
Untersuchung des Wirkmechanismus von FTY720 in einem ...
111
Universitätsklinikum Ulm Klinik für Neurologie Ärztlicher Direktor: Prof. Dr. A. C. Ludolph Untersuchung des Wirkmechanismus von FTY720 in einem Mausmodell der Amyotrophen Lateralsklerose (ALS) Dissertation zur Erlangung des Doktorgrades der Medizin der Medizinischen Fakultät der Universität Ulm Vorgelegt von Ingo Wantzen Ulm, 2016
Transcript of Untersuchung des Wirkmechanismus von FTY720 in einem ...

Universitätsklinikum Ulm
Klinik für Neurologie
Ärztlicher Direktor: Prof. Dr. A. C. Ludolph
Untersuchung des Wirkmechanismus von
FTY720 in einem Mausmodell der Amyotrophen
Lateralsklerose (ALS)
Dissertation zur Erlangung des Doktorgrades der Medizin
der Medizinischen Fakultät
der Universität Ulm
Vorgelegt von Ingo Wantzen
Ulm, 2016

Amtierender Dekan: Prof. Dr. R. Kilian
1. Berichterstatter: Prof. H. Tumani
2. Berichterstatter: Prof. J. Weiss
Tag der Promotion: 13.07.2017

i
Inhaltsverzeichnis
Abkürzungsverzeichnis ......................................................................................................................... ii
1 Einleitung ............................................................................................................................................ 1
1.1 Die antiinflammatorische Wirkung des Immunmodulators FTY720 .............................. 1
1.2 Die vermutete zusätzliche neuroprotektive Wirkung von FTY720 bei MS .................... 3
1.3 Tiermodell der ALS, ein primär neurodegeneratives Krankheitsmodell ............................ 3
1.4 Die Amyotrophe Lateralsklerose und deren Pathomechanismen ...................................... 5
1.5 Neuroprotektion ........................................................................................................................ 10
1.6 Ziel der Arbeit ............................................................................................................................ 10
1.7 Fragestellungen ........................................................................................................................ 11
2 Material und Methoden ................................................................................................................... 12
2.1 Material ...................................................................................................................................... 12
2.2 Methoden ................................................................................................................................... 21
3 Ergebnisse ........................................................................................................................................ 34
3.1 Effekt von FTY720 auf α-Motoneurone von SOD1G93Adl-Tieren ........................................ 34
3.2 Effekt von FTY720 auf proinflammatorische Parameter im ZNS und im Blut von
SOD1G93Adl-Tieren ........................................................................................................................... 40
4 Diskussion ........................................................................................................................................ 71
4.1 Allgemeine Bemerkung zur Diskussion ................................................................................ 71
4.2 Effekt von FTY720 auf α-Motoneurone von SOD1G93Adl-Tieren ........................................ 72
4.3 Effekt von FTY720 auf proinflammatorische Parameter im ZNS und im Blut von
SOD1G93Adl-Tieren ........................................................................................................................... 77
5 Zusammenfassung .......................................................................................................................... 86
6 Literaturverzeichnis ......................................................................................................................... 88
Anhang ................................................................................................................................................. 97
Vortest der Neurofilamente NFH .................................................................................................. 97
Danksagung ................................................................................................................................... 101
Lebenslauf...................................................................................................................................... 103

ii
Abkürzungsverzeichnis
µl Mikroliter
µmol Mikromol
ADL Aktivitäten des täglichen Lebens
ALS Amyotrophe Lateralsklerose
AMPA α-Amino-3-hydroxy-5-methyl-4-isoxazol-
Propionsäure
ANOVA Analysis of variance
Apaf-1 Apoptotic Protease activating Factor 1
APC Antigenpräsentierende Zelle
AS Aminosäure
ATP Adenosin-Triphosphat
bcl-2 B-cell lymphoma 2
BNDF Brain derived neurotrophic Faktor
bp Basenpaar
Ca2+ Calcium
CA3 Cornu Ammonis region 3
CCR7 Rezeptor CC-Chemokine receptor 7
CCS co-exprimierendes Kupferchaperon
cm Zentimeter
CNF ciliary neurotrophic Faktor
Cu/Zn Kupfer/Zink
d Tag
d Durchmesser
DAB 3,3'-Diaminobenzidine
DC Dendritische Zellen
DNA Desoxyribonukleinsäure
EAAT2 Excitatory amino-acid transporter 2
EAE Experimentelle Autoimmun Enzephalomyelitis
EDTA Ethylendiamintetraacetat
EGTA Ethylenglycol-bis(aminoethylether)-N,N,N′,N′-
tetraessigsäure

iii
ELISA Enzyme-Linked Immunosorbent Assay
End Endstadium
ERK Extracellular-signal Regulated Kinases
FACS Fluorescence activated cell sorting oder
Durchflusszytometrie
fALS familiäre Amyotrophe Lateralsklerose
FDA Food Drug Administration
FITC Fluoresceinisothiocyanate
FSC Follicular Stroma cells
FTLD frontotemporal lobar degeneration
FTY720 Fingolimod oder 2-Amino-2-(2-(4-
octylphenyl)ethyl)propan-1,3-diol
FTY720-P FTY720-Phosphat
g Gramm
G37R Glycin 37 Arginin
G59S Glycin 59 Serin
G86R Glycin 86 Arginin
G93A Glycin 93 Alanin
GABA γ-Aminobuttersäure
GFAP Glial fibrillary acidic protein
GLT-1 Glial Glutamate Transporter 1
Glu Glutamat
GLuR2 Glutamate receptor, ionotropic, AMPA
GM1 Monosialotetrahexosylgangliosid
GNDF Glial cell line-derived neurotrophic Faktor
GPCR G-Protein gekoppelter Rezeptor
h Stunde
H2O Wasser
H2O2 Wasserstoffperoxid
HCL Salzsäure
HE Hämatoxylin & Eosin
hSOD humane Superoxiddismutase
Hz Herz
IBA1 ionized calcium-binding adapter molecule 1

iv
IFN Interferon
IGF-1 Insulin-like-growth-factor-1
IL Interleukin
INA α-Internexin
K+ Kalium
KCl Kaliumchlorid
K-EDTA Kalium-EDTA
kg Kilogramm
KG Körpergewicht
KSP lysin-serin-prolin
l Liter
LDH Laktatdehydrogenase
LFA-1 Lymphocyte function-associated antigen 1
LPS Lipopolysacharid
M Molar
MAPK Mitogen-activated protein kinase
mg Milligramm
Mg2+ Magnesium
MgCl2 Magnesiumchlorid
min Minute
ml Milliliter
mm Millimeter
mRNA messenger Ribonukleinsäure
mut mutiert
n Anzahl
NaCl Natriumchlorid
NaOH Natriumhydroxid
NaOH Natriumhydroxid
NFH Neurofilament Heavy Chain
NF-κB der Transkriptionsfaktor: "nuclear factor
'kappa-light-chain-enhancer' of activated B-
cells"
NFL Neurofilament light chain
NMDA N-methyl-D-Aspartat

v
NFM Neurofilament middle chain
Nrf2 nuclear factor erythroid 2–related factor 2
NT Neurotransmitter
O2 Sauerstoff
PBS phosphate buffered saline
PCR Polymerase Kettenreaktion
PE Propidiumbromid
PFA Paraformaldehyd
pH pH-Wert
pmol Picomol
PMSF Phenylmethylsulfonylfluorid
pNPP p-Nitrophenyl Phosphate
prä präsymptomatische Phase
Ptdin-3-K Phosphatidylinositol-3-Kinasen
RANTES Regulated on Activation, Normal T Cell
Expressed and Secreted
Reg.-Nr. Registernummer
ROS Reaktive Sauerstoffspezies
rpm round per minute
RT reverse Transkriptase
S1P Sphingosin-1-phosphat
sALS sporadische Amyotrophe Lateralsklerose
SCID severe combined immunodeficiency
SD Standardabweichung
sec. Sekunde
SOD Superoxid-Dismutase
symp symptomatische Phase
TAE TRIS-Acetat-EDTA-Puffer
Tcm zentrale Gedächtniszellen
TDP-43 TAR DNA‐Binding Protein 43
Tem Effektor Gedächtniszellen
tg transgen
TLR Toll-like Rezeptor
Tn naive T-Zellen

vi
TNF Tumornekrosefaktor
TRIS Tris(hydroxymethyl)-aminomethan
TVA Tierversuchsantrag
Urea Harnstoff
V Volt
VEGF vascular endothelial growth Faktor

1
1 Einleitung
1.1 Die antiinflammatorische Wirkung des Immunmodulators FTY720
FTY720 wird als immunmodulatorische Substanz weltweit zur Behandlung der
Multiplen Sklerose eingesetzt (Scott 2011) und unter dem Namen Gilenya® von der
Firma Novartis vertrieben. Es ist ein Metabolit der Zellmembran, bestehend aus
Sphingomyelin (Kingwell et al. 2010). Es agiert als ein Strukturanalogon für
Sphingosine. Sphingosine und FTY720 werden durch die Sphingosin-Kinasen -1 und
-2 zu Sphingosin-Phosphat (S1P) bzw. zu FTY720-Phosphat in ihre jeweils aktiven
Formen phosphoriliert. Sowohl Spingosin-Phosphat als auch FTY720-Phosphat
binden spezifisch auf Lymphozyten an den Spingosin-1 Phosphat Rezeptor. Durch die
Bindung von phosphoriliertem FTY720 an den S1P 1 -Rezeptor kommt es zu einer
Internalisierung und Degradierung des Rezeptors und zu einer Reduktion des mRNA-
Niveaus (Mehling et al. 2011). Der S1P - Rezeptor ist dadurch nicht mehr länger
verfügbar und der durch den S1P 1 -Rezeptor vermittelte Austritt der Lymphozyten
aus den Lymphknoten in das ZNS wird blockiert. Durch die funktionelle antagonistische
Wirkung von FTY720 wird das Ziel erreicht, die für die Neuronen im Rahmen der
Multiplen Sklerose schädlichen Effekte der T-Zellen vom zentralen Nervensystem
(ZNS) fernzuhalten (Mehling et al. 2011), (Pinschewer et al. 2011).
Es gibt fünf verschiedene S1P-Rezeptor Subtypen. FTY720 bindet an alle Subtypen
(Dev et al. 2008) (Brinkmann et al. 2002). S1P-Rezeptoren gibt es, wie folgt, in
absteigender Menge in verschiedenen Organen: Gehirn > Lunge = Milz > Herz und
Gefäße > Niere. Im ZNS werden S1P-Rezeptoren auf Neuronen, Astrozyten,
Oligodendrozyten und Mikroglia exprimiert. Ihre Funktionen sind sehr vielfältig. In vitro
Studien zeigten, dass S1P-Rezeptoren eine Rolle bei der Migration der Mikroglia und
bei Ausbreitung und Rückzug von Oligodendrozyten spielen. Dem S1P-1-Rezeptor
wird eine Schlüsselrolle bei der Angiogenese und Neurogenese sowie der Regulation
des Transports im Immunsystem zugeschrieben. Studien zeigen, dass eine genetische
Deletion im S1P-1 Rezeptor oder der Sphingosin-Kinasen zu einer Beeinträchtigung
der Neurogenese und des Neuralrohrverschlusses führt (Mizugishi et al. 2005). Es
liegt daher nahe, dass der S1P-1 Rezeptor Zell protektive Effekte ausübt.

2
Der S1P1-Rezeptor wird aber auch in den Lymphknoten exprimiert. Durch funktionelle
Antagonisierung des Rezeptors durch FTY720-Phosphat kommt die, schon in
zahlreichen Studien belegte, antiinflammatorische Wirkung zustande. Das durch den
S1P-1 Rezeptor vermittelte Signal wird über ein G-Protein vermitteltes
Retentionssignal für Lymphozyten, welches durch den CCR7-Rezeptor (CC-
chemokine-receptor 7) erzeugt wird, aufrechterhalten (Pham et al. 2008). FTY720
beeinflusst aber nicht alle Lymphozyten gleichermaßen. Bei den Lymphozyten werden
folgende Typen unterschieden: CCR+ Naive T-Zellen (TN), die noch keinen
Antigenkontakt hatten und CCR+ zentrale T-Gedächtniszellen (TCM), die durch
schwache Antigenstimulation entstehen und keine oder nur wenig Effektorfunktion
haben. TCM zirkulieren zwischen Blut und den sekundären Lymphorganen und warten
dort auf ein spezifisches Antigen. Bei Antigenkontakt wandern die TCM über
hochendotheliale Venulen zurück in den Lymphknoten, um dort das Antigen zu
präsentieren. Daraufhin proliferieren und differenzieren sie sich zu CCR- Effektor T-
Gedächtniszellen (TEM). Dabei geht die Funktion der Expression des CCR7-
Rezeptors und das durch ihn vermittelte Retentionssignal verloren. TEM entstehen
durch hoch intensive Antigenstimulation, rezirkulieren zum Entzündungsherd und
wirken dort als Effektorzelle (Sanna et al. 2006), (Pham et al. 2008).
S1P1-Rezeptoren bewirken vor allem die CCR7 vermittelte Retention von TN und TCM
Zellen, aber nicht von TEM-Zellen. FTY720 wirkt deshalb auch nur auf TN- und TCM-
Zellen.
Alle TCM-Zellsubtypen werden unabhängig von Th1 und Th2-Helfer Zellen von
FTY720 beeinflusst (Mehling et al. 2008). FTY720 hat zwar keinen direkten Einfluss
auf TEM-Zellen im ZNS, aber durch, die Verhinderung des Eintritts von TN und TCM-
Zellen, die sich zu TEM-Zellen differenzieren können, wird auch die Zahl der TEM-
Zellen, die sich im ZNS differenzieren, vermindert (Mehling et al. 2008).
Die antiinflammatorische Wirkung von FTY720 wurde schon in zahlreichen Studien
belegt, eine direkte neuroprotektive Wirkung wird bisher nur vermutet (Soliven et al.
2011).

3
1.2 Die vermutete zusätzliche neuroprotektive Wirkung von FTY720 bei MS
Soliven et al. vermutet, dass der positive Effekt von FTY720 auf die humane MS vor
allem durch die veränderte Lymphozytenzirkulation vermittelt sein muss. Doch nimmt
er an, dass es aufgrund der Anwesenheit von S1P-Rezeptoren im ZNS einen weiteren
möglichen Mechanismus geben könnte. Auf der Basis seiner Ergebnisse in einer MS
Studie, bei der FTY720 und Interferon-β verglichen wurden, vermutet er, dass der
positive Effekt von FTY720 auf die Pathologie bei der MS nur teilweise durch die
antiinflammatorische Wirkung erklärt wird. Er hält einen direkten neuroprotektiven
Effekt für wahrscheinlich (Soliven et al. 2011). Zudem lassen die Ergebnisse anderer
Studien einen direkten neuroprotektiven Effekt von FTY720 vermuten (Di Menna et al.
2013), (Hasegawa et al. 2010), (Wei et al. 2011), (Pardo et al. 2014), (Cipriani et al.
2015), (Norimatsu Y 2012), (Gao et al. 2012). Unter anderem beobachtete Pardo A et
al. in vivo anhand von FTY720 behandelten HD R 6/2 Mäusen (Mausmodell des M.
Huntington) eine verbesserte motorische Funktion und eine verringerte Degeneration
an striatalen Zellen (Pardo et al. 2014).
1.3 Tiermodell der ALS, ein primär neurodegeneratives Krankheitsmodell
Zur Aufklärung von Pathomechanismen und zur Entwicklung von Medikamenten
werden heutzutage Tiermodelle in der biomedizinischen Forschung verwendet. Dabei
handelt sich oft um gentechnisch veränderte Mäuse oder Ratten, die zum Beispiel ein
humanes mutiertes Protein überexprimieren und in Folge Symptome der
entsprechenden humanen Erkrankung entwickeln. Das am häufigsten verwendete und
am besten charakterisierte ALS-Mausmodell ist das transgene SOD1G93A –Maus
modell (Gurney et al. 1994). Diese Art von genetisch definiertem Modellorganismus
ermöglicht es die Pathogenese und eine mögliche Therapie der ALS systematisch zu
untersuchen.
Transgene Mausmodelle der fALS, die das human mutierte SOD1 Gen in sich tragen,
haben zu einem wahren Boom geführt, was die Erforschung der Ursache der ALS
angeht. (Cleveland 1999).
Die Übertragbarkeit der Untersuchungsergebnisse auf den Menschen ist meist nur in
begrenztem Maße möglich (Benatar 2007). Zur wissenschaftlichen Erforschung von
relevanten Pathomechanismen der ALS hat sich das transgene Mausmodell jedoch

4
als sehr nützlich erwiesen (Vargas et al. 2011). Es gibt weitere transgene
Mausmodelle mit anderen Mutationen in der Superoxid-Dismutase 1 wie bspw. G37R,
G86R (Gaudette et al. 2000).
1.3.1 Das transgene SOD1-G93A Mausmodell
Das am häufigsten verwendete und am besten charakterisierte ALS-Mausmodell ist
die transgene high copy SOD1-(G93A)–Maus B6SJL-Tg(SOD1-G93A) 1G/J,
abgekürzt SOD1G93A (Gurney et al. 1994). SOD1G93A-Mäuse entwickeln die
Motoneuronenpathologie und die klinischen Symptome, wie sie bei der ALS üblich sind
(Ripps et al. 1995), (Dal Canto et al. 1994), (Dal Canto et al. 1995), (Dal Canto et al.
1997). In dieser Studie wurde das transgene low copy SOD1 (G93A)-Mausmodell
B6SJL-Tg(SOD1-G93A)dl 1G/J verwendet. Dieses Mausmodell hat einen späteren
Krankheitsbeginn und eine längere Lebensspanne im Vergleich zu den high copy
Tieren. Die Krankheitsprogression entspricht jedoch, der der high copy Tiere. Dies
ermöglicht es, pathologische Veränderungen in Folge über einen längeren Zeitraum
zu studieren. Das Zeitfenster für medikamentöse Interventionen ist größer (Al-
Chalabi et al. 2013). Der Zeitpunkt, an dem die erste ALS-Symptomatik auftritt, ist
direkt abhängig von der Kopienanzahl der transgenen Maus. Diese SOD1G93A low copy
Mäuse zeigten im Alter von ca. 200 Tagen erste Gangveränderungen an den
Hintergliedmaßen. In einem Alter zwischen 230-250 Tagen kommt es zu ein- oder
beidseitigen Lähmungen der Hintergliedmaßen. Im Alter von ca. 265 Tagen erreichen
die Tiere das Endstadium in dem sie aus Tierschutzgründen getötet werden. Das
Endstadium ist wie folgt definiert: Ein Tier hat das Endstadium erreicht wenn es nicht
mehr in der Lage ist sich unverzüglich aufzurichten nachdem man es auf die Seite
gelegt hat.
Mattiazzi et al. demonstrieren in ihrer Studie, dass es sowohl beim Menschen als auch
bei SOD1G93A-Mäusen zu einer oxidativen Schädigung und in Folge zum Absterben
der Motoneurone bei der ALS kommt (Mattiazzi et al. 2002).
Im Gegensatz zur humanen ALS beobachtet man bei den SOD1G93A-Tieren keinen
neuronalen Zelluntergang im Motorkortex. Die Manifestation ist hier primär spinal
lokalisiert (Almer 2003).
Effekte von neuen pharmakologischen Substanzen und Pathomechanismen der ALS
werden anhand des SOD1 G93A-Mausmodells erforscht. Ludolph et al. weisen darauf

5
hin, dass die Reproduzierbarkeit und die Übertragung der Ergebnisse präklinischer
Studien auf den Menschen begrenzt ist (Ludolph et al. 2010).
1.4 Die Amyotrophe Lateralsklerose und deren Pathomechanismen
Die ALS ist seit mehr als 100 Jahren bekannt und wurde das erste Mal von dem
französischen Neurologen Jean Martin Charcot beschrieben. Indem er die Symptome
dokumentierte, histologische Präparate anfertigte und in Ansätzen den
Krankheitsverlauf untersuchte, lieferte er eine bedeutungsvolle Beschreibung der ALS
mit nachhaltiger Wirksamkeit (Ludolph et al. 2000). Die Diagnosestellung erfolgt
heutzutage durch klinische, histologische und elektrophysiologische Untersuchungen.
Validierte Forschungsergebnisse zeigen auf, dass die Kombination zwischen dem
klinischen Verlauf und elektrophysiologischer Untersuchungen unabdingbar sind für
eine genaue Diagnosestellung (Körner et al. 2010).
Bei der Amyotrophen Lateralsklerose handelt es sich um eine neurodegenerative
Erkrankung, die ausschließlich das erste (primärer Kortex) und das zweite Motoneuron
(Hirnstamm und Rückenmark) betrifft. Sie ist mit einer jährlichen Inzidenz von 0,5-
2/100.000 und einer Prävalenz von 2-3/100.000 die häufigste
Motoneuronenerkrankung des Erwachsenenalters (Rowland 1994).
Klinisch manifestiert sich der Untergang der motorischen Nervenzellen auf Grund
unterschiedlicher Degenerationsmuster mit teilweise sehr variabler Symptomatik
(Rowland 1995).
Die spinale Form besteht aus einem dualen System aus schlaffen und spastischen
Lähmungen. Durch die Degeneration des ersten Motoneuron wird die spastische
Lähmung (pseudobulbär) hervorgerufen. Durch die Degeneration des zweiten
Motoneurons zeigt sich meist eine einseitige, an der oberen Extremität beginnende
fortschreitende schlaffe muskuläre Lähmung (Wijesekera et al. 2009).
Bei der bulbären Form ist die Muskulatur, die von den Hirnnerven (V, VII, IX, X, XI,
XII) innerviert wird, betroffen. Das Sprechen, Schlucken und Kauen werden hier
beeinträchtigt. Andere neurologische Fähigkeiten, wie die Sensibilität, kognitive
Fähigkeiten, Hören, Riechen, Schmecken, Sehen und die Blasen- und
Mastdarmfunktion bleiben erhalten. Die Erkrankung endet in 80% der Fälle innerhalb
von 1-5 Jahren durch respiratorisches Versagen tödlich (Almer 2003). Eine
differentialdiagnostische Abgrenzung zu anderen neuromuskulären Erkrankungen

6
erfolgt elektromyographisch, neurographisch, serologisch, histologisch, radiologisch
und klinisch.
1.4.1 Ätiologie
Die Ursache für die ALS ist nur teilweise bekannt. Die Forschung geht mittlerweile von
einer multifaktoriellen Erkrankung aus.
In 90% der Fälle tritt die ALS in der sporadischen Form (sALS) auf. Ungefähr 10%
zeigen eine positive Familienanamnese (fALS). Ein Fünftel davon tragen eine Mutation
(G93A) im Gen der Cu/Zn Superoxiddismutase 1 (Rosen DR 1993).
Bei dieser Mutation kommt es zum Aminosäureaustausch zwischen Glycin und Alanin.
Heute sind über 90 Mutationen im Gen der Superoxid Dismutase 1 bekannt,
beispielsweise G37R, G59S, G86R (Gaudette et al. 2000). Die SOD1 ist ein
ubiquitäres zytoplasmatisches Enzym, das die Konversion des freien
Sauerstoffradikals O2- zu H2O2 katalysiert. Dies wird wiederum mit Hilfe der
Glutathionperoxidase weiter zu H20 umgewandelt. Bei über 80% der fALS-Patienten
bleibt die Mutation unbekannt. Es wird vermutet, dass es zu einem komplexen
Zusammenspiel zwischen Umweltfaktoren und zahlreichen genetischen Loci kommt,
die in ihrer Einzelheit nur einen geringen Einfluss ausüben (Dunkley et al. 2007). Von
Damme hält einen Zusammenhang zwischen Mutationen im SOD1 Gen und der
sporadischen Form für wahrscheinlich (Van Damme et al. 2005).
Weitere Überlegungen zur Ätiopathogenese sind Umweltfaktoren, virale/infektiöse
Genese, Autoimmunerkrankungen, exogene und endogene Neurotoxine; auf
molekularer Ebene unter anderem Apoptose, Beeinträchtigung im axonalen Transport
und die Exzitoxizität.
Es gibt relativ viele epidemiologische Daten, die Hinweise zur Pathogenese der ALS
liefern. Diese können aber nur wenig substantielle Befunde vorweisen.
Zu diesen epidemiologischen Daten gehört unter anderem das Phänomen der
Golfkrieg ALS, bei der ein Anstieg der Inzidenz unter den, in ihr Heimatland
zurückgekehrten Soldaten, zu verzeichnen war. Als Ursache für diese Häufung wurde
in dieser Zeit ein umweltbedingtes oder ein infektiöses Agens vermutet. (Barohn et al.
2002).
Einen großen Fortschritt in der ALS Forschung konnte im Jahre 2006 durch die
Entdeckung des TAR DNA‐Binding Proteins 43 (TDP‐43) gemacht werden
(Neumann et al. 2006), (Arai et al. 2006).

7
1.4.2 Pathomechanismen
„Gain of funtion“ des mutierten SOD1-Proteins
Mit Entdeckung der krankheitsauslösenden humanen SOD1G93A-Mutation bei ALS-
Patienten im Jahr 1993 nahm man an (Rosen et al. 1993), dass die Mutation zu einen
Verlust der enzymatischen Aktivität führt und somit der enzymatische Abbau von
Sauerstoffradikalen gestört ist („Loss of Function-Theorie“), (Bowling et al. 1995),
(Gurney et al. 1994). Bis heute sind über 100 humane SOD1 Mutationen bekannt, die
zum Ausbruch der ALS beim Menschen führen (Conwit 2006), (Al-Chanlabi et al.
2000). Es wurde aber beobachtet, dass in vielen Mutationen die enzymatische Aktivität
der SOD1 unbeeinträchtigt oder sogar erhöht ist. An SOD1 knock-out Mäusen wurde
gezeigt, dass diese keine Motoneuronenerkrankung entwickeln und eine normale
Lebensspanne haben (Gurney et al. 1994), (Wong et al. 1995), (Cleveland et al.
1995). Diese Daten lassen vermuten, dass die verschiedenen Mutationen das Protein
mit einer oder mehreren neuen toxischen Eigenschaften ausstatten, ganz unabhängig
von der enzymatischen SOD1-Aktivität. Dieses Phänomen wird als „Gain of Function“
bezeichnet. Die Art der neugewonnen Eigenschaften des mutierten SOD1-Proteins
bleibt bis heute jedoch unklar (Almer 2003).
Apoptose
Unter Apoptose versteht man den programmierten Zelltod. Sie dient unter anderem
der Beseitigung defekter Zellen, da diese eine Gefahr für den gesamten Organismus
darstellen können (Rassow et al. 2008).
Durch eine Fülle von Daten geht man mittlerweile davon aus, dass am Ende einer
langen Kaskade, die zum SOD1- vermittelten Motoneuronen Untergang führt, die
Apoptose steht (Almer 2003).
Unter anderem konnte in einer Studie dargelegt werden, dass die Überexpression von
bcl-2, einem anti-apoptotischen Protein, zu einer wesentlichen Verzögerung des
Ausbruchs der Erkrankung sowie einer Verlängerung der Lebenszeit der transgenen
Mäuse führt (Kostic et al. 1997). Dies wurde zusätzlich untermauert, als wenig später
die Aktivierung der Kaspasen 1 und 3, zweier wichtiger Schlüsselenzyme, bei der
Auslösung der Apoptosekaskade im Rückenmark von mutierten SOD1G93A-Mäusen
gezeigt werden konnte (Pasinelli et al. 2000).

8
In einer weiteren Studie, die die Apoptosetheorie stützt, wurde der therapeutische
Effekt bei einer intrathekalen Applikation eines Kaspaseninhibitors (zVAD-fmk)
beschrieben (Li et al. 2000). Zudem konnte bei transgenen Mäusen, die eine Mutation
in dem humanen SOD1-Gen überexprimieren, eine vermehrte Expression von
Kaspase 1 schon im präklinischen Stadium nachgewiesen werden (Yoshihara et al.
2002).
Hypothese des oxidativen Stresses
Oxidativer Stress entsteht, wenn die Produktion freier Sauerstoffradikale die Kapazität
der anti-oxidativen Abwehr überschreitet. Äußere Einflüsse (Glutamat oder H2O2),
intrazelluläre Störungen (Gendefekte) und eine verminderte Abbaurate der ROS
können für die Akkumulation von reaktiven Sauerstoffspezies verantwortlich sein. Zu
den reaktiven Sauerstoffspezies gehören Radikale wie das Superoxidradikal, das
Hydroxylradikal, das Hydroperoxyradikal, aber auch nicht-Radikale wie H2O2,
singulärer Sauerstoff und Hypochlorid. Nicht radikalische Verbindungen werden meist
durch bestimmte Reaktionen in Radikale umgewandelt.
Radikale sind definitionsgemäß Atome oder Moleküle, die ein oder mehrere
ungepaarte Elektronen tragen (Halliwell et al. 1989).
Bei vielen Hypothesen, die sich auf die mutierte SOD1 (fALS) beziehen, scheint das
Kupfer Ion eine entscheidende Rolle zu spielen.
Eine Studie stützt sich auf die veränderte Struktur des mutierten SOD1 –Proteins.
Durch das weniger gefaltete Protein tritt das Kupfer Ion hervor und vermittelt oxidative
Prozesse. Ein anderer möglicher Mechanismus könnte auf der Instabilität des
mutierten SOD1-Proteins beruhen, bei der sich das Risiko der Proteolyse und der
Freisetzung ins Zytoplasma erhöht. Zudem konnte man bei fALS-Patienten zweifach
erhöhte Kupfermengen nachweisen (Watanabe et al. 1997). Die zentrale Rolle des
Kupfers in der Toxizität der SOD1 Mutation wird von einer Studie gestützt, bei der
Kupferchelatoren neben Kaspaseinhibitoren, bcl-2 und Vitamin E, zu den potentesten
Substanzen gehören, in vitro den Zelltod von mutierten SOD1 PC12 Zellen
abzuwenden.
Mitochondrien spielen im Zusammenhang mit oxidativen Stress eine wichtige Rolle.
Einerseits sind die Mitochondrien die Kraftwerke der Zelle und verantwortlich für den
Elektronentransport. Andererseits sind sie sehr vulnerabel gegenüber oxidativem
Stress (Packer et al. 1996).

9
Bei Verlust der mitochondrialen Integrität könnte die Verbindung zur Apoptose
stattfinden. Kommt es zur Ruptur in der äußeren Membran der Mitochondrien, wird
Cytochrom C freigesetzt, welches in Folge die Aktivierung der Kaspasen und damit die
Apoptose einleitet (Kroemer et al. 1997).
In neuropathologischen Studien konnte in Mäusen mit der SOD1G93A –Mutation gezeigt
werden, dass die ersten sichtbaren morphologischen Veränderungen, Aufblähungen
und Vakuolenbildungen in Mitochondrien zu finden sind (Wong et al. 1995) (Dal
Canto et al. 1994).
Excitoxizität
Eine der wichtigsten Funktionen ist die Regulation der Glutamatkonzentration über den
EAAT2/GLT-1 Glutamat Transporter im synaptischen Spalt. Glutamat ist ein wichtiger
exzitatorischer Neurotransmitter im zentralen Nervensystem (Almer 2003). Sowohl bei
der fALS und der sALS als auch dem transgenen Mausmodell konnte gezeigt werden,
dass es zum Verlust des EAAT2/GLT-1 Glutamat Transporters kommt (Rothstein et al.
1995), (Fray et al. 1998), (Howland et al. 2002). Dieser Rezeptor ist für ca. 90% des
Abtransports des synaptischen Glutamats im Bereich von Motoneuronen
verantwortlich (Rothstein et al. 1996). Es ist deshalb nicht verwunderlich, dass die
Glutamatkonzentration im synaptischen Spalt bei Verlust des Rezeptors sehr stark
ansteigt und einen toxischen Effekt auf Motoneurone ausübt. Man bezeichnet dieses
Phänomen als Exzitoxizität. Durch die abnorme exzitatorische Stimulation des
Glutamats im synaptischen Spalt (Almer 2003) werden die ionotropen Glutamat-
Rezeptoren AMPA und NMDA auf den Motoneuronen überstimuliert. Der dadurch
resultierende exzessive Ca2+ Einstrom hat Einfluss auf die Integrität der
Mitochondrien, die verdauenden Enzyme (Proteasen, Endonucleasen, NO-
Synthetasen…) und die Ionenströme. Durch den Ca2+-Einstrom in die Mitochondrien
kommt es aufgrund der Akkumulation des Ca2+ zur Erniedrigung des
elektrochemischen Gradienten und damit zu einer verminderten ATP-Synthese. Unter
anderem wird Cytochrom C in das Zytoplasma freigesetzt, das über den „Apoptotic
Protease activating Factor 1“ (Apaf-1) die Apoptosekaskade einleitet (Rassow et al.
2008).
Riluzol reduziert unteranderem die Freisetzung von Glutamat aus den Nervenenden
und ist im Moment das einzig zugelassene Medikament zur Behandlung der ALS,
welches die Rolle der Exzitoxizität bei der ALS festigt.

10
1.5 Neuroprotektion
Unter Neuroprotektion versteht man den Versuch Nervenzellen und Nervenfasern
durch pharmakologische Methoden vor dem Absterben zu bewahren.
Ziel der Neuroprotektion ist es, die zugrundeliegende Ätiologie oder Pathogenese einer
Erkrankung günstig zu beeinflussen und dadurch den Erkrankungsbeginn, die
Symptomatik oder den klinischen Verlauf zu verzögern (Schwarz et al. 2005).
1.6 Ziel der Arbeit
Die antiinflammatorische Wirkung des Immunmodulators FTY720 wurde bereits
in zahlreichen klinischen und präklinischen Studien bei Patienten mit Multipler
Sklerose nachgewiesen. Dabei wurde eine zusätzliche neuroprotektive Wirkung von
FTY720 vermutet, allerdings bisher nur unzureichend untersucht (Soliven et al.
2011) .
Daher lag die Frage nahe, ob FTY720 direkte neuroprotektive Eigenschaften
unabhängig von der antiinflammatorischen Wirkung haben könnte. Um dieser Frage
nachgehen zu können, wurde das Tiermodell der ALS, einem primär
neurodegenerativen Krankheitsmodell, herangezogen. Die ALS ist eine
neurodegenerative Motoneuronenerkrankung, bei der es gegenwärtig keinen kurativen
Therapieansatz gibt. Jedoch konnte durch Riluzol (NMDA-Rezeptorantagonist) das
Überleben um 3-6 Monate verlängert werden (Nirmalananthan et al. 2005).
1.6.1 Teilprojekt 1 Die neuroprotektive Wirkung wurde an Hand von folgenden Parametern evaluiert: Wir
führten eine Quantitative Bestimmung der α-Motoneuronenanzahl, Astrozyten und
Mikroglia im lumbalen Rückenmark durch. Zudem führten wir eine quantitative Analyse
der Neurofilamente Heavy Chain im Hirn-Homogenat durch.

11
1.6.2 Teilprojekt 2
In Teilprojekt 2 sollte die Wirkung auf das Ausmaß der Neuroinflammation im SOD
G93A -Mausmodell der ALS bestimmt werden. Um die Wirkung auf die
Neuroinflammation zu untersuchen wurden folgende Parameter ermittelt:
Quantitative Bestimmung der B- und T-Lymphozyten bzw.CD4+- und CD8+-T-
Lymphozyten im Vollblut und der Milz mittels FACS.
Quantitative Bestimmung der proinflammatorischen (IFN-γ, IL-2) und
antiinflammatorischen (IL-4, IL-10) Zytokine mittels ELISA.
1.7 Fragestellungen
1.7.1 Teilprojekt 1
• Hat FTY720 unabhängig von der antiinflammatorischen Wirkung eine direkte
neuroprotektive Wirkung?
• Welchen Einfluss hat die Dosis auf die Wirkung von FTY720?
• Welchen Einfluss hat das Medikament auf das Überleben der G93A SOD1
Mäuse?
1.7.2 Teilprojekt 2
• Welchen Einfluss hat FTY720 auf das Ausmaß der Neuroinflammation im
Rahmen des SOD1 G93A Mausmodells?
• Welchen Einfluss hat die Dosis auf die Wirkung von FTY720 auf die
Neuroinflammation?

12
2 Material und Methoden
2.1 Material
2.1.1 Die Versuchstiere
In der Studie wurden B6SJL-Tg(SOD1-G93A)dl 1Gur/J Maus verwendet. Die Zuchttiere
wurden bei Jackson Labratory (600 Main Street Bar Harbor, ME USA 04609) gekauft.
Die Versuchstiere stammen aus einer eigenen Nachzucht am Tierforschungszentrum
Ulm. Diese Tiere tragen die humane hemizygote G93A-Mutation im Gen der Cu/Zn-
Superoxid Dismutase (SOD1). Sie sind zurzeit der einzige validierte
Modellorganismus, mit dessen Hilfe die Wirkung verschiedener Substanzen zur
Behandlung neurodegenerativer Erkrankungen verlässlich erfasst werden kann.
Die SOD1 G93A low copy Maus überexprimiert die humane Form der SOD1-Mutation
und zeigt einen ähnlichen Phänotyp wie die humane ALS. Sie zeigt unter anderem
Lähmungen an einem oder mehreren Hintergliedmaßen, Motoneuronenverlust vor
allem auf lumbaler Rückenmarksebene, die Akkumulation von aktivierten Mikroglia,
Astrogliose und die krankheitsprogressive Akkumulation von SOD1-Aggregaten. In der
Studie wurden nur männliche Tiere verwendet, um geschlechtsspezifische
Unterschiede zu vermeiden.
2.1.2 Haltung der Tiere
In Teilprojekt 1 wurden die Mäuse in offenen Regalsystemen in Käfigen vom Typ „M2
Long“ gehalten. Es wurden max. 3 Tiere in einem Käfig gehalten, da bei männlichen
Tieren die Gefahr von Unverträglichkeiten bestand. Wasser und Futter standen ad
libidum zur Verfügung. Den Mäusen wurde Weichholzgranulat als Einstreu und
Zellstoff als Nestbaumaterial im Käfig zur Verfügung gestellt.
Das Teilprojekt 2 unterscheidet sich in der Haltung der Mäuse. Weil das Zytokinprofil
und der Immunstatus der Tiere untersucht werden sollte und Entzündungen durch evtl.
Bisswunden die Ergebnisse beeinflussen würden, wurden die Mäuse einzeln gehalten.

13
2.1.3 Studiendesign
Die präklinische Studie wurde in Kooperation mit der Firma Norvatis, Schweiz
durchgeführt. Die Studie umfasste 2 Teilprojekte.
Das Medikament FTY720 wurde ab dem 35. Lebenstag den Tieren in zwei
unterschiedlichen Dosierungen gewichtsspezifisch mittels Schlundsonde appliziert.
Die Tiere der Negativkontrolle erhielten Wasser über eine Schlundsonde. Die Tötung
der Tiere erfolgte in drei verschiedenen Krankheitsstadien. Präsymptomatisch im Alter
von 70 Tagen, Symptomatisch beim ersten Auftreten von Lähmungen und dem
Endstadium der Erkrankung. Als Kontrolle dienten hier altersgleiche unbehandelte
transgene SOD G93A- Tiere. Die Durchführung der präklinischen Studie erfolgte
gemäß des, vom Regierungspräsidium Tübingen am 06.12.2012 genehmigten,
Tierversuchsantrags Reg.-Nr. 1123.
2.1.3.1 Teilprojekt 1
Transgene SOD1G93A Mäuse
1. Behandlungsgruppe (n=5): Tägliche orale Gabe von 1,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zu einem Alter von 70 Tagen. Die Tiere
wurden in der präsymptomatischen Phase (Tag 70) getötet.
2. Behandlungsgruppe (n=5): Tägliche orale Gabe von 1,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250μl) ab Tag 35 bis zu einem Alter von 130 Tagen. Die Tiere
wurden mit beginnender symptomatischer Phase (ca. Tag 200) getötet.
3. Behandlungsgruppe (n=5): Tägliche orale Gabe von 1,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zum Erreichen des Endstadiums. Die
Tiere wurden mit Erreichen des Endstadiums getötet.
4. Behandlungsgruppe (n=5): Tägliche orale Gabe von 3,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zu einem Alter von 70 Tagen.
Die Tiere wurden in der präsymptomatischen Phase (Tag 70) getötet.
5. Behandlungsgruppe (n=5): Tägliche orale Gabe von 3,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zu einem Alter von 130 Tagen. Die
Tiere wurden mit beginnender symptomatischen Phase (ca. Tag 200) getötet.

14
6. Behandlungsgruppe (n=5): Tägliche orale Gabe von 3,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zum Erreichen des Endstadiums. Die
Tiere wurden mit Erreichen des Endstadiums getötet.
7. Behandlungsgruppe (n=5): Kontrolltiere. Die Tiere wurden im Alter von 65-70
Tagen (Präsymptomatisch) getötet. Die Tiere haben 250μl H2O mittels Schlundsonde
erhalten.
8. Behandlungsgruppe (n=5): Kontrolltiere. Die Tiere wurden im Alter von ca. 200
Tagen (Symptomatisch) getötet. Die Tiere haben 250μl H2O mittels Schlundsonde
erhalten.
9. Behandlungsgruppe (n=5): Kontrolltiere. Die Tiere wurden im Alter von ca.240
Tagen (Endstadium) getötet. Die Tiere haben 250μl H2O mittels Schlundsonde
erhalten.
2.1.3.2 Teilprojekt 2
Teilprojekt 2: Messung des Immunstatus und des Zytokinprofils in vivo
- Es erfolgten Blutabnahmen mittels Punktion aus der Vena facialis an folgenden
Tagen: 40d; 60d; 80d; 100d; 120d; 170-180d; Symptomatische Phase;
Endstadium
- 2 unterschiedliche Dosierungen: 1mg/kg KG und 3mg/kg KG FTY720
- Tötung der Tiere in Endstadium
1. Gruppe (n=7): Tägliche orale Gabe von Trinkwasser mittels Schlundsonde
(Volumen 250 µl) ab Tag 35 bis zum Erreichen des Endstadiums. Die Tiere wurden im
Endstadium getötet.
2. Gruppe (n=7): Tägliche orale Gabe von 1,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zum Erreichen des Endstadiums. Die
Tiere wurden im Endstadium getötet.
3. Gruppe (n=7): Tägliche orale Gabe von 3,0 mg/kg Tag FTY720 mittels
Schlundsonde (Volumen 250 µl) ab Tag 35 bis zum Erreichen des Endstadiums. Die
Tiere wurden im Endstadium getötet.

15
2.1.4 Antikörper
2.1.3.1 ELISA
Zytokine:
MSD® MULTI-SPOT Assay System, Proinflammatory Panel 1 (mouse) Kits
INFγ, IL-2, IL-4, IL-10 von Mesocale
Neurofilamente und Proteinanalyse:
Phosphorylated Neurofilament H (pNF-H) Sandwich ELISA Kit, Chemicon International
(Cat.No. NS170, Lot: 2485687)
2.1.3.2 Immunhistochemie
Rabbit Anti Iba1 Code No. 019-19741 Wako Pure Chemical Industries
Anti-GFAP Antikörper ab 7260 Abcam
goat anti-rabbit IgG-B
(Sekundärantikörper)
sc-2040
SantaCruz Biotechnology, Inc.
Impact Nova Red Kit Cat. No. SK-4800 Vectorlabs
VECTASTAIN ABC KIT Cat. No. PK-4000 Verctorlabs
2.1.3.3 FACS-Analyse
Antikörper Firma
IgG Maus Antikörper Chrom Pure Mouse; FA.
Jackson, no 015-000-003
CD19 PE (B-Zellen) eBioscience #12-0193-81
CD3 FITC (T-Zellen) eBioscience #11-0031-82
CD4 APC (CD4+ T-Zellen) eBioscience #17-0041-82
CD8 PE (CD8+T-Zellen) eBioscience #12-0081-82

16
AK- Färbungen (Verdünnungen alle in FACS-Puffer)
B- und T-Zellen: CD19 PE (1:200) und CD3 FITC (1:50)
CD4 und CD8-Zellen: CD4 APC (1:400) und CD8 PE (1:800)
single stainings:
CD3 FITC (1:50)
CD19 PE (1:200)
CD11b APC (1:400)
ungefärbte Kontrolle (Zellen mit Totfarbstoff)
2.1.5 Puffer, Lösungen und Material
2.1.2.1 SOD PCR
10 × TAE- Puffer(pH 8,3)
- TRIS-Base 0,4M, Roth
- Eisessig 0,2M, Normapure
- EDTA 10mM, Sigma
Primer und DNA-Polymerase Kit, Thermo Scientific ,Ulm
• SOD Maus 42: 5‘- cta ggc cac aga att gaa aga tct-3‘
• SOD Maus 43: 5’-cta ggt gga aat tct agc atc atc-3‘
• SOD Maus 113: 5‘-cat cag ccc taa tcc atc tga-3‘
• SOD Maus 114: 5‘-cgc gac taa caa tca aag tga-3‘
• Arbeitskonzentration der Primer 10µM
2.1.2.2 Immunhistochemie
Objektträger: Menzel-Gläser Superfrost Plus Menzel
Objektträgerkisten VWR
Objektträgermappen VWR
Deckgläser Menzel

17
Färbetröge mit Deckel und Einsatz Brand
Eppendorf Reaktionsgefäße (1,5 ml,0,5 ml) Sarstedt
Rotilabor®-embedding cassettes, POM mit
Deckel L40 x B28 x H6,8mm
Carl Roth GmbH
Rotilabor®-embedding cassettes, Macro,
weiß, Spezialpolymer, L41 x B27,5 x H12mm
Carl Roth GmbH
Peel-A-Way® Disposable Embedding Molds
(S-22), 22 x 22 x 20mm deep
Polyscience Inc.(Cat# 186446A
LOG# 654361)
Impact Nova Red Kit, Vectorlabs, Cat. No. SK-4800
VECTASTAIN ABC KIT, Verctorlabs, Cat. No. PK-4000
1M NaOH, Fluka
1M HCl, Fluka
1×TRIS-HCl-Tween 20 Puffer (pH 7,6), Sigma-Aldrich
- 0,5M Tris-Base
- 0,5% Tween 20
- 1 M HCl
Citratpuffer (pH 8,0)
- 10mM Zitronensäure, Roth
- 10mM Na2 HPO4 * 2H2O, Roth
Bovine Serum Albumin Lösung
- 5g Albumin Fraktion V, GE Healthcare
- 0,25ml Triton X, Sigma
- 100ml Tris-HCl-Puffer (pH 7,6)
Mayers Hämolaun Lösung, Merck Millipore
Eosin 1%, Roth
99,8% Alkohol vergällt 642, VWR
Xylol, Sigma Aldrich
Liquid Blocker-Super PAP Pen, Dako
EUKITT: Xylol lösliches Eindeckmedium, Sigma Aldrich
Kresylviolett 1%, Merck Millipore
Aqua dest: Avium pro VF, Sartorius stedim Biotech

18
2.1.2.3 ELISA
Fa. Sarstedt Biosphere® Filter tips
- 20-300 μl
- 0,5-10 μl
- 0,5-20 μl
- 2,0-20 μl
- 2-100 μl
Quality Pipette tips, 100-1000 μl, Fa Sarstedt
Puffer für Homogenisate
- 10mM TRIS-HCL pH 7,6
- 4M Urea, Sigma-Aldrich
- 1mM EDTA, Sigma
- 1mM EGTA, Sigma-Aldrich
- 0,2 mM PMSF, Sigma-Aldrich
Ampuwa, VWR
2.1.2.4 FACS-Analyse
FCS (Fetales Kälberserum)
- 1×DPBS-CaCl2-MgCl2, Fa. Gibco
FACS-Puffer:
- 1x PBS ohne Ca/Mg, Fa Gibco
- FCS (5%), Fa Gibco
Ammoniumchlorid-Lösung, Fa stemcell technologies, no 07800
EDTA zum Spülen der Kanülen und Spritzen: UltraPure 0,5M EDTA, pH 8,0; Fa. Gibco
no 15575 100ml
Propidiumjodid (PI)-Lösung:
Propidiumjodid, Fa Invitrogen, no P3566
1:2500 Verd. in FACS-Puffer, (UV sensitiv,1-2 Wochen haltbar)

19
Blocking Reagenz:
Maus IgG Antikörper (ChromPure Mouse IgG; Fa Jackson Immuno Research),
no 015-000-003
Verdünnung 1:50 in FACS-Puffer
Material:
S-Monovette, Sarstedt,cat no 05.1167.001
Cell strainer (Nylonnetz 40 µm), BD Falcon, cell strainer REF 352340
Cell strainer (5 ml Polystyrene Round Bottom Tube with cell strainer cap) BD Falcon,
Ref 352235
FACS-Röhrchen: 5ml Polstyrene round bottom tube with cell strainer cap von BD
Falcon, REF 352235
Cell strainer 40µm Nylon von BD Falcon, REF 352340
Lysepuffer: Ammoniumchlorid (NH4Cl) von Fa. Stemcell technologies, no. 07800
Totfarbstoff: Propidiumjodid von Fa. Invitrogen, no. P3566
FACS-Puffer 1×PBS without Ca/Mg + 5% FCS (=Fetales Kälberserum)
2.1.6 Präparation und Lagerung
Forene 100% (V/V) 250ml, Wirkstoff: Isofloran, Abbott
1×PBS, pH=7,4
- 137mM NaCl, Sigma-Aldrich
- 2,7mM KCl, Sigma
- 12mM Na2HPO4 , Roth
Eismaschine, Scotsman AF80
2.1.7 Geräte
Personal Thermocycler, Biometra
Dampfkocher, Braun
Mikrotom RM 2265, Leica
Modulares Einbettsystem mit Kühlplatte, Leica EG1150 C

20
Entwässerungsautomat, Leica
Schüttler stuart® SSL4, Biocote
Vortexer, IKA
96-Well Mikroplate reader,
Kühlzentrifuge Fresco 21, Thermo Scientific
Tischzentrifuge (Biofuge pico), Heraeus instruments
FACS-Gerät, BD FACS Calibur 3607, BD Bioscience
ELISA: Berthold Technologies Twinkle LB 970

21
2.2 Methoden
2.2.1 DNA-Isolierung aus Schwanzspitzenbiopsiematerial
Zur DNA Isolierung wurde jeweils ein ca. 3mm langes Schwanzstück vom Schwanz
der Maus entnommen und in einen Eppendorf Tube gegeben. Um die DNA aus den
Zellen zu lösen wurde zu jeder Probe 230µl Quick Extrakt Puffer dazugegeben. Jede
Probe wurde daraufhin 15 sec gevortext. Anschließend wurde die Probe bei 65°C für
6 Min. erhitzt, im Folgenden wieder 15 Sekunden gevortext und zu Letzt bei 95°C für
2 Min. erhitzt.
2.2.2 Polymerasekettenreaktion
Die PCR dient der Genotypisierung der transgenen G93A-Mäuse. Es handelt sich
hierbei um eine molekularbiologische Methode. Sie erlaubt es in vitro die DNA nach
Belieben stark zu vervielfältigen, damit sie sehr einfach nachzuweisen ist.
Zur Umsetzung dieser Methode werden folgende Komponenten benötigt:
• die DNA-Polymerase, die die einzelnen DNA-Stränge miteinander verbindet.
• ein sog. Primer, der als Startpunkt gilt und an den die Bausteine der DNA (Nucleotide)
angebaut werden.
• ein sog. Template (DNA-Strang), welches als Kopiervorlage dient.
Der PCR-Zyklus besteht aus drei Teilschritten:
Der erste Schritt umfasst die Denaturierung. Bei 95°C werden hier die DNA-Stränge
aufgetrennt. Im zweiten Schritt folgt das „Annealing“, auch Hybridisierung genannt. In
diesem Schritt heften sich die Primer der einzelsträngigen DNA an. Danach folgt die
Elongation (Verlängerung). Die Temperatur wird auf 72°C erhöht, damit die Taq-
Polymerase optimal arbeiten kann. Die verwendete Taq-Polymerase ist hitzestabil. In
den meisten Organismen werden die DNA-Polymerasen bei 95°C zerstört. Doch durch
die Entdeckung des Mikroorganismus „Thermus aquaticus“, welches in heißen Quellen
bei über 100°C gefunden wurde, konnte eine hitzestabile DNA-Polymerase, isoliert
werden. Die Taq-Polymerase beginnt über die OH—Gruppe am 3’Ende des Primers
die Nucleotide aufzufüllen.

22
Tabelle 1: Reagenzien für die SOD-PCR. SOD= Superoxid Dismutase; PCR=
Polymerasekettenreaktion; Master Mix= vorgefertigte gebrauchsbereite Reagenzienmischung; Taq-
Polymerase= thermostabile DNA-Polymerase.
Inhalt Menge Inhalt
DreamTaq Master Mix 50nmol Taq-Polymerase,
dNTP’s, MgCl2, Loading
Buffer
Primer 113 10pmol
Primer 114 10pmol
Primer 42 10pmol
Primer 43 10pmol
Ampuwa 3,5pmol
Tabelle 2: Programm der SOD-PCR. Die Reihenfolge der einzelnen Reaktionsschritte der SOD-PCR
mit Berücksichtigung der Temperatur und Dauer der einzelnen Reaktionsschritte. Diese wird vom
Hersteller Thermoscientific vorgegeben. SOD= Superoxid Dismutase; PCR= Polymerasekettenreaktion.
PCR-Schritte Temperatur Dauer
Initiale Denaturierung 95°C 3min
Denaturierung 95°C 30sec
Annealing 58°C 30sec
Automatisierte Fluoreszenz
Extension
72°C 10min
Finale Extension 72°C 10min
2.2.3 Gelelektrophorese
Aufgrund der negativen elektrischen Ladung der DNA können DNA-Moleküle in einem
konstanten elektrischen Feld aufgetrennt werden. Da die meisten DNA-Moleküle im
Verhältnis zu ihrer Masse annähernd die gleiche Ladung besitzen, verwendet man zur
Auftrennung der DNA Agarose- oder Polyacrylamidgele. Die Gele bestehen aus einem
System von Poren, deren Größe von der jeweiligen Gelkonzentration abhängig ist. Die
Wanderungsgeschwindigkeit der Moleküle verhält sich umgekehrt proportional zu ihrer
Größe. Zur Größenabschätzung der einzelnen DNA-Fragmente trägt man parallel
einen Satz von Restriktionsfragmenten bekannter Größe, einen sogenannten

23
Größenmarker, auf. Über einen Positionsvergleich der Banden kann man die
Fragmentgrößen ermitteln. Dabei ist die Laufstrecke linearer Nukleinsäuren
proportional zum Logarithmus ihres Molekulargewichts.
Durch Zugabe von Ethidiumbromid werden die DNA-Moleküle im Gel sichtbar
gemacht. Ethidiumbromid interkaliert zwischen die einzelnen Basen des DNA-
Doppelstrangs und fluoresziert rot-orange bei Bestrahlung mit ultraviolettem Licht von
254 nm oder 300 nm Wellenlänge. Die Nachweisgrenze von ethidium¬bromid
gefärbten Gelen liegt bei 5 ng je Bande [Mülhardt, 1999].
Zur Bestimmung von DNA-Fragmenten zwischen 800 bp und 5.000 bp wurden 1,0 -
2,0%ige Agarosegele, für Fragmente von 200 bp bis 1.000 bp 2,0 2,5%ige Gele
verwendet. Die Agarose wurde mit 1x-TAE-Puffer, pH 8,0 angesetzt und bis zum
vollständigen Lösen der Agarose erhitzt. Nach Zugabe von 0,5 µg Ethidiumbromid (10
mg/ml) pro 1 ml Gellösung und Abkühlen auf ca. 60°C erfolgte das Gießen der Gele
im Gelschlitten mit einer Dicke von 0,7 mm. Das Probenauftragsvolumen betrug jeweils
10 µl.
Für eine nähere Charakterisierung der aufgetrennten Banden, wurden präparative
Agarosegele verwendet, aus denen die gewünschten Banden mittels eines Skalpells
ausgeschnitten werden. Die Konzentration der präparativen Agarosegele lag bei 0,7%,
die Dicke variierte zwischen 1,0 – 1,5 cm, je nach Probenauftragsvolumen.
Zusammensetzung:
4,0 - 9,0 µl DNA (je nach Konzentration)
1,0 µl 10x-Probenpuffer (pH 7,2)
X µl ddH2O
10,0 µl
Laufbedingungen:
Laufpuffer 1x-TAE, pH 8,0
Spannung 90 V
Zeit 15 - 30 min

24
2.2.4 Gewinnung des zu analysierenden Materials
Die Maus wurde mit Isofloran in Narkose gelegt. Nach ca. 30 sec. wurde durch Kneifen
in die Zehenzwischenräume die Tiefe der Narkose überprüft. Danach erfolgte die
transkardiale Perfusion mittels Herzpunktion.
Für die FACS-Analyse wurden 200µl Vollblut verwendet. Der Rest wurde verwendet
um eine Zytokinanalyse mittels ELISA durchzuführen. Das Blut wurde ab zentrifugiert,
das Zellpellet verworfen und das Plasma bei -80°C schockgefroren.
2.2.5 Präparation, Fixierung und Lagerung des Gewebes
Nach der Blutabnahme wurde die Milz frei präpariert, entnommen und in PBS gelegt.
Eine Hälfte diente der Herstellung eines Homogenats. Sie wurde in einen Kryotube
gegeben und sofort in Trockeneis schockgefroren. Die Lagerung erfolgte bei -80°C.
Die andere Hälfte und der Rest der Maus wurden für 48h in 4% PFA fixiert. Danach
erfolgte die Präparation des Rückenmarks unter Eröffnung des Wirbelkanals. Die
Gehirnhälfte und das Rückenmark wurden in PBS gegeben und in Paraffin eingebettet.
2.2.6 Immunhistochemie
2.2.6.1 Das Prinzip
Die Immunhistologie ermöglicht es in Geweben spezifische Proteine mittels Antikörper
nachzuweisen und anzufärben.
2.2.6.2 Einbetten
Das Einbetten erfolgte in 4 Schritten.
Das Gewebe wurde schrittweise durch Inkubation in Alkohol entwässert. Der Alkohol
wurde dann durch Xylol (Lösungsmittel) ersetzt. Xylol wurde daraufhin durch Paraffin
ersetzt. Das Gewebe wurde in einem Einbettförmchen (Peel-A-Way® Disposable
Embedding Molds) mit flüssigem Paraffin in die gewünschten Position gebracht. Das
Förmchen wurde zum Abkühlen auf eine kalte Platte gestellt.

25
2.2.6.2 Paraffinschnitte
Das in Paraffin eingebettete Gewebe wurde mit dem Mikrotom „EG1150 C“ von Leica
geschnitten. Die Schnittdicke lag bei 7 µm. Bevor mit dem Schneiden begonnen
werden konnte, wurden die Paraffinblöcke für 2 h in die Kühltruhe (-20°C) gelegt.
Um eine gute Anhaftung zu erreichen (Joppien et al. 2011), wurden die Objektträger
mit den aufgezogenen Geweben für 24h in einer Wärmekammer bei 37°C getrocknet.
Die darauffolgende Lagerung erfolgte bei Raumtemperatur.
2.2.7 Nissl-Färbung
Mit der Nissl-Färbung werden sogenannte Nissl-Schollen angefärbt. Dabei handelt es
sich um endoplasmatisches Retikulum (ER) und Zellkerne von Nervenzellen. Das
Prinzip liegt in der elektrostatischen Anlagerung des basischen Farbstoffs
„Kresylviolett“ an die sauren Gruppen der Nucleinsäuren, der Zellkerne und des rauen
ER, die in einem sauren Milieu (PH 4,9) erfolgt.
Das Präparat auf dem Objektträger wurde in Xylol entparaffiniert (Xylol Reihe) und für
je 3min in absteigender Alkoholreihe (100% - 90% - 70%) rehydriert. Im Folgenden
verblieb das Präparat für 5 Min. in destilliertem Wasser. Dann wurde das Präparat 10
Min. in Kresylviolett (1%) wässrig bei Raumtemperatur inkubiert, um danach nochmal
in destilliertem Wasser gespült zu werden. Im folgenden Schritt wurde die Färbung in
96% Ethanol differenziert (ca. 7 Min.). Die Differenzierung wurde durch Gabe des
Präparats in 100% Ethanol unterbrochen und durchlief im Anschluss die Xylol Reihe.
Mit Hilfe eines Deckglases und des Eindeckmediums Eukitt wurde die Färbung und
das Gewebe auf dem Objektträger ausreichend fixiert.
2.2.8 Antikörperfärbungen
2.2.8.1 Prinzip der indirekten Antikörperfärbung
Der Primärantikörper bindet spezifisch an sein Antigen. Um den Primärantikörper
sichtbar zu machen, markiert man ihn mit einem biotinylierten Sekundärantikörper. Bei
der Avidin-Biotin-Komplex Methode reagiert der Avidin-Biotin-Enzymkomplex mit dem
biotinylierten Sekundärantikörper. Hierbei macht man sich die starke Affinität von

26
Avidin gegenüber Biotin zunutze. Aufgrund der vier Bindungsstellen des Avidins ist die
Sensitivität höher als bei der direkten Immunfluoreszenzfärbung, bei der der
Primärantikörper direkt mit einem Enzym markiert wird. Durch Inkubation mit einem
Enzymsubstrat (Impact Nova Red) kommt es dann zur Anfärbung der Antigene
(Joppien et al. 2011).
2.2.8.2 Durchführung
1. Tag
Das Präparat auf dem Objektträger wurde in Xylol entparaffiniert (Xylol Reihe: Xylol 1:
3sec; Xylol 2: 5min; Xylol 3: 5min; Xylol 4: 5min) und in absteigengender Alkoholreihe
(100%: 3sec; 100%: 5min; 96%: 5min; 70%: 5min) rehydriert. Im Folgenden verblieb
das Präparat für 5 Min. in destilliertem Wasser. Je nach Antikörperfärbung erfolgte eine
Vorbehandlung. Die Vorbehandlung dient der Antigendemaskierung. Die Fixierung mit
Formalin führt teilweise zur Entstehung von sog. Cross-links, d.h. zur Quervernetzung
der Proteinstrukturen. Die Erhitzung des Gewebes beim Einbettungsvorgang auf bis
zu 60°C kann zu Konfirmationsänderungen am Epitop des Antigens führen.
Deshalb versucht man durch das Erhitzen in Citratpuffer die Konformation der Epitope
teilweise oder ganz wiederherzustellen (Joppien et al. 2011). Anschließend folgte die
Inkubation in 1% H2O2-Lösung über 20 Minuten. Dies dient der Inaktivierung der
körpereigenen Peroxidasen und vermindert den Background. Nachfolgend wurden die
unspezifischen Bindungsstellen mit ca. 200µl BSA Lösung geblockt. Aufgrund der
Ladungsverteilung und der Konformation anderer Proteine oder der Oberfläche des
Objektträgers kann es zu unspezifischen Bindungen kommen, was wiederum zu einer
störenden Hintergrundfärbung führt (Joppien et al. 2011). Die Inkubation betrug ein
Stunde. Nachdem die Inkubationszeit vorbei war, wurde die BSA Lösung von den
Präparaten abgeklopft und für jeweils 5 Min. in 1×TRIS-HCl Puffer pH (8,0) gelagert
und geschüttelt. Dazwischen wurde der gewechselt. Im nächsten Schritt folgte die
Inkubation mit dem Primärantikörper über Nacht. Bei Raumtemperatur wurde je nach
Antikörperfärbung mit einer unterschiedlicher Verdünnung gearbeitet.
Folgende Verdünnungen wurden verwendet:
- IBA-1: 1:1000 in TRIS-HCl Puffer (pH 8,0)
- GFAP: 1:1000 in TRIS-HCl Puffer (pH 8,0)

27
2. Tag
Am 2. Tag wurde die Antikörperlösung abgeklopft und die Präparate für 3× jeweils 5
Min in 1×TRIS Puffer unter schütteln gewaschen. Anschließend folgte die Inkubation
mit dem Sekundärantikörper in einer 1:200 Verdünnung in 1×TRIS-HCl Puffer (pH 8,0)
über 1h bei Raumtemperatur. Das Präparat wurde danach wieder abgeklopft und für
jeweils 5 Min in TRIS Puffer gewaschen und geschüttelt. Im Folgenden wurde das
Präparat für eine Stunde mit dem ABC-Kit (Avidin+Peroxidase) behandelt. Das
Präparat wurde danach wieder abgeklopft und für jeweils 5min in TRIS Puffer gelagert
und geschüttelt. Dazwischen wurde der TRIS-Puffer gewechselt. Danach folgte die 15-
minütige Inkubation mit Nova Red, dem Substrat der Peroxidase. Das Präparat wurde
dann 5 Min. in destilliertem Wasser gespült. Um den Kontrast zu erhöhen wurde eine
Gegenfärbung mit Hämolaun durchgeführt. Danach wurden die Präparate für 10 Min.
unter fließendem Leitungswasser gespült. Dann erfolgte die Dehydrierung in
aufsteigender Ethanol Reihe (70%: 3 sec; 96%: 5min; 100%: 3sec; 100%: 5min) und
Inkubation in Xylol (Xylol Reihe: Xylol 1: 3sec; Xylol 2: 5min; Xylol 3: 5min; Xylol 4:
5min). Zu guter Letzt wurden die Präparate eingedeckelt. Die Präparate wurden zum
Trocknen 24h in einen Wärmeschrank bei 37°C gegeben.
2.2.9 Auswertung der immunhistochemischen Färbungen
2.2.9.1 Quantitative Bestimmung der Motoneurone
Die Zellkörper wurden mit einer Nissl-Färbung angefärbt. Es wurden alle Nervenzellen
gezählt, die im anterioren lumbalen Rückenmark liegen, einen Durchmesser d > 20µm
haben und in einem definiertem Feld liegen. Das definierte Feld wurde mit Hilfe eines
rechten Winkels konstruiert. Der rechte Winkel wurde so gelegt, dass sein
Scheitelpunkt im Canalis centralis liegt und einer seiner Strahlen durch die Fissura
mediana anterior verläuft.
Frühere Studien haben diese Methode genutzt, um Motoneurone zu identifizieren (4).
Xiaoxing Ma et al. bestätigten, dass der Durchmesser aller mit NeuN angefärbten
Nervenzellen größer als 20µm ist (Ma et al. 2011).

28
2.2.9.2 Bestimmung des Ausmaßes der Astrogliose
Das „Glial fibrillary acidic Protein“ (kurz: GFAP) ist ein Intermediär Filament, das
hochspezifisch auf Astroglia exprimiert wird (Reeves et al. 1989). Mit Hilfe des Anti-
GFAP Antikörpers (Abcam 7260) wurden die Astrozyten markiert und angefärbt. Somit
war es möglich hyperplastische und proliferative Veränderungen der Astroglia zu
beurteilen.
Es wurden alle GFAP-positiven Zellen gezählt, die in einem klar definierten Bereich, in
der grauen Substanz des anterioren Rückenmarks lagen. Das definierte Feld wurde
mit Hilfe eines rechten Winkels konstruiert. Der rechte Winkel wurde so gelegt dass
sein Scheitelpunkt im Canalis centralis liegt und einer seiner Strahlen durch die Fissura
mediana anterior verläuft.
2.2.9.3 Bestimmung des Ausmaßes der Einwanderung der Mikroglia
IBA1 (= ionizing calcium-binding adaptor molecule 1) ist ein Protein, welches
vorwiegend auf Makrophagen/ Mikroglia exprimiert wird. Je stärker diese Zellen
aktiviert werden, desto stärker exprimieren sie IBA1 auf ihrer Oberfläche. Mit Hilfe des
Anti-Iba1, Rabbit von Wako wurden die Mikroglia markiert und angefärbt. So war es
möglich die proliferativen und morphologischen Veränderungen zu beurteilen.
Es wurden alle Iba1-positiven Zellen, wie schon zuvor beschrieben, gezählt.
2.2.3. ELISA
2.2.3.1 Prinzip des ELISA
ELISA bedeutet Enzyme-linked immunosorbent Assay. Es handelt sich um einen
Immunoassay, mit dem Substanzen wie Zytokine in geringen Konzentrationen auf
Grundlage des Antigen-Antikörperprinzipes nachgewiesen werden können. Zur
Bestimmung von Zytokinen und Neurofilamenten (Antigene) wurde der nicht-
kompetive Immunoassay, die sog. Sandwich-Methode eingesetzt. Bei dieser Methode
wird das Antigen von zwei Antikörpern gebunden und ist dadurch sensitiver als die
kompetive Methode. Wie in Abb. 2 gezeigt, bindet der erste Antikörper
(Primärantikörper) an eine feste Phase, die sog. 96-Well-Mikrotiterplatte.

29
Zur quantitativen Messung der Zytokine im Plasma wurde das MSD® MULTI-SPOT
Assay System, Proinflammatory Panel 1 (mouse) Kit verwendet. Die Durchführung
erfolgte nach Herstellerangaben.
2.2.3.2 Vorbereitung der Plasmaproben
Die Plasmaproben wurden gemeinsam aufgetaut und auf 4 Tubes ( Neurofilamente,
Gelsolin, Zytokine) aliquotiert, zwei Tubes mit 3μl; zwei Tubes mit je 20μl.
2.2.3.3 Phosphorilierte Neurofilamte Heavy Chain ELISA
Herstellung des Homogenatpuffers
10mM TRIS-HCl wurden in VE-Wasser gelöst und mit Hilfe einer 1M NaOH-Lösung
der PH-Wert auf 7,2 eingestellt. Alle weiteren Bestandteile (4M Urea, 1mM EDTA, 1mM
EGTA, 0,2 mM PMSF) des Puffers wurden unter Rühren in dem 10mM TRIS-HCl
Puffer gelöst.
Zur quantitativen Analyse der Neurofilamente wurde das Phosphorylated
Neurofilament H (pNF-H) Sandwich ELISA Kit von Chemicon International. Die
Durchführung erfolgte nach Herstellerangaben.
2.2.4 Quantitative Proteinanalyse
Die Quantitative Proteinanalyse der Hirn-Homogenate diente dazu, um die Menge der
Neurofilamente NFH ins Verhältnis zum Gesamtprotein des jeweiligen Gehirngewebes
zu setzen. So war es möglich, unabhängig von der Menge des Gehirngewebes, eine
Aussage über die Quantität der Neurofilamente NFH zu machen. Zur Quantitativen
Proteinanalyse wurde das Quant-it Protein Assay Kit verwendet. Die Durchführung und
Auswertung erfolgte nach Herstellerangaben.
2.2.5 Auswertung der Neurofilamente
Die quantitative Bestimmung der Neurofilamente (heavy Chain) in den Plasmaproben
erfolgte mit Hilfe des ELISA Plate Readers bei einer Wellenlänge von 430 nm. Der

30
ELISA Plate Reader maß die Extinktion und berechnete mit Hilfe der Standardkurve
die Konzentration der Neurofilamente (heavy Chain) in den Wells.
Die Konzentrationen wurden mit dem Faktor 100 multipliziert, um die Verdünnung zu
bereinigen. Dann wurden sie zur Gesamtproteinkonzentration in der jeweiligen
Plasmaprobe ins Verhältnis gesetzt. Sie wurden in der Einheit μg/mg angegeben.
Die statistische Auswertung erfolgte mit Excel.
2.2.6 FTY720-Messung im Plasma und Hirn-Homogenat
Die quantitative Analyse von FTY720 und FTY720-Phospaht wurde von der Norvatis
Pharma GmbH durchgeführt.
2.2.7 FACS-Analyse von Maus-Vollblut und Milz
Vorbereitungen
• Totfarbstoff Propidiumjodid (PI): PI-Lösung und FACS-Puffer frisch ansetzen
• NH4Cl-Lösung auftauen
• Kanüle und Spritze für die Herzpunktion mit 0,5 M EDTA durchspülen
• Verwendung von K-EDTA Monovetten zur Herzpunktion
2.2.7.1 Lyse der Erythrozyten
Ca 300 µl Vollbut (EDTA-Vollblut) werden mit der 5-10 fachen Menge an Lysepuffer
(NH4Cl-Lösung) versetz und 10 min auf Eis inkubiert. Dann erfolgt die Zugabe von 1
ml FACS- Puffer damit die Lysereaktion abgestoppt wird. Anschließend wird die Probe
bei 2200 rpm für 3 min bei 4 °C zentrifugiert; der Überstand wird verworfen du das
Zellpellet wird in 1 ml FACS-Puffer resuspendiert. Dieser Waschschritt wird 2x
wiederholt.
2.2.7.2 Vorbereitung der Milz
Die Milz wird mit Hilfe einer Pipettenspitze oder eines Gummistempels (aus einer
Spritze) und unter Zugabe von 1 ml FACS-Puffer durch einen cell strainer (BD Falcon,
cell strainer REF 352340) das ist eine Art Nylonnetz mit einer Porengröße von 40 µm
in ein 50 ml Falconröhrchen gepresst um das Gewebe zu zerteilen und Einzelzellen zu
erhalten. 300 µl der Zellsuspension (ca 1-2 x 106-Zellen) werden abgenommen und in

31
ein 1,5 ml Eppendorf Reaktionsgefäß überführt und mit FACS-Puffer auf 1 ml
aufgefüllt. Anschließend wird die Probe bei 2200 rpm für 3 min bei 4 °C zentrifugiert;
der Überstand wird verworfen du das Zellpellet wird in 1 ml FACS-Puffer resuspendiert.
Dieser Waschschritt wird 1x wiederholt.
Blocken der Zellen (Blut und Milz)
Nach dem letzten Waschschritt werden die Zellen jeder Probe jeweils in 50 µl Blocking-
Reagenz luftblasenfrei resuspendiert und 15-20 min auf Eis inkubiert. Dieser Schritt
dient der xxx. Anschließend wird die Probe in zwei gleiche Volumina zu je 30 µl
aliquotiert und bei 2200 rpm für 3 min bei 4°c zentrifugiert. Der Überstand wird
verworfen.
2.2.7.3 Antikörper Master-Mix und Antikörper Färbung (Blut und Milz)
Die jeweiligen Antikörper sind mit unterschiedlichen Fluoreszenzfarbstoffen konjugiert
und wurden in den unten stehenden Verdünnungen eingesetzt. Dabei wurden zwei
verschiedene Master-Mixe hergestellt; Master-Mix 1 diente der Färbung von B- und T-
Zellen, Master-Mix 2 diente der Färbung von CD4- und CD8-Zellen
Färbung von B-Zellen: CD3 FITC (1:50) in FACS-Puffer
Färbung von T-Zellen: CD19 PE (1:200) in FACS-Puffer
Färbung von CD4-Zellen: CD4 APC (1:400) in FACS-Puffer
Färbung von CD8-Zellen: CD8 PE-Zellen (1:800) in FACS-Puffer
Das Zellpellet wird in jeweils 100 µl Antikörper-Master-Mix (Master-Mix 1 oder 2)
vorsichtig resuspendiert (ohne zu schäumen) und im Kühlschrank für 60 min inkubiert.
Danach erfolgt die Zugabe von 1 ml FACS-Puffer. Anschließend wird die Probe bei
1800 rpm für 3 min bei 4°C zentrifugiert und der Überstand verworfen. Danach wird
das Zellpellet in 300 µl Totfarbstoff-Lösung resuspendiert um xxx. Die Arbeiten
erfolgen im Dunkeln, da die mit Fluoreszenzfarbstoffen konjugierten Antikörper UV
sensitiv sind.
Kontrollmessungen
Zusätzlich zu den zu messenden Blut- und Milz-Proben wird jeweils die Blutprobe einer
Kontrollmaus (unbehandeltes wildtyp-Tier) gemessen um die Einstellungen des FACS
Gerätes zu überprüfen und gegebenenfalls zu korrigieren. Die Zelllyse und das
Blocken der Zellen erfolgt wie zuvor beschrieben. Im Anschluss an die Zentrifugation
wird die Probe in vier gleiche Volumina zu je 15 µl aliquotiert und bei 2200 rpm für 3

32
min bei 4°c zentrifugiert. Der Überstand wird verworfen. Es werden drei verschiedene
Antikörper Master-Mixe angesetzt. Drei (der vier) Zellpellets werden in 100 µl des
jeweiligen Master-Mixes vorsichtig resuspendiert. Das vierte Zellpellet wird in 250 µl
Totfarbstofflösung resuspendiert.
Master-Mix 1 CD3-FITC (1:50) 100 µl FACS-Puffer 2 µl AK
Master Mix 2 CD19-PE (1:200) 200 µl FACS-Puffer 1 µl AK
Master Mix 3 CD11b-APC (1:400) 400 µl FACS-Puffer 1 µl AK
Vor der FACS-Messung werden alle Proben über einen cell strainer (5 ml Polystyrene
Round Bottom Tube with cell strainer cap, BD Falcon, Ref 352235) pipettiert um,
eventuell vorhandene Zellklumpen, zu entfernen.
2.2.8 Bestimmung des Alters bei Beginn der symptomatischen und Endstadium-
Phase
Zur Überprüfung der Wirksamkeit des Medikaments wurde sowohl bei Eintritt in die
symptomatische Phase als auch in die Endstadium-Phase das Alter der Tiere
bestimmt.
Zur Bestimmung des Eintritts in die Symptomatische Phase wurden folgende Kriterien
verwendet:
• Gewichtsverlust von 3g im Vergleich zum max. Gewicht der jeweiligen Maus
• Wenn man die Maus am Schwanz hochhält, sind die Beine im Allgemeinen im
45° Winkel abgespreizt; bei symptomatischen Tieren sind die Beine nicht mehr
abgespreizt. Sie zittern und hängen unter dem Körper
• Gangveränderungen
Das Endstadium ist, wie folgt, definiert:
• Die Tiere haben das Endstadium erreicht, sobald sie nicht mehr in der Lage
sind, sich unverzüglich aufzurichten, nachdem sie auf die Seite gelegt wurden.
Die Tiere entwickeln Symptome, die der humanen ALS sehr ähnlich sind. Daher
würden sie ohne Euthanasie an einer respiratorischen Insuffizienz sterben.
Aus Gründen des Tierschutzes wurden die Tiere daher getötet, sobald sie das
Endstadium der Erkrankung erreicht hatten.

33
Die Ergebnisse wurden mit einfachen Parametern der deskriptiven Statistik wie
Mittelwert, Standardabweichung analysiert. Die visuelle Darstellung der Ergebnisse
erfolgte in einem Kaplan-Maier Plot.
2.2.9 Statistische Analyse
2.2.9.1 Deskriptive Statistik
Mittelwert und Standardabweichung
In unsere Studie haben wir den Mittelwert und Standartabweichung zur besseren
Veranschaulichung unserer Ergebnisse verwendet.
2.2.9.2 Signifikanztestung
Anova one way und Post Hoc Tests
Der „One-way Analysis of Variance“ (abgekürzt one-way ANOVA) ermöglicht es, drei
oder mehr Behandlungsgruppen miteinander zu vergleichen. Mit diesem Test kann
gezeigt werden, ob ein signifikanter Unterschied zwischen den Behandlungsgruppen
besteht oder nicht. Dabei zeigt dieser Test nicht, zwischen welchen
Behandlungsgruppen ein signifikanter Unterschied besteht. Dafür werden im
Anschluss an den ANOVA sogenannte Post Hoc Tests verwendet.
Post Hoc Tests vergleichen die Unterschiede zwischen den Mittelwerten
verschiedener Behandlungsgruppen, um signifikante Unterschiede zu differenzieren.
In unserer Studie wurden folgende Post Hoc Tests verwendet:
• Least Significant Difference-Test,
• Tukey-Test und
• Scheffé-Test
2.2.10 Spezielle Software
- Word Microsoft Office 2013
- Excel Microsoft Office 2013
- Graph Prism Version 6.07

34
3 Ergebnisse
3.1 Effekt von FTY720 auf α-Motoneurone von SOD1G93Adl-Tieren
Anmerkung für die immunhistochemischen Färbungen:
Unter der Voraussetzung, dass Wildtyptiere keinen altersabhängigen
Motoneuronenverlust sowie Astrogliose und Mikrogliaaktivierung im lumbalen
Rückenmark zeigen, wurden die unbehandelten Wildtyptiere nur zu einem
Alterszeitpunkt (200 Tage) untersucht. Aus Tierschutzgründen waren die transgenen
Tiere der Kontrollgruppe ebenfalls unbehandelt (Verzicht auf eine Applikation von
Wasser mittels Schlundsonde), da nicht davon auszugehen ist, dass eine
Schlundsondenbehandlung Auswirkungen auf die bei der IHC gemessenen Parameter
hat.
3.1.1 Quantitative Bestimmung der α-Motoneurone im lumbalen Rückenmark
Nachgewiesenermaßen zeigen SOD1G93Adl-Tiere im Krankheitsverlauf eine
ausgeprägte Degeneration der α-Motoneurone im lumbalen Rückenmark.
Das Ziel der Nissl-Färbung war es herauszufinden, ob FTY720 im Krankheitsverlauf
des SOD1G93Adl-Mausmodells einen protektiven Effekt auf α-Motoneurone im lumbalen
Rückenmark hat.
Im Vergleich zu Wildtyptieren zeigen unbehandelte SOD1G93Adl- Kontrolltiere bereits in
der präsymptomatischen Phase der Erkrankung, d.h. im Alter von ca. 70 Tagen, einen
Motoneuronenverlust von ca. 20%.
Bei den präsymptomatischen SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag FTY720
behandelt wurden, sind ca. 21 % mehr α-Motoneurone nachweisbar als bei den
unbehandelten SOD1G93Adl- Kontrolltieren. Dies zeigt, dass bei den
präsymptomatischen SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag FTY720 behandelt
wurden, kein Motoneuronenverlust zu beobachten ist.
Dieser Effekt zeigt sich ebenfalls bei den präsymptomatischen SOD1G93Adl-Tieren, die
mit 3,0 mg/kg Tag FTY720 behandelt wurden. Im Vergleich zu den unbehandelten
SOD1G93Adl- Kontrolltieren sind bei den 3,0 mg/kg Tag FTY720 dosierten Tieren ca.
28% mehr α-Motoneurone im lumbalen Rückenmark nachweisbar, so dass auch hier
in der präsymptomatischen Phase kein Motoneuronenverlust aufgetreten ist.

35
In der symptomatischen Phase der Erkrankung ist bei den unbehandelten SOD1G93Adl-
Kontrolltieren im Vergleich zu Wildtyp-Tieren ein Verlust von ca. 57% an α-
Motoneuronen nachweisbar. Im Vergleich zu präsymptomatischen SOD1G93Adl-Tieren
haben symptomatische SOD1G93Adl-Tiere ca. 46 % weniger α-Motoneurone. Die
Behandlung mit 1,0 mg/kg Tag FTY720 führt zu einem geringeren Verlust an α-
Motoneurone.
SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, besitzen in der
symptomatischen Phase 32 % mehr α-Motoneurone als unbehandelte transgene
SOD1G93Adl-Tiere. Die symptomatischen SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag
FTY720 behandelt wurden, haben jedoch ca. 8 % weniger α-Motoneurone als die
unbehandelten SOD1G93Adl- Kontrolltiere. Bei den symptomatischen SOD1G93Adl-
Tieren, die mit der Dosierung 1,0 mg/kg Tag FTY720 behandelt wurden, ist ein
neuroprotektiver Effekt erkennbar. Im Gegensatz dazu zeigt sich bei den höher
dosierten SOD1G93Adl-Tieren kein oder sogar ein neurotoxischer Effekt.
Unbehandelte SOD1G93Adl-Tiere weisen im Vergleich zu Wildtyp-Tieren im Endstadium
der Erkrankung ca. 71% weniger α-Motoneurone auf. Im Gegensatz zu
präsymptomatischen unbehandelten SOD1G93Adl- Kontrolltieren haben unbehandelte
SOD1G93Adl- Kontrolltiere im Endstadium ca. 63 % weniger α-Motoneurone.
Die SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, weisen ca. 33
% weniger α-Motoneurone auf als die im Endstadium unbehandelten SOD1G93Adl-
Kontrolltiere. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY 720 behandelt wurden,
haben im Endstadium ca. 5 % weniger α-Motoneurone als die symptomatischen
unbehandelten SOD1G93Adl- Kontrolltiere. Ein neuroprotektiver Effekt von FTY720 ist
hier in keiner Dosierung erkennbar. Die beobachteten neuroprotektiven Effekte der
präsymptomatischen und symptomatischen Tiere, die mit 1,0 mg/kg Tag FTY720
behandelt wurden, sind jedoch nicht signifikant.
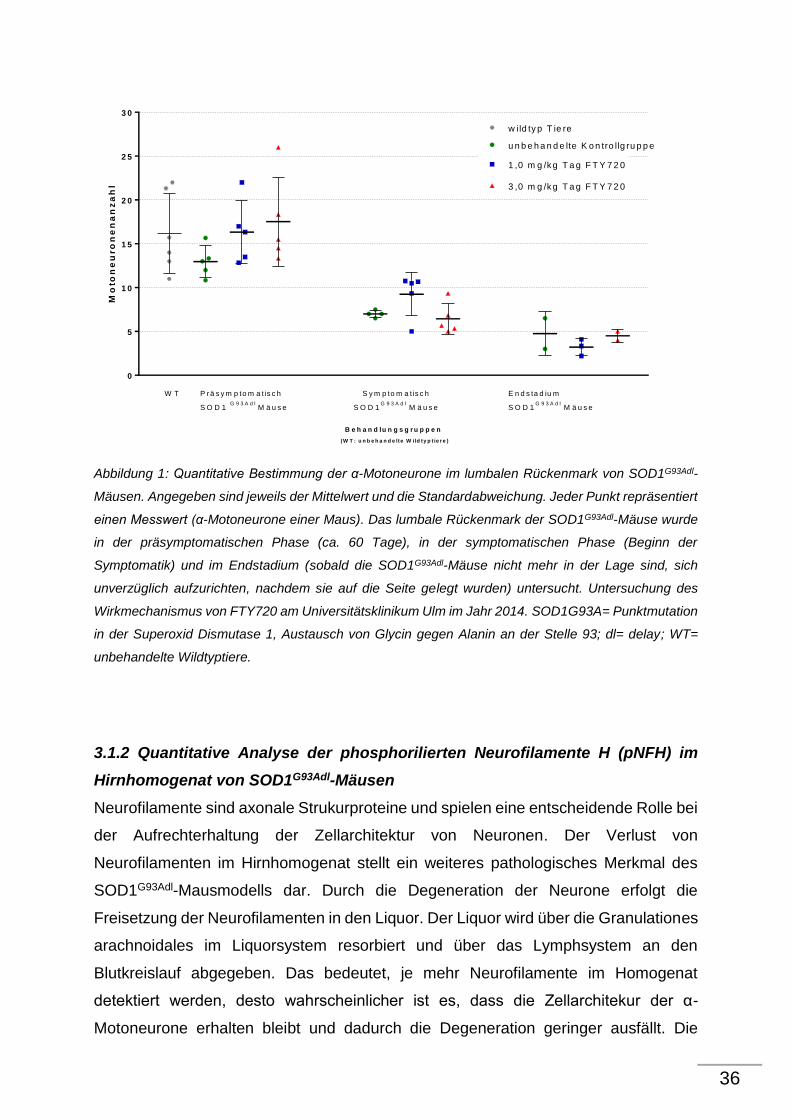
36
0
5
1 0
1 5
2 0
2 5
3 0
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Mo
ton
eu
ro
ne
na
nz
ah
l
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
3 ,0 m g /k g T a g F T Y 7 2 0
W T
Abbildung 1: Quantitative Bestimmung der α-Motoneurone im lumbalen Rückenmark von SOD1G93Adl-
Mäusen. Angegeben sind jeweils der Mittelwert und die Standardabweichung. Jeder Punkt repräsentiert
einen Messwert (α-Motoneurone einer Maus). Das lumbale Rückenmark der SOD1G93Adl-Mäuse wurde
in der präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der
Symptomatik) und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich
unverzüglich aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay; WT=
unbehandelte Wildtyptiere.
3.1.2 Quantitative Analyse der phosphorilierten Neurofilamente H (pNFH) im
Hirnhomogenat von SOD1G93Adl-Mäusen
Neurofilamente sind axonale Strukurproteine und spielen eine entscheidende Rolle bei
der Aufrechterhaltung der Zellarchitektur von Neuronen. Der Verlust von
Neurofilamenten im Hirnhomogenat stellt ein weiteres pathologisches Merkmal des
SOD1G93Adl-Mausmodells dar. Durch die Degeneration der Neurone erfolgt die
Freisetzung der Neurofilamenten in den Liquor. Der Liquor wird über die Granulationes
arachnoidales im Liquorsystem resorbiert und über das Lymphsystem an den
Blutkreislauf abgegeben. Das bedeutet, je mehr Neurofilamente im Homogenat
detektiert werden, desto wahrscheinlicher ist es, dass die Zellarchitekur der α-
Motoneurone erhalten bleibt und dadurch die Degeneration geringer ausfällt. Die

37
quantitative Analyse der phosphorilierten Neurofilamente H (pNFH) im
Hirnhomogenanat wurde mit Hilfe des Sandwich ELISA durchgeführt. pNFH ist neben
NFM und NFL einer der 3 Hauptuntereinheiten von Neurofilamenten.
Ziel war es, einen Effekt von FTY720 auf die pNFHs im Hirnhomogenat im
Krankheitsverlauf des SOD1G93Adl-Mausmodells zu zeigen.
Eine Abnahme der pNFHs bei den unbehandelte SOD1G93Adl-Kontrolltieren gegenüber
den Wildtyp-Tieren ist im Krankheitsverlauf des SOD1G93Adl-Mausmodells nicht zu
beobachten. Damit ist ein Vergleich der mit FTY720 behandelten Therapiegruppen und
den unbehandelten SOD1G93Adl- Kontrolltieren nicht möglich.
0 .0 0 0
0 .0 0 1
0 .0 0 2
0 .0 0 3
0 .0 0 4
0 .0 0 5
0 .0 0 6
0 .0 0 7
NF
H i
n
mo
l/m
l
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
3 ,0 m g /k g T a g F T Y 7 2 0
W T
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 2: Quantitative Analyse der phosphorilierten Neurofilamente H (pNFH) im Hirnhomogenat von
SOD1G93Adl-Mäusen. Angegeben sind jeweils der Mittelwert und die Standardabweichung. Jeder Punkt
repräsentiert einen Messwert (pNFH in μmol/ml einer Maus). Das Hirnhomogenat der SOD1G93Adl-Mäuse
wurde in der präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der
Symptomatik) und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich
unverzüglich aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay; pNFH=
phosphorilierte Neurofilamente Heavy Chain; WT= unbehandelte Wildtyptiere.
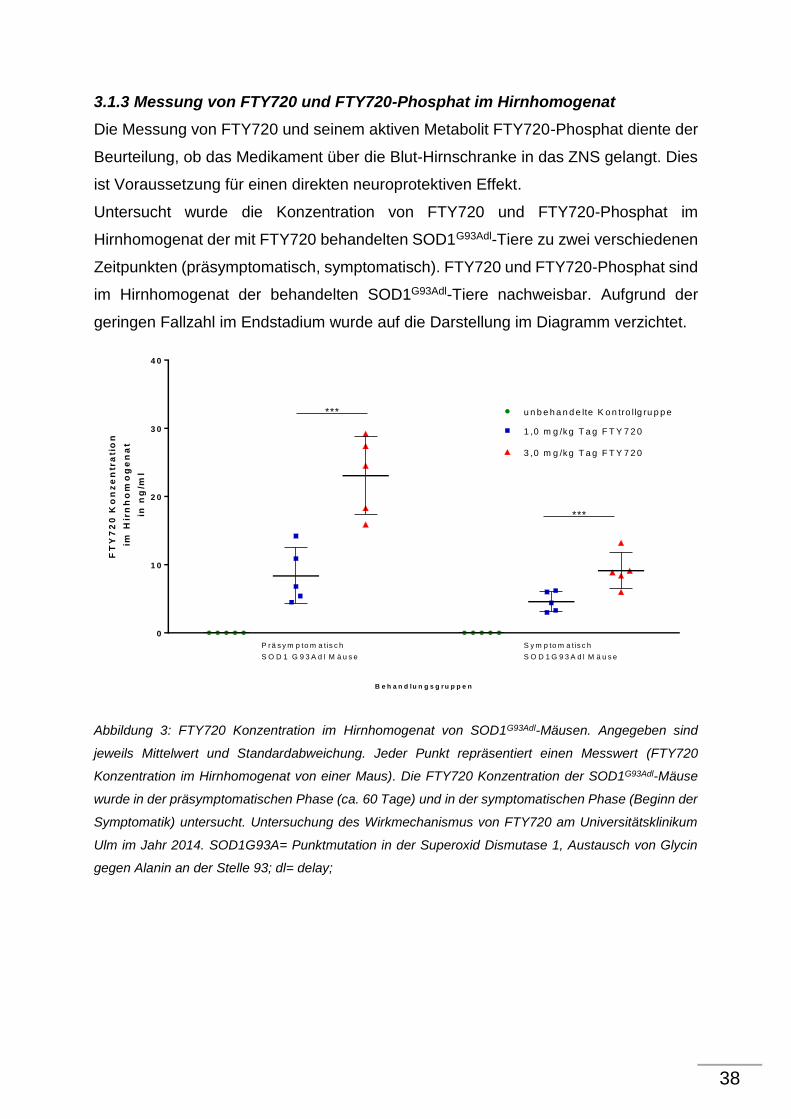
38
3.1.3 Messung von FTY720 und FTY720-Phosphat im Hirnhomogenat
Die Messung von FTY720 und seinem aktiven Metabolit FTY720-Phosphat diente der
Beurteilung, ob das Medikament über die Blut-Hirnschranke in das ZNS gelangt. Dies
ist Voraussetzung für einen direkten neuroprotektiven Effekt.
Untersucht wurde die Konzentration von FTY720 und FTY720-Phosphat im
Hirnhomogenat der mit FTY720 behandelten SOD1G93Adl-Tiere zu zwei verschiedenen
Zeitpunkten (präsymptomatisch, symptomatisch). FTY720 und FTY720-Phosphat sind
im Hirnhomogenat der behandelten SOD1G93Adl-Tiere nachweisbar. Aufgrund der
geringen Fallzahl im Endstadium wurde auf die Darstellung im Diagramm verzichtet.
0
1 0
2 0
3 0
4 0
B e h a n d lu n g s g r u p p e n
FT
Y7
20
Ko
nz
en
tra
tio
n
im
Hir
nh
om
og
en
at
in
ng
/ml
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1 G 9 3 A d l M ä u s e
S y m p to m a tis c h
S O D 1 G 9 3 A d l M ä u s e
3 ,0 m g /k g T a g F T Y 7 2 0
***
***
Abbildung 3: FTY720 Konzentration im Hirnhomogenat von SOD1G93Adl-Mäusen. Angegeben sind
jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (FTY720
Konzentration im Hirnhomogenat von einer Maus). Die FTY720 Konzentration der SOD1G93Adl-Mäuse
wurde in der präsymptomatischen Phase (ca. 60 Tage) und in der symptomatischen Phase (Beginn der
Symptomatik) untersucht. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum
Ulm im Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin
gegen Alanin an der Stelle 93; dl= delay;
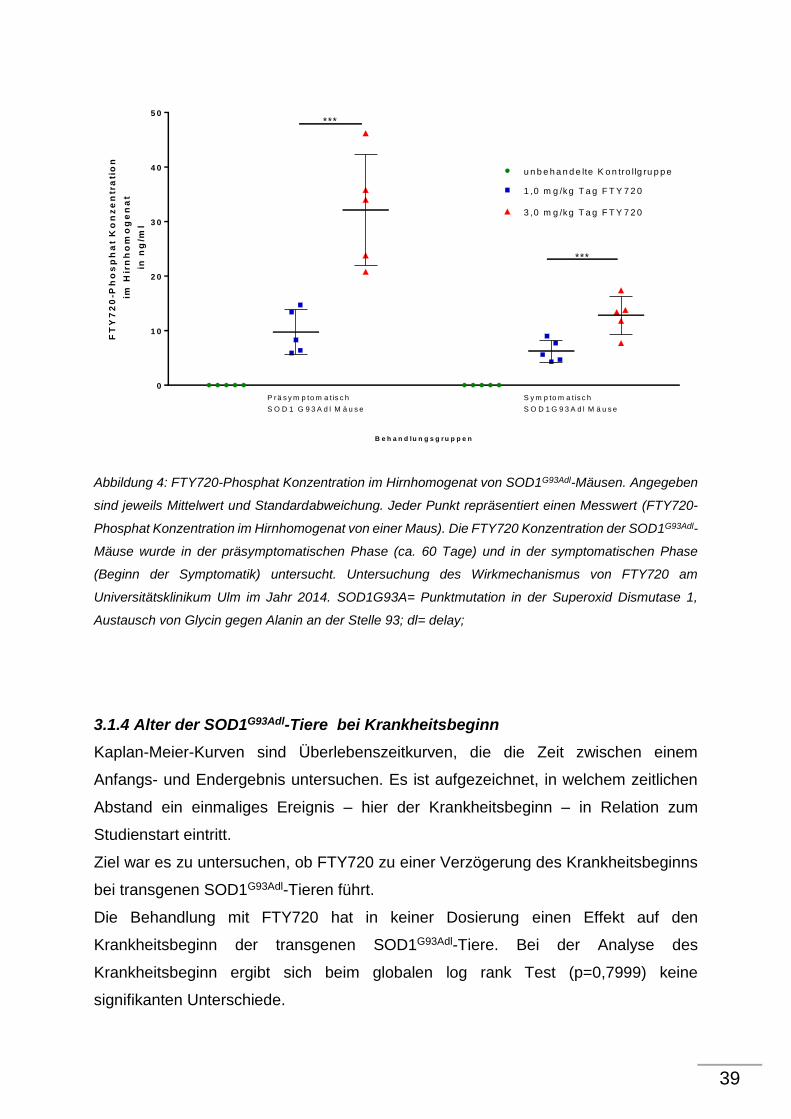
39
0
1 0
2 0
3 0
4 0
5 0F
TY
72
0-P
ho
sp
ha
t K
on
ze
ntr
ati
on
im
Hir
nh
om
og
en
at
in
ng
/ml
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1 G 9 3 A d l M ä u s e
S y m p to m a tis c h
S O D 1 G 9 3 A d l M ä u s e
3 ,0 m g /k g T a g F T Y 7 2 0
B e h a n d lu n g s g r u p p e n
***
***
Abbildung 4: FTY720-Phosphat Konzentration im Hirnhomogenat von SOD1G93Adl-Mäusen. Angegeben
sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (FTY720-
Phosphat Konzentration im Hirnhomogenat von einer Maus). Die FTY720 Konzentration der SOD1G93Adl-
Mäuse wurde in der präsymptomatischen Phase (ca. 60 Tage) und in der symptomatischen Phase
(Beginn der Symptomatik) untersucht. Untersuchung des Wirkmechanismus von FTY720 am
Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1,
Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay;
3.1.4 Alter der SOD1G93Adl-Tiere bei Krankheitsbeginn
Kaplan-Meier-Kurven sind Überlebenszeitkurven, die die Zeit zwischen einem
Anfangs- und Endergebnis untersuchen. Es ist aufgezeichnet, in welchem zeitlichen
Abstand ein einmaliges Ereignis – hier der Krankheitsbeginn – in Relation zum
Studienstart eintritt.
Ziel war es zu untersuchen, ob FTY720 zu einer Verzögerung des Krankheitsbeginns
bei transgenen SOD1G93Adl-Tieren führt.
Die Behandlung mit FTY720 hat in keiner Dosierung einen Effekt auf den
Krankheitsbeginn der transgenen SOD1G93Adl-Tiere. Bei der Analyse des
Krankheitsbeginn ergibt sich beim globalen log rank Test (p=0,7999) keine
signifikanten Unterschiede.

40
Abbildung 5: Alter der SOD1G93Adl-Tiere bei Krankheitsbeginn
3.2 Effekt von FTY720 auf proinflammatorische Parameter im ZNS und im
Blut von SOD1G93Adl-Tieren
Es sollte die Wirkung von FTY720 auf die Neuroinflammation SOD1G93Adl-Mausmodell
untersucht werden. Dafür wurde die Anzahl an aktivierten Astrozyten und Mikroglia im
lumbalen Rückenmark analysiert, die T- und B-Zellen bzw. CD4+ und CD8+ T-Zellen
im Vollblut und Milzhomogenat mittels FACS-Analyse bestimmt und ein Zytokinprofil
mittels ELISA erstellt. FTY720 ist ein Immunmodulator, der als funktioneller Antagonist
den S1P 1 -Rezeptor vermittelten Austritt der Lymphozyten aus den Lymphknoten
in das ZNS sowie in das Blut blockiert (Mehling et al. 2011), (Pinschewer et al. 2011).
3.2.1 Quantitative Bestimmung der Astrozyten (GFAP)
Astrogliose beschreibt einen Zustand der Astrozyten, bei dem diese hypertroph und
hyperplastisch werden. Zudem exprimieren sie in diesem Zustand vermehrt das „Glial
A lte r (T a g e n )
Pro
ze
nt
Kra
nk
he
its
be
gin
n
0 5 0
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
1 5 0 2 0 0 2 5 0 3 0 0
K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0

41
fibrillary acidic protein“, kurz GFAP genannt. Die Astrogliose stellt ein wichtiges
pathologisches Merkmal des SOD1G93Adl-Mausmodells dar. Im Verlauf nimmt sie stetig
zu und korreliert somit im Krankheitsverlauf des SOD1G93Adl-Mausmodells umgekehrt
proportional zu der Anzahl der α-Motoneurone im lumbalen Rückenmark.
Zweck der immunhistochemischen Antikörperfärbung mit dem Anti-GFAP Antikörper
war es, einen potentiellen Effekt des Medikaments auf die Astrogliose im SOD1G93Adl-
Mausmodell zu untersuchen.
Vergleicht man die präsymptomatischen unbehandelten SOD1G93Adl- Kontrolltiere mit
den Wildtyp-Tieren kann in der präsymptomatischen Phase der Erkrankung noch keine
Astrogliose beobachtet werden.
Morphologisch zeigten sich die Astroglia in der präsymptomatischen Phase in allen
Behandlungsgruppen punktförmig und schwach durch den Anti-GFAP Antikörper
angefärbt. Dies spricht auch für einen späteren Beginn der Astrogliose im
Krankheitsverlauf des SOD1G93Adl-Mausmodells.
In der symptomatischen Phase der Erkrankung ist bei den unbehandelten SOD1G93Adl-
Kontrolltieren im Vergleich zu den unbehandelten SOD1G93Adl- Kontrolltieren der
präsymptomatischen Phase eine Zunahme der GFAP positiven Astrozyten von ca. 115
% zu beobachten. In diesem Fall kann man von Astrogliose sprechen. 1,0 mg/kg und
3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere weisen in der symptomatischen
Phase ca. 3-5 % mehr GFAP positive Astrozyten auf, als die unbehandelten
SOD1G93Adl- Kontrolltiere.
Ein Effekt von FTY720 auf die Astrogliose in der symptomatischen Phase zeigt sich
deshalb in keiner Dosierung.
Die unbehandelten transgenen SOD1G93Adl-Tiere weisen im Endstadium im Vergleich
zu symptomatischen unbehandelten SOD1G93Adl- Kontrolltieren 22 % mehr GFAP
positive Astrozyten auf.
1,0 mg/kg und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere zeigen gegenüber
den unbehandelten SOD1G93Adl- Kontrolltieren im Endstadium ca. 20 % weniger GFAP
positive Astrozyten. Eine Reduktion der Astrogliose von FTY720 ist sowohl in der
niedrigen als auch in der hohen Dosierung erkennbar. Aufgrund der geringen Fallzahl
von 2-3 Endstadiumtieren ist die Beobachtung einer verminderten Astrogliose bei den
1,0 mg/kg Tag und 3,0 mg/kg Tag FTY720 behandelten Tieren nicht aussagekräftig.
Die beobachtete Verminderung der Astrogliose bei den behandelten Endstadiumtieren
ist nicht signifikant.
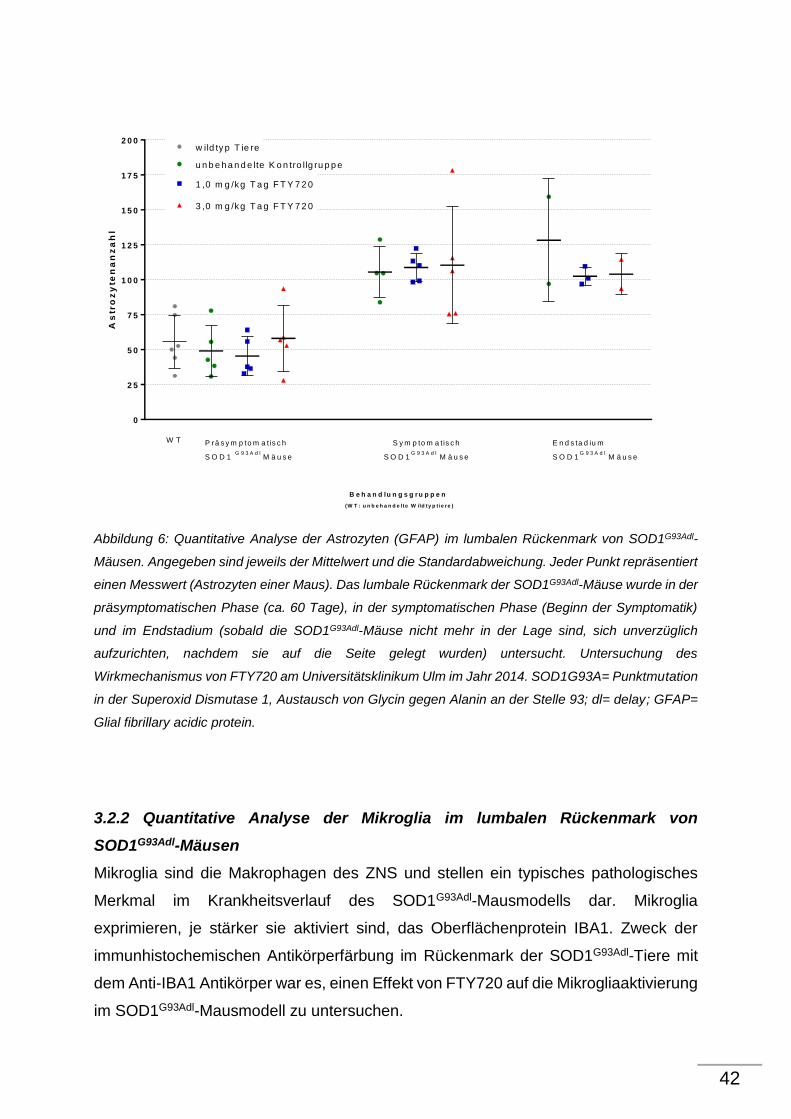
42
0
2 5
5 0
7 5
1 0 0
1 2 5
1 5 0
1 7 5
2 0 0
As
tro
zy
ten
an
za
hl
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
3 ,0 m g /k g T a g F T Y 7 2 0
W T
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 6: Quantitative Analyse der Astrozyten (GFAP) im lumbalen Rückenmark von SOD1G93Adl-
Mäusen. Angegeben sind jeweils der Mittelwert und die Standardabweichung. Jeder Punkt repräsentiert
einen Messwert (Astrozyten einer Maus). Das lumbale Rückenmark der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik)
und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich
aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay; GFAP=
Glial fibrillary acidic protein.
3.2.2 Quantitative Analyse der Mikroglia im lumbalen Rückenmark von
SOD1G93Adl-Mäusen
Mikroglia sind die Makrophagen des ZNS und stellen ein typisches pathologisches
Merkmal im Krankheitsverlauf des SOD1G93Adl-Mausmodells dar. Mikroglia
exprimieren, je stärker sie aktiviert sind, das Oberflächenprotein IBA1. Zweck der
immunhistochemischen Antikörperfärbung im Rückenmark der SOD1G93Adl-Tiere mit
dem Anti-IBA1 Antikörper war es, einen Effekt von FTY720 auf die Mikrogliaaktivierung
im SOD1G93Adl-Mausmodell zu untersuchen.

43
Vergleicht man die präsymptomatischen unbehandelten SOD1G93Adl- Kontrolltiere mit
den Wildtyp-Tieren kann in der präsymptomatischen Phase der Erkrankung noch keine
Mikrogliaaktivierung beobachtet werden.
Morphologisch zeigen sich die Mikroglia in der präsymptomatischen Phase in allen
Behandlungsgruppen eher punktförmig und schwach durch den Anti-IBA1 Antikörper
angefärbt, was zusätzlich für einen späteren Beginn der Mikrogliaaktivierung im
Krankheitsverlauf des SOD1G93Adl-Mausmodells spricht.
In der symptomatischen Phase weisen die unbehandelte SOD1G93Adl- Kontrolltiere
gegenüber unbehandelten SOD1G93Adl- Kontrolltieren der präsymptomatischen Phase
ca. 527 % mehr IBA1-positive Mikroglia auf, so dass man hier von Mikrogliaaktivierung
sprechen kann. Zudem sind die Mikroglia in dieser Phase morphologisch stark
verzweigt und deutlich angefärbt.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere zeigen sich in der
symptomatischen Phase im Vergleich zu den unbehandelten SOD1G93Adl-
Kontrolltieren ca. 5-15 % mehr IBA1-positive Mikroglia.
In der symptomatischen Phase ist eine Verminderung der Mikrogliaaktivierung durch
FTY720 in keiner Dosierung zu sehen.
Im Endstadium weisen die unbehandelten SOD1G93Adl- Kontrolltiere verglichen mit den
unbehandelten SOD1G93Adl- Kontrolltieren in der symptomatischen Phase ca. 54 %
mehr IBA1 positive Mikroglia auf.
Die SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, zeigen
gegenüber unbehandelten SOD1G93Adl- Kontrolltieren im Endstadium ca. 6% weniger
IBA1-positive Mikroglia. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720
behandelt wurden, offenbaren im Endstadium ca. 15 % mehr IBA1-positive Mikroglia
als bei den unbehandelten SOD1G93Adl- Kontrolltieren. Eine geringfügige Reduktion der
Mikrogliaaktivierung ist nur in einer Dosierung von 1,0 mg/kg Tag FTY720 erkennbar.
Aufgrund der geringen Fallzahl von 2-3 Endstadiumtieren ist die Beobachtung einer
Abschwächung der Mikrogliaaktivierung bei den 1,0 mg/kg Tag FTY720 behandelten
Tieren nicht aussagekräftig. Zudem zeigt die Verminderung der Mikrogliaaktivierung
bei den 1,0 mg/kg Tag FTY720 behandelten Tieren im Endstadium keine Signifikanz
auf.

44
0
2 5
5 0
7 5
1 0 0
1 2 5
1 5 0
1 7 5M
icro
gli
aa
nz
ah
l
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
3 ,0 m g /k g T a g F T Y 7 2 0
W T
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 7: Quantitative Analyse der Mikroglia (IBA1) im lumbalen Rückenmark von SOD1G93Adl-
Mäusen . Angegeben sind jeweils der Mittelwert und die Standardabweichung. Jeder Punkt repräsentiert
einen Messwert (Mikroglia einer Maus). Das lumbale Rückenmark der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik)
und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich
aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay; IBA1=
ionized calcium binding adapter molecule 1 .
3.2.3 Quantitative Analyse von T- und B-Zellen bzw. CD4+- und CD8+-T-Zellen im
Vollblut von SODG93Adl-Tieren
Anmerkung zur FACS-Analyse:
Alle Wildtyp-Tiere wurden im Alter von 200 Tagen untersucht.
3.2.3.1 B-Zellen im Vollblut
Die B-Zellen sind Lymphozyten, die zur Gruppe der Leukozyten gehören. Sie stellen
zusammen mit den T-Zellen das adaptive Immunsystem dar. Sie können durch T-
Zellen aktiviert werden und differenzieren sich daraufhin zu Plasmazellen oder
Gedächtniszellen.
Die quantitative FACS-Analyse von B-Zellen im Vollblut diente dem Zweck, den
Einfluss von FTY720 auf B-Zellen im SOD1G93Adl-Mausmodell zu zeigen.

45
Im Vergleich zu Wildtyp-Tieren zeigen unbehandelte SOD1G93Adl-Kontrolltiere in der
präsymptomatischen Phase der Erkrankung ca. 14 % weniger B-Zellen.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere haben ca. 58 -
64% weniger B-Zellen im Blut als unbehandelte transgene SOD1G93Adl-Tiere. Ein
signifikanter Effekt von FTY720 hinsichtlich einer Reduzierung der B-Zellanzahl im
Vollblut der SOD1G93Adl-Tiere in der präsymptomatischen Phase des SOD1G93Adl-
Mausmodells ist in beiden verwendeten Dosierungen erkennbar (p< 0,001).
In der symptomatischen Phase offenbaren die unbehandelten SOD1G93Adl-
Kontrolltiere gegenüber präsymptomatischen unbehandelten SOD1G93Adl-
Kontrolltieren ca. 30 % mehr B-Zellen im Vollblut.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere besitzen in der
symptomatischen Phase, verglichen mit den unbehandelten transgenen SOD1G93Adl-
Tieren, ca. 82-90 % weniger B-Zellen im Vollblut.
Ein signifikanter Effekt von FTY720 bezüglich der Reduzierung der B-Zellanzahl im
Vollblut der SOD1G93Adl-Tiere ist in der symptomatischen Phase unabhängig von der
Dosis erkennbar (p< 0,001).
Im Endstadium zeigen die unbehandelten transgenen SOD1G93Adl-Tiere im Vergleich
zu symptomatischen unbehandelten SOD1G93Adl- Kontrolltieren 30% weniger B-Zellen
im Vollblut.
1,0 mg/kg und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere weisen im
Vergleich zu unbehandelten SOD1G93Adl- Kontrolltieren im Endstadium ca. 90 %
weniger B-Zellen im Vollblut auf. Ein signifikanter Effekt von FTY720 ist hinsichtlich der
Verminderung der B-Zellanzahl im Vollblut von SOD1G93Adl-Tieren in beiden
verwendeten Dosierungen erkennbar (p<0,001). In keinem Krankheitsstadium ist ein
dosisabhängiger Effekt zu sehen.
In der symptomatischen Phase zeigen unbehandelte transgene SOD1G93Adl-Tiere ca.
30% mehr B-Zellen im Vollblut als in der präsymptomatischen Phase sowie im
Endstadium.

46
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0B
-Ze
lle
n i
n%
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
W T
K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
***
***
***
*** ***
***
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 8: Quantitative Analyse der B-Zellen im Vollblut von SOD1G93Adl-Mäusen. Angegeben sind
jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (B-Zellanzahl
einer Maus). Das Vollblut der SOD1G93Adl-Mäuse wurde in der präsymptomatischen Phase (ca. 60 Tage),
in der symptomatischen Phase (Beginn der Symptomatik) und im Endstadium (sobald die SOD1G93Adl-
Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten, nachdem sie auf die Seite gelegt
wurden) untersucht. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im
Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin gegen
Alanin an der Stelle 93; dl= delay;
3.2.3.2 T-Zellen im Vollblut
T-Zellen sind Lymphozyten, die zur Gruppe der Leukozyten gehören. Zusammen mit
den B-Zellen bilden sie das adaptive Immunsystem.
Die quantitative FACS-Analyse von T-Zellen im Vollblut diente dem Zweck, den
Einfluss von FTY720 auf T-Zellen im SOD1G93Adl-Mausmodells zu zeigen.
In der präsymptomatischen Phase besitzen unbehandelte transgene SOD1G93Adl-
Kontrolltiere gegenüber Wildtyp-Tieren ca. 37 % mehr T-Zellen.
1,0 mg/kg und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere haben in der
präsymptomatischen Phase ca. 85-92 % weniger T-Zellen als unbehandelte
SOD1G93Adl-Kontrolltiere. Ein signifikanter Effekt von FTY720 bezüglich der
Verminderung der T-Zellanzahl im Vollblut ist unabhängig von der Dosis erkennbar
(p<0,001).

47
In der symptomatischen Phase offenbaren die unbehandelte SOD1G93Adl- Kontrolltiere
gegenüber präsymptomatischen unbehandelten SOD1G93Adl- Kontrolltieren ungefähr
gleich viel T-Zellen.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere besitzen in der
symptomatischen Phase, verglichen mit den unbehandelten SOD1G93Adl-Kontrolltieren,
ca. 88-91 % weniger T-Zellen im Vollblut. Es zeigt sich in der symptomatischen Phase
dosisunabhängig ein signifikanter Effekt von FTY720 in Bezug auf die Verminderung
der T-Zellen im Vollblut (p<0,001).
Im Endstadium weisen die unbehandelten SOD1G93Adl- Kontrolltiere gegenüber den
unbehandelten SOD1G93Adl- Kontrolltieren der symptomatischen Phase ca. 35 %
weniger T-Zellen auf.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere haben im
Endstadium, verglichen mit unbehandelten SOD1G93Adl-Kontrolltieren, ca. 91-93 %
weniger T-Zellen im Vollblut. Ein signifikanter Effekt von FTY720 in Bezug auf die
Verminderung der T-Zellanzahl im Vollblut ist unabhängig von der Dosis erkennbar
(p<0,001).

48
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
T-Z
ell
en
in
%
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
***
***
***
***
***
***
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 9: Quantitative Analyse der T-Zellen im Vollblut von SOD1G93Adl-Mäusen. Angegeben sind
jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (T-Zellanzahl
einer Maus). Das Vollblut der SOD1G93Adl-Mäuse wurde in der präsymptomatischen Phase (ca. 60 Tage),
in der symptomatischen Phase (Beginn der Symptomatik) und im Endstadium (sobald die SOD1G93Adl-
Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten, nachdem sie auf die Seite gelegt
wurden) untersucht. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im
Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin gegen
Alanin an der Stelle 93; dl= delay;
3.2.3.2 CD4+ T-Zellen im Vollblut
CD4 ist ein Glykoprotein auf der Zelloberfläche der T-Zellen. Es handelt sich um einen
Corerezeptor. Er interagiert gemeinsam mit dem T-Zellrezeptor und dem MHC-Klasse-
II Molekül auf Antigen präsentierenden Zellen. Aktivierte CD4+-T-Helferzellen spielen
eine entscheidende Rolle für die Aktivierung (Lizenzierung) reifer,
antigenpräsentierender dendritischer Zellen (Livingstone et al. 2009), und sie spielen
damit eine entscheidende Rolle bei der Vermittlung einer langanhaltenden
Immunantwort und einer ausgeprägten Inflammation. CD4+ T-Zellen gehören zu den

49
T-Zellen. Die quantitativen FACS-Analyse von CD4+ T-Zellen im Vollblut diente dem
Zweck, den Einfluss von FTY720 auf CD4+ T-Zellen im SOD1G93Adl-Mausmodell zu
zeigen.
In der präsymptomatischen Phase haben unbehandelte SOD1G93Adl- Kontrolltiere
gegenüber Wildtyp-Tieren ca. 60 % mehr CD4+-T-Zellen im Vollblut.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere haben in der
präsymptomatischen Phase ca. 94-95 % weniger CD4+-T-Zellen als unbehandelte
SOD1G93Adl-Kontrolltiere. Ein signifikanter Effekt von FTY720 ist bezüglich der
Verminderung der Anzahl an CD4+ T-Zellen im Vollblut, unabhängig von der Dosis,
erkennbar (p<0,001).
In der symptomatischen Phase haben unbehandelte SOD1G93Adl- Kontrolltiere
gegenüber unbehandelten SOD1G93Adl- Kontrolltieren in der präsymptomatischen
Phase ungefähr 57 % weniger CD4+-T-Zellen.
Die 1,0 mg/kg behandelten SOD1G93Adl-Tiere besitzen wie auch die 3,0 mg/kg Tag FTY
720 behandelte SOD1G93Adl-Tiere, verglichen mit unbehandelten SOD1G93Adl-
Kontrolltieren in der symptomatischen Phase, ca. 86-98 % weniger CD4+ T-Zellen. Es
lässt sich ein signifikanter Effekt von FTY720 hinsichtlich der Verminderung der Anzahl
der CD4+-T-Zellen im Vollblut beobachten (p<0,001).
Im Endstadium haben unbehandelte SOD1G93Adl-Kontrolltiere gegenüber
unbehandelten SOD1G93Adl- Kontrolltieren der symptomatischen Phase ca. 40 % mehr
CD4+-T-Zellen im Vollblut.
Die SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, haben im
Endstadium, verglichen mit den unbehandelten SOD1G93Adl-Kontrolltieren, ca. 97 %
weniger CD4+-T-Zellen. Gleiches gilt auch für SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag
FTY720 behandelt wurden. Ein signifikanter Effekt von FTY720 bezüglich der
Verminderung der Anzahl der CD4+-T-Zellen im Vollblut der behandelten SOD1G93Adl-
Tiere ist unabhängig von der Dosis erkennbar (p<0,001).

50
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0C
D4
+-T
-Ly
mp
ho
zy
ten
in
%
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
***
***
***
*** ***
***
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 10: Quantitative Analyse der CD4+ T-Zellen im Vollblut von SOD1G93Adl-Mäusen. Angegeben
sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (CD4+-T-
Zellanzahl einer Maus). Das Vollblut der SOD1G93Adl-Mäuse wurde in der präsymptomatischen Phase
(ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik) und im Endstadium (sobald
die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten, nachdem sie auf die
Seite gelegt wurden) untersucht. Untersuchung des Wirkmechanismus von FTY720 am
Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1,
Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay;
3.2.3.3 CD8+ T-Zellen im Vollblut
CD8 ist ein transmembranes Glykoprotein, das als Co-Rezeptor mit dem T-
Zellrezeptor agiert. Die Aufgabe der CD8+ zytotoxischen T-Zellen ist das Töten von
Krebszellen, von Virus infizierten Zellen oder anderen beschädigten Zellen. Sie
exprimieren den T-Zellrezeptor, der ein spezifisches Antigen erkennen kann. Antigene
werden innerhalb der Zelle durch das MHC-I-Klasse Molekül gebunden und an der
Zelloberfläche präsentiert. Die CD8+ zytotoxischen T-Zellen binden an das MHC-I-
Klasse Molekül, werden aktiviert und üben ihre zytotoxische Wirkung gegenüber der
Zelle aus. Sie stellen eine Subpopulation der T-Zellen dar und werden deshalb durch
FTY720 beeinflusst. Dabei werden nur Lymphozyten beeinflusst, die den CCR7
Retentionsrezeptor tragen.
Die quantitative FACS-Analyse von CD8+-T-Zellen im Vollblut diente dem Zweck, den
Einfluss von FTY720 auf CD8+T-Zellen im SOD1G93Adl-Mausmodell zu zeigen.

51
Unbehandelte SOD1G93Adl- Kontrolltiere haben in der präsymptomatischen Phase
gegenüber Wildtyp-Tieren ca. 10 % mehr CD8+-T-Zellen im Vollblut. 1,0 mg/kg Tag
FTY720 behandelte SOD1G93Adl-Tiere besitzen wie auch die 3,0 mg/kg Tag FTY 720
behandelten SOD1G93Adl-Tiere ca. 96-97 % weniger CD8+-T-Zellen im Vollblut als
unbehandelte SOD1G93Adl-Kontrolltiere. Unabhängig von der Dosis ist ein signifikanter
Effekt von FTY720 bezüglich der Verminderung der Anzahl der CD8+-T-Zellen im
Vollblut der behandelten SOD1G93Adl-Tiere erkennbar (p<0,001).
Unbehandelte SOD1G93Adl-Kontrolltiere der symptomatischen Phase haben gegenüber
unbehandelten SOD1G93Adl- Kontrolltieren der präsymptomatischen Phase ca. 3 %
mehr CD8+-T-Zellen.
1,0 mg/kg und 3,0 mg/kg Tag FTY 720 behandelte SOD1G93Adl-Tiere besitzen in der
symptomatischen Phase, verglichen mit unbehandelten SOD1G93Adl-Kontrolltieren, ca.
92-93 % weniger CD8+ T-Zellen im Vollblut. Hinsichtlich der Verminderung der CD8+-
T-Zellzahl ist unabhängig von der Dosis ein signifikanter Effekt von FTY720 im Vollblut
der behandelten SOD1G93Adl-Tiere erkennbar (p<0,001).
Im Endstadium weisen unbehandelte SOD1G93Adl- Kontrolltiere gegenüber
unbehandelten SOD1G93Adl- Kontrolltieren in der symptomatischen Phase ca. 27 %
weniger CD8+-T-Zellen auf.
Die SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, haben im
Endstadium, verglichen mit den unbehandelten SOD1G93Adl-Kontrolltieren, ca. 96 %
weniger CD4+-T-Zellen. Gleiches gilt auch für SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag
FTY720 behandelt wurden. Hinsichtlich der Verminderung der Anzahl der CD4+-T-
Zellen im Vollblut von behandelten SOD1G93Adl-Tieren lässt sich ein signifikanter Effekt
von FTY720, unabhängig von der Dosis, beobachten (p<0,001).

52
0
5
1 0
1 5
2 0
2 5
3 0C
D8
+-T
-Ly
mp
ho
zy
ten
in
%
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
***
***
***
***
***
***
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 11: Quantitative Analyse der CD8+ T-Zellen im Vollblut von SOD1G93Adl-Mäusen. Angegeben
sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (CD8+-T-
Zellanzahl einer Maus). Das Vollblut der SOD1G93Adl-Mäuse wurde in der präsymptomatischen Phase
(ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik) und im Endstadium (sobald
die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten, nachdem sie auf die
Seite gelegt wurden) untersucht. Untersuchung des Wirkmechanismus von FTY720 am
Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1,
Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay;
3.2.4 Quantitative Analyse von T- und B-Zellen bzw. CD4+- und CD8+-T-Zellen im
Homogenat der Milz
3.2.4.1 B-Zellen im Milzhomogenat
Die B-Zellen sind Lymphozyten, die zur Gruppe der Leukozyten gehören. Sie stellen
zusammen mit den T-Zellen das adaptive Immunsystem dar. Sie können durch T-
Zellen aktiviert werden und differenzieren sich daraufhin zu Plasmazellen oder
Gedächtniszellen.
Die Milz ist das größte, sekundäre lymphatische Organ und spielt eine wichtige Rolle
in der Abwehr von hämatogenen Infektionen. Wie schon in der Einleitung beschrieben,
ist der Aufbau und die Organisation der Zellen in der Milz sehr komplex. Unter anderem
war es durch die Entdeckung von FTY720 möglich, die Prozesse in der Milz besser
erklären zu können. In der Milz zirkulieren die B-Zellen S1PR1-abhängig. FTY720

53
blockiert in seiner aktivierten Form, dem FTY720-Phosphat, als funktioneller
Antagonist den S1P-Rezeptor 1 (Mehling et al. 2011), (Pinschewer et al. 2011).
Dadurch bleiben die B-Zellen in der Milz konstant oder nehmen sogar zu (Brinkmann
2015).
Die quantitative FACS-Analyse von B-Zellen im Homogenat der Milz diente dem
Zweck, den Einfluss von FTY720 auf B-Zellen im SOD1G93Adl-Mausmodell zu zeigen.
Im Vergleich zu Wildtyp-Tieren weisen unbehandelte SOD1G93Adl- Kontrolltiere in der
präsymptomatischen Phase der Erkrankung, d.h. im Alter von ca. 70 Tagen, ca. 40 %
mehr B-Zellen auf. Dieser Unterschied weist keine Signifikanz auf.
präsymptomatische SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt
wurden, haben ca. 4 % mehr B-Zellen im Milzhomogenat als unbehandelte
SOD1G93Adl-Kontrolltiere. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720
behandelt wurden, besitzen im Vergleich zu unbehandelten SOD1G93Adl-Kontrolltieren
sogar ca. 17 % mehr B-Zellen im Milzhomogenat. Ein dosisabhängiger Effekt von
FTY720 zeigt sich in der präsymptomatischen Phase in Bezug auf den Anstieg der B-
Zellen im Milzhomogenat der SOD1G93Adl-Tiere. Die höhere Dosierung offenbart
hinsichtlich des Anstiegs der B-Zellen im Milzhomogenat der SOD1G93Adl-Tiere einen
stärkeren Effekt.
In der symptomatischen Phase zeigen die unbehandelten SOD1G93Adl- Kontrolltiere
gegenüber unbehandelten SOD1G93Adl- Kontrolltieren der präsymptomatischen Phase
ca. 35 % weniger B-Zellen.
1,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere, besitzen in der symptomatischen
Phase im Gegensatz zu unbehandelten SOD1G93Adl-Kontrolltieren ca. 27 % mehr B-
Zellen im Milzhomogenat. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720
behandelt wurden, haben im Vergleich zu unbehandelten SOD1G93Adl-Kontrolltieren ca.
15 % mehr B-Zellen im Milzhomogenat.
In Bezug auf den Anstieg der B-Zellen im Milzhomogenat der SOD1G93Adl-Tiere ist ein
Effekt von FTY720 in der symptomatischen Phase dosisabhängig erkennbar. Im
Gegensatz zu der präsymptomatischen Phase zeigen 1,0 mg/kg Tag FTY720
behandelte SOD1G93Adl-Tiere bezüglich des Anstieg der B-Zellen im Milzhomogenat
der SOD1G93Adl-Tiere einen stärkeren Effekt.

54
Im Endstadium zeigen die unbehandelten SOD1G93Adl-Kontrolltiere ca. 18 % weniger
B-Zellen im Milzhomogenat, verglichen mit symptomatischen unbehandelten
SOD1G93Adl-Kontrolltieren.
Die 1,0 mg/kg behandelte SOD1G93Adl-Tiere besitzen ebenso wie die 3,0 mg/kg Tag
FTY720 behandelten SOD1G93Adl-Tiere ca. 6-10 % weniger B-Zellen im Milzhomogenat
auf. Ein dosisunabhängiger Effekt von FTY720, hinsichtlich der Verminderung der
Anzahl der B-Zellen im Milzhomogenat der behandelten SOD1G93Adl-Tiere, ist nur
schwach ausgeprägt.
Die beobachteten Effekte von FTY720 in Bezug auf die Erhöhung der B-Zellen in der
präsymptomatischen und symptomatischen Phase sowie der Verminderung der B-
Zellen im Endstadium sind jedoch nicht signifikant.
0
1 0
2 0
3 0
4 0
5 0
6 0
B-Z
ell
en
in
%
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 12: Quantitative Analyse der B-Zellen im Milzhomogenat von SOD1G93Adl-Mäusen.
Angegeben sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert
(B-Zellanzahl einer Maus). Das Milzhomogenat der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik)
und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich
aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay;

55
3.2.4.2 T-Zellen im Milzhomogenat
T-Zellen sind Lymphozyten, die zur Gruppe der Leukozyten gehören.
Durch Blockade dieses Rezeptors kommt es durch FTY720 zur Reduktion der T-Zellen
in der Milz (Brinkmann 2015). Die quantitative FACS-Analyse von T-Zellen im
Homogenat der Milz diente dem Zweck, den Einfluss von FTY720 auf T-Zellen im
SOD1G93Adl-Mausmodell zu zeigen.
In der präsymptomatischen Phase besitzen die unbehandelten SOD1G93Adl-
Kontrolltiere gegenüber Wildtyp-Tieren ca. 52 % mehr T-Zellen im Milzhomogenat.
1,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere besitzen wie auch die 3,0 mg/kg
Tag FTY720 behandelten SOD1G93Adl-Tiere ca. 45-50 % weniger T-Zellen als
unbehandelte SOD1G93Adl-Kontrolltiere. Ein signifikanter Effekt von FTY720 ist
hinsichtlich der Abnahme der T-Zellanzahl im Milzhomogenat der behandelten
SOD1G93Adl-Tiere, unabhängig von der Dosis, erkennbar (p<0,001).
Unbehandelte SOD1G93Adl- Kontrolltiere offenbaren in der symptomatischen Phase
gegenüber präsymptomatischen unbehandelten SOD1G93Adl- Kontrolltieren ca. 18 %
weniger T-Zellen.
1,0 mg/kg Tag FTY720 und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere
besitzen in der symptomatischen Phase, verglichen mit den unbehandelten
SOD1G93Adl- Kontrolltieren, ca. 71-75 % weniger T-Zellen im Milzhomogenat.
Hinsichtlich der Verminderung der T-Zellanzahl im Milzhomogenat bei den
behandelten SOD1G93Adl-Tieren lässt sich ein signifikanter Effekt von FTY720
beobachten (p<0,001).
Unbehandelte SOD1G93Adl-Kontrolltiere im Endstadium weisen gegenüber
symptomatischen unbehandelten SOD1G93Adl- Kontrolltieren ca. 23 % weniger T-Zellen
auf.
Die SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, haben im
Endstadium, verglichen mit den unbehandelten SOD1G93Adl-Kontrolltieren, ca. 45 %
weniger T-Zellen. Im Vergleich zu unbehandelten SOD1G93Adl-Kontrolltieren haben im
Endstadium SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, ca.
60 % weniger T-Zellen. In Bezug auf die Verminderung der T-Zellzahl im
Milzhomogenat der behandelten SOD1G93Adl-Tiere ist ein signifikanter Effekt von
FTY720 zu erkennen (p<0,001). Die höhere Dosierung von 3,0 mg/kg Tag FTY720
zeigt in diesem Zusammenhang den etwas stärkeren Effekt.

56
0
1 0
2 0
3 0
4 0
5 0
6 0
T-Z
ell
en
in
%
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0***
******
***
***
***
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 13: Quantitative Analyse der T-Zellen im Milzhomogenat von SOD1G93Adl-Mäusen.
Angegeben sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert
(T-Zellanzahl einer Maus). Das Milzhomogenat der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik)
und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich
aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay;
3.2.4.3 CD4+ T-Zellen im Milzhomogenat
Wie schon ausführlich beschrieben, gehören CD4+-Zellen zu den T-Zellen.
Durch Blockade des S1P-Rezeptors 1 durch FTY720-Phosphat kommt es zur
Reduktion von CD4+-T-Zellen in der Milz (Brinkmann 2015).
Die quantitative FACS-Analyse von CD4+-T-Zellen im Homogenat der Milz diente dem
Zweck, den Einfluss von FTY720 auf CD4+ T-Zellen im SOD1G93Adl-Mausmodell zu
zeigen.
In der präsymptomatischen Phase besitzen unbehandelte SOD1G93Adl- Kontrolltiere
gegenüber Wildtyp-Tieren ca. 46 % mehr CD4+-T-Zellen im Milzhomogenat. Dieser
Unterschied weist keine Signifikanz auf.
In der präsymptomatischen Phase sind bei den 1,0 mg/kg Tag FTY 720 und 3,0 mg/kg
Tag FTY720 behandelten SOD1G93Adl-Tieren ca. 42-47 % weniger CD4+-T-Zellen im

57
Milzhomogenat nachweisbar als bei unbehandelten SOD1G93Adl-Kontrolltieren.
Unabhängig von der Dosis zeigt sich ein Effekt von FTY720 bezüglich der
Verminderung der T-Zellanzahl im Milzhomogenat der behandelten SOD1G93Adl-Tiere.
In der symptomatischen Phase offenbaren die unbehandelten SOD1G93Adl-
Kontrolltiere gegenüber präsymptomatischen unbehandelten SOD1G93Adl-
Kontrolltieren ca. 8 % mehr CD4+-T-Zellen.
SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, sowie 3,0 mg/kg
Tag FTY720 behandelte SOD1G93Adl-Tiere besitzen in der symptomatischen Phase,
verglichen mit den unbehandelten SOD1G93Adl-Kontrolltieren, ca. 68-72 % weniger
CD4+-T-Zellen im Milzhomogenat. In der symptomatischen Phase ist ein signifikanter
Effekt von FTY720 hinsichtlich der Verminderung der T-Zellzahl im Milzhomogenat
erkennbar (p<0,001).
Im Endstadium weisen unbehandelte SOD1G93Adl-Kontrolltiere gegenüber
unbehandelten SOD1G93Adl- Kontrolltieren in der symptomatischen Phase ca. 30 %
weniger CD4+-T-Zellen im Milzhomogenat auf.
1,0 mg/kg Tag FTY720 und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere
haben im Endstadium verglichen mit unbehandelten SOD1G93Adl- Kontrolltieren, ca. 30-
37 % weniger CD4+ T-Zellen im Milzhomogenat. In Bezug auf die Verminderung der
CD4+ T-Zellanzahl lässt sich ein signifikanter Effekt von FTY720 im Milzhomogenat
von behandelten SOD1G93Adl-Tieren beobachten (p<0,001). Die beobachteten Effekte
in der präsymptomatischen Phase sind jedoch nicht signifikant.

58
0
5
1 0
1 5
2 0
2 5
3 0
CD
4+
-T-L
ym
ph
oz
yte
n i
n %
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
***
***
***
***
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 14: Quantitative Analyse der CD4+ T-Zellen im Milzhomogenat von SOD1G93Adl-Mäusen.
Angegeben sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert
(CD4+T-Zellanzahl einer Maus). Das Milzhomogenat der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik)
und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich
aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay; CD4=
cluster of differentiation 4.
3.2.4.4 CD8+ T-Zellen im Milzhomogenat
Wie schon ausführlich beschrieben, stellen die CD8+-T-Zellen eine Subpopulation der
T-Zellen dar und werden deshalb durch FTY720 beeinflusst.
Durch Blockade des S1P-Rezeptors 1 durch FTY720-Phosphat kommt es zur
Reduktion von CD8+ T-Zellen in der Milz (Brinkmann 2015). Die quantitative FACS-
Analyse von CD8+-T-Zellen im Homogenat der Milz diente dem Zweck, den Einfluss
von FTY720 auf CD8+T-Zellen im SOD1G93Adl-Mausmodells zu zeigen.
In der präsymptomatischen Phase haben unbehandelte SOD1G93Adl-Kontrolltiere
gegenüber Wildtyp-Tieren ca. 10 % mehr CD8+-T-Zellen im Milzhomogenat.
Bei den SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, sind in
der präsymptomatischen Phase ca. 41 % weniger CD8+-T-Zellen nachweisbar als bei

59
unbehandelten SOD1G93Adl-Kontrolltieren. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag
FTY720 behandelt wurden, haben im Vergleich zu unbehandelten SOD1G93Adl-
Kontrolltieren ca. 66 % weniger CD8+-T-Zellen im Milzhomogenat. Ein signifikanter
Effekt von FTY720 hinsichtlich der Verminderung der CD8+-T-Zellanzahl im
Milzhomogenat der behandelten SOD1G93Adl-Tiere ist zu erkennen (p<0,001). Die
höhere Dosierung von FTY720 zeigt dabei, in Bezug auf die Verminderung der CD8+-
T-Zellanzahl, einen stärker ausgeprägten Effekt. Dieser ist aber nicht signifikant.
In der symptomatischen Phase offenbaren die unbehandelten SOD1G93Adl-
Kontrolltiere gegenüber präsymptomatischen unbehandelten SOD1G93Adl-
Kontrolltieren ca. 3 % mehr CD8+-T-Zellen.
Die SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, besitzen in
der symptomatischen Phase, verglichen mit den unbehandelten SOD1G93Adl-
Kontrolltieren, ca. 49 % weniger CD8+-T-Zellen im Milzhomogenat. Die SOD1G93Adl-
Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, offenbaren im Gegensatz zu
unbehandelten SOD1G93Adl-Kontrolltieren ca. 66 % weniger CD8+-T-Zellen . In der
symptomatischen Phase zeigen hinsichtlich der Verminderung der CD8+-T-Zellen im
Milzhomogenat nur die 3,0 mg/kg Tag FTY720 behandelten SOD1G93Adl-Tiere einen
signifikanten Effekt von FTY720 (p<0,05). Zudem ist ein Effekt von FTY720 bezüglich
der Verminderung der CD8+ T-Zellen im Milzhomogenat der mit 1,0 mg/kg Tag
FTY720 behandelten SOD1G93Adl-Tiere zu erkennen.
Unbehandelte SOD1G93Adl- Kontrolltiere im Endstadium gegenüber unbehandelten
SOD1G93Adl- Kontrolltieren in der symptomatischen Phase weisen ca. 21 % weniger
CD8+ T-Zellen im Milzhomogenat auf.
Im Endstadium haben SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt
wurden, verglichen mit unbehandelten SOD1G93Adl-Kontrolltieren, ca. 66 % weniger
CD8+ T-Zellen im Milzhomogenat. Im Vergleich zu unbehandelten SOD1G93Adl-
Kontrolltieren offenbaren im Endstadium SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag
FTY720 behandelt wurden, ca. 52 % weniger CD8+-T-Zellen im Milzhomogenat. In
Bezug auf die Verminderung der CD8+-T-Zellen ist ein signifikanter Effekt von FTY720
im Milzhomogenat der behandelten SOD1G93Adl-Tiere erkennbar (p<0,001). Die
niedrigere Dosierung weist im Vergleich zur höheren Dosierung diesbezüglich einen
stärkeren Effekt auf. Die beobachteten dosisabhängigen Effekte in allen drei
Krankheitsstadien sowie die 1,0 mg/kg Tag FTY 720 behandelten SOD1G93Adl-Tiere in
der symptomatischen Phase sind nicht signifikant.

60
0 .0
2 .5
5 .0
7 .5
1 0 .0
1 2 .5
1 5 .0
1 7 .5
2 0 .0
CD
8+
-T-L
ym
ph
oz
yte
n i
n %
P rä s y m p to m a t is c h
S O D 1G 9 3 A d l
M ä u s e
S y m p to m a tis c h
S O D 1G 9 3 A d l
M ä u s e
E n d s ta d iu m
S O D 1G 9 3 A d l
M ä u s e
W T
w ild ty p T ie re
u n b e h a n d e lte K o n tro llg ru p p e
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
***
******
***
*
B e h a n d lu n g s g r u p p e n
(W T : u n b e h a n d e lte W ild ty p t ie re )
Abbildung 15: Quantitative Analyse der CD8+ T-Zellen im Milzhomogenat von SOD1G93Adl-Mäusen.
Angegeben sind jeweils Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert
(CD8+-T-Zellanzahl einer Maus). Das Milzhomogenat der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage), in der symptomatischen Phase (Beginn der Symptomatik)
und im Endstadium (sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich
aufzurichten, nachdem sie auf die Seite gelegt wurden) untersucht. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. SOD1G93A= Punktmutation
in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der Stelle 93; dl= delay; CD8=
cluster of differentiation 8.
3.2.5 Quantitative Analyse von Zytokinen im Blutplasma von SOD1G93Adl-
Tieren
3.2.5.1 IFN-γ im Plasma
IFN-γ ist ein proinflammatorisches Zytokin und wird unter anderem von Tem-Zellen
(Sallusto et al. 1999), B-Zellen, NK-Zellen und APCs sezerniert (Schroder et al. 2003).
FTY720 blockiert in seiner aktivierten Form, dem FTY720-Phosphat, als funktioneller
Antagonist den S1P-Rezeptor 1 (Mehling et al. 2011), (Pinschewer et al. 2011). Durch

61
Blockade dieses Rezeptors kommt es durch FTY720 zur Reduktion von T-Zellen und
B-Zellen im Vollblut (Brinkmann 2015).
Die quantitative Analyse von IFN-γ mittels Sandwich ELISA diente dem Zweck, einen
möglichen Einfluss von FTY720 auf die Neuroinflammation im SOD1G93Adl-Mausmodell
zu zeigen. IFN-γ wurde im Plasma zu acht verschiedenen Zeitpunkten gemessen (40d,
60d, 80d, 100d, 120d, 170-180d, Symptomatische Phase, Endstadium).
Im Alter von 40-120 Tagen zeigen unbehandelte SOD1G93Adl-Kontrolltiere relativ
niedrige IFN-γ Blutplasmaspiegel zwischen 0,23-0,4 pg/ml IFN-γ im Plasma. Die IFN-
γ Blutplasmaspiegel der behandelten SOD1G93Adl-Tiere liegen im gleichen
Konzentrationsbereich wie die unbehandelten SOD1G93Adl-Kontrolltiere, so dass ein
Einfluss von FTY720 nicht erkennbar ist.
Erst im Alter von 170-180 Tagen kommt es im Vergleich zu den 120 Tage alten
unbehandelten SOD1G93Adl-Kontrolltieren zu einem Anstieg des IFN-γ
Blutplasmaspiegels von ungefähr 330% auf eine Blutplasmakonzentration von 3,35
pg/ml.
In der Gegenüberstellung von unbehandelten SOD1G93Adl-Kontrolltieren mit
SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, weisen die
behandelten SOD1G93Adl-Tiere im Alter von 170-180 Tagen ca. 14 % weniger INF-γ im
Plasma auf.
3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere offenbaren im Vergleich zu
unbehandelten SOD1G93Adl-Kontrolltieren in diesem Zeitraum ca. 80 % weniger IFN-γ
im Plasma.
Ein dosisabhängiger Effekt von FTY720 ist hinsichtlich der Senkung der
Blutplasmakonzentration von IFN-γ zum Zeitpunkt von 170-180 Tagen erkennbar. Die
höhere Dosierung von 3,0 mg/kg Tag FTY720 übt dabei einen stärkeren Effekt aus.
Zum Zeitpunkt der symptomatischen Phase kommt es bei den unbehandelten
SOD1G93Adl-Kontrolltieren im Vergleich zu den 170-180 Tagen alten unbehandelten
SOD1G93Adl-Kontrolltieren zu einem Abfall des INF-γ Blutplasmaspiegels um ca. 67%
auf eine Blutplasmakonzentration von 1,11 pg/ml IFN-γ.
SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, haben verglichen
mit unbehandelten SOD1G93Adl-Kontrolltieren in der symptomatischen Phase ca. 35 %
weniger IFN-γ im Plasma. SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt
wurden, haben im Gegensatz zu unbehandelten SOD1G93Adl-Kontrolltieren in der
symptomatischen Phase ca. 3 % weniger IFN-γ im Plasma. Bei den 1,0 mg/kg Tag

62
FTY720 behandelten SOD1G93Adl-Tieren zeigt sich ein Einfluss von FTY720 bezüglich
der Abnahme des IFN-γ Blutplasmaspiegels. In der höheren Dosierung ist kein Effekt
erkennbar.
Unbehandelte transgene SOD1G93Adl-Tiere im Endstadium haben gemittelt 1,05 pg/ml
IFN-γ im Plasma und zeigen damit ca. 6 % weniger IFN-γ im Plasma auf als
unbehandelte SOD1G93Adl-Kontrolltiere in der symptomatischen Phase.
In der Gegenüberstellung von unbehandelten SOD1G93Adl-Kontrolltieren mit
SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, haben die
behandelten SOD1G93Adl-Tiere im Endstadium ca. 67 % weniger INF-γ im Plasma.
SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, weisen im
Vergleich zu unbehandelten transgenen SOD1G93Adl-Tieren im Endstadium ca. 87 %
weniger IFN-γ im Plasma auf. Im Endstadium lässt sich ein dosisabhängiger Effekt von
FTY720 hinsichtlich der Abnahme des IFN-γ Blutplasmaspiegels beobachten.
Dieser Effekt zeigt sich bei den höher dosierten SOD1G93Adl-Tieren stärker ausgeprägt.
Tabelle 3: IFN-γ wurde im Plasma von SOD1G93Adl-Mäusen zu acht verschiedenen Zeitpunkten
gemessen (40d, 60d, 80d, 100d, 120d, 170-180d, Symptomatische Phase, Endstadium).
Der Mittelwert und die Standardabweichung wurde von allen gemessenen Tieren zu einem Zeitpunkt
bestimmt. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014.
SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der
Stelle 93; dl= delay; IFN= Interferon; d= Tag; symptomatischen Phase= Beginn der Symptomatik;
Endstadium= Sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten,
nachdem sie auf die Seite gelegt wurden.
Mittelwert Standardabweichung Mittelwert Standardabweichung Mittelwert Standardabweichung
Zeit in Tagen (d)
unbehandelte
Kontrollgruppe
(pg/ml)
unbehandelte
Kontrollgruppe
(pg/ml)
1,0 mg/kg
Tag FTY720
(pg/ml)
1,0 mg/kg Tag FTY720
(pg/ml)
3,0 mg/kg
Tag FTY720
(pg/ml)
3,0 mg/kg Tag FTY720
(pg/ml)
40d 0.40 0.23 0.42 0.31 0.41 0.23
60d 0.20 0.18 0.54 0.49 0.31 0.17
80d 0.35 0.08 0.26 0.19 0.27 0.19
100d 0.26 0.09 0.48 0.40 0.42 0.28
120d 0.23 0.19 0.30 0.12 0.33 0.08
170-180d 3.35 2.63 2.87 1.68 0.67 0.23
Symptomatische
Phase 1.11 1.11 0.73 0.63 1.08 0.64
Endstadium 1.05 1.03 0.34 0.43 0.13 0.05

63
3.2.5.2 IL-2 im Plasma
IL-2 ist ein proinflammatorisches Zytokin, das unter anderem von Tcm-Zellen
sezerniert wird (Sallusto et al. 1999). FTY720 blockiert in seiner aktivierten Form, dem
FTY720-Phosphat, als funktioneller Antagonist den S1P-Rezeptor 1 (Mehling et al.
2011), (Pinschewer et al. 2011). Durch Blockade dieses Rezeptors kommt es durch
FTY720 zur Reduktion von T-Zellen im Vollblut (Brinkmann 2015).
Die quantitative Analyse von IL-2 im Plasma mittels Sandwich ELISA diente dem
Zweck, einen möglichen Einfluss von FTY720 auf die Neuroinflammation im
SOD1G93Adl-Mausmodell zu zeigen. IL-2 im Plasma wurde zu acht verschiedenen
Zeitpunkten gemessen (40d, 60d, 80d, 100d, 120d, 170-180d, Symptomatische Phase
und Endstadium).
Im Alter von 40-80 Tagen lassen sich bei den unbehandelten SOD1G93Adl-Kontrolltieren
höhere IL-2 Blutplasmaspiegel beobachten als im Alter von 100d bis zum Endstadium.
Im Alter von 40 Tagen haben unbehandelte SOD1G93Adl-Kontrolltiere gemittelt 1,41
pg/ml IL-2 im Blutplasma. Im Vergleich zu SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag
FTY720 behandelt wurden, besitzen die unbehandelten SOD1G93Adl-Kontrolltiere im
Alter von 40 Tagen ca. 27 % weniger IL-2 im Blutplasma. SOD1G93Adl-Tiere, die mit 3,0
mg/kg Tag FTY720 behandelt wurden, weisen im Gegensatz zu unbehandelten
SOD1G93Adl-Kontrolltieren im Alter von 40 Tagen 60 % weniger IL-2 im Blutplasma auf.
Zum Zeitpunkt 40 Tage ist ein dosisabhängiger Effekt von FTY720 in Bezug auf die
Abnahme von IL-2 im Blutplasma von SOD1G93Adl-Tieren zu erkennen. Die Dosierung
von 3,0 mg/kg Tag FTY720 übt diesbezüglich einen stärkeren Einfluss aus.
Die unbehandelten SOD1G93Adl-Kontrolltiere im Alter von 60 Tagen haben mit 1,46
pg/ml im Vergleich zu 40 Tage alten unbehandelten SOD1G93Adl-Kontrolltieren
ungefähr gleich viel IL-2 im Blutplasma. 60 Tage alte SOD1G93Adl-Tiere, die mit 1,0
mg/kg Tag FTY720 behandelt wurden, weisen, verglichen mit unbehandelten
SOD1G93Adl-Kontrolltieren, ca. 45 % weniger IL-2 im Blutplasma auf. SOD1G93Adl-Tiere,
die mit 3,0 mg/kg Tag FTY720 behandelt wurden, besitzen gegenüber unbehandelten
SOD1G93Adl-Kontrolltieren im Alter von 60 Tagen ca. 75 % weniger IL-2 im Blutplasma.
Bezüglich der Abnahme von IL-2 im Blutplasma ist ein dosisabhängiger Effekt von
FTY720 erkennbar, bei der die höhere Dosierung den stärkeren Effekt ausübt.
Im Alter von 80 Tagen fällt der IL-2 Blutplasmaspiegel von unbehandelten SOD1G93Adl-
Kontrolltieren verglichen mit unbehandelten SOD1G93Adl-Kontrolltieren im Alter von 60

64
Tagen um ca. 17 %. Die 80 Tage alten Tiere zeigen eine IL-2 Blutplasmakonzentration
von ca. 1,21 pg/ml.
SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, haben verglichen
mit unbehandelten SOD1G93Adl-Kontrolltieren im Alter von 80 Tagen ca. 66 % weniger
IL-2 im Blutplasma. 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere besitzen im
Vergleich zu unbehandelten SOD1G93Adl-Kontrolltieren im Alter von 80 Tagen ca. 91
% weniger IL-2 im Plasma. Es stellt sich ein dosisabhängiger Effekt von FTY720
hinsichtlich der Abnahme der IL-2 Blutplasmakonzentration dar. Auch hier zeigt die
höhere Dosierung den stärker ausgeprägten Effekt.
Im Alter von 100 Tagen fällt der IL-2 Blutplasmaspiegel bei unbehandelten SOD1G93Adl-
Kontrolltieren im Vergleich zu 80 Tage alten unbehandelten SOD1G93Adl-Kontrolltieren
um bis zu 57 % auf eine IL-2 Blutplasmakonzentration von 0,52 pg/ml.
In der Gegenüberstellung von unbehandelten SOD1G93Adl-Kontrolltieren mit
SOD1G93Adl-Tieren, die mit 1,0 mg/kg Tag FTY720 behandelt wurden, weisen die
behandelten SOD1G93Adl-Tiere ca. 14 % weniger IL-2 im Plasma auf.
SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, offenbaren
verglichen mit unbehandelten SOD1G93Adl-Kontrolltieren, im Alter von 100 Tagen ca.
83 % weniger IL-2 im Blutplasma.
Auch hier lässt sich hinsichtlich der Abnahme der IL-2 Blutplasmakonzentration ein
dosisabhängiger Effekt von FTY720 beobachten. Die höhere Dosierung übt auch hier
den stärkeren Effekt aus.
Zu den Zeitpunkten 120 Tage, 170-180 Tage und dem Zeitpunkt der symptomatischen
Phase bewegen sich die IL-2 Blutplasmakonzentrationen von unbehandelten
SOD1G93Adl-Kontrolltieren auf einem niedrigen Niveau zwischen 0,22 und 0,54 pg/ml.
1,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere offenbaren zu diesen Zeitpunkten
generell höhere IL-2 Blutplasmaspiegel im Vergleich zu unbehandelten SOD1G93Adl-
Kontrolltieren. Ein Effekt von FTY720 in Bezug auf die Abnahme der IL-2
Blutplasmakonzentration ist deshalb nicht erkennbar.
Zu den Zeitpunkten 120 Tage, 170-180 Tage und zum Zeitpunkt der symptomatischen
Phase zeigt sich bei den 3,0 mg/kg Tag FTY720 behandelten SOD1G93Adl-Tieren im
Vergleich zu 1,0 mg/kg Tag FTY720 behandelten SOD1G93Adl-Tieren ein
gegensätzliches Verhalten in Bezug auf die Änderung der IL-2
Blutplasmakonzentration. Die mit der geringen Dosis behandelten SOD1G93Adl-Tiere

65
zeigten zu diesen Zeitpunkten generell höhere Blutplasmaspiegel im Vergleich zu
unbehandelten SOD1G93Adl-Kontrolltieren.
SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, haben im
Gegensatz zu unbehandelten SOD1G93Adl-Kontrolltieren im Alter von 120 Tagen ca. 58
% weniger IL-2 im Plasma.
SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, zeigen verglichen
mit unbehandelten SOD1G93Adl-Kontrolltieren, im Alter von 170-180 Tagen eine
Abnahme von IL-2 im Blutplasma von ca. 32 %. In der symptomatischen Phase haben
SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt wurden, im Gegensatz zu
unbehandelten SOD1G93Adl-Kontrolltieren in der symptomatischen Phase ca. 97 %
weniger IL-2 im Blutplasma.
Unbehandelte SOD1G93Adl-Kontrolltiere im Endstadium haben gemittelt 0,58 pg/ml IL-
2 im Plasma. Sie besitzen damit im Vergleich zu unbehandelten SOD1G93Adl-
Kontrolltieren in der symptomatischen Phase ca. 164 % mehr IL-2 im Blutplasma.
1,0 mg/kg Tag FTY720 und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere im
Endstadium weisen verglichen mit unbehandelte SOD1G93Adl-Kontrolltieren ca. 64-
90% niedrigere IL-2 Blutplasmaspiegel auf.
Ein Effekt von FTY720 ist hinsichtlich der Abnahme der IL-2 Blutplasmakonzentration
bei den behandelten SOD1G93Adl-Tieren im Endstadium zu erkennen.

66
Tabelle 4: IL-2 wurde im Plasma von SOD1G93Adl-Mäusen zu acht verschiedenen Zeitpunkten gemessen
(40d, 60d, 80d, 100d, 120d, 170-180d, Symptomatische Phase, Endstadium).
Der Mittelwert und die Standardabweichung wurde von allen gemessenen Tieren zu einem Zeitpunkt
bestimmt. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014.
SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der
Stelle 93; dl= delay; IL= Interleukin; d= Tag; symptomatischen Phase= Beginn der Symptomatik;
Endstadium= Sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten,
nachdem sie auf die Seite gelegt wurden.
3.2.5.3 IL-4 im Plasma
IL-4 ist ein antiinflammatorisches Zytokin, welches vor allem von Th2-Zellen sezerniert
wird. Es hemmt Th1-Zellen und Makrophagen sowie die Sekretion von IFN-γ und IL-
12.
Die quantitative Analyse von IL-2 im Plasma mittels Sandwich ELISA diente dem
Zweck, einen möglichen Einfluss von FTY720 auf die Neuroinflammation im
SOD1G93Adl-Mausmodell zu zeigen.
IL-4 wurde im Plasma zu acht verschiedenen Zeitpunkten gemessen (40d, 60d, 80d,
100d, 120d, 170-180d, Symptomatische Phase und Endstadium).
Hinsichtlich der IL-4 Blutplasmakonzentrationen stellen sich im Verlauf der Erkrankung
des SOD1G93Adl-Mausmodells bei den unbehandelten SOD1G93Adl-Kontrolltieren zwei
Konzentrationsgipfel dar. Zum einen zeigen sie sich zu den Zeitpunkten 40d, 60d und
80d, bei dem sich Blutplasmakonzentrationen von 0,41-0,56 pg/ml beobachten lassen
und zum anderen zum Zeitpunkt 170-180 Tage, bei dem eine Blutplasmakonzentration
von 0,79 pg/ml erreicht wird.
Im Alter von 40 Tagen besitzen unbehandelte SOD1G93Adl-Kontrolltiere gemittelt 0,56
pg/ml IL-4 im Blutplasma. SOD1G93Adl-Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt
wurden, haben im Vergleich zu unbehandelten SOD1G93Adl-Kontrolltieren im Alter von
Mittelwert Standardabweichung Mittelwert Standardabweichung Mittelwert Standardabweichung
Zeit in Tagen (d)
unbehandelte
Kontrollgruppe
(pg/ml)
unbehandelte
Kontrollgruppe
(pg/ml)
1,0 mg/kg
Tag FTY720
(pg/ml)
1,0 mg/kg Tag FTY720
(pg/ml)
3,0 mg/kg
Tag FTY720
(pg/ml)
3,0 mg/kg Tag FTY720
(pg/ml)
40d 1.41 0.62 1.03 0.70 0.56 0.13
60d 1.46 0.27 0.81 0.62 0.37 0.13
80d 1.21 0.67 0.41 0.51 0.11 0.06
100d 0.52 0.32 0.44 0.33 0.09 0.07
120d 0.48 0.39 0.87 0.55 0.20 0.04
170-180d 0.54 0.39 0.75 0.15 0.37 0.23
Symptomatische
Phase 0.22 0.23 0.82 0.82 0.01 0.01
Endstadium 0.58 0.79 0.06 0.09 0.21 0.15

67
40 Tagen ca. 58 % weniger IL-4 im Plasma. SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag
FTY720 behandelt wurden, weisen zu unbehandelten SOD1G93Adl-Kontrolltieren im
Alter von 40 Tagen ungefähr gleich viel IL-4 im Blutplasma auf. Bezüglich der Abnahme
von IL-4 zeigt sich ein Effekt bei den 1,0 mg/kg Tag FTY720 behandelten SOD1G93Adl-
Tieren. In der höheren Dosierung ist kein Effekt erkennbar.
Im Alter von 60 Tagen haben unbehandelte SOD1G93Adl-Kontrolltiere gemittelt 0,54
pg/ml IL-4 im Blutplasma. 1,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere wie
auch 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere weisen, verglichen mit
unbehandelten SOD1G93Adl-Kontrolltieren, im Alter von 60 Tagen ca. 38-39 % weniger
IL-4 im Blutplasma auf. Ein Einfluss von FTY720, hinsichtlich der Abnahme von IL-4
im Blutplasma von behandelten SOD1G93Adl-Tieren, lässt sich unabhängig von der
Dosis beobachten.
80 Tage alte unbehandelte SOD1G93Adl-Kontrolltiere zeigen gemittelt 0,41 pg/ml IL-4
im Blutplasma.
Sie offenbaren damit ca. 24 % weniger IL-4 im Blutplasma als im Alter von 60 Tagen.
1,0 mg/kg Tag FTY720 und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere
offenbaren, verglichen mit unbehandelten SOD1G93Adl-Kontrolltieren im Alter von 80
Tagen, ca. 54-61 % weniger IL-4 im Blutplasma. Somit lässt sich ein
dosisunabhängiger Effekt in Bezug auf die Abnahme der IL-4 Konzentration zum
Zeitpunkt 80 Tage darstellen.
Zum Zeitpunkt 100 Tage kommt es bei den unbehandelten SOD1G93Adl-Kontrolltieren
zu einem Abfall der IL-4 Blutplasmakonzentration von 57 % auf einen
Blutplasmaspiegel von 0,18 pg/ml. Im Alter von 120 Tagen fallen die IL-4
Blutplasmakonzentrationen von unbehandelten SOD1G93Adl-Kontrolltieren auf einen
Tiefpunkt von 0,10 pg/ml.
Zu diesen Zeitpunkten ist ein Effekt von FTY720 hinsichtlich der Veränderung des IL-
4 Blutplasmaspiegels aufgrund des sehr niedrigen Niveaus der
Blutplasmakonzentrationen nicht nachvollziehbar.
Der zweite Gipfel der IL-4 Blutplasmakonzentration von unbehandelten SOD1G93Adl-
Kontrolltieren offenbart sich im Alter von 170-180 Tagen mit einem Wert von 0,79 pg/ml
und weist damit einen um ca. 665 % höheren IL-4 Blutplasmaspiegel auf als bei 120
Tage alten unbehandelten SOD1G93Adl-Kontrolltieren.
1,0 mg/kg Tag FTY720 und 3,0 mg/kg Tag FTY720 behandelte SOD1G93Adl-Tiere
besitzen im Alter von 170-180 Tagen ca. 51-55 % weniger IL-4 im Blutplasma.

68
Ein dosisunabhängiger Einfluss von FTY720 ist in Bezug auf die Abnahme der IL-4
Blutplasmakonzentration zu beobachten.
Zu den Zeitpunkten der symptomatischen Phase sowie des Endstadiums fällt die IL-4
Blutplasmakonzentration von unbehandelten SOD1G93Adl-Kontrolltieren erneut auf ein
niedriges Niveau von 0,10-0,28 pg/ml. Aufgrund der niedrigen IL-4
Blutplasmakonzentrationen kann ein Effekt von FTY720 hinsichtlich der Veränderung
des IL-4 Blutplasmaspiegels nicht nachvollzogen werden.
Tabelle 5: IL-4 wurde im Plasma von SOD1G93Adl-Mäusen zu acht verschiedenen Zeitpunkten gemessen
(40d, 60d, 80d, 100d, 120d, 170-180d, Symptomatische Phase, Endstadium).
Der Mittelwert und die Standardabweichung wurde von allen gemessenen Tieren zu einem Zeitpunkt
bestimmt. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014.
SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin gegen Alanin an der
Stelle 93; dl= delay; IL= Interleukin; d= Tag; symptomatischen Phase= Beginn der Symptomatik;
Endstadium= Sobald die SOD1G93Adl-Mäuse nicht mehr in der Lage sind, sich unverzüglich aufzurichten,
nachdem sie auf die Seite gelegt wurden.
Mittelwert Standardabweichung Mittelwert Standardabweichung Mittelwert Standardabweichung
Zeit in Tagen (d)
unbehandelte
Kontrollgruppe
(pg/ml)
unbehandelte
Kontrollgruppe
(pg/ml)
1,0 mg/kg
Tag FTY720
(pg/ml)
1,0 mg/kg Tag FTY720
(pg/ml)
3,0 mg/kg
Tag FTY720
(pg/ml)
3,0 mg/kg Tag FTY720
(pg/ml)
40d 0.56 0.21 0.23 0.23 0.58 0.34
60d 0.54 0.41 0.34 0.22 0.33 0.28
80d 0.41 0.26 0.16 0.20 0.19 0.22
100d 0.18 0.21 0.19 0.30 0.00 0.00
120d 0.10 0.17 0.26 0.18 0.07 0.04
170-180d 0.79 0.55 0.39 0.20 0.36 0.50
Symptomatische
Phase 0.10 0.15 0.02 0.02 0.25 0.19
Endstadium 0.28 0.15 0.01 0.03 0.18 0.14
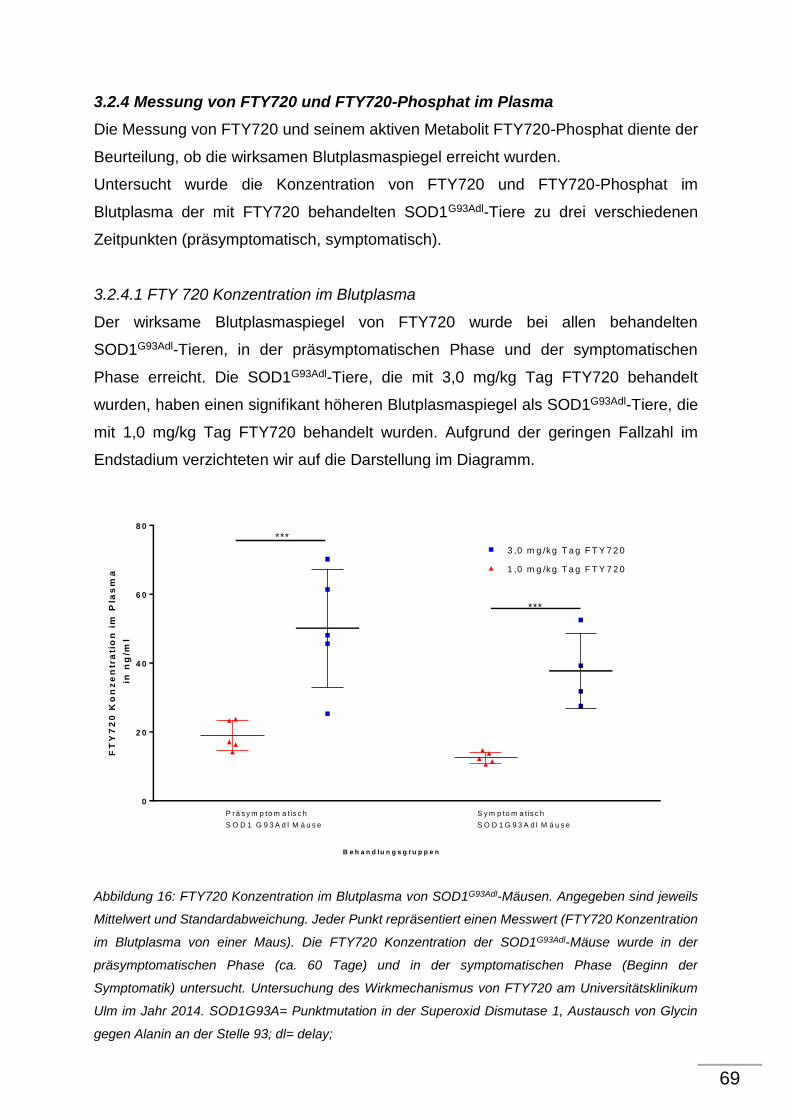
69
3.2.4 Messung von FTY720 und FTY720-Phosphat im Plasma
Die Messung von FTY720 und seinem aktiven Metabolit FTY720-Phosphat diente der
Beurteilung, ob die wirksamen Blutplasmaspiegel erreicht wurden.
Untersucht wurde die Konzentration von FTY720 und FTY720-Phosphat im
Blutplasma der mit FTY720 behandelten SOD1G93Adl-Tiere zu drei verschiedenen
Zeitpunkten (präsymptomatisch, symptomatisch).
3.2.4.1 FTY 720 Konzentration im Blutplasma
Der wirksame Blutplasmaspiegel von FTY720 wurde bei allen behandelten
SOD1G93Adl-Tieren, in der präsymptomatischen Phase und der symptomatischen
Phase erreicht. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720 behandelt
wurden, haben einen signifikant höheren Blutplasmaspiegel als SOD1G93Adl-Tiere, die
mit 1,0 mg/kg Tag FTY720 behandelt wurden. Aufgrund der geringen Fallzahl im
Endstadium verzichteten wir auf die Darstellung im Diagramm.
0
2 0
4 0
6 0
8 0
FT
Y7
20
Ko
nz
en
tra
tio
n i
m P
las
ma
in
ng
/ml
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1 G 9 3 A d l M ä u s e
S y m p to m a tis c h
S O D 1 G 9 3 A d l M ä u s e
***
***
B e h a n d lu n g s g r u p p e n
Abbildung 16: FTY720 Konzentration im Blutplasma von SOD1G93Adl-Mäusen. Angegeben sind jeweils
Mittelwert und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (FTY720 Konzentration
im Blutplasma von einer Maus). Die FTY720 Konzentration der SOD1G93Adl-Mäuse wurde in der
präsymptomatischen Phase (ca. 60 Tage) und in der symptomatischen Phase (Beginn der
Symptomatik) untersucht. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum
Ulm im Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin
gegen Alanin an der Stelle 93; dl= delay;
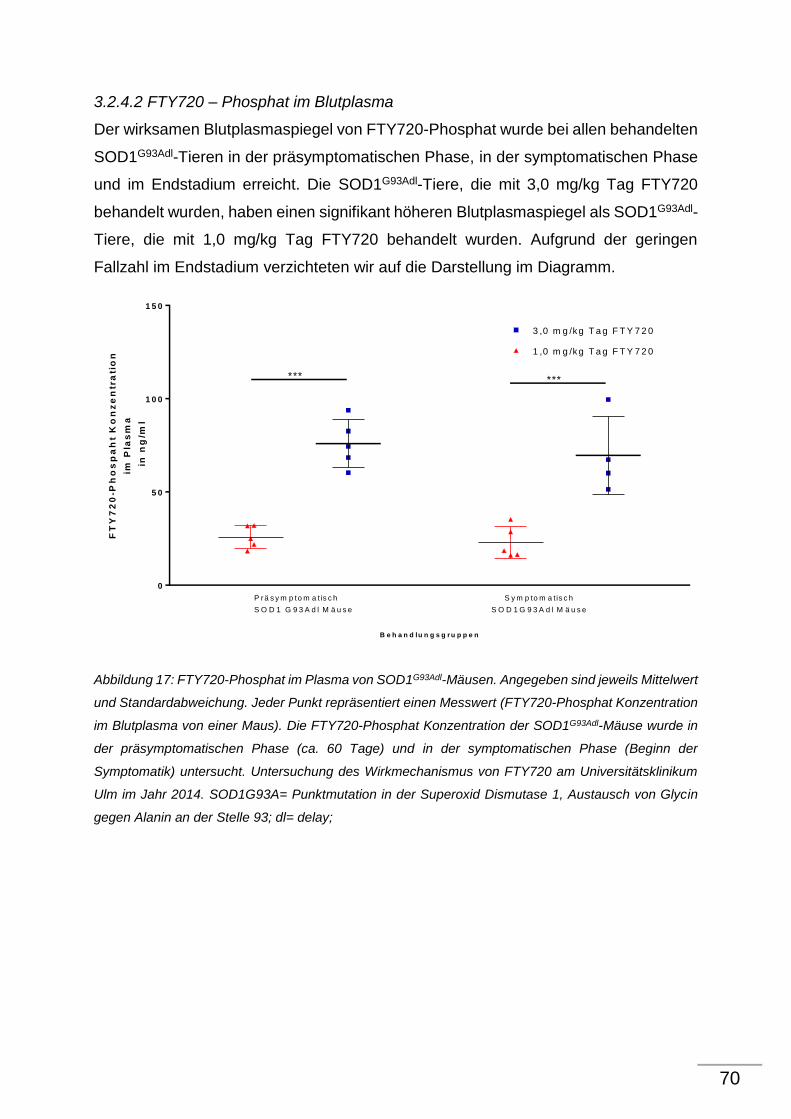
70
3.2.4.2 FTY720 – Phosphat im Blutplasma
Der wirksamen Blutplasmaspiegel von FTY720-Phosphat wurde bei allen behandelten
SOD1G93Adl-Tieren in der präsymptomatischen Phase, in der symptomatischen Phase
und im Endstadium erreicht. Die SOD1G93Adl-Tiere, die mit 3,0 mg/kg Tag FTY720
behandelt wurden, haben einen signifikant höheren Blutplasmaspiegel als SOD1G93Adl-
Tiere, die mit 1,0 mg/kg Tag FTY720 behandelt wurden. Aufgrund der geringen
Fallzahl im Endstadium verzichteten wir auf die Darstellung im Diagramm.
Abbildung 17: FTY720-Phosphat im Plasma von SOD1G93Adl-Mäusen. Angegeben sind jeweils Mittelwert
und Standardabweichung. Jeder Punkt repräsentiert einen Messwert (FTY720-Phosphat Konzentration
im Blutplasma von einer Maus). Die FTY720-Phosphat Konzentration der SOD1G93Adl-Mäuse wurde in
der präsymptomatischen Phase (ca. 60 Tage) und in der symptomatischen Phase (Beginn der
Symptomatik) untersucht. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum
Ulm im Jahr 2014. SOD1G93A= Punktmutation in der Superoxid Dismutase 1, Austausch von Glycin
gegen Alanin an der Stelle 93; dl= delay;
0
5 0
1 0 0
1 5 0
FT
Y7
20
-Ph
os
pa
ht
Ko
nz
en
tra
tio
n
im P
las
ma
in
ng
/ml
1 ,0 m g /k g T a g F T Y 7 2 0
3 ,0 m g /k g T a g F T Y 7 2 0
P rä s y m p to m a t is c h
S O D 1 G 9 3 A d l M ä u s e
S y m p to m a tis c h
S O D 1 G 9 3 A d l M ä u s e
B e h a n d lu n g s g r u p p e n
******

71
4 Diskussion
4.1 Allgemeine Bemerkung zur Diskussion
Das Präparat FTY720 wird unter dem Handelsnamen Gilenya® weltweit zur
verlaufsmodifizierenden Langzeitbehandlung der Multiplen Sklerose eingesetzt (Scott
2011).
Die anti-inflammatorische Wirkung des Immunmodulators FTY720 wurde bereits in
zahlreichen Publikationen belegt. Eine direkte neuroprotektive Wirkung von FTY720
wird vermutet, ist allerdings bisher nur unzureichend untersucht worden (Soliven et al.
2011).
Aus diesem Grund wurde das primär neurodegenerative ALS-SOD1G93Adl
Mausmodell zur Aufklärung des angenommenen neuroprotektiven Wirkmechanismus
von FTY720 herangezogen.
Die SOD1G93Adl-Maus stellt derzeit das am besten etablierte und erforschte
Mausmodell dar (Gurney et al. 1994). Die experimentellen Voraussetzungen und
Bedingungen stimmen mit der humanen ALS soweit als derzeit möglich überein
(Stieber et al. 2000). Es gibt gegenwärtig keinen kurativen Therapieansatz im Rahmen
der ALS. Durch das Präparat Riluzol (NMDA-Rezeptorantagonist) kann das Überleben
der betroffenen Patienten lediglich um 3-6 Monate verlängert werden
(Nirmalananthan et al. 2005), so dass wirksamere Neuroprotektiva dringend benötigt
werden.
In unserer Studie setzten wir FTY720 in den Dosierungen 1,0 mg/kg Tag und 3,0 mg/kg
Tag ein.
Unsere Studie befasste sich mit folgenden Fragestellungen:
• Hat FTY720, unabhängig von der antiinflammatorischen Wirkung, eine direkte
neuroprotektive Wirkung?
• Welchen Einfluss hat die Dosis auf die Wirkung von FTY720?
• Welchen Einfluss hat das Medikament auf den Eintritt in die symptomatische Phase
der SOD1 G93A Mäuse?
• Welchen Einfluss hat FTY720 auf die Neuroinflammation im Rahmen der ALS?

72
4.2 Effekt von FTY720 auf α-Motoneurone von SOD1G93Adl-Tieren
4.2.1 FTY720 hat einen gering ausgeprägten direkten neuroprotektiven Effekt auf
die α-Motoneurone von SOD1G93Adl-Tieren.
In der vorliegenden Studie ist ein gering ausgeprägter neuroprotektiver Effekt von
FTY720 auf die alpha-Motoneurone der SOD1G93Adl-Tiere feststellbar. Dieser zeigt sich
in der präsymptomatischen Phase sowohl in der niedrigen als auch in der höheren
Dosierung (1,0 mg/kg Tag und 3,0 mg/kg Tag FTY720) mit einer positiven Dosis-
Wirkungs-Beziehung. In der symptomatischen Phase der Erkrankung ist der gering
ausgeprägte neuroprotektive Effekt von FTY720 lediglich in der niedrigen Dosierung
von 1,0 mg/kg Tag FTY720 zu erkennen. Bei symptomatischen Tieren, die mit 3,0
mg/kg Tag FTY720 behandelt wurden, ist die Anzahl der Motoneurone vergleichbar
mit der Motoneuronenanzahl bei unbehandelten SOD1G93Adl-Tieren. Dies könnte
eventuell ein Hinweis auf einen neurotoxischen Effekt der höheren Dosierung sein.
Im Endstadium ist ein neuroprotektiver Effekt von FTY720 in keiner Dosis erkennbar.
Die beobachteten neuroprotektiven Effekte sind allerdings sowohl in der
präsymptomatischen Phase (beide Dosierungen) als auch in der symptomatischen
(niedrige Dosis) nicht signifikant.
Die Ergebnisse im Endstadium haben aufgrund der niedrigen Fallzahl von 2-3 Tieren
nur beschränkte Aussagekraft. Die beschriebene Degeneration der α-Motoneurone im
SOD1G93Adl-Mausmodell kann auch in unserer Studie anhand der in den drei Stadien
(präsymptomatisch, symptomatisch, Endstadium) untersuchten unbehandelten
SOD1G93Adl-Kontrolltiere klar nachvollzogen werden.
In einigen früheren Studien gibt es ebenfalls Hinweise auf eine direkte neuroprotektive
Wirkung von FTY720.
Doi Y et al. beobachteten in vitro anhand von kortikalen NeuN-positiven Neuronen aus
Mausembryonen vom Typ C57BL/6J, die mit dem neurotoxischen Amyloid β stimuliert
wurden, dass FTY720-Phosphat über eine Erhöhung der Expression des BNDF eine
Reduzierung der A-β induzierten Neurotoxizität erreicht. Die A-β Neurotoxizität ist ein
pathologisches Merkmal des M. Alzheimer.

73
Auch der M. Alzheimer ist eine neurodegenerative Erkrankung und zeigt unter
anderem mit der Degeneration von Neuronen und einer eher gering ausgeprägten
Neuroinflammation einige Parallelen zur ALS und dem SOD1G93Adl-Mausmodell.
Zudem stellten Doi Y et al. fest, dass durch Bindung des BNDF (Brain derived
neutrophic factor) an den TrKB Rezeptor die sogenannte ERK ½ (extracellular-signal-
regulated kinases) Signalkaskade aktiviert wird und damit eine FTY720-Phosphat
vermittelte Neuroprotektivität bezüglich einer Verminderung der Aβ induzierten
Neurotoxizität vermittelt wird. Der BNDF und sein Rezeptor TrkB werden im ZNS weit
verbreitet exprimiert (Conner et al. 1997), (Yan et al. 1997), (Yan et al. 1997).
Sie zeigten in ihrer Studie auch, dass alle Neurone im ZNS S1P Rezeptoren
exprimieren. Dies macht einen direkten Einfluss von FTY720-Phosphat auf Neurone
wahrscheinlich (Doi et al. 2012).
Pardo A et al. konnten einen neuroprotektiven Effekt von FTY720 in vitro und in vivo
in einem Mausmodell des Morbus Huntingtons darstellen. Die Huntington Erkrankung
gehört, wie auch die ALS, zu den neurodegenerativen Erkrankungen. Sie zeigten in
vitro, dass FTY720 die Apoptose in Zelllinien des M. Huntington signifikant reduziert.
In vivo beobachteten sie die FTY720 induzierte Phosphorylierung von AKT und ERK.
Phosphoryliertes AKT und ERK fördern die Aktivierung von Signalwegen die das
Überleben der Zellen vermitteln.
Zudem offenbarte sich in vivo anhand von FTY720 behandelten HD R 6/2 Mäusen
(Mausmodell des M. Huntington) eine verbesserte motorische Funktion und eine
verringerte Degeneration an striatalen Zellen.
Wie auch schon Doi Y et al in ihrem Versuchsmodell einen Anstieg des BNDF Spiegels
durch FTY720 zeigen konnten, beobachtete dies auch Pardo A et al. bei HD R 6/2
Mäusen. In Kombination mit den beobachteten Effekten in vivo vermuten sie, dass der
neuroprotektive Effekt durch das Neutrophin BNDF vermittelt wird (Pardo et al. 2014).
Wie schon in der Einleitung beschrieben, gibt es einige Hypothesen zur Entstehung
der ALS. Dabei nimmt die Excitoxizität einen wichtigen Stellenwert ein. Das liegt vor
allem daran, dass Riluzol das erste Medikament ist, das gegen die ALS eingesetzt
wird. Es stellt sich die Frage, ob auch FTY720 Einfluss auf die Excitoxizität hat.
Menna L et al. zeigten in vitro, dass FTY720 und sein aktiver Metabolit (FTY 720-
Phosphat) kortikale Nervenzellen gegen den Tod durch die Excitoxizität schützen.
Genauso wie Pardo A et al. beobachteten sie, dass FTY720 die neuroprotektiven

74
Signalwege durch die Phosphorylierung von AKT und ERK aktiviert (Di Menna et al.
2013).
Capriani et al. konnten sowohl in vitro als auch in vivo einen neuroprotektiven Effekt
von FTY720 hinsichtlich der Reduktion von Excitoxizität durch FTY720 aufzeigen. Dies
konnte bei niedrigen Dosierungen von 10nM und 100nM beobachtet werden. Bei der
höheren Dosierung von 1000nM offenbarte sich ein leichter toxischer Effekt
(Cipriani et al. 2015).
Das SOD1G93Adl-Mausmodell ist wie auch die ALS vorwiegend eine neurodegenerative
Erkrankung, weist aber auch eine gering ausgeprägte Neuroinflammation auf. Deshalb
ist es möglich, dass die geringe Neuroprotektivität die FTY720 in unserer Studie zu
vermitteln scheint, durch die antiinflammatorische Wirkung des Medikaments zustande
kommt (Gao et al. 2012).
Die Vergleichbarkeit unterschiedlicher Mausmodelle bezüglich der direkten
Neuroprotektivität ist nur eingeschränkt möglich. Es handelt sich zum einen um andere
Pathomechanismen und zum anderen spielt die Neuroinflammation eine
unterschiedlich ausgeprägte Rolle.
In der vorliegenden Studie beschreiben wir einen gering ausgeprägten direkten
neuroprotektiven Effekt von FTY720 anhand von SOD1G93Adl-Tieren insbesondere in
der präsymptomatischen Phase. In der Vergangenheit konnte ein neuroprotektiver
Effekt von FTY720 in anderen Mausmodellen aufgezeigt werden.
4.2.2 FTY720 geht über die Bluthirnschranke
Die wirksamen Blutplasmaspiegel von FTY720 wurden bei den behandelten
SOD1G93Adl-Tieren in allen drei Krankheitsstadien erreicht. Zudem stellten wir fest,
dass FTY720 dosisabhängig über die Bluthirnschranke gelangt. Schon Meno-Tetang
G et al. beobachteten in ihrer Studie anhand von Ratten, dass FTY720 leicht über die
Bluthirnschranke diffundiert (Meno-Tetang et al. 2006). Bei der Anfertigung des
Hirnhomogenats war eine Trennung von Hirnparenchym und Hirngefäßen aufgrund
der sehr kleinen Strukturen nicht möglich. Dies hat zur Folge, dass sich zu den Zellen
des ZNS auch Endothelzellen im Hirnhomogenat befanden. Da FTY720-Phosphat
auch an S1P-Rezeptoren auf dem Endothel von Gefäßen bindet, muss man sich
vergewissern, dass die FTY720-Phosphat Konzentration im Hirnhomogenat nicht
ganz derjenigen im ZNS der SOD1G93Adl-Tiere entspricht.

75
Im Hinblick auf den Nachweis von FTY720 und FTY720-Phosphat im Hirnhomogenat
der SOD1G93Adl-Tiere ist es möglich, dass FTY720 direkte neuroprotektive Effekte an
α-Motoneuronen ausübt.
4.2.3 FTY720 hat keinen Einfluss auf den Krankheitsbeginn von SOD1G93Adl-
Tieren
FTY720 hat in der vorliegenden Studie keinen Einfluss auf den Krankheitsbeginn von
SOD1G93Adl-Tieren.
Mit dem high copy SOD1G93A-Mausmodell untersuchten wir in einer unserer früheren
nicht publizierten Studien (TVA-Reg 865) den Einfluss von FTY720 hinsichtlich des
Krankheitsbeginns und des Überlebens der SOD1G93A-Tiere.
Auch hier wurden die folgenden Dosierungen 1,0 und 3,0 mg/kg Tag an FTY720 oral,
über den gesamten Applikationszeitraum, mittels Schlundsonde verabreicht. In beiden
Dosierungen wurden die wirksamen Blutplasmaspiegel erreicht. Es konnte dennoch
kein signifikanter lebensverlängernder Effekt der Substanz nachgewiesen werden.
Jedoch konnte bei einer Dosis von 1,0 mg/kg Tag FTY720 in der weiblichen
Untergruppe ein verzögerter Krankheitsbeginn von 5 Tagen im Vergleich zur
unbehandelten Kontrollgruppe festgestellt werden. Dieser positive Effekt blieb im
weiteren Krankheitsverlauf bestehen. Eine um 50% verminderte maximale Aktivität im
Laufrad zeigten diese Tiere erst im Alter von 110 Tagen und damit 4 Tage später als
die unbehandelten Kontrolltiere.
In der Literatur gibt es keine Anhaltspunkte für Effekte von FTY720 hinsichtlich der
Verzögerung des Krankheitsbeginns. Jedoch gibt es eine Studie, die den Einfluss von
FTY720 auf das Überleben untersuchte.
Anhand von HD R 6/2 Mäusen beobachtete Pardo A et al. bei den 1,0 mg/kg Tag
FTY720 behandelten Tieren einen Überlebensvorteil (Pardo et al. 2014).
Die hier vorliegende Studie wurde nicht dafür ausgelegt den Effekt von FTY720
bezüglich einer Verzögerung des Krankheitsbeginns zu untersuchen. Für dieses
Vorhaben hätte man mit anderen Fallzahlen planen müssen.

76
4.2.4 Der Einfluss von FTY720 auf den Abbau des axonalen Zytoskeletts
In der vorliegenden Studie konnte bei den unbehandelten SOD1G93Adl-Kontrolltieren im
Vergleich zu den Wildtyp-Tieren kein Abfall der phosphorylierten Neurofilamente H
(pNFH) im Hirnhomogenat festgestellt werden. Deshalb ist eine Beurteilung des
Effekts von FTY720 bezüglich einer Reduktion der Abnahme an pNFH im
Hirnhomogenat von behandelten SOD1G93Adl-Tieren nicht möglich. Es gibt jedoch
Hinweise, dass FTY720 Einfluss auf die pNFH hat.
Jin S et al. beobachteten anhand eines Mausmodells der Experimentellen
Autoimmunen Encephalomyelitis (EAE), dass 1,0 mg/kg Tag FTY720 den axonalen
Schaden vermindert und damit die Konzentration an pNFH im lumbosakralen
Rückenmark im Vergleich zur Kontrollgruppe steigert (Jin et al. 2014).
Wie bei dem Mausmodell der Multiplen Sklerose kommt es auch beim SOD1G93dl-
Mausmodell aufgrund der Degeneration der Neurone und des axonalen Schadens zur
Freisetzung von pNFH in den Liquor. Über diesen gelangen die pNFH anschließend
ins Plasma.
Zur Feststellung eines Effekts von FTY720 hinsichtlich einer Reduktion der Abnahme
an pNFH im Hirnhomogenat der SOD1G93Adl-Tiere scheint die quantitative Analyse der
pNFH mittels ELISA ungeeignet zu sein.
4.2.5 Schlussfolgerung
Unsere Ergebnisse sprechen eher dafür, dass FTY720 keinen direkten
neuroprotektiven Effekt auf α-Motoneurone im SOD1G93Adl-Mausmodell hat. Soliven et
al. vermuten einen direkten neuroprotektiven Effekt von FTY720 bei der schubförmigen
Multiplen Sklerose (Soliven et al. 2011). Dieser Effekt ist basierend auf unseren
Ergebnissen, eher durch die antiinflammatorische Wirkung des Medikaments
verursacht. Doch es besteht die Möglichkeit, dass FTY720 bei der schubförmigen
Multiplen Sklerose und dem Mausmodell der ALS jeweils unterschiedliche Signalwege
beeinflusst. So wäre es möglich, dass es einen direkten neuroprotektiven Effekt von
FTY720 bei der schubförmigen MS gibt, aber nicht in dem hier untersuchten
Mausmodell der ALS.
In einigen Mausmodellen korreliert die Neuroprotektion von FTY720 mit einer
Erhöhung des BDNF-Spiegels und der Aktivierung der dazu gehörigen Signalkaskade.
Dies hat zur Folge, dass FTY720 im SOD1G93Adl-Mausmodell eventuell gar nicht über

77
diese Signalkaskade wirkt, so dass es auch nicht zu einer Erhöhung des BDNF-
Spiegels kommt.
4.3 Effekt von FTY720 auf proinflammatorische Parameter im ZNS und im
Blut von SOD1G93Adl-Tieren
4.3.1 FTY720 hat keinen Einfluss auf die Astrogliose bei SOD1G93Adl-Tieren
In der präsymptomatischen und symptomatischen Phase der SOD1G93Adl-Tiere im
lumbalen Rückenmark ist kein Effekt von FTY720 in den Dosierungen von 1,0 mg/kg
Tag und 3,0 mg/kg hinsichtlich einer Abnahme der Astrogliose erkennbar.
Bei den Endstadiumtieren ist lediglich ein minimaler Effekt von FTY720 festzustellen.
Dieser hat hier jedoch aufgrund der niedrigeren Fallzahl von 2-3 Endstadiumstieren
nur geringe Aussagekraft.
In der vorliegenden Studie konnte kein Einfluss von FTY720 auf die Astrogliose
gezeigt werden. In Studien aus der Vergangenheit gab es im Rahmen von anderen
Mausmodellen aber Hinweise, dass Effekte von FTY720 hinsichtlich der Reduktion der
Astrogliose beobachtet wurden.
Norimatsu Y et al. stellten anhand von SCID Mäusen, denen eine
Rückenmarkskontusion zugefügt worden war, fest, dass FTY720 in einer Dosierung
von 3,0 mg/kg Tag eine Abnahme der Astrogliose zeigte.
SCID Mäuse besitzen keine T- und B-Zellen. Dadurch fehlt ihnen die Ausbildung eines
adaptiven Immunsystems. Aufgrund dessen vermuteten Norimatsu Y et al., dass
FTY720 die Lymphozytenfunktion nicht mehr beeinflussen konnte und somit
immunsystemunabhängige Effekte des Medikaments zum Vorschein kamen. Somit
nahmen sie an, dass FTY720 nicht immunologische bzw. direkte Effekte auf die Zellen
des ZNS ausübt und damit auch die Astrogliose vermindert (Norimatsu Y 2012).
Im Rahmen einer Studie zu einem Mausmodell der Multiplen Sklerose wurde kürzlich
berichtet, dass phosphoriliertes FTY720 den Verlust von S1P-Rezeptoren auf
Astrozyten zu induzieren scheint. Choi J W et al. gingen davon aus, dass durch den
Funktionsverlust der S1P1 Rezeptoren auf der Oberfläche der Astrozyten die
Wirksamkeit des Medikaments hinsichtlich der Abnahme der Astrogliose erst
zustandekommt (Norimatsu et al. 2012).

78
Anhand von Ratten, an denen ein Status epilepticus mit Hilfe von Lithium/ Pilocarpin
ausgelöst wurde, stellten Gao et al. einen Effekt von FTY720 bezüglich einer Abnahme
der Astrogliose im Hippocampus im Vergleich zu Kontrolltieren fest.
Zudem kam es hierbei auch zu einer verminderten hippocampalen Expression von
proinflammatorischen Zytokinen wie TNFα und IFN-β.
Bei der Epilepsie nimmt anders als bei der ALS die Neuroinflammation eine
Schlüsselrolle ein, so dass eventuell davon auszugehen ist, dass FTY720 seinen
Effekt hinsichtlich der Abnahme der Astrogliose durch eine Verminderung der
proinflammatorischen Zytokine induziert.
Philips T et al. beobachteten in vitro anhand von mutierten SOD1G93A Astrozyten eine
vermehrte Expression von proinflammatorischen Zytokinen (Philips et al. 2011).
Es liegt deshalb nahe, dass FTY720 die proinflammatorischen Zytokine im SOD1G93Adl-
Mausmodell senkt und damit zu einer Abnahme der Astrogliose führt. Dies konnten wir
aber in unserer Studie nicht zeigen.
In der vorliegenden Studie ist zu keinem Zeitpunkt der Erkrankung von SOD1G93Adl-
Tieren ein Effekt von FTY720 auf Astrozyten festzustellen. In anderen Mausmodellen
gab es jedoch Hinweise auf eine FTY720 vermittelte Reduktion der Astrogliose.
4.3.2 FTY720 hat keinen Einfluss auf die Mikrogliaaktivierung im SOD1G93Adl-
Mausmodell
Ein Effekt von FTY720 ist bezüglich der Abnahme der Mikrogliaaktivierung im lumbalen
Rückenmark von SOD1G93Adl-Tieren zu keinem Zeitpunkt feststellbar.
Trotz unserer Ergebnisse gibt es Hinweise, dass FTY720 die Mikrogliaaktivierung
hinsichtlich einer Abnahme beeinflusst. Unter der Behandlung mit 1,0 mg/kg/d FTY720
beobachtete Gao et al. in einem Epilepsiemausmodell eine Verminderung der
Mikrogliaaktivierung im Hippocampus (Gao et al. 2012).
Wir stellten in unserer Studie zwar fest, dass FTY720 keinen Einfluss auf die Abnahme
der Mikrogliaaktivierung hat, doch könnte es durch FTY720 in der vorliegenden Studie
zu einer Mikroglia vermittelten Neuroprotektion der α-Motoneurone gekommen sein.
Anhaltspunkte hierfür zeigte Noda H in vitro im Rahmen des EAE Mausmodells.
FTY720 behandelte Mikroglia übten dabei neuroprotektive Effekte aus, indem sie die
Ausschüttung von proinflammatorischen Zytokinen (TNFα, IL-1β und IL-6) senkten und
die Ausschüttung von neurotrophen Faktoren erhöhten. Schon frühere Studien

79
konnten zeigen, dass Mikroglia neurotrophe Faktoren wie den BNDF und GNDF
freisetzen, welche das neuronale Überleben unterstützen (Mizuno et al. 2004),
(Liang et al. 2010).
FTY720 hat in der vorliegenden Studie keinen Effekt in Bezug auf die Abnahme der
Mikrogliaaktivierung, nimmt aber eventuell Einfluss auf die Mikroglia vermittelte
Ausschüttung von neurotrophen Faktoren und damit auf die Neuroprotektion.
4.3.3 Überprüfung des antiinflammatorischen Effekts von FTY720 im SOD1G93Adl-
Mausmodell
In allen drei Krankheitsstadien senkt FTY720 signifikant und dosisunabhängig die T-
und B-Zellen, sowie CD4+ und CD8+ T-Zellen im Vollblut von SOD1G93Adl-Tieren.
Die antiinflammatorische Wirkung von FTY720 wurde schon in zahlreichen Studien
belegt. Über den S1P1 Rezeptor vermittelt FTY720 ein Retentionssignal, dass die T-
Lymphozyten im Lymphknoten verweilen lässt („LN Homing“) (Pham et al. 2008). Es
kommt zur Lymphopenie (Brinkmann et al. 2002), (Mandala et al. 2002). Betroffen von
diesem Signal sind nur naive T-Zellen (Tn) und zentrale T-Gedächtniszellen, die durch
schwache Antigenstimulation entstehen. Aufrechterhalten wird das Retentionsignal
durch den sogenannten CCR7-Rezeptor. Effektor T-Gedächtniszellen (Tem)
exprimieren diesen Rezeptor nicht und können deshalb auch nicht direkt durch
FTY720 beeinflusst werden.
Mehling et al. beobachteten unter anderem bei FTY-behandelten MS-Patienten eine
Senkung aller Lymphozyten um 61% im Vergleich zum Referenzwert. Die CD8+ T-
Zellen wurden auch reduziert, jedoch nicht im gleichen Ausmaß wie die CD4+ T-
Lymphozyten. Der beschriebene Effekt von FTY720 bezüglich der stärkeren Reduktion
der CD4+ T-Zellen im Vergleich zu CD8+ T-Zellen ist in der vorliegenden Studie nicht
zu erkennen.
Grützke B et al. konnten, wie schon in früheren Studien beschrieben, eine signifikante
Reduktion von B-Zellen im Blut zeigen. In ihrer Studie beschrieben sie die
verschiedenen Subpopulationen der B-Zellen. Sie konnten beobachten, dass FTY720
den Anteil regulatorischer B-Zellen bei FTY-behandelten MS-Patienten signifikant
ansteigen ließ. Im Gegensatz dazu sank der Anteil an naiven (Tn) und zentralen
Gedächtnis T-Zellen (Tcm) leicht.

80
Zudem stellten Grützke B et al. fest, dass Tn- und Tcm-Zellen mehr „lymph node
homing triad“ Moleküle (L-Selectin, CCR7 und LFA-1) als regulatorische B-Zellen
exprimieren (Grützke et al. 2015). Deshalb werden regulatorische B-Zellen durch
FTY720 weniger beeinflusst. Dies könnte in der vorliegenden Studie im Vergleich zu
T-Zellen die geringer ausgeprägte Reduktion der B-Zellanzahl im Vollblut von
SOD1G93Adl-Tieren erklären. Mit unseren Beobachtungen im Vollblut von SOD1G93Adl-
Tieren bestätigen wir die bekannte antiinflammatorische Wirkung von FTY720
hinsichtlich der signifikanten Reduktion von T- und B-Zellen sowie der CD4+ und CD8+
T-Zellen im Vollblut.
4.3.4 Überprüfung des Effekts von FTY720 auf T- und B-Zellen sowie CD4+ und
CD8+ T-Zellen im Milzhomogenat von SOD1G93Adl-Tieren
4.3.4.1 B-Zellen im Milzhomogenat
FTY720 hat in der präsymptomatischen und symptomatischen Phase einen gering
ausgeprägten, dosisabhängigen Effekt hinsichtlich der Zunahme der B-Zellen im
Milzhomogenat von SOD1G93Adl-Tieren. In der präsymptomatischen Phase ist der
beschriebene Effekt bei 3,0 mg/kg Tag FTY720 behandelten SOD1G93Adl-Tieren stärker
ausgeprägt. Im Gegensatz dazu zeigt sich der stärkere Effekt in der symptomatischen
Phase bei den 1,0 mg/kg Tag FTY720 behandelten SOD1G93Adl-Tieren. Im Endstadium
stellen wir in beiden verwendeten Dosierungen von FTY720 eine sehr gering
ausgeprägte Reduktion der B-Zellanzahl im Milzhomogenat der SOD1G93Adl-Tiere fest.
Die beschriebenen Effekte wiesen keine Signifikanz auf.
Mit den follikulären und den MZ B-Zellen beobachteten Arnon und Cyster die
Subpopulationen der B-Zellen in der Milz.
Mit Hilfe der Zwei-Photonen Mikroskopie zeigten sie einen S1P-Rezeptor 1 (S1PR1)
abhängigen Signalweg auf, der die Auswanderung von follikulären B-Zellen aus der
weißen Pulpa steuert. Durch Blockade dieses Signalweges durch FTY720 wurde die
Abgabe der follikulären B-Zellen aus der weißen Pulpa an den venösen Sinus
unterbrochen. Damit wurden keine follikulären B-Zellen an das Blut abgegeben.
Sie stellten sogar fest, dass die B-Zellanzahl in der Milz leicht zunahm. Abhängig vom
S1PR1 pendeln die MZ B-Zellen zwischen der MZ (Marginalzone) und den Follikeln.
Dies wird als „MZ B-Zell Shuttling“ bezeichnet. Die MZ B-Zellen werden bei diesem

81
Mechanismus nicht ans Blut abgegeben. Durch Blockade des S1PR1 durch FTY720
wird deshalb nur dieser Kreislauf unterbrochen. Die MZ B-Zellen verlassen die Milz
deshalb nicht (Arnon et al. 2014). Dieser Mechanismus könnte erklären, weshalb sich
die Zahl der B-Zellen im Milzhomogenat nicht ändert.
In der präsymptomatischen und symptomatischen Phase lässt sich eine leichte
Zunahme der B-Zellen im Milzhomogenat der SOD1G93Adl-Tiere feststellen. Damit
bestätigen wir die Beobachtungen von Arnon und Cyster (Arnon et al. 2014).
Im Endstadium lässt sich das Verhalten hinsichtlich einer leichten Zunahme der B-
Zellen im Milzhomogenat der SOD1G93Adl-Tiere nicht mehr nachvollziehen. Im
Gegenteil, es kommt im Endstadium sogar zu einer leichten Abnahme der B-Zellzahl.
4.3.4.2 T-Zellen sowie CD4+ und CD8+ T-Zellen im Milzhomogenat
In der vorliegenden Studie senkt FTY720 die T-Zellen sowie die CD4+ und CD8+ T-
Zellen im Milzhomogenat von SOD1G93Adl-Tieren signifikant (p<0,001).
Die beobachteten Effekte von FTY720 auf die T-Zellen sowie den CD4+ und CD8+ T-
Zellen sind aus früheren Studien bekannt.
Bei der Reduktion der T-Zellen in der Milz spielt der CCR7 Rezeptor eine
Schlüsselrolle. Er vermittelt über ein G-Protein (Cyster et al. 1995) die Einwanderung
der T-Zellen in die T-Zell Zone der Milz. Der CCR7 ist S1P-Rezeptor 1 abhängig
(Pham et al. 2008). Dabei dient das CCR7 abhängige Chemokin CCL21 als Lockstoff
für T-Zellen. CCL21 wird von FRC exprimiert (Bajenoff et al. 2008). Durch Blockade
des S1P-Rezeptors 1 durch FTY720 wird das Einwanderungssignal des CCR7
Rezeptors aufgehoben. Das hat zur Folge, dass die Zahl der T-Zellen sowie die Zahl
der CD4+ und CD8+ T-Zellen in der Milz abnehmen.
Wir bestätigen damit die Effekte von FTY720 hinsichtlich des beschriebenen
Verhaltens von T-Zellen sowie den CD4+ und CD8+ T-Zellen im Homogenat der Milz
für das SOD1G93Adl-Mausmodell.

82
4.3.5 FTY720 senkt die Freisetzung der proinflammatorischen und
antiinflammatorischen Zytokine
4.3.5.1 IFN-γ im Blutplasma
FTY720 zeigt hinsichtlich der Abnahme des proinflammatorischen Zytokins IFN-γ im
Blutplasma von SOD1G93Adl-Tieren zu den Zeitpunkten 170-180 Tage, in der
symptomatischen Phase und im Endstadium einen dosisabhängigen Effekt. Dabei übt
die höhere Dosierung des Medikaments bei Tieren im Alter von 170-180 Tagen und
bei Tieren im Endstadium den stärkeren Effekt aus. Die niedrigere Dosierung vermittelt
hingegen den stärkeren Effekt in der symptomatischen Phase.
Ein Einfluss von FTY720 zu den Zeitpunkten 40-120 Tage ist nicht festzustellen. Die
erhöhte IFN-γ Blutplasmakonzentration von SOD1G93Adl-Kontrolltieren im Alter von
170-180 Tage bis zum Endstadium interpretieren wir als eine reaktive Inflammation auf
die schon abgelaufene Neurodegeneration bei den α-Motoneuronen.
Tem-Zellen sezernieren unter anderem IFN-γ (Sallusto et al. 1999). Sie werden durch
FTY720 nicht beeinflusst und damit auch nicht die Sekretion von IFN-γ im Blutplasma.
Das Medikament hat zwar keinen direkten Einfluss auf Tem-Zellen im ZNS, aber durch
Verhinderung des Eintritts von Tn- und Tcm-Zellen, die sich zu Tem-Zellen
differenzieren können, wird auch die Zahl der Tem-Zellen vermindert (Mehling et al.
2008). Eine Abnahme der Tem-Zellen könnte eine mögliche Erklärung für die
Verminderung von IFN-γ Blutplasmaspiegel bei FTY720 behandelten SOD1G93Adl-
Tieren sein.
B-Zellen sezernieren ebenfalls IFN-y. Man kann vermuten, dass die in der
vorliegenden Studie beobachtete FTY720 vermittelte Reduktion der B-Zellen eine
Abnahme des IFN-γ Blutplasmaspiegels bewirkt. In der symptomatischen Phase
stellen wir zudem bei SOD1G93Adl-Kontrolltieren eine Zunahme der B-Zellen im Vollblut
fest. Wir nehmen an, dass dies zu dem starken Anstieg der IFN-γ Blutplasmaspiegel
im Alter von 170-180 Tagen führt.
In zwei unabhängigen Studien wurden protektive Effekte von FTY720 in einem
Mausmodell der Arteriosklerose gezeigt. Dabei zeigten sich unter anderem bei den
FTY-behandelten Tieren niedrigere Plasmakonzentrationen für das
proinflammatorische Zytokin IFN-γ (Pinschewer et al. 2000), (Brinkmann et al. 2004).

83
4.3.5.2 IL-2 im Blutplasma
FTY720 senkt das proinflammatorische Zytokin IL-2 im Blutplasma von SOD1G93Adl-
Tieren dosisabhängig zu allen gemessenen Zeitpunkten. Im Blutplasma von
SOD1G93Adl-Kontrolltieren beobachten wir zu den Zeitpunkten 40-80 Tage und im
Endstadium erhöhte IL-2 Blutplasmakonzentrationen. Hinsichtlich der Abnahme der IL-
2 Blutplasmakonzentration bei SOD1G93Adl-Tieren üben beide Dosierungen von
FTY720 zu den Zeitpunkten 40-80 Tage einen Effekt aus. Die höhere Dosierung
vermittelt diesbezüglich den stärkeren Effekt.
Im Zeitraum 100 Tage bis zur symptomatischen Phase ist bei den 1,0 mg/kg Tag
FTY720 behandelten SOD1G93Adl-Tieren hinsichtlich einer Abnahme der IL-2
Blutplasmakonzentration ein Effekt von FTY720 nicht erkennbar. Nur in der Dosierung
von 3,0 mg/kg Tag FTY720 kommt es in diesem Zeitraum bei SOD1G93Adl-Tieren zu
einer Abnahme der IL-2 Blutplasmakonzentration bei SOD1G93Adl-Tieren. Zum
Zeitpunkt Endstadium beobachten wir einen dosisunabhängigen Effekt von FTY720
bezüglich der Abnahme der IL-2 Blutplasmakonzentration.
Tcm-Zellen sezernieren IL-2 (Sallusto et al. 1999). Sie werden wie die Tn-Zellen von
FTY720 beeinflusst, da sie den CCR 7 Rezeptor auf ihrer Zelloberfläche tragen. Dockt
FTY720 an den S1P Rezeptor 1 an, vermittelt der CCR7 Rezeptor über ein G-Protein
(Cyster et al. 1995) die Retention der T-Zellen im Lymphknoten. Somit können Tcm-
Zellen auch kein IL-2 mehr ins Blutplasma freisetzen.
Mehling et al. analysierten die Blutproben von FTY720 behandelten MS Patienten und
stellten fest, dass die Tcm-Zellen die Produktion des Proliferationsfaktors IL-2
vermindern und damit die Proliferationsrate reduzieren. Jedoch beobachteten sie eine
reaktive Produktion von IFN-γ, welche vergleichbar ist mit der der Kontrollgruppen
(unbehandelte MS-Patienten und gesunde Patienten). Es wurde eine Dosierung von
1,25 mg/kg KG FTY720 verwendet (Mehling et al. 2011). Eine reaktive Zunahme der
IFN-γ Blutplasmakonzentration ist, wie sie bei der Multiplen Sklerose beobachtet
wurde, in der vorliegenden Studie nicht erkennbar. Im Gegenteil, es kommt
größtenteils zu einer Abnahme der IFN-γ Blutplasmakonzentration bei behandelten
SOD1G93Adl-Tieren.
Auch Brinkmann et al. beobachteten in vitro anhand von isolierten T-Zellen aus dem
Blut von FTY720-behandelten Transplantations- und MS-Patienten, die dem Tem
Phänotyp entsprachen, eine Reduktion der Sekretion an IL-2 und deren
Proliferationsaktivität. Hier konnte wie in der vorliegenden Studie kein Einfluss von

84
FTY720 hinsichtlich einer Zunahme der IFN-γ Blutplasmakonzentration beobachtet
werden (Brinkmann et al. 2001).
4.3.5.3 IL-4 im Blutplasma
Zudem stellen wir einen dosisabhängigen Effekt von FTY720 hinsichtlich der Abnahme
des antiinflammatorischen Zytokin IL-4 im Blutplasma von behandelten SOD1G93Adl-
Tieren zu den Zeitpunkten 40d, 60d, 80d und 170-180 Tage fest. Die höhere Dosierung
von FTY720 übt diesbezüglich einen stärkeren Effekt aus. Zu den übrigen
gemessenen Zeitpunkten zeigen die SOD1G93Adl-Kontrolltiere sowie die behandelten
SOD1G93Adl-Tiere sehr niedrige IL-2 Blutplasmaspiegel, so dass ein Effekt von FTY720
schwer zu beurteilen ist.
Im Anfangsstadium von SOD1G93A-Tieren findet man vor allem CD4+ T-Zellen und
davon hauptsächlich Th2-Helferzellen, die IL-4 produzieren (Beers et al. 2011),
(Fiala et al. 2010). Dies kann anhand der höheren IL-4 Blutplasmakonzentrationen zu
den Zeitpunkten 40 Tage, 60 Tage und 80 Tage in der vorliegenden Studie
nachvollzogen werden. Inwieweit FTY720 IL-4 im Blutplasma beeinflusst, ist noch nicht
beschrieben worden. Hinsichtlich der Abnahme der naiven CD4+ T-Zellen im Blut von
SOD1G93Adl-Tieren nehmen wir an, dass sich weniger naive CD4+ T-Zellen zu Th2-
Helferzellen differenzieren können. Aufgrund der verminderten Anzahl an Th2-Zellen
könnte auch weniger IL-4 ins Blutplasma freigesetzt werden.
Im Rahmen des SOD1G93Adl-Mausmodells stellen wir fest, dass FTY720 die
proinflammatorischen Zytokine IL-2 und IFN-γ und das antiinflammatorische Zytokin
IL-4 dosisabhängig senkt. Dieser Effekt ist nicht zu allen Zeitpunkten erkennbar, da die
Blutplasmakonzentrationen der untersuchten Interleukine sowohl bei SOD1G93Adl-
Kontrolltieren als auch bei den behandelten SOD1G93Adl-Tieren sehr gering ausgeprägt
ist.
Die Übertragung unserer Ergebnisse auf den Menschen ist nur bedingt möglich und
sinnvoll, da die Reproduzierbarkeit und die Übertragung der Ergebnisse präklinischer
Studien auf den Menschen begrenzt sind.
Zudem ist die Vergleichbarkeit unterschiedlicher Mausmodelle nur eingeschränkt
möglich, da es sich teilweise um andere Pathomechanismen handelt. Diese sind oft
nicht ausreichend erforscht. Auch ist die Gegenüberstellung von in vivo und in vitro

85
Studien nur bedingt möglich. In vitro Studien können systemische, organspezifische
und pharmakologische Aspekte nicht erfassen (Spielmann H 1994).
In Zukunft ist es sicherlich sinnvoll den möglichen direkten neuroprotektiven Effekt von
FTY720 noch einmal im SOD1G93Adl-Mausmodell zu überprüfen. Ob FTY720
insbesondere in der präsymptomatischen Phase mit beiden Dosierungen und in der
symptomatischen Phase nur in einer Dosierung von 1,0 mg/kg Tag neuroprotektiv
wirkt, wie in unserer Studie beschrieben, muss ebenfalls noch genauer erforscht
werden. Dabei stellt sich unter anderem die Frage, in welcher Dosierung sich die
neuroprotektiven und neurotoxischen Effekte die Waage halten.
Durch Messung des BDNF könnte die Frage beantwortet werden, ob FTY720 im
SOD1G93Adl-Mausmodell überlebensfördernde Signalwege beeinflusst.
Um weitere Erkenntnisse über die Neuroprotektivität von FTY720 zu gewinnen, bietet
es sich an, einen möglichen Effekt auf phosphorylierten Neurofilamente H mit einer
suffizienten Methode zu wiederholen.
Im Rahmen der Zytokinmessung sollte in einer neu aufgelegten Studie die Fallzahl der
verschiedenen Behandlungsgruppen angepasst werden, um aussagekräftigere
Ergebnisse zu erhalten.
Es ist außerdem empfehlenswert den Einfluss von FTY720 im Rahmen eines anderen
ALS-Mausmodells zu prüfen.
In Zukunft sollte der Frage nachgegangen werden, auf welche weiteren pathologischen
Prozesse FTY720 im SOD1G93Adl-Mausmodell Einfluss hat. Dafür ist eine
immunhistochemische Färbung der abgelagerten und abnorm gefalteten SOD1
Proteine im Rückenmark sowie eine Messung von Gelsolin, einem proapoptotischen
Protein, im Plasma und Hirn-Homogenat geplant.

86
5 Zusammenfassung
Das Präparat FTY720 wird weltweit zur verlaufsmodifizierenden Langzeitbehandlung
der schubförmigen Multiplen Sklerose eingesetzt und wird unter dem Namen Gilenya®
von der Firma Novartis vertrieben.
Die antiinflammatorische Wirkung des Immunmodulators FTY720 wurde bereits in
zahlreichen Publikationen belegt. Eine direkte neuroprotektive Wirkung von FTY720
wird vermutet, wurde bisher allerdings nur unzureichend untersucht.
Zur Untersuchung dieses Sachverhaltes wurde das primär neurodegenerative
SOD1G93Adl-Mausmodell der Amyotrophen Lateral Sklerose gewählt.
Bei der ALS handelt es sich um eine primär neurodegenerative Erkrankung, die
innerhalb von 3-5 Jahren unweigerlich zum Tode führt. Riluzol ist zurzeit das einzige
zugelassene Medikament zur Behandlung dieser Motoneuron-Krankheit.
In der vorliegenden Studie wurde FTY720 in den Dosierungen 1,0 mg/kg Tag und 3,0
mg/kg Tag verwendet.
Diese Dissertationsarbeit beschäftigte sich mit zwei Fragestellungen. Zum einen sollte
ein möglicher neuroprotektiver Effekt von FTY720 auf die α-Motoneurone erforscht
werden.
Um diesen Sachverhalt zu untersuchen, wurden folgende Methoden verwendet:
• Quantitative Bestimmung der α-Motoneurone im Rückenmark mittels Nissl-Färbung
• FTY720-Messung im Homogenat von SOD1G93Adl-Tieren durch die Firma Norvatis
• Quantitative Analyse von phosphorilierten Neurofilamenten H im Hirnhomogenat von
SOD1G93Adl-Tieren mittels ELISA
• Untersuchung des Effekts von FTY720 in Bezug auf den Krankheitsbeginn von
SOD1G93Adl-Tieren
Die quantitative Analyse der α-Motoneurone im lumbalen Rückenmark von FTY720
behandelten SOD1G93Adl-Tieren ergab einen gering ausgeprägten neuroprotektiven
Effekt. Dieser trat sowohl in der präsymptomatischen als auch in der symptomatischen
Phase auf. In der präsymptomatischen Phase konnte der Effekt in der niedrigen und
hohen Dosierung dosisabhängig beobachtet werden. In der symptomatischen Phase
hingegen zeigte sich dieser nur bei den 1,0 mg/kg Tag FTY720 behandelten
SOD1G93Adl-Tieren. Die beschriebenen Effekte waren nicht signifikant.
Konzentationen von FTY720 und FTY720-Phosphat wurden außerdem im
Hirnhomogenat gemessen, und wir konnten damit zeigen, dass FTY720 über die

87
Bluthirnschranke diffundiert. Ein direkter neuroprotektiver Effekt ist damit zumindest
möglich.
Mit Hilfe der quantitativen Analyse der phosphorylierten Neurofilamente H (pNFH) im
Hirnhomogenat von SOD1G93Adl-Tieren konnte der Krankheitsverlauf der
unbehandelten SOD1G93Adl-Kontrolltiere nicht nachvollzogen werden, so dass eine
abschließende Beurteilung des Effekts von FTY720 hinsichtlich einer Reduktion von
pNFH-Werten bei behandelten SOD1G93Adl-Tieren nicht möglich war.
Einen Einfluss von FTY720 auf den Krankheitsbeginn von SOD1G93Adl-Tieren wurde
nicht beobachtet.
In einigen früheren Studien konnte ein neuroprotektiver Effekt von FTY720 wie
beispielsweise in einem Mausmodell des M. Huntington aufgezeigt werden. Aufgrund
unserer negativen Ergebnisse im ALS-Tiermodell gehen wir aber eher davon aus, dass
FTY720 allgemein seine postulierte neuroprotektive Wirkung bei der multiplen
Sklerose über den antiinflammatorischen Wirkmechanismus vermittelt.
Es stellt sich die Frage, ob FTY720, wie schon in anderen Mausmodellen gezeigt, im
SOD1G93Adl-Mausmodell neuroprotektive Signalwege überhaupt beeinflusst.
Des Weiteren sollte der Effekt von FTY720 auf die inflammatorischen Parameter im
SOD1G93Adl-Mausmodell untersucht werden. Um dieser Fragestellung nachzugehen,
wählten wir folgende Methoden aus:
• Quantitative Bestimmung von Astrozyten im Rückenmark mittels IHC
• Quantitative Bestimmung von Mikroglia im Rückenmark mittels IHC
• Quantitative Analyse der T-und B-Zellen im Blut und Milzhomogenat mittels FACS
• Zytokinmessung im Blutplasma mittels ELISA
• FTY720-Messung im Plasma durch die Firma Norvatis
In der vorliegenden Studie stellten wir keinen Effekt von FTY720 bezüglich einer
Abnahme der Astrogliose und der Mikrogliaaktivierung bei behandelten SOD1G93Adl-
Tieren fest.
Hinsichtlich der Wirkung auf B-und T-Zellen sowie den CD4+ und CD8+ T-Zellen
bestätigten wir die bekannte antiinflammatorische Wirkung von FTY720 im Vollblut und
Milzhomogenat von SOD1G93Adl-Tieren.
Sowohl die proinflammatorischen Zytokine IFN-γ und IL-2 als auch das
antiinflammatorische Zytokin IL-4 im Blutplasma von SOD1G93Adl-Tieren wurden durch
FTY720 dosisabhängig gesenkt.

88
6 Literaturverzeichnis
[1] Al-Chalabi A , Hardiman O. The epidemioogy of ALS: a conspiracy of genes,
environment and time. Nat Rev Neurol. 9: 617–28. (2013).
[2] Al-Chanlabi A , Leigh P N. Recent advances in amyotrophic lateral sclerosis.
Current Opinion in Neurology. 13; 397-405. (2000).
[3] Almer G. Amyotrophe Lateralsklerose: Überlegungen zu Ursprung und
Pathophysiologie der Erkrankung. Journal für Neurologie, Neurochirurgie und
Psychiatrie. 4: 6-12. (2003).
[4] Arai T , Hasegawa M , Akiyama H , Ikeda K , Nonaka T , Mori H , Mann D ,
Tsuchiya K , Yoshida M , Hashizume Y , Oda T. TDP‐43 is a component of ubiquitin‐
positive tau‐negative inclusions in frontotemporal lobar degeneration and amyotrophic
lateral sclerosis. Biochem Biophys Res Commun. 351: 602‐611 . (2006).
[5] Arnon T I , Jason G C. Blood, Sphingosine-1-Phosphate and Lymphocyte
Migration Dynamics in the Spleen. Current Topics in Microbiology and Immunology.
378. (2014).
[6] Bajenoff M , Glaichenhaus N , Germain R N. Fibroblastic reticular cells guide T
lymphocyte entry into and migration within the splenic T cell zone. J Immunol. 181:
3947–3954. (2008).
[7] Barohn R J , Rowland L P. Neurology and gulf war veterans. Neurology. 26,
1484-1485. (2002).
[8] Beers D R , Henkel J S , Zhao W , Wang J , Huang A , Wen S , Liao B , Appel
S H. Endogenous regulatory T lymphocytes ameliorate amyotrophic lateral sclerosis in
mice and correlate with disease progression in patients with amyotrophic lateral
sclerosis. Brain. 134, 1293–1314. (2011).
[9] Benatar M. Lost in translation: treatment trials in the SOD1 mouse and in human
ALS. Neurobiol Dis. 26: 1-13. (2007).
[10] Bowling A C , Barkowski E E , McKenna-Yasek D , Sapp P , Horvitz H R , Beal
M F , Brown R H. Superoxide dismutase concentration and activity in familial
amyotrophic lateral sclerosis. J Neurochem. 64: 2366–9. (1995).
[11] Brinkmann V. personal communication. 2015.
[12] Brinkmann V , Chen S , Feng L , Pinschewer D , Nikolova Z , Hof R. FTY720
alters lymphocyte homing and protects allografts without inducing general
immunosuppression. Transplant Proc. 33: 530-531. (2001).

89
[13] Brinkmann V , Cyster J , Hla T. FTY720: Sphingosine 1-phosphate receptor-1
in the control of lymphocyte egress and endothelial barrier function. Am J Transplant.
4: 1019-1025. (2004).
[14] Brinkmann V , Davis M D , Heise C E , Albert R , Cottens S , Hof R , Bruns C ,
Prieschl E , Baumruker T , Hiestand P , Foster C A , Zollinger M , Lynch K R. The
immune modulator FTY720 targets sphingosine 1-phosphate receptors. J Biol Chem.
277: 21453–21457. (2002).
[15] Cipriani R , Chara J C , Rodríguez-Antigüedad A , Matute C. FTY720 attenuates
excitotoxicity and neuroinflammation. Journal of Neuroinflammation. 12: 86. (2015).
[16] Cleveland D W. From Charcot to SOD1: mechanisms of selective motor neuron
death in ALS. Neuron. 24: 515–520. (1999).
[17] Cleveland D W , Laing N , Hurse P V , Brown R H. Toxic mutants in
Charcot’sclerosis. Nature. 378: 342–3. (1995).
[18] Conner J M , Lauterborn J C , Yan Q , Varon S. Distribution of brain-derived
neurotrophic factor (BDNF) protein and mRNA in the normal adult rat CNS: evidence
for anterograde axonal transport. J Neurosci. 17: 2295–2313. (1997).
[19] Conwit R A. Preventing familial ALS: a clinical trial may be feasible but is an
efficacy trial warranted. Journal of the Neurological Sciences. 251: 1-2. (2006).
[20] Cyster J G , Goodnow C C. Pertussis toxin inhibits migration of B and T
lymphocytes into splenic white pulp cords. J Exp Med. 182: 581–586. (1995).
[21] Dal Canto M C , Gurney M E. Neuropathological changes in two lines of mice
carrying a transgene for mutant human Cu, Zn SOD, and in mice overexpressing wild
type human SOD: a model of familial amyotrophic lateralsclerosis (FALS). Brain Res.
676: 25–40. (1995).
[22] Dal Canto M C , Gurney M E. Development of central nervous system pathology
in a murine transgenic model of human amyotrophic lateral sclerosis. Am J Pathol. 145:
1271–1279. (1994).
[23] Dal Canto M C , Gurney M E. A low expressor line of transgenic mice carrying
a mutant human Cu, Zn superoxide dismutase (SOD1) gene develops pathological
changes that most closely resemble those in human amyotrophic lateral sclerosis. Acta
Neuropathol. 93: 537–550. (1997).
[24] Dev K K , Mullershausen F , Mattes H , Kuhn R R , Bilbe G , Hoyer D , Mir A.
Brain sphingosine-1-phosphate receptors: implication for FTY720 in the treatment
ofmultiple sclerosis. Pharmacol Ther. 117: 77–93. (2008).

90
[25] Di Menna L , Molinaro G , Di Nuzzo L , Riozzi B , Zappulla C , Pozzilli C , Turrini
R , Caraci F , Copani A , Battaglia G , Nicoletti F , Bruno V. Fingolimod protects cultured
cortical neurons against excitotoxic death. Pharmacol Res. 67: 1-9. (2013).
[26] Doi Y , Takeuchi H , Horiuchi H , Hanyu T , Kawanokuchi J , Jin S , Parajuli B ,
Sonobe Y , Mizuno T , Suzumura A. Fingolimod-phosphate attenuates Oligomeric
Amyloid beta–Induced Neurotoxicity via increased Brain-Derived Neurotrophic Factor
Expression in Neurons. PLoS ONE. 8: e61988. (2012).
[27] Dunkley T , Huentelman M J , Craig D W , Pearson J V , Szelinger S , Joshipura
K , Halperin R F , Stamper C , Jensen K R , Letizia D , Hesterlee S H , Pestronk A ,
Levine T , Bertorin T , Graves M C. Whole-genome analysis of sporadic amyotrophic
lateral sclerosis. New Engl J Med. 357: 775-88. (2007).
[28] Fiala M , Chattopadhay M , La Cava A , Tse E , Liu G , Lourenco E , Eskin A ,
Liu P T , Magpantay L , Tse S , Mahanian M , Weitzman R , Tong J , Nguyen C , Cho
T , Koo P , Sayre J , Martinez-Maza O , Rosenthal M J , Wiedau-Pazos M. IL-17A is
increased in the serum and in spinal cord CD8 and mast cells of ALS patients. J
Neuroinflammation. 7: 76. (2010).
[29] Fray A E , Ince P G , Banner S J , Milton I D , Usher P A , Cookson M. The
expression of the glial glutamate transporter protein EAAT2 in motor neurondisease:
an immunohistochemical study. European Journal of Neuroscience. Vol. 10, 8: 2481–
2489. (1998).
[30] Gao F , Liu Y , Li X , Wang Y , Wei D , Jiang W. Fingolimod (FTY720) inhibits
neuroinflammation and attenuates spontaneous convulsions in lithium-pilocarpine
induced status epilepticus in rat model. Pharmacol Biochem Behav. 103: 187–196.
(2012).
[31] Gaudette M , Hirano M , Siddique T. Current status of SOD1 mutations in familial
amyotrophic lateral sclerosis. Amyotrophe Lateral Scler Other Motor Neuron Disord. 1:
83-89. (2000).
[32] Gaudette M , Hirano M , Siddique T. Current status of SOD1 mutations in familial
amyotrophic lateral sclerosis. Amyotrophe Lateral Scler Other Motor Neuron Disord. 1:
83-89. (2000).
[33] Grützke B , Hucke S , Gross C C , Herold M V , Posevitz-Fejfar A , Wildemann
B T , Kieseier B C , Dehmel T , Wiendl H , Klotz L. Fingolimod treatment promotes
regulatory phenotype and function in multiple sclerosis. Annals of Clinical and
Translational Neurology. 2: 119–130. (2015).

91
[34] Gurney M E , Pu H , Chiu A Y , Dal Canto M C , Polchow C Y , Alexander D D ,
Caliendo J , Hentati A , Kwon Y H , Deng H X , Chen W , Zhai P , Sufit R L , Siddique
T. Motor neuron degeneration in mice that express a human Cu, Zn superoxide
dismutase mutation. Science. 264: 1772–1775. (1994).
[35] Halliwell B , Gutteridge J M. Free radicals in Biology and Medicine. Oxford
University Press. 2nd ed. (1989).
[36] Hasegawa Y , Suzuki H , Sozen T , Rolland W , Zhang J H. Activation of
sphingosin-1-phosphat receptor by FTY 720 is neuroprotektive after ischemic stroke in
rats. Stroke. 69: 119-29. (2010).
[37] Howland D S , Liu J , She Y , Goad B , Maragakis N J , Kim B , Erickson J , Kulik
J , DeVito L , Psaltis G , DeGennaro L J , Cleveland D W , Rothstein J D. Focal loss of
the glutamate transporter EAAT2 in a transgenic rat model of SOD1 mutant-mediated
amyotrophic lateral sclerosis (ALS). Proceedings of the National Academy of Sciences
of the United States of America. Vol. 99, 3: 1604–1609. (2002).
[38] Jin S , Takeuchi H , Horiuchi H , Wang H , Kawanokuchi J , Mizuno T , Suzumura
A. Fingolimod ameliorates axonal damage in experimental autoimmune
encephalomyelitis. Clinical and Experimental Neuroimmunology. 5: 315–320. (2014).
[39] Joppien S , Maier S L , Wendling D S. BASICS Experimentelle Doktorarbeit, 1.
Auflage. Elsevier: Urban und Fischer. S. 46-93. 2011.
[40] Kingwell E , Koch M , Leung B , Isserow S , Geddes J , Rieckmann P , Tremlett
H. Cardiotoxicity and other adverse events associated with mitoxantrone treatment for
MS. Neurology. 74: 1822–1826. (2010).
[41] Körner S , Dengler R , Petri S. Diagnostik der Amyotrophen Lateralsklerose.
Journal für Neurologie, Neurochirurgie und Psychiatrie. 11: 1-5. (2010).
[42] Kostic V , Jackson-Lewis V , De Bilbao F , Dubois-Dauphin M , Przedborski S.
Bcl-2: prolonging life in a transgenic mouse model of familial amyotrophic lateral
sclerosis. Science. 277: 559–62. (1997).
[43] Kroemer G , Zamzani N , Susin A. Mitochondrial control of apoptosis. Trends
Immunol. 18: 44–51. (1997).
[44] Liang J , Takeuchi H , Jin S , Noda M , Li H , Doi Y , Kawanokuchi J , Sonobe Y
, Mizono T , Suzumura A. Glutamate induces neurotrophic factor production from
microglia via protein kinase C pathway. Brain Res. 1322: 8-23. (2010).

92
[45] Li M , Ona V , Guegan C , Chen M , Jackson-Lewis V , Andrews L , Olszweski
A J , Stieg P , Lee J P , Pzredborski S , Friedlander R M. Functional role of caspase-1
and caspase-3 in an ALS transgenic mouse model. Science. 288: 335–9. (2000).
[46] Livingstone M A , Wilson E B , Ontiveros F , Wang J C. Unravelling the
mechanisms of help for CD8+ T cell responses. Immunol Res. 45; 209-17. (2009).
[47] Ludolph A C , Bendotti C , Blaugrund E , Chio A , Greensmith L , Loeffler J P ,
Mead R , Niessen H G , Petri S , Pradat P F , Robberecht W , Ruegg M ,
Schwalenstöcker B , Stiller D , van den Berg L , Vieira F , von Horsten S. Guidelines
for preclinical animal research in ALS/MND: A consensus meeting. Amyotroph Lateral
Scler. 11: 38-45. (2010).
[48] Ludolph A C , Dengler R , Zierz S. Amyptrophe Lateralsklerose: Geschichte
Epidemiologie und diagnostische Kriterien. 2. Auflage, 1-16. (2000).
[49] Mandala S , Hajdu R , Bergstrom J , Quackenbush E , Xie J , Milligan J ,
Thornton R , Shei G J , Card D , Keohane C. Alteration of lymphocyte trafficking by
sphingosine-1-phosphate receptor agonists. Science. 296: 346-349. (2002).
[50] Ma X , Peterson R , Turnbull J. Adenylyl Cyclase type 3, a marker of primary
cilia,is reduced in primary cell culture and in lumbar spinal cord in situ in G93A SOD1
mice. BMC Neuroscience. 12: 71. (2011).
[51] Mattiazzi M , D'Aurelio M , Gajewski C D , Martushova K , Kiaei M , Beal M F ,
Manfredi G. Mutated human SOD1 causes dysfunction of oxidative phosphorylation in
mitochondria of transgenic mice. J Biol Chem. 277: 29626-29633. (2002).
[52] Mehling M , Brinkmann V , Antel J , Bar-Or A , Goebels N , Vedrine C , Kristophic
C , Kuhle J , Lindberg R L , Kappos L. FTY720 therapy exerts differential effects on T-
cell subsets in multiple sclerosis. Neurology. 71: 1261–1267. (2008).
[53] Mehling M , Johnson T A , Antel J , Kappos L , Bar-Or A. Clinical immunology
of the sphingosine 1-phosphate receptor modulator fingolimod (FTY720) in multiple
scleoris. Neurology. 76: 20-27. (2011).
[54] Mehling M , Kappos L , Derfuss T. Fingolimod for multiple sclerosis: mechanism
of action, clinical outcomes, and future directions. Curr Neurol Neurosci Rep. 11: 492-
497. (2011).
[55] Meno-Tetang G M , Li H , Mis S , Pyszczynski N , Heining P , Lowe P.
Physiologically based pharmacokinetic modeling of FTY720 (2-amino-2[2-(-4-
octylphenyl)ethyl]propane-1,3-diol hydrochloride) in rats after oral and intravenous
doses. Drug Metab Dispos. 34: 1480-1487. (2006).

93
[56] Mizugishi K , Yamashita T , Olivera A , Miller G F , Spiegel S , Proia R L.
Essential role for sphingosine kinases in neural and vascular development. Mol Cell
Biol. 25: 11113–11121. (2005).
[57] Mizuno T , Kurotani T , Komatsu Y , Kawanokuchi J , Kato H , Mitsuma N ,
Suzumura A. Neuroprotective role of phosphodiesterase inhibitor ibudilast on neuronal
cell death induced by activated microglia. Neuropharmacology. 46: 404-411. (2004).
[58] Neumann M , Sampathu D M , Kwong L K , Truax A C , Micsenyi M C , Chou T
T , Bruce J , Schuck T , Grossman M , Clark C M , McCluskey L F , Miller B L , Masliah
E , Mackenzie I R , Feldman H , Feiden W , Kretzschmar H A , Trojanowski J Q , Lee
V M. Ubiquitinated TDP‐43 in frontotemporal lobar degeneration and amyotrophic
lateral sclerosis. Science. 314: 130‐133. (2006).
[59] Nirmalananthan N , Grennsmith L. Amyotrophic Lateral Sclerosis: Recent
advances and future therapies. Curr Opin Neurol. 18: 712-719. (2005).
[60] Norimatsu Y O T. FTY720 Improves Functional Recovery after Spinal Cord
Injury by Primarily Nonimmunomodulatory Mechanisms. The American Journal of
Pathology. Vol. 180, No. 4. (2012).
[61] Norimatsu Y , Ohmori T , Kimura A , Madoiwa S , Mimuro J , Seichi A , Yatomi
Y , Hoshino Y , Sakata Y. FTY720 Improves Functional Recovery after Spinal Cord
Injury by Primarily Nonimmunomodulatory Mechanisms. The American Journal of
Pathology. No 4. (2012).
[62] Packer M A , Porteous C M , Murphy M P. Superoxide production by
mitochondria in the presence of nitric oxide forms peroxynitrite. Biochem Molec Biol
Intl. 40: 527–34. (1996).
[63] Pardo A , Amico E , Favellato M , Castrataro R , Fucile S , Squitieri F. FTY720
(fingolimod) is a neuroprotective and disease-modifying agent in cellular and mouse
models of Huntington disease. Human Molecular Genetics. 23: 9. (2014).
[64] Pasinelli P , Housewart M K , Brown R H , Cleveland D W. Caspase-1 and -3
are sequentially activated in motor neuron death in Cu/Zn superoxide dismutase
familial ALS. Proc Natl Acad Sci USA. 97: 13901–6. (2000).
[65] Pham T H , Okada T , Matloubian M , Lo C G. S1P1 receptor signaling overrides
retention mediated by Gαi-coupled receptors to promote T-cell egress. Immunity. 28:
122–133. (2008).
[66] Philips T , Robberecht W. Neuroinflammation in amyotrophic lateral sclerosis:
role of glial activation in motor neuron disease. Lancet Neurol. 10: 253–263. (2011).

94
[67] Pinschewer D , Brinkmann V , Merkler D. Impact of sphingosine 1-phosphate
modulation on immune outcomes. Neurology. 76: 15-19. (2011).
[68] Pinschewer D D , Ochsenbein A F , Odermatt B , Brinkmann V , Hengartner H ,
Zinkernagel R M. FTY720 immunosuppression impairs effector T cell peripheral
homing without affecting induction, expansion, and memory. J Immunol. 164:
5761−5770. (2000).
[69] Rassow J , Hauser K , Netzker R , Deutzmann R. Duale Reihe: Biochemie.
Georg thieme Verlag KG. 2008.
[70] Reeves S A , Helman L J , Allison A , Israel M A. Molecular cloning and primary
structure of human glial fibrillary acidic protein. Proc Nat Acad Sci. 86: 5178-5182.
(1989).
[71] Ripps M E , Huntley G W , Hof P R , Morrison J H , Gordon J W. Transgenic
mice expressing an altered murine superoxide dismutase gene provide an animal
model of amyotrophic lateral sclerosis. Proc Natl Acad Sci USA. 92: 689–693. (1995).
[72] Rosen DR S T. Mutations in Cu/Zn superoxide dismutase gene are associated
with familial amyotrophic lateral sclerosis. Nature. 362, 59-62. (1993).
[73] Rosen D R , Siddique T , Patterson D , Figlewicz D A , Sapp P , Hentati A ,
Donaldson D , Goto J , O'Regan J P , Deng H-X , Rahmani Z , Krizus A , Yasek M ,
Cayabyab A , al. e. Mutations in Cu/Zn superoxide dismutase gene are associated with
familial amyotrophic lateral sclerosis. Nature. 362, 59 - 62. (1993).
[74] Rothstein J D , Dykes-Hoberg M , Pardo C A , Bristol L A , Jin L , Kuncl R W ,
Kanai Y , Hediger M A , Wang Y , Schielke J P , Welty D F. Knockout of glutamate
transporters reveals a major role for astroglial transport in excitotoxicity and clearance
of glutamate. Neuron. 16: 675–86. (1996).
[75] Rothstein J D , Van Kammen M , Levey A I , Martin L J , Kuncl R W. Selective
loss of glial glutamate transporter GLT-1 amyotrophic lateral sclerosis. Annals of
Neurology. vol. 38, 1: 73–84. (1995).
[76] Rowland L P. Amyotrophic lateral sclerosis: theories and therapies. AnnNeurol.
35: 129–30. (1994).
[77] Rowland . Hereditary and acquired motor neuron diseases. Williams & Wilkins.
1995.
[78] Sallusto F , Lenig D , Forster R , Lipp M , Lanzavecchia A. Two subsets of
memory T lymphocytes with distinct homing potentials and effector functions. Nature.
401: 708–712. (1999).

95
[79] Sanna M G , Wang S K , Gonzalez-Cabrera P , Don A , Marsolais D , Matheu
M , Wei S H , Parker I , Jo E , Cheng W C , Cahalan M D , Wong C H , Rosen H.
Enhancement of capillary leakage and restoration of lymphocyte egress by a chiral
S1P1 antagonist in vivo. Nature Chem Biol. 2: 434–441. (2006).
[80] Schroder K , Hertzog P J , Ravasi T , Hume D A. Interferon-γ: an overview of
signals, mechanisms and functions. Journal of Leukocyte Biology. Vol. 75, 2: 163-189.
(2003).
[81] Schwarz J , Sharp P S , Dick J R , Greensmith L. The effect of peripheral nerve
injury on disease progression in the SOD1(G93A) mouse model of amyotrophic lateral
sclerosis. Neuroscience. 130: 897–910. (2005).
[82] Scott L J. Fingolimod: a review of its use in the management of relapsing-
remitting. CNS Drugs. 25: 673-698. (2011).
[83] Soliven B , Miron V , Chun J. The neurobiology of sphingosine 1-phosphate
signaling and sphingosine 1-phosphate receptor modulators. Neurology. 76: 9-14.
(2011).
[84] Spielmann H M H. In-Vitro-Methoden in Lehrbuch der Toxicologie, 1. Auflage .
BI-Wissenschaftlicher Verlag. 1994.
[85] Stieber A , Gonatas J O , Gonatas N K. Aggregates of mutant protein appear
progressively in dendrites, in periaxonal processes of oligodendrocytes, and
inneuronal and astrocytic perikarya of miceexpressing the SOD1(G93A) mutation of
familial amyotrophic lateral sclerosis. J Neurol Sci. 177: 114-123. (2000).
[86] Thermoscientific. www.thermo.com/pierce. 2007Abrufbar unterZuletzt
abgerufen .
[87] Van Damme P , Dewil M , Robbrecht W , Van den Bosch L. Excitotoxicity and
Amyotrophic Lateral Sclerosis. Neurodegenerative Dis. 2: 147-159. (2005).
[88] Vargas M R , Johnson D A , Johnson J A. Decreased glutathione accelerates
neurological deficit and mitochondrial pathology in familial ALS-linked hSOD1(G93A)
mice model. Neurobiol Dis. 43: 543-551. (2011).
[89] Watanabe Y , Kono Y , Nanba E , Ohama E , Nakashima K. Instability of
expressed Cu/Zn superoxide dismutase with 2 bp deletion found inamyotrophic lateral
sclerosis. FEBS Lett. 400: 108–12. (1997).
[90] Wei Y , Yemisci M , Kim H H , Yung L M , Shin H K , Hwang S K , Guo S , Qin
T , Alsharif N , Brinkmann V , Liao J K , Lo E H , Waeber C. Fingolimod provides long

96
term protection in rodent models of cerebral ischämia. Annais of Neurology. 69: 119-
29. (2011).
[91] Wijesekera L C , Leigh P N. Amyotrophic lateral sclerosis. Orphanet J Rare. 4:
3. (2009).
[92] Wong P C , Pardo C A , Borchelt D R , Lee M K , Copeland N G , Jenkins N A ,
Sisodia S , Cleveland D W , Price D L. An adverse property of a familial ALS-linked
SOD1 mutation causes motor neuron disease characterized by vacuolar degeneration
of mitochondria. Neuron. 14: 1105–16. (1995).
[93] Yan Q , Matheson C R , Talvenheimo J , Welcher A A , Feinstein S C.
Immunocytochemical localization of TrkB in the central nervous system of the adult rat.
J Comp Neurol. 378: 135–157. (1997).
[94] Yan Q , Rosenfeld R D , Matheson C R , Hawkins N , Lopez O T , Bennett L ,
Welcher A A. Expression of brain-derived neurotrophic factor protein in the adult rat
central nervous system. Neuroscience. 78: 431–448. (1997).
[95] Yoshihara T , Ishigaki S , Yamamoto M , Liang Y , Niwa J , Takeuchi H , Doyu
M , Sobue G. Differential expression of inflammation- and apoptosis-related genes in
spinal cords of a mutant SOD1 transgenic mouse model of familial amyotrophic lateral
sclerosis. J Neurochem. 80: 158–67. (2002).

97
Anhang
Vortest der Neurofilamente NFH
„Spike-and-Recovery“ und „Linearity of Dilution“ Experimente sind wichtige Methoden
zur Validierung und Beurteilung der Genauigkeit von ELISAs (Thermoscientific 2007).
„Spike and Recovery“
Das Spike und Recovery wird gewöhnlich bestimmt, um festzustellen, ob ein
Unterschied durch die Detektion des zu analysierenden Materials zwischen dem
Verdünnungsmittel (Diluent) der Standardkurve und der biologischen „sample matrix“
(biologische Probe +Verdünnungsmittel) besteht.
Dabei wird ein bekannter Betrag an Analyt (Spike) zu dem zu analysierenden Material
hinzugefügt und im Assay gemessen. Verglichen wird der gemessene Wert mit dem
identischen Spike in der Standardkurve. Als Kontrolle dient das zu analysierende
Material + Diluent (Thermoscientific 2007).
„Linearity of Dilution“
Das Linearity of Dilution Experiment sagt etwas über die Genauigkeit der Assay
Ergebnisse bei unterschiedlicher Verdünnung der zu testenden Probe aus.
Die Linearität ist hier definiert als der kalkulierte Betrag des zu analysierenden
Materials basierend auf der Standardkurve. Ist die Linearität über einen weiten Bereich
der Verdünnung vorhanden, dann ist der Assay sehr flexibel in der Verwendung von
verschiedenen Probenspiegeln. Ein zu analysierendes Material mit hohen
Probenspiegeln kann mehrfach verdünnt werden, ohne dass es aus der Standardkurve
herausfällt. Dagegen kann ein zu analysierendes Material mit niedrigen Spiegeln nur
ohne Verdünnung gemessen werden (Thermoscientific 2007).
Durchführung des Vortests:
Zur Herstellung der Verdünnung Probe 1 (Nr. 297) wurden 2,4μl des Homogenats in
117,6μl des ELISA Assay Puffers (1:50) gelöst und 5 Sek. gevortext. Anschließend
wurden aus der einfachen (1:50) Verdünnung 60μl entnommen und in 60μl des ELISA

98
Assay Puffers (1:2) gelöst. Ich einem weiteren Schritt wurden ebenfalls 60μl
entnommen und in 60μl des ELISA Assay Puffers (1:4) gelöst.
Der pNF-H Standard wurde, wie vom Hersteller beschrieben vorbereitet. Aus dem
Tube mit 15ng/ml blieben 20μl für den Vorversuch übrig. Von dieser Menge wurden
10μl entnommen und mit 30μl ELISA Assay Puffer (1:3) auf eine Konzentration von
3,75 ng/ml verdünnt. Anschließend wurden 10μl aus den 40μl der 3,75 ng/ml
Standardlösung herausgenommen und mit 40μl der jeweils unterschiedlich (1, 1:2, 1:4)
verdünnten Proben 1 gelöst und für 5 Sek. gevortext.
Zur Herstellung der Verdünnungsreihe (1:2; 1:4; 1:8; 1:16) von Probe 2 (Nr.
311)wurden 4,8μl des Homogenats in 235,2μl des ELISA Assay Puffers gelöst und 5
Sek. gevortext. Anschließend wurden aus der einfachen (1:50) Verdünnung 120μl
entnommen und in 120μl des ELISA Assay Puffers (1:2) gelöst. In einem weiteren
Schritt wurden ebenfalls 60μl entnommen und in 60μl des ELISA Assay Puffers (1:4)
gelöst. Mit den andern beiden Verdünnungen wurde genauso verfahren.
Bevor die Wells mit den Proben belegt werden konnten, wurden sie mit je 100 μl TBS
hydratisiert und 10 Min. bei Raumtemperatur inkubiert. Dann entfernte man das TBS
und klopfte die Wells trocken.
Anschließend wurden sie, wie im Protokoll dokumentiert, mit je 50μl des Standards
und der Proben 1 + 2 belegt.
Jede Probe wurde doppelt bestimmt. Der Standard und die Proben wurden 1 h bei RT
unter leichtem Schütteln inkubiert. Die Wells wurden während der Inkubationszeit mit
einem Plate Sealer abgedeckt.
Nachdem die Inkubationszeit abgelaufen war, wurden die Wells von den
Probelösungen befreit und ausgeklopft. Es folgte der Waschschritt:
Die Wells wurden mit je 200 μl Waschpuffer gelöst. Nach kurzem Schütteln der
Wellplatte wurden sie erneut ausgeklopft. Dieser Vorgang wurde sechsmal wiederholt.
Infolge wurde jeder Well mit 100 μl des gelösten anti-pNF-H Antikörpers gelöst, und
sie wurden erneut 1 h bei RT unter leichtem Schütteln inkubiert.
Nachdem die Inkubationszeit abgelaufen war, folgte ein weiterer Waschschritt, wie
schon oben beschrieben.
Im folgenden Schritt wurde jeder Well mit 100 μl des gelösten Goat anti-rabbit
polyklonalen Antikörpers, der mit einer Alkaline Phosphatase konjugiert war, benetzt

99
und erneut 1 h bei RT unter leichtem Schütteln inkubiert. Es folgte wieder ein
Waschvorgang.
Anschließend wurden die Wells mit jeweils 200 μl 1×TBS gespült und danach wieder
trocken geklopft.
Infolge gab man dann 100 μl des gelösten 1× pNPP Alkaline Phospatase Substrats in
die Wells.
Die 96-Well Platte wurde daraufhin im Dunkeln ca. 1h bei RT gelagert. Sofort danach
wurden die Wells mit Hilfe des ELISA Plate Readers bei einer Wellenlänge von 430
nm gemessen.
Auswertung des Vortest
Jede Probe wurde doppelt bestimmt und deren gemessene Werte wurden gemittelt.
Recovery
Recovery in %=100× ((Probe mit Spike – Spike)/ Probe ohne Spike)
Tabelle 6: Auswertung des Spike and Recovery Experimentes der pNFH. Gemessen wurde die Probe
2 mit und ohne Spike in den Verdünnungen 1, 1:2, 1:4, sowie der Spike ohne Probe 2. Es wurde
daraufhin der Mittelwert, der Recovery und der Recovery in % ermittelt. Untersuchung des
Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014. Grün= die verwendete
Verdünnung im Experiment pNFH ELISA, pNFH= phosphorilierte Neurofilamente Heavy Chain.
2. Probe
ohne Spike:
1-fach:
2. Probe
ohne Spike:
1:2
2. Probe
ohne Spike:
1:4
2. Probe mit
Spike: 1-
fach:
2. Probe mit
Spike: 1:2
2. Probe mit
Spike: 1:4 Spike
1. Rohdaten 0.88404443 0.53506564 0.31728919 1.6892751 1.6892751 1.26037475 1.08964512
2. Rohdaten 0.98577664 0.61030831 0.53506564 1.83192048 1.66584685 1.8078966 1.3258696
Mittelwert 0.935 0.573 0.426 1.761 1.678 1.534 1.208
Spike 0.553 0.470 0.326
Recovery 0.826 1.105 1.108
Recovery in
% 68.4 91.5 91.7

100
Linearity of Dilution
Tabelle 7: Auswertung des Linearity of Dilution Experiment der pNFH. Gemessen wurde die Probe 2 in
den Verdünnung 1, 1:2, 1:4, 1:8; 1:16. Jede Verdünnungsreihe wurde doppelt bestimmt und der
Mittelwert ermittelt. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im
Jahr 2014. Grün= die verwendete Verdünnung im Experiment pNFH ELISA, pNFH= phosphorilierte
Neurofilamente Heavy Chain.
Abbildung 18: Standardkurve des Vortest pNFH ELISA. pNFH= phosphorilierte Neurofilamente Heavy
Chain. Untersuchung des Wirkmechanismus von FTY720 am Universitätsklinikum Ulm im Jahr 2014.
Probe 2: 1-
fach Probe 2: 1:2 Probe 2: 1:4 Probe 2: 1:8 Probe 2: 1:16
1. Rohdaten 0.784415932 0.591374473 0.370599005 0.299691994 0.162197235
2. Rohdaten 0.884044429 0.591374473 0.479496996 0.424672343 0.213048521
Mittelwert 0.834 0.591 0.425 0.362 0.188

101
Danksagung

102

103
Lebenslauf


















