
Biosynthesis of Vitexin and Isovitexin: Enzymatic Synthesis of the C ...

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
Phosphate Production and Analysis in the Non-Enzymatic Activation of Amino Acids by ATP when Using Hydroxylamine as a Trapping Agent
Dail W . Mullins (Jr.) and James C. Lacey (Jr.)* Division of Molecular Biology, University of Alabama in Birmingham, University Station, Birmingham, Alabama 35294, USA
Z. Naturforsch. 36b, 732-734 (1981); received January 26, 1981 Phosphate Production, Hydroxylamine, ATP
Saygin and Decker [6, 7] have reported that the formation of hydroxamic acids in solutions containing ATP, MgCl2, hydroxylamine and an acidic acceptor (acetic acid or amino acids) is not accompanied by the release of inorganic phosphate from ATP. Using an ion exchange chromatography method developed by Lowenstein [8] to separate the various components of such reaction mixtures, we now show that inorganic phosphate, as well as some pyrophosphate, is produced during incubation at 50 °C. Evidence is also presented to indicate that the failure of Saygin and Decker [6, 7] to detect phosphate is due to a total inhibition of their phosphate assay system by ATP.
We have recently studied [1,2] the non-enzymatic activation of amino acids and simple carboxylic acids by ATP. In these studies we used hydroxyl-amine to trap the activated species because the resulting hydroxamates can be quantitated colori-metrically following complex formation with ferric ions under acid conditions as originally described by Lipmann and Tuttle [3].
In our studies, we assumed that activation pro-ceeded via the formation of an aminoacyl adenylate anhydride intermediate, as suggested by Paecht-Horowitz and Katchalsky [4], or possibly through the production of an aminoacyl phosphate ester, as proposed by Lowenstein and Schatz [5]. Regardless of which mechanism operates, inorganic phosphate would be produced, one mole for each mole of acid activated. However, in similar studies, only using acetate as the principal acid studied, Saygin and Decker [6, 7] reported that the formation of hydrox-amates in such systems cannot proceed by the above mechanism because there is no release of inorganic phosphate from ATP. It was suggested by the workers that the formation of hydroxamates in the presumed activation reaction may simply be the result of catalysis by metal-sugar complexes, rather than through the involvement of the high-energy, phosphodiester bonds of ATP. These reports of Saygin and Decker [6, 7], coupled with further observations of our own which indicated that the reaction systems employing hydroxylamine are
considerably more complex than originally thought, have encouraged us to delve more deeply into the chemistry of these reaction systems.
The principal concern of the present paper is to examine the reaction system with respect to the question of whether or not phosphate is produced. We were somewhat puzzled by Saygin and Decker's report [6, 7] because we had observed that AMP did not facilitate the formation of aminoacyl hydroxa-mates to nearly the extent that A D P and A T P did. Although this fact would not, in itself, rule out only steric involvement of the phosphodiester moiety, it seemed unlikely that inorganic phosphate would not be produced to some extent simply as a consequence of ATP hydrolysis.
In the present study we actually separated the various products (including phosphate) and starting materials of the reaction, using ion exchange chromatography as described by Lowenstein [8]. Because we did find phosphate as a principal product of the reaction, we then questioned why Saygin and Decker [6, 7] had not detected it. Repeating their phosphate assay under the conditions they used, we found that ATP and ADP, in the concentrations employed, completely inhibited the development of color (325 nm). We conclude that they did not detect phosphate because of this interference.
Experimental Reagents
Adenosine 5'-triphosphate (disodium salt). MgCl2 • 6 H2O, phenylalanine and hydroxylamine were purchased from Sigma Chemical Co. and used without further purification. Dowex 1 - X 8 (200 to
2 .0 A B C D
- Pj
" I AMP
r \ ADP
\ \J \ P P j ATP \ 5 10 0 5 10 15 20 25 30 35 40 0 5 10 0 10 20 30 40
Fractions
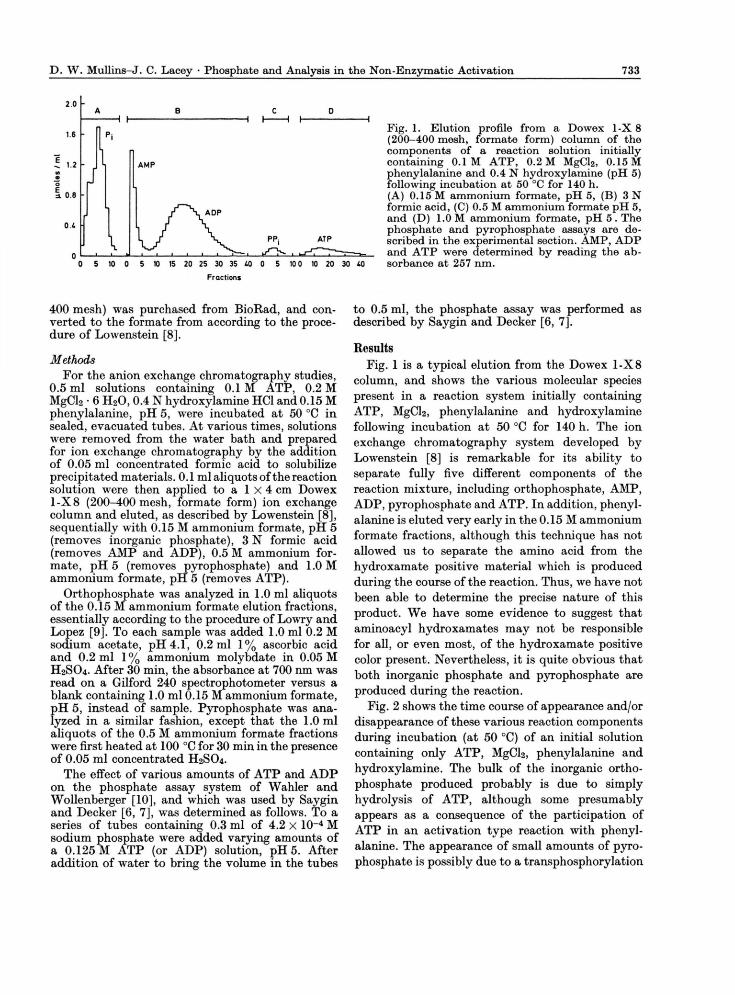
Fig. 1. Elution profile from a Dowex 1-X 8 (200-400 mesh, formate form) column of the components of a reaction solution initially containing 0.1 M ATP, 0.2 M MgCl2, 0.15 M phenylalanine and 0.4 N hydroxylamine (pH 5) following incubation at 50 °C for 140 h. (A) 0.15 M ammonium formate, pH 5, (B) 3 N formic acid, (C) 0.5 M ammonium formate pH 5, and (D) 1.0 M ammonium formate, pH 5 . The phosphate and pyrophosphate assays are de-scribed in the experimental section. ÄMP, ADP and ATP were determined by reading the ab-sorbance at 257 nm.
400 mesh) was purchased from BioRad, and con-verted to the formate from according to the proce-dure o f Lowenstein [8].
Methods For the anion exchange chromatography studies,
0.5 ml solutions containing 0.1 M ATP, 0.2 M MgCl2 • 6 H 2 0 , 0.4 N hydroxylamine HCl and 0.15 M phenylalanine, p H 5, were incubated at 50 °C in sealed, evacuated tubes. At various times, solutions were removed from the water bath and prepared for ion exchange chromatography by the addition of 0.05 ml concentrated formic acid to solubilize precipitated materials. 0.1 ml aliquots of the reaction solution were then applied to a 1 x 4 cm Dowex 1 - X 8 (200-400 mesh, formate form) ion exchange column and eluted, as described by Lowenstein [8], sequentially with 0.15 M ammonium formate, p H 5 (removes inorganic phosphate), 3 N formic acid (removes A M P and ADP) , 0.5 M ammonium for-mate, p H 5 (removes pyrophosphate) and 1.0 M ammonium formate, p H 5 (removes ATP) .
Orthophosphate was analyzed in 1.0 ml aliquots of the 0.15 M ammonium formate elution fractions, essentially according to the procedure of Lowry and Lopez [9J. T o each sample was added 1.0 ml 0.2 M sodium acetate, p H 4 . 1 , 0.2 ml 1 % ascorbic acid and 0.2 ml 1 % ammonium molybdate in 0.05 M H2SO4. After 30 min, the absorbance at 700 nm was read on a Gilford 240 spectrophotometer versus a blank containing 1.0 ml 0.15 M ammonium formate, p H 5, instead of sample. Pyrophosphate was ana-lyzed in a similar fashion, except that the 1.0 ml aliquots of the 0.5 M ammonium formate fractions were first heated at 100 °C for 30 min in the presence o f 0.05 ml concentrated H2S04.
The effect of various amounts of A T P and A D P on the phosphate assay system of Wahler and Wollenberger [10], and which was used by Saygin and Decker [6, 7], was determined as follows. T o a series of tubes containing 0.3 ml of 4.2 x 10 - 4 M sodium phosphate were added varying amounts o f a 0.125 M A T P (or A D P ) solution, p H 5. After addition of water to bring the volume in the tubes
to 0.5 ml, the phosphate assay was performed as described by Saygin and Decker [6, 7].
Results Fig. 1 is a typical elution from the Dowex 1 - X 8
column, and shows the various molecular species present in a reaction system initially containing A T P , MgCl2, phenylalanine and hydroxylamine following incubation at 50 °C for 140 h. The ion exchange chromatography system developed by Lowenstein [8] is remarkable for its ability to separate fully five different components of the reaction mixture, including orthophosphate, AMP, ADP, pyrophosphate and ATP. In addition, phenyl-alanine is eluted very early in the 0.15 M ammonium formate fractions, although this technique has not allowed us to separate the amino acid from the hydroxamate positive material which is produced during the course of the reaction. Thus, we have not been able to determine the precise nature of this product. W e have some evidence to suggest that aminoacyl hydroxamates may not be responsible for all, or even most, of the hydroxamate positive color present. Nevertheless, it is quite obvious that both inorganic phosphate and pyrophosphate are produced during the reaction.
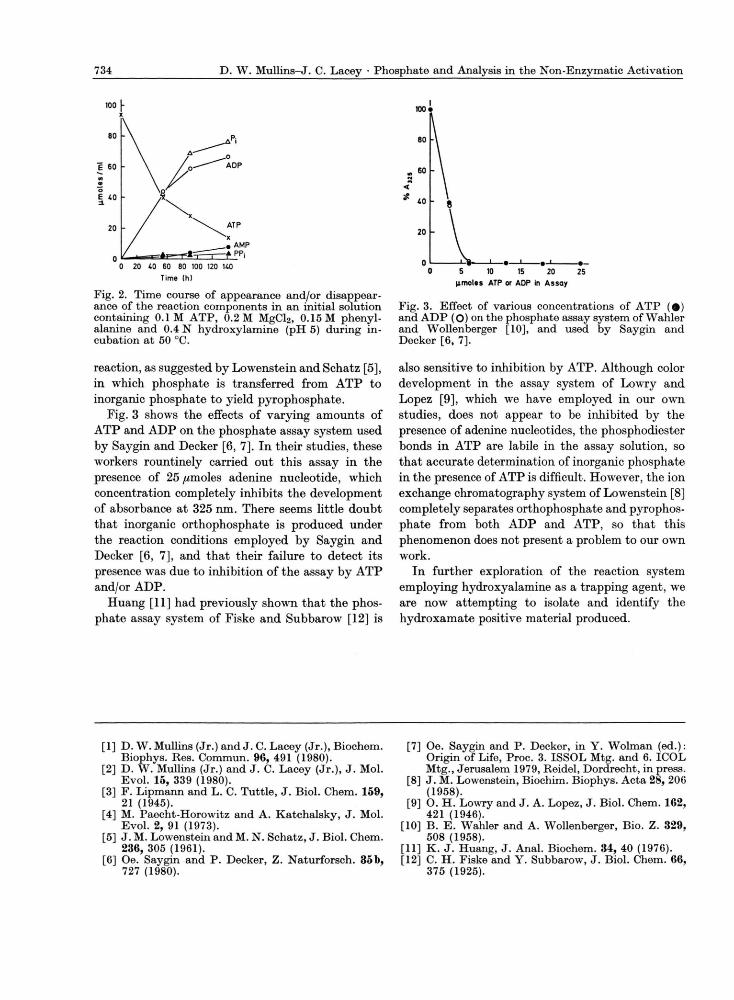
Fig. 2 shows the time course of appearance and/or disappearance of these various reaction components during incubation (at 50 °C) of an initial solution containing only A T P , MgCl2, phenylalanine and hydroxylamine. The bulk of the inorganic ortho-phosphate produced probably is due to simply hydrolysis of A T P , although some presumably appears as a consequence of the participation of A T P in an activation type reaction with phenyl-alanine. The appearance of small amounts of pyro-phosphate is possibly due to a transphosphorylation
Time (h)
Fig. 2. Time course of appearance and/or disappear-ance of the reaction components in an initial solution containing 0.1 M ATP, 0.2 M MgCl2, 0.15 M phenyl-alanine and 0.4 N hydroxylamine (pH 5) during in-cubation at 50 °C.
o 5 10 15 20 25 l imoles ATP or ADP in Assay
Fig. 3. Effect of various concentrations of ATP (•) and ADP (O) on the phosphate assay system of Wähler and Wollenberger [10], and used by Saygin and Decker [6, 7].
reaction, as suggested by Lowenstein and Schatz [5], in which phosphate is transferred from A T P to inorganic phosphate to yield pyrophosphate.
Fig. 3 shows the effects of varying amounts of ATP and A D P on the phosphate assay system used by Saygin and Decker [6, 7]. In their studies, these Avorkers rountinely carried out this assay in the presence of 25 //moles adenine nucleotide, which concentration completely inhibits the development of absorbance at 325 nm. There seems little doubt that inorganic orthophosphate is produced under the reaction conditions employed by Saygin and Decker [6, 7], and that their failure to detect its presence was due to inhibition of the assay by A T P and/or ADP.
Huang [11] had previously shown that the phos-phate assay system of Fiske and Subbarow [12] is
also sensitive to inhibition by ATP. Although color development in the assay system of Lowry and Lopez [9], which we have employed in our own studies, does not appear to be inhibited by the presence of adenine nucleotides, the phosphodiester bonds in ATP are labile in the assay solution, so that accurate determination of inorganic phosphate in the presence of A T P is difficult. However, the ion exchange chromatography system of Lowenstein [8] completely separates orthophosphate and pyrophos-phate from both ADP and ATP, so that this phenomenon does not present a problem to our own work.
In further exploration of the reaction system employing hydroxyalamine as a trapping agent, we are now attempting to isolate and identify the hydroxamate positive material produced.
[1] D. W. Mullins (Jr.) and J. C. Lacey (Jr.), Biochem. Biophys. Res. Commun. 96, 491 (1980).
[2] D. W. Mullins (Jr.) and J. C. Lacey (Jr.), J. Mol. Evol. 15, 339 (1980).
[3] F. Lipmann and L. C. Tuttle, J. Biol. Chem. 159, 21 (1945).
[4] M. Paecht-Horowitz and A. Katchalsky, J. Mol. Evol. 2, 91 (1973).
[5] J. M. Lowenstein and M. N. Schatz, J. Biol. Chem. 236, 305 (1961).
[6] Oe. Saygin and P. Decker, Z. Naturforsch. 35b, 727 (1980).
[7] Oe. Saygin and P. Decker, in Y. Wolman (ed.): Origin of Life, Proc. 3. ISSOL Mtg. and 6. ICOL Mtg., Jerusalem 1979, Reidel, Dordrecht, in press.
[8] J. M. Lowenstein, Biochim. Biophys. Acta 28, 206 (1958).
[9] O. H. Lowry and J. A. Lopez, J. Biol. Chem. 162, 421 (1946).
[10] B. E. Wahler and A. Wollenberger, Bio. Z. 329, 508 (1958).
[11] K. J. Huang, J. Anal. Biochem. 34, 40 (1976). [12] C. H. Fiske and Y. Subbarow, J. Biol. Chem. 66,
375 (1925).


















