Über das diencephal-telencephale neurosekretorische System beim Salamander (Salamandra salamandra...
39
Zeitschrift ffir Zellforschung 89, 371--409 (1968) t~ber das diencephal-telencephale neurosekretorische System beim Salamander (Salamandra salamandra und S. tigrinum)* WOLFGANG ARNOLD Institut fSr Histologie und experimentelle Biologie der Universit~t Miinchen (Direktor: Prof. Dr. med. R. BACH~IA~N) Eingegangen am 11. M/irz 1968 The Dieneephalic-Telencephalic Neurosecretory System in Salamandra salamandra and S. tigrinum Summary. The diencephalic neurosecretory system of Salamandra is examined both histologically and histochemically. Special notice was given to a vessel complex in the front of the 3rd ventricle, which corresponds to the subfornical organ. The results are as follows: 1. The preoptic nucleus is situated in the wall of the 3rd ventricle, caudally to the anterior commissure. Neurosecretory fibers are directed to the infundibulum, the neural lobe, the telencephalon, the posterior commissure and to a vessel complex near the confluence of the lateral ventricles. 2. In the upper infundibulum the neurosecretory fibers have contacts to special vessels of the anterior lobe. 3. Neurosecretory fibers running to the telencephalon terminate close to small caves limited by glial fibers. 4. During embryogenesis the posterior lobe is filled up with neurosecretory material. Parts of the preoptic-hypophyseal tract can be demonstrated at that stage; these are in contact to capillaries of the anterior lobe. 5. Increased secretion into the 3rd ventricle and to the subfornical organ follows to osmotic changes. An accumulation of chromhematoxylin-positive granules in the proximal stump is the result of an experimental interruption of the telencephalic tract near the optic chiasm. 6. The subfornical organ belongs to the septal part of the telencephalon. The internal zone contains a glomerulum-like vessel complex covered by a high ependyma. Small vacuoles are normally found between the capillaries and the ependyma. Laterally astrocytic cndfeet reach the wall of these vessels. From the interhemispheric fissure the subfornical organ is separated by a group of cells consisting of glial and parenchymal cells as well as of CHP-positive cells. 7. Considering its position we believe it to be homologous to the subfornical organ of higher vertebrates. It is possible that in the salamander this organ comprises both, the organum vasculosum laminae terminalis and the subfornical organ. In the brain of the turtle, which is more recent in phylogenesis, these two organs are separated. Zusammen/assung. Das neurosekretorische System des Salamanders wurde unter beson- derer Beriicksichtigung der Einbeziehung eines Gef~florgans am rostralen Ende des 3. Ven- trikels histologisch und histochemisch untersucht. Die Untersuchungen brachten folgende Ergebnisse: 1. Bei Salamandra finder sich der Nucleus praeopticus in der Wand des 3. Ventrikels caudal hinter der Commissura anterior. Von hier ziehen neurosekretorische Fasern zum Infun- dibulum, zur Neurohypophyse, zum Boden des Telencephalons, zur Commissura posterior, ferncr zu einem Gef~iBorgan am Zusammenflufl der beiden Seitenventrikel. * Herrn Dr. med. H. RUDERT und Herrn Dr. med. A. WEINDL bin ich fiir freundliche Unterstiitzung sehr verbunden. Ffir technische Unterstfitzung sei Frl. C. DEGEN, Frl. I. DiiRR, Fr. A. GADACZ und Fr. B. ROTTMANN herzlich gedankt.
-
Upload
wolfgang-arnold -
Category
Documents
-
view
218 -
download
3
Transcript of Über das diencephal-telencephale neurosekretorische System beim Salamander (Salamandra salamandra...

Zeitschrift ffir Zellforschung 89, 371--409 (1968)
t~ber das diencephal-telencephale neurosekretorische System beim Salamander (Salamandra salamandra und S. tigrinum)*
WOLFGANG ARNOLD
Ins t i tu t fSr Histologie und experimentelle Biologie der Universi t~t Miinchen (Direktor: Prof. Dr. med. R. BACH~IA~N)
Eingegangen am 11. M/irz 1968
The Dieneephalic-Telencephalic Neurosecretory System in Salamandra salamandra and S. tigrinum
Summary. The diencephalic neurosecretory system of Salamandra is examined both histologically and histochemically. Special notice was given to a vessel complex in the front of the 3rd ventricle, which corresponds to the subfornical organ. The results are as follows:
1. The preoptic nucleus is s i tuated in the wall of the 3rd ventricle, caudally to the anterior commissure. Neurosecretory fibers are directed to the infundibulum, the neural lobe, the telencephalon, the posterior commissure and to a vessel complex near the confluence of the lateral ventricles.
2. In the upper infundibulum the neurosecretory fibers have contacts to special vessels of the anterior lobe.
3. Neurosecretory fibers running to the telencephalon terminate close to small caves limited by glial fibers.
4. During embryogenesis the posterior lobe is filled up with neurosecretory material. Par ts of the preoptic-hypophyseal t rac t can be demonstra ted at t ha t stage; these are in contact to capillaries of the anterior lobe.
5. Increased secretion into the 3rd ventricle and to the subfornical organ follows to osmotic changes. An accumulat ion of chromhematoxylin-posit ive granules in the proximal s tump is the result of an experimental interruption of the telencephalic t rac t near the optic chiasm.
6. The subfornical organ belongs to the septal par t of the telencephalon. The internal zone contains a glomerulum-like vessel complex covered by a high ependyma. Small vacuoles are normally found between the capillaries and the ependyma. Laterally astrocytic cndfeet reach the wall of these vessels. From the interhemispheric fissure the subfornical organ is separated by a group of cells consisting of glial and parenchymal cells as well as of CHP-positive cells.
7. Considering its position we believe it to be homologous to the subfornical organ of higher vertebrates. I t is possible t ha t in the salamander this organ comprises both, the organum vasculosum laminae terminalis and the subfornical organ. In the brain of the turtle, which is more recent in phylogenesis, these two organs are separated.
Zusammen/assung. Das neurosekretorische System des Salamanders wurde unter beson- derer Beriicksichtigung der Einbeziehung eines Gef~florgans am rostralen Ende des 3. Ven- trikels histologisch und histochemisch untersucht .
Die Untersuchungen brachten folgende Ergebnisse:
1. Bei Salamandra finder sich der Nucleus praeopticus in der Wand des 3. Ventrikels caudal hinter der Commissura anterior. Von hier ziehen neurosekretorische Fasern zum Infun- dibulum, zur Neurohypophyse, zum Boden des Telencephalons, zur Commissura posterior, ferncr zu einem Gef~iBorgan am Zusammenflufl der beiden Seitenventrikel.
* Herrn Dr. med. H. RUDERT und Herrn Dr. med. A. WEINDL bin ich fiir freundliche Unters t i i tzung sehr verbunden. Ffir technische Unterstf i tzung sei Frl. C. DEGEN, Frl. I. DiiRR, Fr. A. GADACZ und Fr. B. ROTTMANN herzlich gedankt.

372 W. ARNOLD:
2. W~hrend ihres Verlaufes durch das distale Infundibulum nehmen die neurosekretori- schen Axone Kontakt mit adenohypophys~iren Gef~]en auf.
3. Im Verlauf des Tractus telencephalicus f~llt eine Beziehung zu Gliakammern am Boden des Telencephalons auf.
4. Bei Embryonen ist der Hypophysenhinterlappen dicht mit Neurosekret gefiillt; der Tractus praeoptico-hypophyseus ist abschnittsweise darstellbar. Eine Beziehung zu adeno- hypophys~ren Gef~Ben ist deutlich.
5. Nach osmotischer Belastung zeigt sich eine vermehrte Sekretabgabe in den 3. Ventrikel und im Subfornikalorgan. Die Durchtrennung des Tractus praeoptico-telencephalicus rostral vor dem Chiasma opticum fiihrt zu einer Stauung des Sekrets an der proximalen Schnittfl~che.
6. Das Subfornikalorgan liegt am Zusammenflul~ der Seitenventrikel in den 3. Ventrikel in der Pars septalis des Telencephalons. Die innere Zone wird von einem glomerulumartigen Gef~il]sinus gebildet, den apikal ein hohes Palisadenependym begrenzt. Vakuolen dr~ngen sich zwischen Gef~Bsinus und Palisadenependym. Lateral ziehen Astrocytenforts~tze mit breiten Fii~chen an die Gef~13schlingen heran. Gegen den Interhemisph~renspalt ist das Organ durch einen Zellschild aus Glia- und Parenchymzellen getrennt. Darunter befinden sich auch sekre- torische Zellen mR Gomori-positiven Forts~tzen.
7. Entsprechend der Lage des Organs sehen wires als ein Homologon des Subfornikal- organs hSherer Wirbeltiere an. Die M5glichkeit wird diskutiert, dab es sich beim Subfornikal- organ des Salamanders um ein Gef~13organ handelt, das in sich Organum vasculosum laminae terminalis und Subfornikalorgan vereinigt. Bei der phylogenetisch hSher stehenden Schild- krSte sind beide Organe vorhanden.
Von al lgemeiner Gi i l t igkei t fiir die gesamte Wirbe l t i e r re ihe is t der enge Zu- sammenhang der sekretor isch t~t igen Kerngeb ie te des H y p o t h a l a m u s mi t der Hypophyse . Bei einer Reihe bisher un te r such te r niederer Wirbe l t i e re zeigt sich zudem, da~ neben dem Trac tus h y p o t h a l a m o - h y p o p h y s e u s zahlreiche neuro- sekretorische Bahnen exist ieren, die das Zwisehenhirn verlassen und ins Telence- phalon, Mesencephalon oder zu den circumventrikul/~ren Organen ziehen. Die Unte r suchungen an Fischen (SCHARR~R, E., 1928 ; HILD, 1950; BARGMANN, 1953 ; ADAM, 1960; ST~RBA, 1961), Amphibien (HILD, 1951; MAZZI, 1953; BOLOO•ARI U. ROTELLA, 1956; L~OAIT, 1957; DIERICKX, 1962; RUDERT, 1965; W]~B]~, 1965), sowie an Reptilien (DIEPEN, 1952; A~ANTHANARYANAN, 1955) weisen auf bemer- kenswer te Unterschiede im Verlauf der neurosekre tor ischen Bahnen hin. Die Ein- beziehung des neurosekre tor i schen Sys tems in den noch wei tgehend u n b e k a n n t e n Funk t ionsk re i s der c i rcumvent r iku l~ren Organe seheint bei Amphibien (BRAAK, 1963; BARAMIKOVA, 1964) ausgepr~gt zu sein. Un te r den ex t r ahypo tha l ami schen Bahnen wurden Verb indungen zum Subforn ika lorgan yon DIERICKX (1962) und RUDERT (1965) beim Frosch beschrieben. I m Anschlul~ an die Unte r suchungen yon RUDERT (1965) lag es nahe, ein weiteres niederes Wirbe l t i e r zu untersuehen, dessen Gehi rnaufbau sich yon dem des Frosches unterscheidet .
I n dieser Arbe i t wird versucht , die Morphologie des normalen und exper imen- tel l be las te ten Gehirns yon Salamandra darzustel len, d .h . die Ver~nderungen im gesamten neurosekre tor ischen Sys tem zu erfassen. Besondere Beaeh tung soll dabe i e inem an der Grenze yon Zwischen- zu Endh i rn gelegenen Organ gewidmet werden, dessen F o r m und Gef~Bversorgung als ein Analogon zum Subforn ika lorgan der S~ugetiere angesehen wird.
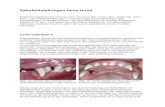
Die Lage des Organs und die Ausdehnung des neurosekre tor isehen Sys tems er l~uter t Abb, 1.

Diencephal-telencephales neurosekretorisches System bei Salamandra 373
T
/ / /
/
\ , \ /
\\ /
D ~ _ p I __
SFO.
SCO
~i, / / -
ucpo .o . ,
/ / - io a
M
\ \ \
\
\ -- -
',\ //:" T " D " M
\
"-~-~ - " : -- -- - " " ' . " - ~ \ r -' ~ ~ . ~ " . , J \~7--.> .<.'.->. : . . . . . . . - : ~ C ~ J .: ', /
J -:-L ."..-:~7~'-,, ~ . . . . .--'~-2.. ...I / / "<,,: ." g r S '" . " ~ L . . ~ ' . " . . )
o . / . / ~ , . . ' �9 .'. �9 ~ ~ X - . . . . ,, ~ . . . . . :! / ~ , , - ~ - ~ \ . . . . ' . . . . . . . . . . . . . . . . . . . ~ . . . ,, .-, �9 . . . / '
b , /
Abb. 1. a Sagit talsehnit t in der Medianebene dureh das Salamandergehirn. T Teleneephalon, D Dieneephalon, M Meseneephalon, SFO Subfornikalorgan, P Paraphyse, C Cerebellum, SCO Subeommissuralorgan, Ch. o. Chiasma faseieulorum optieorum, s.po. Suleus praeoptieus, nuc.po.a, nucleus praeoptieus an~r ior , io ineisura fase. opt., Nh. Neurohypophyse, p.vent. pars ventralis thalami, ca eommissura ant., cp eommissura post., ro reeessus optieus. Die vom nue.po, ausgehenden Bahnen sind dureh Pfeile gekennzeiehnet, b I-Iorizontalsehnitt dureh das Gehirn des Salamanders. Sehnit tebene in HShe der Foramina Monroi. N P Nucleus praeoptieus, Pl Plexus ehor. I Interhemisphiirenspalt , P H Primerdium hippoeampi, grS graue Substanz, wS weiBe Substanz, S V Seitenventrikel, F.o. Faseieulus opt. a neurosekretorisehe Bahn zur I-Iypophyse, b ins Teleneephalon, c zum Subfornikalorgan, d erst mi t b verlaufend,
dann aber rfiekl~ufig zu a; e in den 3. Ventrikel
M a t e r i a l u n d M e t h o d e n
Die Untersuchungen wurden an 120 Amphibien beiderlei Geschlechts der Spezies Sala- mandra maculosa, an 12 Tieren der Untergruppe Ambystoma tigrinum, an 6 einheimisehen FrSschen der Spezies Rana temporaria und vergleichsweise an l%ptil ien (SchildkrSten:

374 W. ARNOLD:
9 Tiere der Gruppe Emys orbicularis, 4 Tiere der Gruppe Testudo graeca und 9 Tiere yon Pseudemys floridana) durchgefiihrt.
Folgende experimentelle Eingriffe wurden an Salamandern durchgefiihrt: in Anlehnung an HILD (1951), E. und H. LEGAIT (I957), RUDERT (1965) und WEBER (1965): a) osmotische Belastung in 1--1,5%iger NaCl-LSsung; in 2,5%iger DextroselSsung. Je drei Salamander wurden fiber 12, 24 und 48 Std in diese hypertonischen LSsungen gesetzt, b) Je fiinf Tiere wurden hypophysektomiert und Each 1, 3 und 7 Tagen getStet, c) Je vier Tiere wurden hypophysektomiert und einen Tag Each dem Eingriff ffir 12, 24 und 48 Std in eine 0,9%ige NaC1-LSsung gesetzt.
Zur An~sthesie wurde den Tieren 1,0--1,5 ml einer 10 % igen w~Brigen LSsung yon Urethan unter die Rfickenhaut injiziert oder sie wurden in eine 1,2 % ige w~Brige AlkohollSsung (BoLz, 1961) gesetzt. Nach Fixierung der auf dem Rficken liegenden Tiere auf einem Wachsbrett wurde der Unterkiefer maximal weir geSffnet und mittels eines Nylonfadens in dieser Stellung gehalten. Nach Li~ngsdurchtrennung der den Oberkiefer bedeckenden Schleimhaut lassen sich am Hinterrand des harten Gaumens bei mikroskopischer Betrachtung die Hypophyse und das Chiasma opticum mit den Nervi optici schnell auffinden. Mit einem Thermokauter wurde das kn5cherne Parasphenoid in der Medianen koaguliert und mit einer Iridektomielanzette seit- lich hochgeklappt. Die Meninx in dieser freien Zone wurde entfernt und die Hypophyse und das distale Infundibulum mit einem Messerchen abgetragen. Gelegentlich auftretende Blu- tungen standen Each leichtem Betupfen mit einem Wattebiiusehchen. Auf eine Schleimhaut- naht konnte verzichtet werden. Die Tiere wurden markiert und zu den Kontrolltieren gesetzt. Adulte Tiere wurden durch Dekapitation getStet und nach ErSffnung der Sch~deldecke und des Gaumens in die FixierlSsung gegeben. Embryonen und Larven wurden direkt in das Fixierungsgemisch eingebracht. Fixierung in: Formalin 1 : 10; Calcium-Formalin 6 % ; neutra- lem Formol 4% (mit Phosphat gepuffert); Gemischen Each BouIN (w 305"), CARNOY (w 226), ELFTMANN (1957), HELLY (w 337), ZENKER (w 336). Die Gewebsstficke wurden in Alkohol entwassert. Weiterbehandlungen: Methylbenzoat, Benzol; Einbettung in Paraplast und Paraf- fin. Schnittdicke 4--10 ~z.
Fiirbungen und histoehemische Reaktionen. Zur Anfarbung von Zellkern und Cytoplasma: Carmalaun Each P. MAYER (w 642); Eisenhamatoxylin nach HANSEN (w 709), nach HEIDEN- HAIN (w 672), Each WEIGERT ( w 677) ; Hamalaun-Eosin nach P. MAYER (w 659) ; Kerneehtrot (w 743) ; Gallaminblau (w 736) ; Gallocyanin (w 734). Zur Darstellung bindegewebiger An- teile: Azanfiirbung Each HEIDENttAIN (w 1489); Massonsche Trichromfiirbung (w 1496); deren Modifikation nach GOLDNER (w 1496, w 1498); Pikrinsi~urefuchsin Each VAN GIESON (w 708); Perjods~iure-Leukofuchsinreaktion Each GRAUMANN (1952). Zur Darstellung yon Gliafasern und Gliazellen: Each HOLZER (w 1848); Each FLEISCHHAUER (1961); um den Verlauf von Nervenfasern darzustellen, Gallocyanin-Chromalaun Each EINARSON (w 1749); Osmium- Zink-Jodid-Methode naeh JABONERO (1962); Nachweis der Ribonueleins~ure Each LJISBERG (1962); Kontrollschnitte wurden mit Ribonuclease (Gewinnung Each BRACHET, 1940) vor- behandelt. Zur Darstellung des Golgiapparates: die Versilberungsmethode nach LASCANO (1959); Osmiummethode Each KOPSCH-KOLATSCHEW (w 1009); ffir die Anf~rbung des Neuro- sekretes dienten die Aldehydthioninreaktion Each PAGET (1959); Chromhamatoxylin-Phloxin Each GOMORI-BARGMANN (PEARSE, 1960); Paraldehydfuchsin nach GABE (1956); Alcianblau- f~rbung Each vorhergehender Oxydation Each ADAMS und SLOPER (1956). Zum Nachweis vermuteter Proteidkomponenten: Histidinnachweis nach BACHMANN und SEITZ (1961); Millonreaktion naeh POLLmTER (GoMoRI, 1952); Pseudoisocyaninreaktion Each SCHIEBLER und SCHIESSLER (1959); fluoreszenzmikroskopischer Nachweis eng benachbarter SOaH- Gruppen des Neurosekrets mit der N,N'-Di~thylpseudoisocyaninchlorid-Methode (STERRA, 1961, 1964); Tyrosinnachweis Each GLENNER und LILLIE (1959). Zum Nachweis yon Kohlen- hydraten: Halesche Eisenbindungsreaktion Each GRAUMANN und CLAUSE (1958); modifizierte Perjods~ure-Leukofuchsinreaktion Each GRAUMANN (1952). Zum Nachweis yon Lipiden: Fettf~rbung Each ROMEm (w 1048), Each LISON mit Sudanschwarz B (w 1050). Zum AusschluB siderogenen Pigmentes: Eisennachweis mit Berlinerblau Each WICKLEIN und FALKENEERG (w 1205). SchlieBlich wurde die Aktivit~t folgender Fermente darzustellen versucht: Acetyl-
* w bezieht sich auf ROMEm (1948).

Diencephal-telencephales neurosekretorisches System bei Salamandra 375
cholinesterase nach KOELLE (1950); Aliesterase nach Go~oRI (1952); Carboanhydrase nach HXUSLi~R (1958); alkalische Phosphatase nach GOMORI (1952); saure Phosphatase nach GOMORI (1952). Zur Kontrolle wurden bei allen Neurosekretnachweisen Schnitte yon K/ilber- hypophysen mituntersucht. - - Die Nomenklatur ist angelehnt an HERRICK (1947).
Befunde
A. Ursprung, Verlau/ und Ziel der sekretorischen Neurone 1. Ursprungszentren
Die Perikarya der sekretorischen Nervcnzellen, deren Gomori-positive Axone in hypothalamischen und cxtrahypothalamischen Regionen angetroffen werden, liegen bei Salamandra maculosa und Ambystoma tigrinum im vordcren markarmen Hypothalamus periventrikul/~r. I t icr bildet der 3. Ventrikel eine rinnenf6rmige Erweiterung, die dorso-ventral durch den Nucleus praeopticus verl/iuft und im ventralen Hypothalamus seitlich in die Recessus praeoptici einmfindet (s. Abb. 1 a). Aberrierende Ganglienzellen finder man gelegcntlich bis in N/ihe der Commissura posterior.
Die Ausdchnung der sekrctorischen Zellen nach der Seite zu ist gering. Caudal vom Commissurenwu]st (Commissura hippocampi, Commissura anterior) finden sich im Hypendym mehrere Lagen sckretorischer Zellen, tells dicht gedr/ingt, teils locker verstreut licgend. Nicht selten kommen solche Ganglienze]len zwischcn den Ependymzellen vor (vgl. K•OWLES, 1967). I m Gomoripr/~parat erkennt man den Nucleus praeopticus schon bei schwacher Vergr61~erung als schwarzblau gef/irbten Bezirk. ~hnliche Ergebnissc bringen die Aldehydthioninf~rbung nach GABs und die Aldehydfuchsinf/irbung.
In wenigen F/illen wurde am rostralen Ende der Seitenventrikel eine kleine, ebenfalls im Hypendym gelegene Ganglienzellgruppe gefunden, die mit den Neurosekretf/irbungen positiv reagiert; doch war der Verlauf der Forts/~tze nicht zu verfolgen.
Die Anzahl der sezernierenden Ganglienzellen des Nucleus praeopticus schwankt insbesondere bei tr/ichtigen Tieren. Die Kerne der Gomori-positiven Zellen besit- zen unterschiedliche Chromatinmengen; meist sind drei Nucleoli vorhanden. Die Kcrnmembran ist glatt und hebt sich deutlich ab. Die Chromatindichte ist ab- h/ingig yon dem Funktionszustand. Das Perikaryon der Ganglienzellen ist nur dort zu erkennen, wo Sekretgranula nachgewicsen werden. Nicht immer ]assen sich die Forts/itzc so eindeutig darstellen wie in Abb. 2. I m Gegensatz zu den neuro- sekretorisch t/itigen Kernarealen mancher S/iuger zeigt der Nucleus praeopticus nicderer Vertebratcn keine vermehrte Vaskularisierung. So liegcn bei Salamandra, wie bei den untersuchtcn Reptilien, die sekretorisch aktiven Ganglienzellen zwi- schen den Zellen des t Iypendyms und nehmen nur gelegentlich Beziehung zu cinem der wenigen Gef/il~e ihrer Region auf. M6glicherweise erkl/irt die unmittel- bare Nachbarschaft des Ventrikels, warum eine vermehrte GefhBversorgung zur Information der osmotischen Blutsituation fiberflfissig ist.
Auffallend ist die hohc Aktivit/it der alkalischen Phosphatase, die fiberall dort, wo Neurosckret vorhanden ist, deut]ich nachzuweisen ist. Bei Amphibien und Reptilien finder man in der Sekretabgabe in den Ventrikel eine Form des Trans- portes, die bei dicser Ticrgruppe einen gro~en Anteil der axonalen Sekretfort- leitung ersetzt.

376 W. ARNOLD:
Abb. 2. Sa. 1/18 ~ Horizontalschnitt durch den Nucleus praeopticus. Fixation nach ELFTMANN (1957); Paraffineinbettung; F~rbung nach GOMORI-BARGMANN. Vergr. 300fach. Anh~ufung
yon Sekret findet man sowohl basal wie apikal zum Ventrikel. V Ventrikel
2. Sekretfortleitung
In der Regel wird nut in einem Zellfortsatz, n~mlich im Neuriten, Gomori- positives Sekret gefunden. Bei unseren Untersuchungsobjekten wurde Neuro- sekret nicht nur in dem yore Ventrikel wegziehenden basalen, sondern auch im apikalen Fortsatz beobachtet. Zwischen den cilienbesetzten Ependymzellen kommen jedoch vereinzelt auch Nervenzellen mit basalen, Gomori-positiv gef/irb- ten Neuriten vor, welche in die Peripherie ziehen. Diese begleiten die ependymalen Tanycytenforts/~tze, durchqueren das Kernareal des Nucleus praeopticus und sind bis an die Membrana limitans externa der Hirnoberfli~che zu verfolgen. Sie enden dort am ~ul~eren Liquorraum mit zarten Ffil~chen.
Bei Emys orbicularis hingegen sind die neurosekretorischen Zellen in zwei Grappen angeord- net. Unter den Ependymzellen liegen in unterschiedlicher Anzahl sekretorisch t~tige, Gomori- sowie Aldehydthionin-positive Ganglienzellen mit apikaler und basaler Sekretion. Davon getrennt durch eine breite Gliazone, die dem Markraum der S~iuger entspricht, finder man in der ,,Rinde" eine weitere Ganglienzellgruppe, die jedoch keine Verbindung zum Ependym aufnimmt.
Die Mehrzahl der Ausl/s der sekretfiihrenden Ganglienzellen zieht in die Peripherie. Anfangs verlaufen sie noch gemeinsam in einem Biindel, teilen sich aber in der weiBen Substanz. Als geschlossenes Fortsatzbfindel lassen sich der Tractus praeoptico-hypophyseus und eine Bahn zum Boden des Telencephalons

Diencephal-telencepha]es neurosekretorisches System bei Salamandra 377
darstellen. Nach anfiinglich horizontalem Verlauf biegen die Fasern in eine vor und in eine hinter der Incisura optica verlaufende Gruppe urn. Die Bahn ins Telencephalon ist dabei faserreicher als die zur Hypophyse ziehende. Allerdings i~ndert ein Teil der Axone wieder seine Verlaufsrichtung und fiberquert dicht unter der Membrana limitans externa die Incisura optica, um sich wieder dem Tractus zur Hypophyse anzuschlie0en (vgl. Abb. l b). Dabei ziehen regelmKl~ig einige Axone zusammen mit dem Tractus opticus in den Thalamus dorsalis, wo sie nicht welter verfolgt werden. Auch in die Periphcrie des Fasciculus opticus ziehende Fasern lassen sich darstellen: wir konnten auf Horizontalserien ein Eindringen yon sekretffihrenden Axonen in das Chiasma verfo]gen (DIERICKX, 1963; Rana). I m Tractus praeoptico-hypophyseus ist bei erwachsenen, unbelasteten Tieren Gomori-positives Sekret oft nur fiber kurze Strecken sichtbar und wird erst wieder in der pars ventralis hypothalami, der Gegend des Tuber cinereum der Sauge- tiere, best~ndig darstellbar. I m proximalen Teil des Infundibulums treffen die konvergierenden Fasern der linken und rechten Seite aufeinander. Beim l~ber- gang vom proximalen ( = caudaler Anteil der Pars ventralis hypothalami) zum distalen Infundibulum beobachteten wir eine starke Sekretstauung in den hier durchziehenden neurosekretorischen Fasern. Dabei kommt es zum l~bertritt yon Granula in das Ventrikellumen des Recessus infundibu]aris. Der Tractus tr i t t auch in direkten Kontak t zu subpialen Gef~]en dieser Gegend. Wo sich Neuro- sekret den Gef~Ben anlagert, hat es nicht mehr die typische Granulastruktur, sondern ein kolloidi~hnliches Aussehen. Welter distal, im Abschnitt der Eminentia mediana (Nomenklatur nach TILN~C, 1936) herrscht das gleiche Bild. Lateral der Eminentia mediana liegen die Fasern des Tractus praeoptico-hypophyseus nut in der Zona interna, wie es bei hSheren Wirbeltieren die Regel ist. Gelegentlich dringen Sekretfasern entlang yon Gef~l~en in die Adenohypophyse ein.
Besondere Verh~ltnisse liegen im distalen Infundibulum vor. Diese Stelle wird yon zahlreichen Autoren als ,,median eminence" bezeichnet. Die Eminentia mediana wird durch die Ansammlung aufgetriebener neurosekretfiihrender Axone gebildet. Beim Salamander stellt sie den infundibul~ren Anteil der Neurohypo- physe dar, in den GefiiBe aus der Adenohypophyse dringen. I m Frontalschnitt erkennt man eine wulstige Anreicherung yon zum Tell erweiterten Neuriten, die selbst beim unbelasteten Tier keine Zona externa mehr erkennen l~Bt. Zahlreiche HerringkSrper liegen im Gewebe verstreut. Erst lateral, gegen die seitliche Begrenzung des Recessus infundibularis hin, kann wieder Zona interna und externa unterschieden werden. Ahnlich wie bei den Myxinoiden und zum Tell bei den Cyclostomen (GouBMAN, 1965) ist ein GroBteil der Oberfl~che der Adenohypo- physe yon einer kr~ftigen Bindegewebsscheide umgeben, welche gegen das Infundi- bulum und die Neurohypophyse bestehen bleibt. Nur an wenigen Stellen wird sie yon gewundenen G e f ~ e n durchbrochen. Die Kapillaren ziehen senkrecht aus der Adenohypophyse an die Scheidewand heran und dringen ein kurzes Stfick in die Pars nervosa ein. Dieser Vorgang ist an der Stauungsstelle beim Eindringen des Tractus in das distale Infundibulum und an der Eminentia mediana zu beob- achten. Hier treten die Gef~l~e durch die bindegewebige Trennwand und verlaufen in spitzem Bogen (Abb. 3a- -c ) oder fiber eine kurze Strecke parallel zur Ober- fl~che durch die Zona externa des Infundibulum, bevor sie wieder senkrecht in die Tiefe dringen. Mit der Masson-, Azan- und Goldnerf~rbung l~I~t sich im

378 W. ARNOLD:
Abb. 3. a Sa. 81/31 ~ Sagittalschnitt durch das distale Infundibulum mit Adenohypophyse; Fixation naeh Botrx~; Paraplasteinbettung; F~rbung nach GOMORI-B~oMAN~. Eine Gomori- positive Membran (M) trennt die Adenohypophyse (A) v o n d e r Zona extema (ze); zi Zona interna; E Ependym. Vergr. 1350fach. b Sa. 66/95 ~ Horizontalschnitt durch die Adeno- hypophyse und Eminentia mediana. Fixation und Weiterbehandlung wie bei a. Vergr. 1250fach. Kapillarschlingen dringen aus der Adenohypophyse in die Eminentia mediana bzw. in die Zona interna vor und nehmen mit den neurosekretorischen Fasern Kontakt auf. k Kapillare. c Schematische Skizze zu den VerhEltnissen im distalen Infundibulum nach einem paramedianen Sagittalschnitt. T.po.hyp. Tractus praeoptico-hypophyseus; iz Innenzone; az Aul]enzone; Sk Sinuskapillaren, die aus der Adenohypophyse Ah bis in die innere Zone reichen; Nh Neurohypophyse; G Gef/~l]e, die den Verlauf des Tractus zur Hypophyse kreuzen
und um die sich das Sekret kappenf6rmig staut. Ri Recessus infundibularis

Diencepha]-telencephales neurosekretorisches System bei Salamandra 379
Gegensatz zu den eigentlichen Hirnkapillaren eine schmale, bindegewebige Scheide hervorheben. Die neurosekretorischen Axone legen sich den Kapillaren dicht an oder enden an ihm mit kSrnigen Auftreibungen. Ein direkter l~bertritt yon Sekret in das Gefg61umen wurde nicht beobachtet. Das Sekret ist bisweilen derart kom- pakt gelagert, dal~ es zu einer kolloid~hnlichen, homogenen Ringbildung um das Gefg6 kommt. Selten treten Neurosekretfasern in die Adenohypophyse fiber (vgl. LEOAIT, 1957).
Dann verlaufen die Axone entlang den Gef~l~en, die aus der Adenohypophyse in die Zona externa oder zurfick ziehen. Ihr weiterer Verlauf ist ]ichtmikroskopisch nieht feststellbar. Gegen die eigentliche Pars nervosa sammelt sich im distalen
)O
T. p o. hyp ~ ~ [ @ " ~ \
\ sk ) , , Ah /
"\ ~ J /
Abb. 3 c
Teil der Pars adenohypophysea des Infundibulums immer mehr grobkSrniges Sekret an, um kontinuierlich in die Neurohypophyse iiberzugehen. Zahlreiche Nervenfasern sind zu HerringkSrpern aufgetrieben. Die Ependymzellen des Reeessus infundibularis sind an vielen Stellen auseinander gerfickt; lichtmikro- skopisch ist keine umschriebene Grenze zwischen Nervengewebe und Ventrikel- lumen siehtbar. Die sekretffihrenden Neuriten dringen knStchenfSrmig zwischen die Ependymzellen, deren Form stark variieren kann. Oft findet man Sekret, das auf dem Ependym zu kolloidAhnlichen Kfigelchen zusammenf]ie6t (vgl. BARG- MA~I~ und HILD, 1951). In der Neurohypophyse selbst ziehen die Neuriten an die zahlreichen GefAl3e; dort lagern sie sich wirbelfSrmig an.
a) Tractus hypothalamo-telencephalicus. Das andere Faserbfindel, welches vor dem Chiasma zum Boden des Telencephalons zieht, scheint zun~chst wesentlich reichhaltiger an sekretfiihrenden Axonen zu sein als der Tractus hypothalamo- hypophyseus. In breitem, gestreutem Verlauf erreichen die Fasern den Boden des Endhirns, wo sie sich zu Bfindeln vereinigen. In allen Schnittrichtungen erkennt man vor der Incisura n. optici eine Stauung der Gomori-blauen Fasern; das gest~ute Sekret bildet mAchtige HerringkSrper. Wie besonders auf Sagittal- und Frontalserien zu erkennen, handelt es sich um rfickl~ufige Faserzfige, die sich (s. oben) wieder dem Tractus zur Hypophyse anschliel3en.

380 W. ARNOLD:
Der weitere Verlauf der telencepha]en Bahn ist nicht klar ersichtlich. Ein Ziel bilden zahlreiche ,,Gliak~mmerchen" am Boden des Endhirns. Dort, wo einzelne neurosekretorische Fasern Kontak t mit ihnen aufnehmen, sind ihre W~nde Gomori- und Aldehydthionin-positiv gef~rbt und zum Teil dicht mit Granula besetzt. Bei st~rkster VergrSBerung erkennt man eine doppelte Wand der Kam- mern, deren ~uBere zumeist den Granulabesatz tr~gtl Das Innere dieser Kammern ist optiseh leer und f~rbt sich auch mit den verschiedenen, auf EiweiB zielenden histoehemischen Methoden nicht an. Lichtmikroskopisch besteht kein Anhalt dafiir, dab die Sekretgranula in das Innere abgegeben werden (Abb. 4). Diese normalerweise m~chtige neurosekretorische Bahn verliert naeh Hypophysektomie, die einen Eingriff in die osmotischen Regelmechanismen darstellt, als erste vor allen anderen extrahypothalamisehen Faserverbindungen ihren Granulagehalt.
Um zu erfahren, ob der Sekretstrom in die Peripherie, also zu den Gliakam- mern und weiter mit dem Tractus olfactorio-peduncularis ins Telencephalon gerichtet sei, nahmen wie bei einigen Tieren in Urethannarkose eine Unterbre- chung dieses Faserzuges vor:
Nach Freilegung der basalen Hirnoberfl~ehe wurde vor der Sehnervenkreu- zung mit einem Irismesser ein etwa 1 m m tiefer Schnitt gesetzt. Nach 24 bzw. 48 Std wurden die Tiere getStet und sofort fixiert. Es ergab sieh, dab die]enigen Axone, welche sieh dem telencephalen Faserzug anschlie~en, an der proximalen Schnittfl~che der Verletzungsstelle m~chtig aufgetrieben waren und das Stau- ungsph~nomen lie~erten, welches HILD (1951) nach Unterbrechung des Tractus hypothalamo-hypophyseus flit Rana esculenta besehrieb. Schon naeh 24 Std beginnen die Axone proximal von der Verletzungsstelle ihre typische Perlketten- konfiguration zu s Man hat den Eindruek, dab die Perlchen zusammen- flieBen. Die Sekretproduktion in den Kerngebieten scheint nicht ver~ndert zu sein. Der Tractus zur Hypophyse zeigt keinerlei Ver~nderung. Am distalen Teil der Sehnittfl~che fehlt das Neurosekret. Bisweilen sind nahe der Schnittstelle hellblaue, stark erweiterte Axonfragmente zu erkennen. Damit ist der SekretfluB in die telencephale Peripherie, den schon BAa~Y (1954, 1960) und LEGAIT (1957) annahmen, beim Salamander best~tigt.
b) Tractus hypothalamo-septalis. In seinem Anfangstefl zwar deutlich aus- gepr~gt, besteht dieses Faserbtindel nur aus wenigen Gomori-blauen Perlschnur- tasern. Nach tI. LEGAIT (1957) ist dieses Faserbiindel bei den Fisehen und Uro- delen rudiment~r angelegt, bei Anuren und VSgeln aber sehr gut ausgebfldet. Es n immt seinen Ursprung vom rostrodorsalen Tell des Nucleus praeoptieus und zieht in kurzem Bogen zum Boden des Zwischenhirns vor das Chiasma. Von den telencephalen Fasern kann man es trennen, da seine Axone weiter medial verlaufen, w~hrend die des Tractus hypothalamo-teleneephalicus an der seitlichen Basis des Zwischenhirns entlangziehen. Ein Teil yon ihnen erreicht den Commissuren- wulst. Nach Durehqueren des Faserfflzes zwischen Commissura hippocampi und Commissura anterior oder, was verh~ltnisms oft zu beobachten ist, nach Uber- wanderung der Commissura hippocampi direkt unter dem bedeckten Ependym (vgl. I~CDV.~T, 1965) verliert sich der GroBteil der Axone zwischen den Ependym- zellen des l~ecessus praecommissuralis. Regelm/~Big tragen die Ependymzellen apical Vakuolen, deren R~nder Gomori-positiv oder Aldehydfuehsin-positiv

Diencephal-telencephales neurosekretorisches System bei Salamandra 381
Abb. 4. Sa. 2/27 ~ Horizontalschnitt am Boden des Telencephalons; Fixation BouI~; Paraffin- einbettung; Chromalaunhiimatoxylinf~rbung nach GOMORI-BARGM.t~N; Vergr. 1250fach. Zahlreiche neurosekretorische Axone des Tractus telencephalicus enden am Boden des End- hirns an Gliakammern. Die W~nde der Kammern sind mit zarten Gomori-positiven Granula besetzt. Gelegentlich finden sich auch dichte, HerringkSrper-~hnliche Auftreibungen an ihren
W~nden. K optiseh leere Kammern, AT/Neurosekretfasern
Abb. 5a--e . Sa. 45/9 g~ Frontalschnitt durch das Subkommissuralorgan und die hintere Kommissur (Cp); Fixation nach BovI~; Paraplasteinbettung; F~rbung nach GO~ORI-BARa- MANN. Vergr. ~ 350fach, b 1250fach, c 1250fach. Neben dem Subcommissuralorgan (SCO) finder man zellul~re Nester mit basalen, Gomori-positiven, biopolaren Forts~tzen (c); Fasern des Tractus praeoptico-habenularis lassen sich beim Durchzug durch die hintere Kommissur
beobaehten (b)

382 W. ARNOLD:
angef/~rbt sind. Ihr Inhalt bleibt dabei optisch leer. Aus Horizontalserien erkennt man, dab es sieh um cytoplasmatische Vorw61bungen des Zelleibes gegen das Ventrikellumen handelt. Gelegentlich sind Fasern zu beobachten, die im Hypen- dym des Recessus praecommissuralis zu einem Gef/~13organ an der rostrMen Begrenzungswand des Recessus ziehen (vgl. S. 389).
c) Tractus praeoptico-habenularis. Eine weitere extrahypothalamische Bahn nimmt ihren Ursprung von den posterodorsalen Zellen des Nucleus praeopticus und folgt dem markarmen Nervenfaserbiindel des Traetus hypothalamo-habe- nularis. W/~hrend W]~B]~R (1965) sie zu den am besten ausgepr/igten extrahypo- thalamisehen Bahnen rechnet, konnten wir sie deutlich und regelm/~Big nur in dem Absehnitt ihres Verlaufs erkennen, wo sie um die Commissura posterior zieht. Diese Fasern verlaufen zum Teil zwischen den hypendymalen Zellen des Subcommissuralorgans; jedoeh sind keine Anzeichen dafiir vorhanden, dab das Subcommissuralorgan von neurosekretorischen Fasern innerviert wird. Abb. 5 zeigt neurosekretbeladene Axone, die dutch den Commissurenfilz ziehen und zum Teil aufgetrieben erseheinen. Davon zu unterscheiden sind die gleichfalls Gomori- positiven basalen Forts/~tze vieler hypendymaler Zellen des Subcommissural- organs. Ihr Sekret ist homogen und sehlierenartig; selbst bei st/~rkster VergrSge- rung ist um den Zellkern keine Granulastruktur zu erkennen. Weiter entfernt vom Organ ist eine Unterscheidung zwischen hypothalamischen Gomori-positiven Axonen und den Forts~tzen der lokalen sekretorischen Elemente des Hypendyms nieht mehr m6glich. Nicht selten liegen augerhalb des subcommissuralen Zell- kranzes vereinzelt Rosetten sekretoriseher Zellelemente, abgesprungene Inseln des speziellen Organependyms. Eine Verbindung des Nucleus praeopticus mit dem Subcommissuralorgan oder mit der Commissura posterior scheint bei niederen Vertebraten keine Seltenheit zu sein.
LEGAIT und GRmNON (1957) beschreiben bei Testudo mauritanica zahlreiche, vom Nucleus praeoptieus stammende Fasern zu GefiiBen dieser Region. Auch bei Salmoniden findet BARAMIKOWA (1964) Gomori-positive Zellforts~tze, die dem Nucleus praeopticus entstammen und zum Subcommissuralorgan ziehen; welter sind sie nicht zu verfolgen.
Ftir die Schlange Thamnophis schildern HrLD (1951) und E. SCnARR]~R (1951) eine ,,neurosekretorisehe Bahn" zur Paraphyse. Neurosekretorisehe Faserver- bindungen zur Epiphyse sind beim Igel (BARGNANN, 1954) und bei einigen Chiropteren (BARRY, 1956) beschrieben worden. Nicht regelm/~Big fanden wir neurosekretfiihrende Fasern, die zum Mittelhirndach, zu Gef/~gen am Boden des Mittelhirns caudal hinter dem Hypophysenstiel (bei Schildkr6ten ein regelm/~giger Befund) und an Gef~ge des Bulbus olfaetorius (WEBwR, 1965) ziehen.
3. Endigung der Axone
Ein groBer Teil der sekrethaltigen Axone endet an den Gef/igen, weitere an den subpialen Gliak/immerchen (s. Abb. 4). Hingegen konnten wir die Endigung der Fasern nieht feststellen, die sieh seheinbar ohne erfagbares Ziel im Gewebe ver- lieren. Letzteres trifft insbesondere fiir solehe Fasern aus dem Traetus praeoptieo- hypophyseus zu, die vor dem Chiasma abseheren und sich verzweigen. W o u n d wie diese Aufzweigungen enden, 1/~gt sieh nieht feststellen (Abb. 6). Einige Perl- sehnurfasern enden im Epithalamus. Die letzten Perlehen schwellen zu kleinen

Diencephal-teleneephales neurosekretorisches System bei Salamandra 383
Herringk6rpern an. Es entsteht ein Gebilde, das aus groBk6rnigen und vielen feink6rnigen Granu]a besteht. Gelegentlich liegen diese Granula um kleine Vakuolen.
B. Morphologie des Sekretes
1. Sekret in den Axonen
Die Morphologie des Neurosekrets in den verschiedenen Abschnitten eines Neurons ist abh~ngig von Funktionszustand und Belastung. Kernnah haben die angef~rbten Neuronenabschnitte ein trichterf6rmiges, gestrecktes Aussehen, in der Peripherie hingegen zeigen sie die bekannten Perlschnurbilder. Im ungef~rbten, entparaffinierten Schnitt erkennt man phasenkontrastmikroskopiseh den Fort- satz mit seinen stark lichtbreehenden W~nden, die sich in regelmiil~igen Abst~nden halbkugelf6rmig ausbuehten. Naeh Anf~rbung mit einer der bekannten Methoden finder man in diesem ,,Rohrsystem" fein verteilt Granula, deren Diehte gegen die Wand hin zunimmt. Gelegentlich sind Teile des Fortsatzes bl~sehenf6rmig erwei- tert oder das Sekret staut sieh zu homogenen Massen an. Dazwischen finden sieh l~bergiinge zu unf6rmigen Herringk6rpern (Abb. 7a). DaB dieses Erseheinungs- bild der Perlschnurfasern nur den augenb]ickliehen Bedingungen zur Zeit der T6tung entspricht, beweist die schnelle •nderung naeh vorausgehender osmoti- seher Belastung oder Hypophysektomie. Ein grol]er Teil der Fasern verliert seine typisehe Struktur. Die vormals runden Perlehen werden in L~ngsrichtung birnen- f6rmig ausgezogen; sie wandeln sich in Bl~schen urn, die ihre Anf/~rbbarkeit verloren haben. Nur an den R~ndern der Axone liegen noeh aufgehellte Sekret- granula. Innerhalb der Einsehnitte, we]ehe die aufgequollenen Per]chen vonein- ander trennen, stellt man homogenes, manchmal ]eicht aufgelockertes Sekret lest, das am distalen P o l d e r Bl~schen besonders intensiv anzufi~rben ist. Bei Tieren, deren T6tung sparer als 24 Std nach dem Eingriff erfolgte, findet man im Verlauf des Tractus praeoptieo-hypophyseus nur mehr vereinzelte Axone in dieser Weise ver~ndert. Sehr eindrucksvoll l~iBt sieh diese Beobachtung in der Chiasmagegend demonstrieren. In der Region trennen sich die beiden Fasermassen zur Hypo- physe und ins Telencephalon. Nach Eingriffen in den Wasserhaushalt kommt es sehr bald zu einer Stagnation, zuerst des telencephalen Sekretabflusses. Schon 12 Std nach einer Hypophysektomie ist diese Bahn mit der Chromalaunhaema- toxylinf~rbung kaum mehr darzustellen.
Zu diesem Zeitpunkt t r i t t im Bereich des Tractus praeoptico-hypophyseus die oben erw~hnte Vakuolisierung auf.
Im proximalen Anteil des durchtrennten Faserbtindels sind die Neuriten vor der Incisionsstelle dureh die St6rung im Sekretabflul~ stark aufgequollen. Die Axone sind unf6rmig, zumeist spindelig aufgetrieben und besitzen Ahnlichkeit mit langgestreekten Herringk6rpern. Ihr Inneres wird yon staubf6rmigen Sekret- k6rnchen erfiillt. Nach weiteren 12 Std sind die Faserstiimpfe mit einem homogen aufgehellten Inhalt angefiillt, in dem keine Granulastruktur mehr zu erkennen ist. Bei Tieren, die erst naeh einigen Tagen get6tet wurden, erscheinen die Faser- st/impfe im Gomoripr~parat r6tlich homogen. Sekretk6rnchen sind nicht mehr vor- handen (Abb. 7 b).
Bei phasenoptiseher Betrachtung an gef~rbten Schnitten, auf denen Fasern des Tractus praeoptico-hypophyseus naeh Uberwanderung des Chiasmas quergetroffen
26 Z. Zellforsch., Bd. 89

384 W. ARNOLD:
Abb. 6. Sa. 82/7 ~ Horizontalschnitt durch den ventralen Hypothalamus caudal des Chiasma. Fixation nach ELFTMANN (1957); Paraplasteinbettung; F~rbung nach GOMORI-B.~.RGMA~. Vergr. 500fach. Sich verzweigender neurosekretorischer Fortsatz. An den Verzweigungsstellen ist das Axon HerringkSrper-~hnlich aufgetrieben. Die genaue Endigungsart dieser Fasern war
nicht festzustellen
Abb. 7. a Sa. 31/21 ~ Horizontalschnitt durch den ventralen Hypothalamus. Fixation BouIN; F/~rbung GOMORI-BARGMANN ; Paraffineinbettung. Vergr. 500fach. Fasern, welche vom Tractus praeoptico-telencephalicus rfickl~ufig sich wieder dem Tractus zur Hypophyse

Diencephal-telencephales neurosekretorisches System bei Salamandra 385
sind, gclingt es, den Inhal t und die Begrenzung der Axone zu unterscheiden. Die unterschiedliche Lichtbrechung l ~ t Axonscheide und Sekret verschieden hell aufleuchten.
2. Sekret in den HcrringkSrpern
Beim Salamander kSnnen Herringk5rper an allen Stcllen des Gehirns bcob- achtct wcrdcn, die durch neurosekretorische Bahnen mit dem Nucleus praeopticus verbunden sind. Ihre GrS~e schwankt yon kleinen spindelfSrmigen Auftreibungen der ,,Perlchen" fiber das ZusammenflieBen mehrerer solcher Auftreibungen bis
Abb. 8. Sa. 15/45 ~ Horizontalschnitt am Boden des Hypothalamus; tr/~chtiges Weibchen. Fixation nach BovIN; Paraffineinbettung; F/irbung nach GOMORI-BARGMANN. Vergr. 1250- fach. Zum HerringkSrper erweiterte Perlschnurfaser mit bl~schenf5rmiger Ausbuehtung B
zu unf6rmigen, blasigen Gebilden. Ihre Dichte ist unterschiedlich. Oft enthalten sie dunkelblaues, dichtes Material. Bei manchen wird das Innere neben zarten K6rnchen yon kleinen Bli~schen, dercn R~inder intensiv tingiert scheinen, aus- geffillt (Abb. 8). Da diese bl~schenhaltigen Herringk6rper neben anderen zweifel- los echten Formen im gleichen Pr~parat vorkommcn, ist ein Artefakt auszu- schliei3en.
AusschlieBlich homogen erscheinende Anreicherungen yon Neurosekret in Gestalt kleiner Herringk6rper finden sich um die GefKSe des Infundibulums, wo diese Gebilde sich kappenf6rmig an der Kapillarwand stauen.
Zahlreiche Herringk6rper entstehen vor dem Chiasma: sie sind als Stauungen aufzufasscn, die dort auftreten, wo sich neurosekretorischc Axonc mit Sehnerven- fasern kreuzen. So entsteht ein Bild spindelf6rmiger Auftreibungen, welche fiber zarte Vcrbindungsstr~nge miteinander gekoppelt sind.
In diesen verschiedenen Erscheinungsformen der Herringk6rper sehen wir Phasen einer durch die Geschwindigkeit der Sekretabgabe bedingten Speicherung.
anschlieflen, kreuzen dabei das dichte System der optischen Projektionsfasern. Dabei bilden sich HerringkSrper. b Sa. 16/191 ~ Sagittalschnitt durch das proximale Infundibulum nach Hypophysektomie. Fixation BouIN; Fs GOMO~Z-BARGMAN~; Paraplasteinbettung. Vergr. 1250fach. Nach Hypophysektomie schwellen die proximalen Faserstfimpfe an. Nach 24~48 Std kommt es zu einem allm~,hlichen Schwund des Neurosekretes in diesem Abschnitt
des Tractus praeoptico-hypophyseus
26*

386 W. ARNOLD :
Bei den perivaskul~rcn, knStchenfSrmigen Auftreibungcn von neurosekretori- schen Axonen handelt es sieh um Vorstufen von HerringkSrpern, welehe bei zu langsamem Abtransport oder zu schnellem Nachsehub an den GefAl]en entstehen. Diese Gebilde kSnnen stark aufgehellt sein. Dies mag der Ausdruck einer struk- turellen Ver~nderung der angeh~uften Sekretgranula im Sinne einer Verflfissigung odor AuflSsung der biologisch wirksamen Substanzen scin (vgl. DuFFY u. MENEFEE, 1965; KOBAYASm etal . , 1966). Zwischen Gef~l~wand und HerringkSrper findet sich ein aufgehellter, unstrukturierter Bereich ; in vivo enth~lt dieser perivascul~re Raum eine dem Blutserum ~hnliehe Flfissigkeit.
C. Das Aultreten neurosekretorischer Verbindungen bei Salamanderembryonen
Salamanderembryonen (S/S 8 ram) wurden nach Entfernung der Eihs in 4%igem neutralem Formalin vor- und im Fixierungsgemisch nach BouIN nach- fixiert. Bei Tieren dieser Entwicklungsstufe lassen sich in den undifferenzierten Zellhaufen des Zwischenhirns noch keine bestimmten Kerngebiete finden. Wie beim erwachsenen Tier kann der Sulcus praeopticus als Orientierungslinie ffir das Aufsuchen des Nucleus praeopticus dienen. Zwar f~rben sich bereits Tefle des Tractus praeoptieo-hypophyseus mit Chromalaun-H~matoxylin und Aldehyd- thionin an, doch ist das zugehSrige Kerngebiet fs noch nicht darstellbar. I m weiteren Verlauf durch das Zwischenhirn nimmt der Sekretgehalt der Fasern offenbar ab. Erst beim Ubertr i t t in die Pars adenohypophysea des Infundibulums treten die gebtindelten Fasern wieder horror. Zarte Granula besetzen die Grenz- membran zu der noch nicht ausgereiften Adenohypophyse. Zahlreiche Blutgef/~l~e treten an diese Grenzmembran heran. Solche Kontaktstellen reagieren besonders stark Gomori-positiv. Daneben beobachtet man, wie neurosekretorische Fasern, zum Toil mit Abweichungen zwischen die Ependymzellen des Infundibulums, zur Pars nervosa der Hypophyse gerichtet sind (Abb. 9). Die Neurohypophyse selbst besteht aus einer einsehichtigen Lage yon Ependymzellen, zwischen und unter denen sich feinkSrniges Neurosekret staut.
In sp~teren Stadien der Gesamtl~nge 2--2,5 cm (die Tiere haben bereits Fadenkiemen) gelingt es auch im Verlaufsabschnitt zwischen Kerngebiet und Infundibulum Gomori-positive Nervenfasern zu finden. Beim Verlauf der Fasern durch den Tractus opticus verdichten sich die neurosekretorischen Axone. Einzelne Neuriten lassen sich fiber die gesamte Strccke zum Infundibulum verfolgen, wobei sic Kontak t zu den Gef/~l~en aufnehmen. Die Neurohypophyse ist in diesem Stadium prall mit Neurosekret geffillt und gut vaskularisiert. I m Larvenstadium verringert sich die Konzcntration des Sekrets am Boden des Zwischenhirns, so dad dieser Toil des Tractus kaum darstellbar ist. Der Sekretgehalt der Neuro- hypophyse bleibt unver~ndert. Mit den angewandten Methoden fArben sich jetzt
Abb. 9. SE. 12/77 Sagittalschnitt durch das Infundibulum mit Adenohypophysenanlage. Fixation ELI~TMANN (1957); F~rbung nach GOMORI-BARGMA~N; Paraplasteinbettung. Vergr. 1250fach. S/S 8 mm. Unter und zwischen den Ependymzellen (E) erkennt man neurosekret- fiihrende Fasern. Die Neurohypophyse (Nh) ist bereits dicht mit Sekret angefiillt. Auch die Basalmembran (B), welche die Adenohypophyse A yon den neuroektodermalen Anteilen des Zwischenhirns trennt, ist besetzt von zarten Gomori-positiven Granula. V Ventrikel, pars infundibularis; K Sinuskapillare aus der Adenohypophyse, die un die Basalmembran zieht;
Er Erythrocyt

Diencephal-telencephales neurosekretorisches System bei Salamandra 387
Abb. 10. Sa. 16/192 9. Sagittalschnitt durch das Subfornikalorgan. Fixation nach ELFTMAN~r (1957); Paraffineinbettung; F/irbung nach GOMORI-BARGMArr162 Vergr. 250fach. Das Organ- zentrum wird yon einem Gef~Bkn~uel gebildet (G), welches gegen den Ventrikel (V) dutch das hohe Palisadenependym getrennt liegt. Zwischen und um die Kapillaren findet sich reichlich Pigment (P). Rostral trennt ein Schild (S) yon Zellen mit nur schmalem Cytoplasmasaum das
Organ yore/ibrigen Hirngewebe

388 W. ARNOLD:
deutlich die Perikarya einiger Ganglienzellen des Nucleus praeopticus an, deren Lokalisation bisher nur durch den Nachweis der alkalischen Phosphatase und die hohe Aktivit~t der unspezifischen Esterase mSglich war.
D. Ver~inderungen des neurosekretorischen Systems bei Salamandern unter experimentellen Bedingungen
1. Befunde nach Hypophysektomie
Bei den meisten hypophysektomierten Salamandern fanden wir neben einer Aufhellung der Sekretpole der Ganglienzellen einen verminderten Chromatin- gehalt der Zellkerne im Nucleus praeopticus. Die proximalen Strecken der Neuriten zur Hypophyse zeigen bereits 12 Std nach dem Eingriff eine deutliche Auf- lockerung ihrer Innenstruktnr. Aus den vormals homogenen oder feingranulierten blauen Perlchen werden trSpfchenfSrmige Gebilde. Der Inhalt der Perlchen ist optisch leer, die R/inder sind fein granuliert und ein Pol, zumeist der proximale, enth/ilt mehr Sekret. Das Bild der Perlschnur bleibt erhalten, doch sind die Perl- chen etwas auseinandergerfickt. Zwischen den einzelnen Perlchen bleibt eine faser- fSrmige Verbindung bestehen. Gegen die Verletzungsstellen bin verlieren die Neu- riten ihren Perlschnurcharakter, quellen gleichm/~Big and massiv auf, ihr Inhalt wird rStlieh und sieht wie ausgelaugt aus. Schon 24 Std nach dem Eingriff ist in den Faserziigen zum Telencephalon kein Sekret mehr zu finden. Keinc Sekret- abnahme zeigt die Verbindung des Kerngebietes zum Ependym des 3. Ventrikels ; sie tritt sogar deutlicher hervor als beim unbelasteten Tier. Diese Tatsaehe 1/~l~t vermuten, dal~ die Unterbrechung des Tractus zur Neuro- und Adenohypophyse zugunsten einer st/irkeren Sekretion in Richtung Ventrikel erfolgt. Das kSnnte auch erld/iren, warum es nach einer L/tsion des Infundibulums nie zu einer mas- siveren Sekretstauung am proximalen Faserstumpf kommt. Die durchtrennten proximalen Neuritenstfimpfe sehen proximal yon der Verletzungsstelle auf- gequollen aus. Es ist anzunehmen, dal3 sie sich nach der Durchtrennung verkiirzen. Aueh naeh liingcr andauernder Unterbrechung des Traetus praeoptico-hypo- physeus konnte keine Regeneration der Faserstfimpfe wahrgenommcn werden. Auch eine subtotale Regeneration der Neurohypophyse (DIERICKX, 1965; Rana temporaria) wurde nicht beobachtet.
Wenige Tage nach dem Eingrfff ist die Neurohypophyse restlos entspeichert; zwischen zarten Gomori-negativen Fasern befinden sich vereinzelt Pituicyten.
2. Befunde nach osmotischer Belastnng
Die Ganglienzellen des Nucleus praeopticus zeigen bei Aufenthalt der Tiere in hypertonischen LSsungen eine vakuolige Erweiterung ihres Zytoplasmas. Das Sekret in Perikaryon und Forts/~tzen sieht grobkSrnig aus. Die Forts/~tze zum Ventrikelependym sind st/irker mit Sekret beladen. Die Verbindungen ins Telence- phalon und in den Epithalamus lassen sich nicht mehr darstellen. Nur das Sub- fornikalorgan enth/ilt vermehrt Gomori-positive Granula. Im Verlauf der Bahn zur Neurohypophyse fallen die beschriebenen vakuoligen Auftreibungen der Perl- schnurfasern auf. Nach mehr als 24 stiindiger osmotischer Belastung sind die Hauptanteile des Faserzuges nicht mehr sichtbar. Die Hypophyse ist stark auf- gehellt, frei yon Herringk5rpern und im Aldehydthioninpr/~parat nur mehr mit ,,staubfSrmigem" Sekret angeffillt. Sowohl die Gef/~$e des Infundibulums als

Diencephal-telencephales neurosekretorisches System bei Salamandra 389
auch der Hypophyse sind erweitert. Herringk6rper werden nur mehr an den Gefi~Ben des Infundibulum gefunden.
E. Das Sub/ornikalorgan vom Salamander
Unter den zahlreichen neurosekretorischen Verbindungen f/ihrt (s. oben) auch eine Bahn vom Hypothalamus zum sog. Subfornikalorgan. Da diese Bahn in einem echten Organ endet, dessen Aufban beim Salamander bisher noch nicht untersucht wurde, beschreiben wires ausffihrlicher.
Bei der Durchsicht der Schnittpr/~parate durch das Salamandergehirn fiel uns ein mit der Basis im Recessus praecommissuralis gelegenes gefi~greiches Organ auf, das nach Lage nnd Gef~6versorgung dem Subfornikalorgan der Wirbeltiere, die einen Fornix besitzen, homolog sein muf.
Friihere Untersuehungen fiber die Morphologie der circumventrikulgren Organe enthalten Beobachtungen, die sich auf eine Einbeziehung dieser Organe in das neurosekretorische System beziehen. E. SCHARRE~ (1951) und HILD (1951) berichten yon einer Nervenfaser- verbindung zwischen dem neurosekretorischen Kernareal des Hypothalamus und der Para- physe. Zu einer ,,besonders reich vaskularisierten Stelle des Ependyms im rostralen Ende des dritten Ventrikels" findet HILD Gomori-positive Faserzfige. Ebenso fanden BARRY (1954, 1960) bei Sgugetieren, E. und H. LEGAIT (1956) bei Sauropsiden, DIERmKX (1962, 1963) und WEBER (1965) bei Amphibien extrahypothalamische neurosekretorische Bahnen zu diesem gef~freichen Organ. Osmotische Belastung bedeutet eine Stimulierung fiir die Zellen der neuro- sekretproduzierenden Kerne. Unter diesen Umst~nden ist zu erwarten, daft auch in die extra- hypothalamischen Faserverbindungen mehr Sekret abgeleitet wird.
WATERMANS (1957) beschreibt das von ihm Subfornikalorgan genannte Gebilde, beim Frosch fiber der vorderen Commissur gelegen, als ein mit einreihigem, hohem Epithcl bedecktes Gefligkn~uel. Einige Gef~fe dringen regelmi~fig bis unter das Ependym. Um die Kapillaren lagern zumeist groge Vakuolen, die in Beziehung zum Ependym treten. Gef~ge aus dem Inter- hemisph~renspalt erreichen das rostrale Ende des Organs. Wir haben daher die Bezeichnung Subfornikalorgan beibehalten, da die Verh~ltnisse beim Salamander in etwa denen beim Frosch ~hneln. Der yon LEGAIT (1942) gepri~gte Begriff des Interventrikularorgans w~re zwar folgerichtiger, doch wird allgemein yon den Autoren, die sich mit diesem Organ oder seinem Homologon besch~ftigen, der Begriff Subfornikalorgan verwendet, um weitere Verwirrung zu vermeiden.
Die Lage des Organs ist beim Salamander durch den Recessns praecommis- suralis und die beiden Foramina Monroi gekennzeichnet. In der dem 3. Ventrikel zugekehrten Wand der Pars ventromedialis des Telencephalons gelegen, dringt es wulstfSrmig in die Lichtung des Recessus vor. Sein Ependym hebt sich im medianen Sagittalschnitt deutlich yon dem sonst flachen Ependym des 3. Ventrikels ab. Bei dieser Schnittrichtung ist im Idealfall auch der Gefi~13zusammenhang mit der Paraphyse nach dorsal und mit den Gefii6en zu erkennen, die aus dem Inter- hemisphgrenspalt in das Organ eindringen. Die Verbindung zur Paraphyse ist bei Embryonen und Larven deutlich ausgeprggt. Horizontalserien, die mit dem Anschnitt der Paraphyse beginnen, lassen kontinuierlich wenigstens ein Gefg6 verfolgen, das unter einer einschichtigen, einreihigen Ependymlage erst waag- recht im Septum ventricnli verli~uft, sich spi~ter in das Ventrikellumen vorzu- wSlben beginnt und so yon dorsal in das Organ einzieht. Aus dem Interhemi- sphi~renspalt erreichen zahlreiche Gef~i6e, begleitet yon zarter Pia mater, den Hilus des Organs. Dazu gelangt yon der Basis des Zwischenhirns ein paariges Gefg6, wahrscheinlich ein Ast der A. ophthalmica aus der A. carotis interna. Strahlenartig treffen die GefgI~e am Hilus des Organs aufeinander und dringen mit kapillaren Schlingen yon rostral ein (Abb. 10).

390
ovl
hy 1 a
W. ARNOLD :
hy
? rsfo
Abb. 11 a--d. Halbschematische Sagittalschnitte durch die Gehirne eines a Primaten (nach AKERT et al., 1961), eines b Kaninchen (nach W~I~DL et al., 1967), einer SchildkrSte c und eines Salamanders d. Der Unterschied des Lageverh~ltnisses von Subfornikalorgan (SFO) und Commissura anterior (c.a.) bei hSheren und niederen Wirbeltieren soll dargestellt werden. AuBerdem fiillt die ~hnlichkeit der Lokalisation zwischen 0rganon vasculosum laminae terminalis {ovlt) des Kaninchen und eines Gef~Borgans an entsprechender Stelle bei der Land- schildkrSte auf. sco Subcommissuralorgan, ep Epiphyse, ~o Psalterium, cc corpus callosum, mi massa intermedia, h habenula, ch chiasma fasiculorum opticorum, hy Neuro- und Adeno-
hypophyse, hc commissura hippocampi, cp Commissura posterior
Auf Sagit talsehnit ten (vgl. Abb. l a) erkennt man die Tiefe des Reeessus praecommissuralis, der vom 3. Ventrikel durch den s~ulenartig in das Cavum ventriculi ragenden vorderen Kommissurenwulst abgegrenzt ist, in dem sich die Commissura hippocampi und die Commissura anterior deutlieh unterseheiden lassen. Die bei S~ugern ffir das Subfornikalorgan so kennzeichnende Beziehung zur Commissura anterior (AKERT, POTTER und ANDERSON, 1961), die selbst bei Frosch und Schildlcr6te noch zu beobachten ist, fehlt hier. Beim Salamander steht die Entwicklung des Gehirns noch auf einer phylogenetisch tieferen Stufe. Das Telencephalon dominiert mit seinem groBen Riechhirnanteil vor dem Zwischen- him. Ers t mit der Aufrichtung der beiden Hemisph~ren, wie sie beim Frosch bereits beginnt, rfieken dann Commissura anterior und Subfornikalorgan dicht anein- ander; das E p e n d y m fiber beiden Bereichen geht ineinander fiber. Der Recessus

Diencephal-telencephales neurosekretorisches System bei Salamandra 391
praecommissuralis wird zu einer winzigen, nur gelegentlich noch auftretenden Ausbuchtung, dem Recessus dorsalis, zusammengedr~ngt. Die beiden vorderen Kommissuren liegen nun in der rostralen Wand des 3. Ventrikels zwischen Lamina terminalis und Lamina supraneuroporica (HALLER voN HALLERSTEI~, 1934, Abb. ll). Somit ergibt sich beim Salamander folgende Lage des Organs: es ragt yon rostral wulstfSrmig in die Lichtung des Recessus praecommissuralis und stellt einen ,,Knotenpunkt" von Kapillaren dar, welche von verschiedenen Gebieten des Gehirns dem Organ zustreben. Kennzeichnend ist zudem ein inniger Kontakt mit Plexusschlingen, die sich dem mit einem Spezialependym ausgestattetem Organ dicht anlegen.
Im Gegensatz zu dem regellosen Aufbau bei zahlreiehen Vertebraten lassen im Subfornikalorgan des Salamanders Gefa~e, Zellen und Vakuolen eine gesetz- m~ige Anordnung erkennen. Im Zentrum des Organs liegen sinusfSrmige Gef~- schlingen mit schmalem, perivaskul~rem Randsaum. Diese gehen aus zahlreichen spiralig gewundenen Gef~l~en hervor, die zusammen mit zartem Bindegewebe dureh den Interhemisph~renspalt in das Organ eindringen. Das Bindegewebe umscheidet den Gef~stamm bis zum ,,Hals" des Kapillarknauels; im Bereich der Schlingen besteht der perivaskul/~re Raum nur aus einer sehr zarten Binde- gewebsscheide zwischen innerer und gu~erer Basalmembran der glomerulum- artigen Kapillare (Abb. 12a, b). An der Spitze des Gef~Skn~uels, wo an manehen Stellen die Kapillaren das Ependym auseinander dr~ngen, trennen nur noch gliaseitige Basalmembran und Ependymplasmalemm den Ventrikelliquor yon der perivaskul/~ren Flfissigkeit.
Um dieses Gefa~system sind Parenehymzellen und Gliazellen angeordnet. Das Organ wird yon einem hochprismatischen, cytoplasmaarmen Ependym bedeckt, das sich vom iibrigen Ventrikelependym stark unterscheidet und beim Salamander nur noch Ahnlichkeit mit dem Spezialependym des Subkommissural- organs besitzt. Besonders eindrucksvoll tritt dieses palisadenartige Ependym auf medianen Sagittalschnitten hervor. Es unterscheidet sich nicht nur in Form und GrSl3e vonder gewShnlichen Ventrikelauskleidung, sondern f~llt schon durch seine intensive Anf/~rbung im Gomoripr~parat auf. Die Ependymzellen liegen hier so eng geschlossen aneinander, dal3 das Bild einer tier dunkelblau gef/~rbten Leiste entsteht (Abb. 10). Mit stark basischen Farbstoffen (Alcianblau, Astrablau und nach Oxydation mit Perameisens/~ure) f/irben sich diese Ependymzellen intensiv blauviolett ; darunter befinden sich immer helle Zellen, die bei Gegenf/~rbung mit Kernechtrot einen rStlichen Farbton annehmen. Feinere Strukturen (Nucleolus, Chromatinzeichnung) kSnnen wegen der starken Affinit/~t der Farbstoffe nicht unterschieden werden. Ober der Palisadenschicht des Ependyms findet sich ein zarter, schlieriger, phloxinroter Belag, der PAS-positiv ist. Gomori-positive Granula werden regelm/~l~ig zwischen den Ependymzellen des Organs gefunden, h/~ufiger jedoch unter dem Ependym. Am Scheitel des Organs wird die Basis dieser Zellen yon Kapillarschlingen begrenzt, die nicht selten die Ependymzellen auseinanderdr/~ngen. H/iufig finder man unter dem Ependym Vakuolen sehr verschiedener GrSBe, in denen offenbar ein starker Innendruck herrscht; dadurch werden die Ependymzellen an ihrer Basis eingebuchtet (Abb. 13). Die seitlichen Wiinde des Organs sind yon einem kubischen Ependym bedeckt. Sie reichen mit langen gestreckten Ausl/iufern tanycytenartig an die /tuBere Basalmembran des

392 W. ARNOLD :
Gef~l~kns wo sie mi t brei ten Ffii~chen enden (Abb. 14). Inne rha lb des Gef~i~- kn~uels, d.h. zwischen den Kapil larschlingen liegen im Zen t rum des Organs zahlreiche grobe K l u m p e n a n d Granula von schwarzbraunen Pigment . Davon
A
~-Ihs-*- I
AZ
Ep
Abb. 12. a Schematische Zeichnung eines Ausschnittes aus dem sagittal geschnittenen Sub- fornikalorgan des Salamanders. Aus dem Interhemisph~renspalt dringen (lhs) arterielle Gef/~Be (A) in das kammartig in den 3. Ventrikel vorspringende Organ ein. Die zellarme (AZ) Aullen- zone ist vonder den Zellschild (Z) enthaltenden Innenzone (IZ) zu unterscheiden. Der Abflu0 des sinusartigen Kapfllarsystems fiihrt zu weitlumigen Gef~l~en, die unter dem Ependym des rostralen Endes des 3. Ventrikels yon der Paraphyse in dorso-ventraler Richtung zum Organ ziehen. Sie stehen wiederum mit Gef/s des InterhemisphKrenspaltes in Verbindung. (Pa paraphys~re Geflil~verbindung, VA ven6ser Abflul~). Unter den Parenchymzellen (PZ) des Zellschildes der Innenzone findet man Gomori-positive Zellen (GPZ), yon denen Perlschnur- fasern (Gs/Gomori-positive Sekretfasern) an die glomerul~ren Schlingen (gS), sowie an die palisadenartigen Ependymzellen (Ep) und die zwischen ihnen gelagerten Vakuolen (V) ziehen. L Ventrikelliquor; Pi Pigmentzellen; g/ Gliafasern; pR perivaskulirer Raum eF extrazellul/ire Fliissigkeit. b Sa. 31/50 ~. Horizontalschnitt dutch den dorsalen Anteil des Organs; Fixation nach BOUIN; zuvor in neutralem gepuffertem Formalin; F~rbung: Azan; Einbettung in Paraplast. Vergr. 300fach. Die Azanf/irbung l~llt die bindegewebige Kompo- nente des Kapillarkn~uels hervortreten. An der Spitze des Organs w61bt sich eine einzelne Kapillarschlinge zwischen den Palisadenzellen gegen das Ventrikellumen (L) vor. Durch den
zarten Bindegewebsmantel scheinen die roten Blutk6rperchen durch

Diencephal-telencephales neurosekretorisehes System bei Salama,wlra 393
Abb. 12b
Abb. 13. Sa. 16/41 $. S~gittalschnitt durch das Subfornikalorgan. Fixation nach ELFTMANN (1957); Paraffineinbettung; F~rbung nach GOMORI-BARGMANN. Vergr. 1250fach. Zwischen die dicht stehenden Palisadenzellen schiebt sich eine Vakuole gegen den Ventrikelliquor (V) vor. Nur ein zarter Zytoplasmasaum trennt die beiden Fliissigkeitsr~ume. E Endothelzellen der
Sinuskapillare

394 W. ARNOLD:
Abb. 14. Sa. 31/40 ~. Horizontalschnitt durch das Subfornikalorgan; Fixation in neutralem, gepuffertem Formol ; Paraplasteinbettung. F~rbung: Aldehydthionin-Hansen. Vergr. 300fach. Vom Ependym der seitlichen Organbegrenzung treten Tanycytenforts~itze an die Gef~gw~inde
heran (T). S Gef~Bstiel aus dem Interhemisph~renspalt. V Ventrikel
zu unterscheiden sind dunkelblaue Gomori-positive Ansammlungen yon Granula, die vorzugsweise entlang den Kapillarschlingen liegen. Die bereits erw~hnten tanycytenart igen Forts~tze nehmen am Aufbau des Gliafflzes tell, der vor allem in der N~he des Gef~i~kni~uels yon blasigen ,,Gliak~mmerchen" aufgelockert wird. Diese optisch leeren R~ume zeigen zumeist einen zarten Gomori- oder Aldehydthionin-positiven Randsaum. Die Gliazellfortss laufen geschl~ngelt zwischen diesen Hohlri~umen hindurch. Subependymal liegen vereinzelt Glia- zellen, deren Forts~tze auch am Aufbau des Gliafilzes beteiligt sind. Eine weitere Gruppe yon Zellen ist um den Gef~l~pol angeordnet und grenzt ihn schildfSrmig gegen den Interhemisph~renspalt ab. In HShe der Basis des Organs dringt yon rostral aus dem Septnm hippocampi beider Hemisph~ren ein schmales Nerven- faserbfindel in das Organ ein, woes sich im Gliafilz aufsplittert.
Somit lassen sich folgende Bauelemente des Organs hervorheben: a) das Gef~Bkn~iuel, bestehend aus Endothelrohr mit Pericyten, einer inneren
und einer iiu~eren Basalmembran. Zwischen den beiden Basalmembranen liegen Adventit ia und Pigmentzellen ;

Diencephal-telencephales neurosekretorisches System bei Salamandra 395
b) die vakuolen/ihnlichen, gliabegrenzten K/~mmerchen fiber der /~uBeren Basalmembran mit ihrem Gomori-positiven Belag;
c) das sich intensiv anf/irbende Palisadenependym;
d) Zellen, welche das Organ rostral gegen den Interhemisph~renspalt abgren- zen und um den Gef/~13pol gelagert sind (Zellsehild).
Es ist nicht mSglich, die schildartig gruppierten Zellen an Hand ihrer Kern- und Zytoplasmastruktur in Parenchym- und Gliazellen zu unterscheiden. Im wesentlichen lassen sich zwei Zelltypen hervorheben. Den zahlenm/~Big grSgten Anteil bilden Zellen mit grol~em ovalen bis runden Zellkern, der zwei bis drei Nucleoli in einem locker angeordneten Chromatin aufweist. Die Kernmembran ist deutlich konturiert. Das Zytoplasma konnte mit keiner der erw/~hnten, dem RN S-(Ni$1-Substanz)-Nachweis dienenden Methoden dargestellt werden. Daneben trifft man kleine, vielgestaltige Zellen an, die reich an Chromatin sind; sic liegen haupts/~chlich am Organhilus. Auch hier ist kein Zytoplasma darstellbar.
Man findet auBer den aufgez~hlten Bauelementen nicht selten Zellen, welche bei ,,Neurosekret"-f/~rbung groSe Ahnlichkeit mit den Ganglienzellen des Nucleus praeopticus besitzen. Nach Eingriffen in den Wasserhaushalt treten sic mit ihren Forts/~tzen besonders hervor. Vorwiegend unter dem Ependym und um das Gef/~ft- kn~uel heben sich Axone mit zarten bis grSberen Granula ab. Typische Perl- schnurgranuherung findet sich vorzugsweise im peripheren Antefl des Axons. Nahe dem Abgang vom Perikaryon ist das Axoplasma gleichm~$ig rein gekSrnt erkennbar (Abb. 15). Am Zelleib selbst, der vakuolig aufgetrieben sein kann, l/~$t sich nur am Sekretpol Zytoplasma erkennen, das bei 1/~ngerer experimenteller Belastung der Tiere stark aufgehellt und im Aldehydthioninpr/~parat homogen erscheint. Die Zellkerne gleichen nach GrSl~e und Anf/~rbbarkeit denen des erst- geschilderten Zelltypus und liegen im basalen Tell des Organs hypendymal oder vor dem Zellschild. Unter osmotischer Belastung nehmen die Vakuolen um die Kapillarschlingen zu. Die Ghafasern werden yon den sich vergrSSernden Vakuolen auseinandergedr/~ngt, anfangs nut um den GefaSsinus herum, nach 1/~ngerer Belastung bis unter das Ependym. Zwischen den einzelnen Vakuolen trifft man Gomori-positive Granula an, die sich ihren R/~ndern anlegen. Zuweilen ballt sich sandf5rmiges Sekret zu runden, kompakten Gebilden zusammen. Nur in wenigen F/~llen 1/il]t sich mit Bestimmtheit eine intrazellul/~re Lage der Vakuolen (RUDERT, 1965; Rana esculenta) feststellen. Eine ErSffnung der Vakuolen in den Ventrikel wurde nicht beobachtet.
Ein Zusammenhang der sekretorischen Zellelemente im Subfornikalorgan mit dem hypothalamischen System konnte nicht festgestellt werden. Dennoch gelan- gen neurosekrethaltige Axone aus dem Hypothalamus fiber den Tractus prae- optico-septalis und den Tractus praeoptico-telencephalicus zur Basis des Organs. Es ist somit nicht zu unterscheiden, welche der feinen Sekretfasern im Organ den ortsst/~ndigen sekretorischen Zellelementen und welche dem Nucleus praeopticus entstammen. Beriicksichtigt man die MSglichkeit, dal] die Gomori-positiven Parenchymzellen des Organs als verlagerter rostraler Anteil des Nucleus prae- opticus zu betrachten sind (KELLY, 1967), SO dfirften die Gomori-blauen Elemente im Subfornikalorgan Teile des hypothalamischen neurosekretorischen Systems verkSrpern.

396 W. ARNOLD:
Abb. 15a u. b. Neurosekret im Ependym des Subfornikalorgans. a Sa. 77/53 9- Horizontal- schnitt dutch das Subfornikalorgan. Fixation nach BovIN; Paraplasteinbettung; Aldehyd- thioninf~irbung. Vergr. 1250fach. Zwischen den Parenchymzellen am Hilus des Organs linden sich stets auch Gomori- oder Aldehydthionin-positive sekretproduzierende Zellen mit typi- schem perlschnurartigem Fortsatz. b Sa. 93/17 9. Horizontalschnitt durch das Subfornikal- organ; Fixation nach EL~'TMANN (1957); Paraffineinbettung; F~rbung mit Aldehydthionin.
Vergr. 1250fach. Aldehydthionin-positive Sekretfaser unter dem hohen Ependym. V Ventrikelliquor
Sehr deutlich li~13t sich die Lage des Subfornikalorgans zur Umgebung mit Hilfe der Reaktion auf alkalische Phosphatase darstellen (Abb. 16). Das Ependym des Subfornikalorgans zeigt im Gegensatz zum benachbarten Ventrikelependym keine Reaktion. Doch weist der Gef~13sinus eine hohe Aktiviti~t auf. Ein weiteres Gefi~fiorgan, das dem Organon vasculosum laminae terminalis der Sduger und Sauropsiden entspr/iche, fehlt beim Salamander. Die Vermutung liegt aber nahe,

Diencephal-telencephales neurosekretorisches Sysf~m bei Salamandra 397
Abb. 16. Sa. 89/33 ~. Horizontalschnitt durch das Subfornikalorgan; Nachweis der alkalischen Phosphatase im nativen Kryostatsehnitt; Eeaktion naeh GOMORI (1952). Vergr. 300fach. Die Gef~13e und eine schild/~hnliehe Parenehymzone um den Stamm des Organs reagieren positiv, w~hrend das Organependym sowie das Zentrum des Gef/iBkniiuels negativ reagieren
da$ es sich bei dem beschr iebenen Organ ebenso um das Gef/i$organ der bei h6heren Wirbe l t i e ren ausgepr/~gten Lamina te rmina l i s hande ln kSnnte, bzw. da$ beide Organe hier noch in e inem einzigen vereinigt sind und erst in der Phylogenese ause inanderr f icken (Abb. 17). Daffir spr icht e inmal die enge Gef/ iSbeziehung zwischen dem Subforn ika lorgan und dem Gef/~$organ der Lamina te rminal i s bei hSheren Wirbel t ie ren , zum anderen die J~hnlichkeit im Aufbau.
Bisher wurde dieses Gefiil3organ aul3er bei S/iugetieren und dem Menschen aueh bei V6geln (DvwRNoY und KORITKG 1963) beschrieben. BRA~_K (1963) berichtet zudem yon einem Organum vasculosum praeopticum bei Chimaera monstrosa, das sehr groBe Ahnlichkeit mit den entspreehenden Bildungen der S/~ugetiere aufweist. Der hahnenkammartige Vorsprung des Gefiil3organs in die Lichtung des Recessus praeopticus veranlal3te dazu, beim Kaninchen (WISLOCKI und KING, 1936) yon einer Crista supraoptiea zu sprechen. Beim Vergleieh mit dem Subfornikalorgan des Salamanders fiillt auf, dab auch hier der OrgankSrper eine ausgepr~gte VorwSlbung des sehmalen Kammes darstellt, der ent.lang der rostralen Wand des 3. Ventrikels in den Reeessus praecommissuralis ragt. Aueh die GefiiBverhaltnisse der Organe stimmen im wesentlichen fiberein. Aus dem Interhemisph~renspalt, der AuSenzone, dringt ein binde- gewebsreicher Keil von rostral in das Parenchym der inneren Zone ein, zahlreiehe ineinander gewundene Kapillarschlingen begleitend.
Bei Chimaera monstrosa erstreckt sich yon ventral, ausgehend yon der Meninx primitiva gefi~Bfiihrendes Bindegewebe in die ependymale Bodenplatte des Reeessus praeoptieus, die wulstfSrmig in den Ventrikel hereinragt. In enger Beziehung zum Gef~Bbaum treten zahlreiche Vakuolen gruppenfSrmig auf, die bis unter das Ependym reiehen, wobei sit die Zellen aus- einanderdr~ngen und abplatten kSnnen. Beim Gef~13organ der Lamina terminalis, beim Organum vasculosum praeopticum und beim Subfornikalorgan des Salamanders sind aueh Gemeinsamkeiten im zellul/~ren Aufbau festzustellen. Die Aktivit~t der alkalischen Phospha- task und unspezifisehen Esterase beim Salamander kaml mit den Befunden am Organum

398 W. ARNOLD:
Abb. 17. Esk. 9/66. Sagittalschnitt durch die Lamina terminalis der europ~ischen Landschild- kr6te. Fixation in Buoinscher L6sung; Parablasteinbettung; Fiirbung nach GO~ORz-BARG- MANY. Vergr. 250fach. Von rostra] dringen schlingenf6rmige Gefiifle in die Lamina terminalis ein und gelangen bis unter das Ependym. Zwischen den Kapillarschlingen und dem Ependym
ziehen regelmi~I3ig neurosekretorische Fasern zur Commissura anterior nach dorsal
vasculosum laminae terminalis (WEINDL, 1965) verglichen werden. Da zum Vergleich auch einige Gehirne yon SchildkrSten untersucht wurden, konnte man erkennen, dal3 bei diesem Reptil bereits ein sog. Subfornikalorgan yon einem Organum vasculosum laminae terminalis unterschieden werden kann.
Der Aufbau der Lamina terminalis mit den yon rostral eindringenden Gef~Bschlingen und dem dariiber ]iegenden Kommissurenkomplex erinnert an die Verh~ltnisse bei manchen Siiuge- tieren (Abb. 17). Vakuolen sind in dem entsprechenden Gef~Borgan bei der SchildkrSte nicht zu beobachten, doch f~llt eine deutliche Beziehung zu neurosekrethaltigen Axonen auf, welche zwischen dem cilienreichen, hochprismatischen Ependym und den Spitzen der ein- dringenden Gef~i3schlingen zur Commissura anterior ziehen. Ihr Ziel scheint das stark aus- gepriigte Subfornikalorgan zu sein. Regelmiiflig stehen beide Organe fiber ein Gefi~fl in Ver- bindung, das rostral yon der Commissura anterior liiuft und mit dem ungeordnet scheinenden Gefiiflsystem des Subfornikalorgans in Verbindung steht. Eine eingehende Untersuchung der beiden Organe bei dieser Reptilienart w~re yon Interesse.

Diencephal-telencephales neurosekretorisches System bei Salamandra 399
Diskussion Unsere Beobachtungen am neurosekretorischen System des Salamanders
entsprechen in den Grundzfigen den Verh/iltnissen bei anderen Amphibien (HrbD, 1950, 1951; MAZZI, 1953, 1954; WILSOn, 1957; KLEIn, 1961 ; B~AMrKOWA, 1964), wonach der Nucleus praeopticus als Bildungsort des Sekrets und die Axone als Transportbahnen gelten, yon denen ein Hauptfaserzug zur Pars nervosa der Hypophyse gelangt. Unsere Befunde an Salamanderembryonen und Larven widersprechen der Hypothese yon DI~P~N (1962), nach der das Sekret ubiquit/ir entstehen soll. Sie stfitzt sich auf die Beobachtung, dab w/ihrend der Ontogenese das Sekret zuerst im Hinterlappen auftrit t und erst zu einem sp/iteren Zeitpunkt in den Perikarya der sekretorischen Kernareale aufzufinden ist (C~RIST, 1962). Entgegen DIEPEN, ENGELHARDT und SMITH-AGREDA (1954) grit heute die Ent- stehung des Neurosekretes im Golgiapparat der Ganglienzellen des Nucleus praeopticus durch elektronenoptische und biochemische Untersuchungen als gesichert (Zusammenfassung bei B ~ M A ~ N , 1967). Mit dem Neurosekretnach- weis nach ST~RBA (1961) gelang es u.a. WEm~R (1965) gleichzeitig mit dem ersten Auftreten fluoreszierenden Neurosekrets in der Hypophyse das Vorhandensein des Sekrets in den Perikarya der hypothalamischen Ganglienzellen beim Sala- mander darzustellen, l~bereinstimmend mit W~BER (1965) konnten wir nach- weisen, da]~ bei Salamanderembryonen yon 8 mm L~nge auBer in der Hypophyse auch schon im Nucleus und vereinzelt im Verlauf des Traetus zur Hypophyse Gomori-positives Neurosekret vorliegt. Wir konnten zudem zeigen, dab bei Embryonen, deren Adenohypophyse noch gar nicht vollst~ndig angelegt war, bereits Gomori-positive Faserzfige zwischen den Ependymzellen des distalen Infundibulums anzutreffen waren. Ahnliche Beobachtungen stellte RINNE (1964) am hypophys~ren Pfortaderkreislauf w~hrend der menschlichen Foetalentwick- lung an. WEBER und BACHMANN (1964) beriehten ebenfalls von einer deutlichen Beziehung neurosekretorischer Neuriten zu portalen Gef~Ben w~hrend der Embryogenese bei der Taube zu einem Zeitpunkt, wo auch in der Neurohypo- physe erstmalig Sekret zu erkennen ist. Der Kontakt zu Gef~Ben der Adeno- hypophyse ist zu diesem Zeitpunkt schon deutlich und die Anhiiufung des Sekretes am neurohypophysenfernen Rand der Gef~Be l~Bt darauf schlieBen, dab das Sekret yon hypothalamischer Seite stammt. Die zeitlich unterschiedliche Dar- stellung des Neurosekretes mit der Fluoreszenzmethode (STERBA, 1964) und mit BA~G~ANNs Chromalaunh~matoxylin beruht darauf, dab erst nach Erreichen eincr bestimmten Konzentrationsschwelle (HILD, 1951) das Sekret mit der letztgenann- ten Methode erfaBbar ist. Nach der Vorschrift yon BOCK (1966) ist jedoch auch mit Chromalaunh~matoxylin Neurosekret wesentlich frfiher als bisher darzu- stellen. Die Ausreifung des Kerngebietes erfolgt beim Salamander in der larvalen Periode. Weite Zfige des Tractus zur Hypophyse sind zuvor schon darstellbar.
Neben den extrahypothalamischen Sekretbahnen des Nucleus praeopticus ist seine Beziehung zum Ventrikel, besonders unter osmotischer Belastung von Interesse. Hinsichtlich einer Gesamtkonzeption yon Neurosekretion in das Liquor- system sei auf das Schema von KI~OWLES (1967) verwiesen. Bei Bu/o vulgaris und Rana esculenta beschreiben BOLOGI~ARI und ROTELLA (1956) neurosekretffihrende Axone zum Ependym des 3. Ventrikels. Die Ganglienzellen liegen im Hypendym und entsenden einen Fortsatz zwischen die Ependymzellen, den anderen in die
27 Z. Zellforsch., Bd. 89

400 W. ARNOLD :
Peripherie. Die Verfasser bezeichnen die zum Ventrikel ziehenden Axone als Dendriten, denen sie die Neuriten zur Hypophyse gegenfiberstellen. Auch DIERICKX (1962, Rana temporaria) nennt solche Fasern Dendriten. In seine Vorstellung fiber den Funktionskreis der osmotischen Regulation bezieht er auch das Subfornikalorgan und die mit diesem in Verbindung stehenden neurosekretori- sehen ,,Dendriten" ein. Die Dendriten enden zwischen den Ependymzellen der pr~optischen Region in engem Kontakt zum Ventrikellumen. Dort laufen sie zu einem EndkSlbchen aus, das mit Neurosekret angeffillt ist. Ahnliche Beschrei- bungen liegen yon SMITH (1951) und WILSON et al. (1957) vor, die jedoch nicht n~her auf eine Deutung oder Definition dieser Fasern eingehen. DIERICKX (1963) riiumt diesen Dendriten rezeptorische Funktion ein. Bei Hyla regilla findet SMOLLER (1965) elektronenmikroskopisch kleine KSlbchen, die zwischen den Ependymzellen in das Lumen des Ventrikels ragen. Ihr Inhalt weist neuro- sekretorische Granula, Mitochondrien und ein endoplasmatisches Retikulum auf. Die Verfasserin sieht darin ein Rezeptorengebilde, dessen Funktion aber nicht allein in der Weitergabe des Milieubefundes besteht, sondern in der Lage sein soll, Neurosekret zu bi]den und in den Liquor abzuleiten. Dagegen beschreibt MURAKAMI (1964) bei Bu/o vulgaris Formosus Forts~tze ventrikelnaher neuro- sekretorischer Ganglienzellen, die an die basalen Teile des Ependyms heran- reichen. Hier dringt Ventrikelliquor in Spalten zwischen die Ependymzellen ein. In diesen Spaltraum reichen die Mehrzahl der in versehiedenen Riehtungen orientierten mikrovilliartigen Ausli~ufer der Ependymzellen hinein. In den dem Extrazellularraum zugekehrten Zellabschnitten der Ependymzellen und der neuro- sekretorischen Zellelemente seien groBe Mengen von mikropinocytotisehen Vesikeln zu finden. Der Autor glaubt dem Verhalten des Extrazellularraumes entnehmen zu kSnnen, dab hier ein besonderer Stoffaustausch stattfindet, konnte aber weder in den Liquor, noch in diesen Raum eine Abgabe yon Neurosekretgranula beob- aehten. Die Tatsache, dab nach einer Hypophysektomie oder bei osmotischer Belastung eine Zunahme des Sekrets in den Fasern in Riehtung Ventrikel erfolgt, bzw. ein Grol~tefl der Fasern unter diesen Bedingungen erst darstellbar wird, ls an einen Mechanismus der Sekretabgabe in den Ventrikel denken. Dieser Weg wfirde unter den genannten Bedingungen eine erhebliche Verkfirzung ffir die Abgabe des Wirkstoffes darstellen, welcher fiber den Liquor ins Blur gelangen kSnnte. Die schlechte Darstellbarkeit dieses subependymalen Sekretfasernetzes unter normalen Bedingungen l~Bt an eine Art von ,,Notventil" denken, das in Strel~situationen genutzt wird. Wie die Drfisenzellen den Impuls zur Sekretabgabe erhalten, ist noch umstritten. Die hohe Aktivitat der Acetylcholinesterase (vgl. KOBAYASHI und FARbTER, 1965) und der unspezifischen Esterase l~l~t eine cholin- ergische Beeinflussung des Systems vermuten.
DaB beim Salamander die MSglichkeit einer Abgabe von Sekret in den Ventrikel besteht, erkennt man im distalen Infundibulum und im Ependym der Neuro- hypophyse selbst. EICHNER (1963) erw~hnt beim Maulwurf den ~Jbertritt von Gomori-positivem Material in den Liquor, ausgehend vonder Eminentia mediana ; auch LEMAmE, ALLEGRINI und MAZER (1961) beobachteten bei hypophysekto- mierten Hunden eine Sekretabgabe in den 3. Ventrikel.
Uber den Regulationsmeehanismus ffir die Produktion des Neurosekretes wissen wir fast gar nichts. Folgende M5glichkeiten sind denkbar: 1. Die Hem-

Diencephal-telencephales neurosekretorisches System bei Salamandra 401
mung oder F6rderung des sekretorischen Neurons dureh cholinerge bzw. adrenerge Neuriten eines vorgeschalteten ( = fibergeordneten) rezeptorischen Neurons. 2. Information des Perikaryons durch Dendriten, die Blut- bzw. Liquormilieu an geeigneten Stetlen, wie z.B. in einem der zirkumventrikul~ren Organe, regi- strieren. 3. Direkte Informationen des sekretorischen Perikaryons dutch engen Kontak t mit einer hypothalamischen Kapillare (ungfinstig wegen dazwischen- geschalteten Astrocyten; vgl. SLUGA und SEITELBERGER, 1967). Zun~chst erschei- nen Mechanismus 1. und 2. am wahrscheinlichsten verwirklicht. Auch im Falle 1. k6nnte das vorgeschaltete Neuron aus einem besonders geeigneten Rezeptorbezirk kommen, z.B. dem Subfornikalorgan.
Sekrethaltige Fasern zum Ependym k6nnen beim Normaltier nicht als Den- driten bezeichnet werden. Unentschieden bleibt die Frage unter experimentellen Bedingungen. Dort k6nnte durch den behinderten Abflul~ der Neuriten zur Hypo- physe Sekret in die Dendriten umgeleitet werden.
Uns fiel beim Salamander eine deutliche Beziehung zwischen Anteilen des Tractus praeoptico-hypophyseus und schlingenartigen Gef~13en der Adenohypo- physe im Infundibulum auf. Nicht nur bei adulten Tieren, sondern bereits beim Embryo findet man einen Kontak t zu den sog. Portalgef~13en (vgl. TROYER, 1965). Die Abhs gewisser adenohypophys~rer Funktionen vom neurosekretori- schen System wurde verschiedentlich er6rtert (vgl. S]~ITZ, 1964). Dies geschieht entweder fiber die Portalgef~l~e oder fiber Gomori-positive Neuriten, die in die Adenohypophyse eindringen.
Allgemein gilt, dal3 es sich bei dem f~rberischen Produkt der hypothalamischen G~nglien- zellen um eine disulfidbrfickenreiche Tr~igersubstanz zweier 0ktapeptide handelt, deren Struk- tur bekarmt ist (VAN DYKE, zit. nach KXaLSON, 1965). Diese sind das Oxytocin und das Vasopressin, Substanzen, deren Struktur sich fiber viele Wirbeltierklassen hinweg sehr kon- stant erh~ilt oder sich nur geringffigig ~ndert (vgl. STEI~BA, 1967). Da die Neurohypophyse als eine Art Vorratskammer dieser beiden Wirkstoffe anzusehen ist, yon wo sie fiber den Blutweg ihre spezifische periphere Wirkung ausl6sen, scheint es zun~chst schwer verst~ndlich, welche Bedeutung dem gleichen morphologischen Produkt ffir die Adenohypophyse zukommt. Beobachtungen nach Exstirpation des Nucleus praeopticus yon Salamanderlarven haben eindeutig die Abh~ngigkeit des Wachstums sowohl des Hinterlappens, als auch des Vorder- lappens gezeigt. Bei Abwesenheit der Gomori-positiven Ganglienzellen im Zwischenhirn kommt es zu einer Atrophie der bereits vorhandenen Neurohypophyse und Wachstums- hemmung mit Mil3bildungen der Adenohypophyse (VOITKWICB und CaELK~, 1964).
Nach Versuchen yon IGARASHI und CANN (1964) beeinflussen Hinterlappen- wirkstoffe das Wachstum yon gonadotropen Drfisenzellen des Vorderlappens; danach enthielte das f~rbbare Produkt der hypothalamischen Neurone zushtzlich einen oder mehrere Faktoren mit dieser Wirkung (vgl. auch ASSENMACH]~R, 1964). DIERICKX (1965) wies bei Rana temporaria Ver~nderungen der Aktiviti~t des Nucleus praeopticus in enger Beziehung zu Wechseln bei der Ovulation, Spermation und Kopulation nach. Nach Enukleation der neurosekretorischen Kerne tr i t t zwar eine normale jahreszeitliche Entwicklung der Gonaden und der sekund~ren Geschlechtsmerkmale ein, die Ovulation jedoch bleibt aus (Dn~RICKX, 1963).
Unter Neurosekret hat man also vorerst eine mit verschiedenen empirischen und histochemischen Methoden erfal~bare Substanz zu verstehen, welche erwie- senermal~en zwei Oktapeptide enthiflt, darfiber hinaus aber noch Wirkstoffe ent- halten kann, welche sowohl fiber die Adenohypophyse gonadotrope (DrEaICKX,
27*

402 W . ARNOLD :
1965; VIERMANN, 1962) als auch ACTH-stimulierende Wirkungen besitzen (MARTINI, 1957; SUOMAL~NV, N, 1957; DE WIED, 1962; BAUMA~Njr., 1965; KIVALO, RINN~ und B]~RGSTRSM, 1964) kSnnten.
Uber die Bedeutung extrahypothalamiseher Faserzfige ist nichts bekannt. Unseren Befunden nach zu schlieBen ist ffir den Teil der telencephalen Bahn, der an den Gliak~mmerchen der Vorderhirnbasis endet, eine sekretorische Funktion anzunehmen. BURY (1954, 1961) und LEGAIT (1957) betraehten s~mtliehe extra- hypothalamischen Gomorifasern als sekretoriseh. In unserem speziellen Fall enden die sekretffihrenden Axone an den Gliak~mmerehen. Eine weitere Aussage fiber das Schieksal des Sekrets im Zusammenhang mit den Gliaki~mmerchen (eventuell Permeabilit~ts~nderung der Membran und Ubertritt) ist liehtmikroskopisch nicht mSglich. Wie erw~hnt, schreibt DIERICKX (1962) diesen Faserzfigen eine rezep- torische Funktion zu, ferner den unter Belastung nachweisbaren Gomorifasern im Chiasma opticum, da sie als Teile des retino-hypothalamischen Systems (BENOIT und ASS~NMACHnR, 1955; KNOCH~, 1960) visuelle Reize an das hypothalamische Zentrum weiterleiten kSnnten. DIERICKX (1962) schlieBt nicht aus, dab sekretori- sche und rezeptorische Funktionen nebeneinander im gleiehen Fasersystem existieren.
Die kennzeichnende Lage des Subfornikalorgans am ZusammenfluB der beiden Seitenventrikel in den 3. Ventrikel und gewisse Grundelemente im Aufbau erlau- ben die Annahme, dab es sich bei diesem Gef~Bkn/~uel im Recessus praeeomis- suralis beim Salamander um ein Homologon des Subfornikalorgans hSherer Wirbeltiere handelt.
Konstant bei allen Wirbeltieren ausgepr/igt ist die auBergewShnhch starke Gef/~Bversorgung mit einem yon Bindegewebselementen erffillten perivaskul~ren Raum, der eine erniedrigte Blut-Hirn-Sehranke (WIsLOCKI und LEDUC, 1952) bedingt. GliSses Grundgewebe umgibt dieses Gef~l~system. Gegen den Ventrikel wird dieses Organ bei den meisten Wirbeltieren yon einem abgeflachten Ependym bedeckt (WATERMANN, 1955). Bei hSheren Wirbeltieren fallen im Organ typische Parenchymzellen mit groBem ehromatinarmen Kern auf, deren ZugehSrigkeit zu Glia- oder Nervenzellen lange umstritten war (WATERMANN, 1955, 1957). Nach neueren Untersuchungen besteht an ihrer neuronalen Herkunft kein Zweifel mehr (HoFER, i965; ANDRES, 1965; WEIN])L, 1965; RUDERT et al., in Vorbereitung). Auch fiber die bei vielen Vertebraten darzustellenden Vakuolen herrschte lange keine fibereinstimmende Meinung. Die bei den meisten S/~ugetieren regellos im Gliagewebe verstreut hegenden Vakuolen entstehen nach elektronenmikro- skopischen Befunden intrazelluliir (RoHR, 1966; Katze).
fi~hnhch wie beim Frosch (RUDERT, 1965) kann beim Salamander ein gewisses Prinzip im Aufbau des Subfornikalorgans festgestellt werden. Zen~rum des Organs ist der kompakte Gef/~Bkomplex, der zu glomerulum/ihnhchen Schlingen geformt und von Vakuolen umgeben ist, an die Ghaforts/ttze grenzen. In Bezie- hung zu dem zarten/iuBeren Bindegewebe und zahlreichen Vakuolen stehen fein granulierte Gomori-blaue Sekretfasern. Nach DIERICKX (1962) stellen sie beson- ders ausgepr/igte Dendriten dar, welche feine osmotische Unterschiede zwischen Liquor und der Fliissigkeit des Perivaskul~rraumes registrieren. Bei dieser Hypo- these bleibt jedoch die Bedeutung der Gomori-positiven Fasern offen. Wir fanden im Subfornikalorgan Gomori-positive Parenchymzellen. AuBer der Produktion

Diencephal-telencephales neurosekretorisehes System bei Salamandra 403
von ttinterlappenhormonen halt ANDRES (1965) aueh die Bereitung adrenerger Transmittersubstanzen in diesem Organ ffir wahrseheinlich. Eine Interpretation dieser Zellen ist nicht mSglich, so lange Ausdehnung und Verlauf ihrer Fortsgtze nicht bestimmt sind und die histoehemische Beschaffenheit der Granula un- geklgrt ist.
Nach den Befunden beim Salamander scheinen die mit blauen Granula besetz- ten Hohlrgume um das Kapillarknguel extrazellul~r zu sein. Im Hypendym und im Ependym des Organs findet man zuweilen kleine intrazellul~r gelegene Vakuo- len, die nur durch ein diinn ausgezogenes Plasmalemm yore Liquor getrennt sind. Sie stehen nicht in Beziehung zu den tiefer gelegenen, extrazellulgren Hohl- rgumen, deren Auslgufer sich in Windungen zwisehen die Ependymzellen schieben, wobei ein wabiges Muster ihrer Anordnung entsteht. Da ihre Wgnde regelmgl~ig mit zarten Gomori- und Aldehydthionin-positiven Granula besetzt sind, kSnnten diese Substanzen Wirkstoffe enthalten, welche die Permeabilit~t der Vakuolen- membranen dahingehend beeinflussen, dab der Durchtritt des histochemisch nicht darstellbaren Vakuoleninhaltes oder Teile desselben in den Liquor ermSglicht wird (vgl. Wirkung des ADH auf die Membranpermeabilits an den Tubuli; SCHWAB, 1959). Der theoretisch mSgliche umgekehrte Weg scheint nicht verwirk. licht ,da das Vordringen der Vakuolenauslgufer gegen das Ventrikellumen eher einen sekretorischen Vorgang andeutet, wobei wohl auch der in den Vakuolen herrschende Druck eine Rolle spielt. Ganz im Gegensatz zu bisherigen Beschrei- bungen steht das hohe, palisadenghnliehe Ependym an der Spitze des Organs. Eine begrenzte Anzahl yon dicht aneinander stehenden tiefblauen Zellen bildet eine kompakte Leiste. Es gleicht etwa dem Ependym des Subkommissuralorgans, ist aber einreihig und enth~lt keinerlei lichtmikroskopiseh sichtbare Fortsatze.
Die Lage des Organs im Recessus praecommissuralis unterhalb und vor der Commissura anterior, sowie seine kammerartige Form erinnert nicht nur an das yon BRAAK (1963) beschriebene Organum vasculosum praeopticum, sondern weist auch gewisse J~hnliehkeit mit dem GefgBorgan der Lamina terminalis h5herer Wirbeltiere auf (DuvER~o:e und KORITX~, 1963; DELL~AN~, 1964; W]~I~DL, 1965). Die Amphibien gehSren zu denjenigen Vertebraten, bei denen in der Ontogenese dureh die Ausstiilpung der Hemisph~ren ein Tell der Lamina supra- neuroporiea, n~mlich die rostrale Begrenzungsplatte (~ der Recessus superior), rostroventral vorgetrieben wird; dadurch entsteht ein gergumiges Teleneephalon medium, dessen Lichtung caudal vom Kommissurenwulst abgegrenzt wird (vgl. vo~ HALL~R, 1934). Eine Lamina terminalis besteht noch nicht ; doch scheint die GefgBanlage an der Stelle, wo spgter die Lamina terminalis zu liegen kommt, bereits angelegt. Dies kommt in dem eigenartigen Gef~Bknguel, ngmlieh dem Sub- fornikalorgan des Salamanders zum Ausdruck. Die bei hSheren Wirbeltieren in der Zweizahl vorhandenen Gefgl3organe scheinen in phylogenetischer Hinsieht yon dem einzigen Gef~Borgan des Salamanders ihren Ursprung zu nehmen.
Literatur
ADAm, H. : Hypophyse und hypothalamo-neurohypophys~res neurosekretorisehes System bei Cyelostomen Myxine fflutinosa und Bdellosto~a stouti. Zool. Anz. 23 Suppl., 157--171 (1960).

404 W. ARNOLD:
ADAMS, C. W. M., and J . C. SLOPER: The thalamic elaboration pituitary principles in man, the rat and dog. Histochemical evidence derived from a performic acid Alcian blue reaction for cystine. J . Endocr. 13, 221--228 (1956).
AKERT, K., N. D. POTTER, and J . W. A~DERSO~: The subfornical organ in mammals. Com- parative and topographical anatomy. J. comp. Neurol. 116, 1--14 (1961).
ANA~THA~ARYA~AN, V. : Nature and distribution of neurosekretory cells of the reptilian brain. Z. Zellforsch. 43, 8--16 (1955).
A~DERS, K. H. : Der Feinbau des Subfornikalorganes vom Hund. Z. Zellforsch. 68, 445--473 (1965).
ASSENMAC~ER, I. : Les regulations hypothalamiques de la fonction gonadotrope. Acta neuroveg. (Wien) 25, 339--382 (1964).
BACU~AN~, Ir U. H. M. SE:Tz: Zur histochemischen Darstellung des Histidin mit Diazonium- salzen. I-Iistochemie 2, 307--312 (1961).
BAR~:KOVA, J . A. : Das hypothalamo-hypophys~re System und die Wanderung der Salmo- niden und Acipenseriden. Biol. Rdsch. 1, 164--173 (1964).
BAaOMA~, W. : Das Zwischenhirn-Hypophysensystem. Berlin-GSttingen-Heidelberg: Sprin- ger 1954.
- - Histologie und mikroskopische Anatomie des Menschen, 6. Aufl. Stuttgart: Georg Thieme 1967.
- - Neurosecretion. International Revue of Cytology, ed. H. H. BOVR~E and J. F. DA~:EI~:, p. 183--201. New York and London: Academic Press 1966.
- - Gehirn und Hypophyse (Schleiden-Vorlesung). Nova Acta Leopoldina, ed. K. MOTHES, N. F., Bd. 34, Nr. 181. Leipzig: Johann Ambrosius Barth 1967.
- - u . W. H:LD: ~ber die Morphologie der neurosekretorischen Verkniipfung yon Hypo- thalamus und Neurohypophyse. Acta anat. (Basel) 8, 264--280 (1949).
- - - - R. ORTMA~ U. Tm H. SC~:EELER: Morphologische und experimentelle Untersuchun- gen fiber das hypothalamisch-hypophys~re System. Acta neuroveg. (Wien) l , 233--271 (1950).
BARRY, J . : Apropos de la notion de "voie neuros6cr~toire". Bull. Soc. Sci. Nancy, N.S. 13, 49--58 (1954).
- - De l'existence de voies neuros~cr6toires hypothalamo-t61enc~phaliques chez la chauve- souris (Rhinolophus/errum equinum) en ~tat d'hibernation. Bull. Soc. Sci. Nancy, N.S. 1 3 , 126--136 (1954).
- - L'hydrenc~phalocrinie de substancesneuro-et glios~cr~toires. Biol. m~d. (Paris) 44, (1955). - - Recherches sur la neuros~cr4tion dienc~phalique chez Talpa europaea. Bull. Soc. Sci.
Nancy 15, 49--58 (1956). - - V o i e s neuros~cr6toires extrahypophysaires et Faction nerveuse centrale des hormons
posthypophysaires. Acta endocr. (Kbh.) 51, 111 (1960). - - , et D. LE~YS: Reeherches sur les relations hypothalamo-pr~hypophysaires chez quelques
mammif~res. Anat. Anz., Erg.-Bd. 109, 456 (1961). BArgAiN jr., J . W. : Effect of hypophysectomy on renal concentrating ability of the rat.
Endocrinology 77, 496--500 (1965). BENOIT, J., et J . ASSE~MACUEa: Le contrSle hypothalamique de l'activit~ pr~hypophysaire
gonadotrope. J . Physiol. (Paris) 47, 427--567 (1955). BooK, R.: ()ber die Darstellbarkeit neurosekretorischer Substanz mit Chromalaun-Gallo-
cyanin im supraoptico-hypophys~ren System beim Hund. Histochemie 6, 362--369 (I966).
- - Zur Darstellbarkeit des Neurosekretes. Verh. Anat. Ges., Erg. zum Bd. 120 (1967) d. Anat. Anz. 139--145.
BOLOONAR:, R., e A. ROTELLA: Sui fenomeni neurosecretori degli Anfibi Anuri. Soc. Peloritana Sci. Fis. Mat. Natur. 2, Fasc. 1, 61--70 (1955/56).
BoLz, W. : Allgemeinnarkose beim Tier. Stuttgart: Ferdinand Enke 1961. BRAAK, H. : Das Ependym der Hirnventrikel yon Chimaera monstrosa (mit besonderer Berfick-
sichtigung des Organon vasculosum praeopticum) Z. Zellforsch. 60, 582--608 (1963). BRAC~RT, G. 1940: in Go~o~:, G.: Microscopic histochemistry. Principles and practice.
Chicago: Chicago University Press 1952.

Diencephal-telencephales neurosekretorisches System bei Salamandra 405
CHRIST, J. : The early changes in the hypophysial neurosecretory fibers after coagulation. I I I . Internat. Conf. on Neurosecretion Bristol 1961, Mem. Soc. Endocrinol. 12 (1962).
])ELLMANN, H. D. : Zur Struktur des Organon vasculosum laminae terminalis des Huhnes. Anat. Anz. l lS , 174--183 (1964).
DIEPEN, R.: Vergleichende anatomische Untersuchungen fiber das Hypophysen-Hypo- thalamussystem bei Amphibien und Reptilien. Anat. Anz., Erg.-Bd. 99, 79--89 (1952).
- - Hypothalamus. In: Handbuch der mikroskopischen Anatomie des Menschen, Bd. 4/VII. Berlin-GSttingen-Heidelberg: Springer 1962.
- - , U. FR. ENGELHARDT: Neurale Ph~nomene im Hypothalamus-Hinterlappensystem. Patho- physiologia diencephalica, p. 122--133. Wien: Springer 1958.
- - - - u. V. SMITmAGR]~DA: Ober Ort und Art der Entstehung des Neurosekrets im supra- optico-hypophys~ren System bei Hund und Katze. Anat. Anz., Erg.-Bd. 101, 276--288 (1954).
D~ERICKX, K. : The dendrites of the preoptic neurosecretory nucleus of Rana temporaria and the osmoreceptors. Arch. int. Pharmacodyn. 140, 708--725 (1962).
- - The dendrites of the preoptic neurosecretory nucleus of Rana temporaria and the osmo- receptors. Naturwissenschaften 49, 405-406 (1962b).
- - The subfornical organ, a spezialized osmoreceptor. Naturwissenschaften 59, 163--164 (1963).
- - Regeneration of the neural lobe of the hypophysis after extirpation of the median eminence in Rana temporaria. Acta anat. (Basel) 60, 181--186 (1965).
- - The origin of the aldehyde-fuchsin-negative nerve fibres of the median eminence of the hypophysis: a gonadotrope centre. Z. Zellforsch. 66, 504--518 (1965).
- - , and F. VA~DESA~DE: The magnocellular preoptic nuclei and the reproduction in Rana temporaria. Z. Zellforsch. 68, 190--193 (1965).
, and D. ROGGE~: The reaction of different areas of the magnocellular preoptic nucleus of Rana temporaria upon progressive dehydration. Z. Zellforsch. 62, 89--101 (1964).
DUFFY, P. E., and M. ME~EFEE : Electron microscopic observations of neurosecrctory granules, nerve and glial fibres and blood vessels in the median eminence of the rabbit. Amer. J . Anat. 117, 215--286 (1965).
DUVERNOY, H., et J. G. KORITK~: Contribution a l'~tude de l'angioarchitecture de l'organe subfornicale. Acta anat. (Basel) 55, 394 (1963).
- - - - Les fibres nerveuses de la lame terminale et leurs rapports avec rorgane vasculaire de la lame terminale. Bull. Ass. Anat. (Nancy) 117, 562--576 (1963).
EICn~ER, D.: Topochemische Untersuchungen am neurosekretorischen Zwischenhirn- Hypophysensystem der Albinoratte unter normalen und experimentellen Bedingungen. Z. Zellforsch. 48, 402--428 (1958).
- - Zur Frage des Neurosekretfibertrittes in den III . Ventrikel beim S/iugetier. Z. mikr.- anat. Forsch. 69, 388--394 (1963).
- - , u. H. T I ~ M A ~ : Elektronenmikroskopische Untersuchungen am Nucleus supraopticus hypothalami der Albinoratte. Bericht des VII. Internat. Anatomenkongr., Wiesbaden 1965, S. 34.
ELFTMANN, H. A. : Chromalaun fixative for the pituitary. Stain. Technol. 32, 25--30 (1957). FLEISCHHAU~R, K.: :Fluoreszenzmikroskopische Untersuchungen fiber den Stofftransport
zwischen Ventrikelliquor und Gehirn. Z. Zellforsch. 62, 6 3 9 ~ 5 4 (1964). - - Untersuchungen am Ependym des Zwischen- und Mittelhirns der LandschildkrSte (Testudo
graeca). Z. Zellforsch. 46, 729--767 (1957). - - Zwei Methoden zur Darstellung der Faserglia in Paraffinschnitten yore Gehirn der Katze.
Zbl. allg. Path. path. Anat. 102, 99 (1961). GABE, M. : Sur quelques applications de la coloration par la fuchsine paraldehyde. Bull. Micr.
appl. Set. II/3, 153--163 (1953). GLE~]~I~, G., and R. LILLIE: Observation on the diazotation coupling reaction for the histo-
chemical demonstration of tyrosine, metal chelation and formazan. J. Histochem. Cyto- chem. 7, 416-421 (1959).
GOMORr, G.: Microscopic histochemistry. Principles and practice. Chicago: Chicago Uni- versity Press 1952.

406 W. ARNOLD:
GORBMAN, A . : Vascular relations between the neurohypophysis and adenohypophysis and the problem of evolution of hypothalamic neuroendocrine control. Arch. Anat. micr. Morph. exp. 54, 163--194 (1965).
GRAVMANN, W., u. W. CLAUSS: Weitere Untersuchungen zur Spezifit~t der histochemischen Polysaccharid-Eisenreaktion. Acta histochem. (Jena) 6, 1--7 (1958).
GUrLL~MI~, R. : On the nature of the hypothalamic substance which controls the secretion of ACTH. Acta neuroveg. (Wien) 28, 58---62 (1962).
HXUSLER, G. : Zur Technik und Spezifit~t des histochemischen Carboanhydrasenachweises im Modellversuch und in Gewebsschnitten von Rattennieren. Histochemie 1, 29--47 (1958).
HAGEN, E . : t~ber die feinere Histologie einiger Abschnitte des Zwischenhirns und der Neuro- hypophyse des Menschen. Aeta anat. (Basel) 16, 367~415 (1952).
- - t?ber die feinere Histologie einiger Abschnitte des Zwischenhirns und der Neurohypophyse. II. Morphologische Ver~nderungen im Zwischenhirn des Hundes nach Pankreatektomie. Acta anat. (Basel) 25, 1--33 (1955).
- - Morphologische Beobachtungen im Hypothalamus und in der Neurohypophyse des Hundes nach Teilli~sion des Infundibulum. Acta anat. (Basel) 81, 193--218 (1957).
HALLER VO~ HALLERSTEIN, V . : Gliederung des Zentralnervensystems. Abschn. Diencephalon, S. 117--141, in: Handbuch der vergleichenden Anatomie der Wirbeltiere, Bd. II , 2. H/~lfte. Berlin u. Wien: Urban & Schwarzenberg 1934.
HERRICK, C. J . : The forebrain of the tigersalamander (Ambystoma tigrinum). Chicago: Chicago University Press 1948.
HERRING, P. T. : The histological appearance of the mammalian pituitary body. Quart. J . exp. Physiol. l , 121--159 (1908).
HIT.D, W. : Zur :Frage der Neurosekretion im Zwischenhirn der Schleie (Tinca vulgaris) und ihrer Beziehung zur Neurohypophyse. Z. Zellforsch. 55, 34--46 (1950).
- - Vergleichende Untersuchungen fiber Neurosekretion im Zwischenhirn yon Amphibien und Reptilien. Z. Anat. Entwickl.-Gesch. 115, 459--479 (1951).
- - Experimentell-morphologische Untersuchungen fiber das Verhalten der ,,Neurosekretori- schen Bahn" nach Hypophysenstieldurchschneidung, Eingriffen in den Wasserhaushalt nnd Belastung der Osmorezeptoren. Virchows Arch. path. Anat. 319, 526--546 (1951).
- - Das Verhalten des neurosekretorischen System nach Hypophysenstieldurchtrennung und die physiologische Bedeutung des Neurosekretes. Acta neuroveg. (Wien) 8, 81--91 (1951).
- - t~ber Neurosekretion im Zwischenhirn des Menschen. Z. Zellforsch. 87, 301--316 (1952). - - Weitere Untersuchungen fiber die Neurosekretion bei Mensch und Hund. Anat. Anz.,
Erg.-Bd. 99, 89--91 (1952). - - Das morphologische, kinetische und endokrinologischeVerhalten yon hypothalamischem
und neurohypophys/irem Gewebe in vitro. Z. Zellforsch. 40, 257--312 (1954). - - , u. G. ZETLER" Experimenteller Beweis ffir die Entstehung der sogenannten Hypophysen-
hinterlappenwirkstoffe im Hypothalamus. Pflfigers Arch. ges. Physiol. 2~7, 169--201 (1953).
HIRO]~OBU, ITG: The receptor in the ventricular wall of the reptilian brain. J. Hirnforsch. 6, 333---338 (1964).
HOFER, H. : Das Subfornikalorgan. In: Primatologia, vol. II , Teil 2/13, 1965. IGARAS~r, M., and S. M. McCAN~: A hypothalamic, follicle stimulating hormone relasing
factor. Endocrinology 74, 446--452 (1964). JAEONERO, V., L. FAERA y R. M. JABON~RO: R6sultados del m6todo del acido osmico-
yoduro de cin para la d6monstracion de los element6s nerviosos perifericos. Trab. Inst. Cajal Invest. biol. LII I , 123, 1--48 (1961).
KARLSO~T, P. : Lehrbuch dcr Biochemie. Stuttgart: Thicme 1966. KELLY, D. E.: Intercolumnar tubercle (subfornical organ) and supraoptic crest. Circum-
ventricular organs. In: Histology and general pathology of the human nervous system (R. ADAMS and HAYMAKER, editors), Springfield, Ill. : Cm C. THOMAS C 26, 1967.
KING, L. S., and G. B. WISLOCKI: The Permeability of the hypophysis and hypothalamus to vital dyes, with a study of the hypophyseal vascular supply. Amer. J . Anat. 58, 421--472 (1936).
KIVALO, E. U .K . , and R. M. A. BERGSTRGEM: Activity of the hypothalamo-neurohypo- physial neurosecretory system of the mouse in continous darkness. Acta neuroveg. (Wien) 2~, 177--186 (1964).

Diencephal-telencephales neurosekretorisches System bei Salamandra 407
KLEIN, A. : t~tude pr61iminaire des cellules neuros6cr6toires des noyaux pr~optiques de Tri- tutus marmoratus. Ann. Endoer. (Paris) 21, 635--649 (1960).
KNOBLOC~, D. v.: Das Subfornikalorgan des dritten Ventrikels in seiner embryonalen und postembryonalen Entwicklung beim Hausschwein (Sus scrofa domesticus). Z. Anat. Entwickl.-Gesch. 106, 379---397 (1937).
KNOCHE, H. : Ursprung, Verlauf und Endigung der retino-hypothalamischen Bahn. Z. Zell- forsch. 51, 588--704 (1960).
K~OWL~S, F.: Neuronal Properties of Neurosecretory cells. Neurosecretion-IV. Internat. Symposium on Neurosecretion, Strasbourg, 25--27 Juillet 1966, ed. F. STUTINSKY. Berlin-Heidelberg-New York: Springer 1967.
KOBAYAS~II, H., and D. S. FARNER: Cholinesterase in the hypothalamo-hypophyseal neuro- secretory system of the White-crowned sparrow, Zonotrichia leucophrys gambelii. Z. Zellforsch. 68, 965--973 (1964).
- - - - Vasculariation of the hypothalamo-hypophysial complex in the White-crowned sparrow, Zonotrichia leucophrys gambelii. Z. Zellforsch. 64, 541--569 (1964).
- - H . I. OATA, H. U E ~ R A , and T. HIRA~O: Electron microscopic and pharmacological studies on the rat median eminence. Z. Zellforsch. 71, 387~405 (1966).
KOELLE, L. : The histochemical differentiation of types of cholinesterases and their localisation in the tissues of the cat. J. Pharmacol. 1O0, 158--179 (1950).
LAGE, C. DA: Innervation neuros6cr6toire de l'ad6nohypophyse chez l'hippocampe. C.R. Ass. Anat. Genes, 41 reunion 361 (1954).
- - Recherches sur le complexe hypophysaire de l'hippocampe. Arch. Anat. mikr. Morph. exp. 47, 401--446 (1958).
LASCANO, E. F. : A new silver method for the Golgi apparatus. Arch. Path. 68, 498--500 (1959).
LE(~AIT, E.: Les organes 6pendymaires du troisibme ventricule. Th6se Univ. Nancy, 1942. - - ~tude histiophysiologiques et exp6rimentale du syst5me hypothalamo-neurohypophysaire
de la poule Rhode Island. Arch. Anat. micr. Morph. exp. 44, 323--331 (1956). - - Les voies 6fferentes des noyaux neuros6cr6toires hypothalamiques chez les oiseaux. C.R.
Soc. Biol. (Paris) 150, 996 (1956). - - , et G. GRmNON: ]~xiste-t-il chez les oiseaux l'homologue de l'organe subfornical du troisi~me
ventricule des mammif~res ? C.R. Ass. Anat. 43, 509--513 (1956). Apropos de la structure et de l ' innervation des organes 6pendymaires du troisi~me
ventricule chez les Batraeiens et ]es Reptiles. C.R. Soc. Biol. (Paris) 150, (2), 1982--1984 (1956).
- - - - Les voies extra-hypophysaires des noyaux neuros6cr6toires hypothalamiques chez les Batraeiens et les Reptiles. Acta anat. (Basel) 30, 429 443 (1957).
- - , et. H. L~GAIT: Recherches sur l'organe subfornical du troisi~me ventricule chez quelques mammif~res. C.R. Ass. Anat. 43, 502--508 (1956).
LEtrSEN, I., and E. LACROlX: Changes in osmolarity in the cerebral ventricles and diuresis. Endocrinology 68, 719--721 (1961).
LIJSB~RG, M. F. : Nissl staining at pH lower than 2. Acta anat. (Basel), Suppl. 44, 1--113 (1962).
MARTINL L. : Neurosecretion and stimulation on the Adenohypophysis. Zweites internat. Symposium fiber Nem'osekretion, Lund 1957, S. 52--55. Berlin-GSttingen-Heidelberg: Springer 1958.
MAzzI, V. : Neurosecrezione e sintesi di proteine. Rend. Accad. naz. Lincei, Ser. VII 4, 214--218 (1948).
- - Rapporti anatomici e funzionali fra ipothalamo e ipofisi. Arch. Zool. ita]., Supp. 38, 53--140 (1953).
- - Alcune osservazioni intorno al sistema neurosecretorio ipothalamo-ipofisario e all'organo sotto-eommissurale nell'ontogenesi di Rana agilis. Monit. zool. ital. 62, 78--82 (1954).
~/[URAKAlVII, lY[.: Elektronenmikroskopisehe Untersuchungen am Nucleus praeopticus der KrSte (Bu/o vulgaris Formosu8). Z. Zellforsch. 63, 208--225 (1964).
- -Elektronenmikroskopisehe Untersuchungen der neurosekretorischen Zellen in Hypo- thalamus der Maus. Z. Zellforsch. 56, 277---299 (1962).
PAGET, G. E.: Aldehydthionin: a stain having similar properties to aldehydfuchsin. Stain Technol. 34, 223--227 (1959).

408 W. ARNOLD:
PEARSE, A. G. E. : I-Iistochemistry, theoretical and applied. London: J. & A. Churchill 1960. PINES, L., u. M. SCH~FTEL: Ist bei den niederen Vertebraten ein Homologon des subfornikalen
Organes der S~iugetiere festzustellen .* Anat. Anz. 67, 203--216 (1929). RANKE, O. : Physiologie des Zentralnervensystems rum Standpunkt der Regulationslehre.
Mfinchen u. Berlin: Urban & Schwarzenberg 1960. RXNNE, U. K. : l~eurosecretory material passing into the hypophysial portal system in the
human infundibulum and its foetal development. Acta neuroveg. (Wien) 25, 310--324 (1964).
- - , and E. K I v ~ o : Maturation of hypothalamic neurosecretion in rat with special reference to the neurosecretory material passing into the hypophysial portal system. Acta neuroveg. (Wien) 27, 112--131 (1965).
RoHR, V. U. : Zum Feinbau des Subfornikalorgans der Katze. I. Z. Zellforsch. 73, 246--271 (1966).
- - Zum Feinbau des Subfornikalorgans der Katze. II. Z. Zellforsch. 75, 11--34 (1966). ROMEIS, B. : Mikroskopische Technik. Mfinchen: Oldenbourg 1948. RUD~R~, H. : Das Subfornikalorgan und seine Beziehungen zum neurosekretorischen System
im Zwischenhirn des Frosches. Z. Zellforsch. 65, 790--804 (1965). SAFF~AN, M., A. V. SCHALLY, M. SEOAL and B. ZIMMER~A~N: Charakterization of the cortico-
tropin releasing factor of the neurohypophysis. Zweites Internat. Symposium fiber Neuro- sekretion, p. 55--59, Lund 1957. Berlin-Gfttingen-Heidelberg: Springer 1958.
SCHARRER, E. : Die Lichtempfindliehkeit blinder Elritzen (Untersuchungen fiber das Zwisehen- h im der Fische I). Z. vergl. Physiol. 7, 1--38 (1928).
- - ~ber sekretorisch t~tige Zellen im Thalamus yon Fundulns heteroclitus L. (Untersuchun- gen fiber das Zwischenhirn der Fische II). Z. vergh Physiol. 11, 767--773 (1930).
- - Ein inkretorisehes Organ im Hypothalamns der Erdkr5te Bu/o vulgaris Laur. ZZ. wiss. Zool. 144, 1--11 (1933).
- - Uber die Beteiligung des Zellkerns an sekretorischen Vorg~ngen in Nervenzellen. Z. Path. 47, 143--151 (1934).
- - A relationship between the paraphysis and paraventrieular nucleus in the garter snake (Thamnophis 8p.) Biol. Bull. 101, 106--113 (1951).
- - Neurosecretion and anterior pituitary in the dog. Experientia (Basel) 16, 264 (1954). - - - - Neurosekretion. In: Handbuch der mikroskopischen Anatomie des Menschen, Abt. 6,
Bd. 5. Berlin-G5ttingen-tteidelberg: Springer 1954. - - , u. B. SCHARREa: ~ber Drfisen-Nervenzellen und neurosekretorische Organe bei Wirbel-
losen und Wirbeltieren. Biol. Rev. 12, 185--216 (1937). - - - - Hormones produced by neurosecretory cells. Recent Progr. Hormone Res. 19, 183--240
(1954). SCHIEBLER, Tr[. H. : Darstellung yon B-Zellen in Pankreasinseln und yon l~eurosekret mit
Pseudoisocyanin. Naturwissenschaften 45, 356--380 (1958). - - , TlZ., u. H. v. BR]~M: lJber jahreszyklische und altersbedingte Ver~nderungen in den
nenrosekretorischen Systemen yon Teleostiern. Naturwissenschaften 45, 450--451 (1958). - - , u. J . HARTMAN~: Experimentelle und histochemische Unt~rsuchungen am neurosekretori-
schen System des Teleostierhypothalamus. Verh. Anat. Ges. Mfinchen 1964, Anat. Anz. 112, 84---92, Erg.-Bd.
- - , u. S. SCHI~.SSLER: ]Dber den Nachweis yon Insulin mit metachromatisch reagierenden Pseudoisocyaninen. Histochemie 1, 445--465 (1959).
SCHWAB, M., u. K. KUEHNS: Die St5rungen des Wasser- und Elektrolytstoffwechsels. Berlin- G5ttingen-Heidelberg: Springer 1959.
Sn~Tz, ]-I. M.: Zur Morphologie und Histochemie des Neurosekrets im Infundibulum des Schweines. J . Hirnforsch. 6, 283--319 (1964).
SLUCA, E . , u. F . SEITELBERGER: gut Ultrastruktur der Kapillaren und ihrer Beziehung zu Nervenzellen im Bereieh der neurosekretorischen Kerne des Hypothalamus. Z. Zellforsch. 78, 303--312 (1967).
SMIT~, S. W. : The correspondence between hypothalamic neurosecretory material and neuro- hypophysial material in vertebrates. Amer. J. Anat. 89, 195--231 (1951).
S~IOLLER, C. : Neuroseeretory processes extending into third ventricle : secretory or sensory ? Science 147, 882--884 (1965).

Dieneephal-telencephales neurosekretorisches System bei Salamandra 409
SI'aANKEL, H.: t~ber die Beziehungen des Plexus des dritten Ventrikels zum subfornikalen Organ bei den Primaten. Naturwissensehaften 47, 383--384 (1960).
STERNA, G. : ~ber eine sehr spezifische Methode zum Nachweis des Neurosekrets. Acta biol. med. germ. 7, 228--231 (1961).
- - Fluoreszenzmikroskopische Untersuchungen fiber die Neurosekretion beim Bachneunauge (Lampetra planeri Bloch). Z. Zellforsch. 55, 763--789 (1961).
- - Grundlagen des histoehemischen nnd biochemischen Nachweises yon Neurosekret (= Tr~- gerprotein der Oxytocine) mit Pseudoisoeyaninen. Acta histochem. (Jena) 17, 268--292 (1964).
SUOMALAINEN, P. : Neurosekretion und Stress. Zweites Internat. Symposium fiber Neuro- sekretion, Lund 1957, S. 26--29. Berlin-GSttingen-Heidelberg: Springer 1958.
TIL~EY, F. : The development and constituents of the human hypophysis. Bull. neurol. Inst. N.Y. 5, 387 (1936).
TROYER, J. R. : Neurosecretion in the hibernating rat. The morphology of the hypothalamo- hypophyseal system. Anat. Rec. 15, 77--92 (1965).
VE~NEY, E. B. : The antidiuretic hormone and the factors, which determine its release. Proe. roy. Soc. B 135, 25--106 (1948).
VIERMA~, B. : Morphologische Beobachtungen im Hypothalamus und in der Neurohypo- physe des Hundes naeh Exstirpation der Keimdrfisen. Acta anat. (Basel) 49, 77--114 (1962).
VOITKWlC~, A. A., and T. K. C~ELK: Effect of extirpation of the embryonic anlages of the neurosecretory-hypophyseal system in amphibians. Dokl. Akad. Nauk SSSR 159, 1183-- 1186 (1964). (Biol. Ber. entnommen).
WAHLERT, G.V.: Molche und Salamander. Kosmos-Ges. der Natuffreunde. Stuttgart: Francksche Verlagshandlung 1966.
WATERMAN~, R . : Zur Morphologie des Subfornikalorganes. Inaug.-Diss. Med. Fakult~it KSln 1955.
- - t~ber das Vorkommen von interstitiellen Vakuolen im Subfornikalorgan. Dtsch. Z. Nerven- heilk. 174, 593--596 (1957).
- - Beitrag zur Frage der Homologie yon Subfornikalorgan und Paraphyse. Acta anat. (Basel) 2 9 , 304--313 (1957).
- - Interventrikularorgan und Trigonum supracommissurale. Anat. Anz. 117, 261--279 (1965). - - , u. G. A. MASSEI~: Eia Vergleich von Subfornikalorgan und Area postrema. Z. Morph.
0kol. Tiere 45, 603--615 (1957). WEBER, W.: Entwicklung und Funktion des neurosekretorischen Systems yon Salamandra
salamandra. Z. Zellforsch. 66, 35--65 (1965). - - , u. H. B):C~MAN~: Zur Entwicklung und Funktion des Zwischenhirn-hypophysensystems
bei Columba livia domestica. Naturwissenschaften 51, 321 (1964). WEI~DL, A. : Zur Morphologie und Histochemie yon Subfornikalorgan. Organum vasculosum
laminae terminalis und Area postrema bei Kaninchen und Ratte. Z. Zellforsch. 67, 740-- 775 (1965).
- - , A. SCn~INK u. R. WETZSTEIN: Der Feinbau des Gef~l]organs der Lamina terminalis beim Kaninchen. I. Die Gef~Be. Z. Zellforsch. 79, 1--48 (1967).
WIED, D. DE: The significance of vasopressin (ADH) for the release of corticotropin from the Adenohypophysis. Acta neuroveg. (Wien) 23, 63--68 (1962).
WILSON, L. D., J. A. WEINBER~, and H. A. B]~R~: The hypothalamic neuroseeretory system of the tree-frog Hyla regilla. J. comp. Neurol. 107, 253--271 (1957).
WISLOCKI, G. B., and E. H. LEDVC: Vital staining of the hematoencephalic barrier by silver nitrate and trypanblue, and cytological comparisons of the neurohypophysis, pineal body, area postrema, intercolumnar tubercle and supraoptic crest. J. comp. Neurol. 96, 371~414 (1952).
WOLFGA~G ARNOLD Institut ffir Histologie und experimentelle Biologie der Universit~t Mtinchen 8 Mfinchen 15, Pettenkoferstr. 11