Praktikum Stoffwechselphysiologie - Uni Ulm Aktuelles ... · Erstabgabe: Enzymkinetik 3 1....
21
Erstabgabe: Enzymkinetik 1 Praktikum Stoffwechselphysiologie WS 2011/2012 Versuch Enzymkinetik Gruppe XY Betreuer: Praktikanten: Versuchstag:
Transcript of Praktikum Stoffwechselphysiologie - Uni Ulm Aktuelles ... · Erstabgabe: Enzymkinetik 3 1....

Erstabgabe: Enzymkinetik
1
Praktikum Stoffwechselphysiologie
WS 2011/2012
Versuch Enzymkinetik
Gruppe XY
Betreuer:
Praktikanten:
Versuchstag:

Erstabgabe: Enzymkinetik
2
Inhaltsverzeichnis 1. Einleitung ............................................................................................................................................ 3
2. Theorie................................................................................................................................................. 3
2.1 Enzyme ...................................................................................................................................... 3
2.1.1 Definition ................................................................................................................................ 3
2.1.2 Aufbau .................................................................................................................................... 3
2.1.3 Katalysestrategien .................................................................................................................. 3
2.1.4 Hauptklassen der Enzyme ...................................................................................................... 4
2.1.5 Beeinflussung der Enzymaktivität .......................................................................................... 4
2.1.6 Enzyminhibitoren ................................................................................................................... 4
2.1.7 Enzymregulation ..................................................................................................................... 5
2.1.8 Enzymkinetik .......................................................................................................................... 6
2.2 Glykolyse und Lactatdehydrogenase (LDH) ................................................................................... 8
2.2.1 Glykolyse................................................................................................................................. 8
2.2.2 Aerobe Umsetzung des Pyruvats............................................................................................ 9
2.2.3 Anaerober Abbau von Pyruvat ............................................................................................... 9
2.2.4 Cori-Zyklus ............................................................................................................................ 10
3. Material und Methoden ..................................................................................................................... 11
3.1 Bestimmung von KM und Vmax; Berechnung der spezifischen Enzymaktivität ........................... 11
3.2 Bestimmung des pH-Optimums .................................................................................................. 12
4. Ergebnisse: ........................................................................................................................................ 13
4.1 Versuch 1 ..................................................................................................................................... 13
4.2 Versuch 2: .................................................................................................................................... 18
5. Diskussion .......................................................................................................................................... 20
5.1 Versuch 1: .................................................................................................................................... 20
5.2 Versuch 2: .................................................................................................................................... 20
6. Quellen .............................................................................................................................................. 21

Erstabgabe: Enzymkinetik
3
1. Einleitung
In diesem Versuch stand die Einführung in die Enzymuntersuchung im Vordergrund. So wurden speziell das pH-Optimum und die Enzymaktivität der Lactatdehydrogenase bestimmt.
2. Theorie
2.1 Enzyme
2.1.1 Definition
Enzyme sind Biokatalysatoren. Sie sind also Makromoleküle, die die Einstellung des chemischen Gleichgewichts einer Reaktion beschleunigen, dabei aber nicht verbraucht werden. Enzyme sind substratspezifisch, das bedeutet, dass sie nur spezielle Moleküle mit einer ganz bestimmten Struktur binden können. Desweiteren sind Enzyme auch wirkspezifisch, was heißt, dass sie das gebundene Substrat nur auf eine Art und Weise umsetzen.
2.1.2 Aufbau
Enzyme sind meist Proteine, also Polypeptidketten aus Aminosäuren (AS), es gibt jedoch auch andere katalytisch aktive Verbindungen, zum Beispiel RNA. Der Ort an dem die Substratumsetzung im Enzym erfolgt heißt aktives Zentrum. Hier befinden sich die Reste der Aminosäuren, die für die Bindung mit dem Substrat wichtig sind. Neben kovalenten Bindungen, werden die Substrate meist über Wasserstoffbrückenbindungen oder Van-der-Waals-Wechselwirkungen am Enzym fixiert. Enzyme können auch einen Nichtproteinanteil kovalent gebunden haben. Dieser wird prosthetische Gruppe genannt und ist maßgeblich an der katalytischen Aktivität des Enzyms beteiligt. Ein Beispiel für ein Enzym mit einer solchen prosthetischen Gruppe ist das Cytochrom C mit seiner Häm-Gruppe (Fe2+-Ion eingeschlossen von einem Porphyrinring). Manche Enzym benötigen für den Umsatz ihres Substrates keinen kovalent gebundenen Nicht-Proteinanteil, sondern sogenannte Co-Faktoren bzw. Co-Substrate. Diese Verbindungen sind nicht permanent an das Enzym gebunden, aber essentiell für dessen katalytische Wirkung. Beispiele für Co-Faktoren und Co-Substraten sind das Coenzym-A und das ATP.
2.1.3 Katalysestrategien
Man unterscheidet grundsätzlich vier Katalysestrategien, die Säure-Base-Katalyse, die Katalyse durch Annäherung, die kovalente Katalyse und die Metall-Ionen vermittelte Katalyse. Bei der Katalyse durch Annäherung werden die beiden Reaktionspartner in der räumlich richtigen Orientierung durch Binden an das Enzym einander näher gebracht, wodurch die Reaktion dann besser ablaufen kann (Bsp.: Nukleosidmonophosphat-Kinase). Bei der Metall-Ionen vermittelten Katalyse wird ein Metall-Ion (z. B.: Zn2+, Fe2+) dazu genutzt beispielsweise ein negativ geladenes Zwischenprodukt zu stabilisieren (Bsp.: Carbonanhydrase). Bei der kovalenten Katalyse, wird das Substrat kovalent an das Enzym gebunden und dadurch umgesetzt (Bsp.: Chymotrypsin). Die Reaktion der Lactatdehydrogenase, die in diesem Versuch genauer untersucht wird ist der Säure-Base-Katalyse zu zuordnen, hierbei wird die Reaktion durch H+- oder OH--Ionen beschleunigt.

Erstabgabe: Enzymkinetik
4
Jedoch nutzen Enzyme meist nicht nur eine Katalysestrategie, sondern setzten ihre Substrate durch eine Kombination mehrerer dieser Katalysetypen um.
2.1.4 Hauptklassen der Enzyme
Die Enzyme werden im Enzyme Coding System (E. C. System) in 6 Hauptklassen und mehrere Unterklassen unterteilt. Die Hauptklassen werden unterschieden in Oxidoreduktasen, die Oxidation und Reduktionen katalysieren. Zu dieser Hauptklasse gehört auch die in diesem Versuch verwendete Lactatdehydrogenase. Dann gibt es noch die Transferasen, die die Übertragung von Gruppen von einem Molekül auf ein anderes katalysieren. Die Hydrolasen spalten Moleküle unter Einbau von Wasser. Die Lyasen katalysieren Eliminierungs- und Additionsreaktionen während die Isomerasen Umlagerungsreaktionen beschleunigen. Die sechste Hauptklasse ist die der Ligasen, die die Verknüpfung von Molekülen unter ATP-Verbrauch katalysieren.
2.1.5 Beeinflussung der Enzymaktivität
Man kann die Enzymaktivität auf ganz verschiedene Arten verändern. Beispielsweise kann man die Temperatur erhöhen bzw. erniedrigen. Denn umso höher die Temperatur ist je schneller bewegen sich die Teilchen. Dies bedeutet dass die Wahrscheinlichkeit, dass ein Substratmolekül auf ein Enzym trifft steigt, weshalb auch die Umsatzrate erhöht wird. Wird die Temperatur jedoch zu stark erhöht, denaturiert das Enzym, also verändert seine Struktur, weshalb dann keine Substratbindung also auch kein Umsatz mehr möglich ist. Wird die Temperatur stark verringert, bewegen sich die Teilchen langsamer, darum sinkt auch die Wahrscheinlichkeit, dass ein Substratmolekül auf ein Enzym trifft, die Umsatzrate sinkt. Aber nicht nur die Temperatur hat einen Einfluss auf die Enzymaktivität auch der pH-Wert spielt dabei eine entscheidende Rolle. Denn die Reste der AS verändern ihre Eigenschaften je nach pH-Wert, dadurch kann sich auch die Struktur des Enzyms umgewandelt werden, wodurch die Umsetzung des Substrates behindert wird. Aus diesem Grund hat jedes Enzym ein Temperatur- und ein pH-Wert-Optimum, bei welchem die höchste und beste Umsatzgeschwindigkeit des Enzyms vorliegt. Desweitern kann auch die Enzymaktivität durch das Vorhandensein von Schwermetallionen oder Inhibitoren herabgesetzt werden. Auch der Körper hat mehrere Mechanismen entwickelt um die Aktivität seiner Enzyme zu regulieren. Beispiele hierfür sind allosterische Effektoren, Isoenzyme, proteolytische Spaltung und noch einiges mehr, worauf später genauer eingegangen wird.
2.1.6 Enzyminhibitoren
Hierbei werden erst einmal zwei Arten der Enzyminhibition unterschieden: die irreversible und die reversible Inhibition. Die irreversible Inhibition kann nicht rückgängig gemacht werden. Das inhibierte Enzym kann seiner Aufgabe nicht mehr nachkommen und muss abgebaut werden. Solche irreversiblen Inhibitoren sind meist Schwermetallionen, wie Quecksilber oder Arsen, oder Penicilline. Bei der reversiblen Inhibition kann das betroffene Enzym wieder in seine aktive Form überführt werden. Auch hier gibt es wieder eine Einteilungen nach der die reversiblen Inhibitoren unterschieden werden können: die kompetitiven und die nicht kompetitiven Inhibitoren. Die kompetitiven Inhibitoren konkurrieren mit den Substratmolekülen um die Bindungsstelle im aktiven Zentrum des Enzyms. Dieser kompetitive Hemmstoff muss also

Erstabgabe: Enzymkinetik
5
eine ähnliche Struktur wie das Substrat aufweisen. Die kompetitive Hemmung kann durch eine Erhöhung der Substratkonzentration abgeschwächt werden. Bei der nicht kompetitiven Inhibition bindet der Hemmstoff außerhalb des aktiven Zentrums am Enzym und verursacht dadurch eine Konformationsänderung des Biokatalysators. Durch die veränderte Struktur kann nun nur noch langsam Substrat falls überhaupt binden, wodurch die Umsatzrate stark sinkt.
2.1.7 Enzymregulation
Wie bereits erwähnt besitzt der Körper viele Mechanismen um die Enzymaktivität zu regulieren. Einerseits kann er die Aktivität der Enzyme mit Hilfe von allosterischen Effektoren verringern. Diese allosterische Effektoren gehören zu den nicht kompetitiven Hemmstoffen und binden außerhalb des aktiven Zentrums des Enzyms. Durch die Bindung eines solchen Effektors kann die Enzymaktivität erhöht oder auch erniedrigt werden. Je nachdem um welches Enzym es sich handelt, führt die Konformationsänderung zur Bindungsmöglichkeit des Substrates oder eben zum Gegenteil. Auch kann das Eintreten des chemischen Gleichgewichtes über Isoenzym beschleunigt werden. Die Isoenzyme besitzen alle eine unterschiedliche Struktur, wodurch sich auch ihr pH-Optimum beispielsweise verändern kann, katalysieren jedoch die gleiche Reaktion. Die Isoenzyme ermöglichen es dem Körper in verschiedenen Geweben bei unterschiedlichen Bedingungen mit den Enzymen zu arbeiten, zu denen das Milieu am besten passt. Auch für die in diesem Versuch verwendete Lactatdehydrogenase besitzt der Körper unterschiedlichste Isoenzyme. Die Lactatdehydrogenase ist ein Tetramer, besteht also aus vier Untereinheiten, die entweder vom Typ M oder H sind. Die Lactatdehydrogenase im Herzen enthält vier Untereinheiten vom Typ H, die Lactatdehydrogenase in der Skelettmuskulatur besteht aus vier M-Untereinheiten. Es gibt auch noch andere Isoenzyme der Lactatdehydrogenase (LDH), die eine Mischung aus M- und H-Untereinheiten aufweisen (LDH-3 im Lungengewebe: 2 H 2 M). Auch ein effektives Mittel des Körpers Enzyme zu aktivieren bzw. zu deaktivieren ist die Interkonversion. Bei diesem Mechanismus der Enzymregulation wird an eine freie OH-Gruppe des Enzyms eine Phosphatgruppe angehängt oder auch davon abgespalten. Dabei ist zu beachten, dass es Enzyme gibt, die in dephosphoryliertem Zustand aktiv sind und welche die erst aktiv werden, wenn sie eine Phosphatgruppe angehängt bekommen. Ebenfalls eine Möglichkeit ein Enzym in eine aktive Konformation zu überführen ist die proteolytische Aktivierung . Hierbei wird wenn das Enzym gebraucht wird ein Teil der Peptidsequenz mit Hilfe von Proteasen abgespalten, wodurch das Enzym endlich die Struktur erhält, die es ihm möglich macht, sein Substrat zu binden und umzusetzen. Diese Art der Enzymaktivierung wird vor allem bei Enzymen des Verdauungstraktes gemacht, damit diese während des Transportes von ihrem Bildungsort zum Wirkort nicht den Organismus selbst abbauen (Selbstverdauung). Der Körper kann die Umsetzungsrate von Substraten nicht nur über Aktivierung und Deaktivierung bereits vorhandener Enzyme steuern, er kann auch die Syntheserate von den Genen für die Enzyme anregen bzw. stoppen über Gen-Induktion bzw. Gen-Repression. Bei der Gen-Induktion führt das Vorhandensein eines bestimmten Effektors zur Aktivierung eines Transkriptionsfaktors und damit zu einer erhöhte Expression von bestimmten Genen,
Erstabgabe: Enzymkinetik
6
die das Enzym codieren. Bei der Gen-Repression zeigt sich der umgekehrte Fall. Hier führt das Vorhandensein eines Repressors zum Stopp der Genexpression von bestimmten Genen.
2.1.8 Enzymkinetik
Unter dem Begriff Enzymkinetik versteht man die Beschreibung eines sich zeitlich ändernden Systems. Vor allem die Theorien von Michaelis und Menten haben auf die Beschreibung großen Einfluss. Diese beiden haben die Michaelis-Menten-Konstante eingeführt, die die Substratkonzentration angibt, bei der das Enzym mit der halbmaximalen Geschwindigkeit arbeitet. Die Theorien von Michaelis und Menten gelten jedoch nur unter zwei Bedingungen. Diese lauten: die Rückreaktion (Reaktion vom Produkt mittels dem Enzym zurück zum Substrat) kann vernachlässigt werden und das immer die gleiche Anzahl von Enzym-Substrat-Komplexen vorhanden ist. Die Michaelis-Menten-Gleichung lautet:
�� = ����∗[�]
���[�]
v0: Anfangsgeschwindigkeit vmax: maximale Geschwindigkeit KM: Michaelis-Menten-Konstante [S]: Substratkonzentration
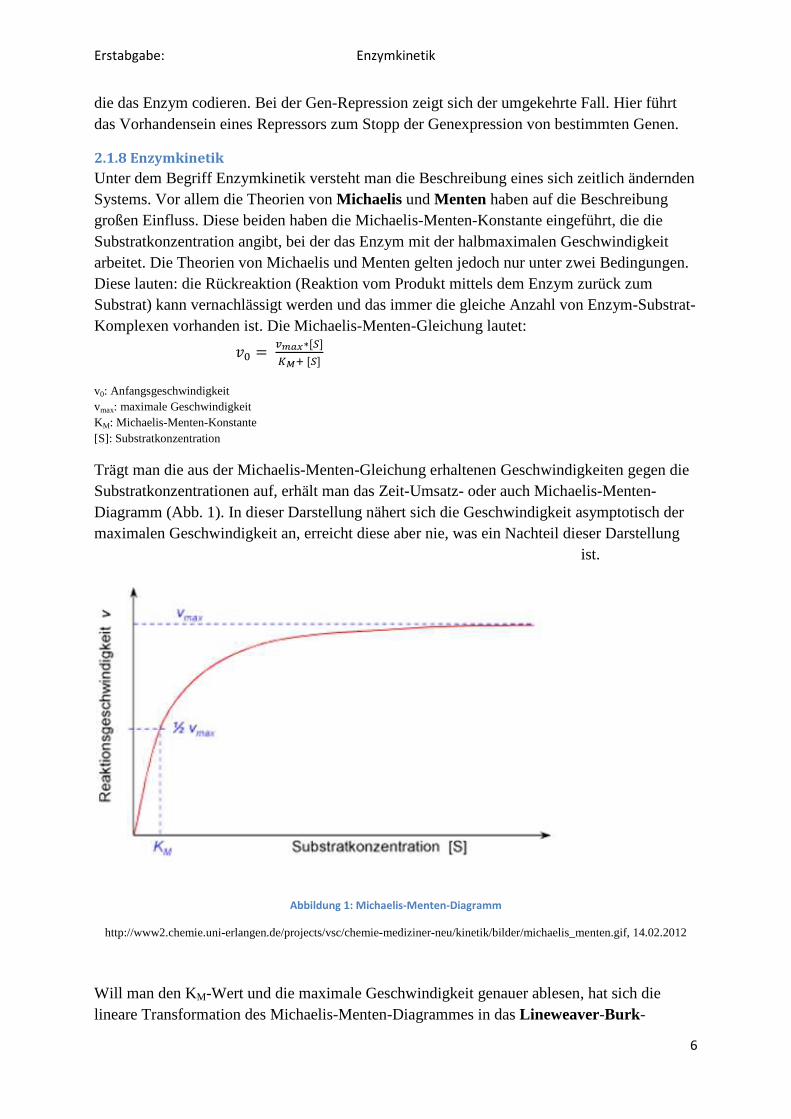
Trägt man die aus der Michaelis-Menten-Gleichung erhaltenen Geschwindigkeiten gegen die Substratkonzentrationen auf, erhält man das Zeit-Umsatz- oder auch Michaelis-Menten-Diagramm (Abb. 1). In dieser Darstellung nähert sich die Geschwindigkeit asymptotisch der maximalen Geschwindigkeit an, erreicht diese aber nie, was ein Nachteil dieser Darstellung
ist.
Abbildung 1: Michaelis-Menten-Diagramm
http://www2.chemie.uni-erlangen.de/projects/vsc/chemie-mediziner-neu/kinetik/bilder/michaelis_menten.gif, 14.02.2012
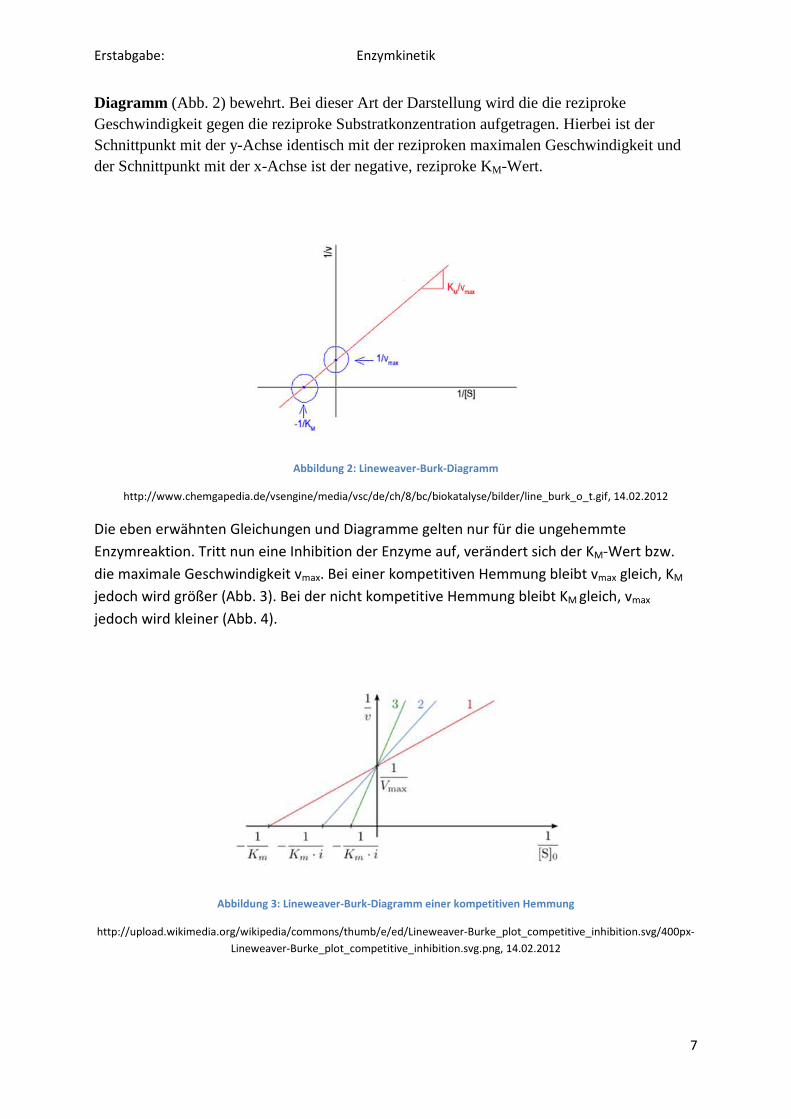
Will man den KM-Wert und die maximale Geschwindigkeit genauer ablesen, hat sich die lineare Transformation des Michaelis-Menten-Diagrammes in das Lineweaver-Burk -
Erstabgabe: Enzymkinetik
7
Diagramm (Abb. 2) bewehrt. Bei dieser Art der Darstellung wird die die reziproke Geschwindigkeit gegen die reziproke Substratkonzentration aufgetragen. Hierbei ist der Schnittpunkt mit der y-Achse identisch mit der reziproken maximalen Geschwindigkeit und der Schnittpunkt mit der x-Achse ist der negative, reziproke KM-Wert.
Abbildung 2: Lineweaver-Burk-Diagramm
http://www.chemgapedia.de/vsengine/media/vsc/de/ch/8/bc/biokatalyse/bilder/line_burk_o_t.gif, 14.02.2012
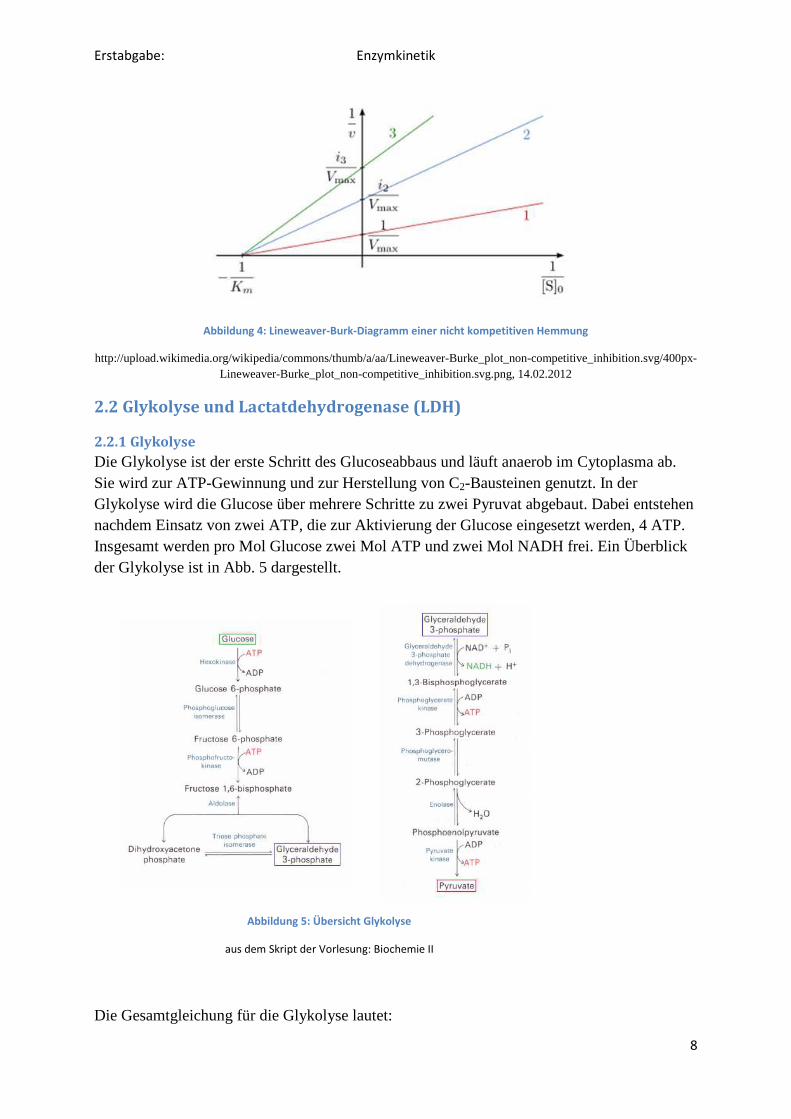
Die eben erwähnten Gleichungen und Diagramme gelten nur für die ungehemmte
Enzymreaktion. Tritt nun eine Inhibition der Enzyme auf, verändert sich der KM-Wert bzw.
die maximale Geschwindigkeit vmax. Bei einer kompetitiven Hemmung bleibt vmax gleich, KM
jedoch wird größer (Abb. 3). Bei der nicht kompetitive Hemmung bleibt KM gleich, vmax
jedoch wird kleiner (Abb. 4).
Abbildung 3: Lineweaver-Burk-Diagramm einer kompetitiven Hemmung
http://upload.wikimedia.org/wikipedia/commons/thumb/e/ed/Lineweaver-Burke_plot_competitive_inhibition.svg/400px-
Lineweaver-Burke_plot_competitive_inhibition.svg.png, 14.02.2012
Erstabgabe: Enzymkinetik
8
Abbildung 4: Lineweaver-Burk-Diagramm einer nicht kompetitiven Hemmung
http://upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lineweaver-Burke_plot_non-competitive_inhibition.svg/400px-Lineweaver-Burke_plot_non-competitive_inhibition.svg.png, 14.02.2012
2.2 Glykolyse und Lactatdehydrogenase (LDH)
2.2.1 Glykolyse
Die Glykolyse ist der erste Schritt des Glucoseabbaus und läuft anaerob im Cytoplasma ab. Sie wird zur ATP-Gewinnung und zur Herstellung von C2-Bausteinen genutzt. In der Glykolyse wird die Glucose über mehrere Schritte zu zwei Pyruvat abgebaut. Dabei entstehen nachdem Einsatz von zwei ATP, die zur Aktivierung der Glucose eingesetzt werden, 4 ATP. Insgesamt werden pro Mol Glucose zwei Mol ATP und zwei Mol NADH frei. Ein Überblick der Glykolyse ist in Abb. 5 dargestellt.
Die Gesamtgleichung für die Glykolyse lautet:
Abbildung 5: Übersicht Glykolyse
aus dem Skript der Vorlesung: Biochemie II

Erstabgabe: Enzymkinetik
9
2.2.2 Aerobe Umsetzung des Pyruvats
Im menschlichen Körper wird das Pyruvat meist aerob zuerst in Acetyl-CoA abgebaut, um dann in den Citratcylus (Abb. 6) eingeschleust zu werden. Hier wird das Acetyl-CoA dann vollständig zu CO2 und den entsprechenden Anzahlen von Reduktionsäquivalenten abgebaut. Die Reduktionsäquivalente (NADH, FADH2) die für den weiteren Glucoseabbau notwendig sind, werden in der Atmungskette unter Aufbau eines Protonengradienten und anschließender ATP-Synthese rückoxidiert. Im aeroben Abbau von Glucose werden so über Glykolyse, Citratcyclus und Atmungskette insgesamt 30 Mol cytosolisches ATP pro Mol Glucose gewonnen.
Abbildung 6: Übersicht über den Citratcyclus
aus dem Skript der Vorlesung: Biochemie II
2.2.3 Anaerober Abbau von Pyruvat
Beim anaeroben Abbau von Pyruvat wird zur Glykolyse kein zusätzliches ATP gewonnen. Der einzige Zweck der Gärung, egal ob alkoholische oder Milchsäuregärung, ist die Rückgewinnung der Reduktionsäquivalente (NADH � NAD+), damit die Glykolyse nicht zum Stillstand kommt. Also wird durch den anaeroben Abbau von Glucose nur zwei ATP gewonnen. Bei der alkoholischen Gärung wird Pyruvat zuerst zu CO2 und Ethanal umgewandelt. Das Ethanal wird schließlich zu Ethanol oxidiert, wobei das NAD+ zurückgewonnen wird. Diese Reaktionen werden von der Pyruvatdecarboxylase und der Alkoholdehydrogenase katalysiert.
Erstabgabe: Enzymkinetik
10
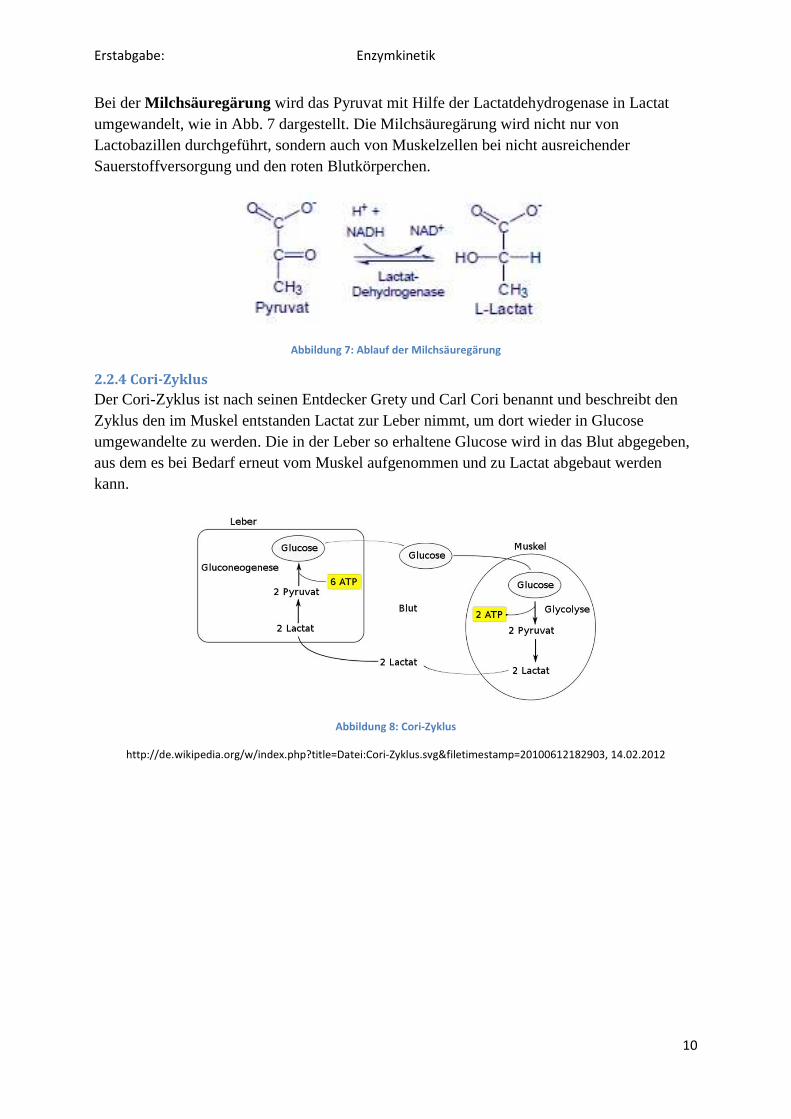
Bei der Milchsäuregärung wird das Pyruvat mit Hilfe der Lactatdehydrogenase in Lactat umgewandelt, wie in Abb. 7 dargestellt. Die Milchsäuregärung wird nicht nur von Lactobazillen durchgeführt, sondern auch von Muskelzellen bei nicht ausreichender Sauerstoffversorgung und den roten Blutkörperchen.
Abbildung 7: Ablauf der Milchsäuregärung
2.2.4 Cori-Zyklus
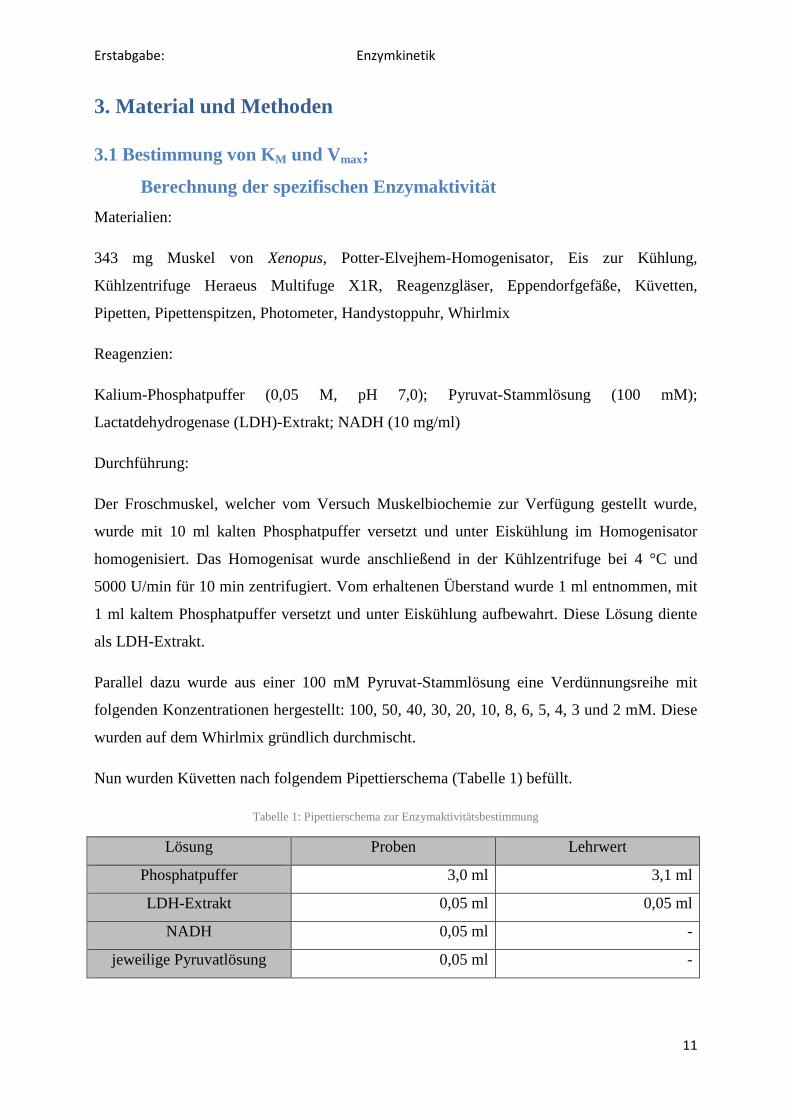
Der Cori-Zyklus ist nach seinen Entdecker Grety und Carl Cori benannt und beschreibt den Zyklus den im Muskel entstanden Lactat zur Leber nimmt, um dort wieder in Glucose umgewandelte zu werden. Die in der Leber so erhaltene Glucose wird in das Blut abgegeben, aus dem es bei Bedarf erneut vom Muskel aufgenommen und zu Lactat abgebaut werden kann.
Abbildung 8: Cori-Zyklus
http://de.wikipedia.org/w/index.php?title=Datei:Cori-Zyklus.svg&filetimestamp=20100612182903, 14.02.2012
Erstabgabe: Enzymkinetik
11
3. Material und Methoden
3.1 Bestimmung von KM und Vmax;
Berechnung der spezifischen Enzymaktivität
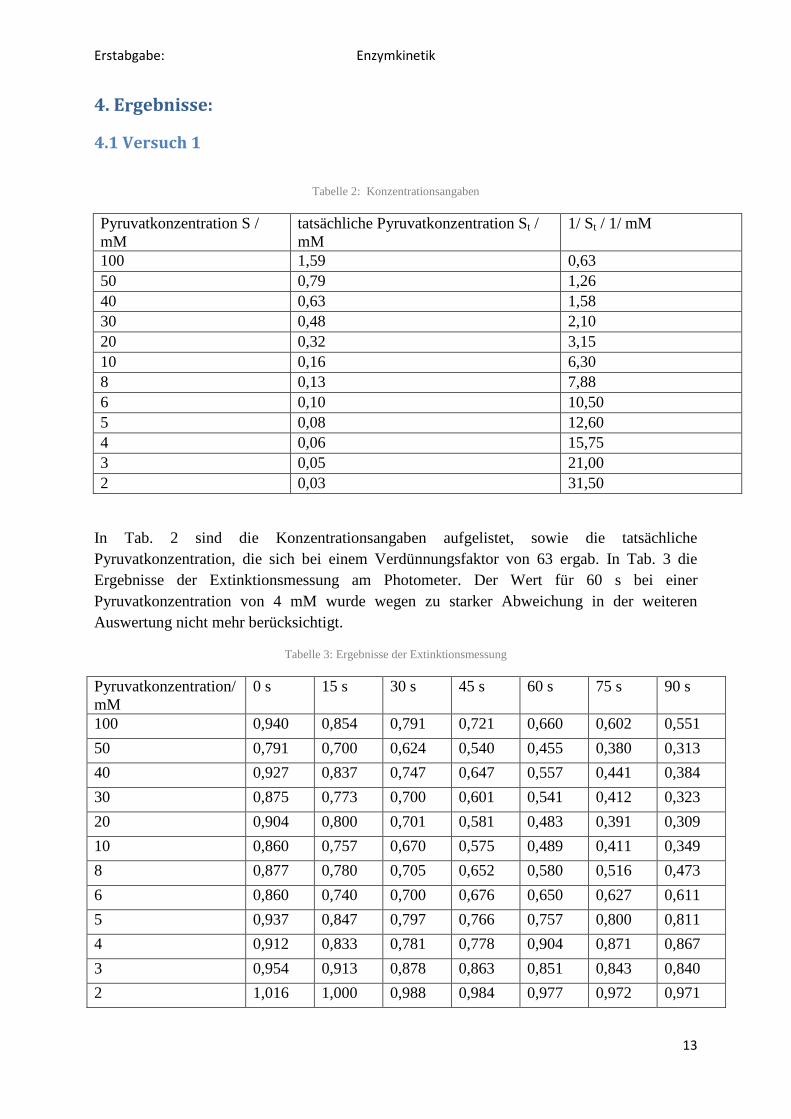
Materialien:
343 mg Muskel von Xenopus, Potter-Elvejhem-Homogenisator, Eis zur Kühlung,
Kühlzentrifuge Heraeus Multifuge X1R, Reagenzgläser, Eppendorfgefäße, Küvetten,
Pipetten, Pipettenspitzen, Photometer, Handystoppuhr, Whirlmix
Reagenzien:
Kalium-Phosphatpuffer (0,05 M, pH 7,0); Pyruvat-Stammlösung (100 mM);
Lactatdehydrogenase (LDH)-Extrakt; NADH (10 mg/ml)
Durchführung:
Der Froschmuskel, welcher vom Versuch Muskelbiochemie zur Verfügung gestellt wurde,
wurde mit 10 ml kalten Phosphatpuffer versetzt und unter Eiskühlung im Homogenisator
homogenisiert. Das Homogenisat wurde anschließend in der Kühlzentrifuge bei 4 °C und
5000 U/min für 10 min zentrifugiert. Vom erhaltenen Überstand wurde 1 ml entnommen, mit
1 ml kaltem Phosphatpuffer versetzt und unter Eiskühlung aufbewahrt. Diese Lösung diente
als LDH-Extrakt.
Parallel dazu wurde aus einer 100 mM Pyruvat-Stammlösung eine Verdünnungsreihe mit
folgenden Konzentrationen hergestellt: 100, 50, 40, 30, 20, 10, 8, 6, 5, 4, 3 und 2 mM. Diese
wurden auf dem Whirlmix gründlich durchmischt.
Nun wurden Küvetten nach folgendem Pipettierschema (Tabelle 1) befüllt.
Tabelle 1: Pipettierschema zur Enzymaktivitätsbestimmung
Lösung Proben Lehrwert
Phosphatpuffer 3,0 ml 3,1 ml
LDH-Extrakt 0,05 ml 0,05 ml
NADH 0,05 ml -
jeweilige Pyruvatlösung 0,05 ml -

Erstabgabe: Enzymkinetik
12
Zuerst wurde die Extinktion der Lehrwertprobe im Photometer bei 340 nm bestimmt und das
Photometer so kalibriert. Danach wurde die Extinktion der einzelnen Proben bei 340 nm
gemessen, wobei zu jeder Probe die jeweilige Pyruvatlösung erst ca. 30 s vor der Messung
zugegeben wurde. Die Lösungen wurden daraufhin für 30 s mit der Pipette gut durchmischt
und anschließend ins Photometer eingesetzt. Die Extinktionen wurden direkt nach dem
Einsetzen, sowie nach 15, 30, 45, 60, 75 und 90 s gemessen. Für die exakte Zeitmessung
wurde eine Handystoppuhr verwendet.
Die Lösungen mit den Pyruvatkonzentrationen 2 und 4 mM wurden je zweimal gemessen.
3.2 Bestimmung des pH-Optimums
Materialien:
Photometer, Küvetten, Pipetten, Pipettenspitzen, Handystoppuhr
Reagenzien:
KH2PO4/K2HPO4-Puffer mit folgenden pH-Werten: pH 4, pH 5, pH 6, pH 7, pH 8, pH 9; 30
mM Pyruvatlösung aus Versuch 1
Durchführung:
Es wurden nun Proben anhand des Schemas aus Versuch 1 hergestellt, jedoch jeweils 2
Proben mit Phosphatpuffern der verschiedenen pH-Werte für eine Doppelbestimmung.
Zu jeder dieser Proben wurde daraufhin 30 s vor jeder Messung 0,05 ml der 30 mM
Pyruvatlösung gegeben, gut mit der Pipette durchmischt und anschließend photometrisch die
Extinktion bei 340 nm bestimmt. Die Extinktion wurde je einmal direkt nach dem Einsetzen
in das Photometer und einmal nach 1 min gemessen.
Erstabgabe: Enzymkinetik
13
4. Ergebnisse:
4.1 Versuch 1
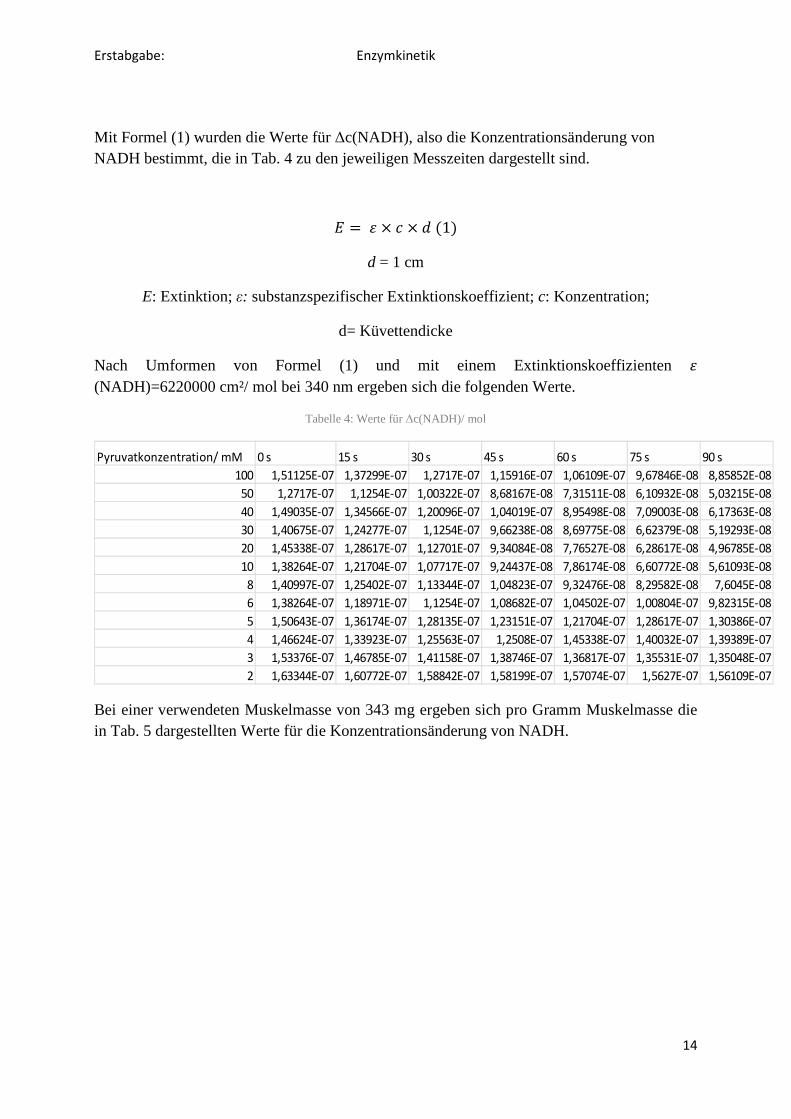
Tabelle 2: Konzentrationsangaben
Pyruvatkonzentration S / mM
tatsächliche Pyruvatkonzentration St / mM
1/ St / 1/ mM
100 1,59 0,63 50 0,79 1,26 40 0,63 1,58 30 0,48 2,10 20 0,32 3,15 10 0,16 6,30 8 0,13 7,88 6 0,10 10,50 5 0,08 12,60 4 0,06 15,75 3 0,05 21,00 2 0,03 31,50
In Tab. 2 sind die Konzentrationsangaben aufgelistet, sowie die tatsächliche Pyruvatkonzentration, die sich bei einem Verdünnungsfaktor von 63 ergab. In Tab. 3 die Ergebnisse der Extinktionsmessung am Photometer. Der Wert für 60 s bei einer Pyruvatkonzentration von 4 mM wurde wegen zu starker Abweichung in der weiteren Auswertung nicht mehr berücksichtigt.
Tabelle 3: Ergebnisse der Extinktionsmessung
Pyruvatkonzentration/ mM
0 s 15 s 30 s 45 s 60 s 75 s 90 s
100 0,940 0,854 0,791 0,721 0,660 0,602 0,551
50 0,791 0,700 0,624 0,540 0,455 0,380 0,313
40 0,927 0,837 0,747 0,647 0,557 0,441 0,384
30 0,875 0,773 0,700 0,601 0,541 0,412 0,323
20 0,904 0,800 0,701 0,581 0,483 0,391 0,309
10 0,860 0,757 0,670 0,575 0,489 0,411 0,349
8 0,877 0,780 0,705 0,652 0,580 0,516 0,473
6 0,860 0,740 0,700 0,676 0,650 0,627 0,611
5 0,937 0,847 0,797 0,766 0,757 0,800 0,811
4 0,912 0,833 0,781 0,778 0,904 0,871 0,867
3 0,954 0,913 0,878 0,863 0,851 0,843 0,840
2 1,016 1,000 0,988 0,984 0,977 0,972 0,971
Erstabgabe: Enzymkinetik
14
Mit Formel (1) wurden die Werte für ∆c(NADH), also die Konzentrationsänderung von NADH bestimmt, die in Tab. 4 zu den jeweiligen Messzeiten dargestellt sind.
� = � × � × �(1)
d = 1 cm
E: Extinktion; ε: substanzspezifischer Extinktionskoeffizient; c: Konzentration;
d= Küvettendicke
Nach Umformen von Formel (1) und mit einem Extinktionskoeffizienten � (NADH)=6220000 cm²/ mol bei 340 nm ergeben sich die folgenden Werte.
Tabelle 4: Werte für ∆c(NADH)/ mol
Bei einer verwendeten Muskelmasse von 343 mg ergeben sich pro Gramm Muskelmasse die in Tab. 5 dargestellten Werte für die Konzentrationsänderung von NADH.
Pyruvatkonzentration/ mM 0 s 15 s 30 s 45 s 60 s 75 s 90 s
100 1,51125E-07 1,37299E-07 1,2717E-07 1,15916E-07 1,06109E-07 9,67846E-08 8,85852E-08
50 1,2717E-07 1,1254E-07 1,00322E-07 8,68167E-08 7,31511E-08 6,10932E-08 5,03215E-08
40 1,49035E-07 1,34566E-07 1,20096E-07 1,04019E-07 8,95498E-08 7,09003E-08 6,17363E-08
30 1,40675E-07 1,24277E-07 1,1254E-07 9,66238E-08 8,69775E-08 6,62379E-08 5,19293E-08
20 1,45338E-07 1,28617E-07 1,12701E-07 9,34084E-08 7,76527E-08 6,28617E-08 4,96785E-08
10 1,38264E-07 1,21704E-07 1,07717E-07 9,24437E-08 7,86174E-08 6,60772E-08 5,61093E-08
8 1,40997E-07 1,25402E-07 1,13344E-07 1,04823E-07 9,32476E-08 8,29582E-08 7,6045E-08
6 1,38264E-07 1,18971E-07 1,1254E-07 1,08682E-07 1,04502E-07 1,00804E-07 9,82315E-08
5 1,50643E-07 1,36174E-07 1,28135E-07 1,23151E-07 1,21704E-07 1,28617E-07 1,30386E-07
4 1,46624E-07 1,33923E-07 1,25563E-07 1,2508E-07 1,45338E-07 1,40032E-07 1,39389E-07
3 1,53376E-07 1,46785E-07 1,41158E-07 1,38746E-07 1,36817E-07 1,35531E-07 1,35048E-07
2 1,63344E-07 1,60772E-07 1,58842E-07 1,58199E-07 1,57074E-07 1,5627E-07 1,56109E-07
Erstabgabe: Enzymkinetik
15
Tabelle 5: Werte für ∆c(NADH) pro Gramm Muskelmasse/ mol/ g
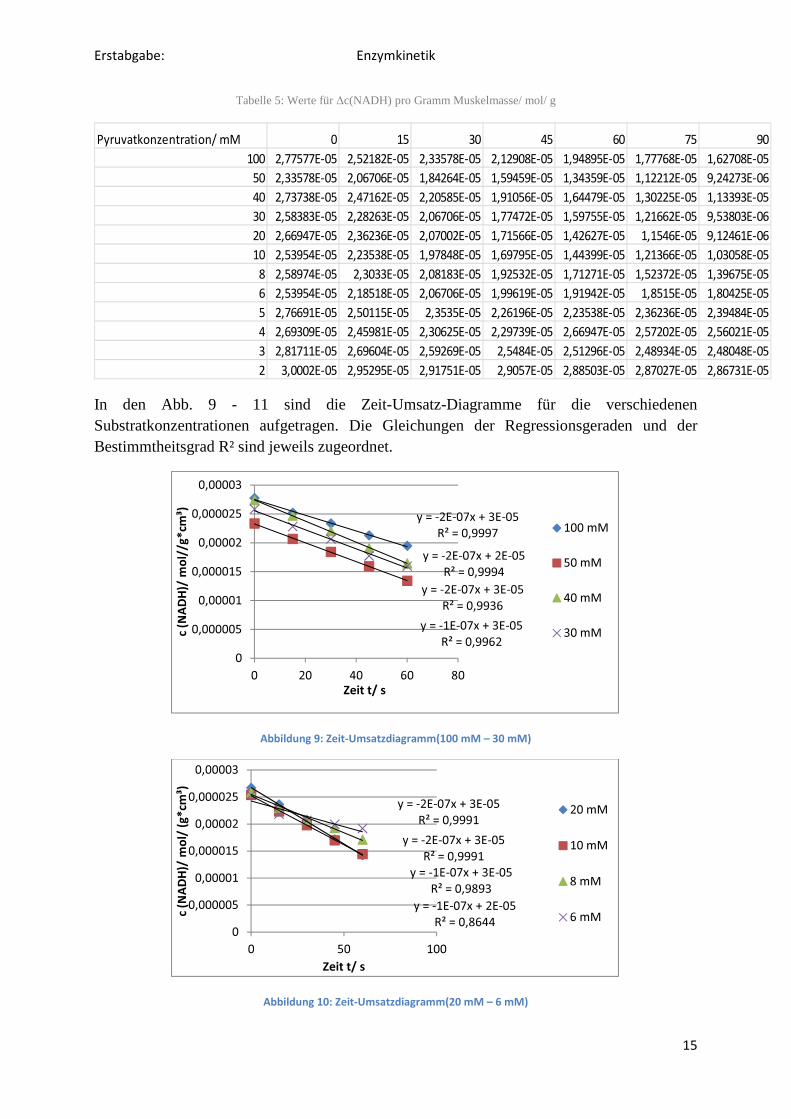
In den Abb. 9 - 11 sind die Zeit-Umsatz-Diagramme für die verschiedenen Substratkonzentrationen aufgetragen. Die Gleichungen der Regressionsgeraden und der Bestimmtheitsgrad R² sind jeweils zugeordnet.
Abbildung 9: Zeit-Umsatzdiagramm(100 mM – 30 mM)
Abbildung 10: Zeit-Umsatzdiagramm(20 mM – 6 mM)
Pyruvatkonzentration/ mM 0 15 30 45 60 75 90
100 2,77577E-05 2,52182E-05 2,33578E-05 2,12908E-05 1,94895E-05 1,77768E-05 1,62708E-05
50 2,33578E-05 2,06706E-05 1,84264E-05 1,59459E-05 1,34359E-05 1,12212E-05 9,24273E-06
40 2,73738E-05 2,47162E-05 2,20585E-05 1,91056E-05 1,64479E-05 1,30225E-05 1,13393E-05
30 2,58383E-05 2,28263E-05 2,06706E-05 1,77472E-05 1,59755E-05 1,21662E-05 9,53803E-06
20 2,66947E-05 2,36236E-05 2,07002E-05 1,71566E-05 1,42627E-05 1,1546E-05 9,12461E-06
10 2,53954E-05 2,23538E-05 1,97848E-05 1,69795E-05 1,44399E-05 1,21366E-05 1,03058E-05
8 2,58974E-05 2,3033E-05 2,08183E-05 1,92532E-05 1,71271E-05 1,52372E-05 1,39675E-05
6 2,53954E-05 2,18518E-05 2,06706E-05 1,99619E-05 1,91942E-05 1,8515E-05 1,80425E-05
5 2,76691E-05 2,50115E-05 2,3535E-05 2,26196E-05 2,23538E-05 2,36236E-05 2,39484E-05
4 2,69309E-05 2,45981E-05 2,30625E-05 2,29739E-05 2,66947E-05 2,57202E-05 2,56021E-05
3 2,81711E-05 2,69604E-05 2,59269E-05 2,5484E-05 2,51296E-05 2,48934E-05 2,48048E-05
2 3,0002E-05 2,95295E-05 2,91751E-05 2,9057E-05 2,88503E-05 2,87027E-05 2,86731E-05
y = -1E-07x + 3E-05
R² = 0,9962
y = -2E-07x + 2E-05
R² = 0,9994
y = -2E-07x + 3E-05
R² = 0,9997
y = -2E-07x + 3E-05
R² = 0,9936
0
0,000005
0,00001
0,000015
0,00002
0,000025
0,00003
0 20 40 60 80
c (N
AD
H)/
mo
l//g
*cm
³)
Zeit t/ s
100 mM
50 mM
40 mM
30 mM
y = -2E-07x + 3E-05
R² = 0,9991
y = -2E-07x + 3E-05
R² = 0,9991
y = -1E-07x + 3E-05
R² = 0,9893
y = -1E-07x + 2E-05
R² = 0,86440
0,000005
0,00001
0,000015
0,00002
0,000025
0,00003
0 50 100
c (N
AD
H)/
mo
l/ (
g*
cm³)
Zeit t/ s
20 mM
10 mM
8 mM
6 mM
Erstabgabe: Enzymkinetik
16
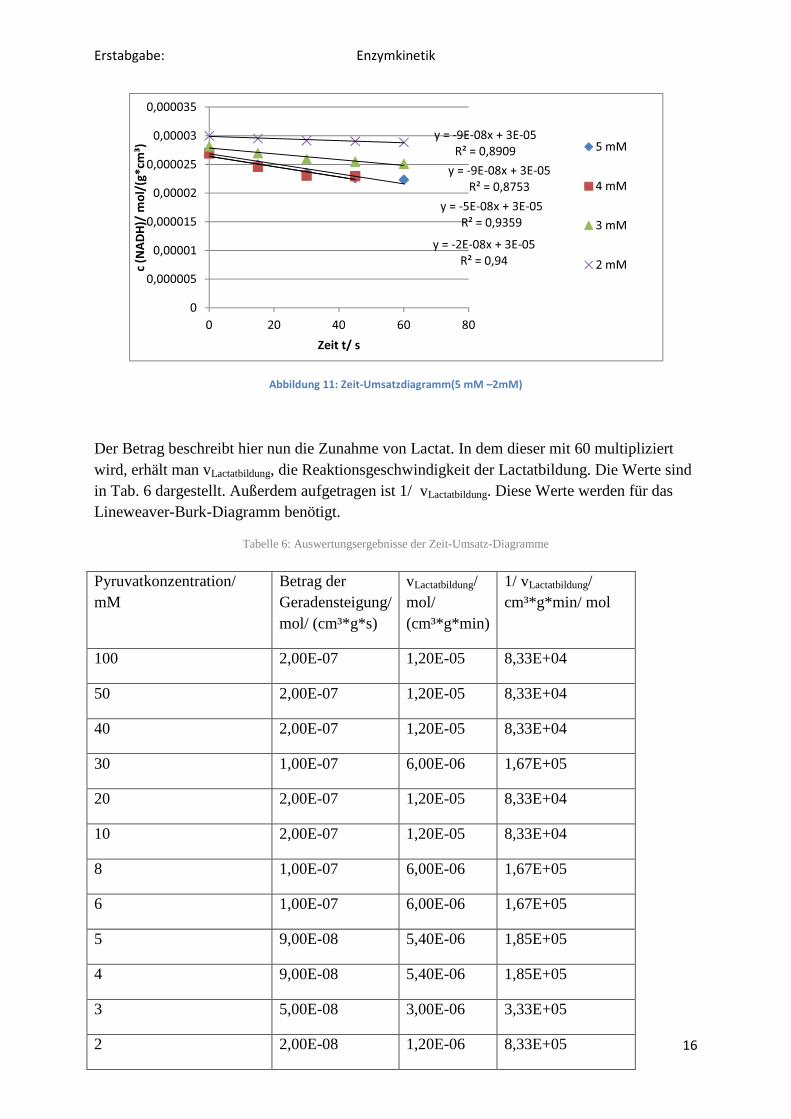
Abbildung 11: Zeit-Umsatzdiagramm(5 mM –2mM)
Der Betrag beschreibt hier nun die Zunahme von Lactat. In dem dieser mit 60 multipliziert wird, erhält man vLactatbildung, die Reaktionsgeschwindigkeit der Lactatbildung. Die Werte sind in Tab. 6 dargestellt. Außerdem aufgetragen ist 1/ vLactatbildung. Diese Werte werden für das Lineweaver-Burk-Diagramm benötigt.
Tabelle 6: Auswertungsergebnisse der Zeit-Umsatz-Diagramme
y = -9E-08x + 3E-05
R² = 0,8909
y = -9E-08x + 3E-05
R² = 0,8753
y = -5E-08x + 3E-05
R² = 0,9359
y = -2E-08x + 3E-05
R² = 0,94
0
0,000005
0,00001
0,000015
0,00002
0,000025
0,00003
0,000035
0 20 40 60 80
c (N
AD
H)/
mo
l/(g
*cm
³)
Zeit t/ s
5 mM
4 mM
3 mM
2 mM
Pyruvatkonzentration/ mM
Betrag der Geradensteigung/ mol/ (cm³*g*s)
vLactatbildung/ mol/ (cm³*g*min)
1/ vLactatbildung/ cm³*g*min/ mol
100 2,00E-07 1,20E-05 8,33E+04
50 2,00E-07 1,20E-05 8,33E+04
40 2,00E-07 1,20E-05 8,33E+04
30 1,00E-07 6,00E-06 1,67E+05
20 2,00E-07 1,20E-05 8,33E+04
10 2,00E-07 1,20E-05 8,33E+04
8 1,00E-07 6,00E-06 1,67E+05
6 1,00E-07 6,00E-06 1,67E+05
5 9,00E-08 5,40E-06 1,85E+05
4 9,00E-08 5,40E-06 1,85E+05
3 5,00E-08 3,00E-06 3,33E+05
2 2,00E-08 1,20E-06 8,33E+05

Erstabgabe: Enzymkinetik
17
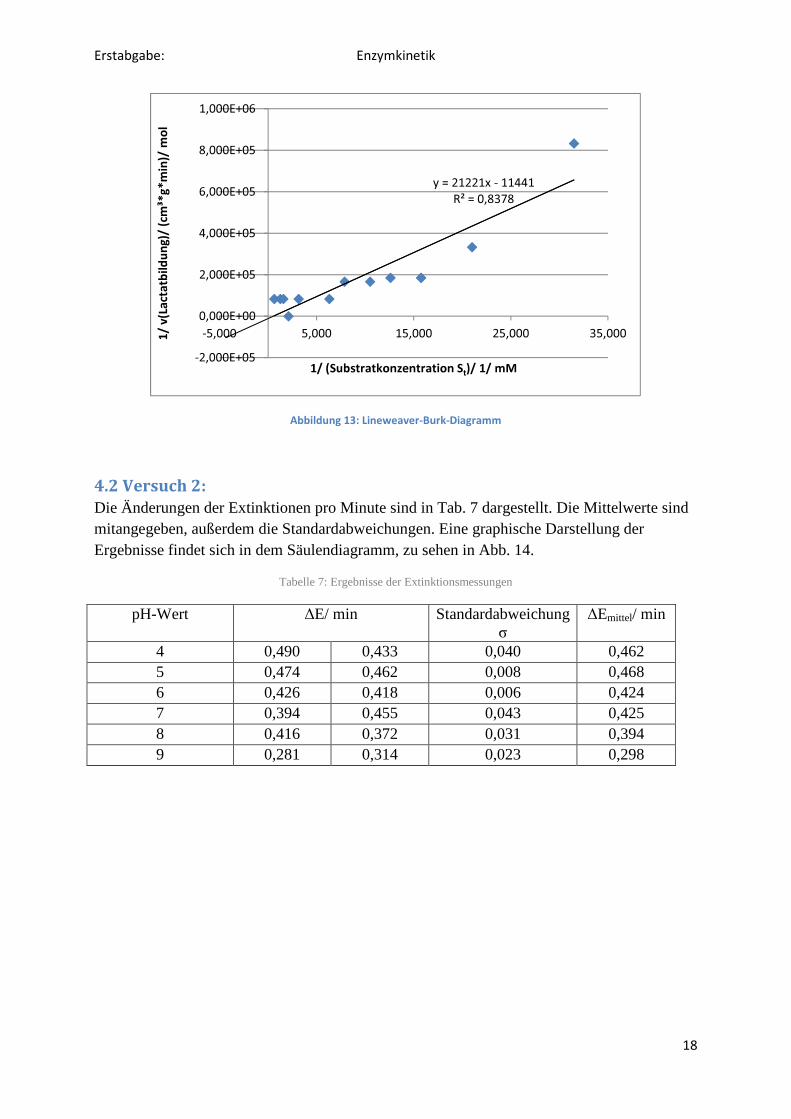
In dem nun vLactatbildung gegen die tatsächliche Substratkonzentration aufgetragen wird, erhält man den näherungsweisen Verlauf einer Michaelis-Menten-Kinetik. Man erkennt eine Maximalgeschwindigkeit vmax=1,2E-05 mol/ (cm³*g*min). Die Michaeliskonstante KM beträgt etwa 0,125 mM. Die graphische Darstellung ist in Abb. 12 zu sehen. In Abb. 13 wurden die Ergebnisse doppeltreziprok aufgetragen. Die Regressionsgerade samt Geradengleichung und Bestimmungsgrad sind dargestellt. Dieses Lineweaver-Burk-Diagramm ist jedoch nicht von Nutzen, da die Regressionsgerade die y-Achse im Negativen schneidet und die x-Achse im Positiven. Es lassen sich aus diesem Diagramm keine Erkenntnisse gewinnen.
Abbildung 12:Darstellung der Kinetik
0,000E+00
2,000E-06
4,000E-06
6,000E-06
8,000E-06
1,000E-05
1,200E-05
1,400E-05
,000 ,500 1,000 1,500 2,000
v(L
act
atb
ild
un
g)/
mo
l/(c
m³*
g*
min
)
Substratkonzentration St/ mM
Erstabgabe: Enzymkinetik
18
Abbildung 13: Lineweaver-Burk-Diagramm
4.2 Versuch 2:
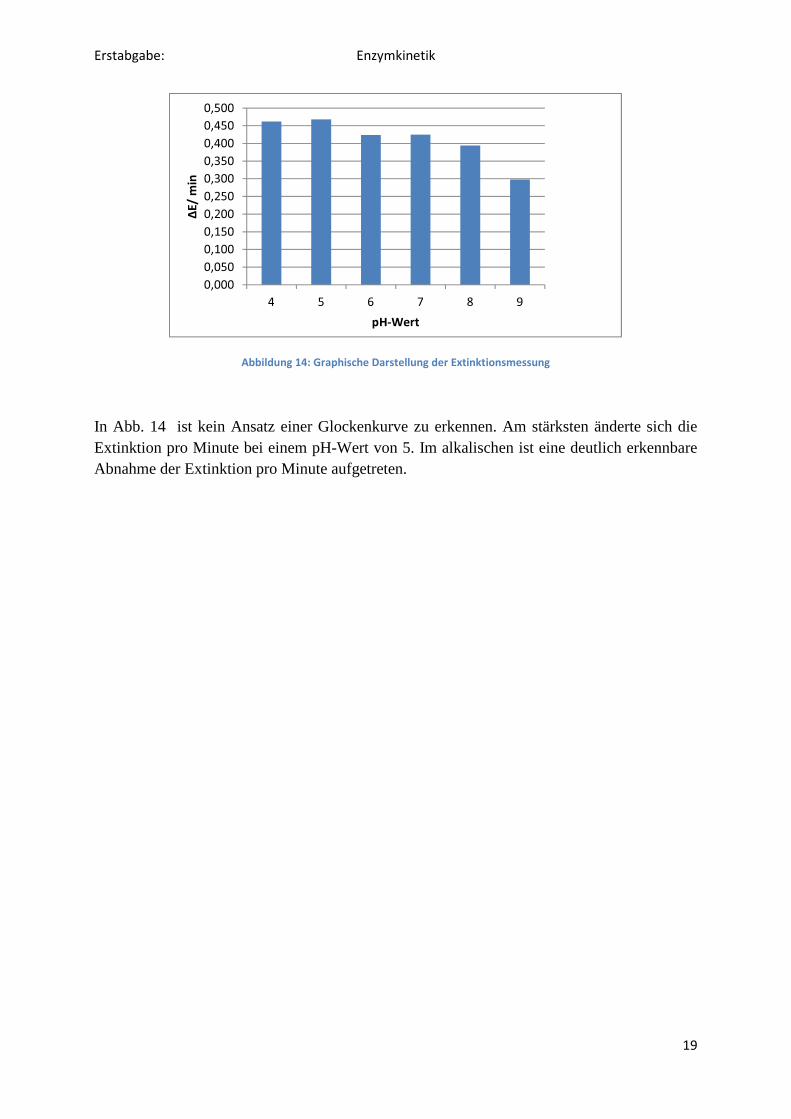
Die Änderungen der Extinktionen pro Minute sind in Tab. 7 dargestellt. Die Mittelwerte sind mitangegeben, außerdem die Standardabweichungen. Eine graphische Darstellung der Ergebnisse findet sich in dem Säulendiagramm, zu sehen in Abb. 14.
Tabelle 7: Ergebnisse der Extinktionsmessungen
pH-Wert ∆E/ min Standardabweichung σ
∆Emittel/ min
4 0,490 0,433 0,040 0,462 5 0,474 0,462 0,008 0,468 6 0,426 0,418 0,006 0,424 7 0,394 0,455 0,043 0,425 8 0,416 0,372 0,031 0,394 9 0,281 0,314 0,023 0,298
y = 21221x - 11441
R² = 0,8378
-2,000E+05
0,000E+00
2,000E+05
4,000E+05
6,000E+05
8,000E+05
1,000E+06
-5,000 5,000 15,000 25,000 35,0001/
v(L
act
atb
ild
un
g)/
(cm
³*g
*m
in)/
mo
l
1/ (Substratkonzentration St)/ 1/ mM
Erstabgabe: Enzymkinetik
19
Abbildung 14: Graphische Darstellung der Extinktionsmessung
In Abb. 14 ist kein Ansatz einer Glockenkurve zu erkennen. Am stärksten änderte sich die Extinktion pro Minute bei einem pH-Wert von 5. Im alkalischen ist eine deutlich erkennbare Abnahme der Extinktion pro Minute aufgetreten.
0,000
0,050
0,100
0,150
0,200
0,250
0,300
0,350
0,400
0,450
0,500
4 5 6 7 8 9
ΔE
/ m
in
pH-Wert

Erstabgabe: Enzymkinetik
20
5. Diskussion
5.1 Versuch 1:
Die in Abb. 12 dargestellte Kinetik zeigt den Verlauf einer Michaelis-Menten-Kinetik. Es muss jedoch berücksichtigt werden, dass ein Wert wegen zu großer Abweichung von den anderen Werten nicht mehr berücksichtigt wurde. Außerdem waren aus dem Lineweaver-Burk-Diagramm keine Werte zu entnehmen, da hier keine sinnvolle Auftragung möglich war. Fehler könnten sich hier bei dem Herstellen der Lösungen ergeben haben. Außerdem ist ein einwandfreies Funktionieren des Photometers nicht immer gewährleistet.
Um eindeutig aussagekräftige Ergebnisse zu erhalten, hätten mehrere Bestimmungen vorgenommen werden müssen. Die in dem Versuch vorgenommene Einzelbestimmung ist somit nur bedingt aussagekräftig. Die diversen Ungenauigkeiten, welche durch Konstruktion und Bedienung der Messinstrumente bestimmt sind, schlagen bei diesen einzelnen Werten zu stark ins Gewicht. Außerdem können die Fehler von Werte zu Wert stark schwanken. Dies erklärt auch, warum nicht alle gemessenen Werte den Erwartungen entsprechen.
5.2 Versuch 2:
Die Ergebnisse entsprachen hier nicht den Erwartungen. Es wurde ein Optimum der Lactat-Dehydrogenase im neutralen Bereich bei pH 7 erwartet. In dem Histogramm, dargestellt in Abb. 14 ist jedoch nur eine deutliche Abnahme im alkalischen Bereich ab pH 9 zu erkennen. Im sauren Bereich ist jedoch entgegen den Erwartungen sogar das Maximum der Enzymaktivität im sauren bei pH 5 aufgetreten. Dort war eine Abnahme der Extinktion pro Minute von 0,468 gemessen worden.
Fehler könnten sich hier wieder am Photometer ergeben haben. Außerdem können nicht korrekt eingestellte Puffer das Problem gewesen sein. Durchaus in Betracht zu ziehen ist, dass die Lösungen vertauscht wurden und somit falsch zugeordnet wurden. Da die Werte im sauren Bereich aber alle deutlich höher als die Erwarteten sind und auch nahe beieinander liegen, würden die Auswirkungen begrenzt sein und nicht zu den Erwarteten Ergebnissen führen.
Die berechneten Standardabweichungen, welche aus Tab. 6 zu entnehmen sind, sind nicht besonders groß, die Werte liegen also nahe beieinander, was ein defektes Photometer als Fehlerquelle somit ausschließt. Maßgebliche Fehlerquelle liegt somit in der Durchführung, die für Anfänger nicht immer fehlerfrei zu bewältigen ist, und in den bereitgestellten Lösungen. Für aussagekräftigere Ergebnisse hätten mehr Bestimmungen vorgenommen werden müssen, darüber hinaus hätte man die Lösungen neu ansetzen können. Dies wurde aus organisatorischen und zeitlichen Gründen jedoch nicht getan.

Erstabgabe: Enzymkinetik
21
6. Quellen
− http://www1.tu-darmstadt.de/fb/ch/Fachgebiete/OC/AKSchmidt/Avimec/UE/Hemmung.html#Irreversibel
− http://de.wikipedia.org/wiki/Cori-Zyklus − Skript der Vorlesung: Biochemie II