
Phänotypische und molekulare Analyse von ... · und feuchtwarmer, strahlungsarmer Witterung...
102
Phänotypische und molekulare Analyse von Kreuzungsnachkommen auf Resistenz gegen Ährenfusariosen bei Triticale (x Triticosecale Wittmack) Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften (Dr. rer. nat.) Fakultät Naturwissenschaften Universität Hohenheim Landessaatzuchtanstalt vorgelegt von Maren Swente Großmann aus Offenbach am Main 2010
-
Upload
phungthien -
Category
Documents
-
view
220 -
download
0
Transcript of Phänotypische und molekulare Analyse von ... · und feuchtwarmer, strahlungsarmer Witterung...

Phänotypische und molekulare Analyse
von Kreuzungsnachkommen auf
Resistenz gegen Ährenfusariosen bei
Triticale (x Triticosecale Wittmack)
Dissertation zur Erlangung des Doktorgrades der
Naturwissenschaften (Dr. rer. nat.)
Fakultät Naturwissenschaften
Universität Hohenheim
Landessaatzuchtanstalt
vorgelegt von
Maren Swente Großmann
aus Offenbach am Main
2010

Dekan: Prof. Dr. rer. nat. Heinz Breer
1. berichtende Person: apl. Prof. Dr. Thomas Miedaner
2. berichtende Person: Prof. Dr. Otmar Spring
Eingereicht am: 30.09.2010
Mündliche Prüfung am: 25.01.2011
Die vorliegende Arbeit wurde am (21.12.2010) von der Fakultät Naturwissenschaften
der Universität Hohenheim als „Dissertation zur Erlangung des Doktorgrades der
Naturwissenschaften“ angenommen.

Für meine Omas

Inhaltsverzeichnis
1. Einleitung .......................................................................................................................... 1
1.1 Bedeutung und Evolution von Triticale .......................................................................... 1
1.2 Ährenfusariosen und deren Bekämpfungsmaßnahmen ................................................. 3
1.3 Reistenz und Resistenzzüchtung .................................................................................. 6
1.4 Molekulargenetische Kartierung .................................................................................... 8
1.5 Kartierung von Resistenz-QTL gegen Ährenfusariosen ................................................. 9
1.6 Zielsetzungen dieser Arbeit ......................................................................................... 11
2. Material und Methoden................................................................................................... 12
2.1 Versuchsüberblick ....................................................................................................... 12
2.2 Pflanzenmaterial ......................................................................................................... 12
2.3 Versuchsstandorte und Design der Feldversuche ....................................................... 13
2.4 Inokulum und Inokulation ............................................................................................ 14
2.5 Symptombonitur und Erfassung agronomischer Merkmale ......................................... 15
2.6 Ernte, Aufarbeitung und Bestimmung des DON-Gehaltes ........................................... 16
2.7 Genotypisierung und Kartierung .................................................................................. 18
DNA-Extraktion ....................................................................................................... 18
PCR, Mikrosatelliten- und DArT-Marker .................................................................. 19
2.8 Statistische Auswertung .............................................................................................. 21
3. Ergebnisse ...................................................................................................................... 23
3.1 Phänotypische Daten .................................................................................................. 23
3.1.1 Kreuzungspopulationen LASKO x ALAMO ......................................................... 23
3.1.1.1 F2:n-Populationen ...................................................................................... 23
3.1.1.2 DH-Population .......................................................................................... 30
3.1.1.3 Vergleich der Populationen ....................................................................... 36
3.1.2 Weitere Populationen ......................................................................................... 37
3.2 Kartierung von zwei Populationen ............................................................................... 40
3.2.1 Genetische Karte LASKO x ALAMO ................................................................... 41
3.2.2 Genetische Karte LASKO x TRIMESTER ........................................................... 42
3.2.3 Integrierte Karte .................................................................................................. 42
3.3 QTL-Analyse von zwei Kreuzungspopulationen .......................................................... 43
4. Diskussion ...................................................................................................................... 48
4.1 Phänotypische Analysen ............................................................................................. 48
4.2 Genotyp-Umwelt-Interaktion ........................................................................................ 52
4.3 Mykotoxine .................................................................................................................. 53

4.4 Genetische Kartierung ................................................................................................ 54
4.5 QTL-Analysen ............................................................................................................. 56
4.6 Schlußfolgerungen und Züchterische Konsequenzen ................................................. 60
5. Zusammenfassung ......................................................................................................... 62
6. Summary ......................................................................................................................... 64
7. Literaturverzeichnis ....................................................................................................... 66
8. Anhang ............................................................................................................................ 76
8.1 Genetische Karten beider Kreuzungspopulationen ..................................................... 76
8.2 LOD-Kurven ................................................................................................................ 82
8.3 Integrierte Karten ........................................................................................................ 85
8.4 Abkürzungsverzeichnis ............................................................................................... 93

1 Einleitung
1. Einleitung
1.1 Bedeutung und Evolution von Triticale
Triticale (x Triticosecale Wittmack) ist eine Kreuzung aus Weizen (Triticum spp.) als Mutter
und Roggen (Secale cereale) als Vater und enthält die Genome des Weizens (AABB oder
AABBDD) und das Genom des Roggens (RR). Wie im Fall der vorliegenden Dissertation,
werden meist hexaploide Triticale-Pflanzen angebaut (2n = 6x = 42, AABBRR; Kuleung et
al., 2004). Erstmalig berichtete der schottische Botaniker A. Stephen Wilson im Jahr 1875
über eine gelungene Bestäubung von Weizen mit Roggenpollen. Es gelang ihm zwei
Pflanzen heranzuziehen, die allerdings steril waren, so dass eine weitere Vermehrung nicht
möglich war (Wilson, 1876). Im Jahr 1883 erzeugte der amerikanische Pflanzenzüchter
Elbert S. Carman eine echte Hybridpflanze aus der Kreuzung aus Weizen und Roggen. Über
den Verbleib der Nachkommen ist nichts bekannt (Oettler, 2005). Dem deutschen
Pflanzenzüchter Wilhelm Rimpau gelang im Jahr 1888 ebenfalls eine fertile Kreuzung. Er
produzierte eine F1-Hybride, welche sogar 12 fertile Nachkommen hatte. Dieser Erfolg wird
auch heute noch in der Genbank in Gatersleben als Triticale Rimpau mit der
Akzessionsnummer TCA 26 instand gehalten (Lelley, 2006). Triticale erhielt den
wissenschaftlichen Namen „Triticosecale rimpaui“ von Ludwig Wittmack im Jahr 1899. Auf
die Empfehlung von Lindschau und Oehler (1935) wurde dann der bis heute gebräuchliche
Name Triticale (x Triticosecale Wittmack) bestimmt (Oettler, 2008).
Es können zwei verschiedene Triticaletypen unterschieden werden, die sich nach Art ihrer
Herstellung und ihres Ausgangsmaterials unterscheiden. Es gibt die sogenannten „primären“
Triticale, die direkt durch die Kreuzung Weizen x Roggen entstehen, und die „sekundären“
Triticale, welche auf einer Kreuzung von Triticale x Triticale beruhen (Oettler, 2008). Für die
Erstellung von primären Triticalen war die Entwicklung von Embryokultur und Anwendung
von Colchizin ein wissenschaftlicher Durchbruch (Blakeslee und Avery, 1937). Hierbei wird
der Embryo etwa 14 Tage nach der Bestäubung mit Hilfe der Embryokultur auf einem
Nährboden herangezogen. Die sich entwickelnde haploide Pflanze wird mit dem Alkaloid der
Herbstzeitlosen Colchicum autumnale L. (Colchizin) behandelt, das die Mitose beeinflusst.
Während die Verdopplung der Chromosomen noch stattfindet, bleibt die anschließende
Aufteilung auf zwei Tochterzellen jedoch aus. So entstehen Pflanzenzellen mit doppeltem
Chromosomensatz (AABBRR). Ohne Colchizinbehandlung käme es zu keiner
Kornausbildung auf der Pflanze, da die F1-Pflanzen mit nur einem Chromosomensatz von
Weizen und Roggen steril wären. Die Colchizinierung führt somit zu amphidiploiden, fertilen
Triticale-Pflanzen. Diese Pflanzen produzieren aufgrund des reinerbigen Zustandes
identische Nachkommen (Hoffmann, 1985). Die anfänglich hergestellten primären Triticale
entsprachen jedoch nicht den erwarteten Leistungen, deshalb wurde ab 1950 vermehrt mit

2 Einleitung
hexaploiden sekundären Triticale gezüchtet. Die ersten, für den Ackerbau zugelassenen
Triticalesorten wurden im Jahr 1968 angebaut und stammten aus Ungarn (Oettler, 2008).
Triticale ist einerseits als Nahrung vorwiegend für Tiere und andererseits als Substrat zur
Produktion von Biogas von wirtschaftlichem Nutzen. Im Jahr 1964 begann die internationale
Organisation CIMMYT (Centro Internacional de Mejoramiento de Maiz y Trigo) mit der
Triticalezüchtung, mit dem Ziel, den Hunger in Entwicklungsländern durch Triticale als neues
Nahrungsmittel zu reduzieren (Oettler, 2008). Bishnoi et al. (1978) fanden heraus, dass
Triticale in Europa einen höheren Viehfutter-Ertrag und Proteingehalt besitzt als
beispielsweise Weizen, Roggen oder Gerste. Der Großteil der Triticaleproduktion wird als
Viehfutter verwendet, vorrangig für Schweine und Geflügel (Lelley, 2006).
Eine der besten Eigenschaften von Triticale ist das hohe Ertragspotenzial. Triticale vereinigt
die robusten Eigenschaften der Winterhärte, Krankheitsresistenz wie auch Trockentoleranz
des Roggens mit den hohen Ertragsleistungen, guten Proteingehalt und
Qualitätseigenschaften des Weizens. Weiterhin kann Triticale, da gegenüber Weizen
besonders stressresistent, sehr gut in rauen Umwelten wachsen, vor allem an Standorten,
an denen weder Weizen noch Roggen vorkommt (Kuleung et al., 2006; Lelley, 2006).
Mittlerweile wird Triticale sehr erfolgreich für die Biogasproduktion eingesetzt. Nach Braun et
al. (2008) hatte Triticale im Jahr 2007 einen Methanertrag von 337 bis 555 m3 pro tvs (tons
volatile solids).
Laut Ernährungs- und Landwirtschaftsorganisation der Vereinten Nationen (FAO) hat sich
die Triticale-Produktion weltweit vom Jahr 1980 bis 2008 erheblich, von 638.042 Tonnen auf
2.381.500 Tonnen, gesteigert. Der durchschnittliche Triticale-Ertrag in Deutschland stieg von
5,41 t ha-1 im Jahr 2007 auf 5,97 t ha-1 im Jahr 2008 an (FAOSTAT, 2010). Im Jahr 2004
gingen in Deutschland sechs von 29 Triticale-Sorten auf polnischen Ursprung zurück (Lelley,
2006). Eine Übersicht zur Triticale-Produktion zeigt Abbildung 1. Auch heute noch ist Polen
das Land mit der größten Triticale-Produktionsmenge. Deutschland ist das Land mit der
zweitgrößten Triticale-Produktionsmenge (2.061.470 t), im Gegensatz zu Tschechien mit
einer Produktionsmenge von 205.513 t (FAOSTAT, 2010).

3 Einleitung
0
500
1000
1500
2000
2500
3000
3500
4000
4500 4.1
47
2.0
61
1.4
50
1.2
41
450
373
276
228
209
206
Pro
du
ktio
n (t x 1
00
0)
Anbaugebiet
Abb. 1: Weltweite Anbaugebiete und deren Triticale-Produktion im Jahr 2007; t = Tonnen; verändert nach FAOSTAT (2010)
Trotz zahlreicher Vorteile erwies sich Triticale als anfällig gegenüber verschiedenen
Krankheitserregern. Neben Blatt- und Spelzenbräune, Halmbasiserkrankungen, Rost und
Mehltau entwickelte sich vor allem der Befall mit Ährenfusariosen zu einem ernst zu
nehmenden Problem (Börner et al., 2009). In den letzten Jahren wurde ein verstärktes
Auftreten von Ährenfusariosen beobachtet (Rodemann, 2005). Bei einer weiteren Zunahme
der Ährenfusariosen muss mit einem Rückgang der Triticale-Anbauflächen gerechnet
werden.
1.2 Ährenfusariosen und deren Bekämpfungsmaßnahmen
Die für Fusarien anfälligsten Getreidearten sind in abnehmender Reihenfolge Mais, Weizen,
Triticale und Hafer. Ährenfusariosen werden durch verschiedene Schimmelpilze verursacht.
In Europa sind überwiegend Fusarium culmorum (F. culmorum), F. graminearum, F. poae,
F. avenaceum, F. sporotrichioides, F. cerealis und F. tricinctum verbreitet (Parry et al., 1995).
Von Miedaner et al. (2001) wird F. culmorum neben F. graminearum als einer der wichtigsten
Vertreter der Gattung Fusarium bei Weizen, Roggen und Triticale beschrieben. Die
natürlichen Populationen von F. culmorum und F. graminearum zeigen zwar eine große
Variation, eine Wirtsspezifität konnte jedoch nicht nachgewiesen werden (Mesterházy, 1995;
Miedaner, 1997)

4 Einleitung
In Abbildung 2 sind die Befallssymptome von F. culmorum an Triticale dargestellt. Es kommt
zunächst an einzelnen Stellen der Ähre zu Ausbleichungen einzelner Ährchen, bis hin zur
typischen rosaroten Verfärbung der kompletten Ähre. Nachfolgend sterben die infizierten
Ährchen ab und der Pilz wächst bis zur Ährenspindel. Durch die Unterbrechung des
Nährstoffstromes in der Ährenspindel stirbt bei anfälligen Sorten meist der gesamte obere
Ährenteil ab, weißlich bis rötlich verfärbte Kümmerkörner sind die Folge.
Abb. 2: Krankheitssymptome bei Triticale nach künstlichem Befall mit Fusarium culmorum
Während von F. culmorum ausschließlich die asexuelle Konidienform bekannt ist, existiert
von F. graminearum darüber hinaus auch die sexuelle Askosporenform Gibberella zeae
(Börner et al., 2009). F. culmorum kann saprophytisch an Pflanzenresten oder auch in Form
langlebiger Chlamydosporen im Boden überdauern. Während feucht-kühler Witterungs-
perioden über mehrere Wochen im Frühjahr entstehen häufig schon die ersten Infektionen
auf Ernteresten des Vorjahres. Die Infektion der Ähren erfolgt ab Anfang Juni, wobei die
Sporen über Regentropfen auf die Ähren übertragen werden. Somit sind kurzstrohige
Getreidepflanzen durch den kürzeren Infektionsweg gefährdeter als langstrohige Pflanzen.
Der Hauptinfektionszeitraum ist die Blüte des Getreides, dabei dienen möglicherweise die
Staubbeutel als Eintrittspforten in die einzelnen Blüten (Prigge, 2004). Bei feucht-warmen
Bedingungen und anfälligen Sorten verbreitet sich nun die Krankheit mit großer
Geschwindigkeit. Askosporen und Konidien können jedoch auch etliche Tage bei
Trockenheit auf den Blättern überleben, wobei die Idealtemperatur für eine Infektion > 15 °C
beträgt. Schon bei einem Temperaturanstieg um 4 °C während der Blüte erhöht sich der
Fusariumbefall, was gleichzeitig mit höheren Ertragsverlusten einhergeht (Brennan et al.,
2005). Grund für die Zunahme von Schadenswahrscheinlichkeit und –ausmaß sind vor allem
die Vorfrucht Mais, in einer Kombination mit nicht wendender, pflugloser Bodenbearbeitung
und feuchtwarmer, strahlungsarmer Witterung während der Getreideblüte mit
Niederschlägen > 3 mm (Beck et al., 1997).

5 Einleitung
Nach Martin und Johnston (1982) kann es aufgrund des Befalls von Ährenfusariosen an
Weizen durch reduzierte Kornzahl pro Ähre, sowie einem verminderten Tausendkorngewicht
zu Ertragsverlusten von bis zu 70 % kommen. Bei Triticale sind keine genauen Angaben
über Ertragsverluste infolge des Fusariumbefalls bekannt. Ährenfusariosen führen neben
Ertrags- und Qualitätsverlusten zur Belastung mit Mykotoxinen, vor allem Deoxynivalenol
(DON), welches das größte Problem des Befalls mit Fusarium darstellt (Buerstmayr et al.,
2003).
Die Mykotoxine lassen sich in verschiedene chemische Stoffgruppen einteilen. Die wichtigste
Klasse ist die der Trichothecene mit ca. 170 bisher bekannten Verbindungen (Offenbächer,
2001), des Weiteren gibt es u. a. noch Zearalenone (Meier, 2003). Mykotoxine sind
bekanntermaßen schädlich für Mensch und Tier. Nachgewiesene Probleme ergeben sich vor
allem in der Schweinehaltung (Dehne, 2001). Sie führen bei Schweinen sowohl zu einem
Rückgang beim Futterverzehr und dadurch zu Gewichtsreduktionen, als auch zu Erbrechen,
Durchfall, Schleimhautentzündungen und Futterverweigerung bis hin zu Totgeburten
(Offenbächer, 2001; Meier, 2003). Zwar sind Rinder und Hühner weniger empfindlich als
Schweine, dennoch sollten auch an sie keine kontaminierten Getreidepartien verfüttert
werden (Kabak et al., 2006). Im Vergleich zur menschlichen Ernährung bestehen die
Futterrationen der Tiere meist zu einem großen Teil aus Getreideprodukten, was das Risiko
einer Vergiftung auch bei nur geringer Kontamination erhöht (D‘Mello et al., 1999). Beim
Menschen konnten bei langfristiger Aufnahme von mit Mykotoxinen belasteten Lebensmitteln
Nierenschäden bis hin zum Nierenversagen ermittelt werden (Rodemann, 1999).
Im Februar 2004 wurden in Deutschland Grenzwerte für Mykotoxine eingeführt. Seit dem 1.
Juli 2006 gelten für die EU definierte Höchstgehalte von Mykotoxinen in Getreide, wobei
Brot, feine Backwaren, Kekse, Getreide-Snacks und Frühstückscerealien maximal
0,5 mg kg-1 DON und Getreidekörner zum direkten Verzehr und verarbeitete
Getreideerzeugnisse zur Herstellung von Säuglings- und Kindernahrung höchstens
0,2 mg kg-1 enthalten dürfen. Bei Futtermitteln für Schweine existiert zwar noch kein Grenz-,
aber ein Orientierungswert, der bei 1 mg kg-1 liegt (Heinrich, 2004). Nach Miedaner und
Schneider (2002) kann der Toxingehalt des Endproduktes durch Verarbeitungsprozesse, wie
Mahlen, Backen und Kochen, reduziert werden, jedoch höchstens um 50 %. Dadurch ist die
gesamte Verarbeitungsindustrie wie die Futtermittel-, Mehl- und Backindustrie bis hin zum
Verbraucher durch die Mykotoxin-Problematik betroffen. Ein weiteres großes Problem bei
der Einhaltung der Grenzwerte liegt in der Art der Probenahme, da einzelne Stichproben
derselben Probe oft stark variieren (Whitaker, 2006).

6 Einleitung
Die Bestimmungen des DON-Gehaltes können mit Hilfe chromatographischer und ELISA-
Verfahren durchgeführt werden. Mit der chromatographischen Methode können bis zu acht
Trichothecene gleichzeitig detektiert werden (Schollenberger et al., 1998). Sie ist aber sehr
zeitintensiv und teuer, ebenso wird ein hoher Laborbedarf benötigt. Das kommerziell
erhältliche ELISA-Testverfahren (enzyme linked immunosorbent assay) bietet dagegen eine
kostengünstigere Alternative. Die mit den ELISA-Verfahren getesteten DON-Gehalte bei
Roggen und Weizen wiesen eine hohe Korrelation zu den mit Gaschromatographie
ermittelten Werten auf (Miedaner et al., 2003). Wenn es sich um größere zu analysierende
Populationen handelt, sind die Kosten des ELISA-Verfahrens jedoch immer noch ziemlich
hoch.
1.3 Resistenz und Resistenzzüchtung
Verschiedene Methoden, den Ährenfusariosen und der Mykotoxin-Problematik entgegen zu
wirken, sind vorhanden, jedoch können bisher keine vollständigen Bekämpfungserfolge
erreicht werden. In engem Zusammenhang mit der zunehmenden Bedeutung von
Ährenfusariosen stehen die ackerbaulichen Entwicklungen, die die Krankheit begünstigen.
Beer (2005) empfiehlt als direkte Maßnahme die Auswahl der gegen Ährenbefall
widerstandsfähigen Sorten und bei hohem Infektionsdruck zusätzlich die gezielte
Anwendung wirksamer Fungizide zur Blütezeit. Fungizide haben nur eine eingeschränkte
Wirkung gegenüber Ährenfusariosen, können demnach Ertragsverluste und
Mykotoxinbelastungen nicht zuverlässig verhindern. Die Bodenbearbeitung spielt ebenfalls
eine wichtige Rolle zur Verminderung des Befalls. Um Erntereste, insbesondere
Maisstoppeln, zu vergraben und somit die Krankheitsübertragung zu verhindern, ist das
Pflügen als wendende Bodenbearbeitung der Mulchsaat und der Direktsaat vorzuziehen.
Auch durch zu starke Verunkrautung kann es aufgrund vermehrter Infektionsquellen zu
einem größeren Fusariumbefall kommen (Jenkinson und Parry, 1994). Der Einsatz von
Fungiziden zur Bekämpfung von Ährenfusariosen gestaltet sich oft sehr schwierig, da die
Zeitspanne für einen wirkungsvollen Fungizideinsatz nur wenige Tage nach optimalen
Infektionsbedingungen während der Blüte beträgt. Überdies ist es in der Praxis aus
technischen Problemen (z.B. durch Regen) nicht immer möglich, die Fungizide innerhalb
eines kurzen Zeitraumes zu applizieren. Da weder eine Krankheitsbekämpfung durch
ackerbauliche Maßnahmen, noch durch Fungizideinsatz ausreichend ist, stellt die Züchtung
resistenter Sorten eine wirksame Alternative dar. Resistenz wird definiert als die genetisch
bedingte Fähigkeit einer Wirtspflanze, die Schäden eines parasitären Befalls zu reduzieren
(Becker, 1993). Bei der Resistenz gegen Ährenfusariosen spielen sowohl morphologische
als auch physiologische Komponenten eine Rolle. Die morphologischen Eigenschaften
umfassen Wuchshöhe, Kompaktheit und Begrannung der Ähre, Offenblütigkeit und

7 Einleitung
Blühdauer, wobei Kurzstrohigkeit und Begrannung den natürlichen Befall mit
Ährenfusariosen unterstützen. Die physiologischen Komponenten beinhalten die Resistenz
gegen das Eindringen des Pilzes, dessen Ausbreitung, die Korninfektion, wie auch die
Resistenz gegen Mykotoxinakkumulation (Miedaner und Schneider, 2002). Es lassen sich
grundsätzlich fünf Resistenztypen hinsichtlich ihrer Anfälligkeit und der Abwehrreaktion,
sowie des visuellen Ährenbefalls und der Belastung mit Mykotoxinen unterscheiden:
Typ I: Resistenz gegenüber dem Eindringen bzw. der Infektion durch den
Schaderreger (Schroeder und Christensen, 1963)
Typ II: Resistenz gegenüber der Ausbreitung des Pathogens im Pflanzengewebe
(Schroeder und Christensen, 1963)
Typ III: Resistenz gegenüber einer Infektion der Körner (Mesterházy, 1995)
Typ IV: Toleranzerscheinungen gegenüber Ährenfusariosen (Mesterházy, 1995)
Typ V: Resistenz gegenüber Toxinen, beispielsweise durch Toxinabbau (Miller,
1985)
Die Züchtung resistenter Sorten ist aufgrund der quantitativen Vererbung und der hohen
Genotyp-Umwelt-Interaktion aufwändig. Bisher sind keine Genotypen bekannt, die
vollkommen befallsfrei sind, obwohl eine große Variation für das Merkmal besteht (Snijders,
1990). Um resistente Sorten zu erhalten, kann man entweder Kreuzungen mit resistenteren
Eltern verwenden, auf die Rekurrente Selektion zurückgreifen oder exotisches Material als
Resistenzträger einkreuzen. Hierzu gibt es verschiedenste Studien, in denen z.B. die
chinesische Sommerweizensorte Sumai 3 oder die brasilianische Sorte Frontana erfolgreich
bei Kreuzungen mit europäischen Weizensorten eingesetzt wurde (Wilde und Miedaner,
2006). Oft besteht jedoch das Problem, dass die Nachkommenschaften keine weiteren
agronomisch wertvollen Eigenschaften besitzen. Bei der Rekurrenten Selektion wird in
mehreren Zyklen auf eine möglichst hohe Resistenz selektiert und die jeweils besten Linien
wieder miteinander kombiniert. Dieser Vorgang wird solange wiederholt, bis ein
zufriedenstellendes Ausmaß an Resistenz erreicht wird, oder die Variation innerhalb einer
Population ausgenützt ist (Miedaner et al., 2000). Die Korrelationen zwischen dem visuellen
Befall und dem DON-Gehalt wurde in verschiedenen Studien als sehr gut beschrieben
(Mesterházy et al., 1999 und Mesterházy 2002; Miedaner et al., 2004; Wilde und Miedaner,
2006).
Für eine aussagekräftige Resistenzbestimmung ist es notwendig, Feldversuche mit
künstlicher Inokulation über möglichst viele Umwelten zu testen. Es lassen sich zwei
Methoden der künstlichen Inokulation unterscheiden. Die Sprühinokulation, die auch in der
vorliegenden Dissertation angewendet wurde, erfolgt ohne mechanische Verletzung, es wird
eine Sporensuspension bei Vollblüte auf die zu untersuchenden Pflanzen gesprüht. Bei
dieser Inokulationsmethode wird nicht zwischen der Typ I- und Typ II-Resistenz
unterschieden, infolge dessen können bei der visuellen Bonitur der Genotypen beide

8 Einleitung
Resistenztypen erfasst werden. Die andere Inokulationsmethode wird durchgeführt, wenn es
vorrangig um die Ausbreitungsresistenz der zu untersuchenden Pflanzen (Typ II) geht. Dazu
wird eine Einzelährcheninokulation vorgenommen und der Anteil infizierter Ährchen zu einem
Zeitpunkt erfasst. Die Züchtung resistenter Sorten benötigt enorme Forschung und einen
hohen Zeitaufwand.
1.4 Molekulargenetische Kartierung
Da Feldversuche mit viel Aufwand verbunden sind, wird nun immer häufiger erfolgreich die
markergestützte Selektion, z.B. anhand von Mikrosatelliten, eingesetzt. Mikrosatelliten sind
kurze DNA-Sequenzabschnitte, in denen ein bestimmtes Nukleotidmuster aus ver-
schiedenen Basenpaaren mehrmals wiederholt wird (simple sequence repeats, SSR). Es
wurden bei Weizen bereits zahlreiche genetische Karten publiziert, die als Referenzkarten
zur Zuordnung zu Chromosomen bei Kartierungen hilfreich sein können (Röder et al., 1998;
Song et al., 2005; Somers et al., 2004). Eine genetische Karte stellt die Anordnung von
Marker-Loci (Position von DNA-Markern auf einem Chromosom) entlang der Chromosomen
eines Genoms dar. Dienliche Genkarten mit SSR-Markern sind u.a. in der Datenbank
GrainGenes (2010) zu finden. SSR-Marker sind also brauchbar für die Genotypisierung, da
die Anzahl der Wiederholungen sich bei verschiedenen Individuen unterscheidet und
deswegen DNA-Fragmente unterschiedlicher Länge hervorbringt. Auf diese Weise können
Polymorphismen in der DNA festgestellt werden. Während der letzten 25 Jahren wurden
zahlreiche Markertechnologien zur Genotypisierung von Nutzpflanzen, wie Restriktions-
Fragment-Längen-Polymorphismus (RFLP), random amplified polymorphic DNA (RAPD),
amplified fragment-length polymorphism (AFLP), simple sequence repeat (SSR) und single
nucleotide polymorphism (SNP) verwendet. Diese Technologien haben jedoch verschiedene
Einschränkungen, Schnelligkeit und einen unterschiedlich großen Marker-Durchsatz. Die
meisten dieser Verfahren haben eine geringe Reproduzierbarkeit, hohe Kosten oder sind von
den DNA-Sequenzinformationen abhängig. Die Diversity Arrays Technology (DArT) bietet
hingegen eine gute Alternative zu den bisherigen Verfahren. DArT basiert auf der DNA-DNA-
Hybridisierung, welche sich die Eigenschaft der Mikroarrays (DNA-Chips) zu Nutze macht
und mit der mehrere tausend Loci gleichzeitig typisiert werden können (Jaccoud et al., 2001).
DArT-Marker erzeugen genetische Fingerabdrücke durch An- bzw. Abwesenheit von DNA-
Fragmenten, die genomische Abbildungen mehrerer Genotypen enthalten und idealerweise
die Vielfältigkeit einer Art abdecken. Die Verfahrensweise zur Herstellung genomischer
Repräsentation wird in Wenzl et al. (2004) beschrieben. Nach Jaccoud et al. (2001) wurde
die DArT-Technologie ursprünglich für Reis entwickelt. Seitdem gibt es Mikroarrays für viele
Pflanzenarten, wie z.B. Gerste, Hafer, Weizen, Roggen und Triticale.

9 Einleitung
Grundlage für die Berechnung einer genetischen Karte ist die Identifizierung von
Rekombinationsereignissen, die mit Hilfe der molekularen Marker ermöglicht wird. Die
Berechnung von Genkarten erfolgt mit Computerprogrammen, wie z.B. JoinMap 4.0 (Van
Ooijen, 2006), in denen die Zuordnung der DNA-Marker zu Kopplungsgruppen, die
Reihenfolge der Marker innerhalb dieser und die genetische Distanz zwischen den Markern
einer Kopplungsgruppe anhand der Rekombinationsfrequenz zwischen den Marker-
genotypen ermittelt wird. Die Abstände zwischen Genorten werden Anhand des
Regressionsverfahrens mit Hilfe vom LOD-Score (LOD = logarithm of odds) berechnet. Der
LOD-Score ist der Logarithmus des Quotienten aus der Kopplungs-Wahrscheinlichkeit geteilt
durch die Wahrscheinlichkeit einer Nicht-Kopplung. Die ermittelte Rekombinationsfrequenz
wird mit Hilfe einer Kartierungsfunktion, z.B. Haldane oder Kosambi (Haldane, 1919;
Kosambi, 1944) in eine genetische Distanz in Centimorgan (cM) umgerechnet.
1.5 Kartierung von Resistenz-QTL gegen Ährenfusariosen
Eine Erweiterung der genetischen Kartierung ist die QTL-Kartierung (quantitative trait loci).
Bei Getreide ist die Resistenz gegen Fusariumbefall quantitativ ausgeprägt. Quantitative
Merkmale werden durch mehrere bis viele Loci kontrolliert, wobei einzelne Loci nur einen
Teil zur Ausprägung des Merkmales beitragen. Molekulare Marker, die eng an jene
Genombereiche gekoppelt sind, die an der Ausprägung der Fusariumresistenz beteiligt sind,
ermöglichen in Zuchtprogrammen die markergestützte Selektion resistenter Genotypen.
Diese Art von Selektion auf DNA-Ebene spart Zeit und ist gut zu handhaben, da die DNA
auch schon aus Keimlingen extrahiert werden kann. Bei Weizen gibt es diverse
Veröffentlichungen mit QTL-Studien zur Resistenz gegen Ährenfusariosen, die in Buerstmayr
et al. (2009) zusammenfassend beschrieben werden.
Die Identifikation von QTL erfordert zunächst eine spaltende Population bezüglich des
Zielmerkmals zur Phänotypisierung und entsprechend dazu polymorphe molekulare Marker
zur Genotypisierung. Die Markerdaten einer Population werden mit der phänotypischen
Varianz des Zielmerkmals innerhalb dieser Population mit Hilfe statistischer Verfahren
verglichen, welche in Computerprogrammen wie z.B. PLABMQTL (Utz, 2009) implementiert
sind. Zusätzlich zur Identifizierung der QTL kann auch der Beitrag einzelner QTL an der
gesamten phänotypischen Ausprägung des Zielmerkmals geschätzt werden. Bestimmte
Rahmenbedingungen, wie z.B. Populationsgröße und -design, Heritabilität der Merkmale,
Markerdichte und Verteilung der QTL über das Genom und Beitrag der einzelnen QTL zur
gesamten genotypischen Varianz tragen zur Präzision der QTL-Kartierung bei (Asins, 2002).
Die QTL-Kartierung wird genauer, je höher die Heritabilität des Merkmales ist. Auch der
Populationstyp beeinflusst die QTL-Analyse. Die erklärte phänotypische Varianz detektierter
QTL kann leicht überschätzt werden, aufgrund von zu geringen Populationsgrößen, wodurch

10 Einleitung
die QTL mit kleineren Effekten nicht erkannt werden (Utz et al., 2000). Der sogenannte
Permutationstest ist eine gute Methode, um sicherzustellen, dass die gefundenen QTL als
signifikant eingestuft werden können (Churchill und Doerge, 1994). Anhand der Verifikation
durch Testsets können bereits detektierte QTL überprüft werden.
In den letzten Jahren wurden bei Weizen verschiedenste Resistenz-QTL für Ährenfusariosen
kartiert. In diversen Studien, u.a. von Waldron et al. (1999), Anderson et al. (2001),
Buerstmayr et al. (2002), Zhou et al. (2002) und Chen et al. (2006) konnte ein QTL, des aus
China stammenden resistenten Genotyps Sumai 3, auf dem Chromosom 3BS detektiert
werden. Dieser erklärte bis zu 60 % der phänotypischen Varianz nach Einzel-
ährcheninokulation (Typ II-Resistenz). Nach der Verwendung der Sprühinokulation konnten
bis zu 20 % der phänotypischen Variation auf diesen QTL zurückgeführt werden (Buerstmayr
et al., 2003; Chen et al., 2006). Ein weiterer QTL wurde auf dem Chromosom 5A identifiziert
(Buerstmayr et al., 2002 und 2003). Dieser hatte einen Effekt auf die Typ I-Resistenz. Die
aus Brasilien stammende, resistente Weizensorte Frontana wurde in mehreren Studien
kartiert, jedoch ohne QTL zu detektieren, die große Effekte hatten (Steiner et al., 2004; Mardi
et al., 2006; Buerstmayr et al., 2009). In der umfangreichen Publikation von Buerstmayr et al.
(2009) wurden Daten von 52 QTL-Studien mit über 100 veröffentlichten QTL für
Fusariumresistenz zusammengefasst. In insgesamt 20 Chromosomen konnten wiederholt
QTL gefunden werden. Es wurden laut dieser Publikation auf allen Weizenchromosomen,
außer auf 7D, QTL detektiert. Bei Triticale wurden bisher weder komplette genetische Karten
publiziert, noch sind QTL-Studien zur Fusariumresistenz oder DON-Gehalt bekannt.

11 Einleitung
1.6 Zielsetzungen dieser Arbeit
Übergeordnetes Ziel der vorliegenden Arbeit ist die QTL-Kartierung. Im Einzelnen soll
folgendes erreicht werden:
Phänotypische Erfassung des mit F. culmorum künstlich inokulierten Fusariumbefalls
von:
a) 250 F2:3 - bzw. F2:4 - Linien und 150 DH-Linien der Kreuzung LASKO x ALAMO
in sechs Umwelten (Jahr x Ort-Kombinationen)
b) je 100 bis 150 F2:4- Linien der Kreuzungen LASKO x VITALIS, LASKO x
TRIMESTER und LUPUS x SW TALENTRO an drei Orten
Beziehung zwischen visuellem Ährenbefall und DON-Gehalt bzw. Kornbonitur des
Erntegutes
QTL-Analysen der Kreuzungsnachkommenschaften von LASKO x ALAMO bzw.
LASKO x TRIMESTER

12 Material und Methoden
2. Material und Methoden
2.1. Versuchsüberblick
In drei Versuchsjahren (2007-2009) wurden verschiedene Triticale-Kreuzungen (resistent x
anfällig) auf Resistenz gegenüber Ährenfusariosen nach künstlicher Inokulation mit Fusarium
culmorum untersucht (Tab.1).
Tab.1: Übersicht über alle in diesem Projekt verwendeten Kreuzungspopulationen; TR=Teilramsch; DH = Doppelhaploide
Kreuzung
(resistent x anfällig)
Generation Anzahl
Linien
Jahr des Feld-
experimentes
Verwendung
LASKO x ALAMO F2:3-/F2:4-TR 250 2007a + 2008b Phänotypisierung
LASKO x ALAMO DH 150 2008 + 2009b QTL-Analyse
LASKO x TRIMESTER F2:4-TR 100 2009 QTL-Analyse
LASKO x VITALIS F2:4-TR 100 2009 Phänotypisierung
LUPUS x SW TALENTRO F2:4-TR 150 2009 Phänotypisierung
a An den Standorten Hohenheim, Bohlingen und Grabau
b An den Standorten Hohenheim, Bohlingen und Ranzin
2.2. Pflanzenmaterial
Das Ausgangsmaterial für dieses Projekt stammt aus Beständen der AG Triticale der
Landessaatzuchtanstalt (Universität Hohenheim, Stuttgart). In vier Populationen wurde
LASKO als resistenter Kreuzungselter gewählt, die Partner waren mittel bis hoch anfällig.
Eine fünfte, unabhängige Population, wurde mit LUPUS als resistenterem mit SW
TALENTRO als anfälligerem Elter aufgebaut.
Die Kreuzungen wurden mit den jeweiligen Eltern durch Handkastration im Gewächshaus
erstellt. Die F1-Generationen wurden im Feld angebaut. Die folgenden F2-Pflanzen
wurden als Einzelpflanzen im Feld angebaut und die Ähren einer Pflanze wurden mit Hilfe
von Papiertüten geselbstet. Von den F2-Pflanzen wurden nun Teilramsche gebildet, die als
F2:3 und F2:4 in den folgenden Jahren als Mikroparzellen angebaut wurden.
Die Herstellung der DH-Pflanzen beruht auf der Methode der Triticale-Mais-Kreuzung, die in
Rogalska et al. (1996) beschrieben wird. Die DH-Herstellung erfolgte mit derselben F1-
Generation, wie die Erstellung der spaltenden Nachkommen der Kreuzung LASKO x
ALAMO. Die Antheren der Pflanze wurden ein bis zwei Tage vor der Blüte entfernt und nach
einigen Tagen mit Hilfe eines Pinsels mit Maispollen bestäubt. Zur Förderung der
Embryonenentwicklung wurde nach einigen Tagen eine Hormonlösung zwischen dem letzten

13 Material und Methoden
Nodium und der Ähre eingespritzt. Die nach 15 bis 16 Tagen gebildeten Embryonen wurden
unter dem Binokular explantiert und auf Nährmedium in Petrischalen übertragen.
Die Embryonen mussten in den ersten ein bis zwei Wochen in vitro im Dunkeln bei 24 °C
kultiviert werden, bevor sie auf Regenerationsmedien in größere Kulturgefäße übertragen
werden konnten und zu kleinen haploiden Pflanzen heranwuchsen. Die neu gebildeten
Pflanzen wurden in Erdsubstrat überführt und im Gewächshaus weiterkultiviert. Nach
weiteren drei bis vier Wochen erfolgte die Colchizinbehandlung. Hierzu wurden die Pflanzen
aus dem Erdsubstrat herausgenommen und Wurzeln bzw. Blätter um ein Drittel gekürzt.
Danach wurden die Wurzeln bei 22 °C bis ca. ein Zentimeter über der Wurzelkrone für fünf
Stunden in eine 0,1 %-ige Colchizinlösung getaucht. Das überschüssige Colchizin wurde
anschließend ausgewaschen und die Pflanzen wieder in Erdsubstrat gepflanzt. Die neu
gebildeten Sprossen erhielten so den identisch verdoppelten Chromosomensatz, waren fertil
und homozygot. Sie wurden im Gewächshaus und eine weitere Generation im Feld
vermehrt.
2.3. Versuchsstandorte und Design der Feldversuche
In allen Versuchsjahren wurden die Feldexperimente aufgrund der großen Bedeutung der
Genotyp x Ort-Interaktion an verschiedenen Standorten in Süd- und Norddeutschland
(Hohenheim, Bohlingen [Saatzucht Dr. Hege] und Grabau [SAKA] im ersten Jahr bzw.
Ranzin/Ostvorpommern [SAKA] in den folgenden zwei Jahren) durchgeführt (Abb.3).
Abb. 3: Übersicht über die Standorte der Feldversuche: Hohenheim bei Stuttgart; Bohlingen bei Radolfzell am Bodensee; Grabau bei Hamburg; Ranzin bei Greifswald

14 Material und Methoden
Die Feldexperimente wurden für alle Kreuzungspopulationen getrennt an den drei
Versuchsstandorten in einer jeweils an die Größe der Population angepassten
unvollständigen Blockanlage ( -Design) mit je drei Wiederholungen angebaut. Die Eltern
wurden in der jeweiligen Kreuzungspopulation in sechsfacher Anzahl mit angebaut.
Zusätzlich wurden in die Blockanlagen die Standards integriert. Alle Prüfglieder wurden in
doppelreihigen Mikroparzellen (0,42 x 1,25 m2) angelegt, dies entspricht einer Fläche von
rund 0,5 m2.
2.4. Inokulum und Inokulation
Die Inokulation erfolgte mit dem F. culmorum - Isolat FC 46, das von Snijders und Perkowski
(1990) als hochagressiv eingestuft wurde. Durch die Fähigkeit, hohe Mengen an DON
zu bilden, stellte es im vorliegenden Projekt einen hohen Infektionsdruck an allen
Versuchsstandorten sicher (Miedaner et al. 2003). Zur Herstellung des Inokulums wurden die
FC 46-Isolate aus der Hohenheimer Stammsammlung (AG Roggen), die in mit sterilem
Wasser gefüllten, autoklavierten Eppendorf-Reaktionsgefäßen im Kühlschrank aufbewahrt
werden, verwendet. Die Isolate wurden auf speziell nährstoffarmen Agar-Platten (SNA-
Platten; Nirenberg, 1981) ausgelegt und etwa eine Woche inkubiert, bis die Platten
vollständig mit Myzel bewachsen waren. Aus diesen myzelbewachsenen Platten wurden
erneut kleine Stücke auf neu gegossene SNA-Platten überimpft. Diese Platten wurden zur
Sporulation einige Tage bei 16 bis 18 °C unter langwelliges UV-Licht gestellt. Anschließend
wurde der Pilz nach der von Bockmann (1962) beschriebenen Methode auf Weizenkörnern
kultiviert. Hierzu wurden Weizenkörner über Nacht in Leitungswasser eingeweicht und
anschließend in Autoklaviersäcke gefüllt und für 40 Minuten bei 121 °C autoklaviert. Die
Säcke wurden über Nacht abgekühlt und am nächsten Tag erneut autoklaviert, um eventuell
vorhandende bakterielle Endosporen abzutöten. Die unter UV-Licht gebildeten Sporen
wurden durch Abschwemmen den Weizensäcken zugefügt. Die Säcke wurden bei
Zimmertemperatur (ca. 20 °C) im Dunkeln inkubiert. Nach einer Inkubationszeit von etwa vier
bis sechs Wochen und einem vollständigen Myzelbewuchs wurden die Weizenkörner in 40 x
60 cm2 große desinfizierte Euro-Schalen überführt, flächig ausgebreitet und mit einer
durchsichtigen Folie abgedeckt. Zur Sporulation wurden sie für ein bis zwei Wochen unter
langwelligem UV-Licht bei 18 bis 20 °C gestellt. Die Weizenkörner sollten anfangs täglich
durchmischt werden, um eine Myzelbildung zu unterdrücken und die Sporulation zu
beschleunigen. Die stufenweise Entfernung der Folie erfolgte, um eine langsame Trocknung
der Weizenkörner und damit die Sporulation zu gewährleisten. Die Sporendichte wurde
anschließend durch Auszählen mit Hilfe einer Zählkammer (Neubauer) bestimmt. Hierzu
wurde ein Gramm Weizenkörner abgewogen, 100 ml Leitungswasser hinzugefügt und für 15
Minuten auf den Schüttler (Edmund Bühler GmbH) gestellt. Die restlichen Weizenkörner

15 Material und Methoden
wurden luftgetrocknet, portionsweise in Autoklaviersäcke abgefüllt und konnten bis zur
weiteren Verwendung bei 6 °C gelagert werden. Die Inokulation erfolgte an allen
Versuchsstandorten zur Vollblüte. Um die Unterschiede der Blühzeitpunkte der jeweiligen
Genotypen auszugleichen, wurden alle Versuche drei bis viermal inokuliert. Die
Sporendichte lag bei 500.000 bis 700.000 Sporen ml-1 Die Ausbringung des Inokulums
erfolgte mit einer Hochfeldspritze (Hege 75) bei bedecktem Himmel oder am
Spätnachmittag, um die Infektion zu unterstützen. In allen drei Versuchsjahren wurden
bereits vor der Blüte präventiv Pflanzenschutzmittel ausgebracht.
2.5. Symptombonitur und Erfassung agronomischer Merkmale
Ein bis zwei Wochen nach der Inokulation erfolgte ab Beginn der Symptombildung eine zwei-
bis fünfmalige Bonitur. Der Ährenbefall wurde im Abstand von drei bis vier Tagen in Prozent
(0 bis 100 %) befallener Ährchen pro Parzelle visuell erfasst (Tab. 2). Aus den Bonituren
wurden die Mittelwerte des jeweiligen Genotyps errechnet, sowie die Gesamtmittelwerte der
einzelnen Versuchsstandorte. Zusätzlich zum Ährenbefall wurde das Datum des
Ährenschiebens (EC 51 bis EC 59; 51 = Anfang Ährenschieben; 59 = Ende Ährenschieben)
bonitiert und die Wuchshöhe (cm) gemessen.
Tab. 2: Prozentuale Bonitur des Ährenbefalls
Visueller Befall aller Ährchen
pro Parzelle (%)
Symptombeschreibung
0 % Kein sichtbarer Befall
5-15 % Beginnende Ausbleichung einzelner Ährchen
20-30 % Ähren bis zu ¼ ausgebleicht
35-45 % An allen Ähren Ausbleichungen vorhanden
50-60 % Ähren bis zur Hälfte ausgebleicht
65-75 % Ähren bis zu ¾ ausgebleicht
80-90 % Über 80% der gesamten Ährenfläche befallen
95-100 % Völlige Ausbleichung aller Ähren
Ergänzend wurde im Jahr 2008 eine Kornbonitur der jeweils zehn Prozent anfälligsten und
resistentesten Genotypen sowie der Standardsorten und Eltern durchgeführt als auch im
Jahr 2009 alle Genotypen der DH-Population LASKO x ALAMO mit zwei Wiederholungen.
Dazu wurde eine Kornprobe von ca. 150 g abgenommen und der visuelle Anteil des
sichtbaren Befalls geschätzt (Abb. 4). Die Befallsnoten hatten die Stufen eins, drei, fünf,
sieben bzw. neun (eins = niedriger Befall; neun = hoher Befall).

16 Material und Methoden
2.6. Ernte, Aufarbeitung und Bestimmung des DON-Gehaltes
Das mit dem Mähdrescher (Wintersteiger) parzellenweise gedroschene Erntegut wurde
getrocknet und anschließend mit Hilfe eines Steigsichters (Pelz) gereinigt. Nach der
Reinigung wurden die Körner mit Hilfe einer Labormühle (Foss 1093 Cyclotec Sample Mill;
Siebporengröße: 1 mm) gemahlen. Von den gemahlenen Proben wurden jeweils fünf Gramm
abgewogen, in kleine Papiertüten überführt und bis zur DON-Gehalts-Bestimmung bei -20 °C
aufbewahrt.
Zweifachbestimmungen der DON-Gehalte der zehn Prozent anfälligsten und resistentesten
Genotypen der F2:n-Linien und der kompletten DH-Linien der Kreuzung LASKO x ALAMO,
sowie die der Standardsorten und Eltern wurden mit Hilfe von ELISA (RIDASCREEN FAST-
DON, R-Biopharm AG, Darmstadt) durchgeführt. Dieser Immunotest ist ein kompetitiver
Enzymimmunoassay zur quantitativen Bestimmung von DON in Getreide. Eine
Mikrotiterplatte ist ausreichend für 96 Messungen inklusive der Standardbestimmungen. Zur
DON-Extraktion wurden fünf Gramm jeder Probe in einen Erlenmayerkolben mit 100 ml
destilliertem Wasser gefüllt und für fünf Minuten bei 14.000 Umdrehungen pro Minute (Upm)
auf einem Rüttler geschüttelt. Von den Proben wird jeweils ein Milliliter abgenommen und in
einem Eppendorf Reaktionsgefäß für fünf Minuten bei 14.000 Upm zentrifugiert. Der
Überstand wird in ein neues Eppendorf Reaktionsgefäß überführt und das Pellet verworfen.
Die Mikrotiterplatte muss ca. 45 Minuten vor der Zugabe des Antikörpers auf
Raumtemperatur gebracht werden. Zunächst werden jeweils 50 µl der Probe in die dafür
Abb. 4: Beispiel von visuell unbefallenen Körnern (links) und mit Fusarium culmorum befallenen Körnern (rechts)

17 Material und Methoden
vorgesehenen Kavitäten pipettiert. Im Anschluss werden 50 µl des Standards in zweifacher
Anzahl in die übrigen Kavitäten pipettiert. Danach werden jeweils 50 µl des
Enzymkonjugates in alle Kavitäten hinzugefügt. Die anschließende anti-DON-
Antikörperlösung wird ebenfalls mit 50 µl in jede Kavität pipettiert und für fünf Minuten
inkubiert. Dabei wird die Platte vorsichtig durch manuelle Bewegung gemischt. Nach der
Inkubationszeit wird die Platte dreimal durch Ausschlagen geleert und mit Waschpuffer
gewaschen. Nachfolgend werden je 100 µl des Substrates Chromogen pipettiert. Die Platte
wird erneut manuell gemischt und für drei Minuten abgedeckt inkubiert. Als letzter Schritt
werden jeweils 100 µl Stopp-Reagenz in jede Kavität gegeben und nach zwei Minuten kann
dann die Messung gestartet werden. Dafür wird die Mikrotiterplatte in einen Photometer
(Sunrise) gestellt und mit Hilfe der Software Magellan (V5.03, TECAN) die DON-
Konzentration photometrisch bei 450 nm in mg kg-1 pro Probe errechnet. In Abbildung 5 ist
ein Beispiel für eine fertige ELISA-Platte dargestellt. Die Grundlage der DON-Gehalts-
Bestimmung ist die Antigen-Antikörper-Reaktion. Das freie und das enzymmarkierte DON
konkurrieren um die DON-Antikörperbindestelle. Zur gleichen Zeit werden die anti-DON-
Antikörper von den Fängerantikörpern gebunden. In den Waschschritten wird nicht-
gebundenes, enzymmarkiertes DON wieder entfernt. Gebundenes Enzymkonjugat wandelt
das zuvor hinzugegebene Chromogen in ein blau gefärbtes Endprodukt um. Die Zugabe des
Stopp-Reagenzes führt zu einem Farbumschlag von blau nach gelb. Die Extinktion der
Lösung ist umgekehrt proportional zur DON-Konzentration in der Probe, d.h. je heller die
Farbe, umso höher die DON-Konzentration in der Probe.
S 1
S 2
S 5
S 3
S 4
S 2
S 1
S 3
S 5
S 4
Abb. 5: Beispiel für eine ELISA-Platte nach Abschluss der Messung; S1=0,00ppm, S2=0,22ppm, S3=0,66ppm, S4=2,0ppm, S5=6,0ppm; je heller die Farbe, umso höher die DON-Konzentration

18 Material und Methoden
2.7. Genotypisierung und Kartierung
DNA-Extraktion
Im Rahmen der vorliegenden Dissertation wurden Pflanzen als Ausgangsmaterial zur
Isolierung genomischer DNA verwendet, die im Jahr zuvor auf dem Feld in Roggen-Isolation
angebaut wurden. Es wurden zwei bis drei Blätter je Genotyp von zwei Wochen alten
Pflanzen abgenommen und anschließend für ein bis zwei Wochen in Silicagel getrocknet.
Das Blattmaterial aller Genotypen wurde in kleinere Stücke geschnitten und für fünf bis
sechs Minuten mit einer Kugelmühle gemahlen. Die Isolierung der DNA basierte auf der
CTAB-Methode (Cetylrimethylammoniumbromid) von Murray und Thompson (1980) in 2 ml-
Eppendorf-Reaktionsgefäßen. Hierzu wurde der Extraktionspuffer (100 mM Tric-HCL; pH
8.0; 50 mM Na2-EDTA, 500 mM NaCl; 1,25 % SDS) auf 60 °C vorgewärmt. Vor der Zugabe
wurde zu 10 ml Puffer 50 mg Natriumbisulfit zugegeben. Zu jeder Probe mit 40 bis 50 mg
gemahlenem Blattmaterial wurde ein Milliliter Extraktionspuffer hinzugegeben und mit dem
Vortex durchmischt, bis zur vollständigen Suspension. Anschließend wurde für etwa eine
Stunde in 60 °C heißem Wasserbad, mit Hilfe des Überkopfschüttlers inkubiert. Nach dem
Auffüllen mit Chloroform/Isomylalkohol (24:1; 670 µl SEVAG) auf das doppelte Volumen
erfolgte eine erneute Inkubation im Überkopfschüttler für 15 bis 20 Minuten. Durch
Zentrifugation (10.000 Upm; 10 min) erfolgte die Phasentrennung und die obere, DNA-
enthaltende, wässrige Phase wurde in ein neues zwei ml Eppendorf Reaktionsgefäß
überführt. Nach dreißigminütigem RNA-Verdau (Zugabe von jeweils fünf bis 10 µl RNAse
(10 mg ml-1)) wurde die DNA durch Zugabe von 0,7 Vol% Isopropanol (ca. 650 µl) bei 20 °C
entweder für 20 Minuten im Überkopfschüttler, oder bei Zimmertemperatur über Nacht
gefällt. Die Proben wurden anschließend für 10 Minuten bei 10.000 Upm zentrifugiert und der
Überstand vorsichtig abgekippt. Die gefällte DNA wurde zwei Reinigungen unterzogen,
wobei für die erste Reinigung jeder Probe 500 µl von 0,2 M Natriumacetat in 76 % Ethanol
zugefügt wurde, für 30 Minuten inkubiert und danach bei 10.000 Upm kurz zentrifugiert
wurde. Der Überstand wurde abgenommen und die Proben für den zweiten
Reinigungsschritt mit 500 µl von 10 mM Ammoniumacetat in 76 % Ethanol versetzt und
erneut kurz bei 10.000 Upm zentrifugiert. Die Waschlösungen wurden nach Zentrifugation
verworfen und die Pellets für 20 bis 30 Minuten getrocknet. Die DNA-Aufnahme erfolgte in
100 bis 200 µl TE-Puffer (10 mM Tris-HCL; pH 8.0; ein mM Na2-EDTA). Am nächsten Tag
wurde die DNA in 100 µl TE-Puffer rückgelöst und deren Konzentration auf einem 0,8 %igem
Agarosegel mittels Standardvergleich ( DNA/HindIII) geschätzt. Die Konzentration wurde
anschließend auf 50 ng µl-1eingestellt und bei 6 °C bis zur weiteren Verwendung gelagert.

19 Material und Methoden
PCR, Mikrosatelliten- und DArT- Marker
Die PCR (polymerase chain reaction) ist eine gängige Methode, um DNA zu vervielfältigen,
wobei mit Hilfe einer hitzestabilen Polymerase und Primern die selektive Amplifikation eines
DNA-Abschnittes erreicht werden kann. Die Amplifikation erfolgt über drei Teilschritte
(Denaturierung, Annealing und Extension), dabei werden die Schritte zyklisch wiederholt und
führen zu einer exponentiellen Anhäufung des gewünschten DNA-Abschnittes. Die PCR-
Methode wurde nach Saiki et al. (1985) durchgeführt.
Die 150 DH-Linien der Kreuzung LASKO x ALAMO wurden mit insgesamt 1197 SSR-
Markern (898 Weizen-Marker, 299 Roggen-Marker) aus verschiedenen Quellen auf
Polymorphismen geprüft (Tab. 3). Die meisten Informationen über SSR-Marker, wie deren
Ursprung, Sequenzen und Referenz-Karten können über die Datenbank GrainGenes (2010)
abgerufen werden.
Tab. 3.: Verwendete SSR-Marker, deren geprüfte Anzahl und Quellen
Primererkennung Anzahl Quellen
BARC 141 Song et al., 2002 und 2005
CFA 24 Sourdille et al., 2003
CFD 43 Sourdille et al., 2003
WMS 523 Röder et al., 1998; M. Ganal, IPK, Gatersleben
WMC 163 Gupta et al., 2002
GDM 4 Pestsova et al., 2000
RMS 61 V. Korzun, KWS-LOCHOW, Bergen
SCM 238 B. Hackauf, JKI, Groß-Lüsewitz
Die Sequenzierung der Amplifikationsprodukte erfolgte nach Sanger et al. (1977) mit dem
ALF-Sequenziergerät (Pharmacia Biotech) bzw. MegaBACE (Amersham Pharmacia). Die
Auswertungen erfolgten mit den jeweiligen Software- Programmen ALF Fragment Analyzer
1.03 bzw. MegaBACE Fragment Profiler 1.2 (Abb. 6). Die anschließenden Marker-
untersuchungen mit den Nachkommen wurden auf die gleiche Weise durchgeführt.

20 Material und Methoden
LASKO
ALAMO
F2-Pool
Fragmentlänge
Fragmentlänge
Fragmentlänge
LASKO
LUPUS
ALAMO
KITARO
SWTalentro
TRIMESTER
VITALIS
Standard 2 bei 227 bpStandard bei 71 bp
Fragmentlänge [bp]
A
B
Abb. 6: Screening auf die Eltern mit Hilfe des MegaBace- (A) und des A.L.F. - Sequenziergerätes (B); Bild A zeigt LASKO, ALAMO und einen F2-Pool; Bild B zeigt beide Eltern und fünf Standardsorten; LASKO und ALAMO sind bei diesem Marker monomorph
Um eine bessere Chromosomenabdeckung für die Kartierung zu erreichen, wurden
zusätzlich zu den SSR-Markern DArT-Marker analysiert. Die Genotypisierung der
genomischen DNA der jeweiligen Eltern, Standardsorten und aller Genotypen beider
Kartierungspopulationen erfolgte durch Diversity Array Technology Pty Ltd in Australien. Es
wurden drei Mikro-Chips (von Weizen, Roggen und Triticale) mit insgesamt 7.300 Markern
für die DH-Population der Kreuzung LASKO x ALAMO und für die Population der Kreuzung
LASKO x TRIMESTER verwendet.

21 Material und Methoden
2.8. Statistische Auswertung
Aus den Daten aller Prüfglieder eines Standortes, die an jedem Standort in dreimaliger
Wiederholung standen, wurde mit Hilfe des Computerprogramms PLABSTAT (Utz, 2007) ein
Gitterausgleich durchgeführt und die dort errechneten adjustierten Gittermittelwerte zur
weiteren Datenauswertung verwendet. Nach der Methode von Anscombe und Tukey (1963)
wurden die Daten auf Ausreißer überprüft, stark abweichende Werte wurden in der
Verrechnung als fehlende Werte behandelt. Basierend auf den adjustierten Gitter-
Mittelwerten wurden mit Hilfe einer Varianzanalyse (ANOVA) die Varianzen über die
Umwelten bzw. über die Jahre analysiert. Die Umwelten wurden als Ort-Jahr-Kombinationen
definiert und nach Cochran und Cox (1957) zusammengefasst. Als Faktoren gingen Umwelt,
Genotyp sowie deren Wechselwirkung ein. Mit den Ergebnissen der Varianzanalysen
wurden die Heritabilitäten (Anteil der genetischen an der phänotypischen Varianz), sowie die
Grenzdifferenzen (GD 5%) für alle Merkmale berechnet.
Die Kartierung der polymorphen Marker erfolgte nach Erstellung einer ABH-Matrix in
Microsoft Excel (2007) und nach einer Überprüfung des 1:1- (DH) bzw. des 1:2:1- (F2:n)
Spaltungsverhältnisses (Chi-Quadrat-Test) der jeweiligen Populationen mit dem
Computerprogramm JoinMap 4.0 (Van Ooijen, 2006). Die genetischen Karten wurden mit der
Kartierungsfunktion von Haldane (1919) erstellt. Die Kopplungsgruppen wurden bei LOD ≥ 3
gebildet und für alle Gruppen wurde ein dritter Anlauf (third run) ermöglicht. Anhand der
veröffentlichten Weizen-Mikrosatelliten-Karten von Röder et al. (1998) und Somers et al.
(2004) und der Roggen-DArT-Marker Karte von Bolibok-Bragoszewska et al. (2009) erfolgte
die Zuordnung der Kopplungsgruppen zu den Chromosomen von Triticale. Die Karten
wurden mit MapChart (Voorips, 2002) zur endgültigen Darstellung bearbeitet. Um eine
dichtere Abdeckung der Chromosomen zu ermöglichen, wurde eine integrierte Karte beider
Populationen erstellt.
Die QTL-Analysen wurden mit dem Computerprogramm PLABMQTL (Utz, 2009)
durchgeführt. Bei der QTL-Verrechnung der F2:4-Population wurden die phänotypischen
Daten Ährenbefall, Ährenschieben und Wuchshöhe eingegeben, bei der DH-Population
wurden zusätzlich zu den genannten Merkmalen die DON-Werte verrechnet. Im „final fit“
kalkuliert PLABMQTL für alle identifizierten QTL die erklärte phänotypische Varianz des
Gesamtmodells. Der LOD-Schwellenwert wurde zuvor auf 3,0 festgelegt, ab dem ein QTL als
signifikant angenommen wurde. Alle QTL wurden durch die Angaben der Genomposition,
der benachbarten Marker, dem Abstand zum nächstliegenden Marker, das Konfidenzintervall
(95%KI), den LOD-Wert und die normalisierte partielle phänotypische Varianz (nR2)
beschrieben. Das 95 %ige KI wurde nach Darvasi und Soller (1997) berechnet. Als „Haupt-

22 Material und Methoden
QTL“ werden im Folgenden alle QTL bezeichnet, die mehr als zehn Prozent der
normalisierten phänotypischen Varianz (nR²) erklären (Semagn et al., 2007). Die
phänotypische Varianz (R2) wurde normalisiert, damit die Summe der detektierten QTL dem
Modell R2 entsprechen (Zhu et al., 2004).
Zusätzlich dazu wurde eine fünf-fache Kreuzvalidierung durchgeführt, um die Aussagekraft
der phänotypischen Varianz der gefundenen QTL zu ermitteln. Hierbei wurden QTL für 80 %
der Individuen verrechnet, um sie dann mit den restlichen 20 % zu validieren. Der Original-
Datensatz (OD) wurde in fünf Untergruppen aufgeteilt, wobei vier Untergruppen den Schätz-
Datensatz (SD) darstellten und die letzte Untergruppe als Test-Datensatz (TD) fungierte. Die
Ergebnisse dieser Datensätze konnten anschließend mit dem Originaldatensatz verglichen
werden.

23 Ergebnisse
3. Ergebnisse
3.1 Phänotypische Daten
Die künstlichen Inokulationen mit F. culmorum aller Versuche sind in den Versuchsjahren
2007 bis 2009 an den Standorten Hohenheim, Singen-Bohlingen, Grabau und Ranzin
erfolgreich verlaufen.
3.1.1 Populationen der Kreuzung LASKO x ALAMO
3.1.1.1 F2:n-Populationen
Die Inokulationen im Jahr 2007 und 2008 der Versuche mit den F2:n-Populationen führten
an allen Standorten zu einem mittleren Befall (Tab. 4). Die Befallsmittelwerte lagen bei den
F2:n-Populationen in den Jahren 2007 und 2008 je nach Standort zwischen 25 und 42 %.
Die Befallswerte im Jahr 2007 waren durchschnittlich höher als im Versuchsjahr 2008.
LASKO, der resistentere Elter zeigte an allen Versuchsstandorten übereinstimmend einen
niedrigeren Ährenbefall als der anfällige Elter ALAMO. Die Standardsorte TRIMESTER
hatte im Mittel über zwei Jahre den höchsten Ährenbefall, wohingegen die Standardsorte
BENETTO den geringsten Ährenbefall aufwies. Die Wuchshöhe lag im Mittel je nach
Versuchsstandort zwischen 98 und 118 cm. LASKO war im Jahr 2007 etwas länger und
früher im Ährenschieben als ALAMO, jedoch im Jahr 2008 in etwa genauso lang und
genauso früh beim Ährenschieben wie ALAMO. Das mittlere Ährenschieben über zwei
Jahre lag bei 57,24 (mittelhohes EC-Stadium am Tag der Bonitur).

24 Ergebnisse
Tab. 4: Mittelwerte des Ährenbefalls, der Wuchshöhe und des Ährenschiebens von LASKO, ALAMO, deren 250 F2:n-Nachkommen und sieben Standardsorten 2007 und 2008
2007 2008
Genotyp HOHa BOH GRA HOH BOH RAN
Ährenbefall (%)
Eltern: LASKO 28,33 29,78 39,90 22,76 21,49 14,54 ALAMO 33,94 43,82 41,70 28,95 24,58 20,63 F2:n (LASKO x ALAMO) 32,56 41,66 41,30 27,32 24,96 18,11 Standards: TRIMESTER 40,06 55,42 45,11 54,08 46,89 17,63 LUPUS 42,03 53,14 42,66 45,79 37,46 10,24 MODUS 31,40 31,96 41,64 52,41 33,00 12,69 SW TALENTRO 34,87 39,46 39,88 39,93 34,22 12,09 KITARO 36,38 40,85 46,54 32,06 28,25 15,50 VITALIS 36,64 41,78 46,25 35,43 24,62 12,33 BENETTO 26,99 26,72 40,68 21,24 22,25 12,56
Wuchshöhe (cm)
Eltern: LASKO 120,13 118,61 114,72 111,20 126,00 117,05 ALAMO 114,96 115,71 106,75 111,43 126,00 117,17 F2:n (LASKO x ALAMO) 113,44 112,93 105,71 109,57 123,97 115,94 Standards: TRIMESTER 101,12 91,48 97,79 96,67 101,67 100,00 LUPUS 108,25 96,97 102,82 93,17 112,50 117,05 MODUS 113,26 118,01 113,35 116,67 116,67 125,00 SW TALENTRO 110,97 92,55 91,24 103,33 113,33 118,33 KITARO 109,16 109,06 101,44 106,67 111,67 110,00 VITALIS 114,67 120,33 102,77 116,67 128,33 123,33 BENETTO 116,53 112,99 105,97 110,00 123,33 121,67
Ährenschieben (EC)b
Eltern: LASKO 53,11 58,22 52,78 59,23 56,00 56,71 ALAMO 59,15 59,00 56,24 58,86 56,00 56,68 F2:n (LASKO x ALAMO) 58,01 58,87
54,71 58,49 56,89 56,46
Standards: TRIMESTER 54,68 57,00 52,25 57,00 53,33 54,67 LUPUS 51,05 54,35 50,94 56,33 57,00 56,23 MODUS 56,94 59,00 51,04 57,00 54,33 55,33 SW TALENTRO 57,07 57,01 53,90 58,33 54,00 57,33 KITARO 57,33 59,00 54,98 58,33 57,00 56,67 VITALIS 59,06 58,32 57,10 58,33 57,67 56,67 BENETTO 56,26 58,35 52,53 59,00 54,33 55,67 a HOH = Hohenheim; BOH = Bohlingen; GRA = Grabau; RAN = Ranzin
b 51 = Beginn Ährenschieben; 59 = Ende Ährenschieben
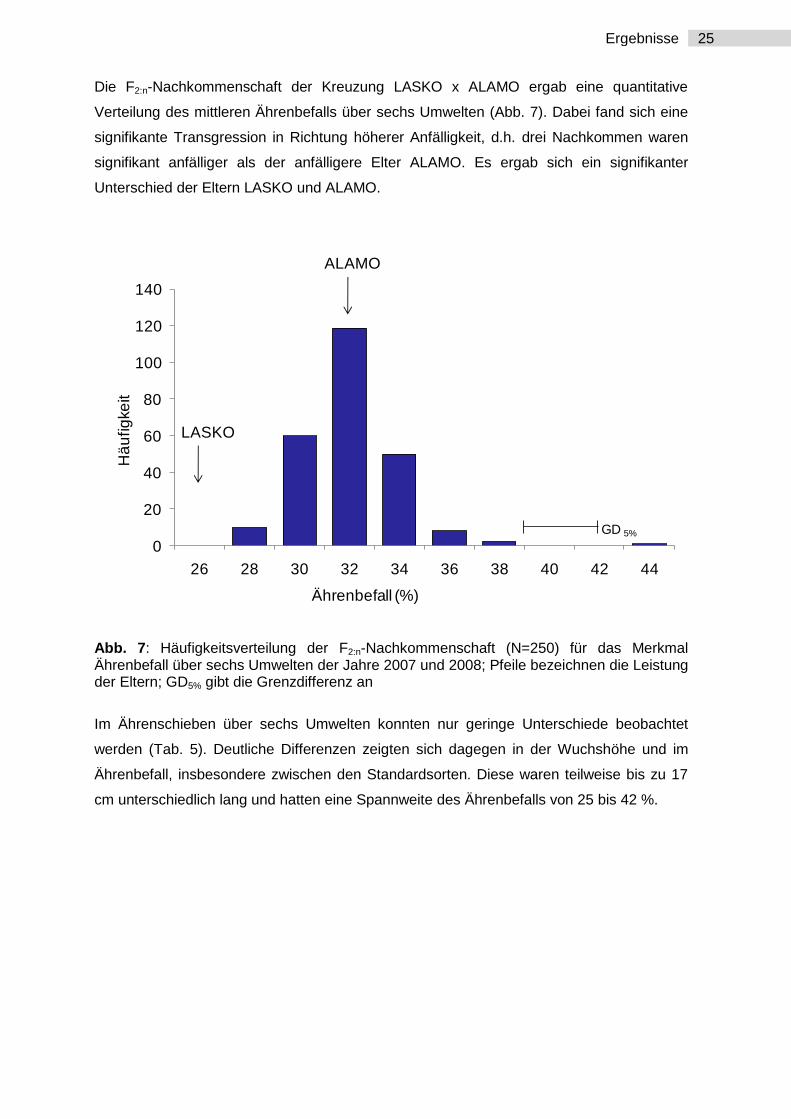
25 Ergebnisse
Die F2:n-Nachkommenschaft der Kreuzung LASKO x ALAMO ergab eine quantitative
Verteilung des mittleren Ährenbefalls über sechs Umwelten (Abb. 7). Dabei fand sich eine
signifikante Transgression in Richtung höherer Anfälligkeit, d.h. drei Nachkommen waren
signifikant anfälliger als der anfälligere Elter ALAMO. Es ergab sich ein signifikanter
Unterschied der Eltern LASKO und ALAMO.
0
20
40
60
80
100
120
140
26 28 30 32 34 36 38 40 42 44
Hä
ufig
ke
it
Ährenbefall (%)
ALAMO
LASKO
GD 5%
Abb. 7: Häufigkeitsverteilung der F2:n-Nachkommenschaft (N=250) für das Merkmal Ährenbefall über sechs Umwelten der Jahre 2007 und 2008; Pfeile bezeichnen die Leistung der Eltern; GD5% gibt die Grenzdifferenz an
Im Ährenschieben über sechs Umwelten konnten nur geringe Unterschiede beobachtet
werden (Tab. 5). Deutliche Differenzen zeigten sich dagegen in der Wuchshöhe und im
Ährenbefall, insbesondere zwischen den Standardsorten. Diese waren teilweise bis zu 17
cm unterschiedlich lang und hatten eine Spannweite des Ährenbefalls von 25 bis 42 %.

26 Ergebnisse
Tab. 5: Mittelwerte des Ährenbefalls, der Wuchshöhe und des Ährenschiebens von LASKO, ALAMO, sieben Standardsorten und deren 250 F2:n-Nachkommenschaften über sechs Umwelten
Genotyp Ährenbefall (%) Wuchshöhe (cm) Ährenschieben (EC)
Eltern:
LASKO 26,13 aa 118,41 54,73
ALAMO 32,27 b 117,26 58,00
F2:n (LASKO x ALAMO) 31,00 113,57 57,29
Standards:
TRIMESTER 43,20 97,96 54,86
LUPUS 38,55 107,21 53,58
MODUS 33,85 117,37 55,63
SWTALENTRO 33,41 104,69 56,28
KITARO 33,26 107,72 57,19
VITALIS 32,83 117,96 57,87
BENETTO 25,07 115,03 56,01 a Unterschiedliche Buchstaben bezeichnen signifikante Unterschiede; GD5% = 3,26; P< 0,05
Zwischen dem Ährenbefall und der Bonitur des Ährenschiebens gab es keine Korrelation (P
> 0,1). Die Korrelation zwischen Wuchshöhe und Ährenbefall der 250 F2:n-Nachkommen-
schaften über sechs Umwelten war mit r = -0,47 signifikant (P< 0,01; Abb. 8). Kürzere
Pflanzen zeigten somit auch einen höheren Befall.
80
85
90
95
100
105
110
115
120
125
130
25 30 35 40 45
Wu
ch
shöh
e (cm
)
Ährenbefall (%)
LASKO ALAMO
GD 5%
rp = -0,47**
rg = -0,5++
Abb. 8: Korrelation zwischen Wuchshöhe und mittlerem Ährenbefall von ALAMO, LASKO (rote Punkte), deren 250 F2:n-Nachkommenschaften (blaue Rauten) und sieben Standardsorten (grüne Dreiecke) über sechs Umwelten; rp= phänotypische Korrelation; rg = genotypische Korrelation

27 Ergebnisse
Der DON-Gehalt (mg kg-1) ausgewählter Genotypen der F2:n-Population, so dass die
ursprüngliche Variation erhalten blieb, wurde über fünf Umwelten bestimmt (Tab. 6). Die
beiden Eltern hatten im Mittel in ihrem DON-Gehalt einen signifikanten Unterschied. Sowohl
im mittleren Ährenbefall, als auch im mittleren DON-Gehalt lag LASKO auf niedrigerem
Niveau als ALAMO. Von allen geprüften Standard-Genotypen hatten BENETTO und
TRIMESTER die geringsten Toxinwerte, MODUS die höchsten. Der Populationsmittelwert
unterschied sich nicht vom Elternmittel.
Tab. 6: Mittelwerte des DON-Gehaltes (mg kg-1) von ALAMO, LASKO, deren 65 ausgewählten F2:n-Nachkommen und sieben Standardsorten an zwei bzw. drei Orten, im Jahr 2007 und 2008
2007 2008
Genotyp HOHa BOH HOH BOH RAN Mittel
Eltern:
LASKO 40,31 22,14 17,33 13,20 21,72 22,94
ALAMO 61,66 55,13 55,10 21,21 25,83 43,79
F2:n (LASKO x ALAMO) 39,89 34,41 35,92 19,00 27,00 31,24
GD5%b 17,32
Standards:
MODUS 56,12 62,06 70,93 81,96 36,88 61,59
KITARO 83,78 74,41 37,21 36,72 50,56 56,54
SW TALENTRO 68,66 85,10 36,17 29,69 32,97 50,52
LUPUS 67,80 52,46 55,97 25,75 42,85 48,97
VITALIS 64,81 48,36 47,62 26,92 32,44 44,03
TRIMESTER 47,89 47,89 48,32 36,04 31,50 42,33
BENETTO 30,60 35,15 37,67 30,72 19,10 30,65 a HOH = Hohenheim; BOH = Bohlingen; RAN = Ranzin
b GD5% gibt die Grenzdifferenz an; P< 0,05
Zwischen DON-Gehalt und Ährenbefall über fünf Umwelten ergab sich eine signifikante
Korrelation von r = 0,65 (P< 0,01; Abb. 9). Generell hatten die Genotypen, die einen
geringeren Ährenbefall hatten auch geringere DON-Gehalte, allerdings war die Streuung
des Ährenbefalls innerhalb derselben DON-Gehalte sehr groß. So hatten BENETTO und
einzelne F2:3-Nachkommen denselben DON-Gehalt von 30 mg kg-1, der Ährenbefall war
aber mit 25 bzw. 45 % signifikant verschieden.

28 Ergebnisse
10
20
30
40
50
60
70
20 25 30 35 40 45
DO
N-G
eh
alt (m
g k
g-1
)
Ährenbefall (%)
ALAMO
LASKO
GD 5%
Abb. 9: Korrelation zwischen dem DON-Gehalt und dem Ährenbefall im Erntegut von ALAMO, LASKO (rote Punkte), sieben Standardsorten (grüne Dreiecke) und deren 65 ausgewählten F2:n-Nachkommenschaften (blaue Rauten) über fünf Umwelten
Für die Kornbonitur wurden dieselben 65 Genotypen, die bereits für die DON-Analyse
ausgewählt wurden, visuell in die Noten eins, drei, fünf, sieben und neun eingeteilt. Die
Korrelationen zwischen der Kornbonitur und dem DON-Gehalt bzw. dem Ährenbefall wurde
gegeneinander aufgetragen (Abb. 10). Es ergaben sich signifikante Korrelationen von
r = 0,6 bzw. r = 0,7 (P< 0,01).

29 Ergebnisse
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8 9
DO
N-G
eh
alt (m
g k
g-1
)
Kornbonitur (1-9)
ALAMO
LASKOGD 5%
10
20
30
40
1 2 3 4 5 6 7 8 9
Äh
ren
be
fall
(%)
Kornbonitur (1-9)
ALAMO
LASKO
GD 5%
A
B
Abb. 10: Korrelation zwischen der Kornbonitur und dem DON-Gehalt (A) bzw. dem Ährenbefall im Erntegut (B) von ALAMO, LASKO (rote Punkte), sieben Standardsorten (grüne Dreiecke) und deren 65 ausgewählten F2:n-Nachkommenschaften (blaue Rauten) über drei Standorte (HOH, BOH, RAN) im Jahr 2008
Die Varianzanalyse über sechs Umwelten zeigte für die Merkmale Ährenbefall,
Ährenschieben, Wuchshöhe und DON-Gehalt hoch signifikante Unterschiede innerhalb der
Population (P < 0,01; Tab. 7). Der Schätzwert der Varianzkomponenten der Genotypen war
für alle Merkmale, ausgenommen der DON-Gehalt, größer als die Varianzkomponenten der
Genotyp-Umwelt-Interaktion. Es zeigten sich mittlere Heritabilitäten. Die höchsten
Heritabilitäten ergaben sich für das Merkmal Wuchshöhe (h2 = 0,89).
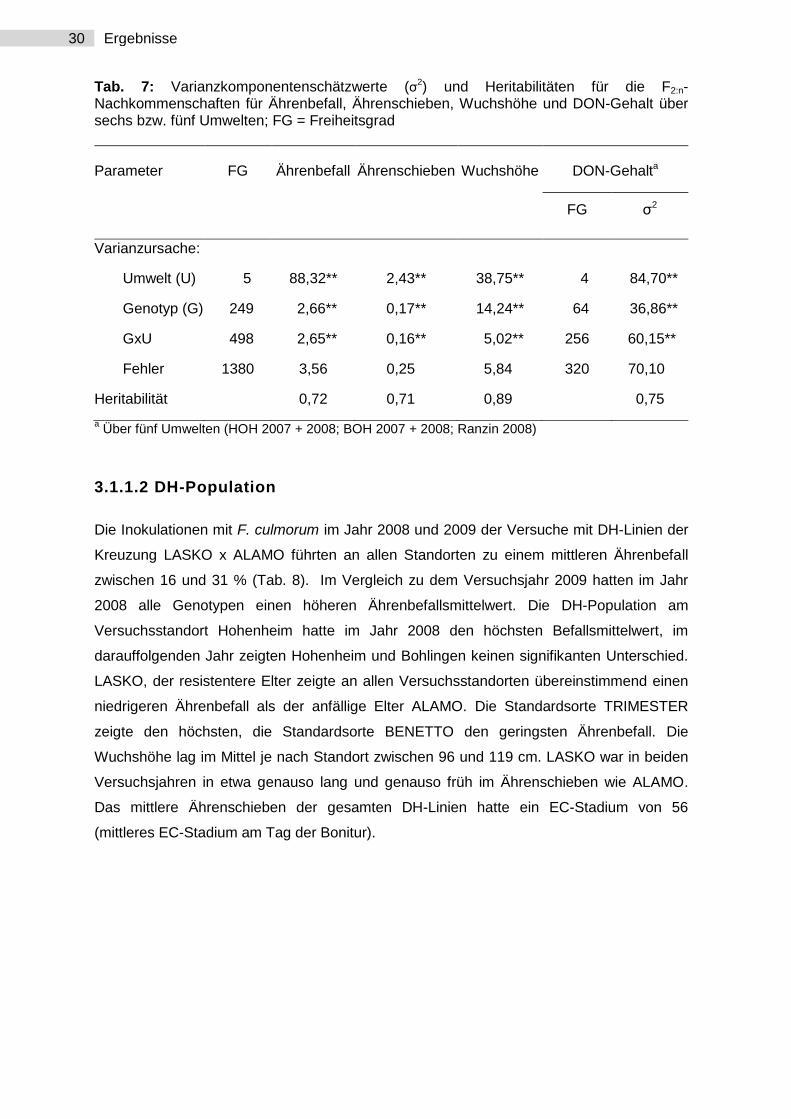
30 Ergebnisse
Tab. 7: Varianzkomponentenschätzwerte (σ2) und Heritabilitäten für die F2:n-Nachkommenschaften für Ährenbefall, Ährenschieben, Wuchshöhe und DON-Gehalt über sechs bzw. fünf Umwelten; FG = Freiheitsgrad
Parameter FG Ährenbefall Ährenschieben Wuchshöhe DON-Gehalta
FG σ2
Varianzursache:
Umwelt (U) 5 88,32** 2,43** 38,75** 4 84,70**
Genotyp (G) 249 2,66** 0,17** 14,24** 64 36,86**
GxU 498 2,65** 0,16** 5,02** 256 60,15**
Fehler 1380 3,56 0,25 5,84 320 70,10
Heritabilität 0,72 0,71 0,89 0,75
a Über fünf Umwelten (HOH 2007 + 2008; BOH 2007 + 2008; Ranzin 2008)
3.1.1.2 DH-Population
Die Inokulationen mit F. culmorum im Jahr 2008 und 2009 der Versuche mit DH-Linien der
Kreuzung LASKO x ALAMO führten an allen Standorten zu einem mittleren Ährenbefall
zwischen 16 und 31 % (Tab. 8). Im Vergleich zu dem Versuchsjahr 2009 hatten im Jahr
2008 alle Genotypen einen höheren Ährenbefallsmittelwert. Die DH-Population am
Versuchsstandort Hohenheim hatte im Jahr 2008 den höchsten Befallsmittelwert, im
darauffolgenden Jahr zeigten Hohenheim und Bohlingen keinen signifikanten Unterschied.
LASKO, der resistentere Elter zeigte an allen Versuchsstandorten übereinstimmend einen
niedrigeren Ährenbefall als der anfällige Elter ALAMO. Die Standardsorte TRIMESTER
zeigte den höchsten, die Standardsorte BENETTO den geringsten Ährenbefall. Die
Wuchshöhe lag im Mittel je nach Standort zwischen 96 und 119 cm. LASKO war in beiden
Versuchsjahren in etwa genauso lang und genauso früh im Ährenschieben wie ALAMO.
Das mittlere Ährenschieben der gesamten DH-Linien hatte ein EC-Stadium von 56
(mittleres EC-Stadium am Tag der Bonitur).

31 Ergebnisse
Tab. 8: Mittelwerte des Ährenbefalls, der Wuchshöhe und Ährenschieben von LASKO, ALAMO, 150 DH-Linien und sieben Standardsorten an drei Orten in den Jahren 2008 und 2009
2008
2009
Genotyp HOHa BOH RAN
HOH BOH RAN
Ährenbefall (%)
Eltern: LASKO 22,76 21,49 14,54 14,74 15,45 7,82 ALAMO 28,95 24,58 20,63 19,22 22,01 14,89 DH-L (LASKO x ALAMO) 26,37 23,77 17,95 18,84 19,87 13,40 Standards: TRIMESTER 54,08 46,89 17,63 21,01 27,08 21,49 SW TALENTRO 39,93 34,22 12,09 24,60 34,32 17,68 LUPUS 45,79 37,46 10,24 22,37 23,42 10,68 MODUS 52,41 33,00 12,69 19,40 19,98 12,00 VITALIS 35,43 24,62 12,23 23,22 24,56 6,86 KITARO BENETTO
32,06 21,24
28,25 22,25
15,50 12,56
20,17 15,85
18,45 18,90
9,66 9,47
Wuchshöhe (cm)
Eltern: LASKO 111,20 126,00 117,05 110,71 120,53 119,15 ALAMO 111,43 126,00 117,17 109,37 119,60 112,67 DH-L (LASKO x ALAMO) 113,71 128,59 118,91 111,50 120,32 113,63 Standards: TRIMESTER 96,67 101,67 100,00 91,99 86,25 102,05 SW TALENTRO 103,33 113,33 118,33 98,02 88,94 102,89 LUPUS 93,17 112,50 117,05 102,40 109,54 108,20 MODUS 116,67 116,67 125,00 103,93 119,07 112,43 VITALIS 116,67 128,33 123,33 108,01 125,57 112,62 KITARO 106,67 111,67 110,00 92,94 106,70 102,77 BENETTO 110,00 123,33 121,67 113,58 120,09 115,40
Ährenschieben (EC)b
Eltern: LASKO 59,23 56,00 56,71 55,55 55,67 56,71 ALAMO 58,86 56,00 56,68 56,45 57,98 56,57 DH-L (LASKO x ALAMO) 58,16 55,53 55,98 56,21 56,33 54,15 Standards: TRIMESTER 57,00 53,33 54,67 55,29 56,89 52,77 SW TALENTRO 58,33 54,00 57,33 55,41 58,35 55,64 LUPUS 56,33 57,00 56,23 54,02 54,59 51,05 MODUS 57,00 54,33 55,33 55,70 56,36 52,78 VITALIS 58,33 57,67 56,67 56,42 57,04 56,52 KITARO 58,33 57,00 56,67 55,86 55,33 55,97 BENETTO 59,00 54,33 55,67 55,98 56,33 53,13 a HOH = Hohenheim; BOH = Bohlingen; GRA = Grabau; RAN = Ranzin
b 51 = Beginn Ährenschieben; 59 = Ende Ährenschieben

32 Ergebnisse
Die DH-Linien der Kreuzung LASKO x ALAMO ergab eine quantitative Verteilung des
mittleren Ährenbefalls über sechs Umwelten (Abb. 11). Der Ährenbefall über sechs
Umwelten hatte eine Spannweite der Genotypen von 14 bis 31 %. LASKO und ALAMO
zeigten einen signifikanten Unterschied im Ährenbefall bei einer Grenzdifferenz von 4,6 im
Mittel. Der Befall der Nachkommen im Mittel über sechs Umwelten lag bei 20,03 %. Es
ergab sich eine signifikante Transgression in Richtung höherer Anfälligkeit, es gab jedoch
auch einen Nachkommen, der einen signifikant geringeren Befallswert hatte als der Elter
LASKO.
0
10
20
30
40
50
60
14 16 18 20 22 24 26 28 30
Hä
ufig
ke
it
Ährenbefall (%)
ALAMO
LASKO
GD 5%
Abb. 11: Häufigkeitsverteilung der DH-Linien über sechs Umwelten; Pfeile bezeichnen die Leistung der Eltern; GD5% gibt die Grenzdifferenz an; P< 0,05
Die Standardsorte TRIMESTER war auffallend kürzer als die anderen Standardsorten und
zeigte den höchsten Ährenbefall (Tab. 9). Im Ährenschieben war LASKO früher als
ALAMO, deren Wuchshöhe zeigte keinen Unterschied. Die Korrelation zwischen dem
Ährenbefall und der Wuchshöhe bzw. dem Ährenschieben der 150 DH-Linien über sechs
Umwelten betrug r = -0,2 bzw. r = 0,16 (P < 0,01).

33 Ergebnisse
Tab. 9: Mittelwerte des Ährenbefalls, der Wuchshöhe und des Ährenschiebens von LASKO, ALAMO, deren 150 DH-Linien, und sieben Standardsorten über sechs Umwelten
Genotyp Ährenbefall (%) Wuchshöhe (cm) Ährenschieben (EC)
Eltern:
LASKO 16,13 117,90 54,61 ALAMO 21,71 117,96 57,47 DH-L 20,03 117,78 56,06
Standards: TRIMESTER 31,45 96,28 55,03 LUPUS 24,99 109,23 54,13 MODUS 24,91 115,84 55,27 SWTALENTRO 27,14 103,87 56,68 KITARO 20,68 104,85 56,47 VITALIS 21,15 119,36 57,12 BENETTO 16,71 117,44 55,73
Die beiden Eltern der DH-Linien der Kreuzung LASKO x ALAMO unterschieden sich im
Mittel in ihrem DON-Gehalt (Tab. 10). Der Elter LASKO lag im DON-Gehalt auf einem
niedrigeren Niveau als ALAMO. Von den Standard-Genotypen hatten BENETTO und
LUPUS die geringsten Toxinwerte, MODUS die höchsten. Bei der Kornbonitur konnte man
im Mittel bei einer Grenzdifferenz von 1,84 einen signifikanten Unterschied der Eltern
feststellen. Mit einer Boniturnote von 8 hatten die Standardsorten MODUS und
TRIMESTER die höchsten, die Standardsorte BENETTO mit 5,7 den geringsten sichtbaren
Befall der Körner.
Tab. 10: Mittelwerte des DON-Gehaltes, des Ährenbefalls und der Kornbonitur von LASKO, ALAMO, deren 150 DH-Linien und sieben Standardsorten an drei Orten, 2009
Genotyp DON-Gehalt (mg kg-1
)
Ährenbefall (%)
Kornbonitur
(1-9)b
HOHa BOH RAN Mittel
Kreuzungspopulation:
LASKO 18,38 6,89 5,66 10,31 12,67 3,28
ALAMO 31,87 22,41 19,10 24,46 18,71 5,06
DH-L (LASKO x ALAMO) 27,87 16,38 13,63 19,29 17,37 4,57
Standards:
MODUS 44,05 45,50 18,37 35,98 17,13 8,33
VITALIS 31,32 39,42 34,74 35,16 18,21 6,33
KITARO 42,68 31,13 18,88 30,90 16,09 6,33
TRIMESTER 39,58 14,50 20,68 24,92 23,19 8,33
SW TALENTRO 25,22 23,51 18,34 22,36 25,53 7,33
LUPUS 24,33 25,18 16,63 22,05 18,82 6,17
BENETTO 18,74 14,82 15,80 16,46 14,74 5,67 a HOH = Hohenheim; BOH = Bohlingen; RAN = Ranzin
b 1 = kein visuell sichtbarer Befall; 9 = höchster visuell sichtbarer Befall

34 Ergebnisse
Für die Korrelation zwischen dem Ährenbefall und dem DON-Gehalt bzw. zwischen der
Kornbonitur der 150 DH-Linien im Jahr 2009 ergaben sich geringe bis mittlere Werte von
r = 0,32 bzw. r = 0,61 (P < 0,01; Abb. 12).
0
5
10
15
20
25
30
35
40
0 5 10 15 20 25 30
DO
N-G
eh
alt (m
g k
g-1
)
Ährenbefall (%)
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8 9
DO
N-G
eh
alt (
mg
kg
-1)
Kornbonitur (1-9)
ALAMO
ALAMO
LASKO
LASKO
GD 5%
GD 5%
A
B
r = 0,32
r = 0,61
Abb 12: Beziehung zwischen der Kornbonitur und dem Ährenbefall im Erntegut (A) bzw. dem DON-Gehalt (B) von ALAMO, LASKO (rote Punkte), deren 150 DH-Linien (blaue Rauten) und sieben Standardsorten (grüne Dreieicke), über drei Standorte (HOH, BOH, RAN) im Jahr 2009

35 Ergebnisse
In der Varianzanalyse für die 150 DH-Linien der Kreuzung LASKO x ALAMO über sechs
Umwelten für die Merkmale Ährenbefall, Ährenschieben, Wuchshöhe und DON-Gehalt
ergaben sich signifikante genotypische Unterschiede innerhalb der Linien (Tab. 11). Der
Schätzwert der Varianzkomponenten der Genotypen war für alle Merkmale größer als die
Varianzkomponenten der Genotyp-Umwelt-Interaktion. Es zeigten sich hohe Heritabilitäten,
der höchste Wert ergab sich für das Merkmal Wuchshöhe (h2 = 0,92).
Tab. 11: Varianzkomponentenschätzwerte (σ2) für die 150 DH-Linien der Kreuzung LASKO x ALAMO für Ährenbefall, Ährenschieben, Wuchshöhe und DON-Gehalt über sechs bzw. drei Umwelten
Parameter FG Ährenbefall Ährenschieben Wuchshöhe DON-Gehalta
FG σ2
Varianzursache:
Umwelt (U) 5 20,74** 1,69** 39,49** 2 58,56**
Genotyp (G) 149 4,46** 0,64** 26,67** 149 21,28**
GxU 745 2,99** 0,43** 8,33** 298 17,60**
Fehler 3198 2,39 0,19 6,04 447 30,44
Heritabilität 0,83 0,86 0,92 0,68
a Für drei Orte im Jahr 2009

36 Ergebnisse
3.1.1.3 Vergleich der Populationen
In Abbildung 13 werden die Häufigkeitsverteilungen der F2:4-Population mit der DH-
Population über drei Orte im Jahr 2008 verglichen. Beide Populationen wiesen eine
Normalverteilung des Ährenbefalls auf. Die DH-Population zeigte im Mittel einen
niedrigeren Ährenbefall. Die Spannweiten des Ährenbefalls von 18 bzw. 21 bis 34 % waren
in etwa gleich.
0
20
40
60
80
100
120
140
16 18 20 22 24 26 28 30 32 34
Hä
ufig
ke
it (A
nza
hl L
inie
n)
Ährenbefall (%)
F2:4
DH
LASKO
ALAMO
GD 5%
Abb. 13: Häufigkeitsverteilung der F2:4-Nachkommenschaften (rot) und der DH-Population (blau) der Kreuzung LASKO x ALAMO im Vergleich für das Merkmal Ährenbefall über drei Orte (Hohenheim, Bohlingen, Ranzin) im Jahr 2008; Pfeile bezeichnen die Leistung der Eltern; GD5% gibt die Grenzdifferenz an
In der Varianzanalyse für die 250 F2:4-Nachkommenschaft und die 150 DH-Linien der
Kreuzung LASKO x ALAMO über drei Orte im Jahr 2009 für das Merkmal Ährenbefall
ergaben sich signifikante genotypische Unterschiede (Tab. 12). Die Varianz-
komponentenschätzwerte der DH-Linien zeigten größere Werte als die der F2:4-
Nachkommenschaften. Es zeigte sich ebenfalls eine höhere Heritabilität.

37 Ergebnisse
Tab. 12: Varianzkomponentenschätzwerte (σ2) für die F2:4-Nachkommenschaften und DH-Linien für Ährenbefall über drei Orte (Hohenheim, Bohlingen, Ranzin) im Jahr 2008, FG = Freiheitsgrad
F2:4 DH-L
Parameter FG Ährenbefall (%) FG Ährenbefall (%)
Varianzursache:
Ort (O) 2 22,86** 2 18,56**
Genotyp (G) 249 1,28** 149 3,93**
GxO 498 1,31** 298 2,45**
Fehler 2328 2,05 2328 2,05
Heritabilität 0,53 0,72
3.1.2 Weitere Populationen
Die Häufigkeitsverteilungen der zwei Kreuzungspopulationen LASKO x TRIMESTER
und LASKO x VITALIS, sowie der unabhängigen Kreuzungspopulation
LUPUS x SWTALENTRO unterschieden sich in ihrer Anfälligkeit (Abb. 14). Die
unabhängige F2:4-Kreuzungspopulation LUPUS x SW TALENTRO entsprach am besten der
Normalverteilung. Mit einem Populationsmittelwert von 16,52 % hatte sie auch den
geringsten Befall. Die Kreuzungspopulation LASKO x VITALIS hatte mit einem
Populationsmittelwert 19,07 % einen etwas höheren Ährenbefall.

38 Ergebnisse
0
10
20
30
40
50
8 10 12 14 16 18 20 22 24 26 28
Hä
ufig
ke
it (A
nza
hl L
inie
n)
Ährenbefall (%)
Lasko x Trimester
0
10
20
30
40
50
8 10 12 14 16 18 20 22 24 26 28
Hä
ufig
ke
it (A
nza
hl L
inie
n)
Ährenbefall (%)
Lasko x Vitalis
0
10
20
30
40
50
8 10 12 14 16 18 20 22 24 26 28
Hä
ufig
ke
it (A
nza
hl L
inie
n)
Ährenbefall (%)
Lupus x SWTalentro
A
C
B
GD 5%
GD 5%
GD 5%
SWTalentro
LASKO
VITALIS
LUPUS
LASKO
TRIMESTER
Abb. 14: Häufigkeitsverteilungen der Populationen LASKO x TRIMESTER (A; N = 100), LASKO x VITALIS (B; N = 100) und LUPUS x SW TALENTRO (C; N = 150) gemittelt über drei Orte; 2009

39 Ergebnisse
Nur in der Kreuzungspopulation LASKO x VITALIS zeigte der resistente Elter LASKO einen
signifikant geringeren Ährenbefall, als der anfällige Elter VITALIS (Tab. 13). Die Mittelwerte
und genotypischen Spannweiten des Ährenbefalls der drei F2:4-Populationen über drei Orte
im Jahr 2009 lagen zwischen 8 und 27 %.
Tab. 13: Mittelwerte und genotypische Spannweiten des Ährenbefalls (%) von drei F2:4-Populationen über drei Orte im Jahr 2009; P1 = LASKO bzw. LUPUS; P2 = TRIMESTER, VITALIS bzw. SW TALENTRO
Eltern Populationen
Population P1 P2 GD5% Mittel Popul. Spannweite GD5%
LASKO x TRIMESTER 14,54 aa 18,09 a 3,76 18,15 11,60-25,12 3,38
LASKO x VITALIS 13,77 a 20,43 b 3,97 19,07 13,27-25,34 3,45
LUPUS x SWTALENTRO 17,10 a 24,13 a 8,74 16,52 8,57-27,08 4,81
a Unterschiedliche Buchstaben bezeichnen signifikante Unterschiede; GD5%=Grenzdifferenz; P< 0,05
In der Varianzanalyse für drei F2:4-Kreuzungspopulationen über drei Orte für das Merkmal
Ährenbefall ergaben sich für die Orte, Genotypen und Genotyp-Ort-Interaktionen hoch
signifikante Unterschiede (Tab. 14). Die Varianzursache Ort zeigte für den Ährenbefall die
höchsten Varianzkomponenten. In allen Kreuzungspopulationen war die Varianz der
Genotyp-Ort-Interaktion kleiner als die des Genotyps. Es zeigten sich hohe Heritabilitäten
zwischen h2 = 0,7 und 0,82.
Tab. 14: Varianzkomponentenschätzwerte für drei F2:4-Populationen für Ährenbefall über drei Orte (HOH, BOH, RAN) im Jahr 2009; FG=Freiheitsgrad
Parameter LASKO x TRIMESTER
LASKO x VITALIS LUPUS x SWTALENTRO
FG Ähren- befall (%)
FG Ähren- befall (%)
FG Ähren- befall (%)
Varianzursache:
Ort (O) 2 18,59** 2 33,73** 2 36,03**
Genotyp (G) 99 6,89** 99 3,57** 149 7,97**
G x O 198 1,72** 198 2,05** 289 5,65**
Fehler 588 2,68 588 2,55 870 3,30
Heritabilität 0,82 0,70 0,73
** Signifikant bei P <0,01

40 Ergebnisse
3.2 Kartierung von zwei Populationen
Die Kreuzungspopulationen LASKO x ALAMO und LASKO x TRIMESTER wurden mit je
898 Weizen- und 299 Roggen-SSR-Markern untersucht. Aus diesen Markern resultierte ein
Polymorphiegrad von 12 bzw. 8 % (Tab. 15). Dieser Polymorphiegrad wurde mit Hilfe von
DArT-Markern leicht erhöht. Der Polymorphiegrad der Kreuzungspopulationen
LASKO x VITALIS und LUPUS x SW TALENTRO war mit 7 % und 10 % noch geringer als
bei den anderen Populationen. Der Polymorphiegrad war somit bei der Population der
Kreuzung LASKO x ALAMO (18 %) am höchsten. Bei beiden kartierten Populationen wurde
durch einen Chi-Quadrat-Test (χ²) bestätigt, dass es bei mehreren Markern zu einer hoch
signifikanten Abweichung des erwarteten Spaltungsverhältnisses kam.
Tab. 15: Polymorphiegrad der Eltern von vier Kreuzungspopulationen bezogen auf Roggen- bzw. Weizen- SSR-Markern (N=1197) und DArT-Markern (N=7300)
Eltern SSR-Marker DArT-Marker
LASKO / ALAMO 12 % 18 %
LASKO / TRIMESTER 8 % 9 %
LASKO / VITALIS 7 % -
LUPUS / SWTALENTRO 10 % -
Die kartierte Gesamtlänge des Genoms betrug bei der Population der Kreuzung LASKO x
ALAMO 1.815 cM und bei der Population der Kreuzung LASKO x TRIMESTER 1.407 cM
(Tab. 16). Für die Kreuzungspopulation LASKO x ALAMO wurden 1328 polymorphe Marker
mit 146 DH-Linien für die Kartierung verwendet. Von diesen Markern konnten jedoch nur
350 Marker kartiert werden, da sehr viele der polymorphen Marker an derselben Stelle des
Chromosoms lagen (Clusterbildung). Bei der Population der Kreuzung LASKO x
TRIMESTER wurden 94 Individuen mit 631 polymorphen Markern kartiert. Davon wurden
310 Marker in die genetische Karte integriert. Es entstand ein durchschnittlicher
Markerabstand von 5,19 bzw. 4,54 cM.
Tab. 16: Übersicht über die Anzahl der polymorphen und kartierten Marker (SSR- und DArT-Marker) der Kreuzungspopulationen LASKO x ALAMO und LASKO x TRIMESTER
Population # Linien # polymorphe Marker
# kartierte Marker
Polymorphe Regionen (cM)
Durchschnittliche Markerdichte (cM)
LASKO x ALAMO 146 1328 350 1.815 5,19
LASKO x TRIMESTER 94 631 310 1.407 4,54

41 Ergebnisse
3.2.1 Genetische Karte LASKO x ALAMO
Die genetische Karte der DH-Population der Kreuzung LASKO x ALAMO umfasst 21
Chromosomen (siehe Anhang; Abb. 17). Das Roggengenom R war mit 151 kartierten
Markern am besten abgedeckt (Tab. 17). Die beiden Weizen-Genome A und B waren mit
95 bzw. 104 Markern in etwa gleich abgedeckt. Von insgesamt 1201 untersuchten SSR-
Markern waren insgesamt 163 Marker polymorph und 102 davon wurden in die genetische
Karte integriert. Die genetische Karte umfasste 52 Kopplungsgruppen, davon wurden 22
Kopplungsgruppen den Chromosomen zugeordnet, wobei Chromosom 2B aus zwei
Kopplungsgruppen bestand. Es konnten alle der 21 Chromosomen des Triticalegenoms
lokalisiert werden. Insgesamt lagen 20 Marker ungekoppelt vor, von denen 10 Marker
anhand der bekannten Kartenposition zugeordnet werden konnten.
Tab. 17: Anzahl kartierter Marker und deren Chromosomenabdeckung der DH-Population LASKO x ALAMO im A-,B- und R-Genom (SSR- und DArT-Marker)
Chromosom A-Genom B-Genom R-Genom
SSR DArT SSR DArT SSR DArT
1 2 3 1 8 1 19
2 6 2 3/3 9/12 6 28
3 6 8 2 0 3 15
4 2 3 6 7 6 23
5 8 6 5 5 2 22
6 7 16 7 10 1 21
7 14 12 11 15 0 4
Summe 45 50 38 66 19 132
In Abbildung 15 ist beispielhaft ein Ausschnitt aus der genetischen Karte der DH-Population
der Kreuzung LASKO x ALAMO dargestellt. Die DArT-Marker werden mit rPt, tPt und wPt
benannt. Exemplarisch für das ganze Genom sind hier die Chromosomen 4A, 4B und 4R
dargestellt. Alle Chromosomen sind relativ dicht abgedeckt, allerdings zeigen die
Weizenchromosomen oft nur kurze polymorphe Bereiche.

42 Ergebnisse
LASKO X ALAMO
1ABS_WMS330.01A_WMS8350.51A_wPt-11679.01A_wPt-628014.81A_wPt-059520.0
Chromosom_1A
2A_wPt-63930.02A_BARC151.62AS_WMS3722.12AB_WMC632.82A_WMS3286.52A_BARC516.02AL_WMS31222.62A_wPt-324423.2
Chromosom_2A
3A_WMS11100.03AL_WMS6383.1
wPt-109229.1tPt-51372629.83AL_WMS15535.7
wPt-34598845.8
3A_wPt-976167.13A_WMS109673.13A_wPt-133976.63A_wPt-439877.73A_WMS43480.83A_wPt-188883.13A_wPt-039887.03AL_WMS122987.6
Chromosom_3A
4AL_BARC1700.04A_WMS9371.84A_wPt-80196.24A_wPt-36386.74A_wPt-944516.7
Chromosom_4A
5A_BARC1410.05A_wPt-97485.95AL_WMS12369.4tPt-51287719.45A_wPt-909419.6
5A_BARC15132.7
5A_CFA216371.85AL_WMS98277.75A_CFA215578.85A_wPt-105283.15A_wPt-509686.15AL_WMS59595.65A_tPt-4184101.6
5AL_WMS291113.3
Chromosom_5A
1B7B_wPt-25400.01B_wPt-31720.91B_WMS11301.81B_wPt-73718.71B_wPt-735916.61A1B_wPt-285917.81B_wPt-030819.31B_wPt-346520.3
1B_tPt-524966.3
Chromosom_1B
2BL_WMS13000.02B_wPt-06940.82B2D_wPt-42234.72B_wPt-91906.42B_wPt-716110.02B_wPt-700411.31B2B_wPt-877613.2rPt-50825930.12B_wPt-442632.62B_WMC36133.62B_wPt-363235.32B_WMS61937.9
Chromosom_2B1
2B_tPt-46020.02B_tPt-16631.72B_wPt-575912.32B_wPt-927415.3tPt-51318817.02B_BARC3519.62B_WMS120323.6tPt-51365925.82B_WMS112835.72B4D_tPt-462737.32B_wPt-769539.3tPt-585341.5
2B_wPt-966898.7wPt-432799.72B_wPt-9402102.9
Chromosom_2B2
WMS2470.03B_WMS1813.5
Chromosom_3B
4B_wPt-52650.0
4B_WMC12522.94B_wPt-104631.1tPt-51365231.84B_wPt-741237.44BL_5ALWMS736b42.74BL_WMS93047.64B_WMS14952.24B_wPt-879652.64BSL_WMS51354.04B_wPt-614955.1
4B_wPt-939375.04AB_WMC61776.8
Chromosom_4B
1R_SCM210.0
rPt-39966234.6rPt-38980336.2
rPt-50712045.7rPt-50839750.5rPt-39031852.3rPt-41076055.1rPt-288857.7rPt-39972365.6rPt-41106366.3rPt-50662968.0rPt-50743869.3rPt-39937874.8rPt-39010379.1rPt-50832586.1rPt-50705988.5rPt-50852095.2rPt-40132297.8rPt-50656198.2rPt-507790103.7
Chromosom_1R
rPt-4111410.0tPt-42692.02R_SCM108.1rPt-5076759.1rPt-40249414.6rPt-39873715.6rPt-41087118.92R_SCM23321.8rPt-100223.32R_RMS120325.3rPt-50619626.02RL_SCM7526.8rPt-38980730.3rPt-40251932.5rPt-50916034.7rPt-50783736.0rPt-38990837.42R_SCM6142.5SCM25444.9tPt-51365746.8rPt-39876148.5rPt-50854349.8rPt-50649660.4rPt-40092362.0rPt-50910566.6rPt-50909667.9rPt-39883174.9rPt-41106679.2rPt-50538592.3rPt-50539193.0tPt-51340794.8rPt-508776102.2rPt-506604103.4rPt-509197107.4
Chromosom_2R
rPt-5053380.0rPt-4023989.9rPt-40026417.2BARC918.2rPt-50951321.5rPt-39042923.6wPt-129125.0tPt-51411427.4tPt-374633.3rPt-085734.4rPt-50617334.9rPt-40250435.93R_RMS102839.0rPt-38970840.0rPt-38945443.4rPt-50789743.9SCM28445.9rPt-50806852.6
Chromosom_3R
SCM2280.0rPt-5085821.4rPt-38994112.9rPt-41122813.8rPt-38927222.1rPt-50516533.2rPt-50789937.6rPt-50645838.6rPt-40156339.94R_SCM6640.84R_RMS118141.9rPt-39043643.2rPt-41093245.6rPt-50717947.8rPt-41076849.0rPt-50807049.4rPt-39878357.1rPt-40082859.4rPt-40181762.0rPt-50670664.44R_SCM33965.6tPt-744866.5rPt-38961067.44R_RMS115468.3tPt-51372971.84R_SCM11674.3rPt-50955275.9rPt-50819979.5rPt-39957581.7
Chromosom_4R
rPt-5061720.0rPt-39974312.9rPt-39946214.9rPt-40239116.4rPt-50967721.6rPt-50558122.3rPt-50674724.4rPt-39880028.75R_RMS126434.4rPt-50966837.9rPt-39966340.5rPt-50890241.6rPt-50889742.4tPt-894044.2rPt-40127551.0rPt-50737351.9rPt-50892552.7rPt-50597357.6rPt-39939758.6rPt-50557759.6
rPt-507326107.65R_SCM280109.2rPt-506066113.9rPt-402334116.1
Chromosom_5R
3.2.2 Genetische Karte LASKO x TRIMESTER
Die genetische Karte der Kreuzungspopulation LASKO x TRIMESTER wurde
ausschließlich mit DArT-Markern erstellt. Von den insgesamt 7300 Markern waren 631
polymorph, davon wurden 310 Marker kartiert. Auch in diesem Fall war das Roggen-
Genom mit 170 kartierten Markern am besten abgedeckt. Die beiden Weizen-Genome
zeigten 61 bzw. 79 kartierte Marker. Die genetische Karte der F2:4-Kreuzungspopulation
LASKO x TRIMESTER umfasst 20 Chromosomen (siehe Anhang, Abb. 18). Sie beinhaltet
insgesamt 56 Kopplungsgruppen, von denen mit Ausnahme von fünf Gruppen alle den 21
Chromosomen des Triticalegenoms zugeordnet werden konnten. Das Chromosom 6B
konnte nicht dargestellt werden.
3.2.3 Integrierte genetische Karte
Die integrierte genetische Karte wurde erstellt, um eine vollständige Abdeckung der
Chromosomen zu erhalten (siehe Anhang, Abb. 22). Es gelang jedoch nur von neun
Chromosomen des Triticalegenoms mit insgesamt 21 Chromosomen eine integrierte Karte
zu erstellen, da zu wenig gemeinsame Marker vorhanden waren. Exemplarisch für die
integrierte Karte ist in Abbildung 16 das Chromosom 3R dargestelllt.
Abb. 15: Ausschnitt
aus der Kartierung
der DH-Population
LASKO x ALAMO mit
SSR- und DArT-
Markern des je-
weiligen 4. Chromo-
soms aller drei
Genome A, B und R;
vollständige Karte
siehe Anhang, Abb.
17.

43 Ergebnisse
rPt-5053380.0
rPt-4023989.9
rPt-40026417.2BARC918.2rPt-50951321.5rPt-39042923.6wPt-129125.0tPt-51411427.4tPt-374633.3rPt-085734.4rPt-50617334.9rPt-40250435.93R_RMS102839.0rPt-38970840.0rPt-38945443.4rPt-50789743.9SCM28445.9
rPt-50806852.6
Chr_3R_LA_x_ALA
rPt-5053380.0
rPt-40239810.1BARC918.7rPt-50951321.5rPt-39042924.2wPt-1291 rPt-50696125.3rPt-50971926.7tPt-51411428.2rPt-50787729.5rPt-50650430.0tPt-374633.9rPt-085734.9rPt-50617335.3rPt-38967636.1rPt-40250436.33R_RMS102839.7rPt-38970841.0rPt-38945444.1rPt-50789744.6rPt-50743445.1rPt-50737445.7rPt-39989746.1rPt-50710546.2rPt-39072446.3rPt-50684746.5SCM28446.6rPt-50841846.7rPt-50860947.2rPt-40116349.2rPt-50806853.2rPt-50522763.9rPt-50780168.7
Int_Chr_3R
rPt-5069610.0rPt-5097191.8rPt-5078775.7rPt-5065046.4rPt-4025048.7rPt-5061739.6rPt-38967612.2rPt-38970816.2rPt-50743418.7rPt-50860919.4rPt-50841819.6rPt-39072420.0rPt-50684720.2rPt-38945420.7rPt-39989721.7rPt-40116322.5rPt-50710523.8rPt-50737424.0
rPt-39001837.7rPt-50522741.9rPt-50757543.1rPt-50780144.8rPt-50656545.0rPt-40129645.6rPt-50645546.1rPt-41124346.5rPt-50696747.1rPt-40173850.3rPt-568855.6rPt-40026461.0
Chr_3R_LA_x_TRI
Das integrierte Chromosom aus beiden Populationen enthält eine größere Marker-Anzahl,
als die Chromosomen der jeweiligen Einzelpopulationen. Die Verteilung der jeweiligen
Marker entspricht größtenteils der vorherigen Anordnung der Marker.
3.3 QTL-Analyse von zwei Kreuzungspopulationen
Für die Kreuzungspopulation LASKO x ALAMO wurden insgesamt sechs QTL für das
Ährenschieben mit einer phänotypisch erklärten Varianz von insgesamt 65,9 %, vier QTL
für die Wuchshöhe (38,1 %), ein QTL für den DON-Gehalt (8,4 %) und vier QTL für den
Ährenbefall (36,9 %) detektiert (Tab. 19). Die Standorte wurden einzeln über zwei Jahre
hinweg verrechnet. Hierbei unterschied sich der Standort Hohenheim von den anderen
beiden Standorten. In Hohenheim konnte nur ein QTL für Ährenschieben und ein QTL für
Ährenbefall gefunden werden. Diese hatten jeweils nur sehr kleine Effekte und waren auf
einem anderen Chromosom zu finden, als an den beiden anderen Standorten.
Abb. 16: Beispiel für
eine Karte des
Chromosoms 3R der
Kreuzungspopulationen
LASKO x ALAMO
(links), Kreuzungs-
population LASKO x
TRIMESTER (rechts)
und integrierte Karte
aus beiden Popul.
(mitte); Verbindungs-
linien markieren die
übereinstimmenden
Marker
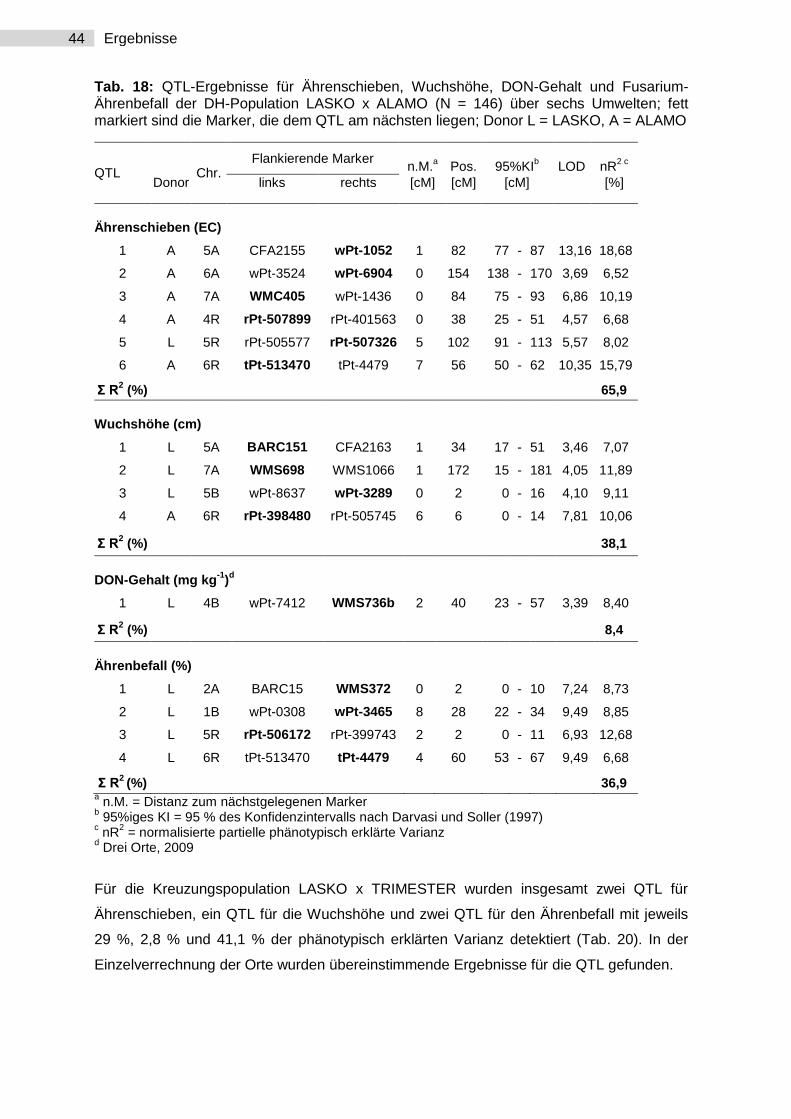
44 Ergebnisse
Tab. 18: QTL-Ergebnisse für Ährenschieben, Wuchshöhe, DON-Gehalt und Fusarium-Ährenbefall der DH-Population LASKO x ALAMO (N = 146) über sechs Umwelten; fett markiert sind die Marker, die dem QTL am nächsten liegen; Donor L = LASKO, A = ALAMO
QTL
Chr. Flankierende Marker
n.M.a Pos. 95%KI
b LOD nR
2 c
Donor links rechts [cM] [cM] [cM] [%]
Ährenschieben (EC)
1 A 5A CFA2155 wPt-1052 1 82 77 - 87 13,16 18,68
2 A 6A wPt-3524 wPt-6904 0 154 138 - 170 3,69 6,52
3 A 7A WMC405 wPt-1436 0 84 75 - 93 6,86 10,19
4 A 4R rPt-507899 rPt-401563 0 38 25 - 51 4,57 6,68
5 L 5R rPt-505577 rPt-507326 5 102 91 - 113 5,57 8,02
6 A 6R tPt-513470 tPt-4479 7 56 50 - 62 10,35 15,79
Σ R2 (%) 65,9
Wuchshöhe (cm)
1 L 5A BARC151 CFA2163 1 34 17 - 51 3,46 7,07
2 L 7A WMS698 WMS1066 1 172 15 - 181 4,05 11,89
3 L 5B wPt-8637 wPt-3289 0 2 0 - 16 4,10 9,11
4 A 6R rPt-398480 rPt-505745 6 6 0 - 14 7,81 10,06
Σ R2 (%) 38,1
DON-Gehalt (mg kg-1
)d
1 L 4B wPt-7412 WMS736b 2 40 23 - 57 3,39 8,40
Σ R2 (%) 8,4
Ährenbefall (%)
1 L 2A BARC15 WMS372 0 2 0 - 10 7,24 8,73
2 L 1B wPt-0308 wPt-3465 8 28 22 - 34 9,49 8,85
3 L 5R rPt-506172 rPt-399743 2 2 0 - 11 6,93 12,68
4 L 6R tPt-513470 tPt-4479 4 60 53 - 67 9,49 6,68
Σ R2 (%) 36,9
a n.M. = Distanz zum nächstgelegenen Marker
b 95%iges KI = 95 % des Konfidenzintervalls nach Darvasi und Soller (1997)
c nR
2 = normalisierte partielle phänotypisch erklärte Varianz
d Drei Orte, 2009
Für die Kreuzungspopulation LASKO x TRIMESTER wurden insgesamt zwei QTL für
Ährenschieben, ein QTL für die Wuchshöhe und zwei QTL für den Ährenbefall mit jeweils
29 %, 2,8 % und 41,1 % der phänotypisch erklärten Varianz detektiert (Tab. 20). In der
Einzelverrechnung der Orte wurden übereinstimmende Ergebnisse für die QTL gefunden.

45 Ergebnisse
Tab. 19: QTL-Ergebnisse für Ährenschieben, Wuchshöhe und Fusarium-Ährenbefall der F2:4-Population LASKO x TRIMESTER (N = 94) über drei Orte, 2009; fett markiert sind die Marker, die dem QTL am nächsten liegen; Donor T = TRIMESTER, Donor L = LASKO
QTL
Chr. Flankierende Marker
n.M.a Pos. 95%KI
b LOD nR
2 c
Donor links rechts [cM] [cM] [cM] [%]
Ährenschieben (EC)
1 T 6A wPt-6904 wPt-5652 5 22 14 - 30 7,62 17,59
2 T 6R rPt-390525 rPt-8205 0 10 0 - 24 4,46 11,39
Σ R2 (%) 29
Wuchshöhe (cm)
1 T 6A rPt-505608 wPt-7445 0 74 57 - 91 3,44 2,77
Σ R2 (%) 2,8
Ährenbefall (%)
1 T 4A wPt-0162 wPt-0763 1 60 50 - 70 6,10 21,07
2 L 6A wPt-5652 wPt-9692 5 34 23 - 45 5,52 20,01
Σ R2 (%)
41,1
a n.M. = Distanz zum nächstgelegenen Marker
b 95%iges KI = 95 % des Konfidenzintervalls nach Darvasi und Soller (1997)
c nR
2 = normalisierte partielle phänotypisch erklärte Varianz
Beide Populationen zeigten im Merkmal Ährenschieben zwei gemeinsame QTL auf den
Chromosomen 6A und 6R. In den anderen Merkmalen wurden keine übereinstimmenden
QTL gefunden. Die zugehörigen LOD-Kurven zur DH-Population sind im Anhang in Abb. 19
bis 21 dargestellt.
In der Kreuzungspopulation LASKO x ALAMO lassen sich 16 Markerklassen je nach
Vorhandensein der resistenten und/oder anfälligen Donor-QTL-Allele unterscheiden (Abb.
17). Es lässt sich eine Tendenz von geringerer Anfälligkeit bei Genotypen mit allen Donor-
QTL-Allelen zu höherer Anfälligkeit bei jenen Genotypen erkennen, die kein Donor-QTL-
Allel besitzen.

46 Ergebnisse
0
5
10
15
20
25
30
Äh
ren
be
fall
(%)
alle 4 QTL
3 QTL
2 QTL
1 QTL
kein QTL
Donor-QTL-Allele 2A
Donor-QTL-Allele 1B
Donor-QTL-Allele 5R
Donor-QTL-Allele 6R
N
+ + + + - + + + - - - - - - + -
+ + - + + + - - - + + + - - - -
+ - + + + - - + + + - - - + - -
+
12
+
7
+
5
-
21
+
16
-
7
+
2
-
7
+
5
-
14
+
9
-
10
+
5
-
3
-
8
-
5
Abb. 17: Übersicht über die Verteilung von 16 Markerklassen mit den resistenten (+) und/oder den anfälligen (-) Donor-QTL-Allelen der Kreuzungspopulation LASKO x ALAMO.
Auch in der Kreuzungspopulation LASKO x TRIMESTER lässt sich bei 4 unterschiedlichen
Markerklassen dieselbe Tendenz von geringerer Anfälligkeit bei Genotypen mit beiden
Donor-QTL-Allelen zu höherer Anfälligkeit bei Genotypen ohne Donor-QTL-Allel erkennen
(Abb. 18).
0
5
10
15
20
25
beide QTL QTL 1 QTL 2 kein QTL
Äh
ren
be
fall
(%)
Donor-QTL-Allele 4A + + - -
Donor-QTL-Allele 6A + - + -
N 14 12 25 9
Abb. 18: Übersicht über die Verteilung von 4 Markerklassen mit den resistenten (+) und/oder den anfälligen (-) Donor-QTL-Allelen der Kreuzungspopulation LASKO x TRIMESTER.

47 Ergebnisse
Um zu sehen, wie aussagekräftig die QTL-Analysen waren, wurde eine „Fünffach-
Kreuzvalidierung“ durchgeführt. Hierbei wurden QTL für 80 % der Individuen verrechnet
und mit den restlichen 20 % validiert. Diese Ergebnisse konnten dann mit dem Original-
Datensatz verglichen werden. Die Kreuzvalidierung zeigte in beiden
Kreuzungspopulationen in dem Schätz-Datensatz eine geringere Anzahl an QTL verglichen
mit dem Original-Datensatz (Tab. 21). Zusätzlich dazu konnte eine verringerte adjustierte
phänotypische Varianz (R2adj.) vom Original-Datensatz zum Test-Datensatz beobachtet
werden.
Tab. 20: Ergebnisse der fünffachen-Kreuzvalidierung mit 30 Durchläufen in zwei Kreuzungspopulationen für Fusarium-Resistenz-QTL
Kreuzungspopulation # Linien # QTL R2adj. (%)b
ODa SD OD SD TD
LASKO x ALAMO 146 4 1 13,21 20,31 8,95
LASKO x TRIMESTER 94 2 1 33,69 36,20 21,55 a OD= Original-Datensatz; SD = Schätz-Datensatz; TD = Test-Datensatz
b Adjustierte phänotypische Varianz bezogen auf alle detektierten QTL je Set

48 Diskussion
4. Diskussion
Die Resistenz von zwei Populationen der Kreuzung LASKO x ALAMO und von drei weiteren
Kreuzungen wurde jeweils über drei Orte und ein bzw. zwei Jahre bei künstlicher Infektion
mit dem F. culmorum Isolat FC 46 geprüft. Bei allen Versuchen konnte eine gute
Differenzierung der Befallswerte beobachtet werden. Eine Infektion des Getreides mit
Fusarium spp. kann zu Verlusten von bis zu 60 % führen (Miedaner et al., 1993; Parry et al.,
1995). Deshalb ist es sehr wichtig, resistente Sorten zu züchten und somit hohe
Ertragsverluste und Mykotoxinkontamination von Getreide zu verringern. Neben der
Resistenz sind jedoch auch andere Eigenschaften der Sorten essentiell. Die geringe
Anfälligkeit einer Sorte bringt nur einen Erfolg, wenn gleichzeitig gute agronomische
Eigenschaften, wie z.B. hohe Erträge bestehen (Miedaner, 1987).
4.1 Phänotypische Analysen Für die vorliegenden Versuche wurde der visuelle Befall aller Ähren pro Parzelle in einer
Skala von 0 bis 100 % durchgeführt. Diese Bonitur hat Vorteile gegenüber der von Miedaner
(1986) und Heinrich (2004) verwendeten Einteilung nach Noten von eins bis neun, da
dadurch eine bessere Differenzierung der Befallswerte ermöglicht wird und zusätzlich zu
einer verminderten Fehlerquelle beiträgt (Büchse et al., 2006). Hierzu ist jedoch zu beachten,
dass die Bonituren an allen Standorten von derselben Person durchgeführt werden sollten,
um eine gleichmäßige Bonitur zu gewährleisten. Es ist wichtig, mehrere Einzelbonituren zu
erfassen, um Unterschiede im Befallsverlauf verschiedener Genotypen aufzeigen zu können
(Mesterházy, 2003). Wird nur eine einzelne Bonitur zu einem festgelegten Zeitpunkt erfasst,
kann es zu Fehlinterpretationen kommen, da für Genotypen mit unterschiedlichem
Blühzeitpunkt und dementsprechend unterschiedlichem Inokulationszeitpunkt im Zeitraum
zwischen der Inokulation und dem Boniturtermin verschiedene Witterungsbedingungen
herrschen können. In der vorliegenden Arbeit wurden alle Genotypen in allen Versuchen drei
bis fünf Mal im Abstand von drei bis vier Tagen bonitiert. Anschließend wurden aus den
Einzelboniturwerten der Mittelwert errechnet und im Weiteren als Merkmal des Ährenbefalls
betrachtet. Um die verschiedenen Termine des Ährenschiebens und die damit
zusammenhängenden unterschiedlichen Blühzeitpunkte innerhalb der Populationen
auszugleichen, wurden drei verschiedene Inokulationstermine veranschlagt. Mit einer
spezifischen Inokulation für jede Linie, könnte man dieses Problem ebenfalls zukünftig
vermeiden, was jedoch mit einem enormen Zeitaufwand und vielen verschiedenen
Inokulationsterminen verbunden wäre. Zwischen dem Ährenschieben und Ährenbefall
ergaben sich keine signifikanten Korrelationen (P > 0,1). Dies deutet darauf hin, dass
erfolgreiche Inokulationen vorgenommen wurden und alle Genotypen zur Vollblüte infiziert

49 Diskussion
wurden. Signifikante Korrelationen zwischen dem Ährenbefall und dem Zeitpunkt des
Ährenschiebens nach Sprühinokulation wurden in zahlreichen Arbeiten festgestellt (Hilton et
al., 1999; Gervais et al., 2003; Somers et al., 2003; Paillard et al., 2004; Schmolke et al.,
2005; Holzapfel, 2009). Bei Liu et al. (2006) hingegen wurde nur in einer von vier Umwelten
eine schwache Korrelation zwischen dem Ährenbefall und dem Zeitpunkt des
Ährenschiebens gefunden. In den Versuchen von Klahr et al. (2007) ergaben sich ebenfalls
nur in einzelnen Umwelten Korrelationen zwischen dem Ährenbefall und dem Zeitpunkt des
Ährenschiebens. Ob zwischen einzelnen Merkmalen Korrelationen auftreten, hängt aber
auch davon ab, wie stark sich die Eltern und somit auch die Nachkommen bezüglich der
erfassten Merkmale unterscheiden.
Die Fusariumbonitur von Triticale ist aufwändiger als die von Weizen, da Triticale eine
schlechtere Symptomausprägung aufweist. Große Unterschiede zeigten sich auch im
Ährentyp, sowohl in der Länge und Farbe der Begrannung, als auch in der Spelzenfarbe
(Furmann et al., 1997). Dadurch ist es sehr schwierig den Befall visuell zu erfassen.
Zusätzlich wird die Bonitur durch den Reifeprozess erheblich erschwert. Sowohl der
Ährenbefall, als auch die Reife führen zur Ausbleichung der Ähren. Für eine gute
Differenzierung sind demnach mehrere frühe Bonituren in kürzeren Abständen sinnvoll.
Während der Bonituren des Merkmals Ährenbefall ergaben sich besondere Bedingungen an
einigen Standorten. In Grabau gab es Probleme mit Lager, wodurch nur zwei Bonituren und
keine ausreichende Differenzierung möglich waren. Dagegen führten die klimatischen
Bedingungen in Ranzin (trocken, heiß) zu schnell abgereiften Pflanzen, wodurch ebenfalls
die Differenzierung erschwert wurde. Demzufolge waren nur drei Bonituren möglich. Die
Standorte Grabau und Ranzin unterschieden sich insgesamt von den anderen Standorten
durch niedrigere Befallswerte. Möglicherweise lag der Unterschied in den Befallswerten der
einzelnen Standorte daran, dass der Krankheitsdruck bei feucht-warmer Witterung höher ist
und somit der Pilz die besseren Bedingungen für seine Entwicklung hat verglichen mit einem
trockenen bzw. heißen Standort. Sowohl Temperaturen von über 25 °C und Feuchteperioden
länger als 24 Stunden, als auch große Temperaturunterschiede begünstigen die
Fusariuminfektion (Brennan et al., 2005). Reinbrecht (2002) zeigte, dass Pflanzen mit
höheren Inkubationstemperaturen weniger offensichtliche Unterschiede und höhere
Befallswerte aufwiesen, als jene Pflanzen mit niedrigeren Inkubationstemperaturen.
Witterungsbedingungen sind in den ersten Stunden nach der Inokulation für die Infektion
entscheidend (Mauler-Machnik und Zahn, 1994; Miedaner et al., 2001; Reinbrecht, 2002).
Die Befallswerte in den vorliegenden Untersuchungen lagen je nach Kreuzungspopulation im
Mittel zwischen 8 und 42 %. Ähnliche Befallswerte ließen sich auch in den Versuchen von
Schmolke (2004) und Wilde et al. (2007) bei Weizen feststellen. Im Vergleich zu den

50 Diskussion
Untersuchungen von Rodemann und Mielke (2007), in denen die Sorte LUPUS über zwei
Jahre an fünf Orten die geringsten Befallswerten erzielte, hatte LUPUS in den dargestellten
Versuchen nur mäßige Befallswerte. LUPUS wurde in der vorliegenden Doktorarbeit für die
Population LUPUS x SW TALENTRO als resistenter Elter ausgewählt. Insgesamt ergab sich
eine gute Differenzierung innerhalb der verschiedenen Kreuzungen. Alle Populationen
erzielten hohe Heritabilitäten und es ergaben sich signifikante genotypische Unterschiede
innerhalb der einzelnen Populationen. Dies deutet auf eine hohe Reproduzierbarkeit der
phänotypischen Daten hin.
Bei den vorliegenden Versuchen wurden Transgressionen in Richtung resistentere und
anfälligere Genotypen in einigen Populationen für das Merkmal Ährenbefall beobachtet. Dies
ist ein Anzeichen dafür, dass Resistenzallele auch vom anfälligen Elter kommen können.
Eine ausgeprägte Transgression zeigte sich in der Kreuzungspopulation LASKO x ALAMO.
Der Elter ALAMO ist im Gegensatz zum Elter LASKO nur mäßig anfällig. Abhängig von der
Rekombination der Resistenz- bzw. Anfälligkeitsallele kann es bei den Nachkommen
entweder zu einer verbesserten Resistenz oder aber auch zu einer erhöhten Anfälligkeit
kommen. Transgressive Spaltungen der Linien in Richtung anfälligere Genotypen, wie auch
eine quantitative Verteilung des Ährenbefalls sind ebenso in den Untersuchungen von
Gervais et al. (2003), Paillard et al. (2004) und Schmolke et al. (2005) zu Weizenlinien nach
Sprühinokulation beobachtet worden.
Verglichen mit kurzstrohigen Genotypen zeigen Pflanzen mit einer langen Halmlänge bei
natürlicher Infektion einen geringeren Befall (Mesterházy, 1995). Es wird vermutet, dass der
Zusammenhang zwischen Wuchshöhe und Befall eine genetische Ursache hat, da
Genotypen mit großer Wuchshöhe auch nach Sprühinokulation einen geringeren Befall
zeigen (Buerstmayr et al., 2000; Gervais et al., 2003; Paillard et al., 2004; Steiner et al.,
2004). Das Merkmal Wuchshöhe wird demnach, wie die Resistenz gegen Ährenfusariosen,
quantitativ vererbt. In europäischen Weizensorten sind verschiedene Verzwergungsgene
(Rht) vorhanden (Holzapfel et al., 2008; Voss et al., 2008). Ein Zusammenhang zwischen
Resistenz und Langstrohigkeit wurde auch in Studien mit Winterweizen von Hartl et al.
(2001) bestätigt. Dort zeigten Sorten mit mittlerer Halmlänge eine hohe Fusariumresistenz.
Hingegen konnte in der Untersuchung von Schneider (2005) weder beim Winterroggen, noch
beim Winterweizen ein enger Zusammenhang zwischen der Ährenbonitur und der
Wuchshöhe beobachtet werden. Auch in den vorliegenden Versuchen wurde zwar eine
signifikante Korrelation zwischen Wuchshöhe und Ährenbefall erkannt, jedoch mit mittleren
Werten (r = -0,47). Wenn man die drei Linien, die deutlich anfälliger waren als die restlichen
Genotypen herausnimmt, ergibt sich nur noch eine phänotypische Korrelation von r = -0,3,
die keine Signifikanz mehr zeigt. Die genetische Korrelation über sechs Umwelten betrug

51 Diskussion
jedoch r = -0,5, was einen Zusammenhang zwischen Wuchshöhe und Ährenbefall bestätigt.
Zur weiteren Überprüfung der Korrelation zwischen Ährenbefall und Wuchshöhe sollten
weitere Triticalekreuzungen in Feldversuchen verglichen werden.
Um eine besseren Einschätzung der Anfälligkeit der einzelnen Genotypen zu gewährleisten,
sollten Kornbonituren hinzugezogen werden (Mesterházy et al., 1999). In den vorliegenden
Versuchen wurden Kornbonituren der Kreuzungspopulation LASKO x ALAMO durchgeführt,
um eine Korrelation zwischen Ährenbefall bzw. DON-Gehalt und Kornbonitur zu erreichen.
Im Vergleich zu den Studien von Mesterházy et al. (1999) konnten in den vorliegenden
Versuchen ähnliche Korrelationen zwischen Kornbonitur und DON-Gehalt gefunden werden
(r = 0,6). Die Korrelationen zwischen der Kornbonitur und dem DON-Gehalt in der Arbeit von
Heinrich (2004) waren deutlich größer (r = 0,8 bzw. r = 0,9). Der Ährenbefall zeigte jedoch
einen geringeren Zusammenhang zur Kornbonitur als in den vorliegenden Versuchen
(r = 0,7).
Das Isolat FC 46 wird nach Snijders und Perkowski (1990) als besonders aggressiv
eingestuft und führt daher zu einem hohen Krankheitsbefall. Wie auch in den vorliegenden
Versuchen wurde es bereits in einigen anderen Untersuchungen erfolgreich bei künstlichen
Inokulationen eingesetzt (Miedaner et al., 2004; Schmolke, 2004; Häberle et al., 2008). Die
Wahl einer geeigneten Inokulumkonzentration basierte auf Erfahrungswerten aus
vorrangegangenen Experimenten mit erfolgreicher künstlicher Inokulation von Roggen
(Miedaner et al., 1993; Miedaner und Perkowski, 1996), Triticale (Maier und Oettler, 1996)
und Weizen (Mesterházy, 1978). Auch in der vorliegenden Doktorarbeit wurden
Inokulumsuspensionen mit 500.000 bis 700.000 Sporen pro ml verwendet, was zu einem
ausreichend hohen Infektionsdruck in allen Umwelten führte. Als Inokulationsmethode wurde
dazu die Sprühinokulation aufgrund einer höheren Heritabilität und eines wesentlich
geringeren Arbeitsaufwandes im Vergleich zur Einzelähreninjektion gewählt (Miedaner et al.,
2003). Auch ist ohne künstliche Inokulation eine effektive Selektion nicht möglich
(Mésterhazy, 2003). Eine Sprühinokulation hat zur Folge, dass morphologische
Eigenschaften einer Pflanze, wie bspw. Begrannung oder Ährentyp weitgehend
ausgeschaltet werden, wodurch es möglich ist, die physiologischen Prozesse der Resistenz
zu nutzen (Lemmens et al., 2004). Die Sprühinokulation erfolgte jeweils für die gesamte
Population. Im Vergleich zu anderen Studien wurden auch in der vorliegenden Arbeit ähnlich
hohe Befalls-Heritabilitäten nach Sprühinokulation festgestellt (Schmolke et al., 2005; Klahr
et al., 2007).

52 Diskussion
Als nützlich erscheinen Vergleiche von Feldversuchen mit künstlicher Inokulation mit denen
von natürlichem Fusariumbefall. Züchterisch gesehen ermöglichen Feldversuche im
Unterschied zu denen im Gewächshaus eine bessere Beurteilung von Umwelteinflüssen wie
beispielsweise Witterung oder Klimaschwankungen. Diese Problematik wird auch in Zeller et
al. (2010) diskutiert, wobei sich Ergebnisse aus Gewächshausversuchen nicht auf Versuche
im Freiland übertragen lassen. Somit sind Freilandversuche wichtig, um natürliche Prozesse
mit einzuschließen.
Eine vollständige Resistenz oder Immunität gegen Ährenfusariosen ist bei Weizensorten und
verwandten Arten bisher nicht gefunden worden (Mielke, 1995). Dies ist ein Hinweis, dass
keine qualitative, sondern eine quantitative Merkmalsausprägung vorliegt. Bei quantitativ
vererbten Merkmalen sind mehrere Gene an der Resistenzausprägung beteiligt (Heinrich,
2004). In den hier dargelegten Ergebnissen zeigte sich eine quantitative Vererbung der
Resistenz gegenüber Ährenfusariosen, wie auch in früheren Untersuchungen von Weizen
(Snijders, 1990; Buerstmayr et al., 1999) und Roggen (Miedaner et al., 1993) beobachtet
worden ist. Die überwiegend additive Vererbung ermöglicht es, das Resistenzniveau über
rekurrente Selektion zu erhöhen (Snijders und Perkowski, 1990).
4.2 Genotyp-Umwelt-Interaktionen
Bei Getreide sind vielfache Untersuchungen von Resistenzmerkmalen mit Genotyp-Umwelt-
Interaktionen beschrieben worden (Miedaner et al., 1993 und 2001; Oettler und Wahle,
2001). Die hohe Umweltabhängigkeit aller an der Resistenz beteiligten Faktoren und
Merkmale zeigte, dass Untersuchungen in mehreren Umwelten durchgeführt werden
müssen, um zuverlässige Aussagen über das Resistenzverhalten eines Genotyps
gegenüber Ährenfusariosen machen zu können, da hohe Genotyp-Umwelt-Interaktionen die
präzise Differenzierung resistenter und anfälliger Genotypen in Feldversuchen verringern.
Auch Becker (1993) wies darauf hin, dass es bei Feldversuchen wichtig ist, in möglichst
vielen Umwelten zu prüfen, um eine hohe Versuchsgenauigkeit zu erhalten. In der
vorliegenden Doktorarbeit wurden Innerhalb des Versuchszeitraums alle Populationen als
randomisierte Gitteranlagen mit unvollständigen Blöcken an drei Standorten in jeweils drei
Wiederholungen angebaut.
Nach Heinrich (2004) und Schneider (2005) ist es vonnöten, die Resistenzprüfung nicht an
Einzelpflanzen, sondern mindestens auf der Basis von Mikroparzellen durchzuführen. Bei
deren Vergleichen zeigte sich, dass eine deutlich bessere Differenzierung der Eltern in der
Parzellenprüfung auftrat als in Einzelpflanzenanordnung. Die Varianz in der
Einzelpflanzenanordnung der Eltern beruhte weitgehend auf Genotyp-Umwelt-Interaktionen.
Dieser bedeutende Einfluss wurde bereits von Miedaner et al., (2001), Oettler und Wahle

53 Diskussion
(2001), Reinbrecht (2002), Klahr et al. (2004), Oettler et al. (2004) und Paillard et al. (2004)
bestätigt. Wie auch in den Untersuchungen von Oettler und Wahle (2001) sind in der
vorliegenden Arbeit signifikante Genotyp-Umwelt-Interaktionen für alle Merkmale beobachtet
worden.
4.3 Mykotoxine
An der Kontamination des Ernteguts mit Mykotoxinen sind mehrere Faktoren, wie der
Infektionszeitpunkt, die Wachstumsdauer des Pilzes, wie auch die Wachstumsbedingungen
beteiligt. Eine Infektion unter kühleren Verhältnissen führt zu erhöhten Bildungsraten an
Trichothecenen. Ebenfalls ist eine erhöhte Luftfeuchtigkeit über 48 Stunden nach Inokulation
für eine höhere DON-Akkumulation verantwortlich (Reinbrecht, 2002). Lacey et al. (1999)
ermittelten bereits 12 Stunden nach der Inokulation in feuchter Umgebung erhöhte DON-
Gehalte bei Weizen. In der vorliegenden Doktorarbeit wurden die DON-Gehalte in zweifacher
Wiederholung pro Genotyp ermittelt. Es ließ sich jedoch feststellen, dass diese Toxinwerte
nicht immer miteinander übereinstimmten. Um eine genauere Toxinbestimmung je Genotyp
zu erzielen, sollte man zukünftig eine größere Anzahl an Wiederholungen durchführen.
Für den DON-Gehalt konnten in der vorliegenden Doktorarbeit zwischen den Genotypen
signifikante Unterschiede (P < 0,01) beobachtet werden, wie bereits Untersuchungen von
Mesterházy et al. (1999) und Miedaner und Schneider (2002) bei Weizen und Roggen
zeigten. In den hier vorliegenden Versuchen wurde im Mittel je nach Standort zwischen 6
und 85 mg kg-1 DON gemessen. Die höchsten DON-Werte wurden am Standort Hohenheim
gemessen. Dies könnte auf die besseren Entwicklungsbedingungen des Pathogens an
diesem Standort zurückzuführen sein. Ähnliche DON-Gehalte wurden auch in den
Untersuchungen mit Triticale von Heinrich (2004) festgestellt.
In den vorliegenden Versuchen traten einige Genotypen auf, bei denen der Ährenbefall
niedriger war, als der der Toxinbestimmung. Für diese Unterschiede zwischen Befallswerten
und DON-Gehalt könnten die sortenabhängigen Symptomausprägungen verantwortlich sein.
Die Korrelationen zwischen DON-Gehalt und Ährenbefall waren mittel bis gering, dennoch
signifikant (r = 0,32 bis 0,65). Dies konnte bei Weizen von Mesterházy und Bartok (1993);
Perkowski et al. (1996) und Liu et al. (1997) bestätigt werden. Bei Triticale wurde in den
Versuchen von Miedaner et al. (2004) ein nur mäßiger Zusammenhang (r = 0,6) zwischen
beiden Merkmalen aufgrund von sehr hohen Genotyp-Umwelt-Interaktionen und eines sehr
hohen Fehlers gefunden. Bei einer Roggenpopulation in den Untersuchungen von Schneider
(2005) ergaben sich ebenfalls nur mäßige Korrelationen von 0,5 bis 0,6. Miedaner und
Schneider (2002) geben an, dass es bei hohen Korrelationen zwischen Ährenbefall und
DON-Gehalt möglich wäre, die Resistenz von Genotypen in frühen spaltenden Populationen

54 Diskussion
ausschließlich mit Hilfe einer Symptombonitur zu erfassen und somit gleichzeitig einen
indirekten Selektionserfolg auf verminderte DON-Akkumulation zu erzielen. Jedoch
verdeutlichen die großen Unterschiede in den Korrelationswerten bei verschiedenen
Getreidearten und Experimenten, dass die Befallshäufigkeit keinen direkten Rückschluss auf
die Toxinmenge zulässt und es bei Resistenzuntersuchungen wichtig ist zusätzlich zur
Befallsbonitur auch eine Mykotoxinbestimmung durchzuführen. Paul et al. (2005) bestätigen
den weiteren Forschungsbedarf zur Beziehung zwischen DON-Gehalt und Fusariumbefall,
da viele Prozesse noch ungeklärt sind. Bei einer hohen Korrelation zwischen DON-Gehalt
und Ährenbefall wäre eine kostengünstigere Variante anhand einer Vorselektion der
Befallswerte eine Möglichkeit. Man sollte in diesem Fall nur die resistenten Genotypen auf
den DON-Gehalt überprüfen.
4.4 Genetische Kartierungen
Die durchgeführten Feldversuche stellten aufgrund der hohen Heritabilitäten eine gute
Grundlage für die anschließende Kartierung und QTL-Analyse dar. Die genetische Karte der
vorliegenden Doktorarbeit ist die erste aus Wintertriticalepopulationen der Kreuzungen
LASKO x ALAMO und LASKO x TRIMESTER. Aus insgesamt 1197 untersuchten SSR-
Markern und 7300 DArT-Markern entstanden zwei genetische Karten mit jeweiligen
Chromosomenlängen von 1815 und 1407 cM. Davon waren 1328 bzw. 631 Marker
polymorph, wovon 350 (LASKO x ALAMO) bzw. 310 (LASKO x TRIMESTER) Marker kartiert
wurden.
Von Roggen wurden bereits diverse genetische Karten veröffentlicht (Devos et al., 1993;
Korzun et al., 1998; Ma et al., 2001; Hackauf und Wehling, 2003; Khlestkina et al., 2004;
Gustafson, 2009). Grundlage dieser genetischen Karten waren RFLP, cDNA-abgeleitete
oder SSR-Marker. Jedoch umfassen diese Karten meist nicht alle Chromosomen (Loarce et
al., 1996). Die Konsensuskarte von Gustafson (2009), die veröffentlichte Karte von Hackauf
et al. (2009) und die umfangreiche genetische Karte von Bolibok-Bragoszewska et al. (2009)
beinhalten hingegen alle sieben Chromosomen mit jeweiligen Gesamtlängen von 779 cM,
124 cM bzw. 3144,6 cM. Das Problem des Erstellens von genetischen Karten bei Roggen
liegt daran, dass nicht genügend Marker vorhanden sind.
Peleg et al. (2008) berichten, dass bei der Kartierung einer Weizenpopulation das B-Genom
60 % besser abgedeckt war, als das A-Genom. Die dort erstellte genetische Karte hatte eine
Chromosomenlänge von 2317 cM für das B-Genom mit einem durchschnittlichen
Markerabstand von 7,5 cM. Auch bei weiteren Studien konnte das Phänomen der ungleichen
Abdeckung bestätigt werden (Peng et al., 2000; Paillard et al., 2004). Ebenfalls trat dies in
der vorliegenden Doktorarbeit auf, das A-Genom war in beiden Kreuzungspopulationen

55 Diskussion
schlechter abgedeckt als das B-Genom. Das A-Genom hatte bei der DH-Population der
Kreuzung LASKO x ALAMO eine Gesamtlänge von 621 cM und einen durchschnittlichen
Marker-Abstand von 6,5 cM. Das B-Genom hingegen hatte eine Gesamtlänge von 617 cM
und einen durchschnittlichen Markerabstand von 5,9 cM. Bei der Population der Kreuzung
LASKO x TRIMESTER wurden beim B-Genom im Unterschied zum A-Genom 18 Marker
mehr kartiert bei einer Gesamtlänge von 530 bzw. 420 cM. Gonzalez et al. (2005) erstellten
eine genetische Karte aus 73 DH-Triticale-Linien, mit einer Gesamtlänge von 2465,4 cM. Bei
dieser Karte konnte, wie bei der in dieser Arbeit erstellten Karte, festgestellt werden, dass
das Roggen-Genom besser abgedeckt war als die beiden Weizengenome. Diese ungleiche
Verteilung liegt zum großen Teil an den Unterschieden der Getreidearten Roggen und
Weizen. Roggen hat als Fremdbefruchter eine hohe Heterozygotie. Durch die größere
Diversität kommt es zu mehr Rekombinationen, zu neuen Gen- und Merkmalskombinationen
und somit auch zu einer höheren Polymorphie. Selbstbefruchter wie Weizen haben hingegen
einen sehr eingeschränkten genetischen Spielraum. Durch züchterische Selektion werden in
der Linienzüchtung bei Weizen manche Bereiche stärker fixiert, was die geringere
Polymorphie im Gegensatz zu Roggen erklären könnte.
Der niedrige Polymorphiegrad der SSR-Marker (7 bis 12 %) führte zu lückenhaften
genetischen Karten in beiden Populationen. Der geringe Polymorphiegrad kann auf die
relativ enge genetische Verwandtschaft der beiden Sorten LASKO und ALAMO zurück
geführt werden. ALAMO entstand aus einer Kreuzungspopulation DAGRO x LASKO. Ein
geringer Polymorphiegrad konnte auch in den Arbeiten mit Weizen von Schmolke (2004) und
Risser (2010) beobachtet werden. Trotz dieser engen genetischen Basis konnte durch
zusätzliche DArT-Marker in beiden Populationen der Polymorphiegrad auf 9 bzw. 18 %
erhöht werden.
Die Lokalisation der SSR-Marker erfolgte mit Hilfe von Referenzkarten nach Röder et al.
(1998) und Somers et al. (2004). Das Problem der Kartierung war jedoch, dass viele Marker
auf dieselbe Centimorgan-Stelle kartierten, die sogenannte Clusterbildung. Dieses
Phänomen wurde bereits bei Röder et al. (1998), Holzapfel (2009) und Risser (2010)
beobachtet. Insbesondere die DArT-Marker neigen zur Clusterbildung, was schon von Akbari
et al. (2006), Semagn et al. (2006) und Peleg et al. (2008) beobachtet wurde. Peleg et al.
(2008) nehmen an, dass die Clusterbildung zum Einen mit mangelnden Rekombinationen
zusammenhängt, zum Anderen deutet es auf genreiche Regionen hin und auf zu viele
Marker bei zu kleinen Populationsgrößen.

56 Diskussion
Aufgrund der geringen Anzahl an verfügbaren Konsensuskarten war eine Zuordnung der
Roggenmarker schwierig, jedoch konnten, basierend auf Bolibok-Bragoszewska et al.
(2009), einige Marker lokalisiert werden. Die Ergebnisse von Akbari et al. (2006)
demonstrieren, dass DArT-Marker für die Genotypisierung besonders für polyploide
Getreidearten mit großen Genomen, wie z.B. Weizen, einen effektiven Einsatz finden. Auch
in Alsop et al. (2010) wurden insgesamt 6144 DArT-Marker auf vier Gerstenpopulationen
getestet, 573 waren polymorph und 31 wurden kartiert. Die Kartenlängen reichten von 710
cM bis zu 988 cM. Die cM-Abstände lagen zwischen 3,90 und 4,47 cM. In der vorliegenden
Untersuchung ergaben sich ähnlich durchschnittliche Markerabstände von 5,19 cM bei der
Kreuzungspopulation LASKO x ALAMO bzw. 4,54 cM bei der Kreuzungspopulation LASKO x
TRIMESTER. Im Gegensatz dazu zeigte Gustafson (2009) wesentlich kleinere
durchschnittliche Markerabstände von 1,6 cM. Dichte genetische Karten können hilfreich für
die Identifikation von molekularen Markern sein, die eine Verknüpfung zu Genen oder QTL
haben können (Varshney et al., 2007). Einschränkend dazu berichtet Asins (2002), dass
Marker, die dichter als 15 cM gekoppelt sind, nur noch eine geringe Verbesserung der QTL-
Schätzung erreichen. Zusätzlich ist die Bestimmung der richtigen Markerreihenfolge auf der
Kopplungsgruppe bei eng gekoppelten Markern deutlich schwieriger (Van Os et al. 2005).
Dies ist jedoch für die Qualität der Kartierung wichtiger als eine hohe Markerdichte (Asins,
2002).
4.5 QTL-Analysen
Diverse Winterweizensorten mit moderaten Resistenzen gegen Ährenfusariosen wurden
bereits für QTL-Studien verwendet (Schmolke et al., 2005; Holzapfel et al., 2008). Nach
Miedaner et al. (2004) könnte die Fusariumresistenz im adaptierten europäischen Genpool
durch das Einkreuzen neuer, exotischer Resistenzquellen gesteigert werden. Zu beachten
sind dabei die agronomischen Merkmale wie z.B. Wuchshöhe, Ährenschieben oder
Lageranfälligkeit, die Anfälligkeit gegenüber anderen Krankheiten und Schädlingen, sowie
der Ertrag eines Genotyps. Bisherige Veröffentlichungen über die Kartierung von
Resistenzgenloci gegen Ährenfusariosen wurden größtenteils im Sommerweizen mit Sumai3
oder mit von Sumai3 abgeleiteten Resistenzdonoren durchgeführt. Der erste Haupt-QTL
Fhb1 für Fusariumresistenz (Qfhs.ndsu-3BS) wurde von Waldron et al. (1999) detektiert und
von Anderson et al. (2001) verifiziert. Er erklärte 24,8 und 41,6 % der phänotypischen
Varianz in zwei Kartierungspopulationen. Eine Übersicht über weitere Resistenz-QTL sind in
Buerstmayr et al. (2009) zusammengefasst.

57 Diskussion
Charakteristisch für eine quantitative Vererbung der Fusariumresistenz sind die, in der
durchgeführten Studie gefundenen, kontinuierlichen Häufigkeitsverteilungen der
Populationen LASKO x ALAMO und LASKO x TRIMESTER. Zurückzuführen ist diese
quantitative Ausprägung auf mehrere Resistenz-QTL mit vergleichsweise geringen
phänotypischen Effekten. Die hier beobachteten Transgressionen verdeutlichen, dass auch
anfällige Eltern Resistenzallele besitzen können. Dies wurde bereits von Waldron et al.
(1999) in einer QTL-Kartierung bestätigt, bei der der anfällige Elter zwei Allele für
Ährenfusarienresistenz trug, die 7,2 bzw. 14,3 % der phänotypischen Varianz erklärten. In
den vorliegenden Untersuchungen war dies in der Kreuzungspopulation LASKO x
TRIMESTER der Fall.
In der Kreuzungspopulation LASKO x ALAMO wurden für Fusarium-Resistenz vier QTL
gefunden, die insgesamt 37 % der phänotypischen Varianz erklärten. In der
Kreuzungspopulation LASKO x TRIMESTER hingegen wurden nur zwei QTL für Fusarium-
Resistenz detektiert. Diese erklärten 41 % der phänotypischen Varianz. Diese
Größenordnung liegt im Bereich bereits veröffentlichter Arbeiten über die QTL-Kartierung
von Fusariumresistenzen im Winterweizen (Buerstmayr et al., 2009). Auch die
molekulargenetischen Studien nach Bai et al. (2000); Buerstmayr et al. (2000) und Anderson
et al. (2001) zeigten, dass QTL für die Resistenz gegen Ährenfusariosen überwiegend
additiv vererbt werden und epistatische Effekte nur bei einzelnen Kreuzungen auftreten
können. Hervorzuheben sind hier zusätzlich die Ergebnisse von Gervais et al. (2003), bei
den in Feldversuchen unter Berücksichtigung von 194 rekombinierten Inzuchtlinien, neun
QTL detektiert wurden, von denen drei über die Umwelt stabil waren, die gemeinsam 30 bis
45 % der Variation erklärten. Dagegen konnten Buerstmayr et al. (2003) bei einer Sumai3-
abgeleiteten Resistenzquelle zwei QTL detektieren, die gemeinsam ca. 50 % der Variation
erklärten. Alle in der vorliegenden Arbeit detektierte QTL für Fusariumresistenz in A- und B-
Genom wurden bereits bei Weizen beschrieben (Buerstmayr et al., 2009).
Es gab keine Kongruenz zwischen den Resistenz-QTL der beiden Kreuzungspopulationen,
obwohl der resistente Elter derselbe war. In der Population LASKO x ALAMO trugen alle
detektierten QTL für Fusariumresistenz das Resistenzallel vom resistenten Elter LASKO.
Dies war in der Kreuzungspopulation LASKO x TRIMESTER nicht der Fall. Hier wurden als
Resistenzdonor sowohl der resistente, als auch der anfällige Elter gefunden. Dies könnte auf
epistatische Effekte mit anderen QTL zurückgeführt werden. Einzelne QTL interagieren so
miteinander, dass sie in einer Population einen ausreichenden Effekt erzielen, um detektiert
zu werden, während in der anderen Population der entsprechende Interaktions-QTL fehlt und
somit kein Effekt erzielt werden kann. Ein weiterer Grund könnte die QTL-Umwelt-Interaktion
darstellen. Dies würde bedeuten, dass unterschiedliche QTL in unterschiedlichen Umwelten

58 Diskussion
aktiv sind bzw. detektiert werden. Bei der Kreuzungspopulation LASKO x ALAMO konnte bei
der Verrechnung an dem Standort Hohenheim über zwei Jahre hinweg nur ein Resistenz-
QTL detektiert werden. Dieser QTL stimmte nicht mit den vier QTL überein, die hingegen an
den beiden anderen Standorten gefunden wurden. Er zeigte nur einen sehr kleinen Effekt.
Des Weiteren könnte der gleiche QTL in beiden Eltern vorhanden sein, was auf eine enge
Kopplung schließen lässt, wodurch sich die Effekte gegenseitig aufheben würden.
Ein weiterer wichtiger Aspekt bei der Bewertung von QTL-Analysen ist die häufig
beobachtete Überlappung von Fusariumresistenz-QTL mit QTL für Merkmale, die einen
Einfluss auf den Verlauf und die Intensität der Infektion haben, wie Wuchshöhe und
Blühzeitpunkt. Zur Diskussion steht hierbei aber, ob ein verminderter Befall durch einen
Locus für eine physiologische Resistenzeigenschaft hervorgerufen wird, oder ob diese
überlappenden QTL-Regionen durch morphologische Eigenschaften bzw. das
Entwicklungsstadium der Pflanzen hervorgerufen wird. In der vorliegenden Arbeit konnten
bei der Kreuzungspopulation LASKO x ALAMO Überlappungen von zwei QTL für Fusarium-
Resistenz mit zwei QTL für den Zeitpunkt des Ährenschiebens auf den Chromosomen 5R
und 6R gefunden werden. Eine ähnliche Überlappung konnte auch Schmolke (2004)
entdecken. In den Fällen einer Überlappung ist jedoch die Größe des tatsächlichen Effektes
der Resistenz-QTL ausschlaggebend. Eine weitere Überlappung wurde in diesen
Untersuchungen in der Kreuzungspopulation LASKO x ALAMO zwischen dem Resistenz-
QTL und dem QTL für Wuchshöhe auf dem Chromosom 6R gefunden. Dabei könnten
pleiotrope Effekte eine Rolle spielen, wobei Veränderungen mehrerer phänotypischer
Merkmale nur durch ein einzelnes Gen hervorgerufen werden. Die zwei weiteren detektierten
QTL für Fusarium-Resistenz auf Chromosomen 2A und 1B wurden unabhängig von
morphologischen Charakteristika, wie Wuchshöhe und Ährenbeschaffenheit vererbt. Des
Weiteren wurde für die Kreuzungspopulation LASKO x ALAMO ein QTL für DON-Gehalt auf
dem Chromosom 4B mit einer phänotypisch erklärten Varianz von 8,4 % detektiert. Der
DON-QTL trägt das Resistenzallel des resistenten Elters LASKO. Horsley et al. (2006)
konnte bei einer Gerstenpopulation ebenfalls einen QTL in drei von vier Umwelten für DON
mit einer Varianz von 9 bis 14 % detektieren. Bei Triticale sind bisher keine QTL für DON
gefunden worden. Der DON-QTL stimmte nicht mit den gefundenen QTL für Resistenz
überein. Hierfür wäre noch weiterer Forschungsbedarf gegeben, um die genauen
physiologischen Zusammenhänge zu analysieren und zu verstehen.
Phänotypische, wie auch markerbasierte Selektion (MAS) sind effektive Methoden in der
Pflanzenzüchtung, durch die Fusariumanfälligkeit und Mykotoxingehalte verringert werden
können. Bisherige theoretische Studien haben gezeigt, dass die MAS in vielen Situationen
höhere Erfolge erzielen kann als die phänotypische Selektion (Moreau et al., 2000).

59 Diskussion
Langfristig könnte durch die MAS auf phänotypische Untersuchungen verzichtet werden.
Dazu müssten jedoch viele bereits detektierte QTL in weiteren Populationen verifiziert
werden, um sie effektiv in schon vorhandenes Material mit guten agronomischen
Eigenschaften einzukreuzen. Denn uneinheitliche QTL-Effekte über diverse
Züchtungspopulationen sind zum jetzigen Zeitpunkt Hauptgründe dafür, dass sie nicht in
MAS Programmen genutzt werden können (Anderson, 2007). Weitere effiziente Marker-
Plattformen könnten in Zukunft zu einer verbesserten Nutzung der MAS in Zuchtprogrammen
beitragen. Obwohl Ährenfusariosen durch die Auswahl der richtigen Sorte vermindert werden
könnten, ist der Anbau resistenter Arten heutzutage aufgrund höherer Ertragsfähigkeit bzw.
geringerer Lagerneigung einiger fusariumanfälliger Weizensorten begrenzt (Miedaner und
Schneider, 2002; Rodemann, 2004). In den letzten Jahren hat jedoch die Züchtung
resistenter Sorten immer größere Bedeutung erlangt, sodass immer weniger anfällige Sorten
auf den Markt gebracht werden. Die Anwendung von MAS im Zusammenhang mit
markergestützter Rückkreuzung ist bei Triticale nicht möglich, da kein exotisches Material
zur Einlagerung von Resistenzen vorhanden ist. Des Weiteren stellt sich die Frage, ob die
bereits detektierten QTL groß genug sind, um solche Methoden bei Triticale anwenden zu
können.
Zur statistischen Validierung müssten Populationen mit mehr als 300 Linien verwendet oder
Linien, die nach gezielter Selektion von Linien mit Rekombinationen in den QTL-Regionen
entwickelt werden (Melchinger et al., 1998; Asins, 2002). Diese Populationsgröße ist ein
wichtiger und oft diskutierter Faktor bei der Ermittlung der Effizienz, wie auch der Präzision
von QTL-Analysen (Schmolke, 2004). Im vorliegenden Versuch traten relativ kleine
Populationsgrößen von 146 (LASKO x ALAMO) bzw. 94 Linien (LASKO x TRIMESTER) auf.
Grundsätzlich ist diese Anzahl an Linien für experimentelle QTL-Analysen nicht
ungewöhnlich, denn mit der Anzahl der Linien steigen auch die Kosten für die Geno- bzw.
Phänotypisierung. Aber selbst bei großen Populationen sind die statistischen Möglichkeiten
zur QTL-Detektion mit kleinen Effekten begrenzt (Utz und Melchinger, 1994).

60 Diskussion
4.6 Schlussfolgerungen und Züchterische Konsequenzen
Die Ergebnisse dieser Arbeit belegen, dass im züchterisch nutzbaren Material von
Wintertriticale genügend genetische Variation für eine ausreichende Selektion auf
Fusariumresistenz vorhanden ist. Die durchgeführten Versuche in der vorliegenden
Doktorarbeit zeigten, dass selbst mit modernen molekularbiologischen Methoden die
Einlagerung einer solchen Resistenz, die auf recht kleinen Effekten mehrerer Gene beruht,
aufwändig ist. So ergibt sich auch in Zukunft ein erhöhter Forschungsbedarf, um die durch
Ährenfusariosen verursachte Schadenshäufigkeit zu verringern. Dazu gehört auch die
markergestützte Züchtung resistenterer Sorten mit zugleich hervorragenden agronomischen
Eigenschaften, denn die Bewertung von QTL-Analysen hängt von vielen Faktoren ab, wie
z.B. der Qualität der genetischen Karte, den phänotypisierten Daten, der Populationsgröße
und der Wahl des statistischen Modells. Hierdurch können bei der QTL-Analyse
verschiedene Effekte ausgelöst werden, die bei der Interpretation der Ergebnisse
berücksichtigt werden müssen.
Die Kopplung zwischen Marker und QTL ist ein wichtiger Aspekt, für die Aussagekraft eines
QTL. Je größer der Abstand zwischen QTL und Marker, umso größer ist die Möglichkeit für
Rekombinationen. Demnach ist die Wahrscheinlichkeit der Beständigkeit eines QTL nur bei
enger Kopplung gegeben. In den vorliegenden Ergebnissen lagen die Abstände von QTL zu
Marker zwischen 0 und 8 cM, wobei nur relativ geringe Effekte vorlagen. Vor einem
wirkungsvollen Einsatz von QTL und der Überprüfung der Ergebnisse von anderen QTL oder
derselben QTL in einem anderen genetischen Hintergrund muss geklärt werden, ob ein
genetischer Zusammenhang zwischen der physiologischen Resistenz und bspw. der
Wuchshöhe besteht. Zusätzlich dazu müssten z.B. Kreuzungen mit anderen Eltern
vorgenommen und geprüft werden, ob die bereits detektierten QTL und deren Effekte
wiedergefunden werden bzw. wie hoch die Effekte wären. Darüber hinaus müssten die
Versuche in anderen Umwelten getestet werden. Weiterführend sollte anderes Material
kartiert werden, um zu sehen, ob bessere Effekte vorhanden sind. Zusätzlich sollte an
markergestützter Selektion und rekurrenten Selektionsprogrammen weitere Forschung
betrieben werden, um Resistenzgene zu kombinieren und die Fusarium- und
Mykotoxinproblematik zu verringern.
Die quantitativ-genetischen Parameter der vorliegenden Doktorarbeit lassen günstige
Voraussetzung für die Resistenzzüchtung im praktischen Zuchtbetrieb erwarten. Basierend
auf den Grundlagen, die diese Arbeit liefert, kann der Einsatz molekularer Marker vor allem
dann als erfolgreich bewertet werden, wenn mit einem QTL-gekoppelten Marker ein hoher
Anteil der genetischen Varianz erklärt werden kann und eine markergestützte Rückkreuzung
mit exotischen Donoren erfolgt. Nachteilig hierbei ist, dass einzelne QTL aber immer nur

61 Diskussion
einen begrenzten Teil der Varianz erklären können und so ein gelegentliches Einschalten
einer phänotypischen Selektion parallel zu markergestützter Selektion empfehlenswert ist,
um einen maximalen Selektionserfolg zu erzielen (Zhou et al., 2003).
Für den Züchter ergibt sich weiterhin die Möglichkeit die Sorten, die über mehrere Umwelten
einen ähnlichen Ährenbefall aufwiesen, wie der resistente Elter LASKO, als Grundlage zu
nehmen, um sie mit anderem Material zu kombinieren. Nach erfolgreicher Aufklärung der
Positionen und Eigenschaften der QTL für eine Resistenz gegen Ährenfusariosen könnten
die Ergebnisse mittels molekularbiologischer Ansätze in angewandte Zuchtprogramme
integriert werden. Anhand der Ergebnisse dieser Doktorarbeit wurde die Basis für weitere
Forschungsansätze gegeben, wodurch sich der Zuchtfortschritt maßgeblich beschleunigen
kann und so zu einer schnelleren Marktreife resistenter Sorten führen könnte.

62 Zusammenfassung
5. Zusammenfassung
Die Bekämpfung des Befalls von Triticale mit Ährenfusariosen ist bisher nur unzureichend
möglich, daher verspricht man sich durch die Resistenzzüchtung erhebliche Verbesserungen
hinsichtlich einer Verringerung des Mykotoxingehaltes. Seit 2006 bestehen vom
Gesetzgeber europaweit strenge Grenzwerte für Deoxynivalenol (DON) in Lebensmitteln.
Die Belastung durch Mykotoxine, die durch den Befall mit Ährenfusariosen verursacht
werden, führen bei Triticale zusätzlich zu hohen Ertragsverlusten. Darüber hinaus verursacht
es in der Fütterung erhebliche Gesundheitsprobleme der Tiere, insbesondere in der
Schweinefütterung, und über die verringerte Futteraufnahme zu finanziellen Verlusten des
Landwirts.
Die Resistenz gegen Ährenfusariosen wird bei Triticale quantitativ mit einer überwiegend
additiven Genwirkung vererbt und ist deshalb nur durch aufwändige Feldprüfungen sicher zu
erfassen. Molekulare Marker beschleunigen den Zuchtgang und ermöglichen die gezielte
Introgression von günstigen QTL. In dieser Doktorarbeit sollten deshalb durch den
innovativen Einsatz molekularer Marker Möglichkeiten zur Erhöhung der Resistenz von
Triticale gegenüber Ährenfusariosen untersucht werden. Ziele waren die Aufklärung der
Mechanismen zur Vererbung von Resistenz mittels mehrortigen und mehrjährigen
Feldversuchen, wie auch die Verringerung der Mykotoxinanhäufung, das Erstellen von
genetischen Karten für zwei Kreuzungspopulationen und die Identifikation von QTL für die
Merkmale Ährenschieben, Wuchshöhe, DON und Fusarium-Resistenz.
Es zeigten sich in allen Populationen signifikante Unterschiede zwischen den einzelnen
Genotypen für die genannten Merkmale. Die Ährenbefallswerte betrugen je nach Standort
und Jahr im Mittel zwischen 8 und 43 %. Die Heritabilitäten waren relativ hoch (0,7 - 0,82).
Die große Bedeutung der Genotyp-Ort-Interaktion zeigte, dass mehrortige und mehrjährige
Prüfungen unbedingt erforderlich sind. Zwischen Ährenbefall und Wuchshöhe bzw.
Ährenschieben (EC-Stadium) ergab sich kein bzw. nur ein mittlerer, jedoch signifikanter
Zusammenhang. Die Korrelation zwischen DON-Gehalt und Ährenbefall lag zwischen r = 0,3
und r = 0,65. Diese Ergebnisse sind zu unpräzise, um eine zuverlässige Vorhersage des
DON-Gehalts anhand des Ährenbefalls zu ermöglichen.
Die Kartierung der Kreuzungspopulationen LASKO x ALAMO und LASKO x TRIMESTER
wurde mit Hilfe von SSR- und DArT-Markern durchgeführt. Anhand der besseren
Chromosomenabdeckung durch DArT-Marker ist der Polymorphiegrad von 7 bis 12 % auf 9
bis 18 % gestiegen. Es wurden genetische Karten mit Gesamtlängen von 1815 bzw. 1407
cM und durchschnittlichen Markerabständen von 5,19 und 4,54 cM erstellt.

63 Zusammenfassung
Bei der QTL-Analyse beider Populationen wurden in der Kreuzungspopulation LASKO x
ALAMO für das Ährenschieben sechs QTL, für die Wuchshöhe vier QTL, für den DON-
Gehalt ein QTL und für die Fusarium-Resistenz vier QTL detektiert. Diese erklärten 8 bis
66 % der phänotypischen Varianz. Für die Population der Kreuzung LASKO x TRIMESTER
wurden jeweils zwei QTL für Ährenschieben, ein QTL für Wuchshöhe und zwei QTL für
Fusarium-Resistenz gefunden, die 3 bis 41 % der phänotypischen Varianz erklärten. Andere
QTL-Studien bei Weizenpopulationen zeigten ähnlich hohe Effekte.
Der Einsatz molekularer Marker zur Erhöhung der Selektionseffizienz dürfte in naher Zukunft
möglich sein, gentechnologische Methoden zur Fusariumresistenz und Toxinverminderung
sind in der Entwicklung. Bei Triticale wurden bisher weder komplette genetische Karten
publiziert, noch sind QTL-Studien zu Fusariumresistenz oder DON bekannt. Die Züchtung
auf Resistenz gegen Ährenfusariosen ist aufwändig, da die Resistenz durch mehrere Gene
vererbt wird und mehrfach im Züchtungsverlauf Resistenzprüfungen mit Inokulation
erforderlich sind. Günstig ist die additive Genwirkungsweise, so dass sich unterschiedliche
Resistenzgene von verschiedenen Eltern kombinieren lassen. Rekurrente
Selektionsprogramme, phänotypische und markerbasierte Selektion sind bereits effektive
Methoden in der Pflanzenzüchtung, um eine weitere Verbesserung der Resistenz gegen
Ährenfusariosen zu züchten und die Mykotoxinbelastung von Triticale zu reduzieren. Die
Ergebnisse dieser Doktorarbeit sind für die weitere Nutzung in der Züchtung resistenter
Sorten sehr vielversprechend.

64 Summary
6. Summary Fusarium head blight (FHB) is an epidemic disease of cereals but disease control still is
insufficiently possible. Resistance breeding can be addressed as one approach to reduce the
mycotoxin contents. Since 2006 the European Union had strict rules governing the values for
Desoxinivalenol (DON) in food products.
The extraordinary loads through mycotoxins in triticale, which are caused by FHB, lead to
large reductions in grain yield and quality. Beyond the fact that it can cause health issues in
animals, especially in pig feed, it may induce financial burdens for farming industries.
Resistance against FHB is inherited quantitatively with mainly additive effects and therefore
is only recorded by complex field experiments. Molecular markers accelerate resistance
breeding and enable specific introgressions of favorable QTL. The main topic of this thesis is
to show the application possibilities of molecular markers for the investigation of FHB
resistance in triticale. In detail this study aims to analyze factors leading to a minimized
mycotoxin accumulation in perennial field trials at several locations. Furthermore genetic
maps of two different crossing populations were compiled and QTL mapping for ear
appearance, plant height, DON content and FHB resistance was implemented.
For all populations significant varieties had been displayed for all characteristics. The
average values for FHB rate were between 8 and 43 % depending on year and location. The
heritability’s showed high values for each population (h2 = 0.7 – 0.82). The substantial
genotype-environment-interaction pointed out the importance of field experiments.
Furthermore no significant correlation was obtained between ear appearance and plant
height. Due to the fact that the correlation factor between DON-content and FHB rate was
very low (r = 0.32) to moderate (r = 0,65). Therefore no assumption about the DON-content
subjected to the FHB rate could be made. The mapping of both populations LASKO x
ALAMO and LASKO x TRIMESTER has been accomplished with SSR- and DArT-markers.
The rate of polymorphism could be increased from 7 - 12 % respectively to 9 - 18 %. Genetic
maps were constructed with length of 1.815 and 1.407 cM. They indicated 5.19 and 4.54 cM
in their average marker distances. Throughout the QTL mapping several QTL were detected
(6 QTL for ear appearance, 4 QTL for plant height, 1 QTL for DON-content and 4 QTL for
FHB rate). These QTL explained 8 to 66 % of the phenotypical variance. In addition crossing
populations of LASKO x TRIMESTER showed 2 QTL for ear appearance, 1 for plant height
and 2 QTL for FHB resistance, which could explain 3 to 41 % of the phenotypical variance.
This was also observed in several other QTL-surveys for wheat.

65 Summary
Prospectively using molecular markers and genetically engineered methods will increase the
research and development of resistant varieties and lead to a reduced mycotoxin
accumulation. So far neither genetic maps nor any QTL studies for FHB resistance or DON-
content in triticale have been published. Due to the fact that resistance is transmitted by
several genes numerous artificial inoculations have to be carried out during breeding.
Additive gene effects are useful to combine several resistance genes from different parents.
Established methods in plant breeding such as recurrent selection, phenotypical and marker-
based selection are successfully in use to reduce FHB- symptoms and to reduce the
mycotoxin value but these methods have to be improved. The results of this thesis are
promising for a superior resistance breeding in the future.

66 Literaturverzeichnis
7. Literaturverzeichnis Anderson, J. A., Stack, R. W., Liu, S., Waldron, B. L., Fjeld, A. D., Coyne, C., Moreno- Sevilla, B., Mitchell Fetch, J., Song, Q. J., Cregan, P. B. und Frohberg, R. C. (2001). DNA markers for Fusarium head blight resistance QTLs in two wheat populations. Theoretical and Applied Genetics 102: 1164-1168.
Anderson, J. A. (2007). Marker-assisted selection for Fusarium head blight resistance in wheat. International Journal of Food Microbiology 119:51–53.
Anscombe, F. J. und Tukey, J. W. (1963). The examination and analysis of residuals. Technometrics 5: 141-160.
Akbari, M., Wenzl, P., Caig, V., Carling, J., Xia, L., Yang, S., Uszynski, G., Mohler, V., Lehmensiek, A., Kuchel, H., Hayden, M. J., Howes, N., Sharp, P., Vaughan, P., Rathmell, B., Huttner, E. und Kilian, A. (2006). Diversity arrays technology (DArT) for highthroughput profiling of the hexaploid wheat genome. Theoretical and Applied Genetics 113: 1409-1420.
Alsop, B. P., Farre, A., Wenzl, P., Wang, J. M., Zhou, M. X., Romagosa, I., Kilian, A. und Steffenson, B. J. (2010). Development of wild barley-derived DArT markers and their integration into a barley consensus map. Molecular Breeding: 1-16.
Asins, M. J. (2002). Present and future of quantitative trait locus analysis in plant breeding. Plant Breeding 121: 281-291.
Bai, G.H., Shaner, G. und Ohm, H. (2000). Inheritance of resistance to Fusarium graminearum in wheat. Theoretical and Applied Genetics 100: 1-8.
Beck, R., Lepschy, J. und Obst, A. (1997). Gefahr aus der Maisstoppel. DLG-Mitteilung 5: 34-38.
Becker, H. (1993). Pflanzenzüchtung. Eugen Ulmer Verlag.
Beer, E. (2005). Arbeitsergebnisse aus der Projektgruppe "Krankheiten im Getreide" der Deutschen Phytomedizinischen Gesellschaft e.V. Gesunde Pflanzen 57: 59-70.
Bishnoi, U. R., Chitapong, P., Hughes, J. und Nishimuta, J. (1978). Quantity and quality of triticale and other small grain silages. Agron Journal 70: 439-441.
Blakeslee, A. F. und Avery, A. G. (1937). Methods of inducing doubling of chromosomes in plant. Journal of Heredity 28: 392-411.
Bockmann, H. (1962). Künstliche Freilandinfektionen mit den Erregern der Fuß- und Ährenkrankheiten des Weizens I. Vorbereitung und Durchführung der Feldinfektion, sowie deren Neben- und Nachwirkungen. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 14: 153-156.
Börner, H., Schlüter, K., und Auermann, J. (2009). Pflanzenkrankheiten und Pflanzenschutz. Springer.
Braun, R., Weiland, P. und Nellinger, A. (2008). Biogas from Energy Crop Digestion. IEA Bioenergy 37: 4-6.
Brennan, J. M., Egan, D., Cooke, B. M. und Doohan, F. M. (2005). Effect of temperature on head blight of wheat caused by Fusarium culmorum and F. graminearum. Plant Pathology 54: 156-160.

67 Literaturverzeichnis
Bolibok-Brągoszewska, H., Heller-Uszyńska, K., Wenzl, P.,Uszyński, G., Kilian, A. und Rakoczy-Trojanowska, M. (2009). DArT markers for the rye genome – genetic diversity and mapping. BMC Genomics.
Büchse, A. und Piepho, H.-P. (2006). Messen, Schätzen, Bonitieren: Konsequenzen des Skalenniveaus für die Auswertung und Interpretation von Versuchsdaten. Beitrag zur DLG-Versuchstechnikertagung Soest.
Buerstmayr, H., Lemmens, M., Fedak, G. und Ruckenbauer, P. (1999). Back-cross reciprocal monosomic analysis of Fusarium head blight resistance in wheat (Triticum aestivum L.). Theoretical and Applied Genetics 98: 76-85.
Buerstmayr, H., Steiner, B., Lemmens, M. und Ruckenbauer, P. (2000). Resistance to Fusarium head blight in two winter wheat crosses: heritability and trait associations. Crop Science 40: 1012-1018.
Buerstmayr, H., Lemmens, M., Hartl, L., Doldi, L., Steiner, B., Stierschneider, M. und Ruckenbauer, P. (2002). Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. I. Resistance to fungal spread (Type II resistance). Theoretical and Applied Genetics 104: 84-91.
Buerstmayr, H., Steiner, B., Hartl, L., Griesser, M., Angerer, A. und Lengauer, D. (2003). Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. II Resistance to fungal penetration and spread. Theoretical and Applied Genetics 107: 503-508.
Buerstmayr, H., Ban, T. und Anderson, J. A. (2009) QTL mapping and marker assisted selection for Fusarium head blight resistance in wheat: a review. Plant Breeding 128: 1-26.
Chen, J., Griffey C., A., Saghai Maroof, M. A., Stromberg, E. L., Biyashev, R. M., Zhao, W., Chappell, M. R., Pridgen, T. H., Dong, Y. und Zeng, Z. (2006). Validation of two major quantitative trait loci for Fusarium head blight resistance in Chinese wheat line W14. Plant Breeding 125: 99-101. Churchill, G. A. und Doerge, R. W. (1994). Empirical threshold values for quantitative trait mapping. Genetics 138: 963-971.
Cochran, W. G. und Cox, G. M. (1957). Experimental designs. Wiley: 553-566.
Daniel, D. (2005). Erstellung von doppelhaploiden Weizenlinien über die Weizen x Mais- Methode. Von http://www.lfl.bayern.de abgerufen.
Darvasi, A. und Soller, M. (1997). A simple method to calculate resolving power and
confidence interval of QTL map location. Behavior Genetics 27: 125-132.
Dehne, H. W. (2001). Fusarium-Befall und Mykotoxinbelastung von Getreide -Ursachen, Auswirkungen, Vermeidungsstrategien. 13. Wissenschaftliche Fachtagung; Bonn 1-3.
Devos, K. M., Milan, T. und Gale, M. D. (1993). Comparative RFLP maps of the homeologous group-2 chromosomes of wheat, rye and barley. Theoretical and Applied Genetics 85: 784-792.
D´Mello, J. P. F., Placinta, C. M. und Macdonald, A. M. C. (1999) Fusarium mycotoxins: a review of global implications for animal health, welfare and productivity. Animal Feed Science and Technology 80:183-205.

68 Literaturverzeichnis
FAOSTAT (2010). Food and Agriculture Organisation of the United Nations. Von http://faostat.fao.org abgerufen.
Furman, B. J., Qualset, C. O., Skovmand, B., Heaton, J. H., Corke, H. und Wesenberg, D. M. (1997). Characterization and analysis of North American triticale genetic resources. Crop Science 37: 1951-1959.
Gervais, L., Dedryver, F., Morlais, J-Y., Bodusseau, V., Negre, S., Bilous, M., Groos, C. und Trottet, M. (2003). Mapping of quantitative trait loci for field resistance to Fusarium head blight in an European winter wheat. Theoretical and Applied Genetics 106: 961-970.
González, J. M., Muñiz, L. M. und Jouve, N. (2005). Mapping of QTLs for androgenetic response based on a molecular genetic map of × Triticosecale Wittmack. Genome 48: 999-1009.
GrainGenes. (2010). GrainGenes 2.0: A Database for Triticeae and Avena. Von http://wheat.pw.usda.gov/ abgerufen.
Gupta, P. K., Balyan, H. S., Edwards, K. J., Isaac, P., Korzun, V., Röder, M., Gautier, M. F., Joudrier, P., Schlatter, A. R., Dubcovsky, J., De la Pena, R. C., Khairallah, M., Penner, G., Hayden, M. J., Sharp, P., Keller, B., Wang, R. C. C., Hardouin, J. P., Jack, P. und Leroy, P. (2002). Genetic mapping of 66 new microsattelite (SSR) loci in bread wheat. Theoretical and Applied Genetics 105: 413-422.
Gustafson J. P., Ma, X-F., Korzun, V. und Snape, J. W. (2009). A consensus map of rye integrating mapping data from five mapping populations Theoretical and Applied Genetics 118: 793–800.
Häberle, J., Schmolke, M., Schweizer, G., Korzun, V., Ebmeyer, E., Zimmermann, G. und Hartl, L. (2008). Effects of Two Major Fusarium Head Blight Resistance QTL Verified in a Winter Wheat Backcross Population. Crop Science 47: 1823-1831.
Hackauf, B. und Wehling, P. (2003). Development of microsatellite markers in rye: map construction. Plant Breeding and Seed Science 48: 143-151.
Hackauf, B., Ruda, S., van der Voort, J. R., Miedaner, T. und Wehling, P. (2009) Comparative mapping of DNA sequences in rye (Secale cereale L.) in relation to the rice genome. Theoretical and Applied Genetics 118: 371-384.
Haldane, J. B. S. (1919). The combination of linkage values and the calculation of distances between loci of linked factors. Journal of Genetics 8: 299-309.
Hartl, L., Wosnitza, A. und Zimmermann, G. (2001). Sortenresistenz wird besser. DLG-Mitteilungen 8: 40-43.
Heinrich, N. (2004). Quantitativ-genetische Untersuchungen zur Vererbung der Resistenz gegen Ährenfusariosen bei Triticale (xTriticosecale Wittmack). Dissertation.
Hilton, A. J., Jenkinson, P., Hollins, T. W. und Parry, D. W. (1999). Relationship between cultivar height and severity of Fusarium ear blight in wheat. Plant Pathology 48: 202-208.
Hoffmann, W., Mudra, A., und Plarre, W. (1985). Lehrbuch der Züchtung landwirtschaftlicher Kulturpflanzen. Paul Parey-Verlag.
Holzapfel, J., Voss, H-H., Miedaner, T., Korzun, V., Häberle, J., Schweizer, G., Mohler, V., Zimmermann, G. und Hartl, L. (2008). Inheritance of resistance to Fusarium head blight in three European winter wheat populations. Theoretical and Applied Genetics 117: 1119-1128.

69 Literaturverzeichnis
Holzapfel, J. (2009). Kartierung von Resistenzloci gegen Ährenfusarium im europäischen Winterweizen unter Berücksichtigung des Kurzstrohgens Rht-D1. Dissertation.
Horsley, R. D., Schmierer, D., Maier, C., Kudrna, D., Urrea, C. A., Steffenson, B. J. und Schwarz, P. B. (2006). Identification of QTLs Associated with Fusarium Head Blight Resistance in Barley Accession CIho 4196. Crop Science 46:145–156.
Jaccoud, D., Peng, K., Feinstein, D. und Kilian, A. (2001). Diversity Arrays: a solid state technology for sequence information independent genotyping. Nucleic Acids Research 29: No. 4 e25
Jenkinson, P. und Parry, D. W. (1994). Isolation of Fusarium species from common broad- leaved weeds and their pathogenicity to winter wheat. Mycological Research; 98 776-780.
Kabak, W., Dobson, A. D. W. und Var, I. (2006). Stategies to prevent mycotoxin contamination of food and animal feed. Critical Reviews in Food Science and Nutrition 46: 593-619.
Klahr, A., Mohler, V., Herz, M., Wenzel, G. und Schwarz, G. (2004). Enhanced power of detection for Fusarium head blight resistance in wheat by means of co-dominant scoring of AFLP and resistance gene analog markers. Molecular Breeding 13: 289-300.
Klahr, A., Zimmermann, G., Wenzel, G. und Mohler V. (2007). Effects of environment, disease progress, plant height and heading date on the detection of QTLs for resistance to Fusarium head blight in an European winter wheat cross. Euphytica 154: 17-28.
Khlestkina, E. K., Than, M. H. M., Pestsova, E. G., Röder, M. S., Malyshev, S. V., Korzun, V. und Börner, A. (2004). Mapping of 99 new microsatellite-derived loci in rye (Secale cereale L.) including 39 expressed sequence tags. Theoretical and Applied Genetics 109: 725-732.
Korzun, V., Röder, M. S., Ganal, M. W., Worland, A. J. und Law, C. N. (1998) Genetic analysis of the dwarfing gene Rht8 in wheat. Part I. Molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.). Theoretical and Applied Genetics 96: 1104-1109.
Kosambi D. D. (1944). The estimation of map distances from recombination values. Annals of Eugenics 12: 172-175.
Kuleung, C., Baenzinger, P. S. und Dweikat, I. (2004). Transferability of SSR markers among wheat, rye and triticale. Theoretical and Applied Genetics 108: 1147-1150.
Kuleung, C., Baenziger, P. S., Kachmann, S. D. und Dweikat, I. (2006). Evaluating the Genetic Diversity of Triticale with Wheat and Rye SSR Markers. Crop Science 46: 1692-1700.
Lacey, J., Bateman, G. L. und Mirocha, C. J. (1999): Effects of infecion time and moisture on development of ear blight and deoxynivalenol production by Fusarium spp. in wheat. Ann.of Applied Biology 134: 277-283.
Lelley, T. (2006). Triticale: A Low-Input Cereal with Untapped Potential. In R. Singh, und P. Jauhar. Genetic Resources, Chromosome Engineering, and Crop Improvement. CRC Press: 395-430.

70 Literaturverzeichnis
Lemmens, M., Buerstmayr, H., Krska, R., Schumacher, R., Grausgruber, H. und Ruckenbauer, P. (2004). The effect of inoculation treatment and long term application of moisture on Fusarium head blight symptoms and deoxynivalenol contamination in wheat grains. European Journal of Plant Pathology 110: 299-308.
Lindschau, M., und Oehler, E. (1935). Untersuchungen am konstant intermediären additiven Rimpau´schen Weizen-Roggenbastard. Der Züchter 7: 228-233.
Liu, W., Langseth, W., Skinnes, H., Elen, O. N. und Sundheim, L. (1997). Comparison of visual head blight ratings, seed infection levels, and deoxinivalenol production for assessment of resistance in cereals inoculated with Fusarium culmorum. European Journal of Plant Pathology 103: 589-595.
Liu, S., Zhang, X., Pumphrey, M. O., Stack, R. W., Gill, B. S. und Anderson, J. A. (2006). Complex microcolinearity among wheat, rice, and barley revealed by fine mapping of the genomic region harbouring a major QTL for resistance to Fusarium head blight in wheat. Functional and Integrative Genomics 6: 83-89.
Loarce Y, Hueros G, Ferrer E (1996). A molecular linkage map of rye. Theoretical and Applied Genetics 93: 1112-1118.
Ma, X.-F., Wanous, M. K., Houchins, K., Rodriguez Milla, M. A., Goicoechea, P. G., Wang, Z., Xie, M. und Gustafson, J. P. (2001). Molecular linkage mapping in rye (Secale cereale L.). Theoretical and Applied Genetics 102: 517-523.
Maier, F. J. und Oettler, G. (1996). Genetic variation for head blight resistance in triticale caused by Fusarium graminearum isolates of different deoxinivalenol production. Euphytica 89: 387-394.
Mardi, M., Pazouki, L., Delvar, H., Kazemi, M. B., Ghareyazie, B., Steiner, B., Nolz, R., Lemmens, M. und Buerstmayr, H. (2006). QTL analysis of resistance to Fusarium head blight in wheat using a ‘Frontana’-derived population. Plant Breeding 125: 313-317. Martin, R. A. und Johnston H. W. (1982). Effects and control of Fusarium diseases of cereal grains in the Atlantic Provinces. The Canadian Journal of Plant Pathology 4: 210-216.
Mauler-Machnik, A. und Zahn, K. (1994). Ährenfusariosen an Weizen- neue Erkenntnisse zur Epidemiologie und zur Bekämpfung mit Folicur ® (Tebucanazole). Pflanzenschutz-Nachrichten Bayer 47: 133-159.
Meier, A. (2003). Zur Bedeutung von Umweltbedingungen und pflanzenbaulichen Maßnahmen auf den Fusarium-Befall und die Mykotoxinbelastung von Weizen. Dissertation.
Melchinger, A. E., Utz, H. F. und Schön, C. (1998). Quantitative trait locus (QTL) mapping using different testers and independent population samples in maize reveals a low power of QTL detection and a large bias in estimates of QTL effects. Genetics 149: 383-403.
Mesterházy, A. (1978). Comparative analysis of artificial inoculation methods with Fusarium spp. on winter wheat varieties. Journal of Phytopathology 93: 12-25.
Mesterházy, A. (1987). Selection of head blight resistant wheats through improved seedling resistance. Plant Breeding 98, 25-36.
Mesterházy, A. und Bartok, T. (1993). Resistance and pathogenicity influencing toxin (DON) contamination of wheat varieties following Fusarium infection. Hodowla Roslin, Aklimatizaca i Nasiennictwo 37: 9-15.

71 Literaturverzeichnis
Mesterházy, A. (1995). Types and components of resistance to Fusarium diseases. Plant Breeding 114: 377-386.
Mesterházy, A., Bartok, T., Mirocha, C. G. und Komoroczy, R. (1999). Nature of wheat resistance to Fusarium head blight and the role of deoxinivalenol for breeding. Plant Breeding 118: 97-110.
Mesterházy, Á. (2003). Breeding wheat for Fusarium head blight resistance in Europe. In: Leonard, K. J. und Bushnell, W. R. (eds.). Fusarium head blight of wheat and barley. APS Press: 211-240.
Miedaner, T. (1986). Entwicklung von Methoden zur Bestimmung der Fusarium-Resistenz in frühen Wachstumsstadien des Weizens. Dissertation.
Miedaner, T., Borchard, D. C. und Geiger, H. H. (1993). Genetic analysis of inbred lines and their crosses for resistance to head blight (Fusarium graminearum, F. culmorum) in winter rye. Euphytica 65: 123-133.
Miedaner, T. und Perkowski, J. (1996). Correlations among Fusarium culmorum head blight resistance, fungal colonization and mycotoxin contents in winter rye. Plant Breeding 115: 347- 351.
Miedaner, T. (1997) Breeding wheat and rye for resistance to Fusarium diseases. Plant Breeding 116:201–220.
Miedaner, T., Reinbrecht, C., Schollenberger, M. und Lauber, U. (2000). Vorbeugende Maßnahmen gegen Befall mit Ährenfusariosen und Mykotoxinbelastung des Getreides. Mühle und Mischfutter 137: 485-489.
Miedaner, T., Reinbrecht, C., Lauber, U., Schollenberger, M. und Geiger, H. H. (2001). Effects of genotype and genotype-environment interaction on deoxinivalenol accumulation and resistance to Fusarium head blight in rye, triticale and wheat. Plant Breeding 120: 97-105.
Miedaner, T. und Schneider, B. (2002). Züchtungsstrategien zur Verringerung von Ährenfusariosen und Mykotoxingehalte bei Getreide. Tagungsband der 13. und 14. Wissenschaftlichen Fachtagung: Fusarium-Befall und Mykotoxinbelastung von Getreide und Schadvernichtung in Ackerböden 55-67.
Miedaner, T., Schneider, B. und Geiger, H. H. (2003). Deoxynivalenol (DON) content and Fusarium head blight resistance in segregating populations of winter rye and winter wheat. Crop Science 43: 519-526.
Miedaner, T., Heinrich, N., Schneider, B., Oettler, G., Rohde, S. und Rabenstein, F. (2004). Estimation of deoxynivalenol (DON) content by symptom rating and exoantigen content for resistance selection in wheat and triticale. Euphytica 139: 123-132.
Mielke, H. (1995). Untersuchungen zur Anfälligkeit verschiedener Weizensorten gegenüber der partiellen Taubährigkeit (Fusarium culmorum (W. G. Sm.) Sacc.). Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 47: 254-262.
Miller, J. D., Young, J. C. und Sampson, R. D. (1985). Deoxynivalenol and Fusarium head blight resistance in spring cereals. Phytopathology 113: 359-367.
Moreau, L., Lemarié, S., Charchosset, A. und Gallais, A. (2000). Economic efficiency of one cycle of marker-assisted selection. Crop Science 40: 329-337.
Murray, M. G. und Thompson, W. F. (1980) Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research 8: 4321-4326.

72 Literaturverzeichnis
Nirenberg,H. I. (1981). A simplified method for identifying Fusarium spp. occurring on wheat. The Canadian Journal of Botany 59: 1599-1609.
Oettler, G. und Wahle, G. (2001). Genotypic and environmental variation of resistance to head blight in triticale inoculated with Fusarium culmorum. Plant Breeding 12: 297-300.
Oettler, G., Heinrich, N. und Miedaner, T. (2004). Estimates of additive and dominance effects for Fusarium head blight resistance of winter triticale. Plant Breeding 123: 525-530.
Oettler, G. (2005). The fortune of botanical curiosity-Triticale: past, present and future. Journal of Agricultural Science 143: 329-346.
Oettler, G. (2008). Triticale, x Triticoselcale Wittmack. Die erste vom Menschen geschaffene Getreideart. In: Röbbelen, G. (Editor). Die Entwicklung der Pflanzenzüchtung in Deutschl. (1908-2008). Vorträge für Pflanzenzüchtung 75: 320-329.
Offenbächer, G. (2001). Zur Analytik von Fusarium-Mykotoxinen. Tagungsband der 13. und 14. Wissenschaftlichen Fachtagung: Fusarium-Befall und Mykotoxinbelastung von Getreide und Schadverdichtung in Ackerböden: 4-13.
Paillard, S., Schnurbusch, T., Tiwari, R., Messmer, M., Winzeler ,M., Keller, B. und Schachermayr, G. (2004). QTL analysis of resistance to Fusarium head blight in Swiss winter wheat (Triticum aestivum L.). Theoretical and Applied Genetics 109: 323-332.
Paul, P. A., Lipps, P. E. und Madden, L. V. (2005). Relationship between visual estimates of Fusarium head blight intensity and deoxynivalenol accumulation in harvested wheat grain: a meta-analysis. Phytopathology 95: 1225-1236.
Peleg, Z., Saranga, Y., Suprunova, T., Ronin, Y., Röder, M. S., Kilian, A., Korol, A. B. und Fahima, T. (2008). High-density genetic map of durum wheat x wild emmer wheat based on SSR and DArT markers. Theoretical and Applied Genetics 117: 103-115.
Peng, J. H., Fahima, T., Röder, M. S., Li, Y. C., Grama, A., Ronin, Y. I., Korol, A. B. und Nevo, E. (2000). Molecular genetic maps in wild emmer wheat, Triticum dicoccoides: genome-wide coverage, massive negative interference, and putative quasi-linkage. Genome Research 10: 1059-1031.
Perkowski, J., Lasocka, I., Stuchlikova, E., Sip, V., Golinski, P. und Bartos, P. (1996). Response of Czech and Slovak wheat varieties to Fusarium culmorum leading to trichothecenes content in grain. Genetica a Slechteni UZPI 32: 73-77.
Parry, D. W., Jenkins, P. und McLeod, L. (1995). Fusarium ear blight (scab) in small grain cereals—a review. Plant Pathology 44: 207-238.
Pestsova, E., Ganal, M. W. und Röder, M. S. (2000). Isolation and mapping of microsattelite
markers specific for the D genome of bread wheat. Genome 43: 689-697.
Prigge, G., Gerhard, M. und Habermayer, J. (2004). Pilzkrankheiten und Schadsymptome im Getreideanbau. Landwirtschaftsverlag.
Reinbrecht, C. (2002). Genetische und physiologische Einflußfaktoren sowie deren Wechlsewirkungen auf die Trichothecenbildung bei Roggen, Trtiticale und Weizen nach Inokulation mit Fusarium culmorum (W.G. Smith) Sacc. Dissertation.
Risser, P. (2010). Mapping of quantitative‐trait loci (QTL) for adult‐plant resistance to Septoria tritici in five wheat populations (Triticum aestivum L.). Dissertation.

73 Literaturverzeichnis
Rodemann, B. (1999). Mykotoxine in Getreide. Forschungsreport für Ernährung, Landwirtschaft und Forsten 2: 6-9.
Rodemann, B. (2004). Untersuchungen zum Auftreten von Ährenfusariosen in Triticale. Mitteilungen der Biologischen Bundesanstalt für Land- und Forstwirtschaft 396: 403.
Rodemann, B. (2005). Fusarium-Resistenzprüfungen bei Triticale. Vortrag im Kolloquium „Triticale-Züchtung und Verwendung― in Braunschweig.
Rodemann, B. und Mielke, H. (2007). Zum Anbau und Pflanzenschutz des Triticale. Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft 409: 5-40.
Röder, M. S., Korzun, V., Wendehake, K., Plaschke. J., Tixier, M. H., Leroy, P. und Ganal, M. M. (1998). A microsatellite map of wheat. Genetics 149: 2007-2023.
Rogalska, S. M., Cybulska-Augustyniak, J. und Kulawinek, A. (1996). Aneuploidy in polish cultivars of winter triticale (x Triticosecale Wittmack). Genetica Polonica 32: 10-15.
Saiki, R. K., Scharf, S., Faloona, F., Mullis, K. B., Horn, G. T., Erlich, H. A., et al. (1985). Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia. Science 230: 1350-1354.
Sanger, F., Nicklen, S., und Coulson, A. R. (1977). DNA-sequencing with chain-terminating inhibitors. Biotechnology 24: 104-108.
Schneider, B. (2005). Aufspaltungsvariation für die Resistenz gegen Ährenfusariosen und den Deoxynivalenolgehalt bei Winterroggen (Secale cereale L.), Wintertriticale (Triticosecale Wittmack) und Winterweizen (Triticum aestivum L.). Dissertation.
Schmolke, M. (2004). Molekulargenetische Charakterisierung und Lokalisierung von Resistenzgenloci gegen Ährenfusariosen bei Winterweizen. Dissertation.
Schmolke, M., Zimmermann, G., Buerstmayr, H., Schweizer, G., Miedaner, T., Korzun, V., Ebmeyer, E. und Hartl, L. (2005). Molecular mapping of Fusarium head blight resistance in the winter wheat population Dream/Lynx. Theoretical and Applied Genetics 111: 747-756.
Schollenberger, M., Lauber, U., Terry-Jara, H., Suchy, S., Drochner, W. und Müller, H. M. (1998). Determination of eight trichothecenes by gas chromatography/mass spectroscopy after sample clean-up by two-stage solid-phase extraction. Journal of Chromatography A 815: 123-132.
Schroeder, H. W. und Christensen, J. J. (1963). Factors affecting resistance of wheat to scab caused by Gibberella zeae. Phytopathology 53: 831-838.
Semagn, K., Bjornstad, A., Skinnes, H., Maroy, A. G., Tarkegne, Y. und William, M. (2006). Distribution of DArT, AFLP and SSR markers in a genetic linkage map of a doubled-haploid hexaploid wheat population. Genome 49: 545-555.
Semagn, K., Skinnes, H., Bjørnstad, Å., Marøy, A. G. und Tarkegne, Y. (2007) Quantitative trait loci controlling Fusarium head blight resistance and low deoxynivalenol content in hexaploid wheat population from Arina and NK93604. Crop Science 47: 294–303.
Snijders, C. H. A. und Perkowski, J. (1990). Effects of head blight caused by Fusarium culmorum on toxin content and weight of wheat kernels. Phytopathology 80: 566-570.
Snijders, C. H. A. (1990). The inheritance of resistance to head blight caused by Fusarium culmorum in winter wheat. Euphytica 50: 11-18.

74 Literaturverzeichnis
Song, Q. J., Shi, J. R., Singh, S., Fickus, E. W., Costa, J. M., Lewis, J., Gill, B. S., Ward, R. und Cregan P. B. (2005). Development and mapping of microsatellite (SSR) markers in wheat. Theoretical and Applied Genetics 110: 550-560.
Somers, D. J., Fedak, G. und Savard, M. (2003). Molecular mapping of novel genes controlling Fusarium head blight resistance and deoxynivalenol accumulation in spring wheat. Genome 46: 555-564.
Somers, D. J., Isaac, P. und Edward, K. (2004). A high-density wheat microsatellite consensus map for bread wheat (Triticum aestivum L.). Theoretical and Applied Genetics 109: 1105-1114.
Sourdille, P., Cadalen, T., Guyomarc´h, H., Snape, J. W., Perretant, M. R., Charmet, G., Boeuf, C., Bernard, S. und Bernard, M. (2003). An update of the Courtout x Chinese Spring intervarietal molecular marker linkage map for the QTL detection of agroniomic traits in wheat. Theoretical and Applied Genetics 106: 530-538.
Steiner, B., Lemmens, M., Griesser, M., Scholz, U., Schondelmaier, J. und Buerstmayr, H. (2004). Molecular mapping of resistance to Fusarium head blight in the spring wheat cultivar Frontana. Theoretical and Applied Genetics 109: 215-224.
Utz, H. F. und Melchinger, A. E. (1994). Comparison of different approaches to interval mapping of quantitative trait loci. In: Van Ooijen, J.W. und Jansen, J.: Proceedings of the Ninth Meeting of the EUCARPIA Section Biometrics in Plant Breeding. CPRO-DLO Wageningen, Niederlande: 195-204.
Utz, H. F., Melchinger, A. E. und Schön, C. C. (2000). Bias and sampling error of the estimation of genotypic variance explained by quantitative trait loci determined from experimental data in maize using cross validation and validation with independent samples. Genetics 154: 1839-1849.
Utz, H. F. (2007). PLABSTAT, a computer programme for statistical analysis of plant breeding experiments, version 3A. Universität Hohenheim, Stuttgart.
Utz, H. F. (2009). PLABMQTL, a computer programme to map QTL, version 0.5.s. Universität Hohenheim, Stuttgart.
Van Ooijen, J. W. (2006). JoinMap® 4.0, Software for the calculation of genetic linkage maps. Plant Research International, Wageningen.
Van Os, H., Stam, P., Visser, R. G. und Van Eck, H. J. (2005). Record: A novel method for ordering loci on a genetic linkage map. Theoretical and Applied Genetics 112: 30-40.
Varshney, R. K., Langridge, P. und Graner, A. (2007). Aplication of genomics to molecular breeding of wheat and barley. Advanced Genetics 58: 121-155.
Voorips, R. E. (2002). MapChart: software for the graphical presentation of linkage maps and QTLs. Journal of Heredity 93: 77-78.
Voss, H.-H., Holzapfel, J., Hartl, L., Korzun, V., Rabenstein, F., Ebmeyer, E., Coester, H., Kempf, H. und Miedaner, T. (2008). Effect of the Rht-D1 dwarfing locus on Fusarium head blight rating in three segregating populations of winter wheat. Plant Breeding 127: 333-339.
Waldron, B. L., Moreno-Sevilla, B., Anderson, J. A., Stack, R. W. und Frohberg, R. C. (1999). RFLP mapping of QTL for Fusarium head blight resistance in wheat. Crop Science 39: 805-811.

75 Literaturverzeichnis
Wenzl, P., Carling, P., Kudrna, D., Jaccoud, D., Huttner, E., Kleinhofs, A. und Kilian, A. (2004). Diversity Arrays Technology (DArT) for whole-genome profiling of barley. PNAS 26: 9915-9920.
Whitaker, T. B. (2006). Sampling foods for mycotoxins. Food Additives and Contaminants 23: 50-61.
Wilde, F. und Miedaner, T. (2006). Selection for Fusarium head blight resistance in early generations reduces the deoxynivalenol (DON) content in grain of winter and spring wheat. Plant Breeding 125: 96-98.
Wilde, F., Korzun, V., Ebmeyer, E., Geiger, H. H. und Miedaner T. (2007). Comparison of phenotypic and marker-based selection for Fusarium head blight resistance and DON content in spring wheat. Molecular Breeding 19: 357-370.
Wilson, A. S. (1876). Wheat and rye hybrids. Transactions and Proceedings of the Botanical Society 12: 286-288.
Wittmack, L. (1899). Gesellschaft Naturforschender Freunde Berlin, Sitzung vom 21. März.
Zeller, S. L., Kalinina, O., Brunner, S., Keller, B. und Schmid, B. (2010). Transgene X environment interactions in genetically modified wheat. Public library of science 5: 1-8.
Zhou, W. C., Kolb, F. L., Bai, G. H., Shaner, G. und Domier, L. L. (2002). Genetic analysis of scab resistance QTL in wheat with microsatellite and AFLP markers. Genome 45: 719-727.
Zhou, W. C., Kolb, F. L., Bai, G. H., Domier, L. L., Boze, L.K. und Smith, N. J. (2003). Validation of a major QTL for scab resistance with SSR markers and use of marker-assisted selection in wheat. Plant Breeding 122: 40-46.
Zhu, S., Rossnagel, B. G. und Kaeppler, H. F. (2004). Genetic analysis of quantitative trait loci for groat protein and oil content in oat. Crop Science 44: 254-260.

76 Anhang
8. Anhang
8.1 Genetische Karten beider Kreuzungspopulationen
LA
SK
O X
AL
AM
O
1A
BS
_W
MS
33
0.0
1A
_W
MS
835
0.5
1A
_w
Pt-
1167
9.0
1A
_w
Pt-
6280
14.8
1A
_w
Pt-
0595
20.0
Ch
r_1A
2A
_w
Pt-
6393
0.0
2A
_B
AR
C15
1.6
2A
S_W
MS
372
2.1
2A
B_W
MC
63
2.8
2A
_W
MS
328
6.5
2A
_B
AR
C5
16.0
2A
L_W
MS
312
22.6
2A
_w
Pt-
3244
23.2
FUS
Ch
r_2A
3A
_W
MS
1110
0.0
3A
L_W
MS
638
3.1
wP
t-1092
29.1
tPt-
513726
29.8
3A
L_W
MS
155
35.7
wP
t-345988
45.8
3A
_w
Pt-
9761
67.1
3A
_W
MS
1096
73.1
3A
_w
Pt-
1339
76.6
3A
_w
Pt-
4398
77.7
3A
_W
MS
434
80.8
3A
_w
Pt-
1888
83.1
3A
_w
Pt-
0398
87.0
3A
L_W
MS
1229
87.6
Ch
r_3A
4A
L_B
AR
C170
0.0
4A
_W
MS
937
1.8
4A
_w
Pt-
8019
6.2
4A
_w
Pt-
3638
6.7
4A
_w
Pt-
9445
16.7
Ch
r_4A
5A
_B
AR
C141
0.0
5A
_w
Pt-
9748
5.9
5A
L_W
MS
1236
9.4
tPt-
512877
19.4
5A
_w
Pt-
9094
19.6
5A
_B
AR
C151
32.7
5A
_C
FA
2163
71.8
5A
L_W
MS
982
77.7
5A
_C
FA
2155
78.8
5A
_w
Pt-
1052
83.1
5A
_w
Pt-
5096
86.1
5A
L_W
MS
595
95.6
5A
_tP
t-4184
101.6
5A
L_W
MS
291
113.3
WUH AES
Ch
r_5A
6A
_w
Pt-
3247
0.0
6A
_w
Pt-
5550
40.2
6A
_W
MC
417
74.6
6A
L_W
MS
1017
83.3
6A
_w
Pt-
5310
91.6
6A
L_W
MS
570
97.7
6A
L_B
AR
C113
106.1
6A
_w
Pt-
2465
107.7
6A
_w
Pt-
7373
108.9
6A
_w
Pt-
2077
110.2
6A
_W
MS
530
111.4
6A
_w
Pt-
7599
113.1
6A
_W
MS
1009
134.5
6A
S_W
MS
1040
145.7
6A
_w
Pt-
3965
147.5
6A
6B
_w
Pt-
7906
147.8
6A
_w
Pt-
7663
149.2
6A
_w
Pt-
3524
153.1
6A
_w
Pt-
6904
153.8
6A
_w
Pt-
9679
154.6
6A
_w
Pt-
7027
154.7
6A
_w
Pt-
5652
155.9
6A
_w
Pt-
4270
178.9
AES
Ch
r_6A
7A
S_W
MS
471
0.0
7A
_w
Pt-
0002
4.7
7A
_w
Pt-
8043
8.2
7A
S_W
MS
681
11.8
7A
_W
MS
635
14.2
7A
_w
Pt-
6824
20.8
7A
_w
Pt-
4835
27.6
7A
_w
Pt-
3523
30.6
7A
_w
Pt-
7785
75.2
7A
_w
Pt-
0321
80.1
WM
C405
83.6
7A
_w
Pt-
1436
84.9
BA
RC
222
86.5
tPt-
513624
89.4
7A
L_W
MS
870
90.6
7A
S_W
MS
913
91.8
7A
_B
AR
C121
97.3
6A
7A
_B
AR
C195
98.7
7A
_tP
t-2230
100.6
WM
S276
106.1
7A
_W
MS
332
131.7
7A
_tP
t-6875
146.3
7A
_W
MS
1126
155.9
7A
_w
Pt-
2260
159.7
7A
L_W
MS
698
170.8
7A
L_W
MS
1066
181.4
AES WUH
Ch
r_7A
Ab
b.
17 :
Ge
ne
tische
Ka
rte
n m
it Q
TL-P
ositio
ne
n f
ür
jew
eili
ge h
om
olo
ge
Ch
rom
osom
en (
Ch
r.)
de
r K
reuzu
ng
spo
pu
lation
LA
SK
O x
AL
AM
O;
Ba
lke
n u
mfa
sse
n d
ie 9
5 %
Ko
nfid
en
zin
terv
alle
für
QT
L;
AE
S =
QT
L f
ür
den
Zeitp
unkt
de
s Ä
hre
nsch
iebe
ns,
WU
H =
QT
L f
ür
Wu
ch
shö
he
, D
ON
=
Q
TL fü
r D
ON
-Ge
ha
lt,
FU
S =
Q
TL fü
r F
usariu
mre
sis
tenz;
Fett
ma
rkie
rt s
ind d
ie M
ark
er,
d
ie d
em
Q
TL am
näch
ste
n lie
ge
n

77 Anhang
LA
SK
O x
AL
AM
O
1B
7B
_w
Pt-
2540
0.0
1B
_w
Pt-
3172
0.9
1B
_W
MS
1130
1.8
1B
_w
Pt-
7371
8.7
1B
_w
Pt-
7359
16.6
1A
1B
_w
Pt-
2859
17.8
1B
_w
Pt-
0308
19.3
1B
_w
Pt-
3465
20.3
1B
_tP
t-5249
66.3
FUS
Ch
r_1
B
2B
L_W
MS
1300
0.0
2B
_w
Pt-
0694
0.8
2B
2D
_w
Pt-
4223
4.7
2B
_w
Pt-
9190
6.4
2B
_w
Pt-
7161
10.0
2B
_w
Pt-
7004
11.3
1B
2B
_w
Pt-
8776
13.2
rPt-
508259
30.1
2B
_w
Pt-
4426
32.6
2B
_W
MC
361
33.6
2B
_w
Pt-
3632
35.3
2B
_W
MS
619
37.9
2B
_tP
t-4602
0.0
2B
_tP
t-1663
1.7
2B
_w
Pt-
5759
12.3
tPt-
513188
17.0
2B
_B
AR
C35
19.6
2B
_W
MS
1203
23.6
tPt-
513659
25.8
2B
_W
MS
1128
35.7
2B
4D
_tP
t-4627
37.3
2B
_w
Pt-
7695
39.3
tPt-
5853
41.5
2B
_w
Pt-
9668
98.7
wP
t-4327
99.7
2B
_w
Pt-
9402
102.9C
hr_
2B
WM
S247
0.0
3B
_W
MS
181
3.5C
hr_
3B
4B
_w
Pt-
5265
0.0
4B
_W
MC
125
22.9
4B
_w
Pt-
1046
31.1
tPt-
513652
31.8
4B
_w
Pt-
7412
37.4
4B
L_5A
LW
MS
736b
42.7
4B
L_W
MS
930
47.6
4B
_W
MS
149
52.2
4B
_w
Pt-
8796
52.6
4B
SL_W
MS
513
54.0
4B
_w
Pt-
6149
55.1
4B
_w
Pt-
9393
75.0
4A
B_W
MC
617
76.8
DON
Ch
r_4
B
5B
_w
Pt-
8637
0.0
5B
_w
Pt-
3289
2.7
5B
_w
Pt-
9569
3.0
5B
_w
Pt-
5120
3.5
5B
_w
Pt-
3461
3.7
5B
_B
AR
C109
4.3
5B
_B
AR
C4
4.6
5B
_W
MS
1180
5.0
5B
_W
MS
197
5.4
5B
L_W
MS
777
71.9
WUH
Ch
r_5
B
wP
t-9572
0.0
6B
_w
Pt-
8239
3.1
6B
_w
Pt-
9990
4.8
6B
_w
Pt-
7954
11.3
6B
_w
Pt-
6674
14.0
6B
L_W
MS
1199
34.8
6B
S_W
MS
825
53.5
6B
_w
Pt-
9195
57.8
6B
_w
Pt-
6247
59.5
6B
_W
MC
756
66.9
6B
_W
MS
644
67.6
6B
_W
MC
473
68.9
6B
_B
AR
C178
94.6
6B
_W
MS
1076
97.6
6B
_w
Pt-
0986
151.6
6B
_w
Pt-
5885
163.8
6B
_w
Pt-
9952
165.3C
hr_
6B
7B
_w
Pt-
7318
0.0
7B
_W
MS
537
5.5
7B
_w
Pt-
8283
7.0
5B
7B
_w
Pt-
4863
13.7
7B
S_W
MS
1184
15.6
7B
S_W
MS
1173
17.6
7B
L_W
MS
983
18.9
7B
_B
AR
C176
21.0
7B
_w
Pt-
11598
24.0
7B
_w
Pt-
8919
25.8
7B
_w
Pt-
2273
33.4
7B
_w
Pt-
9925
44.1
7B
_W
MC
517
47.8
wP
t-11711
52.0
tPt-
513969
52.8
rPt-
411046
53.9
7B
_w
Pt-
5892
58.4
7B
_tP
t-7362
62.7
tPt-
513197
63.9
7B
_W
MS
611
65.6
7B
L_W
MS
577
67.6
7B
_W
MS
238
69.1
7B
L_W
MS
745
70.6
7B
D_
CF
A2040
72.4
wP
t-7546
73.7
7B
_w
Pt-
4875
92.2C
hr_
7B
Fort
setz
ung
der
Ab
bild
un
g 1
7

78 Anhang
LA
SK
O x
AL
AM
O
1R
_S
CM
21
0.0
rPt-
399662
34.6
rPt-
389803
36.2
rPt-
507120
45.7
rPt-
508397
50.5
rPt-
390318
52.3
rPt-
410760
55.1
rPt-
2888
57.7
rPt-
399723
65.6
rPt-
411063
66.3
rPt-
506629
68.0
rPt-
507438
69.3
rPt-
399378
74.8
rPt-
390103
79.1
rPt-
508325
86.1
rPt-
507059
88.5
rPt-
508520
95.2
rPt-
401322
97.8
rPt-
506561
98.2
rPt-
507790
103.7C
hr_
1R
rPt-
411141
0.0
tPt-
4269
2.0
2R
_S
CM
10
8.1
rPt-
507675
9.1
rPt-
402494
14.6
rPt-
398737
15.6
rPt-
410871
18.9
2R
_S
CM
233
21.8
rPt-
1002
23.3
2R
_R
MS
1203
25.3
rPt-
506196
26.0
2R
L_S
CM
75
26.8
rPt-
389807
30.3
rPt-
402519
32.5
rPt-
509160
34.7
rPt-
507837
36.0
rPt-
389908
37.4
2R
_S
CM
61
42.5
SC
M254
44.9
tPt-
513657
46.8
rPt-
398761
48.5
rPt-
508543
49.8
rPt-
506496
60.4
rPt-
400923
62.0
rPt-
509105
66.6
rPt-
509096
67.9
rPt-
398831
74.9
rPt-
411066
79.2
rPt-
505385
92.3
rPt-
505391
93.0
tPt-
513407
94.8
rPt-
508776
102.2
rPt-
506604
103.4
rPt-
509197
107.4C
hr_
2R
rPt-
505338
0.0
rPt-
402398
9.9
rPt-
400264
17.2
BA
RC
918.2
rPt-
509513
21.5
rPt-
390429
23.6
wP
t-1291
25.0
tPt-
514114
27.4
tPt-
3746
33.3
rPt-
0857
34.4
rPt-
506173
34.9
rPt-
402504
35.9
3R
_R
MS
1028
39.0
rPt-
389708
40.0
rPt-
389454
43.4
rPt-
507897
43.9
SC
M284
45.9
rPt-
508068
52.6C
hr_
3R
SC
M228
0.0
rPt-
508582
1.4
rPt-
389941
12.9
rPt-
411228
13.8
rPt-
389272
22.1
rPt-
505165
33.2
rPt-
507899
37.6
rPt-
506458
38.6
rPt-
401563
39.9
4R
_S
CM
66
40.8
4R
_R
MS
1181
41.9
rPt-
390436
43.2
rPt-
410932
45.6
rPt-
507179
47.8
rPt-
410768
49.0
rPt-
508070
49.4
rPt-
398783
57.1
rPt-
400828
59.4
rPt-
401817
62.0
rPt-
506706
64.4
4R
_S
CM
339
65.6
tPt-
7448
66.5
rPt-
389610
67.4
4R
_R
MS
1154
68.3
tPt-
513729
71.8
4R
_S
CM
116
74.3
rPt-
509552
75.9
rPt-
508199
79.5
rPt-
399575
81.7
AES
Ch
r_4
R
rPt-
506172
0.0
rPt-
399743
12.9
rPt-
399462
14.9
rPt-
402391
16.4
rPt-
509677
21.6
rPt-
505581
22.3
rPt-
506747
24.4
rPt-
398800
28.7
5R
_R
MS
1264
34.4
rPt-
509668
37.9
rPt-
399663
40.5
rPt-
508902
41.6
rPt-
508897
42.4
tPt-
8940
44.2
rPt-
401275
51.0
rPt-
507373
51.9
rPt-
508925
52.7
rPt-
505973
57.6
rPt-
399397
58.6
rPt-
505577
59.6
rPt-
507326
107.6
5R
_S
CM
280
109.2
rPt-
506066
113.9
rPt-
402334
116.1
FUS AES
Ch
r_5
R
rPt-
398480
0.0
rPt-
505745
21.5
6R
_S
CM
180
38.4
rPt-
410850
44.2
tPt-
513470
48.8
tPt-
7900
49.7
tPt-
4479
64.8
rPt-
509453
69.3
rPt-
506099
70.4
rPt-
402552
72.9
rPt-
506039
73.7
rPt-
509043
78.1
rPt-
411354
80.6
rPt-
399597
88.9
rPt-
399421
93.2
rPt-
401346
95.8
rPt-
505250
96.3
rPt-
400935
99.0
rPt-
506093
99.8
rPt-
507320
102.5
tPt-
513959
105.0
rPt-
509167
112.0
WUH AES
FUS
Ch
r_6
R
rPt-
400843
0.0
rPt-
400287
0.2
rPt-
507636
1.4
tPt-
514139
3.1C
hr_
7R
Fort
setz
ung
der
Ab
bild
un
g 1
7

79 Anhang
LA
SK
O x
TR
IME
ST
ER
1A
_w
Pt-
0196
0.0
1A
_w
Pt-
4666
0.8
1A
_w
Pt-
2527
2.2
1A
_w
Pt-
6122
3.2
1A
_w
Pt-
6280
21.7
1A
_w
Pt-
3198
24.3
1A
_w
Pt-
6709
24.8C
hr_
1A
2A
_w
Pt-
5839
0.0
2A
_w
Pt-
7626
0.8
2A
_tP
t-5584
4.9C
hr_
2A
3A
_w
Pt-
1353
0.0
3A
_w
Pt-
7890
1.3
3A
_tP
t-6949
6.6
3A
_tP
t-0519
7.0
3A
_w
Pt-
0797
19.4
3A
_w
Pt-
2478
19.5
3A
_w
Pt-
2866
24.9
wP
t-4349
40.1
3A
_w
Pt-
2253
43.0
3A
_w
Pt-
9634
44.9C
hr_
3A
4A
_w
Pt-
1091
0.0
tPt-
513019
1.1
4A
_w
Pt-
9860
44.5
4A
_w
Pt-
0817
45.4
4A
_w
Pt-
0056
48.3
4A
_w
Pt-
9305
51.9
4A
_w
Pt-
6867
55.1
4A
_w
Pt-
0162
58.7
4A
_w
Pt-
0763
80.6
4A
_w
Pt-
1155
81.9
4A
_w
Pt-
4424
82.1
4A
_w
Pt-
3845
83.0
FUS
Ch
r_4
A
5A
_w
Pt-
5096
0.0
wP
t-4106
4.1
wP
t-7804
43.8
tPt-
8506
44.9
tPt-
514325
48.1
5A
_tP
t-6495
55.3
5A
_tP
t-4184
56.1C
hr_
5A
6A
_w
Pt-
1285
0.0
6A
_w
Pt-
9382
2.9
6A
_w
Pt-
4270
7.5
6A
_w
Pt-
6520
10.7
6A
_w
Pt-
6904
16.7
6A
_w
Pt-
5652
28.9
6A
_w
Pt-
9692
46.7
6A
_w
Pt-
9690
48.0
rPt-
505608
72.9
6A
_w
Pt-
7445
74.1
6A
_w
Pt-
3452
104.8
AES
FUS WUH
Ch
r_6
A
7A
_w
Pt-
2199
0.0
7A
_w
Pt-
3059
0.3
7A
_w
Pt-
1441
4.9
7A
_w
Pt-
3523
24.8
7A
_w
Pt-
8149
56.4
7A
_w
Pt-
6967
56.8
7A
_w
Pt-
2523
56.9
wP
t-2056
58.1
7A
_w
Pt-
4835
71.5
7A
_w
Pt-
6447
72.0
7A
_w
Pt-
0321
101.6C
hr_
7A
Ab
b.
18:
Ge
netisch
e K
art
en m
it Q
TL-P
ositio
ne
n fü
r je
we
ilig
e h
om
olo
ge
C
hro
mo
som
en (C
hr.
) d
er
Kre
uzu
ng
spo
pu
latio
n L
AS
KO
x
TR
IME
ST
ER
; B
alk
en u
mfa
ssen
die
95
% K
onfid
en
zin
terv
alle
fü
r Q
TL;
AE
S =
QT
L für
den
Ze
itp
unkt
des Ä
hre
nsch
iebe
ns, W
UH
= Q
TL f
ür
Wu
ch
shö
he
, F
US
= Q
TL f
ür
Fusariu
mre
sis
tenz;
Fe
tt m
ark
iert
sin
d d
ie M
ark
er,
die
dem
QT
L a
m n
äch
ste
n lie
ge
n

80 Anhang
LA
SK
O x
TR
IME
ST
ER
tPt-
513348
0.0
1B
_w
Pt-
1876
0.7
1B
_w
Pt-
4729
1.3
1B
_w
Pt-
5899
2.3
1B
_w
Pt-
8744
21.2
1B
_w
Pt-
1573
28.9
1B
_w
Pt-
1726
29.1
1B
3A
_w
Pt-
6012
29.8
1B
_w
Pt-
0441
29.9
1B
_w
Pt-
1684
30.0
1B
_w
Pt-
3103
30.2
1B
_w
Pt-
1248
37.4
1B
_w
Pt-
6690
41.8
1B
_w
Pt-
8280
43.9C
hr_
1B
2B
_w
Pt-
9859
0.0
2B
_w
Pt-
6706
1.2
2B
_w
Pt-
6311
1.6
2B
_w
Pt-
0100
55.5
2B
_w
Pt-
9274
59.0
tPt-
2971
70.7
2B
_w
Pt-
5374
72.7
tPt-
513473
84.8
tPt-
514208
88.5
2B
6B
_w
Pt-
9423
105.8
2B
_w
Pt-
9402
105.9
2B
_w
Pt-
7619
108.6C
hr_
2B
3B
_w
Pt-
3342
0.0
3B
_w
Pt-
4412
9.5
3B
_w
Pt-
7158
12.6
3B
_w
Pt-
1306
33.2
3B
_w
Pt-
0343
33.6
wP
t-9989
37.2
wP
t-9488
37.3
3B
_w
Pt-
1311
39.8
tPt-
513153
39.9
3B
6B
_w
Pt-
7627
40.0
3B
_w
Pt-
8352
40.2
3B
_w
Pt-
0995
40.3
3B
_w
Pt-
1822
40.4
3B
_w
Pt-
0365
40.6
3B
3D
_w
Pt-
9786
49.9
tPt-
513516
64.2
3B
_w
Pt-
5704
65.4
3B
_w
Pt-
0990
86.8C
hr_
3B
4B
_w
Pt-
1272
0.0
4B
_w
Pt-
4280
0.1
tPt-
513652
0.6
tPt-
513338
1.3
4B
_w
Pt-
2141
62.1C
hr_
4B
5B
_w
Pt-
8637
0.0
5B
_tP
t-8942
6.7
tPt-
0714
7.5
5B
_w
Pt-
6348
53.6
5B
7B
_w
Pt-
5914
53.7
5B
_w
Pt-
5346
53.9
5B
7B
_w
Pt-
5175
54.0
5B
_w
Pt-
5120
58.4
5B
_w
Pt-
1548
78.8
5B
_tP
t-3719
78.9
5B
_tP
t-7183
79.0
5B
_w
Pt-
8393
79.2
5B
_w
Pt-
3457
116.6C
hr_
5B
7B
_w
Pt-
2273
0.0
7B
_w
Pt-
4300
50.4
7B
_w
Pt-
9880
58.4
7B
_w
Pt-
7295
58.7
7B
_w
Pt-
0217
58.9
wP
t-7546
61.4
7B
_w
Pt-
0600
62.7
7B
_w
Pt-
4820
63.8
7B
_w
Pt-
5585
85.7
7B
_w
Pt-
4038
85.9
7B
_w
Pt-
5462
86.6
7B
_w
Pt-
5228
87.2
7B
_w
Pt-
7413
87.3
7B
_w
Pt-
9954
88.3
tPt-
513687
99.6
tPt-
513663
100.6
7B
_tP
t-3700
112.1C
hr_
7B
Fort
setz
ung
der
Ab
bild
un
g 1
8
81 Anhang
LA
SK
O x
TR
IME
ST
ER
rPt-
399
68
40.0
rPt-
509
57
83.6
rPt-
507
91
25.2
rPt-
401
38
59.4
rPt-
401
61
011.9
rPt-
506
39
628.3
tPt-
513
36
128.8
rPt-
389
44
629.3
rPt-
506
13
629.7
rPt-
390
33
830.2
rPt-
254
230.3
rPt-
506
43
130.5
rPt-
410
92
731.2
rPt-
402
05
532.9
tPt-
513
06
733.1
rPt-
399
76
933.8
rPt-
411
06
334.8
rPt-
401
41
036.2
rPt-
505
93
641.1
rPt-
400
05
361.9
rPt-
399
83
163.9C
hr_
1R
rPt-
399
72
10.0
rPt-
411
06
60.6
tPt-
513
24
50.7
tPt-
999
27.8
rPt-
402
48
117.3
rPt-
399
89
518.9
rPt-
507
98
220.8
rPt-
508
95
721.8
rPt-
402
51
922.0
rPt-
100
222.6
rPt-
400
41
022.8
rPt-
402
36
422.9
rPt-
506
17
623.1
rPt-
389
44
123.8
rPt-
507
94
623.9
rPt-
390
53
024.1
tPt-
514
06
924.2
tPt-
513
86
124.5
rPt-
505
93
325.4
rPt-
390
78
926.3
rPt-
507
71
427.4
rPt-
411
47
046.4
rPt-
507
32
357.5
rPt-
506
92
658.8
rPt-
390
28
360.3
tPt-
512
92
962.4
rPt-
410
87
171.9
rPt-
399
82
472.6
rPt-
506
69
774.0
rPt-
505
89
174.6
rPt-
505
48
179.3
rPt-
411
17
481.7
rPt-
507
67
586.6
rPt-
402
36
1104.3C
hr_
2R
rPt-
506
96
10.0
rPt-
509
71
91.8
rPt-
507
87
75.7
rPt-
506
50
46.4
rPt-
402
50
48.7
rPt-
506
17
39.6
rPt-
389
67
612.2
rPt-
389
70
816.2
rPt-
507
43
418.7
rPt-
508
60
919.4
rPt-
508
41
819.6
rPt-
390
72
420.0
rPt-
506
84
720.2
rPt-
389
45
420.7
rPt-
399
89
721.7
rPt-
401
16
322.5
rPt-
507
10
523.8
rPt-
507
37
424.0
rPt-
390
01
837.7
rPt-
505
22
741.9
rPt-
507
57
543.1
rPt-
507
80
144.8
rPt-
506
56
545.0
rPt-
401
29
645.6
rPt-
506
45
546.1
rPt-
411
24
346.5
rPt-
506
96
747.1
rPt-
401
73
850.3
rPt-
568
855.6
rPt-
400
26
461.0C
hr_
3R
rPt-
389
74
00.0
rPt-
509
55
21.0
rPt-
411
32
94.7
rPt-
506
20
78.6
rPt-
507
87
311.3
rPt-
401
64
313.1
rPt-
402
41
113.3
tPt-
513
85
820.6
rPt-
401
82
540.7
rPt-
505
35
241.6
rPt-
400
31
742.2
rPt-
505
77
542.5
rPt-
401
11
742.7
rPt-
402
60
143.1
rPt-
507
49
843.2
rPt-
508
12
443.4
rPt-
507
54
843.7
rPt-
389
45
743.8
rPt-
507
20
244.6C
hr_
4R
tPt-
918
00.0
rPt-
507
66
40.9
tPt-
980
81.6
rPt-
508
83
44.6
rPt-
399
64
110.8
rPt-
508
92
511.1
rPt-
509
55
114.1
rPt-
505
67
915.4
rPt-
411
26
724.7
rPt-
399
95
326.9
rPt-
411
27
027.0
rPt-
398
50
627.6
rPt-
506
11
327.8
rPt-
508
26
628.9
rPt-
398
76
729.2
rPt-
508
08
130.8
rPt-
509
55
046.6
rPt-
390
15
149.6
rPt-
506
17
249.9
rPt-
505
22
051.9
rPt-
390
72
252.8
rPt-
508
56
653.3
rPt-
402
62
553.5
rPt-
505
48
555.2
rPt-
401
97
455.5
rPt-
505
58
156.0
rPt-
509
67
756.1
rPt-
390
09
356.8C
hr_
5R
rPt-
410
85
00.0
rPt-
401
50
51.6
tPt-
790
02.8
tPt-
514
24
46.8
rPt-
390
52
59.9
rPt-
820
513.3
tPt-
513
06
015.7
rPt-
400
93
517.9
rPt-
508
37
132.1
rPt-
509
15
434.8
rPt-
400
60
836.3
rPt-
508
38
837.3
rPt-
399
42
137.4
tPt-
466
937.5
rPt-
401
34
638.1
rPt-
509
39
538.6
rPt-
506
040.3
rPt-
508
84
341.6
rPt-
505
48
856.1
rPt-
400
78
963.4
rPt-
390
25
266.4
AES
Ch
r_6
R
rPt-
399
84
80.0
rPt-
399
84
50.9
rPt-
411
20
51.4
rPt-
399
30
82.9
rPt-
506
34
44.0
rPt-
509
49
74.6
rPt-
061
54.7
rPt-
399
70
55.2
rPt-
509
31
47.3
rPt-
401
36
638.5
rPt-
508
66
941.2
rPt-
399
65
142.6
rPt-
509
06
553.9
rPt-
507
39
856.4
rPt-
509
40
157.5
tPt-
513
78
658.9
rPt-
508
61
659.6C
hr_
7R
Fort
setz
ung
der
Ab
bild
un
g 1
8

82 Anhang
8.2 LOD-Kurven
Chr. 5A
Chr. 6R
Chr. 7A
Chr. 5R
Chr. 6A
Chr. 4R
Abb. 19: Darstellungen der LOD-Kurven (LOD gegenüber cM) für die Kreuzungspopulation LASKO x ALAMO für das Merkmal Ährenschieben für die QTL auf den Chromosomen 5A, 6A, 7A, 4R, 5R und 6R; der LOD-Wert von 3 bezeichnet den Schwellenwert, über dem ein QTL als signifikant angenommen wird (Linie); unter den einzelnen Grafiken ist das jeweilige Chromosom horizontal dargestellt, das Dreieck bezeichnet die Stellen der QTL

83 Anhang
Chr. 5A
Chr. 6RChr. 5B
Chr. 7A
Abb. 20: Darstellungen der LOD-Kurven für die Kreuzungspopulation LASKO x ALAMO für das Merkmal Wuchshöhe für die QTL auf den Chromosomen 5A, 7A, 5B und 6R; der LOD-Wert von 3 bezeichnet den Schwellenwert, über dem ein QTL als signifikant angenommen wird (Linie); unter den einzelnen Grafiken ist das jeweilige Chromosom horizontal dargestellt, das Dreieck bezeichnet die Stellen der QTL

84 Anhang
Chr. 2A
Chr. 6R
Chr. 1B
Chr. 5R
Abb. 21: Darstellungen der LOD-Kurven für die Kreuzungspopulation LASKO x ALAMO für das Merkmal Ährenbefall für die QTL auf den Chromosomen 2A, 1B, 5R und 6R; der LOD-Wert von 3 bezeichnet den Schwellenwert, über dem ein QTL als signifikant angenommen wird (Linie); unter den einzelnen Grafiken ist das jeweilige Chromosom horizontal dargestellt, das Dreieck bezeichnet die Stellen der QTL
´

85 Anhang
8.3. Integrierte Karten
5A_BARC1410.0
5A_wPt-97485.95AL_WMS12369.4
tPt-51287719.45A_wPt-909419.6
5A_BARC15132.7
5A_CFA216371.8
5AL_WMS98277.75A_CFA215578.85A_wPt-105283.15A_wPt-509686.1
5AL_WMS59595.6
5A_tPt-4184101.6
5AL_WMS291113.3
Chr_5A_LA_x_ALA
5AL_WMS2910.0
5AL_WMS59518.7
5A_wPt-509628.05A_wPt-105230.95A_CFA215535.25AL_WMS98236.2
5A_CFA216342.1
5A_BARC15181.2
5A_wPt-909494.4tPt-51287794.5
5AL_WMS1236104.65A_wPt-9748108.1
5A_BARC141114.0
Int_Chr_5A
5A_wPt-50960.0wPt-41064.1
wPt-780443.8tPt-850644.9tPt-51432548.15A_tPt-649555.35A_tPt-418456.1
Chr_5A_LA_x_TRI
Abb. 22: Darstellungen der integrierten Karten der neun Chromosomen 5A, 6A, 5B, 7B, 2R,
3R, 5R und 6R; Kreuzungspopulation LASKO x ALAMO (links), Kreuzungspopulation
LASKO x TRIMESTER (rechts) und integrierte Karte aus beiden Populationen (mitte);
Verbindungslinien markieren die jeweiligen übereinstimmenden Marker

86 Anhang
6A_wPt-32470.0
6A_wPt-555040.2
6A_WMC41774.66AL_WMS101783.36A_wPt-531091.66AL_WMS57097.76AL_BARC113106.16A_wPt-2465107.76A_wPt-7373108.96A_wPt-2077110.26A_WMS530111.46A_wPt-7599113.16A_WMS1009134.56AS_WMS1040145.76A_wPt-3965147.56A6B_wPt-7906147.86A_wPt-7663149.26A_wPt-3524153.16A_wPt-6904153.86A_wPt-9679154.66A_wPt-7027154.76A_wPt-5652155.96A_wPt-4270178.9
Chr_6A_LA_x_ALA
6A_wPt-32470.0
6A_wPt-555040.26A_wPt-345271.86A_WMC41774.66AL_WMS101783.36A_wPt-531091.66AL_WMS57097.76A_wPt-7445102.6rPt-505608103.76AL_BARC113106.16A_wPt-2465107.76A_wPt-7373108.96A_wPt-2077110.26A_WMS530111.46A_wPt-7599113.16A_wPt-9690128.56A_wPt-9692129.96A_WMS1009134.56AS_WMS1040145.76A_wPt-3965147.46A6B_wPt-7906147.86A_wPt-7663149.26A_wPt-3524153.16A_wPt-6904153.46A_wPt-9679154.66A_wPt-7027154.76A_wPt-5652156.76A_wPt-6520169.66A_wPt-4270176.46A_wPt-1285180.1
Int_Chr_6A
6A_wPt-12850.06A_wPt-93822.96A_wPt-42707.56A_wPt-652010.76A_wPt-690416.7
6A_wPt-565228.9
6A_wPt-969246.76A_wPt-969048.0
rPt-50560872.96A_wPt-744574.1
6A_wPt-3452104.8
Chr_6A_LA_x_TRI
Fortsetzung der Abbildung 22

87 Anhang
5B_wPt-86370.05B_wPt-32892.75B_wPt-95693.05B_wPt-51203.55B_wPt-34613.75B_BARC1094.35B_BARC44.65B_WMS11805.05B_WMS1975.4
5BL_WMS77771.9
Chr_5B_LA_x_ALA
5B_wPt-34570.0
5BL_WMS7777.0
5B_tPt-894216.8tPt-071417.3
5B_wPt-839341.45B_tPt-371941.75B_tPt-718341.85B_wPt-154842.05B_wPt-634865.45B7B_wPt-591465.55B_wPt-534665.75B7B_wPt-517565.85B_wPt-512073.15B_wPt-328973.85B_wPt-956974.15B_wPt-346174.65B_BARC475.25B_BARC10975.45B_WMS118075.85B_WMS19776.25B_wPt-863780.7
Int_Chr_5B
5B_wPt-86370.0
5B_tPt-89426.7tPt-07147.5
5B_wPt-634853.65B7B_wPt-591453.75B_wPt-534653.95B7B_wPt-517554.05B_wPt-512058.4
5B_wPt-154878.85B_tPt-371978.95B_tPt-718379.05B_wPt-839379.2
5B_wPt-3457116.6
Chr_5B_LA_x_TRI
Fortsetzung der Abbildung 22

88 Anhang
7B_wPt-73180.07B_WMS5375.57B_wPt-82837.05B7B_wPt-486313.77BS_WMS118415.67BS_WMS117317.67BL_WMS98318.97B_BARC17621.07B_wPt-1159824.07B_wPt-891925.87B_wPt-227333.47B_wPt-992544.17B_WMC51747.8wPt-1171152.0tPt-51396952.8rPt-41104653.97B_wPt-589258.47B_tPt-736262.7tPt-51319763.97B_WMS61165.67BL_WMS57767.67B_WMS23869.17BL_WMS74570.67BD_CFA204072.4wPt-754673.7
7B_wPt-487592.2
Chr_7B_LA_ALA
7B_wPt-48750.07B_wPt-43006.27B_wPt-482014.97B_wPt-988015.07B_wPt-021715.27B_wPt-729515.3wPt-754618.67B_wPt-060019.67BD_CFA204019.97BL_WMS74521.67B_WMS23823.17BL_WMS57724.67B_WMS61126.6tPt-51319728.47B_tPt-736229.57B_wPt-589233.8rPt-41104638.3tPt-51396939.4wPt-1171140.37B_wPt-558541.57B_wPt-403841.77B_wPt-9954 7B_wPt-522843.07B_wPt-546243.17B_wPt-741343.47B_WMC51744.47B_wPt-992548.1tPt-51368755.4tPt-51366356.47B_wPt-227358.87B_wPt-891966.47B_wPt-1159868.37B_BARC17671.27BL_WMS98373.37BS_WMS117374.67BS_WMS118476.75B7B_wPt-486378.57B_wPt-828385.27B_WMS53786.77B_wPt-731892.2
Int_Chr_7B
7B_wPt-22730.0
7B_wPt-430050.47B_wPt-988058.47B_wPt-729558.77B_wPt-021758.9wPt-754661.47B_wPt-060062.77B_wPt-482063.8
7B_wPt-558585.77B_wPt-403885.97B_wPt-546286.67B_wPt-522887.27B_wPt-741387.37B_wPt-995488.3tPt-51368799.6tPt-513663100.6
7B_tPt-3700112.1
Chr_7B_LA_x_TRI
Fortsetzung der Abbildung 22

89 Anhang
rPt-4111410.0tPt-42692.02R_SCM108.1rPt-5076759.1rPt-40249414.6rPt-39873715.6rPt-41087118.92R_SCM23321.8rPt-100223.32R_RMS120325.3rPt-50619626.02RL_SCM7526.8rPt-38980730.3rPt-40251932.5rPt-50916034.7rPt-50783736.0rPt-38990837.42R_SCM6142.5SCM25444.9tPt-51365746.8rPt-39876148.5rPt-50854349.8
rPt-50649660.4rPt-40092362.0rPt-50910566.6rPt-50909667.9
rPt-39883174.9
rPt-41106679.2
rPt-50538592.3rPt-50539193.0tPt-51340794.8
rPt-508776102.2rPt-506604103.4rPt-509197107.4
Chr_2R_LA_x_ALA
rPt-5091970.0rPt-4023613.3rPt-5087764.0rPt-5066044.8tPt-51340712.9rPt-50539114.6rPt-50538515.2rPt-41106630.8rPt-39883132.8rPt-39028334.8rPt-39029236.1rPt-50909639.4rPt-50910540.6rPt-40092344.4rPt-50649646.5rPt-50854356.6tPt-999257.5rPt-39876157.9tPt-51365759.6SCM25461.42R_SCM6163.7rPt-39977467.2rPt-38990869.0rPt-50783770.2rPt-40248170.8tPt-51424370.9rPt-40251973.9rPt-50798274.1rPt-38980775.1rPt-50895775.6rPt-40041076.5rPt-50617676.7rPt-40236476.8rPt-39078977.9tPt-514069 rPt-50794678.0rPt-389441 tPt-51386178.1rPt-39053078.22RL_SCM7578.6rPt-50619679.42R_RMS120380.0rPt-100280.82R_SCM23383.6rPt-39873789.9rPt-40249490.8rPt-50767596.02R_SCM1097.1tPt-4269103.3rPt-411141105.2
Int_Chr_2R
rPt-3997210.0rPt-4110660.6tPt-5132450.7
tPt-99927.8
rPt-40248117.3rPt-39989518.9rPt-50798220.8rPt-50895721.8rPt-40251922.0rPt-100222.6rPt-40041022.8rPt-40236422.9rPt-50617623.1rPt-38944123.8rPt-50794623.9rPt-39053024.1tPt-51406924.2tPt-51386124.5rPt-50593325.4rPt-39078926.3rPt-50771427.4rPt-41147046.4rPt-50732357.5rPt-50692658.8rPt-39028360.3tPt-51292962.4rPt-41087171.9rPt-39982472.6rPt-50669774.0rPt-50589174.6rPt-50548179.3rPt-41117481.7rPt-50767586.6
rPt-402361104.3
Chr_2R_LA_x_TRI
Fortsetzung der Abbildung 22

90 Anhang
rPt-5053380.0
rPt-4023989.9
rPt-40026417.2BARC918.2rPt-50951321.5rPt-39042923.6wPt-129125.0tPt-51411427.4tPt-374633.3rPt-085734.4rPt-50617334.9rPt-40250435.93R_RMS102839.0rPt-38970840.0rPt-38945443.4rPt-50789743.9SCM28445.9
rPt-50806852.6
Chr_3R_LA_x_ALA
rPt-5053380.0
rPt-40239810.1BARC918.7rPt-50951321.5rPt-39042924.2wPt-1291 rPt-50696125.3rPt-50971926.7tPt-51411428.2rPt-50787729.5rPt-50650430.0tPt-374633.9rPt-085734.9rPt-50617335.3rPt-38967636.1rPt-40250436.33R_RMS102839.7rPt-38970841.0rPt-38945444.1rPt-50789744.6rPt-50743445.1rPt-50737445.7rPt-39989746.1rPt-50710546.2rPt-39072446.3rPt-50684746.5SCM28446.6rPt-50841846.7rPt-50860947.2rPt-40116349.2rPt-50806853.2rPt-50522763.9rPt-50780168.7
Int_Chr_3R
rPt-5069610.0rPt-5097191.8rPt-5078775.7rPt-5065046.4rPt-4025048.7rPt-5061739.6rPt-38967612.2rPt-38970816.2rPt-50743418.7rPt-50860919.4rPt-50841819.6rPt-39072420.0rPt-50684720.2rPt-38945420.7rPt-39989721.7rPt-40116322.5rPt-50710523.8rPt-50737424.0
rPt-39001837.7rPt-50522741.9rPt-50757543.1rPt-50780144.8rPt-50656545.0rPt-40129645.6rPt-50645546.1rPt-41124346.5rPt-50696747.1rPt-40173850.3rPt-568855.6rPt-40026461.0
Chr_3R_LA_x_TRI
Fortsetzung der Abbildung 22

91 Anhang
rPt-5061720.0rPt-39974312.9rPt-39946214.9rPt-40239116.4rPt-50967721.6rPt-50558122.3rPt-50674724.4rPt-39880028.75R_RMS126434.4rPt-50966837.9rPt-39966340.5rPt-50890241.6rPt-50889742.4tPt-894044.2rPt-40127551.0rPt-50737351.9rPt-50892552.7rPt-50597357.6rPt-39939758.6rPt-50557759.6
rPt-507326107.65R_SCM280109.2rPt-506066113.9rPt-402334116.1
Chr_5R_LA_x_ALA
rPt-4023340.0rPt-5060662.25R_SCM2806.9rPt-5073268.5rPt-50826625.1rPt-506113 rPt-39850626.0rPt-399953 rPt-41127026.3rPt-41126726.6rPt-39876727.6rPt-50808128.1rPt-50883456.4rPt-50557756.6rPt-50766457.0rPt-39939757.6rPt-50597358.6rPt-50737363.9rPt-50892564.4rPt-40127565.2rPt-50567966.8rPt-39964169.0rPt-50955170.0tPt-894071.9rPt-50889773.7rPt-50890274.6rPt-39966375.6rPt-50966878.25R_RMS126481.6rPt-39880087.3rPt-50955090.1rPt-50674791.5rPt-50558193.9rPt-50967794.4rPt-50548595.7rPt-40197495.9rPt-39009396.5rPt-40239199.3rPt-402625100.6rPt-399462100.8rPt-508566100.9rPt-505220101.0rPt-390722101.1rPt-399743102.8rPt-390151103.2rPt-506172111.6
Int_Chr_5R
tPt-91800.0rPt-5076640.9tPt-98081.6rPt-5088344.6rPt-39964110.8rPt-50892511.1rPt-50955114.1rPt-50567915.4rPt-41126724.7rPt-39995326.9rPt-41127027.0rPt-39850627.6rPt-50611327.8rPt-50826628.9rPt-39876729.2rPt-50808130.8rPt-50955046.6rPt-39015149.6rPt-50617249.9rPt-50522051.9rPt-39072252.8rPt-50856653.3rPt-40262553.5rPt-50548555.2rPt-40197455.5rPt-50558156.0rPt-50967756.1rPt-39009356.8
Chr_5R_LA_x_TRI
Fortsetzung der Abbildung 22

92 Anhang
rPt-3984800.0
rPt-50574521.5
6R_SCM18038.4rPt-41085044.2tPt-51347048.8tPt-790049.7
tPt-447964.8rPt-50945369.3rPt-50609970.4rPt-40255272.9rPt-50603973.7rPt-50904378.1rPt-41135480.6rPt-39959788.9rPt-39942193.2rPt-40134695.8rPt-50525096.3rPt-40093599.0rPt-50609399.8rPt-507320102.5tPt-513959105.0rPt-509167112.0
Chr_6R_LA_x_ALA
rPt-3984800.0rPt-50574520.9rPt-39975521.16R_SCM18037.9rPt-41085045.4tPt-51347048.6tPt-790049.7rPt-40150551.0tPt-51424455.2rPt-39052558.6rPt-39958762.5rPt-820562.6tPt-447963.6tPt-51306065.3rPt-50933365.8rPt-50609967.6rPt-50945368.4rPt-40255271.5rPt-50603972.3rPt-50904376.7rPt-41135477.9rPt-39959788.1rPt-50884389.6rPt-506091.0rPt-39942192.8rPt-40134694.7rPt-50939595.4rPt-50525095.9rPt-50915496.9rPt-50609399.3rPt-507320101.1rPt-400935101.9tPt-513959104.7rPt-509167111.7rPt-505488114.7rPt-399312120.4rPt-507626126.1
Int_Chr_6R
rPt-4108500.0rPt-4015051.6tPt-79002.8tPt-5142446.8rPt-3905259.9rPt-820513.3tPt-51306015.7rPt-40093517.9rPt-50837132.1rPt-50915434.8rPt-40060836.3rPt-50838837.3rPt-39942137.4tPt-466937.5rPt-40134638.1rPt-50939538.6rPt-506040.3rPt-50884341.6rPt-50548856.1rPt-40078963.4rPt-39025266.4
Chr_6R_LA_x_TRI
Fortsetzung der Abbildung 22

93 Anhang
8.4 Abkürzungsverzeichnis
AFLP Amplified fragment length polymorphism
AG Arbeitsgruppe
ANOVA Analysis of variance
BOH Standort Bohlingen
CIMMYT Centro Internacional de Mejoramiento de Maiz y Trigo
cM Centimorgan
CTAB Cetylrimethylammoniumbromid
DArT Diversity Arrays Technology
DH Doppelhaploid
DNA Deoxyribonucleic Acid
DON Deoxynivalenol
EC Stadium des Ährenschiebens
EDTA Ethylendiamintetraacetat
ELISA Enzyme-linked Imunosorbent Assay
F Filialgeneration
FHB Fusarium head blight
GD Grenzdifferenz
GRA Standort Grabau
HOH Standort Hohenheim
LOD Logarithm of Odds
PCR Polymerase Chain Reaction
QTL Quantitative Trait Loci
RADP Random amplified length polymorphism
RAN Standort Ranzin

94 Anhang
RFLP Restriktions-Fragment-Längen-Polymorphismus
RNA Ribonucleic acid
SDS Sodium dodecylsulfate
SEVAG CHCl3/Isoamylalkohol, 24:1 (v/v)
SNA Speziell nährstoffarmer Agar
SNP Single Nucleotide Polymorphism
SSR Simple sequence repeat; Mikrosatellit
TR Teilramsch
tvs Tons volatile solids
Upm Umdrehungen pro Minute

Danksagung Ich möchte mich bei allen bedanken, die bei der Erstellung dieser Arbeit mitgewirkt haben.
Für die Überlassung des Themas, die sehr gute Betreuung und stetige Hilfe möchte ich mich ganz herzlich bei meinem Doktorvater Herrn apl. Prof. Dr. Thomas Miedaner bedanken.
Vielen Dank auch an die Züchter Elmar Weissmann, Günther Wahle und Carsten Reinbrecht für die Bereitstellung der Versuchsflächen und deren freundliche Unterstützung in diesem Projekt.
Ein Dankeschön an Herrn Prof. Dr. Otmar Spring und Herrn apl. Prof. Dr. Klaus Haas vom Institut für Botanik für die Übernahme der Korreferate und die Teilnahme als Prüfer an meiner mündlichen Prüfung.
Vielen Dank an Marion Röder für die Bereitstellung der SSR-Marker.
Sabine Frey-Tippelt, Hans Häge, Justus von Kitlitz, Agnes Rölfing-Finze und Jacek Till danke ich für die freundliche Aufnahme in die Arbeitsgruppe, für die vielen schönen und lustigen gemeinsamen Arbeitstage und für ein stets offenes Ohr. Danke auch für die enorme Hilfe bei der Durchführung der Feldversuche.
Janna, Helke und Manfred Fautz, Manfred Großmann, Sandra Fischer, Horst Leimbeck und Carolin Schneider möchte ich für das Korrekturlesen meiner Arbeit danken.
Für die Hilfe im Labor möchte ich mich besonders bei Eva Bauer, Gunda Brunsbach, Angela Harmsen, Sylvia Kaiser, Rasha Kalih, Bärbel Lieberherr, Barbara Renz, Marika Takacs und Firas Talas bedanken.
Allen fleißigen Praktikanten, meiner Bachelorstudentin und Hiwis, insbesondere Katharina Alheit, Carolina Bogenschütz und Katharina Seibold möchte ich für die Hilfe bei der vielen und manchmal auch anstrengenden, harten Arbeit danken.
Vielen Dank auch an Hr. Utz, der mir bei vielen Fragen zu den Statistikprogrammen zur Seite stand.
Ich möchte Martin Messerschmidt für die schöne Zeit im gemeinsamen Büro danken und für die vielen hilfreichen Verbesserungsvorschläge bei der Korrektur meiner Arbeit.
In sehr guter Erinnerung werde ich die Arbeitsatmosphäre an der LSA in Hohenheim behalten und möchte herzlich allen danken, die dazu beigetragen haben, u.a.: Sankalp Bhosale, Sebastian Bopper, Chrisin Falke, Heike Goßlau, Manje Gowda, Marlen Hübner, Wenxin Liu, Mathias Martin, Hans-Peter Maurer, Bettina Müller, Elena Orsini, Heiko Parzies, Peter Risser, Jana Steinhoff, Delphine Van Ingelandt, Christiane Von der Ohe, Hans-Henning Voss und Tobias Würschum.
Letztlich danke ich meiner Familie und meinen Freunden, die mir in den letzten drei Jahren zur Seite standen, insbesondere meinem Ehemann, Nils, für die stete Unterstützung Durchhaltevermögen zu behalten, um diese Arbeit fertig zu stellen.
Das Forschungsvorhaben wurde vom Bundesministerium für Verbraucherschutz, Ernährung und Landwirtschaft (BMVEL) und der Gemeinschaft zur Förderung der privaten deutschen Pflanzenzüchtung e.V. (GFP) gefördert - vielen Dank.

Lebenslauf
Name: Maren Swente Großmann geb. Fautz
Geburtsdatum, -ort: 12. März 1981, Offenbach am Main
Schulausbildung
1987-1991 Gerhart-Hauptmann-Schule, Dreieich (Grundschule)
1991-1995 Ricarda-Huch-Schule, Dreieich (Gymnasium)
1995-1998 Heinrich-Heine-Schule, Dreieich (Gesamtschule,
Gymnasialzweig)
1998-2000 Weibelfeldschule, Dreieich (gymnasiale Oberstufe)
Abitur; Abschluss im Mai 2000
Studium
2000-2006 Biologie an der Technischen Universität Darmstadt mit den
Fachrichtungen Pflanzenphysiologie, Spezielle Zoologie,
Tierphysiologie
Abschluss: Diplombiologin
Promotion
2007-2010 Promotion an der Universität Hohenheim,
Institut für Agrarwissenschaften, Landessaatzuchtanstalt

Erklärung
Hiermit erkläre ich an Eides statt, dass die vorliegende Arbeit von mir selbst verfasst wurde
und lediglich unter Zuhilfenahme der angegebenen Quellen und Hilfsmittel angefertigt wurde.
Wörtlich oder inhaltlich übernommene Stellen wurden als solche gekennzeichnet.
Die vorliegende Arbeit wurde in gleicher oder ähnlicher Form noch keiner anderen Institution
oder Prüfungsbehörde vorgelegt.
Insbesondere erkläre ich, dass ich nicht früher oder gleichzeitig einen Antrag auf Eröffnung
eines Promotionsverfahrens unter Vorlage der hier eingereichten Dissertation gestellt habe.
Stuttgart, im September 2010
________________
Maren Großmann


















