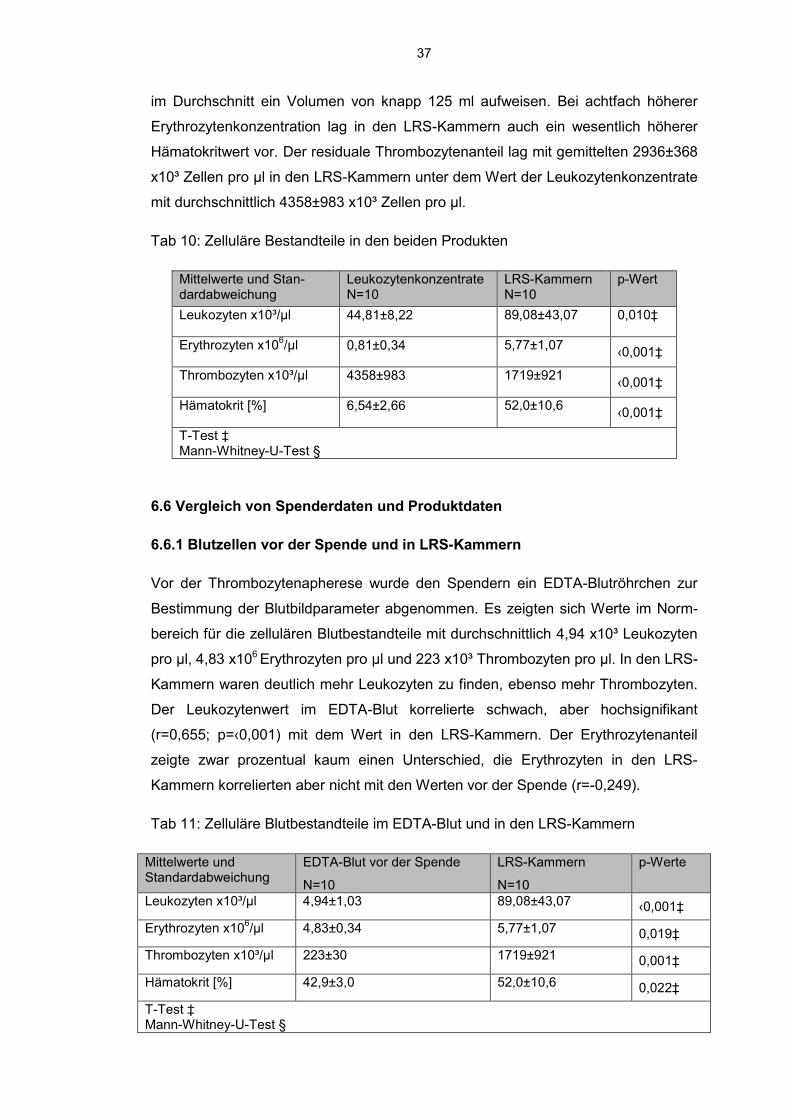
37
im Durchschnitt ein Volumen von knapp 125 ml aufweisen. Bei achtfach höherer
Erythrozytenkonzentration lag in den LRS-Kammern auch ein wesentlich höherer
Hämatokritwert vor. Der residuale Thrombozytenanteil lag mit gemittelten 2936±368
x10³ Zellen pro µl in den LRS-Kammern unter dem Wert der Leukozytenkonzentrate
mit durchschnittlich 4358±983 x10³ Zellen pro µl.
Tab 10: Zelluläre Bestandteile in den beiden Produkten
Mittelwerte und Stan-dardabweichung
Leukozytenkonzentrate N=10
LRS-Kammern N=10
p-Wert
Leukozyten x10³/µl 44,81±8,22 89,08±43,07 0,010‡
Erythrozyten x106/µl 0,81±0,34 5,77±1,07
‹0,001‡
Thrombozyten x10³/µl 4358±983 1719±921 ‹0,001‡
Hämatokrit [%] 6,54±2,66 52,0±10,6 ‹0,001‡
T-Test ‡ Mann-Whitney-U-Test §
6.6 Vergleich von Spenderdaten und Produktdaten
6.6.1 Blutzellen vor der Spende und in LRS-Kammern
Vor der Thrombozytenapherese wurde den Spendern ein EDTA-Blutröhrchen zur
Bestimmung der Blutbildparameter abgenommen. Es zeigten sich Werte im Norm-
bereich für die zellulären Blutbestandteile mit durchschnittlich 4,94 x10³ Leukozyten
pro µl, 4,83 x106 Erythrozyten pro µl und 223 x10³ Thrombozyten pro µl. In den LRS-
Kammern waren deutlich mehr Leukozyten zu finden, ebenso mehr Thrombozyten.
Der Leukozytenwert im EDTA-Blut korrelierte schwach, aber hochsignifikant
(r=0,655; p=‹0,001) mit dem Wert in den LRS-Kammern. Der Erythrozytenanteil
zeigte zwar prozentual kaum einen Unterschied, die Erythrozyten in den LRS-
Kammern korrelierten aber nicht mit den Werten vor der Spende (r=-0,249).
Tab 11: Zelluläre Blutbestandteile im EDTA-Blut und in den LRS-Kammern
Mittelwerte und Standardabweichung
EDTA-Blut vor der Spende
N=10
LRS-Kammern
N=10
p-Werte
Leukozyten x10³/µl 4,94±1,03 89,08±43,07 ‹0,001‡
Erythrozyten x106/µl 4,83±0,34 5,77±1,07 0,019‡
Thrombozyten x10³/µl 223±30 1719±921 0,001‡
Hämatokrit [%] 42,9±3,0 52,0±10,6 0,022‡
T-Test ‡ Mann-Whitney-U-Test §
38
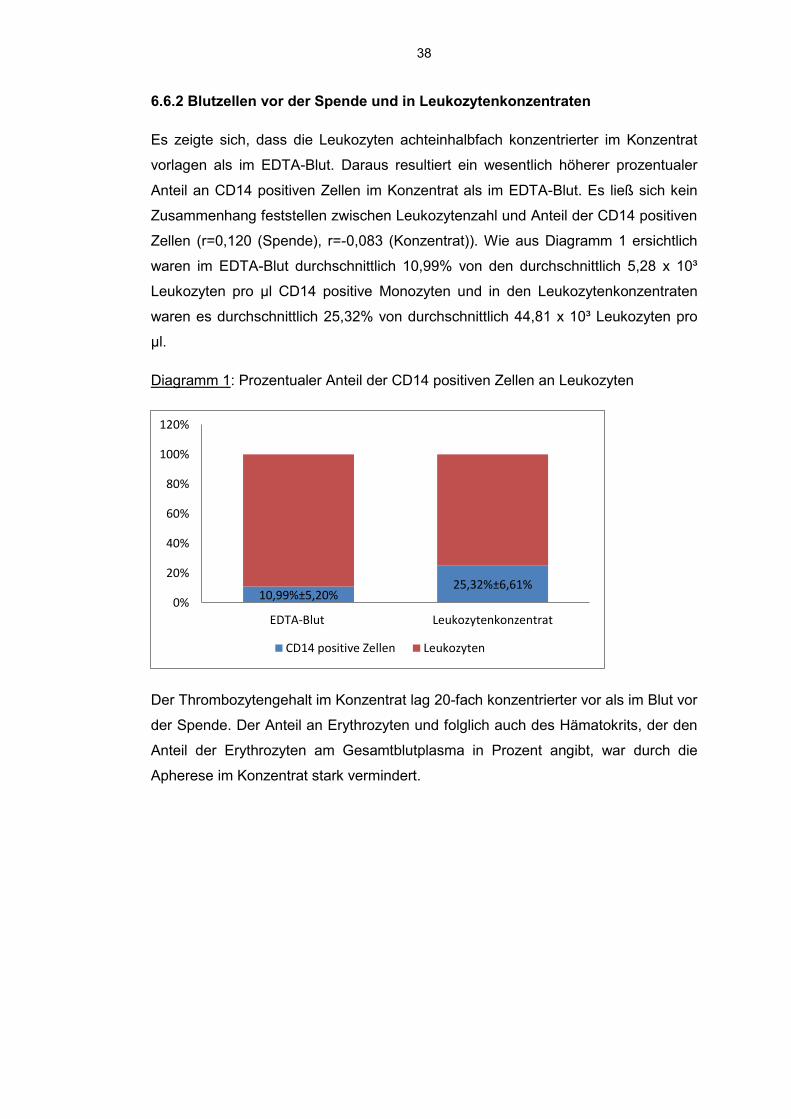
6.6.2 Blutzellen vor der Spende und in Leukozytenkonzentraten
Es zeigte sich, dass die Leukozyten achteinhalbfach konzentrierter im Konzentrat
vorlagen als im EDTA-Blut. Daraus resultiert ein wesentlich höherer prozentualer
Anteil an CD14 positiven Zellen im Konzentrat als im EDTA-Blut. Es ließ sich kein
Zusammenhang feststellen zwischen Leukozytenzahl und Anteil der CD14 positiven
Zellen (r=0,120 (Spende), r=-0,083 (Konzentrat)). Wie aus Diagramm 1 ersichtlich
waren im EDTA-Blut durchschnittlich 10,99% von den durchschnittlich 5,28 x 10³
Leukozyten pro µl CD14 positive Monozyten und in den Leukozytenkonzentraten
waren es durchschnittlich 25,32% von durchschnittlich 44,81 x 10³ Leukozyten pro
µl.
Diagramm 1: Prozentualer Anteil der CD14 positiven Zellen an Leukozyten
Der Thrombozytengehalt im Konzentrat lag 20-fach konzentrierter vor als im Blut vor
der Spende. Der Anteil an Erythrozyten und folglich auch des Hämatokrits, der den
Anteil der Erythrozyten am Gesamtblutplasma in Prozent angibt, war durch die
Apherese im Konzentrat stark vermindert.
10,99%±5,20% 25,32%±6,61%
0%
20%
40%
60%
80%
100%
120%
EDTA-Blut Leukozytenkonzentrat
CD14 positive Zellen Leukozyten
39
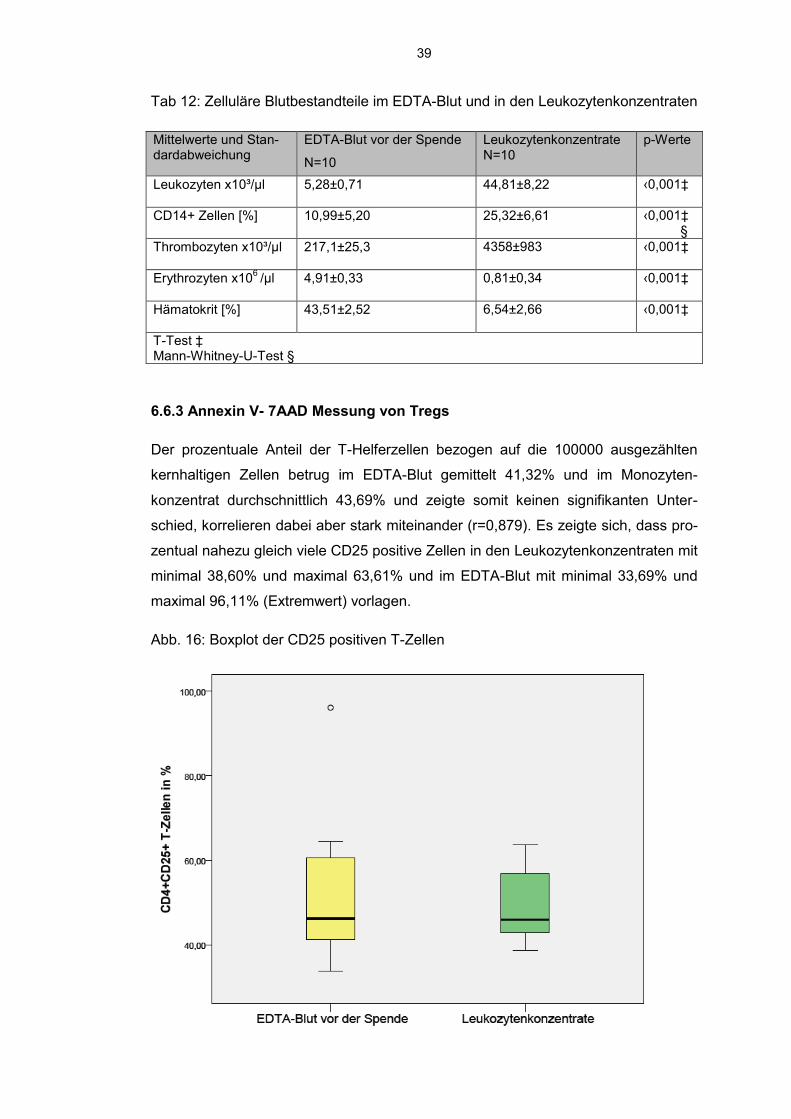
Tab 12: Zelluläre Blutbestandteile im EDTA-Blut und in den Leukozytenkonzentraten
Mittelwerte und Stan-dardabweichung
EDTA-Blut vor der Spende
N=10
Leukozytenkonzentrate N=10
p-Werte
Leukozyten x10³/µl 5,28±0,71 44,81±8,22 ‹0,001‡
CD14+ Zellen [%] 10,99±5,20 25,32±6,61 ‹0,001‡ §
Thrombozyten x10³/µl 217,1±25,3 4358±983 ‹0,001‡
Erythrozyten x106 /µl 4,91±0,33 0,81±0,34 ‹0,001‡
Hämatokrit [%] 43,51±2,52 6,54±2,66 ‹0,001‡
T-Test ‡ Mann-Whitney-U-Test §
6.6.3 Annexin V- 7AAD Messung von Tregs
Der prozentuale Anteil der T-Helferzellen bezogen auf die 100000 ausgezählten
kernhaltigen Zellen betrug im EDTA-Blut gemittelt 41,32% und im Monozyten-
konzentrat durchschnittlich 43,69% und zeigte somit keinen signifikanten Unter-
schied, korrelieren dabei aber stark miteinander (r=0,879). Es zeigte sich, dass pro-
zentual nahezu gleich viele CD25 positive Zellen in den Leukozytenkonzentraten mit
minimal 38,60% und maximal 63,61% und im EDTA-Blut mit minimal 33,69% und
maximal 96,11% (Extremwert) vorlagen.
Abb. 16: Boxplot der CD25 positiven T-Zellen
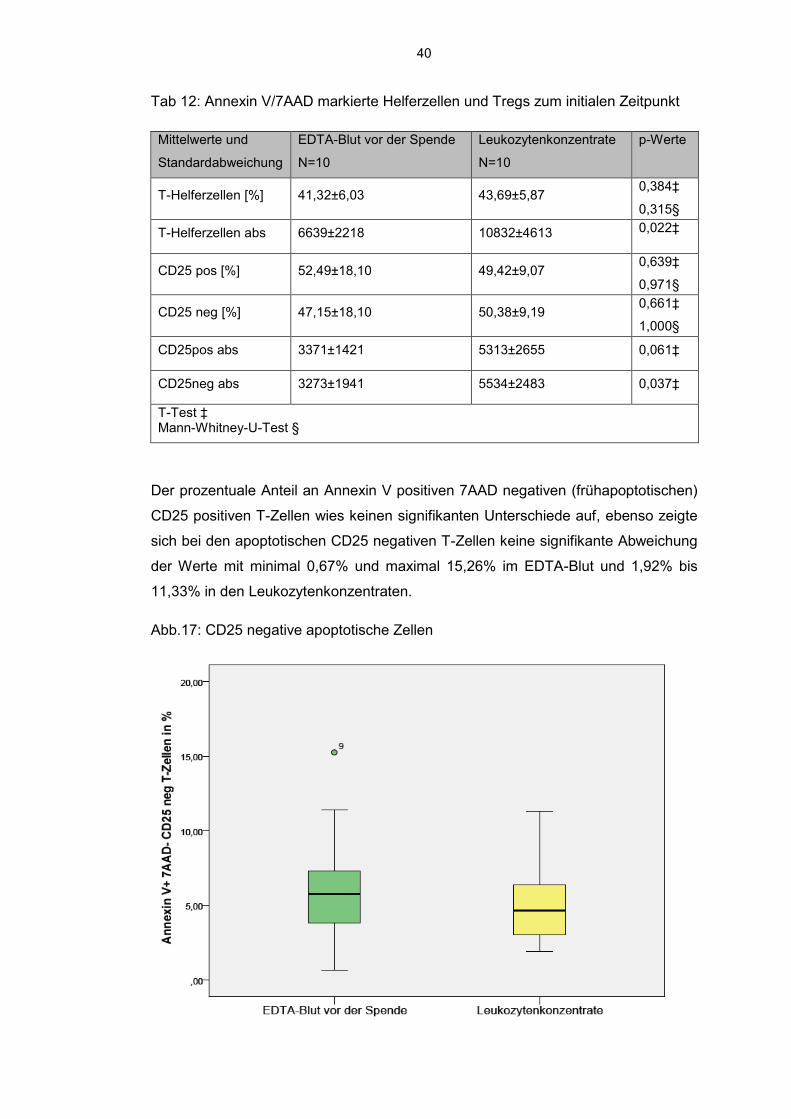
40
Tab 12: Annexin V/7AAD markierte Helferzellen und Tregs zum initialen Zeitpunkt
Mittelwerte und
Standardabweichung
EDTA-Blut vor der Spende
N=10
Leukozytenkonzentrate
N=10
p-Werte
T-Helferzellen [%] 41,32±6,03 43,69±5,87 0,384‡
0,315§
T-Helferzellen abs 6639±2218 10832±4613 0,022‡
CD25 pos [%] 52,49±18,10 49,42±9,07 0,639‡
0,971§
CD25 neg [%] 47,15±18,10 50,38±9,19 0,661‡
1,000§
CD25pos abs 3371±1421 5313±2655 0,061‡
CD25neg abs 3273±1941 5534±2483 0,037‡
T-Test ‡ Mann-Whitney-U-Test §
Der prozentuale Anteil an Annexin V positiven 7AAD negativen (frühapoptotischen)
CD25 positiven T-Zellen wies keinen signifikanten Unterschiede auf, ebenso zeigte
sich bei den apoptotischen CD25 negativen T-Zellen keine signifikante Abweichung
der Werte mit minimal 0,67% und maximal 15,26% im EDTA-Blut und 1,92% bis
11,33% in den Leukozytenkonzentraten.
Abb.17: CD25 negative apoptotische Zellen
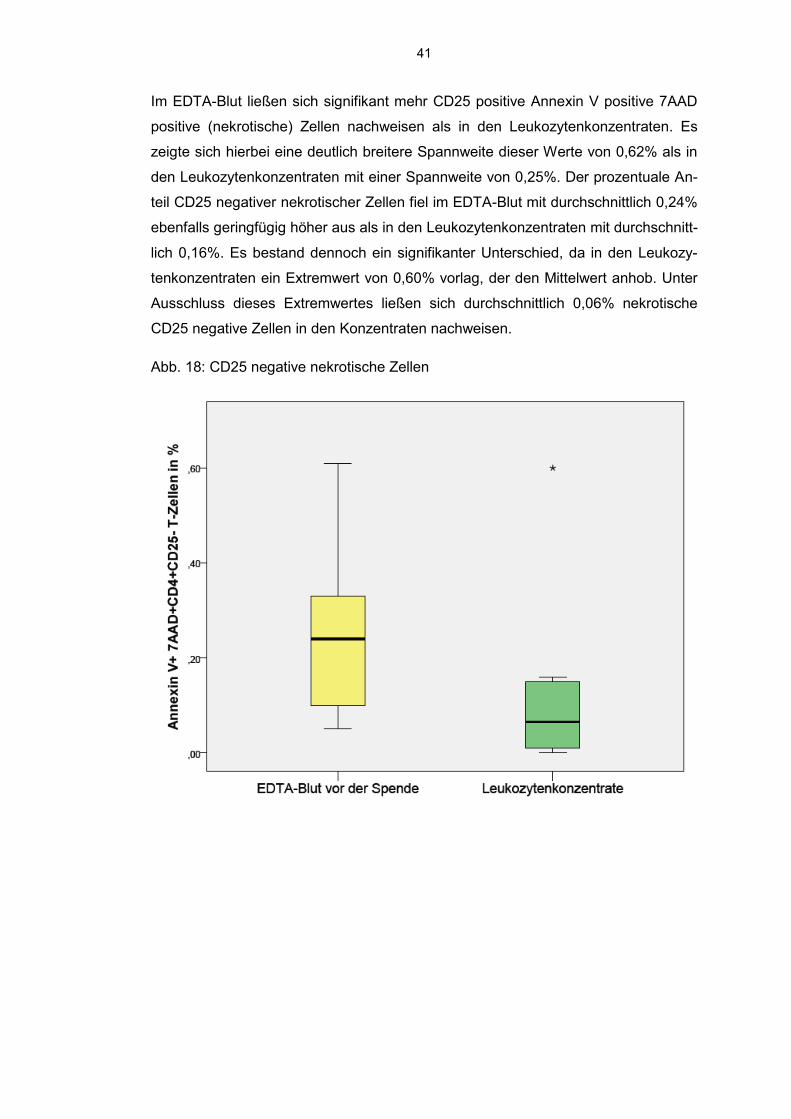
41
Im EDTA-Blut ließen sich signifikant mehr CD25 positive Annexin V positive 7AAD
positive (nekrotische) Zellen nachweisen als in den Leukozytenkonzentraten. Es
zeigte sich hierbei eine deutlich breitere Spannweite dieser Werte von 0,62% als in
den Leukozytenkonzentraten mit einer Spannweite von 0,25%. Der prozentuale An-
teil CD25 negativer nekrotischer Zellen fiel im EDTA-Blut mit durchschnittlich 0,24%
ebenfalls geringfügig höher aus als in den Leukozytenkonzentraten mit durchschnitt-
lich 0,16%. Es bestand dennoch ein signifikanter Unterschied, da in den Leukozy-
tenkonzentraten ein Extremwert von 0,60% vorlag, der den Mittelwert anhob. Unter
Ausschluss dieses Extremwertes ließen sich durchschnittlich 0,06% nekrotische
CD25 negative Zellen in den Konzentraten nachweisen.
Abb. 18: CD25 negative nekrotische Zellen
42
Tab 13: Annexin V/7AAD markierte Tregs zum initialen Zeitpunkt
Mittelwerte und
Standardabweichung
EDTA-Blut vor der Spende
N=10
Leukozytenkonzentrate
N=10
p-Werte
CD25 pos
apoptotisch [%] 9,39±3,59 10,09±2,33 0,611‡
CD25 neg
apoptotisch [%] 6,38±4,19 5,22±3,05 0,490‡
CD25 pos
nekrotisch [%] 0,31±0,20 0,16±0,82 0,041‡
CD25 neg
nekrotisch [%] 0,24±0,17 0,12±0,18
0,140‡
0,035§
T-Test ‡ Mann-Whitney-U-Test §
6.6.4 JC1-Messung von Lymphozyten und Monozyten
Tabelle 14 zeigt die sowohl vitalen als auch apoptotischen Lymphozyten und Mo-
nozyten nach JC1 Markierung. Die vitalen Lymphozyten des EDTA-Blutes zeigten
einen Minimalwert von 86,22% und einen Maximalwert von 98,52% und unterschie-
den sich nicht signifikant von den Werten im Leukozytenkonzentrat, in welchem sich
die Werte über eine Spannbreite von 75,97% bis 96,60% verteilten. Ebenso zeigte
sich bei den vitalen Monozyten, die im EDTA-Blut einen Spannweitenbereich von
6,15% und im Leukozytenkonzentrat einen Bereich von 6,75% umfassen, kein signi-
fikantes Ergebnis.
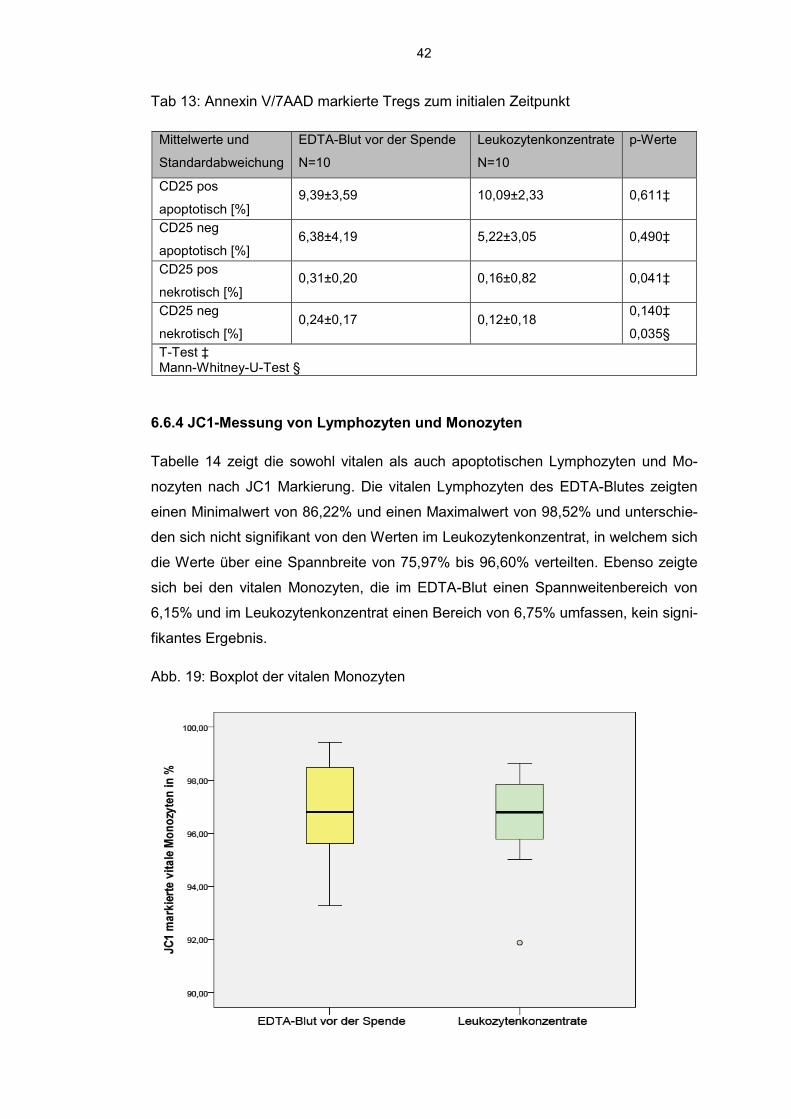
Abb. 19: Boxplot der vitalen Monozyten
43
Dieser Spannweitenbereich blieb auch bei den apoptotischen Monozyten erhalten.
Die apoptotischen Lymphozyten zeigten eine wesentlich größere Streuung der Wer-
te im Leukozytenkonzentrat mit minimal 1,48% und maximal 13,14% apoptotischer
Zellen als im EDTA-Blut mit einem Minimalwert von 0,57% und einem Maximalwert
von 6,72%.
Tab 14: JC1 markierte mononukleäre Zellen zum initialen Zeitpunkt
Mittelwerte und
Standardabweichung
EDTA-Blut vor der Spende
N=10
Leukozytenkonzentrate
N=10
p-Werte
Lymphozyten vital
[%] 93,61±3,71 88,96±6,02 0,055‡
Monozyten vital
[%] 96,83±1,89 96,51±2,00 0,718‡
Lymphozyten apopto-
tisch [%] 6,25±3,58 11,05±6,02 0,047‡
Monozyten apopto-
tisch [%] 3,16±1,88 3,49±2,00 0,702‡
T-Test ‡ Mann-Whitney-U-Test §
6.6.5 BrdU Messung von mononukleären Zellen
Die mit BRDU markierten mononukleären Zellen wurden hinsichtlich vitaler und
apoptotischer Zellen unterschieden. Es zeigte sich eine sehr große Spannweite der
Werte im EDTA-Blut sowohl bei den vitalen mononukleären Zellen mit minimal
65,68% (Extremwert) und maximal 98,80% als auch bei den apoptotischen Zellen
mit einem Minimalwert von 1,35% und einem Maximalwert von 34,32% (Extrem-
wert), während im Leukozytenkonzentrat die Spannweite bei den vitalen Zellen von
96,48% bis 99,63% und bei den apoptotischen Zellen zwischen 0,37% bis 3,42%
lag. Unter Vernachlässigung der Extremwerte ließ sich kein signifikanter Unter-
schied zwischen EDTA-Blut und Leukozytenkonzentraten nachweisen.
44
Tab 15: BRDU markierte mononukleäre Zellen zum initialen Zeitpunkt
Mittelwerte und
Standardabweichung
EDTA-Blut vor der Spende
N=10
Leukozytenkonzentrate
N=10
p-Werte
MNC vital
[%] 90,90±12,40 98,38±1,05
0,089‡
0,007§
MNC apoptotisch
[%] 8,92±12,48 1,61±1,04
0,098‡
0,019§
T-Test ‡ Mann-Whitney-U-Test §
6.7 Darstellung mittels unterschiedlicher Analysen
6.7.1 Annexin V- 7AAD positive Tregs
Der prozentuale Anteil an T-Helferzellen in den Leukozytenkonzentraten betrug
durchschnittlich 43,69±5,87% und in den LRS-Kammern 45,30±6,11%. Davon wur-
de der prozentuale Anteil an regulatorischen T-Zellen ermittelt, die als Oberflä-
chenantigene CD4 positiv und CD25 positiv waren. Die folgenden Diagramme zei-
gen, dass sich die Anteile an regulatorischen T-Zellen in den Leukozytenkonzentra-
ten und in den LRS-Kammern nicht signifikant voneinander unterschieden.
Diagramm 2: Prozentuale Anteile der Tregs zum initialen Zeitpunkt
49,42±9,07% 51,40+16,85%
50,38±9,19% 48,60±16,85%
0%
20%
40%
60%
80%
100%
Leukozytenkonzentrate LRS-Kammern
CD25 positive T-Zellen CD25 negative T-Zellen
45
Diagramm 3: Prozentuale Anteile der Tregs nach 48-stündiger Lagerung
Drei Stunden nach Konzentratgewinnung befanden sich in den Leukozytenkon-
zentraten zwischen 6,30% bis 13,73% apoptotische CD25 positive T-Zellen und
somit hochsignifikant mehr als in den LRS-Kammern mit minimal 3,11% bis maximal
10,37%.
Abb. 20: Boxplot der apoptotischen CD25 positiven Zellen zum initialen Zeitpunkt
41,41±8,0% 49,80±17,44%
58,60±8,00% 50,20±17,44%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Leukozytenkonzentrate LRS-Kammern
CD25 positive T-Zellen CD25 negative T-Zellen
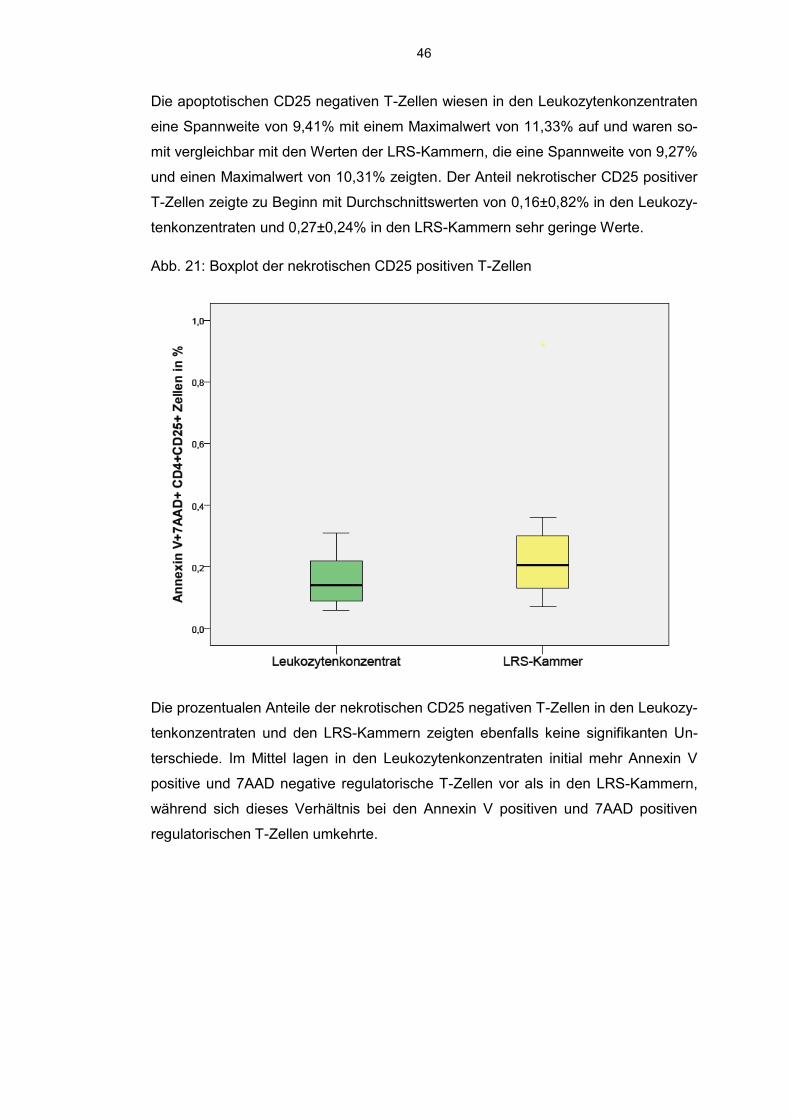
46
Die apoptotischen CD25 negativen T-Zellen wiesen in den Leukozytenkonzentraten
eine Spannweite von 9,41% mit einem Maximalwert von 11,33% auf und waren so-
mit vergleichbar mit den Werten der LRS-Kammern, die eine Spannweite von 9,27%
und einen Maximalwert von 10,31% zeigten. Der Anteil nekrotischer CD25 positiver
T-Zellen zeigte zu Beginn mit Durchschnittswerten von 0,16±0,82% in den Leukozy-
tenkonzentraten und 0,27±0,24% in den LRS-Kammern sehr geringe Werte.
Abb. 21: Boxplot der nekrotischen CD25 positiven T-Zellen
Die prozentualen Anteile der nekrotischen CD25 negativen T-Zellen in den Leukozy-
tenkonzentraten und den LRS-Kammern zeigten ebenfalls keine signifikanten Un-
terschiede. Im Mittel lagen in den Leukozytenkonzentraten initial mehr Annexin V
positive und 7AAD negative regulatorische T-Zellen vor als in den LRS-Kammern,
während sich dieses Verhältnis bei den Annexin V positiven und 7AAD positiven
regulatorischen T-Zellen umkehrte.
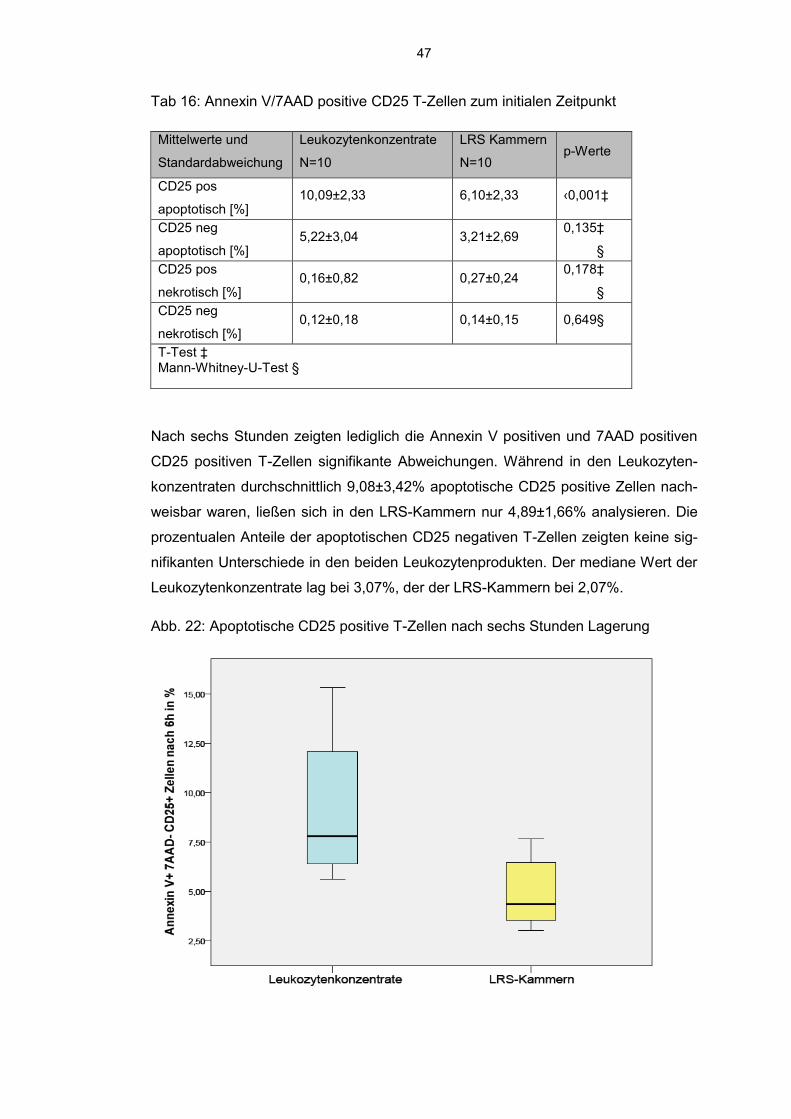
47
Tab 16: Annexin V/7AAD positive CD25 T-Zellen zum initialen Zeitpunkt
Mittelwerte und
Standardabweichung
Leukozytenkonzentrate
N=10
LRS Kammern
N=10 p-Werte
CD25 pos
apoptotisch [%] 10,09±2,33 6,10±2,33 ‹0,001‡
CD25 neg
apoptotisch [%] 5,22±3,04 3,21±2,69
0,135‡
§
CD25 pos
nekrotisch [%] 0,16±0,82 0,27±0,24
0,178‡
§
CD25 neg
nekrotisch [%] 0,12±0,18 0,14±0,15 0,649§
T-Test ‡ Mann-Whitney-U-Test §
Nach sechs Stunden zeigten lediglich die Annexin V positiven und 7AAD positiven
CD25 positiven T-Zellen signifikante Abweichungen. Während in den Leukozyten-
konzentraten durchschnittlich 9,08±3,42% apoptotische CD25 positive Zellen nach-
weisbar waren, ließen sich in den LRS-Kammern nur 4,89±1,66% analysieren. Die
prozentualen Anteile der apoptotischen CD25 negativen T-Zellen zeigten keine sig-
nifikanten Unterschiede in den beiden Leukozytenprodukten. Der mediane Wert der
Leukozytenkonzentrate lag bei 3,07%, der der LRS-Kammern bei 2,07%.
Abb. 22: Apoptotische CD25 positive T-Zellen nach sechs Stunden Lagerung
48
In den prozentualen Anteilen der Annexin V positiven und 7AAD positiven Tregs
konnten keine signifikanten Unterschiede in den beiden Produkten nachgewiesen
werden.
Tab 17: Annexin V/7AAD positive CD25 T- Zellen nach sechs Stunden
Mittelwerte und
Standardabweichung
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
CD25 pos apoptotisch [%] 9,08±3,42 4,89±1,66 0,004‡
CD25 neg apoptotisch [%] 5,18±4,55 2,70±1,17 0,126‡
0,123§
CD25 pos nekrotisch [%] 0,30±0,18 0,25±0,13 0,529§
CD25 neg nekrotisch [%] 0,26±0,37 0,14±0,12 0,853§
T-Test ‡ Mann-Whitney-U-Test §
Nach 24-stündiger Lagerung zeigten sowohl die apoptotischen CD25 positiven als
auch die apoptotischen CD25 negativen T-Zellen in den beiden Zellprodukten ähn-
liche durchschnittliche Werte. An Annexin V positiven und 7AAD positiven CD25
positiven Zellen konnten in den Leukozytenkonzentraten durchschnittlich
2,02±2,74% Zellen nachgewiesen werden, während bei den LRS-Kammern lediglich
im Durchschnitt 0,58±0,40% nekrotische Zellen nachzuweisen waren. Ebenfalls
zeigten sich deutlich mehr nekrotische CD25 negative Zellen in den Leukozyten-
konzentraten mit gemittelt 1,13±1,19% Zellen. In den LRS-Kammern fanden sich
durchschnittlich dreimal weniger nekrotische CD25 negative Zellen. Somit lag der
prozentuale Anteil an Annexin V positiven und 7AAD positiven regulatorischen T-
Zellen in den Leukozytenkonzentraten nach 24-stündiger Lagerung höher als in den
LRS-Kammern.
Tab 18: Annexin V/7AAD markierte CD25 T- Zellen nach 24-stündiger Lagerung
Mittelwerte und
Standardabweichung
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
CD25pos apoptotisch [%] 6,34±1,12 6,40±3,80 0,965‡
CD25neg apoptotisch [%] 3,92±2,46 3,75±2,47 0,221§
CD25pos nekrotisch [%] 2,02±2,74 0,58±0,40 0,047§
CD25neg nekrotisch [%] 1,13±1,19 0,37±0,29 0,079‡
T-Test ‡ Mann-Whitney-U-Test §
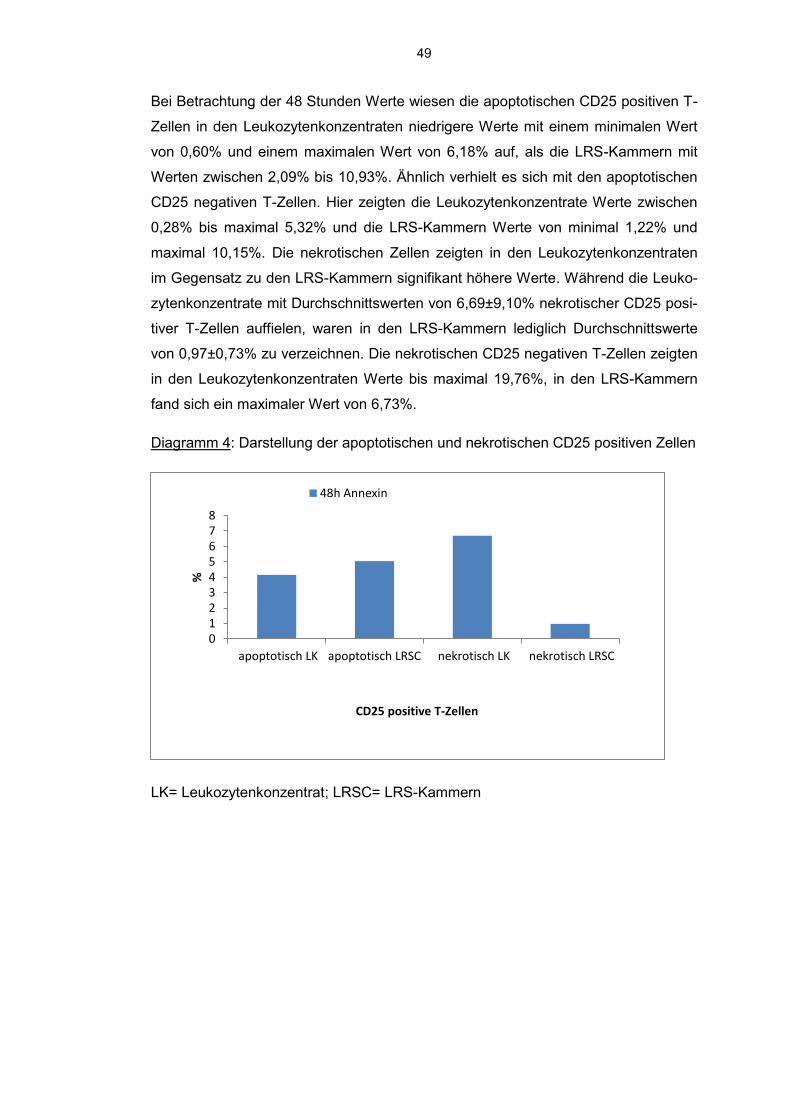
49
Bei Betrachtung der 48 Stunden Werte wiesen die apoptotischen CD25 positiven T-
Zellen in den Leukozytenkonzentraten niedrigere Werte mit einem minimalen Wert
von 0,60% und einem maximalen Wert von 6,18% auf, als die LRS-Kammern mit
Werten zwischen 2,09% bis 10,93%. Ähnlich verhielt es sich mit den apoptotischen
CD25 negativen T-Zellen. Hier zeigten die Leukozytenkonzentrate Werte zwischen
0,28% bis maximal 5,32% und die LRS-Kammern Werte von minimal 1,22% und
maximal 10,15%. Die nekrotischen Zellen zeigten in den Leukozytenkonzentraten
im Gegensatz zu den LRS-Kammern signifikant höhere Werte. Während die Leuko-
zytenkonzentrate mit Durchschnittswerten von 6,69±9,10% nekrotischer CD25 posi-
tiver T-Zellen auffielen, waren in den LRS-Kammern lediglich Durchschnittswerte
von 0,97±0,73% zu verzeichnen. Die nekrotischen CD25 negativen T-Zellen zeigten
in den Leukozytenkonzentraten Werte bis maximal 19,76%, in den LRS-Kammern
fand sich ein maximaler Wert von 6,73%.
Diagramm 4: Darstellung der apoptotischen und nekrotischen CD25 positiven Zellen
LK= Leukozytenkonzentrat; LRSC= LRS-Kammern
012345678
apoptotisch LK apoptotisch LRSC nekrotisch LK nekrotisch LRSC
%
CD25 positive T-Zellen
48h Annexin
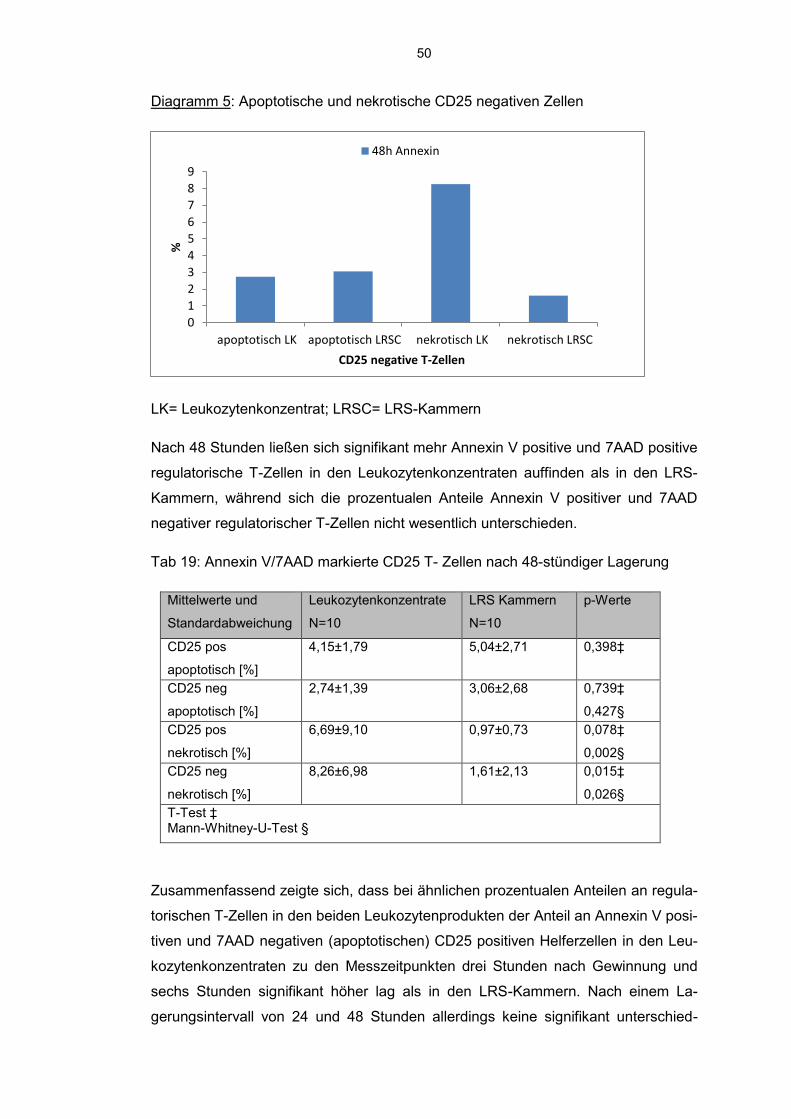
50
Diagramm 5: Apoptotische und nekrotische CD25 negativen Zellen
LK= Leukozytenkonzentrat; LRSC= LRS-Kammern
Nach 48 Stunden ließen sich signifikant mehr Annexin V positive und 7AAD positive
regulatorische T-Zellen in den Leukozytenkonzentraten auffinden als in den LRS-
Kammern, während sich die prozentualen Anteile Annexin V positiver und 7AAD
negativer regulatorischer T-Zellen nicht wesentlich unterschieden.
Tab 19: Annexin V/7AAD markierte CD25 T- Zellen nach 48-stündiger Lagerung
Mittelwerte und
Standardabweichung
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
CD25 pos
apoptotisch [%]
4,15±1,79 5,04±2,71 0,398‡
CD25 neg
apoptotisch [%]
2,74±1,39 3,06±2,68 0,739‡
0,427§
CD25 pos
nekrotisch [%]
6,69±9,10 0,97±0,73 0,078‡
0,002§
CD25 neg
nekrotisch [%]
8,26±6,98 1,61±2,13 0,015‡
0,026§
T-Test ‡ Mann-Whitney-U-Test §
Zusammenfassend zeigte sich, dass bei ähnlichen prozentualen Anteilen an regula-
torischen T-Zellen in den beiden Leukozytenprodukten der Anteil an Annexin V posi-
tiven und 7AAD negativen (apoptotischen) CD25 positiven Helferzellen in den Leu-
kozytenkonzentraten zu den Messzeitpunkten drei Stunden nach Gewinnung und
sechs Stunden signifikant höher lag als in den LRS-Kammern. Nach einem La-
gerungsintervall von 24 und 48 Stunden allerdings keine signifikant unterschied-
0
1
2
3
4
5
6
7
8
9
apoptotisch LK apoptotisch LRSC nekrotisch LK nekrotisch LRSC
%
CD25 negative T-Zellen
48h Annexin
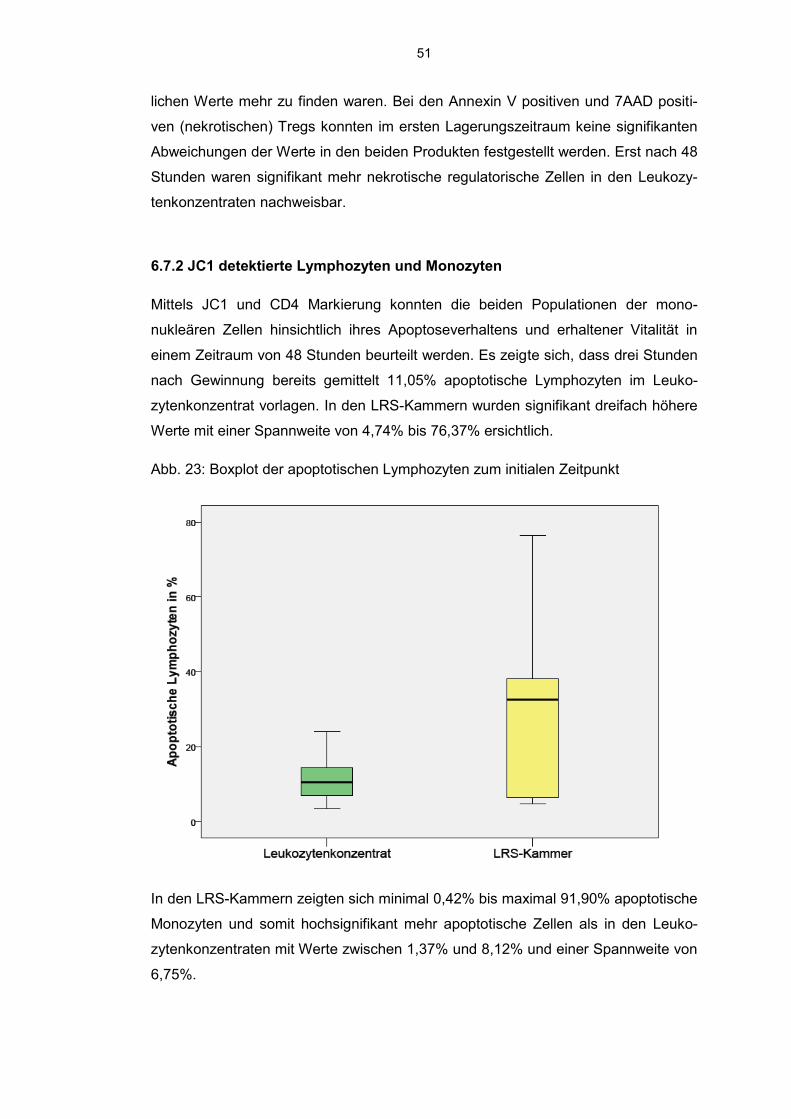
51
lichen Werte mehr zu finden waren. Bei den Annexin V positiven und 7AAD positi-
ven (nekrotischen) Tregs konnten im ersten Lagerungszeitraum keine signifikanten
Abweichungen der Werte in den beiden Produkten festgestellt werden. Erst nach 48
Stunden waren signifikant mehr nekrotische regulatorische Zellen in den Leukozy-
tenkonzentraten nachweisbar.
6.7.2 JC1 detektierte Lymphozyten und Monozyten
Mittels JC1 und CD4 Markierung konnten die beiden Populationen der mono-
nukleären Zellen hinsichtlich ihres Apoptoseverhaltens und erhaltener Vitalität in
einem Zeitraum von 48 Stunden beurteilt werden. Es zeigte sich, dass drei Stunden
nach Gewinnung bereits gemittelt 11,05% apoptotische Lymphozyten im Leuko-
zytenkonzentrat vorlagen. In den LRS-Kammern wurden signifikant dreifach höhere
Werte mit einer Spannweite von 4,74% bis 76,37% ersichtlich.
Abb. 23: Boxplot der apoptotischen Lymphozyten zum initialen Zeitpunkt
In den LRS-Kammern zeigten sich minimal 0,42% bis maximal 91,90% apoptotische
Monozyten und somit hochsignifikant mehr apoptotische Zellen als in den Leuko-
zytenkonzentraten mit Werte zwischen 1,37% und 8,12% und einer Spannweite von
6,75%.
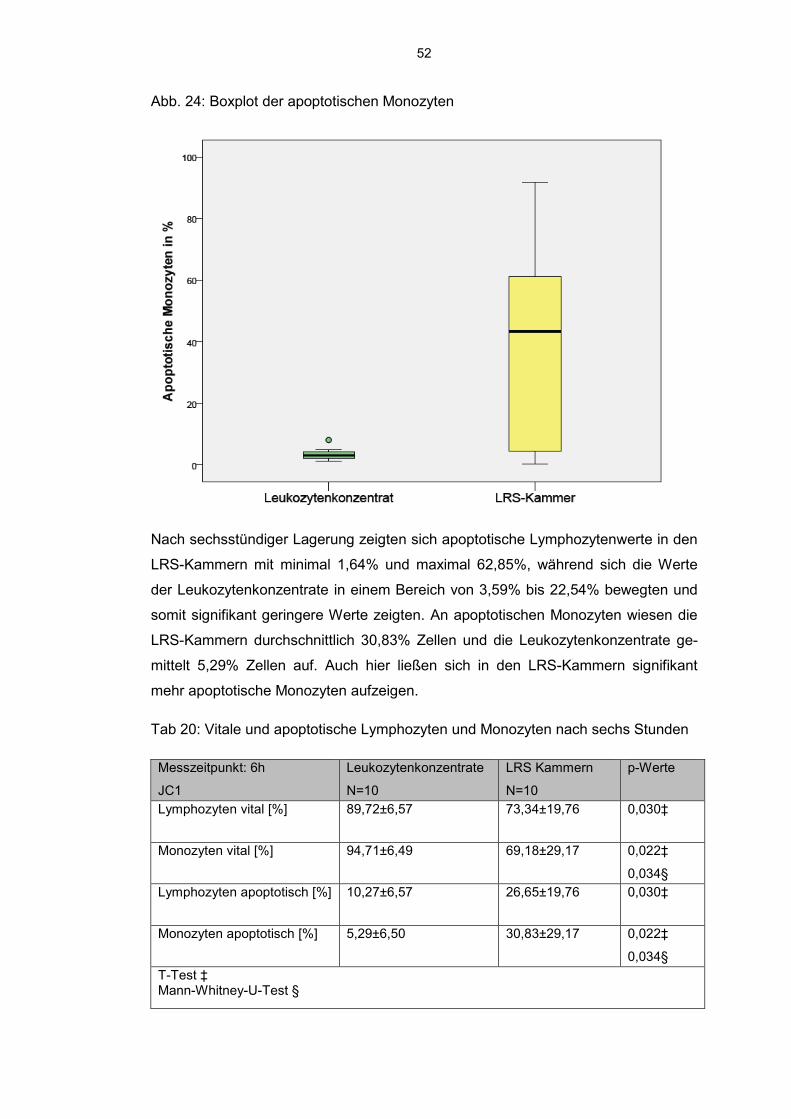
52
Abb. 24: Boxplot der apoptotischen Monozyten
Nach sechsstündiger Lagerung zeigten sich apoptotische Lymphozytenwerte in den
LRS-Kammern mit minimal 1,64% und maximal 62,85%, während sich die Werte
der Leukozytenkonzentrate in einem Bereich von 3,59% bis 22,54% bewegten und
somit signifikant geringere Werte zeigten. An apoptotischen Monozyten wiesen die
LRS-Kammern durchschnittlich 30,83% Zellen und die Leukozytenkonzentrate ge-
mittelt 5,29% Zellen auf. Auch hier ließen sich in den LRS-Kammern signifikant
mehr apoptotische Monozyten aufzeigen.
Tab 20: Vitale und apoptotische Lymphozyten und Monozyten nach sechs Stunden
Messzeitpunkt: 6h
JC1
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
Lymphozyten vital [%] 89,72±6,57 73,34±19,76 0,030‡
Monozyten vital [%] 94,71±6,49 69,18±29,17 0,022‡
0,034§
Lymphozyten apoptotisch [%] 10,27±6,57 26,65±19,76 0,030‡
Monozyten apoptotisch [%] 5,29±6,50 30,83±29,17 0,022‡
0,034§
T-Test ‡ Mann-Whitney-U-Test §
53
Mit längerer Lagerungsperiode wurden keine signifikanten Unterschiede mehr zwi-
schen den beiden untersuchten Zellprodukten ersichtlich. In den Leukozyten-
konzentraten befanden sich nach 24-stündiger Lagerung durchschnittlich 22,04%
apoptotische Lymphozyten, in den LRS-Kammern durchschnittlich 33,52%. Auch
nach weiteren 24 Stunden Lagerung zeigten sich weder in den LRS-Kammern noch
in den Leukozytenkonzentraten signifikante Abweichungen in den Lymphozyten-
werten.
Tab 21: Vitale und apoptotische Lymphozyten und Monozyten nach 48 Stunden
Messzeitpunkt: 48h
JC1
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
Lymphozyten vital [%]
50,28±29,47 66,58±22,91 0,185‡
Monozyten vital [%]
43,53±31,72 67,02±29,09 0,102‡
0,070§
Lymphozyten apoptotisch [%] 49,71±29,48 33,42±22,92 0,185‡
Monozyten apoptotisch [%] 56,44±31,76 32,96±29,09 0,102‡
0,070§
T-Test ‡ Mann-Whitney-U-Test
Die apoptotischen Monozyten verhielten sich ähnlich. In den Leukozytenkonzen-
traten zeigte sich nach 24-stündiger Lagerung eine Spannweite der Werte von
9,13% bis 23,46% und nach längerem Lagerungsintervall Werte von 12,41% bis
97,22%. Die LRS-Kammern wiesen innerhalb der Lagerungszeit Werte von anfangs
6,5% bis 91,95% und am Ende der Lagerung von 7,12% bis 85,30% auf.
Diagramm 6 und 7 zeigen deutlich die prozentualen Unterschiede des apop-
totischen Verhaltens der Lymphozyten und Monozyten. Während der Messzeitpunk-
te initial (drei Stunden nach Gewinnung), sechs Stunden und 24 Stunden lag der
Anteil an apoptotischen Zellen in den Leukozytenkonzentraten wesentlich niedriger
als in den LRS-Kammern. Zum Messzeitpunkt 48 Stunden waren deutlich mehr
apoptotische Zellen in den Leukozytenkonzentraten zu finden als in den LRS-
Kammern.
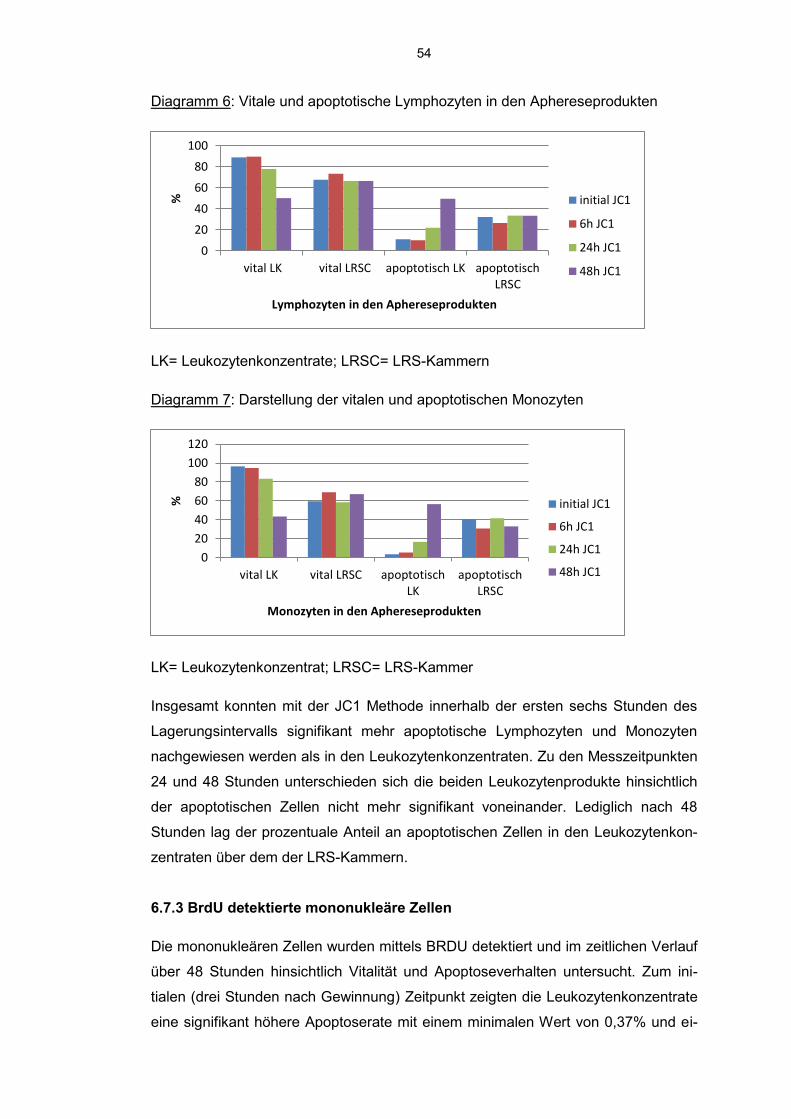
54
Diagramm 6: Vitale und apoptotische Lymphozyten in den Aphereseprodukten
LK= Leukozytenkonzentrate; LRSC= LRS-Kammern
Diagramm 7: Darstellung der vitalen und apoptotischen Monozyten
LK= Leukozytenkonzentrat; LRSC= LRS-Kammer
Insgesamt konnten mit der JC1 Methode innerhalb der ersten sechs Stunden des
Lagerungsintervalls signifikant mehr apoptotische Lymphozyten und Monozyten
nachgewiesen werden als in den Leukozytenkonzentraten. Zu den Messzeitpunkten
24 und 48 Stunden unterschieden sich die beiden Leukozytenprodukte hinsichtlich
der apoptotischen Zellen nicht mehr signifikant voneinander. Lediglich nach 48
Stunden lag der prozentuale Anteil an apoptotischen Zellen in den Leukozytenkon-
zentraten über dem der LRS-Kammern.
6.7.3 BrdU detektierte mononukleäre Zellen
Die mononukleären Zellen wurden mittels BRDU detektiert und im zeitlichen Verlauf
über 48 Stunden hinsichtlich Vitalität und Apoptoseverhalten untersucht. Zum ini-
tialen (drei Stunden nach Gewinnung) Zeitpunkt zeigten die Leukozytenkonzentrate
eine signifikant höhere Apoptoserate mit einem minimalen Wert von 0,37% und ei-
0
20
40
60
80
100
vital LK vital LRSC apoptotisch LK apoptotischLRSC
%
Lymphozyten in den Aphereseprodukten
initial JC1
6h JC1
24h JC1
48h JC1
0
20
40
60
80
100
120
vital LK vital LRSC apoptotischLK
apoptotischLRSC
%
Monozyten in den Aphereseprodukten
initial JC1
6h JC1
24h JC1
48h JC1
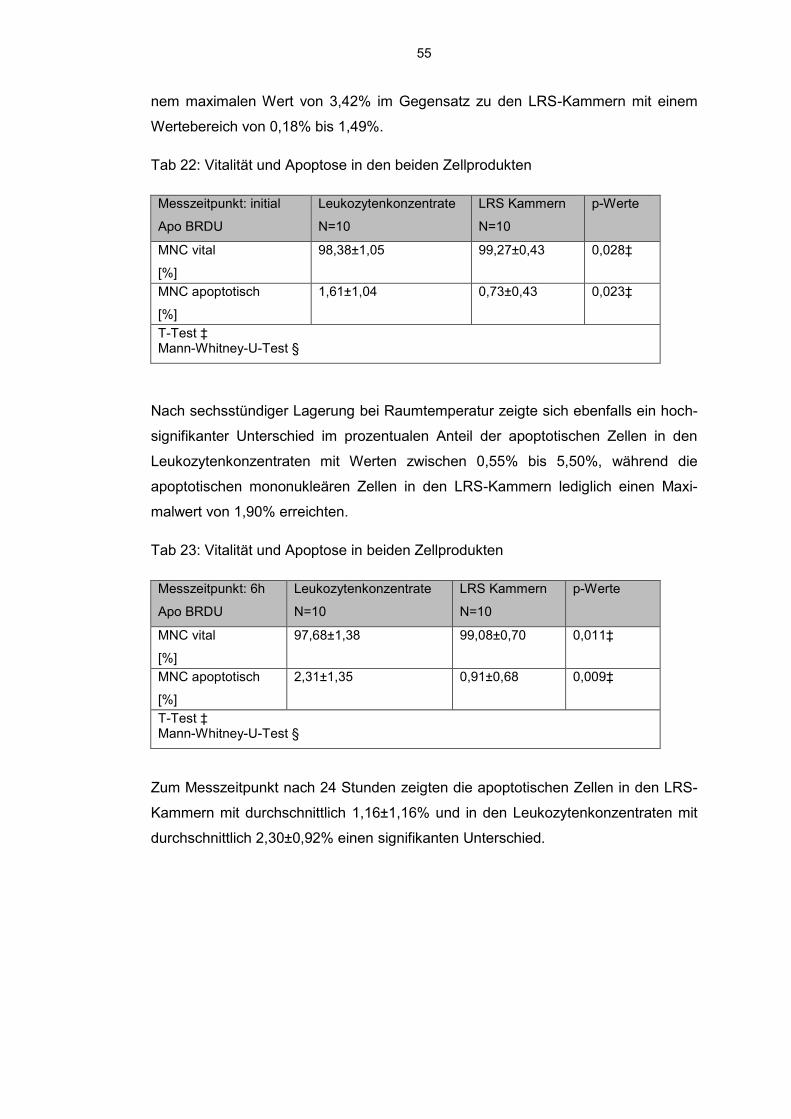
55
nem maximalen Wert von 3,42% im Gegensatz zu den LRS-Kammern mit einem
Wertebereich von 0,18% bis 1,49%.
Tab 22: Vitalität und Apoptose in den beiden Zellprodukten
Messzeitpunkt: initial
Apo BRDU
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
MNC vital
[%]
98,38±1,05 99,27±0,43 0,028‡
MNC apoptotisch
[%]
1,61±1,04 0,73±0,43 0,023‡
T-Test ‡ Mann-Whitney-U-Test §
Nach sechsstündiger Lagerung bei Raumtemperatur zeigte sich ebenfalls ein hoch-
signifikanter Unterschied im prozentualen Anteil der apoptotischen Zellen in den
Leukozytenkonzentraten mit Werten zwischen 0,55% bis 5,50%, während die
apoptotischen mononukleären Zellen in den LRS-Kammern lediglich einen Maxi-
malwert von 1,90% erreichten.
Tab 23: Vitalität und Apoptose in beiden Zellprodukten
Messzeitpunkt: 6h
Apo BRDU
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
MNC vital
[%]
97,68±1,38 99,08±0,70 0,011‡
MNC apoptotisch
[%]
2,31±1,35 0,91±0,68 0,009‡
T-Test ‡ Mann-Whitney-U-Test §
Zum Messzeitpunkt nach 24 Stunden zeigten die apoptotischen Zellen in den LRS-
Kammern mit durchschnittlich 1,16±1,16% und in den Leukozytenkonzentraten mit
durchschnittlich 2,30±0,92% einen signifikanten Unterschied.
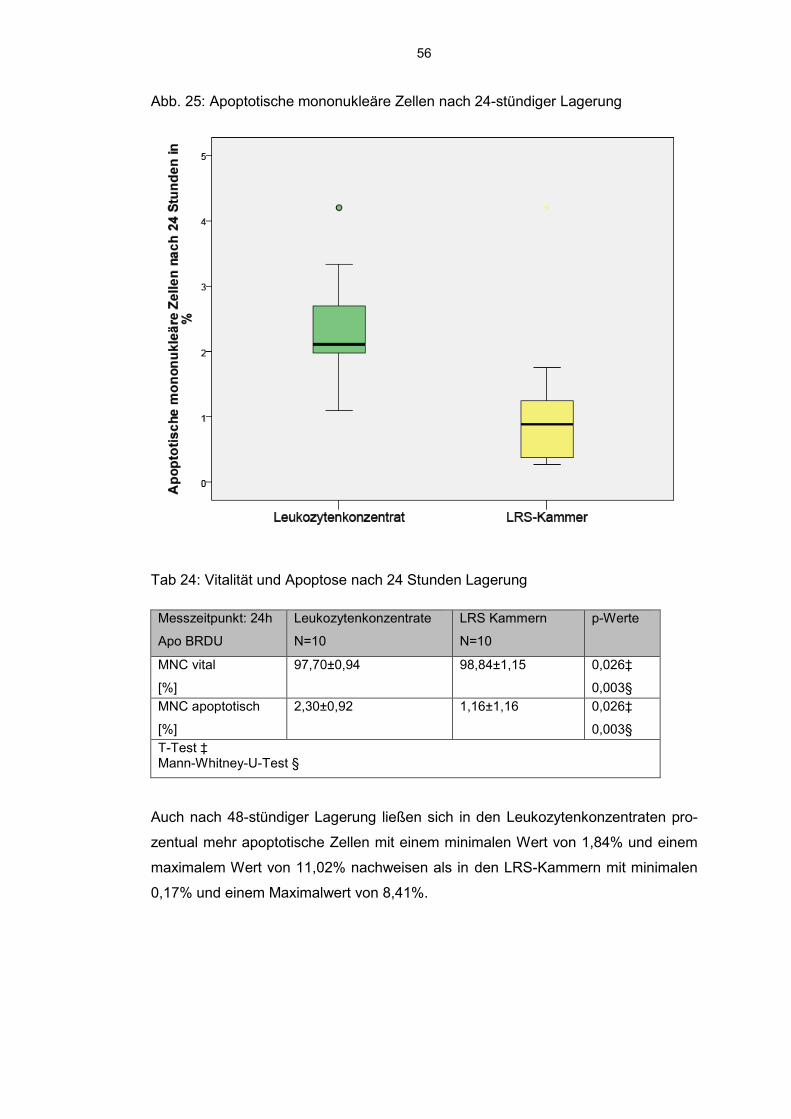
56
Abb. 25: Apoptotische mononukleäre Zellen nach 24-stündiger Lagerung
Tab 24: Vitalität und Apoptose nach 24 Stunden Lagerung
Messzeitpunkt: 24h
Apo BRDU
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
MNC vital
[%]
97,70±0,94 98,84±1,15 0,026‡
0,003§
MNC apoptotisch
[%]
2,30±0,92 1,16±1,16 0,026‡
0,003§
T-Test ‡ Mann-Whitney-U-Test §
Auch nach 48-stündiger Lagerung ließen sich in den Leukozytenkonzentraten pro-
zentual mehr apoptotische Zellen mit einem minimalen Wert von 1,84% und einem
maximalem Wert von 11,02% nachweisen als in den LRS-Kammern mit minimalen
0,17% und einem Maximalwert von 8,41%.
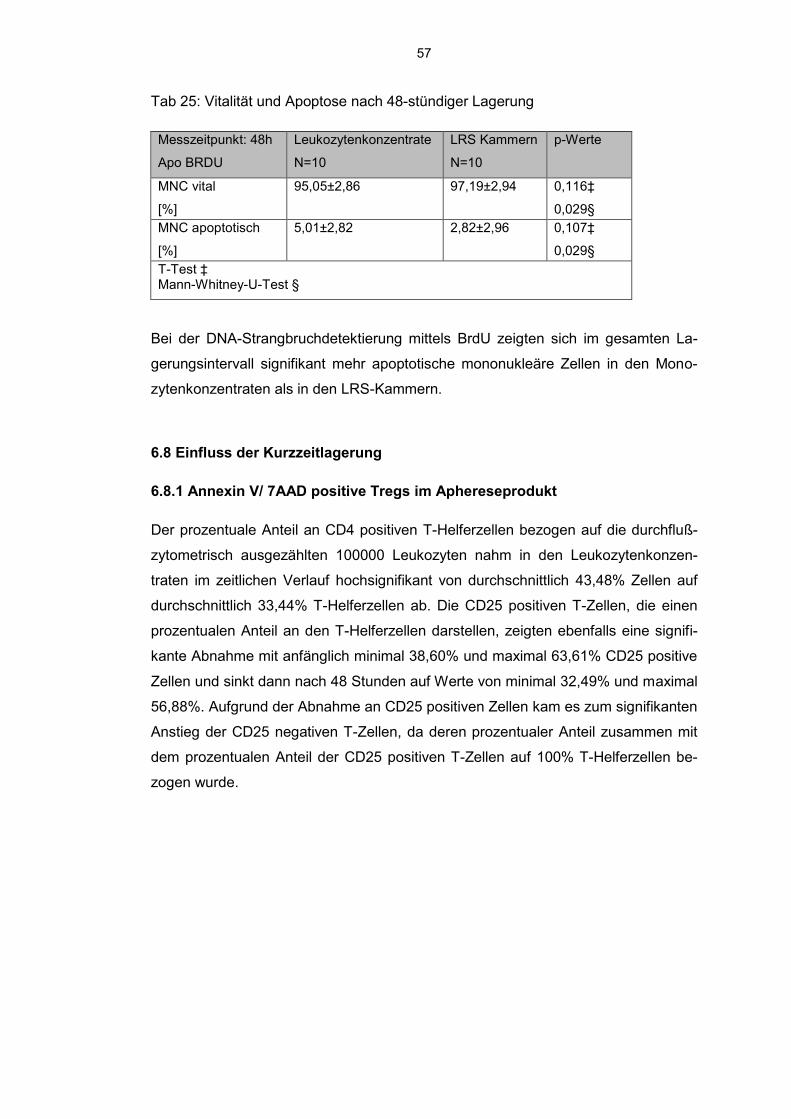
57
Tab 25: Vitalität und Apoptose nach 48-stündiger Lagerung
Messzeitpunkt: 48h
Apo BRDU
Leukozytenkonzentrate
N=10
LRS Kammern
N=10
p-Werte
MNC vital
[%]
95,05±2,86 97,19±2,94 0,116‡
0,029§
MNC apoptotisch
[%]
5,01±2,82 2,82±2,96 0,107‡
0,029§
T-Test ‡ Mann-Whitney-U-Test §
Bei der DNA-Strangbruchdetektierung mittels BrdU zeigten sich im gesamten La-
gerungsintervall signifikant mehr apoptotische mononukleäre Zellen in den Mono-
zytenkonzentraten als in den LRS-Kammern.
6.8 Einfluss der Kurzzeitlagerung
6.8.1 Annexin V/ 7AAD positive Tregs im Aphereseprodukt
Der prozentuale Anteil an CD4 positiven T-Helferzellen bezogen auf die durchfluß-
zytometrisch ausgezählten 100000 Leukozyten nahm in den Leukozytenkonzen-
traten im zeitlichen Verlauf hochsignifikant von durchschnittlich 43,48% Zellen auf
durchschnittlich 33,44% T-Helferzellen ab. Die CD25 positiven T-Zellen, die einen
prozentualen Anteil an den T-Helferzellen darstellen, zeigten ebenfalls eine signifi-
kante Abnahme mit anfänglich minimal 38,60% und maximal 63,61% CD25 positive
Zellen und sinkt dann nach 48 Stunden auf Werte von minimal 32,49% und maximal
56,88%. Aufgrund der Abnahme an CD25 positiven Zellen kam es zum signifikanten
Anstieg der CD25 negativen T-Zellen, da deren prozentualer Anteil zusammen mit
dem prozentualen Anteil der CD25 positiven T-Zellen auf 100% T-Helferzellen be-
zogen wurde.
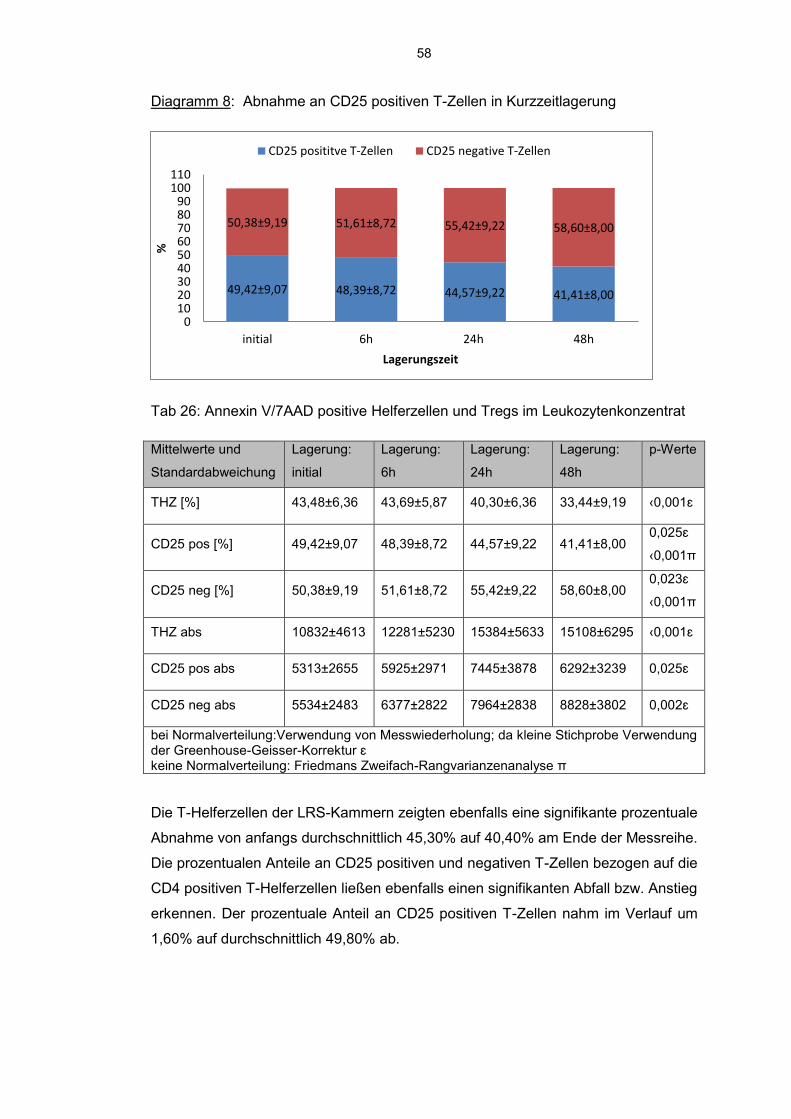
58
Diagramm 8: Abnahme an CD25 positiven T-Zellen in Kurzzeitlagerung
Tab 26: Annexin V/7AAD positive Helferzellen und Tregs im Leukozytenkonzentrat
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
THZ [%] 43,48±6,36 43,69±5,87 40,30±6,36 33,44±9,19 ‹0,001ɛ
CD25 pos [%] 49,42±9,07 48,39±8,72 44,57±9,22 41,41±8,00 0,025ɛ
‹0,001π
CD25 neg [%] 50,38±9,19 51,61±8,72 55,42±9,22 58,60±8,00 0,023ɛ
‹0,001π
THZ abs 10832±4613 12281±5230 15384±5633 15108±6295 ‹0,001ɛ
CD25 pos abs 5313±2655 5925±2971 7445±3878 6292±3239 0,025ɛ
CD25 neg abs 5534±2483 6377±2822 7964±2838 8828±3802 0,002ɛ
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwendung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
Die T-Helferzellen der LRS-Kammern zeigten ebenfalls eine signifikante prozentuale
Abnahme von anfangs durchschnittlich 45,30% auf 40,40% am Ende der Messreihe.
Die prozentualen Anteile an CD25 positiven und negativen T-Zellen bezogen auf die
CD4 positiven T-Helferzellen ließen ebenfalls einen signifikanten Abfall bzw. Anstieg
erkennen. Der prozentuale Anteil an CD25 positiven T-Zellen nahm im Verlauf um
1,60% auf durchschnittlich 49,80% ab.
49,42±9,07 48,39±8,72 44,57±9,22 41,41±8,00
50,38±9,19 51,61±8,72 55,42±9,22 58,60±8,00
0102030405060708090
100110
initial 6h 24h 48h
%
Lagerungszeit
CD25 posititve T-Zellen CD25 negative T-Zellen
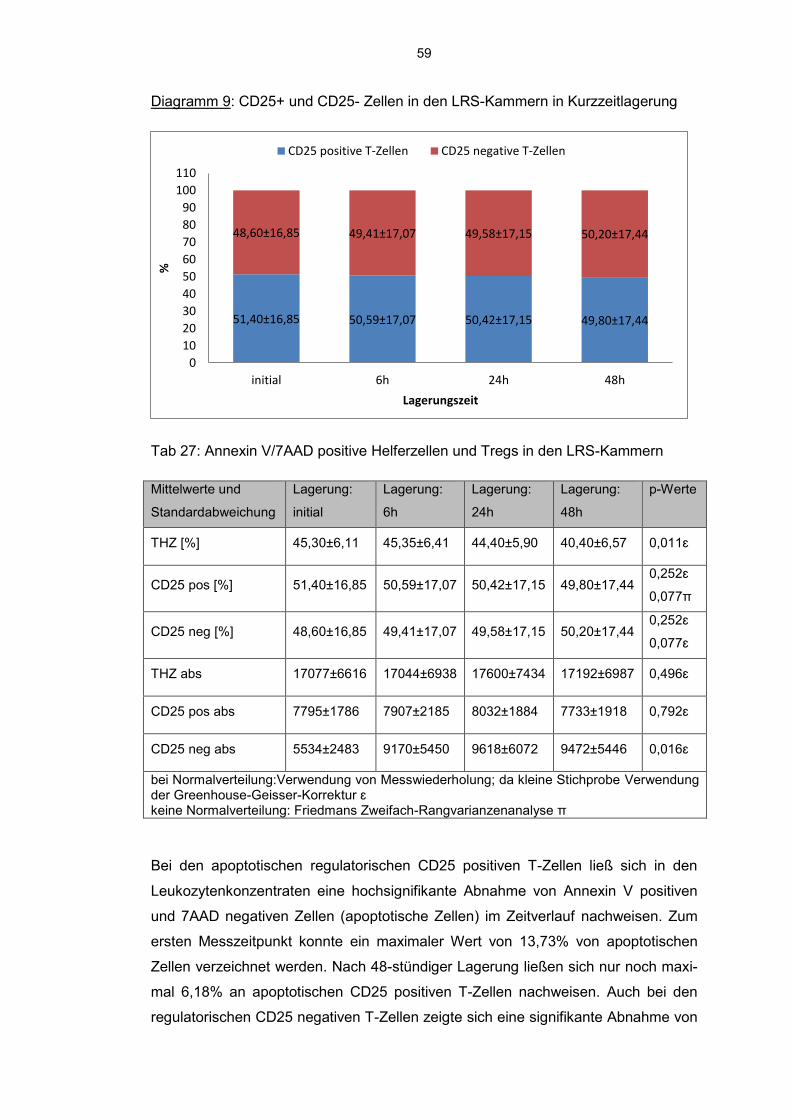
59
Diagramm 9: CD25+ und CD25- Zellen in den LRS-Kammern in Kurzzeitlagerung
Tab 27: Annexin V/7AAD positive Helferzellen und Tregs in den LRS-Kammern
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
THZ [%] 45,30±6,11 45,35±6,41 44,40±5,90 40,40±6,57 0,011ɛ
CD25 pos [%] 51,40±16,85 50,59±17,07 50,42±17,15 49,80±17,44 0,252ɛ
0,077π
CD25 neg [%] 48,60±16,85 49,41±17,07 49,58±17,15 50,20±17,44 0,252ɛ
0,077ɛ
THZ abs 17077±6616 17044±6938 17600±7434 17192±6987 0,496ɛ
CD25 pos abs 7795±1786 7907±2185 8032±1884 7733±1918 0,792ɛ
CD25 neg abs 5534±2483 9170±5450 9618±6072 9472±5446 0,016ɛ
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwendung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
Bei den apoptotischen regulatorischen CD25 positiven T-Zellen ließ sich in den
Leukozytenkonzentraten eine hochsignifikante Abnahme von Annexin V positiven
und 7AAD negativen Zellen (apoptotische Zellen) im Zeitverlauf nachweisen. Zum
ersten Messzeitpunkt konnte ein maximaler Wert von 13,73% von apoptotischen
Zellen verzeichnet werden. Nach 48-stündiger Lagerung ließen sich nur noch maxi-
mal 6,18% an apoptotischen CD25 positiven T-Zellen nachweisen. Auch bei den
regulatorischen CD25 negativen T-Zellen zeigte sich eine signifikante Abnahme von
51,40±16,85 50,59±17,07 50,42±17,15 49,80±17,44
48,60±16,85 49,41±17,07 49,58±17,15 50,20±17,44
0
10
20
30
40
50
60
70
80
90
100
110
initial 6h 24h 48h
%
Lagerungszeit
CD25 positive T-Zellen CD25 negative T-Zellen
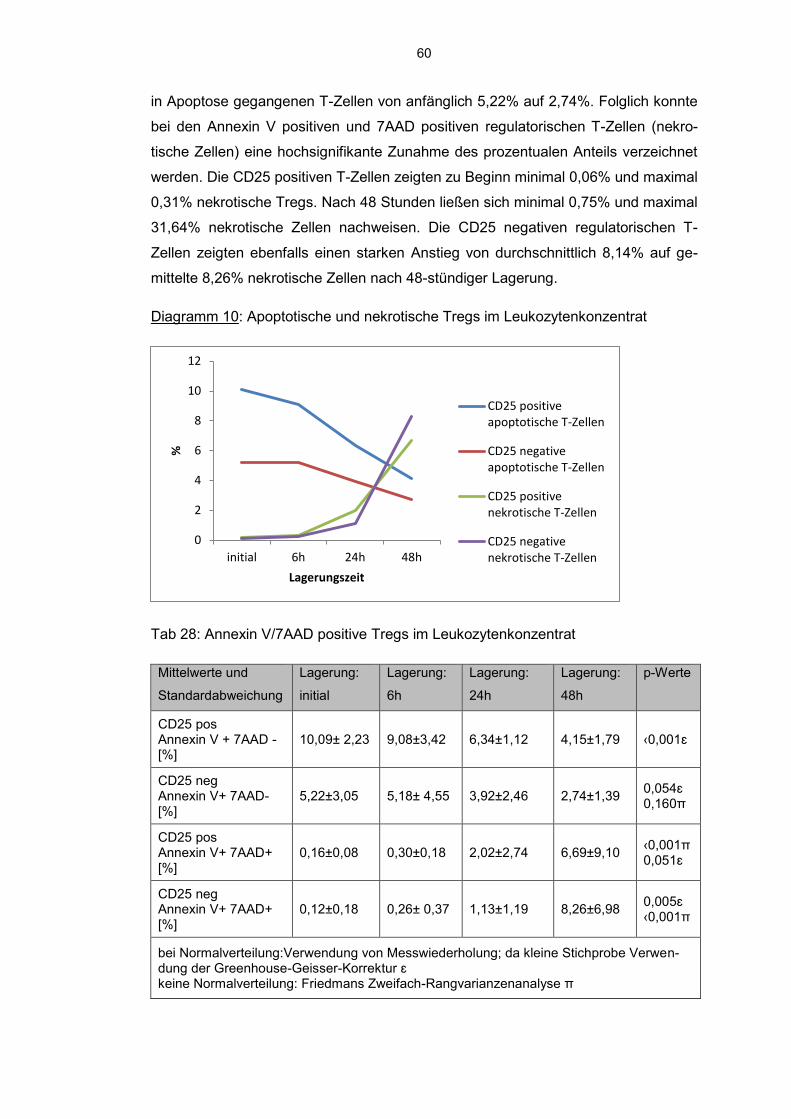
60
in Apoptose gegangenen T-Zellen von anfänglich 5,22% auf 2,74%. Folglich konnte
bei den Annexin V positiven und 7AAD positiven regulatorischen T-Zellen (nekro-
tische Zellen) eine hochsignifikante Zunahme des prozentualen Anteils verzeichnet
werden. Die CD25 positiven T-Zellen zeigten zu Beginn minimal 0,06% und maximal
0,31% nekrotische Tregs. Nach 48 Stunden ließen sich minimal 0,75% und maximal
31,64% nekrotische Zellen nachweisen. Die CD25 negativen regulatorischen T-
Zellen zeigten ebenfalls einen starken Anstieg von durchschnittlich 8,14% auf ge-
mittelte 8,26% nekrotische Zellen nach 48-stündiger Lagerung.
Diagramm 10: Apoptotische und nekrotische Tregs im Leukozytenkonzentrat
Tab 28: Annexin V/7AAD positive Tregs im Leukozytenkonzentrat
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
CD25 pos Annexin V + 7AAD - [%]
10,09± 2,23 9,08±3,42 6,34±1,12 4,15±1,79 ‹0,001ɛ
CD25 neg Annexin V+ 7AAD- [%]
5,22±3,05 5,18± 4,55 3,92±2,46 2,74±1,39 0,054ɛ 0,160π
CD25 pos Annexin V+ 7AAD+ [%]
0,16±0,08 0,30±0,18 2,02±2,74 6,69±9,10 ‹0,001π 0,051ɛ
CD25 neg Annexin V+ 7AAD+ [%]
0,12±0,18 0,26± 0,37 1,13±1,19 8,26±6,98 0,005ɛ ‹0,001π
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
0
2
4
6
8
10
12
initial 6h 24h 48h
%
Lagerungszeit
CD25 positiveapoptotische T-Zellen
CD25 negativeapoptotische T-Zellen
CD25 positivenekrotische T-Zellen
CD25 negativenekrotische T-Zellen
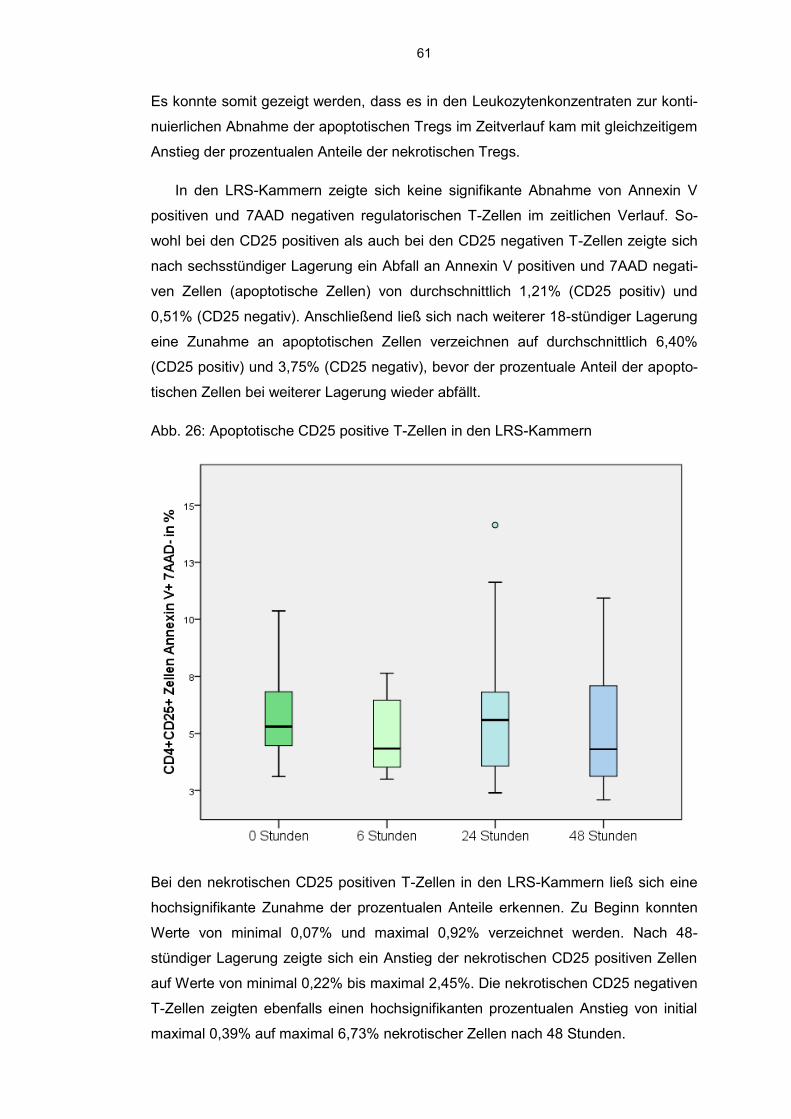
61
Es konnte somit gezeigt werden, dass es in den Leukozytenkonzentraten zur konti-
nuierlichen Abnahme der apoptotischen Tregs im Zeitverlauf kam mit gleichzeitigem
Anstieg der prozentualen Anteile der nekrotischen Tregs.
In den LRS-Kammern zeigte sich keine signifikante Abnahme von Annexin V
positiven und 7AAD negativen regulatorischen T-Zellen im zeitlichen Verlauf. So-
wohl bei den CD25 positiven als auch bei den CD25 negativen T-Zellen zeigte sich
nach sechsstündiger Lagerung ein Abfall an Annexin V positiven und 7AAD negati-
ven Zellen (apoptotische Zellen) von durchschnittlich 1,21% (CD25 positiv) und
0,51% (CD25 negativ). Anschließend ließ sich nach weiterer 18-stündiger Lagerung
eine Zunahme an apoptotischen Zellen verzeichnen auf durchschnittlich 6,40%
(CD25 positiv) und 3,75% (CD25 negativ), bevor der prozentuale Anteil der apopto-
tischen Zellen bei weiterer Lagerung wieder abfällt.
Abb. 26: Apoptotische CD25 positive T-Zellen in den LRS-Kammern
Bei den nekrotischen CD25 positiven T-Zellen in den LRS-Kammern ließ sich eine
hochsignifikante Zunahme der prozentualen Anteile erkennen. Zu Beginn konnten
Werte von minimal 0,07% und maximal 0,92% verzeichnet werden. Nach 48-
stündiger Lagerung zeigte sich ein Anstieg der nekrotischen CD25 positiven Zellen
auf Werte von minimal 0,22% bis maximal 2,45%. Die nekrotischen CD25 negativen
T-Zellen zeigten ebenfalls einen hochsignifikanten prozentualen Anstieg von initial
maximal 0,39% auf maximal 6,73% nekrotischer Zellen nach 48 Stunden.
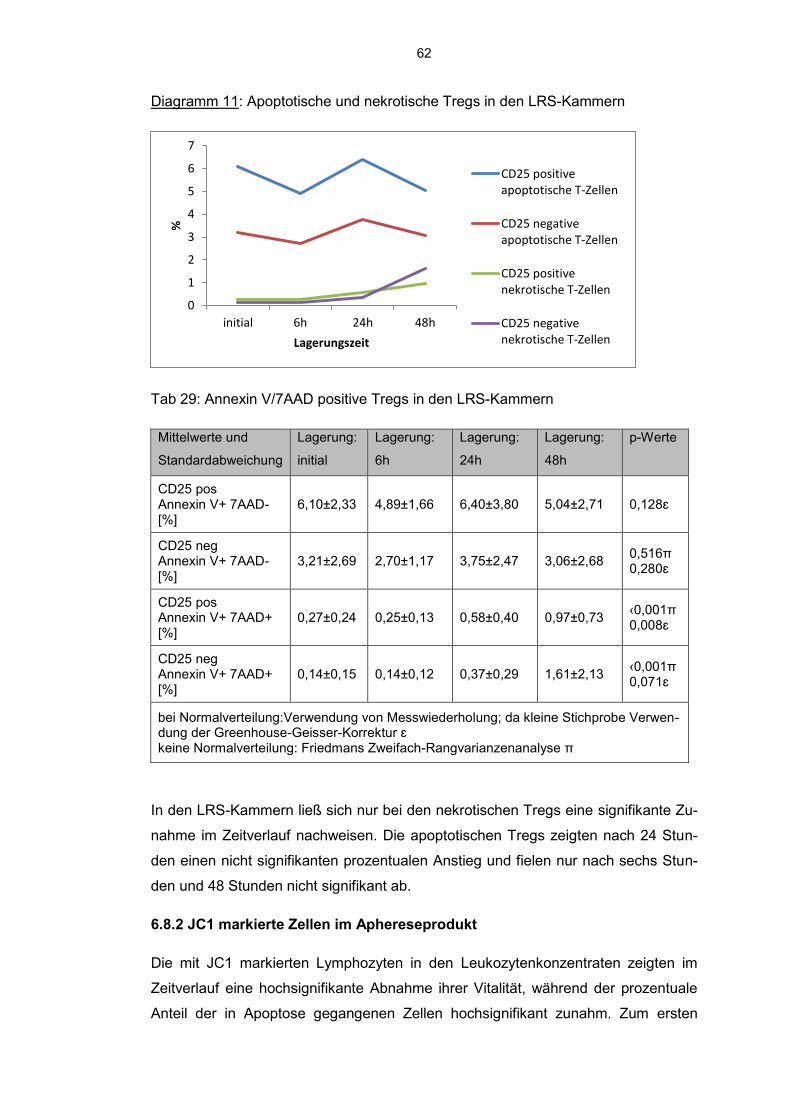
62
Diagramm 11: Apoptotische und nekrotische Tregs in den LRS-Kammern
Tab 29: Annexin V/7AAD positive Tregs in den LRS-Kammern
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
CD25 pos Annexin V+ 7AAD- [%]
6,10±2,33 4,89±1,66 6,40±3,80 5,04±2,71 0,128ɛ
CD25 neg Annexin V+ 7AAD- [%]
3,21±2,69 2,70±1,17 3,75±2,47 3,06±2,68 0,516π 0,280ɛ
CD25 pos Annexin V+ 7AAD+ [%]
0,27±0,24 0,25±0,13 0,58±0,40 0,97±0,73 ‹0,001π 0,008ɛ
CD25 neg Annexin V+ 7AAD+ [%]
0,14±0,15 0,14±0,12 0,37±0,29 1,61±2,13 ‹0,001π 0,071ɛ
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
In den LRS-Kammern ließ sich nur bei den nekrotischen Tregs eine signifikante Zu-
nahme im Zeitverlauf nachweisen. Die apoptotischen Tregs zeigten nach 24 Stun-
den einen nicht signifikanten prozentualen Anstieg und fielen nur nach sechs Stun-
den und 48 Stunden nicht signifikant ab.
6.8.2 JC1 markierte Zellen im Aphereseprodukt
Die mit JC1 markierten Lymphozyten in den Leukozytenkonzentraten zeigten im
Zeitverlauf eine hochsignifikante Abnahme ihrer Vitalität, während der prozentuale
Anteil der in Apoptose gegangenen Zellen hochsignifikant zunahm. Zum ersten
0
1
2
3
4
5
6
7
initial 6h 24h 48h
%
Lagerungszeit
CD25 positiveapoptotische T-Zellen
CD25 negativeapoptotische T-Zellen
CD25 positivenekrotische T-Zellen
CD25 negativenekrotische T-Zellen
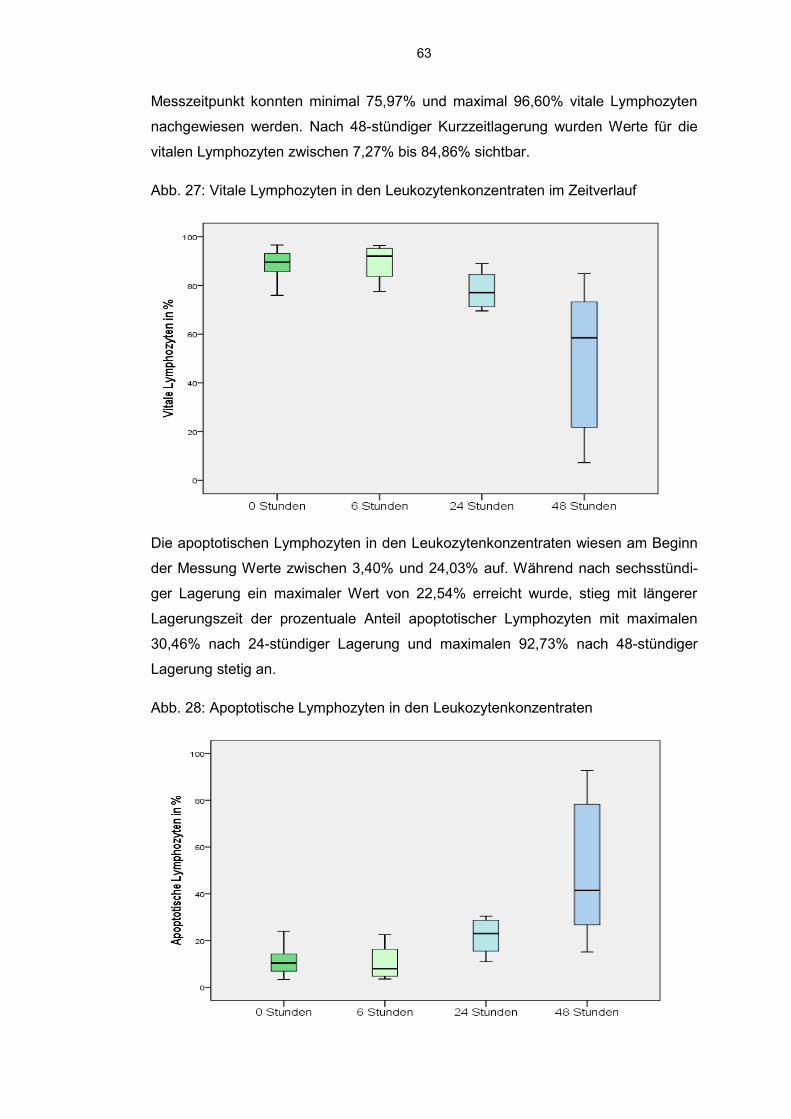
63
Messzeitpunkt konnten minimal 75,97% und maximal 96,60% vitale Lymphozyten
nachgewiesen werden. Nach 48-stündiger Kurzzeitlagerung wurden Werte für die
vitalen Lymphozyten zwischen 7,27% bis 84,86% sichtbar.
Abb. 27: Vitale Lymphozyten in den Leukozytenkonzentraten im Zeitverlauf
Die apoptotischen Lymphozyten in den Leukozytenkonzentraten wiesen am Beginn
der Messung Werte zwischen 3,40% und 24,03% auf. Während nach sechsstündi-
ger Lagerung ein maximaler Wert von 22,54% erreicht wurde, stieg mit längerer
Lagerungszeit der prozentuale Anteil apoptotischer Lymphozyten mit maximalen
30,46% nach 24-stündiger Lagerung und maximalen 92,73% nach 48-stündiger
Lagerung stetig an.
Abb. 28: Apoptotische Lymphozyten in den Leukozytenkonzentraten
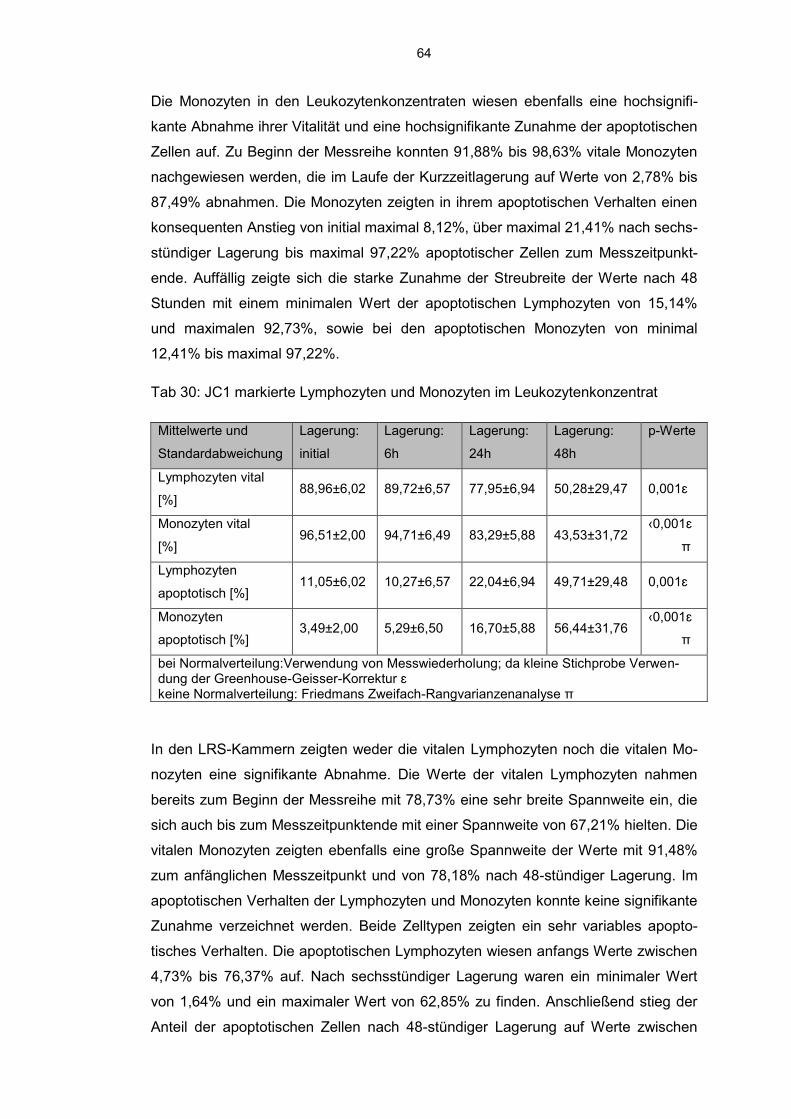
64
Die Monozyten in den Leukozytenkonzentraten wiesen ebenfalls eine hochsignifi-
kante Abnahme ihrer Vitalität und eine hochsignifikante Zunahme der apoptotischen
Zellen auf. Zu Beginn der Messreihe konnten 91,88% bis 98,63% vitale Monozyten
nachgewiesen werden, die im Laufe der Kurzzeitlagerung auf Werte von 2,78% bis
87,49% abnahmen. Die Monozyten zeigten in ihrem apoptotischen Verhalten einen
konsequenten Anstieg von initial maximal 8,12%, über maximal 21,41% nach sechs-
stündiger Lagerung bis maximal 97,22% apoptotischer Zellen zum Messzeitpunkt-
ende. Auffällig zeigte sich die starke Zunahme der Streubreite der Werte nach 48
Stunden mit einem minimalen Wert der apoptotischen Lymphozyten von 15,14%
und maximalen 92,73%, sowie bei den apoptotischen Monozyten von minimal
12,41% bis maximal 97,22%.
Tab 30: JC1 markierte Lymphozyten und Monozyten im Leukozytenkonzentrat
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
Lymphozyten vital
[%] 88,96±6,02 89,72±6,57 77,95±6,94 50,28±29,47 0,001ɛ
Monozyten vital
[%] 96,51±2,00 94,71±6,49 83,29±5,88 43,53±31,72
‹0,001ɛ
π
Lymphozyten
apoptotisch [%] 11,05±6,02 10,27±6,57 22,04±6,94 49,71±29,48 0,001ɛ
Monozyten
apoptotisch [%] 3,49±2,00 5,29±6,50 16,70±5,88 56,44±31,76
‹0,001ɛ
π
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
In den LRS-Kammern zeigten weder die vitalen Lymphozyten noch die vitalen Mo-
nozyten eine signifikante Abnahme. Die Werte der vitalen Lymphozyten nahmen
bereits zum Beginn der Messreihe mit 78,73% eine sehr breite Spannweite ein, die
sich auch bis zum Messzeitpunktende mit einer Spannweite von 67,21% hielten. Die
vitalen Monozyten zeigten ebenfalls eine große Spannweite der Werte mit 91,48%
zum anfänglichen Messzeitpunkt und von 78,18% nach 48-stündiger Lagerung. Im
apoptotischen Verhalten der Lymphozyten und Monozyten konnte keine signifikante
Zunahme verzeichnet werden. Beide Zelltypen zeigten ein sehr variables apopto-
tisches Verhalten. Die apoptotischen Lymphozyten wiesen anfangs Werte zwischen
4,73% bis 76,37% auf. Nach sechsstündiger Lagerung waren ein minimaler Wert
von 1,64% und ein maximaler Wert von 62,85% zu finden. Anschließend stieg der
Anteil der apoptotischen Zellen nach 48-stündiger Lagerung auf Werte zwischen
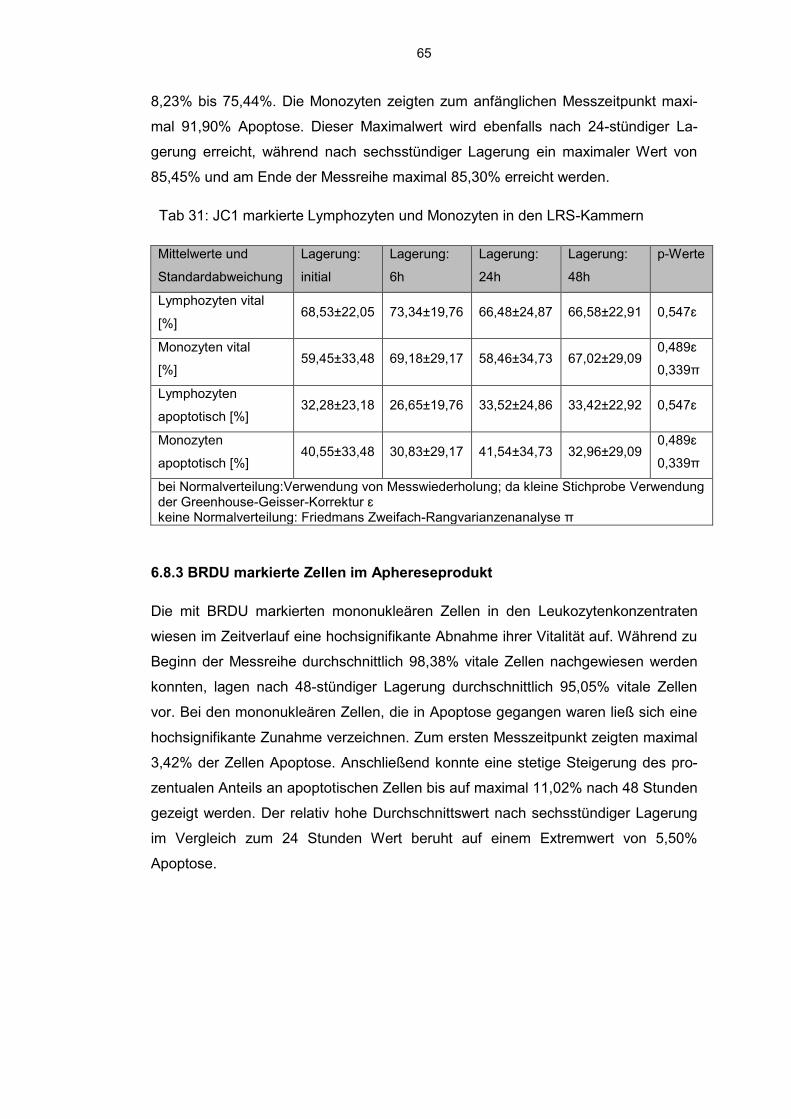
65
8,23% bis 75,44%. Die Monozyten zeigten zum anfänglichen Messzeitpunkt maxi-
mal 91,90% Apoptose. Dieser Maximalwert wird ebenfalls nach 24-stündiger La-
gerung erreicht, während nach sechsstündiger Lagerung ein maximaler Wert von
85,45% und am Ende der Messreihe maximal 85,30% erreicht werden.
Tab 31: JC1 markierte Lymphozyten und Monozyten in den LRS-Kammern
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
Lymphozyten vital
[%] 68,53±22,05 73,34±19,76 66,48±24,87 66,58±22,91 0,547ɛ
Monozyten vital
[%] 59,45±33,48 69,18±29,17 58,46±34,73 67,02±29,09
0,489ɛ
0,339π
Lymphozyten
apoptotisch [%] 32,28±23,18 26,65±19,76 33,52±24,86 33,42±22,92 0,547ɛ
Monozyten
apoptotisch [%] 40,55±33,48 30,83±29,17 41,54±34,73 32,96±29,09
0,489ɛ
0,339π
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwendung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
6.8.3 BRDU markierte Zellen im Aphereseprodukt
Die mit BRDU markierten mononukleären Zellen in den Leukozytenkonzentraten
wiesen im Zeitverlauf eine hochsignifikante Abnahme ihrer Vitalität auf. Während zu
Beginn der Messreihe durchschnittlich 98,38% vitale Zellen nachgewiesen werden
konnten, lagen nach 48-stündiger Lagerung durchschnittlich 95,05% vitale Zellen
vor. Bei den mononukleären Zellen, die in Apoptose gegangen waren ließ sich eine
hochsignifikante Zunahme verzeichnen. Zum ersten Messzeitpunkt zeigten maximal
3,42% der Zellen Apoptose. Anschließend konnte eine stetige Steigerung des pro-
zentualen Anteils an apoptotischen Zellen bis auf maximal 11,02% nach 48 Stunden
gezeigt werden. Der relativ hohe Durchschnittswert nach sechsstündiger Lagerung
im Vergleich zum 24 Stunden Wert beruht auf einem Extremwert von 5,50%
Apoptose.
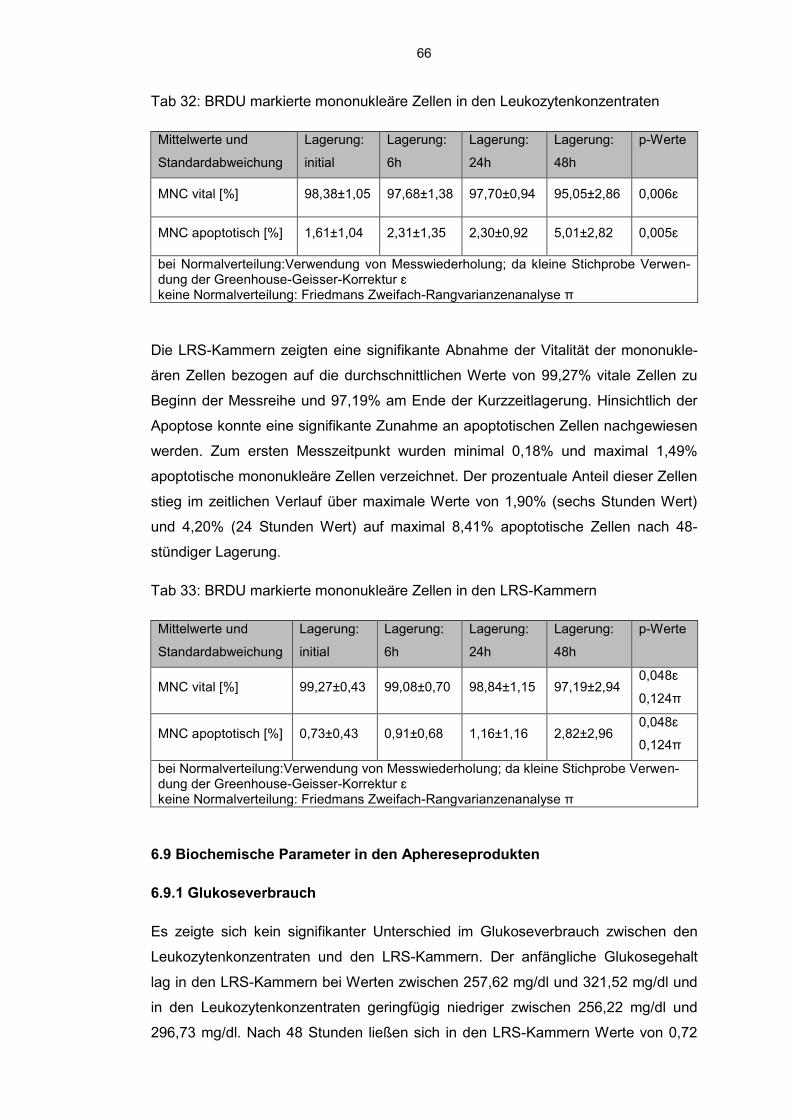
66
Tab 32: BRDU markierte mononukleäre Zellen in den Leukozytenkonzentraten
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
MNC vital [%] 98,38±1,05 97,68±1,38 97,70±0,94 95,05±2,86 0,006ɛ
MNC apoptotisch [%] 1,61±1,04 2,31±1,35 2,30±0,92 5,01±2,82 0,005ɛ
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
Die LRS-Kammern zeigten eine signifikante Abnahme der Vitalität der mononukle-
ären Zellen bezogen auf die durchschnittlichen Werte von 99,27% vitale Zellen zu
Beginn der Messreihe und 97,19% am Ende der Kurzzeitlagerung. Hinsichtlich der
Apoptose konnte eine signifikante Zunahme an apoptotischen Zellen nachgewiesen
werden. Zum ersten Messzeitpunkt wurden minimal 0,18% und maximal 1,49%
apoptotische mononukleäre Zellen verzeichnet. Der prozentuale Anteil dieser Zellen
stieg im zeitlichen Verlauf über maximale Werte von 1,90% (sechs Stunden Wert)
und 4,20% (24 Stunden Wert) auf maximal 8,41% apoptotische Zellen nach 48-
stündiger Lagerung.
Tab 33: BRDU markierte mononukleäre Zellen in den LRS-Kammern
Mittelwerte und
Standardabweichung
Lagerung:
initial
Lagerung:
6h
Lagerung:
24h
Lagerung:
48h
p-Werte
MNC vital [%] 99,27±0,43 99,08±0,70 98,84±1,15 97,19±2,94 0,048ɛ
0,124π
MNC apoptotisch [%] 0,73±0,43 0,91±0,68 1,16±1,16 2,82±2,96 0,048ɛ
0,124π
bei Normalverteilung:Verwendung von Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
6.9 Biochemische Parameter in den Aphereseprodukten
6.9.1 Glukoseverbrauch
Es zeigte sich kein signifikanter Unterschied im Glukoseverbrauch zwischen den
Leukozytenkonzentraten und den LRS-Kammern. Der anfängliche Glukosegehalt
lag in den LRS-Kammern bei Werten zwischen 257,62 mg/dl und 321,52 mg/dl und
in den Leukozytenkonzentraten geringfügig niedriger zwischen 256,22 mg/dl und
296,73 mg/dl. Nach 48 Stunden ließen sich in den LRS-Kammern Werte von 0,72
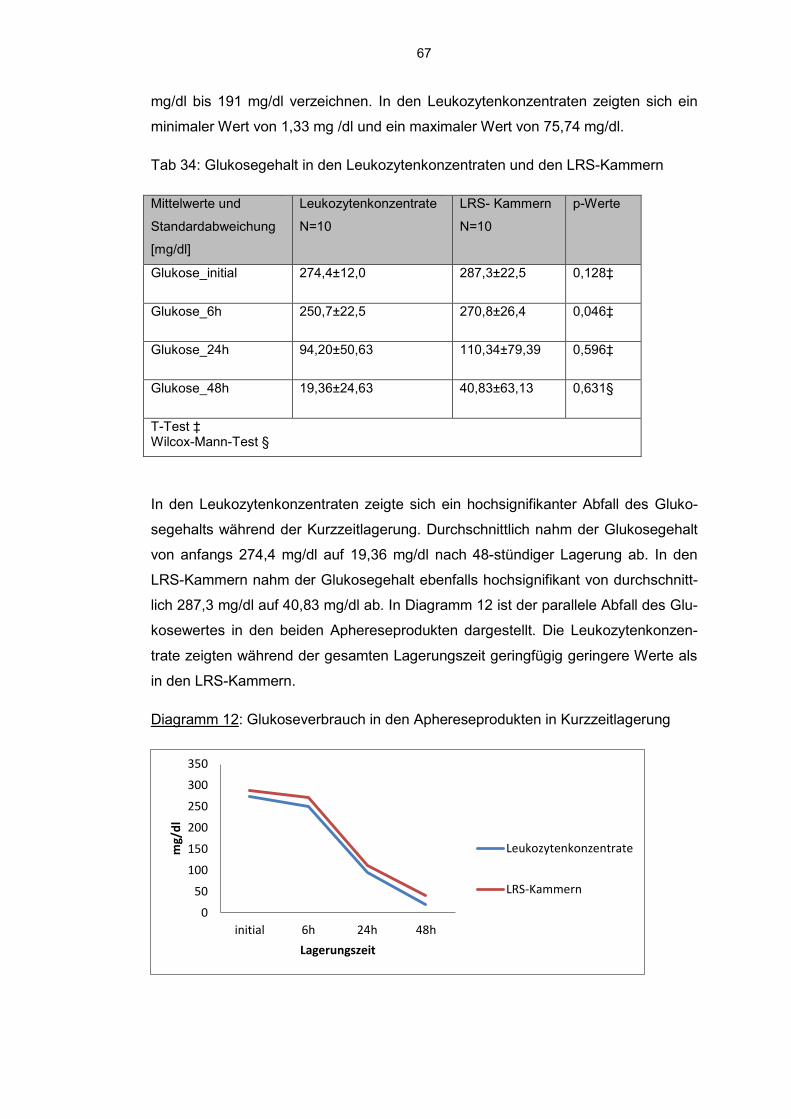
67
mg/dl bis 191 mg/dl verzeichnen. In den Leukozytenkonzentraten zeigten sich ein
minimaler Wert von 1,33 mg /dl und ein maximaler Wert von 75,74 mg/dl.
Tab 34: Glukosegehalt in den Leukozytenkonzentraten und den LRS-Kammern
Mittelwerte und
Standardabweichung
[mg/dl]
Leukozytenkonzentrate
N=10
LRS- Kammern
N=10
p-Werte
Glukose_initial 274,4±12,0 287,3±22,5 0,128‡
Glukose_6h 250,7±22,5 270,8±26,4 0,046‡
Glukose_24h 94,20±50,63 110,34±79,39 0,596‡
Glukose_48h 19,36±24,63 40,83±63,13 0,631§
T-Test ‡ Wilcox-Mann-Test §
In den Leukozytenkonzentraten zeigte sich ein hochsignifikanter Abfall des Gluko-
segehalts während der Kurzzeitlagerung. Durchschnittlich nahm der Glukosegehalt
von anfangs 274,4 mg/dl auf 19,36 mg/dl nach 48-stündiger Lagerung ab. In den
LRS-Kammern nahm der Glukosegehalt ebenfalls hochsignifikant von durchschnitt-
lich 287,3 mg/dl auf 40,83 mg/dl ab. In Diagramm 12 ist der parallele Abfall des Glu-
kosewertes in den beiden Aphereseprodukten dargestellt. Die Leukozytenkonzen-
trate zeigten während der gesamten Lagerungszeit geringfügig geringere Werte als
in den LRS-Kammern.
Diagramm 12: Glukoseverbrauch in den Aphereseprodukten in Kurzzeitlagerung
0
50
100
150
200
250
300
350
initial 6h 24h 48h
mg/
dl
Lagerungszeit
Leukozytenkonzentrate
LRS-Kammern
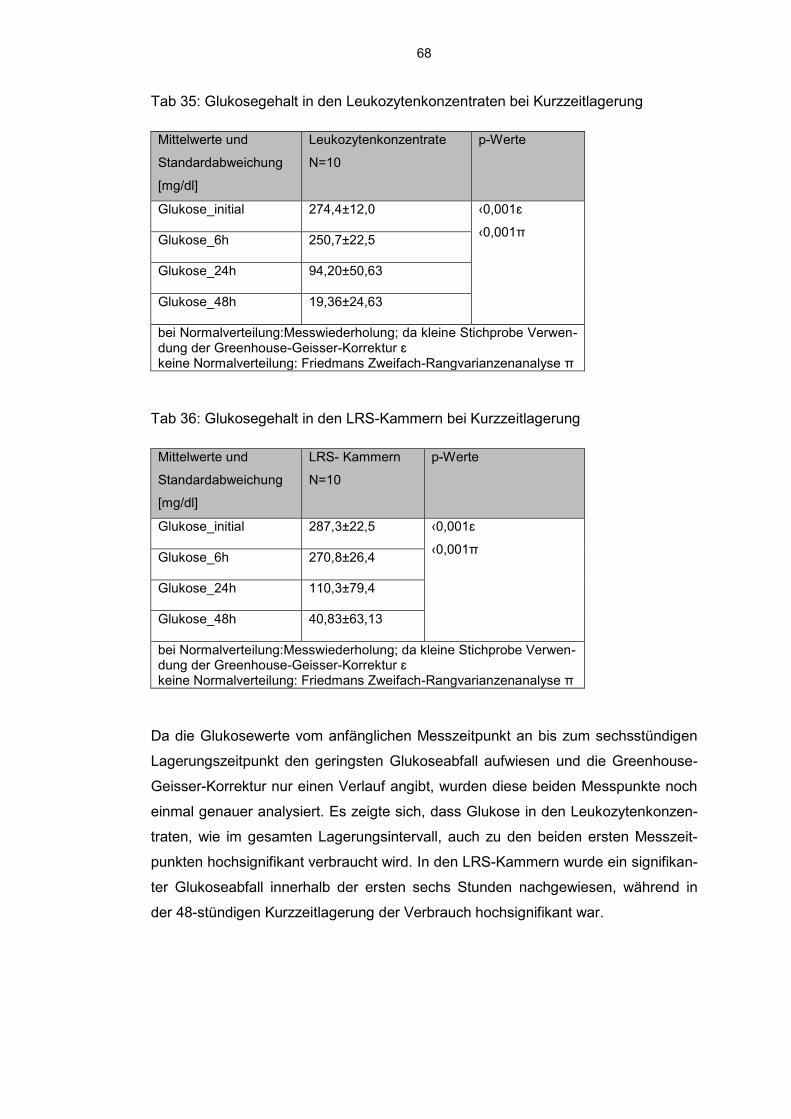
68
Tab 35: Glukosegehalt in den Leukozytenkonzentraten bei Kurzzeitlagerung
Mittelwerte und
Standardabweichung
[mg/dl]
Leukozytenkonzentrate
N=10
p-Werte
Glukose_initial 274,4±12,0 ‹0,001ɛ
‹0,001π Glukose_6h 250,7±22,5
Glukose_24h 94,20±50,63
Glukose_48h 19,36±24,63
bei Normalverteilung:Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
Tab 36: Glukosegehalt in den LRS-Kammern bei Kurzzeitlagerung
Mittelwerte und
Standardabweichung
[mg/dl]
LRS- Kammern
N=10
p-Werte
Glukose_initial 287,3±22,5 ‹0,001ɛ
‹0,001π Glukose_6h 270,8±26,4
Glukose_24h 110,3±79,4
Glukose_48h 40,83±63,13
bei Normalverteilung:Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
Da die Glukosewerte vom anfänglichen Messzeitpunkt an bis zum sechsstündigen
Lagerungszeitpunkt den geringsten Glukoseabfall aufwiesen und die Greenhouse-
Geisser-Korrektur nur einen Verlauf angibt, wurden diese beiden Messpunkte noch
einmal genauer analysiert. Es zeigte sich, dass Glukose in den Leukozytenkonzen-
traten, wie im gesamten Lagerungsintervall, auch zu den beiden ersten Messzeit-
punkten hochsignifikant verbraucht wird. In den LRS-Kammern wurde ein signifikan-
ter Glukoseabfall innerhalb der ersten sechs Stunden nachgewiesen, während in
der 48-stündigen Kurzzeitlagerung der Verbrauch hochsignifikant war.
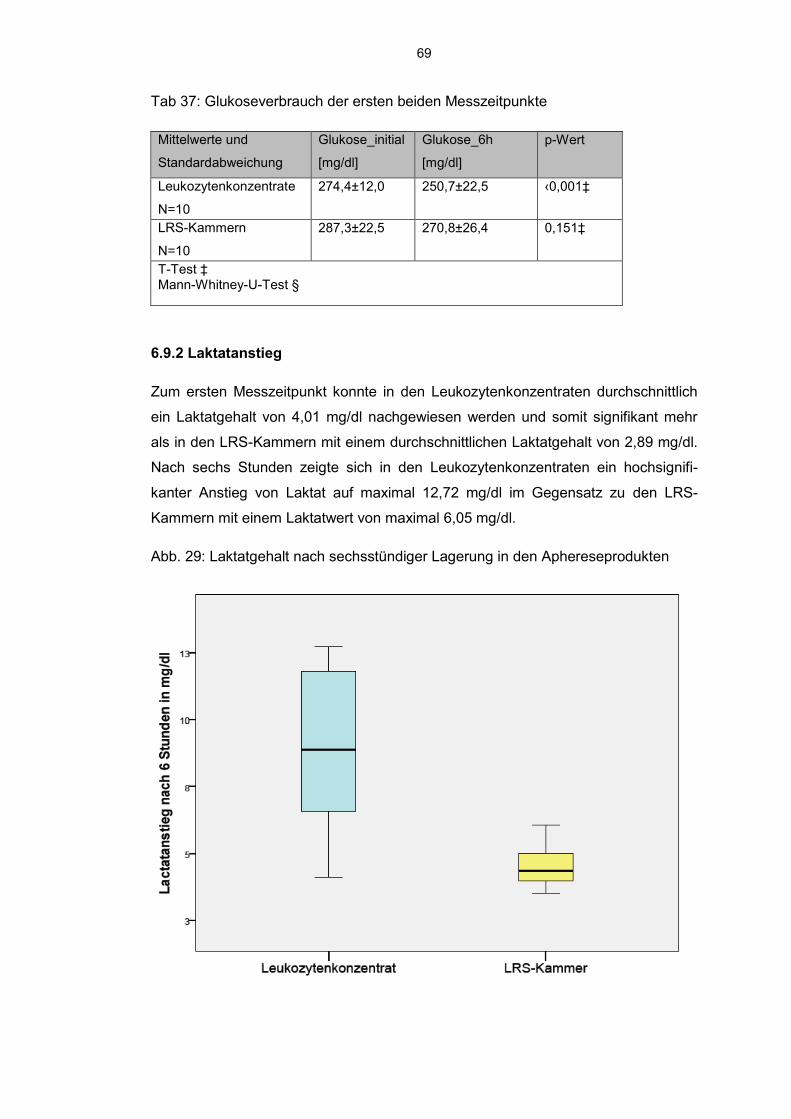
69
Tab 37: Glukoseverbrauch der ersten beiden Messzeitpunkte
Mittelwerte und
Standardabweichung
Glukose_initial
[mg/dl]
Glukose_6h
[mg/dl]
p-Wert
Leukozytenkonzentrate
N=10
274,4±12,0 250,7±22,5 ‹0,001‡
LRS-Kammern
N=10
287,3±22,5 270,8±26,4 0,151‡
T-Test ‡ Mann-Whitney-U-Test §
6.9.2 Laktatanstieg
Zum ersten Messzeitpunkt konnte in den Leukozytenkonzentraten durchschnittlich
ein Laktatgehalt von 4,01 mg/dl nachgewiesen werden und somit signifikant mehr
als in den LRS-Kammern mit einem durchschnittlichen Laktatgehalt von 2,89 mg/dl.
Nach sechs Stunden zeigte sich in den Leukozytenkonzentraten ein hochsignifi-
kanter Anstieg von Laktat auf maximal 12,72 mg/dl im Gegensatz zu den LRS-
Kammern mit einem Laktatwert von maximal 6,05 mg/dl.
Abb. 29: Laktatgehalt nach sechsstündiger Lagerung in den Aphereseprodukten
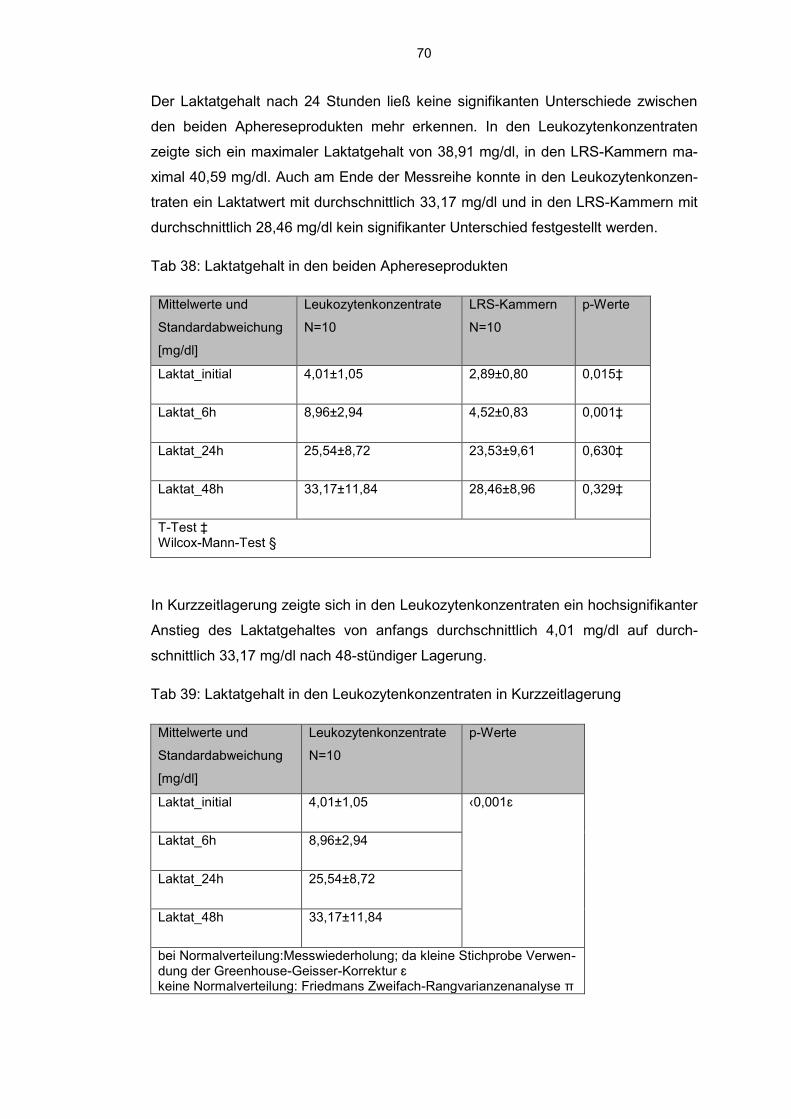
70
Der Laktatgehalt nach 24 Stunden ließ keine signifikanten Unterschiede zwischen
den beiden Aphereseprodukten mehr erkennen. In den Leukozytenkonzentraten
zeigte sich ein maximaler Laktatgehalt von 38,91 mg/dl, in den LRS-Kammern ma-
ximal 40,59 mg/dl. Auch am Ende der Messreihe konnte in den Leukozytenkonzen-
traten ein Laktatwert mit durchschnittlich 33,17 mg/dl und in den LRS-Kammern mit
durchschnittlich 28,46 mg/dl kein signifikanter Unterschied festgestellt werden.
Tab 38: Laktatgehalt in den beiden Aphereseprodukten
Mittelwerte und
Standardabweichung
[mg/dl]
Leukozytenkonzentrate
N=10
LRS-Kammern
N=10
p-Werte
Laktat_initial 4,01±1,05 2,89±0,80 0,015‡
Laktat_6h 8,96±2,94 4,52±0,83 0,001‡
Laktat_24h 25,54±8,72 23,53±9,61 0,630‡
Laktat_48h 33,17±11,84 28,46±8,96 0,329‡
T-Test ‡ Wilcox-Mann-Test §
In Kurzzeitlagerung zeigte sich in den Leukozytenkonzentraten ein hochsignifikanter
Anstieg des Laktatgehaltes von anfangs durchschnittlich 4,01 mg/dl auf durch-
schnittlich 33,17 mg/dl nach 48-stündiger Lagerung.
Tab 39: Laktatgehalt in den Leukozytenkonzentraten in Kurzzeitlagerung
Mittelwerte und
Standardabweichung
[mg/dl]
Leukozytenkonzentrate
N=10
p-Werte
Laktat_initial 4,01±1,05 ‹0,001ɛ
Laktat_6h 8,96±2,94
Laktat_24h 25,54±8,72
Laktat_48h 33,17±11,84
bei Normalverteilung:Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
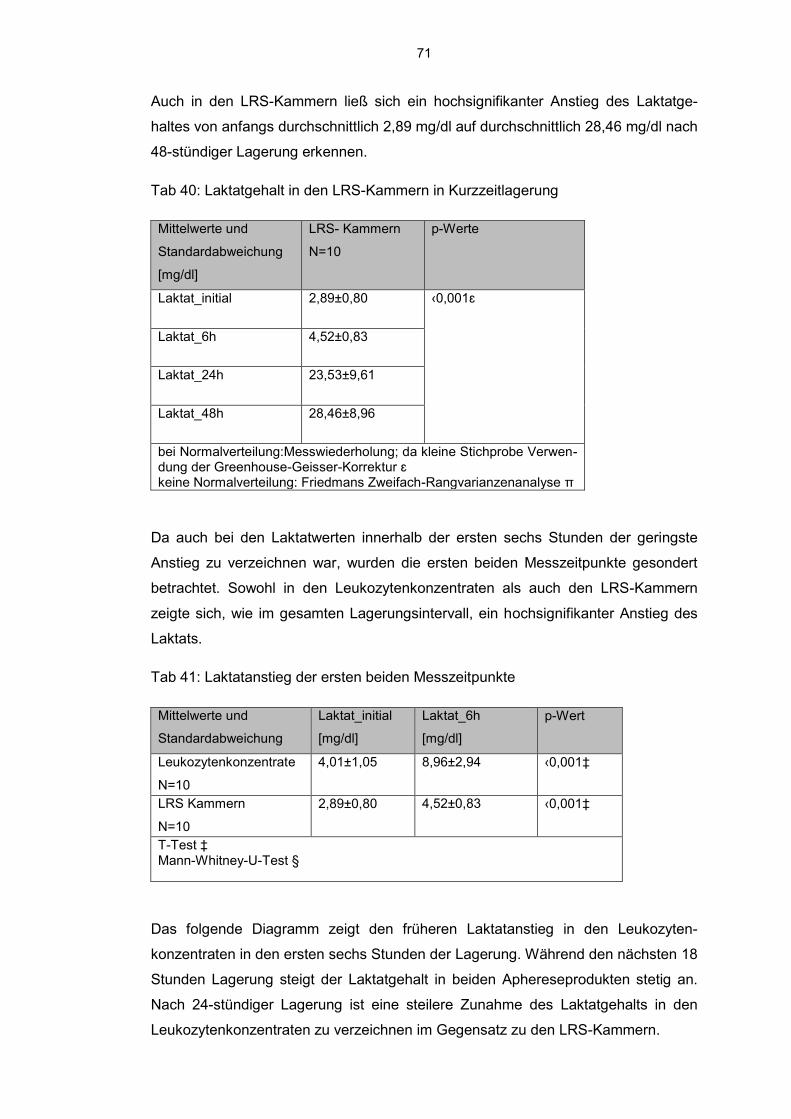
71
Auch in den LRS-Kammern ließ sich ein hochsignifikanter Anstieg des Laktatge-
haltes von anfangs durchschnittlich 2,89 mg/dl auf durchschnittlich 28,46 mg/dl nach
48-stündiger Lagerung erkennen.
Tab 40: Laktatgehalt in den LRS-Kammern in Kurzzeitlagerung
Mittelwerte und
Standardabweichung
[mg/dl]
LRS- Kammern
N=10
p-Werte
Laktat_initial 2,89±0,80 ‹0,001ɛ
Laktat_6h 4,52±0,83
Laktat_24h 23,53±9,61
Laktat_48h 28,46±8,96
bei Normalverteilung:Messwiederholung; da kleine Stichprobe Verwen-dung der Greenhouse-Geisser-Korrektur ɛ keine Normalverteilung: Friedmans Zweifach-Rangvarianzenanalyse π
Da auch bei den Laktatwerten innerhalb der ersten sechs Stunden der geringste
Anstieg zu verzeichnen war, wurden die ersten beiden Messzeitpunkte gesondert
betrachtet. Sowohl in den Leukozytenkonzentraten als auch den LRS-Kammern
zeigte sich, wie im gesamten Lagerungsintervall, ein hochsignifikanter Anstieg des
Laktats.
Tab 41: Laktatanstieg der ersten beiden Messzeitpunkte
Mittelwerte und
Standardabweichung
Laktat_initial
[mg/dl]
Laktat_6h
[mg/dl]
p-Wert
Leukozytenkonzentrate
N=10
4,01±1,05 8,96±2,94 ‹0,001‡
LRS Kammern
N=10
2,89±0,80 4,52±0,83 ‹0,001‡
T-Test ‡ Mann-Whitney-U-Test §
Das folgende Diagramm zeigt den früheren Laktatanstieg in den Leukozyten-
konzentraten in den ersten sechs Stunden der Lagerung. Während den nächsten 18
Stunden Lagerung steigt der Laktatgehalt in beiden Aphereseprodukten stetig an.
Nach 24-stündiger Lagerung ist eine steilere Zunahme des Laktatgehalts in den
Leukozytenkonzentraten zu verzeichnen im Gegensatz zu den LRS-Kammern.
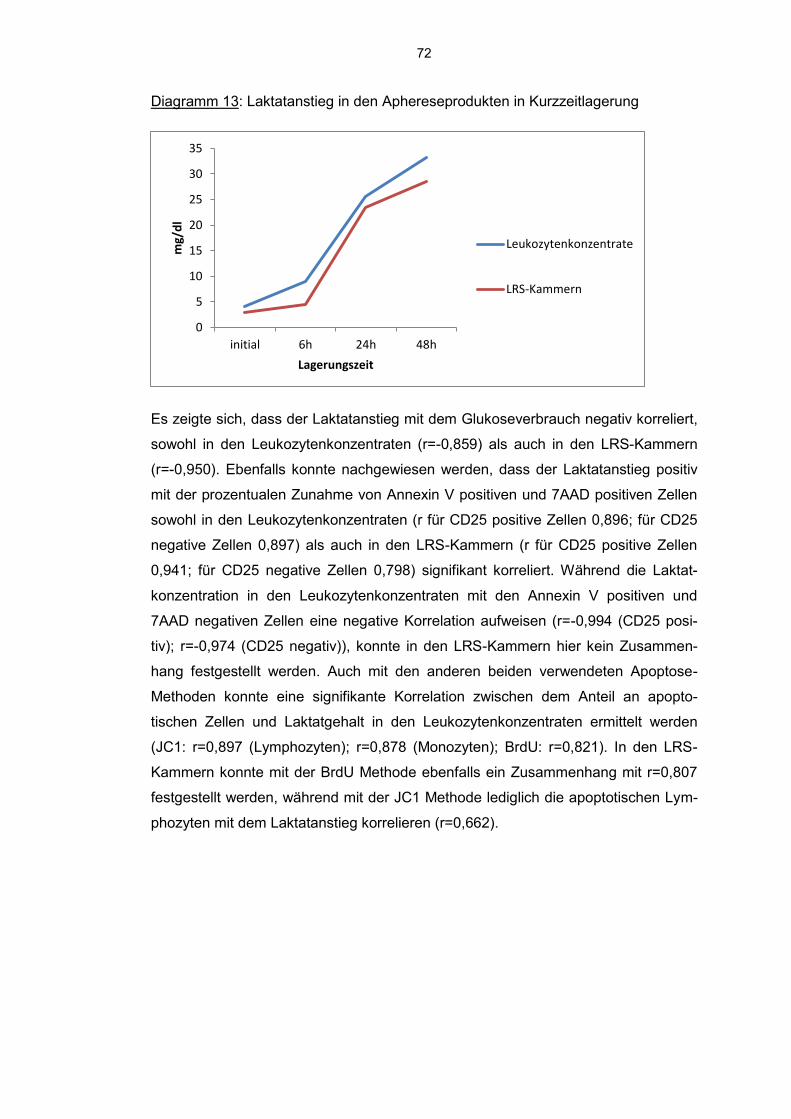
72
Diagramm 13: Laktatanstieg in den Aphereseprodukten in Kurzzeitlagerung
Es zeigte sich, dass der Laktatanstieg mit dem Glukoseverbrauch negativ korreliert,
sowohl in den Leukozytenkonzentraten (r=-0,859) als auch in den LRS-Kammern
(r=-0,950). Ebenfalls konnte nachgewiesen werden, dass der Laktatanstieg positiv
mit der prozentualen Zunahme von Annexin V positiven und 7AAD positiven Zellen
sowohl in den Leukozytenkonzentraten (r für CD25 positive Zellen 0,896; für CD25
negative Zellen 0,897) als auch in den LRS-Kammern (r für CD25 positive Zellen
0,941; für CD25 negative Zellen 0,798) signifikant korreliert. Während die Laktat-
konzentration in den Leukozytenkonzentraten mit den Annexin V positiven und
7AAD negativen Zellen eine negative Korrelation aufweisen (r=-0,994 (CD25 posi-
tiv); r=-0,974 (CD25 negativ)), konnte in den LRS-Kammern hier kein Zusammen-
hang festgestellt werden. Auch mit den anderen beiden verwendeten Apoptose-
Methoden konnte eine signifikante Korrelation zwischen dem Anteil an apopto-
tischen Zellen und Laktatgehalt in den Leukozytenkonzentraten ermittelt werden
(JC1: r=0,897 (Lymphozyten); r=0,878 (Monozyten); BrdU: r=0,821). In den LRS-
Kammern konnte mit der BrdU Methode ebenfalls ein Zusammenhang mit r=0,807
festgestellt werden, während mit der JC1 Methode lediglich die apoptotischen Lym-
phozyten mit dem Laktatanstieg korrelieren (r=0,662).
0
5
10
15
20
25
30
35
initial 6h 24h 48h
mg/
dl
Lagerungszeit
Leukozytenkonzentrate
LRS-Kammern

73
7.Diskussion
Die Leukapherese gilt als die beste Methode, um effektiv Monozyten in hoher Aus-
beute und Reinheit zu gewinnen [73]. Anfang der sechziger Jahre begannen die
ersten Versuche die Zellseparation zu optimieren, um neue Therapieoptionen im
Rahmen der Leukämiebehandlung zu ermöglichen [25]. Körbling und Pflieger rekru-
tierten Ende der 70er Jahre Patienten und freiwillige Spender zur Stammzellaphere-
se und belegten mittels in vitro Kulturverfahren den Erhalt funktionstüchtiger und
proliferationsfähiger Zellen [41]. Durch die Möglichkeit mittels Zytokinen wie G-CSF
(granulocyte colony stimulating factor) hämatopoetische Vorläuferzellen zu mobili-
sieren, konnten Stammzellen in 100fach höherer Ausbeute durch Leukozytapherese
aus dem peripheren Blut gewonnen werden, als ohne Mobilisierung aus dem Kno-
chenmark. Im Laufe der Jahre wurden die unterschiedlichsten Zellseparatoren er-
probt, um eine ausreichende Anzahl mononukleärer Zellen zu sammeln. Daraus
können antigen-präsentierende Zellen generiert werden, die im Rahmen von Im-
muntherapien eingesetzt, das Überleben bei malignen Erkrankungen verbessern
sollen [88].
In dieser Arbeit wurden die mononukleären Zellen, die durch Leukozytapherese
mit dem ComTec Zellseparator gewonnen wurden und aus LRS-Kammern, die bei
der Thrombozytapherese mit dem Trima Accel Zellseparator als Nebenprodukt ent-
stehen, hinsichtlich ihrer Zellqualität und des Zellverlustes während der Kurzzeitla-
gerung untersucht. Die Leukozytenausbeute und der Restzellgehalt sind von den
eingestellten Parametern am Apheresegerät abhängig. Bei der Leukapherese dau-
erte die Separation durchschnittlich 121 Minuten mit einem gemittelten Verbrauch
von 522 ml Acid citrate dextrose Formula A (ACD-A) bei einem prozessierten Blut-
volumen von 4831 ml. Bei der Thrombozytapherese mittels Trima Accel Zellsepara-
tor wurde bei einer Separationsdauer von durchschnittlich 51 Minuten hochsignifi-
kant weniger ACD-A (322 ml) verbraucht bei einem prozessierten Blutvolumen von
2965 ml. Gemäß dem Transfusionsgesetz der Bundesärztekammer darf die Spen-
denfrequenz von maximal 26 Thrombozytapheresen pro Jahr nicht überschritten
werden [83]. Die beschriebene Möglichkeit eines Leukozytenverlustes von 1 bis 5
x109 Zellen pro Liter pro Spende mit daraus resultierendem klinisch bedeutsamem
Immundefizit bei hoher Blutspendefrequenz [80] konnte beim Vergleich des EDTA-
Blutes vor der Monozytenspende mit dem EDTA-Blut vor der Thrombozytenspende
nicht nachgewiesen werden. Die Spender zeigten durchschnittlich nur einen Leuko-
zytenverlust von 0,3 x109 Zellen pro Liter pro Spende.

74
Die Leukozytapherese mittels ComTec Zellseparator lieferte im Konzentrat
durchschnittlich eine Leukozytenkonzentration von 45 x109 Leukozyten pro Liter,
wobei der Zellgehalt bezogen auf das Produktvolumen 5,6 x109 Leukozyten beträgt.
Ähnliche Werte sind in der Literatur zu finden [66; 78]. Eine Studie von Strasser und
Mitarbeiter, die verschiedene Zellseparatoren miteinander verglich, zeigte, dass der
ComTec Zellseparator die höchste Sammeleffizienz aufwies [73]. In den untersuch-
ten Leukozyten-Reduktion-System Kammern (LRSC), die als neue Ressource für
mononukleäre Zellen durch die Entwicklung von leukozyten-reduzierender Filter bei
der Herstellung von Thrombozytenkonzentraten eingesetzt werden [20], enthielten
durchschnittlich eine Leukozytenkonzentration von 89 x109 Leukozyten pro Liter,
wobei der Zellgehalt aufgrund des kleinen Produktvolumens nur bei 0,89 x109 Leu-
kozyten lag.
Die unterschiedliche Herstellung der Leukozytenprodukte verursacht unter-
schiedliche Zellkonzentrationen. So unterschieden sich die Leukozytapheresepro-
dukte und LRS-Kammern nicht nur hochsignifikant in der Leukozytenausbeute, son-
dern auch im Anteil der Restblutzellen. Die durch Leukozytapherese gewonnenen
und untersuchten Leukozytenkonzentrate zeigten 4,4 x1012 Thrombozyten pro Liter
und 0,8 x1012 Erythrozyten pro Liter. Ähnliche Thrombozytenwerte wurden bereits in
der Literatur mit 5,0 x1012 Thrombozyten pro Liter beschrieben [74]. Es ist bekannt,
dass der ComTec Zellseparator im Vergleich zu anderen Zellseparatoren, wie dem
Fenwal Amicus Zellseparator, eine größere Verunreinigung mit Thrombozyten auf-
weist [3; 73]. Als mögliche Ursache hierfür wurde die zusätzliche Gewinnung von
thrombozytenreichem Plasma während der Konzentratherstellung angesehen [76].
Durch die hohe thrombozytäre Verunreinigung könnte es zu einem Verlust an mo-
nonukleären Zellen im Produkt kommen [29; 76], da Thrombozyten mit Monozyten
interagieren und dadurch deren Ausdifferenzierung beeinflussen können [39]. Der
geringe Erythrozytenanteil im Konzentrat war vergleichbar mit den Ergebnissen der
Leukozytapherese mit dem Cobe Spectra Zellseparator [77; 78]. Der Restzellgehalt
in den LRS-Kammern betrug 1,7 x1012 Thrombozyten und 5,8 x1012 Erythrozyten
pro Liter. Die Werte zeigten sich gut reproduzierbar mit den in einer von Strasser
und Mitarbeiter erhobenen Studie von durchschnittlich 2,5 x1012 Thrombozyten und
5,5 x1012 Erythrozyten pro Liter [79]. Im Vergleich mit dem Cobe Spectra Zellsepa-
rator liegt der Anteil an residualen Zellen in den LRS-Kammern bei der Trima Accel
wesentlich höher, allerdings steigt auch die Ausbeute an Leukozyten deutlich [79].
Im Vergleich zu den Leukozytenkonzentraten enthalten die LRS-Kammern anteils-
mäßig fünffach mehr residuale Erythrozyten, bei einem mittleren korpuskulären Vo-
lumen von 80,7 fl in den Leukozytenkonzentraten und 90,1 fl in den LRS-Kammern.

75
Die Zellqualität lässt sich mit unterschiedlichen Methoden zur Apoptosedetektion
untersuchen. Die Arbeit verglich drei unterschiedliche Apoptosemethoden mitein-
ander. Da nicht jede Zelle alle Apoptosestadien durchläuft, ist eine Kombination der
Methoden erforderlich, um sinnvolle Aussagen über die Qualität der Zellen zu erhal-
ten. Die Annexin V Färbung ist eine sehr sensitive und in zahlreichen Studien gut
etablierte Methode zur Apoptosedetektion [56; 84; 86; 89]. In der Frühphase der
Apoptose kommt es zur Umverteilung der Phospholipide in der Plasmamembran, so
dass Phosphatidylserin auf die Außenseite transloziert wird, woran Annexin V spezi-
fisch bindet. Da dieser Prozess auch in nekrotischen Zellen abläuft, wurde die Me-
thode mit der 7AAD Kernfärbung verknüpft, da in apoptotischen Zellen die Plasma-
membran noch intakt ist und 7AAD nicht eindringen kann [86]. Da allerdings auch
spätapoptotische Zellen ihre Zellintegrität meist verlieren, kann mit der Annexin V
Färbung allein nicht zwischen den verschiedenen Apoptosestadien, Frühapoptose,
Spätapoptose und Nekrose unterschieden werden [45]. Eine weitere Methode zur
Messung der Apoptose stellt die Detektion des mitochondrialen Potentials mittels
JC1 Färbung dar. Diese Methode ist zwingend an die noch bestehende Integrität
der Zelle gebunden, da sich die Mitochondrien im Zytoplasma befinden. Nekrotische
Zellen sollten demnach nicht erfasst werden, da es zum Austritt von Zytoplasma und
der Zellorganellen in den Extrazellularraum kommt und dadurch diese Zellen nicht
mehr durchflußzytometrisch messbar wären. Mit der JC 1 Methode wird eine sehr
frühe Phase der Apoptose erfasst. Allerdings ist nicht klar, ob die Änderung des
mitochondrialen Potentials generell auftritt [48; 51]. Es ist auch nicht auszuschlie-
ßen, dass lebende Zellen mit erfasst werden, da in dem Stadium der Veränderung
des mitochondrialen Potentials die Zellintegrität und die Funktion der Plasmamem-
bran noch weitestgehend erhalten sind [19]. Das dritte verwendete Nachweisverfah-
ren ist die DNA-Strangbruch-Markierung mit BrdU (5-Bromo-2´-deoxyuridin), die
sogenannte TUNEL-Methode. Lugli und Mitarbeiter konnten zeigen, dass die DNA-
Fragmentierung im späteren apoptotischen Prozess stattfindet, so dass diese Me-
thode zum Nachweis der Spätapoptose dient [48]. Nekrotische Zellen werden nicht
erfasst, da diese bereits außerhalb des durchflußzytometrischen Messbereichs lie-
gen. Zu bedenken ist hierbei allerdings, dass mit dieser Methode nicht nur apoptoti-
sche Prozesse nachgewiesen werden, sondern auch Zellen, die aus anderen Grün-
den DNA-Strangbrüche aufweisen [22]. In der Literatur existieren einige Arbeiten,
die diese Methoden zur Detektion der unterschiedlichen Apoptosephasen einsetzen
[49; 63; 67].
Mit der Annexin V und 7AAD Methode wurden in dieser Studie speziell CD25
positive T-Helferzellen (regulatorische T-Zellen) hinsichtlich ihres apoptotischen Ver-

76
haltens untersucht. Es ließ sich zu Beginn der Messreihe in den Leukozytenkonzen-
traten bei den CD4 positiven und CD25 positiven T-Zellen eine Apoptoserate von
10% nachweisen und somit hochsignifikant mehr als in den LRS-Kammern mit nur
6%. Amer und Mitarbeiter wiesen nach, dass durch maschinelle Manipulation oxida-
tiver Stress ausgelöst wird und zu einem deutlichen Zellschaden führen kann [4].
Ein Unterschied am Anteil der Annexin V positiven und 7AAD positiven CD25 positi-
ven regulatorischen T-Zellen wurde in den beiden Leukozytenprodukten zu Beginn
nicht ersichtlich (0,2% versus 0,3%). Zum Messzeitpunkt nach 24 Stunden über-
steigt dann der Anteil an Annexin V positiven und 7AAD positiven CD25 positiven
Tregs in den Leukozytenkonzentraten mit 2% den Anteil in den LRS-Kammern mit
0,5% hochsignifikant. Der Unterschied im prozentualen Anteil an Annexin V positi-
ven und 7AAD positiven T-Zellen wird nach 48-stündiger Lagerung noch deutlicher
und zeigt in den Leukozytenkonzentraten hochsignifikant höhere Werte im Vergleich
zu den LRS-Kammern (7% versus 1%). Ein signifikant höherer Anteil an Annexin V
positiven und 7AAD positiven Lymphozyten nach 48-stündiger Lagerung im Leuko-
zytaphereseprodukt im Gegensatz zu den LRS-Kammern wurde bereits beschrie-
ben [87]. In der Studie wurden die unterschiedlichen Herstellungsverfahren der bei-
den Produkte als mögliche Ursache für den signifikant unterschiedlichen Anteil an
lebenden Zellen angesehen [87]. Da die Annexin V und 7AAD Konjugation an die
Zelle davon abhängig ist, wie intakt die Plasmamembran ist, binden nur Zellen mit
Verlust der Membranintegrität Annexin V und 7AAD und weisen so einen irreversib-
len Zellschaden auf. Es ist daher anzunehmen, dass die Zellen bei der Leukozyt-
apherese größeren Schaden nehmen als die Zellen bei der Gewinnung der LRS-
Kammern, da die Zellen der LRS-Kammern schonender gewonnen werden und
nicht so stark wie im Buffy coat komprimiert werden.
Durch die begrenzte Anzahl an Kanälen des verwendeten Durchflußzytometers,
war eine differenzierte Darstellung der Tregs mit der JC 1 Apoptosemessung limi-
tiert, so dass nur der Gesamtanteil an Lymphozyten und Monozyten analysiert wur-
de. Da Monozyten mehr Zellorganellen besitzen als Lymphozyten und somit mehr
JC1 aufnehmen können, ließen sich diese beiden Zellgruppen gut unterscheiden
[15]. Die Leukozytenkonzentrate in dieser Studie zeigten einen höheren Gehalt an
vitalen Lymphozyten (89% versus 69%) und Monozyten (97% versus 60%) als die
LRS-Kammern. Hierfür könnte die längere Separationszeit der Leukozytapherese,
bei der ein größeres Blutvolumen prozessiert wird, verantwortlich sein [12]. Bei der
Apoptosedetektion mittels JC1 Färbung ließ sich bereits zu Beginn mit 32% in den
LRS-Kammern ein dreimal so hoher Anteil an apoptotischen Lymphozyten nachwei-
sen als in den Leukozytenkonzentraten mit 11%. Der Gehalt an apoptotischen Mo-

77
nozyten lag in den LRS-Kammern sogar sechsfach höher vor als in den Leukozy-
tenkonzentraten. Im Vergleich zur Annexin V und 7AAD Methode wären in den LRS-
Kammern weniger apoptotische Zellen zu erwarten gewesen als in den Leukozyten-
konzentraten. Erst nach 48 Stunden ließen sich in den Leukozytenkonzentraten
prozentual mehr apoptotische Zellen mit 50% versus 33% Lymphozyten und 56%
versus 33% Monozyten nachweisen. Da die Verunreinigung mit Erythrozyten in den
LRS-Kammern höher ist, könnte dies als mögliche Ursache für die bereits anfäng-
lich hohe Apoptoserate in Erwägung gezogen werden. Auch andere zelluläre Vor-
gänge, die in der Lage sind das mitochondriale Potential zu verändern, wären denk-
bar. Ebenso sollte daran gedacht werden, dass nicht alle Mitochondrien in den Zel-
len zur selben Zeit depolarisieren [48], so dass sich gewisse apoptotische Unter-
schiede daraus ergeben könnten. Diese Hypothesen sollten Gegenstand weiterer
Untersuchungen sein und überprüft werden.
Mit der DNA-Strangbruchdetektion mittels BrdU konnten, bedingt durch die Limi-
tierung durch das Durchflußzytometers, nur mononukleäre Zellen detektiert werden.
Bereits zu Beginn ließ sich in den Leukozytenkonzentraten eine signifikant höhere
Apoptoserate der mononukleären Zellen im Vergleich zu den LRS-Kammern (2%
versus 1%) nachweisen. Auch nach 48-stündiger Lagerung lagen in den Leukozy-
tenkonzentraten signifikant mehr apoptotische Zellen vor als in den LRS-Kammern.
Der Anteil an apoptotischen Zellen lag hier bei 5% (Leukozytenkonzentraten) und
3% (LRS-Kammern). Da die DNA-Strangbruchdetektion ausschließlich Zellen in der
Spätapoptose erfasst, war entsprechend der Lagerungszeit mit niedrigen Werten für
die apoptotischen Zellen zu rechnen. Die Vermutung liegt nahe, dass noch nicht alle
Zellen in die Spätapoptose eingetreten sind und sich noch keine DNA-Strangbrüche
ereignet hatten. Mit der BrdU Methode zeigte sich wieder, wie bei der Annexin V
und 7AAD Methode, dass in den Leukozytenkonzentraten der Anteil an apopto-
tischen Zellen höher liegt als in den LRS-Kammern. Wie bereits erwähnt könnte
hierfür der durch die Apherese ausgelöste oxidative Stress verantwortlich sein.
Denkbar wäre auch, dass die gewonnenen Leukozytenkonzentrate durch den hohen
Restgehalt an Thrombozyten vermehrt apoptotische Zellen aufweisen. Einige Stu-
dien berichten über eine durch die Apherese bedingte Erhöhung der Stoffwech-
selaktivität der Thrombozyten, welche mit einer gesteigerten Apoptose korrelieren
[5; 7; 35]. Dadurch könnte der Stoffwechsel der Leukozyten beeinträchtigt werden,
so dass es vermehrt zur Zellschädigung kommen könnte [87].
Einen hohen Stellenwert für die Viabilität der Zellen haben die Lagerungs-
bedingungen hinsichtlich Temperatur, Lagerungsmedium und Lagerungszeit, wes-

78
halb viele Studien sich mit deren Optimierung befassen. Hubel und Mitarbeiter zeig-
ten beispielsweise, dass es bei der Lagerung von Plazentarestblut im Lagerungs-
medium über einen Zeitraum von 24 Stunden kaum zu Zellverlust kommt [36].
Garraud und Mitarbeiter wiesen nach, dass es keinen prozentualen Unterschied im
Anteil an T-Helferzellen und an T-Suppressorzellen gab im Vergleich von frischen
mit gelagerten (22°C) Blutprodukten [27]. Diese Studie evaluiert allerdings den spe-
ziellen Einfluss der Lagerung auf die Zellqualität in Leukozytenkonzentraten und
LRS-Kammern, die jeweils in Beutelsystemen aus CO2 und O2 durchlässigem Fluor-
ethylenpropylen-Teflon abgefüllt, bei Raumtemperatur über einen Zeitraum von 48
Stunden aufbewahrt wurden. Sowohl in den Leukozytenkonzentraten als auch in
den LRS-Kammern wurden innerhalb dieses Lagerungsintervalls der Anteil an
apoptotischen Zellen mittels der drei bereits beschriebenen Färbungen zum
Apoptosenachweis bestimmt.
Mit der Annexin V und 7AAD Nachweismethode konnte ein signifikanter Anstieg
an Annexin V positiven und 7AAD positiven regulatorischen T-Helferzellen innerhalb
der 48-stündigen Lagerung in beiden Leukozytenprodukten aufgezeigt werden. Da-
bei zeigte sich ein deutlich höherer Anteil Annexin V positiver und 7AAD positiver
Tregs in den Leukozytenkonzentraten mit maximal 8% im Vergleich zu den LRS-
Kammern mit maximal 2%. Dieser Unterschied scheint, wie bereits erwähnt, in der
schonenderen Gewinnung der LRS-Kammern und des damit verbundenen gering-
eren oxidativen Stresses begründet zu sein. Es zeigte sich, dass besonders nach 24
Stunden Lagerung der Anteil an Annexin V positiven und 7AAD positiven Tregs
stark ansteigt. Dies lässt vermuten, dass in den ersten 24 Stunden die Apoptose
dominiert bevor sie nach 24 Stunden in die Nekrose mündet. Lund und Mitarbeiter
zeigten denselben Effekt in ihrer Studie über die Kultivierung von Monozyten [49].
Auffällig zeigte sich ebenfalls ein wesentlich höherer Anteil an Annexin V positiven
und 7AAD negativen CD25 positiven Tregs mit anfangs 10% im Vergleich zu den
CD25 negativen T-Zellen mit einer initialen Apoptoserate von 5%. Die höhere
Apoptoserate der CD25 positiven Tregs im Vergleich zu den CD25 negativen T-
Zellen bleibt im Zeitverlauf bestehen und zeigt sich sowohl in den Leukozytenkon-
zentraten als auch in den LRS-Kammern. Da einige Studien darüber berichten, dass
CD25 positive T-Zellen die Proliferation von CD25 negativen T-Zellen supprimieren
können [46; 70], könnte dies die Ursache für den deutlich geringeren Anteil an
apoptotischen CD25 negativen T-Zellen sein.
Da bei der JC 1 Methode zur Detektion des mitochondrialen Potentials die Dif-
ferenzierung der T-Zellsubpopulation nicht möglich war, wurde der Gesamtanteil an

79
Lymphozyten und Monozyten analysiert. Auch diese Methode zeigte, dass in den
Leukozytenkonzentraten der Anteil an apoptotischen Zellen nach 24 Stunden La-
gerung stark ansteigt. Zu Beginn der Messreihe lagen für die Lymphozyten bereits
11% apoptotische Zellen vor. Auch die Annexin V und 7AAD Methode zeigte für die
CD25 positiven Tregs zu Beginn eine Apoptoserate von 10%. Der Anteil apopto-
tischer Monozyten lag in den ersten 24 Stunden deutlich unter dem Anteil apopto-
tischer Lymphozyten. Initial konnte nur eine Apoptoserate von knapp 4% nachge-
wiesen werden. Strasser und Mitarbeiter zeigten bereits, dass Lymphozyten im Ge-
gensatz zu Monozyten stabiler sind und den Apheresevorgang besser überstehen.
Auch die unterschiedliche Lebensdauer der Zellen in vivo könnte eine Rolle für die
unterschiedliche Apoptoserate spielen [49]. Möglich wäre auch, dass die Inkuba-
tionstemperatur einen Einfluss hat [82]. Die Untersuchung der apoptotischen Lym-
phozyten und Monozyten in den LRS-Kammern mit der JC 1 Methode zeigte vom
ersten Messzeitpunkt an ein Kontinuum im Anteil apoptotischer Zellen, so dass die-
se Ergebnisse im ersten Moment unplausibel erscheinen, weshalb diese Ergebnisse
zunächst kontrolliert werden müssen.
Limitiert durch die Durchflußzytometrie konnten mit der DNA-Strangbruch-
Markierung mittels des Thymidinanalogons BrdU nur mononukleäre Zellen darge-
stellt werden. Wie bei der Annexin V und 7AAD Methode zeigte die BrdU Methode
in den Leukozytenkonzentraten einen höheren Anteil an apoptotischen Zellen im
Vergleich zu den LRS-Kammern. Der Anteil apoptotischer mononukleärer Zellen lag
aber sowohl in den Leukozytenkonzentraten mit 5% und in den LRS-Kammern mit
knapp 3% nach 48 Stunden Lagerung noch recht niedrig. Da keine Apoptose indu-
zierenden Reagenzien verwendet wurden, ist davon auszugehen, dass das unter-
suchte Zeitintervall nicht ausreichend lang war, um Zellen in der Spätapoptose zu
detektiert. Da mit der JC 1 Methode nach 48-stündiger Lagerung mehr als 50%
apoptotische Zellen nachgewiesen werden konnten, liegt die Vermutung nahe, dass
bei dem Großteil der Zellen die Zellveränderungen der Spätapoptose noch nicht
aufgetreten sind.
Des Weiteren wurden die biochemischen Parameter Glukose und Laktat und
deren Bezug zur Apoptoserate in dieser Arbeit analysiert. In den letzten Jahren be-
schäftigten sich viele Studien [38; 69; 71] mit der optimalen Lagerung und den damit
verbundenen metabolischen Veränderungen der Zellen, da man weiß, dass kleinste
Abweichungen zum Verlust der Überlebensfähigkeit der Zelle führen können. Den
Zellen stehen zwei verschiedene Wege zur Energiegewinnung offen. Die sehr effek-
tive oxidative Verstoffwechselung von Glukose und Fettsäuren zu Adenosintriphos-

80
phat (ATP) und die wenig adäquate anaerobe Glykolyse. Unter hypoxischen Bedin-
gungen versuchen die Zellen deshalb ihren Energiebedarf mittels gesteigerter an-
aerober Glykolyse zu decken, wobei vermehrt Laktat als Stoffwechselprodukt anfällt
[28]. In dieser Studie verzeichneten die Leukozytenkonzentrate einen Glukosever-
brauch von 93% in 48 Stunden und einen achtfachen Anstieg der Laktatkonzentra-
tion. Während des Lagerungszeitraums kam es nach 24 Stunden rapide zum Zuk-
kerverbrauch bzw. Anstieg der Laktatkonzentrationen. Die LRS-Kammern, die unter
den gleichen Bedingungen wie die Leukozytenkonzentrate gelagert waren, zeigten
ebenfalls einen starken Abfall der Glukosekonzentration von 86% und einen neun-
fachen Anstieg der Laktatkonzentration. Auch hier kam es im Zeitintervall von sechs
Stunden bis 24 Stunden zur stärksten Veränderung. Es ist zu vermuten, dass in
dieser Zeitspanne die Zellen eine hohe metabolische Aktivität aufweisen, gekoppelt
an eine deutliche Zunahme an apoptotischen Zellen in diesem Zeitintervall [32]. In
dieser Arbeit wurde bereits mit der Annexin V und 7AAD Methode sowie mit dem JC
1 Nachweisverfahren gezeigt, dass der prozentuale Anteil apoptotischer Zellen nach
24 Stunden stark ansteigt. Auch Kilkson und Mitarbeiter [38] sowie Murphy und Mit-
arbeiter [52] konnten in Studien nachweisen, dass die steigende Laktatproduktion
als Indikator für den Verlust der Viabilität der Zellen anzusehen ist. Diese Hypothese
konnte mit der starken Korrelation zwischen apoptotischen Zellen aller drei verwen-
deter Apoptose-Methoden und Laktatgehalt bestätigt werden. Da ein inverser Bezug
zwischen Annexin V positiver und 7AAD negativer Zellen und Laktatanstieg zu ver-
zeichnen war, ist zu vermuten, dass die Laktatproduktion mit Spätapoptose und
Nekrose von Zellen verbunden ist, da Annexin V positive und 7AAD negative Zellen
noch vollständig intakt sind und nur Annexin V positive und 7AAD positive Zellen,
die einen Verlust der Plasmamembran aufweisen, einen positiven Zusammenhang
aufzeigen. Da eine inverse Korrelation zwischen Glukoseverbrauch und Laktatan-
stieg besteht, kann darauf rückgeschlossen werden, dass mit zunehmendem Gluko-
severbrauch der Anteil an apoptotischen Zellen stetig steigt [74].
In wie weit zusätzliche Analysemethoden, wie die Massenspektrometrie zum
Nachweis molekularer Veränderungen, während der Zelllagerung eingesetzt werden
können, ist derzeit noch unklar. Die Veränderung der Glycerophospholipid-Zu-
sammensetzung der Zellmembran während der Apoptose wurde bereits nachgewie-
sen und mit Verfahren wie der Annexin V und 7AAD Methode dargestellt. Weitere
mögliche Veränderungen, wie der intrazelluläre Ceramid Gehalt, der darüber ent-
scheidet, ob eine Zelle in Apoptose geht oder überlebt [81], könnten massenspek-
trometrisch erfasst werden. Pienimäki-Römer und Mitarbeiter [59] wiesen mittels
Massenspektrometrie eine Verminderung des Sphingolipidspiegels und eine Um-

81
wandlung in Ceramide in gelagerten Thrombozyten nach. Erste massenspek-
trometrische Untersuchungen hinsichtlich des Gehaltes an Glycerophospholipiden,
Sphingolipiden, Acylcarnithinen, biogenen Aminen und Aminosäuren im Lagerungs-
zeitraum von 48 Stunden konnten noch keinen Ausblick auf die zu erwartenden Ver-
änderungen liefern.
Zusammenfassend zeigten die Apoptosemethoden einen höheren Anteil an
apoptotischen Zellen in den Leukozytenkonzentraten als die Zellen, die im Rahmen
der Thrombozytapherese in den LRS-Kammern gewonnen wurden, so dass sich die
Hypothese, dass die Zellen bei der Leukozytapherese einen größeren Schaden
nehmen als bei der Gewinnung durch Thrombozytapherese, zu bestätigen scheint.
Da mit der JC 1 Methode ein hoher prozentualer Anteil an apoptotischen Zellen
auch in den LRS-Kammern gemessen wurde, stellt sich die Frage, welche Apop-
toseereignisse durch die verglichenen Methoden mit welcher Sensitivität tatsächlich
nachgewiesen werden. Die Detektion eines sehr frühen Apoptosestadiums, in dem
sich die Zelle möglicherweise wieder erholen kann, ist hierbei von den späten
Apoptoseereignissen mit nachfolgender Zellnekrose zu unterscheiden. In einer neu-
en Fragestellung sollte daher untersucht werden, ob und in welchem Anteil früh-
apoptotische Zellen, erfasst durch JC-1 (mitochondriales Potential) oder Annexin-V-
positive Zellen, alle Phasen der Apoptose bis zum Zelltod durchlaufen oder nur pas-
sager geschädigt überleben können. Die BrdU Methode zeigte in dieser Studie gute
vergleichbare Daten mit der Annexin V und 7AAD Methode. Der Anteil apoptotischer
Zellen war allerdings gering, so dass davon auszugehen ist, dass ein Großteil der
apoptotischen Zellen außerhalb des durchflußzytometrischen Messfensters lag und
diese apoptotischen Ereignisse somit nicht erfasst wurden. Auf biochemischer
Ebene scheint mit abnehmender Glukosekonzentration und zunehmendem Laktat-
gehalt im gelagerten Produkt die Viabilität der Zellen verloren zu gehen. Besonders
nach 24 Stunden Lagerung steigt der Anteil apoptotischer Zellen rapide an. In der
Studie konnte eine starke Korrelation zwischen steigender Laktatkonzentration und
dem Anteil an apoptotischen Zellen mit den drei verwendeten Apoptose-Methoden
gezeigt werden. Weiterhin könnte die massenspektrometrische Analyse Gegen-
stand neuer Forschungsarbeiten sein, um molekulare Zellveränderungen während
der Zelllagerung spezifizieren zu können, Metabolite in der Zellkultur, die entweder
ab- oder zunehmen zu identifizieren, um die Zellkultur bedarfsgerecht gezielt mit
Stoffen zu versetzen und dadurch die Apoptoserate zu senken und die Zellqualität
weiter zu verbessern.

82
Literaturverzeichnis
1. Ahrends MJ, Morris RG, Wyllie AH. „Apoptosis. The role of the
endonuclease.“ Am J Pathol 136, (1990): 593-608.
2. Albanyan AM, Harrison P, Murphy MF. „Markers of platelet activation and
apoptosis during storage of apheresis- and buffy coat–derived platelet
concentrates for 7 days.“ Transfusion 49, (2009): 108-117.
3. Altuntas F, Kocyigit I, Ozturk A, Kaynar L, Sari I, Oztekin M, Solmaz M,
EserB, Cetin M, Unal A. „Comparison of the Fenwal Amicus and Fresenius
Com.Tec cell separators for autologous peripheral blood progenitor cell
collection.“ Transfus Apher Sci 36, (2007): 159-167.
4. Amer J, Frankenburg S, Fibach E. „Apheresis induces oxidative stress in
blood cells.“ Ther Apher Dial 14, (2010): 166-171.
5. Bakry, R., Sayed, D., Galal, H., und Shaker, S. „Platelet function, activation
and apoptosis during and after apheresis.“ Ther Apher Dial 14, (2010): 457-
464.
6. Banchereau J, Schuler-Thurner B, Palucka AK, et al. „Dendritic cells as
vectors for therapy.“ Cell, (2001): 106:271-274.
7. Barry Op, Praticò D, Lawson JA, FitzGerald GA. „Transcellular Activation of
Platelets and Endothelial Cells by Bioactive Lipids in Platelet Microparticles.“
J.Clin.Invest 99, (1997): 2118–2127.
8. BD Biosciences. „Handbuch FACS Calibur.“ erhältlich unter
www.bdbiosciences.com
9. Berger DP, Engelhardt R, Mertelsmann R. Das Rote Buch. Ecomed
Hamburg; (2010).
10. Bopp T. „Molekulare Mechanismen der Kontrolle von T-Helferzellen durch
natürlich vorkommende CD4+CD25+ regulatorische T-Zellen.“ Dissertation
in Biologie Universität Mainz, (2006).
11. Brosius F. „SPSS 8.“ available from www.molar.unibe.ch/help/statistics/spss,
(1998).
12. Brügel J. „Untersuchung der Effizienz der Herstellung CD14- und CD16-
positiver Monozyten mit den Zellseparatoren COBE Spectra und COM.TEC.“
Dissertation in Humanmedizin Universität Erlangen, (2010).
13. Burgstaler E., Winters J., Pineda A. „Paired comparison of Gambro Trima
Accel versus Baxter Amicus single-needle plateletpheresis.“ Transfusion
Vol.44, (2004): 1612-1619.
14. Chen W, Jin W, Hardegen N, Lei K, Li Li, Marinos N, McGrady G, Wahl SM.
„Conversion of Peripheral CD4+CD25− Naive T Cells to CD4+CD25+

83
Regulatory T Cells by TGF-β Induction of Transcription Factor Foxp3.“ JEM
12, (2003): 1875-1886.
15. Cossarizza A. „Measure of Mitochondrial Membrane Potential (D y) with the
Fluorescent Probe JC1.“ Cytometry CD ROM 3, (1997).
16. Cossarizza A, Baccarani-Contri M, Kalashnikova G, Franceschi C. „A new
method for the cytofluorimetric analysis of mitochondrial membrane potential
using the J-Aggregate forming lipophilic cation 5,5`,6,6`-Tetrachloro-
1,1`,3,3`-Tetraethylbenzimidazolcarbocyanine Iodide (JC1).“ Biochem.
Biophs. Res. Comm 197, (1993): 40-45.
17. Dada A, Rothe G, Schmitz G, Klouche M. „Qualitätskontrollen und rechtliche
Grundlagen bei der Herstellung von autologen und allogenen peripheren
Stammzellpräparaten.“ J Lab Med 26, (2002): 408-417.
18. Darzynkiewicz Z, Huang X, Okafuji M. „Detection of DNA Strand breaks by
Flow and Laser Scanning Cytometry in Studies of Apoptosis and Cell
Proliferiation.“ Biomediacal and Life Sciences, (2006): 607-619.
19. Darzynkiewicz Z, Juan G, Li X, Gorczyca W, Murakami T, Traganos F.
„Cytometry in Cell Necrobiology: Analysis of Apoptosis and Accidental Cell
Death (Necrosis).“ Cytometry 27, (1997): 1-20.
20. Dietz AB, Bulur PA, Emery RL, Winters JL, Epps DE, Zubair AC. „A novel
source of viable peripheral blood mononuclear cells from leukoreduction
system chambers.“ Transfusion 46, (2006): 2083-2089.
21. Directive 2004/23/EC of the European Parliament and of the European
Council. „On setting standards of quality and safety for the donation,
procurement, testing, processing, preservation,storage and distribution of
human tissues and cells.“ Off J Eur Union L102, (2004): 48-58.
22. Ehemann V, Sykora J, Vera-Delgado J, Lange A, Otto HF. „Flow cytometric
detection of spontaneous apoptosis in human breast cancer using the
TUNEL-technique .“ Cancer Letters 194, (2002): 125-131.
23. Elsner F. „Statistische Datenanalyse mit SPSS für Windows.“ available from
www.home.uni-osnabrueck.de/elsner/Skripte/spss.pdf, (2009).
24. Fadok VA, Voelker DR, Cammpbell PA, Cohen JJ, Bratton DL, Henson PM.
„Exposure of phosphatidylserine on the surface of apoptotic lymphocytes
triggers specific recognition and removal by macrophages.“ J Immunol 148,
(1992): 2207-2216.
25. Freireich EJ, Judson G, Levin RH. „Separation and collection of leukocytes.“
Cancer Res 25, (1965): 1516-1520.

84
26. Gallimore AM, Simon AK. „Positive and negative influences of regulatory T-
cells on tumour immunity.“ Oncogene 27, (2008): 5586-93.
27. Garraud O, Moreau Th. „Effect of Blood Storage on Lymphocyte
Subpopulation.“ JIM 75 , (1984): 95-98.
28. Glaser A, Friedlein H, Zingsem J, Zimmermann R, Weisbach V, Ruf A,
Eckstein R. „Storage of single donor platelet concentrates: Paired
comparison of storage as single or double concentrates.“ J Clin Apher 16,
(2001): 148-154.
29. Glaser A, Schuler-Thurner B, Feuerstein B, Zingsem J, Zimmermann R,
Weisbach V, Eckstein R. „Collection of MNCs with the cell separators for
adoptive immunotherapy in patients with stage IV melanoma.“ Transfusion
41, (2002): 117-122.
30. Gorczyca W, Gong J, Darzynkiewicz Z. „Detection of DNA strand breaks in
individual apoptotic cells by the in situ terminal deoxynucleotidyl transferase
and nick translation assays.“ Cancer Research 53, (1993): 1945-51.
31. Graca L, Le Moine A, Lin CY, Fairchild PJ, Cobbold SP, Waldmann H.
„Donor-specific transplantation tolerance: The paradoxical behavior of
CD4+CD25+ T cells.“ PNAS 101, (2004): 10122–10126.
32. Gulliksson H. „Additive solutions for the storage of platelets for transfusion.“
Transfusion, 10 (2000): 257-264.
33. Hentze M.W., Kulozik A.E., Bartram C.R. Molekulare Medizin. de Gruyter
Berlin, 1.Auflage, (2000); 146-147.
34. Hiruma K, Nakayama S, Okuyama Y. „A comparative study of a new, fully
automated procedure and the standard mononuclear cell program using the
Cobe Spectra for peripheral blood stem cell collection.“ Therapeutic
Apheresis and Dialysis, (1998): 2:273-276.
35. Holmsen H. „Platelet metabolism and activation.“ Semin Hematol 22, (1985):
219-240.
36. Hubel A, Carlquist D, Clay M, McCullough J. „Short-term liquid storage of
umbilical cord blood.“ Transfusion 43, (2003): 626-632.
37. Kiefel V. Transfusionsmedizin und Immunhämatologie. Springer Verlag
Berlin, 4.Auflage, (2010).
38. Kilkson H, Holme S, Murphy S,. „Platelet metabolism during storage of
platelet concentrates at 22°C.“ Blood 64, (1984): 406-414.
39. Kissel K, Berber S, Nockher A, Santoso S, Bein G, Hackstein H. „Human
platelets target dendritic cell differentiation and production of
proinflammatory cytokines.“ Transfusion 46, (2006): 818-827.

85
40. Koopman G, Reutelingsperger CP, Kuijten GA, Keehnen RM, Pals ST and
van Oers MH. „Annexin V for flow cytometric detection of phosphatidylserine
expression on B cells undergoing apoptosis.“ Blood 84, (1994): 1415-1420.
41. Körbling M, Ross WM, Pflieger H. „Procurement of human blood stem cells
by continuous-flow-centrifuagtion-further comment.“ Blood 50, (1977): 753-
754.
42. Korczak-Kowalska G, Wierzbicki P, Bocian K, Klosowska D, Niemczyk M,
Wyzgal J, Korecka A, Durlik M, Chmura A, Pączek L, Górski A. „The
Influence of Immuosuppressive Therapy on the Development of
CD4+CD25+TCells After Renal Transplantation.“ Transplantation 39, (2007):
2721-2723.
43. Leaver HA, Schou AC, Rizzo MT, Prowse CV. „Calcium-sensitive
mitochondrial membrane potential in human platelets and intrinsic signals of
cell death.“ Platelets 17, (2006): 368-377.
44. Lewkowicz P, Lewkowicz N, Sasiak A, Tchorzewski H. „Lipopolysaccharide-
Activated CD4+CD25+ T Regulatory Cells Inhibit Neutrophil Function and
Promote Their Apoptosis and Death.“ J Immunol 177, (2006): 7155-7163.
45. Lizard G, Fournel S, Genestier L, Dhedin N, Chaput C, Flacher M, Mutin M,
Panaye G, Revillard JP. „Kinetics of plasma membrane and mitochondrial
alterations in cells undergoing apoptosis.“ Cytometry 21, (1995): 275-283.
46. Longhi MS, Hussain M, Mitry RR, Sunil AR, Mieli-Vergani G, Vergani D, Ma
Y. „Functional Study of CD4+CD25+ Regulatory T Cells in Health and
Autoimmune Hepatitis.“ JIM 176, (2006): 4484-4491.
47. López-Hoyos, M, D Segundo, G Fernández-Fresnedo, MJ Marín, V
González-Martín, und M Arias. „Regulatory T Cells in Renal Transplantation
and Modulation by Immunosuppression.“ Transplantation 88, (2009): 31-39.
48. Lugli E, Troiano L, Ferraresi R, Roat E, Prada N, Nasi M, Pinti M, Cooper
EL, Cossarizza A. „Characterization of Cells with Different Mitochondrial
Membrane Potential During Apoptosis.“ Cytometry Part A 68A, (2005): 28-
35.
49. Lund P, Westvik A, Joø G, Øvstebø R, Haug K, Kierulf P. „Flow cytometric
evaluation of apoptosis, necrosis and recovery when culturing monocytes.“
JIM,(2001): 45-55.
50. Lund PK, Namork E, Brorson SH, Westvik A, Joø G, Øvstebø R, Kierulf P.
„The fate of monocytes during 24 h of culture as revealed by flow cytometry
and electron microscopy.“ JIM 270, (2003): 63-76.

86
51. Ly JD, Grubb DR, Lawen A. „The mitochondrial membrane potential (∆ψm)
in apoptosis; an update.“ Apoptosis 8, (2003): 115-128.
52. Murphy S, Gardner FH. „Platelet storage at 22°C: metabolic, morphologic
and functional studies.“ J Clin Invest 50, 2 (1971): 370-377.
53. Néron S, Dussault N, Racine C. „Whole-blood leukoreduction filters are a
source for cryopreserved cells for phenotypic and functional investigations
on peripheral blood lymphocytes.“ Transfusion 46, (2006): 537-544.
54. Neron S, Thibaut L, Dussault N. „Characterization of mononuclear cells
remaining in the leucoreduction system chambers of apheresis instruments
after routine platelet collection:a new source of viable human blood cells.“
Transfusion, (2007): 47:1042-1049.
55. Neumann J. Immunbiologie. Springer Verlag Berlin, (2008).
56. Overbeeke R, Steffens-Nakken H, Vermes I, Reutelingsperger C, Haanen C.
„Early features of apoptosis detected by four different flow cytrometry
assays.“ Apoptosis 3, (1998): 115-121.
57. Peter W. „Statistik und Datenauswertung.“ available from
www.arbeitszimmer.statistik-peter.at/2010/04/08/analyse-von-wiederholten-
messungen/,(2012).
58. Picker S, Oustianskaia L, Schneider V, Gathof B. „Annexin V Release and
Transmembrane Mitochondrial Potential during Storage of Apheresis-
Derived Platelets Treated for Pathogen Reduction.“ Transfus Med Hemother
37, (2010): 7-12.
59. Pienimaeki-RoemerA, Ruebsaamen K, Boettcher A, Orso E, Scherer M,
Liebisch G, Kilalic D, Ahrens N, Schmitz G. „Stored platelets alter
glycerophospholipid and sphingolipid species, which are differentially
transferred to newly released extracellular vesicles.“ Transfusion, (2012): 1-
15.
60. Preiß J, Dornoff W, Hagmann FG, Schmieder A. Taschenbuch Onkologie
2008/2009. Zuckschwerdt Verlag München, 14. Auflage,(2008).
61. Rasch, Friese, Hofmann, Naumann. „Quantitative Methoden Band 2
2.Auflage.“ Springer Verlag Berlin, (2006).
62. Rump G, Braun R, Jahn UR, Krakowitzky P, Sibrowski W, van Aken H.
„Transfusionsmedizin compact.“ 8-10. Thieme Verlag Stuttgart, (2003).
63. Savitskiy VP, Shman TV, Potapnev MP. „Comparative measurement of
spontaneous apoptosis in pediatric acute leukemia by different techniques .“
Cytometry 56B, (2003): 16-22.

87
64. Schiebler TH, Korf HW. „Anatomie 10.Auflage.“ 141-142. Steinkopff Verlag
Berlin, (2007).
65. Schwarz J. „Methodenberatung der Universität Zürich.“ available from
www.methodenberatung.uzh.ch, (2010).
66. Schwella N, Movassaghi K, Scheding S, Ahrens N, Salama A. „Comparison
of two leukapheresis programs for computerized collection of blood
progenitor cells on a new cell separator.“ Transfusion 43, (2003): 58-64.
67. Sdralia ND, Patmanidi AL, Velentzas AD, Margaritis LH, Baltatzis GE,
Hatzinikolaou DG, Stavridou A. „The mode of lymphoblastoid cell death in
response to gas phase cigarette smoke is dose-dependent.“ Respiratory
Research , (2009): 1-13.
68. Sedlacek HH. „Die Immunabwehr des Menschen.“ 32-35. Skript Verlag
Kühnel, 1.Auflage, Altendorf Schweiz, ( 2009).
69. Seghatchian J, Krailadsiri P. „The Platelet storage lesion.“ Transfusion 11,
(1997): 130-144.
70. Shevach EM. „CD4+CD25+ Suppressor T cells: more questions than
answers.“ Nature 2, (2002): 389-400.
71. Solheim BG, Flesland O, Seghatchian J, Brosstad F. „Clinical implications of
red blood cell and platelet storage lesions:an overview.“ Transfusion 31,
(2004): 185-189.
72. Stoll Christian. „Phänotypische Untersuchungen zur prognostischen
Bedeutung Proliferations- und Apoptose-assozierter Faktoren sowie die
Expression von Adhäsionsmolekülen in Plattenepithelkarzinomen der
Mundhöhle und des Ösophagus.“ Habilitationsschrift Universität Berlin,
(2000).
73. Strasser E, Eckstein R. „Optimization of leukocyte collection and monocyte
isolation for dendritic cell culture.“ Transfusion 24, (2010): 130-139.
74. Strasser E, Keller B, Hendelmeier M, Ringwald J, Zingsem J, Eckstein R.
„Short-term liquid storage of CD14+ monocytes, CD11c+, and CD123+
precursor dendritic cells produced by leukocytapheresis.“ Transfusion 47,
(2007): 1241-12149.
75. Strasser E, Weidinger T, Weiss DR, Strobel J, Zimmermann R, Eckstein R.
„Storage induced apoptosis of peripheral blood mononuclear cells obtained
from leucoreduction system chambers.“ Vox Sanguinis 101, (2011): 106-111.
76. Strasser E., Schremmer M. „Automated CD14+ monocyte collection with the
autoMNC program of the COM.TEC cell seperator.“ Transfusion Volume 47,
(2007): 2297-2304.

88
77. Strasser EF, Berger TG, Weisbach V , Zimmermann R, Ringwald J, Schuler-
Thurner B, Zingsem J, Eckstein R. „Comparison of two apheresis systems
for the collection of CD14+ cells intended to be used in dendritic cell culture.“
Transfusion 43, (2003): 1309-1316.
78. Strasser EF, Hendelmeier M, Weisbach V, Zimmermann R, Ringwald J,
Eckstein R. „CD14+ cell collection in non-cytokine-stimulated donors with the
COM.TEC cell separator.“ Transfusion 46,(2006): 66-73.
79. Strasser EF, Weidinger T, Zimmermann R, Ringwald J, Eckstein R.
„Recovery of white blood cells and platelets from leukoreduction system
chamber of Trima Accel and Cobe Spectra plateletpheresis devices.“
Transfusion, (2007): 1943-1944.
80. Strauss RG. „Effects on donors of repeated leucocyte losses
duringplateletpheresis.“ Journal of Clinical Apheresis 9, (1994): 130-134.
81. Taha Ta, Hannun YA, Obed LM. „Sphingosine Kinase:biochemical and
cellular regulation and role in disease.“ J Biochem Mol Biol 39, (2006): 113-
131.
82. Trahtemberg U, Atallah M, Krispin A. „Calcium, leukocyte cell death and the
use of Annexin V: fatal encounters.“ Apoptosis 12, (2007): 1769-1780.
83. Transfusionsgesetz, Bundesärztekammer. „Richtlinien zur Gewinnung von
Blut und Blutbestandteilen und zur Anwendung von Blutprodukten.“( 2010).
84. van Engeland M, Nieland L, Ramaekers , Schutte B, Reutelingsperger C,.
„Annexin V-Affinity Assay: A Review on an Apoptosis Detection System
Based on Phosphatidylserine Exposure.“ Cytometry 31, (1998): 1-9.
85. Vermes I, Haanen C, Reutelingsperger C. „Flow cytometry of apoptotic cell
death.“ JIM, (2000): 167-190.
86. Vermes I, Haanen C, Steffens-Nakken H, et al. „A novel assay for apoptosis
flow cytometric detection of phosphatidylserine expression on early apoptotic
cells using fluorescein labelled Annexin V.“ JIM, (1995): 39-51.
87. Weidinger T, Keller A, Weiss D, Zimmermann R, Eckstein R, Strasser E.
„Peripheral blood mononuclear cells obtained from leukoreduction system
chambers show better viability than those from leukapheresis.“ Transfusion
51, (2011): 2047-2049.
88. Wiesneth M. „Gewinnung und Präparation von periheren Blutstammzellen.“
In Transfusionsmedizin und Immunhämatologie, von Kiefel V, Springer
Verlag Berlin, 4.Auflage, (2010): 246-256
89. Zhang G, Gurtu V, Kain SR, Yan G. „Early detection of apoptosis using a
fluorescent conjugate of annexin V.“ Biotechniques 23, (1997): 525-531.

89
Anlage 1
Bestandteile der verwendeten Kits:
Annexin V Apoptosis Detection
Kit I
10x Annexin V Binding Buffer
0,5ml PE Annexin V
BD Mitoscreen Kit lyophilisiertes JC-1 (5,5`,6,6`-tetrachloro-
1,1`,3,3`-
tetraethylbenzimidazolcarbocyanine io-
dide)
60ml 10x Assay Buffer
APO-BRDU Kit 0,3ml FITC-labeled Anti-BrdU mAb
30ml PI/RNase Staining Buffer
0,6ml Reaction Buffer
126ml Rinsing Buffer
120ml Wash Buffer
0,48ml Br-DUTP
5ml Negative Control Cells
5ml Positive Control Cells
0,045ml TdT Enzyme

90
Danksagung
Meinen herzlichsten Dank an Herrn Prof. Dr. med. R. Eckstein für die Überlassung
des Themas und der Bereitstellung eines Arbeitsplatzes in seiner Abteilung.
Herrn Prof. Dr. med. E. Strasser danke ich für die hervorragende Betreuung und
Förderung. Seine Hilfestellungen und geduldige Beantwortung aller Fragen haben
besonderen Dank verdient, da sie für das erfolgreiche Gelingen dieser Arbeit unver-
zichtbar waren.
In besonderem Maße möchte ich auch Frau Dr. rer. nat. A. Pfeiffer für ihre Hilfsbe-
reitschaft und Anleitung bei den durchflußzytometrischen Messungen danken, die
ohne sie nicht möglich gewesen wären.
Weiterhin danke ich den Schwestern und MTAs in der Blutspende und im Labor der
Kinderklinik für ihre Hilfe.
Nicht zuletzt danke ich ganz herzlich meiner Familie und meinem Freund für die
stetige Unterstützung und den motivierenden Einfluss.


















