
Histomorphometrische Untersuchung zur gerichteten ...
95
Aus der der Universitätsklinik für Zahn-, Mund- und Kieferheilkunde der Albert-Ludwigs-Universität Freiburg im Breisgau Abteilung Klinik für Mund, Kiefer- und Gesichtschirurgie Histomorphometrische Untersuchung zur gerichteten Nervendistraktion anhand des Nervus facialis der Ratte Inaugural - Dissertation zur Erlangung des Zahnmedizinischen Doktorgrades der Medizinischen Fakultät der Albert-Ludwigs-Universität Freiburg im Breisgau Vorgelegt 2003 von Bettina Maria Kriesche geboren in Villingen-Schwenningen
Transcript of Histomorphometrische Untersuchung zur gerichteten ...

Aus der der Universitätsklinik für Zahn-, Mund- und Kieferheilkunde
der
Albert-Ludwigs-Universität Freiburg im Breisgau
Abteilung Klinik für Mund, Kiefer- und Gesichtschirurgie
Histomorphometrische Untersuchung
zur gerichteten Nervendistraktion
anhand des Nervus facialis der Ratte
Inaugural - Dissertation zur
Erlangung des Zahnmedizinischen Doktorgrades
der Medizinischen Fakultät
der Albert-Ludwigs-Universität
Freiburg im Breisgau
Vorgelegt 2003
von Bettina Maria Kriesche
geboren in Villingen-Schwenningen

Dekan: Prof. Dr. Josef Zentner
1. Gutachter: Prof. Dr. Dr. Nils-Claudius Gellrich
2. Gutachter: Frau PD Dr. Petra Ratka-Krüger
Jahr der Promotion 2004

Meinen Eltern gewidmet

Die vorliegende Arbeit wurde in der Zeit von April 2000 bis März 2003 in der
Klinik und Poliklinik für Zahn-, Mund- und Kieferheilkunde der Albert-Ludwigs-
Universität Freiburg im Breisgau angefertigt.
Mein besonderer Dank gilt Herrn Professor Dr. Dr. N.-C. Gellrich für die Vergabe
des Themas der Dissertation und für seine Unterstützung bei ihrer Durchführung.
Des Weiteren möchte ich mich bei all jenen sehr herzlich bedanken, die mir
während dieser Dissertationsarbeit zur Seite gestanden haben:
Herrn Dr. Dr. M. Nilius danke ich für die Anleitung zur Durchführung der
klinischen Arbeiten,
Frau Hübner für Ihre Hilfe bei der Anfertigung und Aufarbeitung der
histologischen Präparate,
Frau Bächle für Ihren Rat und die Bereitstellung des Computerprogramms
analySIS ®,
Frau Neubacher von der Ruhr-Universität Bochum für ihre Beratung während
der Anfertigung und Aufarbeitung der Präparate,
Herrn Hartmann für seine stetige Hilfe bei PC Problemen
sowie meinen Freunden für Ihr Verständnis und Ihren Beistand.
Ein besonderes Dankeschön möchte ich meiner Mutter für ihre große
Unterstützung und ihr Verständnis aussprechen.

Inhaltsverzeichnis
Inhaltsverzeichnis
1 Einleitung.......................................................................................................1 1.1 Geschichte der traumatischen Schädigung peripherer Nerven...............2 1.2 Anatomische Grundlagen ........................................................................3 1.2.1 Aufbau eines peripheren Nerven ........................................................4 1.2.2 Der Verlauf des Nervus facialis ...........................................................6 1.3 Schädigung des Nervus facialis...............................................................9 1.3.1 Symptome der Facialisparese.............................................................9 1.3.2 Pathophysiologie der Nervenläsion...................................................11 1.3.3 Diagnostik..........................................................................................14 1.4 Möglichkeiten der Rekonstruktion..........................................................15 1.5 Ziel der Arbeit ........................................................................................18
2 Material und Methode ................................................................................19 2.1 Versuchstiere.........................................................................................19 2.2 Materialien .............................................................................................19 2.3 Methoden...............................................................................................21 2.3.1 Operationen.......................................................................................21 2.3.2 Distraktion des Ramus buccalis n. facialis ........................................26 2.3.3 Entnahme der Nervenproben............................................................26 2.3.4 Histologische Aufarbeitung der Präparate ........................................27 2.3.5 Epoxideinbettung ..............................................................................29 2.3.6 Herstellung der Semi-Dünnschnitte ..................................................30 2.3.7 Brechung der Glasmesser ................................................................31 2.3.8 Kontraststeigerung mit para-Phenylendiamin ...................................31 2.3.9 Histomorphometrische Auswertung mittels Lichtmikroskopie ...........32 2.3.10 Histologische Auswertung der Präparate..........................................35 2.3.11 Quantitative Auswertung des Nervenquerschnitts ............................35 2.3.12 Statistische Auswertung....................................................................36
3 Ergebnisse...................................................................................................37 3.1 Histologische Auswertung .....................................................................37 3.2 Auswertung der histomorphometrischen Ergebnisse ............................50 3.2.1 Normalwerte des Ramus buccalis.....................................................50 3.2.2 Der Ramus buccalis nach Distraktion ...............................................51 3.3 Quantitative Auswertung der Axonanzahl .............................................59
4 Diskussion ...................................................................................................62 4.1 Methodenkritik .......................................................................................64 4.2 Morphologische und histomorphometrische Ergebnisse.......................66
5 Zusammenfassung.....................................................................................73
6 Literaturverzeichnis ....................................................................................74
7 Lebenslauf ...................................................................................................90

1
Einleitung
1 Einleitung
In den letzten Jahren hat die Mikronervenchirurgie wesentlich an Bedeutung
zugenommen. Aufgrund der technischen Möglichkeiten und der klinischen
Erfahrung können unter Beachtung der korrekten Indikationsstellung besonders
auf dem Gebiet der Mund-Kiefer- und Gesichtschirurgie gute Ergebnisse erzielt
werden.
Verschiedene Hirnnerven sind im Rahmen der Tumorchirurgie, der
Traumatologie und durch Infektionen gefährdet, so auch der Nervus facialis in
seinem extratemporalen Verlauf. Betroffen können auch sensible Äste des
Nervus trigeminus sein, wie der Nervus alveolaris inferior, der Nervus lingualis
und der Nervus infraorbitalis. Gerade den beiden genannten Trigeminusästen
kommt in der rekonstruktiven Chirurgie klinische Bedeutung zu, da diese auch bei
zahnärztlichen Maßnahmen beschädigt werden können (Girod und al. 1989). Der
Nervus glossopharyngeus, der Nervus vagus und der Nervus phrenicus sind
dagegen selten betroffen (Grobe und Raithel 1978).
Im Gegensatz zu Knochen- und Weichteilverletzungen müssen
Nervenverletzungen nicht unbedingt primär rekonstruiert werden. Da die
Regenerationsfähigkeit von Nerven lange erhalten bleib, besteht die Möglichkeit
einer frühen sekundären Wiederherstellung nach 3-4 Wochen oder einer
sekundären Rekonstruktion nach mehreren Monaten.
In vielen Fällen führen die bisherigen Rekonstruktionsmöglichkeiten dennoch
nicht zu befriedigenden Ergebnissen. Daher ist das Bestreben vieler
Arbeitsgruppen darauf ausgerichtet, neue Methoden zu entwickeln, um ein
besseres funktionelles Ergebnis zu erzielen und den Leidensdruck der Patienten
zu vermindern.

2
Einleitung
1.1 Geschichte der traumatischen Schädigung peripherer Nerven
Die erste Erwähnung einer akuten Nervenverletzung kann etwa 3500 Jahre
zurückdatiert werden und findet sich bereits in der Bibel. In dem bekannten
Bericht des Alten Testaments erlitt Jakob im Kampf mit Gott eine Verletzung des
Nervus ischiadicus und eine traumatische Hüftdislokation (Genesis 32: 23-33)
(Cornwell 2000).
Über erste Versuche, durchtrennte Nervenenden durch einen einfachen
Wundverschluss wieder zusammenzuführen, berichtete Paul von Aegina
(625-690) (Novak et al. 2002).
Als Pionier der chirurgischen Versorgung peripherer Nerven kann Gabriele
Ferrara (1543-1672) angesehen werden. Er war der erste Chirurg, der die
Technik beschrieb, verletzte Nervenenden durch eine Naht zusammenzufügen.
Seine Methode beinhaltete die präzise Identifizierung der Nervenstümpfe und ihr
vorsichtiges Vernähen, wie auch die Desinfektion von Nadel und Faden mit
einem Gemisch aus Rotwein, Rosmarin und Rosenblättern. Im Grundsatz wird
ein ähnliches chirurgisches Vorgehen auch heute noch praktiziert (Artico et al.
1996).
1850 schilderte August Waller die absteigende Nervendegeneration.
Hueter (1871, 1873) stellte das Konzept der epineuralen Nervennaht vor (Sharon
und Fishfield 2002). 1876 erörterte Albert die Überbrückung von Nervendiastasen
durch Transplantation.
Die Erfahrungen mit Kriegsverletzungen während des Ersten und Zweiten
Weltkriegs lieferten wichtige Informationen über die Beurteilung und Behandlung
von Nervenverletzungen (Sharon und Fishfield 2002). Fortschritte in der
Entwicklung neurochirurgischer Instrumente und verbesserte Diagnoseverfahren
haben zu guten Ergebnissen in der Versorgung akuter Nervenverletzungen
geführt (Artico et al. 1996). Allerdings konnte eine entscheidende, klinisch
fassbare Verbesserung der funktionellen Ergebnisse auch in den letzten Jahren
nicht erreicht werden (Trumble et al. 2000).

3
Einleitung
1.2 Anatomische Grundlagen In dieser Arbeit wird die Reaktion eines gesunden peripheren Nervenastes auf
eine moduliert gerichtete und gesteuerte primäre Nervendistraktion untersucht.
Aufgrund seiner günstigen anatomischen Verhältnisse bietet sich der Ramus
buccalis der Ratte für die Durchführung derartige Experimente an.
Bei den höher entwickelten Lebewesen sorgen Nerven für die
Informationsübertragung im Organismus. Die Gesamtheit des Nervengewebes,
welches Reize aufnimmt, sie weiterleitet, im Zentralnervensystem verarbeitet und
eine Antwort in den peripheren Empfängern auslöst, bildet das Nervensystem.
Dieses und die damit verbundenen Organe machen es möglich, rasch auf
unterschiedliche Reize zu reagieren.
Es wird unterschieden zwischen dem Zentralen Nervensystem (Gehirn und
Rückenmark) und dem peripheren Nervensystem (Rückenmarksnerven,
Hirnnerven und periphere Ganglien). In verschiedener Hinsicht ähneln die
Hirnnerven den Spinalnerven des Rückenmarks. Letztere versorgen Rumpf und
Extremitäten, während die Hirnnerven überwiegend den Kopf innervieren.
Im Gegensatz zu den Rückenmarksnerven enthalten die Hirnnerven jedoch nicht
alle nervösen Faserarten in fast gleicher Zusammensetzung, sondern jeweils nur
Fasern aus einzelnen Terminalkernen mit spezieller Funktion.
Der Nervus facialis (siebenter Hirnnerv) versorgt in erster Linie die mimische
Muskulatur des Gesichtes und des Halses. Er enthält jedoch auch
Geschmacksfasern und parasympathische Faserbündel. Diese sind in der Regel
als gesonderter Nerv anzutreffen (Nervus intermedius).
Patienten, die einen permanenten Verlust der Fazialis-Funktion erleiden, haben
nicht nur Funktionsstörungen beim Essen und Sprechen. Vielmehr führt die
Parese der mimischen Muskulatur, die mit dem Verlust der emotionellen
Ausdrucksmotorik und einer morphologischen Entstellung einhergeht, zu nicht zu
unterschätzenden psychischen und psychosozialen Konsequenzen. Der
Rehabilitation dieser Patienten ist somit eine hohe Priorität zuzuordnen.

4
Einleitung
1.2.1 Aufbau eines peripheren Nerven
Ein genaues Verständnis von Aufbau und Physiologie eines peripheren Nerven
ist die Vorbedingung für die chirurgische Rekonstruktion von Nervenverletzungen
(Battista und Lusskin 1986) und unabdingbar für die morphologische Bewertung
von Nervenpräparaten. Die Überprüfung auf Abweichungen der
Nervenmorphologie dient in dieser Arbeit als Grundlage für die Beurteilung einer
vorhandenen Schädigung eines Nerven nach primärer Distraktion.
Eine Nervenzelle lässt sich in Perikaryon (Zellleib), Nervenzellfortsätze,
Dendriten und das Axon (Neurit, Achsenzylinder) gliedern. Mehrere Dendriten
bilden die afferente Komponente der Nervenzelle. Das Axon ist die efferente
Struktur. Es leitet die Erregung in die Peripherie und kann beim Menschen bis zu
1 m lang sein. Im Gegensatz zu den Dendriten, deren Durchmesser variieren,
besitzt es einen konstanten Durchmesser von 15-20 µm.
Sowohl die einzelnen peripheren Nervenfasern als auch die Faszikel, zu denen
sie zusammengefasst sind, und der Nerv in seiner Gesamtheit verlaufen
wellenförmig. So wird bei Bewegung eine geringfügige Verlängerung der
Strukturen möglich, ohne dass eine Überdehnung der Nervenfasern eintritt
(Sunderland 1990).
Je nach Differenzierung der Hülle kann man zwischen markscheidenhaltigen
oder markscheidenlosen Nervenfasern unterscheiden (Kuczynski 1980).
Der untersuchte Nervus facialis enthält wie die meisten peripheren Nerven
markscheidenhaltige Fasern.
Die Markscheidenreifung beginnt durch die Einsenkung eines Axons in die
Vertiefungen von hintereinander angeordneten Schwann´schen-Zellen. In der
entstehenden Rinne lagern sich Membranen der Schwann´schen-Zelle
aneinander und bilden das Mesaxon. Dieses wickelt sich mehrfach um das Axon.
Diese Wicklungen und Verschmelzungen der benachbarten Zellmembrananteile
bilden die Markscheide, welche einen lamellaren Bau aufweist. Die eigentliche
Substanz dieser Schicht, die der elektrischen Isolierung dient, ist das Myelin.
Dieses besteht zu 70% aus Lipiden (Phospholipide, Cholesterin und Glycolipide).

5
Einleitung
Zu 30% ist Myelin aus einer Anzahl verschiedener Proteine zusammengesetzt,
darunter Protein 0 (P0), P2, peripheres Myelinprotein 22 (PMP 22), myelin-
assoziiertes Glycoprotein (MAG) und Connexin 32 (CX32) (Snipes und Suter
1995). Diese Proteine spielen eine wichtige Rolle bei der Bildung, Erhaltung und
Degeneration der Myelinscheiden (Quarles 2002).
Zwischen dem Durchmesser des Axons und der zugehörigen Markscheide
besteht in der Regel eine positive Korrelation (Morell und Norton 1980).
Die Myelinscheide wird außen von einer Hülle aus Zytoplasma umgeben, dem
Neurolemm.
Eine Schwann´sche-Zelle umhüllt das Axon auf einer Länge von 0,2 bis 1,5 mm.
Zwischen benachbarten Zellen befindet sich ein Interzellularspalt, der als
Ranvier´scher-Schnürring bezeichnet wird. In diesen Spalten ist die Isolierung
geringer. Auf diese Weise wird eine saltatorische Erregungsleitung mit erhöhter
Leitgeschwindigkeit ermöglicht.
Durch Bindegewebsstrukturen sind die Nerven untereinander und mit ihrer
Umgebung verbunden.
1) Das Endoneurium ist eine Bindegewebsschicht, die jede Nervenfaser
umgibt und zarte kollagene Fasern, eine geringe Anzahl elastischer
Fasern und retikuläre Fasern enthält. Zur Nervenfaser hin schließt sie
mit einer Basalmembran ab.
2) Das Perineurium ist ein straffes Bindegewebe aus Kollagen- und
dickeren elastischen Fasern. Es schließt mehrere Nervenfasern zu
Faszikeln zusammen. Außerdem dient es als Diffusionsbarriere
zwischen Endo- und Epineuralraum (Martin 1964; Rechtland 1987).
3) Das Epineurium fasst die einzelnen Faszikel zu einem peripheren Nerv
zusammen und vermittelt einen beweglichen Einbau in seine Umgebung.
Der Anteil elastischer Fasern ist relativ gering. Es kann daher davon
ausgegangen werden, dass die Eigenelastizität eines peripheren Nerven primär
auf Kollagenfasern zurückzuführen ist (Tassler et al. 1994).
In der Anordnung der Faszikel können drei Grundtypen unterschieden werden:
monofaszikulär, oligofaszikulär und polyfaszikulär. Der Faszikelanordnung kommt
für die Technik der Nervenkoaptation große Bedeutung zu (Hausamen und
Schmelzeisen 1996).

6
Einleitung
1.2.2 Der Verlauf des Nervus facialis
Ein Vergleich zwischen dem Verlauf des menschlichen Nervus facialis und dem
Verlauf dieses Nerven bei der Ratte soll dargestellt werden.
Der Nervus facialis ist der siebente der 12 Hirnnerven und gehört
entwicklungsgeschichtlich zum zweiten Kiemenbogensegment. Er hat drei
Ursprungskerne, welche die motorischen, sensiblen, sensorischen und
parasympathischen Qualitäten wiederspiegeln.
Der Nucleus originis n. facialis ist der motorische Ursprungskern. Er besteht aus
einem Hauptkern für die mimische Muskulatur und den Musculus stylohyoideus,
sowie aus zwei akzessorischen Kernen.
Nach Szentágothai (1949) ist der Nucleus dorsalis das Kerngebiet des Venter
posterior des Musculus digastricus. Man nimmt an, dass der zweite Kern, der
Nucleus accessorius ventralis, an der Innervation des Musculus stapedius
beteiligt ist (Chouard et al. 1972).
Für die parasympathischen Anteile ist der Nucleus salivatorius superior
verantwortlich. Tränendrüse, Nasen-Gaumen- und Unterzungendrüsen wie auch
Orbita, Mund- und Nasenhöhle werden von diesen Anteilen versorgt.
Die afferenten Geschmacksfasern aus der Chorda Tympani enden im Nucleus
tractus solitarii.
Die sensiblen und parasympathischen Faserbündel bilden den Nervus
intermedius und verlassen den Hirnstamm getrennt vom motorischen
Hauptstamm (Banfai 1976). Der N. facialis verlässt gemeinsam mit dem N.
vestibulocochlearis den Hirnstamm im Kleinhirnbrückenwinkel. Er tritt in den
Meatus accusticus internus ein und erreicht dort den Canalis facialis des
Felsenbeins. Durch eine fast rechtwinklige Biegung an der vorderen
Felsenbeinwand nach dorsal bildet er das Geniculum n. facialis. Bogenförmig
zieht der Nervus facialis um das Tympanon nach kaudal. Es besteht hier eine
enge topographische Beziehung mit dem Sinus sigmoideus (Aslan et al. 2001).

7
Einleitung
Noch im Canalis facialis zweigen vom N. facialis folgende Nerven ab:
der N. stapedius, der den gleichnamigen Muskel versorgt, die Chorda tympani
sowie der Ramus communicans cum plexu tympanico. Dieser nimmt aus dem
Plexus tympanicus des Nervus glossopharyngeus und dem Ramus auricularis
nervi vagi sensible Afferenzen auf. So ist er an der sensiblen Innervation der
Schleimhaut der Paukenhöhle und der Haut des äußeren Gehörgangs beteiligt
(Eshraghi et al. 2002).
Durch das Foramen stylomastoideum verlässt der siebte Hirnnerv den Canalis
facialis. Kurz nach dem Austritt gibt er den N. auricularis posterior nach dorsal ab
und versorgt die Muskeln der inneren Ohrmuschel. Nach kaudal gibt er den
Ramus stylohyoideus und den Ramus digastricus ab, sie versorgen die
gleichnamigen Muskeln.
Der N. facialis tritt in die Parotis ein und bildet dort den Plexus parotideus, der die
Ohrspeicheldrüse in einen oberflächlichen und tiefen Teil teilt. Unter der Fascia
masseterica strahlt er am vorderen Rand des Musculus masseter, in einer
topographischen Schicht liegend, fächerförmig in die oberflächliche mimische
Gesichtsmuskulatur ein (Becker 1999).
Über den Jochbogen ziehen die Rami temporales zum Venter frontalis des
Musculus occipito-frontalis sowie zum Musculus orbicularis oculi. Die Rami
zygomatici versorgen die Umgebung der unteren Lidspalte, der Nasen- und der
oberen Lippenregion.
Der Musculus buccinator wie auch der Musculus orbicularis oris werden von den
Rami buccales innerviert. Parallel zum Unterkiefer verläuft der Ramus marginalis
mandibulae, der die Muskulatur der unteren Mundspalte versorgt. Die oben
genannten Rami bilden Anastomosen mit dem Nervus trigeminus (Paturet 1964).
Eine Innervation des Platysmas erfolgt durch den Ramus colli und den
Nervus transversus colli aus dem Plexus cervicalis, die ebenfalls Anastomosen
bilden.
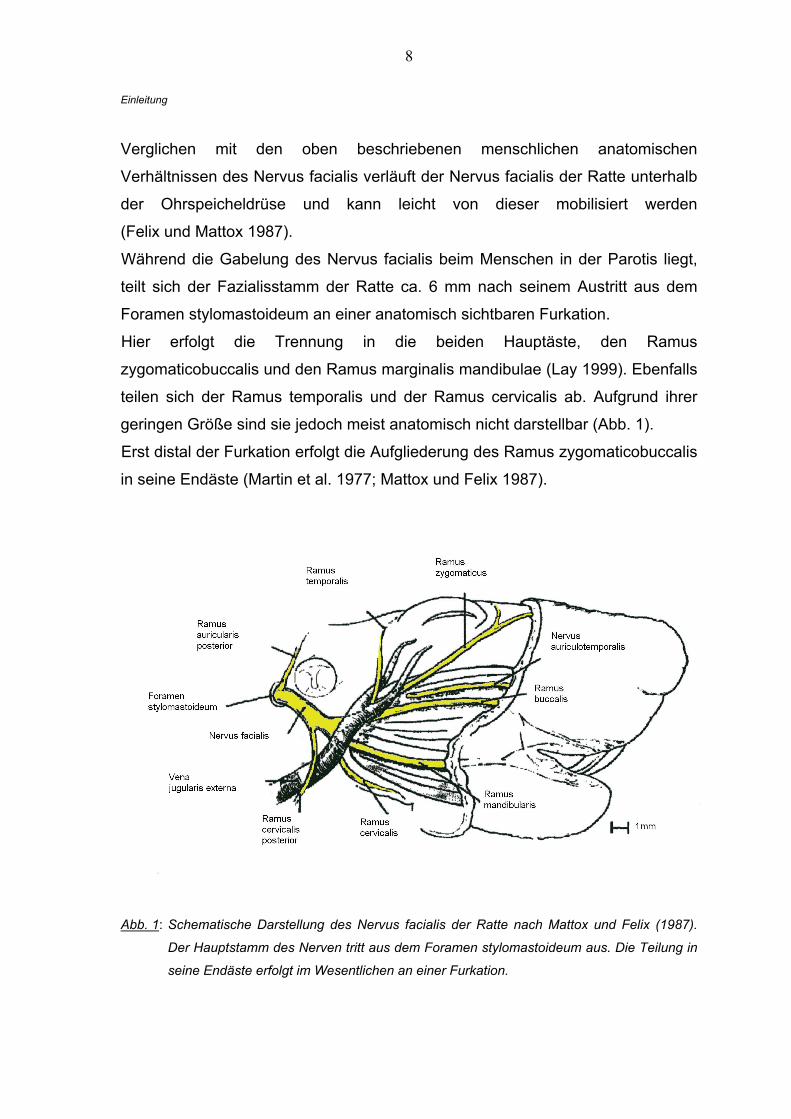
8
Einleitung
Verglichen mit den oben beschriebenen menschlichen anatomischen
Verhältnissen des Nervus facialis verläuft der Nervus facialis der Ratte unterhalb
der Ohrspeicheldrüse und kann leicht von dieser mobilisiert werden
(Felix und Mattox 1987).
Während die Gabelung des Nervus facialis beim Menschen in der Parotis liegt,
teilt sich der Fazialisstamm der Ratte ca. 6 mm nach seinem Austritt aus dem
Foramen stylomastoideum an einer anatomisch sichtbaren Furkation.
Hier erfolgt die Trennung in die beiden Hauptäste, den Ramus
zygomaticobuccalis und den Ramus marginalis mandibulae (Lay 1999). Ebenfalls
teilen sich der Ramus temporalis und der Ramus cervicalis ab. Aufgrund ihrer
geringen Größe sind sie jedoch meist anatomisch nicht darstellbar (Abb. 1).
Erst distal der Furkation erfolgt die Aufgliederung des Ramus zygomaticobuccalis
in seine Endäste (Martin et al. 1977; Mattox und Felix 1987).
Abb. 1: Schematische Darstellung des Nervus facialis der Ratte nach Mattox und Felix (1987).
Der Hauptstamm des Nerven tritt aus dem Foramen stylomastoideum aus. Die Teilung in
seine Endäste erfolgt im Wesentlichen an einer Furkation.

9
Einleitung
1.3 Schädigung des Nervus facialis
Neben der traumatischen Läsion einzelner Fazialis-Äste oder des Fazialis-
Stammes bei tiefreichenden Verletzungen der lateralen Gesichtsregion kommen
hauptsächlich maligne, teilweise aber auch benigne Neoplasien der
Ohrspeicheldrüse mit nachfolgender radikaler Resektion des Tumors als Ursache
eines Funktionsverlustes des Nervus facialis in Frage.
Akustikusneurinome können aufgrund der anatomischen Variabilität des N.
facialis nach einer Resektion ebenfalls zu einer Facialisparese führen (Samii und
Matthies 1997). Auch entzündliche Erkrankungen stellen eine Bedrohung für die
Fazialisfunktion dar. Als Ursachen stehen hier die Borreliose oder der Zoster
oticus (reaktivierte Varizella-Zoster Infektion) im Vordergrund.
Ebenso kommen Hirntumoren oder vaskuläre Prozesse als Grund einer Störung
in Frage. Sie führen zu einer zentralen Störung des Nervus facialis.
1.3.1 Symptome der Facialisparese
Als Leitsymptom entwickelt sich eine Lähmung der mimischen Muskulatur. Diese
betrifft in den meisten Fällen nur eine Gesichtshälfte, nämlich die Seite der
Nervenschädigung. Schmerzen hinter dem Ohr können die ersten Zeichen sein.
Als Folgesymptome zeigen sich (Tab. 1):
• Erweiterte Lidspalte
• Lidspaltendifferenz
• Positives Bell-Phänomen (unvollständiger Augenschluss, beim Versuch des
Lidschlusses wird die physiologische Aufwärtsbewegung des Bulbus oculi
nach oben sichtbar)
• Verstrichene Stirn- und Nasolabialfalte
• Beeinträchtigte Aussprache der betroffenen Patienten
• Verlust der Ausdrucksmotorik
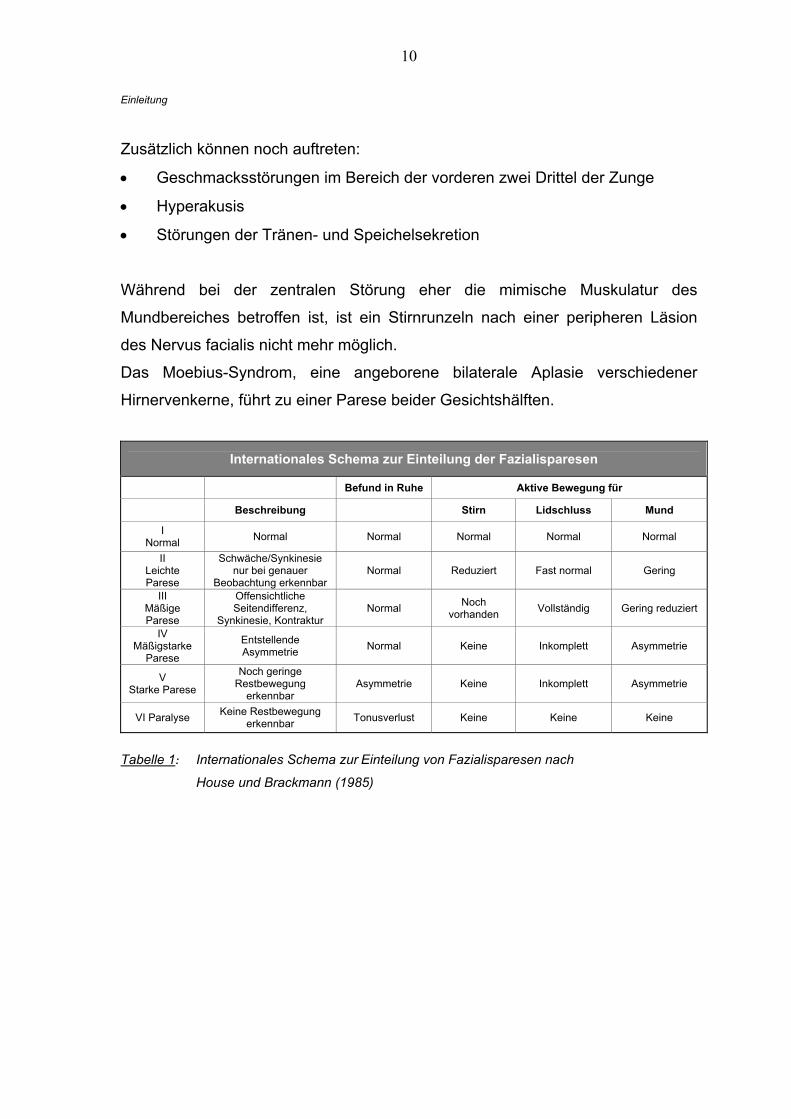
10
Einleitung
Zusätzlich können noch auftreten:
• Geschmacksstörungen im Bereich der vorderen zwei Drittel der Zunge
• Hyperakusis
• Störungen der Tränen- und Speichelsekretion
Während bei der zentralen Störung eher die mimische Muskulatur des
Mundbereiches betroffen ist, ist ein Stirnrunzeln nach einer peripheren Läsion
des Nervus facialis nicht mehr möglich.
Das Moebius-Syndrom, eine angeborene bilaterale Aplasie verschiedener
Hirnervenkerne, führt zu einer Parese beider Gesichtshälften.
Internationales Schema zur Einteilung der Fazialisparesen
Befund in Ruhe Aktive Bewegung für
Beschreibung Stirn Lidschluss Mund
I Normal Normal Normal Normal Normal Normal
II Leichte Parese
Schwäche/Synkinesie nur bei genauer
Beobachtung erkennbar Normal Reduziert Fast normal Gering
III Mäßige Parese
Offensichtliche Seitendifferenz,
Synkinesie, Kontraktur Normal Noch
vorhanden Vollständig Gering reduziert
IV Mäßigstarke
Parese
Entstellende Asymmetrie Normal Keine Inkomplett Asymmetrie
V Starke Parese
Noch geringe Restbewegung
erkennbar Asymmetrie Keine Inkomplett Asymmetrie
VI Paralyse Keine Restbewegung erkennbar Tonusverlust Keine Keine Keine
Tabelle 1: Internationales Schema zur Einteilung von Fazialisparesen nach
House und Brackmann (1985)

11
Einleitung
1.3.2 Pathophysiologie der Nervenläsion
Die Neurapraxie ist durch eine Markscheidenschädigung mit Ödembildung
innerhalb der Markscheide gekennzeichnet. Die Axone bleiben dagegen intakt.
Während die Axonotmesis mit Degeneration sowohl der Markscheide als auch
der Axone einhergeht, die bindegewebigen Mantelstrukturen des Nerven aber
unversehrt sind, charakterisiert das Verletzungsmuster der Neurotmesis eine
vollständige Unterbrechung der Nervkontinuität (Seddon 1943, Menger 1992).
Klinisch sind Axonotmesis und Neurotmesis anfänglich schwer zu differenzieren.
In beiden Fällen liegt eine Parese der beteiligten Muskulatur vor. Das
Elektromyogramm zeigt eine vollständige Unterbrechung der Nervenleitung mit
verlangsamter Muskelkontraktion. Distal der Nervenläsion tritt die von August
Waller 1850 beschriebene sekundäre Degeneration (Waller´sche Degeneration)
auf, bei der es gleichzeitig zu De- und Regenerationsvorgängen kommt
(Fawcett und Keynes 1990).
Während der Degeneration sind histologisch betrachtet Form und Durchmesser
der Myelinscheiden unregelmäßig. Es resultiert eine Quellung und Invagination
der Myelinscheiden mit einer Aufsplitterung der äußeren Lamellen. Binnen einer
Woche zerfallen die Markscheiden. Durch Lysophosphatide erfolgt schließlich die
Auflösung der Axone. Der weitere Prozess beinhaltet die Entmischung und
Verklumpung des Axoplasmas, eine diffuse Granulierung der axonalen
Mitochondrien, die Aufnahme von Markfragmenten in die Schwann´sche Scheide
und das Auftreten von Fettkörnchenzellen. Es kommt zur Markballenbildung
(Lahoda 1988; Tackman 1989).
Eine massive Makrophageninvasion mit CD8-Expression und eine
T-Zellinfiltration schließen sich an (Jander et al. 2001). Die Myelin-Fragmente
werden abgebaut, und es wird ein Milieu geschaffen, in dem der Prozess der
Regeneration stattfinden kann (Kury et al. 2001). Durch Proliferation der
Schwann´schen-Zellen und Bildung feiner Fibrillen vom Axon des proximalen
Nervenstumpfes aus beginnt die Regeneration. Die durchschnittliche
Geschwindigkeit beträgt hierbei 1-2 mm/Tag (Quaan und Bird 1999).

12
Einleitung
Allerdings kann der Regenerationsprozess bei einer Neurotmesis zu einer
ungeordneten Aussprossung der Axone (Neurom) führen, da hier auch die
bindegewebigen Leitstrukturen durchtrennt sind, die sonst als Schiene für die
Wiedereinsprossung dienen.
Im Jahre 1951 wurden Nervenverletzungen von Sunderland detailliert
unterschieden und in fünf Kategorien aufgeteilt:
• Erster Grad: Die Kontinuität des Axons ist erhalten. Es besteht ein
vorübergehender Verlust der Leitfähigkeit am Ort der Schädigung. Es
kommt zu einer Demyelinisierung, nicht aber zur Waller´schen
Degeneration.
• Zweiter Grad: Die Kontinuität des Axons ist unterbrochen, das Endoneurium
und die bindegewebigen Mantelstrukturen des Nerven bleiben erhalten.
Waller´sche Degeneration tritt auf. Distal der Läsion ist eine elektrische
Stimulation der Erfolgsorgane nicht mehr möglich, doch durch
Wiedereinsprossung entlang der endoneuralen Leitschienen kommt es nach
Monaten zu einer völligen Regeneration.
• Dritter Grad: Hierbei handelt es sich um eine intrafaszikuläre Läsion. Die
äußere Struktur der Faszikel ist zwar intakt, aber die Kontinuität von Axon
und Endoneuralscheide ist unterbrochen. So kommt es zum Zerfall der
Axone und zu axonaler Degeneration, wobei Blutungen, Ödeme, Ischämie,
Neuroinflammation und eine Fibrose des Endoneuriums auftreten können.
Durch die Unterbrechung der Endoneuralscheide ist während der
Regeneration die geordnete Wiedereinsprossung der Axone nicht mehr
gewährleistet. Es verbleibt oft ein Verlust der motorischen und/oder
sensorischen Qualität.
• Vierter Grad: Die Struktur der Faszikel ist zerstört. Die äußere Hülle des
Nerven wird lediglich durch unstrukturiertes Gewebe erhalten. Das
traumatisierte Segment wird durch Narbengewebe ersetzt, welches aber
eine Einsprossung neuer Axone verhindert. Die Wahrscheinlichkeit, dass die
Funktion ohne chirurgischen Eingriff wiederkehrt, ist äußerst gering.

13
Einleitung
• Fünfter Grad: Hier ist die Kontinuität der kompletten Nervenbahn
unterbrochen. Das Ereignis dieser Verletzung ist meist auf pathologische
Prozesse zurückzuführen. Aufgrund der großen Unterbrechung tritt ein
kompletter Verlust der sensorischen, motorischen und autonomen
Funktionen auf. Das klinische Bild weist eine Rötung durch Vasodilatation in
dem betroffenen Gebiet auf. Nach wenigen Wochen wirkt die Region infolge
von Vasokonstriktion blass und kühl. Auch das Auftreten trophischer
Störungen der Haut ist möglich. Selbst nach einem chirurgischen Eingriff ist
die völlige Wiederherstellung der Funktion nicht gewährleistet.
Seddon Sunderland Pathologie
Neurapraxie 1 Segmentale Demyelinisierung ,reversibler Leitungsblock, keine axonale Degeneration
Axonotmesis 2 Axonale Degeneration, Endoneurium intakt
Axonotmesis 3 Axonale Degeneration, Zerreißung des Endoneuriums, Perineurium intakt
Axonotmesis 4 Axonale Degeneration, endoneurale und perineurale Zerreißung, Epineurium intakt
Neurotmesis 5 Komplette axonale Degeneration, Zerreißung aller verbindenden Gewebestrukturen
Tabelle 2: Vergleich der Einteilung von Nervenverletzungen zwischen Seddon (1943)
und Sunderland (1951)

14
Einleitung
1.3.3 Diagnostik
Einer der wichtigsten Punkte im Rahmen der klinischen Diagnostik ist die
Befragung des Patienten. Die Kenntnis von dem Zeitpunkt und der Art der
Verletzung haben einen entscheidenden Einfluss auf die therapeutische
Vorgehensweise und die Vorhersagbarkeit des Ergebnisses.
Zu den einfachen Möglichkeiten der Funktionsdiagnostik zählen die Überprüfung
der Hautsensibilität, der motorischen Willkürbewegungen, die Druck- und
Temperaturempfindungen sowie das Tinel-Hoffmann-Zeichen, welches bei
Perkussion eines geschädigten Nerven Schmerzen in dem betroffenen Hautareal
hervorruft. Der Verlust der Wahrnehmung von Bewegung und Lage,
augenscheinliche Muskelatrophien, das Ausbleiben der Schweißsekretion sowie
die Hautfarbe und -beschaffenheit können einen Hinweis auf die Verletzung
motorischer und/ oder sensorischer Nerven geben.
Weitere klinische Untersuchungen werden mittels Neurographie,
Computertomographie (Nemeth und Cigic 1999; Martin-Duverneuil 1999) und
Magnetresonanztomographie durchgeführt. Letztere erscheint bei Läsionen im
extratemporalen Bereich des N. facialis als sehr geeignet (Sutherland et al. 1997;
Kumar et al. 2000). Es können ebenfalls verschiedene elektrophysiologische
Untersuchungen durchgeführt werden, die die Stimulation motorischer Neurone
mit Ableitung evozierter Potentiale (Gellrich et al. 1995), die Elektromyographie
und die Elektroenzephalographie einschließen.

15
Einleitung
1.4 Möglichkeiten der Rekonstruktion
Je nach Ausmaß der Verletzung werden bisher verschiedene
Rekonstruktionsverfahren eingesetzt, die entweder die Kontinuität des Nerven
und seine elektrophysiologische Leitfähigkeit wiederherstellen oder funktionell die
Aufgabe des Nervus facialis übernehmen und ersetzen sollen (Abb. 2). Darüber
hinaus können dynamische Muskeltransplantate der mimischen Muskulatur
eingesetzt werden.
Abb. 2: Vergleich zwischen dem entstandenen Nervendefekt und der Distanz von proximalem
und distalem Ende der Nervenstümpfe. Die Distanz entspricht dem Defekt oder
vergrößert sich, falls die Nervenenden nach der Verletzung auseinander gezogen
worden sind (Mackinnon 1989).
Nur in umschriebenen Fällen gelingt eine End-zu-End-Anastomose. Eine
spannungsfreie Adaptation der Nervenstümpfe ist nur bei einer scharfen
Durchtrennung ohne wesentliche Traumatisierung der Nervenenden möglich.
Sowohl im Rahmen der Traumatologie als auch in der Tumorchirurgie wird sehr
viel öfter die Indikation zur Überbrückung der entstandenen Nervendefekte
gestellt.

16
Einleitung
Um die Nervendiastase zu überbrücken, werden autogene Interponate
unterschiedlicher Herkunft zur Wiederherstellung der Nervenkontinuität
verwendet (Abb. 3 und Abb. 4). Dafür haben sich freie Transplantate aus dem
Plexus cervicalis, dem Nervus suralis oder Ästen des Nervus medianus als
besonders gut geeignet herausgestellt (Hagan 1981; Stephanian et al. 1992;
Haller et al: 1997). Das Verfahren der fazio-fazialen Anastomose mit Interposition
autologer Nervensegmente hat ansprechende funktionelle Ergebnisse gezeigt
(Anderl 1982; Monballiu 1982).
Wenn eine direkte Versorgung des Nervus facialis nicht möglich ist, kann eine
Fremdnervenanastomose zur Rehabilitation der distalen Funktion des Nervus
facialis beitragen. Hierfür kommen verschiedene Spendernerven in Frage
(Hoffmann 1992). Durchgesetzt hat sich vor allem eine Anastomose aus dem
Nervus hypoglossus und dem Nervus facialis (Kunhirio et al. 1991).
Im Gegensatz zu sensiblen Nerven, bei denen eine Rekonstruktion der
Nervenkontinuität zeitlich nicht strikt limitiert ist, sollte die Wiederherstellung der
Nervenkontinuität bei motorischen Nerven innerhalb eines Jahres erfolgen, da
das Erfolgsorgan Muskel ohne Innervation unweigerlich atrophiert und nach
dieser Zeitspanne die Aussichten einer funktionellen Rehabilitation erheblich
sinken.
In Fällen einer länger bestehenden Paralyse des Nervus facialis müssen
umfangreiche Rekonstruktionsmaßnahmen ergriffen werden, die auf eine
Wiederherstellung der muskulären Transposition abzielen. In Frage kommen
hierbei neurovaskulär reanastomosierte Muskeltransplantate des Musculus
gracilis (Guelinckx und Sinsel 1996, Schmelzeisen et al. 1996), des
Musculus latissimus dorsi (Harii et al. 1998), des Musculus masseter und des
Musculus temporalis (Hoffmann 1992) sowie des Musculus rectus abdominis
(Koshima et al. 1997).
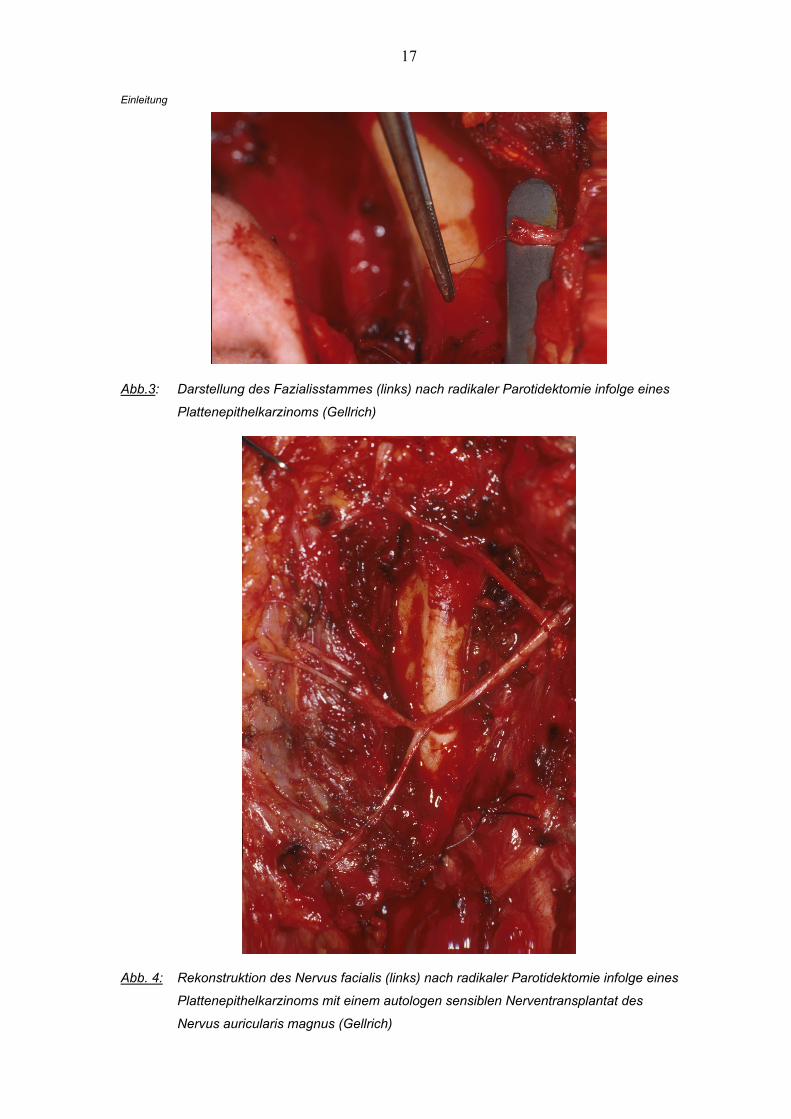
17
Einleitung
Abb.3: Darstellung des Fazialisstammes (links) nach radikaler Parotidektomie infolge eines
Plattenepithelkarzinoms (Gellrich)
Abb. 4: Rekonstruktion des Nervus facialis (links) nach radikaler Parotidektomie infolge eines
Plattenepithelkarzinoms mit einem autologen sensiblen Nerventransplantat des
Nervus auricularis magnus (Gellrich)

18
Ziel
1.5 Ziel der Arbeit
Die vorliegende Arbeit dient der Etablierung eines neuen Modells der gesteuerten
Nervenregeneration mittels primärer Nervendistraktion. Als
Grundlagenuntersuchung ist der Nachweis wichtig, dass ein physiologisch
intakter Nerv oder einzelne Äste eines Nerven in ihrer Länge aktiv verändert
werden können. Untersuchungen der Distraktionsosteogenese haben gezeigt,
dass eine sekundäre Distraktion peripherer Nerven unter definierten Ausmaßen
der Längenveränderung zu einer wenig veränderten Struktur des Nerven führen
(Battison et al. 1992; Block et al. 1993; Hu et al. 2001; Wang et al. 2002).
Auch Verlängerungen mittels Gewebeexpandern zeigten gute Ergebnisse
(Manders et al. 1987; Wood et al. 1991; Hall et al. 1993; Van der Wey et al. 1995;
Van der Wey et al. 1996). Nur eine langsame Dehnung des Nerven mittels
Expander geht ohne einen Funktionsverlust einher, ähnlich dem Verhalten
peripherer Nerven bei langsam wachsenden, benignen Tumoren
(Mackinnon 1989; Anand 1997).
In unserem Modell wird mit einem Zugschraubensystem, welches am Jochbogen
fixiert ist, der Ramus buccalis des Nervus facialis primär distrahiert.
Histologisch aufbereitete Präparate des distrahierten Segments sowie der nicht-
behandelten Gegenseite werden auf lichtmikroskopisch sowohl quantitativ
(anhand von Axonzahl, Myelinscheidendicke und Durchmesser) als auch
qualitativ (im Hinblick auf das Degenerationsverhalten im Vergleich zur gesunden
Seite) ausgewertet.

19
Material und Methode
2 Material und Methode
2.1 Versuchstiere
Die Versuche wurden mit 16 weiblichen Albinoratten der Gattung Wistar
durchgeführt, deren Gewicht zwischen 271g und 437g betrug (durchschnittliches
Gewicht 300g). Das durchschnittliche Alter der Tiere lag bei 1 Jahr. Die Züchtung
und Haltung erfolgte durch das Tierzentrum am Klinikum der Albert-Ludwigs-
Universität Freiburg im Breisgau.
Eine Genehmigung der Experimente durch die Ethik-Kommission und das
Regierungspräsidium Freiburg lag selbstverständlich vor.
2.2 Materialien
Operationsinstrumente und -materialien
Handbohrer mit Hudson-Ansatz Aesculap AG & Co. KG, D-Tuttlingen
Minischraube Synthes GmbH & Co. KG, D-Umkirch
Ethilon® Ethicon, Johnson & Johnson, B-Brüssel
Prolene® Ethicon, Johnson & Johnson, B-Brüssel
Vicryl® Ethicon, Johnson & Johnson, B-Brüssel
Cyanoacyralat-Kleber Renfert GmbH, D-Hilzingen
Chemikalien und Reagenzien
Isopropanol Sigma-Aldrich Chemie GmbH, D-Sinsheim
Methanol Merck KgaA, D-Darmstadt
Ethanol Merck KgaA, D-Darmstadt
Aceton Merck KgaA, D-Darmstadt
3-Aminopropyltriethoxy-Silane Sigma-Aldrich Chemie GmbH, D-Sinsheim
para-Phenylendiamin Sigma-Aldrich Chemie GmbH, D-Sinsheim
Osmiumtetraoxidlösung 2% Merck KgaA, D-Darmstadt
Cacodylat-Puffer Carl Roth GmbH, D-Karlsruhe
Durucrupan ACM® Fluka Chemie AG, CH-Buchs
De Pex®,Brechungsindex 1,52 Serva Electrophoresis GmbH, D-Heidelberg

20
Material und Methode
Geräte
Operationsmikroskop Carl Zeiss Jena GmbH, D-Jena
Mikrotom Supercut 2065 Leica Instruments GmbH, D-Nussloch
7800B Knife Maker LKB-Produkter AB, S-Bromma
Glasstreifen Leica Instruments GmbH, D-Nussloch
Objektträger 76x26 mm Langenbrinck, D-Emmendingen
Probegläschen Kummer, D-Freiburg i.Br
Deckgläser 25x27, Langenbrinck, D-Emmendingen
Inversionsoel Carl Zeiss Jena GmbH, D-Jena
Axioskop Carl Zeiss Jena GmbH, D-Jena
3-Chip-Farbkamera AVT Horn, D-Aalen
Heizplatte Medax, D-Kiel
Computer Software
Axio Vision® 3.0 Carl Zeiss Vision GmbH, D-Eching
analySis® Soft Imaging System GmbH, D-Münster
Excel 2000® Microsoft Corporation, USA-Redmond, WA
SPSS®, Version 10.0 SPSS Inc., USA-Chicago, IL

21
Material und Methode
2.3 Methoden
Albinoratten der Gattung Wistar weisen einen ausreichend dicken Fazialisstamm
auf, dessen Verlauf und Zusammensetzung bezüglich der Faszikel dem der
Menschen sehr ähnlich ist. Während des ersten operativen Eingriffs wurde bei den 16 Albinoratten eine
punktuelle, mikroskopgestützte, perineurale Anschlingung des Ramus buccalis
des Nervus facialis vorgenommen. Die Fixation des Fadens erfolgte an einer
Minischraube (ø 1mm) (Synthes GmbH & Co. KG, D-Umkirch), welche sich im
Jochbeinmassiv befand. Täglich wurde der Nerv insgesamt 10 Tage lang um eine
viertel Umdrehung der Schraube distrahiert. Im Idealfall würde dies einem
täglichen Längengewinn des Nerven von 0,785 mm entsprechen ( π×= dU ).
In einem zweiten Eingriff erfolgten die Entnahme des distrahierten
Nervensegmentes und eines entsprechenden Nervensegmentes der nicht
behandelten Gegenseite zur Kontrolle. Nach der histologischen Aufarbeitung
wurden die Präparate histomorphometrisch mit Hilfe eines Lichtmikroskops
(Axioskop Carl Zeiss Jena GmbH, D-Jena) und mittels der Software Axio Vision®
3.0 (Carl Zeiss Vision GmbH, D-Eching) und analySis®
(Soft Imaging System GmbH, D-Münster) computerassistiert auf Zeichen einer
Degeneration des Nerven nach Distraktion ausgewertet.
2.3.1 Operationen
Die Tiere wurden mit 5,4 Vol% Isofluran auf 6 Liter Sauerstoff für ungefähr 2 min.
in einer Narkosevorrichtung prämediziert. Die Anästhesie erfolgte mit
4 mg Chloralhydrat/100g Körpergewicht, welches intraperitoneal injiziert werden
musste (Gellrich 1995). Bei nicht ausreichender Wirkung des Anästhetikums
wurde entsprechend nachinjiziert.

22
Material und Methode
Chloralhydrat führt zu einer Allgemeinanästhesie der betroffenen Ratte. Dies hat
den Vorteil, dass es im Gegensatz zu Barbituraten, Halotan oder Stickoxiden
weder die Hämodynamik noch den Metabolismus im Kortex negativ beeinflusst
(Ueki et al. 1988). Eine Intubation zur Beatmung des Tieres ist daher nicht
notwendig.
Zur Darstellung des Nervus facialis wurde das Tier für die Dauer des Eingriffs in
einer Mikrohalterung adjustiert (Gellrich 1995). Der Schädel der Ratte wurde
hierzu beidseitig durch einen Metallstift am Meatus accusticus externus in der
stereotaktischen Halterung fixiert. Die folgenden Schritte konnten nun mit Hilfe
des Operationsmikroskops (Carl Zeiss GmbH, D-Jena) durchgeführt werden. Es
wurde immer die linke Gesichtsseite als Distraktionsseite gewählt, während die
rechte Seite als Kontrollseite diente.
Der operative Zugang erfolgte präaurikulär durch eine mediansagittale Inzision.
Die Kopfhaut wurde unter Umgehung einer Orbitotomie abpräpariert, bis sich
distal der Stamm des Nervus facialis mit seinen Aufzweigungen und proximal das
Jochbein mit der dort ansetzen Muskulatur darstellen ließ.
Der aufgesuchte Ramus buccalis wurde nach proximal in seinem Verlauf
freipräpariert, bis sich seine Endaufzweigungen in der mimischen Muskulatur
verloren. Unter Verdrängung der Muskulatur wurde anschließend mit einem
Handbohrer (Aesculap AG & Co. KG, D-Tuttlingen) die Vorbohrung zur
Aufnahme der Distraktionsschraube (Synthes GmbH & Co. KG, D-Umkirch) in
das Jochbeinmassiv durchgeführt. Anschließend wurde die Minischraube mit
einem Durchmesser von 1 mm und einer Länge von 6 mm oder 8 mm mit einem
Mikroschraubenzieher (Handbohrer mit Hudson-Ansatz, Aesculap AG & Co. KG,
D-Tuttlingen) eingebracht. Der ausreichende Halt im Jochbein wurde überprüft.
Darauf folgte die perineurale Anschlingung des Ramus buccalis n. facialis mittels
eines Polyacrylamidfadens (Ethicon, B-Brüssel) der Stärke 10/0 oder 9/0.
Unterschiedliche Nahtmaterialien und Nadelformen waren zuvor erprobt worden,
wobei sich die gebogenen Nadelformen als am geeignetsten erwiesen. Die
beiden Enden der Naht wurden um die Schraube geschlungen und
festgebunden. Eine zusätzliche Fixierung wurde durch Aufbringen eines
Methacrylat-Klebers (Renfert GmbH, D-Hilzingen) mittels einer Minisonde
erreicht.

23
Material und Methode
Der Faden wurde durch Drehen an der Schraube auf Primärspannung gebracht.
Zur Überprüfung der Längenveränderung des Nerven diente eine Markierung in
Form einer Naht mit Vicryl® 4/0 (Ethicon, B-Brüssel), die auf der proximalen Seite
der Anschlingung angebracht war. Der Abstand zum Distraktionsfaden wurde
gemessen und protokolliert (Abb. 5).
Die abpräparierte Gesichtshaut wurde anschließend zurückadaptiert. Mittels einer
Stichinzision wurde die Schraube durch die Gesichtshaut geführt. Der primäre
Wundverschluss erfolgte durch eine Naht mit Vicryl® 4/0.
Anschließend wurden die Tiere den Tierpflegern übergeben, die das Erwachen
und die postoperative Heilungsphase kontrollierten. Im Falle postoperativer
Wundschmerzen erfolgte die Medikation mit Paracetamol oder Acetylsalicylsäure.
Zu jedem Tier wurde ein Operations- und Versuchsprotokoll erstellt (Abb. 6).
Abb. 5: Schematische Darstellung des Zugschraubensystems zur Distraktion des
Ramus buccalis. Die Drehung der Schraube erfolgte täglich um eine viertel
Umdrehung nach rechts.

24
Material und Methode
Abb. 6: Versuchsprotokoll
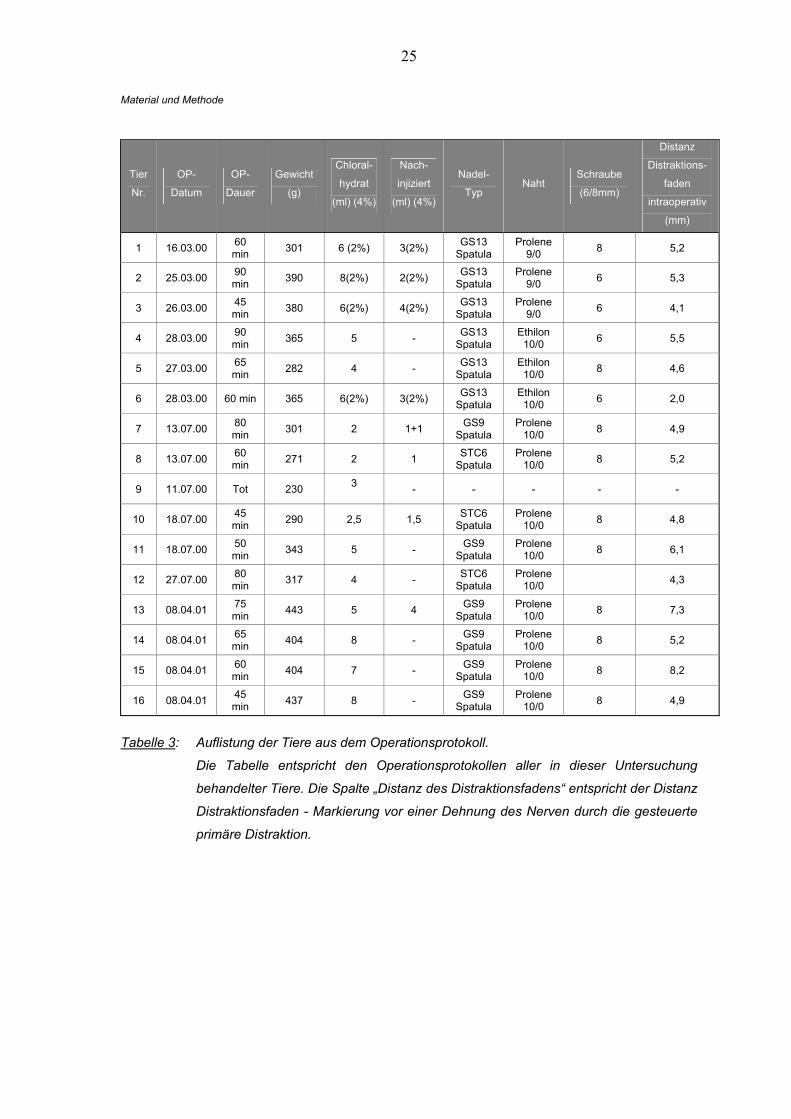
25
Material und Methode
Tier
Nr.
OP-
Datum
OP-
Dauer
Gewicht
(g)
Chloral-
hydrat
(ml) (4%)
Nach-
injiziert
(ml) (4%)
Nadel-
Typ Naht
Schraube
(6/8mm)
Distanz
Distraktions-
faden
intraoperativ
(mm)
1 16.03.00 60 min 301 6 (2%) 3(2%) GS13
Spatula Prolene
9/0 8 5,2
2 25.03.00 90 min 390 8(2%) 2(2%) GS13
Spatula Prolene
9/0 6 5,3
3 26.03.00 45 min 380 6(2%) 4(2%) GS13
Spatula Prolene
9/0 6 4,1
4 28.03.00 90 min 365 5 - GS13
Spatula Ethilon
10/0 6 5,5
5 27.03.00 65 min 282 4 - GS13
Spatula Ethilon
10/0 8 4,6
6 28.03.00 60 min 365 6(2%) 3(2%) GS13 Spatula
Ethilon 10/0 6 2,0
7 13.07.00 80 min 301 2 1+1 GS9
Spatula Prolene
10/0 8 4,9
8 13.07.00 60 min 271 2 1 STC6
Spatula Prolene
10/0 8 5,2
9 11.07.00 Tot 230 3 - - - - -
10 18.07.00 45 min 290 2,5 1,5 STC6
Spatula Prolene
10/0 8 4,8
11 18.07.00 50 min 343 5 - GS9
Spatula Prolene
10/0 8 6,1
12 27.07.00 80 min 317 4 - STC6
Spatula Prolene
10/0 4,3
13 08.04.01 75 min 443 5 4 GS9
Spatula Prolene
10/0 8 7,3
14 08.04.01 65 min 404 8 - GS9
Spatula Prolene
10/0 8 5,2
15 08.04.01 60 min 404 7 - GS9
Spatula Prolene
10/0 8 8,2
16 08.04.01 45 min 437 8 - GS9
Spatula Prolene
10/0 8 4,9
Tabelle 3: Auflistung der Tiere aus dem Operationsprotokoll.
Die Tabelle entspricht den Operationsprotokollen aller in dieser Untersuchung
behandelter Tiere. Die Spalte „Distanz des Distraktionsfadens“ entspricht der Distanz
Distraktionsfaden - Markierung vor einer Dehnung des Nerven durch die gesteuerte
primäre Distraktion.

26
Material und Methode
2.3.2 Distraktion des Ramus buccalis n. facialis
Zur Distraktion des Ramus buccalis war die im Jochbein fixierte Schraube täglich
um eine viertel Umdrehung (0,785 mm) im Uhrzeigersinn zu bewegen (Abb. 5).
Die Dauer der Distraktion wurde auf 10 Tage festgelegt.
Um den Tieren keine zusätzlichen Schmerzen zuzufügen und die Schraube ohne
Störung drehen zu können, wurden die Ratten in einer dafür vorgesehenen
Kammer einer Kurznarkose unterzogen. Die Narkose erfolgte mit Isofluran
(5,4 Vol%), welches mit 6 l/min Sauerstoff in die Narkosekammer geleitet wurde.
Nach ungefähr zwei Minuten waren die Tiere narkotisiert. Anschließend erfolgte
die Drehung der Schraube mittels eines Mikroschraubenziehers. Nachdem die
Ratten aus der Narkose erwacht waren, wurden sie den Tierpflegern übergeben.
2.3.3 Entnahme der Nervenproben
In einer zweiten Operation wurden die Nervenproben entnommen. Die Tötung
der Tiere erfolgte vor der Operation in einer CO 2 -Kammer. Auf der linken Seite
wurde der ehemalige Operationszugang erneut eröffnet. Nach Freilegung des
Ramus buccalis und Aufsuchen der Anschlingung konnte ein 1,5 cm langes
Segment des Nervus facialis entnommen werden. Aufgrund seiner
ausreichenden Länge war später eine Einteilung in je drei Probenstücke möglich.
Die Schraube wurde ebenfalls entfernt.

27
Material und Methode
Zur Entnahme der Kontrollpräparate auf der rechten, nicht-behandelten Seite
wurde präauriculär eine winkelförmige Inzision vorgenommen. Die Gesichtshaut
wurde abpräpariert, bis der Stamm des Nervus facialis dargestellt werden konnte.
Der Ramus buccalis wurde aufgesucht und freipräpariert. Anschließend war auch
hier ein 1,5 cm langes Segment zu entnehmen. Die Proben wurden danach in
Probegläschen mit einer 4%-igen Formalinlösung gelegt und dort bis zur
histologischen Aufarbeitung aufbewahrt. Die Mindestverweildauer betrug 24
Stunden.
2.3.4 Histologische Aufarbeitung der Präparate
Das in 4 % Formaldehydlösung primär fixierte Gewebe wurde mit 0,9 % Natrium-
Chlorid-Lösung zwei Stunden lang gespült. Die Lösung musste währenddessen
dreimal gewechselt werden.
Die Nervensegmente wurden in drei Teile getrennt, um eine genaue
Lokalisierung eventueller Schäden vornehmen zu können. Die Einteilung erfolgte
in ein proximales, mittleres und distales Drittel. Proximal entspricht dabei „in
Richtung Nervenstamm“, während als distal das Nervensegment bezeichnet wird,
welches peripherwärts auf die Distraktionsnaht folgt.
Die proximalen und distalen Anteile wurden etwa auf eine Länge von 0,4 cm
gebracht, während das mittlere Stück, an welchem der Distraktionsfaden fixiert
war, ungefähr eine Länge von 0,7 cm aufwies (Abb. 7).
Für die Kontrastierung und Fixierung des Myelins ist die Methode der Osmierung
sehr geeignet. Durch Osmiumtetraoxid werden ungesättigte Lipide und
ungesättigte Fettsäuren von Proteinen fixiert und durch Oxidation schwarz
eingefärbt.

28
Material und Methode
Abb. 7: Einteilung der Nervensegmente in drei Teilstücke
Die Proben wurden zur Nachfixierung für 3 bis 4 Stunden in eine gekühlte 2%-ige
Osmiumtetraoxidlösung (Merck KgaA, D-Darmstadt) in 6%-iger Cacodylatpuffer
(pH 7,6) (Carl Roth GmbH, D-Karlsruhe), eingelegt worden
(Holländer et al. 1968).
Es erfolgte eine Zwischenspülung mit 2,4%-iger NaCl-Lösung, die während 1½
Stunden dreimal gewechselt wurde. Im Anschluss folgte die Dehydrierung der
Proben in einer aufsteigenden Alkoholreihe, beginnend mit 10%-igem Ethanol.
Die Konzentration wurde alle zehn Minuten um 10% erhöht, bis 100% Ethanol
erreicht waren. Zur gänzlichen Entwässerung verblieben die Proben für eine
Stunde in reinem Ethanol, welches nach 30 Minuten nochmals ausgetauscht
wurde. Die Dehydrierung wurde mit Aceton abgeschlossen, das während einer
Einwirkzeit von 30 Minuten dreimal gewechselt wurde.

29
Material und Methode
2.3.5 Epoxideinbettung
Die Einbettung der Nervenproben erfolgte in einem Epoxyharz
(Bridge et al. 1994). In der vorliegenden Arbeit wurde Durucrupan ACM® (Fluka
Chemie, CH-Buchs) verwendet, welches auf Araldit-Basis hergestellt wird.
Durucrupan-ACM® besteht aus vier Stammlösungen (Stammlösung A:
M-Epoxyharz, Stammlösung B: Härter, Stammlösung C: Beschleuniger,
Stammlösung D: Weichmacher). Diese vier Komponenten ergeben bei korrekter
Anwendung ein sehr hartes Einbettmaterial für histologische Proben. Zunächst
wurden jeweils 25 ml Lösung A und B gut vermischt. Es folgte die Zugabe von 1
ml Stammlösung C. Am Ende wurden noch 0,5 ml Lösung D hinzugegeben.
Die Proben mussten für eine halbe Stunde bei 37°C in der Aralditlösung belassen
werden. Nach Austausch des Araldits wurden die Proben bei 0°C über Nacht im
Kühlschrank gelagert und am nächsten Tag nochmals auf 37°C erwärmt. Darauf
folgten ein weiterer Austausch des Araldits und eine nochmalige Erwärmung auf
37°C.
Nach drei Stunden konnten die Nervenproben entnommen werden. Sie wurden in
beschriftete Silikon-Formen gegeben und darauf mit Araldit ausgegossen.
Anschließend erfolgte die Polymerisation der Proben bei 75°C für 48 Stunden.
Es wurden von jedem Tier sechs Präparate-Blöcke hergestellt, drei der
distrahierten linken Seite und je drei der Kontrollseite.
Sie waren jeweils in ein distales, ein mittleres und ein proximales Drittel eingeteilt
und mit einer numerischen Kennzeichnung versehen. Insgesamt wurden neunzig
Probe-Blöcke weiter verarbeitet (Tab. 4).

30
Material und Methode
Tier Nr. Datum der Entnahme
Proben Nr. Distraktion
Proben Nr. Kontrolle
1 05.04.00 11 12 2 13.04.00 13 14 3 13.04.00 15 16 4 13.04.00 17 18 5 13.04.00 21 22 6 13.04.00 19 20 7 01.08.00 1 2 8 01.08.00 3 4
10 01.08.00 5 6 11 01.08.00 7 8 12 09.08.00 9 10 13 22.04.01 23 24 14 22.04.01 25 26 15 22.04.01 27 28 16 22.04.01 29 30
Tabelle 4: Histologische Nummerierung der Proben nach der Entnahme der Nervensegmente.
Die Einteilung erfolgte nach Distraktionsseite (ungerade Zahlen) und Kontrolle
(gerade Zahlen). Alle Proben wurden in ein proximales, ein mittleres und ein distales
Drittel unterteilt.
Beispiel: Tier Nr. 1, Seite der Distraktion Proben Nr. 11, Segment distal = 11 distal
2.3.6 Herstellung der Semi-Dünnschnitte
Mit Hilfe des Mikrotoms (Leica Instruments, D-Nussloch) wurden Semi-
Dünnschnitte von 1,5 µm Dicke angefertigt (Holländer et al. 1968). Ein Mikrotom
ermöglicht es, mittels eines Glas- oder Diamantmessers von einem Einbettblock
Schnitte in einer vorgegebenen Stärke anzufertigen. Die Epoxidblöcke wurden in
die dafür vorgesehene Halterung eingespannt. Manuell erfolgte das Heranführen
der zukünftigen Schnittfläche an das hier verwendete Glasmesser. Danach wurde
auf maschinelle Bearbeitung umgestellt. Der Schneidevorgang basiert auf einer
Auf- und Abbewegung des Epoxidblocks, der dabei während der absteigenden
Bewegung senkrecht auf die Schneide des Glasmessers trifft. Die mit
3-Aminopropyltriethoxy-Silan (Sigma-Aldrich Chemie GmbH, D-Sinsheim)
beschichteten Objektträger (Langenbrinck, D-Emmendingen) wurden mit je 10
Wassertropfen benetzt. Mittels einer Pinzette wurden die Schnitte auf die
Wassertropfen des Objektträgers gestreckt. Die Trocknung erfolgte auf einer
Heizplatte (Medax, D-Kiel). Pro Probe wurden 5 Objektträger mit je 10 Semi-
Dünnschnitten angefertigt.

31
Material und Methode
2.3.7 Brechung der Glasmesser
Die Herstellung der Glasmesser (Knife Maker, LKB, S-Bromma) erfolgte durch
Brechung. Aus einer Glasstange (Leica Instruments GmbH, D-Nussloch) wurde
durch Ritzung mit einer scharfen Stahlklinge und nachfolgender Brechung durch
Druck erst ein Quadrat hergestellt. In einem Winkel von 45° wurde das Quadrat
ebenfalls zuerst durch Ritzung die Sollbruchstelle markiert und durch den
aufgebrachten Druck gebrochen. Die unscharfe Glashälfte wurde verworfen.
2.3.8 Kontraststeigerung mit para-Phenylendiamin
Der Kontrast durch Osmiumtetraoxid allein ist nicht ausreichend, um Semi-
Dünnschnitte qualitativ und quantitativ auswerten zu können. Es wäre nur eine
Groborientierung möglich.
Mit para-Phenylendiamin (Sigma-Aldrich Chemie GmbH, D-Sinsheim) werden
osmophile Strukturen dargestellt, während basophile Gewebe nicht gezeigt
werden (Böck 1984). So ist es hier möglich, nur das Myelin hervorzuheben. Die
ideale Dicke der Semi-Dünnschnitte für die Färbung liegt zwischen 1,5 und 2,5
µm (Holländer und Vaaland 1968).
Die Objektträger mit den aufgebrachten Präparaten wurden auf einer Heizplatte
(Medax, D-Kiel) bei 37°C getrocknet und anschließend für 5 Minuten in eine 1% -
ige Lösung von para-Phenylendiamin in Methanol und Isopropanol
(Verhältnis 1:1) eingebracht. Es folgte eine kurze Spülung der Präparate in
Methanol und Isopropanol und ihre anschließende Trocknung.
Das Eindecken der Präparate wurde mit De Pex (Serva, D-Heidelberg)
vorgenommen.

32
Material und Methode
2.3.9 Histomorphometrische Auswertung mittels Lichtmikroskopie
Degenerationsvorgänge peripherer Nerven nach einer Verletzung sind in der
Literatur bekannt und vielfach beschrieben. Es gibt definierte histologische
Bewertungskriterien, die beachtet werden müssen. Wichtige Parameter hierfür
sind: die Anzahl und der Durchmesser der Axone, sowie die Myelinscheidendicke
(Zemp et al. 1981; Gillespie und Stein 1983; Bridge et al. 1994;
O´Sullivan et al. 1998; Lindmuth et al. 2002).
Mit einem Lichtmikroskop (Axioskop, Zeiss, D-Jena), an dem eine Kamera
(AVT, D-Aalen) angeschlossen war, wurde das Bild eines Nervenquerschnitts auf
den Computer übertragen. Zur verbesserten Ansicht wurden die Präparate mit
einem Tropfen Dispersionsöl (Carl Zeiss GmbH, D-Jena) bedeckt.
Die Messungen erfolgten mit Hilfe der Computersoftware analySis®
(Soft Imaging, D-Münster).
Der Querschnitt des Nerven wurde bei 200-facher Vergrößerung in vier
Quadranten eingeteilt (Abb. 8). Durch die Quadranten 1 und 3 wurden die
Messungen in der äußeren, durch die Quadranten 2 und 4 in der inneren Hälfte
vorgenommen. Es wurde beurteilt, ob Zentrum und Peripherie unterschiedliche
Reaktionen auf die Distraktion zeigen.
Abb. 8: Einteilung des Gesamtquerschnitts in 4 Quadranten

33
Material und Methode
In 1000-facher Vergrößerung wurde ein Rahmen von 760 x 572 Pixel
(82,8 x 62,3 µm) gesetzt, um einen definierten Quadrantenabschnitt zu erhalten,
der allen Messungen zugrunde liegt (Abb. 9).
Angeschnittene Axone wurden nicht in die Messung einbezogen.
Abb. 9: Ausschnitt des gesetzten Rahmens von 82,3 x 62,3 µm
Die erste Messung bezog sich auf die Myelinscheidendicke. An vier Punkten
wurde die Markscheidendicke in 3-, 6-, 9-, 12- Uhrposition gemessen
(Funktion analySis®: Messen/beliebiger Abstand) und in eine Excel® Tabelle
(Miccrosoft Coperation, USA-Redmond, WA) übertragen (Abb.10).
Abb. 10: Messung der Myelinscheidendicke an vier Punkten der Myelinscheide

34
Material und Methode
Anschließend wurde der Umfang der Myelinscheiden im selben Abschnitt
gemessen (Funktion analySis®: Messen/beliebiger Umfang/ beliebige Fläche)
und ebenfalls in eine Excel® Tabelle übertragen worden (Abb. 11).
Abb. 11: Messung des Umfangs der Myelinscheiden
Um den folgenden festgelegten Quadranten zu erfassen, wurde dieser zunächst
in 200-facher Vergrößerung eingestellt und in die 1000-fache Vergrößerung
übertragen.
Der mittlere Außendurchmesser von Axon und Myelinscheide wurde aus der
gemessenen Fläche errechnet: πAD = x 2 (Abb.12)
Die Entscheidung der mathematischen Berechnung basiert auf der Beobachtung,
dass eine direkte Messung schwierig gewesen wäre, da Axon und Myelinscheide
meist nicht kreisrund sind. Mit den errechneten Werten können Messfehler
vermieden und ein objektiver Vergleich angestellt werden.
Abb. 12: Berechnung des Durchmessers aus der Fläche

35
Material und Methode
2.3.10 Histologische Auswertung der Präparate
Die Betrachtung der histologischen Präparate erfolgte ebenfalls in 1000-facher
Vergrößerung mit Dispersionsöl. Folgende Merkmale wurden als
Degenerationszeichen gewertet:
• Atrophie von Axon und Myelinscheide, Ablösen des Axons von der
Myelinscheide (Abe et al 1996)
• Kapillarverdichtung, dünn myelinisierte große Axone (Bridge et al 1994)
• Fibrosierung des Endoneuriums und weit auseinanderliegende Axone
(Manders et al. 1987)
• Schwellung der Myelinscheiden (Logroscino et al. 1980) und die
Aufsplitterung der Myelinscheiden in ihre Lamellen (Lahoda 1988; Tackman
1989)
• Unregelmäßigkeit der Form von Axon und Myelinscheide (Gillespie und
Stein 1983)
• Abnahme der Anzahl von Axonen (Carter und Linsey 1987; Holland und
Robinson 1990)
2.3.11 Quantitative Auswertung des Nervenquerschnitts
Um die Anzahl der Axone zwischen distrahierter und nicht-distrahierter Seite
vergleichen zu können, wurden die Axone des gesamten Nervenquerschnitts
gezählt.
Über das Axioskop (Zeiss, D-Jena) und der daran angekoppelten
3-Chip-Farbkamera (AVT, D-Aalen) konnten in 200-facher Vergrößerung die
Bilder der Nervenquerschnitte mit Hilfe des Computerprogramms Axio Vision®
(Zeiss Vision, D-Eching) übertragen, ausgedruckt und von Hand ausgezählt
werden.
Die Werte wurden tabellarisch zusammengefasst. Die Kontrollseite des Tieres
Nummer 15 konnte nicht bewertet werden, da der entnommene Faszikel dem
kleinen Ramus zygomaticus mit einer wesentlich geringeren Axonanzahl
entspricht. Somit wurde dieser nicht in den Vergleich miteinbezogen.

36
Material und Methode
2.3.12 Statistische Auswertung
Die Statistische Auswertung der Ergebnisse erfolgte mittels der Software
SPSS® (USA-Chicago, IL)
Durch eine Varianzanalyse für mehr als zwei unabhängigen Stichproben
(einfaktorielle ANOVA) und dem nachfolgenden Test auf Gruppenbildung (nach
Duncan) wurde festgestellt, ob eine Korrelation zwischen den Quadranten
vorliegt. Ebenso wurden mit diesem Verfahren die Unterschiede zwischen den
Sektionen (proximal, mitte, distal) und den Ouadranten (1, 2, 3, 4) der
verschiedenen Seiten verglichen.
Anhand des t-Tests zweier unabhängiger Stichproben
(van der Wey et al. 1996) wurde der Einfluss der Distraktion auf die
Messparameter (Myelinscheidendicke, Umfang, Fläche, mittlerer
Außendurchmesser) im Vergleich zu der Kontrollseite statistisch überprüft. Durch
diesen Test können die Gleichheit der Varianzen (Standardfehler im Quadrat)
und die Irrtumswahrscheinlichkeit p der zu vergleichenden Mittelwerte
(Signifikanz) ermittelt werden. Die Vorrausetzung zur Durchführung eines t-Tests
ist das Vorliegen einer Normalverteilung. Dies war bei allen zu vergleichenden
Proben der Fall.
Die oben genannten Messparameter der distrahierten Seite wurden mit
Parametern der nicht-behandelten Seite verglichen.
Aus den Einzelwerttabellen der gemessenen Parameter wurden in der Computer-
Software Excel® (Microsoft USA, Redmond, WA) das Maximum, das Minimum,
der Mittelwert und die Standardabweichung ermittelt. So war es möglich, den
mittleren Innendurchmesser (Innendurchmesser D= Außendurchmesser D - 2x
Myelinscheidendicke) und die G-Ratio zu berechnen. Die G-Ratio ist ein wichtiger
Faktor für die Nervenleitgeschwindigkeit und beschreibt das Verhältnis zwischen
Axondurchmesser ohne Myelinscheide und Axondurchmesser mit Myelinscheide.
Der optimale theoretische Wert liegt bei 0,6 (Rushton 1951,
De Jong et al. 1991). Aufgrund der Errechnung aus Mittelwerten wird für die
beiden letztgenannten Werte nur eine Tendenz angegeben.

37
Ergebnisse
3 Ergebnisse Von den insgesamt 16 Tieren verstarb ein Tier während der Narkose. Der
Distraktionsfaden des Tieres Nr. 8 war während der Verlängerung des Nerven
gerissen, der Zeitpunkt der Divulsion konnte nicht bestimmt werden.
Anhand von zwei Stichproben wurde der Längengewinn des Ramus buccalis
nach der moduliert gerichteten primären Distraktion überprüft. Diese führte bei
Tier Nr. 7 zu einer Verlängerung von 5,1 mm, bei Tier Nr. 15 zu einer
Verlängerung von 8 mm.
3.1 Histologische Auswertung
Die Anzahl der zu untersuchenden Präparate betrug auf Seiten der Distraktion
N=45 und auf der Kontrollseite N=42. Das Kontrollpräparat des Tieres 16 konnte
nicht in die Auswertung einbezogen werden, da der Faszikel des kleinen Ramus
zygomaticus mit einer sehr geringen Axonanzahl entnommen wurde.
Das erste Präparat mit der ungeraden Probennummer entsprach der distrahierten
Seite des Tieres, darauf folgte mit der geraden Nummerierung die entsprechende
Kontrollseite des Tieres.
Bei zwei Tieren waren die Präparate der distrahierten Seite und ihre
Kontrollseiten völlig degeneriert. Dies ist auf einen Fehler in der Lagerung oder in
der histologischen Aufbereitung der Präparate zurückzuführen.
Das Gesamtergebnis der histologischen Untersuchung erwies, dass in allen
Segmenten (proximal, mitte, distal) degenerative Veränderungen der
Myelinscheiden als Folge der primären Distraktion der Nervus facialis auftraten.
Starke Degenerationserscheinungen waren vermehrt im distalen Drittel der
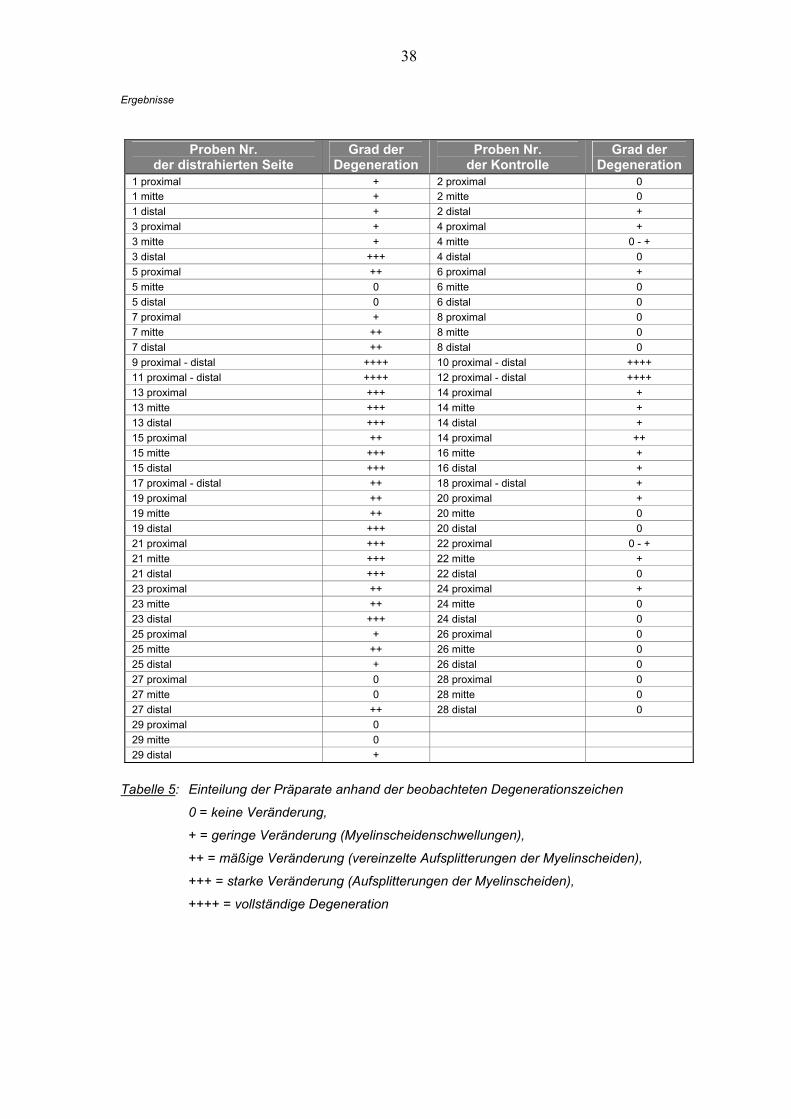
entnommenen Nervenproben zu erkennen (Tab. 5).
38
Ergebnisse
Proben Nr. der distrahierten Seite
Grad der Degeneration
Proben Nr. der Kontrolle
Grad der Degeneration
1 proximal + 2 proximal 0 1 mitte + 2 mitte 0 1 distal + 2 distal + 3 proximal + 4 proximal + 3 mitte + 4 mitte 0 - + 3 distal +++ 4 distal 0 5 proximal ++ 6 proximal + 5 mitte 0 6 mitte 0 5 distal 0 6 distal 0 7 proximal + 8 proximal 0 7 mitte ++ 8 mitte 0 7 distal ++ 8 distal 0 9 proximal - distal ++++ 10 proximal - distal ++++ 11 proximal - distal ++++ 12 proximal - distal ++++ 13 proximal +++ 14 proximal + 13 mitte +++ 14 mitte + 13 distal +++ 14 distal + 15 proximal ++ 14 proximal ++ 15 mitte +++ 16 mitte + 15 distal +++ 16 distal + 17 proximal - distal ++ 18 proximal - distal + 19 proximal ++ 20 proximal + 19 mitte ++ 20 mitte 0 19 distal +++ 20 distal 0 21 proximal +++ 22 proximal 0 - + 21 mitte +++ 22 mitte + 21 distal +++ 22 distal 0 23 proximal ++ 24 proximal + 23 mitte ++ 24 mitte 0 23 distal +++ 24 distal 0 25 proximal + 26 proximal 0 25 mitte ++ 26 mitte 0 25 distal + 26 distal 0 27 proximal 0 28 proximal 0 27 mitte 0 28 mitte 0 27 distal ++ 28 distal 0 29 proximal 0 29 mitte 0 29 distal +
Tabelle 5: Einteilung der Präparate anhand der beobachteten Degenerationszeichen
0 = keine Veränderung,
+ = geringe Veränderung (Myelinscheidenschwellungen),
++ = mäßige Veränderung (vereinzelte Aufsplitterungen der Myelinscheiden),
+++ = starke Veränderung (Aufsplitterungen der Myelinscheiden),
++++ = vollständige Degeneration

39
Ergebnisse
Beschreibende Ergebnisse der Histologie:
Abkürzungen: proximal= prox, mitte= mitt, distal = dist
1prox, 1mitt: Auffällige Zeichen axonaler Degeneration sind nicht zu
beobachten.
1dist: Die Myelinscheiden in diesem Präparat sind häufig längs
angeschnitten in diesen Abschnitten ist die Differenzierung
aufgeschwollener Myelinscheiden schwierig; in den quer
angeschnittenen Bereichen sind vereinzelt aufgeschwollene
Myelinscheiden zu erkennen.
2prox, 2mitt: Myelinscheiden und Axone sind intakt (Abb. 13).
2dist: Der Nerv weist Degenerationszeichen auf, bei einzelnen
Myelinscheiden ist eine Aufspaltung der Myelinlamellen
erkennbar, der Abstand zwischen den Axonen ist etwas
vergrößert.
Abb. 13: Präparat 10 proximal
3prox, 3mitt: Außer verdickten Myelinscheiden können fast keine
Degenerationszeichen festgestellt werden.

40
Ergebnisse
3dist: Besonders im ersten Quadranten zeigen sich Zeichen einer
Degeneration: Formveränderungen der Axone und ihrer
Myelinscheiden, Schwellung und Aufsplitterung in Myelinlamellen
(Abb.14).
Abb. 14: Präparat 3 distal
4prox: Das Präparat ist sehr dunkel eingefärbt, es ist wenig
Degeneration im klassischen Sinn vorhanden. Es sind kleine,
dicht gepackte Axone zu erkennen.
4mitt: Außer geringen Formabweichungen der Myelinscheiden sind
wenige Veränderungen zu beobachten. Einzelne untergegangene
Axone sind in den Quadranten 1 und 2 zu sehen.
4dist: Es fallen keine Degenerationszeichen auf, die Axone sind relativ
regelmäßig geformt, die kleinen Axone haben verhältnismäßig
dünne Myelinscheiden.
5prox: Einzelne Axone fallen durch geschwollene oder aufgesplitterte
Myelinscheiden auf, die Nervenfasern erscheinen sehr klein, eine
Häufung aufgeschwollener Myelinscheiden befindet sich im
Quadranten 3.
5prox, 5dist: Es sind keine Anzeichen von Degeneration zu erkennen.

41
Ergebnisse
6prox, 6mitt: Die äußere Form der Faszikel dieser Präparate ist sehr
unregelmäßig, sonst sind sie intakt. Es sind längs angeschnittene
Myelinscheiden vorhanden.
6dist: Faszikel, Axone und Myelinscheiden sind unversehrt.
7prox-7dist: In diesen Präparaten sind vereinzelt angeschwollene
Myelinscheiden zu sehen.
8prox: Das Präparat ist unauffällig.
8mitt: Längs angeschnittene Myelinscheiden in diesem sonst
unversehrten Querschnitt zu beobachten.
8dist: Vereinzelte Axone zeigen aufgeschwollene Myelinscheiden.
9prox: Die axonale Degeneration ist fortgeschritten, aufgeschwollene
Myelinscheiden und die Aufsplitterung in Myelinlamellen sind bei
den meisten Markscheiden zu sehen. Bei 1/3 der Myelinscheiden
ist keine Dicke messbar.
Abb. 15: Präparat 9 mitte

42
Ergebnisse
9mitt, 9dist: Das Präparat zeigt auffällige Abweichungen, die Myelinscheiden
sind aufgesplittert. Es dominieren zu 2/3 Markballen.
(Abb. 15 und 16).
Abb. 16: Präparat 9 distal
10prox-10dist: Diese Nervenpräparate sind durch starke Degeneration
gekennzeichnet. Zu 50% ist eine Bildung von Markballen sichtbar.
Die restlichen Myelinscheiden sind aufgesplittert, was
wahrscheinlich auf einen Fehler während der Aufbewahrung oder
der Aufbereitung der Präparate zurückzuführen ist (Abb.17).
Abb. 17: Präparat 10 mittel
11prox: Markballen und aufgesplitterte Myelinlamellen sowie
auseinandergerissene Axone mit fibrösem endoneuralen
Zwischengewebe bestimmen das Bild.

43
Ergebnisse
11mitt: Fast alle Myelinscheiden weisen eine Aufsplitterung der Lamellen
auf.
11dist: Die Hälfte der Axone lässt eine Aufsplitterung der Lamellen
erkennen.
12prox-12dist: Wie in Präparat 18 ist wahrscheinlich ein Fehler in der
Verarbeitung oder der Aufbewahrung für das degenerative
Erscheinungsbild verantwortlich (Abb. 18).
Abb. 18: Präparat 12 mitte
13prox: Die Myelinscheiden sind aufgeschwollen, ebenso kann
Markballenbildung beobachtet werden.
13mitt: Nur wenig myelinisierte Axone mit viel Zwischensubstanz sind
dargestellt, wobei die vorhandenen Axone einen intakt sind. Es
fallen viele kleine Nervenfasern auf, deren Myelinscheiden etwas
dünner als im Kontrollpräparat wirken.
13dist: Es sind wenig intakte myelinisierte Axone in großem Abstand
zueinander zu sehen, allerdings befinden sich außerordentlich
kleine Nervenfasern im dritten Quadranten; dies könnte auf einen
Regenerationsprozess hindeuten (Abb. 19).
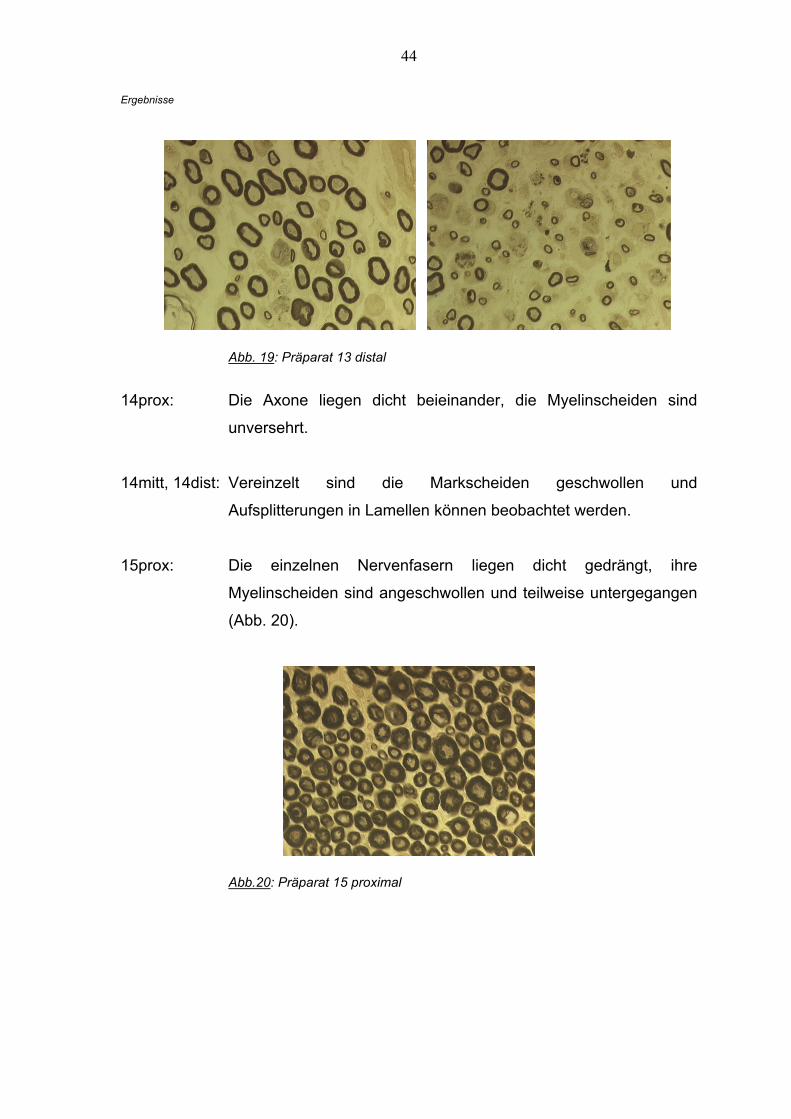
44
Ergebnisse
Abb. 19: Präparat 13 distal
14prox: Die Axone liegen dicht beieinander, die Myelinscheiden sind
unversehrt.
14mitt, 14dist: Vereinzelt sind die Markscheiden geschwollen und
Aufsplitterungen in Lamellen können beobachtet werden.
15prox: Die einzelnen Nervenfasern liegen dicht gedrängt, ihre
Myelinscheiden sind angeschwollen und teilweise untergegangen
(Abb. 20).
Abb.20: Präparat 15 proximal

45
Ergebnisse
15mitt, 15dist: Es sind weit auseinandergerissene Axone zu sehen, und
Markfragmente untergegangener Axone treten in Erscheinung,
das weitere Bild wird durch sehr wenig intakte Axone mit
ausreichender Myelinscheidendicke und gleichmäßiger Form
geprägt (Abb. 21).
Abb. 21: Präparat 15 distal
16prox-16dist: Die Axone liegen dicht beisammen, und es zeigen sich einzelne
Axone mit aufgeschwollenen Myelinscheiden.
17prox-17dist: Die Nervenfasern haben einen etwas größeren Abstand
zueinander als im Kontrollpräparat, die Myelinscheiden wirken
partiell angeschwollen, Schollenbildung der Myelinscheiden
lässt sich erkennen (Abb. 22).
Abb. 22: Präparat 17 proximal

46
Ergebnisse
18prox: Viele Axone sind längs angeschnitten, deutliche
Degenerationszeichen sind erkennbar, die Markscheiden sind
eher kugelförmig geschwollen.
18mitt: Kugelförmige Schwellungen der Markscheiden sind zu
beobachten.
18dist: Die Myelinscheiden sind sehr dünn. Einige zeigen eine
Aufsplitterung der Myelinlamellen.
19prox: Auch in diesem Präparat können viele längs angeschnittene
Axone mit kugelförmiger Degeneration der Myelinscheiden
wahrgenommen werden. Es fallen dichte Abstände der
Nervenfasern zueinander auf.
19mitt, 19dist: Die Axone liegen relativ nahe beieinander; die Markscheiden sind
vereinzelt aufgeschwollen (Abb. 23).
Abb. 23: Präparat 19 distal
20prox-20dist: Vereinzelt sind degenerierte Axone mit aufgeschwollenen
Myelinscheiden zu beobachten.

47
Ergebnisse
21prox: Die Axone liegen dicht beieinander, sie zeigen keine
Formabweichungen, Ödeme der Myelinscheiden und viele kleine
Axone sind sichtbar (Abb. 24).
Abb. 24: Präparat 21 proximal
21mitt: Die Qualität des Präparates ist fraglich, da die meisten
Nervenfasern längs angeschnitten sind; es fallen kugelförmige
Myelinscheidenverdickungen auf (vor allem in den Quadranten 3
und 4).
21dist: Die Anzahl der Axone mit geschwollenen Myelinscheiden hat im
Vergleich zum proximalen und mittleren Drittel dieses
Nervensegmentes zugenommen, kugelförmige
Markscheidenverdickung sowie Aufsplitterung des Myelins zeigen
sich.
22prox-22dist: Das Präparat ist unauffällig, vereinzelt sind aufgeschwollene
Myelinscheiden zu erkennen.
23prox: Die Axone liegen etwas weiter auseinandergedrängt, die
Markscheiden sind sehr dick, doch sie zeigen keine Zeichen von
Aufsplitterung oder Kugelbildung. Leichte Formveränderungen
sind zu bemerken.

48
Ergebnisse
23mitt: Größtenteils sind die Nervenfasern längs getroffen, relativ dicke
Myelinscheiden und verhältnismäßig große Axone fallen auf.
23dist: Die Axone haben eine gleichmäßige Form und wirken etwas
auseinandergedrängt, die Myelinscheiden sind nicht geschwollen.
Im zweiten Quadranten befinden sich auffällig kleine
Nervenfasern, was auf einen die auf Regenerationsprozess
hindeutet (Abb.25).
Abb. 25: Präparat 23 distal
In der Abbildung rechts sind Anzeichen von Regeneration zu sehen.
24prox-24dist: Die Form der Markscheiden ist gleichmäßig, es sind viele kleine
Axone vorhanden.
25prox: Schwellungen der Markscheiden sind vereinzelt zu beobachten.
25mitt, 25dist: Die myelinisierten Axone sind intakt, sie weisen etwas größere
Abstände zueinander auf als in ihrem Kontrollpräparat.
26prox-26dist: Die Axone sind unversehrt, die Myelinscheiden wirken drall, aber
nicht geschwollen.

49
Ergebnisse
27prox-27dist: Die Axone sind in Form und Größe unauffällig, im distalen
Präparat sind vereinzelt Axone mit aufgeschwollenen
Myelinscheiden feststellbar.
28prox-28dist: Es sind fast keine Degenerationszeichen zu sehen, nur Präparat
36 dist zeigt mehr aufgeschwollene Markscheiden als die
Kontrollpräparate 35 prox-dist.
29prox: Es sind keine Verdickungen, aber starke Formabweichungen der
Myelinscheiden in diesem Präparat zu beobachten, die
Nervenfasern liegen dicht gedrängt, und ihre Größe ist gering.
29mitt, 29dist: Es ist keine Waller´sche Degeneration zu erkennen (Abb. 26).
Abb. 26: Präparat 29 distal
50
Ergebnisse
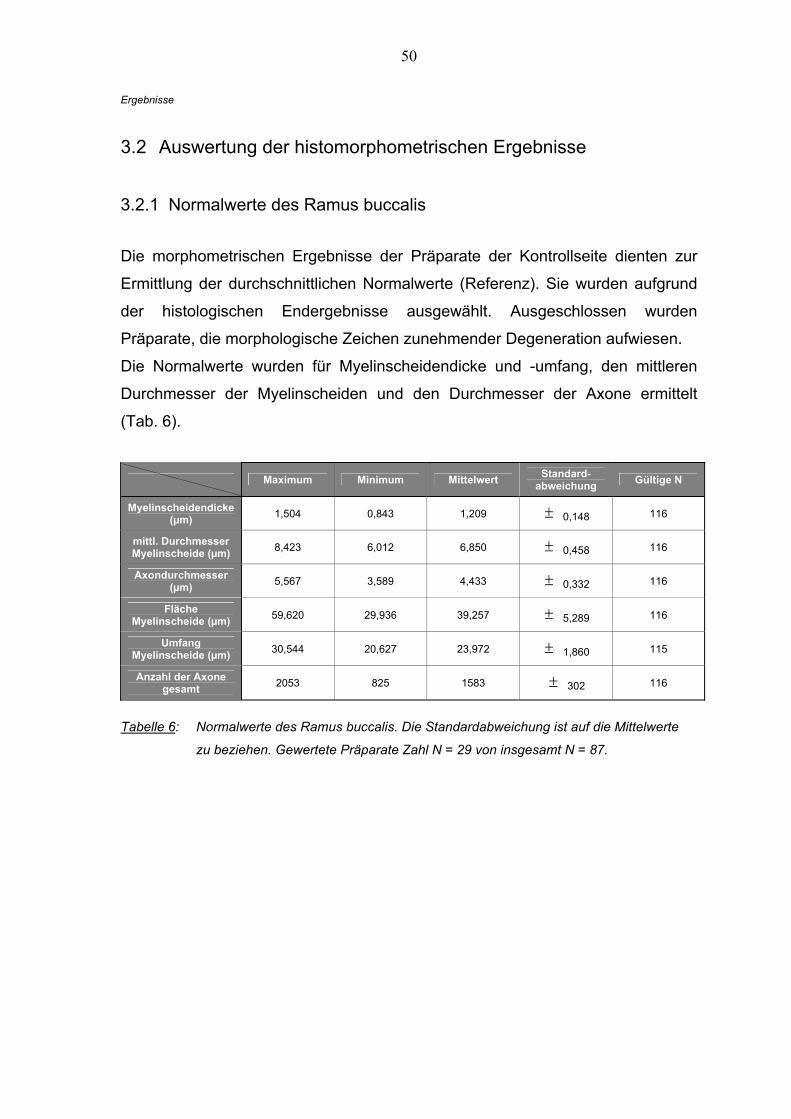
3.2 Auswertung der histomorphometrischen Ergebnisse
3.2.1 Normalwerte des Ramus buccalis
Die morphometrischen Ergebnisse der Präparate der Kontrollseite dienten zur
Ermittlung der durchschnittlichen Normalwerte (Referenz). Sie wurden aufgrund
der histologischen Endergebnisse ausgewählt. Ausgeschlossen wurden
Präparate, die morphologische Zeichen zunehmender Degeneration aufwiesen.
Die Normalwerte wurden für Myelinscheidendicke und -umfang, den mittleren
Durchmesser der Myelinscheiden und den Durchmesser der Axone ermittelt
(Tab. 6).
Maximum Minimum Mittelwert Standard-abweichung Gültige N
Myelinscheidendicke (µm) 1,504 0,843 1,209 ± 0,148 116
mittl. Durchmesser Myelinscheide (µm) 8,423 6,012 6,850 ± 0,458 116
Axondurchmesser (µm) 5,567 3,589 4,433 ± 0,332 116
Fläche Myelinscheide (µm) 59,620 29,936 39,257 ± 5,289 116
Umfang Myelinscheide (µm) 30,544 20,627 23,972 ± 1,860 115
Anzahl der Axone gesamt 2053 825 1583 ± 302 116
Tabelle 6: Normalwerte des Ramus buccalis. Die Standardabweichung ist auf die Mittelwerte
zu beziehen. Gewertete Präparate Zahl N = 29 von insgesamt N = 87.

51
Ergebnisse
3.2.2 Der Ramus buccalis nach Distraktion
Als Grundlage für die Bewertung der Auswirkung der Distraktion auf den Ramus
buccalis wurden bei den statistischen Tests nicht die oben genannten Idealwerte
benutzt. Sie dienen nur zur Übersicht.
Die Untersuchungen basieren auf einem intraindividuellen Seitenvergleich
zwischen distrahierter und Kontrollseite der gemessenen Werte eines jeden
Tieres.
Myelinscheiden
Die Dicke wurde an vier Punkten der Myelinscheiden gemessen. Es standen
117.665 Einzelwerte für den Vergleich zur Verfügung, wobei die Distraktionsseite
60.860 und die Kontrollseite 58.848 Werte enthielten.
Präparat Nr. 29 ist nur in der Gesamtwertung, nicht im Seitenvergleich vertreten.
Der Vergleich innerhalb der Quadranten und Sektionen (einfaktorielle ANOVA)
und der Test auf Untergruppen (Duncan) wurde vor dem t-Test durchgeführt, um
festzustellen, ob eine Korrelation der Quadranten vorliegt. Da dies nicht der Fall
ist, konnte ein Seitenvergleich aller Werte eines Tieres vorgenommen werden.
Das Ergebnis wird aus methodischen Gründen erst später erläutert.
Gesamtwertung:
Der Vergleich der Mittelwerte zeigt einen höchst signifikanten Unterschied
zwischen den beiden Seiten. Dabei ist der Mittelwert auf der distrahierten Seite
geringer als in der Kontrolle.
Es kann die Aussage getroffen werden, dass in der Gesamtuntersuchung die
Myelinscheidendicke auf der distrahierten Seite abnimmt.
Einzelwertung:
In den Vergleichen zwischen Distraktion und Kontrolle eines jeden Tieres stellt
man bei 4 Tieren eine Zunahme der Myelinscheidendicke auf der distrahierten
Seite fest.
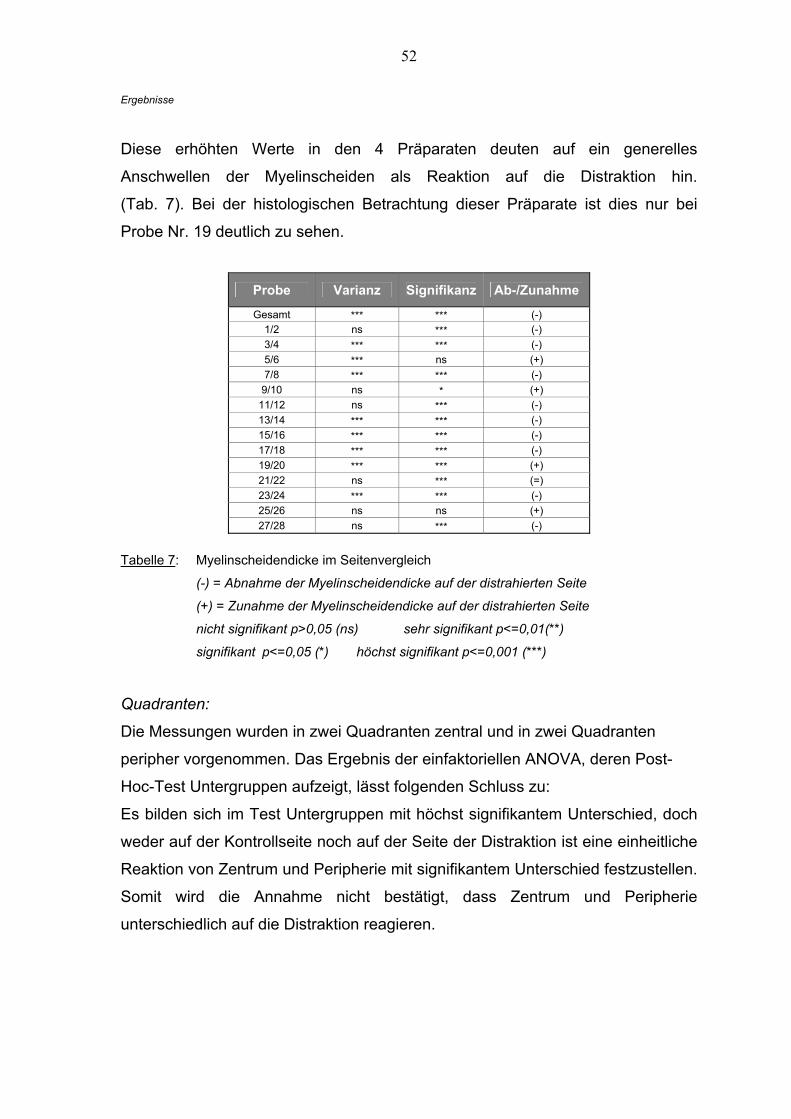
52
Ergebnisse
Diese erhöhten Werte in den 4 Präparaten deuten auf ein generelles
Anschwellen der Myelinscheiden als Reaktion auf die Distraktion hin.
(Tab. 7). Bei der histologischen Betrachtung dieser Präparate ist dies nur bei
Probe Nr. 19 deutlich zu sehen.
Probe Varianz Signifikanz Ab-/Zunahme
Gesamt *** *** (-) 1/2 ns *** (-) 3/4 *** *** (-) 5/6 *** ns (+) 7/8 *** *** (-)
9/10 ns * (+) 11/12 ns *** (-) 13/14 *** *** (-) 15/16 *** *** (-) 17/18 *** *** (-) 19/20 *** *** (+) 21/22 ns *** (=) 23/24 *** *** (-) 25/26 ns ns (+) 27/28 ns *** (-)
Tabelle 7: Myelinscheidendicke im Seitenvergleich
(-) = Abnahme der Myelinscheidendicke auf der distrahierten Seite
(+) = Zunahme der Myelinscheidendicke auf der distrahierten Seite
nicht signifikant p>0,05 (ns) sehr signifikant p<=0,01(**)
signifikant p<=0,05 (*) höchst signifikant p<=0,001 (***)
Quadranten:
Die Messungen wurden in zwei Quadranten zentral und in zwei Quadranten
peripher vorgenommen. Das Ergebnis der einfaktoriellen ANOVA, deren Post-
Hoc-Test Untergruppen aufzeigt, lässt folgenden Schluss zu:
Es bilden sich im Test Untergruppen mit höchst signifikantem Unterschied, doch
weder auf der Kontrollseite noch auf der Seite der Distraktion ist eine einheitliche
Reaktion von Zentrum und Peripherie mit signifikantem Unterschied festzustellen.
Somit wird die Annahme nicht bestätigt, dass Zentrum und Peripherie
unterschiedlich auf die Distraktion reagieren.
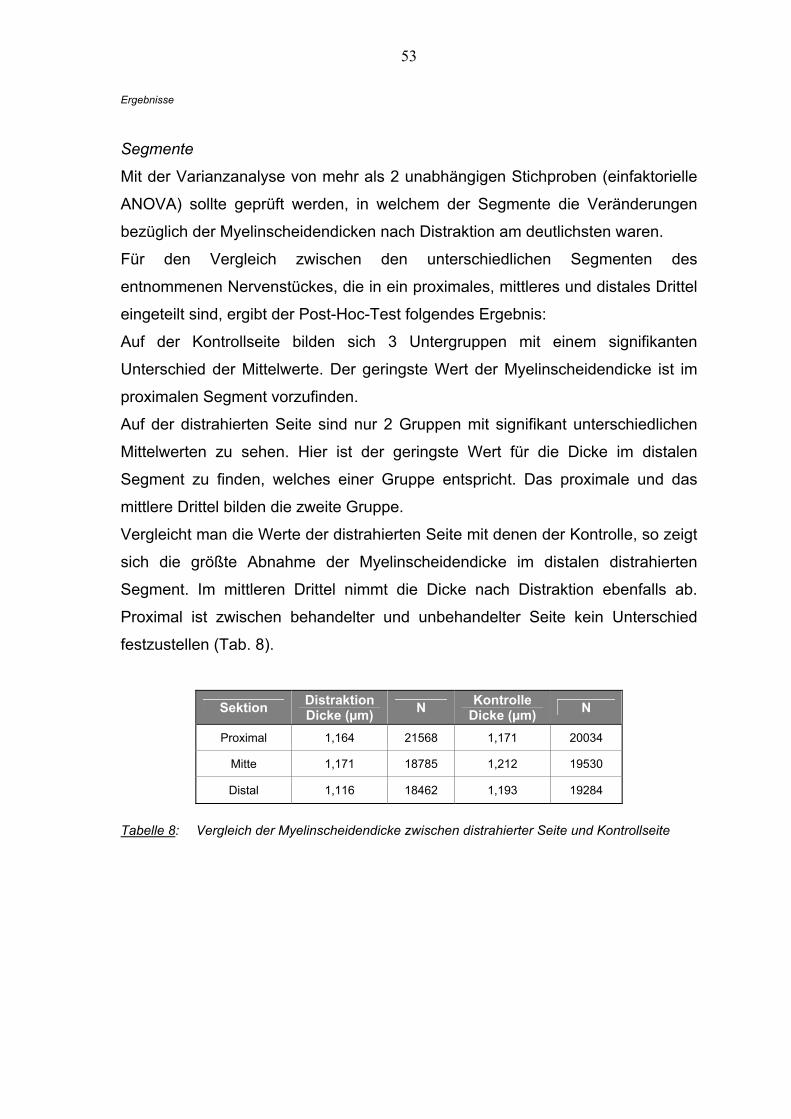
53
Ergebnisse
Segmente
Mit der Varianzanalyse von mehr als 2 unabhängigen Stichproben (einfaktorielle
ANOVA) sollte geprüft werden, in welchem der Segmente die Veränderungen
bezüglich der Myelinscheidendicken nach Distraktion am deutlichsten waren.
Für den Vergleich zwischen den unterschiedlichen Segmenten des
entnommenen Nervenstückes, die in ein proximales, mittleres und distales Drittel
eingeteilt sind, ergibt der Post-Hoc-Test folgendes Ergebnis:
Auf der Kontrollseite bilden sich 3 Untergruppen mit einem signifikanten
Unterschied der Mittelwerte. Der geringste Wert der Myelinscheidendicke ist im
proximalen Segment vorzufinden.
Auf der distrahierten Seite sind nur 2 Gruppen mit signifikant unterschiedlichen
Mittelwerten zu sehen. Hier ist der geringste Wert für die Dicke im distalen
Segment zu finden, welches einer Gruppe entspricht. Das proximale und das
mittlere Drittel bilden die zweite Gruppe.
Vergleicht man die Werte der distrahierten Seite mit denen der Kontrolle, so zeigt
sich die größte Abnahme der Myelinscheidendicke im distalen distrahierten
Segment. Im mittleren Drittel nimmt die Dicke nach Distraktion ebenfalls ab.
Proximal ist zwischen behandelter und unbehandelter Seite kein Unterschied
festzustellen (Tab. 8).
Sektion Distraktion Dicke (µm) N Kontrolle
Dicke (µm) N
Proximal 1,164 21568 1,171 20034
Mitte 1,171 18785 1,212 19530
Distal 1,116 18462 1,193 19284
Tabelle 8: Vergleich der Myelinscheidendicke zwischen distrahierter Seite und Kontrollseite

54
Ergebnisse
Durchmesser
Betrachtet man das Ergebnis des t-Testes für die Gesamtheit des mittleren
Durchmessers, so nimmt dieser auf der distrahierten Seite ab.
Die mittlere Differenz beträgt dabei minus 0,1 µm, wobei sich die Anzahl N der
untersuchten Werte von 15.639 der Kontrollseite auf 16.249 auf der
Distraktionsseite erhöht. Proximal steigt die Anzahl N der zu vergleichenden
Werte am stärksten an, was auf einen Regenerationsprozess durch
ausprossende Axone von proximal zurückzuführen ist.
Bei den einzelnen Seitenvergleichen (N=14) der Tiere, kann in 10 Fällen eine
Abnahme und in 4 Fällen (Proben-Nr.: 11, 23, 27, 33) eine Zunahme des
Durchmessers festgestellt werden (Abb. 27).
Zwischen proximal, medial und distal ist ein höchst signifikanter Unterschied der
Mittelwerte zu erkennen, im distalen Segment nimmt der Durchmesser mit
0,209 µm am meisten ab.
Anhand der Überprüfung der Quadranten kann die Aussage getroffen werden,
dass in der Reaktion auf die Distraktion kein Unterschied zwischen Zentrum und
Peripherie erkennbar ist.
Seite proximal Untersuchte Einzelwerte
N mitte
Untersuchte Einzelwerte
N distal
Untersuchte Einzelwerte
N Distraktion 6,39 5959 6,83 5118 6,51 5172
Kontrolle 6,49 5391 6,80 5044 6,73 5204
Tabelle 9: Vergleich der Mittelwerte des Durchmessers
Anzahl der untersuchten Präparate: Seite der Distraktion N = 90,
Seite der Kontrolle N= 87.

55
Ergebnisse
Abb. 27: Mittelwerte des Durchmessers der Axone mit ihrer Myelinscheide :
Der Unterschied der Werte des mittleren Durchmessers ist
mit p < 0,001 im Seitenvergleich höchst signifikant.
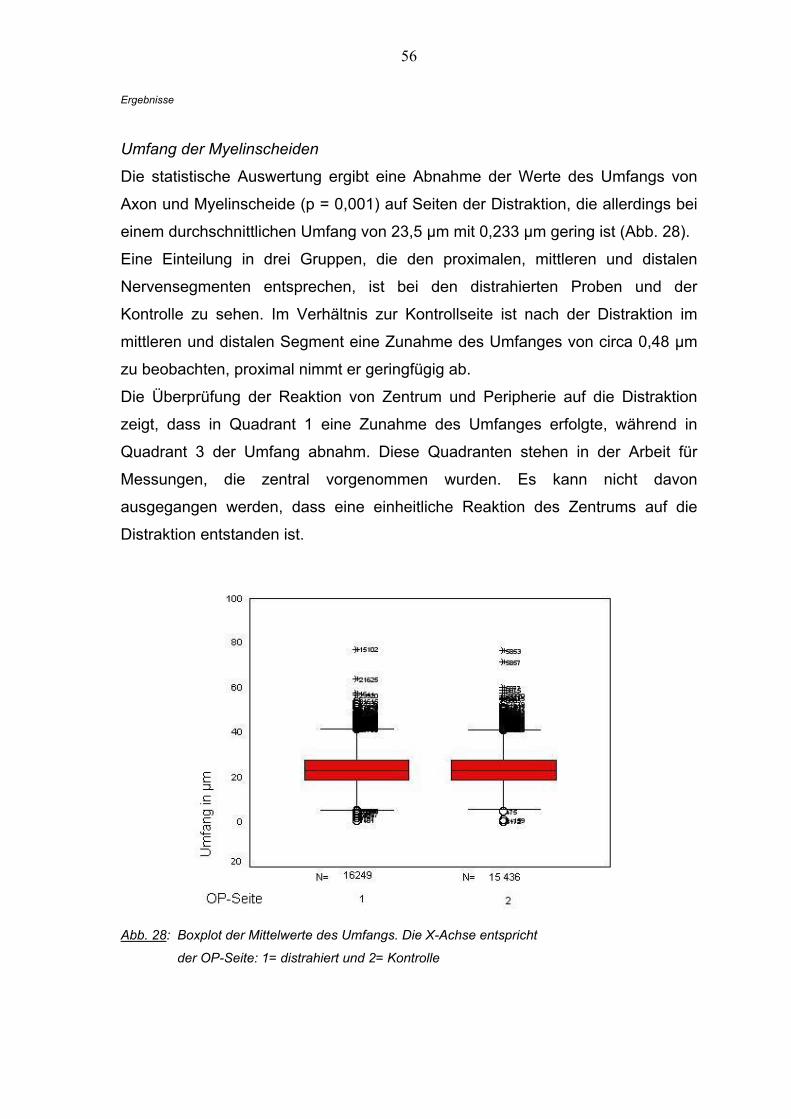
56
Ergebnisse
Umfang der Myelinscheiden
Die statistische Auswertung ergibt eine Abnahme der Werte des Umfangs von
Axon und Myelinscheide (p = 0,001) auf Seiten der Distraktion, die allerdings bei
einem durchschnittlichen Umfang von 23,5 µm mit 0,233 µm gering ist (Abb. 28).
Eine Einteilung in drei Gruppen, die den proximalen, mittleren und distalen
Nervensegmenten entsprechen, ist bei den distrahierten Proben und der
Kontrolle zu sehen. Im Verhältnis zur Kontrollseite ist nach der Distraktion im
mittleren und distalen Segment eine Zunahme des Umfanges von circa 0,48 µm
zu beobachten, proximal nimmt er geringfügig ab.
Die Überprüfung der Reaktion von Zentrum und Peripherie auf die Distraktion
zeigt, dass in Quadrant 1 eine Zunahme des Umfanges erfolgte, während in
Quadrant 3 der Umfang abnahm. Diese Quadranten stehen in der Arbeit für
Messungen, die zentral vorgenommen wurden. Es kann nicht davon
ausgegangen werden, dass eine einheitliche Reaktion des Zentrums auf die
Distraktion entstanden ist.
Abb. 28: Boxplot der Mittelwerte des Umfangs. Die X-Achse entspricht
der OP-Seite: 1= distrahiert und 2= Kontrolle
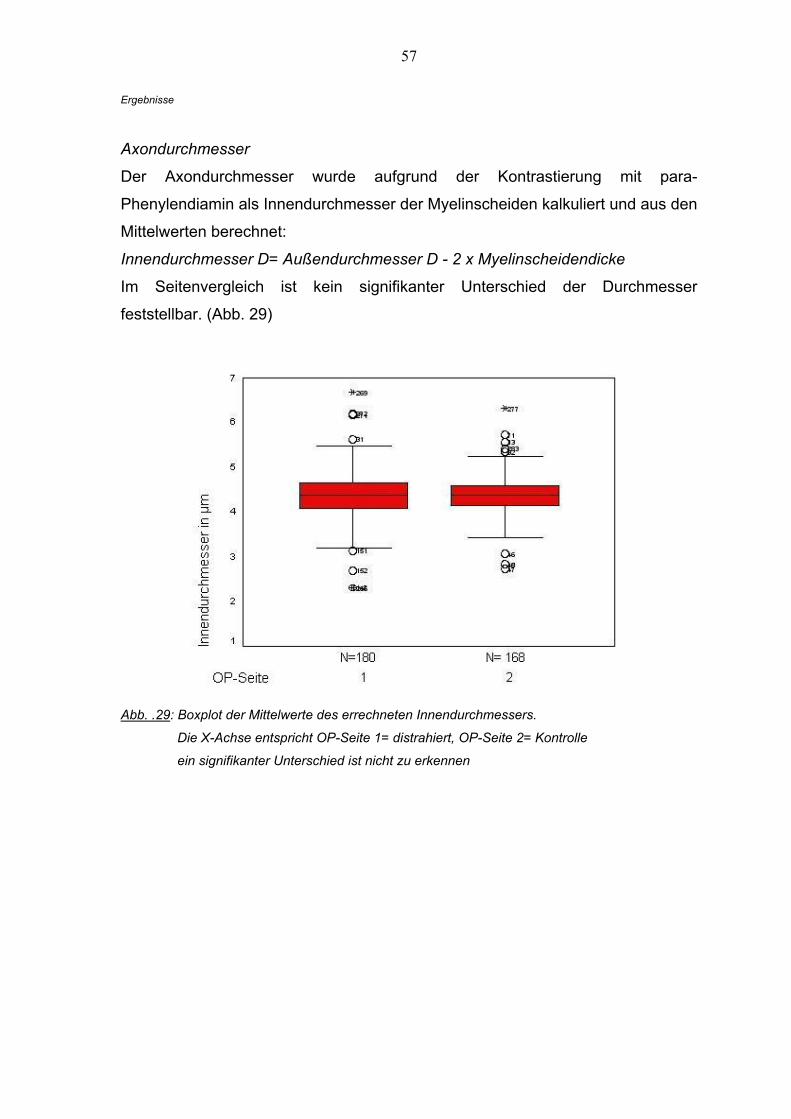
57
Ergebnisse
Axondurchmesser
Der Axondurchmesser wurde aufgrund der Kontrastierung mit para-
Phenylendiamin als Innendurchmesser der Myelinscheiden kalkuliert und aus den
Mittelwerten berechnet:
Innendurchmesser D= Außendurchmesser D - 2 x Myelinscheidendicke
Im Seitenvergleich ist kein signifikanter Unterschied der Durchmesser
feststellbar. (Abb. 29)
Abb. .29: Boxplot der Mittelwerte des errechneten Innendurchmessers.
Die X-Achse entspricht OP-Seite 1= distrahiert, OP-Seite 2= Kontrolle
ein signifikanter Unterschied ist nicht zu erkennen

58
Ergebnisse
G-Ratio
Die G-Ratio ist ein Parameter für die Nervenleitgeschwindigkeit. Sie kann Werte
zwischen 0 und 1 annehmen. Der ideale Wert der G-Ratio liegt bei 0,6. Als
Reaktion auf eine Zunahme der Myelinscheidendicke folgt eine Abnahme der G-
Ratio.
Vergleicht man die Mittelwerte im t-Test, so liegen sie mit 0,642 ± 0,042 auf der
Kontrollseite und 0,650 ± 0,037 auf der distrahierten Seite im idealen Bereich
(Abb. 30). Die Minimal- und Maximalwerte der G-Ratio auf der distrahierten Seite
und der Kontrollseite sind Tabelle 10 zu entnehmen.
Abb. 30: Mittelwerte der G-Ratio: Die meisten Werte für die G-Ratio liegen bei 0,65.
Der Minimalwert liegt bei 12 proximal zu sehen.
OP Seite (li1/re2) Probennummer Wert
Maximum 1 dist 0,73 1
Minimum 15 prox 0,53
Maximum 10 dist 0,78 G-Ratio
2 Minimum 12 prox 0,47
Tabelle 10: Minimal- und Maximalwerte der G-Ratio auf der Distraktions- und Kontrollseite
1= Distraktion, 2= Kontrolle

59
Ergebnisse
3.3 Quantitative Auswertung der Axonanzahl Im Vergleich der Mittelwerte ist eine Abnahme der myelinisierten Axone von
-13,32 auf Seiten der Distraktion zu beobachten (Tab. 11).
Es ergibt sich eine Heterogenität der Varianzen zwischen Seite 1 und 2. Dennoch
erweist sich die Differenz der Mittelwerte als statistisch nicht signifikant.
Proben Nr. gedehnt Anzahl Proben Nr.
Kontrolle Anzahl Unterschied (+ gedehnt mehr als Kontrolle)
1 prox 1721 2 prox 1673 + 48 1 mitte 1810 2 mitte 1962 -152 1 dist 1849 2 dist 1917 - 68 3 prox 1755 4 prox 1511 +244 3 mitte 1494 4 mitte 1764 -270 3 dist 1478 4 dist 1497 - 19 5 prox 1783 6 prox 1390 +393 5 mitte 1759 6 mitte 1373 +386 5 dist 1638 6 dist 1636 + 2 7 prox 1163 8 prox 825 +338 7 mitte 1165 8 mitte 808 +357 7 dist 1082 8 dist 1346 -264 9 prox 1489 10 prox 1744 -255 9 mitte 1458 10 med 1583 -125 9 dist 1012 10 dist 1553 -541
11 prox 1714 12 prox 1737 - 23 11 mitte 1378 12mitte 1448 - 70 11 dist 1387 12 dist 1550 -163 13 prox 1558 14 prox 1407 +151 13 mitte 1448 14 mitte 1483 - 35 13 dist 1451 14 dist 1461 - 10 15 prox 1581 16 prox 1912 -331 15 mitte 1591 16 mitte 1691 -100 15 dist 1562 16 dist 1826 -264 17 prox 1571 18 prox 1506 + 65 17 mitte 1358 18 mitte 1304 + 54 17 dist 1343 18 dist 1168 +175 19 prox 1781 20 prox 1785 - 4 19 mitte 1528 20 mitte 1683 -155 19 dist 1319 20 dist 1801 -482 21 prox 1386 22 prox 1704 -318 21 mitte 1436 22 mitte 1558 -120 21 dist 1373 22 dist 1542 -169 23 prox 1667 24 prox 1002 + 65 23 mitte 1825 24 mitte 977 +848 23 dist 2001 24 dist 1044 +957 25 prox 1758 26 prox 2053 -295 25 mitte 1652 26 mitte 1866 -214 25 dist 1587 26 dist 1664 - 77 27 prox 2011 28 prox 1837 +174 27 mitte 1638 28 mitte 1775 -137 27 dist 1758 28 dist 1503 +255 29 prox 1323 30 prox (191) nicht auszuwerten 29 mitte 1387 30 mitte nicht auszuwerten 29 dist 1276 30 dist nicht auszuwerten
Tabelle 11: Anzahl der Axone im intraindividuellen Seitenvergleich

60
Ergebnisse
Bei einer Gesamtzahl von 45 Vergleichswerten, die 100% entsprechen würden,
ergibt sich aus der Gegenüberstellung von distrahierter und nicht behandelter
Seite in 31,1% eine höhere Axonanzahl auf der distrahierten Seite. In 57,8 % ist
die Anzahl der Axone im Vergleich zur Kontrollseite erniedrigt (Abb. 31). Bei 7,7%
konnte keine Auswertung vorgenommen werden, da Präparat 38 dem Faszikel
des kleinen Ramus zygomaticus entspricht (Anzahl der Axone 191) und so ein
objektiver Vergleich nicht möglich ist.
Abb. 31: Der Vergleich der Mittelwerte bezogen auf die Anzahl der Axone im gesamten
Nervenquerschnitt: Die linke Seite zeigt die Kontrolle, während rechts die Werte der
Distraktion dargestellt sind. Maximal- und Minimalwert entsprechen in der Graphik
UCL (= upper control limit) und LCL (= lower control limit)
TEIL distrahiert Kontrolle
dist Mittelwert 1474,40 1536,29 N 15 14 Standardabweichung 268,89 239,91 Varianz 72302,114 57554,835
med Mittelwert 1528,47 1519,64 N 15 14 Standardabweichung 184,49 325,51 Varianz 34036,124 105959,478
prox Mittelwert 1617,40 1577,57 N 15 14 Standardabweichung 213,90 339,44 Varianz 45751,829 115222,264
Insgesamt Mittelwert 1540,09 1544,50 N 45 42 Standardabweichung 227,92 298,31 Varianz 51946,992 88987,378
Insgesamt ist die Abnahme der Anzahl
myelinisierter Axone mit -4,41 auf der
distrahierten Seite der Tiere gering.
Prüft man die unterschiedlichen Werte
innerhalb der Segmente, erkennt man, dass
eine leichte Abnahme der myelinisierten
Axone im distalen Drittel. In den Proben des
proximalen und mittleren nimmt dagegen
Axonanzahl leicht zu.
Tabelle 12: Betrachtung der Mittelwerte Anzahl von Axonen in einem Nervenquerschnitt
bezüglich der Einteilung in die Segmente poximal, medial und distal.
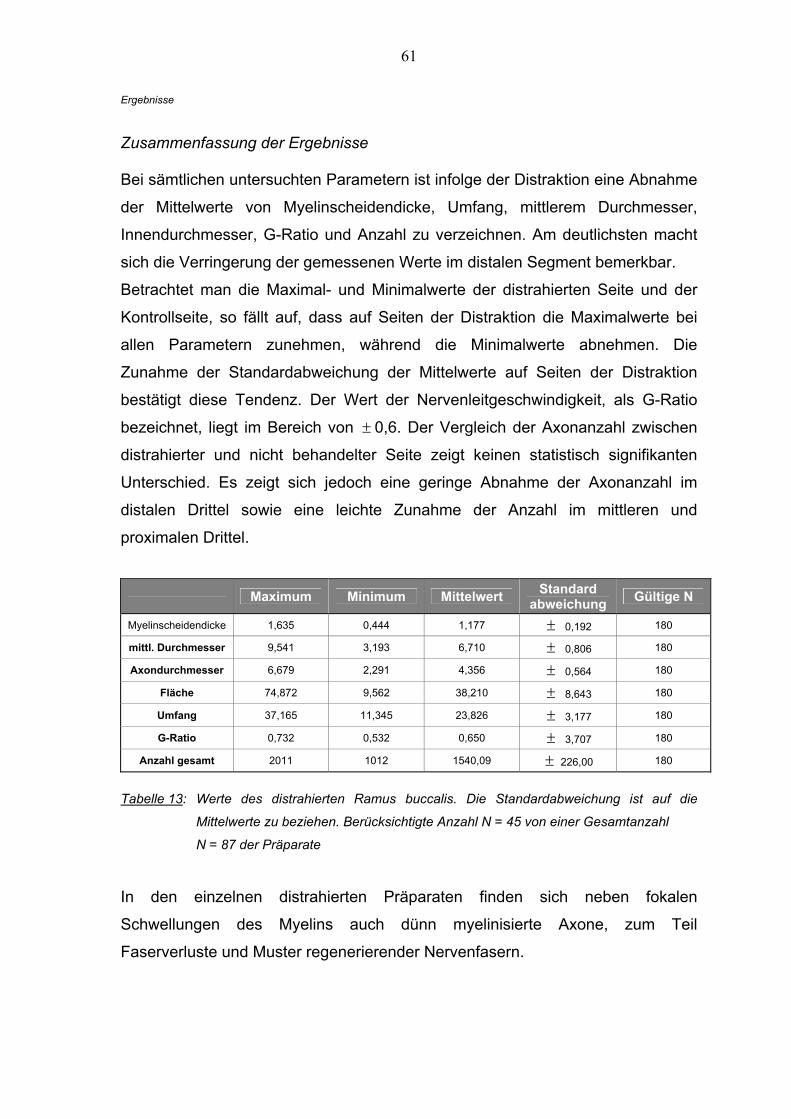
61
Ergebnisse
Zusammenfassung der Ergebnisse
Bei sämtlichen untersuchten Parametern ist infolge der Distraktion eine Abnahme
der Mittelwerte von Myelinscheidendicke, Umfang, mittlerem Durchmesser,
Innendurchmesser, G-Ratio und Anzahl zu verzeichnen. Am deutlichsten macht
sich die Verringerung der gemessenen Werte im distalen Segment bemerkbar.
Betrachtet man die Maximal- und Minimalwerte der distrahierten Seite und der
Kontrollseite, so fällt auf, dass auf Seiten der Distraktion die Maximalwerte bei
allen Parametern zunehmen, während die Minimalwerte abnehmen. Die
Zunahme der Standardabweichung der Mittelwerte auf Seiten der Distraktion
bestätigt diese Tendenz. Der Wert der Nervenleitgeschwindigkeit, als G-Ratio
bezeichnet, liegt im Bereich von ± 0,6. Der Vergleich der Axonanzahl zwischen
distrahierter und nicht behandelter Seite zeigt keinen statistisch signifikanten
Unterschied. Es zeigt sich jedoch eine geringe Abnahme der Axonanzahl im
distalen Drittel sowie eine leichte Zunahme der Anzahl im mittleren und
proximalen Drittel.
Maximum Minimum Mittelwert Standard
abweichung Gültige N
Myelinscheidendicke 1,635 0,444 1,177 ± 0,192 180
mittl. Durchmesser 9,541 3,193 6,710 ± 0,806 180
Axondurchmesser 6,679 2,291 4,356 ± 0,564 180
Fläche 74,872 9,562 38,210 ± 8,643 180
Umfang 37,165 11,345 23,826 ± 3,177 180
G-Ratio 0,732 0,532 0,650 ± 3,707 180
Anzahl gesamt 2011 1012 1540,09 ± 226,00 180
Tabelle 13: Werte des distrahierten Ramus buccalis. Die Standardabweichung ist auf die
Mittelwerte zu beziehen. Berücksichtigte Anzahl N = 45 von einer Gesamtanzahl
N = 87 der Präparate
In den einzelnen distrahierten Präparaten finden sich neben fokalen
Schwellungen des Myelins auch dünn myelinisierte Axone, zum Teil
Faserverluste und Muster regenerierender Nervenfasern.

62
Diskussion
4 Diskussion Zahlreiche Studien beschreiben die Abläufe der sekundären Nervendegeneration
nach einer Verletzung. Die Neurotmesis nach Seddon (1943) oder die
Nervenläsion des Grades vier/fünf nach Sunderland (1951) stellen die
schwerwiegendste Art einer Nervenverletzung dar. Beide Einteilungen stehen für
eine komplette Durchtrennung des peripheren Nerven mit einer Parese der
beteiligten Muskulatur. Im Sinne der Waller´schen Degeneration kommt es zur
Axiolyse und Myelinolyse distal der Nervenläsion (Lundborg 1987).
Schwann´sche-Zellen des distalen Nervenstumpfes proliferieren und bilden das
Haken-Büngener-Band, einen kompakten Strang, der als Leitschiene für die
proliferierenden Nervenfasern dient (Son und Thompson 1995). Experimentelle
Studien belegen, dass die Regeneration des proximalen Stumpfes und die
Degeneration der axonalen Reste zeitlich synchron verlaufen (Fawcett und
Keynes 1990). Die Wachstumsgeschwindigkeit der regenerierenden Axone liegt
dabei zwischen 0,6 mm/Tag und 4,2 mm/Tag. Beim Menschen kann ein
Mittelwert von 1 mm/Tag angenommen werden (Seddon et al. 1943; Jasper
1946; Sunderland 1946; Guth 1956; Isch et al. 1968; Thomas 1988; Seeburger
und Springer 1993; Lewis et al. 1993). Trotz dieser Regenerationsfähigkeit
peripherer Nerven ist die Möglichkeit einer Rückkehr der Funktion ohne
chirurgische Intervention bei einer gravierenden Nervenverletzung nicht möglich.
Eher bilden sich schmerzhafte Neurome, die die zu überbrückende Lücke der
Nervenstümpfe nach einer Resektion noch vergrößern.
Zur Wiederherstellung der Nervenkontinuität besteht die Möglichkeit der
Verwendung autogener Nerventransplantate oder neurovaskular-
reanastomosierter Muskeltransplantate. Im Bereich der Mund-, Kiefer- und
Gesichtschirurgie werden oft sensible Nerven wie der Nervus suralis und der
Nervus auricularis magnus als Ersatz für motorische Nerven genutzt
(Hagen 1981; Stephanian et al. 1992). Transplantate bringen aber unweigerlich
Nachteile mit sich: Es ist ein zweiter operativer Eingriff an der Entnahme- oder
Spenderstelle erforderlich. Eine erhöhte Morbidität im Bereich der
Entnahmeregion ist zu erwarten.

63
Diskussion
Falls eine primäre Reanastomose nicht möglich ist, muss mit einem Ausfall des
vom Spendernerv versorgten Gebiets gerechnet werden.
Ebenso besteht bei Nerven, die mit Interponaten rekonstruiert wurden, das Risiko
einer inadäquaten Funktionsrehabilitation sowie einer aberranten Regeneration
(Mackinnon 1989). Ob unterschiedliche Nervenqualitäten zwischen Spender- und
Empfängernerven dafür verantwortlich sind, ist bisher nicht bekannt. Auch ein
Vorliegen unterschiedlicher Faszikelanordnungen (Sunderland 1990) und die
Tatsache, dass die Nervenregeneration über zwei Anastomosestellen schwieriger
zu erreichen ist (Manders et al. 1987), könnten für einen partiellen oder totalen
Misserfolg verantwortlich sein.
Um diese Nachteile zumindest teilweise zu umgehen, wurden einige
Untersuchungen zur Nervenverlängerung mittels eines Gewebeexpanders
durchgeführt, die erfolgreich waren (Manders et al. 1987; Wood et al. 1991;
Hall et al. 1993; van der Wey et al. 1995; Van der Wey et al. 1996).
Während die genannten Studien also die sekundäre Dehnung des Nerven
behandelten, wurden bislang noch keine morphologischen und
histomorphometrischen Untersuchungen über die primäre Distraktion eines
Nerven durchgeführt. Die vorliegende Arbeit erforscht diese Parameter erstmals.
Als Grundlage für die Überprüfung, ob ein Nerv aktiv in seiner Länge verändert
werden kann, dienten folgende Fragen:
1) Welche morphologischen Abweichungen können beobachtet werden?
2) Treten Veränderungen bezüglich Myelinscheidendicke, -durchmesser und
-umfang nach der Distraktion auf, die als Hinweis auf zu erwartende
Veränderungen der elektrischen Nervenleitgeschwindigkeit dienen
können?
3) Unterscheiden sich Peripherie und Zentrum des Nervenquerschnitts
bezüglich ihrer Reaktion auf die Distraktion?
4) Erfolgt eine Zu- oder Abnahme der Anzahl myelinisierter Axone?
Anhand dieser Punkte wird die Hauptfrage erörtert werden, ob es nach der
gesteuerten primären Distraktion zu einer Degeneration des Nervus facialis
kommt, und wenn ja, in welchem Ausmaß.

64
Diskussion
4.1 Methodenkritik Tiermodell In den Experimenten zu dieser Arbeit wurden ausschließlich weibliche Tiere
benutzt, da Testosteron das Überleben der Motoneurone im Hirnstamm nach
Axotomie der entsprechenden Nerven verstärkt und die Regeneration peripherer
Nerven fördert (Drengler et al. 1997; Kinderman et al. 1998; Bodjanac et al. 2000;
Larowski et al. 2000). Es wurde festgestellt, dass geschlechtsspezifische
Unterschiede bezüglich des Verlustes an Nervenfasern und der Regeneration
nach Nervenverletzungen zu beobachten sind (Yu und McGinnis 2001).
Als klinisch übertragbarer Versuchsansatz wurde zur Distraktion des Nervus
buccalis in unseren Versuchen ein Schraubensystem verwendet (Abb. 5). Mittels
einer perineuralen Anschlingung des Ramus buccalis, deren Fadenenden an
einer am Jochbogen fixierten Schraube befestigt waren, wurde die tägliche
Distraktion vorgenommen. Für die Anwendung dieses Verfahrens spricht, dass
das Perineurium ein straffes Bindegewebe aus Kollagenfasern und dickeren
elastischen Fasern ist. Es schließt mehrere Nervenfasern zu Faszikeln
zusammen und dient außerdem als Diffusionsbarriere zwischen Endo- und
Epineuralraum. (Martin 1964; Rechtland 1987). Durch scharfe Gegenstände
können Hernien peripherer Nerven entstehen. Dieser Mechanismus wird als
perineurales Fenster beschrieben. Sugimoto et al. (2002) untersuchten die
Auswirkungen eines solchen perineuralen Fensters und vermuten, dass
neurologische Symptome und Nervendegeneration eher durch den Austritt der
Nervenfasern aus dem Perineurium hervorgerufen werden, als durch die
Unterbrechung der endoneuralen Homöostase.
Ein Austritt von Nervenfasern wurde bei der Entnahme der Nervenproben in
unseren oben geschilderten Untersuchungen nicht beobachtet und kann nahezu
ausgeschlossen werden. Es ist also davon auszugehen, dass der Einfluss der
Distraktionsnaht auf das Degenerationsverhalten des Ramus buccalis im
Verhältnis zu der eigentlichen Dehnung des Nerven kaum eine Rolle spielte.

65
Diskussion
Histologische und histomorphometrische Methodik
Bei der Herstellung der Präparate wurde darauf geachtet, schnittbedingte
morphologische Veränderungen der zu untersuchenden Parameter durch
senkrecht zur Längsachse des Nervenstammes angelegte Schnitte zu
vermeiden. Dennoch befanden sich aufgrund des wellenförmigen Verlaufes des
Nerven (Sunderland 1990) Längsschnitte in den Präparaten.
Die Präparate wurden mit Osmiumtetraoxid fixiert; weil eine normale Fixierung zu
einer Schrumpfung der wasserreichen Achsenzylinder führen würde.
Die Schnittdicke mit 1,5 µm wurde für diese Untersuchung als ideal angesehen
(Holländer und Vaaland 1968).
Die hier angewandte Kontrastierung mit para-Phenylendiamin zur Erfassung der
wichtigen Parameter hat sich laut vieler vorangegangener Arbeiten
bewährt (Böck 1984; Colman und Stockert 1983; Gellrich et al. 2002 b, 2002 c).
Auf eine elektronenmikroskopische Untersuchung wurde verzichtet, da diese bei
stärkerer Degeneration keine zusätzliche Information liefert, es kann lediglich
unterschieden werden, ob ein Nerv leicht oder gar nicht verletzt wurde (Gellrich et
al. 2002 a).
Die Untersuchung des Axondurchmessers und der G-Ratio wurde als Tendenz
angegeben, weil para-Phenylendiamin nur osmophile Strukturen anfärbt
(Böck 1984). Die Axone können also mit dieser Kontrastierung nicht eindeutig
identifiziert werden.
Die unbehandelte rechte Seite der Tiere diente als Kontrolle.
Aus einer Studie von Mattox und Felix (1987) ist bekannt, dass der Stamm des
Nervus facialis der Ratte zwischen rechter und linker Seite lediglich eine
Seitendifferenz von 2,1% aufweist. Mit 13,7 % ist diese im Ramus buccalis zwar
höher, liegt für die vorliegende Untersuchung aber durchaus in einem Bereich,
mit dem ein Vergleich der Seiten möglich ist.

66
Diskussion
4.2 Morphologische und histomorphometrische Ergebnisse
De- und Regenerationsvorgänge
Zwischen völliger Degeneration und der partiellen Verletzung eines Nerven liegt
ein breites Spektrum degenerativer Vorgänge, abhängig von Schweregrad,
Dauer und Typ der Verletzung (Gellrich et al. 2002 c). Untersuchungen zu der
moduliert gerichteten Quetschung des Nervus opticus ergaben ein lineares
Verhalten zwischen angewandter Kraft und axonaler Degeneration
(Klöcker et al. 2001; Gellrich et al. 2002 b). Erst beim Überschreiten der
kritischen Grenzen ist eine Veränderung der vorhandenen Schädigung nicht
mehr zu beobachten (Gellrich et al. 2002 b, 2002 c). Die Anzahl der
regenerierenden Axone ist ebenfalls von der Art der Verletzung abhängig
(Gutman und Sanders 1943; Shawe 1955; Evans und Murray 1956;
Jenq und Coggeshall 1985). Nach einem Crush (Quetschung) ist die
Wiederaussprossung der Axone weniger ausgeprägt als bei Axotomie mit oder
ohne Nervennaht. Überdies wird der Normalwert der Axonanzahl früher erreicht
(Jenq und Coggeshall 1985). Je größer die Diastase zwischen proximalem und
distalem Nervenstumpf ist, desto mehr nimmt die Ausprossung der Axone in den
distalen Stumpf ab (Jenq und Coggeshall 1985).
Die experimentelle Studie von 6 verschiedenen Verletzungsmustern anhand des
Nervus facialis (Cai et al. 1998) zeigte:
• Freilegung und Kompression des Nerven rufen eine Neurapraxie hervor.
• Quetschung und Dehnung führen zu einer Axonotmesis.
• Axotomie und Nervennaht nach Axotomie fallen unter das Verletzungsmuster
der Neurotmesis. Es entwickelt sich keine völlige Regeneration innerhalb von
sechs Monaten.
Eine wichtige Aufgabe dieser Studie war, zu bestimmen, welchem Schweregrad
die Degenerationserscheinungen zuzuordnen sind. Durch die moduliert
gerichtete Dehnung des Nerven wurde bei den Experimenten versucht, die oben
festgestellte Axonotmesis zu vermeiden.

67
Diskussion
Aus der Abnahme der Myelinscheidendicke in dieser Untersuchung ist zu
schließen, dass eine De- und Remyelinisierung der Axone vorliegt. Dabei waren
in einzelnen Präparaten Ödeme der Myelinscheiden feststellbar. Der Verlust der
Myelinscheidendicke war im distalen Segment am deutlichsten. Ein ähnliches
Ergebnis zeigte die Analyse des mittleren Durchmessers und des Umfangs der
Myelinscheiden. Der Vergleich der Mittelwerte der Axonanzahl weist mit einem
nicht signifikanten Unterschied noch weniger auf eine Axonotmesis hin als die
oben genannten Werte. Erst bei der genauen Beobachtung der Einzelwerte fällt
auf, dass in einigen Präparaten entweder eine erhöhte Zu- oder Abnahme der
Werte vorliegt. Tabelle 12 zeigt in der Gesamtübersicht eine geringe
Verminderung der Anzahl im distalen Segment, während im proximalen und
mittleren Drittel eine leichte Erhöhung der Axonanzahl sichtbar wird. Das Defizit
der Axonanzahl ist auf die Waller´sche Degeneration im distalen Nervenstumpf
zurückzuführen. Unterdessen deutet ein Zuwachs an Axonen auf einen
beginnenden Regenerationsprozess der Nerven von proximal. Zum besseren
Verständnis soll im Folgenden nochmals auf die De- und Regenerationsvorgänge
eingegangen werden.
Eine Unterbrechung der Axone durch Axotomie oder Crush / Stretching führt zu
einer absteigendenden Degeneration. Ein typisches Kennzeichen ist der Zerfall
der Myelinscheiden im distalen Nervenstumpf (Gellrich et al. 2002 c).
Eingewanderte Makrophagen phagozytieren die Axonreste und schaffen so ein
Milieu, in dem der Prozess der Regeneration stattfinden kann (Kury et al. 2001).
Untersuchungen zur Regeneration des Zentralen Nervensystems ergaben, dass
die dafür wesentlichen Informationen im degenerierenden Nervenende peripherer
Nerven zu finden sind, da deren Transplantation auch im Zentralen
Nervensystem eine Regeneration hervorrufen (Stoll und Müller 1999). Dagegen
erzielten Versuche, die Regeneration des Nervus opticus durch fetale
Transplantate aus Strukturen des Zentralen Nervensystems zu fördern, keinen
positiven Effekt auf dessen Regenerationsfähigkeit (Gellrich et al. 1994). Im
Gegensatz zum distalen Nervenstumpf wird das proximale Membranleck
verschlossen, und die Axone bleiben erhalten (Lay 1999).

68
Diskussion
Als „regenerating unit“ (Mackinnon et al.1991) werden aussprossende Axone
bezeichnet, die vom letzten proximalen Ranvier-Knoten entlang der Innenfläche
der Basalmembran innerhalb eines Stranges formierter Schwann´scher-Zellen
(Haken-Büngener-Band) in Richtung des distalen Nervenstumpfes einwachsen.
Eine solche Einheit kann mehrere myelinisierte Axone enthalten. Daraus
resultiert ein Anstieg der Axonanzahl in der Regenerationsphase, der den
Normalwert um ein Vielfaches überschreiten kann. Neuromuskuläre
Verbindungen in der Peripherie werden wiederhergestellt.
Axone, die diese funktionellen Verbindungen nicht aufbauen, werden einem
degenerativen Prozess unterworfen. Es kommt zum „die back"
(Rückwärtssterben), gleichzeitig wird eine weitere Ausprossung der Axone
verhindert (Lay 1999).
Die Hoffnung, durch die moduliert gerichtete Verlängerung (0,79 mm/d) nur das
Verletzungsmuster der Neurapraxie zu erhalten, hat sich in unserem Versuch
nicht erfüllt. In etwa 60% der Präparate musste anhand der Axonanzahlen und
der histologischen Befunde festgestellt werden, dass es sich um eine
Axonotmesis handelt (Tab. 11). Dabei erscheint hier die Einteilung von
Sunderland passender, da ein Verletzungsmuster des Grades II vorliegt. Ähnlich
der Neurapraxie (Grad I) folgt eine vollständige Regeneration (Sunderland 1990).
Drei Nerven (Abb.19, Abb. 21) zeigten deutliche Merkmale einer Regeneration.
Histologisch waren hier extrem kleine Axone erkennbar.
Bei Präparat 23 (Abb. 25) zeigte sich zusätzlich eine deutlich erhöhte Anzahl von
Axonen, was auf eine Wiederaussprossung der Axone von proximal
zurückzuführen ist. Die Entnahme dieser Nerven erfolgte etwa eine Woche nach
Beendigung der Distraktion. Der verhältnismäßig zeitnahe Beginn der
Regeneration mit einer relativ geringen Anzahl an überschießenden Axonen in
den einzelnen Nervenpräparaten sowie der statistisch nicht signifikante
Unterschied zwischen der distrahierten Seite und den Kontrollpräparaten in ihrer
Gesamtheit bestätigt die Annahme einer Verletzung von Grad II nach
Sunderland (1951).

69
Diskussion
Vergleich der Distraktionsarten Mit der Untersuchung der Reaktion des intakten Nervus facialis auf eine aktive
Längenveränderung sollte eine Grundlage geschaffen werden, die es ermöglicht,
das Modell später in den klinischen Einsatz zu übertragen. Die vorliegenden
Ergebnisse werden von vorangegangenen Studien zur Distraktion eines
peripheren Nerven gestützt und sollen mit ihnen verglichen werden.
In Anlehnung an Gavriel A. Ilizarov (1951) wurde die Distraktionsosteogenese
auch in den Bereich der Kiefer- und Gesichtchirurgie übertragen. Im Vordergrund
standen dabei jedoch knöcherne Korrekturen des Unterkiefers und des
zygomatico-maxillären Bereichs. Gerade bei Korrekturen des Unterkiefers spielt
die Toleranz des Nervus alveolaris inferior gegenüber Dehnung eine große Rolle.
Untersuchungen haben gezeigt, dass eine Längenveränderung bei einem
definierten Ausmaß von 1 mm/d zu guten Ergebnissen führt (Battison et al. 1992;
Block et al. 1993; Hu et al. 2001; Wang et al. 2002).
Bei einem Längengewinn des Nervus alveolaris von 7 mm wurden bei 25 % von
insgesamt 381 myelinisierten Axonen Degenerationserscheinungen wie
Demyelinisierung, axonale Schwellung festgestellt. Ebenso erfolgte eine
Verdunkelung des Axonplasmas, die auf eine ischämische Schädigung der
Axone hinweist (Block et al. 1993). Diese Veränderungen können entweder auf
die mechanische Einwirkung oder auf eine Ischämie der Vasa nervorum aufgrund
einer Kompression zurückgeführt werden (Block et al. 1993). Das
Verletzungsmuster entspricht einer Neurapraxie, ähnlich den Nervenläsionen
nach Kompression.
Hu et al. (2001) beobachten nach einer Verlängerung des Nervus alveolaris
inferior mittels Distraktionsosteogenese von 10 mm ebenfalls axonale
Schwellungen, Demyelinisierung, eine Verdunkelung des Axonplasmas und eine
Abnahme der Anzahl von Axonen. Dabei ist das Bild einer Degeneration bei einer
täglichen Distraktion von 1 mm weniger ausgeprägt als bei einer täglichen
Verlängerung von 2 mm.
Durch Gewebeexpander ist die selektive Distraktion peripherer Nerven möglich.
Beobachtungen bestätigen, dass eine Dehnung peripherer Nerven während
eines langsamen benignen Tumorwachstums ohne einen Verlust der Funktion
einhergeht (Mackinnon 1989; Anand 1997).

70
Diskussion
Malis et al. (1995) erzielten bei der Distraktion des Nervus facialis des Schweins
mittels Expander innerhalb von 43 Tagen einen Längengewinn des Nerven von
20 bis 54 mm. Die Dehnung wurde vor und während der Versuche
elektromyographisch kontrolliert. Ein Funktionsverlust konnte nicht festgestellt
werden. Das histologische Bild ergab jedoch eine mittlere bis schwere
Degeneration der betroffenen Nerven wie auch ihrer Kontrollseiten.
Dagegen erzielten Manders et al. (1987) wie auch van der Wey et al. (1995;
1996) einen Längengewinn mit weniger auffälligen histologischen Befunden. Es
werden Myelinscheidenschwellung, dünn myelinisierte Axone oder Axone ohne
Myelinscheide und Axondegeneration beschrieben.
Ya et al. (2000) stellten nach Elongation des Nervus facialis histologisch eine
Läsion des Grades III (Sunderland 1951) fest, wobei, wie bereits von Malis et al.
geschildert, ein Funktionsverlust nicht festzustellen war.
Da in de vorliegenden Untersuchungen keine elektromyographischen Befunde
erhoben wurden, stützt sich der Vergleich nur auf die histologischen Ergebnisse:
Ödeme der Myelinscheiden und dünn myelinisierte Axone waren am häufigsten
zu beobachten. Auch die Aufsplitterungen der Myelinscheiden in Lamellen und
der Verlust von Fasern waren zu sehen (Tab. 5). Die beiden letztgenannten
Beschreibungen stellen ein Zeichen Waller´scher Degeneration dar, was zu der
Schlussfolgerung führt, dass es sich in manchen Präparaten mindestens um eine
Verletzung des Grades II handeln muss.
Die Anzeichen der Degeneration waren trotz allem relativ mild, und die zeitnahe
Regeneration spricht ebenfalls eher für eine Verletzung des Grades II als des
Grades III. (Abb. 17und Abb. 19).
Zwei Tiere wiesen eine komplette Degeneration der Axone und Markscheiden mit
Markballenbildung auf. Da aber die Kontrollseite in gleichem Maße davon
betroffen war, ist anzunehmen, dass ein Fehler in der Aufbereitung der Präparate
dafür verantwortlich war.
Bei der kritischen Betrachtung der soeben geschilderten Resultate zeigt es sich,
dass hier ähnliche histologische Ergebnisse erzielt wurden wie in den
Untersuchungen der oben genannten Studien.

71
Diskussion
Die Berechnung der G-Ratio (Parameter für die Nervenleitgeschwindigkeit) ist
zwar als Tendenz angegeben, weist jedoch mit 0,650 ± 0,037 auf der
distrahierten Seite einen Wert auf, der nahe dem Idealwert von 0,6 liegt. Ein
starker Funktionsverlust kann auf jeden Fall ausgeschlossen werden, da bei
Malis et al. (1995) die Funktion des Nervus facialis trotz extremer histologischer
Veränderung nicht eingeschränkt war.
Aufgrund dieser Feststellungen ist davon auszugehen, dass die moduliert
gerichtete und gesteuerte primäre Nervendistraktion im Vergleich zu anderen
Methoden der Nervenverlängerung keine Nachteile aufweist. Dagegen verspricht
sie den Vorteil, dass eine übermäßige Dehnung der umliegenden Gewebe
vermieden werden kann. Die Methode der primären Distraktion dürfte daher im
klinischen Einsatz ein für den Patienten nutzbarer Ansatz sein, da mit
zunehmendem Alter und bei starker Beanspruchung die Elastizität der Haut und
der Muskelgewebe nachlässt. Eine Überdehnung der Haut und der Muskeln,
welche zu einer ästhetischen Einbuße des sonst erfolgreichen Ergebnisses
führen könnte, tritt bei diesem Behandlungsverfahren nicht ein.

72
Diskussion
Schlussfolgerung
1) Die Verlängerung eines Nerven mittels der gesteuerten primären
Distraktion ist möglich.
2) Es wurden bei der primären Nervendistraktion ähnliche Ergebnisse erzielt
wie in vorrangegangenen Studien, die die Verlängerung eines Nerven
mittels Distraktionsosteogenese oder Gewebeexpandern untersuchten.
Jedoch wird in der vorliegenden Untersuchung das umliegende Gewebe
während der Distraktion nicht beeinträchtigt.
3) Als morphologische Veränderungen werden Schwellungen der
Myelinscheiden, dünn myelinisierte Axone, eine Abnahme der Faseranzahl
und lamellenförmige Aufsplitterungen der Myelinscheiden beobachtet.
4) Der Vergleich der Mittelwerte der gemessenen Parameter deutet auf eine
Abnahme der Myelinscheidendicke, des Durchmessers und des Umfangs
hin. Die stärkste Abnahme der Markscheidendicke und des Durchmessers
erfolgt distal.
5) Peripherie und Zentrum zeigen keine nachweisbar unterschiedliche
Reaktion auf die Distraktion des Ramus buccalis.
6) Im Vergleich der Mittelwerte nimmt die Anzahl der Axone leicht ab, am
auffälligsten im distalen Segment.
Die primäre Distraktion des Ramus buccalis führt bei ca. 60% der distrahierten
Nerven zu einer Verletzung des Grades II nach Sunderland (1951).
Mit einer vollständigen und zeitnahen Regeneration der Verletzung ist zu
rechnen. Dies kann als positives Ergebnis für die Untersuchung gewertet werden.

73
Zusammenfassung
5 Zusammenfassung
Ziel der Arbeit ist die Etablierung eines auf den Menschen übertragbaren Modells
der selektiven, moduliert gerichteten und gesteuerten primären Nervendistraktion
bei der Ratte. Unter Ausnutzung der bei der Nervendegeneration und ihrer
Regeneration bisher gewonnenen Ergebnisse sollten die Technik zur
Durchführung und der Grad der degenerativen Veränderungen des Nerven nach
der moduliert gerichteten Nervenverlängerung erarbeitet werden.
Im Tiermodell wurde bei weiblichen Wistar-Ratten (N=15) mit einem
Zugschraubensystem die Distraktion des Ramus buccalis vorgenommen. Die
unbehandelte Kontralateralseite diente als Kontrolle. Täglich wurde der Nerv
durch die am Jochbogen befestigte Schraube um 0,785 mm gedehnt. Nach 10
Tagen war die Distraktion abgeschlossen. Die Sakrifikation der Tiere erfolgte
nach weiteren 20 Tagen in einer CO 2 -Kammer. Nach der Entnahme des
distrahierten Segmentes und des entsprechenden Nervensegments der
Gegenseite zur Kontrolle folgte die lichtmikroskopische Untersuchung der Proben
auf morphologische Zeichen einer Degeneration des distrahierten Nerven. Mit
Hilfe des Bildanalysesystems analySIS® (Soft Imaging, D-Münster) wurden
Myelinscheidendicke, Durchmesser und Umfang der Myelinscheiden
morphometrisch ermittelt. Die Axonanzahl wurde anhand von Bildern ausgezählt.
Die statistische Auswertung der ermittelten Daten erfolgte durch die
Statistiksoftware SPSS® (SPSS Inc., USA-Chicago, Illinois).
Als wichtigstes Ergebnis ist festzustellen, dass die Verlängerung des Nervus
facialis von 0,79 mm/d mittels der gesteuerten primären Nervendistraktion
möglich ist. Das Resultat der morphologischen und histomorphometrischen
Analyse ergab als Folge der gesteuerten primären Distraktion in 60% der
untersuchten Proben eine Nervenverletzung des Grades II nach Sunderland
(1951). Die restlichen der Präparate wiesen auf eine Verletzung des Grades I hin
(fokale Schwellungen der Myelinscheiden und dünn myelinisierte Axone). Eine
Regeneration der Nerven ist bei diesen Verletzungsmustern zu erwarten.
Auf dem Weg, die bisherigen Rekonstruktionsmöglichkeiten zu ergänzen und zu
verbessern, bietet die hier dargestellte Methode eine überzeugende Alternative,
die zu weiteren Forschungen ermutigt.

74
Literaturverzeichnis
6 Literaturverzeichnis 1. Abe Y, Doi K, Katoh Y, Yamamoto H, Kataoka H, Kwai S (1996) The limit of low speed peripheral nerve elongation, neurological and
circulatory aspects.
J Neurol Sci 140: 61-66
2. Anand VK (1997) Plasticity of the extracranial nerve.
Laryngoscope 107: 1393-1404
3. Anderl H (1982) Cross-face nerve transplantation in facial palsy.
Ann Chir Gynaecol 71: 70-76
4. Artico M, Cervoni L, Nucci F, Giuffre R (1996) Birthday of peripheral nervous system surgery: the contribution of
Gabriele Ferrara (1543-1627).
Neurosurgery 39: 380-382
5. Aslan A, Gotkan C, Okmus M, Tarhan S, Unulu H (2001) Morphometric analysis of anatomical relationship of the facial nerve for mastoid
surgery.
J Laryngol Otol 115: 447-449
6. Banfai P (1976) Die angewandte klinische Anatomie des Nervus Facialis I: Kernsystem,
supranukleäre Verbindung, Peripherie.
HNO 24: 253-264
7. Banfai P (1976) Die angewandte klinische Anatomie des Nervus Facialis II: Die Anastomosen.
HNO 24: 289-294

75
Literaturverzeichnis
8. Battista AF, Lusskin R (1986) The anatomy and physiology of the peripheral nerve.
Foot Ankle 7: 65-70
9. Battison B, Buffoli P, Vigaso A, Brunelli G, Antoinini L The effect of lengthening on nerves.
Ital J Orthop Traumatol 18: 79-86
10. Becker M (1999) Die extratemporalen Äste des Nervus facialis im Schichtaufbau der
Gesichtsweichteile und ihre Schonung beim indirekten Zugang zur seitlichen und
infratemporalen Schädelbasis.
Inaug-Diss, Med. Fak., Rheinische Friedrich-Willhelms-Universität, Bonn
11. Block MS, Daire J, Stover J, Marthews M Changes in the inferior alveolar nerve following mandibular lengthening in the dog
using distraction osteogenesis.
J Oral Maxillofac Surg 51: 652-660
12. Bodjanac WT, Terzis JK, Luizzi FJ (2000) Effects of testosterone on a cross-facial nerve graft model.
J Reconstr Microsurg 16: 449-456
13. Böck P (1984) Der Semidünnschnitt.
Bergmann, München
14. Bridge PM, Ball DJ, Mackinnon SE, Nakao Y, Brandt K, Hunter D, Hertel C (1994)
Nerve crush injuries - a model for axonotmesis.
Exp Neurol 127: 284-290

76
Literaturverzeichnis
15. Cai Z, Yu G, Ma D, Tan J, Yang Z, Zhang X (1998) Experimental studies on traumatic facial nerve injury.
J Laryngol Otol 112: 243-247
16. Chouard CH, Charachon R, Morgon A, Cathala HP (1972) Anatomie, pathologie et chirurgie du nerf facial.
Masson, Paris
17. Carter DA, Linsney SJ (1987) The number of unmyelinated and myelinated axons in normal and regenerated
rat saphenous nerve.
J Neurol Sci 80: 163-171
18. Colman O, Stockert JC (1983) Observations on the osmium tetroxoide-p-phenylendiamine staining method for
semithin sections of nervous tissue.
Acta Histochem 72: 123-125
19. Cornwell R, Radomisli TE (2000) Nerve injury in traumatic dislocation of the hip.
Clin Orthop 377: 84-91
20. De Jong GI, Jansen ASP, Horvath E, Gispen WH, Luiten PGM (1991) Nimodipine effects on cerebral microvessels and sciatic nerve in aging rats.
Neurobiol Aging 13: 73-81
21. Drengler SM, Handa RJ, Jones KJ (1997) Effects of axotomy and testosterone on androgen receptor mRNA expression in
hamster facial motoneurons.
Exp Neurol 146: 374-379

77
Literaturverzeichnis
22. Eshraghi AA, Buchmann CA, Telischi FF (2002) Sensory auricular branch of the facial nerve.
Otol Neurotol 23: 393-396
23. Evans DHL, Murray JG (1956) A study of regeneration in a motor nerve with unimodal with unimodal fibre
diameter distribution.
Anat Rec 126:311-329
24. Fawcett JW, Keynes RJ (1990) Peripheral nerve regeneration.
Annu Rev Neurosci 13: 43-60
25. Gellrich NC, Gellrich MM, Bremerich A (1994) Influence of fetal brain grafts on axotomized retinal ganglion cells.
Int J Oral Maxillofac Surg 23: 403-495
26. Gellrich NC (1995 ) Histomorphologische und elektrophysiologische Untersuchung zur traumatischen
Sehnervschädigung.
Habilitationsschrift, Med. Fak. Ruhr-Universität, Bochum
27. Gellrich NC, Greulich W, Gellrich MM, Eysel U (1995) Flash visual evoked potentials (VEP) and electroretinograms (ERG) in evaluation
of optic nerve damage due to midface and skullbase trauma.
Mund Kiefer Gesichtschir 19: 98-101
28. Gellrich NC, Gutwald R, Makrutzki M (2002 a) Traumatic lesion of the rat optic nerve: A transmission-electron- and light-
microscopic study.
MXj 2002.02.0003.1

78
Literaturverzeichnis
29. Gellrich NC, Schimming R, Zerfowski M, Eysel UT (2002 b) Quantification of histomorphological changes after calibrated crush of the
intraorbital optic nerve in rats.
Br J Ophtalmol 86 (2): 233-237
30. Gellrich NC, Schramm A, Rustenmeyer J, Schön R, Eysel UT (2002 c) Quantification of the neurodegenerative impact on the visual system following
sudden retrobulbar expanding lesion - an experimental model.
J Craniomaxiollofac Surg 30, 230-236
31. Gillespie MJ, Stein RB (1983) The relationship between axon diameter, myelin thickness
and conduction velocity during atrophy of mammalian peripheral nerves.
Brain Res 259: 41-56
32. Girod SC, Neukam FW, Girod B, Reumann K, Semrau H (1989) The fascicular structure of the lingual nerve and the chorda tympani: an anatomic
study.
J Oral Maxillofac Surg 47: 607-609
33. Grobe T, Raithel D (1978) Peripheral nerve injuries complicating extracranial vascular
Surgery.
Thoraxchir Vask Chir 26: 373-376
34. Guelinckx PJ, Sinsel NK (1996) Muscle transplantation for reconstruction of a smile
after facial paralysis , past, present, and future.
Microsurgury 17:391-401
35. Guth L (1956) Regeneration of the mammalian nervous system.
Physiol Rev 36: 441-4478

79
Literaturverzeichnis
36. Gutmann F, Sanders FK (1943) Recovery of fibre numbers and diameters in the regeneration of peripheral
nerves.
J Physiol 101: 489-518
37. Hagan WE (1981) “How I do it” - plastic surgery. Practical suggestions on facial plastic surgery.
Microneural techniques for nerve grafting.
Larngoscope 91: 1759-1766
38. Hall GD, Van der Wey CW, Kung FT, Compton-Allen M (1993) Peripheral nerve elongation with tissue expansion techniques.
J Trauma 34. 401-405
39. Haller JR, Shelton C (1997) Medial antebrachial cutaneous nerve: a new donor graft for
repair of facial defects at the skull base.
Laryngoscope 107: 1048-1052
40. Harii K, Asato H, Yosshimura K, Sugawara Y, Nakatsuka T, Ueda K (1998)
One-stage transfer of the latissimus dorsi muscle for reanimation of a paralyzed
face: a new alternative.
Plast Reconstr Surg 102: 941-950
41. Hausamen JE, Schmelzeisen R (1996) Current principles in microsurgical nerve repair.
Br J Oral Maxillofac Surg 34:143-156
42. Hoffmann WY (1992) Reanimation of the paralyzed face.
Otolaryng Klin North Am 25: 649-667

80
Literaturverzeichnis
43. Holländer H, Vaaland JL (1968) A reliable staining method for semi-thin sections in experimental neuroanatomy.
Brain Res 10: 120-126
44. Holland GR, Robinson PP (1990) The number and size of axons central and peripheral to inferior alveolar
nerve injuries in the cat.
J Anat 173: 129-137
45. House JW, Brackman DE (1985) Facial grading system.
Otolaryn Head Neck Surg 93: 146-147
46. Hu J, Tang Z, Wang D Buckley MJ (2001) Changes in the inferior alveolar nerve after mandibular lengthening with different
rates of distraction.
J Oral Maxillofac Surg 59: 1041-1046
47. Isch F, Isch-Treussard C, Jesel M (1968) EMG findings on reinnervation in peripheral nerve lesions.
EEG Clin Neurophysiol 24: 404-405
48. Jander S, Lausberg F, Stoll G (2001) Differential recruitment of CD8 + macrophages during Wallerian degeneration in
the peripheral and central nervous system.
Brain Pathol 11: 27-38
49. Jenq CB, Coggeneshall RE (1985) Numbers of regenerating axons in parent and tributary peripheral nerves in rat.
Brain Res 326: 27-40
50. Kinderman NB, Harrington CA, Drengler SM Jones KJ (1998) Ribosomal RNA transcriptional activation and processing in hamster facial
motoneurons: effects of axotomy with or without exposure to testosterone
J Comp Neurol 401: 205-216

81
Literaturverzeichnis
51. Klöcker N, Zerfowski M, Gellrich N-C, Bähr M (2001) Morphological and functional analysis of an incomplete CNS fibre tract lesion -
graded crush of the rat optic nerve.
J Neurosci Meth 110 (1-2): 147-153
52. Koshima I, Tsuda K, Hamanaka T, Moriguchi T (1997) One-stage reconstruction of established facial paralysis using a rectus abdominis
muscle transfer.
Plast Reconstr Surg 99: 234-238
53. Kumar A, Mafee MF, Mason T (2000) Value of disorders of the facial nerve.
Top Magn Reson Imaging 11: 38-51
54. Kury P, Stoll G, Müller HW (2001) Molecular mechanisms of cellular interactions in peripheral nerve regeneration.
Curr Opin Neurol14: 635-639
55. Kuczynski K (1980) Histology of the peripheral nerve trunks.
Int Surg 65: 495-498
56. Kanazaki J, Kunihiro T, Ouchi T (1991) Intracranial reconstruction of the facial nerve. Clinical observation.
Acta Otolaryngol Suppl 487: 85-90
57. Lahoda F (1988) Neurologische Untersuchungen in der Praxis-Erkrankungen des
peripheren Nervensystems. 5. Aufl.
Einhorn-Presse-Verlag, Reineck
58. Lay HC (1999) Regeneration des Nervus facialis nach Nervennaht und Nimodipingabe bei der
Ratte: Eine stereologische Untersuchung des peripheren Nerven.
Inaug-Diss, Med. Fak., Universität zu Köln, Köln

82
Literaturverzeichnis
59. Larowski TD, Drengler SM, Tanzer L, Jones KJ (2000) Androgen receptor mRNA regulation in adult male and female hamster facial
motoneurons: effects of axotomy and exogenous androgens.
J Neurobiol 45: 207-214
60. Lewis ME, Neff NT, Contreras PC, Stong DB, Oppenheim RW, Grebow PE, Vaught JL (1993)
Insulin-like growth factor-I: potential for treatment of motoneural disorders.
Exp Neurol 124: 73-88
61. Lindmuth R, Ernzerhof C, Schimrigk K (2002) Comparative morphometry of myelinated fibres in the normal and pathologically
altered human sural and tibial nerve.
Clin Neuropathol 21: 29-34
62. Logroscino CA, Sanguinetti C, Catalano F (1980) Electron microscopic study of the early changes proximal to a constriction in
myelinated nerve.
Int Orthop 4: 121-125
63. Lundborg G (1987) Nerve regeneration and repair.
Acta Orthop Scand 58: 145-169
64. Mackinnon SE (1989) Surgical management of peripheral nerve gap.
Clin Plast Surg 16: 587-603
65. Mackinnon SE, Dellon AL, O´Brien JP (1991) Changes in nerve fibre numbers distal to a nerve repair in rat sciatic nerve model.
Muscle Nerve 14: 1116-1122
66. Malis DJ, Mac Millan JG, Kelly JL, Mc Gath JH (1995) Tissue expansion of the facial nerve in an animal model.
ENT 74:261-270

83
67. Manders EK, Saggers GC, Diaz-Alonso O (1987) Elongation of peripheral nerve and viscera containing smooth muscle.
Clin Plast Surg 14: 551-562
68. Martin KH (1964) Untersuchung über die perineurale Diffusionsbarriere an
gefriergetrockneten Nerven.
Z Zellforsch Mikr Anat 64: 404-415
69. Martin MR, Caddy KWT, Biscoe TJ (1977) Numbers and diameters of motoneurons and myelinated axons in the facial
nucleus and nerve of the albino rat.
J Anat 123: 579-587
70. Martin-Duverneuil N, Lafitte F, Chiras J (1999) Cross-sectional anatomy of the facial nerve.
JBR-BTR 82: 301-305
71. Maschino MC, Giudiceandrea F, Grimaldi M, Cervelli V (1991) Surgical lesions of the facial nerve: anatomy, semilology, and nervous function.
Riv Eur Sci Med Farmacol13: 247-250
72. Mattox DE, Felix H (1987)
Surgical anatomy of the rat facial nerve.
Am J Otol 8: 43-47
73. Menger H (1992) Affektionen peripherer Nerven im Sport.
Dtsch Z Sportmed 1992,43: 44-58
74. Morell P, Norton WT (1980) Myelin.
Sci Am 242: 74-89

84
Literaturverzeichnis
75. Monballiu G (1981) Cross-face nerve grafting in facial paralysis.
Acta Chirg Bel 80:147-154
76. Nagatsu I, Karasawa N, Shimizu n (1978) Early Changes in the structure of rat sciatic nerves by ligation.
Arch Histopatol Jpn 41: 439-451
77. Nemeth E, Cigic T (1999) Recovery of facial nerve function using sural nerve transplantation after its injury
in nerve surgery.
Med Pregl 52: 283-286
78. Novack CB, Mackinnon SE, Baratz ME (2002) Peripheral nerve injuries.
eMedicine: topic 537
79. O´Sullivan KL, Pap SA, Megerian CA, Li Y, Sheffler LR, Smith TW, Lawrence WT (1998) Improved axon diameter and myelin sheath thickness in facial nerve cable
grafts wrapped in temporopartial fascial flaps.
Ann Plast Surg 40: 478-485
80. Paturet G (1964) Traite d´anatomie humaine.
Masson, Paris
81. Quan D, Bird SJ (1999) Nerve conduction studies and electromyography in the evaluation of peripheral
nerve injuries.
J Orthop 12: 45-51

85
Literaturverzeichnis
82. Quarles RH (2002) Myelin sheaths: glycoproteins involved in their formation, maintenance and
degeneration.
Cell Mol Life Sci 59: 1851-1871
83. Rechtland E, Rapoports SI (1987) Regulation of the microenvironment of peripheral nerve: Role of blood nerve
barrier.
Prog Neurobiol 28: 303-343
84. Rushton WAH (1951) A theory of the effects of fibre size in medullated nerve.
J Physiol 115: 101-122
85. Samii M, Matthies C (1997) Management of 1000 vestibular schwannomas (acoustic neuromas): the facial
nerve-preservation and restitution of function.
Neurosurgury 40: 684-695
86. Schmelzeisen R, Neukam FW, Hausamen J-E (1996) Atlas der Mikrochirurgie im Kopf-, Halsbereich.
Hanser, München
87. Seddon HJ (1942) A classification of nerve injuries.
Br Med J 1: 237-239
88. Seddon HJ (1943) Three types of nerve injuries.
Brain 66: 237-388
89. Seddon HJ, Medawar PB, Smith H (1943) Rate of regeneration of peripheral nerves in man.
Neurol 124: 64-72

86
Literaturverzeichnis
90. SeeburgerJL, Springer JE (1993) Experimental rationale for the therapeutic use of neutrophins in amyotrophic
lateral sclerosis.
Neurol 124: 64-72
91. Sharon I, Fishfield C (2002) Acute nerve injury.
eMedicine: topic 2908
92. Snipes GJ, Suter U (1995) Molecular anatomy and genetics of myelin proteins in the peripheral nervous
system.
J Anat 186 (Pt3): 483-494
93. Son YJ, Thompson WJ (1995) Schwann cell processes guide regeneration of peripheral axons.
Neuron 14:125-132
94. Son YJ, Thompson WJ (1995) Nerve sprouting in muscle is induced and guided by processes extended by
Schwann cells.
Neuron 14: 133-141
95. Stephanian E, Sekhar LN, Janecka IP, Hirsch B (1992) Facial nerve repair by interposition nerve graft: results in 22 patients.
Neurosurgery 31: 73-76
96. Stoll G, Müller HW (1999) Nerve injury, axonal degeneration and neural regeneration: insights.
Pathol 9: 313-325
97. Sugimoto Y, Takayama S, Horiuchi Y, Toyama Y (2002) An experimental study on the perineural window.
J Peripher Nerv Sys 7: 104-111

87
Literaturverzeichnis
98. Sunderland S (1946) Course and rate of regeneration of motor fibres following lesions of the radial
nerve.
Arch Neurol Psychiatry 56: 133-157
99. Sunderland S (1951) A classification of the peripheral nerve injuries producing loss of function.
Brain 74: 491-516
100. Sunderland S (1990) The anatomy and physiology of nerve injury.
Muscle Nerve 13: 771-784
101. Sutherland D, Buist R, Dort JC (1997) Magnetic resonance imaging of the extratemporal facial nerve of the rat.
J Otolaryngol 26: 112-115
102. Szentágothai J (1949) Functional representation in the motor trigeminal nucleus.
J Comp Neurol 90: 111-120
103. Tackman W, Richter HP, Stöhr M (1989) Kompressionssyndrome peripherer Nerven.
Springer, Berlin
104. TasslerPL, Dellon AL, Canoun C (1994) Identification of elastic fibres in the peripheral nerve.
J Hand Surg 19: 48-54
105. Thomas PK (1988) Clinical aspects of PNS regeneration.
Adv Neurobiol 49: 9-29

88
Literaturverzeichnis
106. Trumble TE in: Felicano DV, Moore EE, Matox KL (2000) Trauma. 4. Aufl.
McGraw-Hill, New York
107. Ueki M, Linn F, Hossmann KA (1988) Functional activation of cerebral blood flow and Metabolism before and after
global ischemia of rat brain.
J Cereb Blood Flow Metab 8: 486-49
108. Van der Wey LP, GabreelsFesten AA, Merks MH (1995) Peripheral nerve elongation by laser doppler flowmetry controlled expansion,
morphological aspects.
Acta Neuropathol 97: 168-171
109. Van der Wey LP, Polder TW, Stegman DF, Gabreels-Festen AA, Spauwein PH, Feesten FJ (1996) Peripheral nerve elongation by laser doppler flowmentry-monitored expansion:
an experimental basis for future application in the management of peripheral
nerve defects.
Plast Reconstr Surg 97: 568-576
110. Wang XX, Wang X, Li ZL (2002) [Effects of mandibular distraction osteogenesis on the inferior alveolar nerve: an
experimental study in monkeys].
Plast Reconstr Surg 109: 2373-2383
111. Wood RJ, Adson MH, Van Beek AL, Peltier GL, Zubkoff MM, Bubrick MP (1991) Controlled expansion of peripheral nerves: comparison of nerve grafting and
nerve expansion/repair for canine sciatic nerve defects.
J Traumata 31: 686-689

89
Literaturverzeichnis
112. Ya Z, Chen Z, Wang J (2000) Establishment of facial nerve elongation model by string type loading.
Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi 14: 90-92
113. Yu WH, McGinnis MY (2001) Androgen receptors in cranial nerve motor nuclei of male and female rats.
J Neurobiol 46: 1-10
114. Zemp C, Bestetti G, Rossi GL (1981) Morphological and morphometric study of peripheral nerves from rats with
streptozotocin-induced diabetes mellitus.
Acta Neuropathol 53: 99-106

90
Lebenslauf
7 Lebenslauf Persönliche Daten Name: Kriesche Vorname: Bettina Maria Geburtsdatum: 29. Dezember 1971 Geburtsort: Villingen - Schwenningen Familienstand: ledig Eltern: Prof. Dipl.-Ing. Josef Kriesche Dr. Marianne Kriesche Staatsangehörigkeit: deutsch Anschrift: Lehenerstr. 152 79106 Freiburg
Schulische Ausbildung 1978-1982 Grundschule Erbsenlachen VS - Villingen 1982-1991 Gymnasium am Hoptbühl VS - Villingen 1991 Allgemeine Hochschulreife 1991-1992 Freiwilliges Soziales Jahr Städtisches Krankenhaus Überlingen Abteilung für Allgemeine Chirurgie
Hochschulstudium 1993 Studium der Zahnmedizin an der Albert- Ludwigs- Universität, Freiburg i Br. 1999 Staatsexamen Zahnmedizin
Beruflicher Werdegang 2000 Approbation zur Zahnärztin Feb. 2001- Sept. 2001 Ausbildungsassistentin Praxis Dr. F. Grunert, Lörrach Okt. 2001-Juli 2003 Ausbildungsassistentin Praxis Dr. W. Polozcek,
Freiburg Seit Aug. 2003 Assistenzzahnärztin im Zentrum für Zahnmedizin der
Universität Basel, Abteilung für Rekonstruktive Zahnmedizin und Myoarthropathien Klinikvorsteher Prof. Dr. C.P. Marinello


















