
Veränderungen des neuronalen Schaltkreises des ... · Solange die horizontale optokinetische...
173
Veränderungen des neuronalen Schaltkreises des optokinetischen Reflexes und optokinetische Verhaltensdefizite bei albinotischen Frettchen Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Biologie an der Ruhr-Universität Bochum angefertigt am Lehrstuhl für Allgemeine Zoologie und Neurobiologie vorgelegt von Nikolaos Garipis aus Serres (Griechenland) Bochum 2001
Transcript of Veränderungen des neuronalen Schaltkreises des ... · Solange die horizontale optokinetische...

Veränderungen des neuronalen Schaltkreises
des optokinetischen Reflexes und
optokinetische Verhaltensdefizite
bei albinotischen Frettchen
Dissertation
zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie
an der Ruhr-Universität Bochum
angefertigt am
Lehrstuhl für Allgemeine Zoologie und Neurobiologie
vorgelegt von
Nikolaos Garipis
aus Serres (Griechenland)
Bochum 2001

Dissertation eingereicht am: 02. November 2001 Betreuer: Prof. Dr. K.-P. Hoffmann (Lehrstuhl Allgemeine Zoologie und Neurobiologie, Fakultät Biologie) Koreferent: Prof. Dr. R. Necker (Lehrstuhl Tierphysiologie, Fakultät Biologie) Dekan: Prof. Dr. Dr. H. Hatt (Lehrstuhl Zellphysiologie, Fakultät Biologie)

I
Inhaltsverzeichnis
Einleitung ............................................................................. 1
I. Der horizontale optokinetische Reflex........................................................................ 1
II. Funktionelle Anatomie des optokinetischen Systems ............................................... 1
III. Albinismus bei Säugetieren......................................................................................... 3
IV. Neuronaler hOKN-Schaltkreis des Frettchens .......................................................... 4
V. Zielsetzungen und Hypothesen................................................................................... 5
Material und Methoden..................................................... 8
I. Teil I: Verhalten........................................................................................................... 8
A. Versuchstiere..................................................................................................................8
B. Fixierung der Versuchstiere ...........................................................................................8
C. Horizontale optokinetische Reizung ...............................................................................9
D. Registrierung der Augenbewegungen...........................................................................12
E. Analyse und Auswertung des EOGs .............................................................................12
1. Bearbeitung und graphische Darstellung der EOG-Spuren.................................... 12
2. Statistik ................................................................................................................... 14
F. Inaktivierung des Prätektums ......................................................................................14
1. Injektionsmethode und Vorbereitung der Versuchstiere ........................................ 14
2. Muscimol-Injektionen............................................................................................. 17
3. Farb-Injektionen ..................................................................................................... 17
G. Sehtest..........................................................................................................................17
1. Versuchsaufbau und Konditionierung .................................................................... 17
2. Durchführung des Perimetrietests........................................................................... 20
3. Auswertung und graphische Darstellung ................................................................ 21
H. Histologische Untersuchung .........................................................................................21
II. Teil II: Neurophysiologie ........................................................................................... 23

II
A. Anästhesie und Tierpräparation...................................................................................23
B. Extrazelluläre Ableitungen und Suche nach dem NOT ................................................25
C. Elektrische Stimulation................................................................................................26
1. Reizparameter und Erläuterungen .......................................................................... 26
2. Reizexperimente ..................................................................................................... 27
a) Antidrome Reizung der NOT-Neuronen von der IO ........................................... 27
b) Orthodrome Reizung der NOT-Neuronen vom VC ............................................ 28
c) Orthodrome Reizung der NOT-Neuronen vom ON............................................ 28
d) Antidrome Reizung kortikaler Neuronen vom NOT und SC .............................. 28
D. Visuelle Stimulation .....................................................................................................30
1. Visuelle Stimulation der prätektalen Zellen ........................................................... 30
2. Visuelle Stimulation der VC-Neuronen.................................................................. 30
3. ON-OFF-Test von Zellen des Mittelhirns .............................................................. 31
4. Lichtverhältnisse ..................................................................................................... 32
E. Datenaufnahme und Analyse........................................................................................32
1. Richtungsspezifitäts-Indizes ................................................................................... 33
2. ON-OFF-Indizes ..................................................................................................... 34
3. Statistik ................................................................................................................... 35
F. Histologie .....................................................................................................................35
Ergebnisse .......................................................................... 37
I. Teil I: Verhalten......................................................................................................... 37
A. Spontane Augenbewegungen (vor Muscimol-Injektion)...............................................37
B. hOKN-Test (vor Muscimol-Injektion)..........................................................................40
1. Optokinetische Reizung der Albinofrettchen ......................................................... 40
a) Monokulare Ganzfeldreizung .............................................................................. 40
b) Binokulare Ganzfeldreizung ................................................................................ 43
2. Optokinetische Reizung der pigmentierten Frettchen ............................................ 47
a) Monokulare Ganzfeldreizung .............................................................................. 47
b) Binokulare Ganzfeldreizung ................................................................................ 47
C. Inaktivierungsstudie und histologische Auswertung.....................................................49
1. hOKN-Test nach Muscimol-Injektion.................................................................... 49

III
2. Histologische Dokumentation der Injektionsorte ................................................... 53
3. Farb-Injektionen ..................................................................................................... 55
D. Ergebnis des Perimetrietests.........................................................................................56
1. Binokulare Messungen ........................................................................................... 57
2. Monokulare Messungen.......................................................................................... 58
II. Teil II: Neurophysiologie ........................................................................................... 64
A. Histologische Verifikation............................................................................................64
1. NOT-Ableitorte, IO- und VC-Reizorte................................................................... 65
2. NOT- und SC-Reizorte und VC-Ableitorte ............................................................ 69
B. Physiologie ...................................................................................................................72
1. Lokalisation des NOT-Areals und der NOT-Neuronen.......................................... 72
2. Umgebung der NOT-Neuronen .............................................................................. 73
3. Allgemeine Charakterisierung der prätektalen und der NOT-Neuronen................ 74
4. ON/OFF-Eigenschaften von Neuronen im Mittelhirn............................................ 74
a) ON/OFF-Eigenschaften von prätektalen und NOT-Neuronen............................ 74
b) ON/OFF-Eigenschaften von SC-Neuronen......................................................... 75
5. DS-Indizes und antidrome Identifizierung der NOT-Neuronen............................. 76
a) Albinofrettchen.................................................................................................... 78
b) Pigmentierte Frettchen......................................................................................... 82
c) Statistischer Vergleich zwischen den beiden Tiergruppen.................................. 82
6. Orthodrome Reizung der NOT-Neuronen vom VC ............................................... 83
a) Albinofrettchen.................................................................................................... 84
b) Pigmentierte Frettchen......................................................................................... 85
c) Vergleich zwischen den beiden Tiergruppen ...................................................... 86
7. Antidrome Reizung der VC-Neuronen vom NOT und / oder SC .......................... 87
a) Anzahl der untersuchten VC-Neuronen............................................................... 87
(1) Albinofrettchen............................................................................................ 87
(2) Pigmentierte Frettchen ................................................................................ 88
b) Richtungsspezifität und Vorzugsstärke im VC.................................................... 88
(1) Albinofrettchen............................................................................................ 89
(2) Pigmentierte Frettchen ................................................................................ 89
(3) Statistischer Vergleich zwischen den beiden Tiergruppen ......................... 90
c) Anti- und orthodrome Reizlatenzen im VC......................................................... 91

IV
(1) Albinofrettchen............................................................................................ 91
(2) Pigmentierte Frettchen ................................................................................ 92
(3) Statistischer Vergleich zwischen den beiden Tiergruppen ......................... 92
d) Antidrome Reizbarkeit der VC-Neuronen und visuelle Reize ............................ 93
(1) Albinofrettchen............................................................................................ 93
(2) Pigmentierte Frettchen ................................................................................ 94
e) Vorzugsrichtungen der VC-Neuronen................................................................. 94
(1) Albinofrettchen............................................................................................ 94
(2) Pigmentierte Frettchen ................................................................................ 96
f) Vom NOT und SC reizbare VC-Neuronen.......................................................... 98
(1) Albinofrettchen............................................................................................ 98
(2) Pigmentiertes Frettchen............................................................................... 98
(3) Vergleich zwischen den beiden Tiergruppen.............................................. 99
8. Orthodrome Reizung der NOT-Neuronen vom ON............................................... 99
Diskussion........................................................................ 104
I. Teil I: Verhalten....................................................................................................... 104
A. Die Meßmethode ........................................................................................................ 104
1. Wahl des EOGs und EOG-Auswertung ............................................................... 104
2. Sensitivität des unkalibrierten EOGs .................................................................... 105
B. Augendrift oder Spontannystagmus beim Albinofrettchen? ....................................... 106
C. Optokinetische Blindheit des Albinofrettchens ........................................................... 106
1. Reizparameter ....................................................................................................... 107
2. Optokinetische Augenbewegungen bei hypopigmentierten Säugern................... 109
D. Muscimol-Experimente .............................................................................................. 111
1. Die Injektionsmethode .......................................................................................... 111
2. Erste Eingrenzung des okulomotorischen Defektes des Albinofrettchens ........... 113
E. Perimetrietest............................................................................................................. 113
1. Annahmen des Perimetrietests.............................................................................. 113
2. Getestetes Gesichtsfeld ......................................................................................... 115
3. Sehfähigkeit des Albinofrettchens unter binokularer Bedingung......................... 115
4. Sehfähigkeit des Albinofrettchens unter monokularer Bedingung....................... 116

V
5. Der Perimetrietest in Bezug auf das optokinetische System ................................ 117
II. Teil II: Neurophysiologie ......................................................................................... 119
A. Identifizierung der NOT-Neuronen............................................................................ 119
B. Ausmaß des physiologischen Defizits.......................................................................... 121
C. Richtungsunspezifität der NOT-Neuronen und optokinetische Blindheit................... 121
D. Kortikaler Eingang des NOT...................................................................................... 125
1. Einordnung der Experimente in den vorhandenen Wissensstand ......................... 125
2. Stärke und Leitungsgeschwindigkeit des kortikalen NOT-Eingangs ................... 128
3. Informationsgehalt des kortikalen NOT-Eingangs ............................................... 130
4. Ursprung des kortikalen NOT-Eingangs beim Frettchen..................................... 132
E. Retinaler Eingang des NOT........................................................................................ 134
1. Stärke und Leitungsgeschwindigkeit des retinalen NOT-Eingangs ..................... 134
2. Informationsgehalt des retinalen NOT-Eingangs ................................................. 138
F. Das morphologische Korrelat zur Richtungsunspezifität der NOT-Neuronen............ 143
Zusammenfassung.......................................................... 145
1. Teil I: Verhalten.................................................................................................... 145
2. Teil II: Neurophysiologie ..................................................................................... 146
a) Richtungsunspezifität der NOT-Neuronen........................................................ 146
b) Kortikaler Eingang der NOT-Neuronen............................................................ 147
c) Retinaler Eingang der NOT-Neuronen.............................................................. 148
Literaturverzeichnis ....................................................... 150
Danksagung ..................................................................... 163
Curriculum vitae............................................................. 164

VI
Abkürzungsverzeichnis
# Nummer (Nr.)
∅ Durchmesser
° Grad Sehwinkel
°C Grad Celsius
µl Mikroliter
µm Mikrometer
Abb. Abbildung(en)
ALLS anterolaterales laterales suprasylvisches Areal
AMLS anteromediales laterales suprasylvisches Areal
APB 2-Amino-4-phosphonobutyrat
AS ansinater Sulcus
ASG anteriorer sygmoider Gyrus
CB Cerebellum (Kleinhirn)
CC Corpus callosum
ccw gegen den Uhrzeigersinn (engl. counterclockwise)
cd Candela
CGL Corpus geniculatum laterale
CNG coronaler Gyrus
CNS coronaler Sulcus
CRS cruciater Sulcus
cw im Uhrzeigersinn (engl. clockwise)
DS-I Richtungsspezifitäts-Index (Plural: -izes) (engl. direction selectivity index)
EOG Elektrookulogramm
GABA γ-Aminobuttersäure
HC Hypocampus
hOKN horizontaler optokinetischer Nystagmus
hOKR horizontaler optokinetischer Reflex
Hz Hertz
IAL Interaurallinie
IO inferiore Olive (bzw. dorsale Kappe der inferioren Olive)
K Helligkeitskontrast
KG Körpergewicht

VII
LG lateraler Gyrus
LS lateraler Sulcus
LTD Langzeit-Depression (engl. long-term depression)
MΩ Megaohm
MD Medulla
MT mediales temporales Areal
N Anzahl
NOT Nukleus des optischen Traktes
NPH Nukleus präpositus hypoglossi
NRTP Nukleus reticularis tegmenti pontis
ON optischer Nerv
OX optisches Chiasma
PC Personalcomputer
PEG posteriorer ectosylvischer Gyrus
PLLS posteriolaterales laterales suprasylvisches Areal
PMLS posteriomediales laterales suprasylvisches Areal
PSG posteriorer sygmoider Gyrus
Q25 unteres Quartil
Q75 oberes Quartil
RF rezeptives Feld
s.c. subcutan
s.d. Standardabweichung (engl. standard deviation)
SC superiorer Colliculus
SSG suprasylvischer Gyrus
SSS suprasylvischer Sulcus
STS superiorer temporaler Sulcus
TTL Transistor-Transistor-Logik
VC visueller Kortex
VK Vestibulariskerne

Einleitung 1
Einleitung
I. Der horizontale optokinetische Reflex
Der horizontale optokinetische Reflex (hOKR) ist ein basaler Mechanismus zur
Stabilisierung des Bildes der Umwelt auf der Retina. Er wird durch die großflächige retinale
Bildverschiebung in horizontaler Richtung ausgelöst, die bei Betrachtung der sich
bewegenden Umwelt und / oder bei Eigenbewegung hervorgerufen wird. Er wirkt dieser
Bildverschiebung entgegen und ermöglicht damit eine Stabilisierung des Bildes auf der
Retina. Folglich ist er von fundamentaler Bedeutung für Mensch und Tier. Bei anhaltender
Bildverschiebung in einer Richtung wird der hOKR durch sogenannte Rückstellsakkaden in
entgegengesetzter Richtung unterbrochen. Die alternierende Auslösung des hOKR und der
Rückstellsakkaden ist der horizontale optokinetische Nystagmus (hOKN). Der hOKR bildet
die langsamen Folgephasen und die Rückstellsakkaden bilden die schnellen Phasen
des hOKNs.
II. Funktionelle Anatomie des optokinetischen Systems
Bei typischen Afoveaten (Lagomorpha und Rodentia) mit lateralstehenden Augen und
nahezu vollständiger Kreuzung der retinofugalen Fasern findet man folgenden neuronalen
hOKN-Schaltkreis.
Ausgehend von der Retina existiert eine monosynaptische Projektion zum kontralateralen
Nucleus des optischen Traktes (NOT) (Klooster et al., 1983; Pak et al., 1987). Diese
Projektion wird von richtungsspezifischen Ganglienzellen des ON-Typs gebildet (Barlow &
Hill, 1963). Wenn durch eine Augeninjektion von APB (2-Amino-4-phosphonobutyrat) die
ON-Ganglienzellen selektiv inaktiviert werden, kann über das injizierte Auge kein hOKN
mehr ausgelöst werden.
Der NOT ist ein paariger Kern und befindet sich im Prätektum. Dieser Kern nimmt eine
zentrale Stellung zwischen Sensorik und Motorik ein, indem er eine herausragende Rolle bei
der Auslösung des hOKR bzw. der langsamen Phase des hOKN spielt (Kaninchen:
Collewijn, 1975; Ratte: Schmidt et al., 1993; Frettchen: Klauer et al., 1990; Katze: Hoffmann
& Schoppmann, 1981; Tammarwallaby: Ibbotson et al., 1994; Totenkopfaffe: Hoffmann,

Einleitung 2
1985; Rhesusaffe & Langschwanzmakak: Hoffmann et al., 1988; für einen Überblick siehe
Cohen et al., 1992). Innerhalb des NOTs finden sich Interneuronen, die γ-Aminobuttersäure
enthalten (Horn & Hoffmann, 1987; Nunes Cardozo & Van der Want, 1990; Nunes Cardozo
et al., 1991; Van der Want & Nunes Cardozo, 1988), die Synapsen mit den NOT-
Ausgangsneuronen bilden. Die Injektion von Muscimol, einem GABAA-Agonisten, in einen
NOT inaktiviert reversibel die NOT-Ausgangsneuronen. Infolgedessen kann der hOKN nicht
mehr in Vorzugsrichtung des inaktivierten NOTs ausgelöst werden.
Die NOT-Ausgangsneuronen sind richtungsspezifisch für Bewegung eines großflächigen
visuellen Musters. Im linken NOT werden sie richtungsspezifisch aktiviert durch eine
Reizbewegung nach links und die im rechten NOT durch die Reizbewegung nach rechts.
Dieses Schema ist bei allen untersuchten Säugetieren mit intaktem optokinetischen System
bestätigt worden.
Grob spaltet sich der NOT-Ausgang in zwei Projektionszweige auf. Der für die Auslösung
der langsamen Folgephase des hOKN verantwortliche Projektionszweig zieht ipsilateral in
den Hirnstamm zum Nukleus präpositus hypoglossi (NPH) und / oder zum Nukleus
reticularis tegmenti pontis (NRTP; Kaninchen: Maekawa et al., 1981, 1984; Ratte: Cazin et
al., 1980, 1982, 1984; Katze: Magnin et al., 1983, 1989; Watanabe et al., 1993; Primaten:
Mustari et al., 1994; Fuchs et al., 1992), die ihrerseits zu den kontralateralen
Vestibulariskernen projizieren. Von den Vestibulariskernen verläuft der Projektionsweg zu
den Augenmuskelkernen, die schließlich die entsprechenden äußeren Augenmuskeln
innervieren. Der zweite Projektionszweig zieht vom NOT zur ipsilateralen inferioren Olive
(IO) und dient unter Einbeziehung des Kleinhirns und der Vestibulariskerne der
Rekalibirierung des vestibulookulären Nystagmus (Ito et al., 1974; Ito, 1982, 1993). Die
richtungsspezifische Information aus dem NOT ist für beide Projektionen unerläßlich für die
zu erfüllenden Funktionen.
Dieser neuronale Schaltkreis ermöglicht den Afoveaten folgendes optokinetisches Verhalten:
Solange die horizontale optokinetische Reizung binokular erfolgt, zeigen alle bis jetzt
untersuchten, visuell intakten adulten Spezies einen symmetrischen hOKN, d.h. die Güte
(Quotient der Augengeschwindigkeit durch die Reizgeschwindigkeit) der Augenfolge-
bewegung ist für beide horizontalen Reizrichtungen gleich.
Unter monokularer Reizbedingung jedoch zeigen die Afoveaten einen stark asymmetrischen
monokularen hOKN (Kaninchen: Hahnenberger, 1977; Maus: Grüsser-Cornehls & Böhm,
1988; Ratte: Hess et al., 1985; Reber et al., 1991). Die Güte der Augenfolgebewegung ist in
temporonasaler Richtung höher als in nasotemporaler Richtung. Der Grund für diese

Einleitung 3
Asymmetrie liegt darin, daß von einem Auge nur der kontralaterale NOT aktiviert wird, der
die langsame Folgephase des hOKN nur in der temporonasalen Richtung des gereizten
Auges auslösen kann.
Mit zunehmender evolutiver Entwicklung des Kortex, Foveation und Frontalstellung der
Augen treten immer mehr ungekreuzte retinofugale Ganglienzellfasern auf. Damit
einhergehend findet eine Erweiterung des neuronalen Schaltkreises des hOKN statt. Der
NOT bekommt zusätzlich zu seinem direkten Eingang von der kontralateralen Retina auch
einen schwächeren Eingang von der ipsilateralen Retina und einen immer stärker werdenden
binokularen Eingang vom ipsilateralen visuellen Kortex (VC) (Katze: Hoffmann &
Schoppmann, 1975; Berman, 1977; Ventre, 1985; Primaten: Lui et al., 1995).
Parallel zu der Erweiterung des hOKN-Schaltkreises nimmt die Symmetrie des monokularen
hOKN zu (Tauber & Atkin, 1968). Bei Katzen (Markner & Hoffmann, 1985) findet man
einen nahezu symmetrischen und bei Primaten (Affe: Koerner & Schiller, 1972; Mensch:
Van den Berg & Collewijn, 1988) schließlich einen völlig symmetrischen monokularen
hOKN. Der Grund für die Symmetriezunahme liegt darin, daß durch die Erweiterung des
hOKN-Schaltkreises beide Retinae mit jedem NOT verbunden werden und damit die
Aktivierung beider NOTs von einem Auge möglich wird (vgl. Abb. 1A). Beispielsweise
aktiviert das linke Auge den rechten NOT bei Reizbewegung nach rechts (über die direkte
Projektion) und den linken NOT bei Reizbewegung nach links (über die direkte ipsilatrale
Projektion und insbesondere über die indirekte binokulare Projektion vom ipsilateralen VC).
Folglich kann der hOKN über jedes Auge allein in beiden horizontalen Richtungen ausgelöst
werden.
Aus diesen anatomischen und verhaltensphysiologischen Befunden stellt sich die Frage, wie
der Symmetriegrad des monokularen hOKN bei einer Spezies verändert wird, bei der die
Anzahl der ipsilateral verlaufenden retinalen Ganglienzellfasern infolge einer genetischen
Mutation reduziert ist (vgl. Abb. 1B). Solche Mutanten sind Albinos.
III. Albinismus bei Säugetieren
Albinotische Säuger zeigen eine Vielzahl von Fehlbildungen und -funktionen, insbesondere
in ihrem visuellen System. Anatomisch findet man das Fehlen von Pigment in der Haut und /
oder im retinalen Pigmentepithel, was mit einer Hypoplasie der Retina und abnormen
retinofugalen Projektionen einhergeht. Am optischen Chiasma der Albinos kreuzen mehr
Ganglienzellaxone als bei normal-pigmentierten Individuen derselben Art (Jeffery et al.,

Einleitung 4
1997; Ilia & Jeffery 1999; Morgan et al., 1987; Stone et al., 1978b). Die Folge dieser
abnormen Überkreuzung ist eine partiell falsche Abbildung des Gesichtsfeldes auf das
Corpus geniculatum laterale (CGL), die sich bis zum visuellen Kortex fortsetzt. Der visuelle
Kortex versucht zwar die Störung seiner retinotopischen Organisation durch Reorganisation
zu korrigieren, trotzdem bleibt die kortikale Repräsentation der visuellen Umwelt partiell
abnorm (Guillery 1971, 1974, 1990; Apkarian et al., 1991; Sanderson et al., 1974). Die
retinoprätectale Projektion ist ebenfalls von
der Reduktion der ipsilateralen retinofugalen
Fasern betroffen (Maus: Balkema et al., 1981;
Pak et al., 1987; Kaninchen: Klooster et al.,
1983; Frettchen: Morgan et al., 1987; Zhang
& Hoffmann, 1993; Siamesiche Katze:
Guillery, 1974; Ault et al. 1995; Albinokatze:
Creel et al., 1982; Ault et al., 1995; Tiger:
Guillery & Kaas, 1973; Mensch: Guillery et
al., 1975; Apkarian et al., 1991).
IV. Neuronaler hOKN-Schaltkreis des
Frettchens
Der hOKN-Schaltkreis des Iltisses (MUSTELA
PUTORIUS FURO, pigmentiertes Frettchen) ist
hinsichtlich seiner sensorischen NOT-
Eingänge zwischen dem der Ratte und dem
der Katze einzuordnen. Der Frettchen-NOT
erhält einen direkten, starken Eingang vom
kontralateralen Auge, einen direkten,
schwachen Eingang vom ipsilateralen Auge
und einen binokularen Eingang vom
ipsilateralen visuellen Kortex (Abb. 1A)
(Klauer et al., 1990; Zhang & Hoffmann,
1993). Entsprechend zeigt das pigmentierte
Frettchen einen monokularen hOKN, der
wesentlich symmetrischer ist als der
Abb. 1: Vereinfachtes Schema des neuronalen hOKN-Schaltkreises des pigmentierten (A) und albinotischen (B) Frettchens. Die gepunkteten grauen Pfeile in A zeigen den zusätz-lichen indirekten NOT-Eingang, der im Verlaufe der Evolution hinzugekom-men ist und beim Albinofrettchen (B) infolge der Reduktion der ipsilateralen retinofugalen Fasern weggefallen ist. Der Übersichtlichkeit halber sind nur die Projektionen vom linken Auge ein-gezeichnet und die dem NOT nachge-schalteten visuomotorischen Struktu-ren wurden weggelassen.
A pigmentiertes Frettchen
NOT NOT
VC VC
CGL CGL
?
NOT NOT
VC VC
CGL
B albinotisches Frettchen

Einleitung 5
monokulare hOKN der Ratte und in dieser Hinsicht dem monokularen hOKN der Katze
nahekommt (Hein et al., 1990).
Bei der albinotischen Form des Iltisses, dem Albinofrettchen, ist der direkte retinale NOT-
Eingang vom ipsilateralen Auge reduziert (Abb. 1B) (Zhang & Hoffmann, 1993). Diese
Reduktion ist eine Folge der allgemeinen Reduktion der ipsilateral projizierenden zugunsten
der kontralateral projizierenden retinofugalen Fasern, die bei allen höheren albinotischen
Säugern beobachtet wird (Frettchen: Guillery 1971; Huang & Guillery, 1985; Morgan
et al., 1987).
V. Zielsetzungen und Hypothesen
Die vorliegende Arbeit wird methodisch in einen Verhaltensteil und einen
elektrophysiologischen Teil gegliedert. Beide Teile werden getrennt dargestellt und
diskutiert. Es werden folge Fragestellungen bearbeitet:
Teil I: Verhalten
Aufgrund der eingangs beschriebenen Anatomie des neuronalen hOKN-Schaltkreises und
des entsprechenden optokinetischen Verhaltens des pigmentierten Frettchens unter
monokularer Bedingung, scheint das Albinofrettchen ein geeigneter Kandidat zu sein für
Untersuchungen bezüglich der Symmetrie des monokularen hOKN.
• Die erste Hypothese ist, daß die Binokularität des kortikalen NOT-Eingangs beim
Albinofrettchen infolge der Reduktion der ipsilateralen retinofugalen Projektionen
schwächer als beim pigmentierten Frettchen ausgebildet ist. Deshalb müßte der
monokulare hOKN des albinotischen Frettchens asymmetrischer sein als der des
pigmentierten Frettchens. Zur Überprüfung dieser Hypothese werden Augenbewegungs-
messungen unter monokularer Bedingung während optokinetischer Ganzfeldreizung
durchgeführt.
Die Augenbewegungsmessungen entdecken überraschenderweise einen schweren
okulomotorischen Defekt bei den Albinofrettchen. Dieser Befund wirft weitere Fragen auf,
deren Klärung den Ursprung des Verhaltensdefizites funktionell eingrenzen soll:
• Liegt der Grund für das okulomotorische Verhaltensdefizit in einem neuronalen Defekt,
der dem NOT nachgeschaltet ist?
Die Hypothese ist, daß das Verhaltensdefizit der Albinos durch einen motorischen Defekt
verursacht ist. Zur Klärung dieser Frage wird der NOT von Albinofrettchen mit Hilfe von
Muscimol einseitig und reversibel chemisch inaktiviert. Damit wird getestet, ob das

Einleitung 6
künstlich erzwungene Aktivitätsungleichgewicht zwischen dem NOT der injizierten und
dem NOT der nicht-injizierten Hirnseite zu einem Spontannystagmus führt. Das
Auftreten eines Spontannystagmus wird gegen einen dem NOT nachgeschalteten Defekt
sprechen und diese Hypothese widerlegen. Zur Kontrolle wird das Muscimol-Experiment
auch mit einem pigmentierten Frettchen durchgeführt.
• Ist das Albinofrettchen überhaupt in der Lage sich visuell zu orientieren?
Die Hypothese ist, daß das Albinofrettchen aufgrund eines defekten optischen Apparates
oder eines allgemeinen neuronalen Defektes gänzlich blind ist und seine Blindheit
unbemerkt geblieben ist, da es sich besonders gut olfaktorisch orientieren kann
(Apfelbach, 1973; Apfelbach & Wester, 1977) und es immer nur in seiner gewohnten
Umgebung beobachtet wurde. In diesem Zusammenhang wird die Frage gestellt,
inwiefern das zentrale Gesichtsfeld noch funktionell ist, welches auf die temporale
Hemiretina projiziert wird, deren Ausgangsfasern abnormerweise am optischen Chiasma
kreuzen. Zur Beantwortung dieser Fragestellungen werden albinotische und pigmentierte
Frettchen einem Sehtest unterzogen, der dem Perimetrietest von Sherman (1973)
angelehnt ist.
Teil II: Neurophysiologie
Aus der Entdeckung des hOKN-Ausfalls beim Albinofrettchen resultieren folgende Frage-
stellungen, die im elektrophysiologischen Teil der vorliegenden Arbeit bearbeitet werden:
• Findet sich in der Aktivität der NOT-Ausgangsneuronen ein Korrelat zum optokineti-
schen Verhaltensdefizit der Albinofrettchen?
Zur Beantwortung dieser Frage werden an narkotisierten albinotischen und pigmentierten
Frettchen NOT-Neuronen hinsichtlich ihrer Richtungsspezifität elektrophysiologisch
charakterisiert und miteinander verglichen.
Diese Experimente decken ein abnormes physiologisches Verhalten im NOT des
Albinofrettchens auf. Hieraus ergeben sich folgende Fragen und Hypothesen:
• Inwiefern sind der retinale und / oder der kortikale NOT-Eingang für die abnorme
Physiologie des NOTs des Albinofrettchens verantwortlich?
Diese Frage basiert auf der Hypothese, daß beim pigmentierten Frettchen die Retina und
/ oder der Kortex über ihre Projektionen zum NOT die Richtungsspezifität der NOT-
Ausgangsneuronen determinieren und folglich auch für den Verlust der
Richtungsspezifität beim Albinofrettchen verantwortlich sein könnten. Zur Überprüfung
dieser Hypothese wurden beide NOT-Eingänge mit Hilfe antidromer und orthodromer

Einleitung 7
elektrischer Reizung bei albinotischen und pigmentierten Frettchen untersucht und
miteinander verglichen.
• Eine weitere Hypothese ist, daß die Unterentwicklung der Retina des Albinofrettchens
eine Veränderung des relativen Vorkommens an ON-Ganglienzellen und somit auch an
ON-Zellen im superioren Colliculus (SC) bedingt. Deshalb wird das relative
Vorkommen von ON-, ON-OFF und OFF-Neuronen im SC ermittelt und zwischen
albinotischen und pigmentierten Frettchen verglichen. Ein Unterschied im relativen
Anteil dieser Ganglienzelltypen zwischen dem Albinofrettchen und dem pigmentierten
Frettchen würde indirekt einen abnormen retinalen NOT-Eingang beim Albinofrettchen
anzeigen, weil der direkte retinale Eingang des NOTs von ON-Ganglienzellen gebildet
wird.

Material und Methoden 8
Material und Methoden
I. Teil I: Verhalten
A. Versuchstiere
Es wurden insgesamt 36 albinotische und 16 pigmentierte Frettchen (MUSTELA PUTORIUS
FURO) beiderlei Geschlechts im Alter von fünf bis zwölf Monaten verwendet. Die Tiere
stammten aus der Zucht des Instituts für Allgemeine Zoologie und Neurobiologie der Ruhr-
Universität Bochum und wogen zwischen 450 und 1500 g. Die Albinofrettchen besaßen ein
gelblich-weißes Fell und rosa-rot leuchtende Augen, während die pigmentierten Tiere ein
schwarz-braun gemustertes Fell und dunkle, schwarze Augen hatten. Die Tiere waren agil
und zeigten weder Verhaltensstörungen noch irgendwelche augenfälligen Defizite. Alle
Versuche waren von den lokalen Behörden und der Ethikkommission genehmigt und ihre
Durchführung erfolgte nach dem deutschen Tierschutzgesetz und den internationalen
Richtlinien für Tierversuche „Principles and Policies of Animal Use at NIH“ (Investigator´s
Handbook „Using Animal in Intramural Research: Guidelines for Investigators“, NIH,
1985).
B. Fixierung der Versuchstiere
Um die Augenbewegungsmessungen durchführen zu können, war es notwendig die
Frettchen möglichst in eine unbewegliche Körperhaltung zu bringen. Dazu kamen sie in eine
Röhre, deren vorderes Ende einen verengten Durchmesser besaß (vgl. Abb. 2). Für die
unterschiedlich großen Tiere standen drei unterschiedlich große Röhren zur Verfügung, so
daß die Tiere bequem eingespannt werden konnten. Die in der Institutswerkstatt der
Allgemeinen Zoologie und Neurobiologie gebauten Röhren bestanden aus Hartplastik und
hatten die Maße (Länge / Durchmesser) 45 cm / 9 cm, 45 cm / 7,5 cm und 32 cm / 3,0 cm.
Der verengte Durchmesser am vorderen Ende betrug entsprechend 4 cm, 3,5 cm und 3,0 cm,
so daß das Tier seinen außerhalb der Röhre ragenden Kopf nicht durchziehen konnte. Der

Material und Methoden 9
Kopf konnte zusätzlich mit Hilfe eines passend gebauten Kopfgeschirrs an einer
Haltevorrichtung in horizontaler Lage befestigt werden.
C. Horizontale optokinetische Reizung
Die horizontale optokinetische Ganzfeldreizung erfolgte in einer optokinetischen
Reiztrommel (∅ 90 cm, Höhe 100 cm), in die die Röhre mit dem Versuchstier gestellt
wurde. Der Abstand zwischen den Augen des Tiers und der inneren Wand betrug ca. 25 cm.
Die innere Wand der Trommel war mit einem schwarz-weißen Zufallspunktemuster
ausgekleidet, dessen Punktgröße entweder ca. 2,3° (feines Reizmuster) oder ca. 10,7°
Sehwinkel (grobes Reizmuster) betrug. Die Trommel hing an einem Seil senkrecht von der
Decke des Raumes. Mittels eines Elektromotors konnte sie kontinuierlich mit verschiedenen
Winkelgeschwindigkeiten um ihre senkrechte Längsachse im Uhrzeiger- (cw) und gegen den
Uhrzeigersinn (ccw) rotiert werden. Der Innenraum der Trommel war beleuchtet, so daß der
Helligkeitskontrast (K) zwischen den weißen und den schwarzen Punkten ca. 87% betrug
(berechnet nach der Formel [K=(a–b)/(a+b)]×100; a = Leuchtdichte der weißen und b =
Leuchtdichte der schwarzen Punkte). Dabei betrug die Leuchtdichte der schwarzen Punkte
2 cd/m2 und die der weißen Punkte 30 cd/m2 (gemessen mit dem Luminanzmeßgerät Minolta
luminance meter). Durch Klopfen auf die Trommel konnten die Tiere auch akustisch gereizt
und dadurch in einem besonders aufmerksamen Zustand gehalten werden.
Vorrichtung für die Befestigung des Geschirrs: Grundfläche der Stifte: 1,5 cm × 1,5 cm Höhe der beiden Stifte: 3 cm Abstand der beiden Stifte: 5,5 cm
Röhre
optokinetische Trommel
Vorrichtung zur Befestigung des Geschirrs
Abb. 2 skizziert die Röhre, in die die Versuchstiere während der optokinetischen Reizung in die Trommel gebracht wurden. Der Umriß der optokinetischen Trommel ist ebenfalls gezeigt (nicht maßstabsgetreu); der Drehpfeil symbolisiert die Rotationsrichtung im Uhrzeigersinn (cw).

Material und Methoden 10
In dieser Weise wurden die Versuchstiere binokular und / oder monokular (nur linkes Auge
sehend, nur rechtes Auge sehend) mit einem oder mit beiden Reizmustern optokinetisch
gereizt. Der Einsatz von zwei Reizmustern mit unterschiedlichen Punktgrößen war
notwendig, da bekannt ist, daß die Auslösbarkeit des hOKN (insbesondere bei Albinos) von
der Punktgröße des Reizmusters abhängen kann (Sirkin et al., 1985). Die Reizgeschwindig-
keiten betrugen 5, 10, 20, 50 und 100°/s jeweils in cw- und in ccw-Richtung. Dieser
Geschwindigkeitsbereich wurde gewählt, weil aus Hein et al. (1990) bekannt ist, daß die
pigmentierten Frettchen solche Reizgeschwindigkeiten mit einer relativ hohen Folgegüte
beantworten können. Für die monokularen Tests wurde eines der beiden Augen mit einer
lichtundurchlässigen Kappe verdeckt. Die Kappe hatte die Form einer hohlen Halbkugel mit
einem Durchmesser von 1,5 cm und wurde am Kopf des Tiers angebracht.
Ein Versuchstier wurde ein Mal pro Tag in einer Sitzung von ca. je einer Stunde getestet. Die
Punktgröße des Reizmusters wurde nur zwischen verschiedenen Sitzungen geändert.
Innerhalb einer Sitzung wurden mehrere Reizperioden (im Falle der spontanen
Augenbewegungen „Aufnahmeperiode“) von 30 s oder 60 s Dauer durchgeführt. Die
Testbedingungen (Okularität, Reizgeschwindigkeit und Reizrichtung) blieben während einer
Reizperiode konstant und wurden immer nur zwischen den Reizperioden geändert.
Nicht alle Versuchstiere wurden unter allen möglichen Testbedingungen untersucht, so daß
die Anzahl der Tiere, die unter einer bestimmten Bedingung getestet wurde, in vielen Fällen
unter der Gesamtanzahl (33 Albino- und zwölf pigmentierte Frettchen) liegt. Tabelle 1 zeigt
die Anzahl (N) der Tiere, die jeweils mit einem der beiden Muster unter den verschiedenen
Okularitätsbedingungen getestet wurden.
Tab. 1: Anzahl der Versuchstiere, bei denen der hOKN durch Reizung in der Trommel jeweils mit den verschiedenen Reizparametern (Punktgröße des schwarz-weißen Zufallspunktemusters, binokular / monokular) getestet wurde. N = Anzahl der Individuen, die unter den verschiedenen Bedingungen horizontal optokinetisch getestet wurden; X = bei einigen Tieren probeweise getestet, jedoch hier nicht gezeigt, — = nicht getestet.
Reizmuster Okularitätsbedingung N (Albinos) N (Pigm.)
rechts sehend 20 — monokular links sehend 20 —
fein (Punktgröße: 2,3° Sehwinkel) binokular 20 12
rechts sehend X — monokular links sehend X —
grob (Punktgröße: 10,7° Sehwinkel) binokular 33 X

Material und Methoden 11
Albinofrettchen: Insgesamt wurden 33 Albinofrettchen optokinetisch getestet. Von diesen
wurden zunächst 20 Tiere mit dem feinen Reizmuster unter allen drei Okularitäts-
bedingungen (binokular, nur rechtes oder nur linkes Auge sehend) gereizt. Anschließend
wurden alle 33 Albinos mit dem groben Reizmuster binokular und einige auch monokular
gereizt. Die wenigen monokularen Messungen mit dem groben Reizmuster werden hier nicht
vorgestellt, da sie sich in ihrem Ergebnis nicht von den binokularen oder den monokularen
Messungen mit dem feinen Muster unterschieden.
Pigmentierte Frettchen: Insgesamt wurden zwölf pigmentierte Frettchen binokular mit dem
feinen Reizmuster optokinetisch getestet. Monokulare Messungen wurden nicht
durchgeführt.
Kontrollmessungen: Als Kontrolle dienten die spontanten Augenbewegungen in Helligkeit
und in Dunkelheit. Für die Aufnahmen in Helligkeit befanden sich die Tiere in der
stehenden, von innen beleuchteten Trommel und konnten auf das Reizmuster blicken. Die
Augenbewegungen der Albinofrettchen wurden auf diese Weise sowohl binokular als auch
monokular bei eingesetztem feinen Muster und nur binokular bei eingesetztem groben
Muster gemessen. Die Messungen mit den pigmentierten Frettchen erfolgten nur binokular
bei eingesetztem feinen Reizmuster. Während der Aufnahmen in völliger Dunkelheit befand
sich die Röhre mit dem Tier in einem durch eine Holzwand abgetrennten, völlig dunklen (0
cd/m2) Raumabschnitt. Die Kontrollmessungen sollten erstens Referenzwerte liefern und
zweitens bei den Albinofrettchen eventuelle okulomotorische Instabilitäten aufdecken, zumal
bei albinotischen Säugern oft ein Spontannystagmus beobachtet wurde (Collewijn et al,
1978, 1985; Precht & Cazin, 1979).
Planetarium: Einige visuelle Stimulationen mit Albinofrettchen erfolgten nicht mit dem
Zufallspunktemuster in der Trommel, sondern mit einem Muster aus unterschiedlich großen,
zufällig im Raum verteilten projizierten Lichtpunkten („Planetarium“). Die Lichtpunkte
wurden mit Hilfe einer hohlen Metallkugel erzeugt, in derem Inneren sich eine Glühbirne
befand. Die Wand der Kugel war randomisiert durchlöchert, so daß durch das Einschalten
der Glühbirne Lichtpunkte in das gesamte abgedunkelte Labor projiziert wurden. Die Kugel
konnte mit Hilfe eines Motors mit verschiedenen Geschwindigkeiten horizontal in cw- und
ccw-Richtung rotiert werden. Das Ergebnis dieser Reizungen unterschied sich nicht von dem
Ergebnis der Reizungen mit den beiden Zufallspunktemustern und wird im Ergebnisteil nicht
dargestellt, sondern nur in der Diskussion erwähnt.

Material und Methoden 12
D. Registrierung der Augenbewegungen
Die Messungen der horizontalen Augenbewegungen während der optokinetischen Reizung
erfolgte mit Hilfe des Elektrookulogramms (EOG). Als EOG-Ableitelektroden dienten feine
(∅ 0,1 mm), ca. 15 cm lange, lackisolierte Silberdrähte, deren chloriertes 0,5 cm langes Ende
mit Hilfe einer Injektionskanüle unter die Haut an den beiden äußeren Augenwinkeln
eingestochen wurde. Dieses sogenannte bitemporale Abgreifen der corneoretinalen
Potentialdifferenz eignet sich bei Registrierungen von konjugierten Augenbewegungen am
besten (Collewijn, 1999).
Das abgegriffene Potential wurde mit Hilfe eines Verstärkers (Eigenbau) 2000-fach
verstärkt, auf einem Oszillographen dargestellt und gleichzeitig zu einem PC (Deskpro 286,
Compaq) geleitet. Der PC stellte es online in Abhängigkeit von der Zeit mit einer zeitlichen
Auflösung von 100 Hz als EOG-Spur auf dem Monitor dar. Am Ende einer Reizperiode
(Aufnahmeperiode), deren Dauer mit Hilfe des Aufnahmeprogramms auf 30 s oder 60 s
eingestellt war, wurde die EOG-Spur zwecks späterer Offline-Analyse als Datei
abgespeichert.
Es handelt sich hierbei um ein unkalibriertes EOG, welches keine Aussage über die
Amplitude der Augenbewegungen in Grad Sehwinkel erlaubt. Deshalb stellt die y-Achse der
Augenspuren das abgegriffene EOG-Potential in mV dar.
E. Analyse und Auswertung des EOGs
1. Bearbeitung und graphische Darstellung der EOG-Spuren
Die Offline-Analyse der EOG-Spuren bestand zunächst aus dem Zuschneiden. Dabei wurden
diejenigen Teile der EOG-Spur herausgeschnitten, die keinen optokinetisch ausgelösten
Augenbewegungen entsprachen. Solche Abschnitte wurden nicht in die weitere Auswertung
einbezogen. Abbildung 3 zeigt beispielhaft eine EOG-Spur vor (A) und nach (B) dem
Zuschneiden. Anschließend erfolgte die Berechnung der Steigungen, die jeweils zwischen
zwei 10 ms entfernten benachbarten Punkten entlang der übriggebliebenen Spur bestanden.
Die berechneten Steigungen mit der Einheit mV/s entsprechen der relativen Geschwindigkeit
der Augenbewegungen während der EOG-Registrierung. Der Medianwert dieser Steigungen
(Steigungsmedian) ist ein relativer Wert, welcher über den Differentialquotienten der EOG-
Spur ein relatives Maß für die Augengeschwindigkeit ergab.

Material und Methoden 13
Im Falle der Kontrollmessungen wurden nahezu konstante EOG-Potentiale aufgenommen
und infolge dessen haben die allermeisten Steigungsmediane den Wert Null oder einen Wert
nahe bei Null (Steigungsmedian = 0). Im Gegensatz dazu liefern die für den hOKN typischen
Sägezahn-förmigen EOG-Spuren Steigungsmediane ungleich Null. Denn da die langsamen
Folgephasen des hOKN wesentlich mehr Zeit in Anspruch nehmen als die schnellen
Rückstellsakkaden, stammen die meisten Steigungen einer Augenspur von diesen langsamen
Folgephasen und bestimmen somit den Medianwert der Gesamtheit aller Steigungen. Das
Vorzeichen des Steigungsmedians einer EOG-Spur wird definitionsgemäß von der Richtung
des hOKN bestimmt, wobei ein hOKN nach rechts (cw) einen positiven (positive Steigung
der langsamen Phasen) und ein hOKN nach links (ccw) einen negativen Steigungsmedian
(negative Steigung der langsamen Phasen) liefert. Die Tatsache, daß die Rückstellsakkaden
in ihrer Richtung den langsamen Folgephasen entgegengesetzt sind und somit Steigungen
mit entgegengesetztem Vorzeichen liefern, wirkt sich auf das Vorzeichen des Medianwertes
nicht aus.
Von jedem Tier wurde ein Steigungsmedian für jeden getesteten Reizparameter berechnet.
Wenn mehrere EOG-Spuren unter demselben Reizparameter aufgenommen wurden, wurde
das arithmetische Mittel der Steigungsmediane der einzelnen Spuren verwendet.
Anschließend wurde die Häufigkeitsverteilung der Steigungsmediane der verschiedenen
Tiere für jeden Reizparameter (verschiedene Reizgeschwindigkeiten, Kontrollmessungen in
Abb. 3: Beispiele für das Zuschneiden einer EOG-Spur eines pigmentierten Frettchens. A:Vollständige 30-s-lange EOG-Spur vor dem Zuschneiden. Die Aufnahme erfolgte während binokularer optokinetischer Reizung im cw-Richtung mit dem feinen Reizmuster. Die Reizgeschwindigkeit betrug 50°/s. B: Die EOG-Spur aus A nach dem Zuschneiden. In diesem Falle wurde der Spurabschnitt ab 13,8 s herausgeschnitten, weil er nicht optokinetisch ausgelösten Augenbewegungen entspricht. Die ersten 13,8 s zeigen die typische Sägezahn-Form des hOKN und wurden für die Berechnung des Medians zugrunde gelegt, der in diesem Falle +4,15 mV/s beträgt. Abszisse: Zeit; Ordinate: bitemporal abgegriffene corneoretinale Potentialdifferenz.
Zeit [ms]
0 5000 10000 15000 20000 25000 30000
1 mV
A
B

Material und Methoden 14
Helligkeit und in Dunkelheit) berechnet und graphisch dargestellt. Da die errechneten
Steigungsmediane innerhalb des Bereichs ±6 mV/s liegen, zeigen die x-Achsen aller
Histogramme den entsprechenden Achsenabschnitt [–6; +6], wobei er in 0,25 breite Klassen
unterteilt ist. Die y-Achse gibt die absolute Anzahl der Medianwerte, die in die jeweiligen
Klassen fallen. Da für jede Versuchsbedingung ein Medianwert von jedem Tier errechnet
wurde, entspricht die Anzahl der Medianwerte innerhalb einer Graphik der Anzahl der Tiere,
die unter der jeweiligen Bedingung getestet wurden.
2. Statistik
Es wurden verteilungsfreie Tests angewandt. Zum Vergleich der Steigungsmediane, die von
derselben Tiergruppe für verschiedene Reizbedingungen errechnet wurden, wurde der
WILCOXON SIGNED RANK TEST angewandt, dessen Nullhypothese („zwei unterschiedliche
Reizbedingungen haben keinen unterschiedlichen Effekt auf die optokinetischen Augen-
bewegungen derselben Tiergruppe“) bei einem P < 0,05 abgelehnt wurde. Zum Vergleich der
Steigungsmediane der beiden unterschiedlichen, ungleich großen Tiergruppen (albinotische
und pigmentierte Tiere) wurde der MANN-WHITNEY RANK SUM TEST angewandt. Die Null-
hypothese („die Medianwerte der einen Gruppe unterscheiden sich nicht von den Median-
werten der anderen Gruppe“) wurde auch in diesem Falle bei einem P < 0,05 abgelehnt. Ein
P > 0,05 zeigt in beiden Tests, daß kein signifikanter Unterschied zwischen den beiden
miteinander verglichenen Gruppen von Steigungsmedianen existiert.
Zum Vergleich zweier Gruppen von Steigungsmedianen hinsichtlich ihrer Varianz wurde der
LEVENE S HOMOGENEITY OF VARIANCE TEST gerechnet, der bei einem P < 0,05 einen
signifikanten Unterschied anzeigt.
F. Inaktivierung des Prätektums
Im Anschluß an den oben beschriebenen hOKN-Test wurden drei albinotische und ein
pigmentiertes Tier für die NOT-Inaktivierungsstudie mit Muscimol verwendet. Diese Tiere
stellen eine Teilgruppe der im hOKN-Test untersuchten Tiere dar.
1. Injektionsmethode und Vorbereitung der Versuchstiere
Um das Muscimol mehrmals, d.h. an verschiedenen Tagen, an derselben Stelle injizieren zu
können, wurde senkrecht über einem Prätektum eine rostfreie Kanüle (Sterican 26 G x 1",
Außendurchmesser 450 µm, Fa. Braun Melsungen) implantiert (gemäß Fischer et al., 1998;

Material und Methoden 15
vgl. auch Bonaventure et al., 1992), die als Führungsröhrchen (guide tube) für die Injektions-
nadel (Micro-canula G33, 18073-90 FST, Außendurchmesser 203,2 µm; Innendurchmesser
101,6 µm) diente. Am oberen, aus dem Tier ragenden Ende des Führungsröhrchens wurde
zuvor in der Feinmechanikerwerkstatt des Instituts für Allgemeine Zoologie und
Neurobiologie ein Metallgewinde angebracht, so daß es mit einem rostfreien Schraub-
verschluß luftdicht verschlossen werden konnte. In der Zeit zwischen den Injektionen befand
sich im Führungsröhrchen ein passendes Stilett aus rostfreiem Stahl, das eine Verstopfung
durch Hirngewebe und / oder Wundsekrete verhinderte. Die Länge der Injektionsnadel war
so gewählt, daß sie gerade aus dem Führungsröhrchen ragte, wenn sie bis zum Anschlag
eingeführt war. Das hintere Ende der Injektionsnadel war mit einem durchsichtigen, 45 cm
langen Schlauch aus Vinyl (Abimed) verbunden. Der Innendurchmesser des Schlauchs
betrug 0,38 mm und sein Außendurchmesser 2,3 mm. Am anderen Ende des Schlauchs
befand sich eine 5 µl-Hamiltonspritze, mit deren Hilfe zuerst Wasser, dann etwas Luft und
schließlich die Muscimol-Lösung in den Schlauch gesaugt wurde. Der Plastikschlauch war in
1 µl-Abschnitte unterteilt, so daß man das injizierte Volumen an der Bewegung der Luftblase
im Schlauch verfolgen konnte.
Die Implantation des Führungsröhrchens erfolgte unter tiefer Narkose. Hierzu wurden die
Tiere durch die subcutane (s.c.) Injektion eines Ketamin/Xylazin-Gemisches [Ketamin:
40 mg/kg KG (Hostaket, 100 mg/ml), Xylazin: 0,6 mg/kg KG (Rompun, 20 mg/ml)] initial
narkotisiert und in einem stereotaktischen Halter mit Ohrstiften und Beißstange befestigt.
Eine zusätzliche s.c. Injektion von 0,0025 mg Atropinsulfat diente der Kreislaufstabilisie-
rung und Verhinderung von Schleimbildung in der Trachea. Der Kaumuskel, der sich beim
Frettchen über die Hälfte der Schädeloberfläche zieht und an einem Knochenkamm entlang
der Mittellinie des Schädels ansetzt, wurde mit einer s.c. Injektion von 0,5 ml
Prilocainhydrochlorid-Lösung (5,0 mg/ml, Xylonest) zusätzlich lokal betäubt. Danach wurde
er vom Schädelknochen stumpf abgelöst und zur Seite gelegt. Weitere Xylonest-Injektionen
erfolgten an den Druckstellen der Ohrstifte. Die Körpertemperatur wurde für die gesamte
Dauer der Narkose rektal gemessen und mit Hilfe eines Heizkissens und einer Decke auf
37°C (±1°C) gehalten. Da der gesamte Eingriff bis zur Implantation der Kanüle ca. vier bis
sechs Stunden dauerte (hauptsächlich wegen der relativ zeitraubenden Suche nach dem NOT
bzw. Prätektum, siehe unten), mußte der Narkosezustand des Tieres mit zusätzlichen s.c.
Injektionen von jeweils 0,1 ml Ketaminlösung (100 mg/ml; je nach Körpergewicht alle 50
bis 80 Minuten) aufrechterhalten werden.

Material und Methoden 16
Nach einer Kraniotomie über dem Prätektum und dem superioren Colliculus (SC)
[stereotaktische Koordinaten der Schädelöffnung nach Horsley und Clarke: 0 - 3,5 mm
posterior der IAL und 0,5 - 3 mm lateral der Mittellinie; zum Vergleich die Position des
NOT aus Klauer et al., (1990): 1,5 - 2,5 mm anterior und 2,5 - 3,0 mm lateral] wurden in der
Nähe der Schädelöffnung zwei rostfreie Schrauben in den Knochen gesetzt, die später zur
Befestigung des Führungsröhrchens dienten. Die Suche nach dem Prätektum bzw. NOT
erfolgte mittels extrazellulärer Ableitungen unter Verwendung von Wolfram-in-Glas-
Mikroelektroden (unisolierte Spitze ca. 6 µm lang und ca. 1 - 8 µm dick; 2 - 4 MΩ). Dabei
orientierte man sich an der relativen Lage des SC zum Prätektum und der retinotopischen
SC-Karte. Beim pigmentierten Tier konnte der NOT anhand seiner charakteristischen
richtungsspezifischen Antwort auf horizontale optokinetische Stimulation mit einem großen
Zufallspunktemuster aus schwarzen und weißen Punkten eindeutig identifiziert werden (eine
detailierte Beschreibung der Lokalisation des NOT befindet sich im Abschnitt „Material und
Methoden“ des neurophysiologischen Teils dieser Arbeit). Danach wurde die Ableitelektrode
aus dem Hirn gefahren und an derselben Stelle das Führungsröhrchen senkrecht
eingestochen, allerdings nur bis zu einer Tiefe von ca. 1 mm oberhalb der zuvor
identifizierten prätektalen Oberfläche. Dadurch sollte eine unerwünschte mechanische
Beeinträchtigung des darunterliegenden Prätektums bzw. NOT vermieden werden.
Anschließend wurde mit Zahnzement (Technovit 4004) das Führungsröhrchen mit dem
Schädelknochen und den zwei Verankerungsschrauben festgeklebt. Auf diese Weise war das
Führungsröhrchen im Hirn fixiert und erlaubte mehrmals an genau derselben Stelle
Muscimol zu injizieren. Die freigebliebene Schädelöffnung um das Führungsröhrchen wurde
mit blutstillendem Gelatineschwamm (Gelita Tampon) bedeckt. Im letzten Präparationsschritt
wurden der Kaumuskel und die Kopfhaut auf den Schädel um den Schraubverschluß des
Führungsröhrchens gelegt und mit dem (der) entsprechenden Kaumuskel (Kopfhaut) auf der
gegenüberliegenden Seite chirurgisch zusammengenäht. Zuletzt wurde auf die Wundnaht
eine Neomycin/Bacitracin-Salbe (Nebacetin) zwecks Prävention von Infektionen
aufgetragen. Ein bis zwei Stunden nach der letzten Ketamin-Injektion waren die Tiere aus
der Narkose gänzlich aufgewacht und konnten in ihr Gehege zu ihren Artgenossen
freigelassen werden. Sie zeigten keine Anzeichen von Infektionen und ihr Körpergewicht
blieb stabil.

Material und Methoden 17
2. Muscimol-Injektionen
Frühestens 40 Stunden nach der Implantation der Kanüle waren die Tiere bereit für die
Muscimol-Injektionen. Nach dem Einspannen in die Röhre wurde 1 µl einer 0,1%-igen
Muscimol-Lösung (1 mg/ml) (vgl. Fischer et al., 1998) auf das Prätektum mit einer Rate von
ca. 0,5 µl/min injiziert. Danach wurde so schnell wie möglich mit der Registrierung der
Augenbewegungen in Helligkeit (2 - 5 cd/m2), in Dunkelheit (0 cd/m2) und unter
optokinetischer Ganzfeldreizung in der Trommel begonnen. Die Zeitspanne zwischen dem
Injektionsende und der Aufzeichnung der ersten EOG-Spur betrug mindestens zwei und
höchstens vier Minuten.
3. Farb-Injektionen
Zur farblichen Markierung der Injektionsorte wurde nach der letzten Muscimol-Injektion ein
Fluoreszenzfarbstoff injiziert [bei zwei Albinofrettchen jeweils 0,2 µl eines roten
Textmarkerfarbstoffes und beim dritten Albinofrettchen und beim pigmentierten Frettchen
jeweils 0,2 µl einer 2%-igen Lösung Granular Blue (EMS-POLYLOY)]. Unter der Annahme,
daß sich die beiden Farbstoffe in ähnlicher Weise im Hirn ausbreiten wie das Muscimol,
sollte ihre Ausbreitung im bzw. auf dem Mittelhirn ein Bild über das Ausbreitungsgebiet des
injizierten Muscimols liefern. Außerdem sollten diese Injektionen nicht zu einer NOT-
Inaktivierung führen und somit keinen Effekt auf die Okulomotorik haben. Insofern kann
man sie auch als Kontrollinjektionen betrachten.
G. Sehtest
Mit fünf albinotischen und vier pigmentierten Frettchen wurde ein Perimetrietest
durchgeführt. Alle Tiere waren intakt und wurden auch im hOKN-Test eingesetzt. Der Test
sollte überprüfen, zum einen ob sich die Albinofrettchen visuell orientieren können oder
eventuell gänzlich blind sind und zum anderen ob sie ein funktionstüchtiges binokulares
Gesichtsfeld haben. Hierzu dienten die pigmentierten Frettchen als Kontolltiere.
1. Versuchsaufbau und Konditionierung
Der Test ist eine Version des Perimetrietests, der von Sherman (1973) und von Elekessy et
al. (1973) erfolgreich mit Katzen durchgeführt wurde. Das Experiment wurde von zwei
Personen auf einem Tisch (66 cm × 132 cm) durchgeführt. Der Tisch war in sechs 30°-
Sektoren unterteilt, die als L90°-, L60°-, L30°-, R30°-, R60°- und R90°-Sektor benannt

Material und Methoden 18
waren (vgl. Abb. 4A). Der Experimentator war immer auf die Hilfe eines Assistenten
angewiesen und befand sich auf der Seite des Startpunktes X. Sein Assistent stand auf der
gegenüberliegenden Seite des Fixationspunktes F. Vor der Durchführung des Tests wurden
die Versuchstiere unter binokularer Bedingung folgendermaßen konditioniert:
In der ersten Trainingsphase hielt der Experimentator mit seiner linken Hand den Kopf des
Tieres am Startpunkt X so, daß es möglichst geradeaus in 0°-Richtung auf den
Fixationspunkt F blickte. Auf der gegenüberliegenden Seite des Tisches klopfte sein
Assistent mit seinem Finger auf den Fixationspunkt F, um die Aufmerksamkeit des Tieres
auf diesen Ort zu lenken. Daraufhin ließ der Experimentator das Tier los. Wenn das Tier
sofort zum Fixationspunkt F lief, wurde es vom Assistenten anfänglich immer mit einem
Stück Trockenfutter belohnt. Pro Tag wurden die Tiere einmal in einer Sitzung von ca. 30
min Dauer trainiert. Vor jeder Trainingssitzung waren sie mindestens 20 Stunden futter-
depriviert. Nach zwei bis sechs Tagen lernten die Tiere schnell und ohne Umweg zum
Fixationspunkt F zu laufen.
In der zweiten Trainingsphase führte der Experimentator mit seiner rechten Hand
gleichzeitig mit dem Loslassen des Tieres einen visuellen Reiz in einen der 30°-Sektoren
seines Gesichtsfeldes ein. Der visuelle Reiz war eine runde schwarze Scheibe aus Pappe (ca.
∅ 3 cm), die an dem Ende eines feinen 90 cm langen Stabes aus durchsichtigem Plexiglas
befestigt war (vgl. Abb. 4B). Der Reiz wurde schnell von oben nach unten bis kurz über der
Oberfläche des Tisches bewegt und dort angehalten. Dabei wurde darauf geachtet, daß der
Reiz nicht den Tisch berührte, damit das Tier keine akustische Information über die
Reizposition zur Verfügung hatte. Wenn das Tier von seiner 0°-Laufrichtung durch den
visuellen Reiz abgelenkt wurde und zu diesem hinlief, belohnte der Assistent es wieder mit
einem Stück Trockenfutter, das er mit einer 20 cm langen Pinzette genau neben den visuellen
Reiz legte. Wenn das Tier den Reiz ignorierte und geradeaus zum Fixationspunkt F lief,
wurde es ab dieser Trainingsphase nur noch in jedem dritten Falle belohnt. Nach weiteren
ein bis drei Tagen mit jeweils einer solchen 30 minütigen Trainingssitzung hatten alle Tiere
gelernt, sofort und ohne Ablenkung zum visuellen Reiz zu laufen, wenn er präsentiert wurde.
Wurde der Reiz zwischendurch nicht präsentiert, so liefen sie weiterhin geradeaus zum
Fixationspunkt F. In beiden Fällen wurden sie dann vom Assistenten mit Trockenfutter
belohnt. Gemäß dieser Beschreibung wurden das Training und später der Test (siehe unten)
mit zwei albinotischen und zwei pigmentierten Frettchen durchgeführt. Beim Training / Test
der übrigen drei Albinos und zwei pigmentierten Tiere wurde eine Röhre aus durchsichtigem
Plexiglas in Längsrichtung auf den Tisch befestigt, so daß ihr vorderes Ende sich genau am

Material und Methoden 19
Startpunkt X befand (vgl. Abb. 4A). Der Experimentator setzte das Tier am hinteren Ende in
die Röhre und die Aufgabe des Tieres war es, durch die Röhre und dann über den Tisch
geradeaus zum Fixationspunkt F zu laufen. In diesem Falle wurde der visuelle Reiz genau in
dem Augenblick präsentiert, in dem das Tier aus der Röhre kam. Der Reiz wurde immer im
gleichen Abstand zum Startpunkt X, d.h. auf dem ca. 33 cm entfernten Halbkreis (vgl. Abb.
4A) gezeigt. Die Belohnungsregeln wurden nicht geändert.
In Abbildung 4C ist das Gesichtsfeld des Frettchens in seiner gesamten horizontalen Breite
skizziert. Es ist ca. 270° Sehwinkel groß und erstreckt sich von ca. 135° rechts-lateral nach
ca. 135° links-lateral (Law et al., 1988). Die geradeaus gerichtete Blickachse (0°) teilt den

Material und Methoden 20
ca. 76° großen binokularen Anteil (vgl. Law et al., 1988 und Quevedo et al., 1996) des
superioren frontalen Gesichtsfeldes (hell grau schraffiert) symmetrisch in einen linken und in
einen rechten Sektor auf. Aus dem Vergleich zwischen Abbildung 4A und 4C geht hervor,
daß der Perimetrietest nur 180° des Gesichtsfeldes (jeweils 90° auf der linken und rechten
Seite) berücksichtigt. Die äußeren temporalen Gesichtsfeldanteile der beiden Augen
(dunkelgrau schraffiert) wurden nicht untersucht. Für die Fragestellung, insbesondere für die
Frage nach der Funktionstüchtigkeit des binokularen Gesichtsfeldes des Albinofrettchens,
waren die 180° völlig ausreichend.
2. Durchführung des Perimetrietests
Eine Testsitzung dauerte in Abhängigkeit von der Motivation des Tiers ca. 30 bis 60
Minuten, wurde einmal am Tag durchgeführt und bestand aus ca. 40 bis 160 Durchläufen
(Loslaufen des Tiers am Startpunkt X und Präsentation bzw. nicht Präsentation des visuellen
Reizes). Wenn sich das Tier im Falle der Präsentation des visuellen Reizes ohne Ablenkung
schnell und eindeutig dem visuellen Reiz zuwendete und zu ihm hinlief, bekam es vom
Assistenten die Belohnung wie in der Trainingsphase zuvor und der Durchlauf wurde als
positiv beantwortet notiert. In allen anderen Fällen (wie z. B. uneindeutige Zuwendung zum
visuellen Reiz weil über Umweg oder völliges Ignorieren des visuellen Reizes) wurde die
Antwort als negativ bewertet. Die Bewertung erfolgte sowohl vom Experimentator als auch
von seinem Assistenten, die beide unabhängig voneinander das Ergebnis des Durchlaufs
notierten. Die Reihenfolge, nach der der visuelle Reiz entweder in einem der verschiedenen
Sektoren präsentiert wurde, war vom Experimentator zufällig gewählt. Innerhalb eines
Sektors wurde der visuelle Reiz ebenfalls zufällig an verschiedenen Gradpositionen
präsentiert. Zwischen den Durchläufen mit Reizpräsentation wurden auch Durchläufe ohne
Reizpräsentation randomisiert durchgeführt. Die Bewertung dieser Durchläufe zwischen-
durch sollte ein relatives Maß für die Motivation der Tiere liefern, schnell und ohne Umweg
zum Fixationspunkt F zu laufen.
Jedes Tier wurde binokular in mindestens vier und monokular in drei Sitzungen getestet. Für
die monokulare Variante wurde ein Auge abgedeckt. Hierzu diente eine schwarze
lichtundurchlässige Haftschale aus Plastikfolie (0,5 mm dick), die wie eine Kontaktlinse
aufgesezt wurde und den ganzen Augapfel verdeckte. Bei drei albinotischen und allen vier
pigmentierten Frettchen wurde (unter monokularer Bedingung) der 60°-Sektor kontralateral
zum sehenden Auge weiter in zwei 15° große Teilsektoren (30-45°- und 45-60°-Sektor)

Material und Methoden 21
unterteilt (vgl. Abb. 4A). Mit jedem Versuchstier wurden mindestens 350 Durchläufe unter
binokularer und 100 Durchläufe unter monokularer Bedingung durchgeführt.
3. Auswertung und graphische Darstellung
Für jeden Sektor und für das Geradeauslaufen ohne Reizpräsentation wird jeweils die
Gesamtanzahl aller Durchläufe 100% gesetzt. Die Anzahl der positiv beantworteten Durch-
läufe wird tabellarisch in Prozent angegeben. Die Differenz zu 100% entspricht der Anzahl
der negativ beantworteten Durchläufe. Zur graphischen Darstellung wird ein Halbkreis
gewählt, der entsprechend der Einteilung des Tisches bzw. Gesichtsfeldes in Sektoren
eingeteilt ist (vgl. Abb. 4C). Der Radius des Halbkreises entspricht 100% und die Länge des
Pfeils in jedem einzelnen Sektor und in 0°-Richtung repräsentiert die Anzahl der positiv
beantworteten Durchläufe in Prozent. Auch hier gibt die Differenz zu 100% die Anzahl der
negativ beantworteten Durchläufe wieder.
H. Histologische Untersuchung
Die Gehirne der Muscimol-injizierten Tiere wurden zur Dokumentation des Implantations-
ortes der Injektionskanülen histologisch aufgearbeitet. Die Perfusion durch den linken
Herzventrikel erfolgte am Ende des Experimentes unter tiefer Narkose mit 1 Liter 0,9%-iger
NaCl-Lösung gefolgt von 1,5 Liter 4%-igem Paraformaldehyd in 0,1 M Phosphatpuffer (pH
7,4) und Succrose (57,6 g pro Liter Fixierlösung). Nach Öffnung des Schädels und
Entfernung der Dura mater wurde das Gehirn stereotaktisch geblockt und über Nacht in
derselben Fixierlösung nachfixiert. In den nächsten Tagen wurde das Gehirn zwecks
Entwässerung für mindestens 12 h jeweils in einer 10, 20 und 30%-igen Succrose-Lösung bis
zum Absinken aufbewahrt, so daß es anschließend gefahrlos schockgefroren werden konnte
(Isopentan, –70° C). Mit einem Gefriermikrotom (Microm HM 500 OM) wurden 50 µm dicke
Frontalschnitte des Mittelhirns und des darüberliegenden Kortex angefertigt, auf gelatinierte
Objektträger gebracht und für einige Tage in den Trockenschrank gelegt. Nachdem die
Schnitte getrocknet waren, wurden sie Klüver-Barrera- und / oder Nissl-gefärbt und unter
dem Lichtmikroskop (Zeiss Axiolab) untersucht. Auf diese Weise wurde die tatsächliche
Position der zuvor implantierten Injektionskanüle und des Injektionsortes dokumentiert.
Diese Vorgehensweise erlaubt allerdings keine eindeutige Identifizierung des NOT als
eigenständigen Kern, da weder die Nissl-, noch die Myelin-Färbung (Klüver-Barrera) klare
Abgrenzungen zwischen den verschiedenen prätektalen Kernen liefern (vgl. Nunes Cardozo

Material und Methoden 22
& Van der Want, 1987). Aus den elektrophysiologischen Experimenten, die erst im zweiten
Teil dieser Arbeit vorgestellt werden, ist aber bekannt, daß der NOT des Frettchens eine
längliche Form besitzt. Seine rostrocaudale Ausdehnung beträgt bis zu ca. 700 µm und seine
mediolaterale Ausdehnung ca. 200 µm. Er zieht sich in seiner ganzen Länge entlang der
lateralen, rostrocaudal verlaufenden Grenze des SC, der nach rostral hin schmaler wird.
Folglich befindet sich der rostrale Rand des NOT medialer als sein caudales Ende, so daß
seine Längsachse schräg von anteromedial nach posterolateral orientiert ist. Rostraler Rand
des SC und rostraler Rand des NOT liegen beim Frettchen in etwa nebeneinander (vgl. mit
Abbildungen in Zhang & Hoffmann, 1993). Deshalb wurde die leicht erkennbare rostrale
Begrenzung des SC als Landmarke gewählt, von der aus
die Entfernung (∆x) des Injektionsortes (Spitze der
Injektionskanüle) in rostrocaudaler (anterior-posterior)
Richtung angegeben wird. Zur Veranschaulichung der
∆x-Werte dient die Skizze in Abbildung 5. Ein positives
∆x bedeutet, daß der Injektionsort rostral des SC liegt
und gibt somit die kürzeste Entfernung zwischen dem
Injektionsort und dem rostralen Ende des NOT an. Ein
negatives ∆x gibt an, daß der Injektionsort caudal des
rostralen SC-Rands liegt.
Außerdem sollte die lichtmikroskopische Untersuchung
der Hirnschnitte bestätigen, daß der injizierte NOT auf
keinen Fall durch die implantierte Injektionskanüle
mechanisch beschädigt worden war.
SC
∆x =0
+∆x
-∆x
∆x = -1000 µm
NOT
caudal
rostral
Mittellinie
med
ial
lateral
Abb. 5: Skizze zur Erläute-rung der ∆x-Angaben. Nicht maßstabsgetreu. SC = rechter superiorer Colliculus.

Material und Methoden 23
II. Teil II: Neurophysiologie
Für den neurophysiologischen Teil wurden insgesamt 32 adulte Frettchen (MUSTELLA
PUTORIUS FURO) beiderlei Geschlechts im Alter von fünf bis zwölf Monaten mit einem
Körpergewicht von 450 bis 1680 g verwandt. Von den 32 Tieren waren 23 albinotisch und
neun pigmentiert. Vier pigmentierte Tiere wurden bei Marshall Farms (New Rose, NY)
erworben. Alle anderen Tiere waren aus der Zucht des Instituts für Allgemeine Zoologie und
Neurobiologie der Ruhr-Universität Bochum. Die pigmentierten Frettchen hatten ein braun-
schwarzes Fell und dunkle schwarze Augen, während die Albinos ein gelblich-weißes Fell
und rosa-rot leuchtende Augen besaßen. Alle Tiere waren intakt und zeigten keine
Verhaltensstörungen oder andere augenfällige Defizite. Alle Experimente waren von den
lokalen Behörden und der Ethikkommission genehmigt und ihre Durchführung erfolgte nach
dem deutschen Tierschutzgesetz und den internationalen Richtlinien für Tierversuche
„Principles and Policies of Animal Use at NIH“ (Investigator´s Handbook „Using Animal in
Intramural Research: Guidelines for Investigators“, NIH, 1985).
A. Anästhesie und Tierpräparation
Die Tiere wurden durch eine s.c. Injektion eines Ketamin/Xylazin-Gemisches [Ketamin: 40
mg/kg KG (Hostaket, 100 mg/ml), Xylazin: 0,6 mg/kg KG (Rompun, 20 mg/ml)] initial
narkotisiert, nach einem Trachealschnitt intubiert und in einem stereotaktischen Halter
befestigt (Ohrstifte, Beißstange). Ab diesem Zeitpunkt befanden sie sich auf einem
Heizkissen, ihre rektale Temperatur und ihr Puls wurden ständig überwacht und auf
37° ± 1°C bzw. 220 bis 300 / s (Oszilloskop-Modell S51B, Telequipment) gehalten. Die
künstliche Beatmung und die Aufrechterhaltung der Narkose erfolgten gleichzeitig mit
einem 3:1-Gemisch aus N2O und O2, wobei der respiratorische Druck unter 20 hPa und der
CO2-Gehalt der Expirationsluft zwischen 3,3% und 4,2% gehalten wurden (Datex Normocap
CD200 oxy, Hoyer Medizintechnik Bremen). Dem N2O/O2-Gemisch wurde mit Hilfe eines
Halothanverdampfers (Vapor, Drägerwerk Lübeck) je nach Bedarf 0,1 bis 0,3 % Halothan
(Eurim-Pharm) beimischt. Zusätzlich wurde die Aufrechterhaltung des Narkosezustands
durch weitere Ketamin-Gaben von jeweils 0,1 ml unterstützt, die in regelmäßigen Abständen
(2 bis 5 Stunden) s.c. verabreicht wurden.
Zur Kreislaufstabilisierung und Verhinderung von Schleimbildung in der Trachea wurden
0,025 mg Atropinsulfat (Atropinsulfat Braun, 0,5 mg/ml) s.c. injiziert. Mit einer weiteren s.c.
Injektion von 0,3 mg Alcuroniumchlorid (Alloferin, 1 mg/ml) erfolgte die Immobilisierung,

Material und Methoden 24
die im Verlauf des Experimentes durch zusätzliche Injektionen (alle 3 bis 5 Stunden)
aufrechterhalten wurde. Sinn der Immobilisierung war die Verhinderung eines langsamen
Abdriftens der Augen von der Geradeausposition, was trotz Narkose auftreten kann. Dies
erleichterte die visuelle Stimulation in relativ zueinander definierten Bereichen des
Gesichtsfeldes bei der Suche nach dem NOT (siehe unten). Weitere 0,5 ml einer
Prilocainhydrochlorid-Lösung (Xylonest, 5 mg/ml) wurden in die Kaumuskeln über dem
Schädel und in die Muskeln hinter den Ohren zur lokalen Anästhesie vor dem Beginn der
chirurgischen Eingriffe injiziert.
Kraniotomien: Die verschiedenen Ableit- und Reizorte wurden über entsprechende Kranio-
tomien zugänglich gemacht, die mit Hilfe eines Zahnarztbohrers (Chiyoda, Etelna) durchge-
führt wurden. Ihre Position und Größe werden im folgenden durch die Horsley-Clarke-
Koordinaten angegeben:
• Zur Durchführung der elektrophysiologischen Ableitungen und elektrischen Reizungen im
Prätektum diente eine Schädelöffnung im Bereich von -3,5 bis 0 mm anterior und +0,5 bis
+3 mm lateral.
• Eine weitere Schädelöffnung im Bereich von ca. -12 bis -5 mm anterior und +3 bis +10
mm lateral setzte die Oberfläche des ipsilateralen visuellen Kortex (VC) frei (dorsaler
Bereich der Areae 17 und 18, vgl. Law et al., 1988). Diese Öffnung ermöglichte zum einen
die Plazierung zweier Reizelektroden, die zur orthodromen Reizung des ipsilateralen
Prätektums vom VC dienten, und zum anderen die Ableitung von Neuronen des VC, die
vom ispilateralen NOT und in manchen Fällen auch vom ipsilateralen superioren
Colliculus (SC) auf Reizbarkeit getestet wurden.
• Eine kleinere runde Schädelöffnung im Bereich +14 bis +15 mm anterior und ca. +1,5 mm
lateral (ca. 6 mm ∅) diente der Plazierung einer Reizelektrode auf den kontralateralen
optischen Nerv (ON).
Zur Durchführung der elektrischen Reizung der ipsilateralen inferioren Olive war keine
Kraniotomie notwendig. Die IO wurde über das Foramen magnum erreicht, welches zuerst
freigelegt und dann vorsichtig geöffnet wurde, so daß der vierte Hirnventrikel unter dem
Binokular (Zeiss, OMPI 1) deutlich gesehen werden konnte.
Die Flüssigkeitsverluste, die während der im Normalfall 30 bis 40 Stunden andauernden
Experimente unvermeidbar sind, wurden durch mehrere s.c. Injektionen von jeweils 0,5 bis
1,0 ml physiologischer Lösung (alle 2 bis 5 Stunden) ausgeglichen. Alternativ wurde eine
s.c. Infusion mit einer Rate von 2 ml/h für die Dauer von 15 bis 25 Stunden eingesetzt. Die
Corneae wurden durch regelmäßiges Befeuchten mit physiologischer Lösung intakt gehalten.

Material und Methoden 25
B. Extrazelluläre Ableitungen und Suche nach dem NOT
Alle elektrophysiologischen Ableitungen erfolgten extrazellulär. Die eingesetzten Ableit-
elektroden waren Wolfram-in-Glas-Mikroelektroden mit einer unisolierten Spitze von ca.
6 µm Länge (1 - 6 µm ∅) und einem Widerstand von 1,5 bis 4 MΩ. Alle Ableitelektroden
wurden im Institut für Allgemeine Zoologie und Neurobiologie der Ruhr-Universität Bochum
von Herrn H. Korbmacher hergestellt. Die Ableitelektrode war an einem Elektrodenhalter
geklemmt, der mit Hilfe eines Steppermotors (burleigh, angesteuert durch den burleigh 6000
Controller) in 0,5 µm-Schritten vorwärts und rückwärts bewegt werden konnte. Der
Steppermotor war an einem stereotaktischen Manipulator festgeschraubt, der auf dem
seitlichen Holm des stereotaktischen Halters angebracht war. Der Manipulator war in allen
drei Richtungen des Raumes kippbar, so daß die Ableitelektrode in nahezu jedem Winkel ins
Hirngewebe gefahren werden konnte. Als indifferente Elektrode diente eine Metallklammer,
die an dem freigelegten Gewebe auf dem Schädel des Tiers festgeklemmt war.
Die Erfahrung aus Vorexperimenten hatte gezeigt, daß der NOT wegen seiner kleinen
Ausmaße — wenn überhaupt — dann eher zufällig durch rein stereotaktisch plazierte
Penetrationen lokalisiert werden könnte. Deshalb erfolgte die Suche nach dem NOT-Areal
mit Hilfe der retinotopischen Karte des ipsilateralen SC. Bei jedem Tier wurde die
Retinotopie des SC grob umrissen, indem die Ableitelektrode an mehreren Orten senkrecht
in die oberflächlichen SC-Schichten eingestochen (5800 bis 7000 µm unterhalb der
kortikalen Oberfläche) und mit Hilfe einer Handlampe die rezeptiven Felder (RF) auf dem
Tangentialschirm (siehe unten) bestimmt wurden. Dabei wurde der Penetrationsort in 200 bis
300 µm Schritten zum rostrolateralen Bereich des SC hin versetzt, wo das ipsilaterale
inferiore Gesichtsfeld mit einem Azimut von ca. –30° und einer Elevation von ca. –20°
Sehwinkel repräsentiert ist (vgl. Quevedo et al.,1996). Von dieser Position aus wurde dann
die Elektrode 300 bis 500 µm lateral und / oder rostral versetzt, wo sich das NOT-Areal
befindet. Es zeigte sich, daß diese Vorgehensweise schneller und mit weniger Penetrationen
zum NOT führt als rein stereotaktische Penetrationen, die in den NOT zielen.
Zur Ableitung der VC-Zellen befand sich die Elektrode in den meisten Fällen in gekippter
Stellung, um zur Kortexoberfläche senkrechte Penetrationen zu ermöglichen.
Nahezu alle Penetrationsorte wurden unter visueller Kontrolle mit Hilfe des Binokulars
gewählt, um das Durchstechen von oberflächlichen Blutgefäßen zu vermeiden. Um einem
Abbrechen der Ableitelektrode rechtzeitig vorbeugen zu können, wurde die visuelle
Kontrolle erst aufgegeben, nachdem die Elektrode eindeutig die Dura mater passiert hatte.

Material und Methoden 26
C. Elektrische Stimulation
1. Reizparameter und Erläuterungen
Die elektrischen Reizpulse hatten eine Dauer von 0,1 ms und eine maximale Stromstärke von
0,5 mA. Jeder Reizpuls diente gleichzeitig als Trigger für ein Speicheroszilloskop (Tektronix,
Guernsey LTD), auf dessen Monitor der Potentialverlauf an der gerade abgeleiteten Zelle
dargestellt wurde. Die anti- und orthodromen Aktionspotentiale konnten auf diese Weise
vom Monitor abfotografiert werden und ihre Latenzen anhand der horizontalen Zeitskala der
Potentialspur am Oszilloskop abgelesen werden. Alle durch die Reizung ausgelösten
Aktionspotentiale mußten sich deutlich vom Hintergrundrauschen abheben.
Ein Aktionspotential wurde als antidrom definiert, wenn es mehrmals (mindestens viermal)
mit konstanter Amplitude und konstanter Latenz ausgelöst werden konnte. Auf dem Monitor
überlagerten sich dann die Potentialverläufe der antidromen Aktionspotentiale nach jedem
Reizpuls zu hundert Prozent. Die Potentialspuren vor und nach dem antidromen
Aktionspotential hatten einen zufälligen und unterschiedlichen Verlauf. In vielen Fällen
diente ein positiver Kollisionstest als zusätzliches Kriterium für ein antidromes
Aktionspotential (vgl. Fuller & Schlag, 1976).
Da bei einer orthodromen Reizung mindestens eine Synapse zwischen Reizort und abgeleite-
ter Zelle liegt, besitzt ein orthodrom ausgelöstes Aktionspotential eine variable Latenz, d.h.
es tritt im Gegensatz zu einem antidromen Aktionspotential innerhalb eines bestimmten
Latenzbereichs auf. Wenn — wie in der vorliegenden Arbeit — mehrmals gereizt wird,
speichert das Oszilloskop mehrere orthodrome Aktionspotentiale mit unterschiedlichen
Latenzen. Deshalb muß sowohl die kürzeste als auch die längste Latenz notiert werden. Im
Falle der vorliegenden Arbeit war dies ohnehin notwendig, weil jede Reizung des ON oder
des VC eine Mehrfachantwort, d.h. mehrere orthodrome Aktionspotentiale, auslöste. Für die
graphische Darstellung der Häufigkeitsverteilungen der verschiedenen orthodromen
Latenzen wurde nur die jeweils kürzeste Latenz gewertet. Zur graphischen Darstellung des
Zeitfensters, innerhalb dessen die Aktionspotentiale der Mehrfachantworten auftraten, wurde
die Häufigkeitsverteilung der maximalen Latenzdifferenzen (längste Latenz minus kürzeste
Latenz) gewählt.
Bei allen Reizungen dienten kleine Metallklammern, die am freigelegten Gewebe des
Versuchstiers angeklemmt waren, als indifferente Reizelektroden.

Material und Methoden 27
2. Reizexperimente
Von den eingangs erwähnten 32 Versuchstieren (23 albinotische und neu pigmentierte
Frettchen) wurden 16 albinotische und fünf pigmentierte Tiere für die antidrome Reizung der
NOT-Zellen von der ipsilateralen IO und für die orthodrome Reizung der NOT-Zellen vom
ipsilaterlaen VC und vom kontralateralen ON verwendet. Die restlichen sieben albinotischen
und vier pigmentierten Frettchen wurden für die antidrome Reizung des VC eingesetzt.
a) Antidrome Reizung der NOT-Neuronen von der IO
Die antidrome Reizung des Prätektums von der ipsilateralen IO wurde sowohl bei albinoti-
schen als auch bei pigmentierten Frettchen durchgeführt und diente zur Identifizierung der
einzelnen NOT-Neuronen. Von der IO antidrom reizbare Zellen des NOT-Areals wurden als
Zellen definiert, die innerhalb des neuronalen OKN-Schaltkreises visuelle Information für
die Auslösung der langsamen Phase des hOKN weiterleiten (vgl. Hoffmann et al., 1976;
Ballas & Hoffmann, 1985). Bei den Albinofrettchen konnten die NOT-Zellen nur auf diese
Weise eindeutig identifiziert werden, da sie richtungsunspezifisch waren. Deshalb wird im
Falle der Albinofrettchen erst dann von „NOT-Zellen“ gesprochen, wenn sie von der IO
antidrom reizbar waren. Nicht antidrom reizbare Zellen werden „prätektale Zellen“ genannt.
Bei den pigmentierten Frettchen konnten die NOT-Zellen auch ohne antidrome Reizung
anhand ihrer charakteristischen richtungsspezifischen Antworten auf die horizontale
optokinetische Reizung identifiziert werden (siehe unten).
Zur elektrischen Reizung der IO wurden zwei konzentrische Reizelektroden (IO-
Reizelektroden) aus Metalldrähten mit einem Durchmesser von 0,3 mm und einer Länge von
5 cm verwendet. Beide IO-Reizelektroden waren in einem Abstand von ca. 1 mm neben-
einander an einem Elektrodenhalter befestigt und wurden gleichzeitig durch das geöffnete
Foramen magnum 1 mm lateral des Obex bei einer 45°-Neigung nach posterior 4 mm tief
eingestochen (weitere Beschreibungen dieser Vorgehensweise sind in Horn & Hoffmann,
1987 und Telkes et al., 2001 zu finden). An dieser Position wurden sie für die Dauer des
gesamten Experimentes belassen.
Bei den meisten Albinofrettchen wurden die antidrome Reizung von der IO zusammen mit
einer der beiden orthodromen Reizungen (vom VC oder ON, siehe unten) durchgeführt. Bei
einigen Tieren wurden alle drei Reizungen durchgeführt, so daß auch Zellen charakterisiert
wurden, die sowohl antidrom von der IO als auch orthodrom vom VC und vom ON reizbar
waren.

Material und Methoden 28
b) Orthodrome Reizung der NOT-Neuronen vom VC
Die orthodrome Reizung der NOT-Zellen vom ipsilateralen VC wurde bei albinotischen und
pigmentierten Frettchen durchgeführt. Mit dieser Reizung wurden die Stärke und
Leitungsgeschwindigkeit des kortikalen Eingangs der zuvor von der IO antidrom
identifizierten NOT-Zellen festgestellt und zwischen den beiden Tiergruppen verglichen.
Dabei wurden auch weitere Daten aus der Literatur herangezogen, die von Klauer et al.
(1990) an pigmentierten Frettchen ermittelt wurden.
Es wurden die gleichen Reizelektroden wie für die antidrome Reizung der NOT-Neuronen
von der IO verwendet. Diese VC-Reizelektroden wurden ca. 2 mm tief in den VC an einem
Ort plaziert, an dem zuvor mit einer Ableitelektrode visuelle Aktivität mit einem RF im
frontalen Gesichtsfeld lokalisiert werden konnte.
c) Orthodrome Reizung der NOT-Neuronen vom ON
Die orthodrome Reizung der NOT-Zellen vom kontralateralen ON wurde bei den
Albinofrettchen durchgeführt. Damit wurden die Stärke und Leitungsgeschwindigkeit des
retinalen NOT-Eingangs der zuvor von der IO antidrom identifizierten NOT-Neuronen
ermittelt und mit den entsprechenden Werten von pigmentierten Frettchen aus der Literatur
(Klauer et al., 1990) verglichen.
Die Elektrode zur Reizung des ON war eine niederohmige (0,2 bis 0,6 MΩ) Wolfram-in-
Glas-Mikroelektrode (herausragende Spitze ca. 20 µm lang und 1 - 20 µm dick) und erlaubte
auch das Ableiten von Mehrzellaktivität. Dadurch ermöglichte sie die Erkennung des ON
anhand seines elektrischen Summenpontentials, welches durch Beleuchten des
entsprechenden Auges ausgelöst wurde. Nur auf diese Weise war es möglich, die Elektrode
auf den ON zu plazieren ohne ihn zu penetrieren. Bei allen Tieren wurde der ON in einer
Tiefe von 11 bis 12,5 mm unter der Kortexoberfläche lokalisiert. Anschließend wurde die
Elektrode am Schädelknochen befestigt (Technovit 4004) und diente im weiteren Verlauf des
Experimentes als ON-Reizelektrode.
d) Antidrome Reizung kortikaler Neuronen vom NOT und SC
Diese Experimentenreihe wurde sowohl mit albinotischen als auch mit pigmentierten
Frettchen durchgeführt. Es wurden Neuronen des VC auf ihre Reizbarkeit vom ipsilateralen
NOT und bei einigen Tieren auch vom ipsilateralen SC getestet.
Das primäre Ziel dieser Experimente war die antidrome Reizung der VC-Zellen vom NOT
und die anschließende visuelle Charakterisierung dieser Zellen. Vom NOT antidrom reizbare

Material und Methoden 29
VC-Zellen wurden als Zellen definiert, deren Axone die kortikale Projektion zum NOT
bilden. Im Falle der Albinofrettchen wurde der NOT anhand seiner relativen Lage zum SC
und seiner charakteristischen, hohen Spontanrate elektrophysiologisch identifiziert, jedoch
nicht durch das Kriterium „antidrome Reizbarkeit von der IO“. Bei den pigmentierten
Frettchen kam als weiteres, eindeutiges Kriterium die charakteristische richtungsspezifische
Beantwortung der horizontalen optokinetischen Stimulation hinzu.
Da die NOT-Neuronen im Brachium des SC liegen, kann jeder Reizpuls im NOT auch zur
Reizung von kortikalen Fasern führen, die nicht im NOT enden, sondern lediglich am NOT
vorbeiziehen und den SC innervieren. Infolgedessen besteht die Möglichkeit, daß eine vom
NOT antidrom reizbare VC-Zelle in Wirklichkeit nicht zum NOT sondern zum SC projiziert.
Aufgrund dieser Verwechslungsgefahr wurde bei einigen Tieren auch die Reizung des SC
durchgeführt und darauf geachtet, daß die SC-Reizelektrode in retinotopischer Übereinstim-
mung mit der Position der kortikalen rezeptiven Felder plaziert war. Als Zellen der
kortikoprätektalen Projektion wurden in diesem Falle nur solche VC-Zellen definiert, die
ausschließlich vom NOT antidrom reizbar waren.
Als NOT- und SC-Reizelektroden wurden niederohmige Elektroden mit den gleichen
Eigenschaften wie die ON-Reizelektrode benutzt. Mit der NOT-Reizelektrode wurde
zunächst das NOT-Areal in der oben beschriebenen Vorgehensweise lokalisiert. Nachdem
auch die SC-Reizelektrode in den rostralen Pol des SC plaziert war, wurden beide Elektroden
am Schädelknochen festzementiert (Technovit 4004) und dienten für den weiteren Verlauf des
Experimentes als Reizelektroden.
Während der Suche nach VC-Zellen, die vom NOT oder SC antidrom reizbar waren, wurden
auch einige VC-Zellen orthodrom gereizt. Die orthodrome Reizung der VC-Zellen war nicht
der Zweck des Experimentes, denn sie erfolgt nicht über die direkte VC-NOT-Verbindung.
Insofern ist sie ein Nebenprodukt der antidromen Reizung.
Am Ende des Experimetes wurden die NOT- und SC-Reizorte durch elektrolytische
Läsionen markiert, so daß ihre Position in der anschließenden Histologie verifiziert werden
konnte.

Material und Methoden 30
D. Visuelle Stimulation
1. Visuelle Stimulation der prätektalen Zellen
Die visuelle Charakterisierung der prätektalen bzw. NOT-Zellen erfolgte hinsichtlich ihrer
Richtungsspezifität (Vorzugsrichtung und Vorzugsstärke). Die horizontale Richtung ist die
adäquate Reizrichtung der NOT-Zellen innerhalb des hOKN-Schaltkreises. Deshalb erfolgte
die optokinetische Reizung des narkotisierten Tieres, bzw. der zu charakterisierenden Zellen
in horizontaler Richtung.
Die Reizbewegung von kontralateral nach ipsilateral in Bezug auf die abgeleitete Hirnseite
wird als ipsiversive Reizung definiert. Die Reizbewegung in die entgegengesetzte Richtung
wird dann kontraversive Reizung genannt. Eine ipsiversive und eine kontraversive
Reizbewegung definieren eine Reizperiode.
Die optokinetische Reizung erfolgte binokular mit einem schwarz-weißen Zufallspunkte-
muster (Punktgröße: 1,5 oder 3° Sehwinkel), welches mit Hilfe eines Diaprojektors
(KINDERMANN universal) und zweier Spiegel um 90° abgelenkt und auf einem Tangentialschirm
142 cm vor dem Tier projiziert wurde. Die beiden Spiegel dieses Projektionssystems waren
auf senkrecht zueinander stehenden Achsen montiert und gegeneinander beweglich. Das
Reizmuster auf dem Tangentialschirm bewegte sich in horizontaler Richtung bei Drehung
des einen Spiegels und in vertikaler Richtung bei Drehung des anderen Spiegels. Beide
Spiegel wurden durch jeweils ein Galvanometer gedreht, das seinerseits von einem variablen
Phasen-Funktionsgenerator (Modell FG-124, Elektronik Starnberg) unabhängig angesteuert
werden konnte. Die horizontale Bewegung des Reizmusters auf dem Tangentialschirm kam
durch ein Dreieckssignal zustande, welches den einen Spiegel bewegte, während der andere
Spiegel stillstand. Die Amplitude war in der Regel so gewählt, daß sich das Reizmuster
innerhalb des Gesichtsfeldbereiches von ca. –30° (ipsilaterales Gesichtsfeld) bis ca. +70°
Azimut (kontralaterales Gesichtsfeld) bewegte. Dabei war die Ruheposition des stehenden
Spiegels so gewählt, daß sich das Muster von ca. +70° bis ca. –30° Elevation erstreckte.
Durch Einstellung der Frequenz wurde die Reizgeschwindigkeit festgelegt, die in der Regel
20°/s betrug, da bei dieser Geschwindigkeit die Antwort der NOT-Zellen des Frettchens
maximal ist (Klauer et al., 1990).
2. Visuelle Stimulation der VC-Neuronen
Die VC-Zellen wurden hinsichtlich ihrer RF-Position und Richtungsspezifität (Vorzugs-
richtung und eventuell auch Vorzugsstärke) visuell charakterisiert.

Material und Methoden 31
Die Bestimmung der RF erfolgte mit Hilfe der Handlampe und der über die Lautsprecher
(siehe unten) hörbaren Zellaktivität. Mit der Handlampe wurden Lichtpunkte und Licht-
balken unterschiedlicher Größe und Orientierung auf dem Tangentialschirm projiziert und in
verschiedene Richtungen mit unterschiedlichen Geschwindigkeiten bewegt.
Die Richtungsspezifität der VC-Zellen wurde entweder durch Reizung mit Hilfe der
Handlampe oder mit Hilfe des Projektionssystems bestimmt. Die Handlampe erlaubte nur die
Bestimmung der Vorzugsrichtung. Durch die Reizung mit dem Projektionssystem wurde
zusätzlich auch die Vorzugsstärke bestimmt (Berechnung eines Richtungsspezifitäts-Indizes,
siehe unten). Dabei erfolgte die Reizung mit den erwähnten Zufallspunktemustern
(Punktgröße: 1,5° und 3° Sehwinkel), mit einem einzelnen Lichtpunkt (24 cm ∅; ca. 9,7 °
Sehwinkel) und / oder mit einem einzelnen Lichtbalken (47 cm × 4 cm; ca. 19° Sehwinkel ×
ca. 1,6° Sehwinkel). Der Lichtbalken war immer senkrecht zur Bewegungsrichtung
ausgerichtet. Da im VC alle Vorzugsrichtungen repräsentiert sind, wurde sowohl horizontal,
als auch vertikal, als auch genau zwischen diesen beiden Kardinalachsen (insgesamt acht
Richtungen) gereizt. Die Abfolge jeweils einer einzelnen Hin- und Herbewegung des Reizes
definierte eine Reizperiode.
3. ON-OFF-Test von Zellen des Mittelhirns
Die ON/OFF-Eigenschaften wurden sowohl von prätektalen Zellen als auch von SC-Zellen
bestimmt. Die SC-Zellen wurden bei der Suche nach dem NOT abgeleitet (siehe weiter
vorne) und befanden sich alle in den oberen visuellen Schichten des SC. Ziel dieser Unter-
suchung war es, das relative Vorkommen von ON-, OFF- und ON-OFF-Zellen im Mittelhirn
und damit indirekt auch das Vorkommen der entsprechenden retinalen Ganglienzellen zu
ermitteln. In Bezug auf die Richtungsspezifität im NOT sind die ON-Ganglienzellen von
besonderer Interesse, da sie den retinalen Eingang des NOT enthalten. Der Test wurde
entweder mit der Handlampe oder mit einem Blendenverschluß durchgeführt.
Handlampe: Mit Hilfe der Handlampe wurde zunächst die Position des RF der zu
charakterisierenden Zelle auf dem Tangentialschirm lokalisiert. Danach wurde die Zelle mit
der Handlampe visuell gereizt und in Abhängigkeit von ihrer akustisch wahrnehmbaren
Reaktion als eine ON-, OFF- oder ON-OFF-Zelle klassifiziert. Die Reizung erfolgte mit
einem stationären Lichtpunkt, der aus dem RF verschwand und wieder auftauchte
(Verdecken und Aufdecken des Lichtstrahls mit der Hand).
Bei den prätektalen Zellen des Albinofrettchens war diese Vorgehensweise in den
allermeisten Fällen nicht möglich, weil das RF nicht eindeutig bestimmt werden konnte.

Material und Methoden 32
Außerdem kommt bei prätektalen Zellen hinzu, daß ihre RF relativ groß sind und deshalb
schwierig als ganzes mit einem Lichtpunkt zu beleuchten. In diesen Fällen wurde nicht das
RF auf der Tangentialwand visuell gereizt, sondern das ganze kontralaterale Auge
ausgeleuchtet. Beide Varianten mit der Handlampe erlauben lediglich eine qualitative
Charakterisierung der ON/OFF-Eigenschaften.
Blendenverschluß: Zunächst wurde die Position des RF der zu charakterisierenden Zelle auf
dem Tangentialschirm mit Hilfe der Handlampe bestimmt. Anschließend wurde mit Hilfe
des oben erwähnten Projektionssystems ein Lichtpunkt auf das RF projiziert und dort
stationär gehalten. Nun wurde eine verschließbare Blende genau in den Lichtstrahl des
Projektionssystems gesetzt. Durch Öffnen des Blendenverschlusses wurde das RF beleuchtet
(„Licht-An“) und durch Schließen des Verschlusses verschwand der Lichtpunkt aus dem RF
(„Licht-Aus“). Diese Methode erlaubte auch eine quantitative Bestimmung der
Zelleigenschaften, denn das Öffnen und Schließen der Verschlusses wurde durch den oben
erwähnten Phasen-Funktionsgenerator gesteuert, so daß die Zellaktivität in Abhängigkeit des
Öffnungszustands aufgenommen und als PSTH gespeichert werden konnte (siehe unten).
4. Lichtverhältnisse
Der Helligkeitskontrast (K) zwischen den weißen und schwarzen Punkten des Reizmusters
betrug 60% bis 71% [berechnet nach der Formel K=(a-b)/(a+b)×100; a = Leuchtdichte der
weißen und b= Leuchtdichte der schwarzen Punkte]. Dabei betrug die Leuchtdichte der
weißen Punkte 6 bis 10 cd/m2 und die der schwarzen Punkte 1 bis 2,5 cd/m2. Die
Leuchtdichte in cd/m2 wurde mit dem Gerät luminance meter der Firma Minolta gemessen.
Während der visuellen Reizung waren die normale Beleuchtung des Labors ausgeschaltet
und die Fenster verdunkelt. Alle Parameter der Lichtverhältnisse wurden während des
gesamten Experimentes konstant gehalten. Sämtliche Parameter der visuellen Reizung waren
bei albinotischen Frettchen und pigmentierten Frettchen gleich.
E. Datenaufnahme und Analyse
Das abgeleitete Signal wurde konventionell verstärkt und gefiltert und zu einem Window-
Diskriminator geleitet. Am Window-Diskriminator war die obere und untere Schwelle des
Potentialfensters festgelegt, innerhalb dessen die Potentiale als Aktionspotentiale
diskriminiert und als TTL-Pulse über ein Interface einem PC (Deskpro 286, Compaq)
übermittelt wurden. Zu Beginn einer Reizperiode sendete der Phasen-Funktionsgenerator ein

Material und Methoden 33
Triggersignal an das Softwareprogramm (spisa3, programmiert von Herrn Dr. G. J.
Dörrscheidt im Institut für Allgemeine Zoologie und Neurobiologie) des PCs, das daraufhin
mit der Registrierung der TTL-Pulse begann. Die Feuerrate der Zelle [Aktionspotentiale (=
TTL-Pulse) / s] wurde online auf dem Monitor des PCs als Peri-Stimulus-Time-Histogramm
(PSTH) dargestellt und am Ende der Aufnahme als EDV-Datei zur späteren Offline-Analyse
gespeichert. Das PSTH einer Aufnahme beinhaltete in der Regel die Zellaktivität aus 10
Reizperioden. Über einen Zweikanal-Audioverstärker (TFE-Profi-Equalizer, TFE GmbH) und
zwei Lautsprecher waren während der gesamten Dauer der Ableitungen das Rohsignal und /
oder die als Aktionspotentiale diskriminierten TTL-Pulse akustisch wahrnehmbar.
1. Richtungsspezifitäts-Indizes
Zur einheitlichen Bewertung der Richtungsspezifität der verschiedenen Zellen wurde
folgender Richtungsspezifitäts-(DS-)Index (DS-I) definiert:
DS-Indizes der prätektalen Zellen: Da die NOT-Zellen im Normalfall die ipsiversive
Reizung bevorzugen, wurde ihre Zellaktivität während der ipsiversiven Reizbewegung als
Feuerrate(Vorzugsrichtung) definiert. Die Feuerrate(Nullrichtung) ist die Zellaktivität, mit der die
kontraversive Reizung beantwortet wurde. Der Betrag des DS-Is ist ein relatives Maß für die
Vorzugsstärke. Ein DS-I von Null bedeutet, daß eine Zelle richtungsunspezifisch ist, d.h.
keine der beiden horizontalen Bewegungsrichtungen des optokinetischen Reizes bevorzugt.
Da zur visuellen Charakterisierung der prätektalen bzw. NOT-Zellen nur in die beiden
horizontalen Richtungen gereizt wurde, verrät das Vorzeichen des DS-Is auch die Vorzugs-
richtung. Ein positives Vorzeichen bedeutet, daß die ipsiversive Reizung bevorzugt wird. Ein
negativer DS-I zeigt die Bevorzugung der kontraversiven Reizbewegung an, was nicht dem
Normalfall entspricht. Definitionsgemäß kann der Wert eines DS-I nur im Bereich von –1 bis
+1 liegen. Zur graphischen Darstellung der DS-Indizes aus horizontalen Reizungen wurde
ihre Häufigkeitsverteilung im Intervall von –1 bis +1 mit einer Klassenbreite von 0,1
gewählt. Die DS-Is von NOT-Zellen, die von der IO antidrom identifiziert wurden, sind in
allen Abbildungen schraffiert dargestellt.
DS-Indizes der VC-Zellen: Die DS-Indizes der VC-Zellen zeigen nur die Vorzugsstärke an.
Denn da die visuelle Reizung in mehr als zwei Richtungen erfolgte, wurde aus allen
DS-I =Feuerrate(Vorzugsrichtung) – Feuerrate(Nullrichtung)
Feuerrate(Vorzugsrichtung) + Feuerrate(Nullrichtung)

Material und Methoden 34
Reizrichtungen diejenige Richtung als Vorzugsrichtung definiert, die die höchste Feuerrate
auslöste. Infolgedessen kann der DS-Index nur einen Wert zwischen 0 und +1 annehmen.
Die zur Vorzugsrichtung genau entgegengesetzte Richtung wurde als Nullrichtung definiert.
Die Häufigkeitsverteilung der DS-Indizes zeigt das Vorkommen von Richtungsspezifität
(Vorzugsstärke) bei Neuronen des VC an, ungeachtet der präferierten Richtungen.
Zur graphischen Darstellung der Vorzugsrichtungen der VC-Zellen mit DS-Index wurde das
Gesichtsfeld in acht 45°-Sektoren unterteilt. Diese acht Sektoren sind [Aufzählung gegen den
Uhrzeigersinn beginnend bei der Richtung auf die abgeleitete Seite (ipsi)]: (1) Sektor 157,5°
- 202,5° (ipsi), (2) Sektor 202,5° - 247,5° (ipsi-unten), (3) Sektor 247,5 - 292,5° (unten), (4)
Sektor 292,5 - 337,5° (kontra-unten), (5) Sektor 337,5° - 22,5° (kontra), (6) Sektor 22,5° -
67,5° (kontra-oben), (7) Sektor 67,5° - 112,5° (oben) und (8) Sektor 112,5° - 157,5° (ipsi-
oben). Anschließend wurde die Häufigkeitsverteilung der DS-Indizes derjenigen VC-Zellen
berechnet, deren Vorzugsrichtungen in die jeweiligen Sektoren fielen. In allen Abbildungen
sind die DS-Indizes von VC-Zellen, die auf Reizbarkeit vom ipsilateralen NOT getestet
wurden, grau markiert. Die Teilmenge der DS-Indizes von VC-Zellen, die vom NOT
antidrom reizbar waren, sind zusätzlich schraffiert.
2. ON-OFF-Indizes
Zur Bestimmung der ON/OFF-Eigenschaften mit Hilfe des Blendenverschlusses wurden von
jeder Zelle 3 PSTH aufgenommen. Das erste PSTH dokumentiert den eigentlichen ON-OFF-
Test und beinhaltet die Aktivität von 10 Reizperioden, wobei jede Reizperiode aus einer
Beleuchtungsphase („Licht-An“-Phase) und einer Dunkelphase („Licht-Aus“-Phase) von
jeweils 1 s Dauer besteht. Das zweite PSTH dokumentiert die Spontanrate der Zelle,
während ihr RF auf dem Tangentialschirm mit dem Lichtpunkt permanent beleuchtet wurde
(Blendenverschluß im offenen Zustand). Das dritte PSTH dokumentiert die Spontanrate bei
Dunkelheit (Blendenverschluß im geschlossenen Zustand). Zur einheitlichen Bewertung der
ON/OFF-Antworten der verschiedenen Zellen wurde folgender ON-OFF-Index (ONF-I)
definiert:
trans. Feuerrate(AN) = transiente Feuerrate bei „Licht-An“ (Öffnung des Blendenverschlusses) trans. Feuerrate(AUS) = transiente Feuerrate bei „Licht-Aus“ (Schließen des Blendenverschlusses) Spontanrate(AN) = Spontanrate während permanenter Beleuchtung des RF (offener Blendenverschluß) Spontanrate(AUS) = Spontanrate während permanenter Dunkelheit (geschlossener Blendenverschluß)
ONF-I =trans. Feuerrate (AN) – Spontanrate(AN)
(trans. Feuerrate (AN) – Spontanrate(AN)) + (trans. Feuerrate (AUS) – Spotanrage(AUS))× 100

Material und Methoden 35
Zur Definition des ONF-I wurden nur die transienten (phasischen) ON- und OFF-Antworten
auf das Erscheinen und Verschwinden des Lichtpunktes und die Spontanraten während
andauernder Beleuchtung und Nicht-Beleuchtung des RF auf dem Tangentialschirm
berücksichtigt. Der Faktor 100 dient lediglich zur Umrechnung des ONF-I in Prozent. Damit
gibt der ONF-I an, wieviel Prozent die phasische ON-Antwort an der phasischen Gesamt-
antwort (Summe aus phasischer ON- und phasischer OFF-Antwort) ausmacht. Ein ONF-I >
50 % bedeutet, daß die ON-Antwort größer als die OFF-Antwort ist. Ein ONF-I < 50 % zeigt
eine stärkere OFF-Antwort und ein ONF-I = 50 % zeigt gleichstarke phasiche ON- und OFF-
Antworten an. Zur graphischen Darstellung der ONF-Indizes wurde ihre Häufigkeits-
verteilung mit einer Bin-Breite von 5 % gewählt.
Definitionsgemäß werden Zellen mit einem ONF-I < 0,33% als OFF-Zellen und mit einem
ONF-I ≥ 0,66% als ON-Zellen klassifiziert. Wenn 0,33% ≤ ONF-I < 0,66% gilt, werden die
Zellen als ON-OFF-Zellen klassifiziert.
3. Statistik
Da in einigen Fällen aus statistischer Sicht relativ wenige Werte vorhanden sind und / oder
extreme „Ausreißer“ auftreten und / oder asymmetrische Verteilungen vorliegen, wurde als
Mittelwert der DS-Indizes und der Reizlatenzen der Median und als Streuungsparameter das
untere (Q0,25) und obere (Q0,75) Quartil angegeben. Entsprechend wurde zum statistischen
Vergleich zweier Gruppen von DS-Indizes oder Reizlatenzen der nicht-parametrische RANG-
SUMMEN-TEST nach Mann und Whitney angewandt. Die Nullhypothese dieses Testes ist, daß
die Mediane der beiden zu vergleichenden Wertegruppen sich nicht signifikant voneinander
unterscheiden. Sie wurde bei einem P < 0,05 abgelehnt. Zur Überprüfung, ob eine
Normalverteilung vorliegt, wurde der nach Lilliefors modifizierte KOLMOGOROV-SMIRNOV-
TEST angewandt, der bei einem P ≥ 0,05 eine Normalverteilung anzeigt. Um zwei absolute
Häufigkeitsverteilungen miteinander zu vergleichen wurde der χ2-Test angewandt, der bei
einem P < 0,05 einen signifikanten Unterschied anzeigt. Alle Tests wurden mit der
Windows-Version der Statistik-Software SigmaStat 2.0 (SPSS Inc.) berechnet.
F. Histologie
Die Verifikation der richtigen Position der Ableitorte im NOT und der IO-Reizelektroden
war bei den albinotischen Frettchen von besonderer Wichtigkeit, da die NOT-Zellen nur

Material und Methoden 36
anhand ihrer antidromen Reizbarkeit von der IO eindeutig identifiziert werden konnten. Das
Auffinden der NOT-Ableitorte wurde bei einigen albinotischen und pigmentierten Frettchen
durch elektrolytische Läsionen erleichtert, die am letzten NOT-Ableitort durch Applikation
eines Stroms von ±10 µA für die Dauer von jeweils 10 s gesetzt wurden. Die Position der
IO- und VC-Reizelektroden wurde durch ihre dicken Einstichspuren dokumentiert. Die
Position der vom NOT antidrom identifizierten VC-Zellen wurde ebenfalls mit
elektrolytischen Läsionen markiert. In diesem Falle sollte beim Frettchen überprüft werden,
ob sich die gereizten VC-Zellen wie bei der Katze (Schoppmann, 1981) in der kortikalen
Lamina V befinden.
Am Ende der Experimente wurde das Versuchstier eingeschläfert und durch den linken
Herzventrikel mit 1 Liter 0,9%-iger NaCl-Lösung gefolgt von 1,5 Liter 4%-igem Paraform-
aldehyd in 0,1 M Phosphatpuffer (pH 7,4) und Succrose (57,6 g pro Liter Fixierlösung)
perfundiert. Nach erneuter Einspannung des Kopfes in die Stereotaxie, Öffnung des Schädels
und Entfernung der Dura mater wurde das Hirn stereotaktisch geblockt und über Nacht in
derselben Fixierlösung nachfixiert. In den folgenden Tagen wurde es zur Entwässerung für
mindestens 12 Stunden jeweils in einer 10, 20 und 30%-igen Succrose-Lösung bis zum
Absinken aufbewahrt und dann in kaltem Alkohol (Isopentan, –70° C) schockgefroren.
Anschließend wurden mit einem Gefriermikrotom (Microm HM 500 OM) 50 µm dicke
Hirnschnitte angefertigt, auf gelatinierte Objektgläser aufgezogen und für einige Tage in
einem Trockenschrank aufbewahrt. Nachdem die Schnitte trocken genug waren, wurden sie
Klüver-Barrera- und / oder Nissl-gefärbt, in Depex eingedeckelt und unter dem
Lichtmikroskop (Zeiss Axiolab) untersucht.
Folgende Serien von Hirnschnitten wurden angefertigt: a) Frontalschnitte vom Mittelhirn
und dem darüberliegenden Kortex zur Dokumentation der Ableit- und Reizorte im Prätektum
und b) Frontalschnitte vom Occipitalkortex zur Dokumentation der Position der VC-Reiz-
elektroden oder der Ableitorte, an denen zuvor VC-Neuronen vom NOT antidrom gereizt
wurden. c) Von drei Frettchen aus IO-Reizexperimenten wurden auch Sagittalschnitte vom
Hirnstamm zur Dokumentation der Position der IO-Reizelektroden angefertigt. In diesen drei
Fällen wurden die Hirnstämme ungefroren auf eine Metallplatte geklebt und mit einem
Vibratom (Micro-Cut H1200, Bio-Rad) in 100 µm dicke Scheiben geschnitten.
Die Perfusion wurde frühestens 30 Minuten nach der letzten elektrolytischen Läsion
gestartet, damit genügend Zeit für die Bildung einer erkennbaren Gliosis verstreichen
konnte, die das Wiederfinden der Läsionen ermöglichte.

Ergebnisse 37
Ergebnisse
I. Teil I: Verhalten
Als erstes werden die Ergebnisse aus den hOKN-Tests vorgestellt, die vor den Muscimol-
Anwendungen durchgeführt wurden. Beginnend mit der Häufigkeitsverteilung der
Steigungsmediane aus den Kontrollaufnahmen (spontane Augenbewegungen) in
Abbildung 6 folgen die Häufigkeitsverteilungen aus den hOKN-Tests, die unter den
verschiedenen Parametern durchgeführt wurden. Da ursprünglich bei Albinofrettchen ein
asymmetrischerer monokularer hOKN als bei pigmentierten Frettchen postuliert wurde,
erfolgten zuerst die monokularen Reizungen. Deshalb werden zuerst die Ergebnisse der
monokularen Reizungen im einzelnen beschrieben.
Anschließend folgen die Ergebnisse aus der Inaktivierungsstudie (Muscimol-Experimente)
und dem Perimetrietest.
A. Spontane Augenbewegungen (vor Muscimol-Injektion)
Die Registrierung der spontanen Augenbewegungen diente als Kontrolle. Alle EOG-Spuren
der spontanen Augenbewegungen haben eine glatte Form, wenn man von den gelegentlichen
Sakkaden und Kopfbewegungen absieht, die in diesem Zusammenhang nicht von Interesse
sind und deshalb herausgeschnitten wurden. Einige EOG-Spuren verlaufen genau horizontal,
d.h. sie besitzen einen Steigungsmedian von Null. Die meisten EOG-Spuren haben
abschnittsweise Steigungen, die von Null etwas abweichen. Da sie aber während einer
Aufnahmeperiode ihr Vorzeichen zufällig ändern, werden im Durchschnitt gleichlange
Abschnitte mit einer konstanten positiven und einer konstanten negativen Steigung
aufgezeichnet. Dieser zufällige Vorzeichenwechsel führt dazu, daß die Steigungsmediane der
Spuren Null sind oder zumindest sehr nahe um den Wert Null liegen. Eine konstante
Steigung der EOG-Spur bzw. eines Spurabschnitts ist als konstante Augendrift (oder Drift
des EOG-Signals) in horizontaler Richtung zu verstehen.
In Abbildung 6 sind die Häufigkeitsverteilungen der Steigungsmediane der EOG-Spuren der
untersuchten Tiere gezeigt, die unter den verschiedenen Bedingungen der Kontrollmessun-

Ergebnisse 38
gen registriert wurden. In der linken Spalte befinden sich die Ergebnisse der Aufnahmen in
Helligkeit, d.h. die Tiere befanden sich in der stehenden, beleuchteten Trommel und konnten
auf das stationäre Zufallspunktemuster blicken. In der rechten Spalte befinden sich die
entsprechenden Ergebnisse der EOG-Aufnahmen, die in völliger Dunkelheit durchgeführt
wurden. Die oberen vier Zeilen der Abbildung 6 zeigen die Befunde der Albinofrettchen, die
unter den vier Bedingungen („binokular“, „nur rechtes sehend“ und „nur linkes Auge
sehend“ jeweils mit dem feinen Reizmuster und „binokular mit dem groben Reizmuster“)
getestet wurden. In der letzten Zeile der Abbildung 6 befindet sich das Ergebnis der
pigmentierten Tiere, die in Helligkeit nur binokular mit dem feinen Muster gemessen
wurden.
Im Falle der völligen Dunkelheit (rechte Spalte) ist es unerheblich, ob die Aufnahmen
binokular oder monokular durchgeführt wurden. Trotzdem sind auch monokulare
Aufnahmen registriert worden, um einen eventuellen Einfluß der Augenkappe auf die
Okulomotorik aufzudecken.
Die einzelnen Häufigkeitsverteilungen in Abbildung 6 zeigen in allen Fällen, daß sich die
Steigungsmediane der allermeisten Tiere zwischen –0,25 und +0,25 befinden. Der Median
dieser Steigungsmediane ist erwartungsgemäß in jeder Häufigkeitsverteilung Null. Folglich
zeigen die verteilungsfreien Tests (WILCOXON SIGNED RANK TEST und MANN-WHITNEY
RANK SUM TEST) keinen signifikanten Unterschied zwischen jeweils zwei beliebigen
Verteilungen der Abbildung 6 (in allen Fällen P > 0,05).
Vergleicht man jedoch die Steigungsmediane aus den verschiedenen Häufigkeitsverteilungen
hinsichtlich ihrer Varianzen, so stellt man bei den Albinos eine größere Varianz fest als bei
den pigmentierten Tieren. Entsprechend ergibt der LEVENE S HOMOGENEITY OF VARIANCE
TEST einen hoch signifikanten Unterschied (P < 0,0001). Dies gilt sowohl für die Aufnahmen
in Helligkeit (Vergleich der Häufigkeitsverteilungen oben-links und unten-links in Abb. 6),
als auch für die Aufnahmen in Dunkelheit (Häufigkeitsverteilungen oben-rechts und unten-
rechts in Abb. 6).
Die größere Varianz der Albinos zeigt, daß die Steigungsmediane ihrer EOG-Spuren dem
Betrag nach größere Schwankungen aufweisen. Dies bedeutet, daß ihre Augen betragsmäßig
mit signifikant größeren Geschwindigkeiten in horizontaler Richtung abdriften als die Augen
der pigmentierten Frettchen. Damit wird bei den Albinofrettchen eine okulomotorische
Instabilität dokumentiert.

Ergebnisse 39
bei Helligkeit
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
bei Dunkelheit
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
alb
alb
alb
alb
pigm
N
N
N
N
N
N
N
N
N
N
Abb. 6: Häufigkeitsverteilung der Steigungsmediane von EOG-Spuren albinotischer und pigmentierter Frettchen, die in völliger Dunkelheit (rechte Spalte) oder in Helligkeit bei stehender Trommel (linke Spalte) registriert wurden (Messung spontaner Augenbewegungen). In allen Fällen sind die Steigungsmediane um Null verteilt und unterschei-den sich hinsichtlich ihrer Mediane nicht signifikant voneinander. Die Werte der Albinofrettchen variieren jedoch signifikant stärker als die Werte der pigmentierten Tiere. Dieser Befund zeigt die okulo-motorische Instabilität der Albinofrettchen an. Die nebenstehende Legende erläutert die Symbole. N = Anzahl der Medianwerte (entspricht der Anzahl der Tiere).
: EOG-Aufnahmen bei Helligkeit : EOG-Aufnahmen bei Dunkelheit : binokular sehend : monokular, linkes Auge sehend : monokular, rechtes Auge sehend : feines Reizmuster : grobes Reizmuster

Ergebnisse 40
B. hOKN-Test (vor Muscimol-Injektion)
1. Optokinetische Reizung der Albinofrettchen
a) Monokulare Ganzfeldreizung
Feines Reizmuster: Alle EOG-Spuren der Albinofrettchen aus den monokularen Reizungen
mit dem feinen Reizmuster weisen überraschenderweise eine glatte Form und einen nahezu
waagerechten Verlauf auf. Hinsichtlich Form und Verlauf unterscheiden sie sich nicht von
den EOG-Spuren der spontanen Augenbewegungen.
Die Häufigkeitsverteilungen der Steigungsmediane der EOG-Spuren aus diesen monokularen
hOKN-Tests der Albinofrettchen sind in Abbildung 7 (linkes Auge sehend, rechtes verdeckt)
und Abbildung 8 (rechtes Auge sehend, linkes verdeckt) gezeigt. Innerhalb einer Abbildung
zeigen die Graphiken in der linken Spalte das Ergebnis der cw-Reizung und die Graphiken in
der rechten Spalte das Ergebnis der ccw-Reizung (jeweils für die Reizgeschwindigkeiten 5,
10, 20, 50 und 100°/s). Die temporonasale (nasotemporale) Reizung ist für das linke Auge
die cw-Reizung (ccw-Reizung) und für das rechte Auge die ccw-Reizung (cw-Reizung).
Folglich entspricht die Stimulation in der linken Spalte der Abbildung 7 der Stimulation in
der rechten Spalte der Abbildung 8 und vice versa.
Wie Form und Verlauf der EOG-Spuren erwarten lassen, sind ihre Steigungmediane in allen
Fällen Null oder nah um Null verteilt. Für jede Häufigkeitsverteilung in Abbildung 7 und 8
gilt, daß sie sich weder von dem Spontanverhalten unter monokularer Bedingung (vgl. Abb.
6), noch von Null signifikant unterscheidet (WILCOXON SIGNED RANK TEST, in allen Fällen
P > 0,05).
Auch der Vergleich von jeweils zwei nebeneinander liegenden Graphiken in Abbildung 7
oder Abbildung 8 ergibt keinen signifikanten Unterschied (WILCOXON SIGNED RANK TEST,
in allen Fällen P > 0,05). Dies bedeutet, daß es keinen Unterschied zwischen der
monokularen cw- und ccw-Reizung des sehenden Auges gibt.
Damit kann festgehalten werden, daß die erwartete Asymmetrie des hOKN bei monokularer
Reizung mit dem feinen Muster ausbleibt. Nachdem die Funktion der EOG-Meßanordnung
nochmals gründlich überprüft und als funktionstüchtig befunden wurde, zwang sich die
Schlußfolgerung auf, daß die Albinofrettchen unter den getesteten Bedingungen nicht in der
Lage waren, einen monokular ausgelösten hOKN zu generieren.

Ergebnisse 41
100°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24100°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
20°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
10°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s cw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
50°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
20°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
10°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s ccw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
N
N
N
N
N
N
N
N
N
N
Abb. 7: Häufigkeitsverteilung der Steigungsmediane der EOG-Spuren der Albinofrettchen, die während monokularer (nur linkes Auge sehend) optokinetischer Ganzfeldreizung registriert wurden. Die Reizung erfolgte mit dem feinen Reizmuster und mit den Reizgeschwindigkeiten 100, 50, 20, 10 und 5°/s. Linke Spalte: Reizung in cw-Richtung, rechte Spalte: Reizung in ccw-Richtung. In allen Fällen unterscheiden sich die Steigungsmediane aus den einzelnen Häufigkeitsverteilungen nicht signifikant von Null oder von den Steigungsmedianen der einzelnen Häufigkeitsverteilungen in Abbildung 6 (Spontane Augenbewegungen derselben Tiere). Es existiert auch kein signifikanter Unterschied zwischen der cw- und ccw-Reizung. Weitere Erläuterungen im Text. Zur Bedeutung der Symbole siehe Legende in Abbildung 6. N = Anzahl der Steigungsmediane (entspricht der Anzahl der getesteten Tiere).

Ergebnisse 42
100°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24100°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
10°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s cw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
10°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s ccw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
N
N
N
N
N
N
N
N
N
N
Abb. 8: Häufigkeitsverteilung der Steigungsmediane der EOG-Spuren der Albinofrettchen, die während monokularer (nur rechtes Auge sehend) optokinetischer Ganzfeldreizung mit dem feinen Reizmuster registriert wurden. Das Ergebnis unterscheidet sich nicht signifikant vom dem Ergebnis in Abbildung 7, weshalb für beide Abbildungen dieselbe Kommentierung gilt. Weitere Erläuterungen im Text. Zur Bedeutung der Symbole siehe Legende in Abbildung 6.

Ergebnisse 43
b) Binokulare Ganzfeldreizung
Feines Reizmuster: Der Befund aus den monokularen Messungen führte notwendigerweise
zu der Frage, ob die Albinofrettchen auch unter binokularer Ganzfeldreizung einen hOKN-
Ausfall zeigen würden. Deshalb wurden mit denselben Albinofrettechen und demselben
feinen Reizmuster auch binokulare Messungen durchgeführt.
Wie die EOG-Spuren aus der monokularen Reizung, weisen auch alle EOG-Spuren aus der
binokularen Reizung mit dem feinen Reizmuster eine glatte Form und einen nahezu
waagerechten Verlauf auf. Hinsichtlich Form und Verlauf unterscheiden sie sich nicht von
den EOG-Spuren der spontanen Augenbewegungen.
Das Ergebnis dieser Messungen ist in Abbildung 9 präsentiert und zeigt, daß auch bei
binokularer Reizung mit dem feinen Zufallspunktemuster die EOG-Spuren der
Albinofrettchen Steigungsmediane liefern, die alle um den Wert Null liegen. Dies gilt für
alle getesteten Reizgeschwindigkeiten (100, 50, 20, 10 und 5°/s) und Reizrichtungen (cw und
ccw). Alle Steigungsmediane befinden sich im Bereich –0,5 bis +0,5, wobei aber die
allermeisten Werte zwischen –0,25 und +0,25 liegen und damit nah bei Null. Der Median der
Steigungsmediane in der jeder Häufigkeitsverteilung ist Null (vgl. auch Abb. 11A weiter
unten). Vergleicht man die beiden Reizrichtungen miteinander (linke und rechte Spalte der
Abb. 9), so findet man aufgrund der Binokularität erwartungsgemäß keinen Unterschied
(WILCOXON SIGNED RANK TEST, in allen Fällen P > 0,05).
Vergleicht man die Steigungsmediane der Teilabbildungen aus Abb. 9 mit den Steigungs-
medianen aus den binokularen Kontrollmessungen bei Licht oder bei Dunkelheit (Abb. 6), so
findet man ebenfalls keinen signifikanten Unterschied (WILCOXON SIGNED RANK TEST, in
allen Fällen P > 0,05). Das bedeutet, daß sich die EOG-Spuren der binokularen Ganzfeld-
reizung mit dem feinen Reizmuster nicht signifikant von den EOG-Spuren unterscheiden, die
bei stehender Trommel oder in völliger Dunkelheit binokular aufgezeichnet wurden. Auch
unterscheiden sich die einzelnen Häufigkeitsverteilungen nicht signifikant von Null (in allen
Fällen P > 0,05). Deshalb kann festgehalten werden, daß die Albinofrettchen auch bei bino-
kularer optokinetischer Reizung mit dem feinen Reizmuster einen hOKN-Ausfall zeigen.
Nach diesem Befund wurde der Entschluß gefaßt, die binokulare Ganzfeldreizung auch mit
dem groben Zufallspunktemuster durchzuführen, bei sonst konstanten Bedingungen.

Ergebnisse 44
100°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24100°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
10°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s cw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
10°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s ccw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
N
N
N
N
N
N
N
N
N
N
Abb. 9: Häufigkeitsverteilung der Steigungsmediane von EOG-Spuren der Albinofrettchen, die während binokularer optokinetischer Ganzfeldreizung registriert wurden. Die Reizung erfolgte mit dem feinen Reizmuster und den Reizgeschwindig-keiten 100, 50, 20, 10 und 5°/s. Linke Spalte: Reizung in cw-Richtung, rechte Spalte: Reizung in ccw-Richtung. Unter allen Reizbedingungen sind die Werte um Null verteilt. In keinem Fall findet man einen signifikanten Unterschied zu den spontanen Augenbewegungen (vgl. Abb. 6). Weitere Erläuterungen im Text. Zur Bedeutung der Symbole siehe Legende in Abbildung 6. N = Anzahl der Steigungsmediane (entspricht der Anzahl der unter der jeweiligen Reizbedingung getesteten Tiere).

Ergebnisse 45
Grobes Reizmuster: Die EOG-Spuren der Albinofrettchen aus den binokularen Reizungen
mit dem groben Reizmuster unterscheiden sich hinsichtlich ihrer glatten Form weder von den
Spuren aus den binokularen Reizungen mit dem feinen Reizmuster, noch von den Spuren der
spontanen Augenbewegungen. Hinsichtlich ihrer Seigung findet man jedoch zum Teil einen
signifikanten Unterschied zu den Spuren aus der Reizung mit dem feinen Muster. Die EOG-
Spuren aus der Reizung mit dem groben Muster haben größere Steigungsmediane, d.h. die
Reizung mit dem groben Muster hat eine höhere Augengeschwindigkeit ausgelöst. Dieser
Befund wird im weiteren bei der Erläuterung der Abbildung 10 und Abbildung 11
eingehender beschrieben.
In Abbildung 10 werden die Häufigkeitsverteilungen der Steigungsmediane aus den hOKN-
Tests mit dem groben Muster für die verschiedenen Reizgeschwindigkeiten und
Reizrichtungen dargestellt. Mit Ausnahme der Steigungsmediane aus der ccw-Reizung mit
100°/s (Median = –0,05), haben die Steigungsmediane in allen anderen Verteilungen in
Abbildung 10 einen Median von Null. Infolge dessen detektiert der WILCOXON SIGNED
RANK TEST nur bei der ccw-Reizung mit 100°/s einen signifikanten Unterschied zwischen
den Steigungsmedianen aus der Reizung mit dem groben (Abb. 10) und denen aus der
Reizung mit dem feinen (Abb. 9) Reizmuster (P = 0,027). Bei allen anderen
Reizgeschwindigkeiten und Reizrichtungen wird kein signifikanter Unterschied entdeckt (in
allen Fällen P > 0,05).
Es ist jedoch auffällig, daß die Steigungsmediane bei Reizung mit dem groben Muster
(Abb. 10) stärker um Null streuen als bei Reizung mit dem feinen Muster (Abb. 9). Dies
resultiert aus dem oben erwähnten Befund, daß die EOG-Spuren zu größeren
Steigungsmedianen tendieren, wenn die Albinofrettchen mit dem groben Reizmuster gereizt
werden. Außerdem fällt bei Reizung mit dem groben Muster (Abb. 10) auf, daß die
Steigungsmediane — im Gegensatz zu den Steigungsmedianen aus der Reizung mit dem
feinen Muster (Abb. 9) — bei cw-Reizung zu positiven und bei ccw-Reizung zu negativen
Werten tendieren. Der statistische Vergleich (WILCOXON SIGNED RANK TEST) zwischen den
beiden Reizrichtungen (linke und rechte Spalte in Abb. 10) ergibt einen signifikanten
Unterschied bei den Reizgeschwindigkeiten 10, 20 und 100°/s (in allen drei Fällen P < 0,05).
Bei 5 und 50°/s existiert kein signifikanter Unterschied (P = 0,249 und 0,082).
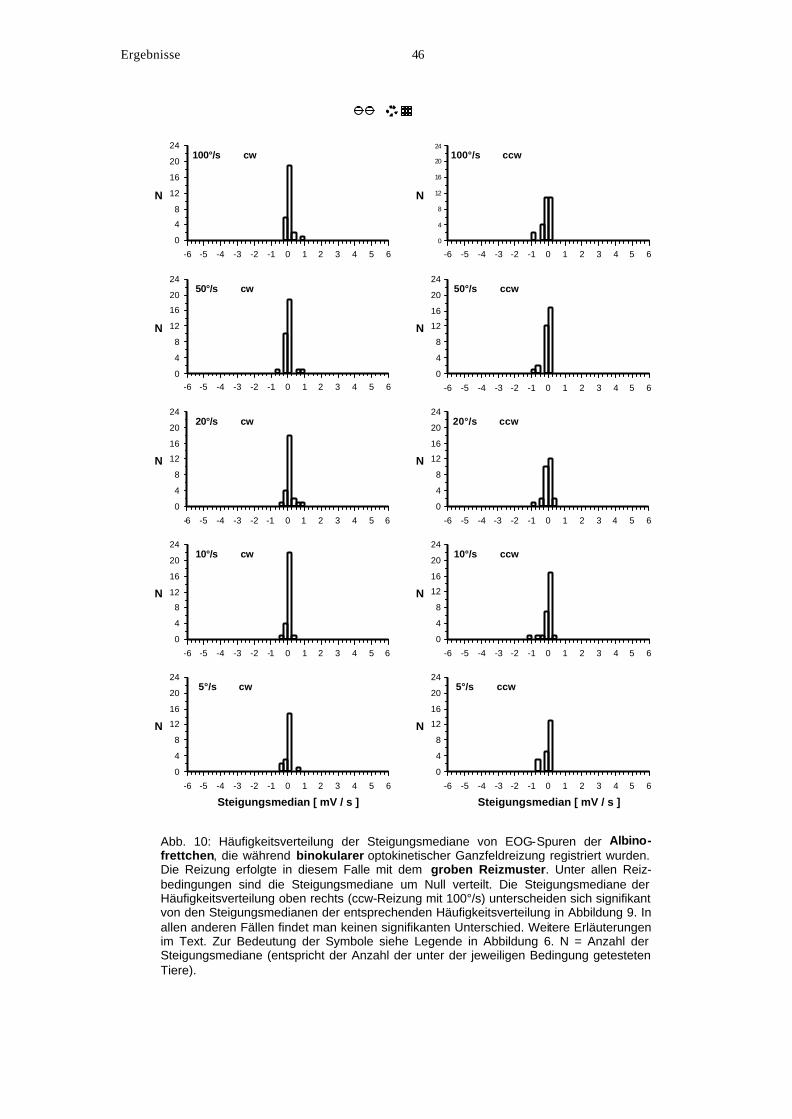
Ergebnisse 46
100°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24100°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
20°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
10°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s cw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
10°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s ccw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
N
N
N
N
N
N
N
N
N
N
Abb. 10: Häufigkeitsverteilung der Steigungsmediane von EOG-Spuren der Albino-frettchen, die während binokularer optokinetischer Ganzfeldreizung registriert wurden. Die Reizung erfolgte in diesem Falle mit dem groben Reizmuster. Unter allen Reiz-bedingungen sind die Steigungsmediane um Null verteilt. Die Steigungsmediane der Häufigkeitsverteilung oben rechts (ccw-Reizung mit 100°/s) unterscheiden sich signifikant von den Steigungsmedianen der entsprechenden Häufigkeitsverteilung in Abbildung 9. In allen anderen Fällen findet man keinen signifikanten Unterschied. Weitere Erläuterungen im Text. Zur Bedeutung der Symbole siehe Legende in Abbildung 6. N = Anzahl der Steigungsmediane (entspricht der Anzahl der unter der jeweiligen Bedingung getesteten Tiere).

Ergebnisse 47
In Abbildung 11 werden der Median, das 10%-, 25%-, 75%- und 90%-Perzentil der
Steigungsmediane aus den binokularen Reizungen mit dem feinen (Abb. 11A, entspricht der
Abb. 9) und groben (Abb. 11B, entspricht der Abb. 10) Reizmuster für jede
Reizgeschwindigkeit und Reizrichtung dargestellt. Der Vergleich zwischen Abbildung 11A
und 11B zeigt und fasst die Befunde besonders deutlich zusammen:
• Die Steigungsmediane in fast allen Häufigkeitsverteilungen in den beiden Abbildungen 9
und 10 haben einen Median von Null. Folglich unterscheiden sich die entsprechenden
Reizungen nicht hinsichtlich ihrer Steigungsmediane (mit Ausnahme der beiden ccw-
Reizungen mit 100°/s).
• Die Mediane der Steigungsmediane streuen mehr bei Reizung mit dem groben Muster.
• Die Mediane der Steigungsmediane tendieren zu positiven Werten bei cw- und zu
negativen Werten bei ccw-Reizung mit dem groben Muster. Diese Tendenz ist bei 10, 20
und 100°/s signifikant und belegt, daß die Albinofrettchen eine sehr schwache Reaktion
auf das grobe Muster zeigen: Die rein zufällige Richtungsänderung der horizontalen
Augendrift, die zuvor bei stehendem Reizmuster (spontane Augenbewegungen)
festgestellt wurde, erfolgt bei Reizung mit dem groben Muster in Abhängigkeit von der
Reizrichtung. Die Augen der Albinos tendieren bei Reizung mit dem groben Muster
dazu, in Reizrichtung abzudriften.
2. Optokinetische Reizung der pigmentierten Frettchen
a) Monokulare Ganzfeldreizung
Zur Erprobung der Methode wurden einige wenige pigmentierte Frettchen mit beiden
Reizmustern monokular getestet. Erwartungsgemäß konnte der monokulare hOKN ausgelöst
werden (vgl. Hein et al., 1990).
b) Binokulare Ganzfeldreizung
Feines Reizmuster: Abbildung 12 zeigt für die Gruppe der pigmentierten Tiere das Ergebnis
aus den binokularen Ganzfeldreizungen mit dem feinen Muster und den verschiedenen Reiz-
geschwindigkeiten. Die EOG-Spuren der pigmentierten Tiere waren Sägezahn-förmig.
Folglich ergaben sich in allen Fällen erwartungsgemäß Steigungsmediane, die je nach
hOKN-Richtung (bzw. Reizrichtung) wesentlich von Null abwichen.

Ergebnisse 48
Die Steigungsmediane in jeder Häufigkeitsverteilung in Abbildung 12 unterscheiden sich
statistisch signifikant sowohl von dem Wert Null als auch von den Steigungsmedianen der
Albinofrettchen aus binokularen Reizungen (Abb. 9 und 10) und auch von den
Steigungsmedianen aus den Messungen der spontanen Augenbewegungen der pigmentierten
Frettchen (vgl. letzte Zeile in Abb. 6; MANN-WHITNEY RANK SUM TEST, in allen Fällen P <
0,05). Vergleicht man die übereinanderstehenden Graphiken innerhalb der linken oder der
rechten Spalte, so fällt auf, daß die Steigungsmediane mit fallender Reizgeschwindigkeit zu
kleineren Beträgen tendieren. Dies zeigt eine Abhängigkeit der Augengeschwindigkeit von
der Reizgeschwindigkeit, die in Abbildung 13 deutlicher dargestellt wird. Die verschiedenen
Abb. 11: Mediane, 10%-, 25%-, 75%- und 90%-Perzentile der Steigungsmediane der Albinofrettchen aus den binokularen Reizungen. Die Häufigkeitsverteilungen dieser Steigungsmediane sind in den Abbildungen 6, 9 und 10 gezeigt. A: Reizung mit dem feinen Muster (entspricht der Abb. 9). B: Reizung mit dem groben Muster (entspricht der Abb. 10). Zusätzlich sind die entsprechenden Werte für die Spontanrate (Abb. 6) bei Licht (L) und bei Dunkelheit (D) angegeben. Mit Ausnahme eines Medians von –0,05 (grobes Reizmuster, 100ccw) sind alle anderen Mediane Null. Die Reizung mit dem groben Reizmuster führte zu einer größeren Streuung der Steigungsmediane (größere 10, 25, 75 und 90%-Perzentile) als die Reizung mit dem feinen Muster. Ein waagerechter Balken unter einer cw- und ccw-Reizung zeigt einen signifikanten Unterschied zwischen den Steigungsmedianen der beiden Reizungen an (WILCOXON SIGNED RANK TEST, P > 0,05). Die nebenstehende Legende erläutert die Position des Medians und der verschiedenen Perzentile.
75%
90%
25%
10%
Median
feines Muster
0 (D) 0 (L) 5cw 5ccw 10cw 10ccw 20cw 20ccw 50cw 50ccw 100cw 100ccwMed
ian
der
Ste
igun
gsm
edia
ne [m
V/s
]
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
grobes Muster
Reizparameter
0 (D) 0 (L) 5cw 5ccw 10cw 10ccw 20cw 20ccw 50cw 50ccw 100cw 100ccwMed
ian
der
Ste
igun
gsm
edia
ne [
mV
/s]
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
A
B

Ergebnisse 49
Reizgeschwindigkeiten sind auf der x-Achse aufgetragen. Die y-Achse zeigt den Median,
das 10%-, 25%-, 75%- und 90%-Perzentil der Steigungsmediane von allen pigmentierten
Tieren für die cw- und ccw-Reizrichtung an.
Aus diesen Befunden lassen sich folgende Schlußfolgerungen ziehen: 1. Die Methode zur
Messung der Augenbewegungen ist in der Lage beim pigmentierten Frettchen einen hOKN
zu detektieren. 2. Die Methode ist auch in der Lage zwischen unterschiedlichen
Antwortstärken (d.h. Geschwindigkeiten der langsamen hOKN-Phasen) zu unterscheiden,
was durch die Geschwindigkeitsabhängigkeit der Steigungsmediane gezeigt ist.
Grobes Reizmuster: Mit dem groben Reizmuster wurden probeweise nur einige wenige
pigmentierte Frettchen getestet. Die Reizwirkung des groben Musters hatte sich nicht von
der des feinen Reizmusters unterschieden, so daß diese Messungen nicht weiter fortgesetzt
wurden und hier nicht dargestellt werden.
C. Inaktivierungsstudie und histologische Auswertung
1. hOKN-Test nach Muscimol-Injektion
Vor den Muscimol-Injektionen zeigten weder die pigmentierten noch die albinotischen
Frettchen einen Spontannystagmus (vgl. Abb. 6), wenngleich bei den albinotischen Frettchen
im Gegensatz zu den pigmentierten Frettchen eine okulomotorische Instabilität gefunden
wurde. Nach den Muscimol-Injektionen wurde bei zwei der drei albinotischen und bei dem
einen pigmentierten Frettchen ein Spontannystagmus beobachtet. In allen Fällen war der
Spontannystagmus schon bei der ersten registrierten EOG-Spur auffällig, die frühestens ca.
zwei Minuten aber spätestens ca. vier Minuten nach der Muscimol-Injektion aufgenommen
wurde. Der Spontannystagmus hatte eine langsame Phase in Richtung auf die nicht-injizierte
Seite, trat sowohl in Dunkelheit als auch bei Helligkeit auf und konnte durch mono- oder
binokulare Ganzfeldreizung in der optokinetischen Trommel nicht sichtbar beeinflußt
werden. Er dauerte mindestens zwei Stunden, war aber am nächsten Tag immer vollständig
verschwunden. Nach dreimaliger Anwendung von Muscimol, meistens an aufeinander-
folgenden Tagen, wurden die Tiere eingeschläfert und perfundiert. Ihre Gehirne wurden
gefärbt und histologisch untersucht. Bei dem dritten albinotischen Tier hatte die Muscimol-
Injektion keinen Effekt auf die okulomotorische Stabilität. Die anschließende histologische
Untersuchung der Schnitte liefert eine mögliche Erklärung dieses Befundes (siehe unten).

Ergebnisse 50
100°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24100°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
10°/s cw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
5°/s cw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
50°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
20°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
12
16
20
24
10°/s ccw
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 60
4
8
1216
20
24
5°/s ccw
Steigungsmedian [ mV / s ]
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
4
8
12
16
20
24
N
N
N
N
N
N
N
N
N
N
Abb. 12: Häufigkeitsverteilung der Steigungsmediane von EOG-Spuren pigmentierter Frettchen, die während binokularer optokinetischer Ganzfeldreizung mit dem feinen Reizmuster registriert wurden. Unter allen Reizbedingungen sind die Steigungsmediane nicht um Null verteilt, sondern signifikant größer (bei cw-Reizung) oder signifikant kleiner (bei ccw-Reizung) als Null. Sie unterscheiden sich ebenfalls signifikant von allen anderen Steigungsmedianen, die in den Abbildungen 6 bis 10 dargestellt sind. Außerdem ist eine Tendenz auffällig, nach der der Betrag der Steigungsmediane mit fallender Reizgeschwindigkeit kleiner wird. Zur Bedeutung der Symbole siehe Legende in Abbildung 6. N = Anzahl der Steigungsmediane (entspricht der Anzahl der unter der jeweiligen Bedingung getesteten Tiere).

Ergebnisse 51
In Abbildung 14 ist der Muscimol-Effekt exemplarisch für ein albinotisches (linke Spalte)
und für das pigmentierte Frettchen (rechte Spalte) dargestellt (Albinofrettchen #1 und
pigmentiertes Frettchen aus Tab. 2). Die einzelnen Teilabbildungen zeigen 5-s-lange
Ausschnitte der EOG-Spuren, die vor und nach der Applikation von Muscimol
aufgenommen wurden. Die binokulare optokinetische Reizung erfolgte in der beleuchteten
Trommel mit dem groben Reizmuster mit einer Reizgeschwindigkeit von 50°/s in
horizontaler cw- und ccw-Richtung. Die Aufnahmen in Dunkelheit erfolgten außerhalb der
Trommel in einem völlig abgedunkelten Raum. Beim Albinofrettchen wurde auf der linken
Seite injiziert, weshalb der Spontannystagmus eine langsame Folgephase nach rechts hat,
während beim pigmentierten Tier die rechte Seite injiziert wurde und der ausgelöste
Spontannystagmus eine langsame Phase nach links zeigt. Bei beiden Tieren wurde der
Spontannystagmus nicht durch die optokinetische Reizung beeinflusst. Einen Tag nach der
Muscimol-Injektion zeigten beide Tiere das gleiche Verhalten wie vor der Inaktivierung,
woraus abgeleitet werden kann, daß die Wirkung des Muscimols abgeklungen und keine
permanente Schädigung entstanden war. Die Farbstoff-Injektion beim Albinofrettchen zeigte
keinen Effekt (Kontrolle).
Abb. 13: Abhängigkeit der Augengeschwindigkeit von der Reizgeschwindigkeit bei den pigmentierten Frettchen während binokularer Ganzfeldreizung mit dem feinen Muster. Definitionsgemäß sind die Steigungsmediane positiv bei Reizung in cw-Richtung und negativ bei Reizung in ccw-Richtung. Entsprechend verhält sich das Vorzeichen ihres Medians. x-Achse: verschiedene Reizgeschwindigkeiten (5, 10, 20, 50, 20 und 100°/s) und Spontanrate bei Dunkelheit [(0°/s (D)] und Licht [(0°/s (D)]. y-Achse: EOG-Median, 10%-, 25%-, 75%- und 90%-Perzentil (vgl. Legende der Abb. 11).
Plot 1
Reizgeschwindigkeit
0°/s 0°/s 5°/s 10°/s 20°/s 30°/s 40°/s 50°/s 60°/s 70°/a 80°/s 90°/s 100°/s
Med
ian
der
Ste
igu
ng
smed
ian
e [ m
V /
s ]
-6
-4
-2
0
2
4
6
cw
ccw
(D) (L)

Ergebnisse 52
binokular, 50°/s albinotisches Frettchen (Muscimol-Injektion links)
pigmentiertes Frettchen (Muscimol-Injektion rechts)
: EOG-Aufnahme bei Helligkeit : EOG-Aufnahme bei Dunkelheit : Reizung in cw-Richtung : Reizung in ccw-Richtung
5 s 5 s
1 mV
vor Muscimol
vor Muscimol
nach Muscimolspontan
Licht
nach Muscimolspontan
Dunkelheit
nach Muscimol
nach Muscimol
nächster Tag
nächster Tag
nach Farbstoffspontan
Licht
nach Farbstoffspontan
Dunkelheit
nach FarbstoffLicht
nach Farbstoff
Licht
0,0
0,10
3,40
-5,00
-4,25
-3,15
4,25
3,80
4,90 -2,70
2,95
0,0
0,0 -4,05
5,60
-3,30
0,10
0,0
-0,10
-0,25
Abb. 14: EOG-Spuren aufgenommen vor und nach einseitiger Inaktivierung des NOT mit Muscimol. Die Zahl oben rechts in jeder Teilabbildung gibt den Steigungsmedian der abgebildeten EOG-Spur an.

Ergebnisse 53
2. Histologische Dokumentation der Injektionsorte
Die implantierten Injektionskanülen hinterließen im Kortex Einstichspuren, die auf den
Hirnschnitten mit bloßen Augen sichtbar waren. Bei den zwei albinotischen und dem einen
pigmentierten Frettchen, die infolge der Muscimol-Injektion einen Spontannystagmus
gezeigt hatten, wurde unter dem Lichtmikroskop sichtbar, daß die Spitzen der Injektions-
kanülen zwar in der Nähe des Prätektums plaziert gewesen waren, das Prätektum selbst aber
nicht berührt bzw. mechanisch zerstört hatten. Dieser Befund spricht für die Reversibilität
des Muscimol-Effekts.
Tabelle 2 listet die implantierten Tiere und das jeweilige Ergebnis der Muscimol- und der
Farb-Injektionen auf. Die Entfernung ∆x gibt in rostrocaudaler (anterior-posterior)
Richtung den Abstand zwischen dem Injektionsort (Spitze der Injektionskanüle) und dem
rostralen Rand des SC (bzw. rostralem Ende des NOT) an. Ein positives (negatives) ∆x
bedeutet, daß der Injektionsort rostral (caudal) des vorderen SC-Rands liegt (vgl. Abb. 5). In
mediolateraler Richtung kann die Entferung zwischen NOT und Injektionsort weniger
genau angegeben werden, da die laterale NOT-Grenze auf den Nissl- und/oder Klüver-
Barrera gefärbten Schnitten nicht erkennbar ist (vgl. Material und Methoden). Unter Zuhilfe-
nahme der Hirnschnitt-Abbildungen aus Zhang & Hoffmann, 1993 läßt sich jedoch
schlußfolgern, daß die Injektionsorte zwischen 0 und maximal 300 µm vom NOT entfernt
waren.
Tab. 2: Ergebnis der Inaktivierungsstudie. Die Muscimol-Injektionen haben bei den Albinos #1 und #2 und dem pigmentierten Frettchen einen Effekt (Spontannystagmus) gezeigt. Bei Albino #3 haben die Muscimol-Injektionen keinen Spontannystagmus ausgelöst. Keine der Farbstoff-Injektionen hat einen Effekt auf die Okulomotorik gehabt. Tier implantierte
Hirnseite Muscimol-
Effekt ∆x Farb-Injektion Effekt der Farb-
Injektion Albino #1 links ja +200 µm roter Marker nein
Albino #2 links ja +50 µm Granular Blue nein
Albino #3 links nein +600 µm roter Marker nein
Pigm. rechts ja –250 µm Granular Blue nein

Ergebnisse 54
Abb. 15: Position der implantierten Injektionskanüle. Es werden zwei schwarz-weiße Fotografien von Nissl-gefärbten frontalen Hirnschnitten eines Albinofrettchens gezeigt. A: Die Schnittebene verläuft durch das rostrale Ende des linken (l-SC) und rechten superioren Colliculus (r-SC). Auf der linken Seite sieht man die Zerstörung des Kortexgewebes, die durch die Kanüle verursacht wurde (umrandet durch die weiße, gestrichelte Linie). B: Mittelpunkt der Kanüle (ca. 450 µm weiter rostral als die Schnittebene in A). Die Zerstörung des Kortex ist auf dieser Schnittebene wesentlich größer und vollständiger. In dieser Schnittebene ist der SC und das NOT-Areal nicht mehr zu sehen. Abkürzungen: l-SC = linker SC, r-SC = rechter SC, HC = Hypocampus, CC = Corpus callosum, NOT = rostrales Ende des NOT-Areals, d = dorsal, v = ventral.
Foto-Nr. 3.15 22.07.1999.aF Schnitt-Nr. 2-26-2
Foto-Nr. 3.7 22.07.1998.aF Schnitt-Nr. 2-25-1
l-SC r-SC
A
B
CC
CC
HC
Kortex
NOT
Kortex
d
v
v
d
HC
1 mm
1 mm
CGL
CGL

Ergebnisse 55
In der Abbildung 15 werden beispielhaft zwei Fotografien von 50 µm dicken Frontalschnit-
ten des Albinofrettchens #2 gezeigt, aus denen sich die Position der implantierten Injektions-
kanüle rekonstruieren läßt. Die Injektionskanüle war 13 Tage in der linken Hirnseite in senk-
rechter Stellung implantiert. Der Hirnschnitt in Abbildung 15A verläuft durch das Mittelhirn
und den darüberliegenden Kortex, in dem sich die Kanüle befand. Die Schnittebene geht
durch den rostralen Bereich des SC, der unten rechts auf dem Foto zu sehen ist. Auf der dor-
salen Seite sind die beiden Hemisphären des Kortex cerebri sichtbar, die durch das Corpus
callosum miteinander verbunden sind. Der helle Pfeil zeigt auf die Position des zu inaktivie-
renden NOT, welcher sich lateral des linken SC befindet. Im Kortex darüber ist kortikales
Gewebe sichtbar, das durch die Injektionskanüle zerstört wurde (weiß gestrichelt umrissener
Bereich). Der Mittelpunkt der Kanüle (bzw. Injektionszentrum) befand sich allerdings nicht
genau in dieser Position, sondern ca. 450 µm weiter rostral, wo die Zerstörung des Kortex
am größten ist (Abb. 15B). Die in Abbildung 15B gezeigte Schnittebene liegt 50 µm (∆x +50
µm) vor der rostralen SC-Grenze, so daß der SC hier nicht mehr zu sehen ist. Der
Injektionsort befindet sich auf der Oberfläche des Mittelhirns direkt unter der Einstichspur.
3. Farb-Injektionen
Aus Tabelle 2 geht hervor, welcher Fluoreszenz-Farbstoff bei welchem Tier injiziert wurde.
Keine der Farb-Injektionen hatte einen Effekt auf die okulomotorische Stabilität. Insofern
sind diese Injektionen auch als Kontrollinjektionen aufzufassen. In der anschließenden
Untersuchung der Hirnschnitte unter dem Fluoreszenz-Mikroskop konnten die Injektionsorte
relativ leicht bestimmt werden, da sie sehr viel stärker gefärbt waren als das umliegende
Gewebe. Dadurch wurde die Lage der Injektionsorte (∆x-Werte) bestätigt, die schon anhand
der Einstichspuren ermittelt wurden.
Granular Blue ist besonders gut vom Gewebe aufgenommen worden, so daß die Injektionen
zu einer weit verbreiteten Färbung des gesamten Mittelhirns geführt haben. Dabei findet man
auch das Prätektum der nicht-injizierten Seite gefärbt. Allerdings ist das Prätektum der
injizierten Seite stärker gefärbt als das der nicht-injizierten Seite. Insgesamt ist die
Fluoreszenz auf der injizierten Seite sehr viel stärker als auf der nicht-injizierten Hirnseite.
Außerdem befindet sich auf der prätektalen Oberfläche der injizierten Seite sehr viel mehr
Granular Blue als auf der gegenüberliegenden Seite. Unter der Annahme, daß sich das
Muscimol in ähnlicher Weise ausgebreitet hat, kann davon ausgegangen werden, daß das
Prätektum der injizierten Seite stärker mit Muscimol durchdrungen und somit auch stärker
inaktiviert wurde als auf der gegenüberliegenden Seite.

Ergebnisse 56
Die Injektionen von roter Textmarkertinte haben zu einer sehr schwachen Färbung der Zellen
des Mittelhirns und speziell des Prätektums geführt. Der meiste Farbstoff befindet auf der
prätektalen Oberfläche und nicht im Gewebe. Das deutet auf eine schlechte Aufnahme des
Farbstoffs durch das Gewebe hin. Bei Albino #1 findet man eine schwache Färbung des Prä-
tektums der injizierten Seite, während bei Albino #3 überhaupt keine Färbung zu sehen ist.
Das völlige Ausbleiben einer Färbung bei Albino #3 zeigt, daß die Injektion der roten
Textmarkertinte gänzlich mißglückt war. Dies könnte auf eine Verstopfung des
Führungsröhrchens zurückzuführen sein, was auch das Ausbleiben des Muscimol-Effektes
bei diesem Tier erklären könnte. Der wahrscheinlichste Grund jedoch für die Wirkungslosig-
keit des Muscimols bei diesem Albino ist die sehr viel größere Entfernung zwischen dem
Injektionsort und dem NOT (∆x = 600 µm).
D. Ergebnis des Perimetrietests
Insgesamt wurden fünf albinotische (Tier #10, #16, #39, #40 und #41) und vier pigmentierte
Frettchen (Tier #33, #35, #37 und #38) unter binokularer und monokularer Bedingung im
Perimetrietest untersucht. Die Ergebnisse werden als Fallbeispiele in den Abbildungen
16 - 20 graphisch dargestellt. Ein horizontaler Balken unter der x-Achse symbolisiert bei
monokularer Bedingung die Abdeckung des entsprechenden Auges. Die genauen Prozent-
sätze positiv beantworteter Durchläufe können den Tabellen 3 und 4 entnommen werden. Da
sich die Daten aus den beiden Testvarianten (mit und ohne Plexiglasröhre, vgl. Material und
Methoden) nicht unterschieden haben, wurden sie zusammengefaßt.
Die beiden Albinofrettchen #10 und #16 (Abb. 16 und Tab. 3) wurden nur mit dem linken
Auge ohne Unterteilung des R60°-Sektors monokular getestet. Alle anderen Frettchen
wurden mit beiden Augen monokular getestet, wobei der 60°-Sektor kontralateral zum
sehenden Auge in die beiden Teilsektoren 30-45° und 45-60° unterteilt wurde.
Alle neun Tiere wurden auch im hOKN-Test eingesetzt. Die fünf Albinos zeigten einen
hOKN-Ausfall. Bei den vier pigmentierten Tieren konnte der hOKN ausgelöst werden.
Bei den verschiedenen Versuchstieren wurde eine ungleich starke Motivation beobachtet,
was sich in den unterschiedlich hohen Trefferquoten in 0°-Richtung (randomisierte Durch-
läufe ohne Reizpräsentation zwischen den Durchläufen mit Reizpräsentation) widerspiegelt.
Die Unterschiede zwischen den verschiedenen Individuen hinsichtlich Kooperationsbereit-
schaft und Lernfähigkeit waren in der Regel schon zu Beginn der Trainingsphasen auffällig.

Ergebnisse 57
1. Binokulare Messungen
Unter binokularen Bedingungen haben beide Tiergruppen in den allermeisten Sektoren 80%,
in einigen Fällen sogar weit über 90% der Durchläufe positiv beantwortet (Tab. 3 und 4 und
obere Reihen der Abb. 16 - 20). Ausnahmen stellen die beiden Albinofrettchen #39 und #41
dar, die im 60°- und 90°-Sektor der rechten Gesichtsfeldhälfte geringere Trefferquoten
erzielten [Tier #39: 72% im R60°- und 52% im R90°-Sektor (vgl. Abb. 17 und Tab. 3); Tier
#41: 65% im R60°- und 69% im R90°-Sektor (vgl. Abb. 18 und Tab. 3)]. Offensichtlich sind
diese beiden Albinofrettchen in ihrer Sehfähigkeit innerhalb des R30°- und R90°-Sektors
gestört. Bei Zusammenbetrachtung aller neun Fallbeispiele (fünf albinotische und vier
pigmentierte Frettchen) ist jedoch die Schlussfolgerung zulässig, daß sich die beiden Tier-
gruppen unter binokularer Bedingung im Perimetrietest nicht voneinander unterscheiden.
Damit kann festgestellt werden, daß sich die Albinofrettchen unter binokularen Bedingungen
genauso gut visuell orientieren können wie ihre pigmentierten Artgenossen.
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°0 20 40 60 80 100020406080100
R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
Albino #10 Albino #16
Abb. 16: Trefferquoten der Albinofrettchen #10 und #16 im Perimetrietest. Unter binokularer Bedingung (obere Reihe) erreichen beide Tiere in allen sechs Gesichtsfeldsektoren eine relativ hohe positive Trefferquote. Unter monokularer Bedingung erreichen sie in den Sektoren innerhalb der Gesichtsfeldhälfte des sehenden Auges eine ähnlich hohe Trefferquote wie in den binokularen Messungen, während die Trefferquoten innerhalb der Sektoren kontralateral zum sehenden Auge unter 5% liegen. Die dunklen Punkte in den Sektoren, auf die die Pfeil-spitzen zeigen, geben die Anteile der positiv beantworteten Durchläufe in % an (vgl. Tabelle 3). Die schwarzen Balken in der unteren Reihe symbolisieren die Abdeckung des rechten Auges. Die Bedeutung der anderen Symbole kann der Legende in Abbildung 6 entnommen werden.

Ergebnisse 58
2. Monokulare Messungen
Unter monokularer Bedingung unterscheiden sich die beiden Tiergruppen ebenfalls nicht,
wenn der visuelle Reiz innerhalb der ipsilateralen Gesichtsfeldhälfte des sehenden Auges
präsentiert wird. Sowohl die Albinos als auch die pigmentierten Frettchen erzielten in diesen
Fällen ähnlich hohe Trefferquoten von über 80%, in vielen Sektoren sogar 100%.
Albino #39 Albino #40
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
Abb. 17: Trefferquoten der Albinofrettchen #39 und #40 im Perimetrietest. Unter binokularer Bedingung erreichen beide Tiere eine relativ hohe positive Trefferquote. Das Tier #39 fällt auf, da es mit dem rechten Auge innerhalb des rechten 60°- und 90°-Sektors besonders niedrige Trefferquoten erzielt. Unter monokularer Bedingung wird in allen Sektoren innerhalb der Gesichtsfeldhälfte des sehenden Auges eine ähnlich hohe Trefferquote erreicht wie in den binokularen Messungen, wobei der Defekt des rechtes Auges des Tieres #39 wieder sichtbar wird. Kontralateral zum sehenden Auge wird eine Trefferquote von maximal 14% erreicht (Tier #40 mit dem rechten Auge im R30°-Sektor). Die einzelnen Trefferquoten können der Tabelle 3 entnommen werden.

Ergebnisse 59
Die Beeinträchtigung im R60°- und R90°-Sektor bei den beiden Albinofrettchen #39 und
#41 (vgl. Abb. 17 und Abb. 18) macht sich auch unter monokularer Bedingung bemerkbar.
Das Albinofrettchen #39 erzielt hier mit dem rechten Auge eine niedrigere Trefferquote als
unter binokularer Bedingung. Im Gegensatz dazu beobachtet man beim Albinofrettchen #41
eine Verbesserung des rechten Auges, wenn das linke Auge abgedeckt ist.
Bei Reizpräsentation im 30°- und 60°-Sektor (bzw. in den beiden Teilsektoren 30-45° und
45-60°) innerhalb der Gesichtsfeldhälfte kontralateral zum sehenden Auge werden
erhebliche Unterschiede zwischen den beiden Tiergruppen sichtbar.
Die höchste Trefferquote in einem 30°-Sektor kontralateral zum sehenden Auge hat unter
den Albinos das Tier #41 (18% mit dem rechten Auge im L30°-Sektor, vgl. Abb. 18). Alle
anderen Albinos erzielten unter monokularer Bedingung in den 30°-Sektoren kontralateral
zum sehenden Auge Trefferquoten von unter 18%, in vielen Fällen von 0%. Die
pigmentierten Frettchen erzielten dagegen eine Trefferquote von mindestens 44% im 30°-
Sektor kontralateral zum sehenden Auge (Tier #33, L30°-Sektor, vgl. Abb. 19). In vielen
Fällen wurde sogar eine wesentlich höhere Trefferquote erreicht, die zwischen 78% (Tier
#37 mit dem linken Auge im R30°-Sektor) und 100% (Tier #38 mit dem rechten Auge im
L30°-Sektor) liegt. Aus diesen Befunden ergibt sich die Schlußfolgerung, daß das frontale
Gesichtsfeld, welches auf die temporale Hemiretina projiziert wird, bei den Albinofrettchen
funktionell unterdrückt ist.
Bei den monokularen Messungen mit den drei Albinofrettchen #39, #40 und #41 und allen
vier pigmentierten Frettchen wurde der 60°-Sektor kontralateral zum sehenden Auge in die
Teilsektoren 30-45° und 45-60° unterteilt. Die Albinofrettchen antworteten, wenn überhaupt,
im 30-45°-Teilsektor positiv [Albino #40 im linken 30-45°-Sektor mit dem rechten Auge
(Abb. 17) und Albino #41 im rechten 30-45°-Teilsektor mit dem linken Auge, Abb. 18]. Im
45-60°-Sektor hat keiner der Albinos auf den visuellen Reiz reagiert. Die pigmentierten
Frettchen zeigten in diesen beiden Teilsektoren folgendes Sehvermögen: Im 30-45°-
Teilsektor wurden immer mehr Duchläufe positiv beantwortet als im 45-60°-Teilsektor.
Dieser Unterschied zwischen den beiden Teilsektoren ist bei den beiden Tieren #35 und #37
besonders groß, bei denen die Trefferquote von mehr als 60% im rechten 30-45°-Teilsektor
steil auf weit unter 50% im rechten 45-60°-Teilsektor fällt. Dieser Befund bestätigt die
Erwartungen, die sich auf elektrophysiologischen Befunden stützen, nach denen sich das
binokulare Gesichtsfeld des Frettchens um ca. 38° in jeder Gesichtsfeldhälfte (Law et al.,
1988; Quevedo et al., 1996) erstreckt (vgl. Abb. 4C). Folglich gehört ungefähr die Hälfte des
30-45°-Sektors zum binokularen Gesichtsfeld und deshalb sollten die pigmentierten

Ergebnisse 60
Frettchen eine höhere Trefferquote in diesem Teilsektor erreichen als im 45-60°-Teilsektor,
der außerhalb des binokularen Gesichtsfeldes liegt und deshalb theoretisch überhaupt nicht
mit dem kontralateralen Auge gesehen werden sollte.
Tab. 3: Ergebnis des Perimetrietests für die Albinofrettchen: Anteil positiv beantworteter Durchläufe in Prozent unter bino- und monokularer Testbedingung. Die Albinos #10 und #16 wurden nur mit dem linken Auge sehend monokular getestet. Die graphische Darstellung der einzelnen Ergebnisse ist in den Abb. 16 - 18 gezeigt. Abkürzungen: bino = binokular, mono = monokular, l = nur linkes Auge sehend, r = nur rechtes Auge sehend.
Albino #10 Albino #16 Albino #39 Albino #40 Albino #41
mono mono mono mono mono bino
l r
bino
l r
bino
l r
bino
l r
bino
l r
L90° 93 100 80 100 92 100 0 98 100 0 91 100 0
45-60° 0 0 0 L60°
30-45° 98 100 94 100
100 100
0
98 100
12
100 100
0
L30° 100 86 96 100 98 97 6 100 100 13 98 100 18
0° 77 100 91 100 76 98 95 93 100 100 96 95 91
R30° 97 7 98 0 98 0 91 100 14 100 93 7 100
30-45° 0 0 4 R60°
45-60° 98 0 96 5
72
0
46 100
0
100 65
0
93
R90° 80 0 91 3 52 0 17 100 0 100 69 0 80
Tab. 4: Ergebnis des Perimetrietests für die pigmentierten Frettchen: Anteil positiv beantworteter Durchläufe in Prozent unter bino- und monokularer Testbedingung mit jedem Auge. Die graphische Darstellung der einzelnen Ergebnisse ist in den Abbildungen 19 und 20 gezeigt. Abkürzungen wie in Tabelle 3.
Pigm. #33 Pigm. #35 Pigm. #37 Pigm. #38
mono mono mono mono bino
l r bino
l r bino
l r bino
l r
L90° 94 100 4 99 92 4 96 94 0 92 100 5
45-60° 31 25 37 42 L60°
30-45° 97 100
44 99 96
61 100 100
75 94 100
78
L30° 98 92 44 100 100 89 100 100 86 99 100 100
0° 98 89 79 97 100 96 100 82 91 98 94 90
R30° 99 49 100 95 80 99 100 78 98 100 88 100
30-45° 15 48 37 43 R60°
45-60° 96
7 100 100
14 100 100
21 100 100
3 100
R90° 90 0 96 99 5 97 82 0 95 100 4 85

Ergebnisse 61
Albino #41
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
Abb. 18: Trefferquoten des Albinofrettchens #41 im Perimetrietest: Dieses Tier zeigt ein ähnliches Verhalten wie die Albinofrettchen #39 und #40. Unter monokularer Bedingung erzielt es mit dem rechten Auge eine maximale Trefferquote von 18% im L30°-Sektor. Die einzelnen Quoten können der Tabelle 3 entnommen werden.

Ergebnisse 62
Pigm. #33
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
Pigm. #35
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
Abb. 19: Trefferquoten der pigmentierten Frettchen #33 und #35 im Perimetrietest: Unter binokularer Bedingung erreichen beide Tiere eine relativ hohe positive Trefferquote. Unter monokularer Bedingung erzielen sie, im Vergleich zu den Albinofrettchen, eine wesentlich höhere Trefferquote innerhalb des 30°- und 60°-Sektors kontralateral zum sehenden Auge. Ihre Sehfähigkeit fällt zwischen dem 30-45°- und dem 45-60°-Teilsektor kontralateral zum sehenden Auge relativ schnell ab. Die einzelnen Quoten können der Tabelle 4 entnommen werden.

Ergebnisse 63
Pigm. #38 Pigm. #37
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
0 20 40 60 80 100020406080100R 90°
R 60°
R 30°
0°
L 30°
L 60°
L 90°
Abb. 20: Trefferquoten der pigmentierten Frettchen #37 und #38 im Perimetrietest: Beide Tiere zeigen qualitativ das gleiche Verhalten wie die pigmentierten Frettchen #33 und #35 (vgl. Abb. 19). Die einzelnen Trefferquoten können der Tabelle 4 entnommen werden.

Ergebnisse 64
II. Teil II: Neurophysiologie
A. Histologische Verifikation
Insgesamt wurden die Gehirne von 10 albinotischen und 5 pigmentierten Frettchen
histologisch aufgearbeitet. Das Ziel der histologischen Auswertung war die Bestätigung der
richtigen Position der verschiedenen Ableit- und Reizorte. Alle elektrophysiologischen
Daten von Tieren, bei denen dieses Ziel nicht erreicht wurde, wurden in der Auswertung
nicht berücksichtigt.
Für alle Experimente kann festgestellt werden, daß die anschließende Auswertung der
Histologie immer das zuvor im elektrophysiologischen Experiment dokumentierte Ergebnis
bestätigte. Dies bedeutet, daß immer die richtige Position des Ableit- und / oder des
Reizortes gefunden werden konnte, wenn zuvor die erwünschten, hier vorgestellten elektro-
physiologischen Daten erhoben werden konnten. In den Fällen, in denen die richtige Position
des Ableit- und / oder Reizortes nicht bestätigt wurde, konnten auch im vorausgegangenen
elektrophysiologischen Experiment keine spezifischen Daten erhoben werden. Im konkreten
Fall bedeutet dies, daß beispielsweise immer dann prätektale Zellen des Albinos von der IO
antidrom gereizt werden konnten, wenn die anschließende Histologie bestätigte, daß
mindestens eine der beiden IO-Reizelektroden auch tatsächlich in der IO plaziert war. In
Bezug auf die NOT-Läsionen kann entsprechend festgestellt werden, daß immer dann von
der IO antidrom reizbare Zellen gefunden werden konnten, wenn anschließend die NOT-
Läsionen tatsächlich im NOT-Areal lokalisiert wurden. Zeigte aber die Histologie, daß beide
Reizelektroden die IO verfehlt hatten oder daß die NOT-Läsion nicht im NOT-Areal plaziert
war, so konnten zuvor auch keine prätektalen Zellen antidrom gereizt werden.
Ein Teil der elektrophysiologischen Daten stammt von Tieren, bei denen keine elektrolyti-
schen NOT-Läsionen gesetzt wurden. In diesen Fällen konnten auf den Hirnschnitten nur
solche Penetrationsspuren identifiziert werden, die in das NOT-Areal führten, jedoch keine
Penetrationsspuren in größerer Entfernung vom NOT-Areal außerhalb des benachbarten SC.
Ein weiterer, kleiner Teil der elektrophysiologischen Daten stammt von Tieren, deren Hirne
nicht histologisch untersucht wurden. In diesen Fällen wird jedoch aufgrund der wieder-
holten Erfahrung mit den histologisch untersuchten Tieren unterstellt, daß die Ableit- und
Reizorte richtig gewesen sein müssen, sonst hätten die entsprechenden elektrophysiolo-
gischen Daten nicht erhoben werden können.

Ergebnisse 65
1. NOT-Ableitorte, IO- und VC-Reizorte
In Tabelle 5 ist die Anzahl der histologisch aufgearbeiteten Gehirne aufgelistet, in denen die
richtige Position der Ableit- und Reizorte dokumentiert wurde. Die Tiere stammten aus der
Experimentenreihe, in der die NOT-Zellen von der ipsilateralen IO antidrom und vom
ipsilateralen VC und kontralateralen ON orthodrom elektrisch gereizt wurden.
Es wurden insgesamt acht Gehirne untersucht. Sechs stammen von albinotischen und zwei
von pigmentierten Frettchen. Bei einem Albinofrettchen wurde die Position des NOT-
Ableitortes durch eine Läsion und bei einem zweiten Albinofrettchen sowohl durch eine
Läsion als auch durch Penetrationsspuren bestätigt. Bei einem anderen Albinofrettchen
wurde keine NOT-Läsion gesetzt und die Identifizierung des NOT-Ableitortes erfolgte nur
durch Penetrationsspuren, die ins NOT-Areal ziehen. Von einem weiteren Albinofrettchen
wurde nur der Hirnstamm histologisch untersucht und die Position der IO-Reizelektroden
bestätigt. Bei den letzten 2 Albinofrettchen wurden Penetrationsspuren entdeckt, die ins
NOT-Areal ziehen. Außerdem wurden bei diesen beiden Tieren die richtige Position der IO-
Reizelektroden und die Position der VC-Reizelektroden dokumentiert. Bei beiden
pigmentierten Frettchen wurde die richtige Position des NOT-Ableitortes durch
elektrolytische Läsionen bestätigt.
Tab. 5: Histologische Untersuchung der Frettchenhirne. Im NOT gefundene elektrolytische Läsionen und Penetrationsspuren verifizieren post mortem die richtige Position der NOT-Ableitorte, d.h. der Orte, an denen prätektale Zellen von der IO antidrom und vom VC und ON orthodrom gereizt wurden. Bei einigen Albinofrettchen wurden keine Läsionen gesetzt. In diesen Fällen konnten Penetrationsspuren entdeckt werden, die in das NOT-Areal ziehen. Albino = albinotische Frettchen, pigm. = pigmentierte Frettchen, N = Anzahl der histologisch untersuchten Hirne albinotischer und pigmentierter Frettchen, Pen-Spuren = Penetrationsspuren, IO = Oliva inferior, VC = visueller Kortex, Σ = Gesamtanzahl der untersuchten Hirne, — = Bestätigung nicht möglich, weil entweder nicht histologisch untersucht oder keine Läsionen gesetzt. Tier N Bestätigung des NOT-Ableitortes Bestätigung der IO-Reizorte
Albino 1 Läsion im NOT —
Albino 1 Läsion und Pen-Spuren im NOT —
Albino 1 Pen-Spuren im NOT —
Albino 1 — Pen-Spuren in der IO
Albino 2 Pen-Spuren im NOT Pen-Spuren in der IO
pigm. 2 Läsion im NOT —
Σ 8 7 3

Ergebnisse 66
Abbildung 21A zeigt die schwarz-weiße Fotografie eines Formol-fixierten Frettchengehirns.
Der rostrale Bereich des Gehirns ist direkt vor dem medialen Ansatz des cruciaten Sulcus
(CRS) abgeschnitten. Die Pfeile zeigen auf die Orte, wo die Penetrationen zur Ableitung des
NOT (a) gesetzt wurden, und wo die VC- (b) und IO-Reizelektroden (c) plaziert waren. Das
Kleinhirn wurde entfernt, um die Einstichorte der IO-Reizelektroden sichtbar werden zu
lassen. Die VC-Reizelektroden waren auf der linken Hirnseite im caudalen Bereich des
lateralen Gyrus plaziert. Gemäß Law et al. (1988) befinden sie sich in Area 17 in der Nähe
der Area 18. Orientiert man sich an den Abbildungen von Redies et al. (1990), so befinden
sie sich genau auf der Grenze der Areae 17 und 18. Bevor die VC-Reizelektroden ca. 2500
µm tief in den Kortex eingefahren wurden, konnte an dieser Stelle die visuelle Aktivität
mehrerer Neuronen abgeleitet werden. Diese VC-Neuronen befanden sich in verschiedenen
Tiefen zwischen 250 und 2500 µm und hatten rezeptive Felder im zentralen Bereich des
Gesichtsfeldes. Die elektrische Reizung an dieser Stelle konnte prätektale Zellen orthodrom
reizen, die auch von der IO antidrom und / oder vom ON orthodrom reizbar waren.
Abbildung 21B ist eine Umrißzeichnung des Frettchengehirns aus der Perspektive von oben
und soll die Lokalisierung der Penetrationsorte aus Abbildung 21A erleichtern.
NOT-Ableitorte: In Abbildung 22 wird die schwarz-weiße Fotografie eines Nissl-gefärbten
Frontalschnittes eines Albinofrettchens gezeigt. Der Schnitt verläuft durch den rostralen SC.
Die Fotografie dokumentiert die Position einer NOT-Läsion, die sich in der „Kerbe“
zwischen dem linken SC (l-SC) und dem linken lateralen Genikulatum (CGL) befindet. Am
Läsionsort wurden NOT-Zellen von der ipsilateralen IO antidrom identifiziert.
Abbildung 23 zeigt die Rekonstruktion des Mittelhirns eines Frettchens. Das Hirn wurde in
diesem Falle von caudal (posterior) nach rostral (anterior) in 50 µm dicke Frontalschnitte
geschnitten. Es sind die Umrisse jedes vierten Schnittes hintereinander angeordnet. Der SC
erstreckt sich auf beiden Hirnseiten in rostrocaudaler Richtung über ca. 3,6 mm und ist als
Hügel erkennbar (Umriß-Nr. 2 bis 18). Auf dem 15. Umriß beginnt seitlich des SC das
Genikulatum, welches nach anterior hin an Volumen gewinnt. Die Sternchen, die sich
seitlich des vorderen SC-Abschnitts in der „Kerbe“ zwischen SC und Genikulatum befinden
(Schnitt-Nr. 14 bis 17), markieren den prätektalen Bereich, in dem die NOT-Läsionen und
NOT-Penetrationsspuren bei den verschiedenen Versuchstieren aus Tabelle 5 gefunden
wurden. Dieser Bereich stimmt mit der in der Literatur angegebenen Position des NOT
eindeutig überein (Zhang & Hoffmann, 1993).
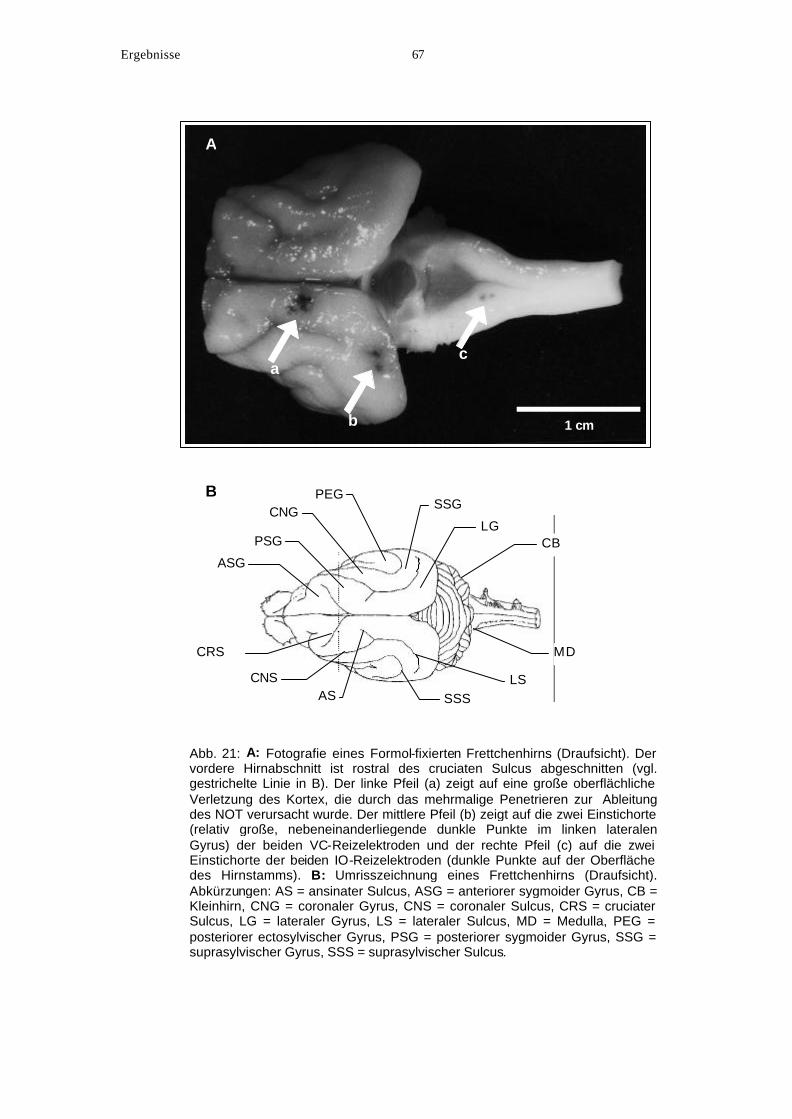
Ergebnisse 67
Abb. 21: A: Fotografie eines Formol-fixierten Frettchenhirns (Draufsicht). Der vordere Hirnabschnitt ist rostral des cruciaten Sulcus abgeschnitten (vgl. gestrichelte Linie in B). Der linke Pfeil (a) zeigt auf eine große oberflächliche Verletzung des Kortex, die durch das mehrmalige Penetrieren zur Ableitung des NOT verursacht wurde. Der mittlere Pfeil (b) zeigt auf die zwei Einstichorte (relativ große, nebeneinanderliegende dunkle Punkte im linken lateralen Gyrus) der beiden VC-Reizelektroden und der rechte Pfeil (c) auf die zwei Einstichorte der beiden IO-Reizelektroden (dunkle Punkte auf der Oberfläche des Hirnstamms). B: Umrisszeichnung eines Frettchenhirns (Draufsicht). Abkürzungen: AS = ansinater Sulcus, ASG = anteriorer sygmoider Gyrus, CB = Kleinhirn, CNG = coronaler Gyrus, CNS = coronaler Sulcus, CRS = cruciater Sulcus, LG = lateraler Gyrus, LS = lateraler Sulcus, MD = Medulla, PEG = posteriorer ectosylvischer Gyrus, PSG = posteriorer sygmoider Gyrus, SSG = suprasylvischer Gyrus, SSS = suprasylvischer Sulcus.
Foto-Nr. 28 05-03-1999.aF
1 cm
a
b
c
A
CRS
ASLS
SSS
ASG
PSG
CNG SSG PEG
MD
CB LG
CNS
B

Ergebnisse 68
Abb. 23: Rekonstruktion des Frettchen-Mittelhirns. Es ist der Umriß jedes viertenHirnschnitts einer 50 µm-Frontalschnitt-Serie dargestellt, so dass der Abstand zwischen zwei Umrissen 200 µm beträgt. Insgesamt sind 4,4 mm in anterior-posterior-Richtung rekonstruiert. Jeder zweite Umriß ist numeriert. Die Sternchen ( ) seitlich des rechten und linken rostralen SC-Abschnitts (SC) markieren den prätektalen Bereich, in dem die NOT-Läsionen (NOT-Ableit- und NOT-Reizorte) bei den verschiedenen Versuchstieren gefunden wurden. Die Position der SC-Läsionen wird durch die schwarzen Punkte ( ) angezeigt. Die gepunkteten Linien markieren die ungefähre Abgrenzung des lateralen Genikulatums (CGL). Der Maßstab bezieht sich nur auf die mediolaterale Ausdehnung der Rekonstruktion.
rostral
caudal
1 mm
SC SC
2
4
6
8
10
12 14
16 18
20
22
CGL CGL
Abb. 22: Schwarz-weiße Fotografie eines 50 µm dicken, Nissl-gefärbten Frontalschnitts durch den rostralen SC-Abschnitt und das linke NOT-Areal eines Albinofrettchens. Zu sehen sind der linke (l-SC) und der rechte SC (r-SC), der darüberliegende Kortex und das caudale Ende des linken lateralen Genikulatums (CGL). Lateral des l-SC wurde eine NOT-Läsion (NOT) gesetzt, die sich in der „Kerbe“ zwischen l-SC und dem linken Genikulatum befindet. Im Kortex über dem NOT-Areal ist ein heller Bereich sichtbar (Pfeil), der durch das wiederholte Penetrieren mit der NOT-Ableitelektrode verursacht wurde. d = dorsal, v = ventral.
Foto-Nr. 2-36 25-07-2000.aF 2,5x-Objektiv
Kortex
l-SC r-SC
NOT
CGL
A
1 mm V
d

Ergebnisse 69
IO-Reizorte: Aus Abbildung 24 geht die Position der IO-Reizelektroden hervor bei einem
Albinofrettchen. Gezeigt wird die schwarz-weiße Fotografie eines 100 µm dicken, Nissl-
gefärbten Sagittalschnittes durch den linken Hirnstamm. Beide IO-Reizelektroden haben
Einstichspuren im Gewebe hinterlassen, die sich makroskopisch erkennen lassen. Die Spuren
erstrecken sich von dorsal nach rostroventral (entsprechend der 45°-Neigung der
Reizelektroden, vgl. Material und Methoden). Die IO erscheint aufgrund ihrer hohen
Zelldichte besonders dunkel. Der Umriß der IO ist gepunktet hervorgehoben. Die rostrale
Penetrationsspur zieht sich durch den rostralen Abschnitt der IO, während die caudale Spur
die IO verfehlt. Bei diesem Tier konnten durch die elektrische Reizung prätektale Zellen
antidrom gereizt und damit als zur IO projizierende NOT-Zellen identifiziert werden.
2. NOT- und SC-Reizorte und VC-Ableitorte
In der Tabelle 6 sind alle Versuchstiere aufgelistet, bei denen die NOT- und SC-Reizorte
durch elektrolytische Läsionen im NOT-Areal und im rostralen Bereich des SC verifiziert
wurden. Die Läsionen im VC dokumentieren die Position der richtungsspezifischen VC-
Abb. 24: Fotografie eines Nissl-gefärbten Sagittalschnitts durch den linken Hirnstamm eines Albinofrettchens. Die beiden Pfeile zeigen auf die Penetrationsspuren (Pfeile) der beiden IO-Reizelektroden (∅ 0,3 mm). Die IO befindet sich im ventralen Abschnitt des Hirnstamms und ist als dunkle, sehr dicht gepackte Zellansammlung mit ovaler Form zu erkennen (gepunkteter Umriß). Abkürzungen: d = dorsal, v = ventral, c = caudal und r = rostral.
Foto-Nr. 5.24 24.02.1999.aF
Hirnschnitt-Nr. 1-8-2 (IO und zwei Einstichspuren)
d
c
v
r
1 mm

Ergebnisse 70
Zellen, die vom NOT antidrom und mit dem Zufallspunktemuster visuell reizbar waren.
Insgesamt wurden sieben Hirne untersucht, die von vier albinotischen und drei pigmentierten
Frettchen stammten.
Tab. 6: Anzahl der histologisch untersuchten Frettchengehirne aus den Reizexperimenten, in denen der NOT und der SC elektrisch gereizt wurden. In allen Fällen befinden sich die „NOT-Läsionen“ im NOT-Areal. Die Läsionen im SC wurden im rostralen Abschnitt des SC lokalisiert und markieren die Repräsentation des zentralen Gesichtsfeldes. Die Läsionen im VC markieren die Position der VC-Zellen, die vom NOT antidrom reizbar waren, und dokumentieren, daß sich diese Neuronen in der Lamina V befinden. Abkürzungen wie in Tabelle 5. Tier N Bestätigung des
NOT-Reizortes Bestätigung des SC-Reizortes
Position der NOT-antidromen VC-Zellen
Albino 1 Läsion im NOT — 2 Läsionen in der Lamina V
pigm. 1 Läsion im NOT — —
Albino 1 Läsion im NOT — 2 Läsionen in der Lamina V
Albino 1 Läsion im NOT Läsion im SC —
Albino 1 Läsion im NOT Läsion im SC Läsion in der Lamina V
pigm. 2 Läsion im NOT Läsion im SC —
Σ 7 7 3 3
NOT- und SC-Reizorte: Die Position der NOT-Läsionen, die die NOT-Reizorte markieren,
unterscheidet sich nicht von der Position der NOT-Läsionen, die die NOT-Ableitorte
markieren (Abb. 22 und 23). Die SC-Läsionen befinden sich in Bezug auf die NOT-Läsionen
weiter medial und weiter caudal. Ihre Position ist in Abbildung 23 durch die beiden dunklen
Punkte angegeben. In diesem Bereich des SC war das frontale Gesichtsfeld repräsentiert.
VC-Ableitorte: Abbildung 25 zeigt eine Läsion im linken visuellen Kortex eines Albino-
frettchens. Zwei VC-Neuronen wurden an diesem Ort vom ipsilateralen NOT antidrom
identifiziert. Beide Neuronen reagierten auf die visuelle Reizung mit dem großflächigen
Zufallspunktemuster richtungsspezifisch. Die Läsion (dunkle Pfeile) befindet sich auf der
ventralen Seite (v) des visuellen Kortex in der Lamina V (helle Pfeile). Die Zuordnung der
Läsion zur Lamina V wurde mit Hilfe der Abbildungen von Rockland, (1985) gemacht.
Gemäß Rockland (1985) und Law et al. (1988) befindet sich die Läsion in der Area 17. Dies
stellt den Normalbefund dar und spricht dafür, daß der VC des Albinofrettchens eine intakte,
zum Mittelhirn projizierende Schicht V besitzt.
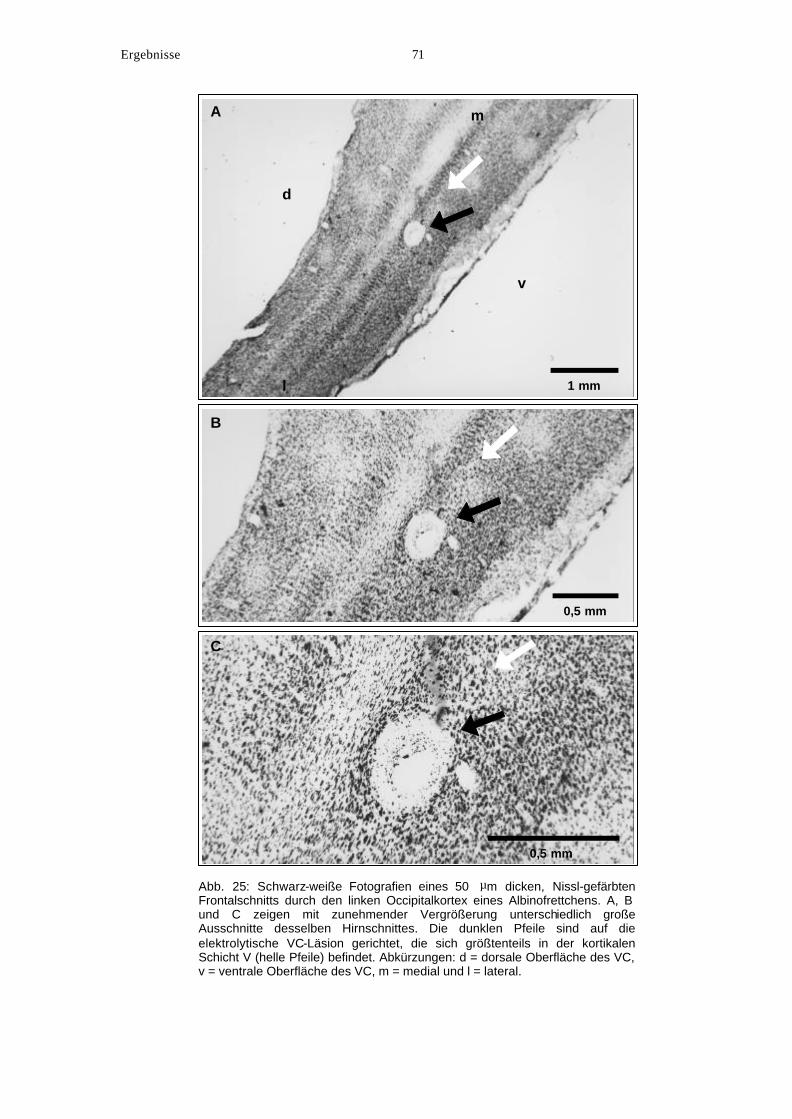
Ergebnisse 71
Abb. 25: Schwarz-weiße Fotografien eines 50 µm dicken, Nissl-gefärbten Frontalschnitts durch den linken Occipitalkortex eines Albinofrettchens. A, B und C zeigen mit zunehmender Vergrößerung unterschiedlich große Ausschnitte desselben Hirnschnittes. Die dunklen Pfeile sind auf die elektrolytische VC-Läsion gerichtet, die sich größtenteils in der kortikalen Schicht V (helle Pfeile) befindet. Abkürzungen: d = dorsale Oberfläche des VC, v = ventrale Oberfläche des VC, m = medial und l = lateral.
Foto-Nr. 2-24 25-07-2000.aF 2,5x-Objektiv
Foto-Nr. 2-26 25-07-2000.aF 10x-Objektiv
Foto-Nr. 2-25 25-07-2000.aF
5x-Objektiv
1 mm
0,5 mm
0,5 mm
A
B
C
m
v
l
d

Ergebnisse 72
B. Physiologie
1. Lokalisation des NOT-Areals und der NOT-Neuronen
Die allermeisten NOT-Zellen wurden sowohl bei den albinotischen als auch bei den pigmen-
tierten Tieren im Bereich von –1 bis –2,5 mm anterior und +2,5 bis +3,0 mm lateral
(Horsley-Clarke-Koordinaten) in einer Tiefe von 6500 bis 7500 µm unter der kortikalen
Oberfläche lokalisiert. Sie befanden sich in einem länglichen Band entlang des
rostrolateralen SC-Randes, welches sich bei den einzelnen Versuchstieren ca. 700 µm in
rostrocaudaler und ca. 200 µm in mediolateraler Richtung erstreckte. Damit wird die von
Klauer et al. (1990) angegebene Position des Frettchen-NOT bestätigt. In Bezug auf die
Oberfläche des Prätektums befanden sich die NOT-Zellen in einer relativ geringen Tiefe von
ca. 100 bis 250 µm, was mit den histologischen Befunden von Zhang & Hoffmann (1993)
und Telkes et al. (2001) übereinstimmt. Aus Abbildung 26 wird die relative Position des
NOT zum benachbarten SC und die relative Position dieser beiden Hirnstrukturen innerhalb
des Horsley-Clarke-Koordinatensystems deutlich. Die y-Achse entspricht der anterior-
posterioren Achse und die x-Achse der mediolateralen Achse des Horsley-Clarke-Systems
(Interaurallinie). Die eingezeichneten Punkte stellen das Penetrationsschema aus einem
konkreten Experiment mit einem Albinofrettchen dar. Innerhalb des NOT-Areals konnten
Zellen von der IO antidrom gereizt werden. Das Dreieckssymbol markiert die elektrolytische
Läsion, die am Ende des Experimentes gesetzt wurde.
Es kam selten vor, daß innerhalb einer Penetration mehrere NOT-Zellen gefunden werden
konnten. Bei den meisten erfolgreichen Penetrationen konnte nur eine NOT-Zelle abgeleitet
werden. Wurde eine Penetration unmittelbar neben einer erfolgreichen Penetration plaziert,
so führte diese Vorgehensweise meistens nicht zum nochmaligen Erfolg. Diese Beobachtun-
gen stehen mit den anatomischen Befunden in Einklang, nach denen die von der ipsilateralen
IO retrograd markierten NOT-Zellen keine dicht gepackten Ansammlungen bilden, sondern
vielmehr ein lockeres großmaschiges Netzwerk (Telkes et al., 2001).
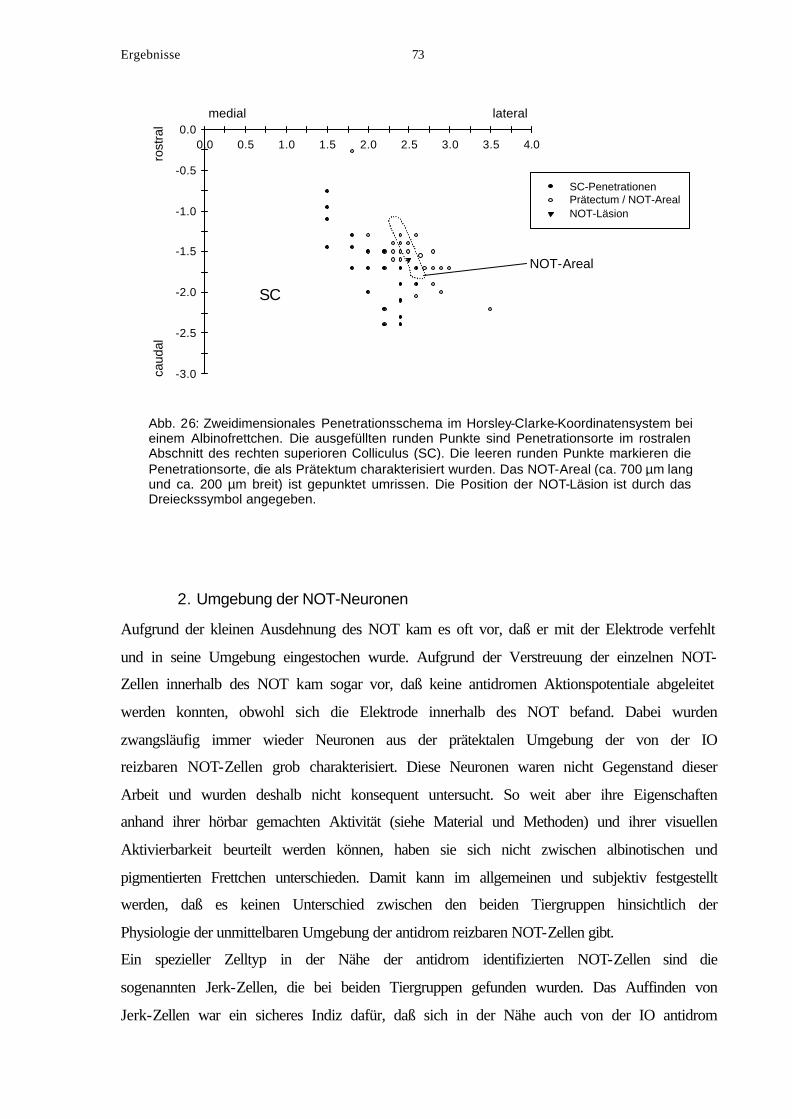
Ergebnisse 73
2. Umgebung der NOT-Neuronen
Aufgrund der kleinen Ausdehnung des NOT kam es oft vor, daß er mit der Elektrode verfehlt
und in seine Umgebung eingestochen wurde. Aufgrund der Verstreuung der einzelnen NOT-
Zellen innerhalb des NOT kam sogar vor, daß keine antidromen Aktionspotentiale abgeleitet
werden konnten, obwohl sich die Elektrode innerhalb des NOT befand. Dabei wurden
zwangsläufig immer wieder Neuronen aus der prätektalen Umgebung der von der IO
reizbaren NOT-Zellen grob charakterisiert. Diese Neuronen waren nicht Gegenstand dieser
Arbeit und wurden deshalb nicht konsequent untersucht. So weit aber ihre Eigenschaften
anhand ihrer hörbar gemachten Aktivität (siehe Material und Methoden) und ihrer visuellen
Aktivierbarkeit beurteilt werden können, haben sie sich nicht zwischen albinotischen und
pigmentierten Frettchen unterschieden. Damit kann im allgemeinen und subjektiv festgestellt
werden, daß es keinen Unterschied zwischen den beiden Tiergruppen hinsichtlich der
Physiologie der unmittelbaren Umgebung der antidrom reizbaren NOT-Zellen gibt.
Ein spezieller Zelltyp in der Nähe der antidrom identifizierten NOT-Zellen sind die
sogenannten Jerk-Zellen, die bei beiden Tiergruppen gefunden wurden. Das Auffinden von
Jerk-Zellen war ein sicheres Indiz dafür, daß sich in der Nähe auch von der IO antidrom
Abb. 26: Zweidimensionales Penetrationsschema im Horsley-Clarke-Koordinatensystem bei einem Albinofrettchen. Die ausgefüllten runden Punkte sind Penetrationsorte im rostralen Abschnitt des rechten superioren Colliculus (SC). Die leeren runden Punkte markieren die Penetrationsorte, die als Prätektum charakterisiert wurden. Das NOT-Areal (ca. 700 µm lang und ca. 200 µm breit) ist gepunktet umrissen. Die Position der NOT-Läsion ist durch das Dreieckssymbol angegeben.
medial lateral
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0ca
udal
ros
tral
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
0.0
SC-PenetrationenPrätectum / NOT-ArealNOT-Läsion
SC
NOT-Areal

Ergebnisse 74
reizbare NOT-Zellen befinden würden. Die Jerk-Zellen befanden sich gelegentlich zwischen,
meistens aber 200 bis 500 µm unterhalb der NOT-Zellen. Damit findet man beim Frettchen
die gleiche relative Position zwischen NOT- und Jerk-Zellen wie bei der Katze (Ballas &
Hoffmann, 1985; Schweigart & Hoffmann, 1992; Schmidt, 1996). Die Jerk-Zellen
beantworteten sehr schnelle ruckartige Bewegungen des Zufallspunktemusters mit einer
kurzen hochfrequenten Salve von Aktionspotentialen. Sie wurden nur in horizontaler
Richtung visuell getestet und beantworteten sowohl die Reizbewegung nach ipsiversiv als
auch nach kontraversiv. Über eine eventuelle, schwache Bevorzugung einer Richtung, die
anhand ihrer hörbaren Antwort nicht detektiert werden konnte, kann keine Aussage gemacht
werden, da ihre Aktivität nicht mit dem PC registriert wurde.
3. Allgemeine Charakterisierung der prätektalen und der NOT-Neuronen
Bei 93 Zellen wurde das Prädikat „diffus visuell“ protokolliert. Die tatsächlich abgeleitete
Anzahl solcher Zellen liegt jedoch wesentlich höher, kann aber nicht angegeben werden, da
nicht alle Zellen diesbezüglich explizit charakterisiert wurden. Solche Zellen reagierten mit
einer Aktivitätsänderung auf die Änderungen der Beleuchtung des gesamten Labors oder des
frontalen Gesichtsfeldes und insbesondere auf die Beleuchtung des kontralateralen Auges
mit Hilfe der Handlampe. Sie reagierten jedoch nicht auf eine Bewegung im Gesichtsfeld des
Tieres, weder auf die Bewegung des Reizmusters, noch auf die Bewegung von Objekten.
Aufgrund dieser Eigenschaften war es nicht möglich, die genaue Ausdehnung ihrer RF auf
dem Tangentialschirm zu bestimmen. Diese Beschreibung gilt auch für fast alle Zellen, die
aufgrund ihrer antidromen Reizbarkeit von der IO als NOT-Zellen definiert wurden.
Im Gegensatz zu den NOT-Zellen des Albinofrettchens reagierten die NOT-Zellen des
pigmentierten Frettchens auf eine großflächige horizontale Bewegung innerhalb ihres
rezeptiven Feldes. Sie wurden sowohl durch die Bewegung des Zufallspunktemusters, als
auch durch die Bewegung von großen Objekten moduliert (vgl. Abb. 28).
4. ON/OFF-Eigenschaften von Neuronen im Mittelhirn
a) ON/OFF-Eigenschaften von prätektalen und NOT-Neuronen
Die Bestimmung der ON/OFF-Eigenschaften von prätektalen Zellen wurde nur bei Albino-
frettchen durchgeführt. Mit Hilfe des Blendenverschlusses wurden zwei prätektale Zellen
charakterisiert, die nicht von der IO reizbar waren. Die ONF-Is dieser beiden Zellen waren
61% und 56%, so daß sie definitionsgemäß (vgl. Material und Methoden) als ON-OFF-

Ergebnisse 75
Zellen angesehen werden können. Durch Beleuchten des kontralateralen Auges mit der
Handlampe oder das Wegnehmen der Beleuchtung (Abdecken des Lichtstrahls) wurden
insgesamt weitere 25 Zellen (100%) des Albinofrettchens als ON-, OFF oder ON-OFF-
Zellen klassifiziert:
• 20% (fünf von 25) dieser Zellen waren ON-Zellen. Sie wurden alle auf Reizbarkeit von der
IO getestet und waren auch alle antidrom reizbar. Damit projizieren alle ON-NOT-Zellen
zur ipsilateralen IO. Eine Zelle wurde auch auf Reizbarkeit vom VC und vom ON getestet
und war in beiden Fällen orthodrom aktivierbar. Die restlichen vier Zellen wurden
diesbezüglich nicht getestet.
• 40% (zehn von 25) waren ON-OFF-Zellen. Acht von diesen wurden auf Reizbarkeit von
der IO untersucht. Sechs waren antidrom reizbar und zwei waren nicht reizbar. Die
restlichen zwei Zellen wurden weder auf Reizbarkeit von der IO, noch vom VC oder ON
untersucht. Damit projizieren die meisten ON-OFF-Zellen des NOT ebenfalls zur
ipsilateralen IO.
• Die übrigen 40% (zehn von 25) der Zellen waren OFF-Zellen. Acht von diesen wurden auf
Reizbarkeit von der IO untersucht. Von diesen acht Zellen war eine Zelle antidrom reizbar
und sieben Zellen waren nicht reizbar. Damit projizieren die meisten OFF-Zellen des NOT
nicht zur IO. Von den sieben nicht zu IO projizierenden Zellen wurden zwei Zellen auch
auf Reizbarkeit vom ON und eine Zelle sowohl auf Reizbarkeit vom ON als auch vom VC
getestet. Alle drei Zellen waren nur vom ON orthodrom reizbar. Die restlichen zwei Zellen
aus der Gruppe der zehn OFF-Zellen wurden weder auf Reizbarkeit von der IO, noch vom
VC oder ON untersucht.
b) ON/OFF-Eigenschaften von SC-Neuronen
Die Bestimmung der ON/OFF-Eigenschaften von Zellen des superioren Colliculus wurde
• sowohl qualitativ als auch quantitativ mit Hilfe des Blendenverschlusses und
• sowohl bei albinotischen als auch bei pigmentierten Frettchen durchgeführt.
Insgesamt wurden 57 SC-Zellen der Albinofrettchen und 25 SC-Zellen der pigmentierten
Frettchen hinsichtlich ihrer ON/OFF-Eigenschaften untersucht.
Von den 57 Zellen der Albinofrettchen wurden acht SC-Zellen nur qualitativ mit Hilfe der
Handlampe untersucht. Davon wurden sieben als ON-OFF-Zellen und eine Zelle wurde als
OFF-Zelle charakterisiert. Die restlichen 49 SC-Zellen der albinotischen und alle 25 SC-
Zellen der pigmentierten Frettchen wurden mit dem Blendenverschluß untersucht, so daß

Ergebnisse 76
ihre ONF-Indizes berechnet werden konnten. Gemäß den festgelegten Klassifizierungs-
kriterien wurden folgende Quoten an ON-, OFF- oder ON-OFF-Zellen gefunden:
• 2% (eine von 49) der SC-Zellen der Albinofrettchen reagierten stärker bei „Licht-An“ als
bei „Licht-Aus“ mit einem ONF-I ≥ 66%. Definitionsgemäß wird diese Zelle als ON-Zelle
klassifiziert. Bei den pigmentierten Frettchen konnte keine einzige SC-Zelle aus der
vorhandenen Stichprobe von 25 Zellen als ON-Zelle eingestuft werden. Da bei den Albino-
frettchen nur 2% ON-Zellen gefunden wurden, scheinen sich die beiden Tiergruppen kaum
voneinander zu unterscheiden.
• 80% (39 von 49) der SC-Zellen der Albinofrettchen und 84% (21von 25) der SC-Zellen der
pigmentierten Frettchen haben einen ONF-Index, der zwischen 33% und 66% liegt. Diese
Zellen werden definitionsgemäß als ON-OFF-Zellen klassifiziert. Damit wird gezeigt, daß
die ON-OFF-Zellen in beiden Tiergruppen in gleichen prozentualen Größenordnungen
vorkommen.
• 18% (neun von 49) der SC-Zellen der Albinofrettchen und 16% (vier von 25) SC-Zellen
der pigmentierten Frettchen haben stärker bei „Licht-Aus“ als bei „Licht-An“ reagiert mit
einem ONF-I < 33%. Definitionsgemäß werden sie als OFF-Zellen eingestuft. Damit findet
man bei beiden Tiergruppen in etwa gleich viele OFF-Zellen.
In Abbildung 27 wird die Häufigkeitsverteilung der 49 ONF-Indizes der Albinofrettchen und
die der 25 ONF-Indizes der pigmentierten Frettchen gezeigt. Die ONF-I der Albinofrettchen
haben einen Median von 48% (Q0,25 = 35%; Q0,75 = 51%) und die der pigmentierten
Frettchen einen Median von 43% (Q0,25 = 40%; Q0,75 = 47%). Der RANG-SUMMEN-TEST
zeigt keinen signifikanten Unterschied zwischen den ONF-Indizes der beiden Tiergruppen
(P = 0,275). Es wird festgestellt, daß die beiden Tiergruppen gleich viele ON-, OFF- und
ON-OFF-Zellen in den oberen Schichten des SC besitzen. Gleichwohl fällt beim
Albinofrettchen eine leichte Tendenz zu mehr bzw. stärkeren OFF-Antworten auf, was sich
im leicht höheren Medianwert widerspiegelt.
5. DS-Indizes und antidrome Identifizierung der NOT-Neuronen
Aufgrund der Eigenschaft „diffus visuell“ konnte an den allermeisten PSTH der NOT-Zellen
des Albinofrettchens keine Richtungspräferenz beobachtet werden. Die Abwesenheit der
Richtungsspezifität stimmte mit dem akustisch unveränderten Geräusch des Rohsignals bzw.
der unveränderten Frequenz des TTL-Signals überein.

Ergebnisse 77
Abbildung 28 zeigt das repräsentative PSTH je einer NOT-Zelle eines albinotischen und
eines pigmentierten Frettchens und veranschaulicht ihr eklatant unterschiedliches
spo ipsi kontra
Albino
Pigmentiert
A
B
Abb. 28: PSTH von jeweils einer NOT-Zelle eines albinotischen (A) und eines pigmentierten (B) Frettchens. Beide Zellen konnten elektrisch von der ipsi-lateralen IO antidrom gereizt werden. Von beiden Zellen wurden 10 Reiz-perioden aufgenommen. A: Die NOT-Zelle des Albinos wird durch die visuelle Reizung nicht moduliert, hat einen DS-Index von Null und wird als richtungs-unspezifisch klassifiziert. B: Im Gegen-satz zur Zelle des Albinofrettchens wird die NOT-Zelle des pigmentierten Frettchens stark moduliert. Sie hat einen DS-I von ca. 0,8 und bevorzugt die normale, d.h. ipsiversive Reizrichtung. y-Achse: Feuerrate in Aktionspotentiale / s; x-Achse: Zeit bzw. Dauer der Reizperioden. Eine Reizperiode dauert insgesamt 5 s [2 s kontraversive (kontra), 2 s ipsiversive Reizung (ipsi) und 1 s Aufnahme der Spontanrate bei stehendem Reizmuster (spo)].
Abb. 27: Häufigkeitsverteilung der ONF-Indizes von 49 SC-Zellen des albinotischen und 25 SC-Zellen des pigmentierten Frettchens. In beiden Tiergruppen werden mehr SC-Zellen mit stärkeren OFF-Antworten (ONF-I < 50%) gefunden. Beim Albino-frettchen beträgt der Median der ONF-Indizes 48% und beim pigmentierten Frettchen 43%. Diese Differenz ist nicht statistisch signifikant (RANG-SUMMEN-TEST, P = 0,275). Die senkrechten, gestrichelten Linien teilen das gesamte Antwortspektrum definitionsgemäß in einen OFF-, ON-OFF- und ON-Bereich.
Albinos
0 10 20 30 40 50 60 70 80 90 100
N
0
2
4
6
8
10
12
14
Pigm.
ONF-I [%]
0 10 20 30 40 50 60 70 80 90 100
N
0
2
4
6
8
10
12
14
N = 49
N = 25
OFF ON-OFF ON
N
N

Ergebnisse 78
Antwortverhalten während horizontaler optokinetischer Reizung. Beide Zellen waren von
der ipsilateralen IO antidrom reizbar. Die NOT-Zelle des Albinofrettchens (Abb. 28A) läßt
sich durch die kontra- (kontra) und ipsiversive (ipsi) Bewegung oder durch das
Stehenbleiben des Reizmusters (spo) nicht beeinflussen, d.h. sie ändert während der ganzen
Reizperiode nicht ihre Spontanrate. Im Gegensatz dazu beobachtet man eine starke
Modulation der NOT-Zelle des pigmentierten Tieres (Abb. 28B). Die Reizbewegung in
Vorzugsrichtung (ipsi) wird von dieser NOT-Zelle mit einer ca. sechsmal höheren Feuerrate
beantwortet als die entgegengesetzte Reizbewegung in Nullrichtung (kontra). Weiterhin ist
auffällig, daß bei Reizung in Nullrichtung ihre Feuerrate unter der Spontanrate liegt, d.h.
diese NOT-Zelle wird durch die kontraversive Reizbewegung inhibiert. Eine solche
Inhibition wurde jedoch nicht bei allen richtungsspezifischen NOT-Zellen des pigmentierten
Frettchens beobachtet.
a) Albinofrettchen
Es wurden insgesamt 198 prätektale Zellen von 16 Albinofrettchen abgeleitet, die aufgrund
ihrer charakteristisch hohen Spontanaktivität und ihrer relativen Lage zum SC und zu Jerk-
Zellen als potentielle NOT-Zellen angesehen werden konnten. Von den 198 Zellen waren
193 Zellen visuell aktivierbar und fünf Zellen nicht. Die Gruppe der 193 visuellen Zellen
beinhaltet die schon erwähnten, explizit als „diffus visuell“ charakterisierten 93 Zellen.
Die ersten 26 der 198 Zellen wurden in frühen Experimenten an vier Tieren abgeleitet, als
noch nicht bekannt war, daß die NOT-Zellen der Albinofrettchen richtungsunspezifisch sind.
Deshalb wurde bei diesen Experimenten noch nicht von der ipsilateralen IO antidrom
gereizt. Da die Erwartung, eine hörbare Richtungsbevorzugung zu finden, nicht erfüllt
wurde, wurden von den meisten dieser richtungsunspezifischen prätektalen Zellen keine
PSTH registriert (Ausnahme fünf Zellen). Die restlichen 172 Zellen wurden auf antidrome
Reizbarkeit von der IO getestet, wobei 58% (99 von 172) Zellen antidrom reizbar und 42%
(73 von 172) Zellen nicht reizbar waren. Von den meisten dieser 172 Zellen existieren
ebenfalls keine PSTH, da die Ableitungen instabil waren.
Von den 99 Zellen, die von der IO antidrom reizbar waren, waren 94 Zellen visuell
aktivierbar. Die übrigen 5% (fünf von 99) Zellen waren die oben erwähnten fünf nicht
visuell aktivierbaren Zellen. Ihre antidromen Latenzen waren: 2,5 ms; 5,5 ms; 1,5 ms; 1,0 ms
und 2,2 ms. Da sie durch Licht nicht gereizt werden konnten, erübrigten sich PSTH-
Aufnahmen. Folglich war auch keine DS-I-Berechnung möglich, so daß diese Zellen nicht in
der Häufigkeitsverteilung der DS-Indizes (siehe unten Abb. 29A) aufgeführt sind.

Ergebnisse 79
Aus der Gesamtgruppe der 198 prätektalen Zellen konnten 42 Zellen ausreichend lange
abgeleitet werden, so daß die Berechnung ihres DS-Indizes möglich war. Diese 42 visuellen
Zellen stammen von zwölf verschiedenen Individuen. Wie schon das PSTH der Beispielzelle
in Abbildung 28A erwarten läßt, haben die meisten dieser 42 Zellen einen DS-Index nahe bei
Null (Median 0,0; Q0,25 = -0,02; Q0,75 = 0,09), was keine oder nur eine sehr schwache
Richtungspräferenz anzeigt. In Abbildung 29A ist die Häufigkeitsverteilung ihrer DS-Indizes
gezeigt. Fünf von diesen 42 prätektalen Zellen wurden nicht auf antidrome Reizbarkeit von
der IO getestet (nt), da sie aus frühen Experimenten stammen, in denen das Kriterium
"antidrome Reizbarkeit von der IO" zur eindeutigen Identifizierung der NOT-Zellen des
Albinofrettchens noch nicht angewandt wurde. Die restlichen 37 Zellen (100%) wurden auf
antidrome Reizbarkeit von der IO getestet, von denen 19% (sieben von 37) nicht reizbar
(IO-) und 81% (30 von 37) reizbar (IO+) waren. Diese 30 Zellen können somit mit
Sicherheit als potentielle Neuronen im neuronalen Schaltkreis des hOKN betrachtet und
definitionsgemäß auch als NOT-Zellen bezeichnet werden. Die DS-Indizes dieser
antidromen NOT-Zellen haben einen Median von 0,00 (Q0,25 = -0,02; Q0,75 = 0,03) und sind
in Abbildung 29A durch die Schraffierung hervorgehoben. Ein DS-Index von Null bedeutet,
daß eine Zelle keine der beiden horizontalen Reizrichtungen bevorzugt und damit
richtungsunspezifisch ist. Die DS-Indizes der fünf nicht getesteten bzw. der sieben
getesteten aber nicht reizbaren prätektalen Zellen unterscheiden sich nicht statistisch
signifikant von den DS-Indizes der 30 antidromen NOT-Zellen (RANG-SUMMEN-TEST,
P = 0,338 bzw. P = 0,194).
Nur zwei antidrom reizbare NOT-Zellen haben einen von Null stark abweichenden DS-I, der
allerdings negativ ist (–0,62 und –0,30). Diese beiden Zellen zeigen also eine Vorzugs-
richtung, die der normalen ipsiversiven Vorzugsrichtung der NOT-Zellen des pigmentierten
Frettchens entgegengesetzt ist. Sie wurden bei demselben Individuum abgeleitet.
In Abbildung 29B ist die Häufigkeitsverteilung der 99 antidromen Latenzen des
Albinofrettchens dargestellt. Sie besitzen einen Median von 2,00 ms (Q0,25 = 1,30 ms, Q0,75 =
2,50 ms) mit einer minimalen Latenz von 0,8 ms und einer maximalen Latenz von 8,5 ms.
Die Latenzen der 30 NOT-Zellen mit DS-Index aus Abbildung 29A sind schraffiert
hervorgehoben und haben einen Median von 2,00 ms (Q0,25 = 1,20 ms, Q0,75 = 2,50 ms) mit
einer minimalen Latenz von 0,9 ms und einer maximalen Latenz von 4,5 ms. Zwischen den
schraffierten und nicht schraffierten Latenzen existiert kein signifikanter Unterschied (RANG-
SUMMEN-TEST, P = 0,351). In Abbildung 30 werden typische, von der IO ausgelöste
antidrome Aktionspotentiale von zwei NOT-Zellen des Albinofrettchens gezeigt. In

Ergebnisse 80
Abbildung 30A sind drei und in Abbildung 30B vier auf den Beginn des Reizpulses (t = 0)
ausgerichtete Potentialspuren dargestellt. Bei beiden NOT-Zellen besitzen die antidromen
Aktionspotentiale in allen Spuren den gleichen Verlauf und die gleiche Latenz (1,2 ms in
Abb. 30A und 2,1 ms in Abb. 30B).
DS-Index
-0.8 -0.6 -0.4 -0.2 0.2 0.4 0.6 0.8-1.0 0.0 1.00
2
4
6
8
10
12
14
16
18
20 24 IO+5 IO- & 8 nt
-0.8 -0.6 -0.4 -0.2 0.2 0.4 0.6 0.8-1.0 0.0 1.0
0
2
4
6
8
10
12
14
16
18
20 30 IO+7 IO- & 5 nt
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
18
20 30 IO+ mit DS-I 69 IO+ ohne DS-I
Latenz [ms]0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
18
20 24 IO+ mit DS-IC D
A B
N N
N N
Abb. 29: Häufigkeitsverteilungen der DS-Indizes (A, C) und antidromen Reizlatenzen von der IO (B, D) von prätektalen bzw. NOT-Zellen des albinotischen (A, B) und pigmentierten (C, D) Frettchens. A: DS-Indizes von insgesamt 42 prätektalen Zellen des Albinofrettchens. Unter den antidromen Zellen fallen zwei Zellen aufgrund ihres besonders stark negativen DS-Index (-0,62 und –0,30) auf. B: 99 IO+ Reizlatenzen des Albinofrettchens. Die 30 Latenzen der Zellen mit DS-I aus A sind schraffiert. C: DS-Indizes von insgesamt 37 NOT-Zellen des pigmentierten Frettchens. D: IO+ Reizlatenzen aus C. Die DS-Indizes in A unterscheiden sich hoch signifikant von den DS-Indizes in C. Die antidromen Reizlatenzen in B unterscheiden sich nicht signifikant von den Latenzen in D. y-Achsen (N): Anzahl der DS-I in A und C, Anzahl der antidromen Latenzen in B und D. Abkürzungen: IO+ = von der IO antidrom reizbar, IO- = von der IO nicht reizbar, nt = nicht auf Reizbarkeit von der IO getestet.

Ergebnisse 81
Abb: 30: Typische antidrome Aktionspotentiale (Pfeile) von der IO, die an zwei NOT-Zellen zweier Albinofrettchen registriert wurden. Die Potentialverläufe wurden mit einem Fotoapparat vom Monitor des Speicheroszilloskops abfotografiert. Die senkrechten, gepunkteten Linien auf der linken Seite markieren den Zeitpunkt der Reizapplikation (Reizbeginn bei t = 0). A: Das antidrome Aktionspotential der ersten Zelle taucht mit einer Latenz von 1,2 ms auf. Gezeigt werden drei auf t = 0 ausgerichtete Potentialspuren. Die Zelle wurde auch auf Reizbarkeit vom kontralateralen ON und vom ipsilateralen VC getestet und war in beiden Fällen orthodrom reizbar (siehe Abb. 31 und 38B). B: Das antidrome Aktionspotential der zweiten Zelle hat eine Latenz von 2,1 ms. Gezeigt werden 4 auf t = 0 ausgerichtete Potentialspuren. Auch diese Zelle wurde auf Reizbarkeit vom kontralateralen ON und vom ipsilateralen VC getestet, war aber in beiden Fällen nicht orthodrom reizbar. Abszisse: Zeitachse; Ordinate: extrazellulär abgeleitetes Zellpotential.
I. Foto-Nr. 10
23+24.02.1999.aF
Pen-Nr. 14 Tiefe 9163
IO-antidromer Spike (2,3 ms)
1 ms
B
t = 0
II. Foto-Nr. 0
23+24.02.1999.aF
Pen-Nr. 11 Tiefe 7426
IO-antidromer Spike (1,2 ms)
1 ms
A
t = 0

Ergebnisse 82
b) Pigmentierte Frettchen
37 NOT-Zellen wurden von fünf pigmentierten Frettchen abgeleitet. Alle diese Zellen
zeigten eine Präferenz für die ipsiversive Bewegung des Zufallspunktemusters. Die
ipsiversive Reizung führte bei allen 37 NOT-Zellen zu einer Erhöhung der Entladungsrate,
während die kontraversive Reizung die meisten Zellen inhibierte oder zumindest nicht
erregte. Entsprechend haben alle NOT-Zellen des pigmentierten Frettchens einen positiven
DS-Index. Dieses Ergebnis steht im Einklang mit den Ergebnissen von Klauer et al. (1990),
die das physiologische Verhalten der NOT-Zellen des pigmentierten Frettchens eingehender
beschrieben.
Abbildung 29C zeigt die Häufigkeitsverteilung der DS-Indizes der 37 charakterisierten
NOT-Zellen. Alle DS-Indizes sind positiv und ihr Median beträgt 0,29 (Q0,25 = 0,19; Q0,75 =
0,40). Von diesen 37 Zellen wurden 29 Zellen (100%) auf antidrome Reizbarkeit von der IO
getestet. 83% (24 von 29) der Zellen waren antidrom reizbar (IO+, schraffierte Zellen) und
der Median ihrer DS-Indizes beträgt 0,28 (Q0,25 = 0,15, Q0,75 = 0,38). Ein DS-I von 0,33
bedeutet, daß die ipsiversive Reizbewegung mit doppelt so vielen Aktionspotentialen
beantwortet wird wie die kontraversive Reizbewegung. Damit kann festgehalten werden, daß
alle zur IO projizierenden NOT-Zellen der pigmentierten Frettchen eindeutig richtungs-
spezifisch sind und die ipsiversive Reizrichtung bevorzugen. Die Häufigkeitsverteilung der
24 antidromen Reizlatenzen ist in Abb. 29D gezeigt. Die Latenzen befinden sich im Bereich
von 1,0 bis 8,2 ms mit einem Median von 1,55 ms (Q0,25 = 1,05 ms, Q0,75 = 3,50 ms).
Die übrigen 17% (fünf von 29) der prätektalen Zellen waren nicht antidrom reizbar (IO-).
Ihre DS-Indizes unterscheiden sich nicht signifikant von den Indizes der 24 IO+ Zellen
(RANG-SUMMEN-TEST, P = 0,933). Die restlichen acht aus der Gruppe der 37 Zellen
stammen aus frühen Experimenten und wurden nicht getestet (nt). Ihre DS-Indizes
unterscheiden sich ebenfalls nicht von den DS-Indizes der 24 IO+ Zellen (RANG-SUMMEN-
TEST, P = 0,147).
c) Statistischer Vergleich zwischen den beiden Tiergruppen
Die 30 DS-Indizes der von der IO antidrom identifizierten NOT-Zellen der Albinofrettchen
(schraffierte Zellen in Abb. 29A) unterscheiden sich hoch signifikant (RANG-SUMMEN-TEST,
P < 0,0001) von den 24 DS-Indizes der richtungsspezifischen NOT-Zellen des pigmentierten
Frettchens, die ebenfalls von der IO antidrom identifiziert wurden (schraffierte DS-I in
Abb. 29C). Somit findet man bei den Albinofrettchen signifikant weniger (fast keine)
richtungsspezifische NOT-Zellen. Ein Vergleich zwischen der Gesamtgruppe von 42 DS-

Ergebnisse 83
Indizes der Albinofrettchen mit den entsprechenden 37 DS-Indizes der pigmentierten
Frettchen ergibt einen ähnlich hoch signifikanten Unterschied (RANG-SUMMEN-TEST, P <
0,0001). Damit wird das physiologische Korrelat zur optokinetischen Blindheit der
Albinofrettchen und zur morphologischen Unreife ihrer NOT-Zellen (Telkes et al., 2001)
nachgewiesen.
Der statistische Vergleich der 99 bzw. 30 antidromen Reizlatenzen des Albinofrettchens
(Abb. 29B) mit den entsprechenden Latenzen des pigmentierten Frettchens (Abb. 29D) zeigt
keinen signifikanten Unterschied hinsichtlich der Mediane an (RANG-SUMMEN-TEST,
P = 0,965 bzw. P 0,610). Es fällt jedoch auf, daß die NOT-Zellen des pigmentierten
Frettchens zu kürzeren Latenzen tendieren als die NOT-Zellen des Albinofrettchens.
6. Orthodrome Reizung der NOT-Neuronen vom VC
Sowohl beim albinotischen als auch beim pigmentierten Frettchen löste die elektrische
Reizung des ipsilateralen VC immer eine Mehrfachantwort der prätektalen bzw. NOT-Zellen
aus. Die Mehrfachantwort bestand in den allermeisten Fällen aus zwei bis drei orthodromen
Aktionspotentialen, wobei die Anzahl der Potentiale bei den einzelnen NOT-Zellen konstant
war. Zwischen dieser qualitativen Beschreibung und der ensprechenden Beschreibung in
Klauer et al. (1990) findet man erwartungsgemäß keinen Unterschied.
Abbildung 31 zeigt beispielhaft das Antwortverhalten einer NOT-Zelle eines
Albinofrettchens auf die elektrische Reizung des VC. Diese Zelle war auch von der IO
antidrom (siehe Abb. 30A) und vom ON orthodrom (vgl. Abb. 38B) reizbar. Gezeigt werden
insgesamt vier Antworten, d.h. vier Potentialspuren, die alle auf den Reizbeginn (t = 0)
ausgerichtet sind. Sie wurden vom Monitor des Speicheroszilloskops abfotografiert. Jede
Spur zeigt eine Mehrfachantwort, die aus 3 orthodromen Aktionspotentialen besteht, so daß
insgesamt zwölf Aktionspotentiale gezeigt werden, die innerhalb eines ca. 5 ms langen
Zeitfensters (ca. 2,2 bis 7,2 ms) auftauchen. Das dritte Aktionspotential jeder
Mehrfachantwort besitzt eine etwas längere Latenz (6,6 bis 7,2 ms) und sondert sich damit
zeitlich ab. Die jeweils ersten beiden Aktionspotentiale der Mehrfachantworten liegen
innerhalb des Zeitfensters 2,2 bis 5,2 ms.

Ergebnisse 84
a) Albinofrettchen
Insgesamt wurden 72 prätektale Zellen von sechs Albinofrettchen auf ihre orthodrome
Reizbarkeit vom VC getestet. Diese sechs Albinos sind eine Teilmenge der zwölf Tiere, von
denen die DS-Indizes in Abbildung 29A ermittelt wurden. 46 Zellen (64%) waren reizbar,
die restlichen 26 (36%) waren nicht reizbar. Die Häufigkeitsverteilung der jeweils kürzesten
Latenzen der reizbaren Zellen ist in Abbildung 32A dargestellt. Nicht alle 72 getesteten
prätektalen Zellen waren jedoch auch von der IO antidrom reizbar, so daß sie mit Sicherheit
als potentielle Neuronen im hOKN angesehen werden können. Aus 49 von der IO antidrom
reizbaren NOT-Zellen des Albinofrettchens, die auch auf orthodrome Reizbarkeit vom
ipsilateralen VC getestet wurden, waren 31 Zellen (63%) orthodrom aktivierbar. Damit ist
gezeigt, daß beim Albinofrettchen 63% der von der IO antidrom identifizierten,
richtungsunspezifischen NOT-Zellen kortikalen Eingang bekommen. Die Gegenrechnung
Abb: 31: Typische orthodrome Aktionspotentiale, die nach Reizung des VC bei demselben NOT-Neuron wie in Abbildung 30A registriert wurden. Gezeigt werden vier Potentialspuren (vier Reizpulse), die alle zeitlich auf den Reizbeginn ausgerichtet sind. Der Reizbeginn (t = 0) ist durch die senkrechte, gepunktete Linie auf der linken Seite markiert. Jeder einzelne Reizpuls löste bei dieser Zelle drei orthodrome Aktionspotentiale aus, die innerhalb eines ca. 5 ms langen Zeitfensters (von ca. 2,2 ms bis ca. 7,2 ms) registriert wurden. Die ersten beiden Aktionspotentiale jeder Mehrfachantwort bilden eine frühe Gruppe (ca. 2,2 – 5,2 ms Latenz) und das dritte Aktionspotential eine späte Gruppe (ca. 6,6 – 7,2 ms Latenz). Diese Zelle war sowohl antidrom von der IO (Abb. 30A) als auch orthodrom vom ON (Abb. 38B) reizbar. Abszisse: Zeit in ms; Ordinate: extrazellulär abgeleitetes Potential.
III. Foto-Nr. 7
23+24.02.1999.aF
Pen-Nr. 11 Tiefe 7426
VC-orthodrome Spikes
1 ms
t = 0

Ergebnisse 85
besagt, daß von den 72 prätektalen Zellen 23 Zellen nicht von der IO reizbar waren. Von
diesen 23 Zellen waren 15 Zellen ebenfalls vom VC aktivierbar. Dies bedeutet, daß 65% (15
von 23) der prätektalen, nicht zur IO projizierenden Zellen ebenfalls einen VC-Eingang
bekommen. Diese 15 Zellen befinden sich innerhalb des NOT-Areals, d.h. zwischen den
verstreuten, von der IO antidrom identifizierbaren NOT-Zellen. In Abbildung 32A sind die
jeweils kürzesten orthodromen Latenzen der 31 von der IO antidrom identifizierten NOT-
Zellen schraffiert hervorgehoben und befinden sich im Bereich von 2,0 bis 7,5 ms mit einem
Median von 3,0 ms (Q0,25 = 2,6 ms; Q0,75 = 4,0 ms).
In Abbildung 32B ist die Häufigkeitsverteilung der Latenzdifferenzen (längste Latenz minus
kürzeste Latenz) derselben NOT-Zellen dargestellt. Es zeigt sich, daß die orthodromen
Aktionspotentiale der Mehrfachantworten sich innerhalb eines maximalen Zeitfensters
befinden, dessen mediane Breite 2,0 ms (Q0,25 = 1,5 ms; Q0,75 = 4,0 ms) betrug. Die
Latenzdifferenzen der 31 von der IO antidrom identifizierten NOT-Zellen sind analog der
Abbildung 32A schraffiert hervorgehoben.
b) Pigmentierte Frettchen
Die orthodrome Reizung des NOT vom VC wurde auch bei einem pigmentierten Frettchen
durchgeführt. Dieses pigmentierte Tier gehörte zu der Gruppe der fünf Tiere, von denen die
DS-Indizes in Abbildung 29C berechnet wurden. Von diesem Tier konnten 14 vom VC
orthodrom reizbare NOT-Zellen abgeleitet werden, die alle richtungsspezifisch und von der
IO antidrom reizbar waren. Demnach sind bei diesem Tier 100% der NOT-Zellen vom VC
orthodrom reizbar. Im Gegensatz zu diesem Befund geben Klauer et al. (1990) einen
entsprechenden Prozentsatz von 65% (33 von 51) an. Dieser Unterschied wird in der
Diskussion aufgegriffen.
In Abbildung 32C sind die kürzesten orthodromen Latenzen der 14 NOT-Zellen im Bereich
von 2,0 bis 5,5 ms verteilt (Median = 3,25 ms; Q0,25 = 3,0 ms; Q0,75 = 4,0 ms; schraffierte
Latenzen). Zusätzlich zu den 14 Latenzen dieses einen pigmentierten Tieres sind weitere 33
orthodrome Latenzen nicht-schraffiert dargestellt. Diese wurden von Klauer et al. (1990)
entnommen und stammen von richtungsspezifischen NOT-Zellen, die nicht auf Reizbarkeit
von der IO getestet wurden. Ihre Häufigkeitsverteilung deckt sich mit der Häufigkeits-
verteilung der in der vorliegenden Arbeit ermittelten 14 Latenzen. Die entsprechenden
Latenzdifferenzen sind in Abbildung 32D verteilt und besitzen einen Median von 2,0 ms
(Q0,25 = 1,5 ms; Q0,75 = 4,0 ms).

Ergebnisse 86
c) Vergleich zwischen den beiden Tiergruppen
Die 31 orthodromen Latenzen vom VC, die an antidrom reizbaren NOT-Zellen des
Albinofrettchens (schraffierte Latenzen in Abb. 32A) ermittelt wurden, unterscheiden sich
nicht signifikant (RANG-SUMMEN-TEST, P = 0,844) von den entsprechenden 14 Latenzen des
pigmentierten Frettchens (schraffierte Latenzen in Abb. 32C).
Abb. 32: Häufigkeitsverteilung der orthodromen Reizlatenzen vom VC (A, C) und der entsprechenden Latenzdifferenzen (B, D) der prätektalen bzw. NOT-Zellen des albinotischen (A, B) und pigmentierten Frettchens (C, D). Die schraffiert markierten Latenzen und Latenzdifferenzen sind von NOT-Zellen, die von der IO antidrom reizbar waren. A: Vom Albinofrettchen waren 46 von 72 getesteten prätektalen Zellen orthodrom reizbar. Die schraffierten 31 Latenzen stammen von antidromen NOT-Zellen und bilden eine Teilmenge aus 49 auf Reizbarkeit von der IO getesteten prätektalen Zellen. B: Häufigkeitsverteilung der maximalen Latenzdifferenzen (längste minus kürzeste Latenz) aus A. C: Von einem pigmentierten Frettchen wurden 14 NOT-Zellen orthodrom vom VC gereizt, die alle auch von der IO antidrom reizbar (IO+) und richtungsspezifisch (DS+) waren. Die übrigen 33 Latenzen in C wurden zu Vergleichszwecken der Literatur entnommen (Klauer et al., 1990). Diese Zellen sind laut Klauer et al. (1990) richtungsspezifische (DS+) NOT-Zellen, die aber nicht auf Reizbarkeit von der IO getestet wurden. D: Häufigkeitsverteilung der maximalen Latenzdifferenzen aus B. y-Achsen: Anzahl der Latenzen in A und C, Anzahl der Latenzdifferenzen in B und D.
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
31 IO+ 15 IO-
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
1631 IO+15 IO-
Latenz [ms]0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
1614 IO+ & DS+33 DS+ (Klauer et al., 1990)
Latenzdifferenz [ms]0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
14 IO+ & DS+ 0 IO-
C D
A B
N N
N N

Ergebnisse 87
Betrachtet man die Verteilung aller 47 orthodromen Latenzen des pigmentierten Frettchens
(Summe aus 14 eigenen und 33 Latenzen aus Klauer et al., 1990) so befinden sie sich im
Bereich von 1 bis 6,5 ms. Ein statistischer Vergleich dieser Gesamtgruppe von 47 Latenzen
mit den 31 Latenzen des Albinofrettchens ist nur mit dem χ2-TEST möglich. Der Grund dafür
ist, daß Klauer et al. (1990) nicht die einzelnen Latenzwerte zur Verfügung stellen, sondern
die absoluten Häufigkeiten. Der χ2-TEST liefert ein P = 0,804 und zeigt damit eine fehlende
Signifikanz für einen Unterschied zwischen den beiden Häufigkeitsverteilungen (schraffierte
Verteilung in Abb. 32A und gesamte Verteilung in Abb. 32C). Somit finden wir bei beiden
Tiergruppen kortikale NOT-Eingänge, die sich hinsichtlich ihrer Leitungsgeschwindigkeit
nicht voneinander unterscheiden.
Zwischen den in Abbildung 32B schraffierten Latenzdifferenzen des albinotischen
Frettchens und den entsprechenden Differenzen des pigmentierten Frettchens in
Abbildung 32D läßt sich ebenfalls kein signifikanter Unterschied feststellen (RANG-
SUMMEN-TEST, P = 0,087).
7. Antidrome Reizung der VC-Neuronen vom NOT und / oder SC
Die rezeptiven Felder der untersuchten VC-Zellen befanden sich im zentralen Gesichtsfeld
der Versuchstiere innerhalb des Bereichs –10° bis +25° Sehwinkel Azimut und +20° bis
–15° Sehwinkel Elevation.
a) Anzahl der untersuchten VC-Neuronen
(1) Albinofrettchen
Insgesamt wurden sieben Albinofrettchen untersucht. Bei einem Albinofrettchen wurde
keine elektrische Reizung durchgeführt. Bei drei Tieren wurde der NOT elektrisch gereizt
und bei den übrigen drei Tieren wurde sowohl der NOT als auch der SC gereizt.
Insgesamt wurden 123 visuelle Zellen des VC untersucht. Von 86 Zellen wurden die DS-
Indizes berechnet (Abb. 33A). Von den restlichen 37 Zellen wurden keine DS-Indizes
berechnet, weil sie entweder nicht lange genug abgeleitet werden konnten oder durch die
Reizung mit der Handlampe eindeutig als richtungsunspezifisch identifiziert wurden.
§ Von den 123 Zellen wurden 78 Zellen auf Reizbarkeit vom NOT getestet und 45 Zellen
nicht getestet. Von den getesteten Zellen waren 30 Zellen (38% von 78) antidrom
(NOTanti+), 20 Zellen (26% von 78) orthodrom (NOTortho+) und 28 Zellen (36% von 78)
gar nicht (NOT–) reizbar.

Ergebnisse 88
§ Von den 78 auf Reizbarkeit vom NOT getesteten Zellen wurden 19 Zellen auch auf Reiz-
barkeit vom SC getestet. Keine der 19 Zellen war antidrom reizbar (SCanti+). Vier (21%
von 19) Zellen waren orthodrom (SCortho+) und 15 Zellen (79% von 19) waren überhaupt
nicht (SC–) vom SC reizbar.
§ Eine Zelle war sowohl vom NOT als auch vom SC orthodrom aktivierbar.
(2) Pigmentierte Frettchen
Insgesamt wurden 63 VC-Zellen von vier pigmentierten Frettchen untersucht (Abb. 33B).
Bei einem der vier pigmentierten Tiere wurde nur der NOT elektrisch gereizt. Von diesem
Tier konnte nur eine VC-Zelle abgeleitet werden. Bei den drei anderen Tieren wurde sowohl
der NOT, als auch der SC elektrisch gereizt. Deshalb wurden fast genau so viele VC-Zellen
auf Reizbarkeit vom NOT wie auch auf Reizbarkeit vom SC getestet. Von 16 VC-Zellen
wurden die DS-Indizes berechnet.
§ Von den 63 Zellen des VC wurden 62 Zellen auf Reizbarkeit vom NOT getestet. 14 Zellen
(23% von 62) waren NOTanti+, fünf Zellen waren (8% von 62) NOTortho+ und 43 Zellen
(69% von 62) NOT–.
§ Von den 62 auf Reizbarkeit vom NOT getesteten VC-Neuronen wurden 61 Neuronen auch
auf Reizbarkeit vom SC getestet. Davon waren elf (18% von 61) SCanti+, drei Zellen (5%
von 61) SCortho+ und 47 Zellen (77% von 61) SC–.
§ Zwei VC-Neuronen waren sowohl vom NOT als auch vom SC antidrom reizbar. Eine Zelle
war von beiden Reizorten orthodrom reizbar.
b) Richtungsspezifität und Vorzugsstärke im VC
Im Rahmen der Untersuchung des kortikalen NOT-Eingangs hinsichtlich seines
Informationsgehaltes sollten zunächst folgende Fragen beantwortet werden:
• Besitzt das Albinofrettchen richtungsspezifische Neuronen im visuellen Kortex?
• Besitzt das Albinofrettchen verhältnismäßig genau so viele richtungsspezifische VC-
Neuronen wie das pigmentierte Frettchen?
• Besitzen beide Tiergruppen relativ gleich viele vom NOT antidrom reizbare VC-Neuronen?
• Haben die richtungsspezifischen VC-Neuronen bei beiden Tiergruppen gleich hohe DS-
Indizes?
• Besitzen beide Tiergruppen richtungsspezifische VC-Neuronen, die sowohl die Bedingung
„antidrome Reizbarkeit vom NOT“ als auch die Bedingung „visuell reizbar mit dem
großflächigen Zufallspunktemuster“ erfüllen?

Ergebnisse 89
Zur Beantwortung dieser Fragen wurden die DS-Indizes der untersuchten VC-Neuronen
benötigt, jedoch nicht ihre Vorzugsrichtungen. Deshalb werden in Abbildung 33 die
Häufigkeitsverteilungen der 86 DS-Indizes des albinotischen (A) und der 16 DS-Indizes des
pigmentierten (B) Frettchens unabhängig von der Vorzugsrichtung gezeigt.
(1) Albinofrettchen
Beim Albinofrettchen sind die 86 DS-Indizes im gesamten Bereich zwischen 0 und 1 verteilt
mit einem Median von 0,36 (Q0,25 = 0,22; Q0,75 = 0,60). 55% (47 von 86) der VC-Zellen
haben einen DS-I ≥ 0,33 (Abb. 33A). Ein DS-I von 0,33 zeigt an, daß die Zelle bei Reizung
in ihrer Vorzugsrichtung um 100% stärker feuert als bei Reizung in ihrer Nullrichtung. Dies
bedeutet, daß 55% der VC-Zellen des Albinofrettchens eine Vorzugsrichtung haben, die sie
mindestens um 50% stärker beantworten als die entgegengesetzte Nullrichtung.
Damit ist die erste Frage beantwortet: Das Albinofrettchen besitzt richtungsspezifische
Zellen im visuellen Kortex.
Von diesen 86 VC-Zellen wurden 41 Zellen auf Reizbarkeit vom ipsilateralen NOT getestet.
Die DS-Indizes dieser 41 Zellen sind in Abbildung 33A grau markiert. Davon waren 32%
(13 von 41) antidrom reizbar (NOT+) und 68% (28 von 41) nicht reizbar (NOT–). Die NOT+
Zellen sind in Abbildung 33A durch die Schraffierung hervorgehoben und befinden sich in
der kortikalen Schicht V, wie die Histologie zeigte (vgl. Abb. 25). Es ist auffällig, daß unter
den NOT+ Zellen auch solche Zellen zu finden sind, die einen relativ hohen DS-I besitzen
(z.B. > 0,2) und folglich eindeutig richtungsspezifisch sind. 54 % (7 von 13) der NOT+
Zellen haben einen DS-I ≥ 0,33.
Damit wird festgestellt, daß beim Albinofrettchen die Projektion der kortikalen Schicht V
auf den NOT unter anderem von richtungsspezifischen Zellen gebildet wird.
(2) Pigmentierte Frettchen
Die 16 DS-Indizes des pigmentierten Frettchens sind zwischen 0 und 0,7 verteilt und haben
einen Median von 0,21 (Q0,25 = 0,09; Q0,75 = 0,34). Für 38% (6 von 16) der DS-Indizes gilt ≥
0,33. Dieser Prozentsatz liegt unter dem korrespondierenden Wert des Albinofrettchens
(55%). Somit findet man in der kleinen Stichprobe beim pigmentierten Frettchen relativ
weniger richtungsspezifische VC-Zellen als beim albinotischen Frettchen.
Alle 16 VC-Zellen des pigmentierten Frettchens sind auf Reizbarkeit vom ipsilateralen NOT
getestet (NOT-getestet), weshalb alle DS-Indizes in Abbildung 33B grau markiert sind.
Davon waren sechs (38% von 16) Zellen antidrom reizbar (NOT+, schraffierte DS-Indizes).

Ergebnisse 90
Somit liegt beim pigmentierten Frettchen der Prozentsatz der vom NOT antidrom reizbaren
VC-Zellen in einer ähnlichen Größenordnung wie beim Albinofrettchen (32% von 41). Aus
dieser Übereinstimmung wird die Schlussfolgerung gezogen, daß sich albinotische und
pigmentierte Frettchen hinsichtlich der Stärke ihrer kortikoprätektalen Projektion nicht
unterscheiden. Damit wird der entsprechende Befund aus der orthodromen Reizung der
NOT-Zellen vom VC bestätigt (siehe oben).
50% (3 von 6) der NOT+ VC-Zellen haben einen DS-I ≥ 0,33. Der analoge Prozentsatz des
Albinofrettchens ist 54%. Dies deutet darauf hin, daß sich die richtungsspezifischen, zum
NOT projizierenden VC-Zellen in beiden Tiergruppen hinsichtlich ihrer Vorzugsstärke nicht
unterscheiden.
(3) Statistischer Vergleich zwischen den beiden Tiergruppen
Die 86 DS-Indizes der Albinofrettchen (Abb. 33A) unterscheiden sich signifikant von den
entsprechenden 16 DS-Indizes der pigmentierten Frettchen in Abbildung 33B (RANG-
Abb. 33: Häufigkeitsverteilung der DS-Indizes von VC-Neuronen des albinotischen (A) und pigmentierten Frettchens (B). A: Die DS-Indizes der 86 VC-Neuronen des Albinofrettchens besitzen einen Median von 0,36. 41 dieser Zellen wurden auf Reizbarkeit vom ipsilateralen NOT getestet, von denen 13 antidrom reizbar waren. B: Beim pigmentierten Frettchen wurde der DS-Index von insgesamt 16 VC-Zellen berechnet (Median = 0,21). Alle 16 Zellen wurden auch auf Reizbarkeit vom NOT getestet. Sechs Zellen waren antidrom reizbar. Die DS-Indizes in A unterscheiden sich statistisch signifikant von den entsprechenden Werten in B (RANG-SUMMEN-TEST, P = 0,013). Vergleicht man nur die schraffierten DS-Indizes miteinander, so findet man beim Albinofrettchen 54% (sieben von 13) und beim pigmentierten Frettchen 50 % (drei von sechs) der DS-Indizes ≥ 0,33. y-Achsen (N): Anzahl der DS-Indizes.
gesamt auf Reizbarkeit vom NOT getestet vom NOT antidrom reizbar
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
2
4
6
8
10
12
14
16
18
20N = 13 / 41 / 86
N
A
DS-Index 0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
0
2
4
6
8
10
12
14
16
18
20N = 6 / 16 / 16
DS-Index
N
B

Ergebnisse 91
SUMMEN-TEST, P = 0,013). Dieser Befund spiegelt sich auch in den unterschiedlich großen
Medianen und Quartilen (siehe oben) wider. Somit kann festgestellt werden, daß die
richtungsspezifischen VC-Zellen des Albinofrettchens stärker eine Richtung bevorzugen als
die entsprechenden Zellen des pigmentierten Frettchens.
c) Anti- und orthodrome Reizlatenzen im VC
Die elektrische Reizung des NOT konnte prinzipiell sowohl zu einer antidromen, als auch zu
einer orthodromen Aktivierung der VC-Zellen führen. Die antidrome Reizung erfolgt
definitionsgemäß über eine direkte Faserverbindung, während die orthodrome Reizung in
diesem Falle über eine Umschaltung im Thalamus erfolgen kann. Bei den einzelnen Zellen
wurde allerdings entweder eine antidrome oder eine orthodrome Aktivierung beobachtet.
Abbildung 34 zeigt beispielhaft das vom NOT antidrom ausgelöste Aktionspontial einer VC-
Zelle eines Albinofrettchens, die positiv auf den Kollisionstest reagierte. Sowohl im oberen
Teil als auch im unteren Teil der Abbildung sind jeweils vier Potentialspuren gezeigt, die
vom Monitor des Speicheroszilloskops abfotografiert wurden. Alle Spuren sind auf den
Reizbeginn (t = 0) ausgerichtet. Die oberen Spuren zeigen zunächst den Kollisionstest: Das
antidrome Aktionspotential ist nicht zu sehen, weil der Reizpuls durch ein spontanes
Aktionspotential (Auslösepotential) mit einer Verzögerung von 1 ms ausgelöst wird. Diese
Verzögerung ist kürzer als die Latenz des antidromen Aktionspotentials (1,3 ms), so daß das
spontane Auslösepotential mit dem antidromen Aktionspotential auf dem Axon kollidiert.
Infolgedessen löschen sich die beiden Aktionspontentiale aus und das antidrome Aktions-
potential kann nicht mehr rechtzeitig an der VC-Ableitelektrode bzw. auf dem Monitor
erscheinen. Als anschließend der Reizpuls extern durch den Experimentator ausgelöst wurde,
blieb die Kollision aus und das antidrome Aktionspotential erschien mit derselben Latenz
und Form wieder (Abb. 34 untere Spuren) wie vor dem Kollisionstest (hier nicht gezeigt).
(1) Albinofrettchen
Insgesamt wurden 30 VC-Zellen des Albinofrettchens vom ipsilateralen NOT antidrom
gereizt. Die Häufigkeitsverteilung der antidromen Latenzen wird in Abbildung 35A gezeigt.
Die kürzeste Latenz ist 1,0 ms und die längste Latenz 8,0 ms. Der Median beträgt 1,4 ms
(Q0,25 = 1,3 ms; Q0,75 = 2,2 ms).
Weitere 20 Neuronen des VC des Albinofrettchens waren vom NOT orthodrom reizbar. In
Abbildung 35B ist die Häufigkeitsverteilung der jeweils kürzesten orthodromen Reizlatenzen

Ergebnisse 92
gezeigt. Die kürzeste Latenz ist 1,0 ms und die längste Latenz 3,5 ms. Der Median beträgt
2,0 ms (Q0,25 = 1,5 ms; Q0,75 = 2,5 ms).
(2) Pigmentierte Frettchen
Beim pigmentierten Frettchen konnten 14 VC-Neuronen vom ipsilateralen NOT antidrom
(Abb. 35C) und 5 VC-Neuronen orthodrom (Abb. 35D) gereizt werden. Die antidromen
Reizlatenzen befinden sich im Bereich zwischen 1,2 ms (kürzeste Latenz) und 2,9 ms
(längste Latenz) mit einem Median von 1,4 ms (Q0,25 = 1,4 ms; Q0,75 = 1,6 ms). Die
orthodromen Latenzen befinden sich zwischen 1,5 und 2,6 ms mit einem Median von 2,3 ms
(Q0,25 = 2,0 ms; Q0,75 = 2,5 ms).
(3) Statistischer Vergleich zwischen den beiden Tiergruppen
Die 30 antidromen Latenzen von VC-Zellen des Albinofrettchens (Abb. 35A) unterscheiden
sich nicht signifikant von den 14 in Abbildung 35C dargestellten Latenzen des pigmentierten
Frettchens (RANG-SUMMEN-TEST, P = 0,554). Damit findet man zwischen albinotischen und
pigmentierten Frettchen keinen Unterschied hinsichtlich der Leitungsgeschwindigkeit der
Abb. 34: Typisches antidromes Aktionspotential einer VC-Zelle des Albino-frettchens nach elektrischer Reizung des NOT und positiver Kollisionstest. Obere Spuren: Der Reizpuls wird durch ein spontanes Aktionspotential (Auslösepotential, Pfeil a) mit einer Verzögerung von 1 ms ausgelöst. Das antidrome Aktionspotential ist nicht zu sehen, weil es mit dem spontanen Aktionspotential kollidiert und sich die beiden Potentiale gegenseitig auslöschen (positiver Kollisionstest). Untere Spuren: Wenn der Reizpuls extern (nicht durch ein Auslösepotential) mit 1 ms Verzögerung ausgelöst wird, kommt keine Kollision zustande und das antidrome Aktionspotential (Pfeil b) erscheint in allen vier Spuren. Abszisse: Zeit in ms; Ordinate: extrazellulär abgeleitetes Potential.
IV. Foto-Nr. 6
24+25.07.2000.aF
Pen-Nr. C1 Tiefe 6366 µm
a) Kollisionstest positiv (antidromer Spike ist weg)
A
1 ms
t = -1 t = 0
a
b

Ergebnisse 93
kortikoprätektalen Fasern. Dies bestätigt den entsprechenden Befund aus der orthodromen
Reizung der NOT-Zellen vom VC (vgl. Abb. 32A mit
Abb. 32C).
Die orthodromen Reizlatenzen vom NOT (Abb. 35B und D) unterscheiden sich ebenfalls
nicht signifikant zwischen den beiden Tiergruppen (RANG-SUMMEN-TEST, P = 0,683).
d) Antidrome Reizbarkeit der VC-Neuronen und visuelle Reize
(1) Albinofrettchen
Von den 30 vom NOT antidrom reizbaren VC-Zellen wurden fünf Zellen nur mit einem
Lichtpunkt (24 cm ∅) visuell gereizt, so daß keine Aussage darüber gemacht werden kann,
Abb. 35: Antidrome (A, C) und orthodrome (B, D) Reizlatenzen von VC-Neuronen des albinotischen (A, B) und des pigmentierten (C, D) Frettchens, die nach elektrischer Reizung des NOT registriert wurden. Als orthodrome Latenzen sind die jeweils kürzesten Latenzen aufgetragen. N = Gesamtanzahl der Latenzen bzw. der vom NOT-Areal aktivierten VC-Zellen. y-Achsen (N): Anzahl der Latenzen.
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
18
20
N = 30
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
18
20
N = 20
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
18
20
N = 14
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
16
18
20
N = 5
C D
A B
N N
N N
Latenz [ms] Latenz [ms]

Ergebnisse 94
ob sie auch mit dem Lichtbalken und Zufallspunktemuster reizbar waren. Weitere 24 Zellen
waren mit dem Lichtbalken und 14 von diesen auch mit dem großen Zufallspunktemuster
visuell reizbar. Von diesen 14 Zellen wurde eine Zelle als richtungsunspezifisch
charakterisiert. Von den restlichen 13 Zellen wurde der DS-Index berechnet (schraffierte DS-
Indizes in Abb. 33A).
(2) Pigmentierte Frettchen
Von den 14 vom NOT antidrom reizbaren VC-Zellen waren neun Zellen nur mit dem
Lichtbalken und fünf Zellen auch mit dem Zufallspunktemuster visuell reizbar. Die sechs in
Abbildung 33B schraffierten DS-Indizes stammen von diesen fünf Zellen und von einer der
neun Zellen.
Damit kann festgestellt werden, daß sowohl die pigmentierten als auch die albinotischen
Frettchen richtungsspezifische VC-Zellen besitzen, die sowohl das Kriterium „antidrome
Reizbarkeit vom NOT“ als auch das Kriterium „visuell reizbar mit dem großflächigen
Zufallspunktemuster“ erfüllen.
e) Vorzugsrichtungen der VC-Neuronen
Unterscheiden sich die beiden Tiergruppen hinsichtlich der Vorzugsrichtungen der zum NOT
projizierenden VC-Neuronen? Zur Beantwortung dieser Frage ist lediglich eine eindeutige
Zuordnung der Vorzugsrichtung in einen der acht Sektoren des Gesichtsfeldes notwendig,
was auch mit der Handlampe ausreichend genau vorgenommen werden konnte. Deshalb
werden hier zusätzlich zu den Vorzugsrichtungen der VC-Zellen mit DS-Index auch die
Vorzugsrichtungen der richtungsspezifischen Zellen, die nur mit der Handlampe untersucht
wurden, berücksichtigt.
(1) Albinofrettchen
In Abbildung 36A sind die 86 DS-Indizes, die in Abbildung 33A unabhängig von der
Vorzugsrichtung dargestellt wurden, nach ihrer Vorzugsrichtung gruppiert. Jede
Teilabbildung zeigt die Häufigkeitsverteilung der DS-Indizes von denjenigen VC-Zellen,
deren Vorzugsrichtung innerhalb eines der acht 45°-Sektoren fällt. Die Position einer
Teilabbildung entspricht der Position des 45°-Sektors (vgl. Material und Methoden). Die DS-
Indizes von VC-Zellen, die auf Reizbarkeit vom NOT getestet wurden, sind weiterhin grau
und die von antidromen (NOT+) Zellen weiterhin schraffiert hervorgehoben. Man findet in
jedem Sektor DS-Indizes, d.h. im VC des Albinofrettchens sind alle Vorzugsrichtungen

Ergebnisse 95
repräsentiert. Innerhalb eines jeden Sektors befinden sich sowohl niedrige als auch höhere
DS-I-Werte, d.h. alle Vorzugsrichtungen werden gleich stark bevorzugt.
0 unten
oben 0
0
0 kontra
0 1
ipsi 5
0
B Abb. 36: Vorzugsrichtungen der VC-Zellen des Albinofrettchens, die auf Reizbarkeit vom ipsilateralen NOT getestet wurden. A: Aufspaltung der 86 DS-Indizes aus Abbildung 33A in die acht Sektoren des Gesichtsfeldes (ipsi, ipsi-unten, unten, kontra-unten, kontra, kontra-oben, oben, ipsi-oben). Jede Teilabbildung stellt die Häufigkeitsverteilung der DS-Indizes von VC-Zellen dar, deren Vorzugsrichtungen innerhalb des entsprechenden Sektors fallen. B: Vorzugs-richtungen der sechs vom NOT antidrom reizbaren Zellen, die nur mit der Handlampe untersucht wurden.
A
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 3 / 4 / 12
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 2 / 7 / 12
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 1 / 5 / 11
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 1 / 3 / 3
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 3 / 6 / 12
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 1 / 10 / 15
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 0 / 1 / 7
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 2 / 5 / 14
gesamt auf Reizbarkeit vom NOT getestet vom NOT antidrom reizbar
ipsi kontra
oben
unten

Ergebnisse 96
Es ist auffällig, daß die 13 schraffierten DS-Indizes sich nicht in einem bestimmten Sektor
anhäufen. Vielmehr findet man in fast allen Sektoren schraffierte DS-Indizes (mit Ausnahme
des Sektors kontra-unten, was aber angesichts der relativ kleinen Datenmenge als Zufall
angesehen werden kann). Diese Verteilung zeigt, daß die Vorzugsrichtungen der vom NOT
antidrom reizbaren VC-Zellen gleichverteilt sind, d.h. daß alle Vorzugsrichtungen unter
diesen VC-Zellen repräsentiert sind.
Innerhalb eines Sektors können die schraffierten DS-Indizes sowohl niedrige als auch höhere
Werte haben. Dieser Befund zeigt, daß die Vorzugsstärken der vom NOT antidrom reizbaren
VC-Zellen ebenfalls gleichverteilt sind, d.h. die zum NOT projizierenden VC-Zellen können
unabhängig von der bevorzugten Richtung sowohl schwach als auch stark
richtungsspezifisch sein.
In Abbildung 36B sind die Vorzugsrichtungen von weiteren sechs vom NOT antidrom
reizbaren (NOTanti+), richtungsspezifischen VC-Zellen (DS) des Albinofrettchens
angegeben. Von diesen Zellen wurden keine DS-Indizes berechnet, so daß keine Aussage
über ihre Vorzugsstärke möglich ist. Ihre Vorzugsrichtung wurde mit der Handlampe
bestimmt. Von den sechs Zellen bevorzugten fünf die ipsiversive (ipsi) Richtung und eine
Zelle die Richtung nach ipsi-unten.
(2) Pigmentierte Frettchen
Die Verteilung der 16 DS-Indizes (vgl. Abb. 33B) der pigmentierten Frettchen in die acht
Sektoren des Gesichtsfeldes ist in Abbildung 37A gezeigt. Unter diesen 16 Zellen findet man
nicht alle Vorzugsrichtungen repräsentiert und mehr als die Häfte der VC-Zellen haben eine
horizontale Vorzugsrichtung. Dies bedeutet jedoch nicht, daß dieser Befund für die gesamte
Zellpopulation des pigmentierten Frettchens repräsentativ ist. Denn von den meisten Zellen
wurden keine Aufnahmen gemacht, wenn aus der Reizung mit der Handlampe hervorging,
daß sie keine horizontale Richtung bevorzugten. Der Grund für die Auswahl der beiden
horizontalen Richtungen lag darin, möglichst viele Zellen mit ipsi- und kontraversiver
Vorzugsrichtung auf antidrome Reizbarkeit vom NOT zu testen. Damit sollte möglichst
schnell ermittelt werden, ob alle zum NOT projizierenden VC-Zellen des pigmentierten
Frettchens nur die ipsiversive Richtung bevorzugen. Das erzielte Ergebnis in Abbildung 37A
erlaubt jedoch keine Aussage, weil die Anzahl der Zellen mit DS-I sehr klein ist.

Ergebnisse 97
1 unten
oben 0
1
2 kontra
1 0
ipsi 1
0
B Abb. 37: Vorzugsrichtungen der VC-Zellen des pigmentierten Frettchens, die auf Reizbarkeit vom ipsilateralen NOT getestet wurden. A: Aufspaltung der 16 DS-Indizes aus Abbildung 33B in die acht Sektoren des Gesichtsfeldes. B: Vorzugs-richtungen der sechs vom NOT antidrom reizbaren (NOTanti+) Zellen, die nur mit der Handlampe untersucht wurden. Darstellungsform und Abkürzungen sind die gleichen wie für das Albinofrettchen in Abbildung 36.
A
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 1 / 10 / 15
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 0 / 0 / 0
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 0 / 0 / 0
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 0 / 0 / 0
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 1 / 6 / 6
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 2 / 4 / 4
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 1 / 2 / 2
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 0 / 1 / 1
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
1
2
3
4
5
N = 2 / 3 / 3
ipsi kontra
oben
unten
gesamt auf Reizbarkeit vom NOT getestet vom NOT antidrom reizbar

Ergebnisse 98
Weitere 29 richtungsspezifische VC-Zellen des pigmentierten Frettchen wurden mit der
Handlampe untersucht. Von diesen wurden 28 Zellen auf Reizbarkeit vom NOT getestet.
Sechs Zellen waren NOTanti+, zwei Zellen waren NOTortho+ und 20 Zellen waren überhaupt
nicht reizbar (NOT-). Von besonderem Interesse sind in diesem Falle die Vorzugsrichtungen
der NOTanti+ Zellen, die in Abbildung 37B dargestellt werden. Von den sechs NOTanti+
Zellen bevorzugte nur eine Zelle die ipsiversive Richtung. Von den übrigen fünf Zellen
bevorzugten zwei Zellen die kontraversive Richtung und jeweils eine Zelle die Richtung
nach unten, nach kontra-oben und nach kontra-unten. Dieser Befund spricht eindeutig gegen
die Hypothese, nach der alle vom NOT antidrom reizbaren VC-Zellen des pigmentierten
Frettchens die ipsiversive Richtung bevorzugen.
Insgesamt kann aus den Abbildungen 37A und B geschlossen werden, daß die
Vorzugsrichtungen der zum NOT projizierenden VC-Zellen des pigmentierten Frettchens,
wie zuvor auch beim Albinofrettchen festgestellt, gleichverteilt sind.
f) Vom NOT und SC reizbare VC-Neuronen
(1) Albinofrettchen
Unter den 19 VC-Zellen, die bei drei Albinofrettchen auf Reizbarkeit sowohl vom SC als
auch vom NOT getestet wurden, befand sich keine einzige Zelle, die von beiden Reizorten
antidrom reizbar war. Eine Zelle war von beiden Orten orthodrom aktivierbar mit einer
Latenz von 2,5 ms nach Reizung des NOT und 3,0 ms nach Reizung des SC. Sie wurde mit
Hilfe der Handlampe als richtungsunspezifische Zelle charaktierisiert.
(2) Pigmentiertes Frettchen
Von den 61 VC-Zellen der drei pigmentierten Frettchen, die auf Reizbarkeit sowohl vom
NOT als auch vom SC getestet wurden, waren 3% (zwei von 61) in beiden Fällen antidrom
reizbar. Die erste Zelle bevorzugte die Richtung nach kontraversiv und besaß eine antidrome
Latenz von 2,7 ms nach Reizung des NOT und eine antidrome Latenz von 3,0 ms nach
Reizung des SC. Ihr DS-Index konnte nicht berechnet werden. Die zweite Zelle bevorzugte
ebenfalls die Richtung nach kontraversiv und besaß einen DS-I von 0,16. Ihre antidromen
Latenzen waren 2,6 ms (nach Reizung des NOT) und 2,7 ms (nach Reizung des SC). Es
erscheint einerseits wahrscheinlich, daß es sich bei diesen beiden VC-Zellen um Zellen
handelt, deren Axone sich aufzweigen und sowohl den NOT als auch den SC innervieren.
Andererseits aber findet man im SC viele richtungsspezifische Neuronen mit einer
kontraversiven Vorzugsrichtung und diese beiden VC-Zellen bevorzugten ebenfalls die

Ergebnisse 99
kontraversive Richtung. Deshalb erscheint es wahrscheinlicher, daß sie nur den SC
innervieren.
Eine dritte VC-Zelle war sowohl vom NOT als auch vom SC orthodrom aktivierbar. Ihre
kürzesten orthodromen Latenzen waren 2,4 ms (nach Reizung des NOT) und 2,5 ms (nach
Reizung des SC). Sie bevorzugte die ipsiversive Reizung mit einem DS-I von 0,64.
(3) Vergleich zwischen den beiden Tiergruppen
Sowohl beim albinotischen als auch beim pigmentierten Frettchen wurden relativ wenige
VC-Zellen abgeleitet, die sowohl vom NOT als auch vom SC reizbar waren. Insofern hätte
ein statistischer Vergleich der Latenzen keine Aussagekraft. Es ist jedoch auffällig, daß die
antidromen und orthodromen Latenzen vom NOT immer etwas kürzer sind als die
entsprechenden Latenzen vom SC. Dies deutet darauf hin, daß sowohl der antidrome Weg
als auch alle möglichen orthodromen Wege zwischen VC und NOT kürzer sind als die
entsprechenden Wege zwischen VC und SC.
8. Orthodrome Reizung der NOT-Neuronen vom ON
Der retinale Eingang des NOT wurde nur beim Albinofrettchen untersucht. Zum Vergleich
mit entsprechenden Daten vom pigmentierten Frettchen werden die Ergebnisse aus Klauer et
al. (1990) herangezogen.
Insgesamt wurden 66 prätektale Zellen von fünf Albinofrettchen auf Reizbarkeit vom
kontralateralen ON getestet. Diese fünf Individuen waren eine Teilmenge aus der Gruppe der
sechs Tiere, bei denen die prätektalen Zellen auch auf Reizbarkeit vom VC getestet wurden.
86% (57 von 66) der Zellen waren vom kontralateralen ON orthodrom reizbar und reagierten
typischerweise mit einer Mehrfachantwort, die aus zwei bis fünf Aktionspotentialen bestand.
Abbildung 38 zeigt beispielhaft zwei typische orthodrome Antworten von zwei NOT-Zellen,
die von unterschiedlichen Albinofrettchen registriert wurden.
In Abbildung 38A werden zwei auf den Zeitpunkt des Reizbeginns (t = 0) ausgerichtete
Spuren von Potentialverläufen gezeigt, die an der ersten Zelle registriert wurden. Beide
Potentialverläufe zeigen zwei orthodrome Aktionspotentiale vom ON mit ca. 1,8 ms und
ca. 3,3 ms Latenz. Diese Zelle wurde auf Reizbarkeit von der ipsilateralen IO getestet und
war nicht antidrom reizbar. Auf Reizbarkeit vom ipsilateralen VC wurde sie nicht getestet.
Ihre relativ kurze orthodrome Latenz ist typisch für die Gruppe prätektaler, nicht zur IO
projizierender Zellen (siehe unten).
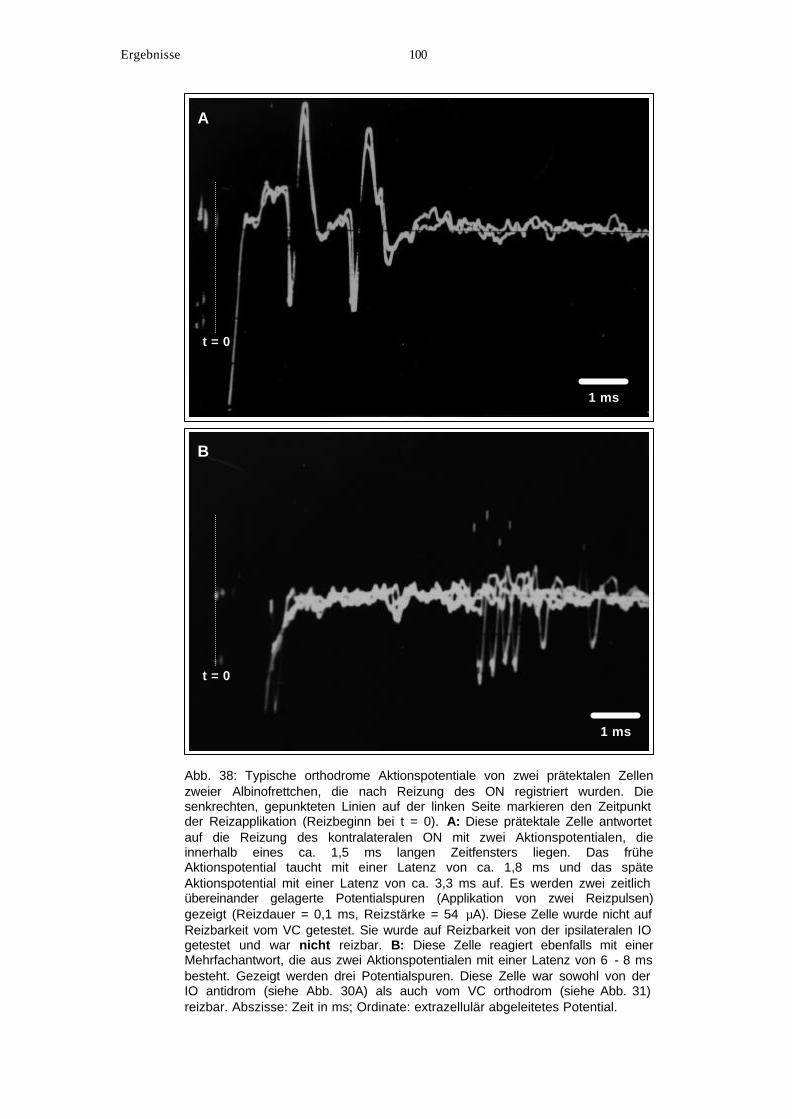
Ergebnisse 100
Abb. 38: Typische orthodrome Aktionspotentiale von zwei prätektalen Zellen zweier Albinofrettchen, die nach Reizung des ON registriert wurden. Die senkrechten, gepunkteten Linien auf der linken Seite markieren den Zeitpunkt der Reizapplikation (Reizbeginn bei t = 0). A: Diese prätektale Zelle antwortet auf die Reizung des kontralateralen ON mit zwei Aktionspotentialen, die innerhalb eines ca. 1,5 ms langen Zeitfensters liegen. Das frühe Aktionspotential taucht mit einer Latenz von ca. 1,8 ms und das späte Aktionspotential mit einer Latenz von ca. 3,3 ms auf. Es werden zwei zeitlich übereinander gelagerte Potentialspuren (Applikation von zwei Reizpulsen) gezeigt (Reizdauer = 0,1 ms, Reizstärke = 54 µA). Diese Zelle wurde nicht auf Reizbarkeit vom VC getestet. Sie wurde auf Reizbarkeit von der ipsilateralen IO getestet und war nicht reizbar. B: Diese Zelle reagiert ebenfalls mit einer Mehrfachantwort, die aus zwei Aktionspotentialen mit einer Latenz von 6 - 8 ms besteht. Gezeigt werden drei Potentialspuren. Diese Zelle war sowohl von der IO antidrom (siehe Abb. 30A) als auch vom VC orthodrom (siehe Abb. 31) reizbar. Abszisse: Zeit in ms; Ordinate: extrazellulär abgeleitetes Potential.
V. Foto-Nr.
1 ms
t = 0
A
VI. Foto-Nr.
1 ms
t = 0
B

Ergebnisse 101
In Abbildung 38B werden drei ebenfalls auf t = 0 ausgerichtete Potentialspuren der zweiten
prätektalen Zelle des Albinofrettchens gezeigt. In diesem Falle besitzen die orthodromen
Aktionspotentiale vom ON eine Latenz von mindestens 6,0 ms. Diese Zelle war von der IO
antidrom reizbar. Ihre relativ lange orthodrome Latenz ist typisch für die zur IO
projizierenden NOT-Zellen (siehe unten).
Die Häufigkeitsverteilung der jeweils kürzesten orthodromen Latenzen der 57 reizbaren
prätektalen Zellen ist in Abbildung 39A gezeigt. Aus der Gesamtmenge der 66 auf
Reizbarkeit vom ON getesteten prätektalen Zellen wurden 60 Zellen auch auf antidrome
Reizbarkeit von der IO getestet. Von den 60 Zellen waren 24 Zellen antidrom reizbar und 36
nicht reizbar. Diese 24 antidromen Zellen, die definitionsgemäß zum hOKN-Schaltkreis
gehören, werden 100% gesetzt. Von diesen 24 Zellen konnten 92% (22) auch vom
kontralateralen ON aktiviert werden. Die entsprechenden Prozentsätze für das pigmentierte
Frettchen lassen sich aus Klauer et al. (1990) entnehmen. Klauer et al. (1990) konnten 90%
(46 von 51) der richtungsspezifischen NOT-Zellen vom optischen Chiasma orthodrom reizen
und folglich 10% nicht reizen.
Desweiteren kann an dieser Stelle festgehalten werden, daß die 24 von der IO antidrom
reizbaren und auf Reizbarkeit vom ON getesteten NOT-Zellen auch auf Reizbarkeit vom VC
getestet wurden. Zwanzig von diesen waren vom VC orthodrom reizbar. Die Schnittmenge
aus beiden orthodromen Reizungen besteht jedoch aus 19 Zellen, d.h. 79% (19 von 24) der
zur IO projizierenden NOT-Zellen des Albinofrettchens waren sowohl vom kontralateralen
ON als auch vom ipsilateralen VC orthodrom aktivierbar. Die gleichzeitige Aktivierbarkeit
einzelner NOT-Zellen vom ON und vom VC war aufgrund von vorausgegangenen
Markierungsexperimenten (Zhang, 1991; Zhang & Hoffmann, 1993) zu erwarten. Denn diese
Experimente zeigen eine beträchtliche räumliche Überlappung von retinalen und kortikalen
Terminalien innerhalb des NOT.
Die 22 Latenzen der von der IO identifizierten NOT-Zellen des Albinofrettchens befinden
sich mit einem Median von 6,0 ms (Q0,25 = 5,0 ms; Q0,75 = 6,5 ms) zwischen 5,0 ms (kürzeste
Latenz) und 8,0 ms (längste Latenz). In Abbildung 39A sind sie durch die Schraffur hervor-
gehoben. Die entsprechenden Latenzen von richtungsspezifischen NOT-Zellen des
pigmentierten Frettchens liegen im Bereich 5,5 bis 8,5 ms (Abb. 39C mit N = 15,
entnommen aus Klauer et al., 1990). Ein Vergleich der 22 Latenzen des Albinofrettchens mit
den 15 Latenzen des pigmentierten Frettchens zeigt keinen signifikanten Unterschied
zwischen den beiden Verteilungen an (χ2-TEST, P = 0,248).
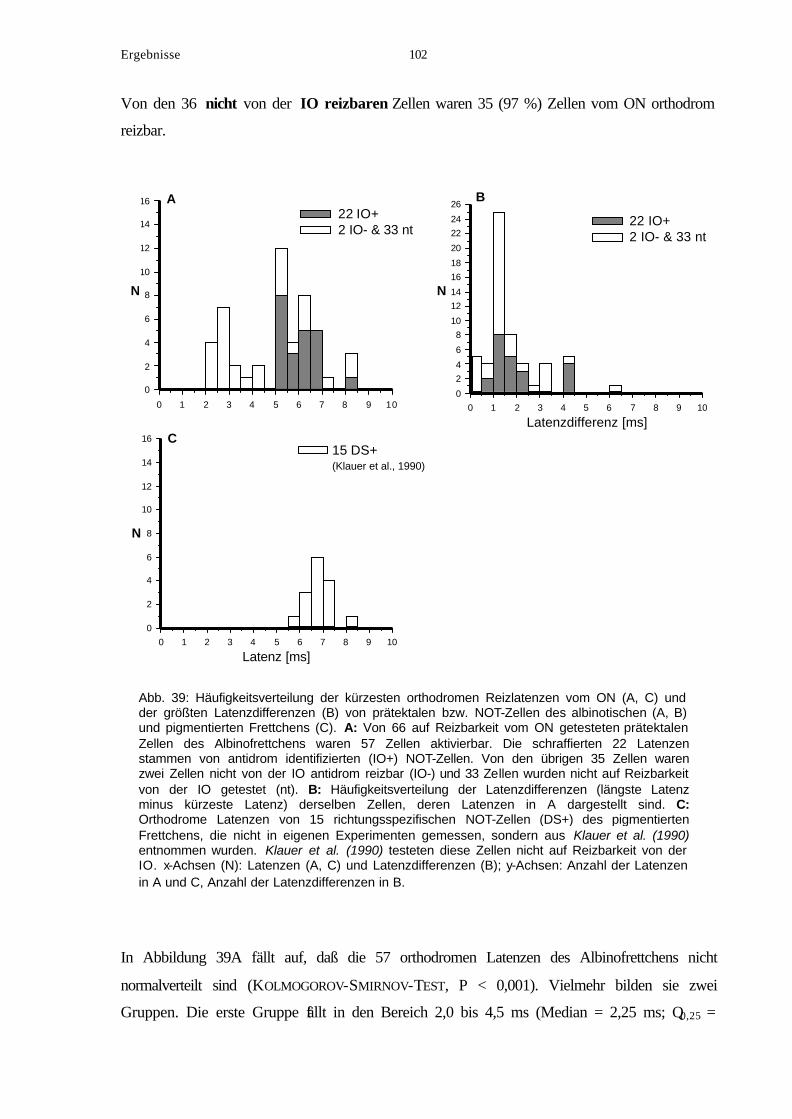
Ergebnisse 102
Von den 36 nicht von der IO reizbaren Zellen waren 35 (97 %) Zellen vom ON orthodrom
reizbar.
In Abbildung 39A fällt auf, daß die 57 orthodromen Latenzen des Albinofrettchens nicht
normalverteilt sind (KOLMOGOROV-SMIRNOV-TEST, P < 0,001). Vielmehr bilden sie zwei
Gruppen. Die erste Gruppe fällt in den Bereich 2,0 bis 4,5 ms (Median = 2,25 ms; Q0,25 =
Abb. 39: Häufigkeitsverteilung der kürzesten orthodromen Reizlatenzen vom ON (A, C) und der größten Latenzdifferenzen (B) von prätektalen bzw. NOT-Zellen des albinotischen (A, B) und pigmentierten Frettchens (C). A: Von 66 auf Reizbarkeit vom ON getesteten prätektalen Zellen des Albinofrettchens waren 57 Zellen aktivierbar. Die schraffierten 22 Latenzen stammen von antidrom identifizierten (IO+) NOT-Zellen. Von den übrigen 35 Zellen waren zwei Zellen nicht von der IO antidrom reizbar (IO-) und 33 Zellen wurden nicht auf Reizbarkeit von der IO getestet (nt). B: Häufigkeitsverteilung der Latenzdifferenzen (längste Latenz minus kürzeste Latenz) derselben Zellen, deren Latenzen in A dargestellt sind. C:Orthodrome Latenzen von 15 richtungsspezifischen NOT-Zellen (DS+) des pigmentierten Frettchens, die nicht in eigenen Experimenten gemessen, sondern aus Klauer et al. (1990)entnommen wurden. Klauer et al. (1990) testeten diese Zellen nicht auf Reizbarkeit von der IO. x-Achsen (N): Latenzen (A, C) und Latenzdifferenzen (B); y-Achsen: Anzahl der Latenzen in A und C, Anzahl der Latenzdifferenzen in B.
Latenz [ms]0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
1615 DS+ (Klauer et al., 1990)
0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
14
1622 IO+2 IO- & 33 nt
C
A
Latenzdifferenz [ms]0 1 2 3 4 5 6 7 8 9 10
0
24
6
810
1214
1618
20
2224
26
22 IO+ 2 IO- & 33 nt
B
N
N N

Ergebnisse 103
1,80 ms; Q0,75 = 2,50 ms) und die zweite Gruppe in den Bereich 5,0 bis 8,5 ms (Median =
6,00 ms; Q0,25 = 5,00 ms; Q0,75 = 6,50 ms). Die in Abbildung 38 gezeigten Spuren der
orthodromen Aktionspontentiale zweier prätektaler Zellen des Albinofrettchnes zeigen
Beispiele für die erste (A) und zweite (B) Latenzgruppe. Die zweite Gruppe wird
hauptsächlich von den erwähnten Latenzen der zur IO projizierenden NOT-Zellen
(schraffierte Latenzen) und einiger prätektaler Zellen gebildet, die entweder nicht zur IO
projizieren oder gar nicht auf Reizbarkeit von der IO getestet wurden. Die Gruppenbildung
läßt die Schlußfolgerung zu, daß es offensichtlich zwei Typen von retinalen
Ganglienzellfasern mit unterschiedlicher Leitungsgeschwindigkeit gibt, die auf prätektale
Neuronen projizieren. Dabei werden die zur IO projizierenden NOT-Zellen nur von Fasern
mit der niedrigeren Leitungsgeschwindigkeit kontaktiert.
Bei der Gruppe der kürzeren Latenzen fällt auf, daß keine einzige Latenz schraffiert ist
(Abb. 39A). Dies bedeutet, daß diese Latenzen an prätektalen Zellen gemessen wurden, die
nicht von der IO antidrom reizbar waren. Hieraus wird offensichtlich, daß der zweite
Ganglienzell-Typ mit den dickeren und deshalb schneller leitenden Axonen nicht auf NOT-
Zellen projiziert, die ihrerseits die IO innervieren.
In Abbildung 39B wird die Häufigkeitsverteilung der Latenzdifferenzen (längste Latenz
minus kürzeste Latenz) derselben Zellen dargestellt. So vermittelt sie einen Eindruck von der
zeitlichen Streuung der Aktionspotentiale der Mehrfachantworten der prätektalen und der
NOT-Zellen. Bei den meisten Zellen findet man eine Latenzdifferenz zwischen 1 und 1,5 ms
(Median 1,3 ms; Q0,25 = 1,0 ms; Q0,75 = 2,0 ms). Die Latenzdifferenzen der von der IO
antidrom identifizierten NOT-Zellen sind schraffiert hervorgehoben.

Diskussion 104
Diskussion
I. Teil I: Verhalten
A. Die Meßmethode
1. Wahl des EOGs und EOG-Auswertung
Die ursprüngliche Hypothese war, daß der monokulare hOKN des Albinofrettchens aufgrund
der Reduktion der ipsilateralen retinofugalen Projektionen asymmetrischer als beim
pigmentierten Frettchen ist. Diese Hypothese ging von der Annahme aus, daß das
Albinofrettchen einen deutlichen hOKN wie auch andere albinotische Säugerspezies machen
kann. Außerdem implizierte der Begriff „asymmetrisch“ einen quantitativen Unterschied.
Aufgrund der Erwartung, daß dieser Unterschied sehr augenfällig und deshalb auch ohne
exakte quantitative, d.h. ohne kalibrierte Augenbewegungsmessungen detektierbar sein
würde, fiel die Entscheidung auf das unkalibrierte EOG, zumal es relativ schnell und einfach
durchführbar ist. Es stellte sich aber sehr schnell heraus, daß Albinofrettchen überhaupt
keinen hOKN zeigen. Von diesem Augenblick an änderte sich zwangsläufig die Intention der
EOG-Messungen. Der Zweck war nicht mehr, eine Asymmetrie des monokularen hOKN zu
beobachten, sondern vielmehr die Auslösbarkeit des hOKN bei den Albinofrettchen zu
testen. Dieser Zweck konnte durch das unkalibrierte EOG erst recht schnell und einfach
erfüllt werden, weshalb die Meßmethode beibehalten wurde.
Zu den allgemeinen Problemen des EOGs (Nicht-Linearität und natürliche Fluktuationen des
zu messenden Potentials, Rauschen und Drift, vgl. Collewijn, 1999) kommt in der
vorliegenden Arbeit hinzu, daß die EOG-Elektroden nicht bei allen Tieren exakt an der
gleichen relativen Position um die Augen plaziert werden konnten. Auch das macht einen
quantitativen Vergleich zwischen den EOG-Spuren zweier Tiere unzulässig. Ein solcher
Vergleich ist selbst zwischen den EOG-Spuren eines Tieres ungenau, wenn sie in
verschiedenen Sitzungen (d.h. an verschiedenen Tagen) registriert wurden, denn die EOG-

Diskussion 105
Elektroden waren nicht permanent implantiert und mussten vor jeder Sitzung neu
eingestochen werden.
Im Bewußtsein der Problematik des unkalibrierten EOG wurde der Entschluß gefaßt, auch
pigmentierte Individuen zu testen, obwohl bekannt war, daß sie sehr wohl optokinetische
Augenbewegungen machen können (Hein et al., 1990). Durch diese Messungen wurden aber
EOG-Spuren von pigmentierten Frettchen mit derselben Methode aufgenommen wie die der
Albinofrettchen. Auf die gleiche Art und Weise konnte Hahnenberger (1977) einen
deutlichen Unterschied zwischen dem monokularen hOKN von albinotischen und dem von
pigmentierten Kaninchen dokumentieren. Dieser Unterschied äußerte sich darin, daß die
albinotischen Kaninchen monokular nur die Reizung in temporonasaler, aber überhaupt nicht
in nasotemporaler Richtung beantworteten. Die pigmentierten Kaninchen beantworteten
beide Richtungen mit unterschiedlicher Stärke. Bei den Frettchen ist der Unterschied
zwischen der albinotischen und der pigmentierten Gruppe noch augenfälliger, denn bei
Albinos bleibt der hOKN komplett aus.
2. Sensitivität des unkalibrierten EOGs
Es hat sich gezeigt, daß das eingesetzte EOG trotz fehlender Kalibrierung in der Lage ist,
auch sehr feine qualitative Unterschiede zu detektieren, wenn genügend statistisch auswert-
bares Datenmaterial zur Verfügung steht. Dies wird durch die Feststellung der okulomotori-
schen Instabilität der Albinofrettchen dokumentiert, die möglich ist, obwohl es für die
Frettchen aufgrund ihres Verhaltenshabitus unmöglich ist, ihre Augen völlig ruhig zu halten.
Infolge dessen liefern sie in den allermeisten Fällen kein völlig konstantes EOG-Potential,
was zu einer gewissen Streuung der Steigungsmediane um den Wert Null führt (vgl. Abb. 6).
Trotz dieser Streuung, die sowohl bei pigmentierten als auch bei albinotischen Tieren
unvermeidbar ist, konnte der signifikante Unterschied in der Varianzgröße festgestellt
werden und damit die okulomotorische Instabilität der Albinos belegt werden.
Darüberhinaus zeigen die Augenbewegungsmessungen der pigmentierten Frettchen, daß das
unkalibrierte EOG unterschiedliche Antwortstärken relativ zueinander in Beziehung setzen
und damit detektieren kann. Dies ist durch die Abhängigkeit der Steigungsmediane von der
Reizgeschwindigkeit in Abbildung 13 dokumentiert. Über die absolute Folgegüte des hOKN
kann aufgrund der Meßmethode keine Aussage gemacht werden. Die Reizgeschwindigkeit
von 50°/s lieferte im Falle der pigmentierten Tiere in der Regel EOG-Spuren mit einem
höheren Steigungsmedian als die Reizgeschwindigkeit von 5°/s, d.h. die Reizgeschwindig-
keit von 50°/s löste — in Übereinstimmung mit Hein et al. (1990) — eine höhere

Diskussion 106
Augengeschwindigkeit aus. Aus Hein et al. (1990) wissen wir aber auch, daß die absolute
Folgegüte des hOKN bei 5°/s Reizgeschwindigkeit wesentlich höher ist als bei 100°/s.
B. Augendrift oder Spontannystagmus beim Albinofrettchen?
Bei albinotischen Formen der Spezies Kaninchen (Collewijn et al., 1978), Ratte (Precht &
Cazin, 1979) und Mensch (Collewijn et al., 1985) wird ein Spontannystagmus beobachtet.
Dieser Spontannystagmus ist aus langsamen Folgephasen und schnellen Rückstellsakkaden
zusammengesetzt, deren Amplitude und Frequenz so hoch sind, daß seine EOG-Spuren eine
— wenn auch ziemlich ungeregelmäßig — gezackte (Sägezahn-)Form haben.
Die Untersuchung der spontanen Augenbewegungen des Frettchens (vgl. Abb. 6) hat bei den
Albinofrettchen ebenfalls eine okulomotorische Instabilität sichtbar werden lassen. Die
Detektion dieser Instabilität war nur durch einen statistischen Vergleich mit den entsprechen-
den Daten vom pigmentierten Frettchen möglich. Denn die „motorische Unruhe“ der Augen
des Albinofrettchens war in ihrer Amplitude Geschwindigkeit und Frequenz so gering, daß
sie mit bloßem Auge nicht am Verlauf der EOG-Spuren erkannt werden konnte. Optisch läßt
sich am Verlauf der EOG-Spuren sicherlich kein Unterschied zwischen dem albinotischen
und dem pigmentierten Frettchen feststellen. Ein weiterer Unterschied zum oben erwähnten
Spontannystagmus der anderen albinotischen Spezies ist, daß an der okulomotorischen
Instabilität der Albinofrettchen keine unterschiedlichen Phasen wie langsame Folgephasen
und schnelle Rückstellsakkaden unterschieden werden können. Aus diesem Grunde wurde
bei der Darstellung dieses Befundes im Ergebnisteil auf den Begriff „Spontannystagmus“
verzichtet und lediglich von einem signifikant stärkeren Abdriften der Augen beim
Albinofrettchen gesprochen.
Die Augendrift des Albinofrettchens führt zu folgender Frage: Wie sind die Informations-
signale beschaffen, die einerseits vom gestörten temporalen Retinabereich (siehe Ergebnis
des Perimetrietests) und andererseits von der vermeintlich intakten nasalen Hemiretina
gleichzeitig denselben kontralateralen NOT erreichen.
C. Optokinetische Blindheit des Albinofrettchens
Während die pigmentierten Frettchen bei optokinetischer Ganzfeldreizung einen eindeutigen
hOKN generieren, der schon in der Publikation von Hein et al. (1990) detaillierter und
quantitativ beschrieben ist, zeigten Albinofrettchen unter den gleichen Reizbedingungen und
Lichtverhältnissen kein als hOKN definierbares Verhalten. Das Kriterium für ein „als hOKN

Diskussion 107
definierbares Verhalten“ war eine reizabhängige Sägezahn-förmige EOG-Spur (bestehend
aus langsamen Folgephasen und schnellen Rückstellsakkaden) mit einem Steigungsmedian,
der wesentlich von Null abweicht, so daß sich auch die Gesamtheit der Steigungsmediane
aus den EOG-Spuren der getesteten Tiere statistisch signifikant von Null unterscheidet. Die
reizabhängige Augendrift der Albinofrettchen, die nur in den binokularen hOKN-Tests mit
dem groben Reizmuster beobachtet wurde und bei den Reizgeschwindigkeiten 10, 20 und
100°/s signifikant war (siehe Abb. 11B), kann nicht als OKN definiert werden. Auch wenn
man sie als eine langsame Folgephase mit einer extrem geringen Folgegüte verstehen will,
muß man feststellen, daß sie nicht durch schnelle Rückstellsakkaden in entgegensetzter
Richtung unterbrochen wird. Folglich erfüllt das Albinofrettchen das obige Kriterium nicht
und wird aus diesem Grunde als „optokinetisch blind“ bezeichnet.
Die unterschiedlichen Reizwirkungen der beiden Reizmuster zeigen, daß das
okulomotorische Verhalten stark von den Reizparametern abhängt. Deshalb sollen zunächst
die Reizparameter in Bezug auf ihre Effektivität diskutiert werden.
1. Reizparameter
Die optokinetische Blindheit des Albinofrettchens erinnert an das Ergebnis von Precht &
Cazin (1979), die ebenfalls einen kompletten Ausfall des hOKN bei der Albinoratte (Wistar)
feststellten. Die optokinetische Ganzfeldreizung hatten sie damals mit einem Muster aus
senkrechten, alternierenden schwarzen und weißen Streifen durchgeführt. Ihre Feststellung
wurde jedoch sechs Jahre später von der gleichen Forschergruppe revidiert (Sirkin et al.,
1985). Die Arbeitsgruppe berichtete nun, daß Ratten des selben albinotischen Stammes
(Wistar) wie auch Albinoratten des Stammes Sprague-Dawley doch einen hOKN machten,
wenn sie mit einem projizierten Lichtpunktemuster (Größe der Lichtpunkte: ca. 10°
Sehwinkel) gereizt würden. In diesem Falle zeigten die Albinoratten im Vergleich zu den
pigmentierten Ratten zwar eine abnorme, doch eindeutige optokinetische Reaktion. Insofern
konnte nicht mehr von einem kompletten Ausfall des hOKN bei Albinoratten die Rede sein.
Sirkin et al. (1985) stellten ebenfalls fest, daß die Auslösbarkeit des hOKN bei ihren
Albinoratten extrem abhängig war vom projizierten Lichtpunktemuster. Denn bei Reizung
mit einem Muster aus gemalten schwarzen Punkten (Punktgröße: 3,6° Sehwinkel) auf
weißem Hintergrund blieb der hOKN bei 13 von 14 Albinoratten im Gegensatz zu den
pigmentierten Ratten komplett aus. Die eine Albinoratte, die die Ausnahme darstellte,
reagierte auf das gemalte Muster erst, nachdem die Lichtintensität herabgesetzt worden war
(siehe unten).

Diskussion 108
Deshalb muß die Frage gestellt werden, ob in der vorliegenden Arbeit die adäquaten
Reizmuster und Lichtverhältnisse gewählt wurden, bzw. ob die Nichtauslösbarkeit des
hOKN auf eine uneffektive Reizung beruhte. Diesbezüglich ist folgendes anzumerken:
• Das uneffektive Reizmuster von Precht & Cazin (1979) war ein vertikal gestreiftes
schwarz-weißes Muster. Im Gegensatz dazu wurden die Frettchen mit Punktemustern
gereizt, die sich bei Sirkin et al. (1985) insgesamt als effektivere Reizmuster
herausgestellt hatten. Insofern sind die in der vorliegenden Arbeit eingesetzten
Reizmuster eher mit den effektiven Reizmustern von Sirkin et al. (1985) vergleichbar.
• Hinsichtlich der Punktgröße unterscheiden sich die in der vorliegenden Arbeit
eingesetzten schwarz-weißen Zufallspunktemuster nicht wesentlich von den beiden
Mustern (Lichtpunktemuster und Muster aus gemalten schwarzen Punkten) von Sirkin et
al. (1985). Die Punkte des groben Zufallspunktemusters (ca. 10,7° Sehwinkel) haben die
gleiche Größe wie die Lichtpunkte des projizierten Musters (ca. 10° Sehwinkel) von
Sirkin et al. (1985). Das feine Zufallspunktemuster (Punktgröße: ca. 2,3° Sehwinkel) ist
mit dem (weniger effektiven) gemalten Punktmuster von Sirkin et al. (1985)
(Punktgröße: ca. 3,6° Sehwinkel) ebenfalls vergleichbar.
• Ein Unterschied zwischen dem groben Reizmuster und dem sehr effektiven Lichtpunkte-
muster von Sirkin et al. (1985) besteht darin, daß das grobe Muster gemalt war und nicht
aus Lichtpunkten bestand. Dieser Unterschied scheint besonders wichtig zu sein, da erst
die Qualität „Licht“punkte das projizierte Muster von Sirkin et al. (1985) zu einem
effektiven Reizmuster machte. Insofern könnte beanstandet werden, daß die optokineti-
sche Blindheit der Albinofrettchen beobachtet wurde, weil die beiden Reizmuster nicht
aus Lichtpunkten bestanden. Um diesem Einwand vorzubeugen, wurden einige
Albinofrettchen auch mit dem Planetarium gereizt. Die optokinetische Reizung mit dem
Planetarium konnte bei den Albinofrettchen, im Gegensatz zu den pigmentierten
Frettchen, ebenfalls keinen hOKN auslösen (in der vorliegenden Arbeit nicht dargestellt).
Dies spricht dafür, daß die Ursache des hOKN-Ausfalls bei den Albinofrettchen nicht in
einer uneffektiven optokinetischen Reizung, sondern in einem neuronalen Defekt zu
suchen ist (siehe Teil II der vorliegenden Arbeit).
• Aus Sirkin et al. (1985) geht hervor, daß auch die Lichtverhältnisse während der opto-
kinetischen Reizung entscheidend für die Auslösbarkeit einer optokinetischen Reaktion
bei Albinoratten sind. Denn alle Albinoratten reagierten auf die Reizung mit dem
Lichtpunktemuster, wenn die mittlere Lichtmenge 3,5 Lumen/Fuß2 (= 37,66 Lumen/m2)

Diskussion 109
betrug. Das auf weißem Hintergrund gemalte Reizmuster aus schwarzen Punkten war bei
gleicher Beleuchtung bei keinem der Tiere effektiv. Dieses gemalte Muster wurde aber
bei einer Albinoratte zu einem effektiven Reizmuster, wenn die Beleuchtung auf ca. 2
Lumen/Fuß2 herabgesetzt wurde. Insofern muß auch die Frage gestellt werden, ob die
Beleuchtung der Albinofrettchen in der optokinetischen Trommel nicht zu stark war. Zur
Zeit kann jedoch kein quantitativer Vergleich zwischen den Lichtverhältnissen bei Sirkin
et al. (1985) und den Lichtverhältnissen bei den Frettchen gemacht werden. Der Grund
hierfür ist, daß die Lichtmengen mit unterschiedlichen Methoden gemessen wurden und
Sirkin et al. (1985) nicht alle Angaben machen, die für eine Umrechnung der einen
Einheit in die andere notwendig sind. Das eigene Meßgerät (Minolta luminance meter)
maß die Leuchtdichte der Musteroberfläche in der Einheit Candela/Fläche, während
Sirkin et al. (1985) die Beleuchtungsstärke in der Einheit Lumen/Fläche angeben.
2. Optokinetische Augenbewegungen bei hypopigmentierten Säugern
Bei den meisten bis jetzt untersuchten albinotischen Säugern lösen Ganzfeldreizungen im
Gegensatz zu den hier untersuchten Albinofrettchen optokinetische Augenbewegungen aus
(Kaninchen: Hahnenberger, 1977; Collewijn, 1978; Ratte: Precht & Cazin, 1979; Lannou et
al., 1982; Sirkin et al., 1985; Mensch: Collewijn et al., 1985). Diese Augenbewegungen sind
zwar im Vergleich zum hOKN der pigmentierten Individuen derselben Art abnorm, können
aber eindeutig als hOKN identifiziert werden.
Ein kompletter hOKN-Ausfall bei optokinetischer Ganzfeldreizung wird in der Literatur nur
von einigen hypopigmentierten Mausmutanten (z.B. die Mutanten albino (C57BL/6J c2J/c2J),
himalayan, pearl und maroon) berichtet (Balkema et al., 1981, 1984; Mangini et al., 1985).
Die Autoren, die aus derselben Arbeitsgruppe stammen, schreiben, daß sie die Mäuse mit
einem Muster aus senkrechten, alternierenden schwarzen und weißen Streifen reizten. Ob
sich ein Muster aus projizierten Lichtpunkten, wie bei den Ratten von Sirkin et al. (1985), als
effektiver herausgestellt hätte, kann hier nicht entschieden werden. Es ist jedoch interessant,
daß von diesen Mausmutanten nur ein Genotyp als albinotisch bezeichnet wird, während die
übrigen Mutanten nicht an völliger Pigmentlosigkeit leiden.
Dieser komplette Ausfall des hOKN bei den hypopigmentierten Mausmutanten, der während
der Ganzfeldstimulation beobachtet wird, tritt nicht auf, wenn nur bestimmte Gesichtsfeld-
bereiche bzw. Retinabereiche optokinetisch stimuliert werden (Teilfeldreizung, restricted
field stimulation). Als Mangini et al. (1985) ihr optokinetisches Reizmuster, mit dem sie bei
der Ganzfeldstimulation überhaupt keine optokinetischen Antworten auslösen konnten, nur

Diskussion 110
im posterioren oder nur im anterioren Gesichtsfeld derselben Mausmutanten bewegten,
konnten sie auf einmal doch optokinetische Reaktionen hervorrufen. Bewegte sich das
Reizmuster nur im posterioren Gesichtsfeld (Reizung der nasalen Retinae), so zeigten die
meisten Mausmutanten einen hOKN, der hinsichtlich seiner Richtung normal war. Wurden
hingegen nur die temporalen Retinae gereizt (Stimulusbewegung nur im anterioren
Gesichtsfeld), so zeigten die meisten Tiere einen invertierten hOKN, d.h. einen hOKN mit
langsamen Phasen gegen die Bewegungsrichtung des Reizmusters. Das gleiche Verhalten
löste die Teilfeldreizung auch bei der Albinoratte (Sirkin et al., 1985) und beim
Albinokaninchen (Collewijn et al., 1978) aus.
Den invertierten hOKN des Albinokaninchens versuchten Winterson & Collewijn (1981) mit
einem Richtungssignal zu begründen, welches von der temporalen Retina anstatt zum ipsi-
lateralen fälschlicherweise zum kontralateralen NOT geleitet werde. Infolge dessen sei für
den NOT dieses Signal ein sog. „invertiertes“ Richtungssignal. Diese Hypothese stellten die
Autoren jedoch unter den beiden Annahmen auf, daß beim pigmentierten Kaninchen erstens
eine retinale Projektion zum ipsilateralen NOT existiert, die infolge der Albinomutation
zugunsten der kontralateralen NOT-Afferenz reduziert ist, und zweitens daß die normale
Vorzugsrichtung der retinalen Ganglienzellen weder verloren geht noch verändert wird.
Unter denselben Annahmen könnte man die gleiche Hypothese auch für den invertierten
hOKN der hypopigmentierten Mausmutanten von Mangini et al. (1985) aufstellen.
Allerdings scheint heute diese Erklärung eher unwahrscheinlich zu sein, denn bis dato ist
eine retinale Projektion zum ipsilateralen NOT weder beim pigmentierten Kaninchen
(Scalia, 1972; Klooster et al., 1983; indirekt auch bei Oyster et al., 1980; Pu & Amthor,
1990) noch bei den pigmentierten Wildtyp-Mäusen (Scalia, 1972; Pak et al., 1987) eindeutig
nachgewiesen. Der Literatur kann entnommen werden, daß sich der anatomische Nachweis
einer ipsilateralen retinoprätektalen Projektion bei Afoveaten — im Gegensatz zu einer
ipsilateralen retinogenikulären Projektion — als besonders schwierig erweist, wie sich bei
der Ratte herausgestellt hat. Der Grund dafür ist wahrscheinlich, daß eine solche Verbindung
— wenn überhaupt existent — viel schwächer ist als bei kortikal höher entwickelten Spezies
wie Frettchen, Katze und Primaten. Die ersten Arbeiten an der Ratte konnten nicht die
geringste Spur einer solchen Projektion nachweisen (z. B. Scalia, 1972; Scalia & Arango,
1979; Perry & Cowey, 1979). Erst später entwickelte, sensitivere Nachweismethoden
konnten die schwache retinoprätektale Projektion bei der Ratte nachweisen (Aarnoutse et al.,
1995; Okada et al., 1991). Angesichts dessen kann die Feststellung, daß der NOT des
Kaninchens und der Maus keine ipsilaterale retinale Afferenz besitzt, immer noch als

Diskussion 111
vorläufig betrachtet werden. Für die Maus jedenfalls könnte man eine schwache retinale
Projektion zum ipsilateralen NOT erwarten, zumal bei ihr eine ipsilaterale retinogenikulo-
kortikale Verbindung nachgewiesen ist, die sogar bei den albinotischen Individuen
hinsichtlich ihrer Stärke reduziert ist (Dräger & Olsen, 1980). Im übrigen handelt es sich bei
den Albinomäusen von Dräger & Olsen (1980) um denselben Albinostamm (C57BL/6J
c2J/c2J), der von Mangini et al. (1985) hinsichtlich der Auslösbarkeit seines hOKN getestet
wurde. Diese Albinomäuse zeigten bei Ganzfeldreizung überhaupt keinen und bei
Teilfeldreizung in Abhängigkeit von den gereizten Retinabereichen einen hinsichtlich seiner
Richtung normalen oder invertierten hOKN (siehe oben).
Für die Albinoratte könnte die oben erwähnte Hypothese zur Begründung des invertierten
Nystagmus relevant sein, unter der Annahme, daß die von der ventrotemporalen Retina
ausgehenden Axone, die bei den pigmentierten Individuen den ipsilateralen NOT innervieren
(Okada et al., 1991), bei den Albinos tatsächlich zum kontralateralen NOT ziehen. Die
Reduktion dieser ipsilateralen Retina-NOT-Verbindung ist allerdings bei Albinoratten noch
nicht nachgewiesen, im Gegensatz zur Reduktion der ipsilateralen retinogenikulären
Projektion (Dreher et al., 1985).
Die in der vorliegenden Arbeit untersuchten Albinofrettchen wurden nur Ganzfeldreizungen
unterzogen. Über das mögliche Resultat einer Teilfeldreizung wird weiter unten bei der
Diskussion des Perimetrietests spekuliert.
D. Muscimol-Experimente
Nach der Entdeckung der optokinetischen Blindheit der Albinofrettchen stellte sich die
Frage, wie das neuronale Korrelat dieser Verhaltensstörung innerhalb des neuronalen
Schaltkreises des hOKN lokalisiert bzw. eingegrenzt werden konnte. Hierzu wurde als erste
Experimentenreihe die NOT-Inaktivierungsstudie durchgeführt.
1. Die Injektionsmethode
Unmittelbar nach einer Muscimol-Injektion trat bei dem einen pigmentierten und bei drei
von vier albinotischen Frettchen tatsächlich ein Spontannystagmus auf, dessen langsame
Phase wie auch bei der Katze (Fischer et al., 1998) und beim Affen (z.B. Cohen et al., 1992;
Yakushin et al., 2000) auf die nicht-injizierte Seite hin gerichtet war. Für das pigmentierte
Frettchen wurde das Auftreten dieses Spontannystagmus erwartet, für die albinotischen Tiere

Diskussion 112
war er jedoch angesichts der Anomalien ihres visuellen und speziell ihres optokinetischen
Systems nicht unbedingt voraussagbar.
Das Auftreten des Spontannystagmus zeigt, daß das Muscimol auch beim Frettchen eine
ähnliche Wirkung wie bei der Katze und beim Affen hat, d.h. ein Aktivitätsungleichgewicht
zwischen dem injizierten und dem nicht-injizierten NOT erzwingt.
Der Befund, daß der Spontannystagmus schon zwei Minuten (wahrscheinlich sogar früher)
nach einer Muscimol-Injektion (1 µl einer 0,1 % Muscimol-Lösung mit einer Rate von ca.
0,5 µl/min) auftrat, spricht zum einen für eine hohe Muscimol-Sensitivität des Frettchen-
NOT, zum anderen für die richtige Plazierung der Injektionen und weiterhin für eine
ausreichende Muscimol-Menge. Fischer et al. (1998) injizierten bei der Katze mit der
gleichen Injektionsmethode ebenfalls 1 µl einer 0,1%-Muscimol-Lösung, allerdings mit einer
Rate von 0,2 µl/min. Ihre niedrigere Injektionsrate ist wahrscheinlich ein Grund dafür, daß
sie den Spontannystagmus meistens erst nach fünf Minuten sehen konnten. In diesem
Zusammenhang muß berücksichtigt werden, daß die absolute Menge zwar gleich, aber die
relative Muscimol-Menge bei der Katze kleiner als beim Frettchen war, denn das Prätektum
der Katze ist größer als das des Frettchens. Dies ist auch eine mögliche Begründung für den
Befund, wonach der induzierte Spontannystagmus bei den Frettchen sowohl in Helligkeit als
auch in Dunkelheit, bei der Katze aber nur in Dunkelheit auftrat. Offensichtlich wurde mit
der gleichen Muscimol-Menge bei den Frettchen eine stärkere Inhibierung des NOT erzielt
als bei der Katze von Fischer et al. (1998). Einen Spontannystagmus in Helligkeit konnten
die Autoren erst mit einer wesentlich größeren Muscimol-Menge auslösen. Für ihre
Fragestellung war aber ein Spontannystagmus in Helligkeit unerwünscht, so daß sie in ihrer
Publikation nur vom Effekt der 1 µl-Muscimol-Injektionen berichten (mündliche Mitteilung
von Fischer et al.,1998).
Andere eher unwahrscheinliche Begründungen des Auftretens des Spontannystagmus sowohl
in Helligkeit als auch in Dunkelheit wären, daß der NOT des Frettchens sensitiver für
Muscimol ist als der NOT der Katze und des Affen und / oder das Frettchen im Gegensatz zu
der Katze und zum Affen keine, wie auch immer beschaffenen Mechanismen besitzt, die den
Spontannystagmus in Helligkeit durch Fixieren von visuellen Reizen unterdrücken können.
Beim Affen z.B. setzen solche Mechanismen auch nach einseitiger Läsion des NOT ein, so
daß die Tiere nur in Dunkelheit einen Spontannystagmus machen (Kato et al., 1986). Dieser
durch die einseitige Läsion des NOT ausgelöste Spontannystagmus hat eine langsame Phase
in Richtung auf die intakte, nicht lädierte Seite hin, die im Falle der Frettchen der nicht-

Diskussion 113
injizierten Hirnseite entspricht. Funktional gesehen entspricht die reversible Inaktivierung
des NOT durch Muscimol einer einseitigen NOT-Läsion.
Um den Effekt einer Muscimol-Injektion ganz sicher reversibel zu halten, war es wichtig,
den NOT der injizierten Seite auf keinen Fall durch das Führungsröhrchen oder durch den
Injektionsdruck mechanisch zu beeinträchtigen. Deshalb sollte das Muscimol nicht „in“ den
NOT, sondern „auf“ den NOT bzw. auf die unmittelbar benachbarte prätektale Oberfläche
injiziert werden. Aus der Position der Spitze der implantierten Injektionskanülen bzw. der
daraus abgeleiteten Injektionsorte konnte bewiesen werden, daß dieses Vorhaben gelungen
ist. Denn es wurde bei keinem der implantierten Tiere eine mechanische Beschädigung des
NOT festgestellt. Dieser anatomische Befund wird durch die vollständige Reversibilität des
Spontannystagmus untermauert.
2. Erste Eingrenzung des okulomotorischen Defektes des Albinofrettchens
Das Resultat der Muscimol-Injektionen erlaubt die erste Eingrenzung des okulomotorischen
Defektes des Albinofrettchens. Denn das Auftreten des Spontannystagmus zeigt, daß das
Albinofrettchen einen dem NOT nachgeschalteten visuomotorischen neuronalen Schaltkreis
besitzt, der seine Funktion für die Generierung eines — wenn auch in diesem Falle künstlich
erzeugten spontanen — Nystagmus wahrnehmen kann. Hieraus wird die Schlußfolgerung
gezogen, daß der gesuchte Defekt nicht dem NOT nachgeschaltet ist. Folglich muß der
Defekt im NOT selbst und / oder in dem dem NOT vorgeschalteten, sensorischen Teil des
neuronalen hOKN-Schaltkreises liegen.
E. Perimetrietest
1. Annahmen des Perimetrietests
Der Perimentrietest macht zwei diskussionswürdige Annahmen (vgl. Shermann, 1973).
Die erste Annahme ist, daß das Frettchen tatsächlich den Fixationspunkt F fixiert, wenn der
visuelle Reiz präsentiert wird. Andernfalls entspricht die Sektoreneinteilung des Tisches
nicht den richtigen, d.h. entsprechenden Gesichtsfeldsektoren des Tieres. Es kann nicht mit
Sicherheit gesagt werden, daß diese Annahme zu hundert Prozent gerechtfertigt und richtig
ist. Vielmehr muß — angesichts der Tatsache, daß die Frettchen recht quirlige und agile
Tiere sind und es deshalb oft schwierig war, sie ruhig festzuhalten — unterstellt werden, daß
der Fixationspunkt F nicht in allen Durchläufen genau fixiert wurde. Deshalb muß eine

Diskussion 114
gewisse Streuung der Blickachse um den Fixationspunkt F angenommen werden. Dies gilt
insbesondere für den Teil der Messungen ohne Plexiglasröhre (vgl. Abb. 4A), in denen die
Versuchstiere vor dem Start am Punkt X vom Experimentator mit der Hand festgehalten
wurden. Dennoch lieferten diese Messungen das gleiche Ergebnis wie die Messungen mit
Plexiglasröhre, für die diese Annahme weniger Relevanz hat. Denn im Falle der Röhre
waren die Tiere völlig ungehindert, kamen sehr schnell aus der Röhre und liefen geradeaus
zum Fixationspunkt F. Folglich sollten sie in diesem Falle eher den Fixationspunkt F fixiert
haben.
Die zweite Annahme ist, daß der in das Gesichtsfeld des Tieres eingeführte Reiz tatsächlich
nur ein visueller Reiz war, d.h. daß das Tier keinen anderen Hinweis hatte, ob und — wenn
`ja´ — wo der Reiz gezeigt werden würde. Solche Hinweise könnten z.B. sein, daß der
Experimentator unbewußt die Tiere unterschiedlich am Startpunkt X festhielt, abhängig
davon, ob er beabsichtigte den Reiz zu präsentieren oder nicht. Ein weiterer Hinweis wäre
die Luftbewegung, die durch die Bewegung des Reizes samt Stab verursacht wurde.
Die Richtigkeit der oben gemachten Annahmen kann nicht mit absoluter Gewißheit
begründet werden, zumal der visuelle Reiz unter monokularen Bedigungen in einigen Fällen
auch in Sektoren gesehen wurde, in denen er theoretisch nicht hätte gesehen werden sollen.
Da wären beispielsweise die wenigen positiv beantworteten Durchläufe im 90°-Sektor
kontralateral zum sehenden Auge (vgl. Tab. 3 und 4), obwohl sie aufgrund der
Augenstellung des Frettchens ganz sicher negativ hätten ausfallen sollen. Solche dennoch
positiv beantwortete Durchläufe können nur erklärt werden, wenn man unterstellt, daß
mindestens eine der oben gemachten Annahmen in diesen Fällen nicht zutraf.
Diese methodische Ungenauigkeit des Perimetrietests kann jedoch im Rahmen seines
Zweckes akzeptiert werden, vor allen Dingen weil die Befunde in Bezug auf die Größe des
binokularen Gesichtsfeldes des Frettchens ziemlich gut den Erwartungen entsprechen. Denn
würde die Blickachse der Versuchstiere um größere Beträge vom Fixationspunkt F
abweichen oder hätten die Frettchen aufgrund der Luftbewegung den Reiz lokalisieren
können, so müßten die monokularen Messungen im Falle der pigmentierten Frettchen ein
größeres binokulares Gesichtsfeld ermittelt haben. Tatsächlich wurde aber ein binokulares
Gesichtsfeld mit einer Azimut-Breite von ca. 70 - 80° Sehwinkel (jeweils 35 - 40° zu jeder
Gesichtsfeldhälfte hin) festgestellt, was ziemlich genau mit der ermittelten Größe aus
elektrophysiologischen Kartierungen des SC (80° Sehwinkel nach Quevedo et al., 1996) und
des primären visuellen Kortex (76° Sehwinkel im superioren Gesichtsfeld nach Law et al.,
1988) übereinstimmt.

Diskussion 115
2. Getestetes Gesichtsfeld
Sämtliche Befunde und Schlußfolgerungen aus dem Perimetrietest beziehen sich
auschließlich auf die horizontale Ausdehnung des Gesichtsfeldes entlang des Azimuts. Der
durchgeführte Test untersucht nicht die vertikale Ausdehnung des Gesichtsfeldes, so daß
diesbezüglich keine Aussagen möglich sind.
3. Sehfähigkeit des Albinofrettchens unter binokularer Bedingung
Der Perimetrietest ist ein weiterer Schritt auf dem Weg zur näheren Eingrenzung des
okulomotorischen Verhaltensdefizites des Albinofrettchens. Die binokularen Messungen
zeigen, daß das Albinofrettchen in der Regel genau so gut wie das pigmentierte Frettchen in
der Lage ist, die Bewegung von kleinen visuellen Reizen in seinem Gesichtsfeld
wahrzunehmen und sich ihnen zuzuwenden. Dies spricht zunächst einmal dafür, daß die
üblichen durch den Albinismus bedingten Beeinträchtigungen des optischen Apparates (z.B.
Herabsetzung der Sehschärfe infolge des erhöhten Streulichtes im Augapfel) beim
Albinofrettchen weniger schwerwiegend sind, zumindest nicht so schlimm, daß dadurch
seine Fähigkeit zur visuellen Wahrnehmung und zu Hinwendereaktionen gestört ist. Deshalb
wird die Schlußfolgerung gezogen, daß seine optokinetische Blindheit nicht durch eventuelle
Unzulänglichkeiten seines optischen Apparates verursacht ist. Dies wiederum spricht dafür,
daß seine optokinetische Blindheit neuronal bedingt ist.
Die geringe Trefferquote, die beim Albinofrettchen # 39 im peripheren Gesichtsfeld des
rechten Auges erzielt wurde, kann nicht begründet werden, weil nicht entschieden werden
kann, ob dieses Verhaltensdefizit durch einen Defekt im optischen Apparat, oder durch einen
Defekt der nasalen Retina oder durch einen zentralnervös gelegenen Defekt verursacht ist.
Eine weitere Schlußfolgerung aus dem Ergebnis der binokularen Messungen ist, daß sich der
Albinismus des Frettchens — erstaunlicherweise trotz der Reduktion der ipsilateralen
retinocolliculären Projektion (Zhang & Hoffmann, 1993) — nicht auf die Retinotopie und
damit auch nicht auf die Funktionstüchtigkeit des SC unter binokularen Bedingungen
auswirkt, denn eine intakte Retinotopie im SC ist für ein visuelles Orientierungsverhalten
wie etwa die Hinwendereaktion auf den visuellen Reiz im Perimetrietest, unbedingt
erforderlich. Diese Schlußfolgerung berücksichtigt auch die Befunde von Quevedo et al.
(1996). Quevedo et al. (1996) fanden mit anatomischen und elektrophysiologischen
Methoden heraus, daß die retinotopische Organisation der oberen visuellen Schichten des SC
bei Albinofrettchen dieselben charakteristischen Merkmale aufweist wie auch bei normal-

Diskussion 116
pigmentierten Individuen. Die beiden Tiergruppen unterscheiden sich weder hinsichtlich der
Größe des im SC repräsentierten Gesichtsfeldes, noch hinsichtlich des Vergrößerungsfaktors
zwischen retinaler Peripherie und Area centralis. Damit liefert der Perimetrietest hinsichtlich
der Größe des Gesichtsfeldes das zu der Elektrophysiologie passende Verhalten. Die
Untersuchungen von Quevedo et al. (1996) wurden im selben Labor und mit Tieren aus
derselben Zucht durchgeführt wie die in der vorliegenden Arbeit.
4. Sehfähigkeit des Albinofrettchens unter monokularer Bedingung
Die monokularen Messungen des Perimetrietests zeigen, daß das Albinofrettchen im
Gegensatz zum pigmentierten Frettchen kein intaktes binokulares Gesichtsfeld besitzt. Über
die Schnauze hinweg ins zentrale aber kontralaterale Gesichtsfeld kann das Albinofrettchen
monokular nicht sehen. Die wahrscheinlichste Begründung dieser Einschränkung des
Gesichtsfeldes eines Auges folgt aus der Reduktion der ipsilateralen retinocolliculären (und
der retinogenikulokortikalen) Projektion (Zhang & Hoffmann, 1993).
Die Aktivierung des SC löst im Normalfall eine Hinwendereaktion nach kontralateral aus,
d.h. die Aktivierung des rechten (linken) SC führt zu einer Reaktion des Tieres nach links
(rechts). Beim Albinofrettchen ziehen die abnormerweise kreuzenden Fasern aus der
temporalen Retina zum kontralateralen, d.h. falschen SC. Wenn diese Fasern eine
funktionelle Projektion bilden und den falschen SC aktivieren würden, so müßten die
Albinos anstatt einer Hinwende- eine Abwendereaktion zeigen. Eine Abwendereaktion
wurde jedoch niemals beobachtet. Folglich muß die Information aus den abnormerweise
kreuzenden Fasern der temporalen Retina im kontralateralen SC funktionell unterdrückt sein.
Die Befunde der monokularen Perimetrietests stimmen mit den Resultaten aus
entsprechenden Perimetriemessungen bei Katzen (Elekessy et al., 1973) überein. Elekessy et
al. (1973) vermassen mit der gleichen Methode das binokulare und monokulare Gesichtsfeld
von Wildtyp- und siamesischen Katzen. Die siamesischen Katzen, die ein albinotisches Allel
besitzen und demnach als „halbe“ Albinos angesehen werden können, weisen wie die
Albinofrettchen eine Reduktion ihrer ipsilateralen retinofugalen Projektionen auf (Guillery,
1974; Stone et al., 1978a). Unter binokularen Bedingungen stellten Elekessy et al. (1973)
zunächst keinen Unterschied zwischen den normalen und den mutierten Katzen fest. Die
Unterschiede wurden erst unter monokularer Bedingung sichtbar, als sich zeigte, daß die

Diskussion 117
Siamesen — im Gegensatz zu den Wildtyp-Katzen — kein intaktes binokulares Gesichtsfeld
haben. Denn monokular konnten sie nicht ins kontralaterale Gesichtfeld sehen, im Gegensatz
zu den normalen Katzen, die mindestens bis 30° in die kontralaterale Gesichtsfeldhälfte
sehen konnten. Elekessy et al. (1973) führten als Ursache für die Suppression dieses
Gesichtsfeldanteils erwartungsgemäß den aberraten Verlauf der Ganglienzellfasern aus der
temporalen Retina auf. Allerdings ließen sie die Frage ungeklärt, warum der komplette
monokulare Gesichtsfeldanteil in der kontralateralen Gesichtsfeldhälfte unterdrückt wurde,
wo doch die ipsilateral verlaufenden retinalen Fasern bei den Siamesen nicht komplett
reduziert sind (Stone et al., 1978b). Für das Albinofrettchen existiert eine solche
Ungewißheit nicht, da die nahezu komplette Unterdrückung des entsprechenden
Gesichtsfeldanteils sehr gut mit der Vollständigkeit der Reduktion der ipsilateralen Fasern
aus der Retina (Zhang & Hoffmann, 1993) in Einklang steht.
An dieser Stelle ist das auffällige Verhalten des pigmentierten Frettchens #33 zu erwähnen,
das unter monokularer Bedingung im 30°-Sektor kontralateral zum sehenden Auge weniger
positive Durchläufe erzielte (siehe Abb. 19 links) als die anderen pigmentierten Frettchen
(vgl. Abb. 19 rechts und Abb. 20). In Bezug auf dieses Verhalten kann dieses pigmentierte
Tier zwischen den anderen pigmentierten und den albinotischen Frettchen eingeordnet
werden. Eine mögliche Erklärung für diesen Befund könnte sein, daß es bezüglich des
Pigment-Gens kein homozygotes Wildtyp-Frettchen war, da es ein relativ helles, blondes
Fell besaß. Die Befunde von Thompson et al. (1991) sprechen zwar gegen diese mögliche
Erklärung, können sie aber nicht ausschließen. Thompson et al. (1991) verglichen die Stärke
der retinofugalen Projektionen zwischen zwei Gruppen von pigmentierten Frettchen
miteinander, die in Bezug auf das Pigment-Gen einerseits homozygot (Wild-Typ) und
andererseits heterozygot (mit einem Albino-Allel) waren. Ihr Befund, nach dem sich die
beiden Tiergruppen bezüglich des Verhältnisses von kreuzenden zu nicht-kreuzenden Fasern
nicht unterscheiden, bezieht sich lediglich auf die Anatomie und schließt einen
physiologischen Unterschied nicht aus.
5. Der Perimetrietest in Bezug auf das optokinetische System
Der Perimetrietest ist nicht spezifisch für das optokinetische System, weil er nicht das
optokinetische Verhalten testet. Da die Hinwendereaktion zum kleinen visuellen Reiz eine
colliculäre und kortikale Leistung ist, beziehen sich die Befunde in erster Linie auf die

Diskussion 118
retinocolliculäre und auf die retinogeniculokortikale Projektion und ihre Funktionstüchtig-
keit, jedoch nicht auf die retinoprätektale Projektion.
Die Befunde der monokularen Perimetriemessungen legen jedoch die Hypothese nahe, daß
die aberrante retinoprätektale Projektion, die zusammen mit den aberranten, funktionell
unterdrückten Anteilen der retinocolliculären und -geniculokortikalen Projektion aus
demselben temporalen Retinabereich entspringt, ebenfalls funktionell unterdrückt sein
müßte. Da nach Zhang & Hoffmann (1993) die ipsilaterale retinoprätektale Projektion des
Albinofrettchens eine viel stärkere Reduktion erfährt (sie ist komplett weg) als die beiden
anderen Projektionen, sollte dies sogar eine massivere Funktionsstörung innerhalb des
optokinetischen Systems verursachen. Diese Erwartung wird durch die in der vorliegenden
Arbeit beschriebene optokinetische Blindheit bestätigt.
Aus der funktionellen Unterdrückung des auf der temporalen Hemiretina abgebildeten
Gesichtsfeldes folgt einerseits die Erwartung, daß eine optokinetische Teilfeldstimulation des
Albinofrettchens keinen invertierten Nystagmus auslösen sollte. Denn zur Auslösung eines
invertierten Nystagmus müßte im unterdrückten Gesichtsfeldbereich (bzw. auf der
temporalen Hemiretina) gereizt werden. Andererseits müßte eine optokinetische Stimulation,
die sich auf das periphere Gesichtsfeld beschränkt, einen normalen Nystagmus auslösen,
denn in diesem Falle würde nur die nasale Hemiretina gereizt, die dem Perimetrietest zufolge
normal ist. Insofern müßte die Teilfeldreizung des Albinofrettchens noch durchgeführt
werden, um bezüglich dieser Fragen Gewißheit zu schaffen.

Diskussion 119
II. Teil II: Neurophysiologie
A. Identifizierung der NOT-Neuronen
Das urprüngliche Ziel der elektrophysiologischen Untersuchungen der vorliegenden Arbeit
war die quantitative und qualitative Erfassung „der NOT-Zellen“, die visuelle Information
für die Auslösung der langsamen Phase des hOKN weiterleiten. Denn bei allen bis jetzt
elektrophysiologisch untersuchten normal-pigmentierten Säugetieren mit intaktem
optokinetischen System sind diese NOT-Zellen richtungsspezifisch mit ipsiversiver
Vorzugsrichtung (Kaninchen: Collewijn, 1975; Ratte: Schmidt et al., 1993; Iltis: Klauer et
al., 1990; Tammarwallaby: Ibbotson et al., 1994; Ibbotson & Mark, 1996; Katze: Hoffmann
& Schoppmann, 1981; Totenkopfaffe: Hoffmann, 1985; Rhesusaffe und Langschwanzmakak:
Hoffmann et al., 1988; Hoffmann & Distler, 1989). Da ihre Richtungsspezifität eine absolute
Notwendigkeit für die Auslösung des hOKN ist, diente sie in den meisten Arbeiten als
hinreichendes Kriterium zu ihrer elektrophysiologischen Identifizierung.
Bei den Albinofrettchen hatte sich jedoch gezeigt, daß das Kriterium „Richtungsspezifität“
unzureichend ist, denn die allermeisten ihrer NOT-Zellen waren nicht richtungsspezifisch
(vgl. Verteilung der DS-Indizes in Abb. 29A). Folglich mußte ein anderes Kriterium
herangezogen werden, das beim Albinofrettchen die Entscheidung ermöglichte, ob eine
abgeleitetete prätektale Zelle tatsächlich eine NOT-Zelle war. Als ein solches Kriterium
wurde die „antidrome Reizbarkeit von der ipsilateralen IO“ gewählt, da bekannt und
allgemein akzeptiert ist, daß NOT-Zellen zur ipsilateralen IO projizieren (Ratte: Teresawa et
al., 1979; Kaninchen: Takeda & Maekawa, 1976; Katze: Hoffmann et al., 1976; Walberg et
al., 1981; Ballas & Hoffmann, 1985; Watanabe et al., 1993; Affe: Sekiya & Kawamura,
1985; Hoffmann et al., 1988; Kato et al., 1995; Büttner-Ennever et al., 1996) und diese
Projektion auch beim Albinofrettchen anatomisch nachgewiesen ist (Zhang & Hoffmann,
1993). Der Nachteil dieser Vorgehensweise ist, daß sie eine gewisse Einschränkung in Bezug
auf die tatsächliche Anzahl der in Frage kommenden Zellen in sich birgt. Denn nicht alle
NOT-Neuronen innerhalb des hOKN-Schaltkreises projizieren zur IO, was durch die
richtungsspezifischen, aber nicht von der IO antidrom reizbaren NOT-Zellen des
pigmentierten Frettchens belegt wird (nicht-schraffierte DS-Indizes in Abb. 29C). Folglich
wird in der vorliegenden Arbeit nicht die Gesamtheit der NOT-Zellen, sondern nur die zur
ipsilateralen IO projizierende Teilmenge betrachtet. Es handelt sich bei diesen NOT-Zellen
um die von Ballas & Hoffmann (1985) bei der Katze als „Bildverschiebungsneuronen“

Diskussion 120
(„retinal slip-neurons“) bezeichnete Zellen, die per Definition auch von der IO antidrom
reizbar sind.
Der große Vorteil des Kriteriums „antidrome Reizbarkeit von der IO“ ist seine hohe
räumliche und funktionelle Selektivität für NOT-Zellen im Sinne des hOKN. Natürlich ist
davon auszugehen, daß die elektrische Reizung der IO auch Zellen und Fasern von
benachbarten Strukturen (z.B. Pyramidenbahn, medialer longitudinaler Fasciculus und
paramedianer retikulärer Nukleus) depolarisiert hat, zumal nicht immer beide IO-
Reizelektroden exakt in der IO plaziert waren (vgl. Abb. 24). Da aber bei keiner dieser
Strukturen ein Eingang vom NOT nachgewiesen ist (Kaninchen: Takeda & Maekawa, 1976;
Ratte: Teresawa et al., 1979; Korb et al., 1989; Frettchen: Zhang, 1991; Zhang & Hoffmann,
1993; Telkes et al., 2001; Katze: Walberg et al., 1981; Magnin et al., 1989; Watanabe et al.,
1993; Schmidt et al., 1995; Affe: Mustari et al., 1994; Büttner-Ennever et al., 1996), muß die
antidrome Reizung der NOT-Zellen von der IO erfolgt sein. Außerdem zeigen die
histologischen Experimente, daß die von der IO retrograd markierbaren NOT-Zellen eine
Ansammlung bilden, die relativ gut von unmittelbar benachbarten Strukturen (SC, andere
prätektale Kerne) abgegrenzt ist. Hieraus resultiert die räumliche Selektivität der antidromen
Reizung für NOT-Zellen. Deshalb kann in der vorliegenden Arbeit unterstellt werden, daß
auch beim Albinofrettchen nur Zellen innerhalb des NOT von der IO antidrom gereizt
wurden. Hierfür sprechen auch die beiden Befunde, daß erstens eine antidrome Reizung der
abgeleiteten prätektalen Zellen unmöglich war, wenn beide Reizelektroden die IO verfehlt
hatten und daß zweitens die Läsionen bei allen erfolgreichen Reizexperimenten tatsächlich
im NOT lokalisiert wurden (vgl. Abb. 22 und 23).
Die grundlegenden physiologischen Eigenschaften der NOT-Zellen des pigmentierten
Frettchens sind nicht Gegenstand dieser Arbeit, da sie schon von Klauer et al. (1990)
ausführlich beschrieben wurden. Deshalb war der Zweck der Ableitung der
richtungsspezifischen NOT-Zellen des pigmentierten Frettchens nicht die Wiederholung der
Datenerhebung von Klauer et al. (1990), sondern die zusätzliche antidrome Identifizierung
dieser Zellen. Diese Vorgehensweise erlaubte folgende exakte Zuordnung: Die von der IO
antidrom identifizierten richtungsunspezifischen NOT-Zellen des Albinofrettchens können
als diejenigen Zellen betrachtet werden, die — hinsichtlich ihrer Lokalisation und
potentiellen Funktion innerhalb des neuronalen hOKN-Schaltkreises — mit den ebenfalls
von der IO antidrom identifizierten, jedoch richtungsspezifischen NOT-Zellen des
pigmentierten Frettchens korrespondieren. Diese Zuordnung ist exakt, weil beim
pigmentierten Frettchen nur solche NOT-Zellen von der IO antidrom reizbar waren, die auch

Diskussion 121
richtungsspezifisch waren und die ipsiversive Richtung bevorzugten. Keine einzige
richtungsunspezifische Zelle innerhalb des NOT war beim pigmentierten Frettchen von der
IO aktivierbar. Der entsprechende Befund bei der Katze wurde von Hoffmann et al. (1976)
und beim Makaken von Hoffmann et al. (1988) festgestellt. Damit ist die oben erwähnte
funktionelle Selektivität der „antidromen Reizung von der IO“ für NOT-Zellen gegeben, die
visuelle Information für die Auslösung der langsamen Phase des hOKN weiterleiten.
Deshalb ist das Kriterium „antidrome Reizbarkeit von der IO“ ein hinreichendes Kriterium
zur physiologischen Identifizierung dieser Zellen.
B. Ausmaß des physiologischen Defizits
An dieser Stelle soll noch einmal darauf hingewiesen werden, daß es — soweit anhand der
qualitativen Charakterisierung der prätektalen Zellen aus der Umgebung der NOT-Zellen
beurteilt werden kann — innerhalb dieser Umgebung keine auffälligen, konsistenten
physiologischen Unterschiede zwischen albinotischen und pigmentierten Tieren gibt. Die
Abwesenheit der Richtungsspezifität der NOT-Zellen des Albinofrettchens ist der einzige
festgestellte Unterschied zu den pigmentierten Frettchen. Inwieweit eine exaktere und
intensivere Untersuchung der prätektalen Zellen doch Unterschiede aufdecken würde, kann
hier nicht vorausgesagt werden. Aufgrund der groben, subjektiven Charakterisierung können
folgende vorläufige Schlußfolgerungen gezogen werden:
Es existiert kein Unterschied zwischen dem albinotischen und pigmentierten Frettchen
• hinsichtlich der Spontanrate der antidrom identifizierten NOT-Zellen und der
benachbarten prätektalen Zellen,
• hinsichtlich der relativen Position der NOT-Zellen zu Jerk-Zellen und
• hinsichtlich der visuellen Aktivierbarkeit der Jerk-Zellen.
C. Richtungsunspezifität der NOT-Neuronen und optokinetische Blindheit
Im vorangestellten Verhaltensteil der vorliegenden Arbeit wurde erstens die optokinetische
Blindheit der Albinofrettchen bei Ganzfeldreizung dokumentiert und zweitens wurden
Beweise dafür erbracht, daß der Grund für die optokinetische Blindheit in einem neuronalen
Defekt zu suchen ist, der entweder dem NOT vorgeschaltet oder im NOT selbst lokalisiert
ist. Die hier vorgestellten elektrophysiologischen Daten von den NOT-Zellen derselben
Albinofrettchen grenzen den gesuchten neuronalen Defekt weiter ein und liefern sogar eine
Begründung für ihre optokinetische Blindheit, denn das Ausbleiben des hOKN kann

Diskussion 122
zunächst als Folge der fehlenden bzw. sehr schwachen Richtungsspezifität der allermeisten
NOT-Zellen betrachtet werden. Dies kann mit Sicherheit behauptet werden, weil der
richtungsspezifische Ausgang des NOT eine notwendige Voraussetzung ist für die adäquate
Aktivierung bzw. Inhibierung der dem NOT nachgeschalteten sensomotorischen und
motorischen Hirnstrukturen. Damit stellt die Richtungsunspezifität der NOT-Zellen das
physiologische Korrelat zur optokinetischen Blindheit der untersuchten Albinofrettchen dar.
Wie in der Diskussion des vorangestellten Verhaltensteils ausführlich erörtert, wurde ein
hOKN-Ausfall schon einmal bei hypopigmentierten Säugetieren (Maus: Mangini et al.,
1985; Ratte: Precht & Cazin, 1979) beschrieben. Lannou et al. (1982) unternahmen den
Versuch, den bei Albinoratten beobachteten hOKN-Ausfall auf neuronaler Ebene mit
elektrophysiologischen Methoden zu erklären. Die Autoren schreiben, daß sie bei ihren
Albinoratten im Gegensatz zu ihren pigmentierten Ratten kein einziges richtungspezifisches
prätektales Neuron extrazellulär ableiten konnten. Hinsichtlich der Aktivierbarkeit durch
„Licht-An“ oder „Licht-Aus“ konnten die Autoren keinen Unterschied zwischen den
prätektalen Zellen der beiden Rattenstämme finden.
Diese Befunde entsprechen genau den hier vorgestellten Befunden, nach denen die NOT-
Zellen des Albinofrettchens richtungsunspezifisch sind und trotz ihrer Richtungsunspezifität
als ON- oder ON-OFF-Zellen klassifiziert werden können. Allerdings muß konstatiert
werden, daß Lannou et al. (1982) „ihre“ sogenannten richtungsunspezifischen prätektalen
Zellen — im Gegensatz zu den hier vorgestellten richtungssunspezifischen NOT-Zellen des
Albinofrettchens — nicht von der IO antidrom identifizierten, sondern nur anhand ihrer
stereotaktischen Koordinaten und den nachträglichen Farbinjektionen in das abgeleitete
prätektale Areal lokalisierten. Aus eigener Erfahrung kann berichtet werden, daß diese
Vorgehensweise sehr unpräzise ist und keine eindeutige Identifizierung von Zellen erlaubt,
weshalb in der vorliegenden Arbeit die aufwändigere aber eindeutige antidrome
Identifizierung von der IO durchgeführt wurde. Deshalb können die richtungsunspezifischen
prätektalen Zellen von Lannou et al. (1982) nicht mit Sicherheit den hier vorgestellten,
richtungsunspezifischen, von der IO antidrom identifizierten NOT-Zellen des
Albinofrettchens gleichgesetzt werden.
Aus der Entdeckung der Richtungsunspezifität der NOT-Zellen des Albinofrettchens
resultiert folgende Frage: Wie kommt die Richtungsspezifität der NOT-Neuronen im
pigmentierten Frettchen zustande und wie geht sie beim Albinofrettchen verloren? Diese
allgemein gehaltene Frage zielt auf den Mechanismus, der der Richtungsspezifität zugrunde

Diskussion 123
liegt und kann aufgrund des in dieser Arbeit erhobenen Datenmaterials nicht endgültig
beantwortet werden. Auf dem Weg zu ihrer Beantwortung soll jedoch zunächst die Frage
nach dem „Wo?“ diskutiert werden: Wo wird die Richtungsspezifität der NOT-Neuronen des
pigmentierten Frettchens determiniert? Die Beantwortung dieser Frage würde gleichzeitig
auch den wahrscheinlichsten Ort nennen, der für den Verlust der Richtungsspezifität beim
Albinofrettchen verantwortlich ist.
Unter Berücksichtigung der im Verhaltensteil gezogenen Schlußfolgerungen, können grund-
sätzlich drei Orte angenommen werden, wo im Normaltier die Richtungsspezifität der NOT-
Neuronen festgelegt werden könnte. Diese drei Orte sind die Ursprungsorte der beiden
sensorischen Haupteingänge des NOT, erstens der VC und zweitens die Retina, und drittens
der NOT selbst. Folglich können alle drei Orte grundsätzlich auch für den Verlust der
Richtungsspezifität beim Albinofrettchen verantwortlich sein:
1. Determination der Richtungsspezifität des NOT im VC bzw. über dem kortikalen NOT-
Eingang: Von Albinos ist bekannt, daß sich die gestörte Retinotopie in ihrer
retinogenikulären Projektion über die genikulokortikale Projektion bis zum VC
fortpflanzt und trotz verschiedener Reorganisationsversuche des VC zu einer abnormen
kortikalen retinotopischen Organisation führt (Kaninchen: Choudhury, 1987; Mink:
Sanderson et al., 1974; Frettchen: Guillery 1971; Huang & Guillery, 1985; Siamesische
Katze: Guillery, 1974, 1990; Albinokatze: Ault et al., 1995; Mensch: Apkarian et al.,
1991; Apkarian & Shallo-Hoffmann, 1991). Hieraus resultierte die Hypothese, daß die
Richtungsspezifität des NOT beim pigmentierten Frettchen im VC bzw. über die
kortikoprätektale Verbindung determiniert wird und infolgedessen der VC bzw. seine
Projektion auf den NOT für den Verlust seiner Richtungsspezifität beim Albinofrettchen
verantwortlich ist. Zur Überprüfung dieser Hypothese wurde der kortikale NOT-Eingang
des Frettchens hinsichtlich seiner Stärke und Leitungsgeschwindigkeit (orthodrome
Reizung der NOT-Zellen vom VC) und seiner Richtungsspezifität (antidrome Reizung
der VC-Zellen vom NOT) untersucht. Dabei wurden die Ableitorte im VC und die
Reizorte im NOT durch elektrolytische Läsionen markiert und verifiziert.
2. Determination der Richtungsspezifität in der Retina bzw. über den retinalen NOT-
Eingang: Der retinale Eingang des NOT könnte physiologisch abnorm sein, zumal von
Albinos bekannt ist, daß sie zum einen eine unterentwickelte / degenerierte Retina haben
(Kaninchen: Oyster et al., 1987; Jeffery et al., 1997; Maus: Jeffery et al., 1997; Ratte:
Webster & Rowe, 1991; Ilia & Jeffery, 1999; Frettchen: Morgan et al., 1987; Jeffery &
Kinsella, 1992; Jeffery et al., 1994; Siamesische Katze: Stone et al., 1978a, 1978b) und

Diskussion 124
daß sie zum anderen Ganglienzellfasern (hauptsächlich aus der temporalen Hemiretina)
besitzen, die am optischen Chiasma fälschlicherweise kreuzen, statt, wie bei den
pigmentierten Formen derselben Art, zur ipsilateralen Hirnseite zu ziehen. Ein solcher
aberranter Verlauf ist bei vielen albinotischen Spezies nachgewiesen und betrifft die
retinoprätektale, die retinogenikuläre und bei manchen Spezies auch die retinocolliculäre
Projektion (Maus: Balkema et al., 1981; Pak et al., 1987; Kaninchen: Klooster et al.,
1983; Frettchen: Morgan et al., 1987; Zhang & Hoffmann, 1993; Siamesiche Katze: Ault
et al. 1995; Albinokatze: Creel et al., 1982; Ault et al., 1995; Tiger: Guillery & Kaas,
1973; Mensch: Guillery et al., 1975). Zur Überprüfung dieser Hypothese wurden die
orthodromen Reizexperimente vom ON durchgeführt. Sie untersuchen beim Albino-
frettchen den retinalen Eingang der NOT-Zellen hinsichtlich seiner Stärke und Leitungs-
geschwindigkeit und erlauben somit einen Vergleich mit den entsprechenden, in der
Literatur vorhandenen Befunden vom pigmentierten Frettchen (Klauer et al., 1990).
3. Determination der Richtungsspezifität im NOT selbst: In diesem Falle müßte man eine
intrinsische Verschaltung im NOT postulieren, die die markante Richtungsspezifität der
NOT-Ausgangsneuronen festlegt. Die Richtungsspezifität dieser Neuronen könnte dann
beim Albinofrettchen verloren gegangen sein, wenn der intrinsiche Schaltkreis durch die
Albinomutation verändert würde. Infolge dessen wäre es aber sehr wahrscheinlich, daß
die NOT-Ausgangsneuronen auch anatomisch verändert wären. Hieraus ergibt sich
folgende Hypothese: Es existiert ein morphologisches Korrelat zur Richtungsunspezifität
der NOT-Zellen des Albinofrettchens. Die Überprüfung dieser Hypothese wurde von
Telkes et al. (2001) mit histologischen Methoden an albinotischen und pigmentierten
Individuen unternommen. Ihre Versuchstiere stammen aus derselben Kolonie, wie die in
dieser Arbeit untersuchten Tiere. Einige ihrer Versuchstiere stellen eine Teilmenge der in
dieser Arbeit verwandten Tiere dar. Ihre Befunde werden weiter unten diskutiert und zu
den physiologischen Befunden der vorliegenden Arbeit in Beziehung gesetzt.
Im folgenden werden alle drei Möglichkeiten in der Reihenfolge ihrer Aufzählung in
gesonderten Abschnitten diskutiert.

Diskussion 125
D. Kortikaler Eingang des NOT
1. Einordnung der Experimente in den vorhandenen Wissensstand
In der Einleitung der vorliegenden Arbeit wurde die Behauptung aufgestellt, daß typische
afoveate Säuger, wie die Maus und das Kaninchen, nur einen subkortikalen hOKN-
Schaltkreis, d.h. keine kortikale Projektion zum NOT, besitzen. Desweiteren wurde erwähnt,
daß mit fortschreitender Foveation, Frontalstellung der Augen und Entwicklung des Kortex
cerebri eine massiver werdende kortikoprätektale Projektion auftritt, die bei Primaten und
insbesondere beim Menschen ihre maximale Stärke erreicht. (Die Konsequenz dieser
Entwicklung ist erstens die Zunahme der Symmetrie des monokularen hOKN und zweitens
die Befähigung zur Beantwortung höherer Reizgeschwindigkeiten.)
An dieser Stelle soll diese Aussage etwas genauer beleuchtet werden, indem die
tatsächlichen Verhältnisse beim Rhesusaffen, bei der Katze, bei der Ratte und schließlich
auch beim Kaninchen vorgestellt werden. Anhand dieser Darstellung soll eine Einordnung
der durchgeführten Untersuchungen in den vorhandenen Wissenstand erfolgen und die
Problematik beschrieben werden, die den Experimenten zugrunde lag.
Affe: Beim Rhesusaffen wurde die kortikofugale Projektion zum NOT mit den gleichen
Methoden wie in der vorliegenden Arbeit untersucht (Hoffmann et al., 1991: orthodrome
Reizung des NOT vom VC und Hoffmann et al., 1992: antidrome Reizung des VC vom
NOT). Mit der orthodromen Reizung wurde zunächst festgestellt, daß der NOT von der Area
V1 und von bewegungssensitiven Arealen im superioren temporalen Sulcus (STS) Eingang
bekommt. Anschließend wurde mittels der antidromen Identifizierung der Zellen im STS
entdeckt, daß sie sich innerhalb des medialen temporalen Areals (MT) in der Lamina V
befinden, rezeptive Felder im zentralen Gesichtsfeld haben und richtungsspezifisch auf
ipsiversive Reizung mit einem großflächigen Zufallspunktemuster reagieren. MT-Zellen mit
anderen Vorzugsrichtungen, die auch gefunden wurden, waren nicht vom NOT antidrom
reizbar. Damit wurde für den Affen eindeutig klargestellt, daß der NOT-Eingang aus dem
STS von richtungsspezifischen Fasern mit überwiegend ipsiversiver Vorzugsrichtung
gebildet wird, und daß alle diese Fasern ihren Ursprung in einem konkreten, näher
abgrenzbaren Areal (MT) haben.
Katze: Die kortikofugale Projektion zum NOT wurde bei der Katze von Schoppmann (1981)
ebenfalls mit Hilfe der antidromen Identifizierung der VC-Zellen vom NOT untersucht.

Diskussion 126
Dabei stellte Schoppmann (1981) fest, daß der kortikale NOT-Eingang von VC-Neuronen
gebildet wird, die in der Lamina V der Areae 17 und 18 liegen. Diese Zellen sind binokulare
Komplex-Zellen und antworten sowohl auf bewegte Lichtbalken als auch auf die Bewegung
großer Zufallspunktemuster. Außerdem zeigen sie ein schärferes Geschwindigkeits-Tuning
und bevorzugen höhere Reizgeschwindigkeiten als die NOT-Zellen. Ihre Vorzugsrichtungen
sind inhomogen, d.h. im Gegensatz zu den NOT-Zellen, die alle eine ipsiversive Vorzugs-
richtung haben, besitzen die kortikalen Zellen auch andere, nicht ipsiversive Vorzugsrichtun-
gen. Deshalb geht Schoppmann (1981) für die Katze davon aus, daß der kortikale NOT-
Eingang nicht die Richtungsspezifität der NOT-Zellen determiniert. Vielmehr befähigt diese
kortikale Projektion die NOT-Neuronen zur Beantwortung höherer Reizgeschwindigkeiten
und zur Generierung eines relativ symmetrischen hOKN unter monokularer optokinetischer
Reizung.
Damit kann für die Katze festgehalten werden, daß der kortikale NOT-Eingang aus Fasern
gebildet wird, die zwar richtungsspezifisch sind, jedoch nicht alle die ipsiversive Richtung
bevorzugen, und daß diese Fasern offensichtlich nicht in einem konkreten kortikalen Areal
lokalisiert, sondern innerhalb der Areae 17 und 18 verstreut sind. Der Versuch von
Schoppmann (1981) bei der Katze mittels der antidromen Reizung vom NOT ein
abgrenzbares kortikales Areal zu finden bzw. zu definieren, das dem Areal MT des Affen
homolog ist, war gescheitert. Die elektrische Reizung der kortikalen Areale 7, 19, 21 und des
suprasylvischen Areals führten zwar zur orthodromen Reizung der NOT-Zellen, allerdings
seltener und weniger verläßlich als die Reizung der Areae 17 und 18. Außerdem waren in
diesen Fällen die orthodromen Latenzen länger als bei Reizung der Areae 17 und 18, was
darauf hindeutet, daß diese Areale nicht monosynaptisch zum NOT projizieren.
Entsprechend ist eine antidrome Aktivierung von VC-Zellen durch Reizung des NOT
außerhalb der Areae 17 und 18 bei der Katze bis heute nicht gelungen.
Die Befunde aus Läsionsexperimenten sprechen jedoch dafür, daß es bei der Katze neben
den Areae 17 und 18 auch weitere kortikale Areale geben muß, die bei der Auslösung und /
oder Steuerung des hOKN beteiligt sind und infolgedessen auch eine Verbindung zum NOT
haben sollten (Malach et al., 1984; Strong et al., 1984; Ventre, 1985 und Tusa et al., 1989).
Aus diesen Experimenten geht hervor, daß die alleinige Zerstörung der Areae 17 und 18
keinen Effekt auf die Güte des hOKN hat, die Zerstörung der suprasylvischen Areale
(AMLS, ALLS, PMLS) jedoch die Güte des hOKN herabsetzt. Unter diesen Arealen scheint
die Läsion von PMLS den größten Effekt auf den hOKN zu haben und wird aus diesem
Grunde als Homologon zu der Area MT des Affen favorisiert.

Diskussion 127
Frettchen: Beim pigmentierten Frettchen zeigten Klauer et al. (1990) zum ersten Mal die
Existenz der kortikoprätektalen Projektion, indem sie die NOT-Zellen orthodrom vom VC
gereizt haben und aus den Latenzen auf eine monosynaptische Verbindung schlossen. Diese
Projektion ist mit der entsprechenden Projektion bei der Katze hinsichtlich ihrer Stärke
vergleichbar. Da auch der bino- und monokulare hOKN des Frettchens vergleichbar mit dem
der Katze ist (Hein et al., 1990), wären im Falle des Frettchens auch ähnliche Verhältnisse
hinsichtlich des Ursprungs des kortikalen NOT-Eingangs zu erwarten.
An diesen Wissenstand schließen die in der vorliegenden Arbeit durchgeführten
Untersuchungen des kortikalen NOT-Eingangs an.
Zuerst wurde die orthodrome Reizung der NOT-Zellen vom VC auch beim Albinofrettchen
durchgeführt. Die Durchführung dieser Experimente wurde als notwendig erachtet, weil sie
den Vergleich zwischen pigmentierten und albinotischen Frettchen hinsichtlich der Stärke
und Leitungsgeschwindigkeit ihrer VC-NOT-Verbindungen erlaubten. Damit sollten die
ersten Indizien ermittelt werden, die bei der Entscheidung helfen sollten, ob die
Richtungsunspezifität des NOT des Albinofrettchens durch den VC verursacht wird. Das
anschließende Vertauschen der Reiz- und Ableitorte (antidrome Reizung der VC-Zellen vom
NOT) sollte weitere, aussagekräftigere Beweise liefern.
Aus diesem Zusammenhang heraus ist die antidrome Identifizierung der VC-Zellen vom
NOT auch als ein Versuch zu verstehen, den Ursprung des kortikalen NOT-Eingangs örtlich
zu lokalisieren und hinsichtlich seiner richtungsspezifischen Eigenschaften zu beschreiben.
Die zu beantwortende Frage in diesem Zusammenhang ist, ob die antidrom reizbaren VC-
Zellen des Frettchens wie im Falle des Rhesusaffen innerhalb eines bestimmten kortikalen
Areals fokusiert oder wie im Falle der Katze verstreut sind. Hieraus resultiert die
Problematik der antidromen Reizung der VC-Zellen, denn die Position der höheren
kortikalen Areale ist zur Zeit beim Frettchen noch unbekannt, was eine gezielte Suche nach
antidromen VC-Zellen erschwert.
Ratte und Kaninchen: Wider Erwarten konnten in elektrophysiologischen Reizexperimenten
kortikale Bereiche (Areae 17, 18 und 18A) gefunden werden, die aller Wahrscheinlichkeit
nach mit der Auslösung und / oder Steuerung des hOKN in Zusammenhang stehen. Schmidt
et al. (1993) konnten bei der narkotisierten Ratte 45% der richtungsspezifischen NOT-
Neuronen vom VC orthodrom reizen und anhand der Latenzen auf eine monosynaptische
VC-NOT-Verbindung schließen. Die anschließende retrograde Markierung der zuvor

Diskussion 128
elektrisch gereizten kortikalen Areale (Injektion von fluoreszierenden Latexpartikeln in den
NOT) bestätigte die VC-NOT-Verbindung.
Pettorossi & Troiani (1983) konnten beim narkotisierten Kaninchen einen kortikalen Bereich
finden, dessen elektrische Reizung 50% der NOT-Neuronen orthodrom aktivierte. Diese
Aktivierung bestand in den meisten Fällen aus einer Mehrfachantwort und ähnelt in dieser
Hinsicht den entsprechenden Befunden beim Frettchen (vgl. Abb. 31). Die Autoren lokali-
sierten diesen kortikalen Bereich dort, wo die Areae 17, 21 und 22 zusammentreffen und
nannten ihn „temporooccipitalen nystagmogenen Kortex“. Beim wachen Kaninchen löste die
elektrische Reizung dieses kortikalen Areals einen Nystagmus aus, der die gleiche Richtung
hatte wie der durch visuelle Reizung des ipsilateralen NOT ausgelöste Nystagmus.
Vergleicht man die Verhältnisse in der dargestellten Reihenfolge (Affe, Katze, Frettchen,
Ratte und Kaninchen) miteinander, so fällt auf, daß bis heute nur beim Affen ein abgrenz-
bares kortikales Areal (MT) beschrieben wurde, welches zum NOT projiziert und eindeutig
nur richtungsspezifische Information mit ipsiversiver Vorzugsrichtung übermittelt. In allen
anderen Spezies konnte ein Homologon zum MT nicht eindeutig identifiziert werden. Bei
der Katze und beim Frettchen haben die in Area 17 und 18 verstreuten antidromen Zellen
nicht alle die ipsiversive Vorzugsrichtung und in dieser Hinsicht sind sich die beiden Spezies
sehr ähnlich. Ein außerhalb der Areae 17 und 18 liegendes Areal, wie z.B. PMLS, konnte bei
der Katze bisher nicht eindeutig vom NOT antidrom identifiziert werden. Ob ein solches
Areal beim Frettchen existiert und auch elektrophysiologisch nachweisbar ist, sollte in dieser
Arbeit überprüft werden und wird in einem gesonderten Abschnitt weiter unten diskutiert.
Die Verhältnisse bei der Ratte und beim Kaninchen scheinen sich ebenfalls sehr zu ähneln.
Die Befunde von Schmidt et al. (1993) und Pettorossi & Troiani (1983) stehen in
Widerspruch zu der anfangs erwähnten, allgemein akzeptierten Hypothese, daß typische
Afoveaten nur einen direkten retinalen NOT-Eingang besitzen und infolgedessen nur einen
asymmetrischen monokularen hOKN generieren können. Angesichts dieser Befunde muß
man diese Hypothese relativieren und die Asymmetrie des monokularen hOKN nicht einzig
und allein durch das Fehlen einer VC-NOT-Verbindung bedingt sehen.
2. Stärke und Leitungsgeschwindigkeit des kortikalen NOT-Eingangs
Beim Albinofrettchen waren 63% (31 von 49) der von der IO antidrom reizbaren NOT-
Zellen vom VC orthodrom reizbar. Dieses Verhältnis liegt im selben Bereich wie das vom
pigmentierten Frettchen, bei dem 65% (33 von 51) der richtungsspezifischen NOT-Zellen

Diskussion 129
von Klauer et al. (1990) entsprechend gereizt wurden. Die Übereinstimmung der beiden
Prozentsätze spricht dafür, daß die beiden Tiergruppen sich hinsichtlich der Stärke ihrer
kortikalen NOT-Eingänge nicht unterscheiden, und daß bei beiden Tiergruppen ähnliche
kortikale Bereiche elektrisch gereizt wurden. Wenn man die Prozentsätze der vom NOT
antidrom reizbaren VC-Zellen miteinander vergleicht, wurden beim Albinofrettchen 32%
und beim pigmentierten Frettchen 38% der VC-Zellen antidrom gereizt. Auch diese
Übereinstimmung spricht dafür, daß die VC-NOT-Verbindung bei beiden Tiergruppen gleich
stark ausgebildet ist.
Weiterhin kann die Übereinstimmung der Prozentsätze von NOT-Zellen, die vom VC
orthodrom reizbar sind, als Indiz angesehen werden, daß die richtungsunspezifischen, von
der IO antidrom identifizierten NOT-Zellen des Albinofrettchens den richtungsspezifischen,
wenn auch nicht von der IO antidrom identifizierten, NOT-Zellen des pigmentierten
Frettchens von Klauer et al. (1900) entsprechen. Unabhängig von diesem Befund wurde
diese Entsprechung bei den in der vorliegenden Arbeit untersuchten NOT-Zellen des
pigmentierten Frettchens in erster Linie durch die antidrome Reizung von der IO eindeutig
sichergestellt.
Aus dem Vergleich der Häufigkeitsverteilungen der orthodromen Latenzen des
Albinofrettchens und des pigmentierten Frettchens (Klauer et al., 1990) ist weiterhin die
Schlußfolgerung zulässig, daß sich die kortikalen NOT-Eingänge der beiden Tiergruppen
auch hinsichtlich ihrer axonalen Leitungsgeschwindigkeit nicht unterscheiden. Denn Klauer
et al. (1990) fanden beim pigmentierten Frettchen orthodrome Latenzen hauptsächlich im
Bereich von 1 bis 6,5 ms mit einem Mittelwert von 3,7 ± 1,5 ms, während die Latenzen vom
Albinofrettchen im Bereich von 2 bis 7,5 ms mit einem Median von 3,0 ms verteilt sind (vgl.
Abb. 32A mit 32C). Für diese Schlußfolgerung sprechen auch die antidromen Latenzen der
VC-Zellen, die sich zwischen den beiden Tiergruppen ebenfalls nicht signifikant
unterscheiden (vgl. Abb. 35A mit 35C).
Die Prozentsätze elektrisch reizbarer Zellen und die Latenzen aus der orthodromen Reizung
der NOT-Zellen vom VC und aus der antidromen Reizung der VC-Zellen vom NOT
sprechen dafür, daß der kortikale NOT-Eingang des Albinofrettchens hinsichtlich seiner
Stärke und Leitungsgeschwindigkeit normal ist. Diese Befunde beweisen jedoch keine
umfassende, lückenlose „Normalität“ des Eingangs, da sie keine Aussage über seinen
Informationsgehalt erlauben. Deshalb wurde anschließend die antidrome Reizung des VC
vom NOT durchgeführt, deren primäres Ziel die visuelle Charakterisierung der vom
ipsilateralen NOT antidrom identifizierten VC-Neuronen war. Erst diese Experimente

Diskussion 130
konnten Aufklärung über den Informationsgehalt des kortikalen NOT-Eingangs leisten und
letztendlich eine Verifikation oder Falsifikation der Hypothese erlauben, nach der der VC die
Richtungsspezifität der NOT-Neuronen beim pigmentierten Frettchen bestimmt und beim
Albinofrettchen auslöscht. Da solche Reizexperimente an pigmentierten Frettchen in der
Literatur nicht beschrieben waren, mußten sie bei beiden Tiergruppen durchgeführt werden.
3. Informationsgehalt des kortikalen NOT-Eingangs
Von der Eigenschaft „Richtungsspezifität“ wurden die beiden Aspekte „Vorzugsstärke“ und
„Vorzugsrichtung“ untersucht und getrennt dargestellt. Es konnte gezeigt werden, daß nicht
alle VC-Neuronen die gleiche Vorzugsstärke besitzen, sondern zwischen totaler Richtungs-
unspezifität (DS-I = 0) und sehr hoher Richtungsspezifität (DS-I > 0,9) verteilt sind (vgl.
Abb. 33). Ab welchem DS-I-Wert man von einer richtungsspezifischen Zelle sprechen kann,
ist rein subjektiv zu entscheiden. So wurde entschieden, daß ein DS-I ≥ 0,33 auf eine
auffällige, unverwechselbare Richtungsspezifität hindeutet. Ein DS-Index von 0,33 bedeutet,
daß die Feuerrate in Vorzugsrichtung gegenüber der Feuerrate in Nullrichtung um 100%
erhöht ist. Damit waren beim Albinofrettchen 55% (47 von 86) und beim pigmentierten
Frettchen 38% (sechs von 16) der untersuchten VC-Zellen eindeutig richtungsspezifisch.
Hieraus folgte dann die Schlußfolgerung, daß das pigmentierte Frettchen prozentual weniger
richtungsspezifische VC-Zellen in der kleinen Stichprobe von 16 Zellen besitzt. Betrachtet
man jedoch nur die vom NOT antidrom reizbaren VC-Zellen, so haben beim Albinofrettchen
54 % (sieben von 13) und beim pigmentierten Frettchen 50 % (drei von sechs) der VC-
Zellen einen DS-I ≥ 0,33. Unter den zum NOT projizierenden VC-Zellen scheint es keinen
Unterschied hinsichtlich ihrer Vorzugsstärke zu geben.
Gibber et al. (2001) fanden in einer Stichprobe von 161 VC-Neuronen des pigmentierten
Frettchens wesentlich mehr (80%) richtungsspezifische Zellen, die ihre Vorzugsrichtung mit
doppelt so vielen Aktionspotentialen beantworteten als ihre Nullrichtung.
Die Eigenschaft „visuelle Reizbarkeit durch das Zufallspunktemuster“ wurde zur Bedingung
gemacht, weil großflächige Reize die adäquaten Reize zur Auslösung des hOKN sind.
Folglich sollten VC-Zellen auf solche Reize reagieren, wenn sie als Teil des hOKN-
Schaltkreises bei der Auslösung und / oder Steuerung des hOKN involviert sind. Die
entsprechenden VC-Zellen der Katze (Schoppmann, 1981) und des Rhesusaffen (Hoffmann
et al., 1992) erfüllen diese Bedingung.
Für diejenigen Neuronen des VC, die auf Reizbarkeit vom SC getestet wurden und nicht
antidrom reizbar waren, kann mit größerer Sicherheit behauptet werden, daß ihre Fasern eher

Diskussion 131
im NOT enden, als am NOT vorbeiziehen, um den rostralen SC zu innervieren, wo das
gleiche frontale Gesichtsfeld repräsentiert ist. Damit erhöht sich wesentlich die Wahrschein-
lichkeit, daß die Fasern dieser richtungsspezifischen Neuronen des VC auf NOT-Zellen
terminieren. Es kann hier aus zwei Gründen nur von einer Wahrscheinlichkeit gesprochen
werden: 1. Die Nichtreizbarkeit der VC-Zellen vom benachbarten SC schließt nicht
hundertprozentig aus, daß die Reizung mit der NOT-Reizelektrode doch über Fasern erfolgt,
die den SC innervieren. 2. Die elektrische Reizung im NOT depolarisiert nicht nur kortikale
Fasern, die NOT-Zellen kontaktieren, sondern auch alle anderen kortikalen Eingänge zu
benachbarten prätektalen Zellen. Dies wäre z.B. bei den Jerk-Zellen durchaus möglich, die
sowohl in der vorliegenden Arbeit beim Frettchen als auch bei der Katze (Ballas &
Hoffmann, 1985; Schweigert & Hoffmann, 1992) in unmittelbarer Nähe zu den NOT-Zellen
gefunden wurden.
Vorbehaltlich dieser methodischen Einschränkung zeigt die antidrome Reizbarkeit von
richtungsspezifischen VC-Neuronen, daß es beim Albinofrettchen NOT-Zellen gibt, die
richtungsunspezifisch sind, obwohl sie einen richtungsspezifischen Eingang vom visuellen
Kortex erhalten. An dieser Stelle könnte man schon zu der Schlußfolgerung verleitet werden,
daß der VC nicht die Richtungsspezifität der NOT-Zellen determiniert und damit die
Untersuchung des kortikalen NOT-Eingangs als abgeschlossen betrachten. Denn unter der
Annahme, daß die Richtungsspezifität des NOT durch seinen kortikalen Eingang
determiniert wird, sollte man erwarten, daß eine NOT-Zelle richtungsspezifisch ist, wenn sie
einen richtungsspezifischen Eingang vom VC erhält, und zwar unabhängig von der Vorzugs-
richtung. Beim Albinofrettchen konnten aber kaum richtungsspezifische NOT-Zellen
gefunden werden, wie aus Abbildung 29A hervorgeht.
Es gibt jedoch eine Möglichkeit, nach der die NOT-Zellen des Albinofrettchens trotz eines
richtungsspezifischen kortikalen Eingangs richtungsunspezifisch sein könnten. Diese
Möglichkeit fällt auf, wenn man die Vorzugsrichtungen der vom NOT antidrom identifizier-
ten VC-Zellen des Albinofrettchens betrachtet (siehe Abb. 36). Denn unter diesen Neuronen
sind wider Erwarten alle Vorzugsrichtungen repräsentiert und damit erscheint folgende
Begründung der Richtungsunspezifität möglich: Die NOT-Zellen des Albinofrettchens
könnten abnormerweise von mehreren VC-Zellen mit unterschiedlichen, d.h.
entgegensetzten Vorzugsrichtungen Eingang bekommen, so daß das Resultat der
synaptischen Integration der übermittelten richtungsspezifischen Signale die
Richtungsunspezifität ist. Wenn dieses Szenario beim Albinofrettchen realisiert wäre und
den abnormen Zustand darstellen würde, so müßte man beim pigmentierten Frettchen

Diskussion 132
erwarten, daß die vom NOT antidrom identifizierbaren VC-Zellen nur die ipsiversive
Vorzugsrichtung oder zumindest nur Vorzugsrichtungen mit einer eindeutig ipsiversiven
Komponente besitzen. Wie erwähnt, ist dies beispielsweise beim Rhesusaffen der Fall, bei
dem nur MT-Zellen mit ipsiversiver Vorzugsrichtung zum NOT projizieren, während
benachbarte MT-Zellen mit anderen Vorzugsrichtungen nicht zum NOT projizieren
(Hoffmann et al., 1992). Erst diese Überlegung machte die Durchführung der entsprechenden
Reizexperimente mit pigmentierten Frettchen notwendig. Wie das Ergebnis aus diesen
Experimenten in Abbildung 37 jedoch zeigt, sind auch beim pigmentierten Frettchen alle
Vorzugsrichtungen unter den richtungsspezifischen, vom NOT antidrom und durch das
Zufallspunktemuster visuell reizbaren VC-Zellen zu finden. Damit wurde beim Frettchen der
von Schoppmann (1981) für die Katze beschriebene Zustand und nicht der von Hoffmann et
al. (1992) für den Rhesusaffen beschriebene Zustand bestätigt.
Der Nachweis, daß sich pigmentierte und albinotische Frettchen in dieser Hinsicht nicht
unterscheiden, bedeutet, daß der Zustand beim Albinofrettchen keinen abnormen, sondern
den normalen Zustand darstellt. Erst diese Feststellung erlaubt die Schlußfolgerung, daß der
VC nicht für den Verlust der Richtungsspezifität beim Albinofrettchen verantwortlich ist
und folglich auch nicht die Richtungsspezifität der NOT-Zellen des pigmentierten Frettchens
determiniert.
4. Ursprung des kortikalen NOT-Eingangs beim Frettchen
Das in dieser Arbeit erhobene Datenmaterial erlaubt keine exakte und vollständige
anatomische Lokalisation des Ursprungs des kortikalen NOT-Eingangs. Es kann festgehalten
werden, daß vom NOT antidrom reizbare VC-Neuronen mit Sicherheit innerhalb der Area 17
liegen. Ferner muß weiter davon ausgegangen werden, daß sich solche VC-Zellen auch in
der Area 18 befinden. Dies kann zwar durch keine VC-Läsion in der Area 18 dokumentiert
werden, allerdings wird es durch die Physiologie einiger VC-Neuronen nahegelegt, die auch
auf schnelle Reizbewegungen reagierten, was eher für die Area 18 typisch ist. Außerdem
wurden einige antidromen VC-Neuronen auch auf der dorsalen Seite des VC abgeleitet (im
Gegensatz zu der in Area 17 befindlichen VC-Läsion, die auf der ventralen Seite des VC
lokalisiert ist, vgl. Abb. 25), was die Wahrscheinlichkeit erhöht, daß sie sich in Area 18
befanden (vgl. Abbildungen von Redies et al., 1990 und Rockland, 1985). Da aber in der
vorliegenden Arbeit weder die Area 17 noch die Area 18 konsequent kartiert wurden, kann
das nicht als bewiesen hingenommen werden.

Diskussion 133
Bei der Suche nach vom NOT antidrom reizbaren VC-Zellen ist weder bei den albinotischen,
noch bei den pigmentierten Frettchen ein Bereich innerhalb des VC aufgefallen, in dem die
reizbaren Zellen besonders häufig zu finden waren. Vielmehr suggeriert die Erfahrung, daß
die vom NOT antidrom reizbaren Zellen nicht fokussiert, sondern verstreut sind.
Andererseits aber war sowohl bei Klauer et al. (1990) als auch in dieser Arbeit auffällig, daß
der Erfolg der orthodromen Reizung der NOT-Zellen vom VC von der Position der
Reizelektroden abhängt. Dies könnte darauf hindeuten, daß die zum NOT projizierenden
VC-Neuronen nicht überall lokalisiert sind. Doch aus diesen Beobachtungen kann kein
spezieller kortikaler Bereich bestimmt werden, da die effektiven Reizorte relativ weit
voneinander entfernt und voneinander getrennt sein können und die VC-Reizelektroden
jedesmal einen relativ großen Bereich des Kortex depolarisieren.
Augrund des vorhandenen Datenmaterials kann nicht ausgeschlossen werden, daß es auch
außerhalb der Area 17 und Area 18 VC-Zellen gibt, die zum NOT projizieren.
Da das Frettchen der Katze sehr ähnlich ist [sowohl hinsichtlich des hier vorgestellten
elektrophysiologischen Datenmaterials als auch hinsichtlich der Güte seines hOKN (Hein et
al., 1990)] würde man erwarten, daß es innerhalb des suprasylvischen Gyrus (SSG) und
suprasylvischen Sulcus (SSS, vgl. Abb. 21B) optokinetisch-relevante Bereiche wie bei der
Katze gibt oder zumindest einen „temporooccipitalen nystagmogenen Kortex“ wie beim
Kaninchen. Inwieweit diese Erwartung zutrifft, müßten zukünftige Untersuchungen zeigen.
In Bezug auf die kortikalen Schichten kann der Ursprung des kortikalen NOT-Eingangs
durchaus genau lokalisiert werden. Denn alle VC-Läsionen, die die Position von
richtungsspezifischen, durch das große Zufallspunktemuster treibbaren VC-Zellen
markierten, wurden in der Lamina V (vgl. Abb. 25) gefunden. Eine entsprechende VC-
Läsion beim pigmentierten Frettchen wurde nicht durchgeführt. Da aber das pigmentierte
Frettchen den Wildtyp repräsentiert und die Lamina V die normale Ausgangsschicht zum
NOT ist (Affe: Hoffmann et al., 1992; Katze: Schoppmann, 1981, Ratte: Schmidt et al.,
1993), gibt es keinen Grund anzunehmen, daß die entsprechenden VC-Zellen des
pigmentierten Frettchens nicht in der Lamina V liegen sollten.
In Bezug auf die Repräsentation des Gesichtsfeldes kann der Ursprung des kortikalen NOT-
Eingangs ebenfalls relativ genau bestimmt werden. Alle vom NOT antidrom reizbaren VC-
Zellen der pigmentierten und albinotischen Frettchen besaßen rezeptive Felder innerhalb des
zentralen Gesichtsfeldes (Azimut: -10° bis +25°, Elevation: +20° bis –15°) und überlappten
mit dem rezeptiven Feld des NOT-Reizortes. Für das pigmentierte Frettchen entsprach dieser
Befund der Erwartung und bestätigt auch den Befund von Klauer et al. (1990), die nur dann

Diskussion 134
eine orthodrome Reizung der NOT-Zellen vom VC durchführen konnten, wenn sich die
rezeptiven Felder des VC und der NOT-Zelle überlappten. Diese Beobachtung wurde auch in
der vergleichbaren Untersuchung bei der Katze von Schoppmann (1981) gemacht und zwingt
zu der Vermutung, daß ein korrekter kortikaler NOT-Eingang eine intakte Retinopie des
projizierenden visuellen Kortex voraussetzt. Deshalb war dieser Befund für das
Albinofrettchen, in Anbetracht seiner gestörten Retinotopie im visuellen Kortex (Guillery,
1971), keine Selbstverständlichkeit.
E. Retinaler Eingang des NOT
Die Retina ist der nächste mögliche Ort, der beim pigmentierten Frettchen für die
Determination der Richtungsspezifität des NOT und beim Albinofrettchen für die Aufhebung
dieser Richtungsspezifität in Frage kommt. Die Wahrscheinlichkeit, daß die Retina eine
solche Funktion erfüllt, erhöhte sich sogar, nachdem in den vorangegangenen Abschnitten
festgestellt wurde, daß der Kortex diese Funktion mit größerer Wahrscheinlichkeit nicht
erfüllt. Desweiteren sind in der Literatur Befunde beschrieben, die mindestens für eine
Prädetermination der Richtungsspezifität des NOT durch die Retina sprechen: zum einen
haben Hoffmann et al. (1995) beim Tamarwallaby die Augenanlage in einer frühen
Entwicklungsphase rotiert und später im adulten Tier festgestellt, daß die Vorzugsrichtung
des NOT entsprechend der Augenrotation mitrotiert hatte, und zum anderen geht aus
Entwicklungsstudien an Katzen hervor, daß erst die retinalen und dann die kortikalen Fasern
in den NOT einwachsen. Solange der NOT nur vom Auge innerviert ist, können die Katzen
nur einen für die Afoveaten typischen asymmetrischen monokularen hOKN machen. In
diesem Stadium ist der NOT aber schon richtungsspezifisch und funktionstüchtig, obwohl er
vom VC noch gar nicht innerviert ist. Einhergehend mit der später erfolgenden Innervation
vom VC erlangt der NOT seine Fähigkeit, höhere Reizgeschwindigkeiten zu beantworten,
und seine Binokularität, die eindeutig mit der Symmetriezunahme des monokularen hOKN
korreliert (Distler & Hoffmann, 1992, 1993).
1. Stärke und Leitungsgeschwindigkeit des retinalen NOT-Eingangs
Die elektrische Reizung der NOT-Zellen vom ON wurde nur beim Albinofrettchen
durchgeführt, da die entsprechenden Daten vom pigmentierten Frettchen in der Literatur
vorlagen (Klauer et al., 1990). Es wurde der kontralaterale ON gereizt, weil der NOT des

Diskussion 135
Albinofrettchens — im Gegensatz zum NOT des pigmentierten Frettchens — ausschließlich
vom kontralateralen Auge direkten retinalen Input bekommt (Zhang & Hoffmann, 1993).
Von 24 antidrom identifizierten NOT-Zellen des Albinofrettchens waren 22 Zellen (92 %)
orthodrom vom kontralateralen ON reizbar. Beim pigmentierten Frettchen konnten Klauer et
al. (1990) 90% (46 von 51) der richtungsspezifischen NOT-Zellen orthodrom vom optischen
Chiasma reizen. Die hohe Übereinstimmung der Prozentsätze legt die Schlußfolgerung nahe,
daß sich die retinalen NOT-Eingänge der beiden Tiergruppen hinsichtlich ihrer Stärke
nicht unterscheiden.
Die hier vorgestellten orthodromen Latenzen (vom ON) der von der IO identifizierten NOT-
Zellen des Albinofrettchens liegen im Bereich 5 bis 8 ms (Median = 6,0 ms; Q0,25 = 5,0 ms;
Q0,75 = 6,5 ms) und unterscheiden sich nicht signifikant von den entsprechenden Latenzen
des pigmentierten Tieres, die im Bereich 5,5 bis 8,5 ms verteilt sind (χ2-TEST, P = 0,248).
Dieses Testergebnis spiegelt sich in der besonders hohen Überlappung auf der x-Achse wider
(vgl. schraffierte Latenzen in Abbildung 39A mit den Latenzen in Abbildung 39C, C
entspricht der Fig. 11B in Klauer et al., 1990). Hieraus wird die Schlußfolgerung gezogen,
daß sich die retinalen NOT-Eingänge der beiden Tiergruppen hinsichtlich ihrer
Fortleitungsgeschwindigkeit nicht unterscheiden. Der Albinismus wirkt sich nicht auf die
Fortleitungsgeschwindigkeit, d.h. auf den Querschnitt der retinoprätektalen W-Fasern aus.
Zwar erscheint beim Albinofrettchen eine leichte Tendenz zu etwas kürzeren Latenzen
möglich. Aufgrund der vorliegenden Datenmaterials ist es aber unmöglich, diese Tendenz als
Folge eines größeren Faserquerschnitts zu interpretieren. Dies könnte — wenn überhaupt —
nur durch einen elektronenmikroskopischen Vergleich der Faserquerschnitte von albinoti-
schen und pigmentierten Frettchen ermöglicht werden. In diesem Zusammenhang ist zu
bedenken, daß die fehlende Richtungsspezifität der NOT-Zellen des Albinofrettchens wahr-
scheinlich dazu verleitete, diese Zellen möglichst oberflächlich im Prätektum zu suchen, da
bekannt ist, daß sich die NOT-Zellen innerhalb des hOKN-Schaltkreises relativ oberflächlich
befinden (Frettchen: Zhang & Hoffmann, 1993; Telkes et al., 2001; Katze: Ballas &
Hoffmann, 1985). Bei der Katze stellten Ballas & Hoffmann (1985) eine hochsignifikante
positive Korrelation zwischen der Tiefe der NOT-Zellen (bezogen auf die Oberfläche des
Prätektums) und ihren orthodromen Latenzen aus elektrischen Reizungen des optischen
Chiasmas (OX) fest. Das heißt, je oberflächlicher die Position der NOT-Zelle war, desto
größer war die Tendenz zu etwas kürzeren OX-Latenzen. Falls es beim Frettchen ebenfalls
eine solche Korrelation gibt, könnte die leichte Tendenz zu etwas kürzeren Latenzen beim
Albinofrettchens durchaus durch die Vorgehensweise bei der Suche bedingt sein.

Diskussion 136
Zusätzlich zur Reizung des kontralateralen ON reizten Klauer et al. (1990) auch am
optischen Chiasma (OX). In diesem Falle fanden sie orthodrome Latenzen (OX-Latenzen)
zwischen 4,3 und 7,0 ms mit einem Mittelwert von 5,42 ms ± 0,66 ms s.d.. In der vorliegen-
den Arbeit wurden beim Albinofrettchen keine OX-Latenzen gemessen. Deshalb können nur
die Latenzen der vom ON ausgelösten orthodromen Aktionspotentiale zum Vergleich
herangezogen werden. Diese Latenzen, die am Albinofrettchen gemessen wurden, sind
erwartungsgemäß etwas länger als die OX-Latenzen des pigmentierten Frettchens, weil die
ON-Reizelektroden ca. 6 mm anterior des optischen Chiasmas plaziert waren. Deshalb
wurde eine Distanz von mindestens 21 mm zwischen ON und NOT zugrunde gelegt, im
Gegensatz zu Klauer et al. (1990), die 15 mm zwischen OX und NOT am perfundierten Hirn
maßen. Berücksichtigt man diese zusätzlichen 6 mm Weges, die von den orthodromen
Aktionspotentialen auf dem Weg zum NOT zusätzlich zurückgelegt werden mußten, so
stehen die orthodromen Latenzen vom ON des Albinofrettchens auch mit den OX-Latenzen
des pigmentierten Frettchens durchaus in Einklang. Gleiche Verhältnisse findet man auch
zwischen den latenzen vom ON und den OX-Latenzen, wenn beide am pigmentierten
Frettchen gemessen werden (Klauer et al., 1990).
Aus dem Abstand zwischen den OX- und ON-Reizorten und dem entsprechenden Latenz-
unterschied berechneten Klauer et al. (1990) die mittlere Fortleitungsgeschwindigkeit der
retinoprätektalen Fasern des pigmentierten Frettchens (5,1 m/s). Da in der vorliegenden
Arbeit nur der ON gereizt wurde, kann keine entsprechende Leitungsgeschwindigkeit für das
Albinofrettchen berechnet werden. Aufgrund der hohen Kongruenz der Latenzverteilungen
(vgl. schraffierte Latenzen in Abb. 39A und Latenzen in Abb. 39C) ist jedoch davon
auszugehen, daß die mittlere Leitungsgeschwindigkeit beim Albinofrettchen im gleichen
Bereich liegt wie beim pigmentierten Frettchen. Folglich kann für das Albinofrettchen, so
wie von Klauer et al. (1990) für das pigmentierte Frettchen, die Schlußfolgerung gezogen
werden, daß der direkte, monosynaptische, retinale NOT-Eingang von langsam-leitenden
Ganglienzellfasern des W-Typs stammt. Diese Schlußfolgerung wird insbesondere durch den
Befund unterstützt, nach dem alle Latenzen vom ON der antidrom identifizierten NOT-
Zellen in die Gruppe der längeren Latenzen fallen (vgl. die zwei Gruppen von Latenzen in
Abb. 39A). Dies bedeutet, daß die retinalen Fasern, die auf die NOT-Zellen projizieren,
langsamer leiten als die Fasern, die benachbarte prätektale Zellen kontaktieren. Für diese
Schlußfolgerung sprechen auch die diesbezüglichen Befunde bei der Katze (Hoffmann &
Schoppmann, 1975; Hoffmann & Stone, 1985), bei der die Axone der W-Zellen ebenfalls die
niedrigste Fortleitungsgeschwindigkeit unter den retinofugalen Fasern besitzen (Stone &

Diskussion 137
Hoffmann, 1972; Stone & Fukuda, 1974) und die von der IO antidrom reizbaren NOT-Zellen
genau von diesem Ganglienzelltyp retinalen Eingang erhalten (Hoffmann et al., 1976). Die
hohe Ähnlichkeit zwischen den Ganglienzellen des Frettchens und der Katze spiegelt sich
auch in der morphologischen Klassifikation der Ganglienzellen wider. Bei der Katze sind die
retinalen Ganglienzellen als α-, β- und γ-Zellen klassifiziert (Boycott & Wässle, 1974). Die
Ganglienzellen des Frettchens werden ebenfalls in drei morphologische Hauptgruppen
eingeteilt, die mit den Gruppen der Katze vergleichbar sind und deshalb als α-, β- und
γ-ähnlich bezeichnet werden (Vitek et al., 1985; Henderson, 1985).
An dieser Stelle werden folgende Fragen diskutiert:
• Warum wurden beim Albinofrettchen zwei Gruppen von orthodromen Latenzen, d.h. zwei
Typen von retinoprätektalen Ganglienzellfasern, gefunden, während Klauer et al. (1990)
beim pigmentierten Frettchen nur eine Gruppe von entsprechenden Latenzen
dokumentieren? Gibt es beim pigmentierten Frettchen keine retinoprätektalen Fasern, die
den schnelleren Fasern des Albinofrettchens entsprechen?
Die wahrscheinlichste Erklärung dieses Unterschieds ist, daß Klauer et al. (1990) aufgrund
ihrer methodischen Vorgehensweise nur den W-Typ detektierten, und daß das pigmentierte
Frettchen ebenfalls die schnelleren Fasern besitzt. Denn sie reizten beim pigmentierten
Frettchen nur die gut isolierten richtungsspezifischen NOT-Neuronen, die genau den von
der IO antidrom identifizierten, richtungsunspezifischen NOT-Neuronen des Albinofrett-
chens entsprechen. Folglich konnten sie nur solche Latenzen vom ON und vom OX finden,
die der Gruppe der längeren Latenzen (vom ON) des Albinos entsprechen. Da nahezu alle
(35 von 36) prätektalen Zellen mit den kurzen Latenzen (vom ON) beim Albinofrettchen
nicht von der IO antidrom reizbar waren, müssten die analogen Zellen beim pigmentierten
Frettchen höchstwahrscheinlich richtungsunspezifisch gewesen sein und deshalb für
Klauer et al. (1990) nicht von Interesse. Diese Interpretation berücksichtigt auch die
Befunde bei der Katze, bei der zwar gemäß der Klassifikation von Stone & Hoffmann
(1972) alle drei Ganglienzell-Typen (W-, X- und Y-Zellen) Axone zum Prätektum bzw.
NOT schicken, die richtungsspezifischen NOT-Zellen im Sinne des hOKN aber nur von
W-Zellen kontaktiert werden (Hoffmann & Schoppmann, 1981; Ballas et al., 1981).
• Gibt es eine andere Möglichkeit, längere orthodrome Latenzen vom ON bei prätektalen
Zellen zu messen, auch wenn sie nicht von W-Ganglienzellen direkt kontaktiert werden
oder vielleicht überhaupt keinen direkten retinalen Eingang erhalten? Diese Frage ist mit
`ja´ zu beantworten, denn es ist durchaus möglich, daß die prätektalen Zellen nicht nur

Diskussion 138
direkt, sondern auch indirekt über das CGL und den Kortex aktiviert werden. In diesem
Falle würde die zeitliche Verzögerung der orthodromen Aktionspotentiale, die durch
mehrere Synapsen und das Zurücklegen des längeren Weges verursacht wird, die
schnellere Fortleitung über Y-Fasern ausgleichen.
• Zu welchem Ganglienzelltyp gehören die zum Prätektum ziehenden Fasern mit der höheren
Leitungsgeschwindigkeit beim Albinofrettchen? Da der W-Typ der Gruppe der längeren
ON-Latenzen zugeordnet wurde, bleiben nur der X- und der Y-Typ, die in Frage kommen
könnten. Die Antwort auf diese Frage muß zur Zeit offen bleiben, denn unglücklicherweise
sind beim Frettchen die verschiedenen Ganglienzelltypen noch nicht physiologisch
eindeutig charakterisiert, so daß die zeitliche Differenz zwischen der Gruppe der längeren
und der Gruppe der kürzeren Latenzen in Abbildung 39A zur Beantwortung dieser Frage
nicht herangezogen werden kann. Möglich erscheinen sowohl die X- als auch die Y-
Fasern. Bei der Katze jedenfalls sind sowohl X- als auch Y-Fasern beschrieben, die in den
NOT ziehen, jedoch nicht auf die richtungsspezifischen NOT-Zellen projizieren (Hoffmann
& Schoppmann, 1981; Hoffmann & Stone, 1985). Unter Berücksichtigung der Tatsache,
daß in unmittelbarer Nähe zu den von der IO antidrom identifizierten NOT-Zellen auch
Jerk-Zellen gefunden wurden, erscheinen die Y-Fasern am wahrscheinlichsten. Denn bei
der Katze bekommen die Jerk-Zellen monosynaptischen retinalen Eingang von schnellen
Y-Fasern (Ballas & Hoffmann, 1985). Dies könnte auch beim Frettchen der Fall sein und
deshalb erscheint es wahrscheinlich, daß die kurzen Latenzen vom ON an Jerk-Zellen
gemessen wurden. Desweiteren liefern Ballas & Hoffmann (1985) Hinweise, die gegen
eine Projektion der Jerk-Zellen zur IO sprechen. Dies paßt zu der eigenen Beobachtung,
nach der keine einzige Jerk-Zelle von der IO gereizt werden konnte.
Bei Betrachtung aller Teilbefunde aus den ON-Reizexperimenten wird die Schlußfolgerung
gezogen, daß die bisherigen Untersuchungen des retinalen NOT-Eingangs in Bezug auf die
Stärke und Leitungsgeschwindigkeit der Retina-NOT-Verbindung keinen Unterschied
zwischen pigmentierten und albinotischen Frettchen aufgedecken. Insoweit scheint beim
Albinofrettchen der direkte NOT-Eingang von der kontralateralen Retina
elektrophysiologisch normal zu sein.
2. Informationsgehalt des retinalen NOT-Eingangs
Es bleibt zur Zeit ungeklärt, ob die retinale Projektion zum NOT beim Albinofrettchen auch
hinsichtlich des Informationsgehaltes normal ist. Denn es gibt weder beim pigmentierten,

Diskussion 139
noch beim albinotischen Frettchen Untersuchungen, die die Klärung dieser Frage ermögli-
chen. Hierzu wäre z. B. — analog zu der antidromen Reizung der VC-Zellen vom NOT —
eine antidrome Reizung der retinalen Ganglienzellen vom NOT notwendig. Die vom NOT
antidrom identifizierten retinalen Ganglienzellen müßten in beiden Tiergruppen
optokinetisch gereizt und hinsichtlich ihrer Richtungsspezifität charakterisiert werden.
Experimente solcher Art wurden bei der Katze von Hoffmann & Stone (1985) durchgeführt.
Dabei konnten aus einer Population von 578 untersuchten Ganglienzellen elf Zellen vom
W-Typ identifiziert werden, die mit größter Wahrscheinlichkeit den retinalen Eingang für
den hOKN vermittelten. Fünf von diesen elf Ganglienzellen waren ON-Zentrum
richtungsspezifische Zellen, die restlichen sechs Zellen hatten uneinheitliche rezeptive
Felder. Beim pigmentierten Frettchen sind bis dato keine richtungsspezifischen, zum NOT
projizierenden retinalen Ganglienzellen beschrieben. Unter der Annahme, daß solche Zellen
existierten, würde sich zwangsläufig die Frage stellen, ob sie auch beim albinotischen
Frettchen präsent und richtungsspezifisch sind. In diesem Falle könnte folgende Hypothese
aufgestellt werden: Die retinalen, zum NOT projizierenden Ganglienzellen, die beim
pigmentierten Frettchen richtungsspezifisch sind, haben beim Albinofrettchen ihre
Richtungsspezifität verloren. Folglich übermitteln sie dem NOT eine abnorme, richtungs-
unspezifische Information und sind deshalb für die Richtungsunspezifität der NOT-
Ausgangsneuronen und als Konsequenz daraus für die optokinetische Blindheit des
Albinofrettchens verantwortlich.
Die Befunde von Knapp et al. (1988) unterstützen die Erkenntnis von Hoffmann & Stone
(1985), daß es sich bei den richtungsspezifischen retinalen W-Ganglienzellen der Katze um
Zellen handelt, die den hOKN antreiben. Knapp et al. (1988) inaktivierten mit einer
Augeninjektion von 2-Amino-4-phosphonobutyrat (APB) selektiv die retinalen ON-Bipolar-
zellen und blockierten damit die ON-Ganglienzellen und die ON-Antworten der
ON-OFF-Ganglienzellen. Bei der intakten Katze zeigte die APB-Injektion zunächst keine
erkennbare Wirkung, höchstwahrscheinlich weil bei dieser Spezies der kortikale Beitrag zur
Auslösung des hOKN ausreichend ist. Doch bei zwei Katzen mit bilateralen kortikalen
Läsionen führte die gleiche Injektion dazu, daß die Versuchstiere mit dem injizierten Auge
keinen monokularen hOKN mehr machen konnten. Dies zeigt bei der Katze, daß die APB-
Blockade der ON-Ganglienzellen die Inaktivierung eines subkortikalen, retinoprätektalen
Pfades zur Folge hat. Beim afoveaten Kanninchen führte das APB auch ohne kortikale
Läsionen zum hOKN-Verlust des injizierten Auges. Dies war zu erwarten, da der hOKN des
Kaninchens im wesentlichen subkortikal vermittelt und durch seine, seit den sechziger

Diskussion 140
Jahren des 20. Jahrhunderts bekannten, richtungsspezifischen ON- und ON-OFF-
Ganglienzellen (Barlow & Hill, 1963; Oyster & Barlow, 1967; Taylor et al., 2000)
kontrolliert wird (Oyster et al., 1972).
Auch beim Frettchen könnten die entsprechenden Experimente mit APB sinnvoll sein. In
diesem Falle müßte nach Injektion von APB in die Retinae von pigmentierten Frettchen der
hOKN gemessen und das physiologische Verhalten der NOT-Zellen abgeleitet werden. Führt
die APB-Injektion zu optokinetischer Blindheit und Richtungsunspezifität der NOT-Neuro-
nen beim pigmentierten Frettchen? Die Antwort auf diese Frage wäre auch eine Aussage
darüber, ob die Richtungsspezifität der NOT-Zellen schon in der Retina determiniert ist.
In diesem Kontext ist die in der vorliegenden Arbeit vorgenommene Charakterisierung der
prätektalen und SC-Zellen hinsichtlich ihrer ON/OFF-Eigenschaften zu sehen. Denn die
APB-Augeninjektionen beim Kaninchen und bei der Katze (Knapp et al., 1988) zeigen, daß
die ON-Ganglienzellen eine entscheidende Rolle bei der Erzeugung von Richtungsspezifität
innerhalb des retinalen Netzwerkes spielen. Folglich müßte jeder Unterschied zwischen
pigmentierten und albinotischen Frettchen hinsichtlich des relativen Vorkommens an ON-
Zellen bzw. ON-Antworten im Mittelhirn ein indirekter Anhaltspunkt über eine mutmaßliche
Anomalie im retinalen Netzwerk des Albinos sein und somit auch hinsichtlich des
neuronalen Substrats der angenommenen Richtungsspezifität in der Retina. Die
Charakterisierung der SC-Zellen wurde bei beiden Tiergruppen durchgeführt und erlaubt
somit einen solchen Vergleich. Es wurde jedoch kein signifikanter Unterschied zwischen
albinotischen und pigmentierten Frettchen entdeckt (RANG-SUMMEN-TEST, P = 0,275).
Insofern hat die Bestimmung der ON/OFF-Eigenschaften im SC keine Hinweise über eine
mutmaßliche Veränderung des retinalen Netzwerks geliefert. Dennoch wird beim Albino-
frettchen eine leichte Tendenz zu mehr ON-Zellen bzw. mehr / stärkeren ON-Antworten
beobachtet, was zu einem etwas größeren Medianwert (48%) der ONF-Indizes führte
(43% beim pigmentierten Frettchen). In diesem Falle könnten immunocytochemische
Methoden, die die ON-Ganglienzellen markieren, eher signifikante Unterschiede zwischen
den beiden Tiergruppen aufdecken.
Ähnlich wie bei der Diskussion des kortikalen NOT-Eingangs, muß auch hier die mögliche
Anordnung der NOT-Eingangsfasern betrachtet werden. Selbst wenn die Richtungsspezifität
des NOT beim pigmentierten Frettchen durch einen richtungsspezifischen retinalen Eingang
determiniert würde und der retinale Eingang auch beim Albinofrettchen richtungsspezifisch
wäre, könnte der Verlust der Richtungsspezifität immer noch durch eine abnorme

Diskussion 141
Anordnung der retinalen NOT-Eingangsfasern bewirkt sein. Man könnte sich beispielsweise
beim pigmentierten Frettchen einen geordneten retinalen Eingang vorstellen, d.h. alle Fasern,
die in einen NOT ziehen, haben die gleiche ipsiversive Vorzugsrichtung wie die NOT-
Ausgangsneuronen. Beim Albinofrettchen könnte dann angenommen werden, daß die
entsprechende Faseranordnung gestört ist. Eine NOT-Zelle könnte gleichzeitig von mehreren
Ganglienzellfasern mit unterschiedlichen, entgegengesetzten Vorzugsrichtungen kontaktiert
werden, so daß das Resultat der synaptischen Integration ihre Richtungsunspezifität ist.
Angesichts der Tatsache, daß beim Albinofrettchen die retinalen Fasern aus der temporalen
Retina abnormerweise zum falschen, d.h. kontralateralen NOT ziehen, erscheint dieses
Szenario durchaus wahrscheinlich. Deshalb stellt sich hier die Frage, ob der Verlust der
Richtungsspezifität der NOT-Zellen des Albinofrettchens durch das gleichzeitige Eintreffen
von normalen und invertierten Richtungssignalen von der Retina verursacht sein könnte?
Diese Frage läßt sich jedoch zur Zeit nicht beantworten, da es keine Informationen über die
Anordnung der retinalen NOT-Eingangsfasern des Frettchens gibt.
In diesem Zusammenhang sind die zwei von der IO antidrom reizbaren NOT-Zellen des
Albinofrettchens zu erwähnen, die eine invertierte Vorzugsrichtung mit einem stark
negativen DS-Index (–0,30 und –0,62; vgl. Abb. 29A) haben. Man könnte sich
beispielsweise vorstellen, daß diese beiden Zellen ausnahmsweise nur ein invertiertes Signal
von der kontralateralen temporalen Hemiretina bekommen und deshalb die invertierte
Vorzugsrichtung haben.
Diese beiden Zellen erinnern an die Befunde von Winterson & Collewijn (1981), die beim
albinotischen Kaninchen unter anderem NOT-Zellen beschreiben, die zwei entgegengesetzte
Vorzugsrichtungen haben. Die Vorzugsrichtung dieser Zellen hängt davon ab, ob sie nur im
anterioren oder nur im posterioren Teil ihres rezeptiven Feldes visuell gereizt werden. Wenn
die visuelle Reizung nur im anterioren Teil des rezeptiven Feldes beschränkt war, bevorzug-
ten die NOT-Zellen die kontraversive Reizrichtung, die im Vergleich zu der Vorzugsrichtung
der meisten NOT-Zellen des pigmentierten Kaninchens invertiert ist. Bei ausschließlicher
Reizung des posterioren Anteils ihres rezeptiven Feldes präferierten dieselben NOT-Zellen
die ipsiversive, d.h. die normale, Reizbewegungsrichtung. Unter den beiden Annahmen, daß
der NOT-Ausgang beim Albinokaninchen nicht gestört ist, sondern nur der NOT-Eingang,
und daß beim pigmentierten Kaninchen eine retinale Projektion zum ipsilateralen NOT
existiert (vgl. jedoch Klooster et al., 1983), die beim Albinokaninchen nach kontralateral
„umgeleitet“ wird, erklären Winterson & Collewijn (1981) diesen Befund durch ein
invertiertes Richtungssignal, das den NOT-Zellen von der Retina übermittelt wird. Fasern

Diskussion 142
aus der temporalen Hemiretina, die im Normalfall (d.h. beim pigmentierten Kaninchen) eine
retinale Bildverschiebung („retinal slip“) auf der temporalen Hemiretina zum ipsilateralen
NOT melden, kreuzen fälschlicherweise am optischen Chiasma des Albinos und übermitteln
das "richtige" Signal zum "falschen" NOT. Dies führt dazu, daß die NOT-Zellen eine
kontraversive Reizbewegung als eine ipsiversive interpretieren und den falschen Nystagmus
auslösen. Auf der Verhaltensebene wird, wie im Verhaltensteil schon diskutiert, der
invertierte Nystagmus beobachtet, wenn das Albinokaninchen nur im frontalen Gesichtsfeld
optokinetisch gereizt wird (Collewijn et al., 1978). Dieselbe Erklärung kann auch für den
invertierten Nystagmus der Albinoratten von Sirkin et al. (1985) und der hypopigmentierten
Mäuse von Mangini et al. (1985) gegeben werden.
Deshalb zwingt sich an dieser Stelle die Frage auf, ob die invertierte Vorzugsrichtung der
beiden NOT-Zellen des Albinofrettchens ebenfalls durch ein invertiertes Richtungssignal
von der Retina verursacht sein könnte? Zur Zeit ist eine eindeutige Beantwortung dieser
Frage nicht möglich. Die bisherigen Befunde sprechen eher gegen die Existenz eines
invertierten Signals:
• Die invertierte Vorzugsrichtung dieser beiden NOT-Zellen wurde gefunden, als die
optokinetische Reizung nicht nur auf das frontale Gesichtsfeld beschränkt war, sondern
sich auch auf weite Teile des temporalen Gesichtfeldes erstreckte.
• Als bei einem anderen Albinofrettchen die optokinetische Stimulation probeweise auf das
frontale (oder temporale) Gesichtsfeld beschränkt wurde, erlangten die von der IO
identifizierten NOT-Zellen keine Richtungsspezifität mit invertierter (oder normaler)
Vorzugsrichtung (unpublizierte Beobachtung). Diese Beobachtung unterstützt auch die
Schlußfolgerung aus dem Ergebnis des monokularen Perimetrietests: Die Schlußfolgerung
war, daß die aberrante Projektion der temporalen Hemiretina zum kontralateralen SC
funktionell unterdrückt ist und deshalb auch die aberrante Projektion zum kontralateralen
NOT ebenfalls funktionell unterdrückt sein sollte. Daraus ergab sich die Spekulation, daß
die optokinetische Reizung der temporalen Hemiretina (Teilfeldstimulation) keinen
invertierten hOKN auslösen sollte. Die entsprechende Spekulation für die Physiologie ist,
daß die optokinetische Reizung der temporalen Hemiretina keine invertierte
Vorzugsrichtung im NOT hervorrufen sollte.
• Hinzu kommt, daß die Verhältnisse beim Kaninchen ohnehin anders sein müssen als beim
Frettchen. Denn ca. 25% der NOT-Zellen haben selbst beim pigmentierten Kaninchen eine
invertierte Vorzugsrichtung (Pettorossi & Troiani, 1983; Maekawa et al., 1984). Beim

Diskussion 143
pigmentierten Frettchen sind bis dato keine NOT-Zellen mit invertierter Vorzugsrichtung
beschrieben.
Abschließend kann zusammenfassend festgestellt werden:
• Es ist nicht nachgewiesen, ob die retinalen Ganglienzellfasern des Frettchens, die den NOT
innervieren, richtungsspezifisch sind. Da die entsprechenden Fasern bzw. Ganglienzellen
bei der Katze und beim Kaninchen richtungsspezifisch sind, würde man jedoch erwarten,
daß sie auch beim Frettchen richtungsspezifisch sind.
• Die Richtungsunspezifität der NOT-Ausgangsneuronen des Albinofrettchens kann nicht
durch einen in Bezug auf die Vorzugsrichtung ungeordneten retinalen Eingang begründet
werden, da diesbezügliche Untersuchungen noch nicht durchgeführt wurden. Unter der
Annahme jedoch, daß der retinale NOT-Eingang richtungsspezifisch ist, erscheint eine
solche Begründung grundsätzlich wegen des aberraten Faserverlaufs aus den temporalen
Hemiretinae sehr wahrscheinlich.
• Die invertierte Vorzugsrichtung der beiden NOT-Zellen des einen Albinofrettchens kann
zur Zeit ebenfalls nicht erklärt werden. Eine Erklärung durch ein invertiertes Signal von
der kontralateralen temporalen Hemiretina erscheint eher unwahrscheinlich, kann aber
nicht ausgeschlossen werden.
• Weitere Untersuchungen sind notwendig, um den Beitrag der Retina zur Determination der
Richtungsspezifität des NOT abschließend beurteilen zu können.
F. Das morphologische Korrelat zur Richtungsunspezifität der NOT-Neuronen
Zu Beginn der Diskussion wurde der NOT selbst als dritter möglicher Ort in Betracht
gezogen, an dem die Richtungsspezifität der NOT-Ausgangsneuronen beim pigmentierten
Frettchen determiniert werden könnte. In diesem Falle müßte die Existenz von speziellen
intrinsischen Verschaltungen im NOT angenommen werden, da sie bis heute nicht
nachgewiesen sind (Horn & Hoffmann, 1987). Beim Albinofrettchen wäre es dann sehr
wahrscheinlich, daß seine richtungsunspezifischen NOT-Ausgangsneuronen auch
anatomisch verändert sind. Eine mögliche Ursache für eine anatomische Veränderung wäre,
daß die NOT-Neuronen während einer kritischen Phase ihrer Reifung zur Entwicklung ihrer
Richtungsspezifität einen spezifischen retinalen und / oder kortikalen Eingang benötigten,
der beim Albinofrettchen möglicherweise nicht vorhanden ist. Aus diesen Überlegungen

Diskussion 144
heraus ergibt sich die Hypothese, daß ein morphologisches Korrelat zur Richtungsunspezifi-
tät der NOT-Zellen des Albinofrettchens existiert.
Die Überprüfung dieser Hypothese wurde von Telkes et al. (2001) mit histologischen
Methoden an gleichaltrigen albinotischen und pigmentierten Frettchen unternommen. Sie
markierten retrograd die NOT-Zellen mit Hilfe von Meerrettichperoxidase- und Biocytin-
Dextran-Injektionen in die ipsilaterale IO. Damit entsprechen die von Telkes et al. (2001)
untersuchten NOT-Zellen genau den sogenannten Bildverschiebungsneuronen, die in der
vorliegenden Arbeit anhand ihrer antidromen Reizbarkeit von der IO identifiziert wurden.
Ihre Versuchstiere stammten aus derselben Kolonie, wie die in der vorliegenden Arbeit
untersuchten Tiere. Einige ihrer Tiere stellen eine Teilmenge der in dieser Arbeit verwandten
Tiere dar. Der von Telkes et al. (2001) durchgeführte morphologische Vergleich zwischen
den NOT-Zellen des albinotischen und pigmentierten Frettchens belegt tatsächlich
signifikante Unterschiede zwischen den beiden Tiergruppen. Die NOT-Zellen des Albino-
frettchens haben signifikant weniger und signifikant kürzere Dendriten als die NOT-Zellen
des pigmentierten Frettchens. Damit einhergehend findet man bei den NOT-Zellen des
Albinofrettchens kein oder ein viel schwächeres dendritisches Netzwerk als bei den NOT-
Zellen des pigmentierten Frettchens.
In diesem Kontext sollten nochmals die Häufigkeitsverteilungen der antidromen Reiz-
latenzen von der IO der NOT-Zellen des albinotischen (Abb. 29B) und pigmentierten
(Abb. 29D) Frettchens betrachtet werden. Der RANG-SUMMEN-TEST zeigte hinsichtlich der
Mediane zwar keinen signifikanten Unterschied zwischen den Reizlatenzen der beiden
Tiergruppen. Beim Albinofrettchen wurde aber eine Tendenz zu längeren Latenzen
beobachtet. Diese Tendenz kann als Indiz dafür angesehen werden, daß die zur IO ziehenden
Axone etwas dünner sind als beim pigmentierten Frettchen. Dies würde einen weiteren
morphologischen Unterschied zwischen den NOT-Neuronen des albinotischen und des
pigmentierten Frettchens bedeuten, der allerdings nur durch elektronenmikroskopische
Untersuchungen eindeutig nachgewiesen werden könnte. Andererseits scheint es keinen
Unterschied zu geben hinsichtlich der Stärke der NOT-IO-Verbindung, denn es konnten bei
beiden Tiergruppen in der gleichen prozentualen Größenordnung prätektale Zellen von der
IO antidrom gereizt werden [81% (30 von 37) bei den albinotischen und 83% (24 von 29) bei
den pigmentierten Frettchen]. Dieser Befund steht wiederum mit dem Befund von Telkes et
al. (2001) in Einklang, wonach sie in beiden Tiergruppen gleichviele NOT-Zellen von der
ipsilateralen IO retrograd markiert haben.

Zusammenfassung 145
Zusammenfassung
1. Teil I: Verhalten
• Die Verhaltensexperimente testeten die Auslösbarkeit des horizontalen optokinetischen
Nystagmus (hOKN) bei albinotischen und pigmentierten Frettchen. Hierzu wurden die
horizontalen Augenbewegungen der Frettchen während optokinetischer Ganzfeldreizung
mittels unkalibrierter Elektrookulographie (EOG) vermessen. Die Reizungen erfolgten in
der optokinetischen Trommel mit zwei unterschiedlichen Zufallspunktemustern
(Punktgröße: 2,3° und 10,7° Sehwinkel) und unterschiedlichen Reizgeschwindigkeiten (5,
10, 20, 50 und 100°/s).
• Das Vermessen der spontanen Augenbewegungen bei stehender Trommel deckte beim
albinotischen, im Gegensatz zum pigmentierten Frettchen, eine okulomotorische
Instabilität auf.
• Die bino- und monokulare optokinetische Reizung mit dem feinen Reizmuster und die
monokulare Reizung mit dem groben Reizmuster konnten bei den Albinofrettchen, im
Gegensatz zu den pigmentierten Frettchen, keinen hOKN auslösen.
• Die binokulare optokinetische Reizung mit dem groben Reizmuster konnte beim
Albinofrettchen ebenfalls keinen hOKN auslösen, im Gegensatz zum pigmentierten
Frettchen. Allerdings wurde festgestellt, daß die Augen des Albinos, die bei fehlender
optokinetischer Reizung in rein zufälliger Richtung abdriften, bei Reizung mit dem
groben Muster dazu tendieren, in Reizrichtung abzudriften.
• Der Perimetrietest belegt, daß sich das Albinofrettchen unter binokularen Bedingungen
genau so gut orientieren kann, wie das pigmentierte Frettchen. Dieser Befund deutet
darauf hin, daß der optische Apparat des Albinofrettchens intakt und somit nicht für das
okulomotorische Verhaltensdefizit verantwortlich ist.
• Aus den monokularen Perimetriemessungen folgt, daß das Albinofrettchen im Gegensatz
zum pigmentierten Frettchen kein binokulares Gesichtsfeld besitzt. Denn jedes Auge des
Albinos kann nur die ipsilaterale Gesichtsfeldhälfte sehen, jedoch nicht den zentralen

Zusammenfassung 146
Anteil der kontralateralen Gesichtsfeldhälfte, der vom entsprechenden Auge des
pigmentierten Frettchens gesehen wird.
• Bei einigen Versuchstieren wurde mit Hilfe eines implantierten Führungsröhrchens
einseitig Muscimol auf das Prätektum in die Nähe des Nukleus des Optischen Traktes
(NOT) injiziert.
• Die Muscimol-Injektionen führten sowohl bei albinotischen als auch bei pigmentierten
Frettchen zu einem reversiblen Spontannystagmus mit einer langsamen Phase in Richtung
auf die nicht-injizierte Hirnseite. Der Spontannystagmus trat in Dunkelheit und bei
Helligkeit auf und konnte durch die horizontale optokinetische Reizung in der Trommel
nicht sichtbar beeinflußt werden. Die anschließende histologische Untersuchung zeigte,
daß der Spontannystagmus nicht auf eine mechanische Zerstörung des NOT
zurückgeführt werden kann. Dies ist eine Schlußfolgerung, für die auch die vollkommene
Reversibilität des Spontannystagmus spricht. Das Muscimol inaktivierte chemisch den
NOT der injizierten Hirnseite und stellte damit ein Aktivitätsungleichgewicht zwischen
den beiden NOTs her. Weiterhin zeigte der Spontannystagmus, daß der Ausfall des
hOKN, der bei den Albinofrettchen vor der Muscimol-Anwendung mit optokinetischer
Ganzfeldreizung festgestellt wurde, nicht durch einen dem NOT nachfolgenden
visuomotorischen Defekt begründet werden kann. Folglich müßte der neuronale Defekt
im NOT selbst und / oder dem NOT vorgeschaltet sein.
• Bei Betrachtung aller Befunde wird die Schlußfolgerung gezogen, daß das albinotische
Frettchen aufgrund eines Defektes im visuellen System optokinetisch blind ist.
2. Teil II: Neurophysiologie
a) Richtungsunspezifität der NOT-Neuronen
Nach der Entdeckung der optokinetischen Blindheit der Albinofrettchen und der
Eingrenzung des zugrundeliegenden Defektes mit Verhaltensexperimenten, stellte sich die
Frage nach einem physiologischen Korrelat dieses Defektes. Da der NOT mit seinen, im
Normalfall richtungsspezifischen Ausgangsneuronen innerhalb des neuronalen hOKN-
Schaltkreises eine grundlegende Rolle spielt, wurde er das Ziel der elektrophysiologischen
Untersuchungen:
• Es kann festgehalten werden, daß die zwölf untersuchten Albinofrettchen — im
Gegensatz zu den fünf pigmentierten Frettchen — signifikant weniger (fast keine)
richtungsspezifische NOT-Neuronen besitzen. Diese Richtungsunspezifität der NOT-

Zusammenfassung 147
Neuronen stellt das physiologische Korrelat der optokinetischen Blindheit der
Albinofrettchen dar. Da diese NOT-Neuronen nicht anhand ihrer Richtungsspezifität
identifiziert werden konnten, wurden sie anhand ihrer antidromen Reizbarkeit von der
ipsilateralen inferioren Olive (IO) identifiziert.
• Die antidromen Reizlatenzen der NOT-Neuronen der beiden Tiergruppen unterscheiden
sich nicht statistisch signifikant voneinander. Es ist jedoch auffällig, daß sie sich beim
pigmentierten Frettchen um einen niedrigeren Wert häufen als beim Albinofrettchen.
b) Kortikaler Eingang der NOT-Neuronen
Das Ziel der Untersuchungen des kortikalen NOT-Eingangs war die Entscheidung, ob der
Kortex bzw. der kortikale NOT-Eingang für den Verlust der Richtungspezifität der NOT-
Zellen des Albinofrettchens verantwortlich ist:
• Beim Albinofrettchen wurden 63% der von der IO antidrom identifizierten NOT-Zellen
orthodrom vom VC gereizt. Der entsprechende Prozentsatz für das pigmentierte Frettchen
kann der Literatur entnommen werden und beträgt 65% (Klauer et al., 1990). Damit sind
bei beiden Tiergruppen prozentual gleichviele NOT-Zellen vom ipsilateralen VC
aktivierbar. Hieraus folgt, daß sich der kortikale NOT-Eingang des Albinofrettchens
hinsichtlich seiner Stärke nicht vom entsprechenden Eingang des pigmentierten
Frettchens unterscheidet. Weiterhin wird aus den gemessenen orthodromen Latenzen
geschlußfolgert, daß sich die beiden Eingänge hinsichtlich ihrer Leitungsgeschwindigkeit
ebenfalls nicht unterscheiden.
• Das Antwortverhalten der NOT-Zellen auf die elektrische Reizung des ipsilateralen VC
ist bei albinotischen und pigmentierten Frettchen gleich. Bei beiden Tiergruppen
antworteten die NOT-Zellen mit zwei bis drei orthodromen Aktionspotentialen auf die
Reizung des VC.
• Im visuellen Kortex des Albinofrettchens findet man richtungsspezifische Neuronen in
der gleichen prozentualen Größenordnung (57% der VC-Zellen mit DS-Index > 0,2) wie
beim pigmentierten Frettchen (55% der VC-Zellen mit DS-Index > 0,2). Hinsichtlich der
Vorzugsstärke der VC-Zellen existiert ebenfalls kein signifikanter Unterschied zwischen
albinotischen und pigmentierten Frettchen.
• Sowohl beim albinotischen als auch beim pigmentierten Frettchen sind unter den
richtungsspezifischen VC-Zellen alle Vorzugsrichtungen gleichmäßig stark repräsentiert.
• Sowohl beim albinotischen als auch beim pigmentierten Frettchen findet man VC-Zellen,
die antidrom vom ipsilateralen NOT reizbar sind. Unter diesen Zellen befinden sich neben

Zusammenfassung 148
richtungsunspezifischen, sowohl schwach richtungsspezifische (niedrige DS-I-Werte) als
auch stark richtungsspezifische (hohe DS-I-Werte) Zellen.
• Die vom NOT antidrom reizbaren VC-Zellen des Albinofrettchens befinden sich in der
kortikalen Schicht V.
• Beim Albinofrettchen waren 32% und beim pigmentierten Frettchen 27% der getesteten
VC-Zellen vom NOT antidrom reizbar. Die gleiche Größenordnung dieser Prozentsätze
unterstützt die obige Schlußfolgerung aus den Befunden der orthodromen Reizung der
NOT-Zellen vom VC, daß sich die kortikalen NOT-Eingänge der beiden Tiergruppen
hinsichtlich ihrer Stärke nicht voneinander unterscheiden.
• Die vom NOT antidromen Reizlatenzen der VC-Zellen des albinotischen Frettchens
unterscheiden sich nicht signifikant von den entsprechenden Latenzen des pigmentierten
Frettchens. Dieser Befund unterstützt die obige Schlußfolgerung, daß sich die kortikalen
NOT-Eingänge der beiden Tiergruppen hinsichtlich ihrer Leitungsgeschwindigkeit
ebenfalls nicht voneinander unterscheiden.
c) Retinaler Eingang der NOT-Neuronen
Der retinale NOT-Eingang wurde bei fünf Albinofrettchen untersucht.
• Der Prozentsatz antidrom identifizierter NOT-Zellen, die vom kontralateralen optischen
Nerv (ON) orthodrom reizbar waren, liegt beim Albinofrettchen mit 92% in der gleichen
Größenordnung wie der Prozentsatz richtungsspezifischer NOT-Zellen des pigmentierten
Frettchens (90%, aus Klauer et al., 1990), die ebenfalls vom ON orthodrom reizbar
waren. Hieraus wird die Schlußfolgerung gezogen, daß sich die retinalen NOT-Eingänge
bei beiden Tiergruppen hinsichtlich ihrer Stärke nicht unterscheiden.
• Die Häufigkeitsverteilungen der orthodromen Reizlatenzen vom ON unterscheiden sich
nicht zwischen dem albinotischen und dem pigmentierten Frettchen. Hieraus folgt die
Schlußfolgerung, daß sich die retinalen NOT-Eingänge der beiden Tiergruppen auch
hinsichtlich ihrer Leitungsgeschwindigkeit nicht unterscheiden.
• Das Antwortverhalten der NOT-Zellen auf die elektrische Reizung des ON läßt keinen
Unterschied zwischen albinotischen und pigmentierten Frettchen erkennen. Bei beiden
Tiergruppen antworteten die NOT-Zellen mit zwei bis fünf orthodromen Aktions-
potentialen auf die Reizung des ON.
• Obwohl die bisherigen elektrophysiologischen Daten vom Albinofrettchen mit den
entsprechenden Daten vom pigmentierten Frettchen übereinstimmen, wird das
physiologische Korrelat der fehlenden Richtungsspezifität der NOT-Zellen des Albino-

Zusammenfassung 149
frettchens in einem retinalen Defekt vermutet. Hier könnten die zum NOT projizierenden
Ganglienzellen ihre Richtungsspezifität verloren haben, oder aber ihre unspezifische
Projektion läßt keine eindeutige ipsiversive Vorzugsrichtung im NOT entstehen.

Literaturverzeichnis 150
Literaturverzeichnis
Aarnoutse, E. J., Van der Want, J. J. L. & Vrensen, G. F. J. M. (1995) Retrograde fluorescent
microsphere tracing of retinal and central afferents to the nucleus of the optic tract in
pigmented and albino rats. Neurosci. Let., 201: 143-146
Apfelbach, R. (1973) Olfactory sign stimulus for prey selection in polecats (Putorius
putorius L.). Z. Tierpsychol., 33: 270-273
Apfelbach, R. & Wester, U. (1977) The quantitative effect of visual and tactile stimuli on the
prey-catching behaviour of ferrets (Putorius furo L.). Behav. Proc., 2: 187-200
Apkarian, P. & Shallo-Hoffmann, J. (1991) VEP projections in congenital nystagmus; VEP
asymmetry in albinism: a comparison study. Invest. Ophthalmol. Vis. Sci., 32: 2653-
2661
Apkarian, P., Eckhardt, P. G. & van Schooneveld, M. J. (1991) Detection of optic pathway
misrouting in the human albino neonate. Neuropediatrics, 22: 211-215
Ault, St. J., Leventhal, A. G., Vitek, D. J. & Creel, D. J. (1995) Abnormal ipsilateral visual
field representation in areas 17 and 18 of hypopigmented cats. J. Comp. Neurol., 354:
181-192
Baker, G. E. & Stryker, M. P. (1990) Retinofugal fibres change conduction velocity and
diameter between the optic nerve and tract in ferrets. Nature, 344: 342-345
Balkema, G. W. Jr., Pinto, L. H., Dräger, U. C. & Vanable, J. W. Jr. (1981) Characterization
of abnormalities in the visual system of the mutant mouse pearl. J. Neurosci. 1: 1320-
1329

Literaturverzeichnis 151
Balkema, G. W. Jr., Mangini, N. J., Pinto, L. H. & Vanable, J. W. Jr. (1984) Visually evoked
eye movements in mouse mutants and inbred strains. Invest. Ophthalmol. Vis. Sci., 25:
795-800
Ballas, I., Hoffmann, K.-P. & Wagner, H.-J. (1981) Retinal projection to the nucleus of the
optic tract in the cat as revealed by retrograde transport of horseradish peroxidase.
Neurosci. Let., 26: 197-202
Ballas, I. & Hoffmann, K.-P. (1985) A correlation between receptive field properties and
morphological structures in the pretectum of the cat. J. Comp. Neurol., 238: 417-128
Barlow, H. B. & Hill, R. M. (1964) Selective sensitivity to direction of movement in
ganglion cells of the rabbit retina. Science, 139: 412-414
Berman, N. (1977) Connections of the pretectum in the cat. J. Comp. Neurol., 174: 227-254
Blanks, R. H. I., Precht, W. & Torigoe, Y. (1983) Afferent projections to the cerebellar
flocculus in the pigmented rat demonstrated by retrograde transport of horseradish
peroxidase. Exp. Brain Res., 52: 293-306
Bonaventure, N., Kim, M. S. & Jardon, B. (1992) Effects on the chicken monocular OKN of
unilateral microinjections of GAGAA antagonist into the mesencephalic structures
responsible for OKN. Exp. Brain Res., 90: 63-71
Boycott, B. B. & Wässle, H. (1974) The morphological types of ganglion cells of the
domestic cat´s retina. J. Physiol., 240: 397-419
Büttner-Ennever, J. A., Cohen, B., Horn, A. K. E., & Reisine, H. (1996) Efferent pathways
of the nucleus of the optic tract in monkey and their role in eye movements. J. Comp.
Neurol., 373: 90-107
Cazin, L., Precht, W. & Lannou, J. (1980) Pathways mediating optokinetic responses of
vestibular nucleus neurons in the rat. Pflügers Arch., 384: 19-29
Cazin, L., Magnin, M. & Lannou, J. (1982) Non-cerebellar visual afferents to the vestibular
nuclei involving the prepositus hypoglossal complex: an autoradiographic study in the
rat. Exp. Brain Res., 48: 309-313

Literaturverzeichnis 152
Cazin, L., Lannou, J. & Precht, W. (1984) An electrophysiological study of pathways
mediating optokinetic responses to the vestibular nucleus in the rat. Exp. Brain Res.,
54: 337-348
Choudhury, B. P. (1987) Visual cortex in the albino rabbit. Exp. Brain Res., 66: 565-571
Cohen, B., Reisine, H., Yokota, J.-I. & Raphan, Th. (1992) The nucleus of the optic tract.
Ann. N. Y. Acad. Sci., 656: 277-296
Collewijn, H. (1975) Direction-selective units in the rabbit´s nucleus of the optic tract. Brain
Res., 100: 489-508
Collewijn, H., Winterson, B. J. & Dubois, M. F. W. (1978) Optokinetic eye movements in
albino rabbits: inversion in anterior visual field. Science, 199: 1351-1353
Collewijn, H., Apkarian, P. & Spekreijse, H. (1985) The oculomotor behaviour of human
albinos. Brain, 108: 1-28
Collewijn, H. (1999) Eye movement recording. in: Carpenter, R. H. S. & Robson, J. G.
(Eds.): Vision Research, A practical guide to laboratory methods: 245-285, Oxford
University Press, Oxford, New York, Tokyo
Creel, D., Hendrickson, A. E. & Leventhal, A. G. (1982) Retinal projections in tyrosinase-
negative albino cats. J. Neurosci., 2: 907-911
Distler, C. & Hoffmann, K.-P. (1992) Early development of the subcortical and cortical
pathway involved in optokinetic nystagmus: the cat as a model for man? Behav. Brain
Res., 49: 69-75
Distler, C. & Hoffmann, K.-P. (1993) Visual receptive field properties in kitten pretectal
nucleus of the optic tract and dorsal terminal nucleus of the accessory optic tract. J.
Neurophysiol., 70: 814-827
Dräger, U. C. & Olsen, J. F. (1980) Origins of crossed and uncrossed retinal projections in
pigmented and albino mice. J. Comp. Neurol., 191: 383-412

Literaturverzeichnis 153
Dreher, B., Sefton, A. J., Ni, S. Y. K. & Nisbett, G. (1985) The morphology, number,
distribution and central projections of class I retinal ganglion cells in albino and
hooded rats. Brain Behav. Evol., 26: 10-48
Elekessy, E. I., Campion, J. E. & Henry, G. H. (1973) Differences between the visual fields
of Siamese and common cats. Vis. Res., 13: 2533-2543
Fischer, W. H., Schmidt, M. & Hoffmann, K.-P. (1998) Saccade-induced activity of dorsal
geniculate nucleus X- and Y-cells during pharmacological inactivation of the cat
pretectum. Vis. Neurosci., 15: 197-210
Fuchs, A. F., Mustari, M. J., Robinson, F. R. & Kaneko, C. R. S. (1992) Visual signals in the
nucleus of the optic tract and their brain stem destinations. Ann. N. Y. Acad. Sci., 656:
266-276
Fuller, J. H. & Schlag, D. (1976) Determination of antidromic excitation by the collision
test: problems of interpretation. Brain Res., 112: 283-298
Gibber, M., Chen, B. & Roerig, B. (2001) Direction selectivity of exitatory and inhibitory
neurons in ferret visual cortex. Neuroreport, 12: 2293-2296
Grüsser-Cornehls, U. & Böhm, P. (1988) Horizontal optokinetic ocular nystagmus in
wildtype (B6CBA+/+) and weaver mutant mice. Exp. Brain Res., 72: 29-36
Guillery, R. W. (1971) An abnormal retinogeniculate projection in the albino ferret (Mustela
furo). Brain Res., 33: 482-485
Guillery, R. W. & Kaas, J. H. (1973) Genetic abnormality of the visual pathways in a
"white" tiger. Science, 180: 1287-1289
Guillery, R.W. (1974) Visual pathways in albinos. Sci. Am., 230: 44-54
Guillery, R. W., Okoro, A. N. & Witkop, C. J. Jr. (1975) Abnormal visual pathways in the
brain of a human albino. Brain Res., 96: 373-377
Guillery, R. W. (1990) Normal and abnormal visual field maps in albinos: central effects of
non-matching maps. Ophthalmic Paediatr. Genet., 11: 177-183

Literaturverzeichnis 154
Hahnenberger, R. W. (1977) Differences in optokinetic nystagmus between albino and
pigmented rabbits. Exp. Eye Res., 25: 9-17
Hein, A., Courjon, J. H., Flandrin, J. M. & Arzi, M. (1990) Optokinetic nystagmus in the
ferret: including selected comparisons with the cat. Exp. Brain Res., 79: 623-632
Henderson, Z. (1985) Distribution of ganglion cells in the retina of adult pigmented ferret.
Brain Res., 358: 221-228
Hess, B. J. M., Precht, W., Reber, A. & Cazin, L. (1985) Horizontal optokinetic ocular
nystagmus in the pigmented rat. Neurosci., 15: 97-107
Hoffmann, K.-P. & Schoppmann, A. (1975) Retinal imput to direction selective cells in the
nucleus tractus opticus of the cat. Brain Res., 99: 359-366
Hoffmann, K.-P., Behrend, K. & Schoppmann, A. (1976) A direct afferent visual pathway
from the nucleus of the optic tract to the inferior olive in the cat. Brain Res. 115: 150-
153
Hoffmann, K.-P. & Schoppmann, A. (1981) A quantitative analysis of the direction-specific
response of neurons in the cat´s nucleus of the optic tract. Exp. Brain Res., 42: 146-157
Hoffmann, K.-P. & Stone, J. (1985) Retinal input to the nucleus of the optic tract of the cat
assessed by antidromic activation of ganglion cells. Exp. Brain Res., 59: 395-403
Hoffmann, K.-P. (1985) Direction selective cells in the nucleus of the optic tract in the
squirrel monkey. Neurosci. Abstr., 11: 1009
Hoffmann, K.-P., Distler, C., Erickson, R. G. & Mader, W. (1988) Physiological and
anatomical identification of the nucleus of the optic tract and dorsal terminal nucleus
of the accessory optic tract in monkeys. Exp. Brain Res., 69: 635-644
Hoffmann, K.-P. & Distler, C. (1989) Quantitative analysis of visual receptive fields of
neurons in the nucleus of the optic tract and dorsal terminal nucleus of the accessory
optic tract in macaque monkey. J. Neurophysiol., 62: 416-428

Literaturverzeichnis 155
Hoffmann, K.-P., Distler, C. & Erickson, R. (1991) Functional projections from striate
cortex and superior temporal sulcus to the nucleus of the optic tract (NOT) and dorsal
terminal nucleus of the accessory optic tract (DTN) of macaque monkeys. J. Comp.
Neurol., 313: 707-724
Hoffmann, K.-P., Distler, C. & Ilg, U. (1992) Callosal and superior temporal sulcus
contributions to receptive field properties in the macaque monkey´s nucleus of the
optic tract and dorsal terminal nucleus of the accessory optic tract. J. Comp. Neurol.,
321: 150-162
Hoffmann, K.-P., Distler, C., Mark, R. F., Marotte, L. R., Henry, G. H. & Ibbotson, M. R.
(1995) Neural and behavioral effects of early eye rotation on the optokinetic system in
the wallaby, Macropus eugenii. J. Neurophysiol., 73: 727-735
Horn, A. K. E. & Hoffmann, K.-P. (1987) Combined GABA-immunocytochemistry and
TMB-HRP histochemistry of prectectal nuclei projecting to the inferior olive in rats,
cats and monkeys. Brain Res., 409: 133-138
Huang, K. & Guillery, R. W. (1985) A demonstration of two distinct geniculocortical
projection patterns in albino ferrets. Dev. Brain Res. 20: 213-220
Ibbotson, M. R., Mark, R. F. & Maddess, T. L. (1994) Spatiotemporal response properties of
direction-selective neurons in the nucleus of the optic tract and dorsal terminal nucleus
of the wallaby, Macropus eugenii. J. Neurophysiol., 72: 2927-2943
Ibbotson, M. R. & Mark, R. F. (1996) Impulse responses distinquish two classes of
directional motion-sensitive neurons in the nucleus of the optic tract. J. Neurophysiol.,
75: 996-1007
Ilia, M. & Jeffery, G. (1999) Retinal mitosis is regulated by Dopa, a melanin precursor that
may influence the time at which cells exit the cell cycle: analysis of patterns of cell
production in pigmented and albino retinae. J. Comp. Neurol., 405: 394-405
Ito, M., Shiida, T., Yagi, N. & Yamamoto, M. (1974) The cerebellar modification of rabbit´s
horizontal vestibulo-ocular reflex induced by sustained head rotation combined with
visual stimulation. Proc. Japan Acad., 50: 85-89

Literaturverzeichnis 156
Ito, M. (1982) Cerebellar control of the vestibulo-ocular reflex — around the flocculus
hypothesis. Ann. Rev. Neurosci., 5: 275-296
Ito, M (1993) Synaptic plasticity in the cerebellar cortex and its role in motor learning. Can.
J. Neurol. Sci., 20, Suppl. 3: 70-74
Jeffery, G. & Kinsella, B. (1992) Translaminar deficits in the retinae of albinos. J. Comp.
Neurol., 326: 637-644
Jeffery, G., Darling, K. & Whitmore, A. (1994) Melanin and the regulation of mammalian
photoreceptor topography. Eur. J. Neurosci., 6: 657-667
Jeffery, G., Brem, G. & Montoliu, Ll. (1997) Correction of retinal abnormalities found in
albinism by introduction of a functional tyrosinase gene in transgenic mice and rabbits.
Dev. Brain Res., 99: 95-102
Kato, I., Harada, K., Hasegawa, T., Igarashi, T., Koike, Y. & Kawasaki, T. (1986) Role of
the nucleus of the optic tract in monkeys in relation to optokinetic nystagmus. Brain
Res., 364: 12-22
Kato, I., Watanabe, S., Sato, S. & Norita, M. (1995) Pretectofugal fibers from the nucleus of
the optic tract in monkeys. Brain Res., 705: 109-117
Klauer, S., Sengpiel, F. & Hoffmann, K.-P. (1990) Visual response properties and afferents
of nucleus of the optic tract in the ferret. Exp. Brain Res., 83: 178-189
Klooster, J., van der Want, J. J. L. & Vrensen, G. (1983) Retinopretectal projections in
albino and pigmented rabbits: an autoradiographic study. Brain Res., 288: 1-12
Knapp, A. G., Ariel, M. & Robinson, F. R. (1988) Analysis of vertebrate eye movements
following intravitreal drug injections. I. Blockade of retinal ON-cells by 2-Amino-4-
Phosphonobutyrate eliminates optokinetic nystagmus. J. Neurophysiol., 60: 1010-1021
Koerner, F. & Schiller, P. H. (1972) The optokinetic response under open and closed loop
conditions in the monkey. Exp. Brain Res., 14: 318-330

Literaturverzeichnis 157
Korp, B. G., Blanks, R. H. I. & Torigoe, Y. (1989) Projections of the nucleus of the optic
tract to the nucleus reticularis tegmenti pontis and prepositus hypoglossi nucleus in the
pigmented rat as demonstrated by anterograde and retrograde transport methods. Vis.
Neurosci., 2: 275-286
Lannou, L., Cazin, L., Precht, W. & Toupet, M. (1982) Optokinetic, vestibular, and
optokinetic-vestibular responses in albino and pigmented rats. Pflügers Archiv, 393:
42-44
Law, M. I., Zahs, K. R. & Stryker, M. P. (1988) Organization of primary visual cortex (Area
17) in the ferret. J. Comp. Neurol., 278: 157-180
Lui, F., Gregory, K. M., Blanks, R. H. I. & Giolli, R. A. (1995) Projections from visual areas
of the cerebral cortex to pretectal nuclear complex, terminal accessory optic nuclei, and
superior colliculus in macaque monkey. J. Comp. Neurol., 363: 439-460
Maekawa, K., Takeda, T. & Kimura, M. (1981) Neural activity of nucleus reticularis
tegmenti pontis – the origin of visual mossy fiber afferents to the cerebellar flocculus
of rabbits. Brain Res., 210: 17-30
Maekawa, K., Takeda, T. & Kimura, M. (1984) Responses of the nucleus of the optic tract
neurons projecting to the nucleus reticularis tegmenti pontis upon optokinetic
stimulation in the rabbit. Neurosci. Res., 2: 1-25
Magnin, M., Courjon, J. H. & Flandrin, J. M. (1983) Possible visual pathways to the cat
vestibular nuclei involving the nucleus prepositus hypoglossi. Exp. Brain Res., 51:
298-303
Magnin, M., Kennedy, H. & Hoffmann, K.-P. (1989) A double labeling investigation of the
pretectal visuo-vestibular pathways. Vis. Neurosci. 3:53-58
Malach, R., Strong, N. P. & van Sluyters, R. C. (1984) Horizontal optokinetic nystagmus in
the cat: effects of long-term monocular deprivation. Dev. Brain Res., 13: 193-205
Mangini, N. J., Vanable J. W. Jr., Williams, M. A. & Pinto, L. H. (1985) The optokinetic
nystagmus and ocular pigmentation of hypopigmented mouse mutants. J. Comp.
Neurol., 241: 191-209

Literaturverzeichnis 158
Markner, C. & Hoffmann, K.-P. (1985) Variability in the effects of monocular deprivation
on the optokinetic reflex of the non-deprived eye in the cat. Exp. Brain Res., 61: 117-
127
Morgan, J. E., Henderson, Z. & Thompson, I. D. (1987) Retinal decussation patterns in
pigmented and albino ferrets. Neurosci., 20: 519-535
Mustari, M. J., Fuchs, A. F., Kaneko, C. R. S. & Robinson, F. R. (1994) Anatomical
connections of the primate pretectal nucleus of the optic tract. J. Comp. Neurol., 349:
111-128
Nunes Cardozo, J. J. & Van der Want, J. J. L. (1987) Synaptic organization of the nucleus of
the optic tract in the rabbit: a combined Golgi-electron microscopic study. J.
Neurocytol., 16: 389-401
Nunes Cardozo, B. & Van der Want, J. (1990) Ultrastructural organization of the retino-
pretecto-olivary pathway in the rabbit: a combined WGA-HRP tracing and GABA
immunocytochemical study. J. Comp. Neurol., 291: 313-327
Nunes Cardozo, B., Buijs, R. & Van der Want, J. (1991) Glutamate-like immunoreactivity in
retinal terminals in the nucleus of the optic tract in rabbits. J. Comp. Neurol., 309: 261-
270
Okada, T., Kato, I., Sato, S., Wanatabe, S. & Takeyama, I. (1991) Retinal ganglion cells
projecting to the nucleus of the optic tract in the rat. Acta Otolaryngol. Suppl., 481:
227-229
Oyster, Cl. W. & Barlow, H. B. (1967) Direction-selective units in rabbit retina: distribution
of preferred directions. Science, 155: 841-842
Oyster, C. W., Takahashi, E. & Collewijn, H. (1972) Direction-selective retinal ganglion
cells and control of optokinetic nystagmus in the rabbit. Vision Res., 12: 183-193
Oyster, C. W., Simpson, J. I., Takahashi, E. S. & Soodak, R. E. (1980) Retinal ganglion cells
projecting to the rabbit accessory optic system. J. Comp. Neurol., 190: 49-61

Literaturverzeichnis 159
Oyster, Cl. W., Takahashi, E. S., Fry, K. R. & Lam, D. M.-K. (1987) Ganglion cell density
in albino and pigmented rabbit retinas labeled with a ganglion cell-specific monoclonal
antibody. Brain Res., 425: 25-33
Pak, M. W., Giolli, R. A., Pinto, L. H., Mangini, N. J., Gregory, K. M. & Vanable, J. W. Jr.
(1987) Retinopretectal and accessory optic projection of normal mice and the OKN-
defective mutant mice beige, beige-J, and pearl. J. Comp. Neurol., 258: 435-446
Perry, V. H. & Cowey, A. (1979) Changes in the retino-fugal pathways following cortical
and tectal lesions in neonatal and adult rats. Exp. Brain Res., 35: 97-108
Pettorossi, V. E. & Troiani, D. (1983) Cortical modulation of the nucleus of the optic tract in
the rabbit. Exp. Neurol., 81: 627-639
Precht, W. & Cazin, L. (1979): Functional deficits in the optokinetic system of albino rats
Exp. Brain Res., 37: 183-186
Pu, M. & Amthor, F. R. (1990) Dendritic morphologies of retinal ganglion cells projecting to
the nucleus of the optic tract in the rabbit. J. Comp. Neurol., 302: 657-674
Quevedo, C., Hoffmann, K.-P., Husemann, R. & Distler, C. (1996) Overrepresentation to the
central visual field in the superior colliculus of the pigmented und albino ferret. Vis.
Neurosci., 13: 627-638
Reber, A., Sarrau, J. M., Carnet, J., Magnin, M. & Stelz, T. (1991) Horizontal optokinetic
nystagmus in unilaterally enucleated pigmented rats: role of the pretectal commissural
fibers. J. Comp. Neurol., 313: 604-612
Redies, C., Diksic, M. and Riml, H. (1990) Functional organization in the ferret visual
cortex: a double-label 2-deoxyglucose study. J. Neurosci., 10: 2791-2803
Rockland, K. S. (1985): Anatomical organization of primary visual cortex (area 17) in the
ferret. J. Comp. Neurol., 241: 225-236
Sanderson, K. J., Guillery, R. W. & Shackelford, R. M. (1974) Congenitally abnormal visual
pathways in mink (Mustela vison) with reduced retinal pigment. J. Comp. Neurol.,
154: 225-248

Literaturverzeichnis 160
Scalia, F. (1972) The termination of retinal axons in the pretectal region of mammals. J.
Comp. Neurol., 145: 223-258
Scalia, F. & Arango, V. (1979) Topographic organization of the projections of the retina to
the pretectal region in the rat. J. Comp. Neurol., 186: 271-292
Schmidt, M., Zhang, H.-Y. & Hoffmann, K.-P. (1993) OKN-related neurons in the rat
nucleus of the optic tract and dorsal terminal nucleus of the accessory optic system
receive a direct cortical input. J. Comp. Neurol., 330: 147-157
Schmidt, M., Schiff, D. & Bentivoglio, M. (1995) Independent efferent populations in the
nucleus of the optic tract: an anatomical and physiological study in rat and cat. J.
Comp. Neurol., 360: 271-285
Schmidt, M. (1996) Neurons in the cat pretectum that project to the dorsal lateral geniculate
nucleus are activated during saccades. J. Neurophysiol., 76: 2907-2918
Schoppmann, A. (1981) Projections from areas 17 and 18 of the visual cortex to the nucleus
of the optic tract. Brain Res., 223: 1-17
Schweigart, G. & Hoffmann, K.-P. (1992) Pretectal jerk neuron activity during saccadic eye
movements and visual stimulations in the cat. Exp. Brain Res., 91: 273-283
Sekiya, H. & Kawamura, K. (1985) An HRP study in the monkey of olivary projections
from the mesodiencephalic structures with particular reference to pretecto-olivary
neurons. Arch. Ital. Biol., 123: 171-183
Sherman, S. M. (1973) Visual field defects in monocularly and binocularly deprived cats.
Brain Res., 49: 25-45
Sirkin, D. W., Hess, B. J. M. & Precht, W. (1985): Optokinetic nystagmus in albino rats
depends on stimulus pattern. Exp. Brain Res., 61: 218-221
Stone, J. & Hoffmann, K.-P. (1972) Very slow-conducting ganglion cells in the cat´s retina:
a major new functional type? Brain Res., 43: 610-616

Literaturverzeichnis 161
Stone, J. & Fukuda, Y. (1974) Properties of cat retinal ganglion cells: a comparison of W-
cells with X- and Y-cells. J. Neurophysiol., 37: 722-748
Stone, J., Rowe, M. H. & Campion, J. E. (1978a) Retinal abnormalities in the siamese cat. J.
Comp. Neurol., 180: 773-782
Stone, J., Campion, J. E. & Leicester, J. (1978b) The nasotemporal division of retina in the
siamese cat. J. Comp. Neurol., 180: 783-798
Strong, N. P., Malach, R., Lee, P. & van Sluyters, R. C. (1984) Horizontal optokinetic
nystagmus in the cat: recovery from cortical lesions. Dev. Brain Res., 13: 179-192
Takeda, T. & Maekawa, K. (1976) The origin of the pretecto-olivary tract. A study using the
horseradish peroxidase method. Brain Res., 117: 319-325
Tauber, E. S. & Atkin, A. (1968) Optomotor responses to monocular stimulation: relation to
visual system organization. Science, 160: 1365-1367
Taylor, W. R., He, S., Levick, W. R. & Vaney, D. I. (2000) Dendritic computation of
direction selectivity by retinal ganglion cells. Science, 289: 2347-2350
Telkes, I., Garipis, N. & Hoffmann, K.-P. (2001) Morphological changes in the neuronal
substrate for the optokinetic reflex in albino ferrets Exp. Brain Res., 140: 345-356
Terasawa, K., Otani, K. & Yamada, J. (1979) Descending pathways of the nucleus of the
optic tract in the rat. Brain Res., 173: 405-417
Thompson, I. D., Jeffery, G., Morgan, J. E. & Baker, G. (1991) Albino gene dosage and
retinal decussation patterns in the pigmented ferret. Vis. Neurosci., 6: 393-398
Thompson, I. D. & Morgan, J. E. (1993) The development of retinal ganglion cell
decussation patterns in postnatal pigmented and albino ferret. Eur. J. Neurosc, 5: 341-
356
Tusa, R. J., Demer, J. L. & Herdman, S. J. (1989) Cortical Areas involved in OKN and VOR
in cats: cortical lesions. J. Neurosci., 9: 1163-1178

Literaturverzeichnis 162
Van den Berg, A. V. & Collewijn, H. (1988) Directional asymmetries of human optokinetic
nystagmus. Exp. Brain Res., 70: 597-604
Van der Want, J. J. L. & Nunes Cardozo, J. J. (1988) GABA immuno-electron microscopic
study of the nucleus of the optic tract in the rabbit. J. Comp. Neurol., 271: 229-242
Ventre, J. (1985) Cortical control of oculomotor functions. I. Optokinetic nystagmus. Behav.
Brain Res., 15: 211-226
Vitek, D. J., Schall, J. D. & Leventhal, A. G. (1985) Morphology, central projections, and
dendritic field orientation of retinal ganglion cells in the ferret. J. Comp. Neurol., 241:
1-11
Walberg, F., Nordby, T., Hoffmann, K.-P. & Holländer, H. (1981) Olivary afferents from the
pretectal nuclei in the cat. Anat. Embryol., 161: 291-304
Watanabe, S., Kato, I., Sato, S. & Norita, M. (1993) Direct projection from the nucleus of
the optic tract to the medial vestibular nucleus in the cat. Neurosci. Res., 17: 325-329
Webster, M. J. & Rowe, M. H. (1991) Disruption of developmental timing in the albino rat
retina. J. Comp. Neurol., 307: 460-474
Winterson, B. J. & Collewijn, H. (1981) Inversion of direction-selectivity to anterior fields in
neurons of nucleus of the optic tract in rabbits with ocular albinism. Brain Res., 220:
31-49
Yakushin, S. B., Reisine, H., Büttner-Ennever, J., Raphan, T. & Cohen, B. (2000) Functions
of the nucleus of the optic tract (NOT). I. Adaptation of the gain of the horizontal
vestibulo-ocular reflex. Exp. Brain Res., 131: 416-432
Zhang, H. Y. (1991) Neuroanatomische Grundlagen des optokinetischen Nystagmus bei
Säugetieren. Dissertation, Ruhr-Universität Bochum
Zhang, H. Y. & Hoffmann, K.-P. (1993) Retinal projections to the pretectum, accessory
optic system and superior colliculus in pigmented and albino ferrets. Eur. J. Neurosci.,
5: 486-500

163
Danksagung
Mein größter Dank gilt meinem Doktorvater Prof. Dr. K.-P. Hoffmann für die Aufgabe
dieses anspruchsvollen und interessanten Themas, für die Aufnahme in das
Graduiertenkolleg KOGNET, ganz besonders aber für die intensive Betreuung bei den
elektrophysiologischen Experimenten und nicht zuletzt für seine Geduld.
Ebenso möchte ich mich beim Graduiertenkolleg KOGNET III für die lehrreichen und
inspirierenden Veranstaltungen bedanken. In diese Danksagung schließe ich die Deutsche
Forschungsgemeinschaft ein, die mich im Rahmen von KOGNET drei Jahre lang finanziell
unterstützte.
Für ihre Unterstützung und Beratung bei der Durchführung und Aufrechterhaltung der
Narkose schulde ich großen Dank Frau PD Dr. C. Distler. Sie hat mir oft aus der Not
geholfen.
Freundlichst bedanke ich mich bei Herrn H. Korbmacher für die kontinuierliche technische
Unterstützung, insbesondere für die Herstellung der Mikroelektroden. Weiterhin bedanke ich
mich für die technische Unterstützung bei Herrn G. Reuter und Herrn B. Thüner aus der
Feinmechanik-Werkstatt und bei Herrn G. Tinney und Herrn H. Marquardt aus der
Elektronik-Werkstatt.
Für die Betreuung der EDV möchte ich mich auch bei Herrn W. Junke bedanken, dem ich
mich besonders verpflichtet fühle.
Desweiteren spreche ich meinen Dank aus an Frau M. Möllmann und S. Krämer für die Hilfe
in der Histologie und an die beiden Tierpfleger, Herrn V. Rostek und Frau M. Schmidt, für
die Betreuung der Frettchen. Für die Assistenz bei der Durchführung des Perimetrietests
danke ich Frau H. Pollmann und insbesondere Frau M. Bronzel, mit der ich das Training der
Tiere optimieren konnte.
Zum Schluß wendet sich mein Dank an alle namentlich nicht genannten Mitarbeiter des
Lehrstuhls Allgemeine Zoologie und Neurobiologie — insbesondere an Herrn M. Klar und
an die Sekretärinnen, Frau M. Vogt, Frau V. Grau und Frau A. Bauswein.
Meinen Zimmergenossen L. Lünenburger, W. Lindner und Dr. D. Kutz danke ich für die
angenehme Arbeitsatmosphäre, die ich besonders zu schätzen wußte.

164
Curriculum vitae
Persönliche Daten
Name: Nikolaos Garipis
Geburtstag: 23.05.1969
Geburtsort: Serres (Griechenland)
Familienstand: verheiratet
Schulausbildung
Sept. 1975 - Jun. 1980 Volksschule in N. Souli (Serres / Griechenland)
Sept. 1980 - Juli 1986 Hauptschule, Bochum; Abschluß: Sekundarabschluß I
Sept. 1986 - Mai. 1989 Städtisches Gymnasium Schiller-Schule, Bochum; Abschluß: Abitur
Akademische Ausbildung
Okt. 1989 - Jan. 1998 Ordentlicher Studierender der Biologie an der Ruhr-Universität
Bochum
Nov. 1996 - Nov. 1997 Diplomarbeit: „Neuronale Grundlagen des veränderten
optokinetischen Reflexes bei albinotischen Säugern“, angefertigt am
Lehrstuhl für Allgemeine Zoologie und Neurobiologie, Ruhr-
Universität Bochum; Betreuer: Prof. Dr. K.-P. Hoffmann
seit Jan. 1998 Doktorand am Lehrstuhl für Allgemeine Zoologie und
Neurobiologie, Ruhr-Universität Bochum
Jan. 1998 - Dez. 2000 Stipendiat im Graduiertenkolleg „Kognition, Gehirn und Neuronale
Netze“ (KOGNET III), Ruhr-Universität Bochum
seit Jan. 2001 Wissenschaftlicher Mitarbeiter am Lehrstuhl für Allgemeine
Zoologie und Neurobiologie, Ruhr-Universität Bochum

















