







Sprachen
Seiten
Rechtliche


MATERIAL UND METHODEN 11
Plastikrohr
Verschluss
Verbindung
zur Stange
Abbildung 2. Sedimentstecher für Sedimentkerne aus
dem Litoral.
3.3 DNS-Extraktion und Aufreinigung
Aus dem Sediment des Bodensees wurde in fünf Ansätzen die Gesamt-DNS extrahiert. Das
Sediment der Ansätze A und B stammte aus dem obersten Zentimeter des Litorals und wurde
für das Erstellen einer Klonbibliothek verwendet. Das Sediment des Ansatzes C stammte
ebenfalls aus dem obersten Zentimeter des Litorals und wurde für eine T-RFLP-Analyse
verwendet. Das Sediment von Ansatz D stammte aus dem Litoral, das Sediment von Ansatz E
aus dem Profundal. Beide Ansätze wurden für eine T-RFLP-Analyse verwendet, wobei in
diesen Ansätzen die Gesamt-DNS aus den Tiefenstufen 0−1 cm und 10−11 cm extrahiert
wurde.
Die DNS-Extraktion wurde in allen Ansätzen mit 1 g Nassgewicht des jeweiligen
Sediments durchgeführt. Dazu wurde das Sediment mit Zirkoniumkügelchen (etwa 0,2 ml;
∅ 0,1 mm), 120 mM Natrium-Phosphat Puffer (800 µl, pH 8) und 10%iger SDS-Lösung
(260 µl; 0,5 M Tris/HCl pH 8,0; 0,1 M NaCl) vermischt und die Zellen mechanisch mit einem
Bead Beater (Fast Prep FP120, Savant Instruments, Inc., Holbrook, NY, USA) (45 s;
6,5 m s−1) aufgeschlossen. Danach wurde der Ansatz abzentrifugiert (12000 rpm, 10 min;
4 °C), der Überstand (0,6−0,8 ml) abgenommen und auf Eis gelagert. Das entnommene
Volumen wurde durch Natrium-Phosphat Puffer ersetzt. Der mechanische Aufschluss wurde

MATERIAL UND METHODEN 12
einmal wiederholt und die beiden Überstände danach vereint. Die einzige Ausnahme bildete
der Ansatz A. Hier wurde 1 g Sediment (Nassgewicht) mit Zirkoniumkügelchen (etwa 0,2 ml;
∅ 0,1 mm), NS Food Lysis Puffer (enthält Guanidinhydrochlorid und Detergenzien)
(Macherey-Nagel, Düren) und Proteinase K vermischt und die Zellen mit einem Bead Beater
mechanisch aufgeschlossen (45 s; 6,5 m s−1).
Die DNS-Aufreinigung wurde mit drei verschiedenen Methoden durchgeführt. Im
Ansatz A wurde das Nucleo Spin Food Kit-Protokoll (Macherey-Nagel, Düren) weiter
durchgeführt. Im Ansatz B erfolgte eine Ausfällung von Proteinen durch Zugabe einer
7,5 M Ammonium-Acetat-Lösung (steril filtriert; 0,2 µm) (DNS : NH4-Ac = 2 : 5, Vol./Vol.).
Dazu wurde das DNS-Extrakt-Ammonium-Acetat-Gemisch für 5 min auf Eis inkubiert.
Anschließend wurde das Gemisch abzentrifugiert (12000 rpm, 5 min, 4 °C),der Überstand
abgenommen und zur DNS-Ausfällung mit Isopropanol (steril filtriert; Raumtemperatur)
vermischt (Überstand : Isopropanol = 1,0 : 0,7; Vol./Vol.). Das Isopropanol-Überstand-
Gemisch wurde danach abzentrifugiert (14000 rpm, 60 min, 4 °C). Daran anschließend wurde
der Überstand dekantiert, das Pellet in 70%igem Ethanol (steril filtriert, −20 °C) gewaschen
und abzentrifugiert (14000 rpm, 5 min, Raumtemperatur). Der Überstand wurde anschließend
durch Pipettieren entfernt. Das Pellet wurde in einer Vakuumzentrifuge (Concentrator 5301,
Eppendorf, Hamburg) getrocknet (ca. 30 min, Raumtemperatur) und in 100 µl EB Puffer
(MiniElute Kit, Qiagen, Hilden) aufgenommen.
In der DNS-Aufreinigung der Ansätze C−E wurde der DNS-Extrakt zur
Proteinausfällung in einem Phase-Lock-Gel-Tube (Eppendorf, 2-ml-Tube) mit 0,5 ml eines
Phenol/Chloroform/Isoamylalkohol-Gemisches (25:24:1) (Sigma, München) vermischt und
anschließend abzentrifugiert (14000 rpm, 5 min, Raumtemperatur). Der Überstand wurde
entnommen und mit einer Isopropanolfällung und anschließender Ethanolreinigung, wie für
Ansatz B beschrieben, aufgereinigt.
Der Anteil an Huminstoffen wurde nach jeder DNS-Aufreinigung mit PVPP-gefüllten
Säulen herabgesetzt. Die genaue Vorgehensweise wurde von Berthelet et al. (1996)
beschrieben.
MATERIAL UND METHODEN 13
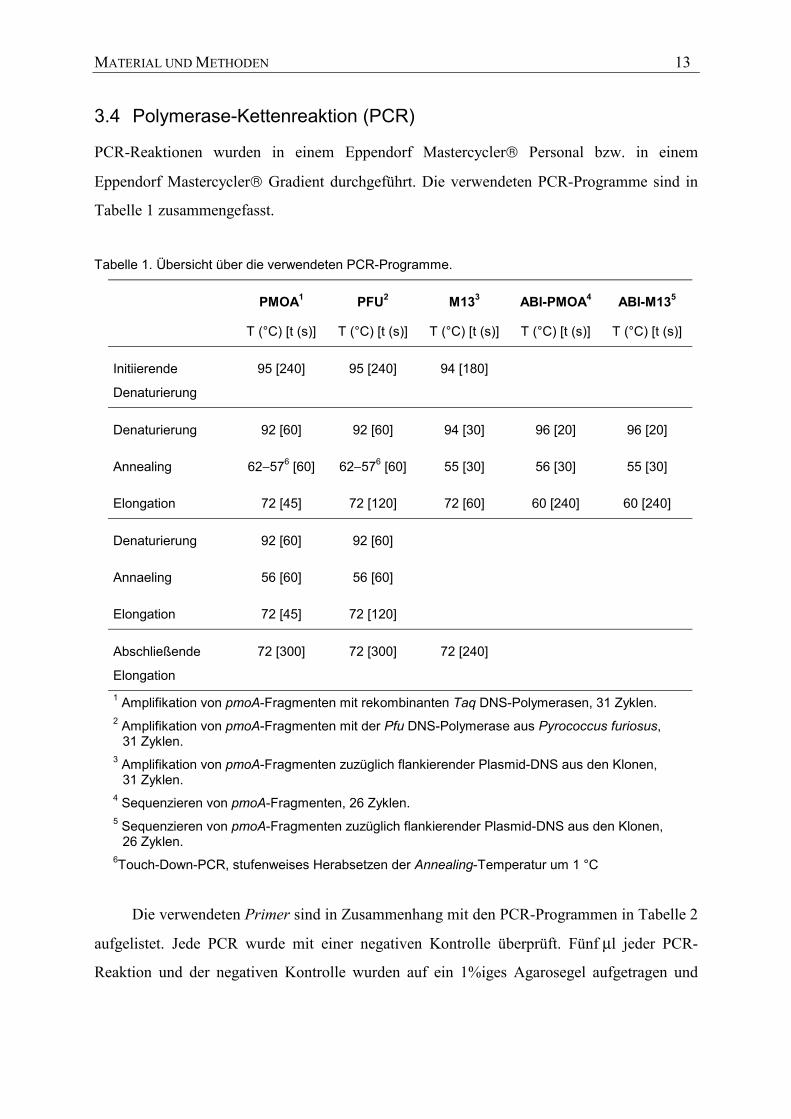
3.4 Polymerase-Kettenreaktion (PCR)
PCR-Reaktionen wurden in einem Eppendorf Mastercycler Personal bzw. in einem
Eppendorf Mastercycler Gradient durchgeführt. Die verwendeten PCR-Programme sind in
Tabelle 1 zusammengefasst.
Tabelle 1. Übersicht über die verwendeten PCR-Programme.
PMOA1
T (°C) [t (s)]
PFU2
T (°C) [t (s)]
M133
T (°C) [t (s)]
ABI-PMOA4
T (°C) [t (s)]
ABI-M135
T (°C) [t (s)]
Initiierende
Denaturierung
95 [240] 95 [240] 94 [180]
Denaturierung 92 [60] 92 [60] 94 [30] 96 [20] 96 [20]
Annealing 62−576 [60] 62−576 [60] 55 [30] 56 [30] 55 [30]
Elongation 72 [45] 72 [120] 72 [60] 60 [240] 60 [240]
Denaturierung 92 [60] 92 [60]
Annaeling 56 [60] 56 [60]
Elongation 72 [45] 72 [120]
Abschließende
Elongation
72 [300] 72 [300] 72 [240]
1 Amplifikation von pmoA-Fragmenten mit rekombinanten Taq DNS-Polymerasen, 31 Zyklen. 2 Amplifikation von pmoA-Fragmenten mit der Pfu DNS-Polymerase aus Pyrococcus furiosus,
31 Zyklen. 3 Amplifikation von pmoA-Fragmenten zuzüglich flankierender Plasmid-DNS aus den Klonen,
31 Zyklen. 4 Sequenzieren von pmoA-Fragmenten, 26 Zyklen. 5 Sequenzieren von pmoA-Fragmenten zuzüglich flankierender Plasmid-DNS aus den Klonen,
26 Zyklen. 6Touch-Down-PCR, stufenweises Herabsetzen der Annealing-Temperatur um 1 °C
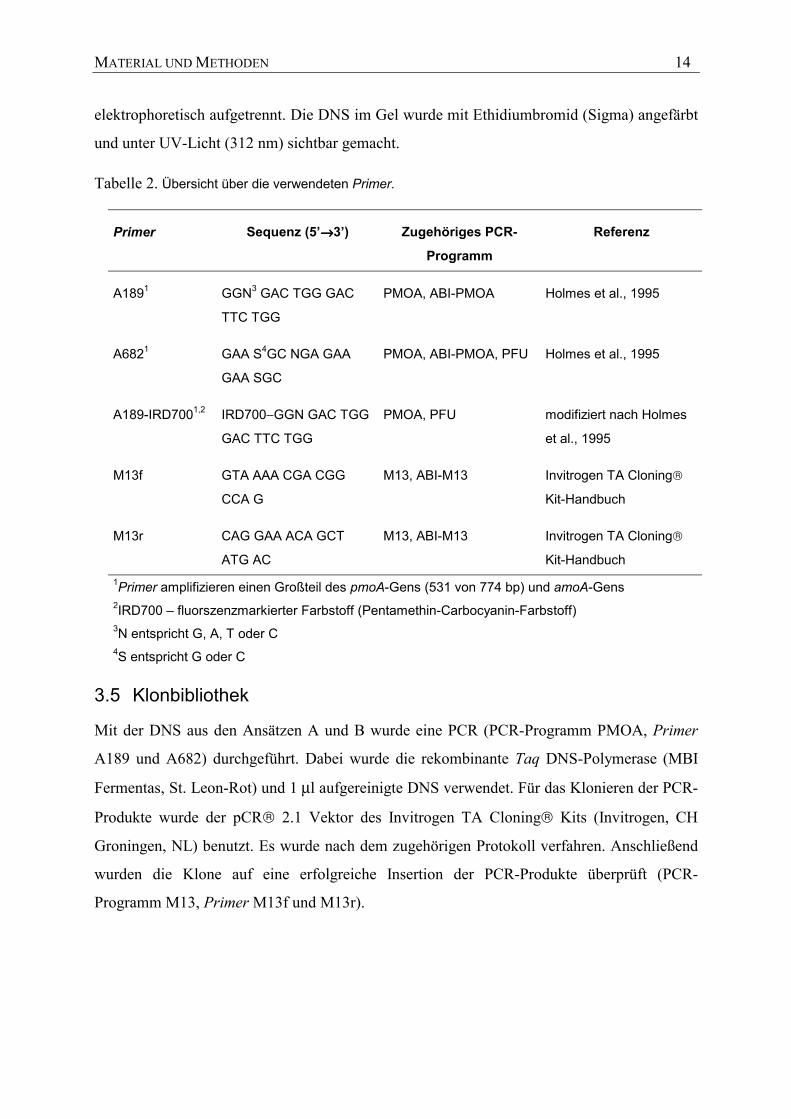
Die verwendeten Primer sind in Zusammenhang mit den PCR-Programmen in Tabelle 2
aufgelistet. Jede PCR wurde mit einer negativen Kontrolle überprüft. Fünf µl jeder PCR-
Reaktion und der negativen Kontrolle wurden auf ein 1%iges Agarosegel aufgetragen und
MATERIAL UND METHODEN 14
elektrophoretisch aufgetrennt. Die DNS im Gel wurde mit Ethidiumbromid (Sigma) angefärbt
und unter UV-Licht (312 nm) sichtbar gemacht.
Tabelle 2. Übersicht über die verwendeten Primer.
Primer Sequenz (5’→→→→3’) Zugehöriges PCR-
Programm
Referenz
A1891 GGN3 GAC TGG GAC
TTC TGG
PMOA, ABI-PMOA Holmes et al., 1995
A6821 GAA S4GC NGA GAA
GAA SGC
PMOA, ABI-PMOA, PFU Holmes et al., 1995
A189-IRD7001,2 IRD700−GGN GAC TGG
GAC TTC TGG
PMOA, PFU modifiziert nach Holmes
et al., 1995
M13f GTA AAA CGA CGG
CCA G
M13, ABI-M13 Invitrogen TA Cloning
Kit-Handbuch
M13r CAG GAA ACA GCT
ATG AC
M13, ABI-M13 Invitrogen TA Cloning
Kit-Handbuch 1Primer amplifizieren einen Großteil des pmoA-Gens (531 von 774 bp) und amoA-Gens 2IRD700 – fluorszenzmarkierter Farbstoff (Pentamethin-Carbocyanin-Farbstoff) 3N entspricht G, A, T oder C 4S entspricht G oder C
3.5 Klonbibliothek
Mit der DNS aus den Ansätzen A und B wurde eine PCR (PCR-Programm PMOA, Primer
A189 und A682) durchgeführt. Dabei wurde die rekombinante Taq DNS-Polymerase (MBI
Fermentas, St. Leon-Rot) und 1 µl aufgereinigte DNS verwendet. Für das Klonieren der PCR-
Produkte wurde der pCR 2.1 Vektor des Invitrogen TA Cloning Kits (Invitrogen, CH
Groningen, NL) benutzt. Es wurde nach dem zugehörigen Protokoll verfahren. Anschließend
wurden die Klone auf eine erfolgreiche Insertion der PCR-Produkte überprüft (PCR-
Programm M13, Primer M13f und M13r).

MATERIAL UND METHODEN 15
3.6 Restriktionsfragment-Längenpolymorphismus (RFLP)
Mit der RFLP-Methode wurden 105 Klone (44 Klone aus Ansatz A, 61 Klone aus Ansatz B)
untersucht. Dazu wurde eine Tooth-Pick-PCR (PCR-Programm M13) mit der rekombinanten
Taq DNS-Polymerase (MBI Fermentas) durchgeführt. Die PCR-Produkte wurden in einem
10-µl-Ansatz im Brutschrank verdaut (16 h, 37 °C). Der 10-µl-Ansatz bestand aus 9 µl PCR-
Produkt, 1 µl Y+/Tango Puffer und 1,5 U MspI (MBI Fermentas). Der Restriktionsverdau
wurde auf ein 4%iges Agarosegel (Nusieve 3:1 Agarose, Biozym, Oldendorf) aufgetragen
und elektrophoretisch aufgetrennt (120 V, 100 min). Die DNS-Fragmente wurden mit
Ethidiumbromid angefärbt und mit UV-Licht sichtbar gemacht. Die Längen der DNS-
Fragmente wurde über einen Vergleich mit einem DNS-Längenstandard (100 bp DNA ladder,
MBI Fermentas) in dem Programm GelScan 5.01 (BioSciTec, Frankfurt/Main) berechnet.
3.7 Sequenzieren
Die pmoA/amoA-Fragmente von 80 Klonen und die pmoA-Fragmente von 13 MOB-Isolaten
wurden sequenziert. Davon wurden die pmoA-Fragmente aller MOB-Isolate und die
pmoA/amoA-Fragmente von 58 der 80 Klone vom 5’-Ende und vom 3’-Ende beginnend
sequenziert. Die pmoA/amoA-Fragmente der restlichen 22 Klone wurden nur von einem Ende
beginnend sequenziert.
Die Sequenzierreaktionen fanden in einem 10-µl-Ansatz statt. Darin enthalten waren
2 µl Big Dye (Applied Biosystems, Foster City, USA), 1 µl Primer (50 mM) und bis zu 2,5 µl
DNS-Template (200−500 ng Plasmid-DNS bzw. 3−10 ng PCR-Produkt). Aufgefüllt wurde
mit sterilem RNAse/DNAse freiem Wasser (Sigma). Die pmoA/amoA-Fragmente der Klone
wurden mit dem PCR-Programm ABI-M13 und die pmoA-Fragmente der MOB-Isolate
wurden mit dem PCR-Programm ABI-PMOA sequenziert.
Im Fall der Klone dienten Plasmide, welche die pmoA/amoA-Fragmente trugen, als
DNS-Template. Die Plasmide wurden mit dem E.Z.N.A. Plasmid Miniprep Kit 1 (PeqLab,
Erlangen) isoliert. Im Fall der MOB-Isolate dienten PCR-Produkte der pmoA-Fragmente als
DNS-Template. Die PCR-Produkte der pmoA-Fragmente wurden aus genomischer DNS der
MOB-Isolate amplifiziert (PCR-Programm PMOA). Die Menge an isolierter Plasmid-DNS
bzw. des PCR-Produkts wurde mit dem Fluoreszenzphotometer Hoefer DyNa Quant 200
und dem fluoreszierenden DNS-Farbstoff H 33258 (Amersham Pharmacia Biotec, Freiburg)
bestimmt.

MATERIAL UND METHODEN 16
Die Ergebnisse der Sequenzier-PCR wurden in der Firma GATC (Konstanz) untersucht
und als ABI-Datei bereitgestellt. Die ABI-Dateien wurden danach mit dem Programm DNA
Star (DNA STAR, Madison, WI, USA) bearbeitet.
Alle Sequenzen wurden einem Chimärentest unterzogen. Dafür wurden die Sequenzen
halbiert und jede Hälfte wurde auf der NCBI-Webseite (GenBank) einem Blast Search
unterzogen. Bei einem Blast Search wird die jeweilige Sequenz mit allen in NCBI und EMBL
veröffentlichen Sequenzen verglichen. Waren die unterschiedlichen Hälften einer Sequenz zu
sehr verschiedenen Sequenzen ähnlich, wurde die jeweilige Sequenz als Chimäre eingestuft.
In den meisten Fällen war die Erkennung von Chimären einfach, weil sie z. B. mit der
Sequenz des Rückwärts-Primers anfingen und aufhörten oder nicht genauso lang waren wie
typische pmoA-Fragmente. Als Chimären eingestufte Sequenzen wurden von weiteren
Analysen ausgeschlossen.
3.8 Phylogenetische Analyse
Die pmoA/amoA-Fragmente der Klone und die pmoA-Fragmente der MOB-Isolate wurden im
Programmpaket ARB mit amoA-Fragmenten und allen pmoA-Fragmenten aus der GenBank-
Datenbank verglichen. Für die phylogenetische Analyse wurden Nukleotidsequenzen in
Aminosäuresequenzen umgewandelt. Damit wurden phylogenetische Bäume mit den in ARB
implementierten Distanz-Modellen Neighbour-Joining und Fitsch-Margoliash und den
implementierten Maximum-Likelihood-Modellen Dayhoff-PAM, Poisson und Jones-Taylor-
Thornton berechnet. Von den Sequenzen dieser Arbeit wurden nur solche verwendet, die von
beiden Enden beginnend sequenziert wurden und untereinander mindestens 1 %
Sequenzunterschied besaßen.
3.9 Rarefaction-Analyse
Die Rarefaction-Analyse sollte nachweisen, ob genügend Klone untersucht wurden, um die
Diversität der pmoA-Gene im Litoralsediment ausreichend zu beschreiben. Dazu wurden alle
pmoA-Sequenzen1 von MOB-Reinkulturen aus der GenBank-Datenbank innerhalb der
einzelnen Gattungen untereinander verglichen. Nach einem Vergleich der maximalen
Sequenzunterschiede innerhalb einer Gattung wurde der Wert 10 % Sequenzunterschied
1 Im Folgenden werden Nukleotidsequenzen als Sequenzen bezeichnet.

MATERIAL UND METHODEN 17
(niedrigster Wert der max. Sequenzunterschiede) festgesetzt, um die pmoA/amoA-Fragmente
der Klone in Gruppen einzuteilen. Mit dieser Gruppeneinteilung wurde analysiert, ob die
Diversität der pmoA-Gene im Litoralsediment ausreichend untersucht wurde. Dazu wurde die
Analytic Rarefaction Software verwendet (Holland, Analytic Rarefaction Software).
3.10 Terminaler Restriktionsfragment-Längenpolymorphismus (T-RFLP)
In der T-RFLP-Analyse wurde die Template-DNS ( Plasmide der Klone, DNS-Extrakte aus
dem Sediment) mit dem PCR-Programm PMOA, der rekombinanten Taq DNS-Polymerase
(Eppendorf) und den Primern A189-IRD700 und A682 amplifiziert. Die PCR wurde mit
einem 1%igen Agarosegel überprüft und anschließend aufkonzentriert, sowie von Salzen,
Primern und Proteinen gereinigt (MiniElute Kit, Qiagen). Die Konzentration des gereinigten
PCR-Produktes wurde anschließend durch einen Vergleich mit einem Menge-Längen-DNS-
Standard (MassRuler DNA Ladder, MBI Fermentas) auf einem 1%igen Agarosegel
bestimmt. Dazu wurde das Software-Programm GelScan 5.01 verwendet. Die Nachweisgrenze
dieser Methode lag bei ca. 1 ng DNS µl−1. Im Anschluss daran wurde das PCR-Produkt (5–50
ng) in einem 20-µl-Ansatz mit dem Restriktionsenzym MspI (3 U) (MBI Fermentas) in einem
Eppendorf Mastercycler Personal verdaut (3 h, 37 °C). Wenn die Konzentration des
aufgereinigten PCR-Produktes unter der Nachweisgrenze lag, wurden 5 bzw. 10 µl PCR-
Produkt im Restriktionsverdau eingesetzt. Im letzten Schritt der T-RFLP-Analyse wurde 5 µl
des verdauten PCR-Produkts mit 3 µl Stop-Solution (LI-COR, Bad Homburg) vermischt und
denaturiert (3 min, 95 °C). Die Proben wurden danach sofort auf Eis gelagert und
anschließend auf ein 5,5%iges Acrylamidgel aufgetragen. Das Acrylamidgel wurde mit dem
Sequenzierer Model 4200 (LI-COR) bei 2000 V, 25 mA, 50 W und 50 °C analysiert. Dabei
wurde der zeitliche Lauf der terminalen Restriktionsfragmente (T-RF’s) an einer definierten
Stelle des Acrylamidgels als TIFF-Datei (8 Bit) aufgenommen. Die TIFF-Datei wurde in dem
Software-Programm GelScan 5.01 analysiert. Die Länge der T-RF’s wurden mit Längen
möglicher T-RF’s von pmoA/amoA-Fragmenten der Klone und veröffentlichen pmoA- und
amoA-Sequenzen (GenBank) in einem Zuordnungsbereich von ± 2 bp verglichen. In
eindeutigen Ausnahmefällen, wie dem ungeschnittenen PCR-Produkt, wurde der
Zuordnungsbereich auf ± 4 bp erhöht.
Abweichungen von diesem Standardprotokoll sind im Folgenden angegeben.

MATERIAL UND METHODEN 18
3.10.1 Modifikation der PCR-Bedingungen
Im Folgenden stellt jeder Unterpunkt ein Experiment mit modifiziertem Standardprotokoll
dar. Es sind nur die Modifikationen angegeben. Als DNS-Template diente 1 ng Plasmid-DNS
von Klon B24.
1. Verschiedene PCR-Zyklen (12, 15, 18, 21, 24, 27 oder 31)
2. PCR mit der Pfu-Polymerase aus Pyrococcus furiosus (MBI Fermentas) und dem PCR-
Programm PFU (10, 11, 12, 15, 18, 21 oder 31 PCR-Zyklen)
3. Verschiedene Verhältnisse von Vorwärts-Primer (A189-IRD700) zu Rückwärts-Primer
(A682) (1:8, 1:4, 1:2, 1:1, 2:1, 4:1 oder 8:1), 20 PCR-Zyklen und 5 ng PCR-Produkt im
Restriktionsverdau.
3.10.2 Modifikation der PCR-Produkt-Aufreinigung
Als DNS-Template diente 1 ng Plasmid-DNS von Klon B24. Zur Aufreinigung wurde der
MiniElute Kit verwendet (Qiagen). Das Protokoll des Herstellers wurde in einem Experiment
folgendermaßen modifiziert: i) das PCR-Produkt wurde zweimal mit dem Waschpuffer
gewaschen, ii) die Bindungssäulen wurden vor jedem Waschschritt 5 min mit Waschpuffer
inkubiert und iii) der Zentrifugationsschritt für das Entfernen des restlichen Ethanols wurde
zweimal durchgeführt.
3.10.3 Modifikationen des Restriktionsverdaus
Im Folgenden stellt jeder Unterpunkt ein Experiment mit modifiziertem Standardprotokoll
dar. Es sind nur die Modifikationen angegeben. Als DNS-Template diente, wenn nicht anders
angegeben, 1 ng Plasmid-DNS von Klon B24.
1. Einsatz verschiedener Enzymmengen [1,5 U; 5,0 U oder 10,0 U MspI (MBI
Fermentas)].
2. Verdau mit MspI von verschiedenen Herstellern [MBI Fermentas , New England
Bioloabs (Frankfurt/Main) und Promega (Mannheim)].
3. Verdau der Klone B24, B60 und B67 mit MspI (MBI Fermentas) vs. Doppelverdau der
Klone B24, B60 und B67 mit AluI/MlsI (MBI Fermentas).
4. Inkubationszeit (3 h oder 16 h)
5. Inkubationsgerät [Brutschrank oder Eppendorf Mastercycler Personal (Eppendorf)]

MATERIAL UND METHODEN 19
3.10.4 Verdau überhängender, einzelsträngiger PCR-Produkte mit Mung Bean
Nuclease
Das DNS-Template von Klon B24 (6 ng Plasmid-DNS) wurde mit dem PCR-Programm PFU,
der Pfu-Polymerase (MBI Fermentas) und den Primern A189-IRD700 und A682 amplifiziert.
Danach wurde das PCR-Produkt (1000 ng DNS) in einem 100-µl-Ansatz mit 5 U Mung Bean
Nuclease (New England Biolabs) verdaut (1 h, 30 °C). Der Verdau wurde anschließend in ein
Phase-Lock-Gel-Tube gegeben, 1:1 (Vol./Vol.) mit einem Phenol/Chloroform/Isoamyl-
alkohol-Gemisch (25:24:1) vermischt und abzentrifugiert (14000 rpm, 5 min, Raum-
temperatur). Danach wurde der Überstand abgenommen, mit dem doppelten Volumen
96%igem Ethanol und dem 0,1fachen Volumen 3 M Natrium-Acetat vermischt und
abzentrifugiert (14000 rpm, 60 min, Raumtemperatur). Anschließend wurde der Überstand
entfernt und das Pellet in einer Vakuumzentrifuge (ca. 30 min, Raumtemperatur) getrocknet.
Das trockene Pellet wurde in 25 µl EB Puffer (MiniElute Kit, Qiagen) aufgenommen. Danach
erfolgte eine zusätzliche Aufreinigung mit dem MiniElute Kit. Anschließend wurden 12,5 ng
DNS mit MspI (MBI Fermentas) verdaut (3 h, 37 °C) und auf dem Sequenzierer Model 4200
analysiert.
3.10.5 T-RFLP-Analyse von Klonen und Klongemischen
In der T-RFLP-Analyse von Klonen wurden folgende Modifikationen des T-RFLP-Protokolls
vorgenommen: das PCR-Programm PMOA wurde mit 20 Zyklen durchgeführt und 5 ng PCR-
Produkt wurden im Restriktionsverdau verwendet.
Die T-RFLP-Analysen von Sedimentproben wurden durch T-RFLP-Experimente mit
Klonen simuliert. Dabei wurden DNS-Templates von verschiedenen Klonen in gleichen
Mengen vermischt. Die Gesamtmenge der eingesetzten Template-DNS betrug 1 ng. Es
wurden Gemische der Klone A8 und B60, A70 und B24, B67 und B60, B24 und B60, und ein
Gemisch aus allen soeben genannten Klonen untersucht.
3.10.6 T-RFLP-Analyse der Sedimente
Die Analyse des Litoral- und Profundalsediments erfolgte ebenfalls nach einem modifizierten
T-RFLP-Protokoll. DNS-Extrakte der Ansätze C−E wurden mit dem FailSafe PCR PreMix
B bzw. C und dem FailSafe PCR Enzyme Mix amplifiziert (Epicentre, Madison, WI, USA).
Maximal 5 ng PCR-Produkt wurden im Restriktionsverdau eingesetzt.

MATERIAL UND METHODEN 20
Alle Fragmente eines T-RF-Profils wurden mit T-RF’s von pmoA/amoA-Sequenzen
dieser Studie und mit T-RF’s veröffentlichter pmoA- und amoA-Sequenzen (GenBank)
verglichen. Dazu wurden alle T-RF’s, die aus primären wie auch aus sekundären
Restriktionsschnittstellen hervorgehen, zum Vergleich herangezogen. Der T-RF-Vergleich
fand im Programmpaket ARB statt.
3.11 Anreicherungskulturen und Reinkulturen
Die DNS aus MOB-Anreicherungskulturen und -Isolaten wurden nach dem Protokoll von
Selenska und Klingmüller (1991) extrahiert und von I. Bussmann zur Verfügung gestellt.
MOB waren zuvor mit Whittenbury-Medium (Whittenbury and Dalton, 1981) und Bussmann-
Medium (Bussmann et al., in Vorbereitung) von I. Bussmann angereichert und isoliert
worden. Die T-RFLP-Analyse der Anreicherungskulturen erfolgte nach dem
Standardprotokoll. Bei den Analysen der Anreicherungskulturen WbC8, WbD6, WbE5,
WbF11, WdC3, WdC4, WdD12, WdF4 und der Gradientenkultur U wurde das T-RFLP-
Protokoll durch Einsatz von 35 PCR-Zyklen modifiziert.

4 ERGEBNISSE
4.1 Diversität der pmoA-Gene im Litoralsediment des Bodensees
4.1.1 Klonbibliothek
Die Diversität der pmoA-Gene im Litoralsediment des Bodensees wurde mit einer pmoA-
Klonbibliothek untersucht. Dazu wurden die DNS-Extrakte A und B aus dem obersten
Zentimeter des Litoralsediments benutzt. Aus beiden Extrakten konnten DNS-Fragmente
(500−600 bp) amplifiziert werden, die der Länge von pmoA-Fragmenten (531 bp)
entsprachen. Zusätzlich wurden ca. 100 bp längere DNS-Fragmente amplifiziert (siehe
Abb. 3). Die PCR-Produkte wurden kloniert und die Klone auf erfolgreiche Insertion der
PCR-Produkte überprüft (siehe Abb. 4). Im Ansatz A enthielten 44 von 87 zufällig
ausgewählten Klonen eine Insertion. Im Ansatz B enthielten 61 von 147 zufällig ausgewählten
Klonen eine Insertion.
M
A B
DNSExtrakt
500 bp
100 bp
1030 bp
M
Insertion– +
500 bp
1030 bp
200 bp
Abbildung 3. PCR mit den DNS-Extrakten A und
B. Die PCR-Produkte wurden für das Erstellen
einer Klonbibliothek benutzt.
Abbildung 4. Überprüfung der Klone auf eine
erfolgreiche Insertion des PCR-Produktes.
4.1.2 RFLP-Analyse und Sequenzierung
Die Insertionen von 105 Klonen (Ansatz A: 44 Klone, Ansatz B: 61 Klone) wurden einer
RFLP-Analyse mit dem Restriktionsenzym MspI unterzogen und nach ihren
Restriktionsmustern gruppiert (siehe Abb. 5). Insgesamt wurden 39 Gruppen (R1−R39)

ERGEBNISSE 22
identifiziert, wovon 29 Gruppen jeweils nur durch einen Klon repräsentiert waren. Vertreter
jeder Gruppe wurden sequenziert, auf Chimären überprüft und mit der in ARB
implementierten Parsimony-Methode vorläufig in einen Stammbaum aus veröffentlichten
pmoA/amoA-Sequenzen eingerechnet (siehe Tab. 3). Die Klonsequenzen der Gruppen
R1−R26 konnten, abgesehen von drei Ausnahmen, als pmoA-Fragmente identifiziert werden.
Zehn Klonsequenzen wurden als Chimären identifiziert, wovon 8 Klonsequenzen jeweils eine
RFLP-Gruppe repräsentierten. Weitere 17 Klonsequenzen (Gruppen R27−R31) waren mit
650 bp wesentlich länger als pmoA/amoA-Fragmente (531 bp). Sequenzierte Vertreter dieser
Gruppen wiesen 83 % Sequenzähnlichkeit zu einem Teil der genomischen DNS von
Methanosarcina acetivorans auf. Mehrere Stopkodons innerhalb dieser Sequenzen und eine
fehlende Zugehörigkeit zu einem Gen (Blast Search) wiesen auf eine nicht kodierende
Sequenz hin.
Um auszuschließen, dass eine RFLP-Gruppe sehr verschiedene pmoA-Fragmente
umfasst, die sich nur in ihrem Restriktionsmuster ähneln, wurde die Mehrzahl der Klone jeder
Gruppe sequenziert. Mit einer Ausnahme waren die Klonsequenzen innerhalb der einzelnen
Gruppen sehr ähnlich; der maximale Sequenzunterschied lag bei 9 %.
M
100 bp
200 bp300 bp500 bp
8 9 15 24 33 36 37 39 40 41
Klone aus Ansatz B
Abbildung 5. Beispiel für die RFLP-Analyse anhand von 10 Klonen des Ansatzes B.

ERGEBNISSE 23
Tabelle 3. Gruppen der RFLP-Analyse und Anzahl der Klone, die einzelnen RFLP-Gruppen zugeordnet
werden konnten. In Klammern ist die Anzahl der sequenzierten Klone pro Gruppe angegeben.
RFLP-Gruppe Anzahl der Klone Zuordnung
R1 22 (11) pmoA-Sequenzen, eine Chimäre
R2 14 (13) pmoA-Sequenzen
R3 10 (9) pmoA-Sequenzen
R4 4 (4) pmoA-Sequenzen
R5 4 (3) pmoA-Sequenzen
R6−R8 2 (2) pmoA-Sequenzen
R9 2 (2) pmoA-Sequenzen, eine Chimäre
R10−R25 1 (1) pmoA-Sequenzen
R26 1 (1) pmoA/amoA-Sequenzen1
R27−R31 insgesamt 17 (7) M. acetivorans2
R32−R39 insgesamt 5 (5) Chimären
1eindeutige Zuordnung zu pmoA- oder amoA-Sequenzen war nicht möglich 2Methanosarcina acetivorans-ähnliche Sequenz
4.1.3 Phylogenetische Analyse
Die als pmoA- bzw. pmoA/amoA-Fragmente identifizierten Klonsequenzen wurden einer
phylogenetischen Analyse unterzogen. Dazu wurde aus den Klonsequenzen und
veröffentlichten pmoA- und amoA-Sequenzen (GenBank) ein phylogenetischer Baum mit dem
Distanz-Modell nach Fitsch-Margoliash berechnet (siehe Abb. 6). Bei einem Vergleich des
berechneten Baumes nach Fitsch-Margoliash mit berechneten Bäumen nach den Maximum-
Likelihood-Modellen Dayhoff-PAM, Poisson und Jones-Taylor-Thornten und dem Distanz-
Modell Neighbour-Joining wurde eine große Übereinstimmung zwischen den Bäumen
festgestellt. Eine Ausnahme war der nach dem Dayhoff-PAM-Modell berechnete Baum; hier
zweigte ein Teil der Typ I-MOB-pmoA-Gene zusammen mit den Typ II-MOB-pmoA-Genen
ab. Alle benutzten Berechnungsmodelle sind im Programmpaket ARB implementiert.

ERGEBNISSE 24
Die pmoA-Klonsequenzen konnten sowohl den Typ I-MOB als auch den Typ II-MOB
zugeordnet werden. Um die einzelnen pmoA-Klonsequenzen miteinander vergleichen zu
können, wurden sie in Gruppen von maximal 10 % Sequenzunterschied eingeteilt. Grundlage
dieser Gruppeneinteilung war der Vergleich der maximalen pmoA-Sequenzunterschiede
innerhalb sieben verschiedener MOB-Gattungen. Dieser Vergleich zeigte, dass sich pmoA-
Fragmente einer Gattung maximal um 10−30 % unterscheiden (siehe Tab. 4). Für die
Gruppeneinteilung wurde der Wert 10 % Sequenzunterschied verwendet, um eine
Unterschätzung der Diverstät auf der Gattungsebene zu vermeiden.
Die pmoA-Klonsequenzen wurden daraufhin in neun Gruppen (B1−B9) eingeteilt.
Sieben Gruppen wurden den Typ I-MOB (B1−B7) und eine Gruppe den Typ II-MOB (B8)
zugeordnet. Eine neunte Gruppe (B9) konnte weder zu pmoA- noch amoA-Genen eindeutig
zugeordnet werden. Die pmoA-Fragmente der Gruppe B1 waren mit pmoA-Genen von
Methylobacter spp. und die pmoA-Fragmente der Gruppe B2 waren mit pmoA-Genen von
Methylococcus spp. am nächsten verwandt. Die Gruppen B3, B4 und B5 konnten zu pmoA-
Klonsequenzen anderer Sediment- und Bodenstudien [marine und limnische Sedimente
(GenBank; Costello und Lidstrom, 1999), Reisfeldboden (Henckel et al., 2001)], aber zu
keiner Reinkultur zugeordnet werden. Die pmoA-Fragmente der Gruppen B6 und B7 konnten
weder zu pmoA-Genen von Reinkulturen, noch zu pmoA-Klonsequenzen anderer
Umweltstudien zugeordnet werden. Hier handelt es sich um bisher nicht beschriebene
Gruppen von pmoA-Genen. Die pmoA-Fragmente der Gruppe B8 waren mit pmoA-Genen von
Methylosinus/Methylocystis spp. am nächsten verwandt.
Eine auf dieser Gruppeneinteilung (10 % Sequenzunterschied) basierende Rarefaction-
Analyse zeigte, dass genügend pmoA-Gene kloniert und untersucht wurden, um die
verschiedenen MOB-Gattungen des Litoralsediments, die sich mit den pmoA/amoA-
spezifischen Primern A189 und A682 erfassen lassen, ausreichend zu beschreiben (siehe
Abb. 7).

ERGEBNISSE 25
Klon pAMC511Klon A55, 79 bp
Klon A67, 531 bp Methylobacter sp. LW12
Methylomonas sp. LW21 Methylomonas sp. LW19
Methylomicrobium buryatense Methylobacter sp. LW14
Methylomicrobium albumKlon FW-50Klon A61
Klon B39Klon B60
Klon A81Klon B9
Klon B13 Methylococcus capsulatus
Klon FW-18Klon B109
Klon A80Klon PS-49Klon B65Klon B63
Methylocaldum gracileKlon A48
Klon B33Klon B37
Klon B77Klon A19
Klon B40Klon B41
Klon M90-P4 Methylothermus HB
Klon A62Klon B75
Klon A78Klon B24
Isolat A Methylocystis sp. SC2 pmoA1Klon B119
Methylocystis sp. KS30Klon B25 Methylosinus sp. LW2
Isolat SWKlon A72Klon A54
Methylocystis sp MKlon B111
Isolat RG Methylosinus sporium SE2
Methylosinus trichosporium strain33-1 Methylocapsa acidophila B2
Methylocystis sp. SC2 pmoA2 Nitrosococcus sp. C-113
Nitrosomonas cryotolerans Nitrosospira multiformis
Nitrosomonas europaeaKlon M90-P69
Klon A70
0.10
B1 (79 bp, 531 bp)
B2, (79 bp)
B3 (79 bp)
B4 (79 bp)B5 (79 bp)
B6 (79 bp)
B7 (79 bp)
B8(244 bp)
B9 (79 bp)
AOB
Typ I-MOB
Typ II-MOB
Abbildung 6. Phylogenetischer Baum aerober, methanotropher Bakterien und Ammonium oxidierender
Bakterien basierend auf abgeleiteten PmoA- und AmoA-Sequenzen (161 Aminosäuren). Der Baum wurde nach
dem Distanz-Modell Fitsch-Margoliash berechnet (Global Rearrangement, 3 Jumbles). Klone dieser Arbeit sind
grau, MOB-Isolate sind grau-unterstrichen und veröffentliche Sequenzen (GenBank) sind schwarz abgebildet.
Die dargestellten Gruppen umfassen Klone dieser Arbeit mit maximal 10% Nukleotidsequenzunterschied.
Hinter den Gruppennamen sind in Klammern die T-RF´s angegeben, die sich aus der primären Schnittstelle
des Restriktionsenzyms MspI ergeben. Im Fall der Gruppe B1 sind die T-RF´s zusätzlich mit den einzelnen
Klonen angegeben. Die Skala zeigt 10 % Aminosäuresequenz-Unterschied an.

ERGEBNISSE 26
Tabelle 4. Maximaler Sequenzunterschied von pmoA-Fragmenten innerhalb verschiedener MOB-
Gattungen.
Gattung Maximaler Sequenzunterschied (%) Länge der pmoA-Fragmente (bp)
Methylosarcina 9,2 ca. 300
Methylomonas 9,8 ca. 500
Methylocaldum 10,1 ca. 500
Methylomicrobium 22,2 ca. 500
Methylobacter 23,9 ca. 500
Methylosinus 26,0 ca. 500
Methylocystis 28,9 ca. 500
Abbildung 7. Rarefaction-Analyse der sequenzierten pmoA-Fragmente der Klone. Zu jedem
berechneten Punkt ist die Standardabweichung aufgetragen.
0123456789
10
0 10 20 30 40 50 60
Sequenzierte pmoA- Fragmente
Neu
e G
rupp
e

ERGEBNISSE 27
Parallel zur Analyse von Sedimentproben aus dem Litoralsediment wurden auch MOB-
Isolate aus dem Litoralsediment auf ihre pmoA-Gene untersucht (siehe Abb. 6). Alle MOB-
Isolate konnten den Typ II-MOB zugeordnet werden. Die Isolate A, G, E, Wd-E3, Wd-C12
und Wd-E12 zeigten in ihren pmoA-Sequenzen 99-100 % Sequenzähnlichkeit zum pmoA1-
Gen von Methylocystis sp. SC2 und wurden unter dem Namen „Isolat A“ zusammengefasst.
Die Isolate SW, 2-8e, 2-10g, WB-E10, WB-D10 und WB-F4 zeigten in ihren pmoA-
Sequenzen 99-100 % Sequenzähnlichkeit zum pmoA-Fragment von Klon B25 und dem pmoA-
Gen von Methylosinus sp. Diese Isolate wurden unter dem Namen „Isolat SW“
zusammengefasst. Das pmoA-Gen von Isolat RG war identisch zum pmoA-Gen von
Methylosinus sporium SE2. In keinem Isolat wurden unterschiedliche pmoA-Kopien gefunden.
4.2 Evaluation der T-RFLP-Methode
Die Methode der T-RFLP beruht auf der Amplifikation einer Ziel-DNS mit einem
fluoreszenzmarkierten Primer und einem anschließenden Verdau des PCR-Produktes durch
ein Restriktionsenzym. Die DNS-Fragmente werden danach in einem Sequenzierer der Größe
nach aufgetrennt. Dabei detektiert der Sequenzierer aufgrund der Fluoreszenzmarkierung nur
das terminale DNS-Fragment des verdauten PCR-Produktes.
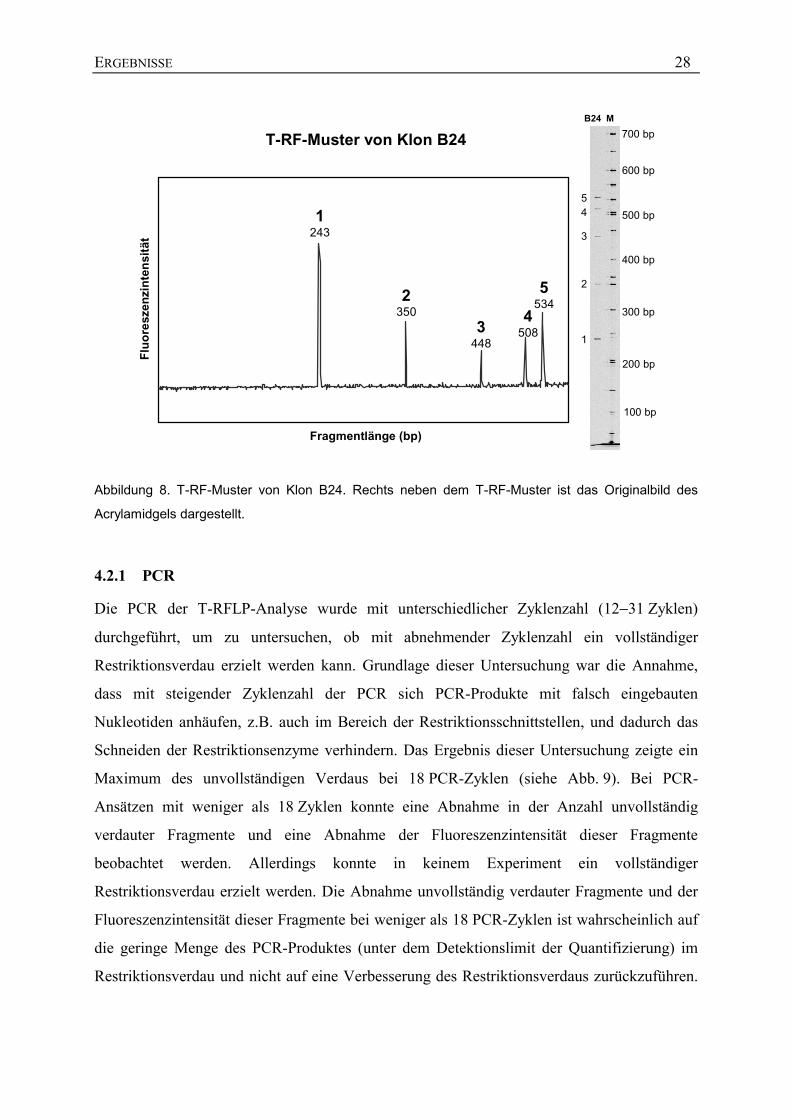
Um die Methode der T-RFLP zu überprüfen, wurden die Klone A8, A70, B24, B60 und
B67 auf ihre Restriktionsfragmentmuster in der T-RFLP untersucht. Theoretisch sollte jeder
Klon durch ein einziges Fragment repräsentiert sein. Die Untersuchung der Klone ergab, dass
Klonsequenzen, die mehrere Schnittstellen aufwiesen, auch durch mehrere Fragmente
repräsentiert waren (siehe Abb. 8). Das kürzeste Fragment entsprach immer der ersten
Schnittstelle des Restriktionsenzyms in der jeweiligen Sequenz und wies immer die stärkste
Fluoreszenzintensität auf. Mit einer Ausnahme konnten allen zusätzlichen Fragmenten
voraussagbare Restriktionsschnittstellen bzw. unverdautes PCR-Produkt zugeordnet werden,
was auf einen unvollständigen Restriktionsverdau hindeutet. Daraufhin wurden die
Einzelschritte der T-RFLP (PCR, Aufreinigung des PCR-Produktes, Restriktionsverdau) auf
mögliche Ursachen des vorliegenden unvollständigen Restriktionsverdaus untersucht.
ERGEBNISSE 28
1243
2350
3448
4508
5534
100 bp
200 bp
300 bp
400 bp
500 bp
600 bp
700 bpT-RF-Muster von Klon B24B24 M
1
2
3
45
Flu
ore
szen
zin
ten
sitä
t
Fragmentlänge (bp)
Abbildung 8. T-RF-Muster von Klon B24. Rechts neben dem T-RF-Muster ist das Originalbild des
Acrylamidgels dargestellt.
4.2.1 PCR
Die PCR der T-RFLP-Analyse wurde mit unterschiedlicher Zyklenzahl (12−31 Zyklen)
durchgeführt, um zu untersuchen, ob mit abnehmender Zyklenzahl ein vollständiger
Restriktionsverdau erzielt werden kann. Grundlage dieser Untersuchung war die Annahme,
dass mit steigender Zyklenzahl der PCR sich PCR-Produkte mit falsch eingebauten
Nukleotiden anhäufen, z.B. auch im Bereich der Restriktionsschnittstellen, und dadurch das
Schneiden der Restriktionsenzyme verhindern. Das Ergebnis dieser Untersuchung zeigte ein
Maximum des unvollständigen Verdaus bei 18 PCR-Zyklen (siehe Abb. 9). Bei PCR-
Ansätzen mit weniger als 18 Zyklen konnte eine Abnahme in der Anzahl unvollständig
verdauter Fragmente und eine Abnahme der Fluoreszenzintensität dieser Fragmente
beobachtet werden. Allerdings konnte in keinem Experiment ein vollständiger
Restriktionsverdau erzielt werden. Die Abnahme unvollständig verdauter Fragmente und der
Fluoreszenzintensität dieser Fragmente bei weniger als 18 PCR-Zyklen ist wahrscheinlich auf
die geringe Menge des PCR-Produktes (unter dem Detektionslimit der Quantifizierung) im
Restriktionsverdau und nicht auf eine Verbesserung des Restriktionsverdaus zurückzuführen.

ERGEBNISSE 29
Bei PCR-Ansätzen mit mehr als 18 Zyklen konnte eine Abnahme der Fluoreszenzintensität
der längsten unvollständig verdauten Fragmente beobachtet werden. Dafür wurde keine
Erklärung gefunden.
M M M
normale Aufreinigung intensive Aufreinigung700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
312721 24181512 312721 24181512
PCR-ZyklenPCR-Zyklen
PCR-Produkt im Verdau (ng)
<15
3827
5050
50 <113
3950
5050
50
Abbildung 9. T-RFLP-Analyse von Klon
B24 mit ansteigender PCR-Zyklenzahl
(PCR-Programm PMOA). Im linken
Ansatz wurde das PCR-Produkt nach
dem Standardprotokoll des MiniElute
Kits aufgereinigt, im rechten Ansatz
wurden zusätzliche Reinigungsschritte
eingeführt.
Die Hypothese der falsch eingebauten Nukleotide wurde daraufhin mit der Pfu DNS-
Polymerase aus Pyrococcus furiosus untersucht. Die Pfu DNS-Polymerase überprüft im
Gegensatz zur rekombinanten Taq DNS-Polymerase ob die richtigen Nukleotide in den neu
synthetisierten DNS-Strang eingebaut wurden (Proofreading-Funktion). Auch in diesem
Ansatz wurden verschiedene PCR-Zyklen (10−31 Zyklen) untersucht. Das Ergebnis zeigte
eine Verschlechterung des unvollständigen Restriktionsverdaus im Vergleich zum Ansatz mit
der rekombinanten Taq DNS-Polymerase. Zusätzlich zu den unvollständig verdauten
Fragmenten traten Fragmente auf, die keiner Restriktionsschnittstelle zugeordnet werden
konnten (siehe Abb. 10).
Eine weitere Ursache des unvollständigen Restriktionsverdaus wurde in der
unterschiedlichen Bindungsaffinität der Primer vermutet. Wenn einer der Primer bei der
vorgegebenen Annaeling-Temperatur viel besser bindet als der andere, könnte es zu einer
Anhäufung einzelsträngiger DNS kommen, die von Restriktionsenzymen nicht geschnitten

ERGEBNISSE 30
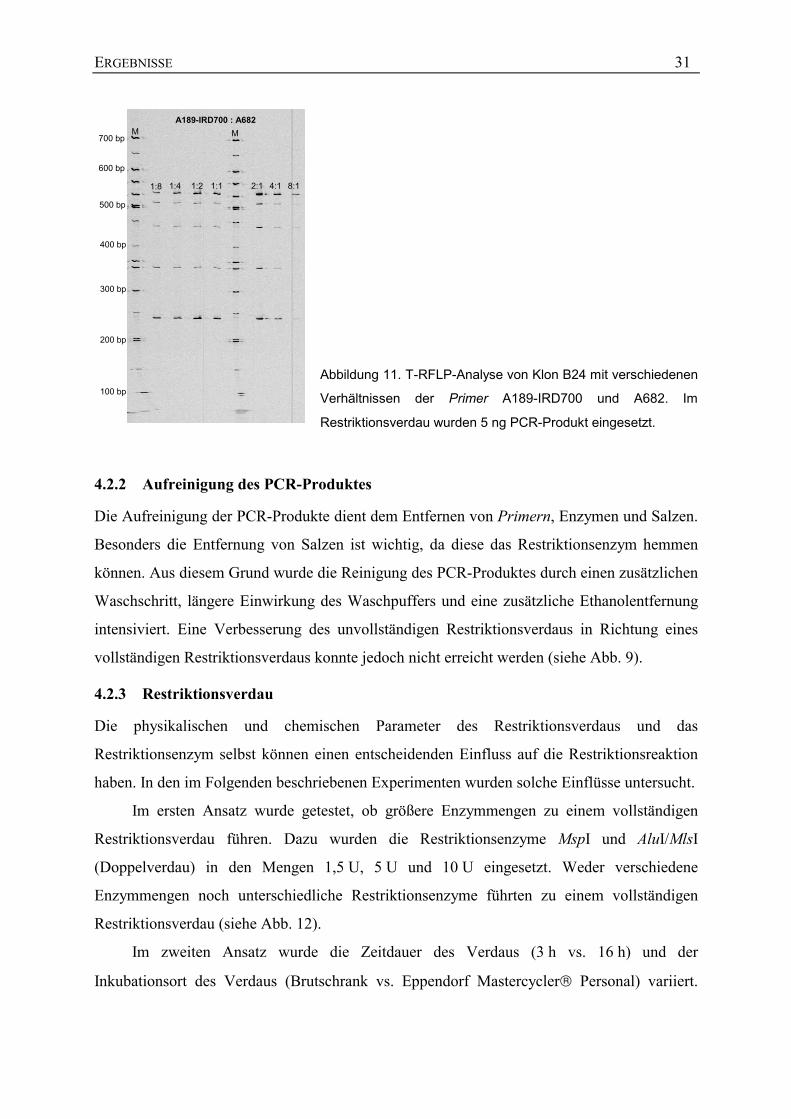
werden kann. Um die unterschiedlichen Bindungsaffinitäten auszugleichen wurden die Primer
in verschiedenen Konzentrationsverhältnissen in der PCR eingesetzt (von 1 : 8 =
0,25 µM : 2,0 µM bis 8 : 1 = 2,0 µM : 0,25 µM). Das Ergebnis zeigte keinen Effekt der
unterschiedlichen Primer-Verhältnisse, außer dass bei einer Zusammensetzung von A189-
IRD700 : A682 = 8 : 1 weniger PCR-Produkt amplifiziert wurde als in den anderen Ansätzen
(siehe Abb. 11).
Allerdings wurde in diesem Experiment gezeigt, dass der Einsatz von 5 ng PCR-Produkt
im Restriktionsverdau im Gegensatz zu 50 ng PCR-Produkt (Experimente mit
unterschiedlichen Zyklenzahlen) das Erscheinen von unspezifischen Fragmenten verhindert.
Die unspezifischen Fragmente sind auf unvollständig amplifizierte PCR-Produkte
zurückzuführen, die zwar nur in geringen Mengen vorkommen, aber sichtbar werden, wenn
eine große Menge verdautes PCR-Produkt auf das Acrylamidgel aufgetragen wird.
M M700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
10 21 31 31 3112 15 1811
PCR-Zyklen
Mung BeanNucleaseVerdau– +
PCR-Produkt im Verdau (ng)<1 <1 <1 <1 50 50 50 50 13
Abbildung 10. T-RFLP-Analyse
von Klon B24 mit der Pfu DNS-
Polymerase. In den letzten
beiden Ansätzen wurden 6 ng
DNS-Template eingestzt. Der
letzte Ansatz wurde mit Mung
Bean Nuclease verdaut. Davor
ist die Kontrolle ohne Mung
Bean Nuclease-Verdau
abgebildet.
ERGEBNISSE 31
M M700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
1:8 1:4 1:2 1:1 2:1 4:1 8:1
A189-IRD700 : A682
Abbildung 11. T-RFLP-Analyse von Klon B24 mit verschiedenen
Verhältnissen der Primer A189-IRD700 und A682. Im
Restriktionsverdau wurden 5 ng PCR-Produkt eingesetzt.
4.2.2 Aufreinigung des PCR-Produktes
Die Aufreinigung der PCR-Produkte dient dem Entfernen von Primern, Enzymen und Salzen.
Besonders die Entfernung von Salzen ist wichtig, da diese das Restriktionsenzym hemmen
können. Aus diesem Grund wurde die Reinigung des PCR-Produktes durch einen zusätzlichen
Waschschritt, längere Einwirkung des Waschpuffers und eine zusätzliche Ethanolentfernung
intensiviert. Eine Verbesserung des unvollständigen Restriktionsverdaus in Richtung eines
vollständigen Restriktionsverdaus konnte jedoch nicht erreicht werden (siehe Abb. 9).
4.2.3 Restriktionsverdau
Die physikalischen und chemischen Parameter des Restriktionsverdaus und das
Restriktionsenzym selbst können einen entscheidenden Einfluss auf die Restriktionsreaktion
haben. In den im Folgenden beschriebenen Experimenten wurden solche Einflüsse untersucht.
Im ersten Ansatz wurde getestet, ob größere Enzymmengen zu einem vollständigen
Restriktionsverdau führen. Dazu wurden die Restriktionsenzyme MspI und AluI/MlsI
(Doppelverdau) in den Mengen 1,5 U, 5 U und 10 U eingesetzt. Weder verschiedene
Enzymmengen noch unterschiedliche Restriktionsenzyme führten zu einem vollständigen
Restriktionsverdau (siehe Abb. 12).
Im zweiten Ansatz wurde die Zeitdauer des Verdaus (3 h vs. 16 h) und der
Inkubationsort des Verdaus (Brutschrank vs. Eppendorf Mastercycler Personal) variiert.

ERGEBNISSE 32
Keine dieser Veränderungen führte zu einer Verbesserung des unvollständigen Restriktions-
verdaus.
M M M M M M
1,5
U M
spI,
Klo
n B
60
5 U
Msp
I,
Klo
n B
60
10 U
Msp
I, K
lon
B60
10 U
Alu
I, 10
U M
lsI,
Klo
n B
24
5 U
Alu
I, 5
U M
lsI,
Klo
n B
24
1,5
U A
luI,
1,5
U M
lsI,
Klo
n B
24
10 U
Alu
I, 10
U M
lsI,
Klo
n B
67
5 U
Alu
I, 5
U M
lsI,
Klo
n B
67
1,5
U A
luI,
1,5
U M
lsI,
Klo
n B
67
B24
B60
B67
nichtverdaut
700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
Abbildung 12. Auswirkungen verschiedener
Enzymmengen auf den Restriktionsverdau mit
MspI bzw. AluI/MlsI. Im Restriktions-verdau
wurden 100 ng PCR-Produkt eingesetzt. In
den Ansätzen mit unverdautem PCR-Produkt
der Klone B24, B60 und B67 sind deutlich
Banden mit schwacher Intensität zu sehen.
Hier handelt es sich höchstwahrscheinlich um
unvollständig amplifiziertes PCR-Produkt.
Im dritten Ansatz wurde das Restriktionsenzym MspI der Hersteller Promega und New
England Bioloabs (NEB) mit dem Restriktionsenzym MspI von MBI verglichen (siehe
Abb. 13). Der Restriktionsverdau mit Promega-MspI resultierte ebenfalls in einem
unvollständigem Restriktionsverdau. Im Vergleich zum Restriktionsverdau mit MBI-MspI
waren jedoch weniger unvollständig geschnittene Fragmente vorhanden, allerdings traten in
unmittelbarer Nähe der Fragmente immer zweite Banden auf. Der Restriktionsverdau mit
NEB-MspI resultierte in einem fast vollständigen Restriktionsverdau. Jedoch konnte auch hier
in unmittelbarer Nähe des Hauptfragments ein zweites Fragment beobachtet werden.
Zusätzlich wies das Hauptfragment starke Intensitätsverluste der Fluoreszenz im Vergleich
zum parallelen Promega-Ansatz auf (gleiche Menge an PCR-Produkt im Restriktionsverdau),
was auf einen starken Verlust an PCR-Produkt während des Restriktionsverdaus schließen
lässt. Der Restriktionsverdau wurde, wie im NEB-Protokoll empfohlen, ohne BSA
durchgeführt, was zu einer Anheftung der DNS an die Reaktionsgefäße und somit zum
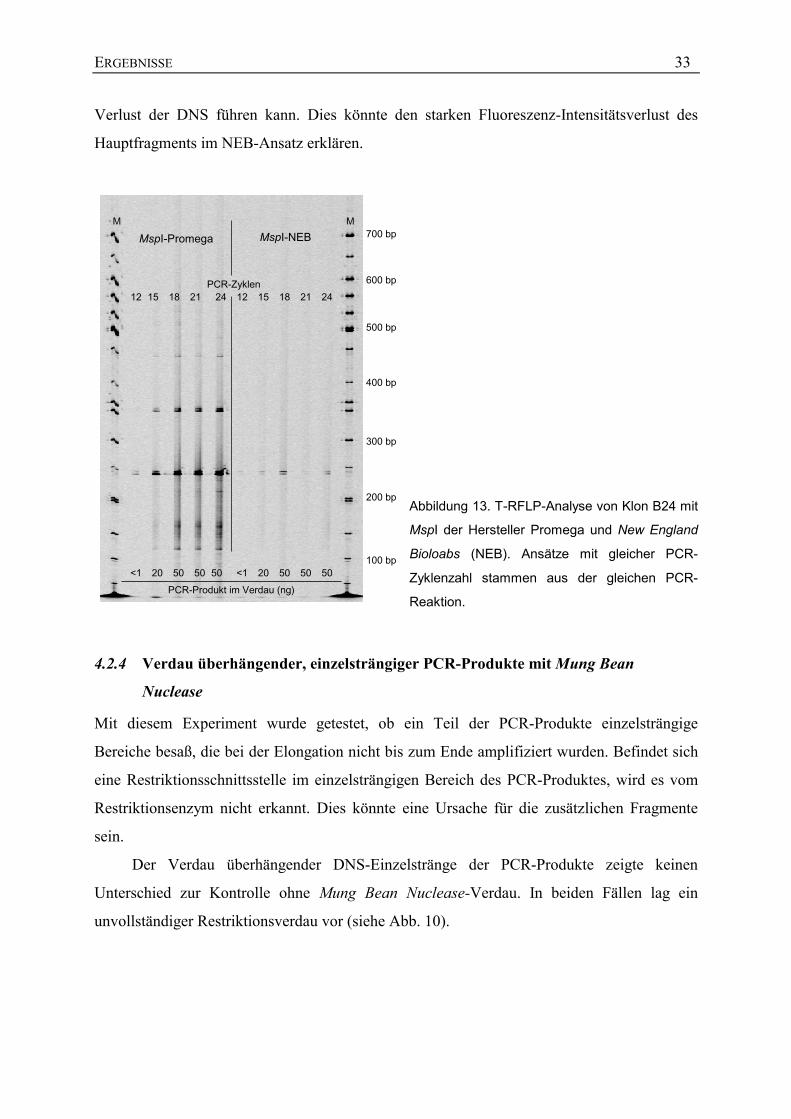
ERGEBNISSE 33
Verlust der DNS führen kann. Dies könnte den starken Fluoreszenz-Intensitätsverlust des
Hauptfragments im NEB-Ansatz erklären.
M M700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
12 15 18 21 24 12 15 18 21 24PCR-Zyklen
MspI-Promega MspI-NEB
PCR-Produkt im Verdau (ng)
<1 <120 2050 50 50 50 50 50
Abbildung 13. T-RFLP-Analyse von Klon B24 mit
MspI der Hersteller Promega und New England
Bioloabs (NEB). Ansätze mit gleicher PCR-
Zyklenzahl stammen aus der gleichen PCR-
Reaktion.
4.2.4 Verdau überhängender, einzelsträngiger PCR-Produkte mit Mung Bean
Nuclease
Mit diesem Experiment wurde getestet, ob ein Teil der PCR-Produkte einzelsträngige
Bereiche besaß, die bei der Elongation nicht bis zum Ende amplifiziert wurden. Befindet sich
eine Restriktionsschnittsstelle im einzelsträngigen Bereich des PCR-Produktes, wird es vom
Restriktionsenzym nicht erkannt. Dies könnte eine Ursache für die zusätzlichen Fragmente
sein.
Der Verdau überhängender DNS-Einzelstränge der PCR-Produkte zeigte keinen
Unterschied zur Kontrolle ohne Mung Bean Nuclease-Verdau. In beiden Fällen lag ein
unvollständiger Restriktionsverdau vor (siehe Abb. 10).
ERGEBNISSE 34
4.2.5 Zusammenfassung der T-RFLP-Evaluation und weiteres Vorgehen
Die Problematik des unvollständigen Restriktionsverdaus der T-RFLP-Methode konnte in
dieser Arbeit nicht gelöst werden. Deshalb wurden im weiteren Vorgehen die
charakteristischen T-RF-Muster einzelner Klone experimentell untersucht. Bei der Analyse
von Sedimentproben wurden die bekannten T-RF-Muster der Klone mit dem T-RF-Profil der
jeweiligen Probe verglichen, um eine gerechtfertigte Zuordnung der einzelnen Fragmente zu
ermöglichen.
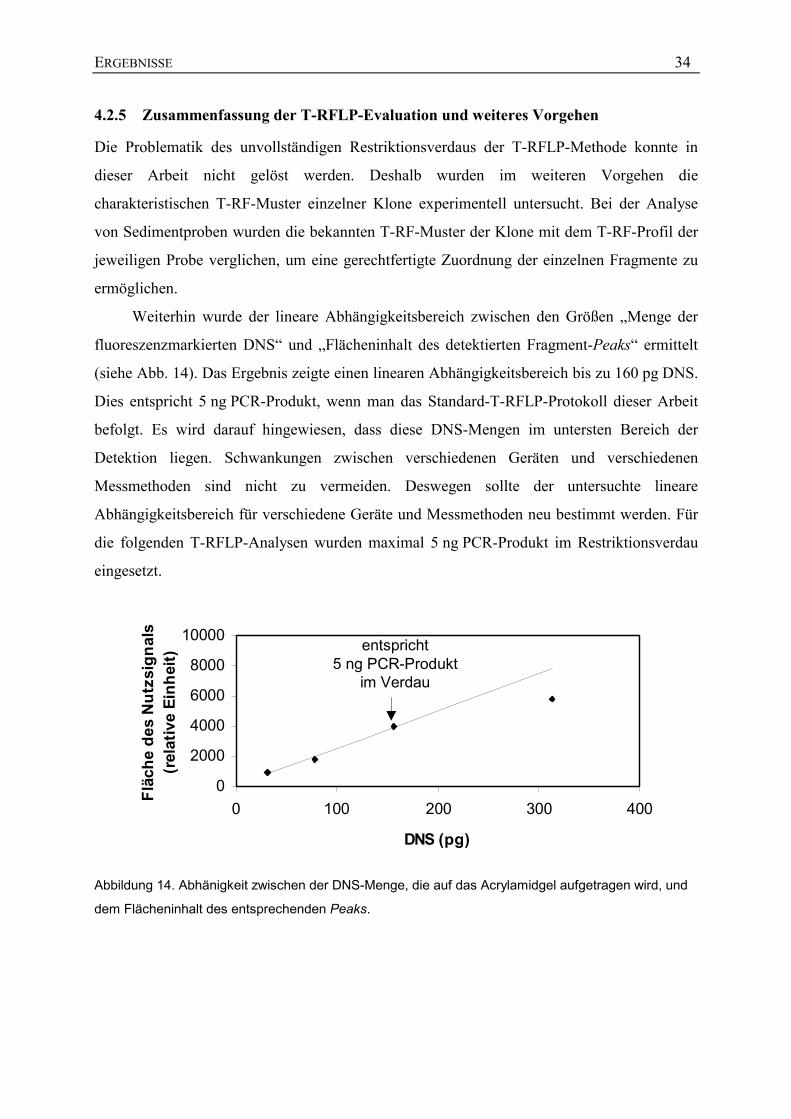
Weiterhin wurde der lineare Abhängigkeitsbereich zwischen den Größen „Menge der
fluoreszenzmarkierten DNS“ und „Flächeninhalt des detektierten Fragment-Peaks“ ermittelt
(siehe Abb. 14). Das Ergebnis zeigte einen linearen Abhängigkeitsbereich bis zu 160 pg DNS.
Dies entspricht 5 ng PCR-Produkt, wenn man das Standard-T-RFLP-Protokoll dieser Arbeit
befolgt. Es wird darauf hingewiesen, dass diese DNS-Mengen im untersten Bereich der
Detektion liegen. Schwankungen zwischen verschiedenen Geräten und verschiedenen
Messmethoden sind nicht zu vermeiden. Deswegen sollte der untersuchte lineare
Abhängigkeitsbereich für verschiedene Geräte und Messmethoden neu bestimmt werden. Für
die folgenden T-RFLP-Analysen wurden maximal 5 ng PCR-Produkt im Restriktionsverdau
eingesetzt.
0
2000
4000
6000
8000
10000
0 100 200 300 400
DNS (pg)
Flä
che
des
Nu
tzsi
gn
als
(rel
ativ
e E
inh
eit)
entspricht5 ng PCR-Produkt
im Verdau
Abbildung 14. Abhänigkeit zwischen der DNS-Menge, die auf das Acrylamidgel aufgetragen wird, und
dem Flächeninhalt des entsprechenden Peaks.

ERGEBNISSE 35
4.3 T-RFLP-Analyse von einzelnen Klonen und Klongemischen
Die Klone B67, B60, B24, A70 und A8 wurden stellvertretend für die Klongruppen B1, B2-
B7, B8, B9 und M.acetivorans-ähnliche Klone auf ihre T-RF-Muster untersucht (siehe
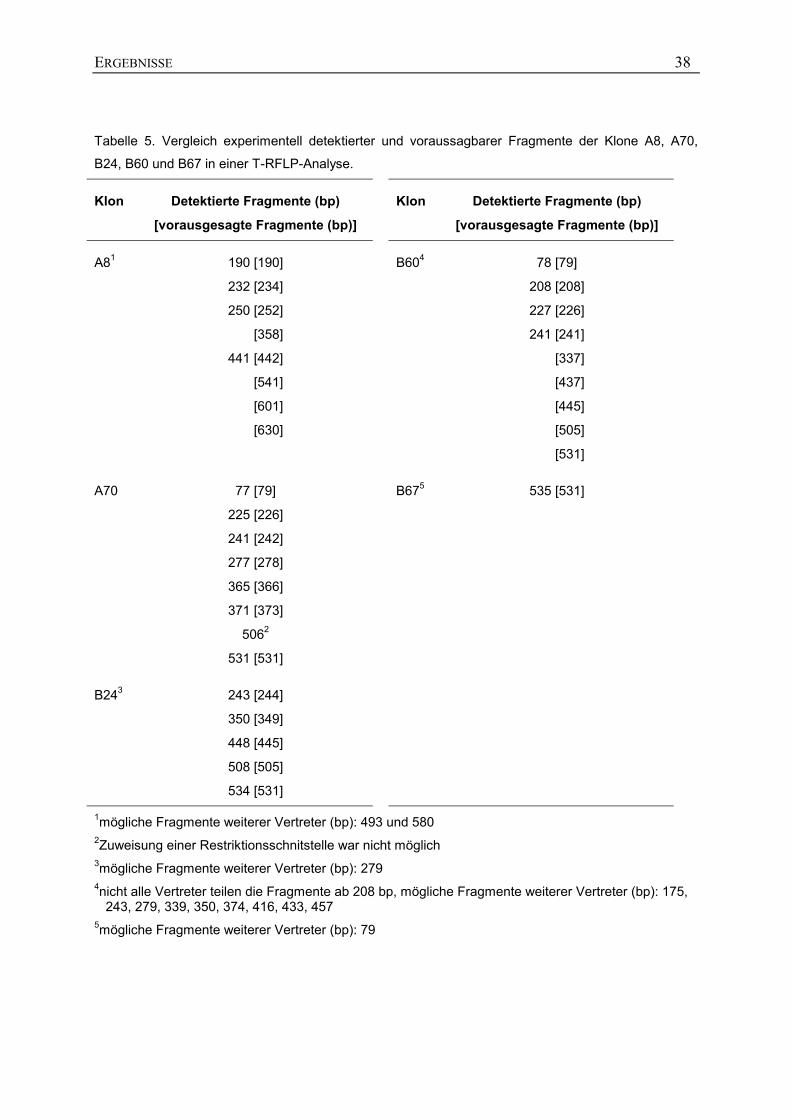
Abb. 15 und Tab. 5). Jedes Fragment der einzelnen T-RF-Profile konnte einer
Restriktionsschnittstelle zugeordnet werden. Die einzige Ausnahme war das Fragment mit
506 bp von Klon A70. Die von beiden Enden sequenzierte Insertion des Klons A70 war an
dieser Stelle mit der Basenabfolge 5’-CCGT-3’ ähnlich zur Restriktionsschnittstelle von MspI
(5’-CCGG-3’).
Mit Hilfe der T-RFLP-Analyse der oben genannten Klone wurde gleichzeitig der
Zuordnungsbereich von experimentell ermittelten Fragmentlängen festgelegt. Dazu wurden
die vorausgesagten Fragmentlängen mit experimentell ermittelten Fragmentlängen verglichen
(siehe Tab. 5), wobei experimentell ermittelte Fragmente mit bis zu 400 bp in einem Bereich
von maximal ± 2 bp, experimentell ermittelte Fragmente mit 400−530 bp in einem Bereich
von maximal ± 3 bp und experimentell ermittelte Fragmente über 530 bp in einem Bereich
von maximal ± 4 bp zugeordnet werden konnten. Auf diesem Ergebnis basierend wurden in
den folgenden T-RFLP-Analysen experimentell ermittelte Fragmente in einem Bereich von
maximal ± 2 bp zu voraussagbaren Fragmenten bekannter pmoA/amoA-Gene zugeordnet. Nur
in eindeutigen Ausnahmefällen, wie dem ungeschnittenen PCR-Produkt wurde der
Zuordnungsbereich auf ± 4 bp erhöht.
Anschließend wurden T-RFLP-Analysen von Sedimentproben mit verschiedenen
Klongemischen aus den oben genannten Klonen simuliert (siehe Abb. 15 und Abb. 16). Dazu
wurden die Template-DNS der Klone in einem 1 : 1-Verhältnis gemischt und einer T-RFLP-
Analyse unterzogen. In der Hälfte der Gemische konnten die eingesetzten Klone in den
jeweiligen T-RF-Profilen wiedergefunden werden und waren durch Fragmente mit ähnlich
starker Fluoreszenzintensität vertreten. In der anderen Hälfte der Gemische war dies nicht der
Fall. Die Fragmente des Klons A8 zeigten immer deutlich geringere Fluoreszenzintensitäten
als die Fragmente anderer Klone und der Klon A70 konnte in keinem Ansatz, in dem er
eingesetzt wurde, eindeutig identifiziert bzw. wiedergefunden werden.

77
22
52
41
27
7
36
53
71
50
65
31
Klo
n A
70
B6
7
53
5K
lon
B6
7
19
0
23
2
25
04
41
Klo
n A
8
78
(B6
0, A70
)
18
9 (
A8
)
20
8 (
B6
0)
22
7 (
B6
0, A70
)2
33
(A
8)
24
5(B
24
)
35
0 (
B2
4)
43
9 (
A8
)44
7 (
B2
4)
50
8 (
B2
4)
53
5(B
67
, B
24
)K
lon
ge
mis
ch
B6
07
8
20
82
41
22
7
Klo
n B
60
24
3
35
0
44
85
085
34
Klo
n B
24
Abbi
ldun
g 15
. T-R
F-M
uste
r der
Klo
ne B
67, B
60, B
24, A
8 un
d A7
0 un
d T-
RF-
Prof
il de
s Kl
onge
mis
ches
aus
B67
, B60
, B2
4 , A
8 un
d A7
0. In
dem
T-R
F-
Prof
il de
s Kl
onge
mis
ches
sin
d di
e er
sten
Fra
gmen
te e
ines
T-R
F-M
uste
rs d
urch
den
fettg
edru
ckte
n N
amen
des
zug
ehör
igen
Klo
ns g
eken
nzei
chne
t.
Nor
mal
ged
ruck
te N
amen
ken
nzei
chne
n Fr
agm
ente
sek
undä
rer S
chni
ttste
llen.
Klo
n A7
0 ko
nnte
in d
em T
-RF-
Prof
il de
s Kl
onge
mis
ches
nic
ht e
inde
utig
zuge
ordn
et w
erde
n un
d is
t des
halb
kur
siv
gedr
uckt
ang
egeb
en.
Frag
men
tläng
e (b
p)
Frag
men
tläng
e (b
p)
Fluoreszenzintensität

ERGEBNISSE 37
190 (A8)
534(B67)A8 und B67
78 (B60)
209 (B60)
227 (B60)
535 (B67)B60 und B67
78 (B60)
209 (B60)
243 (B24)
350 (B24) 447 (B24)
509(B24)
535(B24)
B24 und B60
227 (B60)
244(B24)
350 (B24)
A70 und B24
448 (B24)
507(B24)
534(B24)
Abbildung 16. T-RF-Profile der im Folgenden aufgelisteten Klongemische: i) Klon A8 und Klon B67,
ii) Klon B60 und Klon B67, iii) Klon B24 und Klon B60 und iv) Klon A70 und Klon B24. Die Template-
DNS der Klone wurde in einem 1 : 1−Verhältnis gemischt und einer T-RFLP-Analyse unterzogen. In
den T-RF-Profilen sind die ersten Fragmente eines T-RF-Musters durch den fettgedruckten Namen
des zugehörigen Klons gekennzeichnet. Normal gedruckte Namen kennzeichnen Fragmente
sekundärer Schnittstellen.
Fragmentlänge (bp)
Fluo
resz
enzi
nten
sitä
t
ERGEBNISSE 38
Tabelle 5. Vergleich experimentell detektierter und voraussagbarer Fragmente der Klone A8, A70,
B24, B60 und B67 in einer T-RFLP-Analyse.
Klon Detektierte Fragmente (bp)
[vorausgesagte Fragmente (bp)]
Klon Detektierte Fragmente (bp)
[vorausgesagte Fragmente (bp)]
A81 190 [190]
232 [234]
250 [252]
[358]
441 [442]
[541]
[601]
[630]
B604 78 [79]
208 [208]
227 [226]
241 [241]
[337]
[437]
[445]
[505]
[531]
A70 77 [79]
225 [226]
241 [242]
277 [278]
365 [366]
371 [373]
5062
531 [531]
B675 535 [531]
B243 243 [244]
350 [349]
448 [445]
508 [505]
534 [531]
1mögliche Fragmente weiterer Vertreter (bp): 493 und 580 2Zuweisung einer Restriktionsschnitstelle war nicht möglich 3mögliche Fragmente weiterer Vertreter (bp): 279 4nicht alle Vertreter teilen die Fragmente ab 208 bp, mögliche Fragmente weiterer Vertreter (bp): 175,
243, 279, 339, 350, 374, 416, 433, 457 5mögliche Fragmente weiterer Vertreter (bp): 79

ERGEBNISSE 39
4.4 T-RFLP-Analyse der Bodensee-Sedimente
Mit Hilfe der T-RFLP-Analyse von pmoA-Genen sollten die MOB-Populationen des Litoral-
und Profundalsediments miteinander verglichen werden. Ebenfalls sollte untersucht werden,
ob es im Litoral- und Profundalsediment Unterschiede in der Zusammensetzung der MOB-
Populationen zwischen der aktiven Schicht der aeroben Methanoxidation (inbegriffen in der
Tiefenstufe 0−1 cm) und einer nicht aktiven Schicht der aeroben Methanoxidation
(Tiefenstufe 10−11 cm) gibt. Für die eindeutige Zuordnung von Klongruppen zu den
einzelnen Fragmenten eines T-RF-Profils wurden nur Fragmente, die aus primären
Schnittstellen resultieren, verwendet. Damit konnten drei zusammengefasste Übergruppen der
Klongruppen B1−B9 unterschieden werden: i) Klongruppe B1a (Klon B67), ii) Klongruppen
B1b (Klon A55), Klongruppen B2−B7 und Klongruppe B9 und iii) Klongruppe B8 (siehe
Abb. 6). Fragmente, die aus sekundären Schnittstellen resultieren können, wurden auf der
Basis von Sequenzinformationen der Klongruppen bzw. mit Hilfe der T-RF-Muster der
untersuchten Klone einzelnen Klongruppen zugewiesen.
4.4.1 Das Litoralsediment
Im T-RF-Profil der aktiven Tiefenstufe wurden zehn Fragmente detektiert (siehe Abb. 17).
Die Fragmente mit den größten Fluoreszenzintensitäten wurden der Klongruppe B1a (534 bp,
ungeschnittenes PCR-Produkt) und den Klongruppen B1b−B7 und B9 (78 bp) zugeordnet
oder konnten keiner Klongruppe zugeordnet werden (88 bp). Die Fragmente mit geringerer
Fluoreszenzintensität wurden sekundären Schnittstellen der Klongruppen B2−B7 und B9
zugewiesen (208 bp, 241 bp, 339 bp und 439 bp) oder konnten keiner Klongruppe zugeordnet
werden (53 bp und 74 bp).
Bei dem Vergleich von T-RF-Profilen der aktiven und nicht aktiven Tiefenstufe wurden
keine Unterschiede in der Fragmentzusammensetzung detektiert (siehe Abb. 18 ). Auch der
Vergleich der T-RF-Profile von verschiedenen Probennahmen (aktive Tiefenstufe, Ansatz C
und D) zeigte für alle Proben ein sehr ähnliches Umweltprofil (siehe Abb. 18). Bei dem
parallelen Auftragen des verdauten PCR-Produkts aus Ansatz D konnten ebenfalls keine
Unterschiede festgestellt werden (siehe Abb. 18).

ERGEBNISSE 40
Um auf Replizierbarkeit zu testen, wurde die T-RFLP-Analyse mit denselben DNS-
Extrakten wiederholt. Das Ergebnis der zweiten T-RFLP-Analyse war identisch mit dem
Ergebnis der ersten T-RFLP-Analyse.
Aufgrund eines Experimentierfehlers (zu hohe Konzentration des Laufpuffers für die
Elektrophorese des T-RFLP-Proben) konnten die Fragmente unter 100 bp in der
Vergleichsstudie der Tiefenstufen nicht berechnet werden. Aus diesem Grund wurde nur das
Originalbild des Acrylamidgels in Abbildung 18 dargestellt.
53
74
78
88
208
241
534
339 439
Litoralsediment 0–1 cm
<48
53
57
78
88113
120
208
226318
338
373 439
457
534Profundalsediment 0–1 cm
<48
53
78
87
113
339 374 439242457
533
Profundalsediment 10–11 cm
Abbildung 17. T-RF-Profile des Litoral- und Profundalsediments. Fettgedruckte und unterstrichene
Fragmentlängen kennzeichnen die ersten Fragmente eines zugeordneten T-RF-Musters (78 bp:
Klongruppe B1b, B2−B7 und B9; 533/534 bp: Klongruppe B1a und ungeschnittenes PCR-Produkt).
Normal gedruckte und unterstrichene Fragmentlängen kennzeichnen mögliche sekundäre Fragmente
der T-RF-Muster der Klongruppe B2−B7. Fragmente, die durch normal gedruckte Fragmentlängen
gekennzeichnet sind, konnten nicht zugeordnet werden.
Fluo
resz
enzi
nten
sitä
t
Fragmentlänge (bp)

ERGEBNISSE 41
100 bp
M M700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
M M
M M
460 bp
C D D D
0–1 cm 10–11cm
C D D D
0–1 cm 10–11cm
C D D D
0–1 cm 10–11cm
Abbildung 18. T-RF-Profile der Tiefenstufen 0-1 cm und 10-11 cm des Litoralsediments mit zwei
vergrößerten Bereichen. Die Pfeile deuten auf die detektierten Fragmente. Die Buchstaben C und D
stehen für die Ansätze C und D der DNS-Extraktion, wobei sich beide Ansätze nur in dem Zeitpunkt
ihrer Probennahme unterscheiden (C: 30.08.02, D: 05.09.02).
4.4.2 Das Profundalsediment
Im T-RF-Profil der aktiven Tiefenstufe wurden 15 Fragmente detektiert (siehe Abb. 17). Die
Fragmente mit den größten Fluoreszenzintensitäten wurden der Klongruppe B1a (534 bp,
unverdautes PCR-Produkt) und den Klongruppen B1b−B7 und B9 (78 bp) zugeordnet oder
konnten keiner Klongruppe zugeordnet werden (88 bp und 113 bp). Die Fragmente mit
geringerer Fluoreszenzintensität wurden sekundären Schnittstellen der Klongruppen B2−B7
und B9 zugewiesen (208 bp, 226 bp, 318 bp, 338 bp, 373 bp, 439 bp und 457 bp) oder
konnten keiner Klongruppe zugeordnet werden (<48 bp, 53 bp, 57 bp, 74 bp und 120 bp).
Im T-RF-Profil der nicht aktiven Tiefenstufe wurden 11 Fragmente detektiert (siehe
Abb. 17). Die Fragmente mit den größten Fluoreszenzintensitäten wurden der Klongruppe
B1a (533 bp, unverdautes PCR-Produkt) und den Klongruppen B1b−B7 und B9 (78 bp)
zugeordnet oder konnten keiner Klongruppe zugeordnet werden (<48 bp, 87 bp und 113 bp).

ERGEBNISSE 42
Die Fragmente mit geringerer Fluoreszenzintensität wurden sekundären Schnittstellen der
Klongruppen B2−B7 und B9 zugewiesen (242 bp, 339 bp, 374 bp, 439 bp und 457 bp) oder
konnten keiner Klongruppe zugeordnet werden (53 bp).
Die T-RF-Profile der unterschiedlichen Tiefenstufen hatten ein sehr ähnliches
Fragmentmuster, allerdings konnte ein Teil der Fragmente aus der aktiven Tiefenstufe in der
T-RFLP-Analyse der nicht aktiven Tiefenstufe nicht mehr detektiert werden bzw. die
Fluoreszenzintensität der jeweiligen Fragmente nahm stark ab. Bei diesen Fragmenten
handelte es sich zum größten Teil um Fragmente, die sekundären Schnittstellen der
Klongruppen B2−B7 und B9 zugewiesen wurden und ist auf die Abnahme der
Fluoreszenzintensität des Fragments mit 78 bp, welches der primären Schnittstelle der
Klongruppen B1b−B7 und B9 zugeordnet wurde, zurückzuführen. Die Fragmente mit den
größten Fluoreszenzintensitäten waren in beiden Tiefenstufen dieselben. Die einzige
Ausnahme bildete das Fragments <48 bp, weil seine Fluoreszenzintensität im T-RF-Profil der
nicht aktiven Tiefenstufe deutlich größer war als im T-RF-Profil der aktiven Tiefenstufe.
4.4.3 Vergleich zwischen Litoral- und Profundalsediment
Die T-RFLP-Analyse der aktiven Tiefenstufe des Litoral- und Profundalsediments zeigte eine
teilweise Übereinstimmung, aber auch deutliche Unterschiede in der Zusammensetzung der
T-RF-Profile beider Sedimente. Eine Übereinstimmung gab es in der Zuordnung der
Klongruppen B1a, B1b−B7 und B9 sowie in den Fragmenten mit 53 und 88 bp. Unterschiede
gab es in der Anzahl und Fluoreszenzintensität von Fragmenten, die sekundären Schnittstellen
der Klongruppe B2−B7 und B9 zugewiesen wurden sowie in den Fragmenten mit 113 und
120 bp, die nur in der T-RFLP-Analyse des Profundalsedimentes detektiert wurden (siehe
Abb. 17).
4.5 T-RFLP-Analyse von Anreicherungskulturen
Mit Hilfe der T-RFLP-Analyse von pmoA-Genen sollten die MOB-Populationen in
verschiedenen Anreicherungsansätzen verglichen werden. Dazu wurden folgende
Anreicherungen untersucht: i) Anreicherungen aus dem Litoralsediment des „Litoralgartens“
mit Whittenbury- und Bussmann-Medium (Ansatz L) ii) Anreicherungen aus dem
Litoralsediment vor Meersburg mit Whittenbury- und Bussmann-Medium (Ansatz M) und

ERGEBNISSE 43
iii) Anreicherungskulturen aus dem Litoralsediment „Litoralgarten“ in einem
Gradientensystem aus Sauerstoff und Methan mit Bussmann-Medium.
4.5.1 Anreicherungen in Mikrotiterplatten
Die T-RFLP-Analyse der Anreicherungskulturen in Mikrotiterplatten ist in den Abbildungen
19 und 20 dargestellt. Die Klongruppe B8 wurde für eine genauere Analyse der T-RF-Profile
in die Untergruppen B8a und B8b unterteilt. Die Klongruppe B8a ist durch das T-RF-Muster
von Klon B24 repräsentiert. Die Klongruppe B8b besitzt eine zusätzliche MspI-Schnittstelle
bei 278 bp und ist durch den Klon B25 repräsentiert. Allerdings ist die Klongruppe B8b
derzeit nur postuliert, da der Klon B25 noch nicht experimentell auf sein T-RF-Muster
untersucht wurde. Die Fragmente im Bereich 532−535 bp (ungeschnittenes PCR-Produkt)
können sowohl der Klongruppe B1a, als auch sekundären Schnittstellen der Klongruppen
B8a/B8b zugeordnet werden. Aufgrund der guten Übereinstimmung der Fragmente mit ca.
244 bp, 350 bp, 447 bp und 533 bp mit dem T-RF-Profil des Klons B24 (siehe Abb. 15)
wurden die Fragmente mit 532−535 bp nur den Untergruppen der Klongruppe B8 zugeordnet.
Die Fragmente von T-RF-Profilen der höchsten Verdünnungsstufen aus Ansatz L
wurden Vertretern der Klongruppe B8a zugeordnet oder konnten nicht zugeordnet werden
(<48 bp, 53/54 bp, 68 bp und 87/88 bp). Dabei waren sich die T-RF-Profile der
Anreicherungen mit Whittenbury- und Bussmann-Medium sehr ähnlich.
Die Fragmente von T-RF-Profilen der höchsten Verdünnungsstufen aus Ansatz M
wurden ebenfalls Vertretern der Klongruppen B8a zugeordent oder konnten nicht zugeordnet
werden (318 bp). Auch hier waren sich die T-RF-Profile der Anreicherungen mit
Whittenbury- und Bussmann-Medium sehr ähnlich.

Wh
itte
nb
ury
-Med
ium
Bu
ssm
an
n-M
ed
ium
<4
88
8
24
4(B
8a
,B8
b)
27
8(B
8b
) 35
0(B
8a
,B8b
)44
7(B
8a
,B8b
)
50
8(B
8a
,B8b
)
53
4(B
8a
,B8b
)10
–4
<4
8 54
88
24
4(B
8a
,B8
b)
27
9(B
8b
)
35
0
(B8
a,B
8b
)4
47
(B8
a,B
8b
)
50
7(B
8a
,B8b
)
53
4(B
8a
,B8b
)10
–5
<4
8 53
87
24
4(B
8a
)
35
1(B
8a
)4
47
(B8
a)
50
8
(B8
a)
53
5(B
8a
)
10
–6
<4
8
48 5
45
8
88
24
3(B
8a
,B8
b)
27
8(B
8b
) 35
0(B
8a
,B8b
) 44
7(B
8a
,B8b
)
50
7(B
8a
,B8b
)
53
3(B
8a
,B8b
)10
–4
<4
8 54
88
24
4(B
8a
,B8
b)
27
32
78
(B8
b)
35
0(B
8a
,B8b
) 44
7
(B8
a,B
8b
)
50
8
(B8
a,B
8b
)
53
5(B
8a
,B8b
)10
–5
<4
8
54
68
88
24
3(B
8a
)
35
0(B
8a
)
44
7(B
8a
)
50
8(B
8a
)
53
5(B
8a
)
10
–6
Ab
bild
ung
19. T
-RF-
Prof
ile d
er A
nrei
cher
unge
n in
Mik
rotit
erpl
atte
n au
s de
m L
itora
lsed
imen
t „Li
tora
lgar
ten“
. Die
Ver
dünn
ungs
stuf
en d
er e
inze
lnen
MPN
-
Ansä
tze
sind
in d
en je
wei
ligen
T-R
F-Pr
ofile
n fe
ttged
ruck
t dar
gest
ellt.
Es
konn
ten
die
Klon
grup
pen
B8a
und
B8b
zuge
wie
sen
wer
den,
wob
ei d
ie K
long
rupp
e
B8b
nur
zuge
wie
sen
wur
de,
wen
n da
s Fr
agm
ent
mit
278
bp v
orha
nden
war
. D
ie e
rste
n Fr
agm
ente
der
zug
ewie
sen
T-R
F-M
uste
r si
nd d
urch
den
fettg
edru
ckte
n N
amen
der
jew
eilig
en K
long
rupp
e ge
kenn
zeic
hnet
. Das
Fra
gmen
t mit
533−
535
bp s
tellt
das
ung
esch
nitte
ne P
CR
-Pro
dukt
dar
.
Frag
men
tläng
e (b
p)
Frag
men
tläng
e (b
p)
Fluoreszenzintensität

<4
8 68
86
16
42
43
(B8a
)
34
9(B
8a
)4
46
(B8a
)
53
2(B
8a
)5
06
(B8a
)
10
–4
<4
8
76
(B3
,B9
) 86
22
5(B
3,B
9)
24
3(B
8a
)3
49
(B8a
)
44
6
(B8
a)
50
7
(B8
a)
53
3(B
8a
)
10
–4
24
4(B
8a
)
34
9(B
8a
)
44
6(B
8a
)
50
6(B
8a
)
53
2(B
8a
)
10
–5
24
3(B
8a
)
34
9(B
8a
)4
47
(B8a
)
50
8(B
8a
)
10
–6
<4
81
64
24
3(B
8a
)
34
9(B
8a
)
44
6(B
8a
) 50
6(B
8a
)
10
–4
<4
8
24
4(B
8a
)
87
35
0(B
8a
)4
46
(B8a
)
50
6(B
8a
)5
32
(B8a
)
10
–4
<4
8
87
16
4
24
4(B
8a
)3
49
(B8a
)
44
6(B
8a
)
50
6(B
8a
)5
32
(B8a
)
10
–5
24
4(B
8a
)
31
835
0(B
8a
)
44
7(B
8a
)50
8(B
8a
)
53
4(B
8a
)
10
–6
Wh
itte
nb
ury
-Med
ium
Bu
ssm
an
n-M
ed
ium
Ab
bild
ung
20. T
-RF-
Prof
ile d
er A
nrei
cher
unge
n in
Mik
rotit
erpl
atte
n au
s dem
Lito
rals
edim
ent „
Mee
rsbu
rg“.
Die
Ver
dünn
ungs
stuf
en d
er e
inze
lnen
MPN
-Ans
ätze
sind
in
den
jew
eilig
en T
-RF-
Prof
ilen
fettg
edru
ckt d
arge
stel
lt. D
ie V
erdü
nnun
gsst
ufe
10−4
stam
mt a
us z
wei
par
alle
len
Ans
ätze
n. D
ie e
rste
n Fr
agm
ente
der
zug
ewie
sen
T-R
F-
Mus
ter s
ind
durc
h de
n fe
ttged
ruck
ten
Nam
en d
er je
wei
ligen
Klo
ngru
ppe
geke
nnze
ichn
et.
Das
Fra
gmen
t mit
533−
535
bp st
ellt
das u
nges
chni
ttene
PC
R-P
rodu
kt d
ar.
Frag
men
tläng
e (b
p)
Frag
men
tläng
e (b
p)
Fluoreszenzintensität

ERGEBNISSE 46
4.5.2 Anreicherungen in einem Gradientensystem
Die Analyse der Gradientenkultur U zeigt ein deutlich verschiedenes T-RF-Profil von den
Anreicherungskulturen auf Mikrotiterplatten (siehe Abb. 21). Im T-RF-Profil der
Gradientenkultur U konnten 13 Fragmente detektiert werden. Die Fragmente mit den größten
Fluoreszenzintensitäten wurden den Klongruppen B1b−B7 und B9 (78 bp) und der
sekundären Schnittstelle der Klongruppe B2, B4 und B5 (208 bp) zugeordnet oder konnten
keiner Klongruppe zugeordnet werden (<48 bp, 86 bp, 234 bp). Die Fragmente mit geringerer
Fluoreszenzintensität wurden der Klongruppe B1a (532 bp, ungeschnittenes PCR-Produkt)
und sekundären Schnittstellen der Klongruppe B2−B7 und B9 zugewiesen (226 bp, 241 bp,
337 bp, 373 bp und 439 bp) oder konnten keiner Klongruppe zugeordnet werden (265 bp und
304 bp).
Die Fragmente mit 234 bp, 265 bp und 304 bp wurden im Vergleich zu den
Anreicherungskulturen in Mikrotiterplatten und den untersuchten Umweltproben nur in der
Gradientenkultur gefunden. Sie konnten auch keiner Klongruppe zugeordnet werden. Die
Fragmente mit 337 bp, 373 bp und 439 bp stellen mögliche sekundäre Schnittstellen von
Klonen aus den Gruppen B3−B7 dar. Sie waren ebenfalls in den untersuchten Umweltproben
vorhanden, jedoch nicht in den Anreicherungskulturen in Mikrotiterplatten.

ERGEBNISSE 47
53
74
78
88
208
241
534
339 439
Litoralsediment, 0–1 cm
<48
78
86
208
226
234
241
265 304
337
373439
532
Gradientenkultur U
<48
48
54 58
88 243
278350
447
507 533
Anreicherungskultur, 10–4
Abbildung 21. T-RF-Profil der Gradientenkultur U im Vergleich zu den T-RF-Profilen der aktiven
Schicht des Litoralsediments (inbegriffen in 0−1 cm) und einer Anreicherung in Mikrotiterplatten mit
Bussmann-Medium (MPN-Verdünnungsstufe ist angegeben). Die fettgedruckten und unterstrichenen
Fragmentlängen kennzeichnen die ersten Fragmente eines T-RF-Musters (78 bp: Klongruppen B1b,
B2−B7 und B9; 532 bp: B1a und ungeschnittenes PCR-Produkt). Normal und unterstrichen gedruckte
Fragmentlängen kennzeichnen Fragmente sekundärer Schnittstellen der Klongruppen B2−B7 und B9,
wobei das Fragment mit 208 bp nur den Klongruppen B2, B4 und B5 zugeordnet wurde.

5 DISKUSSION
In dieser Arbeit wurde die Diversität aerober, methanotropher Bakterien (MOB) im
Litoralsediment des Bodensees anhand einer pmoA-Klonbibliothek untersucht. Darauf
aufbauend wurden die MOB-Populationen des Litoral- und Profundalsediments sowie die
Tiefenverteilung der MOB-Populationen innerhalb der jeweiligen Sedimente mit Hilfe der
Fingerprint-Methode T-RFLP verglichen. In einem zweiten Teilprojekt wurde mit einer
T-RFLP-Analyse die Zusammensetzung von MOB-Populationen verschiedener
Anreicherungsansätze aus dem Litoralsediment untereinander und mit einer Probe aus dem
Litoralsediment verglichen. Die verschiedenen Anreicherungsansätze bestanden aus:
i) Anreicherungen in Mikrotiterplatten mit Whittenbury-Medium, ii) Anreicherungen in
Mikrotiterplatten mit Bussmann-Medium und iii) Anreicherungen mit Bussmann-Medium in
einem gegenläufigen Gradientensystem aus Methan und Sauerstoff. Ein drittes Teilprojekt
befasste sich mit der Evaluation der T-RFLP-Methode.
5.1 Diversität der pmoA-Gene im Litoralsediment
Im Litoralsediment des Bodensees wurde mit Hilfe einer pmoA-Klonbibliothek eine große
Diversität2 von pmoA-Genen gefunden. Die sequenzierten pmoA-Insertionen der Klone
zeigten Ähnlichkeit zu sehr unterschiedlichen pmoA-Genen der Typ I- und Typ II-MOB und
waren über die gesamte Bandbreite bekannter pmoA-Gene gleichmäßig verteilt (siehe Abb. 6).
Basierend auf 10 % Sequenzunterschied wurden die pmoA-Fragmente der Klonbibliothek in
sieben Gruppen innerhalb der Typ I-MOB (B1−B7) und in einer Gruppe innerhalb der Typ II-
MOB (B8) zusammengefasst. Eine neunte Gruppe (B9) konnte weder zu pmoA- noch amoA-
Genen eindeutig zugeordnet werden. Da ein Chimärentest des DNS-Fragments der Gruppe B9
negativ ausfiel und DNS-Fragmente mit ähnlichen Zuordnungsschwierigkeiten auch in
Reisfeldböden gefunden wurden (Horz et al., 2001), handelt es sich hier wahrscheinlich um
eine neue Gruppe von MOB oder AOB. Aus dieser Gruppe wurden noch keine Reinkulturen
isoliert, weshalb über ihre ökologische Bedeutung noch keine näheren Aussagen getroffen
werden können. Zusätzlich konnten auch die Klongruppen B6 und B7 keinen nahverwandten
2 Die Diversität wurde an der Anzahl von pmoA-Fragmenten (531 bp), die sich um mindestens
1 % in ihrer Sequenz unterschieden, gemessen.

DISKUSSION 49
pmoA-Genen (maximal 10 % Sequenzunterschied) zugeordnet werden. Sie repräsentieren
möglicherweise noch nicht beschriebene MOB-Arten.
Ein Vergleich mit anderen Umweltstudien zeigte, dass ähnlich große Diversitäten von
pmoA-Genen auch im Profundalsediment von Lake Washington (Costello und Lidstrom,
1999), in Reisfeldern (Horz et al., 2001) und in verschiedenen Böden gefunden wurden
(Bourne et al., 2001). Dies deutet darauf hin, dass in Süßwassersedimenten und Böden eine
merhrere Gattungen umfassende Artenzahl von MOB anzutreffen ist, wobei nicht jedes
unterschiedliche pmoA-Gen eine andere MOB-Art repräsentiert, da sich die pmoA-Kopien
einer MOB-Art zwar meist sehr ähnlich sind (Costello und Lidstrom, 1999; Stolyar et al.,
1999), aber auch unterscheiden können (Dunfield et al., 2002).
Obwohl mit den verwendeten Primern A189 und A682 sowohl pmoA- als auch amoA-
Fragmente amplifiziert werden können (Holmes et al., 1995), wurden in dieser Studie keine
amoA-Gene nachgewiesen. Dieser Befund und nicht detektierbare Ammonium-
Oxidationsraten im Litoralsediment (Bosse et al., 1993) weisen auf eine sehr geringe
Abundanz von Ammonium oxidierenden Bakterien im Litoralsediment des Bodensees hin.
5.2 T-RFLP-Analyse des Litoral- und Profundalsediments
Ein Vergleich der Klonbibliothek mit dem T-RF-Profil der aktiven Tiefenstufe der
Methanoxidation im Litoralsediment (inbegriffen in 0−1 cm) zeigte teilweise eine
Übereinstimmung und teilweise Unterschiede in den detektierten MOB/AOB. Die
Klongruppen B1a , B1b−B7 und B9 wurden sowohl mit der Klonbibliothek nachgewiesen, als
auch in dem T-RF-Profil Fragmenten zugeordnet. Dagegen wurden einerseits die Klongruppe
B8 und M. acetivorans-ähnliche Klone nur in der Klonbibliothek nachgewiesen und
andererseits in dem T-RF-Profil Fragmente mit 53 bp, 74 bp und 88 bp detektiert, die keinen
Klongruppen zugeordnet werden konnten. Entweder sind diese Unterschiede auf die
unterschiedlichen Jahreszeiten der Probennahme zurückzuführen (Klonbibliothek: Winter,
T-RFLP-Analyse: Sommer) oder die eingesetzten Methoden wirken auf unterschiedliche
pmoA/amoA-Gene selektiv.
Bei einem Vergleich der aktiven (0−1 cm) und nicht aktiven (10−11 cm) Tiefenstufe des
Litoralsediments konnten keine nennenswerten Unterschiede zwischen den MOB-
Populationen beider Tiefenstufen festgestellt werden (siehe Abb. 18). Es besteht die
Möglichkeit, dass in der nicht aktiven Tiefenstufe die DNS toter MOB und keine lebenden

DISKUSSION 50
MOB detektiert wurden, da im Bodensee, aufgrund der geringen Eindringtiefe des Sauerstoffs
(Frenzel et al., 1990), die aerobe Methanoxidation auf die obersten Millimeter des Sediments
beschränkt ist. Allerdings sprechen folgende Befunde dagegen: Bosse et al. (1993) haben
potentielle aerobe Methanoxidations-Aktivitäten im Litoralsediment des Bodensees bis auf
5 cm Sedimenttiefe untersucht und nachgewiesen. Zwischen 3 und 5 cm Sedimenttiefe betrug
die Methanabbaurate ca. 75 % der Abbaurate des obersten Zentimeters. Versuche im
Sommervertiefungskurs 2002 am Lehrstuhl für Mikrobielle Ökologie/Limnologie zeigten
ebenfalls potentielle aerobe Methanoxidations-Aktivitäten im Litoralsediment der
Tiefenstufen 2−3 cm, 10−11 cm und 17−18 cm. Zusätzlich wurde bereits gezeigt, dass MOB
über längere Zeiträume hungern können (Roslev and King, 1995), was das Vorkommen
lebender MOB in anoxischen Sedimentschichten erklärt.
In der T-RFLP-Analyse der aktiven und nicht aktiven Tiefenstufe des
Profundalsediments wurden ebenfalls sehr ähnliche MOB-Populationen in beiden
Tiefenstufen detektiert (siehe Abb. 17). Allerdings wurde noch nicht untersucht bzw. gibt es
keine Hinweise darauf, ob in der nicht aktiven Tiefenstufe lebende MOB, DNS toter MOB
oder eine Mischung aus beiden detektiert wurden.
Ein Vergleich der aktiven Tiefenstufe des Litoral- und Profundalsediments zeigte eine
teilweise Übereinstimmung, aber auch deutliche Unterschiede in der Zusammensetzung der
T-RF-Profile beider Sedimente (siehe Abb. 17). In beiden T-RF-Profilen wurden Fragmente
detektiert, die keiner Klongruppe zugeordnet werden konnten und somit entweder von
unbekannten pmoA- oder amoA-Genen stammen. Da es jedoch starke Hinweise auf sehr
geringe Abundanzen von AOB im Bodensee gibt (Bosse et al., 1993; diese Arbeit), weisen die
Unterschiede zwischen den T-RF-Profilen wahrscheinlich auf eine unterschiedliche
Zusammensetzung der MOB-Populationen zwischen dem Litoral- und Profundalsediment hin.
Die unterschiedliche Zusammensetzung der MOB-Populationen zwischen Litoral- und
Profundalsediment können auf folgende Unterschiede in den Umweltbedingungen dieser
Habitate zurückgeführt werden: i) im Litoralsediment wird, unabhängig von der Jahreszeit,
immer mehr Methan produziert als im Profundalsediment (Frenzel et al., 1990; Thebrath et
al., 1993), ii) das Litoralsediment unterliegt einer jahreszeitlichen Temperaturänderung
(Thebrath et al., 1993), während das Profundalsediment über eine konstante Temperatur von
4 °C verfügt (Schulz and Conrad, 1995), iii) das Litoralsediment unterliegt aufgrund der
Photosynthese einer diurnalen Verschiebung der oxisch-anoxischen Grenzschicht (S.

DISKUSSION 51
Gerhardt, A. Brune und B. Schink, nicht veröffentlichte Ergebnisse); im Profundalsediment
existiert diese Dynamik aufgrund einer fehlenden Photosyntheseaktivität nicht und iv) das
Litoralsediment unterliegt sehr wahrscheinlich häufiger und massiver physikalischen
Störungen, wie z.B. Wellenschlag oder Sturmereignissen, als das Profundalsediment.
Zusammengefasst kann das Litoralsediment als ein Habitat angesehen werden, welches
häufiger Störungen und periodischen Änderungen der oxisch-anoxischen Schicht unterliegt,
als das Profundalsediment. Dies könnte die Ursache für die unterschiedliche
Zusammensetzung der MOB-Populationen sein.
Die ähnliche Zusammensetzung von MOB-Populationen über die Tiefe des Sediments
hinweg kann durch zwei Hypothesen erklärt werden. Einerseits ist es möglich, dass MOB im
Laufe der Zeit durch Sedimentation begraben und somit vom Sauerstoff abgeschnitten
werden, jedoch über lange Zeiträume lebensfähig bleiben (1. Hypothese). Andererseits
könnten MOB eine Strategie entwickelt haben, die auf Resuspensions- und
Belüftungsereignissen der anoxischen Sedimentschichten aufbaut (2. Hypothese).
Resuspensionsereignisse können physikalischen oder biologischen Ursprungs sein.
Ursachen physikalisch bedingter Resuspensionsereignisse stellen Stürme, Strömungen,
Durchmischungsereignisse und Wellenschlag dar. Auf das Litoralsediment haben alle
genannten Ereignisse einen Einfluss. Das Profundalsediment vieler Seen wird vor allem in
Folge von Durchmischungsereignissen ein bis mehrere Mal im Jahr gestört. Solche
Störungsereignisse können einige Millimeter bis einige Zentimeter des Sediments
resuspendieren (Wetzel, 2001).
Resuspensionsereignisse biologischen Ursprungs haben vor allem auf die oberen
Zentimeter des Sedimentes einen Effekt. Zum Beispiel wühlen benthische Fische das
Sediment auf, um es nach Nahrung zu durchsuchen (Shormann und Cotner, 1997). Im
Bodensee übernehmen z.B. Brachsen und Kaulbarsche diese Rolle (Prof. Eckmann,
persönliche Kommunikation). Welchen quantitativen Effekt dies auf die Resuspension des
Sedimentes hat, wurde noch nicht untersucht. Auch die im Sediment lebende Fauna hat einen
Effekt auf die stellenweise Belüftung anoxischer Sedimentschichten. Für Larven von
Micropsectra sp. (Chironomidae) konnte gezeigt werden, dass sie im Litoriprofundal
(20−25 m Tiefe) aufgrund ihrer Abundanz das Volumen des oxischen Sedimentes um
ca. 25 % erhöhen im Vergleich zur reinen Diffusion von Sauerstoff in das Sediment. Dieses
erhöhte Sauerstoffangebot bewirken sie durch ihre Lebensweise in nach unten blind

DISKUSSION 52
auslaufenden Röhren (ca. 2,5 cm tief) und ihre rhythmischen Pumpaktivitäten (jede
20−40 min bei 4 °C) (Frenzel, 1990). Dadurch entsteht um die Röhren zeitweise ein
Sauerstoffangebot, das MOB nutzen können. Larven von Micropsectra sp. können einen
immensen Einfluss auf die Belüftung des Sedimentes haben, wenn sie große Abundanzen, wie
im Einfluss des Rheins in den Bodensee (50000 Larven/m2), erreichen. Im Profundal des
Bodensees spielen sie jedoch keine große Rolle (Frenzel, 1990). Allerdings könnten hier
andere Arten der Sediment-Infauna die Rolle der Micropsectra-Larven übernehmen, z.B.
Oligochaeten. Für Oligochaeten, insbesondere die Tubificiden, konnte gezeigt werden, dass
sie die Schichtung der Sedimente durch ihre Aktivitäten verändern. Am häufigsten findet man
sie in 2 bis 4 cm Sedimenttiefe, sie können jedoch auch bis zu 15 cm Sedimenttiefe
vorgefunden werden (Wetzel, 2001).
Der Einfluss solcher Resuspensions- und Belüftungsereignisse erstreckt sich
wahrscheinlich nur auf die obersten Zentimeter des Sediments und nicht auf die untersuchte
Tiefenschicht von 10−11 cm (nicht aktive Tiefenschicht). Zusätzlich kommt hinzu, dass das
Profundalsediment der Tiefenschicht 10−11 cm seit einer viel längeren Zeit anoxischen
Bedingungen ausgesetzt ist als die gleiche Tiefenschicht im Litoralsediment. Trotzdem
konnten sowohl im Litoral- als auch im Profundalsediment ähnliche MOB-Populationen über
die Tiefe hinweg detektiert werden. Die Fähigkeit von MOB über längere Zeiträume ohne
Sauerstoff zu überleben (1. Hypothese) würde ihnen ermöglichen die Zeit des Wartens auf
Sauerstoff (2. Hypothese) zu überbrücken. Beide postulierten Schicksale der MOB in
anoxischen Sedimentschichten könnten miteinander in Verbindung stehen, wobei die
Notwendigkeit ohne Sauerstoff überleben zu können mit der Sedimenttiefe zunimmt und eine
Erklärung für das Vorhandensein von lebenden MOB in einer Sedimenttiefe von 10−11 cm
darstellt. Die Ähnlichkeit der T-RF-Profile in beiden Tiefenstufen deutet darauf hin, dass es
keine speziell angepassten MOB gibt, die anoxische Bedingungen besser überstehen als
andere MOB. Es sollte aber auch berücksichtigt werden, dass die Möglichkeit der Detektion
von DNS toter MOB nicht ausgeschlossen werden konnte, was zu einer gleichzeitigen
Detektion von lebenden, angepassten und toten, nicht angepassten MOB führen kann.

DISKUSSION 53
5.3 Zusammensetzung von MOB-Anreicherungen aus dem
Litoralsediment
Die T-RF-Profile von Anreicherungskulturen in Mikrotiterplatten mit Bussmann- und
Whittenbury-Medium unterschieden sich nur geringfügig. Mit beiden Medien wurde dieselben
MOB angereichert. Geringfügige Unterschiede gab es in den niedrigsten untersuchten
Verdünnungsstufen, in denen die T-RF-Profile der Anreicherungen mit Bussmann-Medium
mehr Fragmente aufwiesen als die T-RF-Profile der Anreicherungen mit Whittenbury-
Medium. Diese zusätzlichen Fragmente konnten mit einer Ausnahme keinem pmoA-Fragment
der Klone zugeordnet werden.
In den höchsten untersuchten Verdünnungsstufen der MOB-Anreicherungen aus dem
Litoralsediment vor Meersburg konnten nur pmoA-Fragmente der Klongruppe B8a (Typ II-
MOB) nachgewiesen werden. In den höchsten untersuchten Verdünnungsstufen der MOB-
Anreicherungen aus dem Litoralsediment des Litoralgartens konnten zusätzlich zu pmoA-
Fragmenten der Klongruppe B8a zwei weitere Fragmente mit großer Fluoreszenzintensität
(<48 bp und 87/88 bp) nachgewiesen werden. Diese Ergebnisse zeigen, dass Anreicherungen
mit Whittenbury- und Bussmann-Medium immer auf MOB der Klongruppe B8 (Typ II-MOB)
selektieren. Dafür sprechen auch die Untersuchungen der MOB-Isolate aus früheren
Anreicherungen mit Whittenburry- und Bussmann-Medium (Quelle: Litoralgarten), die
ebenfalls der Klongruppe B8 zuzuordnen sind. Die Klongruppe B8 wurde allerdings nicht in
dem T-RF-Profil des Litoralsediments (0−1 cm) nachgewiesen, was auf eine generell geringe
Abundanz dieser MOB im Litoralsediment hindeutet.
Zusätzlich wurden in den höchsten untersuchten Verdünnungsstufen aus dem
Litoralgarten Fragmente detektiert (53 bp und 87/88 bp), die auch in dem T-RF-Profil des
Litoralsediments vorhanden waren. Einerseits können diese Fragmente dadurch eindeutig den
MOB zugeordnet werden und andererseits zeigt dies, dass mit Whittenbury- und Bussmann-
Medium auch MOB angereichert werden, die möglicherweise dominierenden MOB-Arten im
Litoralsediment angehören.
Weiterhin wurden mit der T-RFLP-Methode MOB-Anreicherungen aus einem
gegenläufigen Gradientensystem mit Methan und Sauerstoff untersucht. Das T-RF-Profil der
Gradientenkultur U hatte zu dem T-RF-Profil des Litoralsediments größere Ähnlichkeit als die
T-RF-Profile der Anreicherungen in Mikrotiterplatten. Dies zeigt, dass in einem
gegenläufigen Gradientensystem aus Methan und Sauerstoff sich die dominierenden Vertreter

DISKUSSION 54
der natürlichen MOB-Population besser kultivieren lassen als in Anreicherungen in
Mikrotiterplatten. Zusätzlich traten in dem T-RF-Profil der Gradientenkultur Fragmente auf,
die weder im T-RF-Profil des Litoralsediments, noch in den T-RF-Profilen der
Anreicherungskulturen in Mikrotiterplatten festgestellt wurden. Diese Restriktionsfragmente
konnten auch keinen pmoA-Fragmenten von Klonen dieser Studie oder veröffentlichten
pmoA-Sequenzen zugeordnet werden und repräsentieren möglicherweise neue methanotrophe
Bakterien.
5.4 Evaluation der verwendeten Methoden
In dieser Arbeit wurde die Gruppe der MOB mit molekularbiologischen Methoden untersucht.
Im Folgenden werden die eingesetzten Methoden kritisch diskutiert und Vergleiche gezogen,
wenn für eine Methode mehrere Vorgehensweisen praktiziert wurden.
Das Ziel des ersten Teilprojekts dieser Arbeit war es die Diversität von pmoA-Genen
mit einer Klonbibliothek zu erfassen. Das Erstellen einer Klonbibliothek ist eine selektive
Methode, wobei die einzelnen Teilschritte DNS-Extraktion, DNS-Aufreinigung, PCR und
Klonierung alle zur Selektivität beitragen können. Für die DNS-Extraktion wurde ein
mechanischer Aufschluss der Zellen durch einen Bead Beater kombiniert mit einem
chemischen Aufschluss der Zellen durch Detergenzien verwendet. Diese Strategie wurde
gewählt, weil in einem Vergleich von verschiedenen DNS-Extraktions-Methoden, diese
Vorgehensweise den Aufschluss der meisten bakteriellen Zellen zur Folge hatte (92 %) (More
et al., 1994; Miller et al., 1999). Der Nachteil dieser Methode ist, dass Zellen der kleinsten
Größenfraktion (0,3−1,2 µm Länge) durch diesen Aufschluss nicht zerstört werden (More et
al., 1994). Auf einen Lysozymverdau vor dem mechanischen Aufschluss der Zellen wurde
aufgrund einer möglichen Verschlechterung der DNS-Ausbeute verzichtet (Miller et al.,
1999).
In dieser Arbeit wurde die beschriebene DNS-Extraktion in zwei Ansätzen (A und B)
variiert. In Ansatz A wurde der NucleoSpin Food Kit verwendet. Dabei wurden die Zellen
mechanisch durch einen Bead Beater und chemisch durch Guanidinhydrochlorid und
Detergenzien zerstört und die Proteine anschließend durch Proteinase K verdaut. In Ansatz B
wurden die Zellen ebenfalls mechanisch durch einen Bead Beater und chemisch durch
10%iges SDS zerstört. Die Proteine wurden anschließend durch Ammoniumacetat ausgefällt.
Im weiteren Vorgehen wurden aus beiden DNS-Extrakten wie in Material und Methoden

DISKUSSION 55
beschrieben eine Klonbibliothek erstellt, dabei war die Vorgehensweise in beiden Ansätzen
identisch. Aus diesem Grund wurde für einen Vergleich der beiden Extraktionsmethoden das
Vorhandensein von Klonen beider Ansätze in den verschiedenen Klongruppen (Gruppen
B1−B9) als Vergleichsmittel benutzt. In fünf der neun Gruppen waren Klone aus beiden
Ansätzen vertreten. Drei Klongruppen enthielten nur Klone des Ansatzes B (davon bestanden
zwei Gruppen nur aus einem Klon) und eine Klongruppe (bestand aus einem Klon) enthielt
nur einen Klon aus Ansatz A. Der Vergleich beider Methoden zeigte einen großen
Überlappungsbereich der nachgewiesenen pmoA-Gene. Durch den Einsatz beider DNS-
Extraktionsmethoden konnte jedoch eine größere Diversität an pmoA-Genen entdeckt werden.
Für die Reinigung der DNS-Extrakte von PCR-inhibierenden Stoffen, insbesondere
Huminstoffen, wurden PVPP-gefüllte Säulen verwendet. Bei diesem Schritt geht ein Teil der
DNS-Menge verloren. Nach der Aufreinigung wird die DNS als Template in einer PCR
verwendet. Welche Teile der DNS amplifiziert werden und in welchen Mengen ist abhängig
von den PCR-Bedingungen (Annealing-Temperatur, Salzkonzentrationen usw.) und den
verwendeten Primern. Die PCR-Bedingungen bestanden aus dem Standardpuffer für die Taq
DNS-Polymerase von MBI Fermentas und einer Touch-Down-PCR. Die Touch-Down-PCR
besitzt den Vorteil, dass die Annealing-Temperatur in den ersten PCR-Zyklen stufenweise
herabgesetzt wird und somit in den ersten PCR-Zyklen nur sehr spezifische PCR-Produkte
amplifiziert werden, die dadurch in den folgenden PCR-Zyklen eine vergrößerte Menge an
spezifischem DNS-Template gegenüber unspezifischen DNS-Template darstellen.
Gleichzeitig werden durch die Touch-Down-PCR optimale Annaeling-Temperaturen für
unterschiedliche spezifische DNS-Templates gewährleistet.
Als Startoligonukleotide der PCR wurden die pmoA/amoA-spezifischen Primer A189
und A682 benutzt. Für die Amplifikation von pmoA-Fragmenten sind noch zwei weitere
Primer-Paare beschrieben, A189-A650 und A189-mb661 (Costello und Lidstrom, 1999;
Bourne et al., 2001). In einer vergleichenden Studie dieser drei Primer-Paare konnte gezeigt
werden, dass mit jedem Primer-Paar teilweise unterschiedliche pmoA-Fragmente aus DNS-
Extrakten von Böden amplifiziert wurden, wobei die Primer A189-mb661 die größte
Diversität erfassten, mit den Primern A189-A682 mehr amoA- als pmoA-Fragmente
amplifiziert wurden und mit den Primern A189-A650 zum großen Teil andere pmoA-
Fragmente amplifiziert wurden, als mit den Primern A189-mb661 (Bourne et al., 2001). Im
Gegensatz zu dieser Vergleichsstudie wurde in dieser Arbeit mit dem Primer-Paar A189-

DISKUSSION 56
A682 eine große Diversität der pmoA-Gene erfasst und es wurden kein amoA-Fragment
amplifiziert. Zusätzlich kommt hinzu, dass mit den Primer-Paaren A189-A650 und A189-
mb661 kürzere pmoA-Fragmente amplifiziert werden, als mit dem Primer-Paar A189-A682,
was sich negativ auf die phylogenetische Analyse der pmoA-Gene auswirkt. Der Zugewinn an
phylogenetischer Information war ein wichtiger Faktor für den Einsatz des Primer-Paars
A189-A682. Eine zusätzliche Überprüfung der pmoA-Diversität mit den Primer-Paaren A189-
A650 und A189-mb661, z.B. mit einer T-RFLP-Analyse, wäre jedoch sinnvoll und sollte in
zukünftigen Experimenten nachgeholt werden.
Der letzte Teilschritt zum Erstellen einer Klonbibliothek ist das Klonieren selbst. Auch
hier kann eine Selektion bei dem Einbau der PCR-Produkte in die Vektorplasmide und bei der
Aufnahme der Vektorplasmide in kompetente Escherichia coli-Zellen vorkommen. Diese
Selektion erflogt nach dem Zufallsprinzip und ist nicht von der Sequenz des PCR-Produkts
abhängig.
Nach dem Erstellen der Klonbibliothek wurden die Insertionen der Klone einer RFLP-
Analyse unterzogen. Die RFLP-Methode basiert auf einem Fingerprint-Prinzip, welches den
Vorteil der schnellen Analyse von vielen Klonen hat. Die entstehenden Fragmentmuster
erlauben jedoch keine genaue phylogenetische Gruppeneinteilung. Um die Aussagekraft der
RFLP-Methode zu überprüfen, wurden von jeder RFLP-Gruppe mindestens die Hälfte der
Vertreter sequenziert. Innerhalb der RFLP-Gruppen unterschieden sich die Insertionen bis auf
eine Ausnahme um maximal 9 %. Da die pmoA-Fragmente dieser Studie auf maximal 10 %
Sequenzunterschied basierend in Gruppen eingeteilt wurden, hätte eine Sequenzierung von
einem, maximal zwei Vertretern einer RFLP-Gruppe dieselbe Aussagekraft gezeigt.
Der zweite Teil dieser Arbeit wurde mit der T-RFLP-Methode durchgeführt. Die DNS-
Extrakte für die T-RFLP-Analysen wurden mit denselben Methoden, wie die DNS-Extrakte
im Ansatz B für die Klonbibliothek hergestellt. Der einzige Unterschied bestand in der Art der
Proteinfällung, die in der DNS-Extraktion für die T-RFLP-Analyse mit einem
Phenol/Chloroform/Isoamylalkohol-Gemisch (25:24:1) durchgeführt wurde. Diese Variante
der DNS-Extraktion wurde aufgrund der effektiveren DNS-Extrakt-Aufreinigung und der
möglichen Entfernung lipophiler, PCR-inhibierender Substanzen gegenüber dem
Extraktionsprotokoll von Ansatz B bevorzugt. Diese DNS-Extrakte ließen sich jedoch nicht
mit den Standardbedingungen der Taq DNS-Polymerasen von MBI Fermentas oder Eppendorf
amplifizieren. Deshalb wurde das FailSafe PCR PreMix-System von Epicentre (PCR-Puffer

DISKUSSION 57
und DNS-Polymerase) benutzt. Der Nachteil an diesem PCR PreMix ist, dass die
Zusammensetzung der PCR-Puffer dieses Kit-Systems nicht bekannt ist. Ob die Probleme der
Amplifikation auf den unterschiedlichen DNS-Extraktions-Methoden oder auf Unterschieden
in der Beschaffenheit des Wintersediments (Klonbibliothek) und Sommersediments (T-RFLP)
beruhen, wurde noch nicht überprüft.
Die T-RFLP-Methode ist seit einigen Jahren eine etablierte Fingerprint-Methode und
wurde schon in vielen Arbeiten für Diverstätsstudien mit 16S rRNA-Genen (Liu et al., 1997;
Lueders und Friedrich, 2000; Lukow et al., 2000) und funktionellen Genen, wie dem pmoA-
Gen, benutzt (Horz et al., 2000; Horz et al., 2001; Horz et al., 2002). Die Ergebnisse dieser
Arbeit haben gezeigt, dass die Analyse von T-RF-Profilen funktioneller Gene, wie sie bisher
durchgeführt wurde, überarbeitet werden muss. Ein entscheidender Schritt der T-RFLP ist der
Restriktionsverdau des PCR-Produktes. Bei allen auf einem Restriktionsverdau basierenden
Fingerprint-Methoden ging man bisher von einem vollständigem Restriktionsverdau aus. Die
Überprüfung, ob der Restriktionsverdau wirklich vollständig war, wurde vernachlässigt. Die
T-RFLP-Analyse von Klonen dieser Studie zeigte, dass kein vollständiger Restriktionsverdau
der Klonsequenzen erzielt werden konnte, sobald eine Sequenz mehrere
Restriktionsschnittstellen aufwies. Die Ursachen des unvollständigen Restriktionsverdaus
wurden in dieser Arbeit gründlich untersucht, doch weder Modifikationen der PCR-
Bedingungen, Reinheit des PCR-Produktes, Modifikationen der Restriktionsbedingungen,
verschiedene Restriktionsendonukleasen oder Restriktionsendonukleasen verschiedener
Hersteller führten zu einem vollständigem Restriktionsverdau. Eine T-RFLP-Analyse mit
einem alternativen Farbstoff als Fluoreszenzmarkierung [6-Carboxyfluorescein (FAM)] und
eine darauf folgende Analyse in dem ABI Sequenzer 373A führten ebenfalls zu einem
unvollständigen Restriktionsverdau (Arbeitsgruppe von PD M.Friedrich, MPI für
Terrestrische Mikrobiologie).
Bei T-RFLP-Analysen von 16S rRNA-Genen hat man ebenfalls einen unvollständigen
Restriktionsverdau beobachtet (Egert und Friedrich, in Vorbereitung). Hier waren die Ursache
des unvollständigen Restriktionsverdaus unvollständig amplifizierte PCR-Produkte mit
überhängenden Einzelstrangbereichen, in denen das Restriktionsenzym nicht schneiden
konnte. Nach einem Verdau der einzelsträngigen Bereiche der PCR-Produkte mit Mung Bean
Nuclease konnte ein vollständiger Restriktionsverdau erzielt werden. Ein Verdau mit Mung
Bean Nuclease wurde auch in dieser Studie durchgeführt. Allerdings konnte kein vollständiger

DISKUSSION 58
Restriktionsverdau erzielt werden, was auf andere Ursachen, als einzelsträngige Bereiche von
PCR-Produkten, schließen lässt.
Aufgrund des unvollständigen Restriktionsverdaus müssen Fragmente, die aus
sekundären Schnittstellen hervorgehen, in der Interpretation von T-RF-Profilen berücksichtigt
werden. Dies erschwert die Analyse von T-RF-Profilen, weil es zu Überlappungen von
Fragmenten, die aus primären und sekundären Schnittstellen resultieren, kommen kann und
dadurch eine eindeutige Zuordnung von Sequenzen unmöglich macht. Um dies zu vermeiden,
muss ein Restriktionsenzym verwendet werden, welches die zu untersuchenden DNS-
Fragmente so verdaut, dass alle Sequenzgruppen durch unterschiedliche primäre Schnittstellen
repräsentiert sind und es zu keiner Überlappung von primären und sekundären Schnittstellen
unterschiedlicher Sequenzen kommt.
In dieser Studie wurde das Restriktionsenzym MspI verwendet, weil es für die
Klongruppen B1−B9 die geforderten Anforderungen erfüllt. Allerdings konnte mit MspI nur
zwischen drei zusammengefassten Übergruppen der einzelnen Klongruppen unterschieden
werden. Dabei war eine Übergruppe (Klongruppe B1a) durch das ungeschnittene PCR-
Produkt repräsentiert. Dies stellt einen Unsicherheitsfaktor dar, weil die Möglichkeit besteht,
dass PCR-Produkte anderer Klongruppen trotz vorhandener Schnittstellen zum Teil ebenfalls
überhaupt nicht verdaut werden (siehe T-RF-Muster Klon B24, Abb. 15). Zusätzlich wurden
in den T-RF-Profilen der Sedimentanalysen und Anreicherungskulturen Fragmente detektiert,
die keinen Klongruppen zugeordnet werden konnten. Da die Sequenzen, die diese Fragmente
repräsentieren, nicht bekannt sind, ist eine Überlappung von primären Schnittstellen der
Klonsequenzen und sekundären Schnittstellen der unbekannten Sequenzen möglich. Eine
Überlappung von primären Schnittstellen unbekannter Sequenzen mit sekundären
Schnittstellen der Klonsequenzen ist ebenso denkbar.
Mit den gegebenen Bedingungen konnten die verschiedenen MOB-Populationen des
Litoral- und Profundalsediments sowie der Anreicherungskulturen durchaus miteinander
verglichen werden. Jedoch konnten nur Aussagen darüber getroffen werden, inwieweit sich
die T-RF-Profile der einzelnen MOB-Populationen ähneln oder unterscheiden. Worin die
einzelnen Unterschiede in der Zusammensetzung der verschiedenen MOB-Populationen
bestanden, konnte nicht eindeutig nachgewiesen werden. Ebenfalls sollten sich ähnelnde
T-RF-Profile kritisch betrachtet werden, da die Auflösung von MspI zwischen den einzelnen
Klongruppen sehr gering war. Ein in silico-Test aller kommerziell erhältlicher

DISKUSSION 59
Restriktionsenzyme zeigte, dass die Auflösung zwischen den einzelnen Klongruppen durch
einen Doppelverdau mit den Restriktionsenzymen AluI und MlsI verbessert werden kann (fünf
Übergruppen aus den neun Klongruppen). Eine T-RFLP-Analyse mit diesen
Restriktionsenzymen könnte die Ergebnisse dieser Arbeit verifizieren.
Die T-RFLP-Methode stellt eine Fingerprint-Methode dar mit der Bakterien-
populationen unter geringem Zeitaufwand miteinander verglichen werden können. Wie groß
der Informationsgehalt des Vergleichs ist, hängt davon ab, wie gut das eingesetzte
Restriktionsenzym bzw. die eingesetzten Restriktionsenzyme (Doppelverdau) zwischen den
einzelnen Gruppen der jeweiligen Bakterienpopulationen unterscheiden können und wie viele
Gruppen der jeweiligen Bakterienpopulation bekannt sind. Der Informationsgehalt kann durch
einen zweiten T-RFLP-Ansatz mit einem anderen Restriktionsenzym bzw. einer Kombination
von anderen Restriktionsenzymen verbessert werden. Es sollte beachtet werden, dass die
T-RFLP-Methode einer Selektion der DNS-Extraktion und der PCR unterliegt. Die Selektion
der PCR wurde auch in dieser Arbeit untersucht und führte in manchen Fällen zu einer starken
Diskriminierung bzw. zu keiner Amplifikation bestimmter DNS-Templates während der PCR
(siehe Abb. 16). Die Ergebnisse einer T-RFLP-Analyse sollten deshalb immer durch DNS-
Extraktions- und PCR-unabhängige Methoden bestätigt werden, z.B. mit der Fluoreszenz-in-
situ-Hybridisierung (FISH) und durch Kultivierungsansätze. Eine Abschätzung von
Abundanzen verschiedener Bakteriengruppen durch eine quantitative Analyse der T-RF-
Profile wird aufgrund der Selektion durch die DNS-Extraktion und PCR sowie der
Problematik der sekundären Schnittstellen nicht empfohlen. Allerdings könnte man die
quantitative Information der T-RF-Profile für die Berechnung von einem Vergleichsindex [z.
B. Morisita-Index, (Dollhopf et al, 2001)] benutzen, um die Ähnlichkeit bzw. Verschiedenheit
verschiedener T-RF-Profile besser vergleichen zu können. Dies würde eine sinnvolle
Interpretation schwer analysierbarer T-RF-Profile ermöglichen. Zum Beispiel könnte man
untersuchen, wie stark sich die MOB-Populationen zwischen Sommer und Winter
unterscheiden.

DISKUSSION 60
5.5 Ausblick
Im Folgenden wird auf Aspekte dieser Arbeit eingegangen, von denen es sich lohnen würde
sie weiter zu untersuchen bzw. zu ergänzen.
1) Die untersuchte Diversität der pmoA-Gene sollte durch Experimente mit den Primer-
Paaren A189-A650 und A189-mb661 ergänzt werden, z. B. durch einen Vergleich der
verschiedenen Primer-Paare mit einer T-RFLP-Analyse. Wenn sich Unterschiede ergeben,
müssten neue Klonbibliotheken mit den jeweiligen Primer-Paaren konstruiert werden.
2) Die T-RFLP-Analysen der Sedimente sollten durch einen T-RFLP-Ansatz mit den
Restriktionsenzymen AluI und MlsI verifiziert werden.
3) Die T-RFLP-Analysen des Litoral- und Profundalsediments wiesen unter anderem
dominierende Peaks mit 87/88 bp und 113 bp auf, die durch keinen Vertreter der
Klonbibliothek repräsentiert waren. Es wäre von Interesse die von diesen Fragmenten
repräsentierten Sequenzen zu finden. Dazu müsste eine neue Klonbibliothek angelegt
werden und mit der RFLP-Methode gezielt nach Fragmentmustern gesucht werden, die
Fragmente dieser Länge enthalten. Vertreter dieser RFLP-Gruppen müssten sequenziert
und die Sequenzen einer T-RFLP-Analyse unterzogen werden, um einen Vergleich mit den
Fragmenten der Sediment-T-RF-Profile zu ermöglichen.
4) Es wäre ebenfalls von Interesse zu untersuchen, ob die MOB homogen oder heterogen im
Sediment verbreitet sind und ob es Unterschiede in der Zusammensetzung der MOB-
Populationen über die Jahreszeiten hinweg gibt. Dazu könnte man vergleichende Analysen
mit der T-RFLP-Methode durchführen
5) In der Diskussion wurde bereits erwähnt, das Ergebnisse von T-RFLP-Analysen durch
PCR-unabhängige Methoden bestätigt werden sollten. FISH bietet sich als solch eine
Methode an, mit dem Vorteil, dass FISH-Sonden bereits für verschiedene Gruppen von
MOB entwickelt wurden (Eller et al., 2001; Gulledge et al., 2001). Die T-RFLP-Analysen
konnten zeigen, dass sich die MOB-Populationen zwischen 0−1 cm und 10−11 cm
Sedimenttiefe qualitativ nicht unterscheiden. Eine FISH-Analyse könnte diese Ergebnisse
verifizieren und eine Quantifizierung der MOB-Populationen über die Tiefe hinweg
ermöglichen. Weiterhin wäre es von Interesse mit Slurry-Experimenten die potentiellen
Methanabbauraten über die Tiefe des Sediments zu verfolgen. Da man das Alter der
einzelnen Sedimentschichten bestimmen kann, könnte man ermitteln, wie lange MOB-
Populationen ohne Sauerstoff hungern können, ohne zu sterben.

DISKUSSION 61
6) Es gibt viele Hinweise darauf, dass MOB in ihrer natürlichen Umgebung
Störungsereignissen wie auch periodischen Veränderungen ausgesetzt sind. Deshalb wäre
es von Interesse zu erfahren, wie sich solche Ereignisse auf die Aktivität und Physiologie
von MOB auswirken. Eine mögliche Herangehensweise an diese Fragestellung wäre die
Untersuchung definierter MOB-Populationen in künstlichen Gradientensystemen.
Allerdings sind für solche Experimente MOB-Isolate notwendig, die den dominierenden
MOB-Populationen der jeweiligen Sedimente angehören. Die T-RFLP-Analyse der MOB-
Anreicherungskulturen hat gezeigt, dass Gradientenkulturen ein realistischeres Bild der
Sediment-MOB-Populationen wiederspiegeln als Anreicherungskulturen in Mikrotiter-
platten. Solche Gradientenkulturen könnten zu MOB-Isolaten führen, die in Experimenten
mit künstlichen Gradientensystemen verwendet werden könnten.
7) Interessant ist auch die Frage nach der jeweiligen Bedeutung von physikalischen und
biologischen Störungsereignissen für das Sediment. Diese Fragestellung könnte man mit
anderen Arbeitsgruppen, wie der Gewässerphysik oder Fischbiologie zusammen
bearbeiten.

6 ZUSAMMENFASSUNG
Aerobe, methanotrophe Bakterien (MOB) spielen durch ihre Fähigkeit Methan abzubauen
eine wichtige Rolle im globalen Kohlenstoffkreislauf. Sie wirken als biologischer Filter für
das Methan, welches ansonsten in die Atmosphäre entweicht und dort als Treibhausgas
Auswirkungen auf das Klima hat. Aufgrund der großen Menge an Methan, das in
Süßwassersedimenten produziert wird, war es von Interesse die Gruppe der MOB im
Bodenseesediment zu untersuchen.
Diese Arbeit konnte mit Hilfe der pmoA-Gene zeigen, dass im Litoralsediment des
Bodensees eine große Diversität von MOB vorliegt. Aufbauend auf diesem Ergebnis wurden
mit der T-RFLP-Methode die MOB-Populationen zwischen Litoral- und Profundalsediment
verglichen und die Tiefenverteilung von MOB-Populationen zwischen der aktiven Schicht der
aeroben Methanoxidation (inbegriffen in der Tiefenstufe 0−1 cm) und einer nicht aktiven
Schicht der aeroben Methanoxidation (10−11 cm) in den jeweiligen Sedimenten untersucht.
Die MOB-Populationen des Litoral- und Profundalsediments zeigten teilweise
Gemeinsamkeiten und teilweise Unterschiede in ihrer Zusammensetzung. Die Unterschiede in
den MOB-Populationen wurden auf die wesentlich größere Dynamik der Umweltbedingungen
im Litoralsediment im Vergleich zum Profundalsediment zurückgeführt. Sowohl im
Litoralsediment als auch im Profundalsediment waren sich die MOB-Populationen zwischen
den Tiefenstufen der jeweiligen Sedimente sehr ähnlich. Slurry-Experimente zum Messen des
Methanabbaus (Bosse et al., 1993; Sommervertiefungskurs 2002) wiesen potentielle
Methanoxidations-Aktivitäten in anoxischen Sedimentschichten nach. Darauf basierend kann
man davon ausgehen, dass die T-RFLP-Analyse in den verschiedenen Tiefenstufen keine
DNS-Artefakte, sondern lebende MOB detektiert hat.
Eine T-RFLP-Analyse von verschiedenen MOB-Anreicherungsansätzen konnte durch
einen Vergleich mit einer T-RFLP-Analyse des Litoralsediments zeigen, dass sich ein
gegenläufiges Gradientensystem aus Methan und Sauerstoff am besten dazu eignet
dominierende MOB-Arten des Litoralsediments anzureichern.
Für die T-RFLP-Methode als solche konnte gezeigt werden, dass man für die
Interpretation von T-RF-Profilen, nicht nur primäre, sondern auch sekundäre Schnittstellen
der Zielgruppensequenzen berücksichtigen muss. T-RFLP-Analysen sollten deshalb immer
zusammen mit einer Klonbibliothek erstellt und die Klone auf ihre T-RF-Muster untersucht
werden. Nur so wird eine sinnvolle Interpretation von T-RF-Profilen gewährleistet.

7 LITERATURVERZEICHNIS
Bäuerle, E.; Gaedke, U. (1998) Advances in Limnology 53: Lake Constance. Characterization
of an ecosystem in transition (Bäuerle, E.; Gaedke, U.; Eds) Schweizerbart’sche
Verlagsbuchhandlung, Stuttgart. Arch. Hydrobiol.
Berthelet, M.; Whyte, L.G.; Greer, C.W. (1996) Rapid, direct extraction of DNA from soils
for PCR analysis using polyvinylpolypyrrolidone spin columns. FEMS Microbiol. Lett.
138, 17−22.
Bosse, U.; Frenzel, P.; Conrad, R (1993) Inhibition of methane oxidation by ammonium in the
surface layer of a littoral sediment. FEMS Microbiol. Ecol. 13, 123−134.
Bourne, D.G.; McDonald, I.R.; Murrell, J.C. (2001) Comparison of pmoA PCR primer sets as
tools for investigating methanotroph diversity in three danish soils. Appl. Environ.
Microbiol. 67, 3802−3809.
Bussmann, I; Pester, M.; Brune, A., Schink, B. Cultivation and community structure of
methanotrophic bacteria from freshwater sediment. In Vorbereitung.
Cicerone, R.J.; Oremland, R.S. (1988) Biogeochemical aspects of atmospheric methane.
Global Biogeochem. Cycles 2, 299−327.
Costello, A. M.; Lidstrom, M. E. (1999) Molecular characterization of functional and
phylogenetic genes from natural populations of methanotrophs in lake sediments. Appl.
Environ. Microbiol. 65, 5066−5074.
Dedysh, S.N.; Liesack, W.; Khmelenina, V.N.; Suzina, N.E.; Trotsenko, Y.A.; Semrau, J.D.;
Bares, A.M.; Panikov, N.S.; Tiedje, J.M. (2000) Methylocella palustris gen. nov., sp. nov.,
a new methane-oxidizing acidophilic bacterium from peat bogs, representing a novel
subtype of serine-pathway methanotrophs. Int. J. Syst. Evol. Microbiol. 50, 955−969.
Dollhopf, S.L.; Hashsham, S.A., Tiedje, J.M. (2001) Interpreting 16S rDNA T-RFLP Data:
Application of self-organizing maps and principal component analysis to describe
community dynamics and convergence. Microb. Ecol. 42, 495−505
Dunfield, P.F.; Yimga, M.T.; Dedysh, S.N.; Berger, U.; Liesack, W.; Heyer, J. (2002)
Isolation of a Methylocystis strain containing a novel pmoA-like gene. FEMS Microbiol.
Ecol. 41, 17−26.

Literaturverzeichnis 64
Egert, M.; Friedrich, M., Formation of pseudo-T-RFs: a PCR based artifact phenomenon in
T-RFLP analysis. In Vorbereitung.
Eller, G.; Stubner, S.; Frenzel, P. (2001) Group-specific 16S rRNA targeted probes for the
detection of type I and type II methanotrophs by fluorescence in situ hybridisation. FEMS
Microbiol. Lett. 198, 91−97.
Frenzel, P. (1990) The influence of chironomid larvae on sediment oxygen microprofiles.
Arch. Hydrobiol. 119, 427−437.
Frenzel, P.; Thebrath, B.; Conrad, R (1990) Oxidation of methane in the oxic surface layer of
a deep lake sediment (Lake Constance). FEMS Microbiol. Ecol. 73, 149−158.
GenBank-Datenbank. National Center for Biotechnology Information (NCBI).
http://www.ncbi.nlm.nih.gov/
Gulledge, J.; Ahmad, A.; Steudler, P.A.; Pomerantz, W.J.; Cavanaugh, C.M. (2001) Family-
and genus-level 16S rRNA-targeted oligonucleotide probes for ecological studies of
methanotrophic bacteria. Appl. Environ. Microbiol. 67, 4726−4733.
Hanson, R.S.; Hanson, T.S. (1996) Methanotrophic bacteria. Microbiol. Rev. 60, 439−471.
Henckel, T.; Friedrich, M.; Conrad, R. (1999) Molecular analyses of the methane-oxidizing
microbial community in rice field soil by targeting the genes of the 16S rRNA, particulate
methane monooxygenase, and methanol dehydrogenase. Appl. Environ. Microbiol. 65,
1980−1990.
Henckel T.; Jäckel, U.; Conrad R. (2001) Vertical distribution of the methanotrophic
community after drainage of rice field soil. FEMS Microbiol. Ecol. 34, 279−291.
Henckel, T.; Jäckel, U.; Schnell, S. and Conrad, R. (2000) Molecular analyses of novel
methanotrophic communities in forest soil that oxidize atmospheric methane. Appl.
Environ. Microbiol. 66, 1801−1808.
Heyer, J.; Galchenko, V.F.; Dunfield, P.F. (2002) Molecular phylogeny of type II methane-
oxidizing bacteria isolated from various environments. Microbiology 148, 2831−46.
Holland, S.M., University of Georgia, Athens: Analytic Rarefaction Software.
http://www.uga.edu/strata/Software/.html

Literaturverzeichnis 65
Holmes, A.J.; Costello, A.; Lidstrom, M.E.; Murrell, J.C. (1995) Evidence that particulate
methane monooxygenase and ammonia monooxygenase may be evolutionarily related.
FEMS Microbiol. Lett. 132, 203−208.
Horz, H.; Raghubanshi, A.S.; Heyer, J.; Kammann, C.; Conrad, R.; Dunfield, P.R. (2002)
Activity and community structure of methane-oxidising bacteria in a wet meadow soil.
FEMS Microbiol. Ecol. 41, 247−257.
Horz, H.P.; Rotthauwe, J.H.; Lukow, T.; Liesack, W. (2000) Identification of major subgroups
of ammonia-oxidizing bacteria in environmental samples by T-RFLP analysis of amoA
PCR products. J. Microbiol. Methods. 39, 197−204
Horz, H.P.; Yimga, M.T.; Liesack, W. (2001) Detection of methanotroph diversity on roots of
submerged rice plants by molecular retrieval of pmoA, mmoX, mxaF, and 16S rRNA and
ribosomal DNA, including pmoA-based terminal restriction fragment length polymorphism
profiling. Appl. Environ. Microbiol. 67, 4177−4185.
Kajan, R.; Frenzel, P. (1999) The effect of chironomid larvae on production, oxidation and
fluxes of methane in a flooded rice soil. FEMS Microbiol. Ecol. 28, 121−129.
Liu, W.T.; Marsh, T.L.; Cheng, H.; Forney, L.J. (1997) Characterization of microbial diversity
by determining terminal restriction fragment length polymorphisms of genes encoding
16S rRNA. Appl. Environ. Microbiol. 63, 4516−4522.
Ludwig, W; Strunk, O. (1996) ARB: A software environment for sequence data.
http://www.mikro.biologie.tu-muenchen.de/pub/ARB/documentation/arb.ps
Lueders, T.; Friedrich, M. (2000) Archaeal population dynamics during sequential reduction
processes in rice field soil. Appl. Environ. Microbiol. 66, 2732−2742.
Lukow, T.; Dunfield P. F.; Liesack W. (2000) Use of the T-RFLP technique to assess spatial
and temporal changes in the bacterial community structure within an agricultural soil
planted with transgenic and non-transgenic potato plants. FEMS Microbiol. Ecol. 32,
241−247.
Miller, D.N.; Bryant, J.E.; Madsen, E.L.; Ghiorse, W.C. (1999) Evaluation and Optimization
of DNA extraction and purification procedures for soil and sediment samples. Appl.
Environ. Microbiol. 65, 4715−4724.

Literaturverzeichnis 66
Moré, M.I.; Herrick, J.B.; Silva, M.C.; Ghiorse, W.C.; Madsen, E.L. (1994) Quantitative cell
lysis of indigenous microorganisms and rapid extraction of microbial DNA from sediment.
Appl. Environ. Microbiol. 60, 1572−1580.
Müller, G. (1966) Die Sedimentbildung im Bodensee. Naturwissenschaften 53, 237−247.
Müller, G. (1967) Beziehungen zwischen Wasserkörper, Bodensediment und Organismen im
Bodensee. Naturwissenschaften 54, 454−466.
Pacheco-Oliver, M.; McDonald, I.R., Groleau, D.; Murell, J.C.; Miguez, C.B. (2002)
Detection of methanotrophs with highly divergent pmoA genes from arctic soils. FEMS
Microbiol. Lett. 209, 313−319.
Roslev, P.; King, G.M. (1995) Aerobic and anaerobic starvation metabolism in
methanotrophic bacteria. Appl. Environ. Microbiol. 61, 1563−1570.
Schulz, S.; Conrad, R (1995) Effect of algal decomposition on acetate and methane
concentrations in the profundal sediment of a deep lake (Lake Constance). FEMS
Microbiol. Ecol. 16, 251−260.
Schulz, S.; Conrad, R (1996) Influence of temperature on pathways to methane production in
the permanently cold profundal sediment of Lake Constance. FEMS Microbiol. Ecol. 20,
1−14.
Schulz, S.; Matsuyama, H.; Conrad, R (1997) Temperature dependence of methane production
from different precursors in a profundal sediment (Lake Constance). FEMS Microbiol.
Ecol. 22, 207−213.
Selenska, S.; Klingmüller, W. (1991) DNA recovery and direct detection of Tn5 sequences
from soil. Lett. Appl. Microbiol. 13, 21−24.
Shormann, D.E.; Cotner, J.B. (1997) The effects of benthivoures smallmouth buffalo (Ictiobus
bubalus) on water quality and nutrient cycling in a shallow floodplain lake. J.Lake
Reservoir Res. 13, 270−278
Stolyar, S.; Costello, A.M.; Peeples, T.L.; Lidstrom, M.E. (1999) Role of multiple gene copies
in particulate methane monooxygenase activity in the methane-oxidizing bacterium
Methylococcus capsulatus Bath. Microbiology 145, 1235−44.

Literaturverzeichnis 67
Thebrath, B.; Rothfuss, F.; Whiticar, M.J.; Conrad, R (1993) Methane production in littoral
sediment of Lake Constance. FEMS Microbiol. Ecol. 102, 279−289.
Wetzel R.G. (2001) Limnology. Lake and river ecosystems (Wetzel, R.G.; Ed.) 3. edn.
Academic Press, London.
Whittenbury, R. and Dalton, H. (1981) The methylotrophic bacteria in: The Prokaryotes: A
handbook on the biology of bacteria: ecophysiology, isolation, identification, applications.
(Balows, A.; Trüper, H.G.; Dworkin, M.; Harder, W.; Schleifer, K.-H.; Eds.) Vol. I, pp.
894−902. Springer, New York.
Wuebbles, D.J.; Hayhoe, K. (2002) Atmospheric methane and global change. Earth-Science
Reviews 57, 177(210.
Zehnder, A.J.B. and Stumm, W. (1988) Geochemistry and biogeochemistry of anaerobic
habitats in: Biology of Anaerobic Microorganisms (Zehnder, A.J.B., Ed.) pp. 1−38. Wiley,
New York.
Top Related