
3 - ,3, * : - zfn.mpdl.mpg.dezfn.mpdl.mpg.de/data/Reihe_C/48/ZNC-1993-48c-0788.pdfThis work has been...
7
This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution 4.0 International License. Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschung in Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht: Creative Commons Namensnennung 4.0 Lizenz. Photoregulation of Nitrogen Metabolism and Protein Accumulation in the Red Alga Corallina elongata Ellis et Soland F. L. Figueroa Departamento de Ecologia, Facultad de Ciencias, Universidad de Malaga, Campus Universi- tario de Teatinos s/n, E-29071 Malaga, Spain Z. Naturforsch. 48c, 788-794 (1993); received June 15/June 29, 1993 Blue-Light Photoreceptor, Corallina elongata, Glutamine Synthetase, Light Quality, Nitrate Reductase Red and blue light pulses of 5 min applied together with 45 (aM K N 03 stimulated the nitrate uptake and reduction and the assimilation of ammonium in darkness in the red alga Corallina elongata. Nitrate reductase and glutamine synthetase activities were increased in darkness after the application of both red and blue light pulses. Red light produced a dramatic increase in enzyme activities after the first hour in darkness but after 4 h the effect of blue light pulses was greater. The photostimulation of nitrogen metabolism was correlated with light-regulated accumulation of soluble proteins. Nitrogen incorporation, assimilation of ammonium, accu mulation of total proteins and the increment in total intracellular nitrogen was greater in N-limited algae (C :N = 17.3) than in N-sufficient algae (C:N = 10.3) after the application of light pulses and nitrate. Because the stimulant effects of red and blue light on N-metabolism were partially reversed by far-red light, the possible involvement of the photoreversible red/ far-red photoreceptor, phytochrome, is proposed. In addition, the blue-light effect seems to be mediated by a specific B-light photoreceptor besides phytochrome due to the different time course of the response and the extent of the stimulation after blue compared to red light pulses. Introduction It has been proposed that chlorophyll, bili protein and total protein synthesis are controlled by phytochrome, a blue-light photoreceptor and a green-light photoreceptor in various N-limited algae [1-5], The photoregulation of pigment and protein synthesis was related to the photocontrol of nitrogen metabolism [4, 6 ]. The steps of light regulation, however, have still not been com pletely defined. Several steps could be regulated by light quality, namely, (a) uptake of nitrate, (b) nitrate reductase (NR) activity by activation of inactivated enzymes and/or by stimulation of de novo biosynthesis, and (c) assimilation of ammo nium by the stimulation of glutamine synthetase (GS) and glutamate synthase activities (GOGAT). Recently, it has been shown that the main step of photoregulation by phytochrome of soluble protein synthesis in Ulva rigida was the nitrate net uptake and the reduction of nitrate caused by the photostimulation of nitrate reductase activity [6 ]. Coaction of a blue-light photoreceptor and phyto chrome has been shown in these processes as well Verlag der Zeitschrift für Naturforschung, D-72072 Tübingen 0939-5075/93/0900-0788 $01.30/0 as in protein accumulation [3, 6 ]. The greatest effect of red and blue light pulses on protein and amino acid accumulation corresponded to sub strate level; more ammonia was available after red than after blue light pulses for protein synthesis. In the green algae Chlamydomonas reinhardii [7] and Monoraphidium braunii [8 ] blue light activated three key steps in the inorganic nitrogen metabo lism, namely, (a) cell uptake in both nitrate of ni trite, (b) nitrate reductase activity, and (c) biosyn thesis of nitrite reductase. Furthermore, in Chlo rella fusca blue light activated the nitrate specific permease [9], The aim of this work is to investigate the effect of light quality on the inorganic nitrogen me tabolism in algae previously incubated under N-limited (C:N=17.3) and N-sufficient condi tions (C:N = 10.3) in order to relate the possible stimulation of N 0 3~ net uptake and reduction and ammonium assimilation with light-regulated accu mulation of proteins. Red and blue light pulses followed by far-red light were applied in order to define the possible action of phytochrome and a B-light photorecep tor and to avoid the interference of photosynthe sis. Biochemical evidence of the existence of phyto chrome-like protein but not of a B-light photore ceptor has been reported in C. elongata [10, 11].
Transcript of 3 - ,3, * : - zfn.mpdl.mpg.dezfn.mpdl.mpg.de/data/Reihe_C/48/ZNC-1993-48c-0788.pdfThis work has been...

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
Photoregulation of Nitrogen Metabolism and Protein Accumulation in the Red Alga Corallina elongata Ellis et SolandF. L. FigueroaDepartamento de Ecologia, Facultad de Ciencias, Universidad de Malaga, Campus Universi- tario de Teatinos s/n, E-29071 Malaga, SpainZ. Naturforsch. 48c, 788-794 (1993); received June 15/June 29, 1993Blue-Light Photoreceptor, Corallina elongata, Glutamine Synthetase, Light Quality, Nitrate Reductase
Red and blue light pulses of 5 min applied together with 45 (aM K N 0 3 stimulated the nitrate uptake and reduction and the assimilation of ammonium in darkness in the red alga Corallina elongata. Nitrate reductase and glutamine synthetase activities were increased in darkness after the application of both red and blue light pulses. Red light produced a dramatic increase in enzyme activities after the first hour in darkness but after 4 h the effect of blue light pulses was greater. The photostimulation of nitrogen metabolism was correlated with light-regulated accumulation of soluble proteins. Nitrogen incorporation, assimilation of ammonium, accumulation of total proteins and the increment in total intracellular nitrogen was greater in N-limited algae (C :N = 17.3) than in N-sufficient algae (C :N = 10.3) after the application of light pulses and nitrate. Because the stimulant effects of red and blue light on N-metabolism were partially reversed by far-red light, the possible involvement of the photoreversible red/ far-red photoreceptor, phytochrome, is proposed. In addition, the blue-light effect seems to be mediated by a specific B-light photoreceptor besides phytochrome due to the different time course of the response and the extent of the stimulation after blue compared to red light pulses.
Introduction
It has been proposed that chlorophyll, biliprotein and total protein synthesis are controlled by phytochrome, a blue-light photoreceptor and a green-light photoreceptor in various N-limited algae [1-5], The photoregulation of pigment and protein synthesis was related to the photocontrol of nitrogen metabolism [4, 6 ]. The steps of light regulation, however, have still not been completely defined. Several steps could be regulated by light quality, namely, (a) uptake of nitrate, (b) nitrate reductase (NR) activity by activation of inactivated enzymes and/or by stimulation of de novo biosynthesis, and (c) assimilation of ammonium by the stimulation of glutamine synthetase (GS) and glutamate synthase activities (GOGAT).
Recently, it has been shown that the main step of photoregulation by phytochrome of soluble protein synthesis in Ulva rigida was the nitrate net uptake and the reduction of nitrate caused by the photostimulation of nitrate reductase activity [6 ]. Coaction of a blue-light photoreceptor and phytochrome has been shown in these processes as well
Verlag der Zeitschrift für Naturforschung,D-72072 Tübingen0939-5075/93/0900-0788 $01.30/0
as in protein accumulation [3, 6 ]. The greatest effect of red and blue light pulses on protein and amino acid accumulation corresponded to substrate level; more ammonia was available after red than after blue light pulses for protein synthesis. In the green algae Chlamydomonas reinhardii [7] and Monoraphidium braunii [8 ] blue light activated three key steps in the inorganic nitrogen metabolism, namely, (a) cell uptake in both nitrate of nitrite, (b) nitrate reductase activity, and (c) biosynthesis of nitrite reductase. Furthermore, in Chlorella fusca blue light activated the nitrate specific permease [9],
The aim of this work is to investigate the effect of light quality on the inorganic nitrogen metabolism in algae previously incubated under N-limited (C :N =17.3) and N-sufficient conditions (C:N = 10.3) in order to relate the possible stimulation of N 0 3~ net uptake and reduction and ammonium assimilation with light-regulated accumulation of proteins.
Red and blue light pulses followed by far-red light were applied in order to define the possible action of phytochrome and a B-light photoreceptor and to avoid the interference of photosynthesis. Biochemical evidence o f the existence of phytochrome-like protein but not of a B-light photoreceptor has been reported in C. elongata [10, 11].
F. L. Figueroa • Control of N-Metabolism by Light Quality in Corallina 789
Materials and Methods
The red alga Corallina elongata Ellis et Soland was harvested in early spring and autumn from rocky shores along the coast o f Malaga (Mediterranean sea, Spain). Then, they were maintained in seawater containing a low level of nitrate ( 2 |im NCV) at a temperature of 16 °C and a cycle of16 h white light (photon fluence rate:80 |imol m -2
s_1): 8 h darkness. After 3—4 days in these conditions, the algae collected in spring had a C : N level of 10.3, corresponding 263 ± 27 mg C gDW ' 1 and 25.5 ± 2 .2 mg N gDW ' 1 (N-sufficient algae) and the algae collected in autumn reached a C : N level of 17.3 corresponding to 156 ± 13 mg C gDW“' and 9.0 mg N g D W 1 (N-limited algae).
Light treatments and light sources
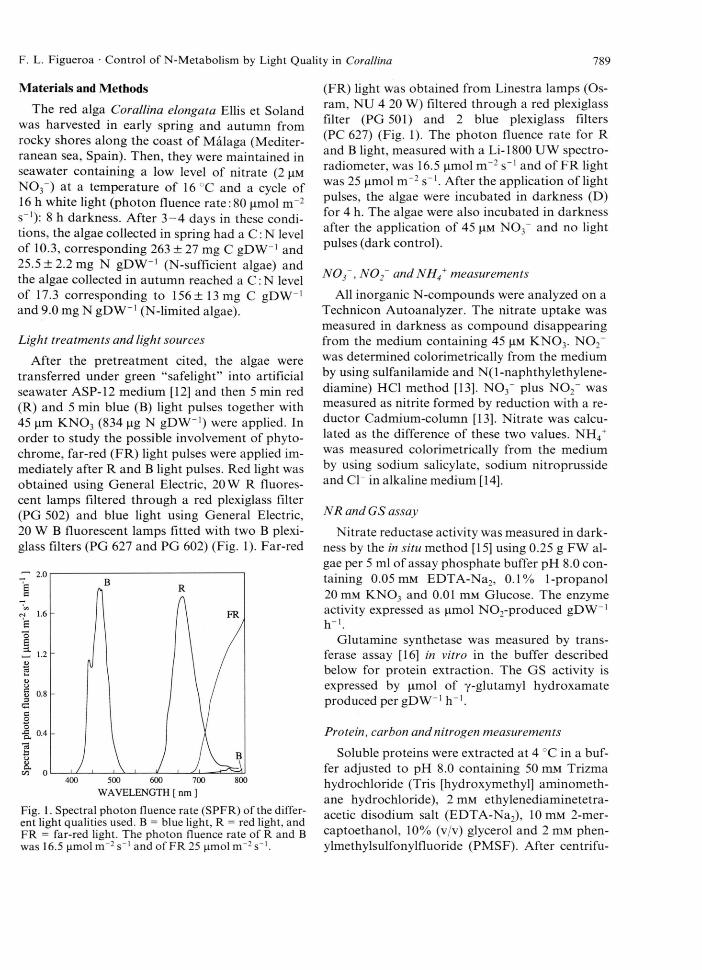
After the pretreatment cited, the algae were transferred under green “safelight” into artificial seawater ASP-12 medium [12] and then 5 min red (R) and 5 min blue (B) light pulses together with 45 nm K N 0 3 (834 jig N gDW -1) were applied. In order to study the possible involvement of phytochrome, far-red (FR) light pulses were applied immediately after R and B light pulses. Red light was obtained using General Electric, 20 W R fluorescent lamps filtered through a red plexiglass filter (PG 502) and blue light using General Electric, 20 W B fluorescent lamps fitted with two B plexiglass filters (PG 627 and PG 602) (Fig. 1). Far-red
WAVELENGTH [ nm ]
Fig. 1. Spectral photon fluence rate (SPFR) of the different light qualities used. B = blue light, R = red light, and FR = far-red light. The photon fluence rate of R and B was 16.5 (imol m ~2 s_1 and of FR 25 |imol m ~2 s_l.
(FR) light was obtained from Linestra lamps (Osram, NU 4 20 W) filtered through a red plexiglass filter (PG 501) and 2 blue plexiglass filters (PC 627) (Fig. 1). The photon fluence rate for R and B light, measured with a Li-1800 UW spectro- radiometer, was 16.5 (imol m ~2 s“ 1 and of FR light was 25 ^mol n r 2 s"1. After the application of light pulses, the algae were incubated in darkness (D) for 4 h. The algae were also incubated in darkness after the application of 45 jim N 0 3~ and no light pulses (dark control).
N O f , NOi~ andN H 4+ measurements
All inorganic N-compounds were analyzed on a Technicon Autoanalyzer. The nitrate uptake was measured in darkness as compound disappearing from the medium containing 45 |im K N 0 3. N 0 2~ was determined colorimetrically from the medium by using sulfanilamide and N(l-naphthylethylene- diamine) HC1 method [13]. N 0 3~ plus NO,~ was measured as nitrite formed by reduction with a re- ductor Cadmium-column [13]. Nitrate was calculated as the difference of these two values. NH4+ was measured colorimetrically from the medium by using sodium salicylate, sodium nitroprusside and C1“ in alkaline medium [14],
NR and GS assay
Nitrate reductase activity was measured in darkness by the in situ method [15] using 0.25 g FW algae per 5 ml of assay phosphate buffer pH 8.0 containing 0.05 m M EDTA-Na2, 0.1% 1-propanol 20 m M K N 0 3 and 0.01 m M Glucose. The enzyme activity expressed as imol NO,-produced gDW “ 1
h-1.Glutamine synthetase was measured by trans
ferase assay [16] in vitro in the buffer described below for protein extraction. The GS activity is expressed by nmol of y-glutamyl hydroxamate produced per gDW -1 h_l.
Protein, carbon and nitrogen measurements
Soluble proteins were extracted at 4 °C in a buffer adjusted to pH 8.0 containing 50 m M Trizma hydrochloride (Tris [hydroxymethyl] aminometh- ane hydrochloride), 2 m M ethylenediaminetetra- acetic disodium salt (EDTA-Na2), 10 m M 2-mer- captoethanol, 10% (v/v) glycerol and 2 m M phen- ylmethylsulfonylfluoride (PMSF). After centrifu-

790 F. L. Figueroa ■ Control of N-Metabolism by Light Quality in Corallina
gation at 10,000 x g for 25 min, the soluble proteins were determined in the supernant spectro- photometrically by the method based on the principle of protein-dye binding by Comassie Blue G-250 [17].
At the end of the dark period the total intracellular C and N contents were estimated by the Per- kin-Elmer Autoanalyzer 240-C.
Statistics
The results are expressed as the mean value ± standard deviation (SD) from 8 samples. 4 replicate samples were collected on each occasion and the experiment was repeated twice. The significance of the means of all data were tested by Fischer’s protected LSD procedure followed by Model 1 one-way ANOVA.
Results and Discussion
Nitrate net uptake was clearly affected by light quality in Corallina elongata in algae previously in
cubated under N-limited conditions (C:N = 17.3) (Fig. 2). Nitrate net uptake at all times was significantly different at 0.05 level in R and B light treatments related to FR light treatment and dark control. FR light reversed partially the effects of R and B light. Throughout the period of darkness, simultaneous uptake and excretion of nitrate were observed (Fig. 2): a rapid initial net uptake in the first 15 min, excretion between 3 0 m in - lh and net uptake in the last 2 h. In the dark control, 32 ± 3.5% of the applied nitrate (45 jim N 0 3 ) disappeared from the medium within 4 h, while, after R and B light pulses, the decrease in nitrate was far greater (65-75 ± 5% of the applied nitrate).
Ammonium net uptake did not occur in dark control and after R + FR and FR light pulses (Fig. 3). NH4+ uptake after 4 h in darkness was significantly greater (p<0.01) after B than after R light treatments. Both R and B effects were partially reversed by far-red light (Fig. 3).
Thus, net nitrate uptake was stimulated by R and B light pulses. Meanwhile, the incorporation
FR
B+FR
B
Fig. 3. Release and uptake of ammonium, expressed as |iM NH4+, in darkness after the same treatment as Fig. 2. The initial level of C : N was 17.3 (N-limited algae). Critical difference values (Fisher PLSD) at 0.05 significance level at 0.15 h (0.57), 0.5 h (0.61), 1 h (0.44), 2 h (0.68), 3 h (0.46), 4 h (0.98). LP = Light pulses.
DarkR+FR
LP + 45 LlM nitrate , .Time m darkness
Fig. 2. Uptake of nitrate expressed as |iM N 0 3~ disappearing from the medium, in darkness, after the application of red (R), blue (B), far-red (FR), R + FR and B + FR light pulses (LP) of 5 min and 45 jim KNO,. The initial level of C :N was 17.3 (N-limited algae). Critical difference values (Fisher PLSD) at 0.05 difference level at 0.15 h (2.80), 0.5 h (2.64), 1 h (3.34), 2 h (2.93), 3 h (2.43), 4 h (2.53). LP = Light pulses.
LP + 45 uM nitrate , , , ,n Time in darkness [ h ]
^ 401 cno£ 30s =*• ™
B+FR
FR
F. L. Figueroa • Control of N-Metabolism by Light Quality in Corallina 791
of the ammonium previously released was mainly stimulated by B-light pulses. High initial net uptake of nitrate and ammonium in various algae, especially after nitrogen starvation, has been reported [18]. In Corallina, nitrate and ammonium seemed to be used simultaneously as in other algae [19, 20]. Because R and B light effects on inorganic nitrogen incorporation were partially reversed, the possible involvement of phytochrome is proposed. The fact that NH4+ uptake in darkness was greater after B than after R light pulses, suggests the involvement of a specific B-light photoreceptor controlling ammonium uptake. The action of only phytochrome in B-light responses is discared because theoretically the level of active phytochrome reached after R is greater (70%) than after B (2 0 %) and more stimulation would be expected after R than after B light pulses; precisely the reverse was found. Different time course after B than after R light is a result that support the proposal on the involvement of a specific B-light photoreceptor. The time course of the responses has been used a criterion to distinguish when the B effect is due to phytochrome or due to specific B-light photoreceptor [21]. In the green alga Ulva rigida R and B light stimulated the nitrate net uptake through phytochrome and blue-light photoreceptor systems [6 ],
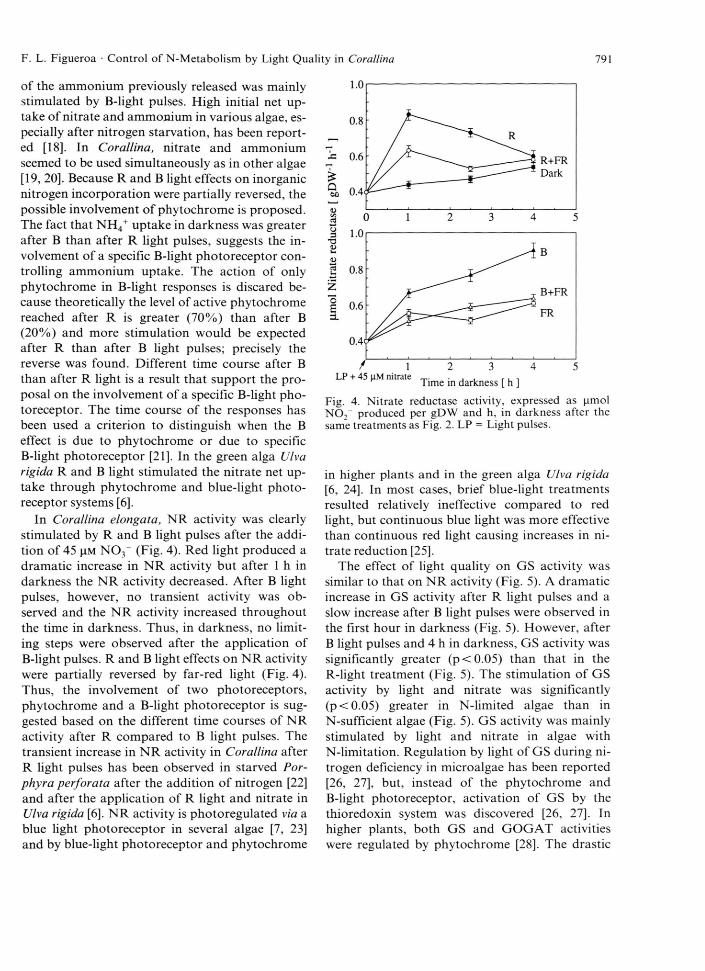
In Corallina elongata, NR activity was clearly stimulated by R and B light pulses after the addition of 45 |im N 0 3~ (Fig. 4). Red light produced a dramatic increase in NR activity but after 1 h in darkness the NR activity decreased. After B light pulses, however, no transient activity was observed and the NR activity increased throughout the time in darkness. Thus, in darkness, no limiting steps were observed after the application of B-light pulses. R and B light effects on NR activity were partially reversed by far-red light (Fig. 4). Thus, the involvement of two photoreceptors, phytochrome and a B-light photoreceptor is suggested based on the different time courses of NR activity after R compared to B light pulses. The transient increase in NR activity in Corallina after R light pulses has been observed in starved Por- phyra perforata after the addition o f nitrogen [2 2 ] and after the application of R light and nitrate in Ulva rigida [6 ], NR activity is photoregulated via a blue light photoreceptor in several algae [7, 23] and by blue-light photoreceptor and phytochrome
Fig. 4. Nitrate reductase activity, expressed as |imol N O r produced per gDW and h, in darkness after the same treatments as Fig. 2. LP = Light pulses.
in higher plants and in the green alga Ulva rigida [6 , 24]. In most cases, brief blue-light treatments resulted relatively ineffective compared to red light, but continuous blue light was more effective than continuous red light causing increases in nitrate reduction [25].
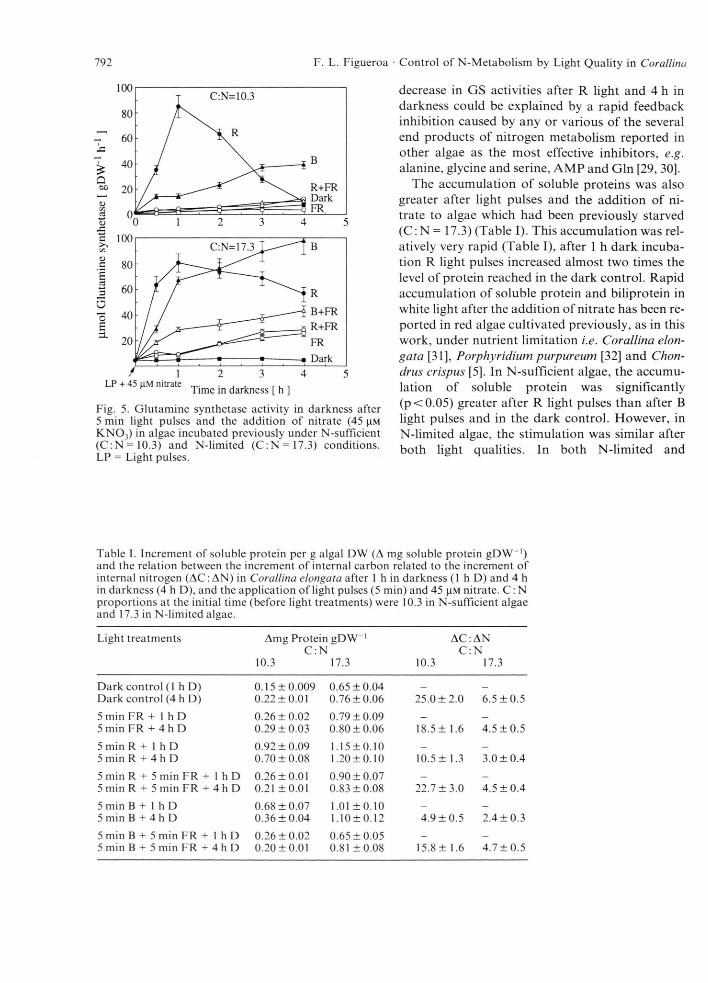
The effect of light quality on GS activity was similar to that on NR activity (Fig. 5). A dramatic increase in GS activity after R light pulses and a slow increase after B light pulses were observed in the first hour in darkness (Fig. 5). However, after B light pulses and 4 h in darkness, GS activity was significantly greater (p<0.05) than that in the R-light treatment (Fig. 5). The stimulation of GS activity by light and nitrate was significantly (p<0.05) greater in N-limited algae than in N-sufficient algae (Fig. 5). GS activity was mainly stimulated by light and nitrate in algae with N-limitation. Regulation by light of GS during nitrogen deficiency in microalgae has been reported [26, 27], but, instead of the phytochrome and B-light photoreceptor, activation of GS by the thioredoxin system was discovered [26, 27]. In higher plants, both GS and GOGAT activities were regulated by phytochrome [28]. The drastic
792 F. L. Figueroa Control of N-Metabolism by Light Quality in Corallina
Fig. 5. Glutamine synthetase activity in darkness after 5 min light pulses and the addition of nitrate (45 K NO j) in algae incubated previously under N-sufficient (C :N = 10 .3 ) and N-limited (C :N = 17 .3 ) conditions. LP = Light pulses.
decrease in GS activities after R light and 4 h in darkness could be explained by a rapid feedback inhibition caused by any or various of the several end products of nitrogen metabolism reported in other algae as the most effective inhibitors, e.g. alanine, glycine and serine, AMP and Gin [29, 30].
The accumulation of soluble proteins was also greater after light pulses and the addition of nitrate to algae which had been previously starved (C : N = 17.3) (Table I). This accumulation was relatively very rapid (Table I), after 1 h dark incubation R light pulses increased almost two times the level of protein reached in the dark control. Rapid accumulation of soluble protein and biliprotein in white light after the addition of nitrate has been reported in red algae cultivated previously, as in this work, under nutrient limitation i.e. Corallina elon- gata [31], Porphyridium purpureum [32] and Chon- drus crispus [5]. In N-sufficient algae, the accumulation of soluble protein was significantly (p < 0.05) greater after R light pulses than after B light pulses and in the dark control. However, in N-limited algae, the stimulation was similar after both light qualities. In both N-limited and
Table I. Increment of soluble protein per g algal DW (A mg soluble protein gDW -1) and the relation between the increment o f internal carbon related to the increment of internal nitrogen (AC: AN) in Corallina elongata after 1 h in darkness (1 h D) and 4 h in darkness (4 h D), and the application of light pulses (5 min) and 45 jaM nitrate. C : N proportions at the initial time (before light treatments) were 10.3 in N-sufficient algae and 17.3 in N-limited algae.
Light treatments Amg Protein gDW 1 A C: ANC :N C : N
10.3 17.3 10.3 17.3
Dark control (1 h D) Dark control (4 h D)
0.15 ±0.009 0.22 ± 0.01
0.65 ±0.04 0.76 ±0.06 25.0 ± 2 .0 6.5 ±0.5
5 min FR + 1 h D 5 min FR + 4 h D
0.26 ± 0.02 0.29 ±0.03
0.79 ±0.09 0.80 ±0.06 18.5 ± 1.6 4.5 ±0 .5
5 min R + 1 h D 5 min R + 4 h D
0.92 ±0.09 0.70 ±0.08
1.15 ± 0.10 1.20 ± 0.10 10.5 ± 1.3 3.0 ± 0 .4
5 min R + 5 min FR + 1 h D 5 min R + 5 min FR + 4 h D
0.26 ± 0.01 0.21 ± 0.01
0.90 ±0.07 0.83 ±0.08 22.7 ±3 .0 4.5 ± 0 .4
5 min B + 1 h D 5 min B + 4 h D
0.68 ±0.07 0.36 ±0.04
1.01 ± 0.10 1.10 ± 0.12 4.9 ±0.5 2.4 ± 0.3
5 min B + 5 min FR + 1 h D 5 min B + 5 min FR + 4 h D
0.26 ± 0.02 0.20 ± 0.01
0.65 ±0.05 0.81 ±0.08 15.8 ± 1.6 4.7 ± 0.5

F. L. Figueroa • Control o f N-M etabolism by Light Quality in Corallina 793
N-sufficient algae FR light pulses reversed partially R and B light stimulant effects on protein synthesis. These results indicate that the action R and B photoreceptors is very complex and it could be affected by the intracellular content of nitrogen.
The relation between the increment of internal carbon related to the increment of internal nitrogen was clearly affected by light quality after 4 h in darkness and the addition o f nitrate (Table I). In both, N-limited and N-sufficient algae, the increment of nitrogen related to carbon was significantly (p < 0.1) greater after R and B light pulses (low AC:AN relation) than in the dark control and after FR light pulses. On the other hand, the relation AC: AN was lower in N-limited algae than in N-sufficient algae in all light treatments (Table I). This indicates that the accumulation of nitrogen related to carbon was greater in algae previously starved than in repleted algae.
The relation between N 0 3~ uptake and reduction and the ammonium assimilation with protein accumulation indicated that the photocontrol of the protein synthesis was produced as a consequence of the photoregulation o f N-metabolism as it has been reported in Ulva [6 ]. The greatest effect of R and B light pulses on protein accumulation
corresponded to substrate level; more ammonia was available after R and B light for protein synthesis than after FR light and in the dark control.
In summary, the involvement of at least two different photoreceptors, namely, phytochrome and a B-light photoreceptor, controlling several steps of the inorganic nitrogen metabolism in the red alga C. elongata: (1) uptake of nitrate, nitrite and ammonium, (2) nitrate reductase activity and (3) glutamine synthetase activity is suggested. The participation of photosynthetic reactions in the control of N-metabolism is excluded because with the light treatments used (short pulses and 4 h in darkness) no differences in photosynthetic rates were observed [6 , 11]. Simultaneously to the action of red/far-red and blue photoreceptor the possible action of other photoreceptors is not discared. More investigations are necessary to define the complex interaction between the different photoreceptors that control N-metabolism.
A cknowledgemen ts
This work was financed by the Spanish Ministry of Education and Science (Project CICYT MAR- 90 0365).
[1] F. Lopez-Figueroa and F. X. Niell, Physiol. Plant 76, 185-193(1989).
[2] F. Lopez-Figueroa, R. Perez, and F. X. Niell, J. Photochem. Photobiol. 4, 185-193 (1989).
[3] F. Lopez-Figueroa and W. Rüdiger, Photochem. Photobiol. 52 (1), 111 -114 (1990).
[4] F. Lopez-Figueroa, Z. Naturforsch. 46c, 542-548(1991).
[5] F. Lopez-Figueroa, Br. Phycol. J. 26 ,383-393 (1991).[6] F. Lopez-Figueroa and W. Rüdiger, J. Phycol. 27,
389-394(1991).[7] M. P. Azuara and P. J. Aparicio, Plant Physiol. 77,
95-98(1985).[8] P. J. Aparicio and M. A. Quinones, Plant Physiol.
95,374-378(1991).[9] F. J. Florencio and J. M. Vega, Phytochemistry 21
(6), 1195-1200(1982).[10] F. Lopez-Figueroa, P. Lindemann, S. E. Braslavsky,
K. Schaffner, H. A. W. Schneider-Poetsch, and W. Rüdiger, Bot. Acta 102, 178-180 (1989).
[11] W. Rüdiger and F. Lopez-Figueroa, Photochem. Photobiol. 55 (6), 949-954 (1992).
[12] H. Iwasaji, J. Phycol. 3, 30-34 (1967).[13] F. D. Snell and T. C. Snell, Colorimetric methods of
analysis, 3rd edition, Van Nostrand, Princeton, New Jersey, Vol. 2, 844 (1949).
[14] G. Slavik and J. J. Macisaac, Deep. Sea. Res. 19, 521-524(1972).
[15] A. Corzo and F. X. Niell, J. Exp. Mar. Biol. Ecol. 146,181-191 (1991).
[16] B. M. Shapiro and E. R. Stadtman, in: Methods Enzymol. (H. Tabor and C. W. Tabor, eds.), Vol. 17, pp. 910-922, Academic Press, New York 1970.
[17] M. M. Bradford, Anal. Biochem. 72 ,248-252(1976).[18] T. E. Thomas and P. J. Harrison, Mar. Biol. (Berl.)
85,269-278(1985).[19] Q. Dortch, J. Exp. Mar. Biol. Ecol. 61, 243-264
(1982).[20] D. H. Turpin, H. G. Weger, R. G. Smith, W. C.
Plaxton, and M. Lin, in: Inorganic nitrogen in plants and microorganisms: uptake and metabolism (W. R. Ullrich, C. Rigano, A. Fuggi, and P. J. Aparicio, eds.), pp. 124-130, Springer-Verlag, Berlin 1990.
[20] A. Corzo and F. X. Niell, Mar. Biol. 112, 223-228(1992).
[21] D. J. Cosgrove, in: Phtomorphogenesis in plants (R. E. Kendrick and G. H. M. Kronenberg, eds.), pp. 341-367, Martinus Nijhoff Publishers, The Netherlands 1986.
[22] T. E. Thomas and P. J. Harrison, J. Exp. Mar. Biol. Ecol. 107, 1-8(1987).
[23] H. Ninnemann, in: Blue light responses: phenomena and occurrence in plants and microorganisms (H. Senger, ed.), pp. 17-29, CRC Press, Boca Raton, Florida 1987.
[24] C. R. Schuster, R. Oelmüller, and H. Mohr, Planta (Berl.) 171, 136-143(1987).

794 F. L. Figueroa • Control of N-Metabolism by Light Quality in Corallina
[25] H. Sasakawa and Y. Yamamoto, Plant Physiol. 63, 1098-1101 (1979).
[26] R. Tischner and A. Schmidt, Plant Physiol. 70, 113 — 116 (1982).
[27] P. J. Syrett and A. M. Peplinska, New Phytol. 109, 289-296(1988).
[28] U. R. Hetsch, R. Oelmüller, S. Schmidt, and H. Mohr, Planta 175, 130-138 (1988).
[29] R. F. Beudeker and F. R. Tabita, Plant Physiol. 75, 516-520(1985).
[30] F. J. Florencio and J. L. Ramos, Biochim. Biophys. Acta 838, 39-48(1985).
[31] J. Vergara and F. X. Niell, J. Phycol. 29, 285-293(1993).
[32] P. Algarra and W. R. Rüdiger, Plant, Cell and Environment 16, 149-159(1993).


















