
Seminar Moderne Anwendung der magnetischen Resonanz · Einleitung Aufklärung der Struktur und der...
27
Seminar Moderne Anwendung der magnetischen Resonanz Vortrag von Jana Koch 14.10.2013 1
Transcript of Seminar Moderne Anwendung der magnetischen Resonanz · Einleitung Aufklärung der Struktur und der...

Seminar Moderne Anwendung der magnetischen Resonanz Vortrag von Jana Koch
14.10.2013 1

Gliederung
Einleitung
Theorie M2-Protein
Protonentransfer
Diskussion
Ergebnisse
Zusammenfassung
Quellen
14.10.2013 2

Einleitung
Aufklärung der Struktur und der funktionellen Dynamik vom pH-sensitiven und protonenselektiven His-37 im M2-Protein gebunden an eine Cholesterin enthaltene Virushülle mit Festkörper NMR
Dient zum besseren Verständnis des Protonen-Leitungs-Mechanismus
14.10.2013 3

Einleitung
Im hohen pH-Bereich verhindern die vier Histidine aus der Kantenvorderseite der π-gestapelten Struktur, eine hydrogengebundene Wasserkette zum führenden Proton
Im niedrigen pH-Bereich
bildet das Imidazol eine ausführliche
Hydrogenbindung zum Wasser
14.10.2013 4

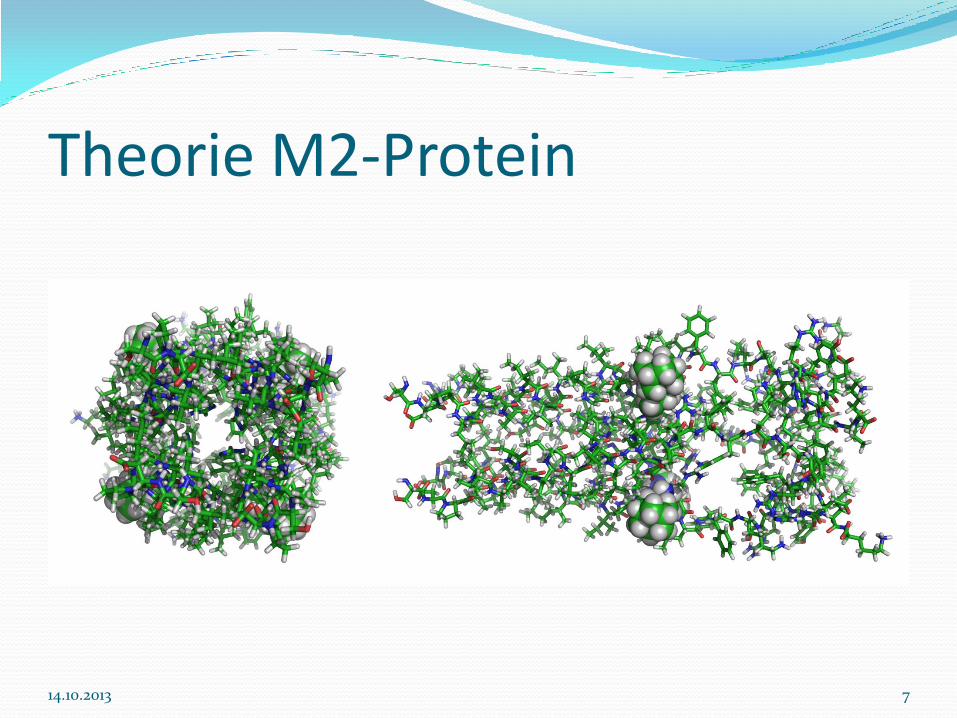
Theorie M2-Protein
M2-Protein ist ein Membranprotein in einer Virushülle
Zählt zum Influenzavirus A
M2-Protein ermöglicht den Zufluss von Protonen in das Virusinnere
M2-Protein ist eines der drei integralen Membranproteine der Influenzavirus-Virion
14.10.2013 5

Theorie M2-Protein
Lysosom niedriger pH-Wert
Anbinden einer Fusionssequenz
Verschmelzung von Virushülle und Membran
Anschließend wird das Virusinnere mit RNA-haltigen Viruskapsiden in das Zytosol freigegeben
14.10.2013 6
Theorie M2-Protein
14.10.2013 7

Protonentransfer
Im synthetischen Material ist das Proton durch Hydrogenbindung an Wasser gebunden
Bindung an titrierbare Heterozyklen
Zum Vergleich: Wasser und titrierbare Protein-Seitenketten werden im biologischen Leitungsmechanismus genutzt
14.10.2013 8

Protonentransfer
M2 Proton bildet einen tetrameren Protonenkanal
Durch Aktivierung bei pH 6 leitet der M2 Kanal 10 bis 10.000 Protonen pro Sekunde
pH-sensitiver und protonenselektiver Rückstand ist dem His-37 zuzuordnen
14.10.2013 9

Diskussion
15N chemische Verschiebung von His-37 in DMPC/DMPG zeigt, dass sich vier His-Doppelschichten bei pKs von 8.2, 8.2, 6.3 und < 5 bilden
Die dritte Protonierung ist verantwortlich für die Kanalaktivierung und den Protonentransfer
14.10.2013 10

Diskussion
Zwei Modelle der Protonenleitung
Verschluss-Modell
Pendel-Modell
Forschungsgruppe beschäftigt sich vorrangig mit dem pH von 8.5 für den geschlossenen Kanal und 4.5 für den leitenden Kanal
14.10.2013 11

Diskussion
15N und 13C chemische Verschiebung von His ist empfindlich gegenüber dem Protonierungszustand und der tautomeren Struktur vom Imidazol
Deprotonierung erhöht 15N chemische Verschiebung von 80 ppm und die chemische Verschiebung von Cγ/Cδ im His-37
14.10.2013 12

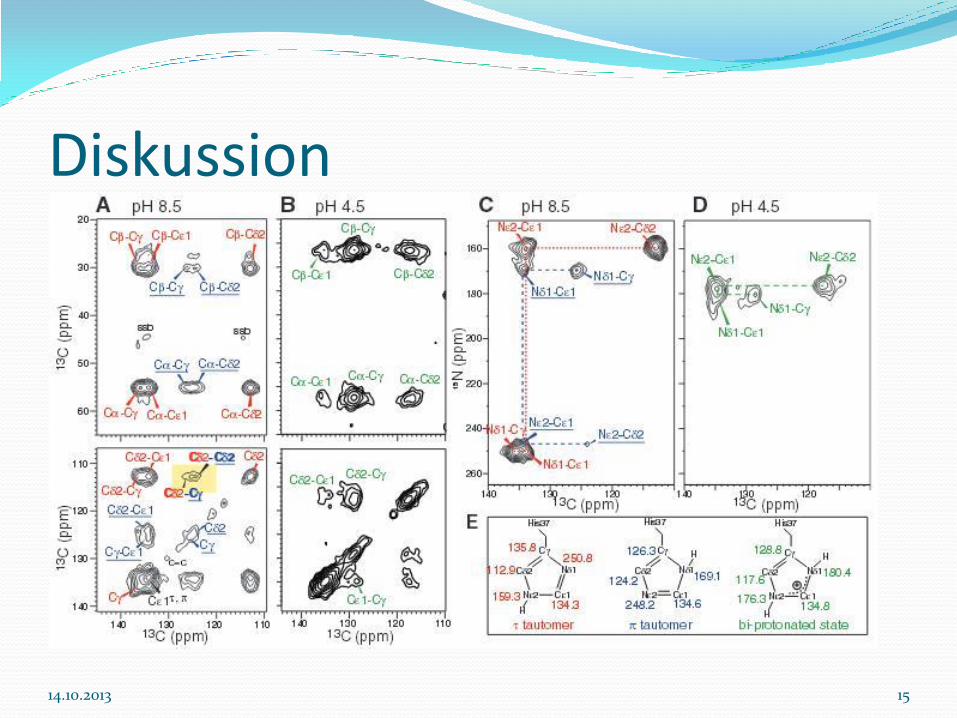
Diskussion
2D 13C-13C und 15N-13C Korrelationsspektren
His-markiert das M2TM
Ausschließlich Signale neutraler Imidazole bei pH 8.5
Nε2-protonierte τ-Tautomer und Nδ1-protonierte π-Tautomer existieren bei einem Verhältnis von 3:1
14.10.2013 13

Diskussion
Die Peaks Cδ2(τ)-Cγ(π) und Cδ2(τ)-Cδ2(π) zeigen, dass beide Tautomere sich in jedem Kanal befinden
Stabilisierung des protonierten Nδ1(π) erfolgt durch Wasserstoffanbindung
Bei pH 4.5 Nδ1 und Nε2 haben Protonen chemische Verschiebung von 170-180 ppm, unprotoniertes Signal beträgt chemische Verschiebung bei 250 ppm
14.10.2013 14
Diskussion
14.10.2013 15

Diskussion
Spiralförmige Helix
des His-37
Entfernung von Cα-Nδ1 schränkt χ2-Verdrehungswinkel ein
Entfernung Cδ2-Nα wirkt auf χ1- und χ2- Winkel
14.10.2013 16

14.10.2013 17

Diskussion
Die Tetradimension hat die Möglichkeit Metallionen zu koordinieren
Inhibieren von Cu2+ im M2-Protein
Das kleinere π-Tautomer kann die vierfache Symmetrie durch das Übernehmen der ττ oder der τ0 Konfiguration des Rotamers annehmen
14.10.2013 18

Ergebnisse
Seitenkette des His-37 bei hohen pH-Werten unbeweglich
Bei niedrigen pH-Werten sollte die Seitenkette dynamisch bleiben
Virenmembran immobilisiert das Protein-Rückgrat (Nα-H und Cα-Hα) und isoliert das Potential der Seitenkettenbewegung
14.10.2013 19

Ergebnisse
Bei pH 8.5 wurde Cγ-Nδ1 der starren Grenze von Cε1-Nδ1 Kopplungen (1.15 kHz und 1.39 Å) zugeordnet und eine starre Grenze der Cδ2-Hδ2 Kopplung (23.9 kHz und 1.08 Å) erhalten
Die bekannte 180° Ringflip-Bewegung wird ausgeschlossen, weil sie wenig Wirkung auf die Cδ2-Hδ2 Kopplung (SCδ2-Hδ2 = 0.94) hat
14.10.2013 20

14.10.2013 21

Ergebnisse
Anstieg der Protonenkonzentration in der Virus-Membran, können Ringflips vorkommen
Bewegung des Ringflips schneller als 104 s, um die Cδ2-Hδ2 Kopplung im Durchschnitt zu erhalten
14.10.2013 22

Ergebnisse
Vorgeschlagen wird Protonenleitungsmechanismus, in dem Imidazol durch Deprotonierung des Cε1-Hε1 eingesetzt wird
Dadurch werden die dauernden Ringflips gewährleistet
Dies erleichtert die Protonenübertragung auf die C-Endwassermoleküle des Imidazols
14.10.2013 23

Ergebnisse
Daten zeigen, dass die höchste Energiebarriere dieses Prozesses, das Imidazol in Bewegung bringt
Dies erleichtert die Protonenleitfähgkeit des temperaturabhängigen M2-Proteins
14.10.2013 24

Zusammenfassung
dynamisches Protonenübertragungs-Modell ist mit der beobachteten Isotop-Wirkung des schweren Wasserstoff im Einklang
H-gebundene Mischketten mit verschiedenen Elementen erkennbar
Die Daten weisen stark darauf hin, dass das His-37 an der Protonenleitung durch das M2-Protein aktiv beteiligt wird
14.10.2013 25

Zusammenfassung
Experimente stellen die Fähigkeit von Festkörper-NMR unter Beweis, um die funktionell wichtige Membran der Protein-Dynamik und der Chemie aufzuklären
14.10.2013 26

Quellen
F. Hu, W. Luo, M. Hong, Science, 2010, 330, 505-508
Supporting Information, Science, 2010, 330
D. Schrama et. al, Nat. Rev. Drug Discov. , 2006, 5, 147-159
14.10.2013 27


















