Entwicklungsbiologie 05 - gobics.degobics.de/lectures/ss07/rv/wimmer/RingWimmer05.pdf · In situ...
12
1 Entwicklungsbiologie 05 Ernst A. Wimmer Abteilung Entwicklungsbiologie Kernäquivalenz • Annahme auf Grund der Gleichverteilung der verdoppelten Chromosomen in der Mitose. • Aber: Wenn jeder Nukleus dem Nukleus der Zygote entspricht, dann sollte jeder Nukleus auch totipotent sein. • Totipotenz: Eigenschaft von Zellen oder Zellkernen, die noch alle Entwicklungsmöglichkeiten besitzen, noch nicht determiniert sind. • Nachweis: Kerntransplantationsexperimente, Klonen • Wenn alle Zellen dieselbe Erbinformation tragen, wie differenzieren sie sich? Differentielle Genexpression Kernäquivalenz: Klonen von Amphibien Spaltung: Genetik - Entwicklungsbiologie • ursprüngliche Genetik (Morgan) noch als Teil der Embryologie (Entwicklungsbiologie) gesehen. • in den 1930ern wurde Genetik eine eigenständige Disziplin. • Zunehmende Feindseligkeit zwischen Genetikern und Embryologen. • Eine genetische Theorie der Entwicklung kann es erst geben wenn geklärt ist: 1. Wie Umweltbedingte Einflüsse die Entwicklung steuern können, z.B. Umweltbedingte Sex-Bestimmung (Temperatur bei Reptilien). 2. Wie Gene die frühen Stadien der Embryonalentwicklung steuern. 3. Wie Chromosomen, die in allen Zellen identisch sind, unterschiedliche und sich verändernde Zellcytoplasmen hervorbringen können, die nötig sind, um die Differenzierung der Zellen zu ermöglichen. Theodor Boveri: „Mendelismus“ Chromosomentheorie der Vererbung • Nicht eine bestimmte Zahl, sondern eine bestimmte Kombination von Chromosomen sind für eine normale Entwicklung notwendig. • „Und dies bedeutet nichts anderes, als daß die einzelnen Chromosomen verschiedene Qualitäten besitzen müssen“ Theodor Boveri: Wechselwirkung zwischen Cytoplasma und Kern • 1910 • „So scheint mir der Fall von Ascaris ein einfaches Paradigma dafür darzustellen, wie die Wechselwirkung von Protoplasma und Kern in der Ontogenese zu denken ist und auf welche Weise aus der äußerst geringen Ungleichartigkeit des Eiprotoplasmas, durch Auslöseeinwirkungen auf den Kern und Rückwirkungen auf das Protoplasma, die schließlich so gewaltigen Verschiedenheiten der entstehenden Zellen hervorgehen können.“ • Grundkonzept der “Differentiellen Genexpression”
Transcript of Entwicklungsbiologie 05 - gobics.degobics.de/lectures/ss07/rv/wimmer/RingWimmer05.pdf · In situ...

1
Entwicklungsbiologie 05Ernst A. Wimmer
Abteilung Entwicklungsbiologie
Kernäquivalenz
• Annahme auf Grund der Gleichverteilung der verdoppelten Chromosomen in der Mitose.
• Aber: Wenn jeder Nukleus dem Nukleus der Zygote entspricht, dann sollte jeder Nukleus auch totipotent sein.
• Totipotenz: Eigenschaft von Zellen oder Zellkernen, die noch alleEntwicklungsmöglichkeiten besitzen, noch nichtdeterminiert sind.
• Nachweis: Kerntransplantationsexperimente, Klonen
• Wenn alle Zellen dieselbeErbinformation tragen, wiedifferenzieren sie sich?
Differentielle Genexpression
Kernäquivalenz: Klonen von Amphibien Spaltung: Genetik - Entwicklungsbiologie• ursprüngliche Genetik (Morgan) noch als Teil der Embryologie
(Entwicklungsbiologie) gesehen.
• in den 1930ern wurde Genetik eine eigenständige Disziplin.
• Zunehmende Feindseligkeit zwischen Genetikern und Embryologen.
• Eine genetische Theorie der Entwicklung kann es erst geben wenn geklärt ist:
1. Wie Umweltbedingte Einflüsse die Entwicklung steuern können,z.B. Umweltbedingte Sex-Bestimmung (Temperatur bei Reptilien).
2. Wie Gene die frühen Stadien der Embryonalentwicklung steuern.
3. Wie Chromosomen, die in allen Zellen identisch sind, unterschiedlicheund sich verändernde Zellcytoplasmen hervorbringen können,die nötig sind, um die Differenzierung der Zellen zu ermöglichen.
Theodor Boveri: „Mendelismus“Chromosomentheorie der Vererbung
• Nicht eine bestimmte Zahl, sondern eine bestimmte Kombination von Chromosomen sind für eine normale Entwicklung notwendig.
• „Und dies bedeutet nichts anderes, als daß die einzelnen Chromosomen verschiedene Qualitäten besitzen müssen“
Theodor Boveri: Wechselwirkung zwischen Cytoplasma und Kern
• 1910• „So scheint mir der Fall von Ascaris ein einfaches
Paradigma dafür darzustellen, wie die Wechselwirkung von Protoplasma und Kern in der Ontogenese zu denken ist und auf welche Weise aus der äußerst geringen Ungleichartigkeit des Eiprotoplasmas, durch Auslöseeinwirkungen auf den Kern und Rückwirkungen auf das Protoplasma, die schließlich so gewaltigen Verschiedenheiten der entstehenden Zellen hervorgehen können.“
• Grundkonzept der “Differentiellen Genexpression”

2
Differentielle Genexpression
• Jeder Zellnukleus enthält das gesamte Genom, das im befruchteten Ei neu zusammengefügt wurde.
• Die nicht genutzten Gene in differenzierten Zellen sind nicht zerstört oder mutiert und behalten grundsätzlich das Potential exprimiert zu werden.
• In jeder Zelle ist nur ein geringer Prozentsatz des Genoms exprimiert. Dies bedeutet, dass der Anteil an synthetisierter RNA in jeder Zelle spezifisch für diesen Zelltyp ist.
Differentielle Genexpression
• Vom Differenzierungszustand abhängige Transkription unterschiedlicher Gene.
• Wird während der Determination programmiert.
Differentielle Genexpression:Nachweis
In situ Hybridisierung
Differentielle Genexpression: Nachweis
Whole mount (Totalpräparate) In situ Hybridisierung (WMISH)
Differentielle Genexpression: Nachweis
In situ Hybridisierung mit radioaktiv markierten Sonden
Differentielle GenexpressionRezeptor-Aktivierung
Protein-Modifikation
Transkriptionsfaktor-Aktivierung
Nukleare Lokalisierung
Chromatin-Dekompaktierung
Transkriptions-Initition und 5‘-Capping
Koppelung: Transkription und RNA-Prozessierung
RNA-Spleißen
3‘-Polyadenylierung
Verpackung der mRNA
mRNA-Export
Translation
Protein-Faltung

3
Moderne Modellorganismen
• Zucht und Handhabbarkeit im Labor.
• Zahl der Nachkommen
• Kurzer Entwicklungszyklus
• Möglichkeit zur Manipulation(Transplantationen, Mikroinjektion, Mutagenese)
• Möglichkeit zur Einzel-Verkreuzung
• Transgenese: Einfügung von rekombinierten Genen
Genetische Modellorganismen
Drosophila melanogasterLebenszyklus
Shockwave: Life Cycle
http://flymove.uni-muenster.de
FLYMOVE
Embryonalentwicklung

4
Genetik der Entwicklungsbiologie
• Systematische Mutagenisierung und Durchmusterung von Fliegenembryonen
• Entdeckung sogenannter Eipolaritätsgene:MaternaleffektgeneEffekt tritt auf bei Nachkommen vonhomozygoten Müttern
z.B. bicoid

5
Morphogengradienten und Musterbildung
• Musterbildung:Etablierung einer klar gegliederten räumlichen Anordnung von Zellaktivitäten.
• Morphogen:Substanz auf die Zellen bei unterschiedlichen Schwellenwerten unterschiedlich reagieren
• Positionsinformation:Lageinformation, durch die Zellen ihre Lage imKeim mitgeteilt bekommen.Diese Information kann von benachbarten Zellenausgehen oder von ferneren Morphogensendern.
Morphogengradientund Musterbildung
Das „Tricolore“-Modell
• Lewis Wolpert, 1978
• Morphogenkonzentrationlegt an jedem Punkt eine exakte Positionsinformation fest.
• Schwellenwertkonzentrationen:Konzentration, oberhalb der Zellen anders reagieren als unterhalb.
Der morphogenetische Gradient von BICOID
Genetik der Entwicklungsbiologie
• Systematische Mutagenisierung und Durchmusterung von Fliegenembryonen
• Entdeckung der zygotischenSegmentierungsgene:
Lückengene (Gap-Gene)
Paar-Regel-Gene
Segmentpolaritätsgene
Drosophila Larve Lückengene (Gap-Gene)• Fehlen mehrerer zusammenhängender
Segmente

6
Lückengene (Gap-Gene)• Expression in einem weiten Bereich des Embryos
hunchback / Krüppel
Paar-Regel-Gene• Fehlen jedes zweiten Segmentäquivalents
Paar-Regel-Gene• Expression in sich wiederholenden Streifen
Segmentpolaritätsgene• Fehlen eines Teilbereichs jeden Segments,
wird durch den verbleibenden Spiegelbildlich ersetzt.
Segmentpolaritätsgene• Expression in einem Teilbereich jeden Segments.
Genetik der Entwicklungsbiologie• Systematische Mutagenisierung und
Durchmusterung von Fliegenembryonen
• Entdeckung der zygotischenSegmentierungsgene:
Lückengene (Gap-Gene)
Paar-Regel-Gene
Segmentpolaritätsgene
führen zur Metamerisierung des Embryos
7
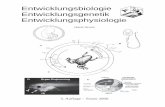
Genetik der Entwicklungsbiologie
• Systematische Mutagenisierung und Durchmusterung von Fliegenembryonen
• Entdeckung der
Homöotischen Selektorgene
führen zur Segmentspezifizierung
Homöotische Transformationen
Antennapedia
Homöotische Transformationen
Ultrabithorax
Expression homöotischeSelektorgene
Edward B.Lewis
Nobelpreis 1995 für Physiologie oder Medizin
“Für ihre Entdeckungen in Bezug auf die genetischeKontrolle der frühen Embryonalentwicklung”
ChristianeNüsslein-Volhard
Eric F.Wieschaus
HomöotischeSelektorgeneund ihre Funktionblieben im Laufe der Evolutionweitgehend unverändert
8
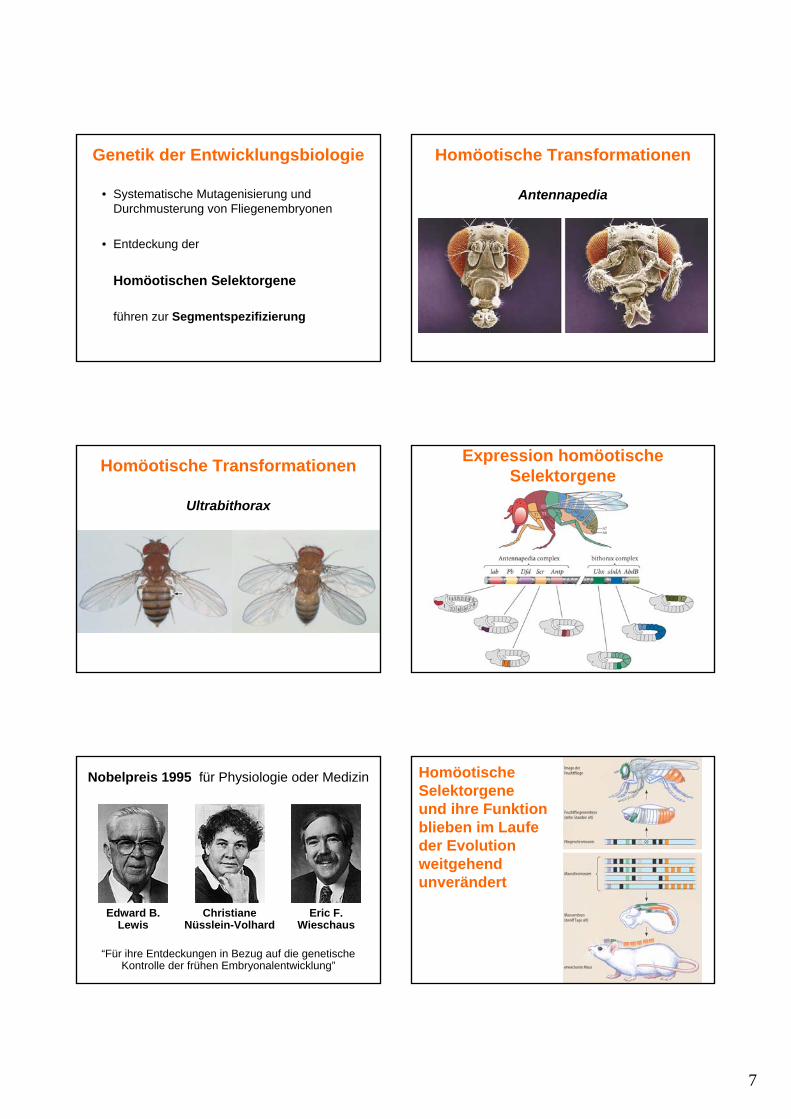
Genetische Modellorganismen Modellorganismus: FadenwurmCaenorhabditis elegans
Zellgenealogie von Caenorhabditis elegans
Zellgenealogie von Caenorhabditis elegansModellorganismus: Caenorhabditis elegans
Eutelie (Zellkonstanz): invariante Anzahl von Zellen (558 somatische Zellen)
Invarianter Zellstammbaum
Genetik: selbstfertilisierender Hermaphrodit
Entdeckung des entwicklungsbiologisch notwendigen Zelltodes:131x programmierter Zelltod (Apoptosis)
9
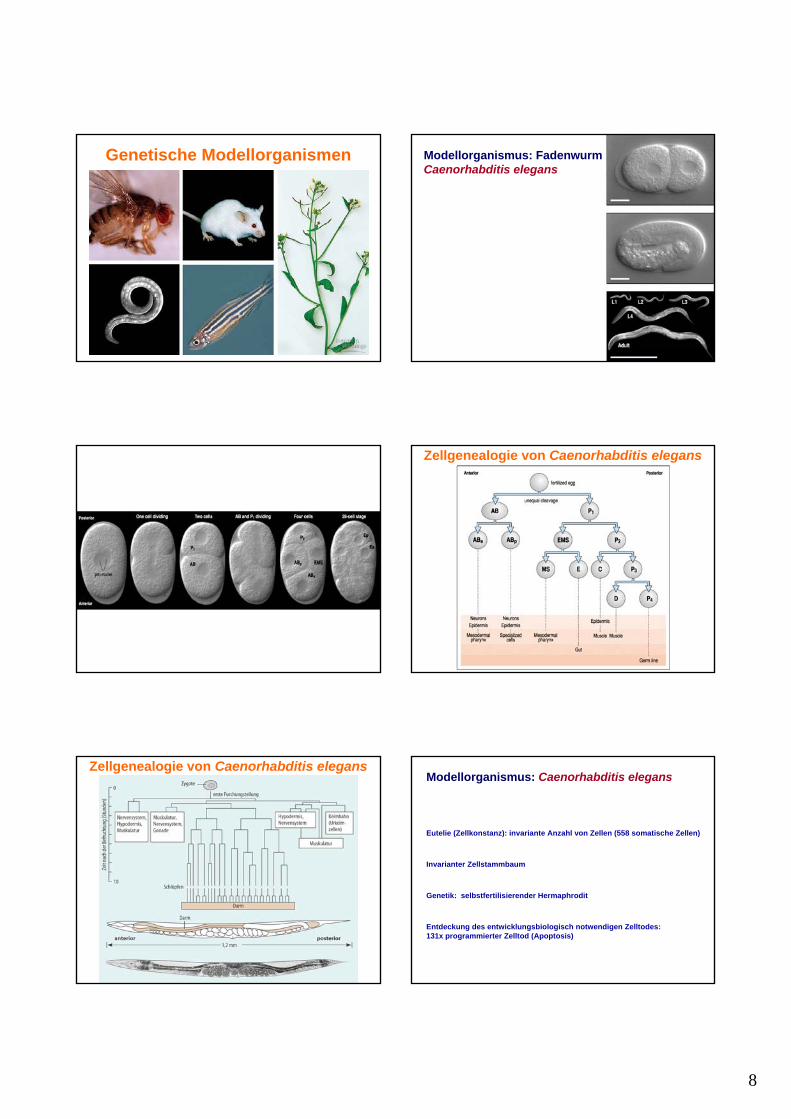
Apoptosis: Programmierter Zelltod
• Zellen durchlaufen genetisches Programm,das zum Selbstmord führt
• Entdeckung bei eutelischem Organismus C. elegans
• Aber auch in anderen Spezies entscheidendbei Entwicklungsprozessen beteiligt:
Gehirnentwicklung
Bildung des Zehen- und Fingerzwischenraums
Sydney Brenner
Nobelpreis 2002 für Physiologie oder Medizin
“Für ihre Entdeckungen in Bezug auf die genetische Kontrolleder Organentwicklung und des programmierten Zelltodes”
H. RobertHorvitz
John E.Sulston
Genetische Modellorganismen Modellorganismus:AckerschmalwandArabidopsis thaliana
1 mm

10
Blüten-ABC
Blüten-ABC
Blüten-ABC Genetische Modellorganismen

11
Modellorganismus: Mus musculus (Maus)
Keine experimentelle Embryologie
Ausgefeilte Genetik
Nähe zum Menschen: Säugetier
Embryonale Stammzellen
Homologe Rekombination: gezielter Gen-Knock-Out möglich
Modellorganismus: Danio rerio (Zebrabärbling)
Experimentelle Embryologie
Genetik
Nähe zum Menschen: Wirbeltier
Morpholinos: gezielter Gen-Knock-Down möglich
Vertebraten Modellorganismen
12
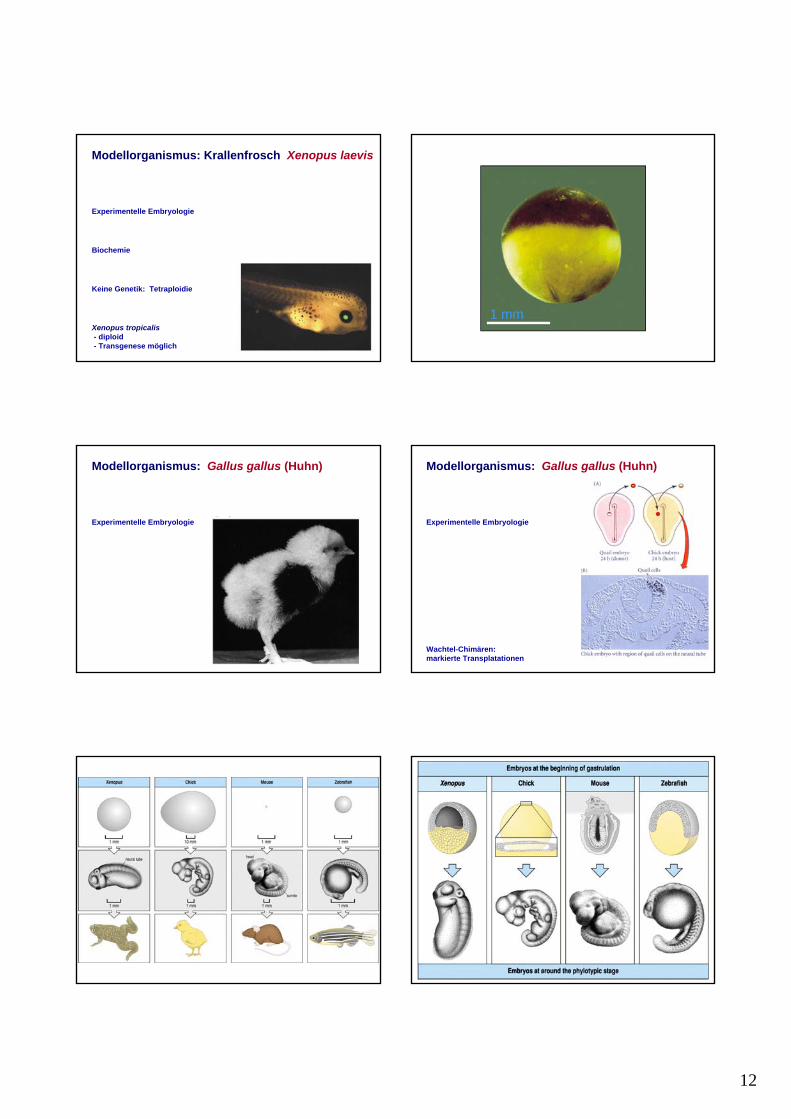
Modellorganismus: Krallenfrosch Xenopus laevis
Experimentelle Embryologie
Biochemie
Keine Genetik: Tetraploidie
Xenopus tropicalis- diploid- Transgenese möglich
1 mm
Modellorganismus: Gallus gallus (Huhn)
Experimentelle Embryologie
Modellorganismus: Gallus gallus (Huhn)
Experimentelle Embryologie
Wachtel-Chimären: markierte Transplatationen