
Experimentelle und Theoretische Untersuchungen Über die Entdifferenzierung und Teilung Pflanzlicher...
36
EXPERIMENTELLE UND THEORETISCHE UNTERSUCHUNGEN OBER DIE ENTDIFFERENZIERUNG UND TEILUNG PFLANZLICHER DAUERZELLEN II. Verschieden gerichtete Permeabilitfitsveriinderungen w~ihrend der Abgliederung kernloser Zellen aus Dauerelementen des Blattes yon ttelodea densa GASP, Von HANS PFEIFFER (Bremen) Mit 12 Figuren Eingegangen am 12. Dezember 1929 Inhaltsiibersicht Einleitende Vorbemerkungen . . . . . . . . . . . . . . . . . 254 I. Absehnitt. DieAbgliederunff kernloserTochterzellen ausDauer- elementen dureh plasmolytisch-deplasmolytisehe Reizung . . . . . . 255 a) Bemerkungen iiber die Eignung der plasmolytisch-de!olasmolytischen Me- rhode zur Induktion yon Zellteilunffen . . . . . . . . . . . . 255 b) Die Beschaffung des Untersuchungsmaterials durch Wiederholung der ]:[ABERLANDT sche~ Versuche . . . . . . . . . . . . . . . . 258 II. Abschnitt. Das Permeabiliti~tsverhalten der entdifferenzierten und der Dauerzellen des Blattrandes und der Spitzhaare yon Helodea densa . . . . . . . . . . . . . . . . . . . . . 263 a) Kritisehe Bespreehung der zur Permeabiliti~tsbestimmung" vorzugsweise geeigneten Methoden . . . . . . . . . . . . . . . . . . 263 b) Oberblick iiber die angestellten Permeabilit~tsversuche . . . . . . 265 Methoclik dot Untersuchungen . , . . . . . . . . . . . . 265 Resultate . . . . . . . . . . . . . . . . . . . . . 267 ILL Absehnitt. Diskussion . . . . . . . . . . . . . . . . . . 272 a) Gesetzmiii~ige Permeabilit~tsverschiebungen bei andern Teilungsprozessen 273 b) Theoretische Untersuehungen fiber den Perraeabilitatsgrad ira IEP des Protoplasmas . . . . . . . . . . . . . . . . . . . . . 277 1. Die Koinzidenz des Permeabiliti~tsminimums mit dem IEP der Plasma- ampholyte . . . . . . . . . . . . . . . . . . . . . 277
-
Upload
hans-pfeiffer -
Category
Documents
-
view
214 -
download
0
Transcript of Experimentelle und Theoretische Untersuchungen Über die Entdifferenzierung und Teilung Pflanzlicher...

EXPERIMENTELLE UND THEORETISCHE UNTERSUCHUNGEN OBER DIE ENTDIFFERENZIERUNG UND TEILUNG PFLANZLICHER
DAUERZELLEN
II. Verschieden gerichtete Permeabilitfitsveriinderungen w~ihrend der Abgliederung kernloser Zellen aus Dauerelementen des Blattes yon
t t e l o d e a d e n s a GASP,
V o n HANS PFEIFFER (Bremen)
Mit 12 Figuren
Eingegangen am 12. Dezember 1929
Inhaltsiibersicht E i n l e i t e n d e V o r b e m e r k u n g e n . . . . . . . . . . . . . . . . . 254
I. Absehnitt. D i e A b g l i e d e r u n f f k e r n l o s e r T o c h t e r z e l l e n a u s D a u e r - e l e m e n t e n dureh plasmolytisch-deplasmolytisehe Reizung . . . . . . 255 a) Bemerkungen iiber die Eignung der plasmolytisch-de!olasmolytischen Me-
rhode zur Induktion yon Zellteilunffen . . . . . . . . . . . . 255 b) Die Beschaffung des Untersuchungsmaterials durch Wiederholung der
]:[ABERLANDT sche~ Versuche . . . . . . . . . . . . . . . . 258 II . Abschnitt. Das P e r m e a b i l i t i ~ t s v e r h a l t e n der e n t d i f f e r e n z i e r t e n
und der D a u e r z e l l e n des Blattrandes und der Spitzhaare yon Helodea densa . . . . . . . . . . . . . . . . . . . . . 263 a) Kritisehe Bespreehung der zur Permeabiliti~tsbestimmung" vorzugsweise
geeigneten Methoden . . . . . . . . . . . . . . . . . . 263 b) Oberblick iiber die angestellten Permeabilit~tsversuche . . . . . . 265
Methoclik dot Untersuchungen . , . . . . . . . . . . . . 265 Resultate . . . . . . . . . . . . . . . . . . . . . 267
ILL Absehnitt. D i s k u s s i o n . . . . . . . . . . . . . . . . . . 272 a) Gesetzmiii~ige Permeabilit~tsverschiebungen bei andern Teilungsprozessen 273 b) Theoretische Untersuehungen fiber den Perraeabilitatsgrad ira IEP des
Protoplasmas . . . . . . . . . . . . . . . . . . . . . 277 1. Die Koinzidenz des Permeabiliti~tsminimums mit dem IEP der Plasma-
ampholyte . . . . . . . . . . . . . . . . . . . . . 277

254 P f e i f f e r
2. Bisherige Versuche der Ermittlung des IEP aus dem Stadium mini- malen P ermeabilitiitsgrades . . . . . . . . . . . . . . . 280
c) Vergleichcnde Auswertung d~r t)ermeabilit~ttsversuche in Hinblick auf die friihere Arbeitshypothese . . . . . . . . . . . . . . . . 281
Z u s a m m e a ~ a s s u n g . . . . . . . . . . . . . . . . . . . . . . 28~ L i t e r a t u r i i b e r s i c h t . . . . . . . . . . . . . . . . . . . . . . 285
Einleitende Vorbemerkungen
Die hier darzustellenden Untersuchungen haben ihren Ausgang genommen yon Beobachtungen HABERLANDTS (1919a, b; 1920), nach welchea eine Teilung yon Dauerzellen durch p l a smo ly t i s chen Reiz hervorgerufen werden kann, wenn die Pflanzen nachher in hypotonische L~sangen iiberfiihrt werden. Allerdings handelt es sich dabei um eine ziemlich ,,primitive" Form der Ze!Iteilungen (H&BERLANDT 1919 a, S. 347; 1919b, S. 732), indem entweder nur Ansi~tze zur Teilung des Karyo- plasmas vorkommen (Coleus Rehneltianus A. BER~., Allium Cepa L.) oder nicht einmal solche mehr festzustellen sind (Helodea densa CAsP~ und canadensis MIG]~.). Well tier Mechanismus der karyoplasmatischen Tel- lung fiir den plasmolytischen Reiz weniger empfindlich ist, kunn in jenen F~llen die Isolierung beider Teilprozesse gelingen (KLEBS 1887, S. 187; HABERLANDT 1919b, S. 732):
Nachdem sich die Deutungsversuche der friiher in dieser Zeitschrift beschriebenen Befunde iiber die Entdifferenzierung phanerogamer Dauer- zellen zu einer Arbeitshypothese (PFEIFFER 19~9e, S. 4~0) verdichtet hatten, mu~te es wertvol! sein, die dort formulierten Ansichten an den allerdings ganz andersartigen E r s c h e i n u n g e n bei der Abg l i ede rung kern lose r Zel len aus k e r n h a l t i g e n D a u e r e l e m e n t e n nachzuprtifen. Daneben aber mufite diese Untersuchung am so eher erfolgen, als sie eine willkommene MSglichkeit zur Durcharbeitung der erst im Entstehen begriffenen Methodik zur E r m i t t l u n g des hypo the t i seh an dem (~eschehen be t e i l i g t en I E P der Protoplasten (PFEIFFER 19~9d) bot, zumal das Untersuchungsmaterial eine besondere Eignung zur Anwendung mehrerer Methoden des Naehweises isoelektrisehen Verhaltens versprach. Von diesen sind zwar auch schon mehrere zur Anwendung gekommen, aber die vorliegenden Ausfiihrungen besch~ftigen sich im wesentliehen mit der im ersten Beitrage (PFEIF:FER 1929e) noeh nieht behandelten Naehweisbarkeit verschiedener Permeabilitht (PFEIFFER 1999 d, S. 1573), zu welcher die Art des Untersuchungsmaterials sieherlieh herausfordert. (~ber die Ergebnisse mit andern Methoden wird spi~ter zu beriehten sein.

Untersuchungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. I I 255
Wir werden also im einftihrenden Abschnitte vor allem die A n- z u c h t yon U n t e r s u c h n n g s m a t e r i a l , das nebeneinander Dauerzellen, entdifferenzierte und bereits geteilte Elemente zeigt, zu schildern haben. Der naehfolgende Tell wird alsdann haupts~tchlich yon den Pe rme- a b i l i t / i t s v e r s u c h e n handeln, aber auch den Stand der anwendbaren M e t h o d i k kritisch zu untersuchen haben. Endlich wird tier letzte Ab- schnitt mit einer Dis ku s s i o n d e r Befunde unter besonderer Bezugnahme auf die aufgestellte Arbeitshypothese schlieBen.
I. A b s c h n i t t
Die Abgliederung kernloser Tochterzellen aus Dauerelementen durch plasmolytisch-deplasmolytische Reizung
a) Bemerkungen fiber die Eignung der plasmolytisch-deplasmolytischen. Methode zur Induktion yon Zellteilungen
Ist auch bekannt, wie die Kernteilung in manchen F~illen durch Plasmolyse aufgehalten werden kann (N]~F~C 1910, S. 266, 270 u. a.), wie tiberhaupt der Eingriff oft oder fast immer die R e s t i t u t i o n des p h y s i o l o g i s c h e n s t a t u s quo a n t e a u s s c h l i e B t (K~s~]~R 1929, S. 9), so ist doch schon mehrfach die Brauchbarkeit der plasmolytisch-deplas- molytisehen Methode zur Abgliederung yon Zellen erkannt worden. Da die yon HABERLAI~DT (1919a, S. 323) tiber die Vorg/inge an plasmoly- sierten Protoplasten mitgeteilten Befuude aus der Literatur in Hinblick auf friihere Erfahrungen speziell bet der p l a s m o l y t i s c h e n I n d u k t i o n yon Z e l l t e i l u n g e n ergiinzt werden k@nen, m6gen einige kurze Hin- weise hier am Platze seth.
Schon ]~LEBS (1887, S. 185) hatte bei Zygnema DE :BARY und Oedogonium LINK in einer Kultur in Zucker-Kongorot eine Teilung yon Zellcn ohne Beteiligung des Kernes und eine fast noch lebhaftere Teilungst~tigkeit in 10% RohrzuckerlSsung bei Euastrum verrucosum EHRBG. konstatiert. Auch in der ausfiihrlicheren Schilderung (1888, S. 550) beschreibt er~ wie er unter Verwendung yon ZuckerlSsungen Teilungen an Mesocarpus HASS., S2irogyra LINK, Cladophora Kt~wz. u. a. ausgel(ist hat. Er stellt bet Oedogonium LINK eine vereinfacht verlaufende Teihng nach vorangegangener Flasmolyse lest und beschreibt das zentrifuffale Vordringen einer einfachen Querwand yon der Peripherie her. Dal3 er auf die gleiche Weise bei Helodea L. C. RICH. nicht zum Ziele gekommen ist, diirfte leicht damit zu erkl~ren sein, alas die dabei verwendete Konzentration des Plas- molytikums zu hoch gewesen ist. Das kann aus seinen eigenen Befunden (1887) an Zygnema-Zellen gefolgert werden, nach welehen mit entspreehender Abnahme des osmoti- schen Druckes des Plasmolytikums anfang's Teilungen ausbleiben (nur Zellhautbildung

~956 Pfe i~fe r
bei 30% Rohrzueker und Lgngenwaehstum bei 25 his 30~ dann erst sp{*rlieh (16%) and schliel~lieh lebhaft (10~ anftreten. 8pgter beschreibt KLEBS (1896, S. 288) noch- reals, wie bei Oedogonium LINK n~eh Plasmolyse uud Regeneration der Plasmahaut eine Teilnng uncl Sehw~rmsporenproduktion hervorgerafen wird. Aueh RE~ARDr (1899) findet an plasmolysierten Oogonien yon 8aj?rotegnia-Fgden eine besonders reiehliehe 0osporenproduktion, und W. ZIMMERMANN (1923, 8. 152) ist es gehngen, Zdlen von S2haeelaria fusea AG. durch Operation im plasmolysierten Zustande trotz ungeteilten Zentriols zur Restitution zu bringen. In derselben Richtnng liegen Befnnde yon PRs (1925), naeh denen in 15 his 20~ SaceharosdSstmg plasrnolysierte Zellert yon Stypo-
Fig. 1. Balantium antarcticum: A normales Prothallium in K~oe-L5sung; B das- selbe naeh vorangegangener Pl~smolyse und jetzt zahlreiehe Adventivsprosse zeigend, 7~/~.
(Nach ISAB~gRO-NAGAI 191~.)
eaulon KOTZ. trotz starker Plasmakontraktionen Kernteilungen erfahren kiSnnen. Dureh SI~mATA (1902) ist bei Monotro3aa u,J,iflora L. nach Vorbehandlung mit 0,3-his 0,b-reel. Traubenzuckerl6sungen der Prozentsatz der unbeDuehtet sieh teilenden Endospermzellen verdoppelt worden. Vermutlieh ist der Befund yon (~IgOSSMANN (1921), dab Glukose- zusatz zur Nghrl~sung die Zellvermohrung und Koloniebildung bei Seenedesmaceen zu f/Srdern vermag, 5hnlieh zu erklgren~ wenn wir uns seine Deutung nicht ungesehen zu eigen machen wollen. Dafiir spricht die Beobaehtung yon ISABURO-NAGM (1914, S. 306, 327), welcher an den untersuehten Prothallien gewisser krten yon Alsophila R. BR.: Balantium KAUL~., Asplenium L., Aspidium Sw., Athyrium ROTH, Ceratopteris BI~ONGN. and Adiantum TouR~. dutch Piasmolys~ und Oberfiihrung in verschied~n konzentrierte

Untersuehungen iibar die Entdifferanziarung u. Teilung pflanzlieher Dauerzellen. I I 257
KNopsehe Lbsungen als eine sehr allgemeine Erseheinung die Adventivsprol~bildung nach vorangegangenen Teilungen und Ausstiilpungen yon Zellen koustatiert hat (Fig. 1). Die Art des Plasmolytikums ist dabei gleichgli]tig (a. a. 0., S. 310). In diesem Zusammen- hange miissen wir aueh an Ergebnisse dureh ~ORTHEIM und L0WI (1909) erinnern, welehe bei Knltur der Pollen yon Philadel2hus L., Amaryllis L. und Tuli~a L. in Rohrzueker- 15sungen geeignater Konzentration Sehl~tuche erhalten haben, in denen ein Tail des Lumens dutch Zellulosamembranen oder gar dureh eehte Querw~nde abgesehlossen ga- wesen sin& Schlieltlich mag auf die Anregung yon Vakuolenteilungen mittels Plasmolyse an den Epidermiszellan der Zwiebelsehuppen yon Allium Cel~v L. (KUSTEI~ 1918, S. 284) hingewiesen warden, wenn wir yon ~hnlich bedingten Teilung'en yon Chromatophoren (E. FJ[EITZ) absehen wollen. Wenn Mr dafiir noehmals auf die Erfolge yon HABERLANDT an ttaarzellen yon Coleus LOinS., Epidermiszellen yon Allium L. (1919a, S. 329, 337) und Blattelementen yon Helodea-Arten (1919b, S. 725) hinweisen, so finden wir wie ]:[ABERLANDT (1919a~ S. 327) naeh dem aug'enbliek]iehen Stande unseres Wissens im ganzen die systamatiseh hbheren Pflanzen (Anthophyten) weniger geeignet zur plasmo- lytischen Induktion yon Zellteilungen ~). Damit wird aueh verst~ndlich, dal~ eigena Be- miihungen, in Vallisneria st~iralis L., MyrioNhyllum L. oder Ceratophyllum L. neue geeignete Untersuehungsobjekte zu finden, bisher keinen Erfolg gehabt haben.
Trotz der ergebnisreiehen Befnnde plasinolytischer Teilungsinduktion mindestens bei systematiseh einfaeher organisierten Pflanzen wollen wir uns abet der S c had l i c h- ke i t des a n g e w e n d e t e n e x p e r i m e n t e l l e n E i n g r i f f e s bewnl~t bleiben, aueh wenn wir dahin geh~rige Stellen der Literatur hier nieht besonders anfiihren. Vor allem kSnnen wir fiir ungeeignete (zu hohe) Konzentrationen des Plasmolytikums die Beob- aehtungen yon KARZEL (1926, S. 581) an dem gleichen Objekte in jeder ttinsicht be- st~tigen (vgl. t:[ABERLANDT 1919a, S. 345; 1920, S. 336).
Es sell bier ferner nieht ganz varschwiegen werden, wie aueh du rch andere e x p e r i m e n t e l l e E i n g r i f f e die A b g l i e d e r u n g k e r n l o s e r Ze l l en g e g l i i c k t ~st. Am bekanntasten sind hier yon einigen Conjugaten und Eiern der Amblystoma Ver- suehe, mit starker Abkiihlung odar tier Einwirkung yon Anaesthetika (J. J. GERASSIMOFF, A. NATHANSOHN~ T. S. ~PAINTER U. a.) ans Ziel zu gelanffen, sowie die Versuche des Zentrifugierens yon Algenf~den (C. VAN WISSELINGH~ C. HOFFMANN) oder solche trauma- tischer Eingriffe an zahlreiehen Eiern (V. JOLLOS, T. 1)/~TERFI: J. SEILER, ~y. FANKHAUSER usw.). Noeh weir st~rkere Unrege]m~l~igkeiten bei der Abgliederung kernloser Zellen an Wurzelspitzen (Vieia Faba L., Allium Cel)a L.) hat sehon B. N/~MEC (1910, S. 11) dureh Behandlung mit Chloralhydrat erzMt. Itierher wiirde auch die yon A. HE1LBRONN dutch Narkotisierung" induzierte Advantivsprogproduktion an Prothallien yon As~vlenium L. geh6ren, die allardings durch ISABURo-NAGAI (1914, S. 305) nieht wieder hervorgerufen worden ist. t~Tbrigens ist eine Absehniirung kernloser Cytoplasmateile aneh bei der als Abfurehung bekannten multiplen Cytoplasmaaufteilung beispielsweise bet Theeamoeben (K. Bf~LA~) und diversen Insekteneiern (3. SEILER), sowia bei der Spermatoeytenteilung yon Algiarien (F. 1V[EYES) bekannt geworden.
1) Im Einklanga mit dieser Erfahrung- steht, dag Fraulein 3{. THIELMANN in ZuekerlSsungen nie Teilung, sondern hgehstens ain Auswaehsen der Epidermis- und Sehliel~zellen beobaehtat hat; vgl. aueh KOSTER (1925, S. 436).
Protoplasma, X 17
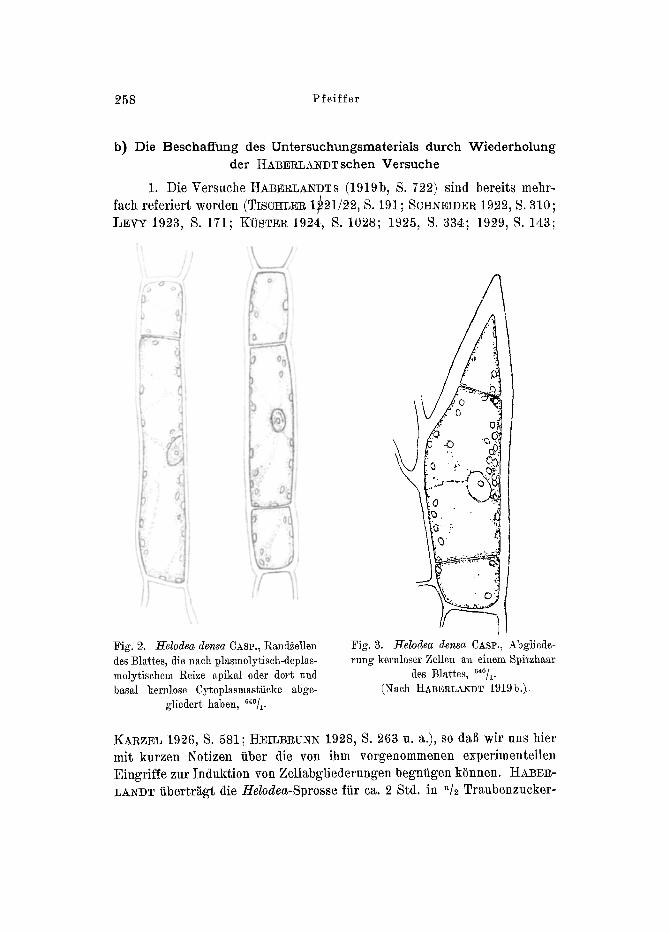
258 Pfeiffer
b) Die Beschaffung des Untersuchungsmaterials dutch Wiederholung der ~ABEI~LAlgDTschen Versuche
1. Die Versuche HABERLANDTS (1919b, S. 722) sind bereits mehr- fach referiert worden (TIscltLER 1~21/22, S. 191; SCHNEIDER 1922, S. 310; LEVY 1923, S. 171; KOSTER 1924, S. 1028; 1925, S. 334; 19297 S. 143;
Fig. 2. Helodea densa CASP., Rand~ellen des Blattes, die nach plasmolytiseh-deplas- molytischem l~eize apikal oder dort und basal kernlose Cytoplasmastiicke abge-
gliedert haben, 6~~
Fig. 3. Helodea densa CASe., Abgliede- rung kernloser Zellen an einem Spitzhaar
des Blattes, 6~~ (Nach HABERLANDT 1919b.).
KARZEI~ 1926, S. 581; HEILBRUNN 1928, S. 263 u. a.), so dal] wir uns hier mit kurzen Notizen fiber die yon ibm vorgenommenen experimenteilen Eingriffe zur Induktion yon ZeUabgliederungen begntigen kSnnen. H ~ E R - LANDT tibertragt die Helodea-Sprosse fiir ca. 2 Std. in "/2 Traubenzucker-

Untersuchung'en fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. I I 259
15sungen und tiberfiihrt dann far 2 Tage in KNoPsche LSsung~) und/oder in Leitungswasser. Nach dieser Frist stellt er in Randzellen (Fig. 2) und besonders in Haarelementen (Fig. 3) die Abgliederung kernlos gebliebener Abschnitte lest. Wegen der dabei leicht eintretenden Uberwucherung mit Algenarten, Diatomeen, Cyanophyceen und Bakterien ist die alleinige Verwendung yon K~oP-LSsung unzweekmi~l~ig. Die zwisehen Mai und Juli bei 18 bis '22 o C durehgefiihrten Versuche ergeben fiir die einzelnen Elemente eine verschiedene Eignung; vo rzuz i ehen sind die e inze l l i gen Blattz~thne. Die dem Blattrande benaehbarten Zellen lassen nicht so oft Teilungen beobaehten, und gegen die Mittelrippe bin nimmt die Er- scheinung weiter ab. Geeignet sind ferner nur kr~tftig vegetierende Sprosse und besonders gut die jtingeren, dem Vegetationspunkt gen~herten Bl~ttter.
2. Die e igene G e s t a l t u n g der Method ik hatte nicht nur die yon I-IABERLANDT gemachten Erfahrungen anzuwenden, sondern auch dartiber hinaus andere Wege zu suchen. Zur Ersparung yon Material ist zuerst versucht worden, an abgeschnittenen Bl~ttern die gleiche Er- scheinung hervorzurufen. Bleibt auch far die wenigen Stunden der plasmolytischen Reizung das Plasmolytikum vor dem Verdunsten bewahrt, so bedarf es doch fiir die mehrt/igige Aufbewahrung der Pri~parate nach der Deplasmolyse besonderer Vorrichtungen zur Vermeidung st~rkerer Verdunstung des Mediums der Bli~tter unter dem Deekglas. Leicht ]~t~t sich das durch Aufbewahrung der Pr~tparate in flaehen, bedeckten (PET•I-) Sehalen, die noch mit befeuchtetem Fliel~papier ausgelegt worden sind, erreiehen, wenn man ferner durch Unterlegen schmaler Korkstreifen oder auf andere Weise die Objekttri~ger hohl hinlegt. Der Vergleich ab- gelSster und am Sprol~ verb]iebener BlOtter ergibt auch unter solchen Mal~nahmen eine betr~ichtliche ~ b e r l e g e n h e i t tier B e h a n d l u n g yon i n t a k t e n Sp ros sen , wenngleich das andere Verfahren keineswegs vSllig unbrauchbar ist. Ftir die sp~tter zu erw~hnenden Permeabilit~tsversuche (S. 265f.) hat abet die geschilderte A u f b e w a h r u n g der B l a t t p r / i p a r a t e wertvolle Hilfe geleistet.
Sodann ist versucht worden, die ftir den gewtinsehten Er[olg opt i - male K o n z e n t r a t i o n des P l a s m o l y t i k u m s (Traubenzueker) heraus- zufinden. Rechnen wir die Zahl der geteilten, ungeteilt gebliebenen and abgestorbenen Haarzellen eines Blattes in Prozent der (zwischen 22 und 33 sehwankenden) Gesamtzahl urn, so erkennen wir am Durchschnitt
1) Sie hat bei ihm in 1 l Wasser enthalten: i g KNOB: je 1/2 g CaSQ und C%(P04)2~ 0,1 g MgS04 und e~ne Spur Eisenchlorid.
17"
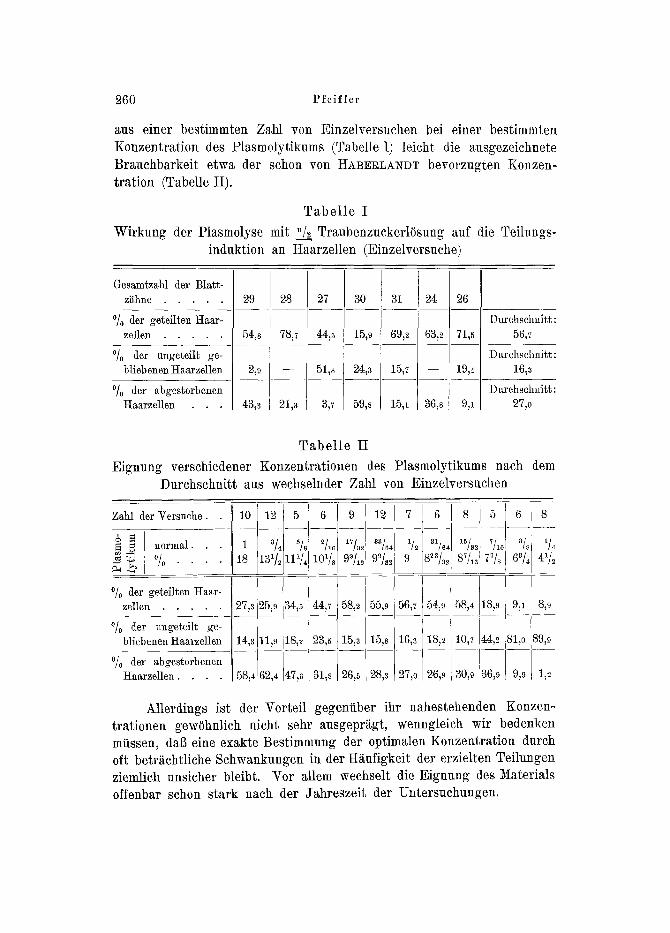
260 Pfeiffer
aus einer bestimmten Zahl yon Einzelversuchen bei einer bestimmten Konzentration des Plasmolytikums (Tabelle I) leicht die ausgezeichnete Brauchbarkeit etwa der schon yon I:IABERLANDT bevorzugten Konzen- tration (Tabelle II).
Tabe l le I
Wirkung der Plasmolyse mit "/--a'-' TraubenzuckerlOsung auf die Teilungs- induktion an Haarzellen (Einzelversuche)
C~esamtzahl der Blatt- zi~hne . . . . .
~ der geteilten ttaar- zellen . . . . .
~ tier ungeteilt ge- bliebenen gaarzellen
~ der abgestorbenen Haarzellen
29
54~s
2,9
43,3
28
_78,~
21,3
2 7 30 _ 31
4 4 , 5 ] 15,~_!_ 69,2
24
63,2
36,s
26
Durchschnitt : 71,5 56,7
Durchschnitt: 19,4 16,s
- " Durchschnitt : 9~11 27,0
Tabel le II
Eignung verschiedener Konzentrationen des Plasmolytikums nach dem Durchschnitt aus wechselnder Zah] von Einzelversuchen
Zahl derVersuche. . 10 12 5 6 9 12 7 // 6 8 5 ' 6
= normal. . .
~ der geteilten gaar- z~]~en . . . . .
~ der ungeteilt .~e- bliebenen Itaarzellen
~ 4er abgestorbenen Haarzellen.
27,3 25,9 34,5
58,4,62,~ ]47~
44 '2
23,5
31,s
58~2 55,9 56,7 54,9
15,3 15,s 16,3 18,2
26~5 28,~ /27,o 26,9
58,4 ~1S~9 9,1
30,9136,9 9 , 9 - - -
8
1/4 41/2
8,9
$9,9
1,2
Allerdings ist der Vorteil gegeniiber ihr nahestehenden Konzen- trationen gewiihnlich nicht sehr ausgepr~gt, wenngleich wir bedenken miissen, daft eine exakte Bestimmung der optimalen Konzentration durch oft betr~tchtliche Schwankungen in der H~ufigkeit der erzielten Teilungen ziemlich unsicher bleibt. Vor allem wechselt die Eignung des Materials offenbar schon stark nach der Jahreszeit tier Untersachungen.

Untersuchungen itber die Entdifferenzierung u. Teilung pi]anzlicher Dauerzellen. II 261
Die Tem p e r a t u r im Versuchsraume ist nicht weiter variiert worden. Doch hat sich gezeigt, dal~ auch niedrigere Temperaturen, als sie yon HA]3ERLANDT angewandt wurden (18--22 o C), durchaus nicht ungeeignet sind, solange sie sich in ertr/~gliehen Grenzen halten. Ebensowenig k0nnen wir eine Modifikation der Versuehe in ttinblick auf die E in - w i r k u n g s z e i t des Plasmolytikums (zwischen 1 und 3 Std.) empfehlen. Durch zu lange Beeinflussung wachst die Zahl der bei dem Eingriff abget5teten unter entsprechender Abnahme der sieh teilenden Zellen, durch zu kurze Einwirkungsdauer nimmt die g~afigkeit der Teilungs- indnktionen wesentlieh ab, w~e denn in vielen F~llen bei zu schwachem Material schon bei einsttindiger Behandlung die erforderliehe Reizschwelle nicht mehr erreieht wird.
Versnche mit a n d e r n P l a s m o ! y t i k a sind nieht welter verfolgt worden, da es ftir die LSsung der gestellten Frage ausreiehend ist, ein mSglichst reiehes Material ftir die n0tigen Permeabilit~tsversuche heran- zuziehen. Von einer Erweiterung der Versuche in dieser Richtung ist vor allem aueh deswegen abgesehen worden, weil, wie tibrigens aueh aus der Literatur iiber die Wirkung versehiedener Plasmolytika (KH2P04, KN03, CaC12 usw.) erkl~rlieh wird, die gebr~uchliehen Salze eine weit gr0t~ere Sch/idigung tier Objekte als die Traubenzuckerl6sungen hervor- rufen, so da6 zwar ebenfalls eine kleine Zahl yon Teitungen an Haar- zellen erzielt wird, tier Prozentsatz der dabei absterbenden Elemente aber rasch in die I-I0he sehnellt.
3. D e s k r i p t i v e A n a t o m i e des Objek tes . - - Seitlich der Mittel- rippe besteht das Blatt tier Helodea densa CASP. aus nur zwei Schichten assimilierender Parenehymzellen, die in tier oberen Lage besonders lang und breit sind. Gegen den Rand hin linden wir als Saum e ine e i nz i ge Schicht yon Zellen, die in der L/~nge etwa die Mitte halten zwisehen den Elementen der oberen und der unteren Lage der iibrigen Lamina, aber schmaler als beide gebaut sind. Mechanisches Gewebe fehlt den Bl~ttern der Helodea densa and wird hOehstens dutch etwas st~rkere Verdickung der Au•enwand ersetzt. Die spitzenw~rts gerichteten, ein- zelligen, dickwandigen und kurzspitzigen B l a t t z ~ h n e sind dutch Aus- waehsen yon Randze]len entstanden und grenzen im Ful]sti~ck, in welchem fast st~ndig der Kern lokalisiert ist, schr/~g oder mehr/minder senkrecht gegen die benachbarten Randzellen.
Bei Behandlung mit 9 % Traubenzuckerl0sung tritt in wenigen Minuten eine P l a s m o l y s e in den Blattz~hnen, den Randzellen und den benachbarten Parenehymelementen ein. Nach 1 bis 3 Std. sind lokale

262 Pfeiffer
Abl6sungen des Protoplasmas ziemlich zur[lckgegangen, die Protoplasten selbst aber starker kontrahiert und abgerundet und fast stets ungeteilt. ttSchstens in den Randzellen wird gelegentlich eine (nicht immer voll- kommene) Aufteilung des Cytoplasmas in meistens zwei nngleiche Teil-
stticke beobachtet. Das Chlorophyll hat sich systro- phisch um den Kern ge- ballt (Fig. 4).
Beim fdbertragen in KNoP-L(isung oder in das ursprtingliche Medium tritt bald eine Deplasmolyse ein, wobei die voriibergehend sistierte S t r ( imungsbe - w e g u n g erneut einsetzt. Etwa zuvor abgelSste Teile des Protoplasten werden wieder aufgenommen oder
verfallen der Nekrose (Fig. 5). Nun aber wird der vakuol~re Anteil der Zelle schon bald von zarten cyto- p l a s m a t i s c h e n Fi~den durchzogen, ohne da6 diese fiir die weitere Dauer ihre Lage beibehalten. Die weitere E,twicklung l~6t eine weitreichende t~ber-
Fig. "4. Helodea densa Fig. 5. Helodea densa einstimmung mit der Tei- CAsp.,inderHaarzelledes CASP., Blattzahn, die lung naeh dem Oedogonium- Blattes eine !systrophen- Nekrose tier bei Deplas- Typus (STASBURGER 1875; artigeBallungdesChloro- molyse nicht mehr ~uf- 1880; 1893, S. 109; vg]. phylls um den~Kern in- genommenen terminalen folgePlasmolyse mit 9% Portion des Cytoplasten aueh TISCHLEI~ 1921/22,
Traubenzucker, 64~ zeigend, 640/, S. 191) erkennen. In dem s p u m o i d e n P l a s m a -
faden, tier sich durch die Vakuole spannt, erfolgt die C e l l u l o s e p r o d u k - t ion sukzedan oder simultan, also unter zentripetalem Fortschreiten vom Rande nach tier Mitte zu oder unter beschleunigter Ausscheidung an der gesamten Randlinie. HABERL~NDT (1919b, S. 725) findet tibrigens

Untersuehungen fiber die Entdifferenzierung u. Teihng pflanzlieher Dauerzellen. II '263
anfangs stets sukzedane Anlage der jugendlichen Zellwand ~hnlich dem Cladophora-Typus (TISCHLER, a. a. 0., S. 184), doch m(igen diese Ver- h~ltnisse auch etwas variieren. ViM seltener, als nach HABERLANDTS Schilderung (S. 7'25) zu erwarten w~re, finder sich dabei eine unvoll- kommene, ,,fraktionierte Querwandbildung" (K~STER 1915, S. 793). Das Material nach ein- bis dreiw6chentlicher Riickiiberftihrung zeigt in der Ausbildung der Q u e r w a n d oft verschiedene Entwieklungsstadien, die zwisehen einer schmalen Ringleiste und nach der Dicke relativ aus- gewachsenen Querw~nden wechseln, nebeneinander (Fig. 6). Ftir die
Fig. 6 . gelodea densa CASP., Partien aus der Wandanlage in Blattz~hnen zufolge naehtrttglicher Plasmolyse in 50 ~ Glyzerin, tells nur eine einzige 0ffnung" Init ver- bindender Plasmabriicke, teils eine grSl~ere Zahl yon Verbindungsbahnen zeigend, 1~s~
(Nach ~:~ABERLANDT 1919b.)
Untersnchung der Wandverhiiltnisse in allen diesen Stadien ist eine er- n e u t e P l a s m o l y s e mit G l y z e r i n sehr anzuraten. Die ausfiihrliche Schilderung HABER~AlVDTS (a. a. 0.) eriibrigt aber hier eine genauere Beschreibung der Befunde.
II. A b s c h n i t t
Das Permeabilitiitsverhalten der entdifferenzierten und der Dauerzellen des Blattrandes und der Spitzhaare von Helodea densa
a) Kritische Besprechung der zur Permeabilittttsbestimmung vorzugsweise geeigneten Methoden
Die recht z a h l r e i c h e n V e r f a h r e n zur Messung des Permeabi]it/~ts- grades yon Protoplasten sind mehrfach zusammengestellt worden, so vor allem yon GRAFE (1924), H0]3ER (1994, S. 406 u. a.), IIJIN 0998, S. 558), REMINGTON (1998, S. 346) und GELI~HOI~N (19~9, S. 4). Ebenso wie die in dieser Zeitsehrift erschienene Arbeit yon REMINGTON bevorzugen wir hier die Einteilung in op ti s che (Farbstoffnaehweis oder Tanninf/~llungen), c h e m i s c h e (analytische Ionenbestimmungen), o s m o t i s e h e (n/imlich

264 P f e i f f e r
mit te ls P lasmolyse) und e l e k t r i s c h e Ver fahren ( B e s t i m m u n g tier e lek-
t r i schen Leitf / ihigkeit) , won denen e t w a der chemisch-ana ly t i sche Nach-
weis a u f g e n o m m e n e r oder a b g e g e b e n e r Ionen (S. 281) oder die opt ische
A u f s u c h a n g e in t r e t ende r und im P r o t o p l a s m a r e a g i e r e n d e r Subs t anzen
oder der das I n h e r e f~rbenden Stoffe als d i r e k t e Methoden den andern
( indirekten) gegen i iberges te l l t werden mSgen. So re ich die Auswah l ist,
so ze igen doch si~mtliche Methoden gewisse Nachtei le , so daft die H e r a n -
z iehung v~rsch iedener yon ihnen ftir die B e w e i s k r a f t der aus den Re-
su l t a t en gefo lger ten Schliisse e r forder l ich wird (GELLHORN 1929, S. 18, 21). Uber die W e r t u n g der e inzelnen Ver fah ren ftir die v e r l a n g t e n Unte r -
suchungsz ie le bes t eh t Ie ider ke ine Einigkei t , so daft dazu einige kr i t i sche
B e m e r k u n g e n hier am P la t ze sind.
Keine der Methoden is t im s t r engen Sinne vol lkommen~ weft sic tells nur bis zu einem gewissen Grade Gin quantitatives Arloeiten ermSglichen, teils keine einwandfreie Abgrenzung gegentiber Adsorption an iiu~ere Zellbestandteile oder gegen- iiber ehemischer oder auderer Nebenwirktmg erlauben. Die meisten indirekten Methodeu berticksiehtigen nut unzureichend die sekundiiren Wirkungen tier Medien auf die in den Zelien aaftretenden Stoffwechselprozesse. Insbesondere gegen 4ie osmot i sehen Ver- f ah ren (W. LEPESCtIKIN, A. TRONDLE, H. FITTING, K. lZfOFLEI~ u. v. a.) ist geltend zu maGhen, alas die Permeabilitat yon Wasser und gelSsten StoffGn nur aus bestimmten Re- aktionen erschlossen wird unter der gleiGhzeitigen Voraussetzung, dag chemische Prozesse, welehe alas Resultat durch Eintritt einer A n a t o n o s e ver~ndern kSnnten, nieht statt- finden. Dabei ist durGh W. S. IL~N (1923; 1928, S. 569) bereits dig MSglichkeit einer derart bewirkten FGhlmessung exakt erwiesen wordGn (vgl. auch: KACZMAREK 1929, S. 255). Er hi~lt daher dig optischen und chemischen ~Iethoden fiir zuverl~tssiger (ILJIN 1928, S. 559). Ferner ist zu bedenken, da6 die plasmolytischen Hethoden mit eiuem hypertonisehen Milieu arbeiten, so daft die Resultate mit ihnen nicht mit unbedingter Sieherheit zu einem Sehlu$ auf gleiches Verhalten unter physiologisehen Bedingungen geeignet zu sein brauehen. Gegen die op t i sehen Ver fah ren (W. PFEFSE~, E. OVEI~- TON~ A. ~ETHE: C. JACOBY: W. RUHLAND: W. STILES~ K. WEIDLING: 1~. COLLANDER: 1~. SCHAEDE usw.) mit Farbstoffen oder mit LSsungen~ welche im Protoplasma einen sicht- baren Niederschlag erkennen lassen, ist eingewendet worden~ daft die an tier Fi~rbungs- i~ltGnsit~tt oder an tier Niedersehlagsquantiti~t gemessene Eintrittsg-esehwindigkeit fremder Substanzen als Gradmesser ungeeignet ist, weil die Rachweisbarkeit der verwendeten kSrperfremden Substauzen yon tier Speieherungsfi~higkeit des Protoplasmas fiir sic weit- gehend abhiingig ist. Ein quantitatives Studium ist auSerdem recht erschwert, well die sichtbar gemachten Niederschl@e und Farbstoffspeicherungen h~ufig nur den Wert quali- tativer Reaktionen besitzen (vgl. aber COLLANDER 1921). Durch die ehemisehen ) ie- thoden (J. ~. JANSE, F. VAN RYSSELBEI~GIIE: F. URANO, G. FAHR, W. J. V. OSTERHOUT, J'. F. ]~c CLENDON, G. EMBDEN, ]~. ADLER, HERMANN LANGE~ B. W. ~J[?JLLER, Ilia. SIEBECK und zahlreiche anderG) wird versucht, ob einder Au~enl~isung zugesetzter Stoff infolffe seiner Diffusion im Innern des Protoplasmas naGhweisbar wird und durch Fortlassen des Stoffes im Medium wieder daraus versehwindet. 8tatt tier Zelle selbst kaan nat~irlieh anGh das ]~ediura untersnGht werden. Dabei ist zwar direkt zwisGhen Impermeabilitiit,

Untersuchungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 265
geringfiigiger Permeabilitiit und Anatonose zu unterseheiden, aber eine Sehwierigkeit erw~chst beispielsweise aus der Mbglichkeit einer Adsorption an der zelluliiren Aui~en- fl~tehe, die hier ebenso wie bei Fa.rbstoffversuchen durch Untersuchung eventueller Gfiltig- keit der Adsorptions-Isotherme ausgeschlossen werden mu$ (H. i~ORAWITZ~ ~. O. ]~[ERZOG, E. ABDERHALDEN, J~. WIECHMANN n.a.). Die e lekt r i schen Best immungen des Ein- trittes yon Substanzen durch Ermittlung yon Leitfi~higkeitsiinderungen (W. ROTH, J. BU~ARSKY, ~R. TANGL, G. 1N T. STEWAR% R. HOBER, A. JOEL, M. GILDEMEISTER~ U. EBBECKE, ~H. BROEMSER, W. J. V. OSTERHOUT, S. C. BROOKS USW.) sch~ieJ]en gewisse Fehlerquellen aller indirekten Vel"fahren, insbesondere die ]~Sglickeit eingetretener Ana- tonose, nicht aus. u allem ist gegen sie angefiihrt worden, dag das ver~tnderte Ver- halten teilweise nieht durch jenes der Zellgrenzsehieht, sondern dutch Auftreten freier Ionen ira Protoplasma oder in tier Zellwand bedingt werden kann. Neben al~en diesen Ver- fahren liegen auch noch solehe fiber die Bestimmung des Gefr ie rpunktes (S. G. I~EOIN), sowie zahlreiche p h y s i o 1 o g i s e h b e g r fin d e t e (0. NASSE, K. I~IROKAWA, Y, HENRI, S. LALOU, V. ~FRANZ, C. SCHWARZ, K. SIEBECK, O. WARBURG ll. v. a.) vor, die im all- gemeinen nur einen bescbri~nkten Anwendungsbereich besitzen and beispielsweise etwa die Bestimmung des diastolisehen HerzstilIstandes (J. LOEB), der Befruehtungsziffern (E. G~LL)t01~), der verminderten Huskelkontraktar (H. VOOEL, E, B. MEreS) oder andere Erseheinunffen betreffen.
Indem die apparative Einriehtunff zu Leitf~,higkeitsbestimmungen 1eider nicht zur Verfiigung steht, mul~ ein derartiges Untersuchungsverfahren hier ausseheiden. Nach den hier angestellten Uberlegungen kommen damit neben dem chemisehen 7Nachweis (S. 281) hauptsiiehlich nur die optisehen ~ethoden in Betracht, aus denen eine m@lichst giinstia'e Auswahl zu treffen war.
b) Uberbl ick fiber die angeste l l ten Permeabi l i t t i t sversuche
M e t h o d i k d e r U n t e r s u c h u n g e n
1. FUr die F a r b s t o f f v e r s u c h e sind vielfach P r@ara t e verschie- dener Provenienz (bei Neutra l rot z. B. : Neutr. Indika tor GR?3]3Y~ER, l~eutr.
Iudikator KAHLBAUM, Neutr. MERCK, Neutr. extra KAttLBAUM) ver-
wendet worden. Die ersten Anf~rbungen s i n d ' a n den ganzen Sprossen
vorgenommen worden, die zuvor in der hier (S. 259) schon beschriebenen
Weise (lurch plasmolyt isch-deplasmolyt isehe Reize zur Indukt ion yon Zellabgliederungen an den bezeichneten Stellen des Blat tes gezwungen
worden waren. Auf solehe Weise erh~lt man, wie im ersten Haapt te i l gezeigt worden ist, mindestens an dan meisten Bliittern neben unver-
/inderten D a u e r z e l l e n , fiir welche j ener Reiz untersehwell ig geblieben ist, auch eiae wechselnde Zahl geeigneter E n t d i f f e r e n z i e r u n g s - u n d
T e i l u n g s s t a d i e n zum Vergleiche ihres Permeabilit/~tsverhaltens. Zur
Untersuchung gelangen abgeschni t tene B1/itter, welche bei sorgf/iltiger Behandlung and Attfbewahrung in den mit befeachtetem Fl ie@apier aus- gelegten PETl~I-Schalen (S. 259) noch Woehen hindurch geniigend lebens-

266 Pfeiffer
f~hig bleiben. Die Beobachtungszeiten, beginnend mit der Einfiihrung in das Plasmolytikum, sind sp~ter einander mehr gen/~hert worden (S. 268).
2. Zur Ermittlung des Permeationsffrades ist zun~chst eine Vital- f~rbung mit 0,05% N e u t r a l r o t l 0 s u n g vorgenommen worden. Nach 1/4- his 3/4-stiindiger Einwirkung auf die Sprosse aller Entwicklungs- stadien (yon erster Plasmolyse und Deplasmolyse his zu mehreren Tagen nach erfolgter Abgliederung der kernlosen Tochterzellen) sind m0glichst gleich besehaffene BlOtter (verglichen nach ihrer Entfernung yore Vege- tationspunkte, nach Gr01~e und Gestalt) abgeschnitten und tells in ~/~o NH3-L6sung, tells in ~/4o NaOH-L0sung iibertragen worden. Dann ist durch Ermittlung des Z e i t p u n k t e s des e r f o l g e n d e n F ~ r b u n g s - u m s c h l a g e s der Baseneintritt gemessen worden (vgl. auch LINSBAUER 1997).
Welter sind gleich beschaffene Sprosse nach dem Vorgange W. PFE~'FERS~ W. I:~UttLAI~DS U. V. a. mit 0,000047 tool. oder 0,01% M e t h y l e n b l a u l 6 s u n g e n (nicht dem technischen Produkt, sondern dem analytisch-reinen Chlorid) bestimmte Zeiten (10, 30 oder 60 Min.) be- handelt wordent), wobei naeh dem Vorschlage COLLANDERS (1921) durch Vergleich mit entspreehenden Farbstoffl6sungen bekannter Konzentra- tion als Medium der Objekte ein Ma6stab fi~r die Menge des e in- g e t r e t e n e n Farbstoffes gewonnen worden ist (vgl. auch PFEIFFER 1997, S. 907).
Im Anschlusse daran sind andere Sprosse gleicher Beschaffenheit eine bis mehrere Stunden mit der gleiehen Konzentration desselben Farb- stoffes behandelt worden, bis eine d e u t l i e h e Blauf /~rbung, aber noch nicht intensivblau gef~rbte F~llungen aufgetreten sind. Neben farbigem Zellsaft werden dabei auch kristallartige, blau tingierte Ausscheidungs- produkte beobachtet. Bei Aufbewahrung in reinem Wasser kann die F~rbung mehrere Tage lang Qhne ersichtliehe Seh/idigung der Zellen erhalten bleiben oder nur wenig abgeschw~cht werden. Dutch Uber- fiihrung der abgesehnittenen BlOtter in 0,01proz. Zitronens~ure ist aber experimentell eine Exosmose des ~ ' a r b s t o f f e s zu erzielen, welehe (ebenso wie die vorangegangene Farbstoffaufnahme) bei Anwendung der
l) Allein auf dieses Verfahren w~ren die weitgehenden Schliisse~ die wir ziehen wollen, nicht erlaubt gewesen; wird doch ,:on YfUM~EL und P~scn~L (1927) festgestellt, dal~ auch yon St~rke ans einer NichtleiterlSsung inehr ~ethylenblau aufgenommen wird als aus einer Salzl~sung: so dal~ eine vergrSSerte Farbstoffa~fnahme nicht sogleich als Permeabilit~tssteigerung gedeutet werden mi~gte. Es ist aber wohl sicher, dal~ im vor- liegenden Falle derartige Befiirchtungen nicht gehegt zu werden brauchen.

Untersuchungen fiber die Entdifferenzierung u. Teilung pttanzlicher Dauerzellen. II 267
COL]~A~D]~Rschen Vergleichsmethode mit Farbstoffl~sungen bekannter Konzentration zum quantitativen Vergleiche des unterschiedlichen Perme- abilit~itsgrades gedient hat.
Endlich sind an entsprechenden Versuchsreihen die B1/itter benaeh- barter Wirtel eines Sprosses mit 10% A t h y l a l k o h o l g e s c h ~ i d i g t worden. Aus der Prozentzahl der in bestimmten Zeiten (1/~, 1/2 , 1 und und 2 Std.) abgestorbenen Haarzellen ergibt sich ein Vergleichsmal~ ftir das EindringungsvermSgen des Zellgiftes. Indem als Kennzeichen ne- krotischer Ver~inderungen die Unf~ihigkei t zu r G l y z e r i n - P l a s m o - lyse gezeigt oder ein anderer Nachweis gefiihrt werden mul~te, kann die Umst~ndliehkeit des Verfahrens neben den nur wenig befriedigenden Ergebnissen zur Entschuldigung dafilr dienen, dab diese Methode nur weniger h/~ufig zur Anwendung gekommen ist.
R e s u l t a t e
1. Verg le i ch des P e r m e a t i o n s g r a d e s d e r Z e l l f o r m e n mi t und ohne K a r y o p l a s m a . - - Von erheblichem Interesse ist zun~ichst wegen tier heute nur erst liickenhaften Kenntnis der physikalischen Eigenschaften kernloser und -haltiger Zellen (KOSTER 1999, S. 48) der Vergleich beider hinsichtlich ihres Permeationsverm~gens.
Dariiber ist durch Untersuchungen yon C. HOFFMANN (1927) an Arten yon Spiro- gyra LINK ermittelt worden, da$ die plasmometrisch (hath K. ]=[0FLER) gewonnenen Werte bei beiden Zellformen fibereinstimmen, sich auch gegenfiber der Beeinflussung der Permeabilitat durch Belichtungs- und Temperaturversehiebungen gleich verhalten [vgl. auch die Angaben yon FR. WEBER (1925, S. 76) gelegentlich einer Besprechung yon KUSTER (1924), sowie LINSBAUE~ (1927, S. 550)]. Diesem Ergebnis steht indessen neben ~lteren Angaben yon J. J. GERASSIMOFF und C. VAN WISSELINGH fiber erh5hten Turgor- druck kernloser Zellen yon Spirogyra LINK die Beobachtung yon BOBILOFF-PREISSER (1917) entgegen, da$ kernlose Pollenschl~uche bei fortgesetztem Wachstume auffallend zu betrKchflicher Wasseraufnahme und deswegen zur Nekrose unter Plasmoptyse neigen. Kt~STEI~ (a. a. 0.), der weitere Angaben nicht verzeichnet, vermag trotzdem die oft kon- statierte ~-eringere Lebensf~ihigkeit kernloser Zellen nicht aus den physikalischen Be- ziehungen zu erkl~ren.
Leider kOnnen die yon uns angestellten Untersuchungen die Beant- wortung der Frage nicht fSrdern, indem ein wesentlicher U n t e r s c h i e d b e i d e r Z e l l f o r m e n h i n s i c h t l i c h des E i n d r i n g e n s von F a r b s t o f f - lOsungen n i c h t g e f u n d e n worden ist, sofern geniigend weir fort- gesehrittene Stadien zum Vergleieh herangezogen worden sind. Da angenommen werden mul3, daI~ die zur Verfiigung stehenden Methodea der Permeabilit~itsermittlang nieht scharf genug sind, um feinere Diffe-

268 Pfeiffer
renzen aufzudecken, eine direkte Beziehung zu den Zielen der vor- liegenden Untersuchungen aul~erdem nicht besteht, ist die Frage vor- l~iufig nicht welter verfolgt worden.
7. Die V a r i a t i o n der A l k a l i p e r m e a b i l i t t i t wl ih rend der E n t d i f f e r e n z i e r n n g und T e i l u n g yon D a u e r z e l l e n . - - Der am Umsehlage der Neutralrotffirbung konstatierte Wechsel des Alkali-Ein- trittes je naeh dem Entwicklungsstadium (S. 759) ergibt fiir n/40 Na0H Werte zwischen 15 und 75 Minuten, fiir "/4o NH.~-L(isnng solche his zu ca. ~/~ Minute. Dieses Ergebnis, da6 die Messungen yon M. M. BROOKS (1973) fiber direkt bestimmte C~-Vertinderungen beim Baseneintritt in den Salt yon Valonia-Zellen ebeuso wie viele andere Angaben der Literatur (vgl. POUs 1978; SMALL 1979, S. 319) best~tigt, zeigt wegen der sehr geringen Unterschiede die geringere Brauchbarkeit yon Nti~ als Versuchsreagens.
Die tabellarische (~bersicht der ersten Gruppen von Na0H-Versuchen lehrt bereits einen z w e i f a c h e n P e r m e a b i l i t ~ i t s w e c h s e l wtthrend der verglichenen Entwicklungsstadien (Tabelie III), indem anscheinend um
Tabe l l e III
Zeit des Fiirbungsumschlages nach Eintritt yon ~1/~o Na0H
Beob achtmlgszeit I des Materials nachStd. 3
Fitrbungsmnschlag I nach iV[in . . . . . 20--25 ca. 20
24
20--25
] 48 ~ 72 168
[ [
20-30135--40 20--25
288
20 -- 25
ca. 79 Std. nach Versuchsbeginn, also um die Zeit der Abgliederung der kernlosen Tochterzellen, eine Ann~herung an ein P e r m e a b i l i t S t s - mi n im um erfaft wird, das zwischen beiderseits erkennbaren Perme- abilit~tsanstiegen gelegen ist. Mit diesem Ergebnis nicht gleieh zu- frieden, sind dann eine Reihe yon Beobaehtungsfolgen in k t i r z e r e n Ze i tabs t i~nden in der Nachbarschaft der kritischen Stadien, beginnend 48 Std. naeh Plasmo]yse (45 Std. naeh Deplasmolyse) der Sprosse, ge- sammelt worden. Da zeigt sich dann aber fiberrasehenderweise (Tab. IV), da6 bei dem nach 77 Std. erreichten Permeabiliti~tsgrade das faktisehe Minimum noch nieht erreicht wird, daft vielmehr das Eindringungsver- mOgen for Alkali noch welter sinkt, bis in einem Stadium zwischen 76 und 84 Std. (vielleicht um ca. 80 Std.) yore Plasmolysebeginne her die Zone m i n i m a l e r P e r m e a b i l i t f i t gelegen sein muff.
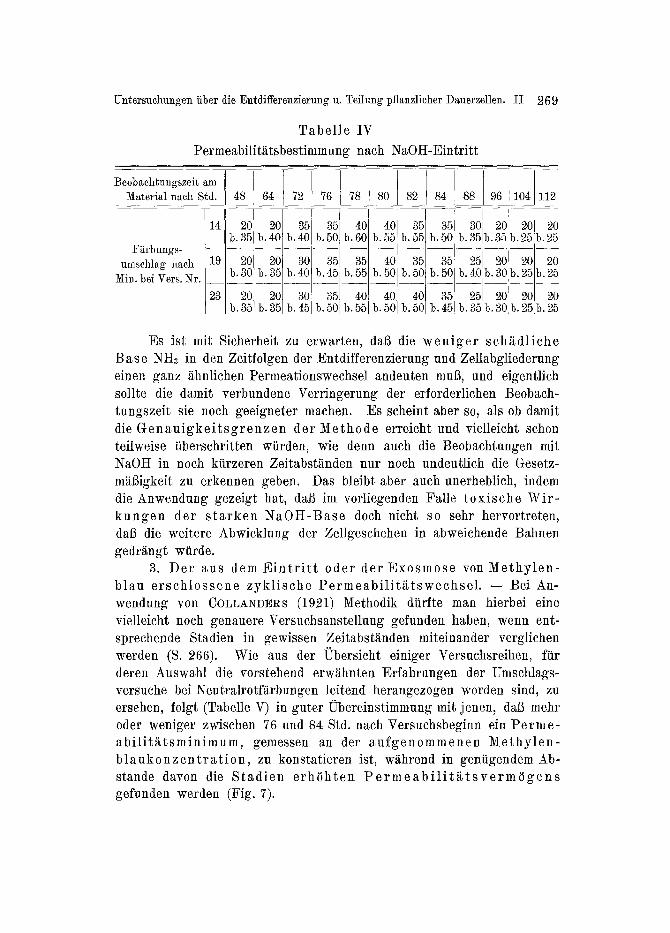
Unr tiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 269
T a b e l l e IV
Permeabilit~ttsbestimmung nach NaOH-Eintritt
Beobachfungszeit am Material nach Std.
14
F~irbnngs- - - umschlag nach 19
Min. bet Vers. ~r. 23
48
20[ b. 35[
u. o ~ 201
b. 35!
64 72 [ 76 78
20 35 35 i 40 b.40 b.40[b.5Ob.60
20 30 35 35 b.35 b.40 b.45 b.55
so l s2 84 8s 961104]112 [
401 351 35 30[ ~0 ~0 .90 b 55 b s~ b ~01 b ~51b 3~ b2.~b 2,~
40 35 3~ ~51 ~0! ~0 ~0 b.50 b.50 b.50 b. 40b. 30b.25b.25 _ _ [ . . . . ; _ _ _ _
4o[ G Es ~st mit Sicherheit zu erwarten, dab die w e n i g e r sch~tdliche
B a s e NH~ in den Zeitfolgen der Entdifferenzierung und Zellabgliederung einen ganz ~thnlichen Permeationswechse] andeuten mull, und eigentlich sollte die damit verbundene Verringerung der erforderlichen Beobach- tungszeit sie noch geeigneter machen. Es scheint aber so, als ob damit die G e n a u i g k e i t s g r e n z e n der M e t h o d e erreicht und vielleicht schon teilweise tiberschritten wiirden, wie denn auch die Beobachtungen mit NaOH in noch kiirzeren Zeitabst~nden nur noch undeutlich die Gesetz- mii6igkeit zu erkennen geben. Das bleibt aber auch unerheblich, indem die Anwendung gezeigt hat, dafl im vorliegenden Falle t o x i s c h e Wir- k u n g e n der s t a r k e n N a 0 t I - B a s e doch nicht so sehr hervortreten, da6 die weitere Abwicklung der Zellgeschehen in abweichende Bahnen gedr~ingt wtirde.
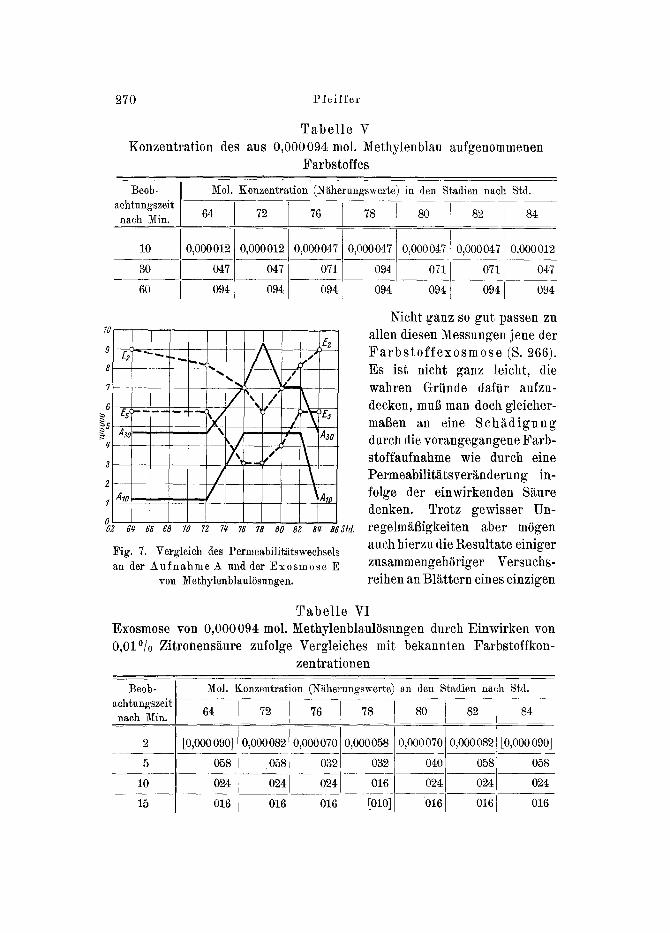
3. Der aus dem E i n t r i t t oder der E x o s m o s e yon M e t h y l e n - b l au e r s c h l o s s e n e z y k l i s c h e P e r m e a b i l i t ~ t t s w e c h s e l . - - Bet An- wendung yon COLLANI)ERS (19~1) Methodik diirfte man hierbei eine vielleicht noch genauere Versuchsanstellung gefunden haben, wenn ent- sprechende Stadien in gewissen Zeitabsti~nden miteinander verglichen werden (S. ~66). Wie aus der tJbersicht einiger Versuchsreihen, far deren Auswahl die vorstehend erw~thnten Erfahrungen der Umschlags- versuche bet Neutralrotfitrbungen leitend herangezogen worden sind, zu ersehen, folgt (Tabelle V) in guter fdbereinstimmung mit jenen, daft mehr oder weniger zwischen 76 und 84 Std. nach Versuchsbeginn ein P e r m e - a b i l i t ~ t s m i n i m u m , gemessen an der a u f g e n o m m e n e n M e t h y l e n - b ] a u k o n z e n t r a t i o n , zu konstatieren ist, w~hrend in geniigendem Ab- stande davon die S t a d i e n e r h S h t e n P e r m e a b i l i t ~ t s v e r m 0 g e n s geflmden werden (Fig. 7).
270 Pfe i f f e r
Tabe l le V Konzentration des aus 0,000094 tool. Methylenblau ~ufgenommenen
Farbstoffes
Beob- h[ol. Konzentratioa (Naherungswerte in den 8tadien nach Std. aehtungszeit nach ~in. 64 72 76 78 80 82 84
10
30
60
0,000012
o94
0,000047 0,000047
- - ~ - 094 L 094
0,000 047 0,000047, 0,000012
071, 047
094 094
7__ ~ .-\ 6 : _ ~ _ _ _ _ ~ . . . . ,
\ - A3~ q ~- J j j ,
3 1 ~ * ' " ~
r A1o _ A1o
0 62 s 66 G8 70 72 7q 7G 78 80 82 8g. 86S/'d.
Fig. 7. Vergleich des Permeabilitiitswechsels an der Aufnahme A und der Exos~lose E
yon MethylenblaulSsungen.
Nicht ganz so gut passen zu allen diesen Messungen jane der F a r b s t o f f e x o s m o s e (S. 266). Es ist nicht ganz leicht, die wahren Griinde dafiir aufzu- decken, muff man doch gleicher- marian an eine S c h ~ d i g u n g dutch die vorangegangene Farb- stoffaufnahme wie dutch eine Permeabilit~ts ver~nderung in- folge der einwirkenden S~ture denken. Trotz gewisser Un- regelm~igkeiten aber m6gen auch hierzu die Resultate einiger zusammengehSriger Versuchs- reihen an Bl~tttern eines einzigen
Tabel le VI Exosmose yon 0,000094 mol. Methylenblaul~isungen durch Einwirken von 0,01% Zitronens~ure zufolge Vergleiches mit bekannten Farbstoffkon-
zentrationen
Beob- Mol. Konzentration (Ni~herungswerte an den Stadien nach Std. achtungszeit nach ~in. 64 72 76 78 80 82 84
[0,000 090]
5 058
10 024
15 016
0,000082 0,000070
024
0,000058
- - o ~ - - ~
0,000070 0,000082
0 4 0 058
024 024
- - ~ ] OlGi
iO,O00 090]
058
024
016

Untersuehungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 271
Sprosses wiedergegeben werden (Tab. u Fig. 7). Aus dem Umstande, daft anf~nglieh eine bessere 0bereinstimmung mit den auf anderen Wegen gewonnenen Messungsergebnissen gefunden wird, mSchte a]s Grund der geringeren Eignung des Verfahrens der S~ureeinf lul~ am wahrschein- lichsten sein.
4. Das z y k l i s e h w e c h s e l n d e A u f n a h m e v e r m ( i g e n fiir Athy l - a lkohol . - - Wie schon aus der Literatur (W. SEIFRIZ) bekannt, verhatten sich die einzelnen Helodea-Pflanzen gegeniiber der vergiftenden Wirkung yon Athylalkohol (S. 267) oft recht verschieden. Die Resistenz w~chst scheinbar (ebenso wie iibrigens der osmotische Wert der Blattzellen) mit einer gewissen Dauer der Kultur. Um vergleichbare Werte zu er- halten, sind an dem betr. Sprosse nach wechselnder Einwirkungszeit des Plasmolytikums und der K~OP- oder audern KulturlSsung nur B lOt t e r b e n a c h b a r t e r W i r t e l verwendet worden. Die Unterschiede sind bei der iiberhaupt z i eml ich g r o b e n Me thode gew(ihnlich verschwindend gering, wenn naeh Verbrauch der BlOtter einer bestimmten Spro~hShe sp~ter die direkt darunter oder dicht dartiber befindlichen zum Ver- gleich benutzt werden. Die (~bereinstimmung mit den vorstehend nach andern Verfahren verzeichneten Messungsergebnisseu ist am deutliehsten bald nach Einwirkung des Zellgiftes (Tabelle VII), indem sich auch bier
Tabe l l e VII
Prozentuelle Anzahl der mit 10% Athylalkohol abgetSteten ttaarzellen
Beobachtungszeit Entwicklungsstadien nach Versuchsstunden nach Std.
64 76 8O 84 92
1~ 40--45 50--60 60--80 50--75 40--45
1~ 60--65 50--70 80--90 60--85 55--65
1 80--90 80--95 9 0 - - 1 0 0 90--95 80--90
2 95--98 ca. 100 ca. 100 95--99 85--95
ein P e r m e a b i l i t ~ t s m i n i m u m etwa im gleichen Stadium wie nach den anderen Untersuchungsverfahren zeigt. Mit der Dauer der Versuchszeit nehmen die Differenzen in der Abtiitungszeit immer welter ab, bis sie schon nach einsttindiger Alkoholeinwirkung ziemlich unkenntlich werden.
5. B e o b a c h t u n g e n fiber g e s e t z m ~ i ~ i g e P e r m e a b i l i t ~ t t s - u n t e r s c h i e d e w ~ h r e n d der Mi tose yon H e l o d e a - K e r n e n . - Zwar

979 Pfeiffer
]iegen die dariiber zu erw~hnenden, nur gelegentlich gesammelten Be- funde nicht gerade im Rahmen der vorgesteckten Untersuchungsziele. Dennoch sollen einige Bemerkungen hier folgen, well daraus die Ver- w a n d t s c h a f t der in diesem Beitrage studierten A b g l i e d e r u n g e n k e r n l o s e r Zel len mit eehten Mitosen (S. 975) besser verdeutlicht wird. Wegen s t a r k e r K u m u l a t i o n der s icher v o r h a n d e n e n Permeab i l i t i~ t sun te r sch iede ist deren Naehweis allerdings ziemlieh erschwert, and so sind entsprechende Resultate anfangs in der Tat iiber- sehen worden. In der beginnenden Telophase ist jedenfalls das Perme- abilit~tsminimum fiir den Eintritt yon hlkalien und den Ein- und Aus- tritt you Farbstoffen schou tiberwunden worden. Aber bei dem sich zur Teilung anschickenden Karyoplasten in den ersten mitotischen Stadien ist eine herabgesetzte Permeabilit~t ebenso wie eine vorangegangene S t e ige rung der Durch l~ss igke i t in den Phasen nach A u f t r e t e n der Spindel doeh mehrfach gefunden worden.
IIL Abschn i t t
Diskussion
Historisch l~flt sich nach einem zeitweiligen TJberwiegen rein mor- phologischer Betrachtungsweise der cytologischeu Erscheinungen eine Richtung erkennen, welche mit dem Riistzeug der physikalischen Chemie in die komplexen physiologischen Gesehehen einzudringen versucht. Wenn wit auch gewissen Mahnungen zur Vorsicht (FR. LEVY, F. WASSER- MANN U. a.) gerne Geh5r sehenken mSchten und uns htiten wollen, mit kolloidchemiscimn Schlagworten bereits eine vollkommene Erkl~rung der Lebensph~tnomene auch ohne die n0tige Bertieksichtigung morphologisch- anatomischer Befunde und tier physikalisch-chemischen Grenzen zu ver- suchen, so brauchen wir uns doch nicht zu versagen, hier die M(iglich- ke i ten einer Deu tung nur mit den Mit te ln der phys ika l i schen and Kol loidchemie aufzuzeigen. Dabei haben wir im vorliegenden Falle gesetzm~Nge Permeabilitlitsverschiebungen einerseits bei Mi t o s e n, anderseits bei den hier studierten E n t d i f f e r e n z i e r u n g s - und Ab- g l i e d e r u n g s p r o z e s s e n an Dauerzellen in Beziehung zu der friiher aufgestellten hrbeitshypothese (PFEIFFER 1929C, S. 417, 419, 490, 499) zu diskutieren. Wegen der Bezugnahme der dahingeh(irigen Vorstel- lungen auf den I E P des P r o t o p l a s m a s werden wir ferner die Be- ziehungen des Permeabilit~tsgrades zu jenem Umladungswerte zu be- sprechen haben.

Untersuchungen fiber die Entdifferenzlernng u. Teflung pflanzlicher Dauerzellen. II 273
a) Gesetzm•fiige Pe rmeab i l i t a t sve r sch iebungen bei ande rn Tei lungsprozessen
Eine bestimmte Per iod iz i t~ t der p h y s i k a l i s c h e n E i g e n s e h a f t e n der p lasmat i schen Kol loidte i lchen parallel mit der Kurve der Ze]l- und Kernteihng ist schon oft dargelegt (DALe0 1928, S. 15, 34, 76 u. a.; HEILBRUNN 1928, S. 255) und vielfaeh auch sehon in Hinbliek auf die Dureh la s s igke i t der Grenzflaehen der Protoplasten diskutiert worden (s. aueh S. 281). So ist die Mlgemeine Annahme einer Permeabilit~tssteigerung mit der Befruehtnng (HOBER 1924, S. 478, 535 nsw.), wenn aueh nicht im strengen Sinn bereits erwiesen, doeh yon hoher Wahrseheinliehkeit (RUNN- STROM 1928, S. 400). Die Erseheinung entsprieht vSllig der Pe rmeab i l i t a t se rhShung bei gesteige~ter Funktion t~itiger Muskeln (G. EMBDEN und E. ADLER) oder andern Pro- zessen periodiseher Funktionssteigerung (Literatur: HOBER 1924, S. 534; REMINGTON 1928, S. 352; (.}ELLHORN 1929, S. 166, 175, u. a.), und auch (}ELLHORN (a. a. 0., S. 174) sehlieJ]t, dM~ der physiologische Erregungsvorgang an tierisehen Zellen gleieh dem experimente]l induzierten bei geeigneter Reizdosierung unabhgngig yon der Qualitat und dem Effekt des Reizes (Bewegung, Sekretion, Entwiekhngsanregung u. a.) regelm~il~ig yon einer Permeabilit~tssteigerung begleitet wird. FAURE-FREMIET (1925, S. 140), der yon den Untersuehungen yon E. P. LYON, G. SPAULDING, A. P. ~ATttEWS~ ~. KONOPACKI, A. R. ~/[00RE, S. R. LILLIE und M. ~ERLANT (1920) die meisten rfieksiehtlieh zykliseher Vari- ationen der Durchlgssigkeit der Protoplasten bei der Entwicklungsanregung zusammen mit eigenen Befunden (1921; 1923) diskutiert 1), findet allerdings die Variationen der Permeabilitgt an sieh teilenden Eizellen nieht immer in g e n f i g e n d e r Para l le le mit dem Fort- sehreiten der Mitosen. Ebenso zeigt naeh Pns (1922 a) eine Zelle yon S2irogyra LINK~ welehe dureh n/~ NaC1 plasmolysiert wird, dauernd denselben Plasmolysegrad wie die sich nicht teilenden Zellen desselben Fadens, woraus geschlossen wird, da] die Perme- abilt~t night wesentlieh erhSht wird. Dal~ der Kolloidzustand der Plasmateilehen schon in der Telophase nieht mehr erheblieh yon jenem ruhender Zellen abweieht, ist iibrigens bier ebenfalls gefunden worden (vgl. S. 272).
Aber schon R. S. LI~LIE (1916, 1918) hat an Eiern yon Arbacia ge-
zeigt, daft die B e f r u c h t u n g zu e r h S h t e r D u r c h l g s s i g k e i t f t i h r t , welche in beschleunigter Schrumpfung in hypertonischen oder Schwellung in hypotonischen LSsungen zum Ausdrucke kommt und nicht e twa mit
verschiedenem osmotischen Drucke erklgrt werden kann. Eine weitere Vert iefung erfahren diese Ergebnisse durch die sehr vielseitigen Unter-
suchungen yon HERLANT (1918; Zusammenfassung: 1920) an dem gleichen Objekt durch Bes t immung der Permeabil i tgt ' far Wasser , Salze, s tarke Basen, Mineralsguren, Fe t t sauren , Saponin, Ammoniak usw. (Fig. 8).
~) Aus diesem Grunde kann hier eine Aufzghlung der raeisten einschlggigen Arbeiten fiiglich unterbleiben, zumal aueh auf die Besprechungen zahlreieher vorliegen~ der Beobachtungen bei P. R. LILLIE (19195, CONKLIN (1924), DALC0 (1928, S. 18) nnd HEILBRUNN (1928, S. 260, 2771 verwiesen werden kann. Trotzdem ist eine ErSrterung einlger besonders wichtiger Ergebnisse sicher wertvoll.
Protoplasma. X 18

274 Pfeiffer
Deutlich ist die Symbas ie des T e i l u n g s r h y t h m u s mit j enem zu- n e h m e n d e r and g e h e m m t e r Permeabi l i t i~ t erkennbar, wobei das Permeabilitiitsmaximum fiir die meisten Substanzen im Stadium der Zell- teilung und w~hrend der Bildung der Diaster liegt, allerdings das Maximum der Wasserdurchl~issigkeit jenem der Salz- und Alkaliperme- abilit~t ein wenig vorausgeht. HERLANT nimmt an, dab das Protoplasma aufzufassen ist als eine Emulsion yon Proteinen and Lipoiden, in welcher das Protein bald die ~ul]ere, zasammenhiingende Phase, bald die disperse Phase des Systems darstellt. Indem jene durch gesteigerte Plasmahaut- permeabilitgt und nur ziemlich hohe Empfindlichkeit gegen lipoidl6sliche Substanzen, letztere durch Hemipermeabilit~t und erhShte Empfindlich-
Fig. 8.
A
/ \ A A
/ / - -
Wa3•ep
Sa/ze
s/arke Basen
Minem/giu pen
f e#sSupen Sapon/ne
, ~ 4 m m o n / a k
Beginn M/He Ende dee Te/lung
Der Permeabilitiitswechsel yon Paraeentrotus-Eiern im Zyklus n~it ihrer ersten Mitose. (Nach HERLANT 1918.)
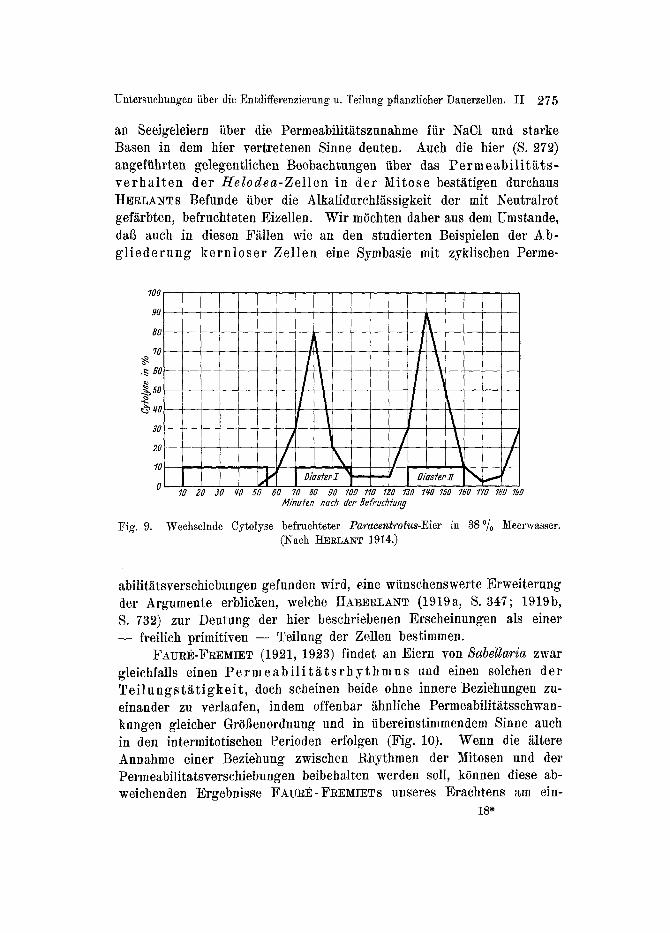
keit in der genannten Richtung eharakterisiert ist, beide Phasen ferner ineinander tiberffibrbar sind, mul~ die E n t w i e k l u n g s a n r e g u n g im wesentlichen da rch den Umsch lag der e inen Emul s ions fo rm in die ande re bed ing t sein. Auf die h(iehst bedeutsamen Untersuchungen yon RU~!I~STROM (19'24) and EPHRUSSI (19~5) fiber Variationen der os- motischen Kriffte und der Permeabilitiit der Plasmagrenzschicht kann hier nur noch hingewiesen werden. Bemerkenswerter sind in diesem Zusammenhange die Befunde HERLANTS fiber die Cy to lyse be f rueh- t e t e r E ie r von Paracentrotus (Fig. 9) beim tlbertragen in Meerwasser, welches zu 6~% mit gew(ihntiehem Wasser verdiinnt wird.
Wean W. RUPPERT (19~4) bei Eiern yon Ascaris eine Variation der E m p f i n d l i c h k e i t gegen U l t r a v i o l e t t b e s t r a h l u n g im Ablaufe der ersten Segmentierungsteilung konstatiert, so l~6t sich dieser Befund ebenso wie entsprechende Beobachtungen yon TCttAHOTINE (1990, 1991)
Untersuehungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 275
an Seeigeleiern iiber die Permeabilit~tszunahme fiir NaC1 und starke Basen in dem hier vertretenen Sinne deuten. Auch die hier (S. ~7~) angefiihrten gelegentlichen Beobachtungen fiber das t )e rmeabi l i t~ t t s - v e r h a l t e n de r Helodea-Ze l l en in der Mi tose best~ttigen durchaus HE,LASTS Befunde tiber die Alkalidurchl~ssigkeit tier mit Neutralrot gef~irbten, befruchteten Eizellen. Wir miichten daher aus dem Umstande, dab auch in diesen F~llen wie an den studierten Beispielen der Ab- g l i e d e r u n g k e r n l o s e r Ze l l en eine Symbasie mit zyktischen Perme-
'~176 t 9o! 1 7o__ / __
.~ aO . . . .
~ 5 0 - -
qg
3g - - / /
Fi~.. 9.
/ f0 20 3g qg 50 50 7g 80 $g lgO 710 12g 73g Iqg 150 160 17g IYg 190
Minuten nach der BefPuchtunq
Weehselnde Cytolyse befruehteter P a r a e e n t r o t u s - E i e r in 38% )/[eerwasser. (Nach HEal,ANT 1914.)
abiliffttsverschiebungen gefunden wird, eine wiinschenswerte Erweiterung der Argumente erblicken, welche HABE•LANT (1919a, S. 347; 1919b, S. 732) zur Deutung der hier beschriebenen Erscheinungen als einer - - freilich primitiven - - Teilung der Zellen bestimmen.
FAURE-FREMmT (1991, 19~3) finder an Eiern yon Sabellaria zwar gleichfalls einen P e r m e a b i l i t ~ t s r h y t h m u s und einen solchen der T e i l u n g s t ~ t i g k e i t , doch scheinen beide ohne innere Beziehungen zu- einander zu verlaufen, indem offenbar ~ihnliche Permeabilit~tsschwan- kungen gleicher Gr(U3enordnung und in tibereinstimmendem Sinne auch in den intermitotischen Perioden erfo]gen (Fig. 10). Wenn die ~ltere Annahme einer Beziehung zwischen Rhythmen der Mitosen und der Permeabilitatsverschiebungen beibehMten werden sol1, k(innen diese ab- weichenden Ergebnisse FAU~E-FREMIETS unseres Erachtens am ein-
18"
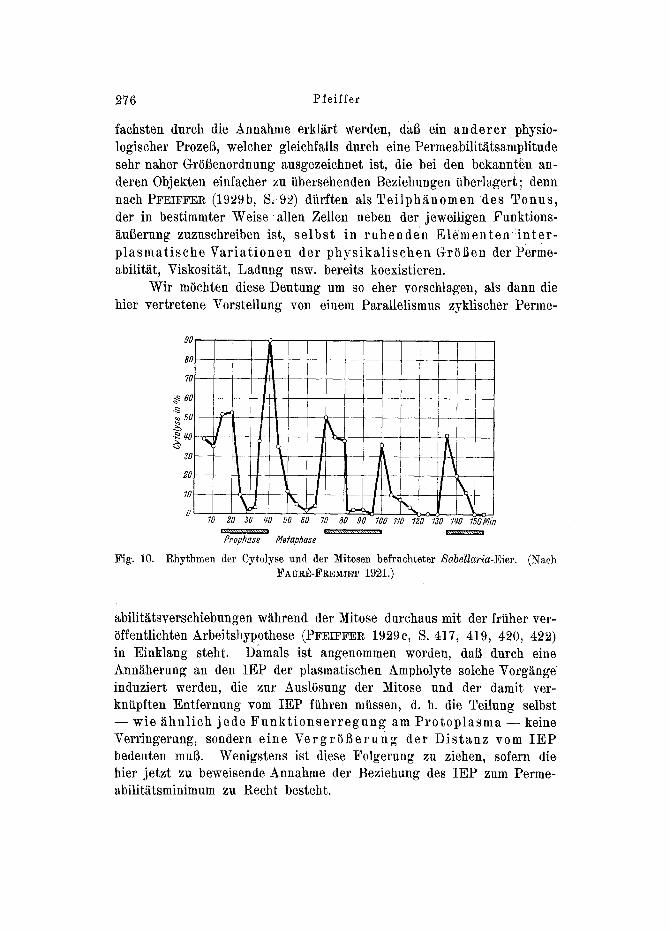
276 Pfeiffcr
fachsten dutch die Annahme erkl~rt werden, da~l ein ande re r physio- logischer Proze~l, welcher gleichfalls durch eine Permeabilit/~tsamptitude sehr naher Gr0flenordnung ausgezeichnet ist, die bei den bekannt~n an- deren Objekten einfacher zu iibersehenden Beziehungen iiberlagert; denn nach PFEIFF]~R (1929b, S, 92) diirften als T e i l p h ~ n o m e n des Tonus , der in bestimmter Weise allen Zellen neben der jeweiligen Funktions- ~ufierung zuzuschreiben ist, se lbs t in r u h e n d e n E l e m e n t e n : inter- p l a s m a t i s c h e Va r i a t i onen der p h y s i k a l i s c h e n C~rSl~en der Perme- abilit~t, Viskosit~t, Ladung usw. bereits koexistieren.
Wir m0chten diese Deutung um so eher vorschlagen, als dann die hier vertretene Vorstellung yon einem Parallelismus zyklischer Perme-
Fig. 10.
90
70
t �84 \ 20 . . . . . ]
L'/ ~ ^
iO 20 30 qO 50 60 70 8D 90 100 710 720 730 7~0 150M/n
Prophase Mefaphase
Rhythmen der Cytolyse und der ]~itosen befruchteter Sabellaria-Eier. (Naeh FAUR~-FREMI]~T 1921.)
abilit~tsverschiebungen w~hrend der Mitose durchaus mit der friiher ver- iiffentlichten Arbeitshypothese (PFEIFFER 1929C, S. 417, 419, 420, 422) in Einklang steht. Damals ist angenommen worden, dab durch eine Ann~herung an den IEP tier plasmatischen h_mpholyte solche u induziert werden, die zur AuslSsung der Mitose und der damit ver- kntipften Entfernung vom IEP fiihren mtissen, d. h. die Teilung selbst - - wie i~hnlich jede F u n k t i o n s e r r e g u n g am P r o t o p l a s m a - - keine Verringerung, sondern eine Verg r ( i l~e rung tier D i s t anz vom I E P bedeuten muir. Wenigstens ist diese Folgerung zu ziehen, sofern die bier jetzt zu beweisende Annahme der Beziehung des IEP zum Perme- abilit~tsminimum zu Recht besteht.

Untersuehungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 277
b) Theoret ische Untersuchungen ilber den Permeabilitgltsgrad im IEP des Protoplasmas
1. Die K o i n z i d e n z des P e r m e a b i l i t / ~ t s m i n i m u m s mi t dem I E P de r P l a s m a a m p h o l y t e
Leider ist nach der Literatur die Auswertung yon Permeabilit~tsmessungen zur Ermittlung einer Ann~herung an den IEP yon Protoplasten deswegen ziemlich ersehwert, weft die herrschenden Ansiehten fiber den Permeabili tKtszustand w~hrend der Entladunff der Ampholyte einander zum Tell diametral gegenfiberstehen. Indem die Beantwortung dieser Frage ffir die Auswertung auch der hier mitgeteilten Resultate (S. 267 f.) yon ausschlaggebender Bedeutung wird, mug eine knappe Darstellung der gegenwgrtigen Sachlaffe hier jetzt erfolgen, zumal die hier angenommene Deutung GISLLHORNs (1929) dessert Monographie zwar wie ein roter Faden durehzieht, aber nicht in der wfinsehenwerten Gesehlossenhei~ im Zusammenhange behandelt w~rd.
1. Zwar folgt aus den Permeabilit/~tstheorien yon F. E. LLOYD (1915) und E. E. FREE (1918), nach welchen Quellungen der dispersen Phase des plasmatischen Kolloidsystems infolge Verringerung der Diffusionswege tier kontinuierlichen Phase die Durchl/~ssigkeit verringern (Entquetlungen umgekehrt den Durchtri t t erleichtern), ftir den IEP zugleich mit dem Quellungsminimum ein Maximum der Permeabilit/~t, und auch die nahe- stehende Theorie yon D. HAYNES (1921) setzt einen maximalen Per- meabilit/~tsgrad far den IEP voraus, indem Salze und Nonelektrolyte im moderierenden Dispersionsmittel des plasmatischen Emulsionskolloids die permeabilit/~t durch Verschiebung der C s ver/~ndern sollen. Wir m0chten dagegen in Ubereinstimmung mit GELLttORN (199,9, S. 3'2, 53, 93, 145, 227 , 945, 366 u. a.) folgern, dab mi t dem Q u e l l u n g s - m i n i m u m in de r N a c h b a r s c h a f t des I E P (PFEIFFER 1999a, S. 14; 1999b, S. 74 u. a.) zugleich e in P e r m e a b i l i t / ~ t s m i n i m u m v e r k n i i p f t ist. Zu dieser Vorstellung gelangen wir aus der auf alle bisherigen Erfahrungen gegrilndeten Annahme, dab m i t s t e i g e n d e m D i s p e r s i t / ~ t s - g r a d e de r P l a s m a k o l l o i d e die P e r m e a b i l i t / ~ t z u n e h m e n mu$ (vgl. auch: KACZMAREK 1999, S. 256).
Zu der gleichen Sehlugfolgerung .gelangen wir aber aueh aus den Versuchen yon 1fOND (1927a; b), nueh welchera bei ~Jbersehreitung des IEP des Globins yon Erythroeyte11, den OSATO (1922) zu pg 8,1 bestimmt hat, d~e elektive Anionen- permeabili t~t in 'e ine Kationendurehlgssigkel t umschlagt (Fig. ll). Die in Rohrzueker suspendierten BlutkSrperchen zeigen namlieh unter Best~tigung der Gesetz- m~l~igkeiten des DO•NAS-G-leiehgewichtes mit zunetmlender Cot t eine wachsende Ver-
Gi ringerung des Quotienten ~-a ffir die Konzentration der Cl' im Innern (Ci) und aul]er-
halb (Ca) und ebenso unter denselben Bedingunffen fiir die S04"-Aufnahme ein Absinken, his ca. bei pH 8 die Aufnahme unterbleibt.
8 mg
7
6
5
q
J
278 Pfeiffer
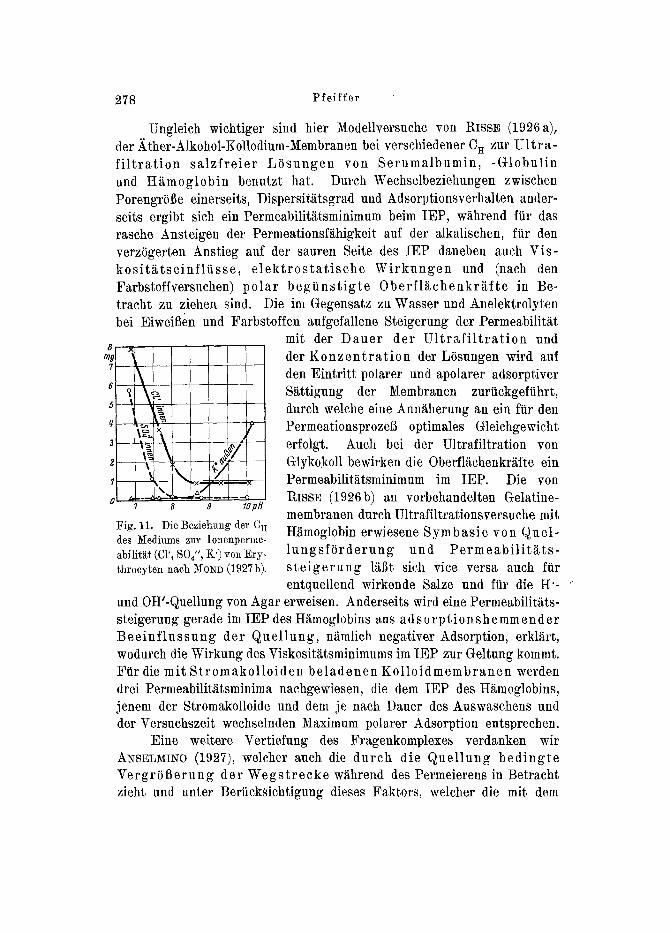
Ungleich wiehtiger sind hier Modellversuche yon RISSE (1926a), der Ather-Alkohol-Kollodium-lVlembranen bei verschiedener C H zur U l t r a - f i l t r a t i o n sa lz f re i e r L S s u n g e n yon Se ruma lbumin , -Globul in und H~moglobin benutzt bat. Durch Wechselbeziehungen zwischen PorengrS~e einerseits, Dispersitiitsgrad und Adsorptionsverhalten ander- seits ergibt sich ein Permeabilit~itsminimum beim IEP, w~hrend ftir das rasche Ansteigen tier Permeationsfi~higkeit auf der alkaliscben, fiir den verzSgerten Anstieg auf der sauren Seite des IEP daneben auch Vis- kos i t~ t s e in f l i i s s e , e l e k t r o s t a t i s c h e W i r k u n g e n und (naeh den Farbstoffversuchen) polar beg i i n s t i g t e Oberf l i~chenkr~f te in Be- tracht zu ziehen sind. Die im Gegensatz zu Wasser und Anelektrolyten bei EiweiBen und Farbstoffen aufgefallene Steigerung der Permeabiliti~t
mit der Dauer der U l t r a f i l t r a t i o n und
0 7 8 9 109//
Fig. 11. Di~ Beziehuug tier C R des Mediums zur Ionenperme- abilit~t (CI'~ SQ" , K') yon Ery- throcyten naeh M0~D (1927b).
der K o n z e n t r a t i o n der L6sungen wird auf den Eintritt polarer und apolarer adsorptiver S~ttigung der Membranen zurtickgeftihrt, dnrch welche eine Ann~herung an ein fiir den PermeationsprozeB optimales Gleichgewicht erfolgt. Auch bei der Ultrafiltration yon Glykokoll bewirken die 0berfl~ehenkriifte ein Permeabilit~tsminimum im IEP. Die yon RIss]~ (19~6b) an vorbehandelten Gelatine- membranen dureh Ultrafiltrationsversuehe mit H~tmoglobin erwiesene Symbas ie yon Quel- l u n g s f O r d e r u n g und P e r m e a b i l i t ~ t s - s t e i g e r u n g li~t sich vice versa auch ftir entquellend wirkende Salze und ftir die H'-
und 0H+-Quellung von Agar erweisen. Anderseits wird eine Permeabilit~ts- steigerung gerade im IEP des H~moglobins aus a d s o r p t i o n s h e m m e n d e r B e e i n f l u s s u n g der Que l lung , niimlich negativer Adsorption, erkli~rt, wodurch die Wirkung des Viskosit~tsminimums im IEP zur Geltung kommt. Ftir die mit S t r o m a k o l l o i d e n be l adenen K o l l o i d m e m b r a n e n werden drei Permeabilit~tsminima nachgewiesen, die dem IEP des H~moglobins, jenem der Stromakolloide und dem je nach Dauer des Auswaschens und tier Versuchszeit wechselnden Maximum polarer Adsorption entsprechen.
Eine weitere Vertiefung des Fragenkomplexes verdanken wir ANSE:5~INO (1927), welcher auch die durch die Que l lung b e d i n g t e VergrO6erung der W e g s t r e c k e w~hrend des Permeierens in Betracht zieht und unter Beriicksichtigung dieses Faktors, welcher die mit dem

Untersuchungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. I I 279
Quellen der Gela t ine anzunehmende Permeabi l i t~ t szunahme tei lweise
he rabse tzen oder ausgle ichen mug, in j e t z t s/~mtlichen F/illen ein Mi -
n i m u m d e r P e r m e a b i l i t ~ t t g e r a d e b e i m I E P erwiesen hat. Noch
widers t re i tende Befunde yon DAVID J. HITC]~COCK (1926) kann er,
unseres E rach t ens mit vollem Rechte: damit erkl~iren, daft bei dessen
ge la t in ier ten Kollodiumfil tern die Quel lung eine P o r e n v e r s t o p f u n g
bewirken mug, w~ihrend an Gela t inemembranen mit der Ausdehnung
ihrer Tei lchen eine P o r e n e r w e i t e r u n g e in t re ten kann. 2. Ganz im Sinne der hicr vertretenen Auffassung einer Koinzidenz von Per-
meabilitiitsminimum mit Quellungsminimmn und Ann~iherung an den IEP steht auch die h~iufige Erfahrung, dal~ quel lend wirkende Salze gleiehzeitig permeabi l i t i i t s - erh(ihenden~ entquellend wirkende vice versa permeabiliti~tsvermindernden Einflul~ zeigen (R. HOBER, G. B. WALLACE: A. R. CUSHNY~ ]~[. OKER-~BLOM~ ]~. BECHHOLD~ J. STIEGLER: 1~. S. LILLIE: W. BRENNER 7 R. CHAMBERS~ W. J. V. 0STERHOUT: ]~[. SAMEC: J. SPEK: E. WERTHEIMER u. V.a.). Sowohl die Qucllung vielcr Hydrogele und Zell- bestandteile als auch die Geschwindigkeit des Eindringens yon Anionen in die Zelle wi~chst in a n t i v a l e n t e r Folget), d. h. in der Reihe
S04 < C1 < Br < J < SCN (BoAs 1928: S. 59; PFEIFFER 1929b, 27 u. ~.). Dasselbe besagt die Regel yon H. KAno (1926): nach welcher die Aufnahme eines Salzes der Kationen-Permeabilitiitsrelhe
K > Na > Li > lVIg > Ba > Ca durch ein bcliebiges anderes gehemmt wird, (lessen Kation in der Reihe weiter rechts steht, and zwar nach Mal]gabe des Abs t andes in der Reihe. Bei gleichem Kation gilt sinngem~g dasselbe fiir die Anioncn naeh der Stellung in dcr Folge
SCN > J > NOa ~ C1 > Tartrat > S04 > Zitrat, d. h. wegen des Umstandes, dal3 beide Reihen zugleich solehe fallender Quel]ungs- beeinflussung darstellen, ist die Symbasie zwischen Que l lungszus t and und Per- meabil i t~i tsgrad grundsiitzlich einleuchtend. Die zweifache Salzwirkung auf das Protoplasms, niimlich entw~ssernd_ bei geringcr Permeabilitiit entsprechend dem osmotischen Drucke tier Salze oder quellend bei gesteigerter Permeabilit~t entsprechend ihrer lyotropen Natur, ist nach PFEIFFER (1929b, S. 73) wohl geeignet, die Diskrepanz zwischen An- gaben verschiedener Autoren, die ungleiches Material verwendet haben, ausreiehend zu erkli~ren.
3. Wie die Auffassung gleichsinniger Verschiebung der Permeabiliti~t und des Quellungsverhaltens zur D e u t u n g mancher Versuchsresu l ta te gccignet crscheint: zeigt GELLIIORN (1929, S. 145), wenn er aus der Parallelitiit tier Kationenreihe der
1) Antivalent ist auch die Anionenfolge ffir (lie Beschleunigung des Plasm olyse- r f ickganges , wie aus ncueren Uutersuchungen an Cyanophyeeen (S. PRi(T, Arch. f. Protistenkd. 52, 142. 1925,) hervorgeht, wclche aber darin attch mit andcrn Objekten iibereinstimmcn (It. FITTING, I:[. KAIt0: A. TRONDLE u.a.). ES ist das zugleich die m y k o t r o p - e l e k t i v e I o n e n o r d n u n g bei dem yon FIL BOAS beschriebenen phyletischen Ionenphfinomen, sowie die Folge der eng mit der Quellung verkaiipften (E. KNOEVENAGEL) Viskos i t i i t szunahme und jene der ebenfal]s damit verbundenen H e r a b s e t z n n g des osmot ischen Druckes yon 0valbuminlSsungen usw. (vgl. 1)FEIFFER: a. a. 0.).
280 Pfe i f fe r
Kaliumkontraktur des in u s e u 1 u S S a r t o r i u s yon R a n a and jener der Quellungswirkung auf die Huskulatnr auf eine Permeabilitiitssteigerung fiir K mit zunehmender Quellunff tier Kolloide in den Zellgrenzschiehten sehliel~t. Endlieh mag noch erw~hnt werden, daft sieh der normale und besonders der akut enter i t i sehe Darm zu~leich stark ge- quollen und erhSht durchlgssig fiir Kristalloide erweist, wghrend dagegen der Darm bei ehroniseher Enter i t i s , da er bereits in bindegewebiger Atrophie begrifEen und folglich wenig qaellbar ist~ einen welt geringeren Stoffdurehtritt ermSglicht, wie wir seit E. ~AYERttOFER und E. ~PRIBRAM wissen, die at~ch auferhalb des 0rganismus an ansgeschnittenen Darmstiicken mittels k~instlieh bewirkter Quellung eine erhShte Durch- l~ssigkeit fiir KCI, NaC1 und Dextrose entspreehend dem Verhalten akuter Enteritis naehgewiesen and damit eine kolloidehemisehe Best~tigunff der klinischen Befunde ]:[. FINKELSTEINs tiber Intoxikationen dureh derart wirkende zuckcrreiehe ~qahrungs- gemische geliefert haben.
2. Bisher ige Versuche der E r m i t t l u n g des I E P aus dem Stadium minimalen P e r m e a b i l i t ~ t s g r a d e s
Trotz seiner innigen Beriihrung auch mit den vorliegendeu Unter- suchungen kann auf diesen Gegenstand nur noch ganz kurz hingewiesen werden. Wie die vorangestellten Betrachtungen zeigen, liegen zahlreiche,
7pH
.... fff ' A _ _
- 3 - - - - I I11 I I
3 q 5 5 o~
-oj5
-1 ,5 i l i l ~ I 3 g " 5 6 7pH
8,5 o
-0,5 - / ,o - / ,5
c
I I 1 [ [ I
3 q 5 6 7pH
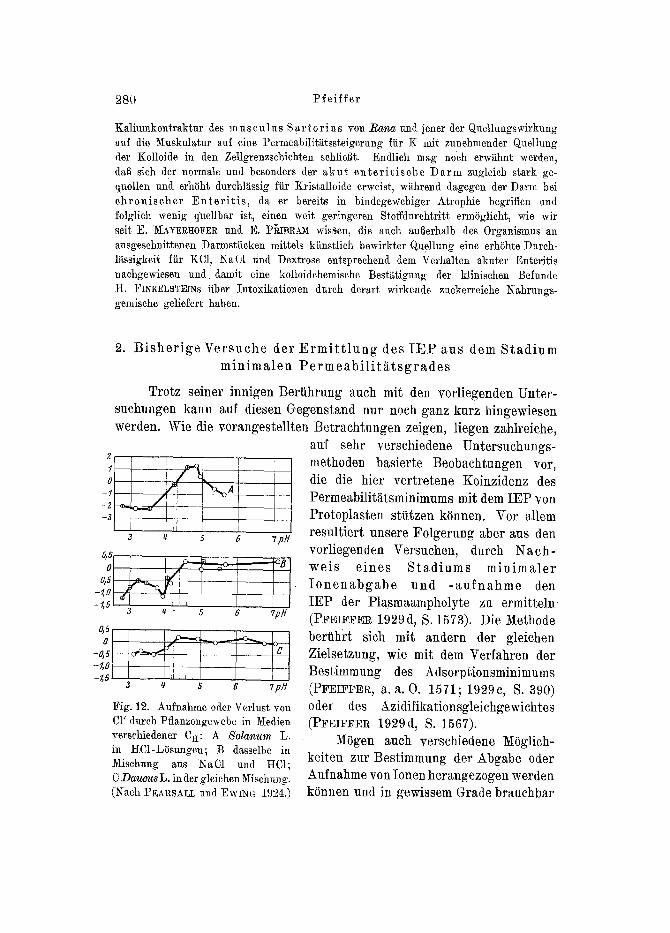
Fig. 12. Aufnahme oder Verlust yon CI' durch Pflanzengewebe in Medien versehiedener CH: A S o l a r i u m L .
in RC1-LSsungen; B dasselbe in ~isehung aus ~aC1 und HC1; C D a u c u s L. in tier gleichen Misehung. (Nach PEARSALL und EwI~G 1924.)
auf sehr verschiedene Untersuchungs- methoden basier te Beobachtungen vor,
die die hier ver t re tene Koinzidenz des
Permeabil i t~tsminimums mit dem I E P yon Protoplas ten st(itzen k~nnen. Vor aliem
resul t ier t unsere Folgerung aber aus den vorliegenden Versuchen, durch N a c h -
w e i s e i n e s S t a d i u m s m i n i m a l e r
I o n e n a b g a b e u n d - a u f n a h m e den I E P der Plasmaampholyte zu e rmi t t e ln (PFETF]PER 1929d, S. 1573). Die Methode
beriihrt sich mit andern der gleichen Zielsetzung, wie mit dem Verfahren der
Best immung des Adsorptionsminimums (PFEIFFER, a. a. 0. 1571; 1929c, S. 390) oder des Azidifikationsgleichgewichtes (PFEIFFEtt 1929d, S. 1567).
MSgen auch verschiedene MOglich- keiten zur Best immung der Abgabe oder Aufnahme yon Ionen herangezogen werden kSnnen und in gewissem Grade brauchbar

Untersuchungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 981
sein,, so ist doch am meisten exakt der c h e m i s c h - a n a l y t i s c h e Nach- w e i s minimaler Ionenpermeabilit~tt. Wegen besonderer Einfachheit der technischen Erfordernisse eignet sich dazu aul~er dem yon D. R. HOAG~Am) gleichfalls geprilften 5 JO/ ;haup t s~ i ch l i ch das C1 +. Hieriiber liegen praktische Erfahrungen au~]er yon I-IOAGLAND haupts~chlich yon PEARSALL und EWING (1924) und yon ROBB~NS und SCOTT (1995) vor. Am meisten durchgearbeitet ist wohl die a r g e n t o m e t r i s c h e F ~ i l l u n g s a n a l y s e mit Ka]iumchromat als Indikator (0. MOHR; vgl. PFErFFER 1999d, S. 1573), wodurch z. B. wertvolle Erfahrungen fiber die Cl'-Aufnahme yon Oeweben yon 3olanum L. und Daucus L. (Fig. 19) gewonnen worden sind, wenn- gleich auch die Bestimmung der E x o s m o s e f a r b i g e n Z e l l s a f t e s in- Iotge Cytolyse (PFmFF~R, a. a. O. 1574) geeignet sein kann.
c} Vergleichende Auswertung der Permeabilit~itsversuche in Hinblick auf die friihere Arbeitshypothese
Wie die wenigen Resultate i~ber g e s e t z m~i~ige Pe rmeab i l i t ~ i t s - v e r ~ n d e r u n g e n im Z u s a m m e n h a n g e mi t M i t o s e n sogleich mit der frtiheren Arbeitshypothese in Einklang zu bringen sind (S. 276), so gilt solches auch ftir die zah]reicheren Beobachtungen t~ber: wechselndes Permeabil[t~itsverhalten bei der A b g ] i e d e r u n g k e r n l o s e r Cyto- p l a s t e n , sofern unsere Ableitungen~ nach denen ftir den IEP ein P6rmeabilitittsminimum zu erwarten ist, zutreffen.
1. Die Tr~iger der physio]ogischen Teilgeschehen, welche den Komplex der Erzeugung kernloser Tochterelemente bilden, sind die Zellen oder ihre Protoplasten, welche aber nicht autonom sind, sondern unter teilweisem Verlust ihrer prospektiven Potenz zu den ,,differenzierten" Elementen der Blattgewebe geworden sind. Es liegen also s p e z i a l i , s i e r t e F u n k t i o n s - ode r D a u e r z e l l e n vor, bei denen Anlagen zu manchen Funktionen, insonderheit zur Teilung des ZellkSrpers, latent geworden sind. Erst durch eine beSondere , , E r r e g u n g " wird eine Teilung iuduziert, wobei dieser Prozef~ in mehr als einer Hinsicht der Ent- wicklungsanregung des tierischen Eies ~ihnelt (F. LEVY, C~. HABERLANDT). Wir kOnnen bier n u n nicht nochmals auf die vielen, oft recht unter- schiedlichen Hypothesen zur Deutung der Teilungsanregung eingehen, zumal S. PRXT dariiber bereits in dieser Zeitschr. (2, 319)refer ier t hat,
"sondern verweisen auf die Ausftihrungen yon FAUR~-FREMI]~T (1925, S. 109, 139 usw.), DALCQ (1998, S. 14, 194, 147 u. a.),, RUi'~I~STROM (1998b) und HEILBRUNN (1998, S. 9 5 5 ) . Schon RUNNSTR0~ (1998b,

282 Pfeiffer
S. 290) vergleicht die Leistungsfahigkeit einer , Q u e l l u n g s t h e o r i e " der Entwieklungserregung (SPEK 19"20, S. 66, 89) mit jener einer in den Grundztigen schon yon M. H. FISCHEt~ und Wo. 0STWALD vor- geschlagenen , ,Koagu la t ions theo r i e " (HEILBRUI~ 1928, S. 260, 280, 285), wobei er noch kritischer ErOrterung der Bezeichnungsweise beider seine der Koagulationshypothese genaherte Einstellung begrtindet. Trotz- dem tritt auch RUNNSTROM fiir eine Hydratotionszunahme der Plasma- kolloide bet der Entwicklungsonregung ein, wobei der Prozef allerdings nur in einer Verschiebung des H y d r a t a t i o n s w a s s e r s im Inne rn der Eize l le zu bestehen braucht. Ahnliches scheint LEV~ (1923, S. 132) bet seiner ,,Analyse des Zellteilungsgeschehens" folgern zu wollen. Er denkt on eine gesetzm~fig verschiedene Empfindlichkeit der Zelle gegen Au6enfaktoren wi~hrend der verschiedenen Phasen der Teilungsphanomene und vermutet in der Hydratationsverschiebung eine Folge lebhofterer Stoffwechselums~tze in der zwischen zwei Teilungsschritten erkennbaren interkinetischen Phase (a. a. O. 170). Indessen bereitet es ouch keine Denkschwierigkeiten, im Kolloidsystem des Protoplasmas gleichfalls die fiir viele heterogene Systeme bekannten, per iodisch ve r l au fenden Prozesse der Grenzflachenbewegungen, der gef6rderten und gehemmten AufRisung ether in die andere Phase und des Auftretens und Absinkens elektromotorischer Kr~fte zwisehen beiden anzunehmen (vgl. tiber die Untersuchungen hauptsachlich yon J. A. DE JONGE, TEMMINCK GROLL, DE BRUIJNE, C. PAAL, W. ttA~T~ANN und C. PH. SLUITE~ die kurze Zusammenfassung bet H~)BER [1924, S. 823]).
2. Es diirfte also die normale Ze l l t e i l uug (ira Anschlufi einer Mitose) ebenso wie die experimentell induzierte Zerschn i i rung des Cytoplasm as (Abgliederung einer kernlosen Tochterzelle) dureh das ge- meinsame Erkl~trungsprinzip ether H y d r a t a t i o n s v e r s c h i e b u n g erfa~t werden. Darin liegt kein Widerspruch zu tier frtiher (PFEIFFER 1929C, S. 417) starker hervorgehobenenVariation des Adso rp t i onsve rha l t en s , m(igen sich beide Erseheinungen doch zugleich mit vielleieht noeh andern teilweise iiberlagern. Wenn wir in diesem Beitrage ein unterschiedliches Permeabilitatsverholten der Protoplasten wahrend verschiedener Stadien der Cytomorphose (i. S. yon M~NOT) als beteiligt an dem ,Teilungs- faktor" (V. JOLLOS) erkannt haben, so werden wir die beschriebenen Versuchsresultate in Best~tigung friiherer Ableitungen (a. o. 0. 419) aueh so deuten k6nnen, daft eine iibersteigerte Hydratation ebenso wie eine besonders starke Permeabilit~tsbegtinstigung dem Funktions- verm(igen der Plasmakolloide gegenli~ufig sein wird (vgl. BOAS 1928,

Untersuchungen iiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. II 283
S. 29), und damit entsprieht der Induktion yon Teilungsprozessen die rtickl/iufige Beeinflussung jenes Faktors. Dann aber bedeutet die T e i l u n g selbst n i ch t e ine A n n ~ h e r u n g an den I E P der P la sma- ampho ly t e , sondern nach weitgehender Verringerung der Distanz gegen jenen Umladungswert ein deutliches Abrticken davon (vgl. PFEIFFER 1929 C, S. 418). Ein aufmerksamer Vergleich der mitgeteilten Befunde tier Per- meabilit/itsmessungen best/itigt sogleich diese Ableitungen.
3. In /ihnlicher Weise l~tl3t sich auch die frtiher vorgetragene Vor- stellung yon dem physiko-chemischen Verhalten der Dauerzellen w/ihrend der E n t d i f f e r e n z i e r u n g (PFEIFI~ER, a. a. O. 420) durch die neuen Ver- suchsresultate belegen. Wenn wir unter Entdifferenzierung die Vor- bereitung zu erneuter Teilungsbereitschaft und zum Riickerwerb der nur noch latent erhaltenen Teilungspotenz begreifen, so gehSren zu den Entdifferenzierungsph~nomenen alle jene, we] eh e ei n e fii r die Tei l u ng u u f ~ h i g e K o l l o i d l a g e zu r e s t i t u i e r e n v e r m S g e n . Unter Ent- differenzierung werden wir sohin die beginnende (oder begonnene?) Abkehr yore w e i t e s t dem I E P g e n ~ h e r t e n S tad ium zu verstehen haben. Im Sinne der in diesem Beitrage haupts~chlich untersuchten ge- setzm~13igen Permeabilit~tsver~tnderungen bedeutet die Entdifferenzierung also /~hnlich wie die Teilung eine Zunahme des vorher minimaleu Per- meabilit/~tsvermSgens der Grenzschiehten der Cytoplasten. Die nahe Verwandtschaft zwischen Entdifferenzierung und Teilung kommt also gut zum Ausdrucke, doch ist die Untersuchung des Permeabilit~ts- verhaltens wenig zur Isolierung der beiden auch zeitlich einander folgenden Prozesse geeignet.
4. Die Untersuchungen yon LEVY (1923), welche er selbst als ein Gegenstiick zu denen yon HABERLANDT (1919a, b; 1920) betrachtet, legen dar, da6 fiir die Beendigung einer Cytoplasmateilung nur die f r i ihes te Einleitung eines Kernteilungsvorganges, event, nur eine Kern- w a n d e r u n g im Cytoplasma, erforderlich zu sein scheint. Bei so welter Begriffsfassung kann letztere sehr wohl schon infolge der tiefgreifenden Ver~tnderungen, welche mit der Plasmolyse und erst rec~t mit der De- plasmolyse verbunden sind (KARzEL 19~6), eintreten. HABERLANDT (1919a, S. 345f; 19~0) m0chte an die Wirksamkeit yon Hormonen denken, deren Konzentration infolge des experimentellen Eingriffes wachsen muB. Mit BoAs (1928, S. 71) werden wir diese Wirkung in- dessen dem yon PFEIFFER (19~9c, S. 4~1) entwiekelten Gedankengange unterzuordnen haben, demzufolge die in dem vorliegenden Beitrage als Begleiterscheinung untersuchte Perm eabi l i t~tsver i~n d erun g den Aus-

284 Pfeiffer
druck der einander gesetzm~flig folgenden Ph~nomene des Ri ickganges u~nd ,der Versti~rkung der Ko l lo idads0 rp t i on darstellt (vgl. auch BOAS 1998, S. 51). l~ber die damit gleichfalls verkntipften , ,auto- k a t a l y t i s c h e n " S o t - G e l - U m w a n d l u n g e n am Kolloidsystem des Pro- toplasmas, die gleichzeitig zu beachten sind~ wird erst in den folgenden Beitr~gen unserer Untersuchungsreihe diskutiert werden kSnnen.
Zusammenfassung 1. NiCht nur bei systematisch niederen Pflanzen, sondern auch bei
manchen Anthophyten, wie bei der untersuchten Helodea densa CASP., ge- tingt durch p l a s m o l y t i s c h e and spi i ter d e p l a s m o l y t i s c h e R e i z u n g eine Indukt~on zur A b g l i e d e r u n g k e r n l 0 s e r Ze l len aus bisherigen Dauerelementen.
2. Diese E n t d i f f e r e n z i e r u n g und T e i l u n g e r s t r e c k e n sich haupts~ichlich auf die einzelligen Haare und auf bestimmte Randzellen des Blattes :and diirfen nicht mit 'gelegentlich ~ durch Plasmolyse hervor- gerafenen Abschntirungen yon Cytoplasmastficken, welche der biekrose verfallen, verwechselt wer@n.
3. In Erg~nzung zu HABERLANDT sind weitere Stiitzen fth" die Deutung der Vorg~nge bei der Abteilung kernloser Gewebelemente als einer - - a l l e rd ings p r im i t i ve ren Form der - - Z e l l t e i l u n g bei- gebracht worden.
4. Unter den zah l r e i chen B e z i e h u n g e n , welche es bei den Erscheinungen der Entdifferenzierung und Teilung zu untersuchen gibt, ist das gesetzm~fig wechselnde P e r m e a b i l i t i ~ t s v e r h a l t e n genauer betrachtet worden,
5. Bei tier kritischen Besprechung der zur Ermittlung des Per- meabilit~tsgrades b r a u c h b a r e n Methoden wird eine fiir das Objekt taugliche Auswah] einfach ausftihrbarer getroffen.
6. Zur Permeabiliti~tsbestimmung werden verschiedene Entwick]ungs- stadien des Versuchsmaterials nach dem A l k a l i e i n t r i t t am F~rbungs- umschlage nach Neutralrottinks nach dem E i n t r e t e n und der Exosmose yon Me thy lenb lau , sowie nach der V e r g i f t u n g durch e i n d r i n g e n d e n A t h y l a l k o h o l verglichen.
7. Wenn auch nicht aile jene Methoden gleich scharf sind, so wird doch mit ausreichender Genauigkeit gefunden, daft mit tier En t - d i f f e r e n z i e r u n g zwar ein P e r m e a b i l i t ~ t s m i n i m u m als Ausdruck der Ann~herung . an den I E P auftritt, aber nicht den Ansto6 zu den Erscheinungen darstellt.

Untcrsuchungen fiber die Entdifferenzierung u. Teilung pflanzlicher Dauerzellen. I I 285
8. Ebenso zeigen Helodea-Zellen in frtihen mi to t i schen Stadien ein Permeabilit~tsminimum, das jedoch njcht die Prozesse e in]eitet.
9. Fiir bekannte Abweichungen yon diesen Beobachtungen wird eine Erkl~rung darin gegeben, daI~ tibereinstimmende Schwankungen in den physikalischen Charakteren als Teilph~nomen des n e b e n dem F u n k t i o n s w e c h s e l a n z u n e h m e n d e n Tonus vermutet werden miissen, die unter nicht genauer bekannten Bedingungen die daneben vorhandene typische Gesetzm~tl~igkeit his zur Unkenntlichkeit iiberlagern.
10. Die beiden Zel l formen mit und ohne Karyoplasma unter- scheiden sich hinsichtlich ihres Permeabilitiitsgrades nicht wesentlich.
11. Es wird diskutiert, wie sich E n t d i f f e r e n z i e r u n g und Te i lung (insbesondere Abgliederung kernloser Cytoplasmastiickei auch bei Betrachtung des Permeabilit~tsverhaltens tier Protoplasten in dem friiher a b g e l e i t e t e n S inge begre i fen lassen, wenn eine Koinzidenz des Permeabilit~ttsminimums mit dem IEP angenommen werden dart.
12. Diese Gesetzmgl3igkeit der B e z i e h u n g des IEP zum Per- meabi l i t~ t smin imum wird dutch viele b i sher ige E r f a h r u n g e n und durch die Methodik des N a c h w e i s e s isoelektrischer Reaktion gesttitzt.
L i t e r a t u r f i b e r s i c h t
1. ANSELMIN0~ K. J., Untersuchungen fiber die Durehliissigkeit kiinsflieher kolloidaler ~[embranen. Biochem. Zei~sehr. 192, 390 (1927).
2. BOAS, FR., Die Pflanze Ms kolloides System (Naturw. u. Landwirtseh. 14:). Freising- Miinchen (F. P. Datterer & Cie.) 1928.
3. BOBILOFF-PRErSSER, W , Zur Physiologic des Pollens. Beih. Bet. Orb., (I)~ 88, 248 (1917).
4. BROOKS~ MATILDA M., Studies on the permeability of living and dead cells~ I I I . The penetration of certain alkalis and ammonium salts into living- and dead cells. Publ. Health Rep. 88, 2074 (1923).
5. COLLANDER, R.: T]ber die Permeabilitiit pflanzlieher Protoplasten ffir Sulfosiiure- farbstoffe. PRtNGSgE[Ms Jahrb. f. wiss. Bet. 60, 354 (1921).
6. CONKLIN, E.~ Ce]i-nlar differentiation, in :EDM. V. COWDRY, General Cytology~ Chicago (Univ. of Chicago Press) 1924, S. 539.
7. DALCQ~ A , Les bases physiologiques de la fgeondation et de la parthgnogen~se. Paris (Les Presses Univers.) 1928.
8. EPHRUSSI, B., Action de ]a chaleur sur l'ceuf d'Oursin, II . Variations de ]a per- mgabilitg aux sels chez l'ceuf vierge. Bull. Instit. 0e~an. n o 458 (1925).
9. - - La r~action ~ la f6condation des oBufs d'Oursin chauffgs (Variations de la perm4a- bilit4 aux se]s). Compt. rend. See. Biol. 93, 1561 (1925).
10. FAUIC~,-FREMIET, E., Variation pgriodique de la sensibilit4 de l~ceuf de Sabellaria alveolata L. aux solva.nts des graisses, ibid. 85, 1051 (1921).

286 Pfe i f f e r
11. FAUR]~-FREMIET, E., Propri~t6s osmotiques de l'ceuf de Sabellaria alveolata. Compt. rend. Soc. Biol. 88, 1028 (1923).
12. - - - - La cin6tique du d6veloppcment. Multiplication cellulaire et croissance. Paris (Les Presses Univers.) 1925.
13. FREE,, E. E., A colloidal hypothesis of protoplasmic permeability. Plant World 21, 141 (1918).
14. GELLHORN, E., Das Permeabilitiitsprob]em (~onogr. a. d. Oes.-Geb. d. Phys. d. Pfl. u. d. T. 16). Berlin (Jul. Springer) 1929.
15. GRAFE, V., ~ethodik der Permeabilitatsbestimmung bei Pflanzenzellen, in E. ABDER- HALDEN. Handb. biol. Arbeitsmeth., (XI), 2, 29 (1924).
16. GR0SSMANN, E., Zellvermehrung and Koloniebildung bei einigen Scenedesmaceen. Intern. Rev. ges. Hydrobiol. u. Hydrogr. 9, 371 (1921).
17. HABERLANDT, Cx., Zur Physiologie der Zellteflung. 3. ~[itt., ]Jber Zellteilungen nach Plasmolyse. Sitz.-Ber. Preu~. Akad. Wiss., math.-phys. K1., 1919, S. 322.
18. - - 4. Mitt., Uber Zellteilungen in Elodea-Blattern nach Plasmolyse. Ibid. 1919, S. 721.
19. - - 5. ~itt.: f~ber das Wesen des plasmolytischen Reizes bei Zellteilungen nach Plasmolyse. Ibid. 1920, S. 323.
20. HAYNES~ D., The action of salts and non-electrolytes upon buffer solutions and amphoteric electrolytes and the relation of these effects to the permeability of cell. Biochem. Journ. 15, 440 (1921).
21. HEILBRUNN, L.V.: The colloid chemistry of protoplasm (Protoplasma-Monogr. 1). Berlin (Gebr. Borntraeger) 1928.
22. HERLANT, 1~[.~ Variations cycliques de la perm6abilit6 ehez l'oBuf activ6. Compt. rend. Soe. Biol. 81, 151 (1918).
23. - - Le cycle de la vie cellulaire chez l'o~uf active. Arch. de Biol. 80, 519 (1920). 24. HITCHCOCK, DAVID J.~ The effect of PH on the permeability of collodion membrane.
Journ. Gen. Phys. 10, 179 (1926). 25. H6BnR, R, Physikalische Chemic der Zclle und der Gewebc, 5. Anti. Leipzig
(W. Engelmann) 1924. 26. HOFFMANN, C., ~ber die Durchlassigkeit kernloser Zellen. Planta ~, 584 (1927). 27. HUMMEL, H. and POSCHEL, J., ]~ber die Zuckerwirkung bei der Guanidinvergiftung
und ihre Bedeutung ffir die Permeabilitatslehre. PFLUGERS Arch. ges. Physiol. 217, 441 (1927).
28. ILJtN~ W. S., Die Permeabilit~t des Plasmas fiir Salze and die Anatonose. Stud. Plant Phys. Labor. Charles Univers. Prague 1, 97 (1923).
29. - - D i e Durchlassigkeit des Protoplasmas, ihre quantitative Bestimmung und ihre Beeinflussung dutch Salze und dureh die Wasserstoffionenkonzentration. Proto- plasma 3, 588 (1928).
30. ISABUI~O-I~AGAI, Physiologische Untersuchungen fiber Farnprothallien. Flora 106, 281 (1914).
31. KACZMAR:EK~ A., Untersuchuugen fiber Plasmo]yse and Deplasmolyse in Abh~ngig- keit yon der Wasserstoffionenkonzentration. Protoplasma 6, 209 (1929).
32. KAH0, It., Das Verhalten der Pflanzenze]le gegen Salze. Ergebn. d. Biol. 1, 380 (1926).
33. KARZEL, R., Uber Nachwirkungen tier Plasmolyse. PRINGSHEIMS Jahrb. f. wiss. Bot. 65, 551 (1926).

Untersuchungen fiber die Entdifferenzierung u. Tcilung pflanzlicher Dauerzellen. I I 287
34. KLEBS, G, Beitr~ge znr Physiolog~ie der Pflanzenzelle. Ber. d. Dtsch. Bot. Ges. 5~ 181 (1887).
35. --Beitr~ge zur Physiologie tier Pflanzenzelle. Unters. Bot. Instit. Tfibingen 2, 489 (lSSS).
36. - - Uber die Fortpflanzungsphysiologic der niederen Organismen, der Protobionten. Spezieller Theft, Die Bedingungen dcr Fortpflanzung bei cinigen Algen und Pilzen. Jena (G. Fischer) 1896.
37. K0STER, E., Zelle und Zellteilung (botan.). ttandwi3rtcrb, d. Naturw. 10, 748 (1915). 38. - - U b e r Vakuolenbildun~ und grobschaumige Protoplasten. Ber. d. Dtsch. Bot.
Ges. 86~ 283 (1918). 39. - - Experimente]le Physiologic der Pflanzenzelle, in E. ABDERttALDEN~ Handb. biol.
Arbeitsmeth. (XI.), 1, 961 (1924). 40. - - Pathologisehe Pflanzenanatomie, 3. Aufl. Jena (G. Fischer) 1925. 41. --Pathologic der Pflanzenzelle, Tell I. Pathologic des Protop]asmas (Proto-
plasma-Monogr. 3). Berlin (Gebr. Borntraeger) 1929. 42. LEVV~ FI~, Untersuehungen fiber abweichende Kern- und Zellteilungsvorg~nge, L
Ubcr heteromorphe Zelten in Hoden yon Amphibien. Ztschr. ges. Anat., (I), 68, 110 (1923).
43. LILLIE, F. R, Problems of fertilization. Chicago i(Univ, of Chicago Press) 1919. 44. LILLIE, R. S.~ Increase of permeability to water following normal and artificial
activation in sea-urchin eggs. Amer. Journ. Physiol. tO, 249 (1916). 45. - - Comparative permeability of fertilized and unfertilized eggs to water. Science 47~
147 (1918). 46. LINSBAUER, K, Weitere Beobaehtungen an SpaltSffnungen. Planta 4:~ 584 (1927). 47. LLOYD, F. E., The behaviour of protoplasm as a colloidal complex. Ycarb.
Carnegie Inst. Washington lt~ 66 (1915). 48. MOND~ R, Umkehr der Anionenpermeabilit~t der roten BlutkSrperchen in eine
elektive Durchl~tssigkeit ffir Kationen bei alkaliseher" Reaktion. Klin. Wochen- schr. 6, 1432 (1927).
49. - -Umkehr der Anionenpermeabilit~t der roten BlutkSrperchen in eine elektive Durchl~ssigkeit ffir Kationen. Ein Beitrag zur Analyse tier Zellmembranen. PFLi)GERS Arch. ges. Physiol. 217, 618 (1927).
50. INI~MEC~ B, Das Problem der Befruchtungsvorg~tnge. Berlin ((kebr. Borntraeger) 1910. 51. OSATO, Srl.~ Der isoelektrische Punkt des Globins. Biochem. Ztschr. 132, 485 (1922). 52. PEARSALL~ W. H. und J. ]~WING, The diffusion of ions from living plant tissues
in relation to protein isoelectric points. New Phytol. 23, 193 (1924). 53. PFEIFFER, H., Uber die l~litwirkung elektro-kapillarer Effekte bei der Vitalfitrbung
pfianzlicher und tierischer Protoplasten. Biol. Ztb. 47~ 201 (1927). 54. - - Der isoelektrische Punkt yon Zellen und Geweben. Biol. Reviews (Cambridge) 4,
1 (1929). 55. - - Elektrizitat und Eiweil~e insbesondere des Zdlplasmas (Wiss. Forsch.-Ber.,
Naturw. Reihe, 21). Dresden u. Leipzig (Th. Steinkepff) 1929. 56. - - Experimentelle and theoretische Untersuehungen fiber die Ent~erenzierung und
Teilun_g pflanzlicher Dauerzellen, I. Der isoelektrische Punkt (IEP) und die aktuelle Azidit~tt yon meristematisierten Zellen. Protoplasma 6, 377 (1929).
57. - - D e r isoelektrische Punkt (IEP) yon Protoplasten und seine Ermittlung. In E. ABDERHALDEN, Handb. biol. Arbeitsmeth., (V), 2/~. 1563(1929).

288 Pfe i f fe r , Unters. fiber d. Entdifferenzicrung u. Teilung pflanzl. Dauerzellen. IT
58. POIJXEVI, A. Die Basenpermeabilit~tt pttanzlicher Zellen. Acta hot. Fennica 4, 1 (1928).
59. PORTltEIM, L. V. and E. LOWI, Untersuchungen fiber die Entwickclungsf~higkeit der PollenkSrner in verschiedenen Medien. 0sterr. Bet. Ztschr. 59, 134 (1909).
60. PI~:T, S., Plasmolyse und Permeabilit~t. Bioch. Ztschr. 128, 557 (1922). 61. - - Weiteres fiber Plasmolyse und Permeabilit~t (Plasmolysis and permeability IV).
V~stnlk kr~l. uSen6 Spol. nauk. Praha 1925, S. 1. 62. REI~ttARDT, M. 0., Plasmolytische Studien zur Kenntnis des Wachstums der Zell-
membran, in Fcstschr. S. SCHWENDENER. Berlin (Gebr. Borntraeger) 1899, S. 425. 63. REMINGTON, ROE E., The high frequency Whcatstone bridge as a tool in cyto-
logical studies; with some observations on the resistance and capacity of the cell of the Beet root. Protoplasma 5, 338 (1928).
64. RISSE: 0., Uber die Durchl~sigkeit yon Kollodium- und EiweiSmembranen ffir einige Ampholyte, I. Der Einfluit der H- und OH-Ioncnkonzcntration. PFLI)GERs Arch. gcs. Physiol. 212, 375 (1926).
65. -- II . Quellungseinflfissc. Ibid. 213, 685 (1926). 66. ROBBINS, W. J. und IEL T. SCOTT, Further studies on isoelectric points of plant
tissue. Journ. Amer. Agr. Rcs. 81, 385 (1925). 67. ]~UN~STRSM, J.: Zur Kenntnis der Zustandsver~ndcrungen der Plasmakolloide bei
der Reifung, Befruchtung und Teilung des Seeigeleies. Acta Zool. 5, 345 (1924). 68. - - Die VerKnderungen der Plasmakolloide bei der Entwicklungsanregung des See-
igelcies, Protoplasma 4, 388 (1928); 5, 201 (1928). 69. RUPPEttT, W., Empfindlichkeits~nderungen des Aseariseies auf verschiedencn Stadien
der Entwicklung gegenfiber der Einwirkung ultravioletter Strah]en. Ztschr. wiss. Zool. 128, 103 (1924).
70. SCHNEIDER, H., Die botanische ~Iikrotechnik. Jena (G. Fischer) 1922. 71. SttIBATA, K., Experimentel]e Studien fiber die Entwicklung des Endosperms bei
Monotropa. Biol. Ctb. 22, 705 (1902). .: 72. S~[ALL, J., Hydrogen-ion concentration in plant cells and tissues (Protoplasma-
Monogr. 2). Berlin (Gebr. Borntraeger) 1929. 73. SPEK, J., Expcrimentclle Beitr~ge zur Kolloidehemie der Zellteilung. Kolloidchem.
Beih. 12, 1 (1920). 74. STRASBUROER, E., Uber Zellbildung und Zellteilung. gena (G. Fischer) 1875. - -
3. Aufl., Jena 1880. 75. -- Uber die Wirkungssph~re der Kerne und die ZellgrSt~e (Histol. Beitr. 5, 95 bis
124). Jena (G. Fischer) 1893. 76. TCHAHOTINE, S.: Action localis~e-des rayons ultra-violets sur le noyau: Compt.
rand. Soc. Biol. 88, 1593 (1920). 77. - - Les ehangements de la perm~abilit6 de ]'ceuf d'Oursin localis6s cxp6rimentalement.
Ibid. 84, 446 (1921). 78. TISCItLER, (~., Allgemeine Pfianzenkaryologie, in K. LINSBAUER, Handb. d. Pfl.-
Anat., I. Abt,, 1. Teil, 2. Berlin (Gebr. BorntrKger) 1921/22. 79. WEBER, FR., Experimentelle Physiologic der Pflanzenzclle. Arch. exper. Zell-
forsch. 2, 67 (1925). 80. Z[H~IERMANN, W., Zytologische Untcrsuchungen an S~haeelaria fusca AG. Ein
Beitrag zur Entwick]ungsphysiologie der Zelle. Ztschr. f. Bot. 15, 113 (1923).
















![Hämodynamische Auswirkungen synthetischer, pflanzlicher ... · weitere endogene Cannabinoide zu identifizieren [24]. Anandamid entsteht aus einer membrangebundenen Vorläuferstufe,](https://static.fdokument.com/doc/165x107/5dd12c40d6be591ccb649235/hmodynamische-auswirkungen-synthetischer-pflanzlicher-weitere-endogene-cannabinoide.jpg)
