
Medizinische Fakultät der Universität Duisburg-Essen · gegen Epilepsie (International League...
54
Medizinische Fakultät der Universität Duisburg-Essen Aus der Abteilung für Neuroanatomie und Molekulare Hirnforschung der Medizinischen Fakultät der Ruhr-Universität Bochum Glia und Epilepsie: Experimentelle Untersuchung klinisch relevanter Antiepileptika an einem in vitro Astrozyten/Mikroglia Entzündungs- Zellkulturmodell Inaugural-Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften in der Medizin durch die Medizinische Fakultät der Universität Duisburg-Essen Vorgelegt von Hannes Dambach aus Bietigheim-Bissingen 2014
-
Upload
truongnguyet -
Category
Documents
-
view
213 -
download
0
Transcript of Medizinische Fakultät der Universität Duisburg-Essen · gegen Epilepsie (International League...

Medizinische Fakultät
der
Universität Duisburg-Essen
Aus der Abteilung für Neuroanatomie und Molekulare Hirnforschung
der Medizinischen Fakultät
der Ruhr-Universität Bochum
Glia und Epilepsie: Experimentelle Untersuchung klinisch relevanter
Antiepileptika an einem in vitro Astrozyten/Mikroglia Entzündungs-
Zellkulturmodell
Inaugural-Dissertation
zur
Erlangung des Doktorgrades der Naturwissenschaften in der Medizin
durch die Medizinische Fakultät
der Universität Duisburg-Essen
Vorgelegt von
Hannes Dambach
aus Bietigheim-Bissingen
2014

2
Dekan: Herr Univ.-Prof. Dr. med. J. Buer
1. Gutachter: Herr Prof. Dr. med. A. Hufnagel
2. Gutachter: Frau Univ.-Prof. Dr. rer. nat. N. Dünker
3. Gutachter: Frau Univ.-Prof. Dr. med. K. Keyvani
Tag der mündlichen Prüfung:
14. Januar 2015

3
Publikation
Dambach H, Hinkerohe D, Prochnow N, Stienen MN, Moinfar Z, Haase CG, Hufnagel A,
Faustmann PM. (2014): Glia and epilepsy: Experimental investigation of antiepileptic drugs
in an astroglia/microglia co-culture model of inflammation. Epilepsia. 55,184-92.

4
Inhaltsverzeichnis
1. Einleitung (S.6)
1.1 Epilepsie ............................................................................................................................................ 6
1.2 Zerebrale Inflammation ..................................................................................................................... 6
1.3 Zusammenhang zwischen Epilepsie und Inflammation .................................................................... 7
1.4 Antiepileptika .................................................................................................................................... 8
1.5 Astrozyten ......................................................................................................................................... 9
1.6 Astrogliale Gap Junctions und Connexin 43 (Cx43) ...................................................................... 10
1.7 Mikroglia ......................................................................................................................................... 10
1.8 Das Astrozyten-Mikroglia Ko-Kultur Model .................................................................................. 11
1.9 Fragestellung der Dissertation ........................................................................................................ 12
2. Material und Methoden (S.15)
2.1 Zellkultur ......................................................................................................................................... 15
2.1.1 Astrozyten-Mikroglia Ko-Kultur.............................................................................................. 15
2.1.2 Inkubation mit den antiepileptisch wirksamen Substanzen ..................................................... 16
2.2 Viabilitätsassay ................................................................................................................................ 17
2.4. Immunocytochemie ........................................................................................................................ 17
2.5 Auswertung der immunocytochemischen Färbungen ..................................................................... 18
2.6 ELISA .............................................................................................................................................. 19
2.7 Western Blot .................................................................................................................................... 21
2.7.1 Zellaufschluss ........................................................................................................................... 21
2.7.2 Konzentrationsbestimmung nach Bradford .............................................................................. 21
2.7.3 SDS-Polyacrylamid-Gelelektrophorese ................................................................................... 22
2.7.4 Western Blot und immunologischer Nachweis der Proteine .................................................... 22
2.8 Statistik ............................................................................................................................................ 23
3. Ergebnisse (S.25)
3.1 Der Einfluss antiepileptischer Substanzen auf die Viabilität primärer M5 und M30
Astrozyten/Mikroglia Ko-Kulturen ....................................................................................................... 25
3.2 Der Einfluss antiepileptischer Substanzen auf die mikrogliale Aktivierung/ Inaktivierung in
primären M5 und M30 Astrozyten/Mikroglia Ko-Kulturen .................................................................. 26

5
3.3 Der Einfluss antiepileptischer Substanzen auf die astrogliale Cx43 Expression in primären M5 und
M30 Astrozyten/Mikroglia Ko-Kulturen .............................................................................................. 29
3.4 Der Einfluss antiepileptischer Substanzen auf die Expression der beiden Zytokine TGF-ß1 und
TNF-α in primären M5 und M30 Astrozyten/Mikroglia Ko-Kulturen .................................................. 30
4. Diskussion (S.32)
4.1 Der Einfluss antiepileptischer Substanzen auf die Viabilität primärer M5 und M30
Astrozyten/Mikroglia Ko-Kulturen ........................................................................................................ 32
4.2. Der Einfluss antiepileptischer Substanzen auf die mikrogliale Aktivierung/ Inaktivierung in
primären M5 und M30 Astrozyten/Mikroglia Ko-Kulturen ................................................................... 33
4.3 Der Einfluss antiepileptischer Substanzen auf die astrogliale Cx43 Expression in primären M5 und
M30 Astrozyten/Mikroglia Ko-Kulturen ................................................................................................ 34
4.4 Der Einfluss antiepileptischer Substanzen auf die Expression der beiden Zytokine TGF-ß1 und TNF-
α in primären M5 und M30 Astrozyten/Mikroglia Ko-Kulturen............................................................ 35
4.5 Klinische Interpretation antiepileptischer Substanzen mit zusätzlichem antiinflammatorischen
Potential bei Patienten mit inflammatorisch bedingten Anfällen ........................................................ 36
5. Zusammenfassung (S.39)
6. Literaturverzeichnis (S.40)
7. Abkürzungsverzeichnis (S.47)
8. Anhang (S.49)
9. Danksagung (S.52)
10. Lebenslauf (S.54)

6
1. Einleitung
1.1 Epilepsie
Epilepsie ist eine der geläufigsten neurologischen Erkrankungen, welche etwa 1 % der
weltweiten Bevölkerung betrifft. Epileptische Anfälle sind gekennzeichnet durch repetitive,
hypersynchrone elektrischen Entladungen von Nervenzellen im Gehirn. Bei etwa einem
Drittel der betroffenen Patienten ist die Behandlung mit derzeitig verfügbaren antiepileptisch
wirksamen Medikamenten nicht kontrollierbar (Seifert et al. 2006). Die Internationale Liga
gegen Epilepsie (International League Against Epilepsy; ILAE) unterscheidet bei
epileptischen Anfällen zwischen generalisierten und fokalen epileptischen Anfällen.
Generalisierte epileptische Anfälle gehen von einem neuronalen Netzwerk aus, bei welchem
beide Großhirnhemisphären beteiligt sind. Diese sogenannten „bilateralen“ Netzwerke
können sowohl kortikale als auch subkortikale Strukturen beinhalten. Hingegen gehen fokale
epileptische Anfälle von einem neuronalen Netzwerk aus, bei welchem lediglich eine
Großhirnhemisphäre beteiligt ist (Berg et al. 2010). Es werden drei zugrunde liegende
Ursachen für epileptische Anfälle behandelt. Epilepsie als Ursache: 1. eines genetischen
Defekts, 2. einer struktureller und metabolischer Veränderungen oder 3. einer bislang nicht
aufgeklärten, unbekannten Ursache.
1.2 Zerebrale Inflammation
Inflammatorische Reaktionen innerhalb des zentralen Nervensystems (ZNS) entstehen durch
eine Reihe unterschiedlicher Erkrankungen. Dazu gehören inflammatorische Erkrankungen,
wie bakterielle und virale Infektionen sowie Multiple Sklerose. Ebenfalls können
Erkrankungen, welche nicht in erster Linie inflammatorischer Natur sind wie Ischämie,
Tumore und Trauma, inflammatorische Reaktionen im ZNS hervorrufen (Walz 2010).
Einerseits schützen inflammatorische Reaktionen das Gehirn vor bakteriellen und viralen
Infektionen. Andererseits können Infektionen des ZNS langanhaltende, neurologische
Spätschäden zur Folge haben. In der Literatur wird unter anderem diskutiert, dass
Gewebeschädigungen aufgrund der inflammatorischen Antwort des Gehirns ernsthaftere
Konsequenzen als die Infektion an sich haben können (Wraith und Nicholson 2012). Lang
andauernde inflammatorische Bedingungen sind gekennzeichnet durch eine Ansammlung,

7
Aktivierung und Proliferation von immunmologisch aktiven Gliazellen wie der Mikroglia. Sie
werden durch ihr biochemisches Umfeld stark beeinflusst und reagieren auf Signale
umliegender Zellen. Unter inflammatorisch veränderten Bedingungen proliferieren Mikroglia
und nehmen einen aktivierten Phenotyp ein (Kettenmann et al. 2011).
1.3 Zusammenhang zwischen Epilepsie und Inflammation
Eine zunehmende Anzahl klinischer als auch experimenteller Studien berichten von einem
Zusammenhang zwischen Epilepsie und einer Inflammation des zentralen Nervensystems
(ZNS) (Vezzani and Granata 2005, Vezzani et al. 2011, Vezzani and Rüegg 2011).
Klinische Erkenntnisse basieren auf der Beobachtung, dass bei Medikament resistenten
Epilepsien eine anti-inflammatorische Therapie bei den Patienten eine Verbesserung bewirkte
(Wirrell et al. 2005, Wheless et al. 2007, Yu et al. 2013). Ein weiterer Hinweis basiert auf
dem erhöhten proinflammatorischen Zytokinlevel in der Zerebrospinalflüssigkeit und im
Serum von chronischen Epilepsie-Patienten nach einem Anfall (Hulkkonen et al. 2004,
Vezzani 2005, Aronica and Crino 2011). Ein Beispiel hierfür ist die erhöht gemessene
Interleukin-6 (IL-6) Konzentration im Liquor und Plasma von Patienten innerhalb von 24
Stunden nach einem tonisch-klonischen Anfall (Peltola et al. 1998). Ein weiterer Hinweis ist
die erhöhte gliale und neuronale Expression inflammatorischer Mediatoren in chirurgisch
entfernten Gehirngewebe von Patienten mit refraktärer Epilepsie (Vezzani and Granata 2005,
Choi et al. 2009, Aronica and Crino 2011, Vezzani 2011, Vezzani and Rüegg 2011, Silveira et
al. 2012). In chirurgisch entfernten Hippocampi von Patienten mit Temporallappenepilepsie
(TLE) konnten ebenfalls Indikatoren für eine Inflammation, wie aktivierte Mikroglia, reaktive
Astrozytose und proinflammatorische Zytokine, nachgewiesen werden (Vezzani et al. 2011).
Experimentelle Untersuchungen in Tiermodellen ergeben ebenfalls Hinweise auf einen
Zusammenhang zwischen Epilepsie und Inflammation: Induzierte epileptische Anfälle führten
zu einer Aktivierung glialer Zellen und folglich erhöhten Anzahl proinflammatorischer
Mediatoren (Vezzani et al. 1999, De Simoni 2000, Voutsine-Porche et al. 2004, Ravizza et al.
2005, Choi and Koh. 2008, Vezzani et al. 2011, Li et al. 2011). Die direkte intrazerebrale
Injektion proinflammatorischer Zytokine oder Lipopolysaccharide (LPS) führte in
Tiermodellen zudem zu einer erhöhten Aktivität von epileptischen Anfällen (Galic et al. 2008,
Auvin et al. 2010). Im Gegensatz führten Zytokinrezeptor Antikörper, wie Interleukin-1
Rezeptorantagonisten, zu einer effektiven antikonvulsiven Aktivität (Vezzani et al. 2000).

8
1.4 Antiepileptika
Der Wirkungsmechanismus der in dieser Dissertationsarbeit verwendeten Antiepileptika
Valproat (VPA), Carbamazepin (CBZ), Phenytoin (PHE) und Gabapentin (GBT) soll in
diesem Abschnitt erläutert werden:
Die therapeutische Wirksamkeit von Antiepileptika zur Senkung der neuronalen Aktivität
kann durch die folgenden Mechanismen erfolgen (Lüllmann et al. 2010):
- Blockade von spannungsabhängigen Na+ -Kanälen (CBZ, VPA, PHE)
- Förderung einer GABAergen Hemmung (GBT, VPA)
- Hemmung eines T-Typ-Ca2+
-Einwärtsstroms
Das Antiepileptikum Carbamazepin (CBZ) wird sowohl für die Behandlung von Epilepsie als
auch neuropathischer Schmerzen und affektiven Funktionsstörungen eingesetzt (Ambrosio et
al. 2002). Es wird bei der Behandlung von Epilepsie-Patienten mit fokalen Anfällen und
sekundär generalisierten tonisch-klonischen Anfällen verwendet (Pohlmann-Eden und
Steinhoff, 2006). Die Wirkung von CBZ erfolgt über die Blockade von spannungsabhängigen
Na+ -Kanälen. Eine Wirksamkeit auf verschiedene Typen anderer Kanäle und Rezeptoren
wird ebenfalls angegeben (Ambrosio et al. 2002).
Valproinsäure (VPS), im englischen durch VPA (valproic acid) abgekürzt, wird für die
Behandlung von Epilepsie und bipolaren Störungen eingesetzt (Monti et al. 2009). Die Salze
der Valproinsäure, die sogenannten Valproate, werden als Antiepileptika für die Behandlung
von Myoklonien, Grand-mal-, generalisierten und fokalen Anfällen verwendet (Pohlmann-
Eden und Steinhoff, 2006). Die antiepileptische Wirkung von VPA erfolgt über Blockade
von spannungsabhängigen Na+ -Kanälen. Außerdem soll VPA zudem die Aktivität des
Neurotransmitters GABA (γ-Aminobuttersäure) verstärken (Monti et al. 2009).
Phenytoin (PHE) wird für die Behandlung von Epilepsie und Herzrhytmusstörungen
eingesetzt (Tunnicliff 1996). Es wird bei der Behandlung von Epilepsie-Patienten mit fokalen
Anfällen, Grand-mal- und komplex-fokalen Anfällen mit Bewusstseinstörungen verwendet
(Pohlmann-Eden und Steinhoff, 2006). Die Wirkung von Phenytoin erfolgt über eine
Blockade von spannungsabhängigen Na+ -Kanälen in der Plasmamembran von Nervenzellen.
Gabapentin (GBT), ein chemisches GABA (γ-Aminobuttersäure) Analog, wird für die
Behandlung von Epilepsie und bei Schmerzen im Rahmen von Neuropathien verabreicht
(Pohlmann-Eden und Steinhoff, 2006). Obwohl GBT ein chemisches GABA Analog ist,

9
bindet es weder an GABAA- noch GABAB-Rezeptoren und wird metabolisch nicht zu GABA
transformiert (Honarmand et al. 2012). Es wird bei Epilepsie-Patienten mit fokalen Anfällen
und großen epileptischen Anfällen eingesetzt (Pohlmann-Eden und Steinhoff, 2006).
Gabapentin bindet an die α2δ Untereinheit spannungsabhängiger Ca2+
-Kanäle und moduliert
den Einstrom von Ca2+
, wodurch es zu einer reduzierten Freisetzung exzitatorischer
Neurotransmitter kommt (Beal et al. 2012).
In der Pathophysiologie der Epilepsie wird der Beteiligung von Gliazellen immer mehr
Bedeutung zugeschrieben (Binder und Steinhäuser 2006, Seifert et al. 2006, Jabs et al. 2008,
Wetherington et al. 2008, Seifert et al. 2010, Devinsky et al. 2013). Astrozyten und Mikroglia
spielen eine wichtige Rolle bei der Initiation und Aufrechterhaltung der zerebralen
Immunantwort sowie der Versorgung der Nervenzellen (Pavone und Cardile 2003,
Kettenmann et al. 2011, Seifert und Steinhäuser 2013). Der Einfluss antiepileptischer
Medikamente auf Gliazellen ist teilweise bekannt, allerdings bleibt der detaillierte
Mechanismus oftmals ungeklärt (Cardile et al. 2001, Pavone und Cardile 2003, Dragunow et
al. 2006, Haghikia et al. 2008, Black et al. 2009, Smith et al. 2010, Andrzejczak 2011,
Gibbons et al. 2011, Stienen et al. 2011, Devinsky et al. 2013).
1.5 Astrozyten
Astrozyten nehmen in etwa 40 % des gesamten Gehirnvolumens ein und liegen in ca. 10 fach
höherer Anzahl als Nervenzellen im humanen Gehirn vor. Ihre Aufgaben im
Zentralnervensystem (ZNS) sind vielseitig. Sie bilden Ausläufer zu den Endothelzellen von
Kapillaren und sind am Aufbau und der Regulation der Blut-Hirn Schranke beteiligt (Nico
und Ribatti 2012). Außerdem umhüllen Astrozyten zentrale neuronale Synapsen und
exprimieren G-Protein gekoppelte Neurotransmitter Rezeptoren sowie Transporter, wodurch
sie direkt auf die Aktivität von Nervenzellen reagieren (Stobart und Anderson 2013). Diese
strukturelle Spezifikation und das breit vernetzte Synzytium ermöglicht es den Astrozyten mit
allen wichtigen Elementen des Zentralnervensystems zu interagieren. In Bezug auf die
neuronale Aktivität wird Astrozyten einerseits eine Versorgungsfunktion als auch effektive
Beseitigungsfunktion von Produkten, welche die Nervenzellen nicht mehr benötigen,
zugeschrieben (Walz 2010).

10
1.6 Astrogliale Gap Junctions und Connexin 43 (Cx43)
Astrozyten bilden ein über Gap-Junction Kanäle gekoppeltes funktionelles Synzytium,
welches für den Erhalt der zerebralen Homöostase essentiell ist (Dermietzel und Spray 1993).
Die interzellulären Kanäle werden durch spezifische Proteine, die Connexine gebildet. Bei
Astrozyten ist Connexin 43 (Cx43) die mit am häufigsten vorkommende Proteinuntereinheit
(Dermietzel et al. 1991; Giaume et al. 1991; Retamal et al. 2007). Insgesamt bilden sechs
Connexine einen Hemikanal (Connexon), welcher aus dem Endoplasmatischen Retikulum zur
Plasmamembran transportiert wird. Die Verknüpfung zweier Connexone benachbarter Zellen,
ermöglicht die Ausbildung eines funktionellen Gap-Junction Kanals zwischen den beiden
Zellen. Die astroglialen Gap-Junction Kanäle ermöglichen einen Austausch von Ionen und
Molekülen zwischen benachbarten Zellen ohne Kontakt zum extrazellulären Milieu.
Neurogliale Interaktionen über Gap Junctions zwischen Gliazellen und Nervenzellen sind
sowohl unter physiologischen als auch pathologischen Konditionen für die neuronale
Aktivität essentiell (Giaume et al. 2013). Veränderungen in der Expression und der Funktion
von Gap-Junction Kanälen sind mit verschiedenen Formen von Nervenkrankheiten assoziiert.
Die zell-spezifische Deletion von Cx43 und Cx30 führte in Mäusen zu einer Generierung
spontaner epileptischer Aktivität (Wallraff et al. 2006). In der Studie von Seifert et al. 2010
wird beschrieben, dass eine verringerte gap junctionale Permeabilität offenbar gegensätzliche
Auswirkungen auf die Exzitabilität zur Folge haben kann: einerseits einen prokonvulsiven
Effekt aufgrund der beeinträchtigten K+
Umverteilung und andererseits einen
antiepileptischen Effekt aufgrund der eingeschränkten neuronalen Energieversorgung. In
weiteren Studien konnte unter inflammatorisch modifizierten Bedingungen bei kultivierten
Astrozyten eine verringerte Cx43 Expression ermittelt werden (Faustmann et. al. 2003,
Hinkerohe et al. 2005). Mikroglia, welche das neuronale Umfeld kontrollieren, sind in der
Lage die astrogliale Zell-Zellkommunikation durch die Produktion einer Vielzahl
proinflammatorischer Zytokine, zu beeinflussen (Aloisi 2001, Faustmann et al. 2003,
Hinkerohe et al. 2005, 2010).
1.7 Mikroglia
Mikroglia, welche oftmals als Makrophagen des Gehirns bezeichnet werden, sind neben
Astrozyten die immunologisch aktiven Zellen des ZNS. Sie migrieren in alle Regionen des
ZNS und nehmen morphologisch einen inaktiven ramifizierten Zustand (resting ramified type

11
RRT) an (Kettenmann et al. 2011). Der Anteil an Mikroglia Zellen im adulten ZNS liegt bei
5-15 % der gesamten glialen Zellpopulation (Walz 2010). Mikroglia tasten ihre territorialen
Domänen ab und kommunizieren über eine große Anzahl an Signalwegen mit Astrozyten,
Oligodendrozyten, Nervenzellen und Zellen des Immunsystems (Kettenmann et al. 2011).
Neuere Studien berichten außerdem, dass Mikroglia, ähnlich wie Astrozyten, an der
Regulation der neuronalen Aktivität beteiligt sind. Diese Erkenntnis beruht unter anderem auf
Studien, welche von einer pathologischen neuronalen Aktivität aufgrund einer Veränderung
der mikroglialen Funktion ausgehen (Béchade et al. 2013). Mikroglia gelten außerdem als
empfindlichste Sensoren für pathologisch veränderte zerebrale Bedingungen. Entdecken
Mikroglia Zellen Hinweise für zerebrale Verletzungen oder Dysfunktionen des
Nervensystems, kommt es zu einem komplexen, mehrstufigen Aktivierungsprozess. In diesem
Prozess wechselt der morphologische Zustand der Mikroglia vom inaktiven ramifizierten
Zustand (resting ramified type RRT) in den aktivierten phagozytierenden Zustand (rounded
phagocytic type RPT). Der morphologische Zustand der Mikroglia ist abhängig vom
immunologischen Status der Zellen (Faustmann et al. 2003; Hinkerohe et al. 2005). Aktivierte
Mikroglia kann zu Verletzungen des Hirngewebes migrieren, dort proliferieren und sowohl
zelluläre Kompartimente als auch komplette Zellen phagozytieren. Im aktivierten Zustand
besitzen Mikroglia außerdem die Fähigkeit eine große Anzahl an verschiedenen Substanzen
zu exprimieren, welche nützlich oder schädlich für umliegende Zellen sein können
(Kettenmann et al. 2011). Die exprimierten Substanzen sind abhängig von dem
unterschiedlichen Status, welchen die Mikroglia im aktivierten Zustand einnehmen können.
Die primären Zustände der aktivierten Mikroglia werden als M1 (klassische Aktivierung) und
M2 (alternative Aktivierung) bezeichnet (Benarroch 2013). Der M1 Zustand ist
gekennzeichnet durch einen phagozytierenden Phenotyp, verbunden mit der Produktion
inflammatorischer Zytokine (IL-1ß, IL-6, IL-23 und TNF-alpha), zytotoxischer Substanzen
(Stickstoffmonoxid), Peroxinitrite und exzitatorischen Aminosäuren. Der M2 Zustand ist
gekennzeichnet durch einen phagozytierenden Phenotyp, bewirkt jedoch eine Produktion
antiinflammatorischer Zytokine (TGF-beta, IL-10) oder unterstützt alternativ die Reperatur
von Gewebe durch die Ausschüttung von Matrixproteinen.
1.8 Das Astrozyten-Mikroglia Ko-Kultur Model
Wie bereits zuvor ausführlich erläutert, kommt es unter inflammatorischen Bedingungen,
verursacht durch zerebrale Infektionen, Verletzungen oder Dysfunktionen, zu einer

12
Aktivierung, Proliferation und Akkumulation immunologisch aktiver Mikroglia. Im
aktivierten Zustand sind sowohl Astrozyten als auch Mikroglia in der Lage eine Vielzahl anti-
/inflammatorischer Substanzen zu exprimieren. Folglich haben sie einen direkten Einfluss auf
benachbarte Nerven- und Gliazellen. Um den direkten Einfluss aktivierter Mikroglia auf die
interzelluläre Kopplungseigenschaften des astrozytären Synzytiums zu untersuchen, etablierte
Faustmann et al. (2003) ein Ko-Kultur Modell. Für diese Studie wurde einerseits eine
Astrozyten Ko-Kultur mit einem Mikroglia Anteil von ungefähr 5 % und von 30 % kultiviert.
Die Astrozyten Kultur mit einem Mikroglia Anteil von 5 % (M5) ist repräsentativ für den
physiologischen und mit einem Anteil von 30 % Mikroglia (M30) für den inflammatorisch
modifizierten Bereich des Gehirns. Basierend auf dem Astrozyten/Mikroglia Ko-Kultur
Modell wurde der Einfluss verschiedener neurologischer Medikamente, pro-/anti-
inflammatorischer Zytokine und anderer Substanzen auf den Mikroglia In-
/Aktivierungszustand sowie den Einfluss auf die astrozytäre Zell-Zell-Kommunikation
untersucht (Faustmann et al. 2003, Hinkerohe et al. 2005, Haghikia et al. 2008, Hinkerohe
2010, Stienen et al. 2011). Im Vergleich zur physiologischen M5 Ko-Kultur ist der Anteil
aktivierter Mikroglia bei der proinflammatorisch modifizierten M30 Ko-Kultur signifikant
erhöht (Faustmann et al. 2003). Des Weiteren konnte bei der M30 Ko-Kultur eine reduzierte,
quantitative Expression des Gap-Junction-Proteins Connexin 43 (Cx43) gemessen werden.
Die reduzierte Cx43 Expression ging zudem mit einem reduzierten Membranruhepotential
(MRP) sowie astrozytären funktionellen Kopplungsfähigkeit einher.
In dieser Dissertationsarbeit wird der direkte Einfluss der vier Antiepileptika VPA, PHE, CBZ
und GBT auf die Gliazellen in dem erläuterten M5/ M30 Astrozyten/Mikroglia Ko-Kultur
Model untersucht. Es wird die gliale Viabilität, Expression des Gap Junction Proteins
Connexin 43, In-/ und Aktivierung von Mikroglia Zellen sowie die Zytokin Expression
untersucht.
1.9 Fragestellung der Dissertation
In etwa 1 % der weltweiten Bevölkerung leiden unter epileptischen Anfällen. Die Ursachen
sind vielfältig: Genetische Prädisposition, traumatische Hirnschädigungen, Hirninfarkt und
Erkrankungen des Zentralnervensystems in Folge einer Meningitis. Die therapeutische
Wirksamkeit von Antiepileptika erfolgt über die Senkung der neuronalen Aktivität. Bei etwa
einem Drittel der betroffenen Patienten ist die Behandlung mit derzeit verfügbaren
Antiepileptika nicht kontrollierbar. Aus diesem Grund beschäftigen sich zunehmend mehr

13
Studien mit neuen Ansätzen. Einer dieser Ansätze, welcher in dieser Dissertation thematisiert
wird, ist der erläuterte Zusammenhang zwischen Epilepsie und einer Inflammation des
zentralen Nervensystems (ZNS). Gliazellen spielen eine wichtige Rolle bei der Initiation und
Aufrechterhaltung der zerebralen Immunantwort sowie der Versorgung der Nervenzellen.
Anhand des beschriebenen Astrozyten/Mikroglia Zellkultur-Modells soll der Einfluss
der alltäglich verwendeten Antiepileptika VPA, PHE, CBZ und GBT auf die Viabilität
von Gliazellen mittels eines Assays zur Bestimmung der Zellviabilität untersucht
werden.
Neurogliale Interaktionen zwischen Gliazellen und Nervenzellen sind für die neuronale
Aktivität essentiell. Gliazellen sind an der Modulation der synaptischen Transmission durch
die Modifikation von Kanälen, Transportern, Rezeptoren und Gap Junction Kanäle beteiligt.
Veränderungen in der Expression und der Funktion von Gap-Junction Kanälen sind mit
verschiedenen Formen von Nervenkrankheiten assoziiert.
Astrozyten bilden ein via Gap-Junction gekoppeltes, funktionelles Synzytium, welches
für den Erhalt der zerebralen Homöostase essentiell ist. Anhand des
Astrozyten/Mikroglia Zellkultur-Modells soll untersucht werden, inwiefern
Antiepileptika die Expression des Gap-Junction Proteins Cx43 beeinflussen können.
Der Zusammenhang zwischen Epilepsie und Inflammation ist durch klinische und
experimentelle Studien erwiesen. Inwiefern etablierte Medikamente gegen Epilepsie
entzündungshemmend oder -fördernd wirken ist jedoch nicht bekannt. Gliazellen sorgen als
Mikroglia für die Entzündungs- und Immunregulation. Unter inflammatorischen Bedingungen
verändert sich der Funktionszustand der Mikroglia von einem inaktiven zu einem aktivierten
Phenotyp. Die mikrogliale Aktivierung gilt somit als eines der wichtigsten Kennzeichen für
eine Entzündungsreaktionen im ZNS und kann mittels immunocytochemischer Färbung
visualisiert werden.
Das in dieser Arbeit verwendete Zellkulturmodell imitiert durch die Ko-Kultur
unterschiedlicher Astroglia- und Mikrogliaanteile normale und entzündliche
Bedingungen im ZNS. Anhand der M5 Ko-Kultur soll mittels immuncytochemischer
Färbung untersucht werden, inwiefern die Antiepileptika VPA, PHE, CBZ und GBT

14
den Aktivierungsgrad der Mikroglia beeinflussen. Hingegen soll anhand der M30 Ko-
Kultur, welche durch eine hohe Anzahl aktivierter Mikroglia gekennzeichnet ist
untersucht werden, inwiefern die Antiepileptika VPA, PHE, CBZ und GBT den
Aktivierungsgrad der Mikroglia herabsetzen und dadurch entzündungshemmende
Eigenschaften aufweisen.
Die beiden Zytokine TGF-beta1 und TNF-alpha sind bei der Regulation von
Entzündungsprozessen im Gehirn beteiligt. Beide Zytokine werden sowohl von Astrozyten als
auch von Mikroglia exprimiert. TGF-beta1 gilt als anti-inflammatorisches Zytokin und TNF-
alpha als proinflammatorisches Zytokin. Die pro- oder antiinflammatorische Wirksamkeit
hängt allerdings von verschiedenen Faktoren wie der Konzentration, der lokalen Verteilung
und dem Einfluss von verschiedenen inhibitorischen Faktoren ab.
Die Bestimmung der Zytokinkonzentrationen von TGF-beta1 und TNF-alpha mittels
Sandwich-ELISA in den Überständen der M5 und M30 Ko-Kulturen nach Inkubation
mit den Antiepileptika VPA, PHE, CBZ und GBT soll zusätzliche Informationen über
die entzündungshemmende oder –fördernde Wirkung der Medikamente auf Gliazellen
ergeben.

15
2. Material und Methoden
2.1 Zellkultur
2.1.1 Astrozyten-Mikroglia Ko-Kultur
Die primären Astrozyten/Mikroglia Ko-Kulturen werden postnatal aus dem Gehirn von
neugeborenen Wistar-Ratten (P0-P2) präpariert. Hierfür wurden die Tiere dekapitiert und der
Schädel mit sterilem Besteck geöffnet. Anschließend wurde das Gehirn vorsichtig entnommen
und in eine durch Eis gekühlte Petrischale mit sterilem PBS (Phosphat-buffered saline)
gelegt. Die Meningen wurden mit zwei sterilen Pinzetten unter Binokular-Kontrolle
abgezogen und der Plexus sowie das Kleinhirn entfernt. Nach erfolgter Präparation wurden
die in PBS gesammelten Hemisphären mechanisch, mit einer 10 ml Pipette durch auf- und
abziehen, zerkleinert und für 30 Minuten bei 37 °C in einer 0,1%igen Trypsin-Lösung (PAA
Laboratories, Pasching, Österreich) im Wasserbad inkubiert. Die verdaute Gehirnmischung
wurde mit PBS auf 20 ml aufgefüllt und bei 500g für 7 Minuten zentrifugiert. Der Überstand
wurde vorsichtig verworfen und das Pellet mit 5 ml DNase I (Serva Electrophoresis,
Heidelberg, Deutschland) 100 µg/ml in DMEM (Dulbecco's modified Eagle's medium) für 5
Minuten bei Raumtemperatur (RT) verdaut. Die Suspension wurde mit DMEM auf 20 ml
aufgefüllt und für 5 Minuten bei 200g zentrifugiert. Der Überstand wurde verworfen und das
Pellet mit 1 ml DMEM pro Hemisphäre resuspendiert. Im Anschluss wurde die
Zellsuspension durch ein steriles Nylon-Netz mit einem Durchmesser von 60 µm filtriert. Das
Filtrat mit den glialen Zellen wurde in Zellkulturflaschen (4 Hemisphären pro 25 cm2
-
Flasche) mit Astrozyten-Medium* ausgesäht und im Wärmeschrank bei 7 % CO2 und 37 °C
kultiviert. Am Folgetag wurden die adhärenten Zellen nochmals gewaschen und frisches
Kulturmedium hinzugegeben. Nach etwa einer Woche waren die Zellen konfluent gewachsen
und konnten für die jeweiligen Experimente ausgesäht werden.
*Inhalt: 87 % DMEM, Invitrogen (Karlsruhe, Deutschland)
10 % Fetales Kälberserum, Biochrom AG (Berlin, Deutschland)
1 % Penicillin/Streptomycin, Invitrogen (Karlsruhe, Deutschland)
1 % Nichtessentielle Aminosäuren, PAA Laboratories (Pasching, Österreich)
1 % Glutamin, PAA Laboratories (Linz, Österreich)

16
Um eine Astrozyten Ko-Kultur mit einem Mikroglia-Anteil von 30 % zu generieren, wird das
Medium der adhärenten Zellen verworfen und mit PBS gewaschen. Da sich die Mikroglia auf
den konfluent gewachsenen Astrozyten absetzen ist es bei der M30 Ko-Kultur wichtig
vorsichtig mit PBS zu waschen, um möglichst wenige Mikroglia abzulösen. Im Vergleich
werden bei der Herstellung einer Astrozyten Ko-Kultur mit einem Mikroglia-Anteil von 5 %
die adhärenten Zellen für 1 Stunde bei 300 rpm unter sterilen Bedingungen bei RT
geschüttelt. Dabei löst sich ein bestimmter Anteil an adhärenten Mikroglia ab und geht in das
Zellkulturmedium über. Das Zellkulturmedium wird anschließend verworfen.
Im weiteren Verlauf wurden die Astrozyten-Mikroglia Ko-Kulturen passagiert und für die
jeweiligen Versuche auf PLL (Poly-L-Lysin) beschichtete Deckgläschen (12 mm) (Assistent,
Sondheim, Deutschland) ausgesäht (60.000 Zellen/Deckgläschen). Alternativ wurden die Ko-
Kulturen auf PLL beschichteten 96 Well-Platten (BD Biosciences, Heidelberg, Deutschland)
mit einer Dichte von 10.000 Zellen/Well ausgesäht.
2.1.2 Inkubation mit den antiepileptisch wirksamen Substanzen
Basierend auf vorherigen Studien mit dem Antiepileptikum Levetiracetam (Haghikia et al.
2008, Stienen et al. 2011) und in Übereinstimmung mit gemessenen Serumkonzentrationen
von Epilepsie Patienten (Fröscher et al. 1999, Grim et al. 2003) werden in dieser Dissertation
klinisch effiziente Konzentrationen der antiepileptischen Substanzen Valproat (VPA),
Carbamazepin (CBZ), Phenytoin (PHE) und Gabapentin (GBT) verwendet. Aufgrund der
Variation gemessener Konzentrationen im Serum, Liquor und Hirngewebe von Epilepsie
Patienten (Schnabel et al. 1996, Mataringa et al. 2002, Rambeck et al. 2006) und um einen
konzentrationsabhängigen Einfluss auf die Ko-Kulturen zu untersuchen, werden folgende
Konzentrationen der Antiepileptika eingesetzt: VPA [50, 100 und 200 µg/ml], CBZ [10, 25,
50 und 100 µg/ml], PHE [10, 25, 50 und 100 µg/ml] und GBT [10, 25, 50 und 100 µg/ml].
Die antiepileptisch wirksamen Substanzen VPA, GBT und PHE (Sigma-Aldrich, Steinheim,
Deutschland) wurden in destilliertem H2O gelöst. CBZ (Sigma-Aldrich, Steinheim,
Deutschland) wurde in DMSO (Dimethyl Sulfoxide) gelöst und mit destilliertem H2O
verdünnt. Die finale Konzentration von DMSO war geringer als 1%. Die Ko-Kulturen wurden
mit VPA, CBZ, PHE und GBT für 24 Stunden im Wärmeschrank bei 7 % CO2 und 37 °C
inkubiert.

17
2.2 Viabilitätsassay
Der konzentrationsabhängige Einfluss der Antiepileptika VPA, CBZ, PHE und GBT auf die
Viabilität und Proliferation der Ko-Kulturen wurde mit dem Cell Proliferation Kit I (Roche
Diagnostics, Mannheim, Deutschland) entsprechend der Beschreibung des Herstellers,
bestimmt. Der Assay basiert auf der Spaltung des gelben Tetrazoliumsalzes MTT (-3-[4,5-
dimethylthiazol-2-yl]-2, 5-diphenyltetrazolium bromide) zu violett gefärbten Formazan
Kristallen, durch Enzyme des Endoplasmatischen Reticulums. Die Verstoffwechselung findet
lediglich in metabolisch aktiven Zellen statt. Die quantitative Ausbeute des violett gefärbten
Formazan korrelierte direkt mit der Anzahl metabolisch aktiver glialer Zellen in den Ko-
Kulturen. Die Absobtion des Farbstoffes wurde mit dem Mikroplatten Lesegerät (Bio-Rad
550, München, Deutschland) bei 570 nm ermittelt.
2.4. Immunocytochemie
Für die Untersuchung der mikroglialen Aktivierung wurden die auf den PLL-beschichteten
Deckgläschen, adhärenten Ko-Kulturen dreimal mit PBS (phosphate buffered saline) für
jeweils 10 Minuten gewaschen und anschließend mit eiskaltem Ethanol (100 %) fixiert.
Nachdem die Zellen fixiert waren, wurde nochmals dreimal mit PBS unter Schütteln
gewaschen und anschließend für 20 Minuten mit PBS + 1 % BSA (bovine serum albumin) +
10 % HS (horse serum) geblockt, um später unspezifische Bindungen der Antikörper zu
vermeiden. Im Anschluss an den Blockschritt wurde der primäre Antikörper in PBS + 1 %
BSA verdünnt und auf die Deckgläschen pipettiert (25 µl/Deckgläschen) und über Nacht bei 4
°C inkubiert. Die Ko-Kulturen wurden mit dem primären mouse anti ED-1 (Serotec,
Düsseldorf, Deutschland) im Verhältnis 1:250 inkubiert. Am Folgetag wurden die Zellen
dreimal mit PBS + 1 % BSA für jeweils 10 Minuten gewaschen und mit dem sekundären
Antikörper Alexa rot 568 anti-mouse (Invitrogen, Karlsruhe, Deutschland) mit einer
Verdünnung von 1:1000 in PBS + 1 % BSA für eine Stunde bei Raumtemperatur (RT)
inkubiert (25 µl/Deckgläschen). Im letzten Schritt wurden die Deckgläschen mit dem
Kernfarbstoff Hoechst 33342 versetzten ProLong-antifade-Kleber (Invitrogen, Karlsruhe,
Deutschland) auf Objekträgern fixiert. Nach 1-2 Tagen konnten die Zellen am
Immunfluoreszenz-Mikroskop (Axiophot Carl Zeiss GmbH, Jena, Deutschland) ausgewertet
werden.

18
In dieser Dissertationsarbeit werden zusätzliche immunocytochemische Färbungen der Ko-
Kulturen sowie histologische Gehirnschnittfärbungen von der Ratte (Kontrolle) mit den
Markern GFAP (Glial fibrillary acidic protein) (Sigma-Aldrich, Steinheim, Deutschland),
NeuN (Neuronal Nuclei) (Millipore, Darmstadt, Deutschland), CNPase (2',3'-cyclic-
nucleotide phosphodiesterase) (Sigma-Aldrich, Steinheim, Deutschland) angefertigt. Diese
Färbungen gelten als Nachweis, dass die Ko-Kulturen, welche aus dem Gehirn von
neugeborenen Wistar-Ratten (P0-P2) gewonnen werden, hautpsächlich aus Astrozyten und
Mikroglia bestehen und keine weiteren Zelltypen vorhanden sind (Details siehe Anhang 7.2).
2.5 Auswertung der immunocytochemischen Färbungen
Die Auswertung der immunocytochemisch gefärbten Astrozyten-Mikroglia Ko-Kulturen
erfolgt visuell am Fluoreszenzmikroskop (Axiophot Carl Zeiss GmbH, Jena). Für die
Bestimmung des prozentualen Mikroglia-Anteils wird das Verhältnis der Gesamtzahl der ED-
1 gefärbten Mikroglia mit der Gesamtzahl durch den Kernfarbstoff Hoechst 33342
angefärbten Kerne insgesamt (Astrozyten und Mikroglia) bestimmt (siehe Anhang 7.1). Es
wurden pro Deckgläschen drei unterschiedliche Sichtfelder ausgezählt (63x Vergrößerung).
Die Einteilung der Mikroglia Phenotypen erfolgt entsprechend Faustmann et al. 2003 in
inaktive ramifizierte Mikroglia (resting ramified type RRT), intermediäre Mikroglia (INT)
und aktive phagozytierende Mikroglia (rounded phagocytic type RPT) (Abb. 2.5). Pro
Deckgläschen wurden jeweils die Gesamtzahl aller Mikroglia von drei unterschiedlichen
Sichtfeldern ausgezählt und in den jeweiligen Funktionszustand eingeteilt (63x
Vergrößerung). Die aktivierten Mikroglia (RPT) sind durch ihre runden Zellkörper ohne
Zellfortsätze deutlich zu erkennen. Im Vergleich zum RPT ist der RRT durch seine langen
Zellfortsätze gekennzeichnet. Die intermediären Mikroglia sind durch kurze Zellfortsätze,
welche den Durchmesser des Soma nicht überschreitet, gekennzeichnet.
Für die M5/M30 Kontrolle als auch für die mit den verschiedenen Antiepileptika inkubierten
Ko-Kulturen wurde der Mittelwert ± SEM für die Anzahl der jeweiligen Phenotypen (RRT,
INT und RPT) aus allen ausgewerteten Sichtfeldern bestimmt. Anschließend wurde der
Mittelwert der einzelnen Mikroglia Phenotypen (RRT und RPT) für die Kontrolle gleich 1
gesetzt und mit dem Mittelwert der Phenotypen der mit Antiepileptika inkubierten Ko-
Kulturen normalisiert.

19
Abb. 2.5. Abbildung entnommen und übersetzt aus (Dambach et al. 2014): Immunocytochemische Färbung
der Mikroglia (rot) mit dem monoklonalen ED-1 Antikörper. Die Zellkerne wurden mit dem Kernfarbstoff DAPI
(blau) visualisiert, um die gesamte Anzahl an Gliazellen zu ermitteln. A. Astrozyten Ko-Kultur mit einem
Mikroglia-Anteil von etwa 30 %, welche in vitro das inflammatorisch betroffene Gewebe des Gehirns
repräsentiert. B. Astrozyten Ko-Kultur mit einem Mikroglia-Anteil von etwa 5 %, welche in vitro das
physiologische Gewebe des Gehirns repräsentiert. C. D. E. Klassifikation der verschiedenen Mikroglia
Phenotypen: Inaktive ramifizierte Mikroglia (resting ramified type, RRT) (C), intermediäre Mikroglia
(intermediate type, INT) (D) und aktive phagozytierende Mikroglia (activated, round phagocytic type, RPT) (E).
2.6 ELISA
Die TGF-beta1 Konzentration in den Zellkultur-Überstanden der Ko-Kulturen wurde mittels
Sandwich-ELISA (Promega, Madison, USA) quantifiziert. Entsprechend der technischen
Beschreibung des Herstellers wurde eine 96-Well Mikrotiterplatte mit dem primären TGF-
beta1 monoklonalen Antikörper (TGF-beta mAB) über Nacht bei 4 °C inkubiert. Am
Folgetag wurde die Mikrotiterplatte mit einem Block-Puffer für 35 Minuten bei 37 °C
blockiert und mit TBST-Puffer (Tris-buffered saline mit Tween 20) ausgewaschen.
Anschließend wurden die zu untersuchenden Proben und eine Standardreihe in die 96-Well
Mikrotiterplatte pipettiert und für 2 Stunden bei RT inkubiert. Nach der Inkubation wurde die

20
Mikrotiterplatte fünfmal mit TBST-Puffer gewaschen und für weitere zwei Stunden bei RT
mit dem sekundären polyklonalen Antikörper (anti-TGF-beta1 pAB) inkubiert. Im Anschluss
wurden je 100 µl TGF-beta1 HRP (horseradish peroxidase) Konjugat in die präparierten
Wells pipettiert, nachdem die Mikrotiterplatte fünfmal mit TBST-Puffer ausgewaschen
wurde. Die Mikrotiterplatte wurde für zwei weitere Stunden bei RT inkubiert. Im Anschluss
an den letzten Waschritt mit TBST-Puffer, wurde die Mikrotiterplatte mit jeweils 100 µl der
vom Hersteller mitgelieferten TMB One Solution pro Well inkubiert. Durch die von dem
Enzym HRP katalysierte Reaktion wird das farblose Substrat TMB (3,3’, 5,5’ –
tetramethylbenziden) in ein blau angefärbtes Produkt umgesetzt. Diese Reaktion wurde nach
15 Minuten durch die Zugabe von 100 µl 1 molarer Salzsäure pro Well gestoppt. Durch den
niedrigen ph-Wert kommt es dabei zu einem Farbumschlag von blau nach gelb. Die optische
Dichte wurde direkt nach Zugabe der Salzsäure mittels eines Mikroplatten-Reader (Bio-Rad
550, München, Deutschland) bei einer Wellenlänge von 450 nm gemessen.
Die TNF-alpha Konzentration in den Zellkultur-Überstanden der Ko-Kulturen wurde mittels
Sandwich-ELISA (R&D Systems, Wiesbaden, Deutschland), entsprechend der Beschreibung
für den Quantikine® Rat TNF-alpha Assay, bestimmt. Die Proben und Standardreihe wurden
auf die mit dem primären TNF-alpha monoklonalen Antikörper (TNF-alpha mAB)
beschichtete 96-Well Mikrotiterplatte pipettiert und für 2 Stunden bei RT inkubiert.
Anschließend wurde die Mikrotiterplatte fünfmal mit dem mitgelieferten Waschpuffer
ausgewaschen und für 2 Stunden bei RT, mit dem TNF-alpha HRP-konjugierten sekundären,
polyklonalen Antikörper, inkubiert. Nach der Inkubation wurde die Mikrotiterplatte fünfmal
gewaschen und mit 100 µl der mitgelieferten Substratlösung pro Well für 30 Minuten bei RT
inkubiert. Die darauf folgende enzymatische Reaktion wurde bereits im vorherigen Abschnitt
beschrieben. Durch die Zugabe von 100 µl Stop Solution wurde die Reaktion gestoppt und die
optische Dichte bei 450 nm mit einer Referenzwellenlänge bis 570 nm mit dem Mikroplatten-
Reader, gemessen.
Die TGF-beta1 und TNF-alpha Konzentrationen der jeweiligen Zellkultur-Überstände
wurden anhand einer Standardkurve aus zwei gleichen Verdünnungsreihen ermittelt. Jedes
Ergebnis entspricht dem Mittelwert ± Standardfehler des Mittelwerts (SEM) als Resultat von
mehreren unabhängig durchgeführten Experimenten.

21
2.7 Western Blot
2.7.1 Zellaufschluss
Für die quantitative Cx43 Proteinbestimmung wurden die Ko-Kulturen, welche in kleinen
Petrischalen (60 x 15 mm) (BD Biosciences, Heidelberg, Deutschland) ausgesäht (300.000
Zellen pro Petrischale) wurden, zuerst mechanisch lysiert. Hierfür wurden 200 µl 1 x
Laemmli Puffer und 4 µl Proteaseinhibitor (Sigma-Aldrich, Steinheim, Deutschland) zu den
Zellen hinzugegeben und mit einem Silikonspatel von den Petrischalen abgekratzt.
Anschließend wurden die Zellen mit einer Spritze mechanisch durch auf- und abziehen lysiert
und die Zellsuspension in 1,5 ml Eppendorf-Tubes überführt.
2.7.2 Konzentrationsbestimmung nach Bradford
Die Proteinbestimmung nach Bradford basiert auf der Verwendung des blauen
Säurefarbstoffes Coomasie-Brilliantblau. In Gegenwart von Proteinen im sauren Milieu
verschiebt sich das Absorptionsmaximum des Coomasie-Brilliantblaus G250 von 465 auf 595
nm. Der Grund für die Verschiebung des Absorptionsmaximums ist vermutlich die
Stabilisierung des Farbstoffes in seiner unprotonierten, anionischen Sulfonat-Form durch
Komplexbildung zwischen Farbstoff und Protein. Für die Erstellung der Eichkurve wird
Rinderserumalbumin (BSA bovine serum albumin) verwendet. Hierfür wurden Proben
unterschiedlicher Proteinkonzentrationen von BSA (1, 2, 4, 6, 8 und 10 µg/µl) mit
destilliertem H2O (799, 798, 796, 794, 793 und 790 µl) und 200 µl Bradford-Standardlösung
(BioRad, München, Deutschland) versetzt. Die Standards wurden innerhalb von 15 Minuten
bei 595 nm in Plastikküvetten (Sarstedt, Nümbrecht, Deutschland) gemessen. Die
Nullkontrolle für die Eichung bestand lediglich aus destilliertem H2O (800 µl) und Bradford-
Standardlösung (200 µl) ohne zugesetzte Proteinlösung. Die BSA-Eichkurve ergibt sich aus
der gemessenen Extinktion, die gegen die bekannte Proteinkonzentration aufgetragen wird.
Von der jeweiligen Zellsuspension wurden 5 µl der Probe mit 795 µl dest. Wasser und 200 µl
Bradford-Reagenz in den Plastikküvetten vermischt und nach 15 Minuten die Extinktion mit
dem Photometer Ultrospec 3000 (GE-Healthcare, München, Deutschland) gemessen.
Anschließend wurde die Konzentration durch Vergleich der ermittelten Extinktion der
unbekannten Proteinprobe mit der Eichkurve ermittelt.

22
2.7.3 SDS-Polyacrylamid-Gelelektrophorese
Die Auftrennung nach der Molekularmasse der Proteine aus den Zellsuspensionen wurde
durch die Gelelektrophorese gewährleistet. Hierbei liegt die Molekularmasse des zu
untersuchenden Gap-Junction Proteins bei 43 Dalton. Die Proben wurden für 5 Minuten auf
95 °C erhitzt, bevor sie auf das Gel aufgetragen wurden. Dadurch kommt es zur Aufspaltung
von Wasserstoff-Brücken und somit zur Auflösung der Sekundär- und Tertiärstruktur der
Proteine. Das Sammel- und Trenngel wird wie folgt zusammengesetzt, wobei der
Radikalstarter APS (Ammoniumperoxodisulfat) (AppliChem, Darmstadt, Deutschland) und
der Katalysator TEMED (Tetramethylethylenediamine) (AppliChem, Darmstadt,
Deutschland), welche zur Polymerisierung des Gels führen, als letztes hinzugegeben werden.
Jeweils 10 µl der Proben sowie 5 µl eines Kaleidoskop-Markers (BioRad, München,
Deutschland) wurden in die Gel-Taschen aufgetragen. Die Elektrophorese wurde bei 100 V
gestartet bis die Proben aus dem Sammelgel in das Trenngel übergegangen sind, anschließend
wurde auf 150 V erhöht.
Trenngel Sammelgel
dest. H2O 5,1 ml 2,9 ml
Tris, pH 8,8 3,2 ml -
Tris, pH 6,8 - 1,25 ml
30 % Acrylamid 4,2 ml 833 μl
10 % SDS 125 μl 50 μl
10 % APS 100 μl 37,5 μl
TEMED 10 μl 7,5 μl
Zusammensetzung für 2 Gele für die SDS-PAGE
2.7.4 Western Blot und immunologischer Nachweis der Proteine
Im Anschluss an die Gelelektrophorese wurden die auf dem Gel separierten Proteine auf ein
Nitrozellulosepapier transferiert. Hierfür wurden die Blotting-Papiere und die
Nitrocellulosemembran (Amersham Pharmacia, Little Chalfont, England) auf das Gel
zurechtgeschnitten und für 10 Minuten in Blotting-Puffer bei 4 °C eingelegt. Bei dem

23
Western-Blot musste die Nitrozellulosemembran zum Plus- und das Gel zum Minuspol
gerichtet werden. Anschließend wurde für eine Stunde bei 90 mA geblottet.
Die Nitrozellulosemembran wurde im Anschluss an den Blot für eine Stunde in 0,5 %
Blocklösung bei RT inkubiert. Im Anschluss wurde die Membran über Nacht bei 4 °C mit
dem primären Antikörper Rabbit anti-Connexin 43 (Invitrogen, Carlsbad, Deutschland) im
Verhältnis von 1:1000 in 0,2 % Blocklösung inkubiert. Am Folgetag wurde dreimal für 20
Minuten mit PBS-Tween gewaschen und für eine Stunde bei RT mit dem sekundären
Antikörper Peroxidase goat anti-rabbit (Jackson ImmunoResearch, Suffold, England) im
Verhältnis von 1:5000 in 0,2 % Blocklösung inkubiert. Anschließend wurde die Membran
dreimal mit PBS-Tween gewaschen. Im Fotolabor wurden 1 ml Peroxidase- und 1 ml
Enhancer-Solution (Thermo Scientific, Bonn, Deutschland) für 2 Minuten direkt auf die
Nitroczellulosemembran pipettiert. Durch Auflegen eines Hyperfilms (Amersham Pharmacia,
Little Chalfont, England) wurden die einzelnen Banden auf der Nitrozellulosemembran
sichtbar gemacht. Die Entwicklung des Hyperfilms erfolgte automatisch mit einer
Röntgenfilm-Entwicklungsmaschine Optimax (PROTEC GmbH, Oberstenfeld, Deutschland).
Der entwickelte Film wurde eingescannt und mit der Quantity-One-Software (Version 4.5.2)
hinsichtlich Dichte, Volumen und Fläche der detektierten Banden ausgewertet.
Im Anschluss an die Detektion der Cx43-Banden wurden die Antikörper von der
Nitrozellulosemembran wieder abgewaschen um eine erneute Markierung mit dem
Referenzprotein ß-Aktin durchzuführen. Hierfür wurde die Nitrozellulosemembran für 35
Minuten in Stripping-Puffer bei 50 °C gelegt und somit die Antikörpermarkierungen gelöst.
Die Membran wurde im Anschluss, wie bereits beschrieben, mit dem Erstantikörper anti-β-
Aktin (Sigma, St. Louis, USA) 1:7500 und mit dem Peroxidase-gekoppelten Zweitantikörper
rabbit anti-mouse IgG (Jackson Immuno Research, West Grove, USA) im Verhältnis von
1:5000 in 0,2 % Blocklösung inkubiert und detektiert.
2.8 Statistik
Die signifikanten Unterschiede in der Viabilität, den Zytokinlevel und den Mikroglia
Phenotypen (RRT und RPT) zwischen der Kontrolle und den mit Antiepileptika inkubierten
Ko-Kulturen (M5 und M30) wurde mittels one-way ANOVA und anschließenden Bonferroni
post-hoc comparison test (GraphPad Software, San Diego, USA) ermittelt. Die signifikanten
Unterschiede des Median der prozentualen Cx43 Expression zwischen der Kontrolle und den
mit Antiepileptika inkubierten Ko-Kulturen wurde mittels one-way ANOVA und

24
anschließenden Kruskal-Wallis test, ermittelt. Die Unterschiede wurden bei p*<0.05,
p**<0.01 und p***<0.001 als signifikant anerkannt.
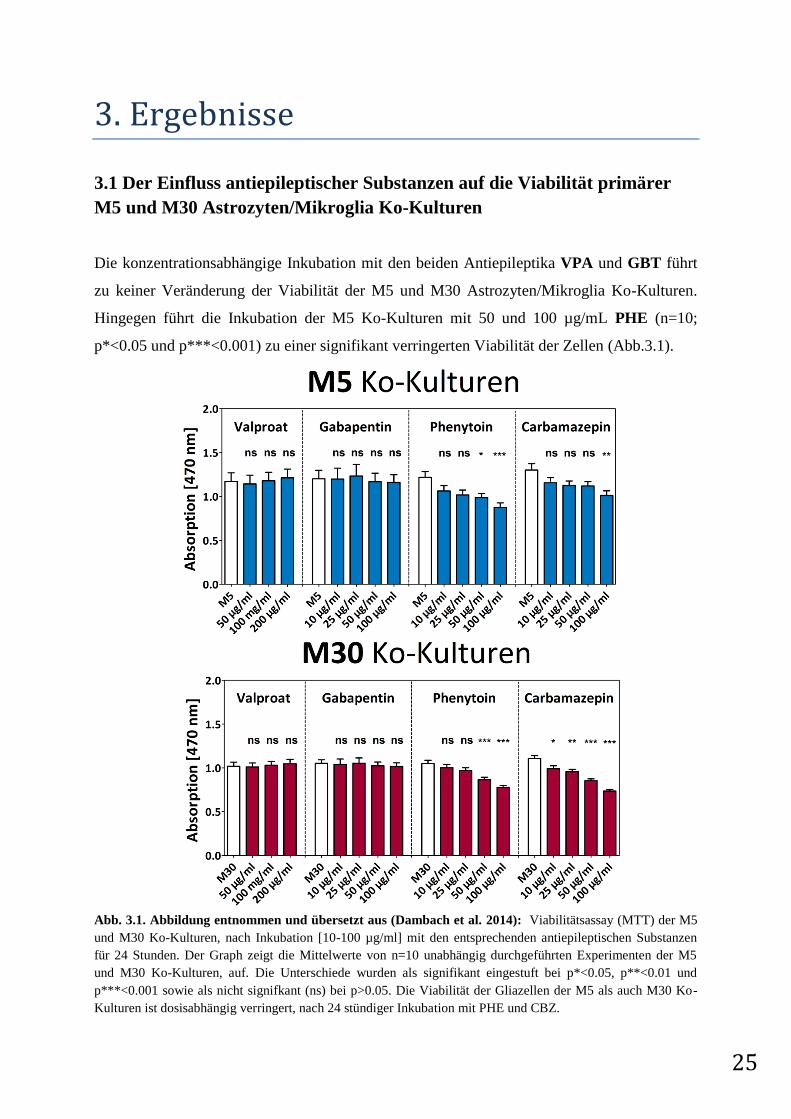
25
3. Ergebnisse
3.1 Der Einfluss antiepileptischer Substanzen auf die Viabilität primärer
M5 und M30 Astrozyten/Mikroglia Ko-Kulturen
Die konzentrationsabhängige Inkubation mit den beiden Antiepileptika VPA und GBT führt
zu keiner Veränderung der Viabilität der M5 und M30 Astrozyten/Mikroglia Ko-Kulturen.
Hingegen führt die Inkubation der M5 Ko-Kulturen mit 50 und 100 µg/mL PHE (n=10;
p*<0.05 und p***<0.001) zu einer signifikant verringerten Viabilität der Zellen (Abb.3.1).
Abb. 3.1. Abbildung entnommen und übersetzt aus (Dambach et al. 2014): Viabilitätsassay (MTT) der M5
und M30 Ko-Kulturen, nach Inkubation [10-100 µg/ml] mit den entsprechenden antiepileptischen Substanzen
für 24 Stunden. Der Graph zeigt die Mittelwerte von n=10 unabhängig durchgeführten Experimenten der M5
und M30 Ko-Kulturen, auf. Die Unterschiede wurden als signifikant eingestuft bei p*<0.05, p**<0.01 und
p***<0.001 sowie als nicht signifkant (ns) bei p>0.05. Die Viabilität der Gliazellen der M5 als auch M30 Ko-
Kulturen ist dosisabhängig verringert, nach 24 stündiger Inkubation mit PHE und CBZ.

26
Eine signifikant verringerte Viabilität kann ebenfalls in den M30 Ko-Kulturen nachgewiesen
werden, nach 24 stündiger Inkubation mit 50 und 100 µg/mL PHE (n=12; p***<0.001 und
p***<0.001). Eine signifikante Verringerung der Metabolisierung des gelben
Tetrazoliumsalzes MTT kann nach Inkubation mit 100 µg/mL (n=10; p**<0.01) CBZ in den
M5 Ko-Kulturen gemessen werden. In den M30 Ko-Kulturen kann eine signifikant
verringerte Viabilität der Zellen nach Inkubation von 10, 25, 50 and 100 µg/mL CBZ (n=12;
p*<0.05, p**<0.001, p***<0.0001 und p***<0.0001) detektiert werden.
3.2 Der Einfluss antiepileptischer Substanzen auf die mikrogliale
Aktivierung/ Inaktivierung in primären M5 und M30 Astrozyten/Mikroglia
Ko-Kulturen
Die Inkubation mit VPA für 24 Stunden führt zu einer signifikanten Aktivierung der
Mikroglia in den physiologischen M5 (Abb. 3.2.a) als auch inflammatorisch modifizierten
M30 (Abb. 3.2.b) Ko-Kulturen. Die Inkubation mit 50 µg/mL VPA führt zu einer
signifikanten Erhöhung der Anzahl des aktivierten Mikroglia Phenotyps (RPT) von 25.00%
(n=66)# auf 62.50% (n=33; p***<0.0001). Entsprechend verringert sich der prozentuale
Anteil des inaktivierten Mikroglia Phenotyps (RRT) signifikant von 42.86% (n=66)#
auf
10.00% (n=33; p***<0.0001). Die signifikante mikrogliale Aktivierung kann ebenfalls nach
Inkubation mit höheren Konzentrationen von VPA (100 und 200 µg/mL) gemessen werden.
Die Inkubation der M30 Ko-Kulturen mit 200 µg/mL VPA führt zu einem signifikant höheren
Anteil an aktivierten Mikroglia (RPT) von 66.67% (n=45)# auf 83.33% (n=24, p**<0.01).
Hingegen ist der prozentuale Anteil an inaktivierten Mikroglia (RRT) bereits ab einer
Konzentration von 50 µg/mL VPA signifikant verringert, im Vergleich zur Kontrolle von
8.62% (n=45)# auf 3.79% (n=24, p*<0.05).
Die konzentrationsabhängige Inkubation der M5 und M30 Ko-Kulturen mit GBT für 24
Stunden bewirkt keine signifikanten Veränderungen der Mikroglia Phenotypen, bei direktem
Vergleich mit der Kontrolle.
Die Inkubation mit PHE bewirkt eine signifikante Mikroglia Aktivierung in den M5, ohne
signifikante Veränderungen in den M30 Ko-Kulturen. Ab einer Konzentration von 50 µg/mL
PHE erhöht sich der prozentuale Anteil des aktivierten Mikroglia Phenotyps (RPT)
signifikant von 24.26% (n=62)# auf 50.00% (n=28, p**<0.01). Dementsprechend verringert
sich der prozentuale Anteil des inaktivierten Mikroglia Phenotyps (RRT) in den M5 Ko-
Kulturen von 41.42% (n=62)# auf 25.00% (n=28, p*<0.05).
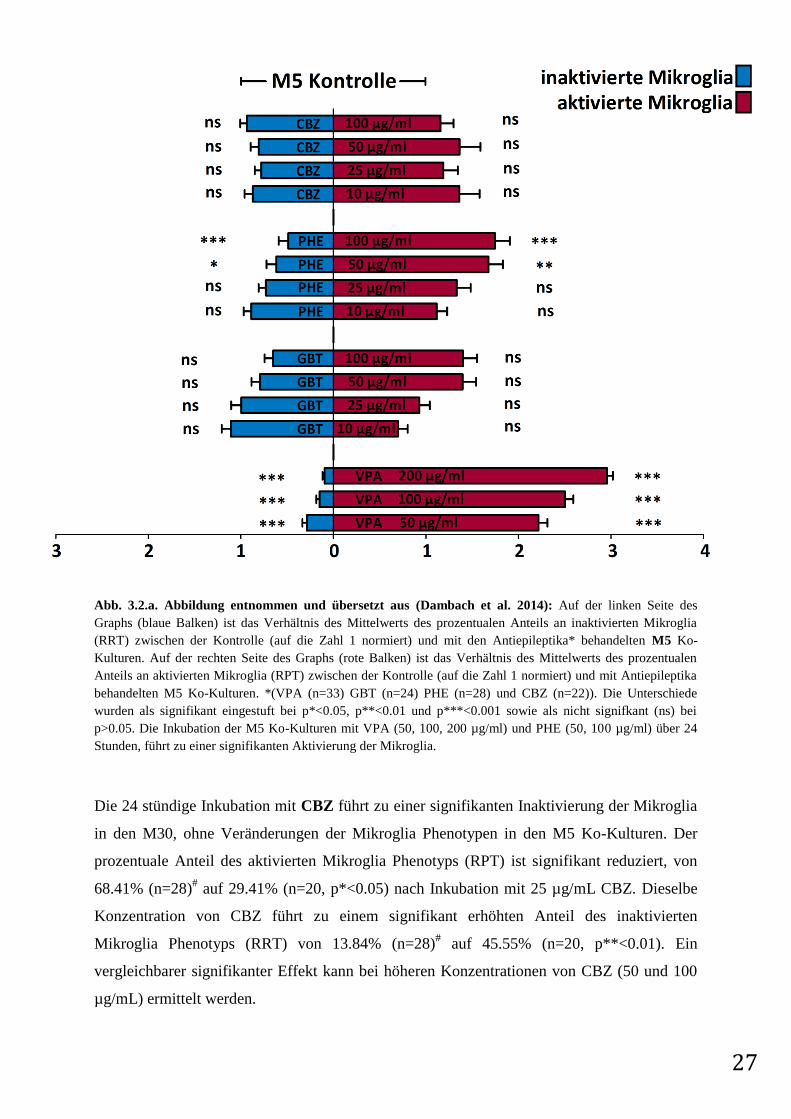
27
Abb. 3.2.a. Abbildung entnommen und übersetzt aus (Dambach et al. 2014): Auf der linken Seite des
Graphs (blaue Balken) ist das Verhältnis des Mittelwerts des prozentualen Anteils an inaktivierten Mikroglia
(RRT) zwischen der Kontrolle (auf die Zahl 1 normiert) und mit den Antiepileptika* behandelten M5 Ko-
Kulturen. Auf der rechten Seite des Graphs (rote Balken) ist das Verhältnis des Mittelwerts des prozentualen
Anteils an aktivierten Mikroglia (RPT) zwischen der Kontrolle (auf die Zahl 1 normiert) und mit Antiepileptika
behandelten M5 Ko-Kulturen. *(VPA (n=33) GBT (n=24) PHE (n=28) und CBZ (n=22)). Die Unterschiede
wurden als signifikant eingestuft bei p*<0.05, p**<0.01 und p***<0.001 sowie als nicht signifkant (ns) bei
p>0.05. Die Inkubation der M5 Ko-Kulturen mit VPA (50, 100, 200 µg/ml) und PHE (50, 100 µg/ml) über 24
Stunden, führt zu einer signifikanten Aktivierung der Mikroglia.
Die 24 stündige Inkubation mit CBZ führt zu einer signifikanten Inaktivierung der Mikroglia
in den M30, ohne Veränderungen der Mikroglia Phenotypen in den M5 Ko-Kulturen. Der
prozentuale Anteil des aktivierten Mikroglia Phenotyps (RPT) ist signifikant reduziert, von
68.41% (n=28)# auf 29.41% (n=20, p*<0.05) nach Inkubation mit 25 µg/mL CBZ. Dieselbe
Konzentration von CBZ führt zu einem signifikant erhöhten Anteil des inaktivierten
Mikroglia Phenotyps (RRT) von 13.84% (n=28)# auf 45.55% (n=20, p**<0.01). Ein
vergleichbarer signifikanter Effekt kann bei höheren Konzentrationen von CBZ (50 und 100
µg/mL) ermittelt werden.

28
Abb. 3.2.b. Abbildung entnommen und übersetzt aus (Dambach et al. 2014): Auf der linken Seite des
Graphs (blaue Balken) ist das Verhältnis des Mittelwerts des prozentualen Anteils an inaktivierten Mikroglia
(RRT) zwischen der Kontrolle (auf die Zahl 1 normiert) und mit den Antiepileptika* behandelten M30 Ko-
Kulturen. Auf der rechten Seite des Graphs (rote Balken) ist das Verhältnis des Mittelwerts des prozentualen
Anteils an aktivierten Mikroglia (RPT) zwischen der Kontrolle (auf die Zahl 1 normiert) und mit Antiepileptika
behandelten M30 Ko-Kulturen. *(VPA (n=24) GBT (n=30) PHE (n=40) und CBZ (n=20)). Die Unterschiede
wurden als signifikant eingestuft bei p*<0.05, p**<0.01 und p***<0.001 sowie als nicht signifkant (ns) bei
p>0.05. Die Inkubation der M30 Ko-Kulturen mit VPA (200 µg/ml) über 24 Stunden, führt zu einer
signifikanten Aktivierung der Mikroglia. Hingegen führt die Inkubation der M30 Ko-Kulturen mit CBZ
(25,50,100 µg/ml) zu einer signifikant verringerten Anzahl aktivierter und erhöhten Anzahl inaktivierter
Mikroglia.

29
3.3 Der Einfluss antiepileptischer Substanzen auf die astrogliale Cx43
Expression in primären M5 und M30 Astrozyten/Mikroglia Ko-Kulturen
Die astrogliale Cx43 Expression in den M5 und M30 Ko-Kulturen wurde mittels quantitativer
Western Blot Analyse untersucht. Die konzentrationsabhängige Inkubation mit allen
untersuchten Antiepileptika VPA, GBT, PHE und CBZ bewirkt keine signifikant veränderte
astrogliale Cx43 Expression sowohl bei den M5 als auch M30 Ko-Kulturen (Abb. 3.3).
Abb. 3.3. Abbildung entnommen und übersetzt aus (Dambach et al. 2014): Die Western Blots zeigen keine
veränderte Cx43 Expression in den Ko-Kulturen, nach Inkubation mit VPA, GBT, PHE und CBZ (n=3) auf.

30
3.4 Der Einfluss antiepileptischer Substanzen auf die Expression der beiden
Zytokine TGF-ß1 und TNF-α in primären M5 und M30
Astrozyten/Mikroglia Ko-Kulturen
Nach 24 stündiger Inkubation mit verschiedenen Konzentrationen der beiden Antiepileptika
VPA und CBZ wurde in den Überständen der M5 als auch M30 Ko-Kulturen keine
veränderten TGF-ß1 und TNF-α Level detektiert (Abb. 3.4).
Die Inkubation mit 100 µg/mL GBT hingegen führt zu einem signifikant erhöhten TGF-ß1
Zytokinlevel in den Überständen der M5 (n=5; p**<0.01) als auch M30 (n=4; p**<0.01) Ko-
Kulturen.
Abb. 3.4. Abbildung entnommen und übersetzt aus (Dambach et al. 2014): Die mittels Sandwich-ELISA
quantitativ gemessene TGF-ß1 (n=4)* und TNF-α (n=3)* Zytokinkonzentration der M5 und M30 Ko-Kultur
Überstande, nach 24 stündiger Inkubation mit den antiepileptischen Substanzen. (n = Anzahl unabhängig
voneinander durchgeführter ELISA). Die Unterschiede wurden als signifikant eingestuft bei p*<0.05, p**<0.01
und p***<0.001 sowie als nicht signifkant (ns) bei p>0.05. Bei undetektierbaren (undetectable, ud) TNF-α Level
in den M30 Ko-Kulturen, einschließlich der Kontrollwerte, sind keine Balken aufgezeigt. Die Inkubation mit
hohen Konzentrationen von GBT als auch PHE führt zu einem signifikant erhöhten TGF-ß1 Level in den M5 als
auch M30 Ko-Kultur Überständen. Schlussendlich führt die 24 stündige Inkubation mit 25 und 100 µg/ml PHE
zu einer signifikant erhöhten TNF-α Zytokinkonzentration in den Überständen der M30 Ko-Kulturen.

31
Des Weiteren führt die Inkubation mit 100 µg/mL PHE zu einem signifikant erhöhten TGF-
ß1 Zytokinlevel in der Überständen der M5 Ko-Kulturen (n=4; p*<0.05). In den M30 Ko-
Kultur Überständen ist die TGF-ß1 Konzentration signifikant erhöht nach Inkubation mit 10,
25, 50 und 100 µg/mL PHE (n=4; p*<0.05, p*<0.05, p**<0.01 und p***<0.0001). Darüber
hinaus führt die Inkubation mit 25 und 100 µg/mL PHE zu einer signifikant erhöhten TNF-α
Zytokinkonzentration (n=3; p**<0.01 und p***<0.0001) in den Überständen der M30 Ko-
Kulturen.

32
3. Diskussion
4.1 Der Einfluss antiepileptischer Substanzen auf die Viabilität
primärer M5 und M30 Astrozyten/Mikroglia Ko-Kulturen
Die Viabilität von primären Astrozyten nach Inkubation mit verschiedenen Konzentrationen
der beiden Antiepileptika GBT und CBZ wurde bereits 2003 von Pavone und Cardile in einer
in vitro Studie untersucht. Sie zeigten eine verringerte Viabilität der Astrozyten nach
Inkubation mit 50 und 100 µg/mL GBT sowie 1, 10, 50 und 100 µg/mL CBZ auf.
Entsprechend der Studie von Pavone und Cardile 2003 führte die Inkubation der M5 Ko-
Kulturen mit 100 µg/mL CBZ ebenfalls zu einer signifikant verringerten Viabilität der
Gliazellen. Im Gegensatz zu der Studie führte die Inkubation mit allen untersuchten
Konzentrationen von GBT in dieser Arbeit bei den M5 Ko-Kulturen zu keiner signifikant
reduzierten Viabilität der Gliazellen. Demnach bewirkt die Ko-Existenz der Mikroglia in den
M5 Ko-Kulturen einen protektiven Effekt auf die Astrozyten, wodurch der zuvor berichtete
Viabilitätverlust nach Inkubation mit GBT verhindert wird. Der genaue Mechanismus dieses
protektiven Effekts der Mikroglia ist allerdings noch zu untersuchen. Letztendlich konnte in
dieser Studie eine konzentrationsabhängige verringerte Viabilität der M5 Ko-Kulturen durch
die Inkubation mit den Antiepileptika CBZ und PHE gemessen werden. Astrozyten sind
verantwortlich für die metabolische und trophische Versorgung der Nervenzellen. Eine
beeinträchtigte Funktion oder Viabilität, ausgelöst durch Antiepileptika, könnte im
ungünstigsten Falle ebenfalls die Versorgung der Nervenzellen bei der Behandlung
beeinträchtigen. Diese Hypothese sollte allerdings durch weitere in vivo Experimente
untersucht werden.
In früheren Untersuchungen mit den inflammatorisch modifizierten M30 Ko-Kulturen im
Vergleich zu den M5 Ko-Kulturen wurde bereits eine Veränderung des
Membranruhepotentials (MRP) als auch der funktionellen Kopplung der Astrozyten ermittelt
(Hinkerohe et al. 2005, Haghikia et al. 2008, Stienen et al. 2011). In dieser Studie bewirkte
die 24 stündige Inkubation mit den beiden Antiepileptika CBZ und PHE eine
konzentrationsabhängig verringerte Viabilität der M30 Ko-Kulturen. Astrozyten reagieren auf
inflammatorische Stimuli (Mediatoren und Zytokine) und verändern verschiedene
Oberflächenrezeptoren, Enzyme, Kanäle sowie intrazelluläre und interzelluläre
Signalmoleküle. Diese astroglialen Veränderungen können verschiedene zelluläre Funktionen
beeinflussen einschließlich die Regulation der zerebralen Homöostase, der Synapsen und

33
neuronalen Funktion sowie der immunologischen und inflammatorischen Aktivität
(Sofroniew 2013). Dieser Prozess ist sehr komplex und hängt von den Stimuli sowie der
zugrunde liegenden pathologischen Veränderung ab. Inwiefern die Verringerung der
Viabilität durch CBZ und PHE unter inflammatorisch modifizierten Bedingungen
Auswirkungen auf die Astrozyten und deren zelluläre Funktion haben, kann in dieser Studie
nicht im Detail geklärt werden. Die signifikant erhöhten TNF-α und TGF-ß1 Zytokinlevel
nach 24 stündiger Inkubation mit PHE deuten jedoch auf einen Einfluss auf die anti-/
inflammatorische Aktivität der Gliazellen hin.
4.2. Der Einfluss antiepileptischer Substanzen auf die mikrogliale
Aktivierung/ Inaktivierung in primären M5 und M30
Astrozyten/Mikroglia Ko-Kulturen
In dieser Studie führte eine 24 stündige Inkubation mit dem Antiepileptikum VPA zu einer
signifikanten dosisabhängigen Aktivierung der Mikroglia Zellen in den physiologischen M5
als auch inflammatorisch modifizierten M30 Ko-Kulturen. Die Inkubation führte jedoch zu
keiner veränderten Viabilität der Zellen. Hingegen zeigen verschiedene Studien eine
verringerte Viabilität sowie eine Caspase-3 vermittelte Apoptose mikroglialer Zellen auf
(Dragunow et al. 2006, Smith et al 2010, Xie et al. 2010 und Gibbons et al. 2011). Die
Koexistenz der Astrozyten, welche ein funktionelles Synzitium ausbilden, verhindert
vermutlich die apoptotisch beschriebene Wirksamkeit von VPA auf die Mikroglia und führt
lediglich zu einer signifikanten Aktivierung der Zellen. Dennoch ist die Aktivierung von
Mikroglia-Zellen ein typisches Merkmal inflammatorisch anhaltender Bedingungen. Im
aktivierten Zustand besitzen Mikroglia die Fähigkeit verschiedene Mediatoren zu
exprimieren, welche nützlich oder schädlich für umliegende Zellen sein können (Kettenmann
et al. 2011). Demnach kann die Aktivierung der Mikroglia-Zellen durch VPA zu einer
Expression diverser Mediatoren führen. Inwiefern die Expression die inflammatorische
Bedingung bei inflammatorisch bedingten epileptischen Anfällen konditioniert, müsste
spezifischer durch eine Bandbreite verschiedener Indikatoren untersucht werden. In dieser
Studie konnte zumindest keine signifikant erhöhte Konzentration des proinflammatorischen
Zytokins TNF-α nach Inkubation mit VPA gemessen werden.
Die konzentrationsabhängige Inkubation mit GBT führte zu keiner Veränderungen der
Mikroglia Phenotypen in den Ko-Kulturen. Bisher gibt es keine repräsentative Studien über
den genauen Wirkungsmechanismus von GBT und der direkte Einfluss auf Mikroglia Zellen

34
ist unerforscht. In dieser Studie konnten wir aufzeigen, dass GBT keinen Einfluss auf die
gliale Viabilität und mikrogliale Aktivierung, ausübt.
Bisher ist über den direkten Einfluss von PHE auf mikrogliale Zellen eher wenig bekannt.
Mikroglia-Zellen exprimieren neben zahlreichen Membranrezeptoren Ionenkanäle wie die
spannungsaktivierten Natriumkanäle. In der Studie von Black et al. (2009) wurde untersucht,
inwiefern aktivierte Mikroglia nach Inkubation mit Lipopolysaccharid (LPS), durch die
Aktivität von Natriumkanälen beeinflusst wird. Die Studie zeigte eine signifikant verringerte
phagozytierende Aktivität der Mikroglia nach der Natriumkanal Blockade durch PHE auf. Bei
den M5 Ko-Kulturen bewirkt die dosisabhängige Inkubation mit PHE eine signifikant erhöhte
Anzahl aktiverter phagozytierender Mikroglia (RPT). Allerdings wird in dieser Studie nicht
überprüft, inwiefern die erhöhte Anzahl aktivierter Mikroglia mit einer erhöhten
phagozytierenden Fähigkeit der Mikroglia einhergehen. Im Gegensatz zu den M5 Ko-
Kulturen bewirkte die Inkubation der M30 Ko-Kulturen keine signifikant erhöhte Anzahl
aktiverter Mikroglia.
Eine anti-inflammatorische Wirksamkeit von CBZ in Ratten wurde unter experimentellen
Bedingungen bereits beschrieben (Bianchi et al. 1995, Chogtu et al. 2011). Ausserdem wird
von einer Reduktion einer inflammatorisch bedingten Hyperalgesie durch dosisabhängige
Applikation von CBZ berichtet (Iwamoto et al. 2011). In dieser Studie bewirkte die
dosisabhängige Inkubation eine signifikante Inaktivierung der Mikroglia in den
inflammatorisch modifizierten M30 Ko-Kulturen. Diese Inaktivierung konnte bei den M5 Ko-
Kulturen nicht ermittelt werden. Die erhöhte Anzahl an inaktivierten Mikroglia (RRT) unter
inflammatorischen Bedingungen (M30) nach Applikation von CBZ könnte eine weitere
antiinflammatorische therapeutische Wirksamkeit von CBZ darstellen. Um diese Vermutung
zu stärken sind jedoch weitere in vivo Untersuchungen notwendig.
4.3 Der Einfluss antiepileptischer Substanzen auf die astrogliale Cx43
Expression in primären M5 und M30 Astrozyten/Mikroglia Ko-
Kulturen
Astrozyten bilden ein via Gap-Junction (Cx 43) gekoppeltes funktionelles Synzytium,
welches für den Erhalt der Homöostase des extrazellulären Milieus von Nervenzellen
essentiell ist (Chew et al. 2010, Pannasch et al 2011). Diese strukturelle Spezifikation der
Astrozyten ermöglicht eine Interaktion und Versorgung der Nervenzellen sowie einen
Abtransport von Substanzen, welche die physiologische neuronale Aktivität einschränken.

35
Dementsprechend können Veränderungen der astrozytären Cx43 Expression mit
Einschränkungen der interzellulären metabolischen und trophischen Versorgung von
Nervenzellen einhergehen. In dieser Studie konnten wir keine signifikanten Veränderungen
der astrozytären Cx43 Expression in den physiologischen M5 Ko-Kulturen nach Inkubation
mit VPA, GBT, PHE sowie CBZ, feststellen.
In neuroinflammatorisch betroffenem Gewebe konnte bereits eine verringerte interzelluläre
gap junctionale Kommunikation von Astrozyten nachgewiesen werden (Karpuk et al. 2011).
In vorangegangenen Studien konnte ausserdem aufgezeigt werden, dass inflammatorisch
modifizierte Konditionen zu einer verringerten Cx43 Expression kultivierter Astrozyten
führen (Faustmann et. al. 2003; Hinkerohe et al. 2005). Aufbauend auf dem M30 Ko-Kultur
Modell wurde das antiinflammatorische Potenzial des Antiepileptikums Levetiracetam (LEV)
untersucht (Haghikia et al. 2008). Es konnte gezeigt werden, dass sich die inflammatorisch
bedingt veränderte astrozytäre Cx43-Expression, die interzelluläre Kopplungsfähigkeit sowie
das Membranruhepotential bei der M30 Ko-Kultur nach LEV Applikation normalisiert. Mit
einer therapeutischen Applikation von 50 µg/ml über 24 Stunden konnte bei den M30 Ko-
Kultur eine erhöhte Cx43-Expression gemessen werden. In dieser Studie bewirkte die
Inkubation mit allen getesteten Antiepileptika keinerlei veränderte Cx43 Expression in den
inflammatorisch modifizierten M30 Ko-Kulturen.
4.4 Der Einfluss antiepileptischer Substanzen auf die Expression der
beiden Zytokine TGF-ß1 und TNF-α in primären M5 und M30
Astrozyten/Mikroglia Ko-Kulturen
Dass Antiepileptika Zytokinlevel beeinflussen können, wurde bereits in in-vitro als auch in-
vivo Experimenten aufgezeigt (Andrzejczak 2011). Erhöhte Level proinflammatorischer
Zytokine, wie beispielsweise TNF-α, stehen unter anderem im Zusammenhang mit einer
Astrogliose und somit beeinträchtigten Bewältigung des extrazellulären Glutamatspiegel
(Choi and Koh. 2008). In dieser Studie konnte aufgezeigt werden, dass die Inkubation mit
allen getesten Antiepileptika zu keinem veränderten TNF-α Zytokinlevel in den M5 Ko-
Kulturen geführt hat.
Das von Gliazellen exprimierte Zytokin TGF-ß1 wird in der Regel im Gehirn als
antiinflammatorisches Zytokin betrachtet. In diesem Zusammenhang wird der glialen TGF-ß1
Expression eine neuroprotektive Rolle zugeschrieben (Doyle et al. 2010 and Caraci et al.
2011). In dieser Studie führte die konzentrationsabhängige Inkubation mit GBT und PHE zu

36
einer erhöhten TGF-ß1 Expression bei den M5 Ko-Kulturen. In Bezug auf die neuroprotektive
Charakteristik von TGF-ß1 ist die erhöhte Expression nach GBT und PHE Applikation, neben
der antikonvulsiven Wirkung, ein positiver Nebeneffekt. Allerdings muss berücksichtigt
werden, dass ständig eine Interaktion zwischen pro- und antiinflammatorischen Zytokinen
besteht. Die pro- und antiinflammatorische Wirksamkeit hängt von verschiedenen Faktoren
wie der Zytokinkonzentration, der lokalen Verteilung der Zytokine sowie dem Einfluss von
verschiedenen inhibitorischen Faktoren ab.
Bei den M30 Ko-Kulturen konnte nach konzentrationsabhängiger Inkubation mit PHE ein
signifikant erhöhtes TNF-α Zytokinlevel gemessen werden. Im Gegensatz konnte bei den M5
Ko-Kulturen keine signifikant erhöhte TNF-α Konzentration gemessen werden. Dieser
Zusammenhang deutet darauf hin, dass die TNF-α Expression hauptsächlich von den
Mikroglia hervorgeht. Das erhöhte TNF-α Zytokinlevel, nach Inkubation der M30 Ko-
Kulturen mit PHE, könnte möglicherweise die inflammatorischen zerebralen Bedingungen
intensivieren. Allerdings scheint dies durch die gleichzeitig erhöhte TGF-ß1 Expression
abgemildert zu werden.
Die Inkubation der inflammatorisch modifizierten M30 Ko-Kulturen mit der höchsten
Konzentration von GBT führte ebenfalls zu einem erhöhten TGF-ß1 Level. Bisher sind keine
vergleichbaren Studien über die gliale Zytokinexpression nach Applilkation von GBT
bekannt. Inwiefern die Expression des antiinflammatorischen und neuroprotektiven Zytokins
einem dauerhaft inflammatorischen Zustandes entgegenwirken könnte, müsste in weiteren
Untersuchungen geklärt werden.
4.5 Klinische Interpretation antiepileptischer Substanzen mit
zusätzlichem antiinflammatorischen Potential bei Patienten mit
inflammatorisch bedingten Anfällen
Epileptische Anfälle als Symptom von Erkrankungen wie einer bakteriellen oder viralen
Meningitis, einer Enzephalopathie oder eines Gehirntumors können ein Merkmal anhaltender
inflammatorischer Bedingungen darstellen. Die glialen Zellkulturen in dieser Arbeit wurden
aus postnatalen Rattenhirnen ohne Neigung zur Epilepsie gewonnen. Die Ergebnisse können
dadurch nicht direkt auf Epilepsie-kranke gliale Zellen übertragen werden, sondern auf den
inflammatorischen Zustand und deren beteiligten Zellen im Gehirn, welcher je nach
Krankheitsbild epileptische Anfälle induzieren kann.

37
Aktivierte Mikroglia-Zellen exprimieren eine Bandbreite molekularer Stoffe, welche den
inflammatorischen Zustand im Gehirn aufrechterhalten. Aus diesem Grund könnte die
signifikante Mikroglia Aktivierung durch VPA und PHE in den Ko-Kulturen, den
andauernden inflammatorischen Zustand bei Patienten unterstützen.
Im Gegensatz führt die Inkubation der inflammatorisch modifizierten M30 Ko-Kulturen mit
CBZ zu einer signifikanten mikroglialen Inaktivierung. Die mikrogliale Inaktivierung bei
anhaltender inflammatorischer Bedigungen könnte eine vorteilhafte Nebenwirkung für
Patienten darstellen, welche mit CBZ behandelt werden. Jedoch müsste im weiteren Verlauf
untersucht werden, inwieweit die mikrogliale Inaktivierung zu einer Verbesserung des
zerebralen inflammatorischen Zustandes bei Patienten beiträgt.
Astrozyten sorgen in einem funktionellen Synzitium für die metabolische und trophische
Versorgung sowie Beseitigung von Produkten der Nervenzellen. Kommt es zu einer Störung
der physiologischen Funktion von Astrozyten sind die in der Zirkumferenz liegenden
Nervenzellen ebenfalls betroffen. Eine verringerte Viabilität von Astrozyten, wie nach der
Applikation hoher Konzentrationen von CBZ und PHE in den Ko-Kulturen, könnte in vivo zu
einer Beeinträchtigung der neuronalen Homöostase führen.
Letztendlich beeinflusst das Antiepileptikum GBT keine der untersuchten Parameter in dieser
Studie. Im Anschluss an diese Arbeit können weitere Untersuchungen an Tiermodellen in-
vivo vorgenommen werden, um die pro-/anti-inflammatorische Wirkungsweise verschiedener
Antiepileptika zu manifestieren.
Bei der Behandlung von epileptischen Anfällen, welche im Zusammenhang mit
entzündlichen Erkrankungen stehen, sollte aufgrund aktueller Erkenntnisse beachtet werden,
dass die Auswahl des Antiepileptikum, Gliazellen und somit die inflammatorischen Prozesse
im Gehirn und ZNS beeinflussen können. Die Zellkulturexperimente in dieser Arbeit zeigen,
dass gebräuchliche Antiepileptika in sehr unterschiedlicher Weise, sowohl
entzündungsverstärkend als auch entzündungshemmend auf Gliazellen wirken.
Aus neurologischer und epileptologischer Sicht wird angeregt, in klinischen Studien die
Auswirkungen gebräuchlicher Antiepileptika nicht nur auf die Schwere und Häufigkeit
epileptischer Anfälle, sondern auch auf entzündliche Reaktionen des ZNS differenziert zu
untersuchen. Die Indikation der jeweiligen Medikation sollte der entzündlichen
Grunderkrankung, die zu einer Epilepsie führt, angepasst werden. Danach würde sich die
Auswahl des Antiepileptikums, z.B. bei der Behandlung einer Epilepsie bei viraler oder
bakterieller Hirnentzündung oder Multipler Sklerose, nicht nur an der Art und Schwere der

38
epileptischen Anfälle, sondern auch an der vorherrschenden entzündlichen Grunderkrankung
orientieren.

39
5. Zusammenfassung
Hintergrund: Die Beteiligung von Gliazellen, wie Astrozyten und Mikroglia, in der
Pathophysiologie der Epilepsie, wird zunehmend anerkannt. Gliazellen spielen eine wichtige Rolle bei
der Initiation und Aufrechterhaltung der zerebralen Immunantwort sowie der metabolischen und
trophischen Versorgung der Nervenzellen. Diverse klinische als auch experimentelle Studien berichten
von einem Zusammenhang zwischen epileptischer Aktivität und einer Inflammation des zentralen
Nervensystems (ZNS). Andauernde inflammatorische Bedingungen im Gehirn sind gekennzeichnet
durch eine Ansammlung, Aktivierung sowie Proliferation von Mikroglia und Astrozyten.
In dieser Arbeit soll der Einfluss alltäglich verwendeter antiepileptischer Substanzen auf die gliale
Viabilität, Expression des Gap Junction Proteins Connexin 43, In-/ Aktivierung von Mikroglia sowie
Zytokin Expression an einem in-vitro Astrozyten/Mikroglia Ko-Kultur Modell, untersucht werden.
Methoden: Eine Astrozyten Kultur (postnatal P0-P2 aus Rattenhirn) wurde mit einem
physiologischen (5%) und einem pathologischen (30%) Anteil Mikroglia, kultiviert. Die Ko-Kulturen
wurden für 24 Stunden mit den Antiepileptika Valproat (VPA), Carbamazepin (CBZ), Phenytoin
(PHE) und Gabapentin (GBT), inkubiert. Die Viabilität und Proliferation wurde mittels Tetrazolium
(MTT) Assay, untersucht. Die Mikroglia-Phänotypen wurden mittels immunocytochemischer
Färbungen, bestimmt. Die astrogliale Connexin 43 (Cx43) Expression wurde quantitativ mittels
Western-Blot und die TGF-beta1 und TNF- α Zytokin Expression mittels Sandwich-ELISA,
ausgewertet.
Ergebnisse: Die dosisabhängige Inkubation der M5 Ko-Kulturen mit PHE und CBZ führt zu einer
signifikant verringerten Viabilität der Zellen. Die Inkubation mit VPA führt zu einer signifikanten
Aktivierung der Mikroglia und die anti-inflammatorische TGF-β1 Zytokinkonzentration war
signifikant erhöht nach der Inkubation mit höheren Konzentrationen von GBT und PHE, in den M5
Ko-Kulturen. Bei den M30 Ko-Kulturen führt die dosisabhängige Inkubation mit PHE zu einer
signifikant verringerten Viabilität der Zellen sowie einer erhöhten TGF-β1 und TNF-α Expression. Die
Inkubation mit CBZ führt zu einer signifikant verringerten Anzahl aktivierter Mikroglia und
gleichzeitig erhöhten Anzahl inaktivierter Mikroglia, sowie einer verringerten Viabilität der Zellen, bei
den M30 Ko-Kulturen.
Schlussfolgerung: Inflammatorische Zustände des ZNS sind charakterisiert durch eine
Beeinträchtigung glialer Zellfunktionen. Die Aktivierung von Mikroglia ist ein typisches Merkmal
inflammatorischer Zustände und wurde ebenfalls durch die Inkubation mit VPA, in den M5 als auch
M30 Ko-Kulturen, induziert. Im aktiven Zustand können Mikroglia in unterschiedlichen Status (M1
oder M2) vorliegen und je nach Status eine unterschiedliche Wirkung (pro- oder antiinflammatorisch)
auf ihre Zirkumferenz ausüben. Eine proinflammatorische Wirkung könnte im Falle einer
inflammatorisch bedingten Epilepsie möglicherweise die inflammatorisch bedingte Grunderkrankung
bei Patienten verschlimmern. Hingegen führt die Inkubation mit CBZ zu einer signifikanten
mikroglialen Inaktivierung in den M30 Ko-Kulturen. Hinsichtlich der entzündlichen
Grunderkrankung, welche zu einer Epilepsie führt, sind Antiepileptika mit einem entzündungs-
hemmenden glialen Potential für Patienten vorteilhaft.

40
6. Literaturverzeichnis
1. Aloisi F. (2001): Immune function of microglia. Glia. 36, 165-79.
2. Ambrósio AF, Soares-Da-Silva P, Carvalho CM, Carvalho AP. (2002): Mechanisms of action
of carbamazepine and its derivatives, oxcarbazepine, BIA 2-093, and BIA 2-024. Neurochem
Res. 27, 121-30.
3. Andrzejczak D. (2011): Epilepsy and pro-inflammatory cytokines. Immunomodulating
properties of antiepileptic drugs. Neurol Neurochir Pol. 45, 275-85.
4. Aronica E, Crino PB. (2011): Inflammation in epilepsy: clinical observations. Epilepsia. 52,
26-32.
5. Auvin S, Mazarati A, Shin D, Sankar R. (2010): Inflammation enhances epileptogenesis in the
developing rat brain. Neurobiol Dis 40, 303-10.
6. Beal B, Moeller-Bertram T, Schilling JM, Wallace MS. (2012): Gabapentin for once-daily
treatment of post-herpetic neuralgia: a review. Clin Interv Aging. 7, 249-55.
7. Béchade C, Cantaut-Belarif Y, Bessis A. (2013): Microglial control of neuronal activity. Front
Cell Neurosci.7, 32.
8. Benarroch EE. (2013): Microglia: Multiple roles in surveillance, circuit shaping, and response
to injury. Neurology. 81, 1079-88.
9. Berg AT, Berkovic SF, Brodie MJ, Buchhalter J, Cross JH, van Emde Boas W, Engel J,
French J, Glauser TA, Mathern GW, Moshé SL, Nordli D, Plouin P, Scheffer IE. (2010):
Revised terminology and concepts for organization of seizures and epilepsies: report of the
ILAE Commission on Classification and Terminology, 2005-2009. Epilepsia 51, 676-685.
10. Bianchi M, Rossoni G, Sacerdote P, Panerai AE, Berti F. (1995): Carbamazepine exerts anti-
inflammatory effects in the rat. Eur J Pharmacol. 294, 71-74.
11. Binder DK, Steinhäuser C. (2006): Functional changes in astroglial cells in epilepsy. Glia. 54,
358-68.
12. Black JA, Liu S, Waxman SG. (2009): Sodium channel activity modulates multiple functions
in microglia. Glia. 57, 1072-1081.
13. Boer K, Jansen F, Nellist M, Redeker S, van den Ouweland AM, Spliet WG, van
Nieuwenhuizen O, Troost D, Crino PB, Aronica E. (2008): Inflammatory processes in cortical
tubers and subependymal giant cell tumors of tuberous sclerosis complex. Epilepsy Res. 78, 7-
21.
14. Buckwalter MS, Wyss-Coray T. (2004): Modelling neuroinflammatory phenotypes in vivo. J
Neuroinflammation. 1, 10.

41
15. Caraci F, Battaglia G, Bruno V, Bosco P, Carbonaro V, Giuffrida ML, Drago F, Sortino MA,
Nicoletti F, Copani A. (2011): TGF-β1 pathway as a new target for neuroprotection in
Alzheimer's disease. CNS Neurosci Ther. 17, 237-49.
16. Cardile V, Pavone A, Renis M, Maci T, Perciavalle V. (2001): Effects of Gabapentin and
Topiramate in primary rat astrocyte cultures. Neuroreport. 12, 1705-1708.
17. Chew SS, Johnson CS, Green CR, Danesh-Meyer HV. (2010): Role of connexin43 in central
nervous system injury. Exp Neurol. 225, 250-261.
18. Chogtu B, Bairy KL, Smitha D, Dhar S, Himabindu P. (2011): Comparison of the efficacy of
carbamazepine, gabapentin and lamotrigine for neuropathic pain in rats. Indian J Pharmacol.
43, 596-598.
19. Choi J, Koh S. (2008): Role of brain inflammation in epileptogenesis. Yonsei Med J. 49, 1-18.
20. Choi J, Nordli DR Jr, Alden TD, DiPatri A Jr, Laux L, Kelley K, Rosenow J, Schuele SU,
Rajaram V, Koh S. (2009): Cellular injury and neuroinflammation in children with chronic
intractable epilepsy. J Neuroinflammation. 6, 38.
21. De Simoni MG, Perego C, Ravizza T, Moneta D, Conti M, Marchesi F, De Luigi A, Garattini
S, Vezzani A. (2000): Inflammatory cytokines and related genes are induced in the rat
hippocampus by limbic status epilepticus. Eur J Neurosci. 12, 2623-33.
22. Dermietzel R, Hertberg EL, Kessler JA, Spray DC. (1991): Gap junctions between cultured
astrocytes: immunocytochemical, molecular, and electrophysiological analysis. J Neurosci. 11,
1421-32.
23. Devinsky O, Vezzani A, Najjar S, De Lanerolle NC, Rogawski MA. (2013): Glia and
epilepsy: excitability and inflammation. Trends Neurosci. 36, 174-84.
24. Doyle KP, Cekanaviciute E, Mamer LE, Buckwalter MS. (2010): TGFβ signaling in the brain
increases with aging and signals to astrocytes and innate immune cells in the weeks after
stroke. J Neuroinflammation. 7, 62.
25. Dragunow M, Greenwood JM, Cameron RE, Narayan PJ, O'Carroll SJ, Pearson AG, Gibbons
HM. (2006): Valproic acid induces caspase 3-mediated apoptosis in microglial cells.
Neuroscience. 140, 1149-1156.
26. Ekdahl CT, Claasen JH, Bonde S, Kokaia Z, Lindvall O. (2003): Inflammation is detrimental
for neurogenesis in adult brain. Proc Natl Acad Sci U S A. 100, 13632-7.
27. Faustmann PM, Haase CG, Romberg S, Hinkerohe D, Szlachta D, Smikalla D, Krause D,
Dermietzel R. (2003): Microglia activation influences dye coupling and Cx43 expression of
the astrocytic network. Glia. 42, 101-8.
28. Friedman A, Dingledine R. (2011): Molecular cascades that mediate the influence of
inflammation on epilepsy. Epilepsia; 52, 33-9.

42
29. Fröscher W, Krämer G, Schmidt D, Stefan H (1999): Serumkonzentration von Antiepileptika
– Praktische Richtlinien zur Messung und sinnvollen Interpretation. Nervenarzt 70, 172–177
30. Galic MA, Riazi K, Heida JG, Mouihate A, Fournier NM, Spencer SJ, Kalynchuk LE, Teskey
GC, Pittman QJ. (2008): Postnatal inflammation increases seizure susceptibility in adult rats. J
Neurosci. 28, 6904-13.
31. Garbelli R, Frassoni C, Condorelli DF, Trovato Salinaro A, Musso N, Medici V, Tassi L,
Bentivoglio M, Spreafico R. (2011): Expression of connexin 43 in the human epileptic and
drug-resistant cerebral cortex. Neurology. 76, 895-902.
32. Giaume C, Fromaget C, el Aoumari A, Cordier J, Glowinski J, Gros D. (1991): Gap junctions
in cultured astrocytes: single-channel currents and characterization of channel-forming
protein. Neuron 6, 133-43.
33. Giaume C, Leybaert L, C Naus C, C Sáez J. (2013): Connexin and pannexin hemichannels in
brain glial cells: properties, pharmacology, and roles. Front Pharmacol. 4, 88
34. Gibbons HM, Smith AM, Teoh HH, Bergin PM, Mee EW, Faull RL, Dragunow M. (2011):
Valproic acid induces microglial dysfunction, not apoptosis, in human glial cultures.
Neurobiol Dis. 41, 96-103.
35. Grim SA, Ryan M, Miles MV, Tang PH, Strawsburg RH, deGrauw TJ, Fakhoury TA,
Baumann RJ. (2003): Correlation of levetiracetam concentrations between serum and saliva.
Ther Drug Monit. 25, 61-6.
36. Haghikia A, Ladage K, Hinkerohe D, Vollmar P, Heupel K, Dermietzel R, Faustmann PM.
(2008): Implications of antiinflammatory properties of the anticonvulsant drug levetiracetam
in astrocytes. J Neurosci Res. 86, 1781-8.
37. Hinkerohe D, Smikalla D, Haghikia A, Heupel K, Haase CG, Dermietzel R, Faustmann PM.
(2005): Effects of cytokines on microglial phenotypes and astroglial coupling in an
inflammatory coculture model. Glia.52, 85-97.
38. Hinkerohe D, Smikalla D, Schoebel A, Haghikia A, Zoidl G, Haase CG, Schlegel U,
Faustmann PM. (2010): Dexamethasone prevents LPS-induced microglial activation and
astroglial impairment in an experimental bacterial meningitis co-culture model. Brain Res. 6,
1329:45-54.
39. Honarmand A, Safavi M, Zare M. (2011): Gabapentin: An update of its pharmacological
properties and therapeutic use in epilepsy. J Res Med Sci.16, 1062-9.
40. Hulkkonen J, Koskikallio E, Rainesalo S, Keränen T, Hurme M, Peltola J. (2004): The
balance of inhibitory and excitatory cytokines is differently regulated in vivo and in vitro
among therapy resistant epilepsy patients. Epilepsy Res. 59, 199-205.

43
41. Ichiyama T, Okada K, Lipton JM, Matsubara T, Hayashi T, Furukawa S. (2000): Sodium
valproate inhibits production of TNF-alpha and IL-6 and activation of NF-kappaB. Brain Res.
857, 246-251.
42. Iwamoto T, Takasugi Y, Higashino H, Ito H, Koga Y, Nakao S. (2011): Antinociceptive
action of carbamazepine on thermal hypersensitive pain at spinal level in a rat model of
adjuvant-induced chronic inflammation. J Anesth. 25, 78-86.
43. Iyer A, Zurolo E, Spliet WG, van Rijen PC, Baayen JC, Gorter JA, Aronica E. (2010):
Evaluation of the innate and adaptive immunity in type I and type II focal cortical dysplasias.
Epilepsia. 51, 1763-73.
44. Jabs R, Seifert G, Steinhäuser C. (2008): Astrocytic function and its alteration in the epileptic
brain. Epilepsia. 49, 3-12.
45. Karpuk N, Burkovetskaya M, Fritz T, Angle A, Kielian T. (2011:) Neuroinflammation leads
to region-dependent alterations in astrocyte gap junction communication and hemichannel
activity. J Neurosci. 31, 414-425.
46. Li G, Bauer S, Nowak M, Norwood B, Tackenberg B, Rosenow F, Knake S, Oertel WH,
Hamer HM. (2011): Cytokines and epilepsy. Seizure. 20, 249-56.
47. Lüllmann H, Mohr K, Hein L. (2010) Pharmakologie und Toxikologie:
Arzneimittelwirkungen verstehen - Medikamente gezielt einsetzen. Auflage: 17. Stuttgart:
Thieme.
48. Maldonado M, Baybis M, Newman D, Kolson DL, Chen W, McKhann G 2nd, Gutmann DH,
Crino PB. (2003): Expression of ICAM-1, TNF-alpha, NF kappa B, and MAP kinase in tubers
of the tuberous sclerosis complex. Neurobiol Dis. 14, 279-90.
49. Mataringa MI, May TW, Rambeck B. (2002): Does lamotrigine influence valproate
concentrations? Ther Drug Monit. 24, 631-6.
50. Monti B, Polazzi E, Contestabile A. (2009): Biochemical, molecular and epigenetic
mechanisms of valproic acid neuroprotection. Curr Mol Pharmacol. 2, 95-109.
51. Nico B, Ribatti D.(2012): Morphofunctional aspects of the blood-brain barrier. Curr Drug
Metab.13, 50-60.
52. Pannasch U, Vargová L, Reingruber J, Ezan P, Holcman D, Giaume C, Syková E, Rouach
N.(2011): Astroglial networks scale synaptic activity and plasticity. Proc Natl Acad Sci U S
A. 108, 8467-72.
53. Pavone A, Cardile V. (2003): An in vitro study of new antiepileptic drugs and astrocytes.
Epilepsia. 44, 34-9.
54. Peltola J, Hurme M, Miettinen A, Keränen T. (1998): Elevated levels of interleukin-6 may
occur in cerebrospinal fluid from patients with recent epileptic seizures. Epilepsy Res.
31(2):129-33.

44
55. Pohlmann-Eden B, Steinhoff BJ. (2009): Antiepileptika verstehen: ein Wegweiser durch den
Medikamenten-Dschungel. 5. Auflage. Stuttgart: Trias Verlag in MVS Medizinvorlage
Stuttgart GmbH & Co. KG.
56. Rambeck B, Jürgens UH, May TW, Pannek HW, Behne F, Ebner A, Gorji A, Straub H,
Speckmann EJ, Pohlmann-Eden B, Löscher W. (2006): Comparison of brain extracellular
fluid, brain tissue, cerebrospinal fluid, and serum concentrations of antiepileptic drugs
measured intraoperatively in patients with intractable epilepsy. Epilepsia 47, 681-94.
57. Ravizza T, Rizzi M, Perego C, Richichi C, Velísková J, Moshé SL, De Simoni MG, Vezzani
A. (2005): Inflammatory response and glia activation in developing rat hippocampus after
status epilepticus. Epilepsia. 46, 113-7.
58. Retamal MA, Froger N, Palacios-Prado N, Ezan P, Sáez PJ, Sáez JC, Giaume C. (2007): Cx43
hemichannels and gap junction channels in astrocytes are regulated oppositely by
proinflammatory cytokines released from activated microglia. J Neurosci. 27, 13781-1392.
59. Rizzi M, Perego C, Aliprandi M, Richichi C, Ravizza T, Colella D, Velískŏvá J, Moshé SL,
De Simoni MG, Vezzani A. (2003): Glia activation and cytokine increase in rat hippocampus
by kainic acid-induced status epilepticus during postnatal development. Neurobiol Dis.14,
494-503.
60. Schnabel R, Rambeck B, May TW, Jürgens U, Lahl R, Villagrán R. (1996): Lack of effect of
histological lesions on the phenytoin and phenobarbital concentrations in the brain cortex of
epileptic patients. J Neurol Sci. 133, 177-82.
61. Seifert G, Carmignoto G, Steinhäuser C. (2010): Astrocyte dysfunction in epilepsy. Brain Res
Rev. 63, 212-21.
62. Seifert G, Steinhäuser C. (2013): Neuron-astrocyte signaling and epilepsy. Exp Neurol. 244,
4-10.
63. Silveira G, de Oliveira AC, Teixeira AL. (2012): Insights into inflammation and epilepsy from
the basic and clinical sciences. J Clin Neurosci. 19, 1071-5.
64. Smith AM, Gibbons HM, Dragunow M. (2010): Valproic acid enhances microglial
phagocytosis of amyloid-beta (1-42). Neuroscience. 169, 505-515.
65. Sofroniew MV. (2013): Multiple Roles for Astrocytes as Effectors of Cytokines and
Inflammatory Mediators. Neuroscientist. 2013 Oct 8. (Epub ahead of print)
66. Stienen MN, Haghikia A, Dambach H, Thöne J, Wiemann M, Gold R, Chan A, Dermietzel R,
Faustmann PM, Hinkerohe D, Prochnow N. (2011): Anti-inflammatory effects of the
anticonvulsant drug levetiracetam on electrophysiological properties of astroglia are mediated
via TGFβ1 regulation. Br J Pharmacol. 162, 491-507.

45
67. Stobart JL, Anderson CM. (2013): Multifunctional role of astrocytes as gatekeepers of
neuronal energy supply. Front Cell Neurosci. 7, 38.
68. Tunnicliff G. (1996): Basis of the antiseizure action of phenytoin. Gen Pharmacol. 27, 1091-7.
69. Verrotti A, Basciani F, Trotta D, Greco R, Morgese G, Chiarelli F. (2001): Effect of
anticonvulsant drugs on interleukins-1, -2 and -6 and monocyte chemoattractant protein-1.
Clin Exp Med. 1, 133-136.
70. Vezzani A. (2005): Inflammation and epilepsy. Epilepsy Curr. 5, 1-6.
71. Vezzani A, Aronica E, Mazarati A, Pittman QJ. (2011): Epilepsy and brain inflammation. Exp
Neurol. 244, 11-21.
72. Vezzani A, Conti M, De Luigi A, Ravizza T, Moneta D, Marchesi F, De Simoni MG. (1999):
Interleukin-1beta immunoreactivity and microglia are enhanced in the rat hippocampus by
focal kainate application: functional evidence for enhancement of electrographic seizures. J
Neurosci. 19, 5054-65.
73. Vezzani A, Granata T. (2005): Brain inflammation in epilepsy: experimental and clinical
evidence. Epilepsia. 46, 1724-43.
74. Vezzani A, Moneta D, Conti M, Richichi C, Ravizza T, De Luigi A, De Simoni MG, Sperk G,
Andell-Jonsson S, Lundkvist J, Iverfeldt K, Bartfai T. (2000): Powerful anticonvulsant action
of IL-1 receptor antagonist on intracerebral injection and astrocytic overexpression in mice.
Proc Natl Acad Sci U S A. 97, 11534-9.
75. Vezzani A, Rüegg S. (2011): The pivotal role of immunity and inflammatory processes in
epilepsy is increasingly recognized: introduction. Epilepsia. 52, 1-4.
76. Voutsinos-Porche B, Koning E, Kaplan H, Ferrandon A, Guenounou M, Nehlig A, Motte J.
(2004): Temporal patterns of the cerebral inflammatory response in the rat lithium-pilocarpine
model of temporal lobe epilepsy. Neurobiol Dis. 17, 385-402.
77. Wallraff A, Köhling R, Heinemann U, Theis M, Willecke K, Steinhäuser C. (2006): The
impact of astrocytic gap junctional coupling on potassium buffering in the hippocampus. J
Neurosci. 26, 5438-47.
78. Walz W. (2010): The Neuronal Environment: Brain Homeostasis in Health and Disease.
Totowa, New Jersey: Contemporary NeuroscienceTM, Human Press Inc.
79. Wetherington J, Serrano G, Dingledine R. (2008): Astrocytes in the epileptic brain. Neuron.
58, 168-78.
80. Wirrell E, Farrell K, Whiting S. (2005): The epileptic encephalopathies of infancy and
childhood. Can J Neurol Sci. 32, 409-18.
81. Wheless JW, Clarke DF, Arzimanoglou A, Carpenter D. (2007): Treatment of pediatric
epilepsy: European expert opinion. Epileptic Disord. 9, 353-412.

46
82. Whitney NP, Eidem TM, Peng H, Huang Y, Zheng JC. (2009): Inflammation mediates
varying effects in neurogenesis: relevance to the pathogenesis of brain injury and
neurodegenerative disorders. J Neurochem; 108, 1343-59.
83. Wood P.L. (2003): Neuroinflammation Mechanisms and Management. Totowa, New Jersey:
Contemporary NeuroscienceTM, Human Press Inc.
84. Wraith DC, Nicholson LB. (2012): The adaptive immune system in diseases of the central
nervous system. J Clin Invest. 122, 1172-9.
85. Xie N, Wang C, Lin Y, Li H, Chen L, Zhang T, Sun Y, Zhang Y, Yin D, Chi Z. (2010): The
role of p38 MAPK in valproic acid induced microglia apoptosis. Neurosci Lett. 482, 51-6.
86. Yang MS, Min KJ, Joe E. (2007): Multiple mechanisms that prevent excessive brain
inflammation. J Neurosci Res. 85, 2298-305.
87. Yu N, Liu H, Di Q. (2013): Modulation of Immunity and the Inflammatory Response: A New
Target for Treating Drug-resistant Epilepsy. Curr Neuropharmacol. 11, 114-27.

47
7. Abku rzungsverzeichnis
Abb. Abbildung
APS Ammoniumpersulfat
Aqua dest. Destilliertes Wasser
BSA Bovine serum albumin
Ca Calcium
CBZ Carbamazepin
CNPase 2',3'-cyclic-nucleotide phosphodiesterase
CO2 Kohlenstoffdioxid
Cx43 Connexin 43
Cx30 Connexin 30
°C Grad Celsius
DAPI 4′,6-Diamidin-2-phenylindol
DMEM Dulbecco's modified Eagle's medium
DMSO Dimethyl Sulfoxide
DNase Desoxyribonuklease
ELISA Enzyme Linked Immunosorbent Assay
GABA γ-Aminobuttersäure
GBT Gabapentin
GFAP Glial fibrillary acidic protein
H2O Wasser
HRP Horseradish peroxidase
IgG Immunglobulin G
IL Interleukin
INT Intermediate type
LEV Levetiracetam
LPS Lipopolysaccharide
mA Milliampere
mAB Monoklonaler Antikörper
M5 Astrozyten Ko-Kultur mit 5 % Mikroglia
M30 Astrozyten Ko-Kultur mit 30 % Mikroglia
ml Mililiter
MRP Membranruhepotential
MTT 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide
mV Milivolt

48
Na Natrium
NEA Nicht-essentielle Aminosäuren
NeuN Neuronal Nuclei
nm Nanometer
ns Nicht Signifikant
P0-P2 Postnatal
pAB Polyklonaler Antikörper
PBS Phosphat-buffered saline
PHE Phenytoin
PLL Poly-L-Lysin
RPT Rounded phagocytic type
rpm Rounds per minute
RRT Resting ramified type
RT Raumtemperatur
SDS Sodiumdodecylsulfat
SEM Standardfehler des Mittelwerts
TBST Tris-buffered saline mit Tween 20
TEMED Tetramethylethylendiamin
TGF-beta Transforming growth factor beta
TLE Temporallappenepilepsie
TMB 3,3',5,5'-Tetramethylbenzidine
TNF-alpha Tumornekrosefaktor-α
TRIS Tris-(hydroxymethyl)-aminomethan
Tween Polyoxyethylen(20)-sorbitan-monolaurat
V Volt
VPA Valproat
VPS Valproinsäure
ZNS Zentrale Nervensystem
μg Mikrogramm
μl Mikroliter
μm Mikrometer

49
8. Anhang
Tabelle 7.1
M5 Ko-Kulturen
M30 Ko-Kulturen
VPA Kontrolle 222.6 ± 13.96 18.1 ± 2.06 195.6 ± 8.82 44.8 ± 3.45 50 ug/ml 200.6 ± 12.64 21.7 ± 1.76 200.4 ± 8.67 43.5 ± 3.37
100 ug/ml 204.5 ± 11.49 23.5 ± 1.99 200.4 ± 9.15 41.6 ± 2.64
200 ug/ml 210.6 ± 12.28 22.9 ± 1.55 199.4 ± 7.52 39.7 ± 2.71
GBT
Kontrolle 132.3 ± 7.99 11.1 ± 1.33 207.8 ± 13.27 41.5 ± 3.14 10 ug/ml 124.1 ± 8.31 10.4 ± 1.79 216.7 ± 12.35 52.2 ± 3.21
25 ug/ml 119.2 ± 8.92 10.2 ± 1.92 215.7 ± 14.24 46.0 ± 2.46
50 ug/ml 127.5 ± 5.35 11.4 ± 1.48 223.5 ± 16.79 43.1 ± 3.55 100 ug/ml 128.3 ± 6.03 11.0 ± 1.37 221.6 ± 15.90 40.0 ± 3.73
PHE
Kontrolle 136.0 ± 6.44 11.9 ± 1.05 209.0 ± 10.53 43.4 ± 3.81 10 ug/ml 118.1 ± 6.87 9.5 ± 1.51 205.0 ± 12.89 47.7 ± 2.89 25 ug/ml 111.2 ± 7.74 8.6 ± 1.48 216.8 ± 12.18 43.9 ± 2.72 50 ug/ml 115.0 ± 7.50 10.8 ± 1.39 218.1 ± 10.37 39.2 ± 2.45
100 ug/ml 113.9 ± 6.76 11.4 ± 1.50 199.3 ± 11.24 35.7 ± 2.43
CBZ
Kontrolle 123.6 ± 8.88 10.3 ± 1.42 238.0 ± 18.09 49.7 ± 4.33 10 ug/ml 114.0 ± 8.31 10.6 ± 1.79 242.0 ± 19.06 46.1 ± 3.07 25 ug/ml 115.6 ± 7.79 10.7 ± 2.02 227.5 ± 13.20 45.9 ± 2.90
50 ug/ml 111.6 ± 10.03 10.5 ± 1.99 179.3 ± 9.25 35.9 ± 2.44 100 ug/ml 98.0 ± 10.70 9.1 ± 1.99 131.0 ± 7.60 20.3 ± 2.34
Gliazellen Mikroglia Gliazellen Mikroglia
Tab. 7.1. Tabelle entnommen und übersetzt aus (Dambach et al. 2014): Die Tabelle zeigt den Mittelwert
± Satndardfehler des Mittelwertes (SEM) der ausgewerteten Anzahl an Glia- und Mikroglia-Zellen der
immunocytochemischen Färbungen der M5* und M30** Ko-Kulturen. *(VPA (n=33) GBT (n=24) PHE
(n=28) und CBZ (n=22)). **(VPA (n=24) GBT (n=30) PHE (n=40) und CBZ (n=20)).
Abbildung 7.2.
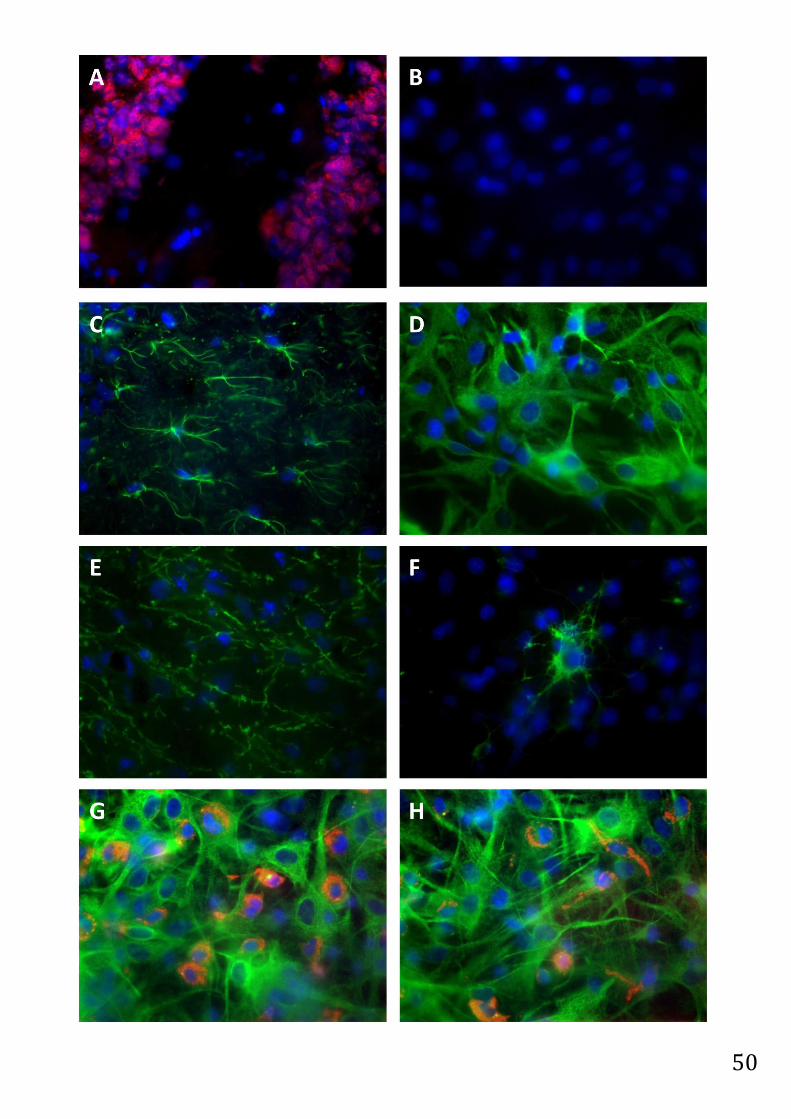
50

51
Abb. 7.2. Abbildung entnommen und übersetzt aus (Dambach et al. 2014):
Neuronen Marker (NeuN)*
A. Positive Kontrolle: Immunohistochemische Färbung von Nervenzellen (rot) in Gehirnschnitten der Ratte mit
dem Marker Anti-NeuN (Millipore, Darmstadt) 1:100 und dem sekundären Antikörper Alexa Fluor 488 (Sigma-
Aldrich, Steinheim) 1:500. Die Zellkerne im Gehirnschnitt wurden zusätzlich mit dem Kernfarbstoff DAPI (4, 6-
diamidino-2-phenyl-indol) (blau) gefärbt, um die gesamte Anzahl an Zellen zu visualisieren. Die dargestellte
Region auf dem Bild zeigt den Hippocampus der Ratte.
B. Ko-Kulturen: Immunocytochemische Färbung der Astrozyten/Mikroglia Ko-Kulturen mit dem neuronalen
(rot) Marker Anti-NeuN (Millipore, Darmstadt) 1:100 und dem sekundären Antikörper Alexa Fluor 488 (Sigma-
Aldrich, Steinheim) 1:500. Die Zellkerne wurden zusätzlich mit dem Kernfarbstoff DAPI (blau) gefärbt, um die
gesamte Anzahl an Zellen zu visualisieren. In den Ko-Kulturen waren keine NeuN positiven Zellen aufzufinden
(n=3).
Astrozyten Marker (GFAP)*
C. Positiv Kontrolle: Immunohistochemische Färbung von Astrozyten (grün) in Gehirnschnitten der Ratte mit
dem Marker Anti-GFAP (Sigma-Aldrich, Steinheim) 1:100 und dem sekundären Antikörper Alexa Fluor 488
(Sigma-Aldrich, Steinheim) 1:500. Die Zellkerne im Gehirnschnitt wurden zusätzlich mit dem Kernfarbstoff
DAPI (blau) gefärbt, um die gesamte Anzahl an Zellen zu visualisieren.
D. Ko-Kulturen: Immunocytochemische Färbung der Astrozyten/Mikroglia Ko-Kulturen mit dem Astrozyten
(grün) Marker Anti-GFAP (Sigma-Aldrich, Steinheim) 1:100 und dem sekundären Antikörper Alexa Fluor 488
(Sigma-Aldrich, Steinheim) 1:500. Die Zellkerne wurden zusätzlich mit dem Kernfarbstoff DAPI (blau) gefärbt,
um die gesamte Anzahl an Zellen zu visualisieren (n=3).
Oligodendrozyten Marker (CNPase)*
E. Positiv Kontrolle: Immunohistochemische Färbung von Oligodendrozyten (grün) in Gehirnschnitten der
Ratte mit dem Marker Anti-CNPase (Sigma-Aldrich, Steinheim) 1:100 und dem sekundären Antikörper Alexa
Fluor 488 (Sigma-Aldrich, Steinheim) 1:500. Die Zellkerne im Gehirnschnitt wurden zusätzlich mit dem
Kernfarbstoff DAPI (blau) gefärbt, um die gesamte Anzahl an Zellen zu visualisieren.
F. Ko-Kulturen: Immunocytochemische Färbung der Astrozyten/Mikroglia Ko-Kulturen mit dem
Oligodendrozyten (grün) Marker Anti-CNPase (Sigma-Aldrich, Steinheim) 1:100 und dem sekundären
Antikörper Alexa Fluor 488 (Sigma-Aldrich, Steinheim) 1:500. Die Zellkerne wurden zusätzlich mit dem
Kernfarbstoff DAPI (blau) gefärbt, um die gesamte Anzahl an Zellen zu visualisieren. In den Ko-Kulturen waren
1,27 % der gesamten Zellanzahl CNPase positive Zellen in insgesamt (n=24) Sichtfeldern aus (n=3) unabhängig
voneinander durchgeführten Färbungen.
Astrozyten (GFAP) und Mikroglia (ED-1) Marker*
G. + H. Ko-Kulturen: Immunocytochemische Färbung der Astrozyten/Mikroglia Ko-Kulturen mit dem
Astrozyten (grün) Marker Anti-GFAP (Sigma-Aldrich, Steinheim) 1:100 und dem sekundären Antikörper Alexa
Fluor 488 (Sigma-Aldrich, Steinheim) 1:500. Zusätzlich wurden die Ko-Kulturen mit dem Mikroglia (rot)
Marker ED-1 (1:250) (Serotec MCA 341R, Eching) und mit dem sekundären Antikörper Alexa Fluor 568
(Invitrogen, Karlsruhe) 1:1000 angefärbt. Die Zellkerne wurden zusätzlich mit dem Kernfarbstoff DAPI (blau)
gefärbt, um die gesamte Anzahl an Zellen zu visualisieren (n=3).
* Alle Färbungen wurden am Immunflourezenz-Mikroskop (Axiophot Carl Zeiss GmbH, Jena) mit einem 63 x
Objektiv ausgewertet.
C D

52
9. Danksagung
Ich möchte mich herzlich bei allen Personen bedanken, welche wissenschaftlich beratend, moralisch
unterstützend und konstruktiv mitwirkend bei dieser Arbeit beteiligt waren:
Herrn Prof. Dr. med. Pedro Faustmann für die unvergleichlich sympathische Unterstützung und
motivierende Betreuung von Beginn meiner Masterarbeit bis zum Ende der Promotion. Ich habe
sowohl wissenschaftlich, medizinisch als auch persönlich viel von ihm gelernt und wäre den Schritt
Richtung Promotion nicht ohne ihn gegangen!
Herrn Prof. Dr. med. Andreas Hufnagel für die Betreuung, Unterstützung und Kooperationen
während dieser Doktorarbeit.
Frau Jun.-Prof. Dr. rer nat. Nora Prochnow für die Möglichkeit meine Doktorarbeit in der Abteilung
für Neuroanatomie erfolgreich zu vollenden, die tollen Kooperationen, die wissenschaftliche
Unterstützung sowie die ersten Einblicke in die medizinische Lehre.
Herrn Dr. med. Daniel Hinkerohe für die tolle, freundschaftliche Unterstützung während meiner
Master- und Doktorarbeit sowie die gemeinsamen Kongressbesuche.
Frau Dr. med. Zahra Moinfar für die aufmunternden wissenschaftlichen und privaten Dialoge sowie
medizinischen Erläuterungen während der Doktorarbeit und die tolle Arbeitsatmosphäre.
Allen Mitarbeitern der Abteilung für Neuroanatomie und Molekulare Hirnforschung, besonders Hans-
Werner Habbes, Sabine Schreiber-Minjoli, Sabine Peukert, Jeanette Wilms, Ulrike Becker und
Monika Birkelbach für die großartige technische und administrative Unterstützung.
Ein Dankeschön auch an meine Kommilitonen mit denen ich das Studium gemeistert habe. Besonders
Jens Christmann, Daniela Damen und Heiner Falkenberg.
Ich möchte mich aus ganzen Herzen bei denjenigen Personen bedanken, welche mein ganzes Leben
immer für mich da waren und mich bedingungslos auf dem Weg Richtung Promotion unterstützt
haben:
Mein allergrößter Dank gilt zweifellos meiner Mutter Hella Dambach, welche mich immer liebevoll
bei allem unterstützt hat was ich bis zu diesem Zeitpunkt erreicht habe und immer für mich da ist. Die
unzähligen aufmunternden Gespräche haben mich immer beruhigt, aufgebaut und vor allem motiviert!
Auch ein herzliches Dankeschön an ihren Lebenspartner Hartmann Rupp.
Meiner lieben Schwester Beate Schieber mit Marco, dem besten Schwager, den ich mir hätte
wünschen können, sowie meinem Neffen Mika und meiner Nichte Roxana. Vielen Dank für die
wunderschönen gemeinsamen Urlaube, Shopping-Touren, Spieleabende, Ablenkungen und
fürsorglichen Ratschläge während meiner Studien- und Promotionszeit!
Meinem großen Bruder Bernd Dambach mit Stefanie, meinen Neffen Robin und Luca sowie meiner
Nichte Jona für die vielen schönen Ablenkungen, Ratschläge und Unterkünfte während meiner
Studien- und Promotionszeit. Besonders herzlich möchte ich meinem Bruder dafür danken, dass er
mich immer bei wichtigen Entscheidungen ermutigt, stärkt und stets fürsorglich unterstützt!
Meinem besten Freund Daniel Rauschenberger für die Ablenkung in stressigen Zeiten. Was wir
schon alles zusammen erlebt haben lässt sich nicht auf einer Seite zusammenfassen.

53
Weitere Danksagung:
Die in dieser Dissertation verwendeten Graphen und Abbildungen aus der unten aufgelisteten
Veröffentlichung* und darin enthaltenen Ergebnisse sind mit der Genehmigung vom 28.4.2014 des
John Wiley & Sons, Inc. Verlages, dem Herausgeber der Fachzeitschrift Epilepsia der
Internationalen Liga gegen Epilepsie (ILAE), genehmigt worden.
*Dambach H, Hinkerohe D, Prochnow N, Stienen MN, Moinfar Z, Haase CG, Hufnagel A,
Faustmann PM. (2014): Glia and epilepsy: Experimental investigation of antiepileptic drugs in an
astroglia/microglia co-culture model of inflammation. Epilepsia. 55,184-92.

54
10. Lebenslauf
Der Lebenslauf ist in der Online-Version aus Gründen des Datenschutzes nicht enthalten.


















