







Sprachen
Seiten
Rechtliche


Charakterisierung und rationale Immobilisierung
von Lipasen in biphasischen Reaktionssystemen
Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der
Rheinisch-Westfälischen Technischen Hochschule Aachen zur Erlangung des
akademischen Grades eines Doktors der Naturwissenschaften genehmigte Dissertation
vorgelegt von
Diplom Biologe
Andreas Buthe
aus Steinfurt
Berichter: Universitätsprofessor Dr.-Ing. Winfried Hartmeier Universitätsprofessor Dr.-Ing. Jochen Büchs
Tag der mündlichen Prüfung: 04.07.2006
Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfügbar.


D 82 (Diss. RWTH Aachen)
Shaker VerlagAachen 2006
Berichte aus der Biotechnologie
Andreas Buthe
Charakterisierung und rationale Immobilisierungvon Lipasen in biphasischen Reaktionssystemen

Bibliografische Information der Deutschen NationalbibliothekDie Deutsche Nationalbibliothek verzeichnet diese Publikation in der DeutschenNationalbibliografie; detaillierte bibliografische Daten sind im Internet überhttp://dnb.d-nb.de abrufbar.
Zugl.: Aachen, Techn. Hochsch., Diss., 2006
Copyright Shaker Verlag 2006Alle Rechte, auch das des auszugsweisen Nachdruckes, der auszugsweisenoder vollständigen Wiedergabe, der Speicherung in Datenverarbeitungs-anlagen und der Übersetzung, vorbehalten.
Printed in Germany.
ISBN-10: 3-8322-5516-8ISBN-13: 978-3-8322-5516-9ISSN 1434-4556
Shaker Verlag GmbH • Postfach 101818 • 52018 AachenTelefon: 02407 / 95 96 - 0 • Telefax: 02407 / 95 96 - 9Internet: www.shaker.de • E-Mail: [email protected]

Vorwort
Diese Arbeit entstand im Rahmen meiner Beschäftigung als wissenschaftlicher Mitarbeiter
am Lehrstuhl für Biotechnologie der RWTH Aachen.
Mein besonderer Dank gilt Herrn Prof. Winfried Hartmeier und Frau Dr. Marion Ansorge-
Schumacher für die Überlassung des interessanten Themas und die exzellente Unterstützung
bei der Durchführung dieser Arbeit. Für die Übernahme des Koreferates danke ich Herrn
Prof. Jochen Büchs, der in fruchtbaren Diskussionen maßgeblich zum Gelingen dieser Arbeit
beitrug. Ebenso ist der Deutschen Forschungsgemeinschaft für die finanzielle Förderung
dieser Arbeit innerhalb des SFB 540 “Modellgestützte experimentelle Analyse kinetischer
Phänomene in mehrphasigen fluiden Reaktionssystemen” zu danken.
Des Weiteren möchte ich mich bei Tobias Recker, Lars Wiemann, Anne van den Wittenboer,
Marco Schlepütz, Mathias Klein, Susanne Dreyer, Meike Beer, Thomas Somrak und Alice
Kapitain herzlich bedanken, die im Rahmen ihrer Diplom- bzw. Bachelorarbeiten meine
Ideen mit unermüdlichem Einsatz und großer Begeisterung umsetzten. Darüber hinaus danke
ich dem Deutschen Akademischen Austauschdienst für die im Rahmen des VIGONI-Projektes
und der „Acciones Integrada“ ermöglichten Auslandsaufenthalte sowie in diesem Zusammen-
hang Dr. Paolo Braiuca, Prof. Andrés Alcantara und Prof. Francesco Molinari. Für die
zahlreichen fachlichen Anregungen und kritischen Diskussionen in inspirierender Atmos-
phäre, die mir die italienische und spanische Lebensart näher brachte, gilt mein besonderer
Dank Fabrizio Sibilla und Dr. Pablo Domínguez de Maria. Natürlich profitierte diese Arbeit
auch von der herzlichen Arbeitsatmosphäre am Lehrstuhl für Biotechnologie, wofür allen
Mitarbeitern herzlich gedankt sei.
Ganz besonders möchte ich meine Dankbarkeit gegenüber meinen Eltern betonen, die in
unvergesslicher Art und Weise die Voraussetzungen für ein erfolgreiches Studium geschaffen
haben!


Verzeichnisse
I
Inhaltsverzeichnis
1. Einleitung ..........................................................................................................................1
1.1 Enzyme – wertvolle Werkzeuge in der Biotechnologie .............................................1
1.2 Lipasen .......................................................................................................................3
1.3 Technische Nutzung von Lipasen ..............................................................................7
1.3.1 Einsatzspektrum .................................................................................................7
1.3.2 Reaktionssysteme für nicht-konventionelle Medien ..........................................9
1.4 Rationaler Einsatz von Biokatalysatoren – Immobilisierung ...................................12
1.4.1 Immobilisierungsverfahren...............................................................................12
1.4.2 Immobilisierung von Lipasen...........................................................................15
1.4.3 Probleme beim technischen Einsatz von Lipase-Immobilisaten ......................17
1.5 Aufgabenstellung......................................................................................................20
2. Material und Methoden .................................................................................................22
2.1 Chemikalien/Enzyme ...............................................................................................22
2.2 Geräte .......................................................................................................................23
2.3 Gaschromatographische Analyse .............................................................................23
2.4 Proteinmengenbestimmung nach BRADFORD [1976] ...............................................24
2.5 Massentransfer im nicht-gelstabilisierten Zweiphasensystem .................................25
2.6 pH-Optimum der lipasenkatalysierten Veresterung .................................................26
2.6.1 Titration der Enzymlösungen ...........................................................................26
2.6.2 Nicht-gelstabilisiertes biphasisches Reaktionssystem......................................26
2.6.3 Reihenuntersuchung weiterer Lipasen..............................................................27
2.7 Einfluss der Grenzfläche auf die lipasenkatalysierte Reaktion ................................27
2.7.1 Alginatkugeln verschiedener Größe .................................................................27
2.7.2 Nicht-gelstabilisiertes biphasisches Reaktionssystem......................................29
2.7.3 Gelstabilisiertes biphasisches Reaktionssystem ...............................................31
2.8 „Molecular modelling“ der Enzymoberfläche..........................................................31
2.9 Immobilisierung in Siliconsphären als Statische Emulsion .....................................32
2.10 Immobilisierung nach der Sol-Gel-Technik .............................................................33
2.11 Bestimmung des Durchmessers der Immobilisate ...................................................34
2.12 Charakterisierung der Statischen Emulsion..............................................................36
2.12.1 Quellungsverhalten...........................................................................................36
2.12.2 Abrieb und Elastizität .......................................................................................37

Verzeichnisse
II
2.12.3 Untersuchungen zum Massentransfer .............................................................. 38
2.12.4 Evaporation der emulgierten wässrigen Phase................................................. 39
2.12.5 Ausbluten ......................................................................................................... 39
2.13 Lipasenkatalysierte Synthese von Octyloctanoat..................................................... 39
2.14 Lipasenkatalysierte Synthese von Fettsäureethylhexylestern .................................. 40
2.15 Lipasenkatalysierte Synthese von Propyllaurat ....................................................... 42
2.16 Lipasenkatalysierte Racematspaltung von Benzoin................................................. 42
3. Ergebnisse und Diskussion............................................................................................ 45
3.1 pH-Optimum der lipasenkatalysierten Estersynthese .............................................. 45
3.1.1 Wahl und Charakterisierung eines geeigneten Untersuchungssystems ........... 46
3.1.2 pH-Optima ....................................................................................................... 49
3.1.3 Einfluss der Propionsäurekonzentration auf das pH-Aktivitätsprofil .............. 51
3.1.3.1 pH-Aktivitätsprofil bei drei verschiedenen Gesamtsäurekonzentrationen .. 51
3.1.3.2 Substratkinetik der protonierten Säure am pH-Optimum ............................ 53
3.1.4 Extrapolation der pH-Optima........................................................................... 55
3.1.5 Mechanistische Erklärung für das pH-Optimum der Veresterung................... 58
3.1.6 Praktische Relevanz ......................................................................................... 61
3.2 Bedeutung der Grenzfläche für die lipasenkatalysierte Reaktion............................ 64
3.2.1 Katalytische Aktivität und die spezifische Grenzfläche .................................. 66
3.2.1.1 Enzym-Alginatkugeln – Variation der spezifischen Grenzfläche................. 66
3.2.1.2 Variable spezifische Grenzfläche................................................................. 68
3.2.1.3 Konstante spezifische Grenzfläche............................................................... 72
3.2.1.4 Variable spezifische Grenzfläche im gelstabilisierten Zweiphasensystem .. 75
3.2.1.5 Einfluss der Wasseraktivität bei variabler spezifischer Grenzfläche .......... 77
3.2.1.6 Hydrolyse von Butylpropionat bei variabler spezifischer Grenzfläche ....... 79
3.2.2 „Molecular Modelling“ der Enzymoberfläche................................................. 81
3.2.3 Thermodynamik der Esterbildung im fluiden Zweiphasensystem .................. 85
3.2.4 Hypothese über die Vorgänge an der Grenzfläche .......................................... 88
3.2.5 Praktische Relevanz ......................................................................................... 90
3.3 Entwicklung einer Immobilisierungstechnik ........................................................... 92
3.3.1 Einfluss von Wasser auf die Produktivität biphasischer Reaktionssysteme .... 93
3.3.2 Erzeugung einer Statischen Emulsion in Silicon ............................................. 96
3.3.3 Aktivität von in Statischer Emulsion immobilisierten Lipasen ....................... 99

III
3.3.3.1 Aktivität der Statischen Emulsion...............................................................100
3.3.3.2 Vergleich mit Sol-Gel-Immobilisaten .........................................................102
3.4 Charakterisierung der Statischen Emulsion............................................................105
3.4.1 Katalytische Stabilität.....................................................................................105
3.4.1.1 Wiederverwendbarkeit................................................................................105
3.4.1.2 Ausbluten ....................................................................................................106
3.4.1.3 Lagerung.....................................................................................................107
3.4.2 Abriebstabilität und Elastizität .......................................................................108
3.4.3 Quellung in Hexan..........................................................................................110
3.4.4 Massentransfer und Verteilung.......................................................................111
3.5 Anwendung der Statischen Emulsion für die Synthese von Fein- und
Bulkchemikalien.................................................................................................................117
3.5.1 Lipase von Thermomyces lanuginosa für die Synthese von Esterölen ..........117
3.5.1.1 Sol-Gel ........................................................................................................119
3.5.1.2 Statische Emulsion......................................................................................120
3.5.1.3 Veresterung von Fettsäuren aus Nebenproduktfraktionen der
Biodieselproduktion....................................................................................................121
3.5.2 Lipase B von Candida antarctica für die Synthese von Propyllaurat............123
3.5.3 Lipase von Pseudomonas stutzeri für die Synthese von S-Benzoinbutyrat....127
4. Zusammenfassung ........................................................................................................134
5. Literatur ........................................................................................................................136

Verzeichnisse
IV
Abkürzungsverzeichnis
BSA Rinderserumalbumin (Bovine serum albumin)
BTMS Butyltrimethoxysilan
c Konzentration
CALA Candida antarctica Lipase A
CALB Candida antarctica Lipase B
CRL Candida rugosa Lipase
Da Dalton
df Filmdicke der Kapillarsäule
E Enantiomerie-Selektivitätsparameter
EC Enzymklasse (enzyme class)
e.e.P Enantiomerenüberschuss Produkt [%]
e.e.S Enantiomerenüberschuss Substrat [%]
FAME Fettsäuremethylester
FFA Freie Fettsäuren
FID Flammenionisationsdetektor
FTIR-ATR Fourier-Transform-Infrarot-abgeschwächte
Totalreflexionspektroskopie
HPLC „High performance liquid chromatography“
ID Innerer Durchmesser der Kapillarsäule
immob. immobilisiert
Km Michaeliskonstante
NF Nebenproduktfraktion der Biodieselproduktion
pKS Säuredissoziationskonstante
Prot. Protein
PSL Pseudomonas stutzeri Lipase
PTMS Propyltrimethoxysilan
PVA Polyvinylalkohol
RI-Detektor Brechungsindexdetektor (refractive index detector)
RT Raumtemperatur
SFB Sonderforschungsbereich
sn “stereospecific number”
THF Tetrahydrofuran

V
TLL Thermomyces lanuginosa Lipase
TMOS Tetramethoxysilan
U Unit (μmol Produkt pro Minute)
Upm Umdrehungen pro Minute
V Volumen


Einleitung
1
1. Einleitung
1.1 Enzyme – wertvolle Werkzeuge in der Biotechnologie
Gegenüber der herkömmlichen chemischen Synthese zeichnen sich biokatalytische Systeme
durch grundlegende Vorteile aus [FABER UND FRANSSEN, 1993; ROZELL, 1998; VAN BEILEN
UND LI, 2002]. Vorrangig zu erwähnen sind die hohe Chemo-, Regio- und Stereoselektivität
sowie die milden Reaktionsbedingungen. Ferner treten Nebenreaktionen, wie Isomerisie-
rungen, Racemisierungen und Epimerisierungen seltener oder gar nicht auf [PATEL, 2001;
SCHOEMAKER ET AL., 2003; BOMMARIUS UND RIEBEL, 2004]. Zusammengenommen können
diese Vorteile die Prozessökonomie positiv beeinflussen, weshalb in den letzten Jahren
Forschungsbemühungen hinsichtlich der Applikation von Biokatalysatoren für industrielle
Biotransformationen intensiviert und bereits für viele chemische Reaktionen biokatalytische
Alternativen gefunden wurden. Als besonders bahnbrechend erwies sich dabei die Anfang der
80er Jahre gewonnene Erkenntnis, dass wasserunlösliche hydrophobe Lösungsmittel ge-
eignete Reaktionsmedien für viele enzymkatalysierte Reaktionen darstellen. Die Biokatalyse
in organischen Lösungsmitteln erlaubt die Umsetzung schwer wasserlöslicher oder in rein
wässriger Umgebung instabiler Verbindungen [ZAKS UND KLIBANOV, 1984; KLIBANOV, 1989;
KVITTINGEN, 1994]. Auch aus weiteren Gründen erweist sich die Anwendung organischer
Lösungsmittel als vorteilhaft: Das thermodynamische Gleichgewicht hydrolytischer Re-
aktionen kann umgekehrt und bisweilen ein positiver Einfluss auf die Thermostabilität,
Substrat-, Regio- und Enantioselektivität verzeichnet werden [FITZPATRICK und KLIBANOV,
1991]. Gleichfalls werden Nebenreaktionen sowie mikrobielle Kontaminationen unterdrückt
und die Produktaufreinigung erleichtert. Insbesondere bei der stereoselektiven Synthese von
Feinchemikalien werden diese Vorteile genutzt [BLASER ET AL., 2001]. Neben organischen
Lösungsmitteln kommen auch andere nicht-konventionelle Medien, wie ionische und über-
kritische Flüssigkeiten, zum Einsatz [GARCÍA ET AL., 2004]. Ebenfalls möglich ist die
Umsetzung flüchtiger Substrate in der Gasphase [LAMARE UND LEGOY, 1995]. Laut
STRAATHOF ET AL. [2002] verdoppelte sich in den letzten 10 Jahren die Zahl biokatalytischer
Prozesse auf industrieller Ebene. Angesichts der Vorteile biokatalytischer Systeme ist auch
weiterhin von hohen Zuwachsraten auszugehen.
Klassische biotechnologische Anwendungsbereiche von Enzymen umfassen die Lebens-
mittel-, Futter-, Papier-, Textil- und Waschmittelindustrie [VAN BEILEN UND LI, 2002], jedoch

Einleitung
2
kam es in den letzten Jahren zu einer starken Verlagerung in die pharmazeutische Industrie.
Das Bewusstsein um die Wirkungsunterschiede von Enantiomeren und der damit verbun-
denen Problematik bei Verabreichung racemischer Pharmazeutika sowie ein im Jahre 1992
vom „Committee for Proprietary Medicinal Products“ und der „US Food and Drugs
Administration“ initiiertes Gesetz, veranlassten die pharmazeutische Industrie, neue chirale
Funktionseinheiten enantiomerenrein zu gewinnen und zu charakterisieren [BREUER ET AL.,
2004]. Verglichen mit anderen Chemikalien werden sie im relativ kleinen Maßstab produziert,
allerdings als Hochwertprodukte. Der stetig wachsende Markt für chirale Pharmaka erreichte
1999 zum ersten Mal ein Volumen von 100 Mrd. Dollar [DRÖGE ET AL., 2001]. Aufgrund
ihrer Enantioselektivität eignen sich Enzyme hervorragend und bisweilen gar ausschließlich
für die Synthese von Enantiomeren bzw. für die präparative Auftrennung von Racematen.
Letzteres gilt nicht nur für Pharmaka, sondern kann in gleicher Weise auch für zahlreiche
Vitamine, Agrochemikalien, Geschmacks- und Geruchsstoffe angeführt werden, die ebenfalls
zunehmend enantiomerenrein vertrieben werden [BLASER ET AL., 2001].
Moderne molekularbiologische Methoden, wie das „Metagenome-Screening“, führen fort-
während zur Entdeckung neuer, für die Biotechnologie interessanter Enzyme [LORENZ ET AL.,
2002]. Von den gegenwärtig mehr als 4000 beschriebenen Enzymen dürften jedoch lediglich
etwa 10 % kommerziell verfügbar sein. Aufgrund fehlender Voraussetzungen werden nur
wenige davon in technischen Prozessen eingesetzt [SCHOMBURG UND STEPHAN, 1997;
BOMMARIUS UND RIEBEL, 2004]. Voraussetzung für den technischen Einsatz von Enzymen ist
eine ausreichende katalytische Aktivität, Spezifität und Stabilität sowie die kostengünstige
Verfügbarkeit. Letzteres kann durch Klonierung gewährleistet werden. Hingegen können un-
zureichende Aktivität bzw. Spezifität gegenüber einem Substrat oder den Reaktionsbe-
dingungen der Überführung eines aussichtsreichen biokatalytischen Prozesses auf die
industrielle Ebene entgegenstehen. Abhilfe schaffen kann die gezielte Anpassung des
Biokatalysators an Substrat und/oder Reaktionsbedingungen durch „Protein-Engineering“
[SCHOEMAKER ET AL., 2003] sowie durch Immobilisierung (Kapitel 1.4)
Auffällig ist, dass die Enzymklasse der Hydrolasen – und hier v.a. Lipasen – im Vergleich zu
anderen Enzymklassen mit einem Anteil von über 40 % die industriellen Biotransformationen
dominiert [STRAATHOF ET AL., 2002]. Hierfür ist nicht nur die vielseitige Einsetzbarkeit der
Lipasen verantwortlich, sondern in erster Linie ihre allgemein hohe Aktivität und Robustheit
[SAXENA ET AL., 1999; JAEGER UND EGGERT, 2002].

3
1.2 Lipasen
Innerhalb der Biotechnologie nehmen Lipasen eine herausragende Stellung ein und haben
mittlerweile Proteasen und Amylasen als die am häufigsten eingesetzten Biokatalysatoren
abgelöst [SAXENA ET AL., 1999; STRAATHOF ET AL., 2002]. Lipasen sind der Gruppe der
Triacylglycerin-Hydrolasen (EC 3.1.1.3) zuzuordnen und kommen als extra- und intrazellu-
läre Enzyme ubiquitär in Tieren, Pflanzen und Mikroorganismen vor [BORNSCHEUER UND
KAZLAUSKAS 1999; SAXENA ET AL., 1999; JAEGER UND EGGERT, 2002]. Hauptaufgabe der
Lipasen in vivo ist die Mobilisierung der Energiereserven, indem sie die Hydrolyse von
Triacylglyceriden in Form von Nahrungs- und Reservefetten zu Glycerin, Mono- und Diacyl-
glyceriden unter Freisetzung von Fettsäuren katalysieren. Die Molekulargewichte von
Lipasen bewegen sich im Bereich von 19 bis 70 kDa. Die pH-Optima der Hydrolyse liegen im
leicht alkalischen bis leicht sauren Bereich, bisweilen können aber auch extremere pH- und
Temperaturoptima beobachtet werden [MISSET ET AL., 1994; NEVES-PETERSEN ET AL., 2001a;
GOPINATH ET AL., 2003; ENUJIUGHA ET AL., 2004]. Bevorzugte Substrate sind langkettige,
schwer wasserlösliche Triacylglyceride, wobei Lipasen ebenfalls in der Lage sind, ein breites
Spektrum an kurzkettigen Substraten umzusetzen [SAXENA ET AL., 1999; JAEGER UND
EGGERT, 2002]. Neben der Lipolyse vermögen die meisten Lipasen auch so genannte
Umesterungen (Interesterifikation, Acidolyse, Alkoholyse) und Veresterungen zu katalysieren
[KVITTINGEN, 1994; SAXENA ET AL., 1999]. Auch die Ausbildung und Spaltung von
Amidbindungen kann durch Lipasen katalysiert werden [GOTOR, 1999]. Im Fall nicht
hydrolytischer Reaktionen erfolgt der Einsatz von Lipasen überwiegend in organischen
Lösungsmitteln, um das thermodynamische Gleichgewicht in Richtung der beabsichtigen
Synthese zu verschieben [REHM ET AL., 1998]. Die hohe Attraktivität von Lipasen für
biotechnologische Anwendungen ist durch ihre hohe Stabilität, insbesondere in organischen
Lösungsmitteln, ihre ausgeprägte Regio- und Stereoselektivität, ihr breites Substratspektrum
sowie die Tatsache, dass sie keine Cofaktoren benötigen, begründet [SAXENA ET AL., 1999;
JAEGER ET AL., 1999].
Ein prägnantes Merkmal vieler Lipasen ist die so genannte Grenzflächenaktivierung. Kenn-
zeichnend hierfür ist der sprunghafte Anstieg der lipolytischen Aktivität, sobald das Substrat
bei Überschreitung seiner Löslichkeit in Wasser nicht mehr homogen gelöst, sondern emul-
giert vorliegt [SARDA UND DESNUELLE, 1958; VERGER UND DE HAAS, 1976; VERGER, 1997].
Erklärt werden kann dieses Phänomen durch das Vorhandensein einer -helikalen Struktur,
dem so genannten Lid, welche das aktive Zentrum bedeckt [BRZOZOWSKI ET AL., 1991;

Einleitung
4
DEREWENDA ET AL.,1992; GROCHULSKI ET AL., 1993; KIM ET AL., 1997]. Der dem aktiven
Zentrum zugewandte Bereich dieser Struktur ist hydrophob, der außen liegende hydrophil.
Infolgedessen wird bei Kontakt mit einer „organisch/wässrigen“ Grenzfläche eine Konforma-
tionsänderung induziert, die den Zugang zum aktiven Zentrum freigibt [GROCHULSKI ET AL.,
1993; SAXENA ET AL., 1999; JAEGER ET AL., 1999]. Dabei wird die lipophile Oberfläche des
Enzyms vergrößert und die Anlagerung an die Grenzfläche erleichtert [VASEL ET AL., 1993].
Das Auftreten einer Grenzflächenaktivierung und das Vorliegen einer Lidstruktur galten lange
Zeit als zuverlässige Kriterien zur Abgrenzung der Lipasen von Esterasen. Da aber in den
letzten Jahren Lipasen identifiziert wurden, die (trotz Lidstruktur) keine Grenzflächen-
aktivierung zeigen, ist diese Differenzierung nicht mehr zulässig. Aus diesem Grund wurde
die unterschiedliche Substratspezifität von Lipasen und Esterasen als Klassifizierungs-
kriterium vorgeschlagen. Lipasen (Triacylglycerol-Hydrolasen, EC 3.1.1.3) weisen maximale
Aktivität gegenüber langkettigen (C 12) Acylglyceriden auf, wohingegen „echte“ Esterasen
(Carboxylester-Hydrolasen, EC 3.1.1.1) vorzugsweise die Ester kurzkettiger Carboxylsäuren
(C 12) hydrolysieren [VERGER, 1997; SCHMID UND VERGER, 1998; JAEGER UND REETZ,
1998].
Abbildung 1.1: 3D-Struktur der Lipase von Candida rugosa (CRL) und der Lipase B von Candida antarctica(CALB); -Helices: rot; -Faltblätter: gelb; die -Faltblätter der -Hydrolase-Faltung sind eingekreist, der „nukleophile Ellbogen“ ist durch einen Pfeil gekennzeichnet; erstellt mit Pymol (Version 0.98 DelanoScientific).
Aufgrund ihrer dreidimensionalen Struktur (Abb. 1.1) werden Lipasen den -Hydrolasen
zugeordnet. Gemeinsames Strukturmerkmal dieser Hydrolasen ist die definierte Anordnung
von mindestens fünf parallelen -Faltblättern, welche durch -Helices verbunden sind [OLLIS
CRL CALB

5
ET AL., 1992; SCHRAG UND CYGLER, 1997]. Dabei weisen sie im aktiven Zentrum eine
katalytische Triade bestehend aus den Aminosäuren Serin, Histidin und Aspartat (bisweilen
auch Glutamat) auf [BRADY ET AL., 1990]. Das nukleophile Serin befindet sich in einer Schlei-
fe zwischen einem der mittleren -Faltblätter und der darauf folgenden -Helix, dem so ge-
nannten „nukleophilen Ellbogen“ (Abb. 1.1), für dessen Ausrichtung das konservierte Penta-
peptid-Konsensusmotiv „Gly-X-Ser-X-Gly“ Voraussetzung ist [SCHRAG UND CYGLER, 1997].
Abbildung 1.2: (A) 3D-Struktur der CALB mit Van-der-Waals-Oberfläche, Vergrößerung des aktiven Zentrums mit Butylpropionat und den Aminosäuren der katalytischen Triade, erstellt mit Pymol (Version 0.98 Delano-Scientific); (B) Reaktionsschema der lipasenkatalysierten Hydrolyse.
Ser 105
His 224
Asp 187
A
B
NNH
HO
OO
NNH
HO
OO
NNH
OO
O R2
O
R1
NNH
HO
OO
O R2
O
H
O
R2
O
R2 O H
O
R2 O R1
O
HisSer
Asp
HisSer
Asp
HisSer
Asp
HisSer
Asp
+
+
+ H2O
- R1OH
- -
- -
+
+
tetraedrisches Intermediat Acyl-Enzym-Komplex
tetraedrisches Intermediat
NNH
HO
OO
NNH
HO
OO
NNH
OO
O R2
O
R1
NNH
HO
OO
O R2
O
H
O
R2
O
R2 O H
O
R2 O R1
O
HisSer
Asp
HisSer
Asp
HisSer
Asp
HisSer
Asp
+
+
+ H2O
- R1OH
- -
- -
+
+
tetraedrisches Intermediat Acyl-Enzym-Komplex
tetraedrisches Intermediat

Einleitung
6
Die Hydroxylgruppe des Serin-Restes führt einen nukleophilen Angriff auf das Carbonyl-C-
Atom der Esterbindung durch, wobei die Nukleophilie durch die Wechselwirkung mit dem
Imidazolring eines benachbarten Histidin-Restes gesteigert wird (Abb. 1.2 B). Der Histidin-
Rest wird dabei durch einen deprotonierten Aspartat-Rest stabilisiert [SAXENA ET AL., 1999].
Aufgrund der besonderen Bedeutung des Serin-Restes für die Funktionalität der Lipasen
werden diese auch als Serin-Hydrolasen bezeichnet. Die eigentliche Hydrolyse erfolgt nach
einem Ping-Pong-Bi-Bi-Mechanismus [LORTIE ET AL., 1993]. Als Folge des nukleophilen
Angriffs kommt es zur Ausbildung eines tetraedrischen Intermediats (Oxyanion). Das dabei
freiwerdende Proton wird vom Imidazolring des Histidins übernommen und das negativ
geladene Intermediat durch die Amidgruppen zweier Aminosäuren in der so genannten
Oxyaniontasche stabilisiert [OLLIS ET AL., 1992]. Danach wird das Proton vom Imidazolring
des Histidins auf den Sauerstoff der Esterbindung übertragen und der Alkohol abgespalten.
Durch Abspaltung des Alkohols vom Intermediat entsteht ein Acyl-Enzym-Komplex, in dem
der Serinrest und die Säurekomponente des Esters kovalent verknüpft sind.
Für die Deacylierung des Enzyms ist ein neuerlicher Angriff eines Nukleophils (Wasser bei
Hydrolyse, Alkohol bei Umesterung) erforderlich. Dabei übernimmt Histidin das Proton des
Wassermoleküls bzw. des Alkohols und das stark nukleophile Hydroxid-Ion greift wiederum
den Carbonylsauerstoff der ans Enzym gebundenen Säurekomponente an (Abb. 1.2). Es bildet
sich ein zweites tetraedrisches Intermediat, welches durch Übertragung des Protons von
Histidin auf den Serin-Sauerstoff anschließend in Enzym und Produkt gespalten wird. Eine
Besonderheit in Zusammenhang mit der Produktfreisetzung wurde erst in jüngerer Zeit
erkannt: NEVES-PETERSEN ET AL. [2001a] stellten eine „Katapult-Theorie“ auf, der zufolge das
aktive Zentrum der meisten Lipasen am pH-Optimum der Reaktion eine negative Netto-
ladung aufweist. Das bei der Hydrolyse synthetisierte Säureanion wird elektrostatisch aus
dem Reaktionszentrum heraus „katapultiert“ – das katalytische Zentrum wird frei und kann
erneut ein Substratmolekül binden. Dennoch ist nach Auffassung mancher Autoren die
Produktfreisetzung der geschwindigkeitsbestimmende Schritt der Reaktion [DEREWENDA ET
AL., 1992; DODSON ET AL., 1992; CAMBILLAU UND VAN TILBEURGH, 1993; BORNSCHEUER UND
KAZLAUSKAS, 1999]. Anderen Autoren zufolge ist die Bildung des ersten tetraedrischen
Intermediats entscheidend [KRAUT, 1977; BEER ET AL., 1996].

7
1.3 Technische Nutzung von Lipasen
1.3.1 Einsatzspektrum
Das technische Einsatzspektrum von Lipasen innerhalb der Biotechnologie ist sehr breit ge-
fächert. Grundsätzlich können hierbei zwei Bereiche abgegrenzt werden: (1) Lipasen als
integraler Bestandteil eines Produktes, um diesem spezifische Eigenschaften zu verleihen
(z.B. Waschmittel, Verdauungspräparate oder Kosmetika) oder (2) Lipasen für die Herstel-
lung von Fein- oder Bulkchemikalien. Die Vielseitigkeit der Lipasen bedingt eine unüber-
schaubare Zahl an konkreten Anwendungen, weshalb nachfolgend nur kurz auf die wich-
tigsten Bereiche eingegangen werden kann.
Waschmittelindustrie
Die Waschmittelindustrie ist wirtschaftlich das bedeutendste Anwendungsfeld von Lipasen.
Etwa 1000 Tonnen an Lipase werden schätzungsweise jährlich zu etwa 13 Billionen Tonnen
Waschmittel hinzugefügt [JAEGER UND REETZ, 1998]. Während des Waschvorgangs hydroly-
sieren die Lipasen Fettverschmutzungen und bewirken eine gute Ablösung der Verschmut-
zung von der Textilfaser [SAXENA ET AL., 1999]. Demnach geht es in der Waschmittel-
industrie weniger um die lipasenkatalysierte Herstellung eines Produktes (mit Ausnahme der
Herstellung bestimmter Detergenzien), sondern vielmehr darum, durch Beimengung von
Lipasen dem Produkt spezifische Eigenschaften zu verleihen.
Lebensmittelindustrie
Unter Einsatz von Lipasen werden in der Lebensmittelindustrie überwiegend Aromastoffe
und Glyceride synthetisiert sowie Fette lysiert bzw. modifiziert [WELSH ET AL., 1990;
MALCATA ET AL., 1990; SAXENA ET AL., 1999; GUNSTONE, 1999]. Fette und Öle (Triglyceride)
sind wichtige Bestandteile von Nahrungsmitteln, deren Nährwert und physikalische Eigen-
schaften durch die Position (sn1, sn2 oder sn3), die Länge und den Sättigungsgrad der
Fettsäuren bestimmt werden. Mit Hilfe von Lipasen lassen sich diese Eigenschaften durch
Austausch bestimmter Fettsäuren gezielt modifizieren [JAEGER UND REETZ, 1998; SCHMID
UND VERGER, 1998; SHARMA ET AL., 2001]. Hingegen dient die lipasenkatalysierte Hydrolyse
von in Molkereiprodukten enthaltenen Fetten meist der Aromaverbesserung [SCHMID UND
VERGER, 1998; SAXENA ET AL., 1999]. Einzig Lipasen kommen aufgrund ihrer besonderen
Eigenschaften und aufgrund lebensmittelrechtlicher Bestimmungen für diese Modifikationen
in Frage.

Einleitung
8
Kosmetikindustrie
Innerhalb der Kosmetikindustrie können Lipasen in Präparaten zum Abbau von Hautfetten
eingesetzt werden. Bedeutsamer ist aber ihre Verwendung zur Herstellung von Fettsäure-
estern, die nach Wasser wichtigste Komponente in vielen kosmetischen Produkten. Dort
dienen sie als Emulgatoren, Aromastoffe [MCCRAE ET AL., 1990; IZUMI ET AL., 1997; PANDEY
ET AL., 1999] oder schützen die Haut vor Austrocknung und halten sie glatt, weich und
elastisch [PANDEY ET AL., 1999; HILLS, 2003]. Gerade im Bereich der Synthese von Fettsäure-
estern kommen die eingangs genannten Vorteile enzymkatalysierter Reaktionen zum Tragen.
Konventionell werden Ester unter extremen Bedingungen (hoher Druck; 160 – 240 °C) und
Verwendung von teilweise toxischen Lösungsmitteln und anorganischen Katalysatoren
synthetisiert – was aufgrund strenger Auflagen zum Schutz des Endverbrauchers problema-
tisch ist [HILLS, 2003]. Demgegenüber erfolgt die lipasenkatalysierte Estersynthese unter
milden Reaktionsbedingungen und zumeist in Abwesenheit von Lösungsmitteln [HILLS,
2003]. Die lipasenkatalysierten Verfahren sind demnach weniger energie- und abwasser-
intensiv. Zusätzlich besteht die Möglichkeit der Produktion von Estern, die chemisch nur
unter großem Aufwand herzustellen sind [THUM, 2004]. Verglichen mit chemisch produzier-
ten Estern verleihen enzymatisch synthetisierte Ester dem Endprodukt eine für den Ver-
braucher ansprechende (helle) Farbe, einen natürlicheren Geruch und der Anteil an nicht
umgesetztem Alkohol und sonstigen Nebenprodukten ist geringer [HILLS, 2003; THUM,
2004].
Chirale Feinchemikalien – Synthesen in der pharmazeutischen und agrochemischen Industrie
Die hier von Lipasen fast ausschließlich in organischen Lösungsmitteln katalysierten stereo-
selektiven Transformationen lassen sich in zwei Grundtypen einteilen: (1) Reaktion mit
prochiralen Substraten und (2) kinetische Racematspaltung von Alkoholen und Säuren bzw.
deren Ester [JAEGER UND REETZ, 1998]. So werden Lipasen zur Produktion der S-Enantiomere
von Naproxen und Ibuprofen (anti-inflammatorische Wirkung) sowie zur Synthese chiraler
Intermediate für die Herstellung von Diltiazem (koronardilatatorische Wirkung) und Taxol
(Mitosehemmer) eingesetzt. [CRAIG UND MARIN, 1998; PANDEY ET AL., 1999; SHARMA ET AL.,
2001]. Die regioselektive Acylierung von Steroiden, Zuckern und Zuckerderivaten stellt ein
weiteres Einsatzbereich für Lipasen dar [SCHMID UND VERGER, 1998; SHARMA ET AL., 2001].
Industrielle Anwendung findet auch die Produktion chiraler Amine [JAEGER und REETZ,
1998; HAEFFNER ET AL., 1998]. Gleichfalls wichtig sind Lipasen bei der enantioselektiven
Synthese von Vorstufen für die Herstellung von Herbiziden und Insektiziden [PANDEY ET AL.,

9
1999; SAXENA ET AL., 1999]. Die stetig wachsende Zahl an Publikationen über den Einsatz
von Lipasen für stereoselektive Biotransformationen spiegelt die große Bedeutung dieses
Anwendungsgebietes wider.
Sonstiges
Ebenfalls wichtig sind Lipasen für die Papier- und Holzindustrie [FARRELL ET AL., 1997],
Gerbereien [MUTHUKUMARAN UND DHAR, 1982] und die Umwelttechnik [DAUBER UND
BOEHNKE, 1993]. Darüber hinaus sind Forschungsbemühungen zu verzeichnen, Lipasen für
die Herstellung biologisch abbaubarer Polymere, Schmiermittel und Hydrauliköle sowie
Biodiesel einzusetzen [LINKO ET AL., 1994 und 1998; HAAS UND SCOTT, 1996; PANDEY ET AL.,
1999; WILLING, 1999; BRENNEIS ET AL., 2002 und 2004].
1.3.2 Reaktionssysteme für nicht-konventionelle Medien
Bei Durchführung biokatalytischer Reaktionen in organischen Lösungsmitteln sind grundsätz-
lich mono- und biphasische Reaktionssysteme zu unterscheiden. Bei Suspension des pulver-
förmigen Enzympräparates oder trockener Immobilisate in einem nicht-wassermischbaren
Lösungsmittel spricht man von monophasischen Reaktionssystemen – im englischen Sprach-
gebrauch findet sich auch die Bezeichnung „microaqueous phase system“ [TWEDDELL ET AL.,
1998]. Lipasen gehören zu den ersten Enzymen, die erfolgreich unter Erhalt ihrer kataly-
tischen Aktivität direkt in derartigen Systemen eingesetzt wurden [ZAKS UND KLIBANOV,
1984; SCHMID UND VERGER, 1998]. In dieser Hinsicht unterscheiden sich Lipasen von den
meisten anderen Enzymen und dies mag zu Recht als Hauptgrund für die Dominanz der
Lipasen in industriellen Biotransformationen angesehen werden. Die überwiegende Zahl der
bisher beschriebenen Enzyme ist in unmittelbarer Umgebung organischer Lösungsmittel nicht
aktiv und wird zumeist auch irreversibel inaktiviert [OGINO AND ISHIKAWA, 2001]. Abhilfe
schaffen kann der Einsatz zweiphasiger Reaktionssysteme, in denen empfindlichere Enzyme
in einer diskontinuierlichen wässrigen Phase homogen gelöst sind, die von einem wasserun-
löslichen organischen Lösungsmittel als kontinuierliche Phase umgeben ist. Das Enzym wird
in der wässrigen Phase vor den inaktivierenden Einflüssen des organischen Lösungsmittels
geschützt. Bisweilen kann aber auch durch Kontakt des Enzyms mit einer Grenzfläche eine
Inaktivierung hervorgerufen werden [GHATORAE ET AL., 1994], weshalb sich dann die Ver-
wendung einer gelstabilisierten wässrigen Phase anbietet [METRANGOLO-RUIZ DE TEMINO ET
AL., 2005]. Ein Spezialfall biphasischer Reaktionssysteme sind so genannte Mikroemul-

Einleitung
10
sionen bzw. reverse Mizellen, bei denen durch Zusatz amphiphiler Moleküle und einem genau
abgestimmten Verhältnis der Volumina von organischer und wässriger Phase eine thermody-
namisch stabile Emulsion geschaffen wird. Die diskontinuierlich vorliegende wässrige Phase
weist dabei Durchmesser von wenigen Nanometern auf, weshalb die Mikroemulsion selbst
ein vollständig homogenes Erscheinungsbild bietet [BALLESTEROS ET AL., 1995; CARVALHO
UND CABRAL, 2000].
Das katalytische Verhalten von Lipasen in mono- und biphasischen Reaktionssystemen wurde
eingehend charakterisiert. Dabei konnten entscheidende Reaktionsparameter, wie die Wasser-
aktivität (Maß für die thermodynamische Verfügbarkeit von Wasser) oder der logPOW-Wert
des Lösungsmittels (dekadischer Logarithmus des Verteilungskoeffizienten in einem Octanol-
Wasser-Gemisch) identifiziert werden. Die katalytische Aktivität hängt zumindest in einpha-
sigen Reaktionssystemen sehr stark von der Wasseraktivität ab [VALIVETY ET AL., 1992 und
1994; KVITTINGEN 1994]. Die Rolle der Wasseraktivität in biphasischen Reaktionssystemen
ist von komplexerer Natur, weshalb dieser Aspekt in Kapitel 3.2.3 aufgegriffen und gesondert
diskutiert wird. In einphasigen Reaktionssystemen kann eine zu hohe Wasseraktivität bei Um-
bzw. Veresterung das thermodynamische Gleichgewicht der Reaktion ungünstig beeinflussen
[SVENSSON ET AL., 1994; KVITTINGEN, 1994]. Eine zu niedrige Wasseraktivität kann sich
jedoch negativ auf das Enzym auswirken. Zur Aufrechterhaltung ihrer Hydrathülle (und somit
ihrer Struktur, Flexibilität und Aktivität) benötigen Lipasen eine bestimmte Menge frei ver-
fügbaren Wassers [AFFLECK ET AL., 1992; ADLERCREUTZ, 1994; BROOS ET AL., 1995; REHM ET
AL., 1998]. Niedrige Wasseraktivitäten und die Verwendung wassermischbarer Lösungsmittel
führen zum Entzug der Hydrathülle und infolgedessen u.U. zu einer irreversiblen Inakti-
vierung. Ein gebräuchliches Auswahlkriterium für Lösungsmittel stellt der log POW-Wert dar.
Es wurde gezeigt, dass Lösungsmittel mit log-POW-Werten > 3 gut für lipasekatalysierte
Reaktionen geeignet sind [REHM ET AL., 1998]. Häufig eingesetzte Lösungsmittel sind u.a.
Hexan, Isooctan, Heptan, Vinylacetat und Toluol [REHM ET AL., 1998]. Neben organischen
Lösungsmitteln kommen für Lipasen jedoch noch weitere nicht-konventionelle Reaktions-
medien, wie ionische und überkritische Flüssigkeiten in Frage [SCHÖFER ET AL., 2001;
GARCÍA ET AL., 2004]. Zudem können Lipasen auch flüchtige Substrate in der Gasphase
umsetzen [LAMARE ET AL., 2001; CAMERON ET AL., 2002]. Auf Prozessebene dominiert aber
bislang der Einsatz organischer Lösungsmittel [GARCÍA ET AL., 2004].

11
Die Verwendung organischer Lösungsmittel in der Enzymtechnologie bringt allerdings auch
einige Nachteile mit sich, die insbesondere bei der industriellen Herstellung von Bulkchemi-
kalien die Wirtschaftlichkeit beeinträchtigen können. So erfordern die Toxizität und der gerin-
ge Flammpunkt organischer Lösungsmittel die Einhaltung kostspieliger Auflagen. Deshalb
wird zunehmend versucht, Enzyme in lösungsmittelfreien Systemen einzusetzen, zumal auf
diese Weise die Produktaufarbeitung erheblich vereinfacht werden kann [BOURG-GARROS ET
AL., 1998]. Dabei kann das Enzym in den reinen Substraten suspendiert oder wässrig gelöst
emulgiert werden, warum es sich prinzipiell auch hier um ein- bzw. zweiphasige Reaktions-
systeme handelt. Allerdings zieht die Gegenwart reiner Substrate bei den meisten Enzymen
eine Inaktivierung nach sich. Einzig Lipasen weisen bislang eine ausreichende Stabilität auf,
weshalb mittlerweile eine Reihe von Prozessen zur lösungsmittelfreien Synthese von Fett-
säureestern beschrieben wurden [CHATTERJEE UND BHATTACHARYYA, 1998; KAIEDA ET AL.,
1999; HILLS, 2003; BRENNEIS ET AL., 2002 UND 2004; THUM, 2004]. Aus Sicherheits-, öko-
nomischen, technologischen und umwelttechnischen Gründen ist davon auszugehen, dass der
Einsatz von Lipasen in lösungsmittelfreien Medien bei Synthese von Bulkchemikalien weiter
an Bedeutung gewinnen wird [BOURG-GARROS ET AL., 1998].
Das vielseitige Potential von Lipasen bedingt zwangsläufig, dass hier genannten Reaktions-
systeme in unzähligen Varianten – etwa durch Immobilisierung (Kapitel 1.4) – zur Anwen-
dung kommen, weshalb obige Darstellung keinesfalls Anspruch auf Vollständigkeit erhebt.
Grundsätzlich stellt sich aber die Frage, ob nun mono- oder biphasische Reaktionssysteme für
die technische Nutzung von Lipasen besser geeignet sind. Für Veresterungen konnte gezeigt
werden, dass unter kinetischen Gesichtspunkten in zweiphasigen Systemen eine höhere Effi-
zienz als in einphasigen Systemen erreicht werden kann [BORZEIX ET AL., 1992; LOUWRIER ET
AL., 1996; TWEDDELL ET AL., 1998]. Als Erklärung wird meist die Agglomeration des pulver-
förmigen Enzyms in monophasischen Systemen angeführt, aufgrund derer nicht alle Enzym-
moleküle gleichartig an der Reaktion partizipieren können. Demgegenüber ist das Enzym in
zweiphasigen Reaktionssystemen homogen in der wässrigen Phase gelöst. Auch wenn diese
Erklärung plausibel erscheint, so muss betont werden, dass sie rein empirisch abgeleitet
wurde und eine ganze Reihe von Fragen unbeantwortet bleibt. Dies ist ein generelles Problem
der Biokatalyse bezogenen Forschung, weshalb die technische Nutzung von Biokatalysatoren
zu einem ganz wesentlichen Anteil auf empirischen Grundlagen beruht [STRAATHOF ET AL.,
2002]. Eine tiefgehende Analyse, z.B. auf Ebene des Enzyms oder unter Berücksichtigung der
Verteilungsphänomenologie und des Massentransfers in biphasischen Reaktionssystemen, ist

Einleitung
12
– wenn überhaupt – nur mit großem Aufwand möglich und unterbleibt zumeist [HEINEMANN
ET AL., 2003].
1.4 Rationaler Einsatz von Biokatalysatoren – Immobilisierung
1.4.1 Immobilisierungsverfahren
Der Etablierung biokatalytischer Prozesse auf industrieller Ebene stehen oft die hohen Kosten
für die verwendeten Enzyme entgegen. Diesem Umstand wird Rechnung getragen, indem ver-
sucht wird, Enzyme wiederholt bzw. kontinuierlich einzusetzen. Dies setzt voraus, dass der
Biokatalysator leicht aus dem Reaktionsmedium abgetrennt werden kann. Wird die Reaktion
in einem wässrigen Reaktionsmedium ausgeführt, liegt der Biokatalysator homogen gelöst
vor und die Abtrennung nach Abschluss der Reaktion ist nur unter großem Aufwand möglich.
Hingegen ist in organischen Reaktionsmedien aufgrund der Unlöslichkeit eine Abtrennung
prinzipiell durch Filtration möglich [SAXENA ET AL., 1999]. Der direkte Einsatz von Enzymen
in organischen Lösungsmitteln kommt allerdings nur dann in Frage, wenn der Biokatalysator
ausreichend stabil ist – eine Bedingung, die nur wenige Enzyme, z.B. Lipasen, erfüllen.
Zudem ist die Abtrennung der feinen Enzympartikel durch Filtration im großtechnischen
Maßstab problematisch. Eine einfache und schonende Abtrennung des Biokatalysators aus
dem Reaktionsmedium ist nicht nur in punkto Recycling des Biokatalysators, sondern auch
aus Sicht der Produktivität wünschenswert. Abhilfe schaffen kann die bewusste Hetero-
genisierung des Reaktionsansatzes, indem der Biokatalysator durch Immobilisierung in eine
leichter handhabbare Form überführt und vor denaturierenden Effekten geschützt wird.
Immobilisierung bezeichnet die Einschränkung der Beweglichkeit von Biokatalysatoren durch
chemische oder physikalische Methoden [HARTMEIER, 1986]. Im Idealfall kann durch Immo-
bilisierung eine für den Biokatalysator optimale Mikroumgebung geschaffen werden, was
mitunter in einer Erhöhung der katalytischen Stabilität oder Aktivität resultiert [REETZ ET AL.,
1996]. Darüber hinaus kann bisweilen ein positiver Einfluss der Immobilisierung auf die
Temperatur- und pH-Charakteristik eines Enzyms beobachtet werden [REETZ ET AL., 1997;
MOHY ELDIN ET AL., 2005].
Da die Immobilisierung von Biokatalysatoren einen wertvollen Beitrag zur Kosteneinsparung
leistet, darf mit einiger Berechtigung geschlussfolgert werden, dass die Immobilisierung eine

13
wesentliche Grundvoraussetzung für den rationalen Einsatz von Biokatalysatoren darstellt. Es
überrascht daher nicht, dass heutzutage ca. 80 % der industriellen Biotransformationsprozesse
unter Einsatz von immobilisierten Biokatalysatoren durchgeführt werden [D’SOUZA, 1999;
TISCHER UND WEDEKIND, 1999].
Die Wahl einer Immobilisierungsmethode setzt stets eine Abstimmung zwischen den
chemischen und technischen Besonderheiten des biokatalytischen Prozesses und den Charak-
teristika des Biokatalysators voraus. Grundsätzlich können zwei unterschiedliche Ansätze zur
Immobilisierung unterschieden werden: (1) Die Fixierung durch Kopplung der Biokata-
lysatoren an bestimmte Träger bzw. durch Quervernetzung untereinander oder (2) die
Immobilisierung durch Membranabtrennung oder Einhüllung in bestimmte Matrizes oder
Mikrokapseln [HARTMEIER, 1986]. Nachfolgend näher vorgestellt werden die Trägerbindung
und die Matrixeinhüllung.
Bei der ionisch/adsorptiven Bindung werden die Biokatalysatoren über ionische und/oder
anders geartete Wechselwirkungen an einen Träger (z.B Aktivkohle, Glas oder DEAE-Cellu-
lose) gebunden [NORIN ET AL., 1988; HARTMEIER, 1986]. Die ionische/adsorptive Bindung ist
die älteste und zugleich einfachste Immobilisierungsmethode. Ihr Hauptvorteil besteht in der
enzymschonenden und preisgünstigen Ausführbarkeit. Nachteilig ist die unzureichende Lang-
zeitstabilität – aufgrund der schwachen Bindungen ist mit einer schleichenden Desorption
(Freisetzung) des Biokatalysators im Prozess zu rechnen. Demgegenüber wird durch kova-
lente Bindung eine sehr viel stabilere Immobilisierung erreicht. Dabei werden die Enzyme
über funktionelle Gruppen der Aminosäuren an geeignete Träger (z.B poröses Glas, Dextran
bzw. Sephadex) fixiert [HARTMEIER, 1986; STARK UND HOLMBERG, 1989; MORENO ET AL.,
1997]. Jedoch besteht die Möglichkeit, dass dabei funktionelle Gruppen von Aminosäuren
innerhalb oder in der Nähe des aktiven Zentrums involviert sind, was in der Regel eine
drastische Beeinträchtigung der katalytischen Aktivität zur Folge hat. Deshalb eignet sich
diese Methode nicht für alle Enzyme [TISCHER UND WEDEKIND, 1999]. Eine Kopplung bzw.
Quervernetzung („Crosslinking“) der Enzyme untereinander führt zur Immobilisierung durch
Bildung unlöslicher Aggregate. Die Verknüpfung beruht auf der Ausbildung kovalenter Bin-
dungen zwischen Enzymen und bifunktionellen Agenzien, z.B. Dialdehyden oder Diisocyana-
ten [HARTMEIER, 1986; GRAY ET AL., 1990]. Durch Einschluss zusätzlicher nichtaktiver Kom-
ponenten („Co-Crosslinking“) können die mechanischen und katalytischen Eigenschaften der

Einleitung
14
Enzymaggregate verbessert werden [HARTMEIER, 1986]. Ähnlich wie bei der kovalenten
Bindung kann es durch die Quervernetzung zu drastischen Aktivitätsverlusten kommen.
Beim Einschluss von Enzymen in polymere Matrizes wird der Biokatalysator in ein drei-
dimensionales Netzwerk, z.B. in Form einer Kugel oder Faser, eingebettet. Aufgrund
optimaler hydromechanischer Eigenschaften bei geringer Abriebtendenz werden sphärische
Strukturen bevorzugt [PRÜßE, 2000]. Der Hauptvorteil dieser Methodik besteht darin, dass der
Biokatalysator, etwa bei Einsatz in organischen Lösungsmitteln, vor inaktivierenden Ein-
flüssen des Reaktionsmediums effektiv geschützt wird. Dieser Vorteil kann durch Kopplung
an Träger nicht erreicht werden. Zudem besteht innerhalb gewisser Grenzen die Möglichkeit,
die Beschaffenheit der Matrix an die Bedürfnisse des Enzyms oder auch an die Erfordernisse
der Reaktion selbst anzupassen [TRIANTAFYLLOU ET AL., 1997; KUNCOVA ET AL., 2003]. Der
Nachteil der Matrixeinhüllung besteht in einem eingeschränkten Massentransfer, da die
Immobilisierungsmatrix als Diffusionsbarriere für Substrat und Produkt zu betrachten ist. In
diesem Zusammenhang sind neben Katalysatordichte, Konzentration und Zusammensetzung
der Immobilisierungsmatrix auch Schichtdicke, Form und Größe der Immobilisate von
Relevanz. Die aus der Matrixeinhüllung resultierende Schutzwirkung kompensiert diesen
Nachteil aber zumeist. Als Materialien für die Matrizes kommen natürliche und synthetische
Polymere in Frage. Natürliche Polymere zeichnen sich im Allgemeinen durch eine hohe
Biokompatibilität aus, weshalb nicht mit inaktivierenden Wechselwirkungen zwischen Enzym
und Matrix zu rechnen ist und die resultierenden Immobilisate im Vergleich zu kovalent
gebundenen Enzymen oftmals höhere Aktivitäten aufweisen [MC MORN UND HUTCHINGS,
2003]. Die Vernetzung der natürlichen Polymere erfolgt entweder durch Kältegelierung, d.h.
durch Abkühlen einer Polymerlösung (Agar, Gellan, Gelatine), oder durch ionotrope Gel-
bildung (Alginat, Carrageenan), bei der divalente Kationen die Polymere miteinander vernet-
zen [ANSORGE-SCHUMACHER ET AL., 2002]. Erhalten werden Hydrogele, die sich vorzüglich
für den Einsatz in organischen Lösungsmitteln eignen. Ferner können auch Hydrogele auf
Basis synthetischer Polymere wie PVA und Polyacrylamid generiert werden [ATIA, 2005].
Für den Einsatz in wässrigen Reaktionsmedien sind Hydrogele weniger prädestiniert, da ihre
Netzwerke zumeist grobmaschig und infolgedessen Enzymverluste und insbesondere Verluste
möglicher Cofaktoren unvermeidlich sind [ANSORGE-SCHUMACHER ET AL., 2002]. Bei Ver-
wendung der Immobilisate in einer hydrophoben Umgebung können diese Verluste minimiert
bzw. vollständig ausgeschlossen werden. In Einzelfällen kommt der Einsatz von Hydrogelen
jedoch nicht in Frage, z.B. bei unzureichendem Massentransfer sehr hydrophober Substrate.

15
Eine Verbesserung des Massentransfers kann in diesem Fall durch Verwendung hydrophober
synthetischer Polymere (z.B. Sol-Gele, Silicone, Polyurethane) erreicht werden [REETZ ET
AL., 1996; FERREIRA-DIAS ET AL., 1999; GILL ET AL., 1999; GILL UND BALLESTEROS, 2000a/b].
Zudem können die Eigenschaften der Matrizes auf Basis synthetischer Polymere, z.B. im
Hinblick auf die Maschenweite oder die Balance zwischen Hydrophilie und Hydrophobie,
besser kontrolliert werden [TRIANTAFYLLOU ET AL., 1997; KUNCOVA ET AL., 2003]. Nachteilig
auf die katalytische Aktivität können sich bei Verwendung synthetischer Polymere unspezi-
fische Reaktionen zwischen Enzym und Monomer während der Polymerisation auswirken.
Dem kann bspw. durch Präpolymerisation begegnet werden [KAWAMOTO UND TANAKA,
2003]. Einige synthetische Polymere bzw. deren Monomere sind toxisch und ihr Einsatz in
sensiblen Bereichen, wie der Lebensmittel- oder Kosmetikindustrie, daher ausgeschlossen.
Für viele Polymere (z.B. Silicon) ist hingegen die Unbedenklichkeit bzw. Biokompatibilität
erwiesen [COMPTON, 1997].
1.4.2 Immobilisierung von Lipasen
Aufgrund der großen Bedeutung von Lipasen für biotechnologische Anwendungen ist es nicht
verwunderlich, dass sämtliche zur Verfügung stehenden Immobilisierungstechniken bereits
zum Einsatz gekommen sind. Einen Überblick über die am häufigsten verwendeten Methoden
und Materialien gibt Tabelle 1.1. Offenkundig ist, dass die Trägerbindung eine sehr wichtige
Rolle bei der Immobilisierung von Lipasen spielt. Es ist deshalb nicht verwunderlich, dass es
sich bei den kommerziell verfügbaren Lipase-Immobilisaten meist um trägergebundene
Lipasen handelt. In diesem Zusammenhang zu erwähnen sind die Präparate Novozym 435
(Novozymes, Dänemark) und Chirazyme L-2 (Biocatalytics, U.S.A.), bei denen die Lipase B
aus Candida antarctica elektrostatisch an einen Träger auf Basis von Polymethylacrylat
gebunden ist und industriell Anwendung findet [KIRK UND CHRISTENSEN, 2002; HILLS, 2003;
THUM, 2004]. Kovalent gebundene oder eingehüllte Lipasen sind zwar auch Forschungs-
gegenstand, haben aber bislang noch keine auch nur annähernd vergleichbare kommerzielle
Verbreitung gefunden.
Neben der Trägerbindung der Lipasen wurde bzw. wird auch an der Einhüllung von Lipasen
in natürliche oder synthetische Polymere geforscht. Eine mögliche Immobilisierungsmethode
ist die Einhüllung von Lipasen in eine Hydrogelmatrix. BETIGERI UND NEAU [2002] unter-
suchten lipasehaltige Hydrogele auf Basis von Gellan, Alginat und Agarose für die Hydrolyse

Einleitung
16
von Triglyceriden in einem wässrigen Reaktionsmedium. Andere Autoren [HERTZBERG ET
AL., 1992; KOCH 1997; DOUMÈCHE, 2002; HEINEMANN, 2003] benutzten Hydrogele als gelsta-
bilisiertes biphasisches Reaktionssystem (Alginatkugeln suspendiert in Hexan) und konnten
die Anwendbarkeit in organischen Lösungsmitteln bestätigen. Allerdings werden natürliche
Polymere bzw. die aus ihnen generierten Hydrogele in der Praxis seltener eingesetzt, da die
umzusetzenden, zumeist hydrophoben Substrate schlecht in die Matrix diffundieren können
[BETIGERI UND NEAU, 2002; KRAJEWSKA, 2004].
Tabelle 1.1: Auswahl der für die Immobilisierung von Lipasen benutzten Methoden und Materialien.
Träger bzw. Matrix Literatur
Ionisc
he/ad
sorpt
ive B
indun
g Acrylglas, Alginat, Celite, Cellulose, Decyl-
chloroacetat-Emulsion, Duolite, silanisiertes
Glas, Glaswolle, Kieselgur, Kunstharze, Poly-
propylen, Polyvinylchlorid, Sephadex, Silica-
Gel, Acrylharz
BRADY ET AL., 1988; ISON ET AL., 1988; NORIN ET AL., 1988;
OMAR ET AL., 1988; PRONK ET AL., 1988; KANG UND RHEE,
1989; GRAY ET AL., 1990; KÉRY ET AL., 1990; WELSH ET AL.,
1990; TAYLOR UND CRAIG, 1991; CAO ET AL., 1992;
VALIVETY ET AL., 1992; DE BACKER UND BARON, 1993;
RUCKENSTEIN UND WANG, 1993; KIERKELS ET AL., 1994;
BYSTROM UND HARTEL, 1994; MARTINS ET AL., 1994
Kova
lente
Bind
ung
Agarose, Amberlite, Cellulose, Chitin,
Chitosan, derivatisiertes Glas, Nylon,
Polyacrylamid, Polyethylenglykol, Polyvinyl-
chlorid, Sepharose, Silicat, Eupergit
OMAR ET AL., 1988; STARK UND HOLMBERG, 1989; KERY ET
AL., 1990; GRAY ET AL., 1990; KHURANA UND HO, 1990;
OTERO ET AL., 1990; SHAW ET AL., 1990; MANJON ET AL.,
1991; MORENO ET AL., 1997
Einh
üllun
g
Alginat, Agarose, Gellan, quervernetzbare
Harzpräpolymere, Membranen, Polyamide,
Polyethylenglykol, Poylpropylenglykol,
Polyurethan, Polyvinylchlorid, Silicon, Sol-
Gel, Phyllosilicate, Organogele mit
Mikroemulsion
RUCKA UND TURKIEWICZ, 1990; DIAS ET AL., 1991; CHEN
UND MCGILL, 1992; CHEN UND YANG, 1992; HERTZBERG ET
AL., 1992; REETZ ET AL., 1996, FERREIRA DIAS ET AL., 1999 ;
GILL ET AL., 1999; STAMATIS AND XENAKIS, 1999; GILL UND
BALLESTEROS, 2000a/b ; ZHOU ET AL., 2001; BETIGERI UND
NEAU, 2002 ; DOUMÈCHE ET AL., 2002; HSU ET AL., 2002;
DELIMITSOU ET AL., 2002 ; RAGHEB ET AL., 2003
Aufgrund der Grenzflächenaktivierung von Lipasen bei der Hydrolyse scheint eine sinnvolle
Anwendung von Lipasen innerhalb einer Gelmatrix ohnehin fragwürdig zu sein, da zu
befürchten ist, dass nur an der Oberfläche befindliche und mit der Grenzfläche in Kontakt
stehende Enzyme aktiv sind. Obwohl für die Estersynthese bislang kaum untersucht, ist auch
dort ein ähnliches Verhalten zu erwarten und nur der mit der Grenzfläche in Kontakt stehende
Bruchteil der Enzymmoleküle entfaltet eine nennenswerte Aktivität [DOUMÈCHE, 2002;
HEINEMANN, 2003]. Auf einer Mikroemulsion basierende Organogele mit im Verhältnis zum
Volumen großer Grenzfläche tragen diesem Umstand Rechnung. Bei diesen Immobilisaten
handelt es sich ebenfalls um Hydrogele, innerhalb derer eine thermodynamisch stabile Mikro-
emulsion eingeschlossen ist [STAMATIS UND XENAKIS, 1999; ZHOU ET AL., 2001; DELIMITSOU
ET AL., 2002]. Aber auch diese Hydrogel-Variante konnte sich bisher nicht durchsetzen. Ver-

17
mutlich können aufgrund der Verteilungskoeffizienten der meist hydrophoben Substrate nur
geringe Konzentrationen im Inneren dieser Hydrogele erreicht werden, so dass der Vorteil der
größeren Grenzfläche nicht zum Tragen kommen kann.
Dieses Problem kann durch Einhüllung der Lipasen in hydrophobe Polymere (z.B. Sol-Gele,
Silicone, Polyurethane) umgangen werden. Durch die hydrophobe Matrix wird nicht nur der
Massentransfer der Substrate und Produkte verbessert, sondern auch eine für die Lipasen
ideale Mikroumgebung geschaffen, erkennbar an einer Aktivitätssteigerung gegenüber dem
nicht immobilisierten Enzym [GILL UND BALLESTEROS, 2000a/b]. So konnten HSU ET AL.
[2002] nach Interkalation von Lipasen in Phyllosilicate und deren Einbettung in eine
hydrophobe Sol-Gel-Matrix eine Aktivitätssteigerung beobachten. Ähnliches gelang GILL ET
AL. [1999] und RAGHEB ET AL. [2003], die Lipasen in Silicon einschlossen sowie FERREIRA-
DIAS ET AL. [1999] bei Einhüllung in Polyurethan. Deutlich höhere Steigerungen wurden
allerdings bei Immobilisierung von Lipasen in den von REETZ ET AL. [1996] entwickelten
alkylsubstituierten Sol-Gelen beobachtet. Aufgrund der erzielten hohen Aktivitätssteige-
rungen und der erwiesenermaßen hohen katalytischen Stabilität stellt die Immobilisierung von
Lipasen in Sol-Gelen gegenwärtig den vielversprechendsten Ansatz dar.
Auch wenn die lipasenkatalysierte Synthese im Vergleich zu konventionellen chemischen
Verfahren nachweislich neue Möglichkeiten eröffnet, muss festgestellt werden, dass eine
gewisse Diskrepanz zwischen der Zahl an in der Literatur beschriebenen und der Zahl an
industriell genutzten biokatalytischen Prozessen besteht. Hauptgrund hierfür mögen die hohen
Kosten der Biokatalysatoren sein, die eine Nutzung über lange Zeiträume und damit eine
effiziente Immobilisierung erfordern [JAEGER UND REETZ, 1998]. Die Entwicklung eines
Immobilisierungsverfahrens für den wirtschaftlichen Einsatz von Lipasen unter industriellen
Gesichtspunkten scheint aber problembehaftet zu sein. Anders ist zumindest nicht zu erklären,
warum trotz der Vielfalt an Immobilisierungsverfahren nur wenige Lipase-Immobilisate kom-
merziell verfügbar sind.
1.4.3 Probleme beim technischen Einsatz von Lipase-Immobilisaten
Im technischen Einsatz werden besondere Anforderungen an ein Immobilisat gestellt. Dies
betrifft u.a. die Größe, die mechanischen Eigenschaften und v.a. die katalytische Aktivität und
Stabilität, wobei stets zu berücksichtigen ist, unter welchen Bedingungen die Immobilisate

Einleitung
18
eingesetzt werden sollen. Es liegt auf der Hand, dass bei einem Einsatz in Rührkesselreaktor
im Vergleich zum Packbettreaktor andere Kriterien bedacht werden müssen. Dabei wird
womöglich mehr Wert auf eine hohe Abriebfestigkeit gelegt, wohingegen im Packbettreaktor
Größe und Kompressibilität der Immobilisate von Interesse sind. Des Weiteren sind Kriterien
wie Langzeitstabilität und Herstellungskosten von großer Bedeutung. Aber auch der Herstel-
lungsprozess selbst muss kritisch betrachtet werden, denn die Verwendung toxischer Hilfs-
stoffe kann die Sicherheit des Endproduktes gefährden und die Eignung einer Immobili-
sierungstechnik grundsätzlich in Frage stellen. Ferner muss die Methode eine hohe Immo-
bilisierungseffizienz im Hinblick auf Enzym- und Materialverluste gewährleisten. Aufgrund
der per se gegebenen hohen Stabilität von Lipasen kann im Vergleich zu anderen Enzymen
davon ausgegangen werden, dass sie über vergleichsweise lange Zeiträume eingesetzt werden
können. Dies wiederum erlaubt den Einsatz aufwändigerer Methoden und teurerer Materialien
für die Immobilisierung, erhebt aber auch sehr hohe Ansprüche, z.B. im Hinblick auf die
Haltbarkeit der Immobilisate.
Zum gegenwärtigen Zeitpunkt kann kein Lipase-Immobilisat allen Anforderungen gerecht
werden. Das kommerziell verfügbare Novozym 435, ein Präparat mit bemerkenswert hoher
Aktivität [BOURG-GARROS ET AL., 1998], wird bereits großtechnisch eingesetzt [HILLS, 2003;
THUM, 2004]. Eine Desorption des Enzyms kann aber nicht ausgeschlossen werden, ebenso
wie die völlig unzureichende Abriebfestigkeit von Novozym 435 dem Einsatz im Rührkessel-
reaktor entgegensteht. Die Aktivität und mechanische Stabilität vieler Hydrogele ist ebenfalls
nicht ausreichend für einen langfristigen technischen Einsatz. Sol-Gele aus alkylsubstituierten
Silanen stellen angesichts der zu erzielenden Aktivitätssteigerungen einen vielversprechenden
Ansatz zur Immobilisierung von Lipasen dar [REETZ ET AL., 1996-2003]. Bei näherer
Betrachtung stellt sich jedoch heraus, dass auch hier die Voraussetzungen für einen tech-
nischen Einsatz nur begrenzt bestehen. Bei der Herstellung wird das giftige Tetramethoxy-
silan (TMOS) verwendet. Eine Gefährdung des Endverbrauchers kann in sensiblen Anwen-
dungsbereichen daher nicht ausgeschlossen werden. Außerdem sind die Immobilisate klein
und spröde, weshalb im Rührkesselreaktor mit einer Desintegration der Immobilisate zu
rechnen ist und deren Rückhaltung somit erschwert wird. Hingegen erscheint eine Anwen-
dung der Sol-Gele im Packbettreaktor aufgrund ihrer geringen Größe und der dadurch
erforderlichen hohen Drücke nicht angeraten. Die kovalente Bindung an Eupergit wird
ebenfalls häufig zur Immobilisierung von Lipasen eingesetzt [IVANOV UND SCHNEIDER, 1997;
BOLLER ET AL., 2002]. Auch wenn der Immobilisierungsgrad und die Stabilität der Immobi-

19
lisierung hoch sind, ist eine Aktivitätssteigerung in der Größenordnung wie bei Sol-Gelen
nicht möglich. Ähnliches gilt bei Immobilisierung durch Einschluss in Silicon und Poly-
urethan – die mechanische Stabilität ist exzellent, wohingegen die Aktivitäten vergleichs-
weise niedrig sind.
Aufgrund der oben skizzierten technischen Probleme wird verständlich, dass die Optimierung
der Immobilisierung von Lipasen bzw. Enzymen im Allgemeinen nach wie vor Gegenstand
intensiver Forschungsaktivitäten ist.

Einleitung
20
1.5 Aufgabenstellung
Die technische Nutzung von Lipasen beruht überwiegend auf rein empirischen Grundlagen,
da in der Forschung der anwendungsbezogene Hintergrund meist im Mittelpunkt steht und
deshalb eine systemimmanente Analyse umgangen wird. Einblicke in das katalytische Verhal-
ten und den Reaktionsmechanismus von Lipasen wurden fast ausschließlich für hydro-
lytische Reaktionen gewonnen. Viele Forscher leiten aus den dortigen Ergebnissen die
voraussichtlichen Gegebenheiten für diverse Reaktionen und Reaktionssysteme ab. Inwieweit
eine Übertragbarkeit gerechtfertigt ist, wird dabei nicht hinterfragt. Gerade bei Betrachtung
der Veresterung in biphasischen Reaktionssystemen ergeben sich aufgrund der Komplexität
dieser Systeme offene Fragen, die in der Fachliteratur bisher nicht ausreichend gewürdigt
wurden. So ist nicht mit hinreichender Genauigkeit bekannt, wo das pH-Optimum der lipasen-
katalysierten Veresterung, ob die protonierte oder deprotonierte Säure das bevorzugte Substrat
ist oder wo die Reaktion in biphasischen Reaktionssystemen stattfindet. Diese Wissenslücken
erschweren eine rationale Prozessoptimierung, insbesondere im Hinblick auf die Immobili-
sierung von Lipasen für den Einsatz in biphasischen Reaktionssystemen.
Im Rahmen dieser Arbeit sollte diesem Mangel durch systematische Untersuchung der Bil-
dung von Butylpropionat aus den Edukten Butanol und Propionsäure in einem gelstabilisier-
ten und einem nicht-gelstabilisierten biphasischen Reaktionssystem entgegengetreten werden.
Die Bedeutung des pH-Wertes für die lipasenkatalysierte Veresterung in biphasischen
Reaktionssystemen sollte für die Lipasen aus Candida rugosa und aus Thermomyces
lanuginosa analysiert werden. Diese beiden Lipasen weisen stark unterschiedliche pH-Optima
bei der Hydrolyse auf [NEVES-PETERSEN ET AL., 2001a], weshalb zunächst erwartet wurde,
dass sich die pH-Optima bei Veresterung in ähnlicher Weise voneinander unterscheiden
sollten. Abschließend sollte der Einfluss der Propionsäurekonzentration auf die Lage der pH-
Optima sowie die katalytische Aktivität beider Lipasen am pH-Optimum untersucht werden.
Um zu klären, ob die Veresterung in biphasischen Reaktionssystemen überwiegend an der
Grenzfläche oder zu einem erheblichen Anteil auch in der wässrigen Phase erfolgt, sollte die
Synthese von Butylpropionat unter Variation der spezifischen Grenzfläche durchgeführt
werden. Es wurde vorab vermutet, dass die Abhängigkeit der Estersynthese von der spezi-
fischen (auf das Volumen der wässrigen Phase bezogenen) Grenzfläche für eine grenzflächen-
aktivierte und eine nicht-grenzflächenaktivierte Lipase unterschiedlich stark ausgeprägt sein
dürfte. Daher sollte das katalytische Verhalten der Lipase von Candida rugosa und der Lipase

21
B von Candida antarctica untersucht und verglichen werden. Letztgenannte Lipase unterliegt
keiner Grenzflächenaktivierung [MARTINELLE ET AL., 1995].
Die gewonnenen Erkenntnisse sollten helfen, eine rationale Basis für die Weiterentwicklung
lipasehaltiger Hydrogele für die Veresterung in organischen Reaktionsmedien zu schaffen.
Vorrangiges Anliegen war es, die Immobilisate an die Erfordernisse der Reaktion unter
Berücksichtigung von Effizienzkriterien wie Immobilisierungsgrad, Ausblutungsverhalten,
katalytische Aktivität und Stabilität in organischen Lösungsmitteln, mechanische Stabilität so-
wie Massentransfer von Substrat und Produkt anzupassen. Eine Abschätzung des Leistungs-
potentials und der Attraktivität für zukünftige Forschungsaktivitäten sollte durch Vergleich
mit dem nach gegenwärtigem Kenntnisstand aussichtsreichsten Verfahren für die Einhüllung
von Lipasen – der Sol-Gel-Technik nach REETZ ET AL. [1996] – erfolgen. Modellreaktion war
hierbei die Veresterung von Octanol und Caprylsäure zu Octyloctanoat.

Material und Methoden
22
2. Material und Methoden
2.1 Chemikalien/Enzyme
BIO-RAD Protein Assay Bio-Rad Laboratories GmbH (München)
BSA (Bovine Serum Albumin) Sigma-Aldrich (Steinheim)
i-Butyltrimethoxysilan ABCR (Karlsruhe)
CaCl2 x 2 H2O Riedel deHæn (Seelze)
Gellan Kelcogel (Hamburg)
n-Hexan Merck (Darmstadt)
Natriumalginat Kelcogel (Hamburg)
Nebenprodukte der Biodieselproduktion Esterco Biofuels Ltd. (St Austell, GB) und
Petrotec GmbH (Borken)
n-Octanol Bayer (Dormagen)
2-Propanol Merck (Darnstadt)
Sylgard® 184 (Komponente A und B) Dow Corning (Wiesbaden)
Syl-Off® Dow Corning (Wiesbaden)
Tributyrin Merck (Darmstadt)
Alle hier nicht aufgeführten Chemikalien wurden von Fluka (Neu-Ulm) bezogen. Tabelle 2.1
listet die in der Arbeit benutzten Enzyme auf.
Tabelle 2.1: Verwendete Enzympräparate, gemessener Proteingehalt sowie kommerzieller Name und Hersteller
Enzym Proteingehalt Kommerzieller Name / Hersteller
Candida antarctica Lipase A (CALA) 1,8 % [v/v] Novozym 735 (Flüssigpräparat) /
Novozymes (Bagsvaerd, Dänemark)
Candida antarctica Lipase B (CALB) 40,5 % [w/w] Chirazyme L-2 lyo / Roche
Diagnostics GmbH (Mannheim)
Candida antarctica Lipase B (CALB) 0,1 -1 % [w/w]* Novozym 435 + Flüssigpräparat /
Novozymes (Bagsvaerd, Dänemark)
Candida rugosa Lipase (CRL) 15,4 % [w/w] L-1754 / Sigma-Aldrich
(Taufkirchen)
Thermomyces lanuginosaLipase (TLL) 35,3 % [w/w] Chirazyme L-8 lyo / Roche
Diagnostics GmbH (Mannheim)
Pseudomonas stutzeriLipase (PSL) 22,6 % [w/w] Jülich Fine Chemicals (Jülich)
Rhizopus arrhizus Esterase 29,0 [w/w] Esterase EL / ASA-Spezialenzyme
(Wolfenbüttel)
* Herstellerangabe

23
2.2 Geräte
Evaporationszentrifuge Speed Vac SC 110 (Savant, Holbrook, USA)
Gaschromatograph HP 5890 Series II (Hewlett Packard, Walbronn)
Gaschromatograph Varian Star 3400 CX (Walnut Creek, USA)
HPLC Beckmann Coulter System Gold (München)
Laborschüttler Vortemp (UniEquip, Martinsried)
Mikroskop (DM RB) DM RB (Leitz, Wetzlar)
Multi-Rührerplatte Variomag Poly 15 (H + P Labortechnik)
Particle Sizer Model 770 (Particle Sizing Systems, Santa
Barbara, USA)
Spektralphotometer UV-160 A (Shimadzu, Kyoto, Japan)
Überkopfschüttler Reax 2 (Heidolph, Schwabach)
2.3 Gaschromatographische Analyse
Durch gaschromatographische Analyse wurden die Substrate und Produkte der verschiedenen
enzymkatalysierten Reaktionen durch zwei- bzw. dreimalige Injektion und anschließende
Mittelwertbildung quantifiziert. Interne Standards waren Decan bzw. Dodecan, als Trägergas
wurde Stickstoff verwendet. Der Nachweis erfolgte mit einem Flammenionisationsdetektor.
Gewählte Betriebsparameter sowie Spezifikationen der Kapillarsäulen sind Tabelle 2.2 zu
entnehmen. Anhand reiner Standards wurden die an der Reaktion beteiligten Edukte und
Produkte identifiziert und quantifiziert. Hinsichtlich Methode d ist anzumerken, dass aufgrund
der komplexen Zusammensetzung des Substratgemisches nicht die individuellen Substanzen
analysiert wurden. Ihr Nachweis erfolgte als Verbindungsklassen, wobei die verschiedenen
Fettsäuren bzw. ihre Ester nach der Länge ihres Kohlenstoffgerüsts klassifiziert wurden. Für
die Analyse der Verbindungsklassen wurden Bedingungen gewählt, die dazu führten, dass die
einer Verbindungsklasse angehörenden Moleküle zur gleichen Zeit von der Säule eluierten.
Material und Methoden
24
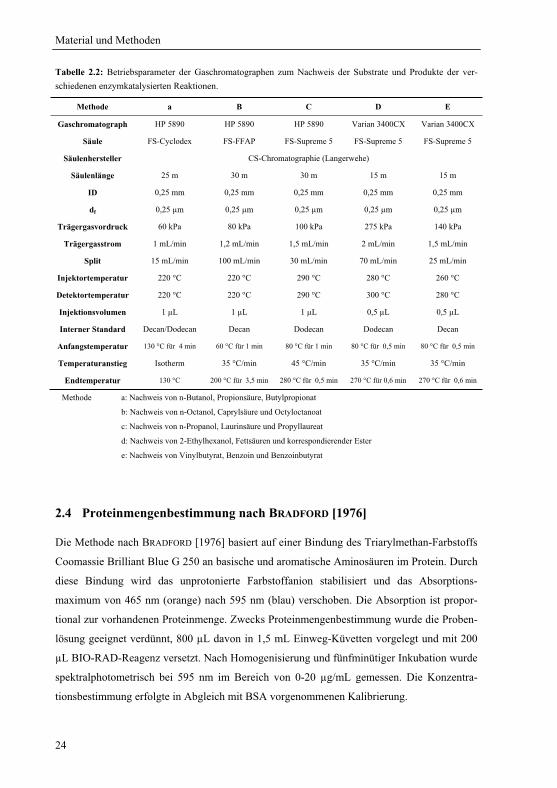
Tabelle 2.2: Betriebsparameter der Gaschromatographen zum Nachweis der Substrate und Produkte der ver-schiedenen enzymkatalysierten Reaktionen.
Methode a B C D E
Gaschromatograph HP 5890 HP 5890 HP 5890 Varian 3400CX Varian 3400CX
Säule FS-Cyclodex FS-FFAP FS-Supreme 5 FS-Supreme 5 FS-Supreme 5
Säulenhersteller CS-Chromatographie (Langerwehe)
Säulenlänge 25 m 30 m 30 m 15 m 15 m
ID 0,25 mm 0,25 mm 0,25 mm 0,25 mm 0,25 mm
df 0,25 μm 0,25 μm 0,25 μm 0,25 μm 0,25 μm
Trägergasvordruck 60 kPa 80 kPa 100 kPa 275 kPa 140 kPa
Trägergasstrom 1 mL/min 1,2 mL/min 1,5 mL/min 2 mL/min 1,5 mL/min
Split 15 mL/min 100 mL/min 30 mL/min 70 mL/min 25 mL/min
Injektortemperatur 220 °C 220 °C 290 °C 280 °C 260 °C
Detektortemperatur 220 °C 220 °C 290 °C 300 °C 280 °C
Injektionsvolumen 1 μL 1 μL 1 μL 0,5 μL 0,5 μL
Interner Standard Decan/Dodecan Decan Dodecan Dodecan Decan
Anfangstemperatur 130 °C für 4 min 60 °C für 1 min 80 °C für 1 min 80 °C für 0,5 min 80 °C für 0,5 min
Temperaturanstieg Isotherm 35 °C/min 45 °C/min 35 °C/min 35 °C/min
Endtemperatur 130 °C 200 °C für 3,5 min 280 °C für 0,5 min 270 °C für 0,6 min 270 °C für 0,6 min
Methode a: Nachweis von n-Butanol, Propionsäure, Butylpropionat
b: Nachweis von n-Octanol, Caprylsäure und Octyloctanoat
c: Nachweis von n-Propanol, Laurinsäure und Propyllaureat
d: Nachweis von 2-Ethylhexanol, Fettsäuren und korrespondierender Ester
e: Nachweis von Vinylbutyrat, Benzoin und Benzoinbutyrat
2.4 Proteinmengenbestimmung nach BRADFORD [1976]
Die Methode nach BRADFORD [1976] basiert auf einer Bindung des Triarylmethan-Farbstoffs
Coomassie Brilliant Blue G 250 an basische und aromatische Aminosäuren im Protein. Durch
diese Bindung wird das unprotonierte Farbstoffanion stabilisiert und das Absorptions-
maximum von 465 nm (orange) nach 595 nm (blau) verschoben. Die Absorption ist propor-
tional zur vorhandenen Proteinmenge. Zwecks Proteinmengenbestimmung wurde die Proben-
lösung geeignet verdünnt, 800 μL davon in 1,5 mL Einweg-Küvetten vorgelegt und mit 200
μL BIO-RAD-Reagenz versetzt. Nach Homogenisierung und fünfminütiger Inkubation wurde
spektralphotometrisch bei 595 nm im Bereich von 0-20 μg/mL gemessen. Die Konzentra-
tionsbestimmung erfolgte in Abgleich mit BSA vorgenommenen Kalibrierung.

25
2.5 Massentransfer im nicht-gelstabilisierten Zweiphasensystem
Das in dieser Arbeit eingesetzte nicht-gelstabilisierte Zweiphasensystem bestand aus einer
verschließbaren Schott-Flasche (Reaktor), in der sich eine wässrige und eine organische Phase
im Volumenverhältnis von 1 : 1 befanden. Beide Phasen wurden durch einen „Zwei-Phasen“-
Magnetrührer (Abb. 2.1) aktiv durchmischt, wobei durch Abstimmung der Füllvolumina (in
Abhängigkeit vom Flaschendurchmesser) die Integrität der Grenzfläche gewahrt blieb.
Gerührt wurde auf einer Multirührerplatte, die ein paralleles Arbeiten mit bis zu 15 Ansätzen
erlaubte. Die Rührerdrehzahl lag stets bei 250 Upm.
Abbildung 2.1: Versuchsaufbau zur Bestimmung des pH-Optimums und der Bedeutung der Grenzfläche im zweiphasigen Reaktionssystem.
Die Geschwindigkeit des Massentransfers der Substrate Butanol und Propionsäure wurde in
einer 25-mL- und einer 250-mL-Schott-Flasche gemessen. Hierfür wurden Konzentrationen
von 50 mmol/L der Substrate und 20 mmol/L Decan als Interner Standard in der organischen
Phase vorgelegt. Durch Entnahme von 75 μL Probe aus der wässrigen und der organischen
Phase in geeigneten Zeitabständen über einen Zeitraum von 200 min wurde die Konzentra-
tionsabnahme dokumentiert. Die Proben der organischen Phasen wurden gaschromatogra-
phisch (Kapitel 2.3, Methode a) analysiert. Die Volumina der wässrigen und organischen
Phase betrugen im 25-mL-Reaktor je 7 mL, im 250-mL-Reaktor je 30 mL. Die pH-abhängige
Verteilung der Propionsäure wurde beispielhaft im 25-mL-Reaktor untersucht. Hierfür
wurden in der wässrigen Phase 150 mmol/L Propionsäure vorgelegt und drei pH-Werte (2,75;
5; 7,5 je ± 0,05) durch Zugabe von HCl (10,18 mol/L) bzw. NaOH (7,5 mol/L) eingestellt.
n-Hexan mit n-Butanol
12 mm
18 mm
Seitenansicht Ansicht von oben
Zweiphasenrührer
Reaktionsgefäß (Schott®-Flasche)
Edelstahl Resopal® Magnetrührer
WässrigePhase mit Lipase und
Propionsäure

Material und Methoden
26
Über einen Zeitraum von 2 h wurden in geeigneten Zeitabständen der wässrigen und der
organischen Phase Proben entnommen.
2.6 pH-Optimum der lipasenkatalysierten Veresterung
2.6.1 Titration der Enzymlösungen
Enzymlösungen der CRL (15 mg/mL) bzw. der TLL (0,2 mg/mL) mit je 150 mmol/L
Propionsäure wurden mit 7,5 M NaOH titriert, um das Puffervermögen des Reaktionssystems
und das Löslichkeitsverhalten beider Lipasen beurteilen zu können. Zu je 5 mL der Enzym-
Propionsäure-Lösungen (initialer pH-Wert 3,0) wurden einige Mikroliter NaOH zugegeben
und die Volumina sowie der resultierende pH-Wert protokolliert. Aufgenommen wurden die
Titrationskurven in einem Bereich von pH 3-11.
2.6.2 Nicht-gelstabilisiertes biphasisches Reaktionssystem
Das pH-abhängige Aktivitätsprofil der CRL und der TLL bei Bildung von Butylpropionat aus
Propionsäure und n-Butanol wurde für drei Propionsäurekonzentrationen bestimmt (75; 150
und 300 mmol/L). Die Lipasen wurden in Lyophilisatkonzentrationen von 15 mg/mL (CRL)
bzw. 0,2 mg/mL (TLL) zusammen mit Propionsäure in deionisiertem Wasser gelöst. Die
erhaltenen Lösungen wurden durch Titration mit HCl (10,18 mol/L) oder NaOH (7,5 mol/L)
auf die gewünschten pH-Werte eingestellt (2,0; 2,75; 3,5; 4,25; 5; 6,25; 7,5 je ± 0,05). Die
zugegebene Menge an Säure bzw. Base wurde notiert, um ggf. die aus der Zugabe resul-
tierende Verdünnung berücksichtigen zu können. Je 7 mL der wässrigen Lösungen wurden in
25 mL fassende und verschließbare Reaktionsgefäße überführt und um die Reaktion zu
starten, mit 7 mL einer Lösung von 150 mmol/L n-Butanol und 20 mmol/L n-Decan (Interner
Standard) in n-Hexan überschichtet. Die beiden Phasen der jeweiligen Reaktionsansätze
wurden mit Hilfe eines Zweiphasenrührers (Abb. 2.2) bei 250 Upm getrennt durchmischt. Die
Reaktion wurde bei RT über einen Zeitraum von bis zu 8 h durchgeführt. Die autokatalytische
Reaktionsrate wurde in enzymfreien Ansätzen der entsprechenden pH-Werte bestimmt. Zu
sechs Zeitpunkten wurden sowohl der wässrigen als auch der organischen Phase je 75 μL
Probe entnommen, um das Phasenverhältnis beizubehalten. Die organische Phase wurde
gaschromatographisch analysiert (Kapitel 2.3, Methode a). Nach Abbruch der Umsetzung

27
wurde der pH-Wert der wässrigen Phase überprüft und mit dem zuvor eingestellten pH-Wert
verglichen. Die initiale Reaktionsrate wurde durch Quantifizierung der Butylpropionat-
Konzentration bestimmt.
2.6.3 Reihenuntersuchung weiterer Lipasen
Die pH-Abhängigkeit der lipasenkatalysierten Veresterung wurde auch für die Lipase A und
die Lipase B aus Candida antarctica (CALA und CALB) sowie für die Lipase aus
Pseudomonas stutzeri (PSL) ermittelt. Dabei wurde die CALB auch in Form von Novozym
435 immobilisiert untersucht. Hierfür wurden 100 μL CALA, 1,4 mg CALB, 250 mg PSL
und 10 mg Novozym 435 in 7 mL der wässrigen Phase vorgelegt. Die wässrige Phase enthielt
50 mmol/L Propionsäure und ihr pH-Wert wurde durch Zugabe von HCl bzw. NaOH auf
2,75, 4,5 und 6,75 eingestellt. Zum Start der Reaktion wurde die wässrige Phase mit 7 mL
Hexan (50 mmol/L Butanol; 20 mmol/L Decan) überschichtet. Die Probenentnahme und
Analyse erfolgte wie in Kapitel 2.6.2 beschrieben. Die Versuchsdauer betrug 10-15 h, die
Experimente wurden im Doppelansatz durchgeführt.
2.7 Einfluss der Grenzfläche auf die lipasenkatalysierte Reaktion
2.7.1 Alginatkugeln verschiedener Größe
Die Herstellung unterschiedlich großer Alginatkugeln erfolgte gemäß der von BUTHE ET AL.
[2004] beschriebenen Methode: Eine Glassäule von 30 cm Länge und einem inneren Durch-
messer von 2,5 cm wurde schrittweise mit 15 mL einer wässrigen Lösung von 2 % (w/v)
CaCl2, 60 mL n-Butanol mit 1 % (w/v) CaCl2, 30 mL n-Butanol und 25 mL n-Hexan gefüllt
(Abb. 2.2). Die unterste Schicht wurde vorsichtig mit einem Magnetrührer durchmischt. Die
Säule mit den geschichteten Lösungen wurde verschlossen und 1 h bei RT belassen. Danach
wurden definierte Volumina einer gekühlten Lösung (4 °C) von 2 % Alginat und Enzym in
deionisiertem Wasser in die obere Hexanphase injiziert. Hierfür wurden Pipettierhilfen mit
abgeschnittenen Pipettenspitzen verwendet. Die eingesetzten Alginatlösungen wiesen Lyo-
philisatkonzentrationen von 10 mg/mL (CRL) bzw. 0,2 mg/mL (CALB) auf. Durch Zugabe
von Hühnerserumalbumin wurden Gesamtproteinkonzentrationen von je 20 mg/mL einge-

Material und Methoden
28
stellt, um nach Absättigung der Interaktionspunkte zwischen Alginat und Protein eine homo-
gene Proteinverteilung zu gewährleisten. Es wurden Kugeln mit Durchmessern von 2, 4, 6
und 8 mm hergestellt. Die erhaltenen Alginatkugeln wurden durch Filtration abgetrennt und
durch zweimaliges Waschen mit Wasser überschüssiges Butanol entfernt. Anschließend
erfolgte die Aushärtung durch Überführung der Kugeln in eine 2%ige wässrige Lösung von
CaCl2 für die Dauer von 1 h. Die Kugeln wurden bis zur weiteren Verwendung in n-Hexan
gelagert. Durchmesser und Rundheit wurden gemäß der in Kapitel 2.11 dargelegten Methodik
bestimmt.
Abb. 2.2: Versuchsaufbau zur Herstellung von Alginatkugeln unterschiedlicher Größen.
Vor Verwendung der Alginatkugeln wurden diese in einer 2%igen CaCl2-Lösung bei pH 3,0
für 20 h äquilibriert. Die Reaktion wurde mit je einer Kugel der verschiedenen Größen in acht
parallelen Ansätzen (für jeden Probenzeitpunkt ein Ansatz) durchgeführt. Verwendet wurden
verschließbare Reaktionsgefäße geeigneter Größe, um eine vollständige Bedeckung der
Kugeln mit der Substratlösung zu gewährleisten. Durch Zugabe einer Hexanlösung mit
50 mmol/L Propionsäure, 50 mmol/L n-Butanol und 20 mmol/L Decan wurde die Reaktion
gestartet. Es wurde stets bei einem Phasenvolumenverhältnis (organisch/wässrig) von 35 : 1
gearbeitet. Die Inkubation erfolgte unter Schütteln bei RT über einen Zeitraum von 70 bis
80 h. In geeigneten Zeitabständen wurden der organischen Phase 75 μL Proben entnommen
und gaschromatographisch analysiert. Über die Quantifizierung der Butylpropionat-Konzen-
tration wurde die initiale Reaktionsrate bestimmt.
n-Hexan
n-Butanol
1 % (w/v) CaCl2 in n-Butanol
2 % (w/v) CaCl2 in H2O
n-Hexan
n-Butanol
1 % (w/v) CaCl2 in n-Butanol
2 % (w/v) CaCl2 in H2O

29
2.7.2 Nicht-gelstabilisiertes biphasisches Reaktionssystem
Als wässrige Phase im nicht-gelstabilisierten zweiphasigen Reaktionssystem diente eine
Enzymlösung mit jeweils definierten Lyophilisatkonzentrationen der CRL (15 mg/mL), der
TLL (0,2 mg/mL) und der Esterase von Rhizopus arrhizus (5 mg/mL). Zudem wurde der
wässrigen Phase Propionsäure in einer Konzentration von 50 mmol/L zugesetzt und der pH-
Wert auf 4,0 eingestellt. Die Reaktionen wurden stets bei einem Phasenvolumenverhältnis
von 1:1 (organisch/wässrig) durchgeführt. Als organische Phase diente Hexan mit 50 mmol/L
n-Butanol und 20 mmol/L n-Decan (Interner Standard). Durch Zugabe der organischen Phase
zur wässrigen Phase wurde die Enzymreaktion gestartet. Die Durchmischung erfolgte mithilfe
von Zweiphasenrührern, wie sie in Kapitel 2.5 beschrieben wurden. Zur Wahrung des Phasen-
volumenverhältnisses wurden beiden Phasen Proben von je 75 μL entnommen, jedoch wurde
nur die organische Phase gaschromatographisch analysiert. Die Versuchsdauer betrug 8 h. Die
initiale Reaktionsrate wurde über die gebildete Menge an Butylpropionat bestimmt. Die spezi-
fische Grenzfläche zwischen den beiden Phasen ergab sich aus dem Verhältnis von Grenz-
fläche zu Phasenvolumen.
A) Variable spezifische Grenzfläche
Durch Verwendung unterschiedlich großer Schott-Flaschen (25-, 50-, 100- und 250-mL),
variierte die Größe der Phasengrenzfläche. Durch Füllen der Flaschen mit definierten
Volumina wurde über die zu messende Füllhöhe und den Innendurchmesser die Größe der
Grenzfläche für mehrere Flaschen einer Größe errechnet und der Mittelwert gebildet. Um zu
gewährleisten, dass der Zweiphasenrührer beide Phasen durchmischt, wurden für die ver-
schiedenen Flaschengrößen geeignete Phasenvolumina gewählt. Diese und die jeweiligen
Größen der Grenzflächen sind in Tabelle 2.3 aufgelistet.
Tabelle 2.3: Phasenvolumina (organisch/wässrig) und Grenzflächengrößen bei variabler Reaktorgröße.
Reaktor-volumen
[mL]
Phasenvolumen (organisch bzw. wässrig)
[mL]
Größe der Grenzfläche
[cm2]
Spezifische Grenzfläche
[cm-1]
25 6 7,59 1,27
50 9 12,66 1,41
100 18 19,71 1,10
250 40 32,67 0,82

Material und Methoden
30
In einer weiteren Versuchsvariante wurden durch Variation der Phasenvolumina in 25-mL-
fassenden Reaktionsgefäßen unterschiedliche spezifische Grenzflächen eingestellt. Die ge-
wählten Phasenvolumina und die daraus resultierenden spezifischen Grenzflächen sind
Tabelle 2.4 zu entnehmen.
Tabelle 2.4: Phasenvolumina und daraus resultierende Grenzflächen bei gleicher Reaktorgröße.
Reaktor-volumen
[mL]
Phasenvolumen (organisch bzw. wässrig)
[mL]
Größe der Grenzfläche
[cm2]
Spezifische Grenzfläche
[cm-1]
25 5 7,59 1,52
25 6,25 7,59 1,21
25 8 7,59 0,94
25 10 7,59 0,76
B) Konstante spezifische Grenzfläche
In einer weiteren Versuchsvariante wurde eine konstante spezifische Grenzfläche von
0,94 cm-1 in Reaktionsgefäßen (Schott-Flaschen) unterschiedlicher Größe (25; 50; 100 und
250 mL) eingestellt, indem diese mit definierten Volumina befüllt wurden. Hierfür wurde der
Innendurchmesser der Reaktionsgefäße individuell vermessen und die erforderlichen Phasen-
volumina berechnet (Tab. 2.5).
Tabelle 2.5: Phasenvolumina und Grenzflächen der verwendeten Reaktoren zur Einstellung einer konstanten spezifischen Phasengrenzfläche.
CRL CALB Reaktor-volumen
[mL]
Spezifische Grenzfläche
[cm-1]Phasenvolumen (org. / wässrig)
[mL]
Größe der Grenzfläche
[cm2]
Phasenvolumen (org. / wässrig)
[mL]
Größe der Grenzfläche
[cm2]
25 0,94 8,0 7,5 8,4 7,9
50 0,94 13,4 12,6 13,5 12,7
100 0,94 21,1 19,8 20,9 19,6
250 0,94 34,0 32,0 35,5 33,3
C) Absenkung der Wasseraktivität
Die Versuchsdurchführung erfolgte analog zu Variante A, wobei jedoch die wässrige Phase
neben 50 mmol/L Propionsäure noch 75 % (w/v) Glucose enthielt. Die Untersuchung be-
schränkte sich auf die CALB (0,2 mg/mL).

31
D) Hydrolyse von Butylpropionat
Der Einfluss der spezifischen Grenzfläche auf die Geschwindigkeit der Hydrolyse von
Butylpropionat im nicht-gelstabilisierten Zweiphasensystem wurde analog zu Variante A
untersucht. Im Gegensatz zur Synthese wurden 50 mmol/L Butylpropionat als Substrat in der
organischen Phase zusammen mit 20 mmol/L Decan (Interner Standard) vorgelegt.
2.7.3 Gelstabilisiertes biphasisches Reaktionssystem
Abweichend zu Variante A aus Kapitel 2.7.2 bestand die wässrige Phase aus einer enzym-
haltigen Gellanschicht. Für deren Herstellung wurde eine Lösung von 3 % (w/v) Gellan durch
Erhitzen auf 90 °C hergestellt und anschließend auf 70 °C temperiert. Parallel wurde eine
Lösung mit 150 mmol/L Propionsäure, 0,4 mg/mL Enzym (CALB) und CaCl2 (10 mmol/L)
auf einen pH-Wert von 4,0 eingestellt und auf 60 °C erhitzt. Beide Lösungen wurden zu
gleichen Teilen vermischt und danach definierte Volumina in 25 mL fassende Reaktions-
gefäße gefüllt (Tabelle 2.3). Nach Aushärtung der Gellanschicht wurde ein dem Volumen der
wässrigen Phase äquivalentes Volumen einer Hexanlösung mit 50 mmol/L n-Butanol und 20
mmol/L n-Decan (Interner Standard) zugegeben und die Ansätze 6 h schüttelnd bei RT
inkubiert. Über diesen Zeitraum verteilt wurden der organischen Phase mehrere Proben von je
75 μL entnommen und gaschromatographisch analysiert.
2.8 „Molecular modelling“ der Enzymoberfläche
Ausgehend von der dreidimensionalen Struktur der CRL in der offenen und der geschlossenen
Konformation und der CALB sowie einer Esterase von Bacillus subtilis sollte softwarege-
stützt die molekulare Oberfläche der Enzymmoleküle charakterisiert werden. Hierfür wurden
die Röntgenstrukturdaten der Enzyme von der „Research Collaboratory for Structural
Bioinfomatics“ (www.rcsb.org) als pdb-Datei heruntergeladen (Tab. 2.6). Die pdb-Dateien
wurden mit Hilfe der Software GREATER (Version 1.2.2) aufbereitet, um anschließend mit
Hilfe der Software GRID (Version 22b) ein dreidimensionales Raumgitter über die Protein-
strukturen zu legen und um für die Knotenpunkte des Raumgitters (Abstand 1 Å) die
physikalischen und chemischen Parameter der Proben „OH2“, „DRY“ und „C3“ zu berechnen
(genauere Erläuterungen in BRAIUCA ET AL., 2006). Die Probe „OH2“ repräsentiert Wasser,
„DRY“ eine nicht näher spezifizierte hydrophobe Probe und „C3“ Methylgruppen. Gewählt

Material und Methoden
32
wurden diese Proben, um für verschiedene Energieniveaus Bereiche in und auf den Enzym-
molekülen zu identifizieren, in denen die Wahrscheinlichkeit, polare bzw. unpolare Moleküle
anzutreffen, erhöht ist. Diese Bereiche auf der Proteinoberfläche konnten mit Hilfe der
Software GVIEW (Version 2.0.1) visualisiert werden. Die Software GREATER, GRID und
GVIEW wurden von der Firma „Molecular Discovery Ltd.“ (www.moldiscovery.com,
London, UK) kostenfrei bezogen.
Tabelle 2.6: Für die Charakterisierung der Enzymoberfläche verwendete pdb-Dateien.
Enzym pdb-Datei
Lipase von Candida rugosa (geschlossen) 1TRH
Lipase von Candida rugosa (offen) 1CRL
Lipase B von Candida antarctica 1TCA
Esterase von Bacillus subtilis 1QE3
2.9 Immobilisierung in Siliconsphären als Statische Emulsion
Die Immobilisierung in Silicon erfolgte unter Verwendung des bei RT polymerisierenden
Zweikomponenten-Siliconkautschuks Sylgard® 184. In einem verschließbaren 15 mL fassen-
den Polypropylengefäß wurden 2 mL Chloroform, 4 g Komponente A ( -divinylterminier-
tes Polydimethylsiloxan) und 0,4 g Komponente B (Copolymer aus Methylhydrosilan und
Dimethylsiloxan) vorgelegt und homogenisiert. Anschließend erfolgte die Zugabe von 1 mL
Enzymlösung. Die zur Immobilisierung eingesetzten Lipasen sowie die zugehörigen Bela-
dungsdichten sind in Tabelle 2.7 aufgeführt. Das Gemisch wurde homogenisiert, der Platin-
Katalysator SylOff® (Bis(1,3-divinyl-1,1,3,3-tetradisiloxan)-platin(0)) zugegeben und danach
das Gemisch für 15-30 min auf einem Überkopfschüttler bei RT bis zu einer erkennbaren
Zunahme der Viskosität inkubiert. Im Anschluss wurde das Gemisch in 150 mL einer auf
45 °C temperierten 4%igen (w/w) PVA-Lösung (10-98 kDa) eingetropft und für 2-3 h
turbulent gerührt. Die Größe der Siliconsphären konnte in Abhängigkeit von der Rührintensi-
tät beeinflusst werden. Die erhaltenen Siliconsphären wurden durch Filtration abgetrennt, mit
deionisiertem Wasser gewaschen und bei RT getrocknet. Je 3 mL der abfiltrierten PVA-
Lösung wurden zur Evaluierung des Immobilisierungsgrades einer Proteinmengenbe-
stimmung nach BRADFORD unterzogen. Abweichend von den Angaben in Kapitel 2.4 wurde
die Kalibrierung mit BSA in einer 4%igen PVA-Lösung vorgenommen. Das Gewicht der
erhaltenen Siliconsphären wurde bestimmt und durch Vergleich mit der Masse der einge-

33
setzten, nicht-flüchtigen Komponenten die Ausbeute bestimmt. Die Siliconsphären wurden
bis zu ihrer weiteren Verwendung bei 4 °C unter Luftabschluss oder in Hexan gelagert. Die
anvisierten Verwendungszwecke sind Tabelle 2.7, nähere Angaben zu den Reaktionen den
Kapiteln 2.13 – 2.16 zu entnehmen.
Tabelle 2.7: Proteinkonzentrationen der zur Herstellung der Statischen Emulsion eingesetzten Proteinlösungen
Enzym Proteinkonzentration [mg/mL]
Verwendung
I 4,5
II 9
III 13,5 CALA
IV 18
Synthese von Octyloctanoat (Kapitel 2.13)
I 16,2
II 22,1
III 33,1
IV 44,2
V 66,2
TLL
VI 88,3
Synthese von Octyloctanoat und
Kühlschmierstoffen (Kapitel 2.13 und 2.14)
I 6
II 12
III 18 CALB
IV 24
Synthese von Propyllaureat (Kapitel 2.15)
I 0,9
II 1,8
III 3,7
IV 5,5
PSL
V 7,3
Racematspaltung von Benzoin
(Kapitel 2.16)
2.10 Immobilisierung nach der Sol-Gel-Technik
Die kommerziell verfügbaren Lipasen wurden in unterschiedlichen Konzentrationen in 0,1 M
Kaliumphosphatpuffer (pH 7,0) gelöst. Die Konzentrationen der für die Immobilisierung ein-
gesetzten Proteinlösungen sind in Tabelle 2.8 aufgeführt. Es wurden je 400 μL der Protein-
lösungen in 2 mL fassende Polypropylengefäße (Fa. Eppendorf) vorgelegt und mit 50 μL NaF
(1 M), 100 μL wässriger PVA-Lösung (15 kDa; 4 % w/v) sowie 100 μL Isopropanol versetzt.
Nach Durchmischung der Ansätze wurden die auf 4 °C temperierten Silan-Monomere nach
absteigender Länge ihrer Alkylgruppen (480 μL i-BTMS, 440 μL PTMS und 148 μL TMOS;

Material und Methoden
34
molares Verhältnis 2,5 : 2,5 : 1) zugegeben und die Ansätze für 10 s auf dem Vortexer ge-
mischt. Die Gefäße wurden daraufhin von Hand geschwenkt und bei beginnender Gelierung
(Erwärmung) 5 min im Eisbad gekühlt. Anschließend wurden sie für 24 h bei RT belassen.
Die erhaltenen Sol-Gele wurden in Petrischalen überführt, für 24 h bei 35 °C unter Normal-
druck getrocknet und anschließend gemörsert. Nach Gewichtsbestimmung wurden die Immo-
bilisate für 2 h bei RT in 10 mL deionisiertem Wasser gewaschen. Der Proteingehalt im
Waschwasser wurde zur Bestimmung des Immobilisierungsgrades herangezogen. Danach
wurden die Immobilisate über eine Glasfritte (D4) abfiltriert und erneut mit deionisiertem
Wasser gewaschen. Die Immobilisate wurden 12 h bei 40 °C unter Normaldruck getrocknet
und anschließend bei RT gelagert. Bei Bedarf wurden auch größere Ansätze unter Multipli-
kation der zuvor genannten Volumina hergestellt. Die Größenbestimmung der Sol-Gel-Immo-
bilisate erfolgte gemäß der in Kapitel 2.11 dargelegten Methodik. Eingesetzt wurden die Sol-
Gel-Immobilisate für die Synthese von Octyloctanoat und Ethylhexyloleat (Kapitel 2.13 und
2.14).
Tabelle 2.8: Proteinnzentrationen der zur Immobilisierung in Sol-Gel eingesetzten Proteinlösungen
Enzym Proteinkonzentration [mg/mL]
I 2,6
II 4,9 TLL
III 7,5
I 4,5
II 8,9 CALA
III 17,8
2.11 Bestimmung des Durchmessers der Immobilisate
Alginatkugeln wurden durch Überführung in eine wässrige Lösung von Methylenblau (0,1 %
w/v) angefärbt und einzeln auf einer mattierten und von unten durchschienenen Plexiglas-
Platte platziert. Seitlich der Kugel waren symmetrisch vier Spiegel angeordnet, um auch die
Seitenansichten evaluieren zu können (Abb. 2.3). Auf diese Weise sollte verhindert werden,
dass durch Auswertung einer ausschließlich zweidimensionalen Ansicht eines abgeflachten
kreisförmigen Immobilisates diesem fälschlicherweise eine perfekte Kugelsymmetrie zuge-
schrieben wird. Die Alginatkugeln und die vier Spiegelbilder wurden mit einer über dem
Versuchsaufbau montierten Digitalkamera fotografiert und das digitale Bild mit der Objekter-

35
kennungssoftware ImageTool (ImageTool für Windows Version 3.0; University of Texas
Health Science Center, USA) im Hinblick auf Rundheit und Durchmesser analysiert. Dabei
wurde stets der Mittelwert aus der direkten Kugelansicht und den vier Projektionen gebildet.
Die Kugelgröße wurde bestimmt als Durchmesser einer perfekten Kreisfläche, welche
dieselbe Fläche wie die analysierte Kugel im zweidimensionalen Bild aufweist. Die genaue
Größenbestimmung war durch Kalibrierung der Objekterkennungssoftware mit Hilfe einer im
experimentellen Aufbau befindlichen Linie bekannter Länge möglich. Die Rundheit der
Kugeln wurde definiert als das Verhältnis der kürzesten zu der senkrecht stehenden längsten
Achse innerhalb der kreisförmigen Fläche. Eine perfekt runde Kugel würde demnach eine
Rundheit von 1 zeigen, wohingegen Werte < 1 eine Abweichung von der Kugelgeometrie
indizieren. Die Genauigkeit der Methode wurde mit Hilfe einer Kugellagerkugel bekannter
Größe überprüft.
Sphärische Silicon-Immobilisate wurden in eine mit einer Lösung von Methylenblau (0,1 %
w/v) gefüllten Petrischale überführt (Volumen ausreichend, um die Kugeln knapp zu
bedecken) und diese auf der von unten durchschienenen Acrylglasplatte platziert (Abb. 2.3).
Abbildung 2.3: Versuchsaufbau zur Bestimmung der Größe und Rundheit kugelförmiger Immobilisate (A:Siliconkugeln; B: Alginatkugeln).
Auch hier wurde durch eine digitale Fotografie mit Hilfe der Objekterkennungssoftware
ImageTool und einer im experimentellen Aufbau befindlichen Linie bekannter Länge der
Durchmesser der Immobilisate bestimmt. Der Durchmesser kleinerer Kugeln (< 1 mm) wurde
hingegen geschätzt. Auf eine Analyse der Rundheit der Silicon-Immobilisate konnte anders
A B
Kameraansicht
Spiegelca. 35 cm
Kamera
Alginatkugel Siliconkugel

Material und Methoden
36
als bei den Alginatkugeln aufgrund des Herstellungsverfahrens (Emulsionspolymerisation)
verzichtet werden.
Die Größenverteilung der Sol-Gel-Immobilisate wurde mit Hilfe eines Systems zur Bestim-
mung der Partikelanzahl und Größenverteilung („Particle Sizer“) ermittelt. Das Funktions-
prinzip des verwendeten Messgerätes beruht auf dem Durchfluss suspendierter Partikel durch
eine vertikale Messzelle. In der Messzelle passieren die Partikel einen senkrecht auf den
Flüssigkeitsstrom treffenden Laserstrahl. Das transmittierte und total reflektierte Licht sowie
die Verweilzeit der Partikel in der Messzelle werden durch Sensoren erfasst. Aus dem Aus-
maß der Totalreflexion des Lichtstrahls ergibt sich der Durchmesser, wohingegen das
Partikelvolumen durch Messung der Transmission und Verweilzeit bestimmt wird. Die nach
Messung erhaltene Größenverteilung gibt Auskunft über die relative Häufigkeit der Partikel
bestimmter Größen an der Gesamtpopulation der gemessenen Partikel. Der durchschnittliche
Durchmesser ergibt sich aus dem Maximum der Größenverteilung. Es wurden jeweils 200 μL
Suspension der zu vermessenden Partikel (100 mg/mL) in das mit 200 mL gefüllte Reservoir
gegeben, die Messzeit betrug 60 s.
2.12 Charakterisierung der Statischen Emulsion
2.12.1 Quellungsverhalten
Zur Untersuchung des Quellungsverhaltens größerer Kugeln (Ø > 1 mm) aus Silicon und
Statischer Emulsion wurden je 20 dieser Kugeln (ohne immobilisiertes Enzym) vor und nach
einstündiger Quellung in Hexan fotografiert. Durchmesser und Volumina der Kugeln wurden
mit der Objekterkennungssoftware ImageTool analysiert und ihre Zunahme nach Quellung
der Kugeln durch Mittelwertbildung ermittelt (Kapitel 2.11).
Zur Analyse des Quellungsverhaltens kleinerer Siliconkugeln (Ø < 1 mm) wurden die Kugeln
in einen Messzylinder gegeben und mit einem Spritzenkolben zusammengedrückt, um die
Räume zwischen den Kugeln zu minimieren (Abb. 2.4). Das Gesamtvolumen der Kugeln
wurde am Messzylinder abgelesen. Anschließend wurde Hexan zu den Kugeln gegeben und
nach einstündigem Quellen das Volumen der gequollenen Kugeln durch erneutes Kompri-
mieren bestimmt. Der Durchmesser des Spritzenkolben wurde reduziert, damit nur größere

37
Partikel (nicht aber Flüssigkeit) komprimiert werden konnten. Die Volumenzunahme ergab
sich aus der Differenz des Kugelvolumens vor und nach Quellung (V2 – V1).
Abbildung 2.4: Versuchsaufbau zur Analyse des Quellungsverhaltens kleiner Siliconsphären (Ø < 1 mm) in Hexan (V1 und V2: Kugelvolumen vor bzw. nach Quellung).
2.12.2 Abrieb und Elastizität
Zur Untersuchung der mechanischen Stabilität wurde eine Siliconkugel (Ø 2-4 mm) auf eine
Waage gelegt und mit einem Stempel komprimiert (Abb. 2.5). Die ausgeübte Gewichtskraft
konnte an der Waage abgelesen werden. Die Verformung der Kugel wurde durch Messung
der von der Druckplatte zurückgelegten Wegstrecke dokumentiert. Die Elastizität bzw.
Plastizität der Kugel wurde durch wiederholtes Komprimieren und Beobachtung dabei
auftretender (evtl. irreversibler) Verformungen abgeschätzt.
Abbildung 2.5: Versuchsaufbau zur Analyse der Elastizität der Siliconsphären (Ø 2-4 mm)
V1
V2V1
V2
Waage
zurückgelegte Wegstrecke
Siliconkugel (Ø 2-4 mm)
motorgetriebener Stempel mit Druckplatte

Material und Methoden
38
Weiterhin wurden die Siliconkugeln auf ihre Abriebfestigkeit hin untersucht. Hierzu wurden
einige große Kugeln (insgesamt ca. 300 mg) mit 8 g Glaskugeln (Ø ca. 0,3 mm) eine Woche
lang turbulent in Hexan geschüttelt und die oberflächliche Beschaffenheit der Kugeln fotogra-
fisch dokumentiert. Außerdem wurde der Gewichtsverlust der Kugeln nach Abrieb bestimmt.
2.12.3 Untersuchungen zum Massentransfer
Es wurden bis zu fünf große Siliconkugeln (Ø 2-3 mm, ohne immobilisiertes Enzym bzw.
wässrige Phase) zu 1 mL einer Lösung der Analyten in Hexan (50 mmol/L Caprylsäure, 50
mmol/L Octanol und 50 mmol/L Octyloctanoat) gegeben und für 2 h bei RT geschüttelt. Von
dieser Lösung wurden 160 μL zu 40 μL Internem Standard (100 mmol/L Decan in Hexan)
gegeben und gaschromatographisch untersucht. Die Siliconkugeln wurden anschließend durch
kurzes Schwenken an Luft oberflächlich getrocknet, in 1 mL reines Hexan überführt und
erneut 2 h bei RT geschüttelt (Abb. 3.38). Von dieser Hexanphase wurden ebenfalls 160 μL
Probe zversetzt mit 40 μL Internem Standard (100 mmol/L Decan in Hexan) zur gaschroma-
tographischen Analyse gegeben. Die in der Probe vorliegenden Molaritäten wurden unter
Berücksichtigung der bei der Probennahme erfolgten Verdünnung (durch Zugabe des Internen
Standards) berechnet. Die Siliconkugeln wurden anschließend über Nacht an Luft getrocknet
und ihre Größe mit der Objekterkennungssoftware ImageTool ermittelt (Kapitel 2.11). Die
Volumenzunahme der Kugeln durch Quellung wurde anhand des zuvor ermittelten Quell-
faktors berechnet und diente der Quantifizierung der Analytkonzentration in den Kugeln.
In einer weiteren Versuchsreihe wurde bei Herstellung der Siliconkugeln (ohne wässrige
Phase) der Mischung der Siliconkomponenten 100 mmol/L Decan beigemengt. Zur Unter-
suchung der invertierten Verteilung wurden drei Siliconkugeln (Ø 2-3 mm) zwei Stunden lang
in 0,5-1 mL Hexan geschüttelt, wovon anschließend 160 μL Probe mit 40 μL Internem
Standard (100 mmol/L Octanol in Hexan) versetzt und gaschromatographisch analysiert
wurden (Abb. 3.41). Für die Untersuchung des Massentransfers wurden fünf Kugeln
(Ø 2-3 mm) in je 0,5 mL Hexan überführt und pro Zeitpunkt (5, 10, 20, 40 und 60 min)
160 μL Probe entnommen, mit 40 μL Internem Standard versetzt und gaschromatographisch
analysiert.
Des Weiteren wurde bei Herstellung der Statischen Emulsion anstelle einer Enzymlösung der
wässrig gelöste pH-Indikator Bromkresolgrün eingesetzt. Nach beendeter Polymerisation

39
wurden die Kugeln in Hexan gequollen. Die Farbveränderung wurde durch Zugabe von
Propionsäure bzw. Butylamin qualitativ untersucht und der Zeitverlauf der Farbänderung
fotografisch dokumentiert.
2.12.4 Evaporation der emulgierten wässrigen Phase
Gewogene Siliconsphären mit emulgierter wässriger Phase wurden in ein Glasgefäß gegeben
und bei 60 °C eine Woche lang bis zur Gewichtskonstanz getrocknet. Anschließend wurde die
Gewichtsdifferenz der Siliconsphären vor und nach Trocknung bestimmt.
Zur Analyse des Einflusses der Trocknungsdauer auf die katalytische Aktivität der Statischen
Emulsion wurden Siliconsphären mit statisch emulgierter CALA und TLL für 1-3 Tage nach
Herstellung an Luft bei RT gelagert und zur Katalyse der Veresterung von Caprylsäure und
Octanol (Kapitel 2.13.1) eingesetzt. Die hierbei ermittelten Aktivitäten wurden in Relation zur
Aktivität frisch eingesetzter Immobilisate derselben Charge gesetzt.
2.12.5 Ausbluten
Für die Analyse des Ausblutens von Protein aus der Statischen Emulsion von CALA und TLL
wurde jeweils 1 g der Siliconsphären mit 5 mL entionisiertem Wasser in einer mit Parafilm
verschlossenen Spritze 5 Tage lang geschüttelt. Das Waschwasser wurde durch Kompression
von den Immobilisaten entfernt und auf den Proteingehalt hin untersucht.
2.13 Lipasenkatalysierte Synthese von Octyloctanoat
Durch Emulsion/Suspension der nativen Enzympräparate und der Immobilisate in Hexan mit
50 mmol/L Caprylsäure, 50 mmol/L n-Octanol und 20 mmol/L Decan wurde die katalytische
Aktivität bei Synthese von Octyloctanoat bestimmt. Sämtliche Reaktionsansätze wurden
schüttelnd bei RT inkubiert. Zu geeigneten Zeitpunkten wurden Proben entnommen und
gaschromatographisch analysiert (Kapitel 2.3, Methode b). Die eingesetzten Mengen an
nativem Enzym und Immobilisaten sowie das Volumen der Substratlösung ist Tabelle 2.9 zu
entnehmen.

Material und Methoden
40
Tabelle 2.9: Bei Synthese von Octyloctanoat eingesetzte Biokatalysatormengen und Substratvolumina.
Enzympräparat Volumen bzw. Einwaage
Volumen der Substratlösung [mL]
CALA 20 μL 3 Nativ
TLL 20 mg 3
CALA 200 mg 3 Statische Emulsion TLL 200 mg 6
CALA 20 und 100 mg 3 Sol-Gel
TLL 20 mg 3
Eingesetzt wurden Immobilisate verschiedener Proteinbeladungsdichten (Tab. 2.7 und 2.8).
Bei Entnahme von Proben aus Reaktionsansätzen mit nativem Enzym wurde dieses vor der
gaschromatographischen Analyse durch Zentrifugation abgetrennt. Um den Einfluss von
Wasser auf die Reaktionskinetik der nativen Enzyme zu untersuchen, wurde weiteren
Ansätzen unterschiedliche Mengen (2,5; 5; 10; 30; 60; 90; 120 % v/v) Kaliumphosphatpuffer
(0,1 mol/L; pH 7,0) zugesetzt.
2.14 Lipasenkatalysierte Synthese von Fettsäureethylhexylestern
Als Modellreaktion für die Synthese von Esterölen diente die Veresterung von Ölsäure mit
2-Ethyl-1-hexanol. Zu diesem Zweck wurden je 250 mg der Statischen Emulsion mit TLL,
250 mg Sol-Gel-Immobilisat mit TLL und 20 mg des lyophilisierten Enzyms in 20 mL
fassende verschließbare Reaktionsgefäße überführt und die Reaktion durch Zugabe von 3 mL
Substratlösung (äquimolare Mengen von 2-Ethyl-1-hexanol und Ölsäure sowie 2,5 % (v/v)
Dodecan als Interner Standard) gestartet. Es wurden Immobilisate unterschiedlicher Enzym-
beladungsdichten eingesetzt (Tab. 2.7 und 2.8). Die Umsetzung erfolgte bei 50 °C unter
vollständiger Durchmischung mittels Rührfisch. Die initiale Reaktionsrate wurde durch Ent-
nahme von 10 μL Probe zu geeigneten Zeitpunkten dokumentiert. Die Proben wurden 1 : 100
mit Hexan verdünnt und gaschromatographisch analysiert (Kapitel 2.3, Methode d). Um den
Einfluss von Wasser abzuschätzen, wurde einigen Ansätzen 1,5 mL deionisiertes Wasser
zugegeben. Sämtliche Versuchsansätze wurden mindestens zweimal durchgeführt. Zudem
wurde durch Zugabe eines organischen Lösungsmittels zu den Versuchsansätzen der Einfluss
von n-Hexan eruiert. Auch hier erfolgte die Bestimmung der initialen Reaktionsrate durch
Entnahme von 10 μL Probe zu geeigneten Zeitpunkten und der Quantfizierung der gebildeten
Menge Ethylhexyloleat.
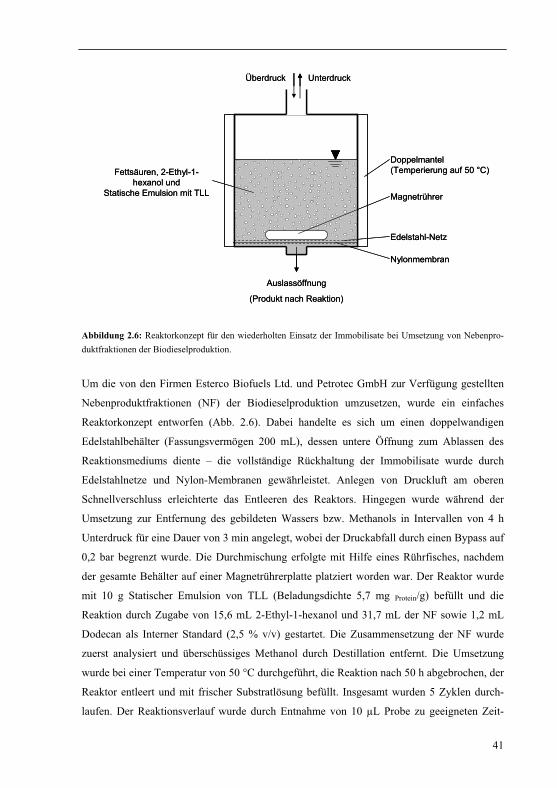
41
Abbildung 2.6: Reaktorkonzept für den wiederholten Einsatz der Immobilisate bei Umsetzung von Nebenpro-duktfraktionen der Biodieselproduktion.
Um die von den Firmen Esterco Biofuels Ltd. und Petrotec GmbH zur Verfügung gestellten
Nebenproduktfraktionen (NF) der Biodieselproduktion umzusetzen, wurde ein einfaches
Reaktorkonzept entworfen (Abb. 2.6). Dabei handelte es sich um einen doppelwandigen
Edelstahlbehälter (Fassungsvermögen 200 mL), dessen untere Öffnung zum Ablassen des
Reaktionsmediums diente – die vollständige Rückhaltung der Immobilisate wurde durch
Edelstahlnetze und Nylon-Membranen gewährleistet. Anlegen von Druckluft am oberen
Schnellverschluss erleichterte das Entleeren des Reaktors. Hingegen wurde während der
Umsetzung zur Entfernung des gebildeten Wassers bzw. Methanols in Intervallen von 4 h
Unterdruck für eine Dauer von 3 min angelegt, wobei der Druckabfall durch einen Bypass auf
0,2 bar begrenzt wurde. Die Durchmischung erfolgte mit Hilfe eines Rührfisches, nachdem
der gesamte Behälter auf einer Magnetrührerplatte platziert worden war. Der Reaktor wurde
mit 10 g Statischer Emulsion von TLL (Beladungsdichte 5,7 mg Protein/g) befüllt und die
Reaktion durch Zugabe von 15,6 mL 2-Ethyl-1-hexanol und 31,7 mL der NF sowie 1,2 mL
Dodecan als Interner Standard (2,5 % v/v) gestartet. Die Zusammensetzung der NF wurde
zuerst analysiert und überschüssiges Methanol durch Destillation entfernt. Die Umsetzung
wurde bei einer Temperatur von 50 °C durchgeführt, die Reaktion nach 50 h abgebrochen, der
Reaktor entleert und mit frischer Substratlösung befüllt. Insgesamt wurden 5 Zyklen durch-
laufen. Der Reaktionsverlauf wurde durch Entnahme von 10 μL Probe zu geeigneten Zeit-
Auslassöffnung
(Produkt nach Reaktion)
Überdruck Unterdruck
Doppelmantel (Temperierung auf 50 °C)
Magnetrührer
Edelstahl-Netz
Nylonmembran
Fettsäuren, 2-Ethyl-1-hexanol und
Statische Emulsion mit TLL
Auslassöffnung
(Produkt nach Reaktion)
Überdruck Unterdruck
Doppelmantel (Temperierung auf 50 °C)
Magnetrührer
Edelstahl-Netz
Nylonmembran
Fettsäuren, 2-Ethyl-1-hexanol und
Statische Emulsion mit TLL

Material und Methoden
42
punkten und anschließender gaschromatographischer Analyse dokumentiert (Kapitel 2.3,
Methode d). Die aufgefangene Produktlösung wurde durch Vakuumdestillation (0,1 bar) bei
100 °C von überschüssigem Alkohol und Wasser befreit. Anschließend wurde das Destillat
mit einem äquivalenten Volumen Ethylamin versetzt, um freie Fettsäuren zu extrahieren.
Nach Zentrifugation (4500 Upm, 4 min) wurde die Oberphase verworfen und der Unterphase
mit einer Evaporationszentrifuge das überschüssige Ethylamin entzogen.
2.15 Lipasenkatalysierte Synthese von Propyllaurat
Zur Bestimmung der katalytischen Aktivität bei Veresterung von n-Propanol und Laurinsäure
wurden in getrennten Reaktionsansätzen 50 μL des nativen CALB-Flüssigpräparates, 50 mg
Statischer Emulsion mit CALB (Beladungsdichten Tab. 2.7) und 2,5 mg Novozym 435 in
verschließbare 10-mL-Reaktionsgefäße aus Glas mit 2 mL Substratlösung (äquimolare Men-
gen n-Propanol und Laurinsäure sowie 2,5 % (v/v) Dodecan als Interner Standard) überführt.
Die Versuchsdauer betrug 4 h, wobei in geeigneten Zeitabständen 10 μL Probe genommen
und mit 990 μL n-Decan verdünnt wurden. Anschließend wurden Edukt- und Produktkonzen-
trationen gaschromatographisch analysiert (Kapitel 2.3, Methode c) und die initiale Reak-
tionsrate berechnet. Die Reaktionstemperatur betrug 60 °C, die Reaktionsansätze wurden mit
Magnetrührern durchmischt. Um den Einfluss von Wasser abzuschätzen, wurde in einer
weiteren Versuchsreihe Reaktionsansätzen mit dem nativen CALB-Flüssigpräparat 1,5 mL
deionisiertes Wasser zugegeben.
2.16 Lipasenkatalysierte Racematspaltung von Benzoin
Es wurden 25 mg und 50 mg der lyophilisierten Lipase von Pseudomonas stutzeri (PSL) in
7,5 mL der Substratlösung suspendiert. Die Substratlösung bestand aus Tetrahydrofuran
(THF) mit 1,084 mol/L Vinylbutyrat, 94,2 mmol/L R/S-Benzoin und 200 mmol/L Decan als
Interner Standard. Die Reaktionsansätze wurden über einen Zeitraum von 50 min inkubiert
und dabei mit Magnetrührern durchmischt. Zudem wurden je 250 mg Siliconsphären mit in
verschiedenen Beladungdichten statisch emulgierter PLS (Tab. 2.7) in 2,5 mL Substratlösung
obiger Zusammensetzung überführt und rührend bei RT über einen Zeitraum von 25 h
inkubiert. Zu geeigneten Zeitpunkten wurden 75 μL Probe entnommen und zunächst gaschro-

43
matographisch analysiert. Die initiale Reaktionsrate wurde über die Quantifizierung der
Benzoinbutyrat-Konzentration ermittelt. Für die Kalibrierung wurden Racemate von Benzoin
und Benzoinbutyrat benutzt, da die reinen Enantiomere als Standards nicht verfügbar waren.
Die Betriebsparameter des Gaschromatographen (Kapitel 2.3, Methode e) wurden so gewählt,
dass jeweils beide Enantiomere von Produkt bzw. Substrat mit der gleichen Retentionszeit
von der Säule eluierten. Anschließend wurden die Proben auf einer chiralen HPLC-Säule
analysiert, um den E-Wert der Reaktion zu berechnen. Die Betriebsparameter der HPLC sind
Tabelle 2.10 zu entnehmen.
Tabelle 2.10: Betriebsparameter der HPLC zur Analyse von R/S-Benzoin und R/S-Benzoinbutyrat.
HPLC-Betriebsparameter
Detektor RI(Refraktometer)
Vorsäule Chiralpak AD-H
0,4 x 1 cm (DAICEL, Japan)
SäuleChiralpak AD-H
0,46 x 25 cm (DAICEL, Japan)
Laufmittel 90 % n-Hexan
10 % i-Propanol (isokratisch)
Temperatur 25 °C
Durchflussrate 0,6 mL/min
Injektionsvolumen 20 μL
R-Benzoinbutyrat 12,06
S-Benzoinbutyrat 16,14
R-Benzoin 29,27
Retentionszeit
[min]
S-Benzoin 39,21
Der von CHEN ET AL. [1982] eingeführte dimensionslose Selektivitätsparameter „E“ entspricht
dem Verhältnis der Geschwindigkeitskonstanten der Reaktionen beider Enantiomere (E =
kR/kS) und ist anders als der Enantiomerenüberschuss unabhängig von Substratkonzentration
und molarer Konversion. Der E-Wert wurde mit Hilfe folgender Formel (1) berechnet:
(1)
mit e.e.P = Enantiomerenüberschuss des Produktes
e.e.S = Enantiomerenüberschuss des Substrates
E = ln [(1 – e.e.S)/(1 + e.e.S/e.e.P)]ln [(1 + e.e.S)/(1 + e.e.S/e.e.P)]E = ln [(1 – e.e.S)/(1 + e.e.S/e.e.P)]ln [(1 + e.e.S)/(1 + e.e.S/e.e.P)]

Material und Methoden
44
Um den Einfluss von Wasser auf die Reaktionskinetik zu untersuchen, wurden 50 mg
lyophilisierter PSL in getrennten Reaktionsansätzen zusammen mit unterschiedlichen Mengen
Wasser (0; 25; 50; 150 μL) in 7,5 mL Substratlösung gegeben und der Reaktionsverlauf durch
Quantifizierung der Konzentrationen von Buttersäure und Benzoinbutyrat dokumentiert. In
einer weiteren Versuchsreihe wurde der Einfluss der Trocknungszeit nach Herstellung der
Siliconsphären auf die Reaktionskinetik untersucht. Zu diesem Zweck wurde eine Statische
Emulsion der höchsten Beladungsdichte (Tab. 2.7) hergestellt und diese in sechs Aliquots
aufgeteilt. 250 mg eines Aliquots wurden unmittelbar in 2,5 mL Substratlösung eingesetzt, die
übrigen Aliquots erst nach einer Expositionsdauer von 3, 6, 16,5, 20 und 25 h bei 40 °C an
Luft. Die Dokumentation des Reaktionsverlaufs erfolgte wie eingangs beschrieben.
Um die Verteilung der Substrate und Produkte dieser Reaktion zu eruieren, wurden 250 mg
Siliconsphären mit enzymfreier wässriger Phase in 1 mL THF suspendiert. Die Substrate
Vinylbutyrat und Benzoin wurden getrennt voneinander in je drei verschiedenen Konzentra-
tionen (50; 75 und 100 mmol/L) in THF vorgelegt. Die Verteilung von Benzoinbutyrat wurde
für zwei Konzentrationen (75 und 100 mmol/L) untersucht. Hierfür wurden Proben vor und
24 h nach Überführung der Siliconsphären in THF genommen und gaschromatographisch
analysiert.

Ergebnisse und Diskussion
45
3. Ergebnisse und Diskussion
3.1 pH-Optimum der lipasenkatalysierten Estersynthese
Enzymmoleküle weisen in der Regel eine Nettoladung auf, die auf funktionelle Gruppen
bestimmter Aminosäurereste zurückgeführt werden kann. In wässrigen Lösungen hängt die
Höhe der Nettoladung von pH-bedingten Änderungen der Dissoziationsgleichgewichte ab
[BISSWANGER, 2000; NEVES-PETERSEN ET AL., 2001a/b]. Da der Dissoziationsgrad einzelner
funktioneller Gruppen sowohl Konformation als auch Interaktion zwischen Enzym und
Substrat beeinflussen kann, ist die Wahl bzw. Identifikation des optimalen pH-Wertes im
Interesse einer hohen Produktivität von großer Bedeutung. Für die lipasenkatalysierte
Veresterung können der Fachliteratur diesbezüglich jedoch kaum verwertbare Informationen
entnommen werden. Viele Wissenschaftler setzen voraus, dass das pH-Optimum der Hydro-
lyse identisch ist mit dem der Umkehrreaktion [GONZÁLEZ-NAVARRO UND BRACO, 1997 und
1998; TWEDDELL ET AL., 1998; DOSANJH UND KAUR, 2002; MIYAKO ET AL., 2003]. Inwieweit
diese Annahme gerechtfertigt ist, wurde dabei nicht hinterfragt. Auch wurde bisher nie die
Frage aufgeworfen, ob die protonierte oder die deprotonierte Säure als Substrat bevorzugt
wird [HEINEMANN, 2003]. Als Folge der gegenwärtig unbefriedigenden Informationslage ist
eine rationale Prozessentwicklung bzw. -optimierung erschwert. Gerade bei Verwendung von
Hydrogelen können die üblichen Puffersysteme bei Zuführung einer Säure einen pH-Abfall
nicht verhindern [HEINEMANN, 2003; DOUMÈCHE, 2002]. Inwieweit weitergehende Maß-
nahmen zur Kontrolle des pH-Wertes angeraten sind, kann erst nach detaillierter Unter-
suchung der pH-Abhängigkeit der lipasenkatalysierten Veresterung beurteilt werden.
Zunächst mag die Feststellung, dass bezüglich des pH-Optimums der lipasenkatalysierten
Estersynthese keine belastbaren Daten verfügbar sind, überraschen. Ein Blick auf die be-
sondere Problematik der Veresterung und ihrer immanenten Komplexität macht es ver-
ständlicher: Normalerweise wird der pH-Wert bei konstanter Substratkonzentration variiert
[BISSWANGER, 2000]. Ist jedoch eine Säure Substrat, muss berücksichtigt werden, dass sich
das Gleichgewicht zwischen deprotonierter und protonierter Säure ebenfalls in Abhängigkeit
vom pH-Wert ändert. Da die deprotonierte Säure jedoch negativ geladen ist, besteht die
Möglichkeit, dass sie im Vergleich zur protonierten Säure anders mit dem aktiven Zentrum
interagiert. Beide Säureformen können daher nicht gleichartig behandelt werden und die pH-

Ergebnisse und Diskussion
46
bedingte Verschiebung des Dissoziationsgleichgewichtes könnte in einer pH-Abhängigkeit
resultieren, die nicht auf enzymspezifischen Effekten, sondern vielmehr auf einer veränderten
Substratkonzentration basiert. Zudem muss die Reaktion in organischen Lösungsmitteln oder
in wässrig-organischen Zweiphasensystemen ausgeführt werden, da aus thermodynamischen
Gründen in rein wässriger Umgebung eine Esterbildung kaum beobachtet werden kann
[KVITTINGEN, 1994]. Bei Suspension des Enzyms im organischen Lösungsmittel stellt sich die
Frage, wie der pH-Wert einzustellen und ggf. zu kontrollieren ist. Zusätzlich muss berück-
sichtigt werden, dass sich im Zuge der Reaktion in unmittelbarer Umgebung des Enzyms eine
distinkte wässrige Phase bildet. Das Vorliegen einer zweiten Phase bedingt jedoch auch, dass
sich Substrat und Produkt in Abhängigkeit von ihren Verteilungskoeffizienten zwischen der
wässrigen und der organischen Phase verteilen. Die Gleichgewichtskonzentrationen hängen
dabei vom Volumenverhältnis zwischen beiden Phasen ab. Erschwerend kommt hinzu, dass
sich die Verteilungskoeffizienten der protonierten und der deprotonierten Säure deutlich
unterscheiden: Die deprotonierte Säure kann nur in der wässrigen Phase, die protonierte Säure
hingegen in beiden Phasen vorliegen – die jeweiligen Konzentrationen in wässriger und
organischer Phase hängen somit ebenfalls vom pH-Wert ab. Die genannten Aspekte beein-
flussen die Enzymkinetik kumulativ und überdecken möglicherweise die intrinsische pH-
Abhängigkeit des Enzyms.
3.1.1 Wahl und Charakterisierung eines geeigneten Untersuchungssystems
Voraussetzung für die Bestimmung des pH-Optimums der Estersynthese ist ein stark verein-
fachtes Untersuchungssystem, das die Kontrolle aller relevanten Faktoren erlaubt. Aus diesem
Grund wurden die anstehenden Untersuchungen in einem an die so genannte Lewis-Zelle
angelehnten Zweiphasensystem durchgeführt (Abb. 2.1). Dabei handelte es sich um einen
Reaktor, in dem die enzymhaltige wässrige Phase mit einer organischen Phase überschichtet
wurde. Um die Ausbildung von Konzentrationsgradienten zu vermeiden, wurden sowohl die
wässrige als auch die organische Phase durchmischt, wobei die Integrität und somit die Größe
der Grenzfläche erhalten blieb. Bisher wurde dieser Reaktortyp eingesetzt, um den Massen-
transfer oder Enzyminaktivierungen in Zweiphasensystemen zu untersuchen [GHATORAE ET
AL., 1994; GARGOURI UND LEGOY, 1997; BAUER ET AL., 2002]. In der anstehenden Unter-
suchung sollten die initialen Reaktionsraten der Lipasen aus Candida rugosa (CRL) und
Thermomyces lanuginosa (TLL) in Abhängigkeit vom pH-Wert bestimmt werden. Ausge-
wählt wurden diese Lipasen, da sie sich in ihren pH-Optima für die Hydrolyse deutlich
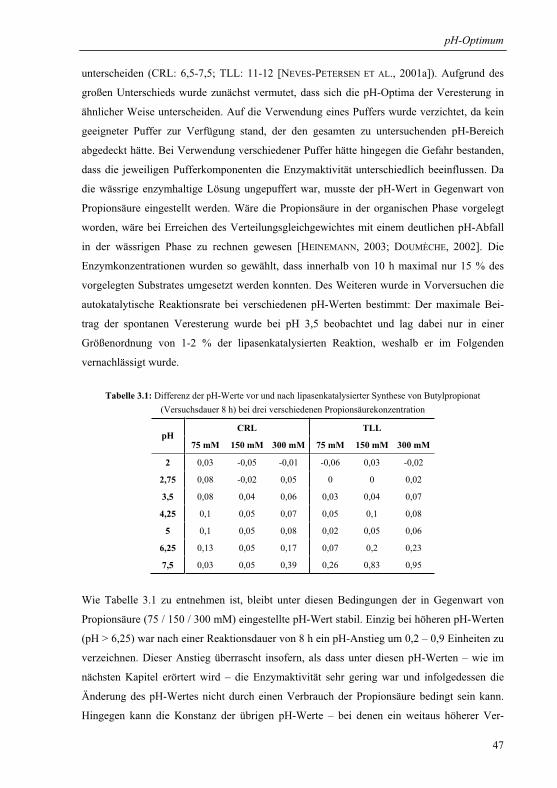
pH-Optimum
47
unterscheiden (CRL: 6,5-7,5; TLL: 11-12 [NEVES-PETERSEN ET AL., 2001a]). Aufgrund des
großen Unterschieds wurde zunächst vermutet, dass sich die pH-Optima der Veresterung in
ähnlicher Weise unterscheiden. Auf die Verwendung eines Puffers wurde verzichtet, da kein
geeigneter Puffer zur Verfügung stand, der den gesamten zu untersuchenden pH-Bereich
abgedeckt hätte. Bei Verwendung verschiedener Puffer hätte hingegen die Gefahr bestanden,
dass die jeweiligen Pufferkomponenten die Enzymaktivität unterschiedlich beeinflussen. Da
die wässrige enzymhaltige Lösung ungepuffert war, musste der pH-Wert in Gegenwart von
Propionsäure eingestellt werden. Wäre die Propionsäure in der organischen Phase vorgelegt
worden, wäre bei Erreichen des Verteilungsgleichgewichtes mit einem deutlichen pH-Abfall
in der wässrigen Phase zu rechnen gewesen [HEINEMANN, 2003; DOUMÈCHE, 2002]. Die
Enzymkonzentrationen wurden so gewählt, dass innerhalb von 10 h maximal nur 15 % des
vorgelegten Substrates umgesetzt werden konnten. Des Weiteren wurde in Vorversuchen die
autokatalytische Reaktionsrate bei verschiedenen pH-Werten bestimmt: Der maximale Bei-
trag der spontanen Veresterung wurde bei pH 3,5 beobachtet und lag dabei nur in einer
Größenordnung von 1-2 % der lipasenkatalysierten Reaktion, weshalb er im Folgenden
vernachlässigt wurde.
Tabelle 3.1: Differenz der pH-Werte vor und nach lipasenkatalysierter Synthese von Butylpropionat (Versuchsdauer 8 h) bei drei verschiedenen Propionsäurekonzentration
CRL TLL pH
75 mM 150 mM 300 mM 75 mM 150 mM 300 mM
2 0,03 -0,05 -0,01 -0,06 0,03 -0,02
2,75 0,08 -0,02 0,05 0 0 0,02
3,5 0,08 0,04 0,06 0,03 0,04 0,07
4,25 0,1 0,05 0,07 0,05 0,1 0,08
5 0,1 0,05 0,08 0,02 0,05 0,06
6,25 0,13 0,05 0,17 0,07 0,2 0,23
7,5 0,03 0,05 0,39 0,26 0,83 0,95
Wie Tabelle 3.1 zu entnehmen ist, bleibt unter diesen Bedingungen der in Gegenwart von
Propionsäure (75 / 150 / 300 mM) eingestellte pH-Wert stabil. Einzig bei höheren pH-Werten
(pH > 6,25) war nach einer Reaktionsdauer von 8 h ein pH-Anstieg um 0,2 – 0,9 Einheiten zu
verzeichnen. Dieser Anstieg überrascht insofern, als dass unter diesen pH-Werten – wie im
nächsten Kapitel erörtert wird – die Enzymaktivität sehr gering war und infolgedessen die
Änderung des pH-Wertes nicht durch einen Verbrauch der Propionsäure bedingt sein kann.
Hingegen kann die Konstanz der übrigen pH-Werte – bei denen ein weitaus höherer Ver-
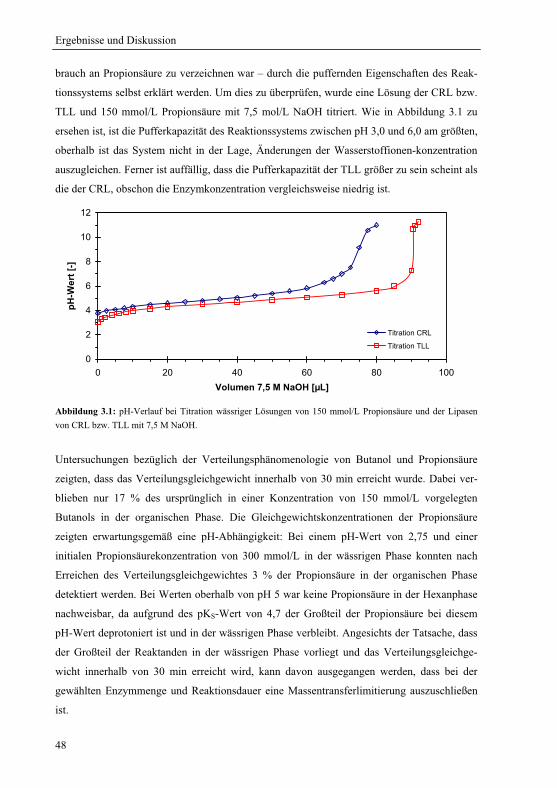
Ergebnisse und Diskussion
48
brauch an Propionsäure zu verzeichnen war – durch die puffernden Eigenschaften des Reak-
tionssystems selbst erklärt werden. Um dies zu überprüfen, wurde eine Lösung der CRL bzw.
TLL und 150 mmol/L Propionsäure mit 7,5 mol/L NaOH titriert. Wie in Abbildung 3.1 zu
ersehen ist, ist die Pufferkapazität des Reaktionssystems zwischen pH 3,0 und 6,0 am größten,
oberhalb ist das System nicht in der Lage, Änderungen der Wasserstoffionen-konzentration
auszugleichen. Ferner ist auffällig, dass die Pufferkapazität der TLL größer zu sein scheint als
die der CRL, obschon die Enzymkonzentration vergleichsweise niedrig ist.
0
2
4
6
8
10
12
0 20 40 60 80 100Volumen 7,5 M NaOH [µL]
pH
-Wer
t [-
]
Titration CRL
Titration TLL
Abbildung 3.1: pH-Verlauf bei Titration wässriger Lösungen von 150 mmol/L Propionsäure und der Lipasen von CRL bzw. TLL mit 7,5 M NaOH.
Untersuchungen bezüglich der Verteilungsphänomenologie von Butanol und Propionsäure
zeigten, dass das Verteilungsgleichgewicht innerhalb von 30 min erreicht wurde. Dabei ver-
blieben nur 17 % des ursprünglich in einer Konzentration von 150 mmol/L vorgelegten
Butanols in der organischen Phase. Die Gleichgewichtskonzentrationen der Propionsäure
zeigten erwartungsgemäß eine pH-Abhängigkeit: Bei einem pH-Wert von 2,75 und einer
initialen Propionsäurekonzentration von 300 mmol/L in der wässrigen Phase konnten nach
Erreichen des Verteilungsgleichgewichtes 3 % der Propionsäure in der organischen Phase
detektiert werden. Bei Werten oberhalb von pH 5 war keine Propionsäure in der Hexanphase
nachweisbar, da aufgrund des pKS-Wert von 4,7 der Großteil der Propionsäure bei diesem
pH-Wert deprotoniert ist und in der wässrigen Phase verbleibt. Angesichts der Tatsache, dass
der Großteil der Reaktanden in der wässrigen Phase vorliegt und das Verteilungsgleichge-
wicht innerhalb von 30 min erreicht wird, kann davon ausgegangen werden, dass bei der
gewählten Enzymmenge und Reaktionsdauer eine Massentransferlimitierung auszuschließen
ist.

pH-Optimum
49
3.1.2 pH-Optima
Überraschenderweise zeigten beide Lipasen ihre maximale katalytische Aktivität bei extrem
niedrigen pH-Werten von 3,5 (CRL) und 4,25 (TLL). Im neutralen bis schwach alkalischen
Bereich konnte für beide Lipasen fast keine Produktbildung nachgewiesen werden. Während
die pH-Optima nahe beieinander liegen, weicht die Form der ermittelten pH-Optimumskurven
deutlich voneinander ab (Abb. 3.2). Für die CRL können im Bereich von + 0,75 pH-Einheiten
um das pH-Optimum noch 80-90 % der maximalen Aktivität erzielt werden, wohingegen die
Aktivität der TLL im gleichen Bereich auf 40 bzw. 20 % absinkt. Die Breite des pH-
Optimums der CRL kann womöglich auf das Vorliegen verschiedener Isoformen im kommer-
ziellen Präparat zurückgeführt werden [DOMÍNGUEZ DE MARÍA ET AL. 2004 und 2006].
Insgesamt sind die gemessenen pH-Optima deutlich niedriger als jene, die für die hydro-
lytische Reaktion in der Literatur genannt werden (6,5 – 7,5 für CRL und 11 – 12 für TLL
[NEVES-PETERSEN ET AL. 2001a]). Auch wenn der Fachliteratur keine präzisen Angaben
hinsichtlich des pH-Optimums der Veresterung entnommen werden können, lässt sich zumin-
dest im Fall der TLL ein Hinweis finden: CROOKS ET AL. [1995] untersuchten die durch die
Lipasen von Thermomyces lanuginosa und Rhizomucor miehei katalysierte Veresterung von
n-Octanol und Decansäure und erzielten für beide Lipasen die höchsten Reaktionsraten bei
pH 6,1, dem niedrigsten in dieser Studie applizierten pH-Wert. Es ist davon auszugehen, dass
eine weitere Absenkung des pH-Wertes eine weitere Steigerung der katalytischen Aktivität
nach sich gezogen hätte. Ähnlichen Beobachtungen machten MANOHAR UND DIVAKAR
[2002]: pH-Werte um pH 6 oder niedriger begünstigen eine Veresterung von m-Kresol und
Essigsäure, während höhere pH-Werte die verwendete Schweine-Pankreas Lipase sogar zu
inaktivieren schienen. In all diesen Arbeiten wurden jedoch keine systematischen Begrün-
dungen für ein Syntheseoptimum bei niedrigen pH-Werten angeführt, die sich mit den unter-
schiedlichen Ladungszuständen der Lipase bei verschiedenen pH-Werten und der damit
veränderten Interaktion von Substrat und Enzym beschäftigten. Erstaunlicherweise scheint es,
dass bisher in keiner Veresterungen betreffenden Untersuchung pH-Werte von weniger als 4,0
eingestellt wurden. Ein Grund hierfür könnte die womöglich allgemein vorherrschende Ver-
mutung sein, dass nur Enzyme aus extremophilen Organismen in Gegenwart derart niedriger
pH-Werte eingesetzt werden können. Dem kann aber entgegengehalten werden, dass die Sta-
bilität der meisten Lipasen nicht allzu stark vom pH-Wert abhängt, wie z.B. von MONTERO ET
AL. [1993] für die CRL im pH-Bereich von 3,0 – 7,0 gezeigt wurde. In diesem Zusammen-
hang muss betont werden, dass die in dieser Arbeit gemessenen Reaktionsraten über die
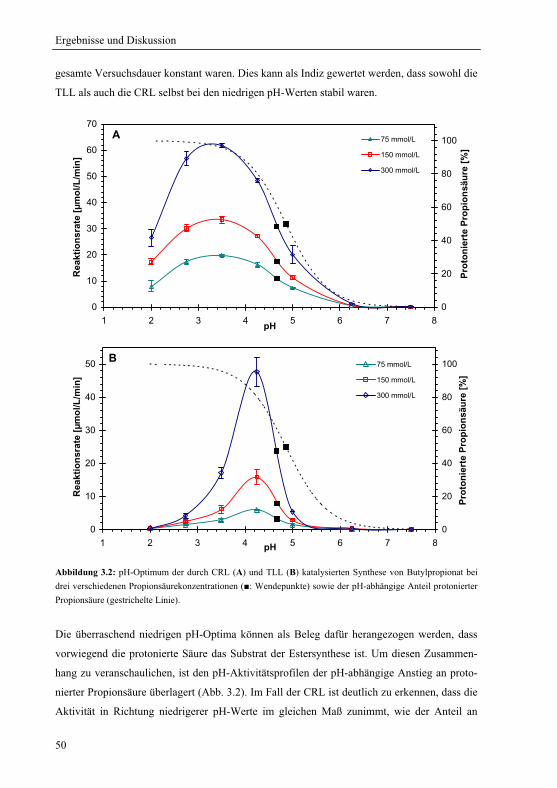
Ergebnisse und Diskussion
50
gesamte Versuchsdauer konstant waren. Dies kann als Indiz gewertet werden, dass sowohl die
TLL als auch die CRL selbst bei den niedrigen pH-Werten stabil waren.
Abbildung 3.2: pH-Optimum der durch CRL (A) und TLL (B) katalysierten Synthese von Butylpropionat bei drei verschiedenen Propionsäurekonzentrationen ( : Wendepunkte) sowie der pH-abhängige Anteil protonierter Propionsäure (gestrichelte Linie).
Die überraschend niedrigen pH-Optima können als Beleg dafür herangezogen werden, dass
vorwiegend die protonierte Säure das Substrat der Estersynthese ist. Um diesen Zusammen-
hang zu veranschaulichen, ist den pH-Aktivitätsprofilen der pH-abhängige Anstieg an proto-
nierter Propionsäure überlagert (Abb. 3.2). Im Fall der CRL ist deutlich zu erkennen, dass die
Aktivität in Richtung niedrigerer pH-Werte im gleichen Maß zunimmt, wie der Anteil an
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8pH
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
0
20
40
60
80
100
Pro
ton
iert
e P
rop
ion
säu
re [
%]
75 mmol/L
150 mmol/L
300 mmol/L
0
10
20
30
40
50
1 2 3 4 5 6 7 8pH
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
0
20
40
60
80
100
Pro
ton
iert
e P
rop
ion
säu
re [
%]
75 mmol/L
150 mmol/L
300 mmol/L
A
B

pH-Optimum
51
protonierter Säure. Ferner fallen die Wendepunkte der Aktivitätsprofile rechts der pH-Optima
beider Lipasen mit dem Wendepunkt der Titrationskurve für die protonierte Säure zusammen.
Die maximale Aktivität der CRL wird bei pH 3,5 beobachtet – bei diesem pH-Wert sind 96 %
der Gesamtsäure protoniert. Der bei weiterer Absenkung des pH-Wertes zu beobachtende
Aktivitätsabfall ist – da die maximale Menge an protonierter Propionsäure verfügbar ist – auf
nicht genauer zu benennende aktivitätsmindernde Effekte zurückzuführen. Ein ähnlicher
Zusammenhang zeigt sich für die TLL, jedoch ist die Übereinstimmung zwischen Zunahme
des Anteils protonierter Säure und dem Aktivitätsanstieg der TLL weniger stark ausgeprägt
als bei der CRL. Die TLL zeigt ihre maximale Aktivität bei einem pH-Wert von 4,25 – bei
diesem pH-Wert sind jedoch nur 80 – 85 % der Propionsäure protoniert. Diese Beobachtung
legt den Schluss nahe, dass die Aktivität der TLL noch von weiteren Faktoren neben der
Verfügbarkeit an protonierter Säure beeinflusst wird, etwa der Protonierung bestimmter funk-
tioneller Gruppen im Enzym selbst. Unterhalb von pH 4,25 ist zudem ein deutlich stärkerer
Abfall der Aktivität zu verzeichnen als bei der CRL – auch das kann nur durch spezifische
aktivitätsmindernde Effekte erklärt werden. Nichtsdestotrotz kann mit einiger Berechtigung
die Aussage getroffen werden, dass die Aktivität beider Lipasen stark mit der Verfügbarkeit
an protonierter Säure korreliert ist und deshalb vermutlich ausschließlich die protonierte Säure
als Substrat für die Estersynthese verwertet wird – eine Behauptung, die in Kapitel 3.1.5
aufgegriffen und mechanistisch untermauert wird.
3.1.3 Einfluss der Propionsäurekonzentration auf das pH-Aktivitätsprofil
3.1.3.1 pH-Aktivitätsprofil bei drei verschiedenen Gesamtsäurekonzentrationen
Um die Frage zu beantworten, ob bei der Estersynthese vorwiegend die protonierte oder
deprotonierte Säure umgesetzt wird, wäre es von Interesse, die Konzentration der protonierten
oder deprotonierten Säurespezies über den gesamten pH-Bereich konstant zu halten. Dies
könnte jedoch nur unter Variation der Gesamtsäurekonzentration erreicht werden, was sich
bei näherer Betrachtung als nicht durchführbar erweist. So beträgt bspw. bei einer Gesamt-
säurekonzentration von 0,3 mol/L und einem pH-Wert von 3,5 die Konzentration protonierter
Propionsäure 0,29 mol/L. Um dieselbe Konzentration bei einem pH-Wert von 6,35 zu
erreichen, müsste die Gesamtsäurekonzentration auf ca. 7,2 mol/L, bei einem pH-Wert von
7,5 sogar auf 123 mol/L angehoben werden. Aus diesem Grund muss bei Interpretation des

Ergebnisse und Diskussion
52
pH-Aktivitätsprofils die pH-abhängige Konzentrationsänderung der beiden Säurespezies
berücksichtigt werden.
Aufgrund der in Kapitel 3.1.2 erzielten Ergebnisse wurde vermutet, dass ausschließlich die
protonierte Säure umgesetzt wird. Folglich darf spekuliert werden, dass bei pH-Werten > pKS
deutlich unterhalb des Km-Wertes agiert wird. Ist dies der Fall, handelt es sich um eine
Reaktion erster Ordnung – eine Erhöhung der Gesamtsäurekonzentration müsste mit einer
proportionalen Erhöhung der Reaktionsgeschwindigkeit einhergehen. Oberhalb des Km-
Wertes wird diese Korrelation abgeschwächt bis hin zu einer Reaktion nullter Ordnung – die
Reaktionsgeschwindigkeit wäre dann unabhängig von der Substratkonzentration. Letztge-
nannte Feststellung gilt natürlich nur dann, wenn keine Substratinhibition besteht. Würden die
pH-Aktivitätsprofile bei verschiedenen Gesamtsäurekonzentrationen ermittelt, wären folgende
Szenarien denkbar:
A: Über den gesamten pH-Bereich wird unterhalb des Km-Wertes operiert und
infolgedessen bei höheren Substratkonzentrationen eine höhere Aktivität gemessen. Die
Lage des pH-Optimums ändert sich dabei nicht.
B: Konstante Lage der pH-Optima – allerdings sind rechts des Optimums deutlich höhere
Aktivitäten feststellbar, da nur hier die Reaktion einer Kinetik erster Ordnung folgt.
C: Mit Erhöhung der Säurekonzentration ist auch eine Verschiebung des pH-Optimums
zu verzeichnen, verursacht 1) durch Substratinhibition und/oder 2) enzymspezifische
Effekte.
Nach Auswertung der Experimente offenbarte sich im untersuchten Konzentrationsbereich für
beide Lipasen Szenario A (Abb. 3.2). Bei Erhöhung der Gesamtsäurekonzentration ist keine
Verschiebung des pH-Optimums und über den gesamten pH-Bereich ein Aktivitätszuwachs
zu verzeichnen. Es ist somit offensichtlich, dass beide Lipasen bei einer Gesamtsäurekonzen-
tration von 75 bis 300 mmol/L im gesamten pH-Bereich unterhalb der maximal möglichen
Umsetzungsgeschwindigkeit arbeiten. Zudem kann in diesem Konzentrationsbereich keine
Substratinhibition festgestellt werden. Nicht vergessen werden darf allerdings, dass in einem
weiter gefassten Konzentrationsbereich das Szenario A fließend in B bzw. C übergehen
könnte. Letztlich handelt es sich bei den im Rahmen dieser Arbeit aufgenommenen pH-
Optima um globale Optima, die sich aus der Summe von enzymspezifischen und aus
unterschiedlichen effektiven Substratkonzentrationen (protonierte Säure) herrührenden Ein-

pH-Optimum
53
flüssen ergeben und deshalb stets abgewogen werden muss, welcher Einfluss dominiert.
Bspw. könnte Szenario C, d.h. eine Verschiebung des pH-Optimums nach rechts bei Erhö-
hung der Gesamtsäurekonzentration, in zwei Fällen beobachtet werden: Im ersten Fall wäre
eine Substratinhibition bei niedrigen pH-Werten der Grund, denn die inhibierende effektive
Säurekonzentration wird dort eher erreicht als bei höheren pH-Werten. Die Konsequenz wäre
ein Abflachen der Kurve mit gleichzeitiger Verschiebung des Optimums nach rechts. Der
zweite Fall setzt voraus, dass enzymspezifische Effekte dominieren, zumindest dann, wenn
die Erhöhung der effektiven Säurekonzentration bei höheren pH-Werten aufgrund einer gün-
stigeren Enzymkonformation mit einer überproportionalen Aktivitätssteigerung im Vergleich
zu niedrigen pH-Werten beantwortet wird. Hier würde der Graph nicht abflachen, sondern ein
nach rechts verschobenes, deutlich höheres Optimum beobachtet werden. Dennoch geben
diese Ergebnisse Anlass zur Untersuchung der Substratkinetik am pH-Optimum, um abzu-
klären, welche Propionsäurekonzentrationen maximal appliziert werden können.
3.1.3.2 Substratkinetik der protonierten Säure am pH-Optimum
Die bisherigen Ergebnisse lassen vermuten, dass bei einer Erhöhung der Propionsäurekon-
zentration über 300 mmol/L eine weitere Steigerung der Reaktionsgeschwindigkeit erreicht
werden könnte. Um weitergehende Aussagen über die Substratkinetik machen zu können,
wurde die Abhängigkeit der Enzymaktivität von der Propionsäurekonzentration bei den zuvor
bestimmten pH-Optima untersucht. Es sei an dieser Stelle einschränkend angemerkt, dass für
eine genauere Beurteilung der Substratkinetik die Konzentration von Butanol als zweites
Substrat ebenfalls hätte variiert werden müssen [BISSWANGER, 2000]. Es ist nicht bekannt, ob
Butanol in der gewählten Konzentration ausreichend verfügbar ist oder sogar schon inhi-
bierend wirkt. Letzteres kann zumindest aufgrund der Arbeit von WEHTJE UND ADLERCREUTZ
[1997] ausgeschlossen werden. Das Hauptaugenmerk war aber auf die Rolle der Propionsäure
gerichtet. Die erzielten Ergebnisse erlauben jedenfalls den Schluss, dass die Verfügbarkeit
von Butanol die Reaktion nicht limitierte. Wie in Abbildung 3.3 zu erkennen ist, konnte für
beide Lipasen die Reaktionsgeschwindigkeit bis zu einer Konzentration 1000 mmol/L
Propionsäure gesteigert werden – oberhalb dieser Konzentration wurde eine Sättigung bzw.
eine deutliche Inhibition beobachtet.
Der anhand der Messdaten mit Hilfe eines Parameterschätzprogrammes (EKI3, Version 1.3,
Physiologisch-Chemisches Institut der Universität Tübingen) ohne Berücksichtigung der Inhi-

Ergebnisse und Diskussion
54
bition für CRL ermittelte Km-Wert lag mit etwa 1600 mmol/L deutlich über den in der Litera-
tur genannten Werten von 100 bis 400 mmol/L Propionsäure [KOCH, 1997; SANDOVAL ET AL.,
2002]. Vermutlich wurde in früheren Arbeiten die Rolle des pH-Wertes unter- bzw. falsch
eingeschätzt, weshalb eine Vergleichbarkeit der Werte ohnehin nicht gegeben ist.
0
20
40
60
80
100
120
140
160
180
200
0 500 1000 1500 2000 2500 3000
Konzentration Propionsäure [mmol/L]
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
TLL
CRL
Abbildung 3.3: Abhängigkeit der Enzymaktivität von der Propionsäurekonzentration bei der durch die CRL (pH 3,5) und die TLL (pH 4,25) katalysierten Synthese von Butylpropionat.
Bei genauerer Betrachtung fällt auf, dass die Inhibition beider Lipasen ein ungewöhnliches
Verhalten zeigt (Abb. 3.3). Es ist eine rapide Aktivitätsabnahme zu verzeichnen und oberhalb
von 2000 bzw. 3000 mmol/L keine Butylpropionatbildung mehr nachweisbar. Üblicherweise
wäre mit einer Abnahme zu rechnen, die sich durch eine asymptotische Annäherung des
Graphen an die x-Achse beschreiben ließe. Es gelang daher weder mit Hilfe klassischer
Methoden graphisch [DIXON, 1972] noch mit geeigneten Inhibitionstermen mathematisch die
Inhibitionskonstante zu bestimmen. Die Ursachen für diese eher ungewöhnliche Art der
Inhibition können derzeit nicht benannt werden.
Anders als bei der CRL zeigt sich bei Auftragung der Aktivität gegen die Substratkon-
zentration für die TLL bis 1000 mmol/L ein sigmoidaler Verlauf (Abb. 3.3). Dies überrascht
ein wenig, da derartige Verläufe v.a. für allosterische Enzyme typisch sind. Für die TLL
wurde jedoch bereits bei hydrolytischen Reaktionen ein sigmoidaler Verlauf beschrieben
[NINI ET AL., 2001]. Basierend auf der Annahme, dass die Lipase als Monomer aktiv ist, sollte
das beobachtete Verhalten mit dem „Slow-Transition-Modell“ zu erklären sein [BISSWANGER,

pH-Optimum
55
2000]. In diesem Modell wird angenommen, dass das Enzym in zwei Zuständen – einer
aktiven und einer weniger aktiven Form – vorliegt, die Umwandlung zwischen beiden Zu-
ständen aber langsam erfolgt. Beide Enzymformen sind in der Lage, Substrat zu binden. Nach
der Substratbindung geht das Enzym in die aktivere Form über, kann aber bei niedrigen
Substratkonzentrationen nach der Produktbildung wieder in die weniger aktive Form zurück-
kehren, ehe das nächste Substratmolekül bindet. Mit zunehmender Substratkonzentration
steigt die Zahl der aktiven Enzymmoleküle und die Zeit bis zur erneuten Bindung eines
Substratmoleküls reicht nicht mehr für die Rückbildung in die weniger aktive Form. Bei
hohen Substratkonzentrationen befinden sich daher alle Enzymmoleküle in der aktiven Form
und das Enzym arbeitet mit höchster Effizienz. Der Unterschied zum bekannten Koopera-
tivitätsverhalten allosterischer Enzyme besteht dabei im hyperbolen Verlauf der Substrat-
bindung an das Enzym, während allosterische Enzyme hier einen sigmoidalen Verlauf zeigen.
Die Aktivität der TLL scheint also einer kinetischen Kooperativität [BISSWANGER, 2000] zu
unterliegen und beruht auf dem Vorhandensein zweier unterschiedlich aktiver Enzymformen.
Diese unterschiedlich aktiven Zustände könnten möglicherweise die offene bzw. geschlossene
Form der Lipase sein. Dass nicht alle Lipasen dieses Verhalten zeigen, liegt vermutlich daran,
dass ein Übergang von der offenen in die geschlossene Konformation bei der TLL vergleichs-
weise langsam erfolgt.
3.1.4 Extrapolation der pH-Optima
Unter der Annahme, dass die protonierte Säure das bevorzugte Substrat der Estersynthese ist,
muss bedacht werden, dass aufgrund der nicht über den gesamten pH-Bereich konstanten
Substratkonzentration, die gemessenen pH-Optima nicht direkt die pH-Abhängigkeit der
Enzymaktvität widerspiegeln. Dies ist in erster Linie – wie bereits erörtert – darauf zurück-
zuführen, dass bei pH-Werten oberhalb des pKS-Wertes die Konzentration an protonierter
Säure stark abnimmt. Dies resultiert zwangsläufig in einer geringeren Aktivität, da nicht alle
Enzymmoleküle mit Substrat abgesättigt werden können. Es liegt auf der Hand, dass bei
höheren Konzentrationen an protonierter Säure eine höhere Aktivität zu verzeichnen sein
sollte. Die Konzentration an protonierter Säure unabhängig vom pH-Wert konstant zu halten,
ist jedoch experimentell nicht möglich, da exorbitant hohe Gesamtsäurekonzentrationen
notwendig wären, um bei hohen pH-Werten die gleiche Menge an protonierter Säure verfüg-
bar zu machen wie bei niedrigen pH-Werten. Dennoch kann ein Einblick in die intrinsische
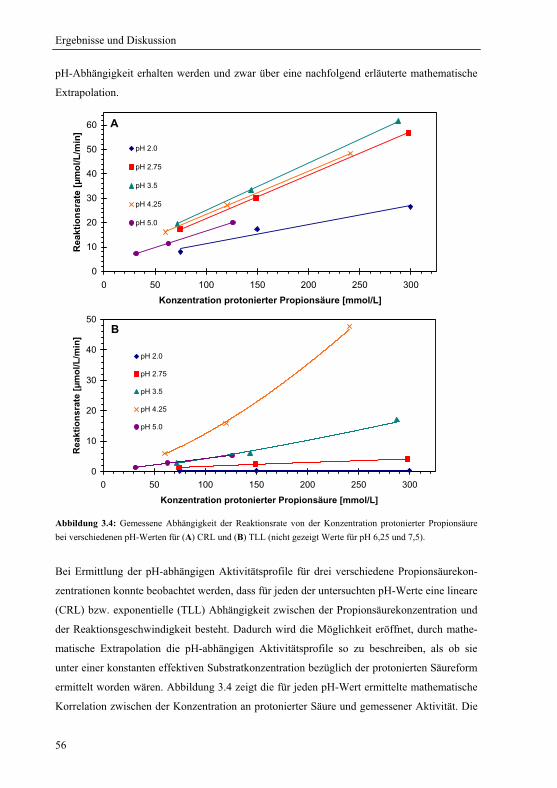
Ergebnisse und Diskussion
56
pH-Abhängigkeit erhalten werden und zwar über eine nachfolgend erläuterte mathematische
Extrapolation.
Abbildung 3.4: Gemessene Abhängigkeit der Reaktionsrate von der Konzentration protonierter Propionsäure bei verschiedenen pH-Werten für (A) CRL und (B) TLL (nicht gezeigt Werte für pH 6,25 und 7,5).
Bei Ermittlung der pH-abhängigen Aktivitätsprofile für drei verschiedene Propionsäurekon-
zentrationen konnte beobachtet werden, dass für jeden der untersuchten pH-Werte eine lineare
(CRL) bzw. exponentielle (TLL) Abhängigkeit zwischen der Propionsäurekonzentration und
der Reaktionsgeschwindigkeit besteht. Dadurch wird die Möglichkeit eröffnet, durch mathe-
matische Extrapolation die pH-abhängigen Aktivitätsprofile so zu beschreiben, als ob sie
unter einer konstanten effektiven Substratkonzentration bezüglich der protonierten Säureform
ermittelt worden wären. Abbildung 3.4 zeigt die für jeden pH-Wert ermittelte mathematische
Korrelation zwischen der Konzentration an protonierter Säure und gemessener Aktivität. Die
0
10
20
30
40
50
60
0 50 100 150 200 250 300Konzentration protonierter Propionsäure [mmol/L]
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
pH 2.0
pH 2.75
pH 3.5
pH 4.25
pH 5.0
0
10
20
30
40
50
0 50 100 150 200 250 300
Konzentration protonierter Propionsäure [mmol/L]
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
pH 2.0
pH 2.75
pH 3.5
pH 4.25
pH 5.0
A
B
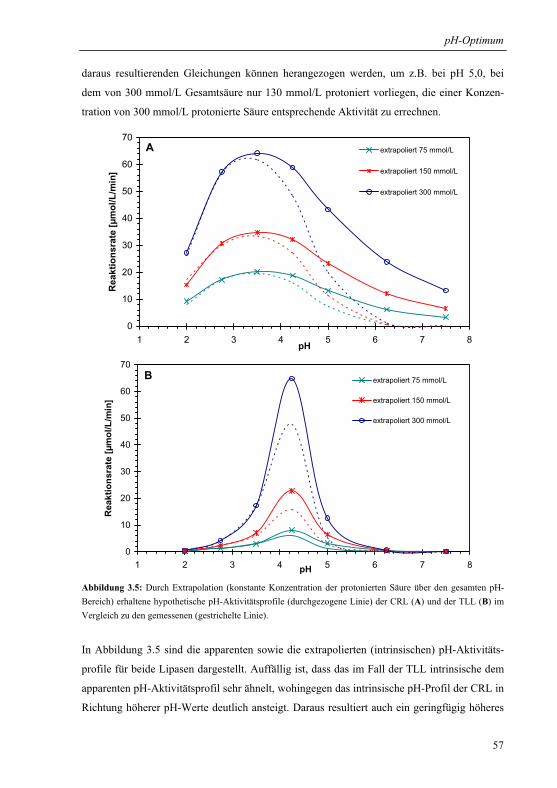
pH-Optimum
57
daraus resultierenden Gleichungen können herangezogen werden, um z.B. bei pH 5,0, bei
dem von 300 mmol/L Gesamtsäure nur 130 mmol/L protoniert vorliegen, die einer Konzen-
tration von 300 mmol/L protonierte Säure entsprechende Aktivität zu errechnen.
Abbildung 3.5: Durch Extrapolation (konstante Konzentration der protonierten Säure über den gesamten pH-Bereich) erhaltene hypothetische pH-Aktivitätsprofile (durchgezogene Linie) der CRL (A) und der TLL (B) im Vergleich zu den gemessenen (gestrichelte Linie).
In Abbildung 3.5 sind die apparenten sowie die extrapolierten (intrinsischen) pH-Aktivitäts-
profile für beide Lipasen dargestellt. Auffällig ist, dass das im Fall der TLL intrinsische dem
apparenten pH-Aktivitätsprofil sehr ähnelt, wohingegen das intrinsische pH-Profil der CRL in
Richtung höherer pH-Werte deutlich ansteigt. Daraus resultiert auch ein geringfügig höheres
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8pH
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
extrapoliert 75 mmol/L
extrapoliert 150 mmol/L
extrapoliert 300 mmol/L
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8pH
Rea
ktio
nsr
ate
[µm
ol/L
/min
]
extrapoliert 75 mmol/L
extrapoliert 150 mmol/L
extrapoliert 300 mmol/L
A
B

Ergebnisse und Diskussion
58
pH-Optimum der CRL nach Extrapolation. Eine derartige Beobachtung kann für die TLL
nicht gemacht werden. Aller Wahrscheinlichkeit nach ist das Aktivitätsprofil der TLL
weniger stark an die Verfügbarkeit der protonierten Säure gebunden ist und hier – wohlge-
merkt nicht bekannte – enzymspezifische Faktoren stärker einwirken.
3.1.5 Mechanistische Erklärung für das pH-Optimum der Veresterung
Bereits mehrfach wurde darauf hingewiesen, dass die erhaltenen Ergebnisse den Schluss nahe
legen, dass die protonierte Säure das bevorzugte Substrat der Estersynthese ist. Es stellt sich
die Frage, inwieweit diese Vermutung mechanistisch untermauert werden kann. In der Arbeit
von NEVES-PETERSEN ET AL. [2001a] wird die Theorie des „elektrostatischen Katapults“
postuliert. Ausgehend von der für viele Lipasen bekannten Tertiärstruktur konnten NEVES-
PETERSEN ET AL. [2001a] das pH-abhängige elektrostatische Potential im aktiven Zentrum mit
Hilfe von computergestützten Titrationskurven ermitteln. Dabei konnten die Autoren zeigen,
dass bei der lipasenkatalysierten Hydrolyse die höchste Aktivität bei pH-Werten (neutral bis
alkalisch) beobachtet werden kann, bei denen der Bereich um das katalytisch aktive Serin ein
negatives elektrostatisches Potential aufweist. Im Zuge der Esterhydrolyse wird eine Carbon-
säure freigesetzt. Die Carbonsäure deprotoniert im neutralen bis alkalischen pH-Bereich und
wird durch das negative elektrostatische Potential im aktiven Zentrum aus diesem herauskata-
pultiert. Zudem steigert die negative Nettoladung im aktiven Zentrum die Nukleophilie und
damit die Reaktivität des katalytischen Serins. Außerdem wird das Substrat durch das elektro-
statische Feld polarisiert, was einen nukleophilen Angriff auf den Carbonylsauerstoff der
Esterbindung erleichtert [NEVES-PETERSEN ET AL., 2001b].
Im Umkehrschluss folgt aus der elektrostatischen Katapulttheorie, dass eine bei neutralen bis
alkalischen pH-Werten negativ geladene Säure nicht in das aktive Zentrum gelangen kann
und somit nicht das Substrat der Estersynthese sein kann. Die der Publikation von NEVES-
PETERSEN ET AL. [2001a] entlehnte Abbildung 3.6 zeigt die in Abhängigkeit vom pH-Wert
ermittelte, im aktiven Zentrum vorherrschende Nettoladung für die CRL und die TLL.
Anhand dieser Abbildung wird deutlich, dass im aktiven Zentrum der CRL bei pH-Werten
oberhalb von 3,0 eine negative Ladung vorherrscht.

pH-Optimum
59
-4
-3
-2
-1
0
1
2
3
2 3 4 5 6 7 8 9 10 11 12 13
pH-Wert [-]
Lad
un
g (
e)
CRL
TLL
Abbildung 3.6: pH-abhängige Ladung im aktiven Zentrum der CRL und der TLL (im Umkreis von 10 Å um das katalytische Serin / geschlossene Konformationen), erstellt nach NEVES-PETERSEN ET AL. [2001a].
Dies könnte eine Erklärung liefern, warum nur die protonierte und damit ungeladene Propion-
säure als Substrat in Frage kommt. Für die CRL folgt daraus, dass sie bei pH-Werten oberhalb
von 6 aufgrund der gegenseitigen elektrostatischen Abstoßung von (fast ausschließlich vor-
handenem) Säureanion und aktivem Zentrum keine oder nur geringe Aktivität zeigen sollte,
während im Bereich um den pKS-Wert der Propionsäure (4,7) die Aktivität aufgrund der stei-
genden effektiven Substratkonzentration stark ansteigen und bei pH-Werten um 3,0 ihr Maxi-
mum erreicht haben sollte – sofern nicht pH-bedingte inaktivierende Effekte auf die Enzym-
konformation überwiegen. Hingegen herrscht im aktiven Zentrum der TLL erst oberhalb von
pH 9,0 eine negative Nettoladung vor. Im Fall der TLL erscheint es somit denkbar, dass auch
die deprotonierte Säure in das aktive Zentrum gelangen und als Substrat für die Estersynthese
fungieren kann. Allerdings haben NEVES-PETERSEN ET AL. [2001a] die pH-abhängige Netto-
ladung im aktiven Zentrum der TLL nur für die geschlossene Konformation ermittelt, da zum
Zeitpunkt der betreffenden Untersuchung die dreidimensionale Struktur der TLL in der
offenen Konformation nicht verfügbar war. Es kann somit nicht ausgeschlossen werden, dass
in der offenen Konformation bereits bei niedrigeren pH-Werten im aktiven Zentrum eine
negative Nettoladung vorliegt und damit der Umsetzung der deprotonierten Säure entgegen-
steht. Derartige Beobachtungen machten NEVES-PETERSEN ET AL. [2001a] zumindest für die
Lipase von Rhizomucor miehei. Die Situation im aktiven Zentrum der TLL kann somit nicht
abschließend geklärt werden. Für den Fall, dass im aktiven Zentrum der TLL auch in der
offenen Konformation keine negative Nettoladung vorherrscht, muss es noch eine andere

Ergebnisse und Diskussion
60
Erklärung geben, um die niedrigen pH-Optima von CRL und TLL bei der Estersynthese zu
erklären.
Eine Schlüsselrolle bei Erklärung der niedrigen pH-Optima wird dem katalytischen Mecha-
nismus bzw. der OH-Gruppe des katalytischen Serin-Restes im aktiven Zentrum zuteil: Für
die Hydrolyse ist bekannt, dass die Nukleophile der OH-Gruppe durch Wechselwirkung mit
dem deprotonierten Imidazolring eines Histidin-Restes gesteigert wird, der seinerseits von
einer deprotonierten Carboxylgruppe eines Aspartat- oder Glutamat-Restes stabilisiert wird.
Bei niedrigen pH-Werten muss jedoch die Funktionalität der katalytischen Triade in Frage
gestellt werden: Ab welchem pH-Wert die Carboxylgruppe des Aspartat- oder Glutamat-
Restes protoniert wird, ist nicht bekannt. Eine mögliche Beeinflussung des Reaktions-
mechanismus durch den pH-Wert wurde bereits von NEVES-PETERSEN ET AL. [2001b] und
PAIVA ET AL. [2000] angedeutet. Die vorliegenden Daten lassen aber keinen definitiven
Schluss darüber zu, ob für die Veresterung neben dem Serin-Rest auch die übrigen Amino-
säurereste der katalytischen Triade essentiell sind. Klar ist, dass die katalytische Wirksamkeit
des Biokatalysators an die Gegenwart eines nukleophilen Restes im aktiven Zentrum ge-
bunden ist. Ist eine organische Säure negativ geladen, so ist die negative Ladung über das
Carbonyl-C-Atom delokalisiert. Für die Ausbildung des Enzym-Acyl-Komplexes ist ein
nukleophiler Angriff auf das Carbonyl-C-Atom der organischen Säure Voraussetzung. Das
einzige hierfür in Frage kommende Nukleophil ist die OH-Gruppe des Serin-Restes, welches
jedoch aufgrund der delokalisierten negativen Ladung nicht stark genug ist. Dies führt zu der
naheliegenden Schlussfolgerung, dass nur die protonierte Säure umgesetzt werden kann und
steht in völliger Übereinstimmung mit den hier erzielten experimentellen Befunden. Diese
Feststellung hätte selbst dann Bestand, wenn das Enzym bei pH-Werten oberhalb des pKS-
Wertes eine womöglich günstigere Konformation aufweisen würde – eine Esterbildung ausge-
hend von der deprotonierten Säure scheint enzymatisch schlichtweg unmöglich zu sein. Im
Endeffekt mag diese Interpretation der Ergebnisse banal anmuten und umso unverständlicher
ist es, dass dieser Aspekt der lipasenkatalysierten Veresterung in der einschlägigen Fachlitera-
tur bisher unberücksichtigt blieb. Hierfür mag die eingangs erwähnte Komplexität der
üblicherweise verwendeten Reaktionssysteme verantwortlich sein und die oft anzutreffende
empirische Herangehensweise an biokatalytische Fragestellungen.

pH-Optimum
61
3.1.6 Praktische Relevanz
Da die meisten organischen Säuren (C2 bis C8) pKS-Werte im Bereich von 4,7 bis 4,9 zeigen,
sollten die pH-Optima auch auf diese Substrate übertragbar sein. Es darf somit basierend auf
den hier erhaltenen Ergebnissen die Empfehlung postuliert werden, dass pH-Werte unterhalb
des pKS-Wertes der zu veresternden Säure die Reaktion begünstigen. Warum in vielen
Arbeiten trotz augenscheinlich eingestellter pH-Werte um 7 bis 8 eine Estersynthese beobach-
tet werden konnte, ist mit dem Massentransfer der in der organischen Phase vorgelegten
Säuren in die wässrige Phase zu erklären [HEINEMANN, 2003; HEINEMANN ET AL., 2003]:
Trotz des mit Hilfe eines Puffers eingestellten pH-Wertes kommt es im Zuge dieses Ein-
stroms zu einem erheblichen pH-Abfall – und zwar in einen für die Reaktion günstigen Be-
reich. In Abhängigkeit von der Säurekonzentration und dem Volumenverhältnis von orga-
nischer und wässriger Phase kommt es dabei zu einer erheblichen Anreicherung der Säure in
der wässrigen Phase. Ein pH-Abfall könnte nur durch Verwendung hochkonzentrierter Puffer
oder spezieller Puffer in der organischen Phase [BLACKWOOD ET AL., 1994] verhindert werden
– mit wahrscheinlich nachteiligen Auswirkungen auf die Enzymaktivität [HEINEMANN, 2003;
DOUMÉCHE, 2002]. Inwieweit der Einsatz eines Puffers Sinn macht, muss daher im Einzelfall
geklärt werden – HEINEMANN [2003] zeigte am Beispiel von in Hexan suspendierten Alginat-
kugeln, dass bei einem Volumenverhältnis von 1 : 50 und einer Propionsäurekonzentration
von 50 mmol/L der pH-Wert in der wässrigen Phase nicht unter Werte von 3,5 fiel – daher
könnte zumindest bei Einsatz der CRL auf die Verwendung eines Puffers verzichtet werden.
In jedem Fall verdient die Kontrolle des pH-Wertes weniger Aufmerksamkeit als zu Beginn
dieser Untersuchungen vermutet wurde. Es erscheint plausibel, dass bei hohen pH-Werten
eine Veresterung aus zwei Gründen nicht stattfinden kann (Abb. 3.7 A):
1) Die negativ geladene Säure kann aufgrund der im aktiven Zentrum vorherrschenden
negativen Nettoladung nicht gebunden werden und
2) die OH-Gruppe des Serins kann keinen nukleophilen Angriff auf das Carbonyl-C-
Atom der negativ geladenen Säure führen.
Diese Einschränkungen werden bei niedrigen pH-Werten aufgehoben (Abb. 3.7 B) – es be-
steht keine elektrostatische Abstoßung mehr zwischen Substrat und aktivem Zentrum und der
Serin-Rest kann wieder als Nukleophil gegenüber der Säure agieren.
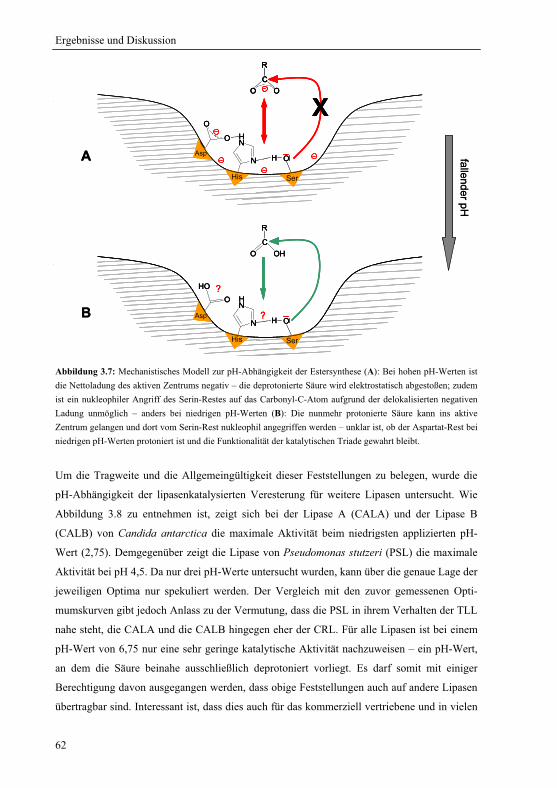
Ergebnisse und Diskussion
62
Abbildung 3.7: Mechanistisches Modell zur pH-Abhängigkeit der Estersynthese (A): Bei hohen pH-Werten ist die Nettoladung des aktiven Zentrums negativ – die deprotonierte Säure wird elektrostatisch abgestoßen; zudem ist ein nukleophiler Angriff des Serin-Restes auf das Carbonyl-C-Atom aufgrund der delokalisierten negativen Ladung unmöglich – anders bei niedrigen pH-Werten (B): Die nunmehr protonierte Säure kann ins aktive Zentrum gelangen und dort vom Serin-Rest nukleophil angegriffen werden – unklar ist, ob der Aspartat-Rest bei niedrigen pH-Werten protoniert ist und die Funktionalität der katalytischen Triade gewahrt bleibt.
Um die Tragweite und die Allgemeingültigkeit dieser Feststellungen zu belegen, wurde die
pH-Abhängigkeit der lipasenkatalysierten Veresterung für weitere Lipasen untersucht. Wie
Abbildung 3.8 zu entnehmen ist, zeigt sich bei der Lipase A (CALA) und der Lipase B
(CALB) von Candida antarctica die maximale Aktivität beim niedrigsten applizierten pH-
Wert (2,75). Demgegenüber zeigt die Lipase von Pseudomonas stutzeri (PSL) die maximale
Aktivität bei pH 4,5. Da nur drei pH-Werte untersucht wurden, kann über die genaue Lage der
jeweiligen Optima nur spekuliert werden. Der Vergleich mit den zuvor gemessenen Opti-
mumskurven gibt jedoch Anlass zu der Vermutung, dass die PSL in ihrem Verhalten der TLL
nahe steht, die CALA und die CALB hingegen eher der CRL. Für alle Lipasen ist bei einem
pH-Wert von 6,75 nur eine sehr geringe katalytische Aktivität nachzuweisen – ein pH-Wert,
an dem die Säure beinahe ausschließlich deprotoniert vorliegt. Es darf somit mit einiger
Berechtigung davon ausgegangen werden, dass obige Feststellungen auch auf andere Lipasen
übertragbar sind. Interessant ist, dass dies auch für das kommerziell vertriebene und in vielen
HN
N OH
HO
O?
Asp
His Ser
?
COHO
R
HN
N OH
O
O
Asp
His Ser
COO
R
X
A
B
fallender pH
HN
N OH
HO
O?
Asp
His Ser
?
COHO
R
HN
N OH
HO
O?
Asp
His Ser
Asp
His Ser
?
COHO
R
HN
N OH
O
O
Asp
His Ser
COO
R
XHN
N OH
O
O
Asp
His Ser
COO
R
COO
R
X
A
B
fallender pH
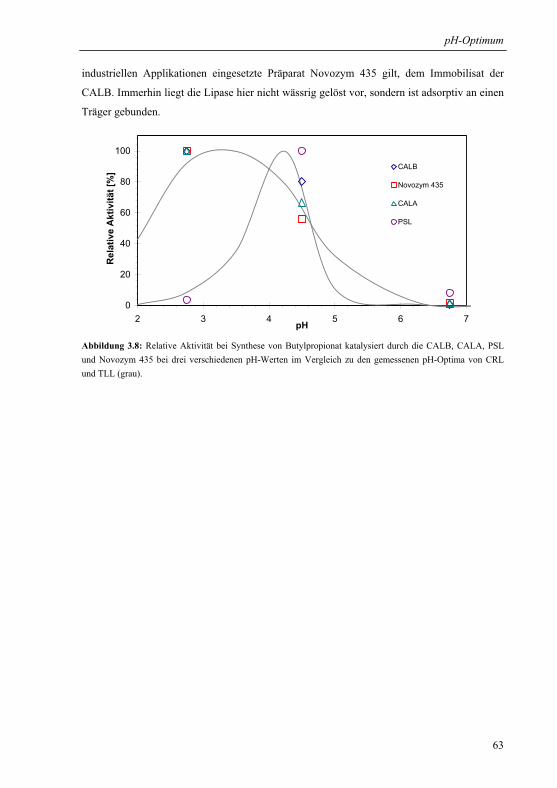
pH-Optimum
63
industriellen Applikationen eingesetzte Präparat Novozym 435 gilt, dem Immobilisat der
CALB. Immerhin liegt die Lipase hier nicht wässrig gelöst vor, sondern ist adsorptiv an einen
Träger gebunden.
Abbildung 3.8: Relative Aktivität bei Synthese von Butylpropionat katalysiert durch die CALB, CALA, PSL und Novozym 435 bei drei verschiedenen pH-Werten im Vergleich zu den gemessenen pH-Optima von CRL und TLL (grau).
0
20
40
60
80
100
2 3 4 5 6 7pH
Rel
ativ
e A
ktiv
ität
[%
]
CALB
Novozym 435
CALA
PSL

Ergebnisse und Diskussion
64
3.2 Bedeutung der Grenzfläche für die lipasenkatalysierte Reaktion
Wie bereits in Kapitel 1.2 dargelegt, muss nach gegenwärtigem Kenntnisstand eine grenz-
flächenaktivierte Lipase in die offene Konformation überführt werden, um Ester effizient
hydrolysieren zu können. Diese Aktivierung setzt einen Kontakt mit einer Grenzfläche
voraus. In der inaktiven Konformation bedeckt eine aus wenigen Aminosäuren bestehende
lidartige Struktur das aktive Zentrum und verwehrt den Zugang der Substrate. Das Auftreten
einer Grenzfläche induziert die Umlagerung dieser Deckelstruktur und damit die für eine
Aktivierung erforderliche Konformationsänderung [BRZOZOWSKI ET AL., 1991; DEREWENDA
ET AL., 1992; GROCHULSKI ET AL., 1993]. Ausgehend von diesen Beobachtungen wird vermu-
tet, dass die hydrolytische Reaktion vornehmlich an der Grenzfläche stattfindet, da die Lipase
in der wässrigen Phase überwiegend in der inaktiven bzw. geschlossenen Konformation vor-
liegt.
Inwieweit diese Betrachtungen auch für die umgekehrte Reaktion – die Estersynthese –
gelten, war bislang nicht Gegenstand einer systematischen Untersuchung. Bisherige Studien
in Zusammenhang mit der Grenzflächenaktivierung beschränkten sich auf hydrolytische
Reaktionen, wo die Grenzfläche durch Emulsion des nur sehr begrenzt wasserlöslichen
Substrates geschaffen wurde. Hingegen ist die Ausgangslage bei Synthesereaktionen eine
andere: Aus thermodynamischen und praktischen Gesichtspunkten wird diese Art der
Umsetzung meist in organischen, nicht-wassermischbaren Lösungsmitteln durchgeführt. Die
Frage, ob ein organisches Lösungsmittel analog zum emulgierten Substrat eine Grenzflächen-
aktivierung herbeiführen kann, wurde so noch nicht gestellt. LOUWRIER ET AL. [1996] konnten
zumindest für die Hydrolyse zeigen, dass das bloße Vorhandensein einer organischen Phase
nicht ausreicht, um eine Aktivierung der Lipase von Rhizomucor miehei zu bewirken. Statt-
dessen scheint hierfür zusätzlich eine wässrige Phase, also eine Grenzfläche vonnöten zu sein.
Bei der Veresterung ist stets eine organische Phase zugegen und der methodisch analoge
Nachweis einer Grenzflächenaktivierung wie bei der Hydrolyse nicht durchführbar.
Erschwerend kommt hinzu, dass für die Synthesereaktion kein universeller Aktivitätstest
etabliert ist [SALIS ET AL., 2003]. Theoretisch könnte zwar die Lipase in lyophilisierter Form
im organischen Lösungsmittel suspendiert und der Wassergehalt sukzessive über den aus der
Reaktion selbst resultierenden Anteil hinaus erhöht werden. Sollte sich dabei aber eine
überproportionale Steigerung der katalytischen Aktivität zeigen, bliebe allerdings unklar, ob
dies auf der Schaffung einer Grenzfläche zurückzuführen oder nur der zunehmenden Solubi-

Bedeutung der Grenzfläche
65
lisierung der Enzymmoleküle geschuldet wäre. Insofern kann diese Strategie nicht herange-
zogen werden, um die Bedeutung der Grenzfläche in Synthesereaktionen aufzuhellen.
In einer Reihe von Studien konnte gezeigt werden, dass unter kinetischen Gesichtspunkten
biphasische gegenüber monophasischen Reaktionssystemen bessere Ergebnisse liefern
[BORZEIX ET AL., 1992; LOUWRIER ET AL., 1996; TWEDDELL ET AL., 1998]. Die Autoren
führten dies darauf zurück, dass die Enzymmoleküle in der wässrigen Phase homogen gelöst
vorliegen. Im Fall monophasischer Systeme hat man es meist mit aus agglomerierten
Enzymmolekülen bestehenden Partikeln zu tun. Infolgedessen können nicht alle Enzymmole-
küle gleichartig an der Reaktion partizipieren. Zudem hat das Auftreten von Wasser als
Nebenprodukt zur Folge, dass die Enzympartikel mit entsprechend negativem Einfluss auf die
katalytische Effizienz verkleben [REETZ ET AL., 1996]. Diese Probleme bestehen bei bipha-
sischen Reaktionssystemen nicht, allerdings wurde dort das Phänomen der Grenzflächen-
aktivierung bzw. die Bedeutung der Grenzfläche für Synthesereaktionen bisher nicht einge-
hend thematisiert, ebensowenig wie auf die besonderen thermodynamischen Aspekte bipha-
sischer Systeme eingegangen wurde.
Gegenwärtig kann nicht gesagt werden, welcher Anteil der katalytischen Gesamtaktivität auf
die an der Grenzfläche befindlichen im Vergleich zu den homogen in der wässrigen Phase
gelösten Enzymmolekülen zurückgeht. Ein erster Aufklärungsansatz wurde von DOUMÈCHE
[2002] durch Anwendung eines hydrophoben lipasespezifischen Inhibitors in einem gelstabi-
lisierten Zweiphasensystem unternommen. Der Inhibitor (n-Dodecyl-p-nitrophenyl-n-hexano-
phosphonat) konnte aufgrund seines Verteilungskoeffizienten nicht in die Gelkugel diffun-
dieren und daher nur die an der Oberfläche der Gelkugel befindlichen Enzymmoleküle
inhibieren. Dabei wurde eine Abnahme der katalytischen Aktivität der CRL auf 15 bis 25 %
beobachtet. Da aber nicht bewiesen werden konnte, ob die gemessene Restaktivität auf die
Enzymaktivität im Inneren der Gelkugel oder auf evtl. nicht inhibierte Enzymmoleküle an der
Oberfläche zurückzuführen war, konnte dies nur als Indiz, nicht aber als Beweis gewertet
werden. Dennoch wird klar, dass der offensichtliche Zusammenhang zwischen den an der
Oberfläche lokalisierten Enzymolekülen und der Produktivität von entscheidender Bedeutung
für die Bewertung der katalytischen Effizienz solcher Systeme ist.
Da seit Mitte der neunziger Jahre bekannt ist, dass nicht alle Lipasen einer Grenzflächen-
aktivierung unterliegen [MARTINELLE ET AL., 1995; VERGER, 1997; SCHMID UND VERGER,

Ergebnisse und Diskussion
66
1998], stellte sich zudem die Frage, ob diese im Vergleich zu grenzflächenaktivierten Lipasen
in biphasischen Reaktionssystemen bei der Estersynthese ein abweichendes katalytisches Ver-
halten zeigen. Aus diesem Grund, und um die Bedeutung der Grenzfläche in biphasischen
Reaktionssystemen beurteilen zu können, wurde im Rahmen dieser Arbeit die Abhängigkeit
der gemessenen Aktivität von der spezifischen auf das Volumen der wässrigen Phase bezoge-
nen Grenzfläche in gelstabilisierten und fluiden biphasischen Reaktionssystemen für eine
grenzflächenaktivierte und eine nicht-grenzflächen-aktivierte Lipase untersucht. Es wurde
davon ausgegangen, dass zu beobachtende Unterschiede im katalytischen Verhalten dazu bei-
tragen können, die Vorgänge an der Grenzfläche besser zu durchschauen. Als nicht-grenz-
flächenaktivierte Lipase diente die Candida antarctica Lipase B (CALB), deren Lid durch
intramolekulare Wechselwirkungen scheinbar dauerhaft geöffnet ist [UPPENBERG ET AL.,
1994]. MARTINELLE ET AL. [1995] bewiesen mit ihrer Arbeit, dass diese Lipase bei der
Hydrolyse tatsächlich keine Grenzflächenaktivierung zeigt und deshalb maximale Aktivität
sowohl gegenüber monomeren, gelösten Substraten als auch an einer Phasengrenzfläche
Wasser/Lipid entfaltet. Als Vertreter der grenzflächenaktivierten Lipasen wurde in Anlehnung
an die Arbeiten von DOUMÈCHE [2002] und HEINEMANN [2003] und aufgrund der im SFB
540 gewählten Modellreaktion die CRL verwendet.
3.2.1 Katalytische Aktivität und die spezifische Grenzfläche
3.2.1.1 Enzym-Alginatkugeln – Variation der spezifischen Grenzfläche
Im gelstabilisierten Zweiphasensystem kann eine Variation der spezifischen Grenzfläche
unter Verwendung unterschiedlich großer Hydrogelkugeln erreicht werden. Da bei konstanten
Phasenvolumenverhältnissen (Kugelvolumen zu Volumen der organischen Phase) gearbeitet
wurde, waren – abgesehen von der spezifischen bzw. volumenbezogenen Grenzfläche – die
direkt zugänglichen reaktionsspezifischen Parameter identisch. Eingesetzt wurden Alginat-
Hydrogelkugeln mit einem Durchmesser von 2 bis 8 mm, die gemäß einer kürzlich entwickel-
ten Methodik hergestellt und vermessen wurden [BUTHE ET AL., 2004]. Kleine Kugeln ver-
fügen im Vergleich zu großen Kugeln über eine größere spezifische Grenzfläche. Unter der
Annahme, dass die Reaktion vorwiegend an der Grenzfläche stattfindet, sollten mit kleineren
Kugeln höhere Produktbildungsraten erzielt werden. Von Interesse hierbei ist, ob eine Korre-
lation zwischen der Größe der spezifischen Grenzfläche und der gemessenen Aktivität beo-
bachtet werden kann und ob sich die grenzflächenaktivierte (CRL) und die nicht-grenz-

Bedeutung der Grenzfläche
67
flächenaktivierte Lipase (CALB) hinsichtlich dieser Korrelation unterscheiden. Grundvoraus-
setzung für die Durchführung dieser Experimente war eine über das gesamte Kugelvolumen
homogene Enzymverteilung. Es ist jedoch bekannt, dass Alginatgele eine inhomogene
Polymerverteilung aufweisen [SKJÅK-BRÆK ET AL., 1989; HEINEMANN ET AL., 2005] und
dementsprechend aufgrund von Interaktionen zwischen Protein und Alginatmatrix bis zu einer
gewissen Konzentration mit einer inhomogenen Enzymverteilung zu rechnen ist. Erst bei
höheren Proteinkonzentrationen stellt sich – wenn alle möglichen Interaktionspunkte abgesät-
tigt sind – eine homogene Enzymverteilung ein [HEINEMANN ET AL., 2002]. Basierend auf
dieser Erkenntnis wurden die Lipasen mit Hühnerserumalbumin coimmobilisiert. Der Einsatz
dieses „Spacerproteins“ war notwendig, da nur so die erforderliche Proteinkonzentration
erreicht werden konnte. Eine Erhöhung der Enzymmenge kam nicht in Frage, da das
Reaktionssystem dann in einen massentransferlimitierten Status überführt worden wäre.
Bei Betrachtung von Abbildung 3.9 wird deutlich, dass sowohl für die grenzflächenaktivierte
(CRL) als auch für die nicht-grenzflächenaktivierte Lipase (CALB) erwartungsgemäß die
höchste Produktbildungsrate für den kleinsten Kugeldurchmesser verzeichnet wurde. Zudem
ist bei Vergleich der relativen Aktivitäten in Abhängigkeit vom Kugeldurchmesser (Abb.
3.10) deutlich zu erkennen, dass sich beide Lipasen diesbezüglich nicht wesentlich unter-
scheiden. Eine Verringerung der spezifischen Grenzfläche wird von beiden Lipasen mit einer
gleichartigen Abnahme der relativen Aktivität beantwortet. HEINEMANN [2003] führte in
diesem Zusammenhang eine Modellsimulation mit dem Resultat durch, dass mutmaßlich nur
etwa 0,002 % der insgesamt in den Alginatkugeln vorhandenen Enzymmoleküle – nämlich
die an der Oberfläche befindlichen – 84 % der Estermoleküle produzieren.
In den Arbeiten von HEINEMANN [2003] und Douméche [2002] wurde ein Zeitbedarf für die
Einstellung des Verteilungsgleichgewichts zwischen Alginatkugel (Ø 3,5 mm) und
umgebender Hexanphase von weniger als 1 h postuliert. Daher wurde, um eine Massen-
transferlimitierung auszuschließen, eine Katalysatormenge gewählt, die innerhalb von 1-2 h
einen Umsatz von maximal 10 % der Substratmenge zuließ. Dennoch könnten die im Ver-
gleich „langen“ Diffusionsstrecken der größeren Kugeln zur Ausbildung markanter Konzen-
trationsprofile über den Kugelquerschnitt führen und die Reaktion infolgedessen im äußeren
Bereich der Kugel schneller ablaufen. Um diese Unwägbarkeiten auszuschließen, wurde die
Fortführung der Untersuchung in einem nicht-gelstabilisierten biphasischen Reaktionssystem
favorisiert, in dem auch die wässrige Phase durchmischt wurde.

Ergebnisse und Diskussion
68
Abbildung 3.9: Synthese von Butylpropionat katalysiert durch in Alginat immobiliserte (A) CALB und (B) CRL bei variabler spezifischer (volumenbezogener) Grenzfläche (unterschiedliche Kugeldurchmesser).
Abbildung 3.10: Relative Aktivität der CRL und CALB bei Synthese von Butylpropionat in Abhängigkeit vom Durchmesser der Alginatkugeln.
3.2.1.2 Variable spezifische Grenzfläche
Der Hauptvorteil des verwendeten und in Kapitel 3.1.1 erläuterten nicht-gelstabilisierten
Zweiphasensystems bestand darin, dass die Größe der spezifischen Grenzfläche leicht variiert
y = 0,0594x
y = 0,0398x
y = 0,0704x
y = 0,0471x
0
5
10
15
20
25
30
35
40
0 100 200 300 400 500Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/L
] 2 mm
4 mm
6 mm
8 mm
y = 0,0025x
y = 0,0023x
y = 0,0018x
y = 0,0015x
0
1
2
3
4
5
6
7
8
0 500 1000 1500 2000 2500 3000Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/L
] 2 mm
4 mm
6 mm
8 mm
0
20
40
60
80
100
2 4 6 8Kugeldurchmesser [mm]
Rel
ativ
e A
ktiv
ität
[%
]
CRL CALB
A
B

Bedeutung der Grenzfläche
69
werden konnte und überdies beide Phasen durchmischt wurden, ohne die Integrität der Grenz-
fläche zu beeinträchtigen. Ähnliche Untersuchungen wurden von TWEDDELL ET AL. [1998] bei
der durch durch die Lipase aus Mucor miehei katalysierten Veresterung von Ölsäure und
Ethanol durchgeführt: Auch hier wurde die wässrige enzymhaltige Phase mit Hexan über-
schichtet, wobei jedoch auf eine aktive Durchmischung der einzelnen Phasen verzichtet wur-
de. TWEDDELL ET AL. [1998] führten die beobachtete Abhängigkeit der Syntheseaktivität von
der Größe der Grenzfläche auf die Geschwindigkeit des Massentransfers zwischen der wäs-
srigen und der organischen Phase zurück, ohne dies näher zu untersuchen. Aufgrund der feh-
lenden Durchmischung beider Phasen und der daraus resultierenden Konzentrationsgradienten
erscheint die dortige Versuchsanordnung ungeeignet, um die eingangs aufgeworfenen Fragen
zu beantworten. Daher wurden beide Phasen durchmischt und die spezifische Grenzfläche
durch unterschiedliche Füllstände variiert (Abb. 3.11). Neben der CRL und der CALB wurde
auch die Esterase aus Rhizopus arrhizus untersucht. Da es sich hier um eine Esterase handelt,
unterliegt sie, wie die CALB, keiner Grenzflächenaktivierung.
Abbildung 3.11: Variation der spezifischen (volumenbezogenen) Grenzfläche durch unterschiedliche Phasen-volumina (Verhältnis 1:1) in gleich großen Reaktionsgefäßen (konstante absolute Grenzfläche), beide Phasen wurden durhmischt.
In Abbildung 3.12 sind die Butylpropionat-Bildungsraten der drei untersuchten Enzyme
wiedergegeben. Die höchste katalytische Aktivität konnte stets für die kleinsten Füllvolumina
und damit für die größte spezifische Grenzfläche beobachtet werden. Dies entspricht den
Beobachtungen, die auch für das gelstabilisierte Reaktionssystem gemacht wurden. Aufgrund
des Versuchsaufbaus und der zuvor vorgenommenen Charakterisierung kann eine Massen-
transferlimitierung ausgeschlossen werden. Bei Auftragung der gemessenen relativen Aktivi-
täten über der spezifischen Grenzfläche (Abb. 3.13) offenbart sich für alle drei Enzyme ein
linearer Zusammenhang. Demzufolge führt eine Verringerung der spezifischen Grenzfläche
zu einer proportionalen Abnahme der katalytischen Aktivität. Diesbezüglich unterscheiden
sich die beiden Lipasen nur minimal.
Hexan mit Butanol (und Decan)
Wässrige Phase mit nativem Enzym
und Propionsäure
Phasengrenzfläche5 mL
1,52 cm-1
6,25 mL
1,21 cm-1
8 mL
0,94 cm-1
10 mL
0,76 cm-1
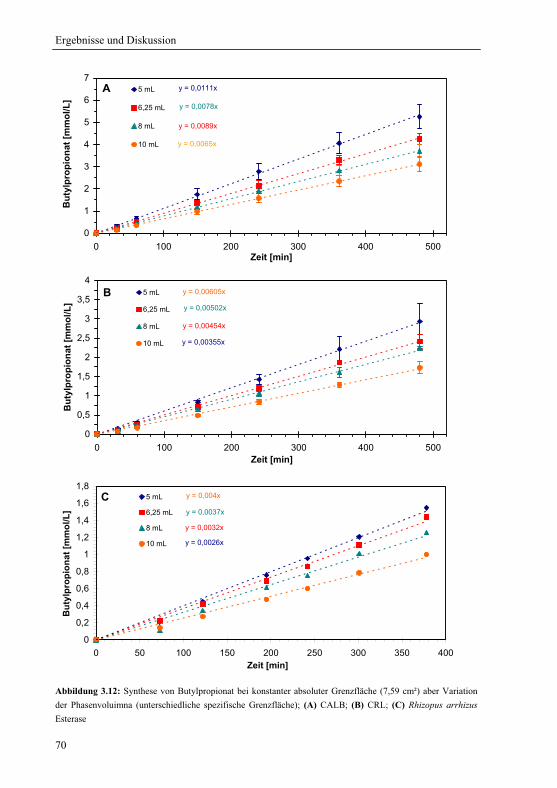
Ergebnisse und Diskussion
70
y = 0,0111x
y = 0,0089x
y = 0,0078x
y = 0,0065x
0
1
2
3
4
5
6
7
0 100 200 300 400 500Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/L
]
5 mL
6,25 mL
8 mL
10 mL
y = 0,00605x
y = 0,00502x
y = 0,00454x
y = 0,00355x
0
0,5
1
1,5
2
2,5
3
3,5
4
0 100 200 300 400 500Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/L
]
5 mL
6,25 mL
8 mL
10 mL
Abbildung 3.12: Synthese von Butylpropionat bei konstanter absoluter Grenzfläche (7,59 cm²) aber Variation der Phasenvoluimna (unterschiedliche spezifische Grenzfläche); (A) CALB; (B) CRL; (C) Rhizopus arrhizusEsterase
A
C
B
y = 0,0026x
y = 0,0032x
y = 0,0037x
y = 0,004x
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
0 50 100 150 200 250 300 350 400Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/L
]
5 mL
6,25 mL
8 mL
10 mL
C

Bedeutung der Grenzfläche
71
y = 0,5136x + 0,2216 R2 = 0,9719
y = 0,5374x + 0,1739 R2 = 0,993
y = 0,4506x + 0,3435 R2 = 0,9401
0,00
0,20
0,40
0,60
0,80
1,00
1,20
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6
Spezifische Grenzfläche [cm-1]
Rel
ativ
e A
ktiv
ität
[ ]
CALB
CRL
Esterase
Abbildung 3.13: Abhängigkeit der relativen Enzymaktivität der CALB, der CRL sowie der Esterase von Rhizopus arrhizus von der spezifischen (volumenbezogenen) Grenzfläche bei Synthese von Butylpropionat.
Hingegen scheint die Abhängigkeit der Aktivität von der spezifischen Grenzfläche für die
Esterase von Rhizopus arrhizus weniger stark ausgeprägt zu sein, erkennbar an der geringeren
Steigung (Abb. 3.13). Nichtsdestotrotz zeigen die Ergebnisse, dass die Veresterung unab-
hängig vom verwendeten Enzym vorwiegend an der Grenzfläche stattfindet. Es muss an
dieser Stelle postuliert werden, dass vermutlich eine bisher unerforschte Phänomenologie an
der Grenzfläche die beobachteten Unterschiede zwischen der Esterase und den beiden Lipasen
bedingt. Ein möglicher Erklärungsansatz wird in Kapitel 3.2.2 entwickelt.

Ergebnisse und Diskussion
72
3.2.1.3 Konstante spezifische Grenzfläche
Um die festgestellte Abhängigkeit der Aktivität von der spezifischen Grenzfläche zu vali-
dieren, wurde in weiteren Versuchsansätzen durch Verwendung von Reaktionsgefäßen ver-
schiedenen Durchmessers und durch abgestimmte Wahl der Füllvolumina eine konstante
spezifische Grenzfläche geschaffen (Abb. 3.14).
Abbildung 3.14: Konstante spezifische (volumenbezogene) Grenzfläche (0,94 cm-1) durch abgestimmte Phasen-volumina (Verhältnis 1:1) in unterschiedlich großen Reaktionsgefäßen (variable absolute Grenzfläche).
Unabhängig von der Reaktorgröße sollten basierend auf der zuvor beobachteten Abhängig-
keit jeweils identische Geschwindigkeiten der Butylpropionatbildung zu beobachten sein. Bei
Betrachtung von Abbildung 3.15 fällt jedoch auf, dass mit zunehmender Größe des Reak-
tionsgefäßes die Aktivität für beide Enzyme sinkt. Dabei kann bei Vergleich der relativen
Aktivitäten, wie Abbildung 3.16 veranschaulicht, für beide Enzyme eine identische Abnahme
beobachtet werden, die demzufolge nicht enzymspezifisch zu sein scheint. Diese Ergebnisse
widersprechen somit den Erwartungen: Die zuvor beobachtete lineare Abhängigkeit der kata-
lytischen Aktivität von der spezifischen Grenzfläche konnte nicht direkt bestätigt werden. Die
hierfür verantwortlichen Faktoren sind nicht bekannt. Eine Massentransferlimitierung wurde
aufgrund von Voruntersuchungen bislang ausgeschlossen, da der Massentransfer unabhängig
von der Behältnisgröße innerhalb von etwa 30 min abgeschlossen war und die durch die
Enzymreaktion hervorgerufenen Konzentrationsänderungen sehr viel langsamer erfolgten
(Abb. 3.17). Da aber stets mit einem zweistufigen Rührwerk gleicher Abmessungen und mit
gleicher Drehzahl in Reaktionsgefäßen unterschiedlicher Geometrie gearbeitet wurde, sind
Unterschiede hinsichtlich Mischzeit und Leistungseintrag kaum zu vermeiden. Im Vorfeld der
Untersuchungen wurde jedoch angenommen, dass die Größenordnung dieser Unterschiede
unerheblich ist.
Hexan mit Butanol (und Decan)
Wässrige Phase mit nativem Enzym
und Propionsäure
Phasengrenzfläche8,0 mL
0,94 cm-1
13,4 mL
0,94 cm-1
34,0 mL
0,94 cm-1
21,1 mL
0,94 cm-1
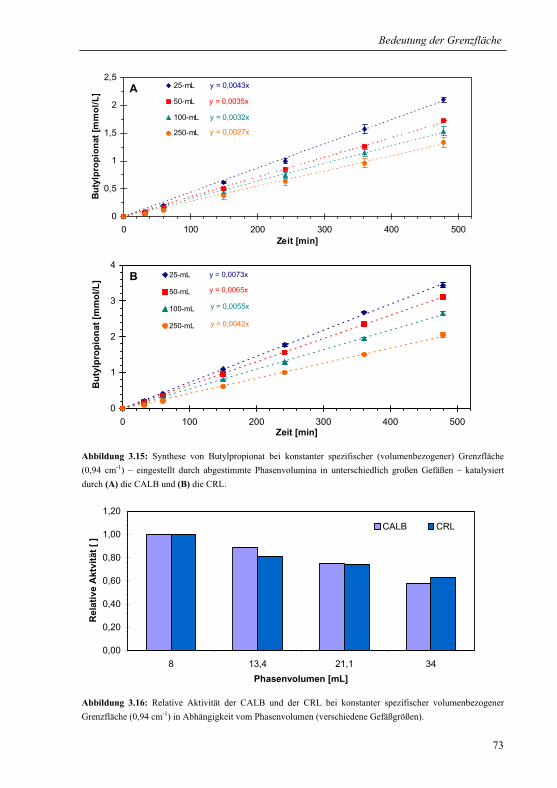
Bedeutung der Grenzfläche
73
y = 0,0043x
y = 0,0035x
y = 0,0032x
y = 0,0027x
0
0,5
1
1,5
2
2,5
0 100 200 300 400 500Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/
L]
25-mL
50-mL
100-mL
250-mL
y = 0,0073x
y = 0,0065x
y = 0,0055x
y = 0,0042x
0
1
2
3
4
0 100 200 300 400 500Zeit [min]
Bu
tylp
rop
ion
at [
mm
ol/L
]
25-mL
50-mL
100-mL
250-mL
Abbildung 3.15: Synthese von Butylpropionat bei konstanter spezifischer (volumenbezogener) Grenzfläche (0,94 cm-1) – eingestellt durch abgestimmte Phasenvolumina in unterschiedlich großen Gefäßen – katalysiert durch (A) die CALB und (B) die CRL.
0,00
0,20
0,40
0,60
0,80
1,00
1,20
8 13,4 21,1 34Phasenvolumen [mL]
Rel
ativ
e A
ktvi
tät
[ ]
CALB CRL
Abbildung 3.16: Relative Aktivität der CALB und der CRL bei konstanter spezifischer volumenbezogener Grenzfläche (0,94 cm-1) in Abhängigkeit vom Phasenvolumen (verschiedene Gefäßgrößen).
A
B
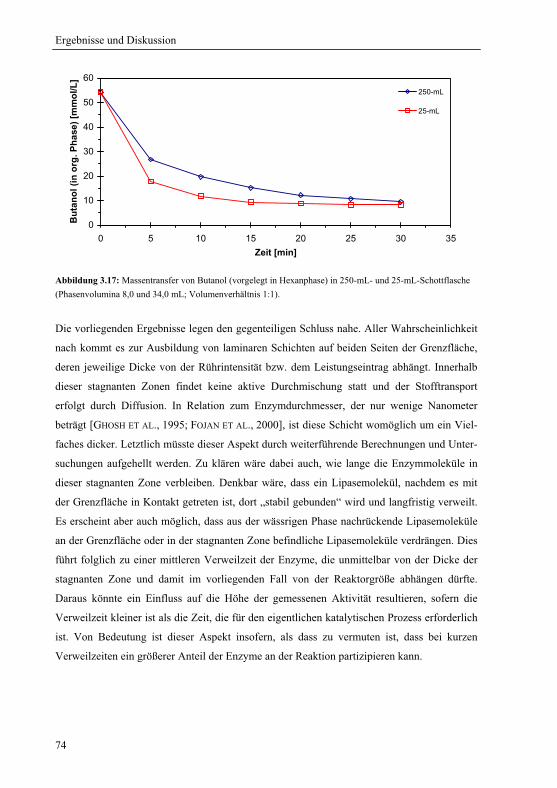
Ergebnisse und Diskussion
74
0
10
20
30
40
50
60
0 5 10 15 20 25 30 35Zeit [min]
Bu
tan
ol (
in o
rg. P
has
e) [
mm
ol/L
]
250-mL
25-mL
Abbildung 3.17: Massentransfer von Butanol (vorgelegt in Hexanphase) in 250-mL- und 25-mL-Schottflasche (Phasenvolumina 8,0 und 34,0 mL; Volumenverhältnis 1:1).
Die vorliegenden Ergebnisse legen den gegenteiligen Schluss nahe. Aller Wahrscheinlichkeit
nach kommt es zur Ausbildung von laminaren Schichten auf beiden Seiten der Grenzfläche,
deren jeweilige Dicke von der Rührintensität bzw. dem Leistungseintrag abhängt. Innerhalb
dieser stagnanten Zonen findet keine aktive Durchmischung statt und der Stofftransport
erfolgt durch Diffusion. In Relation zum Enzymdurchmesser, der nur wenige Nanometer
beträgt [GHOSH ET AL., 1995; FOJAN ET AL., 2000], ist diese Schicht womöglich um ein Viel-
faches dicker. Letztlich müsste dieser Aspekt durch weiterführende Berechnungen und Unter-
suchungen aufgehellt werden. Zu klären wäre dabei auch, wie lange die Enzymmoleküle in
dieser stagnanten Zone verbleiben. Denkbar wäre, dass ein Lipasemolekül, nachdem es mit
der Grenzfläche in Kontakt getreten ist, dort „stabil gebunden“ wird und langfristig verweilt.
Es erscheint aber auch möglich, dass aus der wässrigen Phase nachrückende Lipasemoleküle
an der Grenzfläche oder in der stagnanten Zone befindliche Lipasemoleküle verdrängen. Dies
führt folglich zu einer mittleren Verweilzeit der Enzyme, die unmittelbar von der Dicke der
stagnanten Zone und damit im vorliegenden Fall von der Reaktorgröße abhängen dürfte.
Daraus könnte ein Einfluss auf die Höhe der gemessenen Aktivität resultieren, sofern die
Verweilzeit kleiner ist als die Zeit, die für den eigentlichen katalytischen Prozess erforderlich
ist. Von Bedeutung ist dieser Aspekt insofern, als dass zu vermuten ist, dass bei kurzen
Verweilzeiten ein größerer Anteil der Enzyme an der Reaktion partizipieren kann.

Bedeutung der Grenzfläche
75
3.2.1.4 Variable spezifische Grenzfläche im gelstabilisierten Zweiphasensystem
Unter der Annahme, dass die Reaktion nur an der Grenzfläche erfolgt, dürfte es keinen
Unterschied machen, ob im verwendeten Zweiphasensystem die wässrige Phase gelstabili-
siert ist oder nicht. Die eingesetzten Enzymkonzentrationen waren identisch und damit theore-
tisch auch die Zahl der Enzymmoleküle an der Grenzfläche. Wie Abbildung 3.18 zeigt, ist
sowohl im gelstabilisierten als auch im nicht-gelstabilisierten System eine Abhängigkeit der
Aktivität der CALB von der spezifischen Grenzfläche zu beobachten. Die fehlende Durch-
mischung der gelstabilisierten wässrigen Phase führt zwar unweigerlich zur Ausbildung eines
Konzentrationsgradienten, jedoch dürfte – falls die Reaktion vorwiegend an der Grenzfläche
stattfindet – die Abhängigkeit der Reaktion von der spezifischen Grenzfläche nicht wesentlich
beeinträchtigt werden. Allerdings fällt auf, dass die Aktivität im gelstabilisierten System
erheblich niedriger ist. Mögliche Gründe könnten in evtl. inaktivierenden Einflüssen der
Gellanmatrix zu suchen sein. Da aber Lipasen im Allgemeinen sehr stabil sind und die
Einhüllung in Gellan als schonende Methode gilt [ANSORGE-SCHUMACHER ET AL., 2002],
kommt diese Erklärung nicht in Betracht. Daher muss auf eine andere Erklärung zurückge-
griffen werden.
y = 0,006x + 0,0019 R2 = 0,9915
y = 0,0005x - 7E-05 R2 = 0,9834
0
0,002
0,004
0,006
0,008
0,01
0,012
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6
Spezifische Grenzfläche [cm-1]
Akt
ivit
ät [
mm
ol/L
/min
]
gelstabilisiert
nicht gelstabilisiert
Abbildung 3.18: Abhängigkeit der Aktivität der CALB von der spezifischen (volumenbezogenen) Grenzfläche bei Synthese von Butylpropionat in einem Zweiphasensystem mit gelstabilisierter bzw. mit nicht-gelstabilisierter wässriger Phase.
Im nicht-gelstabilisierten System sind die Enzymmoleküle innerhalb der wässrigen Phase frei
beweglich. Auch wenn die Reaktion beinahe ausschließlich an der Grenzfläche stattfindet,
kann es zu einem Austausch der an der Grenzfläche lokalisierten Enzyme kommen – die
Gesamtzahl der an der Reaktion partizipierenden Enzymmoleküle wäre demnach deutlich

Ergebnisse und Diskussion
76
größer als im gelstabilisierten System (Kapitel 3.2.1.3). Dort ist mit einer sehr viel geringeren
Beweglichkeit der Enzymmoleküle zu rechnen [LONGO ET AL., 1992] und infolgedessen ist
die durchschnittliche Verweilzeit der einzelnen Enzymmoleküle an der Grenzfläche vermut-
lich wesentlich höher. Möglicherweise können die Enzymmoleküle in einer gelstabilisierten
wässrigen Phase auch nicht völlig frei mit der Grenzfläche in Wechselwirkung treten.
Gestützt wird diese Hypothese durch die Arbeit von DICKINSON ET AL. [1988], welche die
Adsorption von Enzymen an Grenzflächen untersuchten. Amphiphile Enzyme wie Lipasen
reichern sich demzufolge an Grenzflächen an – allerdings nur dann, wenn sie in der wässrigen
Phase frei beweglich sind. Dies ist in einer gelstabilisierten Phase nur eingeschränkt möglich.
Die geringere Beweglichkeit und der eingeschränkte Kontakt würden somit die Zahl der an
der Reaktion partizipierenden Enzymmoleküle reduzieren, die Aktivität folglich geringer sein.
Fraglich bleibt, ob es – bei genügend langer Wartezeit – zu einer dem nicht-gelstabilisierten
System vergleichbaren Anreicherung der Enzyme an der Grenzfläche des gelstabilisierten
Systems gekommen wäre und die gemessene Aktivität folglich hätte höher ausfallen können.
y = 0,5368x + 0,1747 R2 = 0,9915
y = 0,7176x - 0,1057 R2 = 0,9834
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6
Spezifische Grenzfläche [cm-1]
Rel
ativ
e A
ktiv
ität
[ ]
gelstabilisiert
nicht gelstabilisiert
Abbildung 3.19: Abhängigkeit der relativen Aktivität der CALB von der spezifischen (volumenbezogenen) Grenzfläche bei Synthese von Butylpropionat in einem Zweiphasensystem mit gelstabilisierter bzw. mit nicht-gelstabilisierter wässriger Phase.
Nachteil der in Abbildung 3.18 gewählten Auftragung ist, dass die aus den verschieden hohen
absoluten Aktivitäten resultierenden Steigungen die Abhängigkeit von der spezifischen
Grenzfläche verfälschen. Diesem Umstand kann durch Auftragung der relativen Aktivitäten
begegnet werden (Abb. 3.19). Dabei fällt auf, dass im gelstabilisierten die Abhängigkeit der
relativen Aktivität von der spezifischen Grenzfläche deutlich höher ist – erkennbar am um
30 % höheren Steigungsfaktor. Dieser Umstand resultiert vermutlich aus denselben Gründen,
die die Aktivitätsminderung im gelstabilisierten Zweiphasensystem bedingen: Die einge-

Bedeutung der Grenzfläche
77
schränkte Beweglichkeit der Enzymmoleküle verhindert einen fortwährenden Austausch mit
Enzymmolekülen aus der wässrigen Phase. Demnach können nur die an der Grenzfläche
fixierten Enzymmoleküle an der Reaktion partizipieren. Im nicht-gelstabilisierten System
hingegen partizipieren durch Austausch beweglicher Enzymmoleküle an der Grenzfläche
insgesamt mehr Enzymmoleküle an der Reaktion, was sich in einer reduzierten Abhängigkeit
von der spezifischen Grenzfläche widerspiegelt.
3.2.1.5 Einfluss der Wasseraktivität bei variabler spezifischer Grenzfläche
Bisherige Untersuchungen zeigten sowohl für die grenzflächenaktivierte als auch für die
nicht-grenzflächenaktivierte Lipase eine identische Abhängigkeit der katalytischen Aktivität
von der spezifischen Grenzfläche. Die Ergebnisse lassen den Schluss zu, dass die Reaktion
fast ausschließlich an der Grenzfläche stattfindet, wofür – wie in Kapitel 3.2.3 eingehend
erörtert wird – in erster Linie thermodynamische Gründe anzuführen sind. Es ist bekannt, dass
in monophasischen Systemen die Esterbildung wasserarme Reaktionsmedien bzw. Wasserak-
tivitäten im Bereich von 0,6 bis 0,8 erfordert [HALLING, 1994; KVITTINGEN, 1994]. In einem
Zweiphasensystem liegt, falls auf eine Kontrolle der Wasseraktivität verzichtet wird, diese
nahe bei eins [HALLING, 1994]. Dabei ist zu berücksichtigen, dass Wasser gegenüber Butanol
ein stärkeres Nukleophil und im Übermaß vorhanden ist. Somit kann nicht ausgeschlossen
werden, dass die thermodynamischen Bedingungen zumindest in der wässrigen Phase grund-
sätzlich einer Esterbildung entgegenstehen. Ziel dieser Versuchsvariante war es, zu über-
prüfen, ob durch eine Absenkung der Wasseraktivität die Abhängigkeit der Aktivität von der
spezifischen Grenzfläche verringert und die Reaktion in der wässrigen Phase verstärkt er-
folgen kann. Eine Absenkung der Wasseraktivität kann durch Lösen einer osmotisch aktiven
Substanz in der wässrigen Phase, durch eine Äquilibrierung über die Dampfphase oder über
spezielle Membrankompartimente, innerhalb derer ein Lösungsmittel oder Gas der gewünsch-
ten Wasseraktivität zirkuliert, vorgenommen werden [KVITTINGEN ET AL., 1992; WEHTJE ET
AL., 1997]. Im verwendeten System kam nur erstgenannte Variante in Frage, wobei allerdings
auf die Verwendung von Salzen verzichtet wurde, da die erforderlichen hohen Salzkonzentra-
tionen die Lipase inaktiviert hätten [KOCH, 1997]. Stattdessen sollte durch Verwendung einer
75%igen (w/v) Glucoselösung die Wasseraktivität im Reaktionsansatz auf etwa 0,8 gesenkt
werden [HALLING, 1994]. Falls durch Absenkung der Wasseraktivität die Reaktion auch in
der wässrigen Phase verstärkt stattfinden kann, müsste die Abhängigkeit der Aktivität der
CALB als nicht grenzflächenaktivierte Lipase von der spezifischen Grenzfläche sinken.
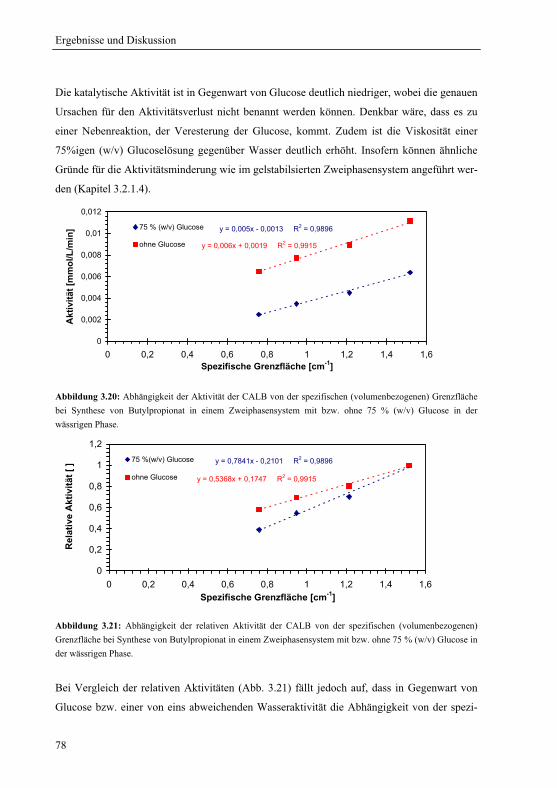
Ergebnisse und Diskussion
78
Die katalytische Aktivität ist in Gegenwart von Glucose deutlich niedriger, wobei die genauen
Ursachen für den Aktivitätsverlust nicht benannt werden können. Denkbar wäre, dass es zu
einer Nebenreaktion, der Veresterung der Glucose, kommt. Zudem ist die Viskosität einer
75%igen (w/v) Glucoselösung gegenüber Wasser deutlich erhöht. Insofern können ähnliche
Gründe für die Aktivitätsminderung wie im gelstabilsierten Zweiphasensystem angeführt wer-
den (Kapitel 3.2.1.4).
y = 0,006x + 0,0019 R2 = 0,9915
y = 0,005x - 0,0013 R2 = 0,9896
0
0,002
0,004
0,006
0,008
0,01
0,012
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6Spezifische Grenzfläche [cm-1]
Akt
ivit
ät [
mm
ol/L
/min
] 75 % (w/v) Glucose
ohne Glucose
Abbildung 3.20: Abhängigkeit der Aktivität der CALB von der spezifischen (volumenbezogenen) Grenzfläche bei Synthese von Butylpropionat in einem Zweiphasensystem mit bzw. ohne 75 % (w/v) Glucose in der wässrigen Phase.
y = 0,5368x + 0,1747 R2 = 0,9915
y = 0,7841x - 0,2101 R2 = 0,9896
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6Spezifische Grenzfläche [cm-1]
Rel
ativ
e A
ktiv
ität
[ ]
75 %(w/v) Glucose
ohne Glucose
Abbildung 3.21: Abhängigkeit der relativen Aktivität der CALB von der spezifischen (volumenbezogenen) Grenzfläche bei Synthese von Butylpropionat in einem Zweiphasensystem mit bzw. ohne 75 % (w/v) Glucose in der wässrigen Phase.
Bei Vergleich der relativen Aktivitäten (Abb. 3.21) fällt jedoch auf, dass in Gegenwart von
Glucose bzw. einer von eins abweichenden Wasseraktivität die Abhängigkeit von der spezi-

Bedeutung der Grenzfläche
79
fischen Grenzfläche erhöht ist. Diese Beobachtung lässt nur den Schluss zu, dass entgegen der
Erwartungen durch Absenken der Wasseraktivität der auf die in der wässrigen Phase gelösten
Enzymmoleküle zurückgehende Anteil an der Gesamtaktivität gesenkt wird. Die hohe
Konzentration an Glucose stellt einen erheblichen Eingriff in das zu untersuchende System
dar. Denkbar wäre auch, dass die hohe Konzentration an Glucose und möglicherweise daraus
resultierende Wechselwirkungen zwischen Glucosemolekülen und Enzymmolekülen den
Zugang der Substrate zum aktiven Zentrum beeinträchtigen. Dennoch ist aufgrund der Ergeb-
nisse wahrscheinlich, dass zumindest in biphasischen Reaktionssystemen die Wasseraktivität
nicht entscheidend ist (Kapitel 3.2.3).
3.2.1.6 Hydrolyse von Butylpropionat bei variabler spezifischer Grenzfläche
Im vorangegangenen Kapitel wurde bereits darauf verwiesen, dass aufgrund widriger thermo-
dynamischer Bedingungen die Reaktionsrate bei der Estersynthese in der wässrigen Phase
erheblich reduziert ist. Bezüglich der Hydrolyse ist aber davon auszugehen, dass die Reaktion
im gesamten Volumen der wässrigen Phase möglich sein sollte. Dies muss jedoch nicht für
eine grenzflächenaktivierte Lipase gelten. Unter der Annahme, dass eine grenzflächen-
aktivierte Lipase nur bei Kontakt mit einer Grenzfläche in der offenen und damit aktiven
Konformation vorliegt, könnte diese Lipase die Hydrolyse nur an der Grenzfläche kataly-
sieren. Hingegen wäre davon auszugehen, dass eine nicht-grenzflächenaktivierte Lipase auch
in der wässrigen Phase katalytisch aktiv ist. Daraus sollte für die nicht-grenzflächenaktivierte
Lipase eine signifikant reduzierte Abhängigkeit der Aktivität von der spezifischen Grenz-
fläche zu konstatieren sein.
Wie Abbildung 3.22 zu entnehmen ist, zeigte sich sowohl für die CALB als auch die CRL
eine gleichgelagerte lineare Abhängigkeit ihrer Aktivität von der spezifischen Grenzfläche für
die Hydrolyse. Demzufolge wird der Ester durch beide Lipasen überwiegend an der Grenz-
fläche hydrolysiert und nicht in der wässrigen Phase. Für die CRL wurde dies erwartet, für die
CALB nicht unbedingt. Aufgrund der etwas höheren Steigung (0,63 cm im Vergleich zu 0,54
cm) ist davon auszugehen, dass die Abhängigkeit der Aktivität der CALB von der spezi-
fischen Grenzfläche sogar etwas stärker ausgeprägt zu sein scheint als die der CRL. Ein
möglicher Grund für diese scheinbare Abhängigkeit von der spezifischen Grenzfläche könnte
in der geringen Löslichkeit von Butylpropionat in der wässrigen Phase bestehen. Dies könnte
dazu geführt haben, dass die Konzentration an Ester in der wässrigen Phase sehr gering war
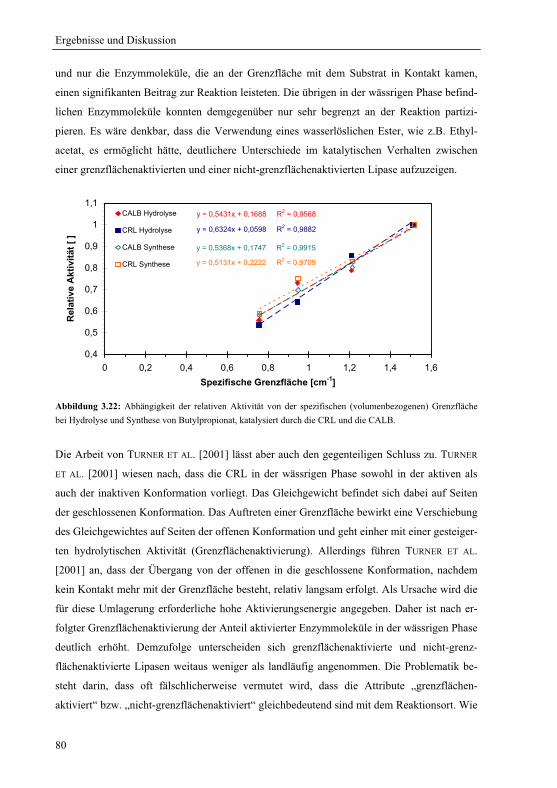
Ergebnisse und Diskussion
80
und nur die Enzymmoleküle, die an der Grenzfläche mit dem Substrat in Kontakt kamen,
einen signifikanten Beitrag zur Reaktion leisteten. Die übrigen in der wässrigen Phase befind-
lichen Enzymmoleküle konnten demgegenüber nur sehr begrenzt an der Reaktion partizi-
pieren. Es wäre denkbar, dass die Verwendung eines wasserlöslichen Ester, wie z.B. Ethyl-
acetat, es ermöglicht hätte, deutlichere Unterschiede im katalytischen Verhalten zwischen
einer grenzflächenaktivierten und einer nicht-grenzflächenaktivierten Lipase aufzuzeigen.
y = 0,6324x + 0,0598 R2 = 0,9882
y = 0,5431x + 0,1688 R2 = 0,9568
y = 0,5131x + 0,2222 R2 = 0,9709
y = 0,5368x + 0,1747 R2 = 0,9915
0,4
0,5
0,6
0,7
0,8
0,9
1
1,1
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6Spezifische Grenzfläche [cm-1]
Rel
ativ
e A
ktiv
ität
[ ]
CALB Hydrolyse
CRL Hydrolyse
CALB Synthese
CRL Synthese
Abbildung 3.22: Abhängigkeit der relativen Aktivität von der spezifischen (volumenbezogenen) Grenzfläche bei Hydrolyse und Synthese von Butylpropionat, katalysiert durch die CRL und die CALB.
Die Arbeit von TURNER ET AL. [2001] lässt aber auch den gegenteiligen Schluss zu. TURNER
ET AL. [2001] wiesen nach, dass die CRL in der wässrigen Phase sowohl in der aktiven als
auch der inaktiven Konformation vorliegt. Das Gleichgewicht befindet sich dabei auf Seiten
der geschlossenen Konformation. Das Auftreten einer Grenzfläche bewirkt eine Verschiebung
des Gleichgewichtes auf Seiten der offenen Konformation und geht einher mit einer gesteiger-
ten hydrolytischen Aktivität (Grenzflächenaktivierung). Allerdings führen TURNER ET AL.
[2001] an, dass der Übergang von der offenen in die geschlossene Konformation, nachdem
kein Kontakt mehr mit der Grenzfläche besteht, relativ langsam erfolgt. Als Ursache wird die
für diese Umlagerung erforderliche hohe Aktivierungsenergie angegeben. Daher ist nach er-
folgter Grenzflächenaktivierung der Anteil aktivierter Enzymmoleküle in der wässrigen Phase
deutlich erhöht. Demzufolge unterscheiden sich grenzflächenaktivierte und nicht-grenz-
flächenaktivierte Lipasen weitaus weniger als landläufig angenommen. Die Problematik be-
steht darin, dass oft fälschlicherweise vermutet wird, dass die Attribute „grenzflächen-
aktiviert“ bzw. „nicht-grenzflächenaktiviert“ gleichbedeutend sind mit dem Reaktionsort. Wie

Bedeutung der Grenzfläche
81
diese Experimente und die Arbeit von TURNER ET AL. [2001] vermuten lassen, ist dem aber
nicht so.
3.2.2 „Molecular Modelling“ der Enzymoberfläche
Wechselwirkungen zwischen Lipasen und Grenzflächen und daraus resultierende Vorgänge
wurden bisher kaum auf molekularer Ebene betrachtet. Einzig JENSSEN ET AL. [2002] und
NOINVILLE ET AL. [2002] führten diesbezüglich Untersuchungen in Zusammenhang mit der
grenzflächeninduzierten Konformationsänderung der Lipase aus Thermomyces lanuginosa
(TLL) durch. NOINVILLE ET AL. [2002] benutzten die FTIR-ATR-Spektroskopie, um die
Adsorption der TLL an die Oberfläche eines hydrophoben Materials zu untersuchen. Dabei
dokumentierten sie die als Folge der Adsorption verstärkte Protonierung von in der Nähe des
Lids bzw. des aktiven Zentrums befindlichen Carboxylgruppen. Folglich wurde postuliert,
dass dieser Bereich bevorzugt mit hydrophoben Oberflächen in Wechselwirkung tritt und
infolgedessen das aktive Zentrum der Grenzfläche zugewandt ist. Zu ähnlichen Rückschlüs-
sen kamen JENSSEN ET AL. [2002], die auf Basis der Elektronendichte durch Röntgenstruktur-
analysen und molekulare Simulationen Aussagen über die Konformation und Orientierung
von an eine Grenzfläche adsorbierten TLL-Molekülen entwickelten. Es stellt sich die Frage,
ob auch für andere Lipasen eine spezifische Orientierung gegenüber der Grenzfläche erwartet
werden darf. Dies wurde des Öfteren angenommen, bisher aber nicht wie von JENSSEN ET AL.
[2002] und NOINVILLE ET AL. [2002] methodisch angegangen. Im Rahmen dieser Arbeit sollte
dieser Aspekt erstmalig in silico über ein „Molecular Modelling“ betrachtet werden.
„Molecular Modelling“ bezeichnet eine softwaregestützte Methode, um experimentelle
Befunde auf einer theoretischen Basis zu erklären [BRAIUCA ET AL., 2006]. Mit Hilfe
hochspezialisierter Software kann ausgehend von der dreidimensionalen Struktur der in dieser
Arbeit untersuchten Enzyme durch Berechnung von physikalischen, chemischen und topo-
logischen Parametern auf den Knotenpunkten eines Raumgitters („GRID-Calculation“) die
molekulare Oberfläche sowie die innere Beschaffenheit der Proteine chemisch charakterisiert
werden. Dabei können Bereiche auf der Proteinoberfläche identifiziert werden, die z.B.
bevorzugt mit polaren oder unpolaren Molekülen interagieren.

Ergebnisse und Diskussion
82
Abbildung 3.23: Superposition der 3D-Struktur der CRL in der offenen und geschlossenen Konformation (A);Visualisierung hydrophober und hydrophiler Bereiche auf der Oberfläche der CRL in der offenen (B) und geschlossenen Konformation (C).
Zunächst wurde die CRL sowohl in der offenen als auch in der geschlossenen Konformation
analysiert (Abb. 3.23). Hellblaue Volumina kennzeichnen hydrophile Bereiche, in denen die
Wahrscheinlichkeit erhöht ist, polare Moleküle, z.B. Wasser, anzutreffen. Gegenteiliges gilt
für die orange-gelben Volumina, dort sind eher unpolare Moleküle zu erwarten. Interessanter-
weise kann im Bereich des aktiven Zentrums ein deutlicher Unterschied zwischen der offenen
und der geschlossenen Konformation erkannt werden. In der offenen Konformation ist die
Ausdehnung der hydrophoben Bereiche durch die vorangegangene Bewegung des -helikalen
Lids gegenüber der geschlossenen Konformation deutlich größer. Ein solches Ergebnis war zu
erwarten, da mehrere Wissenschaftler in Zusammenhang mit Röntgenstrukturanalysen nach-
weisen konnten, dass die hydrophobe Oberfläche in der offenen Konformation deutlich größer
ist [VASEL ET AL., 1993; MARTINELLE ET AL., 1995]. Bisher finden sich jedoch weniger ein-
deutige Informationen, die die Topologie der Proteinmoleküle berücksichtigen. Sowohl für
die CRL als auch die CALB kann topologisch eine Polarität bezüglich hydrophiler und hydro-
phober Bereiche auf der Oberfläche erkannt werden (Abb. 3.24 und 3.25).
B (offen) C (geschlossen)
A
Offene Konformation
Geschlossene Konformation
Hydrophile Bereiche
Hydrophobe Bereiche
Aktives Zentrum
Aktives Zentrum
Lid
B (offen) C (geschlossen)
A
Offene Konformation
Geschlossene Konformation
Hydrophile Bereiche
Hydrophobe Bereiche
Aktives Zentrum
Aktives Zentrum
Lid

Bedeutung der Grenzfläche
83
Abbildung 3.24: Visualisierung hydrophober und hydrophiler Bereiche auf der Oberfläche der CALB.
Abbildung 3.25: Visualisierung hydrophober und hydrophiler Bereiche auf der Oberfläche der CRL.
Die Oberfläche im Bereich des aktiven Zentrums ist überwiegend hydrophob, die distalen
Bereiche demgegenüber hydrophil. Diese Beobachtung legt den Schluss nahe, dass stets der
Bereich um das aktive Zentrum der Grenzfläche zugewandt ist. Zudem kann daraus auch
geschlossen werden, dass es zu einer Anreicherung der Lipasen an der Grenzfläche kommt,
wie im Fall amphiphiler Proteine hinreichend belegt ist [DICKINSON ET AL., 1988; HALLING ET
AL., 1998]. Gerade bei grenzflächenaktivierten Lipasen ist dieser Aspekt von Bedeutung, da
z.B. ihr Lid als eine Art „Anker“ an der Grenzfläche fungieren kann [MARTINELLE ET AL.,
CRL
Aktives Zentrum
Aktives Zentrum
Hydrophobe Bereiche Hydrophile Bereiche
CALB
Aktives Zentrum
Aktives Zentrum
Hydrophobe Bereiche Hydrophile Bereiche

Ergebnisse und Diskussion
84
1995]. Zur Aufklärung des Reaktionsmechanismus wäre es von Bedeutung zu wissen, wie das
aktive Zentrum an der Grenzfläche lokalisiert ist. Aufgrund der Ergebnisse kann davon
ausgegangen werden, dass das aktive Zentrum unmittelbar an der Grenzfläche „andockt“ und
infolgedessen ein direkter Stoffaustausch zwischen aktivem Zentrum und wässriger Phase nur
über die organische Phase erfolgen kann. Leider gibt die Literatur keine Anhaltspunkte zur
Beantwortung dieser Frage.
In Kapitel 3.2.1.2 wurde festgestellt, dass für die Esterase von Rhizopus arrhizus im Ver-
gleich zu den beiden Lipasen (CALB und CRL) die Abhängigkeit der katalytischen Aktivität
von der spezifischen Grenzfläche weniger stark ausgeprägt zu sein scheint. Um dieses
Ergebnis interpretieren zu können, wurde erwogen, die Oberfläche der Esterase analog zu den
beiden Lipasen hinsichtlich hydrophober und hydrophiler Bereiche zu charakterisieren. Aller-
dings ist die dreidimensionale Struktur dieser Esterase bislang nicht publiziert worden. Aus
diesem Grund wurde die Oberfläche der Esterase von Bacillus subtilis charakterisiert.
Abbildung 3.26: Visualisierung hydrophober und hydrophiler Bereiche auf der Oberfläche der Esterase von Bacillus subtilis
Dabei konnte keine topologische Polarität wie bei den Lipasen erkannt werden. Die
hydrophilen und hydrophoben Bereiche scheinen mehr oder weniger gleichmäßig verteilt zu
sein (Abb. 3.26). Zwar ist nicht bekannt, inwieweit die beiden Esterasen miteinander ver-
glichen werden können, doch ist davon auszugehen, dass Esterasen im Vergleich zu Lipasen
auf eine andere Weise mit der Grenzfläche interagieren. Möglicherweise werden Esterasen
weniger stark an der Grenzfläche angereichert und/oder können sich leichter von ihr lösen.
Esterase von Bacillus subtilis
Hydrophobe Bereiche Hydrophile Bereiche

Bedeutung der Grenzfläche
85
Sollte dies zu einer gesteigerten Fluktuation der Enzymmoleküle an der Grenzfläche führen,
könnten insgesamt mehr Enzymmoleküle an der Reaktion partizipieren – erkennbar an einer
geringen Abhängigkeit zwischen Aktivität und spezifischer Grenzfläche (Kapitel 3.2.1.3).
3.2.3 Thermodynamik der Esterbildung im fluiden Zweiphasensystem
Wird in biphasischen Reaktionssystemen auf Maßnahmen zur Kontrolle der Wasseraktivität
verzichtet, so liegt diese unweigerlich nahe 1,0. Bei oberflächlicher Betrachtung erscheint es
zunächst verwunderlich, dass auch unter diesen Bedingungen eine Esterbildung beobachtet
werden kann. Dies lässt aus empirischer Sicht vermuten, dass die lokalen Wasserkonzentra-
tionen ausschlaggebend sind. In der wässrigen Phase ist der Wassergehalt sehr hoch. Im
Gegensatz dazu ist unmittelbar an der Grenzfläche und in der organischen Phase der
Wassergehalt relativ gering. Dies hat – wie auch die bisherigen Ergebnisse nahelegen – zur
Folge, dass die Reaktion bevorzugt an der Grenzfläche stattfinden wird. Thermodynamisch ist
die Esterbildung auch in der organischen Phase möglich. Da aber die Enzymmoleküle nahezu
ausschließlich in der wässrigen Phase gelöst sind, können sie auch nur dort die Reaktion
beschleunigen. Dieser Aspekt erschwert – wie nachfolgend dargelegt – die thermodynamische
Betrachtung des Gesamtsystems, weshalb der Literatur diesbezüglich nur wenig zu entneh-
men ist. Von Interesse wäre aber, wie in dieser Arbeit geschehen, zu eruieren, inwieweit eine
Estersynthese in der wässrigen Phase eines Zweiphasensystems unter thermodynamischen
Gesichtspunkten überhaupt möglich ist.
Die hier untersuchte Bildung von Butylpropionat aus Butanol und Propionsäure erfolgte in
einem zweiphasigen Reaktionssystem. Auch wenn sich in einem solchen System die Konzen-
trationen der Einzelkomponenten in wässriger und organischer Phase im Gleichgewichtszu-
stand unterscheiden, sind die thermodynamischen Aktivitäten in jeder Phase gleich [HALLING,
1994]. Die Beschreibung der Gleichgewichtslage der Reaktion muss daher unter Verwendung
der thermodynamischen Aktivitäten erfolgen (2):
SäureAlkohol
WasserEstera
aaaaK
Ist das Verhältnis der Aktivitätsprodukte größer als Ka, so findet eine Hydrolyse statt, wohin-
gegen im umgekehrten Fall bevorzugt die Estersynthese erfolgt. So ist in einem rein
(2)

Ergebnisse und Diskussion
86
wässrigen System (aw = 1) aufgrund der geringen Löslichkeit des Esters die Esteraktivität
bereits bei sehr niedrigen Konzentrationen nahe 1,0 und die Synthese kommt frühzeitig zum
Erliegen [TWEDDELL ET AL., 1998]. Dies ist mithin auch der Hauptgrund dafür, dass die Ab-
senkung der Wasseraktivität einen eher unwesentlichen Einfluss auf das Reaktionsgleich-
gewicht haben dürfte – die Esteraktivität bleibt hiervon unbeeinflusst. Mit Einführung einer
organischen Phase und der daraus resultierenden Extraktion des Esters sinkt dessen Aktivität,
während die Wasseraktivität nahezu unverändert bleibt [HALLING, 1994; KVITTINGEN, 1994;
TWEDDELL ET AL., 1998]. Die mit Einführung einer organischen Phase eintretende Verschie-
bung der Gleichgewichtslage hin zur Esterbildung beruht demnach nicht auf der Senkung der
Wasseraktivität bzw. des dort niedrigen Wassergehaltes, sondern auf der Senkung der
Esteraktivität. Eine Veränderung des Phasenvolumenverhältnisses wässrig/organisch (vor
Einstellung des Gleichgewichtszustandes) wirkt sich ebenfalls auf die Gleichgewichtslage aus
[HALLING, 1994]. Eine Vergrößerung der organischen Phase erlaubt einen nahezu voll-
ständigen Umsatz der Edukte, da die Konzentration bzw. Aktivität des gebildeten Esters in
der organischen Phase sinkt [BORZEIX ET AL., 1992; TWEDDELL ET AL., 1998].
Sollte die Butylpropionatbildung bevorzugt an der Grenzfläche stattfinden, wie die hier
erzielten Ergebnisse unzweifelhaft nahelegen, erscheint die Verwendung der Gleichge-
wichtskonstanten Ka ungeeignet. Mit ihr wird das System nur als Ganzes beschrieben, jedoch
müssen auf molekularer Ebene lokale Konzentrationen berücksichtigt werden [HALLING,
1994]. Das Verhalten eines Biokatalysators ist demnach abhängig von der Phase, in der er
sich befindet. Daher muss die konzentrationsabhängige Gleichgewichtskonstante Kc für jede
Phase aus Ka abgeleitet werden [HALLING 1994; KVITTINGEN, 1994]. In stark verdünnten
Lösungen ist i konstant (Henry’sches Gesetz) und KX = Kc und daraus folgt:
iXi
KKXX
XXaa
aaK
i
I
XSäureAlkohol
WasserEster
SäureAlkohol
WasserEster
SäureAlkohol
WasserEstera
KomponetederMolenbruchKomponentedertkoeffizienAktivitäts
aWasserEster
SäureAlkoholac KK
KK 1
(3)
(4)

Bedeutung der Grenzfläche
87
Die Aktivitätskoeffizienten sind dabei abhängig vom gewählten Lösungsmittel und bewirken
eine Veränderung der Gleichgewichtskonstanten Kc. Deshalb unterscheidet sich die Lage des
Gleichgewichtes in den jeweiligen Phasen. Zudem ist das Verteilungsverhalten der Kompo-
nenten zwischen zwei verschiedenen Lösungsmitteln (hier Wasser und Hexan) zu berück-
sichtigen, woraus folgt [KVITTINGEN 1994; HALLING, 1994]:
Wasserwobei
KomponentederntskoeffizieVerteilung
i
Hexanii
i
cHexanWasserEster
SäureAlkoholcWasser
ccP
iP
KPP
PPK
Für die untersuchte Veresterung von Butanol und Propionsäure hat dies zur Konsequenz, dass
aufgrund der niedrigen Verteilungskoeffizienten von Butanol, Propionsäure und Wasser
einerseits und des recht hohen Verteilungskoeffizienten von Butylpropionat andererseits die
Gleichgewichtskonstante KcWasser deutlich geringer ist als KcHexan. Somit liegt das Reaktions-
gleichgewicht in der Hexanphase im Vergleich zur wässrigen Phase stärker auf Seiten von
Butylpropionat.
Die von HALLING [1994] getroffene Aussage, das Verhalten eines Biokatalysators sei abhän-
gig von der Phase, in der er sich befindet, muss in einem zweiphasigen Reaktionssystem und
gerade im Fall der Lipasen differenziert betrachtet werden. Der Großteil des Enzyms ist in der
wässrigen Phase gelöst, trägt aber aufgrund der geringeren Gleichgewichtskonstanten KcWasser
nur unwesentlich zur Esterbildung bei. Wird vorausgesetzt, dass die in Kontakt mit der
Grenzfläche stehenden Enzyme der organischen Phase zuzurechnen sind, dann erfolgt hier die
Reaktion aufgrund der größeren Gleichgewichtskonstanten KcHexan schneller. Diese Aussage
kann herangezogen werden, um die experimentell beobachtete Abhängigkeit der Butyl-
propionatbildungsrate von der spezifischen Grenzfläche zu erklären. Das Vorhandensein einer
Grenzfläche bewirkt den Kontakt von Enzym und organischer Phase und die Vergrößerung
der spezifischen Grenzfläche erhöht den Anteil an Enzymmolekülen mit Kontakt zur
organischen Phase. Aus diesem Grund steigt die katalytische Aktivität.
(5)

Ergebnisse und Diskussion
88
3.2.4 Hypothese über die Vorgänge an der Grenzfläche
Die durchgeführten Untersuchungen konnten klar zeigen, dass eine erhebliche Abhängigkeit
der katalytischen Aktivität von der spezifischen Grenzfläche besteht und überdies kein Unter-
schied zwischen grenzflächenaktivierten und nicht-grenzflächenaktivierten Lipasen festzustel-
len ist. Abbildung 3.27A versucht, die in dieser Arbeit getroffenen Feststellungen zusammen-
fassend zu illustrieren. Zugegebenermaßen basiert die Abbildung überwiegend auf einer rein
hypothetischen Grundlage, die aber angesichts der erzielten Ergebnisse einer gewissen Plausi-
bilität nicht entbehrt: Durch Kontakt mit der Grenzfläche wird die grenzflächenaktivierte
Lipase in die aktive Konformation überführt (1). Zu einem gewissen Anteil erfolgt dort die
Reaktion nach Bindung der Substrate aus der organischen Phase und anschließender Frei-
setzug des Produktes (2b und 5). Denkbar wäre aber auch, dass sich die Lipasemoleküle nach
der Aktivierung von der Grenzfläche ablösen und die Bindung der Substrate in der wässrigen
Phase aufgrund der dort vorherrschenden hohen Konzentrationen womöglich bevorzugt
erfolgt (2a) – was zwar spekulativ ist, jedoch aufgrund der erzielten Ergebnisse und der Arbeit
von TURNER ET AL. [2001] durchaus im Bereich des Möglichen liegt. Demzufolge könnte die
Reaktion auch von in der wässrigen Phase befindlichen Enzymmolekülen katalysiert werden
(3a). Allerdings ist dies, wie Reaktionsgleichgewicht und Thermodynamik nahe legen, eher
unwahrscheinlich, da die hohe Nukleophilie von Wasser dem entgegensteht und zudem
möglicherweise gebildete Ester unmittelbar hydrolysiert werden könnten. Sollten dennoch
nennenswerte Mengen an Ester gebildet werden, stellt sich die Frage, ob aufgrund der
geringen Wasserlöslichkeit überhaupt mit einer Freisetzung des Substrats in die wässrige
Phase zu rechnen ist. Wahrscheinlicher ist, dass die Enzymmoleküle nach Substratbindung in
der wässrigen Phase an die Grenzfläche „zurückkehren“ (4) und dort Reaktion sowie
Freisetzung (5) des Produktes direkt in die organische Phase erfolgen.
Prinzipiell können die Feststellungen bezüglich der Substratbindung und Produktfreisetzung
direkt auf nicht-grenzflächenaktivierte Lipasen und Esterasen übertragen werden. Allerdings
sind diese Systeme insgesamt weniger kompliziert, da keine Übergänge zwischen einer
aktiven und einer inaktiven Konformation berücksichtigt werden müssen (Abb. 3.27 B).

Bedeutung der Grenzfläche
89
Abbildung 3.27: Hypothetisches Modell zur Esterbildung in fluiden Zweiphasensystemen mit einer grenz-flächenaktivierten (A) und einer nicht-grenzflächenaktivierten Lipase (B):
1) Anlagerung an die Grenzfläche Überführung in die aktive Konformation 2a) Ablösen der Lipase von der Grenzfläche und Bindung der Substrate aus der wässrigen Phase 2b) Lipase ist stabil an die Grenzfläche gebunden – Bindung der Substrate aus der organischen Phase 3a) Reaktion in wässriger Phase 3b) Reaktion an Grenzfläche 4) Anlagerung an Grenzfläche (folgt auf 3a) 5) Produktfreisetzung an der Grenzfläche in die organische Phase L) Leerlauf – keine Substratbindung und Konformationsänderung der Lipase von aktiv zu inaktiv SZ) stagnante Zone.
Hexan
H2O
4
SZ
Substratbindung
Produktfreisetzung in organische Phase
Substratbindung
2b
Produktfreisetzung in wässrige Phase ?
2a
5
3b
3a
Hexan
H2O
4
SZ
Substratbindung
Produktfreisetzung in organische Phase
Substratbindung
2b
Produktfreisetzung in wässrige Phase ?
2a
5
3b
3a
Hexan
H2O
2a
L
SZ
Substratbindung
Produktfreisetzung in organische Phase
3b
Substratbindung
2b
3a
5
1
4
Produktfreisetzung in wässrige Phase ?
Hexan
H2O
2a
LL
SZ
Substratbindung
Produktfreisetzung in organische Phase
3b
Substratbindung
2b
3a
5
11
4
Produktfreisetzung in wässrige Phase ?
A
B
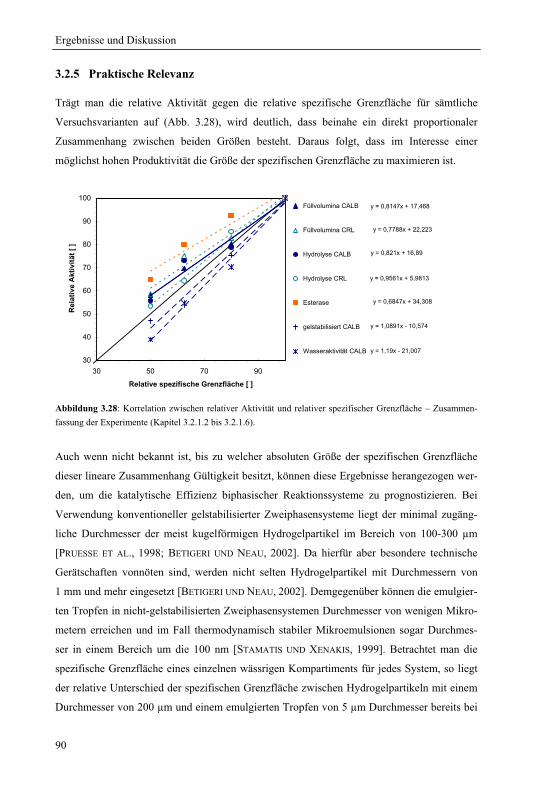
Ergebnisse und Diskussion
90
3.2.5 Praktische Relevanz
Trägt man die relative Aktivität gegen die relative spezifische Grenzfläche für sämtliche
Versuchsvarianten auf (Abb. 3.28), wird deutlich, dass beinahe ein direkt proportionaler
Zusammenhang zwischen beiden Größen besteht. Daraus folgt, dass im Interesse einer
möglichst hohen Produktivität die Größe der spezifischen Grenzfläche zu maximieren ist.
Abbildung 3.28: Korrelation zwischen relativer Aktivität und relativer spezifischer Grenzfläche – Zusammen-fassung der Experimente (Kapitel 3.2.1.2 bis 3.2.1.6).
Auch wenn nicht bekannt ist, bis zu welcher absoluten Größe der spezifischen Grenzfläche
dieser lineare Zusammenhang Gültigkeit besitzt, können diese Ergebnisse herangezogen wer-
den, um die katalytische Effizienz biphasischer Reaktionssysteme zu prognostizieren. Bei
Verwendung konventioneller gelstabilisierter Zweiphasensysteme liegt der minimal zugäng-
liche Durchmesser der meist kugelförmigen Hydrogelpartikel im Bereich von 100-300 μm
[PRUESSE ET AL., 1998; BETIGERI UND NEAU, 2002]. Da hierfür aber besondere technische
Gerätschaften vonnöten sind, werden nicht selten Hydrogelpartikel mit Durchmessern von
1 mm und mehr eingesetzt [BETIGERI UND NEAU, 2002]. Demgegenüber können die emulgier-
ten Tropfen in nicht-gelstabilisierten Zweiphasensystemen Durchmesser von wenigen Mikro-
metern erreichen und im Fall thermodynamisch stabiler Mikroemulsionen sogar Durchmes-
ser in einem Bereich um die 100 nm [STAMATIS UND XENAKIS, 1999]. Betrachtet man die
spezifische Grenzfläche eines einzelnen wässrigen Kompartiments für jedes System, so liegt
der relative Unterschied der spezifischen Grenzfläche zwischen Hydrogelpartikeln mit einem
Durchmesser von 200 μm und einem emulgierten Tropfen von 5 μm Durchmesser bereits bei
y = 1,19x - 21,007
y = 1,0891x - 10,574
y = 0,6847x + 34,308
y = 0,7788x + 22,223
y = 0,8147x + 17,468
y = 0,9561x + 5,9813
y = 0,821x + 16,89
30
40
50
60
70
80
90
100
30 50 70 90
Relative spezifische Grenzfläche [ ]
Rel
ativ
e A
ktiv
ität
[ ]
Füllvolumina CALB
Füllvolumina CRL
Hydrolyse CALB
Hydrolyse CRL
Esterase
gelstabilisiert CALB
Wasseraktivität CALB

Bedeutung der Grenzfläche
91
einem Faktor von 40. Der Unterschied bezüglich der Effizienz dürfte bei ansonsten ver-
gleichbaren Reaktionsbedingungen in einer ähnlichen Größenordnung liegen. Bei der Wahl
einer geeigneten Immobilisierungsvariante muss dies unbedingt Berücksichtigung finden,
auch wenn daraus folgt, dass die Verwendung konventioneller Hydrogele eher nicht in Be-
tracht gezogen werden sollte. Maßgebliches Kriterium bei der Immobilisierung von Lipasen
für den Einsatz in biphasischen Reaktionssystemen muss die Schaffung von Grenzfläche sein,
an bzw. in der die Reaktion stattfinden kann.

Ergebnisse und Diskussion
92
3.3 Entwicklung einer Immobilisierungstechnik
Durch die vorherigen Untersuchungen konnte gezeigt werden, dass bei Einsatz von Lipasen in
biphasischen Reaktionssystemen sich deren katalytische Aktivität vorwiegend an der Grenz-
fläche entfaltet. Folgerichtig sollte bei Immobilisierung von Lipasen eine Maximierung der
spezifischen Grenzfläche angestrebt werden. Die Eignung konventioneller Hydrogele, wie
Alginat oder Chitosan, scheint vor diesem Hintergrund fraglich, da mit ihnen nur Hydrogel-
partikel mit minimalen Durchmessern von 100-300 μm generiert werden können [PRUESSE ET
AL., 1998; BETIGERI UND NEAU, 2002] und dort folglich nur ein Bruchteil der Enzymmoleküle
an der Reaktion partizipieren kann. Hingegen dürfte im Fall thermodynamisch stabiler Mikro-
emulsionen die maximal mögliche spezifische Grenzfläche erreicht sein [CARVALHO UND
CABRAL, 2000]. Eine Immobilisierung derartiger Mikroemulsionen kann in Form so genan-
nter Organogele erfolgen, in welchen die lipasehaltige diskontinuierliche Phase in ein polares
Polymernetzwerk eingebettet ist [STAMATIS UND XENAKIS, 1999]. Ein direkter Kontakt mit
der Grenzfläche ist dabei aber nicht mehr gegeben und infolgedessen ist die katalytische
Effizienz gegenüber den freien Mikroemulsionen vermutlich vermindert. Zudem erfüllen die
mechanischen Eigenschaften der Organogele nicht die Anforderungen an einen technischen
Einsatz. Deshalb darf mit einiger Berechtigung die Aussage getroffen werden, dass gegen-
wärtig keine optimal geeignete Methode zur Immobilisierung von Lipasen in biphasischen
Reaktionssystemen zur Verfügung steht.
Hauptanliegen dieser Arbeit war es, eine adäquate Alternative aufzuzeigen, die den in Kapitel
1.4.3 formulierten Anforderungen an das „ideale Immobilisat“ möglichst nahe kommt und
auch den Ergebnissen von Kapitel 3.2 Rechnung trägt. Aus Gründen, die im Folgenden
herausgearbeitet werden, bestand diese Alternative in der Emulsion einer wässrigen lipase-
haltigen Phase in einem Silicon-Elastomer. Da für die Lipase A von Candida antarctica
(CALA) von BRENNEIS ET AL. [2002 und 2004] bei der Synthese von Esterölen gezeigt wurde,
dass deren katalytische Effizienz durch Wasser deutlich gesteigert werden kann, wurde sie
neben der TLL für die anstehenden Untersuchungen ausgewählt. Anders als den Kapiteln 3.1
und 3.2 zugrunde liegenden Untersuchungen, wurde von der bisherigen durch den SFB 540
vorgegebenen Reaktion – der Synthese von Butylpropionat – Abstand genommen. Haupt-
grund hierfür war, dass es sich bei Butanol und Propionsäure um vergleichsweise hydrophile
Substrate handelt. Im Allgemeinen werden Veresterungen jedoch mit hydrophoberen Sub-
straten durchgeführt. Insofern hätte bei Beibehaltung des bisherigen Reaktionssystems nicht

Entwicklung der Statischen Emulsion
93
ausgeschlossen werden können, dass das Potential des neu entwickelten Verfahrens im Ver-
gleich zu althergebrachten falsch bewertet worden wäre. Leider kann im Gegensatz zur
Hydrolyse für die Veresterung nicht auf einen allgemein etablierten Aktivitätstest zurückge-
griffen werden [SALIS ET AL., 2003]. Nach Durchsicht einschlägiger Literatur schien die
Synthese von Octyloctanoat aus Caprylsäure und Octanol in Hexan durchaus geeignet zu sein.
REETZ ET AL. [1996] und RAGHEB ET AL. [2003] nutzten bspw. eine vergleichbare Reaktion
(Veresterung von Laurinsäure und Octanol in Isooctan) zwecks Charakterisierung der von
ihnen entwickelten Lipase-Immobilisate.
3.3.1 Einfluss von Wasser auf die Produktivität biphasischer Reaktionssysteme
Der in Kapitel 3.1 und 3.2 benutzte Reaktor besitzt aufgrund der geringen spezifischen Grenz-
fläche infolge der fehlenden Durchmischung beider Phasen ineinander keine praktische Rele-
vanz. Regulär basieren biphasische Reaktionssysteme auf durch turbulente Durchmischung
hervorgerufener Emulsion der diskontinuierlichen enzymhaltigen wässrigen Phase in der kon-
tinuierlichen organischen Phase. Allerdings finden sich in der Literatur kaum Angaben zum
optimalen Verhältnis der Volumina von organischer und wässriger Phase. BRENNEIS ET AL.
[2002 und 2004] identifizierten zur Umesterung von Triglyceriden mit 2-Ethyl-1-hexanol
einen optimalen Wassergehalt von 114 % (v/v), wobei auf Verwendung eines Lösungsmittels
verzichtet wurde und infolgedessen die organische Phase durch die Substrate selbst gebildet
wurde. Da die Übertragbarkeit dieses Ergebnisses auf das vorliegende System nicht gegeben
war, wurde die Abhängigkeit der Produktivität vom Wassergehalt in einem Bereich von 0 –
120 % (v/v) für beide Lipasen untersucht.
Wie in Abbildung 3.29 zu erkennen ist, kann ein beträchtlicher Aktivitätsanstieg bei Erhö-
hung des Wasseranteils verzeichnet werden, wobei für beide Lipasen der optimale Wasser-
anteil im Bereich von 90 % zu liegen scheint. Die oberhalb von 90 % beobachtete Abnahme
der Produktivität könnte dadurch bedingt sein, dass die wässrige Phase bei einem Wasser-
anteil von 120 % die kontinuierliche Phase bildet. Bezogen auf die Aktivität, die bei bloßer
Suspension des Lyophilisates in Hexan erzielt wird, kann durch Zugabe von Wasser die der
CALA etwa um den Faktor 14, die der TLL etwa um den Faktor 35 gesteigert werden (Tab.
3.2). Eine genauere Abschätzung ist aufgrund der hohen Standardabweichungen problema-
tisch. Die mangelnde Reproduzierbarkeit ist nicht untypisch für Zweiphasensysteme und lässt
sich vermutlich nicht ohne Weiteres eliminieren. Dennoch konnte eine klare Tendenz erkannt

Ergebnisse und Diskussion
94
werden, die unter Berücksichtigung der in Kapitel 3.2.4 getroffenen Schlussfolgerungen inter-
pretiert werden kann.
0
20
40
60
80
100
120
140
0 2,5 5 10 30 60 90 120
Puffergehalt [%] (v/v)
spez
ifis
che
Akt
ivit
ät [
U/g
Pro
tein
]
0
200
400
600
800
1000
1200
1400
0 2,5 5 10 20 30 60 90 120
Puffergehalt [%] (v/v)
spez
ifis
che
Akt
ivit
ät [
U/g
Pro
tein
]
Abbildung 3.29: Spezifische Aktivitäten bei Veresterung von Caprylsäure und Octanol mit nativer CALA (A)und TLL (B) bei steigendem Wassergehalt (bezogen auf das Volumen der organischen Phase).
Tabelle 3.2: Vergleich der spezifischen Aktivitäten nativer CALA und TLL mit und ohne Puffer.
Spezifische Aktivität [U/g Protein]Enzym
ohne Puffer mit 90 % [v/v]
Puffer
CALA 5,8 81,4
TLL 23,3 877,9
Im Fall lyophilisierter Lipasen war bereits in Kapitel 3.2 postuliert worden, dass bei Suspen-
sion der Enzympartikel nicht alle Lipasen an der Reaktion partizipieren können. Der Vorteil
biphasischer Reaktionssysteme wird in der Regel durch die vollständige Solubilisierung sämt-
licher Lipasemoleküle erklärt [BORZEIX ET AL., 1992; LOUWRIER ET AL., 1996; TWEDDELL ET

Entwicklung der Statischen Emulsion
95
AL., 1998]. In der vorliegenden Untersuchung war die TLL bereits bei einem Wassergehalt
von weniger als 10 % vollständig gelöst, die CALA lag ohnehin als Flüssigpräparat vor. Für
die beobachtete Aktivitätssteigerung muss es demnach noch eine weitergehende Erklärung
geben. Unter der Annahme, dass der mittlere Durchmesser der emulgierten Tropfen im be-
trachteten System weitgehend konstant ist, führt die Steigerung des Wassergehalts nicht zu
einer größeren auf das Volumen der wässrigen Phase bezogenen spezifischen Grenzfläche.
Die absolute Grenzfläche jedoch ist deutlich größer. Da sowohl Enzymkonzentration als auch
Volumen der organischen Phase beibehalten wurden, wird absolut mehr Grenzfläche geschaf-
fen, in der die Reaktion stattfinden kann. Der Anteil aktiver, an der Grenzfläche lokalisierter
bzw. angereicherter Enzyme wäre somit erhöht.
Somit untermauern die in diesem Kapitel erzielten Ergebnisse die Aussage, dass bei Immobi-
lisierung in Zweiphasensystemen eine Maximierung der Grenzfläche anzustreben ist. Wie
zuvor erörtert, sind Hydrogele hierfür weniger gut geeignet – einerseits sind sie zu groß und
andererseits wirkt sich die eingeschränkte Beweglichkeit der Lipasemoleküle innerhalb der
Gelmatrix vermutlich negativ auf die katalytische Aktivität aus (Kapitel 3.2.1.4). Normaler-
weise kann eine maximale Grenzfläche bzw. Emulsion in fluiden Systemen aufgrund der
Koaleszenz nur durch turbulente Durchmischung aufrecht erhalten werden. Daher wurde er-
wogen, die diskontinuierliche wässrige enzymhaltige Phase einer Emulsion zu stabilisieren
und als „Statische Emulsion“ durch Einbettung in eine hydrophobe Matrix in eine handhab-
bare Form zu bringen. Die hydrophobe Matrix kann dabei als stabilisierte organische Phase
aufgefasst werden, die im organischen Lösungsmittel suspendiert wird. Aufgrund der Arbei-
ten von RAGHEB ET AL. [2003] und GILL ET AL. [1999] schienen Silicon-Elastomere hierfür
geeignet (Kapitel 1.4.2). Allerdings wurden dort Silicon-Granulate oder -Blöcke eingesetzt.
Angeraten schien die Verwendung sphärischer Silicon-Elastomere, wie sie mit dem von
HILGERS [2001] entwickelten Solvent-Evaporationsverfahren generiert werden können.
Nachfolgend wird die Erzeugung einer „Statischen Emulsion“ dokumentiert und diese im
Hinblick auf prozessrelevante Eigenschaften charakterisiert. Bei Charakterisierung der kataly-
tischen Aktivität eines Enzym-Immobilisates stellte sich die Frage, ob die mit oder die ohne
Wasser ermittelte Aktivität herangezogen werden sollte. Ist die ohne Wasser ermittelte Akti-
vität maßgebend, fiele die durch Immobilisierung erzielte Aktivitätssteigerung günstiger aus.
Dem ist entgegenzuhalten, dass der wirtschaftliche Nutzen einer Immobilisierungstechnik nur
dann objektiv beurteilt werden kann, wenn die maximal mit dem nativen Präparat zugängliche

Ergebnisse und Diskussion
96
Aktivität – also mit Wasser – herangezogen wird. Dies findet in der Literatur allerdings keine
Berücksichtigung, weshalb stets die ohne Wasserzusatz ermittelte Aktivität des nicht immobi-
lisierten Enzyms zur Charakterisierung der Immobilisate verwendet wird [CROOKS ET AL.,
1995; FURUKAWA ET AL., 2001; HSU ET AL., 2002; KUNCOVA ET AL., 2003; REETZ ET AL.,
2003]. REETZ ET AL. [2003] dokumentierten eine Steigerung der Aktivität der TLL bei Immo-
bilisierung in alkylsubstituiertes Sol-Gel um den Faktor 617. Würde man hingegen die auf der
Zugabe von Wasser beruhende Aktivitätssteigerung (Faktor 35) berücksichtigen, fiele die
Aktivitätssteigerung durch die Sol-Gele erheblich niedriger aus (Faktor 18). Um die Ver-
gleichbarkeit der im Rahmen dieser Arbeit ermittelten Ergebnisse zu den in der Literatur auf-
geführten Daten zu gewährleisten, wurde die native Aktivität ohne Wasser zur Bestimmung
der relativen Aktivität des Immobilisates herangezogen.
3.3.2 Erzeugung einer Statischen Emulsion in Silicon
In Anlehnung an die Arbeit von HILGERS [2001] wurde die Mischung eines bei Raumtem-
peratur polymerisierenden Zweikomponenten-Silicon-Kautschuks zur Erzeugung sphärischer
Silicon-Elastomere in einer PVA-Lösung emulgiert. Silicone umfassen eine Gruppe synthe-
tischer Polymere, in denen Silizium-Atome über Sauerstoff-Atome ketten- und/oder netzartig
verknüpft und die restlichen Valenzen des Siliziums durch Kohlenwasserstoffe abgesättigt
sind. Durch ihre physikalischen und chemischen Eigenschaften (hydrophob, gummielastisch,
temperatur- und oxidationsbeständig) darf vermutet werden, dass sie sich durchaus für die
Immobilisierung von Lipasen und den Einsatz in unkonventionellen Reaktionsmedien eignen.
Der Reaktionsmechanismus der Vernetzung der in dieser Arbeit eingesetzten Präpolymere
(Sylgard® 184, Komponente A: -divinyl-terminiertes Polydimethylsiloxan, Komponente
B: Copolymer von Methylhydrosiloxan und Dimethylsiloxan) kann Abbildung 3.30 entnom-
men werden.
Für die Erzeugung einer statischen Emulsion wurde in der Siliconmischung vor Polymerisa-
tion eine wässrige (enzymhaltige) Phase emulgiert (Kapitel 2.9). Wurde vor Überführung der
Silicon-Wasser-Emulsion in die PVA-Lösung eine angemessene Zeit gewartet, um eine
Präpolymerisation des Silicons zu ermöglichen – erkennbar an einer Zunahme der Viskosität
– kam es in der PVA-Lösung nicht zu einer Entmischung der Silicon-Wasser-Emulsion. Erste
orientierende Versuche zeigten, dass ein Wassergehalt von mehr als 25 % (w/w) im
Silicongemisch die Polymerisation beeinträchtigt, weshalb im Folgenden ein Wassergehalt
von 22,5 % (w/w) eingestellt wurde. Durch lichtmikroskopische Betrachtung war zu

Entwicklung der Statischen Emulsion
97
erkennen, dass auch nach Abschluss der Polymerisation die wässrige Phase weiterhin dis-
kontinuierlich vorlag. Die erzeugten Siliconsphären waren von gummielastischer Konsistenz
und in Abhängigkeit von der Rührintensität konnten Durchmesser bis zu 4 mm erreicht
werden (Abb. 3.31). Bedingt durch das Herstellungsverfahren, waren die Siliconsphären von
augenscheinlich perfekter Rundheit. Für die anstehenden Experimente wurden meist Silicon-
sphären mit Durchmessern im Bereich von 0,5-2 mm hergestellt, wobei bezogen auf den
mittleren Durchmesser eine relativ breite Größenverteilung beobachtet werden konnte.
H2C CH Si O Si O CH CH2 + 2 R O Si HCH3 CH3 CH3 OR1
CH3 CH3 CH3 CH3n
Si
CH2 Si O Si O CH2 CH2 OSi RCH3 CH3 CH3 OR1
CH3 CH3 CH3 CH3n
SiCH2O SiRO R1
CH3
[Pt]
Komponente A Komponente B
R, R1: Siloxan
Abbildung 3.30: Reaktionsschema der platinkatalysierten Vernetzung von dinvinylterminiertem Polydimethyl-siloxan und Methylhydrosilan.
Abbildung 3.31: Siliconsphären mit lipasehaltiger Statischer Emulsion. Der Sphärendurchmesser kann durch die Rührintensität bei der Herstellung kontrolliert werden.
Bezogen auf die eingesetzte Menge der Silicon-Präpolymere und aller übrigen Komponenten
lag die Materialausbeute gewöhnlich zwischen 90 und 95 %. Der Materialverlust wurde durch
5 mm5 mm

Ergebnisse und Diskussion
98
die Adhäsion der zähviskosen präpolymerisierten Silicon-Mischung an den verwendeten
Behältnissen bei Überführung in die PVA-Lösung verursacht. Wurden die vollständig vul-
kanisierten Silicon-Elastomere über einen längeren Zeitraum einer Temperatur von 60 °C aus-
gesetzt, war durch Evaporation des in ihnen enthaltenen Wassers ein Gewichtsverlust festzu-
stellen. Die Höhe des Gewichtsverlusts zeigte an, dass offenbar nur 83,7 +/- 0,3 % des zuvor
in der Mischung der Silicon-Präpolymere vorgelegten Wassers im Elastomer eingeschlossen
wurde. Erklärt werden könnte der Wasserverlust durch Koaleszenz mit der PVA-Lösung
während der Polymerisation des Elastomers. Allerdings konnten weitergehende Untersuchun-
gen zeigen, dass 89 – 96 % des in der wässrigen Phase gelösten Proteins in der Siliconmatrix
immobilisiert wurden und der Proteinverlust somit deutlich geringer ist. Die Diskrepanz
zwischen den Werten kann nur dadurch erklärt werden, dass die gewählten experimentellen
Bedingungen nicht die vollständige Evaporation des Wassers zuließen oder noch unbekannte
Wechselwirkungen zwischen Silicon und Protein, anders als bei Wasser, einer Freisetzung des
Proteins während der Polymerisation entgegenstehen. Da es sich um ein neues Verfahren
handelt, ist davon auszugehen, dass durch systematische Optimierung des Herstellungsprozes-
ses der Materialverlust reduziert und der ohnehin schon hohe Immobilisierungsgrad weiter
erhöht werden kann. Derartige Optimierungbemühungen schienen in diesem Entwicklungs-
stadium jedoch noch nicht angebracht. Anders verhält es sich bezüglich des Durchmessers der
emulgierten Tröpfchen: Angesichts der in Kapitel 3.2 erhaltenen Ergebnisse sollte eine
Minimierung des Durchmessers bzw. eine Maximierung der Grenzfläche unmittelbar einen
positiven Effekt auf die katalytische Aktivität der Immobilisate ausüben.
Zu diesem Zweck wurde eruiert, ob durch Zugabe eines Tensids zur wässrigen Phase der
Durchmesser der emulgierten Tröpfchen reduziert werden kann. Um die Bildung uner-
wünschter Nebenprodukte zu vermeiden, kamen hierfür nur solche Tenside in Frage, die nicht
von Lipasen umgesetzt werden können und keinen negativen Einfluss auf die katalytische
Aktivität haben. Diese Bedingungen wurden von PVA 10-98 und Gummi Arabicum erfüllt.
Zur Abschätzung des Einflusses der Tenside auf den Durchmesser der in Silicon eingehüllten
Tröpfchen wurde wässrig gelöstes Methylenblau in Gegenwart verschiedener Tensidkonzen-
trationen statisch emulgiert und Querschnitte der Siliconsphären mikroskopisch analysiert.
Unter Zuhilfenahme eines Größenstandards konnte der Durchmesser der emulgierten Tröpf-
chen abgeschätzt werden.

Entwicklung der Statischen Emulsion
99
Abbildung 3.32: Wässrige Phase mit Methylenblau in Silicon statisch emulgiert ohne (a), mit 0,5 % (b) und mit 1 % (c) PVA (100fache Vergrößerung).
Wie aus Abbildung 3.32 ersichtlich, streut die Tröpfchengröße um einen Bereich von ca.
5-60 μm, wobei kein signifikanter Unterschied zwischen Ansätzen mit und ohne PVA zu
erkennen ist – auch die Zahl der Tröpfchen scheint identisch. Entsprechende Beobachtungen
wurden bei Verwendung von Gummi Arabicum gemacht. Die ausbleibende Wirkung der
Tenside kann womöglich durch die vergleichsweise hohe Viskosität der Siliconmischung
erklärt werden. Tenside verhindern nur die Koaleszenz der erzeugten Tröpfchen. Es muss
davon ausgegangen werden, dass die bei Herstellung herrschenden hydrodynamischen Scher-
kräfte vermutlich nicht ausreichen, um noch kleinere Tröpfchen zu erzeugen. Daher wurde
auf die Verwendung von Tensiden bei der Erzeugung einer Statischen Emulsion in Silicon
verzichtet.
3.3.3 Aktivität von in Statischer Emulsion immobilisierten Lipasen
Da in Kapitel 3.3.1 gezeigt werden konnte, dass die katalytischen Aktivitäten der Lipase aus
Thermomyces lanuginosa (TLL) und der Lipase A aus Candida antarctica (CALA) in
biphasischen Reaktionssystemen gesteigert werden können, sollte die Eignung der Statischen
Emulsion durch Immobilisierung dieser Lipasen aufgezeigt werden. Zu Beginn der Unter-
suchungen war der Einschluss in alkylsubstituierte Sol-Gele angesichts der nachgewiesenen
Aktivitätssteigerungen die vielversprechendste Variante zur Immobilisierung von Lipasen
(Kapitel 1.4.2). Um das Potential der Statischen Emulsion angemessen bewerten zu können,
empfahl sich daher ein Vergleich mit Sol-Gel-Immobilisaten. Es wurden unterschiedliche
Proteinmengen der CALA und der TLL in Statischer Emulsion und Sol-Gel immobilisiert und
die katalytische Aktivität der Immobilisate in Hexan bei Veresterung von Octanol und
Caprylsäure ermittelt (Kapitel 2.13).
a) b) c)
100 µm 100 µm 100 µm
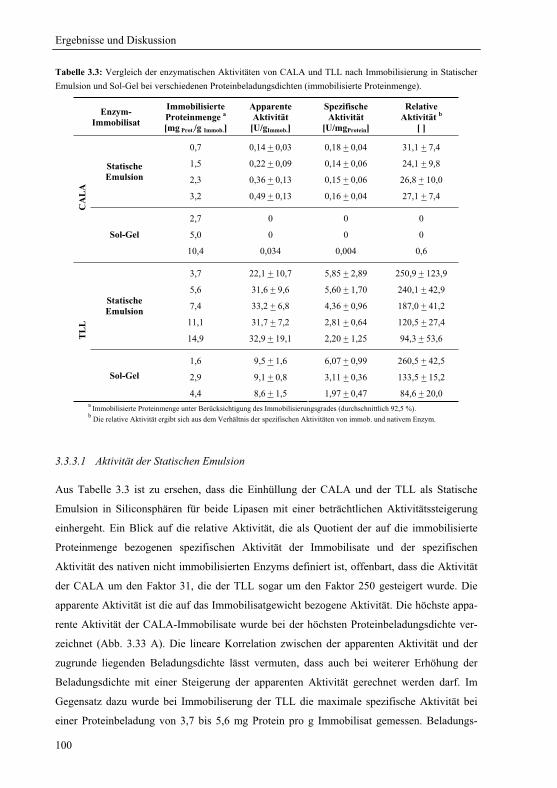
Ergebnisse und Diskussion
100
Tabelle 3.3: Vergleich der enzymatischen Aktivitäten von CALA und TLL nach Immobilisierung in Statischer Emulsion und Sol-Gel bei verschiedenen Proteinbeladungsdichten (immobilisierte Proteinmenge).
Enzym-Immobilisat
Immobilisierte Proteinmenge a
[mg Prot./g Immob.]
ApparenteAktivität
[U/gImmob.]
Spezifische Aktivität
[U/mgProtein]
Relative Aktivität b
[ ]
0,7 0,14 + 0,03 0,18 + 0,04 31,1 + 7,4
1,5 0,22 + 0,09 0,14 + 0,06 24,1 + 9,8
2,3 0,36 + 0,13 0,15 + 0,06 26,8 + 10,0 Statische Emulsion
3,2 0,49 + 0,13 0,16 + 0,04 27,1 + 7,4
2,7 0 0 0
5,0 0 0 0
CA
LA
Sol-Gel
10,4 0,034 0,004 0,6
3,7 22,1 + 10,7 5,85 + 2,89 250,9 + 123,9
5,6 31,6 + 9,6 5,60 + 1,70 240,1 + 42,9
7,4 33,2 + 6,8 4,36 + 0,96 187,0 + 41,2
11,1 31,7 + 7,2 2,81 + 0,64 120,5 + 27,4
Statische Emulsion
14,9 32,9 + 19,1 2,20 + 1,25 94,3 + 53,6
1,6 9,5 + 1,6 6,07 + 0,99 260,5 + 42,5
2,9 9,1 + 0,8 3,11 + 0,36 133,5 + 15,2
TL
L
Sol-Gel
4,4 8,6 + 1,5 1,97 + 0,47 84,6 + 20,0 a Immobilisierte Proteinmenge unter Berücksichtigung des Immobilisierungsgrades (durchschnittlich 92,5 %). b Die relative Aktivität ergibt sich aus dem Verhältnis der spezifischen Aktivitäten von immob. und nativem Enzym.
3.3.3.1 Aktivität der Statischen Emulsion
Aus Tabelle 3.3 ist zu ersehen, dass die Einhüllung der CALA und der TLL als Statische
Emulsion in Siliconsphären für beide Lipasen mit einer beträchtlichen Aktivitätssteigerung
einhergeht. Ein Blick auf die relative Aktivität, die als Quotient der auf die immobilisierte
Proteinmenge bezogenen spezifischen Aktivität der Immobilisate und der spezifischen
Aktivität des nativen nicht immobilisierten Enzyms definiert ist, offenbart, dass die Aktivität
der CALA um den Faktor 31, die der TLL sogar um den Faktor 250 gesteigert wurde. Die
apparente Aktivität ist die auf das Immobilisatgewicht bezogene Aktivität. Die höchste appa-
rente Aktivität der CALA-Immobilisate wurde bei der höchsten Proteinbeladungsdichte ver-
zeichnet (Abb. 3.33 A). Die lineare Korrelation zwischen der apparenten Aktivität und der
zugrunde liegenden Beladungsdichte lässt vermuten, dass auch bei weiterer Erhöhung der
Beladungsdichte mit einer Steigerung der apparenten Aktivität gerechnet werden darf. Im
Gegensatz dazu wurde bei Immobiliserung der TLL die maximale spezifische Aktivität bei
einer Proteinbeladung von 3,7 bis 5,6 mg Protein pro g Immobilisat gemessen. Beladungs-
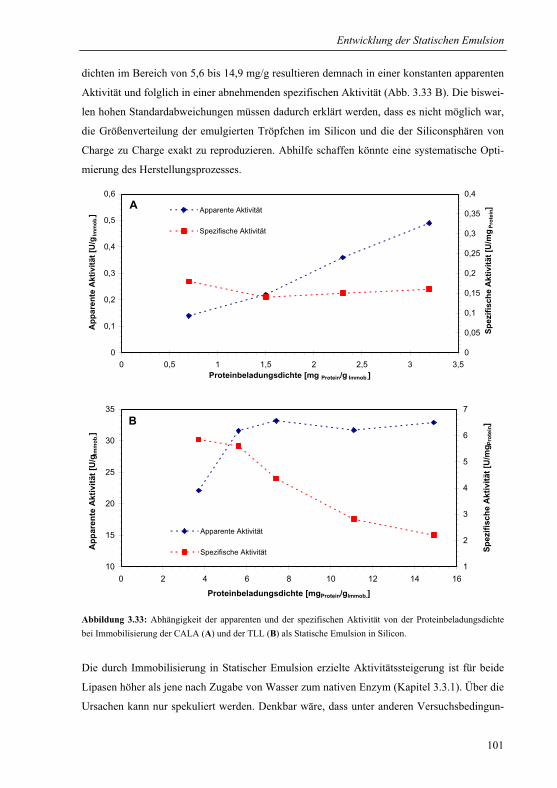
Entwicklung der Statischen Emulsion
101
dichten im Bereich von 5,6 bis 14,9 mg/g resultieren demnach in einer konstanten apparenten
Aktivität und folglich in einer abnehmenden spezifischen Aktivität (Abb. 3.33 B). Die biswei-
len hohen Standardabweichungen müssen dadurch erklärt werden, dass es nicht möglich war,
die Größenverteilung der emulgierten Tröpfchen im Silicon und die der Siliconsphären von
Charge zu Charge exakt zu reproduzieren. Abhilfe schaffen könnte eine systematische Opti-
mierung des Herstellungsprozesses.
0
0,1
0,2
0,3
0,4
0,5
0,6
0 0,5 1 1,5 2 2,5 3 3,5Proteinbeladungsdichte [mg Protein/g Immob.]
Ap
par
ente
Akt
ivit
ät [
U/g
Imm
ob
.]
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Sp
ezif
isch
e A
ktiv
ität
[U
/mg
Pro
tein
]Apparente Aktivität
Spezifische Aktivität
10
15
20
25
30
35
0 2 4 6 8 10 12 14 16
Proteinbeladungsdichte [mgProtein/gImmob.]
Ap
par
ente
Akt
ivit
ät [
U/g
Imm
ob
.]
1
2
3
4
5
6
7
Sp
ezif
isch
e A
ktiv
ität
[U
/mg P
rote
in]
Apparente Aktivität
Spezifische Aktivität
Abbildung 3.33: Abhängigkeit der apparenten und der spezifischen Aktivität von der Proteinbeladungsdichte bei Immobilisierung der CALA (A) und der TLL (B) als Statische Emulsion in Silicon.
Die durch Immobilisierung in Statischer Emulsion erzielte Aktivitätssteigerung ist für beide
Lipasen höher als jene nach Zugabe von Wasser zum nativen Enzym (Kapitel 3.3.1). Über die
Ursachen kann nur spekuliert werden. Denkbar wäre, dass unter anderen Versuchsbedingun-
B
A

Ergebnisse und Diskussion
102
gen auch für die nativen Präparate Aktivitätssteigerungen in der gleichen Größenordnung
erhalten worden wären. Angesichts der Ergebnisse steht fest, dass die Stabilisierung einer dis-
kontinuierlichen wässrigen enzymhaltigen Phase durch Einschluss in eine hydrophobe Matrix
eine durchaus gangbare Strategie zur Immobilisierung von Lipasen darstellt. Auf den ersten
Blick steht dies im Widerspruch zu den Ergebnissen von RAGHEB ET AL. [2003]. Sie immobi-
lisierten die Lipase von Candida rugosa (CRL) direkt in Silicon, wobei dort die Zugabe von
Wasser vor Polymerisation zu deutlich geringeren Aktivitäten führte. Eine elektronenmikro-
skopische Analyse der Matrix ergab jedoch, dass das zuvor suspendierte trockene Enzym-
präparat durch die Zugabe von Wasser agglomerierte [RAGHEB ET AL., 2003]. Das zugegebene
Volumen Wasser reichte nicht aus, um die Lipase homogen zu lösen und in Form emulgierter
Tröpfchen in die Matrix einzubringen. Bei der hier beschriebenen Statischen Emulsion wird
die 20fache Menge Wasser zugegeben und das Enzympräparat homogen gelöst. Infolgedessen
wird eine Grenzfläche geschaffen und die in Kapitel 3.2.4 beschriebene Phänomenologie ist
höchstwahrscheinlich für die Steigerung der katalytischen Aktivität verantwortlich. Belegt
wird dies nicht nur durch die Arbeit von RAGHEB ET AL. [2003], sondern auch durch die
Arbeit von GILL ET AL. [1999]. In beiden Arbeiten konnte durch Dispersion des trockenen
Enzympräparates in einem Silicon-Elastomer keine mit der Statischen Emulsion vergleichbare
Aktivierung erreicht werden. Die niedrigen Aktivitäten mögen auch der Hauptgrund sein,
warum die Immobilisierung von Lipasen in Silicon bisher nur sehr selten untersucht wurde.
Der ansonsten vorteilhaften Matrix Silicon könnte in Form der Statischen Emulsion nunmehr
zum Durchbruch verholfen werden.
3.3.3.2 Vergleich mit Sol-Gel-Immobilisaten
Bei Immobilisierung der CALA und der TLL in alkylsubstituierte Sol-Gele aus TMOS/
PTMS/BTMS wurden weiße pulverförmige Immobilisate erhalten. Der mittlere Partikel-
durchmesser lag im Bereich von 100 μm, wobei die gemessene Partikelgrößenverteilung sehr
breit war (Kapitel 2.11). Die Herstellungsprozedur der Sol-Gele beinhaltet die mechanische
Zerkleinerung der gebildeten spröden Polymere. In Abhängigkeit von der Intensität, kann die
Größenverteilung daher über einen weiten Bereich beeinflusst werden.
Die Immobilsierung der TLL führte zu einer Aktivitätssteigerung gegenüber dem nativen
Enzym um den Faktor 260 (Tab. 3.3). REETZ ET AL. [1996 und 2003] erzielten bei Immobili-
sierung des gleichen Enzyms in TMOS/PTMS eine Aktivitätssteigerung um den Faktor 87,6

Entwicklung der Statischen Emulsion
103
und unter Verwendung von TMOS/BTMS gar um den Faktor 617. Untersucht wurde die
Veresterung von Laurinsäure mit Octanol. Die in dieser Arbeit erzielte maximale Aktivitäts-
steigerung liegt zwischen diesen Werten, was vermutlich weniger auf die unterschiedlichen
Substrate als vielmehr auf die Kombination der Silane PTMS und BTMS zurückzuführen ist.
Die Kombination der Silane war erforderlich, da bei ausschließlicher Verwendung von BTMS
Sol-Gele von klebriger Konsistenz erhalten wurden. Da aber die Hydrophobizität einer
TMOS/PTMS/BTMS-Matrix im Vergleich zu einer reinen TMOS/BTMS-Matrix geringer ist,
sind auch die von REETZ ET AL. [1996] angeführten aktivitätssteigernden Effekte weniger
stark ausgeprägt. Auffällig ist, dass bei Immobilisierung der CALA in Sol-Gel eine drama-
tische Abnahme der katalytischen Aktivität festgestellt wurde (Tab. 3.3). Dieses Ergebnis war
nicht unbedingt zu erwarten, da REETZ ET AL. [1996] bei Immobilisierung der Lipase aus
Candida antarctica zwar eine niedrige, aber signifikante Aktivitätssteigerung beobachteten
(Faktor 3,2). Allerdings spezifizierten REETZ ET AL. [1996] nicht, ob es sich um die Lipase A
oder die Lipase B aus Candida antarctica handelte. Vermutlich benutzten sie eine Mischung
beider Lipasen, wie sie für kommerzielle Lipase-Präparate aus Candida antarctica nicht un-
typisch ist. Denkbar wäre somit, dass die aktivitätssteigernde Wirkung der Sol-Gel-Immobi-
lisierung nur auf die Lipase B entfällt, während die CALA keine oder nur eine sehr geringe
Aktivität innerhalb der Sol-Gel-Matrix entfaltet. Auch GILL ET AL. [1999] stellten fest, dass
die Einhüllung der Lipase aus Candida antarctica in eine hydrophobe Mikroumgebung weni-
ger vorteilhaft ist als bei anderen Lipasen. Leider spezifizierten auch GILL ET AL. [1999] nicht,
um welche der beiden Lipasen es sich handelte. In diesem Zusammenhang ist bemerkenswert,
dass bisher nur sehr wenige Studien zur Immobilisierung der CALA veröffentlicht wurden
[DOMÍNGUEZ DE MARÍA ET AL., 2005]. Es darf davon ausgegangen werden, dass die Ein-
hüllung der CALA als Statische Emulsion in Silicon die bisher am besten geeignete Methode
zur Immobilisierung dieses Enzyms ist und damit völlig neue Perspektiven für dessen Anwen-
dung eröffnet werden.
Bei Immobilisierung der TLL in Sol-Gel war eine Abnahme der relativen Aktivitäten bei
steigenden Beladungsdichten zu erkennen, obschon im Vergleich zur Statischen Emulsion
geringere Proteinmengen immobilisiert wurden. Die apparente Aktivität war unabhängig von
der Beladungsdichte dementsprechend konstant (Tab. 3.3). Diese Ergebnisse können nur
dahingehend interpretiert werden, dass die maximale Enzymbeladung bereits bei sehr
niedrigen Proteinkonzentrationen erreicht wird und eine apparente Aktivität von etwa 9
U/gImmob. das zugängliche Maximum darstellt. REETZ ET AL. [1996] machten bei vergleich-

Ergebnisse und Diskussion
104
baren Beladungsdichten ähnliche Beobachtungen und führten dies nicht nur auf eine Massen-
transferlimitierung innerhalb der Matrix zurück, sondern auch auf eine mögliche Agglomera-
tion der Enzymmoleküle in der Matrix selbst. Demgegenüber konnten in Statischer Emulsion
höhere Proteinbeladungen realisiert und folglich auch eine dreimal höhere apparente Aktivität
(ca. 32 U/gImmob.) erreicht werden. Somit besteht Anlass zu der Vermutung, dass das Ausmaß
der Massentransferlimitierung innerhalb der Statischen Emulsion im Vergleich zur Sol-Gel-
Matrix geringer ist. Allerdings muss darauf verwiesen werden, dass in dieser Arbeit nur die
„erste Generation“ der Sol-Gele untersucht wurde. Durch diverse Weiterentwicklungen im
Bereich der Sol-Gel-Immobilisierung konnten die für die erste Generation üblichen Aktivi-
tätssteigerungen deutlich verbessert werden [FURUKAWA ET AL., 2001; KUNCOVA ET AL.,
2003; REETZ ET AL., 2003; GHANEM UND SCHURIG, 2003]. Es kann daher nicht ausgeschlossen
werden, dass mit der „zweiten Generation“ der Sol-Gele höhere apparente Aktivitäten als mit
der Statischen Emulsion erreicht worden wären. Allerdings wurden bisher noch keine
Anstrengungen zur systematischen Optimierung der Statischen Emulsion unternommen –
weitere Aktivitätssteigerungen scheinen auch hier möglich. Außerdem dürfte das hier ent-
wickelte Verfahren gegenüber der Sol-Gel-Immobilisierung auch aus anderen Gründen über
eine höhere Attraktivität verfügen. Die Siliconsphären sind aufgrund ihrer Größe und ihrer
mechanischen Robustheit leichter handhabbar als Sol-Gele. Ihre Präparation ist weniger
aufwändig und innerhalb weniger Stunden abgeschlossen. Demgegenüber erstreckt sich die
Herstellung der Sol-Gele über zwei Tage und umfasst mehr Verfahrensschritte [REETZ ET AL.,
1996]. Der Hauptvorteil besteht aber darin, dass bei Herstellung keine giftigen Substanzen,
wie TMOS, zum Einsatz kommen und somit die Sicherheit des Endverbrauchers nicht
gefährdet wird: Ein Aspekt der bei einem Einsatz der Immobilisate in der Lebensmittel- oder
kosmetischen Industrie von großer Bedeutung ist [HILLS, 2003].

Charakterisierung der Statischen Emulsion
105
3.4 Charakterisierung der Statischen Emulsion
3.4.1 Katalytische Stabilität
3.4.1.1 Wiederverwendbarkeit
Im Interesse der Prozessökonomie sollte ein Immobilisat ausreichend aktiv und stabil sowie
leicht zu handhaben und ohne Verlust der katalytischen Aktivität wiederholt einsetzbar sein.
Die Abtrennung der Siliconsphären mit statisch emulgierter Lipase aus dem Reaktions-
medium ist vollkommen unproblematisch und ihre mechanische Stabilität ausreichend hoch,
um den Einsatz in Rühr- oder Packbettreaktoren zuzulassen. Noch nicht überprüft wurde, ob
die katalytische Aktivität über mehrere Synthesezyklen hinweg Konstanz zeigt. Daher wurde
in dieser Arbeit die katalytische Stabilität der Statischen Emulsion von CALA und TLL in
gleicher Weise untersucht wie bei REETZ ET AL. [1996].
0
20
40
60
80
100
120
140
1. Zyklus 2. Zyklus 3. Zyklus 4. Zyklus
Rel
ativ
e A
ktiv
ität
[%
]
CALA TLL
Abbildung 3.34: Enzymatische Aktivität von CALA und TLL (statisch in Silicon emulgiert) bei wiederholtem Einsatz zur Veresterung von Caprylsäure und Octanol.
Auch nach vierfachem Einsatz der Immobilisate war bei beiden Lipasen kein Verlust der
ursprünglichen Aktivität zu konstatieren (Abb. 3.34). Im 3. und 4. Reaktionszyklus wurden
relative Aktivitäten deutlich über 100 % gemessen, was vermutlich auf Schwankungen der
Raumtemperatur (20-25 °C) und dem daraus resultierenden Einfluss auf die Reaktionsge-
schwindigkeit zurückgeht.
Die Wiederverwendbarkeit von in Silicon immobilisierten Lipasen wurde auch von RAGHEB
ET AL. [2003] untersucht, wobei nach 60 Synthesezyklen nur eine minimale Abnahme der

Ergebnisse und Diskussion
106
katalytischen Aktivität verzeichnet wurde. Auch GILL ET AL. [1999] konnten den langfristigen
Erhalt der katalytischen Aktivität ihrer Silicon-Lipase-Immobilisate nachweisen. Allerdings
waren die Lipasen sowohl bei RAGHEB ET AL. [2003] als auch bei GILL ET AL. [1999] direkt in
die Siliconmatrix eingebettet, weshalb die Ergebnisse nicht ohne Weiteres auf die Statische
Emulsion zu übertragen sind. Zudem wurde hier nur der vierfache Einsatz der Statischen
Emulsion untersucht, womit eine hohe Langzeitstabilität kaum bewiesen werden kann.
Dennoch liegt der Schluss nahe, dass auch bei wiederholtem Einsatz nicht mit einem nennens-
werten Verlust der katalytischen Aktivität zu rechnen ist. Dies darf als ein weiterer Beleg für
die hervorragende Eignung der Statischen Emulsion in der Enzymtechnologie gewertet
werden.
3.4.1.2 Ausbluten
Voraussetzung für die katalytische Stabilität und die Wiederverwendbarkeit ist die Dauer-
haftigkeit der Immobilisierung. Im Fall einer stabilen Immobilisierung sollte während der
Nutzung keine Freisetzung bzw. kein Ausbluten des immobilisierten Proteins zu beobachten
sein. Um die Stabilität der Immobilisierung in Statischer Emulsion zu überprüfen, wurde eine
definierte Menge der enzymhaltigen Siliconsphären in Wasser überführt und eine Woche
schüttelnd inkubiert. Dabei konnte nur die Freisetzung sehr geringer Proteinmengen nachge-
wiesen werden: 0,75 0,18 % bei CALA und 0,04 0,01 % bei TLL. GILL ET AL. [1999]
beobachteten eine Freisetzung in Höhe von 0,1 bis 0,6 % des immobilisierten Proteins.
RAGHEB ET AL. [2003] beobachteten bei wiederholten Waschschritten ein Ausbluten von bis
zu 1,3 %. Die Freisetzung von Protein aus Silicon-Elastomeren scheint somit vernachlässig-
bar zu sein. Zudem besteht die Möglichkeit, dass die hier nachgewiesenen Proteinmengen auf
oberflächliche, aus dem Herstellungsprozeß herrührende Proteinanhaftungen zurückgehen.
Diese Untersuchungen lassen den Schluss zu, dass die emulgierten Tröpfchen samt Protein
unbeweglich in die Matrix eingebettet sind und die Hydrophobizität des Silicons ein Aus-
bluten der wässrigen Phase samt Protein unter Einsatzbedingungen unwahrscheinlich macht.
Über die Maschenweite des Silicon-Netzwerkes kann keine Aussage gemacht werden. Es ist
aber aufgrund der chemischen Beschaffenheit (Abb. 3.30) anzunehmen, dass sie im Vergleich
zum Enzymdurchmesser klein ist und einen Proteinverlust effektiv verhindert.
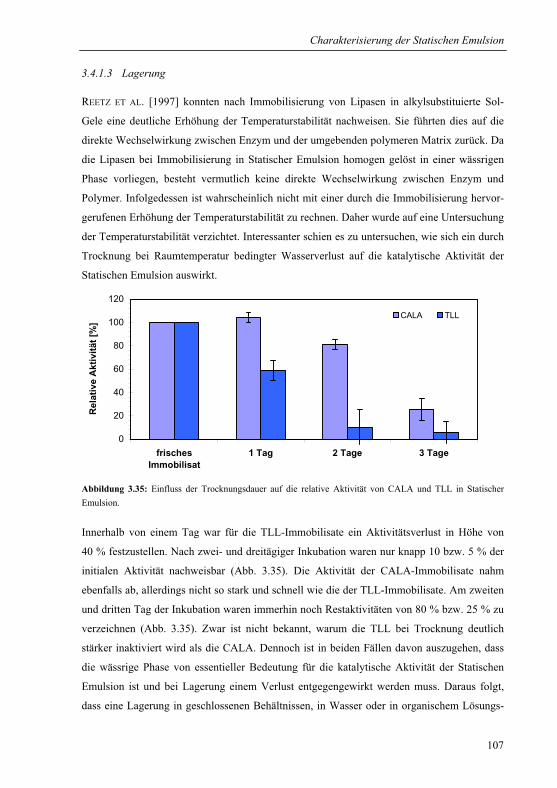
Charakterisierung der Statischen Emulsion
107
3.4.1.3 Lagerung
REETZ ET AL. [1997] konnten nach Immobilisierung von Lipasen in alkylsubstituierte Sol-
Gele eine deutliche Erhöhung der Temperaturstabilität nachweisen. Sie führten dies auf die
direkte Wechselwirkung zwischen Enzym und der umgebenden polymeren Matrix zurück. Da
die Lipasen bei Immobilisierung in Statischer Emulsion homogen gelöst in einer wässrigen
Phase vorliegen, besteht vermutlich keine direkte Wechselwirkung zwischen Enzym und
Polymer. Infolgedessen ist wahrscheinlich nicht mit einer durch die Immobilisierung hervor-
gerufenen Erhöhung der Temperaturstabilität zu rechnen. Daher wurde auf eine Untersuchung
der Temperaturstabilität verzichtet. Interessanter schien es zu untersuchen, wie sich ein durch
Trocknung bei Raumtemperatur bedingter Wasserverlust auf die katalytische Aktivität der
Statischen Emulsion auswirkt.
0
20
40
60
80
100
120
frischesImmobilisat
1 Tag 2 Tage 3 Tage
Rel
ativ
e A
ktiv
ität
[%
]
CALA TLL
Abbildung 3.35: Einfluss der Trocknungsdauer auf die relative Aktivität von CALA und TLL in Statischer Emulsion.
Innerhalb von einem Tag war für die TLL-Immobilisate ein Aktivitätsverlust in Höhe von
40 % festzustellen. Nach zwei- und dreitägiger Inkubation waren nur knapp 10 bzw. 5 % der
initialen Aktivität nachweisbar (Abb. 3.35). Die Aktivität der CALA-Immobilisate nahm
ebenfalls ab, allerdings nicht so stark und schnell wie die der TLL-Immobilisate. Am zweiten
und dritten Tag der Inkubation waren immerhin noch Restaktivitäten von 80 % bzw. 25 % zu
verzeichnen (Abb. 3.35). Zwar ist nicht bekannt, warum die TLL bei Trocknung deutlich
stärker inaktiviert wird als die CALA. Dennoch ist in beiden Fällen davon auszugehen, dass
die wässrige Phase von essentieller Bedeutung für die katalytische Aktivität der Statischen
Emulsion ist und bei Lagerung einem Verlust entgegengewirkt werden muss. Daraus folgt,
dass eine Lagerung in geschlossenen Behältnissen, in Wasser oder in organischem Lösungs-

Ergebnisse und Diskussion
108
mittel erfolgen muss. War diese Bedingung erfüllt, konnte selbst bei Raumtemperatur nach
mehrwöchiger Lagerung kein nennenswerter Aktivitätsverlust beobachtet werden.
3.4.2 Abriebstabilität und Elastizität
Der technische Einsatz von Immobilisaten geht mit hohen mechanischen Belastungen einher.
Die Ursachen für diese Belastungen sind vielfältiger Natur und bestehen neben dem ober-
flächlichen Abrieb in oszillierenden Zug- und Druckbeanspruchungen innerhalb der Immo-
bilisate, hervorgerufen u.a. durch hydrodynamische Scherkräfte oder Kollisionen der Immo-
bilisat-Partikel miteinander oder mit dem Reaktor [DOS SANTOS ET AL., 1997]. Für den tech-
nischen, mitunter sogar wiederholten Einsatz ist es daher bedeutsam, ausreichend stabile
Immobilisate bereitzustellen und Parameter zu definieren, die die mechanischen Eigen-
schaften der Immobilisate klassifizieren. In früheren Arbeiten begnügte man sich mit der
Angabe des Bruchpunktes – dem Verlust der strukturellen Integrität bei Kompression. DOS
SANTOS ET AL. [1997] zeigten jedoch, dass der Bruchpunkt nicht geeignet ist, die Wider-
standsfähigkeit eines Materials gegenüber mechanischen Einflüssen zu kennzeichnen. Daher
waren die Bemühungen der letzten Jahre auf die Entwicklung von Messtechniken gerichtet,
um aussagekräftigere physikalische Kennziffern zu erhalten. Eine derart umfassende zeit- und
materialaufwändige Charakterisierung der Statischen Emulsion war im jetzigen Entwick-
lungsstadium jedoch nicht angebracht, weshalb sich die hier vorgenommene Charakteri-
sierung auf Untersuchungen zur Abriebstabilität und Elastizität beschränkte.
Abbildung 3.36: Siliconkugeln vor und nach einwöchigem Abrieb durch Glaskugeln in Hexan.
Bei Prüfung der Abriebstabilität wurden Siliconsphären (ohne emulgierte wässrige Phase)
zusammen mit Glaskugeln in Hexan turbulent über einen Zeitraum von einer Woche geschüt-
telt, um im Zeitraffer extreme Belastungen, wie sie in der Realität nicht anzutreffen sind, zu
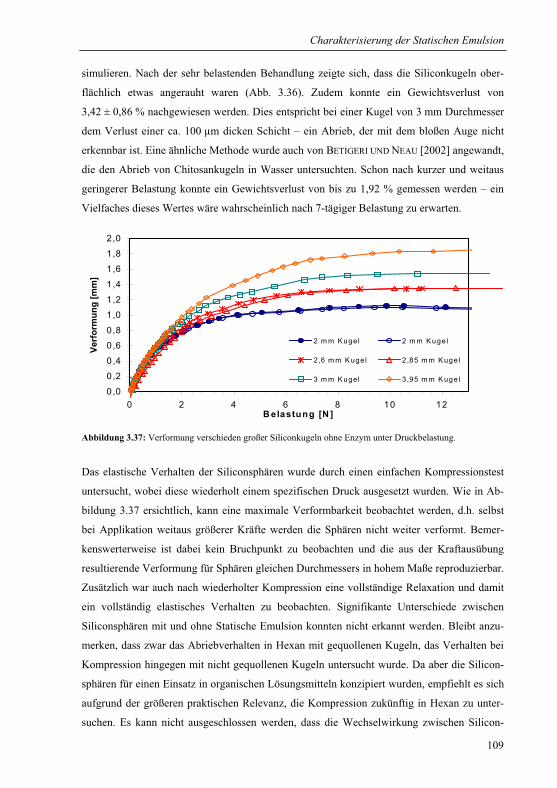
Charakterisierung der Statischen Emulsion
109
simulieren. Nach der sehr belastenden Behandlung zeigte sich, dass die Siliconkugeln ober-
flächlich etwas angerauht waren (Abb. 3.36). Zudem konnte ein Gewichtsverlust von
3,42 ± 0,86 % nachgewiesen werden. Dies entspricht bei einer Kugel von 3 mm Durchmesser
dem Verlust einer ca. 100 μm dicken Schicht – ein Abrieb, der mit dem bloßen Auge nicht
erkennbar ist. Eine ähnliche Methode wurde auch von BETIGERI UND NEAU [2002] angewandt,
die den Abrieb von Chitosankugeln in Wasser untersuchten. Schon nach kurzer und weitaus
geringerer Belastung konnte ein Gewichtsverlust von bis zu 1,92 % gemessen werden – ein
Vielfaches dieses Wertes wäre wahrscheinlich nach 7-tägiger Belastung zu erwarten.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
0 2 4 6 8 10 12B elastung [N ]
Ver
form
un
g [
mm
]
2 m m Kugel 2 m m Kugel
2 ,6 m m K ugel 2 ,85 m m K ugel
3 m m Kugel 3 ,95 m m K ugel
Abbildung 3.37: Verformung verschieden großer Siliconkugeln ohne Enzym unter Druckbelastung.
Das elastische Verhalten der Siliconsphären wurde durch einen einfachen Kompressionstest
untersucht, wobei diese wiederholt einem spezifischen Druck ausgesetzt wurden. Wie in Ab-
bildung 3.37 ersichtlich, kann eine maximale Verformbarkeit beobachtet werden, d.h. selbst
bei Applikation weitaus größerer Kräfte werden die Sphären nicht weiter verformt. Bemer-
kenswerterweise ist dabei kein Bruchpunkt zu beobachten und die aus der Kraftausübung
resultierende Verformung für Sphären gleichen Durchmessers in hohem Maße reproduzierbar.
Zusätzlich war auch nach wiederholter Kompression eine vollständige Relaxation und damit
ein vollständig elastisches Verhalten zu beobachten. Signifikante Unterschiede zwischen
Siliconsphären mit und ohne Statische Emulsion konnten nicht erkannt werden. Bleibt anzu-
merken, dass zwar das Abriebverhalten in Hexan mit gequollenen Kugeln, das Verhalten bei
Kompression hingegen mit nicht gequollenen Kugeln untersucht wurde. Da aber die Silicon-
sphären für einen Einsatz in organischen Lösungsmitteln konzipiert wurden, empfiehlt es sich
aufgrund der größeren praktischen Relevanz, die Kompression zukünftig in Hexan zu unter-
suchen. Es kann nicht ausgeschlossen werden, dass die Wechselwirkung zwischen Silicon-

Ergebnisse und Diskussion
110
Elastomer und Hexan mit Konsequenzen für die mechanischen Eigenschaften behaftet ist.
Dafür spricht auch die Beobachtung, dass nach Quellen in Hexan die Siliconsphären deutlich
härter waren und sich nur unter Ausübung etwas größerer Kräfte verformen ließen. Aber auch
bei Verwendung der Siliconsphären in Hexan konnte zu keinem Zeitpunkt eine Desintegration
beobachtet werden.
Angesichts dieser Ergebnisse ist davon auszugehen, dass Silicon-Elastomere unter mecha-
nischen Gesichtspunkten für den Einsatz in Rührkessel- aber auch Packbettreaktoren hervor-
ragend geeignet sind. In Kombination mit den hohen Aktivitätssteigerungen ist dies ein
weiteres Indiz dafür, dass die Statische Emulsion vielversprechende Perspektiven für den
industriellen Einsatz bietet.
3.4.3 Quellung in Hexan
Bei Überführung des Silicon-Elastomers in Hexan konnte eine beträchtliche Volumenzunah-
me durch Quellung beobachtet werden. Die Quantifizierung dieser Volumenzunahme ist –
wie noch in Kapitel 3.4.4 gezeigt wird – für die Analyse der Verteilung und des Massen-
transfers von Bedeutung. Eine durch organische Lösungsmittel hervorgerufene Quellung von
Silicon ist auch in der Literatur belegt: GILL ET AL. [1999] beobachteten eine Volumen-
zunahme von ca. 60-230 %; RAGHEB ET AL. [2003] hingegen quantifizierten die Quellung in
Isooctan zur Abschätzung des Polymerisationsgrades, nannten diesbezüglich aber keine defi-
nierten Werte. Beiden Literaturstellen sind keine Angaben zur Durchführung der Analyse zu
entnehmen, weshalb eigene Methoden entwickelt wurden. Größere Kugeln ( > 1 mm)
wurden vor und nach Quellung in Hexan fotografiert und anschließend über eine Objekt-
erkennungssoftware die Volumenzunahme errechnet (Kapitel 2.11). Die Quellung kleinerer
Kugeln ( < 1 mm) wurde durch Messen der Volumina in einem Messzylinder vor und nach
der Quellung quantifiziert (Kapitel 2.12.1).
Bei kleinen Kugeln ( bis ca. 2 mm) kam es innerhalb von 3 min zu einer Verdoppelung des
Volumens, bei größeren Kugeln ( ca. 4-5 mm) hingegen erst nach 10 min. Aller Wahr-
scheinlichkeit nach beruht die Volumenzunahme auf der Hydrophobizität der Matrix und der
daraus resultierenden Aufnahme entsprechender Volumina aus der umgebenden organischen
Phase. Die durch hydrophobe Wechselwirkung zwischen den Polymeren sehr kompakte
Matrix wird durch das Einströmen von Hexan vermutlich relaxiert. Der genaue Faktor der

Charakterisierung der Statischen Emulsion
111
Volumenzunahme unterscheidet sich bei Siliconkugeln mit und ohne statisch emulgiertem
Enzym ( > 1 mm) nicht signifikant (2,04 ± 0,11 bzw. 2,01 ± 0,12). Bei kleineren Silicon-
kugeln ( < 1 mm) wurde eine etwas stärkere Volumenzunahme um das 2,5fache ermittelt.
Die somit etwas stärkere Quellung lässt einen geringeren Vernetzungsgrad im Vergleich zu
den großen Sphären vermuten, könnte aber auch auf den verschiedenen Messmethoden
beruhen.
Wie der Arbeit von RAGHEB ET AL. [2003] entnommen werden kann, hängt der Quellungsgrad
direkt vom Vernetzungsgrad der Silicon-Präpolymere ab. Daraus folgt, dass die erhobenen
Daten nur Gültigkeit für die hier verwendeten Siliconsphären haben. Modifikationen im
Herstellungsprozess (z.B. Dauer der Polymerisierung, Verhältnis der Komponenten A und B)
oder Wechsel der verwendeten Siliconkomponenten, erfordern zwangsläufig eine neuerliche
Quantifizierung der Quellung. Insgesamt handelt es sich bei der Quellung um ein vorteilhaftes
Phänomen, da davon auszugehen ist, dass sich die Relaxation des polymeren Silicon-Netz-
werkes günstig auf den Massentransfer auswirkt.
3.4.4 Massentransfer und Verteilung
Die Eignung einer Immobilisierungsmatrix ergibt sich nicht nur aus vorteilhaften mecha-
nischen Eigenschaften, sondern auch aus einer für die Reaktion günstigen Verteilung der
Reaktanden zwischen Matrix und umgebenden Reaktionsmedium sowie einem ausreichend
schnellen Massentransfer in der Matrix selbst. Vereinfacht betrachtet geht es bei Verteilung
und Massentransfer darum, dass das Substrat schnell zum Reaktionsort gelangt und das
Produkt schnell wieder extrahiert werden kann. Anders als bei Hydrogelen ist bei
Verwendung einer hydrophoben Matrix, wie Silicon, mit einer gleichmäßigeren Verteilung
hydrophober Substanzen zwischen Immobilisat und organischer Phase zu rechnen. Die
quantitative Kenntnis dieser Phänomenologie, etwa in Form von Verteilungskoeffizienten und
Diffusionskoeffizienten, wäre hilfreich, um die Eignung einer Immobilisierungsmatrix beur-
teilen zu können. Massentransfer und Verteilung werden üblicherweise durch Überführung
definierter Mengen der Immobilisate in eine Analytlösung und anschließender Quantifizie-
rung der zeitabhängigen Konzentrationsabnahme der Analyten in der kontinuierlichen Phase
(organisches Lösungsmittel) bis zur Gleichgewichtseinstellung untersucht [DE BACKER UND
BARON, 1993; LI ET AL., 1996; MATEUS ET AL., 1999]. Eine direkte Quantifizierung der

Ergebnisse und Diskussion
112
Konzentrationen in der diskontinuierlichen Phase (Immobilisat) ist messtechnisch nicht oder
nur mit großem Aufwand möglich [KUEPPERS ET AL., 2002].
Bisher haben sich nur relativ wenige Studien mit der Immobilisierung von Enzymen in
Silicon befasst [WANG ET AL., 1996; RAGHEB ET AL., 2003; GILL ET AL., 1999; GILL UND
BALLESTEROS, 2000b]. Daher überrascht es nicht, dass Silicon-Elastomere hinsichtlich Vertei-
lung und Massentransfer noch gänzlich uncharakterisiert sind. In dieser Arbeit wurde erst-
malig die Verteilung der Substrate und des Produktes der gewählten Modellreaktion unter-
sucht: Die Analyten wurden in einer definierten Konzentration in Hexan vorgelegt und an-
schließend eine definierte Menge an Siliconkugeln hinzugefügt (Kapitel 2.12.3). Auch nach
2 h konnte in Hexan keine signifikante Abweichung von der ursprünglich eingestellten
Konzentration festgestellt werden. Der Quotient der Analyt-Konzentrationen vor und nach
Gleichgewichtseinstellung lag bei 1,03 ± 0,04 für Octanol und bei 1,00 ± 0,04 für Caprylsäure
und Octyloctanoat. Diese Ergebnisse können einerseits zu der Interpretation führen, dass
zwischen Hexanphase und Siliconsphären keine oder nur eine sehr langsame Verteilung statt-
findet oder aber zu der – wahrscheinlicheren – Interpretation, dass die Analyten zusammen
mit Hexan so gut in das Siliconnetzwerk eindringen können, dass es zu einer völlig gleich-
mäßigen Verteilung in der Matrix und der organischen Phase kommt und somit kein Konzen-
trationsunterschied nachzuweisen ist. Dabei wären die Siliconsphären mit einem porösen
Schwamm zu vergleichen, der sich in Wasser rasch vollsaugt, was angesichts der beträcht-
lichen und schnellen Quellung der Siliconkugeln in Hexan durchaus plausibel erscheint
(Kapitel 3.4.3). Die Volumenzunahme der Siliconsphären bei Quellung in Hexan ist wahr-
scheinlich darauf zurückzuführen, dass eine entsprechende Menge Hexan und die darin
gelösten Analyten vom Elastomer aufgenommen werden.
Um die Hypothese der gleichmäßigen Verteilung der Analyten zu verifizieren, wurde eine
einzelne Siliconkugel (Kugelvomumen VK) nach 2 h aus einer analythaltigen Hexanphase
(Volumen V1; Konzentration c1 = 50 mM) in eine analytfreie Hexanphase (Volumen V3)
überführt (Abb. 3.38). Unter der Annahme, dass durch Quellung ein definiertes Volumen der
analythaltigen Hexanphase von den Siliconsphären aufgenommen wurde (Volumenzunahme
V2), ist davon auszugehen, dass sich die nunmehr in den Siliconsphären befindlichen Ana-
lyten (Konzentration c2) nach Überführung der Sphären in analytfreies Hexan (V3) wiederum
gleichmäßig verteilen und sich dort eine bestimmte Analytkonzentration (c3) einstellt (Abb.
3.38). Träfe die Hypothese der gleichmäßigen Verteilung zu, müssten die Konzentrationen c1

Charakterisierung der Statischen Emulsion
113
und c2 identisch sein und c3 könnte bei Kenntnis der Volumina und dem daraus resultieren-
dem Verdünnungsfaktor (VF) errechnet werden. Sollte die errechnete Konzentration mit der
gemessenen Konzentration c3 übereinstimmen, wäre die Hypothese bestätigt.
Abbildung 3.38: Versuchsablauf bei Analyse der Verteilung von Caprylsäure, Octanol und Octyloctanoat zwi-schen Hexan und Siliconkugeln.
Tabelle 3.4: Verteilung von Caprylsäure, Octanol und Octyloctanoat zwischen der diskontinuierlichen (Silicon) und kontinuierlichen Phase (Hexan) – gemessene und errechnete Konzentratione im Gleichgewicht.
Caprylsäure Octanol Octyloctanoat
V2 [µL] 49,1 24,8
C3 errechnet [mmol/L]* 2,5 1,2
C3 gemessen [mmol/L] 3,1 0,3 3,9 1,2 3,0 0,6 * berechnet unter der Annahme c1 = c2
Wie Tabelle 3.4 zu entnehmen ist, lagen die gemessenen Konzentrationen von Caprylsäure
und Octyloctanoat etwa 20 %, die von Octanol 40 % über der berechneten Konzentration.
Ungenauigkeiten bei Analyse der Größe der Siliconsphären vor und nach Quellung hatten er-
hebliche Auswirkungen auf die Berechnung des von den Sphären aufgenommenen Hexan-
volumens (V2) und der Konzentration (c3), erkennbar an den hohen Standardabweichungen.
Insofern ist es schwierig, die errechneten und gemessenen Konzentrationen miteinander zu
vergleichen. Die Werte liegen in einer ähnlichen Größenordnung, wobei nicht gesagt werden
kann, warum ausgerechnet für Octanol eine deutlich höhere Konzentration gemessen wurde.
Für alle drei Substanzen ist die gemessene Konzentration offenbar höher als die errechnete.
Letzteres kann vermutlich dadurch erklärt werden, dass bei oberflächlicher Trocknung der
Siliconsphären an Luft nur das Hexan an der Kugeloberfläche verdunstet war und erhebliche
Mengen der Analyten außen an der Oberfläche haften blieben. Dies hätte einen höheren, über
das Volumen V2 hinausgehenden Eintrag der Analyten in das zuvor analytfreie Hexan zur
Folge. So entspräche bspw. eine 100 μm dicke Hexanschicht außen auf einer gequollenen
Siliconkugel (VK) in analythaltiges Hexan (c1; V1)
Überführen der gequollenen Kugel (VK + V2; c2) in reines Hexan (V3)
Analyse der org. Phase (c3)

Ergebnisse und Diskussion
114
Siliconsphäre ( 3 mm) einem Volumen von 3,02 μL. Durch Quellung der Siliconsphäre
werden nur 7,21 μL Hexan (V2) aufgenommen. Demzufolge würde eine um 40 % erhöhte
Stoffmenge in das analytfreie Hexan gelangen. Dieser Fehler würde sich je nach Dicke der
Hexanschicht auf der Sphärenoberfläche mehr oder weniger stark auswirken. Da aber über die
Dicke dieser Hexanschicht keine Aussage getroffen werden kann, ist eine genaue Quantifi-
zierung unmöglich. Nichtsdestotrotz legen die erzielten Ergebnisse den Schluss nahe, dass
tatsächlich mit einer gleichmäßigen Verteilung zwischen Siliconsphären und umgebendem
organischen Reaktionsmedium zu rechnen ist.
Abbildung 3.39: Decan-Freisetzung aus Siliconkugeln in reines Hexan.
Im Fall einer Gleichverteilung kann aber die Geschwindigkeit des Massentransfers nicht
durch Konzentrationsabnahme in der umgebenden Hexanphase ermittelt werden. Aus diesem
Grund schien es angebracht, ausreichend hohe und genau definierte Konzentrationen eines
Analyten in den Siliconsphären vorzulegen und die Freisetzung in das umgebende Medium zu
dokumentieren (Abb. 3.39). Zu diesem Zweck wurde bei Herstellung der Siliconsphären dem
Gemisch der Präpolymere Decan in einer Konzentration von 100 mmol/L zugegeben. Bei
einem Kugelvolumen von etwa 10 μL ( 2,7 mm) und einem Volumen der umgebenden
Hexanphase von 500 μL wäre bei gleichmäßiger Verteilung eine Decan-Konzentration von
etwa 2 mmol/L zu erwarten. Nach Gleichgewichtseinstellung konnten jedoch lediglich etwa
35 % des Decans in der organischen Phase nachgewiesen werden (Abb. 3.40). Dies kann nur
dadurch erklärt werden, dass bei Herstellung nicht die gesamte Menge an Decan in die
Siliconsphären gelangte oder eine erhebliche Menge an Decan in den Sphären verblieb. Die
genauen Ursachen hierfür können derzeit nicht benannt werden. Hinsichtlich der geschwin-
digkeitsabhängigen Freisetzung des Decans kann festgehalten werden, dass diese relativ
schnell erfolgte und nach 0,5 h keine Konzentrationsänderung mehr beobachtet werden
konnte. Diese Ergebnisse lassen den Schluss zu, dass der Massentransfer in angemessen
Decanhaltige Siliconkugel in Hexan
Analyse der organischen Phase
Decanhaltige Siliconkugel in Hexan
Analyse der organischen Phase

Charakterisierung der Statischen Emulsion
115
kurzen Zeitspannen erfolgt. Die maximale Konzentrationsänderung erfolgt innerhalb der
ersten 10 min – die gleiche Zeitspanne, innerhalb der die Kugeln quellen.
0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60Zeit [min]
Dec
an [
%]
Abbildung 3.40: Massentransfer von Decan aus Siliconsphären in eine umgebende Hexanphase.
Abbildung 3.41: Farbveränderung von statisch emulgiertem pH-Indikator (Bromkresolgrün) in Siliconkugeln (Querschnitte) in Hexan durch Zugabe von Propionsäure.
Obige Untersuchungen bezüglich Massentransfer und Verteilung beschränkten sich auf die
enzymfreie bzw. wasserfreie Siliconmatrix. Siliconsphären mit statisch emulgierter Lipase
stellen ein komplexeres System dar, da hier Substanzen zunächst von Hexan in die Silicon-
matrix und anschließend in die dort emulgierte wässrige Phase diffundieren müssen. Da aber
der geringe Anteil der wässrigen Phase gegenüber der Siliconmatrix und der Hexanphase zur
Folge gehabt hätte, dass die aus der Verteilung in die wässrige Phase resultierende Konzen-
trationsabnahme relativ gering ausgefallen wäre, wurde auf die Bestimmung der Verteilungs-
koeffizienten verzichtet. Wahrscheinlich hätten diese ohnehin weitestgehend denen einer
typischen Verteilung in einem wässrig-organischen Zweiphasensystem entsprochen. Um
wenigstens die Geschwindigkeit des Massentransfers qualitativ untersuchen zu können, wurde
ein wässrig gelöster pH-Indikator (Bromkresolgrün) statisch in Siliconsphären ( ca. 4-
abnehmender pH-Wert
0 3 6 9 12 15 18
Zeit [min]
abnehmender pH-Wert
0 3 6 9 12 15 18
Zeit [min]

Ergebnisse und Diskussion
116
5 mm) emulgiert. Durch Zugabe von Propionsäure konnte innerhalb von 18 min ein deutlicher
und das gesamte Kugelvolumen umfassender Farbumschlag der Sphären beobachtet werden
(Abbildung 3.41). Durch Zugabe von Butylamin konnte dieser Farbumschlag ähnlich schnell
wieder rückgängig gemacht werden. Aufgrund des großen Durchmessers der untersuchten
Kugeln darf vermutet werden, dass bei Verwendung kleinerer Kugeln der Farbumschlag sehr
viel schneller erfolgt wäre.
In ihrer Gesamtheit lassen die Ergebnisse den Schluss zu, dass die Geschwindigkeit des
Massentransfers relativ hoch ist und die hydrophoben Substanzen sich gleichmäßig in der
organischen Phase und dem Silicon verteilen. Die Eignung dieser Matrix zur Einhüllung von
statisch emulgierten Lipasen darf somit angenommen werden.

Anwendung der Statischen Emulsion
117
3.5 Anwendung der Statischen Emulsion für die Synthese von Fein- und Bulkchemikalien
Die zuvor vorgenommene Charakterisierung der Statischen Emulsion erfolgte anhand einer
einfachen Modellreaktion in organischem Lösungsmittel. Auch wenn die Ergebnisse berech-
tigterweise Anlass zu der Vermutung geben, dass es sich bei der Statischen Emulsion um eine
innovative und äußerst vielversprechende Immobilisierungsmethodik handelt, steht der prak-
tische Beweis noch aus. Weiterführende Untersuchungen sollten klären, inwieweit sich die
Statische Emulsion für die anwendungsorientierte enantioselektiven Synthese von Fein-
chemikalien eignet. Ferner sollte ebenfalls untersucht werden, ob auch die lösungsmittelfreie
Synthese von Bulkchemikalien mit derartigen Immobilisaten möglich ist. Die Synthese von
Bulkchemikalien ist in der Regel mit einer geringen Wertschöpfung verbunden. Aus diesem
Grund ist der kostenintensive Einsatz toxischer und leicht entflammbarer Lösungsmittel
problematisch [BOURG-GARROS ET AL., 1998].
Daher wurde gemeinsam mit Kooperationspartnern des Lehrstuhls für Biotechnologie der
RWTH Aachen, die mit der Entwicklung bzw. Durchführung marktrelevanter enzymkata-
lysierter Reaktionen befasst sind, die diesbezügliche Eignung der Statischen Emulsion über-
prüft. Untersucht wurde die enantioselektive Synthese von Benzoinbutyrat in Tetrahydrofuran
sowie die lösungsmittelfreien Synthesen von Propyllaurat und 2-Ethyl-1-hexyloleat.
3.5.1 Lipase von Thermomyces lanuginosa für die Synthese von Esterölen
Aus der sich abzeichnenden Erschöpfung der Erdöllagerstätten ergibt sich die Notwendigkeit,
mineralölbasierte Produkte zu substituieren: Als prominentes Beispiel hierfür können Fetts-
äuremethylester angeführt werden, die als „Biodiesel“ in Deutschland im Jahr 2005 bereits ein
Marktvolumen von fast 2 Mio. t erreichen dürften [HEIM ET AL., 2005]. Hingegen können
Ester aus Fettsäuren und höhermolekularen Alkoholen (Esteröle) als Lösungsmittel für
Druckertinten, Detergenzien, Metallreiniger und Schmierstoffe Verwendung finden [LINKO ET
AL., 1995; MANG UND DRESEL, 2001]. Neben den ökologischen Vorteilen sind einige der
esterbasierten Produkte unter technologischen Gesichtspunkten ihrem mineralölbasierten
Pendant überlegen [HESSELBACH ET AL., 2003; MANG UND DRESEL, 2001]. Gegenwärtig
erfolgt die Herstellung dieser Fettsäureester überwiegend mit anorganischen Katalysatoren,
wobei in den letzten Jahren verstärkt Forschungsbemühungen bezüglich der lipasenka-

Ergebnisse und Diskussion
118
talysierten Synthese zu verzeichnen sind [LINKO ET AL., 1994-1998; NELSON ET AL., 1996;
SCHUCHARDT ET AL., 1998, HESSELBACH ET AL., 2003; BRENNEIS ET AL., 2004].
An der Bundesanstalt für Materialforschung und -testung (BAM) wurde im Rahmen eines
DBU-Projektes (AZ 13078) ein erfolgreiches Verfahren zur lipasenkatalysierten Bildung von
Verlustschmierstoffe aus Altfetten und 2-Ethyl-1-hexanol entwickelt. Die Umesterung wurde
mit der nicht immobilisierten Lipase A von Candida antarctica (CALA) bei Temperaturen
von 50 °C im Rührkessel ausgeführt [BRENNEIS ET AL., 2002 und 2004; HESSELBACH ET AL.,
2003]. Jedoch war es im Rahmen dieses Verfahrens bislang nicht möglich, freie Fettsäuren
mit 2-Ethyl-1-hexanol zu verestern. Durch direkte Veresterung könnten aber auch alternative
kostengünstige Rohstoffquellen, wie die bei der Biodieselproduktion anfallenden Fettsäuren,
verwendet werden. In erster Linie wird dort im Zuge der Umesterung von Rapsöl mit
Methanol Glycerin als Nebenprodukt gebildet. Die polare Glycerinphase enthält allerdings
nicht unerhebliche Mengen an freien Fettsäuren, die durch Ansäuerung leicht abgetrennt
werden könnten. Durch Veresterung der freien Fettsäuren könnte ein Mehrwert mit ent-
sprechend positiver Auswirkung auf die ökonomische Gesamtbilanz der Biodieselproduktion
geschaffen werden. Ein derartiger Ansatz wurde in der Literatur bislang nicht beschrieben.
Daher wurde im Rahmen dieser Arbeit in Zusammenarbeit mit der BAM an der Entwicklung
eines Verfahrens zur direkten Veresterung von freien Fettsäuren der Biodieselproduktion mit
2-Ethyl-1-hexanol mittels der immobilisierten Lipase von Thermomyces lanuginosa (TLL)
gearbeitet. Die Lipase wurde dabei in Sol-Gel und Statischer Emulsion immobilisiert – als
Modellreaktion zur Charakterisierung der Immobilisate diente im Interesse einer hohen
Reproduzierbarkeit die Bildung von 2-Ethyl-1-hexyloleat aus einer äquimolaren Mischung
von Ölsäure und 2-Ethyl-1-hexanol. Die aus der Nebenproduktfraktion abgetrennten Fett-
säuren stellen ein komplexes Gemisch unterschiedlicher Fettsäuren dar – die genaue Zusam-
mensetzung ist aller Wahrscheinlichkeit nach zu starken Schwankungen unterworfen, um das
notwendige Maß an Reproduzierbarkeit zuzulassen.
Das vorliegende lyophilisierte Präparat der Lipase aus Thermomyces lanuginosa wies für die
Modellreaktion bei 50 °C eine spezifische Aktivität von 0,29 U/mgProt. auf. Die Zugabe von
Wasser (33 % v/v) führte zu einer mehr als doppelt so hohen spezifischen Aktivität von
0,67 U/mgProt.. Es sei an dieser Stelle darauf hingewiesen, dass die in einem enzymfreien
Ansatz ermittelte autokatalytische Reaktionsrate etwa 6 % der enzymkatalysierten Reaktions-
rate betrug. Daher wurden die in den nachfolgenden Untersuchungen gemessenen Reaktions-

Anwendung der Statischen Emulsion
119
raten um die Autokatalyse bereinigt, damit jeweils der auf die Enzymaktivität zurückgehende
Anteil der Esterbildung bestimmt werden konnte.
3.5.1.1 Sol-Gel
Bei Immobilisierung in Sol-Gel konnten durchschnitllich 74 % der vorgelegten Proteinmenge
immobilisiert werden, im Fall der Statischen Emulsion durchschnittlich 95 %. Wie in Tabelle
3.5 zu ersehen, schneidet das Sol-Gel im Vergleich zur Statischen Emulsion hinsichtlich der
apparenten und spezifischen Aktivität deutlich schlechter ab. Bezogen auf die eingesetzte
Menge an Enzym wurde eine relative Aktivität von nur 60 % erreicht. Dies überrascht ein
wenig, da im Rahmen dieser Arbeit (Kapitel 3.3.3.2) bei gleichartiger Immobilisierung des-
selben Enzyms eine Steigerung der Aktivität um den Faktor 260 beobachtet werden konnte.
Allerdings wurde dabei die Veresterung in einem organischen Lösungsmittel untersucht. Die
Anwendung von Sol-Gelen in lösungsmittelfreien Systemen wurde hingegen bisher nicht in
der Literatur beschrieben. Es darf daher spekuliert werden, dass der Massentransfer in die Sol-
Gel-Matrix aufgrund der deutlich höheren Viskosität der Substratmischung eingeschränkt ist.
Nicht nur aus diesem Grund ist von einem Einsatz der Sol-Gele abzusehen, denn wie Ab-
bildung 3.42 veranschaulicht, liegt das anfangs pulverförmige Sol-Gel (Abb. 3.42 A) gegen
Ende der Reaktion als kaum erkennbare amorphe Masse vor (Abb. 3.42 B). Nach Zentri-
fugation des Reaktionsansatzes konnten drei Zonen abgegrenzt werden (Abb. 3.42 C): Unten
sammelte sich das im Zuge der Veresterung gebildete Wasser – an der Phasengrenze ist die
amorphe Sol-Gel-Masse mit dem darüberliegenden Produktgemisch zu erkennen. Die
während der Reaktion auftretenden hydromechanischen Scherkräfte führen vermutlich
aufgrund der Sprödheit der Sol-Gele zu einer fortschreitenden Desintegration. Sol-Gele sind
für ihre spröde Beschaffenheit und der daraus resultierenden schlechten Eignung für den
Einsatz im Bioreaktor bekannt [GILL UND BALLESTEROS, 2000a/b]. Eine Abtrennung der
amorphen Masse dürfte verfahrenstechnisch jedenfalls erhebliche Probleme bereiten. Auf-
grund der geringen Aktivität und der unzureichenden mechanischen Eigenschaften kann die
Eignung des Sol-Gels zumindest mittelfristig ausgeschlossen werden, weshalb keine weiteren
Untersuchungen durchgeführt wurden.

Ergebnisse und Diskussion
120
Abbildung 3.42: Morphologie des Sol-Gels im Reaktionsverlauf A: frisches Sol-Gel im Substratgemisch zu Reaktionsbeginn; B: Sol-Gel während bzw. nach erfolgter Reaktion; C: abzentrifugierter Reaktionsansatz: Unten durch Veresterung gebildetes Wasser, darüberliegend amorphes Sol-Gel und nachfolgend Produktge-misch.
3.5.1.2 Statische Emulsion
Aus Tabelle 3.5 wird ersichtlich, dass im Fall der Statischen Emulsion die spezifische
Aktivität im Vergleich zum nativen Enzym um das Zehnfache gesteigert werden konnte.
Jedoch ist die Aktivitätssteigerung moderat im Vergleich zu der bei Veresterung von Octanol
und Caprylsäure in organischem Lösungsmittel erzielten Aktivitätssteigerung um den Faktor
250 (Kapitel 3.3.3.1). Ferner kann Tabelle 3.5 und Abbildung 3.43 entnommen werden, dass
höhere Beladungsdichten mit einer tendenziell konstanten bzw. nur mit einer geringfügig
gesteigerten apparenten Aktivität einhergehen. Die infolgedessen reduzierten spezifischen
Aktivitäten deuten auf einen limitierten Massentransfer hin. Wie bereits erörtert, kann eine
Steigerung der apparenten Aktivität bei konstanter spezifischer Aktivität nur durch Opti-
mierung der Silicon-Matrix erreicht werden. Gegenwärtig ist eine Enzymbeladungsdichte von
5,7 mg/g zu favorisieren. Dies stimmt sehr gut mit der in Kapitel 3.3.3.1 getroffenen Feststel-
lung überein. Dort lag die ideale Proteinbeladungsdichte zwischen 3,7 und 5,6 mgProt./gImmob..
Oberhalb dieser Beladungsdichte blieb die apparente Aktivität konstant. Andererseits irritiert
in diesem Zusammenhang, dass das Ausmaß der Quellung im Substrat bzw. Produkt deutlich
geringer ausfiel als in Hexan. Bei Inkubation der Statischen Emulsion in einem äquimolaren
Gemisch von Ölsäure und 2-Ethyl-1-hexanol wurde eine Volumenzunahme von nur 6,0 +
3,0 % festgestellt, im Ester in Höhe von 14,0 + 0,7 %. Die geringere Quellung erklärt sich
möglicherweise durch die hohe Viskosität der Substrat- bzw. Produktlösung und könnte zu-

Anwendung der Statischen Emulsion
121
dem das geringere Ausmaß der Aktivitätssteigerung erklären (Kapitel 3.5.2). Allerdings
müsste dann aufgrund des vermutlich erhöhten Massentransferwiderstands bereits bei
niedrigeren Beladungsdichten eine konstante apparente Aktivität beobachtet werden. Eine
genauere Interpretation ist bei der derzeitigen Datenlage nicht möglich.
Tabelle 3.5: Aktivitäten von Sol-Gel und Statischer Emulsion mit TLL (unterschiedliche Beladungsdichten) bei der Synthese von Ethylhexyloleat.
Immobilisierte Proteinmenge a
[mg Prot ./g Immob.]
ApparenteAktivität
[U / g Immob.]
Spezifische Aktivität
[U / mg Protein]
Relative Aktivität
[ ] b
Sol-Gel 1,2 0,2 +/- 0,1 0,17 +/- 0,8 0,59
2,9 8,1 +/- 1,3 2,79 +/- 0,45 9,73
4,4 9,4 +/- 1,0 2,14 +/- 0,23 7,46
5,7 13,1 +/- 1,1 2,30 +/- 0,19 8,02
8,9 13,1 +/- 4,6 1,47 +/- 0,52 5,13
Statische Emulsion
11,7 16,6 +/- 1,4 1,42 +/- 0,12 4,95 a Immobilisierte Proteinmenge unter Berücksichtigung des Immobilisierungsgrades (durchschnittlich 95 %). b Die relative Aktivität ergibt sich aus dem Verhältnis der spezifischen Aktivitäten von immobilisiertem und nativem Enzym.
0
2
4
6
8
10
12
14
16
18
2 4 6 8 10 12Immobilisierte Proteinmenge [mg Protein/g Immobilisat]
Ap
par
ente
Akt
ivit
ät [
U/g
Imm
ob
.]
0
0,5
1
1,5
2
2,5
3
Sp
ezif
isch
e A
ktiv
ität
[U
/mg
Pro
tein
]Apparente Aktivität
Spezifische Aktivität
Abbildung 3.43: Apparente und spezifische Aktivitäten der Statischen Emulsion mit TLL in Abhängigkeit von der Beladungsdichte bei Synthese von Ethylhexyloleat.
3.5.1.3 Veresterung von Fettsäuren aus Nebenproduktfraktionen der Biodieselproduktion
Aus der Nebenproduktfraktion der Biodieselproduktion abgetrennte Fettsäuren wurden von
den Produzenten Esterco Biofuels Ltd. (Cornwall, GB) und PETROTEC GmbH (Borken)
bezogen. Die Charakterisierung der erhaltenen Proben zeigte, dass neben freien Fettsäuren
beträchtliche Mengen an Fettsäuremethylestern (ca. 5 %) und Methanol enthalten waren. Um
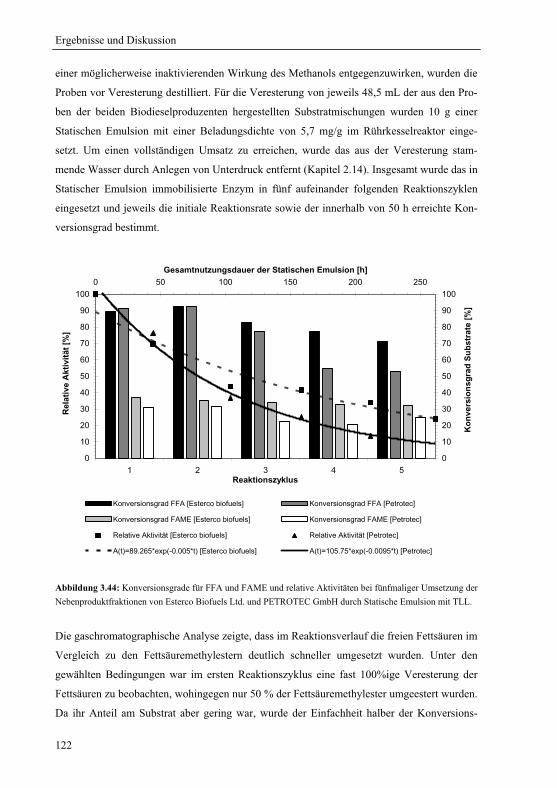
Ergebnisse und Diskussion
122
einer möglicherweise inaktivierenden Wirkung des Methanols entgegenzuwirken, wurden die
Proben vor Veresterung destilliert. Für die Veresterung von jeweils 48,5 mL der aus den Pro-
ben der beiden Biodieselproduzenten hergestellten Substratmischungen wurden 10 g einer
Statischen Emulsion mit einer Beladungsdichte von 5,7 mg/g im Rührkesselreaktor einge-
setzt. Um einen vollständigen Umsatz zu erreichen, wurde das aus der Veresterung stam-
mende Wasser durch Anlegen von Unterdruck entfernt (Kapitel 2.14). Insgesamt wurde das in
Statischer Emulsion immobilisierte Enzym in fünf aufeinander folgenden Reaktionszyklen
eingesetzt und jeweils die initiale Reaktionsrate sowie der innerhalb von 50 h erreichte Kon-
versionsgrad bestimmt.
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5Reaktionszyklus
Rel
ativ
e A
ktiv
ität
[%
]
0
10
20
30
40
50
60
70
80
90
1000 50 100 150 200 250
Gesamtnutzungsdauer der Statischen Emulsion [h]
Ko
nve
rsio
nsg
rad
Su
bst
rate
[%
]
Konversionsgrad FFA [Esterco biofuels] Konversionsgrad FFA [Petrotec]
Konversionsgrad FAME [Esterco biofuels] Konversionsgrad FAME [Petrotec]
Relative Aktivität [Esterco biofuels] Relative Aktivität [Petrotec]
A(t)=89.265*exp(-0.005*t) [Esterco biofuels] A(t)=105.75*exp(-0.0095*t) [Petrotec]
Abbildung 3.44: Konversionsgrade für FFA und FAME und relative Aktivitäten bei fünfmaliger Umsetzung der Nebenproduktfraktionen von Esterco Biofuels Ltd. und PETROTEC GmbH durch Statische Emulsion mit TLL.
Die gaschromatographische Analyse zeigte, dass im Reaktionsverlauf die freien Fettsäuren im
Vergleich zu den Fettsäuremethylestern deutlich schneller umgesetzt wurden. Unter den
gewählten Bedingungen war im ersten Reaktionszyklus eine fast 100%ige Veresterung der
Fettsäuren zu beobachten, wohingegen nur 50 % der Fettsäuremethylester umgeestert wurden.
Da ihr Anteil am Substrat aber gering war, wurde der Einfachheit halber der Konversions-

Anwendung der Statischen Emulsion
123
grad nur aus dem Verhältnis der freien Fettsäuren und der Ethylhexylester berechnet. Wie Ab-
bildung 3.44 veranschaulicht, war bei wiederholtem Einsatz des Immobilisats eine sukzessive
Abnahme der katalytischen Aktivität zu verzeichnen. Daraus erklärt sich auch die Abnahme
der bei einer konstanten Reaktionsdauer von etwa 50 h ermittelten Konversionsgrade – die ge-
wählte Reaktionsdauer reichte bereits nach drei Reaktionszyklen nicht mehr für einen
vollständigen Umsatz des Substrates. Die Abnahme der initialen Reaktionsrate ist vermutlich
in erster Linie auf eine thermische Inaktivierung des bei 50 °C eingesetzten Biokatalysators
zurückzuführen. Inwieweit das Ausmaß dieser Inaktivierung in einer für das Enzym typischen
Größenordnung liegt, konnte nicht geklärt werden, da in der Fachliteratur keine Daten
bezüglich einer vergleichbaren Reaktion zu finden waren. Auffällig ist, dass bei Umsetzung
der von der PETROTEC GmbH bezogenen Fettsäuren die Inaktivierung stärker ausgeprägt
war. Dies deutet darauf hin, dass auch andere Faktoren inaktivierend wirken. Möglicherweise
waren zu große Mengen an Methanol in der Substratmischung enthalten. Weitergehende
Untersuchungen zur Klärung dieses Sachverhalts wurden jedoch nicht durchgeführt.
Die prinzipielle Machbarkeit der Veresterung von aus Biodiesel-Nebenproduktfraktionen
abgetrennten Fettsäuren mittels in Statischer Emulsion immobilisierter Lipase zur Herstellung
von Esterölen konnte aufgezeigt werden. An dieser Stelle bleibt festzuhalten, dass die erreich-
ten Produktivitäten angesichts der erzielten Aktivitätssteigerung zufriedenstellend sind,
jedoch noch zu gering, um einen großtechnischen Einsatz zu ermöglichen. Vermutlich sind
hierfür Massentransferwiderstände in der Matrix verantwortlich, denen durch eine Opti-
mierung der Siliconmatrix begegnet werden kann. Ferner empfiehlt sich eine Reihenunter-
suchung, um möglicherweise besser geeignete Lipasen zu identifizieren.
3.5.2 Lipase B von Candida antarctica für die Synthese von Propyllaurat
Die industrielle, lipasenkatalysierte Produktion von Fettsäureestern wurde in den letzten
Jahren systematisch ausgebaut [HILLS, 2003]. Der Hauptvorteil der enzymkatalysierten Reak-
tion gegenüber konventionellen Verfahren liegt hier in der Unterdrückung unerwünschter
Nebenreaktionen. Dies ist auf die sehr milden Reaktionsbedingungen zurückzuführen und vor
dem Hintergrund der verwendeten Rohstoffe von großer Bedeutung: Ausgangsmaterial sind
pflanzliche Triglyceride mit z.T. hohem Anteil an ungesättigten Fettsäuren, die besonders
anfällig für Nebenreaktionen sind. Hauptbestimmungszweck der bei einem Kooperations-
partner des Lehrstuhls für Biotechnologie der RWTH Aachen produzierten Fettsäureester ist

Ergebnisse und Diskussion
124
ihr Einsatz in Kosmetika, woraus besonders strenge Anforderungen an ihre Qualität bzw.
Reinheit resultieren [HILLS, 2003]. Konventionelle chemische Verfahren sind unweigerlich
mit einer unangenehmen Geruchsentwicklung sowie einer unerwünschten Verfärbung des
Endproduktes assoziiert, die nur durch eine aufwändige Nachbehandlung eliminiert werden
können. Diese kostenintensive Nachbehandlung entfällt bei der lipasenkatalysierten Synthese
und stellt neben dem aufgrund der milden Reaktionsbedingungen erheblich reduzierten
Energiebedarf einen weiteren Vorteil dar [HILLS, 2003; THUM, 2004]. Voraussetzung für die
Etablierung eines unter ökonomischen Gesichtspunkten effizienten Prozesses war die Ver-
wendung immobilisierter Biokatalysatoren [HILLS, 2003].
Der Kooperationspartner verwendet gegenwärtig das hochaktive kommerzielle Präparat
Novozym 435 (Novozymes, Dänemark) mit adsorptiv an einen Träger auf Basis von Poly-
methylacrylat gebundener Lipase B aus Candida antarctica (CALB). Aufgrund der unzu-
reichenden Abriebfestigkeit des Trägermaterials kann der immobilisierte Biokatalysator
bislang nur im Packbettreaktor eingesetzt werden. Zudem bedingt die vergleichsweise
schwache adsorptive Bindung eine fortwährende Desorption der Lipase im operativen
Einsatz. Gerade letzter Aspekt ist von großer Bedeutung, da durch das allergene Potential von
Proteinen deren Gegenwart im Endprodukt die Sicherheit des Endverbrauchers beeinträch-
tigen kann. Aus o.g. Gründen wurde am Lehrstuhl für Biotechnologie der RWTH Aachen auf
Anregung des Kooperationspartners die Eignung der Statischen Emulsion für die lösungsmit-
telfreie Synthese von Propyllaurat aus n-Propanol und Laurinsäure bei 60 °C erprobt.
Ausgehend von einem kommerziell verfügbaren Flüssigpräparat der CALB (Lipozyme) mit
einem Proteingehalt von 2,4 % (w/v) und einer spezifischen Aktivität von 65 U/mg bei 60 °C
bzgl. der Synthese von Propyllaurat wurden Statische Emulsionen mit Beladungsdichten im
Bereich von 1,3 – 5,3 mg/g hergestellt. Dabei wurde durchschnittlich eine Immobilisierungs-
ausbeute von 96 +/- 3,5 % erzielt.
Wie Abbildung 3.45 und Tabelle 3.6 zu entnehmen ist, wird die höchste apparente Aktivität
(142 U/g) bei der höchsten Beladungsdichte gemessen. Hingegen zeichnet sich bei Vergleich
der spezifischen Aktivitäten ein gegenläufiges Bild ab: Die höchste Aktivität (69 U/mg) wird
bei der geringsten Beladungsdichte erzielt. Letztlich muss ein Kompromiss zwischen spezi-
fischer und apparenter Aktivität gefunden werden, der hier bei einer Beladungsdichte im
Bereich von 2,5 – 3,5 mg/g liegen dürfte.
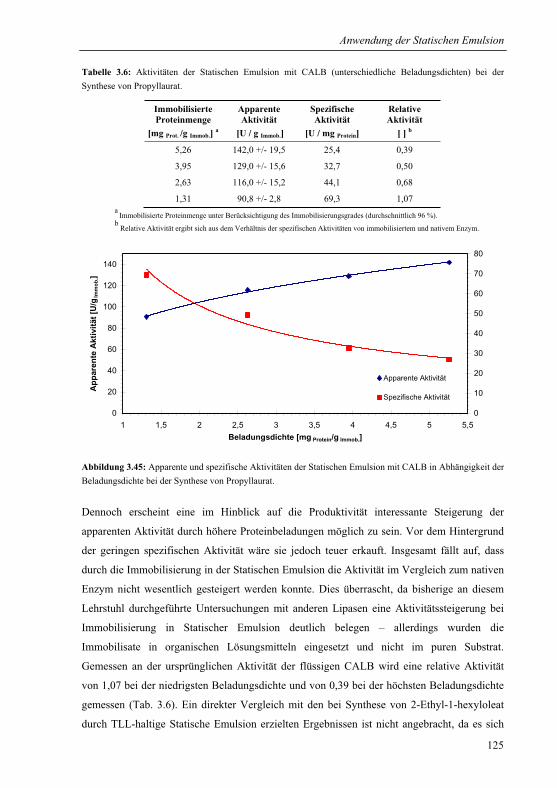
Anwendung der Statischen Emulsion
125
Tabelle 3.6: Aktivitäten der Statischen Emulsion mit CALB (unterschiedliche Beladungsdichten) bei der Synthese von Propyllaurat.
Immobilisierte Proteinmenge
[mg Prot. /g Immob.]a
ApparenteAktivität
[U / g Immob.]
Spezifische Aktivität
[U / mg Protein]
Relative Aktivität
[ ] b
5,26 142,0 +/- 19,5 25,4 0,39
3,95 129,0 +/- 15,6 32,7 0,50
2,63 116,0 +/- 15,2 44,1 0,68
1,31 90,8 +/- 2,8 69,3 1,07 a
Immobilisierte Proteinmenge unter Berücksichtigung des Immobilisierungsgrades (durchschnittlich 96 %). b
Relative Aktivität ergibt sich aus dem Verhältnis der spezifischen Aktivitäten von immobilisiertem und nativem Enzym.
Abbildung 3.45: Apparente und spezifische Aktivitäten der Statischen Emulsion mit CALB in Abhängigkeit der Beladungsdichte bei der Synthese von Propyllaurat.
Dennoch erscheint eine im Hinblick auf die Produktivität interessante Steigerung der
apparenten Aktivität durch höhere Proteinbeladungen möglich zu sein. Vor dem Hintergrund
der geringen spezifischen Aktivität wäre sie jedoch teuer erkauft. Insgesamt fällt auf, dass
durch die Immobilisierung in der Statischen Emulsion die Aktivität im Vergleich zum nativen
Enzym nicht wesentlich gesteigert werden konnte. Dies überrascht, da bisherige an diesem
Lehrstuhl durchgeführte Untersuchungen mit anderen Lipasen eine Aktivitätssteigerung bei
Immobilisierung in Statischer Emulsion deutlich belegen – allerdings wurden die
Immobilisate in organischen Lösungsmitteln eingesetzt und nicht im puren Substrat.
Gemessen an der ursprünglichen Aktivität der flüssigen CALB wird eine relative Aktivität
von 1,07 bei der niedrigsten Beladungsdichte und von 0,39 bei der höchsten Beladungsdichte
gemessen (Tab. 3.6). Ein direkter Vergleich mit den bei Synthese von 2-Ethyl-1-hexyloleat
durch TLL-haltige Statische Emulsion erzielten Ergebnissen ist nicht angebracht, da es sich
0
20
40
60
80
100
120
140
1 1,5 2 2,5 3 3,5 4 4,5 5 5,5Beladungsdichte [mg Protein/g Immob.]
Ap
par
ente
Akt
ivit
ät [
U/g
Imm
ob
.]
0
10
20
30
40
50
60
70
80
Apparente Aktivität
Spezifische Aktivität

Ergebnisse und Diskussion
126
um eine andere Reaktion und ein anderes Enzym handelte. Da aber beide Synthesen in
lösungsmittelfreien Systemen durchgeführt wurden, können zumindest die gleichen Gründe
für die moderaten Aktivitätssteigerungen angeführt werden.
Bei Inkubation der Siliconsphären in einer äquimolaren Mischung der Substrate n-Propanol
und Laurinsäure war eine Volumenzunahme in Höhe von 17 + 5 % zu verzeichnen, bei Inku-
bation in n-Propyllaurat hingegen in Höhe von 45 + 8 %. Die stärkere Quellung im Ester ist
vermutlich durch dessen hydrophoberen Charakter zu erklären, so dass es zu einer stärkeren
Relaxation des polymeren Netzwerkes kommt. Bei Inkubation des Silicon-Elastomers in
Hexan konnte eine Volumenzunahme um mehr als 100 % beobachtet werden (Kapitel 3.4.3),
wobei postuliert wurde, dass sich die Quellung günstig auf den Massentransfer auswirkt. Die
ausgebliebene Aktivitätssteigerung der CALB könnte daher, wie im vorherigen Kapitel bei
der TLL, zum Teil in der deutlich geringeren Quellung des Silicons und einem daraus
resultierenden höheren Diffusionswiderstand begründet sein. Ferner ist die Mischung der
Substrate deutlich viskoser als eine Hexan-Lösung, was sich ebenfalls auf den Massentransfer
auswirken dürfte. Dem könnte bspw. durch eine Verringerung des Vernetzungsgrades in der
Siliconmatrix entgegengewirkt werden.
Festzuhalten bleibt, dass die Immobilisierung der CALB in der Statischen Emulsion und der
nachfolgende Einsatz in einem lösungsmittelfreien Reaktionssystem möglich ist. Die Höhe
der bisher erzielten apparenten Aktivitäten mit 142 U/g ist jedoch zu niedrig, um Novozym
435 (bis zu 10.000 U/g) zu ersetzen. Allerdings weist das verwendete Silicon-Elastomer,
anders als Novozym 435, vorzügliche mechanische Eigenschaften auf – es ist abriebfest und
vollkommen elastisch. Zudem konnte anders als bei Novozym 435 keine Freisetzung von
Protein aus dem Elastomer beobachtet werden. Basierend auf den bisherigen Ergebnissen ist
aber davon auszugehen, dass eine Optimierung möglich ist. Im Mittelpunkt zukünftiger
Bemühungen muss eine Steigerung der apparenten Aktivität durch höhere Enzymbeladungs-
dichten einerseits und durch einen verbesserten Massentransfer andererseits stehen.

Anwendung der Statischen Emulsion
127
3.5.3 Lipase von Pseudomonas stutzeri für die Synthese von S-Benzoinbutyrat
Optisch reine -Hydroxyketone repräsentieren eine wichtige Klasse pharmakologisch
wirksamer Verbindungen, wie etwa das Antidepressivum Bupropion oder die Antibiotika
Olivomycin A und Chromomycin A3 [FANG ET AL., 2002; ROUSH ET AL., 1996]. Chemisch
handelt es sich dabei um homo- oder heterozyklische Benzoin-Derivate, die über die soge-
nannte Benzoin-Kondensation zugänglich sind. Neben traditionellen chemischen Methoden
ist die Bildung chiraler Benzoine auch enzymatisch mit Hilfe Thiamindiphosphat-abhängiger
Enzyme möglich [DEMIR ET AL., 2001; HISCHER ET AL., 2005]. Weitere biokatalytische
Methoden zur Darstellung enantiomerenreiner Benzoine bestehen in der enantioselektiven
Reduktion von -Diketonen oder der lipasenkatalysierten Racematspaltung [AOYAGI ET AL.,
2000].
Am Lehrstuhl für Organische Chemie und Pharmazie der Universidad Complutense de
Madrid – einem Kooperationspartner des Lehrstuhls für Biotechnologie der RWTH Aachen –
wird unter Leitung von Prof. Andrés Alcántara an der Entwicklung eines Verfahrens zur
enantiomerenreinen Darstellung von R-Benzoin und S-Benzoinbutyrat durch lipasenkata-
lysierte Umesterung racemischen Benzoins mit Vinylbutyrat gearbeitet. In Anbetracht der
geringen Löslichkeit von Benzoin und seiner Ester in Wasser sowie den meisten organischen
Lösungsmitteln, wird die Reaktion in Tetrahydrofuran (THF) ausgeführt. Hervorragende
Ergebnisse wurden dort mit der Lipase von Pseudomonas stutzeri (PSL) erzielt [HOYOS ET AL.
2006]. In Zusammenarbeit beider Lehrstühle sollte im Hinblick auf eine spätere industrielle
Anwendung die Eignung der Immobilisierung der PSL in Statischer Emulsion evaluiert
werden. Der Einsatz der Statischen Emulsion in THF stellt im Vergleich zu den bisherigen
Reaktionssystemen einen Sonderfall dar, denn THF ist mit Wasser mischbar. Es ist somit
davon auszugehen, dass die wässrige Phase innerhalb der Statischen Emulsion mit THF ange-
reichert bzw. ein Gleichgewichtszustand hinsichtlich der Zusammensetzung der fluiden Phase
innerhalb und außerhalb der Silicon-Elastomere erreicht wird. Bei mehrfachem Gebrauch der
Immobilisate kann sogar mit einem Austausch der wässrigen Phase gegen THF gerechnet
werden. Dies kann bei Verwendung nicht wassermischbarer Lösungsmittel – wie bei den
übrigen untersuchten Systemen – ausgeschlossen werden.
Bei Herstellung der Statischen Emulsion mit PSL erwies sich die geringe Löslichkeit des
kommerziellen Präparates als problematisch. Obwohl das Präparat nur einen Proteingehalt
von 22,57 % (w/w) aufwies, konnten Konzentrationen oberhalb von 2,5 mg des Lyophilisates

Ergebnisse und Diskussion
128
nicht mehr homogen in Lösung gebracht werden. Es war zudem nicht möglich, die Löslich-
keit durch Variation des pH-Wertes zu beeinflussen. Um dennoch ausreichend hohe Protein-
konzentrationen in der Statischen Emulsion gewährleisten zu können, wurden 50 mg des Fest-
stoffes in Wasser suspendiert. Nach Abtrennung der unlöslichen Bestandteile wies die Lösung
einen Proteingehalt von 7,3 – 8,1 mg/mL auf. Somit konnte die Statische Emulsion mit
Beladungsdichten im Bereich von 0,16 – 1,30 mgProtein/gImmobilisat hergestellt werden. Der
Immobilisierungsgrad lag bei durchschnittlich 91 %. Eine Freisetzung von Protein aus der
Statischen Emulsion in das umgebende Lösungsmittel (THF) war auch nach einwöchiger
Inkubation nicht zu beobachten.
Tabelle 3.7: Aktivitäten der Statischen Emulsion mit PSL (unteschiedliche Beladungsdichten) bei der enantio-selektiven Umesterung von R/S-Benzoin und Vinylbutyrat.
Immobilisierte Proteinmenge
[mg Prot /g Immob.]a
ApparenteAktivität
[U / g Immob.]
Spezifische Aktivität
[U / mg Protein]
Relative Aktivität
[ ] b
E-Wert
[ ]
1,30 0,0531 40,9 0,51 > 200
0,91 0,0906 99,1 1,23 > 200
0,59 0,0413 70,1 0,87 > 200
0,28 0,0372 133,8 1,66 > 200
0,16 0,0119 73,5 0,91 > 200 a Immobilisierte Proteinmenge unter Berücksichtigung des Immobilisierungsgrades (durchschnittlich 91 %). b Relative Aktivität ergibt sich aus dem Verhältnis der spezifischen Aktivitäten von immobilisiertem und nativem Enzym.
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,1
0 0,2 0,4 0,6 0,8 1 1,2 1,4
Immobilisierte Proteinmenge [mg Protein / g Immob.]
Ap
par
ente
Akt
ivit
ät [
U/g
Imm
ob
.]
-0,01
0,01
0,03
0,05
0,07
0,09
0,11
0,13
0,15
Sp
ezif
isch
e A
ktiv
ität
[U
/mg
Pro
tein
]
Apparente Aktivität
Spezifische Aktivität
Abbildung 3.46: Abhängigkeit der spezifischen und apparenten Aktivität der Statischen Emulsion mit PSL in Abhängigkeit von der Beladungsdichte.
Die nach Immobilisierung erzielten spezifischen Aktivitäten lagen im Bereich von 40-130 U/g
(Tab. 3.7 und Abb. 3.46). Im Vergleich zur Aktivität des nicht immobilisierten Enzyms

Anwendung der Statischen Emulsion
129
(80 U/g) konnte demnach nur eine sehr moderate Steigerung der katalytischen Aktivität durch
Immobilisierung verzeichnet werden. Ein Unterschied zwischen nativem und immobilisiertem
Enzym hinsichtlich der sehr hohen Enantioselektivität war nicht festzustellen. Die scheinbar
drastische Aktivitätsminderung für die Statische Emulsion der höchsten Beladungsdichte
(1,3 mg/g) muss differenziert betrachtet werden: Nach gaschromatographischer Analyse
konnten neben den Substraten und S-Benzoinbutyrat abweichend von den übrigen Ansätzen
beträchtliche Mengen an Buttersäure nachgewiesen werden. Die Freisetzung von Buttersäure
kann durch Hydrolyse des Substrates Vinylbutyrat erklärt werden. Es handelt sich dabei um
eine unerwünschte Nebenreaktion und ihr ausschließliches Auftreten bei der höchsten
Beladungsdichte ist weniger auf die höhere Proteinkonzentration als vielmehr auf die
Gegenwart von Wasser zurückzuführen. Obwohl die Statische Emulsion bzw. die Silicon-
sphären direkt nach Herstellung für 24 h bei Raumtemperatur an Luft getrocknet wurden, ist
davon auszugehen, dass zumindest in der Charge mit der höchsten Beladungsdichte der
Wassergehalt nicht auf das für die Reaktion notwendige Maß reduziert werden konnte.
Um diesen Sachverhalt zu überprüfen, wurde zunächst der Einfluss von Wasser auf die
Reaktionskinetik untersucht. Es zeigte sich, dass mit steigendem Wassergehalt die Bildung
von Benzoinbutyrat zugunsten der Freisetzung von Buttersäure unterdrückt wird, demnach
also gegenläufige Reaktionsraten beobachtet werden (Abb. 3.47). Bei einem Wassergehalt
von nur 2 % (v/v) war das eigentliche Zielmolekül dieser Synthese nur in geringen Mengen
nachzuweisen. Ein ähnlicher Wassergehalt wird bei Einsatz der Statischen Emulsion per se
erreicht und damit die Eignung der Statischen Emulsion für diese Synthese in Frage gestellt.
Da aus Voruntersuchungen bekannt war, dass der Wassergehalt in den Siliconsphären durch
Evaporation reduziert werden kann, wurden Siliconsphären mit Statischer Emulsion einer
Charge unterschiedlich lange an Luft getrocknet. Wie Abbildung 3.48 zu entnehmen ist,
konnte bzgl. der Bildung von Benzoinbutyrat eine mit zunehmender Trocknungsdauer
steigende Reaktionsrate verzeichnet werden, wobei das Maximum nach einer Trocknungs-
dauer von 20 h erreicht wurde.
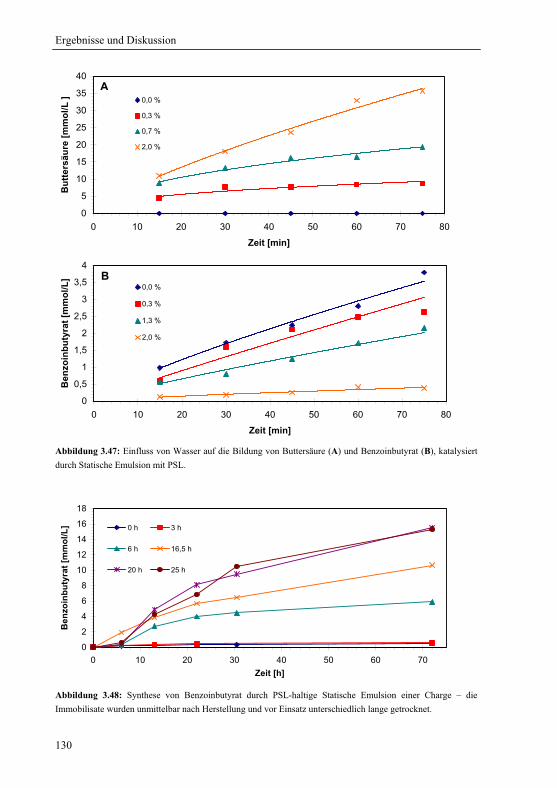
Ergebnisse und Diskussion
130
Abbildung 3.47: Einfluss von Wasser auf die Bildung von Buttersäure (A) und Benzoinbutyrat (B), katalysiert durch Statische Emulsion mit PSL.
Abbildung 3.48: Synthese von Benzoinbutyrat durch PSL-haltige Statische Emulsion einer Charge – die Immobilisate wurden unmittelbar nach Herstellung und vor Einsatz unterschiedlich lange getrocknet.
0
0,5
1
1,5
2
2,5
3
3,5
4
0 10 20 30 40 50 60 70 80
Zeit [min]
Ben
zoin
bu
tyra
t [m
mo
l/L]
0,0 %
0,3 %
1,3 %
2,0 %
0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60 70 80
Zeit [min]
Bu
tter
säu
re [
mm
ol/L
] 0,0 %
0,3 %
0,7 %
2,0 %
0
2
4
6
8
10
12
14
16
18
0 10 20 30 40 50 60 70Zeit [h]
Ben
zoin
bu
tyra
t [m
mo
l/L] 0 h 3 h
6 h 16,5 h
20 h 25 h
A
B

Anwendung der Statischen Emulsion
131
Aufgrund der möglichen Nebenreaktion, der Hydrolyse von Vinylbutyrat, ist die Gegenwart
von Wasser problematisch. Grund hierfür ist die stärkere Nukleophilie von Wasser im Ver-
gleich zu Benzoin, die für die Auflösung des Enzym-Acyl-Intermediats maßgeblich ist. Wie
in Kapitel 3.3.3 und 3.4.1.3 postuliert wurde, ist vermutlich gerade die Gegenwart einer
wässrigen Phase für die bei Veresterung erzielten Aktivitätssteigerungen verantwortlich. Was
jedoch bei Veresterung von Vorteil ist, erweist sich bei Umesterung vermutlich eher als Nach-
teil. Da die Siliconsphären nur etwa 20 h bei RT an Luft getrocknet wurden, ist aufgrund
vorheriger Untersuchungen nicht davon auszugehen, dass das im Silicon-Elastomer emul-
gierte Wasser vollständig entfernt wurde. Erschwerend kommt hinzu, dass das verwendete
Lösungsmittel THF wassermischbar ist und es sich hier nicht um ein biphasisches Reaktions-
system handelt. Es gibt demnach keine definierte Grenzfläche, in der die Reaktion stattfinden
kann, sondern nur enzymhaltige Kavitäten innerhalb der Siliconsphären. Die Reaktion kann
dann vermutlich im gesamten Volumen dieser Kompartimente stattfinden. Dennoch belegen
die hier erzielten Ergebnisse, dass die Umesterung von Vinylbutyrat mit Benzoin auch mit der
in Statischer Emulsion immobilisierten Lipase von Pseudomonas stutzeri katalysiert werden
kann und zwar ohne dass es zur Freisetzung von Buttersäure kommt. Möglicherweise kann
ein gewisser Anteil an Wasser ohne nachteiligen Einfluss auf die Reaktionskinetik toleriert
werden. Ebenfalls denkbar wäre, dass im Fall vollständiger Trocknung der Siliconsphären und
infolgedessen wasserfreien Reaktionsansätzen die Immobilisierung gegenüber dem Einsatz
des nativen Enzyms Vorteile verspricht. Das trockene und daher innerhalb der Kavitäten
präzipitierte Enzym würde nach Einbringung der Siliconsphären in THF suspendiert. Es ist
davon auszugehen, dass die Suspension des Enzyms dabei günstiger ausfällt als bei bloßer
Suspension des pulverförmigen und bisweilen stark agglomerierten Enzyms im Lösungs-
mittel. Somit wären die moderaten Aktivitätssteigerungen zu erklären. Letztlich konnte dieser
Aspekt nicht abschließend geklärt werden.
Für eine Beurteilung der Eignung der Statischen Emulsion muss auch die Verteilung der
Substrate zwischen Silicon-Elastomer und umgebendem Lösungsmittel Berücksichtigung
finden. Ausgehend von den in Kapitel 3.4.4 vorgestellten Ergebnissen sollte auch hier keine
Konzentrationsänderung nach Einbringen der Silicon-Elastomere in das hydrophobe Lösungs-
mittel THF beobachtet werden. Wie die Ergebnisse in Tabelle 3.8 zeigen, ist im Verteilungs-
gleichgewicht im Vergleich zu den Initialkonzentrationen stets eine höhere Konzentration an
R/S-Benzoin, Vinylbutyrat und R/S-Benzoinbutyrat zu verzeichnen. Dies erscheint auf den
ersten Blick nur schwer zu erklären. Es besteht die Möglichkeit, dass während der 24 h Inku-

Ergebnisse und Diskussion
132
bation Lösungsmittel evaporiert. Dies wurde aber selbst nach mehrtägiger Inkubation der
übrigen Reaktionsansätze nicht beobachtet, weshalb eine andere Erklärung gefunden werden
muss. Unter der Annahme, dass das Lösungsmittel THF in die Sphären eindringt und dort zu
einer Relaxation des polymeren Netzwerkes führt, kann die beobachtete Konzentrations-
zunahme nur dadurch erklärt werden, dass für die Substrate und Produkte nicht im gleichen
Maße die Möglichkeit besteht, in das polymere Netzwerk zu diffundieren wie für THF. Bei
genauer Betrachtung fällt auf, dass für Vinylbutyrat die beobachtete Konzentrationszunahme
(< 10 %) geringer ausfällt als für R/S-Benzoin (+ 30-50 %) und R/S-Benzoinbutyrat (+ 40-50
%). Das deutet daraufhin, dass Vinylbutyrat besser in das Silicon-Elastomer diffundieren kann
und erscheint vor dem Hintergrund der geringen Löslichkeit von Benzoin und Benzoinbutyrat
in den meisten organischen Lösungsmitteln durchaus plausibel. Die Höhe der Konzentrations-
zunahme im Fall von Benzoin und Benzoinbutyrat erscheint unter Berücksichtigung der durch
die Quellung des Elastomers hervorgerufenen Volumenänderung ebenfalls plausibel: Es
wurden 500 mg enzymfreier Statischer Emulsion in 2 mL substrat- bzw. produkthaltigem
THF inkubiert – unter der Annahme, dass es zu einer Verdoppelung des Volumens der
Siliconsphären kommt (quantitative Daten wurden nicht erhoben) verringert sich bei alleini-
ger Diffusion von THF in die Siliconsphären der Lösungsmittelgehalt bezüglich Benzoin und
Benzoinbutyrat um ca. 30 %. Dies würde bedeuten, dass nahezu die gesamte im Ansatz
befindliche Menge an R/S-Benzoin und R/S-Benzoinbutyrat außerhalb des Silicon-Elastomers
verbleibt. Genauere Aussagen sind gegenwärtig nicht möglich, u.a. auch deshalb, weil die
GC-Analytik gegenwärtig noch zu ungenau ist, wie die hohen Standardabweichungen für R/S-
Benzoin und R/S-Benzoinbutyrat erkennen lassen (Tab. 3.8). Es empfiehlt sich aber, diesen
Aspekt in zukünftigen Untersuchungen aufzugreifen, da zumindest theoretisch die
Möglichkeit besteht, den offensichtlich doch erheblich limitierten Massentransfer durch
Modifikation der Matrix zu verbessern.
Tabelle 3.8: Gaschromatographisch bestimmte Konzentrationen vor und nach Einstellung des Verteilungsgleich-gewichtes zwischen THF und dem Silicon-Elastomer.
Gemessene Konzentration [mmol/L]
R/S-Benzoin Vinylbutyrat R/S-Benzoinbutyrat
Vor nach Vor Nach vor nach
Gleichgewichtseinstellung
1 100,5 +/- 10,1 147,5 +/- 3,1 114,0 +/- 0,4 121,9 +/- 1,3 69,0 +/- 6,7 95,7 +/- 9,7
2 74,4 +/- 10,6 103,9 +/- 7,3 84,1 +/- 0,2 90,8 +/- 1,1 49,0 +/- 1,9 73,9 +/- 2,4
3 49,7 +/- 2,1 65,1 +/- 5,1 54,9 +/- 0,1 58,2 +/- 1,1 - -

Anwendung der Statischen Emulsion
133
Auch wenn die spezifischen Aktivitäten nach Immobilisierung in Statischer Emulsion in der
gleichen Größenordnung liegen wie für das nicht immobilisierte Enzym, stehen die gegen-
wärtig geringen Beladungsdichten und die folgerichtig geringen apparenten Aktivitäten einer
industriellen Anwendung entgegen. Im Mittelpunkt zukünftiger Bemühungen muss daher die
Steigerung der Aktivität stehen. Zudem müsste auch die mechanische Stabilität der Silicon-
sphären für den Einsatz in THF optimiert werden. Zwar konnte in THF eine Quellung der
Siliconsphären beobachtet werden, die in der gleichen Größenordnung lag wie in Hexan, aber
nach mehrtägiger Inkubation war z.T. eine Desintegration der Siliconsphären zu beobachten.
Es zeigte sich, dass die Siliconsphären in THF ihre Elastizität verloren hatten und mit
moderater Kraftanstrengung zerdrückt werden konnten. Aus derzeit unbekannten Gründen
eignet sich das hier verwendete Siliconelastomer offensichtlich nicht für den Einsatz in THF –
inwieweit dies auch andere Silicone betrifft, kann nicht gesagt werden.
Gleichwohl die hier erzielten Ergebnisse deutlich machen, dass ein baldiger technischer Ein-
satz dieses Systems für die PSL-katalysierte Racematspaltung von R/S-Benzoin nicht in Frage
kommt, kann davon ausgegangen werden, dass es sich auch hier um ein vielversprechendes
Verfahren mit noch erheblichem Optimierungspotential handelt. Ferner belegen die hier
durchgeführten Untersuchungen, dass prinzipiell neben der Veresterung auch Umesterung
und Hydrolyse durch die Statische Emulsion katalysiert werden können.

Zusammenfassung
134
4. Zusammenfassung
Zentrale Aspekte der lipasenkatalysierten Veresterung in biphasischen Reaktionssystemen
sind bis heute nicht abschließend geklärt. Dies gilt insbesondere für das pH-Optimum und die
Frage, ob und inwieweit in biphasischen Systemen die Estersynthese auch in der wässrigen
Phase erfolgen kann. Bei Einsatz lipasehaltiger Hydrogele in organischen Reaktionsmedien ist
die Klärung dieser Sachverhalte für eine rationale Prozessoptimierung von essentieller Bedeu-
tung.
Für die Lipasen von Candida rugosa und Thermomyces lanuginosa konnte erstmalig gezeigt
werden, dass das pH-Optimum der lipasenkatalysierten Veresterung in biphasischen Reak-
tionssystemen mit pH 3,5 bzw. pH 4,25 deutlich unter dem der Hydrolyse liegt. Ein Anstieg
der Konzentration an protonierter Säure ging mit einer Zunahme der katalytischen Aktivität
einher, erklärbar durch den Reaktionsmechanismus: Anders als die protonierte kann die de-
protonierte Säure mit ihrer delokalisierten negativen Ladung im aktiven Zentrum nicht
nukleophil attackiert werden. Die Allgemeingültigkeit und Tragweite dieser Schlussfolgerung
konnte durch eine Reihenuntersuchung weiterer Lipasen bestätigt werden. Der in bipha-
sischen Reaktionssystemen durch die Anreicherung der Säure in der wässrigen Phase hervor-
gerufene pH-Abfall ist demnach weitaus weniger problematisch als gemeinhin vermutet.
Aufwändige Maßnahmen zur Stabilisierung des pH-Wertes sind verzichtbar.
Des Weiteren konnte nachgewiesen werden, dass eine erhebliche Abhängigkeit der kataly-
tischen Aktivität von der spezifischen Grenzfläche besteht. Ein Unterschied zwischen der
grenzflächenaktivierten Lipase von Candida rugosa und der nicht-grenzflächenaktivierten
Lipase B von Candida antarctica konnte nicht festgestellt werden. Eine eingehende Analyse
der Ergebnisse unter Berücksichtigung der besonderen thermodynamischen Gegebenheiten
führte zur Entwicklung einer detaillierten Hypothese über die Vorgänge an der Grenzfläche
bei der lipasenkatalysierten Veresterung. Die Estersynthese findet nahezu ausschließlich an
der Grenzfläche statt, weshalb im Interesse einer hohen Produktivität eine Maximierung der
spezifischen Grenzfläche anzustreben ist. Auf Basis dieser Hypothese wurde die Sinnhaftig-
keit der Verwendung lipasehaltiger Hydrogele in Frage gestellt und ein alternatives Konzept
für die Immobilisierung von Lipasen in biphasischen Reaktionssystemen entwickelt. Die
Innovation bestand darin, dass eine wässrig gelöste Lipase erstmalig innerhalb eines hydro-
phoben Silicon-Elastomers statisch emulgiert wurde. Erhalten wurden sphärische Immobili-

135
sate von gummielastischer Konsistenz. Die katalytischen und mechanischen Eigenschaften
der Statischen Emulsion wurden eingehend charakterisiert. Dabei konnte für mehrere Lipasen
gezeigt werden, dass durch Immobilisierung in der Statischen Emulsion eine Aktivitäts-
steigerung erreicht wurde, wie sie bisher nur bei Immobilisierung in alkylsubstituierten Sol-
Gelen zugänglich war. Gleichzeitig wurde deutlich, dass die Statische Emulsion im Hinblick
auf ihre mechanischen Eigenschaften Hydrogelen und Sol-Gelen deutlich überlegen ist. Die
praktische Anwendbarkeit der lipasehaltigen Statischen Emulsion wurde anhand der Synthese
von Fettsäureethylhexylestern und Propyllaurat in lösungsmittelfreien Reaktionssystemen
sowie anhand der Racematspaltung von Benzoin in Tetrahydrofuran demonstriert. Gerade der
Vergleich mit alkylsubstituierten Sol-Gelen – dem bislang aussichtsreichsten Verfahren zur
Immobilisierung von Lipasen – veranschaulicht, dass es sich bei der neuartigen Statischen
Emulsion um eine Immobilisierungstechnik mit außerordentlichem Potential handelt.

Literatur
136
5. Literatur
ADLERCREUTZ, P. (1994): Enzyme-catalysed lipid modification. Biotechnology and Genetic Engineering Reviews 12: 231-254
AFFLECK R., XU Z.F., SUZAWA V., FOCHT K., CLARK, D.S., DORDICK J.S. (1992): Enzymic catalysis and dynamics in low-water environments. Proceedings of the National Academy of Sciences of the United States of America 89: 1100-1104
ANSORGE-SCHUMACHER M.B., PLEß G., METRANGOLO D., HARTMEIER W. (2002): Beads from natural hydrogels as encapsulation matrices for cofactor-dependent enzymes in organic solvents. Landbauforschung Völkenrode SH 241: 99-102
AOYAGI Y., AGATA N., SHIBATA N., HORIGUCHI M., WILIAMS R.M. (2000): Lipase TL-mediated kinetic resolution of benzoin: facile synthesis of (1R,2S)-erythro-2-amino-1,2-diphenylethanol. TetrahedronLetter 41: 8010-8014
ATIA K.S., EL-BATAL A. (2005): Preparation of glucose oxidase immobilized in different carriers using radiation polymerization. Journal of Chemical Technology and Biotechnology 80: 805-811
BALLESTEROS A., BORNSCHEUER U., CAPEWELL A., COMBES D., CONDORET J.S., KOENIG K., KOLISIS
F.N., MARTY A., MENGUE U., SCHEPER T., XENAKIS A. (1995): Enzymatic reaction in non conventional media. Biocatalysis and Biotransformation 13: 1-42
BAUER M., GEYER R., GRIENGL H., STEINER, W. (2002): The use of Lewis cell to investigate kinetics of an (S)-hydroxynitrile lyase in two-phase systems. Food Technology and Biotechnology 40: 9-19
BEER H.D., WOHLFAHRT G., MCCARTHY J.E., SCHOMBURG D., SCHMID R.D. (1996): Analysis of the catalytic mechanism of a fungal lipase using computer-aided design and structural mutants. ProteinEngineering 9: 507-517
BETIGERI S., NEAU S.H. (2002): Immobilization of lipase using hydrophilic polymers in the form of hydrogel beads. Biomaterials 23: 3627-3636
BISSWANGER, H. (2000): Enzymkinetik. Wiley-VCH, Weinheim 3. Ausgabe ISBN 3-527-30096-1
BLACKWOOD A.D., CURRAN L.J., MOORE B.D., HALLING P.J. (1994): Organic phase buffers’ control biocatalyst activity independent of initial aqueous pH. Biochimica et Biophysica Acta 1206: 161-165
BLASER H.U., SPINDLER F., STUDER M. (2001): Enantioselective catalysis in fine chemicals production. Applied Catalysis A: General 221: 119-143
BOLLER T., MEIER C., MENZLER S. (2002): Eupergit oxirane acrylic beads: How to make enzymes fit for biocatalysis. Organic Process Research and Development 6: 509–519
BOMMARIUS A.S., RIEBEL B. (2004): Biocatalysis: fundamentals and applications. Wiley-VCH, Weinheim ISBN: 3-527-30344-8
BORNSCHEUER U.T., KAZLAUSKAS R.J. (1999): Hydrolases in Organic Synthesis. Wiley-VCH, WeinheimISBN 3-527-31029-0

137
BORZEIX F., MONOT F., VANDECASTEELE J.P. (1992): Strategies for enzymatic esterification in organic solvents: comparison of microaqueous, biphasic, and micellar systems. Enzyme and Microbial Technology 14: 791-797
BOURG-GARROS S., RAZAFINDRAMBOA N., PAVIA A.A. (1998): Optimization of lipase-catalyzed synthesis of (z)-3-hexen-1-yl acetate by direct esterification in hexane and a solvent-free medium. Enzyme and Microbial Technology 22: 240-245
BRADFORD M.M. (1976): A rapid sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye-binding. Analytical Biochemistry 72: 248-254
BRADY C., METCALFE L., SLABOSZEWSKI D., FRANK D. (1988): Lipase immobilized on a hydrophobic, microporous support for the hydrolysis of fats. Journal of the American Oil Chemists’ Society 65: 917-921
BRADY L.E.A. (1990): A serine protease triad forms the catalytic centre of a triacylglycerol lipase. Nature 343: 767-770
BRAIUCA P., EBERT C., BASSO A., LINDA P., GARDOSSI L. (2006): Computational methods to rationalize experimental strategies in biocatalysis. Eingereicht bei Trends in Biotechnology
BRENNEIS R., KÖCHER P., KLEY G., BAECK B., SCHWILLING T. (2002): Untersuchungen zur Herstellung von biologisch abbaubaren Verlustschmierstoffen aus Altfetten. Müll und Abfall 5: 244-249
BRENNEIS R., BAECK B., KLEY G. (2004): Alcoholysis of waste fats with 2-ethyl-1-hexanol using Candida antarctica lipase A in large-scale tests. European Journal of Lipid Science and Technology106: 809-814
BREUER M., DITRICH K., HABICHER T., HAUER E., KEßELER M., STÜRMER R., ZELINSKI T. (2004): Industrial methods for the production of optically active intermediates. Angewandte Chemie International Edition 43: 788-824
BROOS J., VISSER A.J.W.G., ENGBERSEN J.F.J., VERBOOM W., VAN HOEK A., REINHOUDT D.N. (1995): Flexibility of enzymes suspended in organic solvents probed by time-resolved fluorescence anisotropy. Evidence that enzyme activity and enantioselectivity are directly related to enzyme flexibility. Journal of the American Chemical Society 117: 12657-12663
BRZOZOWSKI A.M., DEREWENDA U., DERWENDA Z.S., DODSON G.G., LAWSON D.M., TURKENBURG
J.P., BJORKLING F., JENSEN B.H., PATKAR, S.A., THIM L. (1991): A model for interfacial activation in lipases from the structure of a fungal lipase-inhibitor complex. Nature 351: 491-494
BUTHE A., HARTMEIER W., ANSORGE-SCHUMACHER M.B. (2004): Novel solvent-based method for preparation of alginate beads with improved roundness and predictable size. Journal of Microencapsulation 21: 865-876
BYSTROM C.E., HARTEL R.W. (1994): Evaluation of milk fat fractionation and modification techniques for creating cocoa butter replacers. Lebensmittel-Wissenschaft und -Technologie 27: 142-150
CAMBILLAU C., VAN TILBEURGH H. (1993): Structure of hydrolases – lipases and cellulases. Current Opinion in Structural Biology 3: 885-895
CAMERON P.A., DAVISON B.H., FRYMIER P.D., BARTON J.W. (2002): Direct transesterification of gases by “dry” immobilized lipase. Biotechnology and Bioengineering 78: 251-256

Literatur
138
CAO, S. G., LIU, Z. B., FENG, Y., MA, L., DING, Z. T., CHENG, Y. H. (1992): Esterification and transesterification with immobilized lipase in organic solvent. Applied Biochemistry and Biotechnology 32:1-6
CARVALHO C.M.L., CABRAL J.M.S. (2000): Reverse micelles as reaction media for lipases. Biochimie82: 1063-1085
CHATTERJEE T., BHATTACHARAYYA D.K. (1998): Synthesis of terpene esters by an immobilized lipase in a solvent-free system. Biotechnology Letters 20: 865-868
CHEN C.S., FUJIMOTO Y., GIRDAUKAS G., SIH C.J. (1982): Quantitative analysis of biochemical kinetic resolutions of enantiomers. Journal of the American Chemical Society 104: 7294-7299
CHEN J.P., MCGILL S.D. (1992): Enzymic hydrolysis of triglycerides by Rhizopus delemar immobilized on biomass support particles. Food Biotechnology 6: 1-18
CHEN J.P., YANG B. (1992): Enhancement of release of short-chain fatty acids from milk fat with immobilized microbial lipase. Journal of Food Science 57: 781-782
COMPTON R.A. (1997): Silicone manufacturing for long-term implants. Journal of Long Term Efficiency of Medical Implants 7: 29-54
CRAIG D., MARIN M.L. (1998): An asymmetric synthetic approach to the A-ring of the Taxol family of anti-cancer-compounds. Molecules 3: 64-70
CROOKS G.E., REES G.D., ROBINSON B.H., SVENSSON M.; STEPHENSON G.R. (1995): Comparison of hydrolysis and esterification behaviour of Humicola lanuginosa and Rhizomucor miehei lipases in AOT-stabilized water-in-oil microemulsions: I. Effect of pH and water content on reaction kinetics. Biotechnology and Bioengineering 48: 78-88
DAUBER S.R., BOEHNKE B. (1993): Verfahren zum Betrieb einer Kläranlage nachgeschalteten Anlage für die Weiterverarbeitung von Klärschlamm. Patent DE 4141832C1
DE BACKER L., BARON G. (1993): Effective diffusivity and tortuosity in a porous glass immobilization matrix. Applied Microbiology and Biotechnology 39: 281-284
DELIMITSOU C., ZOUMPANIOTI M., XENAKIS A., STAMATIS H. (2002): Activity and stability studies of Mucor miehei lipase immobilized in novel micromemulsion-based organogels. Biocatalysis and Biotransformation 20: 319-327
DEMIR A.S., HAMAMCI H., SESENOGLU O., AYDOGAN F., CAPANOGLU D., NESLIHANOGLU R. (2001):Simple chemoenzymatic access to enantiopure pharmacologically interesting (R)-2-hydroxypropio-phenones. Tetrahedron: Asymmetry 12: 1953-1956
DEREWENDA U., BRZOZOWSKI A.M., LAWSON D.M., DEREWENDA Z.S. (1992): Catalysis at the interface: The anatomy of a conformational change in a triglyceride lipase. Biochemistry 31: 1532-1541
DIAS S. F., VILAS-BOAS L., CABRAL J. M.S., FONSECA M.M.R. (1991): Production of ethyl butyrate by Candida rugosa lipase immobilized in polyurethane. Biocatalysis 5: 21-34
DICKINSON E., MURRAY B.S., STAINSBY G. (1988): Protein adsorption at air-water and oil-water interfaces. In: Advances in Food Emulsions and Foams, Herausgeber: Dickinson, E.; Stainsby, G., Elsevier Applied Science, London 123-162

139
DIXON M. (1972): The graphical determination of Km and Ki. Biochemical Journal 129: 197-202
DODSON G.G., LAWSON D.M., WINKLER F.K. (1992): Structural and evolutionary relationships in lipase mechanism and activation. Faraday Discussions 93: 95-105
DOMÍNGUEZ DE MARÍA P., XENAKIS A., STAMATIS H., SINISTERRA J.V. (2004) : Unexpected reaction profile observed in the synthesis of propyl laurate when using Candida rugosa lipases immobilized in microemulsion based organogels. Biotechnology Letters 26: 1517-1520
DOMÍNGUEZ DE MARÍA P., CARBONI-OERLEMANS C., TUIN B., BARGEMAN G., VAN DER MEER A.B.,VAN GEMERT R.W. (2005): Biotechnological applications of Candida antarctica lipase A: State-of-the-art. Journal of Molecular Catalysis B: Enzymatic 37: 36-46
DOMÍNGUEZ DE MARÍA P., SÁNCHEZ-MONTERO J.M., SINISTERRA J.V., ALCÁNTARA A.R. (2006): Understanding Candida rugosa lipases: An overview. Biotechnology Advances 24: 180-196
DOSANJH N.S., KAUR J. (2002): Immobilization, stability and esterification studies of a lipase from Bacillus sp. Biotechnology and Applied Biochemistry 36: 7-12
DOS SANTOS V.A.P.M., LEENEN E.J.T.M., RIPPOLL M.M., VAN DER SLUIS C., VAN VLIET T., TRAMPER
J. (1997): Relevance of rheological properties of gel beads for their mechanical stability in bioreactors. Biotechnology and Bioengineering 56: 517-529
DOUMÈCHE, B. (2002): Fundamental phenomenological description and experimental optimisation of gel-stabilized biocatalyst in a two-phase system. Dissertation an der Math.-Nat. Fakultät, RWTH Aachen
DRÖGE M.J., BOS R., QUAX W.J. (2001): Paralogous gene analysis reveals a highly enantioselective 1,2-O-isopropylidenglycerol caprylate esterase of Bacillus subtilis. European Journal of Biochemistry268: 3332-3338
D’SOUZA S.F. (1999): Immobilized enzymes in bioprocess. Current Science 77 (1): 69-80
ENUJIUGHA V.N., THANI F.A., SANNI T.M., ABIGOR R.D. (2004): Lipase activity in dormant seeds of the African oil bean (Pentaclethra macrophylla Benth). Food Chemistry 88: 405-410
FABER K., FRANSSEN M.C.R. (1993): Prospects for the increased application of biocatalysts in organic transformations. Trends in Biotechnology 11: 461-470
FANG Q.K., HAN Z., GROVER P., KESSLER D., SENANAYADE C.H., WALD S.A. (2002): Rapid access to enantiopure bupropion and its major metabolite by stereospecific nucleophilic substitution on an -keto triflate. Tetrahedron Asymmetry 11: 3659-3663
FARRELL R.L., HATA K., WALL M.B. (1997): Solving pitch problems in pulp and paper processes by the use of enzymes of fungi. Advances in Biochemical Engineering/Biotechnology 57: 197-212
FERREIRA-DIAS A., CORREIRA A.C., BAPTISTA F.O. (1999): Activity and batch operational stability of Candida rugosa lipase immobilized in different hydrophilic polyurethane foams during hydrolysis in a biphasic medium. Bioprocess Engineering 21: 517-524
FITZPATRICK P.A., KLIBANOV A.M. (1991): How can the solvent affect enzyme enantioselectivity? Journal of the American Chemical Society 113: 3166-3171
FOJAN P., JONSON P.H., NEVES-PETERSEN M.T., PETERSEN S.B. (2000): What distinguishes an esterase from a lipase: A novel structural approach. Biochimie 82: 1033-1041

Literatur
140
FURUKAWA S., ONO T., IJIMA H., KAWAKAMI K. (2001): Enhancement of activity of sol-gel immobilized lipase in organic media by pretreatment with substrate analogues. Journal of Molecular Catalysis B: Enzymatic 15: 65-70
GARCÍA S., LOURENGO N.M.T., LOUSA D., SEQUEIRA A.F., MIMOSO P., CABRAL J.M.S., AFONSO
C.A.M., BARREIROS S. (2004): A comparative study of biocatalysis in non-conventional solvents: Ionic liquids, supercritical fluids and organic media. Green Chemistry 6: 366-470
GARGOURI M., LEGOY M.D. (1997): The kinetic behaviour of a two-enzyme system in biphasic media: coupling hydrolysis and lipoxygenation. Biochimica et Biophysica Acta 1337: 227-232
GHANEM A., SCHURIG V. (2003): Entrapment of Pseudomonas cepacia lipase with peracetylated -cyclodextrin in sol-gel: application to the kinetic resolution of secondary alcohols. Tetrahedron: Asymmetry 14: 2547-2555
GHATORAE A.S., BELL G., HALLING P.J. (1994): Inactivation of enzymes by organic solvents: New techniques with well-defined interfacial area. Biotechnology and Bioengineering 43: 331-336
GHOSH D., WAWRZAK Z., PLETNEV V.Z., LI N., KAISER R., PANGBORN W., JORNVALL H., ERMAN M.,DUAX W.L. (1995): Structure of uncomplexed and linoleate-bound Candida cylindracea cholesterol esterase. Structure 3: 279-288
GILL I., PASTOR E., BALLESTEROS A. (1999): Lipase-silicone biocomposites: Efficient and versatile immobilized biocatalysts. Journal of the American Chemical Society 131: 9487-9496
GILL I., BALLESTEROS A. (2000a): Bioencapsulation within synthetic polymers (Part I): sol-gel encapsulated biologicals. Trends in Biotechnology 18: 282-296
GILL I., BALLESTEROS A. (2000b): Bioencapsulation within synthetic polymers (Part II): non-sol-gel protein-polymer biocomposites. Trends in Biotechnology 18: 469-479
GONZÁLEZ-NAVARRO H., BRACO L. (1997): Improving lipase activity in solvent-free media by interfacial activation-based molecular bioimprinting. Journal of Molecular Catalysis B: Enzymatic3: 111-119
GONZÁLEZ-NAVARRO H., BRACO L. (1998): Lipase-enhanced activity in flavour ester reactions by trapping enzyme conformers in the presence of interfaces. Biotechnology and Bioengineering 59: 122-127
GOPINATH S.C.B., HILDA A., LAKSHMI PRIYA T., ANNADURAI G., ANBU P. (2003): Purification of lipase from Geotrichum candidum: conditions optimized for enzyme production using Box-Behnken design. World Journal of Microbiology and Biotechnology 19: 681-689
GOTOR V. (1999): Non-conventional hydrolase chemistry: amide and carbamate bond formation catalyzed by lipases. Bioorganic & Medicinal Chemistry 7: 2189-2197
GRAY C.J., NARANG J.S., BARKER S.A. (1990): Immobilization of lipase from Candida cylindraceae and its use in the synthesis of menthol esters by transesterification. Enzyme and Microbial Technology 12: 800-807
GROCHULSKI P., LI Y., SCHRAG J.D., BOUTHILLIER F., SMITH P., HARRISON D., RUBIN, B., CYGLER M.(1993): Insights into interfacial activation from an open structure of Candida rugosa lipase. Journal of Biological Chemistry 268: 12843-12847

141
GUNSTONE F.D. (1999): Enzymes as biocatalysts in the modification of natural lipids. Journal of the Science of Food and Agriculture 79: 1535-1549
HAAS M.J., SCOTT K.M. (1996): Diesel fuel as a solvent for the lipase-catalyzed alcoholysis of triglycerides and phosphatidylcholine. Journal of the American Oil Chemists’ Society 73: 497-504
HAEFFNER F., NORIN T., HULT K. (1998): Molecular modeling of the enantioselectivity in lipase-catalyzed transesterification reactions. Biophysical Journal 74: 1251-1262
HALLING P.J. (1994): Thermodynamic predictions for biocatalysis in nonconventional media: Theory, tests and recommendations for experimental design and analysis. Enzyme and Microbial Technology16: 178-206
HALLING P.J., ROSS A.C., BELL G. (1998): Inactivation of enzymes at the aqueous-organic interface. In: Stability and Stabilization of Biocatalysts, Herausgeber: Ballesteros A., Plou F.J., Iborra J.L., Halling P.J., Elsevier Science B.V., 365-372
HARTMEIER W. (1986): Immobilisierte Biokatalysatoren – Eine Einführung. Springer Verlag, Berlin Heidelberg New York ISBN 3-540-16335-2
HEIM N., BOCKEY D., VON SCHENCK W. (2005): Ölsaaten und Biokraftstoffe. UFOP (Union zur Förderung von Öl und Proteinpflanzen)-Marktinformation 7/05
HEINEMANN M., WAGNER T., DOUMÈCHE B., ANSORGE-SCHUMACHER M., BÜCHS J. (2002): A new approach for the spatially resolved qualitative analysis of the protein distribution in hydrogel beads based on confocal laser scanning microscopy. Biotechnology Letters 24(10): 845-850
HEINEMANN M. (2003): Experimental analysis, modeling and dynamic simulation of thermodynamic and kinetic phenomena in gel-stabilized enzyme carriers. Fortschritt-Berichte. VDI Reihe 3 Nr. 795. VDI-Verlag, Düsseldorf ISBN 3-18-379503-5
HEINEMANN M., KÜMMEL A., GIESEN R., ANSORGE-SCHUMACHER M.B., BÜCHS J. (2003): Experimental and theoretical analysis of phase equilibria in a two-phase system used for biocatalyzed esterifications. Biocatalysis and Biotransformation 21: 115-121
HEINEMANN M., MEINBERG H., BÜCHS J., KOSS H.J., ANSORGE-SCHUMACHER M.B. (2005): Method for quantitative determination of spatial polymer distribution in alginate beads using Raman spectroscopy. Applied Spectroscopy 59: 280-285
HERTZBERG S., KVITTINGEN L., ANTHONSEN T., SKJÅK-BRAEK G. (1992): Alginate as immobilization matrix and stabilizing agent in a two-phase liquid system: application in lipase-catalyzed reactions. Enzyme and Microbial Technology 14: 42-47
HESSELBACH J., HERRMANN C., BOCK R., DETTMER T., KLEY G., KÖCHER P., BRENNEIS R., MEXER-PITTROFF R., FALK O., HANSEN A. (2003): Kühlschmierstoffe aus technischen tierischen Fetten und Altspeisefetten – Herstellung, Technologie und Ökobilanzierung. Transkript Sonderband Biokatalyse 24-27
HILGERS C. (2001): Entwicklung neuer Wirkstofffreisetzungssysteme zur Modifizierung etablierter Medizinprodukte. Dissertation an der Math.-Nat. Fakultät, RWTH Aachen
HILLS G. (2003): Industrial use of lipases to produce fatty acid esters. European Journal of Lipid Science Technology 105: 601-607

Literatur
142
HISCHER T., GOCKE D., FERNANDEZ M., HOYOS P., ALCÁNTARA A.R., SINISTERRA J.V., HARTMEIER
W., ANSORGE-SCHUMACHER M.B. (2005): Stereoselective synthesis of novel benzoins catalysed by benzaldehyde lyase in a gel-stabilised two-phase system. Tetrahedron 61: 7378-7383
HOYOS P., FERNÁNDEZ M., SINISTERRA J.V., ALCÁNTARA A.R. (2006): Dynamic kinetic resolution of benzoines by lipase-metal combo catalysis. Eingereicht bei Tetrahedron Letters
HSU A.-F., JONES K., FOGLIA T.A., MARMER W.N. (2002): Immobilized lipase-catalysed production of alkyl esters of restaurant grease as biodiesel. Biotechnology and Applied Biochemistry 36: 181-186
ISON A.P., DUNNILL P., LILLY M.D. (1988): Effect of solvent concentration on enzyme catalyzed interesterification of fats. Enzyme and Microbial Technology 10: 47-51
IVANOV A.E., SCHNEIDER M.P. (1997): Methods for the immobilization of lipases and their use for ester synthesis. Journal of Molecular Catalysis B: Enzymatic 3: 303-309
IZUMI T., TAMURA F., AKUTSU M., KATOU R., MURAKAMI S. (1997): Enzymic transesterification of 3,7-dimethyl-4,7-octadien-1-ol using lipases. Journal of Chemical Technology and Biotechnology 68: 57-64
JAEGER K.-E., REETZ M.T. (1998): Microbial lipases form versatile tools for biotechnology. Trends in Biotechnology 16: 396-403
JAEGER K.-E., DIJKSTRA B.W., REETZ M.T. (1999): Bacterial biocatalysts: Molecular Biology, three dimensional structures and biotechnological applications of lipases. Annual Review of Microbiology53: 315-351
JAEGER K.-E., EGGERT T. (2002): Lipases for biotechnology. Current Opinion in Biotechnology 13: 390-397
JENSEN M., JENSEN T.R., KJAER K., BJØRNHOLM T., MOURITSEN O.G., PETERS G.H. (2002): Orientation and Conformation of a lipase at an interface studied by molecular dynamics simulations. BiophysicalJournal 83: 98-111
KAIEDA M., SAMUKAWA T., MATSUMOTO T., BAN K., KONDO A., SHIMADA Y., NODA H., NOMATO F.,OHTSUKA K., IZUMOTO E., FUKUDA H. (1999): Biodiesel fuel production from plant oil catalyzed by Rhizopus oryzae Lipase in a water-containing system without an organic solvent. Journal of Bioscience and Bioengineering 88: 627-631
KANG S.T., RHEE J.S. (1989): Characteristics of immobilized lipase-catalyzed hydrolysis of olive oil of high concentration in reverse phase system. Biotechnology and Bioengineering 33: 1469-1476
KAWAMOTO T., TANAKA A. (2003): Entrapment of biocatalysts by prepolymer methods. Food Science and Technology 122: 331-341
KÉRY V., HAPLOVA J., TIHLARIK K., SCHMIDT S. (1990): Factors influencing the activity and thermostability of immobilized porcine pancreatic lipase. Journal of Chemical Technology and Biotechnology 48: 201-207
KHURANA A.L., HO C.T. (1990): Immobilization of porcine-pancreatic lipase on silica support to study lipolysis and reverse hydrolysis reaction. Journal of Liquid Chromatography 13: 715-725
KIERKELS J.G.T., VLEUGELS L.F.W., GELADE E.T.F., VERMEULEN D.P., KAMPHUIS J., WANDREY C.,VAN DEN TWEEL W.J.J. (1994): Pseudomonas fluorescens lipase adsorption and the kinetics of hydrolysis in a dynamic emulsion system. Enzyme and Microbial Technology 16: 513-521

143
KIM K.K., SONG H.K., SHIN D.H., HWANG K.Y., SUB S.W. (1997): The crystal structure of a triacyl-glycerol lipase from Pseudomonas cepacia reveals a highly open conformation in the absence of a bound inhibitor. Structure 5: 173-185
KIRK O., CHRISTENSEN M.W. (2002): Lipases from Candida antarctica: Unique Biocatalysts from a Unique Origin. Organic Process Research & Development 6: 446-451
KLIBANOV A.M. (1989): Enzymatic catalysis in anhydrous organic solvents. Trends in Biochemical Science 14: 141-144
KOCH M. (1997): Hydrogel-eingehüllte Enzyme und deren Einsatz in organischen Lösungsmitteln. Dissertation an der Math.-Nat. Fakultät, RWTH Aachen
KRAJEWSKA B. (2004): Application of chitin- and chitosan-based materials for enzyme immobilizations: a review. Enzyme and Microbial Technology 35: 126-139
KRAUT J. (1977): Serin proteases: Structure and mechanism of catalysis. Annual Review Biochemistry46: 331-358
KUEPPERS M., HEINE C., HAN S., STAPF S., BLUEMICH B. (2002): In situ observation of diffusion and reaction dynamics in gel microreactors by chemically resolved NMR microscopy. Applied Magnetic Resonance 22: 235-246
KUNCOVA G., SZILVA J., HETFLEJS J., SABATA S. (2003): Catalysis in organic solvents with lipase immobilized by sol-gel technique. Journal of Sol-Gel Science and Technology 26: 1183-1187
KVITTINGEN L., SJURSNES B.J., ANTHONSEN T., HALLING P. (1992): Use of salt hydrates to buffer optimal water level during lipase catalyzed synthesis in organic media: a practical procedure for organic chemists. Tetrahedron 48: 2793-2802
KVITTINGEN L. (1994): Some aspects of biocatalysis in organic solvents. Tetrahedron 50: 8253-8274
LAMARE S., LEGOY M. (1995): Working at controlled water activity in a continuous process: the gas/solid system as a solution. Biotechnology and Bioengineering 45: 387-397
LAMARE S., CAILLAUD B., ROULE K., GOUBET I., LEGOY M.D. (2001) : Production of natural esters at the pre-industrial scale by solid/gas biocatalysis. Biocatalysis and Biotransformation 19: 361-377
LI R.H., ALTREUTER D.H., GENTILE F.T. (1996): Transport characterization of hydrogel matrixes for cell encapsulation. Biotechnology and Bioengineering 50: 365-373
LINKO Y.Y., LÄMSÄ M., HUHTALA A., LINKO P. (1994): Lipase-Catalyzed Transesterification of Rapeseed Oil and 2-Ethyl-1-Hexanol. Journal of the American Oil Chemists’ Society 71(12): 1411-1414
LINKO Y.Y., LÄMSÄ M., HUHTALA A., RANTANEN O. (1995): Lipase Biocatalysis in the Production of Esters. JAOCS 72: 1293-1299
LINKO Y.Y., LÄMSÄ M., WU X., UOSUKAINEN E., SEPPÄLÄ J., LINKO P. (1998): Biodegradedable products by lipase biocatalysis. Journal of Biotechnology 66: 41-50
LONGO M.A., NOVELLA I.S., GARCÍA L.A., DÍAZ M. (1992): Diffusion of proteaes in calcium alginate beads. Enzyme and Microbial Technology 14: 586-590

Literatur
144
LORENZ P., LIEBETON K., NIEHAUS F., ECK J. (2002): Screening for novel enzymes for biocatalytic process: accessing the metagenome as a resource of novel functional sequence space. Current Opionion in Biotechnology 13: 572-577
LORTIE R., TRANI M., ERGAN F. (1993): Kinetic Study of the lipase-catalyzed synthesis of triolein. Biotechnology and Bioenineering 41: 1021-1026
LOUWRIER A., DRTINA J.G., KLIBANOV A.M. (1996): On the issue of interfacial activation of lipase in nonaqueous media. Biotechnology and Bioengineering 50: 1-5
MALCATA F.X., REYES H.R., GARCIA H.S., HILL C.G.J., AMUNDSON C.H. (1990): Immobilized lipase reactors for modification of fats and oils. Journal of the American Oil Chemists’ Society 67: 890-910
MANG T., DRESEL W. (2001): Lubricants and Lubrication. Wiley-VCH, Weinheim 1. Auflage ISBN3-527-29536-4
MANJÓN A., IBORRA J.L., AROCAS A. (1991): Short-chain flavor ester synthesis by immobilized lipase in organic media. Biotechnology Letters 13: 339-344
MANOHAR B., DIVAKAR S. (2002): Application of central composite rotatable design to lipase catalysed synthesis of m-cresyl acetate. World Journal of Microbiology and Biotechnology 18: 745-751
MARTINELLE M., HOLMQUIST M., HULT K. (1995): On the interfacial activation of Candida antarcticalipase A and B as compared with Humicola lanuginosa lipase. Biochimica et Biophysica Acta 1258: 272-276
MARTINS J.F., BORGES DE CARVALHO I., CORREA DE SAMPAIO T., BARREIROS S. (1994): Lipase-catalyzed enantioselective esterification of glycidol in supercritical carbon dioxide. Enzyme and Microbial Technology 16: 785-790
MATEUS D.M.R., ALVES S.S., DA FONSECA M.M.R. (1999): Diffusion in cell-free and cell immobilizing -carrageenan gel beads with and without chemical reaction. Biotechnology and Bioengineering 63:
625-631
MCCRAE A., ROEHL E.L., BRAND H.M. (1990): Bio-Ester. Seifen Öle Fette Wachse 116: 201-205
MC MORN P., HUTCHINGS G.J. (2003): Heterogeneous enantioselective catalysts: strategies for the immobilisation of homogeneous catalysts. Chemical Society Reviews 33: 108-122
METRANGOLO RUÌZ DE TEMINIO D., HARTMEIER W., ANSORGE-SCHUMACHER M.B. (2005): Entrapment of the alcohol dehydrogenase from Lactobacillus kefir in polyvinyl alcohol for the synthesis of chiral hydrophobic alcohols in organic solvents. Enzyme Microbial Technology 36: 3-9
MISSET O., GERRITSE G., JAEGER K.-E., WINKLER U., COLSON C., SCHANCK K., LESUISSE E., DARTOIS
V., BLAAUW M., RANSAC S. (1994): The structure-function relationship of the lipases from Pseudomonas aeruginosa and Bacillus subtilis. Protein Engineering 7: 523-529
MIYAKO E., MARUYAMA T., KAMIYA N., GOTO M. (2003): Transport of organic acids through a supported liquid membrane driven by lipase-catalyzed reactions. Journal of Bioscience and Bioengineering 96: 370-374
MOHY ELDIN M.S., HASSAN E.A., ELAASSAR M.R. (2005): -Galactosidase covalent immobilization on the surface of alginate beads and its application in lactose hydrolysis. Deutsche Lebensmittel-Rundschau 101: 255-259

145
MONTERO S., BLANCO A., VIRTO M.D., LANDETA C., AGUD I., SOLOZABAL R., LASCARAY J.M., DE
RENOBALES M., LLAMA M.J., SERRA J.L. (1993): Immobilization of Candida rugosa lipase and some properties of the immobilized enzyme. Enzyme and Microbial Technology 15: 239-247
MORENO J.M., HERNAIZ M.J., SÁNCHEZ-MORENO J.M., SINISTERRA J.V., BUSTOS M.T., SÁNCHEZ M.E.,BELLO J.F. (1997): Covalent immobilization of pure lipase A and B from Candida rugosa. Journal of Molecular Catalysis B: Enzymatic 2: 177-184
MUTHUKUMARAN N., DHAR S.C. (1982): Comparative studies on the degreasing of skins using acid lipase and solvent with preference to the quality of finished leathers. Leather Science 29: 417-424
NELSON L.A., FOGLIA T.A., MARMER W.N. (1996): Lipase-catalyzed production of biodiesel. JAOCS73: 1191-1195
NEVES-PETERSEN M.T., FOJAN P., PETERSEN S.B. (2001a): How do lipases and esterases work: the electrostatic contribution. Journal of Biotechnology 85: 115-147
NEVES-PETERSEN M.T., PETERSEN E.I., FOJAN P., NORONHA M., MADSEN R.G., PETERSEN S.B. (2001b): Engineering the pH-optimum of a triglyceride lipase: from predictions based on electrostatic computations to experimental results. Journal of Biotechnology 87: 225-254
NINI L., SARDA L., COMEAU L.C., BOITARD E., DUBES J.P., CHAHINIAN H. (2001): Lipase-catalyzed hydrolysis of short-chain substrates in solution and in emulsion: a kinetic study. Biochimica et Biophysica Acta 1534: 34-44
NOINVILLE S., REVAULT M., BARON M.-H., TISS A., YAPOUDJIAN S., IVANOVA M., VERGER R. (2002) : Conformational changes and orientation of Humicola lanuginosa lipase on a solid hydrophobic surface : An in situ interface Fourier transform infrared-attenuated total reflection study. BiophysicalJournal 82: 2709-2719
NORIN M., BOUTELJE J., HOLMBERG E., HULT K. (1988): Lipase immobilized by adsorption. Effect of support hydrophobicity on the reaction rate of ester synthesis in cyclohexane. Applied Microbiology and Biotechnology 28: 527-530
OGINO H., ISHIKAWA H. (2001): Enzymes which are stable in the presence of organic solvents. Journalof Bioscience and Bioengineering 91: 109-116.
OLLIS D.L., CHEAH E., CYGLER M., DIJKSTRA B., FROLOW F., FRANKEN S.M., HAREL M., REMINGTON
S.J., SILMAN I., SCHRAG J., SUSSMANN J.L., VERSCHUEREN K.H.G., GOLDMANN A. (1992): The alpha/beta hydrolase fold. Protein Engineering 5: 197-211
OMAR I.C., SAEKI H., NISHIO N., NAGAI S. (1988): Hydrolysis of triglycerides by immobilized thermo-stable lipase from Humicola lanuginosa. Agricultural and Biological Chemistry 52: 99-105
OTERO C., PASTOR E., FERNANDEZ V.M., BALLESTEROS A. (1990): Influence of the support on the reaction course of tributyrin hydrolysis catalyzed by soluble and immobilized lipases. Applied Biochemistry and Biotechnology 23: 237-247
PAIVA A.L., BALCÃO V.M., MALCATA F.X. (2000): Kinetics and mechanisms of reactions catalyzed by immobilized lipases. Enzyme and Microbial Technology 27: 187-204
PANDEY A., BENJAMIN S., SOCCOL C.R., NIGAM P., KRIEGER N., SOCCOL V.T. (1999): The realm of microbial lipases in biotechnology. Biotechnology and Applied Biochemistry 29: 119-131

Literatur
146
PATEL R.N. (2001): Enzymatic synthesis of chiral intermediates for drug development. AdvancedSynthesis & Catalysis 343: 527-546
PRONK W., KERKHOF P.J.A.M., VAN HELDEN C., VAN'T RIET K. (1988): The hydrolysis of triglycerides by immobilized lipase in a hydrophilic membrane reactor. Biotechnology and Bioengineering 32: 512-518
PRUESSE U., FOX B., KIRCHHOFF M., BRUSKE F., BREFORD J., VORLOP K.-D. (1998): New process (jet cutting method) for the production of spherical beads from highly viscous polymer solutions. Chemical Engineering Technology 21: 29-33
PRÜßE U. (2000): Entwicklung, Charakterisierung und Einsatz von Edelmetallkatalysatoren zur Nitratreduktion mit Wasserstoff und Ameisensäure sowie des Strahlschneider-Verfahrens zur Herstellung Polyvinylalkohol-verkapselter Katalysatoren. Landbauforschung Völkenrode Sonderheft214, Braunschweig ISBN 3-933140-36-6
RAGHEB A., BROOK M.A., HRYNYK M. (2003): Highly activated, silicone entrapped, lipase. Chemical Communications 18: 2314-2315
REETZ M.T., ZONTA A., SIMPELKAMP J. (1996): Efficient immobilisation of lipases by entrapment in hydrophobic sol-gel materials. Biotechnol. Bioeng. 49: 527-534
REETZ M.T. (1997): Entrapment of biocatalysts in hydrophobic sol-gel materials for use in organic chemistry. Advanced Materials 9: 943-954
REETZ M.T., TIELMANN P., WIESENHOEFER W., KOENEN W., ZONTA A. (2003): Second generation sol-gel encapsulated lipases: Robust heterogeneous biocatalysts. Advanced Synthesis and Catalysis 345: 717-728
REHM H.-J., REED G., STADLER P. (1998): Biotransformations Vol. I - Water, Organic Solvents and Other Reaction Media. In: Biotechnology - a multi volume comprehensive treatise. Vol. 8a. Wiley-VCH, Weinheim ISBN 3-527-28310-2
ROUSH W.R., BRINER K., KESLER B.S., MURPHY M., GUSTIN D.J.(1996): Studies on the synthesis of aureolic acid antibiotics: Acyloin glycosylation studies. Journal of Organic Chemistry 61: 6098-6099
ROZELL D.J. (1998): Commercial scale biocatalysis: myths and realities. Bioorganic & Medicinal Chemistry 7: 2253-2261
RUCKA M., TURKIEWICZ B. (1990): Ultrafiltration membranes as carriers for lipase immobilization. Enzyme and Microbial Technology 12: 52-55
RUCKA M., TURKIEWICZ B., ZUK J.S. (1990): Polymeric membranes for lipase immobilization. Journal of the American Oil Chemists’ Society 67: 887-889
RUCKENSTEIN E., WANG X. (1993): Lipase immobilized on hydrophobic porous polymer supports prepared by concentrated emulsion polymerization and their activity in the hydrolysis of triacylglycerides. Biotechnology and Bioengineering 42: 821-828
SALIS A., SVENSSON I., MONDUZZI M., SOLINAS V., ADLERCREUTZ P. (2003): The atypical lipase B from Candida antarctica is better adapted for organic media than the typical lipase from Thermomyces lanuginosa. Biochimica et Biophysica Acta 1646: 145-151

147
SANDOVAL G., CONDORET J.S., MONSAN P., MARTY A. (2002): Esterification by immobilized lipase in solvent-free media: Kinetic and thermodynamic arguments. Biotechnology and Bioengineering 78: 313-320
SARDA L., DESNUELLE P. (1958): Action de la lipase pancréatique sur les esters en émulsion. Biochimica et Biophysica Acta 30: 513-521
SAXENA R.K., GOSH P.K., GUPTA R., DAVIDSON W.S., BRADOO S., GULATI R. (1999): Microbial lipases: Potential biocatalysts for the future industry. Current Science 77: 101-116
SCHMID R.D., VERGER R. (1998): Lipases: Interfacial enzymes with attractive applications. Angewandte Chemie - International Edition 37: 1608-1633
SCHOEMAKER H.E., MINK D., WUBBOLTS M.G. (2003): Dispelling the myth-biocatalysis in industrial synthesis. Science 299: 1694-1697
SCHÖFER S.H., KAFTZIK N., WASSERSCHEID P., KRAGL U. (2001): Enzyme catalysis in ionic liquids: Lipase catalyzed kinetic resolution of 1-phenylethanol with improved enantioselectivity. Chemical Communications 5: 425-426
SCHOMBURG D., STEPHAN D. (1997): Class 2.5. – EC 2.1.104, Transferases, Enzyme handbooks Vol. 13 Springer Verlag Berlin, Heidelberg, New York ISBN 3-540-62608-5
SCHRAG J.D., CYGLER M. (1997): Lipases and / hydrolase fold. Methods in Enzymology 284: 85-107
SCHUCHARDT U., SERCHELI R., VARGAS R.M. (1998): Transesterification of vegetable oils: a review. Journal of the Brazilian Chemical Society 9: 199-210
SHARMA R., CHISTI Y., BANERJEE U.C. (2001): Production, purification, characterization and applications of lipases. Biotechnology Advances 19: 627-662
SHAW J.F., CHANG R.C., WANG F.F., WANG Y.J. (1990): Lipolytic activities of a lipase immobilized on six selected supporting materials. Biotechnology and Bioengineering 35:132-137
SKJÅK-BRÆK G., GRASDALEN H., SMIDSRØD O. (1989): Inhomogenous polysaccharide ionic gels. Carbohydrate Polymers 10: 31-54
STAMATIS H., XENAKIS A. (1999): Biocatalysts using microemulsion-based polymers gels containing lipase. Journal of Molecular Catalysis B: Enzymatic 6: 399-406
STARK M.B., HOLMBERG K. (1989): Covalent immobilization of lipase in organic solvents. Biotechnology and Bioengineering 34: 942-950
STRAATHOF A.J.J., PANKE S., SCHMID A. (2002): The production of fine chemicals by biotransformations. Current Opinion in Biotechnology 13: 548-556
SVENSSON I., WEHTJE E., ADLERCREUTZ P., MATTIASSON B. (1994): Effects of water activity on reaction rates and equilibrium positions in enzymic esterifications. Biotechnology and Bioengineering 44: 549-556
TAYLOR F., CRAIG J.C. (1991): Scale-up of flat-plate reactors for enzymic hydrolysis of fats and oils. Enzyme and Microbial Technology 13: 956-959
THUM O. (2004): Enzymatic Production of Care Specialties Based on Fatty Acid Esters. Tensides, Surfactants and Detergents 41: 287-290

Literatur
148
TISCHER W., WEDEKIND F. (1999): Immobilized enzymes: methods and applications. Topics in Current Chemistry 200: 95-126
TRIANTAFYLLOU A.Ö., WANG D., WEHTJE E., ADLERCREUTZ P. (1997): Polyacrylamides as immobilisation supports for use of hydrolytic enzymes in organic media. Biocatalysis and Biotransformation 15: 185-203
TURNER N.A., NEEDS E.C., KHAN J.A., VULFSON E.N. (2001): Analysis of conformational states of Candida rugosa lipase in solution: Implications for mechanisms of interfacial activation and separation of open and closed forms. Biotechnology and Bioengineering 72: 108-118
TWEDDELL R.J., KERMASHA S., COMBES D., MARTY A. (1998): Esterification and interesterification activities of lipases from Rhizopus niveus and Mucor miehei in three different types of organic media: a comparative study. Enzyme and Microbial Technology 22: 439-445
UPPENBERG J., HANSEN M.T., PATKAR S., JONES T.A. (1994): The sequence, crystal structure determination and refinement of two crystal forms of lipase B from Candida antarctica. Structure15: 293-308
VALIVETY R.H., HALLING P.J., PEILOW A.D., MACRAE A.R. (1992): Lipases from different sources vary widely in dependence of catalytic activity on water activity. Biochimica et Biophysica Acta 1122: 143-146
VALIVETY R.H., HALLING P.J., PEILOW A.D., MACRAE A.R. (1994): Relationship between water activity and catalytic activity of lipases in organic media. Journal of Biochemistry 222: 461-466
VAN BEILEN J.B., LI Z. (2002): Enzyme technology: an overview. Current Opinion in Biotechnology 13: 338-344
VASEL B., HECHT H.J., SCHMID R.D., SCHOMBURG D. (1993): 3D-structures of the lipase from Rhizomucor miehei at different temperatures and computer modelling of a complex of the lipase with trilaurylglycerol. Journal of Biotechnology 28: 99-115
VERGER R., DE HAAS G.H. (1976): Interfacial enzyme kinetics of lipolysis. Annual Review of Biophysics and Bioengineering 5: 77-117
VERGER R. (1997): ‘Interfacial activation’ of lipases: Facts and artifacts. Trends in Biotechnology 15: 32-38
WANG H.Y., KOBAYASHI T., SAITOH H., FUJII N. (1996): Porous polydimethylsiloxane membranes for enzyme immobilization. Journal of Applied Polymer Science 60: 2339-2346
WEHTJE E., ADLERCREUTZ P. [1997]: Water activity and substrate concentration effects on lipase activity. Biotechnology and Bioengineering 55: 798-806
WEHTJE E., KAUR J., ADLERCREUTZ P., CHAND S., MATTIASSON A.L. (1997): Water activity control in enzymatic esterification processes. Enzyme and Microbial Technology 21: 502-510
WELSH F.W., WILLIAMS R.E., DAWSON K.H. (1990): Lipase-mediated synthesis of low-molecular-weight flavor esters. Journal of Food Science 55: 1679-1682
WILLING A. (1999): Oleochemical esters. Environmentally compatible raw materials for oils and lubricants from renewable resources. Fett/Lipid 101: 192-198

149
ZAKS A., KLIBANOV A.M. (1984): Enzymatic catalysis in organic media at 100°C. Science 224: 1249-1251
ZHOU G.W., LI G.Z., XU J., SHENG Q. (2001): Kinetic studies of lipase-catalyzed esterification in water-in-oil micromeulsions and the catalytic behaviour of immobilized lipase in MBGs. Colloids and Surface A 194: 41-47

150

151
Lebenslauf
Persönliche Daten:
Name, Vorname: Buthe, Andreas
Geburtsort, -datum: Steinfurt, 03.02.1976
Familienstand: ledig
Ausbildung:
1982-1986 St. Georg Grundschule, Essen-Heisingen
1986-1995 Helmholtz-Gymnasium, Essen-Rüttenscheid
ab WS 1995/96 Studium der Metallurgie und Werkstofftechnik an der RWTH Aachen
ab WS 1996/97 Studium der Biologie an der Ruprecht-Karls-Universität Heidelberg
02/1999 Vordiplom Biologie
ab SS 1999 Studium der Biologie an der RWTH Aachen – Schwerpunkt
Biotechnologie
08/2002 Diplom Biologie
11/2002 – 12/2005 Wissenschaftlicher Mitarbeiter am Lehrstuhl für Biotechnolgie an der
RWTH Aachen
Top Related