
Flexibility of Tropocollagen from Sedimentation and Viscocity · Comparison of experiment.al...
18
C .o llagen Symp .o sium , X (1973) FlexibilityofTropocollagen from Sedimentation andViscocity Hiroyasu Utiyama , Kuniaki Sakato , Kenji Ikehara , Takashi Setsuiye and Michio Kurata 1n s ti tu te f .o r Chemical Research andDepartment 。 f1ndusirialChemistry , Ky .o t .o University SYNOPSIS Sedimentati .o n c .o nstant and int 1 ' insic visc .o sity we 1 ' e measu 1 ' ed .o n purified tr .o p .o collagens ext 1 ' ac ted fr .o m ea 1 ' thw .o rm-cu ticle and la thy 1 ' itic ra tsl 、 in ・ A cartesian diver visC .o meter was used t .o make visc .o sity measu 1 ' ements at small shea 1 ' st 1 ' esses and t .o avoid th 巴 effects .o fsurface f .o1' ces. By c .o mpa 1 ' ing the expe 1 ' imental data with the hyd 1'.o dynamic the .o1' ies .o f w .o1' m- like-c .o il .o fUllman , a value .o f 1300 A has been assigned f .o1' the pe 1 ' sist- ence length .o fthese t 1'.o p .o c .o llagens. Othe 1 ' fact .o1' s which may affect the estimate a 1 ' e discussed. INTRODUCTION -29- Like the a- helix .o f p .o lypeptide and the d .o uble-helix .o fDNA , the t 1 ' iple heli cal st 1 ' uctu 1 ' e .o f c .o llagen is .o ne .o fthe unique c .o nf .o1' mati .o ns .o fbi .o l .o gical mac 1'.o m.o le- cules. 1ts gene 1 ' al featu 1 ' es that a 1 ' e n .o w widely accepted as c .o1'1' ect a 1 ' e th 1 ' ee helical p .o lypeptide chains a 1 ' e w .o und a 1'.o und each .o the 1 ' t .o f .o1' m a th 1 ' ee-st 1 ' anded 1'.o pe- like st 1 ' uctu 1 ' e. Ramachand 1 ' an and Ka 1 ' tha 1) fi 1 ' st pr .o p .o sed this s truc tu 1 ' e .o n the basis .o fthe fact that the 1 ' e is a n .o n-integ 1 ' al sc 1 ' ew axis which 1 ' elates equivalent units by a t 1 ' anslati .o n .o f2.9λand a 1'.o tati .o n .o fapp 1'.o ximately 108" , a 吋 thateach chain , has ev 巴1' ythi 1 ' d residue glycine. 1n additi .o n , the 1 ' e is n .o w little d .o ubtin that this t 1' uctu 1' e is unive 1 ' sal t .o ve 1 ' tebrate and inve 1 ' teb 1 ' ate c .o llagens. The ex- pe 1 ' imental evidences f .o1' this view a 1 ' e the glycine 1 ' esidue c .o ntents a 1 ' e ve 1 ' y cl .o se t .o .o ne thi 1 ' d , and the wide-angle X- 1 ' ay patte 1 ' ns .o fsamples .o1' iented by st 1 ' etching a 1 ' e typical .o f c .o llagens:;) The m.o1' e p 1 ' ecise m.o lecula 1 ' c .o nf .o1' mati .o n has l .o ng been c .o n-
Transcript of Flexibility of Tropocollagen from Sedimentation and Viscocity · Comparison of experiment.al...

C.ollagen Symp.osium, X (1973)
Flexibility of Tropocollagen
from Sedimentation and Viscocity
Hiroyasu Utiyama, Kuniaki Sakato, Kenji Ikehara,
Takashi Setsuiye and Michio Kurata
1n s ti tu te f.o r Chemical Research and Department
。f1ndusirial Chemistry, Ky.ot.o University
SYNOPSIS
Sedimentati.on c.onstant and int 1'insic visc.osity we1'e measu1'ed .on purified
tr.op.ocollagens ext1'ac ted fr.om ea1'thw.orm-cu ticle and la thy 1'itic ra tsl、in・
A cartesian diver visC.ometer was used t.o make visc.osity measu1'ements
at small shea1' st1'esses and t.o avoid th巴 effects.of surface f.o1'ces. By
c.ompa1'ing the expe1'imental data with the hyd1'.odynamic the.o1'ies .of w.o1'm-
like-c.oil .of Ullman, a value .of 1300 A has been assigned f.o1' the pe1'sist-
ence length .of these t1'.op.oc.ollagens. Othe1' fact.o1's which may affect the
estimate a1'e discussed.
INTRODUCTION
-29-
Like the a-helix .of p.olypeptide and the d.ouble-helix .of DNA, the t1'iple heli
cal st1'uctu1'e .of c.ollagen is .one .of the unique c.onf.o1'mati.ons .of bi.ol.ogical mac1'.om.ole-
cules. 1ts gene1'al featu1'es that a1'e n.ow widely accepted as c.o1'1'ect a1'e th1'ee
helical p.olypeptide chains a1'e w.ound a1'.ound each .othe1' t.o f.o1'm a th1'ee-st1'anded 1'.ope-
like st1'uctu1'e. Ramachand1'an and Ka1'tha 1) fi 1's t pr.op.osed this s truc tu1'e .on the
basis .of the fact that the1'e is a n.on-integ1'al sc1'ew axis which 1'elates equivalent
units by a t1'anslati.on .of 2.9λand a 1'.otati.on .of app1'.oximately 108" , a吋 thateach
chain, has ev巴1'y thi1'd residue glycine. 1n additi.on, the1'e is n.ow little d.oubt in
that this t1'uctu1'e is unive1'sal t.o ve1'tebrate and inve1'teb1'ate c.ollagens. The ex-
pe1'imental evidences f.o1' this view a1'e the glycine 1'esidue c.ontents a1'e ve1'y cl.ose
t.o .one thi1'd, and the wide-angle X-1'ay patte1'ns .of samples .o1'iented by st1'etching a1'e
typical .of c.ollagens:;) The m.o1'e p1'ecise m.olecula1' c.onf.o1'mati.on has l.ong been c.on-

Aυ 。。
trovertial due to the limited detail of the X-ray pattern of collagen and the com-
plexity in the amino acid sequence 3-6). 1 t seems tha t the triple helical confor-
mation proposed by Yonath and Traub71, with one NH.. 0 interchain hydrogen bond per
。tripeptide of a hydrogen bond length of 2.86 A, is increasingly favored by the recent
8.9) mvestJgatJons 3)
This structure resembles collagen II type in its mode of hy-
drogen bonding, but the hydrogen-bond length is significantly longer than the values
previously reported.
The size and shape of tropocollagen in solution were established by Boedtker
and Doty in 1956 on collagen extracted from the carp swim bladder tunics (ichthyocol)
from measurements of light-scattering, osmotic pressure, intrinsic viscosity, sedi-
mentation constant, and flow birefringencelO). Comparison of experiment.al results
with a Iight-scattering th巴oryof thin rodsl1
) and hydrodynamic theories of prolate
ellipsoids of revolutionI2-15) lead them to an unequivocal and unified conclusion that tro-
pocollagen consists of rigid, rod-shaped molecules with a diameter and a length of 13.6
A and 3,000 A, respectively. This conclusion stood in remarkable agreement with
findings from X-ray diffraction j) and electron microscopic s tudiesI6). S imilar results
have been repeatedly obtained on numerous oth'er collagens17田:
1t may well be expected, however, that longer tropocollagen will show up some
flexibility, becaus巴 nomatter is completely rigid. The tropocollagen extracted from
the earthworm-cuticle is quite interesting in this respect; the molecular weight is 2
to 6 times as large as the usual vertebrate collagens, but otherwise similar24.251.
If we calculate the particle length, assuming a prolate ellipsoid of revolution, from
the intrinsic viscosity data obtained by Josse and Harrington25J, it will amount to only
about one half of the values expected for rigid rods. This clearly indicates that sol-
ution properties of earthworm-cuticle collagen need to be analyzed in terms of the
wormlike-coil modeI26-V;)
1n treating with this problem, there are two important aspects to be considered
with care. One is experimental in nature and the other is theoretical. First,
viscosity of solutions of very rigid and long particles such as earthworm-cuticle col-
lagen should be considerably subject to the influence of the sh巴ar stress. 1n addi-
tion, viscosity measurements will be badly influenced by the solid surface film formed
at the liquid-air interface. Second, the existing theories of intrinsic viscosity and
sedimentation constant on the wormlike-coil model include hydrodynamic diameter and
friction constant per unit length as the param巴terswhich are no way uniquely deter-
min巴d. Therefore, the persistence length is estimated with greater credence frbm
comparison of hydrodynamic properties of more than one samples of the same species
but of different molecular weight. With respect to the former, we may obtain accu-29)
rate viscosity data with use of a viscometer of cartesian-diver type For the
latter, we may regard earthworm-cuticlp collagen and other vertebrate collagens such

唱
EA
qd
as ratskin collagen as a homologous series, because in these collagens the content
of the glysine residue is very close to one third and the molecular w巴ight of dena-
tured gelatin is also approximately one third of the native tropocollagen.
Thus, in the present investigation we attempt to reinvestigate intrinsic viscosi-
ty and sedimentation constant of tropocollagens extracted from the earthworm-cuticle
and the ratskin in the hope of deducing a universal persistence length.
MATERIALS AND METHODS
Extrαc tionαηd purific日tionof co IIαgen
Earthworm-cuticle collagen and lathyritic ratskin collagen were prepared essen-
tially following the procedures described by Josse and Harr・ington251 and Nimni叫
respectively. The proc巴duresare briefly described here. Lathyritic collagen was
employed simply to use in a later work of renaturation kinetics on a-gelatin.
Earthworms, Phere tma communis s ima, were collec ted locally and s tored f rozen
in a refrigerator until use. After thawing, about 1, 000 earthworms were removed off
cuticles with forceps. The cuticles were minced with scissors, washed thoroughly
with water, and ground with a mortar and pestle until a thick slurry was obtained.
This material was suspended in 100 ml of 0.5 M NaCl and collagen was extracted for
48 hr with stirring. The resuItant viscous suspension was separated from undis-
sol ved material by centrifugation and subject巴d to precipitation twice with saturated
ammoniun sulfa te. The pellet collected by centrifugation was finally dissolved in
30ml of O. 2 M r、JaCl,centrifuged at 105, 000 x g for 1 hr, and th巴 supernatantliquid
collected. This solution was thoroughly dialyzed against 0.2 M NaCl or 0.06 M sodium
acetate buffer, pH4. 8, and stored at 40C.
Rats (Sprague-Dawley) were placed on a diet supplemented with 400 mg of D-peni-
cillamine per kg for 7 days. About 30 g of dorsal skin were cleaned of adhering
tissue and fat. After mincing with scissors into small pieces and washing with water,
they were homogenized for 20 min with 20 volume's of 0.15 M NaCl in Sakuma homoge-
nizer. The homogenized material was suspended in 30 ml of 0.15 M t、JaCl and collagen
was extracted for 48 hr under stirring. Undissolved material was then separated from
the solution by filtering through several layers of cheese cloth and by centrifugation
at 77,000 x g for 2 hr. The crude collagen solution was purifi巴dby precipita tions
with ac巴ticacid and saturated NaCl solution. The precipitate was collected by cen-
trifugation and dissolved in O. 5 M acetic acid with stirr・ing for 24hr. After cen-
trifugation, the supernatant was adjusted to pH 4.8 by addition of 1 M, potassium a-
cetate and thoroughly dialyz巴d against 0.06 M' potassium acetate buffer, pH4. 8, and
10'3 M sodium bisulfite. Sodium bisulfite was added to prevent the formation of

32
free aldehydes叫 Thecollagen solution was finally centrifuged at 105,000 x g for
4 hr and the supernatant collected as the sample solution.
until use.
Conceηtration determination
1t was stored at 40
C
Concentrations were determined as an average of at least thre巴 independentmi-
cro-Kieldahl determinations. A nitrogen content of 18.2% was used for both the
ra tskin and earthworm-cuticle collagens to convert nitrogen analysis to dry weight
of collagen 25.:121.
Intrins icγiscosity
Viscosities of sample liquids were measured in a rota ting cartesian diver vis-{~)
cometer which was described in detail elsewhere" Th巴 rotorused was like a test
tube in shape; 9mm in outer diameter and 50mm in length. A copper ring which
fitted tightly insid巴 the rotor was fixed at about 5 mm from the open end with epoxy
resin. The rotor was submerged upside-down in the test liquid in the stator with
an inner diameter of 10.66 mm. Driving torque was produced through the interaction
of the external rotating magnetic field with the magnetic moment induced by the eddy-
色urrents in the copper ring. The vertical position of the rotor was maintained
constant to 土 0.1 mm by manually regulating pressure inside the rotor while viewing
the rotor through a cathetometer. To achieve the stationary and concentric revo-
lution of the rotor, the copper ring should have b巴enappropria tely fi!ed in advance.
The temperature of the sample liquid could be kept constant to 土1.01 'C by circu-
lating water in the mantle surrounding the stator from a constant temperature bath.
The range of shear stress obtainable by changingthe magnet speed was 0.003 -0.4
dynes・ cm-"forwater at 20 oC. The relative viscosity was estimated according to
the equa tion:
ηr = (P - Pml / (P 0-Pml (1)
where P and P 0 denote the speeds of rotation of the rotor in the collagen solution
and the salt solution used as solvent ,respectively, and P ~ the speed of the magnet. 町1
The relative viscosity could be m巴asuredcorr巴ctly to 士 0.1 %. 1ntrinsic viscosity
was estimated as the coincident intercept of th巴 plotsofη可、/c versus η( the 'ISp' - . ----- '/ sp、
S chulz and B laschke plot ) and in ηrel/ c versus c (the Mead and Fuoss plotl.
Partiα1 specific volume
Densities of solvent and solutions were measured in a measuring-cylinder pycno-
meter of the Lipkin-Davison type. For lathyritic ratskin collagen in 0.06 M potas-
sium buffer, pH 4.8, the partial specific volume was determined as 0.698土 0.005ml/
g. This vaJue is compared to the data reported in the Jiterature: 0.705 for dog-2:{)
fish shark skin collagen in sodium formate, pH 3.8, 20 oC 0.70t and 0.686 in so-
dium phosphate buffer, pH 7.4, and sodium citrate buffer, pH 3.7, respectively, for

qa qa
calf skin collagenl9J
. Density measurements on earthworm-cuticle collagen could not
be made due to shortage of the sample. We therefore used a value of 0.69 ml / g
which was calculated from the amino acid composition25J.
Sedimeηtation constαnt
Sedimentation velocity measurements were made at 20'C in a Beckmaim-Spinco
Mod巴1E ultracentrifuge at a rotor speed of 59,780 rpm. Sedimentation of the
boundary was measured with use of the Schlieren optics, and th巴 Schlierenpatterns
obtain巴d were read on a Nikon universal contour projector, Model V-16, accurate to
土 0.005mm. Sedimentation constant at infinite dilution was estimated from a plot
。f1/ S versus concentration, and the result obtained was converted to that for the
standard condition of water at 20'C by multiplying by (7Jsolvent/ザH20)andby (1予pw)/
(1ー予ρ ) . solvent
S edimenta tion equi lib rium
The short column m巴thodof Van Holde and Baldwin341 was employed for sedim巴nta-
tion eqilibrium measurements on ratskin collagen. The experim巴ntalprocedures were
describ巴d in detail elsewhere35J. Five test solutions of different concentration
were simultaneous¥y centrifuged in a multicell rotor of An-G type with a solution
co¥umn height of approximate¥y 0.17 cm. An inert ¥iquid was not inserted at the cell 361
bottom. The overspeeding t巴chniquedescrib巴d by Hexner et al.~' was used to reduce
the time r巴qUlr巴d for the attainment of巴qui¥ibrium. The attainment of sedimenta-
t10n 巴qui¥ibriumwas assured by plotting against time the horizontal distance between
two Ray¥巴igh fringes s巴paratedas distant as possible on the photograph. The cen-
trifugation was not stopped b巴fore the distance was observed constant at ¥east for 2
hr. Since the ratskin' collagen preparations were fair¥y mon odisperse, the mo¥ecu¥ar
weight and the second viria¥ coefficient can be estimated unambiguous¥y from
M.~~ =M+ B ( c.-c、)/2十higher terms in c and c.. app a b a h (2)
Here M is the molecu¥ar w巴ightof the so¥ute, c a and cb the so¥ute concentrations
in equilibrium at the meniscus and at the bottom, respective¥y, and B is twice the
second viria¥ coefficient. The apparent mo¥ecu¥ar weight M appis defined by
M ム二三三一 2 RT (3)
一 -app c 0 ( 1ーマグ)ω2(ri-r;)
with P" the so¥vent density,ωthe angu¥ar speed of the rotor, r a and r b the radi-
al distances from the center of rotation to the meniscus and bottom of the so¥ution
co¥umn, r巴spectively,R the gas contant, T the absolut巴 temperature,and c 0 the
solute concentration prior to centrifugation. One may obtain, as we did in the pre--1
sent investigation, accurate mo¥ecular weight by simp¥y p¥otting Mapp versus c 0 as in
eq. (5) below, but the s¥ope in this p¥ot does not give the s巴condviria¥ coefficient.

34-
For eathworm-cuticle collagen solutions we employed the Yphantis procedure制,
because the concentration of the collagen preparation was rath巴r low. Centrifuga-
tion was made with the Schlieren optics using an eight-channel centerpiece. The
apparent molecular weight was estimated from
R T 1 dc =一一一←一一一一(-)
app (1-示p")ωrc0 dr r
(4)
whe re r, the radial position whe re the concentra tion remains unchanged upon cen tri-
fugation, is assumed as the midpoint of the liquid column. 1n this method, eq. (2)
is replaced by
M-l= M l十 Bco+・・・ (5)
RESULTS
Experimental results of M app obtained by s巴dimentation equilibrium measure ments
are plotted in Fig. 1 versus the solute concentration prior to centrifugation. Linear
extrapolation of the plotted points to infinite dilution yields the molecular weights
of 3ρ1 土 0.3 x10 and 169 士10 X 10' for ratskin and earthworm-cuticl巴 collagens
resp巴ctively. The former r巴sultstands in very good agreement with the data
of 29.4 X 10' reported by Lewis and Piez231
• The molecular weight of earthworm 23)
4 cuticle collagen may be compared with value of 190 X 10', which was obtained by
25) combining the sedimentation constant and intrinsic viscosity~J' with a βvalue of 3.65
X 10 ぺ
Fig. 2 shows the results of s巴dimentationvelocity measurem巴nts. The plott巴d
points of each collagen follow a straight line, and the intercept at zero solute con-
centration yields S"20, wof 2.82 土 0.05and 5.08士 0.05for ratskin and earthworm
collagens, respectively. The present experim巴ntalresults on earthworm collage口 ar巴
251
contrasted with those of Josse and Harrington ,in which data points in the similar
plot exhibited marked d巴viation from linearity below the concentration of about 0.5 mg
/ ml. They obtained a value of 7.0 for SZO, win 0.2 M NaCl at pH 6. The sedト
mentation constant for ratskin collagen is compared to the value of 3.15, which was
obta.in巴d in citrate buffer, pH 3.7動.
Experimental results of viscosity measurements are shown in Figs. 3 and 4 for
ratskin and earthworm collagens, respectively. Th巴 shear stree used in the meas-
urements was 0.17 dynes / cm -. The plotted points wer巴 extrapolatedto infinite di-
lution so as obtain a common int巴rceptand the same value of Huggins' constant in
the two types .of plot. The intrinsic viscosity of 12.4 dl / g obtained on ratskin
collagen seems to b巴 in satisfactory agreement with a literature value of 13.7 dl / g

4.0
(0
~ 3.5 x '0. a ro
芝
1.0
0.5 O
.
0.5 1.0 1.5 Co (mg/ml)
Fig. 1 Results of sedimentation equilibrium on collagens at
200
C as a plot of reciprocal apparent molecular weight
versus concentration prior to centrifugation. Data
for ratskin collagen (open circles) were obtained in
0.06 M potassium acetate buffer, pH 4.8, with the
standard sedimentation equilibrium method (eq. 3) ,
and those for earthworm collagen (closed circles) 口
0.06 M sodium acetate buffer, pH 4.8, with the short
column method of Yphantis (eq. 4).
-35-

po nべυ
0.3
0.5
0.4
Tの
0.2
o 0.5 1.0 1.5
CONCENTRATION (mg/mt)
Fig. 2 Concent ration dependence of reciprocal sedimentation
constant at 2{)" C for ratskin collagen in 0.06 M po
tassium acetate buffer, pH 4.8 (open circles), and
for earthworm collagen in 0.06 M sodium acetate buf
fer, pH 4.8 (c1osed circles)
~{)J
in citrate buffer, pH 3.1 The intrinsic viscosity of earthworm-cuticle col-
lagen ([η,) = 109 dl/ g} obtained is markedly higher than the 62 dl/ g reported by
Josse and Harrington Z51. The difference should be due, at least partly, to the
e ffec ts 0 f the s hear gradien t in the capill ary viscome te r emp loyed by them. We,
too, attempted to measure the viscosity with a viscometer of Ubbelohde type, but
the reproducibility of the measurement was observed very poor.

巧
tq宅U
O 22
。.. 20 u
‘句、
ra ω h
1 8 .、• .. 1 6 U “、
qhu • $::- 14 c ー
1 2
10 O
0.2
"7sp,。0.4 0.6 0.8 1.0 1.2
• 2 3 4
c (g Id l) x 1 03) •
5 6
Fig. 3 Viscosity results on ratskin collagen at 20"C in 0.06 M
potassium acetate buffer, pH 4.8, as a Schulz and Blasch-
ke plot (open circles) and a Mead and Fuoss plot (c losed
circles). Ext rapolations were made with a Huggins con
stant of 0.58.
All the results obtained are summarized in Table 1.
Table I Experimental results of sedimentation equilibrium,
sedime口tationvelocity, and viscosity at 200
C on
retskin collagen in 0.06 M potassium acetate buffer,
pH 4,8, and on earthworm-cuticle collagen in 0.06 M
sodium acetate buffer, pH 4.8.
CoIlagen sample M XW' S~, 20. w 〔甲 J(dl / g)
Earthworm、cuticle
Ratskin
169:!: 10
30. 1士0.3
5. 19:!: 0.05
2. 82:!:0. 05
109
12.4

-38一
O 170
;150 a. ω
た"
• ど 130GI
p
c
110
O
"'Isp~o 0.2 0.4 0.6
• 234
c (gldl)x103a.
Fig. 4 Viscosity results on earthworm-cuticle collagen at 200 C
in 0.06 M sodium acetate buffer, pH 4.8, as a Schulz and
Blaschke plot (open circlesl and a Mead and Fuoss plot
(closed circles). The extrapolations were made with a
Huggins constant of 0.56.
DISCUSSION
0.8
5 6
The hydrodynamic experimental results obtained are now compared with theory
to estimate th巴 pers is tence length. o f the presen tly available hydrodynamic the-,)9-42) 41. 42)
ories of the wormlil日 coil we employ those of U¥!man .., "', because sedimen-
tation and viscosity were calculated on th巴 same model of the wormlike chain. The
polymer chain was treated as a continuous body and was not divid巴d into segments.
It was assumed that the source of hydrodynamic r巴sistance was on the axis of a

-39-
molecule of finite cross section, and that the hydrodynamic interaction was experi-
enced at the surface of the molecule. The excluded volume effect was not consi-
dered. Th巴 theoreticalresults are given as a function of several variables:λ 。。
(A ), the reciprocal Kuhn statisticallength; L(Al, the contour lengt.h;d(Al, the
hydrodynamic diameter;a, a dimensionless factor defined by /2c/ 2rc7}o・ The Kuhn •
statistical length is twice the persistence length, and c denot巴s the fric tion con-
stant per unit length and 7}0 the solvent viscosity. Since the parameter c cannot
be determtned uniquely by any method, some uncertainty may be introduced in the
estimate of the persistence length.
The recent work of Edwards and Oliver431
is quite instructive in this r巴spect.
They hav巴 shownin the theory of the sedimentation constant on a flexible coil mod-
el, a cylinder with.a random flight axis, that we can dispence with the segmental
friction constant by taking into consideration the non-slip boundary condition for the
liquid-cylinde r su rface. This suggests the importance of the similar theory be
developed for the wormlike coil. In addition, in view of the fact that the ca¥cu-
lation of Edwards and Oliver agrees with that of Kirkwood and Riseman441
in the
non-draining limit, it may be suggested that the U¥lman ca¥culation be applied in the
range where the draining parameter h proportional to c /(12庁 3)lnb770(b:the ef
fective bond lengthl is large. The draining parameter h is given forthe wormlike
coil by (6λL/π)lnQ.
The structural par,.ameters for the tropocol¥agen triple-helical structure are a 。
diameter of 15 A and three residues in 2.9A translation in the axial direction. The
average molecular weight per unit residue is ca¥culated from the amino acid com-
position24.251 as 90 for both ratskin and earthworm col¥agens. Thus we used 93 dal-
tons per angstrom, which gives as the contour lengths of respective collagens 3,200
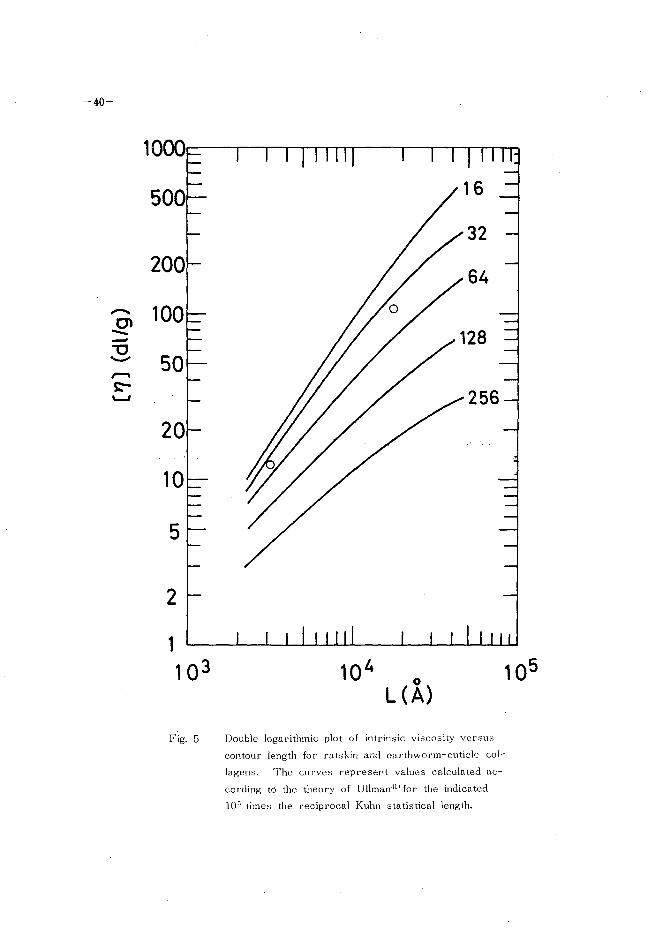
A and 18,000 A. Fig. 5 shows the double logarithmic plot of [ηJ versus L. The
curves drawn in the figure represent the theoretical relations for various values
。f入withα= 5. O. This value ofαwas chosen to obtain for the both collagens 5
a co口sistentvalue ofλ, which is evaluated from Fig. 5 as 42 x 10 On the con-
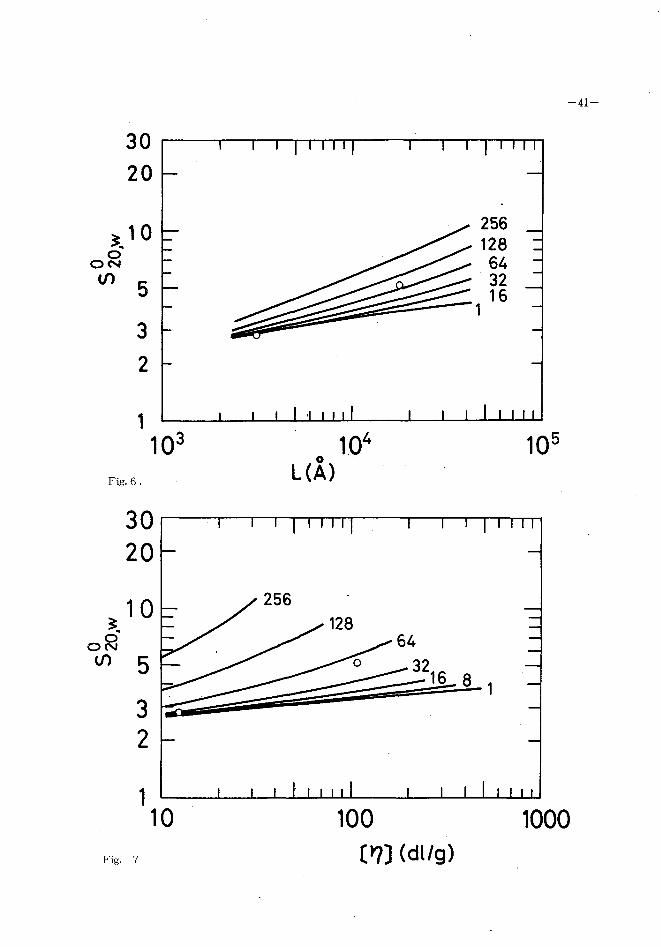
trary, diffe ren t values 0 fλare assigned to the two col¥agens from the analysis of
sedimentation constant as shown in Fig. 6. This -discrepancy may partly originate
from the error in the determination of molec.ular weight, because the more consis-
tent Kuhn statistical length is obtained from the double logarithmic plot of S~~ 20,w
a gains t[η(Fig. 7). However, it is possible as wel¥ that the theoretical values
of sedimentation constant for shorter chains are overestimated in comparison to
those for longer chains. This is suggested from the fact that the comparison of
the same experimental results with the theory of Hearst and Stockmayer yields for -~)
λa value of 38 x 10 common to the two collagens (Fig. 8). The two theories
10∞ -40一
500
200 64
100
50
(四三刀)〔
?
〕 256
20
10
5
2
105
し(久)104 103
Double logarithmic plot of intrinsic viscosity versus
contour length for ratskin a口dearthworm-cuticle col-
lagens. The curves represent values calculated ac-
cording to the theory of Ullman1lJ for the indicated
10ユtimesthe reciprocal Kuhn statistical length.
Fig.5
30
20
ぇ10O
oか4
cf) 5
3
2
103
Fig.6.
30 20
主10oおの 5
3 2
Fig. 7
10
256
104
し(A)
128
100
〔η](dl/g)
256
128 64 32 16
105
1000
41-

-42-
20
1 0
Fig. 6 Double logarithmic' plot of sedimentation constant ver
sus contour length for ratskin and earthworm-cuticle
collagens. The curves represent values calculated ac-42)
cording to the theory of Ullman .~. for the indicated t
10" times the reciprocal Kuhn statistical length.
Fig. 7 Double logarithmic plot of sedimention constant and
intrinsic viscosity of ratskin and earthworm collagens.
For the curves see legends to Figs. 5 and 6.
1250
予三J三F 三J三2蜘5∞
3 I ~岳ョ=1200,.ααm M3
103
Fig.8
104 105
し(A)
Sedime口tatlo口 datareplotted as in Fig. 6. The curves
represent values calculated according to the theory of
Hearst and Stockmayer澗 forthe indicated persistence
length.
differ 0111y il1 whethe1' 01' not the hydrodynamic interaction is allowed withil1 the
bond len引 h b of the chain, and fo1' most values of interest, there is a constant
diffe 1'ence be tween the two calcu la tion s i1' 1'e spec tive 0 f入, L, and a. This sug-
gests, as understood from the discussion above in connection to the f1'iction con-
stant, that the1'e is no 1'eason to p1'efer Ullman calculation to that of Hearst and
Stocl、maye1'. Mo reove 1', sedimen tion cons tan t is quite insens itive to the magni-
tude of the Kuhn statistical length. The1'efo1'e, we tentatively conclude that

qtυ a-
the hydrodynamic properties of ratskin and earthworm collagens are consistent with 。
the wormlike-coil model of a persistence length of 1300 A, which is twice as large
as that for double-helical DNA. From the value so obtained the drainillg parameter
h is calculated as 7.8 and 18.5 for ratskin and earthworm collagens, respectively.
The other factors which should be considered in di,scussing the flexibility of tropo-
collagen are (1) the telopeptide46l at the amino end may assume a structure dif-
ferent from the standard triple helix; (2) the unusuaIly large molecular weight of the
earthworm-cuticle collagen suggests subunits of smaller molecular weight held together
by some other unknown covalent bond. As long as the hydrodynamic proper-
ties are concerned, the experimental data shown above do not clearly indicate the
importance of these possibilities. Needless to say, however, we are unable to ex
clude for earthworm collag巴11 other model conformations such as brokell rods.
ACKNOWLEDGMENTS
lt is our pleasure to thank Professor H. Fujita for a good review of the recent
work of Edwards and Oliver. This investigation was supported in part by the Grant-
in-Aid for Operative'R巴search (8007 and 8(13), the Ministry of Education of Japan
(to H. U.l.
REFERENCES
1) Ramachandran, G. N., and Kartha, G. Nature, 176, 593 (1955)
2 ) Ramachandran, G. N., : in Tγeatse on Collagen, Vol. 1, Academic Press, New York
(1967), p. 170
3) Rich, A., and Crick, F. H. C. : J. Mol. J3iol., 3, 483 (1961)
4) Cowan, P. M., McGavin, S., and North, A. C. T. : Nature, 176, 1062 (1955)
5) Ramachandran, G. N., and Sasisekharan, V. : Biochim. Biophys. Acta, 109, 314 (1965)
6) Ramachandran, G. N., and Chandrasek!祖 国n,R. : Biopoかmers,6, 1649 (1968)
7) Yonath, A., and Traub, W. : J. Mol. Biol., 43, 461 (1969)
8) Traub, W., Yonath, A., and Segal, D. M. : Nature, 221, 914 (1969)
9) Chapman, Go E., Campbell, 1. D., and McLauchlan, K. A. : Ibid, 225, 639 (1970)
10) Boedtker, H., and Doty, P. : J. Am. Che冊 . Soc., 78, 4267 ('1956)
11) Neugebauer, T. : Ann. Physik, ( 5) 42, 509 (1943)
12) Simha, R. : J. Phys. Chem., 44, 25 (1940)
13) Perrin, F. : J. Phys. Radium, ( 7) 5, 33 (1934)
14) Peterlin, A., and Stuart, H. A. Z. Physik, 112, 1, 129 (1939)
15) Scheraga, H. A., Edsall, J. T., and Gadd, J.O., Jr. : J. Chem. Phys., 19, 1101
( 1951)
16) Hall, C. E., and Doty, P. : J. Am. Che叫 . Soc., 80, 1269 (1958)

44
17) Doty, P., and Nishihara. T. : in Recent Advances in Gelatin and Glue Research
(ed. G. Stainsby), Pergamon Press, London (1958), p. 92
18) Nishihara, T., and Doty, P.: Pγoc. Natl. Acad. Sci., 44, 411 (1958)
19) Rice, R. V., Casassa, E. F., Kerwin, R. E., and Maser, M. D. : Arch. Biochem.
Biophys., 105, 409 (1964)
20) Burge, R. E., and Hynes, R. D. : J. Mol. Biol., 1, 155 (1959)
21) Young, E. G., and Lorimer, J. W. : Arch. Biochem. Biophys., 88, 373 (1960)
22) Engel, J., and Beier, G. : Hoppe-Seyler's Z. Physiol. Chem., 334, 201 (1963)
23) Lewis. M. S., and Piez, K. A. : J. Biol. Chem., 239, 3336 (1964)
24) Maser, M. D., and Rice, R. V. Biochim. Biophys. Acta, 63, 255 (1962)
25) Josse, J., and Harrington, W. F. : J. Mol. Biol., 9, 269 (1964)
26) Kratky, 0., and Porod, G. : Rec. Tγav. Chim., 68, 1106 (1949)
27) Daniels, H. E. : Pγoc. Roy. Soc. Edinburgh, 63, 290 (1952)
28) Hermans, J. J., and Ullman, R. : Physica, 18, 951 (1952)
29) Gill, S. J., and Thompson, D. S. Pγoc. Natl. Acad. Sci., 57, 562 (1967)
30) Nimni, M. E. : J. Biol. Chem., 243, 145 (1968)
31) Deshmukh, K., and Nimni, M. E. : Ibid., 244, 1787 (1969)
32) Piez, K. A., Weiss, E., and Lewis, M. S. : Ibid., 235, 1987 (1960)
33) Sugi, N., Tsunashima, Y., Ikehara, K., and Utiyama, H. : Japan. J. Appl. Phys., 11,
No. 10 (1972)
34) Van Holde, K. E., and Baldwin, R. L. : J. Phys. Chem., 62, 734 (1958)
35) Utiyama, H., Tagata, N., and Kurata, M. : Ibid., 73, 1448 (1967) 1969
36) Hexner, P. E., Radford, L. E., and Beams, J. W. : Pγoc. Natl. Acad. Sci., 47, 1848
( 1961)
37) Yphantis, D. A. : Aπ札 N.Y. Acad. SCi., 88, 586 (1960)
38) o rekhovich, V. N., and Shpikiter, V. O. : Biokhi miya, 20, 438 (1955)
39) Hearst. J., and Stockmayer, W. : J. Chem. Phys., 37, 1425 (1962)
40) Ptitsyn, O. B., and Eizner, Yu. E. Vysokomol., Soedin., 3, 1863 (1961)
41) Ullman, R. : J. Chem. Phys., 49, 5486 (1968)
42) Ullman, R. : Ibid., 53, 1734 (1970)
43) Edwards, S. F., and Oliver,加lA. : J. Phys. A: Gen. Phys., 4, 1 (1971)
44) Kirkwood, J. G., and R;'5eman, J. : J. Chys., 16, 565 (1948)
45) Piez, K. A., Eigner, E. A., and Lewis, M. S. Biochemistγy, 2,・58(1963)
46) Rubin, A. L., Drake, M. f'., Davison, P. F., Speakman, P. T., and Schmitt, F. O.・
Ibid., 4, 181 (1965) Kang, A. H., Bornstein, P., and Piez, K. A. : Ibid., 6, 788
(1967)

phυ
daτ
沈降および粘度測定から評価した
トロポコラーゲンのかたさ
内山敬康¥坂戸邦昭,池原健二,節家孝志,倉田道夫
(京都大学化学研究所,京都大学工学部*)
トロポコラーゲン構造の形や大きさについては, Boedtkerと Dotyが ichthyocolコラーゲ
ンを使用して光散乱や流動複屈折などの測定から,長さ 3000A,直径 13.6Aの棒状であるこ
とを見出して以来,他の多くのコラーゲンについて研究が行なわれ,同校の結果が繰り返し得
られてきた O しかし二本鎖 DNAについて見られる様に,分子が長くなると分子全体としては
可携性が表われてくる筈である. ミミズの表皮から抽出されるコラーゲンは,分子量が1.9X
106 にも達し,ゴラーゲン三本鎖構造のかたさについて,より詳細な情報が得られる可能性が
ある.事実 Josseと Harr・Ingtonの得た粘度の結果を回転惰円体の理論と比較して得られる粒
子の長さは,実際の長さの約去にしかならない.この様な粒子は,いわゆる Kratkyと Porod
の wormlike-coil 模型を用いて解析する必要があり,分子のかたさは persistenelengthに
より定量化される.但し,この模型による沈降系数や極限粘度の現在の理論には,流体力学的
半径および単位長あたりの摩擦係数が変数として含まれており,かっこれらは一義的に決めら
れないので,構造が同じで長さの異なる試料の結果を統」的に記述できる様に,二れらの変数
むよび persistencelength が決'JE・される/
試仰としてミミズの表皮およびラット外皮から抽出したコラーゲンを使用し,沈降係数や粘
度の測定には0.06M K acetate buffer, pH 4.8を月Jい200
Cで行った.
分子量の決定は, Spinco E型超遠心機による沈降平衡法によった.但しミミズのコラーゲン
については, Schlieren光学系,および Yphantisのふチャンネルセルを使用し, ラット・コ
ラーゲンについては, Rayleigh干渉光学系を使用する通常の沈降平衡法によった. 得られた
分子量は,夫々 169X104, 30.1 X104
である Ratskinの分子量は,文献値の29.4X104
(Lewis & Piez, J. Biol. Chem., 239, 3336 (1964)) とよく一致していと3が, ミミズの結果
は,粘度と沈降{系数から JosseとHarrington (J. Mol. Biol., 9,269(1964)) が得た190X
104 より 10%程小さい.但し彼らの粘度測定には問題がある.
沈降係数 S020, w は,夫々 2.82士0.05(ratskin ), 5.19土0.05(ミミズ)であった. Josse
と Harr・ingtonがミミズ表皮コラーゲンについて報告している沈降係数の濃度依存性の異常は,
見られなかった.
粘度測定には,気~液界面に形成される膜の影響を除き,かっ低速度勾配で実験を行なうた
め,特別に設計したcartesian-divertYIieの回転粘度計を使用した.得られた結果は, 12.4dl
/ g (ratskin) および109dl/g (ミミズ)で, 後者は通常の毛管粘度計による測定値62dl/g

46-
に比し著しく大きい.
分子軸に沿って lAあたりの分子量を93として ratskinおよびミミズ表皮コラーゲンの
contour lengthは,夫々 3200λ,18,oooAと求められる.粘度の結果をUllmanの理論と比較
すると,分子量に無関係に persistencelength は1250Aとなる.一方沈降係数の結果からも,
大約一致した値1300Aが得られる.但し沈降係数の persistencelength依存性はごくわずか
で,粘度から求められた perslstenc巴 length の方が信頼度は高い.これらはDNAの persis
tence lengthの約 21音である.
以上は,流体力学的にミミズ表皮のコラーゲンと ratskin コラーゲンが完全に同じ構造をも
ち,その相異は長さだけであるという仮定のもとに導かれた結論であることは,注意する必要
があるが,グリシンの含量が共にすに近いこと,変性後の分子量が約すになる事実などは,上
記の仮定を支持するものと考えられる.
イ寸 記
第22回コラーゲン・シンポジウムにおける話題 I細胞内でのトロポコラーゲン構造形成の機構に
に関する物理化学的イメージ、」については,別に発表する予定である.
但し, HCHOにより架橋したコラーゲンを使用したコラーゲン構造再生の動力学的実験に関
する質問への回答は,ここに記載させて項きます.
討 議
御 橋広真氏 の系では回復しません.
Recoveryしなかった fraction の ORDへの (2) 使用したトロポコラーゲンは,全ての分
contributionをどう考えられますか? 子が架橋される様な条件で反応を行ないました.
内山敬康氏 結合した HCHO の平均数は20~100で,また実際
変性ゼラチンのORD と同一で、あると仮定して の架橋数には分布がある筈であります.原理的
います. には,鎖のregistrationさえできればよいので
林 利彦氏 すが,完全な構造再生を行なうには,温度・濃
(1) 37'Cでの回復は如何ですか? 度の領域を適当に定めねばならない筈でありま
(2) HCHOによる cross-linksがどの{立少なく す.
てもよく戻るものですか? (3) 架橋の数,位置が適当であれば, トロポ
(31 Spontaneous renaturationの%の大きさ コラーゲン分子聞の非特異性結合による構造形
と生体内での 3本鎖構造形成の調節との関係は 成のおくれは,問題にならないと考えられ,コ
如何ですか? ラーゲン合成の前駆体 procollagenでは,構造
内山敬康氏 形成の速度は非常に速いものと考えています.
(1) 使用した系でのTmは38.6"cであり,こ


















