
CHE 324 Anorganische Chemie V - chem.uzh.chffffffff-8715-1fc6-ffff-ffffb5f38e... · ACV HS 2012-1....
92
ACV HS 2012-1 Bioanorganik: Metalle in der Biochemie und Biologie Bioanorganik: Metalle in der Biochemie und Biologie CHE 324 Anorganische Chemie V CHE 324 Anorganische Chemie V R. Alberto, E. Freisinger, G. Gasser G. Patzke, R. Sigel, B. Spingler, F. Zelder HS 2012
Transcript of CHE 324 Anorganische Chemie V - chem.uzh.chffffffff-8715-1fc6-ffff-ffffb5f38e... · ACV HS 2012-1....

ACV HS 2012-1
Bioanorganik: Metalle in der Biochemie und BiologieBioanorganik: Metalle in der Biochemie und BiologieCHE 324 Anorganische Chemie VCHE 324 Anorganische Chemie V
R. Alberto, E. Freisinger, G. Gasser G. Patzke, R. Sigel, B. Spingler, F. ZelderHS 2012

ACV HS 2012-2
Übersicht
Woche 1 – 2: R. Alberto; Einführung – Hydrogenase - Nitrogenase
Woche 3 – 4: B. Spingler; DNA / RNA / Cisplatin
Woche 5 – 6: E. Freisinger-Sigel; Metallohomöostasis– Detoxifikation
Woche 7 – 8: G. Patzke; Biomineralisation – Polyoxometallate
Woche 8: Verteilung der Vortragsthemen
Woche 9 – 10: F. Zelder; Tetrapyrrole - Corrine und Corphine
Woche 11 – 12: R. Sigel; ATP Hydrolyse / Polymerase
Woche 13 – 14: Vorträge
CHE324 Anorganische Chemie VCHE324 Anorganische Chemie V

ACV HS 2012-3
W. Kaim, B. Schwederski: Bioanorganische Chemie, Teubner Studienbücher, 2005
S.J. Lippard, J.M. Berg: Principles of Bioinorganic ChemistryUniversity Science Books, 1994 (1995 in German)
H.A.O. Hill, P.J. Sadler, A.J. Thomson (eds.): Metal sites in proteins and models, Springer (series)1999
J.A. Cowan: Inorganic Biochemistry, Wiley-VCH 1997
R.J.P. Williams, J.J.R. Frausto da Silva: The natural selection of the elementsClarendon Press, 1997
Literatur:
CHE324 Anorganische Chemie VCHE324 Anorganische Chemie V

ACV HS 2012-4
1. Die speziellen Eigenschaften von Übergangsmetallkationen
2. Die Selektion der Elemente
3. Ausgewählte Beispiele
3.1. Kupferhaltige O2 Transporter und Aktivierer
3.2. Nitrogenasen / N2 – Fixierung
3.3. Hydrogenasen und Wasserstoff
3.4. Metallfreie Hydrogenase, a fake ?
CHE324 Anorganische Chemie VCHE324 Anorganische Chemie V

ACV HS 2012-5
In biologischen Systemen sind Übergangsmetallkationenessentielle Bestandteile
- Das sogenannte "biologische Periodensystem" gibt Auskunft überessentielle metallische und nicht-metallische Elemente
- Einige dieser Elemente sind echte Spurenelemente, andere kommen in grösseren Mengen vor. Von einigen ist der essentielle Charakter ungewiss.
1. Die speziellen Eigenschaften von Übergangsmetallkationen1. Die speziellen Eigenschaften von Übergangsmetallkationen

ACV HS 2012-6
Rollen von Metallkationen:
- als katalytische Zentren
- in Transportprozessen
- als Strukturbildner
- als Speichermedien
In Prozessen, in denen Metalle involviert sind, genügt die reine Reaktion nicht, Es sind noch andere Charakteristika verlangt, die eben nur Metalle übernehmen können
z. Bsp. Phosphatesterspaltung
Die speziellen Eigenschaften von ÜbergangsmetallkationenDie speziellen Eigenschaften von Übergangsmetallkationen
Metallkationen sind im dem Sinne nicht einmalig,
Viele rein organische Enzyme können einige der prinzipiellen Reaktionstypen katalysieren.

ACV HS 2012-7
high spin low spin
— — — —
— — — — — —
Ionenradien
- high spin / low spin Wechsel durch verändertes Ligandenfeld oder -wechsel
Einfluss auf den Ionenradius rFe2+ (hs) = 78 (65) pm vs rFe2+ (ls) = 61 (55) pm
Einfluss auf Koordinationsgeometrie, Kinetik und Thermodynamik
z. Bsp. [FeIIpor]-Cl vs [FeIIpor]-CO
Die speziellen Eigenschaften von ÜbergangsmetallkationenDie speziellen Eigenschaften von Übergangsmetallkationen
Übergangsmetallkationen haben eine variable d-Elektronenbesetzung
Eigenschaften welche inhärent an diese dx Konfiguration gebunden sind

ACV HS 2012-8
Wechsel der Grösse bei gleicher Oxidationsstufe ist nur bei Übergangsmetallen möglich
Konsequenzen für die Bioanorganik
Variable Anzahl von Liganden und Koordinationsgeometrie
Metall induzierte strukturelle Veränderungen im Proteingerüst
induzierte Aktivität an anderen Stellen im Enzym.
Die speziellen Eigenschaften: IonenradienDie speziellen Eigenschaften: Ionenradien

ACV HS 2012-9
- Redoxpotentiale spielen eine wichtige Rolle, sei das bei reinen Elektronenübertragungsreaktionen oder bei Atomtransferreaktionen (Oxygenasen, Hydrogenasen etc.)
- Eine einmalige Eigenschaft der Übergangsmetallkationen ist die Verfügbarkeit von verschiedenen Oxidationsstufen, welche für wässerige Systeme teilweise recht
ungewöhnlich sein können.
- Da die Grösse der Redoxpotentiale inhärent mit der elektronischen Struktur verbunden ist, können diese einfach beeinflusst werden.
Die speziellen Eigenschaften: RedoxpotentialeDie speziellen Eigenschaften: Redoxpotentiale

ACV HS 2012-10
Art der Ligandeni) HSAB Prinzip, passt der Ligand zur Oxidationsstufe?
ii) Coliganden, nicht vom Makromolekül abgeleitet (i.e. Porphyrine, Corrine...)
Geometrie der Ligandeni) starre Geometrie, der Oxidationsstufe angepasst / nicht angepasst
ii) flexible Geometrieiii) Donor / Akzeptorfähigkeiten
- Metallzentrumi) Mögliche Oxidationsstufenii) Nachbarmetalle (Cluster)
Wie in der Koordinationschemie werden Redoxpotentiale durch verschiedene Faktoren beeinflusst und können für ein bestimmtes Zentrum von
oxidierend bis reduzierend gehen.
Einflussfaktoren:
Die speziellen Eigenschaften: RedoxpotentialeDie speziellen Eigenschaften: Redoxpotentiale

ACV HS 2012-11
Die speziellen Eigenschaften: RedoxpotentialeDie speziellen Eigenschaften: Redoxpotentiale

ACV HS 2012-12
Im Gegensatz zu den p-Elementen: d-Elemente wechseln ihre Bindungspartner leicht
Dies bedeutet jedoch nicht, dass die Bindungen sehr schwach sindsondern nur dass eine kleine Energiebarriere überwunden werden muss.
schneller Ligandaustausch: mechanistisches Prinzip und weniger thermodynamisches
Damit der Ligandaustausch schnell erfolgen kann, muss:
- die LFSE klein sein (im Idealfall 0)
die Geometrie des „Grundzustandes“ nahe beim Übergangszustand sein -(s. entatisches Prinzip später)
der Übergangszustand stabilisiert sein
Die speziellen Eigenschaften: AustauschreaktionenDie speziellen Eigenschaften: Austauschreaktionen
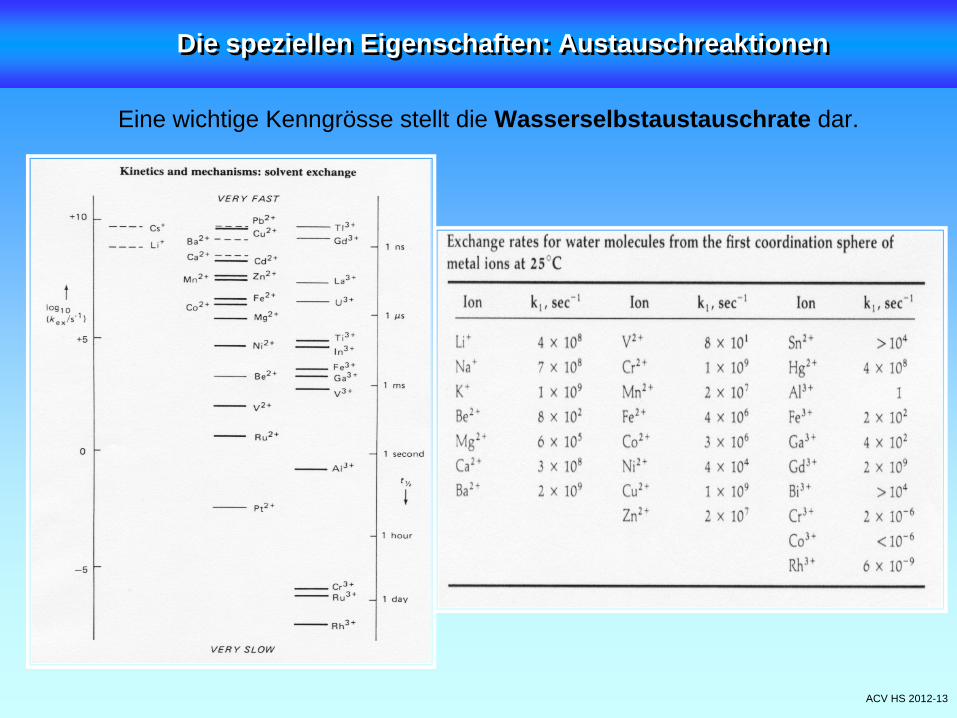
ACV HS 2012-13
Eine wichtige Kenngrösse stellt die Wasserselbstaustauschrate dar.
Die speziellen Eigenschaften: AustauschreaktionenDie speziellen Eigenschaften: Austauschreaktionen

ACV HS 2012-14
- Schnelle biochemische Katalyse: Substrat/Produkt on/off rate muss gross sein.
Typische inerte Kationen werden in der bioanorganischen Chemie selten gefunden.
Die speziellen Eigenschaften: AustauschreaktionenDie speziellen Eigenschaften: Austauschreaktionen
Es sind keine e--Transferreaktionen zwischen Metalloproteinen bekannt, welche nach dem Innensphärenmechanismus ablaufen.
-Elektronentransferreaktionen meistens nach Aussensphärenmechanismus(mechanistisch grosses Interesse)
nieder koordinierte Kationen (KZ
4): assoziativ (Kinetik 2. Ordnung)
höheren Koordinationszahlen KZ
6): dissoziativ (Kinetik 1. Ordnung)

ACV HS 2012-15
Die spezielle elektronische Struktur der Übergangsmetallkationen bewirkt über die Koordination von Substratmolekülen deren Aktivierung.
Diese Aktivierung erfolgt auf vielerlei Art und Weise
Erniedrigung des pKa von koordinierte H2 O resp. Stabilisierung von M-OH
Die speziellen Eigenschaften: Aktivierung kleiner MoleküleDie speziellen Eigenschaften: Aktivierung kleiner Moleküle
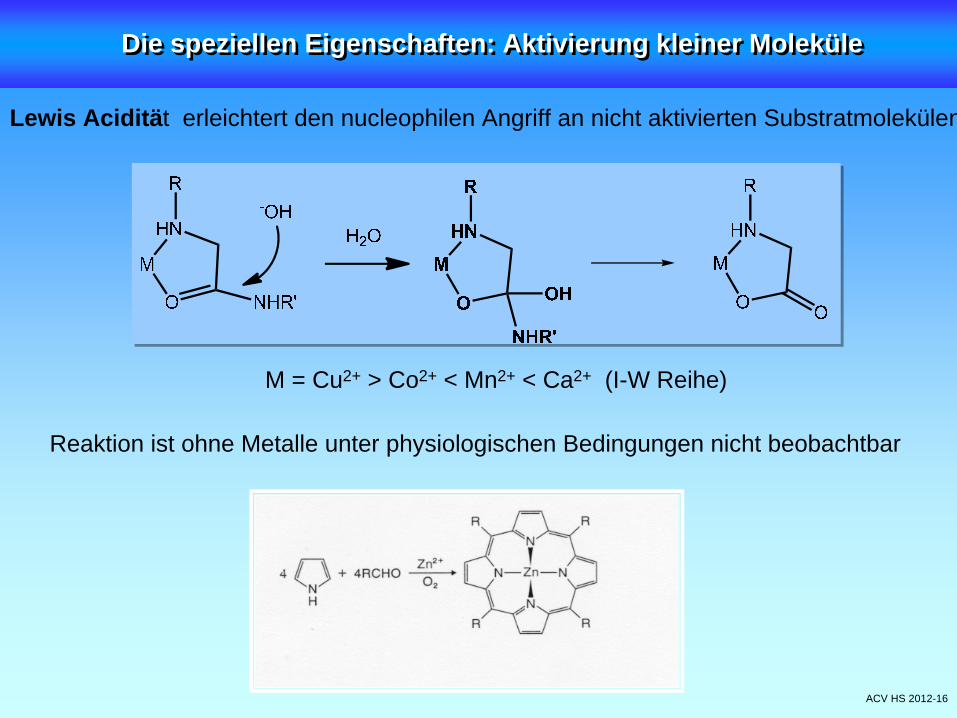
ACV HS 2012-16
Lewis Acidität erleichtert den nucleophilen Angriff an nicht aktivierten Substratmolekülen
Reaktion ist ohne Metalle unter physiologischen Bedingungen nicht beobachtbar
M = Cu2+ > Co2+ < Mn2+ < Ca2+ (I-W Reihe)
Die speziellen Eigenschaften: Aktivierung kleiner MoleküleDie speziellen Eigenschaften: Aktivierung kleiner Moleküle

ACV HS 2012-17
Die meisten Reaktionen mit O2 und organischen Substraten sind verboten und deshalb sehr langsam (Spinmutliplizität)
Durch „Spinübertragung“ auf das Metallzentrum werden sie erlaubt.z. Bsp. die meisten O-Übertragungsreaktionen aus O2
Isomerisierungen bei Vitamin B12
R CO
HR—CH3 + O2 + H2 O
S T S S
2 CuI + O2 2 CuII + O22- + 2 CuI + H2 OR C
O
H
Angeregt T
T D + D S S SSonst S
- Stabilisierung und Generierung von Radikalen
2 H+ + 2 e-
Die speziellen Eigenschaften: Aktivierung kleiner MoleküleDie speziellen Eigenschaften: Aktivierung kleiner Moleküle
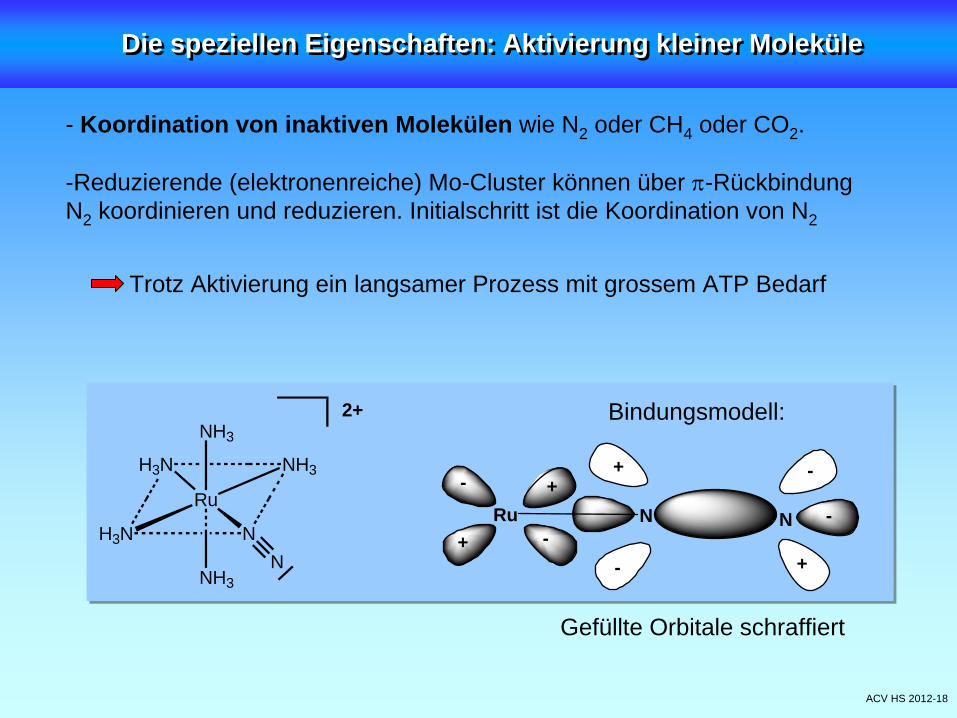
ACV HS 2012-18
- Koordination von inaktiven Molekülen wie N2 oder CH4 oder CO2 .
-Reduzierende (elektronenreiche) Mo-Cluster können über -Rückbindung N2 koordinieren und reduzieren. Initialschritt ist die Koordination von N2
H3N NH3
H3N N
Ru
NH3
NH3N
2+
Ru N N
-
+
-
+
-
- +
+ -
Bindungsmodell:
Gefüllte Orbitale schraffiert
Trotz Aktivierung ein langsamer Prozess mit grossem ATP Bedarf
Die speziellen Eigenschaften: Aktivierung kleiner MoleküleDie speziellen Eigenschaften: Aktivierung kleiner Moleküle

ACV HS 2012-19
Viele Metallkationen nehmen mit sterisch anspruchlosen Liganden eine streng definierteGeometrie ein, in der Regel KZ = 6 oder 4 mit den entsprechenden Geometrien
Wie vorher gesehen, können biologische Makromoleküle einem Metallkation eine Geometrie aufzwingen und damit die Metalleigenschaften bestimmen.
Mn+ + Protein M-Protein inaktiv aktiv
vs Mn+ + Protein M-Protein
inaktiv aktiv
Mn+ + Protein M-Protein inaktiv aktiv
vs Mn+ + Protein M-Protein
inaktiv aktiv
- Der eigentlich aktive Teil ist nicht das Metall sondern das Protein
der umgekehrte Fall wird häufig angetroffen
ohne Metall ist das Enzym (Apoenzym) wirkungslos.
Die speziellen Eigenschaften: KoordinationsgeometrieDie speziellen Eigenschaften: Koordinationsgeometrie

ACV HS 2012-20
Metallkation aktiviert Enzym Kontrollmechanismus über [Mn+]
Steuerung über bevorzugte Liganden oder Koordinationsgeometrie
Anspruchsvolle Koordinationssphären: KoordinationsgeometrieZn2+, Zn-Finger, Genregulation
Anspruchslose Geometrie, variable DonorenCa2+, Calmodulin, second messenger
Die speziellen Eigenschaften: KoordinationsgeometrieDie speziellen Eigenschaften: Koordinationsgeometrie

ACV HS 2012-21

ACV HS 2012-22
1) Metallkationen können variable Koordinationszahlen haben
Anbindung und Aktivierung von Substraten wegen resultierender Lewisacidität
2) Metallkationen können variable Koordinationsgeometrien haben
fine tuning von elektronischen Eigenschaften wie Redoxpotentialen
3) Liganden können mit geringen Strukturveränderungen substituiert werden
schneller Ligandaustausch, schnelle (aber unspezifische) Katalyse
4) Metallkationen können die Grösse (bei gleicher Oxidationszahl)ändern
variable Anzahl Liganden, redox(in)aktiv
5) Metallkationen können über breite Oxidationsstufen variieren
Atomtransferreaktionen, Redoxreaktionen, Elektronenspeicher
Funktion der Metalloenzyme
Die speziellen Eigenschaften: ZusammenfassungDie speziellen Eigenschaften: Zusammenfassung

ACV HS 2012-23
je nach gewünschter Reaktion kann ein bestimmtes Metallzentrum gewählt werden
z. Bsp. schnelle Hydrolyse von Peptiden
Ansprüche:
- Substratanbindung- Lewisacidität ( Acidifizierung von H2 O)
- geringe Strukturänderungen- kein Redox
Lösung: Zn2+!!
Die speziellen Eigenschaften: ZusammenfassungDie speziellen Eigenschaften: Zusammenfassung
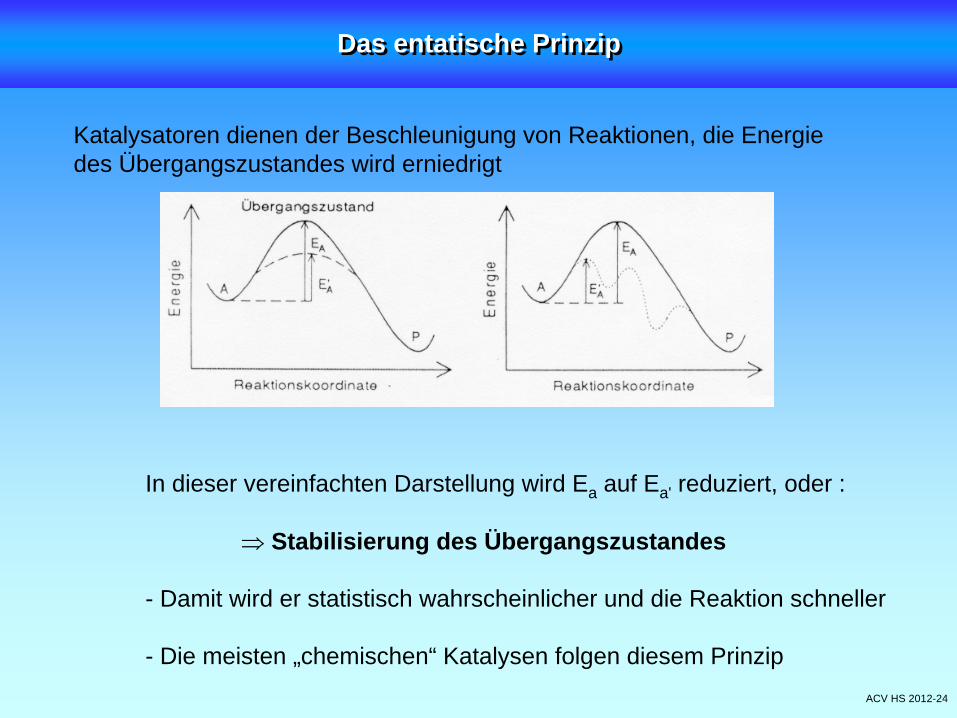
ACV HS 2012-24
In dieser vereinfachten Darstellung wird Ea auf Ea' reduziert, oder :
Stabilisierung des Übergangszustandes
- Damit wird er statistisch wahrscheinlicher und die Reaktion schneller
- Die meisten „chemischen“ Katalysen folgen diesem Prinzip
Katalysatoren dienen der Beschleunigung von Reaktionen, die Energiedes Übergangszustandes wird erniedrigt
Das entatische PrinzipDas entatische Prinzip

ACV HS 2012-25
Möglich, indem im Grundzustand keine reguläre, sondern eine verzerrte und damit
aktivierte Geometrie vorliegt, welche den Übergangszustand schon vorgebildet hat.
Das Protein ist damit in einem gespannten (entatischen) Zustand. Es zwingt
dem Metall eine “nicht optimale“ Koordinationsgeometrie auf.
Das gleiche Ziel wird erreicht, wenn der Grundzustand destabilisiert wird
Das entatische PrinzipDas entatische Prinzip

ACV HS 2012-26
Viele Enzyme zeigen im metalllosen (Apoenzym) und im metallhaltigen Zustand
keine oder nur geringfügige strukturelle Veränderungen auf.
Das entatische PrinzipDas entatische Prinzip

ACV HS 2012-27
Die meisten Reaktionsabläufe welche enzymkatalysiert ablaufen, folgen einem sogenannten Michaelis-Menten Mechanismus
E + S ES P + E
Damit entspricht der Mechanismus in etwa einem Vorgleichgewichtsmechanismus mit Ausnahme, dass E wieder in dieses Ggw eingreift.
k1
k1 'k2
k1 [E][S]k1 ' + k2
k1 ([E0 ] - [ES]) [S]k1 ' + k2
E = Enzym, S = Substrat mit [E] + [ES] = [E0 ]
mit [S] = [S0 ]
[ES] =
[ES] =
Michaelis-Menten KinetikMichaelis-Menten Kinetik

ACV HS 2012-28
variiert also linear mit [E0 ] woraus k2 direkt abgelesen werden kann
k1 [E0 ][S]k1 ' + k2 + k1 [S]
dPdt
k2 [S]KM + [S]
k1 ' + k2
k1
dPdt = k2 [E0 ]
= k [E0 ] mit k =
die Michaeliskonstante ist
nach Umformung folgt
[ES] =
wobei KM =
Substrat in sehr grossem Überschuss: Gleichung reduziert sich auf
oder mit
Michaelis-Menten KinetikMichaelis-Menten Kinetik

ACV HS 2012-29
Die Reaktion ist O. Ordnung in S
k2 heisst maximal turn over Zahl [sec-1] und
kB [E0 ] die maximale Geschwindigkeit der enzymatischenReaktion.
für Verhältnisse [S] << [Km]:
1 1 KMk k2 k2 [S]= +
Plot von 1/k gegen 1/[S] gibt direkt KM (und k2 )
ohne jedoch k1 ' und k1 zu ergeben aus
Michaelis-Menten KinetikMichaelis-Menten Kinetik

ACV HS 2012-30
Auswahl der Elemente für biologische Prozesse: abhängig von mehreren Faktoren
-Die absolute Menge an Element in der Erdkruste oder im Wasser
-Die chemische Form der Elemente
-Die Möglichkeit zur Mobilisierung dieser Elemente
Während der erste Punkt in absoluten Werten mehr oder weniger fixiert ist, ist der 2. und 3. Punkt ganz wesentlich von Umgebungsbedingungen abhängig,
vorwiegend pH und Redoxpotentialen.
Diese Variablen beeinflussten im Laufe der Evolution sowohl die chemische Form wie damit auch die Verfügbarkeit.
2. Die Selektion der Elemente2. Die Selektion der Elemente

ACV HS 2012-31
Während die Elemente der 2. und 3. Übergangsmetalle an sich schon selten und damit schwierig verfügbar sind, sind die
Elemente der 1. Übergangsmetallreihe vergleichsweise häufig.
Die Verfügbarkeit hat sich aber im Laufe der Evolution signifikant verändert
Die Selektion der ElementeDie Selektion der Elemente

ACV HS 2012-32
Im Gegensatz dazu ist heute Fe2+ nicht vorhanden, während Cu2+ und Zn2+ existent sind.
Ein guter Überblick über die Verhältnisse sowie über die Verfügbarkeiten gibt diefolgende Figur, welche die Leichtigkeit darstellt, mit welcher die Elemente aus der
Kruste herausgelöst werden können.
Die Selektion der ElementeDie Selektion der Elemente

ACV HS 2012-33
-Redoxaktivität, welche über die Ligandsphäre getunt werden kann,
-eine Koordinationsgeometrie die stark vom Oxidationszustand abhängig ist
-sehr schnellen Ligand/substrataustausch
- starke Lewisacidität
Aktivierung von Substraten
Tatsächlich finden sich für viele aktiven Eisenenzyme funktionell ähnliche Kupferenzyme
Kupfer als aktives Metall kann Eisen in vielen Reaktionen ersetzen.
Hr: Hemerythrin Fe2+-Fe2+ Fe2+-Fe3+ Fe3+-Fe3+
Hc: Hemocyanin Cu+-Cu+ Cu+-Cu2+ Cu2+-Cu2+
deoxy semi-met met
O2 Transporter
3. Ausgewählte Beispiele: Cu-haltige O2 Transporter und Aktivierer3. Ausgewählte Beispiele: Cu-haltige O2 Transporter und Aktivierer

ACV HS 2012-34
Die verschiedenen Zentren in Cu-Enzymen geben Hinweise auf ihre Funktion
Typ 1 Typ 2
- koordinativ gesättigt- stabile Liganden
- stark tetraedrisch verzerrt
- koordinativ ungesättigt- praktisch planar
- ESR aktiv
CuN N
S-
SCH3
HN NH
(met)
(His) (His)
(Cys-)
- dimeres mit und ohne Brücke- 2 oder 3 histidine
-ESR inaktiv -(antiferromagnetisch gekoppelt)
Typ 3
3. 1. Cu-haltige O2 Transporter und Aktivierer3. 1. Cu-haltige O2 Transporter und Aktivierer

ACV HS 2012-35
O2 Bindung in Typ III Kupferzentren
-staggered imidazoles along Cu……Cu bond
-kein Brückenligand
- praktisch planar
Hämocyanin (Arthropoden und Mollusken) Struktur von deoxy-Hc
(his)3 Cu……Cu(his)3
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer

ACV HS 2012-36
- Im Unterschied zu Hb ist Hc ein teilweise kooperatives System
- Hr und Hc kommen in einigen Wirbellosen vor
-Im Gegensatz zu Hb/Mb, wo O2 im wesentlichen als Neutralmolekülgebunden ist, findet Hc (und Hr) ein 2-Elektronen Redoxprozess statt
- Gebunden ist in jedem Fall O22- wie über Resonanz-Ramanspektroskopie
eindeutig gezeigt werden konnte
Hemocyanin
O2 Bindung zwingt strukturelle Änderung auf.
praktisch planare N2 (O2 )N2 Koordination ein his in quadratisch-planare Anordnung
Basis für Kooperation in den multimeren Hc Proteinen.z. Bsp. trigonal antiprismatisches Hexamer
(bis 48 subunits bekannt)
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer
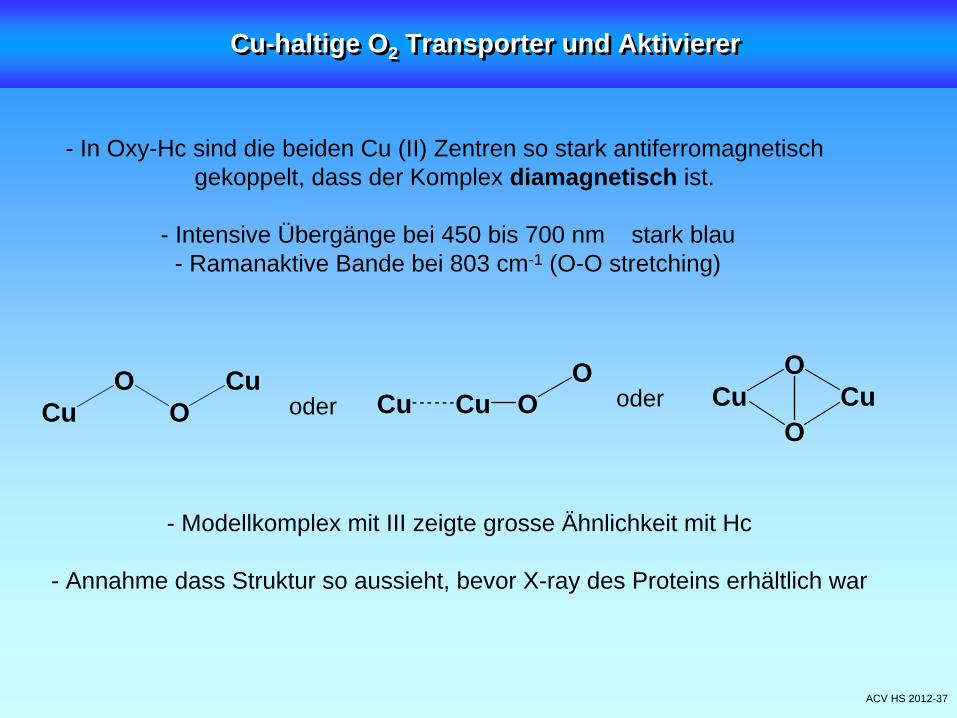
ACV HS 2012-37
- In Oxy-Hc sind die beiden Cu (II) Zentren so stark antiferromagnetischgekoppelt, dass der Komplex diamagnetisch ist.
- Intensive Übergänge bei 450 bis 700 nm stark blau- Ramanaktive Bande bei 803 cm-1 (O-O stretching)
- Modellkomplex mit III zeigte grosse Ähnlichkeit mit Hc
- Annahme dass Struktur so aussieht, bevor X-ray des Proteins erhältlich war
CuO
OCu
Cu Cu OO
oder CuO
CuO
oder
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer

ACV HS 2012-38
-O2 Reduktion und Aktivierung für den Einbau stellt ein Mehrelektronenprozess dar
- Oxidasen koppeln den 1-, 2- oder 4-e- Prozess der Substratoxidation zum 2- oder 4- e-
der Reduktion von O2 zu H2 O2 oder H2 O
- Oxygenasen bauen entweder 1 oder 2 Sauerstoffatome ins Substrat ein
oxidases 2 electron 2 RH + O2 2 R + H2O2RH2 + O2 R + H2O24 RH + O2 4 R + 2 H2O
4 electron 2 RH2 + O2 2 R + 2 H2ORH4 + O2 R + 2 H2O
monooxygenases External(uncoupled)
S + RH2 + O2 SO + R + H2OS' + RH2 + O2 S' + R' + H2O2
Internal SH2 + O2 SO + H2Odioxygenases Intramolecular S + O2 SO2
Intermolecular S + Co + O2 SO + CoO
aRHn = reductant; S = substrate; S' = poor substrate; Co = cofactor
Oxidase and Oxygenase Reactions
Der Übergang von Cu-O2 Transportern zu Oxidasen, Oxygenasen
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer

ACV HS 2012-39
Mechanismus einer Aminoxidase
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer
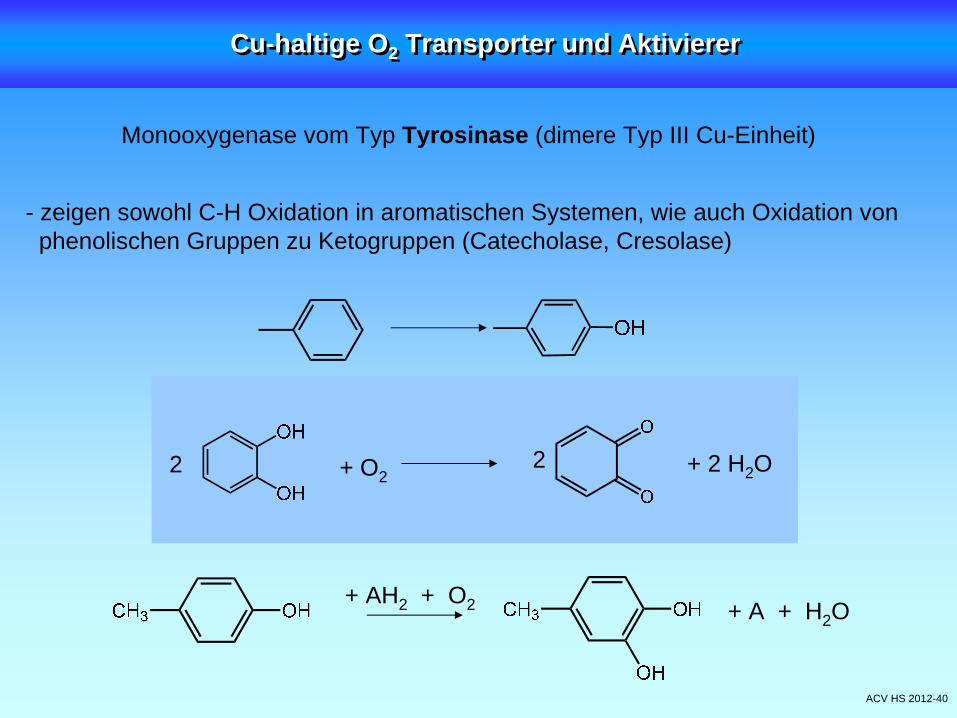
ACV HS 2012-40
- zeigen sowohl C-H Oxidation in aromatischen Systemen, wie auch Oxidation von phenolischen Gruppen zu Ketogruppen (Catecholase, Cresolase)
Monooxygenase vom Typ Tyrosinase (dimere Typ III Cu-Einheit)
+ O22 2 + 2 H2 O
+ AH2 + O2 + A + H2 O
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer

ACV HS 2012-41
OHHO
CuII CuIIN O
N O N
N
O O
CuII CuIIN O
N O N
N
Cu+ Cu+N
N N
N
OHHO
OO
CuIICuIIN N
OH NN
CuII CuIIN
N N
N
OH
OO
O
CuII CuIIN O
N O N
N
OH
+ H2 OH+
2H+
met
oxy-D
oxy
oxy-T
H+
Diphenolasecycle
Monophenolasecycle
deoxymet-D
3H+
+ H2 O2H+
O2
OO
Oxidation von Phenolen zu Chinonen
Mechanismus einer dinuklearen Tyrosinase

ACV HS 2012-42
Es ist eine interessante Parallele zwischen Hc/ Tyrosinase und den dinuklearen Fe-Zentren in Hr und MMO feststellbar
beide haben
- reversible O2 Anbindung
- gemischte Funktionen als Oxidasen/Oxygenasen,
abhängig von Proteinstruktur in welche sie eingebettet sind.
Cu-haltige O2 Transporter und AktiviererCu-haltige O2 Transporter und Aktivierer

ACV HS 2012-43
Begriffe im globalen N2 Kreislauf
-N2 - Fixierung N2 + 3H22NH3
H°R = -46.2 kJ / molS°R = -198 en6° R = -16.7 kJ / mol
N2 + 10H+ + 8e-
eNitrogenas
bar1/C20
'A/K/Fe
bar100/C400
-Nitrifizierung NH4+ + 2O2 NO3
- + H2 O + 2e-
-Denitrifizierung 2NO3- + 12H+ + 10e- N2 + 6H2 O
z.B. Flavobakterien für O2 Gewinnung
2 [NH4 ]+ + H2
Dies ist eine relativ grobe Einteilung, es sind auch N-Verbindungen in anderen
Oxidationsstufen beteiligt z.Bsp. NO2-, NO etc.
3.2. Nitrogenase – N2 Fixierung3.2. Nitrogenase – N2 Fixierung

ACV HS 2012-44
Vor der Einführung des Haber Prozesses, waren Gewitter und vulkanische Aktivitäten die einzigen Zulieferer von fixiertem N2 neben der bakteriellen Tätigkeit.
Heute ist die N2 Fixierung aus menschlichen Prozessenvergleichbar mit natürlichen Kreisläufen.
Gesamt Umsatz N2 :
250 – 300 · 106 t/y = 1 ppm von N2 gesamt
Jedes N-Atom ca. 4000 x transformiert(O2 nur ca. 60 x)
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-45
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-46
N2 wird ausschliesslich durch ....
.....Prokaryonten (Z.B. Azobacter)
.....Rhizobium Knöllchenbakterien
.....Blaualgen....
Bilanzen:Gewitter + 30 ·106 t Dünger + 80 ·106
Biologie + 175 ·106 t Nitrifikation –20 ·106
Denitrifikation – 210 ·106
gesamt ~O
....fixiert und in eigene Aminosäuren etc. eingebaut oder als NH4+ abgegeben.
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-47
NH4+ + 3O2 + 2H2 O 2NO2
- + 4H3 O+
2NO3- + O2 2NO3
-
dann NO3- - NH2
resp. -NH2 CO(NH2 )2 NH3 + CO2
Pflanzen bevorzugen NO2- und nicht NH4
+, deshalb bauen aerobeMikroorganismen NH4
+ über NO2- zu NO3
- unter Energiegewinn ab.
Nitrosomas:
Nitrobacter:
damit ist der Kreislauf wieder geschlossen
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-48
Mikroorganismen: diazotroph
Azobacter vinelandii: aerobClostridium pasteurianum: anaerobKlebsiella pneumoniae: je nachdem
die meisten haben keine agronomische Bedeutung, da freilebend (schwimmend)
nur symbiotische sind von Bedeutung
N2 + 8H+ + 16 Mg ATP + 8 e- 2 NH3 + H2 + 16 MgADP + 16 PO4 3-N2 + 8H+ + 16 Mg ATP + 8 e- 2 NH3 + H2 + 16 MgADP + 16 PO43-
4 Klassen von N2 -Enzymen, 3 sehr ähnlich, eine ist Superoxidabhängig.
in allen Mo-abhängigen Nitrogenasen ist die Struktur stark konserviert
Annahme gilt auch für Rhizobia (agronomisch relevant)
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-49
Die overall Struktur von Nitrogenase aus Azobacter vinelandii
setzt sich aus 2 Komponenten zusammen.Komponente II
Komponente I
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-50
Das Fe-Protein ist eine homodimere Einheit,
wobei der [4Fe-4S] Cluster die beiden Einheiten verbindet.
Das Fe-Protein enthält zusätzlich 2 Bindungsstellen für Nukleotide,
eine auf jeder Einheit.
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-51
Das FeMo-Protein ist ein 2 2 tetramer, welche je einen neuartigen
[8Fe-7S] Cluster (P-Cluster) und einen MoFe Cofaktor enthalten
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-52
Insgesamt stellt sich die Organisation in der Nitrogenase schematisch wie folgt dar:

ACV HS 2012-53
Mechanistische Vorstellung des Transfers von einem Elektron
- Das Fe-Protein scheint nur die Rolle eines Reduktionsmittels zu spielen,
Rolle in Wirklichkeit viel komplexer
- Es finden Wechselwirkungen zwischen Fe-Cluster, Nucleotiden und
Bindungsstelle FeMo-Protein statt
-Substrat (N2 ) Reduktion findet ausschliesslich mit dem Fe-Protein statt
-Der FeMoCo kann reduziert oder oxidiert werden (durch kleine Moleküle)
es findet aber keine N2 -Reduktion ohne die anderen Teile statt
" Driving force" ist nicht der beschränkende Schritt
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-54
Bild der Situation im Fe-Protein
Fe-Protein ist nicht nur Reduktionsmittel
rot: ohne gebundenens Nukleotidblau: mit gebundenem MgADP und [AlF4 ]-
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-55
Wichtige (ungelöste) Fragen:
- Wie ist MgATP Bindung mit e--Transfer und Substrat Reduktion verknüpft?
- Was ist die Rolle des P-Clusters im e- -Transfer?
- Wie bindet das Substrat im FeMoco?
Nukleotid-[4Fe-4S] Cluster: ~15Å
Signal Übertragung durch Konfromationsänderung
Signal Übertragung bis zum eigentlichen Substrat
Switches
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-56
[4Fe-4S]
Geht je zu einem terminalen cystein in [4Fe-4S]
Signifikante Konformationsunterschiedemit / ohne Nukleotid
Nukleotid Bindung wird an [4Fe-4S] weitergegeben
Mechanistische Betrachtungen
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-57
Wichtiger Schritt ist auch die Assoziation von Fe-Protein mit FeMo-Protein
Diese Konformationsänderung wird als Switch I bezeichnet
Gemäss Stöchiometrie muss nach jedem e- Transfer das Fe-Protein
dissozieren und einer neuen Komponente II Platz machen!!!
Rate limiting step
-Switch I ist Kommunikationsweg zwischen Fe-Protein und FeMo-Protein
Postulat für Mechanismus:
MgATP + Fe-Protein
Assoziation
Dissoziation
e--Transfer
HydrolyseMgADP + Fe-Protein
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-58
Der [8Fe-7S] – Cluster – P-Cluster
liegt an der Granzfläche -, -subunit, ~10Å im Inneren verborgen
und ungefähr äquidistant zu [4Fe-4S] und [FeMoCo]
Cluster Struktur:
Zwei Würfel, über ein gemeinsames, sechsfach koordiniertes “S“ verbrückt.
Im Proteingerüst verankert, 2-terminale cys, 2 verbrückende cys pro subunit
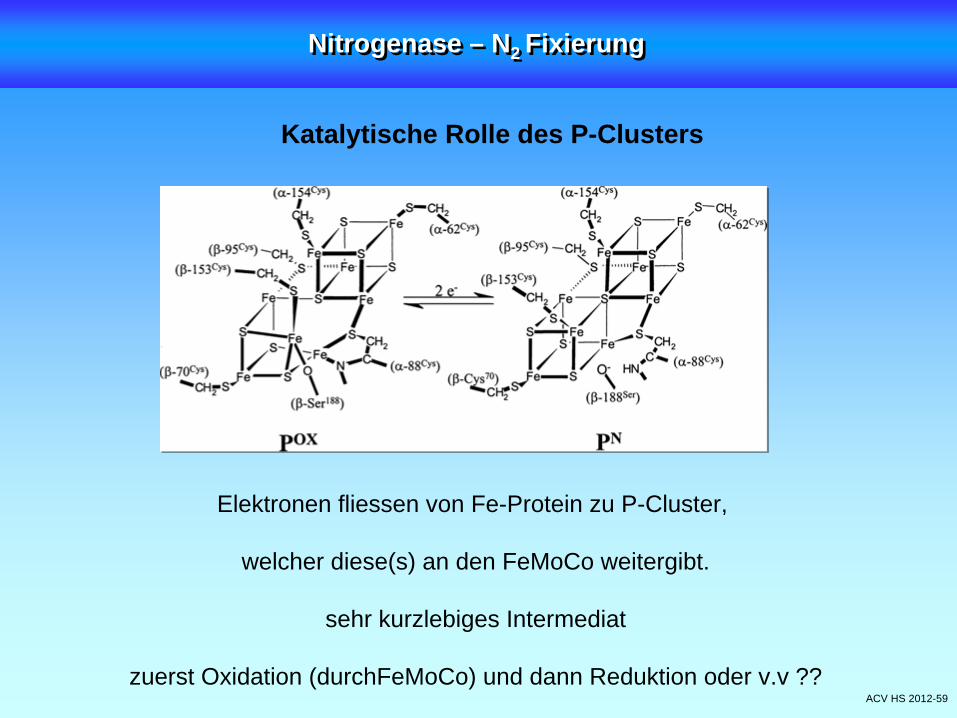
ACV HS 2012-59
Katalytische Rolle des P-Clusters
Elektronen fliessen von Fe-Protein zu P-Cluster,
welcher diese(s) an den FeMoCo weitergibt.
sehr kurzlebiges Intermediat
zuerst Oxidation (durchFeMoCo) und dann Reduktion oder v.v ??
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-60
Elektronentransfer von P-Cluster zu FeMoCo
DistanzDonor - Akzeptor E0 Unterschiede
Natur des “Mediums”Effizienz in biologischem e--Transfer abhängig von:
vier parallele -Helizes gehen von P-Cluster zu FeMoCo,
zwei davon sind direkt über cys in P-Cluster verankert
und führen in die direkte Umgebung von FeMoCo
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-61
Das Herz des Katalysators, der Eisen-Molybdän Cofaktor (FeMoCo)
FeMoCo vollständig in -subunit der Komponente I
aufgebaut aus [4Fe-3S] und [3Fe-3S-Mo], abgeleitet aus würfelartigen Strukturen
homocitrat an Mo gebunden
gegenseitige Enden über cys / his im Proteingerüst verankert
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-62
Mo Koordination, 3S, 2O, 1N
entlang der Proteinachse
Das Herz des Katalysators, der Eisen-Molybdän Cofaktor (FeMoCo)

ACV HS 2012-63
Das Herz des Katalysators, der Eisen-Molybdän Cofaktor (FeMoCo)
In der ursprünglichen Ansicht sind 6 Fe trigonal (!) koordiniert
Eine neuere hochaufgelöste Struktur zeigt aber in der Mitte der
Cluster ein unbekanntes Atom X (N, C, oder O?)
Einsle et al. Science, 2002, 279, 1696
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung
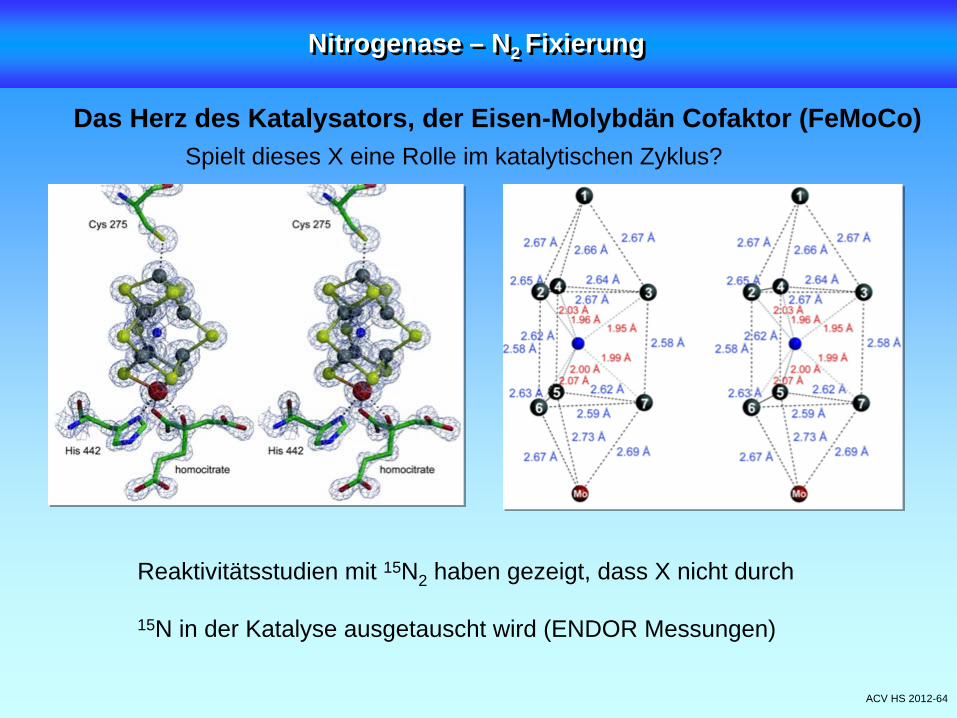
ACV HS 2012-64
Das Herz des Katalysators, der Eisen-Molybdän Cofaktor (FeMoCo)Spielt dieses X eine Rolle im katalytischen Zyklus?
Reaktivitätsstudien mit 15N2 haben gezeigt, dass X nicht durch
15N in der Katalyse ausgetauscht wird (ENDOR Messungen)
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-65
Substrat Bindungsstellen am FeMoCo
Verschiedene Möglichkeiten, aber keine eindeutig verifiziert
- Kleine, olefinische Substrate reagieren mit Nitrogenase und werden
reduziert, z.Bsp. C2 H2 , CS2 , C2 H4 etc
- CO ist ein potenter Inhibitor
Diese kleinen Moleküle verlangen aber nur wenig Elektronen
2 H+ + 2e- H2
C2 H2 + 2 H+ + 2e- C2 H4
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-66
Substrat Bindungsstellen am FeMoCo
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-67
Mechanismus Substratbindung und Reduktion
Thorneley-Lowe stellten ein Modell vor, welches der sukzessiven
Elektronenlokalisation am FeMoCo Rechnung trägt
Dieser Mechanismus ist prinzipiell akzeptiert
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-68
Mechanismus Substratbindung und Reduktion: DFT
Bei der Katalyse wird immer ein H2 frei pro Reduktion von einem N2
auch bei hohen (50 atm) N2 Drucken.
ohne N2 wird H+ sehr effizient zu H2 reduziert
Bei der Katalyse wird immer ein H2 frei pro Reduktion von einem N2
auch bei hohen (50 atm) N2 Drucken.
ohne N2 wird H+ sehr effizient zu H2 reduziert
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-69
Nitrogenase: Mechanismus, nochmaliger Überblick
Nitrogenase – N2 FixierungNitrogenase – N2 Fixierung

ACV HS 2012-70
Die einfachste Reaktion in der Chemie
2H+ + 2e- H2
Benötigt sehr komplexe Enzyme, um sie in Wasser und bei RT durchzuführen
Intensive biologische und chemische Studien werden gemacht, um diese
Reaktionen in Brennstoffzellen oder zur H2 -Produktion zu verwenden.
Wasserstoff Zeitalter
3.3. Hydrogenasen3.3. Hydrogenasen

ACV HS 2012-71
H2 existiert als ortho- und para-Wasserstoff
o-H2 p-H2 G0 = -0.08 kJ/mol
Umwandlung ohne Katalysatoren dauert Jahre
Mit Katalysatoren (paramagnetische Substanzen) Sekunden
durch Absorption an Al2 O3 kann o-H2 in 99% Reinheit gewonnen werden
Hydrogenasen: Einige Eigenschaften von H2 Hydrogenasen: Einige Eigenschaften von H2

ACV HS 2012-72
H2 2H+ + 2 e- Eo = -0.41V (pH=7)
Die einfachste Reaktion der Welt
Ei = +1312 kJ/mol ED = 436 kJ/molEa = -73 kJ/mol pKs = 39 ( entspricht H- = 10-32 M)H+:Hhyd = -1168 kJ/mol H-:Hhyd = -350 kJ/mol
- genügend reduzierendes Potential für chemische / biologische Reaktionen
Koordination von H2 an ein Metall kann pKs bis zu pKs von H2 SO4 reduzieren
Heterogene H2 Spaltung
Hydrogenasen: Einige Eigenschaften von H2 Hydrogenasen: Einige Eigenschaften von H2

ACV HS 2012-73
H2 2H+ + 2 e- Eo = -0.41V (pH=7)
- Genügend stark reduzierendes Potential für chemische Reaktionen
- H-H Bindung sehr stark ~ 400 kJ/mol
- pKa von H2 sehr hoch ~ 35
- Koordination von H2 an ein Metall kann pKa bis um 20 erniedrigen
Heterogene Spaltung
H2 Produktion und Verbrauch in Mikroorganismen: ~ 200 Mio t/y!!!
H2 Herstellung heute: - elektrolytisch (teuer)- Wasser-Gas-Shift Reaktion- aus Methan oder KW
Hydrogenasen: Einige Eigenschaften von H2 Hydrogenasen: Einige Eigenschaften von H2

ACV HS 2012-74
Die biochemische Wassergas-Shift Reaktion
H2 O + CO H2 + CO2 H0 = - 42 kJ/mol
Verschiedene Enzyme sind in die CO Produktion involviert
Das bekannteste ist die CO Dehydrogenase (CODH)
Bacterial roles of H2ases
Methanogenic
Photosynthetic Nitrogen fixing
AcetogenicSulfate reducing
[Fe only] [NiFe] [NiFeSe]H2 production
Activity 10 - 100 x [NiFe]H2ase
Most O2 sensitive H2ase
H2 consumption
Majority of H2ases
H2 affinity 100 x [Fe]H2ase
H2 consumption
Terminal S-Cys Se-Cys
Least O2 sensitive H2ase
D.J. Evans et al. Chem. Soc. Rev. 2003, 32, 268Desulvoibrio desulfuricans Desulvovibrio gigar
HydrogenasenHydrogenasen

ACV HS 2012-75
-methanogenen-acetogenen
-Nitrat- und Sulfatreduzierenden Bakterien-anaeoroben archae Bakterien
-anaeoroben Algen
-methanogenen-acetogenen
-Nitrat- und Sulfatreduzierenden Bakterien-anaeoroben archae Bakterien
-anaeoroben Algen
Produktion aus anaerober Fermentierung von Kohlehydraten, Fetten etc.
H+ als Elektronenempfänger Methanogene als H2 EmpfängerCO2 + H2 CH4 + H2 O
oder Sulfatreduzierer als ultimative Elektronenempfänger
HydrogenasenHydrogenasen
Hydrogenasen erstmals 1931 entdeckt
gefunden in:

ACV HS 2012-761FEH J.W. Peters et al. Science, 1998, 282, 1853
Fe-only Hydrogenase aus Clostridium pasteurianum
eine Stelle an Fe ist frei (weit entferntes Wasser Molekül) kann ..
.. aber leicht durch exogenes CO besetzt werden (1C4A)
vacant site
HydrogenasenHydrogenasen

ACV HS 2012-77
Der proximale Cluster geht über den medialen zum distalen, Abstände ca. 10Å, optimal für e- Leitung sogenannter “hard wire“.
Nahe beim distalen Cluster findet man oft Bindungstasche für Cytochrome
HydrogenasenHydrogenasen

ACV HS 2012-78
Fe-only Hydrogenase aus Clostridium pasteurianum
neuere Erkenntnis
F.A. Armstrong, Curr. Opin. Chem. Biol., 2004, 8, 133M. Frey, ChemBioChem, 2002, 3, 153
D2 O + H2 HOD + HD
Fe-only HydrogenasenFe-only Hydrogenasen

ACV HS 2012-79
Das aktive Zentrum enthält in seiner reduzierten Form ungewöhnliche Einheiten
Fe2 hat eine freie Koordinationsstelle und scheint da mit H2 zu reagieren
Ebenfalls Koordination von CO und CN
Hoch Feld Liganden low spin Fe
Fe-only Hydrogenase aus aus D. desulfuricans (DdHase)
Fe-only HydrogenasenFe-only Hydrogenasen

ACV HS 2012-80
Mechanistische Betrachtungen Möglicher katalytischer Zyklus
H-Cluster = Fe2 -Zentrum, Anbingungsstelle ist 5-fach koordiniertes Fe
Heterolytische H2 -Spaltung
Fe Fe
S
COCN CN
CO
HSH SH
HH2
Cluster
Cys
N
H+
Fe-only HydrogenasenFe-only Hydrogenasen

ACV HS 2012-81
Fe-only Hydrogenase aus Clostridium pasteurianum
Reaktionszentren (in Hydrogenasen) enthalten alle Metallspezifischen Merkmale
- Redoxfähigkeit- Flexibilität in Koordinationssphäre- wechselnde Anzahl Bindungspartner- schneller Substrat- Ligandenaustausch- variable elektronische Struktur - hoher Substrat Durchsatz 9000 H2 /sec-1 (200‘000 l/sec.mol)
Fe-only Hydrogenase Fe/Ni Hydrogenase Überlagerung
Fe-only HydrogenasenFe-only Hydrogenasen

ACV HS 2012-82
Übersicht Struktur
“Fe-only”FeNi
Fe-only Hydrogenase aus aus D. desulfuricans (DdHase)
Fe-only HydrogenasenFe-only Hydrogenasen

ACV HS 2012-83
3[4Fe-4S] Clusteractive site : 2 kerniges Fe-Zentrummw : 46 kDVm (H2 ) : 10000mol/min/mg (mit MV+ als Donor)aerobic (Ausnahme)
CO blockt in allen Fällen die Reaktion
Wichtige Sequenzen sind in allen Fe-only Hydrogenasen konserviert (~80)
Fe-only Hydrogenase aus aus D. desulfuricans (DdHase)
Fe-only HydrogenasenFe-only Hydrogenasen
Einige Eigenschaften

ACV HS 2012-84
Einbindung in biologische Kreisläufe
Relative Grösse und Metallgehalt der verschiedenen Enzyme
Fe-only HydrogenasenFe-only Hydrogenasen
Fe-only Hydrogenase aus aus D. desulfuricans (DdHase)
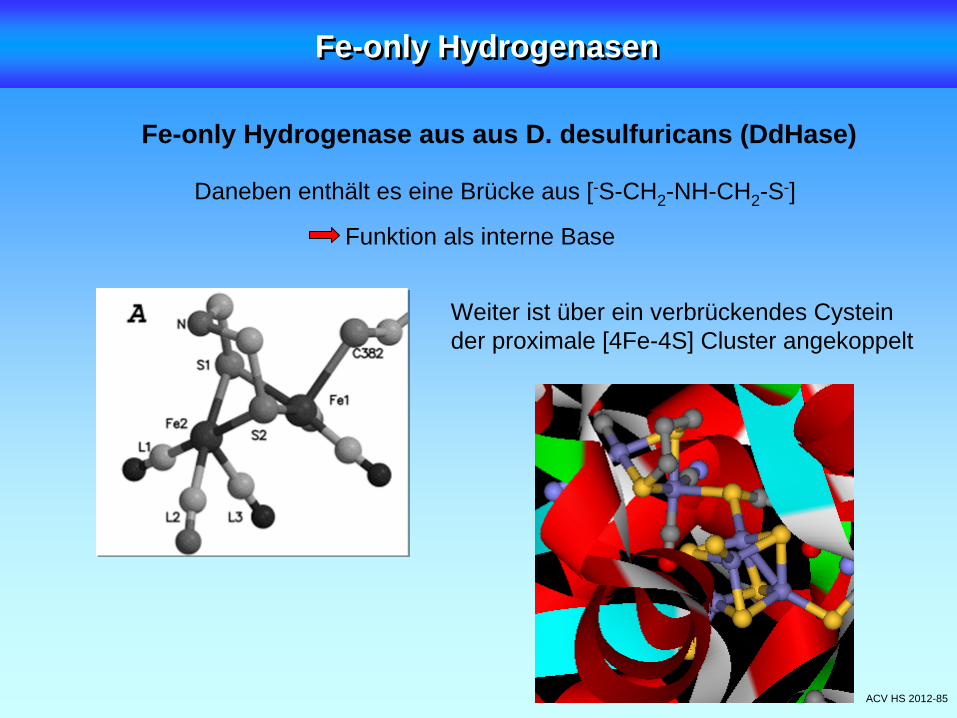
ACV HS 2012-85
Daneben enthält es eine Brücke aus [-S-CH2 -NH-CH2 -S-]
Funktion als interne Base
Weiter ist über ein verbrückendes Cysteinder proximale [4Fe-4S] Cluster angekoppelt
Fe-only HydrogenasenFe-only Hydrogenasen
Fe-only Hydrogenase aus aus D. desulfuricans (DdHase)

ACV HS 2012-86
Enzym aus methanogenen Archaea katalysiert die Reduktion eines Pterins
[HC
H4 MPT]+ + H2
H2 C
H4 MPT + H+ G° = -5.5 kJ/mg
-Diese Reaktion ist ein Intermediärschritt der CH4 Bildung aus CO2 + H2
-Vergleichbar mit der Bildung von Carbokationen in Supersäuren
Metallfreie HydrogenasenMetallfreie Hydrogenasen
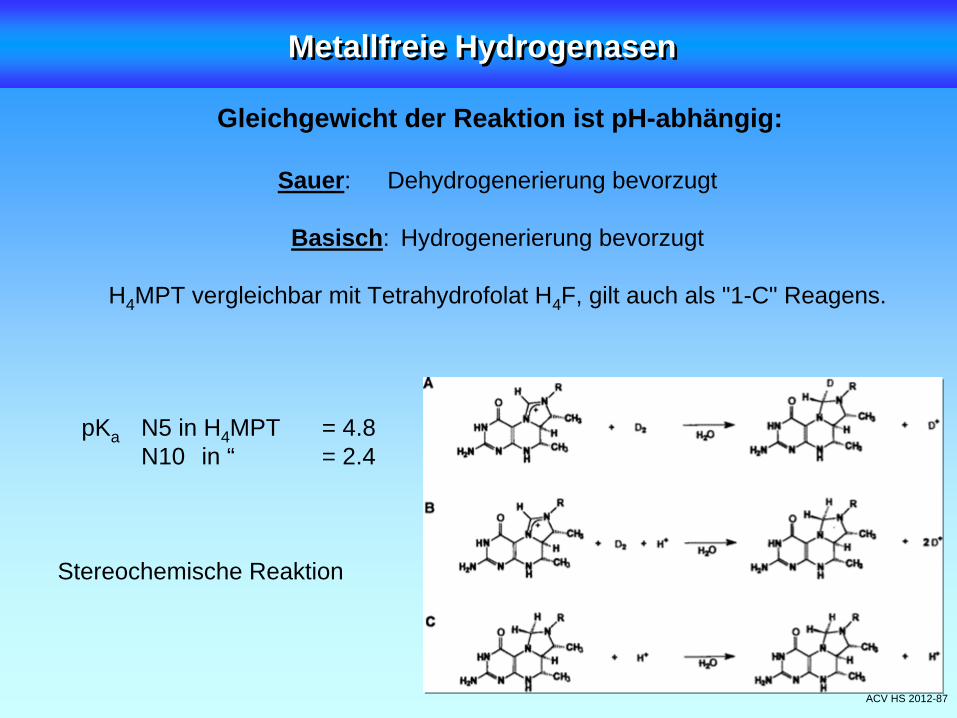
ACV HS 2012-87
Gleichgewicht der Reaktion ist pH-abhängig:
Sauer: Dehydrogenerierung bevorzugt
Basisch: Hydrogenerierung bevorzugt
H4 MPT vergleichbar mit Tetrahydrofolat H4 F, gilt auch als "1-C" Reagens.
pKa N5 in H4 MPT = 4.8N10 in “ = 2.4
Stereochemische Reaktion
Metallfreie HydrogenasenMetallfreie Hydrogenasen

ACV HS 2012-88
- Reaktionsgeschwindigkeit mit D2 und H2 ungefähr gleich
- folgt M-M Kinetik
Km ~ 50 · 10-6M
Vmax ~1000 U/mg
-Kein H2 / H+ Austausch ohne HC
H4 MPT
resp. H2 + D+ HD + H+
H2 + 2D+ D2 + 2H4
para H2 ortho H2
(alle HC
H4 MPT+ abhängig)
Metallfreie HydrogenasenMetallfreie Hydrogenasen

ACV HS 2012-89
Der Mechanismus ist vergleichbar mit Supersäuren
(H3 C)3 C-H + H+(H3C)3--H
H(H3 C)3 C++H2
Das C14 hat ähnliche Eigenschaften
[HC
H4 MPT]+
HN
N+ + H2 H
N
N
H+ H+
Metallfreie HydrogenasenMetallfreie Hydrogenasen

ACV HS 2012-90
Für Carbokation werden Supersäuren gebraucht
im Gegensatz ist die Bildung von H2 aus H2 C
H4 MPT bei pH=7 bereitsexergonisch
spiegelt die Delokalisierung der Ladung und Solvatation
- H2 C-H4 MPT muss so eingerastet sein, dass die Ione-pairs anti-periplanarzum “H-” sind
- Planare Form muss destabilisiert werden, damit Reaktion mit H2 möglich
schliesslich ....
Metallfreie HydrogenasenMetallfreie Hydrogenasen

ACV HS 2012-91
Ein Beispiel aus der Bioanorganik: HydrogenasenEin Beispiel aus der Bioanorganik: Hydrogenasen
.... war alles ein Irrtum und das Enzym eine Fe Hydrogenase
The Crystal Structure of [Fe]-Hydrogenase Reveals the Geometry of the Active Site Science 2008 321, 572.
Allgemeiner Review: Structure–function relationships of anaerobic gas- processing metalloenzymes Nature 2009 460, 814.

ACV HS 2012-92
3. Ein Beispiel aus der Bioanorganik: Hydrogenasen3. Ein Beispiel aus der Bioanorganik: Hydrogenasen
Möglicher Mechanismus der Fe Hydrogenase:
The Crystal Structure of an [Fe]-Hydrogenase-
Substrate Complex Reveals the Framework for H2 Activation Source Angew. Chem. Int. Ed.
2009 48, 6457.


















