
DIE AUGEN DER MOLLUSKEN - techfak.uni-bielefeld.detkaempfe/lehre/UWvision/... · 8 Die Augen der...
19
Seminar Unterwasserbildverarbeitung SoSe 2002 DIE AUGEN DER MOLLUSKEN Hana Boukricha Andrea Finke Michael Bongenberg
Transcript of DIE AUGEN DER MOLLUSKEN - techfak.uni-bielefeld.detkaempfe/lehre/UWvision/... · 8 Die Augen der...

Seminar Unterwasserbildverarbeitung SoSe 2002
DIE AUGEN DER MOLLUSKEN Hana Boukricha Andrea Finke Michael Bongenberg

2
Referat: Unterwasserbildverarbeitung. Stamm Molluska – Klasse Cephalopoda: Die Augen der Molusken.
1. Einführung zur Mollusken (Cephalopoda)
1.1. Arten 1.2. Lebensraum
2. Retina und Photorezeptorenzellen
2.1. Definition 2.2. Rolle 2.3. Photorezeptoren
3. Augen
3.1. Grubeaugen (Nautilus) 3.2. Linseauge: Vergleich mit Wirbeltiere (Octopus)

3
1.1. Mollusken sind Wirbellosetiere, sie wurden auch als Weichtiere genannt. Es gibt zwei Arten von der Klasse Cephalopoda im Stamm Molluska und zwar Octopus (Tintenfische) und Nautilus (Tintenschnecken). Die Cephalopoda sind sehr entwickelte Mollusken und stammen aus dem Cambrian seit 400 Millionen Jahren. Octopus sind Mollusken ohne Schale, Nautilus sind aber mit Schale.
1.2. Die Cephalopoda leben nur im Meer und sind Carnivors. Sie haben sehr
schnelle Bewegungen und sehr entwickelte Augen.
Figur 1: Arten von Cephalopoda
2.1. Die Retina enthält das neuronales Stoff des Auge, denn sie gehört zum Nervensystem. Sie ist durch drei Hauptschichtzellen struktuiert und zwar die Ganglionschicht, die Körnige Schicht und die Photorezeptorenschicht. Diese Schichten enthälten die Ganglionzellen, die zweipolligenzellen, die Horizontalzellen, Stäpchen und Zäpfchen. • Die Zweipolligenzellen befinden sich zwischen den Photorezeptorenzellen
und die Ganglionzellen, es gibt zwei Typen und zwar die Stäbchen Zweipoligenzellen, die die Stäbchen mit Ganglionzellen zusammenbinden und die Zäpfchen Zweipolligenzellen, die die Zäpfchen mit Ganglionzellen zusammenbinden.
• Die Horizontalzellen befinden sich zwischen den Rezeptorenzellen und die Zweipoligenzellen.

4
2.2. Die Retina ermittelt die Lichtinformation zum Gehirn. Die vom Aussen
gekommene Lichtinformation wird durch die Retina (Photorezeptoren) in Nervensignalen übersetzt, dann zum Gehirn in Stufen zugeschickt. Die Neuronalenzellen ermitteln zwischen sich die Lichtinformation, die durch die Photorezeptoren in Nervensignalen übersetzt wird und in Stufen zum Gehirn zugeschickt (in elektrichen Stossen).Das Gehirn bearbeitet das Bild Punkt durch Punkt und Farbe durch Farbe.
Figur 2: Retina
2.3. Wenn das Licht durch das Auge geht und die Retina trifft, wird er durch die
Photorezeptorenzellen zum Gehirn ermittelt, diese Photorezeptoren sitzen an der tiefsten Schicht der Retina und sind Lichtempfindlich. Es gibt zwei Typen von Photorezeptoren:Zäpfchen und Stäbchen. • Stäbchen: diese Zellen sind 95% der Photorezeptorenzellen. Sie
ermöglichen das nächtliche Sehen(man sieht alles grau ) und sind zum Unterschied zwischen Dunkelheit und Licht empfindlich. Sie haben die grösseste Lichtempfindlichkeit und sind zur sehr niedrigen Lichtintensität begabt.
• Zäpfchen: grosse sensorielle Zellen. Sie haben einen Anteil von 5% der Photorezeptorenzellen. Diese Zellen ermöglichen das Tageslicht und sind zum Farben Unterschied sehr empfindlich. Über diese Zellen ist das Sehen sehr prezize, detalliert und sensible zur Bewegungen. Dadurch kommen die meisten Sehsignalen zum Gehirn.

5
Figur 3: Zäpfchen und Stäbchen
Zäpfchen sind die Strukturen, die wie ein Olivenkern aussehen, und die Stäbchen sind dazwischen.
Figur 4: Photorezeptoren

6
3.1. Mollusken sehen keine Farben denn sie haben nur Stäbchen als Photorezeptoren. Grubeaugen (Nautilus) sind Augen ohne linse, ohne Cornea, so dass das Innere Teil des Auge offen zum Meer ist. Sie haben auch mobile Pupille deren Durchmesser zwischen 0,4 und 2,8 mm ist, diese Länge variert durch die Lichtintensität.Diese Augen sind mobil und vertikal orientiert wenn das Tier sich bewegt. Es gibt 4.106 Rezeptoren in der Retina, deren Durchmesser zwischen 5 und 10 µm ist. Sie nehmen aber kein richtiges Bild war (die Augen).
Figur 5: Grubeauge
3.2. Die linseaugen bei Cephalopoda (Octopus) sind sehr änlich mit denen von
Wirbeltiere. Sie haben mehr als 20 Millionen Rezeptoren per Aguen. Das grosseste Auge bei Cephalopoda hat einen 40 cm Durchmesser und 1010 Rezeptoren (bei Menschen sind nur 108).
Figur 6: Linseaugen

7
Die Augen der Mollusken (Stamm Mollusca) Klasse Gastropoda GLIEDERUNG
(1) Allgemeines
1.1 Arten 1.2 Verbreitung 1.3 Lebensraum
(2) Augen
2.1 Augentypen / Aufbau 2.2 Eigenschaften 2.3 Auflösungsvermögen 2.4 Linsentyp
(3) Hautlichtsinn 3.1 Photorezeptoren
(4) Besonderheit
4.1 „Scanner“ – Augen bei Heteropoda

8
Die Augen der Mollusken (Stamm Mollusca) Klasse Gastropoda
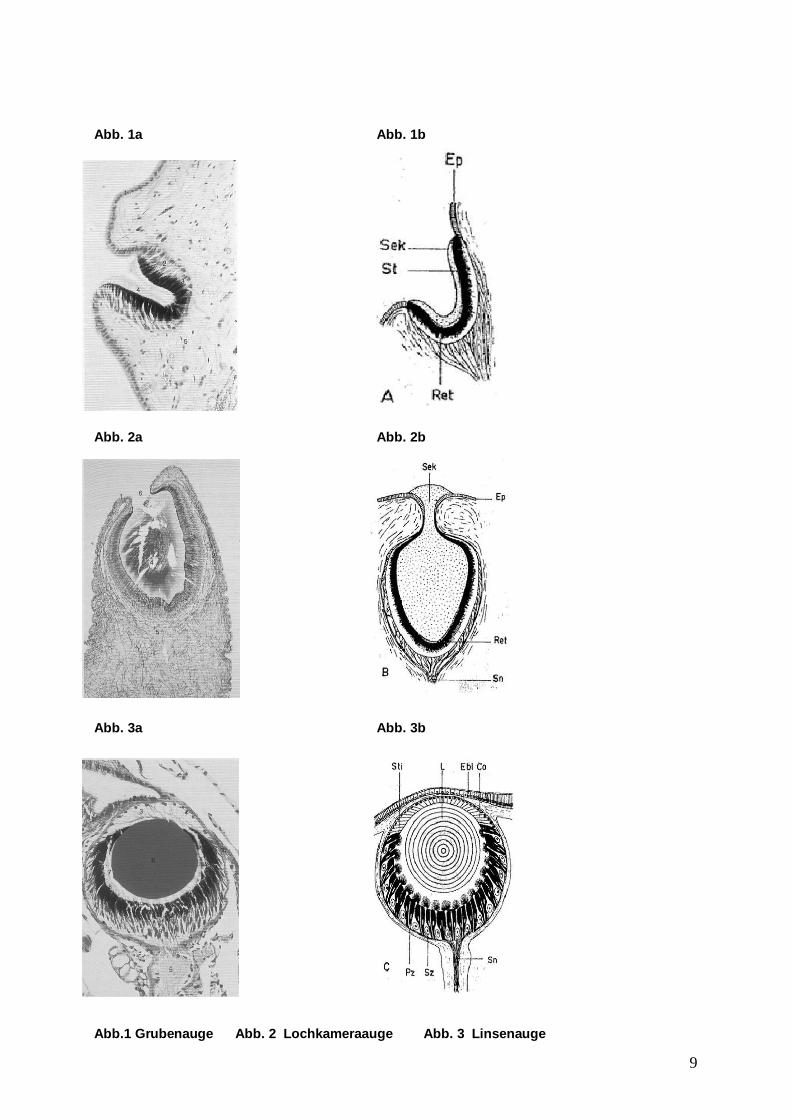
1.1 Die Gastropoda, Bauchfüßer oder Schnecken genannt, gehören zum Stamm der Mollusca (Weichtiere). Diese sind eine Untergruppe der Wirbellosen. Insgesamt gibt es etwa 110.000 verschiedene Arten. 1.2 Gastropoda sind Kosmopoliten. Als einzige Klasse der Mollusken sind sie an Land, in Süßwasser und in den Meeren zu finden, wobei der größte Anteil, nämlich etwa 68.000 Arten im Meer zu finden sind. In Süßwasser leben etwa 10.000 Arten, bei dem Rest handelt es sich um terrestrische Arten. Im Hinblick auf das Thema des Seminars sollen hier primär die marinen Gastropoden betrachtet werden. 1.3 Gastropoden sind vor allem im Küstenbereich und anschließenden Flachwasser zu finden, einige wenige Arten leben jedoch auch in größeren Tiefen. Unter den marinen Gastropoden gibt es sowohl Arten, die im Sande kriechen, als auch Schwimmschnecken und solche, die im Gewirr von Algen leben, zumeist wurmförmige Nacktschnecken. 2.1 Die Mehrzahl der Gastropoden hat ein Paar Augen im Kopf, entweder an der Spitze der Tentakeln (Fühler) oder in Nähe des Celebralganglions. Diese variieren jedoch je nach Art beträchtlich in Größe, Struktur und Anzahl der Rezeptoren. Dieses hat entscheidenden Einfluss auf die optischen Fähigkeiten der jeweiligen Schnecke. Primär lassen sich zwei Typen von Augen unterscheiden: Zum einen die einfachen Gruben- bzw. Lochkameraaugen (z. B. Patella, Haliotis) ohne Linse und die höher entwickelten Linsenaugen, wobei hier die jeweilige Beschaffenheit der Linse ebenfalls noch variiert. Es gibt Linsen, die eine weiche Struktur und einen sehr niedrigen refraktiven Index (Brechungsindex) haben sowie Linsen, die hart und sphärisch sind. Die letztgenannten werden später noch ausführlich behandelt werden.
9
Abb. 1a Abb. 1b
Abb. 2a Abb. 2b
Abb. 3a Abb. 3b
Abb.1 Grubenauge Abb. 2 Lochkameraauge Abb. 3 Linsenauge

10
2.2 Der erstgenannte Typ, nämlich die Augen ohne Linsen in der einfachsten Variante, lassen die Schnecke lediglich Hell und Dunkel unterscheiden und die Lichtrichtung feststellen, die fortentwickelte Version, als Lochkameraauge bezeichnet, entwirft bereits richtige Bilder, wobei diese aber sehr klein sind und von den wenigen Rhabdomeren in eine so geringe Anzahl von Bildpunkten zerlegt werden, dass man nicht von einem richtigen Bildsehen sondern lediglich von einem Bewegungssehen sprechen kann. Formsehen tritt erst bei Linsenaugen auf. Das Formsehen generell bei Schnecken mit Linsenaugen existiert wurde mittels eines einfachen Experiments nachgewiesen. Man ließ die Strandschnecke Littorina (Linsenaugen) auf aufgestellte schwarze Papptäfelchen zukriechen. Diese wurden aus mehreren cm Entfernung angesprochen und durch Änderung der Laufrichtung ohne Berührung umgangen. Generell wird davon ausgegangen, dass Littorina die Fähigkeit besitzt, vertikale und horizontale Strukturen zu unterscheiden. Die Augen der Gastropoden sind extrem lichtempfindlich, die Schwelle liegt teilweise unter 11106 −× Watt/sec. Zum Vergleich: Das menschliche Auge muss bereits einige Minuten dunkeladaptiert sein um eine solche Lichtquelle überhaupt als schwach leuchtenden Punkt zu erkennen. Diese hohe Lichtempfindlichkeit wird auf die große Anzahl von Stäbchen auf der Retina zurückgeführt; bei Pterocera (Strombus) sind es 10.000 auf 1 mm². Eine Besonderheit der visuellen Reaktion ist die Lichtkompassorientierung (Menotaxis), die sowohl bei terrestrischen wie auch marinen Gastropoden zu finden ist. Hier sei wiederum die Littorina beispielhaft angeführt. Diese orientiert sich bei ihren Nahrungsgängen streng nach der Sonne, im Labor reagiert sie auch auf polarisiertes Licht und auf Änderung seiner Schwingungsebene. Der Orientierungswinkel zu den Lichtquellen liegt, soweit bekannt, immer zwischen 45° und 135°. Dieses erklärt sich durch die Augenstruktur der Schnecken, denn einfallendes Licht erreicht ihre Retina nur in einem Winkel von 35°-130°. 2.3 Das Auflösungsvermögen der Gastropoda Augen variiert ebenso stark wie deren Bauart. Ausschlaggebend ist hier die Größe des Auges wie auch die Anzahl der Rezeptoren (Stäbchen). Die Bestimmung des Grades der Auflösung erfolgt mit Hilfe von anatomischen Maßen. Die Berechnung erfolgt durch Dividierung des Abstands zweier Rezeptoren (Stäbchen) durch die Länge des Fokus, bestimmt durch den Abstand des Mittelpunktes der Linse zur Oberfläche der Retina. Dieser Quotient, multipliziert mit 57,3, gibt den Grad der Auflösung an.

11
Abb. 4 Auflösungsvermögen verschiedener Molluskenaugen Spezies Augengröße Auflösung Gastropoda Littorina 155 mµ 4,4° Strombus 1 mm 0,23° Pterotrachea 1250 mµ 0,21° Oxygyrus 540 mµ 0,44° Bivalvia Barbatia 200 mµ 10° Pecten 1 mm 2° Cephalopoda Nautilus 10 mµ 2,3° - 16° Octopus 10 mm 0,02°/1,3 min Biene 1° Mensch 0,008° = 0,5 min
2.4 Die bestentwickelten Augen der Gastropoda verfügen über harte sphärische Linsen, wie sie auch bei den Cephalopoda, den Fischen und überhaupt bei allen Meeresbewohnern, die Augen mit einer Kammer haben , vorkommen. Dieses stellt eine Anpassung an die besonderen Anforderungen des marinen Lebensraumes dar. Das faszinierende an dieser Tatsache ist, dass sich bei den

12
verschiedenen Tieren dieses Sehsystem evolutionär betrachtet völlig unabhängig von einander entwickelt hat. Es ist nicht einmal sicher, ob nicht sogar bei drei Familien der Gastropoda Prosobrachia , die diesen Linsentyp aufweisen, die Entwicklung unabhängig verlaufen ist. Aufgrund dieser Vermutung liegt die These nahe, dass es nur einen einzigen Typ Linsen aus biologischem Material gibt, der den Anforderungen des Unterwassersehens gerecht wird. Die Hornhaut ist - anders als an Land - unter Wasser als Licht brechende Oberfläche unbrauchbar, daher muss die gesamte Brechung allein von der Linse erledigt werden. Der Vorteil einer sphärischen Linse ist, dass sie eine kürzere Fokuslänge aufweist als andere Linsenformen. Doch dies allein würde das Problem nicht lösen, denn eine homogene sphärische Linse würde ernsthafte Abberationen verursachen, die in erster Linie dadurch entstehen, dass das Licht an den äußeren Bereichen zu stark gebrochen wird. Die Natur hat hier eine außergewöhnliche Lösung entwickelt. Statt mit homogenen Linsen sind die schon genannten Meerestiere mit inhomogenen Linsen ausgestattet. Diese haben den Vorteil, dass erstens eintretende Strahlen kontinuierlich und nicht nur an der Oberfläche gebrochen werden und zweitens die Abberationen korrigiert werden, weil die äußeren Bereiche das Licht weniger brechen als die mittleren. Abb. 5 Inhomogene sphärische Linsen
3.1 Gastropoden haben neben den Hauptaugen ein weiteres System, das der optischen Wahrnehmung dient. Dieses wir als Hautlichtsinn bezeichnet. Es handelt sich hier um Photorezeptoren, die entweder über den ganzen Körper verteilt sind oder sich konzentriert an bestimmten Stellen befinden. Während die Hauptaugen primär dem Auffinden und Auswählen des Habitats dienen, lösen die Photorezeptoren den sogenannten Schattenreflex („off response“) aus, der wohl immer eine Schutzreaktion gegen herannahende Feinde darstellt. Die Schnecken reagieren auf

13
eine plötzlichen Wechsel der Lichtintensität (wie in der Natur bei Schatteneinwirkung); ein Abfall von 18 % ist dabei ausreichend, um eine Reaktion auszulösen. Bei einigen Gatropoden (z. B. Onchidium verruculatum) spricht man von Rückenaugen oder augenähnlichen Strukturen am Rücken. Diese Augen haben keine Microvilli sondern statt dessen cilia. Ihre Funktion ist ebenfalls den Schattenreflex auszulösen. Somit kann bei Gastropoden von einem dualen Sehsystem gesprochen werden: Während die Hauptaugen Licht also Helligkeit wahrnehmen, reagieren die anderen Photorezeptoren auf Schatten. Das führt zu der Schlussfolgerung, dass Schnecken zwei unterschiedliche Arten von Rezeptoren mit komplementären Funktionen haben, einfach gesprochen haben sie „an“ und „aus“ Rezeptoren. 4.1 Unter den Gastropoden gibt es eine Gruppe, deren Augen zahlreiche Besonderheiten aufweisen, die Heteropoden. Ein Ursache hierfür mag sein, dass diese Schneckenart als eine der ganz wenigen räuberisch lebt. Die Augen der Heteropoden haben zwar ebenfalls sphärische Linsen, haben aber ansonsten wenig mit denen der anderen Schnecken gemeinsam. Ihre Rezeptoren unterscheiden sich durch zahlreiche cilia. Besonders auffallend ist die Retina, die mehrere hundert Zellen lang, aber nur 3-6 Zellen breite ist. Dies ist einzigartig. Ihre Form ist nicht wie bei anderen Gastropoden ein hemispärischer Halbkreis, sondern ein schmales Band. Man könnte annehmen, dass eine solche Retina nur ein sehr begrenztes Blickfeld bietet, doch dies wäre einem Räuber wenig hilfreich. Er würde wohl verhungern. Die Heteropoden bedienen sich eines Tricks um mit diesen Augen ein großes Sichtfeld abzudecken: ihre Augen vollführen konstante Scan-Bewegungen. Das gesamte Auge bewegt sich um eine Achse durch die Linse und parallel zur Retina, wobei das Sichtfeld der Retina eine Kurve von etwas mehr als 90° beschreibt, das Gebiet unterhalb und seitlich des Tieres abdeckend. Die Bewegungen nach unten sind dabei erheblich schneller (250°/sec.) als die nach oben zurück in die Ruheposition (80°/sec.). Die Augen von Heteropoda stellen eine sehr kompakte und rationelle Realisierung eines visuellen Systems dar. Leider ist es sehr schwer mit Heteropoden zu arbeiten, so dass noch viele Fragen hinsichtlich des Aufbaus ihrer Augen, besonders auch der Rezeptoren, unbeantwortet bleiben.

14
Bivalvia
Das folgende Kapitel beschäftigt sich mit einer Unterart der Mollusken, den sogenannten Bivalvia. Als Bivalvia werden die Lebensformen bezeichnet, die im gängigeren Sprachgebrauch auch als Muscheln bekannt sind. Bisher gibt es in etwa 25.000 bekannte Muschelarten, doch die tatsächliche Anzahl existierender Bivalviaarten liegt laut wissenschaftlicher Schätzungen weit darüber. Insbesondere in den Tiefenregionen der Meere steckt die Artenforschung, nicht nur im Bereich der Bivalvia, noch in den Kinderschuhen. Das liegt unter anderem daran, daß Tiefseeforschung in erster Linie eine Frage des Kapitals ist, da entsprechende Forschungsgeräte, sowie zum Beispiel Tiefensonden, nicht selten einen Anschaffungspreis von mehreren Millionen Dollar haben können, was natürlich viele Forscher abschreckt. Daher sind die Tiefmeere noch immer weitgehend unerforscht, was die dortige Flora und Fauna betrifft. Schätzungen zufolge beläuft sich die Artenanzahl der Bivalvia auf bis zu über 50.000 und sogar noch darüber, je nachdem welcher Quelle man letztendlich seinen Glauben schenkt. Augen der Bivalvia: Fakt ist, Muscheln können ihre Umgebung wahrnehmen. Das hat sicherlich jeder schon mal festgestellt der Schnorcheln war, mit der Hand in die Nähe einer Muschel kam und beobachten konnte, wie sich diese sofort schloss, um sich zu schützen. Die Muschel hat die Hand also scheinbar gesehen. Nun stellt sich die Frage wenn man sich eine „Durchschnittsmuschel“ betrachtet, wo haben diese Tierchen denn überhaupt Augen? Die Augen sitzen bei den Bivalvia am Mantelrand. Das ist jene Region einer Muschel, entlang den äußeren, beiden Rändern ihrer Schale. Dort befinden sich Sinnesorgane wie Fühler und Taster die auf einem Muskelboden sitzen, welcher bei geöffneter Schale leicht hinausgewölbt werden kann, und dort eingebettet befinden sich auch die eben angesprochenen Augen. Die Anzahl der Augen schwankt von Muschelart zu Muschelart sehr stark. Im Schnitt kann man sagen, daß Muscheln zwischen 20-500 Augen besitzen. Allerdings gibt es auch Arten die nahezu blind sind, wie zum Beispiel eine Tiefseemuschel, die allerdings im Gegenzug feinste Fühler von bis zu zwei Metern Länge ausbildet, um dieses Manko scheinbar auszugleichen. Auch gibt es eine Muschel die sich Rote Korallenmuschel nennt, welche knapp an die 5000 Augen zählt. Verwunderlich daran ist die Tatsache, daß diese Muschel nur knapp unter der Wasseroberfläche lebt. Bei den guten Lichtverhältnissen dort ist es bisher noch ungeklärt, wozu diese enorm vielen Augen nützlich sind. Allgemein kann man sagen, daß entgegen der beiden Ausnahmebeispielen eben, die Anzahl der Augen bei den Bivalvia zunimmt, je tiefer sie leben. Das ist leicht erklärlich, zumal das Licht im Wasser zunehmend schlechter wird je tiefer man kommt. Es wird trüber und es kommen Störfaktoren wie umhertreibende Algenfetzen hinzu. Und um sich in diesen Verhältnissen zurechtzufinden, scheinen mehrere Augen von Vorteil zu sein.

15
Mirror Eyes Bis 1965 wurde angenommen, daß Muscheln, genau wie Fische, LensEyes besitzen. Erschüttert wurde diese Annahme in diesem Jahr dann durch einen Forscher namens Land, der sich damit befasste die Augen von Muscheln genauer zu untersuchen. Land führte folgenden Versuch durch: Er untersuchte ein Muschelauge unter einem Mikroskop. Der Versuch war so aufgebaut, das Land mithilfe seines Mikroskops direkt von vorne in das Auge hineinblicken konnte, da er die Netzhaut eines Muschelauges untersuchen wollte. Doch als Land dann schließlich in das Mikroskop blickte sah er nicht wie erwartet die Netzhaut, sondern er sah sich selbst als Spiegelbild im Muschelauge. Dieser Effekt ist bei LensEyes nicht anzutreffen, was Land dann dazu motivierte sich noch mehr mit Muschelaugen zu befassen, da es ja anscheinend doch gravierende, bisher nicht angenommene Unterschiede zum LensEye gab. Seine Forschungen brachten folgende Einsichten:
Das Muschelauge besitzt eine Linse, dahinter zwei Retinas und zusätzlich noch eine Spiegelfläche am Augenhintergrund, die sogenannte Argentea. Dieser Spiegel war es, den Land in seinem Mikroskop gesehen hat. Die beiden Retinas werden unterschieden als eine Distalretina und eine Proximalretina. Die Distalretina liegt vorne im Auge, also hinter der Linse. Die Proximalretina hingegen liegt am Augenhintergrund, fast im Kontakt mit der Spiegelfläche. Mit der Distalretina ist es der Muschel möglich Bilder scharf zu sehen, was folgendermaßen funktioniert: Das Licht fällt durch die Linse in das Auge, wobei der Brechungsindex der Linse allerdings viel zu gering ist, als daß sie das Bild scharf auf eine der beiden Retinas projizieren könnte. Das Bild läge etwa 1-2mm hinter dem Auge, wäre allein die Linse für die Lichtbrechung zuständig. Doch bei den als MirrorEyes bezeichneten Augen der Muschel wird das Licht nach der Brechung durch die Linse noch ein weiteres mal gebrochen, nämlich durch den Spiegel am Augenhintergrund. Das nun zweimal gebrochenen Licht trifft scharf auf die Distalretina des Auges, was es der Muschel ermöglicht mit dieser Retina ein scharfes Bild wahrzunehmen. Auch ein Bewegungssehen ist der Muschel somit möglich.
Jetzt bleibt noch die Frage offen, wofür eine Proximalretina benötigt wird. Das die Proximalretina ein brauchbares, scharfes Bild liefern kann bleibt ausgeschlossen, da das Licht in keiner Weise scharf auf Diese fällt. Doch es gibt Vermutungen, daß die Proximalretina eine erhebliche Rolle spielt, was die Orientierung der Muschel im Wasser angeht. Einige Muscheln können sich fortbewegen. Sie ziehen sich mithilfe eines Fußes über den Meeresgrund, oder sie schwimmen durch schnelles Auf- und Zuklappen ihrer Schale durch das Wasser. Doch eines ist bei allen Muschelarten gleich wenn sie sich bewegen. Sie

16
bewegen sich immer genau zum Licht hin, oder genau vom Licht weg. Die geläufigste und plausibelste Vermutung diesbezüglich ist nun, daß die Proximalretina die Rolle eines Lichtsensors spielt. Die Distalretina könnte diese Information, aus welcher Richtung genau das Licht kommt, im dunklen, trüben Wasser nicht liefern. Allerdings beinhaltet auch diese Vermutung noch ein Problem, denn wenn man sämtliche Rezeptoren aller Proximalretinas einer Muschel aufaddieren würde, käme man auf eine Anzahl von gut 500.000 Rezeptoren. Daraus ergibt sich das sogenannte „SightOverkill“-Problem, denn um lediglich auszumachen aus welcher Richtung das Licht kommt, würden laut Spekulation auch schon ein Dutzend Rezeptoren genügen. Hinzu kommt auch noch die Tatsache, daß jedes Auge ein Sichtfeld von etwa 90° hat. Da die Augen nahe nebeneinander liegen, ergibt sich daraus eine nahezu komplette Sichtfeldüberlagerung. Wozu also diese Reizüberflutung? Annahme: Man geht davon aus, daß dieses Overkill-Problem eine entscheidende Rolle spielt, was die Fehlerreduktion bei der Reizinterpretation angeht. Wie das genau funktioniert, soll anhand eines Beispiels klargemacht werden: Wenn eine Muschel weniger Rezeptoren besäße, zur Vereinfachung in diesem Beispiel angenommen 4, wäre die Fehleranfälligkeit bei einer Reizinterpretation viel größer. Aus folgendem Grund: Im Meerwasser sind diverse Störfaktoren vorhanden, wie zum Beispiel Algenfetzen, Kleinorganismen oder aufgewirbelter Dreck. Dieses alles ist sicherlich für die Muschel keine Bedrohung, doch werfen alle diese Störfaktoren eben auch störende Schatten. Die Muschel muß nun unterscheiden welche Schatten keine Bedrohung bedeuten, oder welche aber eine Bedrohung durch einen Fressfeind ankündigen. Der Schatten eines Fressfeindes ist im Vergleich zu den Störschatten verhältnismäßig groß. Wenn man nun annimmt, das eine Muschel 4 Rezeptoren besäße kann man sich denken, das es schnell zu Fehlinterpretationen kommen könnte, sobald zum Beispiel 4 kleine Algenfetzen genau derart an der Muschel vorbeitreiben, daß die 4 Schatten genau auf die 4 Rezeptoren fallen. Und bei sehr trübem, dreckigem Wasser ist dieser Fall nicht unwahrscheinlich. In der Interpretation der Muschel wären dieses dann allerdings keine 4 kleine, harmlose Schatten, sondern ein gewaltiger Schatten der die gesamte Sicht der Muschel abdecken würde. In dem Glauben ein großer Organismus würde sich nähern würde sich die Muschel völlig grundlos schließen. Ebenso könnte es vorkommen, daß aufgrund mangelnder Interpretation durch zu wenige Rezeptoren ein großer Organismus zu spät oder gar nicht erkannt wird, und die Muschel somit zur Beute wird. Es ist also wichtig für die Muschel, daß sie differenzieren kann welche Schatten harmlos sind und welche Bedrohung verkünden. Und je mehr Rezeptoren sie dafür zur Verfügung hat, desto genauer kann sie die Situation interpretieren und „unwichtige“ Schatten außer Beacht lassen. Ein weiterer Punkt der für eine hohe Zahl von Rezeptoren spricht ist auch folgender: Im tiefen, trüben Wasser werfen Fressfeinde keine scharfkantigen Schatten mehr, und sind somit schwerer auszumachen. Daher kommen den Bivalvia auch hierbei viele Rezeptoren zur genaueren Umgebungsbeobachtung zugute. Das MirrorEye der Bivalvia ist exzellent was die Lichtausnutzung betrifft, allerdings auf Kosten der Sehschärfe. Das Licht passiert auf seinem Weg durch das Auge insgesamt 4 mal die Netzhaut. Zuerst trifft es unscharf auf die Distalretina, dann zweimal auf die Proximalretina (wobei die Proximalretina so dicht an der Argentea sitzt, daß nach der Reflexion durch Diese das Licht unmittelbar dieselbe Stelle der Proximalretina nochmals passiert) und dann erst scharf wieder zurück auf die Distalretina. Die Lichtauskostung ist daher enorm. Eine Muschel hat eine 5 mal bessere Lichtwahrnehmung als vergleichsweise ein Fisch. Allerdings geht die gewonnene

17
Bildhelligkeit auf Kosten der Bildqualität, da sich das scharfe Bild mit dem unscharfen Bild auf der Distalretina vermischt. Viele Details kann eine Muschel daher nicht wahrnehmen, und das Bild verliert wohl stark an Kontrast. Zusammenfassend läßt sich über die Retinas also sagen: Die Distalretina ist zuständig für das scharfe Sehen und das Bewegungssehen, und die Proximalretina dient der Orientierung im Wasser sowie zur rechtzeitigen Feinderkennung auch bei nur unscharfen Schatten. Die Argentea (Spiegelschicht) Die Argentea ist die Spiegelschicht am Augenhintergrund der MirrorEyes der Bivalvia. Sie besteht aus Cytoplasma und aus lichtspiegelnden Guaninkristallen, welche in mehreren Schichten übereinander angelagert sind. Guanin ist der Stoff der immer dann im Tierreich vorkommt, sobald ein Tier, Insekt oder Säugetier, eine spiegelnde Fläche aufweist. Als Beispiel seien hier die Neonfischchen genannt mit ihrem leuchtendem, blauem Seitenstreifen, der glitzernde Staub an den Flügeln einer Motte, oder der blaue Spiegel am Flügel einer Ente. Guanin ist der einzige Stoff den ein Organismus bilden kann, der zu einer Lichtreflexion fähig ist. Außerdem wird Guanin in der Wissenschaft verwendet, um farbselektive Spiegel zu bauen. Für eine Anwendung im Auge allerdings, wie im Falle der Bivalvia, ist Guanin wohl nicht die Ideallösung wenn es darum geht exakte Spiegel zu konstruieren. Es hat nämlich einen entscheidenden Nachteil. Guanin ist kein totalreflektierender Stoff, wie zum Beispiel Metall. Für die Bivalvia bringt das folgenden Nachteil mit sich: Das Licht fällt auf die erste Guaninschicht, wird dort aber nur zum Teil reflektiert. Das unreflektierte Licht durchdringt die erste Guaninschicht, bis es auf die Zweite trifft, wo es dann wieder zum Teil reflektiert wird und so weiter, Schicht für Schicht... Der Nachteil für die Bivalvia ist nun der, daß das Licht welches angenommen erst auf der siebten Guaninschicht reflektiert wurde nicht auf der Stelle der Retina auftrifft wie das Licht desselben Lichtstrahls, welches schon auf der ersten Schicht reflektiert wurde. Je tiefer das Licht in die Guaninschichten eindringt, desto mehr wird es von seinem eigentlich korrektem Treffpunkt auf der Retina versetzt. Das Bild wird dadurch unscharf. Allerdings, da es keinen anderen Stoff zur Lichtspiegelung im Tierreich gibt, muß dieser Fehler wohl oder übel hingenommen werden. Ein effektiverer Stoff um Licht total zu reflektieren, wurde bisher bei keinem Lebewesen gefunden. Die Guaninschichten stellen also etwas wie die bestmögliche Notlösung dar.

18
Augenentwicklung (Schema) Anhand dieser Skizze wird die Evolution des Auges dargestellt. Zu Beginn der Augenevolution stand das Grubenauge, welches noch keine Linse oder Ähnliches besaß. Das Innere des Auges war frei zugänglich und Licht wurde daher beim Einfall nicht gebrochen. Die Augenevolution läuft nun so ab, daß sich die einzelnen Bestandteile des Auges die wir heute kennen, wie etwa die Linse oder die Hornhaut, nacheinander über das Auge „hinüberfalten“. So sieht man bei Abbildung B zum Beispiel wie sich das Auge schließt, da sich das Körperepithel über die Augengrube hinüberwölbt. Es entsteht ein „Lochkameraauge“, welches qualitativ dem Grubenauge schon wesentlich überlegen ist. Bei Abbildung C ist das Auge komplett geschlossen und erste Ansätze der Iris sind zu erkennen. Bei D beginnt sich die Linse zu bilden und auch die Cornea ist im Ansatz zu erkennen. Und bei E letztendlich hat das Auge fast den höchsten Evolutionsstand erreicht. Die Cornea ist nahezu geschlossen und auch sonst weist das Auge alle Bestandteile auf, die typisch für ein uns bekanntes Linsenauge sind. Bei den Mollusken sind alle diese Stadien der Augenevolution anzutreffen.

19
Literaturverzeichnis
(1) M. F. Land, Molluscs (2) Kurt Fiedler/ Johannes Lieder, Mikroskopische Anatomie der Wirbellosen,
Gustav Fischer Verlag 1994 (3) H.-E. Guner, Lehrbuch der speziellen Zoologie, begründet von A. Kaestner,
Band I: Wirbellose Tiere










![HNO UND AUGEN - Dr. Franz Schramm : Arzt für ... · Hordeolum 2 Tage alt PRAXIS PLUS - HNO UND AUGEN ... Microsoft PowerPoint - Augen+HNO 2009.ppt [Kompatibilit.tsmodus] Author:](https://static.fdokument.com/doc/165x107/5b81bf1a7f8b9ae87c8cfd29/hno-und-augen-dr-franz-schramm-arzt-fuer-hordeolum-2-tage-alt-praxis.jpg)







