Dissertation Hornickel Korr - elib.tiho-hannover.de · Bereich Histologie und Embryologie...
207
Transcript of Dissertation Hornickel Korr - elib.tiho-hannover.de · Bereich Histologie und Embryologie...



Bibliografische Informationen der Deutschen Bibliothek
Die Deutsche Bibliothek verzeichnet diese Publikation in der Deutschen Nationalbibliografie; Detaillierte bibliografische Daten sind im Internet über http://dnb.ddb.de abrufbar.
1. Auflage 2009
© 2009 by Verlag: Deutsche Veterinärmedizinische Gesellschaft Service GmbH, Gießen Printed in Germany
ISBN 978-3-941703-20-9
Verlag: DVG Service GmbH Friedrichstraße 17
35392 Gießen 0641/24466
[email protected] www.dvg.net

Tierärztliche Hochschule Hannover
Investigations on the innate immunity
of the esophagus epithelium
of domesticated mammals
INAUGURAL-DISSERTATION
zur Erlangung des Grades einer
DOKTORIN DER VETERINÄRMEDIZIN
-Doctor medicinae veterinariae-
(Dr. med. vet.)
Vorgelegt von
Isabelle Nina Hornickel
aus Hagen
Hannover 2009

Wissenschaftliche Betreuung: Univ.-Prof. Dr. rer. nat. habil. Wilfried Meyer
Stiftung Tierärztliche Hochschule Hannover
Anatomisches Institut
Bereich Histologie und Embryologie
Bischofsholer Damm 15
30173 Hannover
1. Gutachter: Univ.-Prof. Dr. rer. nat. habil. Wilfried Meyer
2. Gutachter: Univ. Prof. Dr. med. vet. habil. M. Hewicker-Trautwein
Tag der mündlichen Prüfung: 8. Mai 2009
Diese Arbeit wurde durch die H. Wilhelm Schaumann Stiftung (Hamburg) finanziell
unterstützt.

Für meine Mama
Gott hat uns nicht gegeben den Geist der Furcht, sondern der Kraft und der Liebe und der Besonnenheit.
(Neues Testament, 2. Timotheus 1, 7)


Table of contents
V
Table of contents
TABLE OF CONTENTS............................................................................................. V
ABBREVIATIONS..................................................................................................... IX
FIGURES .................................................................................................................. XI
TABLES.................................................................................................................. XIII
1 INTRODUCTION ..............................................................................................1
2 LITERATURE...................................................................................................3
2.1 Macroscopic anatomy of the esophagus: Overview .............................................. 3
2.2 Histological structure: Overview.............................................................................. 3
2.3 Innate immunity: General aspects ........................................................................... 8
2.4 Antimicrobial peptides: Overview .......................................................................... 15
2.4.1 ß-defensins ............................................................................................................ 18
2.4.2 Cathelicidin ............................................................................................................ 22
2.4.3 Biological activity of antimicrobial peptides............................................................ 24
2.5 Toll-like receptors .................................................................................................... 25
2.6 C-type lectin receptors (ß-glucan receptors) ........................................................ 28
2.6.1 Collectins and ficolins: General overview .............................................................. 28
2.6.2 Mannan-binding lectin (MBL) ................................................................................. 29
2.6.3 Ficolins................................................................................................................... 30
2.6.4 Structure of collectins and ficolins.......................................................................... 31
2.6.5 Functional features of the collectins and ficolins: Activation of the complement
system.................................................................................................................... 32
2.6.6 Pathogene specificity of collectins and ficolins ...................................................... 35
2.6.7 Dectin-1.................................................................................................................. 39
2.7 Lysozyme.................................................................................................................. 41
2.8 Dendritic cells .......................................................................................................... 42

Table of contents
VI
2.9 Questions addressed in this study ........................................................................ 45
3 MATERIAL AND METHODS......................................................................... 46
3.1 Animals ..................................................................................................................... 46
3.2 Sampling ................................................................................................................... 46
3.3 Tissue Fixation ......................................................................................................... 48
3.3.1 Bouin’s solution (after BÖCK 1989) ....................................................................... 48
3.3.2 Calcium acetate-buffered formalin (after LILLIE and FULLMER 1976).................. 48
3.3.3 HOPE® fixation (after OLERT et al. 2001) .............................................................. 48
3.3.4 Fixation after KARNOVSKY (1967)........................................................................ 49
3.3.5 Liquid nitrogen sampling ........................................................................................ 49
3.4 Paraffin embedding.................................................................................................. 49
3.4.1 Paraffin embedding for samples fixed in Bouin’s solution and Ca-acetate buffered
formalin................................................................................................................... 49
3.4.2 Paraffin embedding HOPE® ................................................................................... 50
3.5 Standard EPON embedding for Karnovsky fixed tissue ...................................... 51
3.6 Sectioning................................................................................................................. 51
3.7 Histological staining ................................................................................................ 52
3.7.1 Hematoxylin-Eosin staining of Bouin and formalin-fixed tissue.............................. 52
3.7.1.1 H.E. staining of HOPE® fixed tissue ............................................................... 53
3.7.2 Trichrome staining (Masson-Goldner) (after BÖCK 1989) ..................................... 55
3.8 Microscopical evaluation ........................................................................................ 56
3.8.1 Light microscopy .................................................................................................... 56
3.8.2 Cryo scanning electron microscopy (Cryo-SEM) ................................................... 56
3.9 Immunohistochemistry............................................................................................ 57
3.9.1 Primary antibodies.................................................................................................. 57
3.9.2 Secondary antibodies............................................................................................. 59
3.9.2.1 Two step indirect method with peroxidase linked secondary antibodies ........ 59
3.9.2.2 Use of a labelled streptavidin biotin Complex (LSBC).................................... 61
3.9.2.3 Immunofluorescence (IF) ............................................................................... 62
3.9.3 Immunohistochemistry: General procedure ........................................................... 64

Table of contents
VII
3.9.4 Controls.................................................................................................................. 66
3.9.5 Preliminary tests for the establishment of the primary antibodies.......................... 66
3.9.5.1 Demasking methods tested............................................................................ 67
3.9.6 Antibody dilutions and visualisation systems applied ............................................ 71
3.9.7 Statistical analyses ................................................................................................ 72
4 RESULTS.......................................................................................................73
4.1 Light and electron microscopical observations ................................................... 73
4.1.1 Demonstration of microorganisms with cryo SEM ................................................. 73
4.1.2 General esophageal structure................................................................................ 77
4.2 Fixation experiment: Influences of different fixation media on IHC results....... 81
4.2.1 ß-defensin 2 and -3 reactions ................................................................................ 81
4.2.2 Cathelicidin reactions............................................................................................. 86
4.2.3 Toll-like receptor 2 reactions.................................................................................. 88
4.2.4 Lysozyme reactions ............................................................................................... 91
4.3 IHC results for the substances of the innate immune system: Species
comparison............................................................................................................... 93
4.3.1 Demonstration of ß-defensin 2............................................................................... 93
4.3.2 Demonstration of ß-defensin 3............................................................................... 97
4.3.3 Demonstration of cathelicidin............................................................................... 102
4.3.4 Demonstration of Toll-like receptor 2 ................................................................... 105
4.3.5 Demonstration of ß-glucan receptors................................................................... 108
4.3.6 Demonstration of lysozyme.................................................................................. 109
4.3.7 Demonstration of Langerhans cells ..................................................................... 112
5 DISCUSSION ...............................................................................................113
5.1 Fixation experiment ............................................................................................... 113
5.1.1 Influences of fixation on structure preservation ................................................... 113
5.1.2 Influences of fixation on immunohistochemical results ........................................ 117
5.2 Defence mechanisms of the mammalian esophagus: Species comparison ... 124
5.2.1 Antimicrobial peptides.......................................................................................... 124
5.2.2 Toll-like receptor 2 ............................................................................................... 129
5.2.3 ß-glucan receptors ............................................................................................... 133
5.2.4 Lysozyme............................................................................................................. 137

Table of contents
VIII
5.2.5 Langerhans cells .................................................................................................. 138
5.3 Microbial colonisation of the esophagus ............................................................ 142
5.4 Conclusions............................................................................................................ 145
6 ZUSAMMENFASSUNG............................................................................... 147
7 SUMMARY .................................................................................................. 150
8 REFERENCES............................................................................................. 152

Abbreviations
IX
Abbreviations
AB Antibody
AP Antimicrobial peptides
APC Antigen presenting cell
BC B-cell
BCR B-cell receptor
ß-GR ß glucan receptor
BSA Bovine serum albumin
CD Cluster of differentiation
CLR C-type lectin receptor
CAP Cationic antimicrobial peptide
CAT Cathelicidin
CFU Colony forming units
CRD Carbohydrate recognition domain
CTL C-type lectin receptor
DC Dendritic cell
FBD Fibrinogen binding domain
Fig. Figure
GalNac N-acetylgalactosamin
GlcNac N-acetyl-D-glucosamin
hBD-2 human ß-defensin 2
hBD-3 human ß-defensin 3
IHC Immunohistochemistry
ITAM immunoreceptor tyrosine-based activation-like motiv
LB Lamellar body
LC Langerhans cell
LN2 Liquid nitrogen
LPS Lipopolysaccharide
m monoclonal
MAC Membrane attacking complex
MCG Membrane coating granules
MDC Myeloid dendritic cell
MHC Major histocompatibility complex
mRNA messenger ribonucleic acid

Abbreviations
X
NF-κB Nuclear factor of kappa light polypeptide gene enhancer
NGS Normal goat serum
NHS Normal horse serum
NRS Normal rabbit serum
NK Natural killer cell
p polyclonal
PAMP Pathogen associated molecular pattern
PBS Phosphate buffered saline
PDC Plasmacytoid dendritic cell
PRR Pathogen recognition receptor
RT-PCR Reverse transcriptase polymerase chain reaction
SEM Scanning electron microscopy
SP-A Surfactant protein A
SP-D Surfactant protein D
Tab. Table
TEM Transmission electron microscopy
TGF Transforming growth factor
TLR Toll-like receptor

Figures
XI
Figures
Fig. 2.1 General histological structure of the mammalian esophagus according to the
great three nutrition groups 7
Fig 2.2: Danger model of innate immunity 12
Fig 2.3: Shia-Matsuzaki-Huang model 21
Fig 3.1: Principle of the two step indirect method 61
Fig 4.1: Microorganisms on the esophagus epithelium of the horse 73
Fig. 4.2: Microorganisms on the esophagus epithelium of the goat 74
Fig. 4.4: Microorganisms on the esophagus epithelium of the rat, surrounding
an excretory duct of a secretory gland 75
Fig 4.5: Microorganisms and mucus covering the esophagus epithelium
of the pig 75
Fig. 4.6: Microorganisms and mucus covering the esophagus epithelium of the
dog 76
Fig. 4.7: Bacteria and fungi without mucus covering on the esophagus epithelium
of the cat 76
Fig. 4.8: Histological structure of the esophagus of the herbivorous horse (left)
and the omnivorous dog (right) 78
Fig. 4.9: Histological overview of the esophagus of the carnivorous cat 78
Fig. 4.10: Semithin sections of the esophagus epithelium of the herbivorous
horse (left) and sheep (right) 79
Fig. 4.11: Semithin section of the esophagus epithelium of the omnivorous
mouse 79
Fig. 4.12: Semithin section of the esophagus epithelium of the carnivorous cat 80
Fig. 4.13: Demonstration of fixation differences for hBD-2 in the equine
esophagus 83
Fig. 4.14: Demonstration of fixation differences for hBD-2 in the
ovine esophagus 84
Fig 4.15: Demonstration of fixation differences for TLR2 in the
equine esophagus 90
Fig 4.16: hBD-2 reaction in the esophagus epithelium of the rat 94
Fig. 4.17: IgG rabbit isotype control for hBD-2 for the esophagus of the rat 94
Fig. 4.18: hBD-2 reaction in the esophagus epithelium of the dog 95
Fig. 4.19: Rabbit IgG isotype controle for hDB-2 in the esophagus of the dog 95

Figures
XII
Fig. 4.20: hBD-2 reaction in the esophagus epithelium of the cat 96
Fig 4.21: Differences in staining intensities in Bouin-fixed samples between the
three nutrition groups for hBD-2 97
Fig 4.22: Demonstration of hBD-3 in the esophagus epithelium of the cattle 98
Fig. 4.23: Demonstration of hBD-3 in the ovine esophagus epithelium 99
Fig. 4.24: Negative control for hBD-3 in the ovine esophagus 99
Fig. 4.25: Positive control for hBD-3 100
Fig. 4.26: Demonstration of hBD-3 in the esophagus epithelium of the pig 100
Fig. 4.27: IgG rabbit isotype control for hBD-3 100
Fig 4.28: Differences in staining intensities in Bouin-fixed samples between
the three nutrition groups for hBD-3 101
Fig. 4.29: Demonstration of CAT in the esophagus epithelium of the horse 103
Fig. 4.30: IgG mouse isotype control for CAT 103
Fig. 4.31: CAT reaction in the ovine esophagus epithelium 104
Fig. 4.32: CAT reaction in the esophagus epithelium of the dog 104
Fig. 4.33: CAT reaction in the esophagus of the cat 104
Fig. 4.34: Negative control for CAT 104
Fig. 4.35: Detection of TLR2 in the esophagus epithelium of the horse 106
Fig. 4.36: Detection of TLR2 in the esophagus epithelium of the dog 106
Fig. 4.37: Detection of TLR2 in the esophagus epithelium of the cat 107
Fig. 4.38: Differences in staining intensities in Bouin-fixed samples between the
three nutrition groups for TLR2 108
Fig. 4.39: Lysozyme reaction in the esophagus epithelium of the mouse 110
Fig. 4.40: IgG rabbit Isotype control for lysozyme 110
Fig. 4.41: Differences in staining intensities in Bouin-fixed samples between
the three nutrition groups for lysozyme 111
Fig. 4.42: Demonstration of langerin + LCs in the esophagus epithelium of
the horse 112

Tables
XIII
Tables
Table 2.1: In vitro antimicrobial activity of antimicrobial peptides in animals 23
Tab 2.2: Comparison of animal collectins and ficolins and their antimicrobial
activity 37
Tab. 3.1: Domesticated mammalian species used 47
Table 3.2: Overview fixation steps HOPE® 50
Tab. 3.3: Primary antibodies 58
Tab. 3.4: Overview of secondary detection systems 63
Tab 3.5: General immunohistochemistry protocol, comparison of
fixation techniques 65
Tab. 3.6: Tested antibody dilutions, demasking methods and
visualisation systems for Bouin and formol-fixed material 69
Tab. 3.7: Overview of tested antibody dilutions and visualisation systems
on HOPE® fixed tissue 70
Tab. 3.8: Antibody dilutions applied for established antibodies 71
Tab. 4.1: IHC results of the ß-defensins 2 reactions according to
the different fixation solutions 84
Tab 4.2: IHC results of the ß-defensins 3 reactions according to
the different fixation solutions 85
Tab 4.3: IHC results of the CAT reactions according to
the different fixation solutions 87
Tab 4.4: IHC results for Toll like receptor 2 according to
the different fixation solutions 90
Tab. 4.5: IHC results for lysozyme according to
the different fixation solutions 92

Introduction
1
1 Introduction
The innate immune mechanisms present in the esophagus epithelium of
domesticated mammals have, until now, not been in the focus of any studies. The
esophagus, as the first part of the digestive tract, has often been neglected in
research. In contrast there is a huge amount of information available on innate
defence mechanisms of the following parts of the gastrointestinal tract (STOKES and
WALY 2006, LOTZ et al. 2007, SCHENK and MUELLER 2008, SINGH et al. 2009).
The only studies available on the immune defence mechanisms of the esophagus
have been conducted in humans (AL YASSIN and TONER 1975, HOPWOOD 1995).
We aim to shed some light on innate immune parameters present in the esophagus
of nine domesticated mammalian species. We grouped the animals in order to get a
more differentiated idea on variations in the expression intensity of such defence
mechanisms, comparing influences of different nutrition types. Thus, we studied
representatives from the groups of herbivores, omnivores and carnivores.
The esophagus epithelium is confronted with mechanical strain, chemical food
additives and pathogens during the transfer of food. Such incriminatory influences
may result in a necessity for fast responding innate defence mechanisms. Therefore,
the first step of our study is to demonstrate microorganisms in the esophagus by
relevant microscopical techniques, such as cryo scanning electron microscopy (cryo
SEM). It is a matter of particular interest to detect such microorganisms in order to
answer the question whether innate defence mechanisms are required or not. The
colonisation of the human intestine with commensal bacteria and the resulting
benefits for the organism have been extensively studied (MACKROWIAK 1982,
ZILBERSTEIN et al. 2007, NEISH 2009). On the contrary, clearly less is known about
commensals in the esophagus.
We will focus on several components of the innate immune system, including
pathogen recognition receptors (PRRs), which are able to identify socalled pathogen
associated molecular patterns (PAMPs). One group of PRRs present on epithelial
cells are Toll-like receptors (AKIRA 2003, CARIO et al. 2002, 2007), which have
already been identified in humans and various mammalian species. For another, the
group of ß-glucan receptors, which are soluble PRRs (UEMURA et al. 2002,

Introduction
2
HOLMSKOV et al. 2003, VAN DE WETERING 2004), are of interest. Furthermore,
we intend to detect antimicrobial peptides (APs). APs occur in several animals and
are produced by different cells, e.g. intestinal cells (GANZ and LEHRER 1998,
BEVINS et al. 1999, SELSTED and OULETTE 2005, LINDE et al. 2008).
Additionally, we want to concentrate on the cellular components of innate immunity.
Cells of the innate immune system ingest foreign antigen, process it and present it on
their surface to trigger a further immune response. Such cells are named antigen
presenting cells (APCs), from which we intend to study a special subgroup, the
Langerhans cells (LCs). Such cells bear similarities with dendritic cells (DCs) and
fulfil a surveillance function in different epithelial tissues (CLARK et al. 2000, DE
CARVALHO et al. 2005). All possible parameters of the innate immune system will
be identified by immunohistochemistry (IHC).
Another aim of this study is to compare three different fixation media and their
influence on the preservation quality of structure and antigens in the esophagus
epithelium. Therefore, all samples collected will be fixed in Bouin´s solution, Ca-
Formol and the just recently established HOPE® solution (OLERT et al. 2001).
Influences on the IHC results obtained will be compared and discussed in detail.

Literature
3
2 Literature
2.1 Macroscopic anatomy of the esophagus: Overview
The digestive tract consists of the oral cavity, pharynx, alimentary channel and
accessory organs. According to NICKEL et al. (2004), the alimentary channel can be
divided into three different sections: Foregut, midgut and hindgut. The foregut
comprises the esophagus as well as the stomach, and the midgut includes the small
intestine (intestinum tenue), whereas the hindgut consists of the caecum and the
large intestine with colon and rectum.
The esophagus is a musculo-membranous tube, which extends from the laryngeal
part of the pharynx to the stomach. It passes through the thorax and runs within the
mediastinum. DE NARDI and RIDDELL (1991) classify the human esophagus to
have four parts, whilst NICKEL et al. (2004) divide the mammalian esophagus into
three parts: A cervical, a thoracic and an abdominal portion.
2.2 Histological structure: Overview
The esophagus is a muscular channel that connects the laryngopharynx with the
stomach and transports liquids along with previously masticated food. It shows the
same histological pattern as the other hollow organs of the digestive tract.
According to LIEBICH (2004), EURELL and FRAPPIER (2006) and SAMUELSON
(2007), the esophagus is composed of four tunics, which are named from the inside
out as follows:
1. Tunica mucosa = mucous membrane or mucosa
2. Tela submucosa (better: Lamina submucosa as part of the T. mucosa,
MEYER, personal communication)
3. Tunica muscularis
4. Tunica serosa (Tunica adventitia outside of the coelomic cavity)
The Tunica mucosa can be subdivided into three layers (LIEBICH 2004, EURELL
and FRAPPIER 2006, SAMUELSON 2007).
The inside layer is the Lamina epithelialis. It is composed of a stratified squamous
epithelium, in which the degree of keratinisation intensity varies between the different

Literature
4
species (EURELL and FRAPPIER 2006, SAMUELSON 2007). Only EURELL and
FRAPPIER (2006) describe the epithelium of cats and dogs as “nonkeratinised” and
that of horses, ruminants and pigs as “keratinised” system. SAMUELSON (2007)
takes also the view that the epithelium of the pig is “nonkeratinised”.
The further characterisation of the epithelium depends on the previously mentioned
definitions. The stratified and squamous epithelium can be subdivided into three
layers (stratum basale, stratum spinosum, stratum superficiale or corneum).
SAMUELSON (2007) emphasises that the term “nonkeratinised” is a misnomer, since
some keratinisation can occur in this tissue.
Unfortunately, LIEBICH (2004) and SAMUELSON (2007) divide the keratinised
stratified squamous epithelium into five parts (stratum basale, stratum spinosum,
stratum granulosum, stratum lucidum and stratum corneum). However, MEYER
(1986) calls attention to the fact that the stratum lucidum is a layer which only occurs
in mechanically strained specific body regions (e.g. foot pad), and cannot be
identified in the stratum corneum from the ultrastructural point of view. Due to own
findings, a modern classification of the lamina epithelialis of the esophagus should be
as following:
1. Stratum basale
2. Stratum spinosum
3. Stratum granulosum
4. Stratum corneum (superficiale)
A detailed description of the structure of a typically keratinised stratified squamous
epithelium is given by several authors (MEYER 1986, 2009; EURELL and
FRAPPIER 2006; WELSCH 2006, SAMUELSON 2007).
The basal layer (stratum basale) consists of a single layer of cuboidal to columnar
cells, which develop basal protrusions with hemidesmosomes that improve the
attachment within the basement membrane and to the lamina propia. The nucleus of
the basal cells is located centrally or apically.
The stratum spinosum consists of one to several layers of cells with a central nucleus
and many desmosomal contacts, whereby cell morphology varies between cuboidal

Literature
5
or flat roundish. This structure results in high resistance to stretching. At the same
time the intercellular spaces between the cells and their processes may provide
conduits for substances secreted by the cells.
The stratum granulosum is characterised by the presence of highly basophil, dense
and amorphous material in the cytoplasm, which can be referred to as keratohyalin
granules. Keratohyalin surrounds the cytofilaments and fills out nearly the entire cell
body. The granules mainly consist of profilaggrin, which is the precursor of filaggrin.
This protein plays an important role in keratin filament aggregation during the
process of keratinisation. Due to their lipid components and the abundance of such
granules, it is often difficult to distinguish the nucleus. The oldest cells of the stratum
granulosum undergo a necrobiotic process. The “typical” keratinisation process
begins already in this layer as a combination of necrosis, water loss and the packing
and release of glycolipids containing granules (membrane coat granules). Afterwards
the cells are flattened and organised in one to several layers (lamellae).
Within the stratum corneum, which may consist of 10 to 50 lamellae, the
keratinocytes now are strongly compressed, and, together with desmosomes and
lipid intercellular deposits, create a tight seal. The lower part of the str. corneum is
directly attached to the stratum granulosum and can be named as stratum corneum
conjunctum. Additionally, MEYER (2009) and SAMUELSON (2007) depict the more
loosely structured outermost layer system of the epithelium as stratum corneum
disconjunctum, in which the seal function has become clearly weaker. The following
process of cell abrasion is termed desquamation.
The second layer of the Tunica mucosa is the Lamina propia. It consists of tightly
interwoven collagen and elastic fibres and many free cells (fibroblasts, fibrocytes,
lymphocytes, etc.). The Lamina propia rests on the third layer of the Tunica mucosa,
the Lamina muscularis mucosae. SAMUELSON (2007) mentions that this layer of the
Lamina mucosa is missing in the dog, and EURELL and FRAPPIER (2006) include
pigs into a species group in which the lamina muscularis mucosae is not present.
The Tela submucosa, better Lamina submucosa (of the Tunica mucosa), consists of
a more or less loose connective tissue containing arteries, veins, large lymph
vessels, nerves (Plexus nervorum submucosus) and seromucous glands (EURELL

Literature
6
and FRAPPIER 2006). Along the full length of the esophagus, seromucous glands
are only present in dogs, whereas in pigs they exist primarily along the cranial half of
the esophagus. The Lamina submucosa normally also contains lymphatic cells,
which in the pig can especially be found around the glands. Herbivorous and
carnivorous species show such gland type only at the pharyngeal – esophageal
junction.
The Tunica muscularis is composed of two layers: an inner circular bundle and an
outer longitudinal one (LIEBICH 2004, EURELL and FRAPPIER 2006, SAMUELSON
2007). The distribution of striated and smooth muscle types within this tunic is
variable according to the species groups.
The authors agree that the Tunica muscularis of dogs consists entirely of smooth
muscle. LIEBICH (2004) and EURELL and FRAPPIER (2006) claim that in the horse
striated muscle tissue can be found in the last third, whereas SAMUELSON (2007)
maintains that the transition area of striated and smooth muscle is located more
caudally. Only EURELL and FRAPPIER (2006) and SAMUELSON (2007) mention
that pigs have mixed smooth and striated muscle tissues in the medium third part of
the esophagus. Herbivorous ruminants show the striated muscles system over the
whole length of the organ. In the cat, the same muscle construction seems possible.
The esophagus is surrounded by a Tunica serosa, containing a one-layered
epithelium. In most species, the serosa can only be found in the thorax. EURELL and
FRAPPIER (2006) specify that a Tunica serosa can be found in the abdominal
section of the equine esophagus, which is very long in comparison to that of other
species. Additionally, a Tunica serosa is present at a very small portion of the feline
and canine esophagus. At the cranial part outside the thorax, the esophagus is
covered by a Tunica adventitia.
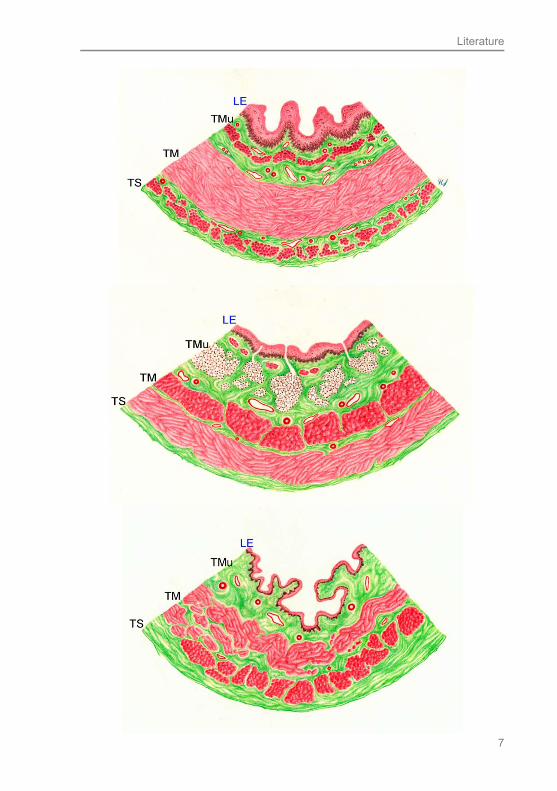
Fig. 2.1: General histological structure of the mammalian esophagus according to the great three nutrition groups Above: horse - herbivorous, central: pig – omnivorous, below: cat – carnivorous; LE = lamina epithelialis, TMu = Tunica mucosa, TM = T. muscularis, TS = T. serosa; drawing by C.I. von Stemm
Literature
7

Literature
8
2.3 Innate immunity: General aspects
There are basically two categories of immunity in mammals: innate immunity and
adaptive immunity.
Innate immunity presents the first line of defence against any pathogenic agent and
is capable of reacting within hours (JANEWAY et al. 2005, TIZARD 2008). The
chemical and cellular immune mechanisms of the innate immune system are evoked,
if an invading microorganism manages to break through the primary physical barrier
of the body (for example the skin).
Innate immunity is based on the fact that microorganisms consist of other
components than the regular cells of the body (TIZARD 2008). JANEWAY (1989)
termed these components on microorganism, which enable the immune system to
recognize them, pathogen-associated molecular patterns (PAMPs). Socalled
“sentinel cells” (macrophages, mast cells and dendritic cells) express receptors on
their surface, which can bind PAMPs on bacteria, fungi, and viruses. These receptors
are called pattern-recognition receptors (PRRs). Different types of PRRs were
developed to identify various microbial molecules. As microorganisms show a wide
variety of structures, cells use PRRs which detect highly conserved molecules found
in a lot of different microorganisms.
A good example of the variety is the composition of cell membranes in bacteria.
Walls of Gram-positive bacteria are mainly made of peptidoglycans and also contain
lipoteichoic acids. Gram-negative bacteria consist of peptidoglycans covered by a
layer of lipopolysaccharide (LPS) and yeasts are covered by a mannan-rich
carbohydrate wall. All these structures present the PAMPs.
The best studied and presumably most important PRRs are the Toll-like receptors
(TLR). TAKEDA et al. (2003), AKIRA (2003) and TIZARD (2008) mention the
existence of 10 different human or mammalian TLRs (TLR1 – TLR10), although
LOTZ et al. (2007) demonstrates 11 TLRs in mice. The various TLRs are capable of
identifying many different ligands of pathogens. For example, TLR2 is able to
recognize peptidoglycan, bacterial lipoprotein, some LPS and necrotic cells, whereas
TLR3 specialised on double-stranded viral DNA. TLRs are expressed by the already
mentioned “sentinel cells”, as well as epithelial cells (KOELLISCH et al. 2005). TLRs
may either be expressed on cell surfaces (TLR2, 4 and, 5) or in cells on endosomal
membranes (TLR3, 7 and, 9) (TIZARD 2008). After TLR activation through

Literature
9
pathogens a signal is passed to the cell triggering the production of numerous
cytokines and antimicrobial peptides (BIRCHLER et al. 2001). Detailed descriptions
about the occurrence and function of TLRs will follow later.
TLRs are of great importance for innate immunity and the recognition of pathogens.
Recently conducted studies indicate that also other, “non-TLR”, PRRs play a pivotal
role in this process. In this context the ß-glucan or C-type lectin-receptor family is
of outstanding interest.
ß-glucans are multibranched glucose polymers. They represent one of the most
important structural cell wall components of bacteria and fungi (KAPTEYN et al.
1995; LIPKE and OVALLE 1998). Structurally, ß-glucans are composed of ß (13)
linkages and additional ß (16) branches. Due to their ability to recognize
pathogens, ß-glucan receptors are members of the PRRs group. Another name for
this receptor group is C-type lectin receptors (CLRs). The CLRs are a large family of
proteins that possess one or more structurally related C-type lectin-like domains in
their carbohydrate recognition domain. The CLRs have been divided into 17 groups
and often mediate fungal binding, uptake and killing (ZELENSKY and GREADY
2005).
One representative of this group of PRRs are the collectins (LU et al. 2002;
HOLMSKOV et al. 2003), which can also be described as soluble patter-recognition
molecules. They form the group III of CLRs (ZELENSKY and GREADY 2005).
Collectins are proteins that recognize certain carbohydrate moieties, and thus belong
to the family of the lectins. Lectins are generally described as proteins capable of
binding carbohydrates (TIZARD 2008). Of this group, mannose-binding lectin (MBL),
surfactant protein A (SP-A), surfactant protein D (SP-D), collectin and conglutinin are
the most important. The collectins belong to the Ca2+-dependent (C-type) lectin
superfamily, and are characterised by the presence of C-type carbohydrate
recognition domain, which enables them to recognize certain sugar moieties on
pathogens (CRD) (WEIS et al. 1998).
Furthermore, another group of PRRs often discussed in the same context as
collectins is the group of the ficolins. Ficolins possess a different type of lectin
domain called the fibrinogen-like binding region (FBG) domain (ICHIJO et al. 1993;
LE et al. 1997). Unlike the collectins, they bind to carbohydrate moieties in a Ca2+

Literature
10
independent manner (LU et al. 2002). Even though the collectins and ficolins bear no
significant similarities in their amino acid sequences, they are often portrayed
together, as they destroy microorganisms via similar effector systems (LU et al. 2002;
HOLMSKOV et al. 2003). Upon recognition of the infectious agent, collectins and
ficolins initiate the lectin pathway of complement activation through attached serine
proteases (MASPs) (HOLMSKOV et al. 2003). A detailed description of this process
will follow later.
Another ß-glucan receptor belonging to group V of the CLRs is Dectin-1. BROWN
and GORDON (2001), HERRE et al. (2004), REID et al. (2004), ZELENSKY and
GREADY (2005), BROWN (2006), KANAZAWA (2007) and TAYLOR et al. (2007)
characterise Dectin-1 as a C-type lectin-like receptor that is capable of recognising
and responding to fungal pathogens, such as Candida species (BROWN and
GORDON 2001; HERRE et al. 2004; REID et al. 2004; BROWN 2006; KANAZAWA
2007; TAYLOR et al. 2007).
One group of effector molecules of the innate immune system are peptides with
antimicrobial activity, thus called antimicrobial peptides (APs). At first scientists
speculated that these peptides were only produced by immune cells, such as
neutrophils and macrophages (GANZ and LEHRER 1994), but very soon studies
demonstrated their production by epithelial cells, for example in the gastrointestinal
or the urinary tract (SCHONWETTER et al. 1995; LEHRER and GANZ 1999). In our
study, we focus on two substances: the defensins and the cathelicidins (CATs), as
these are widely distributed throughout the animal and plant kingdom (GANZ and
LEHRER 1999; LEHRER and GANZ 2002). Antimicrobial peptides are part of the
innate immunity by directly destroying or covering microorganisms (TIZARD 2008).
Moreover, enzymes participate in the first line of defence. An enzyme targeting
microbial structures is lysozyme. It destroys the peptidoglycans of Gram-positive
bacteria by cleaving the bond between N-acetyl glucosamin and N-acetyl muraminic
acid (NIYONSABA and OGAWA 2005; TIZARD 2008).
The body has to cope with several different antigens, which can principally be
allocated to two groups. The first group includes foreign microorganisms, such as
bacteria, that invade the body. Their antigens are called exogenous antigen. In order

Literature
11
to activate cells of the acquired immunity, these antigens need to be processed by
specialised antigen-presenting cells (APCs). One function of the APC is to
incorporate exogenous antigens, process them and then present them on their
surface to lymphocytes (DE CARVALHO 2005).
The second group of antigens that the body has to deal with are endogenous
antigens. These differ from the previously mentioned ones, as they are made in the
body (TIZARD 2008). For example, virus antigen can be found intracellularly, as it
forces the cells to produce new viral proteins, and thus is produced endogenously.
Molecules which present antigen on the cell surface of APCs are called major
histocompatibility complex (MHC). Exogenous and endogenous antigens are
handled via two different types of MHC molecules. The presentation of endogenous
antigen is controlled by MHC I, whereas exogenous antigen is presented by MHC II.
The two MHCs are processed within the APCs in a very different way, and obtain
certain characteristics during this process (JANEWAY et al. 2005, TIZARD 2008).
MHC molecules enable APCs to come into contact with lymphocytes. T-lymphocytes
mainly interact with APCs, whereas B-lymphocytes are usually activated by T-cells.
MHC molecules are not the only receptors expressed on cell surfaces, T- and B- cells
also carry characteristic surface molecules. One unique glycoprotein, which only
exists on T- lymphocytes, is the T-cell receptor (TCR). This immunoglobulin has
similar heterodimer functions as PRRs. Due to additional surface molecules, T cells
can be assigned to two groups: CD4 positive T-lymphocytes and CD8 positive T-
lymphocytes.
CD4+ T-cells fulfil regulatory tasks and are synonymously named T helper cells.
CD8+ T-cells are also called cytotoxic T cells and function by directly killing
pathogenes.
Antigen is presented to the T cell through a MHC molecule. CD4+ cells respond to
MHC II molecules, whereas CD8+ cells only detect the antigen together with a MHC I
molecule (JANEWAY et al. 2005; TIZARD 2008).
The cellular part of innate immunity is mainly conveyed via APCs. APCs are located
in areas with a high density of antigens, thus they are present at epithelial surfaces
and on the mucosa of the alimentary tract and other organs (CLARK et al. 2000; DE
CARVALHO 2005; TIZARD 2008).
Literature
12
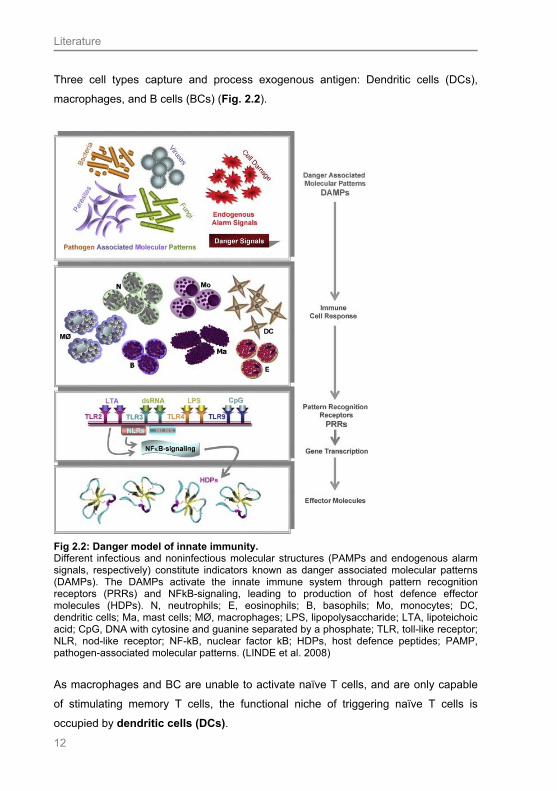
Three cell types capture and process exogenous antigen: Dendritic cells (DCs),
macrophages, and B cells (BCs) (Fig. 2.2).
Fig 2.2: Danger model of innate immunity. Different infectious and noninfectious molecular structures (PAMPs and endogenous alarm signals, respectively) constitute indicators known as danger associated molecular patterns (DAMPs). The DAMPs activate the innate immune system through pattern recognition receptors (PRRs) and NFkB-signaling, leading to production of host defence effector molecules (HDPs). N, neutrophils; E, eosinophils; B, basophils; Mo, monocytes; DC, dendritic cells; Ma, mast cells; MØ, macrophages; LPS, lipopolysaccharide; LTA, lipoteichoic acid; CpG, DNA with cytosine and guanine separated by a phosphate; TLR, toll-like receptor; NLR, nod-like receptor; NF-kB, nuclear factor kB; HDPs, host defence peptides; PAMP, pathogen-associated molecular patterns. (LINDE et al. 2008)
As macrophages and BC are unable to activate naïve T cells, and are only capable
of stimulating memory T cells, the functional niche of triggering naïve T cells is
occupied by dendritic cells (DCs).

Literature
13
The origin of DCs is the bone marrow stem cell and the population of DCs can be
divided into two subpopulations. The myeloid DC (MDCs), which can be found in the
tissues are derived from blood monocytes. In contrast to that, the plasmacytoid DCs
(PDCs) are found in blood and lymphoid organs, and are derived from lymphoid
precursors (CELLA et al. 1997). Furthermore, one can differentiate immature and
mature DCs. Immature DCs are antigen trapping cells which have not interacted with
a pathogen so far (TIZARD 2008). After encountering an antigen, the cells are
activated and mature rapidly or die within days. This activation is triggered by PAMPs
and the TLRs act as PRRs. Hence the DC is now called mature DC, bears an
antigen and migrates to lymph nodes to trigger a T cell response. Upon antigen
presentation via MHCs, expression of co-stimulatory molecules increases at the cell
surface (for example CD86).
In summary, immature DCs are efficient in antigen processing and poor at activation
of naïve T cells, whereas mature DCs loose the capacity to take up proteins but have
the ability to present processed antigen to activated lymphocytes. DCs are the only
cells capable presenting antigen to naïve T cells and triggering their response, whilst
all other APC can only stimulate memory T cells. Therefore DCs occupy a very
unique functional niche in the immune system.
In this study we concentrate on Langerhans cells (LCs), which are immature MDCs
discovered by Paul Langerhans in the year 1868. They have been found in the
epidermis and in the stratified squamous epithelia that line mucous membranes in
humans (AL YASSIN and TONER 1975; DE FRAISSINETTE et al. 1989) and
animals (MEYER 1986) as well as in the esophagus of mice (BOCK 1974; ROWDEN
et al. 1977) and chicken (PEREZ-TORRES et al. 2002).
The hallmark feature of LCs is the presence of typical disc- or cup-shaped
cytoplasmatic organelles termed Birbeck granules (BIRBECK et al. 1961).
LCs remain in an immunological immature state in mucosal tissue and epidermis,
where they can efficiently identify, take up and process antigen, including microbial
antigens. After capturing antigens they emigrate from the epithelial tissue and mature
to professional antigen-presenting cells capable of initiating a primary T-cell mediated
response in the regional lymph node (BANCHEREAU et al. 2000).
As explained above, dendritic cells are characterised by their CD receptors. As we
wanted to detect immature dendritic cells, we tried to identify CD1a positive DCs by

Literature
14
immunohistochemical staining, as various authors (ELDER et al. 1993; CLARK et al.
2000) found out that surface epithelium DCs express CD1a, whilst DCs in non-
epithelial tissue lack CD1a expression. Additionally, we conducted experiments
applying antibodies against langerin, another characteristically cell surface marker of
LCs (ROMANI et al. 2003; MIZUMOTO and TAKASHIMA 2004; NFON et al. 2008).
The presence of LCs in the human esophagus epithelium was reported over 30 years
ago (AL YASSIN 1975; GEBOES et al. 1983; TERRIS and POTET 1995), whereas
later studies could also prove the existence of CD1a positive LCs in the human
esophagus (ZAVALA et al. 2002).
Macrophages can detect microorganisms via TLRs and respond by producing
cytokines (for example interleukins and tumor necrosis factor). Another characteristic
of macrophages is the ability to directly destroy ingested material by lysosomal
proteases and oxidants. Although killing is a helpful ability, it also results in less
antigen being processed and presented on the cell surface by MHCs.
Macrophages are only able to activate sensitised T-cells, as they are unable to
engage in prolonged interactions with T-cells. Thus they cannot activate naïve T-
cells, which have not encountered an antigen yet (JANEWAY et al. 2005; TIZARD
2008).
B-lymphocytes are covered with 200,000 to 500,000 antigen receptors, the socalled
B-cell receptors (BCRs). Each BCR consists of multiple peptide chains and can be
divided into a signalling component and an antigen-binding site. These two
components work closely together, so that the stimulation of a B-lymphocyte results
in the production of antibodies by the cell. Binding of an antigen to a BCR is the
essential first step in triggering a B cell response. However, this is insufficient to
trigger antibody formation, as co-stimulation by T helper cells is necessary. The B cell
captures antigen with the BCR, processes it, and by presenting antigen in MHCs to a
T-cell receives co-stimulation from this cell. This whole process results in the
production of antibodies, which are actually soluble BCRs. Due to the high specificity
of BCRs towards antigens, the BCs play a larger role in the secondary immune
response, when T cells are already sensitised and easier to stimulate.

Literature
15
Consequently B cells have two functions: Responding to antigen by producing
antibodies and acting as professional antigen presenting cell (JANEWAY et al. 2005;
TIZARD 2008).
Even though the esophageal mucosa is the first site after the oral cavity to come into
contact with environmental factors, such as pathogenic microorganisms, chemical
irritants or food additives, there is very little information available about the defence
mechanisms in the esophagus. Most knowledge about the presence of an effective
immunological system stems from studies of the resorptively strongly active intestinal
tract.
In the following chapters, a more detailed review of literature is given concerning the
immune mechanisms which might be involved in the esophagus epithelium.
2.4 Antimicrobial peptides: Overview
Antimicrobial peptides (APs) are some of the most important effector molecules of
innate immunity. Due to their ability to interact with various pathogens in several
ways, they have been the centre of attention for many scientists.
As precise molecular techniques were developed in the past 20 years, the isolation
and identification of different peptides allowed a detailed description of their structure
and functions (GANZ and LEHRER 1994).
Antimicrobial peptides have been isolated from various species. They could not only
be segregated from humans and animals but also from plants (DE FERNANDEZ
CALEYA et al. 1972), fungi and bacteria (GANZ and LEHRER 1999; LEHRER and
GANZ 1999). Numerous authors showed great interest in two types of APs:
Defensins and cathelicidins (CATs). YANG et al. (2002), BROGDEN et al. (2003),
SMET (2005), KELLY et al. (2005); HANCOCK (2006), YANG et al. (2007) and
LINDE et al. (2008) point out that particularly those two groups of peptides are of
growing medical interest, due to their therapeutic potential as endogenous antibiotics
and immune stimulants (YANG et al. 2002). Hence it is hoped that antimicrobial
peptides could be possible alternatives to antibiotics and might be another approach
to diminish the number of antibiotic-resistances. Due to their broad range of
immunomodulatory properties, other fields of application might be found.

Literature
16
Generally APs are small, cationic or anionic and amphipathic peptides, which are
very heterogeneous in length, sequence and structure (SMET 2005). GANZ and
LEHRER (1999) argue that only gene encoded, ribosomally synthesised, polypeptide
antimicrobial substances of mammals with a length of up to 100 amino acid residues
can be defined as APs. Peptides synthesised by fungi and bacteria incorporate
atypical amino-acids and are nonribosomally synthesised.
The Italian Triest Database (at http://www.bbcm.univ.trieste.it/) lists approximately
900 different sequences of APs, which have been identified to this date. The peptides
are put into groups according to similarities in charge, sequence homology, functional
similarity and 3-dimensional structure (NICOLAS and MOR 1995; NISSEN-MEYER
and NES 1997; LEHRER and GANZ 1999). Two categories are distinguished: the
first group are anionic APs and the second one are cationic APs (ELLISON et al.
1985; BROGDEN et al. 2003).
Anionic APs are rich in aspartatic and glutamic acids and have been detected in
cattle, especially in the bronchoalveolar fluid (CAVERLY et al. 2001). They are
effective against Gram-negative and Gram-positive bacteria (BROGDEN et al. 2003),
as they are able to penetrate the outer cell membrane. The peptides attach to
ribosomes and inhibit the ribonuclease activity.
Compared to anionic APs, more attention is paid to cationic APs, especially in
veterinary medical research, as they are very common in domesticated animals.
HANCOCK (2006) classifies cationic AP into four structural groups: α-helical peptides
(for example cathelicidins), ß-sheet peptides with 2-4 disulfid bonds (for example α-
and ß-defensins), loop peptides with one disulfid bridge (for example bactenecin) and
extended peptide structures rich in arginine, glycine, histidine, praline and
tryptophan.
The effect of cationic APs mainly depends on their (tertiary) structure; hence their
structural features are of direct interest (SIMA et al. 2003).
BROGDEN et al. (2003) and TIZARD (2008) depict the defensins as peptides which
are 16-40 residues in size, contain cystein, have two or more disulfide bonds and
form a stabilised ß-sheet structure. α-defensins and ß-defensins are the major
members of this group. They differ in the number of residues (ß-defensins have

Literature
17
slightly more), variation in cystein linkage and in distribution. Whereas α-defensins
have been detected in the azurophil granules in humans and smaller mammalian
species, such as guinea pigs, rabbits, rats and mice (GANZ and LEHRER 1994), ß-
defensins can primarily be found in epithelial cells of humans and a number of
animals. In humans, α-defensin expression in the intestinal tract is highly restricted to
Paneth cells, which are an epithelial cell lineage unique to small intestine. Expression
in other epithelial cell lineages is absent (SELSTED and OUELLETTE 2005). In
contrast to α-defensins, ß-defensins are expressed in enterocytes of the small and
large intestine (O'NEIL et al. 1999). According to AMBATUPUDI and DEANE (2008),
LINDE et al. (2008) and VAN DIJK (2008), ß-defensins are expressed by
domesticated mammals (cattle, goat, sheep, pig, horse, dog, and cat), wild mammals
(e.g. wallabies) and birds.
θ-defensins are the third group of APs and have been identified recently. In contrast
to α- and ß-defensins, which are flat triple-stranded ß sheets, they are double-
stranded circular molecules (BOMAN 2003). As the other defensins are widely
distributed across species, θ-defensins are only traced in granulocytes of Old World
monkeys, such as the rhesus macaque or orang-utans (SELSTED and OUELLETTE
2005).
In cathelicidins (CATs), a determing factor which led to their name is the similarity of
their proregions to cathelin, a 12-kDa protein from porcine leukocytes (ZANETTI et al.
1995; TOMASINSIG and ZANETTI 2005). In a large number of species, CATs could
be detected. Besides in humans, LINDE et al. (2008) sum up that CATs have also
been isolated from the guinea pig, rat, mouse, rabbit (GALLO et al. 1997; IIMURA et
al. 2005), dog, horse (SCOCCHI et al. 1999), sheep (MAHONEY et al. 1995;
HUTTNER et al. 1998), goat (SHAMOVA et al. 1999), pig (ZANETTI et al. 1994) and
birds. Mostly they can be found in the neutrophils of the previously mentioned
animals. Additionally, they have been detected in the testis, inflamed skin (GANZ and
LEHRER 1998), epithelial cells of the colon (HASE et al. 2002) and gingival
epithelium of humans (DALE and FREDERICKS 2005).
The range of microbicidal activity of cathelicidins is highly diverse. They do not only
possess activity against Gram-negative and Gram-positive bacteria (GENNARO and
ZANETTI 2000), but also against fungi (RYAN et al. 1998), and various parasites
(SCHONWETTER et al. 1995).

Literature
18
2.4.1 ß-defensins
ß-defensins were discovered in the early 1990´s as antimicrobial peptides in
epithelial cells of the airways in cattle (DIAMOND et al. 1991). During the last 25
years ß-defensins have also been detected in epithelial surfaces of the skin,
respiratory tract, urogenital tract and gastrointestinal tract. For a long period of time
only two human ß-defensins, ß-defensin-1 and -2 (hBD-1 and -2) were known (GANZ
and LEHRER 1998), however, another two ß-defensins were discovered later on.
Human ß-defensin 3 (hBD-3) has been isolated from human keratinocytes, epithelial
cells of the respiratory tract (HARDER et al. 2001; WEHKAMP et al. 2003) and
recently from the placenta, endometrium, pharynx, and from intestinal epithelial cells
(DHOPLE et al. 2006; SALZMAN et al. 2007; MUKHERJEE et al. 2008). In the
intestinal epithelial cells, the Paneth cells are the major producers of defensins
(DANN and ECKMANN 2007). Moreover, the peptide was identified in stratified
squamous epithelia, such as the epidermis; it was also detected in the epithelial root
sheath of hair follicles and their glands of wild mammals (MEYER et al. 2003;
MEYER and SEEGERS 2004).
In the year 2001, hBD-4 was discovered, and it is proven that it is expressed in
several tissues (testis, gastric antrum, uterus, lung, kidney). hBD-4, -5 and -6 are the
latest three defensins being isolated. hBD-4 and -5 are specifically expressed in
human epididymidis, and hBD-6 was found in Escherichia coli (HUANG et al. 2008).
ß-defensins can either be constitutively expressed or are induced by several
pathogens. SALZMANN et al. (2007) mentioned that in general ß-defensin
expression is inducible at sites of inflammation or infection, whereas DALE and
FREDERICKS (2005) distinguished more precisely between the different groups of ß-
defensins: They pointed out that hBD-1 is constitutively expressed in epithelial cells
in lots of tissues (integument, gut, urinary tract), whereas hBD-2 and -3 is up-
regulated by bacteria and proinflammatory stimuli. A remarkable finding was that
hBD-2 is also expressed in human uninflammed gingival tissue (DALE and
FREDERICKS 2005). The authors suggested that the high level of hBD-2 expression
is a result of the exposure of the tissue to commensal, nonpathogenic bacteria.
Regarding this fact, it can be assumed that this ß-defensin has a normal surveillance
function (DALE and FREDERICKS 2005). The same findings were obtained by

Literature
19
(VORA et al. 2004) for intestinal epithelial cells, as they described hBD-2 production
being induced by activation of TLR through commensal bacteria in the intestine.
Moreover hBD-2 is expressed in human uninflammed skin. It can be localised in the
uppermost layers and the stratum corneum of the epidermis. It has been suggested
that hBD-2 is synthesised and stored in the lamellar bodies of the keratinocytes of
the stratum spinosum and granulosum. Differentiation of the keratinocytes and
barrier disruption leads to release of hBD-2. As a result, hBD-2 can be found in the
intercellular spaces of the stratum corneum (HUH et al. 2002; OREN et al. 2003).
JIA et al. (2000) and GARCIA et al. (2001) independently confirmed or discussed
hBD-3 expression in keratinocytes, as well as in epithelia of the gastrointestinal and
respiratory tract, including the human esophagus epithelium. Additionally it was found
in tonsils, trachea, placenta, adult heart and skeletal muscle.
Due to economic interest, researchers mainly focus on production animals and
studies conducted on companion animals are scarce. LINDE et al. (2008) summed
up the occurrence of ß-defensins in different species.
In cattle, ß-defensin was found in the trachea, lung, tongue, mammary gland and
intestine. ß-defensin has also been detected in the ovine and caprine respiratory and
gastrointestinal tract.
Pigs evoked the interest of scientists due to the fact that the antibiotics, used as
growth promoters in sub-therapeutic levels for many decades, were prohibited in the
European Union in 2006, thus resulting in the necessity for alternative strategies.
VELTHUIZEN et al. (2008) isolated ß-defensin 2 from the porcine intestine and
studied its effect and up-regulation upon infection with different pathogens. They
found out that ß-defensin 2 is expressed in the intestine, and that ß-defensin 2 is
upregulated upon infection with Salmonella typhimurium. Additionally, LINDE et al.
(2008) demonstrated that ß-defensin 2 is also expressed in liver and kidney, though
the peptide is mostly present in the ileum.
Information on innate immune mechanisms in companion animals is rare. Up to now
ß-defensins have only been found in the canine testis (SANG et al. 2005).
As mouse and rat are the most common species used as scientific animal models,
the occurrence of defensins has been intensively studied in these species. Thus,
defensins have been detected in almost every organ (BALS et al. 1999).

Literature
20
Interestingly, the mouse is the only species until now in which expression of ß-
defensin has been reported in the esophagus epithelium (JIA et al. 2000).
ß-defensins interact with pathogens. The microbicidal properties of defensins can be
explained by their ability to interact with a bacterial surface. Membrane disruption
through pore-forming activities is the major mechanism. This mode of action is
described in detail by the Shia-Matsuzaki-Huang model (Fig. 2.3). MATSUZAKI et al.
(1999), YANG et al. (2000) and ZASLOFF et al. (2000) agree on the fact that the
microbicidal activity of defensins is due to their cationic and amphiphilic nature. The
composition of the cell membrane of microorganisms is characterised by an
abundance of negatively charged phospholipids and an absence of cholesterol. This
fact allows defensins to insert themselves into the phospholipid membranes. The
hydrophobic regions are within the lipid membrane and the cationic region is located
on the outside. Thus, a carpet-like structure is formed by defensins on the microbial
surface. Subsequently the contiguity of the microbial membrane is disrupted and
pores are formed. This leads to the complete collapse of the bacterial membrane.
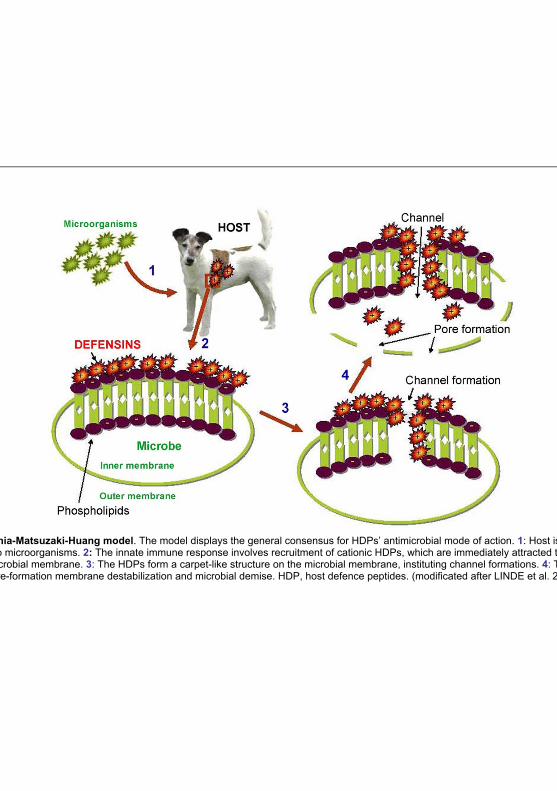
Literature
21
Fig 2.3: Shia-Matsuzaki-Huang model. The model displays the general consensus for HDPs’ antimicrobial mode of action. 1: Host is initially exposed to microorganisms. 2: The innate immune response involves recruitment of cationic HDPs, which are immediately attracted toward the anionic microbial membrane. 3: The HDPs form a carpet-like structure on the microbial membrane, instituting channel formations. 4: The channels lead to pore-formation membrane destabilization and microbial demise. HDP, host defence peptides. (modificated after LINDE et al. 2008)

Literature
22
2.4.2 Cathelicidin
CAT is the second member of the group of antimicrobial peptides, which needs to be
discussed. CATs can be put into the following groups: Peptides with α-helical or
extended helical or loop structure (RAMANATHAN et al. 2002). These groups have
been identified in humans, monkeys, mice, rats, guinea pigs, rabbits, sheep, cattle,
pigs, dogs and horses (RAMANATHAN et al. 2002; BROGDEN et al. 2003; LINDE et
al. 2008). In vitro, the CATs show a wide range of antimicrobial activity against Gram-
positive and Gram-negative bacteria as shown in Tab. 2.1. CATs either kill the
pathogens directly, by disruption of the cytoplasmatic membranes leading to lysis of
the target cell, or bind lipopolysacchrides. The mechanism of killing is similar to that
one of the ß-defensins (compare 2.4.1).
The first product of CAT producing cells is an inactive pro-form. This pro-form is
activated by proteolytic cleavage processes mediated via an elastase in cattle and
pigs (PANYUTICH et al. 1997), and proteinase-3 in humans (SORENSEN et al.
2001).
In horses, cattle, pigs, dogs, rabbits, and mice CAT is synthesised by neutrophils
(MIRGORODSKAYA et al. 1993; SCOCCHI et al. 1999; RAMANATHAN et al. 2002;
SANG et al. 2007). In addition to myeloid derived cells, CAT is expressed in other
cell types. For example, in humans it is present in the testis (MALM et al. 2000) and
in squamous epithelia, as in the oral mucosa and the skin (RAMANATHAN et al.
2002; BRAFF et al. 2005; DALE and FREDERICKS 2005; NIYONSABA et al. 2006).
Concerning the epidermis, it was shown that CAT resides in keratinocytes granules
(MCGs, LBs) of the superficial epidermal layers in humans and mice (BRAFF et al.
2005).
CAT is also present in human airway epithelial cells (BALS et al. 1998). In the human
intestinal tract, CAT was detected in surface epithelial cells of the stomach and
intestine (HASE et al. 2002, 2003). According to the results for the human intestine,
IIMURA et al. (2005) found that CAT is also expressed by murine intestinal cells. The
distribution pattern for CAT in humans was partly corroborated for murine species. In
mice, CAT was constitutively expressed largely in the colon, but was not detected in
the stomach. Only little expression was found in the small intestine. In contrast to
IIMURA et al. (2005), GALLO et al. (1997) detected CAT in the intestine and also in
Literature
23
the stomach of mice. Moreover CAT was found in the testis and spleen of these
animals (GALLO et al. 1997).
With the aim to explore the functional importance of CAT in early intestinal innate
antimicrobial defence, IIMURA et al. (2005) infected CAT knockout mice with a
pathogen bacterium (Citrobacter rodentium). Their results emphasised the
antimicrobial importance of CAT, as it was demonstrated that CAT knockout mice
developed greater surface epithelial cell damage and systemic infection, in
comparison to normal mice.
As described above, the different types of ß-defensins can be either constitutively
expressed or their expression is increased due to microbial pressure. Concerning
CAT, HASE et al. (2002) mentioned its expression in the intestine independent of the
presence of microbiota, although it was modestly upregulated by enteroinvasive
microorganisms. Canine CAT is predominantly expressed in myeloid precursor cells
and to a lower extent in the spleen, liver and testis. Minimal expression was also
found in the kidney, lung and intestinal tissue (SANG et al. 2007).
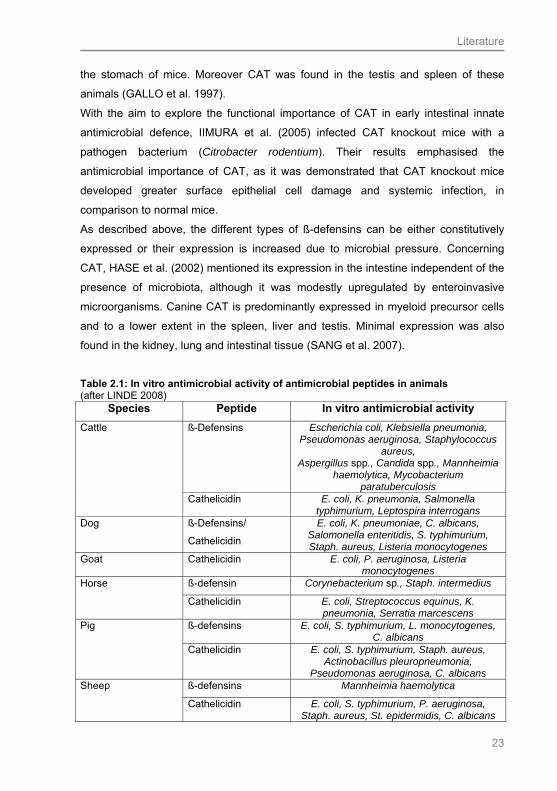
Table 2.1: In vitro antimicrobial activity of antimicrobial peptides in animals (after LINDE 2008)
Species Peptide In vitro antimicrobial activity
ß-Defensins Escherichia coli, Klebsiella pneumonia, Pseudomonas aeruginosa, Staphylococcus
aureus, Aspergillus spp., Candida spp., Mannheimia
haemolytica, Mycobacterium paratuberculosis
Cattle
Cathelicidin E. coli, K. pneumonia, Salmonella typhimurium, Leptospira interrogans
Dog ß-Defensins/
Cathelicidin
E. coli, K. pneumoniae, C. albicans, Salomonella enteritidis, S. typhimurium, Staph. aureus, Listeria monocytogenes
Goat Cathelicidin E. coli, P. aeruginosa, Listeria monocytogenes
ß-defensin Corynebacterium sp., Staph. intermedius Horse
Cathelicidin E. coli, Streptococcus equinus, K. pneumonia, Serratia marcescens
ß-defensins E. coli, S. typhimurium, L. monocytogenes, C. albicans
Pig
Cathelicidin E. coli, S. typhimurium, Staph. aureus, Actinobacillus pleuropneumonia,
Pseudomonas aeruginosa, C. albicans ß-defensins Mannheimia haemolytica Sheep
Cathelicidin E. coli, S. typhimurium, P. aeruginosa, Staph. aureus, St. epidermidis, C. albicans

Literature
24
2.4.3 Biological activity of antimicrobial peptides
Antimicrobial peptides are the frontiers of innate immunity, thus participating in
several modes of action. RAMANATHAN et al. (2002), BROWN (2006) and LINDE et
al. (2008) recapitulate the different functions of APs as further described:
Protective effects of the peptides have been attributed to direct killing of bacteria. The
functional feature of this effect is represented by the Shia-Matsuzaki-Huang model
(Fig. 2.3). However, many of these mechanisms were analysed in vitro, but GANZ
and LEHRER (1998, 1999) state that under physiological salt conditions ß-defensins
have reduced antimicrobial activities. It is likely that the direct antimicrobial effect
occurs mainly on the mucosal epithelia, where concentration of salt is lower.
There is evidence that CAT and ß-defensins show a synergistic effect. NAGAOKA et
al. (2000) found out that in contrast to ß-defensins, CAT is resistant to high salt
conditions. The authors presumed that the various resistances are due to the
different structure of CAT and ß-defensins. They concluded that ß-defensins cannot
function as antimicrobial molecules by themselves. In fact they are working
synergistically with CAT by increasing membrane permeabilisation of target cells.
The second function of APs is their ability to act as immune system modifiers, i.e. as
chemoattractants for immune cells enhancing their function (NIYONSABA et al.
2002). For example, they increase the cytotoxicity of natural killer cells (SCOTT and
HANCOCK 2000). Additionally, they exhibit anti-inflammatory properties by
suppressing bacterial-induced cytokine production and binding lipopolysaccharide
(LPS). As a result, LPS fail to induce tumor necrosis factor production in macrophage
cell lines thereby preventing endotoxemia.
Furthermore, cathelicidins and defensins promote cell proliferation, vasculogenesis
and wound healing (MURPHY et al. 1993; AARBIOU et al. 2004; CHAVAKIS et al.
2004).
APs can even act at the interface of innate and adaptive immunity, as it is supposed
that they support the differentiation of certain cell lineages. For example, BIRAGYN
et al. (2004) demonstrated that ß-defensin 2 simulates dendritic cell maturation and
increases the expression of co-stimulatory molecules (CD40, CD80 and CD86).
Thus, APs can be seen as a connecting link between innate and adaptive immunity.

Literature
25
2.5 Toll-like receptors
Toll-like receptors (TLRs) play a pivotal role in host defence, as they recognize
microbial ligands and are members of the group of PRRs. As already mentioned, 11
mammalian TLRs have been identified. Characteristically they show three common
structural features: a divergent ligand-binding extracellular domain with leucin-rich
repeats, a short trans-membrane region, and a highly homologous cytoplasmatic toll /
interleukin-1 receptor domain, which is essential for the initiation of signalling
cascades (AKIRA 2003).
TLRs have been detected on various cell types. In our study we want to put the focus
on the expression of TLRs on the epithelium of the esophagus. In the literature,
several authors discussed the occurrence of TLRs in a number of organs. Most of the
studies conducted concentrate on TLR expression by human intestinal cells, as many
authors are of the opinion that TLRs play an important role in the pathogenesis of
inflammatory bowel disease (RAKOFF-NAHOUM et al. 2004; VORA et al. 2004;
CARIO and PODOLSKY 2006). Furthermore, the existence of TLRs on human
keratinocytes (KOELLISCH et al. 2005; NAGY et al. 2005; SUMIKAWA et al. 2006;
BUECHAU et al. 2008), the cornea (KUMAR et al. 2006), lung epithelial cells
(BIRCHLER et al. 2001; DROEMANN et al. 2003) and vaginal epithelial cells has
been described (PIVARCSI et al. 2005).
The function of the TLRs as PRRs results in an activation of defence mechanisms of
the cell. As already mentioned previously, one of the first line defence mechanisms is
the production of antimicrobial peptides. Several authors assumed a correlation for
the expression of TLRs and the production of antimicrobial peptides (APs) by the cell
(BIRCHLER et al. 2001; VORA et al. 2004; SUMIKAWA et al. 2006; BUECHAU et al.
2008). BIRCHLER et al. (2001) were the first to mention that TLRs mediate induction
of the synthesis of APs. They found out that human lung epithelial cells constitutively
express TLR2 and produce ß-defensin 2 in response to bacterial lipoprotein.
Subsequently many studies on different epithelial cell types followed.
In the intestine, triggering of TLRs is mainly caused by commensal bacteria, and the
crucial function of the TLRs in the intestine is to provide epithelial homeostasis and
integrity (HOOPER et al. 2001; RAKOFF-NAHOUM et al. 2004).
Not all of the 12 different TLRs seem to be activated under normal steady-state
conditions; moreover TLR2 and TLR4 are in the focus of attention and seem to
contribute to the production of APs (VORA et al. 2004). TLR2 is required for the

Literature
26
recognition of bacterial lipopeptide, and, in combination with TLR6, for the recognition
of peptidoglycan and lipoteichoic acid, which are components of Gram-positive
bacteria (TAKEDA et al. 2003). Additionally, TLR2 is able to detect the yeast cell wall
component zymosan (AKIRA 2003). TLR4 mediates the recognition of
lipopolysaccharides (LPS), found on the outer membrane of Gram-negative bacteria
(AKIRA 2003).
In order to connect the activation of TLRs and the resulting production of APs,
signalling pathways are necessary. Signals from TLRs can be mediated into the cell
via different pathways. The nuclear factor kappa-B (NF-κB) pathway plays the key
role in this manner (VORA et al. 2004). NF-κB is a transcription factor and possesses
an important role in immunity. It is localised in the cell cytoplasm and is hold in its
inactive form through its association with the unphosphorylated IκB proteins, thus
NF-κB cannot move to the nucleus or activate genes. One way to activate the
transcription factor is via triggering TLRs with PAMPs. This results in an alteration of
the shape of the TLR. Subsequently the TLR binds several adaptor molecules, of
which the myeloid differentiation primary response gene 88 (MyD88) is the most
important one; following various kinases are activated. Through their activation in the
last step, the IκB protein is phosphorylated leading to its destruction and the release
of the active NF-κB (O'NEILL 2006).
TLR activation can be a potent stimulus of AP production, but also for the synthesis
of other factors, such as interleukin 6 or tumor necrosis factor. These substances are
involved in cytoprotection and tissue repair in the intestine (RAKOFF-NAHOUM et al.
2004) and other organs, for example in the lung (WARD et al. 2000).
As already mentioned previously, correlation between the occurrence of TLRs on an
epithelium and the resulting production of APs has not only been described for
intestinal epithelium, but also for other organs. The expression of TLR2 and a
resulting production of APs has additionally been observed in human keratinocytes of
the skin (KOELLISCH et al. 2005; NAGY et al. 2005; SUMIKAWA et al. 2006;
BUECHAU et al. 2008), in lung epithelial cells (BIRCHLER et al. 2001), in vaginal
epithelial cells (PIVARCSI et al. 2005) and also in corneal epithelial cells (KUMAR et
al. 2006). Not all APs seem to be triggered by the activation of TLRs, as from most
studies conducted can be concluded that mainly ß-defensin production is
upregulated by the activation of TLR2 or TLR4 (BIRCHLER et al. 2001; VORA et al.

Literature
27
2004; NAGY et al. 2005; KUMAR et al. 2006; SUMIKAWA et al. 2006). Concerning
intestinal epithelial cells, MUKHERJEE et al. (2008) mentioned that hBD-2 is
normally low in cultured epithelial cells and can be induced in response to microbial
influence in a TLR2 dependent manner.
Only for the skin a relationship between TLR2 expression and enhanced cathelicidin
production was shown (BUECHAU et al. 2008).
Most of the observations mentioned beforehand have been obtained from human
epithelial cells, whereas publications about TLRs expression and their functional
relationship to APs for animals are sparse. For the dog, TLR expression has been
reported in different uninfected tissues. Thus, TLR4 was detected in the epithelium of
the lung, small intestine, cornea and renal tubules by performing
immunohistochemistry (WASSEF et al. 2004). mRNA of TLR4 was found in
peripheral blood leukocytes (PBL), in the spleen, stomach and small intestine, and
was moderately expressed in the liver. However, it was not detected in the kidney,
large intestine and skin (ASAHINA et al. 2003). Applying polymerase chain reaction
(PCR), mRNA of TLR2 was found in blood monocytes, lymph nodes, lung, liver,
spleen, bladder, pancreas, small intestine and skin of the dog (ISHII et al. 2006), and
also in canine heart tissue (LINDE et al. 2007). Intriguingly in one study it was
mentioned that a dysregulation of TLR2 and TLR4 may contribute to the
pathogenesis of inflammatory bowel disease (IBD) in dogs, which is characterised by
a chronic inflammation of the small intestine (SWERDLOW et al. 2006).
TLR9 occurrence in canine tissue has been maintained by real-time PCR for lymph
nodes and spleen (HASHIMOTO et al. 2005). The wide spread of TLRs in the dog
might also indicate the occurrence of APs in the mentioned tissues (LINDE et al.
2007).
Descriptions about feline TLRs are made in context with the feline immunodeficiency
virus (FIV), and the authors primarily mention TLR expression on lymphocytes
(IGNACIO et al. 2005). Additionally, mRNA of feline TLR4 was highly expressed in
lung, bladder and peripheral blood monocytes, moderately in kidney, liver, spleen
and large intestine, and with low levels in pancreas and small intestine (ASAHINA et
al. 2003).

Literature
28
Selected bovine and ovine tissues have been tested for the occurrence of various
TLRs. As a result, quantitative real-time PCR confirmed expression of 10 TLRs within
the ovine jejunum, Peyers patches and lymph nodes. Whereas TLRs 3, 5 and 6 were
abundant in the jejunum, all TLRs had been detected in bovine skin; here TLRs 2
and 7 were most abundant (MENZIES and INGHAM 2006).
In cattle, a relationship between ß-defensin 5 production and TLR2 and TLR4
expression in mastitis infected mammary glands has been observed (GOLDAMMER
et al. 2004).
Occurrence of TLR2 in pigs has been verified for epithelial cells lining the
tracheobronchial and intestinal tracts, bile ducts in the liver and renal tubules.
Additionally it was found in the basal cell layer of the epidermis (ALVAREZ et al.
2008). Furthermore, TLR4 mRNA has been detected in tissue samples of the bone
marrow, thymus, lymph node, spleen, brain, liver, kidney and ovary (ALVAREZ et al.
2006).
Information about the expression of TLRs in equine tissue is scarce, although TLR9
was detected in the spleen and lymph nodes (ZHANG et al. 2008). Furthermore,
tissue expression profiles showed that TLR3 was highly expressed in the kidney,
duodenum, spleen and liver, and moderately expressed in bone marrow, lung, and
skin (SANG et al. 2008).
In summary, TLR signalling pathways might be involved in commensal-induced
antimicrobial peptide production, therefore helping to prevent pathogenic bacteria
from break through the epithelial barrier. TLR distribution is consistent with a
surveillance function at entry sites, allowing early detection of microbial invasion.
2.6 C-type lectin receptors (ß-glucan receptors)
2.6.1 Collectins and ficolins: General overview
Collectins and ficolins are pathogen recognition molecules, which bind to
oligosaccharide structures on the surface of microorganisms, subsequently leading to
the killing of bound microorganisms through complement activation and
phagocytosis.

Literature
29
Collectins and ficolins are members of the group of lectins, which are in general
described as carbohydrate-binding proteins. They serve as weapons against
pathogens by being activating by the invaders (FUJITA et al. 2004).
Both groups bear no significant similarity in amino acid sequences. Nevertheless,
they have similar domain organisations and, functionally, both collectins and ficolins,
mediate microbial killing through similar effector systems. This is the reason why both
groups and their representatives are described in one chapter.
2.6.2 Mannan-binding lectin (MBL)
The existence of a molecule with the ability to recognize the yeast cell surface
component was first reported by KAWASAKI et al. (1978). They isolated a molecule
from rabbit liver and named it mannan-binding protein. The human variant of this
molecule is today known as mannan-binding lectin (MBL) or mannose-binding lectin.
MBL is, beside surfactant proteins A and D (SP-A and SP-D), the best studied family
member of the collectins. The greatest amounts of MBL are produced by the liver
and are secreted into the blood (SASTRY and EZEKOWITZ 1993; LU et al. 2002;
WAGNER et al. 2003; VAN DE WETERING et al. 2004). SP-A is mainly synthesized
by alveolar type II cells and unciliated bronchial epithelial cells (Clara cells) in the
lung (MENDELSON et al. 1991; WONG et al. 1996; MADSEN et al. 2003). SP-D was
detected in alveolar type II cells and also the production by Clara cells has been
proven (CROUCH et al. 1992; VOORHOUT et al. 1992). The extrapulmonary
production of both surfactant proteins has been demonstrated in various organs.
Besides epithelial cells of the skin, small intestine, urinary tract, and the collecting
duct of the kidney, even in the esophagus epithelium of human SP-D was detected
by IHC. It was concluded that SP-D is generally present on mucosal surface and is
not restricted to a subset of cells in the lung (MADSEN et al. 2000) . The same is
right for SP-A. PCR on trachea, prostata, pancreas, thymus, salivary glands and the
colon led to positive results, whereas immunohistochemical experiments revealed no
positive results in humans (MADSEN et al. 2003).
Results presented for animals differ from those of humans. SP-A was found in murine
lung, uterus and gastric mucosa (RUBIO et al. 1995; AKIYAMA et al. 2002), in the
porcine Eustachian tube (PAANANEN et al. 1999), in the rabbit middle ear and
maxillar sinus (DUTTON et al. 1999).

Literature
30
In contrast to SP-A and SP-D, the extrahepatic biosynthesis of MBL has not been
reported so frequently for other organs.
In rodents, rabbits, pigs and rhesus monkeys two forms of MBL have been identified,
named as MBL-A, which can be found in serum, and MBL-C, that is detectable in
liver (SASTRY et al. 1991; AGAH et al. 2001). For rodents, RT-PCR showed that the
liver is the major site of expression for both MBL genes. Both were also detected in
the kidney, brain, spleen, and muscle, whereas MBL-A is exclusively expressed in
the testis. MBL-C was solely detected in the small intestine of murine species. These
findings were supported also on the protein level by positive immunohistochemical
reactions (UEMURA et al. 2002; WAGNER et al. 2003). As the non-hepatic MBL-C
expression was highest in the small intestine, the authors concluded that this MBL
type plays a role in the defence against microbial invasion and colonisation in the
intestinal tract. It was also reasoned that MBL might be the counterpart of secretory
IgA of the aquired immune system in the intestine (WAGNER et al. 2003).
In contrast to the above mentioned species, for humans, chimpanzees and chicken
only one form of MBL has been characterised (HANSEN and HOLMSKOV 1998).
Due to the fact that MBL deficiency is one of the most common immune-deficiencies
in humans (TURNER 2003; GUPTA and SUROLIA 2007), the extrahepatic sources
of MBL have been studied extensively in this species; for example mRNA could be
localized in the small intestine and the testis (SEYFARTH et al. 2006).
Also in chicken the occurrence of MBL was corroborated. Likewise it was detected in
extrahepatic organs, for example in the caecal tonsils of healthy chickens (NIELSEN
et al. 1998; LAURSEN and NIELSEN 2000).
2.6.3 Ficolins
Ficolins were originally discovered in the porcine endometrium, where they were
identified as transforming growth factor (TGF)-ß1-binding proteins (ICHIJO et al.
1991). In a broad spectrum of species ficolins were identified, such as in humans
(MATSUSHITA et al. 1996), mouse (FUJIMORI et al. 1998), pig, and hedgehog
(OMORI-SATOH et al. 2000), and the South African clawed toad (Xenopus leavis)
(KAKINUMA et al. 2003).
In humans, three types of ficolins, named H-ficolin, L-ficolin and M-ficolin have
been found. The hepatocytes are the primary source of L-ficolin and H-ficolin
(MATSUSHITA et al. 1996), but the latter substance H is also synthesised by the bile

Literature
31
duct epithelial cells, and in the lung by alveolar type II cells and ciliated bronchial
epithelial cells (AKAIWA et al. 1999).
The third form of ficolins, M-ficolin, is produced by monocytes (HASHIMOTO et al.
1999) and also detectable in the lung (FUJITA et al. 2004). Whereas H-ficolin and L-
ficolin are considered to be serum proteins, this does not apply to M-ficolin. It has
been demonstrated that this lectin is expressed on the surface of peripheral blood
monocytes. These results suggest that M-ficolin plays a role in innate immunity by
acting as a PRR for pathogens on monocytes (FUJITA et al. 2004).
Mice, as well as rats, have two ficolin forms, named ficolin-A and ficolin-B. Ficolin-A
is highly expressed in liver and spleen (FUJIMORI et al. 1998). OHASHI and
ERICKSON (1998) were the first to identify ficolin-B and observed its expression in
bone marrow cells and the spleen. Just recently, ficolin-B was found to be expressed
by peritoneal macrophages (RUNZA et al. 2008).
Also in pigs two forms of ficolins have been identified. They have been named as
porcine ficolin-α and –β (ICHIJO et al. 1993). The expression of ficolin-α mRNA
could be verified for the lung, liver, uterus, spleen and bone marrow (OHASHI and
ERICKSON 1998). Ficolin-β has also be detected in the cattle, whereby tissue
expression was observed in bone marrow and spleen (LILLIE et al. 2005).
2.6.4 Structure of collectins and ficolins
The collectins own their name, due to the fact that their structure is based on
collagenous regions as well as on lectin domains (TURNER 2003). Basically
collectins are built of structural subunits, each composed of three identical
polypeptide chains (timer). The number of trimeric units per collectin molecule varies
among the collectins.
In a single peptide subunit, four structural domains can be distinguished: A N-
terminal cystein rich domain, a collagen domain, a coiled neck domain and a C-type
lectin domain, which represents the carbohydrate recognition domain (CRD)
(TURNER 2003; VAN DE WETERING et al. 2004; GUPTA and SUROLIA 2007).
The ficolins lack the coiled neck domain and possess a different type of lectin
domain, called fibrinogen-like domain (FBG) (LU and LE 1998; FUJITA et al. 2004;
RUNZA et al. 2008). This unique feature resulted in their name “ficolins” (ICHIJO et
al. 1993).

Literature
32
Generally spoken, the structure of collectins and ficolins can be described as follows:
Three polypeptides fold together to form the structural subunit, and three to six of
these subunits join to form a mature protein (FUJITA et al. 2004).
Due to this arrangement, the tertiary structure of MBL and ficolins reminds of a
bouquet of flowers, constructed of octadecamers of six trimeric subunits (LU et al.
2002; HOLMSKOV et al. 2003).
Through the collagen like domain, the collectins and ficolins are associated with
serine proteases (FUJITA et al. 2004; GUPTA and SUROLIA 2007). They are of
great functional importance according the activation of the complement pathway and
the resulting destruction of microorganisms.
2.6.5 Functional features of the collectins and ficolins: Activation of the
complement system
Aiming the elimination of microorganisms, the collectins cooperate with phagocytes
and humoral factors, including the complement.
The complement system is a defence mechanism activated by innate and acquired
immune mechanisms. It consists of serum proteins together with an associated group
of cell membrane proteins. When complement proteins bind to the surface of
microorganisms, enzymatic pathways are activated, and in the next step
microorganisms are destroyed. In healthy animals, these pathways are inactive, but
they can be evoked either by the presence of antibodies on the surface of organisms
or by recognition of the complex carbohydrate moieties on the surface of infectious
agents (TIZARD 2008). The complement system consists of proteins, which are
either labelled numerically with the prefix C (for example C1 or C2) or designated by
letters of the alphabet. The complement proteins are mainly synthesised in the liver
(C3, C6; C8 and B), but some are also made by macrophages (C2, C3, C4, C5, B, D,
and P). A number of the proteins can be found free in the serum, whereas others are
cell-bound.
The complement cascade can be activated through three distinct pathways, which
are named the classical, alternative and lectin pathways.
The activation of the classical pathway requires pre-sensitisation of microorganisms
with antibodies, and is thus part of the acquired immune system. The first component
of this pathway is a multimolecular protein complex called C1, which consists of three

Literature
33
proteins: C1q, C1r and C1s. C1 can be described as a complex of C1q, which is
associated with C1s and C1r. C1q is able to recognize the Fc region of IgG and IgM
immunoglobulins bound to an antigen. Subsequently recognition leads to the
formation of a membrane attack complex (MAC) and microbial lysis. This occurs via
the activation of several complement components and the initiation of a potent
protease (C3 convertase).
In contrast to the classical pathway, both, the alternative and the lectin pathway are
independent of clonal immunity and for that reason parts of innate immunity.
The alternative pathway is triggered when microbial cell walls come into contact
with complement components in the bloodstream. The complement protein C3 plays
a key role within this pathway, as it binds to acceptor groups on many pathogens and
marks them for destruction by immune cells.
C3 spontaneously breaks down into two fragments called C3a and C3b, even under
normal conditions in healthy animals. This process is called tick-over. C3b is
constructed of a highly reactive carbonyl group, which irreversibly binds to the target
cell surface. The target cell is not necessarily a pathogen, such as a bacterium, but
can also be a normal cell of the body. To avoid activation of the alternative pathway
by normal cells, the further activity of C3b is controlled by two factors, named factor H
and I. Factor H interacts with normal cell surfaces and glycoproteins rich in sialic acid
and other neutral or anionic polysaccharides, resulting in the destruction of C3b by
the activation of factor I. Consequently in a healthy individual, factors H and I destroy
C3b as fast as it is generated. Due to the fact that the components of bacterial cell
walls, such as lipopolysaccharides (LPS), lack sialic acid, factor H cannot bind to
C3b. Factor I remains inactive, and the C3b persists. Subsequently the pathogen is
marked by C3b and accessible for recognition by phagocytes, which then destroy the
cell. The process of marking a pathogen, in this case with a complement factor, is
called opsonisation. Furthermore, also the formation of a membrane attack complex
(MAC), which inserts itself into a microbial cell membrane resulting in osmotic lysis of
the pathogen, can be triggered by bound C3b. Binding of C3b leads to activation and
cleaving of several other complement factors, in which factor C5 plays an important
role (JANEWAY et al. 2005; TIZARD 2008).

Literature
34
In our study the lectin pathway is of major interest, as it is activated by MBL and
also by ficolins, which bind to the surface sugar residues on microorganisms (IKEDA
et al. 1987; HOLMSKOV et al. 1994; EPSTEIN et al. 1996; LU 1997; MATSUSHITA
and FUJITA 2001; VAN DE WETERING et al. 2004; PHANEUF et al. 2007).
The discovery that MBL activates complement independently of antibodies was made
in 1987 by IKEDA et al. (1987). This finding was noticed to be a major breakthrough
for the understanding of its role in innate immunity (HOLMSKOV et al. 2003).
The ligands for the collectins are bacterial cell wall components like lipoteichoic acid
of Gram-positive and LPS of Gram-negative bacteria (POLOTSKY et al. 1996;
HOLMSKOV et al. 2003; GUPTA and SUROLIA 2007).
As collectins coat microorganisms, they act as opsonins. Subsequently specific
interaction between the collectins and receptors on phagocytic cells may result in
increased association, uptake and killing of the pathogens (MCNEELY and
COONROD 1994; PIKAAR et al. 1995; OFEK et al. 2001). In case of MBL it either
opsonises microorganisms directly or it can lead to opsonisation through complement
activation and deposit of complement factor C3 (PETERSEN et al. 2001;
HOLMSKOV et al. 2003).
MBL circulates in complex with serine proteases, known as mannan-binding lectin
associated serine proteases (MASPs). Three types of MASPs have been indentified
until now: MASP-1 (MATSUSHITA and FUJITA 1992), MASP-2 (THIEL et al. 1997)
and MASP-3 (DAHL et al. 2001). If MBL binds to the surface of microorganisms,
MASPs are activated to recruit and cleave complement factors. MASP-2 has been
shown to cleave C4 and C2 (THIEL et al. 1997). By the splitting of C4 and C2 a cell-
bound complex called C4bC2b is generated, which is able to activate the C3
convertase. As a result, great amounts of C3b and C3a are created. C3a acts as a
peptide mediator for inflammatory cells, whereas C3b binds the surface of
microorganisms and functions as an opsonin, resulting in cell marking for destruction
through phagocytes. Furthermore, C3b attaches to C5 and the C5 convertase is
generated. It subsequently cleaves C5 into C5a and C5b. C5a also, like C3a, is an
important mediator for inflammatory cells. C5b functions as an activator for the
synthesis of the MAC (JANEWAY et al. 2005; TIZARD 2008). The MASP-1 is
responsible for the direct cleavage of C3 (MATSUSHITA and FUJITA 1995). The
function of MASP-3 still remains unclear, and no relevant substrate for this serine
protease has been identified by now (GÁL et al. 2007).

Literature
35
Also ficolins activate the lectin complement pathway, by association of the molecules
with MASPs (MATSUSHITA et al. 2000; RUNZA et al. 2008).
It needs to be emphasised that the lectin pathway and the classical pathway are not
completely isolated from each other; neither in structure of the activating molecules,
nor in function (FUJITA et al. 2004).
The lectin-MASP complex is structurally and functional equivalent to C1, which is
composed of the serine proteases C1r and C1s and the recognition subcomponent
C1q. This complex represents the first component of the classical complement
pathway.
In the MBL pathway, MASP-2 is the enzyme part, like C1s in the classical pathway,
which cleaves the complement components C4 and C2 to form the C3 convertase
C4bC2b (THIEL 1992; THIEL et al. 1997; FUJITA et al. 2004). Besides, MASP-1 is
capable of cleaving C3 directly (DAHL et al. 2001; ROSSI et al. 2001), with a
resulting activation of the alternative pathway (MATSUSHITA and FUJITA 1995).
By means of structure, C1q and MBL also show great similarity. As already described
previously, MBL is organised in form of a flower bouquet, consisting if six trimeric
subunits. The structure of C1q consists of six globular heads, each connected by a
strand to a central fibril-like region, composed of collagen-like triple-helical structure
(KISHORE and REID 2000).
One difference between MBL and C1q structure results from the fact that MBL is
composed of three identical polypeptide chains, respectively, whereas C1q is
composed of three different chains (FUJITA et al. 2004)
However, it still can be resumed that the overall structure of C1q is similar to that of
MBL and ficolins (HOLMSKOV et al. 2003; VAN DE WETERING et al. 2004).
Furthermore, the structure of MASP family members associated with MBL and
ficolins resembles that of C1r and C1s, as they both consist of six domains,
composed of proteins and proteases (FUJITA et al. 2004).
2.6.6 Pathogene specificity of collectins and ficolins
As already mentioned previously, collectins and ficolins bind to carbohydrates with
their lectin domain. The lectin domain of MBL is the CRB, respectively named FBG in
ficolins.
With its CRD, MBL binds carbohydrates with 3- and 4-hydroxyl groups in the
pyranose ring in the presence of calcium. Thus MBL ligands for MBL are mannose,

Literature
36
glucose, L-fucose, N-acetyl-mannosamine and N-acetylglucosamine (LU et al. 2002;
HOLMSKOV et al. 2003; FUJITA et al. 2004). With this wide range of ligands, MBL is
able to recognize various types of bacteria.
Most carbohydrate structures in animals are terminated by sugars not recognized by
the collectins, e.g., galactose or sialic acid. Mammalian cells do not present the
pathogen-associated molecular patterns characteristically for microorganisms
(HOLMSKOV et al. 2003).
In contrast to collectins, ficolins do not show such a wide variety of sugar binding
capacity. Ficolins mainly bind to N-acetyl-D-glucosamine (GlcNAc) and N-
acetylgalactosamine (GalNAc) (LE et al. 1997, 1998; LU et al. 2002; FUJITA et al.
2004). For humans it has been shown that L-ficolin recognizes cell wall components
of Salmonella typhimurium.
Binding capacities of ficolins and collectins have been discussed controversial
(POLOTSKY et al. 1996; LYNCH et al. 2004). The latter authors observed that L-
ficolin, but not MBL or H-ficolin, specifically binds to lipoteichoic acid, common for
Gram-positive bacterial cell walls. In contrast to this opinion, POLOTZKY et al. (1996)
argues that also MBL shows binding capacity for lipoteichoic acid of Gram-positive
bacteria.
In the literature, various descriptions of the recognition capacity of MBL for different
microorganisms can be found. Generally spoken, common microbial structures
recognized by the collectins are lipopolysaccharides (LPS) and mannan-like high
mannose structures. Due to the fact LPS is a major glycolipid component on the
outer membrane of Gram-negative bacteria, these are ideal ligands for collectins (LU
et al. 2002; HOLMSKOV et al. 2003; VAN DE WETERING et al. 2004; GUPTA and
SUROLIA 2007). Moreover, MBL has been shown to interact with a wide variety of
Gram-positive bacteria, and various types of their cell wall component lipoteichoic
acid (LTC) (POLOTSKY et al. 1996). LU et al (2002) sums up that the wide range of
bacteria recognized by collectins highlights the importance of collectins in innate
immunity.
Even though binding of collectins and ficolins to bacteria has been reviewed
intensively for humans (HOLMSKOV et al. 2003; VAN DE WETERING et al. 2004;
GUPTA and SUROLIA 2007), information about their role in innate immunity of
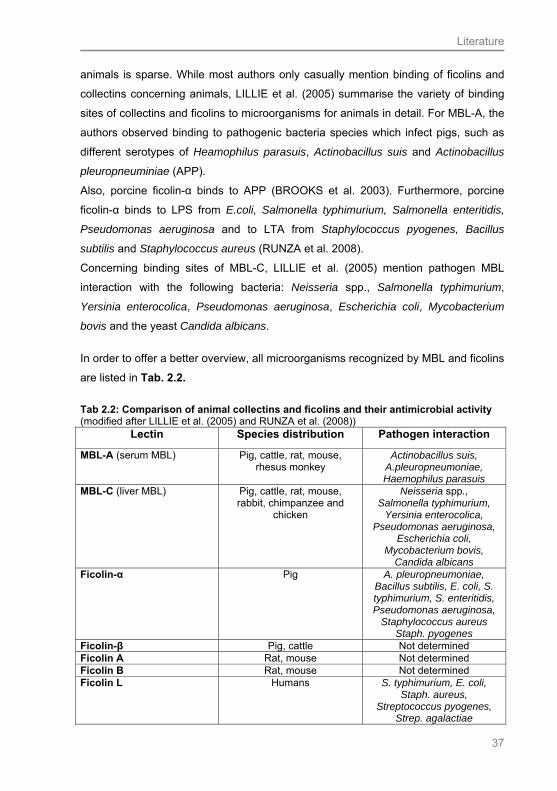
Literature
37
animals is sparse. While most authors only casually mention binding of ficolins and
collectins concerning animals, LILLIE et al. (2005) summarise the variety of binding
sites of collectins and ficolins to microorganisms for animals in detail. For MBL-A, the
authors observed binding to pathogenic bacteria species which infect pigs, such as
different serotypes of Heamophilus parasuis, Actinobacillus suis and Actinobacillus
pleuropneuminiae (APP).
Also, porcine ficolin-α binds to APP (BROOKS et al. 2003). Furthermore, porcine
ficolin-α binds to LPS from E.coli, Salmonella typhimurium, Salmonella enteritidis,
Pseudomonas aeruginosa and to LTA from Staphylococcus pyogenes, Bacillus
subtilis and Staphylococcus aureus (RUNZA et al. 2008).
Concerning binding sites of MBL-C, LILLIE et al. (2005) mention pathogen MBL
interaction with the following bacteria: Neisseria spp., Salmonella typhimurium,
Yersinia enterocolica, Pseudomonas aeruginosa, Escherichia coli, Mycobacterium
bovis and the yeast Candida albicans.
In order to offer a better overview, all microorganisms recognized by MBL and ficolins
are listed in Tab. 2.2.
Tab 2.2: Comparison of animal collectins and ficolins and their antimicrobial activity (modified after LILLIE et al. (2005) and RUNZA et al. (2008))
Lectin Species distribution Pathogen interaction
MBL-A (serum MBL) Pig, cattle, rat, mouse, rhesus monkey
Actinobacillus suis, A.pleuropneumoniae, Haemophilus parasuis
MBL-C (liver MBL) Pig, cattle, rat, mouse, rabbit, chimpanzee and
chicken
Neisseria spp., Salmonella typhimurium,
Yersinia enterocolica, Pseudomonas aeruginosa,
Escherichia coli, Mycobacterium bovis,
Candida albicans Ficolin-α Pig A. pleuropneumoniae,
Bacillus subtilis, E. coli, S. typhimurium, S. enteritidis, Pseudomonas aeruginosa,
Staphylococcus aureus Staph. pyogenes
Ficolin-β Pig, cattle Not determined Ficolin A Rat, mouse Not determined Ficolin B Rat, mouse Not determined Ficolin L Humans S. typhimurium, E. coli,
Staph. aureus, Streptococcus pyogenes,
Strep. agalactiae

Literature
38
For animals, there is still little known about the roles of the several collectins and
ficolins in viral infections. Concerning MBL, no information was available up to now;
only for SP-A and SP-D increased expression has been observed in lambs infected
with parainfluenza virus type 3 (GRUBOR et al. 2004).
In contrast to animals, detailed descriptions of target pathogens, beside bacteria, are
available from several studies for humans.
Thus not only bacteria were recognized by MBL in humans, but also fungi. Especially
the noncapsulated form of Cryptococcus neoformans (SCHELENZ et al. 1995) and
Candida albicans (NETH et al. 2000) were mentioned in this connection.
Regarding infections with viruses, it was an outstanding finding that MBL binds to the
human immunodeficiency virus (HIV) (HAURUM et al. 1993), and as well to influenza
virus (READING et al. 1993). Other collectins, such as SP-A and SP-D, detect
herpes simplex virus and the non-enveloped rotavirus (HOLMSKOV et al. 2003).
Concerning infections with parasites, binding of MBL to Leishmania species in
humans was observed (GREEN et al. 1994). Leishmania is an intracellular parasite,
mainly infecting macrophages, and in case of this infection it is considered that
binding of MBL and covering with complement factors provides no protective effect,
but in fact stimulates the uptake of the parasite via complement receptors on the
surface of macrophages (SANTOS et al. 2001).
In contrast to MBL, for ficolin no other binding sites, except those for bacteria, have
been described in the literature.
The general function of collectins, thus, is to protect the host by recognising PAMPs
on microorganisms and eliciting appropriate responses by activation of multiple
processes of innate immunity (HOLMSKOV et al. 2003; VAN DE WETERING et al.
2004).

Literature
39
2.6.7 Dectin-1
Dectin-1 is a C-type lectin receptor (CTL) and a member of the larger family of
pathogen recognition receptors (HERRE et al. 2004; GANTNER et al. 2005;
TAYLOR et al. 2007). The C-type lectins recognize sugar ligands through the
carbohydrate recognition domain (CRD) in a Ca2+ dependent manner. Dectin-1 is the
most recently characterised ß-glucan receptor as it was identified in humans in 2001
(BROWN and GORDON 2001). In the animal kingdom it was first identified in mice
(RYAN et al. 2002).
Human Dectin-1 differs from the murine receptor, as in humans two major isoforms
exist (WILLMENT et al. 2001; GRUNEBACH et al. 2002). The isoforms are named ß-
glucan receptor-A (ßGR-A) and ß-glucan receptor-B (ßGR-B). Concerning the
structure, they differ by the presence and absence of a stalk region, but functionally
both recognize ß-glucans of fungi (WILLMENT et al. 2001).
Dectin-1 is expressed on a variety of human leukocytes including monocytes,
macrophages, eosinophils, neutrophils and NK cells (STEELE et al. 2003;
GANTNER et al. 2005); DCs as well express Dectin-1. For our study, the fact that
Dectin-1 was detected on Langerhans cells of the human epidermis is of outstanding
interest (ARIIZUMI et al. 2000; YOKOTA et al. 2001; GAVINO et al. 2005), as the
esophagus epithelium bears similarity to the epidermis.
The receptor is also expressed on B cells and a subpopulation of T cells. WILLMENT
et al. (2005) conclude that this cell expression demonstrates that human Dectin- 1 is
not myeloid restricted. In mice, Dectin-1 is detectable on macrophages (WILLMENT
et al. 2005); REID et al. (2004) found out that in this species Dectin-1 is also
expressed on neutrophils, macrophages in the splenic red and white pulp, alveolar
macrophages, Kupffer cells of the liver, macrophages and on DCs in the lamina
propia of gut villi. The authors concluded that this expression pattern is consistent
with its role in pathogen surveillance. Supporting the results in humans, also in the
murine epidermis Dectin-1 positive LCs were detected.
In bovine breeds, mRNA transcripts of Dectin-1 were demonstrated in bone-marrow
cells, monocytes, macrophages, NK-cells and DCs (WILLCOCKS et al. 2006). Also in
sheep, Dectin-1 mRNA was found, as in the spleen, lung, kidney and skin
(NALUBAMBA et al. 2007).

Literature
40
As already mentioned previously, Dectin-1 is a C-type lectin receptor. A characteristic
for this family is the extracellular carbohydrate-recognition domain (CRD). The
cytoplasmatic part of the Dectin-1 receptor contains an immunoreceptor tyrosine-
based activation-like (ITAM-like) motif (BROWN and GORDON 2001; BROWN
2006). Between the extracellular CRD and the cytoplasmatic ITAM a stalk
transmembrane region is located. Functionally the ITAM-region is of great interest.
The ITAM can be phosphorylated and activated by ß-glucan binding in the CRD.
Through the phosphorylation of the cytoplasmatic ITAM motif, Dectin-1 is able to
mediate intracellular signalling, which results in a variety of cellular responses. For
example, the production of several cytokines and chemokines (TNF, IL2, IL6, IL10,
IL-12 and IL23) is triggered and the respiratory burst system is activated (GANTNER
et al. 2003; BROWN 2006; WILLMENT and BROWN 2008).
There is evidence that Dectin-1 collaborates with TLR-2 in recognising
microorganisms. GANTNER et al. (2003) describe how co-expression of Dectin-1
and TLR-2 results in an increasing production of TNF and IL-12. Both receptors
activate the NF-κB signalling way and mediate the production of TNF and IL-12
(DENNEHY and BROWN 2007).
Dectin-1 is capable to recognize fungal cell wall components (BROWN and
GORDON 2001; HERRE et al. 2004; DENNEHY and BROWN 2007; KANAZAWA
2007; TAYLOR et al. 2007). In vitro analysis showed that Dectin-1 possesses the
ability of identifying a wide variety of fungal species, including Aspergillus, Candida,
Coccidioides, Pneumocystis and Saccharomyces (GANTNER et al. 2005; STEELE et
al. 2005; BROWN 2006).
The important role of Dectin-1 in fungal infections in vivo is underlined by the results
of recent studies. All experiments mentioned in the following were accomplished with
mice. STEELE et al. (2005) found out that blockage of Dectin-1 during intratracheal
infection with Aspergillus fumigatus reduced lung inflammatory responses
significantly and increased fungal burdens. Via infecting a Dectin-1 knock-out mice
with Candida albicans, it was proven that lack of Dectin-1 results in an enhanced
susceptibility to systemic Candida albicans infection (TAYLOR et al. 2007).

Literature
41
2.7 Lysozyme
Lysozyme is a peptidoglycan N-acetylmuramoylhydrolase. It is a member of the class
of enzymes which mainly destroys the cell wall of Gram-positive bacteria. In this
connection, lysozyme is capable to cleave the bond between N-acetylglucosamine
and N-acetylmuramic acid of the peptidoglycan in the bacterial cell wall.
NIYONSABA and OGAWA (2005) explain why Gram-positive but not Gram-negative
bacteria are preferred targets of lysozyme: The natural substrate of lysozyme is
peptidoglycan. Gram-positive bacteria are composed of up to 40 layers of
peptidoglycan, whereas Gram-negative only have a single layer of peptidoglycan
surrounded by an outer membrane (MASSCHALCK et al. 2002). As a result, Gram-
negative bacteria are protected by their specific outer membrane, which acts as
barrier preventing contact with lysozyme.
The major cellular sources of lysozyme are neutrophilic granulocytes and
macrophages, where it is detectable in the lysosomes (DUSZYK 2001). Furthermore,
lysozyme is present in various tissues and secretion fluids of glands. So it was
detected in the skin, epithelial cells of the trachea and in Paneth cells of the small
intestine of humans (DUSZYK 2001; NIYONSABA and OGAWA 2005; WEHKAMP et
al. 2006). Concerning expression in Paneth cells, the highest amounts of lysozyme
were found in the duodenum, where it was also expressed by the Brunner`s glands.
In the jejunum and ileum, lysozyme appeared only in smaller amounts (WEHKAMP et
al. 2006).
Analyses of the distribution pattern of lysozyme in the human epidermis revealed that
it is expressed much more in the upper than the basal cells layers (NIYONSABA and
OGAWA 2005). Lysozyme was also identified in the skin of different wild mammals,
including dolphins. Here it was mostly found in cells and intercellular spaces of the
stratum spinosum and str. corneum (MEYER et al. 2003; MEYER and SEEGERS
2004).
Interestingly, lysozyme was also detected in human esophageal submucosal glands
(HOPWOOD et al. 1986). The authors described positive immunohistochemical
reactions for the substance in the duct epithelium cells.
Secretions containing lysozyme are saliva, tears, or urine, and secretions of cervical
glands (REITAMO et al. 1977; NIYONSABA and OGAWA 2005)

Literature
42
2.8 Dendritic cells
As mentioned previously, the ability of DCs to integrate a variety of environmental
signals, like the conflict with microorganisms, either commensal or pathogenic,
together with their ability to respond with an immunological reaction, makes them
cells of particular relevance and importance in the mucosal environment.
Mainly two ways of responding towards an environmental signal are possible in the
mucosal tissue: On one hand, an active defensive response to mucosal pathogens is
feasible, and on the other hand, a “tolerant” response to harmless environmental
agents, such as commensal bacteria and food-derived antigens, is the opposite type
of reaction (HAVERSON and RIFFAULT 2005).
Antigen presentation via human LCs is possible due to their expression of MHC class
II molecules, and upon the expression of CD1 molecules and langerin (SALAMERO
et al. 2001; PENA-CRUZ et al. 2003; HUNGER et al. 2004; NFON et al. 2008).
Beside MHC class molecules, which present peptide antigens towards T-cells, also
the family of the more recently discovered CD1 molecules initialises a T-cell receptor
(TCR) activation. This results in a T-cell defence reaction against other, non-peptide
lipid and glycolipid antigens, which are presented via CD1 molecules on APCs
(PORCELLI and FRANK 1995; SUGITA et al. 2000; PENA-CRUZ et al. 2003;
HUNGER et al. 2004; MOODY et al. 2004; SUGITA et al. 2004; BARRAL and
BRENNER 2007; MORI and DE LIBERO 2008). The CD1 molecules are a lineage of
cell-surface glycoproteins related in structure and evolutionary origin to the MHC-
encoded antigen-presenting molecules (KNOWLES and BODMER 1982; CALABI et
al. 1989).
Due to sequence comparisons, CD1 proteins were classified into two separate
groups over decades: human CD1a, -b and -c form group 1, and the significant more
divergent CD1d defines group 2 (PORCELLI and FRANK 1995). More recently
conducted studies implicate the existence of a third group of CD1 molecules, which is
named CD1e. This molecule has low homology with the other CD1 proteins, and thus
is not included in CD1 group 1 or 2. A supplementary difference to the other CD1
molecules, which are expressed on the cell-surface, is the fact that CD1e is
expressed in DC without reaching the plasma membrane (ANGENIEUX et al. 2000).
CD1 molecules belonging to group 1 mainly present microbe-derived lipid antigens to
CD4 and CD8 positive T cells to clear microbial infection, whereas CD1d activates a

Literature
43
specialised T cell subset known as natural killer cells (BENDELAC et al. 1995;
ROSAT et al. 1999; PENA-CRUZ et al. 2003; HUNGER et al. 2004; MIZUMOTO and
TAKASHIMA 2004).
Although CD1 molecules show homology to MHC molecules, their atomic structures
of the antigen-binding sites and their routes of intracellular trafficking differ from the
ones described for MHC molecules (SUGITA et al. 2004; VAN DEN ELZEN et al.
2005; VAN RHIJN et al. 2005; MORI and DE LIBERO 2008).
The epitopes for CD1a, CD1b and CD1c are located on various cells, as they can be
detected on myeloid dendritic cells, LC and B cells. CD1c is expressed on 20-50% of
the B cells from blood, tonsil and spleen (SMALL et al. 1987; TIZARD 2008) and,
together with CD1b, on dendritic cells in a variety of tissues including the skin, liver,
kidney, lung, and lymphoid organs in humans (PORCELLI and FRANK 1995;
JANEWAY et al. 2005).
CD1a is the specific marker molecule for LCs in the skin, which express this molecule
in great amounts (ELDER et al. 1993; CLARK et al. 2000; PENA-CRUZ et al. 2003;
MIZUMOTO and TAKASHIMA 2004; HUSSEIN 2008). Other authors also allude the
occurrence of CD1a positive DCs in the epithelium of different tissues, such as the
bronchus (TAZI et al. 1993), conjunctiva (YOSHIDA et al. 1997), gingiva
(CRAWFORD et al. 1989), vagina and cervix (MILLER et al. 1992). This tissue
distribution might be due to the fact that antigen-presentation via CD1a plays a
pivotal role at sites constantly exposed to external pathogens (PENA-CRUZ et al.
2003).
Beside Birbeck granules, detectable in LCs and CD1a, expressed as surface
molecule on LCs, langerin (CD207), a C-type lectin, is another characteristical
molecule on LCs (VALLADEAU et al. 2000; ROMANI et al. 2003; MIZUMOTO and
TAKASHIMA 2004), whereby the three molecules show a functional relationship
(VALLADEAU et al. 1999, 2000). The C-type lectin langerin, exclusively expressed
by LCs, recognizes mannose residues via its single carbohydrate recognition domain
(CRD) and captures glycolipid antigens. The complex of glycolipids and langerin is
internalised and delivered to the Birbeck granules inside the LC. In the Birbeck
granules also CD1a, which has been internalised from the cell surface, can be
detected. The interrelation between CD1a and langerin can be explained by the fact,
that CD1a is coupled with lipid moieties, collected by langerin on the cell surface

Literature
44
inside the Birbeck granules. Finally, the loaded CD1a molecule is transported back to
the cell surface to present lipid antigen to T cells (VALLADEAU et al. 2000).
In the literature various statements about the occurrence of DCs and LCs in animals
can be found. Most studies mention LC appearance in the skin (ROMANI et al.
2003). Just recently a study showed the existence of langerin positive LCs in the
porcine epidermis (NFON et al. 2008), corroborating former information (MEYER
1986). In mice it is notable that langerin is also expressed by non-cutaneous, non-
Langerhans cell derived dendritic cells in lymphoid tissue and the lung (DOUILLARD
et al. 2005; SUNG et al. 2006).
Most other reports about the occurrence of LCs in animals concentrate on the
detection of CD1a positive (CD1a +) LCs. For dogs expression of CD1a + LCs in skin
was just recently maintained (VAN BEECK et al. 2008); also in cats, CD1a + LCs
were detected (MARCHAL et al. 1997).
Description of LC cell-like dendritic cells in the intestinal tract of animals, including the
esophagus, can be found very rarely. Only one study confirmed the presence of LCs
in the esophagus of mice, verified by ultrastructural observations (ROWDEN et al.
1977). Furthermore, Langerhans cell-like dendritic cells have been identified at the
mucosal site of the chicken esophagus (PEREZ-TORRES et al. 2002).

Literature
45
2.9 Questions addressed in this study
1. Is the esophagus epithelium of domesticated mammals colonised by
microorganisms?
2. Do any mechanisms of innate immunity exist in the esophagus epithelium of
domesticated mammals?
3. Are the APs, hBD-2, -3 and CAT, produced by the epithelium?
4. Is TLR2 expressed on epithelial cells? Can a co-expression with APs be
demonstrated?
5. Is it possible to detect L-ficolin, MBL and Dectin-1 in the esophagus
epithelium?
6. Is the enzyme lysozyme produced by epithelial cells?
7. Are CD1a(+) and/or langerin(+) LCs detectable in the esophagus of
domesticated mammals?
8. Where can such innate defence mechanisms be identified in the esophagus
epithelium?
9. Do differences exist in the expression intensity of innate defence mechanisms
in herbivorous, omnivorous and carnivorous species?
10. Are structure preservation and IHC results influenced by the three different
fixation media (Bouin`s solution, Ca-Formol, HOPE®) applied?

Material and Methods
46
3 Material and Methods
3.1 Animals
Esophagus samples were collected from 10 domesticated mammalian species. The
three great nutrition groups were considered: Omnivores (pig, dog, mouse, rat),
carnivores (cat) and herbivores (goat, sheep, rabbit, cattle, horse).
One group of samples had already been taken during earlier studies on the
oesophagus epithelium of the mentioned mammals, and was available fixed and
embedded. These samples had been obtained from the Institute for Pharmacology,
Toxicology and Pharmacy of the University of Veterinary Medicine Hannover, the
abattoir in Gleidingen (Hannover) and several practitioners in the surroundings of
Hannover.
The other group of samples for this study was compiled from different other institutes
of the University of Veterinary Medicine Hannover, again the abattoir in Gleidingen
(Hannover) and small animal practitioners in Hannover.
Goat and horse samples were collected with the help of the Institute for Anatomy.
The samples of porcine esophagi were provided from the Institute for Nutrition and
the canine esophagi were supplied from the Institute for Parasitology. Sheep, rabbit,
rat and mouse samples were taken with support of the Institute for Physiolology and
the Institute for Pharmacology, Toxicology and Pharmacy. Feline oesophagi were
collected from a veterinary surgery in Hannover and the Small Animal Clinic of the
University of Veterinary Medicine Hannover; the abattoir in Gleidingen (Hannover)
provided bovine oesophagi.
3.2 Sampling
The esophagus of the different freshly dead animals was opened by careful
dissection. Following, six to seven small pieces (maximum 1 cm x 1 cm) were
excised and immediately transferred into the different fixation solutions.

Material and Methods
47
Normally the samples were taken from the medium part of the esophagus (compare
also SCHOENNAGEL 2005).
Especially for samples fixed in HOPE® and Karnovsky solutions it was important to
control that the samples were not larger than 8 mm x 8 mm for HOPE® and 5 x 5 mm
for Karnovsky fixation, as penetration into the material would have been too slowly or
incomplete.
Tab. 3.1: Domesticated mammalian species used Nutrition type Species Number and gender
Horse 11 (6 geldings, 5 females)
Cattle 4 (females)
Goat 4 (1 male, 3 females)
Herbivores
Sheep 10 (2 males, 8 females)
Dog 9 (2 males; 2 females; 5
subadult males)
Pig 15 (females)
Mouse 9 (3 males, 6 females)
Omnivores
Rat 8 (2 males, 6 females)
Carnivores Cat 8 (4 males, 4 females)

Material and Methods
48
3.3 Tissue Fixation
3.3.1 Bouin’s solution (after BÖCK 1989)
The following chemicals were mixed:
1500 ml of saturated picric acid
+ 500 ml of 37 % formaldehyde (Merck)
+ 100 ml glacial acetic acid 99-100 %
The samples remained in the solution for 48 hours. Afterwards they were washed
several times in 70% ethanol with the addition of some drops of ammonia (conc.) to
remove the picric acid, and stored in 80% ethanol before running through an
ascending concentration of ethanol for embedding.
3.3.2 Calcium acetate-buffered formalin (after LILLIE and FULLMER
1976)
Calcium acetate-buffered formalin is a mixture of the following chemicals:
20 g Ca-acetate monohydrate (Merck)
+ 100 ml 37 % formaldehyde solution (Merck)
+ 900 ml de-ionised water
The samples remained in the solution for at least 24 hours, and later were washed
and stored in 70% ethanol.
3.3.3 HOPE® fixation (after OLERT et al. 2001)
The HOPE® fixation solutions are ready for use, and therefore no mixing of basic
chemicals is necessary. The complete HOPE® protocol is described in Tab. 3.2. Two
different fixation solutions were employed.
1. Incubation in HOPE I solution for 12-72 h at 0-4°C in the refrigerator
2. Removal of HOPE I solution
3. Incubation in 5 ml of an ice cold HOPE II / acetone solution (100 ml
acetone + 100µl HOPE II) for 2 h at 0-2°C on ice in the refrigerator.

Material and Methods
49
3.3.4 Fixation after KARNOVSKY (1967)
In order to prepare semithin sections and for future electron microscopical
observations, esophagus samples of each species were also fixed in Karnovsky`s
solution.
The following substances were mixed:
20 g para-formaldehyde (Merck) were dissolved in 250 ml distilled water
+ 100 ml glutaraldehyde (Merck)
+ 150 ml of a 0.2 molar cacodylate buffer (Serva)
The solution has to be stored at 4°C. The tissue pieces have to be very small (5mm x
5mm) to obtain good fixation and must be left in the solution for at least 24 hours.
3.3.5 Liquid nitrogen sampling
Liquid nitrogen samples were taken in order to analyse the surface of the esophagus
epithelium applying cryo scanning electron microscopy (Cryo-SEM). Therefore, small
pieces (2 x 4 x 8 mm) of the esophagus were transferred into liquid nitrogen (LN2) (-
180 °C) immediately after dissection, and then stored separately in small cavities of a
covered aluminium block, that was kept in LN2 until use.
3.4 Paraffin embedding
3.4.1 Paraffin embedding for samples fixed in Bouin’s solution and Ca-
acetate buffered formalin
Both types of samples were initially cut into small pieces and put into marked plastic
capsules for paraffin embedding. Afterwards, the Bouin-fixed samples were stored in
80 % ethanol and then dehydrated in ascending concentrations of ethanol in order to
facilitate paraffin penetration into the tissue.
In contrast to the Bouin-fixed samples, the formalin-fixed material was stored in the
fixation medium, before it was rinsed in tap water over night and then transferred into
70% ethanol.

Material and Methods
50
The following steps were identical for both sample types.
1. 24 hours 80% ethanol
2. 2 hours 96% ethanol
3. 2 hours isopropanol (abs.)
In order to remove the ethanol from the tissue, the samples in the capsules were
immersed in xylene (Riedel-de Haen) for 2 hours for two times, before they were
transferred to 60°C paraffin I (Paraplast, Sherwood) over night. The next day, the
capsules were stored in paraffin II for 4 hours and for another 4 hours in paraffin III.
In the last step, they were embedded with fresh 60°C paraffin in moulds, before
hardening at 4°C in the refrigerator.
3.4.2 Paraffin embedding HOPE®
After the HOPE II/acetone solution had been discarded, the samples were incubated
with freshly prepared cold acetone over 2 hours. This step was repeated twice, each
incubation lasting 2 hours.
Acetone is a dehydrating solution and removes the HOPE solution from the tissue.
After dehydration, the specimens were directly transferred into low melting paraffin
(52-54°C), which was prepared and stored in a warming cupboard. The first
incubation in paraffin I was carried out at 54°C for 12-16 hours; the second one in
paraffin II required 4 hours. It is important to avoid a trapping of air bubbles in the
paraffin. In the last step, the tissues were embedded in paraffin and the blocks were
stored in the refrigerator to cure until sectioning.
Table 3.2: Overview fixation steps HOPE® Reagent Incubation time
HOPE I 12-72 hours
HOPE II / acetone 2 hours
Acetone 2 hours
Acetone 2 hours
Acetone 2 hours
Low melting paraffin (54°C) I 12-16 hours
Low melting paraffin (54°C) II 4 hours

Material and Methods
51
3.5 Standard EPON embedding for Karnovsky fixed tissue
In comparison to paraffin embedded tissue, less shrinkage and less separation of the
tissue layers in EPON embedded material occurs. The synthetic material is much
harder than paraffin and it is possible to prepare semithin (0.2-1 µm) and ultrathin
slides (about 80 nm), which can be used for transmission electron microscopy (TEM).
For embedding, the tissue samples were dehydrated in ascending concentrations of
ethanol. In the next step, the samples were incubated in propylenoxide for 30 to 60
minutes. Following they were put in a propylenoxide-Epon mixture, and finally in an
absolute Epon mixture for another 30 to 60 minutes. After preparing a fresh Epon
mixture and adding the polymerisation enhancer dimethyl-amino-phenol (DMP-30),
the small samples were blocked in rubber moulds. Polymerisation was carried out in
an incubator at 36°C during the first day; the temperature was increased to 45°C on
the second day and to 60°C on the third day.
3.6 Sectioning
The procedure of sectioning is rather similar for most of the different samples
embedded in different media. After hardening in the refrigerator, the paraffin blocks
were removed from the metal moulds and sectioned with a rotation microtome
(Reichert Jung). The sections were not thicker than 5 µm, for the
immunohistochemical experiments conducted.
For the sections of paraffin embedded material only one water bath was necessary
with a temperature of approximately 50°C to allow stretching; for sections of the
HOPE®-fixed material, however, two water baths were needed. In the latter case, the
sections were collected in the first water bath, which had to be cold water, and the
second water bath needed to be tempered at 35-39°C. One by one section had to be
dipped in the heated water baths and just a short period of stretching is required.
After stretching, the sections were transferred on adhesive slides and dried in an
incubator. HOPE® sections are sensitive against heat, and therefore were dried
overnight at 37°C, whereas the other slides were dried for 3-4 hours in a warming
cupboard (50°C).

Material and Methods
52
The Epon series embedded samples were treated slightly different. In order to
prepare semithin sections (0.5 µm) an ultra-microtome (Ultracut E, Leica) with a
diamond knife was used.
3.7 Histological staining
3.7.1 Hematoxylin-Eosin staining of Bouin and formalin-fixed tissue
In order to evaluate the quality of tissue preservation, all collected samples were
stained in the H.E. and Masson Goldner trichrome stains.
For a good differentiation of nuclei and cytoplasm, the preferred method was H.E..
After this procedure, nuclei are blue and the cytoplasm is pink, the nuclear and cell
boundaries are well defined (LILLIE and FULLMER 1976). The H.E. staining
represents a dichromatic staining, which is composed of the nucleus dye hematoxylin
and the acidic cytoplasm dye eosin (BÖCK 1989). The originally herbal dye
hematoxylin is activated after a process of acid cleaning.
Before starting staining, hemalaun after DELAFIELD had to be prepared. Therefore,
4 g of hematoxylin (Merck) were dissolved in 25 ml absolute ethanol and in the next
step 400 ml of 10% ammonium aluminium sulfate (Merck) were added. After four
days of ripening, 100 ml of glycerin (Merck) and 100 ml of methanol were added.
Over several months, the hemalaun had to oxidize and to be aerated, before it was
ready for use.
To make the ready-to-use cytoplasm dye, an acetic 1 % eosin (Merck) solution with
96 % ethanol was prepared.
For the paraffin embedded material it is necessary to remove the surrounding wax
from the tissue before any other treatment is possible. This process is called
deparaffinisation. Therefore the slides were immersed in xylene two times for 15
minutes, whereby the paraffin was extracted. Subsequently, water had to be brought
back into the tissue to allow staining in an aqueous milieu.
The process of rehydration is achieved with the help of descending concentrations of
ethanol:

Material and Methods
53
1. 2 minutes isopropanol (abs.)
2. 2 minutes 96 % ethanol
3. 2 minutes 80 % ethanol
4. 2 minutes 70 % ethanol
Following the slides were brought in de-ionised water, before staining was started.
The following staining protocol was used (BÖCK 1989):
1. 8 minutes hematoxylin (after DELAFIELD)
2. 10-15 seconds in 0.1 % HCL
3. 15 minutes under running tap water
4. 5 min in Eosin (Annex) in distilled water with 5 drops acetic acid glacial
(Riedel-de Haen)
5. flushing in 70 % ethanol
6. 2 minutes 70 % ethanol
7. 2 minutes 80 % ethanol
8. 2 minutes 90 % ethanol
9. 2 minutes 96 % ethanol
10. 2 minutes ethanol (abs.)
11. 2 minutes isopropanol (abs.)
12. 5 minutes xylene
13. 5 minutes xylene
Finally the slides were covered with the synthetic resin EUKITT® (Kindler).
3.7.1.1 H.E. staining of HOPE® fixed tissue
The HOPE® fixed tissue needs also the procedure of deparaffinisation. This process
differs slightly from that one described for the Bouin and formalin-fixed material.
Deparaffinisation is achieved via an incubation in 60°C isopropanol (abs.), and
rehydration is accomplished in 70% acetone at 2-8°C.
To obtain the ideal conditions for deparaffinisation, cuvettes with isopropanol were
placed in a warming cupboard until they had reached the correct temperature. The
same procedure was conducted for the cuvettes with cold 70% acetone, as they

Material and Methods
54
were stored in the refrigerator until they had achieved the suitable temperature. To
guarantee good deparaffinisation and rehydration, the following protocol was applied:
1. 10 minutes 60°C temperate isopropanol (abs.)
2. 3 minutes twice fresh 60°C isopropanol (abs.)
3. 10 minutes twice fresh 70% cool (2-8°C) acetone
Incubation times in the dyes for H.E. staining are normally shorter for HOPE®-fixed
material than for the other two fixation methods. Another important aspect is to keep
attention to the fact that HOPE®-fixed tissue is sensitive for any influence of acid. For
this reason, no acetic acid was added to the eosin dye. Furthermore, the immersion
times in the ascending concentrations of ethanol had to be different. The following
protocol was used:
1. Flushing in de-ionised water twice
2. 5 minutes de-ionised water
3. 2-4 minutes hemalaun after DELAFIELD
4. short washing in de-ionised water
5. 1-2 minutes under running tap water to perform bluing reaction
6. 2-4 minutes eosin
7. Washing in de-ionised water twice
8. Dipping in 70% isopropanol twice
9. Dipping in ethanol (abs.) twice
10. Incubation for 10 minutes in ethanol (abs.)
11. Brief washing in xylene (Riedel-de Häen)
12. 5 minutes xylene
Finally slides were removed, dip-dried and mounted with Eukitt® (Kindler).

Material and Methods
55
3.7.2 Trichrome staining (Masson-Goldner) (after BÖCK 1989)
This staining is normally used for the demonstration of connective tissue (green),
which can be distinguished from muscle tissue (red).
Only the Bouin and formol fixed samples were stained after Masson-Goldner, as the
acidic substances used are too aggressive for the HOPE® fixed material and no
satisfactory results could be expected.
After deparaffinisation, the tissue was stained as follows:
1. 10 minutes hemalaun (after Delafield)
2. 10 seconds 0.1% HCL
3. 15 minutes running tap water
4. 5 minutes acid-fuchsin-ponceau [0.2 ponceau de xyline (Merck) + 0.1 g acid
fuchsin (Merck) + 300 ml de-ionised water + 0.6 ml acetic acid glacial]
5. 5 minutes 1 % acetic acid
6. 10 minutes phosphotungstic acid-orange-G [10 g phosphotungstic acid (Fluka)
+ 5g orange-G (Fluka) + 250 ml de-ionised water] to bleach the connective
tissue
7. 5 minutes 1 % acetic acid
8. 5 minutes lightgreen [0.5 g lightgreen (Merck) + 250 ml de-ionised water + 0.5
ml acetic acid glacial]
9. 5 minutes 1 % acetic acid
10. 2 minutes 80 % ethanol
11. 2 minutes 96 % ethanol
12. 2 minutes ethanol (abs.)
13. 2 minutes isopropanol (abs.)
14. two times 5 minutes xylene
15. mounting with EUKITT® (Kindler)

Material and Methods
56
3.8 Microscopical evaluation
3.8.1 Light microscopy
After staining with H.E. and/or the Masson-Goldner trichrome procedure, the Bouin,
Ca-Formol and HOPE® fixed esophagus samples were analysed with a light
microscope [Zeiss Photomicroscope II with a digital camera (Olympus DP70)]. After
this inspection for structure quality, it was decided which samples would allow
immunohistochemical experiments.
3.8.2 Cryo scanning electron microscopy (Cryo-SEM)
For analysis, one piece of the esophagus samples stored in LN2 was transferred in
the LN2 reservoir of a cryo sample loading device (Bal-Tec) and carefully mounted in
a specially designed clamping device on top of an object table. By means of a LN2-
cooled vacuum cryo transfer system (cryo shuttle VCT 100, Bal-Tec), in which the
sample was kept in an atmosphere of evaporising nitrogen for about 15 sec
(temperature lower than -180 °C); the loaded object table was rapidly transferred in a
freeze-etch / freeze-fracture unit (MED 020, Bal-Tec). Therein it was placed on a cryo
stage, where it was kept under high vacuum (5 x 10–6 mbar) at a minimum
temperature of -150 °C. After appropriate positioning of the stage, the esophagus
sample was hit once with a blade, thus creating a freeze-fracture from the outermost
to the deeper skin layers. Minor amounts of ice were removed from the sample
surface by gentle sublimation (-120 °C for 5 min, 1.7x10-4 mbar). Subsequently, the
surface of the sample was sputter-coated with gold palladium (40 sec), whereby all
samples were sputtered twice and from opposite directions (40 mA, 2.5 x 10-2 mbar,
argon). Using the shuttle, the coated sample was transferred onto the cryo stage
(Bal-Tec) of an adapted LEO 1430 vp scanning electron microscope (Zeiss) and
inspected at minimum vacuum of 9 x 10-6 mbar with an accelerating voltage of 10 kV.
The findings were documented the help of a digital camera.
For the demonstration of microorganisms on the surface of the esophagus it was
very important to rely on careful and artifact-free tissue processing (see also MEYER
et al, 2007). In this context, cryo SEM was the most suitable approach due to the
following reasons: Biological material, both plant and animal tissue, consist to a large
amount of water, which might occupy up to 98% of total tissue volume. Most types of
tissues, particularly those with hydrated structural elements, like several forms of

Material and Methods
57
connective tissue, are heavily altered under conditions of conventional SEM
preparations that apply alcoholic dehydration and critical point drying (CPD). The
abdication of chemical fixation, dehydration and solvents during cryo SEM
processing means that delicate structures are maintained without shrinkage and
without loss of soluble sample components. The latter feature is especially crucial
studying the localisation and distribution of extra- and intracellular aqueous or
colloidal material, such as, additionally, the mucus cover of the esophagus surface.
In contrast to other methods, which to variable extents are associated with the
generation of structural artifacts, the use of cryo freezing techniques instead of
chemical fixation allows to attain an ultrastructure which is as close as possible to its
natural state.
3.9 Immunohistochemistry
Immunohistochemistry is the localisation of antigens or proteins in tissue sections by
the use of labelled antibodies as specific reagents through antigen-antibody
interactions that are visualised by a colour marker, such as fluorescent dyes,
enzymes, or colloidal gold.
Five important representatives produced by the innate immune system were chosen:
Antimicrobial peptides, ß-glucan receptors, Toll-like receptors, lysozyme and
Langerhans cells. In order to detect the latter groups, different primary antibodies
were used.
As another aim of this thesis, different fixation methods (Bouin`s solution, Ca-
buffered formalin, HOPE® fixation) and their influences on immunohistochemical
reations were compared.
3.9.1 Primary antibodies
For the demonstration of the various parts of the innate immune system of the
esophagus epithelium several primary antibodies were applied (see Tab. 3.3). All
antibodies mentioned were diluted with PBS and 1 % bovine serum albumin (BSA)
as an additive until the suitable functional concentration had been reached. The
antibodies were stored at 4 °C and the PBS / 1% BSA mixture prepared at -20 °C
until use.

Material and Methods
58
Tab. 3.3: Primary antibodies AP: Antimicrobial peptide, ß-GR: ß-glucan receptor, LC: Langerhans cell, m: monoclonal, p: polyklonal
Primary antibody
Origin Specificity Clonality, isotype
Vendor
ß-defensin-2 (AP)
rabbit human, mouse p Biologo,
Kronshagen, Germany
ß-Defensin-3 (AP)
rabbit human, mouse p Biologo,
Kronshagen, Germany
Cathelicidin (LL37/CAP18)
(AP) mouse human
m, IgG1
HyCult Biotechnology,
Sanbio, Beutelsbach,
Germany
CD1a (clon O 10)
(LC) mouse human
m, IgG1
Biologo, Kronshagen,
Germany
Langerin (CD 207)
(LC) rabbit mouse, rat p
Biologo, Kronshagen,
Germany
Dectin-1 (ß-GR)
goat mouse p Biologo,
Kronshagen, Germany
L-Ficolin (clon GN4)
(ß-GR) mouse human
m, IgG1
Biologo, Kronshagen,
Germany
MBL (clon 3E7)
(ß-GR) mouse human
m, IgG1
Biologo, Kronshagen,
Germany
Toll-like-receptor 2
mouse human m,
IgG2a
Biologo, Kronshagen,
Germany
Lysozyme rabbit human p DAKO,
Golstrup, Denmark

Material and Methods
59
3.9.2 Secondary antibodies
As secondary detection systems two different types of visualisation were tested. The
first one is known as direct method. It is a one step staining technique and involves
a labelled antibody (for example FITC labelled). The antibody reacts directly with the
antigen in the tissue. Due to the fact that this method is rather insensitive, as little
amplification signal occurs, the indirect method is the technique of choice. For the
indirect method an unlabelled primary antibody is applied, which reacts with the
tissue antigen. To amplify the signal of the reaction, a labelled secondary antibody is
used (PEARSE 1985). These antibodies can be labelled with various substances. In
our study we applied antibodies labelled with a fluorescent dye (FITC), and
streptavidin-biotin and/or with horseradish peroxidase (HRP).
3.9.2.1 Two step indirect method with peroxidase linked secondary antibodies
To detect primary antibodies produced in the mouse or rabbit, we used the EnVision®
systems (DAKO). This visualisation technique is a two-step indirect staining method
(Fig. 3.1). The secondary antibody is conjugated with a horseradish peroxidase
(HRP) labelled dextran polymer. It is important to use a secondary antibody, which
has a strong affinity to the first antibody. Subsequently, the second antibody must be
directed against the immunoglobulin isotype of the species in which the first antibody
has been raised. For example, a primary antibody raised in the mouse must be
incubated with a secondary antibody raised in the rabbit, reacting as an anti mouse
immunoglobin.
To visualise the reaction, a 3,3`-diaminobenzidine (DAB) chromogen solution is used.
If the peroxidase linked secondary antibody has bound to the primary antibody,
peroxidase reduces DAB and a brownish colour reaction can be observed. As a
catalyst for this reaction, hydrogen peroxide (H2O2) is needed. It is important to notice
that mainly in granulocytes, mast cells and erythrocytes an endogenous peroxidase
activitivity can be observed. The easiest way to block the endogenous peroxidase is
to incubate the samples in an 80 % ethanol solution with H2O2 (197 ml 80 % ethanol
+ 3 ml 30% H2O2) for 30 minutes during the process of rehydration. As the
rehydration of HOPE® samples is accomplished in cold 70 % acetone, a blocking of
the endogenous peroxidase has to be conducted after the step of rehydration
through a 10 minutes incubation in de-ionised water and H2O2 (197 ml de-ionised
water + 3ml H2O2).

Material and Methods
60
Two types of DAKO visualisation systems were applied, depending on the origin of
the primary antibody. EnVision® mouse was used to detect antibodies with mouse
specificity, while EnVision® rabbit was employed to visualise the binding of a primary
anti-rabbit antibody. The kits contained the peroxidase labelled secondary antibody,
DAB and a HRP substrate buffer.
A general immunohistochemical protocol can be found in Tab. 3.5. The visualisation
step for EnVision® is described in detail as following; the procedure is equal for all
three fixing methods:
1. Deparaffinisation, rehydration and blocking of the endogenous peroxidase
2. Washing in phosphate-buffered saline (PBS) or de-ionised water
3. Demasking (3.9.5.1)
4. Washing in PBS
5. Blocking of electrostatic binding sites (3.9.3)
6. Incubation of primary antibody over night at 4°C
7. Washing in PBS
8. Visualisation:
Incubation with secondary peroxidase linked antibody EnVision® rabbit
or mouse over 45 minutes
Washing in PBS to remove excess antibody
Incubation with DAB (40µl DAB + 1000 µl HRP substrate buffer)
over 5 minutes.
9. Rinsing in running tap water and/or PBS
10. Dehydration in ascending concentrations of ethanol
11. Mounting with Eukitt® (Kindler)
Furthermore, another two step indirect visualisation technique was utilised, as not all
primary antibodies used derived from the mouse or rabbit. The antibody to detect
Dectin-1 was extracted from goats, thus a secondary anti-goat antibody was needed.
The antibody applied was peroxidase labelled, similar to the one described above,
but as no visualisation kit was available, the binding of the secondary antibody had to
be visualised with a self-prepared DAB solution. Therefore DAB (Sigma) as a powder
was dissolved in 200 ml PBS and filtered. 14 µl of 30% H2O2 were added and the
slides were incubated in the solution over 10 minutes under constantly stirring.

Material and Methods
61
Fig 3.1.: Principle of the two step indirect method DAB: 3,3`-diaminobenzidine
3.9.2.2 Use of a labelled streptavidin biotin Complex (LSBC)
This method uses the high affinity of avidin for biotin and is described as a three layer
technique. Avidin is a large glycoprotein and can be labelled with peroxidase. Avidin
shows a high electrostatical binding-affinity to the tissue and contains carbohydrate
groups, which show affinity to tissue lectins. These problems were eliminated through
the development of streptavidin, an avidin derived from Streptococcus avidini. Biotin
is a low molecular weight vitamin, which can be conjugated to antibodies.
As already mentioned previously, this method is of a three layer type. The first layer
is the primary antibody. The second layer is a biotinylated secondary antibody,
whereas the third layer is an enzyme-streptavidin conjugate (HRP-streptavidin) to
replace the complex of avidin-biotin peroxidase. The reaction is visualised by
applying the substrate DAB. All substrates mentioned and secondary antibodies were
included in the visualisation kit from BioGenex/DCS.
The blocking of the endogenous peroxidase is conducted as described under 3.9.2.1.
Primary
antibody
Secondary Peroxidase labelled antibody
DAB colour reaction
Epitope

Material and Methods
62
According to the fact that the steps before and after visualisation equal those ones
described before and in Tab. 3.5 the following protocol is restricted to the step of
visualisation:
Incubation with a biotinylated secondary antibody over 40 minutes
Washing in PBS 3x5 minutes to remove excess antibody
Incubation with HRP-streptavidin for 40 minutes
Washing in PBS 3x5 minutes to remove excess enzyme
Visualisation with DAB (250 µl DAB substrate buffer + 2250 µl de-ionised
water + 2 drips liquid DAB + 1 drip H2O2), incubation for 5 minutes
3.9.2.3 Immunofluorescence (IF)
The third visualisation technique used was immunofluorescence. To identify the
primary antibodies, secondary antibodies conjugated with fluorescein isothiocyanate
(FITC) were applied. FITC was first introduced by John L. Riggs in 1960 as a stable
yellow powder and today is available as commercial product (PEARSE 1985). The
principle of IF is that of a relatively fast decreasing light emission of molecules, which
have been activated via the absorption of energetic rays. The light emission is visible
in the fluorescence microscope, where FITC shows a green-yellowish reaction, using
a specific filter [Zeiss Axioskop equipped with an epifluorescence device (FITC filter
combination: BP450-490, FT510, LP520)]. As the reaction intensity decreases very
fast, reaction analysis has to be made immediately and pictures have to be taken.
Different FITC labelled antibodies depending on the origin of the first antibody were
used and are shown in Tab. 3.4. In contrast to the other visualisation systems, FITC
needs no blocking of endogenous peroxidase, as no chromogen is used. The slides
were mounted with Mowiol (Sigma Aldrich), in order to obtain a longer lasting
reaction. The Mowiol solution was self prepared after the following protocol:
1. 2.4 g Mowiol (Sigma Aldrich)
+ 6.0 g glycerol (99.5%) (Sigma)
+ 6 ml aqua dest.
The components were mixed with a magnetic stir bar for 4 hours
2. In the next step, 12 ml of 0.2 M tris-buffer pH 8.5 [for 100 ml: 0.88 g TRIZ
hydrochlorid powder + 1.75 TRIZ base powder (Sigma)] were added.
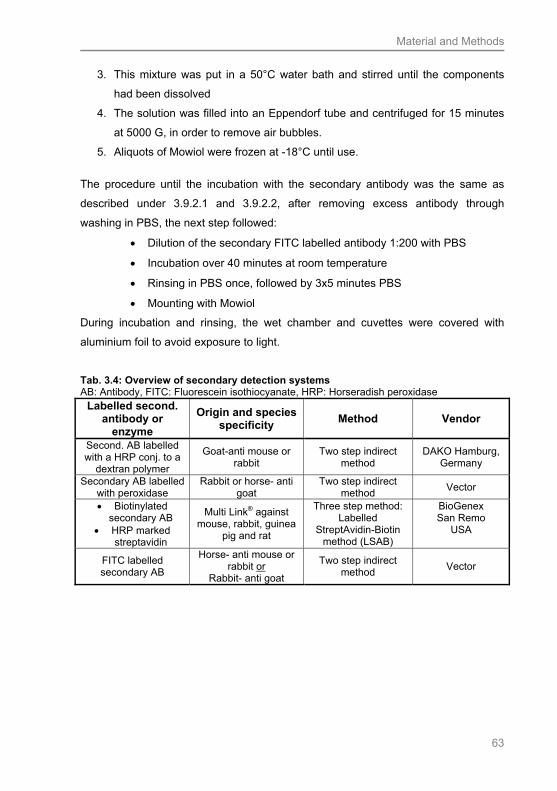
Material and Methods
63
3. This mixture was put in a 50°C water bath and stirred until the components
had been dissolved
4. The solution was filled into an Eppendorf tube and centrifuged for 15 minutes
at 5000 G, in order to remove air bubbles.
5. Aliquots of Mowiol were frozen at -18°C until use.
The procedure until the incubation with the secondary antibody was the same as
described under 3.9.2.1 and 3.9.2.2, after removing excess antibody through
washing in PBS, the next step followed:
Dilution of the secondary FITC labelled antibody 1:200 with PBS
Incubation over 40 minutes at room temperature
Rinsing in PBS once, followed by 3x5 minutes PBS
Mounting with Mowiol
During incubation and rinsing, the wet chamber and cuvettes were covered with
aluminium foil to avoid exposure to light.
Tab. 3.4: Overview of secondary detection systems AB: Antibody, FITC: Fluorescein isothiocyanate, HRP: Horseradish peroxidase
Labelled second. antibody or
enzyme
Origin and species specificity
Method Vendor
Second. AB labelled with a HRP conj. to a
dextran polymer
Goat-anti mouse or rabbit
Two step indirect method
DAKO Hamburg, Germany
Secondary AB labelled with peroxidase
Rabbit or horse- anti goat
Two step indirect method
Vector
Biotinylated secondary AB
HRP marked streptavidin
Multi Link® against mouse, rabbit, guinea
pig and rat
Three step method: Labelled
StreptAvidin-Biotin method (LSAB)
BioGenex San Remo
USA
FITC labelled secondary AB
Horse- anti mouse or rabbit or
Rabbit- anti goat
Two step indirect method
Vector

Material and Methods
64
3.9.3 Immunohistochemistry: General procedure
Before starting any reaction on the paraffin embedded material, deparaffinisation and
rehydration needs to be accomplished as described under 3.7.1 / Tab. 3.5. The
blocking of the endogenous peroxidase activity was already described under 3.9.2.
During fixation, especially in formalin and Bouin-fixed tissue, it occurs that protein-
cross links are formed and methyl-bridges are accumulated during the formaldehyde
reaction with amino acids. This fact might result in a reduced immuno reactivity of the
protein (epitope masking). To decrease the amount of protein cross links, an epitope
demasking might be necessary to avoid wrong negative reactions. Epitope
demasking can also be called antigen retrieval and several techniques can be
applied. In our study, four different types of pre-treatments were tested until the
suitable method for the respective antibody was established.
To reduce background staining due to hydrophobic interactions between the antibody
and tissue, normal serum was put on the slides over 20 minutes before the
incubation with the primary antibody. Proteins of the normal serum bind the various
charges of the tissue, and thus the following primary antibody is able to react
specifically with the tissue. We used normal goat (NGS), normal rabbit (NRS) and
normal horse serum (NHS), depending on the origin of the secondary antibody.
Before use, the normal serum was inactivated in a 56°C water bath for 30 minutes. In
order to avoid background staining, due to the proteins in the normal serum, the
normal serum was diluted 1:500 with PBS (500 µl normal serum + 2000 µl PBS).
According the same reason, the primary antibody was diluted with 1% bovine serum
albumin (BSA) (Sigma).
The tissue was incubated with the diluted primary antibody over night at 4°C to
ensure a good linkage of the primary antibody. Afterwards the reaction was
visualised applying the different detection systems (3.9.2).
To give an overview of the general approach of immunohistochemistry, the
procedure is described for Ca-formol and Bouin-fixed material in comparison to
HOPE® in Tab. 3.5.

Material and Methods
65
Tab 3.5: General immunohistochemistry protocol, comparison of fixation techniques Abs.: Absolute, AR: Antigen retrieval, min: Minutes, IF: immunofluorescence, NHS: Normal horse serum, NGS: Normal goat serum, NRS: Normal rabbit serum, PBS: Phosphate buffered saline, RT: room temperature, TEC: Tris-EDTA-citrate buffer
Step Formol-Ca and Bouin-
fixed tissue HOPE® fixed tissue
Deparaffinisation 2 x10 min. xylene 2 min. isopropanol
2x10 min. 60°C isopropanol
2x3 min 60°C isopropanol
Rehydration + blocking endogenous peroxidase
2 min. ethanol 30 min. 80%
alcohol+H2O2 2 min. 70 % alcohol
2x10 min. 4°C acetone
Washing 3x5 min. PBS 2x3 min. de-ionised water 5 min. de-ionised water
Demasking
TEC-buffer Citrat buffer AR buffer (DAKO) Trypsin
Blocking endogenous peroxidase
10 min. de-ionised water+ H2O2
Washing Short rinse in PBS 3x5 min PBS Blocking electrostatic binding sites
20 min NGS, NRS or NHS
20 min NGS, NRS or NHS
Incubation of primary antibody
Over night at 4°C in suitable dilution
Over night at 4 °C in suitable dilution
Washing 3x5 min. PBS 3x5 min. PBS
Visualisation EnVision® BioGenex® IF
EnVision® BioGenex® IF
Washing
For IF: 1x rinse in PBS, 3x5 min PBS
Others: 5 min PBS, 10 min. running water
For IF: 1x rinse in PBS, 3x5 min PBS
Others: 5 min PBS, 2x10 min PBS
Rehydration
Only necessary for EnVision® and BioGenex®
2 min. 70% alcohol 2 min. 80% alcohol 2 min. abs. alcohol 2 min. isopropanol 2x5 min. xylene
Only necessary for EnVision® and BioGenex® 2x rinse in 70%
isopropanol 2x rinse in ethanol
(abs.) 10 min ethanol (abs.) rinse in xylene 5 min. xylene
Mounting
For EnVision® and BioGenex®:
Eukitt® (Kindler) For IF: Mowiol
For EnVision® and BioGenex®: Eukitt® (Kindler)
For IF: Mowiol

Material and Methods
66
3.9.4 Controls
For all immunohistochemical experiments positive, negative and isotype controls
were attendantly conducted on all animal species studied.
As references for the tissues needed as a positive control the antibody datasheets of
the vendor were used. Positive controls were conducted if the reference tissue
needed was available. Thus, the following positive control tissues were applied:
Murine skin (langerin), equine skin (CD1a, hBD-2 and -3) and equine liver (MBL and
L-ficolin).
To conduct the negative controls, the first antibody was omitted and instead the
tissue was incubated with PBS / 1% BSA.
In order to exclude the possibility of non-specific binding of the Fc-part of the
respective primary antibody, isotype controls were conducted. Therefore, the primary
antibody was omitted and replaced by a IgG mouse (Sigma) or rabbit (Sigma)
antibody depending on the origin of the first antibody. The respective protein
concentrations of the antibodies were considered. This means, that the IgG mouse or
rabbit antibody was applied in the same protein concentration, as the primary
antibody tested.
3.9.5 Preliminary tests for the establishment of the primary antibodies
For the primary antibodies listed in Tab. 3.3, initially immunohistochemical
preliminary tests were conducted. Therefore various demasking methods and
different antibody dilutions were tested and controls were accomplished. The first
preliminary tests on the animal esophagi were conducted on pig, mouse / rat, horse,
cat and goat tissue, in order to get an impression of the reactivity within the different
nutrition types. If the suitable antibody had been established, the
immunohistochemical experiments were performed for all species and esophagi
collected. If it was not possible to establish an antibody during the first preliminary
tests for the species mentioned, the same tests were applied on the sheep, dog and
rabbit esophagus. In the next step, different pre-treatments and visualisation systems
were considered, as described in Tabs. 3.6 and 3.7.

Material and Methods
67
3.9.5.1 Demasking methods tested
Basically two types of antigen retrieval techniques can be distinguished: heat induced
epitope retrieval and proteolyse induced epitop retrieval. In our study we tested
trypsin as proteolyse induced epitope retrieval, whereas the others ones were heat
induced.
Trypsin:
200 ml distilled water were heated in a warming cupboard until it had a temperature
of approximately 37°C. Following 0.2 g of trypsin (Sigma) and 0.2 g calcium-chloride
powder (Merck) were added, and the pH was adjusted to 7.6 with diluted NaOH
(Merck). The solution was transferred to a warm cuvette and the slides were
incubated for 30 minutes in a warming cupboard at 37°C.
Heating pre-treatment with citrate buffer:
To use citrate buffer as a pre-treatment, two stock solutions had to be prepared. The
first one (stock solution A) comprised 0.1 M citric acid. To prepare 0.1 M citric acid,
21g citric acid powder (Riedel-de-Haen) were added to 1000 ml de-ionised water. For
the second stock solution (stock solution B), 0.1 M sodium citrate was prepared. In
this case 29.4 g sodium citrate powder was mixed in 1000 ml de-ionised water.
In the following step, 18 ml of stock solution A were added to 82 ml of stock solution
B and mixed with 1000 ml de-ionised water (pH 6).
Finally the solution was heated to 96-98°C and the slides were incubated in the
solution for 30 minutes.
Heating pre-treatment with Tris-EDTA-citrate buffer (TEC):
900 ml of distilled water and 100 ml TEC stock solution (composition below) were
filled in a beaker. A magnetic stir bar and an empty slide holder were put into the
solution. In the next step, the beaker was put on a heating plate and under constant
stirring the solution was heated up to 96°C. Following, the slide holder with the slides
was placed into the hot solution. The slides were left in the solution for 30 minutes at
96-98°C.
TEC stock solution:
1. 2.5 g Tris-base (MW 121.1) (Sigma)
2. 5.0 g EDTA (Riedel-de-Haen)
3. 3.2 g Tri-Natrium-Citrat (Merck)

Material and Methods
68
4. dissolve in 800 ml de-ionised water
5. adjust to pH 7.8 with NaOH
fill up to 1000 ml with de-ionised water
Heating treatment with DAKO target retrieval solution (TRS):
The TRS from DAKO was tested with pH 6 and pH 9 (TRIS EDTA base). 7 ml of the
TRS stock solution pH 6 or 9 were mixed with 63 ml distilled water (1:10). This
solution was filled into a beaker and heated under constant stirring to 96°C. Than the
slides were put into the hot solution and heated at 96-98°C. It was important to keep
attention to heating time, as the pre-treatment of the slides in the pH 9 solution
should not exceed 20 minutes. Slides in the pH 6 solution were pre-treated for 30
minutes. Finally, a cooling down period of maximum 15 minutes followed.

Material and Methods
69
Tab. 3.6: Tested antibody dilutions, demasking methods and visualising systems for Bouin and formol-fixed material. VS: Visualising system, 1: DAKO EnVision rabbit (two-layer method), 2: DAKO EnVision mouse, 3: DAB as pure substance, 4: BioGenex (LSAB), 5: Immunofluorescence, AB: antibody, TEC: Tris-EDTA buffer
Antibody Antibody dilution, tested demasking methods and visualising systems ß-defensin 2 1:50, 1:100, 1:250, 1:350,
1:450 no pre-treatment, VS: 1
1:250, 1:350 TEC-buffer VS: 1
ß-defensin 3 1:50, 1:100, 1.250, 1:500, 1:1000, 1:1200, 1:1500 no pre-treatment, VS: 1
1:1000, 1:1200, 1:1500 TEC-buffer, VS: 1
Cathelicidin 1:10, 1:20, 1:30, 1:40, 1:50no pre-treatment, VS: 2
1:20, 1:30 TEC-buffer, VS: 2
CD1a 1:10, 1:20, 1:50 no pre-treatment, VS: 2,4 + 5
1:10, 1:20 TEC-buffer, VS: 2, 4 +5
1:10, 1:20 Trypsin VS: 2+4
1:10, 1:20 DAKO TRS pH 6+9 VS: 2+4
Dectin-1 1st AB: 1:10, 1:20 2nd AB: 1:200 No pre-treatment, VS: 3+4
1st AB:1:10, 1:20 2nd AB: 1:200 TEC- buffer , VS: 3+5
1st AB: 1:10, 1:20 2nd AB: 1.200 Trypsin, VS: 3
1:10, 1:20, DAKO TRS pH 6+9 VS: 3+5
1:10, 1:20 citrate buffer VS: 3
Langerin 1:10, 1:20 No pretreatmentVS: 1
1:10, 1:20 TEC-buffer VS: 1
L-Ficolin 1:10, 1:20 No pre-treatment, VS: 2,4+5
1:10, 1:20 TEC-buffer, VS: 2,4+5
1:10, 1:20 Trypsin, VS: 2
1:10, 1:20 DAKO TRS pH 6+9 VS: 2,4+5
1:10, 1:20 citrate buffer, VS: 2
Mannose-binding-lectine
1:10, 1:20 No pe-treatment VS: 2, 4+5
1:10, 1:20 TEC-buffer VS: 2,4+5
1:10, 1:20 Trypsin VS: 2
1:10, 1:20 DAKO TRS pH 6+9 VS: 2+5
1:10, 1:20 Citratebuffer VS: 2
Toll-like-receptor 2 1:10, 1:20 No pre-treatment VS: 2
1:10, 1:20 TEC-buffer VS: 2
1:10, 1:20 Trypsin VS: 2
Lysozyme 1:50, 1:75, 1:100 No pretreatment, VS: 1
1:50, 1:75, 1:100 TEC-buffer ,VS: 1
1:50, 1:75, 1:100 Trypsin, VS: 1
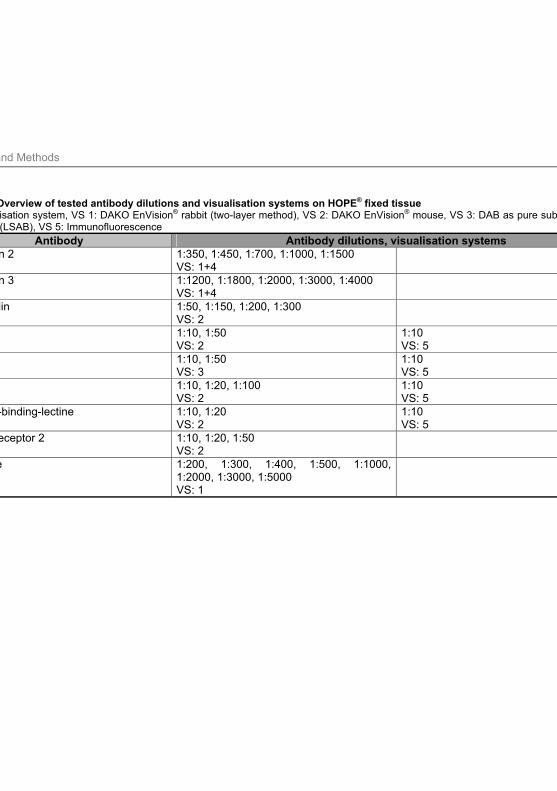
Material and Methods
70
Tab.3.7.: Overview of tested antibody dilutions and visualisation systems on HOPE® fixed tissue VS: Visualisation system, VS 1: DAKO EnVision® rabbit (two-layer method), VS 2: DAKO EnVision® mouse, VS 3: DAB as pure substance, VS 4: BioGenex (LSAB), VS 5: Immunofluorescence
Antibody Antibody dilutions, visualisation systems ß-defensin 2 1:350, 1:450, 1:700, 1:1000, 1:1500
VS: 1+4
ß-defensin 3 1:1200, 1:1800, 1:2000, 1:3000, 1:4000 VS: 1+4
Cathelicidin 1:50, 1:150, 1:200, 1:300 VS: 2
CD1a 1:10, 1:50 VS: 2
1:10 VS: 5
Dectin-1 1:10, 1:50 VS: 3
1:10 VS: 5
L-Ficolin 1:10, 1:20, 1:100 VS: 2
1:10 VS: 5
Mannose-binding-lectine 1:10, 1:20 VS: 2
1:10 VS: 5
Toll-like-receptor 2 1:10, 1:20, 1:50 VS: 2
Lysozyme 1:200, 1:300, 1:400, 1:500, 1:1000, 1:2000, 1:3000, 1:5000 VS: 1
Material and Methods
71
3.9.6 Antibody dilutions and visualisation systems applied
After conducting the preliminary tests it was decided which dilution of the
primary antibody seemed to ensure the best results (compare Tab. 3.8), and
also the advantages or disadvantages of a pre-treatment were considered.
Furthermore, the different results of the visualisation systems were compared
and it was decided which system had to be the suitable one.
As some primary antibodies could not be established in an acceptable way,
experiments were accomplished on the whole animal material only for the
established antibodies. Due to the fact that the reactivity of the established
primary antibodies for lysozyme, ß defensin-2 and -3, cathelicidin and TLR2
could not be improved through application of a pre-treatment, it was decided to
conduct the experiments without pre-treatments.
The two layer visualisation system EnVision® rabbit and EnVision® mouse from
DAKO seemed to be the most suitable visualisation system, because with the
three layer technique of BioGenex, colour intensity was not satisfactory and
false positive reactions on HOPE® fixed material could be observed.
Tab. 3.8: Antibody dilutions applied for established antibodies Dilution with PBS+1% BSA
Antibody Dilution for Bouin and formol-fixed tissue
Dilution for HOPE® fixed tissue
ß defensin-2 1:350 1:1500
ß defensin-3 1:1200 1:1200
Cathelicidin 1:30 1:150
Toll-like receptor 2 1:10 1:20
Langerin (CD207) 1:20 Not conducted
Lysozyme 1:50 1:2000

Material and Methods
72
3.9.7 Statistical analyses
To describe the immunohistochemical results, a semiquantitative scale was
used. The staining intensity was classified using six levels (0: no reaction, 0.5:
very weak reaction, 1: weak reaction, 1.5: mild reaction, 2: medium reaction,
2.5: strong reaction, 3.0: very strong reaction).
For the evaluation of the statistical significance we used the computer program
SAS. Basically two different statistical tests were applied. For analysing the
results found for the various substances and receptors in the three fixation
solutions (Bouin`s solution, Ca-formol, HOPE®), the signed rank test was
applied, as it was considered that the results found did not show normal
distribution. The three different fixation solutions were compared with each
other, considering each of the five layers of the esophagus epithelium. All data
obtained from the experiments were pooled, considering the species and
fixation solution applied. The results were expressed as standard error of mean,
and values of p < 0.05 were regarded as significant difference.
The comparison of the nutrition groups was accomplished applying the
Wilcoxon test. All species were allocated to one nutrition group (herbivorous,
carnivorous, or omnivorous). Regarding all five layers of the epithelium, the
results of the different groups were compared with each other. P-values < 0.05
demonstrated a statistical significant difference between the nutrition groups.

Results
73
4 Results
4.1 Light and electron microscopical observations
4.1.1 Demonstration of microorganisms with cryo SEM
The demonstration of a microbial occupation of the surface of the esophagus
epithelium was realised using cryo SEM. The pictures revealed a one layer
colonisation of the epithelium with bacteria and fungi in each species studied
(Figs. 4.1 to 4.7). The species considered were: Horse, cattle, goat, pig, rat,
dog and cat. Species related differences in the density of microorganisms found
could not be observed.
Fig 4.1 Microorganisms on the esophagus epithelium of the horse Predominantly bacteria (fungus marked as: ►, bacteria marked as: →)
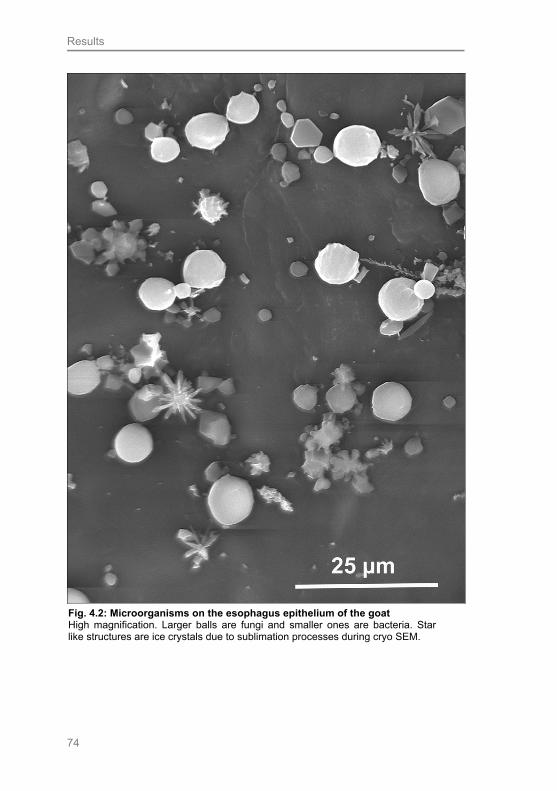
Results
74
Fig. 4.2: Microorganisms on the esophagus epithelium of the goat High magnification. Larger balls are fungi and smaller ones are bacteria. Star like structures are ice crystals due to sublimation processes during cryo SEM.

Results
75
Fig. 4.4: Microorganisms on the esophagus epithelium of the rat, surrounding an excretory duct of a secretory gland Field of bacteria is marked by: →, a fungus is marked by: ►
Fig 4.5: Microorganisms and mucus covering the esophagus epithelium of the pig

Results
76
Fig. 4.6: Microorganisms and mucus covering the esophagus epithelium of the dog Bacterium is marked by: →
Fig. 4.7: Bacteria and fungi without mucus covering on the esophagus epithelium of the cat Bacterium is marked by a: → and field of fungi is marked by a: ►

Results
77
4.1.2 General esophageal structure
In order to compare the structural composition of the esophagus epithelium, all
tissue samples collected were stained with H.E. and Masson-Goldner trichrome
stain. The procedure was the same for all fixation media applied. The HOPE®
fixed samples were only stained in H.E.. The protocol applied slightly differed
from the one used for samples fixed in Ca-formol and Bouin. Before IHC
experiments were started, the integrity of the tissue was evaluated. The best
results regarding structure preservation were obtained from the Bouin-fixed
samples. Almost all samples revealed very good tissue integrity. The HOPE®
fixed samples exhibited a slightly diminished quality of structure preservation. A
separation of the lamina epithelialis and the lamina propia could be observed in
some of these samples. Additionally, in some cases the epithelium rolled up. In
a number of the Ca-formol fixed esophagus samples, tissue preservation was
relatively bad, whereby the different layers (lamina epithelialis, lamina propia
and Tunica muscularis) appeared as completely separated from each other.
The structure of the esophagus epithelium of the species studied was mainly
evaluated using semithin sections stained with toluidine blue. In this connection,
differences between the three nutrition groups could be demonstrated. The
epithelium of the herbivorous species was more prominent, in comparison to
that one of the omnivorous and carnivorous species studied. This fact
concerned all layers, but was most obvious in the stratum corneum. While the
carnivorous cat exhibited a very thin stratum corneum, better named stratum
superficiale, as almost no keratinisation occured in this species, the stratum
corneum of the herbivorous animals was thicker. Furthermore, the corneocytes
of the latter seemed to be organised in dense layers. This structural feature was
most obvious in the equine esophagus epithelium. The stratum corneum of the
omnivorous pig was even more prominent, compared to conditions in the cat. In
contrast to the stratum corneum of the herbivorous species, the pig did not
show such a dense organisation.
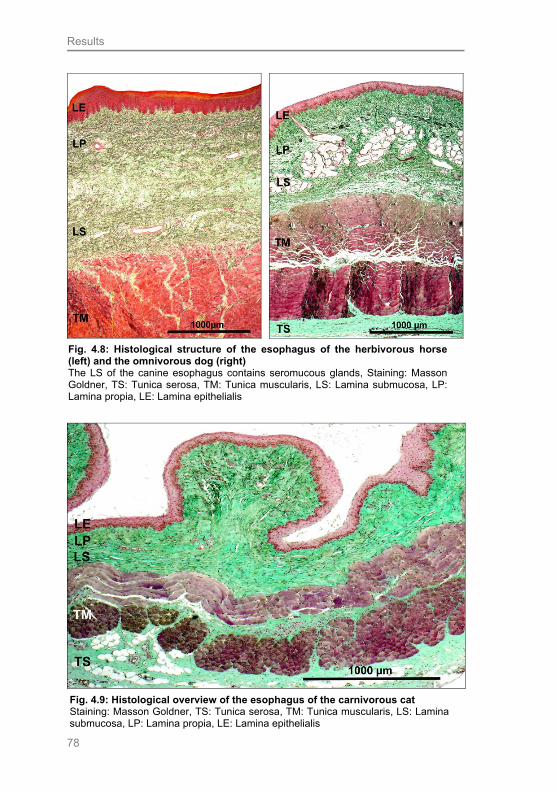
Results
78
Fig. 4.8: Histological structure of the esophagus of the herbivorous horse (left) and the omnivorous dog (right) The LS of the canine esophagus contains seromucous glands, Staining: Masson Goldner, TS: Tunica serosa, TM: Tunica muscularis, LS: Lamina submucosa, LP: Lamina propia, LE: Lamina epithelialis
Fig. 4.9: Histological overview of the esophagus of the carnivorous cat Staining: Masson Goldner, TS: Tunica serosa, TM: Tunica muscularis, LS: Lamina submucosa, LP: Lamina propia, LE: Lamina epithelialis
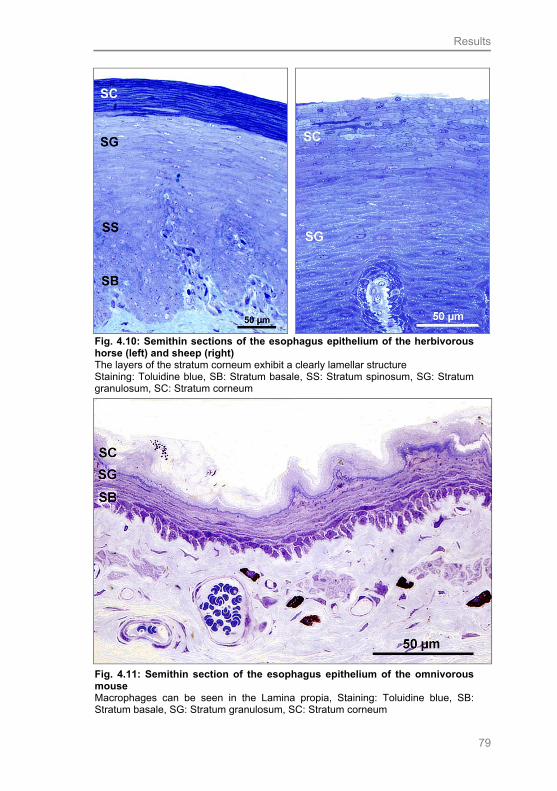
Results
79
Fig. 4.10: Semithin sections of the esophagus epithelium of the herbivorous horse (left) and sheep (right) The layers of the stratum corneum exhibit a clearly lamellar structure Staining: Toluidine blue, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.11: Semithin section of the esophagus epithelium of the omnivorous mouse Macrophages can be seen in the Lamina propia, Staining: Toluidine blue, SB: Stratum basale, SG: Stratum granulosum, SC: Stratum corneum
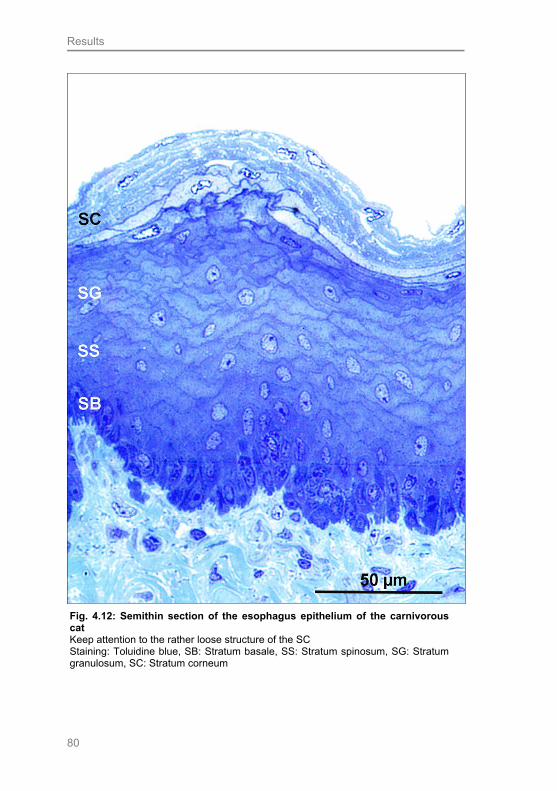
Results
80
Fig. 4.12: Semithin section of the esophagus epithelium of the carnivorous cat Keep attention to the rather loose structure of the SC Staining: Toluidine blue, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
81
4.2 Fixation experiment: Influences of different fixation
media on IHC results
4.2.1 ß-defensin 2 and -3 reactions
Human ß defensins-2 (hBD-2) and human ß defensins-3 (hBD-3) are both
members of the group of antimicrobial peptides, and thus will be discussed
together.
Concerning hBD-2, for the samples fixed in Bouin`s solution and Ca-formol
equal dilutions of the primary antibody were used (hBD-2: 1:350). On the
contrary, higher dilutions were applied for the HOPE® fixed samples, as such
samples exhibited too strong reaction intensities with lower dilutions (hBD-2:
1:1500). Regarding hBD-3, equal antibody dilutions for all three fixation media
were used.
The comparison of the expression patterns of the two hBD types within the
lamina epithelialis of the esophagus revealed several differences as related to
the different fixation solutions. Those differences are shown in detail in Tabs.
4.1 and 4.2, and Figs. 4.13 and 4.14. A statistically relevant variation of the
staining intensity for hBD-2 could be observed primarily in the horse and the
sheep. Intriguingly, the comparison of the results found for Bouin and Ca-formol
fixed samples in the equine esophagus showed significant differences regarding
all layers (p-value < 0.05), except the stratum corneum (Fig. 4.13). Whereas in
the Ca-formol fixed tissue normally no positive reaction was observable in the
layers, an obviously positive reaction could be shown for the Bouin-fixed
samples in all layers. These findings were confirmed for hBD-3 (p-value < 0.05),
apart from the fact that the stratum basale showed a very weak positive reaction
in the Ca-formol fixed samples. Comparing the results for hBD-2 of the Bouin
and HOPE® fixed samples in the equine esophagus revealed a statistically
significant difference for the stratum spinosum (lower and upper layer) (p-value
< 0.05); again the Bouin samples exhibited a stronger reaction. The same fact
was noticed for hBD-3, besides a difference demonstrated in the stratum basale
and granulosum (p-value < 0.05). Concerning the sheep, a statistically relevant
difference for hBD-2 could be demonstrated comparing the results of the Bouin
and Ca-formol fixation as well as comparing the Bouin and HOPE® fixation (p-

Results
82
value < 0.05) (Fig. 4.14). Interestingly, for both comparisons a difference was
shown for the same layers (stratum corneum, stratum granulosum, stratum
basale). Again, the HOPE® fixed samples exhibited a weaker reaction than the
Bouin samples. The results of hBD-3 expression in the sheep comparing Bouin
and Ca-Formol fixed samples equalled the ones described for hBD-2. It is a
remarkable finding, that statistically significant differences observed in the horse
and the sheep could be corroborated for hBD-2 and -3 in the same
localisations. The same finding was made analysing the IHC results of the
porcine esophagus. In this regard, a reaction difference for hBD-2 and -3 was
found for the stratum basale and the str. granulosum (p-value < 0.05)
The reaction comparison between the Ca-formol and HOPE® fixed samples
revealed no statistically relevant differences (p-values > 0.05).
SB SS (l) SS (u) SG0
1
2
3
Ca-FormolBouin`s solution
HOPE
* **
** *
Sta
inin
g in
ten
sit
ies
Fig. 4.13: Demonstration of fixation differences for hBD-2 in the equine esophagus. Shown data: Mean and standard deviation. Applied antibody dilutions for the primary antibody: Bouin`s solution and Ca-Formol: 1:350, HOPE®: 1:1200 (SB: Stratum basale, SS (l): lower Stratum spinosum, SS (u): upper Stratum spinosum, SG: Stratum granulosum, *: p-value < 0.05)
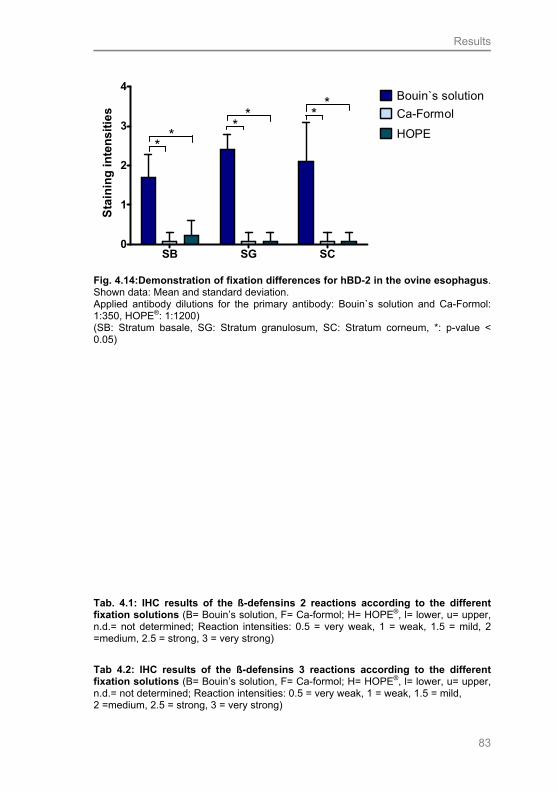
Results
83
SB SG SC0
1
2
3
4
Ca-Formol
Bouin`s solution
HOPE*
*
**
**
Sta
inin
g i
nte
ns
itie
s
Fig. 4.14:Demonstration of fixation differences for hBD-2 in the ovine esophagus. Shown data: Mean and standard deviation. Applied antibody dilutions for the primary antibody: Bouin`s solution and Ca-Formol: 1:350, HOPE®: 1:1200) (SB: Stratum basale, SG: Stratum granulosum, SC: Stratum corneum, *: p-value < 0.05)
Tab. 4.1: IHC results of the ß-defensins 2 reactions according to the different fixation solutions (B= Bouin’s solution, F= Ca-formol; H= HOPE®, l= lower, u= upper, n.d.= not determined; Reaction intensities: 0.5 = very weak, 1 = weak, 1.5 = mild, 2 =medium, 2.5 = strong, 3 = very strong)
Tab 4.2: IHC results of the ß-defensins 3 reactions according to the different fixation solutions (B= Bouin’s solution, F= Ca-formol; H= HOPE®, l= lower, u= upper, n.d.= not determined; Reaction intensities: 0.5 = very weak, 1 = weak, 1.5 = mild, 2 =medium, 2.5 = strong, 3 = very strong)
Results
84
Layers Species
Str. corneum Str. granulosum Str. spinosum (u) Str. spinosum (l) Str. basale
B 0,5 B 1,5 B 1,5 B 2 B 2 F 0 F 0 F 0 F 0 F 0 Horse
H 0 H 1 H 0,5 H 0,5 H 1 B 1,5 B 0 B 0,5 B 0,5 B 2 F 1 F 0 F 0 F 0 F 0,5 Cattle
H 0,5 H 0,5 H 1 H 1 H 1 B 2 B 2,5 B 2 B 2 B 2,5
F 0,5 F 0 F 0 F 0,5 F 0,5 Goat
H 1 H 2,5 H 2 H 1 H 1 B 2 B 2,5 B 0,5 B 0,5 B 2 F 0 F 0 F 0 F 0 F 0
Herbivorous
Sheep
H 0 H 0 H 0 H 0 H 0 B 0,5 B 1,5 B 0 B 0 B 2 F 1,5 F 0 F 0 F 0 F 0,5 Pig H 0 H 1 H 1 H 1,5 H 1
B 2,5 B 0,5 B 1 B 2 B 2,5 F 2 F 0 F 0 F 0 F 0,5 Dog H 0 H 1 H 1 H 1 H 0 B 1 B 1 B 0 B 0,5 B 1,5 F 0 F 0 F 0 F 0 F 0 Mouse H 1 H 1,5 H 1,5 H 1,5 H 1 B 1 B 3 B 2 B 2 B 2,5 F 1 F 1 F 0 F 0 F 0,5
Omnivorous
Rat n.d. n.d n.d n.d n.d. B 3 B 0 B 0 B 1 B 2 F 2 F 0 F 0 F 0 F 1 Carnivorous Cat H 1 H 1 H 1 H 2 H 0,5
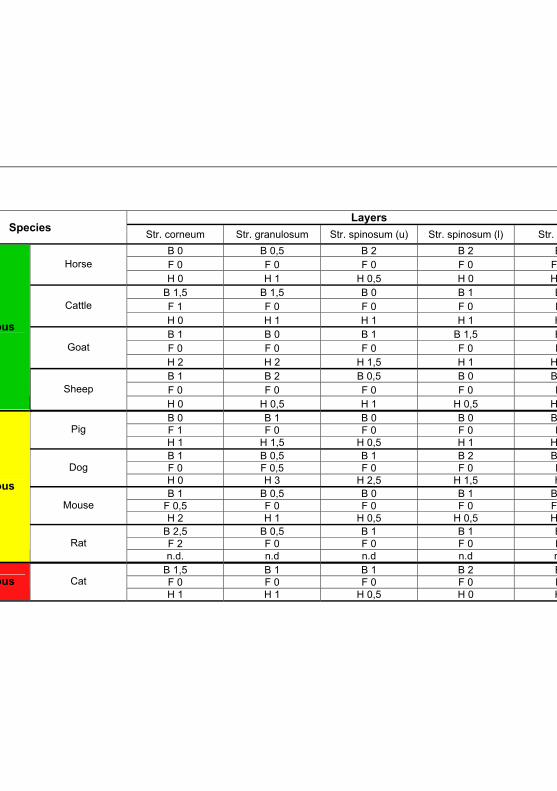
Results
85
Layers Species
Str. corneum Str. granulosum Str. spinosum (u) Str. spinosum (l) Str. basale
B 0 B 0,5 B 2 B 2 B 2 F 0 F 0 F 0 F 0 F 0,5 Horse
H 0 H 1 H 0,5 H 0 H 0,5 B 1,5 B 1,5 B 0 B 1 B 2 F 1 F 0 F 0 F 0 F 0 Cattle
H 0 H 1 H 1 H 1 H 1 B 1 B 0 B 1 B 1,5 B 2 F 0 F 0 F 0 F 0 F 0 Goat
H 2 H 2 H 1,5 H 1 H 0,5 B 1 B 2 B 0,5 B 0 B 1,5 F 0 F 0 F 0 F 0 F 0
Herbivorous
Sheep
H 0 H 0,5 H 1 H 0,5 H 0,5 B 0 B 1 B 0 B 0 B 1,5 F 1 F 0 F 0 F 0 F 0 Pig H 1 H 1,5 H 0,5 H 1 H 0,5 B 1 B 0,5 B 1 B 2 B 2,5 F 0 F 0,5 F 0 F 0 F 0 Dog H 0 H 3 H 2,5 H 1,5 H 0 B 1 B 0,5 B 0 B 1 B 1,5
F 0,5 F 0 F 0 F 0 F 0,5 Mouse H 2 H 1 H 0,5 H 0,5 H 0,5
B 2,5 B 0,5 B 1 B 1 B 2 F 2 F 0 F 0 F 0 F 1
Omnivorous
Rat n.d. n.d n.d n.d n.d.
B 1,5 B 1 B 1 B 2 B 3 F 0 F 0 F 0 F 0 F 0 Carnivorous Cat H 1 H 1 H 0,5 H 0 H 0

Results
86
4.2.2 Cathelicidin reactions
The third member of the group of antimicrobial peptides analysed in this
dissertation was CAT. In this case the following antibody dilutions were used:
1:30 for Bouin and Ca-formol fixed samples and 1:150 for the HOPE® fixed
samples (compare Tab. 3.8), as the staining intensity in the latter was too
intensive in a lower dilution.
Comparable to the results described for the ß defensins, statistically relevant
differences again were observed for the equine, ovine and porcine esophagus
epithelium (compare Tab. 4.3). The differences found for the ovine tissue were
limited to the stratum basale and stratum granulosum (p-value < 0.05). It should
be emphasised, that these findings were also made comparing the Bouin
fixation with the Ca-formol fixation, and comparing Bouin with the HOPE®
fixation. In both combinations, the results varied at the same location.
Furthermore, these results confirmed those ones described for hBD-2 and -3.
The differences mentioned resulted from the fact that in the Ca-formol and
HOPE® fixed sections almost no positive IHC reaction could be observed.
The only statistically significant difference between the fixation methods in the
horse was detected comparing Bouin fixation with Ca-formol fixation, and
concerning the lower stratum basale (p-value < 0.05). The statistical analysis of
the results for the Bouin and HOPE® fixed sections of the porcine esophagus
only revealed a difference for the stratum basale (p-value < 0.05).
The comparison of the Ca-formol and HOPE® results revealed no statistical
disparity (p-values > 0.05).
Tab 4.3: IHC results of the CAT reactions according to the different fixation solutions (B= Bouin’s solution, F= Ca-formol; H= HOPE®, l= lower, u= upper, n.d.= not determined; Reaction intensities: 0.5 = very weak, 1 = weak, 1.5 = mild, 2 = medium, 2.5 = strong, 3 = very strong)
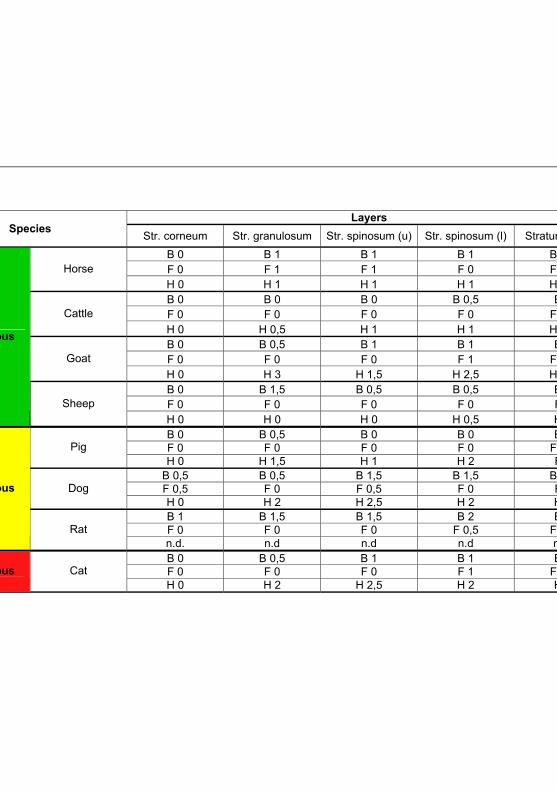
Results
87
Layers Species
Str. corneum Str. granulosum Str. spinosum (u) Str. spinosum (l) Stratum basale
B 0 B 1 B 1 B 1 B 2,5 F 0 F 1 F 1 F 0 F 2 Horse
H 0 H 1 H 1 H 1 H 2 B 0 B 0 B 0 B 0,5 B 2 F 0 F 0 F 0 F 0 F 0,5 Cattle
H 0 H 0,5 H 1 H 1 H 1,5 B 0 B 0,5 B 1 B 1 B 2 F 0 F 0 F 0 F 1 F 1,5 Goat
H 0 H 3 H 1,5 H 2,5 H 2,5 B 0 B 1,5 B 0,5 B 0,5 B 2 F 0 F 0 F 0 F 0 F 0
Herbivorous
Sheep
H 0 H 0 H 0 H 0,5 H 0 B 0 B 0,5 B 0 B 0 B 3 F 0 F 0 F 0 F 0 F 0,5 Pig H 0 H 1,5 H 1 H 2 F 2
B 0,5 B 0,5 B 1,5 B 1,5 B 2,5 F 0,5 F 0 F 0,5 F 0 F 0 Dog H 0 H 2 H 2,5 H 2 H 3 B 1 B 1,5 B 1,5 B 2 B 2 F 0 F 0 F 0 F 0,5 F 0,5
Omnivorous
Rat n.d. n.d n.d n.d n.d. B 0 B 0,5 B 1 B 1 B 2 F 0 F 0 F 0 F 1 F 1,5 Carnivorous Cat H 0 H 2 H 2,5 H 2 H 2

Results
88
4.2.3 Toll-like receptor 2 reactions
Interestingly, differences according to the fixation solutions used were found for
the equine and ovine esophagus epithelium. These findings are comparable to
those described for the APs. In the case of TLR2, statistically significant
differences for the equine esophagus were identified comparing Bouin fixation
with Ca-formol and HOPE® fixations (compare Tab. 4.4 and Fig. 4.15). It is
important to mention, that the HOPE® sections were incubated with an antibody
diluted 1:20, and for the ones of Bouin and Ca-formol fixed tissue a dilution of
1:10 was applied. Such differences were obvious in the stratum basale and the
stratum spinosum, comparing Bouin and Ca-formol fixation (p-value < 0.05).
Bouin-fixed samples exhibited a stronger staining intensity (Fig. 4.15). The
same differences were observed, comparing Bouin with the HOPE® fixation. In
this case, additionally, a significant difference was observed for the stratum
granulosum of the equine esophagus (p-value < 0.05). Reaction staining
conducted on Bouin-fixed tissue produced the strongest staining intensity in all
layers, except the stratum granulosum (compare Tab. 4.4). In the latter layer,
the HOPE® fixation method resulted in a stronger staining. In contrast to the
comparison of Bouin fixation with Ca-formol and HOPE® fixation, the
comparison of Ca-formol with HOPE® fixation only rarely showed relevant
differences for the substance studied. Regarding TLR2, a difference could be
observed for the equine esophagus. This difference was evident in the stratum
granulosum (p-value < 0.05). Intringuingly, in this case HOPE® fixed samples
showed the stronger reaction intensity.
In the ovine esophagus epithelium, a difference was apparent in the stratum
basale comparing Bouin fixation with Ca-formol and HOPE® fixation. These
observations are comparable to those for the APs studied (compare 4.2.1 and
4.2.2).
Additionally, statistically significant differences were identified in the stratum
basale of the caprine and canine esophagus. Regarding the fixation media, the
latter observations were made in the canine esophagus comparing the Bouin-
fixed samples with the Ca-formol fixed ones. The samples fixed in Bouin`s
solution exhibited the stronger reaction. In the caprine esophagus, the
difference was noticeable comparing the Bouin and HOPE ® fixation. Again the
Bouin-fixed samples revealed the stronger staining intensity.
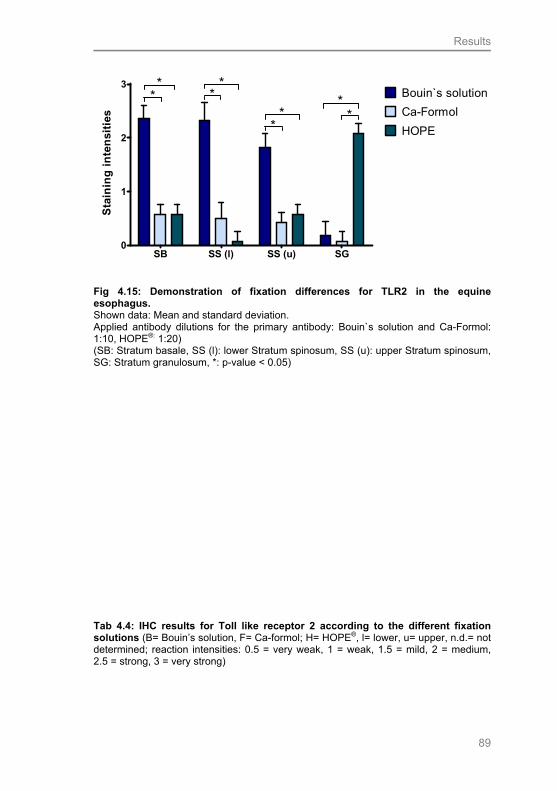
Results
89
SB SS (l) SS (u) SG0
1
2
3
Ca-Formol
Bouin`s solution
HOPE
**
**
**
**
Sta
inin
g i
nte
nsi
ties
Fig 4.15: Demonstration of fixation differences for TLR2 in the equine esophagus. Shown data: Mean and standard deviation. Applied antibody dilutions for the primary antibody: Bouin`s solution and Ca-Formol: 1:10, HOPE®: 1:20) (SB: Stratum basale, SS (l): lower Stratum spinosum, SS (u): upper Stratum spinosum, SG: Stratum granulosum, *: p-value < 0.05)
Tab 4.4: IHC results for Toll like receptor 2 according to the different fixation solutions (B= Bouin’s solution, F= Ca-formol; H= HOPE®, l= lower, u= upper, n.d.= not determined; reaction intensities: 0.5 = very weak, 1 = weak, 1.5 = mild, 2 = medium, 2.5 = strong, 3 = very strong)
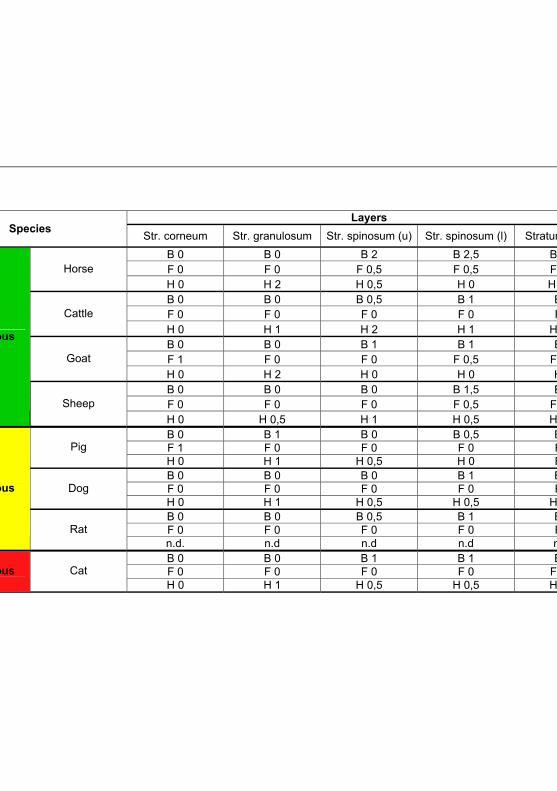
Results
90
Layers Species
Str. corneum Str. granulosum Str. spinosum (u) Str. spinosum (l) Stratum basale
B 0 B 0 B 2 B 2,5 B 2,5 F 0 F 0 F 0,5 F 0,5 F 0,5 Horse
H 0 H 2 H 0,5 H 0 H 0,5 B 0 B 0 B 0,5 B 1 B 2 F 0 F 0 F 0 F 0 F 0 Cattle
H 0 H 1 H 2 H 1 H 0,5 B 0 B 0 B 1 B 1 B 2 F 1 F 0 F 0 F 0,5 F 0,5 Goat
H 0 H 2 H 0 H 0 H 0 B 0 B 0 B 0 B 1,5 B 2 F 0 F 0 F 0 F 0,5 F 0,5
Herbivorous
Sheep
H 0 H 0,5 H 1 H 0,5 H 0,5 B 0 B 1 B 0 B 0,5 B 1 F 1 F 0 F 0 F 0 F 1 Pig H 0 H 1 H 0,5 H 0 F 0 B 0 B 0 B 0 B 1 B 2 F 0 F 0 F 0 F 0 F 0 Dog H 0 H 1 H 0,5 H 0,5 H 0,5 B 0 B 0 B 0,5 B 1 B 2 F 0 F 0 F 0 F 0 F 1
Omnivorous
Rat n.d. n.d n.d n.d n.d. B 0 B 0 B 1 B 1 B 2 F 0 F 0 F 0 F 0 F 0,5 Carnivorous Cat H 0 H 1 H 0,5 H 0,5 H 0,5

Results
91
4.2.4 Lysozyme reactions
The only difference between the fixation solutions used was found for the
equine esophagus (Tab. 4.5). The lysozyme antibody for the Ca-formol and
Bouin fixed samples was applied in a 1:50 dilution and for the HOPE® fixed
ones in a 1:2000 one. In contrast to most of the other substances studied, a
difference was also identified comparing the Ca-formol fixation with the HOPE®
fixation. In this case, a disparity appeared in the upper stratum spinosum (p-
value < 0.05). Surprisingly, the samples fixed in Ca-formol exhibited the
stronger reactions. The comparison of Bouin and HOPE® revealed a difference
between the stratum basale and lower stratum spinosum (p-values < 0.05).
Again, the staining intensity in the HOPE® fixed samples was weaker. For the
Bouin - Ca-formol comparison, a difference was only shown for the stratum
basale (p-value < 0.05), where the Bouin-fixed tissue revealed stronger
reactions. The other species studied revealed no disparities (p-values > 0.05).
Tab. 4.5: IHC results for lysozyme according to the different fixation solutions (B= Bouin’s solution, F= Ca-formol; H= HOPE®, l= lower, u= upper, n.d.= not determined, reaction intensities: 0.5 = very weak, 1 = weak, 1.5 = mild, 2 = medium, 2.5 = strong, 3 = very strong)

Results
92
Layers Species
Str. corneum Str. granulosum Str. spinosum (u) Str. spinosum (l) Str. basale B 0,5 B 1,5 B 1,5 B 1,5 B 2,5 F 0 F 2 F 2 F 1 F 1 Horse
H 0 H 1,5 H 0,5 H 0 H 1 B 0 B 0 B 0 B 1 B 2,5 F 2 F 0,5 F 1,5 F 2 F 1 Cattle
H 0 H 0,5 H 1 H 0,5 H 0,5 B 0 B 1 B 1 B 1 B 1
F 0,5 F 0,5 F 0 F 0 F 0,5 Goat
H 2 H 2 H 0 H 0 H 2 B 0 B 2 B 0 B 0,5 B 1,5 F 1 F 1 F 0,5 F 0,5 F 1
Herbivorous
Sheep
H 0 H 2 H 0,5 H 1 H 1 B 1 B 1 B 0 B 0 B 2
F 0,5 F 0 F 0 F 0,5 F 0,5 Pig H 1 H 1,5 H 1 H 0 H 2 B 0 B 0,5 B 0,5 B 1,5 B 2 F 1 F 1 F 1 F 0 F 2 Dog H 0 H 1,5 H 1 H 1 H 1 B 0 B 0 B 0 B 0 B 2
F 0,5 F 0,5 F 0,5 F 0,5 F 1 Mouse H 1,5 H 2 H 1,5 H 1,5 H 1 B 0 B 0 B 0 B 0 B 2
F 0,5 F 0,5 F 0,5 F 0,5 F 1
Omnivorous
Rat n.d. n.d n.d n.d n.d. B 3 B 0 B 0 B 1 B 2,5 F 1 F 0 F 1,5 F 2 F 2 Carnivorous Cat H 0 H 1 H 1 H 1 H 1,5

Results
93
4.3 IHC results for the substances of the innate immune
system: Species comparison
4.3.1 Demonstration of ß-defensin 2
Due to the fact that the Bouin-fixed esophagus tissues revealed the best structural
and most reliable IHC results, the following descriptions will primarily focus on
observations made after using this fixation medium. However, significant findings
from HOPE® or Ca-Formol fixed tissue concerning the distribution pattern of the
analysed substances will also be mentioned.
As we aimed to compare the occurrence and distribution pattern of the substances
studied between the three important nutrition groups (herbivorous, omnivorous,
carnivorous), the description of the findings will be accomplished focussing on
each group. In order to identify significant differences between the three nutrition
types, statistical tests were applied (compare 3.9.7), and each layer of the
epithelium was considered.
Almost all herbivorous species studied displayed the same distribution and
reaction pattern for hBD-2 within the esophagus epithelium: A strong reaction was
observed in the stratum basale, upwards the staining intensity decreased in the
stratum spinosum but was up-regulated again in the stratum granulosum, except
in the cattle, where no reaction in the stratum granulosum could be found. The
goat exhibited the strongest reaction intensities in all layers. Within the cells, the
positive staining was limited to the cytoplasm. In all herbivorous species, a positive
reaction could be detected in the stratum corneum, even though the horse only
showed a weak staining. The positive staining in the stratum corneum was limited
to the most luminal part, and the reacting granules seemed to cover the epithelium
in the form of a thin layer. The cells of the stratum corneum were completely
negative.
In the omnivorous species, the distribution pattern of hBD-2 was almost the same
as demonstrated for the herbivorous animals. The strongest reactions were found
in the esophagus epithelium of the laboratory rat (Fig. 4.16). In contrast to the

Results
94
other omnivorous species, the dog showed no increase, but a decrease of staining
intensity in the stratum granulosum (Fig. 4.18).
The cat, as a typical carnivorous animal, not surprisingly displayed a distribution
pattern of hBD-2 comparable to that one of the dog, despite the fact that no
staining was observed in the upper stratum spinosum and the stratum granulosum
(Fig. 4.20).
Fig 4.16: hBD-2 reaction in the esophagus epithelium of the rat Very strong reaction intensity in the SG, Bouin fixation SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.17: IgG rabbit isotype control for hBD-2 for the esophagus of the rat The IgG rabbit isotype control was applied in the same protein concentration as the primary antibody, Bouin fixation SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
95
Fig. 4.18: hBD-2 reaction in the esophagus epithelium of the dog Keep attention to the decrease of staining intensity in the SG Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.19: Rabbit IgG isotype control for hDB-2 in the esophagus of the dog The isotype control was applied in the same protein concentration as the primary antibody. Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
96
The comparison of the herbivorous and the omnivorous species revealed no
statistically significant difference in the staining intensity of the esophagus samples
fixed in Bouin`s solution (p-value > 0.05). In contrast, significant differences could
be observed comparing herbivorous and omnivorous species with the carnivorous
cat (Fig. 4.21). Differences in staining intensity were found for the stratum
corneum and the stratum granulosum, comparing herbivorous species and the cat
(p-value < 0.05). In the stratum corneum, the cat displayed a stronger reaction,
whereas almost no reaction was observed in the stratum granulosum of the
esophagus of this species. The comparison of the omnivorous animals with the cat
exhibited a significantly stronger reaction only in the stratum corneum of the cat (p-
value < 0.05).
In contrast to the Bouin-fixed tissue, a difference between the herbivorous and
omnivorous species was observed for the HOPE® fixed samples. Statistically
significant differences were found for the stratum corneum and the lower and
upper stratum spinosum (p-value < 0.05). The omnivorous species studied reacted
slightly stronger. However, no differences were found comparing the herbivorous
Fig. 4.20: hBD-2 reaction in the esophagus epithelium of the cat Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
97
or omnivorous animals with the carnivorous cat (p-value > 0.05), as related to the
Bouin-fixed material.
SG SC0
1
2
3
4
*
* Herbivorous species
Omnivorous species
Carnivorous species
*
Sta
inin
g i
nte
nsi
ties
Fig 4.21: Differences in staining intensities in Bouin-fixed samples between the three nutrition groups for hBD-2. Data shown: Mean and standard deviation of staining intensities. (SB: Stratum basale, SS: Stratum spinosum, *: p- value < 0.05)
4.3.2 Demonstration of ß-defensin 3
Concerning the herbivorous species, the cattle and the sheep expressed almost
the same distribution pattern of hBD-3 (compare Tab 4.2). Therefore, the most
prominent expression of this antimicrobial peptide (AP) was observed in the
stratum basale (Fig. 4.22 and 4.23). Within the stratum spinosum, only for the
lower part of this stratum of the cattle a slightly positive reaction was observed,
whereas the upper part of the stratum spinosum reacted completely negative in
both species. An up-regulation of reaction intensity was demonstrated in the
stratum granulosum of the esophagus epithelium in the cattle and the sheep. The
sheep showed a slightly weaker staining in the stratum corneum. In the cattle, the
reaction intensity was consistently strong within both layers. Similar to the
distribution of hBD-2, a reaction of the cells in the stratum corneum was absent in
all species, but the epithelium surface was covered by a thin layer of granules
reacting positively for hBD-3. In the esophagus epithelium of the horse and the
goat, the hBD-3 reaction was also mostly strong within the stratum basale. This
degree of reaction intensity remained constant in the stratum spinosum of the
horse, and appeared significantly diminished in the stratum granulosum and
stratum corneum. In the last two layers only a weak reaction could be

Results
98
demonstrated. The goat displayed almost the same distribution pattern, albeit of
the fact that in this species no positive reaction staining was found in the stratum
granulosum.
In the omnivorous species, the pig expressed a distribution of hBD-3 similar to the
sheep, as both species exhibited a strongly positive reaction in the stratum basale
and no staining in the stratum spinosum, before the staining was up-regulated
again in the stratum granulosum and remained constant in the stratum corneum
(Fig 4.26). The mouse and the rat displayed a strong reaction in the stratum
basale, a slightly weaker one in the stratum spinosum and stratum granulosum,
before the staining intensity increased again in the stratum corneum. Interestingly,
the reaction in stratum corneum of the rat was the most prominent one of all
species.
The cat and the dog showed comparable results as the reaction intensity
decreased continuously from a very strong one in the stratum basale towards a
weak one in the stratum granulosum; the stratum corneum again displayed a
stronger reaction. Comparing the reaction intensities of the different species, it is
of great interest that the cat showed the strongest reaction intensities of all the
species studied in the stratum basale.
Fig 4.22: Demontration of hBD-3 in the esophagus epithelium of the cattle Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
99
Fig. 4.23: Demonstration of hBD-3 in the ovine esophagus epithelium hBD-3 is up-regulated in the SG Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.24: Negative control for hBD-3 in the ovine esophagus First antibody was omitted and replaced by PBS+1% BSA. Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
100
Fig. 4.25: Positive control for hBD-3 As positive control tissue equine skin was used and the primary antibody was applied in the same concentration as for the reactions on the esophagus tissue (1:2000), a hair follicle (with hair) in the skin is marked by: →, Bouin fixation
Fig. 4.26 Demontration of hBD-3 in the esophagus epithelium of the pig hBD-3 is up-regulated in the SG, SB: Stratum basale, SS: Stratum spinosum, SG: Str granulosum, SC: Str. corneum
Fig. 4.27: IgG rabbit isotype control for hBD-3 Demonstrated for the porcine esophagus, IgG and hBD-3 antibody were applied in the same protein concentrations, for abbr. see Fig. 4.26

Results
101
The comparison of the three nutrition types displayed similar results, as described
for the hBD-2 reactions. Statistical analysis comparing the results for the Bouin-
fixed tissue of the herbivorous and omnivorous animals studied showed no
differences between these two nutrition groups (p-value > 0.05). Only in the
stratum basale of the herbivorous animals and the carnivorous cat a significant
variation in staining intensity could be observed (p-value < 0.05), the cat exhibiting
the strongest reaction (Fig. 4.28). The same situation was found comparing the
omnivorous species with the cat, but the cat, additionally, showed a significantly
stronger reaction in the lower stratum spinosum (p-value < 0.05).
Comparable to the results found for hBD-2 with the HOPE® fixed samples, a
statistical difference for hBD-3 was also observed for the herbivorous and
omnivorous species (p-value < 0.05), even though this variation was only present
in the stratum corneum. However, in contrast to the herbivorous animals, a slightly
positive reaction could be noticed for the omnivorous species in this layer.
SB SS (l)0
1
2
3
4
**
*
Herbivorous species
Omnivorous species
Carnivorous species
Sta
inin
g i
nte
nsi
ties
Fig 4.28: Differences in staining intensities in Bouin-fixed samples between the three nutrition groups for hBD-3. Data shown: Mean and standard deviation of staining intensities. (SB: Stratum basale, SS (l): lower Stratum spinosum, *: p- value < 0.05)

Results
102
4.3.3 Demonstration of cathelicidin
All herbivorous species reacted strongly positively for the AP cathelicidin (CAT) in
the stratum basale. In the horse, the reaction intensity decreased to a weak one in
the stratum spinosum and remained stable up to the stratum granulosum, before it
decreased in the stratum corneum (Fig. 4.29). The goat almost showed the same
reaction pattern, despite the fact that the reaction within the upper stratum
spinosum and the stratum granulosum was very weak (Fig. 4.31). The cattle and
the sheep both displayed a very weak reaction in the lower stratum spinosum,
before the staining disappeared in the upper stratum spinosum. In contrast to the
cattle, the sheep reacted strongly positive for CAT within the stratum granulosum.
The positive reactions were limited to the cytoplasm of the cells. Concerning the
stratum corneum, no positive reaction was demonstrated within the cells covering
the epithelium.
The pig was the species that showed the strongest reaction intensity of all
mammals studied within the stratum basale. Interestingly, in many pigs the
positive reactions were exclusively evident in this layer of the esophagus
epithelium, whereas in the other species the AP again was identified within the
stratum granulosum. Both, the dog and the rat displayed a strong reaction within
the stratum spinosum. In the stratum granulosum of the rat the intensity remained
constantly high, whereas it decreased in this layer of the canine esophagus (Fig.
4.32). Intriguingly, the last two mentioned species were the only ones reacting
positively for CAT within the stratum corneum cells, showing a thin layer of
granules reacting positively for CAT on the epithelium.
The distribution pattern of CAT in the canine epithelium was the same as detected
for the esophagus epithelium of the cat, despite the fact that the latter species
exhibited no positive reaction within the stratum corneum (Figs 4.32 and 4.33).

Results
103
Fig. 4.29: Demonstration of CAT in the esophagus epithelium of the horse The reaction intensity in SB is strong. Bouin fixation, SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.30: IgG mouse isotype control for CAT Demonstrated for the equine esophagus epithelium. Bouin fixation, SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
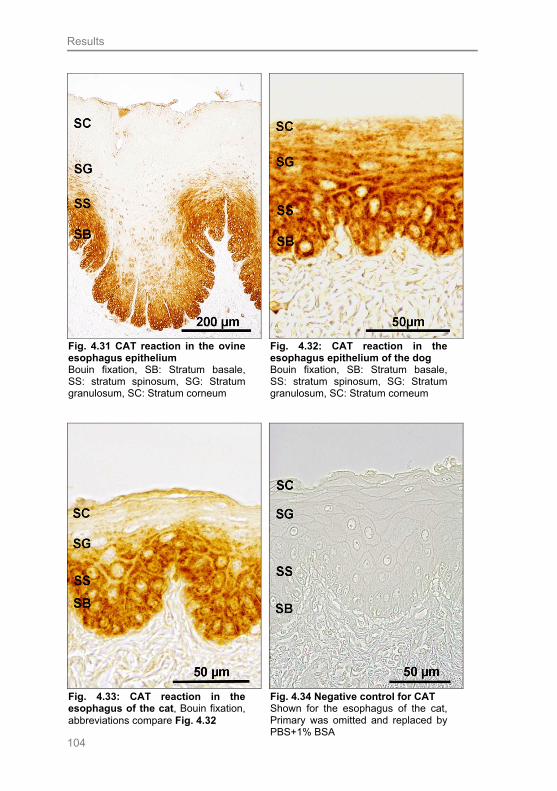
Results
104
Fig. 4.31 CAT reaction in the ovine esophagus epithelium Bouin fixation, SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.32: CAT reaction in the esophagus epithelium of the dog Bouin fixation, SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.33: CAT reaction in the esophagus of the cat, Bouin fixation, abbreviations compare Fig. 4.32
Fig. 4.34 Negative control for CAT Shown for the esophagus of the cat, Primary was omitted and replaced by PBS+1% BSA

Results
105
Similar to the other APs, for CAT no variation between herbivorous and
omnivorous species could be detected in Bouin-fixed samples (p-value > 0.05). In
contrast to the ß-defensins, no difference between the three nutrition types could
be observed for any of the layers.
The reactions found in the HOPE® fixed samples slightly differed from those ones
observed for Bouin-fixed material. Comparing the herbivorous with the omnivorous
animals a significant difference between the nutrition groups was found (p-value <
0.05). This variation was particularly present in the stratum granulosum and the
stratum spinosum. The comparison of the other nutrition types corroborated the
results found for Bouin-fixed tissue. The goat, particularly, showed a very strong
reaction in these layers compared to the omnivorous species.
4.3.4 Demonstration of Toll-like receptor 2
In all herbivorous species studied a strongly positive reaction for TLR2 was
observed in the stratum basale (Fig. 4.35). This reaction slightly diminished
towards the lower stratum spinosum in all the herbivorous animals, nevertheless in
the horse this reduction occurred in the upper stratum spinosum. The more luminal
esophagus layers, the stratum granulosum and the stratum corneum, generally
displayed negative reactions in the herbivorous group.
Resembling the situation in the herbivorous animals, in the omnivorous species a
strong reaction for TLR2 was generally discernable in the stratum basale (Fig.
4.36). Likewise, a reduction of reaction intensity towards the stratum spinosum
was found in all the omnivorous species studied. A significant finding was detected
for the pig, exhibiting a weak reaction within the stratum granulosum, whereas all
the other animals showed no reaction within this layer. It should be emphasised,
that it was not possible to detect TLR2 in the stratum corneum of any of the
species studied.
The positive reaction for TLR2 in the cat was limited to the stratum basale and the
stratum spinosum (Fig. 4.37). Comparable to the results found for the APs, the cat
and the dog displayed the same distribution of reaction patterns for TLR2 within
the esophagus epithelium.
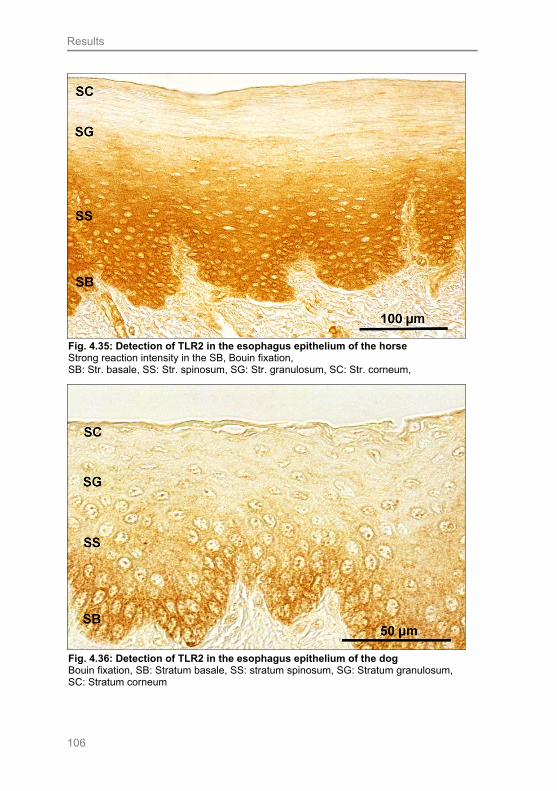
Results
106
Fig. 4.35: Detection of TLR2 in the esophagus epithelium of the horse Strong reaction intensity in the SB, Bouin fixation, SB: Str. basale, SS: Str. spinosum, SG: Str. granulosum, SC: Str. corneum,
Fig. 4.36: Detection of TLR2 in the esophagus epithelium of the dog Bouin fixation, SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum

Results
107
For the Bouin-fixed samples, in four of the five layers of the esophagus epithelium
a statistically significant difference was demonstrated for the herbivorous animals
compared to the omnivorous species (p-value < 0.05) (Fig. 4.38). All layers,
except the stratum corneum, exhibited differences. Primarily the herbivorous
animals showed distinct reactions in the different layers. A stronger reaction in the
omnivorous species was exclusively evident for the stratum granulosum. As
already mentioned, the pig was the only species showing a distinct reaction in this
layer. For the herbivorous and carnivorous species studied a difference was only
found for the lower stratum spinosum (p-value < 0.05), which reacted slightly
stronger in the herbivorous species. The statistical analysis of the omnivorous and
the carnivorous cat revealed a difference between these two nutrition types in the
upper stratum spinosum and stratum granulosum (p-value < 0.05). As the pig was
the only species exhibiting a positive reaction in the stratum granulosum, the cat,
of course, showed the weaker staining intensities. In contrast, the reaction
intensities in the upper stratum spinosum were higher in the cat compared to the
omnivorous species. In contrast, in HOPE® fixed samples no differences between
the three nutrition groups could be observed.
Fig. 4.37: Detection of TLR2 in the esophagus epithelium of the cat Bouin fixation, SB: Stratum basale, SS: stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
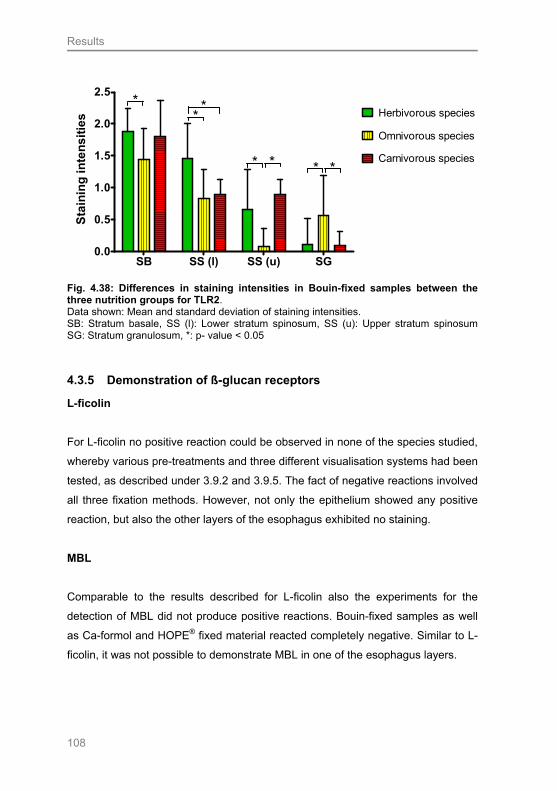
Results
108
SB SS (l) SS (u) SG0.0
0.5
1.0
1.5
2.0
2.5* Herbivorous species
Omnivorous species
Carnivorous species** *
**
*
Sta
inin
g i
nte
ns
itie
s
Fig. 4.38: Differences in staining intensities in Bouin-fixed samples between the three nutrition groups for TLR2. Data shown: Mean and standard deviation of staining intensities. SB: Stratum basale, SS (l): Lower stratum spinosum, SS (u): Upper stratum spinosum SG: Stratum granulosum, *: p- value < 0.05
4.3.5 Demonstration of ß-glucan receptors
L-ficolin
For L-ficolin no positive reaction could be observed in none of the species studied,
whereby various pre-treatments and three different visualisation systems had been
tested, as described under 3.9.2 and 3.9.5. The fact of negative reactions involved
all three fixation methods. However, not only the epithelium showed any positive
reaction, but also the other layers of the esophagus exhibited no staining.
MBL
Comparable to the results described for L-ficolin also the experiments for the
detection of MBL did not produce positive reactions. Bouin-fixed samples as well
as Ca-formol and HOPE® fixed material reacted completely negative. Similar to L-
ficolin, it was not possible to demonstrate MBL in one of the esophagus layers.

Results
109
Dectin-1
Experiments conducted on the esophagus tissue of all studied species did not
show positive results, pertaining to all fixation solutions. None of the esophagus
layers showed a positive reaction.
4.3.6 Demonstration of lysozyme
Appling IHC for lysozyme revealed very strong positive reactions within the
stratum basale of the horse and the cattle. The staining intensity diminished
towards the lower stratum spinosum in both species. The weak reaction in the
cattle disappeared towards all following upper layers. In the horse, the reaction in
the lower stratum spinosum was slightly stronger and continuous up to the stratum
granulosum. A positive reaction within the stratum corneum of the herbivorous
animals was only demonstrable for the horse, which showed a weak reaction. In
the stratum basale of the goat and the sheep, the reaction intensity for lysozyme
was slightly reduced, in comparison to the other herbivorous animals. The goat
expressed the same weak reaction intensity throughout all layers, except for the
stratum corneum where the reaction decreased. Comparable to the distribution
pattern of CAT, the reaction intensity of lysozyme in the sheep again increased in
the stratum granulosum, after it had decreased in the stratum spinosum.
In the mouse and the rat, the positive reaction for lysozyme was limited to the
stratum basale, whereas the other layers showed no positive staining (Fig. 4.39).
A significant positive reaction in the pig was demonstrated in the stratum basale.
The stratum spinosum reacted negatively, the stratum granulosum and stratum
corneum only showed a weakly positive staining. A continuous decrease of
reaction intensity towards the lumen was demonstrated for the esophagus
epithelium of the dog and this species displayed no positive reaction within the
stratum corneum.
In the cat the most prominent reaction was found in the stratum basale and also in
the stratum corneum. It should be emphasised that the cat expressed the
strongest reaction of all species in the stratum corneum. The layers between the
stratum basale and the stratum corneum of the cat generally displayed no positive
reaction.
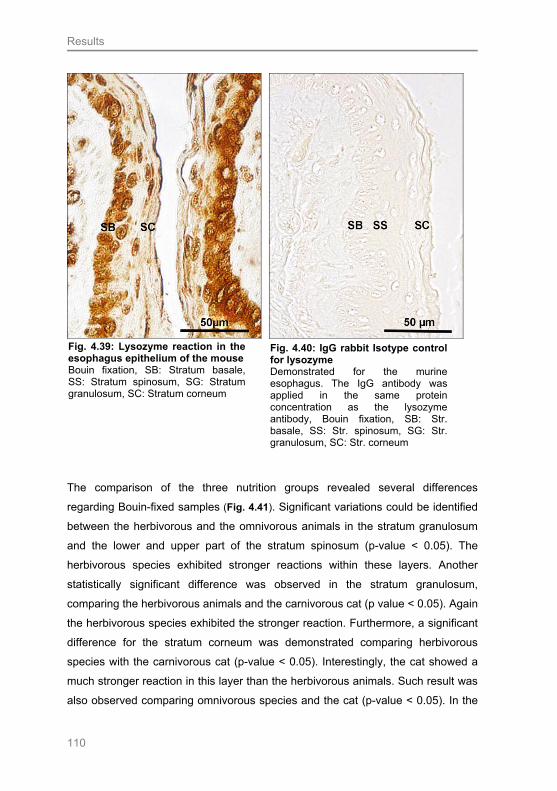
Results
110
The comparison of the three nutrition groups revealed several differences
regarding Bouin-fixed samples (Fig. 4.41). Significant variations could be identified
between the herbivorous and the omnivorous animals in the stratum granulosum
and the lower and upper part of the stratum spinosum (p-value < 0.05). The
herbivorous species exhibited stronger reactions within these layers. Another
statistically significant difference was observed in the stratum granulosum,
comparing the herbivorous animals and the carnivorous cat (p value < 0.05). Again
the herbivorous species exhibited the stronger reaction. Furthermore, a significant
difference for the stratum corneum was demonstrated comparing herbivorous
species with the carnivorous cat (p-value < 0.05). Interestingly, the cat showed a
much stronger reaction in this layer than the herbivorous animals. Such result was
also observed comparing omnivorous species and the cat (p-value < 0.05). In the
Fig. 4.39: Lysozyme reaction in the esophagus epithelium of the mouse Bouin fixation, SB: Stratum basale, SS: Stratum spinosum, SG: Stratum granulosum, SC: Stratum corneum
Fig. 4.40: IgG rabbit Isotype control for lysozyme Demonstrated for the murine esophagus. The IgG antibody was applied in the same protein concentration as the lysozyme antibody, Bouin fixation, SB: Str. basale, SS: Str. spinosum, SG: Str. granulosum, SC: Str. corneum

Results
111
latter, another difference was found for the lower stratum spinosum, again the cat
here displayed a stronger reaction intensity.
The immunohistochemical experiments conducted using the HOPE® fixed samples
brought about results consistent for the three nutrition types. Only in the upper
stratum spinosum a significant difference between the herbivorous and
omnivorous species was detected (p-value < 0.05). The omnivorous species
displayed slightly stronger reactions.
SS (l) SS (u) SG SC0
1
2
3
4
*
* Herbivorous species
Omnivorous species
Carnivorous species
*
**
**
Sta
inin
g i
nte
nsi
ties
Fig. 4.41: Differences in staining intensities in Bouin-fixed samples between the three nutrition groups for lysozyme. Data shown: Mean and standard deviation of staining intensities. SS (l): Lower Stratum spinosum, SS (u): Upper Stratum spinosum SG: Stratum granulosum, SC: Stratum corneum, *: p- value < 0.05

Results
112
4.3.7 Demonstration of Langerhans cells
Langerin
Langerin (+) LCs were sporadically detected in the esophagus of the mouse and
the horse (Fig. 4.42). In both animal species such cells were organised within the
stratum basale. In the other species studied no positive reactions for langerin (+)
LCs could be demonstrated.
CD1a
Applying an antibody against the surface molecule CD1a of Langerhans cells did
not produce positive results. In contrast to langerin, none of the species studied
reacted positively. The results did not differ between the three various fixation
solutions used.
Fig. 4.42: Demontration of langerin + LCs in the esophagus epithelium of the horse A langerin (+) LCs is marked by a: →, Bouin fixation

Discussion
113
5 Discussion
5.1 Fixation experiment
5.1.1 Influences of fixation on structure preservation
One aim of this thesis was to compare three different fixation media and to
evaluate their particular influence on the preservation of the structure of the
esophagus tissue and on the reactions of the IHC experiments performed. The
three fixation media used were: Bouin’s solution (after BÖCK 1989), Calcium
buffered formalin (after LILLIE and FULLMER 1976) (Ca-formol) and the HOPE®
fixation technique (after OLERT 2001). The first two fixation solutions are for the
most part formalin based (additionally Bouin`s medium is supplemented with picric
acid). Due to the fact that formalin is a hazardous substance and bears several
other disadvantages, e.g. masking of antigen epitopes by cross-linking proteins,
new alternatives are more and more in the focus of attention (OLERT et al. 2001;
BUESA 2008). We demonstrated that all samples fixed in Bouin`s solution
revealed an excellent tissue structure. In contrast, approximately 35 % of the
HOPE® fixed samples showed a decrease in the quality of tissue preservation.
This means in detail that tissue integrity was not always guaranteed. For example,
the epithelium of the esophagus was separated from the lamina propia and
revealed distinct clefts. In approximately 60 % of the Ca-formol fixed samples, the
quality of structure preservation was substantially reduced. The integrity of the
epithelium and the lamina propia was destroyed. An ideal fixation should preserve
the original structure of the tissue as good as possible and should be able to
provide an equivalent close to the natural state (BÖCK 1989). This state can be
reached by fast penetration of the fixation fluid into the tissue, thus avoiding
autolysis and make feasible rapid conservation (PEARSE 1985; BÖCK 1989).
Under these circumstances the best preservation of the structure as well as
continuity of the esophagus tissue were obtained using Bouin`s fixation medium.
The HOPE® fixation showed the next best results, followed by the relatively
unsatisfactory results obtained after Ca-formol fixation. OLERT et al. (2001)
claimed good formalin-like structure preservation for HOPE® fixed samples. The

Discussion
114
comparison of our results with the findings of the latter authors emphasises a
clearly controversial state on this matter. Unlike the fixation media we used,
common formalin (not buffered formaldehyde in aqueous solution) was applied in
the latter study. On one hand we can state, that tissue preservation of HOPE® was
even superior to Ca-formol fixation, but, on the other hand, it was not as good as
Bouin fixation. Studies conducted on HOPE® fixed human placenta demonstrated
that this fixation can result in a structural quality that is superior to cryo sections
(BLASCHNITZ et al. 2008). The comparison of HOPE® and formalin fixation was
also subject to other studies. ANDREI et al. (2008) observed that structural
alterations in the human uterine cervix, such as nuclear distortion and epithelial
clefts, were minimal in HOPE® fixed samples. The findings of a study comparing
structure preservation of ovine lung tissue fixed in 10% neutral buffered formalin
(NBF) or Bouin`s solution displayed no differences in the structure preservation
between the two fixatives used (BENAVIDES et al. 2006).
Specimen collection
The process of fixation is influenced by several different factors. The first critical
aspect is specimen collection, whereby several factors have to be taken into
account to obtain representative results. It was important to keep the time period
between tissue removal and its immersion in the fixation solution as short as
possible. In our study, all samples were obtained from freshly dead animals. Due
to the fact that the samples fixed in the solution HOPE® I had to be very small, the
preparation of this material took some time. On the one hand, this fact could
explain the slightly reduced structure quality of the HOPE® fixed samples, on the
other hand, however, the time period between sampling and immersing the tissue
was short (< 15 minutes), and the samples were constantly cooled on an icepack.
As the Bouin and Ca-formol samples were treated identically, the tissue collection
and preparation could not cause the reduced quality of structure preservation of
the Ca-formol fixed samples observed.

Discussion
115
Chemical characteristics of the fixation media
An additional fact, which has to be taken into account, is the chemical composition
of the fixation solutions used. To some extent, Bouin and Ca-formol fixation are
both based on formalin, which belongs to the chemical group of aldehyds. Fixation
with these chemicals is a complex process including a rapid penetration that stops
autolysis, followed by covalent binding and cross-linking. These three parts of
fixation proceed simultaneously, but at very different rates (penetration is 12 times
faster than binding) (BÖCK 1989; BUESA 2008). In contrast to Ca-formol, Bouin`s
solution additionally contains picric acid and glacial acetic acid. Bouin`s solution
has the advantage of penetrating into the tissue more rapidly, and therefore it
produces good tissue structure excluding shrinkage (LILLIE and FULLMER 1976).
This fact is in accordance with our findings, as the use of Bouin`s solution resulted
in the best morphological preservation of all tissue samples collected. The fast
penetration rate of Bouin`s solution is perhaps the most relevant explanation for
our findings. In this context, it is particularly difficult to explain the fixation process
of the HOPE® fixed samples, as the selling company (DCS) offers no detailed
description of the ingredients of the HOPE® I solution (also named protection
solution) and the HOPE® II solution. OLERT et al. (2001), who introduced the
HOPE® fixation into histological research, stated that HOPE® I solution is a
hyperosmolar mixture of different amino acids (10-100 mMol), with a pH of 5.8-6.4,
and works like an immersion fixative. In the second step, the tissue is immersed
into a mixture of HOPE® II and acetone (absolute), followed by an incubation in
acetone (absolute). Acetone acts as a dehydrating agent, according to the
specifications of the manufacturer, and replaces the increasing ethanol
concentrations used for the embedding of Bouin or Ca-formol fixed tissue. The
HOPE® solutions are considered to protect the tissue from protein cross-linking
occurring during dehydration and incubation in the low melting paraffin. From our
point of view, the HOPE® fixation seems to be based mainly on acetone fixation.
Acetone fixates tissue by coagulation of the proteins present. Pure acetone lacks
popularity as routine fixative as it does not preserve tissue structure as good as,
for example, formaldehyde (WERNER et al. 2000). A negative side effect of using
acetone is that it can cause strong shrinkage artefacts, as also confirmed by our
observations. After a histological staining, HOPE® fixed tissue often equally

Discussion
116
displayed shrinkage artefacts, such as the epithelium rolling up. As acetone is a
potent dehydration medium, a solid crust at the periphery of the tissue might be
formed and cause diminished penetration rates of the paraffin. This aspect might
be an explanation for the lower quality of tissue structure of the HOPE® fixed
samples compared to the Bouin fixed samples.
Influence of the temperature
Another critical aspect is the temperature during fixation. Whereas fixation in Ca-
formol and Bouin`s solution are fairly independent of temperature influences and
can be mainly conducted at room temperature, the first steps of the HOPE®
fixation have to be done under constant cooling. In general, low temperatures
retard autolysis, but also decrease the diffusion rate and thus prolong penetration.
In our experience, it can be concluded that it is particularly important to have a
temperature of not more than 4°C during the incubation in HOPE® I and HOPE® II
solutions. Even only slightly higher temperatures resulted in a reduced quality of
tissue preservation.
Influence of the duration of fixation
Another important aspect influencing structure quality is fixation duration. For
formalin based solutions, a time of less than 24 hours generally results in a
mixture of formalin and ethanol fixation. An interruption of formalin fixation before it
is completed will lead to cross-linking only at the tissue periphery. Coagulation
occurs near the centre, caused by the ascending ethanol solutions during tissue
dehydration, or the centre of the tissue sample remains unfixed (WERNER et al.
2000). In our case, the possibility of a fixation period which was too short and
caused reduced tissue quality of the Ca-Formol fixed samples can be neglected,
as we incubated the samples for 30-40 hours. Bouin`s fixative is suitable for a
longer fixation without causing over fixation (BÖCK 1989; MEYER personal
communication); however, tissue fixed in Ca-formol is susceptible for such
problem. Over fixation may lead to excessive cross-linking and cause irreversible
damage of epitopes. Moreover, this damage of epitopes affects the reactions of
IHC experiments (compare 5.1.2) (WERNER et al. 2000; VAN ALSTINE et al.

Discussion
117
2002; BLASCHNITZ et al. 2008). In the HOPE® fixed esophagus tissue, over
fixation did not seem to be much of a problem. However, we incubated the tissue
12-72 hours according to the instruction of the manufacturers. To evaluate the
possible influence of over fixation on the HOPE® fixed samples, further
experiments with varying incubation times are necessary.
According to the structural quality of the esophagus tissues, it can be concluded
that the samples fixed in Bouin`s solution produced the best results followed by
the HOPE® fixed samples. Regarding the latter findings, some tissue damage
might be due to wrong handling of the tissue (tissue pieces may have been too
large). Another reason for the slightly reduced quality of structure preservation
might be the influence of acetone, in particular on the esophagus tissues, as
experiments on human placenta and uterus revealed good tissue preservation
(ANDREI et al. 2008; BLASCHNITZ et al. 2008). The esophagus is composed of
several different tissues types including accurately defined cell structures and
functions. For example, the epithelium contains a great amount of keratins
(SCHOENNAGEL 2005), in contrast to other organs. Thus, the slightly reduced
tissue preservation in our study might be a result of these particular tissue
characteristics. According to the diminished structural quality of Ca-formol fixed
tissue, it might be presumed that the penetration rate of the fluid is rather slow in
comparison to picric acid and acetic acid (glacial) that are added to Bouin`s
solution. As already emphasised earlier (LILLIE and FULLMER 1976; JAMES and
TAS 1984; PEARSE 1985), a mixture of different components is the still the best
way to achieve high quality tissue preservation.
5.1.2 Influences of fixation on immunohistochemical results
One aim of this thesis was to discuss the influence of fixation on the quality of the
IHC results obtained from the esophagus epithelium. In this context we provide the
first comprehensive study on the fixation of the mammalian esophagus, comparing
the use and effects of three different fixation solutions. One central question of our
study was, whether or not the staining intensities of an IHC reaction in the five
layers of the esophagus epithelium differed depending on the fixation media used.
Therefore, the staining intensity was evaluated with a semi quantitative scale
system (0= no reaction up to 3= very strong reaction). This comparison was

Discussion
118
controlled by statistical analyses of the semiquantitative data obtained. The
findings can already be summarised to a first general statement: Our study
demonstrated no general significant differences in the staining intensities between
the fixation media used, even though a difference could be shown in certain layers
of the esophagus epithelium in certain species for some antibodies. Discussing
and evaluating the reasons for these results is nevertheless of great importance.
Although the IHC findings did not differ significantly between the three fixation
media applied, it is, firstly, important to consider the influences of fixation on the
structural preservation of the esophagus tissue (compare 5.1.1). Only samples
with the best preservation quality were taken into the IHC procedure. Such feature
was evaluated after routine H.E. staining, and only tissue sections of high
structural quality were used for the IHC experiments. In this connection, almost all
samples fixed in Bouin`s solution could be taken. In contrast, only approximately
40% of the Ca-formol and 65% of the HOPE® fixed samples were usable. Due to
the great quantity of the samples collected, tissue sections from almost every
animal species used were available for IHC analyses.
Influences on the quantity of antibodies
One astonishing finding of several former studies was that the quantities of the
primary antibodies could be reduced using HOPE® fixed tissue, in comparison to
formalin-fixed tissue. These observations are corroborated by findings of
GOLDMANN et al. (2003) and ANDREI et al. (2008). According to our results, in
most cases it was possible to reduce the required primary antibody amount usally
by 50-80% and for lysozyme even by 97.5% compared to samples fixed in Bouin`s
solution or Ca-formol (compare Tab 3.8). Such feature emphasises economical
advantage, however, the question why HOPE® fixation offers this opportunity has
still to be clarified.
Principles of fixation
In contrast to the two well established fixatives Bouin`s solution and Ca-formol, the
HOPE® fixation solution is not formalin based. Formalin fixation was introduced

Discussion
119
more than 100 years ago by the German medical practitioner F. Blum (1893). He
already demonstrated that formaldehyde forms methylene compounds with amino,
amide and hydroxyl groups, thus affecting the solubility and reactivity of proteins
(BLUM 1893, 1896). Therefore, it can be concluded that the formation of such
methylene cross-links (PUCHTLER and MELOAN 1985) is absent in HOPE® fixed
tissue (OLERT et al. 2001). Nevertheless, the primary characteristic of a fixative is
to stabilise tissue structure in order to protect against shrinking effects during the
use of concentrated organic solvents. OLERT et al. (2001) argue that the direct
application of organic solvents would be an ideal option, as the accumulation of
denaturing events that takes place during fixation could be circumvented. This
technique is rarely applied due to the incalculable negative affects of organic
solvents on the tissue. Although the exact mechanism of the HOPE® fixation still
needs to be elucidated, its aim is to protect the tissue from the influences of the
dehydration solution (acetone) used and to reduce the number of protein cross-
links. For this purpose, the tissue is directly transferred into a special protection
solution (ingredients not listed in detail) to weaken the negative influences of
acetone. Earlier studies already recommended acetone for an optimal retention of
antigenic activity in embedded tissue (KAKU et al. 1983). The authors stated that
the major disadvantage of acetone fixation is a considerable hardening and
shrinkage of the material. Such negative side effects should be prevented in the
HOPE® fixation system via previous incubation in a so-called protection solution.
As a consequence the reduction of protein-cross links in HOPE® fixed specimens
might directly result in improved preservation of antigenicity. In contrast, formalin
fixation results in a loss or decrease of antigenicity, due to methylene cross-links
of reactive sites on proteins making certain epitopes inaccessible for antibodies.
This drawback can be partly overcome by techniques based on heat denaturation
or enzymatic digestion, which facilitate the retrieval of antigens (WERNER et al.
2000; CARRASCO et al. 2004). Some authors argue that this time consuming
procedure could be avoided using the HOPE® fixation, as less antigen masking
occurs (GOLDMANN et al. 2003; ANDREI et al. 2008). Due to the fact that the
established antibodies used in our studies did not require any pre-treatment, we
cannot corroborate or refute these statements concerning the HOPE® fixation. It is
important to emphasise that in our study the attempt to establish some of the
antibodies in the esophagus (MBL, L-ficolin, Dectin-1 and CD1a) remained

Discussion
120
inconclusive after the use of all three fixatives. Due to the fact that the primary
antibodies for MBL and L-ficolin showed positive results on the equine liver
(positive control), such antibodies revealed a cross-reactivity at least for this
species. Hence, it can be concluded that MBL and L-ficolin are not present in the
esophagus epithelium of the horse. Regarding the other species, we cannot totally
refute the existence of the latter ß-glucan receptors, as it is possible that the
applied antibodies exhibit no cross-reactivities. These findings correspond with
observations made by BLASCHITZ et al. (2008), who argued that inadequate
antibodies lacking specificity cannot lead to satisfying IHC results and that the
HOPE® technique will not solve such problems. In conclusion, HOPE® did not
result in a direct advantage concerning the establishment of antibodies in our
study, which might be related to the applied antibodies. Nevertheless, supportive
evidence is provided that the improved preservation of antigenicity of HOPE®
fixation was directly related to the effect that the HOPE® specimens required only
50-2.5% of the amount of primary antibodies in comparison to the other two
fixation solutions. The influence of mild pre-treatments (for example heating in
citrate buffer) on HOPE® fixed esophagus tissue needs to be evaluated in future
experiments.
The main advantage of HOPE® fixation is, as discussed by BLASCHITZ et al.
(2008), that it provides the possibility of applying cryo-compatible antibodies to
paraffin sections. It has been demonstrated that antibodies in the human placenta
immunolocalised their antigens on cryo sections and on HOPE® fixed but not on
formalin-fixed paraffin sections. These results were related to the improved
preservation of soluble proteins in HOPE® fixed samples, in comparison to
samples fixed by the formalin method. The authors emphasised the benefit of the
HOPE® technique for cases when only cryo-compatible antibodies are available. In
our study most of the antibodies applied were established for use on paraffin
sections. Only the antibody designed for the detection of the collectin MBL had
been tested on cryo sections and not on paraffin ones. Our findings could not
corroborate the findings of BLASCHITZ et al. (2008), as no positive reactions
could be demonstrated on HOPE® fixed samples for MBL. The missing cross-
reactivity of the antibody in other species, than the horse, has to be taken into
account. Nevertheless, the HOPE® fixation can be looked upon as an
advantageous tool for fixation when discussing the aspect of saving time. The

Discussion
121
HOPE® protocol is shorter than the protocol used for the other two fixatives, owing
to the fact that the dehydration step (increasing ethanol concentrations) and the
incubation time in paraffin is more time-consuming for the Bouin and Ca-formol
fixed samples. The dehydration step is limited to the incubation in acetone for the
HOPE® fixed samples.
In spite of the possible advantages of the HOPE® fixation procedure, it is of great
importance to discuss the HOPE® fixation more critically. One aspect to be kept in
mind is that the ingredients of the HOPE® solutions are not listed or defined in
detail by the selling company. It is only mentioned that the solutions contain amino
acids (OLERT et al. 2001). An interaction of amino acids with antigens is
conceivable and could result in a change of epitope structure, which could
influence the immunoreactivity of the antigens to be detected. A second critical
feature is that the tissue sections prepared should not be stored for more than 7
days (personal communication DCS) before starting an IHC experiment. During
our IHC experiments, we tried to use freshly prepared tissue sections, as false
negative reactions were observed when using tissue sections prepared eight
weeks earlier. On the contrary, tissue sections of Bouin and Ca-formol tissue could
be used for IHC over years, without any recognisable negative influence on the
immunoreactivity (MEYER personal communication). A possible explanation for
the shorter usability of the HOPE® sections could be a decreased number of
protein cross-links than in formalin-fixed material, yielding a diminished stability of
proteins and tissue structure. Due to this fact it is also important to keep the
sections and paraffin blocks refrigerated until use after HOPE® fixation.
Another fact disqualifying HOPE® from routine diagnostics is the necessity of
refrigerating the samples immersed in HOPE® solution. In contrast, there is no
need to refrigerate formalin-fixed samples.
Differences in reaction intensities
In some cases significant differences between the fixation media could be
determined. Such differences were most obvious in the equine and ovine
esophagus epithelium, revealing a diminished reaction intensity in the HOPE®
fixed samples. A decrease of staining intensity was observed, particularly after

Discussion
122
analysing the results for ß-defensin 2 and 3 and TLR2. As already mentioned
before, in some cases the amount of primary antibody necessary could be
reduced by more than one third for the HOPE® sections, in comparison to the
Bouin and Ca-formol sections. Concerning the ß-defensins, it can be presumed
that this reduction was too high, resulting in a weaker reaction in the equine and
ovine esophagus epithelium compared to the other fixation media. Due to the
great quantity of species studied, it was particularly difficult to determine a suitable
antibody dilution for all species.
Regarding the results found for Ca-formol fixed esophagus tissue in our study, one
possible explanation for the bad quality of structure preservation and
immunoreactivity of some of the Ca-buffered formol fixed tissues could be the high
pH level of the solution. Ca-bufferd formol has a pH level of 7.1 – 7.3 and Bouin`s
solution fixes at a pH level of about 6.0. PUCHTLER and MELOAN (1984)
emphasised that maximum tissue fixation occurs in the pH range 4 to 5.5, no
increase in tissue stabilisation was observed above pH 5.5. They concluded that
the increased amount of formaldehyde bound at higher pH levels only blocks
numerous reactive groups. We can support the latter findings and assume that the
reduced antigenicity of the Ca-formol fixed samples is due to the higher pH-level of
the latter solution. Furthermore, an over fixation of the tissue in Ca-formol could
affect the stability of epitopes and result in diminished immunoreactivity (WERNER
et al. 2000). However, the aspect of over fixation is negligible, as we incubated the
samples for not more than 48 hours.
The positive and negative effects of various fixatives on the IHC based detection
of antigens have been discussed extensively in the literature. Supportive evidence
for our findings that fixation in Bouin`s solution leads to the best structural and IHC
results has been supplied by several authors (BEDOSSA et al. 1987; SMITT et al.
1993; CARRASCO et al. 2004). SMITT et al. (1993) agreed that Bouin`s fixative
obtained the best structural and IHC reactions for fixed human cerebellum in
contrast to formalin. The effect of fixation on ovine lung tissue was studied by
CARRASCO et al. (2004). They demonstrated that Bouin`s solution proved to be
the most suitable fixative for structural and immunohistochemical studies.
BEDOSSA et al. (1987) investigated the influences of fixation on human liver

Discussion
123
tissue and also emphasised that Bouin`s solution was the best fixative for their
studies. Several other authors discussed the negative effect of formalin fixation on
various tissues (PUCHTLER and MELOAN 1985; ARNOLD et al. 1996; VAN
ALSTINE et al. 2002). ARNOLD et al. (1996) found that neutral buffered formalin
was generally the poorest fixative (in comparison to ethanol and Bouin`s solution)
for maintaining antigen recognition by IHC. VAN ALSTINE et al. (2002) discussed
the possible effects of over fixation on tissues fixed in formalin in detail. They
concluded that an incubation exceeding three days results in a substantially
decreased sensitivity of an immunohistochemical test. Our samples were only
fixed for a maximum of 48 hours. In a more recently conducted study, BUESA
(2008) summarised the advantages and disadvantages of formalin fixation. On the
one hand the author argued that formalin fixation has been well established for
more than 100 years and is rapid as well as economic. Furthermore, he stated that
most antibodies are optimised for use on formalin-fixed paraffin embedded tissue
(FFPET), and that cross-links of proteins are reversible. On the other hand, the
author stresses the carcinogenic potential of formalin. From our point of view this
fact is one of the only really important advantages of the HOPE® fixation method,
because its media probably do not contain hazardous ingredients.
We can conclude that the HOPE® fixation technique offers some advantages,
especially concerning the costs of antibodies and time. If the HOPE® protocol is
well established in a laboratory, the preservation of structure is to some extend
comparable to samples fixed in Bouin`s solution, although differentiated structural
information cannot be obtained. In order to choose HOPE® as a serious alternative
to other fixatives, the ingredients of the solutions should be clearly defined to
achieve a critical evaluation of results obtained.

Discussion
124
5.2 Defence mechanisms of the mammalian esophagus:
Species comparison
5.2.1 Antimicrobial peptides
We focused our immunohistochemical studies on several parameters of the innate
immune system, in order to understand whether or not parts of the innate system
participate in the mucosal defence of the mammalian esophagus epithelium.
Although constitutive, this system is activated by the presence of microorganisms
and their products, providing a rational for a potential bio defence strategy (NEISH
2009). Besides our aim to detect receptors of the innate immune system
expressed by epithelial cells (TLRs) or on sentinental cells (LCs), our intent was to
detect substances produced and secreted by the epithelial cells (APs, ß-glucan
receptors, and lysozyme).
Mucosal defence strategies in the human esophagus
Information about esophageal mucosal defence is sparse, even in humans.
Nevertheless, HOPWOOD (1995) demonstrated several interesting aspects of
esophageal defence, e.g. various defence mechanisms can be elicited by reflux
damage. The author argued that chemical damage wrought by proteinases is
countered by cell turnover, intercellular barriers and pH management.
Furthermore, the presence of epithelial growth factor (EGF) is an added defence
feature that acts when ulcers are formed. For our study, the evidence of an
“intercellular barrier” is of particular interest. HOPWOOD (1995) demonstrated that
the intercellular space between the cells in the stratum spinosum was not empty,
as it appeared to be in electron microscopy, but showed the presence of
“mucosubstances”. However, during his studies the characteristics and functions
of this material remained unclear. In contrast to these findings, we confirmed the
presence of APs as intercellular material between the layers of the esophagus
epithelium of the nine mammalian species studied. This might be due to the fact
that the substances accumulate in large amounts between cells of the stratum
spinosum only in the human esophagus (HOPWOOD 1995), but are generally
present in smaller amounts in other layers. In our study we used highly sensitive

Discussion
125
IHC to detect APs, so that it was possible to give a detailed description of their
distribution in the esophagus epithelium.
Additionally, we provide the first artefact-free electron microscopical pictures (cryo-
SEM) of the surface of the esophagus epithelium, thus verifying the existence of a
one-layered microbial colonisation in all species studied. To be able to tolerate and
benefit from these microorganisms, the eukaryotic host must monitor the
microbiota and control their number and composition. One possible way to control
colonisation with commensal and potentially pathogenic bacteria is the production
of APs, which are part of the first line of defence (INAMOTO et al. 2008; NEISH
2009).
As APs are found in various epithelial tissues (DALE and FREDERICKS 2005;
BRAFF and GALLO 2006) and due to their antimicrobial and immunoregulatory
activity, scientists perceive APs as promising candidates for new therapeutic
drugs, also in veterinary medicine (LINDE et al. 2008; SANG and BLECHA 2009).
In accordance with the sites expressing APs, our discussion focuses on two other
organ systems. To start out, an analysis of the AP expression pattern in other
parts of the digestive tract is of interest. As a matter of fact APs are involved in the
control of the physiological microbial flora and mucosal homeostasis (BEVINS et
al. 1999; IIMURA et al. 2005; LOTZ et al. 2007; SALZMAN et al. 2007;
MUKHERJEE et al. 2008; VELDHUIZEN et al. 2008). For example, the production
of APs (mainly α-defensin) in the small intestine of humans is restricted to Paneth
cells located in the intestinal crypts, whereas in the large intestine CAT and ß-
defensins contribute to the antimicrobial barrier (LOTZ et al. 2007). Interestingly
enough, reduced numbers of Paneth cells and a decrease in the α-defensin
production were noted in Crohn`s disease patients (WEHKAMP et al. 2005).
Thereafter the specific construction of the esophagus epithelium has to be taken
into account. In most herbivorous and omnivorous species the esophagus shows a
more or less keratinised stratified squamous epithelium, which is not really
comparable to the one of the epidermis of the skin. The latter organ presents a
reliable mechanical, as well as an immunological barrier to infection, displaying a
considerable capacity of innate immunity. Due to constant attacks of
microorganisms and remarkable mechanical strain, research focussed on the
expression of APs in the epidermis (OREN et al. 2003; BRAFF et al. 2005; ELIAS

Discussion
126
2005; ELIAS and CHOI 2005; BRAFF and GALLO 2006). Whenever the
occurrence of this substance group is discussed, it is of great interest whether the
APs are produced constitutively or inducible at the site of inflammation. BRAFF et
al. (2005) found that CAT is constitutively expressed by neutrophils in the skin and
inducible in keratinocytes in response to infection. In contrast to this, ELIAS and
CHOI (2005) argued that hBD-2 and -3, and CAT are expressed at low levels in
unperturbed skin, but occur in higher levels in healing wounds and in inflammatory
dermatoses. Our results demonstrated that hBD-2 and -3 and CAT are expressed
constitutively in the esophagus epithelium of the species studied. It is
inconceivable that all animals were infected, and thus the constitutive production
of the considered APs is necessary to maintain the homeostasis of the esophagus
epithelium. None of the animal species revealed negative results, only the reaction
intensity differed between the species. Due to our findings we agree with ELIAS
and CHOI (2005) and can transfer their findings demonstrated for the skin to a
certain extent to the esophagus epithelium of the domesticated mammals studied.
Nevertheless, additional experiments analysing the influence of infection by certain
pathogens need to be conducted, whereby the up-regulation of APs could be a
likely effect.
Comparison of expression sites in the esophagus epithelium
In the skin, APs are normally stored in so-called lamellar bodies (LBs) or
membrane coating granules (MCGs), respectively, but were also found in the
intercellular space (OREN et al. 2003; ELIAS and CHOI 2005). While the glycolipid
contents of the LBs are well known, the additional AP contents were a surprise.
OREN et al. (2003) gave a detailed description on the expression sites of hBD-2 in
the epidermis. They performed immunogold labelling and showed that hBD-2 is
predominantly located in the spinous and granular layers. Dense labelling was
demonstrated for the LBs of the keratinocytes and some additional staining was
seen in the intercellular space. In contrast, ELIAS and CHOI (2005) described a
different expression pattern of hBD-2 and additional hBD-3 and CAT in the
epidermis. Conducting IHC, they showed that all three APs were predominantly
localised in the outer epidermis. At this site they appeared in LBs and in the
intercellular spaces of the stratum corneum. The authors argued that the APs are

Discussion
127
perfectly positioned to intercept pathogenic microorganisms when they attempt to
penetrate the epidermis in between the corneocytes. With regard to our own
findings from the esophagus epithelium, we could not corroborate the results of
OREN et al. (2003) and ELIAS and CHOI (2005). One contrasting finding was the
observation in all species and APs studied, that constantly strong to very strong
reaction intensities occurred in the cells of the stratum basale. Due to the fact that
this layer has the highest rate of proliferation, we presume a direct connection to
AP production intensities. With regard to the three nutrition types studied, a
statistically significant difference could be shown for hBD-3 in the stratum basale.
Interestingly enough, the carnivorous cat exhibited a slightly stronger reaction in
comparison to the herbivorous and omnivorous species analysed. The increased
production of hBD-3 in the feline stratum basale could be considered a reaction
due to diminished preventative mechanical protection properties of the esophagus
epithelium. The feline esophageal epithelium is only weakly keratinised, and
therefore cannot function as a reliable mechanical barrier against invading
microorganisms. Thus, the epithelium has to be protected by larger amounts of
APs. Supporting evidence for this theory is provided by the results observed for
the stratum corneum. What is more, statistically relevant differences were found in
this layer when comparing herbivorous and omnivorous species with the
carnivorous cat. This time hBD-2 was expressed in greater amounts in the
carnivorous cat. Moreover, hBD-3 was expressed at a higher level in the cat, even
though the difference between the nutrition groups was not obvious from the
statistical point of view. Consequently, the feline esophagus epithelium is
intensively protected by ß-defensins. Due to its reduced mechanical protection by
the weakly keratinised epithelium and the constant influence of microorganisms, ß-
defensin production is remarkable and has to fulfil a distinct protective task.
Presumably hBD-2 is more effective on the epithelial surface, and hBD-3 functions
in the more basal part of the feline esophagus epithelium. This explanation and the
exact amount of APs produced remains to be examined in future experiments (for
example by the application of densitometry). On the one hand, these observations
are to some extent astonishing, as one would expect the herbivorous species to
exhibit the strongest reactions, due to their rough-textured food and the resulting
high mechanical strain. On the other hand, our findings showed the influence of
other epithelial properties, such as a regulated AP production. A more keratinised

Discussion
128
epithelium protects the mucosa from invading microorganisms mechanically, and a
less keratinised epithelium is protected by an increased production of ß-defensins.
It remains to be elucidated why the reaction intensity of all three APs decreased
from the stratum basale towards the lower and upper stratum spinosum. One
explanation could be that APs are stored in vesicles, comparable to the LBs
(MCGs) of the epidermis (OREN et al. 2003; ELIAS and CHOI 2005), and are not
detectable by the primary antibody, due to the compact packing. The idea of
substance transport in vesicles corresponds with findings of INAMOTO et al.
(2008), who investigated the epithelial response towards the attachment of
indigenous bacteria in the small intestine of the rat. One of their observations was
that defensins and lysozyme are transferred via cellular vesicles into the lumen,
where they fused with the membrane of an invaginated bacterium. However, in
some cases an increase in reaction intensity could be observed towards the
stratum granulosum of the esophagus. Concerning hBD-2, this was true for the
goat, sheep, pig, mouse and rat. Regarding hBD-3, the phenomenon was
demonstrated for cattle, sheep, pigs and mice, and for CAT an increase was
observed in the ovine and porcine esophagus epithelium. Analysing the species
that showed the increase mentioned makes the existence of a correlation between
these results and the nutrition type a realistic feature. All of the species are
herbivorous or omnivorous and because of the enormous mechanical strain on
their esophagus epithelium, in comparison to the esophagus of the carnivorous
cat, APs have to be stored in the stratum granulosum. This “layer” of APs in the
latter stratum probably indicates some kind of barrier function. An intriguing finding
is that such a “barrier” was constantly detectable in the porcine esophagus
epithelium. As shown by the analysis of the structure of the esophagus epithelium,
the pig revealed a rather loose and varying organisation of the stratum corneum.
Such feature may result in an increased penetration of microorganisms. This
specific structural aspect explains the necessity of a protective layer of APs. Due
to the fact that the esophagus epithelium of the cat is protected by a layer of hBD-
2 covering the surface, the storage of APs in the stratum granulosum is not
necessary in this species. For CAT, the strongest reaction intensity was seen in
the pig, mainly in the stratum basale. One explanation could be that the
commensal flora of the porcine esophagus especially triggers this AP production.
LINDE et al. (2008) showed in in-vitro experiments that CATs are produced by the

Discussion
129
porcine organism and demonstrated its high effectiveness against: E. coli,
Salmonella typhimurium, Staphylococcus aureus, Actinobacillus pleuropneumonia,
Pseudomonas aeruginosa, and Candida albicans. It remains to be elucidated via
microbial differentiation, whether these microorganisms also belong to the
commensal flora of the porcine esophagus surface.
LINDE et al. (2008) showed in-vitro activity of equine ß-defensins against the
following microorganisms: Corynebacterium spp. and Staphylococcus intermedius.
These two Gram-positive cocci were not found in the microbial flora of the
esophagus of the horse (MEYER et al. 2009). Furthermore, LINDE et al. (2008)
described in-vitro activity of CAT against the following bacteria: Escherichia coli,
Streptococcus equinus, Klebsiella pneumonia and Serratia marcescens. In
contrast to the afore-mentioned results for ß-defensins, this spectrum of
antimicrobial activity is more consistent with the findings made by MEYER et al.
(2009) concerning the actual microbial status of the equine esophagus. These
authors demonstrated a high occupation with the Gram-negative facultative
anaerobic rods E.coli. However, E.coli is an ubiquitous bacterium and cannot be
made out as specific for the equine species or generally explain the expression of
CAT in the equine esophagus. The results indicate that the characteristic microbial
flora of the esophagus of the different domesticated mammals still needs to be
analysed more intensively.
With regard to our own findings and the literature studied, it can be concluded that
the diverse repertoire of APs is most likely a key factor in allowing the esophagus
mucosal surface to maintain homeostasis concerning the diverse colonising
bacterial populations.
5.2.2 Toll-like receptor 2
Toll-like receptors (TLRs) comprise a class of transmembrane pattern recognition
receptors that play a key role in microbial recognition, induction of antimicrobial
genes and control of adaptive immune responses (JANEWAY and MEDZHITOV
2002). Eleven different TLRs have been identified in mice (LOTZ et al. 2007) and
ten in humans (AKIRA 2003; JANEWAY et al. 2005). One aim of this study was to
provide the first proof of TLR expression in epithelial cells of the esophagus of the
species studied. After an analysis of the literature available, we decided to put our

Discussion
130
focus on TLR2, as several authors emphasised the occurrence of this TLR on
intestinal epithelial cells of the human and murine gut (CARIO and PODOLSKY
2006; CARIO et al. 2007; TIZARD 2008), and on epidermal keratinocytes
(KOELLISCH et al. 2005; LEBRE et al. 2007). For the intestine it was
demonstrated that TLRs are involved in tissue regeneration and inflammation, as
well as in controlling the integrity of the intestinal epithelial barrier (RAKOFF-
NAHOUM et al. 2004; CARIO and PODOLSKY 2006; CARIO et al. 2007).
Regarding the skin, LEBRE et al. (2007) showed that TLRs actively contribute to
the induction of an immune response.
In this dissertation thesis the existence of TLR2 on epithelial cells of the
esophagus was verified by IHC. Thus, we provide the first comprehensive
evidence for the expression of the latter TLR in the nine different species studied.
So far no information concerning the expression of TLRs in the esophagus
epithelium was available from other mammals including humans. In our study we
were able to demonstrate the existence of TLR2 in all species, which indicates a
constitutive expression of TLR2. These observations are in line with the findings of
CARIO et al. (2007) who emphasised the functional relevance of TLR2 to control
tight junction associated intestinal barrier integrity and to balance mucosal
homeostasis. Furthermore, a constitutive expression of TLR2 was observed on
human keratinocytes in the skin (LEBRE et al. 2007). A constitutive expression of
TLRs in cells creating an epithelial barrier against invading microorganisms is
comprehensible as these PRRs enable the organism to continuously screen the
microbial composition, readily recognize potential imbalances and rapidly initiate
innate immune responses.
In regard to the specific locations of TLR expression in the esophagus epithelium,
it could be observed in all species that the staining intensity of TLR2 was very high
in the stratum basale and decreased towards the stratum spinosum. The cells of
the stratum granulosum and the stratum corneum showed no positive reaction.
Only in the porcine esophagus a positive staining was observed in the stratum
granulosum. It appears logical that the stratum corneum cells do not express
TLR2, as it can be assumed that such cells do not possess the full ability to
respond, i.e. by the production of antimicrobial products. The fairly isolated

Discussion
131
expression of TLR2 in the basal part of the epithelium may have a functional
background. It is conjecturable that TLR expression at this site of the esophagus
epithelium is correlated with high production of APs in the stratum basale
(compare 5.3.1). Several authors described a functional relation between the
expression and activation of TLRs and the inducible production of APs via a TLR
dependent pathway (BIRCHLER et al. 2001; VORA et al. 2004; SUMIKAWA et al.
2006; BUECHAU et al. 2008; LINDE et al. 2008). The findings of the latter authors
correspond with our results. It was only possible within the scope of this study to
demonstrate a co-expression of TLR2 and APs, leaving the evaluation of the direct
functional relation of both to further studies. However, we could not accurately
differentiate which AP is induced due to a TLR2 response. BIRCHLER et al.
(2001) argued that only hBD-2 was induced via a TLR2 dependent pathway, whilst
hBD-3 was not inducible via TLR2. Later, the findings of BIRCHLER et al. (2001)
were confirmed for intestinal and epidermal cells (VORA et al. 2004; KUMAR et al.
2006; MUKHERJEE et al. 2008). To our knowledge, only SUMIKAWA et al. (2006)
demonstrated a TLR2 dependent hBD-3 production in human keratinocytes.
Nevertheless, a TLR2 dependent expression of CAT was only demonstrated for
the skin (BUECHAU et al. 2008). It is an interesting finding that TLR2, hBD-2 and -
3 and CAT expression was shown for the stratum granulosum of the porcine
esophagus. This feature might indicate a TLR2 triggered production of the latter
APs in the stratum granulosum. Further studies should concentrate on the
expression of TLR4 in the esophagus epithelium, as this TLR is considered to
participate in the surveillance of the intestinal tract, and co-expression with APs
has already been demonstrated (CARIO et al. 2000; CARIO et al. 2002; VORA et
al. 2004).
Functions of TLR2 in the esophagus epithelium
Besides being responsible for triggering an innate immune response, i.e. induction
of AP production, TLRs are thought to fulfil further tasks to help maintain epithelial
homeostasis.
RAKOFF-NAHOUM et al. (2004) demonstrated the functions of TLR2 by
simulating intestinal injury and inflammation in TLR2 and MyD88 (TLR signalling
protein) knock-out mice. First, they showed a dysregulation of proliferation and

Discussion
132
differentiation in the intestinal epithelium in the absence of TLR signals. TLR
absence resulted in an increased number of proliferating cells. Secondly, the
authors demonstrated a decreased production of cytoprotective and reparative
factors (IL-6 and TNF) in such knock-out mice. IL-6 and TNF play a pivotal role in
protecting the epithelium from injury and in the initiation of repair responses.
Moreover, it was shown that the production of such cytokines is dependent on the
presence of commensals, which stimulate TLRs. It can be assumed that the loss
of commensal-mediated cytoprotection of the intestinal barrier, due to TLR2 /
MyD88 dysfunction, results in the destruction of epithelial homeostasis. After the
breakdown of epithelial integrity normally innocuous bacteria, such as Escherichia
coli, can cross the Tunica mucosa, invade the blood stream, and cause fatal
systemic infection (JANEWAY et al. 2005). Besides functioning as PRRs, TLR2
might also act as regulatory feature of the epithelial integrity in the esophagus
epithelium, i.e. in case of injury. This could be one explanation for our finding that
herbivorous mammals revealed stronger reaction intensities to TLR2, in
comparison to the omnivorous and carnivorous species studied. The epithelium of
the herbivorous species is exposed to higher mechanical stress than that of the
other two nutrition groups. It remains to be elucidated in future experiments, if IL-6
and TNF are also present in the esophagus epithelium of the animal species
studied, in order to support or refute the latter theory.
Since TLRs play a pivotal role in innate immunity and are the front liners in
pathogen recognition, we hypothesised that TLRs are also expressed in the
esophagus epithelium. Furthermore, we aimed to reveal whether a co-existence
with APs occurs, as previously shown for other epithelial surfaces. After
demonstrating a colonisation of the esophagus surface with microorganisms by
cryo SEM, we had strong evidence that PRRs are in fact necessary in the
esophagus epithelium. In this context we were clearly able to demonstrate TLR2
expression in the nine species studied. Moreover, we can support our hypothesis
of a simultaneous expression of TLR2 and the APs hBD-2, -3, and CAT. It has to
be taken into account that TLR2 is stimulated by the commensal bacteria and
induces the production of APs by the epithelial cells. Furthermore, the regulation of
antimicrobial responses by TLR2 activation could prevent over production of
antimicrobial proteins that could interfere with intestinal homeostasis. More

Discussion
133
specific information about the functional relationship of TLR2 and the APs has to
be gained from future experiments, also meaning that the signal pathway of TLR2
(e.g. NF-κB) in the epithelial cells of the esophagus has to be studied in more
detail.
5.2.3 ß-glucan receptors
The ß-glucan receptors studied in this thesis comprise MBL, L-ficolin and Dectin-1
and, like all ß-glucan receptors, belong to the group of lectins. Lectins are able to
recognize carbohydrate moieties, the unique “sugar codes”, on pathogens with
their characteristic lectin binding domain, hence acting as PRRs (LU et al. 2002;
GUPTA and SUROLIA 2007). Such lectin molecules are divided into further
groups. MBL belongs to the group of collectins, L-ficolins is member of the ficolin
group and Dectin-1 is another C-type lectin receptor, specific for recognising
fungal invaders. The structural and functional features of each group have been
extensively described in the literature chapter of this thesis. Therefore we will
waive a discussion of these aspects in detail at this point.
There was substantial evidence that collectins and ficolins are predominantly
found at the interface of body and environment, i.e. at epidermal (MEYER et al.
2008) and mucosal surfaces (DELAY 1999; UEMURA et al. 2002; WAGNER et al.
2003; VAN DE WETERING et al. 2004; LILLIE et al. 2005). Regarding these
findings, we aimed to support or refute the presence of such molecules of innate
immunity in the esophagus epithelium of the nine domesticated mammals studied.
We conducted several IHC experiments to demonstrate ß-glucan receptors in the
esophagus epithelium of the various species. However, all our experiments ended
without a positive result despite the fact that various antigen retrieval techniques
and different visualisation methods were tested. The positive controls on equine
liver revealed a distinct staining for MBL and L-ficolin. Hence, it can be resulted
that at least in this species the latter ß-glucan receptors are not present. In order
to totally refute their occurrence in the other species studied, the cross-reactivity of
the antibodies applied needs also to be elucidated on positive control tissue.
Furthermore, a new fixation technique, the HOPE® fixation, was employed, which
we hypothesised would result in better antigen preservation than the formalin

Discussion
134
based fixations (compare 5.1.2). However, even the putative improvement in
antigen preservation did not bring about any positive results concerning ß-glucan
receptors. On one hand, the results described are not surprising, as to this day no
substantial evidence for the occurrence of the ß-glucan receptors examined in the
esophagus was provided for humans. Only the collectin surfactant protein D (SP-
D) could be shown here by IHC (MADSEN et al. 2000). On the other hand, the
collectin MBL has already been identified in the intestinal tract of mice and pigs.
UEMURA et al. (2002) and WAGNER et al. (2003) demonstrated MBL in the
murine small intestine by IHC and PCR, and DELAY (1999) also proved a positive
reaction for the latter collectin in the intestinal crypts of the pig. The expression on
mucosal surfaces suggests that MBL has a general function in innate immunity in
the gastrointestinal tract (VAN DE WETERING et al. 2004). The results of
UEMURA et al. (2002) actually revealed that MBL, which is secreted from villous
epithelial cells, acts as a humoral immune factor in the intestine, comparable to the
secretory IgA. The latter authors found the maximum expression of MBL in the
jejunum, which they characterised as relatively sterile in comparison to the colon
and argued that MBL, particularly, may be able to contribute to this sterility. These
findings do not correspond with our own observations. Following the
argumentation of UEMURA et al. (2002), one would consider MBL to be
expressed extremely high in the esophagus to maintain sterility. We consider that
a certain “antigenic pressure” is required for the production of this substance.
However, such level is not reached in the esophagus, and thus, MBL is not
produced by the epithelial cells of the esophagus. The esophagus reveals only a
thin layer of microorganisms on its luminal surface (compare 5.4). Consequently,
ß-glucan receptors may not be required to complement the innate defence
mechanisms as a first line of defence in the esophagus. These findings contradict
the theory of other authors, who characterised ß-glucan receptors as molecules of
an early immune response (HOLMSKOV et al. 2003; GUPTA and SUROLIA 2007;
SCHOREY and LAWRENCE 2008). The hypothesis that a ß-glucan receptor
response is only required at sites exposed to a high number of pathogens, is
consistent with recent observations from the integument of marine and freshwater
aquatic mammals, whereby MBL, L-ficolin and Dectin-1 were demonstrated
(MEYER et al. 2008). Staining for L-ficolin was generally the strongest and most
obvious in all layers of the epidermis, except in the stratum basale. Concerning

Discussion
135
MBL, the stratum basale showed a weak reaction. Additionally, positive reactions
for this lectin were observed in macrophages. Positive staining for Dectin-1 was
limited to secretory cells and the epidermis of the capybara. The authors
emphasised that in marine or freshwater ecosystems the skin has to cope with a
multitude of microorganisms. It is a remarkable finding that the sparsely-haired
capybara was the only species exhibiting a positive staining for Dectin-1. This
result indicates that Dectin-1 might be the ß-glucan receptor that is activated due
to increased pathogen pressure. Another reason could be that the skin of the
capybara is strained with a high number of fungi, for which Dectin-1 is the
specialised PRR. In this context, the pathogen specificity of the different ß-glucan
receptors should be taken into consideration. The primer ligand for MBL is LPS,
the major cell wall component of Gram-negative bacteria (HOLMSKOV et al. 2003;
VAN DE WETERING et al. 2004; LILLIE et al. 2005). L-ficolins have a wider range
of pathogen specificity and interact with Gram-negative and positive bacteria
(RUNZA et al. 2008). In contrast, Dectin-1 primarily recognizes the cell wall
components of fungi (WILLMENT and BROWN 2008). MEYER et al. (2009)
isolated bacteria from the equine esophagus. The genera found most frequently
comprised Gram-positive Streptococci, Gram-negative E. coli, Actinobacillus spp,
and Prevotella spp. and yeasts. LILLIE et al. (2005) and RUNZA et al. (2008)
studied the distribution of MBL and ficolins between the species and their
interaction with pathogens in various animals (compare also Tab. 2.2).
Unfortunately, they did not provide information concerning the horse. One finding
corresponding with the microbial flora of the equine esophagus is that the authors
demonstrated antimicrobial activity of MBL and ficolins against Actinobacillus spp.
and Escheriachia coli. The rather low occupation of the equine esophagus with
Candida spp. may indicate that Dectin-1, the ß-glucan receptor specific for fungi, is
rarely required. There are two principal explanations as to why the horse revealed
no positive reaction for any ß-glucan receptors. The first one being that bacterial
antigens are very species specific. Hence, as an example, the comparison of
porcine and equine Actinobacillus spp. is not tenable scientifically. Eventually, the
microbiota of the esophagus of the mammals studied differed considerably from
that recognized by ß-glucan receptors. Further experiments on the composition of
the natural microflora are needed. The second explanation is that the very species
specific genotypes of lectins make it particularly difficult to predict from one

Discussion
136
mammalian species to another (FUJITA et al 2004; LILLIE et al. 2005). As already
described in the literature chapter, various subtypes of lectins are present in the
animal kingdom. The comparison of lectin orthologues from different species
showed that they differ in many aspects (RUNZA et al. 2008). As a result, cross-
reactivity for antibodies specific for humans and animals cannot be assumed.
Further investigations are necessary in order to abandon the theory of a ß-glucan
receptor occurrence in the esophagus of domesticated mammals. Other methods,
e.g. in situ hybridization or PCR, could be applied and/or species specific
antibodies should be designed.
It is important to emphasise the different functional features of the epithelium of
the small and large intestine compared to the esophagus epithelium. In contrast to
the esophagus, the other parts of the intestine fulfil a wider range of secretory and
resorptive tasks (INAMOTO et al. 2008). Regarding this fact, it can be conjectured
that the epithelial cells of the mammalian esophagus are rarely needed for the
production of ß-glucan receptors.
YOKOTA et al. (2001), GANTNER et al. (2003) and DENNEHY and BROWN
(2007) demonstrated a co-expression and collaboration of Dectin-1 and TLR2. The
observation of cooperative pathogen recognition could not be corroborated in our
study. The co-expression of both receptors results in an increased production of
TNF and IL-12. One can regard these two chemokines as not necessary for
ensuring tissue homeostasis in the esophagus epithelium.
Furthermore, Dectin-1 was detected on LCs of the skin (ARIIZUMI et al. 2000;
YOKOTA et al. 2001; GAVINO et al. 2005). Regarding the esophagus, however,
our results do not provide supportive evidence for this theory. In our study
langerin(+) LCs were identified in the equine and murine esophagus (compare
5.3.4). Although yeasts were present, the equine esophagus has not to deal with a
very high fungal burden in comparison to the skin. This may explain the missing
expression of Dectin-1 on LCs of the esophagus epithelium.

Discussion
137
5.2.4 Lysozyme
Lysozyme is an enzymatically active antimicrobial protein, which targets essential
cell wall components of microorganisms. Functionally, lysozyme is a glycosidase
that hydrolyses linkages of peptidoglycan, an important constituent especially of
the cell wall of Gram-positive bacteria. Due to its preference for peptidoglycan,
lysozyme is more effective against this bacterial group (GANZ 2004).
Besides being present in a number of secretes, including tears and saliva,
lysozyme is also actively involved in the mucosal defence of the intestinal tract
(WEHKAMP et al. 2005; INAMOTO et al. 2008; MUKHERJEE et al. 2008).
Considering these recent findings, we tried to detect lysozyme in the esophagus
epithelium of nine different domesticated mammals. Our results revealed positive
reactions in all the species studied. This observation indicates that lysozyme is
constitutively expressed in the mammalian esophagus epithelium. Our findings
correspond with the observation made in microbiologically sterile “germ-free” mice.
These laboratory animals revealed lysozyme expression in the intestine,
independent of microbial signals (HOOPER et al. 2001). However, the role of the
commensal bacteria of the esophagus in the induction of lysozyme production
remains to be elucidated. MEYER et al. (2009) described the microbial flora of the
equine esophagus. Although the bacteria detected were predominantly Gram-
negative, lysozyme still seems to play a pivotal role in the innate immune defence
of the esophagus epithelium. Due to the fact that α-haemoloytic Streptococcus
spp. were also isolated in high numbers, a lysozyme induction is likely.
The distribution pattern of lysozyme varied between the different layers of the
esophagus epithelium. The only observation made in all species was the generally
high expression of the enzyme in the stratum basale. We think that lysozyme is
produced by the cells of the basal layer and then transported to the epithelial
surface. Our findings differ from those made by NIYONSABA and OGAWA (2005),
who described the distribution pattern of lysozyme in the human epidermis. They
found lysozyme expression to be much stronger in the upper cells layers than in
the basal part. In some species, the staining intensity of lysozyme decreased
continuously from the stratum basale towards the stratum corneum, e.g. the horse
or the dog (Tab. 4.5). In others, for example in the cat, the staining persisted in the

Discussion
138
layers between the stratum basale and stratum corneum. Similar observations
have been made in the epidermis of marine mammals (MEYER et al. 2003;
MEYER and SEEGERS 2004). This phenomenon may be explained by a vesicular
transport or storage of the enzyme during epithelial development towards the
lumen. Supportive evidence for this theory is also provided by INAMOTO et al.
(2008), who described a vesicular transport of lysozyme through the intestinal
epithelium of rats. It is possible that the vesicular form of lysozyme was not
detectable by the antibody we applied. In order to corroborate this statement,
further experiments are needed, such as immunogold labelling of the secondary
antibody for TEM analysis.
As expected, in three of the five esophagus layers studied the herbivorous species
exhibited the strongest expression of lysozyme in comparison to the omnivorous
mammals. We reason that this expression pattern is a typical feature of the
esophagus epithelium of the herbivorous species being exposed to high
mechanical strain. Although the epithelium comprises thick layers of corneocytes,
this mechanical protection obviously cannot completely guarantee epithelial
integrity. As no strongly keratinised cells protect the epithelium of the carnivorous
cat, a thin layer of lysozyme covers the epithelium instead. This explains the
statistically relevant increase of lysozyme expression in the feline stratum corneum
in comparison to the herbivorous and omnivorous species.
In conclusion, lysozyme forms an innate chemical shield in the esophagus
epithelium of domesticated mammals, and is another vital player of immunity
against microbial threats.
5.2.5 Langerhans cells
An additional objective of this thesis was to support or refute the fact that cellular
mechanisms participate in innate immune mechanisms of the esophagus
epithelium of the nine different species studied. Supporting evidence concluding
that cells of the innate immune system are also actively involved in the defence of
epithelial surfaces was provided in earlier studies on the esophagus and other
epithelial tissues, such as the epidermis (AL YASSIN and TONER 1975; ZAVALA
et al. 2002; PENA-CRUZ et al. 2003; ROMANI et al. 2003; DE CARVALHO et al.
2005; HUSSEIN 2008; NFON et al. 2008). Most authors focussed their attention

Discussion
139
on two cell subsets of the innate immunity: Dendritic cells (DCs) and T-cells. There
is a strong interaction between these two cell types, i.e., DCs possess antigen-
presenting molecules, which stimulate a T-cell response (PENA-CRUZ et al. 2003;
SUGITA et al. 2004; VAN RHIJN et al. 2005; MORI and DE LIBERO 2008). To
identify the first instances of innate immunity, we concentrated on the detection of
a DC subgroup, the Langerhans cells (LCs). For decades LCs were thought to be
the typical DCs present in stratified squamous epithelia, such as the epidermis.
Due to modern findings, LCs can no longer be regarded as representatives of DCs
in general. Several studies encouraged the view that LCs may possess various
features that make them special and distinguish them from other types of DCs
(ROMANI et al. 2003). Principally there are three characteristical “cell markers” for
LCs: The antigen presenting molecule CD1a, the glycolipid receptor langerin
(CD207), and the tennis racket-shaped intracellular organelles, the Birbeck
granules (VALLEDEAU et al. 2000; ROMANI et al. 2003; MIZUMOTO and
TAKASHIMA 2004). We chose to detect LCs by IHC, using primary antibodies
against the CD1a and the langerin molecule. The advantage of the latter approach
is that this receptor is also expressed on immature LCs (VALLEDEAU et al. 2000).
In contrast CD1a and Birbeck granules are mainly found in activated LCs.
The presence of LCs in the human esophageal epithelium has been reported
previously (AL YASSIN 1975; GEBOES et al. 1983; TERRIS and POTET 1995;
ZAVALA et al. 2002). The authors applied electron microscopy or IHC for
identification. Our results showed a positive staining for langerin(+) LC in the
equine and mouse esophagus epithelium, whereby no positive staining was
observed in any of the nine species studied when using the CD1a antibody.
Several pre-treatments were applied to identify CD1a positive LCs, however, all
the methods tested were inconclusive. Thus, our findings only partly correspond
with the most recent findings of TERRIS and POTET (1995) and ZAVALA et al.
(2002). These authors detected LCs in the esophagus epithelium of humans by
using a CD1a antibody. Similarities can be observed in regard to the location of
LCs: Both of the latter author groups detected LCs in the stratum basale and
spinosum, which is in line with our findings. We could not corroborate the results of
AL YASSIN and TONER (1976), because these authors detected LCs in the
medial and superficial layers (stratum granulosum and str. corneum). Additionally,

Discussion
140
a positive reaction for LCs was demonstrated in the stratum basale of the feline
and equine esophagus by applying the zinciodide-osmium method
(SCHOENNAGEL 2005). Furthermore, SCHOENNAGEL (2005) showed that
CD1a(+) LCs are present in the porcine and ovine esophagus. However, our
observations are only partially in accordance with the findings of this author. A
corresponding finding is that we also observed a positive reaction in the equine
esophagus and detected langerin(+) LCs. The positive reaction for the ovine and
porcine species could not be corroborated.
The theory of a more basal orientation of the LCs seems more conclusive due to
two aspects. First, LCs are predominantly present near T-cells, which are the cells
they stimulate. T-cells are mainly found in the suprabasal cell layers of the
esophagus epithelium (TERRIS and POTET 1995; RESNICK et al. 1999). Second,
similar results for LC distribution were described in the skin, where LCs could be
detected in the basal part of the epidermis in humans (ROMANI et al. 2003), but
also in different other mammals, such as in the normal epidermis of the horse
(HAMADA et al. 1992) and the pig (MEYER 1986) by electron microscopy. In
porcine epidermis, LCs were also found via IHC, with antibodies against langerin
(NFON et al. 2008). In a recently conducted study, LCs were identified in canine
skin by IHC against CD1a surface molecules (VAN BEECK et al. 2008). Infection
correlated occurrence of LCs in the skin was described for the dog. LCs play an
important role in lupus erythematosus, atopic dermatitis (DAY 1996), and in
parasitic diseases such as demodicosis and leishmaniosis (DAY 1997; DE
CARVALHO 2005).
It remains to be elucidated, why some species reacted positive for langerin, while
none revealed a positive staining for CD1a. One theory is that the human specific
antibodies applied simply displayed no cross-reactivity with the studied species.
DE CAVALHO et al. (2005) already stated that monoclonal antibodies directed
against one species do not necessarily exhibit cross-reactivity with other species,
especially as the CD molecules are very species specific. Cells which exhibited
langerin did not simultaneously express CD1a antigen-presenting molecules on
their surface. We have to bear in mind which functional features both receptors
possess: Langerin captures glycolipids, transports these to the Birbeck granules,

Discussion
141
where CD1a is loaded and finally transported to the surface in order to present the
internalised antigen fragments to T-cells (VALLADEAU et al. 2000; HUNGER et al.
2004). This means that langerin(+) / CD1a(-) LCs are capable of internalising
circulating glycolipids with their langerin receptors into the cell. Nevertheless, they
do not behave as APCs, due to the fact they do not express CD1a on their
surface. It is possible that such cells may be able to provide a mechanism to
sequester circulating antigen and reduce its immunogenic potential (MORI and DE
LIBERO 2008). As a result, the antigenic pressure on the mucosal surface is
reduced. In case of increased antigenic pressure such cells eventually mature to
APCs and express CD1a on their surface.
One recent finding which has to be taken into account is the fact that dermal DCs
can express langerin too (NAGAO et al. 2009). Thus, the hypothesis that langerin
is a specific marker for LCs has to be abandoned. With regard to our findings this
means that the cells detected could also belong to the DC subset. NAGAO et al.
(2009) determined that LCs and langerin(+) DCs exhibit variable migratory
capabilities in vitro and repopulate distinct anatomic compartments in the skin at
diverse rates after conditional depletion in vivo. Langerin(+) DCs, in contrast to
LCs, did not require TGFbeta1 for development. Further studies on cell cultures
stimulated with TGFbeta1 have to be conducted, in order to differentiate between
langerin(+) DCs and LCs in the esophagus.
The observations of our study encourage the view that in the esophagus of murine
and equine species LCs also contribute to the first immunological barrier towards
luminal antigens. It remains to be elucidated, if such cellular mechanisms are also
present in other domesticated mammals. Species specific antibodies against the
CD epitopes of various species need to be designed for this approach.

Discussion
142
5.3 Microbial colonisation of the esophagus
Our study povides the first evidence of microbial colonisation on the esophagus
surface of the important domesticated mammals. By applying cryo SEM an
authentic display of the microorganisms covering the surface was achieved
(compare figures at 4.1.1). In the same way it was shown that innate defence
mechanisms are required in the esophagus where the epithelium has to control
and ward off bacterial threats. The epithelial surface revealed an occupation by a
thin layer of bacteria and fungi. Species specific differences in the quantity of the
microbiota could not be determined. In order to quantify the number of
microorganisms further studies are necessary, whereby they should begin by
establishing a standardised method for collecting samples.
The interest in the microbial composition of the regular digestive tract developed in
the 19th century; during the lifetime of Louis Pasteur, who believed that a symbiotic
relationship between man and bacteria was essential for life. The development of
experimental (animal) models allowed the research on this relationship in
absolutely germ-free mice, demonstrating the role of commensal microflora, i.e. a
function in vitamin synthesis (vitamin K and B12), the metabolism of nitrogen
compounds and lipids, and especially the participation as barrier against the
invasion of pathogenic microorganisms (MACKROWIAK 1982; ZILBERSTEIN et
al. 2007). The latter authors studied the microbial flora of the human digestive tract
in healthy volunteers. They collected samples from the mouth, esophagus,
stomach, duodenum, jejunum, ileum, colon, and rectum, and determined the flora
composition. The bacteria most frequently encountered in the human esophagus
were Streptococcus sp. (40%) and Staphylococcus sp. (20%). Furthermore, the
esophagus revealed a colonisation by Corynebacterium sp., Lactobacillus sp. and
Peptococcus sp. (all 10%). The authors claimed that the esophageal microbiota
are mainly transitory, similar to that detected in the mouth. However, they stated
that when alimentary stasis occurs, as for example in neoplasia, there is an
excessive increase in bacterial growth (LAU et al. 1981). In order to determine
whether the microbial colonisation of the esophagus of the species studied in this
thesis is only transitory or also indigenous, it is necessary to begin by
characterising the microbial flora of the oral cavity of every animal. Comparing the
microbial flora of the human esophagus (ZILBERSTEIN et al. 2007) with that of

Discussion
143
the only other species studied, the horse (MEYER et al. 2009), revealed no
similarities. Such finding is not surprising, since humans and horses have
completely different nutrition habits. For a comparison of the human microbiota
with the microflora of an animal species, the porcine digestive tract should be
investigated in detail, as swine are often used as models for humans also due to
similarities regarding the digestive organs.
In contrast to the esophagus, a lot of information is available on the microbial
colonisation of other parts of the human digestive tract. Information about bacterial
communities is expected to provide crucial information how to develop therapies
for various gastrointestinal diseases. Furthermore, scientists hope to generate
knowledge about the next generation of probiotic bacteria as part of functional
food (SINGH et al. 2009). The gut bacterial microbiota has established multiple
mechanisms to influence the eukaryotic host, generally in a beneficial way. The
prokaryotic genomes of the human microbiota encode a spectrum of metabolic
capabilities beyond that of the host genome, making the microbiota an integral
component of human physiology (HOOPER et al. 2001; SCHENK and MUELLER
2008; NEISH 2009). Modulating bacterial numbers and diversity is entrusted to the
gastrointestinal immune system. Although bacteria are distributed throughout the
intestine, the major concentration of microorganisms can be found in the large
intestine (SINGH et al. 2009). The diversity of species encompasses around 400-
500 different bacteria (LOTZ et al. 2007). The small intestine has microbial
communities with 1x108 CFU g-1 ileal content. Through the remainder of the
digestive tract, bacterial concentrations gradually increase, reaching 1x1011 CFU
g-1 (SINGH et al. 2009). Within the 400-500 species mentioned, 30-40 species
make up for 99% of the total population of the intestine (GUARNER 2006). The
results of several studies indicate that five genera are predominant in the large
intestine of humans: Bacteroides, Eubacterium, Bifidobacterium, Peptococcus,
and Fusobacterium (SANDBORN et al. 2003; SINGH et al. 2009). It is important to
note that although immunostimulatory structures recognized by the innate immune
system are generally produced by both, commensal and pathogenic bacteria, the
synthesis of low stimulatory hypoacytylated LPS from Bacteroides might contribute
to homeostasis (WEINTRAUB et al. 1989).

Discussion
144
With regard to our own observations, one difference was most obvious between
the colonisation of the intestinal tract and the esophagus, i.e. the quantity of
microorganisms detected. In contrast to the resorpative intestine, only a thin layer
of microbiota covered the epithelial surface of the esophagus. These findings are
in line with observations of LOTZ et al. (2007) from the human esophagus, which
is covered with only low to very low numbers of bacteria. As the esophagus
mucosa does not fulfil such digestive tasks, as the absorption of nutrients or
synthesis of vitamins, the colonisation with commensal bacteria here seems not
essential for nutrient absorption. As a result, much less bacteria, in comparison to
the intestine, can be observed. Nevertheless, due to the fact that we demonstrated
the existence of pattern recognition receptors in the esophagus epithelium, we
also showed that already a relatively low number of microorganisms is able to
evoke the response of the innate immune system. It is possible that such a low
number of microorganisms maintains homeostasis in the esophagus, whereby an
excessive colonisation with pathogenic bacteria may be avoided.

Discussion
145
5.4 Conclusions
From the results obtained in the current study, it can be concluded that a first-line
of defence mechanisms of the innate immune system contributes to maintaining a
microbial homeostasis at the surface of the esophagus epithelium of domesticated
mammals. The esophageal antimicrobial arsenal encompasses a variety of PRRs
and antimicrobial proteins and peptides.
One of the first authorities for pathogen recognition appears to be TLR2. Most
likely this receptor recognizes sugar moieties on the surface of pathogens, thus
triggering an innate immune response. Apparently one of these responses is the
production of antimicrobial peptides (Aps). We observed expression of ß-defensin
2 and -3 as well as CAT in all species studies. The co-expression of TLR2 and
APs by the epithelial cells was clearly demonstrated for all animals. Comparing the
three different nutrition groups (herbivorous, omnivorous, carnivorous) revealed
differences in the expression pattern of the TLR2 and APs studied. Regarding the
ß-defensins, it could be shown that the weakly keratinised esophagus epithelium
of the carnivorous cat compensates the diminished mechanical protection by an
increased production of ß-defensin 2 and -3. In contrast, the herbivorous species
exhibited a higher expression of TLR2 when compared to the carnivorous and
omnivorous species. We explained this observation with the functional feature of
TLR2 to trigger the production of substances that maintain epithelial integrity
(RAKOFF-NAHOUM et al. 2004). The esophagus epithelium of the herbivorous
species is confronted with enormous mechanical strain, clearly making such a
function necessary.
Another antimicrobial protein contributing to the formation of an innate epithelial
shield in the esophagus epithelium is lysozyme, detectable in the esophagus of all
species studied. In contrast to ß-defensin 2 and -3, lysozyme was expressed in
higher amounts in the herbivorous species studied. Although their epithelium is
protected by an often distinctly keratinised epithelium, the complementary
production of lysozyme seems to be required to protect the epithelium from
invading bacteria, as perforations can occur from the rough plant material (MEYER
et al. 2008, unpublished data).
Since Langerhans cells could only be proven in the murine and equine esophagus
epithelium, their importance for the surveillance of the esophagus epithelium in
healthy animals has to remain open.

Discussion
146
In regard to the fact that ß-glucan receptors could not be identified in the present
study, we believe that these PRRs do not play a key role in the epithelial defence
of the esophagus. It is conjecturable that the microbial colonisation of the
esophagus epithelium, in contrast to that of the epidermis, is too insignificant as to
trigger a ß-glucan receptor response (MEYER et al. 2008). In general, IHC offered
a good possibility to study the presence of innate defence mechanisms in the
esophagus epithelium.
In addition, the application of cryo SEM permitted the first artefact free
demonstration of microbial colonisation of the esophagus. The pictures obtained
revealed a thin layer of bacteria and fungi covering the epithelial surface. This
result can be declared as vital for justifying the necessity of innate defence
mechanisms.
The third main topic of this dissertation was the comparison of three different
fixation media. Our results indicated that the recently established HOPE® fixation
offers a good opportunity to study certain immune parameters in the esophagus
epithelium. Nevertheless, it is no serious alternative to the well established
formalin based Bouin`s solution but could be an alternative to cryosections.
Fixation in the latter medium resulted in the best preservation of tissue structure
and antigens. The frequently applied Ca-formol fixation proved to be the poorest
fixative in regard to preservation quality and maintenance of antigenicity in the
esophagus epithelium.

Zusammenfassung
147
6 Zusammenfassung
Untersuchungen zur angeborenen Immunität des Oesophagusepithels der
Haussäugetiere
Isabelle N. Hornickel
Ziel dieser Dissertation war zu klären, ob das Oesophagusepithel von
Haussäugetieren charakteristische Merkmale der angeborenen Immunität
aufweist. Im Rahmen aktueller Studien am humanen Darm wurde bereits das
Vorhandensein und die Notwendigkeit von Mechanismen der angeborenen
Immunität offenkundig. Die Autoren unterstrichen im Besonderen das
Zusammenspiel zwischen den im Darmtrakt zahlreich vorhandenen kommensalen
Mikroorganismen und einer Aktivierung der angeborenen Immunität, um die
epitheliale Homöostase zu garantieren. Im Rahmen der vorliegenden Dissertation
wurde zunächst der Frage nachgegangen, ob überhaupt eine bemerkenswerte
Besiedlung des Oesophagusepithels mit Mikroorganismen besteht. Im nächsten
Schritt sollte geklärt werden, ob und inwieweit vorhandene Bakterien und Pilze zur
Aktivierung des angeborenen Immunsystems beitragen.
Methodische Grundlage für die vorliegende Studie waren immunhistochemische
Färbungen, die qualitativ und semiquantitativ ausgewertet wurden. Als Material
dienten Oesophagusproben von verschiedenen frischtoten Haussäugetierarten mit
unterschiedlicher Ernährungsweise (Omnivore: Schwein, Hund, Maus, Laborratte,
Karnivore: Katze und herbivore: Ziege, Schaf, Kaninchen, Rind, Pferd). Als
zusätzlicher methodischer Aspekt waren die Einflüsse von drei verschiedenen
Fixationsmedien auf die immunhistochemischen Ergebnisse zu beurteilen.
Verglichen werden sollte die Verwendung von zwei formalinhaltigen Fixantien
(Bouin´sche Lösung und Ca-Acetat Formalin) mit einer neuen, möglicherweise
Antigen schonenden Fixationsmethode namens HOPE®. Nach der Fixation im
jeweiligen Medium wurden die Proben in Paraffin eingebettet. Um eine
artefaktfreie Darstellung der die Oesophagusoberfläche besiedelnden
Mikroorganismen sicher zu stellen, wählten wir die Untersuchung mittels eines

Zusammenfassung
148
Cryo Rasterelektronenmikroskop (Cryo REM). Hierzu wurden die einzelnen
Proben bis zur Untersuchung in flüssigem Stickstoff gelagert.
Die Ergebnisse lieferten den Beweis für die Existenz von Parametern der
angeborenen Immunität im Oesophagusepithel der Haussäugetiere. Es konnte
gezeigt werden, dass dem tierischen Organismus verschiedene Pathogen-
erkennende Rezeptoren zur Verfügung stehen, um eine frühe Antwort des
Immunsystems auf fakultativ pathogene Keime zu aktivieren. Als erste Instanz
einer Pathogenerkennung scheint der Toll-like Rezeptor 2 von besonderer
Bedeutung zu sein, der hauptsächlich von den Zellen des Stratum basale des
Oesophagusepithels exprimiert wurde. Zum weiteren Schutz der epithelialen
Barriere produzierten die Epithelzellen kationische antimikrobielle Peptide.
Innerhalb dieser Studie, wurden dabei Cathelizidin, sowie ß-Defensin 2 und -3
nachgewiesen. Somit konnten Ergebnisse aus Studien an humanen intestinalen
und kutanen Epithelgeweben bestätigt werden, die eine Koexpression von Toll-like
Rezeptoren und antimikrobiellen Peptiden zeigten. Das nur schwach verhornte
Oesophagusepithel der karnivoren Katze wurde vor einer Keiminvasion durch
besonders hohe Mengen an antimikrobiellen Peptiden geschützt, während im
Epithel der herbivoren und omnivoren Spezies eine mehr oder weniger starke
Keratinisierung und ein charakterischer histologischer Aufbau für einen guten
mechanischen Schutz sorgten.
Ein Nachweis der löslichen, Pathogen-erkennenden ß-glucan Rezeptoren auf den
Zellen des Oesophagusepithels der Haussäugetiere war innerhalb dieser Studie
nicht möglich. Auf Grund des Vergleichs mit anderen Arbeiten, z.B. von der Haut
mariner Säuger, lässt sich vermuten, dass die Expression dieser Rezeptoren eine
höhere Belastung mit Mikroorganismen erfordert, als es im Oesophagus der Fall
ist. Weitere Untersuchungen auf diesem Bereich sind also notwendig.
Neben den antimikrobiellen Peptiden konnte außerdem das enzymatisch aktive
Protein Lysozym in allen Spezies nachgewiesen werden, welches somit ebenfalls
einen Beitrag zur Epithelhomöostase liefert.

Zusammenfassung
149
Des Weiteren wurde ein Beweis für das Vorkommen zellulärer Abwehrinstanzen
innerhalb des Oesophagusepithels der Haussäugetiere erbracht. In diesem
Zusammenhang gelang es, Langerhans Zellen beim Pferd und bei der Maus durch
immunhistochemische Methodik zu erfassen.
Die immunhistochemischen Analysen dieser Studie wurden durch die Cryo REM
Darstellung einer dünnen Schicht von Bakterien und Pilzen auf der
Oesophagusoberfläche ergänzt.
Mit der vorliegenden Arbeit konnte außerdem ein Beitrag zur aktuellen Diskussion
über Alternativen zu formalinbasierenden Fixationsmethoden geliefert werden.
Bezüglich dieser Thematik ist festzuhalten, dass die HOPE® Fixierung in der
Grundlagenforschung eine kostensenkende Alternative darstellt, jedoch für eine
Routinediagnostik auf Grund vieler variabler Parameter nicht unbedingt geeignet
erscheint. Eine weitere Schlussfolgerung auf der Basis unserer Resultate ist
daher, dass immunhistochemische Befunde auch immer im Zusammenhang mit
der Fixierungsmethode kritisch diskutiert werden sollten. So zeigte sich in unserer
Studie, dass die immunhistochemischen Färbungsergebnisse von Ca-Formol
fixierten Proben kaum verwertbar waren, hingegen die Fixierung in Bouin`scher
Lösung gute Ergebnisse in allen Bereichen (Strukturerhaltung und
Immunhistochemie) erbrachte.
Zusammenfassend gesehen, liefert die vorliegende Dissertation eine detaillierte
Charakterisierung der Parameter der angeborenen Immunität des
Oesophagusepithels der Haussäugetiere. Die gewonnenen Erkenntnisse könnten
bei der Erforschung humaner Erkrankungen der Speiseröhre, wie zum Beispiel der
Refluxoesophagitis oder des Barrett Oesophagus, besonders im Zusammenhang
mit den vorhandenen Tiermodellen (Maus, Ratte, Schwein) von Nutzen sein.

Summary
150
7 Summary
The aim of this dissertation thesis was to support or refute the hypothesis, whether the
esophagus epithelium of domesticated mammals exhibits characteristical features of
innate immunity or not. Within the scope of recent studies on human intestinal tissue, the
occurrence and necessity of innate immune mechanisms has already been demonstrated.
The authors emphasised that the interaction between the numerously occurring intestinal
microorganisms and the activation of the innate immune system is required to maintain
intestinal homeostasis. In this context, first, we tried to answer the question if it is possible
to show a microbial colonisation of the esophageal surface. Second, it should be clarified
to which extend this colonisation contributes to an activation of innate immune
mechanisms.
The experimental basis of this study was immunohistochemical staining which was
evaluated in a qualitative and semiquantitative matter. The esophagus samples used were
obtained immediately after euthanization from nine species of domesticated mammals of
three nutriton types (omnivores: pig, dog, mouse, rat; carnivores: cat; herbivores: goat,
sheep, rabbit, cattle, horse). An additional feature of this thesis was to analyse the
influence of different fixation media on the immunohistochemical results. Two formalin
based fixation solutions (Bouin`s solution and Ca- acetate formalin) were compared with a
recently introduced, probably more gentle fixation medium, the HOPE® fixation. In order to
guarantee an artefact free demonstration of microorganisms colonising the esophagus
epithelium, we applied cryo scanning electron microscopy (cryo SEM).
We clearly confirmed the existence of several innate immune parameters in the
esophagus epithelium of domesticated mammals. In this context, various PRRs which
trigger a first response of the innate immune system were demonstrated. Toll-like receptor
2 seems to play a pivotal role in this manner. This PRR was primarily expressed on the
epithelial cells of the stratum basale. Furthermore, cationic antimicrobial peptides
contribute to the maintainance of epithelial integrity. Within this study, the antimicrobial
peptides cathelicidin and ß-defensin 2 and 3 could be demonstrated. Hence, we
corroborated results obtained from studies on human intestinal and cutanous tissue,
which showed a co-expresseion of Toll-like receptors and antimicrobial peptides. The less
keratinised esophagus epithelium of the carnivorous cat was protected from exogenous
strain by an extraordinary high amount of antimicrobial peptides. In contrast, in the
epithelium of the herbivorous and omnivorous species a stronger keratinisation and a
characteristical epithelial layer structure guaranteed an effective mechanical barrier.

Summary
151
A detection of the soluble pathogen recognising ß-glucan receptors remained
inconclusive. Based on the comparison with publications, for example from marine
mammals, it can be conjectured that an epithelial exposition to higher amounts of
microorganisms is required to trigger a production of these PRRs. This hypothesis is in
line with the rather low number of microorganisms present in the esophagus of the
animals studied.
Additionally, the enzymatically active protein lysozyme could be demonstrated in the
mammalian esophagus epithelium. Thus, it can be assumed that this protein participates
as another parameter of the innate innate immune mechanisms present in the esophagus
epithelium.
Furthermore, cellular components of the innate immune system were present in the
esophagus epithelium. In this context, we demonstrated langerin (+) Langerhans cells in
basal layers of the murine and equine esophagus.
The immunohistochemical results were complemented by observations made by cryo
SEM. It could be shown for all species studied that the esophagus epithelium is covered
by a thin layer of bacteria and fungi. However, detailed analyses of the microbial
composition for each species have still to be conducted.
Finally, a substantial contribution to the actual discussion concerning the advantage of
formalin alternatives could be provided by the present study. On the one hand, the HOPE®
fixation method offers a rather cheap alternative, as the antibody amounts could be
reduced. On the other hand, an application in routine diagnostic is not advisable, because
of several variable parameters. It can be concluded that immunohistochemical results
always have to be discussed in connection to the fixation method applied. We clearly
demonstrated a diminished immunoreactivity for Ca-formol fixed samples; satisfactory
results were obtained, particularly, from samples fixed in Bouin`s solution.
It can be summarised that the present dissertation offers a detailed characterisation of the
innate immune parameters present in the esophagus epithelium of domesticated
mammals. Our findings might also contribute to the progress of research on human
esophageal illnesses, such as reflux esophagitis or Barrett`s esophagus. In this
connection, the use of already established animal models (mouse, rat, pig) could benefit
from our results to some extent.

References
152
8 References
AARBIOU, J., R. M. VERHOOSEL, S. VAN WETERING, W. I. DE BOER, J. H. VAN KRIEKEN, S. V. LITVINOV, K. F. RABE and P. S. HIEMSTRA (2004): Neutrophil defensins enhance lung epithelial wound closure and mucin gene expression in vitro. American Journal of Respiratory Cellular and Molecular Biology 30, 193-201 AGAH, A., M. C. MONTALTO, K. YOUNG and G. L. STAHL (2001): Isolation, cloning and functional characterization of porcine mannose-binding lectin. Immunology 102, 338-343 AKAIWA, M., Y. YAE, R. SUGIMOTO, S. O. SUZUKI, T. IWAKI, K. IZUHARA and N. HAMASAKI (1999): Hakata antigen, a new member of the ficolin opsonin p35 family, is a novel human lectin secreted into bronchus alveolus and bile. Journal of Histochemistry and Cytochemistry 47, 777-785 AKIRA, S. (2003): Mammalian Toll-like receptors. Current Opinion in Immunology 15, 5-11 AKIYAMA, J., A. HOFFMAN, C. BROWN, L. ALLEN, J. EDMONDSON, F. POULAIN and S. HAWGOOD (2002): Tissue distribution of surfactant proteins A and D in the mouse. Journal of Histochemistry and Cytochemistry 50, 993-996 AL YASSIN, M. T., TONER P.G. (1975): Langerhans cells in the human oesophagus. Journal of Anatomy 122, 435-445 ALVAREZ, B., C. REVILLA, S. CHAMORRO, M. LOPEZ-FRAGA, F. ALONSO, J. DOMINGUEZ and A. EZQUERRA (2006): Molecular cloning, characterization and tissue expression of porcine Toll-like receptor 4. Developmental and Comparative Immunology 30, 345-355 ALVAREZ, B., C. REVILLA, N. DOMENECH, C. PEREZ, P. MARTINEZ, F. ALONSO, A. EZQUERRA and J. DOMIGUEZ (2008): Expression of toll-like receptor 2 (TLR2) in porcine leukocyte subsets and tissues. Veterinary Research 39, 12 AMBATIPUDI, K. and E. M. DEANE (2008): In search of neutrophil granule proteins of the tammar wallaby (Macropus eugenii). Molecular Immunology 45, 690-700

References
153
ANDREI, F., M. NEAGU, M. CEAUSU, A. GEORGESCU, F. VASILESCU, M. MIHAI, D. TERZEA, C. IOSIF, C. DOBREA, E. SIMONA, D. ENE, G. BUTUR and C. ARDELEANU (2008): Advantages of hope fixation for immunohistochemistry. Histopathology 53 (Suppl. 1), 294-295 ANGENIEUX, C., J. SALAMERO, D. FRICKER, J. P. CAZENAVE, B. GOUD, D. HANAU and H. DE LA SALLE (2000): Characterization of CD1e, a third type of CD1 molecule expressed in dendritic cells. Journal of Biological Chemistry 275, 37757-37764 ARIIZUMI, K., G. L. SHEN, S. SHIKANO, S. XU, R. RITTER, T. KUMAMOTO, D. EDELBAUM, A. MORITA, P. R. BERGSTRESSER and A. TAKASHIMA (2000): Identification of a novel, dendritic cell-associated molecule, Dectin-1, by subtractive cDNA cloning. Journal of Biological Chemistry 275, 20157-20167 ARNOLD, M. M., S. SRIVASTAVA, J. FREDENBURGH, C. R. STOCKARD, R. B. MYERS and W. E. GRIZZLE (1996): Effects of fixation and tissue processing on immunohistochemical demonstration of specific antigens. Biotechnic & Histochemistry 71, 224-230 ASAHINA, Y., N. YOSHIOKA, R. KANO, T. MORITOMO and A. HASEGAWA (2003): Full-length cDNA cloning of Toll-like receptor 4 in dogs and cats. Veterinary Immunology and Immunopathology 96, 159-167 BAILEY, G. D. and D. N. LOVE (1991): Oral associated bacterial infection in horses: studies on the normal anaerobic flora from the pharyngeal tonsillar surface and its association with lower respiratory tract and paraoral infections. Veterinary Microbiology 26, 367-379 BALS, R., X. R. WANG, R. L. MEEGALLA, S. WATTLER, D. J. WEINER, M. C. NEHLS and J. M. WILSON (1999): Mouse beta-defensin 3 is an inducible antimicrobial peptide expressed in the epithelia of multiple organs. Infection and Immunity 67, 3542-3547 BALS, R., X. R. WANG, M. ZASLOFF and J. M. WILSON (1998): The peptide antibiotic LL-37/hCAP-18 is expressed in epithelia of the human lung where it has broad antimicrobial activity at the airway surface. Proceedings of the National Academy of Sciences of the United States of America 95, 9541-9546 BANCHEREAU, J., F. BRIERE, C. CAUX, J. DAVOUST, S. LEBECQUE, Y. T. LIU, B. PULENDRAN and K. PALUCKA (2000): Immunobiology of dendritic cells. Annual Review of Immunology 18, 767-811

References
154
BARRAL, D. C. and M. B. BRENNER (2007): CD1 antigen presentation: how it works. Nature Reviews Immunology 7, 929-941 BEDOSSA, P., J. BACCI, G. LEMAIGRE and E. MARTIN (1987): Effects of fixation and processing on the immunohistochemical visualisation of type-I, -III and -IV collagen in paraffin-embedded liver tissue. Histochemistry 88, 85-89 BENAVIDES, J., C. GARCÍA-PARIENTE, D. GELMETTI, M. FUERTES, M. C. FERRERAS, J. F. GARCÍA-MARÍN and V. PÉREZ (2006): Effects of fixative type and fixation time on the detection of Maedi Visna virus by PCR and immunohistochemistry in paraffin-embedded ovine lung samples. Journal of Virological Methods 137, 317-324 BENDELAC, A., O. LANTZ, M. E. QUIMBY, J. W. YEWDELL, J. R. BENNINK and R. R. BRUTKIEWICZ (1995): CD1 recognition by mouse NK1+ T lymphocytes. Science 268, 863-865 BEVINS, C. L., E MARTIN-PORTER AND T GANZ (1999): Defensins and innate host defence of the gatrointestinal tract. Gut 45, 911-915 BIENERT, A., C. P. BARTMANN, J. VERSPOHL and E. DEEGEN (2003): Bacteriological findings for endodontical and apical molar dental diseases in the horse. Deutsche Tierärztliche Wochenschrift 110, 358-361 BIRAGYN, A., P. A. RUFFINI, M. COSCIA, L. K. HARVEY, S. S. NEELAPU, S. BASKAR, J. M. WANG and L. W. KWAK (2004): Chemokine receptor-mediated delivery directs self-tumor antigen efficiently into the class II processing pathway in vitro and induces protective immunity in vivo. Blood 104, 1961-1969 BIRBECK, M. S., A. S. BREATHNACH and J. D. EVERALL (1961): An electron microscope study of basal melanocytes and high-level clear cells (Langerhans cells) in vitiligo. Journal of Investigative Dermatology 37, 51-63 BIRCHLER, T., R. SEIBL, K. BUCHNER, S. LOELIGER, R. SEGER, J. P. HOSSLE, A. AGUZZI and R. P. LAUENER (2001): Human Toll-like receptor 2 mediates induction of the antimicrobial peptide human beta-defensin 2 in response to bacterial lipoprotein. European Journal of Immunology 31, 3131-3137 BLASCHNITZ, A., M. GAUSTER and G. DOHR (2008): Application of cryo-compatible antibodies to human placenta paraffin sections. Histochemistry and Cell Biology 130, 595-599

References
155
BLUM, F. (1893): Der Formaldehyd als Härtungsmittel. Zeitschrift für wissenschaftliche Mikroskopie 10, 314-315 BLUM, F. (1896): Ueber Wesen und Wert der Formaldehydhärtung. Anatomischer Anzeiger 11, 718-727 BOCK, P. (1974): Fine structure of Langerhans cells in the stratified epithelia of the esophagus and stomach of mice. Zeitschrift für Zellforschung und Mikroskopische Anatomie 147, 237-247 BÖCK, P. (1989): Romeis Mikroskopische Technik. 17. Auflage, Urban & Schwarzenberg, München, Wien, Baltimore BOMAN, H. G. (2003): Antibacterial peptides: basic facts and emerging concepts. Journal of Internal Medicine 254, 197-215 BRAFF, M. H., A. DI NARDO and R. L. GALLO (2005): Keratinocytes store the antimicrobial peptide cathelicidin in lamellar bodies. Journal of Investigative Dermatology 124, 394-400 BRAFF, M. H. and R. L. GALLO (2006): Antimicrobial peptides: an essential component of the skin defensive barrier. Current Topics of Microbiology and Immunology 306, 91-110 BRAFF, M. H., M. ZAIOU, J. FIERER, V. NIZET and R. L. GALLO (2005): Keratinocyte production of cathelicidin provides direct activity against bacterial skin pathogens. Infection and Immunity 73, 6771-6781 BROGDEN, K. A., M. ACKERMANN, P. B. MCCRAY and B. F. TACK (2003): Antimicrobial peptides in animals and their role in host defences. International Journal of Antimicrobial Agents 22, 465-478 BROOKS, A. S., J. HAMMERMUELLER, J. P. DELAY and M. A. HAYES (2003): Expression and secretion of ficolin by porcine neutrophils. Biochimica et Biophysica Acta-General Subjects 1624, 36-45 BROWN, G. D. (2006): Dectin-1: a signalling non-TLR pattern-recognition receptor. Nature Reviews Immunology 6, 33-43 BROWN, G. D. and S. GORDON (2001): Immune recognition - A new receptor for beta-glucans. Nature 413, 36-37

References
156
BUECHAU, A. S., J. SCHAUBER, T. HULTSCH, A. STUETZ and R. GALLO (2008): Pimecrolismus enhances TLR2/6-induced expression of antimicrobial peptides in keratinocytes Journal of Investigative Dermatology 128, 2646-2654 BUESA, R. J. (2008): Histology without formalin? Annual Diagnostic Pathology 12, 387-396 CALABI, F., J. M. JARVIS, L. MARTIN and C. MILSTEIN (1989): 2 Classes of Cd1 Genes. European Journal of Immunology 19, 285-292 CARIO, E., D. BROWN, M. MCKEE, K. LYNCH-DEVANEY, G. GERKEN and D. K. PODOLSKY (2002): Commensal-associated molecular patterns induce selective toll-like receptor-trafficking from apical membrane to cytoplasmic compartments in polarized intestinal epithelium. American Journal of Pathology 160, 165-173 CARIO, E., G. GERKEN and D. K. PODOLSKY (2007): Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 132, 1359-1374 CARIO, E. and D. K. PODOLSKY (2006): Toll-like receptor signaling and its relevance to intestinal inflammation. Inflammatory Bowel Disease: Genetics, Barrier Function, Immunologic Mechanisms, and Microbial Pathways 1072, 332-338 CARIO, E., I. M. ROSENBERG, S. L. BRANDWEIN, P. L. BECK, H. C. REINECKER and D. K. PODOLSKY (2000): Lipopolysaccharide activates distinct signaling pathways in intestinal epithelial cell lines expressing Toll-like receptors. Journal of Immunology 164, 966-972 CARRASCO, L., A. NÚÑEZ, P. J. SÁNCHEZ-CORDÓN, M. PEDRERA, M. FERNÁNDEZ DE MARCO, F. J. SALGUERO and J. C. GÓMEZ-VILLAMANDOS (2004): Immunohistochemical detection of the expression of pro-inflammatory cytokines by ovine pulmonary macrophages. Journal of Comparative Pathology 131, 285-293 CAVERLY, J. M., Z. A. RADI, C. B. ANDREASEN, R. A. DIXON, K. A. BROGDEN and M. R. ACKERMANN (2001): Comparison of bronchoalveolar lavage fluid obtained from Mannheimia haemolytica-inoculated calves with and treatment with the selectin without prior inhibitor TBC 1269. American Journal of Veterinary Research 62, 665-672

References
157
CELLA, M., F. SALLUSTO and A. LANZAVECCHIA (1997): Origin, maturation and antigen presenting function of dendritic cells. Current Opinion in Immunology 9, 10-16 CHAVAKIS, T., D. B. CINES, J. S. RHEE, O. D. LIANG, U. SCHUBERT, H. P. HAMMES, A. A. HIGAZI, P. P. NAWROTH, K. T. PREISSNER and K. BDEIR (2004): Regulation of neovascularization by human neutrophil peptides (aleph-defensins): a link between inflammation and angiogenesis. FASEB Journal 18, 1306-1308 CLARK, G. J., N. ANGEL, M. KATO, J. A. LOPEZ, K. MACDONALD, S. VUCKOVIC and D. N. J. HART (2000): The role of dendritic cells in the innate immune system. Microorganisms and Infection 2, 257-272 CRAWFORD, J. M., J. M. KRISKO, G. A. MORRIS and D. A. CHAMBERS (1989): The distribution of Langerhans cells and CD1a antigen in healthy and diseased human gingiva. Regional Immunology 2, 91-97 CROUCH, E., D. PARGHI, S. F. KUAN and A. PERSSON (1992): Surfactant protein-D - subcellular-localization in nonciliated bronchiolar epithelial-cells. American Journal of Physiology 263, L60-L66 DAHL, M. R., S. THIEL, M. MATSUSHITA, T. FUJITA, A. C. WILLIS, T. CHRISTENSEN, T. VORUP-JENSEN and J. C. JENSENIUS (2001): MASP-3 and its association with distinct complexes of the mannan-binding lectin complement activation pathway. Immunity 15, 127-135 DALE, B. A. and L. P. FREDERICKS (2005): Antimicrobial peptides in the oral environment: Expression and function in health and disease. Current Issues in Molecular Biology 7, 119-133 DANN, S. M. and L. ECKMANN (2007): Innate immune defenses in the intestinal tract. Current Opinion in Gastroenterology 23, 115-120 DAY, M. J. (1996): Expression of major histocompatibility complex class II molecules by dermal inflammatory cells, epidermal Langerhans cells and keratinocytes in canine dermatological disease. Journal of Comparative Pathology 115, 317-326 DAY, M. J. (1997): An immunohistochemical study of the lesions of demodicosis in the dog. Journal of Comparative Pathology 116, 203-216

References
158
DE CARVALHO, M. C., BONNEFONT-REBEIX C., RIGAL D., CHABANNE L. (2005): Dendritic cells in different animal species: an overview. Pathologie Biologie 54, 85-93 DE FERNANDEZ CALEYA, R., B. GONZAALEZ-PASCUAL, F. GARCIA-OLMEDO and P. CARBONERO (1972): Susceptibility of phytopathogenic bacteria to wheat purothionins in vitro. Applied Microbiology and Biotechnology 23, 998-1000 DE FRAISSINETTE, A., D. SCHMITT and J. THIVOLET (1989): Langerhans cells of human mucosa. Journal of Dermatology 16, 255-262 DELAY, J. (1999) Biochemical Characterization and Bacterial Binding Functions of Porcine Ficolins. Guelph, University of Guelph, D.V.Sc Thesis. DENNEHY, K. M. and G. D. BROWN (2007): The role of the beta-glucan receptor Dectin-1 in control of fungal infection. Journal of Leukocyte Biology 82, 253-258 DHOPLE, V., A. KRUKEMEYER and A. RAMAMOORTHY (2006): The human beta-defensin-3, an antibacterial peptide with multiple biological functions. Biochimica et Biophysica Acta-Biomembranes 1758, 1499-1512 DIAMOND, G., M. ZASLOFF, H. ECK, M. BRASSEUR, W. L. MALOY and C. L. BEVINS (1991): Tracheal antimicrobial peptide, a cysteine-rich peptide from mammalian tracheal mucosa - peptide isolation and cloning of a cDNA. Proceedings of the National Academy of Sciences of the United States of America 88, 3952-3956 DE NARDI, F. G. and R. H. RIDDELL (1991): The normal esophagus. American Journal of Surgical Pathology 15, 296-309 DOUILLARD, P., P. STOITZNER, C. H. TRIPP, V. CLAIR-MONINOT, S. AIT-YAHIA, A. D. MCLELLAN, A. EGGERT, N. ROMANI and S. SAELAND (2005): Mouse lymphoid tissue contains distinct subsets of Langerin/CD207+dendritic cells, only one of which represents epidermal-derived Langerhans cells. Journal of Investigative Dermatology 125, 983-994 DROEMANN, D., T. GOLDMANN, D. BRANSCHEID, R. CLARK, K. DALHOFF, P. ZABEL and E. VOLLMER (2003): Toll-like receptor 2 is expressed by alveolar epithelial cells type II and macrophages in human lung. Histochemicistry Cell Biology 119, 103-108

References
159
DUSZYK, M. (2001): CFTR and lysozyme secretion in human airway epithelial cells. Pflugers Archiv-European Journal of Physiology 443, S45-S49 DUTTON, J. M., K. GOSS, K. R. KHUBCHANDANI, C. D. SHAH, R. J. SMITH and J. M. SNYDER (1999): Surfactant protein A in rabbit sinus and middle ear mucosa. Annals of Otology Rhinology and Laryngology 108, 915-924 ELDER, J. T., N. J. REYNOLDS, K. D. COOPER, C. E. GRIFFITHS, B. D. HARDAS and P. A. BLEICHER (1993): CD1 gene expression in human skin. Journal of Dermatological Science 6, 206-213 ELIAS, P. M. (2005): Stratum corneum defensive functions: an integrated view. Journal of Investigative Dermatology 125, 183-200 ELIAS, P. M. and E. H. CHOI (2005): Interactions among stratum corneum defensive functions. Experimental Dermatology 14, 719-726 ELLISON, R. T., 3RD, D. BOOSE and F. M. LAFORCE (1985): Isolation of an antibacterial peptide from human lung lavage fluid. The Journal of Infectious Diseases 151, 1123-1129 EPSTEIN, J., Q. EICHBAUM, S. SHERIFF and R. A. B. EZEKOWITZ (1996): The collectins in innate immunity. Current Opinion in Immunology 8, 29-35 EURELL, J. A. and B. L. FRAPPIER (EDS:) (2006) Dellmann`s Textbook of Veteriary Histology. 6th ed. Blackwell Publishing, Ames, IA, pp. 184-185 FUJIMORI, Y., S. HARUMIYA, Y. FUKUMOTO, Y. MIURA, K. YAGASAKI, H. TACHIKAWA and D. FUJIMOTO (1998): Molecular cloning and characterization of mouse ficolin-A. Biochemical and Biophysical Research Communications 244, 796-800 FUJITA, T., M. MATSUSHITA and Y. ENDO (2004): The lectin-complement pathway - its role in innate immunity and evolution. Immunological Reviews 198, 185-202 GÁL, P., L. BARNA, A. KOCSIS and P. ZÁVODSZKY (2007): Serine proteases of the classical and lectin pathways: Similarities and differences. Immunobiology 212, 267-277

References
160
GALLO, R. L., K. J. KIM, M. BERNFIELD, C. A. KOZAK, M. ZANETTI, L. MERLUZZI and R. GENNARO (1997): Identification of CRAMP, a cathelin-related antimicrobial peptide expressed in the embryonic and adult mouse. Journal of Biological Chemistry 272, 13088-13093 GANTNER, B. N., R. M. SIMMONS, S. J. CANAVERA, S. AKIRA and D. M. UNDERHILL (2003): Collaborative induction of inflammatory responses by Dectin-1 and toll-like receptor 2. Journal of Experimental Medicine 197, 1107-1117 GANTNER, B. N., R. M. SIMMONS and D. M. UNDERHILL (2005): Dectin-1 mediates macrophage recognition of Candida albicans yeast but not filaments. Embo Journal 24, 1277-1286 GANZ, T. (2004): Antimicrobial polypeptides. Journal of Leukocyte Biology 75, 34-38 GANZ, T. and R. I. LEHRER (1994): Defensins. Current Opinion in Immunology 6, 584-589 GANZ, T. and R. I. LEHRER (1998): Antimicrobial peptides of vertebrates. Current Opinion in Immunology 10, 41-44 GANZ, T. and R. I. LEHRER (1999): Antibiotic peptides from higher eukaryotes: biology and applications. Molecular Medicine Today 5, 292-297 GARCÍA, J.R., JAUMANN, F., SCHULZ, S., KRAUSE, A., RODRÍGUEZ-JIMÉNEZ, J., FORSSMANN, U., ADERMANN, K., KLÜVER, E., VOGELMEIER, C., BECKER, D., HEDRICH, R., FORSSMANN, W.G. and BALS, R. (2001): Identification of a novel, multifunctional beta-defensin (human beta-defensin 3) with specific antimicrobial activity. Its interaction with plasma membranes of Xenopus oocytes and the induction of macrophage chemoattraction. Cell and Tissue Research 306, 257-264 GAVINO, A. C. P., J. S. CHUNG, K. SATO, K. ARIIZUMI and P. D. CRUZ (2005): Identification and expression profiling of a human C-type lectin, structurally homologous to mouse dectin-2. Experimental Dermatology 14, 281-288 GEBOES, K., C. DEWOLFPEETERS, P. RUTGEERTS, J. JANSSENS, G. VANTRAPPEN and V. DESMET (1983): Lymphocytes and Langerhans Cells in the human esophageal epithelium. Virchows Archiv a-Pathological Anatomy and Histopathology 401, 45-55

References
161
GENNARO, R. and M. ZANETTI (2000): Structural features and biological activities of the cathelicidin-derived antimicrobial peptides. Biopolymers 55, 31-49 GOLDAMMER, T., H. ZERBE, A. MOLENAAR, H. J. SCHUBERTH, R. M. BRUNNER, S. R. KATA and H. M. SEYFERT (2004): Mastitis increases mammary mRNA abundance of beta-defensin 5, toll-like-receptor 2 (TLR2), and TLR4 but not TLR9 in cattle. Clinical and Diagnostic Laboratory Immunology 11, 174-185 GOLDMANN, T., E. VOLLMER and J. GERDES (2003): What's cooking? detection of important biomarkers in HOPE-fixed, paraffin-embedded tissues eliminates the need for antigen retrieval. American Journal of Pathology 163, 2638-2640 GREEN, P. J., T. FEIZI, M. S. STOLL, S. THIEL, A. PRESCOTT and M. J. MCCONVILLE (1994): Recognition of the major cell-surface glycoconjugates of Leishmania parasites by the human serum mannan-binding protein. Molecular and Biochemical Parasitology 66, 319-328 GRUBOR, B., J. M. GALLUP, D. K. MEYERHOLZ, E. C. CROUCH, R. B. EVANS, K. A. BROGDEN, H. D. LEHMKUHL and M. R. ACKERMANN (2004): Enhanced surfactant protein and defensin mRNA levels and reduced viral replication during parainfluenza virus type 3 pneumonia in neonatal lambs. Clinical and Diagnostic Laboratory Immunology 11, 599-607 GRUNEBACH, F., M. M. WECK, J. REICHERT and P. BROSSART (2002): Molecular and functional characterization of human Dectin-1. Experimental Hematology 30, 1309-1315 GUARNER, F. (2006): Enteric flora in health and disease. Digestion 73 Suppl 1, 5-12 GUPTA, G. and A. SUROLIA (2007): Collectins: sentinels of innate immunity. Bioessays 29, 452-464 HAMADA, M., M. TAKECHI and C. ITAKURA (1992): Langerhans' cells in equine cutaneous papillomas and normal skin. Veterinary Pathology 29, 152-160 HANCOCK, R. A. H. G. S. (2006): Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nature biotechnology 24,

References
162
HANSEN, S. and U. HOLMSKOV (1998): Structural aspects of collectins and receptors for collectins. Immunobiology 199, 165-189 HARDER, J., J. BARTELS, E. CHRISTOPHERS and J. M. SCHROEDER (2001): Isolation and characterization of human beta-defensin-3, a novel human inducible peptide antibiotic. Journal of Biological Chemistry 276, 5707-5713 HASE, K., L. ECKMANN, J. D. LEOPARD, N. VARKI and M. F. KAGNOFF (2002): Cell differentiation is a key determinant of cathelicidin LL-37/human cationic antimicrobial protein 18 expression by human colon epithelium. Infection and Immunity 70, 953-963 HASE, K., M. MURAKAMI, M. IIMURA, S. P. COLE, Y. HORIBE, T. OHTAKE, M. OBONYO, R. L. GALLO, L. ECKMANN and M. F. KAGNOFF (2003): Expression of LL-37 by human gastric epithelial cells as a potential host defense mechanism against Helicobacter pylori. Gastroenterology 125, 1613-1625 HASHIMOTO, M., Y. ASAHINA, J. SANO, R. KANO, T. MORITOMO and A. HASEGAWA (2005): Cloning of canine Toll-like receptor 9 and its expression in dog tissues. Veterinary Immunology and Immunopathology 106, 159-163 HASHIMOTO, S., T. SUZUKI, H. Y. DONG, S. NAGAI, N. YAMAZAKI and K. MATSUSHIMA (1999): Serial analysis of gene expression in human monocyte-derived dendritic cells. Blood 94, 845-852 HAURUM, J. S., S. THIEL, I. M. JONES, P. B. FISCHER, S. B. LAURSEN and J. C. JENSENIUS (1993): Complement activation upon binding of mannan-binding protein to HIV envelope glycoproteins. Aids 7, 1307-1313 HAVERSON, K., RIFFAULT S. (2005): Antigen presenting cells in mucosal sites of veterinary species. Veterinary Research 37, 339-358 HERRE, J., S. GORDON and G. D. BROWN (2004): Dectin-1 and its role in the recognition of [beta]-glucans by macrophages. Molecular Immunology 40, 869-876 HOLMSKOV, U., R. MALHOTRA, R. B. SIM and J. C. JENSENIUS (1994): Collectins - collagenous C-type lectins of the innate immune defense system. Immunology Today 15, 67-74

References
163
HOLMSKOV, U., S. THIEL and J. C. JENSENIUS (2003): Collectins and ficolins: Humoral lectins of the innate immune defense. Annual Review of Immunology 21, 547-578 HOOPER, L. V., M. H. WONG, A. THELIN, L. HANSSON, P. C. FALK and J. I. GORDON (2001): Molecular analysis of commensal host-microbial relationships in the intestine. Science 291, 881-884 HOPWOOD, D. (1995): Oesophageal defence mechanisms. Digestion 56 Suppl 1, 5-8 HOPWOOD, D., G. COGHILL and D. S. SANDERS (1986): Human oesophageal submucosal glands. Their detection mucin, enzyme and secretory protein content. Histochemistry 86, 107-112 HUANG, L., C. B. CHING, R. R. JIANG and S. S. J. LEONG (2008): Production of bioactive human beta-defensin 5 and 6 in Escherichia coli by soluble fusion expression. Protein Expression and Purification 61, 168-174 HUH, W. K., T. OONO, Y. SHIRAFUJI, H. AKIYAMA, J. ARATA, M. SAKAGUCHI, N. H. HUH and K. IWATSUKI (2002): Dynamic alteration of human beta-defensin 2 localization from cytoplasm to intercellular space in psoriatic skin. Journal of Molecular Medicine 80, 678-684 HUNGER, R. E., P. A. SIELING, M. T. OCHOA, M. SUGAYA, A. E. BURDICK, T. H. REA, P. J. BRENNAN, J. T. BELISLE, A. BLAUVELT, S. A. PORCELLI and R. L. MODLIN (2004): Langerhans cells utilize CD1a and langerin to efficiently present nonpeptide antigens to T cells. Journal of Clinical Investigation 113, 701-708 HUSSEIN, M. R. (2008): Evaluation of Langerhans' cells in normal and eczematous dermatitis skin by CD1a protein immunohistochemistry: preliminary findings. Journal of Cutaneous Pathology 35, 554-558 HUTTNER, K. M., M. R. LAMBETH, H. R. BURKIN, D. J. BURKIN and T. E. BROAD (1998): Localization and genomic organization of sheep antimicrobial peptide genes. Gene 206, 85-91 ICHIJO, H., U. HELLMAN, C. WERNSTEDT, L. J. GONEZ, L. CLAESSON-WELSH, C. H. HELDIN and K. MIYAZONO (1993): Molecular cloning and characterization of ficolin, a multimeric protein with fibrinogen- and collagen-like domains. Journal of Biological Chemistry 268, 14505-14513

References
164
ICHIJO, H., L. RONNSTRAND, K. MIYAGAWA, H. OHASHI, C. H. HELDIN and K. MIYAZONO (1991): Purification of transforming growth factor-beta 1 binding proteins from porcine uterus membranes. Journal of Biological Chemistry 266, 22459-22464 IGNACIO, G., S. NORDONE, K. E. HOWARD and G. A. DEAN (2005): Toll-like receptor expression in feline lymphoid tissues. Veterinary Immunology and Immunopathology 106, 229-237 IIMURA, M., R. L. GALLO, K. HASE, Y. MIYAMOTO, L. ECKMANN and M. F. KAGNOFF (2005): Cathelicidin mediates innate intestinal defense against colonization with epithelial adherent bacterial pathogens. Journal of Immunology 174, 4901-4907 IKEDA, K., T. SANNOH, N. KAWASAKI, T. KAWASAKI and I. YAMASHINA (1987): Serum lectin with known structure activates complement through the classical pathway. Journal of Biological Chemistry 262, 7451-7454 INAMOTO, T., Y. KAWATA, W. M. QI, K. YAMAMOTO, K. WARITA, J. KAWANO, T. YOKOYAMA, N. HOSHI and H. KITAGAWA (2008): Ultrastructural study on the epithelial response against attachment of indigenous bacteria to epithelial membranes in Peyer`s patches of rat small intestine. Journal of Veterinary Medical Science 70, 235-241 ISHII, M., M. HASHIMOTO, K. OGUMA, R. KANO, T. MORITOMO and A. HASEGAWA (2006): Molecular cloning and tissue expression of canine Toll-like receptor 2 (TLR2). Veterinary Immunology and Immunopathology 110, 87-95 JAMES, J. and J. TAS (1984): Histochemical Protein Staining Methods (Microscopy handbooks 04). Oxford University Press, Royal Microscopy Society, Oxford JANEWAY, C. A. (1989): Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harbor Symposia on Quantitative Biology 54 Pt 1, 1-13 JANEWAY, C. A. and R. MEDZHITOV (2002): Innate immune recognition. Annual Review of Immunology 20, 197-216 JANEWAY, C. A., P. TRAVERS, M. WALPORT and M. J. SHLOMCHIK (2005): Immuno Biology - The Immune System in Health and Disease. 6th edition, Garland Science Taylor & Francis Group, New York, pp. 48-54, 103-169, 203-241

References
165
JIA, H. P., S. A. WOWK, B. C. SCHUTTE, S. K. LEE, A. VIVADO, B. F. TACK, C. L. BEVINS and P. B. MCCRAY (2000): A novel murine beta-defensin expressed in tongue, esophagus, and trachea. Journal of Biological Chemistry 275, 33314-33320 KAKINUMA, Y., Y. ENDO, M. TAKAHASHI, M. NAKATA and M. MATSUSHITA (2003): Molecular cloning and characterization of novel ficolins from Xenopus laevis. Immunogenetics 55, 29-37 KAKU, T., J. K. EKEM, C. LINDAYEN, D. J. BAILEY, A. W. VAN NOSTRAND and E. FARBER (1983): Comparison of formalin- and acetone-fixation for immunohistochemical detection of carcinoembryonic antigen (CEA) and keratin. American Journal of Clinical Pathology 80, 806-815 KANAZAWA, N. (2007): Dendritic cell immunoreceptors: C-type lectin receptors for pattern-recognition and signaling on antigen-presenting cells. Journal of Dermatological Science 45, 77-86 KAPTEYN, J. C., R. C. MONTIJN, G. J. P. DIJKGRAAF, H. VANDENENDE and F. M. KLIS (1995): Covalent association of beta-1,3-glucan with beta-1,6-glucosylated mannoproteins in cell-walls of Candida albicans. Journal of Bacteriology 177, 3788-3792 KARNOVSKY, M.J. (1967): A formaldehyd-glutaraldehyd fixative of high osmolality for use in electron microscopy. Journal of Cell Biology 27, 137A KAWASAKI, T., R. ETOH and I. YAMASHINA (1978): Isolation and characterization of a mannan-binding protein from rabbit liver. Biochemical and Biophysical Research Communication 81, 1018-1024 KISHORE, U. and K. B. M. REID (2000): C1q: structure, function, and receptors. Immunopharmacology 49, 159-170 KNOWLES, R. W. and W. F. BODMER (1982): A monoclonal-antibody recognizing a human thymus leukemia-like antigen associated with beta-2-microglobulin. European Journal of Immunology 12, 676-681 KOELLISCH, G., KALALI B.N., VOELCKER V., WALLICH R., BEHRENDT H., RING J., BAUER S., JAKOB T., MEMPEL M., AND OLLERT M. (2005): Various members of the Toll-like receptor family contibute to the innate immune response of human epidermal keratinocytes. Immunology 114, 531-541

References
166
KUMAR, A., J. ZHANG and F.-S. X. YU (2006): Toll-like receptor 2-mediated expression of [beta]-defensin-2 in human corneal epithelial cells. Microorganisms and Infection 8, 380-389 LAU, W. F., J. WONG, K. H. LAM and G. B. ONG (1981): Oesophageal microbial flora in carcinoma of the oesophagus. Australian and New Zealand Journal of Surgery 51, 52-55 LAURSEN, S. B. and O. L. NIELSEN (2000): Mannan-binding lectin (MBL) in chickens: molecular and functional aspects. Developmental and Comparative Immunology 24, 85-101 LE, Y., S. H. LEE, O. L. KON and J. LU (1998): Human L-ficolin: plasma levels, sugar specificity, and assignment of its lectin activity to the fibrinogen-like (FBG) domain. FEBS Letters 425, 367-370 LE, Y., S. M. TAN, S. H. LEE, O. L. KON and J. LU (1997): Purification and binding properties of a human ficolin-like protein. Journal of Immunological Methods 204, 43-49 LEBRE, M. C., A. M. VAN DER AAR, L. VAN BAARSEN, T. M. VAN CAPEL, J. H. SCHUITEMAKER, M. L. KAPSENBERG and E. C. DE JONG (2007): Human keratinocytes express functional Toll-like receptor 3, 4, 5, and 9. Journal of Investigative Dermatology 127, 331-341 LEHRER, R. I. and T. GANZ (1999): Antimicrobial peptides in mammalian and insect host defence. Current Opinion in Immunology 11, 23-27 LEHRER, R. I. and T. GANZ (2002): Defensins of vertebrate animals. Current Opinion in Immunology 14, 96-102 LIEBICH, H. G. (2004): Funktionelle Histologie: Farbatlas und Kurzlehrbuch der mikroskopischen Anatomie der Haussäugetiere. 4 th ed. Schattauer Verlagsgesellschaft, Stuttgart, New York LILLIE, B. N., A. S. BROOKS, N. D. KEIRSTEAD and M. A. HAYES (2005): Comparative genetics and innate immune functions of collagenous lectins in animals. Veterinary Immunology and Immunopathology 108, 97-110 LILLIE, R. D. and H. M. FULLMER (1976): Histopathologic Technic and Practical Histochemistry. 4.ed., McGraw-Hill Book Company, New York

References
167
LINDE, A., F. BLECHA and T. MELGAREJO (2007): Toll-like receptor (TLR) 2 and TLR4 gene expression in canine heart. American Journal of Animal and Veterinary Sciences 2, 6-10 LINDE, A., C. R. ROSS, E. G. DAVIS, L. DIB, F. BLECHA and T. MELGAREJO (2008): Innate immunity and host defense peptides in veterinary medicine. Journal of Veterinary Internal Medicine 22, 247-265 LIPKE, P. N. and R. OVALLE (1998): Cell wall architecture in yeast: New structure and new challenges. Journal of Bacteriology 180, 3735-3740 LOTZ, M., S. MÉNARD and M. HORNEF (2007): Innate immune recognition on the intestinal mucosa. International Journal of Medical Microbiology 297, 379-392 LU, J. H. (1997): Collectins: Collectors of microorganisms for the innate immune system. Bioessays 19, 509-518 LU, J. H. and Y. LE (1998): Ficolins and the fibrinogen-like domain. Immunobiology 199, 190-199 LU, J. H., C. TEH, U. KISHORE and K. B. M. REID (2002): Collectins and ficolins: sugar pattern recognition molecules of the mammalian innate immune system. Biochimica et Biophysica Acta-General Subjects 1572, 387-400 LYNCH, N. J., S. ROSCHER, T. HARTUNG, S. MORATH, M. MATSUSHITA, D. N. MAENNEL, M. KURAYA, T. FUJITA and W. J. SCHWAEBLE (2004): L-ficolin specifically binds to lipoteichoic acid, a cell wall constituent of Gram-positive bacteria, and activates the lectin pathway of complement. Journal of Immunology 172, 1198-1202 MACKROWIAK, P. A. (1982): The normal microbial flora. National English Journal of Medicine 307, 83-93 MADSEN, J., A. KLIEM, I. TORNOE, K. SKJODT, C. KOCH and U. HOLMSKOV (2000): Localization of lung surfactant protein D on mucosal surfaces in human tissues. Journal of Immunology 164, 5866-5870 MADSEN, J., I. TORNOE, O. NIELSEN, C. KOCH, W. STEINHILBER and U. HOLMSKOV (2003): Expression and localization of lung surfactant protein A in human tissues. American Journal of Respiratory Cell and Molecular Biology 29, 591-597

References
168
MAHONEY, M. M., A. Y. LEE, D. J. BREZINSKI-CALIGURI and K. M. HUTTNER (1995): Molecular analysis of the sheep cathelin family reveals a novel antimicrobial peptide. FEBS Lett 377, 519-522 MALM, J., O. SORENSEN, T. PERSSON, M. FROHM-NILSSON, B. JOHANSSON, A. BJARTELL, H. LILJA, M. STAHLE-BACKDAHL, N. BORREGAARD and A. EGESTEN (2000): The human cationic antimicrobial protein (hCAP-18) is expressed in the epithelium of human epididymis, is present in seminal plasma at high concentrations, and is attached to spermatozoa. Infection and Immunity 68, 4297-4302 MARCHAL, I. S., C. DEZUTTERDAMBUYANT, B. J. WILLETT, J. C. WOO, P. F. MOORE, J. P. MAGNOL, D. SCHMITT and T. MARCHAL (1997): Immunophenotypic characterization of feline Langerhans cells. Veterinary Immunology and Immunopathology 58, 1-16 MASSCHALCK, B., D. DECKERS and C. W. MICHIELS (2002): Lytic and nonlytic mechanism of inactivation of Gram-positive bacteria by lysozyme under atmospheric and high hydrostatic pressure. Journal of Food Protection 65, 1916-1923 MATSUSHITA, M., Y. ENDO and T. FUJITA (2000): Cutting edge: Complement-activating complex of ficolin and mannose-binding lectin-associated serine protease. Journal of Immunology 164, 2281-2284 MATSUSHITA, M., Y. ENDO, S. TAIRA, Y. SATO, T. FUJITA, N. ICHIKAWA, M. NAKATA and T. MIZUOCHI (1996): A novel human serum lectin with collagen- and fibrinogen-like domains that functions as an opsonin. Journal of Biological Chemistry 271, 2448-2454 MATSUSHITA, M. and T. FUJITA (1992): Activation of the classical complement pathway by mannose-binding protein in association with a novel C1s-like serine protease. Journal of Experimental Medicine 176, 1497-1502 MATSUSHITA, M. and T. FUJITA (1995): Cleavage of the 3rd component of complement (C3) by mannose-binding protein-associated serine-protease (Masp) with subsequent complement activation. Immunobiology 194, 443-448 MATSUSHITA, M. and T. FUJITA (2001): Ficolins and the lectin complement pathway. Immunological Reviews 180, 78-85

References
169
MATSUZIKA, K. (1999): Why and how are peptide-lipid interactions utelized for self-defense? Magainin and tachyplesins as archetypes. Biochimica et Biophysica Acta 1482, 1-10 MCNEELY, T. B. and J. D. COONROD (1994): Aggregation and opsonization of type-a but not type-B Haemophilus influenzae by surfactant protein-A. American Journal of Respiratory Cell and Molecular Biology 11, 114-122 MENDELSON, C. R., M. J. ACARREGUI, M. J. ODOM and V. BOGGARAM (1991): Developmental and hormonal regulation of surfactant protein A (SP-A) gene expression in fetal lung. Journal of Developmental Physiology 15, 61-69 MENZIES, M. and A. INGHAM (2006): Identification and expression of Toll-like receptors 1-10 in selected bovine and ovine tissues. Veterinary Immunology and Immunopathology 109, 23-30 MEYER, W. (1986): Die Haut des Schweines. Vergleichende histologische und histochemische Untersuchungen an der Haut von Wildschweinen, Hausschweinen und Kleinschweinen. Schlütersche Verlagsanstalt, Hannover MEYER, W. (2009): Haut und Hautorgane. In: Wissdorf H, Gerhards H, Huskamp B, Deegen E, eds. Praxisorientierte Anatomie des Pferdes. 3rd ed. Schlütersche Verlagsanstalt, Hannover, in press. MEYER, W., J. KLOEPPER and L. G. FLEISCHER (2008): Demonstration of ß-glucan receptors in the skin of aquatic mammals-a preliminary report. Journal of European Wildlife Research 54, 479-486 MEYER, W. and U. SEEGERS (2004): A preliminary approach to epidermal antimicrobial defense in the Delphinidae. Marine Biology 144, 841-844 MEYER, W., U. SEEGERS, J. HERRMANN and A. SCHNAPPER (2003): Further aspects of the general antimicrobial properties of pinniped skin secretions. Diseases of Aquatic Organisms 53, 177-179 MEYER, W., J. KACZA, N. H. ZSCHEMISCH, S. GODYNICKI, J. SEEGER (2007): Observations on the actual structural conditions in the stratum superficiale dermidis of porcine ear skin, with special reference to its use as model for human skin. Annals of Anatomy 189, 143-156.

References
170
MEYER, W., J. KACZA, A. SCHNAPPER, J. VERSPOHL, I. HORNICKEL and J. SEEGER (2009): Microbial occupation of the mammalian esophagus, a first report based on observations from the horse. Cells Tissues Organs (submitted) MILLER, C. J., M. MCCHESNEY and P. F. MOORE (1992): Langerhans cells, macrophages and lymphocyte subsets in the cervix and vagina of rhesus macaques. Laboratory Investigation 67, 628-634 MIRGORODSKAYA, O. A., A. A. SHEVCHENKO, K. O. ABDALLA, I. V. CHERNUSHEVICH, T. A. EGOROV, A. X. MUSOLIAMOV, V. N. KOKRYAKOV and O. V. SHAMOVA (1993): Primary structure of three cationic peptides from porcine neutrophils. Sequence determination by the combined usage of electrospray ionization mass spectrometry and Edman degradation. FEBS Lett 330, 339-342 MIZUMOTO, N. and A. TAKASHIMA (2004): CD1a and langerin: acting as more than Langerhans cell markers. Journal of Clinical Investigation 113, 658-660 MOODY, D. B., D. C. YOUNG, T. Y. CHENG, J. P. ROSAT, C. ROURA-MIR, P. B. O'CONNOR, D. M. ZAJONC, A. WALZ, M. J. MILLER, S. B. LEVERY, I. A. WILSON, C. E. COSTELLO and M. B. BRENNER (2004): T cell activation by lipopeptide antigens. Science 303, 527-531 MORI, L. and G. DE LIBERO (2008): Presentation of lipid antigens to T cells. Immunology Letters 117, 1-8 MUKHERJEE, S., S. VAISHNAVA and L. V. HOOPER (2008): Multi-layered regulation of intestinal antimicrobial defense. Cellular and Molecular Life Sciences 65, 3019-3027 MURPHY, C. J., B. A. FOSTER, M. J. MANNIS, M. E. SELSTED and T. W. REID (1993): Defensins are mitogenic for epithelial cells and fibroblasts. Journal of Cell Physiology 155, 408-413 NAGAO, K., F. GINHOUX, W. W. LEITNER, S. I. MOTEGI, C. L. BENNETT, B. E. CLAUSEN, M. MERAD and M. C. UDEY (2009): Murine epidermal Langerhans cells and langerin-expressing dermal dendritic cells are unrelated and exhibit distinct functions. Proceedings of the National Academy of Science of the U. S. A. 106, 3312-3317

References
171
NAGAOKA, I., S. HIROTA, S. YOMOGIDA, A. OHWADA and M. HIRATA (2000): Synergistic actions of antibacterial neutrophil defensins and cathelicidins. Inflammation Research 49, 73-79 NAGY, I., A. PIVARCSI, A. KORECK, M. SZELL, E. URBAN and L. KEMENY (2005): Distinct strains of Propionibacterium acnes induce selective human beta-defensin-2 and interleukin-8 expression in human keratinocytes through Toll-like receptors. Journal of Investigative Dermatology 124, 931-938 NALUBAMBA, K. S., A. G. GOSSNER, R. G. DALZIEL and J. HOPKINS (2007): Differential expression of pattern recognition receptors in sheep tissues and leukocyte subsets. Veterinary Immunology and Immunopathology 118, 252-262 NEISH, A. S. (2009): Microorganisms in gastrointestinal health and disease. Gastroenterology 136, 65-80 NETH, O., D. L. JACK, A. W. DODDS, H. HOLZEL, N. J. KLEIN and M. W. TURNER (2000): Mannose-binding lectin binds to a range of clinically relevant microorganisms and promotes complement deposition. Infection and Immunity 68, 688-693 NFON, C. K., H. DAWSON, F. N. TOKA and W. T. GOLDE (2008): Langerhans cells in porcine skin. Veterinary Immunology and Immunopathology 126, 236-247 NICKEL, R., A. SCHUMMER, E. SEIFERLE (2004): Lehrbuch der Antomie der Haustiere, Bd. 2: Eingeweide. 9th ed. Parey -Thieme, Stuttgart NICOLAS, P. and A. MOR (1995): Peptides as weapons against microorganisms in the chemical defense system of vertebrates. Annual Reviews of Microbiology 49, 277-304 NIELSEN, O. L., P. H. JORGENSEN, J. HEDEMAND, J. C. JENSENIUS, C. KOCH and S. B. LAURSEN (1998): Immunohistochemical investigation of the tissue distribution of mannan-binding lectin in non-infected and virus-infected chickens. Immunology 94, 122-128 NISSEN-MEYER, J. and I. F. NES (1997): Ribosomally synthesized antimicrobial peptides: their function, structure, biogenesis, and mechanism of action. Archives of Microbiology 167, 67-77

References
172
NIYONSABA, F., K. IWABUCHI, H. MATSUDA, H. OGAWA and I. NAGAOKA (2002): Epithelial cell-derived human beta-defensin-2 acts as a chemotaxin for mast cells through a pertussis toxin-sensitive and phospholipase C-dependent pathway. International Immunology 14, 421-426 NIYONSABA, F., I. NAGAOKA and H. OGAWA (2006): Human defensins and cathelicidins in the skin: Beyond direct antimicrobial properties. Critical Reviews in Immunology 26, 545-575 NIYONSABA, F. and H. OGAWA (2005): Protective roles of the skin against infection: Implication of naturally occurring human antimicrobial agents [beta]-defensins, cathelicidin LL-37 and lysozyme. Journal of Dermatological Science 40, 157-168 O'NEIL, D. A., E. M. PORTER, D. ELEWAUT, G. M. ANDERSON, L. ECKMANN, T. GANZ and M. F. KAGNOFF (1999): Expression and regulation of the human beta-defensins hBD-1 and hBD-2 in intestinal epithelium. Journal of Immunology 163, 6718-6724 O'NEILL, L. A. J. (2006): How Toll-like receptors signal: what we know and what we don't know. Current Opinion in Immunology 18, 3-9 OFEK, I., A. MESIKA, M. KALINA, Y. KEISARI, R. PODSCHUN, H. SAHLY, D. CHANG, D. MCGREGOR and E. CROUCH (2001): Surfactant protein D enhances phagocytosis and killing of unencapsulated phase variants of Klebsiella pneumoniae. Infection and Immunity 69, 24-33 OHASHI, T. and H. P. ERICKSON (1998): Oligomeric structure and tissue distribution of ficolins from mouse, pig and human. Archives of Biochemistry and Biophysics 360, 223-232 OLERT, J., K.-H. WIEDORN, T. GOLDMANN, H. KÜHL, Y. MEHRAEIN, H. SCHERTHAN, F. NIKETEGHAD, E. VOLLMER, A. M. MÜLLER and J. MÜLLER-NAVIA (2001): HOPE fixation: A novel fixing method and paraffin-embedding technique for human soft tissues. Pathology - Research and Practice 197, 823-826 OMORI-SATOH, T., Y. YAMAKAWA and D. MEBS (2000): The antihemorrhagic factor, erinacin, from the European hedgehog (Erinaceus europaeus), a metalloprotease inhibitor of large molecular size possessing ficolin/opsonin P35 lectin domains. Toxicon 38, 1561-1580

References
173
OREN, A., T. GANZ, L. LIU and T. MEERLOO (2003): In human epidermis, beta-defensin 2 is packaged in lamellar bodies. Experimental and Molecular Pathology 74, 180-182 PAANANEN, R., V. GLUMOFF and M. HALLMAN (1999): Surfactant protein A and D expression in the porcine Eustachian tube. FEBS Lett 452, 141-144 PANYUTICH, A., J. S. SHI, P. L. BOUTZ, C. Q. ZHAO and T. GANZ (1997): Porcine polymorphonuclear leukocytes generate extracellular microbicidal activity by elastase-mediated activation of secreted proprotegrins. Infection and Immunity 65, 978-985 PEARSE, A. G. E. (1985): Histochemistry - Theoretical and Applied. 4th ed., Churchill Livingston, Edinburgh PENA-CRUZ, V., S. ITO, C. C. DASCHER, M. B. BRENNER and M. SUGITA (2003): Epidermal Langerhans cells efficiently mediate CD1a-dependent presentation of microbial lipid antigen to T cells. Journal of Investigative Dermatology 121, 1240-1240 PEREZ-TORRES, A., USTARROZ-CANO M., AND MILLAN-ALDACO D. (2002): Langerhans cell-like dendritic cells in the cornea, tongue and oesophagus of the chicken (Gallus gallus). Histochemical Journal 34, 507-515 PETERSEN, S. V., S. THIEL, L. JENSEN, R. STEFFENSEN and J. C. JENSENIUS (2001): An assay for the mannan-binding lectin pathway of complement activation. Journal of Immunological Methods 257, 107-116 PHANEUF, L. R., B. N. LILLIE, M. A. HAYES and P. V. TURNER (2007): Binding of mouse mannan-binding lectins to different bacterial pathogens of mice. Veterinary Immunology and Immunopathology 118, 129-133 PIKAAR, J. C., W. F. VOORHOUT, L. M. G. VANGOLDE, J. VERHOEF, J. A. G. VANSTRIJP and J. F. VANIWAARDEN (1995): Opsonic activities of surfactant protein-a and protein-D in phagocytosis of Gram-negative bacteria by alveolar macrophages. Journal of Infectious Diseases 172, 481-489 PIVARCSI, A., I. NAGY, A. KORECK, K. KIS, A. KENDERESSY-SZABO, M. SZELL, A. DOBOZY and L. KEMENY (2005): Microbial compounds induce the expression of pro-inflammatory cytokines, chemokines and human beta-defensin-2 in vaginal epithelial cells. Microorganisms and Infection 7, 1117-1127

References
174
POLOTSKY, V. Y., W. FISCHER, R. A. B. EZEKOWITZ and K. A. JOINER (1996): Interactions of human mannose-binding protein with lipoteichoic acids. Infection and Immunity 64, 380-383 PORCELLI, S. A. and J. D. FRANK (1995): Advances in Immunology - The CD1 Family: A Third Lineage of Antigen-Presenting Molecules Academic Press, New York PUCHTLER, H. and S. N. MELOAN (1985): On the chemistry of formaldehyde fixation and its effects on immunohistochemical reactions. Histochemistry 82, 201-204 RAKOFF-NAHOUM, S., J. PAGLINO, F. ESLAMI-VARZANEH, S. EDBERG and R. MEDZHITOV (2004): Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 118, 229-241 RAMANATHAN, B., E. G. DAVIS, C. R. ROSS and F. BLECHA (2002): Cathelicidins: microbicidal activity, mechanisms of action, and roles in innate immunity. Microorganisms and Infection 4, 361-372 READING, P. C., C. A. HARTLEY and E. M. ANDERS (1993): A serum mannose-binding lectin mediates complement-dependent lysis of influenza virus-Infected cells. Journal of Leukocyte Biology, 5, 45-45 REID, D. M., M. MONTOYA, P. R. TAYLOR, P. BORROW, S. GORDON, G. D. BROWN and S. Y. C. WONG (2004): Expression of the beta-glucan receptor, Dectin-1, on murine leukocytes in situ correlates with its function in pathogen recognition and reveals potential roles in leukocyte interactions. Journal of Leukocyte Biology 76, 86-94 REITAMO, S., M. KLOCKARS and A.-M. RAESTE (1977): Immunohistochemical identification of lysozyme in the minor salivary glands of man. Archives of Oral Biology 22, 515-519 RESNICK, M. B., Y. FINKELSTEIN, A. WEISSLER, J. LEVY and E. YAKIREVICH (1999): Assessment and diagnostic utility of the cytotoxic T-lymphocyte phenotype using the specific markers granzyme-B and TIA-1 in esophageal mucosal biopsies. Human Pathology 30, 397-402

References
175
ROMANI, N., S. HOLZMANN, C. H. TRIPP, F. KOCH and P. STOITZNER (2003): Langerhans cells - dendritic cells of the epidermis. APMIS 111, 725-740 ROSAT, J. P., E. P. GRANT, E. M. BEEKMAN, C. C. DASCHER, P. A. SIELING, D. FREDERIQUE, R. L. MODLIN, S. A. PORCELLI, S. T. FURLONG and M. B. BRENNER (1999): CD1-restricted microbial lipid antigen-specific recognition found in the CD8(+) alpha beta T cell pool. Journal of Immunology 162, 366-371 ROSSI, V., S. CSEH, I. BALLY, N. M. THIELENS, J. C. JENSENIUS and G. J. ARLAUD (2001): Substrate specificities of recombinant mannan-binding lectin-associated serine proteases-1 and-2. Journal of Biological Chemistry 276, 40880-40887 ROWDEN, G., M. G. LEWIS and A. K. SULLIVAN (1977): Ia antigen expression on human epidermal Langerhans cells. Nature 268, 247-248 RUBIO, S., T. LACAZE-MASMONTEIL, B. CHAILLEY-HEU, A. KAHN, J. R. BOURBON and R. DUCROC (1995): Pulmonary surfactant protein A (SP-A) is expressed by epithelial cells of small and large intestine. Journal of Biological Chemistry 270, 12162-12169 RUNZA, V. L., W. SCHWAEBLE and D. N. MÄNNEL (2008): Ficolins: Novel pattern recognition molecules of the innate immune response. Immunobiology 213, 297-306 RYAN, E. J., A. J. MARSHALL, D. MAGALETTI, H. FLOYD, K. E. DRAVES, N. E. OLSON and E. A. CLARK (2002): Dendritic cell-associated lectin-1: A novel dendritic cell-associated, C-type lectin-like molecule enhances T cell secretion of IL-4. Journal of Immunology 169, 5638-5648 RYAN, L. K., J. RHODES, M. BHAT and G. DIAMOND (1998): Expression of beta-defensin genes in bovine alveolar macrophages. Infection and Immunity 66, 878-881 SALAMERO, J., H. BAUSINGER, A. M. MOMMAAS, D. LIPSKER, F. PROAMER, J. P. CAZENAVE, B. GOUD, H. DE LA SALLE and D. HANAU (2001): CD1a molecules traffic through the early recycling endosomal pathway in human Langerhans cells. Journal of Investigative Dermatology 116, 401-408

References
176
SALZMAN, N. H., M. A. UNDERWOOD and C. L. BEVINS (2007): Paneth cells, defensins, and the commensal microbiota: A hypothesis on intimate interplay at the intestinal mucosa. Seminars in Immunology 19, 70-83 SAMUELSON, D. A. (2007): Textbook of veterinary histolgy. Elasevier, New York, 1st edition SANDBORN, W. J., V. W. FAZIO, B. G. FEAGAN and S. B. HANAUER (2003): AGA technical review on perianal Crohn's disease. Gastroenterology 125, 1508-1530 SANG, Y. and F. BLECHA (2009): Porcine host defense peptides: expanding repertoire and functions. Developmental and Comparative Immunology 33, 334-343 SANG, Y., M. TERESA ORTEGA, K. RUNE, W. XIAU, G. ZHANG, J. L. SOULAGES, G. H. LUSHINGTON, J. FANG, T. D. WILLIAMS, F. BLECHA and T. MELGAREJO (2007): Canine cathelicidin (K9CATH): gene cloning, expression, and biochemical activity of a novel pro-myeloid antimicrobial peptide. Developmental and Comparative Immunology 31, 1278-1296 SANG, Y., J. YANG, C. R. ROSS, R. R. ROWLAND and F. BLECHA (2008): Molecular identification and functional expression of porcine Toll-like receptor (TLR) 3 and TLR7. Veterinary Immunology and Immunopathology 125, 162-167 SANG, Y. M., M. T. ORTEGA, F. BLECHA, O. PRAKASH and T. MELGAREJO (2005): Molecular cloning and characterization of three beta-defensins from canine testes. Infection and Immunity 73, 2611-2620 SANTOS, I. K. F. D., C. H. N. COSTA, H. KRIEGER, M. F. FEITOSA, D. ZURAKOWSKI, B. FARDIN, R. B. B. GOMES, D. L. WEINER, D. A. HARN, R. A. B. EZEKOWITZ and J. E. EPSTEIN (2001): Mannan-binding lectin enhances susceptibility to visceral leishmaniasis. Infection and Immunity 69, 5212-5215 SASTRY, K. and R. ALAN EZEKOWITZ (1993): Collectins: pattern recognition molecules involved in first line host defense. Current Opinion in Immunology 5, 59-66 SASTRY, K., K. ZAHEDI, J. M. LELIAS, A. S. WHITEHEAD and R. A. B. EZEKOWITZ (1991): Molecular characterization of the mouse mannose-binding proteins - the mannose-binding protein-a but not protein-C is an acute phase reactant. Journal of Immunology 147, 692-697

References
177
SCHELENZ, S., R. MALHOTRA, R. B. SIM, U. HOLMSKOV and G. J. BANCROFT (1995): Binding of host collectins to the pathogenic yeast Cryptococcus neoformans - human surfactant protein-D acts as an agglutinin for acapsular yeast-cells. Infection and Immunity 63, 3360-3366 SCHENK, M. and C. MUELLER (2008): The mucosal immune system at the gastrointestinal barrier. Best Practice & Research Clinical Gastroenterology 22, 391-409 SCHOENNAGEL, B. (2005) Vergleichende Untersuchungen zur Struktur und Funktion des Oesophagus-Epithels bei Vertebraten in Bezug zur Ernährungsweise, unter besonderer Berücksichtigung der Haussäugetiere. Hannover, Tierärztliche Hochschule Hannover, Dissertation. SCHONWETTER, B. S., E. D. STOLZENBERG and M. A. ZASLOFF (1995): Epithelial antibiotics induced at sites of inflammation. Science 267, 1645-1648 SCHOREY, J. S. and C. LAWRENCE (2008): The pattern recognition receptor Dectin-1: From fungi to mycobacteria. Current Drug Targets 9, 123-129 SCOCCHI, M., D. BONTEMPO, S. BOSCOLO, L. TOMASINSIG, E. GIULOTTO and M. ZANETTI (1999): Novel cathelicidins in horse leukocytes. FEBS Letters 457, 459-464 SCOTT, M. G. and R. E. W. HANCOCK (2000): Cationic antimicrobial peptides and their multifunctional role in the immune system. Critical Reviews in Immunology 20, 407-431 SELSTED, M. E. and A. J. OUELLETTE (2005): Mammalian defensins in the antimicrobial immune response. Nature Immunology 6, 551-557 SEYFARTH, J., P. GARRED and H. O. MADSEN (2006): Extra-hepatic transcription of the human mannose-binding lectin gene (mbl2) and the MBL-associated serine protease 1-3 genes. Molecular Immunology 43, 962-971 SHAMOVA, O., K. A. BROGDEN, C. ZHAO, T. NGUYEN, V. N. KOKRYAKOV and R. I. LEHRER (1999): Purification and properties of proline-rich antimicrobial peptides from sheep and goat leukocytes. Infection and Immunity 67, 4106-4111

References
178
SIMA, P., I. TREBICHAVSKY and K. SIGLER (2003): Mammalian antibiotic peptides. Folia Microbiologica 48, 123-137 SINGH, V., K. SINGH, S. AMDEKAR, D. D. SINGH, P. TRIPATHI, G. L. SHARMA and H. YADAV (2009): Innate and specific gut-associated immunity and microbial interference. FEMS Immunol Med Microbiol 55, 6-12 SMALL, R. M., S. M. WALDEN and S. J. EWALD (1987): Effects of Ly-5 antibodies on antibody-dependent cell-mediated cytotoxicity (ADCC). Immunology 60, 159-165 SMET, R. C. A. K. D. (2005): Human antimicrobial peptides: defensins, cathelicidins and histatins. Biotechnology Letters 27, 1337-1347 SMITT, P., C. VANDERLOOS, J. DEJONG and D. TROOST (1993): Tissue fixation methods alter the imunohistochemical demonstrability of neurofilament proteins, synaptophysin, and glial fibrillary acidic protein in human cerebellum. Acta Histochemica 95, 13-21 SORENSEN, O. E., P. FOLLIN, A. H. JOHNSEN, J. CALAFAT, G. S. TJABRINGA, P. S. HIEMSTRA and N. BORREGAARD (2001): Human cathelicidin, hCAP-18, is processed to the antimicrobial peptide LL-37 by extracellular cleavage with proteinase 3. Blood 97, 3951-3959 STEELE, C., L. MARRERO, S. SWAIN, A. G. HARMSEN, M. Q. ZHENG, G. D. BROWN, S. GORDON, J. E. SHELLITO and J. K. KOLLS (2003): Alveolar macrophage-mediated killing of Pneumocystis carinii f. sp muris involves molecular recognition by the Dectin-1 beta-glucan receptor. Journal of Experimental Medicine 198, 1677-1688 STEELE, C., R. R. RAPAKA, A. METZ, S. M. POP, D. L. WILLIAMS, S. GORDON, J. K. KOLLS and G. D. BROWN (2005): The beta-glucan receptor Dectin-1 recognizes specific morphologies of Aspergillus fumigatus. Plos Pathogens 1, 323-334 STOKES, C. and N. WALY (2006): Mucosal defence along the gastrointestinal tract of cats and dogs. Veterinary Research 37, 281-293 SUGITA, M., M. CERNADAS and M. B. BRENNER (2004): New insights into pathways for CD1-mediated antigen presentation. Current Opinion in Immunology 16, 90-95

References
179
SUGITA, M., P. J. PETERS and M. B. BRENNER (2000): Pathways for lipid antigen presentation by CD1 molecules: Nowhere for intracellular pathogens to hide. Traffic 1, 295-300 SUMIKAWA, Y., H. ASADA, K. HOSHINO, H. AZUKIZAWA, I. KATAYAMA, S. AKIRA and S. ITAMI (2006): Induction of [beta]-defensin 3 in keratinocytes stimulated by bacterial lipopeptides through toll-like receptor 2. Microorganisms and Infection 8, 1513-1521 SUNG, S. S. J., S. M. FU, C. E. ROSE, F. GASKIN, S. T. JU and S. R. BEATY (2006): A major lung CD103 (alpha(E))-beta(7) integrin-positive epithelial dendritic cell population expressing Langerin and tight junction proteins. Journal of Immunology 176, 2161-2172 SWERDLOW, M. P., D. R. KENNEDY and J. S. KENNEDY (2006): Expression and function of TLR2, TLR5, and Nod2 in primary canine colonic epithelial cells. Veterinary Immunology and Immunopathology 114, 313-319 TAKEDA, K., T. KAISHO and S. AKIRA (2003): Toll-like receptors. Annual Review of Immunology 21, 335-376 TAYLOR, P. R., S. V. TSONI, J. A. WILLMENT, K. M. DENNEHY, M. ROSAS, H. FINDON, K. HAYNES, C. STEELE, M. BOTTO, S. GORDON and G. D. BROWN (2007): Dectin-1 is required for beta-glucan recognition and control of fungal infection. Nature Immunology 8, 31-38 TAZI, A., F. BOUCHONNET, M. GRANDSAIGNE, L. BOUMSELL, A. J. HANCE and P. SOLER (1993): Evidence that granulocyte macrophage-colony-stimulating factor regulates the distribution and differentiated state of dendritic cells/Langerhans cells in human lung and lung cancers. Journal of Clinical Investigation 91, 566-576 TERRIS, B. and F. POTET (1995): Structure and role of Langerhans`cells in the human oesophageal epithelium. Digestion 56, 9-14 THIEL, S. (1992): Mannan-binding protein, a complement activating animal lectin. Immunopharmacology 24, 91-99

References
180
THIEL, S., T. VORUPJENSEN, C. M. STOVER, W. SCHWAEBLE, S. B. LAURSEN, K. POULSEN, A. C. WILLIS, P. EGGLETON, S. HANSEN, U. HOLMSKOV, K. B. REID and J. C. JENSENIUS (1997): A second serine protease associated with mannan-binding lectin that activates complement. Nature 386, 506-510 TIZARD, I. A. (2008): Veterinary Immunology. 8 th ed., Saunders Elsevier, St. Louis, pp. 50-58, 70-75, 81-95 TOMASINSIG, L. and M. ZANETTI (2005): The cathelicidins - Structure, function and evolution. Current Protein & Peptide Science 6, 23-34 TURNER, M. W. (2003): The role of mannose-binding lectin in health and disease. Molecular Immunology 40, 423-429 UEMURA, K., M. SAKA, T. NAKAGAWA, N. KAWASAKI, S. THIEL, J. C. JENSENIUS and T. KAWASAKI (2002): L-MBP is expressed in epithelial cells of mouse small intestine. Journal of Immunology 169, 6945-6950 VALLADEAU, J., V. DUVERT-FRANCES, J. J. PIN, C. DEZUTTER-DAMBUYANT, C. VINCENT, C. MASSACRIER, J. VINCENT, K. YONEDA, J. BANCHEREAU, C. CAUX, J. DAVOUST and S. SAELAND (1999): The monoclonal antibody DCGM4 recognizes Langerin, a protein specific of Langerhans cells, and is rapidly internalized from the cell surface. European Journal of Immunology 29, 2695-2704 VALLADEAU, J., O. RAVEL, C. DEZUTTER-DAMBUYANT, K. MOORE, M. KLEIJMEER, Y. LIU, V. DUVERT-FRANCES, C. VINCENT, D. SCHMITT, J. DAVOUST, C. CAUX, S. LEBECQUE and S. SAELAND (2000): Langerin, a novel C-type lectin specific to Langerhans cells, is an endocytic receptor that induces the formation of Birbeck granules. Immunity 12, 71-81 VAN ALSTINE, W. G., M. POPIRLARCZYK and S. R. ALBREGTS (2002): Effect of formalin fixation on the immunohistochemical detection of PRRS virus antigen in experimentally and naturally infected pigs. Journal of Veterinary Diagnostic Investigation 14, 504-507 VAN BEECK, F. A. L., D. M. ZAJONC, P. F. MOORE, Y. M. SCHLOTTER, F. BROERE, V. RUTTEN, T. WILLEMSE and I. VAN RHIJN (2008): Two canine CD1a proteins are differentially expressed in skin. Immunogenetics 60, 315-324

References
181
VAN DE WETERING, J. K., L. M. G. VAN GOLDE and J. J. BATENBURG (2004): Collectins - Players of the innate immune system. European Journal of Biochemistry 271, 1229-1249 VAN DEN ELZEN, P., S. GARG, L. LEON, M. BRIGL, E. A. LEADBETTER, J. E. GUMPERZ, C. C. DASCHER, T. Y. CHENG, F. M. SACKS, P. A. ILLARIONOV, G. S. BESRA, S. C. KENT, D. B. MOODY and M. B. BRENNER (2005): Apolipoprotein-mediated pathways of lipid antigen presentation. Nature 437, 906-910 VAN DIJK, A., E. J. A. VELDHUIZEN and H. P. HAAGSMAN (2008): Avian defensins. Veterinary Immunology and Immunopathology 124, 1-18 VAN RHIJN, I., D. M. ZAJONC, I. A. WILSON and D. B. MOODY (2005): T-cell activation by lipopeptide antigens. Current Opinion in Immunology 17, 222-229 VELDHUIZEN, E. J. A., M. RIJNDERS, E. A. CLAASSEN, A. VAN DIJK and H. P. HAAGSMAN (2008): Porcine [beta]-defensin 2 displays broad antimicrobial activity against pathogenic intestinal bacteria. Molecular Immunology 45, 386-394 VOORHOUT, W. F., T. VEENENDAAL, Y. KUROKI, Y. OGASAWARA, L. M. G. VANGOLDE and H. J. GEUZE (1992): Immunocytochemical localization of surfactant protein-D (Sp-D) in type-Ii cells, Clara cells, and alveolar macrophages of rat lung. Journal of Histochemistry & Cytochemistry 40, 1589-1597 VORA, P., A. YOUDIM, L. S. THOMAS, M. FUKATA, S. Y. TESFAY, K. LUKASEK, K. S. MICHELSEN, A. WADA, T. HIRAYAMA, M. ARDITI and M. T. ABREU (2004): beta-Defensin-2 expression is regulated by TLR signaling in intestinal epithelial cells. Journal of Immunology 173, 5398-5405 WAGNER, S., N. J. LYNCH, W. WALTER, W. J. SCHWAEBLE and M. LOOS (2003): Differential expression of the murine mannose-binding lectins A and C in lymphoid and nonlymphoid organs and tissues. Journal of Immunology 170, 1462-1465 WARD, N. S., A. B. WAXMAN, R. J. HOMER, L. L. MANTELL, O. EINARSSON, Y. F. DU and J. A. ELIAS (2000): Interleukin-6-induced protection in hyperoxic acute lung injury. American Journal of Respiratory Cell and Molecular Biology 22, 535-542

References
182
WASSEF, A., K. JANARDHAN, J. W. PEARCE and B. SINGH (2004): Toll-like receptor 4 in normal and inflamed lungs and other organs of pig, dog and cattle. Histology and Histopathology 19, 1201-1208 WEHKAMP, J., H. CHU, B. SHEN, R. W. FEATHERS, R. J. KAYS, S. K. LEE and C. L. BEVINS (2006): Paneth cell antimicrobial peptides: Topographical distribution and quantification in human gastrointestinal tissues. FEBS Letters 580, 5344-5350 WEHKAMP, J., N. H. SALZMAN, E. PORTER, S. NUDING, M. WEICHENTHAL, R. E. PETRAS, B. SHEN, E. SCHAEFFELER, M. SCHWAB, R. LINZMEIER, R. W. FEATHERS, H. CHU, H. LIMA, JR., K. FELLERMANN, T. GANZ, E. F. STANGE and C. L. BEVINS (2005): Reduced Paneth cell alpha-defensins in ileal Crohn's disease. Proceedings of the National Academy of Sciences of the U S A 102, 18129-18134 WEHKAMP, J., K. SCHMIDT, K. R. HERRLINGER, S. BAXMANN, S. BEHLING, C. WOHLSCHLAGER, A. C. FELLER, E. F. STANGE and K. FELLERMANN (2003): Defensin pattern in chronic gastritis: HBD-2 is differentially expressed with respect to Helicobacter pylori status. Journal of Clinical Pathology 56, 352-357 WEINTRAUB, A., U. ZAHRINGER, H. W. WOLLENWEBER, U. SEYDEL and E. T. RIETSCHEL (1989): Structural characterization of the lipid A component of Bacteroides fragilis strain NCTC 9343 lipopolysaccharide. European Journal of Biochemistry 183, 425-431 WEIS, W. I., M. E. TAYLOR and K. DRICKAMER (1998): The C-type lectin superfamily in the immune system. Immunological Reviews 163, 19-34 WELSCH, U. (2006): Sobotta - Lehrbuch der Histologie. 2nd ed. Urban & Fischer, München, Jena WERNER, M., A. CHOTT, A. FABIANO and H. BATTIFORA (2000): Effect of formalin tissue fixation and processing on immunohistochemistry. American Journal of Surgical Pathology 24, 1016-1019 WILLCOCKS, S., Y. YAMAKAWA, A. STALKER, T. J. COFFEY, T. GOLDAMMER and D. WERLING (2006): Identification and gene expression of the bovine C-type lectin Dectin-1. Veterinary Immunology and Immunopathology 113, 234-242 WILLMENT, J. A. and G. D. BROWN (2008): C-type lectin receptors in antifungal immunity. Trends in Microbiology 16, 27-32

References
183
WILLMENT, J. A., S. GORDON and G. D. BROWN (2001): Characterisation of the human ß-glucan receptor and its alternatively spliced isoforms. Journal of Biological Chemistry 276, 43818-43823 WILLMENT, J. A., A. J. MARSHALL, D. M. REID, A. WILLIAMS, S. Y. C. WONG, S. GORDON and G. D. BROWN (2005): The human ß-glucan receptor is widely exprssed and functionally equivalent to murine Dectin-1 on primary cells. European Journal of Immunology 25, 1539-1547 WONG, C. J., J. AKIYAMA, L. ALLEN and S. HAWGOOD (1996): Localization and developmental expression of surfactant proteins D and A in the respiratory tract of the mouse. Pediatric Research 39, 930-937 YANG, L., T.M: WEISS, R.I. LEHRER (2000): Crystallization of antimicrobial pores in membranes: Magainin and protegrin. Biophysical Journal 79, 2002-2009 YANG, D., A. BIRAGYN, L. W. KWAK and J. J. OPPENHEIM (2002): Mammalian defensins in immunity: more than just microbicidal. Trends in Immunology 23, 291-296 YOKOTA, K., A. TAKASHIMA, P. R. BERGSTRESSER and K. ARIIZUMI (2001): Identification of a human homologue of the dendritic cell-associated C-type lectin-1, Dectin-1. Gene 272, 51-60 YOSHIDA, A., S. IMAYAMA, S. SUGAI, Y. KAWANO and T. ISHIBASHI (1997): Increased number of IgE positive Langerhans cells in the conjunctiva of patients with atopic dermatitis. Britisch Journal of Ophthalmology 81, 402-406 ZANETTI, M., R. GENNARO and D. ROMEO (1995): Cathelicidins - a novel protein family with a common proregion and a variable C-terminal antimicrobial domain. FEBS Letters 374, 1-5 ZANETTI, M., P. STORICI, A. TOSSI, M. SCOCCHI and R. GENNARO (1994): Molecular cloning and chemical synthesis of a novel antibacterial peptide derived from pig myeloid cells. Journal of Biological Chemistry 269, 7855-7858 ZASLOFF, M. (2002): Antimicrobial peptides of multicellular organisms. Nature 415, 389-395

References
184
ZAVALA, W. D., D. S. DE SIMONE, F. L. SACERDOTE and J. C. CAVICCHIA (2002): Variation in Langerhans cell number and morphology between the upper and lower regions of the human esophageal epithelium. Anatomical Record 268, 360-364 ZELENSKY, A. N. and J. E. GREADY (2005): The C-type lectin-like domain superfamily. FEBS Journal 272, 6179-6217 ZHANG, Y. W., E. G. DAVIS, F. BLECHA and M. J. WILKERSON (2008): Molecular cloning and characterization of equine Toll-like receptor 9. Veterinary Immunology and Immunopathology 124, 209-219 ZILBERSTEIN, B., A. G. QUINTANILHA, M. A. SANTOS, D. PAJECKI, E. G. MOURA, P. R. ALVES, F. MALUF FILHO, J. A. DE SOUZA and J. GAMA-RODRIGUES (2007): Digestive tract microbiota in healthy volunteers. Clinics 62, 47-54

a
Danksagung
Ich danke allen, die mich auf meinem Weg bis an diese Station meines Lebens
begleitet haben.
Meinen besonderen Dank möchte ich meinem lieben Betreuer Prof. Dr. Wilfried
Meyer aussprechen. Durch seine Hilfsbereitschaft, fachlich kompetente
Unterstützung und Förderung und auch persönlichen Beistand, hat er mir viele
Türen geöffnet. Er hat es geschafft, dass ich mich an meine Zeit als seine letzte
Doktorandin sehr gerne zurück erinnern werde. Der „ewige Fibroblast“ und die
gute Zusammenarbeit in ganz Deutschland werden mir in schöner Erinnerung
bleiben.
Ich danke meinen Freundinnen und Kolleginnen Marion Gähle und Doris Walter
(„Mutti“) sehr herzlich für die unglaubliche Unterstützung bei der Bearbeitung
meiner zahlreichen Proben. Darüber hinaus gilt ihnen mein Dank aber ebenso
dafür, dass ich jeden Tag gerne ins Institut gekommen bin und mit ihnen lachen,
weinen und essen (!) konnte.
Weiterhin möchte ich der besten VMTA Praktikantin der Welt, Ellen Baumeister,
für ihre ungeheuer motivierte Mitarbeit an meinem Projekt meine besondere
Anerkennung aussprechen!
Kerstin Rohn danke für ihre hervorragende und schnelle Hilfe bei der EPON
Einbettung und der Herstellung der Semidünnschnitte.
Die wundervollen zeichnerischen Darstellungen der Oesophagusstruktur in
meiner Dissertation verdanke ich der Künstlerin Caren Imme von Stemm.
Bei PD Dr. Johannes Kazca und Prof. Dr. Johannes Seeger möchte ich mich
für die Realisierung und Unterstützung bei unserem Projekt der Cryo REM
Darstellung von Mikroorganismen im Oesophagus bedanken.
PD Dr. Anke Schnapper danke ich für die fachliche Unterstützung, die
technische Hilfe bei Word und viele tolle Gespräche.
Ich bedanke mich bei der H. Wilhelm Schaumann Stiftung für die finanzielle Unter-
stützung dieser Arbeit.

b
Den folgenden Angehörigen der Stiftung Tierärztliche Hochschule Hannover und
Kollegen aus der Kleintierpraxis danke ich für ihre Hilfe bei der Erlangung von
Oesophagusproben: Prof. Dr. Wolfgang Löscher und Dr. Maren Fedowitz für die
Bereitstellung der Rattenproben, Prof. Dr. Andrea Meyer-Lindenberg und Dr.
Miriam Hemmes für die Bereitstellung der Oesophagusproben von Katzen sowie
Prof. Dr. Gerhard Breves, Prof. Dr. Josef Kamphues und Prof. Dr. Thomas
Schnieder und ihren Arbeitsgruppen für die unkomplizierte Kooperation bei der
Gewinnung einer Vielzahl von Oesophagusproben von Kleinsäugern, Schweinen
und Hunden.
Die freundliche, kompetente Hilfe bei der statistischen Auswertung von Dr. Martin
Beyerbach sei ebenfalls dankend anerkannt.
Mein besonderer Dank für die unermüdliche Hilfe bei der sprachlichen Korrektur der
Arbeit gilt meiner Freundin Emily Northrup.
Ich danke meinen „acht Mädels“ für ihren treuen Beistand und viele tolle Momente
neben der Arbeit. Ohne euch wäre das Leben nur halb so lustig und schön!
Meiner besten Freundin, Bernadette Hellmann, danke ich für ihre tiefe Freundschaft.
Wann fahren wir wieder nach Berlin?!?
Meiner Tante und meinem Onkel, Monika und Klaus Hornickel, danke ich für die
Unterstützung bei meinem Studium und dafür, dass sie mir vertrauensvoll ihren süßen
Mops überlassen.
Bei meinem Freund, David Finkenstädt, bedanke ich mich dafür, dass er immer an
mich glaubt und mich bei der Realisierung meiner Ziele unterstützt. Das tiefe Vertrauen
zwischen uns hat mir viel Kraft gegeben und diese Leistung ermöglicht. „Lieben
belebt“…, das sagte schon Goethe, und er hatte Recht.
Der Dank an meine Mama, Karin Hornickel, ist auf Grund ihrer Leistung schwer mit
wenigen Worten zusammenfassen. Ich danke ihr für ihre Liebe, ihren Stolz, ihre
Förderung und für ein Vertrauenvolles Mutter-Tochter Verhältnis in guten und in
schlechten Zeiten. Du hast mir die Werkzeuge an die Hand gegeben, die das
Wahrwerden meiner Träume ermöglicht haben.

c