
Stress- und Schmerzbelastung des Schweines bei … · Ablauf der Cortisoluntersuchungen.....32 7....
90
Inaugural-Dissertation zur Erlangung der Doktorwürde der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Stress- und Schmerzbelastung des Schweines bei Entnahme eines Tracheobronchialabstriches im Vergleich zum Nasentupfer und der Fixierung in der Oberkieferschlinge von Christine Weiß aus Altötting München 2015
-
Upload
truongxuyen -
Category
Documents
-
view
216 -
download
0
Transcript of Stress- und Schmerzbelastung des Schweines bei … · Ablauf der Cortisoluntersuchungen.....32 7....

Inaugural-Dissertation zur Erlangung der Doktorwürde
der Tierärztlichen Fakultät
der Ludwig-Maximilians-Universität München
Stress- und Schmerzbelastung des Schweines bei Entnahme eines
Tracheobronchialabstriches im Vergleich zum Nasentupfer und der
Fixierung in der Oberkieferschlinge
von
Christine Weiß
aus
Altötting
München 2015

Aus dem Zentrum für Klinische Tiermedizin
der Tierärztlichen Fakultät
der Ludwig-Maximilians-Universität München
Lehrstuhl für Krankheiten des Schweines
Arbeit angefertigt unter der Leitung von: Univ.-Prof. Dr. Mathias Ritzmann
Mitbetreuung durch: Dr. Nicole Übel und Dr. Susanne Zöls

Gedruckt mit der Genehmigung der Tierärztlichen Fakultät
der Ludwig-Maximilians-Universität München
Dekan: Univ.-Prof. Dr. Braun
Berichterstatter: Univ.-Prof. Dr. Mathias Ritzmann
Korreferent: Univ.-Prof. Dr. Dr. Michael Erhard
Tag der Promotion: 31. Januar 2015

Meiner Familie, meinen Freunden und den Tieren

Inhaltsverzeichnis 7
INHALTSVERZEICHNIS
I. EINLEITUNG .......................................................................................... 11
II. LITERATURÜBERSICHT .................................................................... 13
1. Diagnostische Mittel ................................................................................. 13
1.1. Gesetzliche Grundlagen ............................................................................. 13
1.2. Diagnostik von Atemwegserkrankungen beim Schwein ........................... 13
1.2.1. Bronchoalveoläre Lavage .......................................................................... 14
1.2.2. Tracheobronchialabstrich ........................................................................... 15
1.2.3. Nasentupfer ................................................................................................ 16
2. Stress und Wohlbefinden ........................................................................ 17
3. Schmerz ..................................................................................................... 18
3.1. Schmerzentstehung, Weiterleitung und Wahrnehmung............................. 19
3.2. Innervation der untersuchten Regionen ..................................................... 20
4. Untersuchte Parameter ............................................................................ 22
4.1. Cortisol ....................................................................................................... 23
4.1.1. Wirkung im Organismus ............................................................................ 23
4.1.2. Transport und Ausscheidung ..................................................................... 24
4.1.3. Speichelcortisol .......................................................................................... 25
4.1.4. Regulation .................................................................................................. 25
4.1.5. Circadiane Rhythmik ................................................................................. 26
4.2. Katecholamine ........................................................................................... 27
4.2.1. Wirkung im Organismus ............................................................................ 28
4.2.2. Bedeutung im Stressgeschehen .................................................................. 28
4.2.3. Regulation .................................................................................................. 29
III. MATERIAL UND METHODEN ........................................................... 31
1. Ziel der Untersuchung ............................................................................. 31
2. Genehmigung des Tierversuchsvorhabens ............................................ 31
3. Studientiere und Haltung ........................................................................ 31
4. Auswahl der Studientiere ........................................................................ 31

8 Inhaltsverzeichnis
5. Gruppeneinteilung ................................................................................... 32
6. Ablauf der Cortisoluntersuchungen ....................................................... 32
7. Ablauf der Katecholaminuntersuchungen ............................................ 34
8. Eingriffe .................................................................................................... 36
8.1. Durchführung eines Tracheobronchialabstriches ....................................... 36
8.2. Fixierung in der Oberkieferschlinge .......................................................... 37
8.3. Entnahme einer Nasentupferprobe ............................................................. 37
9. Statistische Auswertung .......................................................................... 38
IV. ERGEBNISSE .......................................................................................... 40
1. Cortisol ...................................................................................................... 40
1.1. Serumcortisol ............................................................................................. 40
1.2. Speichelcortisol .......................................................................................... 44
2. Katecholamine .......................................................................................... 47
2.1. Noradrenalin............................................................................................... 47
2.2. Adrenalin .................................................................................................... 50
V. DISKUSSION ........................................................................................... 53
1. Cortisol ...................................................................................................... 54
1.1. Kontrollgruppe ........................................................................................... 55
1.2. Fixierung in der Oberkieferschlinge .......................................................... 56
1.3. Nasentupferentnahme ................................................................................. 57
1.4. Tracheobronchialabstrichentnahme ........................................................... 58
1.5. Speichelcortisol .......................................................................................... 60
1.6. AUC ........................................................................................................... 61
2. Katecholamine .......................................................................................... 62
2.1. Kontrollgruppe ........................................................................................... 62
2.2. Fixierung in der Oberkieferschlinge .......................................................... 62
2.3. Nasentupfer- und Tracheobronchialabstrichentnahme .............................. 63
VI. SCHLUSSFOLGERUNG ........................................................................ 67
VII. ZUSAMMENFASSUNG ......................................................................... 69

Inhaltsverzeichnis 9
VIII. SUMMARY .............................................................................................. 71
IX. LITERATURVERZEICHNIS ................................................................ 73
X. ABBILDUNGSVERZEICHNIS ............................................................. 89
XI. TABELLENVERZEICHNIS .................................................................. 90
XII. DANKSAGUNG ....................................................................................... 92

10 Abkürzungsverzeichnis
ABKÜRZUNGSVERZEICHNIS
ACTH Adrenocorticotropes Hormon
Abb. Abbildung
App. Actinobacillus
pleuropneumoniae
AUC area-under-the-curve
ca. circa
CBG Cortisol-bindendes Globulin
cm Zentimeter
CRH Corticotropin-Releasing-
Hormon
h Stunde(n)
HHN Hypothalamus-Hypophysen-
Nebennieren
HWZ Halbwertszeit
l Liter
MESOR midline estimating statistic
of rhythm
M. hyo M. hyopneumoniae
min Minute(n)
ml Milliliter
nmol Nanomol
NNM Nebennierenmark
NNR Nebennierenrinde
pCBG porcines Corticosteroid-
bindendes Globulin
PCR Polymerase-Kettenreaktion
PCV2 porcines Circovirus Typ 2
pg Pikogramm
PGE2 Prostaglandin E2
RL Richtlinie
PRRSV Porcine reproductive and
respiratory syndrome virus
SD Standardabweichung
(standard deviation)
sec Sekunde(n)
SIV Influenza-A-Virus
SEM Standardfehler (standard
error of the mean)
TÄBEO Berufsordnung für die
Tierärzte
TIERSCHG Tierschutzgesetz
U/min Umdrehungen pro Minute

I. Einleitung 11
I. EINLEITUNG
Atemwegsinfektionen sind weltweit in Schweinebeständen verbreitet, führen auf
Grund von Gewebeschädigungen zu Schmerzen bei den betroffenen Tieren und
verursachen hohe wirtschaftliche Verluste (HENKE und ERHARDT, 2001; VAN
ALSTINE, 2012). Die Verbesserung des Tierwohls sowie die Reduzierung von
Antibiotika in der Nutztierhaltung liegen derzeit im Fokus des tierärztlichen und
gesellschaftlichen Interesses. Ein besonderes Augenmerk gilt daher der
Krankheitsprophylaxe durch Impfungen. Dafür bedarf es einer ätiologischen
Diagnose, die es ermöglicht Atemwegsinfektionen gezielt vorzubeugen (GROSSE
BEILAGE et al., 2013). Als Diagnostikmethode bei Atemwegserkrankungen des
Schweines wird in Deutschland seit circa zehn Jahren die Bronchoalveoläre
Lavage (BAL) durchgeführt. Zur Vorbereitung auf die BAL müssen die Schweine
narkotisiert werden (FLASSHOFF, 1996), um eine sichere Fixierung zu
gewährleisten und um dem Schwein die Erstickungsangst zu ersparen, die durch
Einbringen von Flüssigkeit in die Lunge hervorgerufen wird (PAUSENBERGER
et al., 2012). Allerdings birgt die Narkose von Tieren, die unter
Atemwegserkrankungen leiden, ein erhöhtes Narkoserisiko (LENDL und
HENKE, 2011). Des Weiteren müssen die Tiere bis zur Wiedererlangung des
Bewusstseins separat aufgestallt werden. Das Narkoserisiko und der höhere Zeit-
und Kostenaufwand begrenzen folglich die Anzahl der beprobten Tiere
(PAUSENBERGER et al., 2012). Von FABLET et al. (2010) wurde erstmalig ein
Tracheobronchialabstrich, der ohne Narkose durchgeführt wird, zur Gewinnung
von Sekret aus den tiefen Atemwegen beschrieben. Da es keiner Narkose bedarf,
bietet sich diese Methode dafür an eine größere Anzahl von Proben zu sammeln
(PAUSENBERGER et al., 2012). Laut PAUSENBERGER et al. (2012) sind
Tracheobronchialabstrichproben ebenso geeignet wie das Material der BAL um
eine Aussage über die Besiedlung der Lunge mit pathogenen Erregern hinsichtlich
Kolonisationszeitpunkten und -prävalenzen zu ermöglichen. Darüber hinaus sind
Nasentupferproben zum Nachweis von Erregern der oberen Atemwege etabliert
(GROSSE BEILAGE et al., 2013).
In der Verantwortung als Tierarzt und der zunehmenden Bedeutung des
Tierschutzgedankens in der Gesellschaft ist es notwendig Eingriffe am Tier neben
ihrer diagnostischen Eignung auch auf Tiergerechtheit zu überprüfen.

12 I. Einleitung
Ziel dieser Arbeit war die Untersuchung der Stress- und Schmerzbelastung von
Schweinen auf Grundlage der Arbeitshypothese, dass ein
Tracheobronchialabstrich, der ohne Narkose durchgeführt wird, mit einer
größeren Belastung für die Schweine einhergeht als die Entnahme eines
Nasentupfers oder die Fixierung in der Oberkieferschlinge. Als Parameter dienten
Cortisolkonzentrationen aus Serum und Speichel sowie aus dem Blutplasma
ermittelte Noradrenalin- und Adrenalinkonzentrationen.

II. Literaturübersicht 13
II. LITERATURÜBERSICHT
1. Diagnostische Mittel
1.1. Gesetzliche Grundlagen
Die Notwendigkeit und die Anforderungen an eine tierärztliche Diagnose werden
zum einen in der Berufsordnung der Bayerischen Landestierärztekammer und zum
anderen im deutschen Tierschutzgesetz geregelt (TÄBEO, 2011; TIERSCHG,
2013). Die Aufgaben des Tierarztes erstrecken sich über die Bewahrung der
Gesundheit der Tierbestände bis zur Gesunderhaltung des Menschen (TÄBEO,
2011). In § 1(1) steht „Der Tierarzt/die Tierärztin (im folgenden "Tierarzt"
genannt) ist berufen, Leiden und Krankheiten der Tiere zu verhüten, zu lindern
und zu heilen, zur Erhaltung und Entwicklung eines leistungsfähigen
Tierbestandes beizutragen […]“ (TÄBEO, 2011). Auch das deutsche
Tierschutzgesetz beschreibt im Grundsatz, dass es in der Verantwortung des
Menschen für das Tier als Mitgeschöpf liegt, dessen Leben und Wohlbefinden zu
schützen und, dass niemand einem Tier ohne vernünftigen Grund Schmerzen,
Leiden oder Schäden zufügen darf (TIERSCHG, 2013). Der vierte Abschnitt
dieses Gesetzes befasst sich mit Eingriffen an Tieren. Grundsätzlich gilt laut §5
(1), dass an einem Wirbeltier ohne Betäubung ein mit Schmerzen verbundener
Eingriff nicht vorgenommen werden darf (TIERSCHG, 2013). Im Folgenden,
zweiten Abschnitt, werden auch Ausnahmen von dieser Regel gewährt, unter
anderem sofern „bei vergleichbaren Eingriffen am Menschen eine Betäubung in
der Regel unterbleibt oder der mit dem Eingriff verbundene Schmerz
geringfügiger ist als die mit einer Betäubung verbundene Beeinträchtigung des
Befindens des Tieres“ (TIERSCHG, 2013).
1.2. Diagnostik von Atemwegserkrankungen beim Schwein
Große Bedeutung für den Erfolg von diagnostischen Untersuchungen hat die
Auswahl geeigneter Tiere, idealerweise werden akut kranke, unbehandelte Tiere
mit typischer Symptomatik ausgewählt (GROSSE BEILAGE et al., 2013;
HÖLTIG und HENNIG-PAUKA, 2013). Ungünstig auf die Interpretation der
Befunde wirkt sich die Untersuchung chronisch kranker Tiere aus, die zwar
Veränderungen aufweisen, welche aber nicht unmittelbar mit der

14 II. Literaturübersicht
Primärerkrankung in Zusammenhang stehen (GROSSE BEILAGE et al., 2013;
HÖLTIG und HENNIG-PAUKA, 2013). Jeglicher weiterführenden Untersuchung
sollte eine eingehende Anamnese und klinische Allgemeinuntersuchung sowie die
spezielle Untersuchung des Atmungstraktes voraus gehen (GROSSE BEILAGE et
al., 2013).
1.2.1. Bronchoalveoläre Lavage
Seit circa zehn Jahren gehört die Bronchoalveoläre Lavage (BAL) in Deutschland
zu den Diagnostikmethoden bei Atemwegserkrankungen des Schweines
(FLASSHOFF, 1996; NIENHOFF et al., 2006; PALZER, 2006; MOORKAMP et
al., 2008). In der Literatur werden bezüglich der Durchführung verschiedene
Methoden beschrieben (GROSSE BEILAGE et al., 2013). HENNIG-PAUKA et
al. (2008) empfahlen die bronchoskopische Lavage für wissenschaftliche
Untersuchungen, während sich die transtracheale Lavage für den Einsatz in der
Praxis zur bakteriologischen und zytologischen Befunderhebung eignet. Mittels
bronchoskopisch gewonnenener Proben lässt sich die höchste diagnostische
Sensitivität für Genomfragmente von M. hyopneumoniae erzielen (HENNIG-
PAUKA et al., 2008). Die für die bronchoskopischen Methoden benötigten
technischen Hilfsmittel sind nur nach aufwendiger Reinigung und Desinfektion in
unterschiedlichen Schweinebetrieben einsetzbar (GROSSE BEILAGE et al.,
2013). In der Praxis haben sich laut GROSSE BEILAGE et al. (2013) Methoden
ohne Sichtkontrolle mit transoralem, transnasalem oder transtrachealem Zugang
etabliert. Für den transnasalen Zugang wird das Schwein in der
Oberkieferschlinge fixiert und nicht narkotisiert (GROSSE BEILAGE et al.,
2013). Das Einbringen von Flüssigkeit in die Lunge ist aber laut
PAUSENBERGER et al. (2012) aus Tierschutzgründen kritisch zu hinterfragen,
da die Tiere unter Erstickungsangst leiden können. Die Lungenspülung unter
transoralem und transtrachealem Zugang erfordert eine Allgemeinanästhesie
mittels Azaperon und Ketamin (NIENHOFF et al., 2006; GROSSE BEILAGE et
al., 2013). ERHARDT et al. (2011) warnten, dass die Narkose von Tieren mit
Atemwegserkrankungen zu einem erhöhten Narkoserisiko führt, da durch
Einführen des Spülkatheters und durch die zusätzlich eingebrachte Flüssigkeit
eine weitere Verkleinerung der Luftwege mit Zunahme des Atemwiderstandes
und Hypoventilation erfolgt (LENDL und HENKE, 2011). Um narkosebedingte
Todesfälle zu vermeiden, sind schwer erkrankte Schweine von der Lungenspülung

II. Literaturübersicht 15
auszuschließen (GROSSE BEILAGE et al., 2013). Gespült wird mit steriler
physiologischer Kochsalzlösung (FLASSHOFF, 1996). Bei der transtrachealen
Methode ist die Gefahr der Kontamination mit Umgebungskeimen, die nicht aus
der Lunge stammen, am geringsten (HENNIG-PAUKA et al., 2008). Für die
Interpretation der Ergebnisse ist es sinnvoll mehrere Proben bakteriologisch-
kulturell oder molekularbiologisch untersuchen zu lassen (HÖLTIG und
HENNIG-PAUKA, 2013). Außerdem empfiehlt sich eine zusätzliche zytologische
Untersuchung hinsichtlich Entzündungszeichen, da viele bakterielle Erreger auch
in gesunden Schweinen zu finden sind (GROSSE BEILAGE et al., 2013).
1.2.2. Tracheobronchialabstrich
FABLET et al. beschrieben erstmalig im Jahr 2010 die Entnahme von
Tracheobronchialtupfern zur Gewinnung von Sekret aus den tiefen Atemwegen.
Dazu werden die Schweine mit einer Oberkieferschlinge unmittelbar hinter den
Eckzähnen fixiert und ein Maulkeil zum Offenhalten der Maulspalte eingelegt
(FABLET et al., 2010). Im Anschluss wird ein steriler Katheter, wie er zur
trachealen Intubation verwendet wird, bei Einatmung des Tieres in die Luftröhre
vorgeschoben und unter Drehen auf und ab bewegt (FABLET et al., 2010). Zum
Versand ins Labor wird die Probe in 2 ml gepufferter Peptonwasserbouillon
transportiert (FABLET et al., 2010). MAROIS et al. (2007) beschrieben den
Tracheatupfer und die BAL als effektivere Methoden zur Detektion von
M. hyopneumoniae im Vergleich zu Nasen- oder Tonsillentupfern. Zu ähnlichem
Ergebnis kamen FABLET et al. (2010). Auch in ihrer Studie erwiesen sich BAL
und Tracheobronchialtupfer gleichermaßen als sensitivste Methoden zur Diagnose
von M. hyopneumoniae mittels nested-PCR (FABLET et al., 2010).
PAUSENBERGER et al. (2012) führten den Begriff Tracheobronchialabstrich
ein, weil dieser der Methode in der deutschen Sprache am nächsten kommt.
PAUSENBERGER et al. (2013) wiesen aus Tracheobronchialabstrichen mittels
PCR Genomfragmente von M. hyopneumoniae (M. hyo), Influenza-A-Virus
(SIV), Porcines Circovirus Typ 2 (PCV2), Porcines Respiratorisches und
Reproduktives Syndrom Virus (PRRSV), Haemophilus parasuis spp. und
Actinobacillus pleuropneumoniae (App) nach und führten auch eine erfolgreiche
bakteriologische Untersuchungen auf α-hämolysierende Streptococcus spp.,
Pasteurella spp., Bordetella spp. und Keime mit Verdacht auf Haemophilus spp.
durch. Dafür wurde die Spitze des Tracheobronchialabstrichkatheters nach der

16 II. Literaturübersicht
Entnahme abgeschnitten und in ein Röhrchen mit 2 ml phosphatgepufferter
Kochsalzlösung überführt. Davon wurde 1 ml abpipettiert und ins Labor versandt
(PAUSENBERGER et al., 2013). Unter Berücksichtigung einer ausreichend
großen Stichprobenzahl ließ sich nach PAUSENBERGER et al. (2013) eine
Einschätzung zu Kolonisationsdruck, -prävalenz und -zeitpunkt geben. Vorteilhaft
wirkt sich laut HÖLTIG und HENNIG-PAUKA (2013) auch aus, dass das Sekret
von den Tracheobronchialabstrichen im Gegensatz zur BAL unverdünnt
gewonnen wird. Nach PAUSENBERGER et al. (2012) ist die Gewinnung von
Tracheobronchialabstrichen einfach durchzuführen und man benötigt zusätzlich
zur Ausrüstung nur einen Maulkeil (FABLET et al., 2010).
1.2.3. Nasentupfer
Ferner dienen Nasentupfer zur Diagnose von Atemwegserkrankungen deren
Erreger sich direkt auf oder in der Nasenschleimhaut vermehren (GROSSE
BEILAGE et al., 2013). Zur Probengewinnung werden sie in die Nasenhöhle
eingeführt und mehrmals über der Schleimhaut vor- und zurückgedreht um
möglichst viel Zellgewebe aufzunehmen (BUER, 2012). Um Irritationen und
Verletzungen zu vermeiden empfahlen GROSSE BEILAGE et al. (2013) Ferkel
bis zu einem Gewicht von 15 kg auf dem Arm zu halten und zusätzlich noch
durch eine Hand des Probennehmers zu immobilisieren. Tiere ab 15 kg werden
mittels Oberkieferschlinge fixiert (GROSSE BEILAGE et al., 2013). Nasentupfer
sind geeignet zur Untersuchung auf toxinbildende Pasteurella-multocida-Stämme
(SCHOSS und ALT, 1995; HÖLTIG und HENNIG-PAUKA, 2013) sowie zum
Nachweis von Influenza (GOODELL et al., 2013; GROSSE BEILAGE et al.,
2013). Besteht die Infektion mit Influenza-A-Virus bereits länger als zwei Tage,
ist ein Nachweis aus Nasentupfern nicht mehr möglich, daher sollten akut kranke
Tiere in der Fieberphase zur Untersuchung ausgewählt werden (VAN REETH et
al., 2012; GROSSE BEILAGE et al., 2013; HÖLTIG und HENNIG-PAUKA,
2013), die Symptome wie Niesen, Nasenausfluss oder Bindehautentzündung
zeigen (HÖLTIG und HENNIG-PAUKA, 2013). Da sich an einem Nasentupfer
nur sehr wenig Material anheftet, ist nach NATHUES und GROSSE BEILAGE
(2009) ein Nachweisverfahren mit hoher Sensitivität wie nested PCR oder Real-
time-PCR zu wählen. Im Gegensatz dazu sind Nasentupfer laut GROSSE
BEILAGE et al. (2013) für den Nachweis von Infektionen der tiefen Atemwege
nicht geeignet. Im Vergleich zum Nasentupfer erwies sich der

II. Literaturübersicht 17
Tracheobronchialabstrich in einer Studie von FABLET et al. (2010) dreieinhalb
mal sensitiver für den Nachweis von M. hyopneumoniae, da M. hyopneumoniae
sich an das Flimmerepithel anheftet und sich im unteren Respirationstrakt
vermehrt (BLANCHARD et al., 1992; KURTH et al., 2002; MAROIS et al.,
2007; FABLET et al., 2010). Die Aussagekraft der Nasentupferproben
hinsichtlich der festgestellten Erreger ist, je nach Fragestellung, sehr
unterschiedlich, da eine Vielzahl von Erregern auch bei gesunden Tieren Nasen-
und Rachenraum besiedelt (GROSSE BEILAGE et al., 2013; HÖLTIG und
HENNIG-PAUKA, 2013).
2. Stress und Wohlbefinden
VEISSIER und BOISSY (2007) schlussfolgerten, dass Wohlbefinden und Stress
gegensätzliche Konzepte bezüglich des mentalen Zustandes eines Individuums
darstellen, da sich Wohlbefinden und Stress gegenseitig ausschließen. Das
„Stress-System“ reagiert mit einer Stress-Antwort, wenn die Homeostase, ein
komplexes dynamisches Gleichgewicht des Lebewesens, welches internen und
externen Einflüssen, den sogenannten Stressoren unterliegt, gestört wird
(CHROUSOS et al., 1995; CHROUSOS, 2009). Die Stress-Antwort ist
gekennzeichnet durch ein Zusammenspiel des Endokrinen-, des Nerven- und des
Immunsystems (SMITH und VALE, 2006), schließt die Aktivierung der HHN-
Achse sowie des sympathischen Nervensystems ein und hilft dem Tier auf
Veränderung zu reagieren (VEISSIER und BOISSY, 2007). Um das
Wohlbefinden von Nutztieren zu untersuchen wurden viele Studien zur
Basalfunktion der HHN-Achse und den Reaktionen auf Umweltreize durchgeführt
(MORMEDE et al., 2007). Unter anderem führt Frustration zu einer Erhöhung des
Plasmacortisolspiegels und zu Verhaltensänderungen (DANTZER et al., 1980). In
einer Untersuchung von DANTZER et al. (1980) wurden Schweine trainiert
Futter als Belohnung auf eine bestimmte Handlung zu erlangen. Anschließend
wurde Frustration erzeugt indem die Belohnung vorenthalten wurde. Diese
Schweine reagierten mit einem erhöhten Cortisolspiegel und aggressivem
Verhalten gegen ein anderes, begleitendes Tier (DANTZER et al., 1980).
MORMEDE et al. (2007) diskutierten die Grenzen der Messung der Aktivität der
HHN-Achse. Da im Gegensatz zu akuten Stressreizen, die nicht zwingend
schädigend sein müssen, bei moderaten, länger andauernden Belastungen
Cortisolkonzentrationen weniger aussagekräftig sind, ist ein mehrdimensionaler

18 II. Literaturübersicht
Ansatz zur Evaluierung von Wohlbefinden nötig, der Verhaltensbeobachtung,
Produktionsdaten und Krankheitsinzidenzen berücksichtigt (MORMEDE et al.,
2007). Stress steht auch in direktem Zusammenhang mit Angst und Schmerz und
unterliegt gegenseitiger Beeinflussung (HENKE und ERHARDT, 2001). Durch
die Reduktion von Stress und Angst kann die Schmerzschwelle angehoben
werden (HENKE und ERHARDT, 2001).
3. Schmerz
„Schmerz ist ein aversives Gefühl oder eine Sinneswahrnehmung, die mit
tatsächlicher oder potentieller Gewebeschädigung einhergeht“ (BROOM, 2001).
Ähnlich wird Schmerz auch von der International Society for the Study of Pain
(1979) beschrieben, die statt aversiv das Adjektiv „unangenehm“ verwendet. Vom
Schmerz betroffene Individuen können von diesen Erfahrungen lernen, um diese
negativen Situationen in Zukunft zu meiden (BROOM, 2001). MELLOR et al.
(2000) beschrieben, dass die Möglichkeiten der Schmerzwahrnehmung bei allen
Spezies gleich sensitiv sind, sich die Schmerztoleranz hingegen unterscheidet. Sie
differenzierten zwischen den Begriffen „stress“ der mit unterschiedlich großen
Veränderungen physiologischer Zustände einhergeht und „distress“ der den
emotionalen Anteil schmerzhafter Ereignisse sowohl emotionaler als auch
physischer Natur beschreibt (MELLOR et al., 2000). Zusätzlich prägten
MELLOR et al. (2000) den Ausdruck „pain induced distress“ der verdeutlicht,
dass emotionale und physikalische Komponenten untrennbar zusammenwirken
und sich in der Antwort des Organismus auf die schädigende Noxe widerspiegeln
(MELLOR et al., 2000). Schmerz löst viele Veränderungen physiologischer
Parameter und des Verhaltens aus (MELLOR et al., 2000). MELLOR und
STAFFORD (2004) stellten einen Leitfaden zur Beurteilung von
schmerzbedingtem Stress mit folgenden fünf Kriterien zusammen: die zur
Schmerzbeurteilung herangezogenen Parameter müssen bei einem nicht
schmerzhaften Tier absent sein; es muss sichergestellt werden, dass der zu
verwendende Parameter das Maß der zu untersuchenden, einzelnen schmerzhaften
Aktion wiedergibt und nicht nur die Gesamtheit des Schmerzes; die Sicherheit
von Verhaltensparametern soll mit Hilfe physiologischer Parameter überprüft
werden; eine Übertragbarkeit auf andere Tierarten und eine Beeinflussung der
Ergebnisse durch andere Faktoren wie z.B. Geschlecht, Rasse und Alter muss
sorgfältig geprüft werden (MELLOR und STAFFORD, 2004).

II. Literaturübersicht 19
3.1. Schmerzentstehung, Weiterleitung und Wahrnehmung
Die Nozizeption beschreibt die Reizaufnahme durch Nozisensoren, die nervale
Weiterleitung und zentrale Verarbeitung, wohingegen Schmerz laut
SILBERNAGEL und DESPOPOULUS (2012a) einer anschließenden subjektiven
Empfindung entspricht. Schmerzauslösende Reize werden in der Peripherie durch
Nozizeptoren von Kopf und Körper aufgenommen, deren Zellkörper in den
Spinalganglien der Dorsalwurzel des Rückenmarks und den Trigeminalganglien
liegen (JULIUS und BASBAUM, 2001; PFANNKUCHE, 2008; SANN, 2010).
Neben den viszeralen Organen sind Nozizeptoren in fast allen somatischen
Körperbereichen außer dem ZNS vertreten (PFANNKUCHE, 2008).
Nozizeptoren können polymodal durch unterschiedliche Reize (z.B. thermische,
mechanische und chemische) oder seltener unimodal durch einen bestimmten
Reiz erregt werden (SCHOLZ und WOOLF, 2002; BROOKS und TRACEY,
2005). Sogenannte schlafende Nozizeptoren verstärken den Schmerz und werden
nur durch pathologische Zustände erregt (SANN, 2010). Entzündungen,
Gewebezerstörungen und Tumorzellen werden von SCHOLZ und WOOLF
(2002) zur „inflammatory soup“ zusammengefasst, welche die Nozizeptoren
sensibilisiert. Die Nozizeptoren können auf Grund ihrer anatomischen und
funktionellen Unterschiede in Gruppen eingeteilt werden (HENKE und
ERHARDT, 2001; JULIUS und BASBAUM, 2001). Grundsätzlich gilt, je dicker
eine Nervenfaser, desto höher ist deren Leitungsgeschwindigkeit (JULIUS und
BASBAUM, 2001). Aus der Peripherie vermitteln die Nozizeptoren Signale über
die Spinalganglien zum Rückenmark, wo sie über den tractus spinothalamicus
zum Thalamus weitergeleitet werden (LÖSCHER und FREY, 2010) ebenso wie
an sympathische Efferenzen und Motoneurone, die eine reflektorische Antwort
auslösen können (HENKE und ERHARDT, 2001; PFANNKUCHE, 2008). Vom
Thalamus werden Signale zum somatosensorischen Kortex, wo der
Schmerzentstehungsort lokalisiert wird aber auch zum Hirnstamm weitergeleitet,
wodurch Schmerzen Atmung und Kreislauf beeinflussen können (HENKE und
ERHARDT, 2001; LÖSCHER und FREY, 2010; SILBERNAGEL und
DESPOPOULUS, 2012a). Die somatosensorische Hirnrinde ist laut LÖSCHER
und FREY (2010) in Projektionsfelder für unterschiedliche Körperregionen
unterteilt. Die Größe dieser Felder richtet sich nicht nach der abgebildeten
Körperoberfläche sondern nach der Rezeptorendichte des jeweiligen Körperareals
(LÖSCHER und FREY, 2010). Deshalb ist laut LÖSCHER und FREY (2010)

20 II. Literaturübersicht
z.B. das Gesicht, das wichtig für den Taststinn ist, entsprechend größer
dimensioniert. Außerdem besteht eine Verbindung zwischen Thalamus und
Hypophyse, die unter Schmerzeinwirkung endokrin die Freisetzung von ACTH
und β-Endorphin aus dem Hypophysenvorderlappen steuert (HENKE und
ERHARDT, 2001). Die Nozizeption wird über deszendierende Bahnen im
Thalamus und Rückenmark gehemmt (SILBERNAGEL und DESPOPOULUS,
2012a). Laut PFANNKUCHE (2008) dient dies möglicherweise zur
Schmerzunterdrückung in körperlichen Extremsituationen wie Flucht, Kampf oder
Geburt. Die wichtigste Rolle als Transmitter in der körpereigenen
Schmerzunterdrückung spielen endogene Opioide (Enkephaline, Dynorphine,
Endorphine) (PFANNKUCHE, 2008). PHOGAT und PARVIZI (2007)
beschrieben, dass endogene Opioide sowohl physischen als auch psychischen
Stress vermindern.
3.2. Innervation der untersuchten Regionen
Organe und Gewebe reagieren unterschiedlich sensibel auf Schmerzreize
(HENKE und ERHARDT, 2001). Als hoch sensibel gelten für HENKE und
ERHARDT (2001) Zahnpulpa, Cornea, Perianalbereich und die Serosen. Des
Weiteren kann auch die Haut, welche über mechanische, thermische und
chemische Rezeptoren verfügt, intensive Schmerzsignale senden (HENKE und
ERHARDT, 2001). Im Gegensatz dazu sind parenchymatöse Organe weniger
sensibel, jedoch kann eine starke Dehnung zu Schmerzhaftigkeit der serösen
Häute führen (HENKE und ERHARDT, 2001). Eine geringe Schmerzhaftigkeit
weist die Muskulatur auf, die vor allem mit Mechanorezeptoren ausgestattet ist
(HENKE und ERHARDT, 2001). Auch Gelenke und Knochen werden von
HENKE und ERHARDT (2001) als relativ unsensibel beschrieben. Als
schmerzunempfindlich gilt das Gehirn selbst, wohingegen die Meningen
besonders sensibel sind (HENKE und ERHARDT, 2001). Durch ausstrahlenden
Schmerz empfindlicher Gewebe ergibt sich z.B. eine hohe Schmerzhaftigkeit für
den Kopf (HENKE und ERHARDT, 2001). Jedes Körperteil wird auf einem
bestimmten Feld des somatosensorischen Kortex somatotopisch repräsentiert
(SILBERNAGEL und DESPOPOULUS, 2012a). Dies bedeutet, dass benachbarte
Körperregionen auch entsprechend auf den Arealen der Großhirnrinde abgebildet
werden (TREEDE, 2007). Allerdings ist bisher wenig über die somatotopische
Gliederung beim Schwein erforscht (CRANER und RAY, 1991). CRANER und

II. Literaturübersicht 21
RAY (1991) gehen von einer ähnlichen Gliederung der somatosensorischen
Gehirnregion beim Schwein wie der bei Menschen aus. Auffällig ist die
Überrepräsentation des Rostrums und die mehrfachen Projektionsfelder für Maul,
Gesicht und auch Rostrum (CRANER und RAY, 1991). GASTL et al. (2014)
konnten beim Menschen eine eigene, bilaterale Projektion für die nasale Mukosa
darstellen. Der N. trigeminus versorgt über den N. ethmoidalis sensibel die
Schleimhaut des dorsalen Teils der Nasenhöhle und mit dem N. nasalis caudalis
(sensibel) die ventralen Anteile der Nasenhöhle und des Gaumens sowie mit dem
N. infraorbitalis, der vom N. maxillaris abzweigt und ebenfalls vom N. trigeminus
stammt, sensibel die Haut des Nasenrückens und der Nasenseitenwand, den
Nasenhöhleneingang und Nasenvorhof sowie die Oberlippe (BUDRAS und
BUDA, 2002; NICKEL et al., 2004; KÖNIG et al., 2012). Die menschliche
Rachenwand ist laut DE CARLOS et al. (2013) mit verschiedenen Rezeptoren
(ähnlich den Ruffini- und Pacini-Tastkörperchen) ausgestattet, die mechanische
Reize der oberen Atemwege aufnehmen. Diese Rezeptoren werden von
sensorischen Nerven begleitet (DE CARLOS et al., 2013). Der
N. glossopharyngeus (IX der Vagusgruppe) versorgt laut NICKEL et al. (2004)
sensibel das kaudale Drittel der Zunge, die Rachenschleimhaut sowie die
Tonsillen. Die Schleimhaut und Drüsen des Gaumens werden von Ästen des
N. trigeminus sensibel und parasympathisch innerviert (NICKEL et al., 2004)
ebenso wie die Kehlkopfschleimhaut (sensibel) (BUDRAS und BUDA, 2002).
Der N. vagus versorgt sensibel, motorisch und parasympathisch die Eingeweide
von Kopf, Hals, Brust und Bauchhöhle, wo er sich plexusartig aufteilt (BUDRAS
und BUDA, 2002; KÖNIG et al., 2012). Die Lunge wird vom Plexus pulmonalis
versorgt (KÖNIG und LIEBICH, 2012). Dieser stellt ein Nervengeflecht aus
sympathischen Fasern des Ggl. cervicothoracicum dar, das sich mit
parasympathischen Anteilen des N. vagus verbindet (KÖNIG und LIEBICH,
2012). SILBERNAGEL und DESPOPOULUS (2012b) zählten auch die
Irritationsendigungen in der Bronchialschleimhaut zu den Afferenzen der Lunge.
Sie sind sowohl beteilig an einer verstärkten Atmung, aktiviert durch verringertes
Lungenvolumen als auch am Hustenreflex, der durch Staub oder reizende Gase
ausgelöst wird (SILBERNAGEL und DESPOPOULUS, 2012b). Freie, afferente
C-Faserendigungen, die in der Alveolar- und Bronchialwand liegen, sind beim
wachen Tier am Husten beteiligt (CANNING et al., 2004; SILBERNAGEL und
DESPOPOULUS, 2012b). Diese sind laut CANNING et al. (2004) mehr an der

22 II. Literaturübersicht
Signalisierung der Hustennotwendigkeit, als an der Aufrechterhaltung
unkontrollierten Hustens beteiligt.
4. Untersuchte Parameter
Zur Einschätzung von Emotionen und Gefühlen von Tieren ist man auf die
Erhebung messbarer Größen angewiesen, von denen man annimmt, dass sie sich
mit dem Zustand eines Tieres verändern (MORMEDE et al., 2007). Grundsätzlich
sollte die Aktivität eines Individuums beurteilt werden, die unter dem Einfluss
von Schmerzen sowohl verringert, als auch vermehrt sein kann (HENKE und
ERHARDT, 2001). Weitere Informationen bietet auch das äußere
Erscheinungsbild (HENKE und ERHARDT, 2001). Für die Einschätzung des
Temperamentes setzen HENKE und ERHARDT (2001) voraus, dass das normale
Verhalten des Tieres bekannt ist. Akute Schmerzen erzeugen oftmals
Lautäußerungen, wobei unter chronischem Schmerz vorwiegend bei größeren
Tieren (Schwein, Hund, Schaf) Stöhnen oder Zähneknirschen beobachtet werden
kann (HENKE und ERHARDT, 2001). VIÑUELA-FERNÁNDEZ et al. (2007)
gaben für die Einschätzung von Schmerz bei Beutetierspezies zu bedenken, dass
deren Schmerzverhalten vom notwendigen Fluchtverhalten überdeckt wird. Eine
verminderte Futter- und Wasseraufnahme und in der Folge auch reduzierter Harn-
und Kotabsatz stellen laut HENKE und ERHARDT (2001) relativ objektive
Kriterien dar. Über eine Aktivierung des sympathischen Nervensystems und der
Hypophysen-Nebennierenachse kommt es in Folge von Schmerz- und
Belastungssituationen zu einer Katecholamin- und Cortisolausschüttung (VON
BORELL, 2013). In vielen Studien zur Schmerzbeurteilung wurden deshalb durch
Schmerz beeinflusste physiologische Mess-Parameter untersucht (PEERS et al.,
2002; ZÖLS et al., 2006; SCHULZ et al., 2007; ÜBEL, 2011; ZIMMERMANN et
al., 2011). Nach einem akuten, kurzen Schmerzreiz dauert eine vermehrte
Adrenalinausschüttung am kürzesten an (2-5 min), gefolgt von Noradrenalin (45-
60 min) und am längsten wird ACTH und Cortisol in Abhängigkeit von der
Schädigung ausgeschüttet (von 2,5 h bis zu 8 h) (MELLOR et al., 2000;
MELLOR und STAFFORD, 2000; MELLOR et al., 2002; PEERS et al., 2002).
Im Gegensatz zu Cortisol steigen Herzfrequenz und arterieller Blutdruck früher an
(MELLOR et al., 2000; MELLOR und STAFFORD, 2000; MELLOR et al., 2002;
PEERS et al., 2002). ESCRIBANO et al. (2013) beschrieben einen Anstieg von
Chromogranin A im Schweinespeichel nach einer akuten Stresssituation.

II. Literaturübersicht 23
4.1. Cortisol
Glucocorticoide spielen eine wichtige Rolle im Intermediärstoffwechsel (GRECO
und STABENFELDT, 2013). Ferner kann die Glucocorticoidkonzentration als
Antwort auf Stress laut GRECO und STABENFELDT (2013) innerhalb von
Minuten ein mehrfaches ihrer ursprünglichen Konzentration erreichen. Dabei ist
die Höhe des Anstieges proportional zum auslösenden Stress (GRECO und
STABENFELDT, 2013). In einer Studie über die Fixierung von Schweinen in der
Oberkieferschlinge beobachteten ROOZEN et al. (1995) bereits dreieinhalb
Minuten nach Beginn dieser Maßnahme einen Anstieg. Die Aktivierung der
HHN-Achse trägt mit einer verlängerten metabolischen und
antiinflammatorischen Antwort zur Erholung und Heilung des Körpers bei
(MELLOR et al., 2000). Die Aussagekraft von Cortisol zur Evaluierung von
Stress bei Tieren wurde durch viele Studien bestätigt (ROOZEN et al., 1995;
ROSOCHACKI et al., 2000; MELLOR et al., 2002; ZÖLS et al., 2006; ÜBEL,
2011).
4.1.1. Wirkung im Organismus
Die Nebenniere besteht aus zwei Anteilen, dem Nebennierenmark (NNM) und der
umgebenden Nebennierenrinde (NNR) (MÖSTL, 2010). In den Markzellen
werden Adrenalin und Noradrenalin gespeichert, (Näheres dazu siehe
4.2. Katecholamine) in der Zona fasciculata der Nebennierenrinde werden
Glucocorticoide, vor allem Cortisol und Corticosteron, und in der Zona
glomerulosa Mineralocorticoide, hauptsächlich Aldosteron, aus Cholesterol
gebildet (MÖSTL, 2010). Außerdem gibt es eine dritte Schicht, die Zona
reticularis die Sexualsteroide sezerniert (GRECO und STABENFELDT, 2013).
Im Gegensatz zu Vögeln, Mäusen und Ratten, die hauptsächlich Corticosteron
synthetisieren, dominiert bei Schweinen und Wiederkäuern Cortisol (MÖSTL,
2010). Hauptaufgabe der Glucocorticoide ist es den Stoffwechsel zu kontrollieren
und die Gluconeogenese in der Leber zu steuern (GRECO und STABENFELDT,
2013).

24 II. Literaturübersicht
Tabelle 1: Zusammenfassung der wichtigsten Wirkungen von Cortisol nach
MÖSTL (2010) und SILBERNAGEL und DESPOPOULUS (2012d)
Wirkung Folgen
Proteinkatabolismus
Gluconeogenese aus
Aminosäuren
Verhinderung der
Aufnahme von Glucose in
Muskel- und Fettzellen
(GRECO und
STABENFELDT, 2013)
Blutglucosespiegel
steigt:
gesteigerte
Glycogenbildung und
reduzierte
Fettsäuresynthese in
der Leber
Antiinflamatorische
Wirkung
Stabilisierung der Lysosomen; Gehemmte
Lymphokinsynthese und Histaminfreisetzung
Herz und Kreislauf:
positiv inotrope und
vasokonstriktorische
Wirkung
Verstärkung katecholaminerger Effekte;
Cortisol induziert vermehrte Bildung von Adrenalin
im NNM und Angiotensinogen in der Leber
Darüber hinaus entfalten Glucocorticoide eine geringgradige mineralocorticoide
Wirkung und tragen dadurch in geringem Maße zur Aufrechterhaltung des
Flüssigkeits- und Mineralstoffhaushaltes bei (MÖSTL, 2010).
4.1.2. Transport und Ausscheidung
Größtenteils sind Glucocorticoide an Proteine gebunden, welche den Transport
der lipophilen Steroidhormone im Blutplasma ermöglichen und eine Speicherform
darstellen (MÖSTL, 2010; GRECO und STABENFELDT, 2013). 75% des
Cortisols sind an Transcortin (Cortisol-bindendes Globulin = CBG) und 15% an
Albumin gebunden (GRECO und STABENFELDT, 2013). Nur 10% sind laut
GRECO und STABENFELDT (2013) in ungebundener, freier Form im Plasma
gelöst. Nur die ungebundenen, freien Corticoide sind biologisch wirksam
(MÖSTL, 2010). Durch eine Konformationsänderung des CBG wird Cortisol
vermehrt in der Umgebung von Entzündungen freigesetzt (SILBERNAGEL und
DESPOPOULUS, 2012d). GRECO und STABENFELDT (2013) geben die
biologische Halbwertszeit von Cortisol mit 60 min an. In der Leber erfolgt die
Konjugation der Steroidhormone, die dadurch wasserlöslich werden und
größtenteils über den Urin und zum Teil über den Kot ausgeschieden werden

II. Literaturübersicht 25
(MÖSTL, 2010; GRECO und STABENFELDT, 2013). COOK (2012) fasste
unterschiedlichste Medien die sich minimalinvasiv gewinnen lassen und die
indirekte Untersuchung von Stress über Cortisol erlauben in seinem Artikel
zusammen. Im Gegensatz zu Säugetieren, bei denen sich Cortisol im Speichel,
Kot, Urin, Haaren und der Milch nachweisen lässt, ist bei Vögeln auch eine
Untersuchung deren Ausscheidungen, der Federn und Eier möglich (COOK,
2012). Er gibt aber auch zu bedenken, dass man bei der Interpretation der
Ergebnisse beachten muss, dass die jeweiligen Testmedien eine unterschiedliche
Zeitspanne wiedergeben, von wenigen Minuten für Speichelcortisol bis zu
mehreren Wochen für die Bildung von Federn (COOK, 2012).
4.1.3. Speichelcortisol
MERLOT et al. (2011), COOK et al. (1996) und PARROTT und MISSON (1989)
empfahlen Speichelcortisolmessungen zum Nachweis einer Aktivierung der
HHN-Achse durch einen Stressor. Dabei muss sichergestellt sein, dass ein
Vergleich zwischen Basalwerten und Werten nach der Belastung möglich ist
(MERLOT et al., 2011). Lipidlösliche, unkonjugierte Hormone wie Testosteron,
Progesteron und Cortisol gelangen über den intrazellulären Weg in den Speichel
(VINING et al., 1983). Die Konzentrationen dieser Hormone werden durch die
Speichelflussrate nicht beeinflusst und spiegeln den ungebundenen, freien Anteil
im Plasma wider (VINING et al., 1983). Freies Cortisol entsprach laut
SCHÖNREITER et al. (1999) zwischen 11,9 % - 16,4% vom Gesamtcortisol,
wobei im Speichel zwischen 5,9 % und 7,5 % des Gesamtcortisols nachgewiesen
wurden. Die mittlere Speichelcortisolkonzentration sank laut GALLAGHER et al.
(2002) von höheren perinatalen Konzentrationen auf unter 27,6 nmol/l bei
30 Tage alten Ferkeln ab.
4.1.4. Regulation
Die Glucocorticoidsynthese in der Nebennierenrinde wird durch das
Adrenocorticotrope Hormon (ACTH) des Hypophysenvorderlappens gesteuert
(GRECO und STABENFELDT, 2013). Die Ausschüttung des ACTH wird
wiederum von einer höheren Instanz, dem Hypothalamus, über die Höhe des
Corticotropin-Releasing-Hormones (CRH) in einem Feedbacksystem geregelt
(CHROUSOS und GOLD, 1992; GRECO und STABENFELDT, 2013).
Glucocorticoide vermindern am Hypothalamus über eine negative Rückkopplung

26 II. Literaturübersicht
eine weitere Ausschüttung von CRH (GRECO und STABENFELDT, 2013). Das
Feedbacksystem wird zum einen von einem circadianen Rhythmus überlagert und
zum anderen durch Stress beeinflusst (GRECO und STABENFELDT, 2013).
PHOGAT und PARVIZI (2007) zeigten, dass die Beziehung zwischen akutem
Stress und Cortisolantwort auch vom beta-adrenergen System und vom
Opioidsystem reguliert wird. Sie beschrieben, dass eine starke Stimulation des
Beta-Adrenozeptors zu einer vermehrten Freisetzung von Cortisol führt
(PHOGAT und PARVIZI, 2007).
4.1.5. Circadiane Rhythmik
Biologische Rhythmen ermöglichen dem Individuum sich seiner zeitlichen
Umgebung anzupassen (MOHR, 2010). Damit kann der Organismus laut MOHR
(2010) den Funktionszustand eines Organsystems bereits anpassen, bevor die
äußeren Umstände, die eine Veränderung nötig werden lassen, eintreten. Der
Begriff „feedforward coupling“ beschreibt die vorausschauende Anpassung des
Organismus auf das Kommende, die im Gegensatz zu einer Reaktion schon im
Voraus erfolgt (MOHR, 2010). Die Produktion von Glucocorticoiden unterliegt
einem tageszeitabhängigen Rhythmus (GRECO und STABENFELDT, 2013). Die
höchste Konzentration an Glucocorticoiden besteht bei tagaktiven Tieren zu
Sonnenaufgang, kurz vor dem Erwachen (BARNETT et al., 1981). EVANS et al.
(1988) beschrieben einen circadianen Rhythmus 28 Wochen alter Schweine, der
sich durch eine ausgeprägte Spitze am Morgen, eine schwächere Spitze am
Nachmittag und ein Tief am Abend auszeichnete. Für die maximale
Blutcortisolkonzentration gaben KLEMCKE et al. (1989) morgens 121,4 nmol/l
und für das Minimum abends 35,6 nmol/l an. EKKEL et al. (1996) gaben zu
bedenken, dass die Corticosteroidkonzentration zum einen natürlichen
Schwankungen unterliegt und zum anderen eine unterschiedliche Hormonantwort
auf einen identischen Stimulus, abhängig davon erfolgt, wann während des
circadianen Zyklus dieser Reiz erfolgte. Während GALLAGHER et al. (2002)
schon bei sechs bis zehn Tage alten Saugferkeln die Stabilisierung eines Zyklus
dokumentierten und auch EKKEL et al. (1996) bei acht Wochen alten Schweinen
einen ausgeprägten circadianen Rhythmus feststellten, vertraten andere Autoren
die Meinung, dass diese tageszeitabhängige Rhythmik bei drei bzw. neun oder
15 Wochen alten Tieren noch nicht entwickelt ist (RUIS et al., 1997; DE JONG et
al., 2000; SKARLANDTOVA et al., 2011). EKKEL et al. (1997) untersuchten

II. Literaturübersicht 27
den Einfluss von chronischem Stress auf die circadiane Rhythmik sechs und
41 Tage nach Neugruppierung von neun Wochen alten Schweine. Dabei konnten
sie keine Unterschiede bezüglich des MESOR (midline estimating statistic of
rhythm; durchschnittliches Cortisol Level, angepasst an die circadiane Rhythmik)
und der Amplitude des circadianen Rhythmus des Speichelcortisols zwischen
gemischten und ungemischten Gruppen feststellen (EKKEL et al., 1997). Für
beide Gruppen war aber ein Anstieg des Speichelcortisols sowohl bezüglich des
MESOR als auch für die Höhe der Amplitude zu verzeichnen, was
möglicherweise auf das höhere Alter zurückzuführen ist (EKKEL et al., 1997).
Auch EVANS et al. (1988) beschrieben, dass Schweine erst mit der Pubertät einen
erwachsenen circadianen Rhythmus erreichen. Ein deutlicheres Muster entwickelt
sich auch mit zunehmendem Gewicht der Schweine (HILLMANN et al., 2008).
Außerdem entwickelten Schweine, die in einer angereicherten Umwelt gehalten
wurden ab einem Alter von 15 Wochen eine ausgeprägtere circadiane Rhythmik
als solche in reizarmer Umgebung (DE JONG et al., 2000). Chronischer Stress
führt nach MUNSTERHJELM et al. (2010) unter Umständen zum Ausbleiben
eines circadianen Rhythmus.
Ähnliches gilt für die Speichelcortisolkonzentration, die ebenfalls einer
circadianen Rhythmik mit höchsten Konzentrationen am späten Vormittag und
frühen Nachmittag folgt (EKKEL et al., 1996; RUIS et al., 1997). Mit
zunehmendem Alter der Tiere nimmt die basale Cortisolkonzentration ab
(EVANS et al., 1988; RUIS et al., 1997). Dies ist nach KATTESH et al. (1990)
und RUIS et al. (1997) möglicherweise auf die zunehmende Bindung von freiem
Cortisol, welches im Speichel nachgewiesen wird, an CBG zurückzuführen.
4.2. Katecholamine
Katecholamine werden vom Nebennierenmark konstant freigesetzt und
unterstützen den Körper in Belastungssituationen und weniger bei der
Basalregulation (MÖSTL, 2010; GRECO und STABENFELDT, 2013). Ihr
Ausstoß wird in bestimmten Situationen immens gesteigert um dem Tier eine
adäquate, sofortige Anpassung an akute Stresssituationen, die auch als „flight or
fight“ Situationen beschrieben werden zu ermöglichen (GRECO und
STABENFELDT, 2013). Laut GOLDSTEIN (1987) unterstützt das
sympathoadrenerge System den Organismus in Zuständen der Hypovolämie,
Hypoxie und Hypoglycämie sowie bei lebensbedrohlichen Schockzuständen.

28 II. Literaturübersicht
Dabei steigt der Adrenalinspiegel im Gegensatz zur Noradrenalinkonzentration
stark an (GRECO und STABENFELDT, 2013). Noradrenalin ist Transmitter der
postganglionär-sympathischen Neurone, gibt die neuronale sympathische
Aktivität wieder und verändert sich unter solchen Zuständen kaum
(GOLDSTEIN, 1987).
4.2.1. Wirkung im Organismus
Die Adrenozeptoren (vier Haupttypen α1, α2, β1 und β2) sind unterschiedlich
empfindlich für Adrenalin und Noradrenalin (GRECO und STABENFELDT,
2013). Laut MÖSTL (2010) haben β-Rezeptoren eine höhere Affinität zu
Adrenalin und α-Rezeptoren zu Noradrenalin. Mit Einfluss auf das Herz- und
Gefäßsystem, unter dem es zur Steigerung des Herzminutenvolumens und zu
unterschiedlichen Auswirkungen auf den Gefäßtonus kommt, wird grundsätzlich
eine Reduktion der Blutversorgung von Haut, Darm und Niere zu Gunsten der
anderen Organe, die eine wichtigere Rolle in Abwehr- und Fluchtreaktionen
spielen, erreicht (MÖSTL, 2010). Unter Katecholamineinfluss verbessert sich
einerseits die Energieversorgung durch die Erhöhung des Blutzuckerspiegels und
die gesteigerte Fettmobilisation andererseits wird auch das respiratorische System
angeregt (MÖSTL, 2010).
4.2.2. Bedeutung im Stressgeschehen
Durch Prämedikation mit einer anxiolytischen Dosis eines Benzodiazepines
konnte sowohl der Adrenalin- als auch der Corticosteronanstieg nach einem
elektrischen Reiz gemindert werden (DE BOER et al., 1990). Dies weist
daraufhin, dass emotionaler Stress und Angst sowohl zu einer Aktivierung des
Nebennierenmarkes als auch der -rinde führen (DE BOER et al., 1990). ROOZEN
et al. (1995) zeigten bei Sauen nach Halten in der Oberkieferschlinge, dass
Adrenalin und Noradrenalin bereits 0,5 min nach dem Beginn der Fixierung
anstiegen. Bei ROOZEN et al. (1995) sowie bei NEUBERT et al. (1996) erwiesen
sich die Katecholamine als geeignete Parameter zur Detektion von kurzzeitigem
Stress bei Schweinen. NEUBERT et al. (1996) empfahlen die
Katecholaminmessung als beste Möglichkeit den Grad der Erregung und den
Zeitpunkt der körperlichen Anstrengung zu ermitteln. Ähnlich erwiesen sich auch
in den Untersuchungen von ZIMMERMANN (2010) zur Kastration männlicher
Saugferkel unter CO2 Betäubung Adrenalin und Noradrenalin als geeignete

II. Literaturübersicht 29
Parameter. Direkt nach dem Eingriff stiegen die Adrenalinkonzentrationen um das
29 fache und die Noradrenalinkonzentration um das bis zu 81 fache an
(ZIMMERMANN, 2010). Die Aussagekraft der Ergebnisse hängt von der Art des
Stimulus ab. Bei Reizen, die einen länger andauernden Zustand erzeugen, ist der
Beprobungzeitpunkt weniger wichtig als bei kurzfristigen Stimuli, die eine genau
determinierte Probennahme erfordern (HJEMDAHL, 1993). Dafür sollte eine ein
bis zwei minütige „wash-out“ Zeit eingeplant werden, welche die Zeitspanne
berücksichtigt, die Noradrenalin braucht um vom Ort der Freisetzung ins
Blutplasma zu gelangen (HJEMDAHL, 1993). Den Effekt der verzögerten
Noradrenalin Freisetzung und Ausschwemmung ins Blut beobachteten auch
MELLOR et al. (2002) bei Lämmern, die mit einem Gummiring kastriert wurden
und deren Schwänze mittels Gummiring kupiert wurden.
4.2.3. Regulation
Das sympathische Nervensystem wird vom Hypothalamus und den
Kreislaufzentren des ZNS, die wiederum vom limbischen System und dem Cortex
beeinflusst werden, gesteuert (MÖSTL, 2010). Das Nebennierenmark kann laut
MÖSTL (2010) als sympathisches Ganglion betrachtet werden. Katecholamine
(Dopamin, Adrenalin, Noradrenalin) werden in den chromaffinen Zellen des
Nebennierenmarkes gespeichert (AXELROD und REISINE, 1984). Stress führt
über unterschiedlich starken Einfluss von neuronaler Aktivität, Glucocorticoiden
und ACTH zu einer Aktivitätssteigerung der an der Katecholaminsynthese
beteiligten Enzyme (AXELROD und REISINE, 1984). Neben dem
Nebennierenmark kann Noradrenalin auch in anderen chromaffinen Geweben
gebildet werden (MÖSTL, 2010). Einen weiteren Syntheseort für Katecholamine
fanden KVETNANSKY et al. (2012) in den Adipozyten, die vor allem durch
physikalischen und psychischen Stress dazu angeregt werden Katecholamine zu
synthetisieren und einen großen Einfluss auf die Lipolyse und die
Thermoregulation auszuüben vermögen (KVETNANSKY et al., 2009). Zusätzlich
zur Hormonwirkung fungiert Noradrenalin als Neurotransmitter im
Zentralnervensystem und im Sympathikus (MÖSTL, 2010). Es wird von
sympathischen Nervenendigungen ins Blut abgegeben, was die höheren
Plasmaspiegel als für Adrenalin erklärt (MÖSTL, 2010). Noradrenalin gibt im
Wesentlichen die sympathische Aktivität wieder, wohingegen Adrenalin zur
Messung der Sekretion des Nebennierenmarkes dient (AXELROD und REISINE,

30 II. Literaturübersicht
1984). Diese Ansicht teilten auch KVETNANSKY et al. (1979), wonach die
Noradrenalin Antwort auf Immobilisationsstress hauptsächlich aus den
sympathischen Endigungen und zu einem kleineren Teil aus den Nebennieren
stammte. Hingegen wird Adrenalin vorwiegend aus dem Nebennierenmark
freigesetzt (KVETNANSKY et al., 1979). Bereits bei kurzem, sanftem Handling
kann es zu einer starken Freisetzung der Katecholamine kommen, was wiederum
die Schwierigkeit erklärt, Basalwerte dafür festzulegen (KVETNANSKY et al.,
1978). Die Halbwertszeiten sind sehr kurz und variieren zwischen 20 sec und
10 min (MÖSTL, 2010). Eine Inaktivierung erfolgt bei Wiederaufnahme in die
sympathischen Fasern oder die chromaffinen Zellen des NNM durch
enzymatische Inaktivierung oder Abbau (z.B. zu Vanillinmandelsäure) (MÖSTL,
2010).

III. Material und Methoden 31
III. MATERIAL UND METHODEN
Die Studie wurde an der Versuchsstation Thalhausen der Technischen Universität
München durchgeführt. Die Probenentnahmen erstreckten sich von Mai bis
Oktober 2013.
1. Ziel der Untersuchung
Ziel der vorliegenden Untersuchung war es, die Stress- und Schmerzbelastung bei
Entnahme eines Tracheobronchialabstriches mit der eines Nasentupfers und mit
der alleinigen Fixierung in der Oberkieferschlinge beim Schwein zu vergleichen.
Zur Ermittlung der Stressbelastung dienten Cortisolkonzentrationen aus Serum
und Speichel sowie aus dem Blutplasma ermittelte Noradrenalin- und
Adrenalinkonzentrationen.
2. Genehmigung des Tierversuchsvorhabens
Die Genehmigung des Tierversuchsvorhabens wurde gemäß §8 Absatz 1 des
Tierschutzgesetzes bei der Regierung von Oberbayern beantragt und am 27. Mai
2013 erteilt. Das Versuchsvorhaben wird unter dem Aktenzeichen 55.2-1-54-
2532-169-12 geführt.
3. Studientiere und Haltung
Bei den Studientieren handelte es sich um Masthybriden aus der Kreuzung
Deutsche Landrasse x Piétrain. Alle Tiere stammten aus der Versuchsstation
Thalhausen der Technischen Universität München. Sie wurden zusammen mit
ihren Buchtengenossen in den Stallungen der Versuchsstation Thalhausen im
Aufzuchtabteil in Buchten zu je zehn Tieren gehalten. Es wurde
Beschäftigungsmaterial angeboten. Gefüttert wurde Futter aus Eigenproduktion
sowie kommerziell erworbenes Futter ad libitum. Ebenso stand Wasser über
Nippeltränken zur freien Verfügung.
4. Auswahl der Studientiere
Für die Cortisol- und Katecholaminuntersuchungen wurden mindestens 24
Stunden vor einem Studientag klinisch gesunde, weibliche Ferkel in einem Alter
von acht Wochen ausgewählt und im Anschluss nach Gewicht in die vier

32 III. Material und Methoden
nachfolgend beschriebenen Gruppen randomisiert eingeteilt. Die Tiere waren
bereits betriebsbedingt mit einer individuellen Ohrmarke versehen, zusätzlich
wurden sie zur Gruppenzuordnung mit einer Nummer am Rücken markiert. Es
wurde darauf geachtet, dass an jedem Studientag die Anzahl der Tiere pro Gruppe
ausgeglichen war.
5. Gruppeneinteilung
Es wurden vier Gruppen, drei Versuchs- und eine Kontrollgruppe, zu je 23 Tieren
für die Cortisoluntersuchungen und zu je 31 Tieren für die
Katecholaminuntersuchungen gebildet (siehe Tabelle 2). Im Teilversuch zur
Cortisolmessung wurde jeweils ein Tier aus der Gruppe I und IV am Studientag
auf Grund klinischer Erkrankung von der Studie ausgeschlossen sowie im
Teilversuch zur Katecholaminuntersuchung ein Tier der Gruppe II.
Tabelle 2: Gruppeneinteilung Cortisol- und Katecholaminuntersuchungen
Gruppe Eingriff
Anzahl (n)
Studie
Cortisol
Anzahl (n)
Studie
Katecholamine
I TBA Tracheobronchialabstrich 22 31
II OK Oberkieferschlinge 23 30
III NT Nasentupfer 23 31
IV KO Kontrolle 22 31
6. Ablauf der Cortisoluntersuchungen
6.1. Zeitlicher Ablauf
Die im Folgenden beschriebenen Schritte wurden entsprechend der
Gruppenzugehörigkeit der Schweine durchgeführt.
Die acht Wochen alten Tiere wurden möglichst ruhig in den Flatdeckbuchten
gefangen und herausgehoben. Unmittelbar danach erfolgte die erste
Blutprobenentnahme zur Ermittlung der basalen Cortisolkonzentrationen. Im
Hinblick auf die circadianen Schwankungen der Cortisolsekretion wurden alle

III. Material und Methoden 33
Blutproben des Basalwertes zwischen 12:00 Uhr und 12:30 Uhr entnommen. Bei
zehn Schweinen pro Studientag ergab sich dabei ein dreiminütiger Abstand
zwischen den Blutproben. Die Reihenfolge der Tiere wurde für den weiteren
Tagesablauf beibehalten. Direkt nach der Blutentnahme wurde eine Speichelprobe
gewonnen. Im Anschluss wurde das Schwein wieder in seine Bucht zurückgesetzt.
30 min später wurde dasselbe Tier wieder aus seiner Bucht gefangen und seiner
Gruppe entsprechend einer Maßnahme unterzogen (unter 8. Eingriffe
beschrieben). 30 min nach dem jeweiligen Eingriff erfolgte die zweite Blut- und
Speichelprobenentnahme. Tieren der Gruppe IV wurden diese Proben ohne
vorherigen Eingriff entnommen. Zwei weitere Blut- und Speichelproben erfolgten
60 min und 90 min nach dem Eingriff.
Abbildung 1: Zeitplan der Cortisoluntersuchungen
6.2. Blut- und Speichelprobenentnahme
Die Blut- und Speichelprobenentnahme erfolgte stets durch dieselbe
studiendurchführende Person. Die Schweine jeder Gruppe wurden einzeln von
einer Hilfsperson aus der Bucht gehoben und auf dem Rücken liegend auf dem
Schoß einer Hilfsperson fixiert. Zur Blutentnahme aus der Vena jugularis externa
wurden Serummonovetten (Primavette®
V, Kabe Labortechnik GmbH,
Nümbrecht-Elsenroth) und sterile Einmalkanülen der Größe 20 G (Sterican,
B.Braun AG, Melsungen) verwendet. Die Serummonovetten wurden vollständig
mit 7,5 ml Blut gefüllt.
Im Anschluss an die Blutentnahme wurden die Schweine in Brustlage gedreht und
locker auf dem Schoß der assistierenden Person gehalten. Ein mittels
Eingriff
Speichel
Speichel
Speichel
Speichel
Blut
Blut
Blut
Blut
-30 60 90 30 t (min)

34 III. Material und Methoden
Arterienklemme fixierter Speicheltupfer (Salivette®
, SARSTEDT AG & Co,
Nümbrecht-Elsenroth) wurde vorsichtig in die Maulspalte geführt und in
kreisenden Bewegungen am Gaumendach und im Kieferwinkel gedreht, bis er
ausreichend durchfeuchtet war.
6.3. Probenbearbeitung
Unmittelbar nach der Blutentnahme wurden die Serummonovetten in Eiswasser
gekühlt. Die Zentrifugation bei 3000 U/min für die Dauer von 10 min bei einer
Temperatur von 4 °C erfolgte noch am selben Tag in der Klinik für Schweine.
Aus dem gewonnen Serum wurden zwei Aliquots zu je mindestens 0,5 ml
hergestellt, wovon ein Exemplar als Rückstellprobe asserviert wurde.
Die Proben wurden bis zur Bestimmung der Cortisolspiegel für maximal acht Wo-
chen bei -22 °C tiefgefroren.
Die Speichelproben wurden bis zur Weiterbearbeitung am Ende des Studientages
bei Raumtemperatur aufbewahrt. Nach Zentrifugation bei 3000 U/min für die
Dauer von 10 min wurde der gewonnene Speichel in ein Eppendorf Tube (Fa.
Eppendorf AG, 1,5 ml Safe-Lock Tubes, Hamburg) pipettiert und bei -22 °C
tiefgefroren. Die Messung der Cortisolkonzentrationen aus Serum und Speichel
wurde im Labor der Klinik für Schweine in Oberschleißheim vorgenommen.
Dafür wurde das Gerät Elecsys 2010®
(Roche Diagnostics, Mannheim) verwendet.
Die Messung erfolgte mittels Elektrochemilumineszenz-Immunoassay (ECLIA)
mit einem speziellen Messreagenz (Cortisol Elecsys®
, Roche Diagnostics,
Mannheim).
7. Ablauf der Katecholaminuntersuchungen
7.1. Zeitlicher Ablauf
Die im Folgenden beschrieben Schritte wurden entsprechend der
Gruppeneinteilung nacheinander an allen Schweinen durchgeführt. Die acht
Wochen alten Schweine wurden möglichst ruhig in den Flatdeckbuchten gefangen
und herausgehoben. Unmittelbar nach dem Einfangen wurde das Schwein in
Rückenlage fixiert, um die erste Blutprobe zu gewinnen. Direkt danach wurde das
Tier in seine Flatdeckbucht zurückgesetzt. Nach 15 min wurde dasselbe Schwein
erneut möglichst stressarm aus der Bucht genommen und dem der jeweiligen
Gruppe zugeordneten Eingriff unterzogen (unter 8. Eingriffe beschrieben).

III. Material und Methoden 35
Unmittelbar im Anschluss an den Eingriff wurden die Schweine in Rückenlage
fixiert und eine zweite Blutprobe gewonnen. Tieren der Gruppe IV wurde diese
Blutprobe ohne vorher erfolgten Eingriff entnommen.
Abbildung 2: Zeitplan der Katecholaminuntersuchungen
7.2. Blutprobenentnahme
Die Blutprobenentnahme erfolgte entsprechend der für Cortisoluntersuchungen.
Allerdings wurden zur Blutentnahme aus der Vena jugularis externa EDTA-
Plasma-Monovetten (Primavetten®
V EDTA 2,6 ml, Kabe Labortechnik GmbH,
Nümbrecht-Elsenroth) und sterile Einmalkanülen der Größe 20 G (Sterican,
B.Braun AG, Melsungen) verwendet. Die auf 4 °C vorgekühlten EDTA-
Monovetten wurden bis zur Füllgrenze mit Blut gefüllt. Nach der Blutentnahme
wurde das Schwein wieder zurückgesetzt.
7.3. Probenverarbeitung
Sofort nach der Blutprobenentnahme wurden die befüllten EDTA-Primavetten für
maximal fünf Minuten in Eiswasser zwischengelagert bis die Zentrifugation in
einer Kühlzentrifuge (Hettich Mikro 22R, Fa. Hettich Zentrifugen, Tuttlingen) bei
4 °C für die Dauer von 10 min erfolgte. Das gewonnene Plasma wurde jeweils zur
Hälfte in zwei 1 ml fassende, gekühlte kryostabile Gefäße (Corning®
2 ml, Fa.
Corning Incorporated, New York) pipettiert. Diese wurden in einem Behälter mit
flüssigem Stickstoff schockgefroren, zur Klinik für Schweine in Oberschleißheim
transportiert und anschließend bei -80 °C tiefgefroren. Auf Trockeneis wurden die
Proben zum Leibniz-Institut für Nutztierbiologie (FBN) nach Dummerstorf zur
Bestimmung der Katecholaminkonzentrationen verbracht.
Blut
Blut
Eingriff
-15 0 t (min)

36 III. Material und Methoden
7.4. Messsung der Katecholamine
Die Plasmaproben wurden am Leibniz-Institut für Nutztierbiologie (FBN) in
Dummerstorf vom Forschungsbereich Verhaltensphysiologie bearbeitet. Unter
Leitung von Herrn Dr. W. Otten wurden die Adrenalin- und
Noradrenalinkonzentrationen bestimmt. Die Analyse erfolgte durch
Hochdruckflüssigkeitschromatographie mit elektrochemischer Detektion nach
vorheriger Extraktion der Katecholamine aus dem Plasma durch Absorption an
Aluminiumoxid (OTTEN et al., 2013).
8. Eingriffe
8.1. Durchführung eines Tracheobronchialabstriches
Bei den Tieren der Gruppe I wurde ein Tracheobronchialabstrich nach FABLET
et al. (2010) entnommen. Dazu wurden die Schweine möglichst ruhig aus der
Bucht gefangen und in den Gang gehoben. Dann wurden sie stehend zwischen
den Beinen einer Hilfsperson positioniert und der Kopf mit einer
Oberkieferschlinge unmittelbar hinter den Eckzähnen fixiert. Ein Maulkeil wurde
zwischen Ober- und Unterkiefer eingelegt, damit die Tiere den Katheter, der im
Anschluss eingeführt wurde, nicht abbeißen und verschlucken konnten. Der
Katheter aus weichem Kunststoff mit einem Durchmesser von 0,4 cm und 50 cm
Länge (Absaugkatheter CH 12, B.Braun Melsungen AG, Melsungen) wurde
zwischen Drahtschlinge und Gaumen zunächst bis zum ersten Widerstand an der
Epiglottis vorgeschoben. Um den Kehldeckel zu überwinden wurde sorgfältig auf
den Atemrhythmus des Tieres geachtet und der Katheter bei Inspiration mit
leichten Drehbewegungen zügig bis zur Bifurcatio tracheae vorgeschoben. Dies
löste einen Hustenstoß aus. Der Katheter wurde dann sofort wieder
herausgezogen, das Tier aus der Fixation gelöst und in die Bucht zurückgesetzt.
Der Tracheobronchialabstrich wurde immer von derselben studiendurchführenden
Person entnommen.

III. Material und Methoden 37
Abbildung 3: Durchführung eines Tracheobronchialabstriches
8.2. Fixierung in der Oberkieferschlinge
Die Tiere der Gruppe II wurden ruhig in der Bucht gefangen und auf den Gang
gehoben. Anschließend wurden die Schweine für 60 sec von der
studiendurchführenden Person in der Oberkieferschlinge fixiert. Nach Ablauf der
Zeit wurden die Tiere unmittelbar zurückgesetzt.
8.3. Entnahme einer Nasentupferprobe
Bei Tieren der Gruppe III wurde ein Nasentupfer entnommen. Dafür wurden die
Schweine ruhig in der Bucht gefangen und auf den Gang gehoben. Die Schweine
wurden stehend zwischen die Beine einer Hilfsperson und zusätzlich am Kopf
mittels Oberkieferschlinge fixiert. Die studiendurchführende Person führte
vorsichtig einen Nasentupfer nacheinander in beide Nasenlöcher ein. Der Tupfer
wurde jeweils bis zur Hälfte in der Nasenhöhle eingeführt und dabei um ca. 260°
gedreht. Nach Entnahme des Tupfers wurde die Fixation aufgehoben und das
Schwein zurückgesetzt.

38 III. Material und Methoden
9. Statistische Auswertung
Die Ergebnisse der Cortisolmessungen wurden mit Hilfe der Programme IBM
SPSS Statistics 22.0 und Microsoft Office Excel 2010 ausgewertet. Individuelle
Ferkel bildeten die Studieneinheit. Anhand der Messwerte von Serum- und
Speichelcortisol und Katecholaminen wurden jeweils Mittelwerte,
Standardabweichungen und Standardfehler ermittelt. Die Nullhypothese besagt,
dass kein Unterschied zwischen den Studiengruppen hinsichtlich der untersuchten
Parameter besteht.
Die Anwendung des Kolmogorov-Smirnov-Testes mit zusätzlicher
Signifikanzkorrektur nach Lilliefors zeigte, dass weder die Serum- noch die
Speichelcortisolwerte einer Normalverteilung folgten. Beim Gruppenvergleich
wies der Kruskal–Wallis-Test 30 min nach dem Eingriff signifikante Unterschiede
sowohl für Serum- als auch Speichelproben nach. Für diesen Zeitpunkt wurden
daher Vergleiche zwischen den Gruppen (vier Vergleiche: TBA mit NT, NT mit
OK, TBA mit OK und OK mit KO Gruppe) mittels einseitigem Mann-Whitney-
Test berechnet. Um den Typ I-Fehler zu verringern, wurde für die Paarvergleiche
eine Korrektur nach Bonferroni-Holm vorgenommen. Die Signifikanzniveaus
wurden entsprechend geändert und auf 1.25 %, 1.67 %, 2.50 % und 5.00 %
angehoben. Es wurde jeweils mit dem p-Wert der höchsten Signifikanz unter den
Paarvergleichen begonnen (HOLM, 1979).
Um innerhalb einer Gruppe Vergleiche zwischen zwei unterschiedlichen
Beprobungszeitpunkten bilden zu können, wurde der nichtparametrische
Wilcoxon-Test angewendet. Von Interesse waren die Vergleiche zwischen den
Zeitpunkten: basal mit 30 min, basal mit 60 min, 60 min mit 90 min, basal mit
90 min Wert. Die vier Vergleiche wurden nach Bonferroni-Holm korrigiert und
die Signifikanzniveaus, wie oben angegeben, angepasst.
Für Speichel- und Serumcortisol wurde der AUC (Area-under-the-curve), wie bei
BLAND (2009) beschrieben, berechnet. Der Kolmogorov-Smirnov-Test
(zusätzliche Signifikanzkorrektur nach Lilliefors) zeigte eine Normalverteilung
der AUC Daten. Eine anschließende einseitige ANOVA wurde durchgeführt, um
signifikante Unterschiede zwischen den Gruppen deutlich zu machen. Basierend
auf diesen Ergebnissen wurden vier Paarvergleiche mittels t-Test zum
Gruppenvergleich zwischen TBA mit NT, NT mit OK, TBA mit OK und OK mit

III. Material und Methoden 39
KO Gruppe eingeleitet. Auch hierfür wurde eine Bonferroni-Holm Adjustierung
angewendet.
Bei der Katecholaminauswertung wurden Tests analog zur Auswertung der
Cortisolkonzentrationen angewendet. Der Kolmogorov-Smirnov-Test zeigte, dass
die Daten nicht normalverteilt waren. Anschließend wurden signifikante
Unterschiede zwischen den Gruppen (vier Vergleiche: TBA mit NT, NT mit OK,
TBA mit OK und OK mit KO Gruppe) mittels Mann-Whitney-Test berechnet und
die Signifikanzniveaus nach Bonferroni-Holm angepasst. Im Gegensatz dazu
konnte für den Wilcoxon-Test, der zwischen vor und nach dem Eingriff verglich,
auf eine Adjustierung nach Bonferroni-Holm verzichtet werden, da es sich nur um
einen einzelnen Vergleich handelte.

40 IV. Ergebnisse
IV. ERGEBNISSE
1. Cortisol
Die mittleren Cortisolkonzentrationen der vier Gruppen variierten zwischen
55,6 nmol/l (KO) und 63,8 nmol/l (TBA) (siehe Tabelle 3) und unterschieden sich
vor den Eingriffen nicht signifikant voneinander und.
1.1. Serumcortisol
Tabelle 3: Mittlere Cortisolkonzentrationen im Serum in nmol/l vor sowie
30 min, 60 min und 90 min nach dem Studieneingriff, mit Minima, Maxima,
Standardabweichungen (SD), Standardfehler (SEM) und der area-under-the-
curve (AUC) der vier Gruppen
Gruppe n
t (min)
AUC
-30 30 60 90
TBA 22
MW 63,8 105,4 96,6 89,0 10889,6
SD 33,6 36,4 33,3 37,2 3558,8
Min. 16,6 35,1 46,8 34,7 4605,9
Max. 123,9 173,1 160,3 185,3 16806,0
SEM 7,2 7,8 7,1 7,9 758,7
OK 23
MW 62,4 92,2 108,2 81,9 10492,8
SD 19,1 38 60,8 35,3 3709,3
Min. 25,4 30,6 31,4 31,8 4251,5
Max. 111,3 166,8 266,7 183,3 17728,5
SEM 4,0 7,9 12,7 7,4 773,5
NT 23
MW 58,7 109,2 111,6 103,3 11571,9
SD 23,6 50,6 42,8 40,1 4348,7
Min. 26,5 37,4 34,3 37,7 4479,5
Max. 113,4 228,7 219,1 196,8 20999,1
SEM 4,9 10,6 8,9 8,4 906,8
KO 22
MW 55,6 53,2 80,3 70,1 7520,1
SD 27,3 28,5 32,7 18,0 2531,2
Min. 21,8 15,1 32,5 30,0 3490,1
Max. 109,0 134,1 152,4 96,0 13136,7
SEM 5,8 6,1 7,0 3,8 539,7
.

IV. Ergebnisse 41
30 min nach dem Eingriff zeigten alle Gruppen außer der Kontrollgruppe einen
Anstieg der mittleren Cortisolkonzentrationen. Der höchste Wert mit 109,2 nmol/l
war bei der Nasentupfergruppe messbar und entsprach einem signifikanten
Anstieg (p < 0,001) vom Basalniveau um 86 % (1,9 facher Basalwert). Gefolgt
wurde die NT Gruppe von der Tracheobronchialabstrichgruppe mit einer 65 %
Erhöhung (1,7 fach) zum Basalwert (p < 0,001).
Abbildung 4: Verlauf der mittleren Cortisolkonzentrationen (nmol/l) im
Serum der vier Gruppen über die Zeit t (min)
Mit einer mittleren Cortisolkonzentration von 105,4 nmol/l erreichte die
TBA Gruppe 30 min nach dem Eingriff ihren maximalen Wert. Auch die mittlere
Cortisolkonzentration der OK Gruppe lag 30 min nach dem Eingriff mit
92,2 nmol/l signifikant über dem Basallevel (p = 0,002). Dies entsprach einer
1,5 fachen Erhöhung (48 % höher als Basallevel) zur Basalkonzentration. 30 min
nach dem Eingriff unterschieden sich die mittleren Cortisolkonzentrationen der
TBA, NT und OK Gruppe nicht signifikant voneinander (siehe Tabelle 4). Der
mittlere Cortisolspiegel der OK Gruppe war um 73 % signifikant höher als der der
Kontrollgruppe (p = 0,001).
0
20
40
60
80
100
120
140
-30 Eingriff 30 60 90
Cort
isol
in n
mol/
l
Zeit (t) in min
Tracheobronchialtupfer Oberkieferschlinge
Nasentupfer Kontrolle
42 IV. Ergebnisse
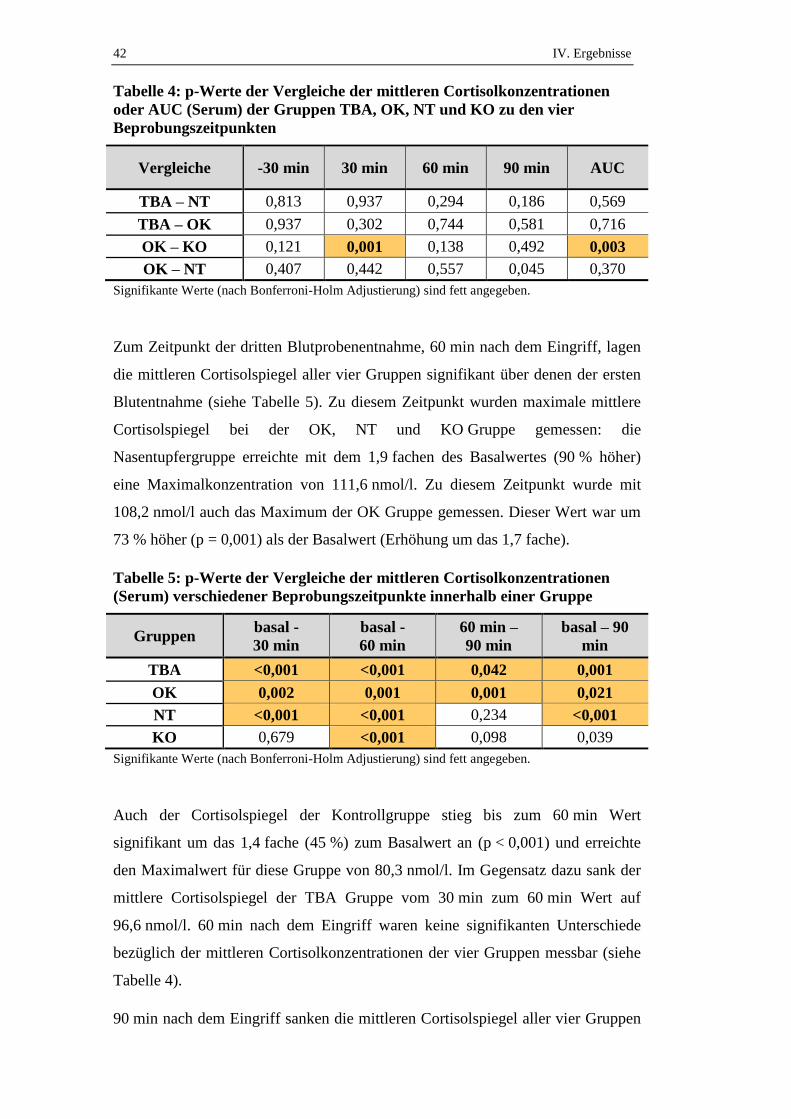
Tabelle 4: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen
oder AUC (Serum) der Gruppen TBA, OK, NT und KO zu den vier
Beprobungszeitpunkten
Vergleiche -30 min 30 min 60 min 90 min AUC
TBA – NT 0,813 0,937 0,294 0,186 0,569
TBA – OK 0,937 0,302 0,744 0,581 0,716
OK – KO 0,121 0,001 0,138 0,492 0,003
OK – NT 0,407 0,442 0,557 0,045 0,370
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
Zum Zeitpunkt der dritten Blutprobenentnahme, 60 min nach dem Eingriff, lagen
die mittleren Cortisolspiegel aller vier Gruppen signifikant über denen der ersten
Blutentnahme (siehe Tabelle 5). Zu diesem Zeitpunkt wurden maximale mittlere
Cortisolspiegel bei der OK, NT und KO Gruppe gemessen: die
Nasentupfergruppe erreichte mit dem 1,9 fachen des Basalwertes (90 % höher)
eine Maximalkonzentration von 111,6 nmol/l. Zu diesem Zeitpunkt wurde mit
108,2 nmol/l auch das Maximum der OK Gruppe gemessen. Dieser Wert war um
73 % höher (p = 0,001) als der Basalwert (Erhöhung um das 1,7 fache).
Tabelle 5: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen
(Serum) verschiedener Beprobungszeitpunkte innerhalb einer Gruppe
Gruppen basal -
30 min
basal -
60 min
60 min –
90 min
basal – 90
min
TBA <0,001 <0,001 0,042 0,001
OK 0,002 0,001 0,001 0,021
NT <0,001 <0,001 0,234 <0,001
KO 0,679 <0,001 0,098 0,039
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
Auch der Cortisolspiegel der Kontrollgruppe stieg bis zum 60 min Wert
signifikant um das 1,4 fache (45 %) zum Basalwert an (p < 0,001) und erreichte
den Maximalwert für diese Gruppe von 80,3 nmol/l. Im Gegensatz dazu sank der
mittlere Cortisolspiegel der TBA Gruppe vom 30 min zum 60 min Wert auf
96,6 nmol/l. 60 min nach dem Eingriff waren keine signifikanten Unterschiede
bezüglich der mittleren Cortisolkonzentrationen der vier Gruppen messbar (siehe
Tabelle 4).
90 min nach dem Eingriff sanken die mittleren Cortisolspiegel aller vier Gruppen

IV. Ergebnisse 43
ab. Die Cortisolspiegel der TBA und der OK Gruppe wiesen im Gegensatz zu
denen der NT und KO Gruppe eine signifikante Reduktion zwischen der 60. und
90. min auf (siehe Tabelle 5). Zum Zeitpunkt der letzten Blutentnahme
unterschieden sich die mittleren Cortisolkonzentrationen der TBA und der NT
Gruppe, der TBA und der OK Gruppe, der OK und der NT Gruppe sowie der OK
und der KO Gruppe nicht signifikant voneinander (siehe Tabelle 4). Verglichen
mit den jeweiligen Basalwerten waren 90 min nach dem Eingriff die mittleren
Cortisolkonzentrationen der TBA um 40 %, der OK um 31 % und der NT Gruppe
um 76 % signifikant erhöht (siehe Tabelle 5). Nur die Kontrollgruppe unterschied
sich zum Zeitpunkt der letzten Blutentnahme nicht signifikant (p = 0,039) vom
Basalwert (26 % höher als Basalwert).
Abbildung 5: Verlauf der mittleren Cortisolkonzentrationen im Serum der
Gruppen OK und KO (AUC)
Die Konzentrations-Zeit-Kurven (Area-under-the-curve (AUC)), welche Dauer
und Intensität der Belastung beschreiben, unterschieden sich zwischen den
Gruppen TBA, OK und NT nicht signifikant. Im Vergleich zwischen der OK und
der KO Gruppe lag der AUC der in der Oberkieferschlinge fixierten Tiere
signifikant (p = 0,003) über jenem der Kontrollgruppe (siehe Abbildung 5).
0
20
40
60
80
100
120
140
-30 Eingriff 30 60 90
Cort
isol
in n
mol/
l
Zeit (t) in min
Oberkieferschlinge (AUC) Kontrolle (AUC)
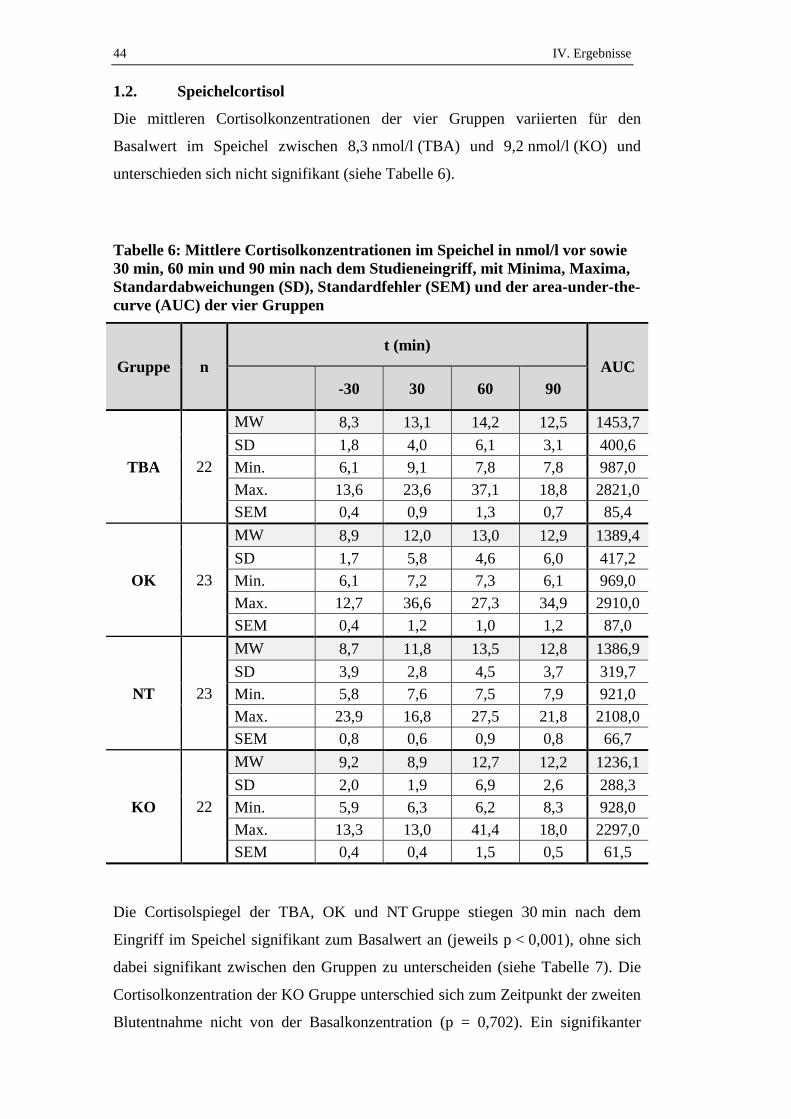
44 IV. Ergebnisse
1.2. Speichelcortisol
Die mittleren Cortisolkonzentrationen der vier Gruppen variierten für den
Basalwert im Speichel zwischen 8,3 nmol/l (TBA) und 9,2 nmol/l (KO) und
unterschieden sich nicht signifikant (siehe Tabelle 6).
Tabelle 6: Mittlere Cortisolkonzentrationen im Speichel in nmol/l vor sowie
30 min, 60 min und 90 min nach dem Studieneingriff, mit Minima, Maxima,
Standardabweichungen (SD), Standardfehler (SEM) und der area-under-the-
curve (AUC) der vier Gruppen
Gruppe n
t (min)
AUC
-30 30 60 90
TBA 22
MW 8,3 13,1 14,2 12,5 1453,7
SD 1,8 4,0 6,1 3,1 400,6
Min. 6,1 9,1 7,8 7,8 987,0
Max. 13,6 23,6 37,1 18,8 2821,0
SEM 0,4 0,9 1,3 0,7 85,4
OK 23
MW 8,9 12,0 13,0 12,9 1389,4
SD 1,7 5,8 4,6 6,0 417,2
Min. 6,1 7,2 7,3 6,1 969,0
Max. 12,7 36,6 27,3 34,9 2910,0
SEM 0,4 1,2 1,0 1,2 87,0
NT 23
MW 8,7 11,8 13,5 12,8 1386,9
SD 3,9 2,8 4,5 3,7 319,7
Min. 5,8 7,6 7,5 7,9 921,0
Max. 23,9 16,8 27,5 21,8 2108,0
SEM 0,8 0,6 0,9 0,8 66,7
KO 22
MW 9,2 8,9 12,7 12,2 1236,1
SD 2,0 1,9 6,9 2,6 288,3
Min. 5,9 6,3 6,2 8,3 928,0
Max. 13,3 13,0 41,4 18,0 2297,0
SEM 0,4 0,4 1,5 0,5 61,5
Die Cortisolspiegel der TBA, OK und NT Gruppe stiegen 30 min nach dem
Eingriff im Speichel signifikant zum Basalwert an (jeweils p < 0,001), ohne sich
dabei signifikant zwischen den Gruppen zu unterscheiden (siehe Tabelle 7). Die
Cortisolkonzentration der KO Gruppe unterschied sich zum Zeitpunkt der zweiten
Blutentnahme nicht von der Basalkonzentration (p = 0,702). Ein signifikanter

IV. Ergebnisse 45
Unterschied ergab sich 30 min nach dem Eingriff zwischen der OK Gruppe und
der Kontrollgruppe (p = 0.001). Der mittlere Cortisolspiegel der OK Gruppe lag
35 % über dem der Kontrollgruppe.
Abbildung 6: Verlauf der mittleren Cortisolkonzentrationen (nmol/l) im
Speichel aller vier Gruppen über die Zeit t (min)
Alle vier Gruppen erreichten ihr Maximum 60 min nach dem Eingriff. Zu diesem
Zeitpunkt zeigte sich bei allen Gruppen ein signifikanter Anstieg zum Basalwert;
dieser betrug bei der OK Gruppe 46 % (1,5 facher Anstieg zum Basalwert), bei
der NT Gruppe 55 % (1,6 facher Anstieg), bei der TBA Gruppe 71 % (1,7 facher
Anstieg) und bei der KO Gruppe 38 % (1,4 facher Anstieg) (siehe Tabelle 8).
Tabelle 7: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen
oder AUC (Speichel) der Gruppen TBA, OK, NT und KO zu den vier
Beprobungszeitpunkten
Vergleiche -30 30 60 90 AUC
TBA - NT 0,468 0,441 0,706 0,986 0,723
TBA - OK 0,184 0,078 0,338 0,657 0,281
OK - KO 0,732 0,001 0,389 0,645 0,126
OK - NT 0,043 0,459 0,740 0,575 0,683
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
5
7
9
11
13
15
-30 Eingriff 30 60 90
Cort
iso
l in
nm
ol/
l
Zeit (t) in min
Tracheobronchialtupfer Oberkieferschlinge
Nasentupfer Kontrolle

46 IV. Ergebnisse
Die mittlere Cortisolkonzentration der TBA Gruppe sank vom 60 min zum 90 min
Wert, im Gegensatz zur OK, NT und KO Gruppe, signifikant ab (p = 0,039).
Weder 60 min noch 90 min nach dem Eingriff wurden zwischen der TBA, NT,
OK und KO Gruppe signifikante Unterschiede festgestellt (siehe Tabelle 7).
90 min nach dem Eingriff lagen die mittleren Speichelcortisolspiegel aller vier
Gruppen signifikant über dem Basallevel (siehe Tabelle 8). Zu diesem Zeitpunkt
wurden zwischen 32 % (KO Gruppe) und 50 % (TBA Gruppe) höhere
Cortisolkonzentrationen als zum Basalwert gemessen.
Tabelle 8: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen
(Speichel) verschiedener Beprobungszeitpunkte innerhalb einer Gruppe
Gruppen basal -
30 min
basal -
60 min
60 min –
90 min
basal –
90 min
TBA <0,001 <0,001 0,039 <0,001
OK <0,001 <0,001 0,771 <0,001
NT <0,001 <0,001 0,468 0,001
KO 0,702 0,001 0,633 <0,001
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
Die AUC der Speichelcortisolkonzentrationen wiesen keine signifikanten
Unterschiede zwischen der TBA, NT, OK und der Kontrollgruppe auf (siehe
Tabelle 7).
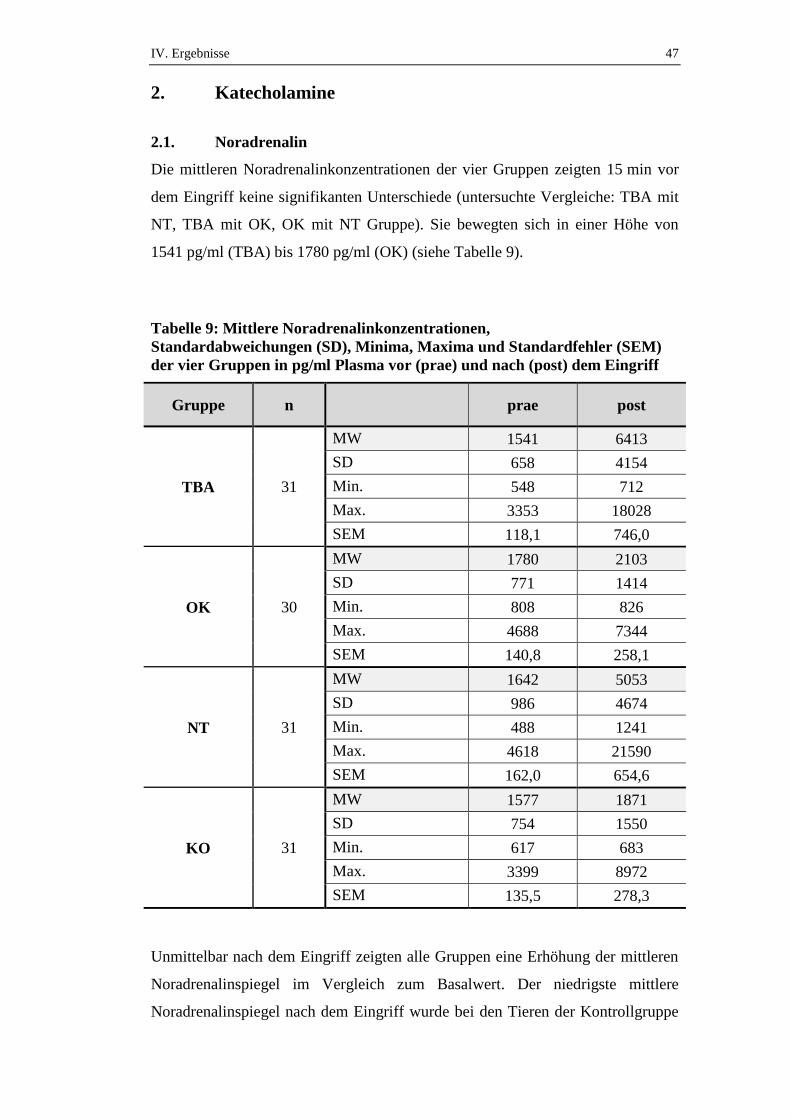
IV. Ergebnisse 47
2. Katecholamine
2.1. Noradrenalin
Die mittleren Noradrenalinkonzentrationen der vier Gruppen zeigten 15 min vor
dem Eingriff keine signifikanten Unterschiede (untersuchte Vergleiche: TBA mit
NT, TBA mit OK, OK mit NT Gruppe). Sie bewegten sich in einer Höhe von
1541 pg/ml (TBA) bis 1780 pg/ml (OK) (siehe Tabelle 9).
Tabelle 9: Mittlere Noradrenalinkonzentrationen,
Standardabweichungen (SD), Minima, Maxima und Standardfehler (SEM)
der vier Gruppen in pg/ml Plasma vor (prae) und nach (post) dem Eingriff
Gruppe n
prae post
TBA 31
MW 1541 6413
SD 658 4154
Min. 548 712
Max. 3353 18028
SEM 118,1 746,0
OK 30
MW 1780 2103
SD 771 1414
Min. 808 826
Max. 4688 7344
SEM 140,8 258,1
NT 31
MW 1642 5053
SD 986 4674
Min. 488 1241
Max. 4618 21590
SEM 162,0 654,6
KO 31
MW 1577 1871
SD 754 1550
Min. 617 683
Max. 3399 8972
SEM 135,5 278,3
Unmittelbar nach dem Eingriff zeigten alle Gruppen eine Erhöhung der mittleren
Noradrenalinspiegel im Vergleich zum Basalwert. Der niedrigste mittlere
Noradrenalinspiegel nach dem Eingriff wurde bei den Tieren der Kontrollgruppe
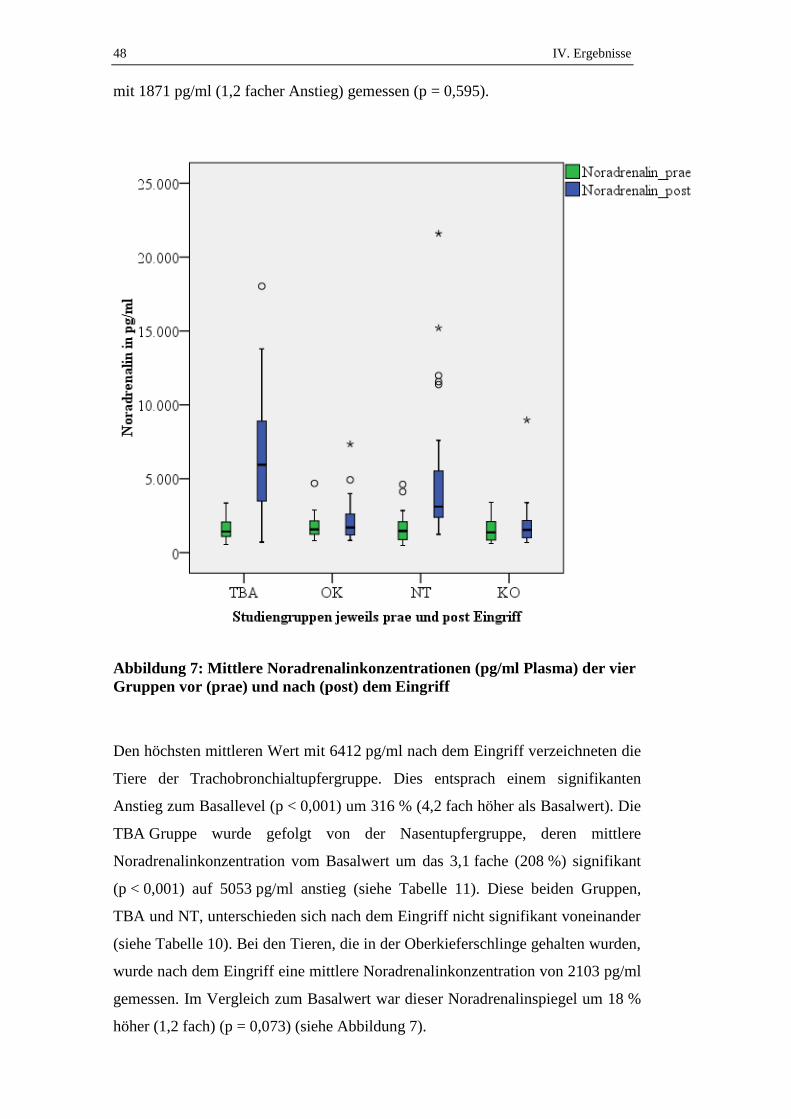
48 IV. Ergebnisse
mit 1871 pg/ml (1,2 facher Anstieg) gemessen (p = 0,595).
Abbildung 7: Mittlere Noradrenalinkonzentrationen (pg/ml Plasma) der vier
Gruppen vor (prae) und nach (post) dem Eingriff
Den höchsten mittleren Wert mit 6412 pg/ml nach dem Eingriff verzeichneten die
Tiere der Trachobronchialtupfergruppe. Dies entsprach einem signifikanten
Anstieg zum Basallevel (p < 0,001) um 316 % (4,2 fach höher als Basalwert). Die
TBA Gruppe wurde gefolgt von der Nasentupfergruppe, deren mittlere
Noradrenalinkonzentration vom Basalwert um das 3,1 fache (208 %) signifikant
(p < 0,001) auf 5053 pg/ml anstieg (siehe Tabelle 11). Diese beiden Gruppen,
TBA und NT, unterschieden sich nach dem Eingriff nicht signifikant voneinander
(siehe Tabelle 10). Bei den Tieren, die in der Oberkieferschlinge gehalten wurden,
wurde nach dem Eingriff eine mittlere Noradrenalinkonzentration von 2103 pg/ml
gemessen. Im Vergleich zum Basalwert war dieser Noradrenalinspiegel um 18 %
höher (1,2 fach) (p = 0,073) (siehe Abbildung 7).

IV. Ergebnisse 49
Tabelle 10: p-Werte der Vergleiche der mittleren
Noradrenalinkonzentrationen der Gruppen TBA, OK, NT und KO zu den
zwei Beprobungszeitpunkten
Vergleiche prae post
TBA – NT 0,955 0,071
TBA – OK 0,236 <0,001
OK – KO 0,280 0,236
OK – NT 0,266 <0,001
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
Nach dem Eingriff überstieg die mittlere Noradrenalinkonzentration der
Tracheobronchialabstrichgruppe (p < 0,001) als auch der Nasentupfergruppe
(p < 0,001) signifikant den Noradrenalinspiegel der Oberkieferschlingengruppe.
Zwischen der mittleren Noradrenalinkonzentration der Tiere, die mit der
Oberkieferschlinge fixiert wurden und den Tieren der Kontrollgruppe wurde kein
signifikanter Unterschied ermittelt (siehe Tabelle 10).
Tabelle 11: p-Werte der Vergleiche der mittleren
Noradrenalinkonzentrationen innerhalb einer Gruppe im Vergleich prae /
post Eingriff
Gruppen prae / post
TBA <0,001
OK 0,073
NT <0,001
KO 0,595
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
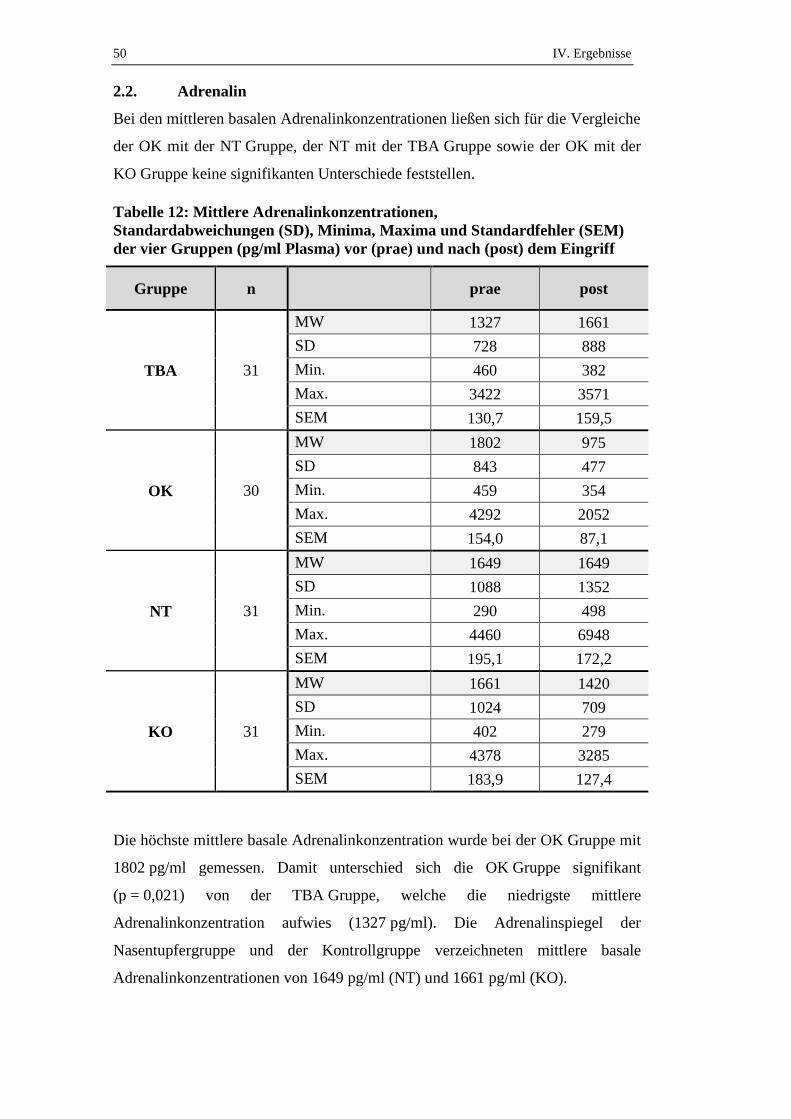
50 IV. Ergebnisse
2.2. Adrenalin
Bei den mittleren basalen Adrenalinkonzentrationen ließen sich für die Vergleiche
der OK mit der NT Gruppe, der NT mit der TBA Gruppe sowie der OK mit der
KO Gruppe keine signifikanten Unterschiede feststellen.
Tabelle 12: Mittlere Adrenalinkonzentrationen,
Standardabweichungen (SD), Minima, Maxima und Standardfehler (SEM)
der vier Gruppen (pg/ml Plasma) vor (prae) und nach (post) dem Eingriff
Gruppe n
prae post
TBA 31
MW 1327 1661
SD 728 888
Min. 460 382
Max. 3422 3571
SEM 130,7 159,5
OK 30
MW 1802 975
SD 843 477
Min. 459 354
Max. 4292 2052
SEM 154,0 87,1
NT 31
MW 1649 1649
SD 1088 1352
Min. 290 498
Max. 4460 6948
SEM 195,1 172,2
KO 31
MW 1661 1420
SD 1024 709
Min. 402 279
Max. 4378 3285
SEM 183,9 127,4
Die höchste mittlere basale Adrenalinkonzentration wurde bei der OK Gruppe mit
1802 pg/ml gemessen. Damit unterschied sich die OK Gruppe signifikant
(p = 0,021) von der TBA Gruppe, welche die niedrigste mittlere
Adrenalinkonzentration aufwies (1327 pg/ml). Die Adrenalinspiegel der
Nasentupfergruppe und der Kontrollgruppe verzeichneten mittlere basale
Adrenalinkonzentrationen von 1649 pg/ml (NT) und 1661 pg/ml (KO).
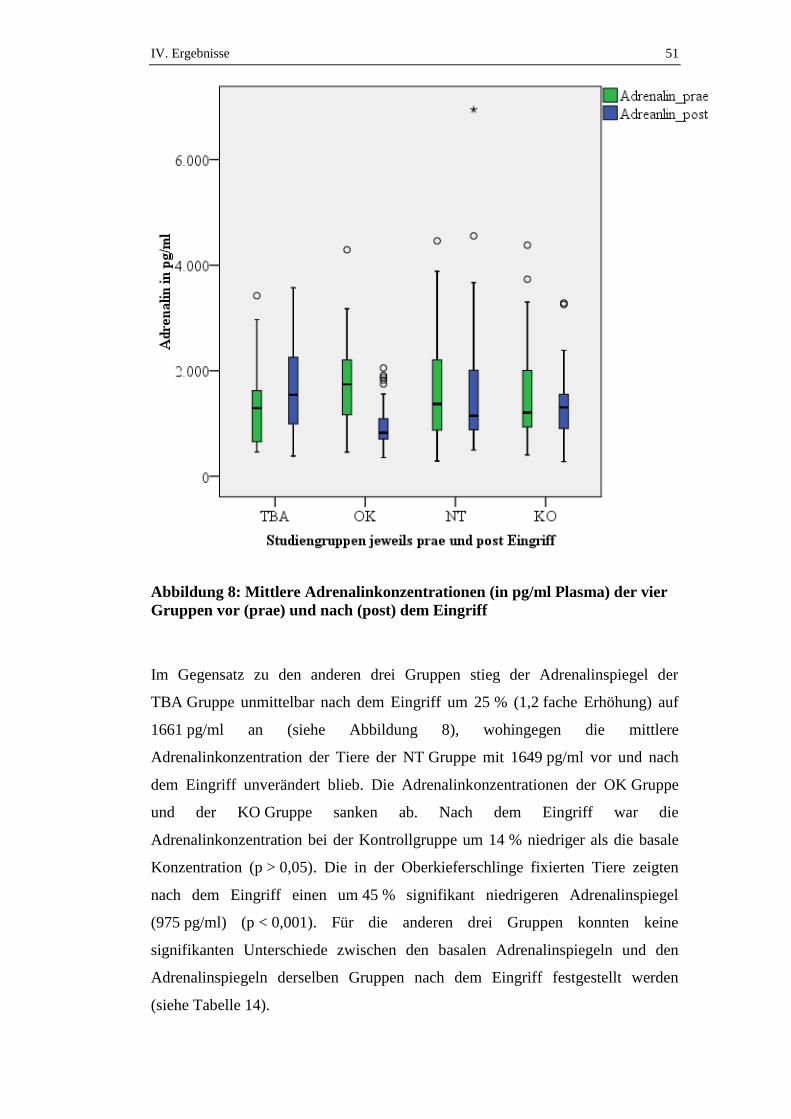
IV. Ergebnisse 51
Abbildung 8: Mittlere Adrenalinkonzentrationen (in pg/ml Plasma) der vier
Gruppen vor (prae) und nach (post) dem Eingriff
Im Gegensatz zu den anderen drei Gruppen stieg der Adrenalinspiegel der
TBA Gruppe unmittelbar nach dem Eingriff um 25 % (1,2 fache Erhöhung) auf
1661 pg/ml an (siehe Abbildung 8), wohingegen die mittlere
Adrenalinkonzentration der Tiere der NT Gruppe mit 1649 pg/ml vor und nach
dem Eingriff unverändert blieb. Die Adrenalinkonzentrationen der OK Gruppe
und der KO Gruppe sanken ab. Nach dem Eingriff war die
Adrenalinkonzentration bei der Kontrollgruppe um 14 % niedriger als die basale
Konzentration (p > 0,05). Die in der Oberkieferschlinge fixierten Tiere zeigten
nach dem Eingriff einen um 45 % signifikant niedrigeren Adrenalinspiegel
(975 pg/ml) (p < 0,001). Für die anderen drei Gruppen konnten keine
signifikanten Unterschiede zwischen den basalen Adrenalinspiegeln und den
Adrenalinspiegeln derselben Gruppen nach dem Eingriff festgestellt werden
(siehe Tabelle 14).

52 IV. Ergebnisse
Tabelle 13: p-Werte der Vergleiche der mittleren Adrenalinkonzentrationen
zwischen den Gruppen TBA, OK, NT und KO zu den zwei
Beprobungszeitpunkten
Vergleiche prae Post
TBA – NT 0,409 0,458
TBA – OK 0,021 0,001
OK – KO 0,253 0,008
OK – NT 0,247 0,007
Signifikante Werte (nach Bonferroni-Holm Adjustierung) sind fett angegeben.
Nach dem Eingriff war die mittlere Adrenalinkonzentration der Tiere der
OK Gruppe im Vergleich zur TBA, NT und KO Gruppe signifikant niedriger
(siehe Tabelle 13). Die Tiere der TBA Gruppe unterschieden sich nicht signifikant
von der NT Gruppe zum zweiten Blutentnahmezeitpunkt (p = 0,458).
Tabelle 14: p-Werte der Vergleiche der mittleren Adrenalinkonzentrationen
innerhalb einer Gruppe im Vergleich prae / post Eingriff
Gruppen prae / post
TBA 0,083
OK <0,001
NT 0,977
KO 0,595

V. Diskussion 53
V. DISKUSSION
In der Verantwortung als Tierarzt und der zunehmenden Bedeutung des
Tierschutzgedankens in der Gesellschaft ist es notwendig, Eingriffe am Tier
neben ihrer diagnostischen Eignung auch auf Tiergerechtheit zu überprüfen. Ziel
der vorliegenden Untersuchung war es, die Belastung bei der Entnahme von
Tracheobronchialabstrichen mit der von Nasentupfern zu vergleichen.
Nasentupfer sind zum Nachweis toxinbildender Pasteurella-multocida-Stämme
(SCHOSS und ALT, 1995; HÖLTIG und HENNIG-PAUKA, 2013) und von
Influenza geeignet (GOODELL et al., 2013; GROSSE BEILAGE et al., 2013).
Die Gewinnung von Sekret durch Entnahme von Tracheobronchialtupfern wurde
erstmalig von FABLET et al. (2010)) beschrieben. Die diagnostischen Ergebnisse
der Tracheobronchialabstriche stimmten mit denen der BAL für die Erreger des
Porcine Respiratory Disease Complex (PRDC) überein (PAUSENBERGER et al.,
2012). Die Bronchoalveoläre Lavage (BAL) wird bereits seit ca. zehn Jahren zur
Diagnostik von Atemwegserkrankungen erfolgreich beim Schwein genutzt. Dafür
werden die Schweine mittels Azaperon / Ketamin-Neuroleptanalgesie narkotisiert
(GROSSE BEILAGE et al., 2013). Die Narkose ist notwendig um den Stress, der
durch das Einbringen von Flüssigkeit in die Lunge für das Tier entsteht zu
vermeiden (PAUSENBERGER et al., 2012). Allerdings ist dadurch kein
Vergleich dieser Methode hinsichtlich Stressbelastung mit der des
Tracheobronchialabstriches, der im wachen Zustand durchgeführt wird, möglich,
da die Belastung für das Tier bei der BAL auf Grund der Narkose selbst und nicht
durch die Manipulation zur BAL entsteht. Dies geht mit Ergebnissen von
SCHULZ (2007) einher, die besagten, dass durch eine Narkose die
Katecholaminausschüttung in Folge einer Fixation verändert wird.
Zum Vergleich der Stressbelastung bei der Entnahme von
Tracheobronchialabstrichen mit der von Nasentupfern, wurden Cortisol- und
Katecholaminkonzentrationen in Blut und Speichel bestimmt. Um Rückschlüsse
auf den Anteil der Stressantwort, der sich durch die dafür notwendige Fixation der
Schweine mit der Oberkieferschlinge ergab, ziehen zu können, wurde eine Gruppe
von Schweinen nur in der Oberkieferschlinge gehalten. Überdies konnten die
Veränderungen die durch wiederholtes Einfangen und Blutprobenentnahme
entstanden mit Hilfe der Kontrollgruppe abgeschätzt werden. Cortisol in Serum

54 V. Diskussion
und Speichel wurde als Parameter zur Ermittlung von akutem Stress der sowohl
durch emotionalen als auch physischen Schmerz ausgelöst wurde, gewählt
(MELLOR et al., 2000). Als weitere Parameter zur Ermittlung der Belastung in
der vorliegenden Studie wurden die Katecholamine Noradrenalin und Adrenalin
gewählt. Sie geben die Aktivität des sympathischen, adrenomedullären Systems
wieder, das vor allem in „fight-fright-flight“ Situationen schnell adaptiert und
damit Gegenwehr oder Flucht ermöglicht (MELLOR et al., 2000).
1. Cortisol
In der vorliegenden Studie wurden zur Bestimmung der Aktivität der HHN-Achse
sowohl Speichel- als auch Serumproben gewonnen und deren
Cortisolkonzentrationen ermittelt. Die Aussagekraft von Cortisol als Parameter
zur Stress- und Schmerzbelastung konnte in vielen Studien belegt werden
(ROOZEN et al., 1995; ZÖLS, 2006; LANGHOFF et al., 2009). Ferner wurde
eine gute Korrelation zwischen Speichel- und Blutcortisolspiegeln sowie die
Eignung von Speichelcortisol zum Nachweis der Aktivierung der HHN-Achse
bereits in mehreren Studien gezeigt (PARROTT et al., 1989; SCHÖNREITER et
al., 1999; MERLOT et al., 2011). Serum- und Speichelcortisol wurden in der
vorliegenden Studie zwar in der gleichen Maßeinheit von nmol/l angegeben, sie
unterschieden sich aber in der Höhe der gemessenen Werte. So wurden für die
Speichelcortisolspiegel Mittelwerte zwischen 8,3 nmol/l und 14,3 nmol/l ermittelt,
für Serumcortisol hingegen mittlere Konzentrationen zwischen 55,6 nmol/l und
111,6 nmol/l. Dies deckt sich mit den Ergebnissen von PARROTT et al. (1989),
die den Basalwert der Speichelcortisolkonzentration in Höhe von 10 % der
Serumcortisolkonzentration angaben. Die Speichelcortisolkonzentration spiegelt
den ungebundenen Anteil des Plasmacortisols wider und wird durch die
Speichelflussrate nicht beeinflusst wird (VINING et al., 1983). Eine wiederholte
Speichelprobenentnahme beeinflusste die Ergebnisse der
Speichelcortisolkonzentration von Aufzuchtschweinen nach COOK et al. (2013)
nicht. Dieser Vorteil konnte allerdings in der vorliegenden Studie nicht genutzt
werden. Um die Anzahl der Versuchstiere gering zu halten wurden demselben
Tier, welches der mehrfachen Blutprobenentnahme ausgesetzt war gleichzeitig
auch Speichelproben entnommen. Diese wiederholten Venenpunktionen stellten
eine unerwünschte zusätzliche Belastung und eine Störvariable dar, die laut
EKKEL et al. (1996) auch ethisch zu diskutieren ist. Ein Nachteil von

V. Diskussion 55
Speichelcortisol im Vergleich zu Serumcortisol als Messgröße für Stress- und
Schmerzbelastung ist die geringere Sensitivität (PARROTT et al., 1989). Darauf
deutet eine Studie von PARROTT et al. (1989) hin, bei der nach ACTH
Stimulation für Plasmacortisol ein maximaler Anstieg um 230 % und für
Speichelcortisol nur um 130 % beobachtet werden konnte. Andererseits stieg in
einer Arbeit von SCHÖNREITER et al. (1999) Speichelcortisol nach Kastration
um 255,7 % und damit deutlich höher als von PARROTT et al. (1989) erwartet.
In die vorliegende Studie wurden nur weibliche Tiere aufgenommen, da nach
RUIS et al. (1997) männliche Schweine im Mittel einen höheren basalen
Speichelcortisolspiegel zeigten, der unter anderem in der vorausgegangen
Kastration begründet liegen könnte. Großer Berücksichtigung erfordert die
circadiane Rhythmik der Cortisolsekretion in der vorliegenden Untersuchung. Im
Gegensatz zu Arbeiten die sich mit der Cortisolantwort nach Saugferkelkastration
beschäftigten (ZÖLS, 2006; SCHULZ, 2007), waren bei den acht Wochen alten
Studientieren alle Eingriffe stets zur selben Tageszeit durchzuführen. Die
Angaben über die Entwicklung eines stabilen tageszeitabhängigen Rhythmus
variierten bei verschiedenen Autoren. Das Alter in dem sich ein
tageszeitabhängiger Rhythmus bildet, wurde zwischen der ersten Lebenswoche
eines Ferkels bis zum Eintritt in die Pubertät angegeben (RUIS et al., 1997;
GALLAGHER et al., 2002) und von EKKEL et al. (1996) bereits für acht
Wochen alte Schweine beobachtet. Um eine additive Beeinflussung der
gemessenen Konzentrationen durch tageszeitabhängige Cortisolausschüttung zu
vermindern, wurde der Beprobungsdurchgang an jedem Studientag mittags zum
tageszeitlichen Cortisol Minimalniveau begonnen (MERLOT et al., 2011).
1.1. Kontrollgruppe
Die Tiere der Kontrollgruppe wurden nur zu den Blutprobenentnahmen
herausgefangen. Zum Zeitpunkt des Eingriffes wurden sie ohne Beunruhigung in
der Bucht belassen. Daraus ergab sich für diese Schweine eine einstündige Pause
zwischen der ersten und zweiten Blutprobe. In dieser Zeit sanken die
Cortisolkonzentrationen der Tiere ab, bei der zweiten Blutprobe wurden wieder
Serum- und Speichelcortisolkonzentrationen, ähnlich dem Basallevel gemessen.
Dies deckt sich mit GRECO und STABENFELDT (2013) die für die biologische
Halbwertszeit von Cortisol 60 min angaben. Maximale Serum- und
Speichelcortisolspiegel wurden in dieser Gruppe zum Zeitpunkt der dritten

56 V. Diskussion
Blutentnahme (60 min-Wert) erreicht, danach sanken die mittleren
Cortisolkonzentrationen trotz dieser weiteren Venenpunktion ab und erreichten
120 min nach der ersten Blutentnahme wieder Basalniveau (kein signifikanter
Unterschied zum Basalwert). Die Cortisolspiegel der Kontrollgruppe dienten als
Maß der Belastung durch eine Blutprobenentnahme aus der Vena jugularis
externa. Dies deckte sich mit der Studie von LANGHOFF (2008), die bei ihren
Handlingstieren bei Durchführung eines ähnlichen Beprobungsplanes eine
Beeinflussung durch Fixation und Blutentnahme beobachtete. Ähnliches wurde
auch in anderen Studien nach wiederholter Venenpunktion beobachtet
(SYLVESTER et al., 1998; MELLOR und STAFFORD, 2000; MELLOR et al.,
2002). SYLVESTER et al. (1998) gewannen von Kälbern einer Kontrollgruppe in
ähnlichem Intervall Blut per Venenpunktion und beobachteten zunächst auch
einen geringgradigen Anstieg der Cortisolkonzentration durch Handling und
Beprobung. Innerhalb einer Stunde erlangte der Cortisolwert wieder Basallevel.
Die kurzfristige Cortisolkonzentrationserhöhung führten sie auf die ungewohnte
Situation zurück (SYLVESTER et al., 1998) an die sich die Tiere gewöhnten
(MELLOR und STAFFORD, 2000; MELLOR et al., 2002). Bezug nehmend auf
diese Studien kann man von einem ähnlichen Effekt in der eigenen Studie
ausgehen.
1.2. Fixierung in der Oberkieferschlinge
Eine Fixierung in der Oberkieferschlinge gewährleistet eine ausreichende
Ruhigstellung von Schweinen zu diagnostischen Maßnahmen (PLONAIT, 2004).
Zur Belastung, die durch diese Zwangsmaßnahme verursacht wird, wurden bereits
Studien mit unterschiedlichen Stressindikatoren durchgeführt. NEUBERT et al.
(1996) zeigten, dass neben der Lactat- die Katecholaminmessung geeignet ist um
den Grad und Zeitpunkt der körperlichen Anstrengung zu bestimmen. Neben
Katecholaminen empfahlen ROOZEN et al. (1995) Beta-Endorphine und Cortisol
als geeigneten Nachweis von kurzzeitigem Stress, ausgelöst durch das Halten der
Schweine in der Oberkieferschlinge. Außerdem zeigten COOK et al. (1996) in
ihrer Studie sowohl die Eignung von Speichelcortisol als Stressparameter als auch
eine starke Korrelation zwischen Serum- und Speichelcortisol. Für diese
Untersuchung wurden die Schweine jeweils fünf Minuten in der
Oberkieferschlinge fixiert und anschließend für 30 min in fünfminütigem Abstand
Blut über einen Venenverweilkatheter und Speichel durch freiwilliges Kauen auf

V. Diskussion 57
einem Wattetupfer gewonnen (COOK et al., 1996). Gegenteilig zur eigenen
Untersuchung, die sowohl maximale mittlere Serum- als auch
Speichelcortisolspiegel 60 min nach Fixierung nachwies, wurden bei COOK et al.
(1996) maximale Cortisolspiegel schon fünf Minuten nach Loslösen aus der
Oberkieferschlinge gemessen. Des Weiteren beruhigten sich die Schweine bei
COOK et al. (1996) bereits nach 30 min und wiesen zu diesem Zeitpunkt zum
Basalwert vergleichbare Serum- und Speichelcortisolspiegel auf. Demgegenüber
sanken die mittleren Serumcortisolspiegel in der eigenen Arbeit erst nach 60 min
ab. Dies stimmte mit den Ergebnissen von MERLOT et al. (2011) überein, die
ebenfalls ein Absinken der Serum- und Speichelcortisolkonzentration 60 min nach
fünfminütiger Fixierung in der Oberkieferschlinge beobachteten. In der
vorliegenden Arbeit erreichten weder die mittleren Serum- noch die
Speichelcortisolkonzentrationen 90 min nach Halten in der Oberkieferschlinge
den Basalwert, entgegen der Beobachtungen von MERLOT et al. (2011), deren
Tiere nach 90 min zum Basalwert vergleichbare mittlere Serum- und
Speichelcortisolkonzentrationen aufwiesen. Allerdings sollte beim Vergleich der
Studien die unterschiedliche Dauer der Fixierung in der Oberkieferschlinge
beachtet werden. Im Gegensatz zur eigenen Arbeit mit einminütiger Fixierung in
der Oberkieferschlinge wurden die Tiere bei COOK et al. (1996) und MERLOT et
al. (2011) für fünf Minuten in der Oberkieferschlinge gehalten. In weiteren
Versuchen zeigten RUSHEN und LADEWIG (1991), dass Schweine, die
15 Minuten in der Oberkieferschlinge gehalten wurden endogene Opioide
freisetzten und dies zu einer vorübergehenden Hypoalgesie führte und die
Stressantwort auf die vorausgegangene Bewegungseinschränkung abmilderte
(RUSHEN und LADEWIG, 1991). Durch diesen Effekt könnte es auch in der
eigenen Studie zu einer Reduzierung der Cortisolfreisetzung nach der Fixierung in
der Oberkieferschlinge gekommen sein. Dies stimmt auch mit den Ergebnissen
von PHOGAT und PARVIZI (2007) überein, in deren Studie es ebenfalls zur
Verminderung der Stressantwort durch endogene Opioide kam.
1.3. Nasentupferentnahme
In der vorliegenden Studie unterschieden sich die mittleren Serum- und
Speichelcortisolkonzentrationen nach Nasentupferentnahme (NT) zu keinem
Zeitpunkt signifikant von denen der TBA und OK Gruppe. Im Gegensatz zu
Tieren bei denen eine Tracheobronchialabstrichentnahme durchgeführt wurde und

58 V. Diskussion
die ihr Maximum 30 min nach dem Eingriff erreichten, stiegen die mittleren
Serum- und Speichelcortisolspiegel der Tiere, bei denen ein Nasentupfer
durchgeführt wurde bis 60 min nach dem Eingriff an. Es gilt zu berücksichtigen,
dass das nozizeptive System in einen peripheren und zentralen Anteil, zu dem
unter anderem der somatosensorische Kortex zählt, gegliedert ist
(PFANNKUCHE, 2008). Bezug nehmend auf GASTL et al. (2014), die für die
nasale Mukosa eine eigenständige Projektion auf dem somatosensorischen Kortex
nachgewiesen haben, kann vermutet werden, dass diese starke Repräsentation
auch in der vorliegenden Studie zu einer länger andauernden Reizantwort führte.
Zusammen mit dem Thalamus ist der somatosensorische Kortex, der einen Teil
der Großhirnrinde darstellt laut ILLES und ALLGAIER (2013) für die bewusste
Schmerzempfindung sowie die Lokalisation und die Registrierung der Stärke von
Schmerzen zuständig. Ferner wirkt sich ein schmerzhafter Stimulus auf das
spinoreticulothalamische System mit einer Erhöhung des Wachzustandes aus
(ILLES und ALLGAIER, 2013). Außerdem führen laut SÖRÖS et al. (2001)
schmerzhafte Reize bereits nach wenigen Minuten zu einer örtlichen
Umstrukturierung der somatosensorischen Großhirnrinde. Es kommt dabei zu
einer Überempfindlichkeit somatotopisch benachbarter Regionen (SÖRÖS et al.,
2001). Durch die Untersuchung der Großhirnrinde konnten CRANER und RAY
(1991) zeigen, dass das Rostrum bei neonaten Schweinen sehr ausgeprägt
somatotopisch repräsentiert und benachbart zur Region von Gesicht und Maul auf
dem Kortex lokalisiert ist. Dies deckt sich mit den Beobachtungen der eigenen
Studie, in der die mittleren Serum- und Speichelcortisolkonzentrationen der NT
und der OK Gruppe bis zum 60 min-Wert anstiegen und für eine Irritation durch
die Eingriffe spricht.
1.4. Tracheobronchialabstrichentnahme
Für die Durchführung des Tracheobronchialabstriches ist ebenso eine Fixierung in
der Oberkieferschlinge nötig wie für die des Nasentupfers. Zu keinem Zeitpunkt
der eigenen Untersuchung unterschieden sich die mittleren Speichel- und
Serumcortisolkonzentrationen der TBA Gruppe weder von der OK noch von der
NT Gruppe signifikant. Bezugnehmend auf die Studie von WIDDICOMBE
(1995) könnte die geringere Sensitivität der Rezeptoren in den unteren
Atemwegen gegen mechanische Reize, verbunden mit einer zunehmenden
Chemosensititvität dafür verantwortlich sein, dass die Stressantwort trotz

V. Diskussion 59
Hustenreiz nicht deutlicher ausfällt als für die Anwendung der Oberkieferschlinge
und des Nasentupfers. Die Ergebnisse deuten darauf hin, dass die Anwendung
eines Tracheobronchialabstriches nach 30 min eine ähnliche Stressreaktion auslöst
wie die Entnahme eines Nasentupfers und es könnte daraus geschlossen werden,
dass der Großteil der Belastung von der Immobilisation herrührt. Denn vergleicht
man die OK Gruppe mit der TBA Gruppe, so konnte zwischen diesen beiden
Gruppen in der vorliegenden Untersuchung 30 min nach dem Eingriff kein
signifikanter Unterschied nachgewiesen werden. Dass eine Immobilisaton für
Schweine mit Stress verbunden ist, wurde auch von LANGHOFF (2008)
nachgewiesen. Demnach waren die mittleren Cortisolkonzentrationen bis zu einer
Stunde nach Fixation der Ferkel im Kastrationsbock signifikant zu den Werten
vor dieser Maßnahme erhöht (LANGHOFF et al., 2009). Allerdings muss die
altersabhängige Reaktion auf Schmerz berücksichtigt werden, nachdem in der
eigenen Studie acht Wochen alte Tiere im Gegensatz zu Saugferkeln
(LANGHOFF, 2008) untersucht wurden. Bezugnehmend auf die Studie von
FANG et al. (2005), die Untersuchungen an zwei, vier und sechs Monate alten
Schweinen durchführten, konnte eine erhöhte Schmerzhaftigkeit bei vier Monate
alten Schweinen nachgewiesen werden. Dies deckt sich mit der Beobachtung von
CARROLL et al. (2006), dass Handling für Schweine mit zunehmendem Alter
belastender wird.
In einer Studie zu Agonisten und Antagonisten des beta-adrenergen Systems
konnte gezeigt werden, dass als Folge der Beta-adrenergen Stimulation der
Cortisolspiegel ansteigt, allerdings durch einen Antagonisten dieses Systems
kaum Einfluss auf die Cortisolbasalsekretion genommen werden kann (PHOGAT
und PARVIZI, 2007). Bei gleichzeitiger Betrachtung der mittleren
Noradrenalinspiegel und der mittleren Speichel- und Serumcortisolspiegel der
TBA Gruppe, die sich vor und nach dem Eingriff jeweils signifikant
unterschieden, ist dieser Effekt nachzuvollziehen. Es wird angenommen, dass eine
Katecholaminfreisetzung nach Tracheobronchialabstrichprobenentnahme eine
vermehrte Cortisolsekretion nach sich zieht. Dies kann analog auf die
Beobachtungen zur Belastung bei Nasentupferprobenentnahmen übertragen
werden. Die Überlegungen zur Freisetzung von endogenen Opioiden bei
Fixierung mit der Oberkieferschlinge sind auch für die TBA Gruppe anzuwenden,
da dabei ebenfalls der Gebrauch der Oberkieferschlinge nötig ist.

60 V. Diskussion
1.5. Speichelcortisol
Der Teil der Speichelcortisolergebnisse, der mit den Serumcortisolergebnissen
übereinstimmt, wurde bereits im Zuge der Diskussion der einzelnen
Studiengruppen erläutert. Abweichungen zwischen den beiden
Untersuchungsmedien werden im Folgenden besprochen.
Alle vier Gruppen erreichten ihre maximale Speichelcortisolkonzentration 60 min
nach dem Eingriff, dagegen wurde die höchste mittlere
Serumcortisolkonzentration der TBA Gruppe schon 30 min nach dem Eingriff
beobachtet. Ferner war zwar ein nummerischer Abfall von der 60 min zur 90 min
Messung der Speichelcortisolwerte zu beobachten, aber außer für die
TBA Gruppe stellte sich kein signifikanter Abfall der Cortisolkonzentrationen
zwischen diesen beiden Beprobungszeitpunkten ein. Überdies kehrte die
Kontrollgruppe bezüglich der mittleren Speichelcortisolkonzentration in der
vorliegenden Studie nicht zum Basalwert zurück wie dies für die mittlere
Serumcortisolkonzentration der Fall war. Dies deckt sich mit der Studie von
PARROTT et al. (1989), die ebenfalls eine verspätete sowie abgeschwächte
Cortisolantwort im Speichel im Vergleich zum Serum beobachteten. Im Hinblick
auf die verzögerte Rückkehr zum Basalwert erzielten BUSHONG et al. (2000)
vergleichbare Ergebnisse wie in der vorliegenden Studie. Ähnlich deren
Untersuchungen, könnte es auch in der eigenen Arbeit zu einem im Serum
festzustellenden Abfall der Gesamtcortisolkonzentration gekommen sein, wovon
aber noch ein relativ großer Anteil nicht an Bindungsproteine gebunden wurde
und somit weiterhin im Speichel gemessen werden konnte (BUSHONG et al.,
2000). BUSHONG et al. (2000) schlussfolgerten, dass Speichelcortisol weniger
einen konstanten prozentualen Anteil vom Gesamtcortisol im Plasma
widerspiegelt, sondern dass die Höhe des im Speichel gemessenen Cortisols mehr
vom ungebundenen Cortisol abhängt als von der Gesamtcortisolkonzentration.
Auch ADCOCK et al. (2007) zeigten, dass zwischen Blutcortisolkonzentration
und porcinem Corticosteroid-bindendem Globulin (pCBG) ein negativer
Zusammenhang besteht. Nach akutem Stress sanken die pCBG, was einen Anstieg
des freien Cortisols, das im Speichel gemessen wurde, zur Folge hatte (ADCOCK
et al., 2007). Auch vermehrter Stress der Muttersauen während der Trächtigkeit
übt einen vermindernden Einfluss auf die Konzentrationen des Cortisol-bindenden
Globulins der Ferkel aus, was zu einem höheren Spiegel von biologisch aktivem

V. Diskussion 61
Cortisol führt (KANITZ et al., 2006).
Entgegen der eigenen Ergebnisse die keinen signifikanten Abfall der
Speichelcortisolwerte von der 60 min zur 90 min Messung, außer für die
TBA Gruppe, dokumentierten, sank der Speichelcortisolspiegel der mit der
Oberkieferschlinge fixierten Tiere bei MERLOT et al. (2011) aber schon nach
30 min ab. Für diese Studie wurden die gleichzeitig erfolgenden Blutproben
jedoch über einen Venenverweilkatheter gewonnen. Dessen Benutzung könnte ein
früheres Absinken der Speichelcortisolkonzentration ermöglichen, da es zu keiner
weiteren Beeinflussung der Tiere durch Venenpunktion kam.
1.6. AUC
Mit Maximalkonzentrationen werden nur Belastungsspitzen wiedergegeben, dies
ist aber laut MELLOR und STAFFORD (2004) ebenso wenig sinnvoll wie allein
die Dauer einer Belastung zu untersuchen. Daher wurden zur besseren Beurteilung
der Gesamtbelastung die gemessenen Konzentrationen einzelner Zeitpunkte
summiert und nach BLAND (2009) die Konzentrations-Zeit-Kurven (AUC)
berechnet. In der vorliegenden Studie wurden keine signifikanten Unterschiede
bezüglich der aus den Serumcortisolwerten errechneten AUC zwischen den
Gruppen TBA, NT und OK ermittelt. Allerdings unterschied sich der AUC der
OK Gruppe für Serumcortisol signifikant von der KO Gruppe. Dieser signifikante
Unterschied zwischen der Oberkieferschlingengruppe und der Kontrollgruppe
ergab sich für einzelne Vergleiche der Beprobungszeitpunkte nur 30 min nach
dem Eingriff. Daraus lässt sich ableiten, dass die Gesamtbelastung für ein Tier
bezogen auf den Parameter Serumcortisol für die Eingriffe
Tracheobronchialabstrichentnahme, Nasentupferentnahme und Fixierung in der
Oberkieferschlinge vergleichbar war und über der Belastung für die
Kontrollgruppe lag. Im Gegensatz dazu unterschieden sich die AUC aus dem
Speichelcortisol zwischen den Gruppen nicht signifikant, obwohl 30 min nach
dem Eingriff, ebenso wie bei der Serumcortisolkonzentration, ein signifikanter
Unterschied zwischen der Kontrollgruppe und der Oberkieferschlingengruppe
bestand. Eine Erklärung dafür könnte sein, dass die Speichelcortisolspiegel länger
erhöht bleiben als die Serumcortisolspiegel (BUSHONG et al., 2000). Dies hätte
bei der Berechnung über die Zeit einen großen Einfluss und verminderte die
Unterschiede zwischen den Gruppen. Auch MERLOT et al. (2011) beschrieben
größere Schwierigkeiten, signifikante Unterschiede bei der Auswertung von

62 V. Diskussion
Speichelcortisol als von Serumcortisol darzustellen, vor allem beim Vergleich
einer Gruppe, der ein stresshafter Reiz zugefügt wurde mit einer Kontrollgruppe.
Dies führten sie auf einen niedrigeren Anstieg für Speichel- als für Serumcortisol
zurück (MERLOT et al., 2011).
2. Katecholamine
Zur Ermittlung der Stress- und Schmerzbelastung wurde Plasma gewonnen und
die Noradrenalin- und Adrenalinkonzentrationen bestimmt. Dass Katecholamine
geeignet sind, um die Belastung von Schweinen zu ermitteln wurde mehrfach in
Studien bestätigt (NEUBERT et al., 1996; LACKNER, 2003; OTTEN et al.,
2004; ZIMMERMANN et al., 2011).
2.1. Kontrollgruppe
In der vorliegenden Studie wurden weder für die mittlere Noradrenalin- noch für
die Adrenalinkonzentration der Kontrollgruppe signifikante Unterschiede
zwischen der ersten und zweiten Blutprobenentnahme beobachtet. Dies spricht für
eine Elimination der Katecholamine, die nach der ersten Blutprobe ausgeschüttet
wurden bis zum Zeitpunkt der zweiten Blutentnahme und stimmt mit MÖSTL
(2010) überein, wonach die Halbwertszeiten der Katecholamine sehr kurz sind
und zwischen 20 sec und 10 min variieren.
2.2. Fixierung in der Oberkieferschlinge
Eine Untersuchung der Stress- und Schmerzbelastung mittels verschiedenen
Parametern, darunter Cortisol und Katecholaminen, wurde bereits von ROOZEN
et al. (1995) und NEUBERT et al. (1996) durchgeführt. Beide untersuchten die
Effekte, die durch eine kurzfristige Fixation von Schweinen mittels
Oberkieferschlinge auftraten und verzeichneten sowohl einen Anstieg der
Katecholamin- als auch der Cortisolkonzentrationen. Dies steht im Gegensatz zu
den eigenen Ergebnissen bezüglich der Noradrenalinkonzentration, die sich nach
der Fixation mittels Oberkieferschlinge nicht signifikant von der
Basalkonzentration unterschied. Ferner lag die mittlere Noradrenalinkonzentration
der OK Gruppe nach dem Eingriff nicht signifikant über der KO Gruppe. Auch
für Adrenalin war kein Anstieg der mittleren Konzentration der OK Gruppe nach
dem Eingriff zu verzeichnen. Der mittlere Adrenalinspiegel lag nach der Fixation
sogar signifikant unter dem Basalwert. Allerdings ging die Bestimmung des

V. Diskussion 63
Basalwertes in der vorliegenden Studie auch mit einer Beeinflussung der Tiere
durch Einfangen, Immobilisierung und Blutentnahme einher. Darum kann der
Basalwert in unserer Studie eher als Referenz- und weniger als Ruhewert
angenommen werden. In einer anderen Studie wurde in zeitlichem Abstand zur
Beprobung ein Venenverweilkatheter platziert, um eine Blutprobenentnahme ohne
Beeinflussung der Katecholaminspiegel zu ermöglichen (ROSOCHACKI et al.,
2000). In der Gruppe, die nur mit der Oberkieferschlinge fixiert wurde stiegen die
Katecholaminkonzentrationen im Gegensatz zu Cortisol, bei dem 30 min nach
dem Eingriff ein signifikanter Unterschied zur Kontrollgruppe zu beobachten war,
nicht an. DE BOER et al. (1990) zeigten ebenfalls, dass erhöhte Cortisolspiegel
nicht immer mit einem Anstieg von Katecholaminen einhergehen. Umgekehrt
bedingen erhöhte Katecholaminspiegel aber eine Aktivierung der HHN-Achse
(DE BOER et al., 1990).
2.3. Nasentupfer- und Tracheobronchialabstrichentnahme
Vergleichbar mit den beschriebenen Cortisolergebnissen, konnte bei der
vorliegenden Untersuchung ein signifikanter Anstieg der
Noradrenalinkonzentration und ein nicht signifikanter Anstieg der
Adrenalinkonzentration nach der Durchführung eines Tracheobronchialabstriches
beobachtet werden. Die mittlere Noradrenalinkonzentration der TBA Gruppe stieg
nach dem Eingriff um das 4,2 fache und für Adrenalin um das 1,2 fache an. Nach
Entnahme eines Nasentupfers erreichte der Noradrenalinspiegel in der
vorliegenden Studie das 3,1 fache und Adrenalin blieb auf dem Niveau des
Basalwertes. Nach dem Eingriff lagen die mittleren Katecholaminspiegel sowohl
der TBA als auch der NT Gruppe signifikant über denen der OK Gruppe. Eine
Erklärung dafür könnte sich in humanmedizinischen Studien finden. Beim
Menschen wurden umso höhere Noradrenalinspiegel beobachtet, desto größer das
Ausmaß des chirurgischen Traumas war (WILMORE et al., 1976; FRIEDRICH et
al., 1999). Die Aktivierung des adrenergen Systems erfolgt durch afferente
Signale vom Ort der Läsion (HALTER et al., 1977). Dies dürfte in der
vorliegenden Studie nur einen sehr geringen Einfluss ausüben, da mit einer
geringgradigen Gewebeschädigung zu rechnen war. Den für die
Tracheobronchialabstriche verwendeten Kathetern hafteten nach Benutzung keine
Gewebeteile oder Blut an. Auch nach der Entnahme der Nasentupfer kam es zu
keinen akuten Blutungen. Die Tupfer zeigten maximal eine rosa Verfärbung, die

64 V. Diskussion
im Rahmen der Diagnostik erwünscht ist und wahrscheinlich mit einer
geringgradigen Schleimhautirritation einherging.
Der mittlere Adrenalinspiegel der Tiere, die in der vorliegenden Untersuchung mit
der Oberkieferschlinge fixiert wurden, nahm nach dieser Zwangsmaßnahme
signifikant ab. Hingegen wurden für die anderen drei Gruppen (TBA, NT, KO)
keine signifikanten Unterschiede zwischen den Adrenalinwerten vor und nach
dem Eingriff beobachtet. Eine mögliche Erklärung könnte darauf beruhen, dass
die Tiere schon bevor die erste Blutentnahme stattfand unterschwellig gestresst
waren (MELLOR et al., 2002). Dagegen spricht aber, dass sich auch die basalen
Adrenalinkonzentrationen in der vorliegenden Studie zwischen den Gruppen
unterschieden. Zwischen der TBA und der OK Gruppe bestand zum Zeitpunkt der
ersten Blutprobenentnahme ein signifikanter Unterschied. Da die Tiere
randomisiert wurden und nach Gruppen gemischt beprobt wurde, ist ein
gruppenspezifischer Stressfaktor vor der ersten Blutentnahme unwahrscheinlich.
Bei der Interpretation der Ergebnisse der eigenen Untersuchung muss aber
beachtet werden, dass die Adrenalinspiegel sich vor und nach dem Eingriff kaum
veränderten und nur ein signifikanter Abfall der Adrenalinkonzentration nach
Fixierung mit der Oberkieferschlinge festgestellt werden konnte. Ferner könnte
sich auch die zeitliche Abfolge der Studienmaßnahmen darauf ausgewirkt haben,
dass die Adrenalinkonzentrationen bei keiner der Gruppen nach dem Eingriff
signifikant anstiegen und für die OK Gruppe nach der Fixierung signifikant
sanken. So wurde die Stressantwort, die der Fixierung in der Oberkieferschlinge
folgte, bei der Ermittlung der Adrenalinkonzentration aus der Blutprobe, die nach
Loslösen des Schweines aus der Fixierung erfolgte, möglicherweise nicht mehr
erfasst. Diese Hypothese deckt sich mit den Ergebnissen von OTTEN et al.
(2004), die eine sehr kurze Plasmahalbwertszeit für Katecholamine beschreiben
und höchste Katecholaminkonzentrationen bereits eine Minute nach Beginn der
Fixierung in der Oberkieferschlinge ermittelten. Nach dem Eingriff lag die
mittlere Noradrenalinkonzentration der TBA Gruppe nummerisch über der
NT Gruppe aber es unterschieden sich weder die Plasmaspiegel von Noradrenalin
noch die von Adrenalin zwischen der TBA und der NT Gruppe signifikant.
Sowohl die Noradrenalinspiegel der Nasentupfergruppe als auch der
Tracheobronchialabstrichgruppe stiegen nach dem Eingriff signifikant zum
Basallevel an. Die geringe Veränderung der Adrenalinspiegel nach den Eingriffen

V. Diskussion 65
wurde auch von MELLOR et al. (2002) bei Lämmern nach Kastration und
Schwanzkupieren per Gummiring beschrieben. Wohingegen die
Noradrenalinspiegel der Lämmer einen Anstieg verzeichneten, den sie auf
ischämischen Schmerz zurückführten (MELLOR et al., 2002). Ähnlich wie in
deren Studie wird auch in der vorliegenden Studie davon ausgegangen, dass die
Erhöhung der Noradrenalinspiegel auf eine periphere Sympathikusaktivität
zurückgeht (MELLOR et al., 2002). Zur Aktivierung des sympathoadrenergen
Systems kommt es in „fight or flight“ Situationen, es unterstützt den Organismus
in Zuständen der Hypovolämie, Hypoxie und Hypoglycämie sowie
lebensbedrohlichen Schockzuständen (GOLDSTEIN, 1987). Dabei steigt der
Adrenalinspiegel im Gegensatz zur Noradrenalinkonzentration stark an, ebenso
wie unter kognitiver Erregung und Angst (DIMSDALE und MOSS, 1980; WARD
et al., 1983; SGOIFO et al., 1996; SILBERNAGEL und DESPOPOULUS,
2012c). Noradrenalin als Transmitter der postganglionär-sympathischen Neurone,
gibt die neuronale sympathische Aktivität wieder und verändert sich unter solchen
Zuständen kaum (GOLDSTEIN, 1987; STARKE, 2013). Diese Ansicht teilten
auch KVETNANSKY et al. (1979) wonach die Noradrenalin-Antwort auf
Immobilisationsstress hauptsächlich aus den sympathischen Endigungen und nur
zu 20% aus den Nebennieren stammt, wohingegen Adrenalin vorwiegend aus dem
Nebennierenmark freigesetzt wird (KVETNANSKY et al., 1979). Aktive
Abwehr- und Wutreaktionen setzten laut DÖCKE und KEMPER (1994) vor allem
Noradrenalin frei. FUKUHARA et al. (1996) stellten steigende
Noradrenalinkonzentrationen auch nach Kältestress fest. Bei der eigenen
Untersuchung lagen die Noradrenalinspiegel der NT als auch der TBA Gruppe
nach dem Eingriff signifikant über denen der OK Gruppe. Gleichzeitig stiegen die
Adrenalinkonzentrationen bei keiner der Gruppen nach dem Eingriff signifikant
an, lediglich ein geringer numerischer Anstieg war für die TBA Gruppe zu
verzeichnen. Bezugnehmend auf die genannten Studien kam es bei der TBA und
NT Gruppe zu einer höheren Freisetzung von Noradrenalin aus den
sympathischen Nervenendigungen in der Peripherie als dies für die OK Gruppe
der Fall war. Dies spricht für eine größere Belastung durch Immobilisationsstress
im Sinne einer Wut- und Abwehrreaktion durch die Probenentnahme mittels
Tracheobronchial- und Nasentupfer als für das Halten in der Oberkieferschlinge.
Möglicherweise wirkte sich die unterschiedliche Fixierung der Tiere insofern aus,
dass die mittleren Katecholaminspiegel sowohl der TBA als auch der NT Gruppe

66 V. Diskussion
nach dem Eingriff signifikant über denen der OK Gruppe lagen. Sowohl die Tiere
der TBA als auch der NT Gruppe wurden zusätzlich zur Fixierung in der
Oberkieferschlinge zwischen den Beinen einer Versuchsperson stehend in der
Flankengegend stabilisiert. Bezugnehmend auf LANGHOFF (2008) die eine
Stressbelastung von Ferkeln in Folge von Immobilisation beschrieb, könnte die
zusätzliche Bewegungseinschränkung der TBA und NT Gruppe nach dem
Eingriff zu den signifikant höheren Katecholaminkonzentrationen dieser beiden
Gruppen als der OK Gruppe geführt haben. Übereinstimmend beobachteten auch
ROOZEN et al. (1995) erhöhte Plasmakonzentrationen von Katecholaminen,
Beta-Endorphin und Cortisol nach Immobilisation. Ferner könnte das Einführen
des Nasentupfers eine Stimulation des N. infraorbitalis, der die Nasenhaut und
den Nasenhöhleneingang sensibel innerviert, hervorrufen sowie zu einer
zusätzlichen Reizung der sensiblen Nasenschleimhaut, die von zwei
unterschiedlichen Ästen des N. trigeminus sensibel versorgt wird, führen
(NICKEL et al., 2004). Ähnliches dürfte für das Einführen des
Tracheobronchialkatheters zutreffen, wodurch es zu Irritationen im Rachen und
der Trachea kommen könnte. Bezugnehmend auf DE CARLOS et al. (2013) ist
die menschliche Rachenwand mit verschiedenen Rezeptoren ausgestattet, die
mechanische Reize der oberen Atemwege aufnehmen. Diese Rezeptoren werden
von sensorischen Nerven begleitet (DE CARLOS et al., 2013). Ferner liegen laut
SILBERNAGEL und DESPOPOULUS (2012b) Irritationsendigungen in der
Bronchialschleimhaut die unter anderem am Hustenreflex beteiligt sind. Der
geringgradige Anstieg der Adrenalinkonzentration der TBA Gruppe nach
Entnahme des Tracheobronchialtupfers könnte auf den psychologischen Anteil
der Belastung hinweisen, wie von DÖCKE und KEMPER (1994) und
DIMSDALE und MOSS (1980) beschrieben. Durch das Einbringen des Katheters
in die Luftröhre wurden die luftleitenden Wege behindert und zum Teil Husten
ausgelöst. Dies könnte zu einer emotionalen Erregung geführt haben. Durch
Erhöhung des Atemwiderstandes lösten ALIUS et al. (2013) auch bei Menschen
Angst und Unwohlsein sowie physiologische Reaktionen aus. Die Ergebnisse der
vorliegenden Studie deuten darauf hin, dass diese Belastung bei den Tieren der
TBA Gruppe vermutlich nur geringgradig ausgeprägt war.

VI. Schlussfolgerung 67
VI. SCHLUSSFOLGERUNG
In der vorliegenden Untersuchung wurde die Belastung bei der Entnahme von
Tracheobronchialabstrichen vergleichend mit der von Nasentupfern untersucht.
Sowohl Cortisol aus Serum als auch aus Speichel bestätigten sich als geeignete
Parameter zur Erfassung von Stress- und Schmerzbelastungen nach
diagnostischen Maßnahmen beim Schwein. Durch den Einschluss einer Gruppe
von Tieren, die in der Oberkieferschlinge gehalten wurden, war eine Aussage über
die Belastung in Folge der Fixierung möglich. Die Konzentrations-Zeit-Kurve
(AUC) ermöglichte in der vorliegenden Studie eine Beschreibung von Intensität
und Dauer der Belastung.
Der Verlauf der Cortisolkonzentrationen nach Entnahme von
Tracheobronchialabstrichen entsprach jenem nach Nasentupfern. Ein
vergleichbarer wenn auch etwas verzögerter Cortisolanstieg wurde durch die
Fixierung mit der Oberkieferschlinge hervorgerufen. Dies deutet darauf hin, dass
die bloße Fixation einen bedeutenden Anteil an der Stressantwort des Tieres trägt.
Außerdem erwies sich die Auswertung der Noradrenalinkonzentrationen als
aussagekräftig. Bezugnehmend auf diese Ergebnisse wurde gezeigt, dass der akute
Stress durch einen Tracheobronchialabstrich vergleichbar mit dem einer
Nasentupferentnahme war und beide eine wesentlich höhere Belastung als die
alleinige Fixierung mit Oberkieferschlinge darstellten. Im Gegensatz dazu waren
die Ergebnisse der Adrenalinkonzentrationen weniger aussagekräftig, was zum
einen an den divergierenden Basalwerten lag und zum anderen an den geringen
Veränderungen der Werte nach dem Eingriff. Die beobachteten Anstiege der
Adrenalinspiegel waren gering und sprechen gegen starken emotionalen Stress
und Angst der Schweine in Folge der durchgeführten Maßnahmen. Dieser
Parameter ist möglicherweise besser zur Evaluierung anderer Belastungen, die mit
Kreislaufbeeinträchtigung einhergehen (GOLDSTEIN, 1987; SILBERNAGEL
und DESPOPOULUS, 2012c) geeignet und erfordert Blutprobenentnahmen
mittels Venenverweilkatheter. Entzündungsmediatoren (Prostaglandine,
Bradykinine etc.) sensibilisieren laut HENKE und ERHARDT (2001) die
Schmerzrezeptoren und erniedrigen die Schmerzschwelle. Dies ist für die
Beprobung von Tieren in der Praxis zu bedenken, denn die zu untersuchenden
Tiere leiden häufig bereits unter einer Vorschädigung, z.B. Atemwegserkrankung.

68 VI. Schlussfolgerung
Darum könnten die Tracheobronchialabstriche und Nasentupfer für erkrankte
Schweine eventuell belastender sein als für gesunde Tiere, die in der vorliegenden
Studie untersucht wurden.
Die vorliegenden Ergebnisse zeigen, dass die Entnahme von
Tracheobronchialabstrichen bei klinisch gesunden Schweinen zu einer Belastung
vergleichbar mit der Entnahme von Nasentupfern führt. Verglichen zur alleinigen
Fixierung mit der Oberkieferschlinge führt sie zu einer deutlich ausgeprägteren
kurzfristigen Stressantwort.

VII. Zusammenfassung 69
VII. ZUSAMMENFASSUNG
Ziel der vorliegenden Untersuchung war es, die Belastung für Schweine bei der
Entnahme eines Tracheobronchialabstriches (TBA) mit der Entnahme eines
Nasentupfers (NT) und mit dem alleinigen Halten in der Oberkieferschlinge (OK)
zu vergleichen. Zur Evaluierung der Stressbelastung dienten
Cortisolkonzentrationen aus Serum- und Speichelproben sowie aus dem
Blutplasma ermittelte Noradrenalin- und Adrenalinkonzentrationen. Hierfür
wurden vier Gruppen, drei Versuchs- und eine Kontrollgruppe (KO), zu je 23
klinisch gesunden Tieren für die Cortisolbestimmungen und zu je 31 klinisch
gesunden Tieren für die Katecholaminbestimmungen untersucht. Von in der
Oberkieferschlinge fixierten Tieren der TBA und NT Gruppe wurde ein
Tracheobronchialabstrich bzw. ein Nasentupfer entnommen. Der
Tracheobronchialkatheter wurde ohne vorherige Narkose der Tiere in die
Luftröhre eingeführt. Bei den Tieren der OK Gruppe erfolgte eine Fixierung in
der Oberkieferschlinge für die Dauer von 60 Sekunden. Im Gegensatz dazu
wurden die Tiere der KO Gruppe zum Zeitpunkt des Eingriffes ohne
Beunruhigung in der Bucht belassen. Im ersten Teilversuch, welcher der
Cortisolbestimmung diente, wurde von jedem Schwein viermal Blut und zum
selben Zeitpunkt Speichel gewonnen: 30 min vor dem Eingriff sowie je 30 min,
60 min und 90 min danach. Die Katecholaminkonzentrationen wurden im zweiten
Teilversuch aus EDTA-Blutproben, die 15 min vor und unmittelbar nach dem
Eingriff entnommen wurden, ermittelt.
Die mittleren Serum- und Speichelcortisolkonzentrationen der Gruppen TBA, NT
und OK stiegen 30 min nach dem Eingriff signifikant zum Basalwert an. Zu
diesem Zeitpunkt war bei mittels Oberkieferschlinge fixierten Tieren (OK) eine
signifikant höhere Cortisolkonzentration als bei Tieren der Kontrollgruppe (KO)
messbar. Ein signifikanter Unterschied der Cortisolkonzentrationen in Serum und
Speichel konnte weder 30 min, 60 min noch 90 min nach den verschiedenen
Eingriffen (TBA, NT und OK) zwischen den Versuchsgruppen ermittelt werden.
Die mittleren Serum- und Speichelcortisolkonzentrationen der drei
Versuchsgruppen (TBA, NT und OK) sanken zum 90 min-Wert ab. In Bezug auf
die Konzentrations-Zeit-Kurven (AUC) unterschieden sich die Serum- und
Speichelcortisolkonzentrationen zwischen den Gruppen TBA, OK und NT nicht

70 VII. Zusammenfassung
signifikant. Die Noradrenalinkonzentrationen stiegen nach
Tracheobronchialabstrich und Nasentupfer signifikant zum Basalwert an, ohne
sich signifikant voneinander zu unterscheiden. Die mittleren
Noradrenalinkonzentrationen der OK Gruppe und der KO Gruppe blieben auf
signifikant niedrigerem Niveau. Der Adrenalinspiegel der TBA Gruppe stieg
unmittelbar nach dem Eingriff geringgradig an, wohingegen die mittlere
Adrenalinkonzentration der Tiere der NT Gruppe vor und nach dem Eingriff
gleich blieb und die der OK Gruppe nach dem Eingriff sank. Die mittleren
Adrenalinkonzentrationen der TBA und NT Gruppe unterschieden sich nach den
Eingriffen nicht signifikant voneinander.
Die Cortisolkonzentrationen nach Durchführung eines Tracheobronchialabstriches
entsprechen denen nach Entnahme eines Nasentupfers. Ein vergleichbarer wenn
auch etwas verzögerter Cortisolanstieg wird durch die Fixierung mit der
Oberkieferschlinge hervorgerufen. Bezugnehmend auf die Noradrenalin-
Ergebnisse der vorliegenden Studie konnte gezeigt werden, dass der kurzfristige
Stress durch die Tracheobronchialabstriche vergleichbar mit der
Nasentupferentnahme ist und beide eine wesentlich höhere Belastung als die
alleinige Fixierung mit der Oberkieferschlinge darstellen. Die insgesamt
beobachteten Veränderungen der Adrenalinspiegel sind gering und sprechen
gegen starken emotionalen Stress und Angst der Schweine nach den
durchgeführten Maßnahmen.

VIII. Summary 71
VIII. SUMMARY
The aim of the present study was to compare stress in pigs resulting from tracheo-
bronchial swabbing compared to nasal swabbing and to fixation in a snare without
sampling. In order to evaluate the stress, cortisol concentrations from serum and
saliva-samples as well as epinephrine and norepinephrine concentrations from
plasma samples have been used. Four groups, three study and one control group,
of clinically healthy animals were formed to measure cortisol (each group n = 23)
and catecholamines (each group n = 31). A tracheo-bronchial swab was taken
from animals of the TBA group (tracheo-bronchial swabbing) during fixation with
a snare. The animals were not anaesthetised for the tracheo-bronchial swabbing.
Animals from the NT group (nasal swabbing) were fixated in the same way, but a
nasal swab was taken. Animals of the OK group (fixation in a snare) were
restrained for 60 sec with a snare without swabbing. The animals of the control
group were left in their pen without any disturbance. In the first part of the study
we looked at cortisol levels in blood and saliva. Blood and saliva samples were
taken simultaneously at the following time points: 30 min before manipulation
and 30 min, 60 min and 90 min afterwards. In the second part of the study we
tested catecholamine levels in plasma samples taken 15 min before and directly
after the manipulation.
30 min after the manipulation, average cortisol levels in serum and saliva in the
study groups (TBA, NT, OK) were significantly higher than the baseline value. At
this point the average cortisol levels of the animals that were only fixated without
swabbing (OK) were significantly higher than those of the animals in the control
group (KO). There was no significant difference in serum or saliva cortisol levels
between the study groups (TBA, NT, OK) at any point after the manipulation. 90
min after the manipulation the average cortisol levels in saliva and serum
decreased in all three study groups (TBA, NT, OK). The concentration-time-
curves (AUC) of the study groups (TBA, NT, OK) did not significantly differ
from each other neither in serum nor in saliva. In both groups where swabs were
taken, either tracheo- bronchial (TBA) or nasal (NT), norepinephrine levels after
the manipulation were significantly elevated compared to the baseline value. They
did not differ significantly between each other though. The average
norepinephrine levels of the groups, where no swabs were taken (OK, KO), were

72 VIII. Summary
significantly lower. We could observe a slight increase in the epinephrine levels
of the TBA group directly after the manipulation, whereas average epinephrine
concentrations of the NT group stayed on a constant level before and after the
manipulation and epinephrine concentrations of the OK group decreased.
However, the average epinephrine levels of the TBA and NT groups after the
manipulation did not significantly differ from each other.
In conclusion, cortisol levels after tracheo-bronchial swabbing are comparable to
that after nasal swabbing. The third study group which was only fixated by snare
(OK) showed the same cortisol increase with a little delay. Concerning the
norepinephrine levels that we measured in our study, we could show that the
short-term stress after tracheo-bronchial swabbing is comparable to the short-term
stress after nasal swabbing. Both manipulations caused significantly higher short-
term stress than fixation with the snare alone. In general, the observed changes in
the epinephrine levels are only small and do not indicate major distress in the
animals after any of the performed manipulations.

IX. Literaturverzeichnis 73
IX. LITERATURVERZEICHNIS
Adcock RJ, Kattesh HG, Roberts MP, Carroll JA, Saxton AM, Kojima CJ.
Temporal relationships between plasma cortisol, corticosteroid-binding globulin
(CBG), and the free cortisol index (FCI) in pigs in response to adrenal stimulation
or suppression. Stress-the International Journal on the Biology of Stress 2007; 10:
305-310.
Alius MG, Pané-Farré CA, Von Leupoldt A, Hamm AO. Induction of dyspnea
evokes increased anxiety and maladaptive breathing in individuals with high
anxiety sensitivity and suffocation fear. Psychophysiology 2013; 50: 488-497.
Axelrod J, Reisine TD. Stress hormones-their interaction and regulation. Science
1984; 224: 452-459.
Barnett JL, Winfield CG, Cronin GM, Makin AW. Effects of photoperiod and
feeding on plasma corticosteroid concentrations and maximum corticosteroid-
binding capacity in pigs. Australian Journal of Biological Sciences 1981; 34: 577-
585.
Blanchard B, Vena MM, Cavalier A, Lelannic J, Gouranton J, Kobisch M.
Electron-microscopic observation of the respiratory-tract of SPF piglets
inoculated with Mycoplasma hyopneumoniae. Veterinary Microbiology 1992; 30:
329-341.
Bland JM. An introduction to medical statistics. 2009. Oxford University Press,
Oxford. 169-171.
Brooks J, Tracey I. Review: From nociception to pain perception: imaging the
spinal and supraspinal pathways. Journal of Anatomy 2005; 207: 19-33.
Broom DM. The evolution of pain. Vlaams Diergeneeskundig Tijdschrift 2001;
70: 17-21.

74 IX. Literaturverzeichnis
Budras K-D, Buda S. Atlas der Anatomie des Rindes. 2002. Schlütersche,
Hannover.
Buer H. Nasentupfer/Trachealspülung. In: NutztierSkills: Arbeitstechniken in der
Großtierpraxis: Rind, Schwein, Ziege. 2012. ed: Palzer A, Buer H. Schattauer,
Stuttgart. 58-59.
Bushong DM, Friend TH, Knabe DA. Salivary and plasma cortisol response to
adrenocorticotropin administration in pigs. Laboratory Animals 2000; 34: 171-
181.
Canning BJ, Mazzone SB, Meeker SN, Mori N, Reynolds SM, Undem BJ.
Identification of the tracheal and laryngeal afferent neurones mediating cough in
anaesthetized guinea-pigs. Journal of Physiology-London 2004; 557: 543-558.
Carroll JA, Berg EL, Strauch TA, Roberts MP, Kattesh HG. Hormonal profiles,
behavioral responses, and short-term growth performance after castration of pigs
at three, six, nine, or twelve days of age. Journal of Animal Science 2006; 84:
1271-1278.
Chrousos GP, Gold PW. The concepts of stress and stress system disorders:
Overview of physical and behavioral homeostasis. Journal of the American
Medical Association 1992; 267: 1244-1252.
Chrousos GP, McCarty R, Pacak K, Cizza G, Sternberg E, Gold PW, Kvetnansky
R. Stress: Basic mechanisms and clinical implications. 1995. New York Academy
of Sciences, New York. 755.
Chrousos GP. Stress and disorders of the stress system. Nature Reviews
Endocrinology 2009; 5: 374-381.

IX. Literaturverzeichnis 75
Cook NJ, Schaefer AL, Lepage P, Jones SM. Salivary vs. serum cortisol for the
assessment of adrenal activity in swine. Canadian Journal of Animal Science
1996; 76: 329-335.
Cook NJ. Review: Minimally invasive sampling media and the measurement of
corticosteroids as biomarkers of stress in animals. Canadian Journal of Animal
Science 2012; 92: 227-259.
Cook NJ, Hayne SM, Rioja-Lang FC, Schaefer AL, Gonyou HW. The collection
of multiple saliva samples from pigs and the effect on adrenocortical activity.
Canadian Journal of Animal Science 2013; 93: 329-333.
Craner SL, Ray RH. Somatosensory cortex of the neonatal pig. 1. Topographic
organization of the primary somatosensory cortex (SI) Journal of Comparative
Neurology 1991; 306: 24-38.
Dantzer R, Arnone M, Mormede P. Effects of frustration on behaviour and plasma
corticosteroid levels in pigs. Physiology & Behavior 1980; 24: 1-4.
De Boer SF, Slangen JL, Van der Gugten J. Plasma catecholamine and
corticosterone levels during active and passive shock-prod avoidance behavior in
rats: effects of chlordiazepoxide. Physiology & Behavior 1990; 47: 1089-1098.
De Carlos F, Cobo J, Macias E, Feito J, Cobo T, Calavia MG, Garcia-Suarez O,
Vega JA. The sensory innervation of the human pharynx: searching for
mechanoreceptors. Integrative Anatomy and Evolutionary Biology 2013; 296:
1735-1746.
De Jong IC, Prelle IT, Van de Burgwal JA, Lambooij E, Korte SM, Blokhuis HJ,
Koolhaas JM. Effects of environmental enrichment on behavioral responses to
novelty, learning, and memory, and the circadian rhythm in cortisol in growing
pigs. Physiology & Behavior 2000; 68: 571-578.

76 IX. Literaturverzeichnis
Dimsdale JE, Moss J. Plasma-catecholamines in stress and exercise. Journal of the
American Medical Association 1980; 243: 340-342.
Döcke F, Kemper A. Nebennierenmark. In: Veterinärmedizinische
Endokrinologie. 1994. ed: Döcke F. Gustav Fischer Verlag, Jena. 357-378.
Ekkel ED, Dieleman SJ, Schouten WGP, Portela A, Cornélissen G, Tielen MJM,
Halberg F. The circadian rhythm of cortisol in the saliva of young pigs.
Physiology & Behavior 1996; 60: 985-989.
Ekkel ED, Savenije B, Schouten WGP, Wiegant VM, Tielen MJM. The effects of
mixing on behavior and circadian parameters of salivary cortisol in pigs.
Physiology & Behavior 1997; 62: 181-184.
Erhardt W, Henke J, Haberstroh J, Baumgartner C, Tacke S. Patienten mit
eingeschränkter respiratorischer Funktion. In: Anästhesie und Analgesie beim
Klein- und Heimtier: mit Exoten, Labortieren, Vögeln, Reptilien, Amphibien und
Fischen. 2011. ed: Erhardt W, Henke J, Baumgartner C. Schattauer, Stuttgart.
485-499.
Escribano D, Soler L, Gutierrez AM, Martinez-Subiela S, Ceron JJ. Measurement
of chromogranin A in porcine saliva: validation of a time-resolved
immunofluorometric assay and evaluation of its application as a marker of acute
stress. Animal 2013; 7: 640-647.
Evans FD, Christopherson RJ, Aherne FX. Development of the circadian rhythm
of cortisol in the gilt from weaning until puberty. Canadian Journal of Animal
Science 1988; 68: 1105-1111.
Fablet C, Marois C, Kobisch M, Madec F, Rose N. Estimation of the sensitivity of
four sampling methods for Mycoplasma hyopneumoniae detection in live pigs
using a Bayesian approach. Veterinary Microbiology 2010; 143: 238-245.

IX. Literaturverzeichnis 77
Fang M, Lorke DE, Li J, Gong X, Yew JCC, Yew DT. Postnatal changes in
functional activities of the pig’s brain: a combined functional magnetic resonance
imaging and immunohistochemical study. Neurosignals 2005; 14: 222-233.
Flasshoff J. Ein praxisrelevantes Verfahren zur frühzeitigen Differenzierung
bakterieller Bronchopneumonieerreger beim Schwein mittels Bronchiallavage
(BAL). Der Praktische Tierarzt 1996; 77: 1020-1024.
Friedrich M, Rixecker D, Friedrich G. Evaluation of stress-related hormones after
surgery. Clinical and experimental obstetrics & gynecology 1999; 26: 71-75.
Fukuhara K, Kvetnansky R, Weise VK, Ohara H, Yoneda R, Goldstein DS, Kopin
IJ. Effects of continuous and intermittent cold (SART) stress on sympathoadrenal
system activity in rats. Journal of Neuroendocrinology 1996; 8: 65-72.
Gallagher NL, Giles LR, Wynn PC. The development of a circadian pattern of
salivary cortisol secretion in the neonatal piglet. Biology of the Neonate 2002; 81:
113-118.
Gastl M, Brunner YF, Wiesmann M, Freiherr J. Depicting the inner and outer
nose: the representation of the nose and the nasal mucosa on the human primary
somatosensory cortex (SI). Human Brain Mapping 2014; 35: 4751-4766.
Goldstein DS. 2-Stress-induced activation of the sympathetic nervous system.
Baillière's Clinical Endocrinology and Metabolism 1987; 1: 253-278.
Goodell CK, Prickett J, Kittawornrat A, Zhou F, Rauh R, Nelson W, O’Connell C,
Burrell A, Wang C, Yoon K-J, Zimmerman JJ. Probability of detecting influenza
A virus subtypes H1N1 and H3N2 in individual pig nasal swabs and pen-based
oral fluid specimens over time. Veterinary Microbiology 2013; 166: 450-460.
Greco D, Stabenfeldt GH. Endocrinology. In: Textbook of Veterinary Physiology.
2013. ed: Cunningham JG. W.B. Saunders Company, Pennsylvania. 359-406.

78 IX. Literaturverzeichnis
Grosse Beilage E, Nathues H, Grummer B, Hartung J, Kamphues J, Kietzmann
M, Rohde J, Spindler B, Weissenböck H. Diagnostik, Prophylaxe und Therapie
von Atemwegserkrankungen in Schweinebeständen. In: Diagnostik und
Gesundheitsmanagement im Schweinebestand: Band 1. 2013. ed: Grosse Beilage
E, Wendt M. Eugen Ulmer, Stuttgart. 200-270.
Halter JB, Pflug AE, Porte D. Mechanism of plasma catecholamine increases
during surgical stress in man. The Journal of Clinical Endocrinology &
Metabolism 1977; 45: 936-944.
Henke J, Erhardt W. Schmerzmanagement bei Klein-und Heimtieren. 2001. Enke
Verlag, Stuttgart. 2-31.
Hennig-Pauka I, Bremerich S, Nienhoff H, Schroder C, Ganter M, Blecha F,
Waldmann KH, Gerlach GF. Methods of lung lavage in pigs - a comparison. Der
Praktische Tierarzt 2008; 89: 505-511.
Hillmann E, Schrader L, Mayer C, Gygax L. Effects of weight, temperature and
behaviour on the circadian rhythm of salivary cortisol in growing pigs. Animal
2008; 2: 405-409.
Hjemdahl P. Plasma catecholamines-analytical challenges and physiological
limitations. Baillière's Clinical Endocrinology and Metabolism 1993; 7: 307-353.
Holm S. A simple sequentially rejective multiple test procedure. Scandinavian
Journal of Statistics 1979; 6: 65-70.
Höltig D, Hennig-Pauka I. Bestandsproblem Atemwegserkrankungen. Der
Praktische Tierarzt 2013; Sonderheft von Der Praktische Tierarzt:
Bestandsbetreuung Rind und Schwein: 40-44.

IX. Literaturverzeichnis 79
Illes P, Allgaier C. Analgetika. In: Allgemeine und spezielle Pharmakologie und
Toxikologie. 2013. ed: Aktories K, Förstermann U, Hofmann FB, Starke K.
Urban & Fischer, 207-213.
Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature 2001; 413:
203-210.
Kanitz E, Otten W, Tuchscherer M. Changes in endocrine and neurochemical
profiles in neonatal pigs prenatally exposed to increased maternal cortisol. Journal
of Endocrinology 2006; 191: 207-220.
Kattesh HG, Charles SF, Baumbach GA, Gillespie BE. Plasma cortisol
distribution in the pig from birth to six weeks of age. Neonatology 1990; 58: 220-
226.
Klemcke HG, Nienaber JA, Hahn GL (1989) Plasma adrenocorticotropic hormone
and cortisol in pigs: effects of time of day on basal and stressor-altered
concentrations. Proceedings of the Society for Experimental Biology and
Medicine. New York. 42-53
König HE, Liebich H-G. Atmungsapparat. In: Anatomie der Haussäugetiere.
2012. ed: König HE, Liebich H-G. Schattauer, Stuttgart. 367-388.
König HE, Misek I, Mülling C, Seeger J, Liebich H-G. Nervensystem. In:
Anatomie der Haussäugetiere. 2012. ed: König HE, Liebich H-G. Schattauer,
Stuttgart. 485-557.
Kurth KT, Hsu T, Snook ER, Thacker EL, Thacker BJ, Minion EC. Use of a
Mycoplasma hyopneumoniae nested polymerase chain reaction test to determine
the optimal sampling sites in swine. Journal of Veterinary Diagnostic
Investigation 2002; 14: 463-469.

80 IX. Literaturverzeichnis
Kvetnansky R, Sun C, Lake C, Thoa NB, Torda T, Kopin IJ. Effect of handling
and forced immobilization on rat plasma levels of epinephrine, norepinephrine,
and dopamine-β-hydroxylase. Endocrinology 1978; 103: 1868-1874.
Kvetnansky R, Weise V, Thoa NB, Kopin IJ. Effects of chronic guanethidine
treatment and adrenal medullectomy on plasma levels of catecholamines and
corticosterone in forcibly immobilized rats. Journal of Pharmacology and
Experimental Therapeutics 1979; 209: 287-291.
Kvetnansky R, Sabban EL, Palkovits M. Catecholaminergic Systems in Stress:
Structural and Molecular Genetic Approaches. Physiological Reviews 2009; 89:
535-606.
Kvetnansky R, Ukropec J, Laukova M, Manz B, Pacak K, Vargovic P. Stress
Stimulates Production of Catecholamines in Rat Adipocytes. Cellular and
Molecular Neurobiology 2012; 32: 801-813.
Lackner A. Untersuchungen zur Schmerzhaftigkeit und der Wundheilung bei der
Kastration männlicher Ferkel zu unterschiedlichen Kastrationszeitpunkten. Diss.
med. vet. 2003. Ludwig-Maximilians-Universität, München
Langhoff R. Untersuchungen über den Einsatz von Schmerzmitteln zur Reduktion
kastrationsbedingter Schmerzen beim Saugferkel. Diss. med. vet. 2008. Ludwig-
Maximilians-Universität, München
Langhoff R, Zöls S, Barz A, Palzer A, Ritzmann M, Heinritzi K. Untersuchungen
über den Einsatz von Schmerzmitteln zur Reduktion kastrationsbedingter
Schmerzen beim Saugferkel. Berliner und Münchener Tierärztliche
Wochenschrift 2009; 122: 325-332.
Lendl C, Henke J. Anästhesien zu diagnostischen und therapeutischen Verfahren.
In: Anästhesie und Analgesie beim Klein- und Heimtier. 2011. ed: Erhardt W,
Henke J, Haberstroh J, Baumgartner C, Tacke S. Schattauer, Stuttgart. 499-515.

IX. Literaturverzeichnis 81
Löscher W, Frey HH. Zentrales Nervensystem (ZNS). In: Physiologie der
Haustiere. 2010. ed: Von Engelhardt W, Breves G. Enke Verlag, Stuttgart. 45-71.
Marois C, Le Carrou J, Kobisch M, Gautier-Bouchardon AV. Isolation of
Mycoplasma hyopneumoniae from different sampling sites in experimentally
infected and contact SPF piglets. Veterinary Microbiology 2007; 120: 96-104.
Mellor D, Cook C, Stafford K. Quantifying some responses to pain as a stressor.
In: The Biology of Animal Stress. 2000. ed: Moberg G, Mench J. CABI
Publishing, Cambridge. 171-198.
Mellor D, Stafford K. Physiological and behavioural assessment of pain in
ruminants: principles and caveats. Alternatives to laboratory animals 2004; 32:
267-271.
Mellor DJ, Stafford KJ. Acute castration and/or tailing distress and its alleviation
in lambs. New Zealand Veterinary Journal 2000; 48: 33-43.
Mellor DJ, Stafford KJ, Todd SE, Lowe TE, Gregory NG, Bruce RA, Ward RN.
A comparison of catecholamine and cortisol responses of young lambs and calves
to painful husbandry procedures. Australian Veterinary Journal 2002; 80: 228-
233.
Merlot E, Mounier AM, Prunier A. Endocrine response of gilts to various
common stressors: a comparison of indicators and methods of analysis.
Physiology & Behavior 2011; 102: 259-265.
Mohr E. Biologische Rhythmen. In: Physiologie der Haustiere. 2010. ed: Von
Engelhardt W, Breves G. Enke Verlag, Stuttgart. 673-677.
Moorkamp L, Nathues H, Spergser J, Tegeler R, Grosse Beilage E. Detection of
respiratory pathogens in porcine lung tissue and lavage fluid. The Veterinary
Journal 2008; 175: 273-275.

82 IX. Literaturverzeichnis
Mormede P, Andanson S, Auperin B, Beerda B, Guemene D, Malmkvist J,
Manteca X, Manteuffel G, Prunet P, van Reenen CG, Richard S, Veissier I.
Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate
animal welfare. Physiology & Behavior 2007; 92: 317-339.
Möstl E. Spezielle Endokrinologie. In: Physiologie der Haustiere. 2010. ed: Von
Engelhardt W, Breves G. Enke Verlag, Stuttgart. 515-579.
Nathues H, Grosse Beilage E. Für Studium und Praxis: Diagnostik und
Bekämpfung der Enzootischen Pneumonie beim Schwein. Tierärztliche Praxis
Ausgabe Großtiere/Nutztiere 2009; 37: 134-141.
Neubert E, Gurtler H, Vallentin G. Effect of restraining growth pigs with snare
restraints on plasma levels of catecholamines, cortisol, insulin and metabolic
parameters. Berliner und Münchener tierärztliche Wochenschrift 1996; 109: 409-
413.
Nickel R, Schummer A, Seiferle E. Peripheres Nervensystem. In: Lehrbuch der
Anatomie der Haustiere. 2004. ed: Nickel R, Schummer A, Seiferle E. Parey,
Stuttgart. 228-384.
Nienhoff H, Staszyk C, Mumme J. Beschreibung eines praktikablen Verfahrens
zur Gewinnung von Lungenspülproben bei Schweinen im schweinehaltenden
Betrieb. Der Praktische Tierarzt 2006; 87: 138-140.
Otten W, Kanitz E, Puppe B, Tuchscherer M, Brussow KP, Nurnberg G,
Stabenow B. Acute and long term effects of chronic intermittent noise stress on
hypothalamic-pituitary-adrenocortical and sympathoadrenomedullary axis in pigs.
Animal Science 2004; 78: 271-283.

IX. Literaturverzeichnis 83
Otten W, Kanitz E, Tuchscherer M, Grabner M, Nurnberg G, Bellmann O, Hennig
U, Rehfeldt C, Metges CC. Effects of low and high protein: carbohydrate ratios in
the diet of pregnant gilts on maternal cortisol concentrations and the
adrenocortical and sympathoadrenal reactivity in their offspring. Journal of
Animal Science 2013; 91: 2680-2692.
Palzer A. Keimspektrum und Erregerassoziationen bei gesunden und an
Pneumonie erkrankten Schweinen. Diss. med. vet. 2006. Ludwig-Maximilians-
Universität, München
Parrott RF, Misson BH. Changes in pig salivary cortisol in response to transport
simulation, food and water deprivation, and mixing. British Veterinary Journal
1989; 145: 501-505.
Parrott RF, Misson BH, Baldwin BA. Salivary cortisol in pigs following
adrenocorticotropic hormone stimulation-comparison with plasma-levels. British
Veterinary Journal 1989; 145: 362-366.
Pausenberger A, Mues G, Nienhoff H, Strutzberg-Minder K. Vergleich der
Untersuchungsergebnisse von bronchoalveolären Lavageproben mit
Bronchialtupferproben (Tracheobronchialabstrichproben) mittels PCR in einem
Ferkelerzeugerbetrieb. Der Praktische Tierarzt 2012; 93: 440-447.
Pausenberger A, Elsbecker C, Wilkes M, Beckmann H. Studie zur kulturell-
bakteriologischen Untersuchung von Bronchialabstrichproben und Vergleich mit
PCR-Ergebnissen. Der Praktische Tierarzt 2013; 94: 912-917.
Peers A, Mellor DJ, Wintour EM, Dodic M. Blood pressure, heart rate, hormonal
and other acute responses to rubber-ring castration and tail docking of lambs. New
Zealand Veterinary Journal 2002; 50: 56-62.
Pfannkuche H. Nozizeption und Schmerz: neurophysiologische Grundlagen.
Tierärztliche Praxis Ausgabe Kleintiere 2008; 36: 5-11.

84 IX. Literaturverzeichnis
Phogat JB, Parvizi N. Beta-adrenergic and opioidergic modulation of cortisol
secretion in response to acute stress. Experimental and clinical endocrinology &
diabetes: official journal, German Society of Endocrinology and German Diabetes
Association 2007; 115: 354-359.
Plonait H. Umgang mit Schweinen, Zwangsmaßnahmen und
Operationsvorbereitung. In: Lehrbuch der Schweinekrankheiten, 4 edn. 2004. ed:
Waldmann KH, Wendt M. Parey Verlag, Stuttgart. 39-49.
Roozen AW, Tsuma VT, Magnusson U. Effects of short-term restraint stress on
plasma concentrations of catecholamines, beta-endorphin, and cortisol in gilts.
American journal of veterinary research 1995; 56: 1225-1227.
Rosochacki SJ, Piekarzewska AB, Poloszynowicz J, Sakowski T. The influence of
restraint immobilization stress on the concentration of bioamines and cortisol in
plasma of Pietrain and Duroc pigs. Journal of veterinary medicine A 2000; 47:
231-242.
Ruis MAW, Te Brake JHA, Engel B, Ekkel ED, Buist WG, Blokhuis HJ,
Koolhaas JM. The Circadian Rhythm of Salivary Cortisol in Growing Pigs:
Effects of Age, Gender, and Stress. Physiology & Behavior 1997; 62: 623-630.
Rushen J, Ladewig J. Stress induced hypoalgesia and opioid inhibition of pigs
responses to restraint. Physiology & Behavior 1991; 50: 1093-1096.
Sann H. Nozizeption und Schmerz. In: Physiologie der Haustiere. 2010. ed: Von
Engelhardt W, Breves G. Enke Verlag, Stuttgart. 75-79.
Scholz J, Woolf CJ. Can we conquer pain? Nature Neuroscience 2002; 5: 1062-
1067.

IX. Literaturverzeichnis 85
Schönreiter S, Huber H, Lohmüller V, Zanella AJ, Unshelm J, Henke J, Erhardt
W. Salivary cortisol as a stress parameter in piglets. Tierärztliche Praxis Ausgabe
Großtiere/Nutztiere 1999; 27: 175-179.
Schoss P, Alt M. Are nasal swabs for swine appropriate for the diagnosis of
bacterial pneumonia agents? Deutsche Tierärztliche Wochenschrift 1995; 102:
427-430.
Schulz C, Ritzmann M, Palzer A, Otten W, Heinritzi K. Verlauf der Noradrenalin-
und Adrenalinkonzentrationen vor und nach der Kastration von Saugferkeln mit
und ohne Isofluran-Narkose. Deutsche Tierärztliche Wochenschrift 2007; 114:
454-459.
Schulz C. Auswirkung einer Isofluran-Inhalationsnarkose auf den
Kastrationsstress und die postoperativen Kastrationsschmerzen von Ferkeln. Diss.
med. vet. 2007. Ludwig-Maximilians-Universität, München
Sgoifo A, De Boer SF, Haller J, Koolhaas JM. Individual Differences in Plasma
Catecholamine and Corticosterone Stress Responses of Wild-Type Rats:
Relationship With Aggression. Physiology & Behavior 1996; 60: 1403-1407.
Silbernagel S, Despopoulus A. Zentralnervensystem und Sinne. In: Taschenatlas
Physiologie. 2012a. ed: Silbernagel S, Despopoulus A. Thieme, Stuttgart. 328-
394.
Silbernagel S, Despopoulus A. Atmung. In: Taschenatlas Physiologie. 2012b. ed:
Silbernagel S, Despopoulus A. Thieme, Stuttgart. 112-144.
Silbernagel S, Despopoulus A. Vegetatives Nervensystem. In: Taschenatlas
Physiologie. 2012c. ed: Silbernagel S, Despopoulus A. Thieme, Stuttgart. 82-92.

86 IX. Literaturverzeichnis
Silbernagel S, Despopoulus A. Hormone, Reproduktion. In: Taschenatlas
Physiologie. 2012d. ed: Silbernagel S, Despopoulus A. Thieme, Stuttgart. 280-
328.
Skarlandtova H, Bicikova M, Neuzil P, Mlcek M, Hrachovina V, Svoboda T,
Medova E, Kudlicka J, Dohnalova A, Havranek S, Kazihnitkova H, Macova L,
Kittnar O. Is there circadian variation in cortisol levels in young sows in heart
catheterization? Prague medical report 2011; 112: 184-192.
Smith SM, Vale WW. The role of the hypothalamic-pituitary-adrenal axis in
neuroendocrine responses to stress. Dialogues in Clinical Neuroscience 2006; 8:
383-395.
Sörös P, Knecht S, Bantel C, Imai T, Wüsten R, Pantev C, Lütkenhöner B, Bürkle
H, Henningsen H. Functional reorganization of the human primary somatosensory
cortex after acute pain demonstrated by magnetoencephalography. Neuroscience
Letters 2001; 298: 195-198.
Starke K. Grundlagen der Pharmakologie des Nervensystems. In: Allgemeine und
Spezielle Pharmakologie und Toxikologie. 2013. ed: Aktories K, Förstermann U,
Hofmann FB, Starke K. Urban&Fischer, München. 105-135.
Sylvester SP, Stafford KJ, Mellor DJ, Bruce RA, Ward RN. Acute cortisol
responses of calves to four methods of dehorning by amputation. Australian
Veterinary Journal 1998; 76: 123-126.
TäBeO (2011) Berufsordnung für die Tierärzte in Bayern. Bayerische
Landestierärztekammer, DTBl 1986, (zuletzt geändert am 01. Januar 2011 (DTBl.
1/2011, S. 88 f.)). 867 ff.
TierSchG (2013) Tierschutzgesetz, "Tierschutzgesetz in der Fassung der
Bekanntmachung vom 18. Mai 2006 (BGBl. I S. 1206, 1313), das zuletzt durch
Artikel 4 Absatz 90 des Gesetzes vom 7. August 2013 (BGBl. I S. 3154) geändert
worden ist" edn. Bundesministerium der Justiz und für Verbraucherschutz, BGBl

IX. Literaturverzeichnis 87
Treede R-D. Das somatosensorische System. In: Physiologie des Menschen. 2007.
ed: Schmidt RF, Lang F. Springer, Heidelberg. 296-323.
Übel N. Untersuchungen zur Schmerzreduktion bei zootechnischen Eingriffen an
Saugferkeln. Diss. med. vet. 2011. Ludwig-Maximilians-Universität, München
Van Alstine WG. Respiratory Diseases. In: Diseases of Swine, 10th edn. 2012. ed:
Zimmerman JJ, Karriker LA, Ramirez A, Schwartz KJ, Stevenson GW. Wiley-
Blackwell, Chichester. 348-363.
Van Reeth K, Brown IH, Olsen CW. Influenza Virus. In: Diseases of swine, 10
edn. 2012. ed: Zimmerman JJ, Karriker LA, Ramirez A, Schwartz KJ, Stevenson
GJ. Wiley-Blackwell, Chichester. 557-572.
Veissier I, Boissy A. Stress and welfare: Two complementary concepts that are
intrinsically related to the animal's point of view. Physiology & Behavior 2007;
92: 429-433.
Vining RF, McGinley RA, Symons RG. Hormones in saliva: mode of entry and
consequent implications for clinical interpretation. Clinical Chemistry 1983; 29:
1752-1756.
Viñuela-Fernández I, Jones E, Welsh EM, Fleetwood-Walker SM. Pain
mechanisms and their implication for the management of pain in farm and
companion animals. The Veterinary Journal 2007; 174: 227-239.
Von Borell E. Schmerzen bei Schweinen - Beurteilung, Vermeidung und
Verminderung. Tierärztliche Umschau 2013; 68: 227-230.
Ward MM, Mefford IN, Parker SD, Chesney MA, Taylor CB, Keegan DL,
Barchas JD. Epinephrine and norepinephrine responses in continuously collected
human-plasma to a series of stressors. Psychosomatic Medicine 1983; 45: 471-
486.

88 IX. Literaturverzeichnis
Widdicombe J. Neurophysiology of the cough reflex. European Respiratory
Journal 1995; 8: 1193-1202.
Wilmore DW, Long JM, Mason AD, Pruitt BA. Stress in surgical patients as a
neurophysiologic reflex response. Surgery Gynecology & Obstetrics 1976; 142:
257-269.
Zimmermann S. Untersuchungen zur Wirkung der Betäubung mittels
Kohlendioxid bei der Kastration männlicher Saugferkel. Diss. med. vet. 2010.
Ludwig-Maximilians-Universität, München
Zimmermann S, Zöls S, Otten W, Palzer A, Ritzmann M, Heinritzi K. Evaluation
of carbon dioxide anaesthesia for the castration of male suckling piglets by stress
hormone concentrations, behaviour and clinical factors. Berliner und Münchener
Tierärztliche Wochenschrift 2011; 124: 368-375.
Zöls S, Ritzmann M, Heinritzi K. Einfluss von Schmerzmitteln bei der Kastration
männlicher Ferkel. Berliner und Münchener Tierärztliche Wochenschrift 2006;
119: 193-196.
Zöls S. Möglichkeiten der Schmerzreduzierung bei der Kastration männlicher
Saugferkel. Diss. med. vet. 2006. Ludwig-Maximilians-Universität, München

X. Abbildungsverzeichnis 89
X. ABBILDUNGSVERZEICHNIS
Abbildung 1: Zeitplan der Cortisoluntersuchungen ............................................. 33
Abbildung 2: Zeitplan der Katecholaminuntersuchungen .................................... 35
Abbildung 3: Durchführung eines Tracheobronchialabstriches .......................... 37
Abbildung 4: Verlauf der mittleren Cortisolkonzentrationen (nmol/l) im Serum der
vier Gruppen über die Zeit t (min) ........................................................................ 41
Abbildung 5: Verlauf der mittleren Cortisolkonzentrationen im Serum der
Gruppen OK und KO (AUC) ................................................................................. 43
Abbildung 6: Verlauf der mittleren Cortisolkonzentrationen (nmol/l) im Speichel
aller vier Gruppen über die Zeit t (min)................................................................ 45
Abbildung 7: Mittlere Noradrenalinkonzentrationen (pg/ml Plasma) der vier
Gruppen vor (prae) und nach (post) dem Eingriff ................................................ 48
Abbildung 8: Mittlere Adrenalinkonzentrationen (in pg/ml Plasma) der vier
Gruppen vor (prae) und nach (post) dem Eingriff ................................................ 51

90 XI. Tabellenverzeichnis
XI. TABELLENVERZEICHNIS
Tabelle 1: Zusammenfassung der wichtigsten Wirkungen von Cortisol nach
MÖSTL (2010) und SILBERNAGEL und DESPOPOULUS (2012d) .................... 24
Tabelle 2: Gruppeneinteilung Cortisol- und Katecholaminuntersuchungen ........ 32
Tabelle 3: Mittlere Cortisolkonzentrationen im Serum in nmol/l vor sowie 30 min,
60 min und 90 min nach dem Studieneingriff, mit Minima, Maxima,
Standardabweichungen (SD), Standardfehler (SEM) und der area-under-the-
curve (AUC) der vier Gruppen .............................................................................. 40
Tabelle 4: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen oder
AUC (Serum) der Gruppen TBA, OK, NT und KO zu den vier
Beprobungszeitpunkten .......................................................................................... 42
Tabelle 5: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen (Serum)
verschiedener Beprobungszeitpunkte innerhalb einer Gruppe ............................. 42
Tabelle 6: Mittlere Cortisolkonzentrationen im Speichel in nmol/l vor sowie
30 min, 60 min und 90 min nach dem Studieneingriff, mit Minima, Maxima,
Standardabweichungen (SD), Standardfehler (SEM) und der area-under-the-
curve (AUC) der vier Gruppen .............................................................................. 44
Tabelle 7: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen oder
AUC (Speichel) der Gruppen TBA, OK, NT und KO zu den vier
Beprobungszeitpunkten .......................................................................................... 45
Tabelle 8: p-Werte der Vergleiche der mittleren Cortisolkonzentrationen
(Speichel) verschiedener Beprobungszeitpunkte innerhalb einer Gruppe ............ 46
Tabelle 9: Mittlere Noradrenalinkonzentrationen, Standardabweichungen (SD),
Minima, Maxima und Standardfehler (SEM) der vier Gruppen in pg/ml Plasma
vor (prae) und nach (post) dem Eingriff ................................................................ 47
Tabelle 10: p-Werte der Vergleiche der mittleren Noradrenalinkonzentrationen
der Gruppen TBA, OK, NT und KO zu den zwei Beprobungszeitpunkten............. 49
Tabelle 11: p-Werte der Vergleiche der mittleren Noradrenalinkonzentrationen
innerhalb einer Gruppe im Vergleich prae / post Eingriff .................................... 49
Tabelle 12: Mittlere Adrenalinkonzentrationen, Standardabweichungen (SD),
Minima, Maxima und Standardfehler (SEM) der vier Gruppen (pg/ml Plasma) vor
(prae) und nach (post) dem Eingriff ...................................................................... 50

XI. Tabellenverzeichnis 91
Tabelle 13: p-Werte der Vergleiche der mittleren Adrenalinkonzentrationen
zwischen den Gruppen TBA, OK, NT und KO zu den zwei Beprobungszeitpunkten
............................................................................................................................... 52
Tabelle 14: p-Werte der Vergleiche der mittleren Adrenalinkonzentrationen
innerhalb einer Gruppe im Vergleich prae / post Eingriff .................................... 52

92 XII. Danksagung
XII. DANKSAGUNG
Mein besonderer Dank gilt Prof. Dr. Mathias Ritzmann für die Bereitstellung
dieses interessanten Themas. Sie waren jederzeit ein ausgezeichneter Doktorvater
und ein hervorragender Chef in der Klinik für Schweine.
Ich freue mich auf zwei wunderbare und lehrreiche Jahre an der Klinik für
Schweine zurückschauen zu dürfen. Ein ganz besonderes Dankeschön richte ich
an meine beiden Betreuerinnen Dr. Nicole Übel und Dr. Susanne Zöls, die mich
stets unterstützt haben und jederzeit Rat wussten. Vielen Dank für eure guten
Anregungen, unermüdlichen Korrekturen und die Lösung sämtlicher
Computerprobleme. Liebe Dr. Jasmin Stark und Dr. Regina Hausleitner
Dankeschön für das gemeinsame Zähmen des Elecsys und vielen Dank an Dr.
Julia Stadler und Dr. Matthias Eddicks für die Beantwortung aller fachlichen
Fragen und euren umfassenden Rat.
Den besten Mitdoktoranden, auch den bereits Ausgeschiedenen, möchte ich
besonders danken. Ohne euch und die Rotationsstudenten, denen ich an dieser
Stelle ebenfalls herzlich danke, wäre diese Arbeit nicht möglich gewesen. Wir
Doktoranden waren eine super Truppe. Es hat wahnsinnig viel Spaß gemacht mit
euch so viele lehrreiche und lustige Tage und Ausfahrten zu verbringen sowie in
unangenehmeren Situationen Aufbau und Halt zu spüren. Außerdem konnte ich
mein Schweinemast- und Grillwissen erweitern!
Ich danke vielmals auch Konrad und allen anderen Mitarbeitern des
Versuchsgutes Thalhausen für die unkomplizierte und hilfsbereite
Zusammenarbeit sowie Frau Dr. Sauter-Louis und Herrn Dr. Reese für die
wertvolle statistische Beratung.
Gedankt sei auch Frau Dr. Pausenberger für ihre Impulse zur Durchführung und
ihre wertvollen Anregungen vor allem zu Beginn dieser Arbeit.
Ebenso danke ich Herrn Dr. Otten und seinem Laborteam vom FBN Dummerstorf
für die hilfreiche Beratung, die gute Zusammenarbeit und das Auswerten der
Katecholaminproben.
Dr. Matthias Huber und der gesamten Praxis danke ich für die Unterstützung und
das große Verständnis von zum Teil recht kurzfristigen Planänderungen.
Bedanken möchte ich mich auch für die ideelle und finanzielle Unterstützung
durch die Promotionsförderung des Cusanuswerkes, die entscheidend zum Erfolg
dieser Arbeit beigetragen und mich persönlich weitergebracht hat.
Meinen Freunden aus München und daheim, die mich zum Teil auch aus
Hamburg unterstützen, meinen Mitbewohnern der alten und neuen WG sowie
meinen Studienfreundinnen, Lisa mit dir hatte ich das Glück auch noch die
Promotion gemeinsam zu bestreiten, danke ich ganz besonders. Vielen Dank für
eure Freundschaft, ihr seid immer für mich da, hört zu und habt mich stets er- und
aufgemuntert!
Meiner Familie, die mir diesen Weg ermöglicht und mich in jeder Hinsicht
gefördert hat (angefangen bei Care Paketen im Studium) danke ich von Herzen.
Danke für eure bedingungslose Unterstützung und, dass ihr mir immer die
Sicherheit vermittelt, dass ich mich auf euch verlassen kann!


















