Diplomarbeit Dopamin und Persönlichkeit - uni-trier.de · Dieser Metabolit lässt sich im Liquor...
113
Universität Trier Studiengang Diplom-Psychologie Diplomarbeit Dopamin und Persönlichkeit Überprüfung des Zusammenhangs zwischen basaler dopaminerger Aktivität und Extraversion oder Spontaneität vorgelegt von: Sonja Römer Gutachter und Betreuer: PD Dr. Dirk Hagemann Prof. Dr. Dieter Bartussek Trier, im März 2005
Transcript of Diplomarbeit Dopamin und Persönlichkeit - uni-trier.de · Dieser Metabolit lässt sich im Liquor...

Universität Trier
Studiengang Diplom-Psychologie
Diplomarbeit
Dopamin und Persönlichkeit
Überprüfung des Zusammenhangs
zwischen basaler dopaminerger Aktivität
und Extraversion oder Spontaneität
vorgelegt von:
Sonja Römer
Gutachter und Betreuer:
PD Dr. Dirk Hagemann
Prof. Dr. Dieter Bartussek
Trier, im März 2005

Dopamin und Persönlichkeit II
Inhaltsverzeichnis Inhaltsverzeichnis.................................................................................................................................II
Abbildungsverzeichnis....................................................................................................................... V
Tabellenverzeichnis............................................................................................................................ VI
Zusammenfassung ............................................................................................................................VII
0 Einleitung...................................................................................................................................... 1
1 Theoretischer Hintergrund......................................................................................................... 3
1.1 Dopamin............................................................................................................................... 3
1.1.1 Struktur, Synthese und Inaktivierung ......................................................................... 3
1.1.2 Dopamin-Rezeptoren..................................................................................................... 4
1.1.3 Beeinflussung der Abläufe an der dopaminergen Synapse ..................................... 5
1.1.4 Dopaminerge Zellgruppen im Gehirn und Projektionen ......................................... 7
1.1.4.1 Dopaminerge Projektionen des Mesencephalons ............................................. 8
1.1.4.2 Dopaminerge Projektionen des Diencephalons............................................... 10
1.1.4.3 Dopaminerge Zellen im Telencepalon.............................................................. 10
1.1.5 Dopamin aus biopsychologischer Sicht..................................................................... 10
1.1.5.1 Die Bedeutung von Dopamin für Psychomotorik und Kognition................ 11
1.1.5.2 Die Bedeutung von Dopamin für Motivation und Emotion ......................... 12
1.2 Spontaner Lidschlag ......................................................................................................... 16
1.2.1 Unterscheidung des spontanen Blinks von anderen Lidbewegungen ................. 16
1.2.2 Physiologie und Anatomie der Blinkgenerierung ................................................... 17
1.2.3 Die Blinkrate als Indikator für zentrale dopaminerge Aktivität............................ 18
1.2.4 Die Blinkrate als psychophysisches Maß .................................................................. 21
1.2.5 Möglichkeiten zur Erfassung von Blinks .................................................................. 23
1.3 Biologische Persönlichkeitstheorien............................................................................... 24
1.3.1 Die Neurobiologie der Persönlichkeitsstruktur nach R. A. Depue ....................... 24
1.3.1.1 Ein Persönlichkeitseigenschaftsmodell............................................................. 24
1.3.1.2 Ein Verhaltensförderungssystem ...................................................................... 27
1.3.1.3 Ein neurobiologisches Förderungsmodell........................................................ 28

Dopamin und Persönlichkeit III
1.3.1.4 Interindividuelle Unterschiede in der dopaminergen Funktion................... 32
1.3.1.5 Ein Schwellenmodell der Verhaltensförderung .............................................. 34
1.3.1.6 Die Interaktion von Extraversion mit Constraint............................................ 36
1.3.1.7 Befunde zur Theorie ............................................................................................ 38
1.3.2 Die allgemeine biosoziale Theorie der Persönlichkeit von C. R. Cloninger......... 39
1.3.2.1 Persönlichkeitsdimensionen............................................................................... 40
1.3.2.2 Monoaminerge Systeme und entsprechende Verhaltenssysteme................. 43
1.3.2.3 Funktionelle Interaktion der neuroadaptiven Systeme.................................. 45
1.3.2.4 Befunde zur Theorie ............................................................................................ 48
1.3.3 Fazit zur basalen dopaminergen Aktivität ............................................................... 50
1.4 Hypothesen........................................................................................................................ 51
2 Methoden.................................................................................................................................... 53
2.1 Versuchspersonen............................................................................................................. 53
2.2 Fragebögen......................................................................................................................... 54
2.2.1 Eysenck Personality Questionnaire - Revised .......................................................... 54
2.2.2 Tridimensional Personality Questionnaire ............................................................... 54
2.2.3 Zusammenhang von EPQ-R und TPQ ...................................................................... 55
2.3 Versuchsablauf .................................................................................................................. 56
2.4 Elektrooculogramm .......................................................................................................... 57
2.4.1 Erfassung des Elektrooculogramms .......................................................................... 57
2.4.2 Identifizierung der Blinks............................................................................................ 58
2.5 Berechnung und Beschreibung der Fragebogendaten................................................. 60
2.6 Berechnung und Beschreibung der Blinkraten............................................................. 61
2.7 Datenanalyse...................................................................................................................... 62
2.7.1 Latent-State-Trait-Analyse der Blinkrate .................................................................. 63
2.7.1.1 Strukturgleichungsmodelle ................................................................................ 63
2.7.2 Korrelationsanalyse...................................................................................................... 71
3 Ergebnisse ................................................................................................................................... 73
3.1 LST-Analyse....................................................................................................................... 73
3.1.1 Trait-Modell................................................................................................................... 73
3.1.2 Latent-State-Trait-Modell ............................................................................................ 74

Dopamin und Persönlichkeit IV
3.1.3 LST-Parameter............................................................................................................... 75
3.1.4 Stabilitätsmodell ........................................................................................................... 75
3.1.5 Fazit zur LST-Analyse.................................................................................................. 77
3.2 Korrelationsanalyse .......................................................................................................... 77
4 Diskussion................................................................................................................................... 80
4.1 LST-Analyse....................................................................................................................... 80
4.2 Korrelationsanalyse .......................................................................................................... 82
4.2.1 Der Zusammenhang zwischen Blinkrate und Extraversion oder Spontaneität .. 82
4.2.1.1 Physiologische Grundaktivität vs. Reaktivität ................................................ 83
4.2.1.2 Die Single-Trait-Single-Transmitter-Hypothese.............................................. 85
4.2.1.3 Die Stichprobe ...................................................................................................... 86
4.2.2 Ein Zusammenhang zwischen Blinkrate und Belohnungsabhängigkeit.............. 86
4.2.3 Überlegungen für zukünftige Untersuchungen....................................................... 90
4.2.3.1 Basale physiologische Aktivität und Reaktivität............................................. 90
4.2.3.2 Neurotransmitter ................................................................................................. 91
4.2.3.3 Indikatoren............................................................................................................ 91
4.2.3.4 Persönlichkeitsdimensionen............................................................................... 91
5 Literaturverzeichnis .................................................................................................................. 93
Anhang............................................................................................................................................... 101

Dopamin und Persönlichkeit V
Abbildungsverzeichnis Abbildung 1 Schematische Darstellung einer dopaminergen Synapse im Gehirn mit
möglichen Angriffspunkten verschiedener Medikamente (nach
Kugele(2000) und Thompson (1994)). ..................................................................... 6
Abbildung 2 Übersicht über das mesotelencephale DA-System................................................ 9
Abbildung 3 Persönlichkeitseigenschaftsmodell (nach Depue & Collins, 1999).................... 26
Abbildung 4 Verhaltensförderungsmodell (nach Depue & Collins, 1999). ............................ 28
Abbildung 5 Schematische Darstellung des MOC-Netzwerks (nach Depue & Collins,
1999). .......................................................................................................................... 30
Abbildung 6 Neurobiologisches Förderungsmodell (nach Depue & Collins, 1999).............. 31
Abbildung 7 Ein psychobiologisches Schwellenmodell der Verhaltensförderung
(nach Depue, 1995). .................................................................................................. 35
Abbildung 8 Interaktion der agency-Komponente von Extraversion mit nicht-
affektivem Constraint (nach Depue (1996) und Depue & Lenzenweger
(2001))......................................................................................................................... 38
Abbildung 9 Interaktion der Persönlichkeitsdimensionen novelty seeking und harm
avoidance (nach Cloninger, 1987). ........................................................................... 46
Abbildung 10 Interaktion der Persönlichkeitsdimensionen novelty sseking und reward
dependence (nach Cloninger, 1987).......................................................................... 46
Abbildung 11 3-fach-Interaktion von Persönlichkeit und monoaminerger Übertragung
(nach Cloninger, 1986) ............................................................................................. 47
Abbildung 12 Drei spontane Blinks im VEOG-Kanal. ................................................................. 59
Abbildung 13 Trait-Modell. ............................................................................................................. 65
Abbildung 14 State-Trait-Modell. ................................................................................................... 66
Abbildung 15 Stabilitätsmodell. ...................................................................................................... 68

Dopamin und Persönlichkeit VI
Tabellenverzeichnis Tabelle 1 Lokalisierung und Nomenklatur dopaminerger Zellgruppen im Gehirn (nach
Dahlström & Fuxe, 1964). .......................................................................................... 8
Tabelle 2 Drei Hauptgehirnsysteme und damit verbundene Lern- und
Persönlichkeitscharakteristika (nach Cloninger, 1987, 1988) ............................. 41
Tabelle 3 Liberalisiertes State-Trait-Modell........................................................................... 74
Tabelle 4 Parameterschätzung des liberalisierten State-Trait-Modells ............................. 74
Tabelle 5 LST-Parameter des liberalisierten State-Trait-Modells ....................................... 75
Tabelle 6 Liberalisiertes State-Trait-Stabilitätsmodell.......................................................... 76
Tabelle 7 Parameterschätzung des liberalisierten State-Trait-Stabilitätsmodells ............ 76
Tabelle 8 Bivariate Korrelationen zwischen der Blinkrate und den Skalen des EPQ-R.. 77
Tabelle 9 Bivariate Korrelationen zwischen der Blinkrate und den Skalen des TPQ...... 78
Tabelle 10 Adjustierung des Signifikanzniveaus für die Korrelationen der Blinkrate
mit den Haupt-Skalen des EPQ-R und des TPQ durch eine sequenzielle
Bonferoni-Korrektur (Holm, 1979)......................................................................... 79
Tabelle A1 Mittelwerte und Standardabweichungen der Fragebogensummenwerte ..... 101
Tabelle A2 Mittelwerte und Standardabweichungen der Itemmittelwertscores.............. 102
Tabelle A3: Mittelwerte, Standardabweichungen und Spannweiten der Blinkrate .......... 102
Tabelle B1: Split-Half-Reliabilität und zeitliche Stabilität (Retest-Reliabiliät) der
EPQ-R-Skalen.......................................................................................................... 103
Tabelle B2: Split-Half-Reliabilität und zeitliche Stabilität (Retest-Reliabilität) der
TPQ-Skalen.............................................................................................................. 104
Tabelle B3: Split-Half-Reliabilität und zeitliche Stabilität (Retest-Reliabilität) der
Blinkrate................................................................................................................... 104
Tabelle C1: Interkorrelationen der Skalen des EPQ-R mit denen des TPQ ........................ 105

Dopamin und Persönlichkeit VII
Zusammenfassung In der vorliegenden Arbeit sollte der Zusammenhang zwischen der basalen zentralen
dopaminergen Aktivität und bestimmten Traits untersucht werden. Basierend auf den
Persönlichkeitstheorien von Depue (z.B. Depue & Collins, 1999) und Cloninger (1986, 1987,
1988) wurde ein Zusammenhang zwischen der zentralen dopaminergen Grundaktivität und
den Persönlichkeitsdimensionen Extraversion und Spontaneität erwartet.
Die Blinkrate im Ruhezustand diente als Indikator für die zentrale dopaminerge
Grundaktivität und die Persönlichkeit wurde mit dem Eysenck Personality Questionnaire -
Revised (EPQ-R) von Eysenck, Eysenck und Barrett (1985a) und dem Tridimensional
Personality Questionnaire (TPQ) von Cloninger (1989) erfasst.
Der erwartete Zusammenhang zwischen der Blinkrate und den Fragebogenskalen
Extraversion (EPQ-R) und Spontaneität (TPQ) konnte nicht festgestellt werden. Stattdessen
zeigte sich ein inverser Zusammenhang der Blinkrate mit der Skala Belohnungsabhängigkeit
(TPQ) und der Lügenskala (EPQ-R).
Möglicherweise spiegelt sich eine unterschiedliche Ausprägung auf den interessierenden
Persönlichkeitsdimensionen weniger in der basalen dopaminergen Aktivität als in der
Reaktivität des dopaminergen Systems wider.
Außerdem scheint die Vorstellung, dass ein Trait durch einen einzigen Neurotransmitter
bestimmt wird, nicht haltbar zu sein. So haben die Transmitter Dopamin und Serotonin
vermutlich einen Einfluss auf den Trait Belohnungsabhängigkeit.

Einleitung 1
0 Einleitung Nachdem sich die Persönlichkeitsforschung lange Zeit vornehmlich auf Beschreibungen der
Persönlichkeitsstruktur beschränkte, die auf subjektiven Instrumenten zur Erfassung von
Traits basierten, ist dank technischer und methodischer Entwicklungen in den
Neurowissenschaften (z.B. funktionale bildgebende Verfahren oder Fortschritte in der
Molekulargenetik) eine biopsychologische Herangehensweise möglich.
Einer der ersten biologisch orientierten Persönlichkeits-Theoretiker war Eysenck, der das
aufsteigende retikuläre Aktivierungssystem (ARAS) als neuroanatomisches Substrat der
Persönlichkeitsdimension Extraversion vorschlug und postulierte, dass Introvertierte im
Vergleich zu Extravertierten ein höheres generelles Aktivierungsniveau (arousal) bezüglich
der kortikoretikulären Schleife aufweisen (Eysenck, 1967; Eysenck, 1994). In neueren
Persönlichkeits-Theorien werden dopaminerge Mechanismen als potentielles Substrat von
impulsivitätsnahen Persönlichkeitsdimensionen (z.B. Extraversion bzw. Psychotizismus,
positive emotionality, novelty seeking oder sensation seeking) diskutiert.
Ziel der vorliegenden Arbeit ist es nun zu untersuchen, ob es einen Zusammenhang
zwischen der zentralen dopaminergen Aktivität und bestimmten
Persönlichkeitsdimensionen gibt. Dabei wird nicht, wie in vielen Studien, die Reaktivität des
dopaminergen Systems, sondern die dopaminerge Grundaktivität im Ruhezustand erfasst. Bei
der Stichprobe handelt es sich (soweit erfassbar) um psychisch und physisch unauffällige
Personen und nicht, wie in einigen anderen Untersuchungen, um Personengruppen mit
bestimmten Auffälligkeiten (z.B. Morbus Parkinson, Epilepsie, Schizophrenie, affektive
Störungen, Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung oder kriminelle
Vergangenheit).
In der vorliegenden Untersuchung werden anhand von Fragebögen (Eysenck Personality
Questionnaire - Revised / EPQ-R und Tridimensional Personality Questionnaire / TPQ) u.a.
die Persönlichkeitsdimensionen Extraversion und Spontaneität erfasst und mit einem Maß
der zentralen dopaminergen Aktivität (Blinkrate) in Zusammenhang gebracht. Zwei
biologisch orientierte Persönlichkeitstheorien, die den Zusammenhang zwischen Dopamin

Einleitung 2
und Extraversion bzw. novelty seeking (Spontaneität) aufgreifen, werden im Verlauf dieser
Arbeit exemplarisch vorgestellt.
Um die neuropsychologischen Hintergründe dieser Theorien besser verstehen zu können,
wird zu Beginn der Arbeit ein Überblick über die unterschiedlichen Facetten des
Neurotransmitters Dopamin gegeben (Kapitel 1.1.). Anschließend wird der Zusammenhang
zwischen Dopamin und der spontanen Blinkrate erläutert, die in der vorliegenden
Untersuchung als Indikator der zentralen dopaminergen Aktivität herangezogen wird
(Kapitel 1.2). Nachdem vorbereitend die biologischen Grundlagen des Neurotransmitters
Dopamin abgehandelt wurden, werden schließlich die Persönlichkeitstheorien von R.A.
Depue und C.R. Cloninger beschrieben, bei denen Dopamin mit den
Persönlichkeitsdimensionen Extraversion bzw. novelty seeking in Verbindung gebracht wird
(Kapitel 1.3). Abschließend werden entsprechende Hypothesen extrahiert (Kapitel 1.4), die
im Verlauf dieser Arbeit überprüft werden.
Nach einer Beschreibung der eingesetzten Methoden (Kapitel 2) und der entsprechenden
Ergebnisse (Kapitel 3) werden die Befunde abschließend diskutiert (Kapitel 4).

Theoretischer Hintergrund 3
1 Theoretischer Hintergrund
1.1 Dopamin
Innerhalb dieser Arbeit werden exemplarisch zwei biologisch orientierte
Persönlichkeitstheorien vorgestellt, die unter anderem dopaminerge Aktivität als
neurobiologisches Korrelat bestimmter Persönlichkeitseigenschaften heranziehen. Um die
Hintergründe dieser Theorien besser verstehen zu können, sollen im Folgenden sowohl
Erkenntnisse über die neurochemischen und -anatomischen Eigenschaften von Dopamin
bzw. dopaminerger Neurone als auch die psychologische Bedeutung von Dopamin
vorgestellt werden.
Einleitend wird zunächst über grundlegende Eigenschaften von Dopamin als
Neurotransmitters informiert. Als Überblick werden die Werke von Köhler (2001) und Elbert
und Rockstroh (1990) empfohlen.
1.1.1 Struktur, Synthese und Inaktivierung
Der Neurotransmitter Dopamin (DA) besitzt eine Amino-Gruppe (-NH2) und zählt damit zu
den Monoaminen bzw. biogenen Aminen, denen im Vergleich zu Aminosäuren die
Carboxyl-Gruppe (-COOH) fehlt. Als Monoamin mit einer Catechol-Gruppe (Benzolring mit
zwei Hydroxyl-Gruppen) kann DA als Catecholamin genauer spezifiziert werden.
L-Tyrosin, das mit der Nahrung aufgenommen wird oder durch Umwandlung der
essentiellen Aminosäure L-Phenylalanin entsteht, stellt den Ausgangspunkt für die
Biosynthese von DA dar. Durch Hydroxylierung mit Hilfe des Enzyms Tyrosin-Hydroxylase
entsteht der DA-Vorläufer L-Dopa, der eine zusätzliche Hydroxyl-Gruppe (-OH) besitzt.
Durch anschließende Decarboxylierung mittels Dopa-Decarboxylase entsteht DA, das
aufgrund der jetzt fehlenden Carboxyl-Gruppe nicht mehr liquorgängig ist, d.h. es passiert
nicht die Blut-Hirn-Schranke.
Nach der Biosynthese im Zellkörper findet ein axonaler Transport des fertigen Transmitters
in die synaptischen Endknöpfchen statt, wo dieser bis zur Ausschüttung in Vesikeln
gespeichert wird.

Theoretischer Hintergrund 4
Die Inaktivierung von DA vollzieht sich im Wesentlichen durch Wiederaufnahme in die
präsynaptische Zelle (Reuptake) mittels Carrier-Proteine, welche die Transmittermoleküle
binden und in die Zelle zurückschleusen (Köhler, 2001). In der präsynaptischen Endigung
als auch in der postsynaptischen Zelle baut das Enzym Monoaminoxidase (MAO)
überschüssige Mengen von DA zu Dihydroxyphenylessigsäure (DOPAC) ab. Ein weiteres
inaktivierendes Enzym in den postsynaptischen Zellen ist die Catechol-O-Methyltransferase
(COMT), die DA zu Methoxytyramin (MT) umwandelt (Elbert & Rockstroh, 1990;
Thompson, 1994). Weiterhin können DOPAC und Methoxytyramin durch COMT bzw. MAO
zu Homovanillinsäure (HVA) metabolisiert werden. Dieser Metabolit lässt sich im Liquor
und teilweise auch in Blut und Urin nachweisen, so dass die Konzentration als (nicht
unumstrittenes) Maß des synaptischen Abbaus und der an den Synapsen verfügbaren
Transmittermengen betrachtet werden kann (Köhler, 2001).
Psychopharmaka und Drogen können in die Synthese und Inaktivierung von DA eingreifen
(siehe Kapitel 1.1.3).
1.1.2 Dopamin-Rezeptoren
Die Wirkung von DA auf eine nachgeschaltete Zelle hängt vom Rezeptor in der
postsynaptischen Membran ab.
Generell werden D1-Rezeptoren und D2-Rezeptoren unterschieden (Kebabian & Calne, 1979).
Neueren Studien (Civelli, Bunzow, Grandy, Zhou & van Tol, 1991; Grandy & Civelli, 1992;
Sibley & Monsma Jr., 1992) zufolge gibt es zwei Untertypen D1-artiger Rezeptoren (D1 und
D5) und drei Untertypen D2-artiger Rezeptoren (D2, D3 und D4).
D1-artige Rezeptoren wirken exzitatorisch und sind fast ausschließlich an der
postsynaptischen Membran lokalisiert. Nach der Bindung des Transmitters an einen solchen
Rezeptor wird eine second-messenger-Kaskade ausgelöst. Über ein G-Protein wird das
Enzym Adenylatcyclase aktiviert, das ATP in cAMP umwandelt. cAMP-abhängige
Proteinkinasen phosphorylieren die Ionenkanäle, was zur Aktivierungsänderung dieser
Kanäle führt (Derrfuß, Krämer, Kugele & Kugler, 2000). Außerdem können auch chemische
Reaktionen innerhalb der Zelle oder sogar im genetischen Material im Zellkern ausgelöst
werden (Thompson, 1994).

Theoretischer Hintergrund 5
D2-artige Rezeptoren wirken inhibitorisch und kommen sowohl in der prä- als auch in der
postsynaptischen Membran vor. Die inhibierende Wirkung dieses Rezeptortyps besteht
einerseits in der Hemmung der cAMP-Bildung und andererseits in der Öffnung der Kalium-
Kanäle in der postsynaptischen Membran über second-messenger-Kaskaden. Der erhöhte
Kalium-Einstrom in die Zelle führt zu einer Hyperpolarisation, welche die
Auftretenswahrscheinlichkeit einer Erregungsweiterleitung durch ein Aktionspotential
vermindert.
D2-Rezeptoren in der präsynaptischen Membran kommen nur im nigrostriären und
mesolimbischen DA-System vor und werden als Autorezeptoren bezeichnet. Sie regulieren
die DA-Ausschüttung über eine negative Rückkopplungsschleife, indem das Enzym
Tyrosinhydroxylase gehemmt wird, sobald DA-Moleküle an diese Rezeptoren binden
(Derrfuß et al., 2000).
Substanzen, die einen Rezeptor besetzen können, aber nicht die Öffnung von Ionenkanälen
bewirken, werden Rezeptorblocker genannt, da sie den Rezeptor gegenüber
konkurrierenden Transmittermolekülen blockieren. So können DA-Rezeptoren durch
Neuroleptika (Antipsychotika) blockiert werden, die entgegengesetzt wirken bzw. die
Wirkung von DA aufheben. Aus diesem Grund werden solche Substanzen auch
Antagonisten (Widersacher) genannt. Dagegen verhalten sich Substanzen, die an einen
Rezeptor binden und eine ähnliche Wirkung ausüben, wie weitere Liganden und werden als
Agonisten bezeichnet (Köhler, 2001). Agonisten und Antagonisten binden mit
unterschiedlicher Stärke an die verschiedenen Rezeptortypen.
Einige Beispiele werden im nächsten Abschnitt beschrieben.
1.1.3 Beeinflussung der Abläufe an der dopaminergen Synapse
Nachdem die normalen Abläufe an dopaminergen Synapsen vorgestellt wurden, werden im
Folgenden mögliche Angriffspunkte verschiedener Psychopharmaka und Drogen
veranschaulicht (Abbildung 1) und aufgelistet, welche die synaptische Übertragung
dopaminerger Nervenzellen beeinträchtigen können (Köhler, 2001; Kugele, 2000; Thompson,
1994).

Theoretischer Hintergrund 6
Abbildung 1 Schematische Darstellung einer dopaminergen Synapse im Gehirn mit möglichen Angriffspunkten verschiedener Medikamente (nach Kugele (2000) und Thompson (1994)). DA: Dopamin, MAO: Monoaminoxidase, COMT: Catechol-o-Methyltransferase, DOPAC: Dihydroxyphenylessigsäure, MT: Methoxytyramin, HVA: Homovanillinsäure.
• Stimulation der DA-Produktion durch liquorgängige Präkursoren (z.B. DA-Vorläufer L-
Dopa bei Morbus Parkinson)
• Hemmung der DA-Produktion durch Diät (tyrosinarme Kost) oder Blockade von
Tyrosinhydroxylase durch α-Methyltyrosin oder verminderte DA-Synthese bei
chronischer Amphetamin-Verabreichung (negative Rückkopplung über Autorezeptoren)
• Beeinflussung der Aufnahme und Speicherung von DA durch Reserpin, das die Vesikel
in den präsynaptischen Endknöpfchen auflöst

Theoretischer Hintergrund 7
• Stimulation der DA-Ausschüttung durch Amphetamine (Beschleunigung der
Vesikelentleerung in den synaptischen Spalt) oder Autorezeptorblockade (schaltet
negative Rückkopplung aus)
• Hemmung der DA-Ausschüttung, z.B. durch γ-Hydroxybuttersäure, welche
Aktionspotentiale in der präsynaptischen Zelle hemmt oder Autorezeptorstimulation
(negative Rückkopplung)
• Verstärkung der Transmittereffekte am Rezeptor durch einen zweiten Liganden
(z.B. Apomorphin, Bromocriptin oder Pergolid als DA-Agonisten, die hauptsächlich an
D1- aber auch an D2-Rezeptoren binden) oder Erhöhung der Rezeptorempfindlichkeit
(Sensibilisierung)
• Schwächung der Transmitterwirkung am Rezeptor durch Neuroleptika-Blockade, wie
z.B. durch Phenothiazine (z.B. Chlorpromazin), Butyrophenonderivate (z.B. Haloperidol)
und einige atypische Neuroleptika als D2-Rezeptorantagonisten und atypische
Neuroleptika (z.B. Clozapin) als D4-Rezeptorantagonisten
• Verstärkung der nachgeschalteten Signalinduktion, z.B. durch Blockade der
Phosphodiesterase, welche die second-messenger-Wirkung beendet (z.B. durch Koffein
oder Sildenafil als Phosphodiesterasehemmer)
• Hemmung der nachgeschalteten Signalinduktion durch Hemmung der second-
messenger-Bildung
• Verhinderung der Inaktivierung durch Hemmung des Abbaus im präsynaptischen
Neuron oder Blockade des Abbauenzyms (Pargylin und Iproniazid sind z.B. MAO-
Hemmer und Tropolon hemmt COMT) oder Reuptake-Hemmung (Amphetamin und
Cocain blockieren DA-Carrier-Proteine)
• Beschleunigung der Inaktivierung, z.B. durch Reserpin und Abbauenzyme
Der Einsatz von Substanzen, die in das dopaminerge System eingreifen, gibt Aufschluss über
bestimmte Funktionen, die durch DA moduliert werden. Der weitreichende Einfluss des
Neurotransmitters wird im nächsten Abschnitt deutlich.
1.1.4 Dopaminerge Zellgruppen im Gehirn und Projektionen
Im Säugetiergehirn werden nach Dahlström und Fuxe (1964) 15 Areale (A1-A15)
unterschieden, in denen catecholaminerge Zellen und Fasern vorkommen. Tabelle 1 gibt

Theoretischer Hintergrund 8
einen Überblick über die Lokalisierung und Nomenklatur dopaminerger Zellgruppen
(Dahlström & Fuxe, 1964).
Tabelle 1 Lokalisierung und Nomenklatur dopaminerger Zellgruppen im Gehirn (nach Dahlström & Fuxe, 1964).
Lokalisierung Dopaminerge Zellgruppen Abkürzung
Mesencephalon Area tegmentalis lateralis Substantia nigra (SN), pars compacta Area tegmentalis ventralis (VTA)
A8 A9 A10
Diencephalon Nucleus infundibularis
A11 A12 A13 A14
Telencephalon Bulbus olfactorius A15
Die nächsten drei Abschnitte, gegliedert nach der Lokalisation der dopaminergen
Zellkörper, sollen einen groben Eindruck darüber vermitteln, in welche Hirnregionen sich
die Axone der genannten Zellgruppen ausdehnen.
1.1.4.1 Dopaminerge Projektionen des Mesencephalons
Nieuwenhuys (1985) fasst die gesamte Vorderhirnprojektion als mesotelencephales DA-
System zusammen, das sich grob in zwei bzw. drei Subsysteme aufgliedern lässt.
Mesostriatale Dopamin-Bahnen
Mesostriatale Projektionen gehen von A8, A9 und A10 aus und enden im Striatum (Nucleus
caudatus und Putamen), im Nucleus accumbens (NAS) und im Globus pallidus.
Dieses DA-System bildet die Grundlage für die Fähigkeit, Motorprogramme willkürlich zu
ändern und ist somit an der Regulation von komplexem Verhalten beteiligt. Speziell die
Aktivierung des NAS bewirkt verstärkte lokomotorische Aktivität. Außerdem wird die
verlangsamte Motorik und Muskelstarre bei Morbus Parkinson auf den fortschreitenden
Verlust dopaminerger Neurone dieses Systems (v.a. in der SN) zurückgeführt (siehe Kapitel
1.1.5).

Theoretischer Hintergrund 9
Mesolimbocorticale Dopamin-Bahnen
Da es keine scharfe Abgrenzung mesolimbischer und mesocorticaler DA-Bahnen gibt,
werden diese Projektionen häufig zusammengefasst und als mesolimbisches (Birbaumer &
Schmidt, 1996; Nieuwenhuys, 1985; Nieuwenhuys, Voogd, von Huijzen & Lange, 1980; Pinel,
1990) oder mesolimbocorticales (Schwarting, 1997) System bezeichnet.
Mesolimbische Projektionen gehen von A10 und vom medialsten Teil der SN (A9) aus und
führen zum Bulbus olfactorius, zum Nucleus olfactorius anterior, zur Substantia perforata
anterior, zum Nucleus septi lateralis, zum Nucleus interstitialis striae terminalis und zur
Amygdala.
Es wird angenommen, dass dieses DA-System an der Pathophysiologie der Schizophrenie
(v.a. an der produktiven Symptomatik) beteiligt ist (siehe Kapitel 1.1.5).
Mesocorticale Projektionen führen ausgehend von A10 und vom medialen Teil der SN (A9)
zum medialen Teil des Frontallappens, zum Cortex präpiriformis, piriformis und
entorhinalis und zum anterioren cingulären Cortex.
Abbildung 2 Übersicht über das mesotelencephale DA-System (vereinfachte schematische Darstellung). (1) mesostriatales System, (2) mesolimbisches System, (3) mesocorticales System; A8: area tegmentalis lateralis, A9: substantia nigra pars compacta, A10: area tegmentalis ventralis.
(2)
(1)
PallidumGyrus cinguli
Nucleus accumbens
Tuberculum olfactorium
Caudatum Putamen
Amygdala
A9
A10 frontaler Cortex
A8
(3)

Theoretischer Hintergrund 10
1.1.4.2 Dopaminerge Projektionen des Diencephalons
Neben den drei mesotelencephalen Projektionen werden außerdem vier efferente Systeme
unterschieden, die dopaminergen Zellgruppen (A11-A14) des Diencephalons entspringen.
Tubero-infundibulare Projektionen gehen hauptsächlich von A12 (Nucleus infundibularis)
aus und führen zur Eminentia mediana. Viele dieser Axone verlaufen weiter zum Lobus
posterior hypophyseos. Es wird angenommen, dass DA von dort zum
Hypophysenvorderlappen transportiert wird, um die Prolactin-Ausschüttung zu hemmen.
Während die Prolactin-Hemmung über diese Projektion in verschiedenen Untersuchungen
als Nachweis für dopaminerge Aktivierung herangezogen wird (z.B. Depue et al., 1994), sind
die incerto-hypothalamische Projektion, die diencephalo-septale Projektion und die
hypothalamo-spinale Projektion im Zusammenhang der vorliegenden Arbeit zu
vernachlässigen und werden hier nicht genauer vorgestellt.
1.1.4.3 Dopaminerge Zellen im Telencepalon
Die wahrscheinlich einzige dopaminerge Zellgruppe (A15) im Telencephalon liegt im
äußeren Bereich des Bulbus olfactorius und besteht aus Interneuronen, die Teil eines
größeren Zellverbands sind.
Nachdem die vorangegangenen Abschnitte eine Einführung in die biologischen Grundlagen
des Neurotransmitters DA bieten sollten, werden nun psychologisch relevante Aspekte von
DA vorgestellt.
1.1.5 Dopamin aus biopsychologischer Sicht
Der Neurotransmitter DA wird mit einigen Verhaltensfunktionen, wie Psychomotorik,
Aufmerksamkeit, Lernen und Motivation in Verbindung gebracht. Da die Herleitung des
Zusammenhangs der Blinkrate mit DA auf der Untersuchung einiger dieser Aspekte beruht
und auch die später vorgestellten Persönlichkeitstheorien auf Studien aus diesen Bereichen
basieren, soll dieses Kapitel einen Überblick über die Funktionen vermitteln, an denen DA
beteiligt ist.

Theoretischer Hintergrund 11
1.1.5.1 Die Bedeutung von Dopamin für Psychomotorik und Kognition
Einige Aspekte der Bedeutung dieses Neurotransmitters werden bei Störungen oder
Ausfällen des dopaminergen Systems deutlich. Das eindeutigste Beispiel ist die Parkinson-
Erkrankung. Hierbei handelt es sich um eine neurodegenerative Erkrankung, bei der vor
allem die dopaminerge Innervation des Neostriatums (Caudatum und Putamen) verloren
geht, was sich in Bewegungs- und Haltungsstörungen äußert. Um die Symptome (Akinese,
Rigor und Tremor) zumindest vorübergehend zu beheben, werden L-Dopa (liquorgängiger
DA-Vorläufer) oder DA-Rezeptoragonisten therapeutisch eingesetzt (Schwarting, 1997).
Auch bei Depression, Autismus, hyperkinetischem Syndrom oder Schizophrenie wird eine
dopaminerge Dysfunktion in Betracht gezogen. So erweisen sich DA-Agonisten, wie z.B.
Apomorphin, bei Depression als therapeutisch wirksam (Kugele, 2000) und DA-
Antagonisten, wie z.B. Chlorpromazin (Neuroleptikum), werden zur Behandlung
schizophrener Störungen eingesetzt, was primär eine Besserung der positiven Symptome
(formale Denkstörungen, Halluzinationen, Wahnvorstellungen und
Leistungseinschränkungen) bewirkt (Birbaumer & Schmidt, 1996; Davison & Neale, 2002).
Amphetamine dagegen verschlimmern positive Symptome aber bessern negative Symptome
(Apathie, Alogie, Anhedonie, flacher Affekt, Asozialität und Aufmerksamkeitsprobleme).
Die DA-Theorie der Schizophrenie erklärt diese Befunde mit einer geringen Aktivität der
DA-Neuronen im präfrontalen Cortex und einer damit zusammenhängenden Überaktivität
im mesolimbischen DA-System (Davison & Neale, 2002). Amphetamine in hohen Dosen
können aber auch bei nicht-schizophrenen Personen motorische Stereotypien und
psychotisches Verhalten hervorrufen (Amphetamin-Psychose). In geringen Dosen führen
Amphetamine hingegen zur Verbesserung der Aufmerksamkeitsleistung (Birbaumer &
Schmidt, 1996). In diesem Sinne werden viele Kinder mit Aufmerksamkeitsdefizit-
/Hyperaktivitätsstörung z.B. mit dem prominenten Amphetamin-Derivat Ritalin behandelt.
In diesem Abschnitt wurden Befunde zu Krankheiten und Störungen vorgestellt, denen eine
dopaminerge Dysfunktion zugrunde liegt. Diese weisen darauf hin, dass dopaminerge
Mechanismen für die Funktion von Motorik und Kognition (insbesondere Aufmerksamkeit)
bedeutsam sind. Im Folgenden wird erläutert, dass DA auch bei motivationalen und
emotionalen Prozessen eine Rolle spielt.

Theoretischer Hintergrund 12
1.1.5.2 Die Bedeutung von Dopamin für Motivation und Emotion
Da bestimmte Traits, die in dieser Arbeit mit DA in Verbindung gebracht werden sollen,
bestimmte motivationale bzw. emotionale Ausprägungen beim Menschen darstellen, sollen
diese Aspekte an dieser Stelle genauer betrachtet werden.
Einige Annahmen über die Bedeutung von DA für Motivation und Emotion wurden aus
Experimenten zur belohnenden elektrischen Hirnreizung (intracranielle Selbststimulation /
ICSS) abgeleitet. Dabei lernen Tiere (meist Ratten), sich durch operantes Verhalten (z.B.
Hebeldrücken) in bestimmten Hirnregionen elektrisch selbst zu reizen, wobei dies sogar
dazu führen kann, dass sie primäre Verstärker, wie Futter oder Sexualpartner,
vernachlässigen. Die elektrische Stimulation ist besonders bei den dopaminergen
Zellkörpern der VTA und dem Bereich der aufsteigenden DA-Fasern auf der Ebene des
lateralen Hypothalamus effektiv. Die direkte Beteiligung von DA an der belohnenden
Hirnreizung wurde anhand pharmakologischer Studien nachgewiesen, die zeigen, dass die
ICSS durch Blockade von DA-Rezeptoren unterbunden werden kann, ohne dass die
Fähigkeit zur Ausführung der operanten Aufgabe beeinträchtigt ist (Schwarting, 1997). Die
Wirkung der DA-Blockade wird also nicht auf eine motorische Unfähigkeit zurückgeführt,
sondern auf ein Ausbleiben positiver Verstärkung in den Gebieten, die stark auf die ICSS
reagieren. Experimentelle Befunde deuten darauf hin, dass die dopaminerge
Signalübertragung und Interaktion sensorischer und motorischer Eingänge im NAS
stattfinden und, dass positive Verstärker (Nahrung, Sexualpartner, ICSS und Suchtdrogen)
mit erhöhter Aktivität in diesem System assoziiert sind (Chen, 1993; Damsma, Pfaus,
Wenkstren, Phillips & Fibinger, 1992; Di Chiara, 1995; Fibinger, 1993; Wise, 1996). Vor
diesem Hintergrund wurde DA lange mit Hirnmechanismen von Belohnung und
Verstärkung sowie Hedonie bzw. Anhedonie in Verbindung gebracht (Schwarting, 1997).
Weitere Befunde lassen allerdings daran zweifeln, dass dopaminerge Neurone als Substrat
von Verstärkung oder ihrer emotionalen, hedonistischen Komponente verstanden werden
kann. So zeigte sich, dass statt dem angenommenen dopaminergen System ein absteigendes
nicht-dopaminerges System durch die ICSS angeregt wird (Huston, 1982), welches mit
ersterem anatomisch eng verbunden ist. Des Weiteren konnten frühere Befunde, bei denen

Theoretischer Hintergrund 13
neuroleptische Rezeptorblockade Amphetamin-induzierte Euphorie verhindert, nicht
bestätigt werden (Brauer & De Wit, 1997). Dies zeigt, dass positive Verstärkung zwar durch
erhöhte DA-Ausschüttung ausgelöst, aber nicht durch DA-Blockade verhindert werden
kann. Beide Befunde legen nahe, dass DA weniger eine direkte als eine modulierende
Wirkung hat.
Schultz et al. (1997) stellten bei der Registrierung der elektrischen Aktivität dopaminerger
Neurone des VTA und der SN bei Affen fest, dass es nur auf unvorhersehbare Belohnungen
eine Reaktion gab (vergleichbar mit frühen Stadien eines Konditionierungsexperiments).
Wenn eine Belohnung erwartet wurde (vergleichbar mit späteren Stadien eines
Konditionierungsexperiments) erfolgte eine Aktivitätszunahme dopaminerger Neurone nur
auf den konditionierten Reiz hin, der die Belohnung ankündigte (Schultz, Dayan &
Montague, 1997). Außerdem erhöht sich die elektrische Aktivität der Zielneurone im NAS
bei ankündigenden, konditionierten Reizen oder Belohnungen, wobei diese Neurone aber
ihre verstärkerabhängige Aktivitätserhöhung beizubehalten scheinen (Schwarting, 1997).
Diese Befunde deuten darauf hin, dass dopaminerge Aktivität das Erlernen von
Verhaltenskontingenzen steuert, und die Zielneurone eher bei der Aufrechterhaltung von
Verhalten eine Rolle spielen (Schwarting, 1997).
Untersuchungen zum Zusammenhang zwischen dopaminerger Aktivität und natürlich
motiviertem Verhalten weisen darauf hin, dass die DA-Ausschüttung während der
Vorbereitung auf eine Belohnung (Annäherung an einen Sexualpartner oder Futtersuche)
ansteigt. Während dies die Auffassung stützt, dass DA eine spezifische Rolle bei der
Anreizmotivation spielt (Robbins & Everitt, 1996), wird oft aber auch ein weiterer Anstieg
der DA-Ausschüttung während der konsumatorischen Phase (Kopulation oder Fressen)
berichtet (Fiorino, Coury & Phillips, 1997).
Einige Autoren (z.B. Robinson & Berridge, 1993) gehen von zwei Komponenten der
Belohnung aus. Es wird zwischen einer eher affektiven oder hedonistischen Komponente,
die als liking (Mögen) bezeichnet wird, und einer appetitiven oder Anreizkomponente, die
als wanting (Wollen / Verlangen) bezeichnet wird, unterschieden. Die Anreizkomponente soll
DA-abhängig sein, wobei das dopaminerge System im NAS eine Transformation von

Theoretischer Hintergrund 14
perzeptiven Signalen vermitteln soll, die zur Anreizhervorhebung (Anreizsalienz) führt.
Dabei sollen konditionierte Reize (sekundäre Verstärker / Anreize) zu gerichteter
Aufmerksamkeit, Orientierung und Annäherung führen und instrumentelle Reaktionen und
kognitive Strategien in Richtung Ziel antreiben (Schwarting, 1997).
Mit der Anreiztheorie lässt sich auch Suchtverhalten erklären. Suchtdrogen erhöhen die
dopaminerge Signaltransmission im NAS, dessen wesentliche Funktion darin zu bestehen
scheint, neutralen Stimuli eine positive Bewertung zu geben, wodurch sie selbst attraktiv
(wanted) werden. Wiederholte Drogeneinnahme kann zu Neuroadaptationen in diesem
System führen, die in einer dauerhaften Sensibilisierung für die mit dem Drogengebrauch
assoziierten Reize resultieren kann, wobei deren Anreizcharakter erhöht wird (Robinson &
Berridge, 1993).
Da dopaminerge Aktivität auch bei aversiven Reizen (z.B. Stressoren) und bei
Vermeidungsverhalten eine Rolle spielt (Salamone, Cousins & Snyder, 1997), wird DA in
einigen neueren Theorien (Le Moal & Simon, 1991) eher eine generelle
verhaltensregulatorische Funktion zugesprochen, wobei der Transmitter, vor allem im NAS,
den Einfluss corticaler und limbischer Afferenzen auf komplexe Aspekte motorischen
Verhaltens moduliert. So soll erhöhte DA-Aktivität zu effektiver Vermeidung aversiver
Reize bzw. Annäherung an positive Reize führen, während dopaminerge Über- oder
Unterfunktion bewirkt, dass aversive Ereignisse weniger effektiv vermieden und positive
Ereignisse weniger effektiv aufgesucht werden.
In diesem Sinne wird das dopaminerge System als Schnittstelle zwischen Motivation und
Handlung aufgefasst, die erfahrungsabhängig die Beziehungen zwischen Input (Reize) und
Output (Verhalten) kontrolliert. Sensorik und Valenz (positiv, negativ), sowie Richtung der
resultierenden Aktion (Annäherung, Vermeidung), werden von anderen Hirnstrukturen und
Neurotransmittern vermittelt (Schwarting, 1997).
Zusammenfassend kann festgehalten werden, dass DA mittlerweile weniger mit der
affektiven (Emotion) als mit der Anreizkomponente (Motivation) von Belohnung in
Verbindung gebracht wird. Dopaminerge Aktivität scheint für das Erlernen von
Verhaltenskontingenzen eine Rolle zu spielen, was eine Grundlage für Anreizhervorhebung

Theoretischer Hintergrund 15
darstellt. Nachdem dopaminerge Mechanismen nicht nur mit Annäherung sondern auch mit
Vermeidung in Verbindung gebracht wurden, wird DA eine eher generelle
verhaltensregulatorische Funktion zugeschrieben. Das dopaminerge System kann als
Schnittstelle zwischen Motivation und Handlung aufgefasst werden, wobei DA eine
modulierende Wirkung zugeschrieben wird.

Theoretischer Hintergrund 16
1.2 Spontaner Lidschlag
Da der spontane Lidschlag (endogener Blink; im Folgenden auch spontaner Blink oder
einfach Blink genannt) in dieser Arbeit als behaviorales Maß der zentralen dopaminergen
Aktivität genutzt wird, soll diese Art der Lidbewegung in den folgenden Abschnitten
genauer beschrieben werden. Dazu gehört sowohl die Unterscheidung von reflektorischen
und willkürlichen Lidschlägen, die für die Erhebung der Blinkrate (BR, Frequenz der
spontanen Blinks; bevorzugte Maßeinheit: Blinks/Minute, bpm) wichtig ist, als auch die
Generatoren und Mechanismen, die der Produktion spontaner Blinks zugrunde liegen. Es
gibt Hinweise auf eine zentrale Kontrolle der Blinkproduktion und es werden Befunde
vorgestellt, die die Annahme stützen, dass die BR durch dopaminerge Systeme reguliert
wird und damit die dopaminerge Aktivität im Gehirn widerspiegelt. Schließlich werden
verschiedene Bedingungen vorgestellt, unter denen die BR innerhalb einer Person variieren
kann.
1.2.1 Unterscheidung des spontanen Blinks von anderen
Lidbewegungen
Der spontane Lidschlag ist ein behaviorales Phänomen, das bei den meisten Wirbeltieren
und allen Säugetieren beobachtbar ist und bezeichnet das schnelle Schließen und
Wiederöffnen des Augenlids in der Abwesenheit identifizierbarer äußerer Stimulation
(Blount, 1928). Solche Lidbewegungen sind also unabhängig von Licht, Hitze oder
Feuchtigkeit und treten auch bei cornealer Anästhesie sowie bei Blinden auf (Ponder &
Kennedy, 1928).
Der reflektorische Lidschlag (Reflexblink), der mit einer Schreckreaktion einhergeht, tritt
zwar auch unwillkürlich und unbewusst auf, stellt aber im Gegensatz zum spontanen Blink
eine Schutzreaktion dar, die durch potentiell schädigende äußere Reize ausgelöst wird.
Außerdem kann der spontane Blink auch vom bewusst ausgeführten willkürlichen Blink
unterschieden werden, der auf eigenen Willen hin oder nach Aufforderung ausgeführt
werden kann (Bentivoglio, Bressman, Cassetta, Caretta, Tonali & Albanese, 1997; Galley,
2001; Karson, 1989; Stern, Walrath & Goldstein, 1984; von Cramon, 1980).

Theoretischer Hintergrund 17
1.2.2 Physiologie und Anatomie der Blinkgenerierung
Unwillkürliche Blinks beruhen im Großen und Ganzen auf der Kontraktion des palpebralen
Teils des musculus orbicularis oculi, der durch den VII. Hirnnerv (nervus facialis) innerviert
wird und den Lidschluss bewirkt. Die Kontraktion des musculus levator palpebrae, der vom
III. Hirnnerv (nervus oculomotorius) innerviert wird, führt anschließend wieder zur
Lidöffnung (Galley, 2001; Karson, 1989; von Cramon, 1980).
Im Gegensatz zum Reflexblink ist der spontane Blink nicht von der Funktionsfähigkeit
optischer, akustischer und trigeminaler Afferenzen abhängig und bleibt bei einer
Durchtrennung der Hirnnerven III, IV und VI unbeeinflusst (von Cramon, 1980). Ponder und
Kennedy (1928) schließen daraus, dass Blinkbewegungen der Augen zentral in einer Region
generiert werden, die mehr oder weniger regelmäßige Impulsentladungen durch den VII.
Hirnnerv verursacht. Sie postulieren einen zentral kontrollierten Blinkmechanismus und
nehmen an, dass die Basalganglien dabei eine Rolle spielen, da der fast vollständige Ausfall
von Blinks eines der auffälligsten Merkmale der postencephalitischen Parkinson-Erkrankung
(siehe Kapitel 1.1.5) ist (Stevens, 1978; von Cramon, 1980).
Karson (1989) vertritt die Annahme, dass spontane Blinks durch ein Nervensystem
kontrolliert werden, das in der pontinen reticulären formation entspringt. SN und Colliculus
superior sollen die Blinkproduktion fördern, während Cerebellum und okzipitalem Cortex
inhibitorische Modulation zugesprochen wird.
Von Cramon (1980) bezieht sich auf die Befunde verschiedener Forschergruppen, um die
Frage nach der zentralen Innervation der spontanen Lidbewegungen zu beantworten. Dabei
werden der Schläfenlappen, dass anteriore Cingulum und die Amygdala genannt, wobei das
Tegmentum eine Zwischenstation darzustellen scheint.
Auch wenn die physiologischen und neuroanatomischen Einzelheiten der Kontrolle des
Lidschlags durch höhere Zentren noch weitgehend ungeklärt sind (Galley, 2001; von
Cramon, 1980), fällt auf, dass einige Gehirnstrukturen in Erwägung gezogen werden, die
unter dopaminerger Kontrolle stehen (siehe Kapitel 1.1.4).
Der folgende Abschnitt soll begründen, dass die Blinkrate als nicht-invasiver Schätzer der
zentralen dopaminergen Aktivität herangezogen werden kann.

Theoretischer Hintergrund 18
1.2.3 Die Blinkrate als Indikator für zentrale dopaminerge
Aktivität
Beobachtungen von Abnormalitäten der BR bei Patienten mit neuropsychiatrischen
Störungen, denen eine dopaminerge Dysregulation zugrunde liegt, führen zur Annahme
eines positiven Zusammenhangs zwischen der BR und der dopaminergen Aktivität im
Zentralnervensystem (z.B. Stevens, 1978; Karson, 1983).
So dokumentiert Stevens (1978) eine erhöhte BR bei medikationsfreien Patienten mit
Schizophrenie (siehe Kapitel 1.1.5) und bei Patienten mit Spätdyskinesie
(Bewegungsstörung; meist infolge neuroleptischer Langzeitmedikation). Da die
untersuchten Patienten früher schon neuroleptisch behandelt wurden ist allerdings unklar,
ob dies ein Teil der Pathologie von Schizophrenie oder ein Residuum der Behandlung oder
ihres Absetzens repräsentiert. Außerdem berichtet Stevens (1978) eine erniedrigte BR bei
Patienten mit postencephalitischem Parkinsonismus. Dieselbe Wirkung haben Phenothiazine
und Butyrophenone (Neuroleptika; siehe Kapitel 1.1.3), die auf Dauer das extrapyramidale
Syndrom (Störung automatischer Bewegungsabläufe und der Regulation des Muskeltonus)
hervorrufen können. Stevens (1978) fasst diese Beobachtungen als Indizien dafür auf, dass
die BR zumindest teilweise durch zentrale dopaminerge Aktivität vermittelt wird und stützt
diese Vermutung durch ein eigenes Experiment. Dabei wurden bei Katzen durch
experimentelle Aktivierung des zentralen dopaminergen Systems u.a. paroxysmale Blinks
hervorgerufen. Stevens (1978) äußert die Erwartung, dass Hypersensibilität von DA-
Rezeptoren des NAS, des tuberculum olfactorium und anderen Endpunkten des
mesolimbischen DA-Systems Augenbewegungen sowie die hypothalamische und limbische
Funktion beeinflussen.
Karson (1979) berichtet eine normale BR bei neuroleptisch naiven Schizophrenie-Patienten
und schließt daraus, dass die erhöhte BR bei behandelten Patienten wahrscheinlich durch die
Medikamente verursacht werden (Karson, 1979). Er stützt diese Vermutung durch ein
Experiment mit Affen, die eine dosisabhängige BR auf Apomorphin-Verabreichung vor und
nach einer Haloperidol-Behandlung aufweisen. Der DA-Rezeptoragonist Apomorphin
bewirkt einen Anstieg der BR, der durch Neuroleptika-Blockade der entsprechenden DA-
Rezeptoren mittels Haloperidol verhindert wird. Es zeigte sich 7 Tage nach Absetzen der

Theoretischer Hintergrund 19
Behandlung keine Änderung der Baseline-BR aber eine erhöhte BR als Reaktion auf niedrig
dosiertes Apomorphin (Karson, Staub, Kleinman & Wyatt, 1981b). Als Mechanismus für
diesen Effekt könnten eine neuroleptisch induzierte postsynaptische Hypersensibilisierung
zentraler DA-Rezeptoren oder Effekte an retinalen DA-Rezeptoren in Frage kommen
(Karson et al., 1981b). Carlsson (1975) berichtet, dass niedrig dosiertes Apomorphin
bevorzugt auf präsynaptische DA-Rezeptoren wirkt (Carlsson, 1975), was wiederum
bedeuten könnte, dass Haloperidol mit präsynaptischen DA-Rezeptoren interagiert, welche
die BR verändern. Schließlich könnte es aber auch sein, dass wiederholte Apomorphin-
Verabreichung die Sensibilität für Apomorphin erhöht oder, dass Haloperidol und
Apomorphin indirekt, über andere Neurotransmitter, eine Änderung der BR bewirken
(Carlsson, 1975).
Laut Karson (1989) unterliegt die Produktion spontaner Blinks einem Nervennetzwerk, das
sich von der Medulla durch den Cortex ausweitet und in dem cholinerge und dopaminerge
Neurotransmission eine wesentliche Rolle spielt. Neuropharmakologische Experimente
liefern Hinweise auf eine neurochemische Kontrolle der Blinkproduktion, die durch
dopaminerge, cholinerge und GABAerge Systeme des Hirnstamms ausgeübt werden. Dabei
zeigt sich, dass dopaminerge Aktivität direkt mit der BR korreliert und Agonismus der
beiden anderen Neurotransmittersysteme die BR hemmen können (Karson, 1989).
Unter Bezug auf die reduzierte BR bei Parkinsonismus postuliert Karson (1989) eine
„Dopamin-Hypothese“ der BR. Diese basiert auf den neuroanatomischen (pathologische
Veränderungen in der SN) und neurochemischen (Verminderung striatalen DAs)
Grundlagen der Parkinson-Erkrankung. Die Hypothese besagt, dass die BR positiv mit
striataler DA-Aktivität korreliert ist und wird durch pharmakologische Experimente mit
subhumanen Primaten unterstützt. So bewirken DA-Agonisten, wie Apomorphin und
Bromocriptin (siehe Kapitel 1.1.3), akute und dramatische Anstiege der BR. Dieser Effekt
wird durch Sulpirid (Neuroleptikum; spezifischer D2-Rezeptorantagonist) blockiert (Karson,
1983). Auch klinische Studien können zur Unterstützung der Hypothese herangezogen
werden (Karson, Kleinman, Berman, Phelps, Wise, De Lisi & Jeste, 1983). Medikationsfreie
Schizophrenie-Patienten hatten eine erhöhte mittlere BR (31 vs. 23 bpm bei Normalen), die
durch neuroleptische Behandlung normalisiert wurde. Diese hemmende Wirkung von
Neuroleptika auf die BR ist allerdings bei nicht-erkrankten Versuchspersonen, die im

Theoretischer Hintergrund 20
Vergleich zu medikationsfreien Schizophrenie-Patienten keine erhöhte BR haben, unklar.
Parkinson-Patienten mit Levodopa-induzierter Dyskinesie zeigten eine doppelt so hohe BR
(21 bpm) wie andere Parkinson-Patienten (11 bpm), während die symptomatischsten der
nicht-dyskinetischen Patienten eine sehr geringe Rate hatten (3 bpm). Diese Beobachtungen
legen nahe, die BR bei Parkinson-Patienten als Marker für den Schweregrad oder den Einsatz
Levodopa-induzierter Dyskinesie zu nutzen und auch drogeninduzierte Veränderungen der
BR bei Schizophrenie-Patienten als Marker für den Anschlag der Behandlung
heranzuziehen.
Experimente von Elsworth et al. (1991) mit Affen erhellen die Art und Weise, wie die BR
durch DA-Rezeptoren reguliert wird. Dihydrexidin (vollständiger D1-Agonist) produzierte
einen schnellen und dosisabhängigen Anstieg der BR. Dieser Effekt wurde durch vorherige
Gabe des spezifischen D1-Antagonisten SCH 23390 vollständig verhindert, war aber
unbeeinflusst durch vorherige Gabe des spezifischen D2-Antagonisten Remoxipirid.
Behandlung mit dem spezifischen D2-Agonisten, (+)-4-propyl-9-hydroxynaphthoxazin
(PHNO), führte zu einem schnellen und dosisabhängigen Anstieg der BR, der durch
Vorbehandlung mit Remoxipirid zunichte gemacht wurde, aber durch Vorbehandlung mit
SCH 23390 nicht beeinflusst wurde.
Diese Befunde weisen darauf hin, dass die spontane BR bei Primaten durch D1- und D2-DA-
Rezeptoren reguliert werden kann, wobei die Rezeptorsubtypen die BR zwar in die gleiche
Richtung aber voneinander unabhängig zu beeinflussen scheinen (Elsworth, Lawrence, Roth,
Taylor, Mailman, Nichols, Lewis & Redmond Jr., 1991).
Zusammenfassend kann festgehalten werden, dass bei klinischen und pharmakologischen
Studien an Tieren und an Patienten mit Schizophrenie oder Parkinson-Erkrankung ein
positiver Zusammenhang zwischen der BR und zentraler dopaminerger Aktivität ersichtlich
ist, so dass auf diesem Hintergrund die BR in dieser Arbeit als Indikator herangezogen wird.
Im nächsten Abschnitt werden weitere Hinweise darauf angeführt, dass die BR nicht nur
interindividuell sondern auch intraindividuell verschieden sein kann, was beim Einsatz der
Blinkrate als Schätzer für die zentrale dopaminerge Aktivität nicht außer Acht gelassen
werden darf.

Theoretischer Hintergrund 21
1.2.4 Die Blinkrate als psychophysisches Maß
Die BR geht bei Geburt gegen Null, steigt aber bis zur Adoleszenz ständig an, bis ein Level
erreicht ist, welches über das Erwachsenenalter hinweg beibehalten wird (Bentivoglio et al.,
1997). Nach Karson et al. (1981a) beträgt die BR eines normalen Erwachsenen etwa 20 bpm
(Karson, Berman, Donnelly, Mendelson, Kleinman & Wyatt, 1981a). Von Cramon (1980)
berichtet eine mittlere BR von 22,8 bpm mit einer Spannweite von 15-36 bpm und einer
Streuung von 10,8 bpm. Die maximale Blinkfrequenz lag zwischen 40,2 und 72 bpm.
Es zeigt sich selbst unter konstanten Umgebungsbedingungen eine erhebliche
interindividuelle als auch intraindividuelle Varianz der mittleren BR (von Cramon, 1980)
und weder physiologische Bedürfnisse des ocularen Systems, wie die Aufrechterhaltung des
cornealen Tränenfilms oder das Wegschwemmen von Fremdkörpern aus dem Auge, noch
indirekte (Schreckreaktion) oder direkte (physische Schädigung) defensive Bedürfnisse
können diese Variabilität erklären (Galley, 2001; Karson, 1983; Stern et al., 1984). Neben den
Hinweisen darauf, dass die großen interindividuellen Unterschiede in der BR auch als
Ausdruck der dopaminergen Aktivierung gedeutet werden dürfen (siehe Kapitel 1.2.3),
legen verschiedene Befunde nahe, dass die BR den Einfluss zentralnervöser Prozesse
widerspiegelt (Blount, 1928; Ponder & Kennedy, 1928).
Von Cramon (1980) beschreibt Lidbewegungen als Teil der mimischen
Ausdrucksbewegungen und geht daher von einer engen Verknüpfung mit psychischen
Prozessen aus. So scheinen kognitive Faktoren als auch organismische Variablen Einfluss auf
die BR zu haben (Galley, 2001; Karson, 1983; Stern et al., 1984; von Cramon, 1980).
Bentivoglio (1997) berichtet z.B., dass die BR bei 67% der normalen Versuchspersonen ein
charakteristisches Muster aufweist. Und zwar ist die mittlere BR im Vergleich zum
Ruhezustand (17 bpm) beim lauten Lesen erniedrigt (etwa 4,5 bpm) und bei Konversation
erhöht (26 bpm). Ähnliche Ergebnisse finden sich auch bei von Cramon (1980), der auch
Untersuchungen anderer Autoren heranzieht, die für einen wesentlichen Einfluss visueller
Aufmerksamkeit auf die BR sprechen. So zeigt sich z.B. bei einem Tracking-Test (Drew,
1951), dass die BR in dem Maß abnimmt, wie die Notwendigkeit für eine detaillierte visuelle
Beobachtung zunimmt, und auch nach Fixation (Kennard & Glaser, 1964) zeigt sich eine
Hemmung der spontanen Blinks, wobei die BR unmittelbar nach Entzug der visuellen

Theoretischer Hintergrund 22
Aufmerksamkeit auf den Ausgangswert oder sogar auf höhere Werte ansteigt. Allerdings
stellt sich auch bei nicht-visuellen Aufgaben (z.B. Irrgarten- oder auditorische Tracking-
Aufgabe) eine verminderte BR ein, was nahe legt, dass Aufmerksamkeit allgemein zu einer
Reduktion der BR führt, die proportional zur Aufgabenschwierigkeit ist (Stern et al., 1984).
Mit dieser Annahme stimmen auch Befunde einer erhöhten BR bei Zuständen mangelnder
oder nachlassender Aufmerksamkeit, wie Ermüdung, Monotonie, Langeweile oder
Geübtheit überein (Galley, 2001).
Als nicht unangezweifelte Erklärung für die hohe BR bei Konversation nimmt von Cramon
(1980) einen Zusammenhang zwischen lidmotorischer und sprechmotorischer Aktivität an
und belegt dies mit einer Untersuchung, bei der die BR nicht bei der Aufgabenlösung
sondern erst bei der lauten Verbalisierung der Antwort ansteigt. Befunde einer erniedrigten
BR beim lauten Lesen werden dann so gedeutet, dass die Blinkhemmung durch visuelle
Aufmerksamkeit wirksamer ist, als die simultane Erleichterung durch sprechmotorische
Aktivität.
Stern (1984) geht eher davon aus, dass eine höhere allgemeine Aktivierung mit erhöhter BR
einhergeht und zieht neben indirekten Beweisen, bei denen von der Aufgabenanforderung
auf das arousal geschlossen wird, auch eine Studie (von Gille, Otto & Ullsperger, 1977) heran,
bei der eine EEG-Analyse und subjektive Berichte darauf hinweisen, dass der Effekt eher
allgemein ist.
Von Cramon (1980) bezeichnet die BR als Maß für allgemeine Erregbarkeit im Sinne von
Affektivität und macht Angaben über affektive Zustände mit sehr hoher oder sehr niedriger
Blinkfrequenz. So liegt die mittlere BR bei Nervosität, Aufregung, Erschrecken,
Überraschung, Wut, Ärger oder dem Gefühl der Bedrohung über 48 bpm, während bei
Angstzuständen im Mittel weniger als 3 bpm gezählt wurden. Die Wirkung von Angst auf
die BR ist aber laut Stern (1984) noch nicht geklärt.
Insgesamt weisen die angeführten Befunde darauf hin, dass die BR bei (psycho-) motorischer
oder genereller Aktivierung bzw. erhöhtem arousal ansteigt und bei nach außen gerichteter
Aufmerksamkeit unterdrückt wird, was bei einer Nutzung der BR als Indikator
berücksichtigt werden muss.

Theoretischer Hintergrund 23
Im Folgenden wird ein Überblick über Methoden gegeben, mit denen die spontanen Blinks
erfasst werden können.
1.2.5 Möglichkeiten zur Erfassung von Blinks
Zur Erfassung von Blinks kommen mehrere Methoden in Frage (z.B. Stern; 1984). Zu den
einfachsten Methoden gehört das Auszählen der Blinks direkt während der Untersuchung
oder nachträglich von einer Videoaufnahme. Zu den komplizierteren Methoden zählt die
Erfassung mit einem Potentiometer, wobei ein Faden am Lid befestigt wird. Außerdem gibt
es mechanische Erfassungsmethoden durch Hebelsysteme, die am Lid befestigt werden oder
optische Methoden mit reflektierenden Spiegeln. Schließlich gibt es auch die Möglichkeit der
elektromyographischen Bestimmung der Aktivität des Orbicularis oculi oder
photoelektrische Techniken, die sclerale Reflexionen vom geöffneten Auge erfassen können.
Die am weitesten verbreitete Methode zur Erfassung von Blinks, ist das Elektrooculogramm
(EOG), das die Änderung der elektrischen Potentiale zwischen Cornea und Retina registriert,
wenn die Lidbewegung erfolgt. Da das EOG auch bei Tieren mit entfernter Retina
funktioniert, vermutet Stern (1984), dass das Potential zwischen Cornea und Fundus
aufgezeichnet wird. Dabei verhält sich das Lid wie ein gleitender Widerstand, der das
bestehende corneo-fundale Potential verändert.
Da die Cornea im Vergleich zum Fundus positiv geladen ist, bildet das Auge einen
elektrischen Dipol (Birbaumer & Schmidt, 1996), dessen elektrisches Feld mit vertikal
gegenüberliegenden Hautelektroden aufgezeichnet werden kann (VEOG). Je nachdem,
welche der Elektroden infra- oder supra-orbital angebracht wird und ob sich das Lid auf
oder ab bewegt, wird eine verstärkte Positivierung oder Negativierung registriert. Wenn
keine Lid- oder Augenbewegungen ausgeführt werden, sollte sich das erfasste
Bestandspotential nicht verändern.
In der vorliegenden Untersuchung wurde zur Erfassung der Blinks das eben beschriebene
EOG eingesetzt.
Zur Begründung der Erwartung eines Zusammenhangs zwischen DA und Persönlichkeit
werden im nächsten großen Abschnitt zwei biologisch orientierte Persönlichkeitstheorien
beschieben, die sich mit diesem Thema befassen.

Theoretischer Hintergrund 24
1.3 Biologische Persönlichkeitstheorien
Im Folgenden werden beispielhaft zwei Persönlichkeitstheorien vorgestellt, für welche die
zentrale dopaminerge Aktivität eine Rolle spielt. Da in der vorliegenden Arbeit die Traits
„Extraversion“ und „Spontaneität“ von Interesse sind, werden an dieser Stelle Theorien
beschrieben, die sich auf diese Traits beziehen.
Zunächst wird die Theorie von Depue umfangreich beschrieben, die sich mit der
neurobiologischen Grundlage der Persönlichkeitsdimension Extraversion befasst. Dann wird
die biosoziale Theorie von Cloninger umrissen, wobei der Schwerpunkt auf die
Persönlichkeitsdimension novelty seeking gelegt wird.
1.3.1 Die Neurobiologie der Persönlichkeitsstruktur nach R. A.
Depue
Depue (z.B. Depue & Collins, 1999) greift einen Trait auf, den er übergreifend als
Extraversion bezeichnet und stellt Parallelen zu einem Verhaltensförderungssystem bei
Tieren her; beides soll mit Anreizmotivation verbunden sein. Integrierend postuliert er ein
neurobiologisches Förderungsmodell, das dem Neurotransmitter DA eine modulierende
Funktion bezüglich Verhalten zuschreibt, das durch Anreize gefördert wird. Ein
psychobiologisches Schwellenmodell der Verhaltensförderung soll einen Zusammenhang
zwischen individuellen Unterschieden in der dopaminergen Übertragung und
unterschiedlichen Reaktionen auf Anreizstimuli erklären.
Als Erstes wird die Persönlichkeitsdimension Extraversion aus Depue´s Sicht beschrieben
und ein entsprechendes Persönlichkeitseigenschaftsmodell vorgestellt.
1.3.1.1 Ein Persönlichkeitseigenschaftsmodell
Trotz unterschiedlicher Benennung findet sich in fast jeder Persönlichkeitstaxonomie ein
emotionaler Trait höherer Ordnung, der die Auslösung von positivem Affekt, Begehren und
Anreizmotivation durch Belohnungssignale einschließt. Obwohl Depue et al. (1994)
aufgrund der emotionalen Aspekte dieses Traits die Bezeichnung „positive Emotionalität“
vorziehen, übernehmen Depue und Collins (1999) den Begriff „Extraversion“, da dieser eine

Theoretischer Hintergrund 25
lange Geschichte hat und bei Traitpsychologen bekannt ist.
Depue und Collins (1999) unterscheiden zwei zentrale Merkmale von Extraversion. Dazu
gehört Impulsivität, die sich aus der Interaktion von Extraversion mit Constraint1 ergibt, und
interpersonelles Engagement. Letzteres besteht wiederum aus zwei Komponenten:
Anschluss (affiliation) äußert sich in der Wertschätzung und dem Genießen
zwischenmenschlicher Bindung und einer warmherzigen und liebevollen Art.
Wirkung (agency) äußert sich in sozialer Dominanz, Bestimmtheit, Angeberei, dem Genießen
einer Führungsrolle und einem Sinn für die Stärke, Ziele zu erreichen. Diese Komponente
stellt eher eine allgemeine motivationale Disposition dar, die Dominanz, Ehrgeiz, Herrschaft,
Wirkungskraft und Leistung einschließt.
Basierend auf Annahmen von Robinson und Berridge (1993) gehen Depue und Collins (1999)
davon aus, dass Extraversion auf positiver Anreizmotivation beruht.
Robinson und Berridge (1993) vermuten, dass die Auseinandersetzung mit
unkonditionierten Anreizstimuli das neuronale Substrat von Freude aktiviert. Das Erleben
von Freude wird durch klassisches assoziatives Lernen mit dem neutralen Reizkontext
verbunden, so dass dieser zu einem konditionierten Anreizstimulus wird und von da an
antizipatorisch Freude und Anreizmotivation auslösen kann. Die Anreizstimuli (oder ihre
zentrale Repräsentation) werden nach ihrer Intensität oder Salienz verschlüsselt, wobei den
Reizen ein motivationaler Wert zugeschrieben wird, der den Einfluss der Anreizmotivation
auf emotionale und Verhaltensreaktionen angibt.
Eine spätere Exposition zu Anreizstimuli (oder die Aktivierung ihrer zentralen
Repräsentationen) löst dann einen Zustand der Anreizmotivation aus, der
Annäherungsverhalten in Richtung Ziel fördert und steuert. Ein solcher Zustand ist beim
Menschen mit starkem positivem Affekt verbunden, der sich in Verlangen, Wollen (wanting),
Erregung, Enthusiasmus, Energie, Stärke und Selbstwirksamkeit äußert. Dies tritt zwar
typischerweise gleichzeitig mit Gefühlen der Freude und des Mögens (liking) auf, ist aber
davon zu unterscheiden (siehe Kapitel 1.1.5).
1 Einer der Hauptfaktoren der Persönlichkeit wird u.a. Psychotizismus oder Constraint genannt. Constraint bezeichnet laut Depue (1996) eine nicht-affektive Verhaltensdimension, welche die Leichtigkeit widerspiegelt, mit der Verhalten durch kontrollierende Reize ausgelöst wird.

Theoretischer Hintergrund 26
Aufbauend auf diesen Annahmen ziehen Depue und Collins (1999) in Betracht, dass die
Variation im Prozess der Verschlüsselung von Anreizsalienz die Basis interindividueller
Unterschiede in der Häufigkeit und Intensität von Anreizmotivation ist und weitergefasst
die Hauptquelle interindividueller Unterschiede in Extraversion darstellt.
Depue und Collins (1999) beschreiben ein Persönlichkeitseigenschaftsmodell (Abbildung 3),
bei dem Extraversion einen Trait höherer Ordnung (eher allgemeines Verhaltensmuster)
darstellt, der von korrelierten Traits geringerer Ordnung (emotionale und
Verhaltensmerkmale) abgeleitet wird. Dabei unterliegt jeder Trait niedrigerer Ordnung den
gleichen Prozessen wie Extraversion. Während einige der Traits geringerer Ordnung (wie
z.B. positiver Affekt und Aktivierung) diese Prozesse direkt repräsentieren, zeigen andere
ihren Einfluss in verschiedenen Kontexten. So wird soziale Stärke in einem kompetitiven
Umfeld, Leistung beim Erreichen von Langzeitzielen und Anschluss in sozialen Situationen
sichtbar.
ANREIZ- STIMULI
sozial sexuell Futter Sicherheit Langzeitziele verhinderte Belohnung
ZUGRUNDE- LIEGENDE PROZESSE
Aktivität-Energie
Anreizbelohnungs-motivation
affektive (Enthusiasmus, Verlangen, Stärke,
positive Stimmung)
komplexe Kognition
TRAIT HÖHERER ORDNUNG
Extraversion
ALLGEMEINES VERHALTENS- MUSTER
Annäherung, Exploration, Engagement
TRAITS NIEDRIGERER ORDNUNG
Aktivierung Leistung soziale Stärke Anschluss positiver Affekt
Abbildung 3 Persönlichkeitseigenschaftsmodell (nach Depue & Collins, 1999)
Depue und Collins (1999) schreiben Extraversion einen zentralen Mechanismus zu, der für
eine fördernde Modulation aller zugrundeliegenden Prozesse sorgen soll. Indem dieser

Theoretischer Hintergrund 27
modulierende Einfluss die Prozesse verbindet und ihre Intensität gewichtet, ergeben sich
variierende Ausprägungen der Förderung behavioraler Annäherung. Spezifische
Verhaltensformen, die an verschiedene Kontexte (z.B. sozial oder leistungsbezogen)
gebunden sind, manifestieren sich in Traits geringerer Ordnung und werden vermutlich
durch spezifische Verhaltenssysteme vermittelt.
Im nächsten Abschnitt wird ein solches System beschrieben, das speziell für die Förderung
von Verhalten zuständig sein soll und sich sehr gut mit dem
Persönlichkeitseigenschaftsmodell deckt.
1.3.1.2 Ein Verhaltensförderungssystem
Für Säugetiere wird ein allgemeines Verhaltenssystem angenommen, das nach Depue und
Collins (1999) zugrundeliegende Prozesse und Verhaltensweisen einschließt, die mit
Extraversion übereinstimmen. Diesem emotionalen System wird zugeschrieben, dass es sich
entwickelt hat, um Vorwärtsbewegung und Suchverhalten zu motivieren, um wiederum die
Bedürfnisse eines Tieres (z.B. nach Futter oder einem Sexualpartner) zu befriedigen. Es wird
durch verschiedenste positiv belohnende Reize aktiviert und soll dazu dienen, das Tier mit
solchen Reizen in Kontakt zu bringen, falls sich diese nicht in unmittelbarer Nähe befinden.
Collins und Depue (1992) nennen dieses System behavioral facilitation system (BFS, frei
übersetzt: Verhaltensförderungssystem).
Primäre und konditionierte Anreizstimuli lösen drei zentrale Prozesse aus, die das BFS und
positive Emotionalität (bzw. Extraversion) gemeinsam haben (Collins & Depue, 1992; Depue,
1995; Depue, Luciana, Arbisi, Collins & Leon, 1994):
• Anreizmotivation als innerer, subjektiver Zustand
• Einleitung lokomotorischer Aktivität zur Unterstützung der Zielerreichung
• Kognitive Prozesse, da aktive Zielsuche die Auseinandersetzung mit der Umwelt, und
somit auch die Notwendigkeit der Bewertung derselbigen, steigert
Das BFS kann somit als dynamisches System angesehen werden, das die emotionale
Bewertung von Anreizstimuli beeinflusst (Collins & Depue, 1992).
Depue und Collins (1999) definieren dieses System als behaviorale Annäherung, die auf
Anreizmotivation beruht, und stellen ein Verhaltensförderungsmodell auf (Abbildung 4). In
diesem Modell wird Annäherungsverhalten von verschiedenen zugrundeliegenden

Theoretischer Hintergrund 28
Prozessen (Bewegungseinleitung, Anreizmotivation, positiver Affekt, kognitive Prozesse)
unterstützt, die Verhalten auf ein belohnendes Ziel ausrichten. Die gemeinsame Aktivierung
dieser Prozesse soll auf einem zentralen Mechanismus höherer Ordnung (Förderung)
basieren.
ANREIZ- STIMULI
sozial sexuell Futter Sicherheit Langzeitziele verhinderte Belohnung
ZUGRUNDE- LIEGENDE PROZESSE
Bewegungs-einleitung
Anreizbelohnungs-motivation
affektive (Neugier, Verlangen, Macht)
komplexe Kognition
ZENTRAL MODULIERENDER MECHANISMUS
Förderung
ALLGEMEINES VERHALTENS- MUSTER
Annäherung, Exploration, Engagement
SPEZIFISCHE VERHALTENS-MUSTER
Erregung Futter-
beschaffung sexuelle
Annäherung sozialer
Wettkampf Anschluss
zielgerichtete Aggression
Abbildung 4 Verhaltensförderungsmodell (nach Depue & Collins, 1999).
Des Weiteren wird ein Modell beschrieben, das die neurobiologische Grundlage der zuvor
beschriebenen Persönlichkeitsdimension und dem entsprechenden Verhaltenssystem
darstellen soll.
1.3.1.3 Ein neurobiologisches Förderungsmodell
Depue und Collins (1999) gehen davon aus, dass die zuvor beschriebene Persönlichkeits-
und Verhaltensebene höherer Ordnung (Extraversion, besonders die agency-Komponente,
und Verhaltensförderung) eng mit Anreizmotivation verbunden sind und postulieren für
beide eine gemeinsame neurobiologische Grundlage.
Sie geben einen Überblick über relevante Literatur aus der Tierforschung, die nahe legt, dass
Verhaltensförderung mit funktionellen Eigenschaften zweier aufsteigender Projektionen des
dopaminergen Systems (ausgehend vom VTA) verbunden ist (z.B. Le Moal & Simon, 1991),

Theoretischer Hintergrund 29
und ordnen Verhaltensförderung innerhalb eines umfassenden neuroanatomischen
Netzwerks ein, das mit Anreizmotivation zielgerichteten Verhaltens zusammenhängt.
Mesolimbische DA-Projektionen tragen zur Förderung von Anreizmotivation und der
Einleitung von Vorwärtsbewegung bei, während mesocorticale DA-Projektionen an der
Förderung komplexer Funktionen beteiligt sind, die adaptives zielgerichtetes Verhalten mit
wechselnden Umweltkontingenzen verbinden (Depue, 1995; Depue et al., 1994).
Das dopaminerge System wird als Modulator höherer Ordnung eines neurobiologischen
Netzwerks in Betracht gezogen, das Verhaltensfunktionen integriert, die mit Extraversion
verbunden sind. Depue und Collins (1999) betonen, dass sie nicht von einer vermittelnden
Funktion von DA ausgehen, da DA allgemein neuronale Prozesse fördert, die zielgerichtetes
Verhalten unterstützen und die funktionalen Effekte weitgehend von den Endstrukturen des
dopaminergen Systems abhängen. DA wird also keine spezifische sondern eine eher
allgemeine und übergreifende Wirkung auf verschiedene Effektoren zugeschrieben.
Bei der Förderung von Anreizmotivation scheint die VTA-DA-Projektion zum NAS eine
entscheidende Rolle zu spielen, was darauf schließen lässt, dass dieser eine Stelle der
Integration von Anreizinformation darstellt (siehe Kapitel 1.1.5). Besonders die caudo-
mediale Schalen(shell)-Region des NAS (NASshell) ist ein Hauptpunkt der Konvergenz
motivationaler Information. Verschiedene limbische Strukturen, die stark miteinander
verbunden sind, übermitteln unterschiedliche spezifische Informationen über den salienten
Anreizkontext (über glutaminerge exzitatorische Afferenzen) zu VTA-DA-Neuronen und
Neuronen des NAS, und tragen so zur Reaktion auf konditionierte Anreize bei (Depue &
Collins, 1999; Depue & Lenzenweger, 2001):
1. Der basolaterale Komplex der Amygdala assoziiert diskrete explizite Reize mit
Verstärkung.
2. Regionen, welche die erweiterte Amygdala einschließen, integrieren
verstärkungsbezogene Information, Reiz-Belohnungs-Assoziationen und Motivation.
3. Der Hippocampus assoziiert räumliche und kontextuelle Beziehungen zwischen
Umgebungsreizen mit Verstärkung.
4. Der präfrontale mediale orbitale Cortex (MOC) bildet eine zusammengefügte Struktur
appetitiver und aversiver Verhaltenskontingenzen der Umgebung, was einen Vergleich

Theoretischer Hintergrund 30
der Valenz und des Ausmaßes von Ergebniserwartungen, die mit verschiedenen
möglichen Reaktionsstrategien assoziiert sind, ermöglicht.
Ausgehend von Arbeiten anderer Autoren schlagen Depue und Collins (1999) ein
corticolimbisch-striatal-thalamisches Netzwerk vor (Abbildung 5), das für die Modulation
von Anreizmotivation zielgerichteten Verhaltens zuständig sein soll und diese reguliert. Da
Ursprungs- und Endregion dieses Netzwerks im präfrontalen medialen orbitalen Cortex
(genauer: MOC 13) liegen, sprechen sie von einem MOC-Netzwerk, das aus drei
wesentlichen Komponenten besteht (Depue & Collins, 1999; Depue & Lenzenweger, 2001):
• Der Antriebskreis integriert, bewahrt und aktualisiert Information zur Verschlüsselung
der Intensität oder Salienz von Anreizstimuli, um einen entsprechenden Zustand der
Anreizmotivation herzustellen, der zum motorischen System übertragen werden kann.
Dieser Kreis umfasst den Nucleus accumbens (NASshell), einen Teil des ventralen
Pallidums (VPm) und dopaminerge Projektionen aus der VTA (VTA-DA-Projektionen).
• Das VTA-DA-Projektionssystem fördert sowohl die neuronale Integration, die im
Antriebskreis auftritt, sowie die Integration innerhalb allgemeinerer
Netzwerkinteraktionen.
• Das MOC 13 (Brodmann-Areal) ist für die Regulation höherer Ordnung von
Netzwerkprozessen zuständig.
Abbildung 5 Schematische Darstellung des MOC-Netzwerks (nach Depue & Collins, 1999). MOC 13: Brodmann-Areal, MD: mediodorsaler Kern des Thalamus, NASshell: Schalenregion des Nucleus accumbens, VPm: Teil des ventralen Pallidums, VTA-DA-Zellen: dopaminerge Zellen des ventralen tegmentalen Areals; durchgehende Pfeile verbinden Strukturen des zentralen Antriebskreises.
MOC 13
VPm NASshell
Erweiterte Amygdala
Basolateraler Komplex der Amygdala
Hippocampus
MD
VTA-DA-Zellen

Theoretischer Hintergrund 31
Depue und Collins (1999) stellen zusammenfassend ein neurobiologisches
Förderungsmodell vor (Abbildung 6), in dem VTA-DA-Projektionen zu Strukturen des
MOC-Netzwerks das neuronale Substrat für Extraversion und Verhaltensförderung
darstellen. Dabei sorgen NAS und MOC für die Aktivierung von DA-Neuronen des VTA
(VTA-DA-Neuronen), die weit projizieren, um Prozesse innerhalb von Strukturen des MOC-
Netzwerks zu fördern.
ANREIZ- STIMULI
sozial sexuell Futter Sicherheit Langzeitziele verhinderte Belohnung
medialer orbitaler Cortex parallele corticale Netzwerke VPm NASshell BL-E Amygdala Hippocampus
NEURONALE STRUKTUREN & PROZESSE DES MOC-NETZWERKS
Bewegungs-einleitung
Anreizbelohnungs-motivation
affektive (Neugier, Verlangen, Macht)
komplexe Kognition
ZENTRAL MODULIERENDER MECHANISMUS
VTA-Dopamin-Förderung
ALLGEMEINES VERHALTENS- MUSTER
Annäherung, Exploration, Engagement
SPEZIFISCHE VERHALTENS-MUSTER
Erregung Futter-
beschaffung sexuelle
Annäherung sozialer
Wettkampf Anschluss
zielgerichtete Aggression
Abbildung 6 Neurobiologisches Förderungsmodell (nach Depue & Collins, 1999). MOC: medialer orbitaler Cortex, VPm: Teil des ventralen Pallidums, NASshell: Schalenregion des Nucleus accumbens, BL-E-Amygdala: basolaterale und erweiterte Amygdala, VTA-Dopamin-Förderung: Förderung neuronaler Prozesse moduliert durch dopaminerge Neuronen des ventralen tegmentalen Areals.
Die Verhaltensförderung umfasst verschiedene Muster der dopaminergen Innervierung aus
dem VTA (Depue & Collins, 1999; siehe auch Abbildung 5):
• Die gegenseitige Innervierung zwischen VTA-DA-Neuronen und Neuronen der
NASshell fördert Anreizmotivation und in Zusammenarbeit mit der Kern(core)-Region
des NAS (NAScore), die Einleitung von Bewegung.

Theoretischer Hintergrund 32
• Dopaminerge Efferenzen des VTA zum VPm modulieren die Zellfeuerraten und können
sowohl die Einleitung appetitiver motorischer Aktivität fördern als auch die
Rückprojektion von Information über Efferenzen vom VPm zum VTA. Dies unterstützt
die zeitliche Aufrechterhaltung eines Zustandes der Anreizmotivation innerhalb des
Antriebskreises.
• Es existiert, ausgehend vom VTA, eine dichte dopaminerge Innervierung der
basolateralen und zentralen amygdaloiden Kerne (BL-E Amygdala), die jeweils die
NASshell innervieren.
Im nächsten Abschnitt werden Einflüsse auf die Neurobiologie beschrieben, die zu
interindividuellen Unterschieden führen, welche sich schließlich auch in der Persönlichkeit
eines Menschen widerspiegeln sollen.
1.3.1.4 Interindividuelle Unterschiede in der dopaminergen Funktion
Genetische und Umweltfaktoren können die Entwicklung neurobehavioraler Systeme
beeinflussen. Collins und Depue (1992) unterscheiden drei Prozesse, die sich während der
Entwicklung neuronaler Systeme überschneiden und gemeinsam zur Regulation der
Neuroarchitektur durch DA beitragen.
Genotypgetriebene (genotyp-driven) Prozesse beeinflussen die Grundstruktur und -funktion
von Nervenbeständen während der embryonalen Entwicklung.
Erfahrungserwartende (experience-expectant) Prozesse finden innerhalb sensibler Phasen der
Gehirnentwicklung statt, die Umwelterfahrungen vorausgehen, und beinhalten eine
weitreichende Überproduktion neuronaler Synapsen. Durch selektive Erhaltung und
Stärkung einer Teilmenge dieser Synapsen werden während der Exposition zu
phylogenetisch vorhersagbaren Formen der Umwelterfahrung, speziestypische Muster der
neuronalen Zellarchitektur eingerichtet. Zeitpunkt und Ort der synaptischen
Überproduktion werden durch genotypische Einflüsse bestimmt, während die funktionalen
Beziehungen in Reaktion auf Umwelterfahrungen variieren.
Erfahrungsabhängige (experience-dependent) Prozesse modifizieren die neuronale
Zellarchitektur, um Umwelterfahrungen zu verschlüsseln. Diese sind für jedes Individuum
einzigartig und damit phylogenetisch unvorhersehbar und genotypisch unbeeinflusst.

Theoretischer Hintergrund 33
Solche Prozesse beinhalten eine lokal begrenzte Synapsenproduktion, die während der
Verschlüsselung von Information aus bedeutsamer Erfahrung eingeleitet wird.
Für diese Arbeit spielt vor allem die heterosynaptische Plastizität im NAS und dem VTA
eine Rolle. Ähnlich einer Langzeitpotenzierung werden dabei synaptische Kontakte von
Neuronen des NAS mit aktiveren corticalen und limbischen Efferenzen durch DA gestärkt.
Dadurch sollen die Verbindungen zwischen dem Eingang des salienten Anreizkontextes und
Anreizprozessen in der NASshell gestärkt werden. Der Eingang von Kontextinformation zu
DA-Neuronen des VTA und Neuronen des NAS veranlassen die Auslösung von
Anreizmotivation während DA kontextbezogene glutaminerge Verbindungen im VTA und
NAS fördert (Collins & Depue, 1992; Depue & Collins, 1999; Depue & Lenzenweger, 2001).
Die Variation im Ausdruck wirkenden (agentic) Affekts und Verhaltens wird großenteils den
Effekten stabiler individueller Unterschiede im Ausmaß, mit dem die VTA-DA-Funktion die
Verbindungen salienten kontextbezogenen Eingangs zu VTA-DA-Neuronen und Neuronen
des NAS fördert, zugeschrieben (Depue & Collins, 1999; Depue et al., 1994). Depue und
Lenzenweger (2001) vermuten, dass es sowohl Effekte interindividueller Unterschiede in der
Funktion von VTA-DA auf die Förderung der Kontextverbindungen im VTA und NAS als
auch Variationen in der dopaminergen Förderung bei späterem Auftreten des
entsprechenden Kontextes sind, die zusammen zu Unterschieden in wirkender (agentic)
Extraversion beitragen.
Nach Collins und Depue (1992) sind interindividuelle Unterschiede in der Anzahl von VTA-
DA-Neuronen das Ergebnis genotypgetriebener Prozesse. Demnach soll ein Individuum mit
relativ großer Zellzahl die strukturelle Kapazität besitzen, während erfahrungserwartender
sensibler Phasen hohe DA-Mengen an Endigungen von VTA-Projektionen auszuschütten.
Falls ein ausreichendes Aktivitätslevel in den VTA-Zellen anhält, wären solche Individuen
prädisponiert, eine große Anzahl synaptischer Kontakte in BFS-Strukturen zu stärken, und
dadurch zu erhalten. Collins und Depue (1992) verweisen auf empirische Befunde bei
gezüchteten Mäusestämmen, bei denen sowohl die Quantität spontaner explorativer
Bewegung als auch Amphetamin-induzierter Bewegung mit der Anzahl der VTA-DA-
Neuronen, mit der relativen Innervationsdichte dopaminerger Endigungen und mit dem

Theoretischer Hintergrund 34
DA-Gehalt im NAS positiv zusammenhängen (Fink & Reis, 1981; Oades, 1985; Sved, Baker &
Reis, 1984, 1985). Dies legt laut Collins und Depue (1992) nahe, dass ein Individuum mit
reicher genetischer Ausstattung an DA-Zellen die Umwelt aktiv explorieren würde, um
belohnende Reize zu suchen. Dieses funktionelle Ergebnis sollte nicht auftreten, wenn die
Umwelterfahrung arm an Belohnung wäre. Wahrscheinlich entsteht eine starke funktionale
Kapazität im dopaminergen System (ausgehend vom VTA), emotionale Reaktionen auf
Signale der Belohnung zu motivieren und zu steuern. Diese Grundlage der BFS-Reaktivität
wäre gegenüber zukünftigen Modifikationen weitgehend resistent.
Bei voranschreitender Entwicklung des neuronalen Systems, könnten erfahrungsabhängige
Prozesse für einen zusätzlichen Anstieg der synaptischen Verbindungen in BFS-Strukturen
sorgen, da eine Prädisposition, sich mit potentiell belohnenden Reizen auseinanderzusetzen,
häufige Forderungen nach zusätzlichen Synapsen in den Endgebieten von VTA-DA-
Projektionen zur Folge hätten. So kann z.B. Hypersensibilisierung (siehe auch Kapitel 1.2.3)
nach wiederholter Verabreichung von DA-Agonisten (Robinson, 1988) als
erfahrungsabhängiger Prozess interpretiert werden (Depue, 1995). Bis zum
Erwachsenenalter würde die extensive synaptische Verzweigung innerhalb des BFS-
Netzwerks Reaktionen des dopaminergen Systems (ausgehend vom VTA) auf Signale der
Belohnung konsistent verstärken. Dementsprechend würde das Individuum ein hohes
stabiles Niveau der BFS-Reaktivität entwickeln (Collins & Depue, 1992).
Zusammenfassend kann festgehalten werden, dass im Laufe des Lebens verschiedene
Prozesse zu interindividuellen Unterschieden in der dopaminergen Übertragung führen.
Diese Unterschiede sind mit Variation in der Verschlüsselung der Salienz positiver
Anreizkontexte und mit der Fähigkeit dieser Kontexte, den Anlass für belohnendes
zielgerichtetes Verhalten auszulösen oder zu bestimmen, verbunden.
Im Folgenden wird ein Modell vorgestellt, das die Auswirkungen solcher interindividueller
neurobiologischer Unterschiede auf das Verhalten darstellen soll.
1.3.1.5 Ein Schwellenmodell der Verhaltensförderung
Depue (1995) beschreibt ein psychobiologisches Schwellenmodell, das die Effekte
interindividueller Unterschiede in der dopaminergen Übertragung auf durch Anreize
gefördertes Verhalten erklären soll (Abbildung 7).

Theoretischer Hintergrund 35
Abbildung 7 Ein psychobiologisches Schwellenmodell der Verhaltensförderung (nach Depue, 1995). A und B repräsentieren beispielhaft zwei Individuen mit niedriger bzw. hoher DA-Rezeptoraktivierung und dementsprechend enger bzw. weiter Spannweite effektiver Anreizstimuli.
Die Minimumschwelle repräsentiert ein zentrales neuronales System, das die äußeren und
inneren Faktoren, die zur Reaktionsförderung beitragen, gewichtet. Das Ausmaß der
Anreizstimulation und das Niveau der postsynaptischen DA-Rezeptoraktivierung
beeinflussen die Schwelle am stärksten. Die Beziehung zwischen diesen beiden
interagierenden Variablen wird als Austauschfunktion dargestellt, so dass sich eine
Diagonale ergibt, die den Minimumschwellenwert der Reaktionsförderung darstellt. Die
Spannweite effektiver Anreizstimuli wird als Funktion des Niveaus der dopaminergen
Aktivierung dargestellt. Durch Sensibilisierung kann sich die DA-Ausschüttung im NAS
steigern, wodurch die Schwelle der Reaktionsförderung über die gesamte Spannweite
effektiver Anreizstimuli herabgesetzt werden kann. Auch der Neurotransmitter Serotonin (5-
Hydroxytryptamin, 5-HT) moduliert die Wahrscheinlichkeit der dopaminergen
Reaktionsförderung über die gesamte Spannweite des Ausmaßes des Anreizstimulus

Theoretischer Hintergrund 36
(Depue, 1995, 1996; Depue & Collins, 1999; Depue, Collins & Luciana, 1996; Depue &
Lenzenweger, 2001).
Mit Hilfe der hypothetischen Individuen A und B (mit geringer bzw. starker
Traitausprägung) lassen sich aus dem Schwellenmodell Verhaltensvorhersagen ableiten, die
Implikationen für die Konzeptualisierung von Extraversion haben (Depue, 1995; Depue &
Collins, 1999):
• Die Ausprägung der Zustands-DA-Reaktion für einen Anreizstimulus wird bei B im
Mittel stärker sein als bei A.
• Der Unterschied im Ausmaß subjektiver Erfahrungen zwischen A und B kann zu
Variation in gleichzeitiger Verschlüsselung der Anreizintensität oder -salienz eines
Stimulus beitragen.
• Traitunterschiede in der dopaminergen Übertragung können sich auf die Spannweite
effektiver (fördernder) Reize auswirken.
• Die breitere Spannweite effektiver Anreize für B legt nahe, dass B im Mittel häufiger die
Auslösung von Annäherungsverhalten und intensivere positive emotionale und
motivationale Gefühle, die mit Extraversion assoziiert sind, erfahren wird.
• Traitunterschiede sowohl in der Anreizverschlüsselung als auch in der Spannweite
effektiver Anreizstimuli können interagieren, um individuelle Variation in der Dynamik
behavioraler Auseinandersetzung mit der Umwelt herzustellen.
• Variation in der dopaminergen Förderung hängt mit Verhaltensflexibilität zusammen,
wenn eine Änderung motorischer, affektiver und kognitiver Reaktionsmuster aufgrund
äußerer Umstände erforderlich ist.
Im nächsten Abschnitt wird der Einfluss der neurobiologische Grundlage eines anderen
Traits auf die Reaktionsschwelle in Betracht gezogen.
1.3.1.6 Die Interaktion von Extraversion mit Constraint
Depue und Lenzenweger (2001) gehen davon aus, dass nicht-affektiver Constraint den Trait
entlang des neurobiologischen Konstrukts der Reaktionsschwelle darstellt. Constraint soll
den Einfluss aller Schwellenmodulatoren beinhalten und weist einen allgemeinen Einfluss
auf Verhalten auf, das nicht offensichtlich mit einem spezifischen neurobehavioral-

Theoretischer Hintergrund 37
motivationalen System assoziiert ist. Neurotransmitter mit inhibitorischem Einfluss sind
besonders bedeutsame Modulatoren der Reaktionsschwelle und tragen wahrscheinlich zu
einem großen Teil zur Varianz nicht-affektiven Constraints bei. Einer dieser
Neurotransmitter ist 5-HT, welches eine substantielle modulierende Rolle für eine
allgemeine neurobiologische Funktion spielt, die verschiedene Formen motivierten
Verhaltens beeinträchtigt (siehe auch Abbildung 7).
Laut Depue und Lenzenweger (2001) wird eine reduzierte Funktion serotonerger
Übertragung häufig mit Verhaltensinstabilität verbunden. Diese äußert sich als Labilität und
verstärkt sich mit ansteigenden fördernden Einflüssen auf die Reaktionsauslösung. Der
qualitative Gehalt instabilen Verhaltens hängt schließlich von dem neurobehavioral-
motivationalen System (oder affektivem Trait) ab, das gerade aktiv ist.
Abbildung 8 stellt die Interaktion von nicht-affektivem Constraint mit wirkender (agentic)
Extraversion dar, wobei 5-HT als inhibitorischer Modulator vieler DA-geförderter
Verhaltensweisen fungiert (Depue, 1996; Depue & Lenzenweger, 2001).
Die Interaktion dieser beiden Traits bildet eine diagonale Dimension der Verhaltensstabilität,
die gleichermaßen für affektive, kognitive, interpersonelle, motorische und Anreizprozesse
gilt, wobei die Diagonale den Verlauf der größten Stabilitätsvarianz (von Labilität bis
Rigidität) repräsentiert (Depue & Lenzenweger, 2001).
Niedrige 5-HT-Werte gehen mit einer niedrigen Schwelle der Förderung behavioraler oder
emotionaler Reaktion einher. Hohe 5-HT-Werte erfordern dagegen einen viel stärkeren
Reizeingang, um sich über den höheren Schwelleneffekt hinwegzusetzen und damit eine
Verhaltensreaktion zu fördern. Die Leichtigkeit, mit der ein Individuum emotionale
Reaktionen ausdrückt, und das Ausmaß, mit dem die emotionale Reaktion zwischen
verschiedenen Verhaltensreaktionen abwechselt (Stabilität) sind gemeinsame Funktionen der
relativen Stärke der modulierenden dopaminergen und serotonergen Einflüsse auf die
Schwelle der behavioral-emotionalen Förderung (Depue, 1996).

Theoretischer Hintergrund 38
Abbildung 8 Interaktion der agency-Komponente von Extraversion mit nicht-affektivem Constraint (nach Depue (1996) Depue & Lenzenweger (2001)).
Nachdem Depue´s neurobiologische Theorie umfassend dargestellt wurde, wird
anschließend eine Untersuchung vorgestellt, mit der Depue seine eigenen Überlegungen
bestätigt.
1.3.1.7 Befunde zur Theorie
Depue et al. (1994) prüften den Zusammenhang interindividueller Unterschiede in der DA-
D2-Reaktivität und Extraversion (gemessen als positive Emotionalität). Bei
Versuchspersonen, die auf der Dimension „positive Emotionalität“ weit verteilt waren,
wurden die Effekte von Bromocriptin auf zwei Indizes zentraler dopaminerger Aktivität
erfasst. Da die Prolactin-Ausschüttung und die spontane BR durch verschiedene
dopaminerge Projektionssysteme innerviert werden, dient ihre Erfassung als Replikation

Theoretischer Hintergrund 39
eines Zusammenhangs von DA und Extraversion (positiver Emotionalität) innerhalb der
Studie (Depue et al., 1994). Diese Befunde wurden mit der gleichen Methode an einer
größeren Stichprobe repliziert (Depue, 1995, 1996).
Es besteht ein hoher signifikanter Zusammenhang zwischen den DA-Indikatoren und dem
Maß für Extraversion, während dieselben Indikatoren mit anderen Traits, wie z.B.
Constraint, fast zu Null korrelieren. Depue und Collins (1999) sehen darin die Bestätigung
einer DA-Dimension, die sich in der Extraversions-Dimension widerspiegelt. Dass der
gefundene Zusammenhang positiv ist, spricht laut Depue et al. (1994) gegen die Auslegung,
dass geringe dopaminerge Funktion, welche erhöhte postsynaptische Rezeptorsensibilität
erzeugt, invers mit dem Extraversions-Maß zusammenhängt (siehe Cloninger, 1999; Kapitel
1.3.2.2).
Depue und Collins (1999) vermuten, dass die hohe Erblichkeit von Extraversion mit
genetischen Einflüssen auf DA-Zellgruppen zusammenhängt und schließen aus den
konsistent starken und spezifischen Zusammenhängen zwischen DA und Extraversion, dass
die emotional-motivationalen Funktionen von DA, die aus der Tierforschung abgeleitet
wurden, auch für den Menschen gelten könnten.
Im Anschluss an die Theorie von Depue soll nun eine weitere Persönlichkeitstheorie
vorgestellt werden, die Annahmen über die biologische Grundlage von Traits, macht. Geht
man davon aus, dass die Persönlichkeitsdimension novelty seeking ein Äquivalent zur
Dimension Extraversion darstellt, sollten beide Traits auf dasselbe neurobiologische bzw.
Verhaltenssystem zurückzuführen sein. Demnach wird die Dimension novelty seeking im
Weiteren genauer betrachtet.
1.3.2 Die allgemeine biosoziale Theorie der Persönlichkeit von C.
R. Cloninger
Cloninger (1986, 1987, 1988) versucht die unterschiedlichen Schwerpunkte bekannter
Persönlichkeitstheorien in einem Entwicklungsmodell zu integrieren, das neurobiologische,
psychosoziale und Lernprozesse berücksichtigt. Er definiert drei unabhängige Dimensionen
der Persönlichkeit und bringt diese mit erblicher Variation von Reaktionsmustern auf
spezifische Arten von Umweltreizen in Verbindung. Des Weiteren vermutet er, dass die

Theoretischer Hintergrund 40
Variation in jeder der Persönlichkeitsdimensionen stark mit der Aktivität in einem
spezifischen zentralen monoaminergen System korreliert ist. Cloninger betont aber auch,
dass Lern- und soziale Einflüsse für die Richtung der Persönlichkeitsentwicklung ebenso
wichtig sind, wie biologische und genetische Einflüsse (Cloninger, 1986, 1987, 1988).
Im Folgenden sollen die Annahmen der Theorie genauer vorgestellt werden, wobei zunächst
ein Überblick über alle von Cloninger postulierten Persönlichkeitsdimensionen gegeben
werden soll. Die Persönlichkeitsdimension novelty seeking (bzw. Spontaneität in der
deutschen Version des TPQ), die im weiteren Verlauf dieser Arbeit von besonderem
Interesse ist, wird dabei vordergründig behandelt.
1.3.2.1 Persönlichkeitsdimensionen
Cloninger (1986, 1987) geht von drei unabhängigen, erblichen Persönlichkeitsdimensionen
aus, deren jeweilige genetische Variation einer Normalverteilung folgt, wobei die meisten
Menschen dem Mittelwert entsprechen:
• Die Dimension novelty seeking (NS bzw. Spontaneität, SP) beruht auf einer erblichen
Tendenz zu äußerst erhebendem Gefühl oder Aufregung als Reaktion auf neue Reize
oder Hinweise auf potentielle Belohnung oder potentielles Entkommen vor einer Strafe.
Dies führt zu häufiger erkundender Aktivität auf der Suche nach potentiellen
Belohnungen oder aktiver Vermeidung von Monotonie und potentieller Bestrafung.
• Die Dimension harm avoidance (HA bzw. Risikovermeidung, RV) beruht auf einer
erblichen Tendenz intensiv auf aversive Reize zu reagieren, wobei gelernt wird
Verhalten zu unterdrücken, um Bestrafung, Neuheit und frustrierende Nicht-Belohnung
zu vermeiden.
• Die Dimension reward dependence (RD bzw. Belohnungsabhängigkeit, BA) beruht auf
einer erblichen Tendenz, intensiv auf Belohnungssignale (besonders verbale Signale für
soziale Zustimmung, Gefühl und Beistand) zu reagieren und Verhalten, das vorher mit
Belohnungen oder mit Entkommen vor Bestrafung verbunden wurde, beizubehalten
oder dessen Extinktion zu widerstehen.
Cloninger (1986, 1988) räumt ein, dass die genotypische Struktur der Persönlichkeit (Erbe),
aufgrund lebenslanger Lernprozesse und sozialer Einflüsse (Umwelt), nicht mit der
phänotypischen Struktur (beobachtbares Verhalten) übereinstimmen muss.
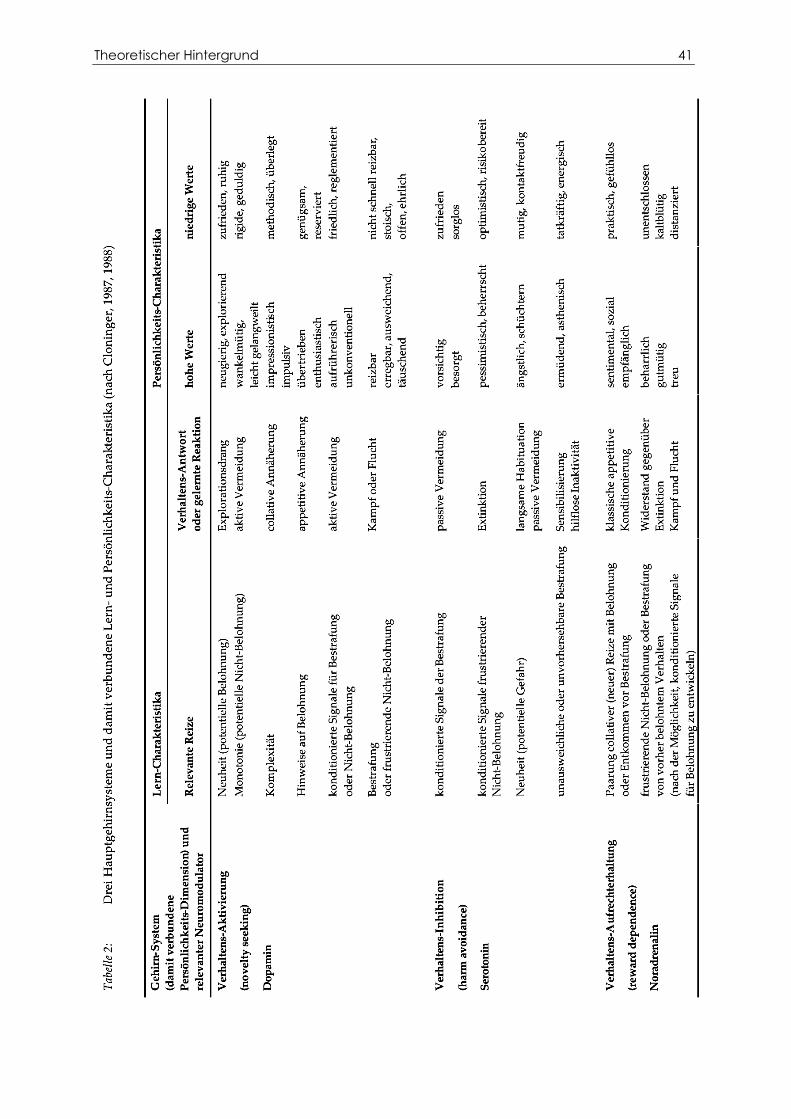
Theoretischer Hintergrund 41

Theoretischer Hintergrund 42
Zur Erfassung der definierten Dimensionen und der Überprüfung der theoretischen
Vorhersagen entwarf Cloninger (1989) u.a. den TPQ (Tridimensional Personality
Questionnaire)2. Dieser Selbstbeurteilungsfragebogen beinhaltet Ratings für die drei
Hauptdimensionen, welche sich aus jeweils vier Subskalen zusammensetzen, die rational auf
der Basis der Reiz-Reaktions-Untergruppen (siehe Tabelle 2) definiert werden.
Für die Dimension novelty seeking ergeben sich folgende Subskalen:
• „Sensationslust vs. stoische Ruhe“ (NS 1)
stellt ein Maß für das Streben nach Erkundung von Neuheit und die aktive Vermeidung
von Monotonie dar. Neuheit wird als potentielle Belohnung gedeutet und Monotonie als
potentielle Nicht-Belohnung. Personen mit hohen Werten auf dieser Skala sind
neugierig, erforschend, wankelmütig und leicht gelangweilt.
• „Impulsivität vs. Reflektion“ (NS 2)
stellt ein Maß für die collative3 Annäherung an Komplexität dar. Impulsivität äußert sich
in der Vernachlässigung von Details und Reflektion in methodischer Überlegung.
Personen mit hohen Werten auf dieser Skala sind leicht zu beeindrucken und impulsiv.
• „Extravaganz vs. Zurückhaltung“ (NS 3)
stellt ein Maß für die aktive Reaktion auf Hinweise auf Belohnung dar. Diese äußert sich
in appetitiver Annäherung an potentiell belohnende Reize. Personen mit hohen Werten
auf dieser Skala sind überschwänglich und enthusiastisch.
• „Flexibilität vs. Rigidität“ (NS 4)
stellt ein Maß für die aktive Reaktion auf Bestrafung oder Nicht-Belohnung oder ihren
Signalen dar. Konditionierte Signale für Bestrafung oder Nicht-Belohnung gehen mit
aktiver Vermeidung einher. Personen mit hohen Werten in diesem Bereich sind
aufrührerisch und unkonventionell. Bestrafung oder frustrierende Belohnung selbst
2 Da Cloninger´s Persönlichkeitsdimensionen in der vorliegenden Untersuchung mit der deutschen Fassung des TPQ erfasst wurden, werden im Folgenden in Bezug auf die vorliegende Untersuchung auch die deutschen Begriffe für die Dimensionen genutzt. Bei allgemeinen Aussagen über Cloninger´s Persönlichkeitsdimensionen, sollen die ursprünglichen Begriffe verwandt werden. 3 Nach Berlyne (1960) werden Orientierungsreaktionen durch Neuheit, Unerwartetheit und Komplexität ausgelöst. Reize, die solche Erfahrungen beinhalten, werden als collative (vergleichende) Reize bezeichnet, weil sie kritische Vergleiche erfordern, um ihre Bedeutsamkeit zu beurteilen.

Theoretischer Hintergrund 43
gehen mit Kampf oder Flucht einher. Personen mit hohen Werten in diesem Bereich sind
reizbar, erregbar, ausweichend und täuschend.
Insgesamt werden Personen, die überdurchschnittliche Werte für novelty seeking und
durchschnittliche Werte für die anderen beiden Dimensionen aufweisen, als impulsiv,
erkundend, wankelmütig, reizbar, leicht erregbar, übertrieben und aufrührerisch
charakterisiert. Sie beschäftigen sich bereitwillig mit neuen Interessen und Aktivitäten und
neigen dazu Details zu vernachlässigen und schnell abgelenkt oder gelangweilt zu sein.
Außerdem sind sie leicht zur Vorbereitung auf Kampf oder Flucht zu bewegen.
Personen mit unterdurchschnittlichen Werten für novelty seeking und durchschnittlichen
Werte für die anderen Dimensionen werden als nachdenklich, rigide, zuverlässig, stoisch,
ausgeglichen, genügsam, gesittet und beharrlich beschrieben. Sie wenden sich nur zögerlich
neuen Interessen zu, verlieren sich oft im Detail und treffen Entscheidungen erst nach langer
Überlegung.
Nachdem nun die verschiedenen Persönlichkeitsdimensionen bekannt sind, werden als
Nächstes die entsprechenden Systeme vorgestellt, die diesen zugrunde liegen sollen.
1.3.2.2 Monoaminerge Systeme und entsprechende Verhaltenssysteme
Studien des menschlichen Zentralnervensystems weisen darauf hin, dass es spezifische
Nervenbahnen oder -netzwerke gibt, die in die Reaktion auf Bestrafung, Belohnung oder
Neuheit involviert sind. Cloninger (1986, 1987, 1988) vermutet, dass die Variation in jeder
der drei Persönlichkeitsdimensionen stark mit der Variation in einer der drei spezifischen
monoaminergen Systeme im Gehirn korreliert ist und, dass jeder Dimension ein bestimmtes
zentrales Verhaltenssystem zugrunde liegt.
So nimmt er an, dass novelty seeking auf einem „Anreiz-“ oder Verhaltensaktivierungssystem
basiert und mit geringer basaler dopaminerger Aktivität zusammenhängt. Harm avoidance
soll im Gegensatz dazu ein „Bestrafungs-“ oder Verhaltensinhibitionssystem repräsentieren
und mit hoher serotonerger Aktivität verbunden sein. Reward dependence soll schließlich auf
ein „Belohnungs-“ oder Verhaltensaufrechterhaltungssystem zurückgehen und mit geringer
basaler noradrenerger Aktivität assoziiert sein.

Theoretischer Hintergrund 44
Da Verhaltensaktivierung mit Exploration neuer Umweltreize, Suche nach und Annäherung
an potentielle Belohnungen, aktiver Vermeidung von Monotonie oder potentieller
Bestrafung und Entkommen vor Bestrafung verbunden ist, geht Cloninger (1987) davon aus,
dass die Ausprägung der Dimension novelty seeking die Variation im „Anreiz“- oder
Verhaltensaktivierungssystem des Gehirns widerspiegelt und zitiert hierzu Quellen
(Routtenberg, 1978; Stellar & Stellar, 1985; Wise, 1980), die angeben, dass dopaminerge
Zellkörper im Mittelhirn an der Funktion des Verhaltensaktivierungssystems beteiligt sind.
Nach Cloninger (1999) ist die dopaminerge Aktivierung von D2-Rezeptoren stark an der
positiven Verstärkung von belohntem Verhalten sowie der subjektiven Erfahrung von
Vergnügen beteiligt (Cloninger, 1999) und niedrige Grundfeuerraten dopaminerger Neurone
werden mit postsynaptischer Hypersensibilität verbunden. Folglich wird erwartet, dass
Individuen mit niedrigen spontanen dopaminergen Entladungsraten außergewöhnlichen
Nervenkitzel brauchen und suchen, um sich zu erfreuen, aber bei entsprechender
Stimulation aufregendere und erhebendere postsynaptische Reaktionen erleben.
Bei Individuen mit hoher Ausprägung auf der Dimension novelty seeking soll der extreme
Anstieg von Vergnügen als Reaktion auf Neuheit die Geschwindigkeit erhöhen, mit der sie
operant konditionierte Verhaltensantworten auf Belohnungen zeigen. Niedrige
hedonistische Grundstimmung führt allerdings zu geringer Ausdauer bei Monotonie und
aversiver Stimulation, was sich in schnellem und häufigem aktivem Vermeidungs- oder
Fluchtverhalten äußert.
Im Gegensatz dazu sollen die dopaminergen Neurone des Mesencephalons bei Individuen
mit geringer Ausprägung auf der Dimension novelty seeking eher hohe spontane Feuerraten
aufweisen. Solche Individuen sollen unabhängig von ihren Umständen relativ glücklich und
zufrieden sein. Aufgrund verhältnismäßiger postsynaptischer Hyposensibilität werden
geringere Antworten auf dopaminerge Aktivierung über ihre hohe Grundaktivität erwartet.
Eine konditionierte Reaktion auf Belohnungen soll sich daher langsamer einstellen,
außergewöhnliche Stimulation soll eher weniger aufgesucht werden, aber die Ausdauer bei
Monotonie und aversiver Stimulation soll größer sein (Cloninger, 1988).

Theoretischer Hintergrund 45
Die eben vorgestellten Systeme, auf welche die Traits zurückgeführt werden, sind nicht ganz
unabhängig voneinander. Im Folgenden soll auf Zusammenhänge zwischen diesen
Systemen hingewiesen werden.
1.3.2.3 Funktionelle Interaktion der neuroadaptiven Systeme
Cloninger (1986) betont, dass die verschiedenen neurobiologischen Dimensionen
untereinander interagieren, um integrierte Reaktionsmuster auf Bestrafung, Belohnung und
Neuheit bereitzustellen, die wiederum durch sozial beeinflusste Muster kognitiver
Bewertung in verschiedenen Situationen modifiziert werden.
Die Systeme, die für Aufrechterhaltung und Aktivierung von Verhalten zuständig sind
arbeiten laut Cloninger (1987) zusammen, indem das dopaminerge Aktivierungssystem
Annäherung und beginnende Aneignung von belohntem Verhalten beeinflusst und das
noradrenerge System die Extinktionsrate von vorher belohntem Verhalten beeinflusst.
Im Vergleich dazu stellt das Verhaltensaufrechterhaltungssystem eher ein Gegengewicht
zum Verhaltensinhibitionssystem dar, da beide gegensätzliche Wirkungen auf die
Extinktionsrate als Reaktion auf Bestrafung oder frustrierende Nicht-Belohnung haben. So
führt z.B. erhöhte serotonerge Aktivität zu Beginn einer Bestrafung zur Hemmung
noradrenerger Neurone im Locus coeruleus (Crow & Deakin, 1985).
Ähnlich arbeitet das Verhaltensaktivierungssystem in entgegengesetzter Richtung zum
Verhaltensinhibitionssystem, da jedes dieser Systeme gegensätzliche Wirkungen auf die
Annäherung an unbekannte oder unsichere Situationen und andere neue Reize hat und
serotonerge Neuronen nigrostriatale dopaminerge Neuronen hemmen.
Die biogenetische Prädisposition zu den Reiz-Reaktions-Charakteristika jedes dieser drei
Systeme scheint in jedem Individuum unabhängig vorgegeben zu sein, aber die Systeme
stehen funktional miteinander in Beziehung. Auf diese Weise ergibt sich eine breite Variation
persönlicher Reaktionscharakteristika, die bei sozialem Verhalten in verschiedenen
Situationen beobachtet werden kann. Die gleiche Reaktionsebene auf einer Dimension kann
sich also, abhängig von der Reaktionsebene auf den anderen Dimensionen, unterschiedlich
ausdrücken.
Abbildung 9 und 10 sollen die Interaktion der Dimension novelty seeking mit den anderen
beiden Dimensionen veranschaulichen.

Theoretischer Hintergrund 46
NS hoch impulsiv erregbar erforschend reizbar
wankelmütig extravagant
gefahrsuchend aggressiv konkurrierend hypothym überaktiv neurotisch ungeduldig leicht bekümmert gesprächig widersprüchlich / unentschlossen extrovertiert
HA zufrieden, sorglos vorsichtig, besorgt HA niedrig ungehemmt, energisch ermüdend, gehemmt hoch
gelassenheitsuchend
hyperthym passiv fröhlich unbestimmt standhaft / stur inaktiv prahlerisch / übertrieben selbstsicher geduldig ruhig
introvertiert
nachdenklich stoisch rigide träge loyal rigide genügsam NS niedrig
Abbildung 9 Interaktion der Persönlichkeitsdimensionen novelty seeking (NS) und harm avoidance (HA) (nach Cloninger, 1987).
NS hoch impulsiv erregbar erforschend reizbar
wankelmütig extravagant
tolerant aufmerksamkeitssuchend opportunistisch nachgiebig gegen sich selbst auf geschickte Weise einnehmend leidenschaftlich kaltblütig bereit, etwas zu tun unsicher eingebildet/ narzisstisch unkonventionell einfallsreich / aufnehmend
RD distanziert ehrgeizig RD niedrig rational sentimental hoch
zurückgezogenheitsuchend autoritär zurückhaltend gewissenhaft unparteiisch aufrichtig / natürlich bescheiden auf herzliche Weise direkt einfallslos traditionell
nachdenklich loyal genügsam nicht leicht reizbar rigide stoisch NS niedrig
Abbildung 10 Interaktion der Persönlichkeitsdimensionen novelty sseking (NS) und reward dependence (RD) (nach Cloninger, 1987).

Theoretischer Hintergrund 47
Da durchschnittliche Werte am häufigsten vorkommen, wird die Beschreibung von
Verhalten, das mit jeder Dimension verbunden ist, durch die Charakteristika, die anfänglich
erwähnt wurden dominiert. Aber es wird erwartet, dass mindestens eine dieser
Kombinationen extremer Werte etwa so häufig auftritt, wie Abweichungen auf nur einer
Dimension.
Abbildung 11 gibt einen Überblick über den Gesamtzusammenhang der drei
Persönlichkeitsdimensionen und monoaminerger Neurotransmission.
Abbildung 11 3-fach-Interaktion von Persönlichkeit und monoaminerger Übertragung (nach Cloninger, 1986)
Die Eckpunkte in Abbildung 11 deuten an, dass zwischen einer besonders starken
Ausprägung auf der Dimension novelty seeking und Merkmalen antisozialer, histrionischer,

Theoretischer Hintergrund 48
passiv-aggressiver und explosiver Persönlichkeitsstörung Parallelen gezogen werden
können (Cloninger, 1987).
Cloninger (1988) weist darauf hin, dass es keine direkten linearen Beziehungen zwischen
einem einzelnen Neurotransmitter und einem spezifischen Verhalten gibt, und dass jedes
der Systeme und deren Interaktion zusätzlich auch durch andere Neurotransmitter, -Peptide
und Hormone beeinflusst werden kann.
Auch für Cloninger´s Theorie wird abschließend eine Untersuchung vorgestellt, welche
Cloninger´s Annahmen weitestgehend stützt.
1.3.2.4 Befunde zur Theorie
Zur Überprüfung von Cloninger´s Theorie scheint keine Studie zu existieren, welche die
Blinkrate als Indikator für die dopaminerge Aktivität heranzieht. Stattdessen werden hier
Untersuchungen vorgestellt, die andere Indikatoren, wie die Prolactin-Hemmung oder die
Wachstumshormon(GH)-Konzentration, nutzen.
In einer Studie von Gerra et al. (2000) wurde der TPQ herangezogen um zu überprüfen, ob
monoaminerge Funktionen mit Traits bei gesunden Personen verbunden sind.
Die Aktivierung des dopaminergen Systems mit dem DA-Agonisten Bromocriptin führte
wie erwartet zu einem Anstieg der GH-Konzentration und Prolactin-Hemmung. Für die
Dimension NS ergaben sich ein positiver Zusammenhang mit der GH-Konzentration sowie
ein negativer Zusammenhang mit der Prolactin-Konzentration.
Die Aktivierung des serotonergen Systems mit dem 5-HT-Agonisten Fenfluramin führte wie
erwartet zu einem Anstieg der Prolactin-Konzentration und des Plasma-Cortisol-Spiegels.
Für die Dimension HA ergab sich ein positiver Zusammenhang sowohl mit der Prolactin-
Konzentration als auch mit dem Cortisol-Spiegel.
Die Aktivierung des noradrenergen Systems mit dem Noradrenalin(NA)-Agonisten
Clonidin führte wie erwartet zu einem Anstieg der GH-Konzentration. Obwohl für die
Dimension RD eine negative Korrelation mit der GH-Konzentration und kein
Zusammenhang mit der Prolactin-Konzentration erwartet wurden, ergaben sich
(vergleichbar mit dem Ergebnis für NS) ein positiver Zusammenhang mit der GH-

Theoretischer Hintergrund 49
Konzentration sowie ein negativer Zusammenhang mit der Prolactin-Konzentration (Gerra,
Zaimovic, Timpano, Zambelli, Delsignore & Brambilla, 2000).
Bis auf die Ergebnisse für RD stimmen die Befunde mit der Theorie von Cloninger überein.
Gerra et al. (2000) hatten keine Erklärung für den entgegengesetzten Zusammenhang von
RD mit der GH-Konzentration und ziehen in Betracht, dass RD möglicherweise mit NS in
Verbindung steht und ebenfalls durch die dopaminerge Funktion beeinflusst wird.
Als Gedankenstütze zur Hypothesengenerierung für die vorliegende Untersuchung sollen
im nächsten Abschnitt die wichtigsten Punkte der zuvor beschriebenen
Persönlichkeitstheorien erinnert werden.

Theoretischer Hintergrund 50
1.3.3 Fazit zur basalen dopaminergen Aktivität
Obwohl sich die vorher beschriebenen Persönlichkeitstheorien vordergründig auf die
Reaktivität des dopaminergen Systems beziehen, soll als Überleitung zu den Hypothesen der
vorliegenden Untersuchung noch einmal kurz zusammengefasst werden, was für
Hypothesen über die basale dopaminerge Aktivität relevant ist.
Depue zieht interindividuelle Unterschiede in der Anzahl von VTA-DA-Neuronen als
genetische Grundlage für die Ausprägung von Extraversion in Betracht. Man könnte zwar
bei einer großen Anzahl von DA-Neuronen, die zu einer positiven Ausprägung von
Extraversion prädisponieren soll, auf eine mögliche hohe dopaminerge Grundaktivität
schließen, aber dazu macht Depue keine konkreten Aussagen. Stattdessen postuliert er eine
hohe Reaktivität eines Verhaltensförderungssystems, das sich im Kontakt mit
Umwelterfahrungen entwickelt und dem eine dopaminerge Moderation zugrunde liegt.
Cloninger hingegen postuliert geringe spontane Feuerraten mesencephaler dopaminerger
Neurone bei hoher Ausprägung auf der Dimension novelty seeking. Dies spricht für eine
geringe dopaminerge Grundaktivität, die aber laut Cloninger durch postsynaptische
Rezeptorhypersensibilität kompensiert wird.
Zusammenfassend kann festgehalten werden, dass sowohl Depue als auch Cloninger, trotz
unterschiedlicher theoretischer Begründung, von einer hohen Reaktivität des dopaminergen
Systems auf Signale der Belohnung ausgehen. Da Depue Cloninger´s Annahme der
postsynaptischen Hypersensibilität bezweifelt (Depue et al., 1994), wird daraus abgeleitet,
dass Depue im Gegensatz zu Cloninger einen positiven Zusammenhang zwischen der
basalen Aktivität und der Reaktivität in Betracht zieht.
Diese unterschiedlichen Annahmen zur basalen dopaminergen Aktivität können mit Hilfe
der vorliegenden Untersuchung geprüft werden. Da die basale dopaminerge Aktivität aus
der Blinkrate im Ruhezustand abgeleitet wird, soll im Rahmen dieser Arbeit mit Hilfe der
Latent-State-Trait-Theorie außerdem untersucht werden, ob die Blinkrate den Ansprüchen
an ein Traitmaß genügt.

Theoretischer Hintergrund 51
1.4 Hypothesen
Ausgehend vom theoretischen Hintergrund werden an dieser Stelle Hypothesen generiert
und die entsprechenden Operationalisierungen vorgestellt.
Hypothese A:
Da die Blinkrate hier als Indikator der zentralen dopaminergen Aktivität eingesetzt wird,
muss abgesichert werden, dass diese ein geeignetes Trait-Maß darstellt.
Es wird demnach erwartet, dass die Blinkrate zuverlässig eine zeitlich stabile und
transsituativ konsistente Verhaltensdisposition (Trait) widerspiegelt. Folglich sollten sich
einerseits ausreichende Reliabilitäten ergeben, und andererseits sollte die Erstellung eines
Strukturgleichungsmodells möglich sein, bei dem der latente Trait einen bedeutsamen Teil
der Varianz der Blinkrate aufklärt. Außerdem wird eine hohe Stabilität der Blinkrate bzw.
des zugrunde liegenden latenten Traits erwartet.
Hypothese B:
Depue und Cloninger postulieren in ihren Persönlichkeitstheorien einen Zusammenhang
zwischen zentraler dopaminerger Aktivität und Extraversion bzw. Spontaneität. In dieser
Arbeit soll untersucht werden, ob sich ein Zusammenhang zwischen diesen beiden
Persönlichkeitsdimensionen und der basalen zentralen dopaminergen Aktivität feststellen
lässt. Dazu wird die Blinkrate im Ruhezustand als Maß für die zentrale dopaminerge
Grundaktivität herangezogen und die relevanten Persönlichkeitsdimensionen werden mit
Hilfe der Fragebögen EPQ-R bzw. TPQ erfasst. Nachdem aus den beiden vorgestellten
Persönlichkeitstheorien eher gegensätzliche Erwartungen über die Richtung des zu
untersuchenden Zusammenhangs abgeleitet werden können, soll in dieser Arbeit überprüft
werden, ob sich ein bedeutsamer Zusammenhang finden lässt oder nicht. Wird ein
bedeutsamer Zusammenhang gefunden, wird dadurch entweder die eine oder die andere
Theorie gestützt.
Zusätzlich soll explorativ untersucht werden, ob ein Zusammenhang zwischen der basalen
zentralen dopaminergen Aktivität und anderen Skalen des EPQ-R (Psychotizismus,
Neurotizismus und Lügenskala) bzw. des TPQ (Risikovermeidung und

Theoretischer Hintergrund 52
Belohnungsabhängigkeit) besteht. Cloninger zufolge wird diesbezüglich kein signifikanter
Zusammenhang erwartet.

Theoretischer Hintergrund 53
2 Methoden In diesem Abschnitt sollen die Versuchspersonen, die eingesetzten Fragebögen und der
Versuchsablauf vorgestellt werden. Außerdem findet sich hier eine Einführung in die
Methode der Elektrooculographie, die hier der Erfassung der Blinks dient. Nach Angaben
zur Berechnung und Beschreibung der Fragebogendaten und Blinkraten folgt abschließend
eine Erklärung zur Latent-State-Trait-Analyse der Blinkrate und der Korrelationsanalyse der
Blinkrate mit den Fragebögen.
2.1 Versuchspersonen
Insgesamt nahmen an der Untersuchung 63 Versuchspersonen teil, die per Aushang und
elektronischer Mail an der Universität Trier akquiriert wurden. Die Teilnehmer bekamen als
Thema der Untersuchung die „Verarbeitung visueller Reize im Gehirn“ mitgeteilt, und für
eine zuverlässige Teilnahme wurde eine Belohnung von 200 DM ausgesetzt. Vor
Untersuchungsbeginn wurden die Versuchspersonen auf die Möglichkeit hingewiesen, diese
jederzeit ohne Angabe von Gründen abbrechen zu können. Zusätzlich wurde eine
schriftliche Zustimmung zur Teilnahme eingeholt. Während eine Versuchsperson aufgrund
ihres auffälligen Verhaltens von der Untersuchung ausgeschlossen wurde, wurden die Daten
zweier weiterer Teilnehmer wegen einem schwerwiegenden technischen Fehler bzw. einem
extrem unregelmäßigen EOG nicht in der Analyse berücksichtigt.
Somit bestand die Stichprobe aus 60 Versuchspersonen, wobei es sich um 30 Studentinnen
(durchschnittliches Alter: 23 Jahre, SD 3.4 Jahre, Spannweite 19-34) und 30 Studenten
(durchschnittliches Alter: 25 Jahre, SD 3.2 Jahre, Spannweite 21-36) der Universität Trier
handelte. Alle Versuchspersonen erwiesen sich nach dem Edinburgh Inventory (Oldfield,
1971) als Rechtshänder und keine der Versuchspersonen trug Kontaktlinsen. Sehschwächen
wurden gegebenenfalls durch eine Brille kompensiert.

Theoretischer Hintergrund 54
2.2 Fragebögen
Im Laufe der Datenerhebung wurden die Versuchspersonen gebeten einige Fragebögen
auszufüllen. Für die vorliegende Arbeit sind der Eysenck Personality Questionnaire -
Revised (Eysenck, Eysenck & Barrett, 1985a) und der Tridimensional Personality
Questionnaire (Cloninger, 1989) relevant.
2.2.1 Eysenck Personality Questionnaire - Revised
Die Versuchspersonen bearbeiteten die deutsche Version (Ruch, 1999) des Eysenck
Personality Questionnaire - Revised (EPQ-R; Eysenck et al., 1985), die sich aus 102 dichotom
gestuften Items zusammensetzt. Der EPQ-R (Ruch, 1999) erfasst die
Persönlichkeitsdimensionen Extraversion, Neurotizismus und Psychotizismus und umfasst
außerdem eine Lügenskala.
Die Dimension Extraversion (E; Summenscore 20 positiv und 3 negativ verschlüsselter
Items) ist für die vorliegende Arbeit von besonderem Interesse. Personen mit hohen Werten
auf dieser Skala sollen gesellig, spontan und impulsiv sein.
Die interne Konsistenz und Reliabilität nach Cronbach´s α (Ruch, 1999: E: .86-.87; P: .81-.84;
N: .85-.87; L: .81-.83; Eysenck et al., 1985: E: .85-.90; P: .76-.78; N: .85-.88; L: .79-.82) kann
insgesamt als ausreichend gut und für die Extraversions-Skala sogar als sehr gut betrachtet
werden.
Neben einer guten Validierung und Herleitung von E über objektive Tests und Ratings
(Amelang & Bartussek, 1997) kann auch die zeitliche Stabilität von E bei vier- bis
sechswöchigem Test-Retest-Intervall als gut beurteilt werden (Ruch, 1999: E: .85-.93; P: .79-
.93; N: .86-.89; L: .81-.86 vs. Intervall von einem Monat bei Eysenck & Eysenck, 1975: E: .87-
.90; P: .71-.83; N: .80-.89; L: .86).
2.2.2 Tridimensional Personality Questionnaire
Des Weiteren bearbeiteten die Versuchspersonen die deutsche Version (Defeu, Kuhn &
Schmidt, 1995) des Tridimensional Personality Questionnaire (Cloninger, 1989). Der TPQ
(Defeu et al., 1995) misst die drei Persönlichkeitsdimensionen Spontaneität (SP, novelty

Theoretischer Hintergrund 55
seeking), Belohnungsabhängigkeit (BA, reward dependence) und Risikovermeidung (RV, harm
avoidance). Diese werden über 98 Items mit Verhaltensaussagen und einem dichotomen
Antwortformat erfasst. Die Spontaneitäts-Skala, die für die vorliegende Arbeit inhaltlich
bedeutsam ist, besteht aus 34 Items (Summenscore 14 positiv und 20 negativ gepolter Items)
und steht für spontanes, freudiges Zugehen auf neue Reizkonstellationen. Die Skala
Risikovermeidung erfasst die Tendenz, neue aversiv antizipierte Reize zu vermeiden und die
Skala Belohnungsabhängigkeit beschreibt die tendenzielle Abhängigkeit von sozialen
Kontakten (Defeu et al., 1995; siehe auch Kapitel 1.3.2.1)
Die Reliabilität und interne Konsistenz nach Cronbach´s α lag für SP in der deutschen
Stichprobe bei .79 (vor und bei .81 nach Entfernung einzelner Items aufgrund mangelnder
Trennschärfe) bei .88 (.89) für RV und bei .69 (Subskala a: .71; Subskala b: .60) für BA4 (Defeu
et al., 1995; zum Vgl. Cloninger, 1987 und Cloninger, Przybeck & Svarakic, 1991: NS: .68-.77,
HA: .77-.85, BA: .55-.69).
Die Validität wurde anhand der Skaleninterkorrelationen, mit Hilfe des Krankeitsbildes
Alkoholismus und durch eine Faktorenanalyse überprüft und stellte sich für SP und RV als
gut heraus. Bei der faktorenanalytischen Überprüfung der Subskalen (mit den verbliebenen
Items) bestätigte sich die Problematik von BA, was eine Vierdimensionalität (SP, RV, BAa,
Bab) des TPQ nahe legt (Defeu et al., 1995). Die zeitliche Stabilität mittels Test-Retest-
Korrelationen lag bei einem sechsmonatigen Intervall für SP bei .88, für RV bei .73, für BAa
bei .67 und für BAb bei .76 (Defeu et al., 1995; zum Vergleich bei Cloninger, 1987; Cloninger,
Przybeck, & Svarakic, 1991: NS: .76, RV: .79, BA: .70).
2.2.3 Zusammenhang von EPQ-R und TPQ
Da sowohl Depue als auch Cloninger davon ausgehen, dass den Persönlichkeitsdimensionen
Extraversion bzw. Spontaneität dopaminerge Mechanismen zugrunde liegen, wird erwartet,
dass beide Skalen ähnliche Aspekte der Persönlichkeit erfassen, welche dieselbe biologische
4 Im Fall von BA wurden 17 der ursprünglich 30 Items aus der Skala entfernt, die danach BAa (vorwiegend Items aus den Subskalen 3 und 4) genannt wird. Von den 17 eliminierten Items bilden 9 die Skala BAb (vorwiegend Items aus den Subskalen 1 und 2). In der vorliegenden Untersuchung wurden allerdings alle 30 Items berücksichtigt, um eine Hauptskala bzw. 4 Subskalen zu berechnen.

Theoretischer Hintergrund 56
Grundlage haben. Diese Ähnlichkeit sollte sich in einer hohen Korrelation der Extraversions-
Skala des EPQ-R mit der Spontaneitäts-Skala des TPQ widerspiegeln.
Cloninger (1988) berichtet substantielle Korrelationen der Extraversions-Skala des EPQ
(Eysenck & Eysenck, 1975) mit den Skalen novelty seeking (.32) und harm avoidance (-.60) des
TPQ (englische Version). Außerdem ergaben sich substantielle Korrelationen der
Neurotizismus-Skala (EPQ) mit allen Skalen des TPQ (NS: .28; HA: .55; RD: .26), aber keine
bedeutsamen Zusammenhänge zwischen der Psychotizismus-Skala (EPQ) und einer der
Skalen des TPQ. Des Weiteren korrelierte die Skala novelty seeking substantiell mit der Skala
harm avoidance (-.20) und letztere wiederum mit der Skala reward dependence (-.19).(Cloninger,
1988).
Weyers et al. (1995) berichten substantielle Korrelationen der Skala novelty seeking (deutsche
Übersetzung des TPQ) mit den Skalen Extraversion (.60) und Psychotizismus (.52) sowie der
Lügenskala (-.36) des EPQ-R. Außerdem zeigten sich substantielle Korrelationen der Skala
Risikovermeidung mit den Skalen Extraversion (-.40) und Neurotizismus (.71) und marginal
bedeutsame Zusammenhänge zwischen der Skala Belohnungsabhängigkeit und den Skalen
Extraversion (.32) und Neurotizismus (.26) des EPQ-R. Die Skala Spontaneität korrelierte mit
der Skala Risikovermeidung zu -.24 und mit der Skala Belohnungsabhängigkeit zu .16,
während letztere zu .14 mit der Skala Risikovermeidung korrelierte (Weyers, Krebs & Janke,
1995).
Fasst man diese Befunde zusammen, wird neben einem positiven Zusammenhang zwischen
Extraversion und Spontaneität auch ein inverser Zusammenhang zwischen
Risikovermeidung und sowohl Extraversion als auch Spontaneität erwartet.
2.3 Versuchsablauf
Die Datenerhebung war auf vier Messzeitpunkte verteilt, die in etwa einmonatigem Abstand
stattfanden, und folgte immer dem gleichen Ablauf:
Zuerst wurden das Ruhe-EEG und das EOG der betreffenden Versuchsperson erfasst. Dazu
wurde die Versuchsperson in einer elektrisch abgeschirmten, schalldichten und
klimatisierten Versuchskabine für die EEG- und EOG-Messung vorbereitet. Dort erhielt die
Versuchsperson auch eine schriftliche Instruktion zum angemessenen Verhalten während

Theoretischer Hintergrund 57
der EEG- bzw. EOG-Ableitung. EEG und EOG wurden dann während 12 einminütiger
Ruheperioden mit je einer Minute Pause nach jeweils 4 Minuten aufgezeichnet. Die Hälfte
der Zeit war die Versuchsperson instruiert, die Augen zu schließen. So ergaben sich 6
Messperioden mit der Instruktion „Augen auf“ (A) und 6 Messperioden mit der Instruktion
„Augen zu“ (Z), die mit Hilfe zweier ausbalancierter Reihenfolgen (AZZAZZAZZA und
ZAAZAAZAAZ) über die vier Messzeitpunkte systematisch variiert wurden.
Dann wurden emotionale Ratings auf zuvor dargebotene Filme und Bilder erhoben, wobei
EEG und EOG während der visuellen Stimulation aufgezeichnet wurden. Dieser Teil der
Untersuchung spielt aber bei der vorliegenden Arbeit keine Rolle.
Abschließend hatten die Versuchspersonen verschiedene Fragebögen auszufüllen, die in
einer für jede Person und jeden Messzeitpunkt randomisierten Reihenfolge vorgelegt
wurden. Während alle Versuchspersonen an jedem Messzeitpunkt den EPQ-R (Eysenck et
al., 1985a) vorgelegt bekamen, wurde der TPQ (Cloninger, 1989) nur von 31
Versuchspersonen an den Messzeitpunkten 3 und 4 ausgefüllt.
Zu Beginn des ersten Messzeitpunktes bearbeiteten die Versuchspersonen zusätzlich einen
Händigkeitsfragebogen (Oldfield, 1971) und unterschrieben eine Einverständniserklärung
zur Teilnahme an der Untersuchung. Am Ende des letzten Messzeitpunktes wurden die
Versuchspersonen über Ziele und Hypothesen der Untersuchung aufgeklärt und bekamen
die Aufwandsentschädigung von 200 DM ausgezahlt.
2.4 Elektrooculogramm
Zunächst wird erläutert, unter welchen Bedingungen das Electrooculogramm abgeleitet
wurde, um dann genauer darauf einzugehen, welche der Potentialverschiebungen als
spontane Blinks gedeutet wurden.
2.4.1 Erfassung des Elektrooculogramms
Neben der Kontrolle von Augenartefakten im EEG diente das Elektrooculogramm (EOG)
hier der Erfassung von spontanen Blinks. Dazu wurde das EOG horizontal (HEOG) und
vertikal (VEOG) mit gesinterten Silber-Silberchlorid-Elektroden bipolar abgeleitet. Für das
horizontale EOG wurden die Elektroden beidseitig am Epicanthus befestigt und für das

Theoretischer Hintergrund 58
vertikale EOG infraorbital und supraorbital auf einer Linie mit der Pupille des rechten
Auges.
Genau wie das EEG wurde auch das EOG mit zwei 32-Kanal-Verstärkern bei einer
Eingangsimpedanz von 10 MΩ und mit Wechselspannungsverstärkung (alternating current,
AC; Rappelsberger, 1980) abgeleitet. Das analoge Signal wurde bei einem Hochpass-Filter
mit einer Grenzfrequenz von 0.3 Hz (-6 dB/Oktave) und einem Tiefpass-Filter mit einer
Grenzfrequenz von 40 Hz (-12 dB/Oktave) extrahiert. Einstreuungen aus dem
Wechselstromnetz (50 Hz) wurden zusätzlich mit einem 50-Hz-Kerbfilter unterdrückt.
Schließlich wurde das Signal bei einer Abtastfrequenz von 200 Hz (Nyquist-Frequenz: 100
Hz) mit einem Analog-Digital-(AD)-Wandler digitalisiert und auf Festplatte gespeichert.
Diese Datenaufzeichnung erfolgte auf einem PC (CPU: Pentium-166 MHz; RAM: 32 MB;
Festplatte: 4 GB; Betriebssystem: DOS 6.22) mit dem Programm Scan (Version 3.0; Modul:
Aquire; Neuroscan, Inc.).
2.4.2 Identifizierung der Blinks
Aufgrund eines vermuteten Zusammenhangs der Blinkrate mit Aktivierungs- und
Aufmerksamkeitsprozessen (siehe Kapitel 1.2.4), wurden nur die Blinks berücksichtigt, die in
der Ruheperiode auftraten; die „Augen zu“-Strecken wurden dabei ausgeschlossen. So
wurden letztendlich mit Hilfe des Programms Brain Vision Analyser (Version 1.03, Brain
Products GmbH) pro Versuchsperson und Messzeitpunkt sechs Minuten des EOGs
ausgewertet.
Abbildung 12 zeigt, dass die Blinks im VEOG-Kanal als hochamplitudige, kurzzeitige und
im Ruhezustand als relativ regelmäßig auftretende Ausschläge zu erkennen sind. Diese sind
meist gut von vertikalen Augenbewegungen zu unterscheiden, die ebenso erfasst werden
und im VEOG als rechteckige Potentialverschiebungen abgebildet werden.
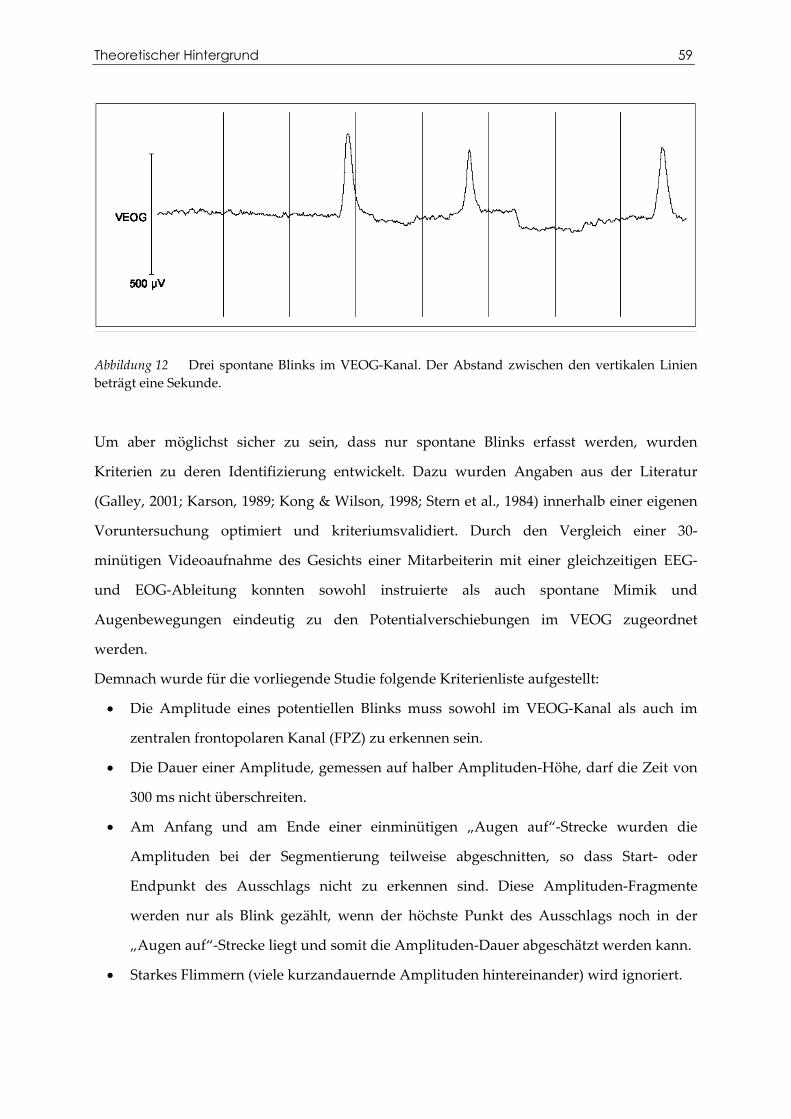
Theoretischer Hintergrund 59
Abbildung 12 Drei spontane Blinks im VEOG-Kanal. Der Abstand zwischen den vertikalen Linien beträgt eine Sekunde.
Um aber möglichst sicher zu sein, dass nur spontane Blinks erfasst werden, wurden
Kriterien zu deren Identifizierung entwickelt. Dazu wurden Angaben aus der Literatur
(Galley, 2001; Karson, 1989; Kong & Wilson, 1998; Stern et al., 1984) innerhalb einer eigenen
Voruntersuchung optimiert und kriteriumsvalidiert. Durch den Vergleich einer 30-
minütigen Videoaufnahme des Gesichts einer Mitarbeiterin mit einer gleichzeitigen EEG-
und EOG-Ableitung konnten sowohl instruierte als auch spontane Mimik und
Augenbewegungen eindeutig zu den Potentialverschiebungen im VEOG zugeordnet
werden.
Demnach wurde für die vorliegende Studie folgende Kriterienliste aufgestellt:
• Die Amplitude eines potentiellen Blinks muss sowohl im VEOG-Kanal als auch im
zentralen frontopolaren Kanal (FPZ) zu erkennen sein.
• Die Dauer einer Amplitude, gemessen auf halber Amplituden-Höhe, darf die Zeit von
300 ms nicht überschreiten.
• Am Anfang und am Ende einer einminütigen „Augen auf“-Strecke wurden die
Amplituden bei der Segmentierung teilweise abgeschnitten, so dass Start- oder
Endpunkt des Ausschlags nicht zu erkennen sind. Diese Amplituden-Fragmente
werden nur als Blink gezählt, wenn der höchste Punkt des Ausschlags noch in der
„Augen auf“-Strecke liegt und somit die Amplituden-Dauer abgeschätzt werden kann.
• Starkes Flimmern (viele kurzandauernde Amplituden hintereinander) wird ignoriert.

Theoretischer Hintergrund 60
• Da Amplitudenhöhe und -form interindividuell sehr unterschiedlich sind, müssen
zuerst alle VEOG-Strecken einer Versuchsperson gesichtet werden, um sich einen
Eindruck dieser beiden Charakteristika zu verschaffen, so dass anschließend besser
beurteilt werden kann, ob eine Amplitude als Blink registriert wird oder nicht.
Dank der Standardisierung der Blink-Kriterien, konnte das Auszählen der Blinks auf zwei
unabhängige Auswerterinnen aufgeteilt werden. Nach einer Übungsphase ergab sich eine
sehr gute Interrater-Reliabilität von .99 mit einer Mittelwertsdifferenz von nur 0,25 bpm.
2.5 Berechnung und Beschreibung der
Fragebogendaten
Um die Vergleichbarkeit mit anderen Studien zu gewährleisten, wurden als erstes für jeden
Messzeitpunkt die in den Testmanualen beschriebenen Summenscores positiv und negativ
gepolter Items berechnet. Mittelwerte und Standardabweichungen für die Skalen des EPQ-R
und des TPQ finden sich in Tabelle A1 bzw. A2 in Anhang A und zeigen keine
nennenswerten Auffälligkeiten.
Da nicht bearbeitete Items bei der Auswertung ein Problem darstellen aber im Mittel nicht
ins Gewicht fallen, wurde in den folgenden Analysen mit den Mittelungen der einzelnen
Items gerechnet. Die Deskriptivstatistik für die Mittelung über jeden einzelnen
Messzeitpunkt, sowie für einen über alle Messzeitpunkte gemittelten Gesamtwert, findet
sich in Tabelle A2 in Anhang A.
Die Reliabilitäten der Fragebögen können in Tabelle B1 und B2 in Anhang A nachgelesen
werden. Für jede Skala des EPQ-R und des TPQ wurden mittels odd-even Methode pro
Messzeitpunkt zwei Testhälften gebildet (ungerade Items: odd, gerade Items: even), anhand
derer die interne Konsistenz als split-half-Reliabilität (Korrelation der beiden Testhälften)
bestimmt werden konnte. Der Fehler, der durch die halbierte Itemanzahl auftritt, wurde
nach der Spearman-Brown-Formel korrigiert. Damit ergaben sich sehr gute korrigierte split-
half-Reliabilitäten für E (.90-.92), N (.89-.91) und RV (.88-.92) sowie eine gute Reliabilität für
SP (.80-.88) und eher geringe Reliabilitäten für P (.66-.73), L (.67-.77.) und BA (.74-.75).

Theoretischer Hintergrund 61
Weiterhin wurde zur Bestimmung der zeitlichen Stabilität als Retest-Korrelation für jeden
einzelnen Messzeitpunkt ein Gesamtwert errechnet. Die Korrelation zwischen dem Aggregat
(Mittelwert) von Messzeitpunkt 1 und 2 und dem Aggregat von Messzeitpunkt 3 und 4 soll
zusätzlich Aufschluss über die zeitliche Stabilität der Werte geben. Die Retest-Korrelationen
für ein Intervall von einem Monat bis hin zu drei Monaten ergaben eine gute zeitliche
Stabilität für E (.77-.93) und befriedigende oder nur geringe Stabilitäten für N (.73-.86), P (.64-
.80) und L (.57-.80). Die Korrelation der Aggregate führte allerdings zu einer sehr guten (E:
.93, N: .91) bis guten (P: .81, L: .84) Stabilität. Da der TPQ nur an zwei Messzeitpunkten
eingesetzt wurde, konnte nur eine einfache Retest-Korrelation mit dem Intervall von einem
Monat ermittelt werden. So ergab sich eine sehr gute zeitliche Stabilität für SP (.92) und eine
befriedigende bis gute Stabilität für BA (.80) und RV (.88).
Abschließend wurden die Interkorrelationen der Fragebögen berechnet. Diese können in
Tabelle C1 in Anhang C nachgeschlagen werden.
Es ergaben sich substantielle Korrelationen der Spontaneitäts-Skala mit den Skalen
Extraversion (.53), Psychotizismus (.38) und Risikovermeidung (-.37). Außerdem ergaben
sich substantielle Korrelationen der Skala Risikovermeidung mit den Skalen Extraversion
(-.67) und Neurotizismus (.65), sowie eine positive Korrelation der Skala
Belohnungsabhängigkeit mit der Extraversions-Skala (.42).
Die Erwartung eines positiven Zusammenhangs der Skalen Extraversion und Spontaneität
und eines inversen Zusammenhangs der Skalen Extraversion und Spontaneität mit der Skala
Risikovermeidung stimmen mit den Befunden von Cloninger (1988) und Weyers et al. (1995)
überein. Die Zusammenhänge sowohl zwischen Spontaneität und Psychotizismus als auch
zwischen Extraversion und Belohnungsabhängigkeit sind mit denen der Untersuchung von
Weyers et al. (1995) vergleichbar.
2.6 Berechnung und Beschreibung der Blinkraten
Zur Vergleichbarkeit mit anderen Studien wurden die Blinks für jede Versuchsperson über
alle Messzeitpunkte aufsummiert und in Blinks pro Minute (bpm) umgerechnet. Die
Deskriptivstatistik (Mittelwerte, Standardabweichungen und Spannweiten) wurde sowohl
für jeden Messzeitpunkt einzeln als auch über alle Messzeitpunkte hinweg bestimmt und

Theoretischer Hintergrund 62
findet sich in Tabelle A3 in Anhang A. Die durchschnittliche Blink-Rate (gemittelt über alle
Messzeitpunkte: 15 bpm) ist mit dem Befund von Bentivoglio et al. (1997; 17 bpm im Ruhe-
Zustand) zu vergleichen.
Zur Feststellung der split-half-Reliabilität der Blinkraten wurden die aufsummierten Blinks
der ungeraden Minuten (odd) mit den aufsummierten Blinks der geraden Minuten (even)
korreliert. Eine abschließende Korrektur mit der Spearman-Brown-Formel ergab sehr gute
Werte (rodd-even ≥ .92). Die zeitliche Stabilität (Retest-Korrelation über ein Intervall von ein bis
drei Monaten) lag zwischen .58 und .80. Die Korrelation des Aggregats (Mittelwert) von
Messzeitpunkt 1 und 2 mit dem Aggregat von Messzeitpunkt 3 und 4 ergibt allerdings eine
noch ausreichende Stabilität von .76. Die Ergebnisse der Reliabilitäten sind in Tabelle B3 in
Anhang A abgebildet.
Für die weitere Analyse mit linearen Strukturgleichungsmodellen wurden außerdem Schiefe
und Exzess der Blinkrate ermittelt. So sollten gegebenenfalls substantielle Abweichungen
von der Normalverteilung aufgedeckt werden. Mit der Schiefe von 0.96 und einem Exzess
von 0.61 kann nach den Kriterien von West, Finch & Curran (1995)5 von einer
Normalverteilung ausgegangen werden, so dass eine Latent-State-Trait-Analyse
durchgeführt werden kann.
2.7 Datenanalyse
Dieses Kapitel beinhaltet zum einen eine Einführung in die Grundlagen der Latent-State-
Trait-Theorie, auf deren Basis untersucht werden soll, ob die Blinkrate ein geeignetes Trait-
Maß darstellt. Zum anderen wird eine kurze Beschreibung darüber gegeben, wie festgestellt
werden soll, ob es einen Zusammenhang zwischen der zentralen dopaminergen
Grundaktivität und den ausgewählten Persönlichkeitsdimensionen gibt.
5 Die univariate Normalverteilung kann mittels Schiefe und Exzess bewertet werden. West et al. (1995) schlagen vor, dass bei einer Schiefe ≥ 2 und einem Exzess ≥ 7 eine substantielle Abweichung vorliegt. Wird eine univariate Normalverteiltheit ausgeschlossen, kann die Variable auch nicht multivariat normalverteilt sein.

Theoretischer Hintergrund 63
2.7.1 Latent-State-Trait-Analyse der Blinkrate
Da die Blinkrate als Indikator für die basale zentrale dopaminerge Aktivität herangezogen
wird, die mit bestimmten Traits zusammenhängen soll, wird in der vorliegenden Arbeit die
Latent-State-Trait(LST)-Analyse eingesetzt um zu überprüfen, ob die Blinkrate ein
ausreichend gutes Trait-Maß darstellt.
Ein Trait ist eine zeitlich stabile und über verschiedene Situationen konsistente
Verhaltensdisposition, die sich nach der Klassischen Testtheorie (KTT) auch als wahrer Wert
beschreiben lässt. Dabei postuliert die KTT, dass ein Messwert nicht genau dem wahren
Wert (true score) entspricht sondern einen Messfehler einschließt.
Als Erweiterung der KTT legt die Latent-State-Trait(LST)-Theorie (Steyer, Ferring & Schmitt,
1992) den wahren Wert als einen latenten Zustand (true state) einer Person aus und
postuliert außerdem, dass dieser sich wiederum in einen latenten Trait und ein latentes
State-Residuum aufgliedern lässt. Dabei stellt der latente Trait den transsituativ konsistenten
Effekt einer Person dar (Konsistenz) und das latente State-Residuum den
messgelegenheitsspezifischen Effekt der Situation und der Person-Situation-Interaktion
(Spezifität).
Wenn der latente Trait mit mindestens zwei Indikatoren (manifeste / beobachtete Variablen)
und in mindestens zwei Messgelegenheiten erfasst wurde, kann die LST-Theorie in ein
Strukturgleichungsmodell überführt werden.
2.7.1.1 Strukturgleichungsmodelle
Ein Strukturgleichungsmodell stellt den Zusammenhang zwischen verschiedenen latenten
Variablen dar, die direkt oder indirekt Einfluss auf die manifesten Variablen haben. Es bietet
damit die Grundlage dafür, die beobachteten Varianzen oder Kovarianzen in hypothetische
Varianzquellen zu zerlegen, die dann genauer bestimmt werden können. Jedem dieser
Einflüsse (Messfehler, latenter Zustand, State-Residuum und Trait) kann ein Pfadkoeffizient
(bei einem gerichteten Zusammenhang: Regressionsgewicht) zugeordnet werden, der die
Stärke des Einflusses angibt.
Es kann außerdem eine latente Variable spezifiziert werden, die auf der Verschiedenheit der
Indikatoren beruht. Diese latente Variable wird Methodenfaktor genannt und sollte über die
Messzeitpunkte hinweg stabil sein. Da es sich bei den hier eingesetzten Indikatoren um die

Theoretischer Hintergrund 64
Summe der Blinks in den ungeraden (odd: Indikator 1) bzw. geraden (even: Indikator 2)
Minuten der EOG-Ableitung handelt, wird nicht von einem grundlegend verschiedenen
Einfluss auf die beobachtete Variable ausgegangen, so dass der Methodenfaktor für die
vorliegende Untersuchung nicht berücksichtigt wird.
Je nach Theorie darüber, welche Einflüsse auf die manifesten Variablen einwirken, können
unterschiedliche Strukturgleichungsmodelle konstruiert werden. Dabei sind sparsamere
Modelle zu bevorzugen, d.h. es sollten so viele freie Modellparameter (Pfadkoeffizienten
und Varianzen latenter Variablen) wie nötig und so wenige wie möglich eingeführt werden.
Außerdem sollte eine Fixierung von Parametern (Normierung von Pfadkoeffizienten oder
Standardisierung von Varianzen latenter Variablen) erwogen werden.
Zunächst wurden also maximal restriktive Modelle aufgestellt, bei denen die Restriktionen
sukzessive aufgehoben wurden, wenn das Modell in dieser Form nicht angenommen wurde.
Konnte ein Modell selbst bei maximaler Liberalisierung nicht akzeptiert werden, wurde es
abgelehnt.
In den nächsten Unterpunkten werden Strukturgleichungsmodelle vorgestellt, die für die
vorliegende Arbeit von Interesse sind.
Trait-Modell
Das sparsamste Modell ist das Trait-Modell. Hier wird davon ausgegangen, dass sich die
Varianz jeder manifesten Variable zu jedem Messzeitpunkt nur durch den Einfluss des
Traits, der hier den wahren Wert darstellt, und eines Messfehlers erklären lässt (Abbildung
13). Die Pfadkoeffizienten sollen für jeden Indikator an jedem Messzeitpunkt die Stärke des
jeweiligen Einflusses (wahrer Wert und Messfehler) auf die manifeste Variable angeben.
Zu Beginn der Untersuchung gab es folgende Restriktionen:
• Gleichheit der Fehlervarianzen für alle Indikatoren und zu jeder Messgelegenheit
• Normierung aller Pfadkoeffizienten
Die Zerlegung der manifesten Varianz ermöglicht nach der LST-Theorie die Berechnung
verschiedener Parameter (Reliabilität / Konsistenz), die jeweils einen bestimmten Anteil an
der manifesten Varianz beschreiben. Der Anteil der Varianz der manifesten Variable, der auf

Theoretischer Hintergrund 65
die Varianz des wahren Wertes der Person zurückzuführen ist, kann als Reliabilität der
Indikatoren aufgefasst. Da in diesem Modell (abgesehen vom Messfehler) keine situations-
oder methodenspezifischen Effekte erwartet werden, stellt dieser Varianzanteil auch
gleichzeitig die Konsistenz dar.
Abbildung 13 Trait-Modell. Die manifesten Variablen y (erste Ziffer: Messzeitpunkt, zweite Ziffer: Indikator) setzten sich aus den Einflüssen der Messfehler e (erste Ziffer: Messzeitpunkt, zweite Ziffer: Indikator) und des Traits zusammen. Die Pfeile stehen für Regressionsgewichte, welche die Stärke der jeweiligen Einflüsse angeben.
Latent-State-Trait-Modell
Bei einem State-Trait-Modell wird angenommen, dass beide Indikatoren innerhalb einer
Messgelegenheit denselben latenten Zustand messen und beide Indikatoren in beiden
Messgelegenheiten denselben latenten Trait messen.
Die Varianz der manifesten Variablen kann dann auf einen Messfehler und einen wahren
Wert zurückgeführt werden, der sich wiederum aus einem messzeitpunktspezifischen
e11
e42
e41
e12
e21
e22
e32
e31
y11
y12
y21
y22
y31
y32
y41
y42
Trait

Theoretischer Hintergrund 66
Faktor (State-Residuum) und einem Trait-Faktor zusammensetzt (Abbildung 14). Die
Pfadkoeffizienten sollen zum Einen für jeden Indikator an jedem Messzeitpunkt die Stärke
des jeweiligen Einflusses (wahrer Wert und Messfehler) auf die manifeste Variable angeben
und zum Anderen die Stärke des jeweiligen Einflusses (State-Residuen und Trait) auf den
wahren Wert.
Abbildung 14 State-Trait-Modell. Die manifesten Variablen y (erste Ziffer: Messzeitpunkt, zweite Ziffer: Indikator) setzten sich aus den Einflüssen der Messfehler e (erste Ziffer: Messzeitpunkt, zweite Ziffer: Indikator) und des wahren Zustands (State 1 bis 4) einer Person zusammen. Der wahre Zustand einer Person gliedert sich wiederum in den Einfluss der State-Residuen (messzeitpunktspezifischer Einfluss; Res 1 bis 4) und des Traits auf. Die Pfeile stehen für Regressionsgewichte, welche die Stärke der jeweiligen Einflüsse angeben.
Zu Beginn der Untersuchung gab es folgende Restriktionen:
• Gleichheit der Fehlervarianzen für alle Indikatoren und zu jeder Messgelegenheit
• Gleichheit der Varianzen der State-Residuen
• Normierung aller Pfadkoeffizienten
Trait
e11
e42
e41
e12
e21
e22
e32
e31
y11
y12
y21
y22
y31
y32
y41
y42
State 1
State 2
State 3
State 4
Res 1
Res 2
Res 3
Res 4

Theoretischer Hintergrund 67
Die Zerlegung der manifesten Varianz ermöglicht nach der LST-Theorie die Berechnung
verschiedener Parameter (Reliabilität, Konsistenz und Spezifität), die jeweils einen
bestimmten Anteil an der manifesten Varianz beschreiben. Der Anteil der Varianz der
manifesten Variable, der auf die Varianz des wahren Wertes der Person zurückzuführen ist,
kann als Reliabilität der Indikatoren aufgefasst werden, die sich auch additiv aus dem Anteil
des Traits an der manifesten Varianz (Konsistenz) und dem Anteil des State-Residuums an
der manifesten Varianz (Spezifität) bestimmen lässt.
Stabilitätsmodell
Neben der herkömmlichen Bestimmung der zeitlichen differentiellen Stabilität mittels
Retestkorrelationen (Kapitel 2.6) einzelner Messzeitpunkte wird die Stabilität innerhalb
dieser Arbeit zusätzlich mit Hilfe eines State-Trait-Stabilitätsmodells (Hagemann, 1999;
Hagemann, Naumann, Thayer & Bartussek, 2002; Abbildung 15) untersucht. Dies hat den
Vorteil, dass Messfehler und Situationseinflüsse explizit berücksichtigt werden.
Zur Stabilitätsbestimmung werden innerhalb eines State-Trait-Modells zwei latente Traits
anhand derselben Indikatoren bestimmt. Einer der Traits wird über die ersten beiden
Messzeitpunkte bestimmt, der andere über die letzten beiden Messzeitpunkte. Die Retest-
Korrelation zwischen diesen beiden Traits stellt einen zuverlässigen Schätzer der
differentiellen zeitlichen Stabilität des latenten Traits dar und taucht auch im Modell als
zusätzlicher Pfadkoeffizient auf. Die Korrelation zwischen den Aggregaten der ersten und
letzten beiden Messzeitpunkte (siehe Kapitel 2.6) kann als Pendant zu diesem Verfahren
verstanden werden.

Theoretischer Hintergrund 68
Abbildung 15 Stabilitätsmodell. Die manifesten Variablen y (erste Ziffer: Messzeitpunkt, zweite Ziffer: Indikator) setzen sich aus den Einflüssen der Messfehler e (erste Ziffer: Messzeitpunkt, zweite Ziffer: Indikator) und des wahren Zustands (State 1 bis 4) einer Person zusammen. Der wahre Zustand einer Person gliedert sich wiederum in den Einfluss der State-Residuen (messzeitpunktspezifischer Einfluss; Res 1 bis 4) und der latenten Traits (Trait 1: Trait aus den Messzeitpunkten 1 und 2; Trait 2: Trait aus den Messzeitpunkten 3 und 4) auf. Die Pfeile stehen für Regressionsgewichte, welche die Stärke der jeweiligen Einflüsse angeben. Der Doppelpfeil stellt die Korrelation zwischen Trait 1 und Trait 2 dar, die als Schätzer für die Stabilität des zugrunde liegenden Traits herangezogen wird.
Modellevaluation
Die konstruierten Strukturgleichungsmodelle können mit verschiedenen Methoden bewertet
und gegeneinander getestet werden (Hu & Bentler, 1995; Hagemann, 2002). An dieser Stelle
werden bestimmte Verfahren herausgegriffen und genauer dargestellt.
Zur Schätzung der Modellparameter können verschiedene Diskrepanzfunktionen verwendet
werden, die ein inverses Maß für die Ähnlichkeit zwischen der empirisch beobachteten
Kovarianzmatrix (der manifesten Variablen) und einer implizierten Kovarianzmatrix (der
Trait 1
e11
e42
e41
e12
e21
e22
e32
e31
y11
y12
y21
y22
y31
y32
y41
y42
State 1
State 2
State 3
State 4
Res 1
Res 2
Res 3
Res 4
Trait 2

Theoretischer Hintergrund 69
geschätzten Modellparameter) darstellen. Über einen iterativen Algorithmus sollen
Modellparameter gefunden werden, die am wenigsten von der empirischen Vorgabe
abweichen.
Aufgrund der relativ kleinen Stichprobe von 60 Versuchspersonen wurde in der
vorliegenden Untersuchung die Funktion der generalized least squares (GLS) eingesetzt, da sie
von der Skalierung der manifesten Variable unabhängig ist und auch für Stichproben mit
weniger als 2500 Versuchspersonen geeignet ist. Die Voraussetzungen zur Anwendung
dieses Verfahrens (Intervallskala und multivariate Normalverteilung der manifesten
Variablen in der Population) sind, soweit überprüfbar, erfüllt (siehe Kapitel 2.6).
Die Modellparameter, die durch die Minimierung der Diskrepanzfunktion ermittelt wurden,
sind best guesses unter der Annahme, dass das Modell in der Population gültig ist. Da das
Minimum der Diskrepanzfunktion praktisch nie den Wert 0 annimmt, muss sichergestellt
werden, ob diese Abweichung auf Stichproben- bzw. Schätzfehlern beruht oder ob das
Modell falsch ist. Das Minimum der Diskrepanzfunktion dient hierbei als statistisches Maß
für die Modellgültigkeit. Je größer der Wert, desto unwahrscheinlicher ist es, dass die
Diskrepanz zwischen den Matrizen nur zufällig zustande gekommen ist. Mit Hilfe einer
Prüfgröße, die vom Minimum der Diskrepanzfunktion und der Stichprobengröße abhängt,
und die sich bei Verwendung von GLS mit steigender Stichprobengröße asymptotisch einer
χ²-Verteilung annähert, kann die Vereinbarkeit des Modells mit den Daten (Nullhypothese,
Minimum der Diskrepanzfunktion = 0) getestet werden. Wird der Test nicht signifikant kann
das Modell beibehalten werden, da angenommen wird, dass die Diskrepanz zwischen
Modell und Daten nur durch zufällige Stichprobenfehler erklärt werden kann. Bei
Signifikanz wird das Modell (mit einer Irrtumswahrscheinlichkeit von α = .05) verworfen, da
es unwahrscheinlich ist, dass die Diskrepanz zufällig ist. Da die Annahme der
Nullhypothese mit einer unbekannten Irrtumswahrscheinlichkeit β verbunden ist und die
Annahme der Alternativhypothese stark von der Stichprobengröße abhängt, wurden im
Falle einer Modellannahme zur Bewertung zusätzliche Indikatoren der Passungsgüte
(goodness of fit indices) angegeben, durch welche die Modellpassung quantifiziert wird.

Theoretischer Hintergrund 70
Der comparative fit index (CFI) basiert auf dem standardisierten Nonzentralitätsparameter (λ)
der χ²-Verteilung und kann Werte zwischen 0 und 1 (perfekte Passung) annehmen. Werte
über .90 werden als sehr gute Passung und Werte über .80 als akzeptable Passung
interpretiert. Der CFI ist unabhängig von der Stichprobe, hat einen geringen Standardfehler,
bildet Fehlspezifikationen des Modells präzise ab und ist gegenüber Verletzungen der
Normalverteilungsannahme robust. Da der CFI den Nachteil hat sparsame Modelle zu
benachteiligen, wird außerdem ein zweiter Indikator herangezogen.
Der root mean square error of approximation (RMSEA) relativiert λ an der Anzahl der
Feiheitsgrade, wodurch komplexere Modelle „bestraft“ werden (parsimonious fit index). Der
Wertebereich des RMSEA liegt ebenfalls zwischen 0 (perfekte Passung) und 1, wobei Werte
von 0.05 und kleiner eine gute und Werte von 0.08 und kleiner eine akzeptable Passung
anzeigen. Werte, die größer als 0.10 ausfallen, werden als inakzeptable Passung interpretiert.
Für den RMSEA lässt sich eine Stichprobenkennwerteverteilung erstellen, welche die
Bestimmung eines Konfidenzintervall zulässt. Dies ermöglicht außerdem einen
Signifikanztest. Die Nullhypothese stellt dabei die Annahme dar, dass der empirisch
gefundene RMSEA einen Wert von 0.05 annimmt oder kleiner ist (closeness of fit).
Wurde ein Modell anhand der beschriebenen Kriterien akzeptiert, wurde abschließend jeder
geschätzte Modellparameter auf Signifikanz getestet. Mit Hilfe des critical ratio (CR) wurde
überprüft, ob ein Modellparameter in der Population einem Wert von 0 entspricht
(Nullhypothese) oder nicht. Der CR gibt das Verhältnis des geschätzten Parameters zum
entsprechenden geschätzten Standardfehler an und lässt sich im Falle der Nullhypothese
und einer multivariat normalverteilten manifesten Variable als Standardnormalverteilung
darstellen. Somit kann die Nullhypothese verworfen werden, wenn der CR nicht in ein
Intervall von ± 1.96 fällt. In diesem Fall weicht der Modellparameter bei einem
Signifikanzniveau von α = 0.05 signifikant von Null ab.
An dieser Stelle kann zusammengefasst werden, dass die Modelle durch eine Kombination
von Modelltest (χ²-Statistik) und Passungsindikatoren (CFI und RMSEA inklusive
Signifikanztest) anhand empirischer Daten bewertet werden. Die Signifikanz einzelner
Parameter wird mit Hilfe des CR beurteilt.

Theoretischer Hintergrund 71
Die Analyse der Strukturgleichungsmodelle wurde mit dem Programm AMOS (Version 3.6,
SmallWaters Corporation; Arbuckle, 1996) unter SPSS für Windows (Version 7.5.2G, SPSS
Inc.) durchgeführt.
2.7.2 Korrelationsanalyse
Um festzustellen, ob es einen bedeutsamen Zusammenhang zwischen der basalen zentralen
dopaminergen Aktivität und bestimmten Traits gibt, wurden für alle Skalen des EPQ-R und
des TPQ bivariate Korrelationen (nach Pearson) mit der Blinkrate berechnet. Die
Korrelationsanalyse wurde mit SPSS für Windows (Version 11.5, SPSS Inc.) durchgeführt,
und da die Richtung des erwarteten Zusammenhangs nach Depue und Cloninger
unterschiedlich ist, wurde eine zweiseitige Testung vorgenommen.
Mit dem Programm GPOWER (Faul & Erdfelder, 1992) konnte festgestellt werden, dass mit
den Daten von 60 Versuchspersonen, die den EPQ-R bearbeitet hatten, einem
Signifikanzniveau von α = .05 und einer Teststärke von 1-β = .90 ein Populationseffekt von r²
= .15 (r = .39) gefunden werden kann, sofern es ihn gibt. Dieser Effekt ist als mittel bis hoch
einzuschätzen (Cohen, 1992). Wird eine geringere Teststärke von 1-β =.80 akzeptiert, kann
unter denselben Bedingungen auch ein eher mittlerer Populationseffekt von r² = .12 (r = .35)
gefunden werden.
Da keine Erfahrungswerte für die empirische Effektgröße vorliegen, soll demnach eine
Korrelation der Blinkrate mit dem EPQ-R von r ≥ .35 als bedeutsam erachtet werden. Das
bedeutet, dass bei einer Korrelation, die nicht signifikant wird, die Nullhypothese (kein
Zusammenhang) interpretiert werden darf.
Mit den Daten von nur 31 Versuchspersonen, die den TPQ bearbeitet hatten, einem
Signifikanzniveau von α = .05 und einer Teststärke von 1-β = .90 kann ein großer
Populationseffekt von r² = .27 (r = .52) gefunden werden, sofern es ihn gibt. Wird eine
geringere Teststärke von 1-β =.80 akzeptiert, kann unter denselben Bedingungen ein
ebenfalls großer Populationseffekt von r² = .21 (r = .46) gefunden werden.
Da auch in diesem Fall keine Erfahrungswerte für einen großen empirischen Effekt sprechen,
und die Teststärke nicht ausreicht einen mittleren Effekt aufzudecken, so dass nicht
signifikante Ergebnisse nicht als Bestätigung der Nullhypothese interpretiert werden
können, wird zur genaueren Beschreibung jedes Korrelationskoeffizienten ein

Theoretischer Hintergrund 72
Konfidenzintervall angegeben. Dieses grenzt den Bereich ein, in dem der wahre Wert liegt,
der den jeweiligen Koeffizienten erzeugt hat. Je größer dieser Bereich ausfällt, desto
ungenauer ist die Schätzung. Schließt dieser Bereich den Wert 0 ein, ist dies ein Hinweis
darauf, dass es keinen Zusammenhang gibt (Bortz, 1999).
Um einer Kumulierung des α-Fehlers zu begegnen, wurden die Signifikanzniveaus für die
jeweiligen Korrelationen durch eine sequenzielle Bonferoni-Korrektur (Holm, 1979)
adjustiert.
Statistische Hypothese:
Es wird eine bedeutsame bivariate Korrelation zwischen der Blinkrate im Ruhezustand und
den Fragebogenskalen Extraversion des EPQ-R bzw. Spontaneität des TPQ erwartet.

Ergebnisse 73
3 Ergebnisse In diesem Kapitel werden zuerst die Ergebnisse der LST-Analyse berichtet (Kapitel 3.1).
Diese umfassen sowohl die Bestimmung des passendsten LST-Modells einschließlich der
Berechnung der entsprechenden LST-Parameter als auch die Bestimmung der zeitlichen
Stabilität mit Hilfe eines Stabilitätsmodells.
Dann folgen die Ergebnisse der Korrelationsanalyse (Kapitel 3.2), die zum Einen den
Zusammenhang zwischen der Blinkrate und den Skalen Extraversion und Spontaneität
betreffen und zum Anderen den Zusammenhang der Blinkrate mit den übrigen
Fragebogenskalen.
3.1 LST-Analyse
Die LST-Analyse wurde in der vorliegenden Arbeit eingesetzt, um festzustellen, ob sich die
Blinkrate als Trait-Maß eignet. Dazu wurden Strukturgleichungsmodelle aufgestellt, welche
die Einflüsse verschiedener latenter Variablen auf die manifesten Variablen darstellen. In die
Analyse gingen die Werte von je zwei Indikatoren an insgesamt vier Messzeitpunkten ein.
Die Indikatoren wurden durch Aufteilung der Blinks mit Hilfe der odd-even-Methode
gebildet. Nach der Testung mehrerer Modelle wurden für das passendste Modell die LST-
Parameter (Reliabilität, Spezififtät und Konsistenz) berechnet. Diese geben an, wie viel
Varianz durch die jeweiligen Einflussgrößen erklärt werden kann.
Des Weiteren diente die LST-Analyse der Einschätzung der zeitlichen Stabilität des
zugrunde liegenden Traits. Das Ergebnis des Stabilitätsmodells kann dem Stabilitätsmaß der
Korrelation der aggregierten Messzeitpunkte 1 und 2 mit dem Aggregat der Messzeitpunkte
3 und 4 vergleichend gegenübergestellt werden.
3.1.1 Trait-Modell
Das Trait-Modell konnte selbst nach maximaler Liberalisierung (Freigabe aller
Fehlervarianzen und Pfadkoeffizienten) nicht akzeptiert werden (χ²(df = 32) = 71,91; p = .00)
und wurde somit von weiteren Analysen ausgeschlossen.

Ergebnisse 74
3.1.2 Latent-State-Trait-Modell
Nachdem ein restriktives Modell kein akzeptables Resultat erbrachte, konnte das State-Trait-
Modell nach Freigabe einer Fehlervarianz akzeptiert werden (χ²(df = 32) = 34,44; p = .352)
und zeigte eine sehr gute Passung (CFI = .96; RMSEA = .04; pRMSEA = .57; Tabelle 3).
Tabelle 3 Liberalisiertes State-Trait-Modell
Liberalisierte Parameter χ2-Statistik Passungsgüte
χ2 df p CFI RMSEA pRMSEA
Var (e11) 34,440 32 0,352 0,964 0,036 0,569
Anmerkung: Die Werte beziehen sich auf ein State-Trait-Modell, bei dem die Fehlervarianz des ersten Indikators zur ersten Messgelegenheit (Var (e11)) freigegeben wurde (liberalisierter Parameter). Alle Pfadkoeffizienten waren auf 1 normiert. Dargestellt sind die χ2-Statistik (χ2-Wert; Freiheitsgrade: df; p-Level) und die Indikatoren der Passungsgüte (CFI; RMSEA incl. p-Level: pRMSEA).
Außerdem konnte mit Hilfe des CR gezeigt werden, dass sich alle geschätzten
Modellparameter bei einem Signifikanzniveau von α = .05 bedeutsam von Null
unterscheiden (Tabelle 4).
Tabelle 4 Parameterschätzung des liberalisierten State-Trait-Modells
manifeste Varianz Var (y11) Var (y12), Var (y21), Var (y22), Var (y31), Var (y32), Var (y41), Var (y42)
68,930 72,211
Fehlervarianz Var (e11) Var (e12), Var (e21), Var (e22), Var (e31), Var (e32), Var (e41), Var (e42)
3,855 7,135
CR 1,981 8,454
State-Residuums-Varianz Var (sr) 14,938 CR = 6,263
Trait-Varianz Var (t) 50,137 CR = 4,594
Anmerkung: Die Werte beziehen sich auf ein State-Trait-Modell, bei dem die Fehlervarianz des ersten Indikators zur ersten Messgelegenheit (Var (e11)) freigegeben wurde. Dargestellt sind die geschätzten Modellparameter des i-ten Messzeitpunktes und k-ten Indikators (Varianz der manifesten Variablen: Var (yik)), die Varianz der Messfehler des i-ten Messzeitpunktes und k-ten Indikators (Varianz der Messfehler: Var (eik)), die Varianz der State-Residuen: Var (s1, s2, s3, s4), die Varianz des Traits: Var (t) und deren critical ratio (CR).

Ergebnisse 75
3.1.3 LST-Parameter
Nach der Entscheidung, welches Modell die empirischen Daten am besten widerspiegelt,
wurden die LST-Parameter für das entsprechende Modell berechnet. Da das Trait-Modell
nicht akzeptiert wurde und auch ein restriktives State-Trait-Modell keine akzeptable Lösung
darstellte, wurden Reliabilität, Spezifität und Konsistenz für das liberalisierte State-Trait-
Modell berechnet (Tabelle 5).
Tabelle 5 LST-Parameter des liberalisierten State-Trait-Modells
REL SISPE KONS
Var (y12)- Var (y42) .90 .21 .69
Var (y11) .94 .22 .73
Anmerkung: Die Werte beziehen sich auf ein State-Trait-Modell, bei dem die Fehlervarianz des ersten Indikators zur ersten Messgelegenheit (Var (e11)) freigegeben wurde. Dargestellt sind die LST-Parameter der Reliabilität (REL), Situationsspezifität (SISPE) und Konsistenz (KONS) für beide Indikatoren zu allen vier Messzeitpunkten.
Da die Fehlervarianz für Indikator 1 am ersten Messzeitpunkt freigegeben wurde, ergeben
sich dementsprechend andere Werte. Insgesamt ließ sich eine sehr gute Reliabilität (.90-.94),
und eine ausreichende Konsistenz (.69-.73) feststellen. Der Einfluss der Situation (Spezifität)
ist dagegen zwar vergleichsweise gering (.21-.22), unterscheidet sich aber dem CR zufolge
bedeutsam von Null.
3.1.4 Stabilitätsmodell
Da sich kein akzeptables Trait-Modell konstruieren ließ, wurde die differentielle zeitliche
Stabilität anhand eines State-Trait-Stabilitätsmodells ermittelt. Nach Freigabe der
Fehlervarianzen des ersten Messzeitpunktes konnte das Modell akzeptiert werden (χ²(df =
29) = 28,36; p = .50) und zeigte eine perfekte Passung (CFI = 1.00; RMSEA = .00; pRMSEA = .70;
Tabelle 6).
Außerdem konnte mit Hilfe des CR gezeigt werden, dass sich die geschätzten
Modellparameter der State-Residuums-Varianz, der Trait-Varianz und der Trait-Kovarianz
bei einem Signifikanzniveau von α = .05 bedeutsam von Null unterscheiden. Nur der CR

Ergebnisse 76
einer Fehlervarianz fiel in den Bereich von ± 1,96 und wurde somit als nicht signifikant
erachtet (Tabelle 7).
Tabelle 6 Liberalisiertes State-Trait-Stabilitätsmodell
Liberalisierte Parameter χ2-Statistik Passungsgüte
χ2 df p CFI RMSEA pRMSEA
Var (e11), Var (e12) 28,355 29 0,499 1,000 0,000 0,695
Anmerkung: Die Werte beziehen sich auf ein State-Trait-Stabilitätsmodell, bei dem die Fehlervarianzen beider Indikatoren zur ersten Messgelegenheit (Var (e11) undnVar (e12)) freigegeben wurden (liberalisierte Parameter). Alle Pfadkoeffizienten waren auf 1 normiert. Dargestellt sind die χ2-Statistik (χ2-Wert; Freiheitsgrade: df; p-Level) und die Indikatoren der Passungsgüte (CFI; RMSEA incl. p-Level: pRMSEA).
Tabelle 7 Parameterschätzung des liberalisierten State-Trait-Stabilitätsmodells
Manifeste Varianz Var (y11) Var (y12) Var (y21), Var (y22) Var (y31), Var (y32), Var (y41), Var (y42)
66,540 69,672 69,736 88,574
Fehlervarianz Var (e11) Var (e12) Var (e21), Var (e22), Var (e31), Var (e32), Var (e41), Var (e42)
3,940 7,072 7,136
CR 1,323 2,244 8,187
State-Residuums-Varianz Var (sr) 12,953 CR = 4,956
Trait-Varianz Var (t1) 49,647 CR = 4,420
Var (t2) 68,485 CR = 4,598
Trait-Kovarianz Kov (t1,t2) 51,395 CR = 4,487
Trait-Korrelation r (t1,t2) .88
Anmerkung: Die Werte beziehen sich auf ein State-Trait-Stabilitätsmodell, bei dem die Fehlervarianzen beider Indikatoren zur ersten Messgelegenheit (Var (e11) und Var (e12)) freigegeben wurden. Dargestellt sind die geschätzten Modellparameter des i-ten Messzeitpunktes und k-ten Indikators (Varianz der manifesten Variablen: Var (yik)), die Varianz der Messfehler des i-ten Messzeitpunktes und k-ten Indikators (Varianz der Messfehler: Var (eik)), die Varianz der State-Residuen: Var (s1, s2, s3, s4), die Varianz der Traits: Var (t1,t2), die Kovarianz der Traits: Kov (t1,t2) und deren Critical Ratio (CR). Zusätzlich wird hier auch die Korrelation zwischen den beiden Traits (r (t1,t2)) angegeben.
Aus den Schätzern der Varianzen der beiden Traits und deren Kovarianz kann die
Korrelation zwischen den beiden Traits berechnet werden, die wiederum einen Schätzer für

Ergebnisse 77
die zeitliche Stabilität darstellt. Es ergab sich eine zeitliche Stabilität von r = .88, die im
Rahmen der vorliegenden Arbeit als gut betrachtet wird und welche die Retest-Korrelation
der Blinkrate der aggregierten Messzeitpunkte (.76; siehe Kapitel 2.6 und Tabelle A6 in
Anhang A) übertrifft.
3.1.5 Fazit zur LST-Analyse
Zusammenfassend kann festgehalten werden, dass die Blinkrate ein gutes Trait-Maß
darstellt. Es konnte ein liberalisiertes State-Trait-Modell konstruiert werden, für das ein
bedeutsamer Anteil an Trait-Varianz ermittelt wurde. Außerdem konnte eine sehr gute
Reliabilität und eine ausreichende Konsistenz feststellen werden. Abschließend konnte mit
Hilfe eines liberalisierten State-Trait-Modells auch eine gute zeitliche Stabilität ermittelt
werden.
3.2 Korrelationsanalyse
Mit Hilfe von bivariaten Korrelationen sollte überprüft werden, ob ein Zusammenhang
zwischen der Blinkrate im Ruhezustand und den Skalen des EPQ-R (Tabelle 7) oder des TPQ
(Tabelle 8) besteht. Dabei waren vor allem die Extraversions-Skala des EPQ-R und die
Spontaneitäts-Skala des TPQ von Interesse.
Keine der Korrelationen der Blinkrate mit den Persönlichkeitsdimensionen Extraversion und
Spontaneität wich signifikant vom Wert 0 ab und die großen Konfidenzintervalle weisen auf
eine ungenaue Schätzung hin.
Tabelle 8 Bivariate Korrelationen zwischen der Blinkrate und den Skalen des EPQ-R.
Fragebogenskalen Blinkrate Konfidenzintervall 95%
Extraversion (E) .01 -.25 - .27
Psychotizismus (P) -.13 -.37 - .13
Neurotizismus (N) .01 -.25 - .27
Lügenskala (L) .26 * .01 - .48
Anmerkung: In die Korrelationen gehen die Daten von 60 Versuchspersonen ein. * Die Korrelation ist auf dem Niveau von .05 (2-seitig) signifikant. Das Konfidenzintervall gilt für α = .05.
Ergebnisse 78
Für den Zusammenhang zwischen der Blinkrate und Extraversion kann, aufgrund einer
ausreichenden Teststärke, mit 95%iger Sicherheit ausgeschlossen werden, dass in der
Population ein Effekt von r² ≥ .12 (r ≥ .35) existiert.
Tabelle 9 Bivariate Korrelationen zwischen der Blinkrate und den Skalen des TPQ.
Anmerkung: SP1 - SP4 kennzeichnen die Subskalen der Persönlichkeitsdimension Spontaneität. RV1 - RV4 kennzeichnen die Subskalen der Persönlichkeitsdimension Risikovermeidung. BA1 - BA4 kennzeichnen die Subskalen der Persönlichkeitsdimension Belohnungsabghängigkeit. In die Korrelationen gehen die Daten von 31 Versuchspersonen ein. ** Die Korrelation ist auf dem Niveau von .01 (2-seitig) signifikant. * Die Korrelation ist auf dem Niveau von .05 (2-seitig) signifikant. Das Konfidenzintervall gilt für α = .05.
Für den Zusammenhang zwischen der Blinkrate und Spontaneität reicht die Teststärke nur
aus um auszuschließen, dass in der Population ein Effekt von r² ≥ .21 (r ≥ .46) existiert.
Da die Teststärke zu gering ist um einen mittleren oder kleinen Effekt zuverlässig zu finden,
kann aber auch die Nullhypothese nicht interpretiert werden.
Fragebogenskalen Blinkrate Konfidenzintervall 95%
Spontaneität (SP) -.09 -.43 - .27
SP 1 -.19 -.51 - .18
SP 2 -.24 -.55 - .13
SP 3 .19 -.18 - .51
SP 4 -.04 -.39 - .32
Risikovermeidung (RV) .13 -.24 - .47
RV 1 .07 -.29 - .42
RV 2 -.01 -.36 - .36
RV 3 .03 -.33 - .38
RV 4 .28 -.08 - .58
Belohnungsabhängigkeit (BA) -.55 ** -.76 - -.24
BA 1 -.51 ** -.73 - -.19
BA 2 -.33 -.61 - .03
BA 3 -.42 * -.68 - -.08
BA 4 -.14 -.47 - .23

Ergebnisse 79
Um bei den einzelnen Korrelationen der Blinkrate mit den Fragebogenskalen eine
Kumulierung des α-Fehlers zu berücksichtigen, wurde eine sequenzielle Bonferoni-
Korrektur (Holm, 1979) durchgeführt, die den α-Fehler für jede einzelne Korrelation
adjustiert (Tabelle 9).
Tabelle 10 Adjustierung des Signifikanzniveaus für die Korrelationen der Blinkrate mit den Haupt-Skalen des EPQ-R und des TPQ durch eine sequenzielle Bonferoni-Korrektur (Holm, 1979).
Fragebogenskalen n pn m-(n-1) α´n = α / (m-(n-1)) pn < α´n
Belohnungsabhängigkeit (BA)
Lügenskala (L)
Psychotizismus (P)
Risikovermeidung (RV)
Spontaneität (SP)
Neurotizismus (N)
Extraversion (E)
1
2
3
4
5
6
7
.001
.044
.314
.484
.619
.947
.959
7
6
5
4
3
2
1
.0071
.0083
*
n. s.
Anmerkung: n: Rangplatz der Fragebogenskalen in aufsteigender Reihenfolge. pn: Wahrscheinlichkeitslevel in aufsteigender Reihenfolge. m: Anzahl der p-Werte. α: Signifikanzniveau von 5%. α´n: adjustierter α-Wert der jeweiligen Fragebogenskala. pn < α´n: Beurteilung der Signifikanz.
Die explorativen Korrelationen der Blinkrate mit den übrigen Skalen der beiden Fragebögen
ergaben signifikante Korrelationen für die Skala Belohnungsabhängigkeit des TPQ (r = .55, α
= .01) und für die Lügenskala des EPQ-R (r = .26, α = .05). Bei einem konservativeren
adjustierten Signifikanzniveau konnte nur die Korrelation der Blinkrate mit der Skala
Belohnungsabhängigkeit als signifikant erachtet werden. Es kann festgehalten werden, dass
in der Population mit einer 94%igen Sicherheit ein großer Effekt von r² ≥ .30 (r ≥ .55) existiert.

Diskussion 80
4 Diskussion Einige neuere Persönlichkeitstheorien befassen sich mit der biologischen Grundlage der
verschiedenen Traits. Depue und Cloninger ziehen in diesem Zusammenhang in Betracht,
dass der Neurotransmitter Dopamin für die Ausprägung der Persönlichkeitsdimensionen
Extraversion bzw. novelty seeking eine Rolle spielt.
Aussagen über den Zusammenhang dieser Traits mit Indikatoren der dopaminergen
Aktivität werden bevorzugt aus der Reaktivität des dopaminergen Systems auf
experimentell induzierte Aktivierung abgeleitet.
In der vorliegenden Arbeit wurde die basale zentrale dopaminerge Aktivität mit Hilfe der
Blinkrate im Ruhezustand geschätzt und mit den Persönlichkeitsskalen des EPQ-R und des
TPQ in Verbindung gebracht. Es wurde erwartet, dass die Blinkrate ein ausreichend gutes
Trait-Maß darstellt, und den vorgestellten Persönlichkeitstheorien zufolge sollte sich ein
Zusammenhang zwischen der Blinkrate und den Skalen Extraversion und Spontaneität
zeigen.
Zunächst werden die Ergebnisse der LST-Analyse diskutiert, die Aufschluss darüber geben
sollte, ob die Blinkrate als Trait-Maß eingesetzt werden kann. Im Anschluss daran wird das
Ergebnis der Korrelationen der Blinkrate mit den Fragebogenskalen diskutiert. Dabei wird
zuerst der Zusammenhang mit den Skalen Extraversion und Spontaneität besprochen und
dann die Ergebnisse der explorativen Tests mit den übrigen Skalen des EPQ-R und des TPQ.
Abschließend werden verschiedene Überlegungen für zukünftige Untersuchungen
vorgestellt.
4.1 LST-Analyse
Die vorliegende Arbeit hatte zum Ziel zu untersuchen, ob es einen Zusammenhang zwischen
der zentralen dopaminergen Grundaktivität und bestimmten Persönlichkeitsdimensionen
gibt. Um die basale zentrale dopaminerge Aktivität bei den psychisch und physisch
unauffälligen Versuchspersonen zu erfassen, wurde mit Hilfe des Electrooculogramms die
Frequenz der spontanen Blinks ermittelt, welche die dopaminerge Aktivität widerspiegeln
soll.

Diskussion 81
Zur Absicherung, dass das Substrat der hier untersuchten zentralen dopaminergen Aktivität
- die Blinkrate - ein ausreichend gutes Trait-Maß darstellt (Hypothese A), sollte ein
akzeptables Strukturgleichungsmodell gefunden werden, bei dem ein latenter Trait einen
bedeutsamen Anteil der Varianz der Blinkrate aufklärt. Außerdem wurden eine
ausreichende Reliabilität und eine hohe Stabilität der Blinkrate bzw. des zugrunde liegenden
Traits erwartet.
Da die Blinkrate, wie schon in Kapitel 1.2.4 erwähnt, mit kognitiven Faktoren und affektiven
Zuständen assoziiert zu sein scheint, ist es nicht erstaunlich, dass die empirischen Daten
nicht mit einem reinen Trait-Modell vereinbar waren.
So berichtet Galley (2001) eine erhöhte Blinkrate bei Zuständen mangelnder oder
nachlassender Aufmerksamkeit, wie Ermüdung, Monotonie, Langeweile oder Geübtheit,
und Stern (1984) postuliert einen positiven Zusammenhang allgemeiner Aktivierung mit der
Blinkrate. Von Cramon (1980) verbindet die Blinkrate mit allgemeiner Erregbarkeit im Sinne
von Affektivität und berichtet eine stark erhöhte Blinkrate bei affektiven Zuständen wie
Nervosität, Aufregung, Erschrecken, Überraschung, Wut, Ärger oder dem Gefühl der
Bedrohung, während die Blinkrate bei Angst stark absinken soll.
Trotz Standardisierung und Homogenisierung der Untersuchungsbedingungen im Labor,
wodurch die situative Varianz gering gehalten werden sollte, konnten solche Faktoren nicht
zuverlässig kontrolliert werden. Schließlich hat der Versuchsleiter keine Kenntnis über den
situativen Kontext, aus dem die Versuchsperson in die Untersuchung kommt, und auch die
Ruhebedingung, unter der das EOG abgeleitet wurde, sagt nichts über das aktuelle arousal
einer Versuchsperson aus. Davon abgesehen kann die ungewohnte Umgebung im Labor
auch für eine erhöhte Aufmerksamkeit gesorgt haben, welche ebenfalls die Blinkrate
beeinflussen soll. So berichtet auch von Cramon (1980) sowohl eine erhebliche
interindividuelle als auch intraindividuelle Varianz der mittleren Blinkrate, selbst unter
konstanten Umgebungsbedingungen, und deutet die interindividuellen Unterschiede als
Ausdruck der dopaminergen Aktivierung (Kapitel 1.2.3).
Daher wurde zusätzlich ein State-Trait-Modell getestet, das neben dem transsituativ
konsistenten Effekt einer Person (Konsistenz) auch den beschriebenen
messgelegenheitsspezifischen Effekt der Situation und der Person-Situation-Interaktion
(Spezifität) berücksichtigt. Es konnte ein liberalisiertes State-Trait-Modell konstruiert

Diskussion 82
werden, für das ein bedeutsamer Anteil an Trait-Varianz ermittelt wurde. Außerdem konnte
eine sehr gute Reliabilität und eine ausreichende Konsistenz festgestellt werden. Die
Blinkrate erfüllt auch die Forderung an ein gutes Trait-Maß, dass die Personeneffekte
wesentlich größer sein sollten als die situativen Effekte. Abschließend konnte mit Hilfe eines
liberalisierten State-Trait-Modells auch eine gute zeitliche Stabilität ermittelt werden.
Zusammenfassend kann festgehalten werden, dass die Blinkrate zuverlässig eine zeitlich
stabile und transsituativ konsistente Verhaltensdisposition widerspiegelt und damit als
gutes Trait-Maß eingeschätzt werden kann.
4.2 Korrelationsanalyse
In diesem Abschnitt werden zunächst die Ergebnisse zur Hypothese B diskutiert. Dabei wird
der Unterschied zwischen physiologischer Grundaktivität und Reaktivität aufgegriffen, und
es werden Überlegungen zur Single-Trait-Single-Transmitter-Hypothese und zur Stichprobe
an sich angestellt.
Des Weiteren werden die Ergebnisse der explorativen Tests mit den übrigen Skalen des
EPQ-R und des TPQ besprochen, wobei vor allem die Korrelation der Blinkrate mit der Skala
Belohnungsabhängigkeit in den Vordergrund rückt. In diesem Zusammenhang wird die
Problematik der genannten Skala angesprochen und es werden Spekulationen über die
Bedeutung dieses Ergebnisses angestellt.
Abschließend werden Überlegungen für zukünftige Untersuchungen dargestellt.
4.2.1 Der Zusammenhang zwischen Blinkrate und Extraversion
oder Spontaneität
Die Persönlichkeit der Versuchspersonen wurde mit den Fragebögen EPQ-R und TPQ erfasst
und mit der Blinkrate in Zusammenhang gebracht. Dabei wurde erwartet, dass speziell die
Persönlichkeitsdimensionen Extraversion und Spontaneität bedeutsam mit der Blinkrate
korrelieren (Hypothese B).
Dass bei einer Teststärke von ca. 80% kein Zusammenhang zwischen der basalen zentralen
dopaminergen Aktivität und der Persönlichkeitsdimension Extraversion gefunden wurde,

Diskussion 83
deutet darauf hin, dass es entweder keinen solchen Effekt gibt oder dieser unbedeutend
klein (r² ≥ .12; r ≥ .35) ausfällt.
Für die Persönlichkeitsdimension Spontaneität war die Teststärke zu gering, um einen
mittleren oder kleinen Effekt zuverlässig aufzudecken. Aus demselben Grund darf aber auch
nicht daraus geschlossen werden, dass es keinen Zusammenhang zwischen der basalen
zentralen dopaminergen Aktivität und der besagten Persönlichkeitsdimension gibt.
Im Folgenden werden einige Punkte angeführt, die zur Erklärung der beschriebenen
Ergebnisse beitragen sollen. Dazu werden Studien herangezogen, die zeigen wie wichtig es
ist, zwischen der physiologischen Grundaktivität und der vorübergehenden Reaktivität auf
Stimulation zu unterscheiden. Des Weiteren wird die Problematik der Sichtweise
angesprochen, dass einem einzigen Trait nur ein bestimmter Neurotransmitter zugrunde
liegen soll (Single-Trait-Single-Transmitter-Hypothese). Abschließend wird die
Beschaffenheit der Versuchspersonenstichprobe kritisch bewertet.
4.2.1.1 Physiologische Grundaktivität vs. Reaktivität
Die Ergebnisse der vorliegenden Untersuchung können als Erweiterung einer Liste von
Studien angesehen werden, die zeigen konnten, dass sich Introvertierte und Extravertierte
im basalen Niveau physiologischer Aktivität nicht unterscheiden (Stelmack, 1990; Stelmack
& Geen, 1992).
Dazu gehören z.B. Studien, bei denen festgestellt wurde, dass es vor einer Stimulation
zwischen Introvertierten und Extrovertierten keinen Unterschied in der Hautleitfähigkeit
gibt (z.B. Davis & Cowles, 1988; Mangan & O´Gorman, 1969). Dies gilt besonders für
Umstände, die ein geringes Erregungspotentials bieten. Des Weiteren wurden unter
Bedingungen, die wenig arousal auslösen, oder während Schlaf auch selten Unterschiede in
Maßen des EEGs festgestellt (Gale, 1981).
In denselben Studien, die Eysenck´s arousal-Hypothese testeten, konnte aber bestätigt
werden, dass sich Introvertierte im Vergleich zu Extravertierten durch eine stärkere
Erregbarkeit oder physiologische Reaktion auf Stimulation auszeichnen.
Auch Rammsayer (2000), der speziell den Zusammenhang zwischen dopaminergen
Mechanismen und Extraversion untersucht, geht davon aus, dass sich Introvertierte und

Diskussion 84
Extrovertierte nicht im absoluten Niveau zentralnervöser Aktivität unterscheiden, postuliert
aber „Unterschiede in der behavioralen Sensitivität, mit der Intro- und Extravertierte auf
Abweichungen vom physiologischen Niveau der dopaminergen Aktivierung reagieren“
(Rammsayer, 2000, S. 218). Dabei sollen Introvertierte im Vergleich zu Extravertierten eine
höhere Reaktivität zeigen (Rammsayer, 2000). Diese Annahme wird durch zwei Experimente
(Rammsayer & Netter, 1993; Rammsayer, 1998) gestützt, bei denen die Auswirkungen einer
pharmakologischen Manipulation der zentralen dopaminergen Aktivität auf die Reaktions-
und Ausführungszeit bei einer motorischen Aufgabe untersucht wurde.
In einem dieser Experimente (Rammsayer & Netter, 1993) wird den Versuchspersonen der
Tyrosinhydroxylasehemmer α-Methyl-para-Tyrosin (AMPT) verabreicht, welcher die DA-
Synthese unterbricht. Die Ausprägung der Versuchspersonen auf der
Persönlichkeitsdimension Extraversion wurde mit dem Eysenck Personality Inventory (EPI;
Eysenck & Eysenck, 1964) erfasst. Vor der AMPT-Applikation unterschieden sich Intro- und
Extravertierte nicht in der Konzentration des freien DAs und des DA-Metaboliten DOPAC
im Urin. Dasselbe galt für die Placebo-Bedingung. Auch bezüglich der Abnahme der
DOPAC-Konzentration nach der pharmakologischen Manipulation konnte kein Unterschied
zwischen Intro- und Extravertierten festgestellt werden. Extraversionsabhängige
Verhaltenseffekte zeigten sich erst in den Reaktionszeitdaten. Dabei reagierten Introvertierte
im Vergleich zu einer Placebo-Kontrollgruppe signifikant langsamer, während die
Manipulation des dopaminergen Systems auf Extrovertierte keinen Einfluss zu haben schien.
Fasst man die Befunde zur Untersuchung des Zusammenhangs der basalen
psychophysiologischen Aktivität mit der Persönlichkeitsdimension Extraversion zusammen,
ist es nicht erstaunlich, dass sich diesbezüglich auch in der vorliegenden Untersuchung keine
signifikanten Ergebnisse finden lassen.
Die Befunde deuten darauf hin, dass die Untersuchung des Zusammenhangs der Reaktivität
des dopaminergen Systems mit bestimmten Traits vorzuziehen ist.
Untersuchungen mit elektrophysiologischen Maßen weisen z.B. darauf hin, dass
Stimulanzien des Zentralnervensystems (z.B. Koffein oder Nikotin) systematische
Unterschiede in der Reaktion auf eine solche Stimulation nach sich ziehen können. So zeigen

Diskussion 85
Introvertierte im Vergleich zu Extravertierten nach der Einnahme von Koffein und bei
mittlerer Stimulusintensität im Mittel eine höhere Hautleitfähigkeit (Smith, 1983).
Auch die stützenden Befunde zu den Persönlichkeitstheorien von Depue und Cloninger
(Depue et al., 1994; Gerra et al., 2000) untersuchten den Zusammenhang zwischen Dopamin
und Persönlichkeit über eine Provokation des dopaminergen Systems mit Hilfe von DA-
Agonisten. Da unklar ist, durch welche anderen Faktoren, in welchem Maße und in welcher
Weise die peripheren Indikatoren der zentralen dopaminergen Aktivität (z.B. die Blinkrate
oder die Prolactin-Hemmung) beeinflusst werden, begründet Depue et al. (1994) den Einsatz
eines direkten DA-Agonisten damit, dass dadurch Fehlervarianz reduziert werden soll.
Die Blinkproduktion wird z.B. möglicherweise auch durch cholinerge und GABAerge
Systeme des Hirnstamms kontrolliert (Karson, 1989), wobei diese Neurotransmittersysteme
die Blinkrate hemmen können. Auch die Prolactin-Sekretion wird beispielsweise durch DA
tonisch inhibiert, während 5-HT als Gegenspieler wirkt.
4.2.1.2 Die Single-Trait-Single-Transmitter-Hypothese
Genauso wie die peripheren DA-Indikatoren können auch die untersuchten Traits einem
unbekannten Einfluss anderer Neurotransmitter unterliegen.
Depue beschreibt in seiner Theorie z.B. ausführlich den tonisch inhibitorischen Einfluss von
5-HT auf die Schwelle der dopaminergen Förderung. Weiterhin zieht er auch den Beitrag
von GABA, Opiaten, der Substanz P und Neurotensin in Betracht (Depue & Lenzenweger,
2001).
Auch Cloninger (1988) weist darauf hin, dass es keine direkten linearen Beziehungen
zwischen einem einzelnen Neurotransmitter und einem spezifischen Verhalten gibt, und
dass jedes der Systeme und deren Interaktion zusätzlich auch durch andere
Neurotransmitter, -Peptide und Hormone beeinflusst werden kann.
So könnte die sehr einfache Vorstellung, dass ein Trait wie Extraversion oder Spontaneität
nur auf der Wirkung eines einzigen Transmitters beruht, ebenfalls dazu beitragen, dass der
Zusammenhang zwischen Dopamin und den besagten Persönlichkeitsdimensionen in der
vorliegenden Arbeit nicht festgestellt werden konnte.
Da die Realität höchstwahrscheinlich komplexer ist, was auch die Autoren der vorgestellten
Persönlichkeitstheorien nicht bestreiten, werden die Untersuchungen zum Beitrag einzelner

Diskussion 86
Transmitter als Schritt in die richtige Richtung gewertet, der Aufschluss über vermutlich
kompliziertere Zusammenhänge geben soll.
4.2.1.3 Die Stichprobe
Eine weitere Erklärung dafür, dass in der vorliegenden Untersuchung kein bedeutsamer
Zusammenhang zwischen DA und den Persönlichkeitsdimensionen Extraversion und
Spontaneität gefunden wurde, könnte die Zusammensetzung der Stichprobe sein.
Depue et al. (1994) stellte seine Versuchpersonen im Verhältnis zu verschiedenen potentiell
interagierenden Traits zusammen; d.h. er wählte nur die Versuchspersonen aus, die
gleichzeitig hohe Werte für das Extraversions-Maß und niedrige Werte für Constraint hatten
oder gleichzeitig hohe Werte für Constraint und niedrige Werte für das Extraversions-Maß.
Die Stichprobe der vorliegenden Untersuchung filterte nicht bestimmte Versuchspersonen
heraus. Es zeigten sich keine nennenswerten Auffälligkeiten in den Persönlichkeitstests und
auch die Deskriptivstatistik der Blinkrate fiel nicht weiter aus dem Rahmen. Vielleicht bietet
eine Stichprobe, die keine Extremwerte auf den betrachteten Skalen liefert, nicht die nötige
Varianz, um einen bedeutsamen Effekt dopaminerger Aktivität auf die Persönlichkeit zu
hervorzubringen, d.h. möglicherweise ist der Effekt innerhalb einer „normalen“
studentischen Stichprobe zu gering, um ihm Bedeutung zuzumessen.
4.2.2 Ein Zusammenhang zwischen Blinkrate und
Belohnungsabhängigkeit
Explorativ wurde auch der Zusammenhang der übrigen Persönlichkeitsdimensionen des
EPQ-R (Psychotizismus, Neurotizismus und Lügenskala) und des TPQ (Risikovermeidung
und Belohnungsabhängigkeit) mit der basalen zentralen dopaminergen Aktivität untersucht.
Dabei zeigte sich wider Erwarten ein signifikanter Zusammenhang der
Persönlichkeitsdimension Belohnungsabhängigkeit des TPQ mit der Blinkrate.
Die Korrelation der Lügenskala des EPQ-R mit der Blinkrate könnte, nach der sequenziellen
Bonferoni-Korrektur zu urteilen, auch zufällig Zustande gekommen sein. Davon abgesehen
war die Stichprobengröße dieser Untersuchung nicht darauf ausgelegt, einen Effekt von
r < .35 mit ausreichender Teststärke zu finden.

Diskussion 87
Wie schon im Methodenteil dieser Arbeit angeschnitten wurde, ist die Interpretation der
Skalen für Belohnungsabhängigkeit problematisch. So legte die Reliabilitätsberechnung und
eine anschließende Faktorenanalyse im Rahmen der Prüfung der Gütekriterien der
deutschen Version des TPQ (Defeu et al., 1995) nahe, dass eine Aufteilung dieser Dimension
in zwei Skalen sinnvoll sein könnte. Diese neu zusammengestellten unabhängigen Skalen
wurden von Defeu et al. (1995) „soziale Orientiertheit“ (BAa) und „emotionale
Abhängigkeit“ (BAb) genannt. Auch Cloninger (1993) beschreibt die Aufnahme einer
unabhängigen vierten Dimension, die er persistence nennt. Im Gegensatz zu Defeu et al.
(1995) wertet Cloninger dabei nur die Subskala BA2 zu einer Hauptskala auf. Diese misst
Ausdauer trotz Frustration und Ermüdung und ist angeblich nicht mit anderen Aspekten
der Belohnungsabhängigkeit korreliert.
Bei der Bestimmung der konvergenten Validität an einer Stichprobe von Alkoholabhängigen
sollten die Skalen BAa und BAb nach Defeu et al. (1995) Variablen prädizieren, die mit
sozialer Bestätigung assoziiert sind. Hier können Parallelen zur Lügenskala des EPQ-R
gezogen werden, welche die Tendenz misst, mit der Versuchspersonen im Sinne sozialer
Erwünschtheit antworten oder die Tendenz, in diesem Zusammenhang bestimmte
Merkmale zu verschweigen oder hervorzuheben.
Während sich für die Skala BAb keine entsprechenden Variablen finden ließen, konnten für
die Skala BAa positive Zusammenhänge mit einer Skala für „soziales Trinken“ und für
„Behandlungsbereitschaft“ gefunden werden und ein negativer Zusammenhang mit
„alkoholbedingten Verkehrsdelikten in der Anamnese“.
Im Gegensatz zu den Skalen der Belohnungsabhängigkeit sollte die Skala Spontaneität laut
Defeu et al. (1995) Variablen vorhersagen, die soziale Auffälligkeiten repräsentieren. Es
zeigten sich Zusammenhänge mit „Partnerproblemen wegen des Trinkens“, „Straftaten in
der Anamnese“, „Beteiligung an Schlägereien in der Anamnese“ und dem Konsum von
„harten Drogen“.
Vorausgesetzt, dass es tatsächlich einen Zusammenhang zwischen der
Persönlichkeitsdimension Spontaneität und dem Maß für die zentrale dopaminerge Aktivität
gibt, könnte die inverse Korrelation der Skala Belohnungsabhängigkeit mit der Blinkrate,
trotz (oder vielleicht wegen) der fraglichen Zusammenstellung dieser

Diskussion 88
Persönlichkeitsdimension, darauf hindeuten, dass möglicherweise Aspekte der Dimension
Spontaneität erfasst werden, die einen Zusammenhang mit dopaminergen Mechanismen
besser zu reflektieren scheint, als es die Dimension Spontaneität selbst kann.
Eine Studie, welche die von Cloninger postulierten Zusammenhänge zwischen Spontaneität
und dopaminerger Aktivität zum Einen und zwischen Risikovermeidung und serotonerger
Aktivität zum Anderen stützt (Gerra et al., 2000), liefert möglicherweise auch einen Beitrag
zur eben formulierten Annahme, dass Belohnungsabhängigkeit eher mit dopaminergen
Mechanismen assoziiert sein könnte. Neben den erwarteten Ergebnissen für die beiden
anderen Persönlichkeitsdimensionen ergab sich für die Dimension reward dependence ein
positiver Zusammenhang mit der GH-Konzentration (erwartet wurde ein negativer
Zusammenhang) und ein negativer Zusammenhang mit der Prolactin-Konzentration
(erwartet wurde kein Zusammenhang; siehe Kapitel 1.3.2.4). Diese Ergebnisse entsprechen
den erwarteten Zusammenhängen von novelty seeking mit dopaminergen Mechanismen.
Gerra et al. (2000) ziehen aufgrund dieser Befunde in Betracht, dass RD möglicherweise mit
NS in Verbindung steht und ebenfalls durch die dopaminerge Funktion beeinflusst wird.
Neben der Untersuchung von Gerra et al. (2000) gibt es außerdem eine Studie (Mulder &
Joyce, 2002), die ebenfalls als Hinweis auf einen Zusammenhang von reward dependence mit
dopaminerger und möglicherweise auch serotonerger Aktivität in Verbindung bringt.
Mulder & Joyce (2002) untersuchten den Zusammenhang zwischen Temperament- und
Verhaltensmaßen anhand einer Fenfluramin-induzierten Prolactin-Reaktion bei depressiven
Männern. Fenfluramin ist ein 5-HT-Agonist und sollte somit die Prolactin-Sekretion erhöhen.
Die Persönlichkeit der Versuchspersonen wurde u.a. mit dem TPQ und dem EPQ (Eysenck
& Eysenck, 1975) erfasst. Wider Erwarten konnte kein Zusammenhang zwischen der
Dimension Risikovermeidung des TPQ, die nach Cloninger durch 5-HT moduliert werden
soll, und der Prolactin-Reaktion festgestellt werden. Stattdessen ergab sich ein positiver
Zusammenhang der Prolactin-Reaktion mit der Skala Belohnungsabhängigkeit des TPQ und
mit der Lügenskala des EPQ. Die Autoren der eben beschriebenen Studie zogen in Betracht,
dass dieses Ergebnis aufgrund der zahlreich durchgeführten Korrelationen nur zufällig sein
könnte.

Diskussion 89
Wenn man davon ausgeht, dass 5-HT (als Gegenspieler von DA) der tonisch hemmenden
Wirkung von DA auf die Prolactin-Sekretion entgegenwirkt, können die Ergebnisse der
Studie von Mulder und Joyce (positive Korrelationen mit BA und LS) mit denen der
vorliegenden Untersuchung (negative Korrelationen mit BA und LS) verglichen werden. Es
könnte also möglich sein, dass die Effekte in beiden Studien nicht zufällig sind. Somit könnte
man vermuten, dass geringe basale dopaminerge Aktivität (hier indiziert durch eine geringe
Blinkrate im Ruhezustand) und hohe serotonerge Reaktivität (bei Mulder & Joyce indiziert
durch eine erhöhte Prolactin-Sekretion als Reaktion auf einen 5-HT-Agonisten) den Trait
reward dependence fördern.
Ginge man bei 5-HT ähnlich wie bei DA von einer postsynaptischen Hypersensibilität und
einer damit verbundenen geringen basalen serotonergen Aktivität aus, würde eine
Interaktion sowohl geringer basaler dopaminerger als auch serotonerger Aktivität und
erhöhten Werten für die Skala Belohnungsabhängigkeit (sollte laut Cloninger mit geringer
basaler noradrenerger Aktivität zusammenhängen) auf eine Persönlichkeitsausprägung
hindeuten, die mit einer histrionischen Persönlichkeit vergleichbar wäre. Hier zeigt sich eine
Überschneidung der Persönlichkeitsdimensionen novelty seeking und reward dependence, da
Cloninger (1987) hohe Werte für beide Dimensionen jeweils u.a. mit einer histrionischen
Persönlichkeitsstörung in Verbindung bringt. Entsprechende Reaktionsmuster für die
histrionische Persönlichkeit, welche Cloninger (1987) auf niedrige basale Werte aller drei
Transmittersysteme zurückführt, sind Impulsivität, Leichtgläubigkeit, emotionale
Empfindlichkeit und Narzissmus (Cloninger, 1987). Laut Cloninger (1987) stimmt dies mit
den Beschreibungen histrionischer oder hysterischer Persönlichkeit im DSM-III und ICD
überein. Zu den narzisstischen Eigenschaften gehören dabei Schauspielerei, Suche nach
Aufmerksamkeit, Nachgiebigkeit gegenüber sich selbst und Eitelkeit. Empfindlichkeit und
emotionale Abhängigkeit sind durch Abhängigkeit, Suche nach Bestätigung, Wärme und
Charme gekennzeichnet, und impulsiv-aggressive Eigenschaften sind Verlangen nach
Aktivität und Aufregung, wütende Ausbrüche und suizidales Verhalten (Horowitz, nach
Cloninger, 1987). Gerade die selbstdarstellerischen Aspekte der histrionischen Persönlichkeit
könnten auch durch die Lügenskala des EPQ-R erfasst worden sein, und die Korrelation der
Skala Belohnungsabhängigkeit mit der Extraversions-Skala könnte auf einen
Zusammenhang zwischen dem Belohnungsaspekt sozialer Bindungen bei reward dependence

Diskussion 90
mit dem Bedürfnis nach Anschluss bei Extraversion zurückzuführen sein. Die Annahme
Cloninger´s (1987), dass die Systeme, die für Aufrechterhaltung (Hintergrund von reward
dependence) und Aktivierung (Hintergrund von novelty seeking) von Verhalten zuständig sind,
zusammenarbeiten (Kapitel 1.3.2.3), legt auch nahe, dass eine enge Verbindung zwischen
den Persönlichkeitsdimensionen novelty seeking und reward dependence besteht.
Ein weiterer Hinweis darauf, dass DA und 5-HT mit der Persönlichkeitsdimension reward
dependence assoziiert sein könnten, liefert eine vorläufige Studie zur genetischen Grundlage
von Traits (Benjamin, Ebstein & Lesch, 1998). Benjamin et al. (1998) fanden, neben einem
Zusammenhang zwischen dem DA-D4-Rezeptor-Gen und der Persönlichkeitsdimension
novelty seeking des TPQ, Hinweise auf eine Interaktion zwischen dem DA-D4-Rezeptor-Gen
und dem 5-HT-2C-Rezeptor-Gen mit einem Effekt auf die Persönlichkeitsdimension reward
dependence des TPQ. Die Ausprägung dieser Dimension war nämlich bei einer Substitution
von Cystein zu Serin bei Aminosäure 23 um etwa 10% reduziert. Dieser Effekt kam bei
gleichzeitigem Auftreten der selteneren 5-HT-2C- und DA-D4-Rezeptor-Allele stärker zur
Geltung.
Die Ergebnisse der vorliegenden Untersuchung könnten als Hinweis darauf interpretiert
werden, dass die Zusammenstellung der Skalen des TPQ nicht optimal gelungen ist und /
oder, dass die postulierten Persönlichkeitsdimensionen jeweils durch mehr als einen
Neurotransmitter beeinflusst werden.
4.2.3 Überlegungen für zukünftige Untersuchungen
Im Folgenden werden einige Punkte angesprochen, die bei zukünftigen Untersuchungen
beachtet werden könnten. Diese betreffen sowohl die physiologische Reaktivität als auch die
zu untersuchenden Neurotransmitter und deren Indikatoren sowie die beobachteten
Persönlichkeitsdimensionen.
4.2.3.1 Basale physiologische Aktivität und Reaktivität
Da im Verlauf der Diskussion schon erklärt wurde, dass sich persönlichkeitsabhängige
Effekte eher in der Reaktion auf Stimulation zeigten als in einer Ruhebedingung, könnte das
zu untersuchende Transmittersystem in zukünftigen Untersuchungen nach einer Erfassung

Diskussion 91
der Grundaktivität zusätzlich manipuliert werden. Um die Konfundierung mit anderen
Variablen möglichst gering zu halten, sollte die Manipulation möglichst spezifisch sein. Im
Fall von DA und Extraversion oder Spontaneität könnte das dopaminerge System künstlich
durch stimulierende Substanzen oder entsprechende Anreiz-Situationen angeregt werden.
Außerdem könnte getestet werden, ob sich sowohl bei Hemmung als auch bei Stimulation
des entsprechenden Systems Unterschiede in der Reaktivität feststellen lassen.
4.2.3.2 Neurotransmitter
Des Weiteren wurde angemerkt, dass eine Single-Trait-Single-Transmitter-Hypothese zu
einfach ist und möglicherweise mehrere verschiedene Modulatoren interagieren.
Zuckerman (1996) postuliert z.B. eine Kombination aus niedrigen 5-HT-Werten, hohen DA-
Werten und niedrigen Noradrenalin-Werten für eine hohe Ausprägung des Traits sensation
seeking, der mit der mit dem Trait novelty seeking verwandt sein soll.
In diesem Sinne sollte angestrebt werden, auch Interaktionen zwischen verschiedenen
Transmittersystemen zu erfassen. Dazu würde es Sinn machen, verschiedene in Frage
kommende Neurotransmitter (zusätzlich zu DA z.B. auch 5-HT und Noradrenalin) zu
untersuchen, indem, soweit möglich, entsprechende Indikatoren gleichzeitig eingesetzt
werden.
4.2.3.3 Indikatoren
Da fraglich ist, ob verschiedene Indikatoren für einen Neurotransmitter funktionell
äquivalent sind, sollten innerhalb einer Untersuchung auch mehrere Indikatoren für einen
Transmitter herangezogen werden, die dann miteinander verglichen werden können. Für
Dopamin könnten z.B. neben der Blinkrate und der Prolactin-Hemmung auch die Sekretion
von Wachstumshormon oder die DA-Metaboliten (DOPAC oder HVA) im Urin erfasst
werden.
4.2.3.4 Persönlichkeitsdimensionen
Laut Depue und Collins (1999) gibt es in fast jeder Persönlichkeitstaxonomie einen
impulsivitätsnahen Trait. Dazu können neben Extraversion und Spontaneität z.B. auch die
Persönlichkeitsdimensionen sensation seeking (Zuckerman, 1979), Impulsivität (impulsiveness)
und Abenteuerlust (venturesomeness) (Eysenck, Pearson, Easting & Allsopp, 1985b)

Diskussion 92
Psychotizismus (Eysenck et al., 1985a) oder positive Emotionalität (Tellegen & Waller, 1996)
gezählt werden.
Da sich diese ähnlichen Traits meist trotzdem in gewissen Aspekten unterscheiden, wäre es
interessant möglichst viele dieser Dimensionen zu erfassen, um feststellen zu können, ob
sich einer dieser Traits vielleicht besser eignet, um einen Zusammenhang zwischen DA und
Persönlichkeit zu finden.
Auch wenn es in dieser Arbeit nicht gelungen ist, einen Zusammenhang zwischen der
basalen zentralen dopaminergen Aktivität und den Persönlichkeitsdimensionen
Extraversion bzw. Spontaneität zu finden, ergaben jedoch explorative Tests mit den übrigen
Persönlichkeitsdimensionen ein überraschendes Resultat. Es scheint, als hätten sowohl die
dopaminerge als auch die serotonerge Aktivität Einfluss auf die Persönlichkeitsdimension
reward dependence.
Es wäre interessant zu sehen, ob sich der Zusammenhang zwischen der Blinkrate im
Ruhezustand und der Persönlichkeitsdimension Belohnungsabhängigkeit auch unter den
verschiedenen Verbesserungsvorschlägen replizieren ließe.

Literaturverzeichnis 93
5 Literaturverzeichnis Amelang, M. & Bartussek, D. (1997). Differentielle Psychologie und Persönlichkeitsforschung.
Stuttgart: Kohlhammer. Benjamin, J., Ebstein, R. P. & Lesch, K.-P. (1998). Genes for personality traits: implications for
psychopathology. International Journal of Neuropsychopharmacology, 1, 153-168. Bentivoglio, A. R., Bressman, S. B., Cassetta, E., Caretta, D., Tonali, P. & Albanese, A. (1997).
Analysis of blink rate patterns in normal subjects. Movement Disorders, 12(6), 1028-1034. Berlyne, D. E. (1960). Conflict, Arousal and Curiosity. New York: McGraw-Hill. Birbaumer, N. & Schmidt, R. F. (1996). Biologische Psychologie. Berlin: Springer-Verlag. Blount, W. P. (1928). Studies of the movements of the eyelids of animals: Blinking. Quarterly
Journal of Experimental Physiology, 18, 111-125. Bortz, J. (1999). Statistik für Sozialwissenschaftler. Berlin: Springer-Verlag. Brauer, L. H. & De Wit, H. (1997). High dose pimozide does not block amphetamine-induced
euphoria in normal volunteers. Pharmacology Biochemistry and Behavior, 56, 265-272. Carlsson, A. (1975). Receptor-mediated control of dopamine metabolism in pre- and
postsynaptic receptors. In: E. Usdin and W. E. Bunney, Jr., Pre- and Postsynaptic Receptors,(pp. 49-65). New York: Marcel Dekker.
Chen, J. (1993). Dopaminergic mechanisms and brain reward. Seminars in the Neurosciences, 5,
315-320. Civelli, O., Bunzow, J. R., Grandy, D. K., Zhou, Q. Y. & van Tol, H. M. (1991). Molecular
biology of the dopamine receptors. European Journal of Pharmacology, 207, 277-286. Cloninger, C. R. (1986). A Unified Biosocial Theory of Personality and its Role in the
Development of Anxiety States. Psychiatric Developments, 3, 167-226. Cloninger, C. R. (1987). A systematic method for clinical description and classification of
personality variants. Archives of General Psychiatry, 44, 573-588. Cloninger, C. R. (1988). A Unified Biosocial Theory of Personality and its Role in the
Development of Anxiety States: A reply to Commentaries. Psychiatric Developments, 2, 83-120.

Literaturverzeichnis 94
Cloninger, C. R. (1999). Personality and Psychopathology: Neuroadaptive Processes. Washington D.C.: American Psychiatric Press.
Cloninger, C. R., Przybeck, T. R. & Svrakic, D. M. (1991) The tridimensional personality
questionnaire: U.S. normative data. Psychological Reports, 69, 1047-1057. Cloninger, C. R., Svrakic, D. M. & Przybeck, T. R. (1993). A psychobiological model of
temperament and character. Archives of General Psychiatry, 50(12), 975-990. Cohen, J. (1992). A Power Primer. Psychological Bulletin, 112(1), 155-159. Collins, P. F. & Depue, R. A. (1992). A Neurobehavioral Systems Approach to Developmental
Psychophathology: Implications for Disorders of Affect. In: D. C. S. L. Toth, Developmental perspectives on depression,(pp. 29-101). New York: University of Rochester.
Crow, T. J. & Deakin, J. F. W. (1985). Neurohumoral transmission, behavior, and mental
disorder. In: M. Shepherd, Handbook of Psychiatry,5: The Scientific Foundations of Psychiatry (pp. 137-182). Cambridge: Cambridge University Press.
Dahlström, A. & Fuxe, K. (1964). Evidence for the existence of monoamine-containing
neurons in the central nervous system. I Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiologica Scandinavica, 62(232), 1-55.
Damsma, G., Pfaus, J. G., Wenkstren, D., Phillips, A. G. & Fibinger, H. C. (1992). Sexual
behavior increases dopamine transmission in the nucleus accumbens and striatum of male rats: comparison with novelty and locomotion. Behavioral Neuroscience, 106, 181-191.
Davis, C. & Cowles, M. (1988). A laboratory study of temperament and arousal: A test of
Gale´s hypothesis. Journal of Research in Personality, 22, 101-116. Davison, G. C. & Neale, J. M. (2002). Klinische Psychologie. Weinheim: Psychologie Verlags
Union. Defeu, P., Kuhn, S. & Schmidt, L. G. (1995). Prüfung der Gütekriterien einer deutschen
Version des ʺTridemsional Personality Questionnaire (TPQ)ʺ von Cloninger bei Alkoholabhängigen - Validity and Reliabiltiy of a German Version of Cloninger´s ʺTridimensional Personality Questionnaire (TPQ) by Alcoholics. Sucht, 41(6), 395-407.
Depue, R. A. (1995). Neurobehavioral factors in personality and depression. European Journal
of Personality, 9, 423-439. Depue, R. A. (1996). A neurobehavioral framework for the structure of personality and
emotion: Implications for personality disorders. In: J. F. Clarkin and M. F. Lenzenweger, Major theories of personality disorder,(pp. 347-390). New York: The Guilford Press.

Literaturverzeichnis 95
Depue, R. A. & Collins, P. F. (1999). Neurobiology of the structure of personality: Dopamine, facilitation of incentive motivation, and extraversion. Behavioral and Brain Science, 22, 491-569.
Depue, R. A., Collins, P. F. & Luciana, M. (1996). A model of neurobiology-environment
interaction in developmental psychopathology. In: M. F. Lenzenweger and J. J. Haugaard, Frontiers of developmental psychopathology,(pp. 44-77). Oxford: Oxford University Press.
Depue, R. A. & Lenzenweger, M. F. (2001). A neurobehavioral dimensional model. In: W. J.
Livesley, Handbook of personality disorders,(pp. New York: The Guilford Press. Depue, R. A., Luciana, M., Arbisi, P., Collins, P. F. & Leon, A. (1994). Dopamine and the
structure of personality: Relation of agonist-induced dopamine activity to positive emotionality. Journal of Personality and Social Psychology, 67(3), 485-498.
Derrfuß, J., Krämer, S., Kugele, O. & Kugler, R. (2000). Transmittersysteme und ihre
Bedeutung für unser Verhalten. In: M. Pritzel, Dopamin und seine Bedeutung für unser Verhalten,(pp. 44-51). Hamburg: Kovac.
Di Chiara, G. (1995). Psychobiology of the role of dopamine in drug-abuse and addiction.
Neuroscience Research Communications, 17, 133-143. Drew, G. C. (1951). Variations in reflex blink-rate during visual-motor tasks. Quarterly Journal
of Experimental Psychology, 3, 73-88. Elbert, T. & Rockstroh, B. (1990). Psychopharmakologie. Berlin: Springer-Verlag. Elsworth, J. D., Lawrence, M. S., Roth, R. H., Taylor, J. R., Mailman, R. B., Nichols, D. E.,
Lewis, M. H. & Redmond Jr., E. R. (1991). D-sub 1 and D-sub 2 dopamine receptors independently regulate spontaneous blink rate in the vervet monkey. Journal of Pharmacology and Experimental Therapeutics, 259(2), 595-600.
Eysenck, H. J. (1967). The biological basis of personality. Springfield: Thomas. Eysenck, H. J. (1994). Personality: Biological foundations. In: P. A. Vernon, The
neuropsychology of individual differences,(pp. 151-207). San Diego: Academic Press. Eysenck, H. J. & Eysenck, S. B. G. (1964). The Eysenck Personality Inventory. London:
University of London Press. Eysenck, H. J. & Eysenck, S. B. G. (1975). Manual of the Eysenck Personality Questionnaire.
London: Hodder & Stoughton. Eysenck, S. B. G., Eysenck, H. J. & Barrett, P. (1985a). A revised version of the psychoticism
scale. Personality and Individual Differences, 6, 21-29.

Literaturverzeichnis 96
Eysenck, S. B. G., Pearson, P. R., Easting, G. & Allsopp, J. F. (1985b). Age norms for impulsiveness, venturesomeness and empathy in adults. Personality and Individual Differences, 6, 613-619.
Fibinger, H. C. (1993). Mesolimbic dopamine: An analysis of its role in motivated behavior.
Seminars in the Neurosciences, 5, 321-327. Fink, J. S. & Reis, D. J. (1981). Genetic variations in midbrain dopamine cell numbers: Parallel
with differences in responses to dopaminergic agonists and in naturalistic behaviors mediated by dopaminergic systems. Brain Research, 222, 335-349.
Fiorino, D. F., Coury, A. & Phillips, A. G. (1997). Dynamic changes in nucleus accumbens
dopamine efflux during the Coolidge effect in male rats. The Journal of Neuroscience, 17(12), 4849-4855.
Gale, A. (1981). EEG studies of extraversion-introversion. In: R. Lynn, Dimensions of
personality: Papers in honour of H. J. Eysenck,(pp. Oxford: Pergamon. Galley, N. (2001). Physiologische Grundlagen, Meßmethoden und Indikatorfunktionen der
okulomotorischen Aktivität. In: F. Roesler, Grundlagen und Methoden der Psychophysiologie,(pp. 237-316). Göttingen: Hogrefe.
Gerra, G., Zaimovic, A., Timpano, M., Zambelli, U., Delsignore, R. & Brambilla, F. (2000).
Neuroendocrine correlates of temperamental traits in humans. Psychoneuroendocrinology, 25, 479-496.
Grandy, D. K. & Civelli, O. (1992). G-protein-coupled receptors: The new dopamine receptor
subtypes. Current Opinion in Neurobiology, 2(3), 275-281. Hagemann, D. (1999). Tonische Asymmetrien corticaler Aktiviertheit und affektive Dispositionen:
Eine empirische Untersuchung latenter State- und Traitvariablen. Berlin: Logos Verlag. Hagemann, D. (2002). Strukturgleichungsmodelle: Ein Skript zur Vorlesung ʺForschungsmethoden
IIʺ. Trier: Universität Trier. Hagemann, D., Naumann, E., Thayer, J. & Bartussek, D. (2002). Does resting EEG asymmetry
reflect a trait? An application of latent state-trait theory. Journal of Personality and Social Psychology, 82, 619-641.
Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal
of Statistics, 6, 65-70. Hu, L. T. & Bentler, P. M. (1995). Evaluating model fit. In: R. H. Hoyle, Structural equation
modeling. Concepts, issues, and applications,(pp. 76-99). Thousand Oaks: Sage Publications.

Literaturverzeichnis 97
Huston, J. P. (1982). Searching for the neural mechanism of reinforcement (of ʺstampingingʺ). In: B. G. Hoebel and D. Novin, The Neural Basis of Feeding and Reward,(pp. 75-83). Brunswick: Haer Institute.
Karson, C. N. (1979). Oculomotor signs in a psychiatric population: A preliminary report.
American Journal of Psychiatry, 136(8), 1057-1060. Karson, C. N. (1983). Spontaneous eye-blink rates and dopaminergic systems. Brain, 106, 643-
653. Karson, C. N. (1989). Blinking. Bulletin de la Societé de belge d´ophtalmologie, 237, 443-457. Karson, C. N., Berman, K. F., Donnelly, E. F., Mendelson, W. B., Kleinman, J. E. & Wyatt, R. J.
(1981a). Speaking, thinking and blinking. Psychiatry Research, 5, 243-246. Karson, C. N., Kleinman, J. E., Berman, K. F., Phelps, B. H., Wise, C. D., De Lisi, L. E. & Jeste,
D. V. (1983). An inverse correlation between spontaneous eye-blink rate and platelet monoamine oxidase activity. British Journal of Psychiatry, 142, 43-46.
Karson, C. N., Staub, R. A., Kleinman, J. E. & Wyatt, R. J. (1981b). Blink Rates And Receptor
Supersensitivity. Neuropharmacology, 20, 91-93. Kebabian, J. W. & Calne, D. B. (1979). Multiple receptors for dopamine. Nature, 277, 93-96. Kennard, D. W. & Glaser, G. H. (1964). An analysis of eyelid movements. Journal of Nervous
and Mental Disease, 139, 31-48. Köhler, T. (2001). Biopsychologie - Ein Lehrbuch. Stuttgart: Kohlhammer. Kong, X. & Wilson, G. F. (1998). A new EOG-based eyeblink detection algorithm. Behavior
Research Methods, Instruments & Computers, 30(4), 713-719. Kugele, O. (2000). Psychopharmaka und Neuroleptika. In: M. Pritzel, Dopamin und seine
Bedeutung für unser Verhalten,(pp. 89-121). Hamburg: Kovac. Le Moal, M. & Simon, H. (1991). Mesocorticolimbic dopaminergic network: functional and
regulatory roles. Physiological Reviews, 71, 155-234. Mangan, G. L. & O´Gorman, J. G. (1969). Initial amplitude and rate of habituation of
orienting reaction in relation to extraversion and neuroticism. Journal of Experimental Research in Personality, 3, 275-282.
Mulder, R. T. & Joyce, P. R. (2002). Relationship of temperament and behaviour measures to
the prolactin response to fenfluramine in depressed men. Psychiatry Research, 109, 221-228. Nieuwenhuys, R. (1985). Chemoarchitecture of the brain. Berlin: Springer-Verlag.

Literaturverzeichnis 98
Nieuwenhuys, R., Voogd, J., von Huijzen, C. & Lange, W. (1980). Das Zentralnervensystem des Menschen. Berlin, Heidelberg: Springer-Verlag.
Oades, R. D. (1985). The role of noradrenaline in tuning and dopamine in switching between
signals in the CNS. Neuroscience & Biobehavioral Reviews, 9, 261-282. Oldfield, R. C. (1971). The assessment and analysis of handedness: Edinburgh inventory.
Neuropsychologia, 9, 97-113. Pinel, J. P. J. (1990). Biopsychologie. Toronto: Allyn and Bacon. Ponder, E. & Kennedy, W. P. (1928). On the act of blinking. Quarterly Journal of Experimental
Physiology, 18, 89-119. Rammsayer, T. (2000). Dopaminerge Mechanismen und Extraversion. Ergebnisse und
Überlegungen zu einer ʺDopamin-Hypothese der Extraversionʺ. Zeitschrift für Differentielle und Diagnostische Psychologie, 21(3), 218-225.
Rammsayer, T. & Netter, P. V., Wolfgang H. (1993). A neurochemical model underlying
differences in reaction times between introverts and extraverts. Personality and Individual Differences, 14(5), 701-712.
Rammsayer, T. H. (1998). Extraversion and dopamine. Individual differences in response to
changes in dopaminergic activity as a possible biological basis of extraversion. European Psychologist, 3(1), 37-50.
Rappelsberger, P. (1980). Verstärkung von EEG-Signalen: Was passiert am
Verstärkereingang und was sollte eine EEG-Assistentin darüber wissen? Das EEG-Labor, 2, 87-103.
Robbins, T. W. & Everitt, B. J. (1996). Neurobehavioural mechanisms of reward and
motivation. Current Opinion in Neurobiology, 6, 228-236. Robinson, T. E. (1988). Stimulant drugs and stress: Factors influencing individual differences
in the susceptibility to sensitization. In: P. Kalivas and C. Barnes, Sensitization in the nervous system,(pp. 145-174). Caldwell: Telford Press.
Robinson, T. E. & Berridge, K. C. (1993). The neural basis of drug craving: an incentive-
sensitization theory of addiction. Brain Research Review, 18, 247-291. Routtenberg, A. (1978). The reward system of the brain. Scientific American, 239(5), 154-164. Ruch, W. (1999). Die revidierte Fassung des Eysenck Personaltiy Questionnaire und die
Konstruktion des deutschen EPQ-R bzw. EPQ-RK / The Eysenck Personality Questionnaire-Revised and the construction of the German standard and short versions. Zeitschrift für Differentielle und Diagnostische Psychologie, 20(1), 1-24.

Literaturverzeichnis 99
Salamone, J. D., Cousins, M. & Snyder, B. J. (1997). Behavioral functions of nucleus accumbens dopamine: Empirical and conceptual problems with the anhedonia hypothesis. Neuroscience & Biobehavioral Reviews, 21, 341-359.
Schultz, W., Dayan, P. & Montague, P. R. (1997). A neural substrate of prediction and
reward. Science, 275, 341-359. Schwarting, R. K. W. (1997). Zur Neurochemie des Verhaltens: Dopamin und Motivation.
Psychologische Rundschau, 48, 211-223. Sibley, D. R. & Monsma Jr., F. J. (1992). Molecular biology of dopamine receptors. Trends in
Pharmacological Sciences, 13, 61-69. Smith, B. D. (1983). Extraversion and electrodermal activity: Arousability and the inverted-U.
Personality and Individual Differences, 4, 411-420. Stellar, J. R. & Stellar, E. (1985). The Neurobiology of Motivation and Reward. New York:
Springer-Verlag. Stelmack, R. M. (1990). Biological Bases of Extraversion: Psychophysiological Evidence.
Journal of Personality, 58(1), 293-310. Stelmack, R. M. & Geen, R. G. (1992). The psychophysiology of extraversion. In: A. Gale and
M. W. Eysenck, Handbook of individual differences: Biological perspectives,(pp. 227-254). New York: Wiley.
Stern, J. A., Walrath, L. C. & Goldstein, R. (1984). The endogenous eyeblink. Psychophysiology,
21(1), 22-33. Stevens, J. R. (1978). Eye blink and schizophrenia: Psychosis or tardive dyskinesia? American
Journal of Psychiatry, 135(2), 223-226. Steyer, R., Ferring, D. & Schmitt, M. (1992). States and traits in psychological assessment.
European Journal of Psychological Assessment, 8, 79-98. Sved, A. F., Baker, H. A. & Reis, D. J. (1984). Dopamine synthesis in inbred mouse strains
which differ in number of dopamine neurons. Brain Research, 303, 261-266. Sved, A. F., Baker, H. A. & Reis, D. J. (1985). Number of dopamine neurons predicts prolactin
levels in two inbred mouse strains. Experientia, 41, 644-646. Tellegen, A. & Waller, N. G. (1996). Exploring personality through test construction:
Development of the multidimensional personality questionnaire. In: S. Briggs and J. Cheek, Personality measures: Development and evaluation,(pp. Greenwich: Jai Press.

Literaturverzeichnis 100
Thompson, R. F. (1994). Das Gehirn - Von der Nervenzelle zur Verhaltenssteuerung. Heidelberg: Spektrum.
von Cramon, D. (1980). Die spontanen Lidbewegungen: Ein Spiegel psychomotorischer
Aktivität? Medizinische Psychologie, 6(1/2), 123-139. von Gille, H. G., Otto, O. & Ullsperger, G. (1977). Aktivierungsabhängigkeit,
Anforderungsspezifität und Zeitverhalten physiologischer und psychologischer Variablen bei gestuften Leistungsanforderungen im Rahmen des Konzentrations-Leistungs-Tests. Zeitschrift für Psychologie, 185, 347-413.
West, S. G., Finch, J. F. & Curran, P. J. (1995). Structural equation models with nonnormal
variables. In R. H. Hoyle, Structural equation modeling. Concepts, issues, and applications (pp. 56-75). Thousand Oaks: Sage Publications.
Weyers, P., Krebs, H. & Janke, W. (1995). Reliability and construct validity of the German
version of Cloninger´s Tridimensional Personality Questionnaire. Personality and Individual Differences, 19(6), 853-861.
Wise, R. A. (1980). Action of drugs of abuse on brain reward systems. Pharmacology
Biochemistry and Behavior, 13, 213-223. Wise, R. A. (1996). Neurobiology of addiction. Current Opinion in Neurobiology, 6, 243-251. Zuckerman, M. (1979). Sensation-seeking: Beyond the optimal level of arousal. Hillsdayle:
Erlbaum. Zuckerman, M. (1996). Serotonin and Dopamine as Mediators of Sensation Seeking Behavior.
Neuropsychobiology, 34, 155-165.
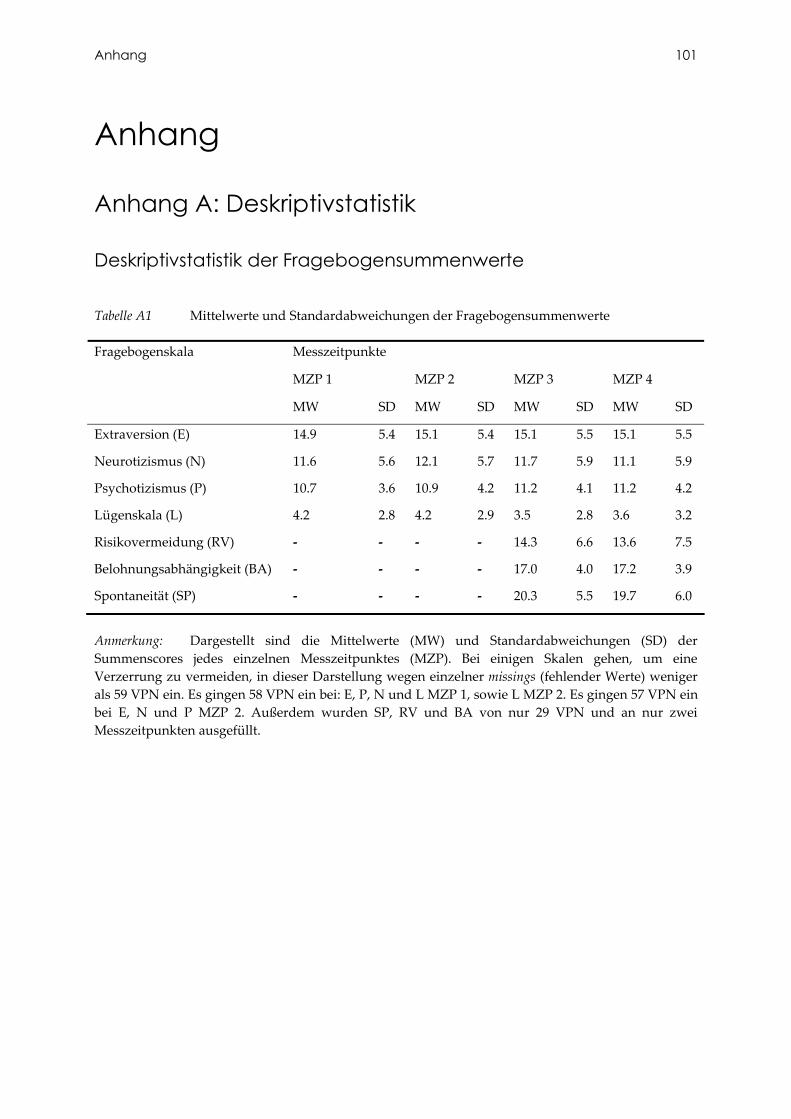
Anhang 101
Anhang
Anhang A: Deskriptivstatistik
Deskriptivstatistik der Fragebogensummenwerte
Tabelle A1 Mittelwerte und Standardabweichungen der Fragebogensummenwerte
Messzeitpunkte
MZP 1 MZP 2 MZP 3 MZP 4
Fragebogenskala
MW SD MW SD MW SD MW SD
Extraversion (E) 14.9 5.4 15.1 5.4 15.1 5.5 15.1 5.5
Neurotizismus (N) 11.6 5.6 12.1 5.7 11.7 5.9 11.1 5.9
Psychotizismus (P) 10.7 3.6 10.9 4.2 11.2 4.1 11.2 4.2
Lügenskala (L) 4.2 2.8 4.2 2.9 3.5 2.8 3.6 3.2
Risikovermeidung (RV) - - - - 14.3 6.6 13.6 7.5
Belohnungsabhängigkeit (BA) - - - - 17.0 4.0 17.2 3.9
Spontaneität (SP) - - - - 20.3 5.5 19.7 6.0
Anmerkung: Dargestellt sind die Mittelwerte (MW) und Standardabweichungen (SD) der Summenscores jedes einzelnen Messzeitpunktes (MZP). Bei einigen Skalen gehen, um eine Verzerrung zu vermeiden, in dieser Darstellung wegen einzelner missings (fehlender Werte) weniger als 59 VPN ein. Es gingen 58 VPN ein bei: E, P, N und L MZP 1, sowie L MZP 2. Es gingen 57 VPN ein bei E, N und P MZP 2. Außerdem wurden SP, RV und BA von nur 29 VPN und an nur zwei Messzeitpunkten ausgefüllt.

Anhang 102
Deskriptivstatistik der Fragebogenmittelwerte
Tabelle A2 Mittelwerte und Standardabweichungen der Itemmittelwertscores
Fragebogenskalen MZP 1-4 MZP 1 MZP 2 MZP 3 MZP 4
MW SD MW SD MW SD MW SD MW SD
Extraversion 0.65 0.23 0.65 0.24 0.66 0.24 0.65 0.24 0.65 0.25
Psychotizismus 0.34 0.11 0.34 0.11 0.34 0.13 0.34 0.13 0.35 0.13
Neurotizismus 0.45 0.21 0.45 0.21 0.47 0.23 0.46 0.24 0.44 0.23
Lügenskala 0.18 0.12 0.19 0.12 0.19 0.13 0.16 0.13 0.16 0.15
Spontaneität 0.58 0.17 - - - - 0.59 0.17 0.56 0.18
Belohnungsabhängigkeit 0.56 0.13 - - - - 0.56 0.14 0.57 0.13
Risikovermeidung 0.41 0.2 - - - - 0.42 0.19 0.4 0.22
Anmerkung: Dargestellt sind die Mittelwerte (MW) und Standardabweichungen (SD) der Itemmittelwertsfragebogenscores der einzelnen Messzeitpunke (MZP) und der über alle vier Messzeitpunkte gemittelten Itemmittelwertscores. Den Skalen Spontaneität, Belohnungsabhängigkeit und Risikovermeidung liegen Daten von 31, den übrigen Skalen Daten von 60 VPn zugrunde. Die Skalen Spontaneität, Belohnungsabhängigkeit und Risikovermeidung wurden außerdem an nur zwei Messzeitpunkten erhoben.
Deskriptivstatistik der Blinkrate
Tabelle A3: Mittelwerte, Standardabweichungen und Spannweiten der Blinkrate
Blinkraten MW SD SPW
MZP 1 14 9,6 2 - 40
MZP 2 14 8,5 3 - 41
MZP 3 15 10,2 3 - 49
MZP 4 15 9,4 1 - 39
MZP1-4 15 8,2 3 - 36
Anmerkung: Dargestellt sind Mittelwerte (MW), Standardabweichungen (SD) und Spannweiten (SPW) der Blinkraten jedes einzelnen Messzeitpunktes (MZP) und der über alle 4 Messzeitpunkte gemittelten Blinkraten (MZP1-4).

Anhang 103
Anhang B: Reliabilitäten
Reliabilitäten der Fragbögen
Tabelle B1: Split-Half-Reliabilität und zeitliche Stabilität (Retest-Reliabiliät) der EPQ-R-Skalen
Skala split-half Retest Aggregation
MZP rodd-even rodd-even-SB MZP 1 MZP 2 MZP 3 r12-34
Extraversion 1 .83 .90 .93
2 .86 .92 .93
3 .83 .90 .79 .80
4 .82 .90 .92 .92 .77
Psychotizismus 1 .54 .70 .81
2 .57 .72 .74
3 .57 .73 .66 .64
4 .50 .66 .80 .75 .75
Neurotizismus 1 .84 .91 .91
2 .83 .91 .75
3 .83 .91 .73 .81
4 .81 .89 .86 .79 .78
Lügenskala 1 .52 .68 .84
2 .50 .67 .64
3 .62 .77 .57 .72
4 .59 .74 .73 .80 .74
Anmerkung: Die Werte rodd-even und rodd-even-SB (Spearman-Brown korrigiert) sind die split-half-Reliabilitäten und geben die Korrelation zwischen den mit der odd-even-Methode gebildeten Testhälften an. Die Retest-Reliabilität ist in Form von Korrelationen zwischen den verschiedenen Messzeitpunkten (MZP) angegeben. Die Werte r12-34 stellen die Stabilität dar, die sich als Korrelation zwischen dem Aggregat der Meßzeitpunkte 1 und 2 und dem Aggregat der Meßzeitpunkte 3 und 4 bestimmen läßt.

Anhang 104
Tabelle B2: Split-Half-Reliabilität und zeitliche Stabilität (Retest-Reliabilität) der TPQ-Skalen
Skala split-half Retest
MZP rodd-even rodd-even-SB MZP 3
Spontaneität 3 .67 .80
4 .79 .88 .92
Belohnungsabhängigkeit 3 .59 .74
4 .61 .75 .80
Risikovermeidung 3 .79 .88
4 .84 .92 .88
Anmerkung: Die Werte rodd-even und rodd-even-SB (Spearman-Brown korrigiert) sind die split-half-Reliabilitäten und geben die Korrelation zwischen den mit der odd-even-Methode gebildeten Testhälften an. In der letzten Spalte wird die Retest-Reliabilität in Form einer Korrelation zwischen den Messzeitpunkten (MZP) angegeben.
Reliabilitäten der Blinkrate
Tabelle B3: Split-Half-Reliabilität und zeitliche Stabilität (Retest-Reliabilität) der Blinkrate
MZP split-half Retest Aggregation
rodd-even rodd-even-SB MZP 1 MZP 2 MZP 3 r12-34
1 .94 .97 .76
2 .85 .92 .61
3 .90 .95 .63 .72
4 .91 .95 .58 .67 .80
Anmerkung: Die Werte rodd-even und rodd-even-SB (Spearman-Brown korrigiert) sind die split-half-Reliabilitäten und geben die Korrelation zwischen den mit der odd-even-Methode gebildeten Testhälften an. Die Retest-Reliabilität ist in Form von Korrelationen zwischen den verschiedenen Messzeitpunkten (MZP) angegeben. Der Wert r12-34 stellt die Stabilität dar, die sich als Korrelation zwischen dem Aggregat der Meßzeitpunkte 1 und 2 und dem Aggregat der Meßzeitpunkte 3 und 4 bestimmen lässt.

Anhang 105
Anhang C: Interkorrelationen der Fragebögen
Tabelle C1: Interkorrelationen der Skalen des EPQ-R mit denen des TPQ
Fragebogenskala E P N L SP RV BA
Extraversion (E)
Psychotizismus (P) .02
Neurotizismus (N) -.32* .04
Lügenskala (L) .18 -.25 -.29*
Spontaneität (SP) .53** .38* -.30 -.02
Risikovermeidung (RV) -.67** .19 .65** -.27 -.37*
Belohnungsabhängigkeit (BA) .42* .02 .08 .22 .12 -.26
Anmerkung: Korrelationen mit einem Stern (*) sind auf dem .05-Niveau signifikant und mit zwei Sternen (**) auf dem .01-Niveau.

Anhang 106
Hiermit versichere ich, dass ich diese Arbeit selbständig verfasst und keine anderen als die
angegebenen Quellen und Hilfsmittel benutzt habe.
Trier, im März 2005