
Exkretion und Osmoregulation Version 2 - Uni Ulm Aktuelles · Grundlage der Osmose ist das...
21
Universität Ulm Praktikum Stoffwechselphysiologie WS 2011/2012 Versuch Exkretion und Osmoregulation Gruppe XY Betreuer: Praktikanten: Versuchstag:
Transcript of Exkretion und Osmoregulation Version 2 - Uni Ulm Aktuelles · Grundlage der Osmose ist das...

Universität Ulm
Praktikum Stoffwechselphysiologie
WS 2011/2012
Versuch Exkretion und Osmoregulation
Gruppe XY
Betreuer:
Praktikanten:
Versuchstag:

2
Inhalt 1. Einleitung ............................................................................................................................................ 3
2. Theorie ................................................................................................................................................ 3
2.1 Osmose .............................................................................................................................................. 3
2.1.1 Begrifflichkeiten ......................................................................................................................... 3
2.1.2 Donnan-Verteilung ..................................................................................................................... 4
2.2 Transportvorgänge ............................................................................................................................. 4
2.3 Exkretionsorgane ............................................................................................................................... 5
2.3.1 Säugerniere ................................................................................................................................. 5
a) Glomeruläre Filtration ............................................................................................................. 6
b) Tubuläre Sekretion .................................................................................................................. 6
2.3.2 Pulsierende Vakuole ................................................................................................................... 7
2.3.3 Protonephridium ......................................................................................................................... 7
2.3.4 Metanephridium ......................................................................................................................... 7
2.3.5 Malpighische Gefäße .................................................................................................................. 8
2.4 Wasserregulation ............................................................................................................................... 8
2.4.1 ADH (Antidiuretisches Hormon) ............................................................................................... 8
2.4.2 Renin-Angiotensin-Aldosteron-System (RAAS) ....................................................................... 8
2.5 Exkretionsarten .................................................................................................................................. 9
2.6 Clearence ........................................................................................................................................... 9
2.7 Enzymatischer Test ........................................................................................................................... 9
2.8 Ornithin-Harnstoff-Zyklus .............................................................................................................. 10
3. Material und Methoden ..................................................................................................................... 11
3.1 Versuch Osmoregulation und Ionenregulation ............................................................................ 11
3.2 Versuch Osmoregulation/ Exkretion ........................................................................................... 12
4. Ergebnisse, Auswertung und Diskussion .......................................................................................... 13
4.1 Versuch Osmoregulation und Ionenregulation ............................................................................ 13
4.2 Versuch Exkretion ....................................................................................................................... 16
5. Quellen .............................................................................................................................................. 21

3
1. Einleitung
Bei den Versuchen zur Osmoregulation wurde die Fähigkeit verschiedener Tiere zur
Regulierung ihrer Osmolarität in verschieden osmolaren Außenmedien untersucht. Für viele
Tiere ist eine korrekte Einstellung ihrer Osmolarität überlebenswichtig, daher ist es
interessant zu wissen inwiefern bzw. in welchem Bereich sie dazu fähig sind.
Im Exkretionsversuch wurde u.a. gesammelter menschlicher Urin auf seinen Harnsäuregehalt
hin untersucht. Hintergrund ist, dass verschiedene Tiergruppen verschiedene Mechanismen
zur Stickstoffausscheidung verwenden. Harnsäure ist neben anderen Stoffen eine Möglichkeit
überflüssigen Stickstoff loszuwerden.
Die Versuche drehen sich also rund um osmotische Vorgänge und Exkretionsorgane. Diesen
Aspekten widmet sich auch der folgende Theorieteil.
2. Theorie
2.1 Osmose Die Osmose beschreibt einen passiven Konzentrationsausgleich zwischen zwei ungleich
konzentrierten wässrigen Lösungen, die durch eine semipermeable Membran getrennt sind. Es
wandert hierbei H2O und nicht die gelösten Teilchen.
Grundlage der Osmose ist das Phänomen der Diffusion. Es beschreibt zufällige Wanderungen
von Teilchen, die durch Braun’sche Molekularbewegungen ausgelöst werden. Im statistischen
Mittel wandern die Teilchen dabei von Regionen höherer Konzentration zu Regionen
niedrigerer Konzentration. Dies geschieht so lange bis ein Konzentrationsgleichgewicht
herrscht.
2.1.1 Begrifflichkeiten Von einer hypoosmotischen Lösung spricht man, wenn die Lösung eine niedrigere
Salzkonzentration aufweist, als eine Vergleichslösung. Eine Lösung ist hyperosmotisch, wenn
ihre Salzkonzentration höher ist, als in der Vergleichslösung. Sind beide Salzkonzentrationen
gleich, so liegen isoosmotische Lösungen vor.

4
Biomembranen sind jedoch nie nur für Wasser durchlässig, sondern immer auch für andere
Teilchen. Daher sollte man bei lebenden Systemen von der effektiven Osmolarität, der
Tonizität, sprechen. Die Tonizität bezieht außer dem Lösungsmittel eben auch andere
Teilchen mit ein, welche die Membran passieren können.
Eine isotone Lösung besitzt das gleiche effektive osmotische Potential wie eine
Vergleichslösung. Hypo- und Hypertonizität sind analog definiert.
Organismen können homoiosmotisch oder poikilosmotisch sein. Osmoregulierer sind
homoiosmotisch, der osmotische Wert ihrer Körperflüssigkeiten wird unabhängig vom
Außenmedium auf konstantem Niveau gehalten. Diese Regulation ist ein aktiver Prozess, der
Energie in Form von ATP verbraucht. Als poikilosmotisch werden Osmokonformer
bezeichnet, sie sind isoosmotisch zum Außenmedium. Stoffwechselvorgänge können deshalb
bei Osmokonformern oft nicht mit höchster Effizienz ablaufen. Grund ist, dass die für
Stoffwechselvorgänge benötigten Katalysatoren, die Enzyme, Osmolaritätsoptima besitzen,
bei denen sie am besten funktionieren. Da Osmokonformer Osmolaritätsschwankungen
unterworfen sind, befinden sich ihre Enzyme meist nicht in der Lösung mit der perfekten
Salzkonzentration. Die Enzyme arbeiten dann langsamer als bei ihrem Optimum und es bleibt
ein gewisses Potential ungenutzt.
Zuletzt wird zwischen euryhalinen und stenohalinen Organismen unterschieden. Euryhaline
Organismen haben einen weiten osmotischen Toleranzbereich, stenohaline einen eher engen
osmotischen Toleranzbereich.
2.1.2 Donnan-Verteilung Die auf Ionen bezogene Donnan-Verteilung beschreibt die Gleichgewichtseinstellung von
zwei Lösungen mit unterschiedlichen gelösten Ionen in einem Zwei-Kammer-System mit
semipermeabler Membran. Die Membran ist zwar für das Lösungsmittel, aber nicht für die
Ionen durchlässig. Das führt dazu, dass sich ein osmotischer Druck, sowie eine
Potentialdifferenz einstellen.
2.2 Transportvorgänge Transport über Membranen kann entweder aktiv oder passiv ablaufen.
Eine Biomembran besteht aus einer Phospholipiddoppelschicht. Bei den Phospholipiden
stehen einem nach außen zeigenden, hydrophilen Kopf mit einer Phosphatgruppe zwei nach
innen zeigende, hydrophobe Alkylschwänze entgegen. Dank dieser hydrophoben Schwänze

5
stellt eine Biomembran eine starke Barriere für polare Teilchen wie Ionen dar. Die
Lipiddoppelschicht ist mit Proteinen bestückt, welche diese teilweise sogar durchstoßen und
als Kanalproteine fungieren können. Sogenannte Glykoproteine, die nach außen zeigen,
können als Erkennungsmerkmale dienen.
Der passive Transport über Membranen findet an Carrierproteinen oder Ionenkanälen statt.
Die Teilchen wandern hier aufgrund der Diffusion in Richtung eines Konzentrationsgefälles.
Aktiver Transport wird unterschieden in Uniport, bei dem nur ein Stoff transportiert wird und
in Cotransport, bei dem ein Stoff aktiv transportiert wird und ein zweiter passiv mitgezogen
werden kann. Es gibt wiederum zwei Arten des Cotransports. Beim Symport werden beide
Stoffe in die gleiche Richtung transportiert, beim Antiport werden sie in entgegengesetzte
Richtungen transportiert.
2.3 Exkretionsorgane
2.3.1 Säugerniere Die Niere von Säugetieren besteht aus zwei Schichten, der äußeren Rinde (Cortex) und dem
inneren Mark (Medulla). Beide sind von der Nierenkapsel umschlossen.
Die Funktionseinheiten der Niere sind die Nephrone (Abb. 1). Im Glomerulus, welcher von
der Bowman-Kapsel umschlossen wird, wird das Blut gefiltert. Das Filtrat wandert durch den
proximalen Tubulus, die Henle-Schleife und den distalen Tubulus bis ins Sammelrohr.

6
Abb. 1: Nephron (Quelle: www.urology-textbook.com/kidney-nephron.html)
a) Glomeruläre Filtration Im Glomerulus, einem Kapillarenknäuel findet eine Ultrafiltration statt. Aufgrund des
Blutdrucks wird hier das Blut durch die Kapillarwände gepresst, wobei die größeren
Blutzellen zurückgehalten werden. Nachdem eine für Proteine undurchlässige Basalmembran
passiert wurde, muss das Filtrat zum Schluss noch durch eine mit Podocyten bedeckte
Kapselwand, durch die nur niedermolekulare Stoffe und Peptide hindurchkommen.
Die Filtration ist von drei Faktoren abhängig:
- vom hydrostatischen Druck zwischen dem Kapillarlumen und der Bowman-Kapsel
- vom kolloidosmotischen Druck, der der Filtration entgegenwirkt
- von den Siebeigenschaften der Membranen
b) Tubuläre Sekretion Im proximalen Tubulus werden zuerst Giftstoffe wie Ammoniak sezerniert, Nährstoffe, K+,
NaCl und H2O werden dagegen resorbiert. Dazu wird beispielsweise Na+ aktiv transportiert,
woraufhin Cl- passiv nachfolgt und das Wasser ebenfalls passiv durch Osmose nachzieht. Zur
pH-Regulation wird H+ sezerniert, die Puffersubstanz HCO3- (Hydrogencarbonat) wird ins
Interstitium zurücktransportiert.
Aufgrund der starken Hypertonizität des Interstitiums wird im absteigenden Ast der Henle-
Schleife passiv H2O reabsorbiert, dadurch wird der Urin stark aufkonzentriert. Im

7
aufsteigenden Ast der Henle-Schleife wird NaCl resorbiert. Zuerst geschieht dies passiv, da
schon viel Wasser hinausdiffundiert ist, im oberen Teil des Astes muss der Transport jedoch
aktiv passieren. Das verdünnt den Urin wieder. In der Henle-Schleife wirkt ein
Gegenstromprinzip. Da beide Schenkel über die interstitielle Flüssigkeit miteinander in
Verbindung stehen beeinflussen sie sich gegenseitig. Eine hohe Effektivität ist die Folge.
Im distalen Tubulus findet neben weiterer pH-Regulation eine K+-Sekretion, sowie die
nochmalige aktive Wiederaufnahme von NaCl und H2O statt.
Danach kommt der Urin im Sammelrohr an, wo ihm weiterhin aktiv NaCl und H2O entzogen
wird.
2.3.2 Pulsierende Vakuole Pulsierende oder kontraktile Vakuolen sind Ausscheidungsorgane von Einzellern wie
Euglena. Sie transportieren Wasser nach außen, daher kommen sie fast nur bei Organismen
vor, die hyperosmotisch zu ihrem Außenmedium sind. Ein von Protonenpumpen geschaffener
Protonengradient über der Vakuolenmembran führt zu einem Einstrom von Ionen in die
Vakuole. Dadurch ist diese hyperosmotisch zum Cytoplasma, was einen Wassereinstrom zur
Folge hat. Ist die Vakuole mit Wasser gefüllt, so fusioniert sie mit der Zellmembran und gibt
ihren Inhalt an das Außenmedium ab.
2.3.3 Protonephridium Protonephridien kommen z.B. bei Plattwürmern vor. Sie sind verzweigte Röhrensysteme an
deren blinden Enden jeweils eine Reusengeißelzelle sitzt. Hier erzeugt eine Wimpernflamme,
welche aus Cilienbündeln besteht, durch ständige schlagende Bewegungen einen
Flüssigkeitsstrom in Richtung des Röhrensystems. Dies erzeugt einen Unterdruck, wodurch
Gewebsflüssigkeit in die jeweilige Röhre einströmt. Dabei wird die Flüssigkeit von
Reusenstäben, die von Podocyten ausgebildet werden, ultrafiltriert. Der so gebildete
Primärharn wird nun im Laufe des Röhrensystems weiter verarbeitet. Alle Stoffe, die der
Körper noch gebrauchen kann werden resorbiert. Der Sekundärharn wird nun ausgeschieden.
Dafür enden die Röhren in Nephridialporen, die mit dem Außenmedium in Verbindung
stehen.
2.3.4 Metanephridium Ringelwürmer, Weichtiere und Krebstiere enthalten diese Exkretionsorgane. Die
Metanephridien bestehen aus Kanälen, die über einen Wimperntrichter mit der sekundären
Leibeshöhle, dem Coelom, verbunden sind und wie bei Protonephridien zur Außenseite des

8
Organismus führen. Am Coelomepithel des Wimperntrichters befinden sich Podocyten,
welche Blutgefäße umgeben. Die Ultrafiltration findet hier an der Basalmembran der
Blutgefäße, getrieben durch den Blutdruck, statt. Analog zu den Protonephridien entsteht hier
der Primärharn, welcher in den Kanälen noch einer Resorption von nützlichen Stoffen
unterzogen und dann als Sekundärharn ausgeschieden wird.
2.3.5 Malpighische Gefäße Hierbei handelt es sich um die Exkretionsorgane von Gliederfüßern (Arthropoden) wie den
Insekten. Es sind viele Ausstülpungen, die an einer Seite blind in der Leibeshöhle enden und
an der anderen Seite in den Darm münden. Anstatt einer Filtration finden hier aktive
Transportprozesse statt. Aus der Hämolymphe wird zunächst K+ und Na+ in die Gefäße
transportiert. Wasser und Harnsäure strömen nach. Nützliche Stoffe, wie Aminosäuren,
Zucker und bestimmte Ionen, werden an das Interstitium abgegeben. Der so gebildete Harn
strömt in den Darm. Im Enddarm werden Ionen aktiv in die Hämolymphe gepumpt, denen
Wasser passiv nachfolgt. Aufgrund dessen spart das Lebewesen viel Wasser ein.
2.4 Wasserregulation
2.4.1 ADH (Antidiuretisches Hormon) Der Elektrolytwert im Blut wird ständig von speziellen Rezeptoren registriert. Als Regler in
diesem Regelkreis fungiert der Hypothalamus, welcher das Hormon bei erhöhter
Blutosmolarität ausschüttet. Dies bewirkt einen Durstreiz, sowie eine Vasokonstriktion, was
den Blutdruck erhöht. Außerdem führt die ADH-Ausschüttung zum Einbau von zusätzlichen
Aquaporinen in die Membranen der Sammelrohre. Dadurch kann mehr Wasser durch die
Membranen transportiert und damit resorbiert werden. All dies führt zu einer Erhöhung der
Wasserkonzentration im Blut, dessen Osmolarität damit sinkt. Folglich wird weniger ADH
ausgeschüttet womit sich der Regelkreis schließt.
2.4.2 Renin-Angiotensin-Aldosteron-System (RAAS) Wenn Blutdruckrezeptoren einen zu geringen Blutdruck feststellen, dann wird Renin
ausgeschüttet. Dieses wandelt Angiotensinogen in Angiotensin 1 um, welches durch das
Enzym ACE in Angiotensin 2 umgewandelt wird. Das Angiotensin 2 führt zu einer
Vasokonstriktion und zu einer Ausschüttung von ADH, dessen Auswirkungen oben
beschrieben wurden.

9
2.5 Exkretionsarten Es gibt drei verschiedene Möglichkeiten, wie ein Tier Stickstoff ausscheiden kann:
- ammoniotele Tiere exkretieren Ammoniak (Abb. 2), dies sind hauptsächlich
aquatische Tiere
- ureotele Tiere exkretieren Harnstoff (Abb. 3), Amphibien und Säugetiere gehören
dazu
- uricotele Tiere verwenden Harnsäure (Abb. 4) zur Stickstoffausscheidung, z.B.
Insekten und Vögel
Abb. 2: Ammoniak (Quelle:
http://de.wikipedia.org/wiki/Ammoniak)
Abb. 3: Harnstoff (Quelle:
http://de.wikipedia.org/wiki/Harnstoff)
Abb.4: Harnsäure (Quelle:
http://de.wikipedia.org/wiki/Harns%C3%A4ure)
2.6 Clearence Die Clearence ist ein Maß für die Ausscheidungsfähigkeit, also die Nierenfunktion. Sie steht
für die Blutplasma- oder Serummenge, die beim Durchfluss durch die Nieren pro Minute
vollständig von Harnstoff befreit wird.
[S]U = Substratkonzentration im Urin; VU = Volumen des Urins pro Minute;
[S]P = Substratkonzentration im Blutplasma
2.7 Enzymatischer Test Die Aktivität von Enzymen kann im Spektralphotometer bestimmt werden. Das Prinzip ist,
dass das Enzymsubstrat und das Enzymprodukt Licht einer bestimmten Wellenlänge
unterschiedlich stark absorbieren. Diese Absorption bzw. Extinktion kann gemessen werden.
Mit dem Lambert-Beer’schen-Gesetz kann dann die Stoffkonzentration bestimmt werden:

10
E = Extinktion; ε = stoffspezifischer Extinktionskoeffizient;
c = Konzentration der absorbierenden Substanz; d = Weg, den das Licht zurücklegt
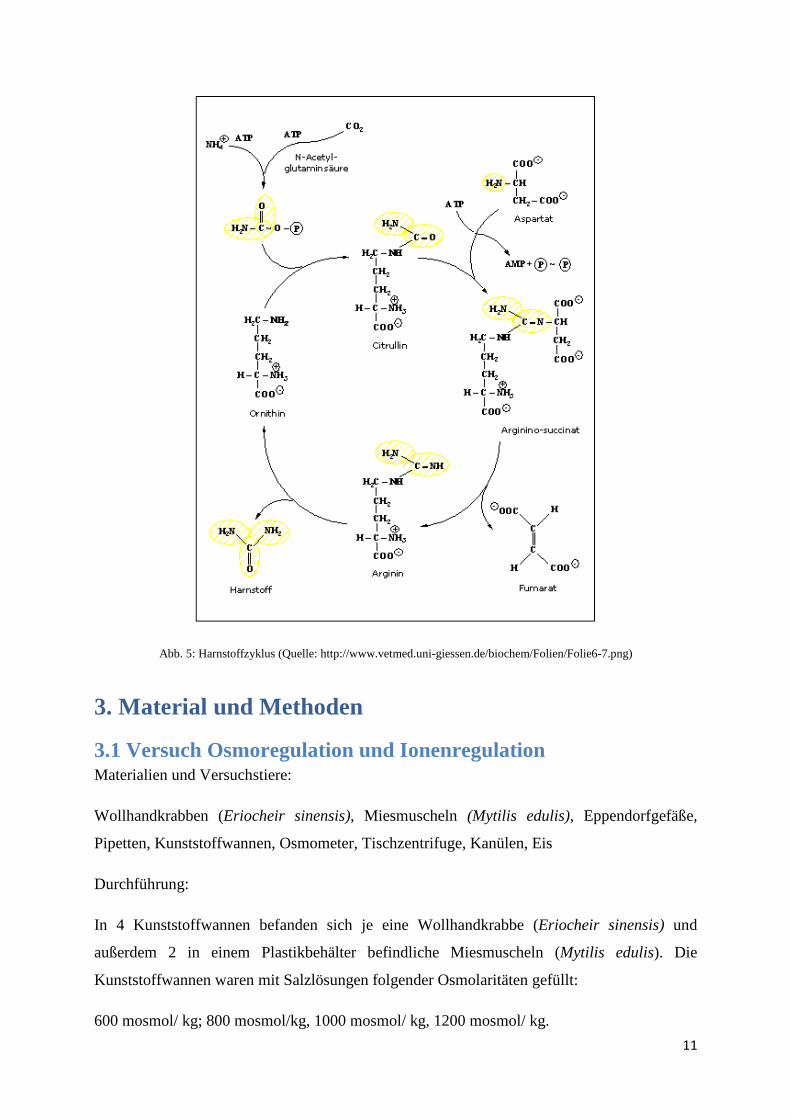
2.8 Ornithin-Harnstoff-Zyklus Der Harnstoffzyklus (Abb. 5) wandelt NH4
+ oder Aminogruppen aus dem Abbau von
stickstoffhaltigen Molekülen in Harnstoff um. Er läuft in der Leber ab.
Durch ATP-Verbrauch wird in den Mitochondrien aus dem Ammonium mit CO2
Carbamylphosphat hergestellt. Dieses wird durch eine Transferase auf Ornithin übertragen. Es
entsteht Citrullin, welches durch Mitochondrienmembranen nach außen ins Cytosol
transportiert werden kann. Durch weiteren ATP-Verbrauch wird Aspartat auf das Citrullin
übertragen, was zu Argininosuccinat führt. Eine Lyase spaltet dieses in Arginin und Fumarat.
Fumarat geht in den Citratzyklus ein, Arginin wird durch eine Arginase in Ornithin und
Harnstoff gespalten. Durch den Rücktransport von Ornithin in die Mitochondrien kann der
Zyklus von vorne beginnen.
11
Abb. 5: Harnstoffzyklus (Quelle: http://www.vetmed.uni-giessen.de/biochem/Folien/Folie6-7.png)
3. Material und Methoden
3.1 Versuch Osmoregulation und Ionenregulation Materialien und Versuchstiere:
Wollhandkrabben (Eriocheir sinensis), Miesmuscheln (Mytilis edulis), Eppendorfgefäße,
Pipetten, Kunststoffwannen, Osmometer, Tischzentrifuge, Kanülen, Eis
Durchführung:
In 4 Kunststoffwannen befanden sich je eine Wollhandkrabbe (Eriocheir sinensis) und
außerdem 2 in einem Plastikbehälter befindliche Miesmuscheln (Mytilis edulis). Die
Kunststoffwannen waren mit Salzlösungen folgender Osmolaritäten gefüllt:
600 mosmol/ kg; 800 mosmol/kg, 1000 mosmol/ kg, 1200 mosmol/ kg.

12
Von den Salzlösungen, dem Haltewasser der Krabben, wurden je 2 Proben von jeweils 100 µl
entnommen und von diesen Proben die Osmolaritäten mit dem Osmometer bestimmt.
Die Krabben wurden nacheinander aus ihrem Becken genommen, mit einer Kanüle am
Gelenk in den Körper eingestochen und dann an dieser mit einer Pipette 20 µl Hämolymphe
entnommen. Die Hämolymphe wurde dann mit 80 µl destilliertem Wasser verdünnt, kurz
zentrifugiert und darauf mit dem Überstand die Osmolarität am Osmometer bestimmt.
Die Miesmuscheln wurden nacheinander aus den Becken genommen und dann mit einem
Spatel angebohrt, um mit einer Mikropipette 20 µl Hämolymphe entnehmen zu können. Diese
wurde mit 80 µl destilliertem Wasser verdünnt, kurz zentrifugiert und mit dem Überstand die
Osmolarität am Osmometer bestimmt.
In einem Diagramm wurden die Werte der Hämolymphe gegen die Werte des Haltungswasser
aufgetragen.
3.2 Versuch Osmoregulation/ Exkretion Materialien:
Kot von Boa Constrictor, Urin des Menschen, Reagenzgläser, Photometer, Pipetten, Falcons,
Homogenisator/Poter, Feinwaage, Eis, Zentrifuge, Wasserbad
Reagenzien:
Reagenzlösung: Phosphatpuffer, TOOS, Fettalkoholpolyglykolether, Ascorbatoxidase
Startlösung: Phosphatpuffer, Kaliumhexacyanoferrat, 4-Aminophenazon, Uricase, Peroxidase
Harnsäurestandard: Harnsäure 0,06 mg/ml = 0,36 mM
Durchführung:
Es wurden 22 g weißer Schlangenkot und 24 g schwarzer Schlangenkot abgewogen, jeweils
mit 10 ml demineralisiertem Wasser verdünnt, mit dem Poter homogenisiert und darauf kurz
abzentrifugiert. Der Überstand des Homogenisats des weißen Kots wurde erneut verdünnt.
Von dieser Flüssigkeit wurden 50 µl zu 1000 µl demineralisiertem Wasser gegeben. Von der
Urinprobe wurden 100 µl zu 900 µl demineralisiertem Wasser gegeben.
13
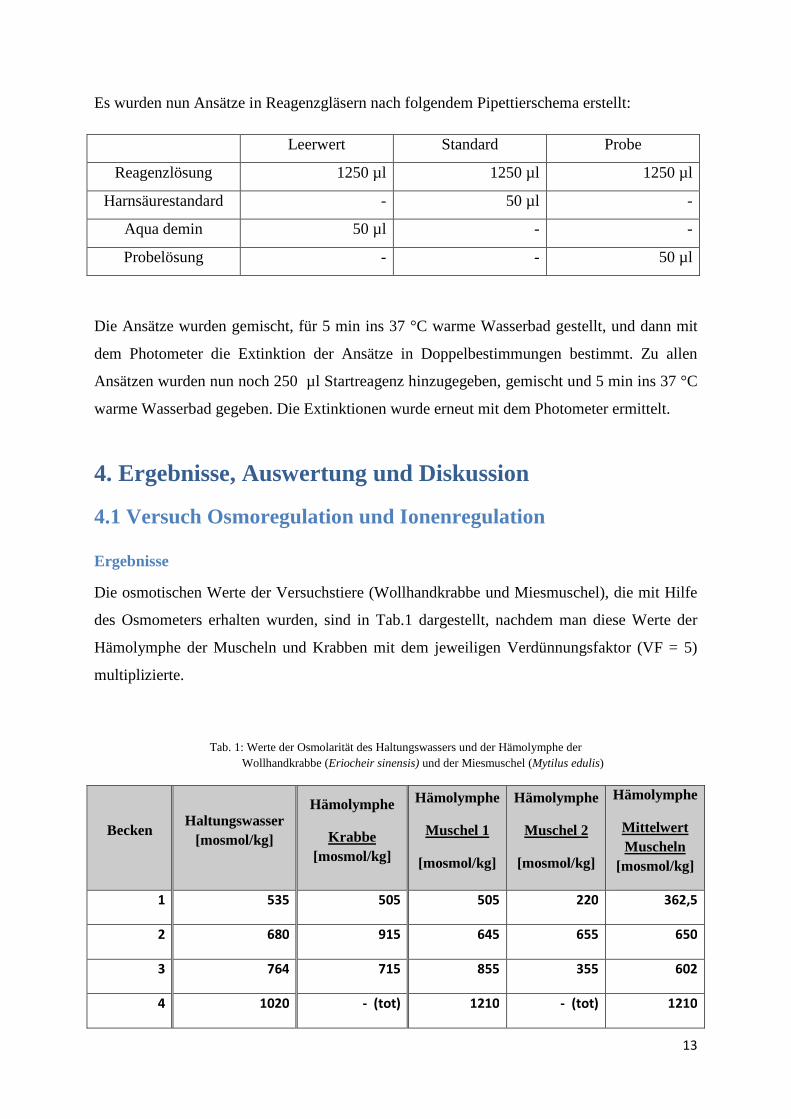
Es wurden nun Ansätze in Reagenzgläsern nach folgendem Pipettierschema erstellt:
Leerwert Standard Probe
Reagenzlösung 1250 µl 1250 µl 1250 µl
Harnsäurestandard - 50 µl -
Aqua demin 50 µl - -
Probelösung - - 50 µl
Die Ansätze wurden gemischt, für 5 min ins 37 °C warme Wasserbad gestellt, und dann mit
dem Photometer die Extinktion der Ansätze in Doppelbestimmungen bestimmt. Zu allen
Ansätzen wurden nun noch 250 µl Startreagenz hinzugegeben, gemischt und 5 min ins 37 °C
warme Wasserbad gegeben. Die Extinktionen wurde erneut mit dem Photometer ermittelt.
4. Ergebnisse, Auswertung und Diskussion
4.1 Versuch Osmoregulation und Ionenregulation Ergebnisse
Die osmotischen Werte der Versuchstiere (Wollhandkrabbe und Miesmuschel), die mit Hilfe
des Osmometers erhalten wurden, sind in Tab.1 dargestellt, nachdem man diese Werte der
Hämolymphe der Muscheln und Krabben mit dem jeweiligen Verdünnungsfaktor (VF = 5)
multiplizierte.
Tab. 1: Werte der Osmolarität des Haltungswassers und der Hämolymphe der Wollhandkrabbe (Eriocheir sinensis) und der Miesmuschel (Mytilus edulis)
Becken Haltungswasser
[mosmol/kg]
Hämolymphe
Krabbe [mosmol/kg]
Hämolymphe
Muschel 1
[mosmol/kg]
Hämolymphe
Muschel 2
[mosmol/kg]
Hämolymphe
Mittelwert Muscheln
[mosmol/kg]
1 535 505 505 220 362,5
2 680 915 645 655 650
3 764 715 855 355 602
4 1020 - (tot) 1210 - (tot) 1210
14
Auswertung
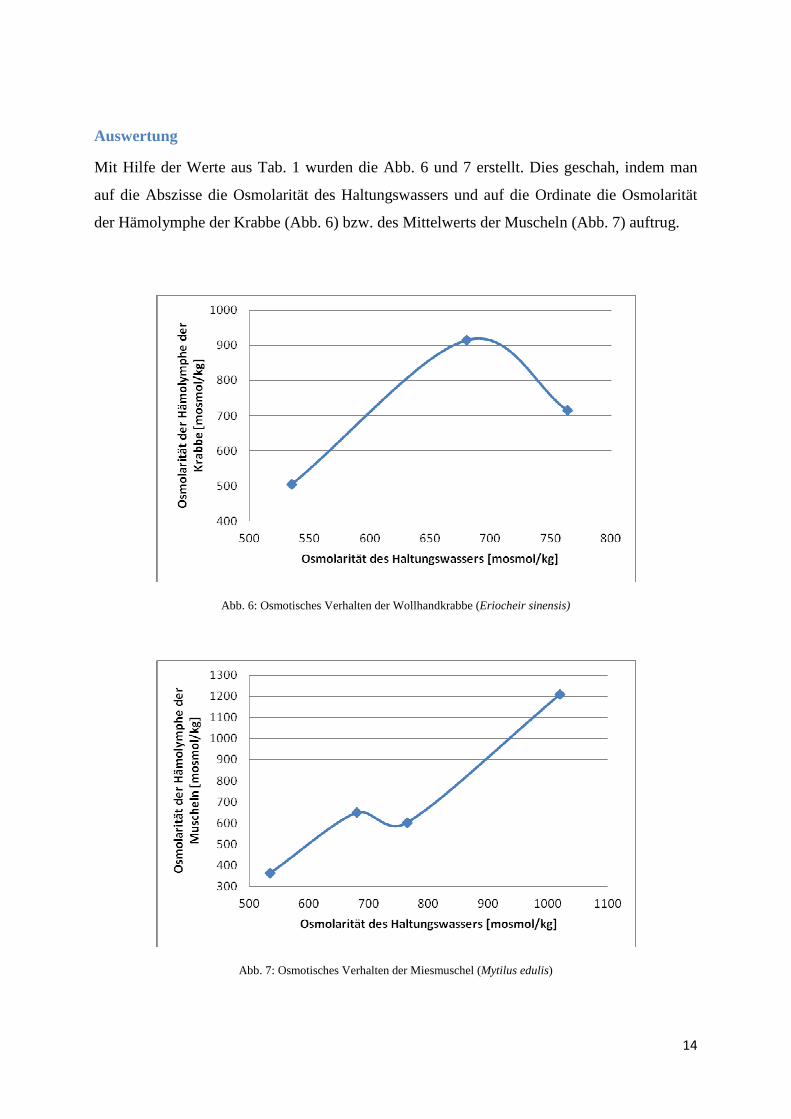
Mit Hilfe der Werte aus Tab. 1 wurden die Abb. 6 und 7 erstellt. Dies geschah, indem man
auf die Abszisse die Osmolarität des Haltungswassers und auf die Ordinate die Osmolarität
der Hämolymphe der Krabbe (Abb. 6) bzw. des Mittelwerts der Muscheln (Abb. 7) auftrug.
Abb. 6: Osmotisches Verhalten der Wollhandkrabbe (Eriocheir sinensis)
Abb. 7: Osmotisches Verhalten der Miesmuschel (Mytilus edulis)

15
Diskussion
In Tab. 1 kann man erkennen, dass die Werte der Osmolarität der Hämolymphe der
Wollhandkrabben stetig mit denen des Haltungswassers steigen. Sie sind sogar höher als die
der Hämolymphe der Miesmuscheln. Bei dem Wert des 2. Haltungsbeckens ist die
Osmolartität des Haltungswasser (680 mosmol/kg) sogar um einiges niedriger als die der
Hämolymphe der Krabbe (915 mosmol/kg). Hierbei muss es sich eindeutig um einen
Ausreißer handeln, was man auch in der Abb. 6 sieht. Denn in Abbildung 6 hat die Kurve
genau an dieser Stelle ein Maximum, um danach wieder steil abzufallen, was nicht
nachvollziehbar ist. Denn eigentlich sollte die Kurve in Abb. 6 nur eine sehr flache Gerade
sein, die erst bei sehr starken Osmolaritätswerten des Haltungswassers stärker ansteigen
sollte. Denn die Wollhandkrabben gehören zu den eingeschränkten Osmoregulieren und sind
euryhalin, was bedeutet, dass sie die Osmolarität ihrer Hämolymphe aktiv regulieren und
diesen Wert auch stabil halten können, wenn sich die Osmolarität des umgebenden Wasser
stark erhöht. Dies ist jedoch bei den hier erhaltenen Werten, wie sie in Tabelle 1 und in
Abbildung 6 dargestellt sind nicht der Fall. Gründe für das unerwartete Ergebnis sind sehr
wahrscheinlich Pipettierfehler. Denn bereits bei der Entnahme der Probe der Hämolymphe
aus den Versuchstieren gab es große Probleme, wie dass die gewünschte Menge an
Probenflüssigkeit nicht erhalten wurde oder starke Verunreinigungen in die Mikropipette
gelangten. Auch hat das Osmometer bei manchen Messungen einen Fehler angezeigt, da nicht
genug Lösung in den Probetubes vorhanden war, sodass noch Wasser nachgefüllt werden
musste und so die Verdünnung nicht mehr exakt nachvollziehbar war. Auch etwas seltsam ist,
dass die Wohlhandkrabbe des 4. Beckens einen Osmolarität des Haltungswassers von 1020
mosmol/kg nicht überlebt hat, denn als euryhaline Tiere sollten sie ein relativ breites
Salztoleranzspektrum besitzen. Jedoch können auch noch andere Gründe, wie Verletzungen
durch zum Beispiel andere Wollhandkrabben oder eine Krankheit Ursache für den Tot sein.
Sieht man sich nun die osmotischen Werte der Miesmuschel in Tab. 1 oder auch Abb. 7 an, so
erkennt man, dass die Osmolarität der Hämolymphe der Muscheln aus Becken 2 und 3 sehr
nahe beieinander liegen (der osmotische Wert der Hämolymphe der Muschel aus Becken 3 ist
um 48 mosmol/kg geringer als der aus Becken 2). Dies entspricht jedoch nicht unseren
Erwartungen, denn bei der Miesmuschel handelt es sich um einen Osmokonformer, dessen
Osmolarität der Hämolymphe sich der des Haltungswassers anpasst. Dies bedeutet, dass wenn
der osmotische Wert des Umgebungswassers steigt, dass auch die Osmolarität der
Hämolymphe des Versuchstiers ansteigen sollte. Aus diesem Grund hätte man eine lineare

16
Steigung bei den osmotischen Werten der Hämolymphe der Miesmuschel erwartet. In Abb. 7
ist eine solche Steigung auch gut zwischen dem 1. und dem 2. Wert, sowie dem 3. und dem 4.
Wert zu erkennen. Wahrscheinlich handelt es sich bei dem 3. Wert um einen Ausreißer, was
auch die starken Unterschiede der Osmolarität der Hämolymphe zwischen den beiden
Muscheln aus dem gleichen Becken vermuten lassen. Diese Verschiedenheit kann durch
ungenaues Pipettieren oder zu starke Verunreinigung der Probe bei der Entnahme zum
Beispiel durch Gewebsstückchen erklärt werden, da sich dadurch die Verdünnung und damit
auch die Osmolarität verändern kann. Damit sind auch die zwei stark unterschiedlichen
Werte der Hämolymphe der Muscheln aus Becken 1 zu erklären. Hierbei wirkt sich die
Differenz nicht so stark auf das Diagramm aus wie bei den Werten aus Becken 3, da die
Werte jedes Mal unter denen von Becken 2 liegen.
4.2 Versuch Exkretion Ergebnisse
Die in Tab. 2 enthaltenen Werte und Daten wurden entweder mit Hilfe von Messgeräten oder
über Berechnungen ermittelt.
Das Gesamtvolumen des von einer Versuchsteilnehmerin über einen Tag lang gesammelten
Urins betrug: Vges(Urin) = 1,3 l
17
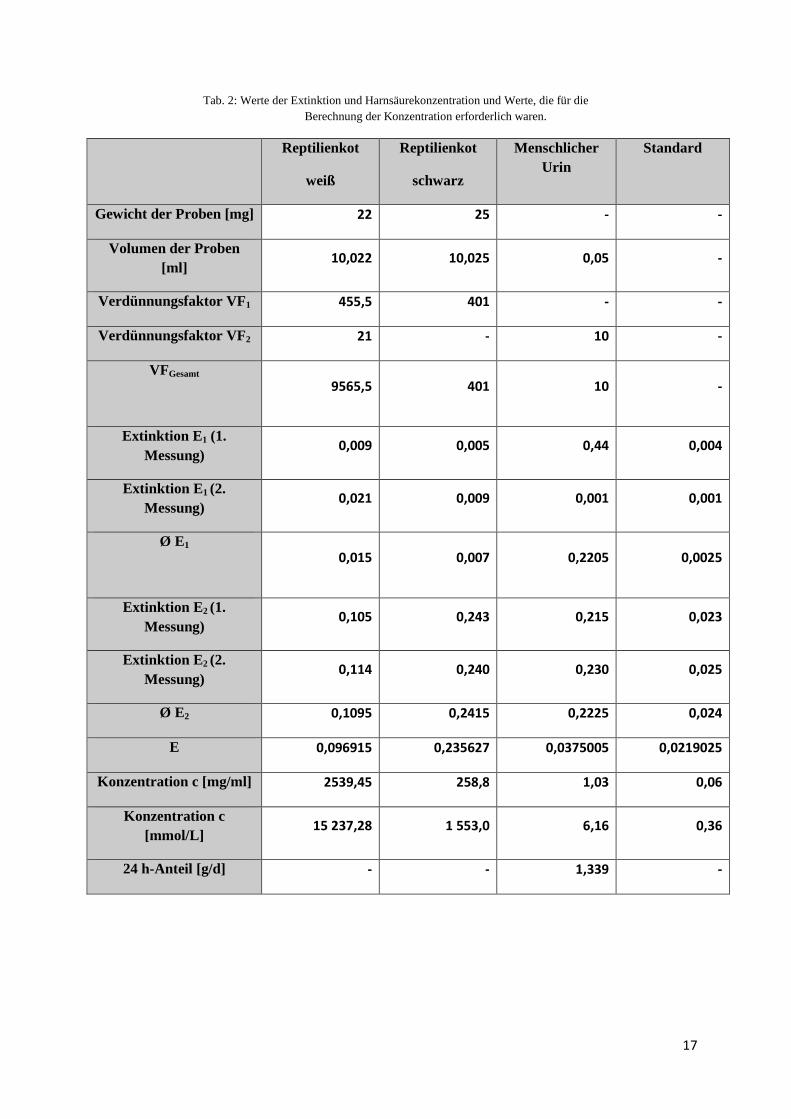
Tab. 2: Werte der Extinktion und Harnsäurekonzentration und Werte, die für die Berechnung der Konzentration erforderlich waren.
Reptilienkot
weiß
Reptilienkot
schwarz
Menschlicher Urin
Standard
Gewicht der Proben [mg] 22 25 - -
Volumen der Proben [ml]
10,022 10,025 0,05 -
Verdünnungsfaktor VF1 455,5 401 - -
Verdünnungsfaktor VF2 21 - 10 -
VFGesamt
9565,5 401 10 -
Extinktion E 1 (1. Messung)
0,009 0,005 0,44 0,004
Extinktion E 1 (2. Messung)
0,021 0,009 0,001 0,001
Ø E1
0,015 0,007 0,2205 0,0025
Extinktion E 2 (1. Messung)
0,105 0,243 0,215 0,023
Extinktion E 2 (2. Messung)
0,114 0,240 0,230 0,025
Ø E2 0,1095 0,2415 0,2225 0,024
E 0,096915 0,235627 0,0375005 0,0219025
Konzentration c [mg/ml] 2539,45 258,8 1,03 0,06
Konzentration c [mmol/L]
15 237,28 1 553,0 6,16 0,36
24 h-Anteil [g/d] - - 1,339 -

18
Auswertung
Der Verdünnungsfaktor VF1 des weißen und schwarzen Reptilienkotes war mit Hilfe der
Formel 1 zu berechnen:
VF1 = (1)
Der Verdünnungsfaktor VF2 wurde im Skript des „Tierphysiologischen Praktikums,
Stoffwechselphysiologie, Versuch Exkretion“ in der Versuchsanleitung angegeben.
Der Gesamtverdünnungsfaktor VFGesamt wurde durch Formel 2 erhalten:
VFGesamt = VF1 * VF2 (2)
Die Durchschnittsextinktion ØE wurde durch die Mittelwertbildung errechnet.
Wollte man nun die Extinktion der Probe E berechnen, musste man mit Formel 3 arbeiten,
wobei es sich bei dem Faktor 0,839 um den Extinktionskoeffizienten ε handelt.
E = ØE2 – (0,839 · ØE1) (3)
Um schließlich die Konzentration der Harnsäure in den Proben zu erhalten, nutzte man
Formel 4:
cProbe = (EProbe ·cStandard ·VFGesamt)/EStandard (4)
Die Konzentration des Standards cStandard in mg/ml wie auch in mmol/l war im bereits oben
genannten Skript enthalten.

19
Der 24 h-Anteil der Harnsäure in g/d im Urin des Menschen wurde mit Hilfe von Formel 5
berechnet:
24 h-Anteil = c [g/l] * Vges(Urin) [l] (5)
Hierbei war zu beachten, dass mit dem Wert der Konzentration der Harnsäure in mg/ml
gerechnet werden konnte, da dies derselbe Wert ist wie in g/l.
Diskussion
In Tab. 2 kann man erkennen, dass die Harnsäurekonzentration im weißen Kot der Boa
constrictor am höchsten ist, ungefähr eine Zehnerpotenz höher als die des schwarzen Kots
dieser Schlange. Die geringste Konzentration der Harnsäure finden wir im menschlichen Urin.
Dies entspricht auch den Erwartungen, da der weiße Kot des Reptils mit dem Urin von
terrestrischen Tieren verglichen werden kann. Reptilien und somit auch die Boa constrictor
sind uricotele Lebewesen, die Stickstoffverbindungen als Harnsäure ausscheiden. Der Kot ist
fest bis breiig, da Harnsäure schwer wasserlöslich ist. Da der weiße Kot also zur Exkretion
von Harnsäure dient ist es klar, dass dieser auch die größte Konzentration daran aufweisen
muss, was auch der Fall ist.
Bei dem schwarze Kot handelt es sich um die Exkremente der Boa constrictor, also die
Ausscheidungen des Darms. Hier sollten eigentlich keine oder nur sehr geringe Mengen von
Harnsäure enthalten sein, da dieser Kot eben nicht der Ausscheidung von Harnsäure dient.
Die doch relativ hohe Konzentration von 258,8 mg/ml (siehe Tab. 2) ist dadurch zu erklären,
dass der weiße und der schwarze Kot nicht getrennt aufbewahrt wurden und dadurch beim
Abwiegen nicht nur schwarzer Kot sondern auch unbemerkt etwas weißer Kot mit in die
Probe des dunklen Exkrementes kam. Ein weiterer Grund dafür ist wahrscheinlich auch, dass
Repitilien eine Kloake besitzen. In dieser Kloake wir sowohl der Darminhalt (schwarzer Kot)
wie auch der Exkretionsstoff (weißer Kot) ausgeschieden, wodurch sich diese beide
Exkremente mit einander leicht vermischen konnten und somit auch der schwarze Kot mit
Harnsäure „verunreinigt“ werden konnte.

20
Aus Tab. 2 kann ebenfalls herausgelesen werden, dass die Konzentration von Harnsäure im
menschlichen Urin sehr gering ist (1,03 mg/ml. Auch dies war zu erwarten, da der Mensch ein
ureoteles Lebewesen ist, Stickstoff also als Harnstoff ausscheidet. Der geringe
Harnsäureanteil ist durch den Purinabbau zu erklären. Die Purinkörper, zum Beispiel Guanin
oder Adenin, sind stickstoffhaltig und können vom Körper nur bis zur Harnsäure abgebaut
werden. Der Grund warum wir Harnsäure auch im menschlichen Urin finden ist, dass der
Körper zelleigene oder mit der Nahrung aufgenommene Purine aus DNA oder RNA oder
auch aus Energieträgern wie GTP und ATP abbaut. Wie aus dem Skript des
Tierphysiologischen Praktikums des Versuches Exkretion zu entnehmen ist, liegt der
Normalwert des 24 h-Anteils der Harnsäure im menschlichen Urin zwischen 0,25 und 0,75
g/d. In Tab.2 kann man jedoch erkennen, dass bei der Versuchsperson der Normalwert
eindeutig überschritten wurde (1,339 g/d). Dies könnte daran liegen, dass diese Person an dem
Tag, als sie den 24 h-Urin sammelte eine erhöhte Menge an Fleisch- und Wurstwaren zu sich
genommen oder eventuell auch viel Alkohol getrunken hatte.
Auch muss man hier bei der Diskussion der Ergebnisse beachten, dass die starken
Schwankungen der Extinktionswerte, vor allem bei den beiden Messungen der
Extinktionswerte E1, zu einer leichten Verfälschung der Werte führen, da hier mit dem
Mittelwert weiter gerechnet wurde. Dies ist besonders bei den E1-Werten des menschlichen
Urins und auch bei den E1-Werten des weißen Kots der Boa constrictor der Fall (siehe Tab.
2). Die unterschiedlichen Extinktionswerte sind durch ungenaues Pipettieren und
Verunreinigungen der Küvetten zu erklären.

21
5. Quellen
− http://www.boa-
constrictors.com/de/Wissenswertes_zur_Boa_constrictor/haltung_der_boas/fuetterung/wi
e_oft_fuettern
− http://www.med4you.at/laborbefunde/lbef_harnsaeure.htm
− Skript des Tierphysiologischen Praktikums, Stoffwechselphysiologie, Versuch:
„Exkretion und Osmoregulation“, der Universität Ulm
− www.wikipedia.org


















