
Skript zum Kernblock „Forstbotanik und Baumphysiologie II ...
30
Skript zum Kernblock „Forstbotanik und Baumphysiologie II“ (430 a) Forstbotanischer Teil I N H A L T S V E R Z E I C H N I S 1. Die Sprossachse 1 1.1 Anatomie der Sprossachse 1 1.1.1 Primärer Bau 1 1.1.2 Das Kambium 2 1.1.3 Leitgewebe 3 1.1.3.1 Xylem 3 1.1.3.2 Sekundäres Dickenwachstum 10 1.1.3.3 Reaktionsholz 12 1.1.3.5 Organisationsstufen des Hydrosystems 14 1.1.3.6 Phloem 14 1.1.3.7 Abschlussgewebe 17 1.2 Morphologie der Sprossachse 19 1.2.1 Lang- und Kurztrieb 19 1.2.2 Weiterwachen der Jahrestriebe 20 1.2.3 Aufbau des Sprosssystems 20 1.2.4 Besonderheiten des Knospentreibens 20 1.2.5 Seitensprossbildung 21 1.2.6 Baumgestalt 21 2. Das Blatt 21 2.1 Morphologie 21 2.2 Blattstellung 22 2.3 Anatomie 22 3. Die Wurzel 24 3.1 Anatomie 24 3.1.1 primäre Struktur 24 3.1.2 Sekundäre Bindungen 26 3.2 Morphologie 28 3.2.1 Wurzelsprosse 28 3.2.2 Wurzelmetamorphosen 28 3.2.2.1 Speicher-Wurzeln 28 3.2.2.2 Stützwurzeln 28 3.2.2.3 Haft- und Kletterwurzeln 28 3.2.2.4 Atemwurzeln 28 3.2.2.5 Zug- und Ankerwurzeln 28 3.2.3 Mykorrhiza 28 3.2.4 Wurzelsysteme 29
Transcript of Skript zum Kernblock „Forstbotanik und Baumphysiologie II ...

Skript zum Kernblock „Forstbotanik und Baumphysiologie II“(430 a) Forstbotanischer Teil
I N H A L T S V E R Z E I C H N I S
1. Die Sprossachse 1
1.1 Anatomie der Sprossachse 1 1.1.1 Primärer Bau 1 1.1.2 Das Kambium 2 1.1.3 Leitgewebe 3 1.1.3.1 Xylem 3 1.1.3.2 Sekundäres Dickenwachstum 10 1.1.3.3 Reaktionsholz 12 1.1.3.5 Organisationsstufen des Hydrosystems 14 1.1.3.6 Phloem 14 1.1.3.7 Abschlussgewebe 17 1.2 Morphologie der Sprossachse 19 1.2.1 Lang- und Kurztrieb 19 1.2.2 Weiterwachen der Jahrestriebe 20 1.2.3 Aufbau des Sprosssystems 20 1.2.4 Besonderheiten des Knospentreibens 20 1.2.5 Seitensprossbildung 21 1.2.6 Baumgestalt 21 2. Das Blatt 21 2.1 Morphologie 21 2.2 Blattstellung 22 2.3 Anatomie 22 3. Die Wurzel 24 3.1 Anatomie 24 3.1.1 primäre Struktur 24 3.1.2 Sekundäre Bindungen 26 3.2 Morphologie 28 3.2.1 Wurzelsprosse 28 3.2.2 Wurzelmetamorphosen 28 3.2.2.1 Speicher-Wurzeln 28 3.2.2.2 Stützwurzeln 28 3.2.2.3 Haft- und Kletterwurzeln 28 3.2.2.4 Atemwurzeln 28 3.2.2.5 Zug- und Ankerwurzeln 28 3.2.3 Mykorrhiza 28 3.2.4 Wurzelsysteme 29

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 1 -
1. Die Sprossachse Die äußere Struktur eines Baumes spiegelt den Bau eines Kormus wieder: eine Sproßachse, bestehend aus Stamm und Ästen, die Blätter und Infloreszenzen tragen, sowie die Wurzel. Gegenüber krautigen Pflanzen zeigen sie zusätzlich Verholzung und Dickenwachstum - nach dem Verzweigungsschema können neben Bäumen (akrotone Verzweigungen) noch Sträucher (basitone Verzweigungen) unterschieden werden.
1.1 Anatomie der Sprossachse
1.1.1 Primärer Bau Die innere Gliederung, der Gewebeaufbau, im jungen (primären) Zustand ist von der frühen Differenzierung aus den embryonalen Zellen des Vegetationskegels ableitbar. Im Zentrum bildet sich das Markmeristem, dieses ist ringförmig umgeben vom Prokambium, vom Rindenmeristem und außen vom Dermatogen.
Vegetationskegel an der Spitze einer Sprossachse
An der Basis gliedern sich die embryonalen Zellen in vier Bildungsgewebe D: Dermatogen; RM: Rindenmeristem; PK: Prokambium; MM: Markmeristem Aus dem Markmeristem entwickelt sich das zentrale Mark, dessen Zellen ein interzellularenreiches Parenchym darstellen, das häufig von Exkretzellen durchsetzt ist. Im Zentrum entsteht ein
Kanal (Markhöhle), dessen Zellen zur Peripherie hin etwas kleiner sind (Markscheide) . Aus den ringförmig angeordneten Zellen des Prokambiums differenzieren sich vorwiegend durch Längsteilung langgestreckte Zellen mit geringem Querschnitt des Protophloems und -xylems. Die kambiumartigen Zellen liegen zwischen den anfangs bündelartigen Strängen des Phloems und Xylems (offenes Leitbündel), erst danach schließen sie sich rund zu einem Zylindermantel. Dieser umschließt das Mark, er selbst ist von einem Sklerenchymring umgeben. Dieser der Leitung von wäßrigen Lösungen dienende Komplex ist der Zentralzylinder. Mit Beginn der Tätigkeit des Kambiums ist der Primärzustand beendet. Aus dem Rindenmeristem entsteht die primäre Rinde, ein interzellularenreiches Parenchym, häufig mit Chlorolasten, das nach innen teilweise mit einer Endodermis abschließt (innere Abschlußschicht). Eine äußere Abschlußschicht, die Epidermis, entsteht aus dem Dermatogen. In dieser Funktion können auch äußere Schichten der prim. Rinde hinzutreten (Hypodermis). Sie enthält keine Chloroplasten und ist so lange funktionsfähig, wie das Organ selbst, dessen Abschluss sie bildet. Im Laufe der weiteren Differenzierung treten häufige Zellteilungen und -streckung auf, so daß z.B. Zellen des Protophloems und -xylems zerreißen und bald durch dauerhaftere ersetzt werden. Das Wachstum beruht anfangs vermehrt auf der Vermehrung von Parenchymzellen und ihrer Streckung im Bereich über und unter den Knoten, bis dann stärker die Teilungstätigkeit des Kambiums für das Dickenwachstum verantwortlich ist .

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 2 -
Entwicklung der jungen Sprossachse eines Baumes aus Braun, 1980)
1.1.2 Das Kambium Das Kambium ist verantwortlich für die Bildung sekundärer Gewebe. Dadurch wird der Umfang vergrößert (sek. Dickenwachstum) und damit gleichzeitig Leit- und Festigungsgewebe vermehrt sowie die Speicherkapazität erhöht. Der gestreckte, meißelförmig zugespitzte Zelltyp des Kambiums (fusiforme Initialen) gliedert axiale Zelltypen ab: Tracheiden, Gefäße, Siebzellen, Siebröhren, Fasern
usw. Parenchymatisches Strahlgewebe wird von kleineren, isodiametrischen Kambiumzellen gebildet. Die vom Kambium durch tangentiale Wandbildung nach innen abgegebenen Zellen bilden den Gewebekomplex des Holzes (Xylem), nach außen wird der Bast (Phloem) gebildet. Holzelemente werden in größerer Zahl produziert, so daß der Kambiumring immer weiter nach außen geschoben wird: Er paßt sich dieser Umfangvergrößerung durch Teilungen der Kambiumzellen in radialer Richtung an (Dilatation). Dabei entsteht, wie häufig bei tropischen Bäumen zu sehen, ein etagiertes Kambium und ein ebensolcher Bau (Stockwerksbau) der daraus abgeleiteten Gewebe. Nicht so hoch entwickelte Angiospermen und vor allem Gymnospermen zeigen bei dem Erweiterungswachstum des Kambiums pseudotransversale Teilung: Die Kambiumzelle teilt sich quer, durch Spitzenwachstum schieben sich die neuen Zellenden aneinander vorbei. Dadurch entsteht ein Kambium mit unregelmäßig verzahnten Zellen, das keinerlei vertikale Schichtung auch in den daraus entstehenden Geweben erkennen lässt.

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 3 -
oben: sich pseudotransversal teilende Kambiumzelle, daneben verzahnt aufgebautes Holz der Esche (Fraxinus excelsior)
unten: Teilung einer Kambiumzelle in radialer Längsrichtung, Stockwerkbau des Holzes einer tropischen Fabacea. (nach Braun, 1970)
Die Teilungsaktivität des Kambiums ist abhängig von inneren Faktoren (vegetative und generative Phase) so wie von der klimatischen Rhythmik entsprechend der geografischen Breite. Im Frühjahr wird die Aktivität eingeleitet durch eine
radiale Streckung. Als Folge sind die Zellen dünnwandig, ein solcher Stamm lässt sich leichter schälen. Die Teilungen beginnen bei zerstreutporigen Hölzern mit dem Knospentreiben, bei ringporigen schon davor.
1.1.3 Leitgewebe Die Pflanze besitzt aus anatomischer Sicht zwei Transportsysteme: das Phloem hauptsächlich zur Assimilatleitung, das Xylem hauptsächlich zur Wasserleitung. Beide Bereiche erscheinen als Gewebekomplexe, deren Zellen in unterschiedlicher Zusammensetzung verschiedene Gewebe bilden.
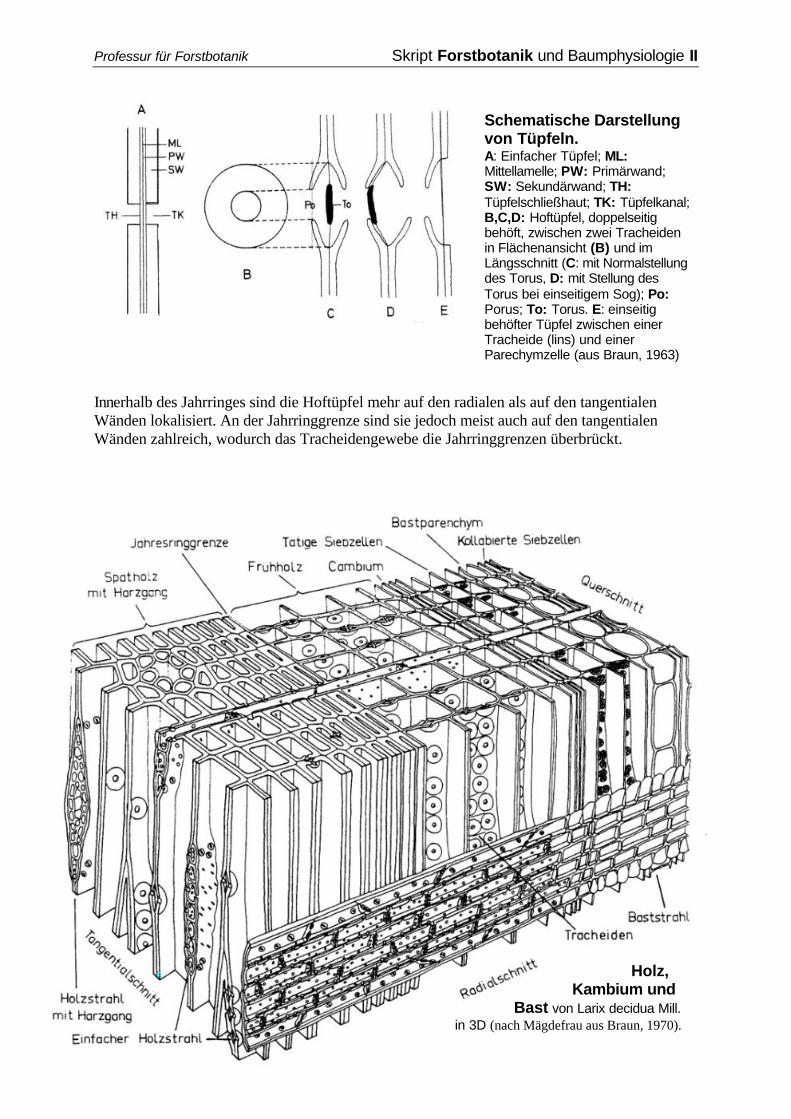
1.1.3.1 Xylem Das Xylem besteht prinzipiell aus den leitenden Elementen, den Gefäßen und den Tracheiden, den Stützelementen in Form von Holzfasern sowie dem axialen und radialen Parenchym. Zellen gleichen Baues und gleicher Funktion bilden Gewebe, wobei nicht alle Zellen im funktionsfähigen Zustand lebend sind. Tracheidengewebe Tracheiden sind langestreckte Zellen, deren gemeinsame Wände von vielen Tüpfelpaaren (Hoftüpfel) durchbrochen sind. Dabei handelt es sich um Kanäle, die vom Zelllumen zur Mittellamelle und von dort in die benachbarte Zelle reichen. Der Kanal kann durch Veränderung der Druckverhältnisse in den zugehörigen Tracheiden mittels des an der Mittellamelle "aufgehängten“ Torus verschlossen werden, indem er sich an einen von der Sekundärwand gebildeten Porus des Hofes legt. Der Porus der Tracheidenhoftüpfel ist kreisförmig. Der Bau der Tüpfel ist abhängig von der Art der benachbarten Zellen. Bei Pinus sind die Tüpfel zwischen einer Tracheide und einer Holzparenchymzelle z.B. einseitig behöfte große Fenstertüpfel ohne Torus.
Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 4 -
Schematische Darstellung von Tüpfeln. A: Einfacher Tüpfel; ML: Mittellamelle; PW: Primärwand; SW: Sekundärwand; TH: Tüpfelschließhaut; TK: Tüpfelkanal; B,C,D: Hoftüpfel, doppelseitig behöft, zwischen zwei Tracheiden in Flächenansicht (B) und im Längsschnitt (C: mit Normalstellung des Torus, D: mit Stellung des Torus bei einseitigem Sog); Po: Porus; To: Torus. E: einseitig behöfter Tüpfel zwischen einer Tracheide (lins) und einer Parechymzelle (aus Braun, 1963)
Innerhalb des Jahrringes sind die Hoftüpfel mehr auf den radialen als auf den tangentialen Wänden lokalisiert. An der Jahrringgrenze sind sie jedoch meist auch auf den tangentialen Wänden zahlreich, wodurch das Tracheidengewebe die Jahrringgrenzen überbrückt.
Holz, Kambium und
Bast von Larix decidua Mill. in 3D (nach Mägdefrau aus Braun, 1970).

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 5 -
Tracheiden haben durch ihre relativ dicken Wände zusätzlich Stützfunktion. Färbungen mit einem Farbstoff im Xylemwasser, die die wasserführenden Elemente anzeigen, zeigen eine Wasserführung des Tracheidengewebes über 10- 20 Jahresringe. Bei den rezenten Gymnospermen finden sich fast ausschließlich Tracheiden, während bei Angiospermen Gefäße hinzukommen
Laubholz. Auschnitt aus dem Holzkörper der Birke an der Grenze zur Rinde in dreidimensionaler Darstellung (nach Mägdefrau)

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 6 -
Gefäße Gefäße sind lange Röhren, die durch Fusion aus Einzelzellen (Gefäßglieder) entstanden sind. Ihre Querwände sind dabei immer vollständig oder teilweise aufgelöst, das Wasser kann frei hindurchfließen. Die Querwände sind schräg bis quer angeordnet und entweder vielfach perforiert oder fast vollständig aufgelöst (zusammengesetzte oder einfache Perforationsplatte). Die Durchbrechungen können leiterförmig (Betula, Magnolia), in Reihen nebeneinander (Liriodendron) oder in diagonalen Reihen (Salix) angeordnet sein. Die Längswände sind ebenfalls getüpfelt und weisen teilweise zum Schutz vor dem Kollabieren (Unterdruck) Sekundärwandauflagerungen auf in Form von Ringen, Schrauben und Netzen. Das
ausdifferenzierte Gefäß ist eine tote Röhre, die bis zu einem Durchmesser von 100µ m als mikropor und darüber bis 400µ m als makropor bezeichnet wird. Es gibt Gehölze, die nur mikropore Gefäße über den Jahrring zerstreut besitzen (zerstreut mikropor, z.B. Buche und viele andere Baumarten des kühl-gemäßigten Klimas) , andere haben makropore Gefäße im Früh- und mikropore im Spätholz (ringporig, cyclopor, z.B. Castanea und andere vor allem aus den wärmeren Lagen des kühl-gemäßigten Klimas und aus mediterranen Klimaten) und eine dritte Gruppe hat zerstreut makropores Holz, die u.a. in den heißen Tieflagen der Tropen beheimatet ist. Gefäße verlaufen häufig in Gruppen zu mehreren in axialer Richtung, wobei sie aber aus dieser Gruppe abzweigen können und sich anderen anschließen. Das so entstehende Gefäßnetz ist dichter bei mikroporen Arten. Bei einigen Bäumen werden Jahrringgrenzen von Gefäßen nicht überbrückt (z.B. Buche), bei anderen zeigen die Gefäße des Spät- und Frühholzes verschiedener Jahrringe starken Tüpfelkontakt (z.B. Populus). Die Länge einzelner Gefäße kann mehrere Meter erreichen.
Grundformen von Zellen,die das axial gerichtete Holzaufbauen. T = Tracheide; G = Gefäße mit vielfacher (leiterförmiger) und einfacher Durchbrechung der Endwände; HF = Holzfasern, links unseptiert und tot, rechts septiert (oft auch unseptiert) und lebend. PZ = Parechymzelle, nachträglich septiert (nach Braun, 1970)

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 7 -
Mikroskopische Querschnittsbilder aus dem Holz verschiedener Laubbäume. Oben links: Fagus sylvatica L.: homogenes, tracheidenreiches Grundgewebe mit zerstreut mikroporen Gefäßen, deren Gefäßnetze noch nicht die Jahresringgrenzen direkt überbrücken („jahresringeigene Gefäßnetze“) Oben rechts: Quercus robur L.: heterogenes Grundgewebe aus Tracheiden- und Holzfaserkomplexen bestehend. Gefäße (inmitten der Tracheidenkomplexe) cyclopor angeordnet mit makroporen Gefäßen im Früh- und mikroporen Gefäßen im Spätholz. Unten links: Afzelia bipindensis Harms: homogenes Holzfaser-Grundgewebe mit zerstreut-makroporen Gefäßen; um die Gefäße zwischen den Holzfasern interfibrilläres Parenchym. Unten rechts: Rhamnus cathartica L.: heterogenes Grundgewebe aus Tracheiden- und Holzfaserkomplexen wie bei Quercus (oben re), nur mit mikroporen Gefäßen inmitten der Tracheidenkomplexe. T = Tracheiden, HF = Holzfaserkomplexe, JG = Jahresringgrenzen, BHOST = breiter Holzstrahl, BA: Bast

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 8 -
Holzfasern Das Holzfasergewebe dient hauptsächlich der Festigung. Die Zellen sind langgestreckt, zugespitzt, haben relativ dicke Wände, die stark verholzt sind. Überwiegend in den radialen Wänden besitzen sie wenige einfache Tüpfel (ohne Hof), die schräg-spaltenförmig erscheinen; sie können später tot sein, ihr enges Lumen ist dann luftgefüllt. In unmittelbarer Nachbarschaft zu Gefäßen können sie auch als Wasserspeicher dienen. Bleiben sie lebend (z.B. bei tropischen Bäumen), dienen sie als Speicher für Stärke und Fette. Übergänge zu kürzeren, weitlumigeren und stärker getüpfelten Zellen heißen Fasertracheiden. Xylemparenchym Parenchymzellen im Holz sind lebende Zellen mit z.T. dicken, teilweise verholzten Wänden, die einzeln oder in kleinen Gruppen am Bau z.B. von Leitgeweben beteiligt sind. Ihre Aufgabe ist dort der vermittelnde Transport auf kurzen Strecken zwischen verschiedenenGeweben sowie die Speicherung verschiedenster Stoffe. Nach der Anordnung der Zellgruppen unterschiedet man axiales (von Prokambium oder Kambium) oder radiales (nur vom Kambium gebildet, Strahlen) Parenchym. Strahlen stellen die parenchymatische Verbindung über die Jahrringgrenze dar. Die im primären Zustand angelegten reichen bis zum zentralen Mark (Marktstrahlen), alle anderen (kürzeren, sekundären Holzstrahlen) wurden später angelegt. Auf gleicher Höhe eines Holzstrahls wird nach außen ein Baststrahl im Phloem gebildet. Die Differenzierung zu einem sekundären Strahl aus einer fusiformen Kambiumzelle beginnt mit einer inäqualen Teilung, wobei die kleinere (plasmareichere) Zelle zur Strahlinitiale wird.
Die ersten Zellen des Strahls sind meist stehend und einreihig. Erst später durch weitere Teilungen entstehende Zellen sind liegende (radial ausgerichtete) Zellen. Neue Strahlen werden erst angelegt, wenn durch das Dickenwachstum zwei benachbarte Strahlen eine (artspezifische) zu große Entfernung voneinander erreicht haben. Durch Querteilung der Initialen kann der Strahl vielzellreihig erhöht werden; er muss nicht einreihig in der Breite bleiben: häufig sind 3 - 10 Zellreihen. Diese Verbreiterung ist in der Mitte des Strahls am stärksten, so dass der Strahl im

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 9 -
Tangentialschnitt spindelförmig erscheint. Durch Ausfall von Fusiforminitialen können Holzstrahlen sekundär zusammenrücken und den Eindruck eines breiten Strahls machen (zusammengesetzter Strahl bzw. Fusionsstrahlen, Carpinus bzw. Quercus). Ein hoher und breiter Strahl kann umgekehrt auch durch Einschub von nicht parenchymachtischen Kambiumabkömmlingen wieder aufgespalten werden ( Fagus ) . Bei Gymnospermen bestehen die Holzstrahlen meist aus liegenden einschichtigen Parenchymzellen. Sie besitzen im Kreuzungsbereich mit den axialen Tracheiden mehrere kleine Tüpfel oder einen großen sog. Fenstertüpfel. Diese Strahlen heißen Kontaktstrahlen. Zu den Parenchymzellen treten noch radial verlaufende Tracheiden, wodurch der Strahl z. T. heterogen wird. Bei Laubbäumen finden sich in ein- und mehrschichtigen Strahlen nur Parenchymzellen ( homogen) , die liegend (radial) oder stehend (axial) ausgerichtet sein können. Sind beide Zelltypen in einem Strahl vorhanden, wird auch dieser als heterogen bezeichnet. Kontaktzellen mit Gefäßen sind ebenfalls reich getüpfelt, ob sie stehend oder liegend sind; die liegende Zelle ist jedoch durch ihre radiale Streckung besser für den Radialtransport geeignet. Vor allem bei tropischen Bäumen wird der Strahl mit stehenden Zellen begonnen, später werden dann im mittleren Bereich liegende Zellen gebildet. Bei Laubbäumen können jedoch Strahlzellen auftreten, die im Bereich von Gefäßen keinen Kontakt über Tüpfel aufnehmen (Isolationszellen). Man findet sie nur unter liegenden Zellen im sekundären Stadium. Solche Strahlen sind in Bezug auf den Kontakt heterogen: Isolationszellen sind prädestiniert für den Radialtransport, Kontaktzellen für den Quertransport über die Gefäße (z.B. Salix und Populus).
Einschichtiger Kontakt - Isolationsstrahl von Salix im Folgestadium. A: Radial-, B: Tangentialansicht. G = Gefäß, KOZ = Kontaktzellen; ISZ = Isolationszellen; IZ = Interzellulargänge; HF = Holzfasern. (nach Kny, aus Braun, 1970) Reine Isolationsstrahlen sind relativ selten (Carpinus, Corylus, Alnus außer A.viridis und Quercus); sie haben mitunter auch keinen räumlichen Kontakt
zu Gefäßen und kommen in den fraglichen Bäumen auch neben Kontakt-Isolations-Strahlen vor. An Hand der Holzstrahlen ist eine übersichtliche anatomische Systematik der Holzgewächse möglich, indem in die drei Organisationsstufen Kontakt-, Kontakt-Isolations- und Isolationsstufe gegliedert wird. Weiterhin kann in die Reihen „ nur stehende“, „ stehende und liegende“. und „nur liegende“ Zellen sowie jeweils die Unterreihen „nur einschichtige“ und „nur mehrschichtige“ Strahlen differenziert werden. In histologisch-funktioneller Betrachtung erkennt man eine strenge Zuordnung speziell der axialen Parenchyme zu bestimmten Elementen des Leitgewebes. Einzelne Zellen oder kleine Gruppen von zwei bis drei nebeneinaner liegenden Zellsträngen liegen diffus verteilt

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 10 -
zwischen den Tracheiden (paratracheidales Parenchym). Ihre Speicherstoffe, die sie aus dem Bast über die Holzstrahlen erhalten, können sie mittels einer reichen Tüpfelung in der Mobilisierungsphase im Frühjahr in die Tracheiden abgeben. Größere Mengen Parenchym in Enzelsträngen oder in größeren Gruppen, vor allem bei tropischen Bäumen, können sich im Holzfasergewebe befinden (interfibrilläres Parenchym) . Zusammen mit lebenden Holzfasern stellt es ein Speichersystem dar. Treten Parenchymzellen in
Begleitung von Gefäßen auf, handelt es sich um paratracheales Kontaktparenchym. Es sind Einzelstränge oder einschichtige Lagen um die Gefäße, wobei diese aber nur teilweise umscheidet sein können (Viertel-, Halb- oder auch Vollscheiden). Bei makroporen Hölzern sind sie am verbreitetsten. Neben der Speicherung und wieder Abgabe in die Gefäße (intensive Tüpfelung zu den Gefäßen hin) sollen sie auch Luftembolien in den Gefäßen verhindern.
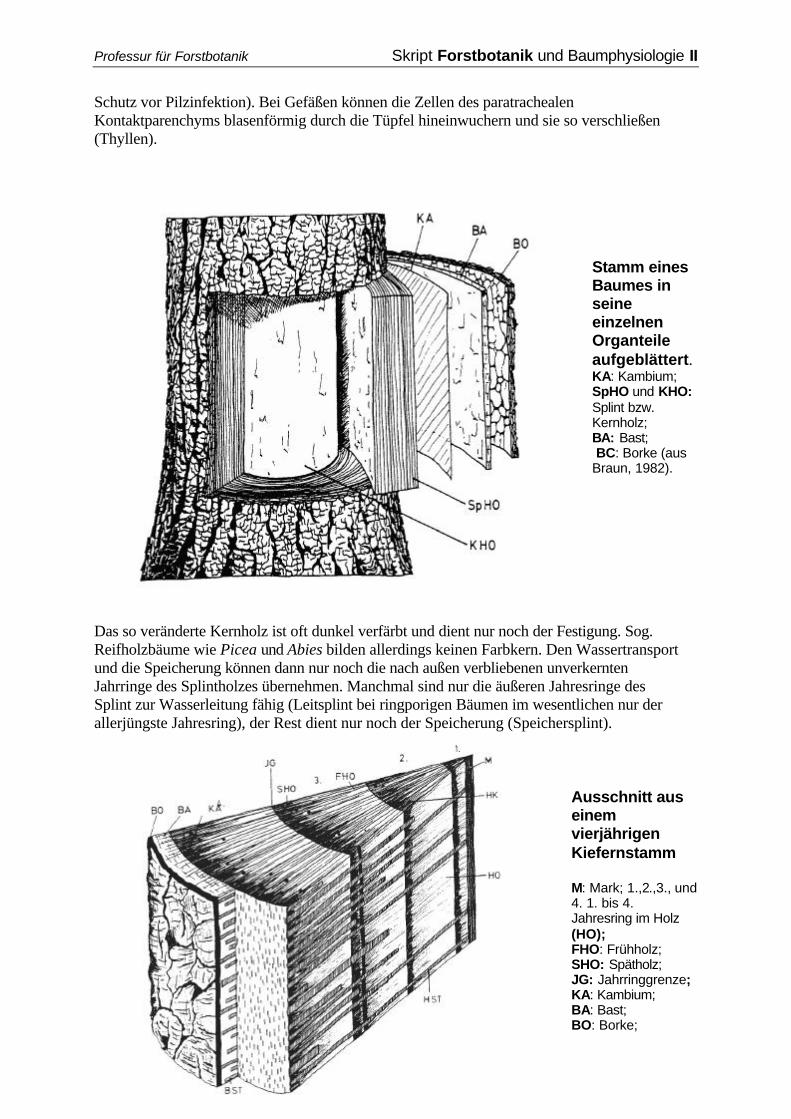
1.1.3.2 Sekundäres Dickenwachstum Durch kambiale Aktivität, speziell der Produktion immer neuer (jährlicher) Ringe von Xylem, kommt es zu einer Umfangvergrößerung des Holzkörpers, der sich das Kambium durch Weitenwachstum anpasst, auf das es aber auch durch die Bildung neuer Strahlen reagiert. Auf diese Weise findet im Holzkörper ein spannungsfreies Wachstum statt, während außerhalb des Kambiums schon eher Spannungen auftreten. Das zentrale Mark gerät in immer größere Entfernung zur Peripherie, seine Protoplasten sterben ab, die Wände verdicken und lignifizieren sich z.T.. Dem Holzteil kommen drei Funktionen zu: Wasserleitung, Speicherung, Festigung. Die Funktionen der Wasserleitung und Speicherung werden nur eine gewisse Anzahl von Jahren aufrechterhalten. In artspezifisch unterschiedlicher Ausdehnung zeigen viele Hölzer im Zentrum des Baumes eine Verkernung: Reservestoffe werden abtransportiert, sekundäre Pflanzenstoffe wie Harze, Gerb- und Farbstoffe werden in die Zellwände eingelagert, das Wasser wird entzogen, die Zellen sterben ab. Bei vielen Gymnospermen verschließen zusätzlich die Tori der Hoftüpfel die Pori, die Tüpfelschließhäute werden lignifiziert (besserer
Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 11 -
Schutz vor Pilzinfektion). Bei Gefäßen können die Zellen des paratrachealen Kontaktparenchyms blasenförmig durch die Tüpfel hineinwuchern und sie so verschließen (Thyllen).
Stamm eines Baumes in seine einzelnen Organteile aufgeblättert. KA: Kambium; SpHO und KHO: Splint bzw. Kernholz; BA: Bast; BC: Borke (aus Braun, 1982).
Das so veränderte Kernholz ist oft dunkel verfärbt und dient nur noch der Festigung. Sog. Reifholzbäume wie Picea und Abies bilden allerdings keinen Farbkern. Den Wassertransport und die Speicherung können dann nur noch die nach außen verbliebenen unverkernten Jahrringe des Splintholzes übernehmen. Manchmal sind nur die äußeren Jahresringe des Splint zur Wasserleitung fähig (Leitsplint bei ringporigen Bäumen im wesentlichen nur der allerjüngste Jahresring), der Rest dient nur noch der Speicherung (Speichersplint).
Ausschnitt aus einem vierjährigen Kiefernstamm
M: Mark; 1.,2.,3., und 4. 1. bis 4. Jahresring im Holz (HO); FHO: Frühholz; SHO: Spätholz; JG: Jahrringgrenze; KA: Kambium; BA: Bast; BO: Borke;

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 12 -
HST: Holzstrahlen; BST: Baststrahlen; HK: Harzkanäle (aus Braun, 1982)
1.1.3.3 Reaktionsholz Bei einseitiger Beanspruchung, z.B. das Gewicht weitausladender Äste oder Winddruck, werden im wachsenden Holz besondere Strukturen gebildet. Bei Gymnospermen auf der Zweigunterseite sowie auf der Leeseite Druckholz, das dichter und ligninreicher ist. Die Tracheiden sind kürzer, dickwandig und stark verholzt. Bei Laubbäumen wird umgekehrt die Oberseite reagieren (sog. Zugholz); das Holz ist schwach lignifiziert, seine Faserzellen bilden gelatinöse, cellulosereiche Wände. 1.1.3.4 Organisationsstufen des Hydrosystems Das Hydrosystem des Stammes läßt aus funktionell-histologischer Sicht mehrere Organisationsstufen erkennen: I. Tracheiden-Stufe Sie ist charakteristisch für Gymnospermen: Ein Tracheidengewebe, die Jahrringe erbrückend, eitet zusammen mit dem paratracheidalen Parenchym Wasser in alle Richtungen; es hat auch zugleich Festigungsfunktion. II. Tracheiden - Gefäß - Stufe Sie stellt die erste Angiospermen-Stufe dar: In das Tracheidengewebe sind hier jahresringeigene Gefäßnetze eingestreut, es besteht enger Tüpfelkontakt. Über die Jahresgrenze hinweg wird Wasser nur zwischen Tracheiden geleitet. III. Eingeschränkte Tracheiden-Gefäßstufe Die Hälfte des Xylems ist gebaut wie Stufe II, die andere Hälfte wird von jetzt hinzukommenden, nicht an der Wasserfernleitung beteiligten Holzfasern, die einen separaten Komplex darstellen, gebildet. Die Festigung beruht hier auf den Tracheiden und den Holzfasern. IV.Gefäß-Holzfaser-Stufe Hier dominiert ein Holzfasergrundgewebe, die Wasserleitung wird allein vom Gefäßnetz besorgt, das über die Jahresgrenze hinweg greift. Neben der Festigung können die Holzfasern durch die enge Nachbarschaft zu den Gefäßen auch als Wasserspeicher dienen. V. Gefäß-Stufe Die letzte Stufe ist ähnlich gebaut wie Stufe IV, doch sind hier alle Gefäße von Vollscheiden aus paratrachealem Parenchym umgeben, was zu einer Isolierung gegenüber den Holzfasern führt. Die Holzfasern fallen also als Wasserspeicher aus, sie dienen der Festigung und bilden im toten Zustand ein pneumatisches System. Lebende Holzfasern dienen neben ausgedehnten Verbänden von interfibrillärem Parenchym der Speicherung von Reservestoffen.

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 13 -
Die Organsation des Hydrosystems der Holzpflanzen in 5 Organisationsstufen und 3 Organisationsreihen Histologische Bilder der Bautypen schematisiert: Fläche mit schwarzen Vierecken = Tracheiden-Grundgewebe. Schwarze Flächen = Holzfaser-Grundgewebe. Kreise oder Kreisverbände = Gefäße bzw. Gefäßgruppen. Fein gepunktete Flächen = Gewebe aus lebenden Holzfasern. Gestreifte Zonen um makropore Gefäße = paratracheales Kontaktparenchym als Viertel-, Halb- und Vollscheiden. Fluoreszenzbilder auf der rechten Seite der Acer- Reihe: Orte der Wasserleitung leuchten durch das dem Transpirationsstrom zugegebene Berberinsulfat hell auf. I. Stufe = Tracheiden-
Stufe mit dem Gvmnospermen-Bautyp. II. Stufe = Tracheiden-Gefäß-Stufe. Links: Fagus sylvatica-Bautyp im weitesten Sinne mit Makrofluoreszenzbild; Mitte: Castanea sativa-Bautyp; rechts: Tristania suaveolens-Bautypim weitesten Sinne. III. Stufe = Eingeschränkte Tracheiden-Gefäß-Stufe, in der jetzt auch Holzfaser-Komplexe auftreten, die Luft führen. Links: Rhamnus carthartica-Bautyp mit Makrofluoreszenzbild; Mitte: Quercus robur-Bautyp: rechts: Lithocarpus molucca-Bautyp. IV. Stufe = Gefäß-Holzfaser-Stufe, in der keine Tracheiden mehr vorkommen und das Holzfaser-Grundgewebe z.T. als Wasserspeicher dient. Links: Aesculus hippocastanum-Bautyp mit Makrofluoreszenzbild (dazu gehörenauch Populus und Salix); Mitte: Ulmus laevis-Bautyp (im Spätholz treten noch Tracheiden-Gewebe auf) rechts: Aucoumea klaineana-Bautyp. V. Stufe = Gefäß-Stufe. Links: Acer pseudoplatanus-Bautyp mit paratrachealen Vollscheiden aus lebenden Holzfasern (mit Makrofluoreszenzbild); Mitte: Fraxinus excelsior-Bautyp; rechts: Albizzia odoratissima-Bautyp. Acer-Reihe: Mikropore Gehölze der gemäßigten Klimazone. Fraxinus-Reihe: Zyklopore Gehölze der wärmeliebenden Auwälder und der mediterranen Zone mit der Tendenz von unten nach oben die makroporen Frühholzgefäße immer vollständiger mit Scheiden aus paratrachealem Kontaktparenchym zu umgeben. Albizzia-Reihe: Makropore Gehölze der (Sub-)Tropen, ebenfalls mit der Tendenz die makroporen Gefäße immer vollständiger mit paratrachealem Kontaktparenchym zu umscheiden. (aus Braun), 1970

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 14 -
1.1.3.5 Organisationsreihen des Hydrosystems Wird in die Betrachtung der Bautypen der jeweiligen Organisations-Stufe die Größe und Anordnung der Gefäße mit einbezogen, lassen sich zusätzlich in den Stufen II bis V drei Reihen unterscheiden. In der I. Stufe ist nur die erste Reihe, die Acer-Reihe , mit mikroporen Hölzern zu finden. In der II. Stufe gehören zu dieser Reihe zahlreiche Holzgewächse der kühl-gemäßigten Klimazone des Fagus-Bautyps wie auch Alnus, Platanus, Prunus, Betula, Carpinus, Corylus, Sorbus und Tilia, in der III. Stufe der Rhamnus-Bautyp, in der IV. Stufe der Aesculus-Bautyp mit Salix und Populus bis hin zu dem Acer-Bautyp in der V. Stufe. Die Fraxinus - Reihe mit makroporen und mikroporen Gefäßen in cycloporer Anordnung wird in der II. Stufe durch den Castanea-, in der III. Stufe durch den Quercus-, in der IV. Stufe durch den Ulmus- und in der V. Stufe durch den Fraxinus-Bautyp repräsentiert. Während in der 1. Reihe die Anforderungen an das Hydrosystem durch Transpirationsgeschwindigkeiten von unter 6 m/h nicht sehr hoch sind, müssen die Gefäße der Bäume der 2. Reihe mit wärmeliebenden, spätaustreibenden Vertretern z.B. der mediterranen Klimazone Geschwindigkeiten von bis zu 40 m/h im Transpirationsstrom im Frühjahr bewältigen. Um Luftembolien zu verhindern, sind entsprechend die Parenchymscheiden um die Gefäße in aufsteigender Richtung in den Stufen zunehmend vollständiger vorhanden, bis hin zu Vollscheiden auch bei mikroporen Gefäßen von Fraxinus. Hartlaubgewächse dieses Klimas fallen mit ihren stark herabgesetzten Transpirationströmen von um 1 m/h aus dieser Gruppe heraus: Sie gehören durch den ausschließlichen Besitz mikroporer Gefäße zur Acer-Reihe . Zur macroporen Albizzia-Reihe gehören in jeder Stufe Vertreter der (sub-) tropischen Klimazone, in deren Hydrosystem zeitweise hohe Fließgeschwindigkeit herrscht. Entsprechend der Ausdehnung des Kernholzes und des Speichersplints ist das Hydrosystem unterschiedlich lange funktionsfähig. Am längsten ist es in der Acer-Reihe tätig: Durchschnittlich 20 Jahresringe leiten das Wasser, wobei 10% auf die äußersten entfällt. Die kürzeste Lebensdauer haben die funktionierenden Gefäße in der Fraxinus-Reihe. Hier leiten nur die äußeren Jahresringe, wobei der letzte bis zu 75% der gesamten Wassermenge transportieren kann. Im Frühjahr kann das Spätholz des letzten Jahres noch leiten, bis der neue Jahrring die gesamte Leitung übernimmt (z.B. Quercus robur).
1.3.6 Phloem Das Phloem (Bast) stellt ein ähnlich komplexes Gewebe dar wie das Xylem. Da seine Hauptfunktion, der Transport der Assimilate aus den Blättern, nur jeweils in den jüngsten Leitelementen des Phloems funktioniert, werden die älteren Schichten allmählich nach außen abgestoßen - das Phloem nimmt also nicht in dem Maße in der Dicke zu wie das Xylem. Die wichtigsten Zellen des Phloems sind die Siebelemente, axiale und radiale Parenchyme sowie Sklerenchymfasern. Siebzellen, Siebröhren Die Siebzelle ist die ursprünglichere Leitbahn im Bast. Sie ist schmal, spitz zulaufend und besitzt Siebfelder meist an den Längswänden, die eine Anhäufung von Poren ( erweiterte Plasmodesmen) darstellen , durch die jeweils ein Plasmaschlauch zieht. In Siebzellen ist der Kern degeneriert und der Tonoplast aufgelöst, so daß Cytoplasma und Vakuolensaft sich zu einer weniger viskosen Lösung vermischen. Im Unterschied zu den bei Farnen und Gymnospermen vorkommenden Siebzellen sind die Siebröhren vor allem der Dicotylen durch Fusion einzelner Siebröhrenglieder entstanden. In den häufig schräg stehenden Querwänden sind auffällige grobporigere Siebplatten ausgebildet, deren Poren ebenfalls am Vegetationsende durch Auflagerung von Callose verschlossen werden. Die ausdifferenzierte Siebröhre ist kernlos; sie ist durch eine inäquale Teilung aus einer Mutterzelle zusammen mit einer Geleitzelle entstanden.

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 15 -
Siebröhrenzellen (Siebröhrenglieder) mit Geleitzellen (und Parenchymzellen). Links schrägstehend Siebporenplatten mit mehreren Siebporenfeldern. (Nach Kollmann aus Lehrb.Bot., 1978) Geleitzellen, Strasburger-Zellen Geleitzellen können durch anschließende Querteilungen zu mehreren die Siebröhren begleiten: Sie besitzen ein dichtes Plasma, eine kleine Vakuole, einen großen (oft endopolyploiden) Kern, viele Mitochondrien und große primäre Tüpfelfelder zur Siebröhre hin. Sie gelten als die Steuerzellen für die kernlosen Siebröhren. Bei Gymnospermen (Pinaceae) übernehmen diese Funktion die sog. „Strasburger-Zellen“:. Es sind hoch stoffwechselaktive, stehende Zellen des Baststrahls, die ihn oben und unten flankieren, mit reichem Tüpfelkontakt zu den Siebzellen. Bei anderen Familien (z.B. Cupressaceae) wird die Steuerung durch axiale, eiweißreiche Phloemparenchymzellen ( „albuminous cells“ ) übernommen. Geleitzellen als auch Strasburgerzellen sterben mit ihren Siebröhren bzw. -zellen am Vegetationsende ab. Phloemparenchym und -fasern Im primären Phloem findet sich nur axiales Parenchym, das als ein- bis zweischichtiges
Band tangential im Jahresring verläuft. Im sekundären Phloem kommt noch radiales Parenchym der Baststrahlen hinzu. Als Festigungselemente findet man im Bast noch Bastfasern (Sklerenchymfasern) . Wenn sie bei Gymnospermen vorhanden sind, dann nur im sek. Phloem. Bei Pinaceen fehlen sie ganz, dafür erfahren die älteren Siebzellen sekundäre Verdickungen. Bei den dicotylen Gehölzen finden sich im primären und sekundären Phloem Fasern, die entweder in tangentialen Bändern sich mit Siebröhren abwechseln, einzeln zwischen anderen Phloemelementen oder in Gruppen zerstreut vorkommen .

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 16 -
Bast der Gymnospermen Der Gewebekomplex des Basts erscheint bei Gymnospermen in zwei Bautypen. Der Larix-Typ (hier auch Abies, Picea, Pinus und Pseudotsuga) zeigt ein mehrschichtiges Siebzellgewebe, das von einem Band Parenchymzellen durchzogen wird. Die Baststrahlen sind stark heterogen: Zellen in der Mitte des Strahls (liegend) sind zu den axial laufenden Parenchymzellen über Tüpfel verbunden, mit den Siebzellen jedoch nicht. Stehende Zellen an den Rändern des Baststrahls haben Tüpfelkontakte zu den Siebzellen (Strasburger-Zellen) oder zu axialen Parenchymzellen. Assimilat- oder Reservestofftransport aus den Siebzellen ist nur über die darunterliegenden Strahlzellen möglich. Die axialen Parenchymzellen können im älteren Bast (mit abgestorbenen und kollabierten Sieb- und Strasburger-Zellen) nur über diese liegenden Strahlzellen versorgt werden.
Ausschnitte aus dem Bast von Larix (Radialansichten). A, B und C zeigen den Kreuzungsbereich zwischen den axiallaufenden Bastgeweben und einem radiallaufenden Baststrahl (BST). Zone 1: Jüngster Bast, gerade vom Kambium gebildet, noch nicht in voller Funktion. Zone 2: Junger Bast-Jahresring in voller Funktion (Leitbast). Zone 3: Speicherbast, nur noch die Parenchymzellen in Funktion. Zu Bild A: Die stehenden Strasburger-Zellen (SBZ) stehen unmittelbar vor den Siebzellen (SZ) und sind mit diesen durch Siebporen verbunden. Die übrigen Zellen des Baststrahles, nämlich die stehenden Parenchymzellen zwischen den Strasburger-Zellen in der oberen und unteren Reihe und die liegenden Parenchymzellen in der Mitte des Baststrahles (PZ),
haben nur Tüpfelkontakt zu den axiallaufenden Parenchymzellen. Zu Bild B: Im Leitbast (Zone 2) werden aus den kernlosen Siebzellen organische Stoffe durch die großkernigen Strasburger-Zellen in den Baststrahl eingeschleust (Strasburger-Zellen = Steuerungs- und Schleusenzellen). Die Stellen und Zellen hoher Stofftransportleistungen weisen starke Enzymaktivitäten auf und sind schwarz gezeichnet. Zu Bild C: Speicherung von Stärke (große, schwarze Punkte) nur in den Parenchymzellen des Bastes. In die axiallaufenden Parenchymzellen gelangen die organischen Stoffe nur via Baststrahlen, was besonders im Speicherbast deutlich wird, wo die Siebzellen und die dazugehörenden Strasburger-Zellen (Zone 3) bereits ausgefallen sind. (aus Braun 1980.)

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 17 -
Beim Chamaecyparis-Typ der Cupressaceae findet man eine recht regelmäßige Bänderung durch ein Abwechseln einschichtiger Lagen von Siebzellen, Parenchymzellen, wieder Siebzellen und Bastfasern. Die homogenen Strahlen bestehen nur aus liegenden Zellen. Zellen des axialen Parenchyms übernehmen die Rolle der Strasburger-Zellen. Bast der Angiospermen Bei ursprünglichen Angiospermen kann noch der einschichtige Typ der Cupressaceen vorkommen. Verbreitet ist jedoch derWechsel mehrzelliger Schichten etwa mit dem Muster Siebröhren /Parenchymzellen/Bastfasern/Parenchymzellen (Robinia) oder recht breite Lagen von Siebröhren/Bastfasern/Kristallzellen/Parenchymzellen (Tilia) . Bei manchen ist eine deutlichere Jahresringgliederung durch Anlegen vorwiegend von Siebröhren im Frühbast und überwiegend Parenchymzellen im Spätbast. Breite Baststrahlen können teilweise sklerotisieren, indem sich ihre Zellen in dickwandige, verholzende Steinzellen umwandeln (Eiche). Bei Fagus werden diese Strahlen bereits im jungen Bast zu Sklerose-Baststrahlen.
Der ausgewachsene Bast (Speicherbast) kann durch spätes Wachstum, ausgehend von axialem Parenchym, auf tangentiale Zugspannung, ausgelöst durch das sek. Dickenwachstum reagieren. Auch diese Zellen können sklerotisieren (Fagus, Abies alba). Dilatation ist aber auch möglich durch sukzessive Verbreitung der Baststrahlen (Tilia).
1.1.3.7 Die Abschlußgewebe Epidermis Die Epidermis als primäres Abschlußgewebe kann vor allem bei Pflanzen mit sekundärem Dickenwachstum nicht auf Dauer einen Schutz vor äußeren Einflüssen bieten. Sie reißt auf und muß nun durch neugebildetes Gewebe ersetzt werden. Periderm Ein solches sekundäres Abschlußgewebe ist das Periderm. Seine Bildung wird eingeleitet durch Reembryonalisierung parenchymatischer Rindenzellen zu einem einschichtigen, parallel zur Oberfläche ausgerichteten Korkkambium (Phellogen) . Von ihm werden nach

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 18 -
außen Korkzellen des Kork (Phellem), nach innen das Phelloderm gebildet. Die Korkzellen sterben bald ab und bilden interzellularenfreie Reihen. Die Phellodermzellen bleiben aktiv und vermitteln zu den innen anschließenden, ebenfalls lebenden Rindenparenchymzellen. Während der Differenzierung der Korkzellen werden sie von Suberin (veresterte polymere Hydroxyfettsäuren) ausgekleidet, die abwechselnde Lamellen mit Wachsen bilden. Durch Einlagerung von Gerbstoffen werden diese Zellen zusätzlich resistent. In stark zusammengedrückter Form ist Kork ein guter Wärmeisolator, benetzbar, aber wasserundurchlässig.
Borke Häufig bleibt es nicht bei der Anlage des ersten Periderm (Initialperiderm) , vielmehr entstehen in tieferen Schichten der primären Rinde und des sekundären Phloems Folgemeristeme, die zu einer Borkenbildung führen. Meist werden sie zunächst zusammenhanglos an verschiedenen Stellen des Stammumfangs angelegt, so daß sich isolierte schuppenförmige Schichten bilden. Durch diese Folgemeristeme werden Bereiche der primären und sekundären Rinde von der Stoffversorgung abgeschnitten und sterben ab. Die so entstehende Borke ist ein Mischgewebe aus Peridermschichten und eingeschlossenen Resten der primären Rinde und des Phloems. Die Art der Anlage des Folgemeristems entscheidet über die Art der gebildeten Borke (Schuppen- und Ringelborke). Meist werden die äußersten Schichten abgestoßen, so dass die Borke in der Dicke konstant bleibt.

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 19 -
Häufig sind Periderme, vor allem wenn die Borke nicht stark rissig ist oder die Folgeperiderme sich nicht voneinander lösen, luftdicht, so daß ein Gasaustausch nicht möglich ist. Die regelmäßige Struktur des Phellems kann jedoch unterbrochen werden durch eine Anhäufung lockerer Zellen, die den Gasdurchtritt erlauben, indem ihre großen Interzellularen in direktem Kontakt zu denen des Rindengewebes stehen. Diese Lenticellen haben ein eigenes Phellogen, das ständig Zellen nach außen abgibt, die schließlich absterben und verwittern; die Abgabe von Phellodermzellen nach innen in größerer Zahl als das normale umgebende Phellogen bewirkt die Erhebung der Lentizelle über die Oberfläche des Periderms.
Histologischer Bau einer Lenticelle
e: Epidermis; K: Korkgewebe; ph: Phelloderm; f: Füllzellen
1.2 Morphologie der Sproßachse
Zur Gliederung der Sproßachse eines Kormus in Nodien und Internodien kommen bei Holzpflanzen noch einige morphologische Besonderheiten hinzu, die sie deutlich von anderen Pflanzen abgrenzt. Eine zusätzliche Gliederung wird durch die Ausbildung von Jahrestrieben erreicht, die durch Sproßverkettung Sproßsysteme aufbauen.
1.2.1 Lang- und Kurztrieb Der Jahrestrieb zeigt häufig eine Gliederung in Langtrieb und Kurztrieb, wobei sich beim Kurztrieb die Internodien nicht strecken und dadurch die Nodien so dicht stehen, dass ihre Blätter eine wirtelige Stellung vortäuschen. Der Langtrieb der Nadelbäume trägt wesentlich

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 20 -
mehr Blätter als der der Laubbäume, ihre Internodien sind kürzer. Kurztriebe bei Laubbäumen tragen häufig Blüten (viele Obstgehölze), während sie bei Coniferen in einem sehr kurzen Trieb mehrere Nadeln tragen (Larix, Cedrus, Pinus) . Während Lang- und Kurztriebe vegetative Triebe darstellen, wird in der reproduktiven Phase der Blütentrieb gebildet: Die Winterknospe treibt zur Blüte oder Infloreszenz, manchmal beginnend mit einem vegetativen Trieb (Aesculus) oder sofort zum fertilen (Kätzchen, Coniferenzapfen) Trieb.
1.2.2 Weiterwachsen der Jahrestriebe Beim monopodialen Wachstum wird am Vegetationsende an der Sproßspitze eine (häufig größere) Endknospe wie bei allen Coniferen und vielen jungen Dicotylen gebildet. Eine echte Endknospe ist nicht in der Achsel eines Blattes entstanden, sie trägt also an ihrer Basis keine Blattnarbe. Im Gegensatz zu diesem Monopodium geht beim Sympodium die Endknospe schon in der Anlage zugrunde, die oberste Achselknospe setzt den Trieb fort. Entsteht er nur aus einer Knospe, entsteht ein Monochasium (Aesculus, Paulownia) , bei zwei Knospen entsteht ein Dichasium (Syringa) , ganz in Analogie zu Infloreszenzen
I. Monopodial und II. sympodial monochasial wachsende Bäume. Co: Cotyledonen, 1 - 11: oberes Ende der Jahrestriebe, III: Strauchartiger Wuchs, 1 - 4: Triebgenerationen (nach Rauh aus Troll, 1973)
1.2.3 Aufbau des Sproßsystems Beim Aufbau des Sproßsystems kommt es nun sehr darauf an, welche Achselknospen gefördert werden, auch in Abhängigkeit von der Blattstellung. Coniferen bilden nur in einer begrenzten Anzahl von Nadelachseln Knospen (die Nadelzahl ist zu groß), Laubgehölze hingegen legen in jeder Blattachsel eine Knospe an. Trotzdem wird nicht jede angelegte Knospe im Frühjahr austreiben, sondern, hormonell gesteuert, nur in bestimmten Förderungszonen, z.B. an der Sproßspitze des Jahrestriebes (Akrotonie durch Apikaldominanz) , in seiner Mitte (Mesotonie, z.B. Cotoneaster) oder an seiner Basis (Basitonie, z.B. Sträucher). Die Förderung besteht in der Größe der angelegten Knospe, in der Frühzeitigkeit und Intensität des Triebs und in der Bereitschaft zur Fortsetzung dieses Triebs im nächsten Jahr.
1.2.4 Besonderheiten des Knospentreibens bei Holzgewächsen Bäume und auch Sträucher bilden als mehrjährige Gewächse an beblätterten Trieben Knospen, die erst nach einer Winterruhe im nächsten Frühjahr austreiben können (retardiertes Knospentreiben). Bei Larix bilden sich in den Achseln einzelner Nadeln des Langtriebs im

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 21 -
nächsten Jahr Kurztriebe. Daneben ist aber auch proleptisches Knospentreiben bekannt, bei dem die Knospe noch im Jahr der Anlage treibt (z.B. bei Pinus Kurztriebe in der Achsel von Schuppenblättern, oder Johannistriebe).
1.2.5 Seitensproßbildung Normalerweise ist die Stellung der Seitenzweige eine Folge der Blattstellung. Häufig findet man an der Hauptachse eine disperse (in alle Richtungen) Stellung der Seitenachsen, an den Nebenachsen jedoch eine distiche (zweizeilig) Verzweigung (Fagus, Carpinus, Ulmus, Tilia). Seitenknospen, die nicht zum Austrieb kommen, können jahrzehntelang als schlafende Knospen (Proventivknospen) am Sproß erhalten bleiben. Entweder sie werden allmählich überwuchert (Tiefknospen), oder durch vermehrte Holzbildung unter ihnen an der Oberfläche gehalten (Hochknospen). Neben diesen axillären Seitensproßentstehungen sind auch solche aus nicht-axillären Knospenanlagen bekannt. Es handelt sich dabei um embryonale Keime, die ebenso an der Wurzel (Wurzelbrut bei Robinia und Ulmus) vorkommen können. Diese Adventivknospen werden sekundär, z.B. im Bedarfsfall angelegt und können aus undifferenziertem Kallusgewebe nach Verletzungen, aus Kambiumderivaten oder reembryonalisierten Parenchymzellen entstehen. Zusammen mit den proventiven Anlagen gelingt es diesen Pflanzen, nach einem Rückschnitt aus älteren Achsen auszuschlagen (z.B. Platane).
1.2.6 Baumgestalt Unter der Einwirkung der bei Holzgewächsen wirksamen Besonderheiten eines Jahrestriebes, des retardierten Knospentreibens, des monopodialen Wachstums (bei manchen wenigstens in der Jungendphase) sowie der Akrotonie, bei der innerhalb des Jahrestriebes die spitzennahen Knospen gefördert werden, kommt eine typische Baumarchitektur zustande. Das Ergebnis aller Faktoren ist ein bestimmtes Verhältnis von Haupt- zu Nebenachsen. In aller Regel wird die Hauptachse gefördert (Monokormie). Im Alter der Bäume beobachtet man eine Reduktion der Zahl treibender Knospen sowie einen größeren Ablaufwinkel der Seitenäste 1. Ordnung als 90 Grad.
Nadelbäume zeigen überwiegend eine strenge Akrotonie sowie eine Anhäufung von Seitenästen zu Beginn des Jahrestriebes (Quirläste). Laubbäume hingegen zeigen häufig eine Gliederung in einen monokormen Stamm und eine polykorme Krone, bei den Langtrieben strecken sich die Internodien stärker, die Zahl der Nodien pro Jahrestrieb ist geringer und der Ablaufwinkel der Äste 1. Ordnung ist stets spitz. In der Jugend der Laubbäume wird nur jeweils der oberste Trieb aufeinanderfolgender Jahre zeitweise gefördert (übgreifende Akrotonie), bis dann im Alter von 5- 10 Jahren vorwiegend eine hypotone ( auf der Zweigunterseite) Seitenzweigförderung eintritt. Die im Freistand der Bäume auftretende Baumarchitektur wird im eng gepflanzten Verband eher der Jugendform entsprechen.
2. Das Blatt
2.1 Morphologie Blätter entstehen exogen aus den peripheren Zellschichten des Scheitelmeristems (Tunica) bei Angiospermen. Sehr früh bildet sich eine Einschnürung an einem Höcker. Der Teil zum Sproß hin (Unterblatt) entwickelt sich zum Blattgrund und evtl. zu Nebenblättern, der nach außen weisende Teil (Oberblatt) zum Blattstiel und zur -spreite (Larnina). Basale und intercalare Meristeme des jungen Blattes vergrößern es bis zum fertigen Blatt und entscheiden

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 22 -
über die endgültige Gestalt, ob es geteilt oder ungeteilt, ganzrandig oder gelappt, glattrandig oder gezähnt bzw. gesägt wird. Das normalerweise flächig gebaute Blatt ist gut angepaßt an die Funktion des Gasaustausches und der Tanspiration. In Abhängigkeit von anderen Funktionen können Blätter in Abweichung von der flächigen Form auftreten, wobei der Blattcharakter nicht immer äußerlich zu erkennen ist. Ist die Blattspreite reduziert, übernimmt aber der Blattstiel oder der -grund ihre Funktion, handelt es sich um Phyllodien (Robinia heterophylla) , bei Kakteen und teilweise Robinia pseudoacacia sind die Blätter in Dornen umgewandelt, rankende Pflanzen haben teilweise zu Ranken umgewandelte Blätter, während bei Zwiebeln der Blattgrund fleischig verdickt ist.
2.2 Blattstellung Blätter inserieren an Knoten (Nodien). Steht an einem Knoten nur ein Blatt, handelt es sich um wechselständige Blätter (z.B. Sorbus-Arten). Stehen zwei Blätter im Nodium, spricht man von gegenständiger Blattstellung ( z .B. Fraxinus) . Ist die Ansatzstelle der Blätter aufeinanderfolgender Nodien dabei um 90° versetzt (auf „Lücke“ stehend), liegt dekussierte Beblätterung vor (z.B. Lamiaceae). Befinden sich mehrere Blätter im Nodium, ist die Blattstellung wirtelig (z.B. Waldmeister). Am jungen Sproß sind nicht alle Blätter gleich gestaltet. Zuerst werden stets die Keimblätter gebildet (Cotyledonen): Sie sind meist einfach gebaut; nach ihrer Zahl unterscheidet man Monocotyle (ein), Dicotyle ( zwei) und Gymnospermen (meist mehrere Keimblätter).
Bei der hypogäischen Keimung (z.B. Eiche) sind die Cotyledonen dick, kohlenhydratreich, im Boden bleibend und kaum ergrünend. Pflanzen mit epigäischer Keimung besitzen Cotyledonen mit normaler Blattspreite, über dem Boden erscheinend und bald ergrünend. Als nächstes werden Laubblätter gebildet: Die ersten (Primärblätter) können einfacher gestaltet sein als die Folgeblätter.
2.3 Anatomie Die meisten Laubblätter sind dorsiventral gebaut: Ober- und Unterseite sind voneinander unterschieden. Der äußerste Abschluß besteht aus einer einschichtigen Epidermis, deren Zellen lückenlos miteinander verzahnt sind, keine Chloroplasten besitzen und nach außen einen dünnen Film bestehend aus Cutin und Wachsen, die Cuticula, ausscheiden. Zwischen der Epidermis der Blattober- und Blattunterseite liegt das Mesophyll, das nach oben in das Palisaden-, nach unten in das Schwammparenchym differenziert ist. Sonnenblätter (in der Baumkrone oben und außen) können längere oder mehrschichtige Palisadenzellen als Schattenblätter besitzen. Die chloroplastenreicheren Palisadenzellen sind langgestreckt und senkrecht zur Oberfläche ausgerichtet, die Zellen des Schwammparenchyms sind unregelmäßig gestaltet und weisen große Interzellularräume auf, die über Spaltöffnungen (Stomata) mit der Außenluft in Verbindung stehen. Spaltöffnungen bestehen aus zwei Schließzellen, die eine Öffnung bilden können, sowie den oft zum Spaltöffnungsapparat gehörenden Nebenzellen. Die Schließzellen besitzen unterschiedlich verdickte Zellwände sowie stets Chlorplasten. Stomata befinden sich meist auf der Blattunterseite ( hypostomatische Blätter) Die Aderung des Blattes (parallel bzw. bogig verlaufend bei monocotylen, netzartig bei dicotylen Pflanzen) entspricht dem Verlauf der Leitbündel. Sie bestehen aus einem Xylem- und einem Phloemteil, bei Hauptadern befindet sich dazwischen ein Kambium (außer bei monokotylen Pflanzen). Entsprechend der Anordnung im Sproß zeigt beim in das Blatt abbiegenden Leitbündel das Phloem zur Blattunterseite. Bis in die feinsten Verzweigungen sind die Leitbündel von Parenchymscheiden umgeben. Zusätzlich werden die stärkeren oft von Sklerenchymfasersträngen begleitet .

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 23 -
Querschnitt durch das bifaciale Blatt der Buche (Fagus sylvatica)
ep: Epidermis; sp: Spaltöffnungen; pl: Palisaden-, s: Schwammparenchym; k: Kristallzellen; L: Leitbündel Das Nadelblatt der
Coniferen zeigt im anatomischen Bau häufig keine deutlichen Unterschiede von Ober- und Unterseite (äquifazial). Die Epidermiszellen sind dickwandig und englumig mit einer dicken Cuticula, darunter liegen ein bis zwei Schichten Hypodermiszellen. Spaltöffnungen sind bis in die Hypodermis eingesenkt, darunter liegt das Assimilationsgewebe. Im Inneren der Nadel, eingebettet in ein Transfusionsgewebe mit einer Endodermis nach außen weisend, liegt der Zentralzylinder mit dem Leitbündel. Die mehrjährigen Nadeln der Coniferen müssen jedes Jahr mindestens die leitenden Elemente des Phloems erneuern. Sie besitzen deshalb im Leitbündel ebenfalls ein Kambium. Zum Ausgleich der verkleinerten Oberfläche der Nadel als Schutz vor schneller Austrocknung (xeromorpher Bau) besitzen die assimilierenden Zellen teilweise Wandausstülpungen nach innen zur Vergrößerung der Oberfläche, an denen Chloroplasten sitzen (Armpalisadenzellen). Zusätzlich werden Coniferennadeln von einer wechselnden Zahl von Harzbehältern längs durchzogen.
Querschnitt durch das Nadelblatt der Waldkiefer (Pinus sylvestris) Als Beispiel für ein xeromorph gebautes Blatt. E: Epidermis; Sp: Spaltöffnung; AP: Assimilatiosparenchym; ED: Endodermis; LB: Leitbündel; TFG: Transfusionsgewebe; HK: Harzkanal (aus Braun, 1980)

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 24 -
3. Wurzel Der Sproß und die Wurzel bilden ein einheitliches Organ - die Achse, nur jeweils im unterschiedlichenRaum. Abweichend vom Stamm besitzt die Wurzel keine Gliederung in Nodien und Internodien und trägt auch keine Blätter, Seitenwurzeln werden endogen angelegt.
3.1 Anatomie
3.1.1 primäre Struktur Im Gegensatz zum Sproß befindet sich das Apikalmeristem der Wurzel subterminal unter der Wurzelhaube (Kalyptra). So wie das Meristem ständig Zellen nachhinten abgibt, ergänzt es fortlaufend nach vorn die Kalyptra, die durch das Verschleimen ihrer Zellen als Schutzgewebe fungiert . Im primären Zustand findet man dicht unter der Initialzellgruppe eine Gliederung von außen nach innen: 1.Dermatogen, aus der die Rhizodermis wird, 2. das Meristem der primären
Rinde (Periblem), aus der das Rindenparenchym entsteht und 3. das Meristem des künftigen Leitgewebes (Plerom) im Zentrum. Spitze einer jungen Wurzel RHD: Rhizodermis; PE: Periblem; PL: Plerom; D: Dermatogen; KLG: Kalyptrogen; KL: Kalyptra; SK: Stärkekörner (nach Lehr.Bot., 1978)
Rhizodermis In Analogie zur Epidermis bildet die äußerste Zellschicht die Rhizodermis, die aber hier endogen unter der Kalyptra angelegt wird und erst weiter hinten an die Oberfläche der Wurzel gelangt. Sie besitzt keine Cuticula und anfangs zarte Zellwände, die sich erst später verdicken und verholzen. Dicht hinter der Wurzelspitze können die Rhizodermiszellen zu Wurzelhaaren zur Oberflächenvergrößerung auswachsen. Häufig wachsen nur bestimmte (Trichoblasten) aus, die in einem bestimmten Abstand zueinander stehen. Sie sind nur wenige Stunden oder Tage lebensfähig. Die Rhizodermis selbst wird nach relativ kurzer Lebensdauer von einer Exodermis abgelöst, die aus einer bis wenigen Schichten des darunterliegenden Rindenparenchyms besteht, dessen Zellen durch Suberinlamellen undurchlässig werden und

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 25 -
durch Sekundärwände verdickt und z.T. verholzt werden. Die Exodermis gilt als primäres Abschlußgewebe . Primäre Rinde Das Rindenparenchym ist interzellularenreich, was bei Wasser und Sumpfpflanzen zu einem besonders ausgedehnten Gewebe führt (Aerenchym). Nach innen ist es abgeschlossen durch die einschichtige Endodermis, deren radiale Wände durch die mikroskopisch sichtbaren
Caspary-Streifen (hydrophob durch Lignin- und Suberineinlagerungen) für wässrige Lösungen unpassierbar sind. Querschnitt durch eine junge Wurzel RHD: Rhizodermis mit Wurzelhaaren; WR: Wurzelrinde; ED: Endodermis mit Casparystreifen; PKA: Perikambium; I und II: Wasseinstrom (Pfeile) durch den Symplasten bzw. durch den Apoplasten bis zur Endodermis (aus Braun, 1982)
Zentralzylinder Funktionell gehört also die Endodermis schon zum Leitsystem. Dieses ist noch umgeben von einer ein- bzw. zweischichtigen ( Dicotyle bzw. Gymnospermen) Zellage, die im primären Zustand als Perikambium, ausdifferenziert als Perizykel bezeichnet wird. Das primäre Leitsystem der Wurzel ist radiär gebaut: im Querschnitt strahlenförmiges Xylem, in den Winkeln nach außen das Phloem. Anfangs befindet sich Kambium nur in den Winkeln zwischen Xylem und Phloem. Im Übergang zum sekundären Zustand wird das Kambium sich zum Ring schließen, ausgehend vom Perikambium über den Xylemstrahlen. Anschließend produziert das Kambium auf dem ganzen Umfang Xylem und Phloem entsprechend dem Sproß; auch hier folgt ein sekundäres Dickenwachstum. Das Xylem der Wurzel ist nicht so einheitlich gebaut wie das des Stammes: Hier findet man eine größere Variation entsprechend den ökologischen Gegebenheiten. Das Wurzelholz der Coniferen hat jedoch einen kaum variierbaren Bau, während bei Laubbäumen eine größere Vielfalt vorliegt. Flachstreichende Wurzeln sind auf feuchten Standorten gefäßreicher, auf trockenen sind es die tiefreichenden. Der Verankerung dienende Wurzeln besitzen viele Faserzellen, andere mit mehr Speicherfunktion sind parenchymreich. Allgemein sind Wurzeln parenchymreicher als der Sproß, sie besitzen aber weniger Holzstrahlen. Echte Jahresringe sind nur in den, jahreszeitlichen Schwankungen ausgesetzten, flachstreichenden Wurzeln erkennbar. Seitenwurzeln werden endogen angelegt. Dazu reembryonalisieren Zellen des Perizykels, meist über den Xylemstrahlen, und beginnen mit dem Teilungswachstum für die junge Wurzel, die die Rindenschichten durchstoßen muss.

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 26 -
Entstehung einer Seitenwurzel PKA: Perikambium; WR: Wurzelrinde; PH: Phloemteil; X: vierstrahliger Xylemteil (aus Braun, 1980)
3.1.2 Sekundäre Bildungen Periderm Durch das Dickenwachstum der Wurzel kann die Exodermis nicht dauerhaft als Abschlußgewebe dienen; sie reißt auf und muss durch ein sekundäres Abschlußgewebe (Wurzelperiderm) ersetzt werden. Im Unterschied zum Spross wird hier immer die Entwicklung in tieferen Schichten beginnen, nämlich im Perizykel.
Sekundäres Dickenwachstum einer Wurzel. A: primärer Zustand mit fünfstrahligem, zentralem Leitbündel; B: das Kambium (punktiert) mit noch sternförmiger Figur; C: Aufreißen der Rhizodermis, der primären Rinde einschließlich der Endodermis durch die stärker werdenden sekundären Gewebe. (Lehrbuch Botanik, 1978) Bei Laubbäumen gibt es keinen qualitativen Unterschied im Periderm vom Stamm und Wurzel, in der Wurzel ist es jedoch wesentlich weniger mächtig. Bei Coniferen hingegen gibt es Unterschiede zu den Laubgehölzen als auch zwischen Sproß und Wurzel. Der hier gebildete Kork (z.B. Picea abies, Pinus sylvestris) besteht aus drei verschiedenen Schichten: aus Steinkork mit dicken, verholzten, aber nicht verkorkten Wänden, aus Schwammkork mit dünnen, unverholzten und auch unverkorkten Wänden sowie aus den völlig verkorkten, dünnwandigen Zellen des Phlobaphenkorks. Im Spross sind die Korke meist jeweils 4-schichtig. Werden Teile des Korks abgestoßen, reißt er an der dünnwandigsten Stelle

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 27 -
(Schwammkork) ab und der darunterliegende, gegen Wasserverlust und Infektionen resistentere dunkle Phlobaphenkork liegt außen. In der Wurzel ist nur der Steinkork mehrschichtig, die beiden anderen einschichtig. Hier wird beim Aufreißen der Schwammkorkschicht auch die eine Phlobaphenkorkschicht zerstört, so daß der helle und gegen mechanische Verletzungen besser Schutz bietende Steinkork im intakten Periderm außen liegt. Er hat zudem den Vorteil, dass er stark quellfähig ist und so wohl eine Wasseraufnahme ermöglicht, auf der anderen Seite aber leichter von Mikroorganismen abgebaut werden kann.

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 28 -
3.2 Morphologie
3.2.1 Wurzelsprosse Bei manchen Bäumen (Robinia, Ulmus) sind die Wurzeln in der Lage, Sprosse zu bilden, deren Apikalmeristem aus dem Perizykel entsteht wie Seitenwurzeln oder aus dem Phellogen des Periderms. Häufig tritt dieses Phänomen bei flach streichenden (plagiotropen) Wurzeln an der Oberseite auf.
3.2.2 Wurzelmetamorphosen
Als Metamorphose wird eine morphologische Umgestaltung mit gleichzeitiger Änderung der Funktion verstanden.
3.2.2.1 Speicher-Wurzeln Bei Speicherwurzeln führt eine Gewebevermehrung z . B. des Parenchyms innerhalb oder außerhalb des Leitgewebes oder ein perikambiales Dickenwachstum mit der zusätzlichen Produktion von Xylem- und Phloemringen zur Verdickung der Wurzel mit der Möglichkeit der vermehrten Speicherung von Wasser oder Reservestoffen.
3.2.2.2 Stützwurzeln
Sproßbürtige, weitausladende Stütz oder Stelzwurzeln können schon im Jugendstadium angelegt werden (Mangroven, z. B. Rhizophora)
3.2.2.3 Haft- und Kletterwurzeln Bei Lianen und Epiphyten kommen Haft- und Kletterwurzeln vor, die geotrop neutral, aber phototrop negativ (dem Substrat zugewandt) reagieren. Sie können ein Hindernis rankenartig umschlingen und sich mit Wurzelhaaren verankern.
3.2.2.4 Atemwurzeln Aerenchymreiche, geotrop negative Wurzeln über sauerstoffarmen Sumpfboden heißen Atemwurzeln. Man findet sie bei Mangroven, Sumpfzypressen und einigen Palmen. In ihnen steht ein ausgedehntes Interzellularen-System über Lenticellen oder Rindenrisse mit der Außenluft in Verbindung.
3.2.2.5 Zug- und Ankerwurzeln Normalerweise verankern die Wurzeln eine Pflanze durch eine reiche Verzweigung sowie durch die Verflechtung der Wurzelhaare um die Bodenpartikel. Es gibt jedoch Wurzeln, die durch ihre Verkürzung die Pflanze in den Boden „ziehen“. Diese Kontraktion kann durch Erweiterung von Parenchymzellen unter gleichzeitiger Verkürzung oder durch Kollabieren von Parenchym in bestimmten Abschnitten zustande kommen .
3.2.3 Mykorrhiza Bei den meisten Waldbäumen zeigen die jungen Wurzeln eine Symbiose mit bestimmten Pilzen. Einzelne Pilzhyphen ersetzen dabei die Wurzelhaare. Bei der ektrotrophen Mykorrhiza befindet sich der Pilz interzellulär in der Rinde, ohne in die Rindenzellen einzudringen (bei unseren meisten Waldbäumen), bei den endotrophen Formen leben die Pilzhyphen intrazellulär (VA-Mykorrhiza bei krautigen Pflanzen und vielen tropischen Bäumen, bei unseren Waldbäumen seltener, z.B.Taxus). Die Affinität beider Partner bzw. ihre Abhängigkeit voneinander kann unterschiedlich stark sein. Obligat ist die Mykorrhiza bei

Professur für Forstbotanik Skript Forstbotanik und Baumphysiologie II
- 29 -
Kiefer, Fichte, Lärche, Eiche und Hainbuche (stark mykotrophe), fakultativ bei Tilia, Populus, Salix, Acer und Fraxinus (schwach mykotrophe Bäume).
3.2.4 Wurzelsysteme Nach der Verbreitung der Wurzel im Boden lassen sich mehrere Typen unterscheiden. Der Pfahlwurzeltyp ist ein einachsiges Wurzelsystem. Zu einer dominierenden Hauptwurzel kommen einige mehr oder weniger kräftige Seitenwurzeln, die sich jeweils bald in viele Feinwurzeln verzweigen. Hierher zählen Abies alba, Pinus sylvestris, Quercus robur und Q.petraea. Als ein mehrachsiges System gilt das Herzwurzelsystem, wie es bei Larix decidua, Fagus, Pseudotsuga, Betula pendula und Carpinus vorkommt. Ohne eine Trennung in horizontal und vertikal verlaufende Wurzeln gehen mehrere Hauptseitenwurzeln schräg bis steil abwärts, ein oder zwei kräftige Vertikalwurzeln reichen tiefer. Ebenfalls mehrachsig ist das Senkerwurzelsystem: Von vorherrschend horizontal streichenden kräftigen Wurzeln, die ein tellerartiges Horizontalwurzelsystem bilden, oder aus dem Wurzelstock, entspringen später meist schwächere Senkerwurzeln ( Picea abies, Fraxinus excelsior , Populus tremula). Nicht immer gelingt eine eindeutige Zuordnung. So besitzt Acer pseudoplatanus ein Mittelding zwischen einem Herz- und Senkerwurzelsystem.
Wurzelsysteme von Waldbäumen Links: Pfahlwurzelzeiger; Mitte: Senkwurzelsysem; Rechts: Herzwurzelsytem (aus Braun, 1982)


















