
Untersuchungen zum „Quorum Sensing“ (QS) cyanobakterieller ... Orszag_Diplomarbeit.pdf ·...
124
Untersuchungen zum „Quorum Sensing“ (QS) cyanobakterieller Begleitbakterien und des Cyanobakteriums Stamm Flo 1 Diplomarbeit vorgelegt dem Fachbereich 2 (Biologie/Chemie) an der Universität Bremen von Peter Orszag Mai 2008 Betreuender Gutachter: Prof. Dr. Ulrich Fischer Zweiter Gutachter: PD Dr. Katarzyna A. Palinska
Transcript of Untersuchungen zum „Quorum Sensing“ (QS) cyanobakterieller ... Orszag_Diplomarbeit.pdf ·...

Untersuchungen zum „Quorum Sensing“ (QS) cyanobakterieller Begleitbakterien und des
Cyanobakteriums Stamm Flo 1
Diplomarbeit
vorgelegt dem
Fachbereich 2 (Biologie/Chemie)
an der Universität Bremen
von
Peter Orszag
Mai 2008
Betreuender Gutachter: Prof. Dr. Ulrich Fischer
Zweiter Gutachter: PD Dr. Katarzyna A. Palinska

Danksagung
Danksagung
An erster Stelle möchte ich mich bei Herrn Prof. Dr. Ulrich Fischer für die
Bereitstellung meines Arbeitsplatzes sowie die Diskussions- und Hilfsbereitschaft bei
der Anfertigung meiner Diplomarbeit bedanken.
Bei Frau PD Dr. Katarzyna A. Palinska möchte ich mich für ihr Interesse und für die
Begutachtung meiner Diplomarbeit bedanken.
Ein großes Dankeschön gilt Dr. Birgit Heyduck-Söller, die mich hervorragend betreut
hat und immer Zeit sowie viel Geduld hatte.
Meine ehemaligen und gegenwärtigen Kollegen der Arbeitsgruppe danke ich für die
angenehme Arbeitsatmosphäre, ihre Unterstützung und stete Hilfsbereitschaft sowie
für die „Verpflegung“ durch Kekse und leckere Kuchen.
Ein besonderer Dank gilt meinen Eltern, die mich über die gesamte Zeit des Studiums
unterstützt haben.

Inhaltsverzeichnis
Inhaltsverzeichnis
Abkürzungsverzeichnis ............................................................................................ i
Nomenklatur für aufgeführte AHL-Signalmoleküle .............................................. iii
1. Einleitung .............................................................................................................. 1
1.1 Zell-Zell-Kommunikation („Quorum Sensing“)............................................................................ 1
1.2 „Quorum Sensing“ bei Vibrio fischeri als grundlegendes Modell einer AHL-abhängigen Genregulation bei Gram-negativen Bakterien............................................................................. 4
1.3 N-Acyl-L-Homoserinlactone bei Gram-negativen Bakterien ...................................................... 6
1.4 AHL-Biosensoren............................................................................................................................ 7
1.5 Interspezifische Kommunikation bei Bakterien........................................................................... 7
1.6 „Quorum Quenching“ (QQ): Inhibierung der Zell-Zell-Kommunikation.................................... 9
1.7 Auswahl an Mikroorganismen als AHL-Produzenten ............................................................... 10
1.8 Ziel der Arbeit................................................................................................................................ 11
2. Material und Methoden....................................................................................... 12
2.1 Verwendete Bakterienstämme und deren Herkunft .................................................................. 12
2.2 Verwendete Kulturmedien ........................................................................................................... 13 2.2.1 ASNIII/2-Medium........................................................................................................................ 13 2.2.2 LB(Luria Bertani)-Medium........................................................................................................ 14
2.2.2.1 LB(Luria Bertani)-Weichagar........................................................................................... 15 2.2.3 Leuchtbakterien-Medium.......................................................................................................... 15 2.2.4 ABG-Medium............................................................................................................................ 16
2.2.4.1 ABG-Weichagar............................................................................................................... 17
2.3 Pufferlösungen.............................................................................................................................. 17
2.4 Lagerung der Bakterienstämme.................................................................................................. 19
2.5 Voranzucht der zu untersuchenden Bakterienstämme ............................................................ 19 2.5.1 Voranzucht der heterotrophen Stämme Bo 53-33 und Bo 10-09 ............................................ 19 2.5.2 Voranzucht der Cyanobakterienstämme Flo 1, Bo 10 und Bo 53............................................ 20
2.6 Untersuchungen zum Wachstumsverhalten der heterotrophen Stämme Bo 53-33 und Bo 10-09......................................................................................................................................... 20
2.6.1 Untersuchungen zum Wachstumsverhalten in ungepuffertem ASNIII/2-Medium im Dunkeln .. 21 2.6.2 Untersuchungen zum Wachstumsverhalten bei unterschiedlichen pH-Werten im Dunkeln.... 21 2.6.3 Untersuchungen zum Wachstumsverhalten bei einem Photonenfluss von 5 µE m-2 s-1 PAR. 22 2.6.4 pH-Wert-Bestimmung in den Kulturüberständen ..................................................................... 22
2.7 Nachweis von N-Acyl-L-Homoserinlactonen ............................................................................. 23 2.7.1 Verwendete Sensorbakterienstämme zum Nachweis von AHL-Signalmolekülen................... 23
2.7.1.1 Chromobacterium violaceum CV026............................................................................... 23 2.7.1.2 Agrobacterium tumefaciens NTL4 (pZLR4) .................................................................... 24

Inhaltsverzeichnis
2.7.2 Kultivierungsbedingungen und Inkubationsdauer der Versuchskulturen zur Untersuchung von AHL-Signalmolekülen........................................................................................................ 25
2.7.2.1 Kultivierungsbedingungen der AHL-Versuchskulturen mit den heterotrophen Bakterienstämmen Bo 53-33 und Bo 10-09 .................................................................... 25
2.7.2.2 Anzucht und Kultivierungsbedingungen der AHL-Versuchskulturen mit dem Cyanobakterium Stamm Flo 1......................................................................................... 26
2.7.3 Extraktion von N-Acyl-L-Homoserinlactonen der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie des Cyanobakteriums Stamm Flo 1........................................................ 27
2.7.3.1 Herstellung zellfreier Kulturüberstände ........................................................................... 27 2.7.3.2 Azidifizierung von Kulturüberständen.............................................................................. 27 2.7.3.3 Extraktion von AHL-Molekülen aus Kulturüberständen................................................... 28 2.7.3.4 Extraktion von AHL-Molekülen aus Bakterienzellen ....................................................... 28
2.7.4 Dünnschichtchromatographische Auftrennung von AHL-Signalmolekülen ............................. 29 2.7.5 Visualisierung der DC-Signale ................................................................................................. 30
2.7.5.1 Anzucht der Indikatorbakterien und weiterer Referenzstämme ...................................... 30 2.7.5.2 Überschichtung der DC-Platten mit Indikatorbakterien ................................................... 31
2.7.6 Auswertung und Dokumentation der DC-Platten ..................................................................... 31 2.7.7 Agarplattentests mit Sensorbakterien zum Nachweis AHL-produzierender Bakterien............ 32
2.7.7.1 Agarplattentest nach RAVN und Mitarbeitern (2001)...................................................... 33 2.7.7.2 Agarplattentest nach FARRAND und Mitarbeitern (2002) .............................................. 33 2.7.7.3 Agarplattentest mit Chromobacterium violaceum CV026 zum Nachweis von AHL-
Signalmolekülen nach MCCLEAN und Mitarbeitern (1997)............................................ 34
2.8 Molekularbiologische Untersuchungen ..................................................................................... 35 2.8.1 Extraktion der chromosomalen DNA........................................................................................ 35 2.8.2 Bestimmung der DNA-Konzentration....................................................................................... 37 2.8.3 Polymerase-Kettenreaktion (PCR)........................................................................................... 37 2.8.4 Gelelektrophoretische Auftrennung der PCR-Produkte........................................................... 39 2.8.5 Elution und Aufreinigung von PCR-Produkten......................................................................... 40 2.8.6 Sequenzierung der 16S-rDNA und des gyrB-Gens ................................................................. 40 2.8.7 Erstellung von phylogenetischen Stammbäumen.................................................................... 40
2.9 Herkunft der verwendeten Chemikalien ..................................................................................... 41
3. Ergebnisse .......................................................................................................... 42
3.1 Verwendete Puffersysteme.......................................................................................................... 42
3.2 Wachstumsverhalten der heterotrophen Stämme Bo 53-33 und Bo 10-09 bei unterschiedlichen pH-Werten ..................................................................................................... 43
3.2.1 pH-Werte der Zellkulturüberstände.......................................................................................... 46
3.3 Wachstumsverhalten der heterotrophen Stämme Bo 53-33 und Bo 10-09 bei einem Photonenfluss von 5 µE m-2 s-1 PAR........................................................................................... 47
3.3.1 pH-Werte der Zellkulturüberstände.......................................................................................... 49
3.4 Screeningtests zum Nachweis AHL-produzierender Bakterien............................................... 50 3.4.1 Nachweis von AHL-Molekülen mit Chromobacterium violaceum CV026 nach MCCLEAN
und Mitarbeitern (1997)............................................................................................................ 50 3.4.2 Nachweis AHL-produzierender Bakterien nach RAVN und Mitarbeitern (2001) ..................... 51 3.4.3 Nachweis AHL-produzierender Bakterien nach FARRAND und Mitarbeitern (2002).............. 52

Inhaltsverzeichnis
3.5 Untersuchungen des Einflusses verschiedener Kultivierungsbedingungen auf die AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie durch das Cyanobakterium Stamm Flo 1 .................................................................................. 54
3.5.1 Einfluss des pH-Wertes auf die AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie durch das Cyanobakterium Stamm Flo 1 ................................ 54
3.5.1.1 Azidifizierung von Kulturüberständen der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 ................................................................................................................... 57
3.5.2 Einfluss von Licht auf die Produktion von AHL-Molekülen durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09............................................................................... 61
3.5.3 Einfluss der Temperatur und der Kohlenstoffquelle auf die Produktion von AHL-Molekülen durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 ....................................... 63
3.5.4 Produktion von AHL-Molekülen durch das Cyanobakterium Stamm Flo 1.............................. 66 3.5.5 Intrazelluläre AHL-Untersuchungen mit den heterotrophen Bakterienstämmen Bo 53-33
und Bo 10-09 sowie dem Cyanobakterium Stamm Flo 1 ........................................................ 67
3.6 Auswertung der molekularbiologischen Untersuchungen ...................................................... 69 3.6.1 Extraktion der chromosomalen DNA........................................................................................ 69 3.6.2 Amplifikation der 16S-rDNA ..................................................................................................... 69 3.6.3 Erstellung von phylogenetischen Stammbäumen auf Basis der 16S-rDNA-Sequenz............ 69 3.6.4 Amplifikation des gyrB-Gens.................................................................................................... 73 3.6.5 Internal Transcribed Spacer (ITS)............................................................................................ 73
4. Diskussion........................................................................................................... 75
4.1 Wachstumsverhalten der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 unter verschiedenen Kultivierungsbedingungen ............................................................................... 75
4.1.1 Verwendete Puffersysteme...................................................................................................... 75 4.1.2 Wachstumsverhalten bei unterschiedlichen pH-Werten .......................................................... 76 4.1.3 Wachstumsverhalten unter Lichteinfluss ................................................................................. 78
4.2 „Quorum Sensing“(QS) bei verschiedenen Gram-negativen Bakterien................................. 79 4.2.1 AHL-Nachweis mit bakteriellen Biosensoren ........................................................................... 80 4.2.2 Screeningtests zum Nachweis AHL-produzierender Bakterien ............................................... 81 4.2.3 Extraktion und Nachweis von N-Acyl-L-Homoserinlactonen ................................................... 82 4.2.4 AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 unter
verschiedenen Kultivierungsbedingungen ............................................................................... 84 4.2.4.1 Einfluss des pH-Wertes auf die AHL-Signalmoleküle ..................................................... 84 4.2.4.2 Einfluss der Temperatur auf die AHL-Signalmoleküle .................................................... 87 4.2.4.3 Einfluss von Licht und Nährstoffen auf die Produktion von AHL-Signalmolekülen......... 88
4.2.5 Identifizierung der AHL-Signalmoleküle der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie des Cyanobakteriums Stamm Flo 1............................................................... 89
4.3 Molekularbiologie ......................................................................................................................... 96
5. Ausblick............................................................................................................... 98
6. Zusammenfassung ........................................................................................... 100
7. Anhang .............................................................................................................. 103
8. Literatur ............................................................................................................. 105

Abkürzungsverzeichnis
Abkürzungsverzeichnis
Abb. Abbildung AI Autoinducer Aqua bidest. doppelt destilliertes Wasser AHL(s) N-Acyl-L-Homoserinlacton(e) ASN Artificial Seawater Nutrient Bchl a Bakteriochlorophyll a BLAST basic local alignment search tool Bp Basenpaare CFB Cytophaga-Flavobacterium-Bacteroides DC Dünnschichtchromatographie DNA Desoxyribonukleinsäure dNTP Desoxynukleosidtriphosphat et al. et alteri ERG Eppendorfreaktionsgefäß FE Fleischextrakt g Gramm g Erdbeschleunigung GC-MS Gaschromatographie-Massenspektrometrie GFP Grün fluoreszierendes Protein h Stunde(n) HPLC High Performance Liquid Chromatography HPLC-MS High Performance Liquid Chromatography-Massen-
spektrometrie HSL(s) Homoserinlacton(e) ITS internal transcribed spacer (interne transkribierte Spacer) Kap. Kapitel l Liter LB Luria Bertani lt. laut M Molar mg Milligramm µg Mikrogramm µl Mikroliter µm Mikrometer
i

Abkürzungsverzeichnis
min Minute(n) ml Milliliter mM Millimolar MPI Max Planck-Institut n Anzahl N-Acyl-HSL(s) N-Acyl-L-Homoserinlacton(e) NCBI National Center for Biotechnology Information NJ Neighbour-Joining NK Negativkontrolle nm Nanometer O.D. optische Dichte p.A. per analyse (zur Analyse) PAR Photosynthetically Active Radiation (photosynthetisch
aktive Strahlung PCR Polymerase Chain Reaction (Polymerasekettenreaktion) Probennr. Probennummer QQ „Quorum Quenching” QS „Quorum Sensing” rDNA ribosomale Desoxyribonukleinsäure Rf-Wert retention factor (Retentionsfaktor) rpm rotation per minute (Rotation pro Minute) RT Raumtemperatur sec Sekunde(n) Tab. Tabelle TMM Trace-Metal-Mix Tris Trishydroxymethylaminomethan U Unit (µmol/min) UFT Zentrum für Umweltforschung und nachhaltige Techno-
logien ÜK Übernachtkultur UV Ultraviolett V Volt v/v (volume per volume) Volumen pro Volumen w/v (weight per volume) Gewicht pro Volumen X-Gal 5-Bromo-4-Chloro-3-Indolyl-β-D-Galactopyranosid 2D-PAGE zweidimensionale Polyacrylamid-Gelelektrophorese
ii

Nomenklatur für AHL-Signalmoleküle
Nomenklatur für aufgeführte AHL-Signalmoleküle
C4-HSL N-Butanoyl-L-Homoserinlacton (BHL)
C6-HSL N-Hexanoyl-L-Homoserinlacton (HHL)
C7-HSL N-Heptanoyl-L-Homoserinlacton
C8-HSL N-Octanoyl-L-Homoserinlacton (OHL)
3-oxo-C4-HSL N-(3-oxo-Butanoyl)-L-Homoserinlacton (OBHL)
3-oxo-C6-HSL N-(3-oxo-Hexanoyl)-L-Homoserinlacton (OHHL)
3-oxo-C8-HSL N-(3-oxo-Octanoyl)-L-Homoserinlacton (OOHL)
3-oxo-C12-HSL N-(3-oxo-Dodecanoyl)-L-Homoserinlacton (OdDHL)
3-hydroxy-C4-HSL N-(3-hydroxy-Butanoyl)-L-Homoserinlacton (HBHL)
3-hydroxy-C6-HSL N-(3-hydroxy-Hexanoyl)-L-Homoserinlacton (HHHL)
3-hydroxy-C8-HSL N-(3-hydroxy-Octanoyl)-L-Homoserinlacton (HOHL)
3-hydroxy-C12-HSL N-(3-hydroxy-Dodecanoyl)-L-Homoserinlacton (HdDHL)
iii

1. Einleitung
1. Einleitung
Lange Zeit bestand die Auffassung, dass Bakterien unabhängig voneinander in strikt
solitärer Lebensweise existieren. Bereits vor 65 Jahren lieferten jedoch
Untersuchungen an marinen Leuchtbakterien erste Hinweise auf ein multizelluläres
Verhalten von Bakterien (DOUDOROFF, 1942). Die Fähigkeit zur interzellulären
Kommunikation bildet hierbei die Voraussetzung für eine derartige mikrobielle
Organisationsform (BEN JACOB et al., 2004; PARSEK und GREENBERG, 2005).
Die Zell-Zell-Kommunikation wurde erstmals bei dem Gram-positiven Bakterium
Streptococcus pneumoniae (TOMASZ, 1965) und den marinen Gram-negativen
Bakterien Vibrio fischeri und Vibrio harveyi (NEALSON et al., 1970; NEALSON und
HASTINGS, 1979) beschrieben, wobei chemische Signale der Kommunikation und
der Koordination von Gruppenaktivitäten dienen. Das Phänomen einer interzellulären
Kommunikation bei Bakterien wurde erst Jahrzehnte später anerkannt und
Kommunikation zwischen Individuen nicht mehr einzig höheren Organismen oder
spezialisierten Bakterien zugeschrieben. Anfang der 1990er Jahre gewann dieses
Arbeitsgebiet aufgrund von Untersuchungen an pflanzen- und humanpathogenen
Gram-negativen Bakterien (GAMBELLO und IGLEWSKI, 1991; BAINTON et al.,
1992; Fuqua et al., 1994) zunehmend an Bedeutung. Mittlerweile wird die Zell-Zell-
Kommunikation als die wichtigste intrazelluläre Signalreaktion zwischen Bakterien
verstanden (SZENTHE und PAGE, 2003).
1.1 Zell-Zell-Kommunikation („Quorum Sensing“) In der Literatur werden Kommunikationssysteme, mit Hilfe derer Bakterien die eigene
Populationsdichte wahrnehmen und in Abhängigkeit von dieser physiologische
Aktivitäten regulieren können, als „Quorum Sensing“-Systeme zusammengefasst.
Dabei stellt die kleinste Einheit, welche zu Zelldichte-abhängigen Reaktionen
befähigt ist, per Definition das Quorum dar (FUQUA et al., 1994; GEISENBERGER,
2000). Zur Ermittlung der Zellpopulationsdichte verwenden Mikroorganismen eine
große Vielfalt an niedermolekularen Substanzen, die unterschiedliche chemische
Strukturen aufweisen (siehe Abb. 1.1) (DONG und ZHANG, 2005; WILLIAMS, 2007).
Diese als „Autoinducer“ bezeichneten Signalmoleküle sind meistens sehr
artspezifisch (SMITH et al., 2004) und werden von den Bakterien zur Koppelung der
Expression bestimmter Gene an die Populationsdichte eingesetzt, welches eine
1

1. Einleitung
synchrone Koordination biologischer Aktivitäten innerhalb einer lokalen Population
ermöglicht (CHA et al., 1998; WILLIAMS, 2007). Das Wahrnehmen der eigenen
Populationsdichte wird dabei durch die Messung der Signalmolekül-Konzentration
ermöglicht, da die Konzentration der produzierten und in die Umwelt freigesetzten
Signalmoleküle mit Zunahme der Bakterienpopulation proportional ansteigt.
Mittlerweile wurden verschiedene QS-Systeme beschrieben, welche sich in der
Verwendung der Signalmoleküle und regulatorischen Proteine unterscheiden. Das
LuxR/LuxI-QS-System mit N-Acyl-L-Homoserinlactonen (AHLs) als Signalmolekülen
wird von den Gram-negativen Bakterien verwendet (FUQUA et al., 2001;
WHITEHEAD et al., 2001). Weitere QS-Systeme vereinigen Komponenten Gram-
negativer und Gram-positiver QS-Systeme, wie für Vibrio harveyi beschrieben
(READING und SPERANDIO, 2006). Dieses marine Bakterium verwendet zwei QS-
Systeme, ein AHL-QS-System zur intraspezifischen Kommunikation (BASSLER et
al., 1994) und ein weiteres System zur interspezifischen Kommunikation (SURETTE
und BASSLER, 1998; FEDERLE und BASSLER, 2003), an dem ein
Furanosylboratdiester (borhaltiges Furanon) als „Autoinducer“ („Autoinducer II“(AI-2),
siehe Abb. 1.1) beteiligt ist (CHEN et al., 2002). Genomanalysen belegten, dass das
für die AI-2-Synthese benötigte LuxS-Enzym unter den Bakterien weitverbreitet ist
(SUN et al., 2004). Für die mittlerweile weiteren nachgewiesenen Gram-positiven
und Gram-negativen Bakterien mit AI-2-Signalmolekülen (LuxS/AI-2-QS-System)
wird ebenfalls angenommen, dass die universellen Signalmoleküle eine
interspezifische Kommunikation ermöglichen (WATERS und BASSLER, 2005).
Die am häufigsten verwendeten Signalmoleküle zur interzellulären Kommunikation
unter den Gram-positiven Bakterien sind lineare, z.T. zyklische Oligopeptide (AIP =
autoinducing peptides) (KLEEREBEZEM et al., 1997; MILLER und BASSLER, 2001;
CAMILLI und BASSLER, 2006), aber es wurden auch γ-Butyrolactone bei
Streptomyceten und AI-2-Signalmoleküle bei z.B. Streptococcus pneumoniae,
S. anginosus und Staphylococcus aureus nachgewiesen (MILLER und BASSLER,
2001; AHMED et al., 2007, WILLIAMS, 2007) (siehe Abb. 1.1).
2

1. Einleitung
halogeniertes Furanon
Al-2
Abb. 1.1: Zusammenstellung verschiedener, repräsentativer QS-Signalmoleküle und deren Strukturen. 3-oxo-C6-HSL, N-3-oxo-hexanoyl-L-homoserine lactone; A-Factor, 2-isocapryloyl-3-hydroxymethyl-γ-butyrolactone; Pseudomonas quinolone signal (PQS), 2-heptyl-3-hydroxy-4(1H)-quinolone; HHQ, 2-heptyl-4(1H)-quinolone; DSF (“diffusible factor“), 11-methyl-2-dodecenoic acid; 3OH-PAME, hydroxyl-palmitic acid methyl ester; AIP-1, staphylococcal autoinducing peptide 1; DPD (AI-2-Precursor-Molekül), 4,5 dihydroxy-2,3-pentanedione; AI-2 (Autoinducer-2), furanosyl borate diester; halogeniertes Furanon aus Delisea pulchra. Strukturen zusammengestellt aus Angaben von WILLIAMS (2007) und FROMMBERGER (2005).
In den letzten Jahren wurde in vielen Gram-negativen Bakterienspezies,
überwiegend Proteobakterien, und in einigen Gram-positiven Bakterienspezies QS-
Systeme identifiziert, welche zum Informationsaustausch über den Zustand der
Gesamtpopulation befähigt sind. Damit bestätigte sich, dass „Quorum Sensing“ ein
weitverbreiteter Mechanismus der Genregulation in Bakterien ist (READING und
SPERANDIO, 2006; CAMILLI und BASSLER, 2006). Die Angaben über QS-Systeme
bei Gram-negativen Bakterien gehen in den Publikationen weit auseinander.
Berichten SHIN und Mitarbeiter (2007) von lediglich über 30 Spezies, konnten alleine
CASE und Mitarbeiter (2008) bei nahezu 50 Spezies (aus etwa 30 Gattungen) QS-
Systeme dokumentieren. Dabei werden für weit mehr Bakterienspezies AHL-QS-
Systeme angenommen.
Bereits 1996 wurde aber am Beispiel der Meeresalge Delisea pulchra aufgezeigt,
dass QS-Komponenten (z.B. halogenierte Furanone, siehe Abb. 1.1) auch in
Eukaryoten vorkommen (GIVSKOV et al., 1996), welche z.B. bakterielle QS-Systeme
durch eine Degradation des LuxR-Rezeptors inhibieren (MANEFIELD et al., 2002).
Die Alge wird folglich kaum von bakteriellen Biofilmen besiedelt.
3

1. Einleitung
„Quorum Sensing“ stellt ein Regulationssystem mit großem Wirkungsbereich dar,
welches eine Reaktion auf sich ändernde Umweltbedingungen ermöglicht bzw. der
Bakterienpopulation einen erheblichen Vorteil verschaffen kann (CASE et al., 2008).
QS kann dabei eine Vielzahl physiologischer Prozesse und phänotypischer
Veränderungen der Bakterienzelle regulieren. Diese Prozesse schließen die
Antibiotika-Produktion, die bakterielle Konjugation, die Produktion von
Virulenzfaktoren, die Sporenbildung, die Lumineszenz, die Biosynthese von
Exopolysacchariden, die Produktion von Sekundärmetaboliten und Exoenzymen, die
Regulation von Zellfunktionen in der stationären Wachstumsphase, die Motilität und
das Schwärmen, die Biofilmbildung und die Zellaggregation ein (WITHERS et al.,
2001; WATERS und BASSLER, 2005; WILLIAMS, 2007; DIGGLE et al., 2007a).
Dementsprechend kommt dem bakteriellen QS z.B. in der Medizin hinsichtlich neuer
Therapieansätze eine entscheidende Bedeutung zu, da die Ausprägung wichtiger
Virulenzfaktoren sowie die Antibiotikaproduktion bei einer Reihe von Organismen,
wie dem humanpathogenen Bakterium Pseudomonas aeruginosa, unter QS-
Kontrolle stehen (BJARNSHOLT und GIVSKOV, 2007).
1.2 „Quorum Sensing“ bei Vibrio fischeri als grundlegendes Modell einer AHL-abhängigen Genregulation bei Gram-negativen Bakterien
Das marine Leuchtbakterium Vibrio (Photobacterium) fischeri lebt in der Natur im
freien Meerwasser oder als Symbiont in Lichtorganen von Fischen und Weichtieren,
in denen das Bakterium zu sehr hohen Zelldichten anwächst (1010 - 1011 Zellen/ml)
und die Lumineszenz ausgelöst (BOETTCHER und RUBY, 1995). Der
Wirtsorganismus zieht Nutzen aus der Lichtemission und stellt den Bakterien
innerhalb der Lichtorgane im Gegenzug Nährstoffe zur Verfügung. Im freien
Meerwasser liegen lediglich geringe Zelldichten vor, wodurch eine Lumineszenz nicht
induziert und keine Energie in dieser nährstoffarmen Umgebung für eine unnötige
Lichtproduktion verbraucht wird (NEALSON und HASTINGS, 1979; LEE und RUBY,
1992).
Bei V. fischeri sind niedermolekulare Verbindungen („Autoinducer“) für diese
Zelldichte-abhängige Regulation der Lumineszenz verantwortlich (EBERHARD et al.,
1981). Die acylierten Derivate des Homoserinlactons (bei V. fischeri: 3-oxo-C6-HSL)
werden dabei von dem LuxI-Protein, einer AHL-Synthase, konstitutiv synthetisiert
(EBERHARD et al., 1981; ENGEBRECHT und SILVERMAN, 1984) und diffundieren
4

1. Einleitung
frei durch biologische Membranen und die Zellwand (KAPLAN und GREENBERG,
1985) (siehe Abb. 1.2 A). Aufgrund der freien Diffusion werden die Signalmoleküle
auch außerhalb der Bakterienzelle lokal akkumuliert. Dementsprechend steigt die
intrazelluläre und (lokale) extrazelluläre Konzentration dieser „Autoinducer“ bei
Zunahme der Zellpopulationsdichte proportional. Die AHL-Signalmoleküle binden bei
einer kritischen Schwellenwert-Konzentration (treshold concentration) als Ligand an
das LuxR-Protein (Transkriptionsaktivator) im Cytoplasma mit nachfolgender
Aktivierung der Transkription des luxICDABE-Operons durch Bindung des LuxR-
„Autoinducer“-Komplexes an die Promotorregion (ENGEBRECHT et al., 1983) (siehe
Abb. 1.2 B). Diese als lux-Boxen bezeichneten speziellen Promotorsequenzen sind
20 bp große invertierte, repetitive Sequenzen (ZHANG et al., 2002a). Neben den für
die Lumineszenz benötigten Genen kodiert das lux-Operon das luxI-Gen. Eine
gesteigerte Transkription der Lumineszenz-Gene führt somit durch eine positive
„feedback“-Regulation zu einer Verstärkung der eigenen Signalmolekül-Produktion
(Autoinduktion) (EBERHARD et al., 1981) (siehe Abb. 1.2 B). Studien an V. fischeri
belegen, dass das LuxR/LuxI-QS-System zudem weitere, nicht für die Lumineszenz
erforderliche Gene reguliert (QIN et al., 2007).
Abb. 1.2: Zelldichte-abhängige QS-Regulation der Lumineszenz bei Vibrio fischeri. A: Bei niedrigen Zelldichten werden die Lumineszenz-Gene luxCDABE aufgrund einer niedrigen Konzentration des Signalmoleküls (3-oxo-C6-HSL) lediglich schwach transkribiert. B: Bei hohen Zelldichten bindet das Signalmolekül bei Erreichen einer Schwellenwert-Konzentration als Ligand an das LuxR. Die gesteigerte Transkription der luxCDABE-Gene führt zur Lichtemission sowe zur Verstärkung der Signalmolekül-Produktion. Abbildung nach WHITEHEAD (2001), verändert nach FROMMBERGER (2005)
5

1. Einleitung
1.3 N-Acyl-L-Homoserinlactone bei Gram-negativen Bakterien In vielen Gram-negativen Bakterien (vor allem bei α-, β- und γ-Proteobakterien)
wurden mittlerweile LuxR/LuxI-homologe QS-Systeme nachgewiesen. Diese auf
molekularer Ebene am besten verstandenen QS-Systeme setzen sich aus
regulatorischen LuxI- und LuxR-homologen Proteinen zusammen, wobei die LuxI-
homologen Proteine die N-Acyl-L-Homoserinlactone (AHLs) als „Quorum
Sensing“(QS)-Signalmoleküle synthetisieren (CASE et al., 2008). Die Stoffklasse der
N-Acyl-L-Homoserinlactone zeichnet sich durch einen Homoserinlactonring und einer
N-acylierten Seitenkette aus, verbunden über eine Amid-Bindung. Die N-Acyl-
Seitenketten unterscheiden sich in Länge (zwischen 4 - 18 C-Atomen), Sättigung
sowie durch eine eventuelle Substitution am C3-Atom (3-Oxo- oder 3-Hydroxygruppe)
(WHITEHEAD et al., 2001; WILLIAMS, 2007) (siehe Abb. 1.3). Dieses bedingt eine
beträchtliche strukturelle Diversität von AHL-Molekülen.
Abb. 1.3: Struktur von AHL-Signalmolekülen. AHL, N-Acyl-L-Homoserinlacton; 3-oxo-AHL, N-3-oxo-Acyl-L-Homoserinlacton; 3-hydroxy-AHL, N-3-hydroxy-Acyl-L-Homoserinlacton. R = C1 bis C15. . Die Acyl-Seitenketten können zudem eine oder mehrere Doppelbindungen aufweisen. 3-oxo-C12-HSL, N-3-oxo-Dodecanoyl-L-Homoserinlacton. Strukturen zusammengestellt aus Angaben von WILLIAMS (2007)
S-Adenosyl-L-Methionin (SAM) und acyliertes acyl carrier protein (acyl-ACP),
letzteres ein Zwischenprodukt der Fettsäurebiosynthese, sind die Substrate der LuxI-
homologen Enzyme (AHL-Synthasen) für die AHL-Synthese. Der
Homoserinlactonring der N-Acyl-HSLs wird dabei aus dem S-Adenosyl-L-Methionin
(SAM) synthetisiert, die N-Acyl-Seitenkette der AHL-Moleküle aus den Acyl-Resten
der acyl-ACP-Moleküle (SCHAEFER et al., 1996).
6

1. Einleitung
1.4 AHL-Biosensoren Die Identifizierung einer großen Anzahl von AHL-QS-Systemen in Bakterien wurde
durch die Entwicklung und den Einsatz bakterieller AHL-Biosensoren bedeutend
vorangebracht, welche auch heutzutage in AHL-Untersuchungen Anwendung finden.
Diese bakteriellen Biosensoren produzieren selber keine AHL-Signalmoleküle,
zeichnen sich aber durch eine phänotypische Nachweisreaktion in Gegenwart
bestimmter N-Acyl-HSLs aus. Diese Reaktion beruht auf dem Vorhandensein eines
LuxR-homologen Proteins, welches die Aktivierung eines bestimmten Reportergens
induziert (z.B. Lumineszenz, β-Galaktosidase, GFP, Violacein-Pigment) (WAGNER-
DÖBLER et al., 2005; STEINDLER und VENTURI, 2007). Das jeweilige Protein aus
der LuxR-Familie bestimmt für jeden AHL-Biosensor die AHL-Spezifität, kann aber
auch in manchen Fällen (bei höheren Konzentrationen) durch nahverwandte AHLs
angesprochen werden. Dementsprechend gibt es gentechnisch konstruierte
Biosensoren, welche auf AHL-Moleküle mit kurz- und mittellangen Acyl-Seitenketten
(z.B. Chromobacterium violaceum CV026: MCCLEAN et al., 1997), mit langen Acyl-
Seitenketten (z.B. Pseudomonas aeruginosa PAO1 (M71LZ): DONG et al., 2005),
aber auch auf ein breites Spektrum von AHL-Molekülen (z.B. Agrobacterium
tumefaciens NTL4 (pZLR4): FARRAND et al., 2002) reagieren. Andere Biosensoren
reagieren hingegen nur auf bestimmte Modifikationen in der Acyl-Seitenkette, wie der
3-hydroxy-AHL detektierende Biosensor Pseudomonas fluorescens 1855 (pSF105,
pSF107) (KHAN et al., 2005). Neben Modifikation und Länge der Acyl-Seitenkette ist
die AHL-Konzentration für eine Aktivierung der jeweiligen Reportergene von großer
Bedeutung (Sensitivität) (STEINDLER und VENTURI, 2007).
1.5 Interspezifische Kommunikation bei Bakterien QS-Signalmoleküle sind größtenteils sehr art-spezifisch (SMITH et al., 2004). Die
N-Acyl-L-Homoserinlactone der Gram-negativen Bakterien weisen eine große
strukturelle Ähnlichkeit auf, werden aber dessen ungeachtet mit einer hohen
Spezifität von ihrem jeweiligen LuxR-homologen Protein erkannt (FEDERLE und
BASSLER, 2003; WAGNER-DÖBLER et al., 2005). Aufgrund dieser Spezifität
zwischen LuxR-Homologen und N-Acyl-HSL-Molekülen wurde das bakterielle AHL-
QS-System vor allem als ein intraspezifisches Kommunikationssystem verstanden
(FEDERLE und BASSLER, 2003; READING und SPERANDIO, 2006). Mittlerweile
sind einige LuxR-homologe Proteine bekannt, welche mehr als ein AHL-
7

1. Einleitung
Signalmolekül erkennen und in erster Linie an einer interspezifischen Kommunikation
beteiligt sind (READING und SPERANDIO, 2006). Eine Interspezies-Kommunikation
ist dabei nicht immer zu einem beiderseitigen Vorteil. Mögliche Interaktionen sind
dabei neben einer Aktivierung des LuxR-homologen Regulatorproteins des Weiteren
eine Blockierung der AHL-Bindungsstelle durch Spezies-fremde AHL-Moleküle. In
diesem Zusammenhang wurde für Chromobacterium violaceum eine Inhibierung der
Produktion von Violacein durch langkettige AHL-Moleküle (C10 - C14-Seitenketten)
beschrieben (MCCLEAN et al., 1997).
Ein Beispiel interspezifischer Kommunikation ist die zwischen den opportunistisch
pathogenen Bakterien Pseudomonas aeruginosa und Burkholderia cepacia, welche
die Lungen von Patienten mit Cystischer Fibrose co-infizieren und gemischte
Biofilme ausbilden. In diesem Fall aktivieren die synthetisierten AHL-Moleküle von
P. aeruginosa das Pathogenese regulierende QS-System bei B. cepacia, wodurch
B. cepacia Nutzen aus Spezies-fremden AHLs zieht (RIEDEL et al., 2001). Die LuxR-
homologen Proteine der beiden Spezies weisen eine hohe Verwandtschaft
zueinander auf, welches evolutionär auf einen lateralen Gentransfer schließen lässt,
resultierend in einer erleichterten Interspezies-Kommunikation (CASE et al., 2008).
Das marine Bakterium Vibrio harveyi existiert in natürlichen Habitaten in gemischten
Populationen, bestehend aus verschiedenen Bakterienspezies (MILLER und
BASSLER, 2001). Das interspezifische QS-System mit dem „Autoinducer II“(AI-2)-
Signalmolekül ermöglicht es V. harveyi neben der eigenen Populationsdichte zudem
die der anderen AI-2-synthetisierenden Bakterienspezies in ihrer lokalen Umgebung
wahrzunehmen (SURETTE und BASSLER, 1998; FEDERLE und BASSLER, 2003).
Die Fähigkeit einer Verhaltensänderung in Abhängigkeit von den
Populationsverhältnissen in der Gesamtpopulation beruht bei diversen
Bakterienspezies auf der Erkennung verschiedener AI-2-Signalmoleküle, ausgehend
vom 4,5-dihydroxy-2,3-pentanedione (DPD, siehe Abb. 1.1) (CHEN et al., 2002;
MILLER et al., 2004).
Untersuchungen an gemischten Bakterienpopulationen mit verschiedenen
Bakterienspezies zeigten, dass die AI-2-Produktion durch Escherichia coli in anderen
Bakterienspezies QS-gesteuerte Vorgänge bereits bei niedrigen Zelldichten
einleitete. Ein Abbau der AI-2-Signalmoleküle durch E. coli verursachte indes bei
benachbarten Bakterienspezies eine zu geringe Einschätzung der eigenen
8

1. Einleitung
Populationsdichte, wodurch QS-gesteuerte Prozesse nicht eingeleitet oder nicht
beendet wurden (XAVIER und BASSLER, 2005).
Neben der bakteriellen Interspezies-Kommunikation wurden auch Interaktionen
zwischen Bakterien und höheren Organismen beschrieben (inter-kingdom). Dabei
können höhere Organismen bakterielle QS-Systeme über sogenannte AHL-Mimetika
stimulieren, wobei Assoziationen mit Bakterien z.B. beim Menschen zur Erhaltung
seiner Darmflora dienen könnten (CAMILLI und BASSLER, 2006). In Reispflanzen
(Oryza sativa) wurden AHL-Mimetika nachgewiesen (DEGRASSI et al., 2007), deren
biologische Bedeutung bisweilen noch nicht erfasst wurde, aber vermutlich im
Zusammenhang mit Reis-pathogenen Burkholderia-Spezies stehen.
1.6 „Quorum Quenching“ (QQ): Inhibierung der Zell-Zell-Kommunikation Die Hemmung der interzellulären Kommunikation („Quorum Quenching“) stellt
womöglich einen wichtigen Mechanismus in der Regulation des bakteriellen QS, aber
auch bei der Interaktion von verschiedenen Mikroorganismen sowie von
Mikroorganismen und höheren Organismen dar. In diesem Zusammenhang
übernimmt wahrscheinlich der Abbau von Signalmolekülen durch „Quorum
Quenching“-Enzyme (als eine Möglichkeit des QQ) eine wichtige Funktion (DONG
und ZHANG, 2005). AHL-degradierende Enzyme, AHL-Lactonasen, wurden
beispielsweise bei Bacillus-Spezies (z.B. DONG et al., 2000) und bei Agrobacterium
tumefaciens (ZHANG et al., 2002b), AHL-Acylasen bei Pseudomonas aeruginosa
(HUANG et al., 2003) und einem Anabaena-Stamm (ROMERO et al., 2008)
nachgewiesen. Ein Abbau bzw. eine Inaktivierung von Signalmolekülen ermöglicht
wahrscheinlich, wie für A. tumefaciens beschrieben (ZHANG et al., 2004), das
Abschalten der QS-regulierten Genexpression. Des Weiteren wird für einige
Bakterienspezies eine Inhibierung der interzellulären Kommunikation konkurrierender
und pathogener Bakterienspezies vermutet. In vielen Fällen regulieren die
bakteriellen QS-Systeme die Expression von Virulenz-Genen sowie die Antibiotika-
Produktion. Die in höheren Organismen detektierten AHL-degradierenden Enzyme
besitzen ebenfalls eine inhibierende Funktion gegen pflanzen- bzw.
humanpathogene QS-Systeme (DONG und ZHANG, 2005). Dementsprechend kann
die bakterielle Virulenz deutlich herabgesetzt werden. Neben QS-Inhibitoren wird den
AHL-degradierenden Enzymen eine Rolle bei der medizinischen Bekämpfung von
humanpathogenen Bakterien zugesprochen (HASELTINE und ARNOLD, 2008).
9

1. Einleitung
1.7 Auswahl an Mikroorganismen als AHL-Produzenten In der vorliegenden Arbeit wurde eine Auswahl von marinen Gram-negativen
Bakterienstämmen für „Quorum Sensing“-Untersuchungen eingesetzt, welche zuvor
auf Grundlage von Voruntersuchungen (SCHOMAKER, 2007) als AHL-Produzenten
beschrieben worden waren.
Der marine heterotrophe Bakterienstamm Bo 53-33 wurde aus der nicht-axenischen
Cyanobakterienkultur Stamm Bo 53 (Nodularia harveyana) isoliert und nach 16S-
rDNA-Sequenzierung der Gattung Porphyrobacter zugeordnet (HUBE, 2008), welche
erstmals von FUERST und Mitarbeitern (1993) beschrieben wurde. Porphyrobacter-
Spezies sind aerobe anoxygen phototrophe Bakterien und gehören zur α-4-
Untergruppe der Proteobakterien (YURKOV und BEATTY, 1998).
Der marine heterotrophe Bakterienstamm Bo 10-09 wurde aus der nicht-axenischen
Cyanobakterienkultur von Oscillatoria brevis Stamm Bo 10 isoliert und nach
Sequenzierung der 16S-rDNA als Vertreter der Gattung Muricauda (Gruppe:
Cytophaga-Flavobacteria-Bacteroides, Familie: Flavobacteriaceae) identifiziert
(HUBE, 2008). Der Typenstamm Muricauda aquimarina ist ein Gram-negatives,
nicht-sporenbildendes, leicht halophiles Stäbchen (YOON et al., 2005). Das Genus
Muricauda wurde erstmals von BRUNS und Mitarbeitern (2001) beschrieben.
Weiterhin wurde das Cyanobakterium Stamm Flo 1 für QS-Untersuchungen
hinzugezogen. Dieser Cyanobakterienstamm gehört zur Ordnung Oscillatoriales und
wurde ursprünglich aufgrund rein morphologischer Kriterien als Oscillatoria limnetica
bezeichnet. Durch molekularbiologische Untersuchungen konnte SCHRÜBBERS
(2007) aber eindeutig aufzeigen, dass Stamm Flo 1 eine Spezies der Gattung
Geitlerinema ist.
Cyanobakterien leben in engen assoziierten Lebensgemeinschaften mit
heterotrophen Bakterien (STEVENSON und WATERBURY, 2006). Cyanobakterien
stellen den heterotrophen Bakterien im Austausch gegen remineralsierte Nährstoffe
labile Substrate zur Verfügung (TUOMAINEN et al., 2006). Durch Fluoreszenz-Insitu-
Hybridisierung (FISH) wurden Porphyrobacter Stamm Bo 53-33 und Muricauda
Stamm Bo 10-09 ebenfalls als Teil der heterotrophen Begleitflora des Geitlerinema-
Stammes Flo 1 detektiert (HUBE, 2008, persönliche Mitteilung). In diesem
Zusammenhang stellt sich die Frage, in wieweit die assoziierten
Lebensgemeinschaften interspezifisch über (AHL)-Signalmoleküle miteinander
kommunizieren und über QS-gesteuerte Prozesse beeinflusst werden.
10

1. Einleitung
1.8 Ziel der Arbeit In der vorliegenden Diplomarbeit sollte eine dünnschichtchromatographische
Methode mit Biosensoren zum Nachweis von AHL-Molekülen etabliert werden. Des
Weiteren sollte die AHL-Produktion in Abhängigkeit vom pH-Wert und der
Wachstumsphase für die heterotrophen Bakterien Porphyrobacter Stamm Bo 53-33
und Muricauda Stamm Bo 10-09 sowie für das Cyanobakterium Stamm Flo 1
untersucht werden. Eine Untersuchung des Lichteinflusses sowie weiterer
Einflussfaktoren wie Temperatur und Kohlenstoffquelle auf die AHL-Produktion der
heterotrophen Bakterien sollte zudem durchgeführt werden. Aussagen zum
Wachstumsverhalten der heterotrophen Bakterien sollten mittels physiologischer
Untersuchungen getroffen werden.
Ferner sollte durch die Anwendung molekularbiologischer Methoden wie 16S-rDNA-
Sequenzierung, gyrB-Gen-Analyse und ITS-Amplifikation überprüft werden, ob es
sich bei den aufgrund morphologischer Kriterien klassifizierten Cyanobakterien
Nodularia harveyana Stamm Bo 53 und Oscillatoria brevis Stamm Bo 10 tatsächlich
um diese Spezies handelt.
11

2. Material und Methoden
2. Material und Methoden
2.1 Verwendete Bakterienstämme und deren Herkunft Die in der vorliegenden Arbeit verwendeten Bakterienstämme sowie deren Herkunft
sind in den Tabellen 2.1 - 2.3 zusammengefasst. Hierbei erfolgte eine Unterteilung in
die zu untersuchenden Bakterienstämme (siehe Tab. 2.1 und Tab. 2.2) sowie in die
für den Nachweis von AHL-Signalmolekülen erforderlichen Stämme und
Referenzstämme (siehe Tab. 2.3).
Die Cyanobakterienstämme (siehe Tab. 2.2) wurden aus der Stammsammlung der
Abteilung Marine Mikrobiologie (UFT, Universität Bremen) entnommen. Die Stämme
Bo 53 und Bo 10 stammen ursprünglich von einem marinen Standort an der Ostsee
(Boiensdorf, Stammbezeichnung Bo), Stamm Flo 1 stammt aus einem
Mangrovenwäldchen in Key Biscayne (Florida (USA), Stammbezeichnung Flo). Die
heterotrophen Bakterienstämme (siehe Tab. 2.1) wurden von Dipl.-Biologin A. Hube
(Abt. Marine Mikrobiologie, UFT, Universität Bremen) zur Verfügung gestellt
(siehe Kap. 1.7).
Tab. 2.1: Liste der ausgewählten heterotrophen Bakterien und ihre Identifizierung nach HUBE (2008)
Stammbezeichnung Identifizierung nach Sequenzierung
Bo 10-09 Muricauda sp.
Bo 53-33 Porphyrobacter sp.
Tab. 2.2: Übersicht der ausgewählten Cyanobakterienstämme aus der Stammsammlung und deren Stammbezeichnung sowie ihre taxonomische Einordnung und ihre Abteilungszugehörigkeit nach RIPPKA und Mitarbeitern (1979)
Stammbezeichnung
Ordnung Oscillatoriales (Unterabteilung III)
Geitlerinema sp. a) Flo 1Oscillatoria brevis Bo 10
Ordnung Nostocales (Unterabteilung IV)
Nodularia harveyana Bo 53
Stamm und Abteilungszugehörigkeit(RIPPKA et al., 1979)
a) reklassifiziert (SCHRÜBBERS, 2007)
12
2. Material und Methoden
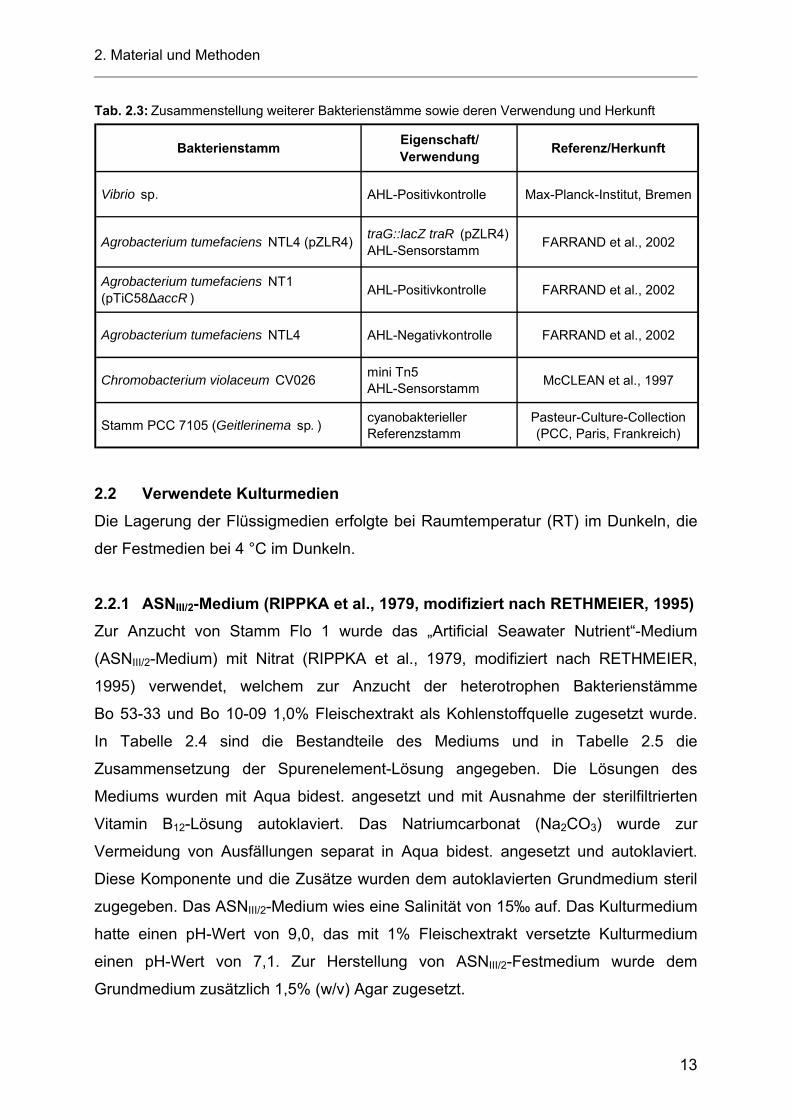
Tab. 2.3: Zusammenstellung weiterer Bakterienstämme sowie deren Verwendung und Herkunft
Bakterienstamm Eigenschaft/Verwendung Referenz/Herkunft
Vibrio sp. AHL-Positivkontrolle Max-Planck-Institut, Bremen
Agrobacterium tumefaciens NTL4 (pZLR4) traG::lacZ traR (pZLR4) AHL-Sensorstamm FARRAND et al., 2002
Agrobacterium tumefaciens NT1 (pTiC58∆accR ) AHL-Positivkontrolle FARRAND et al., 2002
Agrobacterium tumefaciens NTL4 AHL-Negativkontrolle FARRAND et al., 2002
Chromobacterium violaceum CV026 mini Tn5 AHL-Sensorstamm McCLEAN et al., 1997
Stamm PCC 7105 (Geitlerinema sp. ) cyanobakterieller Referenzstamm
Pasteur-Culture-Collection(PCC, Paris, Frankreich)
2.2 Verwendete Kulturmedien Die Lagerung der Flüssigmedien erfolgte bei Raumtemperatur (RT) im Dunkeln, die
der Festmedien bei 4 °C im Dunkeln.
2.2.1 ASNIII/2-Medium (RIPPKA et al., 1979, modifiziert nach RETHMEIER, 1995) Zur Anzucht von Stamm Flo 1 wurde das „Artificial Seawater Nutrient“-Medium
(ASNIII/2-Medium) mit Nitrat (RIPPKA et al., 1979, modifiziert nach RETHMEIER,
1995) verwendet, welchem zur Anzucht der heterotrophen Bakterienstämme
Bo 53-33 und Bo 10-09 1,0% Fleischextrakt als Kohlenstoffquelle zugesetzt wurde.
In Tabelle 2.4 sind die Bestandteile des Mediums und in Tabelle 2.5 die
Zusammensetzung der Spurenelement-Lösung angegeben. Die Lösungen des
Mediums wurden mit Aqua bidest. angesetzt und mit Ausnahme der sterilfiltrierten
Vitamin B12-Lösung autoklaviert. Das Natriumcarbonat (Na2CO3) wurde zur
Vermeidung von Ausfällungen separat in Aqua bidest. angesetzt und autoklaviert.
Diese Komponente und die Zusätze wurden dem autoklavierten Grundmedium steril
zugegeben. Das ASNIII/2-Medium wies eine Salinität von 15‰ auf. Das Kulturmedium
hatte einen pH-Wert von 9,0, das mit 1% Fleischextrakt versetzte Kulturmedium
einen pH-Wert von 7,1. Zur Herstellung von ASNIII/2-Festmedium wurde dem
Grundmedium zusätzlich 1,5% (w/v) Agar zugesetzt.
13

2. Material und Methoden
Tab. 2.4: Zusammensetzung des ASNIII/2-Mediums (RIPPKA et al., 1979, modifiziert nach RETHMEIER, 1995)
Komponenten g/l oder ml/lGrundmedium
NaCl 12,5 gMgCl2 x 6 H2O 1,0 gKCl 0,25 gNaNO3 0,75 gMgSO4 x 7 H2O 1,75 g CaCl2 x 2 H2O 0,25 gFleischextrakta) 10,0 gAqua bidest. ad 900 ml Na2CO3 0,12 g/100 ml
ZusätzeKH2PO4 x 3 H2O-Stammlösung (4 g/l) 2,50 mlFe-NH4-Citrat-Lösung (6 g/l) 0,25 mlVitamin B12-Stammlösungb) (1 mg/100 ml) 0,50 mlSpurenelement-Lösung (Trace Metal Mix) 0,50 ml
a) Anzucht der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 b) sterilfiltriert
Tab. 2.5: Zusammensetzung der Spurenelement-Lösung (Trace Metall Mix) (RIPPKA et al., 1979, modifiziert nach RETHMEIER, 1995)
Komponenten g/lNa3-Citrat x 2 H2O 3,0Na2-EDTA x 2 H2O 5,5H3BO3 2,86MnCl2 x 4 H2O 1,81ZnSO4 0,222Na2MoO4 x 2 H2O 0,318CuSO4 x 5 H2O 0,079Co(NO3)2 x 6 H2O 0,0494Aqua bidest. ad 1000 ml
2.2.2 LB(Luria Bertani)-Medium (SAMBROOK et al., 1989) Die Anzucht von Chromobacterium violaceum CV026 erfolgte im LB-Flüssigmedium
(SAMBROOK et al., 1989). Die Zusammensetzung des Mediums ist in Tabelle 2.6
dargestellt. Vor dem Autoklavieren wurde der pH-Wert durch Zugabe von 1 M HCl
bzw. 1 M NaOH auf 7,0 eingestellt (Sartorius, PB-11). Durch Zugabe von 1,5% (w/v)
Agar wurde LB-Festmedium hergestellt.
14

2. Material und Methoden
Tab. 2.6: Zusammensetzung des LB-Flüssigmediums (SAMBROOK et al., 1989)
Komponenten g/lTryptone 10,0Hefeextrakt 5,0NaCl 10,0Aqua bidest. ad 1000 ml
2.2.2.1 LB(Luria Bertani)-Weichagar (SAMBROOK et al., 1989) In Tabelle 2.7 sind die Medienkomponenten des LB-Weichagars (SAMBROOK et al.,
1989) zusammengefasst, welcher nach Zugabe von 50 ml Sensorkultur (siehe
Kap. 2.7.5.2) bei 150 ml Gesamtvolumen 0,75% (w/v) Agar enthält.
Tab. 2.7: Zusammensetzung des LB-Weichagars (SAMBROOK et al., 1989)
Komponenten gAgar 1,12Tryptone 1,0Hefeextrakt 0,5NaCl 1,0Aqua bidest. ad 100 ml
2.2.3 Leuchtbakterien-Medium (Angaben lt. MPI, Bremen) Die Anzucht von Vibrio sp. erfolgte im Leuchtbakterien-Medium (Angaben lt. MPI,
Bremen). Die Medienkomponenten wurden getrennt in Aqua bidest. angesetzt und
anschließend autoklaviert. Die Zusammensetzung des Mediums ist in Tabelle 2.8
dargestellt.
Tab. 2.8: Zusammensetzung des Leuchtbakterien-Mediums (Angaben lt. MPI, Bremen)
Komponenten g/lCaCl2 x 2 H2O 1Glycerin 3Hefeextrakt 3KCI 0,5MgSO4 x 7 H2O 9NaCl 20Pepton 5Aqua bidest. 1000 ml
15
2. Material und Methoden
2.2.4 ABG-Medium (CHILTON et al., 1974) Die Anzucht der Agrobacterium tumefaciens-Stämme erfolgte im ABG-Medium
(CHILTON et al., 1974). In Tabelle 2.9 ist die Zusammensetzung des ABG-Mediums,
in den Tabellen 2.10 und 2.11 die der 20x AB-Salze und des 20x AB-Puffers
dargestellt. Vor dem Autoklavieren der 20x AB-Puffer-Stammlösung wurde der pH-
Wert durch Zugabe von 1 M HCl bzw. 1 M NaOH auf 7,0 eingestellt (Sartorius
PB-11). Der Puffer, die Salze und die Glucoselösung wurden dem autoklavierten
Aqua bidest. steril zugegeben. ABG-Festmedium wurde durch Zugabe von 1,5 %
(w/v) Agar hergestellt.
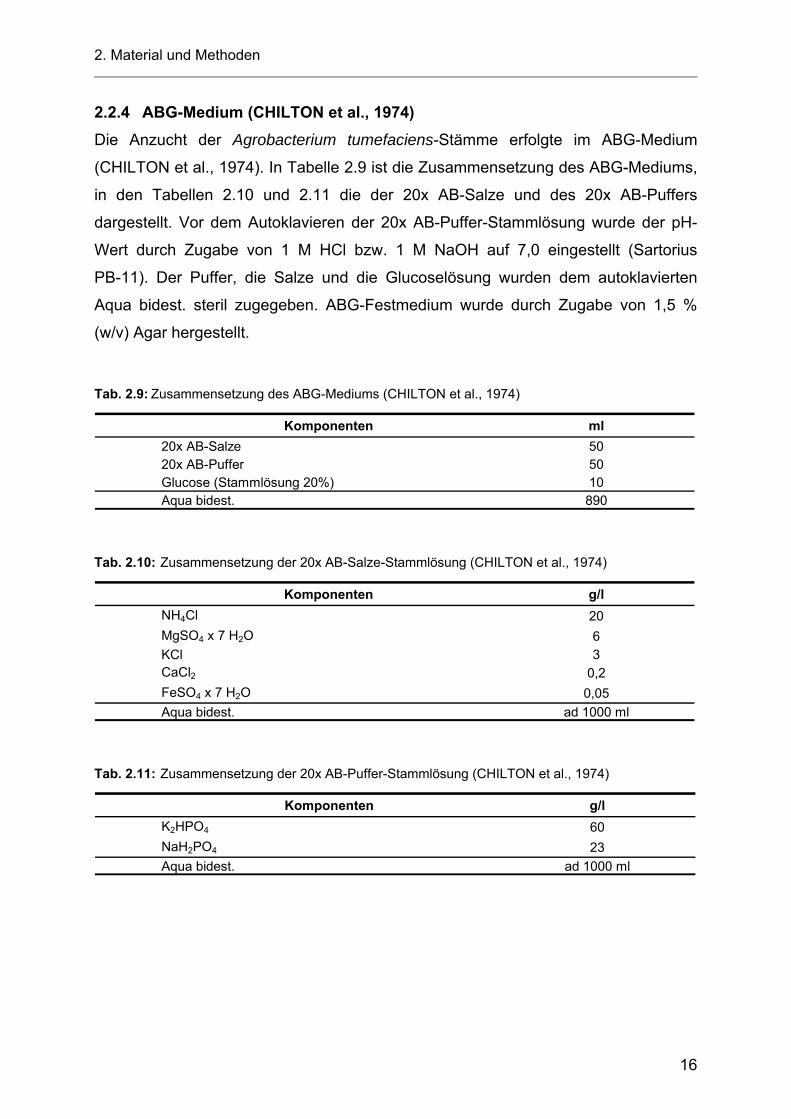
Tab. 2.9: Zusammensetzung des ABG-Mediums (CHILTON et al., 1974)
Komponenten ml20x AB-Salze 5020x AB-Puffer 50Glucose (Stammlösung 20%) 10Aqua bidest. 890
Tab. 2.10: Zusammensetzung der 20x AB-Salze-Stammlösung (CHILTON et al., 1974)
Komponenten g/lNH4Cl 20MgSO4 x 7 H2O 6KCl 3CaCl2 0,2FeSO4 x 7 H2O 0,05Aqua bidest. ad 1000 ml
Tab. 2.11: Zusammensetzung der 20x AB-Puffer-Stammlösung (CHILTON et al., 1974)
Komponenten g/lK2HPO4 60NaH2PO4 23Aqua bidest. ad 1000 ml
16

2. Material und Methoden
2.2.4.1 ABG-Weichagar (CHILTON et al., 1974) In Tabelle 2.12 sind die Medienkomponenten des ABG-Weichagars (CHILTON et al.,
1974) zusammengefasst, welcher nach Zugabe von 50 ml Sensorkultur (siehe
Kap. 2.7.5.2) bei 150 ml Gesamtvolumen 0,75% (w/v) Agar enthält. Die
Zusammensetzung der 20x AB-Salze und des 20x AB-Puffers ist in Kapitel 2.2.4,
Tabelle 2.10 und 2.11 dargestellt. Der Puffer, die Salze und die Glucoselösung
wurden der autoklavierten Agar-Aqua bidest.-Mischung steril zugegeben.
Tab. 2.12: Zusammensetzung des ABG-Weichagar (CHILTON et al., 1974)
Komponenten g oder mlAgar 1,12 gAqua bidest. 89 ml20x AB-Salze 5 ml20x AB-Puffer 5 mlGlucose (Stammlsg. 20 %) 1 ml
2.3 Pufferlösungen Für die Pufferung von Kulturmedien wurden in der vorliegenden Arbeit verschiedene
Pufferlösungen (DAWSON et al. 1986) verwendet, deren Zusammensetzungen in
der nachfolgenden Tabelle 2.13 zusammengefasst sind. Dabei wird jeweils die
Pufferzusammensetzung für ein Endvolumen von 100 ml angegeben. Zur
Herstellung der verschiedenen Pufferlösungen wurden die Komponenten abhängig
von dem anzusetzenden pH-Wert in einem bestimmten Verhältnis miteinander
gemischt und nachfolgend gegebenfalls mit Aqua bidest. auf 100 ml aufgefüllt.
17
2. Material und Methoden
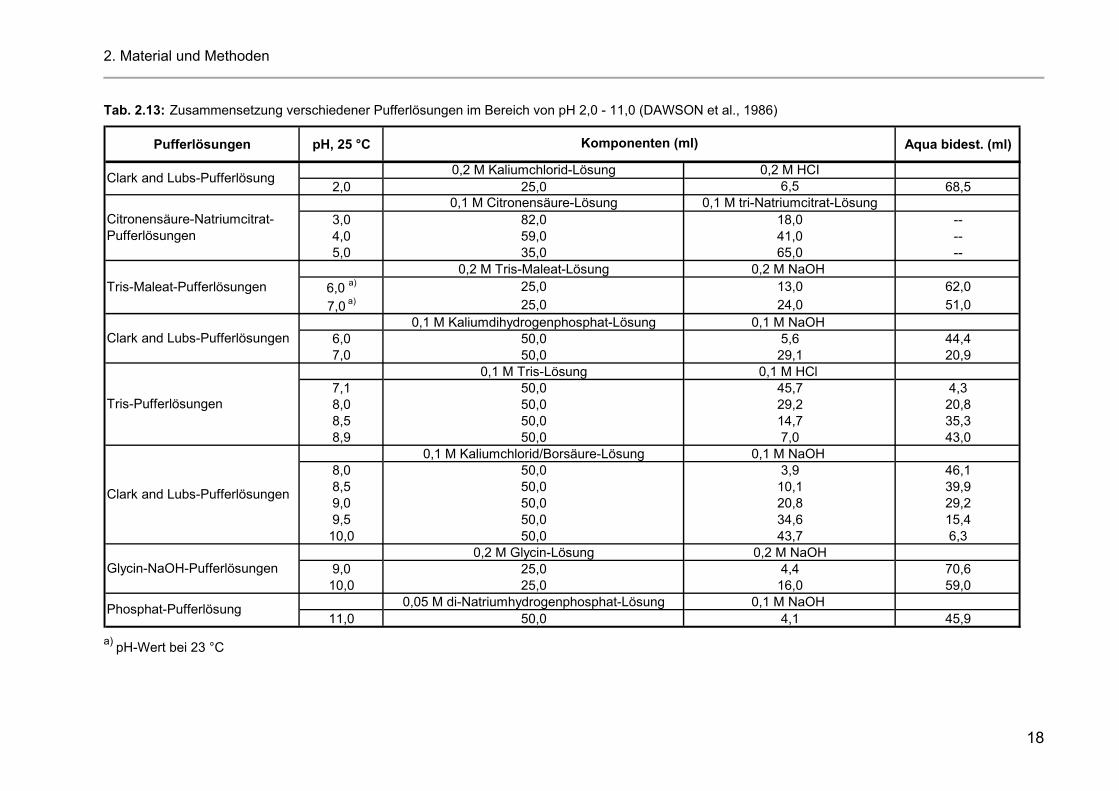
Tab. 2.13: Zusammensetzung verschiedener Pufferlösungen im Bereich von pH 2,0 - 11,0 (DAWSON et al., 1986)
Pufferlösungen pH, 25 °C Aqua bidest. (ml)
0,2 M Kaliumchlorid-Lösung 0,2 M HCI2,0 25,0 6,5 68,5
0,1 M Citronensäure-Lösung 0,1 M tri-Natriumcitrat-Lösung3,0 82,0 18,0 --4,0 59,0 41,0 --5,0 35,0 65,0 --
0,2 M Tris-Maleat-Lösung 0,2 M NaOH 6,0 a) 25,0 13,0 62,07,0 a) 25,0 24,0 51,0
0,1 M Kaliumdihydrogenphosphat-Lösung 0,1 M NaOH 6,0 50,0 5,6 44,47,0 50,0 29,1 20,9
0,1 M Tris-Lösung 0,1 M HCl 7,1 50,0 45,7 4,38,0 50,0 29,2 20,88,5 50,0 14,7 35,38,9 50,0 7,0 43,0
0,1 M Kaliumchlorid/Borsäure-Lösung 0,1 M NaOH 8,0 50,0 3,9 46,18,5 50,0 10,1 39,99,0 50,0 20,8 29,29,5 50,0 34,6 15,410,0 50,0 43,7 6,3
0,2 M Glycin-Lösung 0,2 M NaOH9,0 25,0 4,4 70,610,0 25,0 16,0 59,0
0,05 M di-Natriumhydrogenphosphat-Lösung 0,1 M NaOH 11,0 50,0 4,1 45,9
Tris-Pufferlösungen
Clark and Lubs-Pufferlösungen
Glycin-NaOH-Pufferlösungen
Phosphat-Pufferlösung
Komponenten (ml)
Tris-Maleat-Pufferlösungen
Clark and Lubs-Pufferlösungen
Citronensäure-Natriumcitrat-Pufferlösungen
Clark and Lubs-Pufferlösung
a) pH-Wert bei 23 °C
18

2. Material und Methoden
2.4 Lagerung der Bakterienstämme Zur langfristigen Lagerung der heterotrophen Bakterien wurden Glycerinstocks
angelegt. Die Bakterienstämme wurden hierzu im jeweiligen Medium angezogen und
in sterilen Eppendorfreaktionsgefäßen (ERG) in Glycerin (Bakterienkultur und steriles
87%iges Glycerin in 4:1 Volumenanteilen) bei -80 °C eingefroren. Für alle
nachfolgenden Untersuchungen wurde mit einer Impföse oder einer
Eppendorfpipette steril Bakterienmaterial entnommen und in das jeweilige
Flüssigmedium überführt. Die Kontrolle der Reinheit erfolgte über die jeweiligen
Agarplatten für das Bakterium mittels Drei-Ösen-Ausstrich und Inkubation bei 28 °C
im Brutschrank (Heraeus Instruments, kelvitron® t) unter aeroben Bedingungen für
2 - 4 Tage.
Parallel zu den Glycerinstocks wurden die heterotrophen Bakterienstämme auf den
jeweiligen festen Nährmedien ausgestrichen und bei 28 °C für 4 - 5 Tage inkubiert.
Anschließend wurden die Agarplatten im Kühlraum bei 4 °C gelagert und bildeten ein
zusätzliches Bakterienmateriallager.
2.5 Voranzucht der zu untersuchenden Bakterienstämme
2.5.1 Voranzucht der heterotrophen Stämme Bo 53-33 und Bo 10-09 Die Anzucht der Vorkulturen erfolgte unter sterilen Bedingungen in mit 20 ml ASNIII/2-
Medium (siehe Kap. 2.2.1) gefüllten 50 ml Erlenmeyerkolben. Vorkulturen wurden
stets mit Bakterienmaterial aus den angelegten Glycerinstocks angeimpft (siehe
Kap. 2.4) und für 16 - 24 h bei 21 °C in einem Schüttelwasserbad (Julabo SW 21) bei
120 rpm unter aeroben Bedingungen inkubiert (= Übernachtkultur (ÜK)).
Zur Untersuchung des Wachstumsverhaltens (siehe Kap. 2.6) als auch zur
Untersuchung auf AHL-Moleküle (siehe Kap. 2.7.2.1) wurden die Vorkulturen mit
einer O.D.600nm von 0,1 angesetzt, wobei diese aus einer Übernachtkultur (ÜK)
inokuliert wurden. Die Bakterienstämme wurden, wenn nicht anders beschrieben, für
12 - 16 h bei 21 °C in einem Schüttelwasserbad (Julabo SW 21) bei 120 rpm unter
aeroben Bedingungen im Dunkeln inkubiert.
19

2. Material und Methoden
2.5.2 Voranzucht der Cyanobakterienstämme Flo 1, Bo 10 und Bo 53 Die Hälterung der Flo 1-Kulturen erfolgte bei einer konstanten Temperatur von 21 °C
im Brutraum mit einem konstanten Photonenfluss von 1 µE m-2 s-1 PAR
(Photosynthetic Active Radiation). Die Kulturen wurden in einem Abstand von 4 - 5
Wochen in frisches ASNIII/2-Medium (siehe Kap. 2.2.1) überführt. Das Überimpfen
erfolgte unter sterilen Bedingungen in mit 40 ml Kulturmedium gefüllten 100 ml
Erlenmeyerkolben bzw. in mit 100 ml Kulturmedium gefüllten 250 ml
Erlenmeyerkolben.
Vor dem eigentlichen Versuch wurden Vorkulturen angesetzt, die an die jeweiligen
Versuchsbedingungen für 3 - 4 Wochen adaptiert wurden.
Die Voranzucht des Cyanobakteriums Stamm Bo 10 wurde von Dipl.-Biologin
A. Hube, die des Cyanobakteriums Bo 53 von Dr. B. Heyduck-Söller durchgeführt.
2.6 Untersuchungen zum Wachstumsverhalten der heterotrophen Stämme Bo 53-33 und Bo 10-09
Zur Untersuchung des Wachstumsverhaltens der Stämme Bo 10-09 und Bo 53-33
wurden Wachstumskurven mit ungepufferten Kulturmedien sowie bei verschiedenen
pH-Werten generiert und das pH-Minimum, -Maximum und -Optimum der
heterotrophen Bakterien ermittelt. Des Weiteren wurden die Kulturen bei einer
Photonenflussdichte von 5 µE m-2 s-1 PAR (Photosynthetic Active Radiation) inkubiert
und Wachstumskurven aufgenommen. Da die Hälterung der Cyanobakterien in der
Stammsammlung der Abt. Marine Mikrobiologie bei einer konstanten Temperatur von
21 °C erfolgt, wurde diese Temperatur auch zur Anzucht der Bakterienstämme
Bo 10-09 und Bo 53-33 gewählt, um eine AHL-Produktion der ansonsten in
Assoziation lebenden heterotrophen Bakterien bezüglich einer interspezifischen
Kommunikation unter identischen Inkubationstemperaturen zu untersuchen.
Die Wachstumsmessungen wurden in der vorliegenden Arbeit über O.D.-Messungen
bei einer Wellenlänge von 600 nm mittels eines Spektralphotometers (biochrom Libra
S12) durchgeführt. Die photometrische Messung erfolgte in Plastikküvetten gegen
das jeweilige unbeimpfte Kulturmedium als Blindprobe. Für jeden Versuchsansatz
wurde eine Doppelbestimmung (n = 2) durchgeführt mit anschließender Berechnung
der Mittelwerte. Der Abbruch des Versuches erfolgte in der stationären Phase der
Kulturen. Während und nach Beendigung der Wachstumsmessungen wurde der pH-
Wert der Kulturüberstände kontrolliert (siehe Kap. 2.6.4).
20

2. Material und Methoden
2.6.1 Untersuchungen zum Wachstumsverhalten in ungepuffertem ASNIII/2-Medium im Dunkeln
Die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 wurden, wie in Kapitel
2.5.1 beschrieben, im ASNIII/2-Medium inkubiert. Nach Ermittlung der O.D.600nm der
Vorkultur wurden die ungepufferten Versuchskulturen mit einer Ausgangs-O.D.600nm
von 0,1 inokuliert und bei 21 °C im Dunkeln in einem Schüttelwasserbad (Julabo SW
21) bei 120 rpm im Doppelansatz inkubiert. Die Untersuchungen zum
Wachstumsverhalten wurden in 50 ml Erlenmeyerkolben mit 25 ml Endvolumen
durchgeführt. Die photometrische Messung erfolgte im Abstand von 4 h (siehe
Kap. 2.6).
2.6.2 Untersuchungen zum Wachstumsverhalten bei unterschiedlichen pH-Werten im Dunkeln
Zur Ermittlung des pH-Minimums, -Optimums und -Maximums der heterotrophen
Testorganismen Stamm Bo 53-33 und Stamm Bo 10-09 wurde das Wachstum in
gepuffertem ASNIII/2-Medium in Schritten von einer pH-Einheit im Bereich pH 2,0 -
10,0 getestet. Eine Ausnahme bildete der pH-Wert von 8,5. Die für diese
Versuchsreihe verwendeten Pufferlösungen sind in Tabelle 2.14 zusammengetragen
und wurden in Vorversuchen ausgewählt (siehe Kap. 2.3). Die Komponenten des
Grundmediums (siehe Kap. 2.2.1, Tab. 2.4) wurden dazu direkt dem jeweiligen
Puffer zugesetzt und autoklaviert. Das Natriumcarbonat (Na2CO3) wurde ungelöst
autoklaviert und dem autoklavierten Grundmedium zugegeben. Der pH-Wert wurde
nach Fertigstellung des gepufferten Kulturmediums kontrolliert und gegebenenfalls
durch Zugabe von 1 M HCl bzw. 1 M NaOH auf den jeweiligen pH-Wert eingestellt
(Sartorius PB-11). Mit steigendem pH-Wert der gepufferten Kulturmedien (pH-
Bereich von 8,5 - 10,0) konnte eine zunehmende Trübung des Mediums durch
Ausfall von Salzen (z.B. Mg2+- und Ca2+-Ionen) festgestellt werden.
21

2. Material und Methoden
Tab. 2.14: Verwendete Pufferlösungen im Bereich von pH 2 - 10 (siehe Kap. 2.3, Tab. 2.13) zur Anzucht der heterotrophen Bakterienstämme Bo 10-09 und Bo 53-33
Verwendete Pufferlösungen pH-Wert
Clark and Lubs-Pufferlösung pH 2,0
Citronensäure-Natriumcitrat-Pufferlösung pH 3,0 - 5,0
Clark and Lubs-Pufferlösung pH 6,0
Tris-Maleat-Pufferlösung pH 7,0
Tris-HCl-Pufferlösung pH 8,0 und pH 8,5
Glycin-NaOH-Pufferlösung pH 9,0 und pH 10,0
Die gepufferten Versuchsansätze wurden aus einer Vorkultur (siehe Kap. 2.5.1) mit
einer O.D.600nm von 0,1 inokuliert und anschließend, wie in Kapitel 2.6.1 beschrieben,
inkubiert. Die Anzucht der heterotrophen Bakterien wurde in 50 ml Erlenmeyerkolben
mit 25 ml Endvolumen durchgeführt. Die photometrische Messung erfolgte im
Abstand von 8 h bzw. 16 h (siehe Kap. 2.6).
2.6.3 Untersuchungen zum Wachstumsverhalten bei einem Photonenfluss von 5 µE m-2 s-1 PAR
Zur Untersuchung des Lichteinflusses auf das Wachstum der heterotrophen
Bakterien wurden ungepufferte Versuchsansätze und Versuchsansätze beim
jeweiligen pH-Optimum des heterotrophen Bakteriums bei einem Photonenfluss von
5 µE m-2 s-1 PAR auf einem Inkubationsschüttler (Heidolph UNIMAX 2010) kultiviert
(siehe Kap. 2.6.1). Die Vorkulturen (siehe Kap. 2.5.1) wurden ebenfalls bei einem
Photonenfluss von 5 µE m-2 s-1 PAR auf dem gleichen Inkubationsschüttler
angezogen. Die Anzucht der heterotrophen Bakterien wurde in 50 ml
Erlenmeyerkolben mit 25 ml Endvolumen durchgeführt. Die photometrische Messung
erfolgte im Abstand von 8 h bzw. 16 h (siehe Kap. 2.6).
2.6.4 pH-Wert-Bestimmung in den Kulturüberständen Der pH-Wert der Versuchskulturen wurde parallel zu den photometrischen
Messungen sowie nach Beendigung der Wachstumsversuche bestimmt. Hierzu
wurden jeweils 200 µl Zellkultur aus den Parallelansätzen für 1 min bei 13.000 rpm
zentrifugiert (Heraeus Instruments, Biofuge pico) und der pH-Wert des Überstandes
mit einem pH-Streifen (Macherey-Nagel; pH 6,0 - 10,0) ermittelt. Die abschließende
22

2. Material und Methoden
pH-Bestimmung in den Kulturüberständen erfolgte nach Zentrifugation (Beckman,
AvantiTM J-25 Centrifuge) der gesamten Zellkultur bei 13.000 rpm für 10 min mittels
einer pH-Elektrode, gekoppelt an einem pH-Meter (Sartorius PB-11). Es wurde eine
Doppelbestimmung (n = 2) mit anschließender Berechnung der Mittelwerte
durchgeführt.
2.7 Nachweis von N-Acyl-L-Homoserinlactonen
2.7.1 Verwendete Sensorbakterienstämme zum Nachweis von AHL-Signalmolekülen
Die Detektion von AHL-Signalmolekülen erfolgte mit Hilfe der AHL-Sensorbakterien
Chromobacterium violaceum CV026 bzw. Agrobacterium tumefaciens NTL4
(pZLR4). Diese Indikatorbakterien synthetisieren durch gentechnische
Veränderungen keine AHL-Signalmoleküle, sind aber in der Lage diese
nachzuweisen (siehe Kap. 2.7.1.1 und 2.7.1.2). Die Sensorbakterien werden
nachfolgend näher beschrieben.
2.7.1.1 Chromobacterium violaceum CV026 (MCCLEAN et al., 1997) In Abhängigkeit von der Zelldichte wird in C. violaceum die Produktion von Violacein
reguliert. Dieses nicht-wasserlösliche, violette Pigment besitzt antimikrobielle
Eigenschaften. Die Doppelmutante C. violaceum CV026 wurde durch miniTn5
Transposonmutagenese generiert. Infolge der Insertion wurde die AHL-Produktion
unterbrochen, gekoppelt mit einem Verlust der Pigmentierung. Der C. violaceum
Wildtypstamm produziert N-hexanoyl-L-Homoserinlacton (C6-HSL) als einziges AHL-
Signalmolekül, demzufolge der Stamm C. violaceum CV026 besonders sensitiv auf
dieses Signalmolekül mit Violaceinproduktion reagiert. Der Sensorstamm eignet sich
besonders für den Nachweis von kurzkettigen, unsubstituierten AHL-Signalmolekülen
(vor allem C6-HSL) als Biosensor, wird hingegen von längerkettigen N-Acyl-L-
Homoserinlactonen (Acyl-Seitenketten länger als C8) sowie von N-Acyl-HSLs mit
3-Hydroxy-Substitution nicht zur Violaceinproduktion angeregt (STEINDLER und
VENTURI, 2007). AHL-Signalmoleküle mit einer C8-, C10-, C12- und C14-Seitenkette
sind jedoch fähig, bei vorheriger Zugabe von C6-HSL oder 3-oxo-C6-HSL in
angemessener Konzentration, die Pigmentierung der Mutante zu inhibieren.
23

2. Material und Methoden
Hierdurch kann mit diesem AHL-Biosensor ebenfalls ein Nachweis längerkettiger
AHLs erzielt werden.
2.7.1.2 Agrobacterium tumefaciens NTL4 (pZLR4) (FARRAND et al., 2002; SZENTHE und PAGE, 2003)
Der Wildtypstamm von A. tumefaciens ist ein pflanzenpathogenes Bakterium,
welches in seinen Wirtspflanzen Wurzeltumorwachstum induziert. Der Prozess der
Tumorbildung wird durch die Übertragung der T-DNA (oncogenic DNA) und ihrer
Integration in das Pflanzenchromosom verursacht. Die T-DNA ist ein kleiner
Abschnitt des Ti-Plasmids in A. tumefaciens. Auf dem Ti-Plasmid sind weitere Gene,
z.B. für den Transfer der T-DNA in die Wirtspflanze (vir-Gene) und den konjugativen
Transfer des Ti-Plasmids zwischen Stämmen von A. tumefaciens (tra-Gene),
lokalisiert. Die für einen konjugativen Transfer des Ti-Plamids nötigen Gene sind
dabei in einer Regulationseinheit (tra-Regulon) zusammengefasst. Bei
A. tumefaciens wird dieser Transfer über „Quorum Sensing“ gesteuert, wobei die
LuxI/LuxR-homologen Proteine TraI und TraR des A. tumefaciens-QS-Systems die
Zelldichte-abhängige Expression des tra-Regulons (tra-Operon, trb-Operon, traM)
regulieren. TraI ist eine Autoinducer-Synthase und synthetisiert das AHL-
Signalmolekül 3-oxo-C8-HSL. Das Protein TraR stellt den entsprechenden
Transkriptionsaktivator dar. Bei hoher Zelldichte und ebenfalls hoher intrazellulärer
N-Acyl-HSL-Konzentration bindet TraR das HSL-Molekül und aktiviert sowohl die
Transkription der tra-Gene als auch die Expression des traI-Gens, letzteres
resultierend in einer positiven Feedback-Schleife.
Der AHL-Sensorstamm A. tumefaciens NTL4 (pZLR4) besitzt kein Ti-Plasmid, aber
ein rekombinantes Plasmid (pZLR4). Dieses enthält Inserts von dem Plasmid pTIC58
mit traR und einem tra-Operon, welches eine traG::lacZ-Fusion aufweist. Neben
diesen Genen sind die Gene für Gentamycin- und Carbenicillin-Resistenz auf dem
Vektor kodiert, wobei Gentamycin das bestgeeignete Antibiotikum zur Selektion
darstellt. Der Stamm ist eine AHL-Synthase-Mutante und somit nicht in der Lage,
eigene AHL-Moleküle zu synthetisieren. Folglich reagiert der Stamm aufgrund der
traG::lacZ-Fusion erst bei Anwesenheit geeigneter exogener AHL-Moleküle über die
Aktivierung des TraR-Proteins mit der Expression der lacZ-kodierten
β-Galaktosidase. TraR aktiviert dabei die Transkription des traG-Gens, welches
jedoch durch die Insertion des lacZ-Gens nicht exprimiert wird. Dementsprechend
24

2. Material und Methoden
kann die Anwesenheit entsprechender AHL-Moleküle (bei ausreichender
Konzentration) über die β-Galaktosidase-Aktivität durch Umsetzung des Substrates
X-Gal zu einem hellblauen Farbstoff nachgewiesen werden.
Der A. tumefaciens-Sensorstamm detektiert ein breites Spektrum an AHL-
Signalmolekülen und weist unter den bisweilen konstruierten Biosensoren gegenüber
diesen Komponenten zudem mitunter die höchste Sensitivität auf (STEINDLER und
VENTURI, 2007). Entsprechend des eigenen QS-Systems zeigt der Indikatorstamm
eine höhere Sensitivität gegenüber 3-oxo-substituierten AHL-Signalmolekülen mit
C4- bis C12-Seitenketten, vor allem gegenüber 3-oxo-C8-HSL. Weiter eignet sich der
Sensorstamm, mit Ausnahme von C4-HSL, zum Nachweis unsubstituierter AHL-
Moleküle sowie von 3-hydroxy-Derivaten mit C6-, C8- und C10-Seitenketten
(STEINDLER und VENTURI, 2007).
Weitere A. tumefaciens-Stämme, welche in der vorliegenden Arbeit Verwendung
fanden, waren der A. tumefaciens-Stamm NTL4 und NT1 (pTIC58∆accR). Stamm
NTL4 fehlt ein Ti-Plasmid und ist demzufolge nicht fähig, AHL-Moleküle zu
synthetisieren. Neben dieser AHL-Negativkontrolle wurde der Stamm
NT1 (pTIC58∆accR) als AHL-Positivkontrolle eingesetzt, da dieser aufgrund eines
veränderten Ti-Plasmids (trac) konstitutiv die tra-Gene exprimiert und folglich auch
konstitutiv N-Acyl-HSLs synthetisiert.
2.7.2 Kultivierungsbedingungen und Inkubationsdauer der Versuchskulturen zur Untersuchung von AHL-Signalmolekülen
2.7.2.1 Kultivierungsbedingungen der AHL-Versuchskulturen mit den heterotrophen Bakterienstämmen Bo 53-33 und Bo 10-09
Zur Untersuchung der AHL-Produktion in Abhängigkeit von verschiedenen
Parametern variierten die Kultivierungsbedingungen für die Stämme Bo 53-33 und
Bo 10-09. Für die Extraktion von AHL-Molekülen (siehe Kap. 2.7.3) wurden die
Bakterienstämme in 500 ml Erlenmeyerkolben mit 250 ml Endvolumen angezogen.
Die Inokulation und Anzucht der jeweiligen Versuchsansätze wurden in den
jeweiligen unten genannten Kapiteln zur Untersuchung des Wachstumsverhaltens
der heterotrophen Bakterienstämme beschrieben.
Zur Ermittlung des pH-Wert-Einflusses auf die AHL-Produktion wurden die AHL-
Gehalte bei dem jeweiligen pH-Minimum, pH-Optimum und pH-Maximum der
25

2. Material und Methoden
heterotrophen Bakterien im Dunkeln bei 21 °C verglichen (siehe Kap. 2.6.2). Des
Weiteren wurden ungepufferte Versuchsansätze im Dunkeln (siehe Kap. 2.6.1) und
bei einer Photonenflussdichte von 5 µE m-2 s-1 PAR (siehe Kap. 2.6.3) bei 21 °C
kultiviert und auf Signalmoleküle untersucht. Die Produktion von AHL-
Signalmolekülen wurde in der stationären Wachstumsphase, für die ungepufferten
und pH-optimalen Ansätze zudem in der logarithmischen Wachstumsphase
untersucht. Die generierten Wachstumskurven (siehe Kap. 3.2 und 3.3) dienten der
Anzucht als Referenz. Zur Überprüfung einer Korrelation des Zellwachstums
zwischen 250 ml und 25 ml Versuchsansätzen, wurde das Wachstum der Kulturen
stichprobenartig durch photometrische Messungen der O.D.600nm kontrolliert. Ferner
war die AHL-Produktion in Abhängigkeit von der Inkubationstemperatur und von der
Kohlenstoffquelle von Interesse. Hierzu wurden Versuchsansätze mit ungepuffertem
ASNIII/2-Medium (siehe Kap. 2.2.1) bei 28 °C im Dunkeln und zur Untersuchung des
Einflusses der Kohlenstoffquelle auf die AHL-Produktion im ASNIII/2-Medium mit 0,1%
Fleischextrakt bei 21 °C im Dunkeln kultiviert. Die Inokulation und Anzucht der
Versuchskulturen erfolgte wie in Kapitel 2.6.1 beschrieben, wobei die Vorkulturen für
die 28 °C Inkubation (Temperaturversuch) bei gleicher Temperatur angezogen
wurden. Die jeweilige Inkubationsdauer der Versuchskulturen ist im Anhang als
Tabelle 7.1 hinterlegt.
2.7.2.2 Anzucht und Kultivierungsbedingungen der AHL-Versuchskulturen mit dem Cyanobakterium Stamm Flo 1
Zur Untersuchung der AHL-Produktion durch Stamm Flo 1 wurden Versuchsansätze
mit ungepuffertem Kulturmedium (siehe Kap. 2.2.1) sowie gepufferten Kulturmedien
(pH 7,0, 8,0, 8,5 und 9,0) bei einer konstanten Temperatur von 21 °C und einem
konstanten Photonenfluss von 5 µE m-2 s-1 PAR für 14 Tage (logarithmische Phase)
bzw. 21 Tage (stationäre Phase) kultiviert. Die Inkubationsdauer wurde auf
Grundlage der von SCHRÜBBERS (2007) erzielten Ergebnisse gewählt. Die
eingesetzten Vorkulturen wurden 3 - 4 Wochen an die Wachstumsbedingungen
adaptiert (siehe Kap. 2.5.2) und anschließend bei 8.000 rpm für 10 min zentrifugiert
(Beckman, AvantiTM J-25 Centrifuge). Der Kulturüberstand wurde verworfen, die
cyanobakterielle Biomasse vereinigt und homogenisiert. Mit Hilfe einer
Analysenwaage (Sartorius, MC 1 Research RC 210P) wurden jeweils 1000 mg
Homogenat in sterile ERG eingewogen und als Inokulum für die 250 ml AHL-
26

2. Material und Methoden
Versuchsansätze eingesetzt. Die Weiterbehandlung der Ansätze nach Beendigung
der Anzucht ist in Kapitel 2.7.3 beschrieben.
Für die Versuchsreihe wurden für pH 7,0 und 8,0 Tris-HCl-Pufferlösungen, für pH 8,5
und 9,0 Clark and Lubs-Pufferlösungen verwendet (siehe Kap. 2.3). Das Ansetzen
der gepufferten Kulturmedien erfolgte wie in Kapitel 2.6.2 beschrieben. Nach
Fertigstellung des gepufferten Kulturmediums wurde der pH-Wert kontrolliert und
gegebenenfalls durch Zugabe von 1 M HCl bzw. 1 M NaOH auf den jeweiligen pH-
Wert eingestellt (Sartorius PB-11).
2.7.3 Extraktion von N-Acyl-L-Homoserinlactonen der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie des Cyanobakteriums Stamm Flo 1
2.7.3.1 Herstellung zellfreier Kulturüberstände Nach erfolgter Inkubation der 250 ml Zellkulturen der heterotrophen
Bakterienstämme Bo 10-09 und Bo 53-33 (siehe Kap. 2.7.2.1) sowie des
Cyanobakteriums Stamm Flo 1 (siehe Kap. 2.7.2.2) wurden diese bei 10.000 xg für
30 min bei Raumtemperatur (RT) zentrifugiert (Beckman, AvantiTM J-25 Centrifuge)
und der Zellkulturüberstand anschließend sterilfiltriert (sterilfiltriert; 0,2 µm
Porengröße). Die bakterielle Biomasse wurde entweder bei -20 °C gelagert oder
sofort für intrazelluläre AHL-Untersuchungen weiterverwendet (siehe Kap. 2.7.3.4).
2.7.3.2 Azidifizierung von Kulturüberständen (YATES et al., 2002) Eine Azidifizierung von ungepufferten Kulturüberständen der heterotrophen Stämme
Bo 53-33 und Bo 10-09 sowie des cyanobakteriellen Stammes Flo 1 wurde nach
YATES und Mitarbeitern (2002) durchgeführt. Nach Gewinnung von zellfreien
Überständen (siehe Kap. 2.7.3.1) wurden diese mit konzentrierter HCl auf einen pH-
Wert < 2 eingestellt und anschließend für 24 h bei 21 °C inkubiert. Die
Weiterbehandlung erfolgte wie in Kapitel 2.7.3.3 beschrieben.
27

2. Material und Methoden
2.7.3.3 Extraktion von AHL-Molekülen aus Kulturüberständen (GEISENBERGER, 2000)
Die Extraktion von AHL-Signalmolekülen der heterotrophen Bakterienstämme
Bo 53-33 und Bo 10-09 sowie des Cyanobakteriums Stamm Flo 1 erfolgte aus
250 ml Zellkulturüberstand nach GEISENBERGER (2000). Hierzu wurde zellfreier
Kulturüberstand (siehe Kap. 2.7.3.1) in einen 1000 ml Scheidetrichter vorgelegt,
zweimal mit 100 ml Dichlormethan versetzt und für 10 min durch kräftiges Schütteln
gemischt. Ein ausreichender Druckausgleich wurde durch zwischenzeitliche
Belüftung des Scheidetrichters gewährleistet. Nach deutlicher Phasentrennung
wurde die untere Dichlormethan-Phase in einem 250 ml Erlenmeyerkolben
aufgefangen. Restliches Wasser in den vereinigten Dichlormethanextrakten wurde
vollständig durch Zugabe von wasserfreien Natriumsulfat und anschließendem
Rühren entfernt. Nachfolgend wurden die Extrakte gefiltert (Sartorius 3 hw) und im
Rotationsevaporator (Heidolph VV2000) bei einer Wasserbadtemperatur von
40 - 42 °C ohne Anlegen eines Vakuums eingetrocknet. Alternativ wurden die
gefilterten Extrakte unter dem Abzug offen stehen gelassen, wodurch diese
eindampften. Die Aufnahme des Rückstandes erfolgte in 250 µl Ethylacetat (versetzt
mit 0,01% (v/v) Essigsäure). Die Lagerung der Proben erfolgte bei -20 °C.
Vibrio sp. wurde für einen positiven Extraktionsnachweis bzw. als positive AHL-
Kontrolle im Leuchtbakterien-Medium (siehe Kap. 2.2.3) bei 4 °C im Dunkeln
angezogen, wobei Bakterienmaterial aus einer 24 h-Übernachtkultur (ÜK) als
Inokulum verwendet wurde. Die ÜK wurde mit Bakterienmaterial aus den angelegten
Glycerinstocks angeimpft (siehe Kap. 2.4). Die Kultivierung wurde nach Auftreten
deutlicher Lumineszenz abgebrochen.
2.7.3.4 Extraktion von AHL-Molekülen aus Bakterienzellen Für den Nachweis von intrazellulären AHL-Signalmolekülen wurde die AHL-
Extraktion aus Bakterienzellen nach BYERS und Mitarbeitern (2002) in abgeänderter
Form durchgeführt. Nach Zentrifugation einer 250 ml Zellkultur (siehe Kap. 2.7.3.1)
wurde das Zellpellet vollständig vom Überstand befreit und die gesamte bakterielle
Biomasse in 40 ml 4 °C kaltem Ethanol resuspendiert und anschließend für 72 h bei
4 °C inkubiert. Nach erfolgter Inkubation wurde die Zellsuspension für 5 min bei
8.000 rpm und Raumtemperatur (RT) zentrifugiert, 30 ml der organischen Phase
ohne Aufnahme von Zellsediment entnommen und diese in einem
28

2. Material und Methoden
Rotationsevaporator (Heidolph VV2000) bei RT mit Anlegen eines Vakuums
eingetrocknet. Der Rückstand wurde in 1 - 1,5 ml 10 mM Ammoniumacetat (pH 6,5)
aufgenommen, erneut mit Hilfe des Rotationsevaporators auf ein Endvolumen von
250 µl eingeengt und anschließend bei -20 °C gelagert.
2.7.4 Dünnschichtchromatographische Auftrennung von AHL-Signal-molekülen
Die Dünnschichtchromatographie ist eine chromatographisches Trennmethode, bei
der ein Lösungsmittel oder eine Lösung eines Substanzgemisches (mobile Phase)
über ein Sorptionsmittel (stationäre Phase), meistens Silicagel (Kieselgel), geleitet
wird (LOTTSPEICH und ZORBAS, 1998). Bei dieser Adsorptionschromatographie
befindet sich letzteres als dünne Schicht auf einem Träger. Die in der mobilen Phase
gelösten Substanzen haben unterschiedliche Affinität zur mobilen bzw. stationären
Phase, resultierend in unterschiedlichen Laufstrecken der einzelnen gelösten
Substanzen. Bei der in der vorliegenden Arbeit verwendeten Umkehrphasen-
Chromatographie (reversed phases chromatography) kommt es durch eine unpolare,
chemisch modifizierte stationäre Phase (Kieselgel lipophilisiert mit unpolaren C18-
Alkylresten) zu einer verbesserten Auftrennung hydrophober Substanzen in einem
stark polaren Laufmittelgemisch (mobile Phase), da unpolare Substanzen stärker
zurückgehalten werden (STADLBAUER, 2006).
Die Auftrennung der N-Acyl-L-Homoserinlacton-Extrakte wurde mit Umkehrphasen-
Dünnschichtchromatographie-Platten (RP-18 W/UV F254s, 20x20 cm, Macherey-
Nagel, Düren, Deutschland) nach Geisenberger (2000) mit geringfügiger Modifikation
durchgeführt. Die Ethylacetat- und Ammoniumacetatlösungen (siehe Kap. 2.7.3.3
und 2.7.3.4) wurden entlang einer Auftragslinie in einem Abstand von 2,0 cm auf die
DC-Platte aufgetragen. Der Abstand vom unteren Rand und von den beiden
Seitenrändern der DC-Platten betrug dabei 2,0 cm. Das Probenvolumen der
Ethylacetat- bzw. Ammoniumacetatlösungen variierte in Abhängigkeit von dem
Testorganismus und dem verwendeten Biosensor zwischen 2,0 µl und 100 µl, wobei
die Proben parallel zur Auftragung mit einem kalten Luftstrom eingetrocknet wurden.
Für die dünnschichtchromatographische Auftrennung der Extrakte wurden 110 ml
Laufmittelmischung aus Methanol und Aqua bidest (60:40 (v/v)) verwendet (ca.
0,7 cm Höhe) und die Seitenwände der DC-Kammer anschließend mit Filterpapier
(Sartorius 3 hw) ausgekleidet. Die DC-Platten wurden bei vollständiger Sättigung der
29

2. Material und Methoden
Kammer (24 h) in diese gestellt. Nach ca. vierstündigem Lauf lag die
Lösungsmittelfront ca. 2 cm unterhalb des oberen DC-Plattenrandes, und die DC-
Platten wurden aus der Kammer genommen. Nach Markierung der exakten Höhe der
Lösungsmittelfront erfolgte die Trocknung der DC-Platten mit kalter Luft eines Föns
für 1 h unter dem Abzug. Die DC-Platten konnten, eingewickelt in Frischhaltefolie,
zur Aufbewahrung bei -20 °C gelagert werden.
2.7.5 Visualisierung der DC-Signale
2.7.5.1 Anzucht der Indikatorbakterien und weiterer Referenzstämme Vorkulturen wurden stets mit Bakterienmaterial aus den angelegten Glycerinstocks
angeimpft (siehe Kap. 2.4). Die Voranzucht erfolgte in 50 ml, die Anzucht der
Versuchskulturen in 250 ml Erlenmeyerkolben auf einem Inkubationsschüttler (New
Brunswick Scientific innovaTM 4000 Incubator Shaker).
Eine A. tumefaciens-Vorkultur wurde als Übernachtkultur (ÜK) in 5 ml ABG-Medium
(siehe Kap. 2.2.4) bei 28 °C und 200 rpm angezogen. Als ÜK wurden Kulturen
definiert, welche 16 - 24 h inkubiert wurden. Die Vorkultur wurde anschließend mit
45 ml Kulturmedium versetzt und unter gleichbleibenden Bedingungen bis in die
spät-exponentielle Phase angezogen. Alternativ konnten 50 ml Kulturmedium mit
1 ml Vorkultur inokuliert werden. Die Anzucht erfolgte bei 28 °C und 200 rpm für
24 h. C. violaceum wurde zur Voranzucht als Übernachtkultur (ÜK) bei 30 °C und
120 rpm in 5 ml LB-Medium (siehe Kap. 2.2.2) angezogen. Anschließend wurden
50 ml LB-Medium mit 1 ml Vorkultur angeimpft und bei 30 °C und 120 rpm für 24 h
inkubiert.
Die Vorkulturen der verwendeten bakteriellen Biosensoren wurden unter
Selektionsdruck durch Zugabe verschiedener Antibiotika kultiviert. Hierzu wurde das
LB-Medium mit 50 µg/ml Kanamycin, das ABG-Medium mit 30 µg/ml Gentamycin
und 100 µg/ml Carbenicillin versetzt.
Die Anzucht der weiteren A. tumefaciens-Stämme NTL4 und NT1 (pTIC58∆accR)
erfolgte ebenfalls als ÜK in 5 ml ABG-Medium bei 28 °C und 200 rpm.
30

2. Material und Methoden
2.7.5.2 Überschichtung der DC-Platten mit Indikatorbakterien Zur Detektion der AHL-Signalmoleküle auf der DC-Platte wurde diese mit 150 ml
Indikatorbakterienkultur in Weichagar überschichtet. Hierzu wurden die DC-Platten in
einen Kunststoffrahmen, ausgekleidet mit Klarschichtfolie, eingesetzt. 50 ml
Biosensorkultur A. tumefaciens NTL4 (pZLR4) bzw. C. violaceum CV026 wurden mit
100 ml ABG-Weichagar (siehe Kap. 2.2.4.1) bzw. LB-Weichagar (siehe Kap. 2.2.2.1)
bei einer Temperatur von ca. 42 °C versetzt und ohne Erzeugung von Luftblasen
gemischt. Vor Zugabe der A. tumefaciens-Kultur wurden dem ABG-Weichagar bei
einer Temperatur < 50 °C 90 µg/ml X-Gal (5-Bromo-4-Chloro-3-Indolyl-β-D-
Galactopyranosid) zugesetzt, resultierend in einer Endkonzentration von 60 µg/ml.
Das X-Gal wurde als Stammlösung in N,N-Dimethylformamid (20 mg/ml) angesetzt
und bei - 20 °C gelagert. Nach Überschichtung der DC-Platten und Verfestigung des
Agars wurden die DC-Platten in einer luftdichten Kammer, ausgekleidet mit feuchtem
Küchenpapier, bei 28 °C (Heraeus Instruments, kelvitron® t) für 48 h inkubiert.
Durch Auftragung der AHL-Extrakte auf überschichtete Festagarplatten ohne den
jeweiligen Biosensor, wie in Kapitel 2.7.7.2 beschrieben, wurden Negativkontrollen
durchgeführt. Eine Extraktion des unbeimpften Kulturmediums (siehe Kap. 2.7.3.3)
und anschließender Auftragung auf A. tumefaciens-Indikatorplatten (siehe
Kap. 2.7.7.2) diente als weitere Negativkontrolle.
2.7.6 Auswertung und Dokumentation der DC-Platten Die Auswertung der überschichteten DC-Platten wurde nach einer Bebrütung bei
28 °C für 48 h durchgeführt. Aufgetrennte AHL-Moleküle konnten je nach
verwendetem Biosensor anhand violetter Spots bei C. violaceum CV026 bzw.
hellblauer Spots durch die β-Galaktosidase-Aktiviät von A. tumefaciens NTL4
(pZLR4) lokalisiert und identifiziert werden. Die Intensität und Größe der DC-Signale
konnte bei längerer Inkubationszeit leicht zunehmen. Durch einen Größenvergleich
der Flecken konnten halbquantitative Aussagen gemacht werden.
Als Referenz wurden synthetische AHL-Moleküle (C4-HSL, C6-HSL und C8-HSL),
gelöst in Ethylacetat (versetzt mit 0,01% (v/v) Essigsäure), auf die DC-Platten
aufgetragen. Die DC-Spots wurden mit den synthetischen Referenzmolekülen
(Standards) verglichen und somit eine Eingrenzung an potentiellen AHLs bzw. eine
Identifizierung von AHLs erreicht. Die Referenzmoleküle dienten des Weiteren dazu,
die Nachweisgrenzen des jeweiligen Sensorsystems einzuschätzen. Die Auftragung
31

2. Material und Methoden
der synthetischen AHLs wurde so bemessen, dass in etwa vergleichbare
Signalintensitäten erhalten werden sollten. Die aufgetragenen AHL-Mengen für die
verschiedenen Biosensoren sind in Tabelle 2.15 dargestellt und wurden durch
Agarplattentests (siehe Kap. 2.7.7.3 und 3.4.1) und dünnschichtchromatographische
Voruntersuchungen ermittelt.
Tab. 2.15: Eingesetzte Stoffmengen der synthetischen AHL-Moleküle zum dünnschichtchromato-graphischen Nachweis unter Verwendung der Biosensoren A. tumefaciens NTL4 (pZLR4) und C. violaceum CV026
AHL-Molekül A. tumefaciens NTL4 (pZLR4) C. violaceum CV026
eingesetzte Stoffmenge (in mol) eingesetzte Stoffmenge (in mol)C4-HSL 1,5 x 10-9 bis 2 x 10-9 1 x 10-8
C6-HSL 1,5 x 10-9 1 x 10-9
C8-HSL 2,5 x 10-10 3 x 10-9
Aufgrund einer begrenzten Anzahl zur Verfügung stehender Referenzmoleküle
wurden zudem die Rf-Werte (retention factor) der DC-Signale bestimmt. Ein Abgleich
der Rf-Werte der DC-Signale mit Literaturwerten zur Identifizierung der AHL-
Signalmoleküle konnte jedoch nicht durchgeführt werden, da die ermittelten Rf-Werte
der Referenzmoleküle nicht mit den bekannten Rf-Werten (SHAW et al., 1997)
übereinstimmten Der Rf-Wert errechnet sich als Quotient aus Laufstrecke der
Substanz zur Laufstrecke des Laufmittels vom Startpunkt aus (LOTTSPEICH und
ZORBAS, 1998).
Für die Dokumentation erfolgten alle Aufnahmen von DC-Platten in der vorliegenden
Arbeit mittels Digitalkamera.
2.7.7 Agarplattentests mit Sensorbakterien zum Nachweis AHL-produzierender Bakterien
Zum schnellen Nachweis AHL-produzierender Bakterien wurden verschiedene
Screeningverfahren in einer Voruntersuchung getestet (siehe Kap. 2.7.7.1 und
2.7.7.2), wobei ein Nachweis über die Biosensoren Agrobacterium tumefaciens NTL4
(pZLR4) und Chromobacterium violaceum CV026 (siehe Kap. 2.7.1) erfolgen sollte.
Des Weiteren wurde ein Agarplattentest nach MCCLEAN und Mitarbeitern (1997)
durchgeführt (siehe Kap. 2.7.7.3). Alle Aufnahmen von Agarplatten in der
vorliegenden Arbeit erfolgten mittels Digitalkamera.
32

2. Material und Methoden
2.7.7.1 Agarplattentest nach RAVN und Mitarbeitern (2001) Zur Detektion von AHL-produzierenden Bakterien wurde ein Agarplattentest nach
RAVN und Mitarbeitern (2001) durchgeführt. Anstelle von LB-Agarplatten (siehe Kap.
2.2.2) wurden ASNIII/2-Agarplatten (siehe Kap. 2.2.1) verwendet, da die zu testenden
Bakterienstämme aufgrund zu geringer Salinität des LB-Mediums nicht auf diesem
Agarmedium wuchsen. Für einen Nachweis mit dem Indikatorstamm A. tumefaciens
NTL4 (pZLR4) wurde das Festmedium zusätzlich nach dem Autoklavieren bei einer
Temperatur < 50 °C mit 50 µg/ml X-Gal versetzt. 50 µl einer 24 h-Übernachtkultur
des auf AHL zu testenden Bakterienstammes (siehe Kap. 2.5.1) wurden auf einer
Agarplatte mit einer Impföse als Bakterienstreifen ausplattiert. In einem geringen
Abstand zum Testorganismus wurden anschließend parallel 50 µl des jeweiligen
Sensorstammes aus einer ÜK (siehe Kap. 2.7.5.1) auf der Agarplatte ausgestrichen.
Das jeweilige Sensorbakterium wurde als Negativkontrolle eingesetzt, wobei bei
Verwendung des Sensorstammes A. tumefaciens NTL4 (pZLR4) zudem der
Ti-plasmidlose Stamm A. tumefaciens NTL4 als Negativkontrolle verwendet werden
konnte. Zur Untersuchung einer Temperatur-abhängigen Produktion von AHL-
Signalmolekülen erfolgte die Inkubation bei 21 °C (Brutraum), 28 °C und 37 °C
(Heraeus Instruments, kelvitron® t) für 24 h. Ein AHL-Nachweis wurde durch eine
violette Pigmentierung der Mutante C. violaceum CV026 bzw. einer Blaufärbung des
Indikatorstammes A. tumefaciens NTL4 (pZLR4) erbracht. Eine Bewertung der AHL-
Produktion wurde über die Intensität der Färbung vorgenommen.
2.7.7.2 Agarplattentest nach FARRAND und Mitarbeitern (2002) Zum weiteren Nachweis AHL-produzierender Bakterien wurde ein Screeningtest
nach FARRAND und Mitarbeitern (2002) durchgeführt. Zur Herstellung der
verwendeten A. tumefaciens-Indikatorplatten wurden ABG-Agarplatten (siehe
Kap. 2.2.4) mit jeweils 4 - 5 ml ABG-Weichagar (siehe Kap. 2.2.4.1) überschichtet,
welcher zuvor mit A. tumefaciens NTL4 (pZLR4) versetzt worden war. Die Anzucht
des Sensorstammes erfolgte wie in Kapitel 2.7.5.1, das Versetzen der Kultur mit dem
Weichagar wie in Kapitel 2.7.5.2 beschrieben. Die Fest- und Weichagar enthielten
zusätzlich 50 µg/ml X-Gal, welches nach dem Autoklavieren bei einer Temperatur
< 50 °C hinzugesetzt wurde. Nach Verfestigung des Weichagars wurden 30 µl einer
24 h-Übernachtkultur des zu untersuchenden heterotrophen Bakterienstammes
(siehe Kap. 2.5.1) oder 30 µl Kulturüberstand einer ungepufferten 14 Tage bzw.
33

2. Material und Methoden
21 Tage alten Geitlerinema Flo 1-Kultur (siehe Kap. 2.7.2.2) auf die A. tumefaciens-
Indikatorplatte aufgetragen. Eine ÜK des AHL-produzierenden Stammes
A. tumefaciens NT1 (pTiC58∆accR) (siehe Kap. 2.7.5.1) wurde mit gleichem
Auftragungsvolumen als Positivkontrolle eingesetzt. Überschichtete Agarplatten ohne
Biosensor, auf denen die Proben aufgetragen wurden, dienten als Negativkontrollen.
Zusätzliche Negativkontrollen mit A. tumefaciens-Indikatorplatten und unbeimpftem
Kulturmedium als Probe sollten eine Aktivierung des AHL-Reporters durch im
Medium enthaltene Komponenten ausschließen. Die Bebrütung der Indikatorplatten
und Negativkontrollen erfolgte bei 28 °C (Heraeus Instruments, kelvitron® t) für 24 h.
Bei der Auswertung der Indikatorplatten wurde eine diffuse blaue Zone im Bereich
der Auftragung als Nachweis einer AHL-Produktion gewertet.
2.7.7.3 Agarplattentest mit Chromobacterium violaceum CV026 zum Nachweis von AHL-Signalmolekülen nach MCCLEAN und Mitarbeitern (1997)
Zum Nachweis von kurzkettigen AHLs mit dem Sensorbakterium C. violaceum
CV026 wurde der Agarplattentest nach MCCLEAN und Mitarbeitern (1997)
durchgeführt. Als Kontrolle wurden synthetische AHL-Moleküle eingesetzt, wobei die
Sensitivitäten (Nachweisgrenzen) sowie Spezifitäten von Stamm C. violaceum
CV026 gegenüber bestimmten AHL-Molekülen untersucht wurden.
Zur Herstellung von C. violaceum-Indikatorplatten wurden LB-Agarplatten (siehe
Kap. 2.2.2) mit jeweils 4 - 5 ml LB-Weichagar (siehe Kap. 2.2.2.1) überschichtet,
versetzt mit C. violaceum CV026. Die Anzucht des Indikatorbakteriums erfolgte wie
in Kapitel 2.7.5.1, das Versetzen der C. violaceum CV026-Kultur mit dem Weichagar
wie in Kapitel 2.7.5.2 beschrieben. Nach Verfestigung des Weichagars wurden in
einem Abstand von ca. 2 cm steril Löcher (Ø 6 mm) in den Agar gestanzt.
In die Löcher wurden 5 x 10-9 mol und 5 x 10-8 mol synthetische AHL-Moleküle
(C4-HSL, C6-HSL und C8-HSL), gelöst in Ethylacetat (versetzt mit 0,01% (v/v)
Essigsäure), bzw. je 10 µl AHL-Extrakt (siehe Kap. 2.7.3.3) pipettiert und mit
LB-Medium (siehe Kap. 2.2.2) auf ein Endvolumen von 50 µl aufgefüllt. Als
Negativkontrolle wurden 10 µl Ethylacetat mit 40 µl LB-Medium versetzt und
eingesetzt. Die Agarplatten wurden 48 h bei 30 °C im Brutschrank (Heraeus
Instruments, kelvitron® t) inkubiert. Eine violette Pigmentierung des bakteriellen
Teppichs um die Auftragungspunkte wurde als ein Nachweis für intakte AHL-
34

2. Material und Methoden
Moleküle gewertet, wobei sich die AHL-Konzentration in der Spotgröße
widerspiegelte.
2.8 Molekularbiologische Untersuchungen In der vorliegenden Arbeit wurden des Weiteren die Cyanobakterienstämme Bo 53
und Bo 10 molekularbiologisch untersucht. Bei beiden Mikroorganismen wurden die
16S-rDNA und als weitere Möglichkeit zur Einordnung auf molekularer Ebene das
Gen gyrB mittels PCR amplifiziert und anschließend sequenziert. Zudem erfolgte die
Amplifikation der ITS (internal transcribed spacer) von Stamm Bo 10. Diese nicht-
kodierenden intergenischen Regionen sind zwischen den 16S-rRNA-, 23S-rRNA-
und 5S-rRNA-Genen lokalisiert. Die Anzahl und Größe der amplifizierten ITS-
Regionen stellen einen zusätzlichen taxonomischen Marker zur phylogenetischen
Einordnung von Mikroorganismen dar (ITEMAN et al., 2002).
Für die heterotrophen Bakterienstämme Bo 10-09 und Bo 53-33 (HUBE, 2008) sowie
für das Cyanobakterium Stamm Flo 1 (SCHRÜBBERS, 2007) wurden in diesem
Zusammenhang von den genannten Autoren bereits 16S-rDNA-Sequenzierungen mit
anschließender phylogenetischer Einordnung durchgeführt. Ergänzend sollte in der
vorliegenden Arbeit eine Amplifikation der ITS von Stamm Flo 1 sowie eine
Amplifikation und Sequenzierung des gyrB-Gens der Stämme Bo 53-33 und
Bo 10-09 durchgeführt werden.
Auf Basis der erstellten Sequenzen sollten über die Erstellung von phylogenetischen
Stammbäumen Verwandtschaftsbeziehungen aufgezeigt werden. Stamm Bo 53
wurde freundlicherweise von Dr. B. Heyduck-Söller, Stamm Bo 10 von Dipl.-Biologin
A. Hube zur Verfügung gestellt. Chromosomale DNA vom Stamm Flo 1 und vom
Referenzstamm Geitlerinema PCC 7105 wurde von Dipl.-Biologen J. Schrübbers für
die Untersuchungen überlassen.
2.8.1 Extraktion der chromosomalen DNA nach NEILAN (1995) und SCHENK (1996)
Für die DNA-Extraktion nach NEILAN (1995) und SCHENK (1996) (modifiziert nach
HEYDUCK-SÖLLER, 2003) wurden 8 - 10 Tage alte Cyanobakterienkulturen von
Stamm Bo 53 und Stamm Bo 10 verwendet, die bei konstanten 21 °C und einer
konstanten Photonenflussdichte von 1 µE m-2 s-1 PAR gehältert wurden. Die Anzucht
der heterotrophen Stämme Bo 53-33 und Bo 10-09 erfolgte als Übernachtkultur bei
35

2. Material und Methoden
einer konstanten Temperatur von 21 °C und 120 rpm im Schüttelwasserbad (Julabo
SW 21) (siehe Kap. 2.5.1).
1 ml der jeweiligen Kultur wurde für 5 min bei 8.000 rpm zentrifugiert (Heraeus
Instruments, Biofuge pico). Der Überstand wurde verworfen, die Kultur anschließend
mit 1 ml STE-Puffer (100 mM NaCl; 10 mM Tris-HCl, pH 8,0; 1 mM Na2EDTA, pH
8,0) versetzt und wie oben beschrieben erneut zentrifugiert. Nach Dekantierung des
Überstandes wurde erneut 1 ml STE-Puffer auf den Ansatz pipettiert, wobei
cyanobakterielle Proben zum Ablösen der heterotrophen Begleitflora in einem
Ultraschallbad (Elma™, Transsonic Digital) bei 20 °C für 10 min bei maximaler
Stärke beschallt wurden. Die Ultraschallbad-Behandlung wurde bei nicht der DNA-
Extraktion aus den heterotrophen Bakterien durchgeführt. Nach Zentrifugation (5 min
und 8.000 rpm) und Dekantierung des Überstandes wurde nochmals 1 ml STE-Puffer
hinzugesetzt, anschließend zentrifugiert (siehe oben) und der Überstand vollständig
entfernt. Nachfolgend wurde der Ansatz mit 400 µl STE-Puffer überschichtet sowie
mit autoklavierten Glasperlen (Ø 0,1 - 0,2 µm; Fa. BIOmatik GmbH) im gleichen
Volumenanteil (des jeweiligen Pellets) und mit 1 ml 75 °C heißem Phenol versetzt.
Nach Vortexen auf einem Whirl-Mixer für 40 - 60 sec und anschließender Kühlung
auf Eis erfolgte zur Trennung von Phenol- und wässriger Phase eine Zentrifugation
(Beckman GS-15R) für 20 min bei 4 °C und 12.000 rpm. Die DNA-enthaltende obere
wässrige Phase wurde in ein steriles ERG überführt und das Volumen bestimmt.
Anschließend wurden 2 µl Poly A RNA (10 mg/ml) zugegeben. Zur Entfernung von
eventuell noch vorhandenen Proteinen wurde 1 Volumenanteil
Chloroform/Isoamylalkohol (24:1 (v/v)) hinzugesetzt, anschließend gevortext und für
5 min bei 12.000 rpm zentrifugiert. Ein Großteil der oberen wässrigen Phase (350 µl)
wurde in ein steriles ERG überführt und mit 0,1 Volumenanteil 3 M Natriumacetat
(pH 5,2) versetzt. Zur Fällung der DNA erfolgte die Überschichtung mit
0,8 Volumenanteilen eisgekühltem Isopropanol, anschließender Lagerung bei -80 °C
für 20 min und Zentrifugation bei 12.000 rpm für 30 min. Nach Dekantierung des
Überstandes wurde das Pellet zur vollständigen Entfernung von eventuell
vorhandenen Salzen mit 1 ml frisch angesetztem 70%igem Ethanol versetzt und
nach Durchmischung für 10 min bei 12.000 rpm zentrifugiert. Nach Entfernung des
Überstandes erfolgte die Zugabe von 1 ml Ethanol abs. und eine erneute
Zentrifugation (s.o.). Das vom Überstand befreite DNA-Pellet wurde bei
Raumtemperatur (RT) zum Trocknen für 1 h stehengelassen. Abhängig von der
36

2. Material und Methoden
DNA-Menge wurde das Pellet in 20 - 40 µl TE-Puffer (10 mM Tris-HCl, pH 8,0; 1 mM
Na2EDTA, pH 8,0) über Nacht bei RT resuspendiert. Die DNA wurde im Dunkeln bei
4 °C gelagert.
2.8.2 Bestimmung der DNA-Konzentration Die Konzentration und die Reinheit der extrahierten DNA wurden mit Hilfe eines
Spektralphotometers (Peqlab Biotechnologie, NanoDrop ND-1000) photometrisch bei
Wellenlängen von 260 und 280 nm ermittelt. Zur Quantifizierung der DNA wurde die
Absorption der eingesetzten DNA-Lösung bei einer Wellenlänge von 260 nm
gemessen. Die Reinheitsbestimmung der DNA erfolgte über das Verhältnis der
Absorptionen bei 260 und 280 nm (A260nm/A280nm).
2.8.3 Polymerase-Kettenreaktion (PCR) Zur Amplifikation der 16S-rDNA, des gyrB-Gens und der ITS (siehe Kap. 2.8) wurde
eine Standard-PCR durchgeführt. Hierzu wurde standardmäßig jeweils ein PCR-
Ansatz mit 50 µl Endvolumen pipettiert, dessen Zusammensetzung in Tabelle 2.16
dargestellt ist. Für die PCR wurde ein Vielfaches der angegebenen Mengen in Form
eines Mastermix (ohne DNA-Template) angesetzt und auf die PCR-Reaktionsgefäße
verteilt. Die Sequenzen der verwendeten Primer zur Amplifikation und
Sequenzierung der 16S-rDNA, des gyrB-Gens und der ITS sind in Tabelle 2.17
zusammengefasst.
37
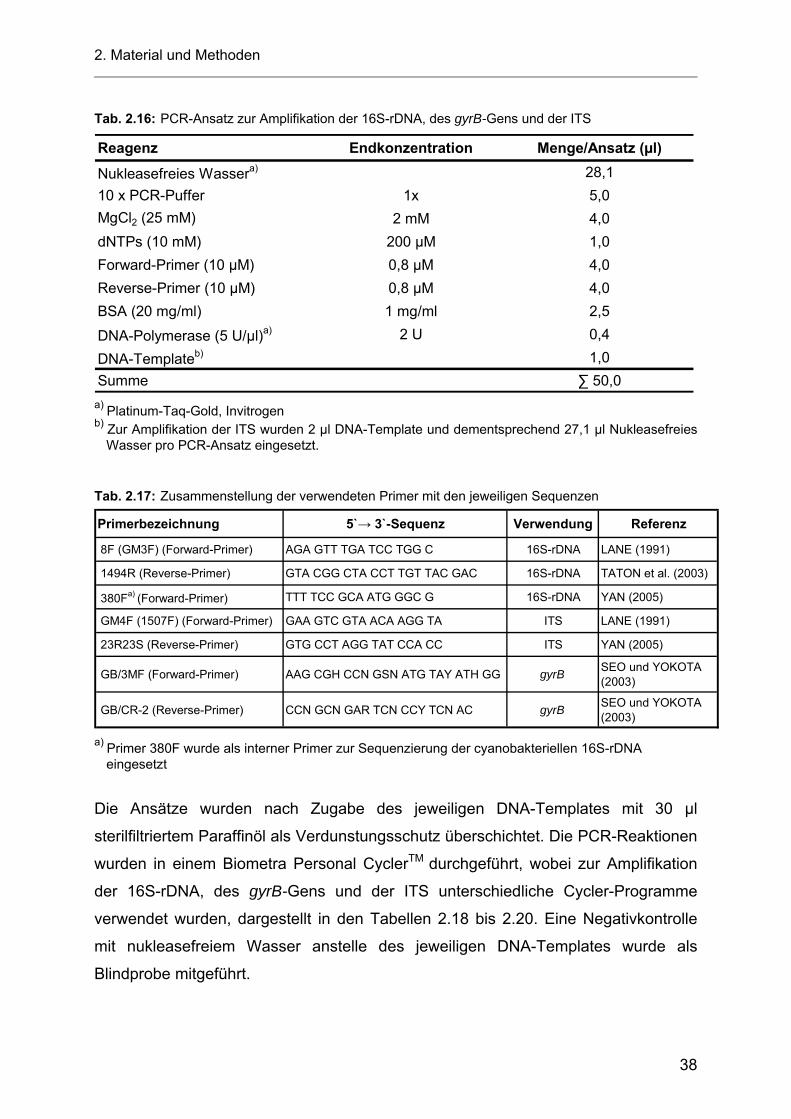
2. Material und Methoden
Tab. 2.16: PCR-Ansatz zur Amplifikation der 16S-rDNA, des gyrB-Gens und der ITS
Reagenz Endkonzentration Menge/Ansatz (µl)
Nukleasefreies Wassera) 28,110 x PCR-Puffer 1x 5,0MgCl2 (25 mM) 2 mM 4,0dNTPs (10 mM) 200 µM 1,0Forward-Primer (10 µM) 0,8 µM 4,0Reverse-Primer (10 µM) 0,8 µM 4,0BSA (20 mg/ml) 1 mg/ml 2,5
DNA-Polymerase (5 U/µl)a) 2 U 0,4
DNA-Templateb) 1,0Summe ∑ 50,0
a) Platinum-Taq-Gold, Invitrogen b) Zur Amplifikation der ITS wurden 2 µl DNA-Template und dementsprechend 27,1 µl Nukleasefreies
Wasser pro PCR-Ansatz eingesetzt.
Tab. 2.17: Zusammenstellung der verwendeten Primer mit den jeweiligen Sequenzen
Primerbezeichnung 5`→ 3`-Sequenz Verwendung Referenz
8F (GM3F) (Forward-Primer) AGA GTT TGA TCC TGG C 16S-rDNA LANE (1991)
1494R (Reverse-Primer) GTA CGG CTA CCT TGT TAC GAC 16S-rDNA TATON et al. (2003)
380Fa) (Forward-Primer) TTT TCC GCA ATG GGC G 16S-rDNA YAN (2005)
GM4F (1507F) (Forward-Primer) GAA GTC GTA ACA AGG TA ITS LANE (1991)
23R23S (Reverse-Primer) GTG CCT AGG TAT CCA CC ITS YAN (2005)
GB/3MF (Forward-Primer) AAG CGH CCN GSN ATG TAY ATH GG gyrB SEO und YOKOTA(2003)
GB/CR-2 (Reverse-Primer) CCN GCN GAR TCN CCY TCN AC gyrB SEO und YOKOTA(2003)
a) Primer 380F wurde als interner Primer zur Sequenzierung der cyanobakteriellen 16S-rDNA eingesetzt
Die Ansätze wurden nach Zugabe des jeweiligen DNA-Templates mit 30 µl
sterilfiltriertem Paraffinöl als Verdunstungsschutz überschichtet. Die PCR-Reaktionen
wurden in einem Biometra Personal CyclerTM durchgeführt, wobei zur Amplifikation
der 16S-rDNA, des gyrB-Gens und der ITS unterschiedliche Cycler-Programme
verwendet wurden, dargestellt in den Tabellen 2.18 bis 2.20. Eine Negativkontrolle
mit nukleasefreiem Wasser anstelle des jeweiligen DNA-Templates wurde als
Blindprobe mitgeführt.
38
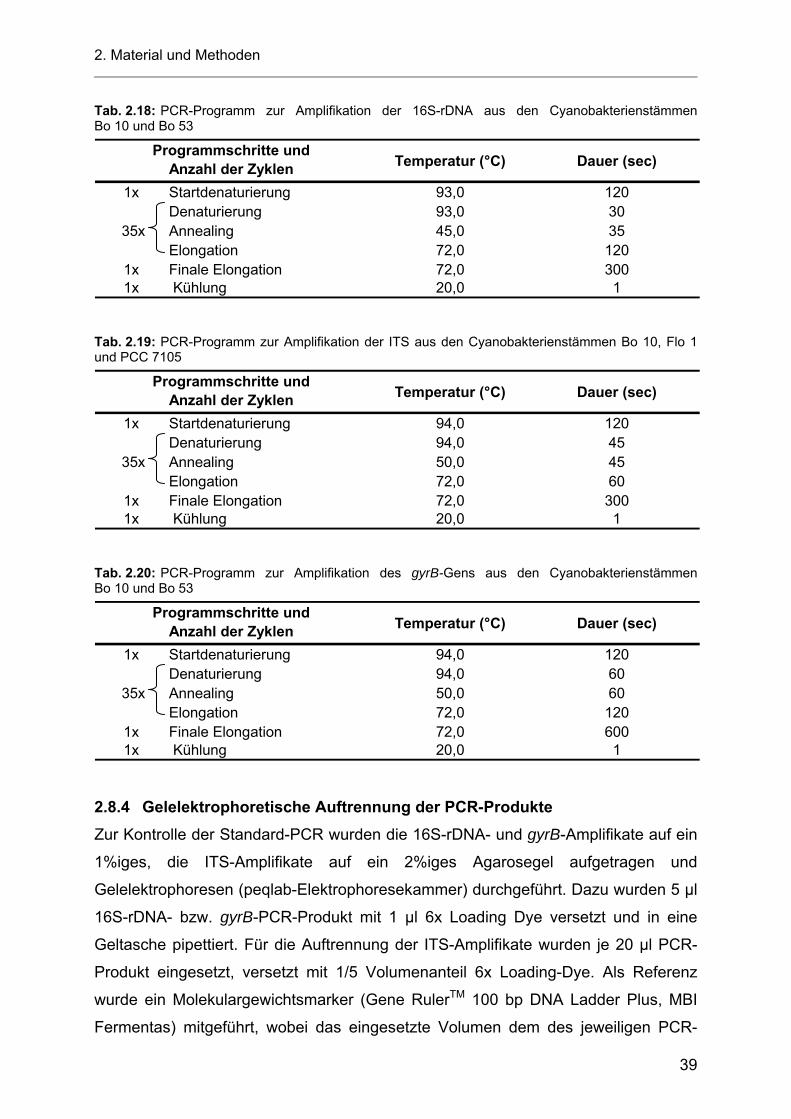
2. Material und Methoden
Tab. 2.18: PCR-Programm zur Amplifikation der 16S-rDNA aus den Cyanobakterienstämmen Bo 10 und Bo 53
Temperatur (°C) Dauer (sec)
1x Startdenaturierung 93,0 120Denaturierung 93,0 30
35x Annealing 45,0 35Elongation 72,0 120
1x Finale Elongation 72,0 3001x Kühlung 20,0 1
Programmschritte undAnzahl der Zyklen
Tab. 2.19: PCR-Programm zur Amplifikation der ITS aus den Cyanobakterienstämmen Bo 10, Flo 1 und PCC 7105
Temperatur (°C) Dauer (sec)
1x Startdenaturierung 94,0 120Denaturierung 94,0 45
35x Annealing 50,0 45Elongation 72,0 60
1x Finale Elongation 72,0 3001x Kühlung 20,0 1
Programmschritte undAnzahl der Zyklen
Tab. 2.20: PCR-Programm zur Amplifikation des gyrB-Gens aus den Cyanobakterienstämmen Bo 10 und Bo 53
Temperatur (°C) Dauer (sec)
1x Startdenaturierung 94,0 120Denaturierung 94,0 60
35x Annealing 50,0 60Elongation 72,0 120
1x Finale Elongation 72,0 6001x Kühlung 20,0 1
Programmschritte undAnzahl der Zyklen
2.8.4 Gelelektrophoretische Auftrennung der PCR-Produkte Zur Kontrolle der Standard-PCR wurden die 16S-rDNA- und gyrB-Amplifikate auf ein
1%iges, die ITS-Amplifikate auf ein 2%iges Agarosegel aufgetragen und
Gelelektrophoresen (peqlab-Elektrophoresekammer) durchgeführt. Dazu wurden 5 µl
16S-rDNA- bzw. gyrB-PCR-Produkt mit 1 µl 6x Loading Dye versetzt und in eine
Geltasche pipettiert. Für die Auftrennung der ITS-Amplifikate wurden je 20 µl PCR-
Produkt eingesetzt, versetzt mit 1/5 Volumenanteil 6x Loading-Dye. Als Referenz
wurde ein Molekulargewichtsmarker (Gene RulerTM 100 bp DNA Ladder Plus, MBI
Fermentas) mitgeführt, wobei das eingesetzte Volumen dem des jeweiligen PCR-
39

2. Material und Methoden
Produktes entsprach. Bei einer angelegten Spannung von 70 V (Power Pac 300,
Biorad) betrug die Laufzeit der 16S-rDNA-Produkte und der gyrB-Amplifikate ca. 1 h,
die der ITS-Produkte ca. 3 h bei 50 V Spannung. Nach anschließender Gelfärbung
für 15 min in 0,1%iger Ethidiumbromid-Lösung erfolgte die Auswertung am
Transilluminator (Fluco-Link, Biometra) unter UV-Licht bei 254 nm. Die 16S-rDNA-
PCR-Produkte bzw. gyrB-PCR-Produkte sollten bei 1500 bp bzw. 1200 bp eine
deutliche Bande aufweisen, während die Bandenanzahl und die Fragmentgröße der
ITS-Amplifikate variieren konnten.
Die Amplifikate der 16S-rDNA von Stamm Bo 10 wurden durch Extraktion (siehe
Kap. 2.8.5) der 1500 bp-Bande aus dem Gel erhalten, da das PCR-Produkt mehrere
Banden im Gel aufwies. Hierzu erfolgte eine Auftrennung des gesamten PCR-
Produkts in einem 2%igen Agarosegel in mehreren Gelläufen. Die Laufzeit betrug
10 h bei einer angelegten Spannung von 50 V.
2.8.5 Elution und Aufreinigung von PCR-Produkten Die Aufreinigung von PCR-Produkten wurde mittels des „Qiagen QIAquick PCR
Purification“-Kits (Qiagen) nach Herstellerangaben durchgeführt. Die DNA-Eluierung
erfolgte in 40 µl EB-Puffer.
Die Elution und Aufreinigung von PCR-Produkten aus Agarosegelen erfolgte unter
Verwendung des „QIAquick Gel Extraktion“-Kits (Qiagen) nach Angaben des
Herstellers. Die DNA wurde in 40 µl EB-Puffer eluiert.
Die Konzentration an PCR-Produkt wurde mit Hilfe eines Spektralphotometers
(Peqlab Biotechnologie, NanoDrop ND-1000) bestimmt (siehe Kap. 2.8.2).
2.8.6 Sequenzierung der 16S-rDNA und des gyrB-Gens Die Sequenzierung der 16S-rDNA und des gyrB-Gens mittels aufgereinigter PCR-
Produkte wurde von der Firma GATC Biotech AG (Konstanz) unter Verwendung der
in Kapitel 2.8.3, Tabelle 2.17 aufgeführten Primer durchgeführt.
2.8.7 Erstellung von phylogenetischen Stammbäumen Das Alignment der Teilsequenzen wurde mit dem Programm ChromasPro
durchgeführt. Die generierten Sequenzen der Cyanobakterienstämme Bo 53 und
Bo 10 wurden unter Anwendung von „nucleotide BLAST” mit den gegenwärtig
hinterlegten Sequenzen der öffentlichen NCBI-Datenbank
40

2. Material und Methoden
(www.ncbi.nlm.nih.gov/blast/Blast.cgi; Stand: März 2008) verglichen. Basierend auf
den erzielten Ergebnissen erfolgte unter Abgleichung mit Sequenzen von
Referenzstämmen mit den größten Homologien aus der NCBI-Datenbank die
Erstellung eines phylogenetischen Stammbaumes. Das Alignment der ausgewählten
Sequenzen sowie das Zuschneiden der Alignmentsequenzen wurden mit den
Programmen Bioedit und GeneDoc durchgeführt. Die phylogenetischen
Stammbäume wurden mit der “Neighbor-Joining”-Methode (NJ) mit dem Programm
MEGA (Version 4.0) konstruiert. Die Berechnung der evolutionären Distanzen
zwischen den Sequenzen erfolgte nach der Methode von JUKES und CANTOR
(1969). Die evolutionäre Distanz ergbt sich aus dem Verhältnis zwischen
verschiedenen Basen zweier gleichlanger Sequenzen zu allen Basen der Sequenz.
Eine statistische Absicherung der Topologie der phylogenetischen Stammbäume
erfolgte durch 1000 Wiederholungen der Bootstrap-Analysen.
2.9 Herkunft der verwendeten Chemikalien Die in der vorliegenden Arbeit verwendeten Chemikalien wiesen p.A.-Qualität oder
HPLC-Grade auf und wurden von den folgenden Firmen bezogen: Acros Organics
(Geel, Belgien), AppliChem (Darmstadt, Deutschland), Fluka (Buchs, Schweiz),
Invitrogen (Karlsruhe, Deutschland), Jannsen Chimica (Geel, Belgien), MBI
Fermentas (St. Leon-Rot, Deutschland), Merck (Darmstadt, Deutschland), Qiagen
(Hilden, Deutschland), Riedel de Haën (Seelze, Deutschland), Roche (Mannheim,
Deutschland), Roth (Karlsruhe, Deutschland), Serva (Heidelberg, Deutschland),
Sigma-Aldrich (Steinheim, Deutschland) und VWR international (Leuven, Belgien).
41

3. Ergebnisse
3. Ergebnisse
3.1 Verwendete Puffersysteme Für die pH-Untersuchungen der Bakterienstämme Bo 53-33, Bo 10-09 und Flo 1
wurde eine große Bandbreite an Puffersystemen getestet (siehe Kap. 2.3), bevor die
in den generierten Wachstumskurven (siehe Kap. 3.2) und zur AHL-Untersuchung
(siehe Kap. 3.5.1) eingesetzten Puffer zum Einsatz kamen.
Erhebliche Schwierigkeiten traten vor allem durch den Ausfall von Salzen (z.B. Mg2+-
und Ca2+-Ionen) in pH-gepufferten ASNIII/2-Medien und durch starke pH-Wert-
Verschiebungen dieser (von dem jeweiligen eingestellten pH-Wert) nach
Autoklavieren auf, die ein Ansetzen von gepufferten Kulturmedien mit höheren pH-
Werten stark erschwerten (pH 8,5 - 10,0) bzw. unmöglich machten (pH 11,0). Die
Trübung des Nährmediums und die pH-Abweichungen nahmen dabei sukzessive mit
steigendem pH-Wert zu, wobei vor allem die mit Fleischextrakt versetzten
Kulturmedien zu Ausfällungen neigten. In diesem Zusammenhang waren
insbesondere die Clark & Lubs-Pufferlösungen im Bereich von pH 8,0 - 10,0
aufgrund oben genannter Probleme zur Kultivierung der heterotrophen
Bakterienstämme Bo 53-33 und Bo 10-09 lediglich bedingt verwendbar (pH 8,0 und
8,5) bzw. völlig ungeeignet gewesen (pH 9,0 und 10,0). Untersuchungen zum
Wachstumsverhalten mit Clark & Lubs-gepufferten Medien (pH 8,0 und 8,5) zeigten
bei Stamm Bo 53-33 kein und bei Stamm Bo 10-09 ein nur schwaches Wachstum
(Daten nicht gezeigt). Neben einem durch Mineralsalz-Ausfällungen bedingten
Nährstoffentzug könnte ebenfalls ein hemmender Einfluss des Puffers ein Grund für
ein nur schwaches Wachstum der heterotrophen Bakterien sein. Die letztlich
verwendeten Tris-HCl- und Glycin-Puffer in diesem pH-Bereich bedingten im
Kulturmedium ebenfalls leichte bis mittlere Ausfällungen und pH-Verschiebungen
nach dem Autoklavieren, welche aber tolerierbar waren. Der geforderte pH-Wert
wurde gegebenenfalls nachträglich durch Zugabe von 1 M HCl bzw. 1 M NaOH
eingestellt. Starke Ausfällungen in den Kulturmedien wurden zudem bei Verwendung
des Tris-HCl-Puffers pH 8,9 und des Clark & Lubs-Puffers pH 7,0 (Phosphat-Puffer)
beobachtet und erwiesen sich somit neben dem Tris-Maleat-Puffer pH 6,0 als
ungeeignet, bei dessen Anwesenheit kein Wachstum der heterotrophen Bakterien
erfolgte.
42

3. Ergebnisse
Weitere Probleme traten mit pH-Wert-Verschiebungen während des Wachstums auf,
die anhand der pH-Endwerte der Versuchskulturen deutlich werden (siehe Kap. 3.2.1
und 3.3.1). Durch die Erhöhung der Molarität und somit der Pufferkapazität (150 mM)
wurde versucht, diesem entgegenzuwirken, welches sich jedoch überwiegend
negativ auf ein Wachstum der Bakterien auswirkte (Daten nicht gezeigt) und zudem
mit stärkeren Ausfällungen im Nährmedium ab pH 8,5 einherging.
Das Ansetzen von getrennt autoklavierten Stammlösungen bestimmter ASNIII/2-
Komponenten (CaCl2 x 2 H2O, MgCl2 x 6 H2O und MgSO4 x 7 H2O), welche nach
Autoklavieren des restlichen gepufferten Grundmediums diesem bei RT hinzugesetzt
wurden, bewirkte sowohl bei schwächer als auch stärker gepufferten Kulturmedien
oberhalb von pH 8,0 hinsichtlich einer Minimierung der Ausfällungen keine Abhilfe.
3.2 Wachstumsverhalten der heterotrophen Stämme Bo 53-33 und Bo 10-09 bei unterschiedlichen pH-Werten
Zur Ermittlung des pH-Toleranzbereiches (pH-Minimum und pH-Maximum) und pH-
Optimums für die heterotrophen Stämme Bo 53-33 und Bo 10-09 wurde das
Wachstum im pH-Bereich von 2,0 - 10,0 untersucht (siehe Kap. 2.6.2). Als Referenz
wurden ungepufferte Versuchsansätze inkubiert (siehe Kap. 2.6.1). Wachstum wurde
definiert als Zunahme in der O.D.600nm von mindestens 0,1 nach 24 h Inkubation.
Gemäß dieser Definition konnte im Bereich von pH 2,0 - 5,0 sowie bei pH 10,0 kein
Wachstum der Stämme Bo 53-33 und Bo 10-09 beobachtet werden (Daten nicht
gezeigt). Die Ergebnisse zum Wachstumsverhalten der heterotrophen Stämme
Bo 53-33 und Bo 10-09 im Bereich von pH 6,0 - 9,0 sowie in ungepuffertem ASNIII/2-
Medium sind in den Abbildungen 3.1 und 3.3 dargestellt, wobei die Ergebnisse durch
Versuchswiederholungen abgesichert wurden (Daten nicht gezeigt).
43
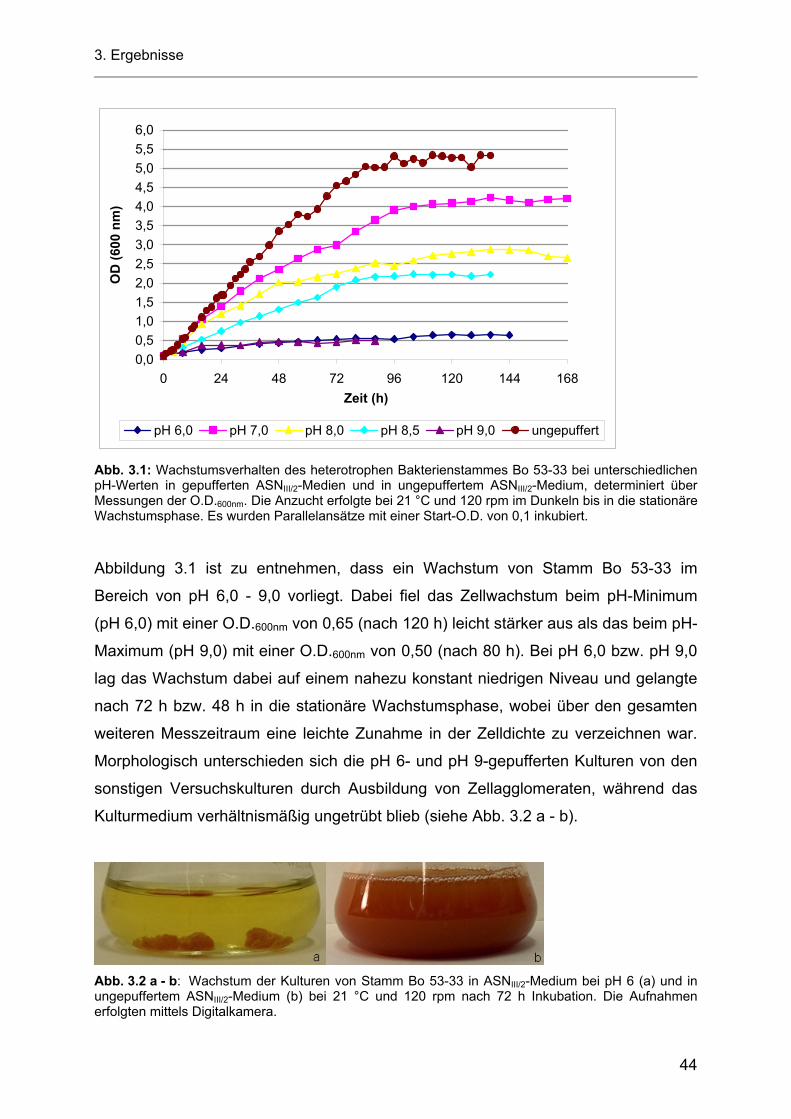
3. Ergebnisse
0,00,51,01,52,02,53,03,54,04,55,05,56,0
0 24 48 72 96 120 144 168Zeit (h)
OD
(600
nm
)
pH 6,0 pH 7,0 pH 8,0 pH 8,5 pH 9,0 ungepuffert
Abb. 3.1: Wachstumsverhalten des heterotrophen Bakterienstammes Bo 53-33 bei unterschiedlichen pH-Werten in gepufferten ASNIII/2-Medien und in ungepuffertem ASNIII/2-Medium, determiniert über Messungen der O.D.600nm. Die Anzucht erfolgte bei 21 °C und 120 rpm im Dunkeln bis in die stationäre Wachstumsphase. Es wurden Parallelansätze mit einer Start-O.D. von 0,1 inkubiert.
Abbildung 3.1 ist zu entnehmen, dass ein Wachstum von Stamm Bo 53-33 im
Bereich von pH 6,0 - 9,0 vorliegt. Dabei fiel das Zellwachstum beim pH-Minimum
(pH 6,0) mit einer O.D.600nm von 0,65 (nach 120 h) leicht stärker aus als das beim pH-
Maximum (pH 9,0) mit einer O.D.600nm von 0,50 (nach 80 h). Bei pH 6,0 bzw. pH 9,0
lag das Wachstum dabei auf einem nahezu konstant niedrigen Niveau und gelangte
nach 72 h bzw. 48 h in die stationäre Wachstumsphase, wobei über den gesamten
weiteren Messzeitraum eine leichte Zunahme in der Zelldichte zu verzeichnen war.
Morphologisch unterschieden sich die pH 6- und pH 9-gepufferten Kulturen von den
sonstigen Versuchskulturen durch Ausbildung von Zellagglomeraten, während das
Kulturmedium verhältnismäßig ungetrübt blieb (siehe Abb. 3.2 a - b).
Abb. 3.2 a - b: Wachstum der Kulturen von Stamm Bo 53-33 in ASNIII/2-Medium bei pH 6 (a) und in ungepuffertem ASNIII/2-Medium (b) bei 21 °C und 120 rpm nach 72 h Inkubation. Die Aufnahmen erfolgten mittels Digitalkamera.
44
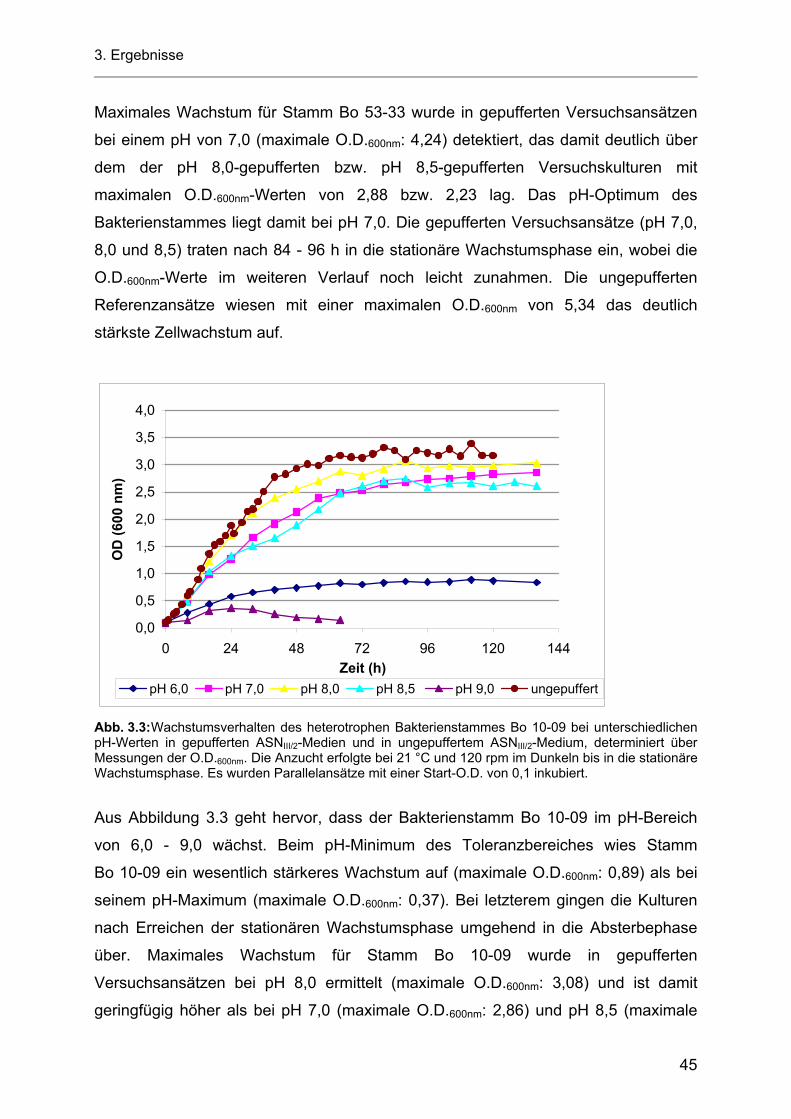
3. Ergebnisse
Maximales Wachstum für Stamm Bo 53-33 wurde in gepufferten Versuchsansätzen
bei einem pH von 7,0 (maximale O.D.600nm: 4,24) detektiert, das damit deutlich über
dem der pH 8,0-gepufferten bzw. pH 8,5-gepufferten Versuchskulturen mit
maximalen O.D.600nm-Werten von 2,88 bzw. 2,23 lag. Das pH-Optimum des
Bakterienstammes liegt damit bei pH 7,0. Die gepufferten Versuchsansätze (pH 7,0,
8,0 und 8,5) traten nach 84 - 96 h in die stationäre Wachstumsphase ein, wobei die
O.D.600nm-Werte im weiteren Verlauf noch leicht zunahmen. Die ungepufferten
Referenzansätze wiesen mit einer maximalen O.D.600nm von 5,34 das deutlich
stärkste Zellwachstum auf.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
0 24 48 72 96 120 144Zeit (h)
OD
(600
nm
)
pH 6,0 pH 7,0 pH 8,0 pH 8,5 pH 9,0 ungepuffert
Abb. 3.3: Wachstumsverhalten des heterotrophen Bakterienstammes Bo 10-09 bei unterschiedlichen pH-Werten in gepufferten ASNIII/2-Medien und in ungepuffertem ASNIII/2-Medium, determiniert über Messungen der O.D.600nm. Die Anzucht erfolgte bei 21 °C und 120 rpm im Dunkeln bis in die stationäre Wachstumsphase. Es wurden Parallelansätze mit einer Start-O.D. von 0,1 inkubiert.
Aus Abbildung 3.3 geht hervor, dass der Bakterienstamm Bo 10-09 im pH-Bereich
von 6,0 - 9,0 wächst. Beim pH-Minimum des Toleranzbereiches wies Stamm
Bo 10-09 ein wesentlich stärkeres Wachstum auf (maximale O.D.600nm: 0,89) als bei
seinem pH-Maximum (maximale O.D.600nm: 0,37). Bei letzterem gingen die Kulturen
nach Erreichen der stationären Wachstumsphase umgehend in die Absterbephase
über. Maximales Wachstum für Stamm Bo 10-09 wurde in gepufferten
Versuchsansätzen bei pH 8,0 ermittelt (maximale O.D.600nm: 3,08) und ist damit
geringfügig höher als bei pH 7,0 (maximale O.D.600nm: 2,86) und pH 8,5 (maximale
45

3. Ergebnisse
O.D.600nm: 2,75). Die gepufferten Kulturen (pH 7,0, 8,0 und 8,5) erreichten nach etwa
64 h die stationäre Wachstumsphase, gefolgt von einer weiteren leichten Zunahme
der Zelldichten. Die ungepufferten Referenzkulturen wiesen mit einer maximalen
O.D.600nm von 3,39 ein leicht stärkeres Wachstum als die gepufferten Kulturen auf.
Die Referenzansätze traten nach etwa 48 h in die stationäre Wachstumsphase ein.
3.2.1 pH-Werte der Zellkulturüberstände Zu den photometrischen Messungen zum Wachstumsverhalten erfolgte parallel eine
pH-Messung der Kulturüberstände mittels pH-Indikatorpapier (Macherey-Nagel;
pH 6,0 - 10,0) (siehe Kap. 2.6.4) (Daten nicht gezeigt).
Nach Abschluss der Untersuchung zum Wachstumsverhalten der heterotrophen
Bakterienstämme Bo 53-33 und Bo 10-09 wurden die pH-Endwerte der
Kulturüberstände mittels einer pH-Elektrode, gekoppelt an einem pH-Meter,
bestimmt. Die ermittelten pH-Endwerte sind in Tabelle 3.1 zusammengefasst.
Tab. 3.1: pH-Werte von unbeimpften Kulturmedien und Kulturüberständen der Stämme Bo 53-33 und Bo 10-09 nach Inkubation. Ungepuffertes ASNIII/2-Medium und gepuffertes ASNIII/2-Medium mit unterschiedlichen pH-Werten diente als Kulturmedium. Die Inkubation erfolgte bis in die stationäre Wachstumsphase (siehe Kap. 3.2) in einem Schüttelwasserbad (120 rpm) bei 21 °C im Dunkeln. n = 2
pH-Wert des Kulturmediums vor Inkubation
pH-Wert des Kulturüberstandesvon Bo 53-33 nach Inkubation
pH-Wert des Kulturüberstandesvon Bo 10-09 nach Inkubation
pH 7,1 (ungepuffert) pH 8,36 pH 8,37pH 6,0 pH 6,32 pH 6,27pH 7,0 pH 8,18 pH 8,31pH 8,0 pH 8,25 pH 8,40pH 8,5 pH 8,50 pH 8,55pH 9,0 pH 8,65 pH 8,80
In Tabelle 3.1 sind die pH-Werte vor und nach Inkubation der Versuchsansätze der
Stämme Bo 53-33 und Bo 10-09 dargestellt.
Bei beiden Bakterienstämmen ist, abgesehen von pH 8,5 und 9,0, ein Anstieg des
pH-Wertes sowohl in den ungepufferten als auch in den gepufferten
Versuchskulturen zu verzeichnen. Eine kontinuierliche Alkalisierung der Überstande
konnte dabei für die ungepufferten Versuchsansätze bereits nach 8 h Inkubation, für
die gepufferten Versuchsansätze hingegen erst nach 40 - 48 h Inkubation festgestellt
werden (Daten nicht gezeigt).
46
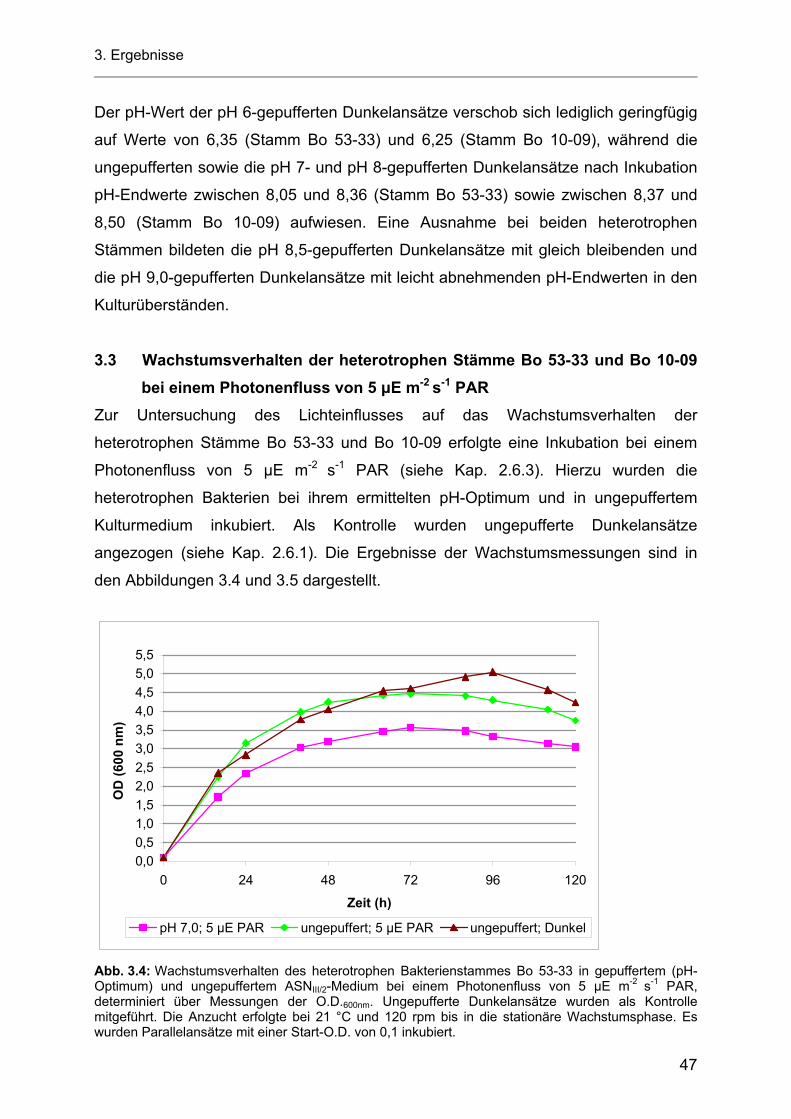
3. Ergebnisse
Der pH-Wert der pH 6-gepufferten Dunkelansätze verschob sich lediglich geringfügig
auf Werte von 6,35 (Stamm Bo 53-33) und 6,25 (Stamm Bo 10-09), während die
ungepufferten sowie die pH 7- und pH 8-gepufferten Dunkelansätze nach Inkubation
pH-Endwerte zwischen 8,05 und 8,36 (Stamm Bo 53-33) sowie zwischen 8,37 und
8,50 (Stamm Bo 10-09) aufwiesen. Eine Ausnahme bei beiden heterotrophen
Stämmen bildeten die pH 8,5-gepufferten Dunkelansätze mit gleich bleibenden und
die pH 9,0-gepufferten Dunkelansätze mit leicht abnehmenden pH-Endwerten in den
Kulturüberständen.
3.3 Wachstumsverhalten der heterotrophen Stämme Bo 53-33 und Bo 10-09 bei einem Photonenfluss von 5 µE m-2 s-1 PAR
Zur Untersuchung des Lichteinflusses auf das Wachstumsverhalten der
heterotrophen Stämme Bo 53-33 und Bo 10-09 erfolgte eine Inkubation bei einem
Photonenfluss von 5 µE m-2 s-1 PAR (siehe Kap. 2.6.3). Hierzu wurden die
heterotrophen Bakterien bei ihrem ermittelten pH-Optimum und in ungepuffertem
Kulturmedium inkubiert. Als Kontrolle wurden ungepufferte Dunkelansätze
angezogen (siehe Kap. 2.6.1). Die Ergebnisse der Wachstumsmessungen sind in
den Abbildungen 3.4 und 3.5 dargestellt.
0,00,51,01,52,02,53,03,54,04,55,05,5
0 24 48 72 96 1
Zeit (h)
OD
(600
nm
)
20
pH 7,0; 5 µE PAR ungepuffert; 5 µE PAR ungepuffert; Dunkel
Abb. 3.4: Wachstumsverhalten des heterotrophen Bakterienstammes Bo 53-33 in gepuffertem (pH-Optimum) und ungepuffertem ASNIII/2-Medium bei einem Photonenfluss von 5 µE m-2 s-1 PAR, determiniert über Messungen der O.D.600nm. Ungepufferte Dunkelansätze wurden als Kontrolle mitgeführt. Die Anzucht erfolgte bei 21 °C und 120 rpm bis in die stationäre Wachstumsphase. Es wurden Parallelansätze mit einer Start-O.D. von 0,1 inkubiert.
47
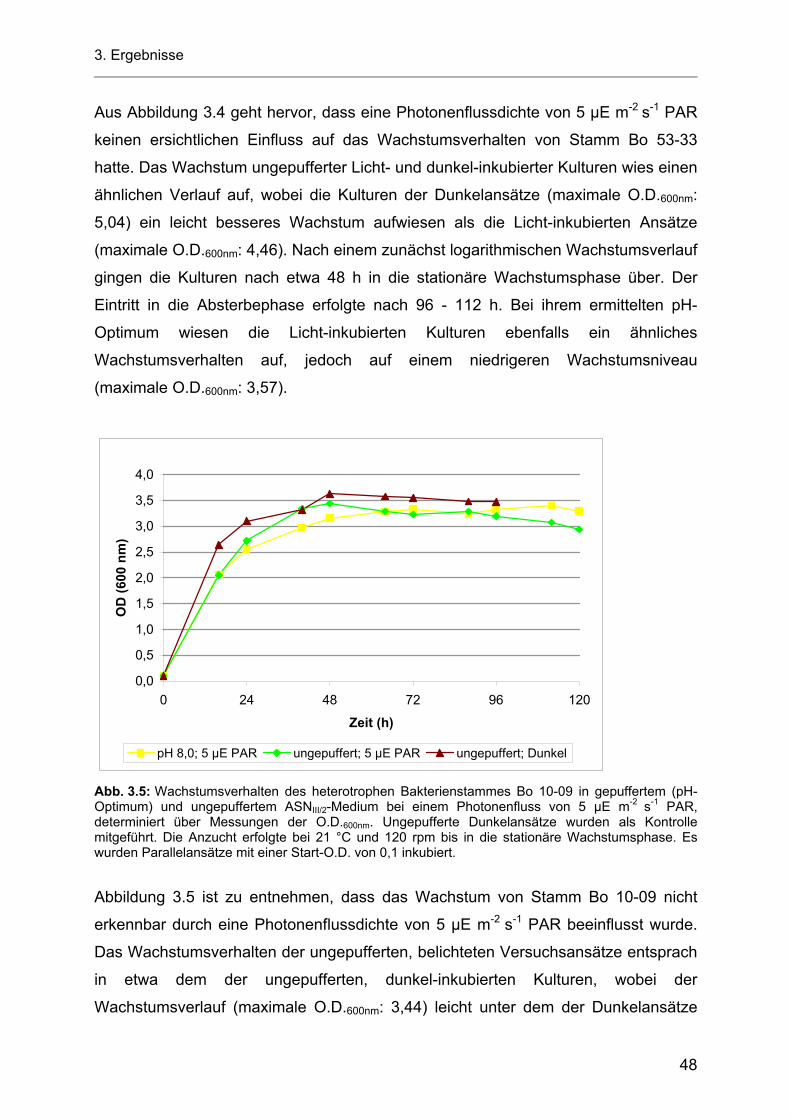
3. Ergebnisse
Aus Abbildung 3.4 geht hervor, dass eine Photonenflussdichte von 5 µE m-2 s-1 PAR
keinen ersichtlichen Einfluss auf das Wachstumsverhalten von Stamm Bo 53-33
hatte. Das Wachstum ungepufferter Licht- und dunkel-inkubierter Kulturen wies einen
ähnlichen Verlauf auf, wobei die Kulturen der Dunkelansätze (maximale O.D.600nm:
5,04) ein leicht besseres Wachstum aufwiesen als die Licht-inkubierten Ansätze
(maximale O.D.600nm: 4,46). Nach einem zunächst logarithmischen Wachstumsverlauf
gingen die Kulturen nach etwa 48 h in die stationäre Wachstumsphase über. Der
Eintritt in die Absterbephase erfolgte nach 96 - 112 h. Bei ihrem ermittelten pH-
Optimum wiesen die Licht-inkubierten Kulturen ebenfalls ein ähnliches
Wachstumsverhalten auf, jedoch auf einem niedrigeren Wachstumsniveau
(maximale O.D.600nm: 3,57).
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
0 24 48 72 96 1
Zeit (h)
OD
(600
nm
)
20
pH 8,0; 5 µE PAR ungepuffert; 5 µE PAR ungepuffert; Dunkel
Abb. 3.5: Wachstumsverhalten des heterotrophen Bakterienstammes Bo 10-09 in gepuffertem (pH-Optimum) und ungepuffertem ASNIII/2-Medium bei einem Photonenfluss von 5 µE m-2 s-1 PAR, determiniert über Messungen der O.D.600nm. Ungepufferte Dunkelansätze wurden als Kontrolle mitgeführt. Die Anzucht erfolgte bei 21 °C und 120 rpm bis in die stationäre Wachstumsphase. Es wurden Parallelansätze mit einer Start-O.D. von 0,1 inkubiert.
Abbildung 3.5 ist zu entnehmen, dass das Wachstum von Stamm Bo 10-09 nicht
erkennbar durch eine Photonenflussdichte von 5 µE m-2 s-1 PAR beeinflusst wurde.
Das Wachstumsverhalten der ungepufferten, belichteten Versuchsansätze entsprach
in etwa dem der ungepufferten, dunkel-inkubierten Kulturen, wobei der
Wachstumsverlauf (maximale O.D.600nm: 3,44) leicht unter dem der Dunkelansätze
48

3. Ergebnisse
(maximale O.D.600nm: 3,63) lag. Die bei einem Photonenfluss von 5 µE m-2 s-1 PAR
und pH-Optimum inkubierten Kulturen wiesen den gleichen Wachstumsverlauf auf
wie die ungepufferten Versuchsansätze, lediglich auf leicht geringerem
Wachstumsniveau (maximale O.D.600nm: 3,40).
3.3.1 pH-Werte der Zellkulturüberstände Zu den photometrischen Messungen zum Wachstumsverhalten wurden parallel die
pH-Werte der Kulturüberstände mittels pH-Indikatorpapier (Macherey-Nagel; pH 6,0 -
10,0) bestimmt (siehe Kap. 2.6.4) (Daten nicht gezeigt).
Nach Abschluss der Wachstumsversuche erfolgte eine Bestimmung der pH-
Endwerte der Kulturüberstände mittels einer pH-Elektrode, gekoppelt an einem pH-
Meter. Die ermittelten pH-Endwerte sind in Tabelle 3.2 zusammengefasst.
Tab. 3.2: pH-Werte von unbeimpften Kulturmedien und Kulturüberständen der Stämme Bo 53-33 und Bo 10-09 nach Inkubation. Ungepuffertes ASNIII/2-Medium und gepuffertes ASNIII/2-Medium mit optimalem pH-Wert für den jeweiligen Stamm diente als Kulturmedium. Die Inkubation erfolgte bis in die stationäre Wachstumsphase (siehe Kap. 3.3) auf einem Rotationsschüttler (120 rpm) bei 21 °C und einem Photonenfluss von 5 µE m-2 s-1 PAR oder im Dunkeln. n = 2
pH-Wert des Kulturmediumsvor Inkubation
pH-Wert des Kulturüberstandesvon Bo 53-33 nach Inkubation
pH-Wert des Kulturüberstandes von Bo 10-09 nach Inkubation
pH 7,1 (ungepuffert) pH 8,78 pH 8,78 pH 7,0 (gepuffert) pH 8,50 -- pH 8,0 (gepuffert) -- pH 8,78 pH 7,1 (ungepuffert) pH 8,80a) pH 8,70a)
a) nach Inkubation im Dunkeln
In Tabelle 3.2 sind die pH-Werte von Kulturmedien vor und nach Inkubation der
Versuchsansätze der Stämme Bo 53-33 und Bo 10-09 dargestellt.
Bei beiden Bakterienstämmen konnte eine deutliche Alkalisierung der ungepufferten
sowie gepufferten Versuchsansätze nach der Inkubation festgestellt werden. Hierbei
wiesen die ungepufferten Kulturüberstände beider Stämme, unabhängig von einer
Licht- oder Dunkelinkubation, pH-Endwerte zwischen 8,7 - 8,8 auf. Bei ungepufferten
Kulturen konnte bereits nach 8 h, bei gepufferten Kulturen hingegen erst nach
32 - 40 h Inkubation eine pH-Verschiebung von dem Ausgangswert beobachtet
werden (Daten nicht gezeigt).
49
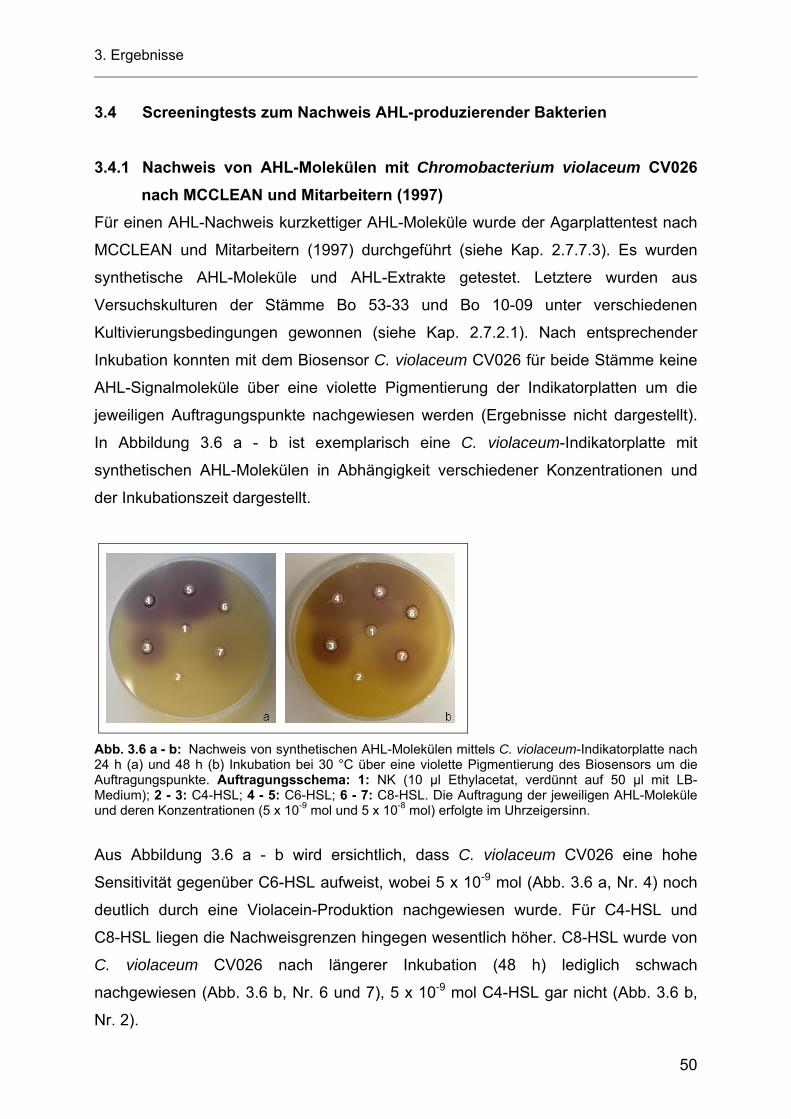
3. Ergebnisse
3.4 Screeningtests zum Nachweis AHL-produzierender Bakterien
3.4.1 Nachweis von AHL-Molekülen mit Chromobacterium violaceum CV026 nach MCCLEAN und Mitarbeitern (1997)
Für einen AHL-Nachweis kurzkettiger AHL-Moleküle wurde der Agarplattentest nach
MCCLEAN und Mitarbeitern (1997) durchgeführt (siehe Kap. 2.7.7.3). Es wurden
synthetische AHL-Moleküle und AHL-Extrakte getestet. Letztere wurden aus
Versuchskulturen der Stämme Bo 53-33 und Bo 10-09 unter verschiedenen
Kultivierungsbedingungen gewonnen (siehe Kap. 2.7.2.1). Nach entsprechender
Inkubation konnten mit dem Biosensor C. violaceum CV026 für beide Stämme keine
AHL-Signalmoleküle über eine violette Pigmentierung der Indikatorplatten um die
jeweiligen Auftragungspunkte nachgewiesen werden (Ergebnisse nicht dargestellt).
In Abbildung 3.6 a - b ist exemplarisch eine C. violaceum-Indikatorplatte mit
synthetischen AHL-Molekülen in Abhängigkeit verschiedener Konzentrationen und
der Inkubationszeit dargestellt.
Abb. 3.6 a - b: Nachweis von synthetischen AHL-Molekülen mittels C. violaceum-Indikatorplatte nach 24 h (a) und 48 h (b) Inkubation bei 30 °C über eine violette Pigmentierung des Biosensors um die Auftragungspunkte. Auftragungsschema: 1: NK (10 µl Ethylacetat, verdünnt auf 50 µl mit LB-Medium); 2 - 3: C4-HSL; 4 - 5: C6-HSL; 6 - 7: C8-HSL. Die Auftragung der jeweiligen AHL-Moleküle und deren Konzentrationen (5 x 10-9 mol und 5 x 10-8 mol) erfolgte im Uhrzeigersinn.
Aus Abbildung 3.6 a - b wird ersichtlich, dass C. violaceum CV026 eine hohe
Sensitivität gegenüber C6-HSL aufweist, wobei 5 x 10-9 mol (Abb. 3.6 a, Nr. 4) noch
deutlich durch eine Violacein-Produktion nachgewiesen wurde. Für C4-HSL und
C8-HSL liegen die Nachweisgrenzen hingegen wesentlich höher. C8-HSL wurde von
C. violaceum CV026 nach längerer Inkubation (48 h) lediglich schwach
nachgewiesen (Abb. 3.6 b, Nr. 6 und 7), 5 x 10-9 mol C4-HSL gar nicht (Abb. 3.6 b,
Nr. 2).
50
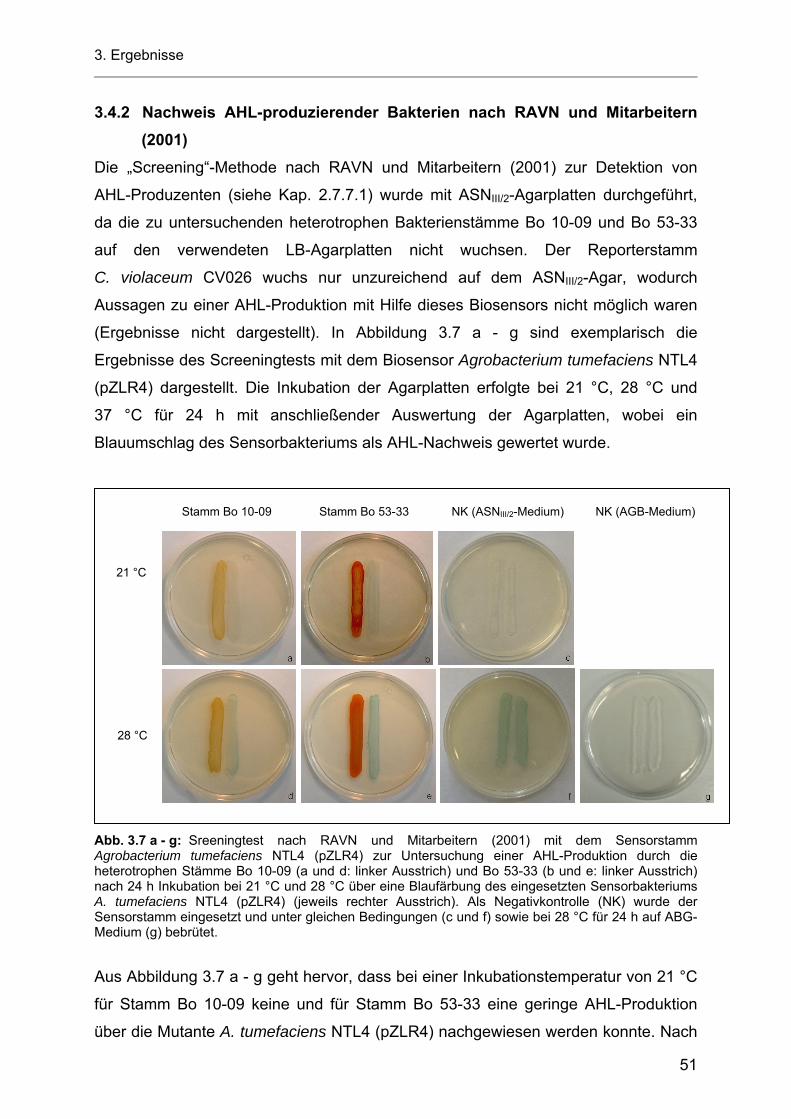
3. Ergebnisse
3.4.2 Nachweis AHL-produzierender Bakterien nach RAVN und Mitarbeitern (2001)
Die „Screening“-Methode nach RAVN und Mitarbeitern (2001) zur Detektion von
AHL-Produzenten (siehe Kap. 2.7.7.1) wurde mit ASNIII/2-Agarplatten durchgeführt,
da die zu untersuchenden heterotrophen Bakterienstämme Bo 10-09 und Bo 53-33
auf den verwendeten LB-Agarplatten nicht wuchsen. Der Reporterstamm
C. violaceum CV026 wuchs nur unzureichend auf dem ASNIII/2-Agar, wodurch
Aussagen zu einer AHL-Produktion mit Hilfe dieses Biosensors nicht möglich waren
(Ergebnisse nicht dargestellt). In Abbildung 3.7 a - g sind exemplarisch die
Ergebnisse des Screeningtests mit dem Biosensor Agrobacterium tumefaciens NTL4
(pZLR4) dargestellt. Die Inkubation der Agarplatten erfolgte bei 21 °C, 28 °C und
37 °C für 24 h mit anschließender Auswertung der Agarplatten, wobei ein
Blauumschlag des Sensorbakteriums als AHL-Nachweis gewertet wurde.
Stamm Bo 10-09 Stamm Bo 53-33 NK (ASNIII/2-Medium) NK (AGB-Medium)
21 °C
28 °C
Abb. 3.7 a - g: Sreeningtest nach RAVN und Mitarbeitern (2001) mit dem Sensorstamm Agrobacterium tumefaciens NTL4 (pZLR4) zur Untersuchung einer AHL-Produktion durch die heterotrophen Stämme Bo 10-09 (a und d: linker Ausstrich) und Bo 53-33 (b und e: linker Ausstrich) nach 24 h Inkubation bei 21 °C und 28 °C über eine Blaufärbung des eingesetzten Sensorbakteriums A. tumefaciens NTL4 (pZLR4) (jeweils rechter Ausstrich). Als Negativkontrolle (NK) wurde der Sensorstamm eingesetzt und unter gleichen Bedingungen (c und f) sowie bei 28 °C für 24 h auf ABG-Medium (g) bebrütet.
Aus Abbildung 3.7 a - g geht hervor, dass bei einer Inkubationstemperatur von 21 °C
für Stamm Bo 10-09 keine und für Stamm Bo 53-33 eine geringe AHL-Produktion
über die Mutante A. tumefaciens NTL4 (pZLR4) nachgewiesen werden konnte. Nach
51

3. Ergebnisse
Inkubation bei 28 °C wies der Indikatorstamm auf den ASNIII/2-Agarplatten mit Stamm
Bo 53-33 und Stamm Bo 10-09, einschließlich der Negativkontrolle (Abb. 3.7 f), eine
Blaufärbung auf. Diese fiel in Gegenwart von Stamm Bo 10-09 wesentlich schwächer
aus als in Gegenwart von Stamm Bo 53-33. Aussagen über eine AHL-Produktion
waren aufgrund der positiven Negativkontrolle nicht bzw. kaum möglich.
Versuchswiederholungen bestätigten die Ergebnisse.
Eine weitere Bebrütung bei der jeweiligen Inkubationstemperatur führte bei allen
ASNIII/2-Agarplatten entweder zu einem Auftreten einer Blaufärbung des Biosensors
oder zu einer weiteren Signalverstärkung (Ergebnisse nicht dargestellt). Bei einer
Inkubationtemperatur von 37 °C wies der Sensorstamm nur ein unzureichendes
Wachstum auf, wodurch eine Auswertung unmöglich wurde (Ergebnisse nicht
dargestellt).
Ein Ausstrich des Biosensors auf ABG-Festagar als NK zeigte nach 24 h (Abb. 3.7 g)
und 48 h Inkubation (Ergebnisse nicht dargestellt) bei 28 °C hingegen keine
Blaufärbung auf.
Weiter konnte in Gegenwart von X-Gal oftmals beobachtet werden, dass der
ansonsten gelb-ockrige Stamm Bo 10-09 eine deutliche Grünblau-Färbung aufzeigte.
3.4.3 Nachweis AHL-produzierender Bakterien nach FARRAND und Mitarbeitern (2002)
Zur Detektion von AHL-Produzenten wurde eine weitere „Screening“-Methode nach
FARRAND und Mitarbeitern (2002) durchgeführt (siehe Kap. 2.7.7.2), da ein
eindeutiger Nachweis nach RAVN und Mitarbeitern (2001) nicht erbracht wurde. In
diesem Screeningtest wurde die AHL-Produktion der zu testenden heterotrophen
Stämme Bo 10-09 und Bo 53-33 sowie des Cyanobakteriums Stamm Flo 1 bei 21 °C
untersucht. In Abbildung 3.8 a - c sind die A. tumefaciens-Indikatorplatten nach 24 h
Bebrütung bei 28 °C dargestellt.
52
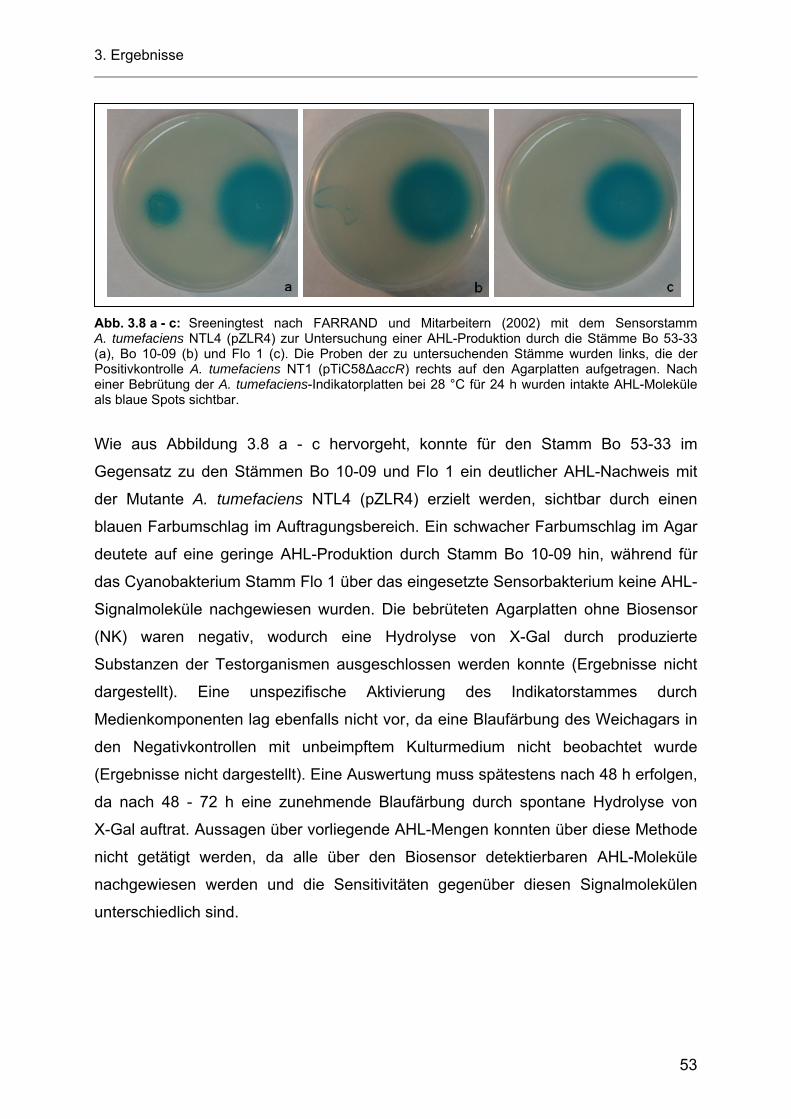
3. Ergebnisse
Abb. 3.8 a - c: Sreeningtest nach FARRAND und Mitarbeitern (2002) mit dem Sensorstamm A. tumefaciens NTL4 (pZLR4) zur Untersuchung einer AHL-Produktion durch die Stämme Bo 53-33 (a), Bo 10-09 (b) und Flo 1 (c). Die Proben der zu untersuchenden Stämme wurden links, die der Positivkontrolle A. tumefaciens NT1 (pTiC58∆accR) rechts auf den Agarplatten aufgetragen. Nach einer Bebrütung der A. tumefaciens-Indikatorplatten bei 28 °C für 24 h wurden intakte AHL-Moleküle als blaue Spots sichtbar.
Wie aus Abbildung 3.8 a - c hervorgeht, konnte für den Stamm Bo 53-33 im
Gegensatz zu den Stämmen Bo 10-09 und Flo 1 ein deutlicher AHL-Nachweis mit
der Mutante A. tumefaciens NTL4 (pZLR4) erzielt werden, sichtbar durch einen
blauen Farbumschlag im Auftragungsbereich. Ein schwacher Farbumschlag im Agar
deutete auf eine geringe AHL-Produktion durch Stamm Bo 10-09 hin, während für
das Cyanobakterium Stamm Flo 1 über das eingesetzte Sensorbakterium keine AHL-
Signalmoleküle nachgewiesen wurden. Die bebrüteten Agarplatten ohne Biosensor
(NK) waren negativ, wodurch eine Hydrolyse von X-Gal durch produzierte
Substanzen der Testorganismen ausgeschlossen werden konnte (Ergebnisse nicht
dargestellt). Eine unspezifische Aktivierung des Indikatorstammes durch
Medienkomponenten lag ebenfalls nicht vor, da eine Blaufärbung des Weichagars in
den Negativkontrollen mit unbeimpftem Kulturmedium nicht beobachtet wurde
(Ergebnisse nicht dargestellt). Eine Auswertung muss spätestens nach 48 h erfolgen,
da nach 48 - 72 h eine zunehmende Blaufärbung durch spontane Hydrolyse von
X-Gal auftrat. Aussagen über vorliegende AHL-Mengen konnten über diese Methode
nicht getätigt werden, da alle über den Biosensor detektierbaren AHL-Moleküle
nachgewiesen werden und die Sensitivitäten gegenüber diesen Signalmolekülen
unterschiedlich sind.
53

3. Ergebnisse
3.5 Untersuchungen des Einflusses verschiedener Kultivierungsbedingungen auf die AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie durch das Cyanobakterium Stamm Flo 1
Die Produktion von AHL-Signalmolekülen wurde unter verschiedenen
Kultivierungsbedingungen untersucht (siehe Kap. 2.7.2).
Zum Nachweis der verschiedenen AHL-Signalmoleküle wurde nach AHL-Extraktion
aus 250 ml Zellkulturüberständen (siehe Kap. 2.7.3.3) bzw. bakterieller Biomasse
(siehe Kap. 2.7.3.4) eine Umkehrphasen-Dünnschichtchromatographie (siehe
Kap. 2.7.4) mit anschließender Überschichtung mit dem Agrobacterium- bzw.
Chromobacterium-Sensor (siehe Kap. 2.7.5.2) zur Visualisierung der AHL-Moleküle
durchgeführt. Eine Identifikation der Signalmoleküle erfolgte, soweit möglich, mit Hilfe
von synthetischen AHL-Referenzmolekülen (siehe Kap. 2.7.6, Tab. 2.15). Aufgrund
der eingesetzten Standardmoleküle konnte, wie in Referenzarbeiten ebenso
(GEISENBERGER, 2000; YATES et al., 2002), auf eine Auswertung über die
ermittelten Rf-Werte verzichtet werden.
Für die DC-Untersuchungen mit dem Agrobacterium-Sensorstamm konnte eine
Hydrolyse von X-Gal durch extrahierte Substanzen (z.B. Enzyme) sowie durch eine
unspezifische Aktivierung des Biosensors durch das Ausbleiben einer Blaufärbung in
den Negativkontrollen nicht festgestellt werden (siehe Kap. 2.7.5.2) (Ergebnisse nicht
gezeigt).
3.5.1 Einfluss des pH-Wertes auf die AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie durch das Cyanobakterium Stamm Flo 1
Zur Ermittlung des pH-Wert-Einflusses auf die Signalmolekül-Produktion wurden die
AHL-Gehalte in den Kulturüberständen gepufferter Versuchskulturen miteinander
verglichen. Für die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 erfolgte
die Untersuchung bei ihrem jeweiligen pH-Minimum, -Optimum und -Maximum (siehe
Kap. 2.7.2.1), ermittelt aus den zuvor generierten Wachstumskurven (siehe
Kap. 3.2). Der pH-Wert 8,5 war das zunächst ermittelte pH-Maximum der
heterotrophen Stämme und fand daher ebenfalls in den AHL-Untersuchungen
Eingang. Eine Kultivierung unter pH-optimalen Bedingungen erfolgte bis in die
logarithmische bzw. stationäre Wachstumsphase. Die weiteren gepufferten
54
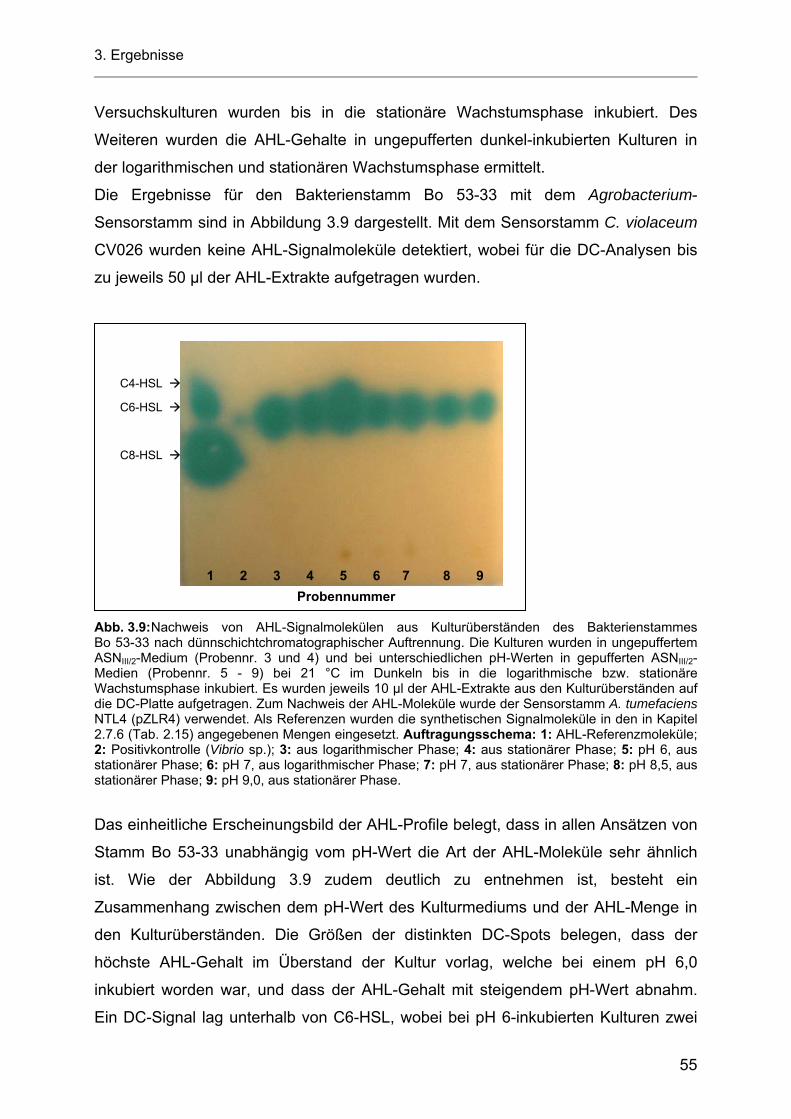
3. Ergebnisse
Versuchskulturen wurden bis in die stationäre Wachstumsphase inkubiert. Des
Weiteren wurden die AHL-Gehalte in ungepufferten dunkel-inkubierten Kulturen in
der logarithmischen und stationären Wachstumsphase ermittelt.
Die Ergebnisse für den Bakterienstamm Bo 53-33 mit dem Agrobacterium-
Sensorstamm sind in Abbildung 3.9 dargestellt. Mit dem Sensorstamm C. violaceum
CV026 wurden keine AHL-Signalmoleküle detektiert, wobei für die DC-Analysen bis
zu jeweils 50 µl der AHL-Extrakte aufgetragen wurden.
C4-HSL
C6-HSL
C8-HSL
1 2 3 4 5 6 7 8 9 Probennummer
Abb. 3.9: Nachweis von AHL-Signalmolekülen aus Kulturüberständen des Bakterienstammes Bo 53-33 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden in ungepuffertem ASNIII/2-Medium (Probennr. 3 und 4) und bei unterschiedlichen pH-Werten in gepufferten ASNIII/2-Medien (Probennr. 5 - 9) bei 21 °C im Dunkeln bis in die logarithmische bzw. stationäre Wachstumsphase inkubiert. Es wurden jeweils 10 µl der AHL-Extrakte aus den Kulturüberständen auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle; 2: Positivkontrolle (Vibrio sp.); 3: aus logarithmischer Phase; 4: aus stationärer Phase; 5: pH 6, aus stationärer Phase; 6: pH 7, aus logarithmischer Phase; 7: pH 7, aus stationärer Phase; 8: pH 8,5, aus stationärer Phase; 9: pH 9,0, aus stationärer Phase.
Das einheitliche Erscheinungsbild der AHL-Profile belegt, dass in allen Ansätzen von
Stamm Bo 53-33 unabhängig vom pH-Wert die Art der AHL-Moleküle sehr ähnlich
ist. Wie der Abbildung 3.9 zudem deutlich zu entnehmen ist, besteht ein
Zusammenhang zwischen dem pH-Wert des Kulturmediums und der AHL-Menge in
den Kulturüberständen. Die Größen der distinkten DC-Spots belegen, dass der
höchste AHL-Gehalt im Überstand der Kultur vorlag, welche bei einem pH 6,0
inkubiert worden war, und dass der AHL-Gehalt mit steigendem pH-Wert abnahm.
Ein DC-Signal lag unterhalb von C6-HSL, wobei bei pH 6-inkubierten Kulturen zwei
55

3. Ergebnisse
AHL-Moleküle detektiert wurden (Probennr. 5). Das zweite DC-Signal lag auf Höhe
von C4-HSL. Signifikante Unterschiede der AHL-Gehalte in den Kulturüberständen
von Stamm Bo 53-33 in der logarithmischen und stationären Wachstumsphase
wurden nicht beobachtet, wobei der Gehalt an AHL-Molekülen in der logarithmischen
Wachstumsphase (Probennr. 3) leicht höher war als in der stationären
Wachstumsphase (Probennr. 4). Die AHL-Positivkontrolle Vibrio sp. wies im
Kulturüberstand AHL-Signalmoleküle auf (Probennr. 2).
Für den Bakterienstamm Bo 10-09 wurde mit dem Agrobacterium-Sensorstamm DC-
Signale detektiert (Abb. 3.10). Mit dem Sensorstamm C. violaceum CV026 wurde
kein AHL-Nachweis erzielt, wobei bis zu jeweils 50 µl der AHL-Extrakte aufgetragen
wurden (Ergebnisse nicht dargestellt).
C4-HSL C6-HSL
C8-HSL
1 2 3 Probennummer
Abb. 3.10: Nachweis von AHL-Signalmolekülen aus Kulturüberständen des Bakterienstammes Bo 10-09 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden bei pH 6 in gepuffertem ASNIII/2-Medium bis in die stationäre Wachstumsphase bzw. in ungepuffertem ASNIII/2-Medium bis in die logarithmische Wachstumsphase bei 21 °C im Dunkeln inkubiert. Es wurden jeweils 30 µl der AHL-Extrakte aus den Kulturüberständen auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle; 2: pH 6, aus stationärer Wachstumsphase; 3: aus logarithmischer Wachstumsphase.
In Abbildung 3.10 sind die AHL-Nachweise für Stamm Bo 10-09 dargestellt. Im
Überstand einer bei pH 6 kultivierten Kultur (Probennr. 2) wurden mindestens zwei
AHL-Signalmoleküle nachgewiesen. In Überständen aus Bo 10-09-Kulturen, welche
beim pH-Optimum (pH 8,0) bis in die logarithmische bzw. stationäre
56

3. Ergebnisse
Wachstumsphase sowie beim pH-Maximum (pH 9,0) bis in die stationäre
Wachstumsphase kultiviert wurden, konnten keine AHL-Signalmoleküle detektiert
werden (Ergebnisse nicht dargestellt). Des Weiteren wurde in Überständen
ungepufferter Kulturen in der logarithmischen Wachstumsphase ein AHL-Nachweis
erzielt (Probennr. 3), jedoch nicht in der stationären Wachstumsphase (Ergebnis
nicht dargestellt).
Bei Stamm Flo 1 erfolgte die Untersuchung auf AHL-Moleküle bei pH-Werten von
7,0, 8,0, 8,5 und 9,0 nach 14 bzw. 21 Tagen Kultivierung bei einem Photonenfluss
von 5 µE m-2 s-1 PAR (siehe Kap. 2.7.2.2). Für die DC-Analysen wurden jeweils 50 µl
der AHL-Extrakte eingesetzt. Ein AHL-Nachweis wurde mit keinem Indikatorstamm
erzielt.
3.5.1.1 Azidifizierung von Kulturüberständen der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09
Im Anschluss an die pH-Untersuchungen wurde eine Azidifizierung von
ungepufferten Kulturüberständen der Stämme Bo 53-33 und Bo 10-09 nach YATES
und Mitarbeitern (2002) durchgeführt (siehe Kap. 2.7.3.2). Eine Kultivierung der
ungepufferten Dunkelansätze erfolgte bis in die logarithmische bzw. stationäre
Wachstumsphase (siehe Kap. 2.7.2.1). Die Ergebnisse des Azidifizierungsversuches
für den Stamm Bo 53-33 mit dem Agrobacterium-Sensorstamm sind in Abbildung
3.11 a dargestellt, wobei ein Vergleich zwischen ungepufferten azidifizierten und
nicht-azidifizierten Kulturüberständen vorgenommen wurde. In der DC-Analyse mit
dem Chromobacterium-Sensor wurden lediglich die azidifizierten Kulturüberstände
untersucht (Abb. 3.11 b).
57
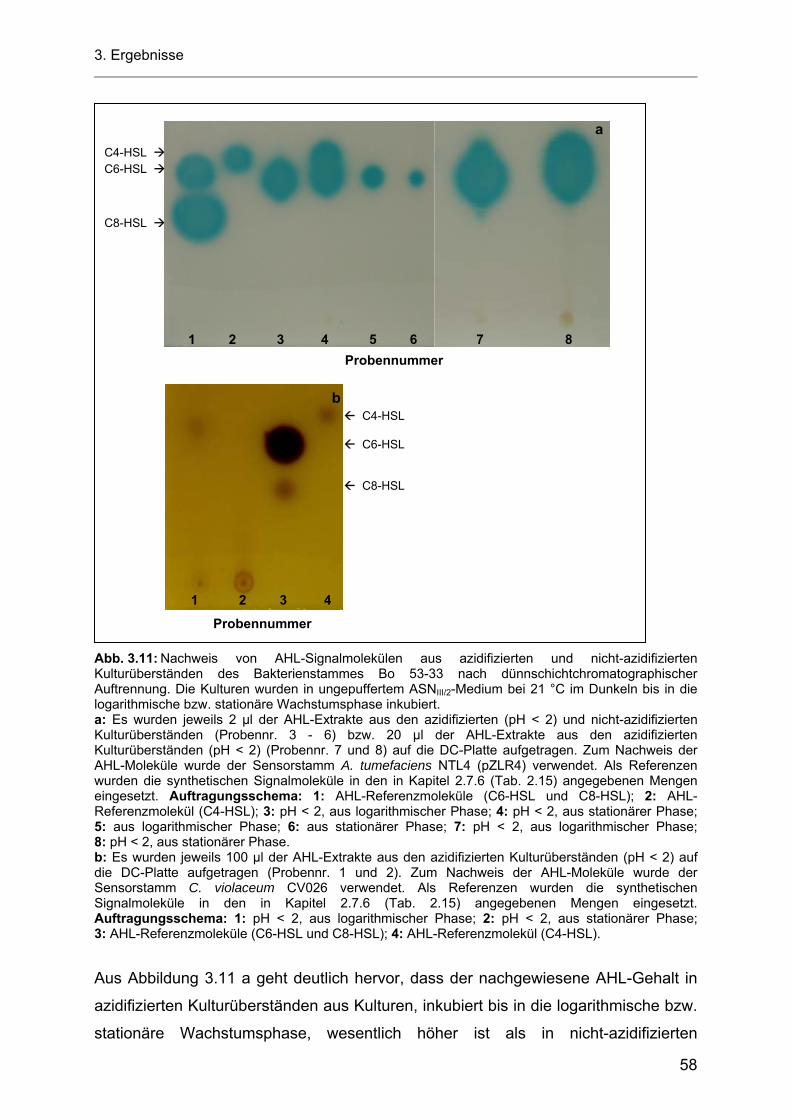
3. Ergebnisse
a C4-HSL C6-HSL
C8-HSL
1 2 3 4 5 6 7 8 Probennummer
C4-HSL b
C6-HSL
C8-HSL
1 2 3 4
Probennummer
Abb. 3.11: Nachweis von AHL-Signalmolekülen aus azidifizierten und nicht-azidifizierten Kulturüberständen des Bakterienstammes Bo 53-33 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden in ungepuffertem ASNIII/2-Medium bei 21 °C im Dunkeln bis in die logarithmische bzw. stationäre Wachstumsphase inkubiert. a: Es wurden jeweils 2 µl der AHL-Extrakte aus den azidifizierten (pH < 2) und nicht-azidifizierten Kulturüberständen (Probennr. 3 - 6) bzw. 20 µl der AHL-Extrakte aus den azidifizierten Kulturüberständen (pH < 2) (Probennr. 7 und 8) auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 2: AHL-Referenzmolekül (C4-HSL); 3: pH < 2, aus logarithmischer Phase; 4: pH < 2, aus stationärer Phase; 5: aus logarithmischer Phase; 6: aus stationärer Phase; 7: pH < 2, aus logarithmischer Phase; 8: pH < 2, aus stationärer Phase. b: Es wurden jeweils 100 µl der AHL-Extrakte aus den azidifizierten Kulturüberständen (pH < 2) auf die DC-Platte aufgetragen (Probennr. 1 und 2). Zum Nachweis der AHL-Moleküle wurde der Sensorstamm C. violaceum CV026 verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: pH < 2, aus logarithmischer Phase; 2: pH < 2, aus stationärer Phase; 3: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 4: AHL-Referenzmolekül (C4-HSL).
Aus Abbildung 3.11 a geht deutlich hervor, dass der nachgewiesene AHL-Gehalt in
azidifizierten Kulturüberständen aus Kulturen, inkubiert bis in die logarithmische bzw.
stationäre Wachstumsphase, wesentlich höher ist als in nicht-azidifizierten
58

3. Ergebnisse
Kulturüberständen. In der logarithmischen Wachstumsphase wurde ein AHL-Molekül
detektiert (DC-Signal unterhalb von C6-HSL) (Probennr. 3). Im Vergleich zur
stationären Wachstumsphase (Probennr. 4) lagen in beiden Wuchsphasen in etwa
gleiche AHL-Mengen vor. Ein weiteres AHL-Molekül wurde in der stationären
Wachstumsphase, dünnschichtchromatographisch auf der Höhe von C4-HSL,
nachgewiesen. Bei einem Auftragungsvolumen von 20 µl AHL-Extrakt konnte zudem
in der logarithmischen Wachstumsphase von Stamm Bo 53-33 noch ein drittes AHL-
Molekül in sehr geringer Menge (DC-Signal auf Höhe von C8-HSL) detektiert werden
(Probennr. 7).
Mit dem Sensorstamm C. violaceum CV026 wurde im azidifizierten Kulturüberstand
der Kultur, welche bis in die logarithmische Wachstumsphase inkubiert worden war,
ein kurzkettiges AHL-Molekül (DC-Spot etwas unterhalb von C4-HSL) nachgewiesen
(Abb. 3.11 b, Probennr. 1).
Ein Vergleich zwischen ungepufferten azidifizierten und nicht-azidifizierten
Kulturüberständen von Stamm Bo 10-09 mit dem Agrobacterium-Sensorstamm ist in
Abbildung 3.12 a dargestellt. Für die DC-Analysen mit dem Chromobacterium-
Sensor wurden lediglich AHL-Extrakte aufgetragen, welche aus azidifizierten
Kulturüberständen gewonnen wurden (Abb. 3.12 b).
59
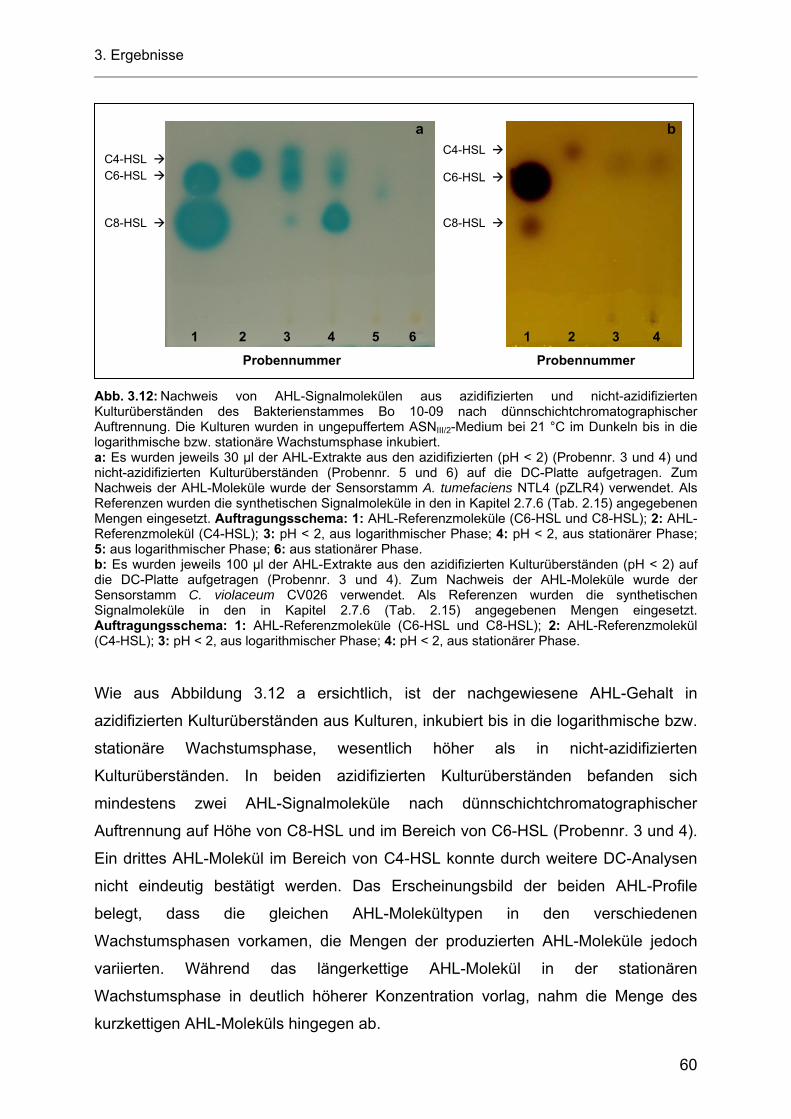
3. Ergebnisse
Abb. 3.12: Nachweis von AHL-Signalmolekülen aus azidifizierten und nicht-azidifizierten Kulturüberständen des Bakterienstammes Bo 10-09 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden in ungepuffertem ASNIII/2-Medium bei 21 °C im Dunkeln bis in die logarithmische bzw. stationäre Wachstumsphase inkubiert. a: Es wurden jeweils 30 µl der AHL-Extrakte aus den azidifizierten (pH < 2) (Probennr. 3 und 4) und nicht-azidifizierten Kulturüberständen (Probennr. 5 und 6) auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 2: AHL-Referenzmolekül (C4-HSL); 3: pH < 2, aus logarithmischer Phase; 4: pH < 2, aus stationärer Phase; 5: aus logarithmischer Phase; 6: aus stationärer Phase. b: Es wurden jeweils 100 µl der AHL-Extrakte aus den azidifizierten Kulturüberständen (pH < 2) auf die DC-Platte aufgetragen (Probennr. 3 und 4). Zum Nachweis der AHL-Moleküle wurde der Sensorstamm C. violaceum CV026 verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 2: AHL-Referenzmolekül (C4-HSL); 3: pH < 2, aus logarithmischer Phase; 4: pH < 2, aus stationärer Phase.
Wie aus Abbildung 3.12 a ersichtlich, ist der nachgewiesene AHL-Gehalt in
azidifizierten Kulturüberständen aus Kulturen, inkubiert bis in die logarithmische bzw.
stationäre Wachstumsphase, wesentlich höher als in nicht-azidifizierten
Kulturüberständen. In beiden azidifizierten Kulturüberständen befanden sich
mindestens zwei AHL-Signalmoleküle nach dünnschichtchromatographischer
Auftrennung auf Höhe von C8-HSL und im Bereich von C6-HSL (Probennr. 3 und 4).
Ein drittes AHL-Molekül im Bereich von C4-HSL konnte durch weitere DC-Analysen
nicht eindeutig bestätigt werden. Das Erscheinungsbild der beiden AHL-Profile
belegt, dass die gleichen AHL-Molekültypen in den verschiedenen
Wachstumsphasen vorkamen, die Mengen der produzierten AHL-Moleküle jedoch
variierten. Während das längerkettige AHL-Molekül in der stationären
Wachstumsphase in deutlich höherer Konzentration vorlag, nahm die Menge des
kurzkettigen AHL-Moleküls hingegen ab.
a b C4-HSL
C4-HSL C6-HSL C6-HSL
C8-HSL C8-HSL
1 2 3 4 5 6 1 2 3 4
Probennummer Probennummer
60

3. Ergebnisse
Mit dem Sensorstamm C. violaceum CV026 wurde in den azidifizierten
Kulturüberständen der Kulturen, welche bis in die logarithmische bzw. stationäre
Wachstumsphase inkubiert worden waren, jeweils ein kurzkettiges AHL-
Signalmolekül (DC-Spot lag geringfügig unterhalb von C4-HSL) nachgewiesen
(Abb. 3.12 b, Probennr. 3 und 4).
3.5.2 Einfluss von Licht auf die Produktion von AHL-Molekülen durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09
Zur Ermittlung des Lichteinflusses auf die Signalmolekül-Produktion in den
heterotrophen Bakterienstämmen wurden die AHL-Gehalte in den Kulturüberständen
Licht-inkubierter und dunkel-inkubierter Kulturen, kultiviert bis in die logarithmische
bzw. stationäre Wachstumsphase, miteinander verglichen (siehe Kap. 2.7.2.1).
Die Ergebnisse für den Bakterienstamm Bo 53-33 mit dem Agrobacterium-
Sensorstamm sind in Abbildung 3.13 dargestellt. DC-Vorversuche mit dem
Chromobacterium-Sensorstamm hatten ergeben, dass trotz großer Auftragsvolumina
von bis zu 100 µl nur sehr schwache AHL-Nachweise mit Stamm Bo 53-33 zu
verzeichnen waren. Daher wurde auf eine weitere Untersuchung mit diesem
Sensorstamm verzichtet.
61

3. Ergebnisse
C4-HSL C6-HSL
C8-HSL
1 2 3 4 5 6
Probennummer
Abb. 3.13: Nachweis von AHL-Signalmolekülen aus Kulturüberständen des Bakterienstammes Bo 53-33 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden bei einem Photonenfluss von 5 µE m-2 s-1 PAR (Probennr. 3 und 4) bzw. im Dunkeln (Probennr. 5 und 6) in ungepuffertem ASNIII/2-Medium bei 21 °C bis in die logarithmische bzw. stationäre Wachstumsphase inkubiert. Es wurden jeweils 10 µl der AHL-Extrakte aus den Kulturüberständen auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 2: AHL-Referenzmolekül (C4-HSL); 3: 5 µE m-2 s-1 PAR, aus logarithmischer Phase; 4: 5 µE m-2 s-1 PAR, aus stationärer Phase; 5: dunkel, aus logarithmischer Phase; 6: dunkel, aus stationärer Phase.
Das einheitliche Erscheinungsbild der AHL-Profile in Abbildung 3.13 bestätigt, dass
die Art der AHL-Moleküle in den Licht-inkubierten und dunkel-inkubierten
Versuchsansätzen sehr ähnlich ist. In allen AHL-Extrakten wurde ein Signalmolekül
detektiert (DC-Spot lag geringfügig unterhalb von C6-HSL). Ein weiteres AHL-
Molekül (DC-Spot lag auf Höhe von C4-HSL) wurde eindeutig in der stationären
Wachstumsphase der Licht-inkubierten Versuchskultur nachgewiesen (Probennr. 4).
Des Weiteren belegen die Spotgrößen, dass in den Überständen der Licht-
inkubierten Versuchsansätze im Vergleich zu den jeweiligen Überständen der
Dunkelansätze aus der logarithmischen bzw. stationären Wachstumsphase keine
signifikanten Unterschiede in den AHL-Konzentrationen vorlagen.
Für den Bakterienstamm Bo 10-09 betrug das Auftragungsvolumen der AHL-Extrakte
jeweils 30 µl. Die Licht-inkubierten Versuchsansätze von Stamm Bo 10-09 wiesen
das gleiche AHL-Profil auf wie die dunkel-inkubierten Ansätze (siehe Kap. 3.5.1,
Abb. 3.10, Probennr. 3). Demnach wurde in der logarithmischen Wachstumsphase
Licht-inkubierter Versuchsansätze in ähnlicher Konzentration ein AHL-Molekül
62

3. Ergebnisse
nachgewiesen. In der stationären Wachstumsphase konnte kein AHL-Nachweis
erzielt werden.
3.5.3 Einfluss der Temperatur und der Kohlenstoffquelle auf die Produktion von AHL-Molekülen durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09
Die in Assoziation mit Cyanobakterien lebenden heterotrophen Bakterien existieren
unter Mangelbedingungen. Eine Auswirkung von verschiedenen
Nährstoffbedingungen auf die Produktion von AHL-Molekülen bei den zu
untersuchenden heterotrophen Bakterien ist dementsprechend von Interesse.
Daher wurden die Bakterienstämme im ASNIII/2-Medium mit nur 0,1% Fleischextrakt
bis zur stationären Phase angezogen (siehe Kap. 2.7.2.1). Die unter diesen
Bedingungen produzierten AHL-Moleküle wurden nach Extraktion mit den AHL-
Molekülen verglichen, welche aus Kulturen mit dem standardmäßig verwendeten
ASNIII/2-Medium mit 1% Fleischextrakt extrahiert worden waren. Zur Detektion der
AHL-Moleküle wurde der Agrobacterium-Sensorstamm eingesetzt. Die Ergebnisse
sind in Abbildung 3.14 dargestellt.
63

3. Ergebnisse
C6-HSL C4-HSL
C8-HSL
1 2 3 4 5 6
Probennummer
Abb. 3.14: Nachweis von AHL-Signalmolekülen aus Kulturüberständen der Bakterienstämme Bo 53-33 und Bo 10-09 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden in 0,1% FE versetztem bzw. standardmäßig verwendetem (1% FE) ASNIII/2-Medium bei 21 °C im Dunkeln bis in die stationäre Wachstumsphase inkubiert. Es wurden jeweils 10 µl (Stamm Bo 53-33) bzw. jeweils 40 µl (Stamm Bo 10-09) der AHL-Extrakte aus den Kulturüberständen auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: Stamm Bo 53-33, 1% FE; 2: Stamm Bo 53-33, 0,1% FE; 3: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 4: AHL-Referenzmolekül (C4-HSL); 5: Stamm Bo 10-09, 1% FE; 6: Stamm Bo 10-09, 0,1% FE. FE = Fleischextrakt.
Aus Abbildung 3.14 wird ersichtlich, dass der mit 0,1% Fleischextrakt kultivierte
Bo 53-33-Ansatz (Probennr. 2) eine leicht stärkere AHL-Produktion aufwies als der
mit 1% Fleischextrakt kultivierte (Probennr. 1), erkennbar durch den größeren DC-
Spotdurchmesser (DC-Spot geringfügig unterhalb von C6-HSL).
Ein schwacher AHL-Nachweis konnte für den Stamm Bo 10-09 in einer
Versuchskultur, kultiviert mit 0,1% Fleischextrakt, erzielt werden
(dünnschichtchromatographisch geringfügig unterhalb von C6-HSL) (Probennr. 6).
Mit dem Agrobacterium-Sensorstamm konnten indes im Überstand einer Kultur,
kultiviert mit 1% Fleischextrakt, keine AHL-Moleküle detektiert werden (Probennr. 5).
Zur Feststellung eines Temperatureinflusses auf die AHL-Menge in Überständen
bzw. auf die AHL-Produktion an sich wurden in einer weiteren Untersuchung
Versuchsansätze der Stämme Bo 10-09 und Bo 53-33 bei 28 °C bis in die stationäre
Wachstumsphase hin inkubiert (siehe Kap. 2.7.2.1). Anschließend wurden die AHL-
Gehalte der Überstände mit denen von Kulturen verglichen, welche bei 21 °C bis in
die stationäre Wachstumsphase inkubiert worden waren. Die Ergebnisse für den
64

3. Ergebnisse
Bakterienstamm Bo 53-33 unter Verwendung des Agrobacterium-Sensorstammes
sind in Abbildung 3.15 dargestellt.
Im Kulturüberstand von Stamm Bo 10-09 konnte auch bei einer
Inkubationstemperatur von 28 °C in der stationären Wachstumsphase kein AHL-
Nachweis erzielt werden (Ergebnisse nicht dargestellt). Für den Stamm Bo 10-09
betrug das Auftragungsvolumen der AHL-Extrakte jeweils 30 µl.
C4-HSL
C6-HSL
C8-HSL
1 2 3 4 Probennummer
Abb. 3.15: Nachweis von AHL-Signalmolekülen aus Kulturüberständen des Bakterienstammes Bo 53-33 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden in ungepuffertem ASNIII/2-Medium bei 21 °C (Probennr. 1) bzw. 28 °C (Probennr. 2) im Dunkeln bis in die stationäre Wachstumsphase inkubiert. Es wurden jeweils 10 µl der AHL-Extrakte aus den Kulturüberständen auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: 21 °C, aus stationärer Wachstumsphase; 2: 28 °C, aus stationärer Wachstumsphase; 3: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 4: AHL-Referenzmolekül (C4-HSL).
Wie aus Abbildung 3.15 hervorgeht, wurden nach einer Kultivierung von Stamm
Bo 53-33 bei 28 °C bis in die stationäre Wachstumsphase zwei Signalmoleküle
detektiert (Probennr. 2). Bei einem Vergleich der Spotdurchmesser (DC-Spot
geringfügig unterhalb von C6-HSL) wird deutlich, dass der bei 28 °C kultivierte
Versuchsansatz eine wesentlich geringere AHL-Menge aufwies als der
Versuchsansatz, welcher bei 21 °C inkubiert wurde (siehe Probennr. 1 und 2).
65

3. Ergebnisse
3.5.4 Produktion von AHL-Molekülen durch das Cyanobakterium Stamm Flo 1 Eine Anzucht von Stamm Flo 1 in ungepufferten Kulturmedien für 14 bzw. 21 Tage
bei einem Photonenfluss von 5 µE m-2 s-1 PAR (siehe Kap. 2.7.2.2) und
anschließender Untersuchung von azidifizierten und nicht-azidifizierten
Kulturüberständen auf AHL-Moleküle mit dem Agrobacterium-Sensorstamm ergab
die in Abbildung 3.16 dargestellten Ergebnisse. Des Weiteren wurden AHL-Moleküle
aus 1000 ml Überstand extrahiert, wobei die Versuchskulturen zuvor für 8 Wochen
unter gleichen Kultivierungsbedingungen angezogen worden waren.
C4-HSL C6-HSL
C8-HSL
1 2 3 4 5 6 7 Probennummer
Abb. 3.16: Nachweis von AHL-Signalmolekülen aus Kulturüberständen des cyanobakteriellen Stammes Flo 1 nach dünnschichtchromatographischer Auftrennung. Die Kulturen wurden in ungepuffertem ASNIII/2-Medium bei 21 °C für 14 bzw. 21 Tage (Probennr. 3 - 6) bzw. für 8 Wochen (Probennr. 7) bei einem Photonenfluss von 5 µE m-2 s-1 PAR inkubiert. Es wurden jeweils 50 µl der AHL-Extrakte aus den azidifizierten (pH < 2) und nicht-azidifizierten 250 ml Kulturüberständen (Probennr. 3 - 6) bzw. 50 µl AHL-Extrakt aus einem nicht-azidifizierten 1000 ml Kulturüberstand (Probennr. 7) auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 2: AHL-Referenzmolekül (C4-HSL); 3: pH < 2, nach 14 Tagen; 4: pH < 2, nach 21 Tagen; 5: nach 14 Tagen; 6: nach 21 Tagen; 7: nach 8 Wochen.
Wie aus Abbildung 3.16 hervorgeht, wurden nach 14- bzw. 21-tägiger Kultivierung
der ungepufferten Flo 1-Kulturen in den Überständen AHL-Moleküle nachgewiesen
(Probennr. 5 und 6). Die kleinen Durchmesser der einzelnen DC-Spots weisen auf
einen geringen AHL-Gehalt in den Überständen hin. Es wurden zwei AHL-
Molekültypen (DC-Signale auf Höhe von C8-HSL und C6-HSL) mit ähnlichen
Konzentrationen in beiden Überständen gefunden. Eine Azidifizierung der
66

3. Ergebnisse
Kulturüberstände führte bei der 21 Tage lang inkubierten Flo 1-Kultur zu einem
stärkeren Nachweis des längerkettigen AHL-Moleküls (DC-Spot auf Höhe von C8-
HSL) (Probennr. 4). Aufgrund einer fehlerhaft durchgeführten Azidifizierung des
Überstandes der Flo 1-Kultur, angezogen für 14 Tage, konnten bezüglich einer AHL-
Produktion in dieser Kultur keine gesicherten Aussagen getätigt werden
(Probennr. 3).
In 1000 ml Überstand aus Flo 1-Kulturen, welche für 8 Wochen inkubiert wurden,
konnten drei AHL-Moleküle detektiert werden. Neben einem deutlich stärkeren Signal
für das längerkettige AHL-Molekül auf Höhe von C8-HSL war ein weiterer DC-Spot
auf Höhe von C4-HSL ein Beleg dafür, dass noch ein drittes AHL-Molekül im
Überstand von Flo 1-Kulturen vorkommt (Probennr. 7).
3.5.5 Intrazelluläre AHL-Untersuchungen mit den heterotrophen Bakterienstämmen Bo 53-33 und Bo 10-09 sowie dem Cyanobakterium Stamm Flo 1
Sowohl die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 als auch der
Cyanobakterienstamm Flo 1 wurden intrazellulär auf AHL-Moleküle hin untersucht.
Der Stamm Flo 1 wurde dazu für 14 bzw. 21 Tage in ungepuffertem Nährmedium bei
einer Photonenflussdichte von 5 µE m-2 s-1 PAR kultiviert (siehe Kap. 2.7.2.2) Die
heterotrophen Stämme wurden, wie bereits in Kapitel 3.5.1 beschrieben, in
gepufferten und ungepufferten Nährmedien bis in die logarithmische bzw. stationäre
Wachstumsphase angezogen. Die Aufarbeitung der bakteriellen Biomasse wurde
nach BYERS und Mitarbeitern (2002) in veränderter Form durchgeführt (siehe
Kap. 2.7.3.4).
Für den heterotrophen Bakterienstamm Bo 10-09 wurde bei Auftragungsvolumina
zwischen 10 - 60 µl der Ammoniumacetat-Extrakte für keinen Versuchsansatz ein
intrazellulärer AHL-Nachweis mit dem Agrobacterium-Sensorstamm erzielt. Die
Ergebnisse der analytischen DC für den Stamm Bo 53-33 und Stamm Flo 1 sind in
Abbildung 3.17 dargestellt, wobei der AHL-Nachweis mit dem Agrobacterium-Sensor
erfolgte.
67
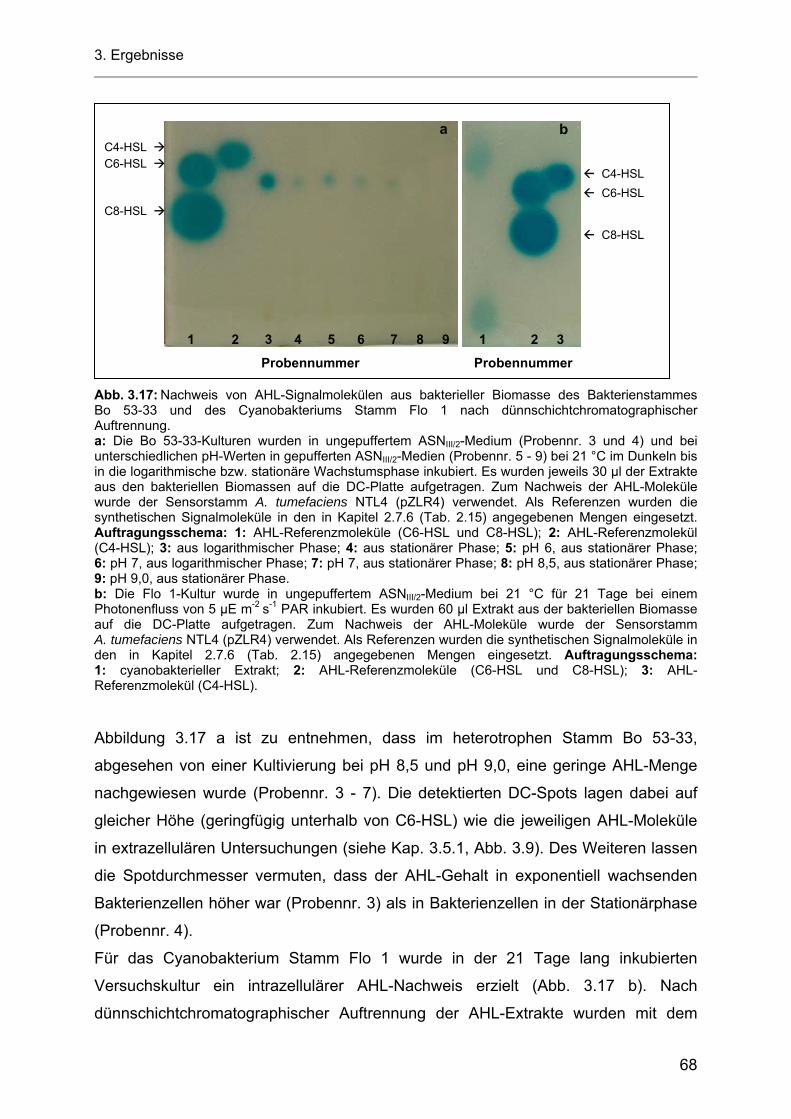
3. Ergebnisse
Abb. 3.17: Nachweis von AHL-Signalmolekülen aus bakterieller Biomasse des Bakterienstammes Bo 53-33 und des Cyanobakteriums Stamm Flo 1 nach dünnschichtchromatographischer Auftrennung. a: Die Bo 53-33-Kulturen wurden in ungepuffertem ASNIII/2-Medium (Probennr. 3 und 4) und bei unterschiedlichen pH-Werten in gepufferten ASNIII/2-Medien (Probennr. 5 - 9) bei 21 °C im Dunkeln bis in die logarithmische bzw. stationäre Wachstumsphase inkubiert. Es wurden jeweils 30 µl der Extrakte aus den bakteriellen Biomassen auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 2: AHL-Referenzmolekül (C4-HSL); 3: aus logarithmischer Phase; 4: aus stationärer Phase; 5: pH 6, aus stationärer Phase; 6: pH 7, aus logarithmischer Phase; 7: pH 7, aus stationärer Phase; 8: pH 8,5, aus stationärer Phase; 9: pH 9,0, aus stationärer Phase. b: Die Flo 1-Kultur wurde in ungepuffertem ASNIII/2-Medium bei 21 °C für 21 Tage bei einem Photonenfluss von 5 µE m-2 s-1 PAR inkubiert. Es wurden 60 µl Extrakt aus der bakteriellen Biomasse auf die DC-Platte aufgetragen. Zum Nachweis der AHL-Moleküle wurde der Sensorstamm A. tumefaciens NTL4 (pZLR4) verwendet. Als Referenzen wurden die synthetischen Signalmoleküle in den in Kapitel 2.7.6 (Tab. 2.15) angegebenen Mengen eingesetzt. Auftragungsschema: 1: cyanobakterieller Extrakt; 2: AHL-Referenzmoleküle (C6-HSL und C8-HSL); 3: AHL-Referenzmolekül (C4-HSL).
Abbildung 3.17 a ist zu entnehmen, dass im heterotrophen Stamm Bo 53-33,
abgesehen von einer Kultivierung bei pH 8,5 und pH 9,0, eine geringe AHL-Menge
nachgewiesen wurde (Probennr. 3 - 7). Die detektierten DC-Spots lagen dabei auf
gleicher Höhe (geringfügig unterhalb von C6-HSL) wie die jeweiligen AHL-Moleküle
in extrazellulären Untersuchungen (siehe Kap. 3.5.1, Abb. 3.9). Des Weiteren lassen
die Spotdurchmesser vermuten, dass der AHL-Gehalt in exponentiell wachsenden
Bakterienzellen höher war (Probennr. 3) als in Bakterienzellen in der Stationärphase
(Probennr. 4).
Für das Cyanobakterium Stamm Flo 1 wurde in der 21 Tage lang inkubierten
Versuchskultur ein intrazellulärer AHL-Nachweis erzielt (Abb. 3.17 b). Nach
dünnschichtchromatographischer Auftrennung der AHL-Extrakte wurden mit dem
a
b C4-HSL C6-HSL
C4-HSL C6-HSL
C8-HSL
C8-HSL
1 2 3 4 5 6 7 8 9 1 2 3
Probennummer Probennummer
68

3. Ergebnisse
Agrobacterium-Sensor zwei DC-Spots erhalten. Ein DC-Signal lag auf Höhe der
Probenauftragung, das andere geringfügig oberhalb von C4-HSL (Abb. 3.17 b,
Probennr. 1).
3.6 Auswertung der molekularbiologischen Untersuchungen
3.6.1 Extraktion der chromosomalen DNA Aus den Cyanobakterienstämmen Bo 10 und Bo 53 sowie den heterotrophen
Stämmen Bo 53-33 und Bo 10-09 wurde deren chromosomale DNA extrahiert (siehe
Kap. 2.8.1) und bei 4 °C im Dunkeln gelagert. DNA-Menge und Reinheit der DNA
wurden mit einem Spektralphotometer (Peqlab Biotechnologie, NanoDrop ND-1000)
ermittelt (siehe Kap. 2.8.2). Die DNA-Ausbeuten sind in Tabelle 3.3
zusammengefasst.
Tab. 3.3: Ermittelte DNA-Ausbeute nach DNA-Extraktion aus den Cyanobakterien- stämmen Bo 10 und Bo 53 sowie den heterotrophen Stämmen Bo 53-33 und Bo 10-09
Bakterienstamm DNA-Ausbeute [ng/µl]
Bo 10 (Cyanobakterium) 740 Bo 53 (Cyanobakterium) 1153 Bo 10-09 (heterotrophes Bakterium) 5975 Bo 53-33 (heterotrophes Bakterium) 6014
3.6.2 Amplifikation der 16S-rDNA Durch die Amplifikation der 16S-rDNA von Cyanobakterium Stamm Bo 53 mittels
einer Standard-PCR (siehe Kap. 2.8.3) wurde ein ca. 1500 bp großes DNA-Fragment
erhalten (Gelbild nicht gezeigt). Für das Cyanobakterium Stamm Bo 10 wurde neben
einer ca. 1500 bp großen Bande noch weitere 1 - 2 Banden im Agarosegel detektiert.
Eine Separation der 16S-rDNA-Amplifikate aus dem Gel erfolgte durch Extraktion
(siehe Kap. 2.8.5).
3.6.3 Erstellung von phylogenetischen Stammbäumen auf Basis der 16S-rDNA-Sequenz
Die ermittelten 16S-rDNA-Sequenzen der Cyanobakterienstämme Bo 53 (1333 bp)
und Bo 10 (1336 bp) sind nach Alignment im Anhang dargestellt (siehe Tab. 7.2 und
Tab. 7.3). Die Sequenzen wurden mit Sequenzen aus der öffentlichen Datenbank
69
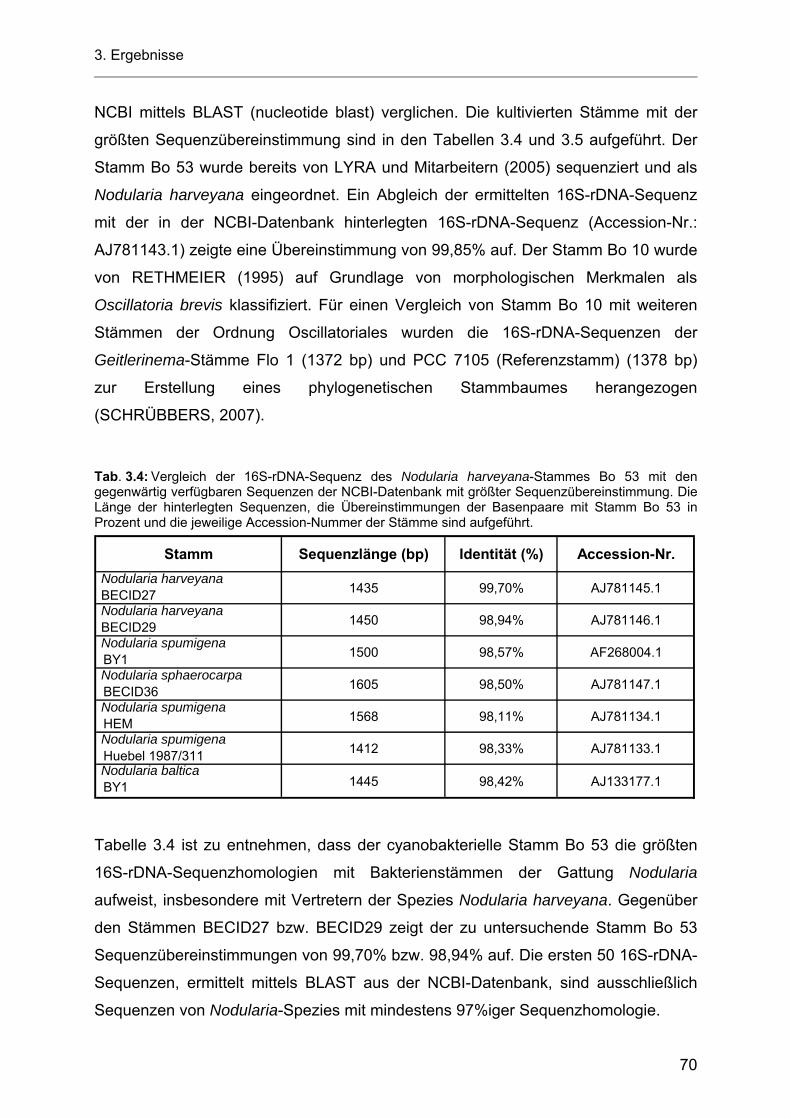
3. Ergebnisse
NCBI mittels BLAST (nucleotide blast) verglichen. Die kultivierten Stämme mit der
größten Sequenzübereinstimmung sind in den Tabellen 3.4 und 3.5 aufgeführt. Der
Stamm Bo 53 wurde bereits von LYRA und Mitarbeitern (2005) sequenziert und als
Nodularia harveyana eingeordnet. Ein Abgleich der ermittelten 16S-rDNA-Sequenz
mit der in der NCBI-Datenbank hinterlegten 16S-rDNA-Sequenz (Accession-Nr.:
AJ781143.1) zeigte eine Übereinstimmung von 99,85% auf. Der Stamm Bo 10 wurde
von RETHMEIER (1995) auf Grundlage von morphologischen Merkmalen als
Oscillatoria brevis klassifiziert. Für einen Vergleich von Stamm Bo 10 mit weiteren
Stämmen der Ordnung Oscillatoriales wurden die 16S-rDNA-Sequenzen der
Geitlerinema-Stämme Flo 1 (1372 bp) und PCC 7105 (Referenzstamm) (1378 bp)
zur Erstellung eines phylogenetischen Stammbaumes herangezogen
(SCHRÜBBERS, 2007).
Tab. 3.4: Vergleich der 16S-rDNA-Sequenz des Nodularia harveyana-Stammes Bo 53 mit den gegenwärtig verfügbaren Sequenzen der NCBI-Datenbank mit größter Sequenzübereinstimmung. Die Länge der hinterlegten Sequenzen, die Übereinstimmungen der Basenpaare mit Stamm Bo 53 in Prozent und die jeweilige Accession-Nummer der Stämme sind aufgeführt.
Stamm Sequenzlänge (bp) Identität (%) Accession-Nr. Nodularia harveyana BECID27 1435 99,70% AJ781145.1
Nodularia harveyana BECID29 1450 98,94% AJ781146.1
Nodularia spumigena BY1 1500 98,57% AF268004.1
Nodularia sphaerocarpa BECID36 1605 98,50% AJ781147.1
Nodularia spumigena HEM 1568 98,11% AJ781134.1
Nodularia spumigena Huebel 1987/311 1412 98,33% AJ781133.1 Nodularia baltica BY1 1445 98,42% AJ133177.1
Tabelle 3.4 ist zu entnehmen, dass der cyanobakterielle Stamm Bo 53 die größten
16S-rDNA-Sequenzhomologien mit Bakterienstämmen der Gattung Nodularia
aufweist, insbesondere mit Vertretern der Spezies Nodularia harveyana. Gegenüber
den Stämmen BECID27 bzw. BECID29 zeigt der zu untersuchende Stamm Bo 53
Sequenzübereinstimmungen von 99,70% bzw. 98,94% auf. Die ersten 50 16S-rDNA-
Sequenzen, ermittelt mittels BLAST aus der NCBI-Datenbank, sind ausschließlich
Sequenzen von Nodularia-Spezies mit mindestens 97%iger Sequenzhomologie.
70

3. Ergebnisse
Auf Basis der 16S-rDNA-Sequenz von Stamm Bo 53 und naheverwandten Stämmen
(siehe Tab. 3.4) erfolgte nach der „Neighbor-Joining“-Methode und dem Model von
JUKES und CANTOR (1969) die Konstruktion eines phylogenetischen
Stammbaumes (siehe Kap. 2.8.7) (siehe Abb. 3.18).
Nodularia spumigena HEM (AJ781134.1)
Nodularia spumigena Huebel 1987/311 (AJ781133.1)
Nodularia spumigena BY1 (AF268004.1)
Nodularia baltica BY1 (AJ133177.1)
Nodularia sphaerocarpa BECID36 (AJ781147.1)
Nodularia harveyana BECID29 (AJ781146.1)
Nodularia harveyana BECID27 (AJ781145.1)
Nodularia harveyana Bo 53
Gloeobacter violaceus PCC 8105 (AF132791.1)
100
71
99
86
67
0.01
Abb. 3.18: Phylogenetischer Stammbaum nach der „Neighbor-Joining“-Methode und dem Model von JUKES und CANTOR (1969) auf Grundlage der 16S-rDNA-Sequenzen von Stamm Bo 53 und naheverwandten Nodularia-Spezies. Gloeobacter violaceus PCC 8105 (AF132791.1) wurde als Außengruppe verwendet. Die Accession-Nummern der Referenz-Sequenzen sind in Klammern angegeben. An den Knotenpunkten des Stammbaumes sind die Bootstrap-Werte in Prozent dargestellt (1000 Wiederholungen). Skalierung: Anzahl an Substitutionen pro Nukleotid-Position.
Abbildung 3.18 zeigt, dass Stamm Bo 53 insbesondere eine enge
Verwandtschaftsbeziehung mit dem Nodularia harveyana-Stamm BECID27 aufweist.
Zu den weiteren Nodularia-Spezies ergab sich ein abgegrenztes Cluster. Der Stamm
Gloeobacter violaceus PCC 8105 grenzt sich als verwendete Außengruppe deutlich
von den Nodularia-Spezies ab.
Tab. 3.5: Vergleich der 16S-rDNA-Sequenz des Stammes Bo 10 mit den gegenwärtig verfügbaren Sequenzen der NCBI-Datenbank mit größter Sequenzübereinstimmung. Die Länge der hinterlegten Sequenzen, die Übereinstimmungen der Basenpaare mit Stamm Bo 10 in Prozent und die jeweilige Accession-Nummer der Stämme sind aufgeführt.
Stamm Sequenzlänge (bp) Identität (%) Accession-Nr.
Phormidium pseudopristleyi ANT.ACEV5.3 1463 99,32% AY493600.1
Phormidium lumbricale UTCC 476 1353 98,64% AF218375.1
Phormidium cf. terebriformis KR2003/25 1348 98,42% AY575936.1
Oscillatoria acuminata 1434 98,12% AB039014.1
71

3. Ergebnisse
Tabelle 3.5 ist zu entnehmen, dass Stamm Bo 10 die nahesten
Verwandtschaftsbeziehungen zu Vertretern der Gattung Phormidium und hier die
größte Übereinstimmung in der 16S-rDNA-Sequenz mit Phormidium pseudopristleyi
Stamm ANT.ACEV5.3 (99,32%) aufweist.
Auf Basis der 16S-rDNA-Sequenz von Stamm Bo 10 und naheverwandten Stämmen
(siehe Tab. 3.5) sowie der 16S-rDNA-Sequenz der Stämme Flo 1 und PCC 7105
erfolgte nach der „Neighbor-Joining“-Methode und dem Model von JUKES und
CANTOR (1969) die Konstruktion eines phylogenetischen Stammbaumes (siehe
Kap. 2.8.7) (siehe Abb. 3.19).
Phormidium cf. terebriformis KR2003/25 (AY575936.1)
Oscillatoria acuminata (AB039014.1)
Oscillatoria brevis Bo 10
Phormidium pseudopristleyi ANT.ACEV5.3 (AY493600.1)
Phormidium lumbricale UTCC 476 (AF218375.1)
Geitlerinema sp. PCC 7105
Geitlerinema sp. Flo 1
Gloeobacter violaceus PCC 8105 (AF132791.1)
10078
66
100
100
0.01
Abb. 3.19: Phylogenetischer Stammbaum nach der „Neighbor-Joining“-Methode und dem Model von JUKES und CANTOR (1969) auf Grundlage der 16S-rDNA-Sequenzen von Stamm Bo 10, naheverwandten Bakterienspezies sowie der Stämme Flo 1 und PCC 7105. Gloeobacter violaceus PCC 8105 (AF132791.1) wurde als Außengruppe verwendet. Die Accession-Nummern der Referenz-Sequenzen sind in Klammern angegeben. An den Knotenpunkten des Stammbaumes sind die Bootstrap-Werte in Prozent dargestellt (1000 Wiederholungen). Skalierung: Anzahl an Substitutionen pro Nukleotid-Position.
Aus Abbildung 3.19 geht hervor, dass Stamm Bo 10 zwar
Verwandtschaftsbeziehungen mit verschiedenen Phormidium-Spezies und einer
Oscillatoria-Spezies aufweist, aber ein eigenes Cluster bildet. Die beiden
Geitlerinema-Stämme Flo 1 und PCC 7105 grenzen sich als Cluster, wie auch der
als Außengruppe verwendete Stamm Gloeobacter violaceus PCC 8105, deutlich von
den Vertretern der Gattungen Phormidium und Oscillatoria ab.
72

3. Ergebnisse
3.6.4 Amplifikation des gyrB-Gens
Durch die Amplifikation des gyrB-Gens der Cyanobakterienstämme Bo 53 und Bo 10
mittels einer Standard-PCR (siehe Kap. 2.8.3) wurde ein ca. 1200 bp großes DNA-
Fragment erhalten (Gelbild nicht gezeigt). Die Sequenzen des gyrB-Gens der
Cyanobakterienstämme Bo 53 und Bo 10 konnten für eine Erstellung von
phylogenetischen Stammbäumen auf Basis der gyrB-Sequenz aber nicht verwandt
werden. Für die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 konnten
nach einer Standard-PCR mit den gyrB-spezifischen Primern keine ca. 1200 bp
großen Banden im Agarosegel detektiert werden.
3.6.5 Internal Transcribed Spacer (ITS) Nach Amplifikation der ITS der cyanobakteriellen Stämme Bo 10, Flo 1 und PCC
7105 (siehe Kap. 2.8.3) wurden die PCR-Produkte gelelektrophoretisch aufgetrennt
(siehe Kap. 2.8.4). Das Gelbild ist in Abbildung 3.20 dargestellt.
AuM
A
F
P
F
Proben bp
− 3000 − 1500 − 1000 − 700 − 500 − 400
− 100
bb. 3.20: ITS-Bandenmuster der Cyanobakterienstämme Bo 10 (Probe 1 und 2), Flo 1 (Probe 3 nd 4) und PCC 7105 (Probe 5 und 6) nach Fragmentauftrennung in einem 2%igen Agarosegel. : Gene RulerTM 100 bp DNA Ladder Plus; NK: Negativkontrolle ohne DNA-Template.
bbildung 3.20 zeigt das Gelbild der ITS-Amplifikate der cyanobakteriellen Stämme
lo 1, PCC 7105 und Bo 10. Für die beiden Geitlerinema-Stämme Flo 1 und
CC 7105 konnte im Bereich von 600 bp - 700 bp eine Bande dokumentiert werden.
ür Stamm Bo 10 wurden im Bereich von 500 bp - 700 bp drei Banden detektiert. Die
73

3. Ergebnisse
DNA-Fragmente wiesen Größen von ~500 bp, ~650 bp und ~700 bp auf. Das
unterschiedliche Bandenmuster bzw. die unterschiedliche Anzahl von amplifizierten
ITS-Regionen bei Stamm Bo 10 gegenüber den Geitlerinema-Stämmen zeigt auf,
dass Stamm Bo 10 nicht der Gattung Geitlerinema zugeordnet werden kann.
74

4. Diskussion
4. Diskussion
Im Mittelpunkt der vorliegenden Diplomarbeit standen sowohl der Nachweis von
AHL-Signalmolekülen bei den heterotrophen Bakterien Porphyrobacter Stamm
Bo 53-33 und Muricauda Stamm Bo 10-09 sowie dem Cyanobakterium Geitlerinema
Stamm Flo 1 als auch die Abhängigkeit der Signalmolekül-Produktion der oben
genannten Mikroorganismen bei verschiedenen Kultivierungsbedingungen.
Neben den Untersuchungen zum Wachstumsverhalten der heterotrophen
Bakterienstämme Bo 53-33 und Bo 10-09 werden des Weiteren in diesem Kapitel die
Ergebnisse der molekularbiologischen Untersuchungen zur phylogenetischen
Einordnung der Cyanobakterienstämme Bo 53 und Bo 10 diskutiert.
4.1 Wachstumsverhalten der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 unter verschiedenen Kultivierungsbedingungen
4.1.1 Verwendete Puffersysteme Zur Untersuchung des Wachstumsverhaltens der Bakterienstämme Bo 53-33 und
Bo 10-09 in Kulturmedien mit verschiedenen pH-Werten mussten zur Abdeckung des
pH-Bereiches von pH 2,0 - 10,0 verschiedene Puffersysteme eingesetzt werden
(siehe Kap. 2.6.2, Tab. 2.14). Jedes Puffersystem beeinflusst das Kulturmedium und
die im Nährmedium wachsenden Bakterien unterschiedlich. Folglich zeigten
bestimmte Puffersysteme einen inhibierenden Einfluss auf das Wachstum der
Bakterien und konnten nicht weiter verwendet werden (siehe Kap. 3.1). So hatte die
im Clark and Lubs(C&L)-Puffer enthaltene Borsäure wahrscheinlich einen toxischen
Einfluss auf die Bakterien (MORTIMER, 2007). Des Weiteren können organischen
Puffer-Komponenten (Citronensäure, Maleat und Glycin) als potentielle
Kohlenstoffquelle Einfluss auf das Wachstumverhalten bei einem bestimmten pH-
Wert nehmen. Wesentliche Probleme bei der Pufferung von mineralischen
Kulturmedien zeichnen sich vor allem im alkalischen Bereich ab, indem Kationen
(z.B. Mg2+- und Ca2+-Ionen) stark polarisierend auf Anionen wirken und kovalente
Bindungen eingehen. Dabei liegen starke Tendenzen zur Bildung von unlöslichen
Komplex-Ionen vor (MORTIMER, 2007). Diese Interaktionen von Puffer und
Kulturmedium sowie von verschiedenen Komponenten des Kulturmediums bei hohen
75

4. Diskussion
pH-Werten resultieren in Ausfällungen und gehen mit einem starken Nährstoffentzug
im Kulturmedium einher.
Die gewählten Puffersysteme stellen dementsprechend einen Kompromiss zwischen
der Pufferkapazität des jeweiligen Puffers sowie dessen Verträglichkeit für die
Testorganismen und den mit steigendem pH-Wert zunehmenden Mineralsalz-
Ausfällungen dar.
4.1.2 Wachstumsverhalten bei unterschiedlichen pH-Werten Mikroorganismen können extreme Umgebungen besiedeln. Das mikrobielle
Wachstum kann dabei durch die Azidität und Alkalität eines Lebensraumes stark
beeinflusst werden (MADIGAN et al., 2001). Aufgrund ihres pH-Toleranzbereiches
können Mikroorganismen in verschiedene Gruppen unterteilt werden. Der Großteil
toleriert dabei als Neutrophile einen pH-Bereich von 5,0 bis 8,5 (zitiert aus RIUS und
LOREN, 1998). Azidophile wachsen in einem sauren pH-Bereich, wobei mikrobielles
Wachstum auch bei sehr niedrigen pH-Werten (pH < 3) vorliegt und oftmals selber
Ursache für die Azidität der Umgebung ist (z.B. bakterielle Oxidation von
Sulfidmineralien) (HALLBERG und JOHNSON, 2001). Alkaliphile können ein
optimales Wachstum bei pH-Werten oberhalb von 9,0, oft zwischen 10,0 - 12,0
aufweisen, wachsen aber nicht oder lediglich langsam bei neutralem pH-Wert
(HORIKOSHI, 1999).
Der heterotrophe Bakterienstamm Porphyrobacter Bo 53-33 wächst in einem pH-
Bereich von 6,0 - 9,0 (siehe Kap. 3.2). Das beste Wachstum konnte bei pH 7,0
festgestellt werden, was nahezu dem standardmäßig eingesetzten ASNIII/2-Medium
(pH 7,1) entspricht. Ein geringes Wachstum wurde bei pH-Werten von 6,0 und 9,0
verzeichnet. Bei diesen pH-Werten bildete der Porphyrobacter-Stamm,
wahrscheinlich zur Schaffung eines eigenen pH-Milieus, deutlich abgegrenzte
Zellagglomerate aus. Aufgrund des relativ großen pH-Toleranzbereiches sowie des
pH-Optimums ist Porphyrobacter Stamm Bo 53-33 als pH-tolerantes, neutrophiles
Bakterium einzuordnen, was sich mit dem natürlichen Habitat mariner Bakterien
deckt (pH 7,8 - 8,2: HAWLEY, 1981).
Im Genus Porphyrobacter, mit mittlerweile 7 klassifizierten Stämmen auf Spezies-
Ebene, wurden identische pH-Toleranzbereiche (6,0 - 9,0) bei den Vertretern
P. sanguineus (HIRAISHI et al., 2002) und P. tepidarius (HANADA et al., 1997)
ermittelt, mit einem langsamen Wachstum bei pH-Werten von 6,0 und 9,0. Einen
76

4. Diskussion
deutlich größeren pH-Toleranzbereich von 5,5 - 10,0 weist dagegen das vor kurzem
beschriebene Bakterium P. meromictius auf (RATHGEBER et al., 2007), für aerobe
anoxygen phototrophe Bakterien (α-4-Unterklasse der Proteobakterien) der bisweilen
maximal tolerierbare pH-Bereich. Das potentielle Wachstum dieses Bakteriums in
diesem großen pH-Bereich ist wahrscheinlich auf dessen natürliche Umgebung,
einem stark Natriumsulfat-haltigen Gewässer, zurückzuführen. Der optimale pH-Wert
für das Wachstum einiger Porphyrobacter-Spezies wurde im Bereich 7,0 - 8,0
detektiert, wobei zwischen den einzelnen Spezies geringfügige Unterschiede
bestehen (HIRAISHI et al., 2002; RAINEY et al., 2003; YOON et al., 2004, 2006)
Der Bakterienstamm Muricauda Bo 10-09 ist aufgrund des pH-Toleranzbereiches
von 6,0 - 9,0, mit einem schwachen Wachstum bei pH 9,0 und einem ermittelten pH-
Optimum bei 8,0 (siehe Kap. 3.2), ebenfalls als neutrophiles Bakterium einzuordnen.
Weitere Untersuchungen mit einem anderen Puffersystem (Tris-HCl-Puffer, pH 7,1)
zeigten auf, dass das pH-Optimum für Stamm Bo 10-09 wahrscheinlich im Bereich
von pH 7,0 liegt (Daten nicht gezeigt).
Das Genus Muricauda (Flavobacteriaceae) ist der Ordnung Cytophagales zugehörig.
Deren Vertreter sind ubiquitär anzutreffen und bilden in marinen Gewässern und
Umgebungen die größte Bakteriengruppe (BRUNS et al., 2001). GLÖCKNER und
Mitarbeiter (1999) konnten dabei in allen untersuchten marinen Proben sowie
Süßwasser-Proben Bakterien der Ordnung Cytophagales nachweisen.
Für die bisher bekannten Arten der Gattung Muricauda (M. ruestringensis,
M. flavescens und M. aquimarina) wurden pH-Optima im Bereich von 6,5 - 7,5 bzw.
um 7,0 dokumentiert (BRUNS et al., 2001; YOON et al., 2005). Mit einem Wachstum
im Bereich von pH 6,0 - 8,0 wurde für das Bakterium M. ruestringensis ein relativ
kleiner pH-Toleranzbereich festgestellt (BRUNS et al., 2001), während für die beiden
anderen Spezies der Gattung noch ein schwaches Wachstum bei einem pH-Wert
von 5,0 nachgewiesen wurde (YOON et al., 2005).
Die lange Kultivierungszeit der heterotrophen Stämme Bo 53-33 und Bo 10-09 bis
zum Eintritt in die stationäre Wachstumsphase ist bei einer gewählten
Inkubationstemperatur von 21 °C primär auf die Anzuchtsbedingungen
zurückzuführen. Ein optimales Wachstum wurde für die Vertreter der Gattung
Porphyrobacter im Bereich von 30 - 37 °C (YOON et al., 2004, 2006) bzw. weit
77

4. Diskussion
oberhalb dessen (HANADA et al., 1997; RAINEY et al., 2003) festgestellt. Für die
von YOON und Mitarbeitern (2005) beschriebene Spezies Muricauda aquimarina
wurde ein Temperatur-Optimum von 30 - 37 °C berichtet.
Das Wachstum der heterotrophen Bakterienstämme war überwiegend durch eine
deutliche Alkalisierung der gepufferten und ungepufferten Kulturmedien
gekennzeichnet (siehe Kap. 3.2.1). Diese pH-Verschiebungen sind wahrscheinlich in
erster Linie auf die Freisetzung von Ammonium-Ionen sowie Kohlendioxid aus
Proteinen und Peptiden des im ASNIII/2-Medium enthaltenen Fleischextraktes
zurückzuführen. Eine längerfristige Pufferung von Versuchskulturen ist demnach bei
Verwendung eines Vollmediums nicht möglich, da zu viele Stoffwechselreaktionen
die Pufferkapazität reduzieren. Des Weiteren kann eine aktive pH-Wert-Einstellung
durch die heterotrophen Bakterien vermutet werden, da bei pH 8,5- bzw. pH 9,0-
gepufferten Kulturen der pH-Wert konstant blieb bzw. sich leicht erniedrigte.
4.1.3 Wachstumsverhalten unter Lichteinfluss Die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 wurden aus nicht-
axenischen Cyanobakterienkulturen isoliert (HUBE, 2008) und können in engen
assoziierten Lebensgemeinschaften mit Cyanobakterien auftreten (TUOMAINEN et
al., 2006; STEVENSON und WATERBURY, 2006). Cyanobakterien nutzen als
photosynthetische Bakterien das Licht als primäre Energiequelle zur CO2-Fixierung
(FUCHS, 2007). Die Porphyrobacter-Spezies (α-Proteobakterien) zeichnen sich
durch die Synthese von Photopigmenten (Bakteriochlorophyll a (Bchl a)) und von
großen Mengen an Carotinoiden aus (YURKOV und BEATTY, 1998). Im Gegensatz
zu den anaeroben anoxygenen phototrophen Proteobakterien besitzen diese
Bakterien aber nicht die Fähigkeit, die Photopigmente zur anoxygenen
Photosynthese für ein phototrophes Wachstum unter anaeroben Bedingungen zu
nutzen (YURKOV und BEATTY, 1998; RATHGEBER et al., 2004) und wurden daher
als aerobe anoxygene phototrophe Bakterien beschrieben. Die
membrangebundenen Photopigmente und akzessorischen Proteine dienen dabei
dem Aufbau eines elektrochemischen Membranpotentials, wobei letztlich ATP-
Äquivalente aus der protonenmotorischen Kraft generiert werden (WAGNER-
DÖBLER und BIEBL, 2006). Geringe Lichtintensitäten führen bei aeroben
anoxygenen Phototrophen zu einer vollständigen Inhibierung der Bchl a-Synthese
(WAGNER und BIEBL, 2006). Dabei wirken bereits Lichtintensitäten von
78

4. Diskussion
20 µE m-2 s-1 PAR auf die Photopigment-Synthese stark inhibierend (zitiert aus
YURKOV und BEATTY, 1998).
Der Porphyrobacter-Stamm Bo 53-33 zeigte bei einem Photonenfluss von
5 µE m-2 s-1 PAR kein sichtlich verändertes Wachstum gegenüber einer
Dunkelinkubation auf (siehe Kap. 3.3). Dementsprechend hatte das Licht als
zusätzliche Energiequelle anscheinend keinen stimulierenden Einfluss auf das
heterotrophe Wachstum der Versuchskulturen. Ein hemmender Einfluss konnte
ebenso nicht festgestellt werden, da die Carotinoide wahrscheinlich einen Schutz vor
photooxidativen Schaden vermitteln und toxische Sauerstoffderivate quenchen
(MADIGAN et al., 2001; WAGNER und BIEBL, 2006). Für den Stamm Bo 10-09
wurde ebenfalls kein verändertes Wachstumsverhalten bei einem Photonenfluss von
5 µE m-2 s-1 PAR festgestellt (siehe Kap. 3.3). Ein Vorkommen von Carotinoiden
wurde in Studien mit verschiedenen Muricauda-Spezies nicht dokumentiert, sind
aber aufgrund eines großen Vorkommens in nicht photosynthetisch aktiven Bakterien
(YURKOV und BEATTY, 1998) auch für den Stamm Bo 10-09 nicht auszuschließen.
4.2 „Quorum Sensing“(QS) bei verschiedenen Gram-negativen Bakterien Interzelluläre Kommunikationsysteme werden in vielen Bakterien zur Regulation der
Expression bestimmter Gene und dementsprechend zur Steuerung physiologischer
Aktivitäten in Abhängigkeit von der eigenen Populationsdichte verwendet. Diese
Anpassung von Aktivitäten können dabei in einer sich verändernden Umgebung
einen deutlichen Vorteil bringen (SZENTHE und PAGE, 2003; CASE et al., 2008)
und z.B. andere Bakterienspezies in einer ökologischen Nische verdrängen. Unter
Gram-negativen Bakterien ist dabei ein AHL-abhängiges „Quorum Sensing“ ein weit
verbreitetes Kommunikationssystem (FUQUA et al., 2001). In den letzten Jahren
wurde zunehmend in Gram-negativen Bakterien zudem ein weiteres QS-System
nachgewiesen, in dem Furanon-Derivate als QS-Signalmoleküle agieren (z.B.
Yersinia pestis: BOBROV et al., 2007) und wahrscheinlich überwiegend an einer
interspezifischen Kommunikation beteiligt sind (FEDERLE und BASSLER, 2003).
79

4. Diskussion
4.2.1 AHL-Nachweis mit bakteriellen Biosensoren Eine Möglichkeit zum Nachweis verschiedener AHL-Signalmoleküle ist der Einsatz
von bakteriellen Biosensoren. Die chemischen Strukturunterschiede der N-Acyl-
Seitenkette der AHL-Moleküle führen bei den einzelnen Biosensoren zu
unterschiedlichen Sensitivitäten gegenüber den verschiedenen AHL-Molekülen,
wobei viele Biosensoren lediglich eine kleine Bandbreite an AHL-Molekülen
detektieren (STEINDLER und VENTURI, 2007). Der Nachweis ist dementsprechend
auf die AHL-Moleküle beschränkt, auf welche der jeweilige Biosensor ab einer
bestimmten AHL-Konzentration reagiert (SHAW et al., 1997). Generell nimmt die
Affinität gegenüber einem AHL-Molekül dabei in der Stärke ab, je mehr sich die
Seitenkette des Moleküls von der des ursprünglich eigenen AHL-Signalmoleküls
unterscheidet. Die große AHL-Diversität bedingt dementsprechend zur Abdeckung
eines breiten AHL-Spektrums den Einsatz mehrerer Biosensoren, welche auf
verschiedene AHL-Moleküle reagieren. Eine AHL-Produktion kann von den
Kultivierungsbedingungen und dem Nährmedium abhängen (FARRAND et al., 2002)
bzw. durch bestimmte Umweltbedingungen reguliert werden (MOHAMMED et al.,
2008) und liegt folglich unter bestimmten Laborbedingungen vielleicht gar nicht vor.
Demzufolge schließt kein AHL-Nachweis die Produktion von AHL-Signalmolekülen
bzw. eines bestimmten AHL-Moleküls im jeweiligen untersuchten Bakterienstamm
grundsätzlich nicht aus.
Die in der vorliegenden Arbeit eingesetzten AHL-Biosensoren Agrobacterium
tumefaciens NTL4 (pZLR4) und Chromobacterium violaceum CV026 weisen
aufgrund ihres eigenen natürlichen AHL-QS-Systems unterschiedliche Sensitivitäten
gegenüber AHL-Signalmolekülen auf. In dünnschichtchromatographischen
Voruntersuchungen und Agarplattentests wurden Spezifitäten und Sensitivitäten der
AHL-Sensoren gegenüber zur Verfügung stehenden synthetischen
Referenzmolekülen (C4-HSL, C6-HSL und C8-HSL) ermittelt. Der Biosensor
C. violaceum CV026 zeigte mit Abstand die höchste Sensitivität gegenüber C6-HSL,
gefolgt von C8-HSL und C4-HSL (siehe Kap. 2.7.6 und 3.4.1). Der A. tumefaciens-
Sensorstamm wies eine sehr hohe Affinität für C8-HSL auf (siehe Kap. 2.7.6).
Gegenüber C6-HSL war der Sensorstamm leicht sensitiver als gegenüber C4-HSL.
Ähnliche Ergebnisse erzielten MCCLEAN und Mitarbeiter (1997) für C. violaceum
CV026 sowie FARRAND und Mitarbeiter (2002) für A. tumefaciens NTL4 (pZLR4). Im
Vergleich mit C. violaceum CV026 wies der Agrobacterium-Sensorstamm wesentlich
80

4. Diskussion
höhere Sensitivitäten gegenüber C8-HSL und C4-HSL und gleiche Affinität
gegenüber C6-HSL auf. Aufgrund der hohen nachgewiesenen Sensitivitäten und des
dokumentierten breiten Nachweisspektrums (STEINDLER und VENTURI, 2007)
erfolgten die weiteren Untersuchungen überwiegend mit dem Agrobacterium-
Sensorstamm.
4.2.2 Screeningtests zum Nachweis AHL-produzierender Bakterien Zum schnellen Nachweis von AHL-produzierenden Bakterien fanden zwei Methoden
in der vorliegenden Arbeit Anwendung. Im Screeningverfahren nach FARRAND und
Mitarbeitern (2002) wurden Bakterienkulturen oder Kulturüberstände direkt auf AHL-
Signalmoleküle untersucht, während RAVN und Mitarbeiter (2001) ausschließlich
Bakterienkulturen einsetzten. Die Methoden ermöglichen jedoch keine
Unterscheidung zwischen verschiedenen AHL-Molekülen. Des Weiteren können die
AHL-Signalmoleküle in den Bakterienkulturen bzw. Kulturüberständen aufgrund einer
fehlenden Aufkonzentrierung für eine Detektion in nicht ausreichender Menge
vorliegen.
In beiden AHL-Screeningsystemen konnte für Stamm Bo 53-33 ein AHL-Nachweis
unter Verwendung des Agrobacterium-Sensors erzielt werden (siehe Kap. 3.4.2 und
3.4.3). Ein schwacher Nachweis auf AHL-Moleküle konnte für Stamm Bo 10-09
lediglich über die Methode von FARRAND und Mitarbeitern (2002) dokumentiert
werden, während eine AHL-Produktion durch das Cyanobakterium Stamm Flo 1 nicht
nachgewiesen wurde.
Im Screeningtest nach RAVN und Mitarbeitern (2001) stellte das verwendete
Nährmedium einen limitierenden Faktor für diesen Assay dar, da sowohl das zu
testende Bakterium als auch der Biosensor auf diesem wachsen müssen. Ein
schwaches Wachstum, z.B. aufgrund unterschiedlicher Mineralsalz- bzw.
Salinitätsansprüche der Bakterien, schränkten Aussagen über eine AHL-Produktion
ein. Ein Kompromiss wurde mit dem ASNIII/2-Medium erzielt, wodurch aber auf eine
AHL-Untersuchung mit dem Biosensor C. violaceum CV026 verzichtet werden
musste. Die stärkeren Nachweisreaktionen über den Screeningtest nach FARRAND
und Mitarbeitern (2002) im Vergleich zum zweiten Assay zeigten die wesentlich
höhere Sensitivität dieses Assays gegenüber QS-Signalmolekülen auf. Eine in
mehreren Untersuchungen aufgetretene Blaugrün-Färbung des Bakterienstammes
Bo 10-09 im Screeningverfahren nach RAVN und Mitarbeitern (2001) ließ annehmen,
81

4. Diskussion
dass dieser X-Gal wahrscheinlich enzymatisch spalten kann. Neben dieser durch
Testorganismen enzymatisch bedingten X-Gal-Hydrolyse wurde des Weiteren von
FARRAND und Mitarbeitern (2002) von einer möglichen schwachen Aktivierung des
Agrobacterium-Biosensors durch Medienkomponenten berichtet, welche zu falsch-
positiven Ergebnissen führen kann. Eine unspezifische Aktivierung des Sensors
durch ASNIII/2-Medienkomponenten könnte auch Ursache für die Blaufärbung des
Sensorstammes bei der Nachweismethode von RAVN und Mitarbeitern (2001) in den
Negativkontrollen gewesen sein (siehe Kap. 3.4.2). Unspezifische
Nachweisreaktionen verdeutlichen, dass bei Untersuchungen mit Biosensoren eine
Durchführung von Negativkontrollen essentiell ist. Eine mögliche spontane Hydrolyse
von X-Gal erfordert zudem zur Vermeidung von Fehlinterpretationen eine
Auswertung beider Assays nach 24 h.
4.2.3 Extraktion und Nachweis von N-Acyl-L-Homoserinlactonen Die Extraktion von AHL-Signalmolekülen aus Kulturüberständen erfolgte in der
vorliegenden Arbeit mittels einer Dichlormethanextraktion nach GEISENBERGER
(2000) (siehe Kap. 2.7.3.3). Die aufwendige Herstellung von AHL-Extrakten war mit
einer Anzucht großer Mengen bakterieller Biomasse verbunden, da für die Extraktion
große Volumina an Kulturüberständen zur Ausschüttelung der AHL-Moleküle mit
dem organischen Lösungsmittel benötigt werden (WAGNER-DÖBLER et al., 2005).
FROMMBERGER (2005) bestimmte die Extraktionsausbeute mit Dichlormethan mit
etwa 60%. Nachfolgend wurde die organische Phase eingeengt. Problematisch
erwies sich hierbei nach Eintrocknung der organischen Phase die vollständige
Aufnahme der Rückstände in einem relativ geringen Volumen an flüchtigem
Ethylacetat. Aufnahmen in unterschiedlichen Ethylacetatvolumen in Abhängigkeit von
der Menge des Rückstandes würden einen AHL-Vergleich zwischen den jeweiligen
Kulturüberständen nicht mehr zulassen. Andere Studien verwendeten Ethylacetat als
Extraktionsmittel (CHA et al., 1998; RAVN et al., 2001; GRAM et al., 2002), wobei
RAVN und Mitarbeiter (2001) erstmals durch Azidifizierung des Ethylacetats die
Effizienz der AHL-Extraktion (75%) deutlich gegenüber nicht-azidifiziertem
Ethylacetat (35%) steigern konnten. Dabei wurde eine abnehmende Polarität der
AHL-Moleküle durch eine Protonierung in der N-Acyl-Seitenkette vermutet,
resultierend in einer leichteren AHL-Extraktion. Aus diesem Grund wurde in der
vorliegenden Arbeit eine Ansäuerung des Ethylacetats mit 0,01% (v/v) Essigsäure
82

4. Diskussion
durchgeführt, um eine maximale Aufnahme des Rückstandes bzw. der extrahierten
AHL-Moleküle zu erreichen.
Die in der vorliegenden Arbeit verwendete Methode zum Nachweis verschiedener
AHL-Moleküle geht ursprünglich auf SHAW und Mitarbeiter (1997) zurück. Dabei
erfolgte im Anschluss an eine Umkehrphasen-Dünnschichtchromatographie zur
Auftrennung der AHL-Moleküle die Detektion der Signalmoleküle mit bakteriellen
Reportersystemen.
In DC-Voruntersuchungen konnte festgestellt werden, dass mit Agrobacterium-
Sensorkulturen in der spät-exponentiellen Wachstumsphase, gekoppelt mit einer
guten Belüftung (200 rpm) während der Anzucht, die besten AHL-Nachweise erzielt
wurden. Des Weiteren ist das spezielle ABG-Kulturmedium für einen optimalen
Nachweis essentiell. Mit LB(Luria Bertani)-Medium konnten diese Ergebnisse nicht
auf diesem Niveau reproduziert werden. Eine unspezifische Aktivierung des
Biosensors durch LB-Medienkomponenten wurde zudem nicht sicher
ausgeschlossen. Ein weiterer Vorteil des ABG-Mediums ist eine Detektion blauer
DC-Spots auf einem hellen Hintergrund. Für eine optimale Visualisierung der AHL-
Moleküle wurde hierzu das X-Gal in N,N-Dimethylformamid gelöst.
Im Laufe der Untersuchungen zeichnete sich ab, dass der angewandten DC-Analytik
sowohl für physiologische Untersuchungen als auch für quantitative Aussagen
Grenzen gesetzt sind, da viele Parameter Einfluss auf die
dünnschichtchromatographischen Ergebnisse nehmen können. Hierbei spielen vor
allem die Herstellung der AHL-Extrakte, die Anzucht des Biosensors, die
Überschichtung der DC-Platten und deren Inkubation eine wesentliche Rolle. Des
Weiteren können Biosensoren geringe AHL-Mengenunterschiede in Form von
geringen Intensitätsabnahmen der DC-Spots kaum detektieren, und AHL-Reporter
sind zudem nicht fähig, die strukturelle Diversität von AHL-Molekülen nachzuweisen
(WAGNER-DÖBLER et al., 2005; CASE et al., 2008). Dementsprechend konnten nur
Tendenzen aufgezeigt, absolute Aussagen aber nicht getroffen werden.
83

4. Diskussion
4.2.4 AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 unter verschiedenen Kultivierungsbedingungen
Im Rahmen der vorliegenden Arbeit sollte der Einfluss verschiedener
Kultivierungsbedingungen auf die AHL-Molekül-Produktion bei verschiedenen Gram-
negativen Bakterien mittels dünnschichtchromatographischer Methode untersucht
werden. Im Mittelpunkt der Untersuchungen standen dabei die AHL-Produktion und
das Vorkommen verschiedener AHL-Molekültypen in Abhängigkeit vom pH-Wert der
Kulturmedien.
QS ist ein Zelldichte-abhängiger Prozess. Die Konzentration der produzierten und in
die Umwelt freigesetzten Signalmoleküle steigt mit Zunahme der Bakterienpopulation
proportional an (CHA et al., 1998; WILLIAMS, 2007). In der vorliegenden Arbeit
wurden oftmals Proben miteinander verglichen, die aus Kulturen mit
unterschiedlichen Zelldichten gewonnen wurden. Weitere vergleichende AHL-
Untersuchungen sollten dementsprechend mit Kulturen gleicher Populationsdichte
durchgeführt werden, um den Einfluss von verschiedenen Kultivierungsbedingungen
besser bewerten zu können.
4.2.4.1 Einfluss des pH-Wertes auf die AHL-Signalmoleküle Die dünnschichtchromatographischen Ergebnisse zeigten, dass ein unmittelbarer
Zusammenhang zwischen dem pH-Wert des Kulturmediums und der AHL-Menge in
den Zellkulturüberständen besteht (siehe Kap. 3.5.1). Dabei nahm die über den
Agrobacterium-Sensorstamm nachgewiesene AHL-Menge in Kulturüberständen von
Stamm Bo 53-33 mit zunehmendem pH-Wert von pH 6,0 - 8,5 kontinuierlich ab und
wies bei pH 8,5 und 9,0 in etwa gleich große AHL-Mengen auf. Für den
heterotrophen Stamm Bo 10-09 konnte der gleiche Zusammenhang festgestellt
werden, wobei für einen Nachweis von AHL-Molekülen wesentlich größere Mengen
an AHL-Extrakt eingesetzt werden mussten und lediglich in den pH 6-gepufferten
Kulturen Signalmoleküle nachgewiesen wurden. Aufgrund der sehr schwachen AHL-
Nachweise bei Stamm Bo 10-09 erwies sich dieser mit der zur Verfügung stehenden
Methode für weitere physiologische Untersuchungen als ungeeignet. Die Ursachen
einer schwachen Nachweisreaktion könnten in einer fehlenden Sensitivität des
Biosensors gegenüber den AHL-Molekülen und in einer für eine AHL-Produktion
nicht gegebenen Kultivierungsbedingung liegen.
84

4. Diskussion
YATES und Mitarbeiter (2002) führten diesen Befund in Studien mit Yersinia
pseudotuberculosis und Pseudomonas aeruginosa auf eine nicht-enzymatische
Hydrolyse der AHL-Moleküle unter basischen Anzuchtsbedingungen zurück. Die
Hydrolyse der Esterbindung des Homoserinlactonrings führt dabei zu einer
Inaktivierung und somit zu einem Funktionsverlust des jeweiligen N-Acyl-L-
Homoserinlactons. Ein weiterer Angriffspunkt ist die Amidbindung zwischen dem
Homoserinlacton und der N-Acyl-Seitenkette. Die degradierten AHL-Moleküle
werden von dem LuxR-homologen Protein des jeweiligen AHL-Biosensors nicht mehr
erkannt, und ein AHL-Nachweis bleibt aus (STEINDLER und VENTURI, 2007).
In Anbetracht einer pH-abhängigen Instabilität der AHL-Moleküle könnte ein
kontinuierlich zunehmender pH-Wert während der Anzucht zudem Grund für die
leicht niedrigeren nachgewiesenen AHL-Mengen in der stationären
Wachstumsphase gegenüber der logarithmischen Wachstumsphase ungepufferter
Kulturen von Stamm Bo 53-33 sein. Damit ließe sich ebenfalls ein AHL-Nachweis in
der logarithmischen Wachstumsphase und das Fehlen von AHL-Molekülen in der
stationären Wachstumsphase in ungepufferten Kulturen von Stamm Bo 10-09
erklären. Eine Abnahme des AHL-Gehaltes in der stationären Wachstumsphase
durch eine nicht-enzymatische, pH-abhängige Spaltung von AHL-Molekülen wurde in
einer Studie von BYERS und Mitarbeitern (2002) bei Erwinia carotovora
dokumentiert.
Nach einer Azidifizierung ungepufferter Kulturüberstände der Stämme Bo 53-33 und
Bo 10-09 konnte der nachgewiesene AHL-Gehalt für beide Stämme deutlich
gegenüber nicht-azidifizierten Kulturüberständen erhöht werden (siehe Kap. 3.5.1.1),
da die inaktiven AHL-Signalmoleküle durch einen reversiblen Ringschluss des
Homoserins in ihre aktive Struktur überführt wurden und somit über AHL-
Biosensoren nachgewiesen werden konnten (YATES et al., 2002). Der
Azidifizierungsversuch und die vorangegangene pH-Wert-Untersuchung (siehe Kap.
3.5.1) zeigten insbesondere beim Stamm Bo 53-33 auf, dass, wie von YATES und
Mitarbeitern (2002) dokumentiert, ein Zusammenhang zwischen der Stabilität des
Moleküls und der Länge der N-Acyl-Seitenkette besteht. Dementsprechend wurde
ein weiteres, kurzkettigeres AHL-Molekül lediglich in azidifizierten und pH 6-
gepufferten Kulturüberständen über den Agrobacterium-Sensorstamm deutlich
nachgewiesen, während ein längerkettiges AHL-Molekül auch bei höheren pH-
Werten detektiert wurde.
85

4. Diskussion
Bei der Expression QS-gesteuerter Gene verstärken die QS-Signalmoleküle
gleichzeitig ihre eigene Signalmolekül-Produktion (Autoinduktion) (EBERHARD et al.,
1981). Folglich wären in AHL-stabilen Umgebungen (sauer bis leicht-alkalisch) ein
kontinuierlicher Anstieg der AHL-Konzentration und damit eine fortwährende
Genexpression zu erwarten. Eine AHL-Akkumulation in AHL-stabilen Umgebungen
wurde nicht beobachtet (LEADBETTER und GREENBERG, 2000), was auf einen
enzymatischen AHL-Abbau schließen ließ. In den letzten Jahren wurden verstärkt
AHL-degradierende Enzyme, AHL-Lactonasen und AHL-Acylasen, in verschiedenen
Bakterienspezies nachgewiesen, welche wahrscheinlich in der Regulation von QS-
gesteuerten Prozessen eine wichtige Rolle spielen (DONG und ZHANG, 2005).
ZHANG und Mitarbeiter (2002b, 2004) konnten einen schnellen Abbau von 3-oxo-
C8-HSL in der stationären Wachstumsphase von A. tumefaciens auf eine AHL-
Lactonase (AttM) zurückführen. Die Expression dieses Enzyms wird dabei durch
Hunger- und Stresssignale induziert und führte zu einer Beendigung des QS-
gesteuerten konjugativen Ti-Plasmid-Transfers.
Die hohen pH-Werte der Kulturüberstände nach Inkubation der Stämme Bo 53-33
und Bo 10-09 lassen Rückschlüsse auf eine enzymatische AHL-Degradation nicht
zu. Der Azidifizierungsversuch zeigte aber auf, dass die nach Azidifizierung wieder
intakten AHL-Moleküle zuvor nicht durch AHL-Acylasen gespalten werden konnten,
da diese enzymatische Reaktion an der Amidbindung des Moleküls irreversibel ist
(ROMERO et al., 2008).
Aufgrund einer freien Diffusion der Signalmoleküle durch biologische Membranen
und die Zellwand steigt die Konzentration der AHL-Moleküle proportional mit
Zunahme der Populationsdichte intrazellulär und extrazellulär gleichermaßen an
(KAPLAN und GREENBERG, 1985). Dementsprechend wurde in den
Versuchsansätzen mit Stamm Bo 53-33 intrazellulär aufgrund geringerer extrahierter
Volumina an Biomasse gegenüber den Kulturüberständen eine wesentlich kleinere
AHL-Menge nachgewiesen als extrazellulär (siehe Kap. 3.5.5). Für Stamm Bo 10-09
konnte kein AHL-Nachweis erzielt werden. Der ausgebliebene AHL-Nachweis bei
höheren pH-Werten (pH 8,5 und 9,0) in Versuchsansätzen mit Stamm Bo 53-33 ist
wahrscheinlich auf nicht-enzymatisch degradierte AHL-Moleküle zurückzuführen. Der
höhere nachgewiesene AHL-Gehalt in der logarithmischen Wachstumsphase kann
ebenfalls durch eine nicht-enzymatische, pH-abhängige Hydrolyse in der
Stationärphase erklärt werden. Eine enzymatische Degradation von AHL-Molekülen
86

4. Diskussion
ist in der Stationärphase aber nicht ausgeschlossen. Über eine QS-regulierte
Expression von Genen in verschiedenen Wuchsphasen wurde z.B. für Pseudomonas
aeruginosa berichtet (SCHUSTER et al., 2003).
4.2.4.2 Einfluss der Temperatur auf die AHL-Signalmoleküle Die Temperatur hat generell einen wesentlichen Einfluss auf die Stabilität der
gebildeten AHL-Moleküle. Bei höheren Temperaturen wie 30 °C (YATES et al., 2002)
bzw. 37 °C (BYERS et al., 2002) konnte bereits bei mild-alkalischen Bedingungen
(oberhalb von pH 7,0 bis pH 8,0) eine deutliche Instabilität der AHL-Moleküle
dokumentiert werden, wobei oberhalb von ~ pH 8,2 keine AHL-Moleküle mehr
nachgewiesen wurden. Folglich wird die Verwendung von Furanon-Derivaten als QS-
Signalmoleküle beim Gram-negativen Bakterium Escherichia coli auf die im Darm
herrschenden Bedingungen (pH 7,6; 37 °C) zurückgeführt (SCHAUDER et al., 2001).
Dem gegenüber wird bei GRAM und Mitarbeitern (2002) für marine Roseobacter-
Stämme bei einer Anzuchtstemperatur von 15 °C erst oberhalb eines pH-Wertes von
8,0 auf eine AHL-Instabilität hingewiesen. Die AHL-Stabilität ist bei niedrigeren
Temperaturen um ein Vielfaches erhöht (BYERS et al., 2002), was für eine
interzelluläre Kommunikation mariner, Gram-negativer Bakterien über AHL-
Signalmoleküle hinsichtlich der pH-Werte mariner Gewässer (Meerwasser, pH 7,8 -
8,2: HAWLEY, 1981) essentiell ist. Eine Kommunikation ist demnach auch noch bei
höheren pH-Werten über AHL-Moleküle möglich.
Bei Stamm Bo 53-33 konnten bei einer Inkubationstemperatur von 21 °C noch bei
pH-Endwerten von ~8,5 AHL-Signalmoleküle nachgewiesen werden (siehe
Kap. 3.5.1). Bei einem Vergleich von Kulturüberständen ungepufferter
Versuchsansätze von Stamm Bo 53-33 wurde ein geringerer AHL-Gehalt nach 28 °C
Inkubation gegenüber einer 21 °C-Inkubation für den Bakterienstamm festgestellt
(siehe Kap. 3.5.3).
Physiologische Experimente sollten zukünftig im pH-neutralen Bereich und bei
niedrigen Inkubationstemperaturen durchgeführt werden bzw., wenn dieses nicht
möglich ist, mit einer anschließenden Azidifizierung der Kulturüberstände gekoppelt
werden.
87

4. Diskussion
4.2.4.3 Einfluss von Licht und Nährstoffen auf die Produktion von AHL-Signalmolekülen
Die Expression von QS-regulierten Genen bei Bakterien kann neben der
Populationsdichte von einer Vielzahl von Umweltstimuli (Stressinduktoren) abhängen
bzw. beeinflusst werden, z.B. der Verfügbarkeit von Nährstoffen (WITHERS et al.,
2001). Ein in Wechselbeziehung mit äußeren Umwelteinflüssen stehendes QS,
involviert in phänotypischen Veränderungen der Bakterienzelle, erhöht dabei für eine
Bakterienzelle die Wahrscheinlichkeit z.B. unter nährstoffarmen Bedingungen oder
anderen Stressbedingungen zu überleben. DAVIES und Mitarbeiter (1998) konnten
in diesem Zusammenhang aufzeigen, dass die Biofilmbildung sowohl vom QS als
auch von externen Umweltstimuli (z.B. pH-Wert und Nährstoffe) abhängig ist.
Studien an Escherichia coli zeigten eine unterschiedliche AI-2-Signalmolekül-
Produktion in Abhängigkeit von umweltbedingten Stressfaktoren auf, was auf die
Fähigkeit einer stressbedingten Anpassung durch QS an sich ändernde
Umweltbedingungen schließen ließ (DE LISA et al., 2001). AHMED und Mitarbeiter
(2007) konnten bei Streptococcus anginosus ebenfalls einen Zusammenhang
zwischen der AI-2-Signalmolekül-Produktion und antibiotischem Stress aufzeigen.
Die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 wurden in den in der
vorliegenden Arbeit durchgeführten Untersuchungen möglichen Stressfaktoren
ausgesetzt.
Bei einer Erhöhung der Photonenflussdichte von 0 auf 5 µE m-2 s-1 PAR wurden
dabei keine signifikanten Unterschiede in der AHL-Menge gegenüber einer
Dunkelinkubation der Kulturen nachgewiesen (siehe Kap. 3.5.2). Folglich hat ein
Photonenfluss von 5 µE m-2 s-1 PAR keinen Einfluss auf die AHL-Produktion bzw. auf
QS-gesteuerte Vorgänge bei den heterotrophen Bakterien. Des Weiteren steht
anscheinend eine Hemmung der Bchl a-Synthese bei Stamm Bo 53-33 bei geringen
Lichtintensitäten ebenfalls in keinem Zusammenhang mit „Quorum Sensing“.
Durch eine deutliche Reduzierung der Kohlenstoffquelle im verwendeten
Nährmedium sollten die Kulturen einem Nährstoffmangel ausgesetzt werden. Unter
den nährstofflimitierten Bedingungen (0,1% Fleischextrakt) konnte im Vergleich zu
den standardmäßigen Nährstoffbedingungen (1% Fleischextrakt) ein leicht stärkerer
AHL-Nachweis mit dem Agrobacterium-Sensorstamm für Stamm Bo 53-33
dokumentiert werden (siehe Kap. 3.5.3). Mit dem standardmäßig verwendeten
Nährmedium blieb ein AHL-Nachweis bei Stamm Bo 10-09 in den Kulturüberständen
88

4. Diskussion
aus, unter nährstofflimitierten Bedingungen wurde hingegen ein geringer AHL-Gehalt
in den Überständen nachgewiesen. Die Ergebnisse lassen vermuten, dass das
Nährmedium einen Einfluss auf die AHL-Produktion durch die heterotrophen Stämme
hat, also eine Abhängigkeit der AHL-Menge von der Verfügbarkeit der
Kohlenstoffquelle vorliegt.
QS ist an einer Vielzahl von phänotypischen Veränderungen und physiologischen
Anpassungen beteiligt (WILLIAMS, 2007), welche insbesondere bei
nährstofflimitierten Bedingungen und Stressbedingungen eine wesentliche Rolle
spielen (WITHERS et al., 2001). Die leicht erhöhten AHL-Gehalte könnten mit einer
verstärkten Expression QS-regulierter Gene zusammenhängen, welche bei einer
geringen Verfügbarkeit von Nährstoffen eine Adaptation an sich ändernde
Umweltbedingungen ermöglichen. Ein Einfluss von Nährmedien bzw. der
Medienzusammensetzung auf die Expression von QS-regulierten Genen wurde für
Pseudomonas aeruginosa beschrieben (WAGNER et al., 2003). In Vollmedien
konnte dabei eine deutliche Senkung der Expression von QS-regulierenden Genen
beobachtet werden. TEPLITSKI und Mitarbeiter (2003) zeigten ebenfalls eine starke
Abhängigkeit des produzierten AHL-Profils von dem Nährmedium auf. Dennoch
konnten QS-Mutanten in LB(Luria Bertani)-Medium zu höheren Zelldichten
heranwachsen als QS-fähige Wildtypstämme von Pseudomonas aeruginosa, da die
QS-regulierte Produktion von diversen Exoprodukten neben den QS-
Signalmolekülen in diesem Nährmedium lediglich einen unnötigen Energieverbrauch
darstellten (DIGGLE et al., 2007b).
4.2.5 Identifizierung der AHL-Signalmoleküle der heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 sowie des Cyanobakteriums Stamm Flo 1
Die Identifizierung der AHL-Moleküle erfolgte nach dünnschichtchromatographischer
Auftrennung durch einen Vergleich mit synthetischen Referenzmolekülen. Die
beschränkte Anzahl kommerzieller Standardmoleküle decken bei weitem nicht die
Bandbreite der in der Umwelt vorkommenden AHL-Signalmoleküle ab und ließen
oftmals eine Zuordnung nicht zu. Eine Zuordnung der DC-Signale über einen
Abgleich der Rf-Werte mit Rf-Werten aus der Literatur konnte nicht durchgeführt
werden, da die Referenzmoleküle von der Literatur abweichende Rf-Werte
aufzeigten. Dieses ist mit großer Wahrscheinlichkeit auf unterschiedlich verwendete
89

4. Diskussion
DC-Platten zurückzuführen. Ein abweichendes Laufverhalten hatte zur Folge, dass
sich die Anzahl der potentiellen AHL-Moleküle für einen detektierten DC-Spot
erhöhte. Des Weiteren weisen 3-oxo- und 3-hydroxy-Derivate mit gleicher
Seitenkettenlänge das gleiche dünnschichtchromatographische Laufverhalten auf, so
dass bei Auftreten beider Derivate eine Überlagerung der Signalmoleküle vorliegt
(SHAW et al., 1997).
Viele terrestrische Gram-negative Bakterien produzieren AHL-Moleküle. Im
Gegensatz dazu ist über das Vorkommen von AHL-Molekülen bei marinen Bakterien,
abgesehen von der Zelldichte-abhängigen Regulation der Lumineszenz bei Vibrio
fischeri und weiteren Vibrio-Spezies, noch relativ wenig bekannt (WAGNER-
DÖBLER et al., 2005; MOHAMMED et al., 2008). Bei marinen heterotrophen
Bakterien aus der Ostsee konnten BRUNS und Mitarbeiter (2002) einen
stimulierenden Effekt von AHL-Signalmolekülen auf die Kultivierung von
Laborkulturen aufzeigen. Es wurde daher eine wesentliche Rolle der AHLs bei der
Anpassung der marinen Bakterien an die Laborbedingungen vermutet. Des Weiteren
wurden AHL-Moleküle (C6-HSL, C8-HSL und 3-oxo-Derivate) bei Untersuchungen
mit einem Agrobacterium-Sensor bei 3 Roseobacter-Isolaten (Roseobacter-
Ruegeria-Untergruppe der α-Proteobakterien) aus “Marine Snow“ nachgewiesen
(GRAM et al., 2002). Eine AHL-Produktion in marinen Schwämmen durch assoziierte
Bakterien wurde ebenfalls auf einen Vertreter der Roseobacter-Ruegeria-
Untergruppe zurückgeführt (TAYLOR et al., 2004). Eine weite Verbreitung von AHL-
QS-Systemen bei marinen α-Proteobakterien, insbesondere aus der Roseobacter-
Gruppe, wurde durch Studien an bakteriellen Isolaten aus verschiedenen marinen
Habitaten (WAGNER-DÖBLER et al., 2005) und an marinen Schwämmen
(MOHAMMED et al., 2008) aufgezeigt, während letztere Arbeitsgruppe zudem für
zwei Isolate der Gattung Erythrobacter eine AHL-Produktion dokumentierte. Durch
Einsatz von verschiedenen bakteriellen Biosensoren konnten MOHAMMED und
Mitarbeiter (2008) für die α-Proteobakterien ein breites AHL-Spektrum, WAGNER-
DÖBLER und Mitarbeiter (2005) bei zusätzlicher Verwendung der GC-MS-Analyse
hingegen überwiegend langkettige AHL-Moleküle (C14, C16 und C18) nachweisen.
C8-HSL wurde zudem von letztgenannter Arbeitsgruppe bei zwei aeroben
anoxygenen phototrophen Bakterien der Gattungen Dinoroseobacter und
Roseobacter detektiert.
90

4. Diskussion
Das weitverbreitete Vorkommen von QS-Systemen in marinen α-Proteobakterien
weist auf eine große Bedeutung dieser Regulationssysteme zur Anpassung der
Organismen in ihren ökologischen Nischen hin. Für einige Proteobakterien wurde
dabei eine QS-regulierte Biofilmbildung vermutet, welche z.B. der Kolonisierung von
pflanzlichen und tierischen Oberflächen oder zur Bildung von “Marine Snow“ dient
(WAGNER-DÖBLER et al., 2005). Des Weiteren wurde z.B. für die aus
Dinoflagellaten isolierten Proteobakterien angenommen, dass QS in einer Regulation
symbiotischer Interaktionen involviert ist. Eine wesentliche Rolle wird den AHL-
Signalmolekülen bei Assoziationen zwischen Proteobakterien und höheren marinen
Organismen zugewiesen, wobei AHL-QS-Systeme wahrscheinlich der Etablierung
und Aufrechterhaltung solcher Assoziationen dienen. Die erhöhte
Nährstoffverfügbarkeit durch das Schwammgewebe ermöglicht dabei im Gegensatz
zum relativ nährstoffarmen Meerwasser das Erreichen hoher Populationsdichten und
somit das Einsetzen QS-regulierter Prozesse (MOHAMMED et al., 2008). Die
Bedeutung des QS in Proteobakterien zeigten CASE und Mitarbeiter (2008) in
Untersuchungen an einer Vielzahl bakterieller Genome auf, die ausschließlich für
eine große Anzahl von Proteobakterien luxI- und luxR-homologe Gene nachwiesen.
Für das heterotrophe marine α-Proteobakterium Porphyrobacter Stamm Bo 53-33,
nahe verwandt mit der Roseobacter-Gruppe, konnten mit dem Agrobacterium-
Sensorstamm zwei AHL-Moleküle nachgewiesen werden (siehe Kap. 3.5.1.1). Nach
Abgleich anhand mitgeführter Referenzmoleküle wurde ein AHL-Molekül mit kurzer
Seitenkette (C4 - C6) und ein weiteres mit kurzer bis mittellanger Seitenkette (C6 -
C8) bestätigt. Damit zeigt der Stamm Bo 53-33 im Vergleich zu anderen AHL-
produzierenden α-Proteobakterien (oftmals 4 - 6 verschiedene AHL-Moleküle)
(MOHAMMED et al., 2008) ein relativ einfaches AHL-Profil auf. Die nachgewiesenen
DC-Spots wiesen eine runde, distinkte Form auf und unterschieden sich damit von
den „tailing spots“, welche von 3-oxo-Derivaten gebildet werden (SHAW et al., 1997).
Da C6-HSL-Moleküle über den Chromobacterium-Sensorstamm nicht nachgewiesen
wurden, kann dieses AHL-Molekül für Stamm Bo 53-33 zudem mit großer
Wahrscheinlichkeit ausgeschlossen werden. Ein zusätzlicher Vergleich mit
Referenzdaten (SHAW et al., 1997; GEISENBERGER, 2000) lässt für das
längerkettige Molekül auf C7-HSL oder 3-hydroxy-C8-HSL und für das kurzkettige
Molekül auf C4-HSL oder 3-hydroxy-C6-HSL schließen. Diese AHL-Moleküle sind mit
91

4. Diskussion
Ausnahme von C4-HSL bei AHL-produzierenden Bakterienspezies selten
vorzufinden. Das 3-hydroxy-C8-HSL wurde bisweilen in Pseudomonas fluorescence
(SHAW et al., 1997) und Photobacterium phosphoreum (FLODGAARD et al., 2005)
detektiert.
Die Azidifizierung der Kulturüberstände zeigte des Weiteren auf, dass das
längerkettigere AHL-Molekül in etwa gleich großen Mengen in der logarithmischen
und stationären Wachstumsphase vorlag. Die geringen Auftragungsvolumina in den
DC-Analysen sprechen für eine hohe Sensitivität des Agrobacterium-Sensorstammes
gegenüber diesem AHL-Molekül. Für das kurzkettige AHL-Molekül konnte ein
deutlicher Nachweis mit dem Agrobacterium-Sensorstamm in der stationären
Wachstumsphase gezeigt werden. In der logarithmischen Wachstumsphase lag das
Signalmolekül hingegen lediglich in geringen Mengen vor. Dieses Molekül konnte
durch den C. violaceum-Sensorstamm weder bestätigt noch ausgeschlossen
werden, da ein Nachweis eines kurzkettigen AHL-Moleküls lediglich in der
logarithmischen Wachstumsphase dokumentiert wurde.
QS könnte bei Porphyrobacter Stamm Bo 53-33 vielleicht, wie bereits für
verschiedene Roseobacter-Stämme vermutet (GRAM et al., 2002), bei hohen
Zelldichten die Produktion von Exoenzymen, eine Antibiotika-Produktion oder eine
Biofilmbildung regulieren. Da die Aggregation bei verschiedenen Bakterienspezies
ein QS-regulierter Prozess ist (WILLIAMS, 2007), könnte die beobachtete Bildung
von Zellagglomeraten bei einem pH-Wert von 6,0 (siehe Kap. 3.2) vielleicht ebenfalls
auf eine AHL-abhängige Regulation zurückgeführt werden. Bei niedrigen Zelldichten
bzw. für Bakterien in einer freilebenden, planktonischen Form sind diese
phänotypischen Veränderungen hingegen kaum von Vorteil für die Population bzw.
das einzelne Bakterium und mit einem unnötigen Verbrauch an Energie verbunden.
Der heterotrophe Bakterienstamm Bo 10-09 wurde nach 16S-rDNA-Sequenzierung
der Cytophaga-Flavobacterium-Bacteroides-Gruppe (CFB-Gruppe) zugeordnet
(HUBE, 2008). In dieser Gruppe sind bisweilen kaum AHL-Produzenten bekannt.
BRUNS und Mitarbeiter (2003) konnten in Kultivierungsversuchen keinen
stimulierenden Einfluss auf das Wachstum von Isolaten der CFB-Gruppe durch
synthetische AHL-Moleküle (C4-HSL und 3-oxo-C6-HSL) dokumentieren. In diesem
Zusammenhang konnten GRAM und Mitarbeiter (2002) ebenfalls bei keinem
kultivierten Isolat der CFB-Gruppe einen AHL-Nachweis über einen Agrobacterium-
92

4. Diskussion
Sensor erzielen, obwohl Bakterien der CFB-Gruppe neben den Proteobakterien eine
dominierende Gruppe in marinen Partikeln darstellen (GRAM et al., 2002). Das
Erreichen hoher Zellpopulationsdichten könnte das Einleiten QS-regulierter Prozesse
ermöglichen. Lediglich WAGNER-DÖBLER und Mitarbeiter (2005) dokumentierten
für einen Flavobacterium-Stamm einen positiven AHL-Nachweis mit einem
Biosensor, sensitiv für kurzkettige AHL-Moleküle. DOBRETSOV und Mitarbeiter
(2007) konnten jedoch durch Einsatz sogenannter QS-Blocker (Derivate des
Furanons) aufzeigen, dass die Signalmoleküle bei Vertretern der CFB-Gruppe eine
Erniedrigung der Zelldichten in bakteriellen Biofilmen auslösten bzw. eine Bildung
von Biofilmen inhibierten und dementsprechend Einfluss auf die Zusammensetzung
bakterieller Gemeinschaften nehmen. Dieses lässt schlussfolgern, dass AHL-
Signalmoleküle bzw. QS-gesteuerte Prozesse in der CFB-Gruppe keine Ausnahmen
darstellen.
In der vorliegenden Arbeit konnten mit dem Agrobacterium-Sensorstamm für
Muricauda Stamm Bo 10-09 mehrere AHL-Signalmoleküle detektiert werden. Dieses
erforderte aber eine Azidifizierung der Kulturüberstände und ein Auftragen sehr
hoher Volumina an AHL-Extrakten für die DC-Analysen (siehe Kap. 3.5.1.1). Dieses
lässt sich entweder auf eine geringe AHL-Produktion durch Stamm Bo 10-09 oder
auf eine geringe Sensitivität des Biosensors gegenüber diesen Molekülen
zurückführen. Nach Abgleich anhand der mitgeführten Referenzmoleküle wurde ein
nachgewiesenes AHL-Molekül als C8-HSL identifiziert. Neben diesem Molekül
konnten wahrscheinlich zwei weitere kurzkettigere AHL-Moleküle mit C6 - C8- bzw.
C4-Seitenketten für den Stamm Bo 10-09 dokumentiert werden, wobei das Fehlen
distinkter DC-Spots eine Zuordnung über einen Abgleich mit den Standards nur
bedingt zuließ. Der wesentlich höhere Gehalt an potentiellen C8-HSL in der
Stationärphase ist auf die höheren Zelldichten in dieser Wuchsphase
zurückzuführen, die Abnahme an kurzkettigen AHL-Molekülen in dieser Phase ist
jedoch zu hinterfragen. Eine QS-Regulation in verschiedenen Wuchsphasen kann für
Stamm Bo 10-09 nur vermutet werden. Ein über den Chromobacterium-
Sensorstamm nachgewiesenes kurzkettiges AHL-Molekül scheint nach Abgleich mit
den Referenzmolekülen keines der AHL-Moleküle zu entsprechen, welche mit dem
Agrobacterium-Sensor nachgewiesen wurden.
93

4. Diskussion
Cyanobakterien stehen im Gegensatz zu den heterotrophen Bakterien kaum im
Fokus von AHL-Untersuchungen. Dennoch konnten AHL-Moleküle in natürlichen
mikrobiellen Gemeinschaften nachgewiesen werden, in denen Cyanobakterien
auftreten bzw. diese sogar dominieren (mikrobielle Matten, cyanobakterielle Blüten)
(MCLEAN et al., 1997; BRAUN und BACHOFEN, 2004). Einen Zusammenhang
zwischen einer Microcystin-Synthese bei Cyanobakterien und AHL-Molekülen
konnten BRAUN und BACHOFEN (2004) jedoch nicht eindeutig aufzeigen.
ROMERO und Mitarbeiter (2006) dokumentierten eine AHL-Produktion durch den
axenischen Anabaena sp.-Stamm PCC 7120, wobei ausschließlich in der
cyanobakteriellen Biomasse AHL-Moleküle detektiert wurden. Es wurde basierend
auf einer Studie von YOON und GOLDEN (1998) angenommen, dass die AHL-
Moleküle (überwiegend 3-hydroxy-C12-HSL) an der Heterocystenbildung oder an
Prozessen der Stickstofffixierung beteiligt sind. Ein Ausbleiben von AHL-Molekülen in
der stationären Wuchsphase führten ROMERO und Mitarbeiter (2008)
wahrscheinlich auf eine enzymatische Degradation durch die AHL-Acylase AiiC
(auto-inducer inhibitor from Cyanobacteria) zurück. Des Weiteren wird zur
Vermeidung von Interferenzen eine intrazelluläre Degradation Spezies-fremder AHL-
Moleküle durch AiiC angenommen.
Bei Untersuchungen cyanobakterieller Kulturen von Stamm Flo 1 konnten zwei sehr
schwache AHL-Nachweise in Kulturüberständen nach 14 bzw. 21 Tagen Inkubation
mit dem Agrobacterium-Sensorstamm dokumentiert werden, wobei ähnliche AHL-
Gehalte in beiden Wuchsphasen vorlagen (siehe Kap. 1.1.1). Nach Abgleich mit
Referenzmolekülen wurde ein längerkettiges AHL-Molekül als C8-HSL und ein
substituiertes oder nicht-substituiertes AHL-Molekül mit einer C6- - C8-Seitenkette
identifiziert. Eine Azidifizierung von Stamm Flo 1-Kulturüberständen führte zu einem
stärkeren Nachweis des potentiellen C8-HSL und des kurzkettigeren Moleküls durch
Rückführung in die aktive Form der AHL-Moleküle (YATES et al., 2002). Die
geringen AHL-Gehalte ließen sich teilweise durch den hohen pH-Wert des
standardmäßig verwendeten Kulturmediums und der Flo 1-Kulturen (pH 9,0)
erklären. Ein zu geringer Einsatz cyanobakterieller Biomasse kann zudem nicht
ausgeschlossen werden. Durch eine Aufkonzentrierung von AHL-Molekülen aus
größeren Volumina an Kulturüberständen von Stamm Flo 1 wurde ein deutlich
stärkerer Nachweis für das potentielle C8-HSL erzielt und zudem ein drittes AHL-
Molekül, vermutlich C4-HSL, nachgewiesen. Die Inkubation von Flo 1-Kulturen für
94

4. Diskussion
8 Wochen und die Verwendung von nicht-axenischen Kulturen lassen jedoch
vermuten, dass die AHL-Produktion nicht auf Stamm Flo 1 sondern auf heterotrophe
Begleitorganismen zurückzuführen ist. Der cyanobakterielle Detritus bildet dabei
vielleicht die Grundlage für ein stärkeres heterotrophes Wachstum und das Erreichen
höherer Zelldichten der heterotrophen Bakterien. Gestützt könnte diese Annahme
durch intrazelluläre, aber nicht gesicherte AHL-Nachweise einer
21 Tage lang inkubierten Flo 1-Kultur werden (siehe Kap. 3.5.5). Nach Abgleich mit
den Referenzmolekülen und in Anbetracht des AHL-Nachweisspektrums von
Agrobacterium tumefaciens NTL4 (pZLR4) (FARRAND et al., 2002) weisen die DC-
Signale auf ein langkettiges AHL-Molekül (mindestens eine C12-Seitenkette) und ein
kurzkettiges AHL-Molekül, vermutlich 3-oxo-C4-HSL oder 3-hydroxy-C4-HSL, hin.
Das Ausbleiben dieser AHL-Moleküle in den Kulturüberständen von Stamm Flo 1
entspräche den bereits geschilderten Beobachtungen von ROMERO und
Mitarbeitern (2006, 2008) bei einem Anabaena sp.-Stamm.
Aussagen über mögliche Interspezies-Kommunikationen in assozzierten
Lebensgemeinschaften von Cyanobakterien und heterotrophen Bakterien können
anhand der erzielten Ergebnisse kaum getroffen werden. Zum einen bringt der
Nachweis von AHL-Molekülen in Überständen aus nicht-axenischen
Cyanobakterienkulturen keinen Aufschluss über den eigentlichen AHL-Produzenten
und zum anderen sind die untersuchten heterotrophen Stämme nicht wesentlicher
Bestandteil der heterotrophen Begleitflora von Stamm Flo 1 (HUBE, 2008,
persönliche Mitteilung). Dennoch ist vorstellbar, dass die produzierten
Signalmoleküle heterotropher Bakterien Einfluss auf physiologische Prozesse von
Stamm Flo 1 nehmen können (unidirektionale Interspezies-Kommunikation). CASE
und Mitarbeiter (2008) konnten bei einer Vielzahl proteobakterieller Genome
unvollständige QS-Systeme (Fehlen des luxI-homologen Gens) bzw. eine höhere
Anzahl von luxR-Homologen gegenüber luxI-Homologen dokumentieren. Diese
„zusätzlichen“ LuxR-homologen Proteine ermöglichen Bakterien (z.B. Brucella
melitensis, Salmonella typhimurium und Escherichia coli) Spezies-fremde QS-
Signale wahrzunehmen und in Abhängigkeit dieser Signalmoleküle physiologische
Prozesse zu modulieren. Eine soziale Intelligenz wurde unter Bakterien auf diesem
Niveau bis vor kurzem noch nicht vermutet.
Die hohen pH-Werte in Kulturen von Stamm Flo 1 (SCHRÜBBERS, 2007) sprechen
indes gegen eine interspezifische Kommunikation über AHL-Moleküle aufgrund der
95

4. Diskussion
Instabilität der Moleküle im alkalischen Bereich. Des Weiteren erreichen heterotrophe
Bakterienspezies in (vitalen) nicht-axenischen Cyanobakterienkulturen lediglich
geringe Zelldichten. QS-gesteuerte Prozesse werden dementsprechend aufgrund
einer zu niedrigen „Autoinducer“-Konzentration nicht eingeleitet.
4.3 Molekularbiologie Die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 wurden aus nicht-
axenischen Cyanobakterienkulturen isoliert. Diese Cyanobakterienstämme Bo 53
und Bo 10 wurden in der vorliegenden Arbeit zur phylogenetischen Einordnung
molekularbiologisch untersucht und auf eine bereits vorgenommene taxonomische
Einordnung überprüft.
Basierte die Klassifizierung von Cyanobakterien hauptsächlich auf morphologischen
Kriterien, sind mittlerweile neben phänotypischen, physiologischen sowie
biochemischen Analysen insbesondere molekularbiologische grundlegend für eine
taxonomische Einordnung von Cyanobakterien und dem Aufzeigen evolutionärer
Verbindungen. Zur Identifikation von Bakterien und Erstellung von phylogenetischen
Stammbäumen wird standardmäßig die 16S-rDNA-Sequenz der Mikroorganismen
herangezogen (SEO und YOKOTA, 2003).
Die 16S-rDNA-Analyse des Cyanobakterienstammes Bo 53 belegte eine
Übereinstimmung von 99,85% mit der bereits in der NCBI-Datenbank hinterlegten
16S-rDNA-Sequenz des Stammes Nodularia harveyana Bo 53 (LYRA et al., 2005).
16S-rDNA-Sequenzen mit einer Homologie von mindestens 97% weisen
Organismen gewöhnlich einer Spezies zu (STACKEBRANDT und GÖBEL, 1994).
Eine Einordnung auf Speziesebene sollte über eine DNA-DNA-Hybridisierung
abgesichert werden, da die 16S-rDNA aufgrund zu geringer Sequenz-Variabilität
hierfür unzureichend ist (FOX et al., 1992; STACKEBRANDT und GÖBEL, 1994).
Eine Verwandtschaftsbeziehung auf Speziesebene setzt dabei eine DNA-
Reassoziation von mindestens 70% voraus (WAYNE et al., 1987). In diesem
Zusammenhang empfehlen STACKEBRANDT und EBERS (2006), basierend auf
publizierten Daten von 16S-rDNA-Analysen und DNA-DNA-Hybridisierungen, eine
Revidierung der Spezies-Grenze auf 98,7 - 99% bzw. definieren diesen Bereich als
denjenigen, welcher DNA-DNA-Hybridisierungsversuche zur Einordnung auf
Spezies-Ebene zwingend erforderlich macht. Demzufolge kann Stamm Bo 53 mit
einer Sequenzhomologie von 98,94 bzw. 99,70 % zu Stämmen von Nodularia
96

4. Diskussion
harveyana dieser Spezies zugeordnet werden (siehe Kap. 3.6.3). Die beiden in sehr
enger Verwandtschaftsbeziehung stehenden Stämme Bo 53 und BECID27 bilden
dabei in dem phylogenetischen Stammbaum ein abgegrenztes Cluster. Eine
Zuweisung auf Speziesebene von Stamm Bo 53 sollte dennoch über eine DNA-DNA-
Hybridisierung zwischen Stamm Bo 53 und Nodularia harveyana Stamm BECID27
bestätigt werden.
Generell werden Unterschiede von 5 - 7% (93 - 95% Identität) in den 16S-rDNA-
Sequenzen zur Unterstützung für neue Gattungen herangezogen (MADIGAN et al.,
2001). Der Cyanobakterienstamm Bo 10 wurde ursprünglich ausschließlich auf
Grundlage morphologischer Merkmale als Oscillatoria brevis klassifiziert
(RETHMEIER, 1995). Die aufgezeigten nahen Verwandtschaftsbeziehungen zu
diversen Phormidium-Spezies über 16S-rDNA-Analysen machen jedoch eine
Einordnung in die Gattung Phormidium wahrscheinlich, obwohl auch 98%
Sequenzhomologie mit einer Oscillatoria-Spezies vorlagen (siehe Kap. 3.6.3). Der
phylogenetische Stammbaum zeigte indes keine nahe Verwandtschaftsbeziehung
von Stamm Bo 10 zu den Geitlerinema-Stämmen Flo 1 und PCC 7105 auf.
Unterstützend zur 16S-rDNA-Sequenzanalyse sollten weitere molekulare Marker wie
das Gen gyrB und die ITS (internal transcribed spacer) untersucht werden. Die ITS-
Sequenzen sind zwischen den rRNA-Genen (16S-rRNA-, 23S-rRNA- und 5S-rRNA-
Gen) lokalisiert (ITEMAN et al., 2000), wobei molekularbiologische Untersuchungen
sich überwiegend auf die intergenische 16S-rRNA-23S-rRNA-Region beschränken.
Anzahl und Größe der ITS-Regionen stellen einen zusätzlichen taxonomischen
Marker für die phylogenetische Klassifizierung cyanobakterieller Stämme dar
(ITEMAN et al., 2002). Ein identisches ITS-Bandenmuster bei nicht-verwandten
Organismen ermöglicht jedoch keine Unterscheidung. Die im Vergleich zu den
Geitlerinema-Stämmen unterschiedliche Anzahl von ITS-Amplifikaten bei Stamm
Bo 10 schloss eine Einordnung in die Gattung Geitlerinema aus (siehe Kap. 3.6.5).
Eine ITS-Sequenzanalyse kann des Weiteren eine Unterscheidung von
Cyanobakterienstämmen auf intra- oder interspezifischer Ebene ermöglichen
(TATON et al., 2003). Die ITS-Sequenz weist dabei gegenüber funktionellen Genen
(z.B. das 16S-rRNA-Gen) eine wesentlich höhere molekulare Evolutionsrate auf
(LEBLOND-BOURGET et al., 1996).
97

5. Ausblick
5. Ausblick
In der vorliegenden Arbeit sind einige Fragen bezüglich des „Quorum Sensing“ bei
Gram-negativen Bakterien offen geblieben, deren Beantwortung teilweise weiterer
Methoden bedarf. Die erzielten Ergebnisse führten zu einer Vielzahl neuer
Fragestellungen und bieten gleichermaßen die Grundlage für weitere QS-
Untersuchungen.
Eine AHL-Produktion in Stamm Flo 1-Kulturen konnte nicht gesichert auf das
Cyanobakterium zurückgeführt werden. Fortführende QS-Untersuchungen an Stamm
Flo 1 sollten mit axenischen Kulturen und ausschließlich intrazellulär durchgeführt
werden. Mit dem Hintergrund der sehr schwachen AHL-Nachweise in Flo 1-Kulturen
ist des Weiteren eine Anzucht im größeren Maßstab erforderlich.
Unter den heterotrophen Bakterien zeichnet sich insbesondere der Porphyrobacter-
Stamm aufgrund der deutlichen AHL-Nachweise für weitere QS-Untersuchungen
aus. Die erzielten Ergebnisse bauen nahezu ausschließlich auf Grundlage eines
Biosensors (Agrobacterium-Sensorstamm) auf. Dementsprechend sollten zusätzlich
spezifischere Biosensoren wie Pseudomonas fluorescens 1855 (pSF105, pSF107)
eingesetzt werden. Dieser Sensorstamm reagiert ausschließlich auf
3-hydroxy-Derivate, welche bei Stamm Bo 53-33 vermutet wurden. Des Weiteren
sollte bezüglich einer Interspezies-Kommunikation in assoziierten
Lebensgemeinschaften von Cyanobakterien und heterotrophen Bakterien eine
Untersuchung auf AI-2-Signalmoleküle erfolgen.
Die Bandbreite der Verfahren zur Untersuchung von AHL-Signalmolekülen ist relativ
eingeschränkt. In den letzten Jahren fanden aber verstärkt MS-gekoppelte
Kapillartechniken in der AHL-Analytik Anwendung. Zur Identifizierung und
strukturchemischen Charakterisierung von AHL-Signalmolekülen sind analytische
Verfahren wie die hochauflösende GC-MS erforderlich, deren Detektionsgrenzen im
nmolaren bis pmolaren Bereich liegen. Alternativ ist eine Strukturanalyse auch mit
der HPLC-MS möglich, wobei das Trennverfahren, die Sensitivität und die
strukturelle Aufklärung nicht dem Niveau der GC-MS entspricht.
Neben der AHL-Produktion ist eine Hinterfragung der AHL-QS-regulierten
Phänotypen bzw. physiologischen Prozesse von großem Interesse. In diesem
Zusammenhang könnte eine Möglichkeit in der Koppelung von
Kultivierungsversuchen mit molekularbiologischen Analysen liegen. Die
98

5. Ausblick
Heterogenität der AHL-Synthase-Gene ermöglicht aber keine Entwicklung von PCR-
Standardprotokollen zur direkten Bestimmung der AHL-Produktionsfähigkeit bei
Mikroorganismen auf molekularer Ebene. Ein Vergleich der Proteinmuster in
verschiedenen Wuchsphasen der Bakterien mittels 2D-PAGE könnte einen Ansatz
bieten, wobei Rückschlüsse auf QS-regulierte Proteine möglich wären. Auf
Transkriptionsebene wurde des Weiteren eine Expression QS-gesteuerter Gene über
eine unterschiedlich stark exprimierte mRNA aufgezeigt (SHIN et al., 2007).
Für eine gesicherte Einordnung des Cyanobakterienstammes Bo 10 in das Genus
Phormidium sollten unterstützend zu den bisherigen phylogenetischen
Untersuchungen eine Sequenzanalyse der ITS-Regionen, eine nochmalige gyrB-
Gen-Analyse und eine Analyse des Fettsäuremusters erfolgen.
99

6. Zusammenfassung
6. Zusammenfassung
1. Die heterotrophen Bakterienstämme Bo 53-33 und Bo 10-09 wachsen in einem
pH-Bereich von 6,0 - 9,0 und können daher als neutrophile Bakterien beschrieben
werden. Der zum Wachstum optimale pH-Wert liegt für Stamm Bo 53-33 bei 7,0 und
für Stamm Bo 10-09 bei 8,0. Bei einem Photonenfluss von 5 µE m-2 s-1 PAR wiesen
die heterotrophen Bakterienstämme kein verändertes Wachstumsverhalten
gegenüber einer Dunkelinkubation auf. Die gepufferten und ungepufferten
Versuchskulturen waren nach Beendigung der Inkubation durch eine Alkalisierung
des Kulturmediums gekennzeichnet.
2. Der Agrobacterium-Sensorstamm erwies sich im Vergleich zum Chromobacterium-
Sensorstamm aufgrund deutlich höherer Sensitivitäten gegenüber den synthetischen
Referenzmolekülen C8-HSL und C4-HSL für den Nachweis von AHL-
Signalmolekülen als wesentlich geeigneter.
3. Die Screeningtests mit dem Agrobacterium-Sensorstamm zum Nachweis AHL-
produzierender Bakterien wiesen Stamm Bo 53-33 als deutlichen AHL-Produzenten
aus. Für den Stamm Bo 10-09 wurde ein schwacher AHL-Nachweis erzielt, für das
Cyanobakterium Stamm Flo 1 wurden hingegen keine AHL-Signalmoleküle
nachgewiesen.
4. Auf Grundlage bereits bestehender Methoden wurde eine dünnschicht-
chromatographische Methode zum Nachweis von AHL-Signalmolekülen mit den
Biosensor Agrobacterium tumefaciens NTL4 (pZLR4) etabliert.
5. Mit zunehmendem pH-Wert (von pH 6,0 bis 9,0) oder bei einer
Temperaturerhöhung von 21 °C auf 28 °C nahm die mit dem Agrobacterium-
Sensorstamm nachgewiesene AHL-Menge in Kulturüberständen von Stamm
Bo 53-33 ab. Bei Stamm Bo 10-09 konnte in gepufferten Versuchskulturen lediglich
bei pH 6,0 ein AHL-Nachweis mit dem Agrobacterium-Sensorstamm erzielt werden.
Die Azidifizierung (pH < 2) von Kulturüberständen der Gram-negativen Bakterien
führte zu einem deutlich stärkeren Nachweis an AHL-Molekülen. Die Stabilität der
AHL-Signalmoleküle ist demnach abhängig vom pH-Wert des Kulturmediums und
100

6. Zusammenfassung
der Temperatur. Der Azidifizierungsversuch konnte insbesondere für Stamm
Bo 53-33 zeigen, dass sich die AHL-Stabilität wahrscheinlich mit abnehmender Acyl-
Seitenkettenlänge verringert.
6. Ein Photonenfluss von 5 µE m-2 s-1 PAR hatte keinen nachweisbaren Einfluss auf
die AHL-Produktion durch die heterotrophen Bakterienstämme Bo 53-33 und
Bo 10-09. Eine Reduzierung der Kohlenstoffquelle im Kulturmedium führte
wahrscheinlich zu einer veränderten AHL-Produktion durch die heterotrophen
Bakterienstämme.
7. Ein intrazellulärer AHL-Nachweis für die heterotrophen Bakterien wurde bei
Stamm Bo 53-33 erzielt, wobei das detektierte AHL-Molekül
dünnschichtchromatographisch identisch ist mit einem nachgewiesenen AHL-Molekül
in den Kulturüberständen. Bei Stamm Bo 10-09 konnten intrazellulär keine AHL-
Moleküle nachgewiesen werden.
8. Für Stamm Bo 53-33 konnte ein AHL-Signalmolekül mit kurzer Acyl-Seitenkette
(C4 - C6) und ein weiteres mit kurzer bis mittellanger Acyl-Seitenkette (C6 - C8)
dokumentiert werden. Für den schwachen AHL-Produzenten Stamm Bo 10-09
wurden neben C8-HSL vermutlich zwei weitere AHL-Signalmoleküle mit einer
C6 - C8-Seitenkette bzw. C4-Seitenkette nachgewiesen. 3-oxo-Derivate konnten
aufgrund fehlender „tailing spots“ der DC-Signale ausgeschlossen werden.
9. In Kulturüberständen des Cyanobakteriums Stamm Flo 1 wurden sehr geringe
AHL-Mengen mit dem Agrobacterium-Sensorstamm nachgewiesen, wobei vermutlich
drei verschiedene Signalmoleküle mit kurzer bzw. mittellanger Acyl-Seitenkette
vorlagen. Die Ergebnisse konnten durch zwei intrazellulär erzielte AHL-Nachweise
nicht bestätigt werden. Der Stamm Flo 1 konnte nicht gesichert als AHL-Produzent
beschrieben werden, da eine AHL-Produktion durch heterotrophe Begleitflora (nicht-
axenische Kulturen) nicht ausgeschlossen werden kann.
101

6. Zusammenfassung
10. Die 16S-rDNA-Sequenz des Cyanobakterienstammes Bo 53 zeigte durch
Vergleich mit Referenzsequenzen aus der NCBI-Datenbank eine Sequenzhomologie
von 99,70% mit Nodularia harveyana Stamm BECID27 auf. Die bereits
vorgenommene Einordnung zu Nodularia harveyana konnte damit bestätigt werden.
11. Der Cyanobakterienstamm Bo 10 wurde bisher auf Grundlage von
morphologischen Merkmalen als Oscillatoria brevis klassifiziert (RETHMEIER, 1995).
Nach 16S-rDNA-Analyse wurden die größten Übereinstimmungen mit Vertretern der
Gattung Phormidium und hier mit Phormidium pseudopristleyi Stamm ANT.ACEV5.3
(99,32%) dokumentiert. Folglich kann Stamm Bo 10 der Gattung Phormidium
zugeordnet werden. Nach einer Amplifikation der ITS konnte für Stamm Bo 10
gegenüber den Geitlerinema-Stämmen Flo 1 und PCC 7105 eine höhere Anzahl an
ITS-Regionen dokumentiert werden, welches eine Einordnung in das Genus
Geitlerinema ausschloss.
102
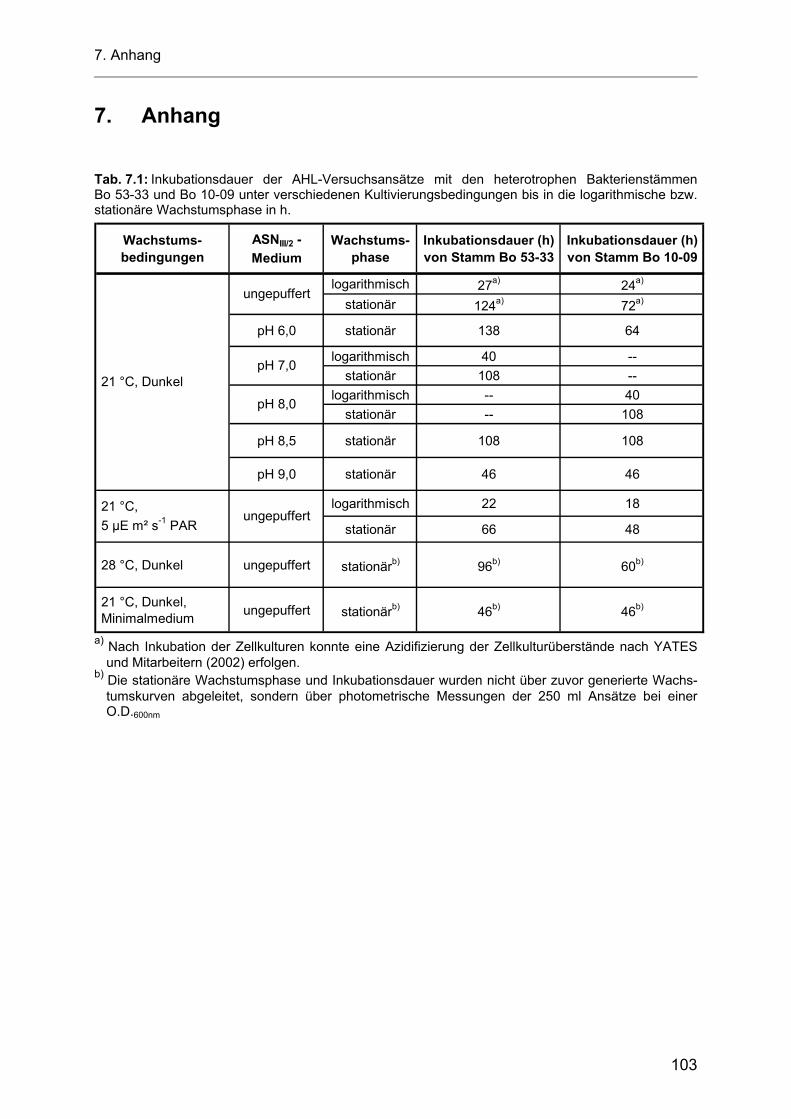
7. Anhang
7. Anhang
Tab. 7.1: Inkubationsdauer der AHL-Versuchsansätze mit den heterotrophen Bakterienstämmen Bo 53-33 und Bo 10-09 unter verschiedenen Kultivierungsbedingungen bis in die logarithmische bzw. stationäre Wachstumsphase in h.
Wachstums-bedingungen
ASNIII/2 - Medium
Wachstums-phase
Inkubationsdauer (h)von Stamm Bo 53-33
Inkubationsdauer (h)von Stamm Bo 10-09
logarithmisch 27a) 24a)
stationär 124a) 72a)
pH 6,0 stationär 138 64
logarithmisch 40 --stationär 108 --
logarithmisch -- 40stationär -- 108
pH 8,5 stationär 108 108
pH 9,0 stationär 46 46
logarithmisch 22 18
stationär 66 48
28 °C, Dunkel ungepuffert stationärb) 96b) 60b)
21 °C, Dunkel, Minimalmedium ungepuffert stationärb) 46b) 46b)
21 °C, Dunkel
21 °C, 5 µE m² s-1 PAR
pH 7,0
pH 8,0
ungepuffert
ungepuffert
a) Nach Inkubation der Zellkulturen konnte eine Azidifizierung der Zellkulturüberstände nach YATES und Mitarbeitern (2002) erfolgen.
b) Die stationäre Wachstumsphase und Inkubationsdauer wurden nicht über zuvor generierte Wachs-tumskurven abgeleitet, sondern über photometrische Messungen der 250 ml Ansätze bei einer O.D.600nm
103

7. Anhang
Tab. 7.2: Sequenzausschnitt der 16S-rDNA des Nodularia harveyana-Stammes Bo 53 (1333 bp)
AGTAACGCGTGAGAATCTAGCTTCAGGTCGGGGACAACCACGGGAAACTGTGGCTAATACCGGATATGCC GAGAGGTGAAAGGCTTGCTGCCTGAAGATGAGCTCGCGTCTGATTAGCTAGTTGGTGTGGTAAGAGCGCA CCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGGACTGAGACACGGCCCAGACT CCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAATACCGCGTGAGGG AGGAAGGCTCTTGGGTTGTAAACCTCTTTTCTCAGGGAAGAAAAAAATGACGGTACCTGAGGAATAAGCA TCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGATGCAAGCGTTATCCGGAATGATTGGGCGTA AAGGGTCCGCAGGTGGCCATGTAAGTCTGCTGTTAAAGAATCTAGCTCAACTAGATAAAAGCAGTGGAAA CTACACGGCTAGAGTGCGTTCGGGGTAGAGGGAATTCCTGGTGTAGCGGTGAAATGCGTAGATATCAGGA AGAACACCAGTGGCGAAGGCGCTCTACTAGGCCGCAACTGACACTGAGGGACGAAAGCTAGGGGAGCGAA TGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGATACTAGGCGTGGCTTGTATCGACCCGAGC CGTGCCGTAGCTAACGCGTTAAGTATCCCGCCTGGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATT GACGGGGGCCCGCACAAGCGGTGGAGTATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCAAGACT TGACATGTCGCGAATCTTTCTGAAAGGAGAGAGTGCCTTAGGGAGCGCGAACACAGGTGGTGCATGGCTG TCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGTTTTTAGTTGCCAG CATTAAGTTGGGCACTCTAGAGAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAGTCAG CATGCCCCTTACGTCTTGGGCTACACACGTACTACAATGCTACGGACAAAGGGCAGCTACACAGCAATGT GATGCAAATCTCAGAAACCGTAGCTCAGTTCAGATCGCAGGCTGCAACTCGCCTGCGTGAAGTAGGAATC GCTAGTAATTGCAGGTCAGCATACTGCAGTGAATTCGTTCCCGGGCCTTGTACACACCGCCCGTCACACC ATGGAAGCTGATCACGCCCGAAGTCGTTACCCCAACTTTTAGGAGAGGGGGATGCCGAAG
Tab. 7.3: Sequenzausschnitt der 16S-rDNA der Oscillatoria brevis Stamm Bo 10 (1336 bp)
GGGTGAGTAACACGTGAGAATCTGCCTCCAGGTCGGGGACAACAGCGGGAAACTGCTGCTAATACCCGAT GTGCCTAAGGGTGAAAGATTAATTGCCTGGAGATGAGCTCGCGTCCGATTAGCTAGTTGGTAGAGTAAAA GCCTACCAAGGCTCCGATCGGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCC AGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACCGCGT GGGGGATGAAGGCTCTTGGGTTGTAAACCCCTTTTCTCAGGGAAGAATAACTGACGGTACCTGAGGAATA AGCATCGGCTAACTCCGTGCCAGCAGCCGCGGTAAGACGGAGGATGCAAGCGTTATCCGGAATTATTGGG CGTAAAGCGTTCGTAGGTGGCTGTTCAAGTCTGCCGTTAAAGACCGGGGCTTAACTCCGGAAAAACTGTG GAAACTGAACAGCTAGAGTATGGTAGGGGTAAGGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATC GGGAAGAACACCGGTGGCGAAGGCGCCTTACTGGGCCATAACTGACACTGAGGGACGAAAGCTAGGGGAG CGAAAGGGATTAGATACCCCTGTAGTCCTAGCTGTAAACGATGGATACTAGGTGTTGCCCGTATCGACCC GGGCAGTGCCGTAGCTAACGCGTTAAGTATCCCGCCTGGGGAGTACGCACGCAAGTGTGAAACTCAAAGG AATTGACGGGGGCCCGCACAAGCGGTGGAGTATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCAG GGCTTGACATGTCCAGAATCTTGGGGAAACTTGAGAGTGCCTTCGGGAGCTGGAACACAGGTGGTGCATG GCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGTCCTTAGTTG CCATCATTAAGTTGGGCACTTTAGGGAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAG TCAGCATGCCCCTTACGTCCTGGGCTACACACGTACTACAATGGGAAGGACAGAGGGTAGCAAGCGCGCG AGTGCAAGCCAATCCCATAAACCTTTCCTCAGTTCAGATTGCAGGCTGCAACTCGCCTGCATGAAGGCGG AATCGCTAGTAATCGCAGGTCAGCATACTGCGGTGAATACGTTCCCGGGCCTTGTACACCCCGCCCGTCA CACCATGGAAGTTGGCCATGCCCGAAGTCATTACCCTAACCGCAAGGAGGGGGATGCCGAAGGCAGGCTA GTACGT
104

8. Literatur
8. Literatur
AHMED NA, PETERSEN FC, SCHEIE AA (2007) AI-2 quorum sensing affects antibiotic susceptibility in Streptococcus anginosus. J Antimicrob Chemother 60: 49-53
BAINTON NJ, STEAD P, CHHABRA SR, BYCROFT BW, SALMOND GP, STEWART GS, WILLIAMS P (1992) N-(3-oxohexanoyl)-L-homoserine lactone regulates carbapenem antibiotic production in Erwinia carotovora. Biochem J 288 (Pt 3): 997-1004
BASSLER BL, WRIGHT M, SILVERMAN MR (1994) Multiple signalling systems controlling expression of luminescence in Vibrio harveyi: sequence and function of genes encoding a second sensory pathway. Mol Microbiol 13: 273-286
BEN JACOB E, BECKER I, SHAPIRA Y, LEVINE H (2004) Bacterial linguistic communication and social intelligence. Trends Microbiol 12: 366-372
BJARNSHOLT T, GIVSKOV M (2007) Quorum-sensing blockade as a strategy for enhancing host defences against bacterial pathogens. Philos Trans R Soc Lond B Biol Sci 362: 1213-1222
BOBROV AG, BEARDEN SW, FETHERSTON JD, KHWEEK AA, PARRISH KD, PERRY RD (2007) Functional quorum sensing systems affect biofilm formation and protein expression in Yersinia pestis. Adv Exp Med Biol 603: 178-191
BOETTCHER KJ, RUBY EG (1995) Detection and quantification of Vibrio fischeri autoinducer from symbiotic squid light organs. J Bacteriol 177: 1053-1058
BRAUN E, BACHOFEN R (2004) Homoserine-lactones and microcystin in cyanobacterial assemblages in Swiss lakes. Hydrobiologia 522: 271-280
BRUNS A, CYPIONKA H, OVERMANN J (2002) Cyclic AMP and acyl homoserine lactones increase the cultivation efficiency of heterotrophic bacteria from the central Baltic Sea. Appl Environ Microbiol 68: 3978-3987
BRUNS A, NUBEL U, CYPIONKA H, OVERMANN J (2003) Effect of signal compounds and incubation conditions on the culturability of freshwater bacterioplankton. Appl Environ Microbiol 69: 1980-1989
BRUNS A, ROHDE M, BERTHE-CORTI L (2001) Muricauda ruestringensis gen. nov., sp. nov., a facultatively anaerobic, appendaged bacterium from German North Sea intertidal sediment. Int J Syst Evol Microbiol 51: 1997-2006
BYERS JT, LUCAS C, SALMOND GP, WELCH M (2002) Nonenzymatic turnover of an Erwinia carotovora quorum-sensing signaling molecule. J Bacteriol 184: 1163-1171
105

8. Literatur
CAMILLI A, BASSLER BL (2006) Bacterial small-molecule signaling pathways. Science 311: 1113-1116
CASE RJ, LABBATE M, KJELLEBERG S (2008) AHL-driven quorum-sensing circuits: their frequency and function among the Proteobacteria. ISME J 2: 345-349
CHA C, GAO P, CHEN YC, SHAW PD, FARRAND SK (1998) Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Mol Plant Microbe Interact 11: 1119-1129
CHEN X, SCHAUDER S, POTIER N, VAN DORSSELAER A, PELCZER I, BASSLER BL, HUGHSON FM (2002) Structural identification of a bacterial quorum-sensing signal containing boron. Nature 415: 545-549
CHILTON MD, CURRIER TC, FARRAND SK, BENDICH AJ, GORDON MP, NESTER EW (1974) Agrobacterium tumefaciens DNA and PS8 bacteriophage DNA not detected in crown gall tumors. Proc Natl Acad Sci USA 71: 3672-3676
DAVIES DG, PARSEK MR, PEARSON JP, IGLEWSKI BH, COSTERTON JW, GREENBERG EP (1998) The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280: 295-298
DAWSON RMC, ELLIOTT DC, ELLIOTT WH, JONES KM (1986) Data for Biochemical Research. 3. Auflage. Oxford University Press, Oxford, Great Britain
DEGRASSI G, DEVESCOVI G, SOLIS R, STEINDLER L, VENTURI V (2007) Oryza sativa rice plants contain molecules that activate different quorum-sensing N-acyl homoserine lactone biosensors and are sensitive to the specific AiiA lactonase. FEMS Microbiol Lett 269: 213-220
DE LISA MP, VALDES JJ, BENTLEY WE (2001) Mapping stress-induced changes in autoinducer AI-2 production in chemostat-cultivated Escherichia coli K-12. J Bacteriol 183: 2918-2928
DIGGLE SP, CRUSZ SA, CAMARA M (2007a) Quorum sensing. Curr Biol 17: 907-910
DIGGLE SP, GRIFFIN AS, CAMPBELL GS, WEST SA (2007b) Cooperation and conflict in quorum-sensing bacterial populations. Nature 450: 411-414
DOBRETSOV S, DAHMS HU, YILI H, WAHL M, QIAN PY (2007) The effect of quorum-sensing blockers on the formation of marine microbial communities and larval attachment. FEMS Microbiol Ecol 60: 177-188
DONG YH, XU JL, LI XZ, ZHANG LH (2000) AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc Natl Acad Sci USA 97: 3526-3531
106

8. Literatur
DONG YH, ZHANG LH (2005) Quorum sensing and quorum-quenching enzymes. J Microbiol 43 Spec No: 101-109
DONG YH, ZHANG XF, SOO HM, GREENBERG EP, ZHANG LH (2005) The two-component response regulator PprB modulates quorum-sensing signal production and global gene expression in Pseudomonas aeruginosa. Mol Microbiol 56: 1287-1301
DOUDOROFF M (1942) Studies on the Luminous Bacteria: I. Nutritional Requirements of Some Species, with Special Reference to Methionine. J Bacteriol 44: 451-459
EBERHARD A, BURLINGAME AL, EBERHARD C, KENYON GL, NEALSON KH, OPPENHEIMER NJ (1981) Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 20: 2444-2449
ENGEBRECHT J, NEALSON K, SILVERMAN M (1983) Bacterial bioluminescence: isolation and genetic analysis of functions from Vibrio fischeri. Cell 32: 773-781
ENGEBRECHT J, SILVERMAN M (1984) Identification of genes and gene products necessary for bacterial bioluminescence. Proc Natl Acad Sci USA 81: 4154-4158
FARRAND SK, QIN Y, OGER P (2002) Quorum-sensing system of Agrobacterium plasmids: analysis and utility. Methods Enzymol 358: 452-484
FEDERLE MJ, BASSLER BL (2003) Interspecies communication in bacteria. J Clin Invest 112: 1291-1299
FLODGAARD LR, DALGAARD P, ANDERSEN JB, NIELSEN KF, GIVSKOV M, GRAM L (2005) Nonbioluminescent strains of Photobacterium phosphoreum produce the cell-to-cell communication signal N-(3-hydroxyoctanoyl)homoserine lactone. Appl Environ Microbiol 71: 2113-2120
FOX GE, WISOTZKEY JD, JURTSHUK PJ (1992) How close is close: 16S rRNA sequence identity may not be sufficient to guarantee species identity. Int J Sys Bacteriol 42: 166-170
FROMMBERGER M (2005) Entwicklung von Methoden zur Analyse von N-Acyl-Homoserinlactonen durch Kapillartrenntechniken und Massenspektrometrie. Dissertation, TU München, Deutschland
FUCHS G (2007) Allgemeine Mikrobiologie. Thieme Verlag, Deutschland
FUERST JA, HAWKINS JA, HOLMES A, SLY LI, MOORE CJ, STACKEBRANDT E (1993) Porphyrobacter neustonensis gen. nov., sp. nov., an aerobic bacteriochlorophyll-synthesizing budding bacterium from fresh water. Int J Syst Bacteriol 43: 125-134
107

8. Literatur
FUQUA C, PARSEK MR, GREENBERG EP (2001) Regulation of gene expression by cell-to-cell communication: acyl-homoserine lactone quorum sensing. Annu Rev Genet 35: 439-468
FUQUA WC, WINANS SC, GREENBERG EP (1994) Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J Bacteriol 176: 269-275
GAMBELLO MJ, IGLEWSKI BH (1991) Cloning and characterization of the Pseudomonas aeruginosa lasR gene, a transcriptional activator of elastase expression. J Bacteriol 173: 3000-3009
GEISENBERGER O (2000) Nachweis, Verbreitung und Bedeutung von N-Acyl-homoserinlactonen bei Gram-negativen Bakterien. Dissertation, TU München, Deutschland
GIVSKOV M, DE NYS R, MANEFIELD M, GRAM L, MAXIMILIEN R, EBERL L, MOLIN S, STEINBERG PD, KJELLEBERG S (1996) Eukaryotic interference with homoserine lactone-mediated prokaryotic signalling. J Bacteriol 178: 6618-6622
GLÖCKNER FO, FUCHS BM, AMANN R (1999) Bacterioplankton compositions of lakes and oceans: a first comparison based on fluorescence in situ hybridization. Appl Environ Microbiol 65: 3721-3726
GRAM L, GROSSART HP, SCHLINGLOFF A, KIORBOE T (2002) Possible quorum sensing in marine snow bacteria: production of acylated homoserine lactones by Roseobacter strains isolated from marine snow. Appl Environ Microbiol 68: 4111-4116
HALLBERG KB, JOHNSON DB (2001) Biodiversity of acidophilic prokaryotes. Adv Appl Microbiol 49: 37-84
HANADA S, KAWASE Y, HIRAISHI A, TAKAICHI S, MATSUURA K, SHIMADA K, NAGASHIMA KV (1997) Porphyrobacter tepidarius sp. nov., a moderately thermophilic aerobic photosynthetic bacterium isolated from a hot spring. Int J Syst Bacteriol 47: 408-413
HASELTINE EL, ARNOLD FH (2008) Implications of rewiring bacterial quorum sensing. Appl Environ Microbiol 74: 437-445
HAWLEY GG (1981) The condensed chemical dictionary, 10th ed. Van Nostrand Reinhold Company, New York, N.Y., USA
HEYDUCK-SÖLLER B (2003) Untersuchungen zur Charakterisierung von marinen Cyanobakterien der Gattung Anabaena und ihrer Licht- und UV-absorbierenden Substanzen. Dissertation, Universität Bremen, Deutschland
108

8. Literatur
HIRAISHI A, YONEMITSU Y, MATSUSHITA M, SHIN YK, KURAISHI H, KAWAHARA K (2002) Characterization of Porphyrobacter sanguineus sp. nov., an aerobic bacteriochlorophyll-containing bacterium capable of degrading biphenyl and dibenzofuran. Arch Microbiol 178: 45-52
HORIKOSHI K (1999) Alkaliphiles: some applications of their products for biotechnology. Microbiol Mol Biol Rev 63: 735-750
HUANG JJ, HAN JI, ZHANG LH, LEADBETTER JR (2003) Utilization of acyl-homoserine lactone quorum signals for growth by a soil pseudomonad and Pseudomonas aeruginosa PAO1. Appl Environ Microbiol 69: 5941-5949
HUBE A (2008) Molekularbiologische und physiologische Untersuchungen an heterotrophen Begleitbakterien mariner filamentöser Cyanobakterien. Dissertation, Universität Bremen, Deutschland
ITEMAN I, RIPPKA R, TANDEAU DE MARSAC N, HERDMAN M (2000) Comparison of conserved structural and regulatory domains within divergent 16S rRNA-23S rRNA spacer sequences of cyanobacteria. Microbiol 146 (Pt 6): 1275-1286
ITEMAN I, RIPPKA R, TANDEAU DE MARSAC N, HERDMAN M (2002) rDNA analyses of planktonic heterocystous cyanobacteria, including members of the genera Anabaenopsis and Cyanospira. Microbiol 148: 481-496
JUKES TH, CANTOR CR (1969) Evolution of protein molecules. In: Mammalian Protein Metabolism (Munro HN, ed.). Academic Press, New York, USA
KAPLAN HB, GREENBERG EP (1985) Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system. J Bacteriol 163: 1210-1214
KHAN SR, MAVRODI DV, JOG GJ, SUGA H, THOMASHOW LS, FARRAND SK (2005) Activation of the phz operon of Pseudomonas fluorescens 2-79 requires the LuxR homolog PhzR, N-(3-OH-hexanoyl)-L-homoserine lactone produced by the LuxI homolog PhzI, and a cis-acting phz box. J Bacteriol 187: 6517-6527
KLEEREBEZEM M, QUADRI LE, KUIPERS OP, DE VOS WM (1997) Quorum sensing by peptide pheromones and two-component signal-transduction systems in Gram-positive bacteria. Mol Microbiol 24: 895-904
LANE DJ (1991) 16S/23S rRNA sequencing. In: STACKEBRANDT E, GOODFELLOW M (ed.), Nucleic acid techniques in bacterial systematics. Wiley, New York: 115-175
LEADBETTER JR, GREENBERG EP (2000) Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J Bacteriol 182: 6921-6926
LEBLOND-BOURGET N, PHILIPPE H, MANGIN I, DECARIS B (1996) 16S rRNA and 16S to 23S internal transcribed spacer sequence analyses reveal inter- and intraspecific Bifidobacterium phylogeny. Int J Syst Bacteriol 46: 102-111
109

8. Literatur
LEE KH, RUBY EG (1992) Detection of the Light Organ Symbiont, Vibrio fischeri, in Hawaiian Seawater by Using lux Gene Probes. Appl Environ Microbiol 58: 942-947
LOTTSPEICH F, ZORBAS H (1998) Bioanalytik. Spektrum Akademischer Verlag, Heidelberg, Germany
LYRA C, LAAMANEN M, LEHTIMAKI JM, SURAKKA A, SIVONEN K (2005) Benthic cyanobacteria of the genus Nodularia are non-toxic, without gas vacuoles, able to glide and genetically more diverse than planktonic Nodularia. Int J Syst Evol Microbiol 55: 555-568
MADIGAN MT, MARTINKO JM, PARKER J (2001) Brock Mikrobiologie, 2. korrigierter Nachdruck 2003. Spektrum Akademischer Verlag, Heidelberg, Germany
MANEFIELD M, RASMUSSEN TB, HENZTER M, ANDERSEN JB, STEINBERG P, KJELLEBERG S, GIVSKOV M (2002) Halogenated furanones inhibit quorum sensing through accelerated LuxR turnover. Microbiol 148: 1119-1127
MCCLEAN KH, WINSON MK, FISH L, TAYLOR A, CHHABRA SR, CAMARA M, DAYKIN M, LAMB JH, SWIFT S, BYCROFT BW, STEWART GS, WILLIAMS P (1997) Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiol 143 (Pt 12): 3703-3711
MCLEAN RJ, WHITELEY M, STICKLER DJ, FUQUA WC (1997) Evidence of autoinducer activity in naturally occurring biofilms. FEMS Microbiol Lett 154: 259-263
MILLER MB, BASSLER BL (2001) Quorum sensing in bacteria. Annu Rev Microbiol 55: 165-199
MILLER ST, XAVIER KB, CAMPAGNA SR, TAGA ME, SEMMELHACK MF, BASSLER BL, HUGHSON FM (2004) Salmonella typhimurium recognizes a chemically distinct form of the bacterial quorum-sensing signal AI-2. Mol Cell 15: 677-687
MOHAMED NM, CICIRELLI EM, KAN J, CHEN F, FUQUA C, HILL RT (2008) Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ Microbiol 10: 75-86
MORTIMER CE (2007) Chemie: das Basiswissen der Chemie, 9. Auflage. Georg Thieme Verlag, Stuttgart, Germany
NEALSON KH, HASTINGS JW (1979) Bacterial bioluminescence: its control and ecological significance. Microbiol Rev 43: 496-518
NEALSON KH, PLATT T, HASTINGS JW (1970) Cellular control of the synthesis and activity of the bacterial luminescent system. J Bacteriol 104: 313-322
110

8. Literatur
NEILAN BA (1995) Identification and Phylogenetic Analysis of Toxigenic Cyanobacteria by Multiplex Randomly Amplified Polymorphic DNA PCR. Appl Environ Microbiol 61: 2286-2291
PARSEK MR, GREENBERG EP (2005) Sociomicrobiology: the connections between quorum sensing and biofilms. Trends Microbiol 13: 27-33
QIN N, CALLAHAN SM, DUNLAP PV, STEVENS AM (2007) Analysis of LuxR regulon gene expression during quorum sensing in Vibrio fischeri. J Bacteriol 189: 4127-4134
RAINEY FA, SILVA J, NOBRE MF, SILVA MT, DA COSTA MS (2003) Porphyrobacter cryptus sp. nov., a novel slightly thermophilic, aerobic, bacteriochlorophyll a-containing species. Int J Syst Evol Microbiol 53: 35-41
RATHGEBER C, BEATTY JT, YURKOV V (2004) Aerobic phototrophic bacteria: new evidence for the diversity, ecological importance and applied potential of this previously overlooked group. Photosynth Res 81: 113–128
RATHGEBER C, YURKOVA N, STACKEBRANDT E, SCHUMANN P, HUMPHREY E, BEATTY JT, YURKOV V (2007) Porphyrobacter meromictius sp. nov., an appendaged bacterium, that produces Bacteriochlorophyll a. Curr Microbiol 55: 356-361
RAVN L, CHRISTENSEN AB, MOLIN S, GIVSKOV M, GRAM L (2001) Methods for detecting acylated homoserine lactones produced by Gram-negative bacteria and their application in studies of AHL-production kinetics. J Microbiol Methods 44: 239-251
READING NC, SPERANDIO V (2006) Quorum sensing: the many languages of bacteria. FEMS Microbiol Lett 254: 1-11
RETHMEIER J (1995) Untersuchungen zur Ökologie und zum Mechanismus der Sulfidadaptation mariner Cyanobakterien der Ostsee. Dissertation, Universität Bremen, Deutschland
RIEDEL K, HENTZER M, GEISENBERGER O, HUBER B, STEIDLE A, WU H, HOIBY N, GIVSKOV M, MOLIN S, EBERL L (2001) N-acylhomoserine-lactone-mediated communication between Pseudomonas aeruginosa and Burkholderia cepacia in mixed biofilms. Microbiol 147: 3249-3262
RIPPKA R, DERUELLES J, WATERBURY JB, HERDMAN M, STANIER RY (1979) Generic assignment, strain histories and properties of pure cultures of cyanobacteria. J Gen Microbiol 111: 1-61
RIUS N, LOREN JG (1998) Buffering capacity and membrane H+ conductance of neutrophilic and alkalophilic gram-positive bacteria. Appl Environ Microbiol 64: 1344-1349
111

8. Literatur
ROMERO M, DIGGLE SP, CAMARA M, FÁBREGAS J, OTERO A (2006) AHL-Type quorum sensing signals in Nostoc PCC7120. Tagungsband vom "12th International Symposium on Phototrophic Prokaryotes"
ROMERO M, DIGGLE SP, HEEB S, CAMARA M, OTERO A (2008) Quorum quenching activity in Anabaena sp. PCC 7120: identification of AiiC, a novel AHL-acylase. FEMS Microbiol Lett 280: 73-80
SAMBROOK J, Fritsch EF, MANIATIS T (1989) Molecular Cloning: a Laboratory Manual, 2nd edn. New York: Cold Spring Habor Laboratory Press.
SCHAEFER AL, VAL DL, HANZELKA BL, CRONAN JE, JR., GREENBERG EP (1996) Generation of cell-to-cell signals in quorum sensing: acyl homoserine lactone synthase activity of a purified Vibrio fischeri LuxI protein. Proc Natl Acad Sci USA 93: 9505-9509
SCHAUDER S, SHOKAT K, SURETTE MG, BASSLER BL (2001) The LuxS family of bacterial autoinducers: biosynthesis of a novel quorum-sensing signal molecule. Mol Microbiol 41: 463-476
SCHENK A (1996) Untersuchungen zur Identifizierung unizellulärer mariner Cyanobakterien der Ostsee auf molekularbiologischer Ebene unter besonderer Berücksichtigung der PCR-Technik. Diplomarbeit, Universität Oldenburg, Deutschland
SCHOMAKER N (2007) Nachweis von N-Acyl-L-homoserinlactonen bei Cyanobakterien und heterotrophen Bakterien. Projektarbeit, Universität Bremen, Deutschland
SCHRÜBBERS J (2007) Charakterisierung des filamentösen Cyanobakteriums Stamm Flo1. Diplomarbeit, Universität Bremen, Deutschland
SCHUSTER M, LOSTROH CP, OGI T, GREENBERG EP (2003) Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: a transcriptome analysis. J Bacteriol 185: 2066-2079
SEO PS, YOKOTA A (2003) The phylogenetic relationships of cyanobacteria inferred from 16S rRNA, gyrB, rpoC1 and rpoD1 gene sequences. J Gen Appl Microbiol 49: 191-203
SHAW PD, PING G, DALY SL, CHA C, CRONAN JE, JR., RINEHART KL, FARRAND SK (1997) Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc Natl Acad Sci USA 94: 6036-6041
SHIN NR, LEE DY, YOO HS (2007) Identification of quorum sensing-related regulons in Vibrio vulnificus by two-dimensional gel electrophoresis and differentially displayed reverse transcriptase PCR. FEMS Immunol Med Microbiol 50: 94-103
SMITH JL, FRATAMICO PM, NOVAK JS (2004) Quorum sensing: a primer for food microbiologists. J Food Prot 67: 1053-1070
112

8. Literatur
STACKEBRANDT E, EBERS J (2006) Taxonomic parameters revisited: tarnished gold standards. Microbiol Today nov 06: 152-155
STACKEBRANDT E, GOEBEL BM (1994) Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology Int J Syst Bacteriol 44: 846-849
STADLBAUER W (2006) Chromatographische Trennverfahren in der Organischen Chemie: Dünnschichtchromatographie. Vorlesungsskriptum (SS 2006), Institut für Chemie, Karl-Franzens-Universität Graz, Austria
STEINDLER L, VENTURI V (2007) Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol Lett 266: 1-9
STEVENSON BS, WATERBURY JB (2006) Isolation and identification of an epibiotic bacterium associated with heterocystous Anabaena cells. Biol Bull 210: 73-77
SUN J, DANIEL R, WAGNER-DÖBLER I, ZENG AP (2004) Is autoinducer-2 a universal signal for interspecies communication: a comparative genomic and phylogenetic analysis of the synthesis and signal transduction pathways. BMC Evol Biol 4: 36
SURETTE MG, BASSLER BL (1998) Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc Natl Acad Sci USA 95: 7046-7050
SZENTHE A, PAGE WJ (2003) Quorum Sensing in Agrobacterium tumefaciens using N-oxo-Acyl-homoserine Lactone chemical signal. In: O'DONNELL MA (ed) Tested studies for laboratory teaching, Volume 24, Pages 145-152. Proceedings of the 24th Workshop/Conference of the Association for Biology Laboratory Education
TATON A, GRUBISIC S, BRAMBILLA E, DE WIT R, WILMOTTE A (2003) Cyanobacterial diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): a morphological and molecular approach. Appl Environ Microbiol 69: 5157-5169
TAYLOR MW, SCHUPP PJ, BAILLIE HJ, CHARLTON TS, DE NYS R, KJELLEBERG S, STEINBERG PD (2004) Evidence for acyl homoserine lactone signal production in bacteria associated with marine sponges. Appl Environ Microbiol 70: 4387-4389
TEPLITSKI M, EBERHARD A, GRONQUIST MR, GAO M, ROBINSON JB, BAUER WD (2003) Chemical identification of N-acyl homoserine lactone quorum-sensing signals produced by Sinorhizobium meliloti strains in defined medium. Arch Microbiol 180: 494-497
TOMASZ A (1965) Control of the competent state in Pneumococcus by a hormone-like cell product: an example for a new type of regulatory mechanism in bacteria. Nature 208: 155-159
113

8. Literatur
TUOMAINEN J, HIETANEN S, KUPARINEN J, MARTIKAINEN PJ, SERVOMAA K (2006) Community structure of the bacteria associated with Nodularia sp. (Cyanobacteria) aggregates in the Baltic Sea. Microb Ecol 52: 513-522
WAGNER VE, BUSHNELL D, PASSADOR L, BROOKS AI, IGLEWSKI BH (2003) Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J Bacteriol 185: 2080-2095
WAGNER-DÖBLER I, BIEBL H (2006) Environmental biology of the marine Roseobacter lineage. Annu Rev Microbiol 60: 255-280
WAGNER-DÖBLER I, THIEL V, EBERL L, ALLGAIER M, BODOR A, MEYER S, EBNER S, HENNIG A, PUKALL R, SCHULZ S (2005) Discovery of complex mixtures of novel long-chain quorum sensing signals in free-living and host-associated marine alphaproteobacteria. Chembiochem 6: 2195-2206
WATERS CM, BASSLER BL (2005) Quorum sensing: cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol 21: 319-346
WAYNE LG, BRENNER DJ, COLWELL RR, GRIMONT PAD, KANDLER O, KRICHEVSKY MI, MOORE LH, MOORE WEC, MURRAY RGE, STACKEBRANDT E, STARR MP, TRÜPER HG (1987) Report of the ad hoc comittee on reconciliation of approaches to bacterial systematics. Int J Syst Bacteriol 37: 463-464
WHITEHEAD NA, BARNARD AM, SLATER H, SIMPSON NJ, SALMOND GP (2001) Quorum-sensing in Gram-negative bacteria. FEMS Microbiol Rev 25: 365-404
WILLIAMS P (2007) Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiol 153: 3923-3938
WITHERS H, SWIFT S, WILLIAMS P (2001) Quorum sensing as an integral component of gene regulatory networks in Gram-negative bacteria. Curr Opin Microbiol 4: 186-193
XAVIER KB, BASSLER BL (2005) Interference with AI-2-mediated bacterial cell-cell communication. Nature 437: 750-753
YAN S (2005) Morphological description and molecular characterization of Subsection IV (Nostocales) of cyanobacteria. Report of Labrotation III in Marmic, M.Sc./Ph.D. program at the University of Bremen, Germany
YATES EA, PHILIPP B, BUCKLEY C, ATKINSON S, CHHABRA SR, SOCKETT RE, GOLDNER M, DESSAUX Y, CAMARA M, SMITH H, WILLIAMS P (2002) N-acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect Immun 70: 5635-5646
YOON HS, GOLDEN JW (1998) Heterocyst pattern formation controlled by a diffusible peptide. Science 282: 935-938
114

8. Literatur
YOON JH, KANG SJ, LEE MH, OH HW, OH TK (2006) Porphyrobacter dokdonensis sp. nov., isolated from sea water. Int J Syst Evol Microbiol 56: 1079-1083
YOON JH, LEE MH, OH TK (2004) Porphyrobacter donghaensis sp. nov., isolated from sea water of the East Sea in Korea. Int J Syst Evol Microbiol 54: 2231-2235
YOON JH, LEE MH, OH TK, PARK YH (2005) Muricauda flavescens sp. nov. and Muricauda aquimarina sp. nov., isolated from a salt lake near Hwajinpo Beach of the East Sea in Korea, and emended description of the genus Muricauda. Int J Syst Evol Microbiol 55: 1015-1019
YURKOV VV, BEATTY JT (1998) Aerobic anoxygenic phototrophic bacteria. Microbiol Mol Biol Rev 62: 695-724
ZHANG HB, WANG C, ZHANG LH (2004) The quormone degradation system of Agrobacterium tumefaciens is regulated by starvation signal and stress alarmone (p)ppGpp. Mol Microbiol 52: 1389-1401
ZHANG HB, WANG LH, ZHANG LH (2002b) Genetic control of quorum-sensing signal turnover in Agrobacterium tumefaciens. Proc Natl Acad Sci USA 99: 4638-4643
ZHANG RG, PAPPAS T, BRACE JL, MILLER PC, OULMASSOV T, MOLYNEAUX JM, ANDERSON JC, BASHKIN JK, WINANS SC, JOACHIMIAK A (2002a) Structure of a bacterial quorum-sensing transcription factor complexed with pheromone and DNA. Nature 417: 971-974
115

Erklärung gemäß § 21, Absatz 6 der Diplomprüfungsordnung für den Studiengang Biologie (vom 7. Dezember 1994) Hiermit erkläre ich, dass ich die vorliegende Arbeit selbstständig verfasst und dabei
keine anderen als die angegebenen Quellen und Hilfsmittel verwendet habe.
Bremen, im April 2008 ____________________________
(Peter Orszag)
116


















